This eBook is for the use of anyone anywhere at no cost and with almost no restrictions whatsoever. You may copy it, give it away or re-use it under the terms of the Project Gutenberg License included with this eBook or online at www.gutenberg.org
Title: Text Book of Biology, Part 1: Vertebrata
Author: H. G. Wells
Release Date: June 8, 2007 [eBook #21781]
Language: English
Character set encoding: ISO-8859-1
***START OF THE PROJECT GUTENBERG EBOOK TEXT BOOK OF BIOLOGY, PART 1: VERTEBRATA***
[Second Edition only text] and -First Edition only text,- and alsowhere more than just a sentence is added or removed. Other things to notice is how some words are spelt or punctuated differently throughout the book, such as;
{Lines from Second Edition only.} and {Lines from First Edition only.}
Blood VesselsI've tried to keep these as close to the original book as possible.
Blood-Vessels
Bloodvessels
In the year 1884 I was invited to give tuition by correspondence, in Biology. Although disposed at the time to ridicule the idea of imparting instruction in natural science by letter, I gladly accepted the opportunity thus afforded me of ascertaining for myself what could and could not be accomplished in that direction. Anyone familiar with the scope of biological enquiry, and the methods of biological instruction, will not need to be reminded that it is only by the most rigorous employment of precise directions for observation, that any good results are to be looked for at the hand of the elementary student. True to this principle, I determined to issue to my correspondence pupils rigid instructions, and to demand in return faithful annotated drawings of facts observed in their usage. In the case of two among the few students who passed through my hands, the result far exceeded my most sanguine anticipations. The notes sent in by one of them-- a man working at a distance, alone and unaided-- far excelled those wrung from many a student placed under the most favourable surroundings; and their promise for the future has been fulfilled to the utmost, the individual in question being now a recognised investigator. It thus became clear that, not-with-standing the complex conditions of work in the biological field, tuition by correspondence would suffice to awaken the latent abilities of a naturally qualified enquirer. The average members of a University Correspondence Class will be found neither better nor worse than those of any other, and they may therefore pass unnoticed; if however, the correspondence system of tuition may furnish the means of arousing a latent aptitude, when the possibilities of other methods of approach are excluded-- and in so doing, of elevating the individual to that position for which he was by nature qualified, ensuring him the introduction to the one sphere of labour for which he was born-- it will have created its own defence, and have merited the confidence of all right-thinking people. The plucking of one such brand from the burning is ample compensation for the energy expended on any number of average dullards, who but require to be left alone to find their natural level.
Mr. Wells' little book is avowedly written for examination purposes, and in conformity with the requirements of the now familiar "type system" of teaching. Recent attempts have been made to depreciate this. While affording a discipline in detailed observation and manipulation second to that of no other branch of learning, it provides for that "deduction" and "verification" by which all science has been built up; and this appears to me ample justification for its retention, as the most rational system which can be to-day adopted. Evidence that its alleged shortcomings are due rather to defective handling than to any inherent weakness of its own, would not be difficult to produce. Although rigid in its discipline, it admits of commentatorial treatment which, while heightening the interest of the student, is calculated to stimulate alike his ambition and his imagination. That the sister sciences of Botany and Zoology fall under one discipline, is expressed in the English usage of the term "Biology." Experience has shown that the best work in either department has been produced by those who have acquired on all-round knowledge of at least the elementary stages of both; and, that the advanced morphologist and physiologist are alike the better for a familiarity with the principles-- not to say with the progressive advancement-- of each other's domain, is to-day undeniable. These and other allied considerations, render it advisable that the elementary facts of morphology and physiology should be presented to the beginner side by side-- a principle too frequently neglected in books which, like this one, are specially written for the biological neophyte. Although the student is the wiser for the actual observation of the fact of nature, he becomes the better only when able to apply them, as for example, by the judicious construction of elementary generalizations, such as are introduced into the pages of this work. So long as these generalizations, regarded as first attempts to deduce "laws" in the form of "generalized statement of facts based observation," are properly introduced into an elementary text-book, intended for the isolated worker cut off from the lecture room, their intercalation is both healthy and desirable.
Mr. Wells has kept these precepts constantly in mind in the preparation of his work, and in the formulation of his plans for its future extension, thereby enhancing the value of the book itself, and at the same time, discouraging the system of pure cram, which is alien to the discipline of biological science.
G. B. Howes
Royal College of Science,
South Kensington;
November 30, 1892.
No method of studying-- more especially when the objects of study are tangible things-- can rival that prosecuted under the direction and in the constant presence of a teacher who has also a living and vivid knowledge of the matter which he handles with the student. In the ideal world there is a plentiful supply of such teachers, and easy access to their teaching, but in this real world only a favoured few enjoy these advantages. Through causes that cannot be discussed here, a vast number of solitary workers are scattered through the country, to whom sustained help in this form is impossible, or possible only in days stolen from a needed vacation; and to such students especially does this book appeal, as well as to those more fortunate learners who are within reach of orderly instruction, but anxious to save their teachers' patience and their own time by some preliminary work.
One of the most manifest disadvantages of book-work, under the conditions of the solitary worker, is the rigidity of its expressions; if the exact meaning is doubtful, he can not ask a question. This has been kept in view throughout; the writer has, above all, sought to be explicit-- has, saving over-sights, used no uncommon or technical term without a definition or a clear indication of its meaning.
In this study of Biology, the perception and memory of form is a very important factor indeed. Every student should draw sketches of his dissections, and accustom himself to copying book diagrams, in order to train his eye to perception of details he might otherwise disregard. The drawing required is within the reach of all; but for those who are very inexperienced, tracing figures is a useful preliminary exercise.
By the time the student has read the "Circulation of the Rabbit" (Sections 34 to 49), he will be ready to begin dissection. It is possible to hunt to death even such a sound educational maxim as the "thing before the name," and we are persuaded, by a considerable experience, that dissection before some such preparatory reading is altogether a mistake. At the end of the book is a syllabus (with suggestions) for practical work, originally drawn up by the writer for his own private use with the evening classes of the University Tutorial College-- classes of students working mainly in their spare time for the London examination, and at an enormous disadvantage, as regards the number of hours available, in comparison with the leisurely students of a University laboratory. This syllabus may, perhaps by itself, serve a useful purpose in some cases, but in this essential part of the study the presence of some experienced overlooker to advise, warn, and correct, is at first almost indispensable.
A few words may, perhaps be said with respect to the design of this volume. It is manifestly modelled upon the syllabus of the Intermediate Examination in Science of London University. That syllabus, as at present constituted, appears to me to afford considerable scope for fairly efficient biological study. The four types dealt with in this book are extremely convenient for developing the methods of comparative anatomy and morphological embryology. Without any extensive reference to related organisms, these four forms, and especially the three vertebrata, may be made to explain and illustrate one another in a way that cannot fail to be educational in the truest sense. After dealing with the rabbit, therefore, as an organic mechanism, our sections upon the frog and dog-fish, and upon development, are simply statements of differences, and a commentary, as it were, upon the anatomy of the mammalian type. In the concluding chapter, a few suggestions of the most elementary ideas of it is hoped to make this first part of our biological course complete in itself, and of some real and permanent value to the student. And the writer is convinced that not only is a constant insistence upon resemblances and differences, and their import, intellectually the most valuable, but also the most interesting, and therefore the easiest, way of studying animal anatomy. That chaotic and breathless cramming of terms misunderstood, tabulated statements, formulated "tips," and lists of names, in which so many students, in spite of advice, waste their youth is, I sincerely hope, as impossible with this book as it is useless for the purposes of a London candidate. On the other hand, our chief endeavour has been to render the matter of the book clear, connected, progressive, and easily assimilable. In the second part Plants, Unicellular Organisms, and Invertebrata will be dealt with, in a wider and less detailed view of the entire biological province.
{Lines from First Edition only.}
-In this volume, we study four organisms, and chiefly in their relation to each other; in the next, we shall study a number of organisms largely in relation to their environment. In this part our key note is the evidence of inheritance; in our second part it will be of adaptation to circumstances.-
This book will speedily, under the scrutiny of the critical reader, reveal abundant weakness. For these the author claims the full credit. For whatever merit it may posses, he must however, acknowledge his profound indebtedness to his former teacher, Professor Howes. Not only has the writer enjoyed in the past the privilege of Professor Howes' instruction and example, but he has, during the preparation of this work, received the readiest help, advise, and encouragement from him-- assistance as generous as it was unmerited, and as unaffected as it was valuable.
{Lines from Second Edition only.}
[The publication of a second and revised edition of this Part affords the author an opportunity of expressing his sense of the general kindliness of his reviewers, and the help they have him in improving this maiden effort. To no one is there vouchsafed such a facility in the discovery of errors in a book as to its author, so soon as it has passed beyond his power of correction. Hence the general tone of encouragement (and in some cases the decided approval) of the members of this termination to a period of considerable remorse and apprehension.]
I have been able through their counsel, and the experience I have had while using this book in teaching, to correct several printer's errors and to alter various ambiguous or misleading expressions, as well as to bring the book up to date again in one or two particulars.
My thanks are particularly due to my friend Miss Robbins, who has very kindly redrawn the occasionally rather blottesque figures of the first edition. Not only have these plates gained immensely in grace and accuracy, but the lettering is now distinct-- an improvement that any student who has had to hunt my reference letters in the first edition will at once appreciate.
H. G. Wells
November, 1892. {First Edition.}
December, 1893. {Second Edition.}
Section 1. It is unnecessary to enter upon a description of the appearance of this familiar type, but it is not perhaps superfluous, as we proceed to consider its anatomy, to call attention to one or two points in its external, or externally apparent structure. Most of our readers know that it belongs to that one of two primary animal divisions which is called the vertebrata, and that the distinctive feature which place it in this division is the possession of a spinal column or backbone, really a series of small ring-like bones, the vertebrae (Figure 1 v.b.) strung together, as it were, on the main nerve axis, the spinal cord (Figure 1 s.c.). This spinal column can be felt along the neck and back to the tail. This tail is small, tilted up, and conspicuously white beneath, and it serves as a "recognition mark" to guide the young when, during feeding, an alarm is given and a bolt is made for the burrows. In those more primitive (older and simpler-fashioned) vertebrata, the fishes, the tail is much large and far more important, as compared with the rest of the body, than it is in most of the air-inhabiting vertebrates. In the former it is invariably a great muscular mass to propel the body forward; in the latter it may disappear, as in the frog, be simply a feather-bearing stump, as in the pigeon, a fly flicker, as in the cow or horse, a fur cape in squirrel, or be otherwise reduced and modified to meet special requirements.
Section 2. At the fore end, or as English zoologists prefer to say, anterior end, of the vertebral column of the rabbit, is of course the skull, containing the anterior portion of the nerve axis, the brain (Figure 1 br.). Between the head and what is called "the body," in the more restricted sense of the word, is the neck. The neck gives freedom of movement to the head, enables the animal to look this way and that, to turn its ears about to determine the direction of a sound, and to perform endless motions in connexion with biting and so forth easily. We may note that in types which swim through the water, the neck dose not appear-- in the fish and frog, for instance-- and the head simply widens out as one passes back to the body. The high resistance offered by water necessitates this tendency to a cigar or ship outline, just as it has determined the cigar shape of the ordinary fish torpedo.
Section 3. In the body of the rabbit, as examined from the outside, we can make out by feeling two distinct regions, just as we might in the body of a man; anteriorly a bony cage, having the ribs at the sides, a rod-like bone in the front, the sternum (Figure 1 -st.-, [stm.]), and the backbone behind, and called the chest or thorax; and posteriorly a part called the abdomen, which has no bony protection over its belly, or ventral surface. These parts together with the neck constitute the trunk. As a consequence of these things, in the backbone of the rabbit there are four regions: the neck, or cervical part, consisting of seven vertebrae, the thoracic part of twelve joined to ribs, the abdominal (also called the lumbar) region of seven without ribs, and the tail or caudal of about fifteen. Between the lumbar and caudal come four vertebrae, the sacral, which tend to run together into a bony mass as the animal grows old, and which form a firm attachment for the base of the hind limb.
Section 4. The thorax and abdomen are separated by a partition, the diaphragm (Figure 1 dia.). This structure is distinctive of that class of the vertebrata called mammals, and which includes man, most of the larger and commoner land animals, and whales and manatee. We shall find later that it is essentially connected with the perfection of the air breathing to which this group has attained. Another characteristic shared by all mammals, and by no other creature, is the presence of hair. In birds we have an equally characteristic cover in the feathers, the frog is naked, and the fishes we find either naked skins or scales.
Section 5. The short strong fore limbs are adapted to the burrowing habit, and have five digits; the hind limbs are very much longer and muscular, enable the animal to progress rapidly by short leaps, and they have four toes. If the student thinks it worth while to attempt to remember the number of digits-- it is the fault of examiners if any value dose attach to such intrinsically valueless facts-- he should associate the number 54 (5 in front, 4 behind) with the rabbit, and observe that with the frog the reverse is the case.
Section 6. We may note here the meaning of certain terms we shall be constantly employing. The head end of the rabbit is anterior, the tail end posterior, the backbone side of the body-- the upper side in life-- is dorsal, the breast and belly side, the lower side of the animal, is ventral. If we imagine the rabbit sawn asunder, as it were, by a plane passing through the head and tail, that would be the median plane, and parts on either side of it are lateral, and left or right according as they lie to the animal's left or right. In a limb, or in the internal organs, the part nearest the central organ, or axis, is proximal, the more remote or terminal parts are distal. For instance, the mouth is anteriorly placed, the tongue on its ventral wall; the tongue is median, the eyes are lateral, and the fingers are distal to the elbow. The student must accustom himself to these words, and avoid, in his descriptions, the use of such terms as "above," "below," "outside," which vary with the position in which we conceive the animal placed.
Section 7. So much for the general form; we may note a few facts of general knowledge, in connection with the rabbit's life-activity. In a day of the rabbit's life a considerable amount of work is done-- the animal runs hither and thither, for instance; in other words, a certain mass of matter is moved through space, and for that we know force must be exerted. Whence comes the force?
Section 8. We find the rabbit occupies a considerable amount of its time in taking in vegetable matter, consisting chiefly of more or less complex combustible and unstable organic compounds. It is a pure vegetarian, and a remarkably moderate drinker. Some but only a small proportion, of the vegetable matter it eats, leaves its body comparatively unchanged, in little pellets, the faeces, in the process of defaecation. For the rest we have to account.
Section 9. We find, also, that the rabbit breathes air into its lungs, which is returned to the atmosphere with a lessened amount of oxygen, and the addition of a perceptible amount of carbon dioxide. The rabbit also throws off, or excretes, a fluid, the urine, which consists of water with a certain partially oxydised substance containing nitrogen, and called urea, and other less important salts. The organs within the body, by which the urine is separated, are called the kidneys.
Section 10. Repeating these facts in other words, the rabbit takes into its body complex and unstable organic compounds containing nitrogen, carbon, hydrogen, a certain amount of oxygen, a small quantity of sulphur, and still smaller amounts of other elements. It also breathes in oxygen.
Section 11. It returns a certain rejected part of its food comparatively unchanged. Besides this, it returns carbon dioxide and water, which are completely oxydised, and very simple and stable bodies, and urea-- a less completely oxydised compound, but a very simple one compared with the food constituents.
Section 12. Now the chemist tells us that when a stable body is formed, or when an unstable compound decomposes into simpler stable ones, force is evolved. The oxydation of carbon, for instance, in the fireplace, is the formation of the stable compound called carbon dioxide, and light and heat are evolved. The explosion of dynamite, again is the decomposition of an unstable compound. Hence, we begin to perceive that force-- the vital force-- which keeps the rabbit moving, is supplied by the decomposition and partial oxydation of compounds continued in its food, to carbon dioxide, water, urea, and smaller quantities of other substances.
Section 13. This is the roughest statement of the case possible, but it will give the general idea underlying our next chapters. We shall consider how the food enters the body and is taken up into the system, how it is conveyed to the muscles in the limbs, to the nerve centres, and to wherever work is done, to be there decomposed and partially oxydised, and finally how the products of its activity-- the katastases, of which the three principal are carbon dioxide, water, and urea-- are removed from the body.
Section 14. There are one or two comparatively modern terms that we may note here. This decomposition of unstable chemical compounds, releasing energy, is called kataboly. A reverse process, which has a less conspicuous part in our first view of the animal's life action, by which unstable compounds are built up and energy stored, is called anaboly. The katastases are the products of kataboly.
Section 15. In an ordinary animal, locomotion and other activity predominate over nutritive processes, which fact we may express, in the terms just given, by saying that kataboly prevails over anaboly. An animal, as we have just explained, is an apparatus for the decomposition and partial oxydation of certain compounds, and these are obtained either directly or indirectly-- through other animals, in the case of meat-eaters-- from the vegetable kingdom. As the student will learn early in his botanical reading, the typical plant has, in its green colouring matter, chlorophyll, a trap to catch the radiating energy of the sun, and to accomplish, by the absorption of that energy, the synthesis (building up) of those organic compounds which the animal destroys. The typical plant is, on whole, passive and synthetic, or anabolic; the typical animal, active and katabolic; and the excess of kataboly over anaboly in the animal is compensated for by the anabolic work stored up, as it were, by the plant, which is, directly or indirectly, the animal's food.
Section 16. Figure 1 represents the general anatomy of the rabbit, but is especially intended to show the alimentary (= food) canal, shortened to a certain extent, and with the proportions altered, in order to avoid any confusing complications. It is evidently simply a coiled tube-- coiled for the sake of packing-- with occasional dilatations, and with one side-shunt, the caecum (cae.), into which the food enters, and is returned to the main line, after probably absorbent action, imperfectly understood at present. A spiral fold in this cul-de-sac {bottom-of-sack}, which is marked externally by constrictions, has a directive influence on the circulation of its contents. The student should sketch Figure 1 once or twice, and make himself familiar with the order and names of the parts before proceeding. We have, in succession, the mouth (M.), separated from the nasal passage (Na.) above the palate; the pharynx (ph.), where the right and left nasal passages open by the posterior nares into the mouth; the oesophagus (oes.); the bag-like stomach, its left (Section 6) end being called the cardiac (cd.st.), and its right the pyloric end (py.); the U-shaped duodenum (ddnm.) and the very long and greatly coiled ileum (il.). The duodenum and ileum together form the small intestine; and the ileum is dilated at its distal end into a thick-walled sacculus rotundus (s.r.), beyond which point comes the large intestine. The colon (co.) and rectum (r.) continue the main line of the alimentary canal; but, at the beginning of the large intestine, there is also inserted a great side-shunt, the caecum (cae.), ending blindly in a fleshy vermiform appendix (v.ap.). The figure will indicate how the parts are related better than any verbal description can. Between the coiling alimentary tube and the body walls is a space, into which the student cuts when he begins dissecting; this is the peritoneal cavity (pt.). A thin, transparent membrane, the mesentery, holds the intestines in place, and binds them to the dorsal wall of this peritoneal space.
Section 17. The food stuffs of an animal, the unstable compounds destined ultimately to be worked into its life, and to leave it again in the form of katastases (Section 13), fall into two main divisions. The first of these includes the non-nitrogenous food stuffs, containing either carbon together with hydrogen and oxygen in the proportion of H2O (the carbo-hydrates), or carbon and hydrogen without oxygen (the hydrocarbons). The second division consists of the nitrogenous materials, containing also carbon, hydrogen, a certain amount of oxygen, sulphur, and possibly other elements. Among the carbohydrates, the commonest are starch and cellulose, which are insoluble bodies, and sugar, which is soluble. The hydrocarbons, fats, oils, and so on, form a comparatively small proportion of the rabbit's diet; the proverb of "oil and water" will remind the student that these are insoluble. The nitrogenous bodies have their type in the albumen of an egg; and muscle substance and the less modified living "protoplasm" of plants, a considerable proportion of the substance of seeds, bulbs, and so on, are albuminous bodies, or proteids. These also are insoluble bodies, or when soluble, will not diffuse easily through animal membranes.
Section 18. Now the essential problem which the digestive canal of the rabbit solves is to get these insoluble, or quasi-insoluble, bodies into its blood and system. They have to pass somehow into the circulation through the walls of the alimentary canal. In order that a compound should diffuse through a membrane, it must be both soluble and diffusible, and therefore an essential preliminary to the absorption of nutritive matter is its conversion into a diffusible soluble form. This is effected by certain fluids, formed either by the walls of the alimentary canal or by certain organs called glands, which open by ducts into it; all these fluids contain small quantities of organic compounds of the class called ferments, and these are the active agents in the change. The soluble form of the carbohydrates is sugar; proteids can be changed into the, of course, chemically equivalent but soluble and diffusible the peptones; and fats and oils undergo a more complicated, but finally similar change.
Section 19. We shall discuss the structure and action of -a gland- [glands] a little more fully in a subsequent chapter. Here we will simply say that they are organs forming each its characteristic fluid or secretion, and sending it by a conduit, the duct, to the point where its presence is required. The saliva in our mouths, tears, and perspiration, are examples of the secretions of glands.
Section 20. In the month of the rabbit the food is acted upon by the teeth and saliva. The saliva contains ptyalin, a ferment converting starch into sugar, and it also serves to moisten the food as it is ground up by the cheek teeth. It does not act on fat to any appreciable extent. The teeth of the rabbit are shown in Figure XVIII., Sheet 4. The incisor teeth in front, two pairs above and one pair below (i.), are simply employed in grasping the food; the cheek teeth-- the premolars (pm.) and molars (m.) behind-- triturate the food by a complicated motion over each. Their crowns are flat for this purpose, with harder ridges running across them.
Section 21. This grinding up of the food in the mouth invariably occurs in herbivorous animals, where there is a considerable amount of starch and comparatively little hydrocarbon in the food. By finely dividing the food, it ensures its intimate contact with the digestive ferment, ptyalin. In such meat-eaters as the cat and dog, where little starchy matter and much fat is taken, the saliva is, of course, of less importance, and this mastication does not occur. The cheek teeth of a dog ({Section 91}), and more so of a cat, are sharp, and used for gnawing off fragments of food, which are swallowed at once. Between the incisors and premolars of a dog come the characteristic biting teeth, or canines, absent in the rabbit.
Section 22. The student will probably ask why the cheek teeth, which are all similar in appearance, are divided into premolars and molars. The rabbit has a set of milk molars-- a milk dentition-- which are followed by the permanent teeth, just as in man. Those cheek teeth of the second set, which have predecessors in the first series, are called premolars; the ones posterior to these are the molars.
Section 23. After mastication, the food is worked by the tongue and cheeks into a saliva-soaked "bolus" and swallowed. The passage down the oesophagus is called deglutition. In the stomach it comes under the influence of the gastric juice, formed in little glandular pits in the stomach wall-- the gastric (Figure VIII. Sheet 3) and pyloric glands. This fluid is distinctly acid, its acidity being due to about one-tenth per cent {of a hundred} of hydrochloric acid, and it therefore stops any further action of the ptyalin, which can act only on neutral or slightly alkaline fluids. The gastric juice does not act on carbo-hydrates or hydrocarbons to any very noticeable degree. Its essential property is the conversion of proteids into peptones, and the ferment by which this is effected is called pepsin. Milk contains a peculiar soluble proteid, called casein, which is precipitated by a special ferment, the rennet-ferment, and the insoluble proteid, the curd, thus obtained is then acted on by the pepsin. In the manufacture of cheese, the rennetferment obtained, from the stomach of a calf is used to curdle the milk.
Section 24. After the food has undergone digestion in the stomach it passes into the duodenum, the U-shaped loop of intestine immediately succeeding the stomach. The duodenum is separated from the stomach by a ring-like muscular valve, the pylorus; this valve belongs to the class of muscles called sphincters, which, under ordinary circumstances, are closed, but which relax to open the circular central aperture. The valve at the anus, which retains the faeces, is another instance of a sphincter.
Section 25. The food at this stage is called chyme; it is an acid and soup-like fluid-- acid through the influence of the gastric juice. The temperature of the animal's body is sufficiently high to keep most of the fat in the food melted and floating in oily drops; much of the starch, has been changed to sugar, and the solid proteids to soluble peptones, but many fragments of material still float unchanged.
Section 26. It meets now with the bile, a greenish fluid secreted by that large and conspicuous gland the liver. The bile is not simply a digestive secretion, like the saliva or the gastric juice; it contains matters destined to mix in, and after a certain amount of change to be passed out of the body with, the faeces; among these substances, of which some portion is doubtless excretory, are compounds containing sulphur-- the bile salts. There is also a colouring matter, bili verdin, which may possibly also be excretory. If the student will compare Sections 10 and 11, he will notice that in those paragraphs no account is taken of the sulphur among the katastases, the account does not balance, and he will at once see that here probably is the missing item on the outgoing side. The bile, through the presence of these salts, is strongly alkaline, and so stops the action of the gastric juice, and prepares for that of the pancreas, which can act only in an alkaline medium. The fermentive action of the bile is trifling; it dissolves fats, to a certain extent, and is antiseptic, that is, it prevents putrefaction to which the chyme might be liable; it also seems to act as a natural purgative.
Section 27. The bile, as we shall see later, is by no means the sole product of the liver.
Section 28. The pancreatic juice, the secretion of the pancreas is remarkable as acting on all the food stuffs that have not already become soluble. It emulsifies fats, that is, it breaks, the drops up into extremely small globules, forming a milky fluid, and it furthermore has a fermentive action upon them; it splits them up into fatty acids, and the soluble body glycerine. The fatty acids combine with alkaline substances (Section 26) to form bodies which belong to the chemical group of Soaps, and which are soluble also. The pancreatic juice also attacks any proteids that have escaped the gastric juice, and converts them into peptones, and any residual starch into sugar. Hence by this stage, in the duodenum, all the food constituents noticed in Section 17 are changed into soluble forms. There are probably, three distinct ferments in the pancreatic juice acting respectively on starch, fat, and proteid, but they have not been isolated, and the term pancreatin is sometimes used to suggest the three together.
Section 29. A succus entericus, a saliva-like fluid secreted by numerous small glands in the intestine wall (Brunner's glands, Lieberkuhnian follicles), probably aids, to an unknown but comparatively small extent, in the digestive processes.
Section 30. The walls of the whole of the small intestine are engaged in the absorption of the soluble results of digestion. In the duodenum, especially, small processes, the villi project into the cavity, and being, like the small hairs of velvet pile, and as thickly set, give its inner coat a velvety appearance. In a villus we find (Figure IX., Sheet 3) a series of small blood-vessels and with it another vessel called a lacteal. The lacteals run together into larger and larger branches until they form a main trunk, the thoracic duct, which opens into the blood circulation at a point near the heart; but of this we shall speak further later. They contain, after a meal, a fluid called chyle.
Section 31. Emulsified fats pass into the chyle. Water and diffusible salts certainly pass into the vein. The course taken by the peptones is uncertain, but Professor Foster favours the chyle in the case of the rabbit-- the student should read his Text-book of Physiology, Part 2, Chapter 1, Section 11, if interested in the further discussion of this question.
Section 32. The processes that occur in the remaining portions of the alimentary canal are imperfectly understood. The caecum is so large in the rabbit that it must almost certainly be of considerable importance. In carnivorous animals it may be so much reduced as to be practically absent. An important factor in the diet of the herbivorous animals, and one absent from the food of the carnivora, is that carbohydrate, the building material of all green-meat- [food], cellulose, and there is some ground for thinking that the caecum is probably a region of special fermentive action upon it. The pancreatic juice, it may be noted, exercises a slight digestive activity upon this substance.
Section 33. Water is most largely absorbed in the large intestine, and in it the rejected (mainly insoluble) portion of the food gradually acquires its dark colour and other faecal characteristics.
Section 34. The next thing to consider is the distribution of the food material absorbed through the walls of the alimentary canal to the living and active parts of the body. This is one of the functions of the series of structures-- heart and blood-vessels, called the circulation, circulatory system, or vascular system. It is not the only function. The blood also carries the oxygen from the lungs to the various parts where work is done and kataboly occurs, and it carries away the katastases to the points where they are excreted-- the carbon dioxide and some water to the lungs, water and urea to the kidneys, sulphur compounds of some kind to the liver.
Section 35. The blood (Figure 4, Sheet 2) is not homogeneous; under the low power of the microscope it may be seen to consist of--
(1.) a clear fluid, the plasma, in which float--
(2.) a few transparent colourless bodies of indefinite and changing shape, and having a central brighter portion, the nucleus with a still brighter dot therein the nucleolus-- the white corpuscles (w.c.), and
(3.) flat round discs, without a nucleus, the red corpuscles (r.c.), greatly more numerous than the white.
Section 36. The chyle of the lacteals passes, as we have said, by the thoracic duct directly into the circulation. It enters the left vena cava superior (l.v.c.s.) near where this joins the jugular vein (ex.j.) (see Figure 1, Sheet 2, th.d.) and goes on at once with the rest of the blood to the heart. The small veins of the villi, however, which also help suck up the soluble nutritive material, are not directly continuous with the other body veins, the systemic veins; they belong to a special system, and, running together into larger and larger branches, form the lieno gastric (l.g.v.) and mesenteric (m.v.) veins, which unite to form the portal vein (p.v.) which enters the liver (l.v.) and there breaks up again into smaller and smaller branches. The very finest ramifications of this spreading network are called the (liver) capillaries, and these again unite to form at last the hepatic vein (h.v.) which enters the vena cava inferior (v.c.i.), a median vessel, running directly to the heart. This capillary network in the liver is probably connected with changes requisite before the recently absorbed materials can enter the general blood current.
Section 37. The student has probably already heard the terms vein and artery employed. In the rabbit a vein is a vessel bringing blood towards the heart, while an artery is a vessel conducting it away. Veins are thin-walled, and therefore flabby, a conspicuous purple when full of blood, and when empty through bleeding and collapsed sometimes difficult to make out in dissection. They are formed by the union of lesser factors. The portal breaks up into lesser branches within the liver. Arteries have thick muscular and elastic walls, thick enough to prevent the blood showing through, and are therefore pale pink or white and keep their round shape.
Section 38. The heart of the rabbit is divided by partitions into four chambers: two upper thin-walled ones, the auricles (au.), and two lower ones, both, and especially the left, with very muscular walls, the ventricles (vn.). The right ventricle (r.vn.) and auricle (r.au.) communicate, and the left ventricle (l.vn.) and auricle (l.au.).
Section 39. The blood coming from all parts of the body, partly robbed of its oxygen and containing much carbon dioxide and other katastases, enters the right auricle of the heart through three great veins, the median vena cava inferior from the posterior parts of the body, and the paired venae cavae superiores from the anterior. With the beating of the heart, described below, it is forced into the right ventricle and from there through the pulmonary artery (p.a.) seen in the figure passing under the loop of the aorta (ao.) to the lungs.
Section 40. The lungs (lg. Figure 1, Sheet 1) are moulded to the shape of the thoracic cavity and heart; they communicate with the pharynx by the trachea (tr. in Figure 1, Sheet 1) or windpipe, and are made up of a tissue of continually branching and diminishing air-tubes, which end at last in small air-sacs, the alveoli. The final branches of the pulmonary arteries, the lung capillaries, lie in the walls of these air-sacs, and are separated from the air by an extremely thin membrane through which the oxygen diffuses into, and the carbon dioxide escapes from, the blood.
Section 41. The mechanism of respiration will be understood by reference to Figure 3, Sheet 2. It will be noted, in dissecting that the lungs have shrunk away from the walls of the thorax; this collapse occurs directly an aperture is made in the thorax wall, and is in part due to their extreme elasticity. In life the cavity of the thorax forms an air-tight box, between which and the lungs is a slight space, the pleural cavity (pl.c.) lined by a moist membrane, which is also reflected, over the lungs. The thorax wall is muscular and bony, and resists the atmospheric pressure on its outer side, so that the lungs before this is cut through are kept distended to the size of the thoracic cavity by the pressure of the air within them. In inspiration (or breathing-in) the ribs are raised by the external intercostal (Anglice, between-ribs, e.i.c.m.) and other allied muscles, and the diaphragm (dia.) contracts and becomes flatter; the air is consequently sucked, in as the lungs follow the movement of the thorax wall. In expiration the intercostals and diaphragm relax and allow the elastic recoil of the lungs to come into play. The thoracic wall is simultaneously depressed by the muscles of the abdominal area, the diaphragm thrust forwards, as the result of the displacement and compression of the alimentary viscera thus brought about. (r.r.r. in the Figure mark ribs.)
Section 42. The oxygen and carbon dioxide are not carried in exactly the same way by the blood. The student will know from his chemical reading that neither of these gases is very soluble, but carbon dioxide is sufficiently so in an alkaline fluid to be conveyed by the liquid plasma. The oxygen however, needs a special portative mechanism in the colouring matter of the red corpuscles, the haemoglobin, with which it combines weakly to form oxy-haemoglobin of a bright red colour, and decomposing easily in the capillaries (the finest vessels between the arteries and veins), to release the oxygen again. The same compound occurs in all true vertebrata, and in the blood-fluid of the worm; in the crayfish a similar substance, haemocyanin, which when oxygenated is blue, and when deoxydized colourless, discharges the same function.
Section 43. The blood returns from the lungs to the left auricle (l.au.) by the pulmonary veins, hidden in the Figure by the heart, passes thence to the thick-walled left ventricle (l.vn.), and on into the aorta (ao.).
Section 44. The beating of the heart is, of course, a succession of contractions and expansions of its muscular wall. The contraction, or systole, commences at the base of the venae cavae and passes to the auricles, driving the blood before it into the ventricles, which then contract sharply and drive it on into the aorta or pulmonary artery; a pause and then a dilatation, the diastole follows. The flow of the blood is determined in one direction by the various valves of the heart. No valves occur in the opening of the superior cavae but an imperfect one, the Eustachian valve, protects the inferior cava; the direction of the heart's contraction prevents any excessive back-flow into the veins, and the onward, tendency is encouraged by the suck of the diastole of the ventricles. Between the left ventricle and auricle is a valve made up of two flaps of skin, the mitral valve, the edges of the flaps being connected with the walls of the ventricle through the intermediation of small muscular threads, the chordae tendinae, which stretch across its cavity to little muscular pillars, the papillary muscles; these attachments prevent the mitral valve from flapping back into the auricle, and as the blood flows into and accumulates in the ventricle it gets behind the flaps of the valve and presses its edges together. When the systole of the ventricle occurs, the increased, tension of the blood only closes the aperture the tighter, and the current passes on into the aorta, where we find three watch-pocket valves, with the pocket turned away from the heart, which are also closed and tightened by any attempt at regurgitation (back-flow). A similar process occurs on the right side of the heart, but here, instead of a mitral valve of two flaps between auricle and ventricle, we have a tricuspid valve with three. The thickness of the muscular walls, in view of the lesser distance through which it has to force the blood, -are- [is] less for the right ventricle than the left.
Section 45. The following are the chief branches of the aorta. The student should be able to follow them with certainty in dissection; they are all displayed in the Figure; but it must not be imagined for a moment that familiarity with this diagram will obviate the necessity for the practical work; (in.) is the innominate artery; it forks into (s.cl.a.) the right subclavian, and (r.c.c.) the right common carotid. Each carotid splits at the angle of the jaw into an internal and an external branch. The left common carotid, (l.c.c.) arises from the base of the innominate,* (l.s.cl.a.) the left subclavian, directly from the aorta. The aorta now curves round to the dorsal middle line, and runs down as seen in Figure 1, Sheet 1 (d.ao.) and Figure 1, Sheet 2 (d.ao.). Small branches are given off to the ribs, and then comes the median coeliac (coe.a.) to the stomach and spleen, the median superior mesenteric (s.mes.a.) to the main portion of the intestine, and the inferior mesenteric (p.m.a.) to the rectum. Note that no veins to the inferior vena cava correspond to these arteries-- the blood they supply going back by the portal vein (p.v.). The paired renal arteries (r.a.) supply the kidneys, and the common iliacs (c.il.a.) the hind legs, splitting into the internal iliacs (i.il.a.) and the femoral (f.).
{Lines from Second Edition only.}
[The student should note that the only arteries in the middle line are those supplying the alimentary canal.]
{Lines from First Edition only.}
* -The figure is inaccurate, and represents the left common carotid as arising from the aortic arch.-
Section 46. The distribution of the veins of the rabbit has only a superficial parallelism with arteries. The chief factors of vena cava inferior are the hepatic vein (h.v.), which receives the liver blood, the renal veins (r.v.), from the kidneys, the ilaeo-lumbar, from the abdominal wall, and the external (e.il.v.) and internal ilias (i.il.v.); with the exception of the renal veins none of these run side by side with arteries. The superior cavae (r. and l.v.c.s.) are formed by the union of internal (i.j.) and external jugular (e.j.) veins with a subclavian (s.cl.v.) from the fore limb. The term pre-caval vein is sometimes used for superior cava. The attention, of the student is called to the small azygos vein (az.) running into the right vena cava superior, and forming the only asymmetrical (not-balancing) feature of the veins in front of the heart; it brings blood back from the ribs of the thorax wall, and is of interest mainly because it answers to an enormous main vessel, the right post-cardinal sinus, in fishes. There are spermatic arteries and veins (s.v. and a.) to the genital organs. All these vessels should be patiently dissected out by the student, and drawn.
Section 47. Between the final branches of the arteries and the first fine factors of the veins, and joining them, come the systemic capillaries. These smallest and ultimate ramifications of the circulation penetrate every living part of the animal, so that if we could isolate the vascular system we should have the complete form of the rabbit in a closely-meshed network. It is in the capillaries that the exchange of gases occurs and that nutritive material passes out to the tissues and katastases in from them; they are the essential factor in the circulatory system of the mammal-- veins, arteries, and heart simply exist to remove and replace their contents. The details of the branching of the pulmonary artery and the pulmonary veins need not detain us now.
Section 48. Summarising the course of the circulation, starting from the right ventricle, we have-- pulmonary artery, pulmonary capillaries, pulmonary vein, left auricle, left ventricle, aorta, arteries, and systemic capillaries. After this, from all parts except the spleen and alimentary canal, the blood returns to systemic veins, superior or inferior cavae, right auricle, and right ventricle. The blood from the stomach spleen, and intestines however, passes via {through} the portal vein to the liver capillaries and then through the hepatic vein to inferior cava, and so on. Material leaves the blood to be excreted in lungs, kidneys, by the skin (as perspiration), and elsewhere. New material enters most conspicuously;
(a) by the portal veins portal veins and
(b) by the thoracic duct and left superior cava.
Section 49. The following table summarises what we have learnt up to the present of the physiology of the Rabbit, considered as a mechanism using up food and oxygen and disengaging energy:--
-Air_ {Nitrogen... returned unchanged.}
{Oxygen... through Pulmonary Vein to--} {see 3.}
-Food_ {Carbo-Hydrates (Starch, Sugar, Cellulose.)} Sugar.
{Protein.} {Peptones.}
{Fat (little in Rabbit.)} {Glycerine, and fatty acids in soups.}
{Rejected matter got rid of in Defaecation.}
1a. {Chyle in Lacteals going via {through} Thoracic Duct and Left
Superior Cava to--} {see 2.}
1b. {Veins of Villi--}
{Portal Vein--}
{Liver--}
{Hepatic Vein and Inferior Cava to--} {see 2.}
2. {Right side of heart; then to lungs, and then to--} {see 3.}
3. {Left side of heart; whence to Systemic Arteries and Capillaries.}
4. {The tissues and -Kataboly_.}
5. {Urea (?Liver) Kidney and Sweat Glands}
{CO2} {Lungs}
{H2O} {Lungs, Kidney, Sweat Glands}
{Other Substances} {Mainly by [Kidney,] Liver and Alimentary Canal}
Section 50. We have thus seen how the nutritive material is taken into the animal's system and distributed over its body, and incidentally, we have noted how the resultant products of the creature's activity are removed. The essence of the whole process, as we have already stated, is the decomposition and partial oxydation of certain complex chemical compounds to water, carbon dioxide, a low nitrogenous body, which finally takes the form of urea, and other substances. We may now go on to a more detailed study, the microscopic study, or histology, of the tissues in which metaboly and kataboly occur, but before we do this it will be convenient to glance for a moment at another of our animal types-- the Amoeba, the lowest as the rabbit is the highest, in our series.
Section 51. This is shown in Figure III., Sheet 3, as it would appear under the low power of the microscope. We have a mass of a clear, transparent, greyish substance called protoplasm, granular in places, and with a clearer border; within this is a denser portion called the nucleus, or endoplast (n.), which, under the microscope, by transmitted light, appear brighter, and within that a still denser spot, the nucleolus (ns.) or endoplastule. The protoplasm is more or less extensively excavated by fluid spaces, vacuoles; one clearer circular space or vacuole, which is invariably present, appears at intervals, enlarges gradually, and then vanishes abruptly, to reappear after a brief interval; this is called the contractile vacuole (c.v.). The amoeba is constantly changing its shape, whence its older name of the Proteus animalcule, thrusting out masses of its substance in one direction, and withdrawing from another, and hence slowly creeping about. These thrust-out parts, in its outline, are called pseudopodia (ps.). By means of them it gradually creeps round and encloses its food. Little particles of nutritive matter are usually to be detected in the homogeneous protoplasm of its body; commonly these are surrounded by a drop of water taken in with them, and the drop of water is then called a food vacuole. The process of taking in food is called ingestion. The amoeba, in all probability, performs essentially the same chemical process as we have summarised in Sections 10, 11, 12; it ingests food, digests it in the food vacuoles and builds it up into its body protoplasm, to undergo kataboly and furnish the force of its motion-- the contractile vacuole, is probably respiratory and perhaps excretory, accumulating and then, by its "systole" (compare Section 44), forcing out of its body, the water, carbon dioxide, urea, and other katastases, which are formed concomitantly with its activity. The amoeba reproduces itself in the simplest way; the nucleus occasionally divides into two portions and a widening fissure in the protoplasm of the animal's body separates one from the other. It is impossible to say that one is the parent cell, and the other the offspring; the amoeba we merely perceive, was one and is now two. It is curious to note, therefore, that the amoeba is, in a sense, immortal-- that the living nucleus of one of these minute creatures that we examine to-day under a microscope may have conceivably drawn, out an unbroken thread of life since the remotest epochs of the world's history. Although no sexual intercourse can be observed, there is reason to believe that a process of supposed "cannabalism," in which a larger amoeba may occasionally engulph a smaller one, is really a conjugative reproductive process, and followed by increased vitality and division.
Section 52. Now if the student will compare Section 35, he will see that in the white blood corpuscles we have a very remarkable resemblance to the amoeba; the contractile vacuole is absent, but we have the protoplasmic body, the nucleus and nucleolus, and those creeping fluctuations of shape through the thrusting out and withdrawal of pseudopodia, which constitute "amoeboid" motion. They also multiply, in the same way, by division.
Section 53. It is not only in the white corpuscle of the blood that we find this resemblance; in all the firmer parts of the body we find, on microscopic examination, similar little blebs of protoplasm, and at an early stage of development the young rabbit is simply one mass of these protoplasmic bodies. Their division and multiplication is an essential condition, of growth. Through an unfortunate accident, these protoplasmic blebs, which constitute the living basis of the animal body, have come to be styled "cells," though the term "corpuscles" is far more appropriate.
Section 54. The word is "cell" suggests something enclosed by firm and definite walls, and it was first employed in vegetable histology. Unlike the typical cells of animals, the cells of most plants are not naked protoplasm, but protoplasm enclosed in a wall of substance (cell wall) called cellulose. The presence of this cellulose cell wall, and the consequent necessity of feeding entirely upon liquids and gases that soak through it instead of being able to ingest a portion of solid food is indeed, the primary distinction between the vegetable and the animal kingdoms, as ordinarily considered.
Section 55. Throughout life, millions of these cells retain their primary characters, and constitute the white corpuscles of blood, "phagocytes," and connective tissue corpuscles; others again, engage in the formation of material round themselves, and lie, in such cases, as gristle and bone, embedded in the substance they have formed; others again, undergo great changes in form and internal structure, and become permanently modified into, for instance, nerve fibres and muscle substance. The various substances arising in this way through the activity of cells are called tissues, the building materials of that complex thing, the animal body. Since such a creature as the rabbit is formed through the co-operation of a vast multitude of cells, it is called multicellular; the amoeba, on the other hand, is unicellular. The rabbit may be thus regarded as a vast community of amoeboid creatures and their products.
Section 56. Figure IV., Sheet 3 represents, diagrammatically, embryonic tissue, of which, to begin with, the whole animal consists. The cells are all living, capable of dividing and similar, but as development proceeds, they differentiate, some take on one kind of duty (function), and some another, like boys taking to different trades on leaving school, and wide differences in structure and interdependence become apparent.
Section 57. It is convenient to divide tissues into three classes, though the divisions are by no means clearly marked, nor have they any scientific value. The first of these comprises tissues composed wholly, or with the exception of an almost imperceptible cementing substance, of cells; the second division includes the skeletal tissues, the tissue of mesentery, and the connective and basement tissue of most of the organs, tissues which, generally speaking, consist of a matrix or embedding substance, formed by the cells and outside of them, as well as the cells themselves; and, thirdly, muscular and nervous tissue. We shall study the former two in this chapter, and defer the third division until later.
Section 58. The outer layer of the skin (the epidermis), the inmost lining of the alimentary canal, the lining of the body cavity, and the inner linings of blood-vessels, glands, and various ducts constitute our first division. The general name for such tissues is epithelium. When the cells are more or less flattened, they form squamous epithelium (Figure VI.) such as we find lining the inside of a man's cheek (from which the cells sq.ep. were taken) or covering the mesentery of various types-- sq.end. are from the mesentery (Section 16) of a frog. A short cylindroidal form of cell makes up columnar epithelium, seen typically in the cells covering the villi of the duodenum (Figure V.). This epithelium of the villi has the outer border curiously striated, and this is usually spoken of as leading towards "ciliated" epithelium, to be described immediately. The epithelium of the epidermis is stratified-- that is to say, has many thicknesses of cells; the deeper layers are alive and dividing (stratum mucosum), while the more superficial are increasingly flattened and drier as the surface is approached (stratum corneum) and are continually being rubbed off and replaced from below.
Section 59. In the branching air-tubes of the lung, the central canal of the spinal cord, and in the ureters of the rabbit, and in most other types, in various organs, we find ciliated epithelium (Figure VII.). This is columnar or cubical in form, and with the free edge curiously modified and beset with a number of hair-like processes, the cilia, by which, during the life of the cell, a waving motion is sustained in one direction. This motion assists in maintaining a current in the contents of ducts which are lined with this tissue. The motion is independent of the general life of the animal, so long as the constituent cell still lives, and so it is easy for the student to witness it himself with a microscope having a 1/4-inch or 1/6-inch objective. Very fine cilia may be seen by gently scraping the roof of a frog's mouth (the cells figured are from this source), or the gill of a recently killed mussel, and mounting at once in water, or, better, in a very weak solution of common salt.
Section 60. The lining of glands is secretory epithelium; the cells are usually cubical or polygonal (8, g.ep.), and they display in the most characteristic form what is called metabolism. Anaboly (see Section 14) we have defined, as a chemical change in an upward direction-- less stable and more complex compounds are built up in the processes of vegetable and animal activity towards protoplasm; kataboly is a chemical running down; metaboly is a more general term, covering all vital chemical changes. The products of the action of a glandular epithelium are metabolic products, material derived from the blood is worked, up within the cell, not necessarily with conspicuous gain or loss of energy, and discharged into the gland space. The most striking case of this action is in the "goblet cells" that are found among the villi; these are simply glands of one cell, unicellular glands, and in Figure V. we see three stages in their action: at g.c.1 material (secretion) is seen forming in the cell, at g.c.2 it approaches the outer border, and at g.c.3 it has been discharged, leaving a hollowed cell. Usually however, the escape of secreted matter is not so conspicuous, and the gland-cells are collected as the lining of pits, simple, as in the gastric, pyloric, and Lieberkuhnian glands (Figure VIII., Sections 23, 29), or branching like a tree or a bunch of grapes (Figure r.g.), as in Brunner's glands (Section 29) the pancreas, and the salivary glands. The salivary glands, we may mention, are a pair internal to the posterior ventral angle of the jaw, the sub-maxillary; a pair anterior to these, the sub-lingual; a pair posterior to the jaw beneath the ear, the parotid, and a pair beneath the eye, the infra orbital.
Section 61. The liver is the most complicated gland in the body (Figure X.). The bile duct (b.d.) branches again and again, and ends at last in the final pits, the lobuli (lb.), which are lined with secretory epithelium, and tightly packed, and squeeze each other into polygonal forms. The blood supply from which the bile would appear to be mainly extracted, is brought by the portal vein, but this blood is altogether unfit for the nutrition of the liver tissue; for this latter purpose a branch of the coeliac artery, the hepatic serves. Hence in the tissue of the liver we have, branching and interweaving among the lobuli, the small branches of the bile duct (b.d.), which carries away the bile formed, the portal vein (p.v.), the hepatic artery (h.a.), and the hepatic vein (h.v.). (Compare Section 45.) Figure X.b shows a lobule; the portal vein and the artery ramify round the lobules-- are inter-lobular, that is (inter, between); the hepatic vein begins in the middle of the lobules (intra-lobular), and receives their blood. (Compare X.a.) Besides its function in the manufacture of the excretory, digestive, and auxiliary bile, the liver performs other duties. It appears to act as an inspector of the assimilation material brought in by the portal vein. The villi, for instance, will absorb arsenic, but this is arrested and thrown down in the liver. A third function is the formation of what would seem to be a store of carbo-hydrate, glycogen, mainly it would appear, from the sugar in the portal vein, though also, very probably, from nitrogenous material, though this may occur only under exceptional conditions. Finally, the nitrogenous katastases, formed in the working of muscle and nerve, and returned by them to the blood for excretion, are not at that stage in the form of urea. Whatever form they assume, they undergo a further metabolism into urea before leaving the body, and the presence of considerable quantities of this latter substance in the liver suggests this as a fourth function of this organ-- the elaboration of urea.
Section 62. Similar from a physiological point of view, to the secretory glands which form the digestive fluids are those which furnish lubricating fluids, the lachrymal gland, and Harderian glands in the orbit internally to the eye, and posterior and anterior to it respectively, the sebaceous glands (oil glands) connected with the hair, and the anal and perineal glands. The secretions of excretory glands are removed from the body; chief among them are the sweat glands and kidneys. The sweat glands are microscopic tubular glands, terminating internally in a small coil (Figure VIII. s.g.) and are scattered thickly over the body, the water of their secretion being constantly removed by evaporation, and the small percentage of salt and urea remaining to accumulate as dirt, and the chief reasonable excuse for washing. The kidney structure is shown diagrammatically in Figure 5, Sheet 7. A great number of branching and straight looped, tubuli (little tubes) converge on an open space, the pelvis. Towards the outer layers (cortex) of the kidney, these tubuli terminate in little dilatations into which tangled knots of blood-vessels project: the dilatations are called Bowman's capsules (B.c.), and each coil of bloodvessel a glomerulus (gl.). In the capsules, water is drained from the blood; in the tubuli, urea and other salts in the urine are secreted from a branching network of vessels.
Section 63. In all the epithelial tissues that we have considered we have one feature in common: they are cells, each equivalent to the amoeba, that have taken on special duties-- in a word, they are specialists. The amoeba is Jack of all trades and a free lance; the protective epidermal cell, the current-making ciliated cell, the bile or urea-making secretory cell, is master of one trade, and a soldier in a vast and wonderfully organized host. We will now consider our second kind of cell in this organization, the cell of which the especial aim is the building round it of a tissue.
Section 64. The simplest variety in this group is hyaline (i.e. glassy) cartilage (gristle). In this the formative cells (the cartilage corpuscles) are enjellied in a clear structureless matrix (Figure XII.), consisting entirely of organic compounds accumulated by their activity. Immediately round the cell lies a capsule of newer material. Some of the cells have recently divided (1); others have done so less recently, and there has been time for the interpolation of matrix, as at 2. In this way the tissue grows and is repaired. A thin layer of connective tissue (see below), the perichondrium, clothes the cartilaginous structure.
Section 65. Connective tissue (Figure XIII) is a general name for a group of tissues of very variable character. It is usually described as consisting typically in the mammals of three chief elements felted together; of comparatively unmodified corpuscles (c.c.), more or less amoeboid, and of fibres which are elongated, altered, and distorted cells. The fibres are of two kinds: yellow, branching, and highly elastic (y.e.f.), in consequence of which they fall into sinuous lines in a preparation, and white and inelastic ones (w.i.f.), lying in parallel bundles. Where the latter element is entirely dominant, the connective tissue is tendon, found especially at the point of attachment of muscles to the parts they work. Some elastic ligaments are almost purely yellow fibrous tissue. A loose interweaving of the three elements is areolar tissue, the chief fabric of mesentery, membrane, and the dermis (beneath the epidermis). With muscle it is the material of the walls of the alimentary canal and bloodvessels, and generally it enters into, binds together, and holds in place other tissue. The connective tissue of fishes displays the differentiation of fibres in a far less distinct manner.
Section 66. Through connective tissues wander the phagocytes, cells that are difficult to distinguish, if really distinct, from the white blood corpuscles. These cells possess a remarkable freedom; they show an initiative of their own, and seem endowed with a subordinate individuality. They occur in great numbers in a tissue called, botryoidal tissue (Figure XIV.), which occurs especially in masses and patches along the course of the alimentary canal, in its walls. The tonsils, swellings on either side of the throat, are such masses, and aggregates occur as visible patches, the Peyer's patches, on the ileum. It also constitutes the mass of the vermiform appendix and the wall of the sacculus rotundus; and in the young animal the "thymus gland," ventral to the heart, and less entirely, the "thyroid gland," ventral to the larynx, are similar structures, which are reduced or disappear as development proceeds. It is evident that in these two latter cases the term "gland" is somewhat of a misnomer. The matrix of botryoidal tissue is a network of stretched and hollowed connective tissue cells-- it is not a secretion, as cartilage matrix appears to be. During digestion, the phagocytes prowl into the intestine, and ingest and devour bacteria, that might otherwise give rise to disease. In inflammation, we may note here, they converge from all directions upon the point wounded or irritated. They appear to be the active agents in all processes of absorption (see osteoclasts under bone), and for instance, migrate into and devour the tissue of the tadpole's tail, during its metamorphosis to the adult frog.
Section 67. Within the connective tissue cells fat drops may be formed, as in Figure XV. Adipose tissue is simply connective tissue loaded with fat-distended cells. The tissue is, of course, a store form of hydro-carbon (Section 17) provided against the possible misadventure of starvation. With the exception of some hybernating animals, such store forms would seem to be of accidental importance only among animals, whereas among plants they are of invariable and necessary occurrence.
Section 68. We now come to Bone, a tissue confined to the vertebrata, and typically shown only in the higher types. As we descend in the scale from birds and mammals to lizards, amphibia (frogs and toads) and fish, we find cartilage continually more important, and the bony constituent of the skeleton correspondingly less so. In such a type as the dog-fish, the skeleton is entirely cartilaginous, bone only occurs in connection with the animal's scales; it must have been in connection with scales that bone first appeared in the vertebrate sub-kingdom. In the frog we have a cartilaginous skeleton overlaid by numerous bony scutes (shield-like plates) which, when the student comes to study that type, he will perceive are equivalent to the bony parts of such scales as occur in the dog-fish, sunk inward, and plating over the cartilage; and in the frog the cartilage also is itself, in a few places, replaced by bony tissue. In the adult rabbit these two kinds of bone, the bone overlying what was originally cartilage (membrane bone), and the bone replacing the cartilage (cartilage bone) have, between them, practically superseded the cartilage altogether. The structure of the most characteristic kind of bone will be understood by reference to Figure XVI. It is a simplified diagram of the transverse section of such a bone as the thigh bone. M.C. is the central marrow cavity, H.v., H.v. are cross sections of small bloodvessels, the Haversian vessels running more or less longitudinally through, the bone in canals, the Haversian canals. Arranged round these vessels are circles of the formative elements, the bone corpuscles or osteoblasts (b.c.) each embedded in bony matrix in a little bed, the lacuna, and communicating one with another by fine processes through canaliculi in the matrix, which processes are only to be seen clearly in decalcified bone (See Section 70). The osteoblasts are arranged in concentric series, and the matrix is therefore in concentric layers, or lamellae (c.l.). Without and within the zone of Haversian systems are (o.l. and i.l.), the outer and inner lamellae. The bone is surrounded by connective tissue, the periosteum. In addition to this compact bone, there is a lighter and looser variety in which spicules and bars of bony tissue are loosely interwoven. Many flat bones, the bones of the skull, for instance, consist of this spongy bone, plated (as an electro spoon is plated) with compact bone.
Section 69. Among the bony bars and spicules of spongy bone occurs the red marrow-- which must not be confused with the yellow marrow, the fatty substance in the central cavity of long bones. In this red marrow are numerous large colourless cells, which appear to form within their substance and then liberate red blood corpuscles. This occurs especially in the spongy bone within the ribs.
Section 70. The matrix of bone differs from that of cartilage or of most other tissues in consisting chiefly of inorganic salts. The chief of these is calcium phosphate, with which much smaller quantities of calcium carbonate, and magnesium phosphate and carbonate occur. These inorganic salts can be removed by immersion of the bone in weak hydrochloric acid, and a flexible network of connecting tissue, Haversian vessels, bone corpuscles, and their processes remains. This is decalcified bone alluded to above.
Section 71. In the very young rabbit, the limb bones, vertebral column, and many of the skull bones are simply plates and bars of cartilage; the future membrane bones, however are planned out in connective tissue. The development of the latter is simple, the connective tissue corpuscles functioning by a simple change of product as osteoblast. The development of the cartilage bones, however, is more complicated. Figure XVII., represents, in a diagrammatic way, the stages in the conversion of a cartilaginous bar to bone. To begin with, the previously sporadically-arranged (scattered anyhow) corpuscles (u.c.c.) are gathered into groups in single file, or in other words, into "columnar" groups (as at c.c.). The matrix becomes clouded with inorganic salts of lime, and it is then said to be calcified. This calcified cartilage then undergoes absorption-- it must not be imagined for a moment that bone is calcified cartilage. Simultaneous with the formation of the cavities (s.) due to this absorption, connective tissue (p.c.i.) from the surrounding perichondrium (p.c.) grows into the ossifying* bar. It is from this connective tissue that the osteoblasts (o.b.) arise, and bone is built up. Throughout life a bone is continually being absorbed and reformed by the activity of the osteoblasts. An osteoblast engaged in the absorption instead of the formation of bone is called an osteoclast.
* The formation of bone is called ossification. To ossify is to become bony.
Section 72. The great thing to notice about this is that cartilage does not become bone, but is eaten into and ousted by it; the osteoblasts and osteoclasts replace entirely the cartilage corpuscles, and are not derived from them.
Section 73. We may mention here the structure of the spleen (Figure 1, Sheet 1). It consists of a connective tissue and muscular coating, with an internal soft matrix much resembling botryoidal tissue, traversed by fibrous trabeculae (= beams, planks) containing blood-vessels, and the whole organ is gorged with blood, particularly after meals. The consideration of its function the student may conveniently defer for the present.
Section 74. Here also, we may notice the lymphatics, a series of small vessels which return the overflow of the blood serum from the capillaries, in the nutrition of the tissues in all parts of the body, to the thoracic duct (see Section 36), and the general circulation. At intervals their course is interrupted by gland-like dilatations, the lymphatic glands, in which masses of rapidly dividing and growing (proliferating) cells occur, of which, doubtless, many are detached and become, first "lymph corpuscles," and, when they reach the veins, white blood corpuscles.
Section 75. We are now in a position to study the rabbit's skeleton. We strongly recommend the student to do this with the actual bones at hand-- they may be cleared very easily in a well-boiled rabbit. This recommendation may appear superfluous to some readers, but, as a matter of fact, the marked proclivity of the average schoolmaster for mere book-work has put such a stamp on study, that, in nine cases out of ten, a student, unless he is expressly instructed to the contrary, will go to the tortuous, and possibly inexact, descriptions of a book for a knowledge of things that lie at his very finger-tips. We have not written, this chapter to give a complete knowledge of the skeleton, but simply as an aid in the actual examination of the bones.
Section 76. We may take the skeleton under five headings. There is the central axis, the chain of little bones, the vertebrae, threaded on the spinal cord (see Figure 1 and Section 1); the thorax, the box enclosed by ribs and sternum; the fore-limb and bones connected with it (pectoral girdle and limb), and the hind-limb and bones connected with it (pelvic girdle). Finally there is the skull, but following the London University syllabus, we shall substitute the skull of the dog for of that of the rabbit, as more typically mammalian (Section 4).
Section 77. In Section 3 (which the student should refer to) we have a division of the vertebrae into four varieties. Of these most representative is the thoracic. A thoracic vertebra (Figure 4, Sheet 5, T.V.) consists of a central bony mass, the body or centrum (b.), from which there arises dorsally an arch, the neural arch (n.a.), completed by a keystone, the neural spine (n.s.); and coming off laterally from the arch is the transverse process (tr.p.). Looking at the vertebra sideways, we see that the arch is notched, for the exit of nerves. Jointed to the thoracic vertebrae on either side are the ribs (r.). Each rib has a process, the tuberculum, going up to articulate with the transverse process, and one, the capitulum articulating between the bodies of two contiguous vertebrae. The facets for the articulation of the capitulum are indicated in the side view by shading. At either end of the body of a vertebra of a young rabbit are bony caps, the epiphyses (ep.), separated from the body by a plane of unossified cartilage (indicated, by the dots). These epiphyses to the vertebral bodies occur only among mammals, and are even absent in some cases within the class. In the adult rabbit they have ossified continuously with the rest of the body.
Section 78. A cervical vertebra (C.V.) seems, upon cursory inspection, to have no rib. The transverse processes differ from those of thoracic series in having a perforation, the vertebrarterial canal, through which the vertebral artery runs up the neck. A study of the development of these bones shows that the part marked f.r. ossifies separately from the rest of the transverse process; and the form of the equivalent structures in certain peculiar lower mammals and in reptiles leaves no doubt that f.r. is really an abbreviated rib; fused up with the transverse process and body. The two anterior cervical vertebrae are peculiar. The first (at.) is called the Atlas-- the figure shows the anterior view-- and has great articular faces for the condyles (Section 86) of the skull, and a deficient centrum. The next is the axis, and it is distinguished by an odontoid peg (od.p.), which fits into the space where the body of the atlas is deficient. In development the centrum of the axis ossifies from one centre, and the odontoid, peg from another, which at that time occupies the position of centrum of the atlas. So that it would seem that the atlas is a vertebra minus a centrum, and the axis is a vertebra plus a centrum, added at the expense of the atlas.
Section 79. The lumbar vertebrae (l.v.) are larger, and have cleft transverse processes, each giving rise to an ascending limb, the metapophyses, and a descending one. The latter (generally spoken of as the transverse processes) point steeply downward, and are considerably longer than those of thoracic series. The sacral vertebrae (s.v.) have great flattened transverse processes for articulation with the ilia. The caudal vertebrae (c.v.) are gradually reduced to the mere elongated centra, as we proceed, towards the tip of the tail.
Section 80. All the vertebrae join with their adjacent fellows through the intermediation of certain intervertebral pads, and also articulate by small processes at either end at the upper side of the arch, the zygapophyses. The normals to the polished facets of these point, in the case of the anterior zygapophyses, up and in (mnemonic: ant-up-in), and in the case of the posterior, down and out. The student should make this, and the other features of vertebrae, out upon actual specimens.
Section 81. The thorax is bounded dorsally by the vertebral column, and ventrally by the sternum. The sternum consists of segments, the sternebrae (st.); anteriorly there is a bony manubrium (mb.), posteriorly a thin cartilaginous plate, the xiphisternum (xi.). Seven pairs of ribs articulate by cartilaginous ends (sternal ribs) with the sternum directly, as indicated in the figure; five (false) ribs are joined, to each other and to the seventh, and not to the sternum directly. The last four ribs have no tuberculum (Section 77).
Section 82. The fore-limb (pectoral limb) consists of an upper arm bone, the humerus (hum.) the distal end of which is deeply excavated by the olecranon fossa (o.f.) as indicated by the dotted lines; of two bones, the ulna (u.) and radius (r.) which are firmly bound by ligament in the position of the figure (i.e., with the palm of the hand downward, "prone"); of a number of small bones (carpalia), the carpus (c.); of a series of metacarpals (mc.); and of three digits (= fingers) each, except the first, or pollex, of three small bones-- the phalanges, only the proximal of which appear in the figure. The ulna has a hook-like head, the olecranon (o.) which distinguishes it easily from the distally thickened radius. The limb is attached to the body through the intermediation of the shoulder-blade (scapula, sc.) a flattened bone with a median external ridge with a hook-like termination, the acromion (acr.). There is also a process overhanging the glenoid cavity (g.) wherein the humerus articulates, which process is called coracoid (co.); it is ossified from two separate centres, and represents a very considerable bone in the bird, reptile, and frog. Along the dorsal edge of the scapula of the rabbit is unossified cartilage, which is called the supra-scapula (s.sc.). In man there runs from the acromion to the manubrium of the sternum a bone, the collar-bone or clavicle. This is represented by a very imperfectly ossified rudiment in the rabbit. The scapula and clavicle, the bones of the body connected with the fore-limb, are frequently styled the pectoral girdle, or shoulder-girdle; this name of girdle will appear less of a misnomer when lower vertebrate types are studied.
Section 83. The hind limb and its body bones-- pelvic limb and girdle-- are shown in Figure 2. The limb skeleton corresponds closely with that of the fore-limb. The femur (fe.) answers to the humerus, and is to be distinguished from it by the greater distinctness of its proximal head (hd.) and by the absence of an olecranon fossa from its distal end. The tibia (ti = the radius) is fused for the distal half of its length with the fibula (fb. = ulna). A tarsus (tarsalia) equals the carpus.* Two of the proximal tarsalia may be noted: one working like a pulley under the tibia, is the astragalus (as.); one forming the bony support of the heel, is the calcaneum (ca.). There is a series of metatarsals, and then come four digits of three phalanges each.
* Such a resemblance as exists between one vertebra and another in the rabbit, or between the humerus and the femur, is called serial homology; the two things correspond with each other to the extent of imperfect reduplication. "Homology" simply is commonly used to indicate the resemblance between any two structures in different animals, in origin and position as regards other parts. Thus the heart of the rabbit and of the frog are homologous structures, corresponding in position, and resembling each other much as two memory sketches of one picture might do.
Section 84. The pelvic girdle differs from the pectoral in most land vertebrata in being articulated with the vertebral column. This difference does not exist in fishes. It consist in the rabbit of four bones; the ilium (i.), the ischium (is.), the pubis (pb.), and the small cotyloid bone-- the first two and the latter one meeting in the acetabular fossa (ac.) in which the head of the femur works. The pubes and ischia are fused along the mid-ventral line. Many morphologists regard, the ilium as equivalent to, that is, strictly corresponding in its relation, to the scapula, the pubis to the cartilaginous substratum of the clavicle, and the ischium to the coracoid.
Section 85. These bones will be studied at the greatest advantage when dissected out from a boiled rabbit. Prepared and wired skeletons, disarticulated skeletons, plates of figures, and written descriptions are in succession more tedious and less satisfactory ways to a real comprehension, of this matter. This chapter directs the student's attention to the most important points in the study of the skeleton, but it is in no way intended to mitigate the necessity of practical work. It is a guide simply.
Section 86. The mammalian skull will be better understood after the study of that of some lower vertebrate. We shall describe its main features now, but their meaning will be much clearer after the lower type is read. Our figures are of Canis. In section (Figure VI., Sheet 6), we perceive a brain case (cranium) opening behind by a large aperture, the foramen magnum (F.M.). In front of this is an extensive passage, the nasal passage (E.N. to P.N.) which is divided from the mouth by a bony floor, the palate, and which opens into the pharynx behind at the posterior nares (P.N.) and to the exterior by the anterior or external nares (E.N.). It is divided into right and left passages by a middle partition, the nasal septum. Outside the skull, on its wings, is a flask-like bone, the bulla tympani (b. in Figures 2 and 3), protecting the middle ear, and from above this there passes an arch, the cheek bone (ju. in Figures 1, 2, and 3), to the upper jaw, forming in front the bony lower protection of the cavity containing the eye, the orbit. The cheek arch, nasal passage, and jaws, form collectively the "facial apparatus," as distinguished from the cranium, and the whole skull is sometimes referred to as, the "cranio-facial apparatus." Two eminences for articulation with the atlas vertebra, the condyles (c.), lie one on each side of the lower boundary of the foramen magnum.
Section 87. The floor of the cranium consists of a series of cartilage bones, the basi-occipital (b.o.), basi-sphenoid (b.sp.), pre-sphenoid (p.sp.), and in front, the ethmoid (eth.), which sends down a median plate, not shown, in the figure, to form the nasal septum between right and left nasal passages. Like extended wings on either side of the basi-occipital are the ex-occipital (e.o.) (the bone is marked in Figure 4, but the letters are a little obscured by shading). Similarly the ali-sphenoids (a.s.), are wings to the basi-, and the orbito-sphenoids (o.s.), to the pre-sphenoid bone (p.sp.). Between the ex-occipital and ali-sphenoid there is wedged in a bone, the periotic (p.o.) containing the internal ear (Section 115). Above the foramen magnum the median supra-occipital bone completes what is called the occipital arch. A pair of parietals (pa.) come above the ali-sphenoids, and a pair of frontals (f.) above the orbito-sphenoids. At the side the brain case is still incomplete, and here the squamosal (sq.) enters into its wall. In the external view (Figure 3) the bulla hides the periotic bone from without. The student should examine all four figures for these bones before proceeding.
Section 88. The outer edge of the upper jaw and the cheek arch are made up of three paired bones. First comes the premaxilla (p.m.) (not p.m.1 or p.m.4), containing in the dog, the three incisors of either side. Then comes the maxilla, bearing the rest of the teeth.* The jugal or malar (ju.) reaches over from the maxilla to meet a zygomatic process (= connecting outgrowth) (z.p.) of the squamosal bone.
* In the dog a sabre-like canine (c.), four premolars (p.m.1 and p.m.4) and two molars (m.1 and m.2).
Section 89. In the under view of the skull (Figure 2) it will be seen that the maxilla sends in a plate to form the front part of the hard palate. Behind, the hard palate is completed by the pair of palatine bones (pal.), which conceal much of the pre- and orbito-sphenoid in the ventral view, and which run back as ridges to terminate in two small angular bones, the pterygoids (pt.) which we shall find represent much more important structures in the lower vertebrata.
Section 90. The pre-maxillae and maxillae bound the sides of the nasal passage, and it is completed above by a pair of splints, the nasals. Along the floor of the nasal passage, on the middle line, lies a splint of bone formed by the coalescence of two halves. It embraces in a V-like groove the mesethmoid (nasal septum) above, and lies on the palate.
{Lines from First Edition only.}
-Its position is indicated by a heavy black line in 4, and it is
called, the vomer bone (vo.).-
{Lines from Second Edition only.}
[In the frog it is represented by two laterally situated bones. This is
the vomer bone (vo.).]
The nasal passages are partially blocked by foliated bony outgrowths, from the inner aspect of their walls, which in life are covered with mucous membrane, and increase the surface sensitive to smell. The ethmoid ends in the ethmo-turbinal (e.t.); the nasal, the naso-turbinal (n.t.); and the maxilla, the maxillo-turbinal (m.t.). In the anterior corner of the orbit there is a bone, the lachrymal (lc. Figure 1), which is hidden by the maxilla in the side view of the skull.
Section 91. The lower jaw (mandible) is one continuous bone in the mammal. Three incisors bite against the three of the upper jaw. Then comes a canine, four premolars, and three molars, the first of which is blade-like (sectorial tooth), and bites against the similar sectorial tooth (last premolar) of the upper jaw. The third molar is small. The arrangement of tooth is indicated in the following dental formula:-- I. 3.3/3.3, C. 1.1/1.1, P.M. 4.4/4.4, M. 2.2/3.3
Section 92. Attached just behind the bulla above, and passing round on either side of the throat to meet at the base of the tongue, is the hyoid apparatus (Figure 6). The stylohyal (s.h.), epihyal (e.h.), and ceratohyal (c.h.) form the anterior cornu of the hyoid. The body of the hyoid (b.h.) forms a basis for the tongue. The posterior coruna (t.h.) of the hyoid are also called the thyrohyals.
Section 93. The following table presents these bones in something like their relative positions. A closer approximation to the state of the case will be reached if the student will imagine the maxilla raised up so as to overlie and hide the palatine and presphenoid, the squamosal similarly overlying the periotic bone, and the jugal reaching between them. Membrane bones are distinguished by capital letters.
-Cranium_
-Nasal_ (paired), Ethmoid Bone (median), -Vomer_
-Upper Jaw_
-Frontal_ (paired), -Lachrymal_ (paired), Orbito-sphenoid (paired),
Pre-sphenoid (median), Eye
-Parietal_ (Paired), Ali-sphenoid (paired), Basi-sphenoid (median)*,
Periotic Bone (paired)
-Bulla_ (paired)
Supra-occipital (median), Ex-occipital (paired), Basi-occipital (median)
-Pre-Maxilla_ (paired)
Palatine (paired)
Pterygoid (paired)
-Lower Jaw_
-Maxilla_ (paired)
-Jugal_ (paired)
-Squamosal_ (paired)*In this table the small bones of the ear are simply indicated by an asterisk.
Section 94. Hidden by the bulla, and just external to the periotic bone, are the auditory ossicles, the incus, malleus, os orbiculare, and stapes. These will be more explicitly treated when we discuss the ear.
Section 95. When we come to the study of the nerves, we shall revert to the skull, and treat of its perforations. The student should not fail, before proceeding, to copy and recopy our figures, and to make himself quite familiar with them, and he should also obtain and handle an actual skull. For all practical purposes the skull of a sheep or cat will be almost as useful as that of the dog.
Section 96. We have, in the skeleton, a complicated apparatus of parts hinged and movable upon one another; the agent moving these parts is the same agent that we find in the heart walls propelling the blood through the circulation, in the alimentary canal squeezing the food along its course, and universally in the body where motion occurs, except in the case of the creeping phagocytes, and the ciliary waving of ciliated epithelium. This agent is muscle. We have, in muscular tissue, a very wide departure from the structure of the primordial cell; to use a common biological expression, a very great amount of modification (= differentiation). Sheet 7 represents the simpler kind of muscular tissue, unstriated muscle, in which the cell character is still fairly obvious. The cells are fusiform (spindle-shaped), have a distinct nucleus and faint longitudinal striations (striations along their length), but no transverse striations.
Section 97. In striated muscle extensive modifications mask the cell character. Under a 1/4 inch objective, transverse striations of the fibres are also distinctly visible, and under a much higher power we discern in a fibre (Sheet 7) transverse columns of rod-like sarcous elements (s.e.), the columns separated by lines of dots, the membranes of Krause (k.m.), and nuclei (n.), flattened and separated into portions, and lying, in some cases, close to the sarcolemma (sc.) the connective tissue enclosing the fibre, in others scattered throughout the substance of the fibre. The figure shows the fibre ruptured, in order to display the sarcolemma; e.p. is the end plate of a nerve (n.v.), and fb. are the fibrillae into which a fibre may be teased.
Section 98. In the heart we have an intermediate kind of muscle cardiac muscle (Figure 2), in which the muscle fibres branch; there is apparently no sarcolemma, and the undivided nuclei lie in the centre of the cell.
Section 99. Unstriated muscle is sometimes called involuntary, and striated, voluntary muscle; but there is really not the connexion with the will that these terms suggest. We have just mentioned that the heart-muscle is striated, but who can alter the beating of the heart by force of will? And the striated muscles of the limbs perform, endless involuntary acts. It would seem that unstriated muscle contracts slowly, and we find it especially among the viscera; in the intestine for instance, where it controls that "peristaltic" movement which pushes the food forward. Voluntary muscle, on the other hand, has a sharp contraction. The muscle of the slow-moving snails, slugs, and mussels is unstriated; all the muscle of the active insects and crustacea (crabs, lobsters, and crayfish) is striated. Still if the student bears the exception of the heart in mind, and considers muscles as "voluntary" that his will can reach, the terms voluntary and involuntary will serve to give him an idea of the distribution of these two types of muscle in his own body, and in that of the rabbit.
Section 100. Muscular contraction, and generally all activity in the body is accompanied by kataboly. The medium by which these katabolic changes are set going and controlled is the nervous system. The nervous system holds the whole body together in one harmonious whole; it is the governing organization of the multicellular community (Section 55), and the supreme head of the government resides in the brain, and is called the mind. But just as in a political state only the most important and most exceptional duties are performed by the imperial body, and minor matters and questions of routine are referred to boards and local authorities, so the mind takes cognisance only of a few of the higher concerns of the animal, and a large amount of the work of the nervous system goes on insensibly, in a perfectly automatic way-- even much that occurs in the brain.
Section 101. The primary elements in the tissue of the nervous system are three; nerve fibres, which are simply conducting threads, telegraph wires; ganglion cells, which are the officials of the system; and neuroglia, a fine variety of connective tissue which holds these other elements together, and may also possibly exercise a function in affecting impressions. A message along a nerve to a ganglion cell is an afferent impression, from a cell to a muscle or other external end is an efferent impression. The passage of an impression may be defined as a flash of kataboly along the nerve, and so every feeling, thought, and determination involves the formation of a certain quantity of katastases, and the necessity for air and nutrition.
Section 102. Unlike telegraph wires, to which they are often compared, nervous fibres usually convey impressions only in one direction, either centrally (afferent or sensory nerve fibres), or outwardly (efferent or motor nerve fibres). But the so-called motor nerve fibres include not only those that set muscles in motion, but those that excite secretion, check impulsive movements, and govern nutrition.
Section 103. Figure 7, Sheet 8, shows the typical structure of nervous tissues. The nerve fibres there figured are bound together by endoneurium into small ropes, the nerves, encased in perineurium. There is always a grey axis cylinder (a.c.), which may (in medullated nerves), or may not (in non-medullated or grey nerves) have a medullary sheath (s.S.) interrupted at intervals by the nodes of Ranvier (n.R.). Nuclei (n.) at intervals under the sheath indicate the cells from which nerve fibres are derived by a process of elongation. The nerves of invertebrata, where they possess nerves, are mostly grey, and so are those of the sympathetic system of vertebrata, to be presently described, g.c., g.c. are ganglion cells; they may have many hair-like processes, usually running into continuity with the axis cylinders of nerve fibres, in which case they are called multi-polar cells, or they may be uni- or bi-polar.
Section 104. The simplest example of the action of the nervous system is reflex action. For instance, when the foot of a frog, or the hand of a soundly sleeping person, is tickled very gently, the limb is moved away from the irritation, without any mental action, and entirely without will being exercised. And when we go from light into darkness, the pupil of the eye enlarges, without any direct consciousness of the change of its shape on our part. Similarly, the presence or food in the pharynx initiates a series of movements-- swallowing, the digestive movements, and so on-- which in health are entirely beyond our mental scope.
Section 105. A vast amount of our activities are reflex, and in such action an efferent stimulus follows an afferent promptly and quite mechanically. It is only where efferent stimuli do not immediately become entirely transmuted into outwardly moving impulses that mental action comes in and an animal feels. There appears to be a direct relation between sensation and motion. For instance, the shrieks and other instinctive violent motions produced by pain, "shunt off" a certain amount of nervous impression that would otherwise register itself as additional painful sensation. Similarly most women and children understand the comfort of a "good cry," and its benefit in shifting off a disagreeable mental state.
Section 106. The mind receives and stores impressions, and these accumulated experiences are the basis of memory, comparison, imagination, thought, and apparently spontaneous will. Voluntary actions differ from reflex by the interposition of this previously stored factor. For instance, when a frog sees a small object in front of him, that may or may not be an edible insect, the direct visual impression does not directly determine his subsequent action. It revives a number of previous experiences, an image already stored of similar insects and associated with painful or pleasurable gustatory experiences. With these arise an emotional effect of desire or repulsion which, passes into action of capture or the reverse.
Section 107. Voluntary actions may, by constant repetition, become quasi-reflex in character. The intellectual phase is abbreviated away. Habits are once voluntary and deliberated actions becoming mechanical in this way, and slipping out of the sphere of mind. For instance, many of the detailed movements of writing and walking are performed without any attention to the details. An excessive concentration of the attention upon one thing leads to absent-mindedness, and to its consequent absurdities of inappropriate, because imperfectly acquired, reflexes.
Section 108. This fluctuating scope of mind should be remembered, more especially when we are considering the probable mental states of the lower animals. An habitual or reflex action may have all the outward appearance of deliberate adjustment. We cannot tell in any particular case how far the mental comes in, or whether it comes in at all. Seeing that in our own case consciousness does not enter into our commonest and most necessary actions, into breathing and digestion, for instance, and scarcely at all in the details of such acts as walking and talking we might infer that nature was economical in its use, and that in the case of such an animal as the Rabbit, which follows a very limited routine, and in which scarcely any versatility in emergencies is evident, it must be relatively inconsiderable. Perhaps after all, pain is not scattered so needlessly and lavishly throughout the world as the enemies of the vivisectionist would have us believe.
Section 109. A little more attention must now be given to the detailed anatomy of the peripheral and central nerve ends. A nerve, as we have pointed out, terminates centrally in some ganglion cell, either in a ganglion or in the spinal cord or brain; peripherally there is a much greater variety of ending. We may have tactile (touch) ends of various kinds, and the similar olfactory and gustatory end organs; or the nerve may conduct efferent impressions, and terminate in a gland which it excites to secretion, in a muscle end-plate, or in fact, anywhere, where kataboly can be set going and energy disengaged. We may now briefly advert to the receptive nerve ends.
Section 110. Many sensory nerves, doubtless, terminate in fine ends among the tissues. There are also special touch corpuscles, ovoid bodies, around which a nerve twines, or within which it terminates.
Section 111. The eye (Figure 8) has a tough, dense, outer coat, the sclerotic (sc.), within which is a highly vascular and internally pigmented layer, the choroid, upon which the percipient nervous layer, the retina (r.) rests. The chief chamber of the eye is filled with a transparent jelly, the vitreous humour (v.h.). In front of the eye, the white sclerotic passes into the transparent cornea (c.). The epidermis is continued over the outer face of this as a thin, transparent epithelium. The choroid coat is continued in front by a ring-shaped muscle, the iris (ir.) the coloured portion of the eyes. This iris enlarges or contracts its central aperture (the black pupil) by reflex action, as the amount of light diminishes or increases. Immediately behind this curtain is the crystalline lens (l.), the curvature of the anterior face or which is controlled by the ciliary muscle (c.m.). In front of the lens is the aqueous humour (a.h.). The description of the action of this apparatus involves the explanation of several of the elementary principles of optics, and will be found by the student in any text-book of that subject. Here it would have no very instructive bearing, either on general physiological considerations or upon anatomical fact.
Section 112. The structure of the retina demands fuller notice. Figure 9 shows an enlarged, diagram of a small portion of this, the percipient part of the eye. The optic nerve (o.n. in Figure 8) enters the eye at a spot called the blind spot (B.S.), and the nerve fibres spread thence over the inner retinal surface. From this layer of nerve fibres (o.n. in Figure 9) threads run outward, through certain clear and granular layers, to an outermost stratum of little rods (r.) and fusiform bodies called cones (c.), lying side by side. The whole of the retina consists of quite transparent matter, and it is this outermost layer of rods and cones (r. and c.) that receives and records the visual impression. This turning of the recipient ends away from the light is hardly what one would at first expect-- it seems such a roundabout arrangement-- but it obtains in all vertebrata, and it is a striking point of comparison with the ordinary invertebrate eye.
Section 113. We may pause to call the student's attention to a little point in the physiology of nerves, very happily illustrated here. The function of a nerve fibre is the conduction of impressions pure and simple; the light radiates through the fibrous layer of the retina without producing the slightest impression, and at the blind spot, where the rods and cones are absent, and the nerve fibres are gathered together, no visual impressions are recorded. If there is any doubt as to the existence of a blind spot in the retinal picture, the proof is easy. Let the reader shut his left eye, and regard these two asterisks, fixing his gaze intently upon the left-hand one of them.
At a distance of three or four inches from the paper, both spots will be focussed on his retina, the left one in the centre of vision, and the right one at some spot internal to this, and he will see them both distinctly. Now, if he withdraws his head slowly, the right spot will of course appear to approach the left, and at a distance of ten or twelve inches it will, in its approach, pass over the blind spot and vanish, to reappear as he continues to move his head away from the paper. The function of nerve fibres is simply conduction, and the nature of the impressions they convey is entirely determined by the nature of their distal and proximal terminations.
Section 114. Certain small muscles in the orbit (eye-socket) move the eye, and by their action contribute to our perception of the relative position of objects. There is a leash of four muscles rising from a spot behind the exit of the optic nerve from the cranium to the upper, under, anterior, and posterior sides of the eyeball. These are the superior, inferior, anterior, and posterior recti. Running from the front of the orbit obliquely to the underside of the eyeball is the inferior oblique muscle. Corresponding to it above is a superior oblique. A lachrymal gland lies in the postero-inferior angle of the orbit, and a Handerian gland in the corresponding position in front. In addition to the upper and lower eyelids of the human subject, the rabbit has a third, the nictitating lid, in the anterior corner of the eye.
Section 115. The ear (Sheet VII.) consists of an essential organ of hearing, and of certain superadded parts. The essential part is called the internal ear, and is represented in all the true vertebrata (i.e., excluding the lancelet and its allies). In the lower forms it is a hollow membranous structure, embedded in a mass of cartilage, the otic capsule; in the mammal the latter is entirely ossified, to form the periotic bone. The internal ear consists of a central sac, from which three semicircular canals spring. The planes of the three canals are mutually at right angles; two are vertical, the anterior and posterior (p.v.c.) vertical canals, and one is horizontal, the horizontal canal (h.c.). There are dilatations, called ampullae, at the anterior base of the anterior, and at the posterior base of the posterior and horizontal canals. Indirectly connected with the main sac is a spirally-twisted portion, resembling a snail shell in form, the cochlea. This last part is distinctive of the mammalia, but the rest of the internal ear is represented in all vertebrata, with one or two exceptions. The whole of the labyrinth is membranous, and contains a fluid, the endolymph; between the membranous wall of the labyrinth and the enclosing bone is a space containing the perilymph. Strange as it may appear at first, the entire lining of the internal ear is, at an early stage, continuous with the general epidermis of the animal. It grows in just as a gland might grow in, and is finally cut off from the exterior; but a considerable relic of this former communication remains as a thin, vertical blind tube (not shown in the figure), the ductus endolymphaticus.
Section 116. The eighth nerve runs from the brain case (Cr.), into the periotic bone, and is distributed to the several portions of this labyrinth. In an ordinary fish this internal ear is the sole auditory organ we should find; the sound-waves would travel through the water to the elastic cranium and so reach and affect the nerves. But in all air-frequenting animals this original plan of an ear has to be added to, to fit it to the much fainter sound vibrations of the compressible and far less elastic air. A "receiving apparatus" is needed, and is supplied by the ear-drum, middle ear, or tympanic cavity (T.). In the mammal there is also a collecting ear trumpet (the ear commonly so-called), the external ear, and external auditory meatus (e.a.m.). A tightly stretched membrane, the tympanic membrane, separates this from the drum. A chain of small bones, the malleus (m.), the incus (i.), the os orbiculare (o.or.), a very small bone, and a stirrup-shaped stapes, swing across the tympanum, from the tympanic membrane to the internal ear. At two points the bony investment of this last is incomplete-- at the fenestra rotunda (f.r.), and at the fenestra ovalis, (f.o.), into which latter the end of the stapes fits, and so communicates the sound vibrations of the tympanic membrane to the endolymph. A passage, the Eustachian tube, communicates between the tympanic cavity and the pharynx (Ph.), and serves to equalize the pressure on either side of the drum-head. A comparative study of the ears of the vertebrata brings to light the fact that, as we descend in the animal scale, the four ear ossicles are replaced by large bones and cartilages connected with the jaw, and the drum and Eustachian tube by a gill slit. We have, in fact, in the ear, as the student will perceive in the sequel, an essentially aquatic auditory organ, added to and patched up to fit the new needs of a life out of water.
Section 117. The impressions of smell are conducted through the first nerve to the brain, and are first received by special hair-bearing cells in the olfactory mucous membrane of the upper part of the nasal passage. The sense of taste has a special nerve in the ninth, the fibres of which terminate in special cells and cell aggregates in the little papillae (velvet pile-like processes) that cover the tongue.
Section 118. At an early stage in development, the brain of a mammal consists of a linear arrangement of three hollow vesicles (Figure 5, Sheet VIII., 1, 2, and 3), which are the fore-, mid-, and hind-brain respectively. The cavities in these in these vesicles are continuous with a hollow running through the spinal cord. On the dorsal side of the fore-brain is a structure to be dealt with more fully later, the pineal gland (p.g.), while on its under surface is the pituitary body (pt.).
Section 119. The lower figure of (5) shows, in a diagrammatic manner, the derivation of the adult brain from this primitive state. From the fore-brain vesicle, a hollow outgrowth on either side gives rises to the (paired) cerebral hemisphere (c.h.), which is prolonged forward as the olfactory lobe (o.l.). From the fore-brain the retina of the eye and the optic nerve also originate as an, at first, hollow outgrowth (op.). The roof of the mid-brain is also thickened, and bulges up to form two pairs of thickenings, the corpora quadrigemina, (c.q.). The hind-brain sends up in front a median outgrowth, which develops lateral wings, the cerebellum (cbm.), behind which the remainder of the hind-brain is called the medulla oblongata, and passes without any very definite demarcation into the spinal cord.
Section 120. Figure 1 is a corresponding figure of the actual state of affairs in the adult. The brain is seen in median vertical section. (ch.) is the right cerebral hemisphere, an inflated vesicle, which, in the mammal-- but not in our lower types-- reaches back over the rest of the fore-brain, and also over the mid-brain, and hides these and the pineal gland in the dorsal view of the brain (Figure 2). The hollow of the hemisphere on either side communicates with the third ventricle, the original cavity of the fore-brain (1 in Figure 5), by an aperture (the foramen of Monro), indicated by a black arrow (f.M.). Besides their original communication through the intermediation of the fore-brain, the hemispheres are also united above its roof by a broad bridge of fibre, the corpus callosum (c.c.), which is distinctive of the mammalian animals. The original fore-brain vesicle has its lateral walls thickened to form the optic thalami (o.th.), between which a middle commissure, (m.c.), absent in lower types, stretches like a great beam across the third ventricle. The original fore-brain is often called the thalamencephalon, the hemisphere, the prosencephalon, the olfactory lobes, the rhinencephalon.
Section 121. The parts of mid-brain (mesencephalon) will be easily recognised. Its cavity is in the adult mammal called the iter; its floor is differentiated into bundles of fibres, the crura cerebri (c.cb.), figured also in Figure 4.
Section 122. The cerebellum (metencephalon) consists of a central mass, the vermis (v.cbm.), and it also has lateral lobes (l.l.), prolonged into flocculi (f.cbm.), which last are -em-bedded in pits, [in] the periotic bone, and on that account render the extraction of the brain from the cranium far more difficult than it would otherwise be. The roof of the hind-brain, before and behind the cerebellum, consists of extremely thin plates of nervous matter. Its floor is greatly thickened to form the mass of the medulla, and in front a great transverse track of fibres is specialized, the pons Varolii (p.V.). Its cavity is called, the fourth ventricle.
Section 123. Figure 2 gives a dorsal view of the rabbit's brain; a horizontal slice has been taken at the level of the corpus callosum. The lateral ventricle (i.e., the hollows of the hemisphere) is not yet opened. A lower cut (Figure 3) exposes this (V.L.). The level of these slices is approximately indicated in Figure 1 by the lines A and B. This latter figure will repay careful examination. The arrow, ar., plunges into the third ventricle, behind the great middle commissure (m.c.), and the barb is supposed to lie under the roof of the mid-brain, the corpora quadrigemina (c.q.). The position of ar. is also indicated in Figure 1. Before reading on, the beginner should stop a while here; he should carefully copy or trace our figures and, putting the book aside, name the parts, and he should then recopy, on an enlarged scale, and finally draw from memory, correct, and again draw. By doing this before the brain is dissected a considerable saving of time is possible.
Section 124. Proceeding from the brain are twelve pairs of cranial nerves. From the fore-brain spring two pairs, which differ from the rest of the cranial nerves in being, first of all, hollow outgrowths of the brain-- the others are from the beginning solid. The first nerve is the olfactory lobe, which sends numerous filaments through the ethmoid bone to the olfactory organ. The second is the optic nerve, the visual sensory nerve.
Section 125. The mid-brain gives rise to only one nerve, the third, which supplies all the small muscles of the eye (see Section 114), except the superior oblique and external rectus.
Section 126. The remainder of the nerves spring from the hind-brain. The fourth pair supply the superior obliques, and the sixth the external recti; so that III., IV., and VI. are alike purely motor nerves, small and distributed, to the orbit. The fifth nerve, the trigeminal, is a much larger and more important one; it is a mixed nerve, having three main branches, of which the first two are chiefly sensory, the third almost entirely motor; it lies deeply in the orbit. V1 (see Sheet 9) runs up over the recti behind the eyeball, it is the ophthalmic branch; V2, the maxillary branch, runs deeply under the eyeball and emerges in front of the malar, and V3, the mandibular branch, runs down on the inner side of the jaw-bone to the jaw muscles and tongue.
Section 127. If the student will now recur to the figures of the dog's skull (Sheet 6), he will see certain apertures indicated in the cranial wall. Of these, o.f. is the optic foramen for the exit of nerve II., perforating the orbito-sphenoid. Behind this there comes an irregular aperture, (f.l.a.), the foramen lacerum anterius, giving exit to III., IV., VI., and V1. V2 emerges from the foramen rotundum, and V3 from the foramen ovale, two apertures uniting behind a bony screen.* Just in front of the bulla is a foramen lacerum medium (f.l.M.), through which no nerve passes.
* In the rabbit's skull f.l. anterius, the foramen rotundum, and foramen ovale are not distinct, and there are two condylar foramina instead of one, through each of which, a moiety of XII. passes.
Section 128. The eighth nerve (auditory) is purely sensory, the nerve of the special sense of hearing; it runs into the periotic bone, and breaks up on the labyrinth. The seventh nerve (facial) is almost entirely motor; it passes through the periotic anterior to VIII., and emerges by the stylo-mastoid foramen (s.m.f.) behind the bulla, to run outside the great jaw muscle across the cheek immediately under the skin (Figure 1).
Section 129. The ninth (glossopharyngeal) nerve is chiefly sensory; it is the special nerve of taste, and is distributed to the tongue. The tenth nerve (vagus) arises by a number of roots, and passes out of the skull, together with IX and XI, by the foramen lacerum -posterium- [posterius] (f.l.p.). It is a conspicuous white nerve, and runs down the neck by the side of the common carotid artery. It sends a superior laryngeal branch (Xa) to the larynx. The left vagus passes ventral to the aortic arch, and sends a branch (l.x.b.) under this along the trachea to the larynx-- the recurrent laryngeal nerve. The corresponding nerve on the right (r.x.b.) loops under the subclavian artery. The main vagus, after this branching, passes behind the heart to the oesophagus and along it to the stomach. XI., the spinal accessory, supplies certain of the neck nerves. XII., the hypoglossal, runs out of the skull by the condylar foramen (c.f.), is motor, crosses the roots of XI., X., and IX., passes ventral to the carotid, and breaks up among the muscles of the tongue and neck.
Section 130. Of the functions of the several parts of the brain there is still very considerable doubt. With disease or willful destruction of the cerebral tissue the personal initiative is affected-- the animal becomes more distinctly a mechanism; the cerebellum is probably concerned in the coordination of muscular movements; and the medulla is a centre for the higher and more complicated respiratory reflexes, yawning, coughing, and so on. The great majority of reflex actions centre, however, in the spinal cord, and do not affect the brain.
Section 131. A cross section of the spinal cord is shown in Figure 6, Sheet 8. It is a cylinder, almost bisected by a dorsal (d.f.) and a ventral (v.f.) fissure. Through its centre runs a central canal (c.c.), continuous with the brain ventricles, and lined by ciliated epithelium. The spinal cord consists of an outer portion, mainly of nervous fibres, the white matter, and of inner, ganglionated, and more highly vascular grey matter. (In the cerebrum the grey matter is external, and the white internal.) The cord, like the brain, is surrounded by a vascular fibrous investment, and protected from concussion by a serous fluid. The nerves which emerge from the vertebral column between the vertebrae, arise, unlike the cranial nerves, by two roots. The dorsal of these, the sensory root (d.n.), has a swelling upon it, the dorsal ganglion, and-- by experiments upon living animals-- has been shown to contain only afferent fibres; the ventral, the motor root, is without a ganglion, and entirely or mainly motor. The two unite outside the cord, and thereafter the spinal nerves are both sensory and motor.
Section 132. Besides the great mass of brain and spinal cord (cerebro-spinal axis), there is, on either side of the dorsal wall of the body cavity, a sympathetic nervous chain. The nerve fibres of this system, like the nerve fibres of invertebrates, are non-medullated. It may be seen as a greyish thread running close by the common carotid in the neck (sym., Figure 1); it then runs over the heads of the ribs in the thorax and close beside the dorsal aorta in the abdominal region. In the anterior region of the neck it dilates to form a superior cervical ganglion, and opposite the first rib it forms an inferior cervical ganglion. Thence, backwards, there is a ganglion on each sympathetic chain opposite each spinal nerve, and the two exchange fibres through a thread, the ramus communicans. To the sympathetic chain is delegated much of the routine work of reflex control of the bloodvessels and other viscera, which would otherwise fall upon the spinal cord.
Section 133. There are eight cervical (spinal) nerves, one in front of the atlas, and one behind each of the cervical vertebrae. The last four and the first thoracic (spinal) contribute to a leash of nerves running out to the fore limb, the brachial plexus (plexus, literally network, but here meaning a plaited cord). The fourth cervical also sends down a phrenic nerve (p.n., Figure 1), along by the external jugular vein and the superior caval vein to the diaphragm. The last three lumbar and the sacral nerves form a sacral plexus, supplying the hind limb.
Section 134. From the sympathetic in the hinder region of the thorax a nerve, the great splanchnic nerve, arises, and runs, back to a ganglionated nervous network, just behind the coeliac artery, into which the vagus also enters; this is the coeliac ganglion, and together with a similar superior mesenteric ganglion around the corresponding artery, makes up a subsidiary visceral nervous network, the solar plexus. A similar and smaller nervous tangle, bearing an inferior mesenteric ganglion, lies near the inferior mesenteric artery.
Section 135. Finally, we may note the pineal gland and the pituitary body, as remarkable appendages above and below the thalamencephalon. Their function, if they have a function, is altogether unknown. Probably, they are inherited from ancestors to whom they were of value. Such structures are called reduced or vestigial structures, and among other instances are the clavicles of the rabbit, the hair on human limbs, the little pulpy nodule in the corner of the human eye, representing the rabbit's third eyelid, and the caudal vertebrae at the end of the human spinal column. In certain lowly reptiles, in the lampreys, and especially in a peculiar New Zealand lizard, the pineal gland has the most convincing resemblance to an eye, both in its general build and in the microscopic structure of its elements; and it seems now more than probable that this little vascular pimple in our brains is a relic of a third and median eye possessed by ancestral vertebrata. The pituitary body is probably equivalent to a ciliated pit we shall describe in the lancelet (Amphioxus).
Section 136. We have now really completed our survey of the individual animal's mechanism. But no animal that was merely complete in itself would be long sanctioned by nature. For an animal species to survive, there must evidently, also, be proper provision for the production of young, and the preservation of the species as well as of the individual. Hence in an animal's physiology and psychology we meet with a vast amount of unselfish provision, and its structure and happiness are more essentially dependent on the good of its kind than on its narrow personal advantage. The mammalia probably owe their present dominant position in the animal kingdom to the exceptional sacrifices made by them for their young. Instead of laying eggs and abandoning them before or soon after hatching, the females retain the eggs within their bodies until the development of the young is complete, and thereafter associate with them for the purposes of nourishment, protection, and education. In the matter of the tail, for instance, already noted, the individual rabbit incurs the disadvantage of conspicuousness for the rear, in order to further the safety of the young.
Section 137. The female organs of reproduction are shown in Sheet 10. The essential organ is the ovary (ov.), in which the ova (eggs) are formed. Figure 3 gives an enlarged and still more diagrammatic rendering of the ovary. There is a supporting ground mass, or stroma, into which numerous bloodvessels and nerves enter and break up. The ova appear first as small cells in the external substance of the ovary (as at 1), and move inward (2 and 3), surrounded by a number of sister cells, which afford them nourishment. At (4) an ovum with its surrounding group of cells is more distinct and near the centre of the ovary; a fluid is appearing within the ovisac as the development proceeds. (5) is a much more mature ovisac or Graafian follicle.
Section 138. The ovum (ov.), is now large, and its nucleus and nucleolus (the germinal vesicle and spot) are very distinct. The wall of the follicle consists, in the mammal, of several layers of cells, the membrana granulosa (or "granulosa" simply); the ovum lies on its outer side embedded in a mass of cells, discus proligerus, separated from actual contact with the ovum by a zona pellucida. The ripening follicle moves to the surface of the ovary and bursts, the ovum falls into the body cavity. In Figure 2, a ripe Graafian follicle (G.F.), projects upon the ovary.
Section 139. The liberated ovum is caught up by the funnel-shaped opening of the Fallopian tube, which passes without any very conspicuous demarcation into the cornu uteri (c.ut.) of its side; the two uterine cornua meeting together in the middle line form the vagina (V.), which runs out into a vestibule (vb.) opening between tumid lips to the exterior. The urinary bladder (ur.b.) also opens into the vestibule, and receives the two ureters from the kidney.
Section 140. In the male we find, in the position of the female uterus, a uterus masculinus (u.m.). The essential sexual organ is the testis (T.), a compact mass of coiling tubuli, which opens by a number of ducts, the vasa efferentia, into a looser and softer epididymis (ep.), which sends the sexual product onward through a vas deferens (v.d.), to open at the base of the uterus masculinus. The urinary bladder and ureters correspond with those of the female, and the common urogenital duct (= vestibule), the urethra, is prolonged into an erectile penis (P.) surrounded by a fold of skin, the prepuce. A prostate gland (pr.), contributes to the male sexual fluid. The character of the essential male element, the spermatozoon, the general nature of the reproductive process, will be conveniently deferred until the chapters upon development are reached.
Section 141. The following facts of classificatory importance may now be considered, but their full force will be better appreciated after the study of other vertebrate types. They are such as come prominently forward in the comparison of the rabbit with other organisms.
Section 142. In the first place, the rabbit is a metazoon, one of the metazoa, i.e., a multicellular organism, as compared with the amoeba, which belongs to the protozoa or one-cell animals (Section 55). In the next place, it is externally bilaterally symmetrical, its parts balance, and where, in its internal anatomy, it departs from this symmetry (as in the case of the aorta, the stomach and intestines, and the kidneys), the departure has an appearance of being the results of partial reductions and distortions of an originally quite symmetrical plan. And the facts of development strengthen this idea; in the very earliest stages we have paired aortic arches, of which, the left only remains, a straight alimentary canal, and less asymmetrical kidneys. In the vast majority of animals the same bilateral symmetry is to be seen, but in the star-fish and sea-urchins, and in the jelly-fish, corals, sea anemones, and hydra, the general form of the animal is, instead, arranged round a centre, like a star and its rays, and the symmetry is called radial.
Section 143. We also see in various organs of the rabbit, and especially in the case of the limbs and vertebral column, what is called metameric segmentation, that is, a repetition of parts, one behind the other, along the axis of the body. Thus the bodies and arches of the vertebrae repeat each other, and so do the spinal nerves. The renal organ of the rabbit, some time before birth, displays a metameric arrangement of its parts; but this disappears, as development proceeds, into the compact kidney of the adult. But the metameric segmentation in the rabbit's organism is not nearly so marked as that of an earthworm, for instance, which is visibly a chain of rings. If the student wants a perfect figure of metameric segmentation he should think of a train of precisely similar carriages, or a string of beads. One bead, one carriage, one vertebra, would be a metamere.
Section 144. In contrast to metameric segmentation is the antimeric repetition of radial symmetry (Section 142), in which each ray of the star is called an antimere. It is possible to have bilateral symmetry without a metameric arrangement of parts, as in the mussel and the cuttle-fish; but metameric segmentation without complete or reduced bilateral symmetry does not occur.
Section 145. We are now in a position to appreciate the fact that the old and more popularly know division of animals into vertebrata and invertebrata scarcely represents the facts of the case, that the primary division should be into protozoa and metazoa, and that the vertebrata are one of several groups of metazoa with a fundamental bilateral symmetry and imperfect metameric segmentation.
The rabbit is one of the vertebrata, and, in common with all the other animals collected under this head, it has--
(a) A skeletal axis (the vertebral column) between its central nervous system and its body cavity. In the adult rabbit this consists of a chain of vertebrae, but in the embryo (i.e., the young rabbit before birth) it is represented by a continuous chord, the notochord, and it remains as such in some of the lowest vertebrata throughout life. In other words, in these lower vertebrata, the vertebral axis is not metameric.
(b) A dorsal and -Tubular_ nervous axis. (Section 131, the central canal)
(c) It has, though in the embryo only, certain slits between the throat and the exterior, like the gill slits of a fish. Such slits are-- with one or two remarkable exceptions outside the sub-kingdom-- distinctly vertebrate features, and remain, of course, in fishes throughout life.
The presence of true cartilage and bone mark a vertebrate, but vertebrata occur in which -these tissues- [bone] -are- [is] absent.
Section 146. The rabbit shares the following features with all the vertebrata, except the true fishes, which do not possess any of them--
(a) Lungs (but many fish have a swimming bladder which answers to the lungs in its anatomical relations.)
(b) Limbs which consist of a proximal joint of one bone an intermediate part of two, and a distal portion which has five digits, or is evidently a reduced form of the five-digit limb.*
(c) The absence of a median fin supported by fin rays.**
* The frog shows indications of a sixth digit.
** The frog's tadpole has a median fin, but no fin rays.
Section 147. The rabbit shares the following features with all the vertebrata above the fishes and amphibia (= frogs, toads, newts, and etc.)--
(a) Absence of gills (not gill slits, note) at any stage in development.
(b) An amnion, and
(c) An allantois in development.
The meaning of (b) and (c) we shall explain to the student in the chapters on embryology. We simply mention them here to render our table complete.
Section 148. The rabbit shares with all mammals, and differs from all other vertebrata (i.e., birds, reptiles, amphibia, and fishes), in having--
(a) Hair.
(b) A diaphragm.
(c) Only one aortic arch, and that on the left side of the body.
(d) Its young born alive. (But two very reptile-like mammals of Australia, the duck-billed platypus and the echidna, lay eggs, and certain fish and reptiles bear living young.)
(e) Epiphyses to its vertebral -centre- [centra].*
(f) The cerebral hemispheres covering the mid-brain.
(g) Corpora quadrigemina instead of bigemina.
[(h) A corpus callosum.]
[(i) A spirally coiled cochlea to the internal ear.]
[(In respect to h and i also, the echidna and platypus are scarcely mammalian.)]
* But certain mammals have no such epiphyses.
Section 149. The rabbit, together with the hares and conies, rats and mice, voles, squirrels, beavers, cavies, guineapigs is included in that order of the class of mammals which is called the rodentia, and is distinguished by the character of the incisor teeth from other orders of the class.
[Many of the above questions were actually set at London University Examinations in Biology.] {In Both Editions.}
Section 1. We will now study the adult anatomy of the frog, and throughout we shall make constant comparisons with that of the rabbit. In the rabbit we have a distinctly land-loving, burrowing animal; it eats purely vegetable food, and drinks but little. In the frog we have a mainly insectivorous type, living much in the water. This involves the moister skin, the shorter alimentary canal, and the abbreviated neck (Rabbit, Section 2) of the frog; the tail is absent-- in a fish it would do the work the frog accomplishes with his hind legs-- and the apertures which are posterior in the rabbit, run together into one dorsal opening, the cloaca. There is, of course (Rabbit, Section 4), no hair the skin is smooth, and an external ear is also absent. The remarkable looseness of the frog's skin is due to great lymph spaces between it and the body wall.
Section 2. If we now compare the general anatomy of the frog (vide Sheet 11) with that of the rabbit, we notice that the diaphragm is absent (Rabbit, Section 4), and the body cavity, or coelom, is, with the exception of the small bag of the pericardium round the heart, one continuous space. The forked tongue is attached in front of the lower jaw, and can be flicked out and back with great rapidity in the capture of the small insects upon which the frog lives. The posterior nares open into the front of the mouth-- there is no long nasal chamber, and no palate, and there is no long trachea between the epiglottis and the lungs. The oesophagus is less distinct, and passes gradually, so far as external appearances go, into the bag-like stomach, which is much less inflated and transverse than that of the rabbit. The duodenum is not a U-shaped loop, but makes one together with the stomach; the pancreas lies between it and the stomach, and is more compact than the rabbit's. There is no separate pancreatic duct, but the bile duct runs through the pancreas, and receives a series of ducts from that gland as it does so. The ileum is shorter, there is no sacculus rotundus, and the large intestine has no caecum, none of the characteristic sacculations of the rabbit's colon, and does not loop back to the stomach before the rectum section commences. The anus opens not upon the exterior, but into a cloacal chamber. The urinary and genital ducts open separately into this cloaca, and dorsally and posteriorly to the anus. The so-called urinary bladder is ventral to the intestine, in a position answering to that of the rabbit, but it has no connection with the ureters, and it is two-horned.
Section 3. The spleen is a small, round body, not so intimately bound to the stomach as in the rabbit, but in essentially the same position.
Section 4. Much that we knew of the physiology of the frog is arrived at mainly by inferences from our mammalian knowledge. Its histology is essentially similar. Ciliated epithelium is commoner and occurs more abundantly than in the rabbit, in the roof of the mouth for instance, and its red blood corpuscles are much larger, oval, and nucleated.
Section 5. The lungs of the frog are bag-like; shelves and spongy partitions project into their cavities, but this structure is much simpler than that of the rabbit's lung, in which the branching bronchi, the imperfect cartilaginous rings supporting them, alveoli, arteries and veins, form together a quasi-solid mass.
Section 6. The mechanism of respiration is fundamentally different from that of the mammal. The method is as follows:-- The frog opens its anterior nares, and depresses the floor of the mouth, which therefore fills with air. The anterior nares are then closed, and the floor mouth rises and forces the air into the lungs-- the frog, therefore, swallows its air rather than inhales it. The respiratory instrument of the rabbit is a suction pump, while that of the frog is a "buccal force pump."
Section 7. The heart is not quadrilocular (i.e., of four chambers), but trilocular (of three), and two structures, not seen in Lepus, the truncus arteriosus and the sinus venosus, into the latter of which the venous blood runs before entering the right auricle, are to be noted. The single ventricle is blocked with bars of tissue that render its interior, not an open cavity, but a spongy mass. Figure 2, Sheet 11, shows the heart opened; l.au. and r.au. are the left and right auricles respectively; the truncus arteriosus is seen to be imperfectly divided by a great longitudino-spiral valve (l.s.v.); p.c. is the pulmo-cutaneous artery -going to the lungs- [supplying skin and lungs]; d.ao., the dorsal aorta [furnishing the supply of the body and limbs]; and c.a. the carotid artery going to the head; all of which vessels (compare Figure 1) are paired.
Section 8. It might be inferred from this that pure and impure blood mix in the ventricle, and that a blood of uniform quality flows to lungs, head, and extremities; but this is not so. The spongy nature of the ventricle sufficiently retards this mixing. It will be noted that the opening of pulmonary arteries lies nearest to the heart, next come the aortic and carotid arches, which have a common opening at A. Furthermore, at c.g.l. [the carotid artery, repeatedly divides to form a close meshwork of arterioles, the carotid gland, forming a sponge-like plug in this vessel.] is a spongy mass of matter, the carotid gland inserted upon the carotid. Hence the pulmonary arteries yawn nearest for the blood, and, being short, wide vessels, present the least resistance to the first rush of blood-- mainly venous blood for the right auricle. As they fill up, the back resistance in them becomes equal, and then greater, than the resistance at A, and the rush of blood, now of a mixed quality passes through that aperture. It selects the dorsal aorta, because the carotid arch, plugged by the carotid gland, offers the greater resistance. Presently, however, the back resistance of the filled dorsal aorta rises above this, and the last flow of blood, from the ventricular systole-- almost purely oxygenated blood for the left auricle-- goes on towards the head.
Section 9. At the carotid gland the carotid artery splits into -an- [a] -external carotid- [lingual] (e.c.), and a deeper internal carotid. The dorsal aorta passes round on each side of the oesophagus, as indicated by the dotted lines in Figure 2, Sheet 11, and meets its fellow dorsal to the liver. Each arch gives off subclavian arteries to the limbs, and the left, immediately before meeting the right, gives off the coeliaco-mesenteric artery [to the alimentary canal]. This origin of the coeliaco-mesenteric artery a little to the left, is the only asymmetry (want of balance) in the arterial system of the frog, as contrasted with the very extensive asymmetry of the great vessels near the heart of the rabbit. [Posteriorly the dorsal aorta forks into two common iliac arteries (right and left) supplying the hind limbs.]
Section 10. Figure 3 gives a side view of the frog, to display the circulation.
{Lines from Second Edition only.}
[The venous return to the heart, as in the rabbit, is by paired venae cavae anteriores and by a single vena cava inferior. The factors of the anterior cava on either side are an external jugular (ex.j.) an innominate vein (in.v.) and subclavian (scl.v.). The last receives not only the brachial vein (b.v.) from the fore limb, but also a large vein bringing blood for the skin, the cutaneous (p.v.). The innominate vein has also two chief factors, the internal jugular (l.i.j.v.) and the subscapular (s.s.v.). The blood returns from each hind limb by a sciatic (l.sc.) or femoral (f.m.) vein, and either passes to a renal portal vein (l.r.p.), which breaks into capillaries in the kidney, or by a paired pelvic vein (l.p.v. in Figures 1 and 3) which meets its fellow in the middle line to form the anterior abdominal vein (a.ab.v.) going forward and uniting with the (median) portal vein (p.v.) to enter the liver.]
-The vessels are named in the references to the figure, which should be carefully copied and mastered. Here we need only- [Comparing with the rabbit, we would especially] call attention to the fact that the vena cava inferior extends posteriorly only to the kidney, and that there is a renal portal system. The blood from the hind limbs either flows by the anterior abdominal vein to the portal vein and liver, or it passes by the renal portal vein to the kidney. There the vein breaks up, and we find in the frog's kidney, just as we find in the frog's and rabbit's liver, a triple system of (a) nutritive arterial, (b) afferent* venous and (c) efferent** venous vessels.
* a, ad = to;
** e, ex = out of.
{This Section missing from Second Edition.}
-Section 11. It is not very improbable that the kidney of the frog
shares, or performs, some of the functions of the rabbit's liver, or
parallel duties, in addition to the simply excretory function. Since
specialization of cells must be mainly the relatively excessive
exaggeration of some one of the general properties of the
undifferentiated cell, it is not a difficult thing to imagine a gradual
transition, as we move from one organism to another, of the functions
of glands and other cellular organs. It is probable that the mammalian
kidney is, physiologically, a much less important (though still quite
essential) organ than the structures which correspond to it in position
and development in the lower vertebrate types.-
Section 12. The lymphatic system is extensively developed in the frog, but, in the place of a complete system of distinctly organized vessels, there are great lymph sinuses (compare Section 1). In Figure 5, Sheet 12, the position of two lymph hearts (l.h., l.h.) which pump lymph into the adjacent veins, is shown.
Section 13. The skull of the frog will repay a full treatment, and will be dealt with by itself later. The vertebral column (Sheet 12) consists of nine vertebrae, the centra of which have faces, not flat, but hollow in front (pro-coelous), and evidently without epiphyses (compare the Rabbit). The anterior is sometimes called the atlas, but it is evidently not the homologue of the atlas of the rabbit, since the first spinal nerve has a corresponding distribution to the twelfth cranial of the mammal, and since, therefore, it is probable that the mammalian skull = the frog's skull + one (or more) vertebrae incorporated with it. Posteriorly the vertebral column terminates in the urostyle, a calcified unsegmented rod. The vertebrae have transverse processes, but no ribs.
Section 14. The fore-limb (Figure 6, Sheet 12) consists of an upper segment of one bone, the humerus, as in the rabbit; a middle section, the radius and ulna, fused here into one bone, and not, as in the mammalian type, separable; of a carpus, and of five digits, of which the fourth is the longest. The shoulder girdle is more important and complete than that of the higher type. There is a scapula (sc.) with an unossified cartilaginous supra-scapula (s.sc.); the anterior border of the scapula answers to the acromion. On the ventral side a cartilaginous rod, embraced by the clavicle (cl.) (a membrane bone in this type), runs to the sternum, and answers to the clavicle of the rabbit. In the place of the rabbit's coracoid process, is a coracoid bone (co.), which reaches from the glenoid cavity to the sternum; it is hidden on the right side of Figure 6, which is a dorsal view of the shoulder girdle. There is a pre-omosternum (o.st.) and a post-omosternum, sometimes termed a xiphisternum (x.).
Section 15. Figure 7 shows the pelvic girdle and limb of the frog. There is a femur (f.); tibia and fibula (t. and f.) are completely fused; the proximal bones of the tarsus, the astragalus (as.), and calcaneum (cal.) are elongated, there are five long digits, and in the calcar (c.) an indication of a sixth. With considerable modifications of form, the three leading constituents of the rabbit's pelvic girdle occur in relatively identical positions. The greatly elongated ilium (il.) articulates with the single (compare Rabbit) sacral vertebra (s.v. in Figure 5). The ischium (is.) is relatively smaller than in the rabbit, and the pubis (pu.) is a ventral wedge of unossified cartilage. The shape of the pelvic girdle of the frog is a wide departure from that found among related forms. In connection with the leaping habit, the ilia are greatly elongated, and the pubes and ischia much reduced. Generally throughout the air-frequenting vertebrata, we find the same arrangement of these three bones, usually in the form of an inverted. Y-- the ilium above, the ischium and pubis below, and the acetabulum at the junction of the three.
Section 16. The uro-genital organs of the frog, and especially those of the male, correspond with embryonic stages of the rabbit. In this sex the testes (T., Sheet 13) lie in the body cavity, and are white bodies usually dappled with black pigment. Vasa efferentia (v.e.) run to the internal border of the anterior part of the kidney, which answers, therefore, to the rabbit's epididymis. The hinder part of the kidney is the predominant renal organ. There is a common uro-genital duct, into which a seminal vesicle, which is especially large in early spring, opens. This is the permanent condition of the frog. In the rabbit, for urogenital duct, we have ureter and vas deferens; the testes and that anterior part of the primitive kidney, the epididymis, shift back into the scrotal sacs, and the ureters shift round the rectum and establish a direct connection with the bladder, carrying the genital ducts looped over them. The oviducts of the female do not fuse distally to form a median vagina as they do in the rabbit. In front of the genital organ in both sexes is a corpus adiposum (c.ad.), which acts as a fat store, and is peculiar to the frogs and toads. The distal end of the oviduct of the female is in the breeding season (early March) enormously distended with ova, and the ovaries become then the mere vestiges of their former selves. The distal end of the oviduct is, therefore, not unfrequently styled the uterus. There is no penis in the male, fertilisation of the ova occurring as they are squeezed out of the female by the embracing fore limbs of the male. The male has a pad, black in winter, shown in Figure 1, which is closely pressed against the ventral surface of the female in copulation, and which serves as a ready means of distinguishing the sex.
Section 17. The spinal cord has a general similarity to that of the rabbit; the ratio of its size to that of the brain is larger, and the nerves number ten pairs altogether. The first of these (sp. 1, in Figure 2, Sheet -12- ) {First Edition error.} [13] corresponds in distribution with the rabbit's hypoglossal nerve, a point we shall refer to again when we speak of the skull. The second and third constitute the brachial plexus. The last three form the sciatic plexus going to the hind limb.
Section 18. The same essential parts are to be found in the brain of both frog and rabbit, but in the former the adult is not so widely modified from the primitive condition as in the latter. The fore-brain consists of a thalamencephalon (th.c. and 1), which is exposed in the dorsal view of the brain, and which has no middle commissure. The cerebral hemispheres (c.h.) are not convoluted, do not extend back to cover parts behind them, as they do in the rabbit, and are not connected above the roof of the thalamencephalon by a corpus callosum. Moreover, the parts usually regarded, as the olfactory lobes (rh.) fuse in the middle line. The mid-brain gives rise to the third nerve, and has the optic lobes on its dorsal side, but these are hollow, and they are not subdivided by a transverse groove into corpora quadrigemina, as in the rabbit. In the hind-brain the cerebellum is a mere band of tissue without lateral lobes or flocculi, and the medulla gives origin only to nerves four to ten; there is no eleventh nerve, and the hypoglossal is the first spinal-- from which it has been assumed that the rabbit's medulla equals that of the frog, plus a portion of the spinal cord incorporated with it. The hypoglossal is very distinctly seen on opening the skin beneath the hyoid plate.
Section 19. The first, second, third, and fourth cranial nerves of the frog correspond with those of the rabbit in origin and distribution. So do five, six and eight. The seventh nerve forks over the ear-drum-- the larger branch emerging behind it and running superficially, as shown in Figure 4. There is also a deeper palatine branch of VII. (P.) running under V2 and V3 below the orbit, and to be seen together with V1 and V2 after removal of the eyeball. The ninth nerve similarly forks over the first branchial slit of the tadpole, and evidence of the fork remains in the frog. It is seen curving round anterior to the hypoglossal nerve, and lying rather deeper in dissection. The vagus (tenth) nerve is distributed to heart, lungs, and viscera-- in the tadpole it also sends for forking branches over the second, third, and fourth branchial slits. It lies deeper than IX., and internal to the veins, and runs close beside the cutaneous artery. Most of these nerves are easily dissected and no student should rest satisfied until he has actually seen them.
Section 20. The sympathetic chain is closely connected with the aorta. It is, of course, paired, and is easily found in dissection by lifting the dorsal aorta and looking at its mesentery. In the presence of ganglia corresponding to the spinal nerves, and of rami communicantes, it resembles that of the rabbit.
Section 21. The whole of this chapter is simply a concise comparison, of frog and rabbit. In addition to reading it, the student should very carefully follow the annotations to the figures, and should copy and recopy these side by side with the corresponding diagrams of the other types.
Section 22. We have already given a description of the mammalian skull, and we have stated where the origin of the several bones was in membrane, and where in cartilage; but a more complete comprehension of the mammalian skull becomes possible with the handling of a lower type. We propose now, first to give some short account of the development and structure of the skull of the frog, and then to show briefly how its development and adult arrangement demonstrate the mammalian skull to be a fundamentally similar structure, complicated and disguised by further development and re-adjustment.
Section 23. Figure 1,I. Sheet 14, shows a dorsal view of a young tadpole cranium; the brain has been removed, and it is seen that it was supported simply upon two cartilaginous rods, the trabeculae cranii (tr.c.). Behind these trabeculae comes the notochord (n.c.), and around its anterior extremity is a paired tract of cartilage, the parachordals (p.c.). These structures, underlying the skull, are all that appear[s] at first of the brain box. In front, and separate from the cranium, are the nasal organs (n.c.); the eyes lie laterally to the trabeculae, and laterally to the parachordals are two tracts of cartilage enclosing the internal ear, the otic capsules.
Section 24. Figure 1, II., is a more advanced, phase of the same structures. The trabeculae have met in front and sent forward a median (c.t.) and lateral parts (a.o.) to support the nasal organs. They have also flattened, out very considerably, and have sent up walls on either side of the brain to meet above it and form an incomplete roof (t.) over it. The parachordals have similarly grown up round, the hind-brain and formed a complete ring, the roof of which is indicated, by b. Further, the otic capsules are fusing with the brain-case. With certain differences of form these elements-- the trabeculae, the parachordals, and the otic capsules, are also the first formed structures of the mammalian cranium.
Section 25. In Figures 1,I. and II., there appears beneath the eye a bar of cartilage (p.p.), the palato-pterygoid cartilage, which is also to be seen from the side in Figures 8,I. and III. It will be learnt from these latter that this bar is joined in front to the cranium behind the nasal organ, and behind to the otic capsule. The cartilaginous bar from the palato-pterygoid to the otic capsule is called the quadrate, and at the point of junction, at the postero-ventral angle of the palato-pterygoid, articulates with the cartilaginous bar which is destined to form the substratum of the lower jaw-- Meckel's cartilage (M.c. in Figure 8,I.).
Section 26. Figure 2 shows a dorsal view of these structures in a young frog. The parts corresponding to these in 1,II. will be easily made out, but now ossification has set in at various points of this cartilaginous cranium. In front of the otic capsule is the paired pro-otic bone (p.o.); behind it at the sides of the parachordal ring is the paired ex-occipital (e.o.); in front of the cranium box, and behind the nasal capsules, is a ring of bone, the (median, but originally paired) sphenethmoid (s.e.). -A paired ossification appears in the palato-pterygoid cartilage the pterygoid bone (pt.), while- A splint of bone, the quadrato-jugal, appears at the angle of articulation with the lower jaw. These are all the cartilage bones that appear in the cranium and upper jaw of the frog.
Section 27. But another series of bones, developed first chiefly in dermal connective tissue, and coming to plate over the cranium of cartilage, are not shown in Figure 2. They are, however, in Figure 3. These membrane bones are: along the dorsal middle line, the parieto-frontals (p.f.), originally two pairs of bones which fuse in development, and the nasals (na.). Round the edge of the jaw, and bearing the teeth, are pre-maxillae (p.m.), and maxillae (mx.), and overlying the quadrate cartilage and lateral to the otic capsules are the T-shaped squamosal bones (sq.). In the ventral view of the skull (Figure 4) we see a pair of vomers (vo.) bearing teeth, a pair of palatines (pal.), [and a pair of pterygoids (pt.)] (which [palatines and pterygoids, we may note,] unlike those of the rabbit, are -stated to be- membrane bones), and a great median dagger-shaped para-sphenoid (p.sp.). These two Figures, and 5, which shows the same bones in side view, should be carefully mastered before the student proceeds with this chapter. The cartilage bones are distinguished from membrane bones by cross-shading.
Section 28. Turning now to Figure 8,I., we have a side view of a tadpole's skull. On the ventral side of the head is a series of vertical cartilaginous bars, the visceral arches supporting the walls of the tadpole's gill slits. The first of these is called the hyoid arch (c.h.), and the four following this, the first (br.1), second, third, and fourth (br.4), branchial arches. Altogether there are four gill slits and between the hyoid arch and the jaw arch, as it is called (= Meckel's cartilage + the palato-pterygoid), is "an imperforate slit," which becomes the ear-drum.* The frog no longer breathes by gills, but by lungs, and the gills are lost, the gill slits closed, and the branchial arches consequently much reduced. Figures 8, II., and 8, III., show stages in this reduction. The hyoid arch becomes attached, to the otic capsule, and its median ventral plate, including also the vestiges of the first, second, and fourth branchial arches, is called the hyoid apparatus. In Figure 5, the apparatus is seen from the side; c.h. is called the (right) anterior cornu** of the hyoid. The function of the hyoid apparatus in the frog is to furnish, a basis of attachment to the tongue muscles; it remains cartilaginous, with the exception of the relic of one branchial arch, which ossifies as the thyro-hyal (Figure 7 th.h.). It will be noted that, as development proceeds, the angle of the jaw swings backward, and the hyoid apparatus, shifts relatively forward. These changes of position are indicated in Figure 8, III., by little arrow-heads.
* We may note here that, comparing the ear of the frog with that of the rabbit, there is no external ear. There is, moreover, no bulla supporting the middle ear, and the tympanic membrane stretches between the squamosal in front and the anterior cornu of the hyoid behind. A rod-like columella auris replaces the chain of ear ossicles, and may, or may not, answer to the stapes alone, or even possibly to the entire series. In the internal ear there is no cochlea, and the otic mass is largely cartilaginous instead of entirely bony.
Section 29. Before proceeding to the comparison of the mammalian skull with this, we would strongly recommend the student thoroughly to master this portion of the work, and in no way can he do this more thoroughly and quickly than by taking a parboiled frog, picking off the skin, muscle, and connective tissue from its skull, and making out the various bones with the help of our diagrams.
Section 30. Figure 9 represents, in the most diagrammatic way, the main changes in form of the essential constituents of the cranio-facial apparatus, as we pass from the amphibian to the mammalian skull. F. is the frog from the side and behind; b.c. is the brain-case, o.c. the otic capsule, e. the eye, n.c. the nasal capsule, p.p. the palato-pterygoid cartilage, mx. the maxillary membrane bones, sq. the squamosal, and mb. the mandible. The student should compare with Figure 5, and convince himself that he appreciates the diagrammatic rendering of these parts. Now all the distinctive differences in form, from this of the dog's skull (D.), are reducible to two primary causes--
(1) The brain is enormously larger, and the brain-case is vastly
inflated, so that--
(a) the otic capsule becomes embedded in the brain-case wall;
(2) The maxilla anteriorly and the palatine posteriorly send down palatine plates that grow in to form the bony palate, cutting off a nasal passage (n.p.) from the mouth cavity (m.p.), and carrying the posterior nares from the front part of the mouth, as they are in the frog, to the pharynx. Hence the vomers of the dog lie, not in the ceiling of the mouth, but in the floor of this nasal passage.
(b) the palato-pterygoid rod lies completely underneath the brain-case instead of laterally to it;
(c) the squamosal tilts down and in, instead of down and out, and the lower jaw articulates with its outer surface instead of below its inner, and, moreover, with the enormous distention of the brain-case it comes about that the squamosal is incorporated with its wall.
Section 31. The quadrate cartilage of the frog is superseded by the squamosal as the suspensorium of the lower jaw. It is greatly reduced, therefore; but it is not entirely absent. In the young mammal, a quadrate cartilage can be traced, connected with the palato-pterygoid cartilage, and articulating with Meckel's cartilage. Its position is, of course, beneath the squamosal, and just outside the otic capsule. As development proceeds, the increase in size of the quadrate, does not keep pace with that of the skull structures. It loses its connection with the palato-pterygoid, and apparently ossifies as a small ossicle-- the incus of the middle ear. A small nodule of cartilage, cut off from the proximal end of Meckel's cartilage, becomes the malleus. The stapes would appear to be derived from the hyoid arch. Hence these small bones seem to be the relics of the discarded jaw suspensorium of the frog utilized in a new function. Considerable doubt, however, attaches to this interpretation-- doubt that, if anything, is gaining ground.
Section 32. The tympanic bulla of the dog is not indicated in Diagram 9, and it would appear to be a new structure (neomorph), not represented in the frog.
Section 33. Besides these great differences in form, there are important differences in the amount and distribution of centres of ossification of the skull of frog and mammal. There is no parasphenoid in the mammal*; and, instead, a complete series of ossifications, the median-, basi-, and pre-sphenoids, and the lateral ali- and orbito-sphenoids occur. The points can be rendered much more luminously in a diagram than in the text, and we would counsel the student to compare this very carefully with that of the Rabbit.
* Faint vestigial indications occur in the developing skulls of some insectivora.
Section 34.
-Cranium_
-Nasal_ (paired), -Vomer_ (paired)
-Upper Jaw_
-Fronto-Parietal_, Sphenethmoid Bone (median), Eye, Pro-otic Bone,
Otic Cartilage, Ex-occipital (paired)
-Para-sphenoid Bone_
-Pre-Maxilla_ (paired), -Palatine_ (paired), Pterygoid (paired),
<-Lower Jaw_
-Squamosal_, Quadrate Cartilage {To 1.}
-Maxilla_
1. Quadrato-Jugal
Mento-meckelian, -Dentary_, -Articulare- [-Angulo Splenial_]
Section 35. -Points especially- [Additional points] to be noticed are:
(1) The otic capsule (= periotic bone) of the dog ossifies from a number of centres, one of which is equivalent to the frog's prootic.
(2) The several constituents of the lower jaw are not to be distinguished in the adult mammal.
(3) The frog has no lachrymal bone.
Section 36. We are now in a position to notice, without any danger of misconception, what is called the segmental theory of the skull. Older anatomists, working from adult structure only, conceived the idea that the brain-case of the mammal represented three inflated vertebrae. The most anterior had the pre-sphenoid for its body, the orbito-sphenoids for its neural processes, and the arch was completed above by the frontals (frontal segment). Similarly, the basi-sphenoids, ali-sphenoids, and parietals formed a second arch (parietal segment), and the ex-, basi-, and supra-occipitals a third (occipital segment). If this were correct, in the frog, which is a more primitive rendering of the vertebrate plan, we should find the vertebral characters more distinct. But, as a matter of fact, as the student will perceive, frontal segment, parietal segment, and occipital segment, can no longer be traced; and the mode of origin from trabeculae and para-chordals show very clearly the falsity of this view. The vertebrate cranium is entirely different in nature from vertebrae. The origin of the parietals and frontals as paired bones in membrane reinforces this conclusion.
Section 37. But as certainly as we have no such metameric segmentation, as this older view implies, in the brain-case of the frog, so quite as certainly is metameric segmentation evident in its branchial arches. We have the four gill slits of the tadpole and their bars repeating one another; the hyoid bar in front of these is evidently of a similar nature; and that the ear drum is derived from an imperforate gill slit is enforced by the presence of an open slit (the spiracle) in the rays and dog-fish in an entirely equivalent position. Does the mouth answer to a further pair of gill slits, and is the jaw arch (palato-pterygoid + Meckel's cartilage) equivalent to the arches that come behind it? This question has been asked, and answered in the affirmative, by many morphologists, but not by any means by all. The cranial nerves have a curious similarity of arrangement with regard to the gill slits and the mouth; the fifth nerve forks over the mouth, the seventh forks over the ear drum, the ninth, in the tadpole and fish, forks over the first branchial slit, and the tenth is, as it were, a leash of nerves, each forking over one of the remaining gill slits. But this matter will be more intelligible when the student has worked over a fish type, and need not detain us any further now.
Section 38. See also Section 13 again, in which is the suggestion that the occipital part of the skull is possibly a fusion of vertebrae, a new view with much in its favour, and obviously an entirely different one from the old "segmental" view of the entire skull, discussed in Section 36.
[All these questions were actually set at London University Examinations.] {In Both Editions.}
Section 1. In the dog-fish we have a far more antique type of structure than in any of the forms we have hitherto considered. Forms closely related to it occur among the earliest remains of vertebrata that are to be found in the geological record. Since the immeasurably remote Silurian period, sharks and dog-fish have probably remained without any essential changes of condition, and consequently without any essential changes of structure, down to the present day. Then, as now, they dominated the seas. They probably branched off from the other vertebrata before bone had become abundant in the inner skeleton, which is consequently in their case cartilaginous, with occasional "calcification" and no distinct bones at all. Unlike the majority of fish, they possess no swimming bladder-- the precursor of the lungs; but in many other respects, notably in the uro-genital organs, they have, in common with the higher vertebrata, preserved features which may have been disguised or lost in the perfecting of such modern and specialized fish as, for instance, the cod, salmon, or herring.
Section 2. Comparing the general build, of a dog-fish with that of a rabbit, we notice the absence of a distinct neck, and the general conical form; the presence of a large tail, as considerable, at first, in diameter as the hind portion of the body, and of the first importance in progression, in which function the four paddle-shaped limbs, the lateral fins, simply co-operate with the median fin along the back for the purpose of steering; and, as a consequence of the size of the tail, we note also the ventral position of the apertures of the body. The anus, and urinary and genital ducts unite in one common chamber, the cloaca. Behind the head, and in front of the fore fin (pectoral fin), are five gill slits (g.s.) leading from the pharynx to the exterior. Just behind the eye is a smaller and more dorsal opening of the same kind, the spiracle (sp.). On the under side of the head, in front of the mouth, is the nasal aperture (olf.), the opening of the nasal sac, which, unlike the corresponding organ of the air-frequenting vertebrata, has no internal narial opening. There is, however, a groove running from olf. to the corner of the mouth, and this, closing, in the vertebrate types that live in air and are exposed to incessant evaporation of their lubricating secretions, constitutes the primitive nasal passage. The limbs are undifferentiated into upper, lower, and digital portions, and are simply jointed, flattened expansions.
Section 3. The skin of the dog-fish is closely set with pointed tooth-like scales, the placoid scales, and these are continued over the lips into the mouth as teeth. Each scale consists of a base of true bone, with a little tubercle of a harder substance, dentine, capped by a still denser covering, the enamel. The enamel is derived from the outer layer of the embryonic dog-fish, the epiblast, which also gives rise to the epidermis; while the dentine and bony base arise in the underlying mesoblast, the dermis. A mammalian tooth has essentially the same structure: an outer coat of enamel, derived from epiblast, overlies a mass of dentine, resting on bone, but the dentine is excavated internally, to form a pulpcavity containing blood-vessels and nerves. Most land animals, however, have teeth only in their mouths, and have lost altogether the external teeth which constitute the armour of the dog-fish. Besides the teeth there perhaps remain relics of the placoid scales in the anatomy of the higher vertebrata, in the membrane bones. How placoid scales may have given rise to these structures will be understood by considering such a bone as the vomer of the frog. This bone lies on the roof of the frog's mouth, and bears a number of denticles, and altogether there is a very strong resemblance in it to a number of placoid scales the bony bases of which have become confluent. In the salamander, behind the teeth-bearing vomers comes a similar toothed parasphenoid bone. The same bone occurs in a corresponding position in the frog, but without teeth. In some tailed amphibians the vomers and splenials are known to arise by the fusion of small denticles. These facts seem to point to stages in the fusion of placoid bases, and their withdrawal from the surface to become incorporated with the cranial apparatus as membrane bones, a process entirely completed in the mammalian type.
Section 4. The alimentary canal of the dog-fish, is a simple tube thrown into a Z shape. The mouth is rough with denticles, and has a fleshy immovable tongue on its floor. In the position of the Eustachian tube there is a passage, the spiracle (sp.), running out to the exterior just external to the cartilage containing the ear. The pharynx communicates with the exterior through five gill slits (g.s.), and has, of course, no glottis or other lung opening. There is a wide oesophagus passing into a U-shaped stomach (st.), having, like the rabbit's, the spleen (sp.) on its outer curvature. There is no coiling small intestine, but the short portion, receiving the bile duct (b.d.) and duct of the pancreas (pan.), is called the duodenum (d'dum.). The liver has large left (L.lv.) and right lobes, and a median lobe (M.lv.), in which the gall bladder (g.bl.) is embedded. The next segment of the intestine is fusiform, containing a spiral valve (Figure 4), the shelf of which points steeply forward; it is sometimes called the colon (co.). It is absorptive in function and probably represents morphologically, as it does physiologically, the greater portion of the small intestine. A rectal gland (r.g.) opens from the dorsal side into the final portion of the canal (rectum).
Section 5. The circulation presents, in many respects, an approximation to the state of affairs in the developing embryos of the higher types. The heart (Figure 3, Sheet -14- {Error in First Edition} [16]) is roughly, Z shape, and transmits only venous blood. It lies in a cavity, the pericardial cavity (P.c.c.), cut off by a partition from the general coelome. At one point this partition is imperfect, and the two spaces communicate through a pericardio-peritoneal canal (p.p.c.), which is also indicated by an arrow (p.p.) in the position and direction in which the student, when dissecting, should thrust his "seeker," in Figure 1 Sheet 15. A sinus venosus (s.v. in Figure 3, Sheet 16) receives the venous trunks, and carries the blood through a valve into the baggy and transversely extended -auricle- [atrium] (au.), whence it passes into the muscular ventricle (Vn.), and thence into the truncus arteriosus. This truncus consists of two parts: the first, the conus or pylangium (c.a.), muscular, contractile, and containing a series of valves; the second, the bulbus or synangium (b.a.), without valves and pulsatile. In the rabbit both sinus and truncus are absent, or merged in the adjacent parts of the heart.
Section 6. From the bulbus there branch, on either side, four arterial trunks, the first of which forks, so that altogether there are five afferent branchials (a.br.) taking blood to be aerated in the gills, here highly vascular filamentary outgrowths of the internal walls of the gill slits.
{Lines from Second Edition only.}
[There are altogether nine vascular outgrowths (demi-branchs), one on each wall of each gill slit except the last, on the hind wall of which there is none. (In the spiracle is a miniature demibranch, the pseudo-branch. This suggests that the spiracle is really a somewhat modified gill slit.)]
Four efferent branchials (e.br.) carry the aerated blood on to the dorsal aorta (d.ao.). A carotid artery runs forward to the head, and a hypo-branchial artery supplies the ventral side of the pharyngeal region. There are sub-clavian, coeliac, mesenteric, and pelvic arteries, and the dorsal aorta is continued through the length of the tail as the caudal artery (Cd.A.).
Section 7. A caudal vein (Cd.V.), bringing blood back from the tail, splits behind the kidneys (K.), and forms the paired renal portal veins (r.p.v.), breaking up into a capillary system in the renal organ. A portal vein brings blood from the intestines to the liver.
Section 8. Instead of being tubular vessels, the chief veins of the dog-fish are, in many cases, irregular baggy sinuses. Three main venous trunks flow into the sinus venosus. In the median line from behind comes the hepatic sinus (H.S.); and laterally, from a dorsal direction, the Cuvierian sinuses (C.S.) enter it. These, as the student will presently perceive, are the equivalents of the rabbit's superior cavae. They receive, near their confluence with the sinus venosus, the inferior jugular vein (I.J.V.). At their dorsal origin, they are formed by the meeting of the anterior (A.C.S.) and posterior (P.C.S.) cardinal sinuses. The anterior cardinal sinus -is, roughly, the equivalent of the internal jugular vein-, lies along dorsal to the gill slits (g.s.), and receives an orbital sinus from the eye. The posterior cardinal sinus receives a sub-clavian vein (s.c.v.) and a lateral vein (L.V.), and fuses posteriorly with its fellow in the middle line. This median fusion is a departure from the normal fish type. It must not be confused with the inferior cava, which is not found in the dog-fish, the [right] posterior cardinals representing the rabbit's azygos vein. A simplified diagram of the circulation of a fish is given in Figure 2, Sheet 16, and this should be carefully compared with the corresponding small figure given of the vascular system of our other types.
{Lines from Second Edition only.}
[The blood of the dog-fish resembles that of the frog.]
Section 9. The internal skeleton, as we have said, is entirely cartilaginous, and only those parts which are pre-formed in cartilage in the skeletons of the higher types are represented here. The spinal column consists of two types of vertebrae, the trunk, bearing short, distinct, horizontally-projecting ribs (r.), and the caudal. The diagrams of Figure 5 [(Sheet 18)] are to illustrate the structure of the centrum of a dog-fish vertebra; C is a side view, D a horizontal median section, A and B are transverse sections at the points indicated by -B and A- [A and B] respectively in Figure C. -(By an unfortunate slip of the pen in the figure, A was substituted for B; section A corresponds to line B, and vice versa.)- The vertebrae are hollowed out both anteriorly and posteriorly (amphi-coelous), and a jelly-like notochord runs through the entire length of the vertebral column, being constricted at the centres of the centra, and dilated between them. The neural arch above the centrum, and containing the spinal cord, is made up of neural plates (n.p.), and interneural plates (i.n.p.), completed above by a median neural spine (n.s.). In the caudal region, instead of ribs projecting outwardly, there are haemal processes, inclined downwards and meeting below, forming an arch, the haemal arch, containing the caudal artery and vein-- the vein ventral to the artery-- and resembling the neural arch, which contains the spinal cord above, in shape and size.
Section 10. The pectoral limb and girdle (Figure 4, Sheet 16) have only a very vague resemblance to the corresponding structures in the rabbit. The girdle (g.) is a transverse bar lying ventral to the pericardial wall, and sending up a portion (sc.), dorsal to the attachment of the limb, which answers to the scapula and supra-scapula of the forms above the fish. Three main cartilages, named respectively the pro- (p.p.), meso- (m.p.), and meta-pterygium, form the base of the limb. With these, smaller cartilaginous plates, rods, and nodules articulate, and form a flattened skeletal support for the fin.
Section 11. The pelvic girdle and limb (Figure 2, Sheet 15) are similar in structure, but the pro-pterygium and meso-pterygium are absent, and the cartilage answering to the meta-pterygium goes by the name of the basi-pterygium. In the male, but not in the female, the pelvic fins are united behind the cloaca, and there are two stiff grooved copulatory organs, the claspers (cl. in Figure 1), which have a cartilaginous support (cl.c.). These claspers form the readiest means of determining the sex of a specimen before dissection.
Section 12. The skull consists of a cartilaginous cranium, and of jaw and visceral arches. The cranium persists throughout life, in what closely resembles a transitory embryonic condition of the higher types. There is a nasal capsule (na.c.), a brain case proper, and lateral otic (auditory) capsules (ot.c.) containing the internal ear. (This should be compared with the frog's embryonic skull.) The upper jaw has a great bar of cartilage, the palato-pterygoid, as its sole support; the arch of premaxilla, maxilla, jugal, and squamosal-- all membrane bones-- is, of course, not represented. In the frog this bar of cartilage is joined directly to the otic capsule by a quadrate portion, but this is only doubtfully represented in the dog-fish by a nodule of cartilage in the pre-spiracular ligament (p.s.). The lower jaw is supported, by Meckel's cartilage (M.C.). The hyoid arch consists of two main masses of cartilage, the hyomandibular (h.m.), and the ceratohyal (c.h.); the former of these is tilted slightly forward, so that the gill slit between it and the jaw arch is obliterated below, and the cartilage comes to serve as the intermediary in the suspension of the jaw from the otic mass. There are five branchia[l] arches, made up pharyngo-, epi- and cerato-branchials, and the ventral elements fuse in the middle line to form a common plate of cartilage. Outside these arches are certain small cartilages, the extra branchials (ex.b.) which, together with certain small labials by the nostrils and at the sides of the gape, probably represent structures of considerably greater importance in that still more primitive fish, the lamprey. The deep groove figured lateral to the otic capsule is the connecting line of the orbital and anterior cardinal sinuses; the outline of the anterior cardinal sinus in this figure and in Figure 1 is roughly indicated by a dotted line.
Section 13. Figure 3a is a rough diagram of the internal ear-- the only auditory structure of our type (compare Rabbit, Sheet 7). To dissect out the auditory labyrinth without injury is a difficult performance, but its structure may be made out very satisfactorily by paring away successive slices of the otic mass. Such a section is shown by Figure 3b; through the translucent hyaline cartilage the utriculus and horizontal canal can be darkly seen. The ductus endolymphaticus (vide Rabbit) is indicated by a dotted line in our figure. It is situated internal to the right-angle between the two vertical canals, and reaches to the surface of the otic capsule.
Section 14. The brain shows the three primary vesicles much more distinctly than do our higher types. The fore-brain has large laterally separated olfactory lobes (rh.), there are relatively small "hemispheres" (pr.c.), the stalk of the pineal gland tilts forward, and the gland itself is much nearer the surface, being embedded in the cartilage of the brain case, and the pituitary body is relatively very large, and has lateral vascular lobes on either side. Following the usual interpretation of the parts, we find optic lobes (op.l.) as the roof of the mid-brain, and behind a very large, median, hollow, tongue-shaped cerebellum (c.b.). The medulla is large, and certain lateral restiform tracts (r.t.) therein, which also occur in the higher types, are here exceptionally conspicuous.
Section 15. The dog-fish has ten pairs of cranial nerves, corresponding to the anterior ten of the rabbit very closely, when we allow for the modification the latter has suffered through the conversion of some part of the spiracular cleft to an eardrum, and the obliteration of the post-hyoid branchial slits.
The first and second nerves are really brain lobes, and nerves of the special senses of smell and sight respectively.
The third (oculomotor), the fourth (patheticus), and the sixth (abducens) are distributed to exactly the same muscles of the eyeball as they are in the rabbit.
The fifth nerve, has, in the dog-fish, as in the rabbit, three chief branches. V.2 and V.3 fork over the mouth just as they do in the mammal; V.1 passes out of the cranium by a separate and more dorsal opening, and runs along a groove along the dorsal internal wall of the orbit, immediately beneath a similar branch of VII., which is not distinct in the rabbit. The grooves are shown in the figure of the cranium, Sheet 18; the joint nerve thus compounded of V. and VII. is called the ophthalmic (oph.). It is distributed to the skin above the nose and orbit. When the student commences to dissect the head of a dog-fish he notices over the dorsal surface of the snout an exudation of a yellowish jelly-like substance, and on removing the tough skin over this region and over the centre of the skull he finds, lying beneath it, a quantity of coiling simple tubuli full of such yellowish matter. These tubuli open on the surface by small pores, and the nerves terminate in hair-like extremities in their lining. These sense tubes are peculiar to aquatic forms; allied structures are found over the head and along a lateral line (see below) in the tadpole, but when the frog emerges from the water they are lost. They, doubtless, indicate some unknown sense entirely beyond our experience, and either only possible or only necessary when the animal is submerged.
In addition to the ophthalmic moiety mentioned above, the seventh nerve has a vidian branch (vid.) running over the roof of the mouth, and besides this its main branches fork over the spiracle, just as V. forks over the mouth, and as IX. and X. fork over gill clefts. This nerve in the rabbit is evidently considerably modified from this more primitive condition.
The eighth is the auditory nerve, as in the rabbit.
The ninth nerve forks over the first branchial cleft.
The tenth nerve is easily exposed by cutting down through the body wall muscles over the gill clefts, into the anterior cardinal sinus (A.C.S.). It gives off (a) branches forking over the posterior four gill slits, (b) a great lateral nerve running inward, and back through the body-wall muscle, and connected with a line of sense organs similar to those in the head, the lateral line, and (c) a visceral nerve curving round to the oesophagus and stomach. In dissection it becomes very evident that the tenth nerve is really a leash of nerves, each one equivalent to the ninth.
We may here call the attention of the reader to the fact of the singular resemblance of V., VII., IX., and the factors of X. That each has a ventral fork, we have already noticed. Each also (?IX.) has a dorsal constituent connected with the sense organs of the skin. The vidian branch of VII., however, is not evidently represented in the others.
Section 16. The coelom of the dog-fish is peculiar-- among the types we treat of-- in the possession of two direct communications with the exterior, in addition to the customary indirect way through the oviduct. These are the abdominal pores (a.p.) on either side of the cloaca in either sex. They can always be readily demonstrated by probing out from the body cavity, in the direction indicated by the arrow (a.p.) in Figure 1, Sheet 15. They probably serve to equalize the internal and external pressure of the fish as it changes its depth in the water, just as the Eustachian tubes equalize the pressure on either side of the mammal's tympanic membrane.
Section 17. The musculature of the dog-fish body is cut into V-shaped segments, the point of the V being directed forward. The segments alternate with the vertebrae, and are called myomeres. Such a segmentation is evident, though less marked, in the body wall muscles of the frog, and in the abdominal musculature of the rabbit and other mammals it is still to be traced.
Section 18. The uro-genital organs of the female dog-fish (Figure 1, Sheet 17) consist of an unpaired ovary (ov.), paired oviducts (o.d.), enlarged at one point to form an oviducal gland (o.d.g.), kidneys (k.), with ureters (ur.) uniting to form a urinary sinus (u.s.) opening into the cloaca by a median urinary papilla separate from the oviducal openings. The eggs contain much yolk, and, like those of the fowl, are very large; like the fowl, too, one of the ovaries is suppressed, and it is the right ovary that alone remains. The two oviducts meet in front of the liver ventral to the oesophagus, and have there a common opening by which the ova are received after being shed into the body cavity. The eggs receive an oblong horny case in the oviduct; in the figure such a case is figured as distending the duct at e. The testes of the male (T. in Figure 2) are partially confluent in the middle line. They communicate through vasa efferentia (v.e.) with the modified anterior part of the kidney, the epididymis (ep.), from which the vas deferens (v.) runs to the median uro-genital sinus (u.g.s.), into which the ureters (ur.) also open. The silvery peritoneum (lining of the body cavity) covers over the reddish kidneys, and hides them in dissection.
Section 19. Figure 3, Sheet 17, is a generalized diagram of the uro-genital organs in the vertebrata; M.L. is the middle line of the body, G. is the genital organ, Pr. is the pronephros, or fore kidney, a structure which is never developed in the dog-fish, but which has functional importance in the tadpole and cod, and appears as a transitory rudiment in the chick. A duct, which is often spoken of as the pronephric duct (p.d.), and which we have figured under that name, is always developed. Anteriorly it opens into the body cavity. It is also called the Mullerian duct, and in the great majority of vertebrata it becomes the oviduct, uniting with its fellow, in the case of the dog-fish, ventral to the oesophagus. In the male it usually disappears; the uterus masculinus of the rabbit is still very generally regarded as a vestige of it. Kolliker has shown, however, that this interpretation is improbable. Ms. is the mesonephros, some or all of which becomes the epididymis in the male of types possessing that organ, and is connected with G. by the vasa efferentia. Mt., the metanephros, is, in -actual fact- [the frog], indistinguishably continuous with Ms., and is the functional kidney, its duct (metanephric duct) being either undifferentiated from the mesonephric (as is the case with the frog) or largely split off from it, as in the dog-fish, to form the ureter.
Section 20. The correspondence of the male organs of the dog-fish with those of the rabbit, will be more evident if the student imagine--
(a) the testes, vasa efferentia, and epididymis of each side to shift posteriorly until they reach a position on either side of the cloaca; and
(b) The uro-genital apertures, instead of meeting dorsally and posteriorly to the anus, to shift round that opening and meet anteriorly and ventrically to it.
Section 21. This completes our survey of this type. Except where we
have specified differences, the general plan of its anatomy follows the
lines of the other vertebrate types described.
Section 1. We find in Amphioxus the essential vertebrate features reduced to their simplest expression and, in addition, somewhat distorted. There are wide differences from that vertebrate plan with which the reader may now be considered familiar. There are no limbs. There is an unbroken fin along the median dorsal line and coming round along the ventral middle line for about half the animal's length. But two lowly vertebrates, the hag-fish and lamprey, have no limbs and a continuous fin. There is, as we shall see more clearly, a structure, the respiratory atrium, not apparently represented in the true vertebrate types, at least in their adult stages. There is no distinct heart, only a debateable brain, quite without the typical division into three primary vesicles, no skull, no structures whatever of cartilage or bone, no genital ducts, no kidneys at all resembling those of the vertebrata, no pancreas, no spleen; apparently no sympathetic chain, no paired sense organs, eyes, ears, or nasal sacs, in all of which points we have striking differences from all true vertebrata; and such a characteristic vertebrate peculiarity as the pineal gland we can only say is represented very doubtfully by the eye spot.
Section 2. The vertebral column is devoid of vertebrae; it is throughout life a rod of gelatinous tissue, the notochord (Figure 1, n.c.), surrounded by a cellular sheath. Such a rod is precursor to the vertebral column in the true vertebrates, but, except in such lowly forms as the lamprey, is usually replaced, partially (e.g., dog-fish) or wholly (as in the rabbit) by at first cartilaginous vertebrae whose bodies are derived from its sheath. Further, while in all true vertebrata the notochord of the developing young reaches anteriorly at most to the mid-brain, and is there at its termination enclosed by the middle portion of the skull, in Amphioxus it reaches far in front of the anterior extremity of the nervous system, to the end of the animal's body.* On this account the following classification is sometimes made of those animals which have a notochord:--
-Chordata_ (= Vertebrata, as used by Lankester).
1. Having the notochord reaching in front of the brain.
Cephalochorda = Amphioxus.
2. Having the notochord reaching anteriorly to the mid-brain, a brain of three primary vesicles and a skull.
Craniata = all "true vertebrata": fishes, amphibia, reptiles, birds, and mammals (Vertebrata of Balfour).
3. Having the notochord confined to the tail.
Urochorda = the ascidians, or sea-squirts, certain forms of life only recently recognised as relatives of the vertebrata.
* The anterior end of the notochord in the developing rabbit or dog lies where the middle of the basisphenoid bone is destined to be.
Section 3. Figure 1, Sheet 19, shows the general anatomy of Amphioxus. We recognise four important points of resemblance to the earlier phases of the higher and the permanent structure of the lower members of the vertebrata, and it is these that justify the inclusion of amphioxus in this volume. In the first place there is the--
-Notochord_.
In the next, just above it (at s.c.) we find--
-A Dorsal Tubular Nervous Axis_,
the spinal cord. Thirdly, the pharynx (ph.) is perforated by--
-Respiratory Slits_,
though these, instead of being straight slashes, are modified from a U-shape [slant very much forward and are much more numerous than in any true vertebrate.]. -And-, Fourthly, there is, as we shall see, a--
-Vertebrate Type of Circulation_.
[And finally the body-wall muscles are divided into--]
[-Myomers_.]
Section 4. The alimentary canal of Amphioxus commences with an "oral cavity," not represented in our vertebrata, surrounded by a number of cirri, or tentacles, supported by a horny substance which seems to be chitin, a common skeletal material among invertebrates. A velum (v.) forms a curtain, perforated by the mouth and by two smaller hyoidean apertures, between the oral cavity and the pharynx (ph.). "Pharynx" is here used in a wider sense than in the true vertebrata; it reaches back close to the liver, and is therefore equivalent to pharynx + oesophagus + a portion or all of the stomach. The [so-called] hyoidean apertures are not equivalent to the similarly-named parts of the vertebrata. Behind the pharynx the intestine (int.) runs straight out to the anus (an.), which opens not in the middle line, as one might expect, but in the left side! The liver lies usually on the creature's right, and instead of being a compact gland, is simply bag-like.
Section 5. The circulation is peculiarly reduced (Figure 2). The cardiac aorta (c.ao.) lies along the ventral side of the pharynx, and sends branches up along the complete bars between the gill slits. There is no -distinct- heart, but the whole of the cardiac aorta is contractile, and at the bases of the aortic arches that run up the bars there are contractile dilatations that assist in the propulsion of the blood. Dorsal to the pharynx, as in fishes, there is a pair of dorsal aorta (d.ao.) that unite above the liver (compare the frog, for instance), and thence run backward as a median dorsal aorta (d.ao.'). A portal vein (p.v.) bring blood back from the intestine (and apparently from the whole posterior portion of the animal) to the liver. Thence hepatic veins (hep.) take it to the cardiac aorta.
{Lines from First Edition only.}
-When we remember that in the embryonic vertebrate the heart is at first a straight tube, this circulation appears even more strikingly vertebrate in its character than before.-
Section 6. The coelom, or body cavity, of Amphioxus lies, of course, as in the vertebrata, between the intestinal wall and the body walls, and, just as in the vertebrata, it is largely reduced where gill slits occur. But matters are rather complicated by the presence of an atrial cavity round the pharynx, which is not certainly represented in the vertebrata, and which the student is at first apt to call the body cavity, although it is entirely distinct and different from that space. The mutual relation of the two will become apparent after a study of Figures 10, 11, 12 (Sheet 21). Figure 10 gives diagrammatically a section of a very young stage of Amphioxus; P is the pharynx portion of the alimentary canal, coe. is the coelom surrounding it at this stage here as elsewhere; mt.c. are certain lymph spaces, the metapleural canals, between which a small invagination (i.e., a pushing-in), at., of the outer epidermis occurs; n.c. is the notochord, and s.c. the spinal cord. The gill slits, by which P. communicates with the exterior, are not shown. Next Figure 11 shows the invagination (at.) pushing its way in, and cut off from the exterior by a meeting of the body wall below. Note that at. is a portion of the animal's exterior thus embraced by its body, and that its lining is therefore of the same material as the external integument. In Figure 12, at. is developing upward, so that the true body hangs into it. Now imagine the gill slits perforated, as shown by the double-headed arrow in Figure 12. Figure 3, on Sheet 20, is a less diagrammatic representation of a cross-section of the pharyngeal region (vide Figure 1, Sheet 19). The student should compare Figure 3, Sheet 20, and Figure 12, Sheet 21. The atrium and metapleural canals are easily recognised in both. In Figure 3 the coelom is much cut up by the gill slits, and we have remaining of it (a) the dorsal coelomic canals (d.c.c.) and (b) the branchial canals (br.c.) in the bars between the slits. The atrial cavity remains open to the exterior at one point, the atrial pore (at.p.).
Section 7. The method of examining cross-sections is an extremely convenient one in the study of such a type as Amphioxus. The student should very carefully go over and copy the six sections on Sheet 20, comparing Figure 1 as he goes. He should do this before reading what follows. One little matter must be borne in mind. These figures are merely intended to convey the great structural ideas, and they are considerably simplified; they must not be regarded as a substitute for the examination of microscopic sections. [He will notice a number of rounded masses from the body wall. The] -For instance, the body-wall- muscles of Amphioxus are arranged in bundles bent sharply in an arrow shape, the point forward. -A number of these bundles are cut in any one section, and so the even shading of our diagrams, if they professed to be anything more than diagrams, should be broken up into masses.- These -bundles, we may mention-, are called myomeres, and they are indicated in Figure 1 by lines pointing acutely forward. [Several are consequently cut in any transverse section (Sheet 20), and these are the rounded masses he sees.] Similar myomeres, similarly situated, are found in fish, behind the head, and, less obviously, they occur with diminishing importance as the scale of the vertebrata is ascended.
Section 8. If we compare the nervous system of amphioxus with that of any vertebrate, we find at once a number of striking differences. In the first place, the skeletal covering of it, the cranium and the neural arches of vertebrae, are represented only by a greatly simplified connective tissue. In the next, a simple and slight anterior dilatation alone represents the brain. A patch of black pigment anterior to this (e.s.) may or may not be what its name implies an eye-spot. There is a ciliated funnel, c.f. (Figure 1, Sheet 19), opening on the left side, which has been assumed to be olfactory in its functions, and in the mouth chamber a ciliated pit (c.p.), which may, or may not, be an organ of taste. The ventral fissure of the spinal cord is absent. The dorsal nerves are without ganglia, and do not come off in pairs, but alternately, one to the left, then one to the right, one to the left, one to the right, and so on. The ventral nerves are very short, more numerous than the dorsal, and never unite with these latter to form mixed nerves.
The student will observe that here, just as in the case of the ciliated funnel and anus, the Amphioxus is not strictly symmetrical, but twisted, as it were, and so departs from the general rule of at least external bilateral symmetry obtaining among the vertebrates. It habitually lies on one side in the mud of the sea bottom, and it is probable that this external asymmetry is due to this habit, so that too much classificatory importance must not be attached to it. The soles and other related fish, for instance, are twisted and asymmetrical, through a similar specific habit, to such an extent that both eyes lie on one side of the animal.
Section 9. No kidney on the vertebrate pattern is found, but the following structures have, among others, been suggested as renal organs:--
(a) Certain canals, the brown tubes of Lankester (b.t.L., Figure 2, Sheet 19), a pair of pigmented tubes opening into the atrium at the hind end of the pharynx, lying forward along by the dorsal coelomic canals, and having an internal opening also.
(b) Certain tubuli described by Weiss as situated in a series along the upper corners of the atrial cavity, and communicating, after the fashion, of the "nephridia" of the earthworm, with the coelom and with the exterior (or, rather, with that portion of the animal's exterior enclosed in by the atrial wall; compare Section 6).
(c) The general epithelial lining of the atrium.
The reproductive organs (Figure 4, Sheet 20, g.) are masses of cells situated in an isolated part of the coelom in the atrial folds, and, having no ducts, their contents must escape into the atrium by rupture of the body-wall. Thence they escape either by gill-slits, pharynx and mouth, or, more generally, through the atrial pore. The animals, like all the vertebrata, are dioecious, i.e., male or female.
Section 10. The endostyle (end.), in Figures 3 and 4, is a ciliated path or groove on the under side of the pharynx, which is generally supposed to represent the thyroid gland of vertebrates. The vertebrate thyroid, early in development, is certainly an open and long narrow groove in the ventral side of the pharynx. The hyper-pharyngeal groove (h.p.) has been in the past compared to the pituitary body, but there is little doubt now that this structure is represented by the ciliated pit.
Section 11. The student is advised to revise this chapter before proceeding, and to schedule carefully the anatomical features under the headings of (1.) distinctly vertebrate characters, (2.) characters contrasting with the normal vertebrate structure, (3.) facts of doubtful import, with the suggestions given in the text written against them.
Section 12. The development of amphioxus, studied completely, is at once one of the most alluring and difficult tasks in the way of the zoologist; but certain of its earlier and most obvious fasts may very conveniently be taken into consideration now.
Section 13. The phenomena of the extrusion of polar bodies and fertilization are treated of later, and will, therefore, not be considered now. We will start our description with an egg-cell, which has escaped, of course, since there are no genital ducts, by rupture of the parent, has been fertilized by the male element, and is about to develop into a young amphioxus. It is simply a single cell, with some power of amoeboid motion, a single nucleus and nucleolus; and in amphioxus its protoplasm is clear and transparent. Frequently ova are loaded with granules of food store (yolk), which enable the young animal to go far with its development before it is hatched and has to begin fending for itself. Such an ovum as that of our present type, however being devoid of such yolk (alecithal = without yolk), necessitates a very early start in life, and, for reasons too complicated to state fully here, the development in such a case is considered particularly instructive and primitive by zoologists.
Section 14. The first thing to be seen in the developing cell is a deepening circular groove (Figure 1, Sheet 21), which divides the ovum into two parts. Another groove then cuts at right angles to this subdividing the two into four (Figure 2). Another groove, at right angles to both the former, follows, making the four eight (Figure 3). And so subdivision goes on. The whole process is called segmentation or cleavage.
Section 15. At the end of segmentation we get a hollow sphere of small cells, the cells separating from one another centrally and enclosing a cavity as the process proceeds. This is the blastosphere, shown diagrammatically in Figure 4, and of which an internal view, rather truer to the facts of the case as regards shape, is given as Figure 5. The central cavity is the segmentation cavity (s.c.).
Section 16. Invagination follows (Figure 6). In this process a portion of the blastosphere wall is the tucked into the rest, as indicated by the arrow, so that a two-layered sack is formed. The space ar. is the archenteron, the primordial intestine, and its mouth is called, the blastopore (bp.). The outer layer of this double-walled sac is called the epiblast. For the present we will give the inner lining no special term. The young amphioxus has, at this stage, which is called the gastrula stage, a curious parallelism with such a lowly form as the Hydra of our ditches. This latter creature, like the gastrula, consists essentially of two layers of cells, an outer protective and sensory layer, and an inner digestive one; it has a primordial intestine, or archenteron, and its mouth is sometimes regarded as being a blastopore. All animals that have little yolk, and start early in life for themselves, pass through a gastrula stage, substantially the same as this of amphioxus.
Section 17. The anus is perforated later near the region occupied at this stage by the blastopore. Hence the anterior end of the future amphioxus, the head end, is pointing towards the Figure 6, and the letters ep. are marked on the side which will be dorsal.
Section 18. Figure 7 i. is a dorsal view of the gastrula at a somewhat later stage, and here indications of distinctly vertebrate relationships already appear. Figure 7 ii. is a cross-section, its position, being shown by cross-lines in 7 i. and 6. Note first that the epiblast along the mid-dorsal line is sinking in to form what is called the neural plate (n.p.), and simultaneously on either side of it rise the neural folds (n.f.). Now, at Figure 8, a slightly later stage is represented, and at 9 i. the inturned part is separated from the general external epiblast as the spinal cord. The remainder of the epiblast constitutes the epidermis.
Section 19. Reverting to Figure 7 ii., along the dorsal side of the archenteron a thickening of its wall appears, and is gradually pinched off from it to form a cellular rod, lying along under the nervous axis and above the intestine. This is the notochord (compare Figures 8 and 9).
Section 20. Finally, we note two series of buds of cells, one on either side of the archenteron in Figure 7 ii. In 8 these buds have become hollow vesicles, growing out from it, the coelomic pouches. They are further developed in 9; and in 9 ii., which is a diagrammatic figure, they are indicated by dotted lines. They finally appear to (? entirely) obliterate the segmentation cavity-- they certainly do so throughout the body; and their cavities are in time cut off from the mesenteron, by the gradual constriction of their openings. In this way the coelom (body cavity) arises as a series of hollow "archenteric" outgrowths, and ms. becomes the alimentary canal. mt.c., the metapleural canals, probably arise subsequently to, and independently of, the general coelomic space, by a splitting in the body-wall substance.
Section 21. Hence, in considering the structure of amphioxus, we have three series of cells from which its tissues are developed:--
1. The epiblast.
2. Walls of the coelomic pouches, which form (a) an inner lining to the epiblast, (b) an outer coating to the hypoblast, and (c) the mesentery (m.), by which the intestine is supported. This is the mesoblast.
3. The lining of the mesenteron, or hypoblast.
From the epiblast the epidermis (not the dermis), the nervous system (including the nerves), and the sensory part of all sense organs are derived. From the mesoblast the muscles, the dermis genital and excretory organs, circulatory fluid and apparatus, any skeletal structures; and all connective tissue are derived. The mass of the body is thus evidently made of mesoblast. The hypoblast is the lining of the intestine and of the glands which open into it; and the material of the notochord is also regarded, as hypoblast.
Section 22. Figure 9 ii. shows all the essential points of the structure of amphioxus. Epiblast is indicated by a line of dashes, mesoblast by dots, and hypoblast, dark or black. The true mouth is formed late by a tucking-in of epiblast, the stomodaeum (s.d.), which meets and fuses with the hypoblast, and is then perforated. The position of this mouth is at the velum. The formation of the atrium has been described. The metapleural folds run forward in front of the velum, as the epipleurs (ep. in Sections 1 and 2), and form an oral hood (b.c.), around which the tentacles appear, and which is evidently not equivalent to the vertebrate mouth cavity, but in front of and outside it. The anus is formed by a tucking in, the proctodaeum, similar to the stomodaeum.
Section 23. The formation of the respiratory slits is complicated, and difficult to describe, but, since investigators have still to render its meaning apparent, it need not detain the elementary student.*
* See Balfour's Embryology, Volume 2, and Quarterly Journal of Microscopical Science March, 1891.
Section 1. We have now to consider how the body of the frog is built up out of the egg cell, but previously to doing so we must revert to the reproductive organs of our type.
Section 2. In the testes of the male is found an intricate network of tubuli, the lining of which is, of course, an epithelium. The cells of this epithelium have their internal borders differentiated into spermatozoa, which, at a subsequent stage, are liberated. A spermatozoon (Figure 3, Sheet 13, sp.) is a rod-shaped cell containing a nucleus; in fact, consisting chiefly of nucleus, with a tail, the flagellum, which is vibratile, and forces the spermatozoon, forward by its lashing. The spermatozoa float in a fluid which is the joint product of the testes, anterior part of the kidney, and perhaps the prostate glands.
Section 3. In the ovary, the ova are formed, and grow to a considerable size. They are nucleated cells, the nucleus going by the special name of the germinal vesicle and the nucleolus the germinal spot. The ova prey upon the adjacent cells as they develop. The protoplasm of the ovum, except at that part of the surface where the germinal vesicle lies, is packed with a great amount of food material, the yolk granules. This yolk is non-living inert matter. An ovum such as this, in which the protoplasm is concentrated towards one pole, is called telolecithal.
Section 4. After the ovum has finished its growth, and elaborated the yolk within itself, a peculiar change occurs in the small area free from yolk-- the animal pole, in which the germinal vesicle lies. This germinal vesicle divides, and one moiety is budded off from the ovum. The ovum has, in fact, undergone cell division into a very large cell containing most of its substance, and a small protoplasmic pimple surrounding half of its nucleus. The disproportion is so great between the two cells, that the phenomenon does not at first suggest the idea of cell division, and it is usually described as the extrusion of the first polar body. There follows a second and similar small cell, behind the first, the second polar body. Since the nucleus of the ovum has divided twice, it is evident that the nucleus remaining now in the ovum is a quarter of the original nucleus. Very little protoplasm is given off with the polar bodies; they play no further part in development, but simply drop off and disappear. Not only in the frog's ovum, but in all vertebrata, two polar bodies are given off in this way before the sexual process occurs. Their exact meaning has been widely discussed. It is fairly evident that some material is removed from the nucleus, which would be detrimental to further developments, and the point debated is what is the precise nature of this excreted material. This burning question we can scarcely deal with here.
Section 5. But here we may point out that in all cells the function of the nucleus appears to be to determine growth and division. It is the centre of directive energy in the cell.
Section 6. Fertilization is effected by a spermatozoon meeting with the ovum. It fuses with it, its nucleus becoming the male pro-nucleus. This and the female pro-nucleus, left after the extrusion of the polar cells, move towards each other, and unite to form the first segmentation nucleus.
Section 7. The ovum next begins to divide. A furrow cutting deeper and deeper divides it into two; another follows at right angles to this, making the two four, and another equatorial furrow cuts off the animal pole from the yolk or vegetative pole. (See Sheet 22, Figures 1, 2, and 3.) And so segmentation (= cleavage) proceeds, and, at last, a hollow sphere, the blastosphere (Figure 4) is formed, with a segmentation cavity (s.c.). But, because of the presence of the yolk at the vegetative pole of ovum, and of the mechanical resistance it offers to the force of segmentation, the protoplasm there is not nearly so finely divided-- the cells, that is to say, are much larger than at the animal pole. The blastosphere of the frog is like what the blastosphere of amphioxus would be, if the future hypoblast cells were enormously larger through their protoplasm being diluted with yolk.
Section 8. The next phase of development has an equally curious resemblance to and difference from what occurs in the case of the ova of animals which do not contain yolk. In such types (e.g., amphioxus) a part of the blastosphere wall is tucked into the rest, and a gastrula formed by this process of invagination. In the frog (Figure 5) there is a tucking-in, but the part that should lie within the gastrula, the yolk-containing cells, are far larger than the epiblast (ep.) which should, form the outer layer of cells. Hence the epiblast can only by continual growth accommodate what it must embrace, and the process of tucking-in is accompanied by one of growth of the epiblast, as shown by the unbarbed arrow, over the yolk. This stage is called the gastrula stage; ar. is the cavity of the gastrula, the archenteron; b.p. is its opening or blastopore. Such a gastrula, formed mainly by overgrowth of the epiblast, is called an epibolic gastrula, as distinguished from the invaginate gastrula of amphioxus. The difference is evidently entirely due to the presence of yolk, and the consequent modification of invagination in the former case.
Section 9. Comparing the two gastrulas, it is not difficult to see that if we imagine the ventral wall of the archenteron of amphioxus to have its cells enormously enlarged through the mixing of yolk with their protoplasm, we should have a gastrula essentially like that of the frog.
Section 10. Figure 6 shows a slightly later ovum than Figure 5, seen from the dorsal side. b.p. is the blastopore. In front of that appears a groove, the neural groove, bordered on either side by a ridge, the neural fold (n.f.). This is seen in section in Figure 7; s.c. is the neural groove; n.f., as before, the neural fold. The neural folds ultimately bend over and meet above, so that s.c. becomes a canal, and is finally separated from the epiblast to form the spinal cord. Below the neural groove a thickening of the dorsal wall of the archenteron appears, and is pinched off to form a longitudinal rod, the precursor of the vertebral column, the notochord, shown in Figure 7 (n.c.), as imperfectly pinched off.
Section 11. Simultaneously, on either side of the notochord appear a series of solid masses of cells, derived mainly by cell division from the cells of the wall of the archenteron, and filling up and obliterating the segmentation cavity. These masses increase in number by the addition of fresh ones behind, during development, and are visible in the dorsal view as brick-like masses, the mesoblastic somites or proto-vertebrae (Figure 6, i., ii., iii.). In Figure 7, these masses are indicated by dotting. In such a primitive type as amphioxus these mesoblastic -somites- [masses] contain a cavity, destined to be the future body cavity, from the first. In the frog, the cavity is not at first apparent; the mesoblast at first seems quite solid, but subsequently what is called the splitting of the mesoblast occurs, and the body cavity (b.c. in Figure 7) appears. The outer mesoblast, lying immediately under the epiblast, constitutes the substance of the somatopleur, and from it will be formed the dermis, the muscles of the body wall, almost all the cartilage and bone of the skeleton, the substance of the limbs, the kidneys, genital organs, heart and bloodvessels, and, in short, everything between the dermis and the coelom, except the nervous system and nerves, and the notochord. The inner mesoblast, the mass of the splanchnopleur, will form the muscle and connective tissue of the wall of the alimentary canal, and the binding substance of the liver and other glands that open into the canal.
Section 12. Figure 8 is one which we reproduce, with the necessary changes in each plate of embryological figures given in this book, so that the student will find it a convenient, one for the purpose of comparison. The lines of dashes, in all cases, signify -epiblast- [hypoblast] , the unbroken black line is -hypoblast-, [epiblast] dotting shows mesoblast, and the shaded rod (n.c.) is the notochord. c.s. is the spinal cord; br.1, br.2, br.3 are the three primary vesicles which constitute the brain, and which form fore, mid, and hind brain respectively. I. is the intestine and Y. the yolk cells that at this early stage constitute its ventral wall.
Section 13. Figure 9 gives a similar diagram of a later stage, but here the blastopore is closed. An epiblastic tucking-in at st., the stomodaeum pre-figures the mouth; pr., the proctodaeum, is a similar posterior invagination which will become the anus. Y., the yolk, is evidently much absorbed. Figure 10 is a young tadpole, seen from the side. The still unabsorbed yolk in the ventral wall of the mesentery gives the creature a big belly. Its mouth is suctorial at this stage, and behind it is a sucker (s.) by which the larvae attach themselves to floating reeds and wood, as shown in the three black figures below.
Section 14. We may now consider the development of the different organs slightly more in detail, though much of this has already been approached. The nervous system, before the closure of the neural groove, has three anterior dilatations, the fore-, mid-, and hind-brains, the first of which gives rise by hollow outgrowths to two pairs of lateral structures, the hemispheres and the optic vesicles. The latter give rise to the retina and optic nerve as described in {Development} Section 40.
Section 15. The hypoblastic notochord is early embraced by a mesoblastic sheath derived from the protovertebrae. This becomes truly cartilaginous, and at regular intervals is alternately thicker and thinner, compressing the notochord at the thicker parts. Hence the notochord has a beaded form within this, at first, continuous cartilaginous sheath. This sheath is soon cut into a series of vertebral bodies by jointings appearing through the points where the cartilage is thickest and the notochord most constricted. Hence what remains of the notochord lies within the vertebral bodies in the frog; while in a cartilaginous fish, such as the dog-fish, or in the embryonic rabbit, the lines of separation appear where the notochord is thickest, and it comes to lie between hollow-faced vertebrae. Cartilaginous neural arches and spines, formed outside the notochordal sheath, enclose the spinal cord in an arcade. The final phase is ossification. As the tadpole approaches the frog stage the vertebral column in the tail is rapidly absorbed, and its vestiges appear in the adult as the urostyle.
Section 16. The development of the skull is entirely dissimilar to that of the vertebral column. It is shown on Figures 1 and 8, Sheet 14; and in the section devoted to the frog's skull a very complete account of the process is given. The process of ossification is described under the histology of the Rabbit.
Section 17. The origin of the circulatory and respiratory organs is of especial interest in the frog. In the tadpole we have essentially the necessities and organization of the fish; in the adult frog we have a clear exposition of the structure of pigeon and rabbit. The tadpole has, at first, a straight tubular heart, burrowed out in somatic mesoblast, and produced forward into a truncus arteriosus. From this arise four afferent branchial arteries, running up along the sides of the four branchial arches, and supplying gills. They unite above on either side in paired hyper-branchial arteries, which meet behind dorsal to the liver, to form a median dorsal aorta. Internal and external carotid arteries supply the head. These four afferent branchial arches are equivalent to the first four of the five vessels of the dog-fish. At first, the paired gills are three in number, external, and tree-like, covered by epiblast (Figures 10 and 11, e.g.), and not to be compared to fish gills in structure, or in fact -with- [to] any other gills within the limits of the vertebrata. Subsequently (hypoblastic) internal gills (int.g., Figure 12), strictly homologous with the gills of a fish, appear. Then a flap of skin outside the hyoid arch grows back to cover over the gills; this is the operculum (op. in Figures 11 and 12, Sheet 22), and it finally encloses them in a gill chamber, open only by a pore on the left, which resembles in structure and physiological meaning, but differs evidently very widely in development, from the amphioxus atrium. At this time, the lungs are developing as paired hollow outgrowths on the ventral side of the throat (Figure 12, L.). As the limbs develop, and the tail dwindles, the gill chamber is obliterated. The capillary interruptions of the gills on the branchial arches (aortic arches) are also obliterated. The carotid gland occupies the position of the first of these in the adult. The front branchial arch here, as in all higher vertebrata, becomes the carotid arch; the lingual represents the base of a pre-branchial vessel; the second branchial becomes the aortic arch. The fourth loses its connection with the dorsal aorta, and sends a branch to the developing lung, which becomes the pulmonary artery. The third disappears. A somewhat different account to this is still found in some text-books of the fate of this third branchial arch. Balfour would appear to have been of opinion that it gave rise to the cutaneous artery, and that the third and fourth vessels coalesced to form the pulmocutaneous, the fourth arch moving forward so as to arise from the base of the third; and most elementary works follow him. This opinion was strengthened by the fact that in the higher types (reptiles, birds, and mammals) no fourth branchial arch was observed, and the apparent third, becomes the pulmonary. But it has since been shown that a transitory third arch appears and disappears in these types.
Section 18. The origin of the renal organ and duct has very considerable controversial interest.* In Figure 13, Sheet 22, a diagrammatic cross-section, of an embryo is shown. I. is the intestine, coe. the coelom, s.c. the spinal cord; n.c. the notochord, surrounded by n.s., the notochordal sheath, ao. is the dorsal aorta. In the masses of somatic mesoblast on either side, a longitudinal canal appears, which, in the torpedo, a fish related to the dog-fish, and in the rabbit, and possibly in all other cases, is epiblastic in origin. This is the segmental duct, which persists, apparently, as the Wolffian duct (W.D.). Ventral to this appears a parallel canal, the Mullerian duct (M.D.), which is often described as being split off from the segmental duct, but which is, very probably, an independent structure in the frog. A number of tubuli, at first metamerically arranged, now appear, each opening, on the one hand, into the coelom by a ciliated mouth, the nephrostome (n.s.), and on the other into the segmental duct. These tubuli are the segmental tubes or nephridia. There grows out from the aorta, towards each, a bunch, of bloodvessels, the glomerulus (compare Section 62, Rabbit). These tubuli ultimately become, in part, the renal tubuli, so that the primitive kidney stretches, at first, along the length of the body cavity from the region, of the gill-slits backward. The anterior part of the kidney, called the pronephros, disappears in the later larval stages. Internal to the kidney on either side there has appeared a longitudinal ridge, the genital ridge (g.r.), which gives rise to testes or ovary, as the case may be.
* In the discussion whether the vertebrata have arisen from some ancestral type, like the earthworm, metamerically segmented, and of fairly high organization, or from a much lower form, possibly even from a coelenterate. Such a discussion is entirely outside the scope of the book, though its mention is necessary to explain the importance given to these organs.
Section 19. The student should now compare the figures on Sheet 17. In the male, tubular connections are established between the testes and the middle part of the primitive kidney (mesonephros). These connections are the vasa efferentia (v.e.), and the mesonephros is now equivalent to the epididymis of the rabbit. The Wolffian duct is the urogenital duct of the adult, and the Mullerian duct is entirely absorbed, or remains, more or less, in exceptional cases.
In the female, the Mullerian duct increases greatly in length-- so that at sexual maturity its white coils appear thicker and longer than the intestine-- and becomes the oviduct; the Wolffian duct is the ureter, and the mesonephros is not perverted in function from its primary renal duty.
Section 20. Tabulating these facts--
In the adult male:Pronephros disappears.
In the adult female:
The Mullerian duct (? = pronephric duct) disappears.
Mesonephros = Epididymis; its duct, the urogenital.
Metanephros and duct, not clearly marked off from
Mesonephros.
(Compare Dog-fish, Section 19.)
Pronephros disappears.
The Mullerian duct, the oviduct.
Mesonephros and Metanephros, the kidney, and their unseparated ducts, the ureters.
Section 21. Hermaphrodism (i.e., cases of common sex) is occasionally found among frogs; the testis produces ova in places, and the Mullerian duct is retained and functional. The ciliated nephrostomata remain open to a late stage of development in the frog, and in many amphibia throughout life. Their connection with the renal tubuli is, however, lost.
Section 22. The alimentary canal is, at first, a straight tube. Its disproportionate increase in length throws it into a spiral in the tadpole (int. Figure 11), and accounts for its coiling in the frog. The liver and other digestive glands are first formed, like the lungs, as hollow outgrowths, and their lining is therefore hypoblastic. The greatest relative length of intestine is found in the tadpole, which, being a purely vegetable feeder, must needs effect the maximum amount of preparatory change in its food.
Section 23. The frog has an ovum with a moderate allowance of yolk, but the quantity is only sufficient to start the little animal a part of its way towards the adult state. The fowl, on the contrary, has an enormous ovum, gorged excessively, with yolk, and as a consequence the chick is almost perfected when it is hatched. The so-called yolk, the yellow of an egg, is the ovum proper; around that is a coating of white albumen, in a shell membrane and a shell. At either end of the yolk (Figure 1, y.) twisted strands of albuminous matter, the chalazae (ch.) keep the yolk in place. The animal pole is a small grey protoplasmic area, the germinal area (g.a.), on the yolk.
Section 24. We pointed out that the presence of the yolk in the frog's egg led to a difference in the size of the cells at the animal and vegetable poles. The late F.M. Balfour, borrowing a mathematical technicality, suggested that the rate of segmentation in any part of an ovum varies inversely with the amount of yolk. In the fowl's egg, except just at the germinal area, the active protoplasm is at a minimum, the inert yolk at a maximum; the ratio of yolk to protoplasm is practically infinity, and the yolk therefore does not segment at all. The yolk has diluted the active protoplasm so much as to render its influence inappreciable. The germinal area segments, and lies upon the yolk which has defeated the efforts of its small mingling of protoplasm to divide. Such a type of segmentation in which only part of the ovum segments is called meroblastic. If we compare this with the typical blastosphere of the lower type, we see that it is, as it were, flattened out on the yolk. This stage is shown in section in the lower figure of Figure 1. b.d., the blastoderm, is from this point of view, a part of the ripped and flattened blastosphere, spread out on the yolk; s.c. is the segmentation cavity, and y. the yolk.
Section 25. There is no open invagination of an archenteron in the fowl, as in the frog--, the gastrula, like the blastosphere, stage is also masked. But, in the hinder region of the germinal area, a thick mass of cells, grows inward and forward, and, appearing in the dorsal view of the egg as a white streak, is called the primitive streak (p.s.). By a comparison of the figures of frog and fowl the student will easily perceive the complete correspondence of the position of this with the blastopore of the frog. The relation of the two will be easily understood if we compare the fowl's archenteron to a glove-finger under pressure-- its cavity is obliterated-- and the frog's to the glove-finger blown out. The tension of the protoplasm, straining over the enormous yolk, answers to the pressure. The gastrula in the fowl is solid. The primitive streak is, in fact, the scar of a closed blastopore. As we should expect from this view of its homology, at the primitive streak, the three embryonic layers are continuous and indistinguishable (Figure 2). Elsewhere in the blastoderm they are distinctly separate. Just as the yolk cells of the frog form the ventral wall of the intestine, so nuclei appear along the upper side of the yolk of the fowl, where some protoplasm still exists, and give rise to the ventral hypoblastic cells. By conceiving a gradually increasing amount of yolk in the hypoblastic cells in the ventral side of the archenteron, the substantial identity of the gastrula stage in the three types, which at first appear so strikingly different, will be perceived. Carry Figures 4 and 5 of the frog one step further by increasing the size of the shaded yolk and leaving it unsegmented, and instead of ar. in 5 show a solid mass of cells, and the condition of things in the fowl would at once be rendered.
Section 26. Figure 3a of the fowl will conveniently serve for comparison with Figure 7 of the frog. The inturning of the medullary groove is entirely similar in the two cases. The mesoblast appears as solid mesoblastic somites. In the section above Figure 4 this layer is shown as having split into somatopleur (so.) and splanchnopleur (spch.). Figure 3 answers to Figure 6 of the frog, and Figure 4 is a later stage, in which the medullary groove is beginning to close at its middle part. The clear club-shaped area around the embryo (a.p.) is the area pellucida; the larger area without this is the area opaca (a.o.), in which the first bloodvessels arise by a running together and a specialization of cells. The entire germinal area grows steadily at its edges to creep over and enclose the yolk.
Section 27. So far, the essential differences between the development of fowl and frog, the meroblastic segmentation, absence of a typical gastrula, and the primitive streak, seem comprehensible on the theory that such differences are due to the presence of an enormous amount of yolk. Another difference that appears later is that, while the tadpole has an efficient pronephros, the fowl, which has no larval (free imperfect) stages in its life history, has the merest indication of such a structure.
Section 28. Another striking contrast, due to, or connected with, this plethora of yolk, is the differentiation of a yolk sac (= umbilical vesicle) and the development of two new structures, the amnion and allantois, in the fowl. If the student will compare Figure 10 of the frog, he will see that the developing tadpole encloses in its abdomen all the yolk provided for it. This is a physical impossibility in the fowl. In the fowl (Figure 2, Sheet 24) the enormous yolk (Y.) lies outside of the embryo, and, as the cells of the germinal area grow slowly over it, umbilical bloodvessels are developed to absorb and carry the material to the embryo. In the case of an embryo sinking in upon, as it absorbs, this mass of nutritive material, a necessity for some respiratory structure is evident. From the hinder end of the fowl's intestine, in a position corresponding to the so-called, urinary bladder of the frog, a solid outgrowth, the allantois, which speedily becomes hollow, appears. Early stages are shown in Figures 1 and 2, Sheet 24 (al.); while the same thing is shown more diagrammatically on Sheet 23, Figure 6 (all.). This becomes at last a great hollow sac, which is applied closely to the porous shell, and the extent of which will be appreciated by looking at Figure 5, Sheet 24, where the allantois is shaded. Allantoic bloodvessels ramify thickly over its walls, and aeration occurs through the permeable shell.
Section 29. The nature of the amnion will be understood by following Figures 4b, 5, and 6 on Sheet 23. The three embryonic layers are indicated by broken lines, dots, and black lines, just as they are in the frog diagrams. Not only is the embryo slowly pinched off from the yolk sac (y.s.), but, as the yolk is absorbed beneath it, and it grows in size, it sinks into the space thus made, the extra-embryonic somatopleur and epiblast rise up round it as two folds, which are seen closing in 5, and closed in 6, over the dorsal side of the young chick. In this way a cavity, a., lined by epiblast, and called the amniotic cavity, is formed. Dorsal to this, in 6, comes a space lined by somatic mesoblast, and continuous with p.p., the pleuro-peritoneal cavity, or body cavity of the embryo. Outside this, again, is a layer, of somatopleur internally and epiblast externally, the false amnion (f.a.), which is continuous with the serous membrane (s.m.) enclosing the rest of the egg. The student should, carefully copy these diagrams, with coloured pencils or inks for the different layers, and should compare them with the more realistic renderings of Figures 2, 5, and 8, Sheet 24.
Section 30. The heart in the fowl appears first as a pair of vessels, which unite to form a straight trunk in the median line, as the flattened-out embryo closes in from the yolk. The way in which this straight trunk is thrown, first of all, into the S shape of the fish heart, and then gradually assumes the adult form, is indicated roughly by Figure 3. In one respect the development of the heart does not follow the lines one would expect. Since, between the fish and the higher form comes the condition of such an animal as the frog, in which the auricles are divided, while there is only one ventricle, we might expect a stage in which the developing chick's heart would have one ventricle, and a septum between the auricles. But, as a matter of fact, the ventricles in fowl and rabbit are separated first, and the separation of the auricles follows, and is barely complete at birth.
Section 31. Two vitelline veins from the yolk sac (v.v.) flow into the heart from behind, as shown in Figure 1. A later more complete and more diagrammatic figure of the circulation is seen in Figure 7. At first there are two anterior cardinal (a.c.), and two posterior cardinal veins (p.c.) uniting to form Cuvierian sinuses (c.s.) that open into the heart just as in the dog-fish. But later the inferior cava is developed and extends backward, the posterior cardinals atrophy, the Cuvierian sinuses become the superior cavae, and the anterior cardinals the internal jugular veins. The vitelline veins (v.v.) flow, at first, uninterruptedly through the liver to the inferior cava, but, as development proceeds, a capillary system is established in the liver, and the through communication, the ductus venosus, is reduced-- at last-- completely. Bearing in mind that the yolk is outside the body in the fowl and inside it in the frog, the vitelline veins of the former have a considerable resemblance in position, and in their relation to the portal vein, to a portion of the single anterior abdominal vein. Blood is taken out to the allantois, however, by the arteries of the latter type.
Section 32. Five aortic arches are generally stated to appear altogether in the fowl, but not simultaneously. The first two, the mandibular and the hyoid vascular arches, early disappear, and are not comparable to any in the frog. The third is the first branchial arch, and, like the corresponding arch in the frog, forms the carotid artery; the second branchial is the aortic arch; and what has hitherto been regarded as the third (the fifth arch, i.e.) the pulmonary artery. A transitory arch, it is now known, however, appears between the second branchial and the last, and it is therefore the fourth branchial arch which is the pulmonary, just as it is in the frog.
Section 33. Blood, it may be mentioned, first appears in the area vasculosa, the outer portion of the area opaca. Embryonic cells send out processes, and so become multipolar; the processes of adjacent cells coalesce. The nucleus divides, and empty spaces appear in the substance of each of the cells.
In this way, the cavities of the smaller vessels and capillaries are formed, and the products of the internal divisions of the cells become the corpuscles within the vessels. The red blood corpuscles of the rabbit, it may be added, are nucleated for a considerable portion of embryonic life. Larger vessels and the heart are burrowed, as it were, out of masses of mesoblast cells. The course of the blood in the embryo is by the veins to the right auricle, thence through the imperfection of the auricular septum already alluded to, into the left auricle. Then the left ventricle, aortic arches (for the future pulmonary artery is in communication by a part presently blocked, the ductus arterious, with the systemic aorta), arteries, capillaries, veins. The liver capillary system and the pulmonary system only become inserted upon the circulation at a comparatively late stage.
Section 34. With the exception of the reduction of the pronephros, what has been said of the development of the frog's nervous system, renal and reproductive organs, and skeleton, applies sufficiently to the fowl for our present purposes. The entire separation of Wolffian and Mullerian ducts from the very beginning of development is here beyond all question (vide Section 18). But the notochord in the fowl is not so distinctly connected with the hypoblast, and so distinct from the mesoblast, as it is in the lower type, and no gills, internal or external, are ever developed. The gill slits occur with a modification due to the slitting and flattening out of the embryo, already insisted upon; for, whereas in the tadpole they may be described as perforations, in the fowl they appear as four notches between ingrowing processes that are endeavouring to meet in the middle line.
Section 35. The early development of the rabbit is apt to puzzle students a little at first. We have an ovum practically free from yolk (alecithal), and, therefore, we find it dividing completely and almost equally. We naturally assume, from what we have learnt, that the next stages will be the formation of a hollow blastosphere, invagination, a gastrula forming mesoblast by hollow outgrowths from the archenteron, and so on. There is no yolk here to substitute epiboly (Section 9) for invagination, nor to obliterate the archenteron and the blastopore through its pressure.
Yet none of these things we have anticipated occur!
We find solid mesoblastic somites, we find primitive streak, allantois and amnion, features we have just been explaining as the consequence of an excess of yolk in the egg. We even find a yolk sac with no yolk in it.
Section 36. A solid mass of cells is formed at the beginning, called a morula, Figure 1. In this we are able to distinguish rather smaller outer layer cells (o.l.c.), and rather larger inner layer cells (i.l.c.), but these cells, in their later development, do not answer at all to the two primitive layers of the gastrula, and the name of Van Beneden's blastopore (V.B.b.), for a point where the outer layer of cells is incomplete over the inner, only commemorates the authorship of a misnomer. The uniformity, or agreement, in the development of our other vertebrate types is apparently departed from here.
Section 37. As the egg develops, however, we are astonished to find an increasing resemblance to that of the fowl. A split occurs at one point between outer layer and inner layer cells, and the space resulting (Y in Figure 2) is filled by an increasing amount of fluid, and rapidly enlarges, so that presently we have the state of affairs shown in 3, in which the inner layer cells are gathered together at one point on the surface of the ovum, and constitute the germinal area. If, with Hubrecht, we regard the outer layer cells as an egg membrane, there is a curious parallelism between this egg and the fowl's the fluid Y representing the yolk; and the inner layer cells the cells of the fowl's germinal area.
At any rate, the subsequent development goes far to justify such a view. The inner cells split into epi-, meso-, and hypo-blast, like the blastoderm in the fowl; there is a primitive streak and no blastopore; an amnion arises; the yolk sac, small and full of serous fluid, is cut off just as the enormous yolk of the fowl is cut off; and an allantois arises in the same way. There is no need to give special diagrams-- Figures 3, 4b, 5, and 6 of the fowl will do in all respects, except proportion, for the development of the rabbit. The differences are such as we may account for, not on the supposition that the rabbit's ovum never had any yolk, but that an abundant yolk has been withdrawn from it. The nutrition of the embryo by yolk has been superseded by some better method. The supposition that the rabbit is descended from ancestors which, like the birds and reptiles, laid eggs with huge quantities of yolk, meets every circumstance of the case.
Section 38. But the allantois and yolk sac of the rabbit, though they correspond in development, differ entirely in function from the similar organs of the fowl. The yolk sac is of the very smallest nutritive value; instead of being the sole source of food, its contents scarcely avail the young rabbit at all as nourishment. Its presence in development is difficult to account for except on the supposition, that it was once of far greater importance. At an early stage, the outgrowing allantois, pushing in front of it the serous membrane, is closely applied to the lining of the mother's uterus. The maternal uterus and the embryonic allantois send out finger-like processes into each other which interlock, and the tissue between the abundant bloodvessels in them thins down to such an extent that nutritive material, peptones and carbohydrates, and oxygen also, diffuse freely through it from mother to foetus,* and carbon dioxide, water, and urea from the foetus to the mother. The structure thus formed by the union of the wall of the maternal uterus, allantois, and the intermediate structures is called the placenta. Through its intermediation, the young rabbit becomes, as it were, rooted and parasitic on the mother, and utilizes her organs for its own alimentation, respiration, and excretion. It gives off CO2, H2O, and urea, by the placenta, and it receives O and elaborated food material through the same organ. This is the better method that has superseded the yolk.
Section 39. In its later development, the general facts already enunciated with regard to the organs of frog and fowl hold, and where frog and fowl are stated to differ, the rabbit follows the fowl. In the circulation the left fourth vascular arch (second branchial) gives rise to the aortic arch; in the right the corresponding arch disappears, except so much of it as remains as the innominate artery. The azygos vein (Chapter 3) -is a vestige of- [is derived from] the right posterior cardinal sinus. Both pulmonary arteries in the rabbit are derived from the left sixth vascular arch (= fourth branchial). Compare Section 32. The allantois altogether disappears in the adult fowl; in the adult mammal a portion of its hollow stalk remains as the urinary bladder, and the point where it left the body is marked by the umbilicus or navel. The umbilical arteries become the small hypogastric arteries on either side of the urinary bladder. There is no trace of a pronephros at all in the rabbit.
Section 40. We may note here the development of the eye. This is shown in Figure 4, Sheet 24. A hollow cup-shaped vesicle from the brain grows out towards an at first hollow cellular ingrowth from the epidermis. The cavity within the wall of the cup derived from the brain is obliterated, [and the stalk withers,] the cup becomes the retina, and -its stalk- [thence fibres grow back to the brain to form] the optic nerve. The cellular ingrowth is the lens. The remainder of the eye-structures are of mesoblastic origin, except the superficial epithelium of the cornea. The retinal cup is not complete at first along the ventral line, so that the rim of the cup, viewed as in Figure 1, r., is horseshoe shaped. -Hence the optic nerve differs from other nerves in being primitively hollow.- In all other sense organs, as, for instance, the olfactory sacs and the ears, the percipient epithelium is derived, from the epiblast directly, and not indirectly through the nervous system. These remarks apply to all vertebrate types.
Section 41. The supposition, that the general characters of the rabbit's ovum were stamped upon it as an heritage from a period when the ancestors of the mammals were egg-laying reptiles, is strengthened by the fact that the two lowest and most reptile-like of all the mammalia, the duck-billed platypus and the echidna, have been shown to depart from the distinctive mammalian character, and to lay eggs. And, in further confirmation of this supposition, we find, in tracing the mammals and reptiles back through the geological record, that in the Permian and Triassic rocks there occur central forms which combine, in a most remarkable way, reptilian and mammalian characteristics.
Section 42. In conclusion, we would earnestly recommend the student to see more of embryological fact than what is given him here. It is seeing and thinking, much more than reading, which will enable him to clothe the bare terms and phrases of embryology with coherent knowledge. In Howes' Atlas of Biology there is a much fuller series of figures of the frog's development than can be given here, and they are drawn by an abler hand than mine can pretend to be. There is also an Atlas d'Embryologie, by Mathias Duval, that makes the study of the fowl's development entertaining and altogether delightful. Such complete series as these are, from the nature of the case, impossible with the rabbit. Many students who take up the subject of biology do so only as an accessory to more extended work in other departments of science. To such, practical work in embryology is either altogether impossible, or only possibly to a very limited extent. The time it will consume is much greater, and the intellectual result is likely to be far less than the study of such plates as we have named.
Section 43. We have now considered our types, both from the standpoint of adult anatomy and from embryological data; and we have seen through the vertebrate series a common structure underlying wide diversity in external appearance and detailed anatomy. We have seen a certain intermediateness of structure in the frog, as compared with the rabbit and dog-fish, notably in the skull and skeleton, in the circulation, in the ear, and in the reduced myomeres; and we have seen that the rabbit passes in these respects, and in others, through dog-fish- and frog-like stages in its development, and this alone would be quite sufficient to suggest that the similarities of structure are due to other causes than a primordial adaptation to certain conditions of life.
Section 44. It has been suggested by very excellent people that these resemblances are due to some unexplained necessity of adherence to type, as though, the power that they assume created these animals originally, as they are now, coupled creative ability with a plentiful lack of ideas, and so perforce repeated itself with impotent variations. On the other hand, we have the supposition that these are "family likenesses," and the marks of a common ancestry. This is the opinion now accepted by all zoologists of repute.
Section 45. It must not be for a moment imagined that it is implied that rabbits are descended from frogs, or frogs from dog-fish, but that these three forms are remote cousins, derived from some ancient and far simpler progenitor. But since both rabbit and frog pass through phases like the adult condition of the dog-fish, it seems probable that the dog-fish has remained more like the primordial form than these two, and similarly, the frog than the rabbit.
Section 46. Hence we may infer that the mammals were the last of the three groups, of which we have taken types, to appear upon the earth, and that the fishes preceded, the amphibia. Workers in an entirely independent province, that of palaeontology, completely endorse this supposition. The first Vertebrata to appear in the fossil history of the world are fishes; fish spines and placoid scales (compare dog-fish) appear in the Ordovician rocks. In the coal measures come the amphibia; and in the Permo-triassic strata, reptile-like mammals. In the Devonian rocks, which come between the Silurian and the coal measures, we find very plentiful remains of certain fish called the dipnoi, of which group three genera still survive; they display, in numberless features of their anatomy, transitional characters between true fish and amphibia. Similarly, in the Permian come mammal-like reptiles, that point also downward to the amphibia. We find, therefore, the story told by the ovum written also in the rocks.
Section 47. Now, when this fact of a common ancestry is considered, it becomes necessary to explain how this gradual change of animal forms may have been brought about.
Section 48. Two subcontrary propositions hold of the young of any animal. It resembles in many points its parent. It differs in many points from its parent. The general scheme of structure and the greater lines of feature are parental, inherited; there are also novel and unique details that mark the individual. The first fact is the law of inheritance; the second, of variation.
Section 49. Now the parent or parents, since they live and breed, must be more or less, but sufficiently, adapted to their conditions of living-- more or less fitted to the needs of life. The variation in the young animal will be one of three kinds: it will fit the animal still better to the conditions under which its kind live, or it will be a change for the worse, or it is possible to imagine that the variation-- as in the colour variations of domesticated cats-- will affect its prospects in life very little. In the first case, the probability is that the new animal will get on in life, and breed, and multiply above the average; in the second, it is probable that, in the competition for food and other amenities of life, the disadvantage, whatever it is, under which the animal suffers will shorten its career, and abbreviate the tale of its offspring; while, in the third case, an average career may be expected. Hence, disregarding accidents, which may be eliminated from the problem by taking many cases, there is a continual tendency among the members of a species of animals in favour of the proportionate increase of the individuals most completely adapted to the conditions under which the species lives. That is, while the conditions remain unchanged, the animals, considered as one group, are continually more highly perfected to live under those conditions. And under changed conditions the specific form will also change.
Section 50. The idea of this process of change may be perhaps rendered more vivid by giving an imaginary concrete instance of its working. In the jungles of India, which preserve a state of things which has existed for immemorial years, we find the tiger, his stripes simulating jungle reeds, his noiseless approach learnt from nature in countless millions of lessons of success and failure, his perfectly powerful claws and execution methods; and, living in the same jungle, and with him as one of the conditions of life, are small deer, alert, swift, light of build, inconspicuous of colour, sharp of hearing, keen-eyed, keen-scented-- because any downward variation from these attributes means swift and certain death. To capture the deer is a condition, of the tiger's life, to escape the tiger a condition of the deer's; and they play a great contest under these conditions, with life as the stake. The most alert deer almost always escape; the least so, perish.
Section 51. But conditions may alter. For instance, while most of these deer still live in the jungle with tigers, over a considerable area of their habitat, some change may be at work that thins the jungle, destroys the tigers in it, and brings in, let us say, wolves, as an enemy to the deer, instead of tigers. Now, against the wolves, which do not creep, but hunt noisily, and which do not spring suddenly upon prey, but follow by scent, and run it down in packs, keen eyes, sharp ears, acute perceptions, will be far less important than endurance in running. The deer, under the new conditions, will need coarser and more powerful limbs, and a larger chest; it will be an advantage to be rough and big, instead, of frail and inconspicuous, and the ears and eyes need not be so large. The old refinements will mean weakness and death; any variation along the line of size and coarseness will be advantageous. Slight and delicate deer will be continually being killed, rougher and stronger deer continually escaping. And so gradually, under the new circumstances, if they are not sufficient to exterminate the species, the finer characteristics will be eliminated, and a new variety of our old jungle deer will arise, and, if the separation and contrast of the conditions is sufficiently great and permanent, we may, at last, in the course of ages, get a new kind of deer specifically different in its limbs, body, sense organs, colour, and instincts, from the deer that live in the jungle. And these latter will, on their side, be still continually more perfected to the jungle life they are leading.
Section 52. Take a wider range of time and vaster changes of condition than this, and it becomes possible to imagine how the social cattle-- with their united front against an enemy, fierce onslaught, and their general adaptation to prairie life-- have differentiated from the ancestors of the slight and timid deer; how the patient camel, with his storage hump, water storage, and feet padded against hot sand, has been moulded by the necessity of desert life from the same ancestral form. And so we may work back, and link these forms, and other purely vegetarian feeders, with remoter cousins, the ancestral hogs. Working in this way, we presently get a glimpse of a possible yet remoter connection of all these hoofed and mainly vegetarian animals, with certain "central types" that carry us across to the omnivorous, and, in some cases, almost entirely vegetarian bears, and to the great and prosperous family of clawed, meat-eaters. And thus we elucidate, at last, a thread of blood relationship between the, at present, strongly contrasted and antagonistic deer and tiger, and passing thence into still wider generalizations, it would be possible to connect the rabbit playing in the sunshine, with the frog in the ditch, the dog-fish in the sea-waters and the lancelet in the sand. For the transition from dog-fish to rabbit differs from the transition from one species of deer to another only in magnitude: it is an affair of vast epochs instead merely of thousands of years.
Section 53. It would, however, be beyond the design of this book to carry our demonstration of the credibility of a common ancestry of animals still further back. But we may point out here that it is not a theory, based merely upon one set of facts, but one singularly rich in confirmation. We can construct, on purely anatomical grounds, a theoretical pedigree. Now the independent study of embryology suggests exactly the same pedigree, and the entirely independent testimony of palaeontology is precisely in harmony with the already confirmed theory arrived at in this way.
Section 54. It is in the demonstration of this wonderful unity in life, only the more confirmed the more exhaustive our analysis becomes, that the educational value and human interest of biology chiefly lies. In the place of disconnected species of animals, arbitrarily created, and a belief in the settled inexplicable, the student finds an enlightening realization of uniform and active causes beneath an apparent diversity. And the world is not made and dead like a cardboard model or a child's toy, but a living equilibrium; and every day and every hour, every living thing is being weighed in the balance and found sufficient or wanting.
Our little book is the merest beginning in zoology; we have stated one or two groups of facts and made one or two suggestions. The great things of the science of Darwin, Huxley, Wallace, and Balfour remain mainly untold. In the book of nature there are written, for instance, the triumphs of survival, the tragedy of death and extinction, the tragi-comedy of degradation and inheritance, the gruesome lesson of parasitism, and the political satire of colonial organisms. Zoology is, indeed, a philosophy and a literature to those who can read its symbols. In the contemplation of beauty of form and of mechanical beauty, and in the intellectual delight of tracing and elucidating relationships and criticising appearances, there is also for many a great reward in zoological study. With an increasing knowledge of the facts of the form of life, there gradually appears to the student the realization of an entire unity shaped out by their countless, and often beautiful, diversity. And at last, in the place of the manifoldness of a fair or a marine store, the student of science perceives the infinite variety of one consistent and comprehensive Being-- a realization to which no other study leads him at present so surely.
To the student who feels inclined to amplify this brief outline of Vertebrate Anatomy, we may mention the following books: Wiedersheim's and Parker's Vertebrates, Huxley's Anatomy of the Vertebrata, Flower's Osteology of the Mammalia, Wallace's Distribution, Nicholson and Lyddeker's Palaeontology (Volume 2), the summaries in Rolleston's Forms of Animal Life (where a bibliography will be found), and Balfour's Embryology. But reading without practical work is a dull and unprofitable method of study.
[All these questions were actually set at London University Examinations.]
{In Both Editions.}
[Most of these questions were actually set at the Biological Examinations of London University.]
{In Both Editions.}
Students preparing for examinations are frequently troubled by "comparison" questions. Tabulation is often recommended, but we are inclined to favour a rather more flexible plan of marking off differences and resemblances. In tabulation a considerable loss of time is occasioned by writing down the features of both the things compared, and this is a serious consideration for the examinee. We advise him therefore, first, if he possibly can, to draw side by side and in corresponding positions the two things under consideration, and then, going over them in a methodical way, to state simply the difference between each homologous part. We append as examples three test answers actually submitted (with figures) in "Correspondence" work:--
1. Compare the brain of the frog with that of the rabbit.
In the frog's fore-brain--The olfactory lobes are fused in the middle line.
In the mid-brain--
There is no corpus callosum, nor is there a middle commissure to the third ventricle.
The cerebral hemispheres are not convoluted, and, looked at from the dorsal aspect, do not hide the thalamencephalon and mid-brain.
The pineal gland lies in the cranial wall and not deeply between the hemispheres, and its stalk is longer and tilts forward.The optic lobes are two, instead of being corpora quadrigemina, and hollow.
In the hind-brain--The cerebellum is a very small transverse band, and has no lateral parts.
The medulla is relatively larger.
There are no spinal accessory nor hypoglossal nerves to the brain.
2. Compare the vertebrae of dog-fish, rabbit, and frog.
The centra of the dog-fish are -opistho- [amphi]-coelous (i.e., hollow at either end).
The centra of the rabbit are flat-faced.
The centra of the frog are procoelous (hollow in front).
The notochord persists between the centra in the dog-fish and rabbit, within the centra in frog.
The centra of the rabbit have epiphyses, absent in the dogfish and frog.
The transverse processes of the rabbit typically bear ribs. Short ribs occur in the dog-fish, but their homology with those of the rabbit is doubtful. The frog has no ribs.
The interneural plates are peculiar to the dog-fish in this comparison.
3. Compare the skull of the dog with that of the frog.
The Brain Case--Of the frog is a cylindrical box, from which the otic capsules project conspicuously on either side. It contains only two ossifications in its cartilaginous substance (the sphen-ethmoid and the ex-occipital), being protected by the membrane bones, the parieto-frontals above and the parasphenoid below.
Jaw Suspension--
In the mammal it is enormously inflated, and the otic capsules are imbedded in its wall. There are supra- and basi- as well as ex-occipital bones; the para-sphenoid is (? entirely) gone, and its place is taken by the basi- and pre-sphenoids, and the lateral walls contain fresh paired ossifications, the ali- and orbito-sphenoids-- all cartilage bones. The sphenethmoid is perhaps represented in part by the ethmoid.
As a result of the inflation of the brain-case, the squamosal, which slopes downward and outward in the frog, and overlies the cartilaginous suspensorium (quadrate cartilage), has become a constituent of the brain-case wall, and slopes downwardly and in.The point of attachment of the jaw has shifted outward, and the original suspensorial cartilage (the quadrate) has taken on a new and minor function as the incus of the middle ear-- the squamosal superseding it as the suspensory part.
Lower Jaw--Distinct bones in the frog; one mass in the dog.
Otic Capsule--Position as specified. One centre of ossification in the frog forming pro-otic; several fuse together and form periotic of the dog.
Palate--
There is no bulla and no external ear in the frog.In the frog the posterior nares open into the front of the mouth. In the dog the maxillae and palatines send plates down and in (the palatine plates) to cut off a nasal passage from the rest of the buccal chamber, and carry the posterior nares back to the pharynx, thus cutting the vomers off from the mouth roof.
The pterygoids in the dog are much reduced, and do not reach back to the suspensorium.
The frog has no lachrymal bone.
We would impress upon the student at the outset the importance of some preliminary reading before dissection is undertaken. No one would dream of attempting to explore a deserted city without some previous study of maps and guide-books, but we find again and again students undertaking to explore the complicated anatomy of a vertebrated animal without the slightest, or only the slightest, preparatory reading. This is entirely a mistake. A student should be familiar with the nomenclature of the structures he contemplates examining, he should have some idea of their mutual relations and functions, or his attention will inevitably be diverted by the difficulty of new names and physiological questionings to the neglect of his dissection, and that careful observation of form and mutual position which is the essential object of dissection. On the other hand, it is equally necessary-- perhaps more so-- to warn students against the bookish fallacy, and to assure them of the absolute impossibility of realizing biological facts from reading alone. Practical work can alone confirm and complete the knowledge to which the text-book is the guide. In scientific teaching it may sometimes be convenient for the thought to precede the thing, but until the thing has been dealt with the knowledge gained is an unsatisfactory and unstable possession.
For such dissection as the subject-matter of this book requires, the following appliances will be needed:--
(a) Two or three scalpels of various sizes.
(b) Scissors, which must taper gradually, have straight blades, and be pointed at the ends, and which must bite right up to the tips (or they are useless). Two pairs, small and large, are advisable.
(c) Forceps, which must hold firmly, and meet truly at the points.
(d) Two needles set in wooden handles.
(e) An ordinary watchmaker's eye-glass is very helpful, but not indispensable.
(f) A dissecting dish-- an ordinary pie dish will do-- into which melted paraffin wax has been poured, to the depth of, say, three-quarters of an inch, and allowed to solidify. (This wax may be blackened by mixture with lampblack. If the wax floats up at any time, it can, of course, be remelted. Or it may be loaded with lead.)
(g) A rough table or board (for the rabbit and dog-fish).
(h) Blanket pins, and ordinary pins.
(i) A pickle or other wide-mouthed jar, and some common, methylated spirit.
(j) A microscope, with low power of 1 inch or 1/2 inch, and high power 1/6 inch or 1/4 inch. Glass slips and cover glasses, and a bottle of very weak (1 per cent.) solution of salt.
Animals for dissection may be obtained from the recognised dealers, who usually advertise in such scientific periodicals as Nature, Natural Science, and Knowledge. Sinel (naturalist, Jersey) is the most satisfactory dealer in dog-fish in our experience; Bolton (Malvern) will supply Amphioxus through the post; frogs and rabbits may be obtained anywhere. The tame variety of rabbit is quite satisfactory for the purpose of dissection.
The following notes may possibly be of some use to the student; they follow the lines of work arranged by the author for the evening classes of the University Tutorial College, classes considerably restricted as regards time, when compared with ordinary laboratory workers. Most of the sections below occupied about three hours, but for a student working alone they are more likely to take four or five, and even then it is not probable that they will be so satisfactory as if performed under skilled supervision. There are many points extremely difficult to convey verbally which are elucidated at once by actual demonstration upon a specimen. Each of these dissections should be repeated, and it is well if a different condition of the type is selected for the repetition-- an old one if the first specimen was immature, a female if the first was a male.
May be killed by chloroform, or potassium cyanide, or drowned. It may also be readily suffocated with house-hold gas. It should be killed immediately before use, as otherwise the gastric juice attacks the wall of the stomach, and the dissection is, in consequence, rendered extremely disagreeable. A very young rabbit is unsatisfactory as regards the genitalia, but otherwise there is no objection to a little one, and it has this advantage-- that it may be immersed more conveniently under water, in a large pie dish, for purposes of fine dissection. The external features of the animal should be examined: eyelids, whiskers and teeth, toes, anus, perineal space on either side of the same, urogenital opening, and position of the ribs, vertebral column, and limb girdles beneath the skin should be made out. Then the animal should be pinned out through the legs, the ventral surface uppermost, the skin opened up along the middle line from pelvic girdle to symphyses of jaw; separated from the body wall below by means of the handle of a scalpel, and turned back; and then the abdominal wall should be cut into and two flaps pinned back to expose its contents. Note the xiphisternum. The caecum and colon will be recognised (Section 16); the stomach, the right and left central, and left lateral lobes of the liver will probably be apparent; and the urinary bladder (especially if distended) in the middle line behind. Without any further dissection, but simply by turning the parts over, all the structures of the abdomen in Figure 1, Sheet 1, will be identified. Seek especially for and note particularly, the gall bladder, bile duct, and portal vein, pancreatic duct, sacculus rotundus, vermiform appendix, ureters (by pulling urinary bladder forward), genital ducts (looping over ureters), spleen, kidneys, and adrenals. The vena cava inferior is seen dorsally. The genital duct guides the student to the genital gland; if the subject is a male, the testes may be exposed by dissection, or by pulling the vas deferens gently the scrotal sac will be turned inside out, and the testes brought into view. The ovary lies exposed without dissection posterior to the kidney. Examine all this carefully, and make small sketches of points of interest-- the duodenal loop and the pyloric end of the stomach, for instance; the meeting of colon, caecum, and sacculus rotundus again; or the urinary bladder and adjacent parts. Note the dorsal aorta and vena cava and their connexions behind. (Compare figure of circulation.) Cut through pelvic girdle, and remove one hind leg, to see bladder and genital ducts better (compare Sheet 10). Wash away any blood that may flow. Turn all the intestines over to the animal's right, and see the dorsal aorta and vena cava inferior of the abdomen, the inferior mesenteric artery, and the spermatic (or ovarian) artery (compare, of course, with figure in book). In front, immediately dorsal to the spleen, is a variable quantity of lymphoidal tissue, which must be very carefully cleared to see the superior mesenteric and coeliac arteries. Separate Spigelian lobe from stomach, and look for vagus nerve descending by oesophagus, solar plexus around the superior mesenteric artery, and thrown up very distinctly by the purple vena cava inferior beneath, and the splanchnic nerve. To see the abdominal sympathetic behind, gently remove the peritoneum that lies on either side of the aorta; blood-vessels will be seen running in between the vertebral bodies, and the sympathetic chain, with its ganglia, made out very distinctly, as it runs across them longitudinally. Now cut oesophagus just in front of stomach, and cut the rectum, cut through the mesentery supporting the intestine, and remove and unravel alimentary canal; cut open, wash out, and examine caecum and stomach. Bleeding to a considerable extent is inevitable, chiefly from the portal vein. The liver had better remain if the same rabbit is to serve for the second dissection.
Second Dissection.-- Skin front of thorax and neck. Note subclavian veins running out to fore limbs-- avoid cutting these. Cut through ribs and remove front of thorax, to expose its contents; cut up middle line of neck, and clear off small muscle bands, to expose bloodvessels; pick away carefully whatever is left of thymus gland; make out structure of heart and blood-vessels, as described, in Chapter 3; note larynx and trachea. Now proceed to the examination of the nerves of this region. See phrenic nerve, by vena cava inferior, and between heart and lungs, and sympathetic, running over the heads of the ribs. By the common carotids will be found the large white vagus nerve, the greyish sympathetic, and a small branch of X., the depressor. Make out branches of X. named in text. The big white cervical spinal nerves will be evident dorsally. Clear forward into the angle between the jaw and the bulla tympani, to see XII. and XI.; IX. will be found, lying deeper, dorsal to the carotid artery and body of the hyoid. Compare with figure given of this. Skin the cheek, and see VII. running over it. Cut through malar and remove it; cut through lower jaw-bone and turn it back, to see the third branch of the fifth nerve on its inner side; examine the muscles of eyeball, and remove it, to expose the first and second branches of V.-- the latter is especially deep within orbit. Remove, open, wash out, and examine the heart. Shave off the dorsal wall of cranium, to expose hemispheres of brain, and then put the head in strong spirit for a week or so. With a second rabbit, this dissection may advantageously be varied by removing the lower jaw, cutting -up- [through] soft palate, and observing openings of the Eustachian tubes. [The tonsils (on the ventral side of the soft palate) must not confused with these.] The heart should also be cut out, washed out and examined (Compare Sections 38, 44.)
Third Dissection.-- (Before this is performed the mammalian skull should have been studied and examined.) Take the head of a rabbit, the brain of which has been hardened by spirit, and carefully remove cranium; be particularly careful in picking away the periotic bone, on account of the flocculi of cerebellum. It is difficult to avoid injury to the pituitary body embedded in the basisphenoid bone. Examine with the help of Sheet 8. Make the sections there indicated.
May be killed by drowning in dilute methylated spirit, or by chloroform. Take a recently-killed frog, and examine a drop of its blood, spread out on a glass slip, under the microscope; compare it with your own. Before using the high power, put a cover glass over the object, of course. Scrape the roof of the mouth of the frog gently, to obtain ciliated epithelium; and mount in very weak salt solution-- the cilia will still be active. Squamous epithelium may be seen by the student similarly scraping the interior of his own cheek. Take a piece of muscle from one of the frog's limbs, tease out with needles upon a glass slip, and examine. To see the striations clearly, the high power will be needed. Compare a piece of muscle from the wall of the alimentary canal. Similarly examine nerve and connective tissue.
First Dissection.-- Pin out the frog in a dissecting dish, ventral surface uppermost, and cover with water. Open up the skin along the mid-ventral line. Note the large sub-cutaneous lymph spaces, the pelvic and pectoral girdles, and the anterior abdominal vein. Cut into the body cavity on one side of this latter, cut across in front of where the vein dips down to liver, and peel the body wall away from it. The xiphisternum will probably be cut in this operation. In early spring the females are greatly distended with ova, and the greater portion of the ovary may, with advantage, be removed. The oviduct is dead white then, and larger and much more in evidence than the (pinkish) intestine even. Turn over the viscera, and compare with Sheet 11; one lung is often found greatly inflated, and then projects back into the body cavity; the stomach is, in some cases, pushed forward and hidden behind the shoulder girdle. Observe the allantoic bladder, the spleen, gall bladder, portal vein, and pancreas. By squeezing the gall bladder gently, the bile duct will be injected with bile, and will be apparent if the stomach is turned over. The oesophagus, just in front of the stomach, should be cut through, and the rectum, and the mesentery and alimentary canal supported by it, removed. This will expose the urogenital organs. (Vide Figures given.) These vary greatly, especially in the females, at different seasons. The condition figured would be seen in late autumn, or winter. In spring females are often found copulating with males, and then the ovary itself is inconspicuous, while the lower part of the oviduct is enormously distended with ova, so as to be mistaken sometimes for the ovary by those who fail to note that the ova are enclosed by a thin semi-transparent skin (wall of oviduct). The vena cava inferior is seen between the kidneys and the renal portal vein beside the ureter. Cutting through the mesentery supporting the kidney laterally, the dorsal aorta is exposed, and on either side of it the sympathetic chain and rami communicantes, often tinged with black pigment. This black pigment is a frequent but variable feature of the frog's anatomy, and usually dapples or blackens the testes, and also sometimes darkens the otherwise pale pink arteries. Behind the kidneys the sciatic plexus also becomes visible. Careful drawings should be made. Cut off the head of the frog, shave off top of brain case, and put the head in strong spirit.
Second Dissection.-- A fresh frog is required. Pin out under water as before, and open up body cavity. Now carefully remove the muscle from the ventral portion of the shoulder girdle, to expose the clavicles and coracoids. Cut away xiphisternum, and then cut through clavicles and coracoids on either side, and remove ventral part of shoulder girdle, to expose the heart. Open out the cut portions of body wall and pin. The veins going towards the heart should now, with a little examination, be evident. Make out the external jugular, the innominate, and its two branches, and the pulmo-cutaneous and vena cava superior. Clear by carefully picking away any shreds of semi-transparent tissue. Make out, by feeling, the position of the hyoid body, and of its anterior cornua. Note the hypoglossal nerve (first spinal) running ventral to this, and the ninth cranial nerve, running parallel to it but dorsal to the hyoid-- hidden therefore by the hyoid, and reappearing in front. The vagus may also be made out less distinctly, running "postero-ventrally" towards the heart. By clearing the muscle by the ramus of the jaw, VII. may be seen, and the third branch of V., running across the jaw at about the middle of its length. Pick off the thin transparent pericardium from the heart very carefully, and proceed to cut away all the veins made out. The truncus arteriosus may then be followed up as it branches. Note all the branches shown in the figures in this book. The precise position of the vessels will vary to a certain extent with the attitude in which the frog is pinned. The cutaneous artery will prevent the student following up the aortic arch until it is cut; then the arch may be followed round until it meets its fellow to form the dorsal aorta. Note the sympathetic again. Make careful drawings of all this. Cut off lower jaw, and note posterior nares and Eustachian openings. If time allows, remove the heart, and examine by cutting open and washing. (Compare, Section 44) Remove eyeball, to see the first and second branches of the fifth nerve, and the Vidian (i.e. palatial) branch of the seventh.
Third Dissection.-- Read the account of the frog's skull carefully. Take the head of a recently killed frog and drop into boiling water for a minute. Then pick off, very carefully, muscle, connective tissue, nerves, and etc., to clear the cranio-facial apparatus; examine the bones, compare with figures given in this book, and draw. Take the head, which has been in spirit a fortnight or so, pick away cranium, and compare brain with figures given. Examine ventricles, by taking sections, after drawings have been made.
First Dissection.-- Examine external characters, nasal grooves-- no internal nares-- fins, spiracle, scales passing over lips, and cloaca. Cut off tail below the cloacal opening. The males are distinguished by the large claspers along the inner edge of the pelvic fin. Open up body cavity. Usually this is in a terrible mess in the fish supplied by dealers, through the post-mortem digestion of the stomach. Wash out all this under a stream of water from a tap or water-bottle. Frequently the testes are washed out of the male in this operation and ova from the loose ovaries in the female. Now compare with figure given in this book, allowing for the collapse of the stomach, if it has occurred. Cut through the oesophagus and rectum, and remove alimentary canal from body; cut open and wash out the intestine, and examine spiral valve. Now make a careful examination of the cloaca and its apertures, and dissect away the peritoneum hiding the kidney. In the female find the opening of the oviducts in front of the liver. Remove liver, and cut off body now behind pectoral fin. Before throwing tail and hinder part of body away, note the myotomes of body wall, the notochord and vertebral body, neural canal, and, in the tail, the haemal canal. [(See {Section 9 the Dog-fish})]
{Lines from First Edition only.}
-The relation of the vertebral bodies to the notochord may be very well seen by taking successive slices, about one-tenth of an inch thick, through the vertebral body. The cartilage is hard and semi-transparent, the notochord jelly-like, least at the centres of the centra, and at a maximum intervertebrally.-
[The notochord is a soft jelly.] Cut away the ventral part of the pectoral girdle, to open pericardium. With a seeker, make out the pericardio peritoneal opening. Cut into the sinus venous, and run seekers into the Cuvierian and hepatic sinuses. [Cut open the Cuvierian and posterior cardinal sinuses, and run seekers into their affluents.] Dissect along the truncus arteriosus to afferent branchials. [Cut away the heart and oesophagus; run a seeker up the dorsal aorta and cut along it from the ventral side to subclavian and efferent branchial arteries.] Skin the top of the head. Note, while doing this, the yellow, jelly-like sense-tubuli beneath the skin. Shave off top of brain-case, and leave the head in spirit for a week or so.
Second Dissection.-- Place the head with the ventral side downward, skin all the dorsal surface as yet unskinned. Refer to book for precise position of the anterior cardinal sinus, and then cut down through body wall into this just over gill slits. The tenth nerve will become visible, with its "slit" branches athwart the floor of the sinus. Clear to make this more evident, and make out its lateral line and visceral branches, and the ninth nerve.
{Lines from Second Edition only.}
[The pharyngo-branchials may be felt beneath the sinus. Run a seeker from the dorsal aorta to the efferent branchials.]
Proceed now to orbit, and, without any dissection beyond the removal of skin, make out recti and oblique muscles of eyeball, and the optic, third and fourth nerves. Cut through these structures carefully and remove, exposing nerves seven, and five, as described and figured in the text. Examine the otic capsule by taking successive slices through it to show the labyrinth of the ear. -Remove the dorsal wall of the skull to obtain a dorsal view of brain. If this is sufficiently hard, examine it; if not, return it to spirit for a more convenient occasion.- [Examine brain.]
Two specimens of this type should be obtained. It should be examined entire by the naked eye and with the low power of the microscope. Immersion, in glycerine will render it more transparent; or it may be cleared with oil of cloves, put up temporarily in that, or permanently in Canada balsam. One specimen should then be pinned out in the dissecting dish, ventral side uppermost, and the atrium opened to expose liver and pharynx. A part of the pharynx may be examined with the low power to see the form of the gill slits. The second specimen should be soaked in turpentine for some time, and then dropped into melted paraffin wax. Transverse sections may then be cut with a razor, the paraffin wax removed from these by solution in turpentine, the turpentine in its turn dissolved out by alcohol, and the sections, after immersion in oil of cloves, may be transferred to Canada balsam for examination and preservation. This work should not be attempted until some practical histological work has been done in botany, and it may be altogether avoided by the purchase of stained and mounted sections.
Laboratory work in this portion of the science is not usually undertaken by elementary students of biology, but the reader will probably find it helpful, in the realization of the facts given in this book, to look out for frog spawn, in February and March, and to catch and examine tadpoles of various sizes. A small dissecting dish may be made by pouring melted paraffin wax into one of those shallow china pots chemists use for cold-cream, and tadpoles may be pinned out with entomologists' pins and dissected with needles. But this is a work of supererogation. Partially incubated hen's eggs may be obtained at a small cost almost anywhere, and the later stages profitably examined and dissected under warm water. For a clear understanding of the allantois and amnion, this last is almost indispensable. A few microscopic slides of sections of embryonic chicks should also be compared with our rough diagrams.
|
Figure 1. Main facts of the Rabbit's Anatomy (diagrammatic). an., anus. a.ao., arch of the aorta. au., auricle. a.r., ad-renal body. br., brain. b.d., bile duct. brch., bronchus. cd.st., cardiac end of stomach. co., colon. cae., caecum. ddnm., duodenum. d.ao., dorsal aorta. dia., diaphragm. ep., epiglottis. g.d., genital duct (either sex). il., ileum. in.art., innominate artery. k., kidney. lg., lung. lv., liver. l., larynx. l.s.c., [l.c.c.] left common carotid artery. m., mouth. na., nasal passage. oes., oesophagus. p.v., pyloric valve. p.d., pancreatic duct. pt., peritoneal cavity. r., rectum. st., stomach. [stm., sternum.] s.r., sacculus rotundus. s.c., spinal cord. tr., trachea. ur., ureter. ur.b., urinary bladder. v.b., a vertebral body. v.ap., vermiform appendix. v.v., [v.p.] velum palatium. v., ventricle of heart. v.c.i., vena cava inferior. Figure 2. The Liver (diagrammatic). g.b., the gall bladder. r.l., r.c., l.l.., l.c., right lateral and central, and left lateral and central, lobes respectively. sp., the Spigelian lobe (fits into angle of stomach and oesophagus). |

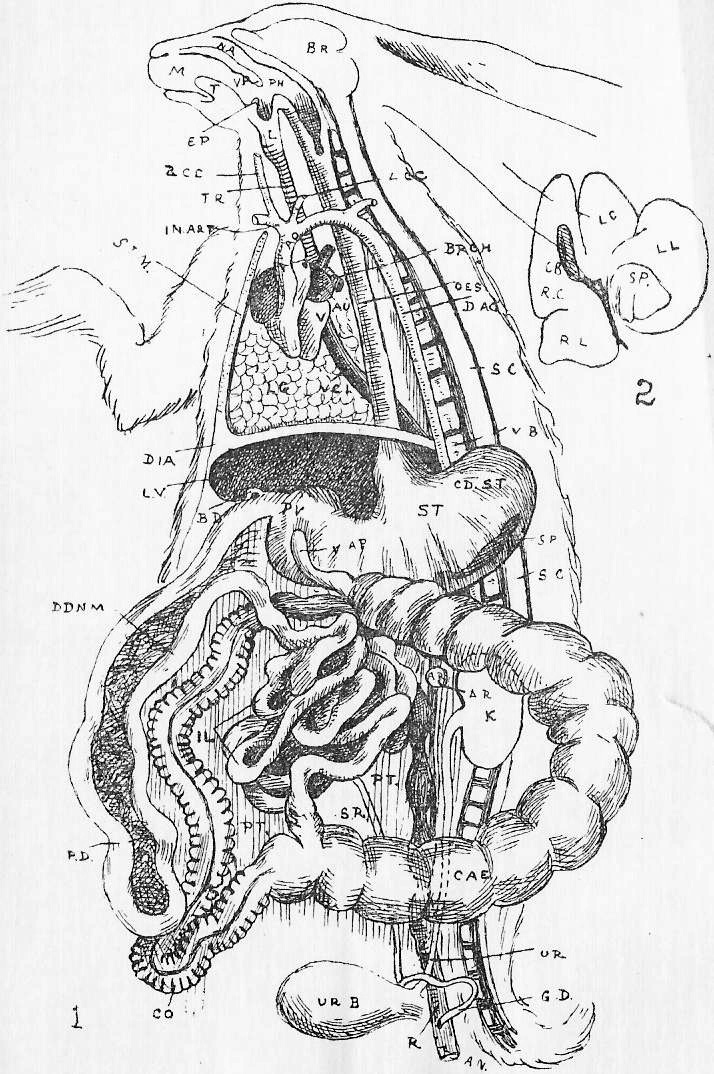
|
Figure 1. The Rabbit's Circulation (see footnote to Section 45). (Throughout l. indicates left, r. right. Vessels without r. or l. prefixed are median.) -[* The figure is inaccurate at one point; l.c.c. should spring from the base of inn. See Sheet 9.]- {First Edition only text} ao.a., aortic arch. au., auricle. az.v., (p.c. in Figure 2), azygos vein. c.c., common carotid. c.il.a., common iliac artery. coe.a., coeliac artery. d.ao., dorsal aorta. e.il.v., external iliac vein. e.ju., external jugular vein. f., femoral artery. h.v., hepatic vein. inn., innominate artery. in.j., internal jugular vein. i.il.a., internal iliac artery. i.il.v., internal iliac vein. k., kidney. lv., liver. l.g.v., lienogastric vein (portal). m.v., mesenteric (portal system). p.m.a., posterior mesenteric artery. p.v., main portal vein. p.a. pulmonary artery. r., rectum. r.a., renal artery. r.v., renal vein. s.v., and a., spermatic (or ovarian) vein and artery (to genital organ). s.mes.a., superior mesenteric artery. s.-cl.a., subclavian artery. s.-cl.v., subclavian vein. v.c.s., vena cava superior. v.c.i., vena cava inferior. v. or vn., ventricle. Figure 2. Figure of Circulation (simplified) illustrating certain points in development to be referred to later. Figure 3. Respiration. See text, Section 41. Figure 4. Blood. See text, Section 35. |
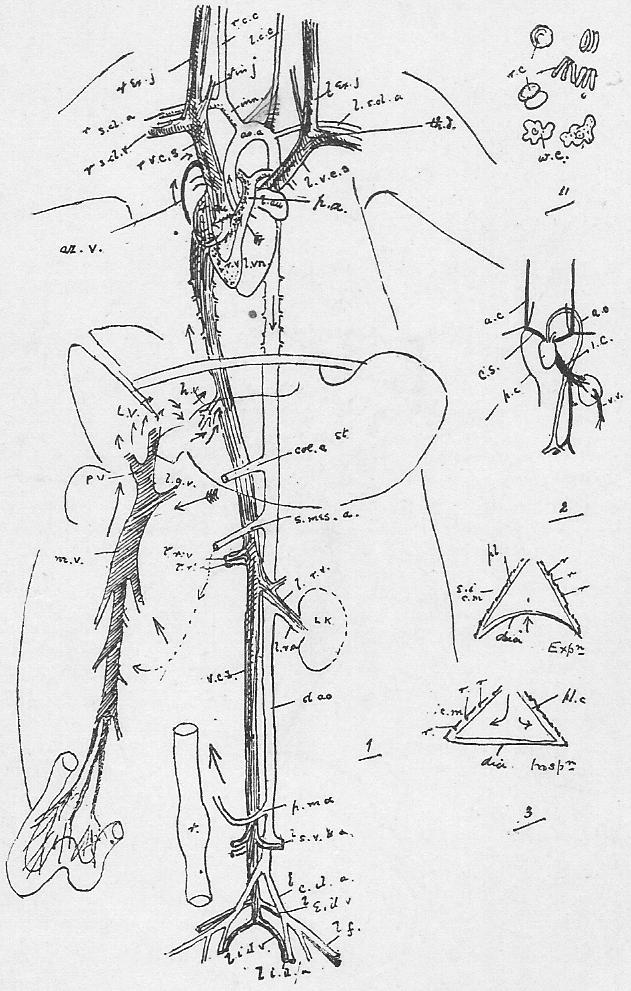
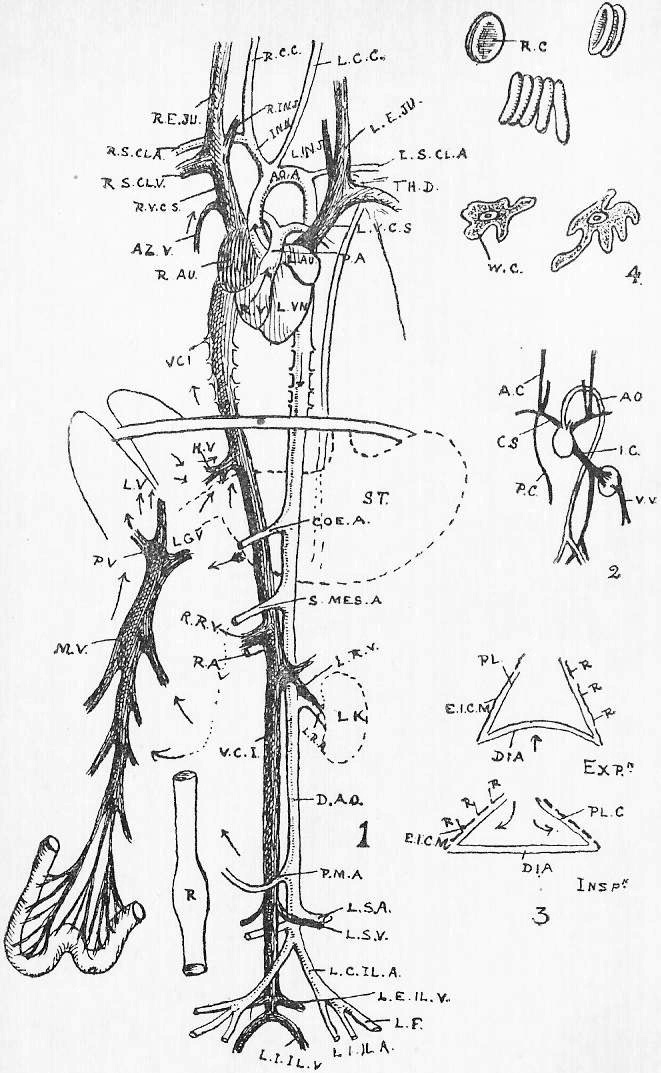
|
{No numbers I., or II.} Figure III. An amoeba.-- n., nucleus. ns., nucleolus. c.v., contractile vacuole. Figure IV. Embryonic tissue from the blastoderm of a chick. Figure V. Columnar epithelium.-- g.c.1, g.c.2, g.c.3, successive phases in the development of a goblet cell. Figure VI. g.end., is geminating endothelium; the cells divide and apparently drop off to become white corpuscles in the lymph current. sq.end., squamous endothelium from the mesentery. sq.ep., squamous epithelium (from the mucous membrane within the cheek). st., are opening (stomata) communicating between the lymphatics in the mesentery and the peritoneal (coelomic) space. Figure VII. Ciliated epithelium from the roof of the frog's mouth. Figure VIII. Forms of glands.-- g.ep., is a gastric gland from the stomach; trs., below, is cross section. This is one of the simplest types of gland. s.g., a sweat gland, is also a simple tube, but convoluted below. r.g., is a racemose gland, such as the pancreas, Brunner's or the salivary glands. The kidney, we shall see later, is simply an aggregate of branching tubuli (Sheet 7). Figure IX. A duodenal villus.-- lac., the lacteal. v., the vein. Figure X.a. Diagram of liver structure.-- b.d., the inter-lobular bile duct. h.a., the hepatic artery, bringing blood to oxygenate and nourish the liver tissue, and similarly distributed. h.v., the hepatic vein taking blood from the liver to the heart, its twigs commencing in the lobuli (intra-lobular). lb. lb., lobuli. p.v., the portal vein bringing blood, from which substances are to be elaborated, into the liver, and breaking up between the lobuli (inter-lobular). Figure X.b. A diagram of the appearance of an injected liver lobule as seen in section under the microscope. |
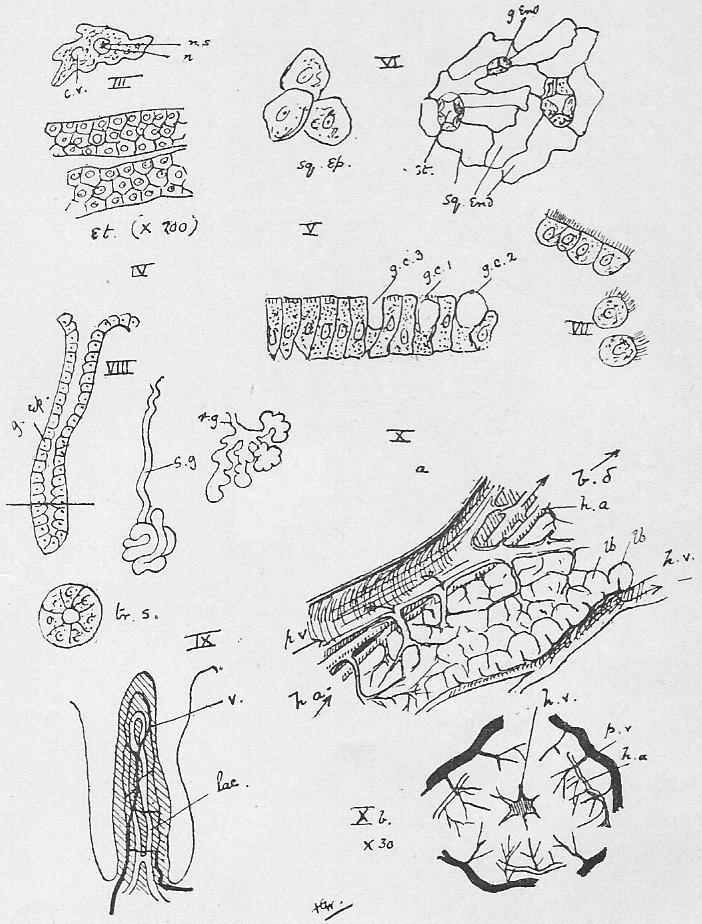
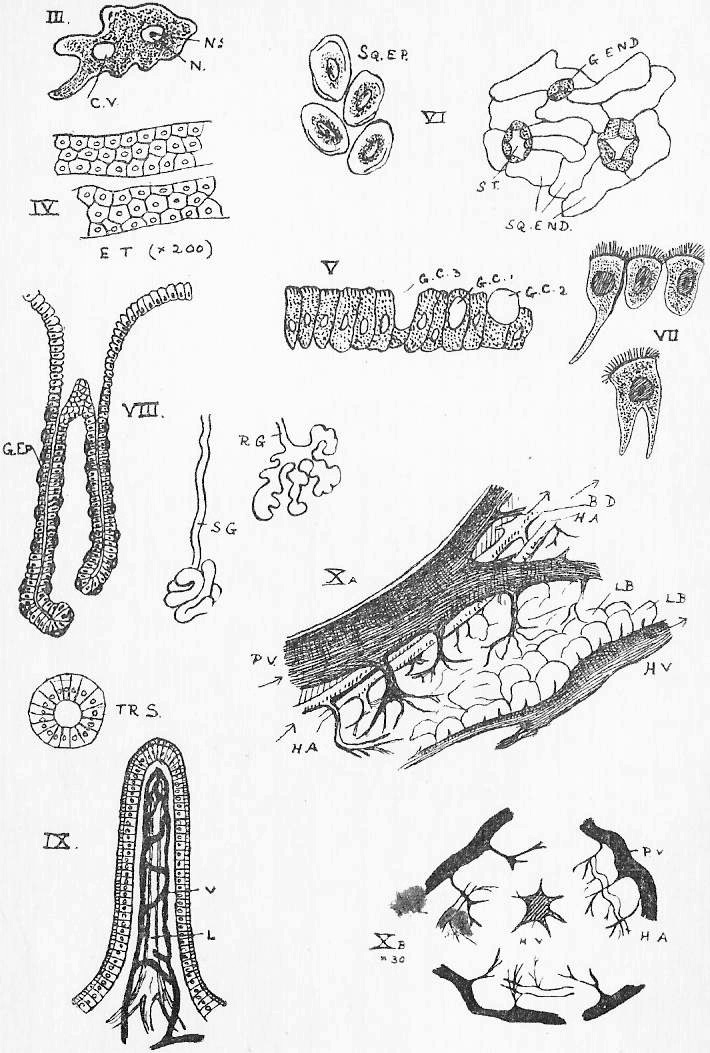
|
Figure XI. A blood capillary. White corpuscles are migrating through the walls into the tissues (compare Section 66). Figure XII. Hyaline cartilage (Section 64). Figure XIII. c.c., connective tissue corpuscle. w.i.f., white inelastic fibres. y.e.f., yellow elastic fibres. Figure XIV. Botryoidal tissue (Section 66). Figure XV. Development of a fat drop.-- f.d., fat drop, in a connective tissue corpuscle; c.c., in the formation of adipose tissue (Section 67). Figure XVI. Diagrammic cross section of a long bone.-- b.c., bone corpuscle in a lacuna. H.v., Haversian vessel (in the Haversian canal) surrounded by concentric lamellae of bone, c.l., and together with these and zones of bone corpuscles, called a Haversian system. i.l., inner lamellae. m.c., medullary canal full of yellow marrow. o.l., outer lamellae. p.o., periosteum. Figure XVII. To illustrate bone development (Section 71). Figure XVIII. Dentition of rabbit, incisors 2/1, canine 0/0, premolar 3/2, molar 3/3. |
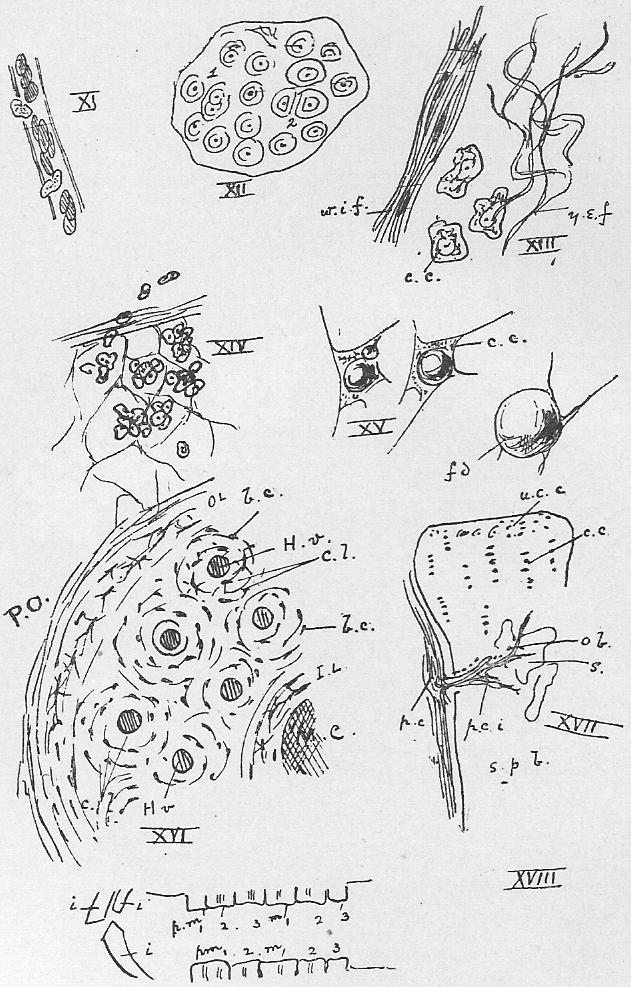
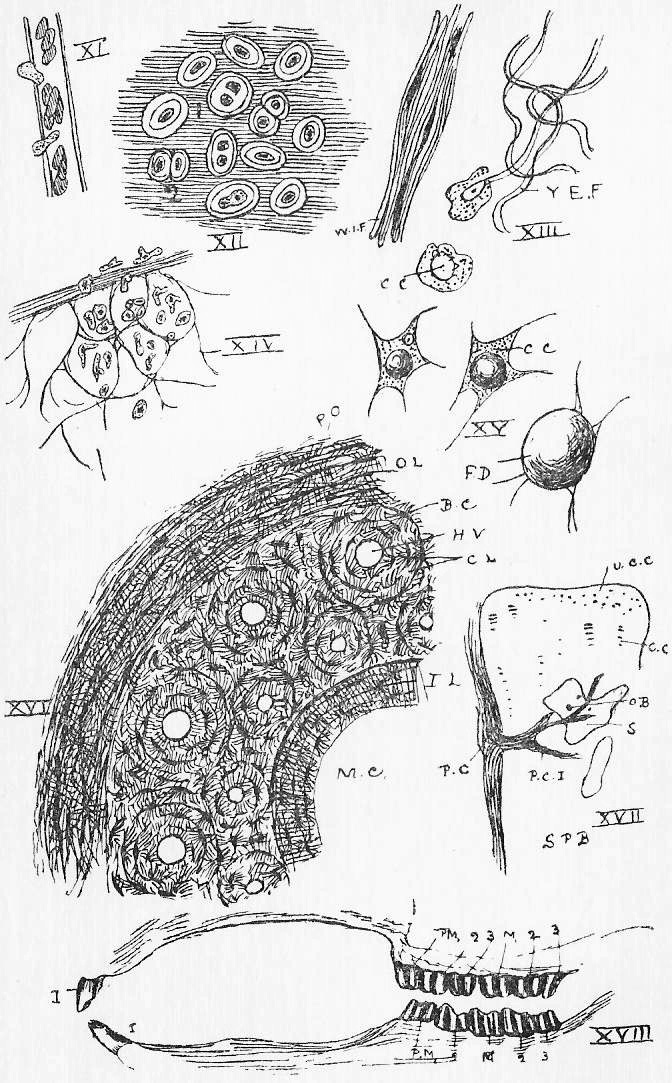
|
D and D' show the fore and hind limbs, to illustrate their homology. D is in the embryonic position. The radius and tibia are, at an early stage in development, on the anterior edge of their respective limbs; the ulna and fibula, posterior; the former are spoken of as preaxial in position, the latter as postaxial. But in the adult the humerus is twisted so that the proximal end of the radius lies at the outer side of the elbow, whence it crosses the ulna, so that its distal end is inside, while the femur is also twisted round, so that the entire tibia is internal. Figures 1 and 2. -Limbs.-- a.c., acetabulum. acr., acromion. as., astragulus. c., carpus. ca., calcaneum. co., coracoid. [coty., cotyloid bone.] fb., fibula. fe., femur. g., glenoid cavity (for head of humerus). hd., head of femur. hum., humerus. i., ilium. is., ischium. m.c., meta-carpals. na., navicular. o., olecranon process of ulna. o.f., olfactory fossa. pb., pubis. r., radius. u., ulna. Figure 3. -Sternum.-- Mb., manubrium. r1., r2., and etc., sternal ribs. st., sternebrae. xi., Xiphisternum. Figure 4. Vertebrae.-- At., Atlas. Ax., axis. c., [b.] centrum. C.V., caudal vertebra. c.v., [Cer.V.] cervical vertebra. ep., epiphysis. f.r., fused rib (in cervical vertebrae). L.V., Lumbar vertebra. m., metapophysis (of lumbar vertebra). n.a., neural arch. n.s., neural spine. r., rib. S.V., sacral vertebra. [T.V., Thoracic.] tr.p., transverse process. v.a.c., vertebrarterial canal. z., zygapophysis. |
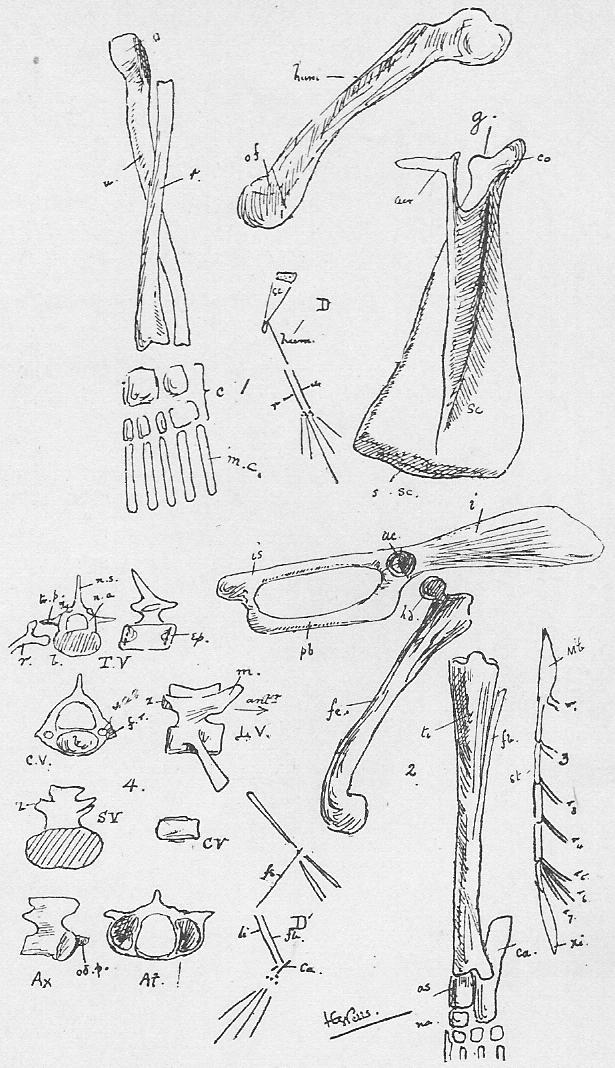
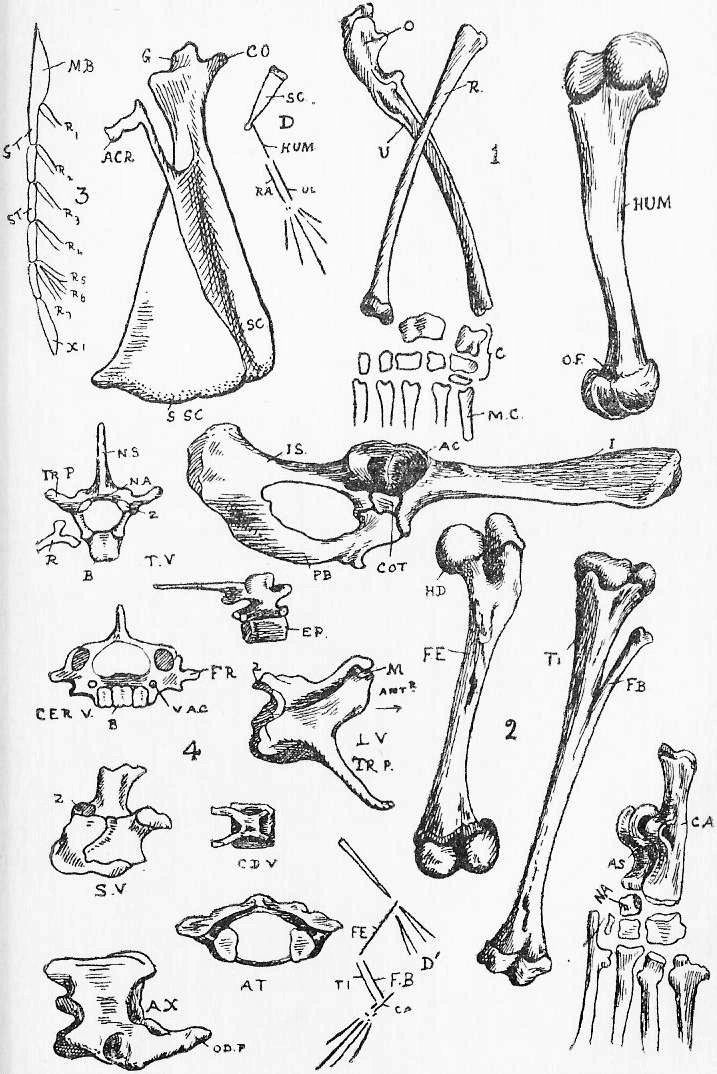
|
a.n., anterior nares. a.s., ali-sphenoid. b.h., body of the hyoid. b.o., basi-occipital. b.sp., basi-sphenoid. c., condyle of the skull. {c.1, c.4, canines.} c.f., condylar foramen (for XII.). c.h., cerato-hyal. E.f., Eustachian foramen. e.h., epihal. -e.n., or a.n., the anterior nares.- e.o., exoccipital. eth., ethmoid. e.t., ethmo-turbinal. f., frontal. f.l.a., foramen lacerum anterius. f.l.m., foramen lacerum medium. f.l.p., foramen lacerum posterius (for IX., X., XI.). F.M., or f.m., foramen magnum. f.o., foramen ovale. f.r., foramen rotundum. {i., incisors.} ju., jugal. m., molars. m.t., maxillo-turbinal. mx., maxilla. na., nasal. n.t., nasal turbinal. o.f., optic foramen. o.s., orbito-sphenoid. p., or pal., palatine. pa., parietal. p.m., pre-maxilla. p.m.1, p.m.4, premolars. p.n., posterior nares. p.sp., pre-sphenoid. pt., pterygoid. s.h., stylo-hyal. s.m.f., stylo-mastoid foramen (for VII.). s.o., supra-occipital. sq., squamosal. s.t., sectorial tooth. t.h. thyro-hyal. vo., -black line indicating position of- vomer. z.p., zygomatic process of squamosal. |
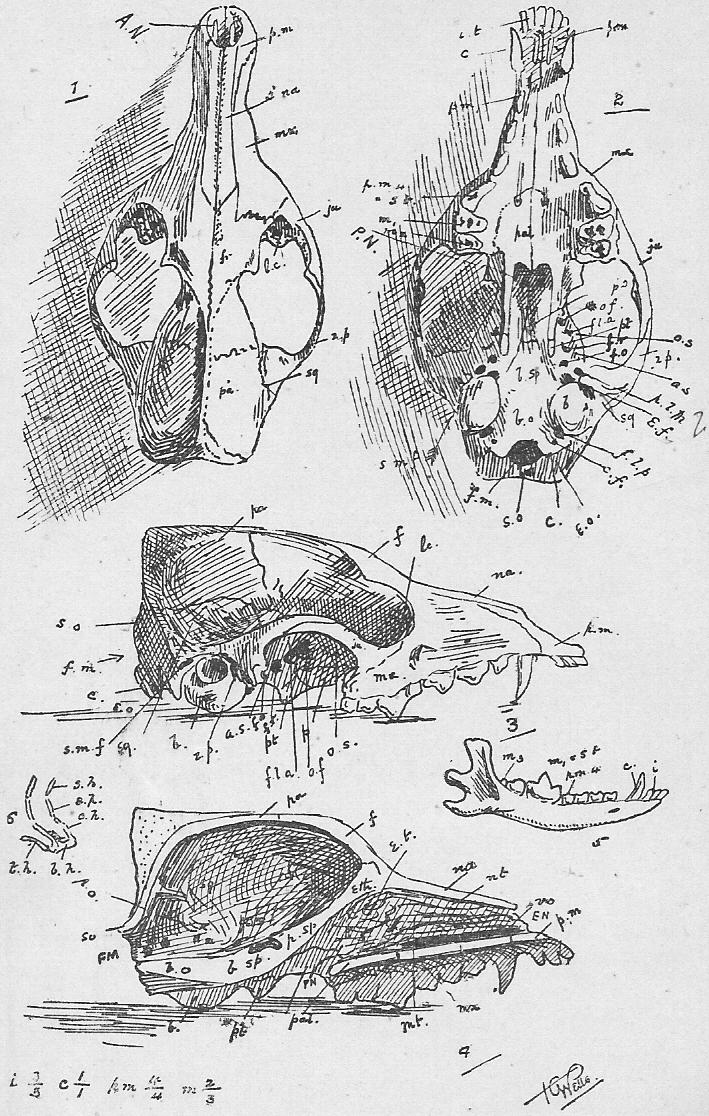
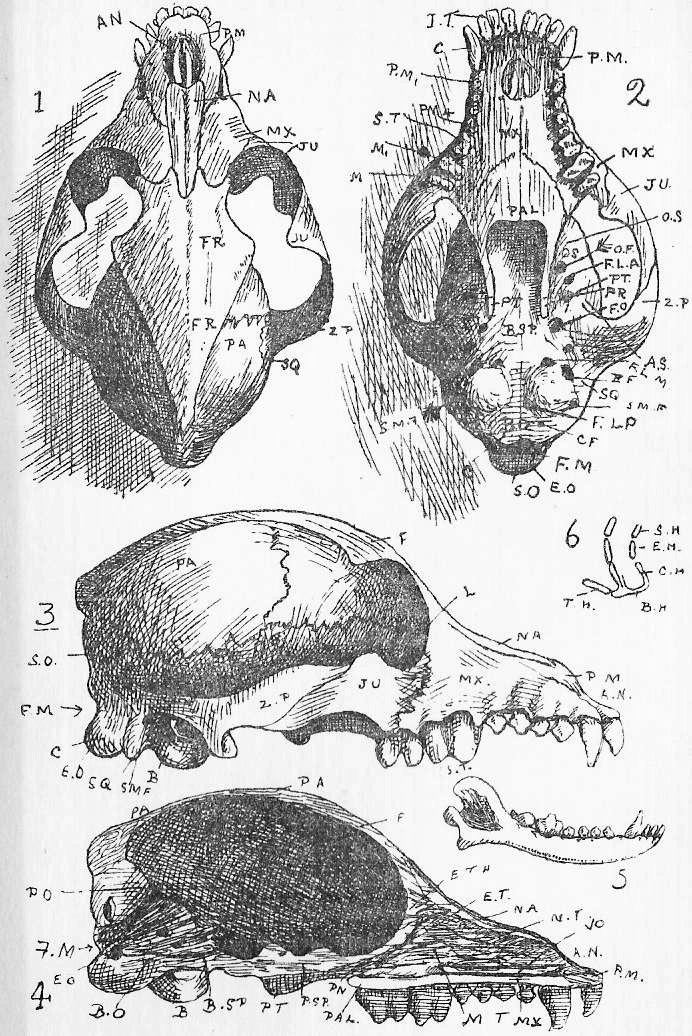
|
Figure 1. Striated muscle fibre (of the Rabbit), ruptured to show sarcolemma. e.p., its end plate. K.m., membrane of Krause. n., nucleus. nv., nerve. sc., sarcolemma. s.e., sarcous elements. Figure 2. Cardiac muscle. Figure 3. Unstriated muscle fibres. Figure 4. Diagram of the Skin. b.v., blood vessel. d., areolar tissue of the dermis (mesoblastic). s.c., stratum corneum, and s.m., stratum mucosum of the epidermis. s.g., sweat gland. t.c., tactile corpuscle. Figure 5. To illustrate Kidney structure.-- a.b.v., and e.b.v., afferent and efferent blood-vessels, of which the latter go to break up upon the tubli. B.c., one of Bowman's capsules of the cortex; ur.t., the uriniferous tubule running from it into the medulla, where it loops and branches; around it branches a blood-vessel, of which the latter go to break up upon the tubuli. c., cortex. g., glomerulus, a knot of blood-vessels in the capsule. m., medulla. p., pelvis. ur., ureter. The water of the urine is probably filtered off in the capsule, the urea and other salts secreted by the tubuli. {No Figure 6.} Figure 7. The Auditory structures of the Rabbit (diagram). See text, Section 115. Figure 8. The Eye (diagram). See text, Section 111. Figure 9. The Retina (diagram). See text, Section 112. |
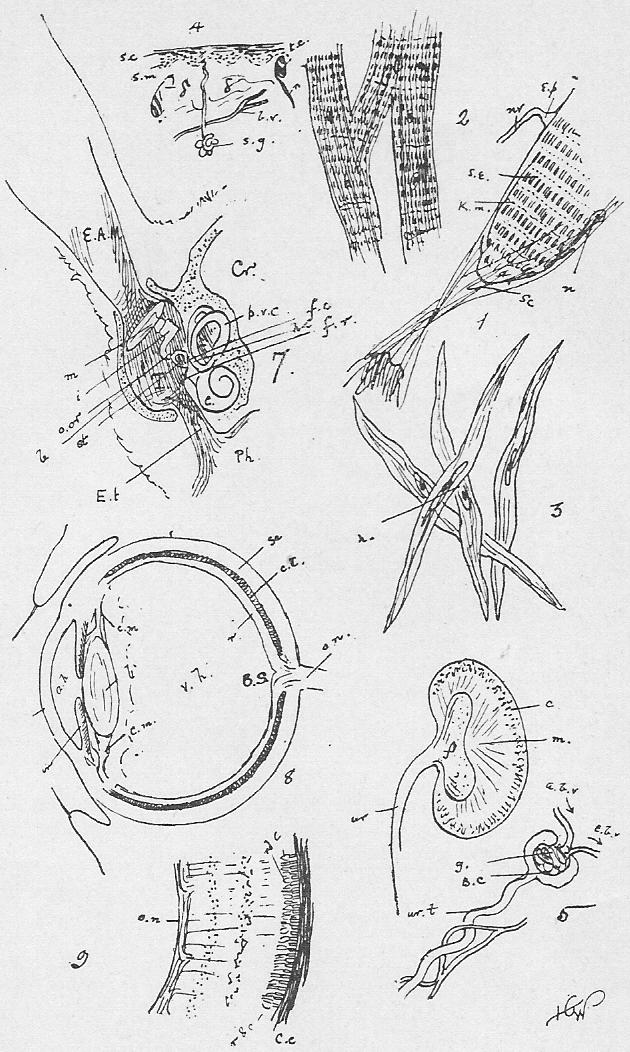
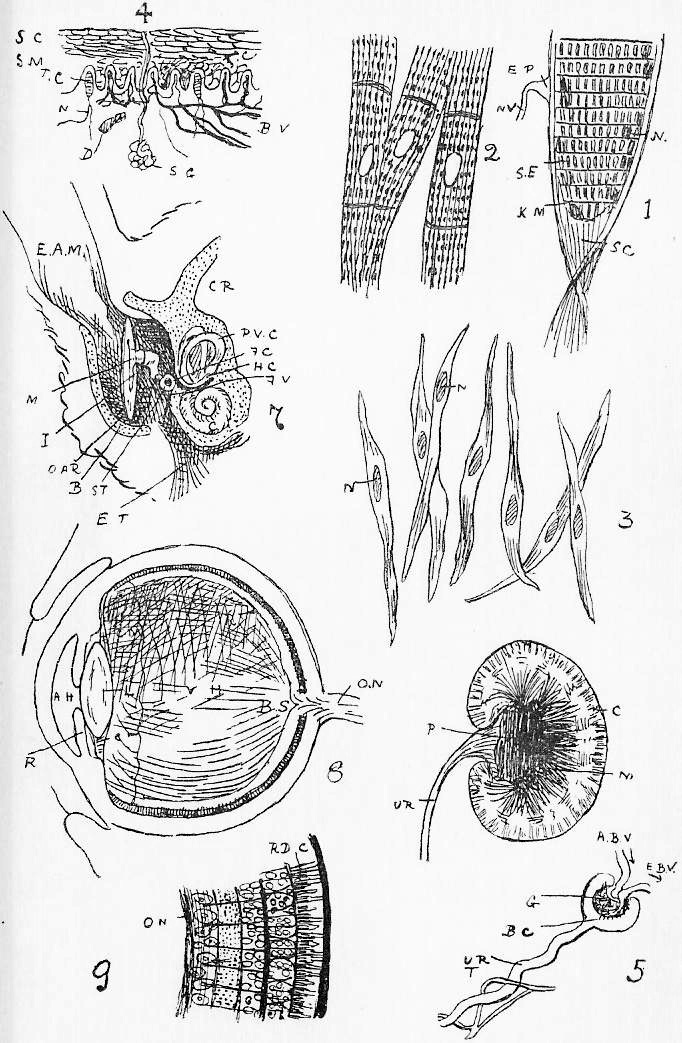
|
{Figures 1-5.} ar., arrow in the iter. a.c., the anterior commissure, a thickening of the anterior wall of the third ventricle. c.c., corpus callosum. c. cb., crura cerebri. c.h., cerebral hemispheres. c.q., corpora quadrigemina. f.cbm. (right), flocculus of the cerebellum. l.h., left cerebral hemisphere (=ch.). l.l., lateral lobe of cerebellum. m.c., middle commissure. m.o., medulla oblongata. op., optic nerve. o.l., olfactory lobe. o.th., (right), optic thalamus. p.c., posterior commissure (thickening of postero-dorsal wall of the third ventricle). p.g., pineal gland. pt., pituitary body. p.V., pons Varolii. s.c., thin roof of the fourth ventricle. v.cbm., vermis of cerebrum. v.l., lateral ventricle. {Figure 4.} Nerves.-- I., Olfactory. II., Optic. III., Oculo-motor. IV., Patheticus. V., Trigeminal. VI., Abducens. VII., Facial (portio dura). VIII., Auditory (portio mollis). IX., Gustatory (glossopharyngeal. X., Pneumogastric or vagus. XI., Spinal Accessory. XII., Hypoglossal. Figure 6. The Spinal Cord in section.-- c.c., the central canal. d.f., the dorsal fissure. d.n., the dorsal nerve root; g., its ganglion. v.f., the ventral fissure. v.n., the ventral nerve root. Note that in Figure 1 the central canal is continuous with the fourth ventricle. Figure 7. Histological elements.-- g.c., multipolar ganglion cell. n., nucleus of a medullated nerve. a.c., its axis fibre. s.S., (sheath of Schwann), medullary sheath interrupted at intervals by n.R., the nodes of Ranvier. n.m.f., a non-medullated fibre. |
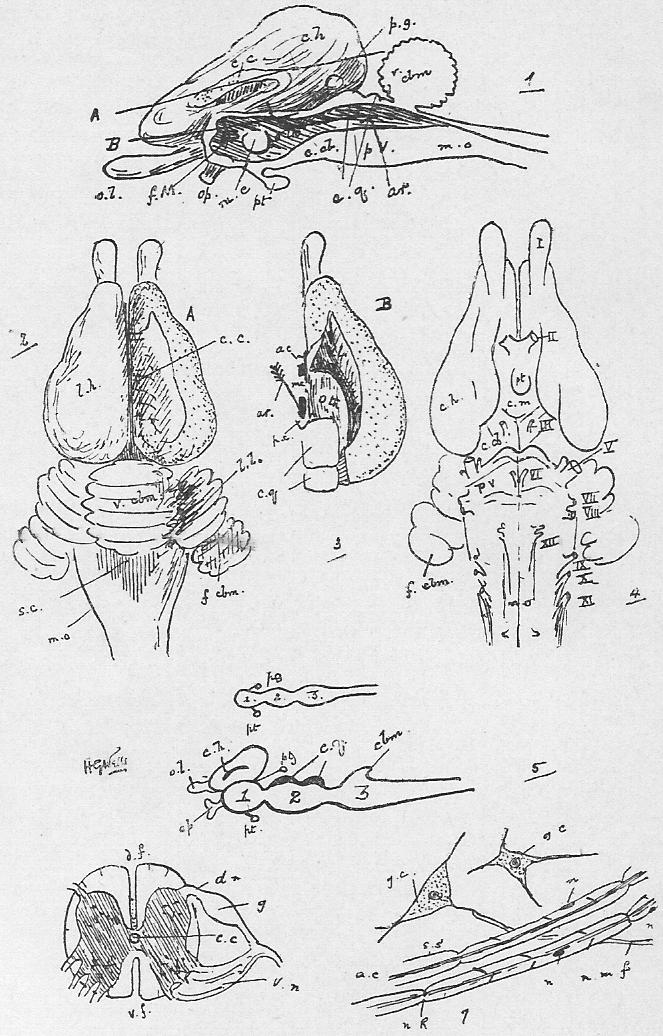
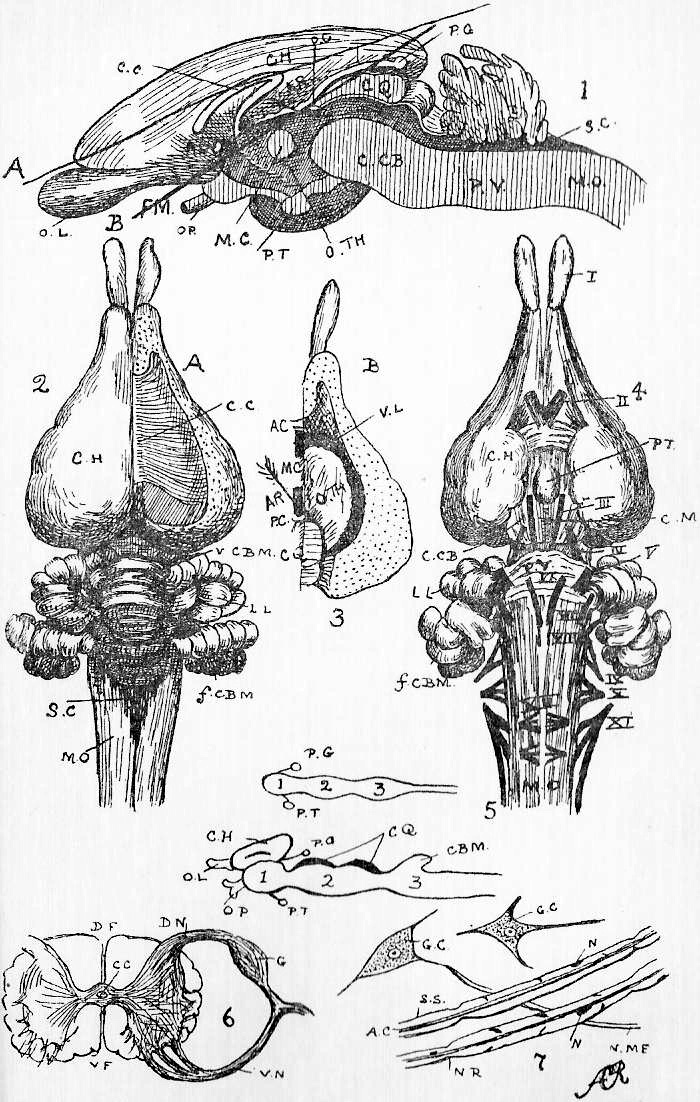
|
Figure I. Rough sketch of dissection of the neck from the left ventral aspect.-- The bands of muscle between hyoid, mandible, and sternum, and the thymus gland carefully cleared. lr., is the larynx, and b., the balla. s.m.g., the right sub-maxillary gland (the left has been removed). The nerves are numbered. l.r.l.n., [r.r.l.n.] is the left recurrent laryngeal looping under that solid connection between the pulmonary artery (p.a.) and ao., the aortic arch, which was an open tube in the embryo, the ductus arteriosus. hy., is the hyoid with its posterior cornua. ph.n., is the phrenic nerve. r.r.l.n., [l.r.l.n.] is the right recurrent looping under the sub-clavian. s.c.g., is the super or cervical ganglion of the sympathetic (sym.); s.l.n., is the left superior laryngeal, and g. the left depressor branch of x. z., is the ramus descendens noni of the twelfth nerve. In early development the heart lay just beneath the pharynx in the position of the larynx (compare Dog-fish and Frog); as the neck elongated, the heart shifted back with its vessels, and so the long loop of the recurrent laryngeal comes to be drawn out in this singular way. Figure II. Diagram of orbit to show V.1 orbit-nasal, V.2 the maxillary, and V.3 the mandibular branch of V. In order to show these in dissection, the malar must be cut away, and the eye and glands of the orbit removed. s.r., e.r. [p.r.], i.r., and a.r., cut ends of the superior, external (or posterior), inferior, and anterior (or internal) recti muscles. s.o., and i.o., the superior and inferior obliques. Figure III. General diagram of the Rabbit's cranial nerves. Figure IV. Rough sketch of dissection of the nerves and blood-vessels dorsal to stomach.-- The stomach turned over to the animal's right, the Spigelian liver lobe cleared from the oesophagus, the mesentery supporting spleen and hiding solar plexus picked off, and the mesentery hiding sympathetic cleared. coe.art., coeliac artery, and s.m.a., superior mesenteric artery. coe.g. coeliac, and s.m.g., superior mesenteric ganglion. The two together form the solar plexus. l.abd.sym., left abdominal sympathetic (in the actual dissection, the right would also be visible). l.a.r., left adrenal. l.sp.n., left splanchnic nerve. r.art., renal artery. r.v., renal vein. st., the stomach, and sp., the spleen. x., the vagus on oes., the oesophagus. |
 >
>
|
Figure 1. The Male. Figure 2. The Female Organs. (The symbols below the figures indicate the sex.) pb., is the pubic symphysis [which has been] cut through. R., the rectum, with r.g., the rectal gland, and a., the anus. t., the tail. r.ur., the right ureter. l.ur., the left ureter. ur.b., the urinary bladder. In the Male ep., the epididymis. P., the penis. pp., the prepuce. scr., the scrotal sac, containing these; r.v.d., the right vas deferens. T., is the testis. u.m., the uterus masculinus. In the Female c.ut, the left cornu uteri. F.t., the left Fallopian tube. ov., is the ovary, with a Graafian follicle, G.F. V., the vagina. v.b., the vestibule. Figure 3. Diagram of ovary with stages in the development of a Graafian follicle 1, 2, 3, 4, 5, see text, Section 137. The arrow indicates the changes in position of the developing follicles. |
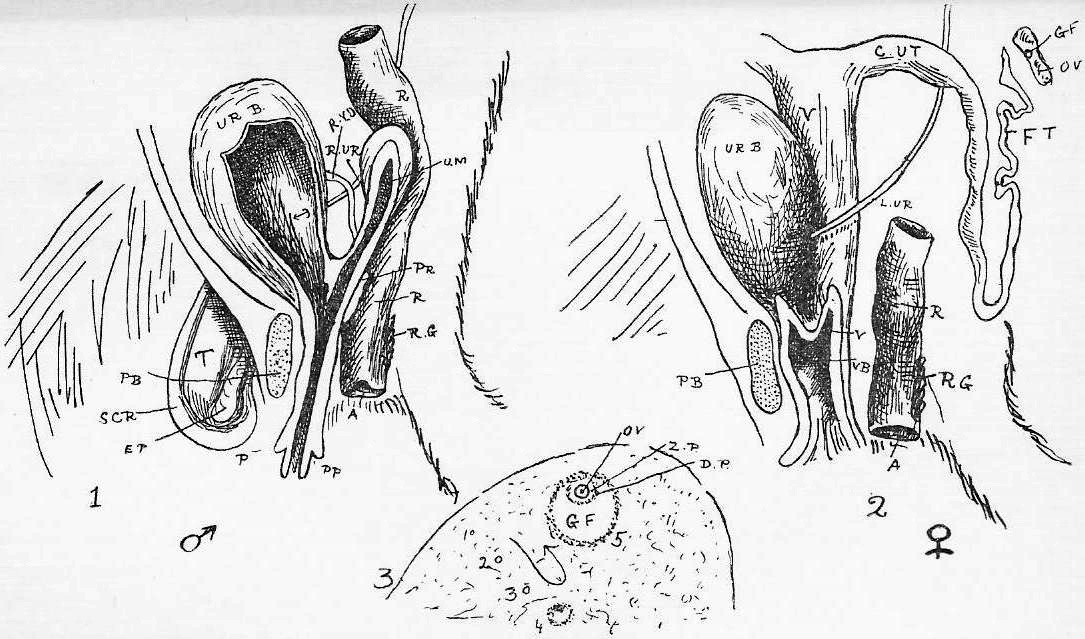
|
Figure 1. General dissection of Frog (male). Figure 2. The heart and great vessels laid open. Figure 3. The circulatory system from the side. Figure 4. Blood. {n., nucleus.} r.c., red corpuscle (oval and nucleated). w.c., white corpuscle Small figure of Frog in left-hand corner is to show position of heel, h. Reference Letters. all.b., allantoic bladder (= urinary bladder). c.ad., corpus adiposum. cl.c., cut end of the right clavicle. d., duodenum. g.b., gall bladder. il., ileum. k., kidney. l.au., left auricle. l.g., lung. l.int., large intestine. l.s.v., longitudino-spiral valve. L.v., Liv., liver. pan., pancreas. r.au., right auricle. sp., spleen. st., stomach. T., testis. t.a., truncus arteriosus. ur., urogenital duct. v., ventricle of heart. Arteries (white). ao., aorta. c.a., carotid arch. c.g. [c.gl.], carotid gland. coe., coeliac. cu., -and pa.",- cutaneous. d.ao., dorsal aorta. e.c., lingual artery. [i.c., internal carotid.] l.a.a., left aortic arch. pa., and p., pulmonary. p.c. [p.cu.], pulmo-cutaneous. r.a.a., right aortic arch. [s.cl., sub-clavian.] t.a., truncus arteriosus. Veins of the Caval System -(black)-. b.v., brachial (from fore limb). e.j., external jugular. h.v., hepatic vein. i.j., internal jugular. [in.v., innominate vein.] l.v.c.s., left vena cava superior. p.v., cutaneous vein. [s.cl.v., sub-clavian vein] s.s.r., sub-scapular vein. v.c.i., vena cava inferior. Veins of the Portal and Renal Portal Systems -(shaded)-. a.ad., and a.ab.v., anterior abdominal vein. b.v., and p.v., united are called the sub-clavian vein. l.fm., left femoral. l.p., left pelvic. l.r.p., (and r.p.) left renal portal. l.sc., left sciatic. p.v., portal vein. -(The anterior abdominal is coloured black in Figure 1.)- The cutaneous artery in the above figures is turned back. In dissection it will be found to lie over and hide the dorsal-ward sweep of the aortic arch. |
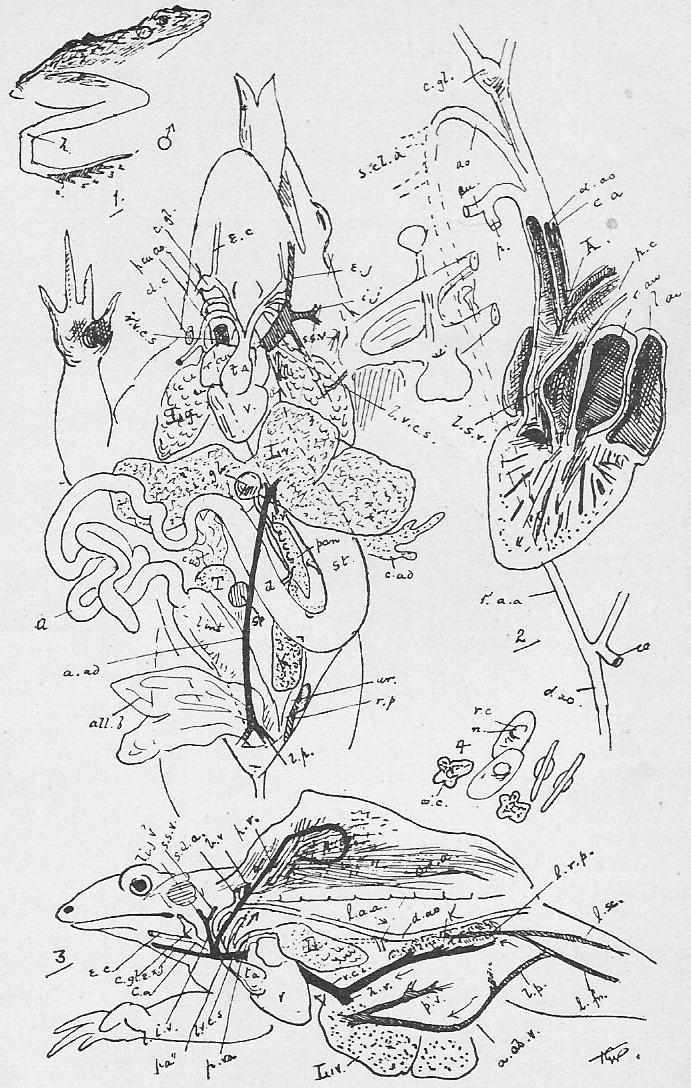

|
Figure 1. Upper view of the Frog's brain. Figure 2. Under view of the same. Figure 3. The same-- median section. Figure 4. The distribution of the Frog's nerves. Compare Sheet 9, Figure III. The shaded part in 4 is the -otic capsule- [tympanum]. The hyoid apparatus is roughly represented in black to show its relation to IX. (dorsal to it) and sp. 1 (ventral). Compare {nerves} IX and XII in Sheet 9. The nerves are numbered. cb., the cerebellum. c.h., cerebral hemispheres. f.t., filum terminale. g.tr., ganglion on the fifth nerve. l.t., lamina terminalis. mb., mid-brain. md., medulla oblongata. o.l., optic lobes. pin., pineal gland. pit., pituitary body. r.h., olfactory lobes (rhinencephalon). th.c., thalamencephalon. sp.1, first spinal nerve. sp.2, 3, brachial plexus to fore limb. Figure 5. The spinal column (and pelvic girdle) of the Frog. Figure 5b. Vertebrae. Figure 6. The pectoral girdle and limb, dorsal view. Figure 7. The pelvic girdle and right limb from the side. (l.h. shows the position of the right lymph hearts-- they are paired.) as., astragalus. b., body. c., calcar (?= a sixth digit). cal., calcaneum. cl., clavicle overlying a procoracoid cartilage. co., coracoid. f., fibula. [FE., femur.] h., humerus. il., ilium. is., ischium. o.st., omosternum. pu., pubis. r., radius. sc., scapula. s.sc., supra-scapula. s.v., sacral vertebra. t., tibia. t.p., transverse process. ul., ulna. ur., urostyle. x., xiphisternum. z., zygapophysis. 1, 2, and etc., first, second, and etc., digits. D. and D'. are simplified diagrams of the limbs for comparison with the similar ones of the Rabbit. In each girdle we have a dorsal ossification (scapula, ilium) and two ventral parts (pubis and procoracoid cartilage, ischium and coracoid), and at the meeting-place of the three in each case the proximal bone of the limb (humerus, femur) articulates. |
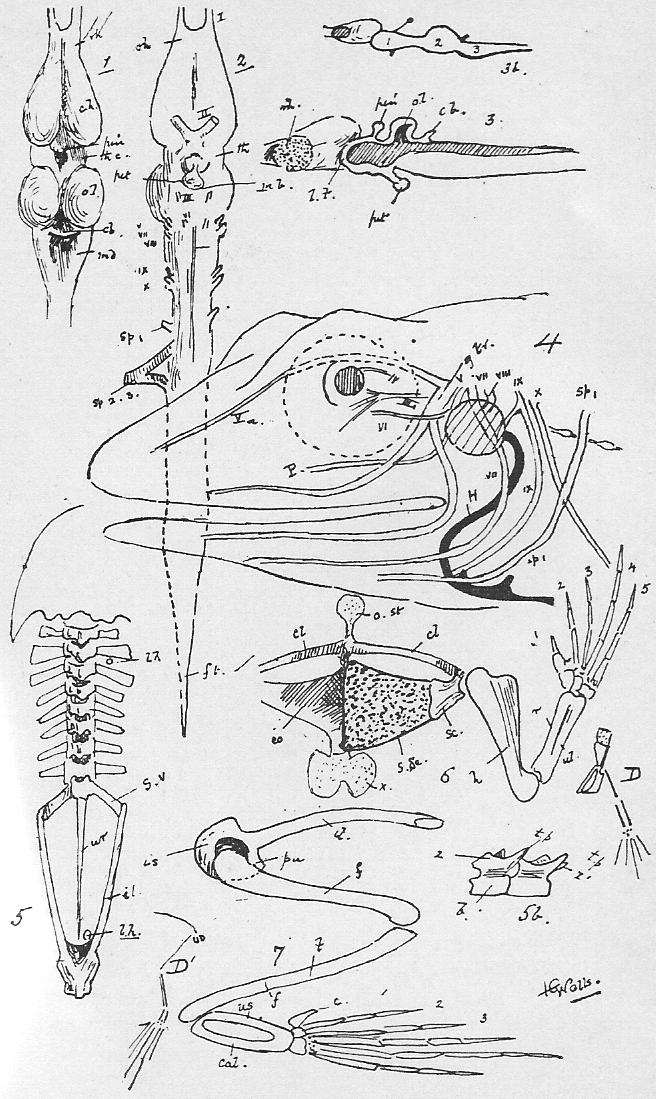
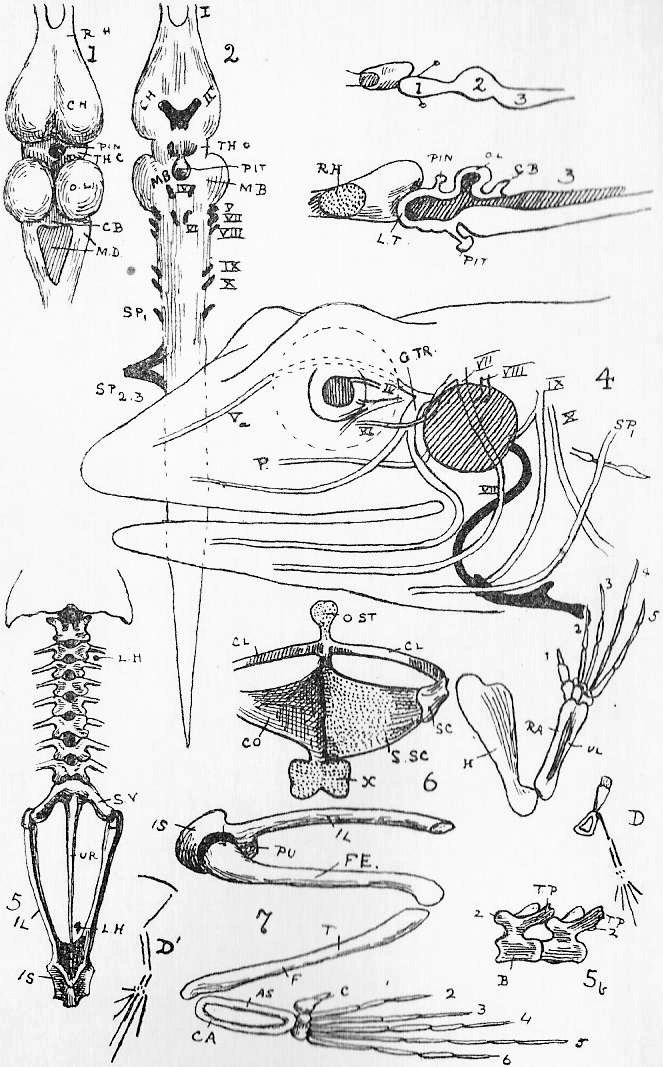
|
Figure 1. The Male. Figure 2. The Female. The oviduct removed on the animal's left, and the ovary on its right. Organs common to both sexes.-- al.b., allantoic bladder. c.ad., corpus adiposum. cl., cloaca. int., intestine. K., kidney. lg., (dotted outline of) lung. oes., oesophagus. r.p.v., renal portal vein. st., stomach. In the Male.-- T., testis. v.e., vasa efferentia. u.g.d., urogenital duct. p., prostate gland. In the Female.-- adr., adrenal. f.t., fallopian tube (anterior part of oviduct). * its opening. o.d., oviduct (letters on [the opening] -uterine portion-). ov., ovary. ur., ureter. (This would be the condition about midwinter.) In March o.d. will be either enormously distended with eggs, or large, flabby, and empty, and ov. will be small and brownish, without any large eggs; the ovary gradually recovers its size through the summer. Figure 3. Spermatozoa attached to the parent cell (g.e.) from the lining epithelium of the testis, and one free. fl., the flagellum. |

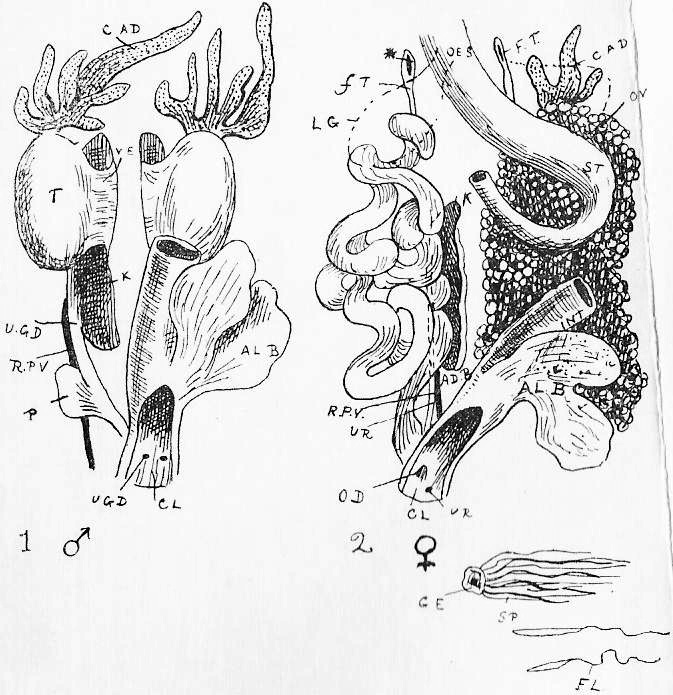
|
Figure 1. I., II., early and late stages of the Tadpole's chindrocranium. Diagrammatic. Figure 2. Dorsal view of a young Frog's cranium-- the membrane bones removed. Diagrammatic. Figures 3 and 4. Dorsal and ventral views, respectively, of the Frog's skull-- the lower jaw removed. Figure 5. Side view of the Frog's skull. Figure 6. Median section of the brain case. Figure 7. The hyoid apparatus. Figure 8. I., II., III., progressive stages of the Tadpole's skull from the side. After W. K. Parker. Figure 9. F., side and hind views of the Frog's skull. D., the same of the Dog. Roughly diagrammatic. N.B.-- In all cartilage is dotted, cartilage bone cross-barred, and membrane bone, white. In Figure 4, pt., should be cross-barred; and in 5, th.h. plain. a.c., anterior cornu of hyoid [(= CH.)] -not lettered, in {Figure} 5-. a.o., antorbital cartilage. ar., angulo-splenial -(On Frog Section 34, for Articulare read -Angulo-Splenial_)-. -b., parachordal part of brain box-. b.c., brain case. b.h., body of hyoid. b.r., branchial arches. CH = a.c. c.t., cornua trabeculi. d., dentary. e., eye. E.N., external nares. e.o., exoccipital bone. f., fenestra (membranous part of cranial wall). -f.p., fronto-parietal.- h.m., hyomandibular cleft = Eustachian tube and ear drum. mb., mandible. [M.C., Meckel's Cartilage.] m.mk., mento-Meckelian bone. m.p., mouth passage. mx., maxilla. n.c., notochord. n.o., nasal organ. n.p., nasal passage. ot., or o.c., otic (auditory) capsule. pal., palatine bone. PAL., hard palate of Mammal. p.c., parachordal. p.f., [parieto-frontal] -see f.p.- p.m., premaxilla. P.N., internal nares. p.o., prootic bone. p.p., palato-pterygoid cartilage. psph., parasphenoid bone. pt., pterygoid bone. q., quadrate cartilage. q.j., quadrato-jugal. s.e., sphenethmoid bone. sq., squamosal. t., trabecular part of brain box. t.c., trabecula. th.h., thyrohyal. |
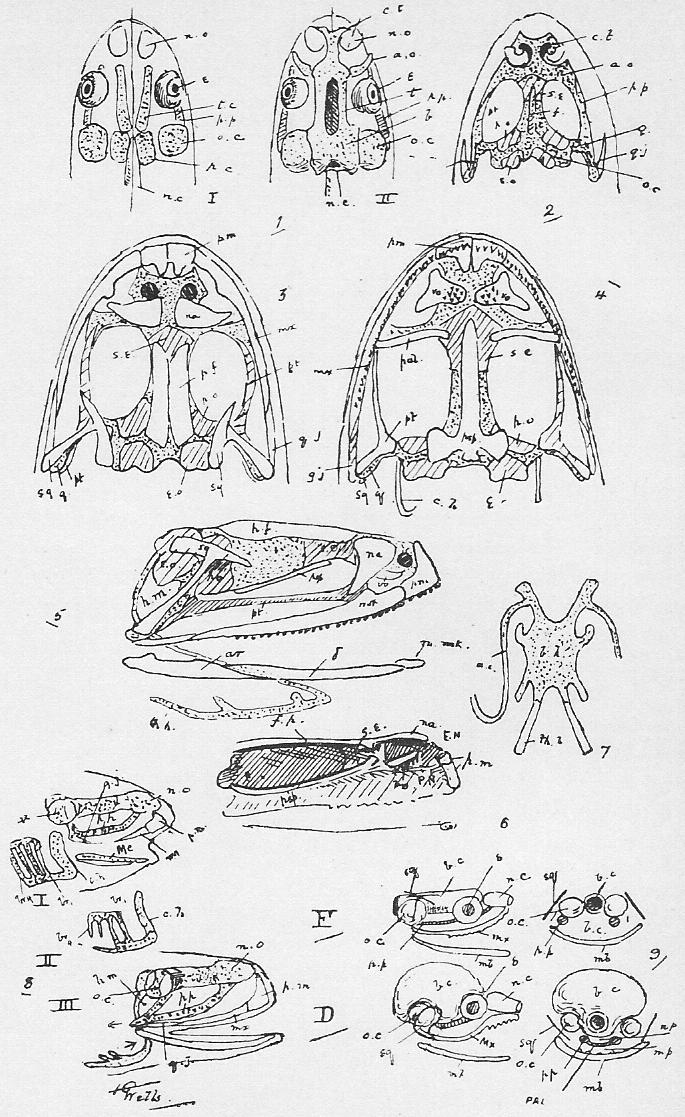
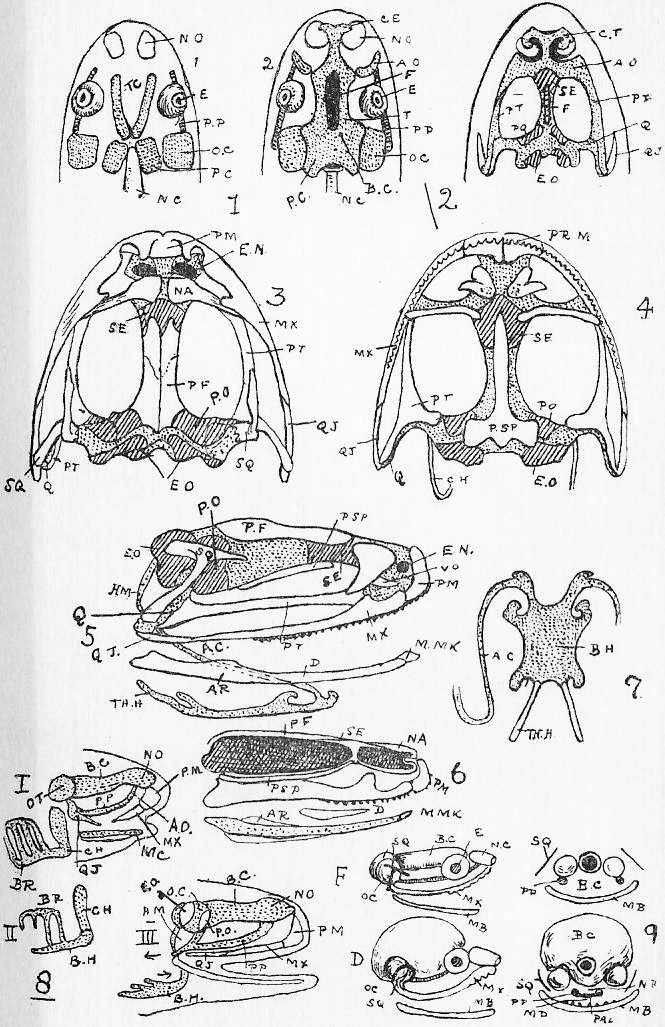
|
Figure 1. Dissection of -Male- [Female] Dog-Fish to show alimentary canal, the pericardium also being opened and the cloaca slit up. [Above is also seen the dorsal view of the head.] Figure 2. The pelvic girdle and fin skeleton [of a male]. {No Figure 3, in First Edition.} Figure 4. The spiral valve in the colon. {Figure 3, in Second Edition.} a.p., abdominal pore. aur., -auricle- [atrium] of heart. b.d., bile duct. b.pt., basi-pterygium. -cl., clasper.- cl.c., -its- [the] supporting cartilage [of the clasper]. co., colon. d'dnm., duodenum. e., the eye. g.bl., gall bladder. g.s., gill slits. L.Lv., left lobe of liver. M.Lv., middle lobe of liver. olf., olfactory opening. [pan., pancreas.] pcd., pericardial wall. pel.g., the pelvic girdle. p.p., arrow through pericardio-peritoneal canal. r.g., rectal gland. [R.Liv., right lobe.] sp., spiracle. spl., spleen. st., the stomach. s.v., sinus venosus. u.g.p., uro-genital pore. v., ventricle. |
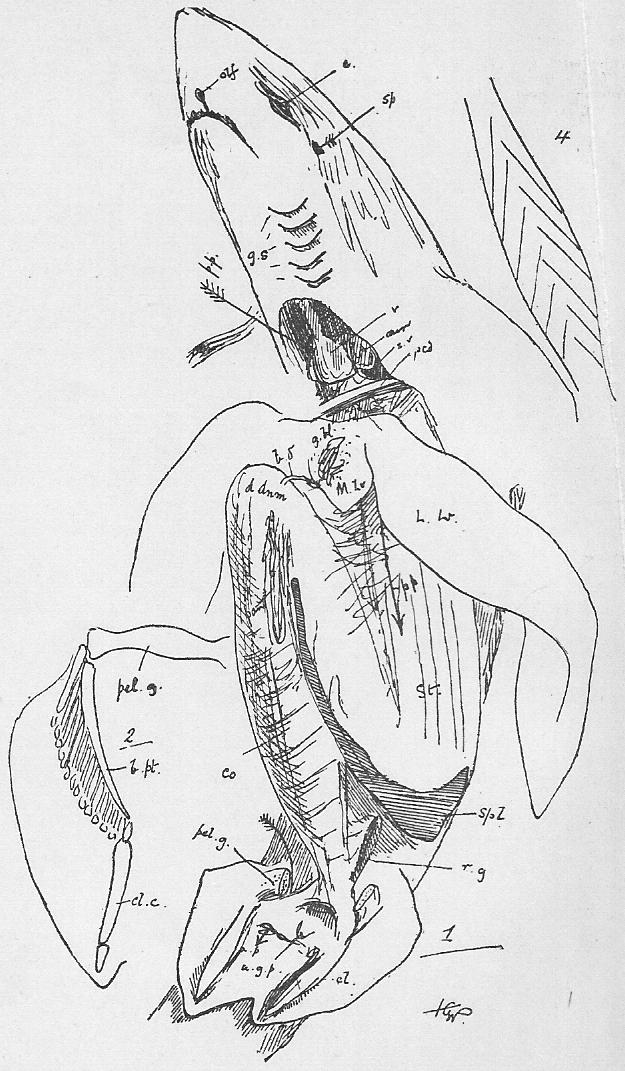
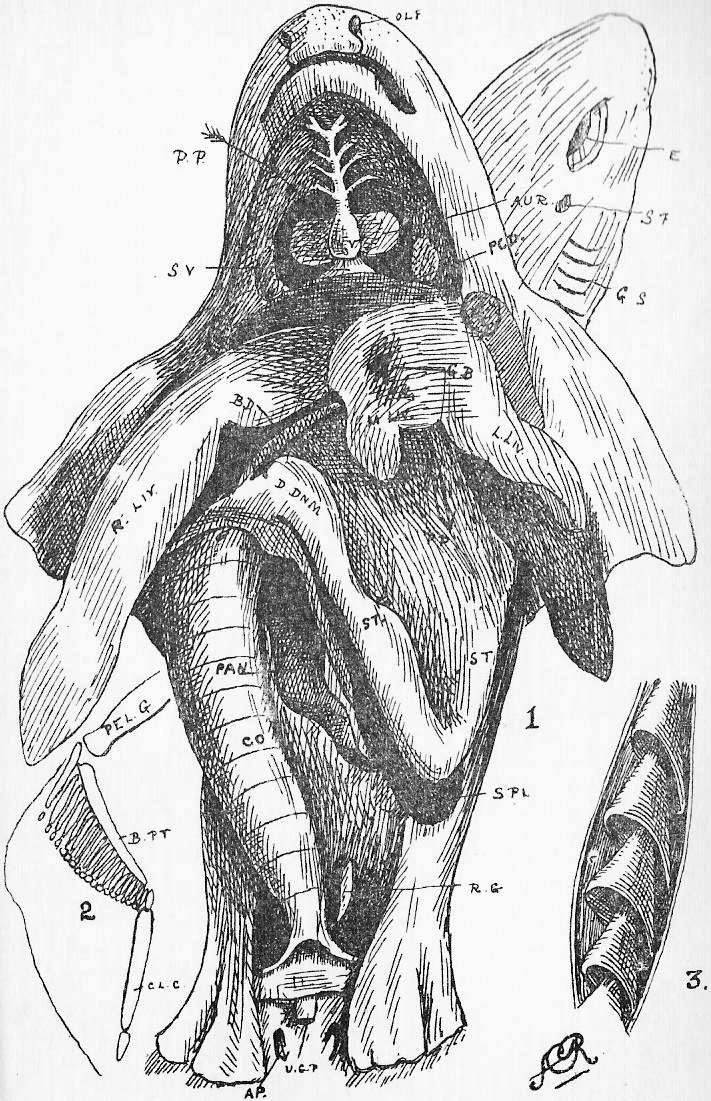
|
Figure 1. Circulation of the Dog-Fish. Figure 2. Simplified and more typical fish circulation, in which the posterior cardinals have not coalesced in the median line. The Cuvierian veins = the vena cava superior of the higher type; the posterior cardinal is represented by the azygos vein in the Rabbit. Compare Sheet 24, Figure 7, and Sheet 2, Figure 2. Figure 3. Side view of the pericardium. a.br., afferent branchial artery. a.c.s., anterior cardinal sinus (= internal jugular vein). au., atrium (auricle) (= the two auricles of higher forms). b.a., bulbus arteriosus. c.a., conus arterious. cd. a., caudal artery. cd.v., caudal vein. c.s., Cuvierian sinus. d.a., dorsal aorta. E., eye. e.br., efferent branchial arteries. g.s., in position of gill slits. h.br.a., hypobranchial artery. H.S., hepatic sinus. [i.j.s., inferior jugular sinus (= external jugular vein).] K., kidney. L.V., lateral vein. [oe.s., ventral wall of oesophagus.] P.C.C., pericardial cavity. P.C.S., posterior cardinal sinus. p.p.c., pericardio-peritoneal canal. P.V., portal vein. r.p.v., reno-portal vein. s.c.v., subclavian vein. Vn., ventricle. -v.s.v., inferior (= external) jugular vein-. Figure 4. Skeleton of pectoral limb, and girdle.-- g., the girdle (also in Figure 3). m.p., meso-pterygium. mt.p., meta-pterygium. p.p., pro-pterygium. sc., its dorsal portion. |
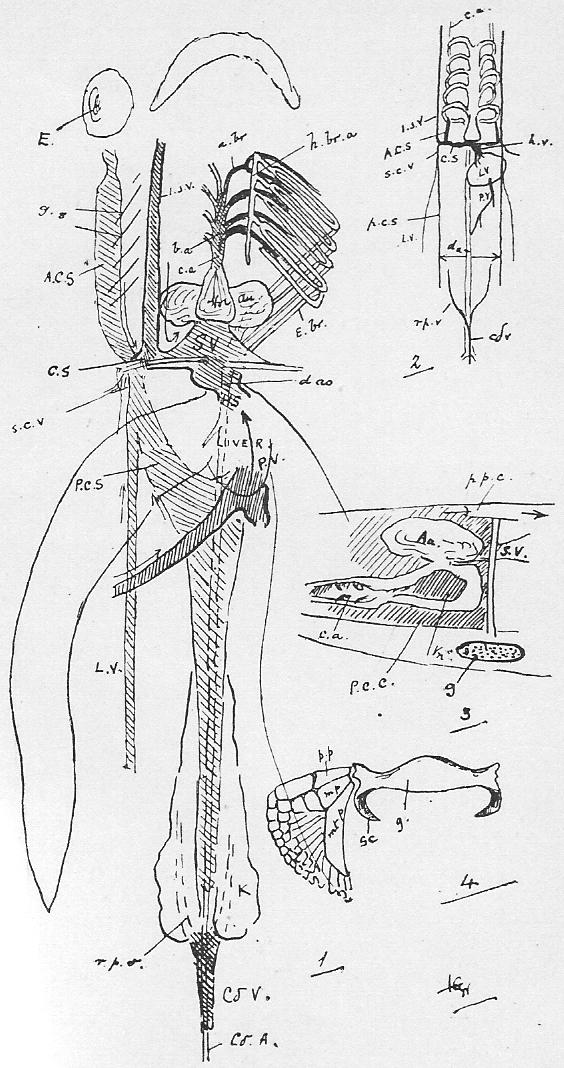
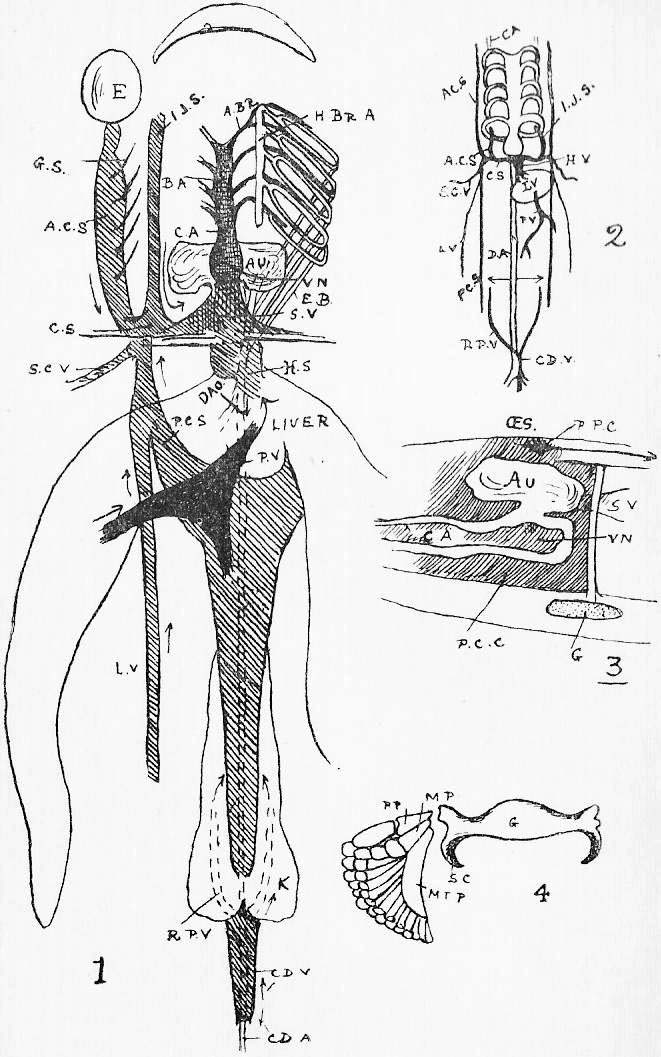
|
Figure 1. The Female, the oviduct of the left side cut away, -and an egg case in the oviduct.- Figure 2. The Male. The rectum is removed in both cases, and the silvery peritoneum dissected off from the kidneys. Figure 3. A generalized diagram of the uro-genital organs.-- All references in text. Ms., the mesonephros, is the epididymis in the male, and is reduced in the female; Ms.d., its duct, is the vas deferens in the male, and persists only as the urinary receptacle in the female. Mt. and Mt.d., the metanephros and metanephric duct, become the functional kidney and ureter in both sexes. G. is the gonad (reproductive gland), and M.L. the animal's middle line (median plane). -Ps.-, [Pr.,] the pronephros, is never developed in the Dog-fish; P.d., its supposed duct, is the oviduct of the female, and is suppressed in the male. |
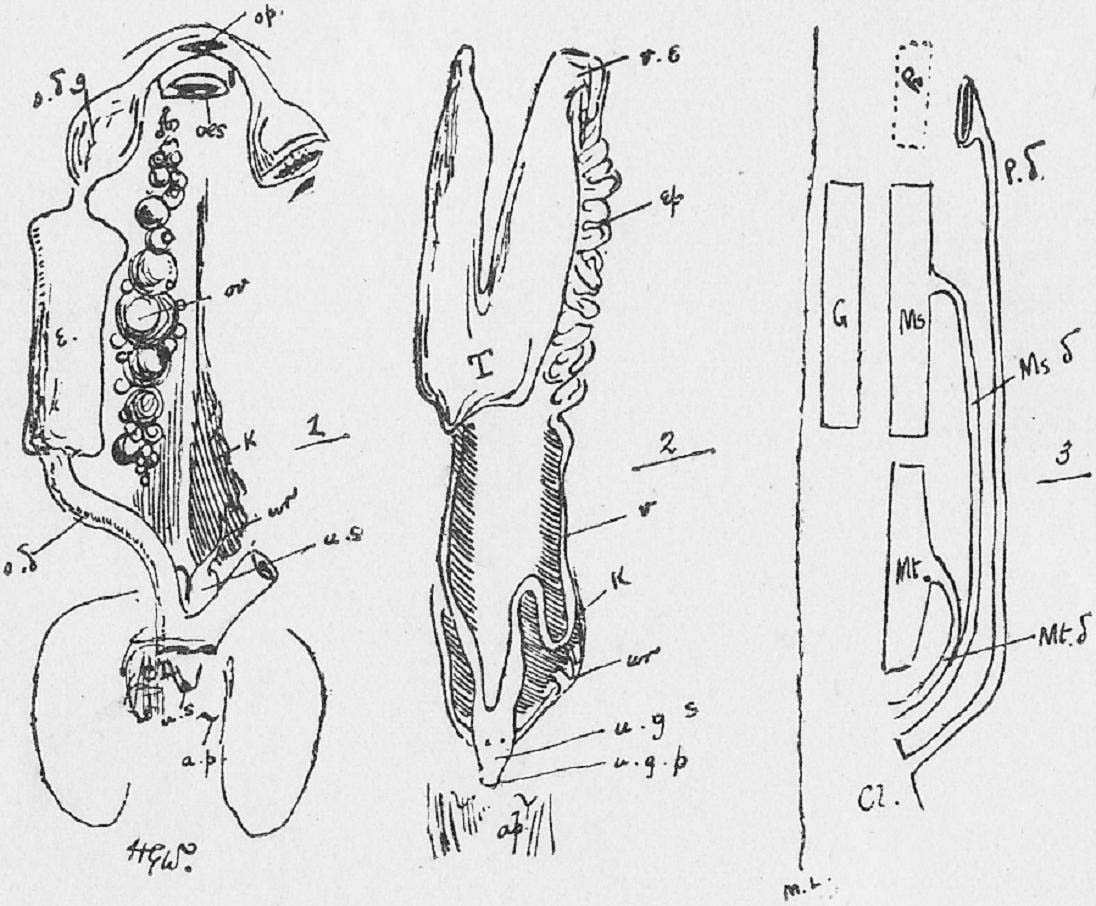
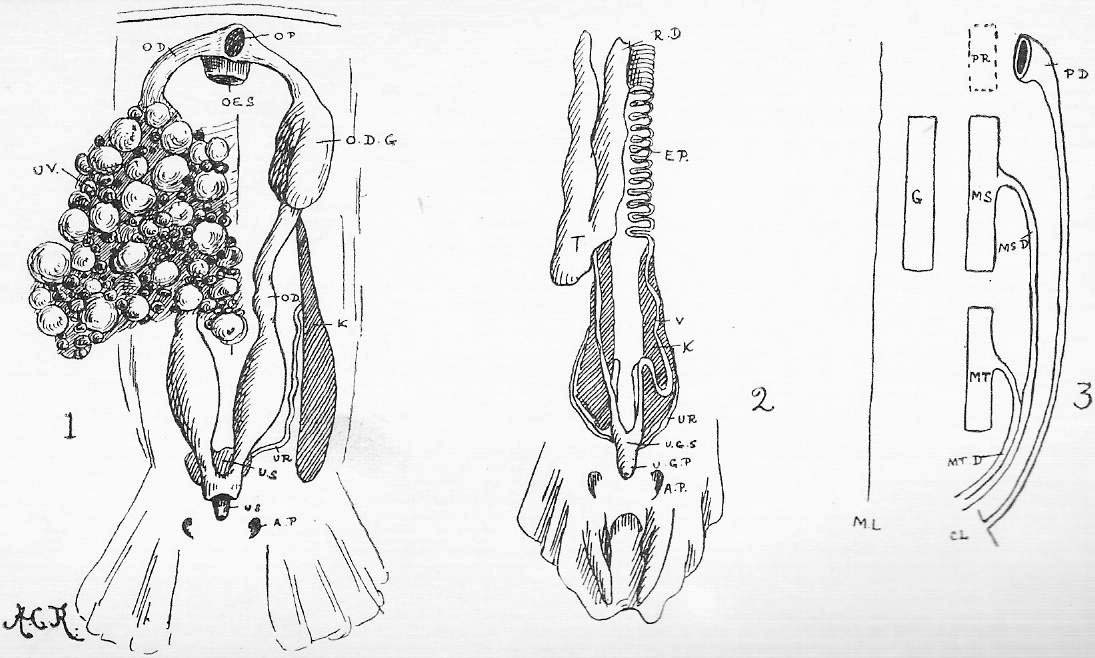
|
Figure 1. The Dog-Fish Brain, dorsal view. Figure 2. Median section of the same. To the right a more diagrammatic figure. The nerves are numbered:-- [BR1, BR2, BR3, BR4 branches of X forking over the second to the fifth gillslit.] cb., cerebellum. h.s.c., horizontal semi-circular canal of ear, exposed by the slicing down of the otic mass. [LAT., lateral-line branch of X.] m.o., medulla oblongata. oph., ophthalmic nerve (V.1+VII.1). op.l., optic lobe. pit., pituitary body. pr.c., prosencephalon (cerebral hemisphere). rh., olfactory lobe (rhinencephalon). r.t., -its- restiform tracts [of medulla]. -st-. [S.P.G.], stalk of the pineal gland. th., thalamencephalon. th.c., thalamencephalon. -ut., the utriculus, seen through the semi-transparent cartilage-. Vid., the Vidian branch of VII. [Visc., visceral branch of X.] Figure 3. Diagram of the ear of a fish. The structure of this is easily made out by clearing otic capsule and cutting slices of the cartilage in the Dog-Fish (e.g., Figure 1, h.s.c.). amp., their ampullae. a.v.c., p.v.c., h.c., anterior, posterior, horizontal canal respectively. [amp., the ampullae.] d.e., the ductus endo-lymphaticus. -sac., the sacculus; c., a small outgrowth of the latter, corresponding to the rabbit's cochlea-. -ut., the utriculus-. Figure 4. The cranium and branchial bars of a Dog-Fish. The groove in the otic capsule connects the orbital and anterior cardinal sinuses. A.C.S., position of the anterior cardinal sinus (dotted outline). c., the vertebral centra. c.b., the cerato-branchial. c.h., the cerato-hyal. e.b., epi-branchial. ex.b., extra-branchial. h.M., the hyo-mandibular. i.n.p., inter-neural plate. M.C., Meckel's (lower jaw) bar. Na.C., the nasal capsule. n.p., neural plate. n.s., neural spine. Ot.C., the otic capsule. ph.b., the pharyngo-branchial. P.pt., the palato-pterygoid bar (upper jaw bar). p.s., pre-spiracular ligament, containing a cartilaginous nodule. r., rib. sp., the position of the spiracle. Figure 5. Diagrams of a vertebral centrum.-- For reference letters, see text (Section 9). {No Figure 6, in First Edition.} [Figure 6. Diagram for comparison with Figure III., Sheet 9.] |
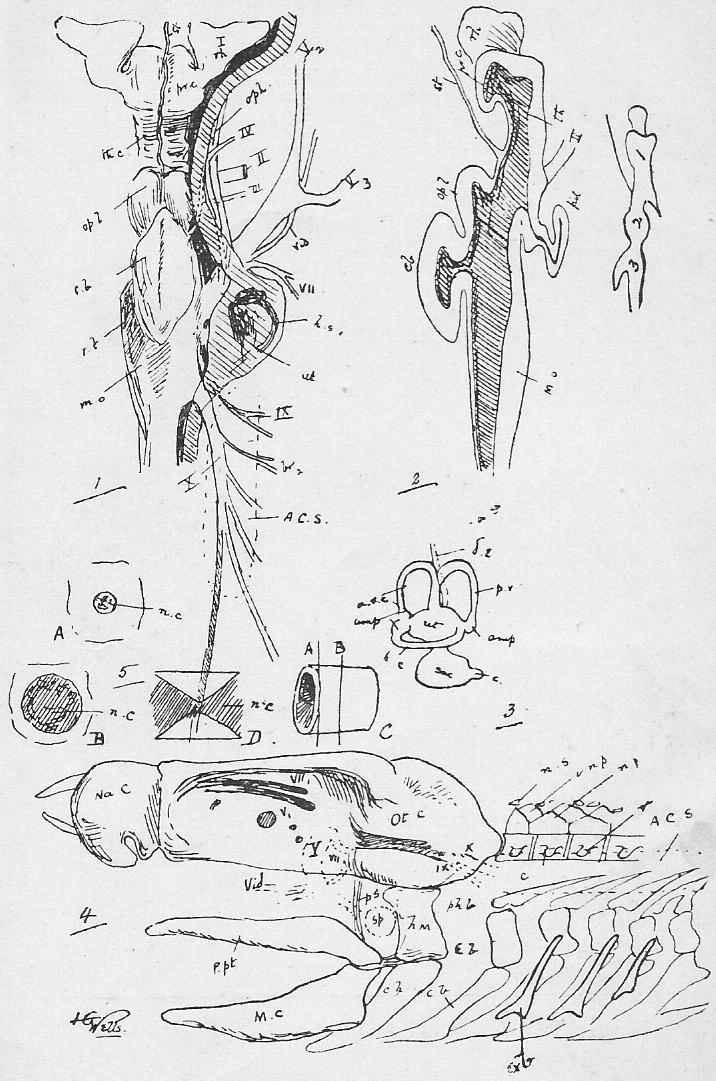
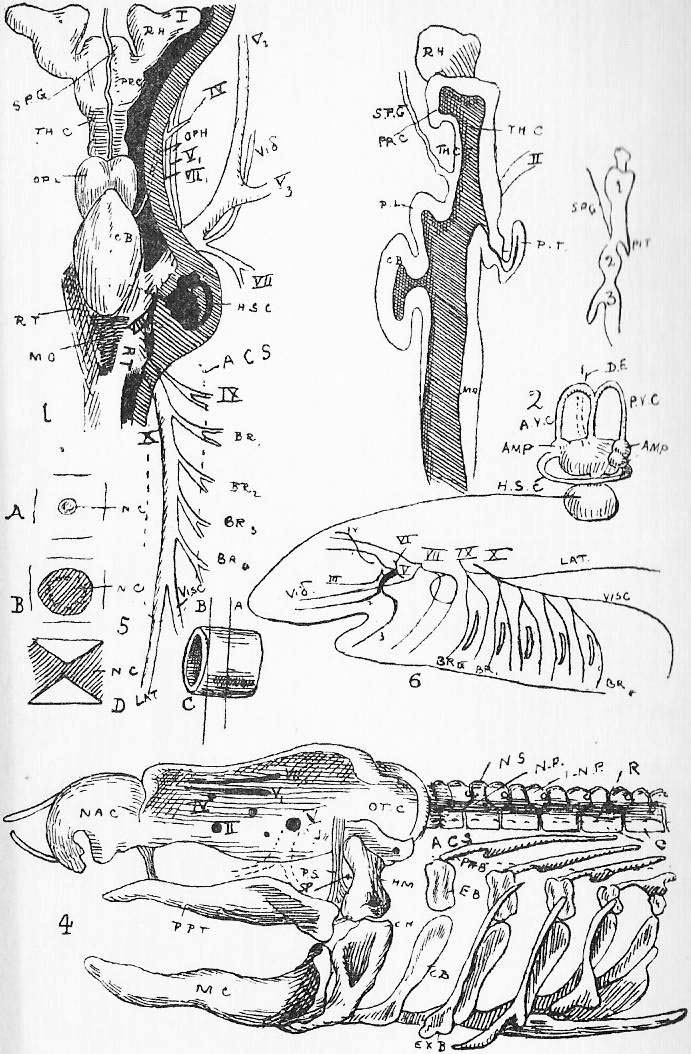
|
Figure 1. Amphioxus, seen from the right side. a----b shows the natural size. The animal is supposed to be clarified, and mounted in some highly refracting medium, so that it is practically transparent; I., II., III., and etc., refer to the section figured on Sheet 20. Figure 2. Amphioxus, General Dissection. (Slightly altered from a figure by Professor E. R. Lankester.) The ventral atrial wall is removed. The pharynx cut away from the dorsal body-wall, and with the true ventral body-wall turned over to the (animal's) right. The arrow a., a., passes through anus to intestine; b., b., is thrust through the atrial pore to the atrial cavity. Note coe., the body cavity. References to the two figures. an., anus. at., atrial cavity. at.w., atrial wall. at.p., atrial pore. a.d., anterior dilatata of nervous system. b.w., body-wall. b.t.L., brown tubes of Lankester. c.f., ciliated funnel. coe., coelome. c.ao., cardiac aorta. d.ao., dorsal aorta (paired). d.ao'., dorsal aorta median. g., gonads (male or female genital gland). hep., hepatic vein. in., intestine. i.w., intestine wall. lv., liver. m.f., median fin. n.c., notochord. p.v., portal vein. ph., pharynx. -p.s.-, [e.s.] pigment spot ("eye spot"). s.c., spinal cord. |
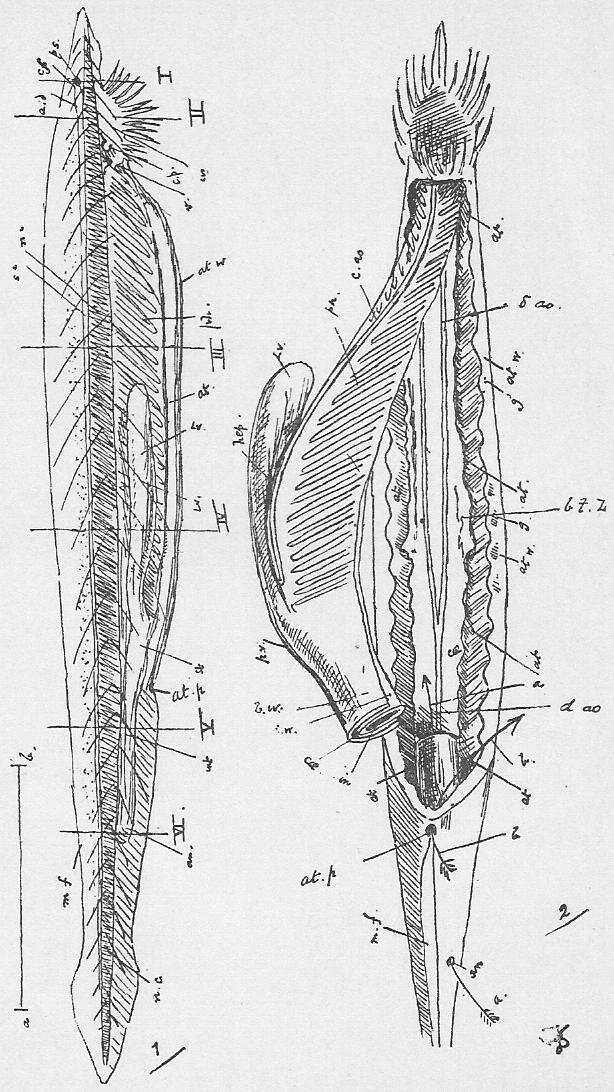
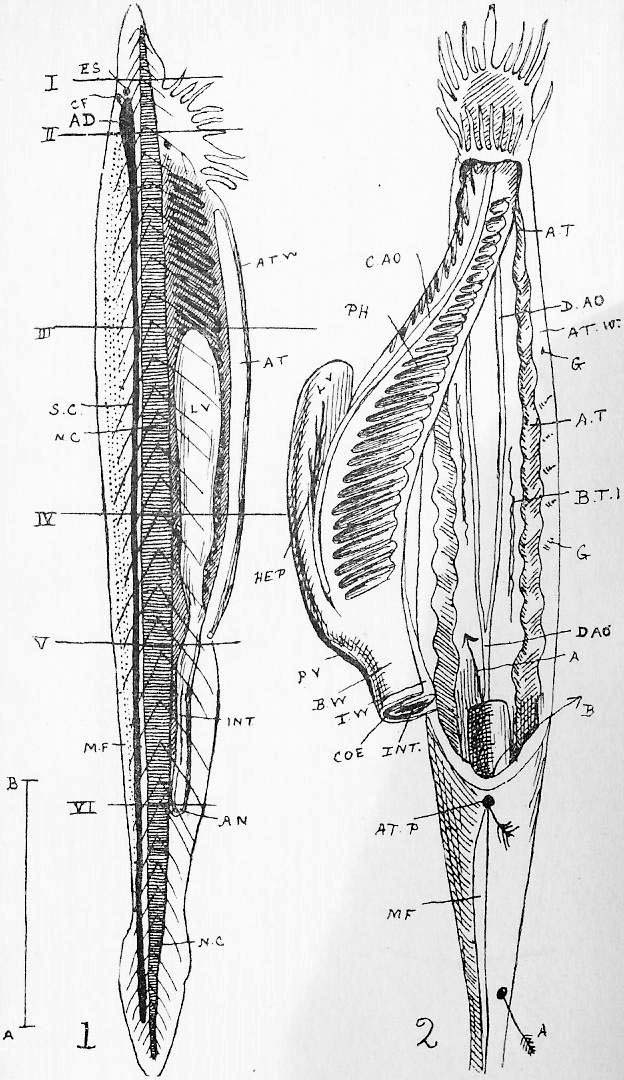
|
The Roman numerals indicate the corresponding region in Figure 1, Sheet 19. The lettering is identical; but note, in addition; br.c., branchial canal. c.f., ciliated funnel. d.c.c., dorsal coelomic canal. end., endostyle. ep., epipleur. e.s., eye spot. h.p., hypopharyngeal grove. h.vn., for hepatic vein. o.c., oral cavity (or hood). |
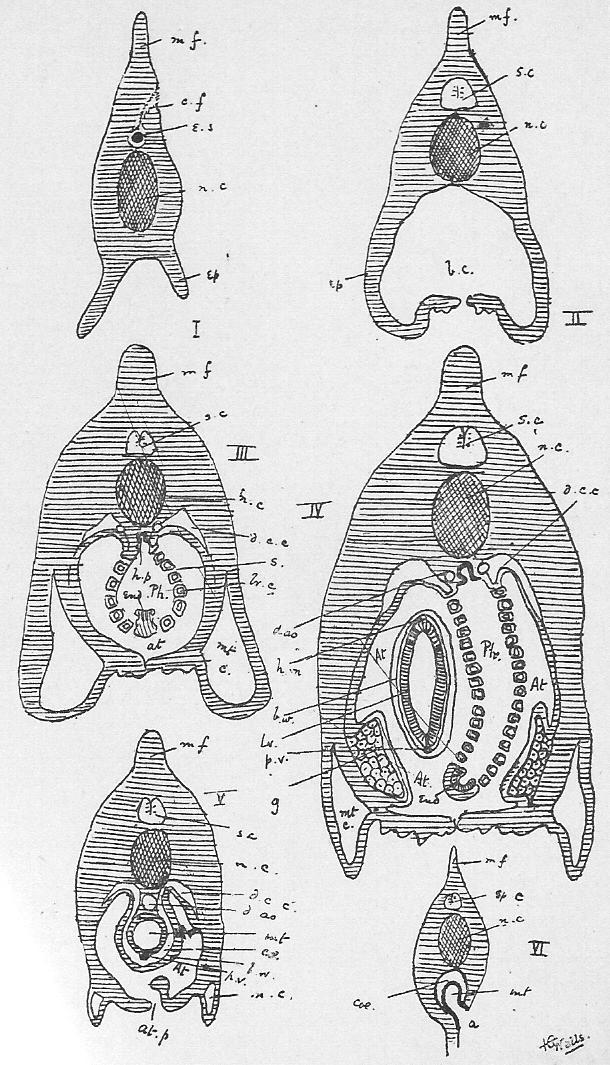
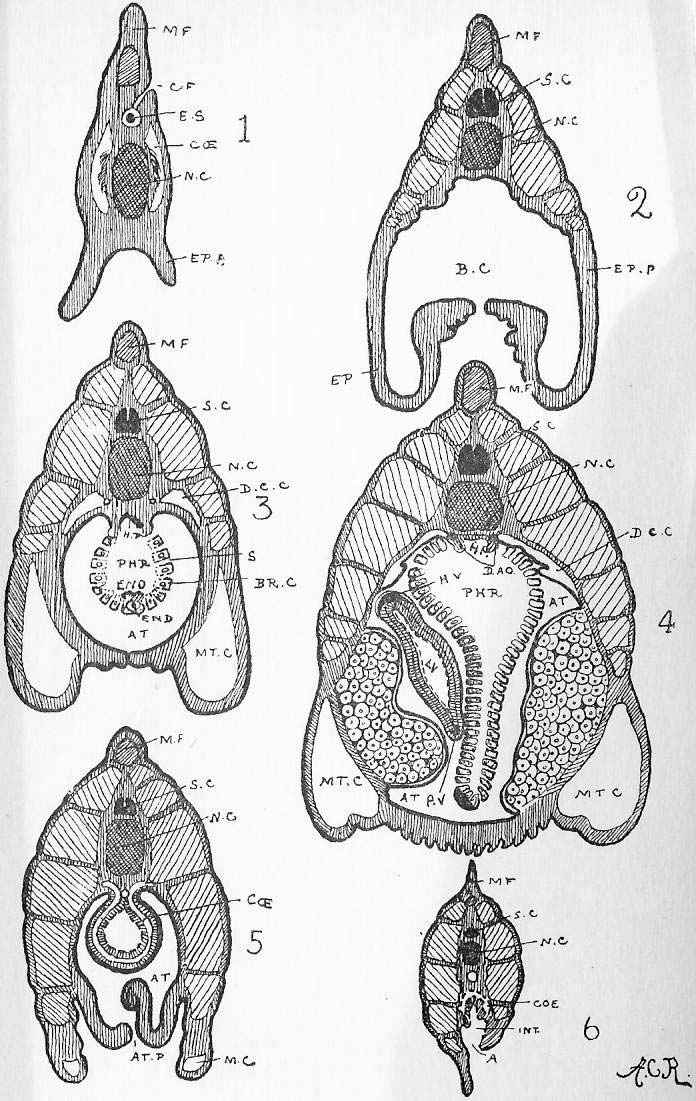
|
Figures 1, 2, 3, 4. Phases in segmentation. Figure 5. The blastosphere. Figure 6. The gastrula in section, anterior end to the right. Figure 7. i. Dorsal view post gastrula stage. Figure 7. ii. Diagrammatic section of the same in the position indicated by the transverse line in 7, i. Figure 8. Diagrammatic section of a later stage. coe.p., the coelomic pouches. n.c., the notochord. n.p., the neural plate. Figure 9.i. Still later section. Figure 9. ii. Diagrammatic view of late embryo. Figures 10, 11, 12 illustrate the formation of the atrium as a median ventral invagination, at. |
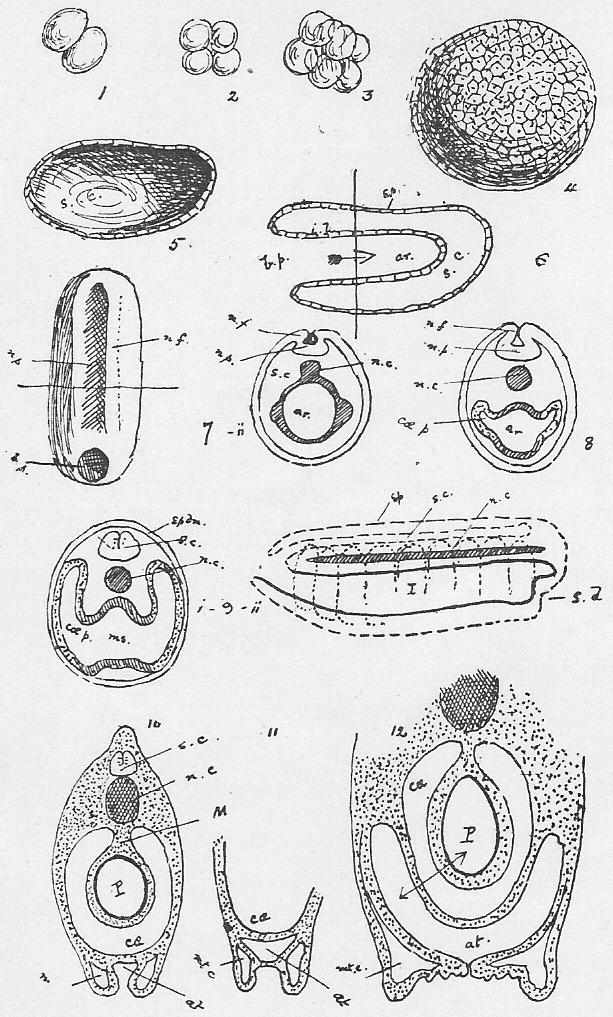
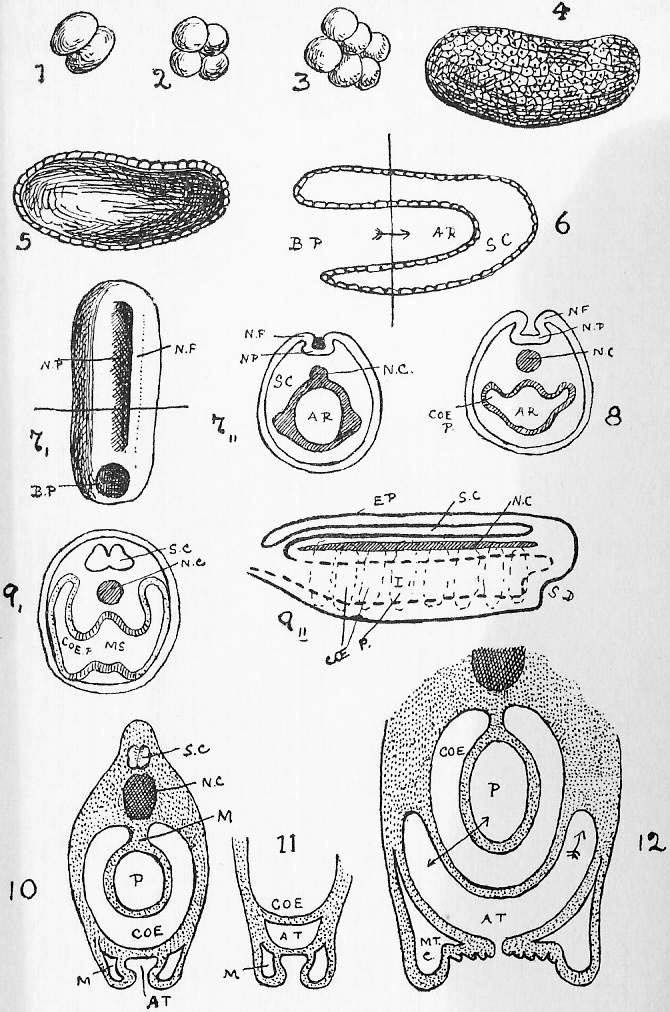
|
Figures 1, 2, 3. Stages in segmentation (compare 1, 2 ,3 of {Sheet 21} Amphioxus). Figure 4. Blastosphere stage (compare 5, Amphioxus). This, on a smaller scale. The cells on the ventral side are so much larger because distended with yolk. Figure 5. Gastrula stage in section (compare 6, Amphioxus). The Frog on a smaller scale than Amphioxus. Figure 6. Dorsal view of gastrula (compare 7, Amphioxus). Figure 7. Part of a transverse section of developing tadpole, corresponding to Figure 8 of Amphioxus. Figures 8 and 9. Diagrammatic longitudinal sections of tadpoles (compare 9. ii. of Amphioxus). Y. represents a mass of yolk cells. Figure 10. Side view of young tadpole, showing external gills (e.g.) and suckers (s.). Note the ventral bulging due to the yolk. Figure 11. Ventral view of a later tadpole. op., the operculum. int., coiling intestine. Figure 12. Head of still later tadpole in horizontal section to show atrial chamber formed by operculum. int.g., internal gills. L., developing lungs. Figure 13. Diagrammatic cross-section of the mid-dorsal part of an embryonic vertebrate. ao., aorta. B.C., Bowman's capsule. coe., coelom. d.g., ganglion on dorsal root of spinal nerve. gl., -its branch- [arteriole] to form glomerulus. g.r., genital ridge. I., intestine. M.D., Mullerian duct. ns. [nst.], nephrostome. n.c., notochord; -n.s.-, [n.sh.] its sheath. s.c., neural canal. W.D., Wolffian duct. |
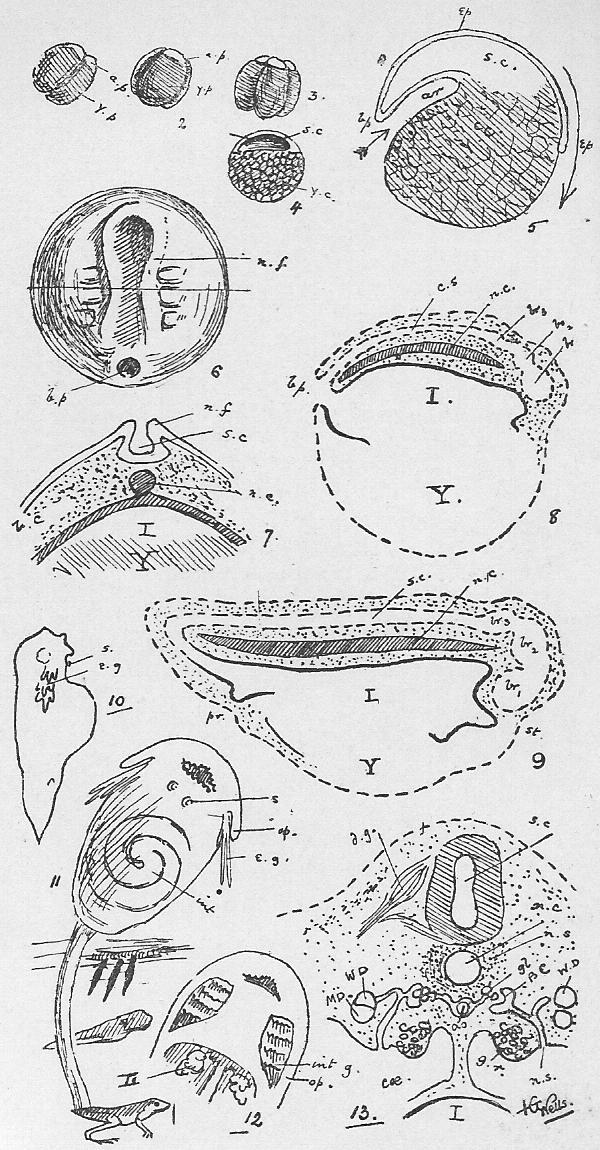
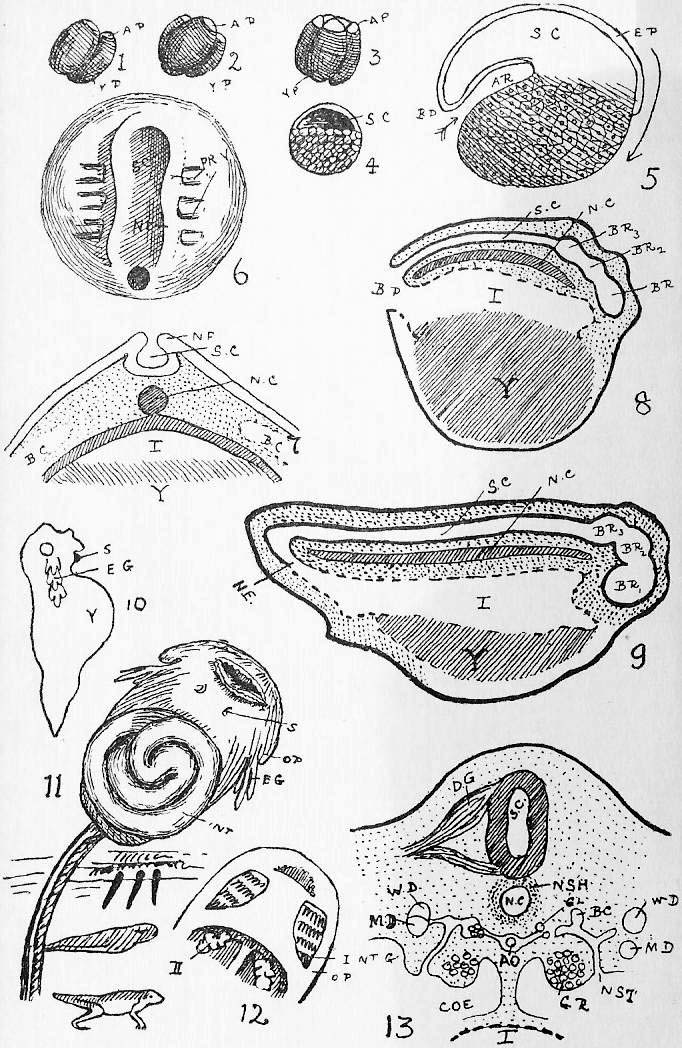
|
Figure 1. Diagram of the early ovum. The section below is a small portion of the blastodermic area. b.d., blastoderm. y., the undivided yolk. s.c., the segmentation between the blastoderm and yolk. Compare s.c. in {Sheet} 22, {Figure} 4. Figure 2. Area pellucida about the sixteenth hour. The figure below is the central part of the section indicated by the transverse line, and showing the primitive streak (p.s.). Figure 3. Area pellucida about the twenty-first hour. Two sections through a and b below. Figure 4. About the twenty-fifth hour; surface view; longitudinal section to right and transverse above. Figure 4b. Diagrammatic rendering of same stage (compare Figure 9 of Frog and 9.ii. Amphioxus). This will be most clearly understood if the reader look at Sheet 22, {Figure} 9, and imagine Y. enormously increased, and the embryo sinking into it. Epiblast, ep., -line of dashes- [black line]. Mesoblast, dotted. Hypoblast, -black- [line of dashes]. pp., the pleuro-peritoneal cavity. Figure 5 and 6 illustrate formation of amnion (a.) and allantois (all.). 6 is about the fourth day. |
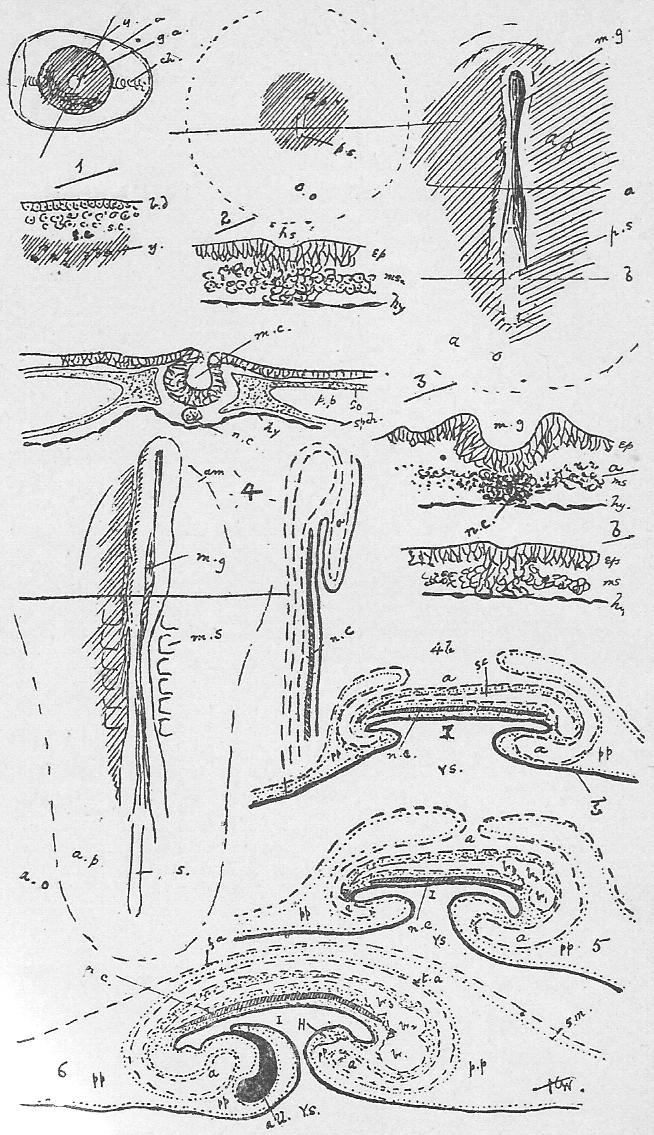
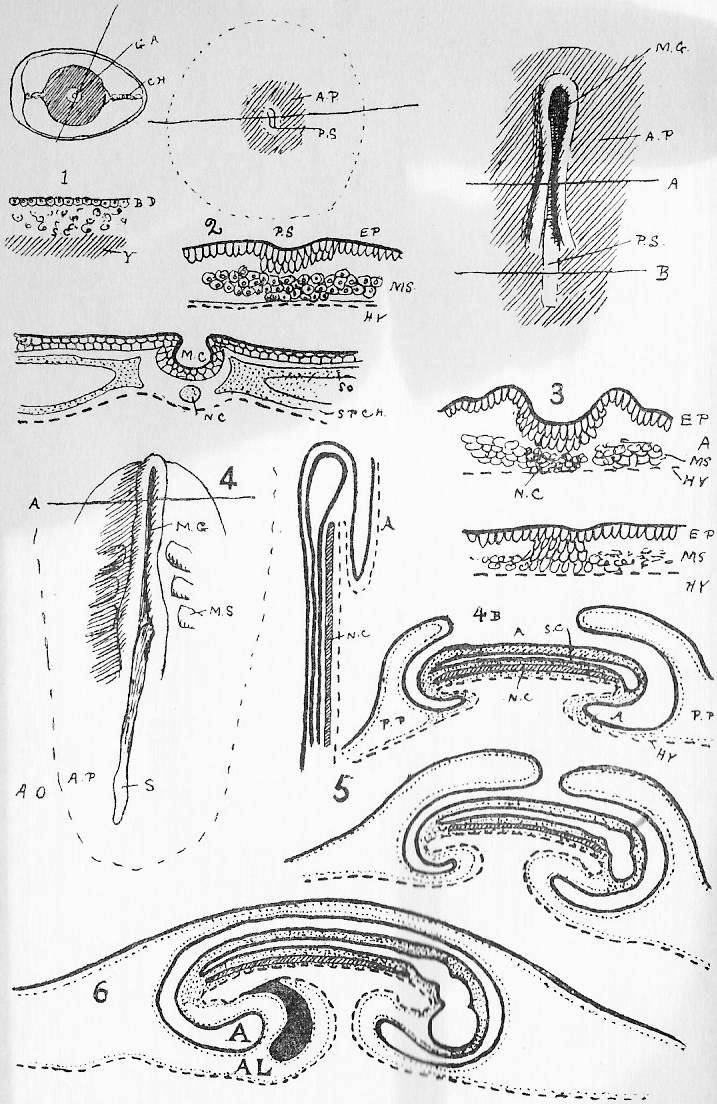
|
Figure 1. Chick about the -fifth- [third] day. At this stage the chick lies on its left side in the yolk. [For lettering of blood vessels, see (7) below.] i., the intestine. u.v., the yolk sac. v.v., the vitelline veins. al., the allantois. Figure 2. Chick about sixth day. Figure 3. Development of heart. Figure 4. Development of the eye. Figure 5. Chick about the sixteenth day. A.M. is the amnion surrounding the embryo. Note particularly how the allantois (al.) has spread over surface of shell and how the yolk sac is shrivelled. Figure 6. Figures to illustrate the relative function and importance of allantois and yolk sac in bird and mammal. In the fowl, however, the blood-vessels of the allantois also probably absorb the albumen of the egg, and may excrete urea into the egg-space. Figure 7. Simplified figure of the embryonic circulation, for comparison with the similar figures annexed to Dog-Fish and Rabbit. {Lines from Second Edition only.} [A.C., anterior cardinal. Ao., Aorta. Br4, sixth aortic arch (fourth branchial). C.S. Cuvierian sinus. H., the heart. I.C., inferior cava. P.C., posterior cardinal vein. Tr.A., truncus arteriosus. v.v., vitelline vein.] Figure 8. Chick on the nineteenth day. |
***END OF THE PROJECT GUTENBERG EBOOK TEXT BOOK OF BIOLOGY, PART 1: VERTEBRATA***
******* This file should be named 21781-h.txt or 21781-h.zip *******
This and all associated files of various formats will be found in:
http://www.gutenberg.org/2/1/7/8/21781
Updated editions will replace the previous one--the old editions will be renamed.
Creating the works from public domain print editions means that no one owns a United States copyright in these works, so the Foundation (and you!) can copy and distribute it in the United States without permission and without paying copyright royalties. Special rules, set forth in the General Terms of Use part of this license, apply to copying and distributing Project Gutenberg-tm electronic works to protect the PROJECT GUTENBERG-tm concept and trademark. Project Gutenberg is a registered trademark, and may not be used if you charge for the eBooks, unless you receive specific permission. If you do not charge anything for copies of this eBook, complying with the rules is very easy. You may use this eBook for nearly any purpose such as creation of derivative works, reports, performances and research. They may be modified and printed and given away--you may do practically ANYTHING with public domain eBooks. Redistribution is subject to the trademark license, especially commercial redistribution.
*** START: FULL LICENSE ***
THE FULL PROJECT GUTENBERG LICENSE
PLEASE READ THIS BEFORE YOU DISTRIBUTE OR USE THIS WORK
To protect the Project Gutenberg-tm mission of promoting the free
distribution of electronic works, by using or distributing this work
(or any other work associated in any way with the phrase "Project
Gutenberg"), you agree to comply with all the terms of the Full Project
Gutenberg-tm License (available with this file or online at
http://www.gutenberg.org/license).
Section 1. General Terms of Use and Redistributing Project Gutenberg-tm
electronic works
1.A. By reading or using any part of this Project Gutenberg-tm
electronic work, you indicate that you have read, understand, agree to
and accept all the terms of this license and intellectual property
(trademark/copyright) agreement. If you do not agree to abide by all
the terms of this agreement, you must cease using and return or destroy
all copies of Project Gutenberg-tm electronic works in your possession.
If you paid a fee for obtaining a copy of or access to a Project
Gutenberg-tm electronic work and you do not agree to be bound by the
terms of this agreement, you may obtain a refund from the person or
entity to whom you paid the fee as set forth in paragraph 1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be
used on or associated in any way with an electronic work by people who
agree to be bound by the terms of this agreement. There are a few
things that you can do with most Project Gutenberg-tm electronic works
even without complying with the full terms of this agreement. See
paragraph 1.C below. There are a lot of things you can do with Project
Gutenberg-tm electronic works if you follow the terms of this agreement
and help preserve free future access to Project Gutenberg-tm electronic
works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the Foundation"
or PGLAF), owns a compilation copyright in the collection of Project
Gutenberg-tm electronic works. Nearly all the individual works in the
collection are in the public domain in the United States. If an
individual work is in the public domain in the United States and you are
located in the United States, we do not claim a right to prevent you from
copying, distributing, performing, displaying or creating derivative
works based on the work as long as all references to Project Gutenberg
are removed. Of course, we hope that you will support the Project
Gutenberg-tm mission of promoting free access to electronic works by
freely sharing Project Gutenberg-tm works in compliance with the terms of
this agreement for keeping the Project Gutenberg-tm name associated with
the work. You can easily comply with the terms of this agreement by
keeping this work in the same format with its attached full Project
Gutenberg-tm License when you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern
what you can do with this work. Copyright laws in most countries are in
a constant state of change. If you are outside the United States, check
the laws of your country in addition to the terms of this agreement
before downloading, copying, displaying, performing, distributing or
creating derivative works based on this work or any other Project
Gutenberg-tm work. The Foundation makes no representations concerning
the copyright status of any work in any country outside the United
States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other immediate
access to, the full Project Gutenberg-tm License must appear prominently
whenever any copy of a Project Gutenberg-tm work (any work on which the
phrase "Project Gutenberg" appears, or with which the phrase "Project
Gutenberg" is associated) is accessed, displayed, performed, viewed,
copied or distributed:
This eBook is for the use of anyone anywhere at no cost and with
almost no restrictions whatsoever. You may copy it, give it away or
re-use it under the terms of the Project Gutenberg License included
with this eBook or online at www.gutenberg.org
1.E.2. If an individual Project Gutenberg-tm electronic work is derived
from the public domain (does not contain a notice indicating that it is
posted with permission of the copyright holder), the work can be copied
and distributed to anyone in the United States without paying any fees
or charges. If you are redistributing or providing access to a work
with the phrase "Project Gutenberg" associated with or appearing on the
work, you must comply either with the requirements of paragraphs 1.E.1
through 1.E.7 or obtain permission for the use of the work and the
Project Gutenberg-tm trademark as set forth in paragraphs 1.E.8 or
1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted
with the permission of the copyright holder, your use and distribution
must comply with both paragraphs 1.E.1 through 1.E.7 and any additional
terms imposed by the copyright holder. Additional terms will be linked
to the Project Gutenberg-tm License for all works posted with the
permission of the copyright holder found at the beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm
License terms from this work, or any files containing a part of this
work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this
electronic work, or any part of this electronic work, without
prominently displaying the sentence set forth in paragraph 1.E.1 with
active links or immediate access to the full terms of the Project
Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary,
compressed, marked up, nonproprietary or proprietary form, including any
word processing or hypertext form. However, if you provide access to or
distribute copies of a Project Gutenberg-tm work in a format other than
"Plain Vanilla ASCII" or other format used in the official version
posted on the official Project Gutenberg-tm web site (www.gutenberg.org),
you must, at no additional cost, fee or expense to the user, provide a
copy, a means of exporting a copy, or a means of obtaining a copy upon
request, of the work in its original "Plain Vanilla ASCII" or other
form. Any alternate format must include the full Project Gutenberg-tm
License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying,
performing, copying or distributing any Project Gutenberg-tm works
unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing
access to or distributing Project Gutenberg-tm electronic works provided
that
- You pay a royalty fee of 20% of the gross profits you derive from
the use of Project Gutenberg-tm works calculated using the method
you already use to calculate your applicable taxes. The fee is
owed to the owner of the Project Gutenberg-tm trademark, but he
has agreed to donate royalties under this paragraph to the
Project Gutenberg Literary Archive Foundation. Royalty payments
must be paid within 60 days following each date on which you
prepare (or are legally required to prepare) your periodic tax
returns. Royalty payments should be clearly marked as such and
sent to the Project Gutenberg Literary Archive Foundation at the
address specified in Section 4, "Information about donations to
the Project Gutenberg Literary Archive Foundation."
- You provide a full refund of any money paid by a user who notifies
you in writing (or by e-mail) within 30 days of receipt that s/he
does not agree to the terms of the full Project Gutenberg-tm
License. You must require such a user to return or
destroy all copies of the works possessed in a physical medium
and discontinue all use of and all access to other copies of
Project Gutenberg-tm works.
- You provide, in accordance with paragraph 1.F.3, a full refund of any
money paid for a work or a replacement copy, if a defect in the
electronic work is discovered and reported to you within 90 days
of receipt of the work.
- You comply with all other terms of this agreement for free
distribution of Project Gutenberg-tm works.
1.E.9. If you wish to charge a fee or distribute a Project Gutenberg-tm
electronic work or group of works on different terms than are set
forth in this agreement, you must obtain permission in writing from
both the Project Gutenberg Literary Archive Foundation and Michael
Hart, the owner of the Project Gutenberg-tm trademark. Contact the
Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable
effort to identify, do copyright research on, transcribe and proofread
public domain works in creating the Project Gutenberg-tm
collection. Despite these efforts, Project Gutenberg-tm electronic
works, and the medium on which they may be stored, may contain
"Defects," such as, but not limited to, incomplete, inaccurate or
corrupt data, transcription errors, a copyright or other intellectual
property infringement, a defective or damaged disk or other medium, a
computer virus, or computer codes that damage or cannot be read by
your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right
of Replacement or Refund" described in paragraph 1.F.3, the Project
Gutenberg Literary Archive Foundation, the owner of the Project
Gutenberg-tm trademark, and any other party distributing a Project
Gutenberg-tm electronic work under this agreement, disclaim all
liability to you for damages, costs and expenses, including legal
fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT
LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE
PROVIDED IN PARAGRAPH F3. YOU AGREE THAT THE FOUNDATION, THE
TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE
LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR
INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH
DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a
defect in this electronic work within 90 days of receiving it, you can
receive a refund of the money (if any) you paid for it by sending a
written explanation to the person you received the work from. If you
received the work on a physical medium, you must return the medium with
your written explanation. The person or entity that provided you with
the defective work may elect to provide a replacement copy in lieu of a
refund. If you received the work electronically, the person or entity
providing it to you may choose to give you a second opportunity to
receive the work electronically in lieu of a refund. If the second copy
is also defective, you may demand a refund in writing without further
opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth
in paragraph 1.F.3, this work is provided to you 'AS-IS,' WITH NO OTHER
WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT LIMITED TO
WARRANTIES OF MERCHANTIBILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied
warranties or the exclusion or limitation of certain types of damages.
If any disclaimer or limitation set forth in this agreement violates the
law of the state applicable to this agreement, the agreement shall be
interpreted to make the maximum disclaimer or limitation permitted by
the applicable state law. The invalidity or unenforceability of any
provision of this agreement shall not void the remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the
trademark owner, any agent or employee of the Foundation, anyone
providing copies of Project Gutenberg-tm electronic works in accordance
with this agreement, and any volunteers associated with the production,
promotion and distribution of Project Gutenberg-tm electronic works,
harmless from all liability, costs and expenses, including legal fees,
that arise directly or indirectly from any of the following which you do
or cause to occur: (a) distribution of this or any Project Gutenberg-tm
work, (b) alteration, modification, or additions or deletions to any
Project Gutenberg-tm work, and (c) any Defect you cause.
Section 2. Information about the Mission of Project Gutenberg-tm
Project Gutenberg-tm is synonymous with the free distribution of
electronic works in formats readable by the widest variety of computers
including obsolete, old, middle-aged and new computers. It exists
because of the efforts of hundreds of volunteers and donations from
people in all walks of life.
Volunteers and financial support to provide volunteers with the
assistance they need, is critical to reaching Project Gutenberg-tm's
goals and ensuring that the Project Gutenberg-tm collection will
remain freely available for generations to come. In 2001, the Project
Gutenberg Literary Archive Foundation was created to provide a secure
and permanent future for Project Gutenberg-tm and future generations.
To learn more about the Project Gutenberg Literary Archive Foundation
and how your efforts and donations can help, see Sections 3 and 4
and the Foundation web page at http://www.gutenberg.org/fundraising/pglaf.
Section 3. Information about the Project Gutenberg Literary Archive
Foundation
The Project Gutenberg Literary Archive Foundation is a non profit
501(c)(3) educational corporation organized under the laws of the
state of Mississippi and granted tax exempt status by the Internal
Revenue Service. The Foundation's EIN or federal tax identification
number is 64-6221541. Contributions to the Project Gutenberg
Literary Archive Foundation are tax deductible to the full extent
permitted by U.S. federal laws and your state's laws.
The Foundation's principal office is located at 4557 Melan Dr. S.
Fairbanks, AK, 99712., but its volunteers and employees are scattered
throughout numerous locations. Its business office is located at
809 North 1500 West, Salt Lake City, UT 84116, (801) 596-1887, email
business@pglaf.org. Email contact links and up to date contact
information can be found at the Foundation's web site and official
page at http://www.gutenberg.org/about/contact
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
gbnewby@pglaf.org
Section 4. Information about Donations to the Project Gutenberg
Literary Archive Foundation
Project Gutenberg-tm depends upon and cannot survive without wide
spread public support and donations to carry out its mission of
increasing the number of public domain and licensed works that can be
freely distributed in machine readable form accessible by the widest
array of equipment including outdated equipment. Many small donations
($1 to $5,000) are particularly important to maintaining tax exempt
status with the IRS.
The Foundation is committed to complying with the laws regulating
charities and charitable donations in all 50 states of the United
States. Compliance requirements are not uniform and it takes a
considerable effort, much paperwork and many fees to meet and keep up
with these requirements. We do not solicit donations in locations
where we have not received written confirmation of compliance. To
SEND DONATIONS or determine the status of compliance for any
particular state visit http://www.gutenberg.org/fundraising/pglaf
While we cannot and do not solicit contributions from states where we
have not met the solicitation requirements, we know of no prohibition
against accepting unsolicited donations from donors in such states who
approach us with offers to donate.
International donations are gratefully accepted, but we cannot make
any statements concerning tax treatment of donations received from
outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation
methods and addresses. Donations are accepted in a number of other
ways including checks, online payments and credit card donations.
To donate, please visit: http://www.gutenberg.org/fundraising/donate
Section 5. General Information About Project Gutenberg-tm electronic
works.
Professor Michael S. Hart is the originator of the Project Gutenberg-tm
concept of a library of electronic works that could be freely shared
with anyone. For thirty years, he produced and distributed Project
Gutenberg-tm eBooks with only a loose network of volunteer support.
Project Gutenberg-tm eBooks are often created from several printed
editions, all of which are confirmed as Public Domain in the U.S.
unless a copyright notice is included. Thus, we do not necessarily
keep eBooks in compliance with any particular paper edition.
Each eBook is in a subdirectory of the same number as the eBook's
eBook number, often in several formats including plain vanilla ASCII,
compressed (zipped), HTML and others.
Corrected EDITIONS of our eBooks replace the old file and take over
the old filename and etext number. The replaced older file is renamed.
VERSIONS based on separate sources are treated as new eBooks receiving
new filenames and etext numbers.
Most people start at our Web site which has the main PG search facility:
http://www.gutenberg.org
This Web site includes information about Project Gutenberg-tm,
including how to make donations to the Project Gutenberg Literary
Archive Foundation, how to help produce our new eBooks, and how to
subscribe to our email newsletter to hear about new eBooks.
EBooks posted prior to November 2003, with eBook numbers BELOW #10000,
are filed in directories based on their release date. If you want to
download any of these eBooks directly, rather than using the regular
search system you may utilize the following addresses and just
download by the etext year.
http://www.gutenberg.org/dirs/etext06/
(Or /etext 05, 04, 03, 02, 01, 00, 99,
98, 97, 96, 95, 94, 93, 92, 92, 91 or 90)
EBooks posted since November 2003, with etext numbers OVER #10000, are
filed in a different way. The year of a release date is no longer part
of the directory path. The path is based on the etext number (which is
identical to the filename). The path to the file is made up of single
digits corresponding to all but the last digit in the filename. For
example an eBook of filename 10234 would be found at:
http://www.gutenberg.org/dirs/1/0/2/3/10234
or filename 24689 would be found at:
http://www.gutenberg.org/dirs/2/4/6/8/24689
An alternative method of locating eBooks:
http://www.gutenberg.org/dirs/GUTINDEX.ALL
*** END: FULL LICENSE ***