The Project Gutenberg eBook, Report on the Radiolaria Collected by H.M.S.
Challenger During the Years 1873-1876, First Part: Porulosa (Spumellaria
and Acantharia), by Ernst Haeckel
| Transcriber's note: |
Some typographical errors in the printed
work have been corrected. The corrected text is underscored in red
like
this. Hover the cursor over the marked text and the explanation
should appear. The Addenda & Errata (Second Part, pp. 1763-4)
have been applied and underscored in this way. |
REPORT
ON THE
SCIENTIFIC
RESULTS
OF THE
VOYAGE OF H.M.S.
CHALLENGER
DURING THE YEARS 1873-76
UNDER THE COMMAND
OF
Captain
GEORGE S. NARES, R.N., F.R.S.
AND THE LATE
Captain
FRANK TOURLE THOMSON, R.N.
PREPARED UNDER THE
SUPERINTENDENCE OF
THE LATE
Sir C. WYVILLE THOMSON, Knt.,
F.R.S., &c.
REGIUS PROFESSOR OF NATURAL
HISTORY IN THE UNIVERSITY OF EDINBURGH
DIRECTOR OF THE CIVILIAN
SCIENTIFIC STAFF ON BOARD
AND NOW OF
JOHN MURRAY
ONE OF THE NATURALISTS OF THE
EXPEDITION
Zoology—Vol. XVIII.
FIRST PART
Published by Order of Her Majesty's
Government
PRINTED FOR HER MAJESTY'S
STATIONARY OFFICE
AND SOLD BY
LONDON:—EYRE &
SPOTTISWOODE, EAST HARDING STREET, FETTER LANE
EDINBURGH:—ADAM &
CHARLES BLACK
DUBLIN:—HODGES, FIGGIS,
& CO.
1887
Price (in Two Parts, with
a Volume of Plates) £5, 10s.
CONTENTS.
Report on the Radiolaria collected
by H.M.S. Challenger during the years
1873-1876.
By Ernst Haeckel, M.D., Ph.D., Professor of Zoology in the
University of Jena.
FIRST PART.—PORULOSA.
(SPUMELLARIA AND ACANTHARIA.)
EDITORIAL NOTES.
The Report on the Radiolaria by Professor Ernst Haeckel of Jena
occupies the whole of the present Volume, the text being bound up in Two Separate Parts and the
Plates in a Third Part. The Report forms Part XL. of the Zoological Series of Reports on the
Scientific Results of the Expedition, and is the largest single Report of the series which has up
to this time been published.
The Manuscript of the Systematic Part was written by Professor Haeckel in the English language,
and was received by me in instalments on the 12th August 1884, 13th July and 4th December 1885,
and 3rd June 1886. The Introduction was written in German and was translated into the English
language by Mr. W. E. Hoyle of the Challenger Editorial Staff; the German text being received in
instalments between the 15th July 1886, and the 25th January 1887.
The Challenger Naturalists found the representatives of this group of animals to be universally
distributed throughout ocean waters, and their dead remains to be nearly equally widely
distributed over the floor of the ocean, the relative abundance and the species differing,
however, with change of locality, and their abundance or variety being intimately connected with
some of the most interesting and intricate problems of general oceanography.
It was a fortunate circumstance that so distinguished a Naturalist, with such an intimate
knowledge of the Radiolaria, should have been willing to undertake the
laborious examination and description of the extensive collections made during the Expedition.
Professor Haeckel has devoted ten years of his life to this work, and this Report sets forth the
results of his labours, on the conclusion of which he will be congratulated by all Naturalists.
The entire literature of the Radiolaria (from 1834 to 1884) is completely
recorded, and the older species (both living and fossil) redescribed, so that the Report is a
complete Monograph, which will be an invaluable aid to all future Investigators.
|John Murray.
Challenger Office, 32 Queen Street,
Edinburgh, 1st February 1887.
THE
VOYAGE OF H.M.S.
CHALLENGER.
ZOOLOGY.
Report on the Radiolaria collected by H.M.S. Challenger
during the Years 1873-76. By Ernst Haeckel, M.D., Ph.D., Professor of
Zoology in the University of Jena.
PREFACE.
The significance of the Radiolaria in regard to the relations of life in the ocean has been
increased in a most unexpected manner by the discoveries of the Challenger. Large swarms of these
delicate Rhizopoda were found not only at the surface of the open ocean but also in its different
bathymetrical zones. Thousands of new species make up the wonderful Radiolarian ooze, which covers
large areas of the deep-sea bed, and was brought up from abysses of from 2000 to 4000 fathoms by
the sounding machine of the Challenger. They open a new world to morphological investigation.
When ten years ago (in the autumn of 1876) I accepted the enticing invitation of Sir Wyville
Thomson to undertake the investigation of these microscopic creatures, I hoped to be able to
accomplish the task with some degree of completeness within a period of from three to five years,
but the further my investigations proceeded the more immeasurable seemed the range of forms, like
the boundless firmament of stars. I soon found myself compelled to decide between making a
detailed study of a selection of special forms or giving as complete a survey as possible of the
varied forms of the whole class; and I decided upon the latter course, having regard both to the
general plan of the Challenger Reports, and to the interests of our acquaintance with the class as
a whole. I must, however, confess at the close of my work that my original intention is far from
having been fulfilled. The extraordinary extent and varied difficulties of the undertaking must
excuse the many deficiencies.
The special examination of the Challenger collection was for the most part completed in the
summer of 1881; I collected its results in my Entwurf eines Radiolarien-Systems auf Grund von
Studien der Challenger-Radiolarien (Jenaische Zeitschr. f. Naturw., Bd. xv., 1881). Since the
manuscript of this preliminary communication was completed only a few days before my departure for
Ceylon, and since I was unable to correct the proofs myself, several errors have crept into the
Prodromus Systematis Radiolarium included in it. These have been corrected in the following more
extensive working out of it. Even at that time I had distinguished 630 genera and more than 2000
species; but on the revision of these, which I undertook immediately on my return from India, this
number was considerably increased. The total number of forms here described amounts to 739 genera
and 4318 species; of these 3508 are new, as against 810 previously described. In spite of this
large number, however, and in spite of the astonishing variety of the new and marvellous forms,
the riches of the Challenger collection are by no means exhausted. A careful and patient worker
who would devote a second decade to the work, would probably increase the number of new forms
(especially of the smaller ones) by more than a thousand; but for a really complete examination,
the lifetime of one man would not suffice.
The richest source of the Challenger material is the Radiolarian ooze of the central Pacific
Ocean (Stations 265 to 274). This remarkable deep-sea mud consists for the greater part of
well-preserved siliceous shells of Polycystina (Spumellaria and Nassellaria). Not less important, however, especially for the study of the Acantharia and Phæodaria, are the wonderful preparations
stained with carmine and mounted in Canada balsam on the spot by Dr. John Murray. One such
preparation (e.g., from Station 271) often contains twenty or thirty, sometimes even fifty
new species. In many of these preparations the individual parts of the unicellular organism are so
well preserved that they show clearly the characteristic peculiarities of the legions and orders.
Since the material for these preparations was taken with the tow-net, not only from the surface of
the sea but also from different bathymetrical zones, it furnishes valuable conclusions regarding
the chorology, as well as the physiology and morphology of the group. For many new discoveries I
am indebted to the study of such preparations, of which I have examined about a thousand from 168
different Stations (compare § 240). In addition to these about 100 bottles
were handed to me, containing partly bottom-deposits, partly tow-net gatherings.
Sir Wyville Thomson, who directed the investigations of the Challenger with so much devotion,
and only partly saw its results, has laid me under a deep debt of obligation; not less is this the
case, however, with his successor, Dr. John Murray. I am especially indebted to both gentlemen for
the freedom they have allowed me in the carrying out of my work, and especially for the permission
to include a description of all known Radiolaria in the Challenger Report, which has thus become a
second edition many times enlarged of my Monograph published in 1862. Since all previous
literature of the subject has been consulted and critically revised, it is hoped that this Report
will form a useful foundation for future investigations. All names of sufficiently described
Radiolaria published during the first half century of our knowledge of the class (from 1834 to
1884), are inserted in alphabetical order in the index at the end of this work.
In addition to the treasures of the Challenger, my own collection of Radiolaria has yielded
many new forms whose description is here included. On my journeys to the Mediterranean (an account
of which is given in the introduction to my Monograph of the Medusæ), I have given special
attention to these delicate microscopic organisms for more than thirty years. Besides the various
points on the Mediterranean, the Atlantic Ocean at the Canaries (in the winter of 1866-67) yielded
many interesting new forms; whilst my voyage across the Indian Ocean, from Aden to Bombay, in
November 1881, thence to Ceylon and back by Socotra in March 1882, was still more productive. In
particular, some extended excursions which I had the opportunity of making from Belligemma and
Matura (at the southern extremity of Ceylon) gave me an insight into the rich treasures of the
Indian Ocean.
Most important, however, as regards the knowledge of the Indian Radiolaria, are the collections
which Captain Heinrich Rabbe of Bremen has so beautifully preserved during his many voyages
through that region. In the neighbourhood of Madagascar and the Cocos Islands more especially, and
also in the Sunda Archipelago, he met with large swarms of Radiolaria, among which were many new
and remarkable forms. These were of special value for completing the chorology, and the more so
since the course of the Challenger in the Indian Ocean lay very far to the southwards. I will
therefore take this opportunity of repeating my best thanks to Captain Rabbe for the friendly
donation of his valuable collection.
The Radiolarian fauna of the North Atlantic Ocean, which was previously but little known and
only slightly increased by the investigations of the Challenger, received a valuable increase from
the interesting collections made by Dr. John Murray on various expeditions to the Færöe Islands
(on the "Knight Errant" in 1880 and on the "Triton" in 1882). A large number of new Radiolaria
were captured in the Færöe Channel, partly at the surface of the Gulf Stream, partly at various
depths, and the proof was thus furnished that at certain points in the North Atlantic Ocean
Radiolaria are very richly developed. I am further indebted to Dr. John Murray for the free use of
this important material as well as for much other assistance in the carrying out of my work.
Another rich source of Radiolaria I found in the alimentary canal of pelagic animals from all
seas. Medusæ, Siphonophoræ, Salpæ, Pteropoda, Heteropoda, Crustacea, &c., which live partly at
the surface of the sea and partly at various depths, and swallow large masses of Radiolaria, often
contain numbers of their shells well-preserved in their intestine. The alimentary canal of Fishes
and Cephalopods too, which live upon these pelagic animal frequently contains considerable
quantities of siliceous shells; and another newly discovered source has been found in the
coprolites of the Jurassic period, which consist largely of Radiolarian skeletons.
In the investigation of this complicated system of organisms, I have endeavoured on the one
hand to give accurately the forms and dimensions of the species observed, and on the other hand to
present a survey of the relationships of the different genera and families; and in this I have
striven especially to combine the phylogenetic aims of the natural system with the essentially
artificial divisions of a practical classification. Being, however, a conscientious supporter of
the theory of descent, I can of course lay no stress upon the value of the categories, which are
here distinguished as Legions, Orders, Families, Genera, &c. All these artificial systematic
grades I regard as of merely relative value; and from the same cause I attach no importance to the
distinction of all the species here described; many of them are probably only developmental
stages, and like my predecessors I have determined their boundaries on subjective grounds. In the
systematic working out of so much material one always runs the risk of doing either too much or
too little in the way of creating species; but in the light of the theory of descent this danger
is of no consequence.
In the carrying out of this extensive task the friendly aid of Dr. Reinhold Teuscher of Jena
was of the greatest benefit to me; at my request he was at the trouble of making a large number of
accurate drawings with the camera lucida, and he also undertook a long series, amounting to some
8000, accurate micrometric measurements, which were of the greatest value in the attempt to settle
the important question of the constancy of the various species; I have alluded to this in a note
at the conclusion of the Report (p. 1760).
My best thanks are due to Dr. Teuscher for the patient and careful manner in which he discharged
these tedious tasks.
The figures of new species of Radiolaria (about 1600 in number) which appear in the
atlas of one hundred and forty plates accompanying this Report, were nearly all drawn with the
camera lucida, partly by Mr. Adolph Giltsch and partly by myself. The names of the genera which
appear at the bottom of the plates have in many cases been changed since they were printed off, as
may be seen from the explanations which accompany them. Had it been possible to complete the
examination of the material before the plates were commenced this might have been avoided, and in
many cases a better selection of figures might have been made. All the drawings have been made
upon the stone by the practised hand of Mr. Adolph Giltsch, in his usual masterly manner, and his
lithographic work, which has lasted fully ten years, is the more valuable since he has himself
microscopically studied the greater part of the species figured. The fact that the atlas presents
so full a picture of the marvellous wealth of form of the Radiolaria is especially due to his
lively interest in the work, to his unwearying care, and to his morphological acuteness. May it be
the means of inducing many naturalists to study more deeply this inexhaustible kingdom of
microscopic life, whose endless variety of wonderful forms justifies the saying—Natura in
minimis maxima.
CONTENTS.
| FIRST PART. |
| GENERAL INTRODUCTION— |
PAGE |
| I. |
Anatomical Section (§§ 1-140), |
i |
|
Chapter |
I. |
The Unicellular Organism, |
i |
|
" |
II. |
The Central Capsule, |
xxiv |
|
" |
III. |
The Extracapsulum, |
li |
|
" |
IV. |
The Skeleton, |
lxviii |
| II. |
Biogenetical Section (§§ 141-200), |
xciii |
|
Chapter |
V. |
Ontogeny (Individual Development), |
xciii |
|
" |
VI. |
Phylogeny (Genealogical Development), |
ci |
| III. |
Physiological Section (§§ 201-225), |
cxxviii |
|
Chapter |
VII. |
Vegetative Functions, |
cxxviii |
|
" |
VIII. |
Animal Functions, |
cxl |
| IV. |
Chorological Section (§§ 226-250), |
cxlvi |
|
Chapter |
IX. |
Geographical Distribution, |
cxlvi |
|
" |
X. |
Geological Distribution, |
clxiv |
| V. |
Bibliographical Section (§§
251-254), |
clxxvi |
| SYSTEMATIC PART, |
1 |
| I. |
Subclass PORULOSA, |
6 |
|
Legion I. SPUMELLARIA vel PERIPYLEA, |
6 |
|
Order |
1. |
Colloidea, |
10 |
|
" |
2. |
Beloidea, |
28 |
|
" |
3. |
Sphæroidea, |
50 |
|
" |
4. |
Prunoidea, |
284 |
|
" |
5. |
Discoidea, |
402 |
|
" |
6. |
Larcoidea, |
599 |
|
Legion II. ACANTHARIA vel ACTIPYLEA, |
716 |
|
Order |
7. |
Actinelida, |
728 |
|
" |
8. |
Acanthonida, |
740 |
|
" |
9. |
Sphærophracta, |
795 |
|
" |
10. |
Prunophracta, |
859 |
|
SECOND PART.
|
| II. |
Subclass OSCULOSA, |
889 |
|
Legion III. NASSELLARIA vel MONOPYLEA, |
889 |
|
Order |
11. |
Nassoidea, |
895 |
|
" |
12. |
Plectoidea, |
898 |
|
" |
13. |
Stephoidea, |
931 |
|
" |
14. |
Spyroidea, |
1015 |
|
" |
15. |
Botryodea, |
1103 |
|
" |
16. |
Cyrtoidea, |
1126 |
|
Legion IV. PHÆODARIA vel CANNOPYLEA, |
1521 |
|
Order |
17. |
Phæocystina, |
1542 |
|
" |
18. |
Phæosphæria, |
1590 |
|
" |
19. |
Phæogromia, |
1642 |
|
" |
20. |
Phæoconchia, |
1710 |
| Note on the Dimensions and
Measurements, |
1760 |
| ADDENDA, |
1761 |
| ERRATA, |
1763 |
| INDEX, |
1765 |
{i}
GENERAL INTRODUCTION.
ANATOMICAL SECTION.
A SKETCH OF OUR KNOWLEDGE OF THE ORGANISATION OF THE RADIOLARIA IN THE YEAR
1884.
Chapter I.—THE UNICELLULAR ORGANISM.
(§§ 1-50.)
1. Definition of the Radiolaria.—Radiolaria are marine
Rhizopoda, whose unicellular body always consists of two main portions, separated by a membrane;
an inner Central capsule (with one or more nuclei) and an Extracapsulum (the
external calymma, which has no nucleus, and the pseudopodia); the endoplasm of the former and the
exoplasm of the latter are connected by openings in the capsule-membrane. The central capsule is
partly the general central organ of the Radiolarian cell, partly the special organ of
reproduction, since its intracapsular protoplasm, along with the nuclei embedded in it, serves for
the formation of flagellate spores. The extracapsulum is partly the general organ for intercourse
with the outer world (by means of the pseudopodia), partly the special organ of protection
(calymma) and nutrition (sarcomatrix). The majority of Radiolaria develop also a skeleton for
support and protection, which presents the utmost variety of form, and is generally composed of
silica, sometimes of an organic substance (acanthin). The Radiolarian cell usually leads an
isolated existence (Monozoa vel Monocyttaria); only in a small minority (of one legion) are
the unicellular organisms united in colonies or cœnobia (Polyzoa vel
Polycyttaria).
The extent of the Radiolaria, as limited by the above definition, which I have
made as compact as possible, differs in several important respects from that allowed to the
group by all previous diagnoses. The shortest expression of its scope might perhaps
be:—Rhizopoda with central capsule and calymma; for the most important character of the
Radiolaria, and that by which they are distinguished from all other Rhizopoda, is the
differentiation of the unicellular body into two principal parts of equal importance and their
separation by a constant capsule-membrane.
2. The Two Subclasses of the Radiolaria.—The systematic catalogue of the
Radiolaria, which forms the second part of this Report, and is brought up to the year {ii}1884, contains 20 orders, 85 families, 739 genera, and 4318
species. The consideration that but a small proportion of the ocean his yet been investigated
renders it likely, however, that even this large number does not include the half of the recent
species. The great progress which our knowledge of the organisation of the Radiolaria has made, by
means of comparative study, renders it possible to arrange this enormous mass of forms in four
main divisions or legions, and these are again related in pairs, so that two divisions of the
highest rank or subclasses are constituted, the Porulosa (or Holotrypasta) and
Osculosa (or Merotrypasta).
The division of the Radiolaria into two subclasses and four legions (or principal
orders), I sought to establish in 1883 in a communication on the Orders of the Radiolaria
(Sitzb. Jena Gesellsch. Med. u. Naturwiss., February 16, 1883). As a believer in the theory of
descent, I regard all the systematic arrangements of specialists as artificial, and all their
divisions as subjective abstractions, and hence I shall be guided in the establishment of such
groups as subclasses, legions, orders, &c., by purely practical considerations, especially
by the desire to give as ready a survey as possible of the complex multitude of forms (compare
§§ 154 to 156).
3. Porulosa or Holotrypasta.—The subclass Porulosa or Holotrypasta
includes the two legions, Peripylea or Spumellaria, and Actipylea or Acantharia, which agree in the following constant and important
characters:—(1) The Central Capsule is primitively a sphere, and retains this homaxon
form in the majority of the species. (2) The Membrane of the central capsule is everywhere
perforated by very numerous minute pores, but possesses no larger principal aperture (osculum).
(3) The Pseudopodia radiate in all directions and in great numbers from the central
capsule, passing through its pores. (4) The Equilibrium of the floating unicellular body is
in most Porulosa pantostatic (indifferent) or polystatic (plural-stable), since a vertical axis is
either absent, or, if present, has its two poles similarly constituted. (5) The
Ground-forms of the skeleton are therefore almost always either spherotypic or
isopolar-monaxon, very rarely zygotypic. The two legions of the Porulosa are distinguished mainly
by the skeleton of the Spumellaria (or Peripylea)
being siliceous, never centrogenous, nor composed of acanthin, whilst in the Acantharia (or Actipylea) it is always centrogenous and
made up of acanthin; hence in the former the nucleus is always central, in the latter always
excentric.
4. Osculosa or Merotrypasta.—The subclass Osculosa or Merotrypasta
includes the two legions Monopylea or Nassellaria,
and Cannopylea or Phæodaria, which agree in the
following constant and important characters:—(1) The Central Capsule is originally monaxon
(ovoid or spheroidal) and retains this ground-form in most of the species. (2) The Membrane
of the central capsule possesses a single large principal aperture (osculum) at the basal pole of
the vertical main axis. (3) The Pseudopodia radiate from a stream of sarcode which passes
out from the central capsule only on one side, namely, through the principal aperture. (4) The
Equilibrium of the floating body is {iii}monostatic or unistable, since the two poles of the principal axis are
always more or less different from each other. (5) The Ground-forms of the skeleton are,
therefore, for the most part grammotypic (centraxon) or zygotypic (centroplan), rarely
spherotypic. The two legions of the Osculosa are distinguished chiefly by the principal opening
(osculum) being closed by a porous plate (porochora with its podoconus) in the Nassellaria (or Monopylea), and by a radiate cover
(operculum with its astropyle) in the Phæodaria (or Cannopylea).
5. The four Legions of Radiolaria.—The four principal groups of
Radiolaria, to which we have given the name "legions," are natural units, since the most important
peculiarities in the structure of the central capsule are quite constant within the limits of the
same legion, and since all the forms in the same legion may be traced without violence to the same
phylogenetic stem. The four legions are, however, related to each other, in so far as they all
exhibit those characters which distinguish the Radiolaria from other Protista. The two which
compose the Porulosa (§ 3) seem somewhat more nearly related to each other
than to the two which make up the Osculosa (§ 4). When, however, the attempt
is made to bring them all into a phylogenetic relationship, it undoubtedly appears that the Spumellaria (or Peripylea) are the primitive stem, out
of which the other three have been developed as independent branches. All three have been derived,
probably independently, from the most ancient stem-form of the Spumellaria, the spherical Actissa.
6. Peripylea or Spumellaria.—Those Radiolaria which we call "Peripylea" on account of the constitution of their central capsule, or "Spumellaria" on account of the nature of their skeleton, are separated from the
other three legions of the class by the combination of the following constant
characters:—(1) The Membrane of the central capsule is single and evenly perforated
all over by innumerable fine pore-canals, but without any larger principal opening (osculum). (2)
The Nucleus always lies centrally in the Spumellaria monozoa and is
serotinous, for it divides only at a later period into the nuclei of the spores; in the Spumellaria polyzoa it is precocious, and divides early into many small nuclei.
(3) The Pseudopodia are exceedingly numerous and distributed evenly over the whole surface
of the central capsule. (4) The Calymma contains no phæodium. (5) The Skeleton is
seldom wanting, is never centrogenous, and is always siliceous. (6) The Ground-form of the
central capsule is originally spherical (often modified); that of the skeleton is also spherical
or, in the majority of cases, derived in different ways from the sphere.
7. Actipylea or Acantharia.—These Radiolaria which we call "Actipylea" on account of the constitution of their central capsule, or "Acantharia" from the formation of their skeleton, are separated from the other
three legions by the combination of the following constant characters:—(1) The
Membrane of the central capsule is single and {iv}perforated by numerous fine pore-canals, which are regularly distributed in
series or groups, but without a larger principal opening (osculum). (2) The Nucleus is
always excentric and generally precocious, since it divides early by a peculiar process of budding
into numerous small nuclei. (3) The Pseudopodia are very numerous and distributed regularly
in groups (or series united into a network). (4) The Calymma contains no phæodium. (5) The
Skeleton is generally present, always centrogenous, and composed of acanthin. (6) The
Ground-form of the central capsule is originally spherical (often modified), that of the
skeleton polyaxon (often modified).
8. Monopylea or Nassellaria.—Those Radiolaria which we call "Monopylea" from the formation of their central capsule, or "Nassellaria" from the nature of their skeleton, are distinguished from the other
three legions of the class by the combination of the following constant characters:—(1) The
Membrane of the central capsule is single, and has only one large principal opening
(osculum) at the basal pole of the vertical main axis; this osculum is closed by a perforated lid
(porochora or operculum porosum) from which there arises within the central capsule a peculiar
cone of threads or pseudopodia (podoconus). (2) The Nucleus is usually excentric and is
always serotinous, since it only divides at a comparatively late period into spore-nuclei. (3) The
Pseudopodia are not very numerous and arise by division of a single stem or bundle of
threads of sarcode, which issues from the porochora. (4) The Calymma contains no phæodium.
(5) The Skeleton (very rarely absent) is never centrogenous, but always extracapsular and
siliceous. (6) The Ground-form of the central capsule is always monaxon (with a vertical
allopolar main axis), originally ovoid, often modified; that of the skeleton is also generally
monaxon, often modified (triradial or bilateral).
9. Cannopylea or Phæodaria.—Those Radiolaria which we call "Cannopylea" from the constitution of their central capsule, or "Phæodaria" on account of their peculiar phæodium, are distinguished from the
other three legions by the combination of the following characters:—(1) The Membrane
of the central capsule is double, consisting of a strong outer and delicate inner capsule, and has
only one principal opening (osculum) at the basal pole of the vertical main axis; this osculum is
closed by a radiate cover (astropyle or operculum radiatum), from the centre of which arises an
external tubular spout (proboscis). Occasionally a few small accessory openings (parapylæ) are
present besides the principal opening. (2) The Nucleus lies centrally or subcentrally in
the capsule (in the vertical main axis), and is serotinous, inasmuch as it only divides at a late
period into spore-nuclei. (3) The Pseudopodia are usually very numerous and arise from a
thick sarcomatrix, formed by the spreading out of a thick stem of sarcode, which issues from the
astropyle. (4) The Calymma always contains a phæodium or peculiar voluminous excentric mass
of pigment. (5) The Skeleton (very rarely absent) is never centrogenous, always
extracapsular and formed of a silicate of carbon. (6) The {v}Ground-form of the central capsule is always monaxon (with a vertical
allopolar main axis) and generally spheroidal; that of the skeleton is very varied.
10. Synopsis of the Subclasses and Legions:—
| First Subclass. |
Second Subclass. |
|
Porulosa vel Holotrypasta.
Central capsule originally spherical, without osculum or principal opening,
with innumerable fine pores.
|
Osculosa vel Merotrypasta.
Central capsule originally monaxon, with an osculum at the basal pole of the
vertical main axis.
|
Legion I.
Spumellaria.
(Peripylea). |
Legion II.
Acantharia.
(Actipylea). |
Legion III.
Nassellaria.
(Monopylea). |
Legion IV.
Phæodaria.
(Cannopylea). |
| Central capsule originally spherical, homaxon. |
Central capsule originally spherical, homaxon. |
Central capsule originally ovoid, monaxon. |
Central capsule always spheroidal, monaxon. |
Capsule-membrane single,
pores innumerable, distributed all over. |
Capsule-membrane single,
pores numerous, regularly distributed. |
Capsule-membrane single,
a porous area (porochora) at the oral pole of the main axis. |
Capsule-membrane always double,
an astropyle (with radiate operculum) at the oral pole of the main axis. |
| Nucleus central, originally spherical (usually dividing late). |
Nucleus excentric, (usually dividing early). |
Nucleus excentric, near the aboral pole (dividing late). |
Nucleus always spheroidal, in the main axis (dividing late). |
| Skeleton absent or siliceous, never centrogenous. |
Skeleton always of acanthin, always centrogenous. |
Skeleton siliceous, usually monaxon, extracapsular. |
Skeleton of a silicate, always extracapsular. |
| Calymma always without phæodium. |
Calymma always without phæodium. |
Calymma always without phæodium. |
Calymma always with phæodium. |
| First Subclass. |
|
Porulosa vel Holotrypasta.
Central capsule originally spherical, without osculum or principal opening,
with innumerable fine pores.
|
Legion I.
Spumellaria.
(Peripylea). |
Legion II.
Acantharia.
(Actipylea). |
| Central capsule originally spherical, homaxon. |
Central capsule originally spherical, homaxon. |
| Capsule-membrane single, pores innumerable, distributed all over. |
Capsule-membrane single, pores numerous, regularly distributed. |
| Nucleus central, originally spherical (usually dividing late). |
Nucleus excentric, (usually dividing early). |
| Skeleton absent or siliceous, never centrogenous. |
Skeleton always of acanthin, always centrogenous. |
| Calymma always without phæodium. |
Calymma always without phæodium. |
| Second Subclass. |
|
Osculosa vel Merotrypasta.
Central capsule originally monaxon, with an osculum at the basal pole of the
vertical main axis.
|
Legion III.
Nassellaria.
(Monopylea). |
Legion IV.
Phæodaria.
(Cannopylea). |
| Central capsule originally ovoid, monaxon. |
Central capsule always spheroidal, monaxon. |
| Capsule-membrane single, a porous area (porochora) at the oral pole of
the main axis. |
Capsule-membrane always double, an astropyle (with radiate operculum) at the oral
pole of the main axis. |
| Nucleus excentric, near the aboral pole (dividing late). |
Nucleus always spheroidal, in the main axis (dividing late). |
| Skeleton siliceous, usually monaxon, extracapsular. |
Skeleton of a silicate, always extracapsular. |
| Calymma always without phæodium. |
Calymma always with phæodium. |
11. Individuality of the Radiolaria.—Like other Protozoa the Radiolaria are
unicellular organisms, the whole fully developed organisation of which falls under the category of
a single cell, both morphologically and physiologically. Since this view is based upon the
composition of the individual body out of two different morphological elements, nucleus and
protoplasm, it is at once justified in the case of the majority of Radiolaria, in which the
plasmatic body encloses only a single nucleus (the so-called "Binnen-Bläschen"); such is the case
in all the Spumellaria monozoa, Nassellaria and
Phæodaria. This aspect of the case might appear doubtful in those
Radiolaria in which the simple primary cell-nucleus divides early into numerous small secondary
nuclei, as is the case in the Spumellaria polyzoa and most Acantharia. Strictly speaking, the multinucleate central capsule should in such
cases be regarded as a syncytium; but since the individual unity of the unicellular organism is as
clearly defined in these precocious multinuclear Radiolaria as in the ordinary serotinous forms,
the former must be considered unicellular Rhizopods just as are the latter. This mode of regarding
{vi}the case is the more necessary, inasmuch as the early
division of the nucleus has no further influence upon the organisation. Just as in many other
classes of the Protista there are monozootic (solitary) and polyzootic (social) forms, so also in
the Radiolaria there are in addition to the ordinary monozootic or monobious forms certain
families in which colonies or cœnobia are formed by the association of individuals; this
distinction may be expressed by the terms "Monocyttaria" and "Polycyttaria."
The unicellular nature of the Radiolaria was first established by Richard Hertwig
in 1879 (L. N. 33),[1] and brought
into conformity with our present histiological knowledge and the new reform of the cell-theory.
Huxley, however, who was in 1851 the first to examine living Radiolaria accurately, declared
Thalassicolla nucleata to be a unicellular Protozoon, and the individual central capsules
of Sphærozoum punctatum to be cells, but, owing to the then condition of the cell-theory,
he was unable to give a conclusive demonstration of this view. Later, when Johannes Müller in
1858 and myself in 1862 recognised the peculiar "yellow cells" which occur in large numbers in
many Radiolaria as true nucleated cells, it appeared impossible any longer to maintain the
unicellular nature of the Radiolaria; also the great complication which I showed to exist in the
structure of Thalassicolla appeared to contradict it. Only after Cienkowski (1871) and
Brandt (1881) had shown that the "yellow cells" do not belong to the Radiolarian organism, but
are symbiotic unicellular algæ, was it possible to revive and demonstrate anew the unicellular
nature of the Radiolaria.
12. Morphological Individuality.—From the morphological standpoint the
individuality of the unicellular elementary organism is obvious in the ordinary solitary
Radiolaria (Monobia), and is to be so regarded that the whole body with all its constituent parts,
and not merely the central capsule, is to be regarded as a cell. Naturally the xanthellæ or
yellow cells (§§ 76, 90), which as independent algæ
live in symbiosis with many Radiolaria, must be excluded. The unicellular organisation of the
Radiolaria is further to be distinguished from that of the other Protista, inasmuch as an internal
membrane (capsule-membrane) separates the central (medullary) from the peripheral (cortical)
portion. In the cœnobia of the social Radiolaria (or Polycyttaria), the morphological
individuality persists only as regards the medullary portions of the aggregated cells (the
individual central capsules), while the cortical portions fuse completely to form a common
extracapsulum. Hence in these Spumellaria polyzoa two different stages of
morphological individuality must be distinguished, the Cell as a Morphon of the first
stage, and the Cœnobium as a Morphon of the second stage.
13. Physiological Individuality.—From the physiological standpoint also the
individuality of the unicellular organism is immediately obvious in the case of the ordinary
solitary Radiolaria (Monobia); as in other Protista it fulfils all the functions of life by itself
alone. This physiological individuality of the monobious Radiolarian cell is furthermore not
influenced by the xanthellæ, which live as independent algæ in symbiosis with many Radiolaria;
even though these often by the production of starch assist in the {vii}nourishment of the Radiolaria, yet they are by no means indispensable to
them. On the other hand, the physiological individuality offers more complicated relations in the
social Radiolaria (Polycyttaria) which live united in colonies or cœnobia. Here the actual
Bion (or the fully developed physiological individual) is not represented by the individual
cells, but by the whole multicellular cœnobium, which in each species has a definite form
and size. In these cœnobia, which are usually spherical or cylindrical jelly-like masses,
several millimeters in diameter, numerous cells are so intimately united that only their medullary
portions (the central capsule with the endoplasm) remain independent; the cortical portions
(calymma and exoplasm) on the contrary uniting into a common extracapsulum. This discharges, as a
whole, the functions of locomotion, sensation, and inception of nutriment, while the separate
central capsules act in the main only as reproductive organs (forming spores) and partly also as
the central organs of metastasis (digestion). Each cœnobium may also be regarded as a
polycyttarium, i.e., a "multicellular Radiolarian," whose numerous central capsules
represent so many sporangia or spore-capsules.
On this head compare the section in my monograph of 1862 (L. N. 16), entitled Die Organisation der Radiolarien-Colonien; Polyzoen oder
Polycyttarien? (pp. 116 to 126); and also
R. Hertwig, Zur Histologie der Radiolarien, 1876 (L. N. 26, p. 23).
14. Monocyttaria and Polycyttaria.—In the majority of the Radiolaria each
unicellular organism passes its individual life in an isolated condition (as a Monocyttarium).
Only in a part of the Spumellaria numerous unicellular individuals are
united into societies which are regarded as cœnobia or colonies (Polycyttaria). This is the
case in three different families belonging to the Peripylea, in the
Collozoida (without a skeleton, Pl. 3),
the Sphærozoida (with a Beloid skeleton, Pl. 4),
and the Collosphærida (with a Sphæroid skeleton, Pls. 5-8). All three
families of Polycyttaria (or social Radiolaria), agree in their mode of forming colonies, since
the central capsules of the social individuals remain separate and lie in a common jelly-like
mass, which is formed by the fusion of their extracapsulum. The chief part of the voluminous
colonies, which attain a diameter of several millimetres (sometimes more than 1 cm.), and are
generally spherical, ellipsoidal or cylindrical, consists therefore of the jelly-like calymma, and
this is penetrated by a sarcoplegma, to whose meshes all the individual organisms contribute by
means of the pseudopodia, which radiate from their sarcomatrix. A further peculiarity in which the
social Spumellaria differ from the solitary consists in the fact that the
former are precocious and the latter serotinous in the division of the nucleus (§ 64). Whilst in the solitary or monozootic Spumellaria
the middle of the central capsule is occupied by the simple nucleus, and this divides only at a
late period (immediately before the formation of spores) into the numerous spore nuclei, in the
colonial or polyzootic Spumellaria this division takes place very early,
and the middle of each central capsule is usually occupied by an oil-globule.
{viii}
The colonial Radiolaria were described as early as the year 1834 by Meyen, the
first investigator of the class, under the name Sphærozoum, and, as Palmellaria,
compared with the gelatinous colonies of the Nostochineæ. The first accurate observations upon
their structure were, however, made in 1851 by Huxley, who described examples of all three
families under the name Thalassicolla punctata. More extended, however, were the
investigations of Johannes Müller, who in his fundamental work (1858) divided the whole class
Radiolaria into Solitaria and Polyzoa. The Radiolaria solitaria he divided
into Thalassicolla, Polycystina and Acanthometra, the Radiolaria polyzoa into Sphærozoa
(without a shell) and Collosphæra (with a shell). The most accurate delineation of the
Polycyttaria was given by Hertwig in his beautiful memoir, Zur Histologie der Radiolarien
(1876). Quite recently, however (1886), since the completion of my manuscript upon the
Challenger Radiolaria, a very complete Monograph of the Polycyttaria has appeared by Karl
Brandt, Die colonie-bildenden Radiolarien (Sphærozoen) des Golfes von Neapel und der
angrenzenden Meeres-Abschnitte (276 pp., 8 pls., Berlin). It contains in particular most
valuable contributions to the physiology and histology.
15. The Central Capsule and Extracapsulum.—The special peculiarity of the
unicellular Radiolarian organism, by which it is clearly distinguished from all other Rhizopoda
(and indeed from most other Protista), is its differentiation into two separate chief
constituents, the central capsule and extracapsulum, and the formation of a special membrane which
separates them. This, the capsule-membrane, is not to be compared with an ordinary cell-membrane,
as an external layer, but rather to be regarded as an internal differentiated product. The
extracapsulum or external (cortical) portion of the body is in most Radiolaria more voluminous
than the central capsule or inner (medullary) portion. The exoplasm of the former (the cortical or
extracapsular protoplasm) is emphatically different from the endoplasm of the latter (the
medullary or intracapsular protoplasm). Besides the most important vital processes are distributed
by division of labour so completely between them that they appear most distinctly co-ordinated.
The central capsule is on the one hand the general central organ of the "cell-soul" for the
discharge of its sensory and motor functions (comparable to a ganglion-cell), on the other hand
the special organ of reproduction (sporangium). The extracapsulum, also, is not less significant,
since on the one hand its calymma acts as a protecting envelope to the central capsule, as a
support to the pseudopodia, and a foundation for the skeleton or a matrix for the development of
the shell, and on the other hand its pseudopodia are of the utmost importance as peripheral organs
of movement and sensation as well as of nutrition and respiration. The central capsule and
the extracapsulum are therefore to be regarded both morphologically and physiologically as
the two characteristic co-ordinated principal parts of the unicellular Radiolarian
organism.
In most of the more modern delineations of the Radiolaria the central capsule is
regarded as the "cell proper" and its membrane as the "cell-wall." The following facts are
opposed to the correctness of this interpretation:—1. In most Radiolaria the exoplasm is
clearly different from {ix}the endoplasm, and the
former is more voluminous than the latter. 2. In all Radiolaria the division of labour is so
carried out between the central capsule and the extracapsulum, that the physiological
significance and independence of both principal parts of the cell is almost equally great. 3. It
is only in the Acantharia that the formation of the skeleton takes place
within the central capsule; in all the other three legions it is quite independent of it.
16. The Malacoma and Skeleton.—Whilst the division of the unicellular
organism into central capsule and extracapsulum is undoubtedly the most important character of the
Radiolarian organism, the development of a skeleton of peculiar and most varied form is of very
striking significance. This skeleton is always a secondary product of the cell, but is
always anatomically so independent, and so clearly marked off from the soft parts or malacoma,
that it seems advisable to regard both separately in a general morphological survey. The skeleton
stands in a different relation to each of the two principal constituents of the malacoma. Only in
the Acantharia is it centrogenous and developed from the central capsule
outwards. In the other three legions the skeleton never arises in the centre of the capsule; in
the Nassellaria and Phæodaria it is always
extracapsular; in the Spumellaria it is also outside the central capsule
originally, but afterwards becomes often surrounded by it, and finally lies in most cases partly
within and partly without the central capsule. The chemical basis of the skeleton in the Acantharia is the curious acanthin (an organic substance allied to chitin), in
the Phæodaria a silicate of carbon, and in the Nassellaria and Spumellaria silica.
17. Ground-Forms of the Radiolaria (Promorphology).—The ground-forms of the
Radiolaria exhibit a greater variety than those of any other class in the organic world, greater
indeed than is to be found in all the remaining groups together. For every conceivable ground-form
which can be defined in the system of promorphology is actually present in the Radiolaria; their
skeleton exhibits, as it were, in material existence, certain geometrical ground-forms which are
found in no other organisms. The cause of this unexampled richness in different forms lies chiefly
in the static relations of the Radiolaria, which swim freely in the sea, partly also in the
peculiar plasticity of their protoplasm and the material of their skeletons.
Regarding the general system of ground-forms compare my Generelle Morphologie
(1866, Bd. i. pp. 375-552; Bd. iv., Allgemeine Grundformenlehre). The ground-forms there
proposed and systematically defined have, however, found but little acceptance (chiefly, no
doubt, owing to the difficult and complicated nomenclature); but having now, twenty years after
their publication, anew carefully revised and critically studied them, I can find no sufficient
reason for abandoning the principles there adopted. On the contrary the study of the Challenger
Radiolaria during the last ten years, with its incomparable wealth of forms, has only confirmed
the accuracy of my system of ground-forms. The customary treatment of these in zoological and
botanical handbooks (such as those of Claus and Sachs) is quite insufficient.
{x}
18. The Principal Groups of Geometrical Ground-Forms.—The great
variety of the geometrical ground-forms which are actually realised in the variously shaped bodies
of the Radiolaria, renders it desirable to classify these in as small a number as possible of
principal groups and a larger number of subdivisions. As extensive principal groups four at least
must be distinguished; the Centrostigma or Sphærotypic, the Centraxonia or
Grammotypic, the Centroplana or Zygotypic, and the Acentrica or Atypic. The natural
centre of the body, about which all its parts are regularly arranged, is in the first group a
point (stigma), in the second a straight line (principal axis), in the third a plane (sagittal
plane), in the fourth a centre is of course wanting.
19. The Centrostigma or Sphærotypic Ground-Forms.—The first group of
geometrical ground-forms, here distinguished as sphærotypic or the centrostigma, is undoubtedly
the most important among the Radiolaria, inasmuch as if these be considered monophyletic, it must
be the original one from which all the other ground-forms have been derived. The common character
of all these sphærotypic ground-forms is that their natural centre is a point (stigma); thus there
is no single principal axis (or protaxon) such as is characteristic of the two following groups.
The sphærotypic ground-forms are subdivided into two important smaller groups, the spheres
(Homaxonia) and the endospherical polyhedra (Polyaxonia). The spherical ground-forms, fully
developed in the central capsule and calymma of Actissa and the Sphæroidea as well as in many Acantharia, present no
different axes; all possible axes passing through the centre of the body are equal (Homaxonia). In
the endospherical polyhedra, on the contrary, numerous axes (three at least) may be distinguished,
which are precisely equal to each other and different from all the remaining axes (Polyaxonia). If
the extremities of these axes, or the poles, which are all equidistant from the common centre, be
united by straight lines, a polyhedral figure is produced whose angles all lie in the surface of
the sphere. According as the poles of the axes are at equal, subequal, or at different distances
from each other, we may divide the endospherical polyhedra into regular, subregular and irregular.
(See Gener. Morphol., Bd. i. pp. 404-416.)
20. The Centraxonia or Grammotypic Ground-Forms.—The second principal
group of organic ground-forms, here called grammotypic or centraxonia, is characterised by the
fact that a straight line (gramma) or a single principal axis (protaxon) forms the natural centre
of the body. This important and extensive group is divided into two subgroups, those with one axis
(Monaxonia) and those with crossed axes (Stauraxonia); in the latter different secondary
transverse or cross-axes may be distinguished, but not in the former. In the Monaxonia, therefore,
every transverse section of the body perpendicular to the principal axis is a circle, in the
Stauraxonia, on the contrary, a polygon. The Monaxonia are further subdivided into two groups, in
one of which the two poles of the principal axis {xi}are
equal and similar (Isopolar), in the other of which they are different (Allopolar); in the former
the two halves of the body, which are separated by the equatorial plane (or the largest transverse
plane, perpendicular to the principal axis), are equal, in the latter unequal. Among the isopolar
uniaxial ground-forms (Monaxonia isopola) may be mentioned the ellipsoidal, spheroidal,
lenticular, &c.; to the allopolar uniaxial forms (Monaxonia allopola) belong the conical,
hemispherical, ovoid, &c. In the same way the pyramidal ground-forms with crossed axes are
divisible into two groups, according as the two poles of the principal axis are equal or not. The
ground-form of the former is the double pyramid, that of the latter the single pyramid. Both the
double and the single pyramids may again be subdivided, each into two important lesser groups, the
regular and the amphithect. In the first division the equatorial plane of the double and the basal
plane of the single pyramid is a regular polygon (square, &c.), whilst in the other division
it is an elongated or amphithect polygon (rhombus, &c.); the crossed axes are equal in the
former, unequal in the latter. (See Gener. Morphol., Bd. i. pp. 416-494.)
21. The Centroplana or Zygotypic Ground-Forms.—The third principal
group of ground-forms includes those which are bilaterally symmetrical in the ordinary sense, or
zeugitic or zygotypic; the natural centre of their body is a plane. These forms are the only ones
in which the distinction between right and left is possible, since their body is divided by the
median plane (planum sagittale) into two symmetrical halves (right and left). In all these
zeugites the position of every part is determined by three axes perpendicular to each other, and
of these three dimensive axes two are allopolar, one is isopolar. The two unlike poles of the
principal (or longitudinal) axis are the oral and aboral, the two unlike poles of the sagittal (or
vertical) axis are the dorsal and ventral; the two similar poles of the frontal (or transverse)
axis, however, are the right and left. This important group of zeugitic or bilateral forms may
also be divided into two clearly distinct lesser groups, the Amphipleura and the
Zygopleura. In the Amphipleura (or bilaterally radial ground-forms) the "radial two-sided"
body is produced by modification of a regular pyramid (as Spatangus from Echinus),
and hence is composed of several (not less than three) antimeres. In the Zygopleura (or
bilaterally symmetrical ground-forms) on the other hand, the bodies consist of two antimeres (as
in all the higher animals, Vertebrata, Arthropoda, &c.). (See Gener. Morphol., Bd. i. pp.
495-527.)
22. The Acentrica or Atypic Ground-Forms.—Among the acentrica or
anaxonia are included all those ground-forms which are absolutely irregular, and in which neither
a definite centre nor constant axes can be distinguished (e.g., most Sponges). These quite
irregular ground-forms are very rare among the Radiolaria, but nevertheless there may be referred
to them the amœboid central capsule of some Colloidea
(Collodastrum, p. 27, Pl. 3, figs. 4, 5)
among the Spumellaria, the irregular shells of many Collosphærida {xii}(Pl. 8,
fig. 2), and the absolutely irregular shells of the Phorticida and Soreumida among the Larcoidea. (See Gener. Morphol., Bd. i. p. 400.)
23. The Subsidiary Groups of Geometrical Ground-Forms.—The four natural principal
groups of ground-forms, which have just been defined according to the nature of the centre of
their bodies, may be divided again into numerous subsidiary groups, defined by the relations of
the constant axes and the two poles of each axis, as well as by the number of the axes and the
differentiation of the secondary with respect to the principal axis. The most important of these
subsidiary groups into which the principal ones are immediately divided are the
following:—(1) The Centrostigma (or sphærotypic) are divided into spheres (Homaxonia)
and endospherical polyhedra (Polyaxonia). (2) The Centraxonia (or grammotypic) into
uniaxial (Monaxonia) and those with crossed axes (Stauraxonia); among the former of these may be
distinguished the isopolar (phacotypic) and the allopolar (conotypic); among the latter the double
and single pyramids. (3) The Centroplana (or bilaterals) are divided into amphipleura (or
bilaterally radial) and zygopleura (or bilaterally symmetrical). (4) The Acentrica (or
Anaxonia) or absolutely irregular ground-forms, present no special subdivisions.
For a complete system of the geometrical ground-forms and their relation to
promorphological classification, see Gener. Morphol., Bd. i. pp. 555-558.
24. The Spherical or Homaxon Ground-Form.—The spherical is the only
absolutely regular ground-form, since only in it are all axes which pass through the centre equal;
it is very often realised among the Radiolaria, especially in the Spumellaria and Acantharia, where it furnishes the
common original ground-form, but it is often to be seen in the shells of many Phæodaria (in most Phæosphæria); on the other hand, it
is never found among the Nassellaria. Geometrical spheres, in the strict
sense of the term, are only to be found among the Spumellaria and Acantharia, namely, in the central capsule of many Collodaria (Pls. 1,
2) and all
Sphæroidea (Pls. 11-30) as well as
many Acanthometra and Acanthophracta (Pls. 128-138).
Nevertheless, speaking generally, one includes those central capsules and skeletons which have
been distinguished here as endospherical polyhedra. (On these ground-forms see Gener. Morphol.,
Bd. i. pp. 404-406.)
25. The Endospherical Polyhedral Ground-Form.—The endospherical
polyhedron or polyaxon ground-form naturally follows the spherical or homaxon. Under it are
included all polyhedra whose angles fall in the surface of a sphere; this ground-form is
especially common among the Spumellaria, especially in the shells of Sphæroidea, but is also found among the Acantharia
(especially in the Astrolophida and Sphærophracta), as well as among the
Phæosphæria (in most genera of the Orosphærida, Sagosphærida, and
Aulosphærida). Strictly speaking, all those lattice-shells which have {xiii}been incorrectly called "spherical" belong to this category, for they
are none of them true spheres in the geometrical sense (like the central capsules of the Sphæroidea), but rather endospherical polyhedra, whose angles are indicated by
the nodal points of the lattice shell, or the radial spines which spring from them. These
endospherical polyhedra may be divided into three groups, the regular, subregular, and irregular.
Of regular polyhedra, properly so-called, it may be shown geometrically that only five can
exist, namely, the regular tetrahedron, cube, octahedron, dodecahedron, and icosahedron. All these
are actually manifested among the Radiolaria, although but seldom. Much more common are the
subregular endospherical polyhedra, e.g., spherical lattice-shells with regular
hexagonal meshes of equal size; they are never exactly equal nor perfectly regular, but the
divergences are so insignificant that they escape superficial observation (Pl. 20, figs. 3, 4;
Pl. 26,
figs. 1-3). On the contrary in the irregular endospherical polyhedra the meshes of the
lattice-sphere are more or less different in size and often in form also (Pl. 28, figs. 4, 8;
Pl. 30,
figs. 4, 6). The five truly regular polyhedra require separate notice on account of their
importance. (See Gener. Morphol., Bd. i. p. 406.)
26. The Regular Icosahedral Ground-Form.—The ground-form whose
geometrical type is the regular icosahedron (bounded by twenty equilateral triangles) is rarely
exemplified, but it occurs among the Phæodaria in the Circoporid genus
Circogonia (Pl. 117,
fig. 1), and also in certain Aulosphærida, but, apparently, only as an accidental variation
(e.g., Aulosphæra icosahedra). Furthermore, this ground-form may also be assumed to
occur in those Sphæroidea whose spherical lattice-shells bear twelve
equidistant radial spines (e.g., many species of Acanthosphæra, Heliosphæra,
and other Astrosphærida); the basal points of these spines indicate the twelve angles of the
regular icosahedron. (See on this head Gener. Morphol., Bd. i. p. 411.)
27. The Regular Dodecahedral Ground-Form.—The ground-form whose
geometrical type is the regular dodecahedron (or pentagonal dodecahedron), bounded by twelve
equilateral and equiangular pentagons, is very rarely found perfectly developed, as in
Circorrhegma dodecahedra (Pl. 117,
fig. 2). This form is by no means so common among the Radiolaria as in the pollen grains of plants
(e.g., Buchholzia maritima, Fumaria spicata, Polygonum amphibium,
&c.). It can, however, be regarded as present in all those Sphæroidea
whose spherical lattice-shells bear twenty equal and equidistant radial spines (e.g., many
species of Acanthosphæra, Heliosphæra, and other Astrosphærida); the basal points of
these spines mark out the twenty angles of the regular pentagonal dodecahedron. (See Gener.
Morphol., Bd. i. p. 412.)
28. The Regular Octahedral Ground-Form.—The ground-form whose
geometrical type is the regular octahedron (bounded by eight equilateral triangles), commonly
appears among the Spumellaria in the family Cubosphærida (p. 169, Pls. 21-25). In {xiv}these Sphæroidea the typical
ground-form is usually indicated by six equal radial spines, which are opposed to each other in
pairs, and lie in three similar axes perpendicular to each other; these are the three axes of the
tesseral crystallographic system; one of them is vertical, whilst the other two cross each other
at right angles in its centre. Occasionally, too, the spherical form of the lattice-shell passes
over into that of the regular octahedron (Pl. 22,
figs. 8, 10). The same form recurs in Circoporus (Pl. 117, fig. 6)
among the Phæodaria. In the vegetable kingdom it is exhibited by the
antheridia of Chara. It is not found in the Nassellaria and Acantharia. (See Gener. Morphol., Bd. i. p. 412.)
29. The Regular Cubic Ground-Form.—The ground-form whose geometrical
type is that of a die or cube, is actually presented in a very striking manner by various
Radiolaria. Among the Spumellaria it occurs in certain Sphæroidea, e.g., in the Astrosphærid genera Centrocubus and
Octodendron (Pl. 18, figs. 1-3);
in these the central medullary shell is a complete cube, bounded by six equal squares, from the
eight angles of which eight equal radial spines project. This form can also be regarded as present
in those Sphæroidea whose spherical lattice-shell bears eight equal and
equidistant radial spines (many Astrosphærida). Besides these the cubic ground-form is to be seen
in certain Nassellaria of the family Tympanida, especially in
Lithocubus (Pl. 82,
fig. 12; Pl. 94,
fig. 13), in many species of Acrocubus, Microcubus, &c.; the twelve bars of its
lattice-skeleton correspond often exactly to the edges of the cube. (See Gener. Morphol., Bd. i.
p. 413.)
30. The Regular Tetrahedral Ground-Form.—The ground-form whose
geometrical type is the regular tetrahedron, bounded by four equilateral triangles, occurs less
frequently in the Radiolaria than the other four regular polyhedra. Among the Spumellaria it is found in the Beloidea, and especially
in those members of the Thalassosphærida and Sphærozoida whose spicules bear four equal branches,
diverging at equal angles from a common centre. Precisely the same structure is seen also among
the Nassellaria in some Plectoidea, as in
Tetraplagia among the Plagonida, and Tetraplecta among the Plectanida. The skeleton
of both these genera consists of four equal rods, which radiate at equal angles from a common
centre, just as do the axes of the regular tetrahedron. The tetrahedral form of these Plectoidea is the more important and interesting since on the one hand it is
related to the similar spicular form of the Beloidea, and on the other
perhaps furnishes the starting point from which Cortina among the Nassellaria may be derived (Plagoniscus, Plectaniscus). (See
Gener. Morphol., Bd. i. p. 415.)
31. The Isopolar-Monaxon or Phacotypic Ground-Form.—The isopolar
uniaxial or phacotypic ground-form is characterised by the possession of a vertical main axis with
{xv}equal poles, whilst no transverse axes are
differentiated. All horizontal planes which cut the axis at right angles are circles, and increase
in size from the poles towards the equator. The most important ground-forms of this group are the
phacoid (the lens or oblate spheroid) and the ellipsoid (or prolate spheroid).
Phacoids (or geometrical lenses with blunt margins) are very often presented by the central
capsules of the Discoidea and of many Acantharia
(Quadrilonchida and Hexalaspida), but the lattice-shells of many Spumellaria and Acantharia exhibit the same form, as
also do a few Phæodaria (e.g., Aulophacus). True geometrical
ellipsoids are seen in the central capsules of many Prunoidea among the
Spumellaria, and of many Amphilonchida and Belonaspida among the Acantharia. Furthermore, the lattice shells of many species of these groups
retain the same essential form, e.g., many Ellipsida, Druppulida, and Spongurida (Pls.
13-17, and 39), as well as most Belonaspida. (See Gener. Morphol., Bd. i. p. 422.)
32. Allopolar-Monaxon or Conotypic Ground-Form.—The allopolar uniaxial
or conotypic ground-form is characterised by the possession of a vertical main axis whose two
poles are unlike, while no transverse axes are differentiated. All horizontal planes cutting the
main axis at right angles are circles, and decrease more rapidly from the largest plane towards
the basal than towards the apical pole. The most important ground-forms of this group are the
ovoid, the cone, and the hemisphere. They often occur (and in geometrical perfection) in the
egg-shaped central capsule and podoconus of the Nassellaria, as well as in
the shells of several groups of this legion, particularly in the Cyrtocalpida or Monocyrtida
eradiata (Pl. 51,
figs. 10-13), and in many Stichocyrtida eradiata; furthermore, they are also seen among the Phæodaria, e.g., certain Challengerida (Pl. 99, figs.
19-22). (See Gener. Morphol., Bd. i. p. 426.)
33. The Regular Dipyramidal or Quadrilonchial Ground-Form.—The
ground-forms whose geometrical type is the regular double pyramid are characterised by a vertical
main axis which possesses equal poles, and which is crossed at its centre by several equal
transverse axes. The horizontal equatorial plane is therefore a regular polygon, and divides the
body into two equal regular pyramids. The simplest and commonest form of this group is the
quadratic octahedron, the ground-form of the quadratic crystallographic system; its equatorial
plane is a square. This regular dipyramidal ground-form occurs among the Spumellaria in the shells of the Staurosphærida as well as of many Discoidea, in which several equidistant radial spines or arms lie in the
quadratic equatorial plane of the body, and project from the margin of the lenticular disc
(e.g., Sethostaurus, Pl. 31;
Histiastrum, Pl. 46,
&c.). It is, however, among the Acantharia that the most important
part is played by this ground-form (and especially by the quadratic octahedron); it forms the
basis of all those Acanthometra and Acanthophracta in which twenty radial spines are disposed according to the
Müllerian Law, and in which {xvi}the four equatorial
spines are of equal dimensions (Icosacantha). (See Gener. Morphol., Bd. i. p. 436-446.)
34. The Amphithect Dipyramidal or Lentelliptical Ground-Forms.—The
ground-forms whose geometrical type is the lenticular or "triaxial" ellipsoid, may also be
designated amphithect double pyramids; they are characterised by the possession of a vertical main
axis which has similar poles, and is crossed at its middle by two transverse axes, unequal but
isopolar. The horizontal equatorial plane of the body is therefore an amphithect or elongated
polygon (a rhombus in the simplest case possible), and divides the whole body into two equal
amphithect pyramids. The simplest and commonest form of this group is the rhombic octahedron,
which is also the ground-form of the rhombic crystallographic system. It plays an important part
in those Acantharia in which twenty radial spines are disposed according
to the Müllerian Law, but in which the two pairs of equatorial spines are unequal (different
geotomical and hydrotomical axes, see p. 719); to this category belong the
Amphilonchida (Pl. 132),
Belonaspida (Pl. 136),
Hexalaspida (Pl. 139),
and Diploconida (Pl. 140).
A form essentially identical obtains also among the Spumellaria in the
majority of the Larcoidea, both in their triaxial lattice-shells, and in
their lentelliptical central capsules, which present geometrically accurate triaxial ellipsoids,
with three unequal isopolar axes at right angles to each other. (See Gener. Morphol., Bd. i. p.
446-452.)
35. The Regular Pyramidal Ground-Forms.—The ground-forms whose
geometrical type is the regular pyramid, and which are the most conspicuous in the Medusæ, Polyps,
Corals, and regular Echinoderms (the Radiata of earlier authors), are almost confined among the
Radiolaria to the legion Nassellaria; they occur, however, in the great
majority of these, and especially in those families which may be classed together as "Cyrtoidea triradiata et multiradiata." Strictly speaking, however, almost all
these Nassellaria, at all events in their origin, are bilateral or
dipleuric, since the primary sagittal ring with its characteristic apophyses marks out the
sagittal median plane, and further, since the three feet of the basal tripod are usually divided
into an unpaired dorsal (pes caudalis) and two paired ventral or lateral (pedes pectorales, dexter
et sinister). On the other hand, it is noteworthy, firstly, that among the primitive Plectoidea there are perfectly regular radial forms, without any indication of
an original bilateral symmetry, and secondly, that similar forms are also very common among the
Cyrtoidea, probably as secondary radial forms, developed from primitive
bilateral ones. Similar cases also occur in certain Phæodaria
(e.g., the Medusettida and Tuscarorida, Pls. 100, 120), but they
are entirely wanting among the Acantharia and Spumellaria. The multiradial Nassellaria have arisen
from the triradial by the interpolation of three, six, nine, or more interradial and adradial
secondary apophyses between the three primary perradial ones. (See Gener. Morphol., Bd. i. pp.
459-874.)
{xvii}
36. The Amphithect Pyramidal Ground-Forms.—The ground-forms whose
geometrical type is the amphithect pyramid, are distinguished from the regular pyramidal forms,
just discussed, chiefly by the form of the basal plane, which is not a regular, but an amphithect
or elongated polygon (in the simplest case a rhombus). Hence in this case the allopolar main axis of the body is
crossed by two transverse axes which are isopolar and at right angles, but are unequal; they
cannot, however be distinguished as sagittal and frontal axes as is the case in the zeugites. In
the animal as well as in the vegetable kingdom, an important part is played by this ground-form,
e.g., in the Ctenophora, where it is the rhombic pyramid. Among the Radiolaria it is not
common, though it is clearly expressed among the Nassellaria in a number
of Stephoidea (Stephanida and Tympanida), as well as in many Spyroidea (e.g., the bipedal Zygospirida). It is very accurately
developed among the Phæodaria in the bivalved Phæoconchia (Pls. 121-128), where
the two valves of the shell (dorsal and ventral) are generally exactly alike, their median keels
corresponding to the poles of the sagittal axis. In the slit between the two valves lie the two
secondary openings (right and left) of the tripylean central capsule, corresponding to the two
poles of the frontal axis, and the main axis stands perpendicularly to both these, its oral pole
being indicated by the astropyle, or principal aperture. (See Gener. Morphol., Bd. i. pp.
479-494.)
37. The Amphipleural Ground-Forms.—By the term amphipleural
ground-forms are to be understood those usually defined as "bilaterally radial"; their geometrical
type is a half amphithect pyramid. The best known examples of this form in the animal kingdom are
the bilateral five-rayed Echinoderms (Spatangus, Clypeaster), in the vegetable
kingdom the symmetrical five-rayed flowers (Viola, Trifolium). The three dimensive
axes have the same relation as in the zygopleura, to be next discussed, and which also resemble
them in being divisible only by one plane (the sagittal median plane) into two equal halves. They
differ, however, the amphipleural body not being made up of two antimeres, but of at least three
pairs of antimeres (or three parameres), being therefore primitively radial. Hence each of the
symmetrical halves of the body contains more than one antimere. Among the Radiolaria this form
does not occur in the Spumellaria, Acantharia, or
Phæodaria; it is very common, however, among the Nassellaria; many Cyrtoidea multiradiata and Spyroidea multiradiata show this bilaterally radial ground-form, inasmuch as
the body consists of two symmetrical halves, and is also composed of numerous (usually three, six,
nine, or more) radial parameres. In the multiradiate Dicyrtida and Tricyrtida the cephalis (the
first joint) is usually bilateral, whilst the thorax (the second joint) is multiradial. (See
Gener. Morphol., Bd. i. pp. 495-506.)
38. The Zygopleural Ground-Forms.—As zygopleural or dipleural
ground-forms, as opposed to the amphipleural, are classed those zeugites or centroplana which are
known {xviii}as "bilaterally symmetrical" in the
strictest sense of the term. This is the most important ground-form in the animal kingdom,
inasmuch as it obtains almost exclusively among the higher animals (Vertebrata, Articulata,
Mollusca, Vermes). The body consists of only two antimeres, which correspond to the two
symmetrical halves of the body. Of the three dimensive axes two are allopolar, one isopolar; the
oral pole of the longitudinal main axis is different from the aboral; the dorsal pole of the
sagittal axis is different from the ventral; but the right pole of the frontal axis is equal to
the left. The right antimere is usually precisely similar to the left (Eudipleura), more rarely it
is slightly dissimilar or asymmetrical (Dysdipleura). Among the Radiolaria this ground-form is
entirely wanting in the Porulosa or Holotrypasta (Spumellaria and Acantharia), but on the contrary it is very common in the Osculosa or
Merotrypasta (Nassellaria and Phæodaria). In the
Nassellaria it is of special importance, for the typical Cortina
(the combination of the primary sagittal ring with the basal tripod) exhibits the zygopleural
ground-form clearly sketched out; indeed it is usually clearly seen even in the sagittal ring
itself, for its ventral segment is more strongly curved than the dorsal; its basal (or oral) pole
is always different from the apical (or aboral). Of the three feet of the basal tripod the
unpaired (caudal) one is directed dorsally and backwards, the two paired (pectoral) ones ventrally
and forwards. The majority of the Nassellaria may be regarded as
modifications of this original ground-form. Its relation to the primitively triradiate tripod
presents a still unsolved problem, and the numerous relations of the zygopleural to the
multiradiate ground-forms in the Nassellaria are exceedingly complicated.
The zygopleural ground-form is less widely distributed among the Phæodaria, though it is very characteristically developed in the rich and varied
group of Challengerida (Pl. 99).
(See Gener. Morphol., Bd. i. pp. 507-527.)
39. Synopsis of the Geometrical Ground-Forms:—
Principal Groups of
Ground-Forms. |
|
Subsidiary Groups of
Ground-Forms. |
|
Geometrical Type. |
|
Examples. |
|
I. Centrostigma.
The geometrical centre of the body is a point. Main axis wanting.
|
 |
I. Homaxonia.
All axes equal
|
 |
1. Sphere, |
 |
Central capsule of the Sphæroidea and of many
Acantharia. |
|
II. Polyaxonia.
Endospherical polyhedra. All the angles of the body lie on the surface of a
sphere. Numerous isopolar axes.
|
 |
2. Endospherical polyhedron, |
|
Lattice-spheres of the Sphæroidea, Sphærophracta, and Phæosphæria. |
| 3. Icosahedron, |
Circogonia. |
| 4. Dodecahedron, |
Circorrhegma. |
| 5. Octahedron, |
Cubosphærida, Circoporus. |
| 6. Cube, |
Centrocubus, Lithocubus, &c. |
| 7. Tetrahedron, |
Tetraplagia, Tetraplecta, &c. |
| {xix}
II. Centraxonia.
The geometrical centre of the body is a straight line (the vertical main axis).
Constant transverse axes (perpendicular to the main axis) are wanting in the
Monaxonia (which have circular transverse sections); on the contrary they are differentiated
in the Stauraxonia (which have polygonal transverse sections).
|
 |
III. Monaxonia.
Uniaxial ground-forms or centraxonia without transverse axes. The transverse
planes (perpendicular to the main axis) are circles.
|
 |
8. Monaxonia isopola.
(Spheroids and ellipsoids; both poles of the main axis similar.)
|
 |
Central capsule and lattice-shell of of many Discoidea (lenses) and Prunoidea (ellipsoids),
Belonaspida, &c. |
|
9. Monaxonia allopola.
(Cone, ovoid and hemisphere; the two poles of the axis dissimilar.)
|
|
Central capsule and lattice-shell of many Nassellaria, especially the Cyrtoidea eradiata
(Cyrtocalpida, &c.). |
|
IV. Stauraxonia.
Pyramidal ground-forms or centraxonia with transverse axes. The transverse
planes (perpendicular to the main axis) are either regular or amphithect polygons.
|
|
10. Dipyramides regulares.
(Quadratic octahedron, or quadrilonchial forms and regular double
pyramids.)
|
|
Acantharia with twenty radial spines, the four
equatorial being equal. Multiradial Discoidea and
Staurosphærida. |
|
11. Dipyramides amphithectæ.
(Rhombic octahedron, lentellipsoid, and amphithect double pyramids.)
|
|
Acantharia with twenty radial spines, whose
four equatorial spines are unequal but paired. Many Larcoidea. |
|
12. Pyramides regulares.
(Regular pyramids.)
|
|
Many Nassellaria (triradial and multiradial).
Medusettida and Tuscarorida. |
|
13. Pyramides amphithectæ.
(Rhombic pyramids.)
|
|
Phæoconchia. Bipedal Spyroidea and Stephoidea. |
|
III. Centroplana.
The geometrical centre of the body is a plane (the sagittal plane).
|
 |
V. Bilateralia (or Zeugita).
Bilateral forms in the general sense, with right and left halves.
|
|
14. Amphipleura
(Bilaterally radial ground-form.)
|
|
Many Cyrtoidea and Spyroidea multiradiata. |
|
15. Zygopleura.
(Bilaterally symmetrical ground-form.)
|
|
Most Nassellaria (primitively at least), many
Challengerida. |
|
IV. Acentra.
There is no geometrical centre.
|
|
VI. Anaxonia.
No definite axes can be determined.
|
|
16. Irregularia.
(Absolutely irregular ground-forms.)
|
|
Collodastrum, Collosphæra, Phorticida, Soreumida. |
| Principal Groups of Ground-Forms. |
|
Subsidiary Groups of Ground-Forms. |
|
Geometrical Type. |
|
Examples. |
|
I. Centrostigma.
The geometrical centre of the body is a point. Main axis wanting.
|
|
|
I. Homaxonia.
All axes equal.
|
|
|
|
|
|
|
1. Sphere, |
|
|
|
|
|
|
|
Central capsule of the Sphæroidea and of many Acantharia. |
|
|
II. Polyaxonia.
Endospherical polyhedra. All the angles of the body lie on the surface of a
sphere. Numerous isopolar axes.
|
|
|
|
|
|
|
2. Endospherical polyhedron, |
|
|
|
|
|
|
Lattice-spheres of the Sphæroidea, Sphærophracta, and Phæosphæria. |
|
|
|
|
3. Icosahedron, |
|
|
|
|
|
|
Circogonia. |
|
|
|
|
4. Dodecahedron, |
|
|
|
|
|
|
Circorrhegma. |
|
|
|
|
5. Octahedron, |
|
|
|
|
|
|
Cubosphærida, Circoporus. |
|
|
|
|
6. Cube, |
|
|
|
|
|
|
Centrocubus, Lithocubus, &c. |
|
|
|
|
7. Tetrahedron, |
|
|
|
|
|
Tetraplagia, Tetraplecta, &c. |
|
II. Centraxonia.
The geometrical centre of the body is a straight line (the vertical main axis).
Constant transverse axes (perpendicular to the main axis) are wanting in the
Monaxonia (which have circular transverse sections); on the contrary they are differentiated
in the Stauraxonia (which have polygonal transverse sections).
|
|
|
III. Monaxonia.
Uniaxial ground-forms or centraxonia without transverse axes. The transverse
planes (perpendicular to the main axis) are circles.
|
|
|
|
|
|
|
8. Monaxonia isopola.
(Spheroids and ellipsoids; both poles of the main axis similar.)
|
|
|
|
|
|
|
|
Central capsule and lattice-shell of of many Discoidea (lenses)
and Prunoidea (ellipsoids), Belonaspida, &c. |
|
|
|
|
9. Monaxonia allopola.
(Cone, ovoid and hemisphere; the two poles of the axis dissimilar.)
|
|
|
|
|
|
|
Central capsule and lattice-shell of many Nassellaria, especially
the Cyrtoidea eradiata (Cyrtocalpida, &c.). |
|
|
IV. Stauraxonia.
Pyramidal ground-forms or centraxonia with transverse axes. The transverse
planes (perpendicular to the main axis) are either regular or amphithect polygons.
|
|
|
|
|
|
|
10. Dipyramides regulares.
(Quadratic octahedron, or quadrilonchial forms and regular double
pyramids.)
|
|
|
|
|
|
|
Acantharia with twenty radial spines, the four equatorial being
equal. Multiradial Discoidea and Staurosphærida. |
|
|
|
|
11. Dipyramides amphithectæ.
(Rhombic octahedron, lentellipsoid, and amphithect double pyramids.)
|
|
|
|
|
|
|
Acantharia with twenty radial spines, whose four equatorial spines
are unequal but paired. Many Larcoidea. |
|
|
|
|
12. Pyramides regulares.
(Regular pyramids.)
|
|
|
|
|
|
|
Many Nassellaria (triradial and multiradial). Medusettida and
Tuscarorida. |
|
|
|
|
13. Pyramides amphithectæ.
(Rhombic pyramids.)
|
|
|
|
|
|
Phæoconchia. Bipedal Spyroidea and Stephoidea. |
|
III. Centroplana.
The geometrical centre of the body is a plane (the sagittal plane).
Constant transverse axes (perpendicular to the main axis) are wanting in the
Monaxonia (which have circular transverse sections); on the contrary they are differentiated
in the Stauraxonia (which have polygonal transverse sections).
|
|
|
V. Bilateralia (or Zeugita).
Bilateral forms in the general sense, with right and left halves.
|
|
|
|
|
|
|
14. Amphipleura
(Bilaterally radial ground-form.)
|
|
|
|
|
|
|
Many Cyrtoidea and Spyroidea
multiradiata. |
|
|
|
|
15. Zygopleura.
(Bilaterally symmetrical ground-form.)
|
|
|
|
|
|
Most Nassellaria (primitively at least), many Challengerida. |
|
IV. Acentra.
There is no geometrical centre.
|
|
|
VI. Anaxonia.
No definite axes can be determined.
|
|
|
|
|
|
|
16. Irregularia.
(Absolutely irregular ground-forms.)
|
|
|
|
|
|
Collodastrum, Collosphæra, Phorticida, Soreumida. |
40. Mechanical Causes of the Geometrical Ground-Forms.—The great
variety of ground-forms exhibited by the Radiolaria is of special interest, since in most
instances their causes admit of recognition, and since they are so intimately related to each
other that even in the remaining cases the assumption that they have arisen by purely mechanical
causæ efficientes seems justified. In this respect the first rank is taken by statical
conditions, especially the indifferent or stable equilibrium of the whole organism, which floats
freely in the water. With regard to these fundamental statical relations, three principal groups
of ground-forms may be distinguished, pantostatic, polystatic, and monostatic.
41. Pantostatic Ground-Forms.—By pantostatic or indifferently stable
ground-forms are meant those in which the centre of gravity coincides with the centre of the body,
so that they are in equilibrium in any given position. Strictly speaking, the only form which
possesses perfectly indifferent equilibrium is the sphere, that being the only truly homaxon and
perfectly regular form. Nevertheless, in a somewhat wider sense many Polyaxonia, especially the
endospherical polyhedra with very numerous sides, may be {xx}included in this category. Such indifferently stable bodies are found among
the Spumellaria in many Collodaria and Sphæroidea, as well as in the Astrolophida among the Acantharia. On the contrary they are entirely wanting among the Nassellaria and Phæodaria, since their central capsule
constantly presents a main axis with a differentiated basal pole, and determines the position of
stable equilibrium.
42. Polystatic Ground-Forms.—Those ground forms are defined as
polystatic or multistable in which the body is in equilibrium in several different positions
(though not in an infinite number). The number of these positions is usually twice as many as that
of the constant equal isopolar axes exhibited by the form. Hence the regular polyhedra have as
many positions of equilibrium as they have angles or sides, the icosahedron twenty, dodecahedron
twelve, octahedron eight, cube six, tetrahedron four. The isopolar monaxon ground-forms (lens,
ellipsoid, cylinder) and the diplopyramidal ground forms (quadrilonchial and lentelliptical) have
two positions of stable equilibrium, since the two poles of the vertical axis are equal and
similar and the body is divided into equal halves by the equatorial plane. This is the case in
many Spumellaria (especially Discoidea, Prunoidea, and Larcoidea), as well as in the great
majority of Acantharia. Perhaps the same holds good also in certain Nassellaria (e.g., isopolar Tympanida) and Phæodaria (e.g., isopolar Phæosphæria), though
here unistable equilibrium appears to be necessitated by the constant main axis of the central
capsule and the differentiated basal pole of the main axis.
43. Monostatic Ground-Forms.—Those ground-forms are classed as
monostatic or unistable in which the body is in equilibrium only in one position, since the centre
of gravity of the body lies in a constant vertical axis below its centre. This fixed position is
only rarely and exceptionally found among the Spumellaria (e.g., in
Xiphostylus, Sphærostylus, Lithomespilus, Lithapium) and among the
Acantharia (e.g., in Zygostaurus and Amphibelone). On
the contrary it is quite usual among the Nassellaria and Phæodaria (with but few exceptions); for here a vertical main axis, with a
differentiated basal pole, is determined even by the formation of the central capsule, and usually
also by the corresponding structure of the skeleton. Among the Nassellaria
this basal pole, with the porochora of the central capsule, appears always to be the lower; as
also in most Phæogromia among the Phæodaria. In
the peculiar bivalved Phæoconchia, on the other hand, the basal pole with
the cannopyle is directed upwards; as also in the Challengerida and Tuscarorida. The Phæosphæria and Phæocystina are probably to a large
extent polystatic. In general unistable equilibrium may be assumed in the following categories of
ground-forms:—(1) Allopolar monaxon (conical and ovoid); (2) pyramidal (regular and
amphithect); (3) Centroplana (amphipleura and zygopleura); (4) Anaxonia.
{xxi}
44. Principal Axes.—From the foregoing consideration of the statical
conditions and their direct causal connection with the geometrical ground-forms of the Radiolaria,
the great mechanical significance of the differentiation of definite axes in these unicellular
free-swimming organisms will be manifest. The most important of these is the primary main axis
(axis principalis, or protaxon), which in all cases has a vertical direction. It is wanting in the
Centrostigma (spheres and endospherical polyhedra), and in the Anaxonia (acentra). It is isopolar
in the phacotypic forms (Monaxonia isopola), and in the double pyramids (Stauraxonia isopola). It
is allopolar in all monastatic ground-forms, in the conotypic forms (Monaxonia allopola), pyramids
(Stauraxonia allopola), and the Centroplana (or bilateral forms).
45. Secondary or Transverse Axes.—In contrast to the vertical main
axis all the other constant axes differentiated in the body may be called "secondary axes," or
"transverse axes," since they cross the former at definite points. All ground-forms whose vertical
axis is crossed by a fixed number of such axes at definite angles may be called "Stauraxonia."
They are divided into two groups, double pyramids and single pyramids; in the former the two poles
of the main axis (or the two halves of the body separated by the equatorial plane) are similar
(Stauraxonia homopola), in the latter dissimilar (Stauraxonia heteropola). If all the secondary
axes be equal, the stauraxon ground-form is regularly radial. If some of them be unequal they are
arranged in certain relations towards two primary transverse axes, perpendicular to each other, to
which all the other secondary axes are subsidiary; the ground-forms are then either amphithect or
bilateral. The two primary transverse axes, which may also be designated "ideal transverse axes"
(euthyni), divide the vertical main axis in its centre; one of them is the sagittal, the other the
frontal. These three dimensive axes give the factors which accurately determine the ground-form
and the dimensions in most Radiolaria; the vertical main axis determines the length (principal
axis); one horizontal transverse axis determines the thickness (sagittal axis), and the other the
breadth (frontal axis). Those ground-forms in which the transverse axes are isopolar are termed
"amphithect," and those in which the one (frontal or lateral) is isopolar and the other (sagittal
or dorso-ventral) is allopolar, are termed "bilateral," or better "zeugitic."
46. Primary and Secondary Ground-Forms.—The geometrical sphere must be
regarded as the original ground-form of the Radiolaria; it being understood that its monophyletic
derivation from a single stem-form, Actissa, is correct. The simplest forms of
Actissa (Procyttarium, Pl. 1,
fig. 1) are in fact geometrically perfect spheres; indeed even the individual parts which
compose their unicellular bodies (nucleolus, nucleus, central capsule and calymma) are concentric
spheres. But in addition the central capsules of most other Spumellaria,
especially the Sphæroidea, as well as of many Acantharia {xxii}are true spheres.
Furthermore the simple or concentrically composed lattice-spheres of Sphæroidea, Sphærophracta, and Phæosphæria may be regarded as spheres, although strictly speaking they are
endospherical polyhedra. From the primary spherical form of the Radiolaria all other secondary
forms may be derived in the following order:—1. By the development of a main axis the
Monaxonia arise. 2. By the development of transverse axes the Stauraxonia arise. 3. In both groups
(Monaxonia and Stauraxonia) the two poles (or upper and lower halves of the body) are at first
similar (Isopola). 4. By differentiation in the two poles or halves of the body (distinction
between the basal pole and the apical) the forms with different poles (Allopola) arise. 5. The
transverse axes of the Stauraxonia are at first equal (regular pyramids and double pyramids). 6.
By differentiation in the transverse axes (distinction between the sagittal and the frontal axis)
the amphithect pyramids and double pyramids arise. 7. From the amphithect pyramids the Amphipleura
arise by differentiation of both poles of the sagittal axis. 8. The zygopleural ground-form
appears last, as the simplest form of the Amphipleura.
47. The Ground-Forms of the Spumellaria.—The Spumellaria, being the oldest and most primitive Radiolaria, have for the most
part either indifferent or multistable equilibrium; e.g., all Colloidea and Beloidea which have a spherical central
capsule, and also most Sphæroidea. Among these primitive Centrostigma
true spheres and endospherical polyhedra are represented in the utmost variety, and the regular
polyhedra in particular. By the development of a vertical main axis these Centrostigma have also
given rise to very numerous Centraxonia, which are usually isopolar, very rarely allopolar.
Sometimes they are Monaxonia (circular in transverse section), sometimes Stauraxonia (polygonal in
transverse section). The vertical main axis is longer in the Prunoidea,
shorter in the Discoidea than any of the other axes. The Larcoidea are distinguished by their lentelliptical or triaxial ellipsoid form;
the three different but isopolar axes corresponding with those of the rhombic octahedron; but even among the Sphæroidea, Prunoidea, and Discoidea, this form is sometimes produced by the differentiation of two
different transverse axes at right angles to each other. Whilst these ground-forms (Centraxonia
and Centrostigma) occur in the utmost variety among the Spumellaria, the
centroplanar (or true bilateral) ground-form is entirely wanting.
48. The Ground-Forms of Acantharia.—In the small family Astrolophida,
which contains the most archaic forms of the legion (Actinelius, Astrolophus), the
Acantharia show a direct relation to the most primitive Spumellaria (Actissa), and like these have indifferent equilibrium; their
central capsule is a sphere, their calymma an endospherical polyhedron, whose angles are indicated
by the distal ends of the numerous {xxiii}equal radial
spines. In the great majority of Acantharia, however (all Acanthonida and Acanthophracta), twenty radial spines
are present, regularly distributed, according to Müller's icosacanthan law, in five parallel
circles, each containing four crossed spines (p. 717). Usually the twenty
spines are equal, and the ground-form is the quadratic octahedron, or a regular double pyramid
with sixteen sides. But in some groups (the Amphilonchida and Prunophracta) two opposite
equatorial spines are much more strongly developed than the other eighteen, and therefore the
hydrotomical axis in the equatorial plane is larger than the geotomical axis (p. 719); the isopolar stauraxonian form passes over into the allopolar, and the
ground-form becomes the rhombic octahedron or the amphithect double pyramid (compare §§ 33 and 34, and p. 720). The
centroplanar ground-form is entirely wanting in the Acantharia.
49. The Ground-Forms of the Nassellaria.—The Nassellaria all possess monostatic ground-forms, inasmuch as by the very
structure of their monopylean central capsule a vertical main axis is necessitated, whose basal
pole occupies the porochora. The same arrangement is also for the most part clearly recognisable
in the corresponding structure of the skeleton, which is generally either centraxon or
centroplanar. Among their manifold skeletal forms different larger groups of ground-forms may be
recognised according as the vertical allopolar main axis is crossed by differentiated transverse
axes or not (Stauraxonia or Monaxonia); the former are either triradial or multiradial. The
triradial, with three lateral or terminal radial apophyses, constitute the greater part of the
Nassellaria, and have probably been derived originally from the triradial
Plectoidea (Triplagia, Triplecta); a more careful
examination, however (especially with reference to the structure of the cortinar septum), reveals
the fact that the ground-form is not strictly regularly pyramidal (with three equal radii), but
amphipleural (with two paired ventral and one unpaired dorsal radius), and that it usually passes
over into a distinctly zygopleural form. The same holds true of the multiradial Nassellaria, where for the most part three interradial or six adradial
(sometimes more) apophyses are intercalated between the three primary perradial ones; sometimes
here also the ground-form is a quite regular hexagonal or nonagonal pyramid, but usually it is
more or less amphithect or amphipleural. Among the eradial Nassellaria,
which have no radial apophyses, the ground-form is sometimes allopolar monaxon (conical, ovoid,
hemispherical, &c.), sometimes amphithect pyramidal (even in the simplest Stephanida,
Archicircus, &c.), or sometimes distinctly zygopleural or bilateral (many Plectellaria).
50. The Ground-Forms of the Phæodaria.—The Phæodaria agree with the Nassellaria in the possession
of a primitively centraxon ground-form, and like them are monostatic, since a vertical main axis
whose basal pole passes through the astropyle is present, owing to the characteristic structure of
their cannopylean central capsule. In {xxiv}the great
majority of Phæodaria the spheroidal central capsule also possesses a pair
of parapylæ near the opposite apical pole of the main axis (Tripylea), and these determine (as the
right and left secondary openings) an isopolar frontal axis. Hence, strictly speaking, in most
Phæodaria the central capsule has the geometrical ground-form of the
amphithect pyramid (as in the Ctenophora), with an allopolar vertical main axis, and two unequal,
but isopolar, horizontal transverse axes. In many Phæodaria the skeleton
also has this amphithect pyramidal ground-form, e.g., the bivalved Phæoconchia and part of the Phæogromia. On the
contrary, in the rest of the Phæodaria the skeleton exhibits very various
geometrical ground-forms, independent of that of the central capsule. In the Phæosphæria it forms preferably spheres or endospherical polyhedra, as also in
the Castanellida and Circoporida among the Phæogromia; among the
Circoporida there are also seen with remarkable distinctness the regular polyhedra (especially the
dodecahedron and icosahedron). Isopolar monaxonia are found among the Aulosphærida
(Aulatractus) and Orosphærida; allopolar monaxonia among the Challengerida
(Lithogromia). The Medusettida and Tuscarorida show various forms of regular pyramids
(allopolar Stauraxonia); and finally, the Challengerida are for the most part centroplanar or
bilateral. Thus the Phæodaria present a great wealth of different
geometrical ground-forms in the development of their skeleton, not in that of their central
capsule.
Chapter II.—THE CENTRAL CAPSULE.
51. Components of the Central Capsule.—In all Radiolaria without exception, at
some period of life or other, the central portion of the soft body is separated from the
peripheral portion by an independent, anatomically recognisable membrane; this membrane with all
its contents is designated the central capsule, and is the peculiar central organ of the
unicellular body, which distinguishes the Radiolaria most clearly from the other Rhizopoda. In the
great majority of the Radiolaria the volume of the central capsule is less than that of the
surrounding peripheral soft body which we place in opposition to it as "extracapsulum." The
"capsule-membrane," which separates these two constituents, arises very early in most Radiolaria,
and persists throughout their whole life. In some species, however, the membrane only appears
later, immediately before the formation of the spores, and hence is absent for a considerable
period. Regarded as a whole, then, the capsule consists of the following parts:—(1) the
capsule-membrane; (2) the enclosed endoplasm, or intracapsular protoplasm; (3) the nucleus. But in
addition, many other non-essential structures may be enclosed in the central capsule, especially
hyaline spheres (vacuoles), fatty spheres, pigment granules, crystals, &c.
The central capsule was first described in my Monograph in 1862 (pp. 69-82) as
the most characteristic component of the Radiolarian organism, and distinguished from the whole
extracapsular {xxv}soft body. The fact that it has
recently been reported as absent by various authors is due to their having observed young or
unripe specimens, before the formation of the spores. In some species of Polycyttaria and Acantharia the membrane persists
only a very short time.
52. The Primary Form of the Central Capsule.—The form of the central
capsule is originally a geometrical sphere; and if in accordance with our monophyletic hypothesis
all Radiolaria are to be derived from one common stem-form (Actissa, see p. 12), then the central capsule of this common stem-form must be regarded as
perfectly spherical (Procyttarium, p. 13, Pl. 1, fig. 1).
Since, further, the enclosed nucleus and the surrounding calymma of this primitive archaic form
must also be spheres, and since the nucleus lies in the centre of the body, and the protoplasm is
evenly distributed between it and the membrane, it follows that no axes or excentrically
differentiated parts are to be distinguished in this most primitive Radiolarian. Rather in the
primary central capsule all parts are concentrically and evenly arranged round its centre. This
primary spherical form becomes modified in most Radiolaria into various secondary ground-forms,
which are correlated partly with the structure of the capsule itself, and partly also with the
development of openings in its membrane. In general the ground-form of the central capsule is
polyaxon in the Porulosa (Spumellaria and Acantharia); but in the Osculosa centraxon forms are more frequently observed;
in the Nassellaria the ovoid (allopolar monaxon) form is predominant, and
in the Phæodaria the rhomboid or amphithect pyramid. In these latter, the
astropyle indicates the basal pole of the vertical main axis, whilst the two parapylæ (right and
left) mark the poles of the frontal transverse axis. In the Nassellaria
the centre of the porochora corresponds with the basal pole of the main axis, whilst no transverse
axes are originally present.
53. The Secondary Forms of the Central Capsule.—The original purely
spherical form of the central capsule persists only in the minority of the Radiolaria, namely, the
greater part of the Spumellaria and Acantharia; it
passes over into various other secondary forms in the majority of the class, in the whole of the
Nassellaria and Phæodaria, and in a considerable
portion of the Spumellaria and Acantharia. These
secondary or derived forms may be divided into two quite distinct groups, which may be designated
endometamorphic and exometamorphic; in the former the cause of the divergence of the secondary
form from the sphere lies in the internal structure of the central capsule; in the latter it lies
in the external influence exerted by the growth of the skeleton. Obviously the former series of
modifications is more significant than the latter.
54. The Endometamorphic Forms of the Central Capsule.—The secondary forms of the
central capsule, which are due to internal causes connected with its growth, are as follows:—
{xxvi}
A. The Ellipsoidal Central Capsule, with one axis elongated, so that it becomes the
vertical main axis of the body.
a. Among the Spumellaria, Actiprunum (p. 14), Colloprunum (p. 25, Pl. 3, fig. 9),
most Prunoidea (p. 288).
b. Among the Acantharia, many Amphilonchida (p. 782, Pl. 132,
figs. 2, 6), and Belonaspida (p. 861).
c. Among the Nassellaria, many Plectoidea (p. 905,
Pl. 91,
figs. 5, 9), Stephoidea (p. 937, Pl. 81, fig.
16), Monocyrtida (Pl. 51,
fig. 3), &c.
B. The Cylindrical Central Capsule, with considerable elongation of the
vertical main axis, which is several times as long as the horizontal transverse axis.
a. Amongst the Spumellaria, Collophidium (p. 26, Pl. 3,
figs. 1-3) and many Prunoidea (Spongurus, &c.).
b. Among the Acantharia, some
Amphilonchida.
C. The Discoidal, Spheroidal, or Lenticular Central Capsule, with one axis shorter
than the others, which becomes the vertical main axis.
a. Among the Spumellaria, Actidiscus (p. 15), Collodiscus (p. 27), and the large group
Discoidea (p. 408).
b. Among the Acantharia, many Quadrilonchida (p. 768, Pl. 131),
and most Hexalaspida (p. 874).
c. Among the Nassellaria, certain Stephoidea and Cyrtoidea.
d. Among the great legion Phæodaria the
spheroidal central capsule is almost always more or less flattened in the direction of the
main axis (p. 1525,
Pls. 101-128).
D. The Lentelliptical Central Capsule (or triaxial ellipsoid), with three unequal but
isopolar axes at right angles to each other, the sections in all three dimensions of space being
ellipses.
a. Among the Spumellaria, Actilarcus and the large
group Larcoidea (p. 604).
b. Among the Acantharia, certain Amphilonchida
and Belonaspida.
E. The Polymorphic, Amœboid or Irregular Central Capsule.
a. Among the Spumellaria, Collodastrum
(p. 28, Pl. 3,
figs. 4, 5), and some Larcoidea.
55. The Exometamorphic Forms of the Central Capsule.—The secondary forms of the
central capsule, which are brought about by external causes, chiefly dependent on the formation of
the skeleton, are very various and in many cases devoid of special interest; in other instances,
on the contrary, they are of great importance, because of the clear relation of cause and effect
which can be traced between the development of the skeleton and of the capsule. The most important
phenomena to be recorded in this connection are as follows:—
{xxvii}
I. Spumellaria.—(A) In many of the Sphæroidea, the central capsule of which is originally enclosed by a simple
lattice-sphere, it puts out protrusions through the meshes of the shell, thus forming
club-shaped processes, corresponding in number with the meshes of the lattice (Pl. 11, figs. 1,
5; Pl. 20,
fig. 1a; Pl. 27,
fig. 3, &c.). The whole surface of the spherical capsule may thus be covered with numerous
independent radial clubs of equal size, but usually they unite again outside the shell to form a
simple sphere with smooth surface. (B) In many Prunoidea whose
originally ellipsoidal body has become cylindrical by the marked prolongation of the main axis,
the central capsule is divided by a series of constrictions into segments, which correspond with
the annular constrictions of the skeleton (Pls. 39, 40). (C) In
most Discoidea whose lentiform or discoidal shell develops radial arms
at its margin, the central capsule sends out processes into these arms, and adapts itself to the
stellate form of the skeleton (p. 409, Pl. 43, fig. 15;
Pl. 47,
&c.) (D) In many Larcoidea whose growth is originally
lentelliptical, but later spiral or irregular, the central capsule follows the mode of growth
and develops irregular protuberances.
II. Acantharia.—Whilst the central capsule of most
Acantharia retains its primitive spherical form, in a minority of the
group it passes over into various secondary forms, which are directly determined by the growth
of the skeleton; especially common are lappet or club-shaped prominences which follow the larger
radial spines. Hence the central capsule may assume the form of a violin, with two lobes
corresponding to the two poles of the elongated main axis, as in many Amphilonchida (p. 782, Pl. 132,
fig. 10), and the Diploconida (p. 884, Pl. 140). On the
other hand the central capsule becomes cruciform, with four lobes disposed at right angles, as
in Lithoptera and other Quadrilonchida (p. 768, Pl. 131, fig.
10, &c.).
III. Nassellaria.—The primitive ellipsoid or ovoid
form of the central capsule persists only in a few Nassellaria, such as
the simplest and most archaic forms, the Nassellida, many Plectoidea,
Stephoidea, Monocyrtida, &c. In the great majority of the Nassellaria, on the contrary, the ellipsoid or ovoid form passes over into a
secondary form which is usually characterised by the presence of lobes, and is obviously
dependent upon the previous development of the skeleton. In many Stephoidea and Spyroidea (probably the majority), a
bilobed central capsule is formed (with symmetrically equal right and left lobes), since the
primary vertical sagittal ring interferes with the growth in the median plane (Pl. 90, figs.
7-10). In other {xxviii}Spyroidea, on the contrary, and the majority of the Cyrtoidea, the central capsule forms at its basis rounded lobes, which
protrude and hang down from the meshes of the cortinar plate; and since this latter has usually
three or four large pores, the capsule similarly develops three or four processes (Pl. 53, fig. 19;
Pl. 55,
figs. 4-11; Pl. 59,
figs. 4-13; Pl. 60,
figs. 3-7; Pl. 65,
fig. 1).
56. The Membrane of the Central Capsule.—The capsule-membrane or envelope of the
central capsule is both morphologically and physiologically one of the most important parts of the
Radiolarian body, for it separates its two main constituents, the capsule with its nucleus and
endoplasm and the extracapsulum with the calymma and exoplasm. The capsule-membrane is invariably
present at some time or other during the life of the organism, even though in a few species it may
persist only for a short time. It is characterised in general by its power of resistance to
chemical and physical reagents, and appears to be related to the elastic tissues or perhaps even
more to the chitinous substances. Its thickness is usually less than 0.0001, though in certain
groups it ranges between 0.001 and 0.002, and in many of the larger Radiolaria (such as Collida
and Phæodaria) it may attain a thickness of 0.003 to 0.006 or more. In the
three legions Spumellaria, Acantharia, and Nassellaria the capsule-membrane is single, while in the Phæodaria it is always double, being composed of a firm outer and a delicate
inner membrane, which are in contact at only few points. Usually it is quite structureless, except
for its apertures; the thicker membrane showing occasionally a fine concentric lamination. In
certain large Colloidea (e.g., Thalassicolla, Pl. 1, fig.
5b) the membrane is covered on the inner surface by a network of polygonal ridges, and in
some large Phæodaria with remarkable small curved rods (Pl. 114, fig. 13).
In all Radiolaria the membrane is perforated by definite openings or pores, through which the
intracapsular and extracapsular protoplasm are in direct communication. These openings (or
"pylae") show very characteristic and constant differences in the four legions, which have given
rise to the names—Peripylea, Actipylea,
Monopylea, Cannopylea.
The capsule-membrane was first indicated as the most important and absolutely
constant component of all Radiolaria, and as the differential character of the class, in my
Monograph (1862, pp. 69-71). The careful investigations of R. Hertwig have confirmed this view
and at the same time have yielded the most important conclusions regarding the nature and
systematic significance of the openings in the capsule (op. cit., 1879, pp. 105-107). On
the contrary, Karl Brandt has recently propounded the theory that the capsule-membrane is by no
means a constant part of the Radiolarian organism, but is lacking in certain species of
Collozoum and Sphærozoum (1881, p. 392). This contradiction is explained by the
fact that in some Collodaria and Acanthometra
the formation of the central capsule takes place much later than in the other Radiolaria, in
some {xxix}species indeed only just prior to the
development of the swarm spores. I have recognised the presence of it in all species which I
have investigated (more than a thousand), and even in those in which Brandt denies its
existence. It is often very delicate and may easily be overlooked, especially when the contents
of the capsule are colourless, but in all cases by the prudent application of staining fluids
and other reagents its presence may be demonstrated. Even in those cases in which the contour of
the capsule was not visible, and its contents appeared to pass without definite boundary into
the matrix of the extracapsulum, it was possible by the use of appropriate stains or reagents,
which would not penetrate the capsule, or of those solvents which were capable of dissolving its
contents and of causing it to swell up like a distended bladder, to recognise the existence of
the membrane. Those Radiolaria in which it is truly absent are young animals of species in which
the membrane is only formed immediately before sporification, and persists but for a short time
(e.g., species of Collozoum, Sphærozoum, Acanthometra,
Acanthochiasma, &c.).
57. The Capsule-Openings of the Peripylea (or Spumellaria).—The capsule-membrane
of the Peripylea is generally perforated by extremely fine and numerous
pores, which are distributed at equal distances over the whole surface, and are precisely alike in
all parts of the capsule. Hence the Spumellaria may be called
"Holotrypasta" or "Porulosa"; they agree with the Actipylea in being
devoid of an osculum or operculum; they are distinguished from the latter group mainly in that
their pores are equally distributed over the whole surface of the capsule, whilst in the Actipylea the pores are disposed in definite groups or lines, separated by large
imporous areas.
The central capsule of the Spumellaria, with its
innumerable fine and evenly distributed pores, must be regarded as the primitive arrangement,
from which the different central capsules of the three other legions have been developed. The
central capsule of the Actipylea has been derived from that of the Peripylea by reduction in the number of the pores and their distribution in
definite, regularly disposed areas in the membrane. The central capsule of the Osculosa is
characterised by the formation of a special main-aperture (osculum) at the basal pole, which is
closed in the Monopylea by the porochora, and in the Cannopylea by the astropyle; the remaining pores, with the exception of the
accessory openings of many Cannopylea, remain undeveloped in both these
legions. In the same way Hertwig regards the central capsule of the Peripylea as the primitive form (1879, L. N. 33, p.
107).
58. The Capsule-Openings of the Actipylea (or Acantharia).—The capsule-membrane of
the Actipylea is perforated by very numerous fine pores, which are
regularly distributed over the surface of the central capsule, and separated by imporous
intervals. Hence the Acantharia belong to the "Holotrypasta" or
"Porulosa"; they have neither osculum nor operculum, and agree in this particular with the Peripylea; but they are separated from these latter chiefly by the fact that
their pores are much less numerous, and marked off into regularly arranged groups or lines by
imporous intervals. In the Peripylea, on the contrary, the pores are much
more numerous and are evenly distributed over the whole surface of the capsule.
{xxx}
The central capsule of the Acantharia has hitherto been
for the most part confounded with that of the Spumellaria, and no clear
distinction has been drawn in this respect between the two legions of the Porulosa. Hertwig, who
in 1879 first discovered the remarkably different structure of the Osculosa (Nassellaria and Phæodaria), recognised no distinction
between the structure of the capsules in the Peripylea and Actipylea (his Acanthometrea), and supposed that in both these legions "very
fine pores were evenly distributed in large numbers over the capsule-membrane" (loc.
cit., p. 106). I have, however, during the last few years convinced myself, by the careful
comparative investigation of numerous Acantharia, that in this respect
they are quite distinct from the Spumellaria (with perhaps the exception
of the Astrolophida, which are nearly related to the primitive Actissa). The number of
pores in the Actipylea is usually very much smaller than in the Peripylea, and they are regularly arranged in groups.
59. The Capsule-Openings of the Monopylea (or Nassellaria.)—The capsule-membrane
of the Monopylea always possesses a single large main-opening, an osculum,
which lies at the basal pole of the main axis, and is closed by a circular perforated lid
(operculum porosum). When seen from the surface this lid appears as a clearly defined porous area
(porochora or area porosa), and forms the horizontal base of a peculiar cone, which stands
vertically in the interior of the capsule and may be designated the "thread-cone" (podoconus). The
Nassellaria may hence be termed "Merotrypasta" or "Osculosa," like the
Cannopylea; the structure and significance of the circular lid
(operculum), which closes the main-opening (osculum) is, however, quite different in the two
legions. Whilst the lid of the Cannopylea (astropyle) is solid, traversed
by radial ribs, and only perforated in its centre by a short tube (proboscis), in the Monopylea the operculum (porochora) is always perforated by numerous vertical
fine pores, and is in connection with the peculiar internal "pseudopodial cone" (podoconus, Pl. 51, figs. 5,
13; Pl. 81,
fig. 16; Pl. 91,
fig. 5; Pl. 98,
fig. 13). The pores are separated by small vertical, highly refractive rods (opercular rhabdillæ);
these become intensely stained by carmine, and are either evenly distributed over the surface of
the porochora or arranged in definite groups. The outer or distal end of each rod is rounded,
sometimes thickened like a club or split into lobes; the inner or proximal end is usually pointed,
and stands in connection with a myophane thread of the podoconus (see § 79).
The primary circular form of the porochora, in which the opercular rhabdillæ are evenly
distributed in a horizontal plane, undergoes various secondary modifications in many Nassellaria. The triradial structure of the skeleton, which characterises the
majority of the legion, causes a splitting of the base of the central capsule into three or four
lobes; this division also affects the porochora, which lies in the centre of the base, so that the
rhabdillæ become arranged in three or four equal circles. If, however, the lobes of the central
capsule become larger and protrude through the three or four collar pores of the cortinar septum,
the central porochora may separate entirely into three or four elongated tracts, which lie on the
axial side of the magnified lobes; the rhabdillæ are then arranged over the whole surface of {xxxi}these tracts, on the outer aspect of which run the
longitudinal myophane fibrillæ of the podoconus (compare §§ 79 and 99).
The porous area of the Monopylea was first described by
Hertwig in 1879, and shown to be the characteristic main-opening of the central capsule in
various families belonging to this legion (L. N. 33, pp. 71, 73, 83, 106,
Taf. vii., viii.). According to his view "the capsule-membrane in the porous area becomes
thickened around each pore into a rod, perforated by a canal," and the intracapsular protoplasm
passes outwards through these fine canals (loc. cit., p. 106). I am not able to share
this interpretation, but think rather that I have convinced myself by the examination of some
living Nassellaria, and of many well-stained and preserved preparations
in the Challenger collection, that the rods are solid, specially modified portions of the
capsular wall, and that the protoplasm does not pass through them but through pores which lie
between them.
60. The Capsule-Openings of the Cannopylea (or Phæodaria).—The capsule-membrane of
the Cannopylea always possesses only a single large main-opening or
osculum, which lies at the basal pole of the vertical main axis, and is closed by a circular
radiated lid (operculum radiatum). This operculum appears, when seen from the surface, as a
sharply defined stellate area (astropyle), from the middle of which arises a shorter or longer
cylindrical tube, the proboscis. Hence the Phæodaria, like the Monopylea, belong to the "Merotrypasta" or "Osculosa"; the structure and
significance of the circular operculum, which closes the main-opening (osculum), are, however,
quite different in the two legions. Whilst the operculum of the Monopylea
(porochora) is perforated by numerous fine vertical pores, and connected with the peculiar
internal pseudopodial cone (podoconus), this structure is entirely wanting in the Cannopylea, and instead of it there is a solid operculum, with radial ribs which
originate at the base of its central tubular mouth; this tube (proboscis) is cylindrical, often
conical at the base, of very variable length and with a round aperture at either end. In spite of
the great difference which the various families of Cannopylea exhibit in
the formation of their skeleton and its appendages, the constitution of this characteristic
stellate main-opening (astropyle) is always essentially the same; both the stellate operculum
itself, and the proboscis which rises from its centre, show only slight differences in the various
groups. In addition to this large main-opening most Phæodaria possess
several small accessory openings (parapylæ); and usually two of these are present, placed
symmetrically right and left of the aboral pole of the main axis and in the frontal plane (Pl. 101, figs. 2,
6, 10; Pl. 104,
figs. 1, 2a). Sometimes there are more numerous accessory openings (three to six or more)
regularly arranged, as in the two peculiar families, Circoporida and Tuscarorida; occasionally
also there is only a single parapyle, at the aboral pole of the main axis (e.g., in
Tuscaridium). The parapylæ seem to be quite absent in the families Challengerida,
Medusettida, Castanellida, and perhaps also in other Phæodaria. The form
and structure of the small accessory openings appear to be always the same. The {xxxii}outer capsule-membrane is elevated in the form of a
short cylindrical tube or "apertural ring" (collare paraboscidis), the external margin of which
bends inwards, and at the base of the ring passes over into the delicate internal capsule
membrane. Upon this apertural ring is situated a longer or shorter "apertural cone" (paraboscis),
which is a tubular, cylindrical or conical, prolongation of the membrane, open externally.
The peculiar capsule-openings of the Phæodaria were first
discovered and carefully described by Hertwig in 1879 (L. N. 33, pp. 95,
107). He found in all the six genera which he examined three openings, a main-opening at
the basal pole of the main axis and two accessory openings, one on either side of the apical
pole; hence he named the whole group "Tripylea." This name, however, is
not applicable to the numerous Phæodaria mentioned above, which have
only a main opening without any accessory openings, nor to those genera in which the number of
the latter is variable. I have, therefore, replaced Hertwig's designation by the term "Cannopylea," which has reference to the peculiar tubular form of the opening.
This I find much more developed in many Phæodaria than Hertwig has
represented, and I must also, in certain particulars, dissent from his delineation of the minute
structure, although this is in the main remarkably accurate.
61. The Nucleus.—The nucleus, enclosed in the central capsule of all
Radiolaria, behaves in every respect like a true cell-nucleus, and thus lies at the base of the
now universal opinion, that the whole Radiolarian organism, in spite of its varied development and
remarkable variations, is unicellular and remains throughout life a true individual cell. This
important theory is not invalidated by the fact that the nucleus undergoes peculiar modifications
in many groups, and in certain groups presents appearances seldom or never seen elsewhere.
62. Uninuclear and Multinuclear Radiolaria (Monocaryotic and
Polycaryotic).—All Radiolaria present two different conditions in respect of the
behaviour of the nucleus, since in their young stages they are uninuclear (monocaryotic),
and in later stages multinuclear (polycaryotic). This is readily explained by the fact that
each individual Radiolarian is developed from a simple unicellular swarm-spore, and that
afterwards, before the formation of swarm-spores, the single nucleus divides into many small
nuclei. Thus in the Radiolaria the nucleus is pre-eminently the organ of reproduction and
inheritance. The division of the originally single nucleus into many small nuclei may take
place, however, at very different periods, so that the Radiolaria may be divided in this respect
into precocious and serotinous.
63. Serotinous and Precocious Radiolaria.—In the great majority of the
Radiolaria the division of the nucleus takes place only at a late period, a short time or even
immediately before the process of spore formation; it then breaks up rapidly into numerous small
nuclei (always more than one hundred, sometimes many thousands), and each of these {xxxiii}either becomes itself the nucleus of a swarm-spore,
or by repeated division gives rise to a group of spore-nuclei. All those Radiolaria which are
uninuclear during the greater part of their existence, and in which the process of division is
late, and takes place rapidly, are called "serotinous" or late-dividing forms. To this category
belong all Phæodaria and Nassellaria, as well as
all the solitary or monozoic Spumellaria and some Acantharia. On the other hand, the name "precocious," or early dividing, is
applied to those Radiolaria in which the division of the nucleus takes place very early, and in
which, therefore, the cell is multinuclear during the greater part of its existence. This is the
case in all the social or polyzootic Radiolaria (Polycyttaria, Pls. 3-8), and also in
the great majority of the Acantharia, both Acanthometra and Acanthophracta. In the last two
groups, however, there are numerous exceptions, and these are seen in remarkably large species,
characterised by the great size of the central capsule. From a phylogenetic point of view, the
conclusion is allowable that the precocious forms are secondary, and have arisen by adaptive
modification from the primitive serotinous stem. In the Polycyttaria (or social Spumellaria, i.e., the three families Collozoida, Sphærozoida, and
Collosphærida), the cause of the adaptation lies most probably in the formation of the colony
itself, for all these three families are so closely related to three corresponding families of
serotinous, monozootic Radiolaria (Thalassicollida, Thalassosphærida, Ethmosphærida), that certain
species of the latter are hardly to be distinguished from isolated individuals of the former.
Perhaps the remarkable formation of the large central oil-globule, which particularly
characterises the Polycyttaria, is the prime cause of their early nuclear division. In the Acantharia the cause is most likely to be found in the characteristic
centrogenous development of their acanthin skeleton, whose radial bars first of all appear
in the centre of the capsule. Hence arises directly the excentric position of the nucleus, which
in the archaic stem of Acantharia (Actissa?) was probably central.
In any case, but little weight is to be laid upon the precocious division of the nucleus in the
Acantharia in general, inasmuch as in certain species (both Acanthometra and Acanthophracta) the more usual
serotinous division persists.
64. Central and Excentric Nuclei.—The position of the nucleus in the
interior of the central capsule was no doubt primitively central, and this situation in the
geometrical centre of the original spherical central capsule has been accurately retained in all
monozootic Spumellaria; in the polyzootic families of this legion
(Polycyttaria), on the contrary, it is obscured by the precocious division of the nucleus. In the
other three legions, which may be phylogenetically derived from the Spumellaria, the position of the nucleus is rarely central, but usually
excentric, or at most subcentral. In the Acantharia (both Acanthometra and Acanthophracta) the central position
of the nucleus is at once excluded by the constantly centrogenous development of the skeleton; the
nucleus is therefore always excentric, and may lie at either side; it usually {xxxiv}divides very early into numerous separate nuclei,
which are usually distributed in the peripheral portions of the central capsule. In the Nassellaria the development of the porochora, and of the podoconus which stands
upon it, brings about the formation of a vertical axis, and in consequence the central capsule
assumes a monaxon form (usually ovoid or conical); the nucleus then lies in the main axis, but
excentrically between the apex of the podoconus and the aboral pole. In many Nassellaria, however, especially when the podoconus is so large that its apex
approaches the aboral pole of the central capsule, the nucleus is pressed to one side and lies
quite excentrically. The Phæodaria exhibit a different arrangement; the
large spheroidal nucleus is always subcentral, so that its main axis corresponds with that of the
concentric spheroidal central capsule; but since the astropyle always occupies the oral pole of
the latter, and since the distance of the nucleus from this pole is always somewhat different from
its distance from the other, it follows that, strictly speaking, the nucleus never lies accurately
in the geometrical centre.
65. Homogeneous and Allogeneous Nuclei.—The nucleus of the Radiolaria
not only exhibits a similar structure and composition, and suffers similar modifications to those
which are found to occur in the case of other cell-nuclei, but also to some extent shows very
peculiar developmental forms, which are seldom or never found in other cells. In the first place
the nuclei may be divided into homogeneous and allogeneous, the former are structureless and
consist of a uniform mass of nuclein, whilst the latter are composed of different substances and
show various structural relations. Homogeneous nuclei, whose whole mass is uniform and
exhibits no structural differentiation, are probably always to be found in the swarm-spores; in
the fully developed Radiolarian body they are found only in the first legion, Spumellaria, and that both in many Monozoa (especially small Sphæroidea and Prunoidea) and in the Polyzoa (or
Polycyttaria). The whole mass of these homogeneous nuclei, which are usually spherical or
ellipsoidal, consists of uniform, perfectly clear and transparent nuclein, and becomes evenly
stained by carmine, hæmatoxyline, &c. They may be readily distinguished by these means from
the clear vacuoles or "hyaline vesicles," which are evenly distributed in the endoplasm of many
Radiolaria, and may be confused with the former. Allogeneous nuclei, which are always
composed of different parts and often show complicated structural relations, are found developed
in the great majority of Radiolaria. The most important differentiation exhibited by these
secondary forms is the separation of the nuclear mass into a firm nuclear substance (caryoplasm)
and a fluid nuclear juice (caryolymph). In addition in each nucleus a nucleolus is visible, and
often several or many may be seen (see §§ 67 to 70).
66. The Form of the Nucleus.—The nucleus of the Radiolaria shows greater
variations in form and structure than are to be found in the majority of cell-nuclei; {xxxv}exception must, however, be made in the case of many
animal ovicells, which, in their peculiar form and composition, often recall large Radiolarian
nuclei. With respect to the external shape two main forms may be distinguished, as primary and
secondary. The primary form of the Radiolarian nucleus is the sphere; it occurs not only in
most swarm-spores, but also in most adult forms belonging to the legion Spumellaria, and in individual instances in other groups; indeed the nuclei of
most Spumellaria, as also the concentric central capsules in which they
lie, are true geometrical spheres. The secondary forms of the nucleus are found in the
majority of adult Radiolaria, and arise from the primary spherical forms in various ways, either
by the elongation or contraction of one axis, or by the formation of apophyses or processes. The
most important of these secondary forms are as follows:—
1. Ellipsoidal nuclei, arising by elongation of one principal axis; very common among
the Nassellaria, as well as in many Prunoidea
and Larcoidea among the Spumellaria; also in
several Acantharia.
2. Discoidal nuclei, arising by contraction of one principal axis, sometimes
lenticular or spheroidal, biconvex, sometimes shaped like a disc or coin; especially common in
the Discoidea among the Spumellaria, also in
some Acantharia; the large nucleus of the Phæodaria is always spheroidal or almost spherical, with a slightly shortened
main axis.
3. Stellate nuclei, spherical, and armed with evenly distributed radial club-shaped or
conical processes; rare but very characteristic, especially in the two large Thalassicollida
Thalassopila (Pl. 1,
fig. 3), and Thalassophysa (Monogr. d. Radiol., Taf. i.); also in some Sphærellaria (Pl. 11,
fig. 5).
4. Amœboid nuclei, with unequal processes irregularly arranged, in certain
irregular forms of Spumellaria and Acantharia.
5. Lobate nuclei, with several (usually two or three) large ovoid or
pyriform lobes, which protrude into corresponding larger lobes of the central capsule, in many
Nassellaria, especially the multiarticulate Cyrtoidea (Pl. 59,
figs. 12, 13). The budding nucleus of the Acantharia is also lobate (Pl.
129,
figs. 6-11).
67. The Nucleus of the Peripylea.—The nucleus of the Spumellaria or Peripylea shows in certain groups a very
primitive arrangement, indeed the archaic structure from which the various forms of nuclei of
other Radiolaria may be derived; but on the other hand, in other groups it exhibits very peculiar
and remarkable differentiations. In the first place it may be noted that the monozootic or
solitary Spumellaria usually possess a single serotinous nucleus, which
only divides into numerous swarm-spores at a late period; {xxxvi}whilst, on the contrary, the polyzootic colonial Spumellaria (or Polycyttaria) are uninuclear only in the young state (Pl. 3, fig. 12), and
speedily present numerous small homogeneous nuclei, which have arisen by precocious division of a
single nucleus; these are usually spherical and 0.008 to 0.012 mm. in diameter. The serotinous
nucleus of the monozootic Spumellaria, in many divisions of this large
legion, and especially in the simply constituted Sphæroidea, is a
homogeneous sphere of nuclein, lying in the middle of the central capsule. In many other cases it
assumes the form of a spherical vesicle ("Binnen-Bläschen"), whose fluid or semi-fluid contents
are enclosed by a more or less firm membrane. This vesicle often contains a single central
spherical nucleolus (Pl. 1,
figs. 1l, 4l), but sometimes a variable number of small excentric nucleoli (Pl. 1, figs.
1a, 2a). The nuclear membrane is often somewhat thick, presenting a double contour,
and in such cases may even exhibit a fine radial striation, the expression of minute pores (Pl. 1, fig.
2a). In the colossal nuclei (as much as 1 to 2 mm. in diameter) of certain large
Thalassicollida the nucleolus presents a very remarkable form, becoming stellate by the protrusion
of processes, which may again branch in a dendritic fashion (as in the common Thalassicolla
nucleata), or it may develop into a very long cylindrical thread, which is disposed in
serpentine coils, and in Thalassophysa pelagica passes into the different cæcal processes
of the stellate nucleus. In many Sphæroidea, whose skeleton is composed
of numerous concentric lattice spheres, the small central spherical nucleus lies at first within
the innermost of these (the medullary shell); but afterwards it grows through the meshes of the
lattice-work, and the radiating club-shaped processes thus formed (Pl. 11, fig. 5)
unite with each other outside the medullary shell, and form an external nuclear sphere which
completely encloses the latter. In the Polysphærida (with several concentric lattice-shells) and
in the Spongosphærida (with spongy lattice-spheres), this process may be several times repeated,
so that eventually the central spherical nucleus attains considerable dimensions, and encloses two
or more concentric lattice-shells with their radial connecting rods. The nuclear membrane is in
these cases usually penetrated by radial bars, which connect the outermost of the enclosed shells
with the remaining cortical shells which surround the central capsule. The same remarkable
arrangement is also very common among the Discoidea. The small spherical
primary nucleus is in such instances immediately surrounded by the innermost earliest developed
lattice-shell, around which the concentric rings are subsequently deposited; it then grows out
through the meshes, and the processes fuse outside the ring to form a homogeneous lentiform
nucleus (Pl. 43,
fig. 15). The same process recurs in certain Prunoidea and Larcoidea, whilst in other Spumellaria of these groups
(e.g., Pylonida) the lobate processes of the nucleus remain free.
Both the simple serotinous nucleus of the monozootic Spumellaria, and the numerous precocious nuclei of the Polycyttaria, were
first described in my Monograph in 1862, the former as the "endocyst" ("Binnen-Bläschen"), the
latter as "spherical transparent vesicles" ("Kugelige {xxxvii}wasserhelle Bläschen"). I was in error, however, in regarding the
latter as identical with the so-called "hyaline spherules" in the central capsule of many
Monozoa, which rather belong to the category of intracapsular vacuoles (see § 72). The credit of recognising, by the aid of the modern methods of staining,
the distinctness of these two structures, which may readily be mistaken for each other, and of
demonstrating the true nature both of the serotinous and precocious nuclei, belongs to Richard
Hertwig (1879, L. N. 33).
68. The Nucleus of the Actipylea.—The nucleus of the Acantharia or Actipylea shows very peculiar relations in
respect of structure and division, particularly special forms of lobular budding, which belong to
the characteristic peculiarities of this singular legion, and are not found among other
Radiolaria. The position of the nucleus is always excentric, even in the youngest Acantharia, for the centrogeneous formation of the skeleton, the constant
development of the earliest radial portions of it in the middle of the central capsule, forces the
nucleus from its normal central position. The majority of the Acantharia,
like most Polycyttaria, are precocious, the primary nucleus early dividing into numerous small
nuclei (see note A below). Nevertheless there are many exceptions to this rule in different
families, e.g., Stauracantha, Xiphacantha, Phatnacantha, and
Pristacantha among the Acanthometra, and Stauraspis,
Echinaspis, Dodecaspis, and Phatnaspis among the Acanthophracta. In these instances the primary nucleus remains for a long time
as a simple excentric ellipsoidal or irregularly round body, even in the fully developed stage,
and only at a very late period (sometimes just before the formation of the spores) divides into
many small nuclei. Since this serotinous division of the nucleus takes place in different genera
of very various groups, it can only be decided by further investigations how widely it is spread
among the Acantharia, and upon what circumstances it is dependent (see
note B). The division of the nucleus appears to be precocious in the majority of this legion, and
a number of small nuclei appear to be early formed by a peculiar process of budding; in most fully
developed Acantharia these are disposed in one or two layers under the
surface of the central capsule, but if their numbers increase to any considerable extent, the
whole space between the skeletal rods becomes filled with small nuclei; sometimes these are
homogeneous, sometimes vesicular, 0.002 to 0.012 mm. in diameter; usually they are spherical and
have a small nucleolus (compare Pl. 129,
figs. 6-11, and note C).
A. The numerous nuclei, which are to be found in the central capsule of most mature Acantharia, were first described in my Monograph (1862) as "spherical,
transparent vesicles, provided with a small dark granule" (p. 374, Taf. xv. figs. 2, 5; Taf.
xvi. figs. 2, 4; Taf. xxi. fig. 7, &c.). Their more minute constitution and peculiar origin
were first accurately delineated by R. Hertwig (1879, loc. cit., pp. 11-24, Taf.
i-iii.).
B. The fact that in a number of Acantharia the nucleus does not
divide early as in the majority of the legion, but only at a later period, was first observed by
R. Hertwig in a species of Acanthometra (Xiphacantha serrata),
and a species of Acanthophracta (Phatnaspis {xxxviii}mülleri = Haliommatidium mülleri) (loc. cit.,
pp. 11 and 27). This serotinous division of the nucleus seems, however, to be rather widely
spread in both sublegions of the Acantharia; I have found, not only in
the forms above mentioned, but also in several others belonging to different genera, a single
large excentric nucleus, even in those individuals in which the skeleton was fully
developed.
C. The peculiar mode of nuclear budding, by which these small nuclei arise,
appears to proceed in the following manner (Pl. 129). The
vesicular primary nucleus, which, in consequence of the centrogeneous development of the
skeleton protrudes as it grows into irregular lobes (Pl. 129, fig.
9), assumes a peculiar concavo-convex form, sometimes that of a hood or dish, sometimes that of
a kidney or sausage. The convex surface is apposed to the capsule-membrane, while the concave is
turned towards the central star of the skeleton (fig. 6). There is now formed at the centre of
the convex surface of the strong, doubly-contoured, nuclear membrane, a flask-shaped
invagination with a narrow neck and expanded base; the membrane now becomes disposed in peculiar
folds, which at the narrow aperture of invagination appear as folds, but on the expanded body of
the flask take the form of concentric rings, laid closely side by side (Pl. 129, fig.
10). The convex bottom of the flask, which is directed towards the concave proximal side of the
nucleus, becomes again invaginated by a central conical apophysis of the enlarged nucleolus,
which is situated between them. Usually the nucleolus has already become flattened into a
lentiform shape, and upon its distal face a conical apophysis has been developed, which is
divisible into a darker proximal and clearer distal portion. The tip of the latter appears to be
in direct connection with the nuclear membrane at the centre of the base of the flask-shaped
invagination (figs. 6, 10). At this stage of development the nucleus of the Acantharia generally presents the characteristic form of a hood-shaped,
concavo-convex vesicle, whose radial axis is also the axis of the flask-shaped distal
invagination, and of the depressed conical nucleolus, which lies between the latter and the
concave side of the nucleus. After this peculiar invagination has persisted for some time in
connection with the enlarged nucleolus, both disappear, and then a remarkable growth of lobular
processes takes place on the concave proximal side of the hood or kidney-shaped nucleus; from
four to eight knobs of unequal size usually appear, and their thickened wall encloses a variable
number of small of nucleoli; these are at first few but afterwards more numerous (fig. 7).
Subsequently these knobs or lobes become completely separated by constriction from the original
central mass of the nucleus, and appear as so many separate independent "sausage-shaped bodies"
in the hollow central capsule (fig. 8). Each of the bodies now appears, and at first on its
convex aspect, to form a large number of small nucleoli, which either separate by constriction
from it or become free by its breaking up and lie in numbers in the central capsule. Finally the
buds or lobes of the nucleus break up entirely into such nucleoli, which are evenly distributed
in the central capsule, and become the nuclei of the swarm-spores (fig. 11). Compare R. Hertwig,
L. N. 33, Taf. i.-iii. pp. 19-25.
69. The Nucleus of the Monopylea.—The nucleus of the mature forms of the Nassellaria or Monopylea is generally simple or lobate,
homogeneous or vesicular and excentric, and appears only to divide into numerous small
nuclei just before the formation of the spores. Nevertheless I have sometimes, though not often,
seen in representatives of very various families of the Monopylea, the
central capsule filled with many small spherical homogeneous nuclei (Pl. 53, fig. 19).
Hence all the families of this legion appear to be serotinous, their simple primitive nucleus
persisting for a long period. It {xxxix}is commonly
placed excentrically, and most usually in the apical or aboral portion of the central capsule,
either between its apex and the podoconus, or quite excentrically on the dorsal aspect. The simple
nucleus of the Nassellaria usually appears to be vesicular and to possess
a somewhat firm membrane, clear contents, and a rather large, dark coloured nucleolus. In many
Nassellaria the nucleus is spherical or ellipsoidal (Pl. 53, fig. 11);
whilst in many Stephoidea and Spyroidea, where
the central capsule is constricted by the sagittal ring and divided into two symmetrical lateral
lobes, the nucleus partakes of the same mode of growth and appears in the middle of the capsule as
a transversely placed ellipsoid or even as a short cylinder (Pl. 90, figs. 7,
9). The most remarkable modification in the form of the nucleus is to be found in the
multi-articulate Cyrtoidea. Here it is usually enclosed in the cephalis
and is spherical, ellipsoidal or spheroidal, often flattened almost into a disc. If now the
central capsule increase greatly in size and put forth three or four clavate lobes which hang down
through the pores of the cortinar septum into the thorax (or even into the succeeding joints), the
nucleus usually undergoes similar modification, and three or four finger-like apophyses are
developed from its base, which project into the corresponding lobes of the central capsule (Pl. 59, figs. 4,
12, 13).
The numerous small, spherical, homogeneous nuclei which are to be found in the
central capsules of those Nassellaria, which are ripe and about to
develop spores, were described in 1862 in my Monograph, as "numerous, small, transparent,
spherical cells" in the case of various Cyrtoidea (Arachnocorys,
Lithomelissa, Eucecryphalus, Eucyrtidium, &c.) (loc. cit., pp.
302, 305, 309, 321, &c.), and I find them of the same form and dimensions, but deeply
stained with carmine in many preparations in the Challenger collection. R. Hertwig has
delineated them very accurately in the case of Tridictyopus (1879, loc. cit., p.
84, Taf. vii. fig. 3). He was also the first to recognise the uninucleate condition of the Nassellaria, which is much more frequently observed than the serotinous
multinucleate condition, and he described very clearly the peculiar lobed nuclei which arise in
Cyrtoidea, owing to the protrusion of the nucleus through the cortinar
septum (loc. cit., p. 85, Taf. viii. figs. 3-8).
70. The Nucleus of the Cannopylea.—The nucleus presents the same remarkable
structures in all species of the Phæodaria or Cannopylea which have been examined, and closely resembles the germinal vesicle
of an amphibian ovum, being a large spherical or spheroidal vesicle with numerous nucleoli. Its
diameter usually amounts to half or two-thirds, sometimes even three-quarters, that of the central
capsule. The vertical main axis of the latter is also that of the nucleus, which usually lies
somewhat nearer to the aboral pole. The nucleus is generally rather more strongly compressed in
the direction of the main axis than the capsule itself. The membrane of the vesicular nucleus is
thin, but firm, and encloses a clear or finely granular mass of nuclein. The number and size of
the contained nucleoli are variable even in one and the same species, and stand in inverse ratio
to each other, an obvious result of the gradual process of division. Commonly {xl}from twenty to fifty roundish or spherical, strongly
refracting nucleoli, are present; more rarely there are several hundred very small ones.
Sometimes the nucleus is penetrated by fine trabeculæ, in whose meshes lie the nucleoli (Pl. 101, fig. 2).
In certain nuclei, which contained a few large nucleoli, these were of irregular form, probably
the result of amœboid movements (Pl. 101,
fig. 1). In the formation of spores in the Cannopylea, the nucleus
apparently becomes dissolved, and its numerous nucleoli develop directly into the nuclei or
mother-nuclei, which produce the nuclei of the flagellate spores. Furthermore, many Phæodaria seem to multiply by simple cell-division, since very commonly
(especially in the Phæocystina and Phæoconchia)
two large nuclei (right and left), may be met with in one central capsule; sometimes also a single
large nucleus, in which a sagittal constriction marks the commencing division of the capsule (Pl.
101, figs.
2, 36; Pl. 104,
fig. 3; Pl. 124,
fig. 6, &c.).
The large nucleus of the Phæodaria was first described in
my Monograph in 1862, in the case of Aulacantha (p. 263), Aulosphæra (p. 359), and
Cœlodendrum (p. 361), as a "large, spherical, thin-walled endocyst," from 0.1 to
0.2 mm. in diameter. More detailed descriptions, especially with respect to the behaviour of the
nucleoli were given by R. Hertwig in 1879 (L. N. 33, p. 97).
71. The Endoplasm or Intracapsular Protoplasm.—In all Radiolaria the
intracapsular protoplasm, which, for the sake of brevity, may be termed "endoplasm," constitutes
originally, and especially in the earliest stages, the only important content of the central
capsule, except the nucleus. In certain Spumellaria and Nassellaria, of simple structure and of small dimensions, this condition
persists for a long period, and the endoplasm then appears as a homogeneous, colourless, turbid or
finely granular, mucous, semi-solid mass, which cannot be distinguished from the ordinary
undifferentiated protoplasm of young cells; no definite structure, and in particular, no fibrillar
network, can be discovered in it even by the use of the customary reagents. In the great majority
of the Radiolaria, however, this primitive homogeneous condition of the endoplasm is very
transient, and it soon undergoes definite modifications, becoming differentiated into separate
parts or producing new constituent contents. Such products of the internal protoplasm are in
particular hyaline spheres (vacuoles and alveoles), oil-globules, pigment-bodies, crystals,
&c. The most important of the differentiations which take place in the endoplasm is that into
an internal, granular, medullary substance and an external, fibrillar, cortical
substance; although the various legions behave somewhat differently in this respect (§§ 77-80).
72. Intracapsular Hyaline Spheres.—The central capsule of very many Radiolaria
contains in its endoplasm numerous spherical bodies of varying size, which consist of watery or
albuminous fluid, and have previously been regarded as nuclei, or described as products of the
internal protoplasm, under various names, such as "spherical transparent {xli}vesicles" (see note A, below), "albumen spheres" (see B), "gelatinous
spheres" (see C), "alveolar cells" (see D), &c. Some of these spheres are perfectly
transparent, structureless and of varying refractive power, producing the impression of drops of
fluid; others contain various formed constituents, such as oil-globules, fat-granules,
pigment-granules, concretions, crystals, &c. From a morphological point of view they may all
be divided into two categories, membraneless vacuoles and vesicular alveoles. The vacuoles
are simple spherical drops of fluid or of gelatinous material, devoid of a special envelope, but
immediately surrounded by the endoplasm. The alveoles, on the other hand, are true vesicles
with a thin spherical envelope, enclosing a drop of fluid or jelly. This envelope is commonly very
thin, homogeneous, and often scarcely discernible, so that in practice a sharp line of demarcation
cannot be drawn between alveoles and vacuoles; the former are usually somewhat larger than the
latter. The fact is, nevertheless, certain that the hyaline spheres, which may be isolated on
rupturing the central capsule of many Radiolaria, in certain cases, particularly in large species,
possess a clear, anatomically demonstrable membrane, whilst in others no such appearance is
presented. It may be assumed that the vesicular alveoles are developed from the drop-like
vacuoles by increase in size, and by the precipitation of a delicate envelope from the endoplasm.
The character common to all these hyaline spheres, whether vacuoles or alveoles, is found in their
aqueous, not adipose, constitution, and in their clear transparent appearance, which allows of no
structure (the above-mentioned contained bodies excepted) being recognised. Their refractive power
and consistency vary somewhat, and probably their chemical constitution still more. Sometimes they
are strongly refractive and shining, and sometimes feebly refractive and pale; their consistency
shows all intermediate stages between a thin fluid, which readily disappears in water, and a firm,
insoluble jelly. As regards their chemical composition (which is probably very variable), the
hyaline spheres may be best divided into two groups, the organic and inorganic. The inorganic
hyaline spheres are simple drops of saline solution without any carbonaceous constituent; the
organic, on the other hand, contain a small quantity of organic matter dissolved in the
watery fluid, and may be either albuminous or gelatinous spheres. The formed contents which are
commonly present are of very various natures, usually small fat-granules, more rarely larger
fat-granules or pigment-granules, sometimes concretions or crystals. In many groups, especially
among the large Phæodaria and Collodaria, the
numerous hyaline spheres are remarkable for their equal size and even distribution throughout the
endoplasm (Pl. 1,
figs. 1, 4; Pl. 104,
fig. 2, &c.). In some genera belonging to the Thalassicollida the alveoles are of enormous
size (Pl. 1,
figs. 2, 3); they then become flattened by mutual pressure into polyhedra and distend the central
capsule to unusual dimensions (in Physematium and Thalassolampe 8 to 12 mm.).
A. The "spherical hyaline vesicles," which I described in my Monograph (1862, p. 71)
as among the most important and constant contents of the central capsule, are partly vacuoles,
{xlii}partly homogeneous nuclei. Most recent
investigators, Bütschli in particular (1882, L. N. 41), have pointed out and
rightly criticised this confusion. The criticism might, however, have been more justly expressed
by stating that, in the preparation of my Monograph (1859-1862), I did not make use of modern
methods of demonstrating the nucleus by staining fluids, which were quite unknown at the time,
and only discovered a decade later. In fact, without the aid of such reagents, it is quite
impossible to distinguish between the various "spherical transparent vesicles," of which those
found in the central capsule of the Phæodaria and many monozootic Collodaria are simple vacuoles lying in the endoplasm, whilst, on the other
hand, those of the Polycyttaria and many other Radiolaria are true homogeneous nuclei. For not
only are the general appearance of the small clear spheres, their refractive power, and regular
distribution in the endoplasm quite similar, but they are also of much the same size, for the
diameter ranges from 0.005 to 0.015 mm., being generally between 0.008 and 0.012 mm. In addition
to this there is generally in each hyaline sphere a dark brightly shining granule, which, in the
case of the vacuole, is simply a fat-granule, whilst in the case of the nucleus, it is a true
nucleolus. The small hyaline spheres in the young uninucleate capsules of the Polycyttaria are
simple vacuoles (Pl. 3,
fig. 12), whilst in the ripe multinucleate capsules they are true nuclei (Pl. 3, figs. 3, 8,
9), and it is quite impossible to discriminate between these two conditions without the use of
reagents. This has been expressly recognised by R. Hertwig, who has the merit of having been the
first to clearly distinguish, by the aid of staining fluids, between these two different
constituents (1879, L. N. 33, p. 108).
B. The "albumen spheres," which were first observed by A. Schneider in 1858 in the
common cosmopolitan Thalassicolla nucleata (L. N. 13, p. 40), and
which appear to occur in only a few other Thalassicollida, are distinguished from the ordinary
hyaline spheres of about the same size by their higher refractive power and by certain
albuminoid reactions, especially the coagulation of a membranous envelope under the influence of
certain reagents (see my Monograph, p. 250, and Hertwig, L. N. 26, 1876, p.
46). They often enclose various formed contents, and require further investigation.
C. The gelatinous spheres of various sizes, found in the endoplasm of the Radiolaria,
agree in their reactions (especially in staining by certain reagents) with the common
extracapsular jelly of the calymma, and are hence distinguishable both from the true
(coagulable) "albumen sphere," and from the ordinary watery vacuoles.
D. The alveoles, which are only accurately known in the case of certain
large monozootic Collodaria, but which also seem to occur in the
central capsule of other remarkably large Radiolaria, were described in my Monograph in the case
of Thalassolampe margarodes and Physematium mülleri, under the name "intracapsular
alveolar cells" (1862, pp. 77, 254, 257). They are not, however, true nucleated cells, and the
body described as a nucleus is not such in reality. Nevertheless these large hyaline spheres do
possess a special envelope, as I have recently convinced myself by the examination of ruptured
central capsules of Thalassolampe maxima, Thalassopila cladococcus, and
Physematium atlanticum (Pl. 1,
figs. 2, 3). The central capsule of these Collodaria becomes distended
to most unusual dimensions (2 to 12 mm. in diameter) by the great development of these large
hyaline vesicles, each of which measure from 0.1 to 0.5 mm. in diameter.
73. The Intracapsular Fat-Globules.—Fat is present in the central
capsule of all Radiolaria in larger or smaller quantities, and generally appears in the form of
very {xliii}numerous, small, spherical granules, which
are either distributed evenly in the endoplasm (as an emulsion) or enclosed in the vacuoles; the
latter, in particular, is the case in most Phæodaria, perhaps generally.
In this group each vacuole contains as a rule a single dark, shining fat-granule, and sometimes
also an irregular bunch composed of from two to five or more granules. In addition to these small
fat-granules (granula adiposa) which are always present, the central capsule of many
Radiolaria contains also larger fat-globules (globuli adiposi). These appear to be
generally wanting in the Phæodaria, and are on the whole rare in the Acantharia; whilst, on the contrary, they are very common in the Nassellaria and Spumellaria. The Polycyttaria or social
Radiolaria are as a rule distinguished by the possession of a single large central oil-globule,
which lies in the centre of the central capsule, and is on an average about one-third of it in
diameter (Pl. 3,
figs. 4, 5). This is absent, however, in those young capsules of the Polycyttaria in which the
primary nucleus is centrally situated (Pl. 3,
fig. 12). Those species of Polycyttaria whose central capsule reaches a considerable size, often
enclose numerous oil-globules, and in Collophidium (species of Collozoum with an
elongated cylindrical capsule, Pl. 3,
figs. 1, 3) the axis of each capsule is occupied by a row of numerous oil-globules. In the
monozootic Spumellaria, in which the nucleus is always centrally situated,
the large oil-globules are, of course, excentric, being in apposition to the inner surface of the
capsule-membrane (Pl. 1,
fig. 3; Pl. 2,
figs. 2, 5). In the Discoidea the oil-globules, which are often present
in large numbers, form elegant concentric rings around the central nucleus, and in those species
with segmented arms, there are one or more transverse rows in each segment (Pl. 43, fig. 15).
In the Nassellaria the number and distribution of the oil-globules are
dependent upon the form of the central capsule. When this is simple, without lobes, and ovoid or
conical, they generally lie in its aboral half above the podoconus (Pl. 51, figs. 5,
13; Pl. 97,
fig. 1). When, on the contrary, the basal portion of the capsule sends out three or four dependent
processes (as in the majority of the Cyrtoidea), a large globule may
generally be seen in the swollen distal part of each conical or ovoid lobe (Pl. 53, fig. 19;
Pl. 60,
figs. 4-7). In many Stephoidea and Spyroidea,
whose central capsule is separated into two lateral portions by the constriction corresponding to
the sagittal ring, each of these contains either a single large globule or a group of small ones
(Pl. 90,
figs. 7, 10). These oil-globules are usually colourless and highly refractive; rarely they are
yellow or brown, sometimes rose-coloured, or an intense blood-red (e.g., in
Thalassophysa sanguinolenta) or even orange (in Physematium mülleri). In many Spumellaria, and particularly in the Polycyttaria, an albuminous substratum may
be recognised in them, which is sometimes disposed in layers, and after extraction of the fat
presents the appearance of a laminated sphere. The physiological significance of the oil-globules
is twofold; in the first place they tend to diminish the specific gravity of the organism; in the
second they may be utilised as a reserve store {xliv}of
nutriment. In the latter respect they are of special importance in the process of spore-formation,
each flagellate spore usually containing a fat-granule.
74. The Intracapsular Pigment-Bodies.—In the majority of Radiolaria when observed
alive, the central capsule is coloured, only in the minority is it colourless. The colour is
never diffuse, but always due to the formation of definite pigment granules or vesicles, which are
sometimes distributed evenly throughout the endoplasm, sometimes aggregated in the central or
peripheral regions. Their form may be either spherical, irregularly rounded, or polyhedral. They
vary much in dimensions, but in most cases are immeasurably small, and appear under a high
magnifying power as fine dust; occasionally, however, their diameter may amount to from 0.001 to
0.005 or more. The chemical constitution of the intracapsular pigment is unknown in most
Radiolaria, and is probably very various. In many instances the pigment-granules consist of fat,
in others not. The commonest colours are yellow, red, and brown; violet and blue are rare, and
green still rarer. Sometimes a definite tone of colour prevails throughout a whole group, and may
then be attributed to inheritance, e.g., red is found in most Sphæroidea, and blue in the Polycyttaria (see note A). One colour is almost
always constant in the members of the same species. True pigment-cells, belonging to the
Radiolarian organism, do not occur within the central capsule. The peculiar yellow cells which are
found in the central capsule of many Acantharia are symbiotic xanthellæ
(see § 76).
A. The number of Radiolaria whose pigment has been examined in the living state,
is too small to allow of any general conclusions being drawn. Regarding the different colours
known, see my Monograph, L. N. 16, p. 76.
75. The Intracapsular Crystals.—The crystals found in the central
capsule of many Radiolaria may be divided into two groups, of very different significance; small
crystals, which are very widely distributed, and large crystals, which occur in only a few genera.
The small crystals may also be termed "spore-crystals," since each swarm-spore often
contains such a crystal. They are rod-like or spindle-shaped, and consist of an organic substance
which probably serves as a reserve of nutriment for the developing spores. Such spore-crystals
have been observed in numerous Spumellaria and Acantharia belonging to various families, and are probably present throughout
the two legions which make up the Porulosa. On the other hand, they have not been noticed in the
Osculosa (Nassellaria and Phæodaria), the few
swarm-spores belonging to these groups which have been observed not exhibiting any crystals. The
large crystals, which occur in small numbers in the endoplasm, have hitherto only been
observed in a few species of Spumellaria, belonging to the Polycyttaria.
They were first noticed in the common Collosphæra huxleyi, and regarded as cœlestin.
They are also found in the central capsule of many other Collosphærida, e.g.,
Buccinosphæra (Pl. 5,
figs. 11, 12). Crystal-masses, crystal-sheaves, or spherical masses of radiating acicular crystals
are enclosed in {xlv}the vacuoles or "albumen globules"
of Thalassicola nucleata and other Thalassicollida, as well as in the central capsule of
Cœlographis and some other Phæodaria (Pl. 127, figs.
4-7). All these large crystals are probably to be regarded as excretory products.
75A. The Intracapsular Concrements.—Concretions,
either mineral or organic, of varying form and constitution, are to be found in the endoplasm of
Radiolaria belonging to very different families. They are most abundant and multiform in
Thalassicolla nucleata, being usually circular or elliptical discs, which are
concentrically laminated and highly refractive, resembling starch-grains. Among them twin forms
may frequently be observed, as though the concrements were in process of division (see note A).
Similar amyloid concretions are to be seen in the central capsule of different Spumellaria and Nassellaria, e.g., in
Cephalospyris triangulata (Pl. 96,
fig. 28). Violin-shaped, highly refractive concrements have been observed in the central capsule
of numerous Spumellaria, Nassellaria, and Acantharia, e.g., Thalassosphæra, Spongosphæra,
Plegmosphæra, Cyrtocalpis, Peripyramis, Botryocella, &c. (see note
B). The chemical constitution of these concrements is insufficiently known.
A. The amyloid concretions of Thalassicolla nucleata have been described in detail in
my Monograph (pp. 80, 250, Taf. iii. figs. 2, 3), and by R. Hertwig in the Histologie der
Radiolarien (1876, p. 47, Taf. iii. figs. 9-13).
B. The violin-shaped concretions of Thalassosphæra bifurca have been
figured in my Monograph (pp. 80, 261, Taf. xii. fig. 1).
76. The Intracapsular Xanthellæ.—The xanthellæ, zooxanthellæ, or symbiotic "yellow
cells" are found within the central capsule only in the Acantharia, whilst
in other Radiolaria they only occur in the extracapsulum. They are most frequent in the Acanthometra, rarer in the Acanthophracta, but even in
the former they are often wanting. Their number is very variable, but usually small, from ten to
thirty in one capsule. They lie for the most part immediately below the capsule membrane, in the
cortical layer of the endoplasm. The form of the yellow cells is either spherical or ellipsoidal,
often also spheroidal or even lentiform. The diameter varies from 0.01 to 0.03 mm. They possess a
distinct membrane and an excentric nucleus, and contain numerous yellow pigment-granules in the
endoplasm. This yellow pigment dissolves in mineral acids to form a green fluid, and in other
respects also behaves somewhat differently from the yellow pigment in the extracapsular yellow
cells of the Spumellaria and Nassellaria. In both
cases, however, the xanthellæ are not integral portions of the organism, but unicellular algae,
living as parasites or symbiontes in the body.
A. The yellow cells in the central capsule of the Acantharia were first observed by Joh. Müller (L. N. 12,
pp. 14, 47). In my Monograph I described them at greater length, and indicated their differences
from the extracapsular yellow cells of other Radiolaria (L. N. 16, pp. 77,
86). Since then, R. Hertwig has demonstrated their cellular nature (L. N. 33, pp. 12, 113), and still more recently {xlvi}Brandt has given further accurate information regarding their
occurrence, constitution, and physiological significance (L. N. 39, ii.
Art., p. 235, figs. 62-73).
77. The Endoplasm of the Peripylea.—The intracapsular protoplasm of the Spumellaria or Peripylea is usually distinguished by a
more or less complete radial arrangement, which does not occur in the same form in other
Radiolaria; it may be regarded as characteristic of this legion, for it probably occurs in all the
species at some period of life or other, and stands in a direct causal relationship with the
typical structure of the capsule-membrane in all the "Peripylea" (see note
A). For as this is commonly perforated by very numerous pores distributed at equal intervals over
the whole surface of the capsule, and since a communication between the intra- and extracapsular
sarcode takes place through these, the radiate structure of the endoplasm may be readily explained
as due to the influence of radial currents which take place continuously or intermittently in the
endoplasm. This radiate structure is most obvious when the endoplasm contains no secondary
products or only an insignificant amount of these, and thus appears colourless and almost
homogeneous, or only finely granular. Under these circumstances, an optical section of the central
capsule usually reveals a distinct radial striation; numerous narrow, straight, dark streaks
alternating regularly with still narrower clear ones; the latter consist of homogeneous, the
former of more or less granular protoplasm (Pl. 20, fig.
1a). Often there may be distinguished in each darker streak a single straight row of
strongly refracting (fat?) granules, sometimes several such rows. Occasionally the whole endoplasm
becomes divided up into a number of large "radial wedges," club-shaped, conical or pyramidal
masses of granular protoplasm, separated by clear divisions of hyaline plasma (e.g., in
Actissa radiata, p. 14, where in the optical section of the central
capsule, between the membrane and the nucleus, twenty-five dark radial wedges of equal size were
separated by thick clear partitions of hyaline protoplasm). In the majority of the Spumellaria this radial striation is partially or entirely concealed by the
formation of pigment or of other products. Very often it is only visible in the cortical layer,
which lies immediately below the capsule-membrane (Pl. 1, figs. 1, 3).
The remarkable "centripetal cones" which characterise the Thalassicollid genus Physematium,
and were formerly described as "centripetal cell-groups," are probably a special development of
these cortical radial wedges; they are conical cortical bodies, regularly distributed on the inner
surface of the membrane of the central capsule, and disposed with the apex turned towards the
centre (see note B). More rarely than in the cortical layer, a similar radial structure is to be
found in the innermost medullary layer immediately surrounding the nucleus. Here the endoplasm
sometimes breaks up into fine radial threads, which are anatomically separable and hang down from
the free nucleus as thin processes (see note C). In some cases it is also possible to isolate
radial rods from the cortical layer of teased out central capsules.
{xlvii}
A. The radial structure of the endoplasm was first described in my Monograph (1862, p. 74),
though R. Hertwig (1879, p. 112) was the first to indicate its typical significance in the case
of the Peripylea, and to demonstrate its causal relation with the radial
currents in the central capsule of this legion. More recent investigations have led me to the
conviction that this phenomenon is more widespread, and often more strongly developed, than was
formerly imagined, and that it is probably one of the typical characters of all Spumellaria (at least of the Monozoa).
B. The centripetal cones of Physematium, which have hitherto been known only in these
colossal Thalassosphærida, were fully described in my Monograph under the name "conical
centripetal cell-groups"; by their first discoverer, A. Schneider (L. N. 13), they were termed "nests," and compared with the "nests" (central capsules)
of the Polycyttaria. In the Physematium mülleri of the Mediterranean (hitherto only observed by
Schneider and myself at Messina) it appeared as though each centripetal cone were composed of a
group of from three to nine (usually four or five) slender wedge-shaped cells, whose common
centripetal apex was produced into a radial thread of sarcode (L. N. 16, p.
258, Taf. iii. fig. 7). Since then (1866) I have observed at Lanzerote, in the Canary Islands, a
nearly related form, which I take to be Physematium atlanticum, Meyen. In this, however,
the "centripetal cell-groups" were wanting, and the whole cortical layer of the endoplasm was
cleft into numerous radial portions, each enclosing a nucleus (probably the mother-cells of
flagellate spores, see p. 35).
C. The radial fibres of the medullary endoplasm which cling to an extracted
nucleus have been observed by Hertwig in certain Sphæroidea
(Diplosphæra, Arachnosphæra) (L. N. 33, p. 40).
78. The Endoplasm of the Actipylea.—The intracapsular protoplasm of the Acantharia or Actipylea is often distinguished by a
partial or complete radial arrangement like that of the Peripylea, but
differing in the number, size, form, and distribution of the radial portions into which the
endoplasm is differentiated. For since the pores of the capsule membrane are distributed at equal
distances all over the surface in the Spumellaria, whilst in the Acantharia they are arranged in definite groups, and since the number and
arrangement of the pores has a direct influence upon the internal currents of the endoplasm, it
follows that the radial structure in the latter legion must be very different from that in the
former. In addition to this there must not be forgotten the important influence which the early
centrogenous formation of the skeletal rods exercises upon the disposition and growth of the
intracapsular structures. Hence the endoplasm of the Acantharia does not
separate into innumerable thin, closely packed radial wedges or cortical radial rods, but into a
small number of large pyramidal portions between which run the radially disposed heterogeneous
portions of the contents of the capsule, viz., the radial bars of acanthin and the peculiar
intracapsular "axial threads." As a direct consequence of the regular disposition of these
heterogeneous radial portions, which is often characteristic of the various families of the Acantharia, a corresponding differentiation of the endoplasm is brought about;
it divides into a number of conical or pyramidal portions (radial pyramids), whose bases rest upon
the capsule-membrane and whose apices are directed towards the centre of {xlviii}the capsule (the central star of the skeleton). These radial
pyramids are, however, but rarely visible, being usually more or less concealed by a dark
pigment.
The differentiations of the endoplasm in the central capsule of the Actipylea have been but little investigated, but they appear to vary somewhat
in the different groups of this legion. In all Acantharia in which the
twenty radial bars are regularly arranged according to the Müllerian law (see p. 717) and in which axial threads constant in number and disposition run
between them from the central star to the capsule-membrane, it obviously follows that the
endoplasm must be divided into more or less distinct radial pyramids, and this must the case
whether these take the form of continuous tracts or of actually separable portions. The regular
polygonal figures, often seen on the surface of the central capsule (with special distinctness
in Acanthometron elasticum and Acanthometron pellucidum) separated by a network of
granular threads, are the bases of such radial pyramids (see Hertwig, L. N. 43, p. 12, Taf. i. figs. 1-7).
79. The Endoplasm of the Monopylea.—The intracapsular protoplasm of the Nassellaria or Monopylea is distinguished from that of
any of the other three legions by the development of a quite peculiar fibrillar structure, the
axial "pseudopodial cone," which may shortly be termed the "podoconus" (foot-cone). Since this is
in direct correlation with the peculiar structure of the capsular opening, the large "porochora,"
which is situated at the basal pole of the main axis, it is quite as characteristic of the legion
as the latter itself (see note A). The podoconus is primitively a vertical regular cone whose
circular base occupies the horizontal porochora or "basal porous area" of the central capsule,
while its vertical axis coincides with that of the latter. The apex of the cone, usually somewhat
rounded off, is therefore directed towards the aboral or apical pole of the central capsule and
separated from it by a larger or smaller interval. In this interval the nucleus originally lies
(as in Pl. 51,
fig. 13; Pl. 98,
fig. 13); but it is usually displaced subsequently and lies excentrically. The cone is of very
variable height; on an average its vertical height is about equal to the diameter of its
horizontal base; these dimensions are, however, dependent upon the form of the central capsule;
the height being greater in slender ovoid or conical capsules, and less in depressed sphæroidal or
discoidal ones, than the diameter of the base. The podoconus consists of differentiated endoplasm,
which becomes more deeply stained by carmine and offers greater resistance to solvents than the
surrounding finely granular protoplasm. The apex, especially, becomes very intensely stained. It
always exhibits a very characteristic fine but distinct striation, numerous straight radial lines
diverging from the apex of the cone towards the base. The number of these striæ appears to
correspond with that of the vertical rods in the porochora, and each of these latter stands
apparently in direct communication with the basal end of an apical stria (§ 59). These threads are probably differentiated constant contractile threads of
endoplasm, or even myophanes, comparable with the contractile cortical threads of the Cannopylea and the permanent axial threads of the Actipylea. The numerous modifications, {xlix}undergone by the form and contents of the central capsule in the
different groups of Monopylea, especially those due to the formation of
the skeleton, are not without influence upon the podoconus. The most important divergencies from
the above described primary form are the following:—(1) The vertical axial cone becomes
oblique, its axis inclining in the sagittal plane and approaching either the dorsal or the ventral
wall of the capsule; the cause of this appears to be usually the excentric development of the
growing nucleus or the formation of a large oil-globule. (2) The smooth mantle of the podoconus
becomes divided by three longitudinal furrows into three equal prominent ridges, which correspond
to three circular lobes in the porochora; the cause of this basal triradial lobular formation lies
probably in the triradial development of the skeleton in many Nassellaria
or in the cortinar structure of the collar septum. (3) The simple podoconus splits into three or
four elongated lobes, which eventually become almost completely separated and correspond to the
lobes of the central capsule, in the axial wall of which they lie as longitudinally striated
bands. The behaviour of these bands justifies the hypothesis that the podoconus is a muscular
differentiated portion of the endoplasm and is composed of myophane fibrillæ, whose contraction
determines the opening of the central capsule.
A. The podoconus of the Monopylea was first described by
R. Hertwig in 1879, and recognised as a characteristic component of the central capsule in the
most various groups of this legion (in Plectoidea, Stephoidea, Spyroidea, and Cyrtoidea; see his figures, loc. cit., Taf. vii., viii., and the
description, pp. 71, 73, 83, 106). Hertwig called it the "pseudopodial cone," and regarded it as
a conical process of the capsule-membrane, which is developed from this latter and projects from
the porous area into the interior of the central capsule; "it is penetrated by fine canals which
arise at the apex of the cone, diverge towards the base, and terminate there in the rods of the
pseudopodial area. The intracapsular protoplasm penetrates at the apex of the pseudopodial cone
into its fine canals, runs along them and emerges from the rods of the porous area in the form
of slender threads" (loc. cit., p. 19). I cannot agree with this view of Hertwig,
although I have been able to confirm the accuracy of his description by my own observations upon
numerous excellently stained and preserved preparations in the Challenger collection. As I have
proved by numerous teased out preparations, and as Hertwig himself correctly states, "the cone
is more readily detached from the membrane than from the protoplasm, when the capsule is teased"
(loc. cit., p. 73). Hence I regard the podoconus not as a differentiated portion of the
capsule-membrane but as endoplasm, and believe that it is composed of myophanes or "contractile
muscular fibrils" in the same manner as the cortical layer of the Cannopylea. Probably the contraction of these fibrils serves to raise the
opercular rods and hence to allow the exit of the endoplasm through the pores which lie between
these opercular rhabdillae (compare § 59).
80. The Endoplasm of the Cannopylea.—The intracapsular protoplasm of the Phæodaria or Cannopylea is distinguished from that of
the other three legions by several characteristic peculiarities, which are very important, since
they stand in causal relation to the typical structure of the capsule-membrane and in particular
of its {l}remarkable aperture. In the case of many and
perhaps of all Phæodaria the endoplasm is differentiated into a granular
medullary and a thin fibrillar cortical layer, the former of which usually encloses numerous small
vacuoles, while the latter contains muscular fibrillæ. In the voluminous central capsule of large
Phæodaria the whole cortical layer of the endoplasm, which lies
immediately below the delicate inner capsule-membrane, sometimes appears delicately and regularly
striated, and most distinctly so under the apertures, towards the centre of each of which the dark
striæ are radially directed (see note A, below). These striæ are probably contractile muscular
fibrillæ; or "myophanes," by whose contraction the openings are voluntarily widened. In the
Tripylea this fibrillar star is much more strongly developed under the astropyle (the main
opening) than under the parapylæ (or accessory openings); and probably the peculiar radial
structure of the operculum of the former is due to the stronger development of these radial
fibrils (being their impression). In many Phæodaria, indeed, the fine
myophane fibrils are only visible under the apertures, whilst in others they form a continuous
fibrillar cortical layer on the whole inner surface of the inner capsule-membrane; the fine
fibrillæ run meridionally from one pole of the main axis to the other; perhaps the whole central
capsule may change its form in consequence of their contractions. The medullary portion of the
endoplasm, which lies below this thin cortical layer, is usually finely granular in the Phæodaria, and permeated by numerous spherical vacuoles, which are noteworthy
from their equal size and regular distribution. Each clear vacuole usually contains a dark shining
fat-granule, more rarely a group of such granules (see note B). Compare § 60, and Pl. 101,
figs. 1-3; Pl. 104,
figs. 1, 2; Pl. 111,
fig. 2; Pl. 128,
fig. 2, &c.
A. The fine fibrillæ in the cortical layer of the endoplasm were first described by Hertwig
in 1879 (L. N. 33, p. 98, Taf x. figs. 6-10). He found them, however, only
below the three openings in the capsule of the Tripylea, where they form three stellate groups
of fibrils. I find them very clearly shown, and with especial distinctness, under the astropyle
in most Phæodaria of which I have had the opportunity of examining
well-stained and preserved central capsules. In many cases, also, the striation is not confined
to the apertures, but spreads over the whole cortical layer. Perhaps this constitutes in all
Phæodaria a thin myophane-sheet, whose contractile fibrils run from one
pole of the main axis to the other and cause by their contraction changes in the form of the
spheroidal central capsule.
B. The granular medullary portion of the endoplasm of the Phæodaria, with its numerous clear spherical vacuoles, was first described in
my Monograph (1862), in the case of Aulacantha (p. 263), Aulosphæra (p. 359), and
Cœlodendrum (p. 361) as a "finely granular, mucous substance (intracapsular
sarcode), packed more or less closely with clear spherical vesicles from 0.005 to 0.015 mm. in
diameter, each of which contains one or two, rarely three, dark shining granules." That these
clear spheres are true vacuoles was first clearly proved by Hertwig (L. N. 33, p. 98). As a rule all the vacuoles of the same central capsule are of equal
size (generally from 0.008 to 0.012 mm. in diameter), and are distributed at equal intervals
throughout the finely granular endoplasm.
{li}
Chapter III.—THE EXTRACAPSULUM.
(§§ 81-100).
81. The Components of the Extracapsulum.—The extracapsulum or
extracapsular malacoma, under which name are included all those parts of the soft body which lie
outside the central capsule, consists of the following constant, and important
constituents:—(1) The calymma or extracapsular jelly-veil; (2) the sarcomatrix
or layer of exoplasm immediately surrounding the membrane of the central capsule; (3) the
sarcodictyum or network of exoplasm, covering the surface of the calymma; (4) the
pseudopodia or radial fibres of exoplasm, which may again be subdivided into intracalymmar
pseudopodia, uniting the sarcomatrix and sarcodictyum, and extracalymmar pseudopodia, radiating
freely into the water outside the calymma.
82. The Calymma.—The calymma or extracapsular jelly-veil of the Radiolaria is
always the most voluminous portion of the extracapsulum, and in spite of its simple structureless
constitution is of great morphological and physiological importance. In all Radiolaria this
gelatinous mantle completely surrounds the central capsule, but is separated from its outer
surface by a continuous, though thin, layer of exoplasm, the sarcomatrix. The pseudopodia
radiating from the latter pierce the calymma, form the sarcodictyum at its surface, and radiate
from its nodal points freely into the surrounding water. The calymma is rarely visible in living
freshly captured Radiolaria, examined in sea-water, for its gelatinous substance is perfectly
hyaline, colourless and pellucid, and possesses the same refractive index as sea-water; but when
the object is removed from this fluid and transferred to carmine solution or some other colouring
matter, the extent and figure of the calymma become apparent, for the staining fluid does not at
first penetrate into the gelatinous material. When this has taken place, however (after a longer
or shorter time), and the gelatinous material has become coloured, its form and size may be
observed by the converse experiment; the object is transferred once more to water and the outlines
of the calymma become as clear as those of the central capsule. The same is the case with dead
specimens in which the sticky surface of the calymma has become covered with dust.
The jelly-veil of the Radiolaria was recognised even by the earliest observers of
the group, Meyen (1834), and Huxley (1851), and compared with that of the Palmellaria; the
former noticed it in Physematium and Sphærozoum (L. N. 1, p.
283), and the latter in Thalassicolla and Collosphæra (L. N. 5,
p. 433). In all these Spumellaria, both in the monozootic
Thalassicolla and in the polyzootic Sphærozoum and Collosphæra, the calymma
is very voluminous and filled with large alveoli. Meyen called them "muco-gelatinous masses, in
the interior of which are contained small equal-sized vesicles"; Huxley likewise found clear
vesicles in the jelly and compared them with Dujardin's vacuoles. Johannes Müller observed the
jelly-veil in many different Radiolaria, in particular in the Acanthometra, first discovered by him, but erroneously believed that it only
originated {lii}after death by liquefaction of the
sarcode (L. N. 12, p. 6). This mistake is, however, easy to understand,
since in living Radiolaria the calymma is usually invisible on account of its perfect
transparency, whilst in dead specimens it is usually quite distinct on account of the dust
clinging to its adhesive surface. I myself believed that the formation of the voluminous hyaline
jelly-veil was only partially due to liquefaction after death, but that it was to some extent
present in the living organism and that it might vanish and subsequently reappear by means of
imbibition (L. N. 16, pp. 109, 110). R. Hertwig was the first to
demonstrate, in 1879, that the jelly-veil is constantly present in living Radiolaria, that it
forms the basis of the extracapsular malacoma and surrounds the central capsule as a second
protective sheath (L. N. 33, p. 114).
83. The Structure of the Calymma.—The extracapsular jelly-veil appears
structureless in most Radiolaria, inasmuch as it represents a homogeneous pellucid excretion of
the exoplasm and contains neither fibres nor other formed structures. In some groups, however,
definite structural characters become secondarily developed. The most common and striking of these
is the formation of alveoles, which takes place in the extracapsulum (see § 86). In consequence of this the calymma assumes a remarkable frothy consistency
and appears to be composed of large, clear, thin-walled vesicles; this is especially the case in
the Collodaria (Colloidea, Pls. 1, 3, and Beloidea, Pls. 2,
4), and in
many large Phæodaria, especially among the Phæocystina (Phæodinida and Cannorrhaphida, Pl. 101, and
Aulacanthida, Pls. 102-104). More
rarely the calymma is not permeated by vacuoles, but there appear in it fine striæ parallel to the
surface as though it were composed of thin concentric laminæ like an onion; perhaps these are the
expressions of a different quantity of water in the various layers. In the calymma of many
Radiolaria thin, straight, radial lines are to be seen, which are probably pseudopodia, and not to
be attributed to any structural modification, or they may be slender canals which serve for the
exit of the pseudopodia. On the outer surface of the calymma of different Radiolaria, and
especially in the Acantharia, a peculiar network of fibres is to be found,
composed of polygonal meshes, like elastic fibres, probably due to a local thickening of the
jelly. These polygonal meshes are often very regularly distributed between the radial spines of
the Acanthometra, and stand in a definite relation to them. The fibres
which form the meshes are often rather strong, resembling elastic fibres, as above-mentioned, and
either simple or composed of bundles of very fine fibrillæ (L. N. 33, p. 15,
Taf. i. fig. 1, Taf. ii. fig. 4).
84. The Consistency of the Calymma.—The gelatinous material of which
the calymma of the Radiolaria consists is a pellucid mass, rich in water and usually quite hyaline
and structureless; its consistency is very variable. In the majority of the Radiolaria it may
perhaps be about equal to that of the jelly which composes the umbrella of most Medusæ; but as in
these latter it may vary between very wide extremes, constituting on the one hand a very soft
jelly-mantle, offering but little {liii}resistance to
mechanical influences and almost disintegrating under the eyes of the observer, and on the other
hand forming a firm gelatinous shell, comparable to cartilage in hardness, elasticity, and power
of mechanical resistance. In many Radiolaria of large dimensions with an alveolar calymma
(especially in numerous Collodaria and Phæodaria)
this may be split by means of dissecting needles and the central capsule extracted like the stone
from a cherry, and then it is easy to ascertain that the firmness and elasticity of this
jelly-veil are not less than those of a cherry. The different degrees of consistency in the
various Radiolaria may be dependent either upon the relative amount of water which they contain,
or upon qualitative or quantitative variations in the organic substance of which the jelly
consists. Great importance is to be attached to the considerable consistency of the calymma,
because it furnishes the indispensable groundwork for the deposition of many parts of the skeleton
and particularly of the lattice-shells.
85. The Primary and Secondary Calymma.—In most Radiolaria the external
form and volume of the calymma are different at different stages of growth, and this difference is
mainly dependent upon the development of the skeleton. Hence it is advisable to distinguish in
general the primary from the secondary calymma. The primary calymma is in the great
majority of Radiolaria a perfect sphere, in the middle of which lies the concentric central
capsule; on the surface of this gelatinous plate the primary spherical lattice-shell is secreted
in most Spumellaria and Acanthophracta, as well
as in those Phæodaria which possess a spherical shell; in the remaining
Phæodaria also and in the Nassellaria, where the
lattice-shell is not spherical but monaxon, it is secreted on the surface of the primary calymma.
This takes place at a definite time, very important in the development of the Radiolarian, which
for the sake of brevity we shall term the "lorication-period." Since the firm surface of
the primary calymma furnishes the necessary foundation for the deposition of the primary
lattice-shell, it is of the greatest mechanical significance in all shell-bearing Radiolaria. The
secondary calymma arises only after the lorication-period by further growth of the
primitive jelly-mantle and in the fully developed Radiolarian usually encloses wholly or partially
the external parts of the skeleton, in consequence of which it assumes the most various forms.
Very often the secondary calymma is polyhedral, being stretched between the radial spines of the
skeleton, the distal ends of the latter then forming the fixed points of the gelatinous
polyhedron.
86. The Extracapsular Vacuoles and Alveoles.—The calymma of the Radiolaria usually
appears completely homogeneous and hyaline without any structure; sometimes it encloses numerous
clear vesicles, vacuoles or alveoles, and then assumes a frothy appearance, the expression of a
more or less distinct alveolar structure. {liv}The clear
vesicles to which this is due are either spherical, or polyhedral from mutual pressure, and like
the similar ones in the central capsule may be divided into membraneless vacuoles and vesicular
alveoles. The vacuoles are simple drops of fluid, without a special envelope, and
immediately surrounded by the gelatinous substance of the calymma, in which they appear as simple
cavities. The alveoles on the contrary are true vesicles, with a thin envelope, which
encloses a drop of fluid or a globule of jelly; in the latter case its contents are different in
refracting power and amount of contained water from the substance of the surrounding calymma. A
sharp boundary between the membraneless vacuoles and the vesicular alveoles cannot be drawn in the
case of the extracapsular hyaline spheres any more than in the intracapsular; the envelope of the
alveoles is sometimes very distinct and even anatomically separable, whilst at other times it is
very thin and scarcely recognisable; it may occasionally arise and disappear within a very short
time (see note A). There is no doubt that in the calymma as in the central capsule the vesicular
alveoles are secondary products, which have arisen from the vacuoles by the secretion of an
enveloping membrane. This membrane is either a delicate sheath of exoplasm, or a firmer and more
resistant skin, distinct from the exoplasm, and probably an excretion from it (e.g., Pl. 4, figs. 2, 3).
In many cases the outer surface even of the vacuoles is covered by a network of pseudopodia, which
form a sarcoplegma similar to a fenestrated alveolar membrane. The colourless pellucid fluid in
the vacuoles and alveoles is usually simple sea-water, more rarely it contains a small quantity of
albumen ("albumen-spheres") or jelly ("gelatinous spheres"). The size of these spheres is very
variable. Quite small vacuoles may be found in the calymma of many Radiolaria. Large vacuoles, on
the other hand, producing the appearance of an alveolar structure, are confined to but few groups,
to a part of the Spumellaria (Colloidea, Beloidea, and a few Sphæroidea), and to the Phæocystina (Phæodaria with incomplete skeleton);
besides they occur only rarely in individual genera, e.g., Nassella among the
skeletonless Nassellaria. Since the volume of the calymma is much
increased by the development of vacuoles, and the power of mechanical resistance is at the same
time much increased, the fact is explained that the vacuoles occur mainly in Radiolaria which have
no skeleton or only an incomplete one (see note B). Among the monozootic Collodaria the alveolar structure is especially well developed in the following
genera; Thalassicolla (Pl. 1,
figs. 4, 5), Thalassophysa, Thalassoplancta, Lampoxanthium (Pl. 2, figs. 1, 2);
among the Phæodaria in most genera of the Phæodinida, Cannorrhaphida and
Aulacanthida (Pls. 101-104), and
probably also in other voluminous Phæodaria (e.g., Phæosphæria). The alveoles or vacuoles in the calymma of these large Radiolaria
lie usually in several layers, one above another, and increase in size from within outwards. The
Polycyttaria or social Radiolaria (the three families Collozoida, Sphærozoida and Collosphærida)
without exception have an alveolar structure, and the special form of {lv}their colonies or cœnobia is to a great extent determined by the
development, number, size and arrangement of the alveoles in their calymma (compare Pls. 3-8). In these
cases there is not unfrequently developed a large central alveole (see note C) whose thickened
wall encloses a globe of jelly and serves as the central support of the whole colony (Pl. 5, fig. 1).
Still more striking, however, is the arrangement of certain Polycyttaria, where each individual of
the colony (or each central capsule with its calymma) is enclosed in a large alveole, whose firm
wall often attains considerable thickness (Pl. 4,
figs. 2, 3). The whole colony then appears as an aggregate of numerous cells, each of which
possesses two envelopes, the inner central capsule and the outer alveolar membrane; between these
lies in the Collosphærida the siliceous lattice-shell (Pl. 6, fig. 2).
These pericapsular alveoles may be regarded as an outer cell-wall more correctly than the membrane
of the central capsule itself, but the arrangement may also be compared to the temporary
encystation of other Protista (see note D).
A. The extracapsular vacuoles in the calymma were first observed in 1851 by Huxley, in
Thalassicolla and Sphærozoum, and compared with Dujardin's sarcode vacuoles (L. N.
5). Afterwards J. Müller noticed that generally these "large clear vesicles
are covered by a fine membrane," and hence he called them "alveoles" (L. N. 12, pp. 3, 7, &c.). In my Monograph I have described them more in detail as
"extracapsular alveoles" (1862, p. 88, Tafs. i.-iii. xxxii.-xxxv.). Ever since then the point
has been debated whether these clear spaces are simple vacuoles in the sense of Huxley or
vesicular alveoles as stated by J. Müller. This contention is unnecessary, for both varieties
are present, and often no sharp line can be drawn between them. R. Hertwig has recently come to
the conclusion that they are as a rule "membraneless vacuoles," but that they "sometimes become
surrounded by a special envelope" (L. N. 33, p. 31). He even succeeded "in
extracting from a Collosphæra the large vesicle which lies in the centre of many colonies
and removing its covering of central capsules and jelly."
B. The mechanical importance of the alveolar structure, which certainly increases the
elasticity and mechanical resistance of the voluminous calymma, has not yet been sufficiently
realised; in the case of those Radiolaria which have no skeleton, or at all events no
lattice-shell, it may take the place of this as a protective envelope. Furthermore, by taking in
and giving out water it may discharge a hydrostatic function, causing the organism to rise or
sink in the water.
C. The large central alveole found in the colonies of many Polycyttaria (especially
Collosphærida) and first described in my Monograph (Taf. xxxiv. fig. 1), has since then been
observed by Hertwig, Bütschli, and other investigators, and recognised as the "central support
of the whole colony, surrounded by a delicate membrane" (compare L. N. 33,
p. 31, and L. N. 41, p. 436). In a colony of Trypanosphæra
transformata (Pl. 5,
fig. 1), which I observed living while in Ceylon in 1881, the membrane of the large central
alveole was surrounded by a firm network of sarcoplegma, and could be mechanically isolated from
the central jelly-sphere which it enclosed.
D. The pericapsular alveoles, figured in Pl. 4, figs. 2, 3,
from a Sphærozoum, and in Pl. 6,
fig. 2, from a Siphonosphæra, were very well preserved in some preparations in the
Challenger collection; perhaps their development coincides with the formation of spores, and may
be regarded as an encystation.
{lvi}
87. The Extracapsular Fat-Globules.—Fat is probably as widely
distributed in the exoplasm as in the endoplasm of the Radiolaria; a considerable proportion of
the small, dark, highly refractive granules appear to consist of fat; most likely they are for the
most part direct products of metastasis. These widely-spread granules, which are sometimes
coloured, and which by their passive motion produce the phenomenon of granular circulation in the
exoplasm, are not the only fatty structures in the extracapsulum; larger globules sometimes occur.
In certain large Collodaria (e.g., Thalassicolla melacapsa,
Pl. 1, fig.
5; Thalassophysa sanguinolenta, &c.) radial series of oil-globules are found in the
calymma, especially in its proximal portion; in others the central capsule is surrounded by a
layer of oil-globules (situated in the sarcomatrix). In the Phæodaria a
part of the phæodium appears to consist of fat-globules.
88. The Extracapsular Pigment.—The formation of colouring matters in the
extracapsulum is on the whole rare in the Radiolaria, apart from the "yellow cells" (see § 91) and from the peculiar phæodium of the Phæodaria,
which will be separately treated of in the next paragraph. Considerable masses of extracapsular
pigment, usually black or blue, rarely brown or red, are found only in a few Radiolaria belonging
to the first three legions; most often in the Spumellaria. Some large
Collodaria, e.g., the common Thalassicolla nucleata and a
few other species of this genus (Pl. 1,
fig. 4), are characterised by a rich deposit of black or blue pigment in the sarcomatrix and in
the proximal portion of the calymma. Brown pigment is deposited in the calymma of many Sphæroidea and Discoidea, as well as of some Nassellaria (Cystidium, Tridictyopus, &c.). In a part of the
Acantharia red pigment granules are thickly strewn in the sarcoplegma and
pass along the free pseudopodia, as for example in Actinelius purpureus and
Acanthostaurus purpurascens. The composition and significance of these extracapsular
pigments are not completely known.
On the extracapsular pigment of Thalassicolla nucleata, compare my
Monograph, pp. 87, 251. On the red extracapsular pigment-granules of the Acantharia, see L. N. 19, pp. 345, 364, &c.
89. The Phæodium of the Phæodaria.—The Phæodaria, which
are distinguished from the other three legions of Radiolaria by the double membrane of the central
capsule, and the peculiar structure of the main-opening (astropyle), differ also in other points,
the most important of which is the constant presence of a voluminous mass of extracapsular
pigment. This possesses a peculiar constitution and special significance, and is not to be
confounded with the extracapsular pigment-granules of other Radiolaria (e.g.,
Thalassicolla), and hence it has been distinguished by the name "Phæodium," and the
individual granules which compose it as "Phæodella" (see note A). The phæodium is always excentric
in position relatively to the central capsule, of which it {lvii}surrounds the oral half in the form of a voluminous concavo-convex cap,
hiding the astropyle at its basal pole so completely that the latter is rarely visible until the
phæodium has been removed (Pls. 99-104; Pl. 115, fig. 8;
Pl. 123,
&c.). The central capsule is generally almost completely embedded in the phæodium, so that
only its aboral pole (with the two parapylæ in the Tripylea) projects. In
the Phæogromia, in which the lattice-shell possesses a special opening
and the central capsule lies excentrically in the aboral position of its interior, the phæodium
occupies the oral aspect, between the capsule and the aperture (Pls. 99, 100, 118-120, &c.).
In the peculiar family Cœlographida (Pls. 126-128) a special
receptacle (galea with its rhinocanna) for the phæodium is developed outside the bivalve shell,
within which the central capsule lies. The proboscis, which in all Phæodaria arises from the centre of the astropyle, lies in the vertical axis of
the phæodium and is entirely surrounded by it. The volume of the phæodium in the majority of the
Phæodaria may be said to be about as great as that of the central capsule,
although in some species it is considerably larger. Its colour is always dark, usually between
green and brown, commonly olive-green or blackish-brown, rarely reddish-brown or black. The
phæodellæ or pigment-granules which make up the greater part of the phæodium (see note B) are
irregular in form and unequal in size and show no definite structure; usually they are spherical
or ellipsoidal, and exhibit fine parallel striæ which run transversely or obliquely (Pl. 101, fig. 3,
6, 10; Pl. 103,
fig. 1, &c.). Between the larger granules is usually found a thick dust-like mass of
innumerable very small grains. The physiological significance of this peculiar phæodium is still
unknown, but is probably considerable, judging from its large size and especially from its
constant topographical relation to the astropyle; the latter consideration would lead to the
supposition that it plays an important part in the nutrition and metastasis of the Phæodaria (see note C).
A. The phæodium of Aulacantha, Thalassoplancta, and Cœlodendrum
was first described in 1862, in my Monograph, as an excentric extracapsular mass of pigment of
blackish-brown or olive-green colour (pp. 87, 262, 264, 361, Taf. ii. iii. xxxii.). Since then
John Murray, who investigated many living Phæodaria during the
Challenger expedition, has shown its general distribution in this legion (Proc. Roy. Soc. Lond.,
vol. xxiv. p. 536, 1876). From the constancy of its presence I gave the legion the name Phæodaria in 1879 (L. N. 34).
B. With regard to the special composition of the phæodium and the constitution of the
phæodellæ, see the general description of the Phæodaria, pp. 1533-1537.
C. Perhaps the phæodellæ are to some extent symbiontes with the Phæodaria; the xanthellæ present in most other Radiolaria are absent in this
legion.
90. The Extracapsular Xanthellæ.—Xanthellæ or Zooxanthellæ, symbiotic "yellow
cells," are very commonly found in the extracapsulum of the Radiolaria, especially in many Spumellaria and Nassellaria; whilst in the Acantharia similar yellow cells usually only occur within the central capsule,
and in the Phæodaria their {lviii}presence has not been certainly demonstrated. The extracapsular
Xanthellæ are found most abundantly in the Collodaria, both in the
monozootic Thalassicollida and in the polyzootic Sphærozoida. They occur in smaller numbers in the
Sphærellaria, and in many divisions of the latter they seem to be
entirely absent. Also it sometimes happens that, though present in large numbers in some Spumellaria, they are entirely absent in others nearly related to them; indeed,
this has also been observed in the case of different individuals of the same species. This fact
alone is sufficient to show that the Xanthellæ are not an integral part of the Radiolarian
organism (as was formerly believed) but parasites or more correctly symbiontes, which live as
inhabitants of the calymma. More recent investigations have shown, that besides the yellow
pigment-grains they contain starch or an amyloid substance, that is to say, vegetable reserve
materials, that their thin envelope contains cellulose, and that their yellow colouring-matter
resembles chlorophyll and is related to that of the Diatomaceæ ("Diatomin"). Hence they are now
generally regarded as unicellular Algæ, nearly related to those which occur as symbiontes in other
marine animals (Exuviella, &c.). The starch, which they develop with the formation of
oxygen, may serve as nutriment to the Radiolaria, while the carbonic acid yielded by the latter is
also beneficial to the Xanthellæ. The form of the Xanthellæ is usually spherical and elliptical,
often also sphæroidal or discoidal. Their diameter is usually between 0.008 and 0.012 mm., rarely
more or less. The differences exhibited by Xanthellæ which live in different groups of Radiolaria
demand further investigation, which will perhaps lead to the establishment of several species of
the genus Zooxanthella. At present Zooxanthella extracapsularis, in the calymma of
Spumellaria and Nassellaria, may be clearly
distinguished from Zooxanthella intracapsularis, in the central capsule of the Acantharia.
The "yellow cells" were first described in 1851 by Huxley, in the Collodaria, and afterwards by J. Müller (1858) in many Spumellaria and Nassellaria. In my Monograph (1862,
pp. 84-87) I gave a detailed account of their structure and increase by division, and laid
special emphasis on the fact that they are the only elements in the Radiolarian organism which
"are undoubtedly cells in the strict histological sense of the word." Afterwards, in my
Beiträge zur Plastiden-Theorie, I showed the constant presence of "starch in the yellow cells of
the Radiolaria" (1870, L. N. 21). Shortly afterwards Cienkowski observed
that the yellow cells live independently and reproduce themselves after the death of the
Radiolaria, and in consequence first put forth the hypothesis that they do not belong to the
Radiolarian organism, but that they are unicellular Algæ parasitic upon it (1871, L. N. 22). This view was ten years later more fully established by Karl Brandt, and
elucidated by comparison with the symbiosis of the gonidia of Algæ, and the hyphæ of Fungi in
the formation of Lichens, which had in the meantime become known (1881, L. N. 38). Brandt gave this unicellular yellow Alga the name Zooxanthella
nutricola, and afterwards gave fuller details regarding its remarkable vital relations (L.
N. 39). Patrick Geddes, who named it Philozoon, supplemented this
account and showed experimentally that it gives off oxygen under the influence of sun-light
(1882, L. N. 42, 43). In consequence {lix}of this there is no doubt that all Xanthellæ (the
Zooxanthella extracapsularis of Spumellaria and Nassellaria, and the Zooxanthella intracapsularis of the Acantharia, and possibly also the Zooxanthella phæodaris of the Phæodaria) do not originally belong to the Radiolarian organism, as was
believed up to the time of Cienkowski, but penetrate actively into it from without, or are taken
in passively by means of the pseudopodia. In any case their symbiosis, when they are associated
with the Radiolarian cell in large numbers, may be of great advantage to both parties, since the
metastasis of the Xanthella is vegetable, that of the Radiolarian animal in character. In any
case their symbiosis is to a large extent accidental, by no means as necessary as in the case of
the Lichens. See on these points in addition to Brandt and Geddes (loc. cit.) also Geza
Enz, Das Consortial-Verhältniss von Algen und Thieren, Biol. Centralbl., Bd. ii. No. 15, 1883,
Oskar Hertwig, Die Symbiose oder das Genossenschaftsleben im Thierreich, Jena, 1883, and
Bütschli, Die Radiolarien, in Bronn's Klass. u. Ord. d. Thierreichs, 1882 (L. N. 41, pp. 456-462).
91. The Exoplasm or Extracapsular Protoplasm.—The extracapsular
protoplasm, which may be shortly termed the "exoplasm" (or ectosarc), is primitively in all
Radiolaria (and especially in their earliest development stages) the only important constituent of
the extracapsulum, besides the calymma. Although the extracapsular and intracapsular protoplasm of
the Radiolaria are everywhere in direct communication, and although the openings in the membrane
of the central capsule bring about an interchange between them, still the two portions of sarcode
show certain constant and characteristic differences, which are due to the physiological division
of labour between the central and peripheral parts of the body and their corresponding
morphological differentiation. The extracapsular, like the intracapsular, protoplasm is originally
homogeneous, but may afterwards become differentiated in various ways, producing the special
constituents of the extracapsulum. Such "external protoplasmic products" are vacuoles,
pigment-bodies, &c. More important, however, are the topographically different sections into
which the exoplasm may be divided according to its relations to the central capsule and the
calymma. In this respect the following parts may be generally distinguished—(1) the
Sarcomatrix, or fundamental layer of the exoplasm, which surrounds the central capsule as a
continuous sheath of sarcode and separates it from the calymma; (2) the Sarcoplegma, an
irregular network of the exoplasm, which spreads throughout the gelatinous material of the
calymma; (3) the Sarcodictyum or network of sarcode on the outer surface of the calymma;
and (4) the Pseudopodia, which project outwards from the latter and radiate into the
water.
92. The Sarcomatrix.—The sarcomatrix, being "the fundamental layer of the
pseudopodia" (or "matrix of the exoplasm"), constitutes the proximal innermost section of the
extracapsular sarcode, and in all Radiolaria forms a thin continuous mucous layer, which covers
the whole outer surface of the central capsule and separates it from the surrounding calymma (see
note A, below). The sarcomatrix communicates internally {lx}through the openings of the central capsule with the endoplasm, whilst
externally the pseudopodia or mucous threads arise from it, which by their union form the
sarcoplegma. The sarcomatrix is only interrupted in the Spumellaria and
Acantharia by those parts of the skeleton which perforate the membrane of
the central capsule. In all Nassellaria and Phæodaria, as in the Collodaria, it appears as a
perfectly continuous sarcode-envelope of the central capsule. Its thickness is variable; in
general it is most strongly developed in the Spumellaria and Phæodaria, less so in the Nassellaria, and is thinnest
in the Acantharia. The thickness seems, however, to vary even in one and
the same individual, the difference depending partly upon the different stages of development and
partly upon nutritional conditions. After abundant inception of nutriment the thin protoplasmic
layer of the matrix is thickened and turbid, rich in granules and irregular masses, which are
probably due to enclosed but only half-digested food; xanthellæ also, as well as foreign bodies
taken up with the nutriment, such as frustules of Diatoms and shells of smaller Radiolaria, and of
pelagic infusoria, larvæ, &c., are often, especially in large individuals, aggregated in
considerable quantities in the matrix. After long fasting, on the contrary, this is poor in these
enclosed bodies and in granules; it then forms a thin colourless more or less hyaline mucous
coating to the central capsule. From a physiological standpoint the sarcomatrix is to be regarded
as the central organ of the extracapsulum, and as of pre-eminent significance. Probably it
is not only the most important organ for the nutrition of the Radiolaria (especially for digestion
and assimilation in particular), but perhaps is also the central organ of perception. On the other
hand the sarcomatrix belongs to those components of the Radiolarian organism which take no part in
the formation of the skeleton.
A. The sarcomatrix was first described in my Monograph in 1862 (p. 110) as the
"Mutterboden der Pseudopodien," possessing a pre-eminent physiological importance. Compare also
my paper on the sarcode elements of the Rhizopoda (Zeitschr. f. wiss. Zool., Bd. xv. p. 342,
1865).
93. The Sarcoplegma.—By the name sarcoplegma, as distinguished from
the remaining extracapsular sarcode, is understood the intracalymmar web of exoplasm or
"ectosarcode network," which ramifies within the gelatinous mass of the calymma. Internally it is
in direct connection with the continuous sheath (sarcomatrix), which encloses the central capsule,
whilst externally it is in contact with the superficial sarcode network (sarcodictyum) which
surrounds the calymma. The configuration of this exoplasmic web, which penetrates the jelly-veil
in all directions, is exceedingly variable; in most Radiolaria it is extremely irregular in form,
like the protoplasmic network in the ground-substance of many kinds of connective tissue. In some
groups, however, it assumes a rather regular shape which it appears to retain (e.g., in
many Acantharia). It must be assumed also that in those instances where
the consistency {lxi}of the calymma approaches that of
cartilage, the tracks of the exoplasmic threads remain constant, but accurate observations are
wanting as to how far the configuration of the sarcoplegma is constant or variable in the
different groups, as well as regarding its peculiar behaviour in those Radiolaria whose calymma is
characterised by the formation of vacuoles or alveoles (see § 86). Usually
it envelops the larger alveoles in the form of a reticulate veil. In many Collodaria the exoplasm is aggregated at certain points of the intracalymmar
web, so that large balls or amœboid bodies appear to be distributed between the alveoles,
e.g., in Thalassophysa pelagica and Thalassicolla melacapsa (Pl. 1, figs. 4, 5).
The sarcoplegma is metamorphosed directly into silex in the Radiolaria spongiosa, or those genera
which possess a spongy cortical skeleton, and were formerly known as Spongurida; to this category
belong the Spongosphærida (Pl. 18)
and Spongodiscida (Pl. 47)
as well as certain Nassellaria and Phæodaria. The
single siliceous spicules, which are irregularly interwoven to form the spongy web, are to be
regarded as the silicified threads of the intracalymmar sarcode network. From a physiological
point of view the sarcoplegma is of importance both for the nutrition and motion of the
Radiolaria, since it brings the sarcomatrix and the sarcodictyum, with the pseudopodia which
radiate from it, into direct communication.
94. The Sarcodictyum.—The sarcodictyum may be defined as the
extracalymmar network of exoplasm, and is a reticular covering which lies upon the outer surface
of the gelatinous calymma. Internally, the sarcodictyum is in direct communication with the
sarcoplegma, or the web of exoplasmic threads which ramifies in the gelatinous substance of the
calymma; externally, on the other hand, the pseudopodia radiate freely from it; thus its relation
to these is similar to that which the sarcomatrix bears to the roots of the sarcoplegma. Relations
similar to those which have led to the separation of the primary from the secondary calymma,
induce us to distinguish also a primary and secondary sarcodictyum. The original or primary
sarcodictyum ramifies over the surface of the original or primary calymma, and like this is of
pre-eminent importance in the formation of the primary lattice-shell; if we regard the surface of
the primary calymma as the indispensable foundation for the deposition of this latter, then the
primary sarcodictyum furnishes the material from which it is developed: silex in the Spumellaria and Nassellaria, a silicate of carbon in the
Phæodaria, and acanthin in the Acantharia. It may
indeed be said that the primary lattice-shell of the Radiolaria arises by a direct chemical
metamorphosis of the primary sarcodictyum, by a chemical precipitation of the dissolved skeletal
material (silex, silicate, or acanthin), which was stored up in the exoplasm of the sarcodictyum.
Hence a deduction from the special conformation of the former to that of the latter is
permissible. The particular form of the primary lattice-sphere with its regular or irregular
meshes is due to the corresponding form of the primary sarcodictyum; both regular and irregular
forms of this {lxii}commonly occurring. The form of the
regular sarcodictyum with circular or regular polygonal, usually hexagonal, meshes is
constantly maintained during the formation of the regular lattice-shells (e.g., Pl. 12, figs. 5-10;
Pl. 52,
figs. 8-20; Pl. 96,
figs. 2-6; Pl. 113,
figs. 1-6). The form of the irregular sarcodictyum, on the other hand, with irregular
polygonal or roundish meshes, persists during the development of the irregular lattice-shells
(e.g., Pls. 29,
70, 97, 106). All this
is true also of the secondary sarcodictyum, or the exoplasmic network which ramifies over
the surface of the secondary calymma. The secondary lattice-shells, which are deposited on the
surface of the latter, retain the configuration of the secondary sarcodictyum, by the chemical
metamorphosis of which they have originated; this is the case in many Spumellaria which develop several concentric lattice-shells (Pl. 29), in some
Nassellaria (Pl. 54,
fig. 5), in the Phractopeltida among the Acantharia (Pl. 133), and in
the double-shelled Phæodaria, Cannosphærida, and part of the
Cœlodendrida and Cœlographida (Pls. 112, 121, 128). In those
Radiolaria which form no lattice-shell whatever, the conformation of the sarcodictyum is usually
irregular, with meshes of irregular form and unequal size; sometimes, however, they seem to be
very regular, as in many Acanthometra (Pl. 129, fig.
4).
95. The Pseudopodia.—On the whole the pseudopodia or thread-like processes of the
exoplasm exhibit in the Radiolaria the same characteristic peculiarities as in all true Rhizopoda;
they are usually very numerous, long and thin, flexible and sensitive filaments of sarcode, which
show the peculiar phenomena of granular movement. Their physiological significance is in several
respects very great, for they serve as active organs for the inception of nutriment, for
locomotion, sensation, and the formation of the skeleton (see note A, below). The presence of a
calymma, however, which distinguishes the Radiolaria from the other Rhizopoda, brings about
certain modifications in the behaviour of the pseudopodia. If in general all the threads, which
arise from the sarcomatrix or fundamental layer and radiate outwards, be called "pseudopodia,"
then that part of them which is included in the gelatinous substance of the calymma and forms the
sarcoplegma may be termed the "collopodia" (or intracalymmar pseudopodia), and the remaining
portion, which passes outwards from the sarcodictyum freely into the water, may be described as
"astropodia" (or extracalymmar pseudopodia). In many Radiolaria these two portions present some
differences in morphological and physiological respects, and certain distinctions are probably
generally present (see note B). Apart from this universal differentiation in the different groups
of the Radiolaria, specially modified forms of pseudopodia may be recognised as the axopodia and
myxopodia of the Acantharia (see § 95, A), and the
sarcode-flagellum of certain Spumellaria (see note C).
A. The pseudopodia of the Radiolaria have been so fully described in my Monograph, in 1862,
both morphologically and physiologically, that I need only refer to the account there given
{lxiii}(pp. 89-127); for supplementary observations
see R. Hertwig (1879, L. N. 33, p. 117) and Bütschli (1882, L. N. 41, pp. 437-445).
B. The Astropodia, or free radiating pseudopodia, are in many Radiolaria more or less
clearly distinguishable from the collopodia, which form the sarcoplegma within the calymma; how
far these distinctions depend upon a permanent differentiation (especially in the Acantharia and Phæodaria) needs further
investigation.
C. The sarcode-flagellum (perhaps better termed axoflagellum) was
first described in my Monograph (1862, p. 115) in the case of various Discoidea (Taf. xxviii. figs. 5, 8; Taf. xxx. fig. 1). Hertwig has given a
substantially similar account of the organ in some other Discoidea (L.
N. 33, p. 67, Taf. vi. figs. 10, 11); probably this peculiar structure is
confined to the order Discoidea among the Spumellaria, but is widely distributed within its limits. The axoflagellum is
a thick cylindrical thread of sarcode, finely striated and pointed towards its free end. It
always lies in the equatorial plane of the discoidal body, and always unpaired in one of its
axes; in the triradiate Discoidea it is in the axis of the unpaired
principal arm and opposite to it (Pl. 43,
fig. 15). In the Ommatodiscida (p. 500, Pl. 48, figs. 8,
19, 20) the axoflagellum probably passes out through the peculiar marginal ostium of the shell.
Perhaps it is always connected with the central nucleus by intracapsular axial fibres, and is to
be regarded as a specially differentiated bundle of pseudopodia (or axopodia?).
95A. The Myxopodia and Axopodia.—The two forms of
pseudopodia which we distinguish as myxopodia and axopodia differ markedly from each other both
morphologically and physiologically. The myxopodia, or ordinary free pseudopodia, which are
found in large numbers in all Radiolaria, and constitute their most important peripheral organs,
are simple homogeneous exoplasmic threads, which arise from the sarcodictyum or extracalymmar
sarcode network, and radiate freely into the water; here they may branch and combine by
anastomosis to form a changeable network, but they never contain an axial thread. The
axopodia, on the other hand, are differentiated pseudopodia, which consist of a firm radial
thread, and a soft covering of exoplasm; they penetrate the whole calymma in a radial direction
and project freely from its surface, and generally (if not always) they are produced inwards to
the middle of the central capsule, perforating its membrane; their proximal end is lost in a dark
central heap of granules. Such axopodia are at present known with certainty only in the Acantharia, where they are widely, and perhaps universally, distributed. Their
development in this legion probably stands in direct causal relation to the peculiar structure of
the central capsule and the centrogenous formation of the skeleton. Since the radial skeletal rods
of the Acanthometra possess originally a thin coating of protoplasm, it
may be said that the centrogenous axopodia of this group became differentiated in two ways, the
firm axial threads of one section remaining very thin and covered by protoplasm, whilst those of
the other section became metamorphosed into radial bars of acanthin. This hypothesis acquires more
probability from the regular distribution and arrangement of the axopodia in the Acantharia; they usually stand at fixed intervals {lxiv}between the radial bars, singly or in groups; sometimes their number
seems to be not greater than that of the bars, whilst in other cases a circlet or group of
axopodia corresponds to each radial bar. Perhaps their fine axial thread consists of acanthin. At
all events the axopodia are constant organs (probably sensory, like the "palpocils") and not
retractile like the movable myxopodia.
The axial threads in the pseudopodia of the Acanthometra
were first discovered by R. Hertwig, who accurately described their peculiar structure and
arrangement (L. N. 33, pp. 16, 117).
96. The Myophriscs of the Acanthometra.—The Acanthometra
are characterised by a very peculiar differentiation of the exoplasm, namely, by the formation of
myophriscs or contractile threads from the sarcodictyum. In most (and perhaps in all) Acantharia of this order each radial bar is surrounded by a circlet of such
contractile threads, which was first described as a "ciliary corona" (see note A, below). The
number of contractile threads in each circlet usually amounts to from ten to twenty, rarely being
more than thirty and less than eight; it often appears to be constant in the individual species
(see note B). In the living state the myophriscs are long, thin filaments, the pointed distal end
of which is inserted into the radial bar, whilst the thicker proximal end is attached to the
surface of the calymma, which is elevated round the base of each rod into the form of a gelatinous
cone or skeletal sheath (see note C). Probably the myophriscs lie on the outer surface of the
apical portion of this gelatinous cone, and are hence to be regarded as exoplasmic threads
differentiated from the sarcodictyum. Sometimes, however (as in Acanthochiasma), they fuse
into a contractile membrane and form the envelope of a cone, whose interior is occupied by a
gelatinous papilla of the calymma. On mechanical irritation the myophriscs contract rapidly and
suddenly, like muscle-fibrillæ, becoming at the same time thicker, and hence are very different
from pseudopodia. Their distal point of insertion being fixed to the firm acanthin rod, they raise
by their contraction the skeletal sheath, to which their bases are attached or in the surface of
which they lie. The result of their contraction is therefore a distention and increase in volume
of the calymma, with which is no doubt connected an inception of water into the gelatinous mass,
and hence a diminution in its specific gravity. Probably the Acanthometra
contract their myophriscs voluntarily when they wish to rise in the water; when these relax the
calymma collapses owing to its elasticity, water is then expelled and the specific gravity
increases. From a physiological point of view, then, the myophriscs are to be regarded as a
hydrostatic apparatus, morphologically as myophanes or muscular fibrillæ, such as also occur in
the intracapsular protoplasm (see §§ 77-80). On more
violent irritation and after the death of the Acanthometra the myophriscs
separate from the radial bars and remain attached to the distal ends of the conical gelatinous
sheaths as free "ciliary coronas." At the same time, {lxv}they melt into short, thick, hyaline rods, the so-called "gelatinous
cilia." The myophriscs are found only in the order Acanthometra, and are
wanting in the Acanthophracta, as well as in the other three legions of
Radiolaria.
A. The "ciliary coronas" on the skeletal rods of dead Acanthometra were first described by the discoverer of this order, Johannes
Müller, and referred to as "the stumps of the contracted, thickened threads" (L. N. 12, p. 11, Taf. xi.).
B. The "number of the gelatinous cilia" I found constant in certain species of Acanthometra, and stated in my Monograph (L. N. 16, p.
115) "that here is to be found the first differentiation of the diffuse sarcode into definite
organs of regular definite number, size, and position, which deserve the name tentacles rather
than pseudopodia."
C. The nature of the myophriscs as fibrillæ allied to muscles was first
discovered by R. Hertwig, who described them as "structures of peculiar nature," under the name
of "contractile threads," and pointed out in detail their histological and physiological
peculiarities (L. N. 33, pp. 16-19, Taf. i.).
97. The Exoplasm of the Peripylea.—The extracapsular protoplasm of the
Spumellaria or Peripylea is in communication with
the intracapsular sarcode by the innumerable fine pores of the capsule-membrane, and like these
pores is evenly distributed over the whole surface. The sarcomatrix which immediately surrounds
the central capsule is moderately strong, and sends out innumerable long, thin pseudopodia, which
probably correspond to the pores of the membrane. Their number is markedly greater in the Spumellaria than in the other three legions. The ramifications and
communications which the radiating fibres of the sarcomatrix undergo within the calymma,
apparently present the most manifold variations, so that the sarcoplegma or intracalymmar network
thus formed has very diverse forms. On the surface of the calymma the exoplasmic threads
constitute a variously disposed sarcodictyum, a regular or irregular exoplasmic network, by the
silicification of which a primary lattice-shell arises in the majority of the Spumellaria. The free ends of the pseudopodia, which arise from this
extracalymmar network and radiate out into the water, appear in most Spumellaria to be relatively short, but exceedingly numerous. Specially modified
pseudopodia and axial threads in particular do not seem to occur in this legion. Perhaps, however,
among the latter may be reckoned the remarkable pseudopodia which combine to form the sarcode
flagellum in many Discoidea (and perhaps in other Spumellaria). This axoflagellum is a particularly strong thread of sarcode,
arising from a definite point in the central capsule; it is cylindrical or slenderly conical in
form, much longer, stronger, and more contractile than the ordinary pseudopodia; it contracts in a
serpentine fashion on mechanical irritation and seems to originate by the fusion of a bundle of
pseudopodia (compare § 95, C).
98. The Exoplasm of the Actipylea.—The extracapsular protoplasm of the
Acantharia or Actipylea differs in several
important respects from that of other {lxvi}Radiolaria,
and appears to undergo more significant differentiations than that of the three other legions.
Since the pores in the wall of the central capsule are not distributed evenly and at equal
intervals over its whole surface (as in the Peripylea), but rather exhibit
a regular disposition in groups at unequal intervals, the number of projecting pseudopodia is much
less and the law of their arrangement different from that which obtains in the Peripylea (§ 58). In many and probably in all Acantharia they are divided into two groups, those which arise from the centre
of the capsule and possess firm axial threads, and those which have not these characters (compare
§ 95, A). The axopodia, or stiff pseudopodia with axial threads, arise from
the centre of the capsule, are present in much smaller numbers than the soft and flexible
myxopodia, and are regularly disposed between the radial bars of acanthin, usually so that they
are as far removed from them as possible, i.e., in the centre between each three or four
bars; these latter may indeed be regarded as strongly developed axial threads, which have become
changed into acanthin (§ 95, A). The soft myxopodia, or pseudopodia without
axial threads, are much more numerous than the others, and arise from the sarcodictyum or
exoplasmic network which ramifies over the surface of the calymma. Their number and arrangement
seem, however, in many (if not in all) Acantharia to be regular and not to
possess the extraordinary variability seen in the other three legions. In many Acanthometra the sarcodictyum exhibits a symmetrical conformation, with regular
or subregular, polygonal (mostly hexagonal) meshes, and generally the stronger threads of the
sarcodictyum secrete a firm, homogeneous or fibrillar, striated substance, which forms a network
of ridges on the surface of the calymma. In the Acanthophracta the place
of this is taken by the acanthin network of the primary lattice-shell. The axopodia of the Acanthometra are usually about as long as the radial spines between which they
stand; their stiff axial thread is surrounded by a soft sheath of protoplasm, communicating with
the thin sarcomatrix which surrounds the central capsule. Numerous branches pass into the calymma
from the exoplasmic sheath of the axial threads, and form by their interweaving a loose
sarcoplegma. The most peculiar differentiated products of the exoplasm of the Acantharia, however, are the myophane fibrillæ of the Acanthometra, which have already been described under the name of myophriscs (§
96).
99. The Exoplasm of the Monopylea.—The extracapsular protoplasm of the
Nassellaria or Monopylea arises only from the
porochora, or the intracapsular podoconus, the oral base of which is formed by this porous area.
The pseudopodia or protoplasmic threads which pass through the pores of the latter, united into a
bundle, are not very numerous (in most Nassellaria probably between thirty
and ninety), and unite just outside it to form a thick discoid sarcomatrix; this covers the
porochora completely below, and spreads out in the form of a thin envelope of exoplasm over the
whole {lxvii}surface of the central capsule; at the
apical portion of the latter the sarcomatrix is often so thin that it can only be recognised by
the aid of reagents; it separates the membrane of the central capsule from the surrounding
calymma. The pseudopodia, which penetrate the latter and by loose anastomoses from a wide-meshed
sarcoplegma within it, are usually not very numerous. The greater part of them radiate in a bunch
downwards from the basal disc of the sarcomatrix, and a smaller number arise from the thinner
envelope which covers the remainder of the central capsule (Pl. 51, fig. 13;
Pl. 65, fig.
1; Pl. 81,
fig. 16). On the outer surface of the calymma the collopodia, which have passed through it, unite
to form the sarcodictyum, and through the silicification of this the primary lattice-shell arises
in the great majority of the Nassellaria. From the surface of the
sarcodictyum arise the astropodia, or free pseudopodia which radiate outwards into the water.
Their number in most Monopylea is relatively small, but their length
appears to be very great.
100. The Exoplasm of the Cannopylea.—The extracapsular protoplasm of
the Phæodaria or Cannopylea is much better
developed as regards volume than in the other three legions, and is connected with the
intracapsular sarcode by only a few apertures in the capsule-membrane. In most Phæodaria three of these are present, the astropyle or main-opening at the oral
pole of the main axis, and the two lateral parapylæ or accessory openings on either side of the
aboral pole (§ 60). In several families the latter appear to be wanting,
whilst in others their number is increased; these families have not yet, however, been observed
during life. The protoplasm projects both from the oral main-opening and from the two aboral
accessory openings in the form of a thick cylindrical rod; the tube into which each opening is
produced in many Phæodaria (longer in the case of the astropyle, shorter
in the parapylæ) being regarded as an excretion from this protoplasmic cylinder. The sarcode
threads within the tube appear like a bundle of fibrils, either quite hyaline or finely striated.
After issuing from the mouth of the aperture they pass over into a thick sarcomatrix, which
surrounds the central capsule entirely and separates it from the enclosing calymma. In the
neighbourhood of the basal astropyle the sarcomatrix is usually swollen into a thick lenticular
disc, which is in direct contact with the peculiar phæodium of this legion (§ 89). The pseudopodia, which radiate from the sarcomatrix, and form by
anastomosis a wide-meshed sarcoplegma within the calymma, are usually not very numerous in the
Phæodaria, but are very strong. Sometimes two stronger bundles of
collopodia may be distinguished at the two poles of the main axis, an oral bundle (in the
direction of the proboscis of the astropyle) and an aboral bundle (at the opposite pole between
the parapylæ). The collopodia of the sarcoplegma unite at the surface of the calymma into a
regular or irregular sarcodictyum, which, in most Phæodaria produces by
the secretion of a peculiar silicate the primary lattice-shell. {lxviii}The free astropodia, which pass outwards from the sarcodictyum into
the water, are in most Phæodaria very numerous (Pl. 101, fig. 10).
Since, however, only a few species of this great legion have been observed in a living state,
their pseudopodia require further accurate examination.
Chapter IV.—THE SKELETON.
(§§ 101-140).
101. The Significance of the Skeleton.—The skeleton of the Radiolaria
is developed in such exceedingly manifold and various shapes, and exhibits at the same time such
wonderful regularity and delicacy in its adjustments, that in both these respects the present
group of Protista excels all other classes of the organic world. For, in spite of the fact that
the Radiolarian organism always remains merely a single cell, it shows the potentiality of the
highest complexity to which the process of skeleton formation can be brought by a single cell. All
that has been brought to pass in this direction by single tissue-cells of animals and plants does
not attain the extremely high stage of development of the Radiolaria. Only very few Rhizopoda of
this very rich and varied class fail to exhibit the power of forming this firm supporting and
protecting organ—indeed, only ten of the seven hundred and thirty-nine genera which are
enrolled in the list of the Challenger collection, namely, six genera of Spumellaria (five Thalassicollida, Actissa, Thalassolampe,
Thalassopila, Thalassicolla, Thalassophysa, Pl. 1, and one genus
of Collozoida, Collozoum, Pl. 3),
and in addition two genera of Nassellaria (the Nassellida,
Cystidium and Nassella, Pl. 91,
fig. 1), and two genera of Phæodaria (the Phæodinida, Phæocolla and
Phæodina, Pl. 101,
figs. 1, 2). These skeletonless forms of Radiolaria are, however, of extreme interest, since they
include the original stem-forms of the whole class as well as of its four legions. All Radiolaria
which form skeletons have originated from soft and skeletonless stem-forms by adaptation, and that
polyphyletically, for the skeletal types of the four legions have been developed independently of
each other (§ 108).
102. The Chemical Peculiarities of the Skeleton.—The chemical composition of the
skeleton shows very marked variations in the different legions of the Radiolaria. The two legions
Spumellaria and Nassellaria (united formerly as
"Polycystina") form their skeleton of pure silica (see note A, below); the legion Phæodaria of a silicate of carbon (see note B), and the Acantharia of a peculiar organic substance—acanthin (see note C). This
explains the well-known fact that the deposits of fossil Radiolaria (or Polycystine marls) are
composed exclusively of the skeletons of Spumellaria and Nassellaria, those of the Acantharia and Phæodaria being entirely absent (in the case of the last group, however,
exception must be made in favour of the Dictyochida, or those Phæodaria
{lxix}whose skeleton is made up of isolated scattered
tangential siliceous fragments). The enormous deposits of Radiolarian skeletons in the deep sea of
today, which constitute the Radiolarian ooze, consist, like the fossil Polycystine marls, almost
exclusively of the shells of Spumellaria and Nassellaria, though here the acanthin skeletons of the Acantharia may be present in very small numbers, and the silicate skeletons of
the Phæodaria, which offer more resistance to the solvent action of
sea-water, somewhat more abundantly. Calcareous skeletons do not occur in the Radiolaria (see note
D).
A. The pure siliceous skeletons of the Polycystina were first recognised in 1833 by Ehrenberg
in chalky marls (L. N. 2, p. 117). Since the two legions Acantharia and Phæodaria were entirely unknown to
Ehrenberg, his name Polycystina has reference only to the Spumellaria
and Nassellaria.
B. The silicate skeleton of the Phæodaria was formerly taken by me
for a purely siliceous one. When I described the first Phæodaria in my
Monograph in 1862, I was only acquainted with five genera and seven species, whilst the number
of Phæodaria here described from the Challenger amounts to eighty-four
genera and four hundred and sixty-five species. In the vast majority of these (though not in
all) the skeleton becomes more or less intensely stained by carmine, and is also more or less
charred at a red heat, in some even becoming of a blackish-brown. In many Phæodaria, furthermore, the hollow skeletal tubes are destroyed by the
continued action of heat. They are also, for the most part, strongly acted upon, or even
destroyed by boiling caustic alkalis, whilst boiling mineral acids have no effect upon them. The
best method of cleaning the skeletons of Phæodaria from their soft parts
is to heat them in concentrated sulphuric acid, and then add a drop of fuming nitric acid; in
this they are not dissolved even on prolonged heating. From these facts it would appear that the
skeletons of the Phæodaria consist of a compound of organic substance
and silica, or a "carbonic silicate." The more intimate composition yet remains to be
discovered, as also the manifold differences which the various families of Phæodaria seem to show in respect of its composition. The small skeletal
fragments of the Dictyochida (the only remains of Phæodaria which occur
as fossils) appear to consist of pure silica.
C. The acanthin skeleton of the Acantharia was first described as
such in my Monograph (1862, pp. 30-32). Johannes Müller, the discoverer of this legion, took
them for siliceous skeletons and defined the Acanthometra as
"Radiolaria without lattice-shell, but with siliceous radial spines" (L. N. 12, p. 46). I formerly supposed that the acanthin skeletons in some of the
Acantharia were partially or wholly metamorphosed into siliceous
skeletons, but, according to the investigations of R. Hertwig, this does not appear to be the
case; he showed that the skeletons of the most varied Acanthometra and
Acanthophracta are completely dissolved under the longer or shorter
action of acids, and supposes that in all Acantharia, without exception,
the skeleton is composed of acanthin (1879, L. N. 33, p. 120). Quite
recently Brandt has found that the acanthin spines dissolve not only in acids, alkalis, and
"liquor conservativus" (as I had shown), but also in solutions of carbonate of soda (1 per
cent.), and even of common salt (10 to 20 per cent.); he concludes from this that they consist
of an albuminoid substance (vitellin) (L. N. 38, p. 400). I am unable to
share this view, for I have never been able to see some of the most important reactions of
albumen in any of the skeletons which I have examined, such for example as the xanthoproteic
reaction, the red coloration with Millon's test, &c. They do not become {lxx}yellow either with nitric acid or with iodine. In
dilute mineral acids they dissolve more rapidly than in concentrated. My usual method of
cleansing the skeleton of Acantharia (which has been practised with the
same result on thousands of specimens) consists in heating the preparation in a small volume of
concentrated sulphuric acid and then adding a drop of fuming nitric acid; all other constituents
(the whole central capsule and the calymma) are thus very rapidly destroyed; the skeleton
remains quite uninjured and withstands the combined action of the mineral acids for a longer or
shorter time, though on prolonged heating it also is dissolved. I do not therefore regard
acanthin as an albuminous substance, but as one related to chitin.
D. Calcareous skeletons have not been certainly demonstrated in the Radiolaria,
and probably do not occur. Sir Wyville Thomson in his Atlantic (1877, L. N. 31, vol. i. p. 233, fig. 51) described under the name Calcaromma
calcarea, a Radiolarian which contained scattered in its calymma numerous calcareous
corpuscles "resembling the rowels of spurs." These are identical with the "toothed bodies,
recalling crystal balls," which Johannes Müller figured in the Mediterranean Thalassicolla
morum so early as 1858, and compared with the "siliceous asterisks of Tethya" (L. N.
12, p. 28, Taf. vii. figs. 1, 2). I formerly regarded these peculiar
calcareous corpuscles, whose solubility in mineral acids I had observed, as spicules of a
Thalassicollid, and hence described the species in my Monograph as Thalassosphæra morum
(L. N. 16, p. 260). I have, however, seen reason to change my view, and am
now led to suppose that those peculiar calcareous corpuscles, which may be named
"Calcastrella," are not formed by the Radiolarian itself, but are foreign bodies which
have been accidentally incorporated into the calymma of a Thalassicollid (Actissa). These
corpuscles occur, often in large numbers, in many preparations in the Challenger collection, and
in the calymma of other Radiolaria, chiefly Discoidea, hence it would
appear that they are foreign bodies taken up by the pseudopodia and carried into the calymma by
the circulation of the sarcode. The Radiolaria which Sir Wyville Thomson figured as
Calcaromma calcarea, and Müller as Thalassicolla morum, I regard as species of
Actissa (see p. 13), perhaps Actissa radiata of the Pacific,
and Actissa primordialis of the Mediterranean (compare the description of the
Thalassosphærida of the Challenger collection, pp. 30, 31).
103. The Physical Properties of the Skeleton.—The skeletons of all
Radiolaria are characterised pre-eminently by a high degree of firmness, which fits them to
serve as protective and supporting apparatus. This is obvious in the case of the pure siliceous
shells of the Polycystina; but the acanthin framework of the Acantharia
also possesses a degree of stiffness but little inferior, whilst the silicate skeletons of the
Phæodaria seem on the whole to be not so firm. The hollow skeletal tubes
of the last-named, which are filled with gelatinous material, are very brittle on account of the
delicacy of their walls. Their elasticity also is very small, whilst that of the acanthin
spines is considerable. The thin long needles of many Acantharia are very
elastic, as are also the bristle-like siliceous spicules of many Spumellaria. The refractive power of the skeleton in the various legions
is very different, depending upon the chemical constitution. The siliceous skeleton of the
Polycystina (Spumellaria and Nassellaria) and the
silicate skeleton of the Phæodaria have the same refractive index as
glycerine, and hence become invisible when mounted in that fluid; they then become visible only on
addition of {lxxi}water, and are clearer in proportion
to the quantity of water which is added. The refractive index of acanthin is, however, very
different from that of glycerine, so that the skeletons of Acantharia are
readily visible when mounted in this fluid. In water, the skeletons of all Radiolaria appear about
equally refractive, as also in Canada balsam. The substance of the skeleton appears almost
entirely hyaline, colourless, and transparent. Very rarely it is faintly coloured (in some Acantharia). A cloudy opaque constitution is seen in some Phæodaria (especially in the "porcellanous shells" of Tuscarorida and
Circoporida, Pls. 100,
114-117); when
dried, these appear by reflected light milky-white or yellowish-white; the cause of this opacity
lies partly in the peculiar "cement-like structure" of these porcellanous shells, partly in their
fine porosity, and the minute air-bubbles contained in their thick walls.
104. The Elementary Structure of the Skeleton.—The general
constitution of the skeleton—or more accurately expressed, of the morphological elements of
which the skeleton consists—is of such a nature that it may be termed structureless. Both
the organic acanthin skeletons of the Acantharia and the silicate
skeletons of the Phæodaria, as well as the inorganic siliceous skeletons
of the Spumellaria and Nassellaria, appear under
the microscope perfectly homogeneous, transparent, colourless, and crystalline. Only very rarely
do they show traces of a concentric striation, which arises from the deposition of the skeletal
substance in layers; as, for example, the thick spines of some Phæodaria
(Pls. 105-107, &c.).
Some of the Phæodaria, however, form an exception to this rule, inasmuch
as their partially tubular skeletal elements possess a remarkable porcellanous structure. In the
tubular or Cannoid skeleton, which occurs in most Cannopylea, the lumen of
the thin-walled flinty tube is filled with jelly, and frequently a thin siliceous thread runs in
its axis, and is connected with the wall by transverse threads (§§ 127, 139). The elementary structure of the opaque porcellanous shells, which
distinguish the two families Circoporida (Pls. 114-117) and
Tuscarorida (Pl. 100),
is quite peculiar. Numerous fine siliceous spicules lie scattered irregularly in a finely granular
or porous matrix.
105. Complete and Incomplete Lattice-Shells.—In the great majority of Radiolaria
(in all four legions) the skeleton has the form of a delicate lattice-shell or a receptacle in
which the central capsule is enclosed. In a small minority, however, this is not the case. The
skeleton then consists only of isolated rigid pieces (radial or tangential spicules), or of a
simple ring (sagittal ring of the Stephoidea), or of a basal tripod with
or without a loose tissue of trabeculæ, &c. (Plectoidea); the central
capsule is then not surrounded by a special latticed receptacle, but only rests upon the skeletal
trabeculæ. According to these different arrangements, two principal groups or sublegions may be
distinguished in each legion, of which one set (Cataphracta) are characterised by a complete {lxxii}lattice-shell, whilst the others (Aphracta) are
without it. The Radiolaria aphracta, then, or Radiolaria without a
complete skeleton, are the Collodaria (p. 9), the
Acanthometra (p. 725), the Plectellaria (p. 895),
and the Phæocystina (p. 1543). On the other
hand, the Radiolaria cataphracta, or Radiolaria with a complete skeleton,
are the Sphærellaria (p. 49), the Acanthophracta (p. 791), the Cyrtellaria (p. 1015),
and the Phæocoscina (p. 1590).
Upon this basis the first subdivision of the Radiolaria was made by Johannes
Müller, who recognised three groups:—"I. Thalassicolla, without receptacle, naked
or with spicules; II. Polycystina, with a siliceous receptacle; III. Acanthometra,
without receptacle, but with siliceous radial spines" (L. N. 12, p. 16).
106. The Ectolithia and Entolithia (Extracapsular and Intracapsular
Skeletons).—The relation of the skeleton to the central capsule in the Radiolaria is
very various in many respects; in the first instance two great groups, Ectolithia and
Entolithia (see note A), may be distinguished topographically by mere external observation;
in the former the skeleton lies entirely outside the central capsule; in the latter, partially at
all events, within it. The Ectolithia, with a completely extracapsular skeleton, include
all Nassellaria and Phæodaria, as well as a great
part of the Spumellaria (all Collodaria and the
most archaic forms of Sphærellaria); the Entolithia, on the other
hand, in which the skeleton lies partly within, partly without the central capsule, include all
Acantharia and the majority of the Spumellaria
(most Sphærellaria, see note B).
A. The difference between Ectolithia and Entolithia was applied in my Monograph in 1862 (p.
222) to separate the Monocyttaria into two main groups. The arrangement was, however, quite
artificial, being contrary to the natural relations of the larger groups, as was shown seventeen
years later by the discovery of the different structural relations of the central capsule.
B. Among the Acantharia, which all possess primitively an
intracapsular and centrogenous skeleton, the remarkable Cenocapsa (Pl. 133, fig.
11), seems to furnish the single exception; in it the skeleton consists of a simple spherical
shell which encloses the concentric central capsule. The exception is, however, only apparent;
the twenty perspinal pores of the shell show that they were originally in connection with twenty
centrogenous acanthin spines, and that those have disappeared by retrograde metamorphosis.
107. Perigenous and Centrogenous Skeletons.—Much more important than
the topographical relation of the skeleton to the central capsule, according to which the
Ectolithia and Entolithia are separated from each other (§ 106), is the
original development of the skeleton within or without the central capsule, which gives rise to
the distinction between perigenous and centrogenous skeletons. Centrogenous skeletons are
found only in the Acantharia, which are further distinguished from all
other Radiolaria by their skeleton being formed of acanthin; in all Acantharia the formation of the skeleton begins in the middle of the central
capsule, from which twenty (the number is inconstant only in the {lxxiii}small group Actinelida) radial spines are
centrifugally developed. The three other legions, on the contrary, possess on the whole a
perigenous skeleton, which originally develops outside the central capsule and never
in its middle. In the Nassellaria and Phæodaria
the skeleton retains this extracapsular position, as also in the Beloidea
and part of the Sphærellaria among the Spumellaria; in the great majority of the latter, however, the primary
perigenous skeleton is subsequently enveloped by the growing central capsule, so that it lies
partially within it (§ 109).
108. Polyphyletic Origin of the Skeleton.—The skeleton of the
Radiolaria has undoubtedly originated polyphyletically, for it is impossible to derive its
manifold varieties from a single ground-form, or to regard them as modifications of one type. It
is much more probable that the different skeletonless Radiolaria have entered upon different ways
of skeleton formation quite independently of each other. At the outset it is quite clear that the
skeletons of the four legions have originated independently of each other. Further, it is
certain that within the legion of the Spumellaria the Beloid skeletons of
the Collodaria are not connected with the Sphæroid skeletons of the Sphærellaria and the forms derived from them (see § 109). In the same way the skeletons of the Phæodaria
are polyphyletic; probably in this legion the Beloid, Sphæroid, Cyrtoid, and Conchoid skeletons
have been developed quite independently (see § 112). In the Nassellaria, on the other hand, it is possible that all the skeletal forms are
to be derived monophyletically from a single simple primitive form (either the sagittal ring or
basal tripod?) (see § 111). Still more probable is it that the Acantharia have arisen monophyletically, for all the forms of their acanthin
skeleton may be derived without violence from Actinelius (see § 110).
109. The Skeleton of the Spumellaria.—The skeletons of the Spumellaria or Peripylea consist of silica, and are very
different and of independent origin in the two orders of this legion. The first order, Collodaria, have either no skeleton whatever (Colloidea, p. 10, Pls. 1, 3), or their
skeleton is Beloid, a loose extracapsular envelope of spicules, consisting of numerous
unconnected portions; the separate parts are usually disposed tangentially, either as simple or
compound siliceous spicules (Beloidea, p. 28, Pls.
2, 4). The second
order of Spumellaria, on the other hand (Sphærellaria, p. 49), develops a siliceous lattice-shell
which consists of a single piece, and is remarkable for the extraordinary variety of its forms
(pp. 50-715, Pls. 5-50). To this
order belong not less than three hundred genera and seventeen hundred species of the Challenger
Radiolaria (that is, about two-fifths of all the genera and species). In spite of this extreme
richness in different forms this large group must be regarded as monophyletic, since all
its forms may be quite naturally derived from a common stem-form, a simple lattice-sphere
(Cenosphæra, p. 61, Pl. 2). The
twenty-eight families of Sphærellaria may be distributed in four
suborders, among which the Sphæroidea constitute the {lxxiv}stem-forms, since they retain the original spherical
shape (Pls. 5-8, 11-30). In the
other three suborders a vertical main axis is developed, which in Prunoidea is longer, in Discoidea shorter than the
other axes of the shell. Hence the shell of the Prunoidea (p. 284, Pls. 13,
bis, 17,
39, 40) is
ellipsoidal or cylindrical, that of the Discoidea, on the other hand,
lenticular or discoidal (p. 402, Pls. 31-38, 41-48). Finally,
the shell of the fourth suborder, Larcoidea, is lentelliptical; it has
the ground-form of a triaxial ellipsoid, and is characterised by the possession of three unequal
dimensive axes, or three isopolar axes of different lengths perpendicular to each other (p. 599, Pls. 9,
10, 49, 50).
110. The Skeleton of the Acantharia.—The skeletons of the Acantharia or Actipylea are distinguished from those of
all other Radiolaria by two very important peculiarities; in the first place, they consist not of
silica but of a peculiar organic substance, Acanthin, and secondly, their development is
centrogenous, numerous radial spines or acanthin spicules being formed which are united in the
middle of the central capsule. Hence the Acantharia are the only
Radiolaria in which the skeleton originates from the first in the middle of the central capsule.
The number of radial spines is primitively indefinite, variable, and often considerable (more than
a hundred), but in the great majority it is limited to twenty. In accordance with this the legion
may be divided into two orders, the more archaic small group Adelacantha, with an indefinite
number of spines, and the more recent group, Icosacantha, which has been developed from them and
possesses twenty regularly disposed spines; of the three hundred and seventy-two species of Acantharia which have been hitherto described, about five per cent. belong to
the former, about ninety-five per cent. to the latter division (see note A, below). The numerous
genera of Icosacantha may then be again divided into two suborders, of which the Acanthonida (p. 740, Pls. 130-132) produce
no complete lattice-shell, and thus agree with the Actinelida, with which
they may be united as Acanthometra in the broader sense (or Acantharia without a lattice-shell). The Acanthophracta, on the other hand (p. 791, Pls. 133-140), produce
a complete lattice-shell, usually by means of two opposite or four crossed transverse processes,
which arise from each radial spine and unite with each other (see note B, below). In most Acanthophracta the lattice-shell remains single; only in the Phractopeltida
does it consist of two concentric lattice-spheres (p. 847, Pl. 133, figs.
1-6). Furthermore, the whole order Acanthophracta may be subdivided into
two suborders according to the different ground-form of the lattice-shell; this remains spherical
in the Sphærophracta (the three families Sphærocapsida, Dorataspida,
Phractopeltida, Pls. 133-138). On the
other hand, it assumes another form in the Prunophracta; it becomes
ellipsoidal in the Belonaspida (Pl. 136,
figs. 6-9), discoidal or lentiform in the Hexalaspida (Pl. 139); and
finally takes the shape of a double cone in the Diploconida (Pl. 140).
{lxxv}
A. The group Adelacantha consists only of the suborder Actinelida,
with the three families Astrolophida, Litholophida, and Chiastolida (p. 728, Pl. 129,
figs. 1-3); the number of the radial spines is very different and variable, sometimes only from
ten to sixteen, but usually from thirty to fifty, and often more than one hundred; they are
generally irregularly distributed, and not as in the second main division. This latter, the
Icosacantha, always possesses twenty radial spines, which are regularly disposed
according to a constant law, the so-called "Müllerian" or "Icosacanthan" law; the twenty spines
are always so placed between the poles of a spineless axis that they form five zones each of
four spines; the four spines of each zone are equidistant from each other, and also from the
same pole, and alternate with those of the neighbouring zones, so that the whole twenty lie in
four meridian planes, which cut out an angle of 45° (compare pp. 717-722, Pls. 130-140). In
spite of the manifold variations in form which are developed in the Icosacantha, they may all be
derived from a common stem-form, Acanthometron (p. 742), since the
law of distribution of the twenty spines is constantly inherited.
B. An exception is found in the peculiar family Sphærocapsida (p. 797, Pl. 133,
figs. 7-11; Pl. 135,
figs. 6-10). Here the shell is composed of innumerable small, perforated plates, which arise on
the surface of the calymma independently of the spines.
111. The Skeleton of the Nassellaria.—The skeletons of the Nassellaria or Monopylea consist of silica, and are
never composed of separate portions, but constitute always a single continuous piece. The
ground-form is originally monaxon, corresponding to that of the central capsule, with a constant
difference between the two poles of the vertical main axis. The ground-form is never spherical or
polyaxon as in the lattice-shells of the Spumellaria, and the skeleton
never consists of hollow tubes, as in the Phæodaria. The legion Nassellaria may be divided into two orders; in the Plectellaria (three suborders Nassoidea, Plectoidea, Stephoidea) the skeleton does not form a
complete lattice-shell; in the Cyrtellaria, on the other hand, which are
derived from these, the siliceous skeleton forms a complete lattice-shell enclosing the central
capsule. The number of forms thus developed is astonishingly great, so that among the Nassellaria no less than two hundred and seventy-four genera and sixteen hundred
and eighty-seven species may be distinguished, almost as many as in the Sphærellaria. In spite of this great variety of forms the legion Monopylea is probably monophyletic; at least all the different skeletal forms
may be derived from three elements which are combined in the most manifold fashion; (1) the
sagittal ring, a simple siliceous ring, which lies vertically in the sagittal plane of the
body, encircles the central capsule and comes into contact with it at the basal pole of the main
axis (§ 124); (2) the basal or oral tripod, composed of three
diverging radial spines, which meet in the middle of the basal pole of the central capsule (or in
the centre of the porochora) (§ 125); (3) the cephalis, or
lattice-head, a simple ovoid or subspherical lattice-shell, which encloses the central capsule and
stands in connection with it at the basal pole of its main axis. Any one of these three important
structural elements of the Nassellarian skeleton may possibly be the
starting-point {lxxvi}for all the remaining forms of
the Monopylea; the great difficulty in their phylogenetic derivation lies
in the facts that, on the one hand, any one of the three elements may alone constitute the
skeleton, and on the other hand, in the great majority of the legion, two or three are united
together (compare §§ 182-185).
112. The Skeleton of the Phæodaria.—The skeleton of the Phæodaria or Cannopylea is always extracapsular, usually
consists of a silicate of carbon (more rarely of pure silica), and in the majority of the legion
is composed of hollow cylindrical tubes, whose siliceous wall is very thin, and whose lumen is
filled with gelatinous material (§ 127). The manifold and remarkable
skeletal forms occurring in this legion are not monophyletic, since they cannot be derived from a
common stem-form; they are, on the contrary, polyphyletic, various skeletonless Phæodaria (Phæodinida) have independently acquired
skeletons of different form and composition. The legion Phæodaria can be
subdivided into four orders, the skeletons of which present the following important
distinctions:—(1) The Phæocystina possess only incomplete Beloid
skeletons (§ 115), composed of many separate pieces, sometimes tangentially
(Cannorrhaphida, Pl. 101),
sometimes radially arranged (Aulacanthida, Pls. 102-105). (2) The
Phæosphæria form Sphæroid skeletons (§ 116),
usually only a simple lattice-shell without special aperture (Pls. 106-111); two
concentric shells united by radial bars occur only in the Cannosphærida (Pl. 112). (3) The
Phæogromia are distinguished by the formation of a simple Cyrtoid
skeleton (§ 123) resembling that of the Monocyrtida; the monothalamus
lattice-shell is usually ovoid or helmet-shaped, more rarely polyhedral or almost spherical; a
vertical main axis can always be distinguished, at the basal pole of which is an aperture usually
armed with teeth or spines (Pls. 99,
100, 113-120). (4) The
Phæoconchia are distinguished from all other Radiolaria by the possession
of a bivalved shell like that of the Conchifera; the two valves of this Conchoid skeleton must be
distinguished as dorsal and ventral, as in the Brachiopoda (Pls. 121-128). The
fifteen families of Phæodaria which are arranged in the four orders just
mentioned, present such great differences among themselves, that the skeleton must be regarded as
probably polyphyletic even within the limits of each order.
113. Types of Skeletal Formation.—No less than twelve different
principal forms may be distinguished as morphological types of the formation of the skeleton in
the Radiolaria; some of these are peculiar to a single legion or even to a smaller group; but
sometimes the same form occurs in several legions. Some types occur only in an isolated manner,
independently of the others, but most exist in various combinations with other types. Of the
twelve described below the Conchoid and Cannoid occur only in the Phæodaria; the Plectoid and Circoid only in the Nassellaria; the Astroid only in the Acantharia; the
remaining seven types are found in several legions in the same form and hence are
polyphyletic.
{lxxvii}
114. The Astroid Skeleton.—Under the name "Astroid" we place the
peculiar star-shaped skeletons of the Acantharia in opposition to those of
all other Radiolaria, for they are separated from them not only fundamentally by reason of the
chemical nature of their substance (Acanthin, § 102), but also by their
centrogenous origin, and the resulting stellate form (Pls. 129-140). The
Acantharia are the only Radiolaria in which the skeleton arises within the
central capsule by the formation of numerous rays or radial spines of acanthin which project on
all sides from the centre. Originally these are united at this point, their conical or pyramidal
points meeting and being supported one upon another. In the great majority of Acantharia this loose apposition is constant, so that when the soft parts are
destroyed the skeleton falls to pieces. Only in a few forms in this legion are the central ends of
the spines fused so that the whole skeleton forms a connected star (Astrolithium). The
small group Chiastolida (or Acanthochiasmida) is characterised by the fact that the two rays which
are opposite to one another in each axis unite and form a diametral bar. The skeleton is almost
always composed of twenty radial spines, which are regularly disposed (Icosacantha), only in the
small primitive group Actinelida is the number variable (Adelacantha, §
110).
115. The Beloid Skeleton.—As Beloid or spicular skeletons are grouped
together all those which consist of several disconnected portions; these always lie outside the
central capsule, either within the calymma or on its surface. Such extracapsular Beloid skeletons
are entirely wanting in the Acantharia and Nassellaria; they occur only in the Beloidea among the
Spumellaria, and in the Phæocystina among the
Phæodaria; the individual Beloid portions of the former are solid, those
of the latter hollow. In both groups the simplest forms of the separate portions are simple
unbranched needles (Thalassosphæra, Thalassoplancta, Physematium,
Belonozoum, among the Spumellaria; Cannobelos and
Cannorrhaphis among the Phæodaria); usually these spicules are
disposed tangentially over the surface of the calymma. Among the Beloidea
branched spicules occur more commonly than these simple ones; they are either stellate (with many
rays united in a centre) or twin-like, with a tangential bar, from each pole of which two or three
(seldom more) radial branches project (Pls. 2,
4). Among the
Phæodaria the subfamily Dictyochida is characterised by the annular shape
of its Beloid portions, either simple rings, or hat-shaped or pyramidal bodies with a latticed cap
over the ring (Pl. 101,
figs. 3-14; Pl. 114,
figs. 7-13). The family Aulacanthida among the Phæodaria, alone possesses
hollow radial tubes, which penetrate the whole calymma, and project distally over its
surface, whilst their proximal ends rest upon the surface of the central capsule. Although in
these cases the enclosed proximal end is always simple, the free distal end develops the most
various processes in adaptation to its prehensile functions (Pls. 102-105).
{lxxviii}
116. The Sphæroid Skeletons or Lattice-Spheres.—The "lattice-spheres"
or sphæroid skeletons are the simplest and most primitive forms of lattice shells, and are widely
distributed in the three legions Spumellaria, Acantharia, and Phæodaria, whilst they are entirely
wanting in the Nassellaria. The round lattice-shell is either a true
sphere in the geometrical sense, or an endospherical polyhedron, i.e., a polyhedron, all
whose angles lie in the surface of a sphere (§ 25). In general,
primary and secondary lattice-spheres may be distinguished, of which the former are
secreted on the outer surface of the primary, the latter on that of the secondary calymma (§ 85). Furthermore, simple and compound lattice-spheres may be
distinguished, the latter of which consist of two or more concentric lattice-spheres firmly united
by radial bars; in such cases the innermost lattice-sphere is always to be regarded as the oldest
or primary, all the succeeding ones as secondary, and the outermost as the youngest (§ 129). The simple lattice-spheres are usually to be regarded as primary; they
may, however, occasionally be secondary, in which case the primary shell, originally enclosed, has
been lost by degeneration (as, for example, in the case of the Aulosphærida and some Sphærellaria).
117. The Lattice-Spheres of the Spumellaria.—The lattice-spheres or
Sphæroid skeletons of the Spumellaria exhibit in spite of their simple
type of structure, an extraordinary variety in the formation of the lattice-work and radial
apophyses, so that in the systematic portion of this work no less than one hundred and seven
genera and six hundred and fifty species are distinguished; these are united in one suborder, the
Sphæroidea (pp. 50-284, Pls.
5-8, 11-30). It may be
divided into two main divisions, the Monosphærida with a single primary lattice-sphere
(Pls. 12-14, 21, 26, 27), and
Pliosphærida (or Sphæroidea concentrica) whose skeleton consists of two or more concentric
lattice-spheres united by radial bars. The latter are subdivided into Dyosphærida with two
concentric lattice-spheres (Pls. 16,
19, 20, 22, 28);
Triosphærida, with three spheres (Pls. 17,
24, 29);
Tetrasphærida, with four (Pls. 23,
30);
Polysphærida, with five or more (Pls. 15,
23); and
Spongosphærida, with spongy lattice-spheres (Pls. 18, 25). A special
group is made up of the simple lattice-spheres of the social Collosphærida (or Sphæroidea polyzoa)
(Pls. 5-8); these are
usually more or less irregular, and characterised by the development of peculiar tubular
processes; the latter are generally wanting in the Sphæroidea monozoa, whose lattice-shell is very
regularly formed. This distinction is interesting and important, inasmuch as the regular
lattice-spheres are explained by the independent development of the free-swimming Monozoa, whilst
the irregular spheres are due to the mutual dependence of the social Polyzoa.
118. The Lattice-Spheres of the Acantharia.—The lattice-shells or
Sphæroid skeletons of the Acantharia are immediately distinguishable from
those of all other Radiolaria by their centrogenous development and the central union of the
radial spines by which they are supported; the only exception is furnished by the remarkable genus
Cenocapsa {lxxix}(Pl. 133, fig. 11),
in which the radial spines are absent, not primitively, however, but in consequence of
degeneration; for the twenty cross-shaped perspinal pores, originally due to the twenty radial
spines, are still present. In the most nearly allied genera, Porocapsa (Pl. 133, fig. 7)
and Cannocapsa (Pl. 133,
fig. 8), the proximal part of the twenty radial spines is still present, while their distal
portion has degenerated; hence in this case they do not stand in direct communication with the
spherical shell. On the other hand, this primitive connection persists in the genera
Astrocapsa (Pl. 133,
figs. 9, 10), and Sphærocapsa (Pl. 135,
figs. 6-10). The five genera just mentioned form the peculiar family Sphærocapsida (pp. 795-802); the spherical shell is in these cases
composed of very numerous small plates disposed like a pavement, each plate or aglet being
perforated by a pore canal; in addition to which there are twenty larger (perspinal) pores (or
twenty cross-shaped groups each of four aspinal pores) at those important points where primitively
the twenty radial spines penetrate the calymma. This peculiar porous "pavement shell" has probably
been developed (independently of the twenty radial spines) upon the calymma of the Acanthonida (Acanthonia, p. 749) by the action of
the sarcodictyum; it has, therefore, quite a different morphological significance from the
spherical lattice-shell of the Dorataspida, which is composed of tangential apophyses of the
twenty Acanthonid spines (pp. 802-847, Pls. 134-138). Each
radial spine here forms either two opposite or four crossed transverse processes, and since their
branches spread over the surface of the spherical calymma and are united suturally at their
extremities, the peculiar lattice-sphere of the Dorataspida arises. This extensive family is again
divided into two subfamilies:—the Diporaspida (Pls. 137, 138) possess
always only two opposite apophyses, and form by the union of their branches two opposite primary
apertures or aspinal meshes. The Tessaraspida, on the other hand (Pls. 135, 138), have
always four crossed transverse processes, and form by their union four primary aspinal meshes.
From the Diporaspida are probably to be derived the Phractopeltida (p. 847,
Pl. 133,
figs. 1-6), the only Acantharia which possess a double lattice-sphere;
their double concentric spherical shell may be compared with that of the Dyosphærida.
119. The Lattice-Spheres of the Phæodaria.—The lattice-spheres or
Sphæroid skeletons of the Phæodaria, which are generally developed quite
regularly, though occasionally in a modified form, fall in the order Phæosphæria into two groups of very different structure, each of which includes
two families. The first group (Phæosphæria inarticulata) contains the families Orosphærida
(Pls. 106,
107) and
Sagosphærida (Pl. 108);
the lattice-work of the former consists of irregular polygonal meshes and very coarse, partially
hollow trabeculæ; in the latter, on the other hand, it consists of triangular meshes and very
slender filiform trabeculæ; in both families the whole sphæroid skeleton forms a single
unsegmented piece as in most Sphæroidea. In the second group of {lxxx}Phæosphæria (Phæosphæria
articulata), on the other hand, the lattice-sphere is segmented in quite a peculiar manner,
and composed of hollow cylindrical tangential tubes, which are separated by astral septa at the
nodal points of the network; this remarkable structure characterises the two families,
Aulosphærida (Pls. 109-111) and
Cannosphærida (Pl. 112);
the segmented lattice-sphere of the former is simple and hollow; while that of the latter is
connected by centripetal radial tubes with a simple concentric inner shell, which is sometimes
solid, sometimes latticed, and provided with a main-opening corresponding to the astropyle of the
enclosed central capsule. Since in the Aulosphærida also, hollow centripetal radial tubes project
from the segmented lattice-sphere, it is possible that they have been derived from the
Cannosphærida by the loss of the primitive internal shell. A special peculiarity of many Phæosphæria (Oroscena, Sagoscena, Auloscena, &c.)
consists in the fact that the whole surface of the lattice-sphere is regularly covered with
pyramidal or tent-shaped prominences (Pl. 106,
fig. 4; Pl. 108,
fig. 1; Pl. 110,
fig. 1). A simple lattice-sphere quite similar to that of most Monosphærida also constitutes the
skeleton of the Castanellida (Pl. 113),
but since it possesses a special main-opening, it must be referred promorphologically to the
Cyrtoid shells of the Phæogromia.
120. The Prunoid Skeleton or Lattice-Ellipsoid.—The
"lattice-ellipsoids" or Prunoid skeletons have arisen from the lattice-spheres or Sphæroid
skeletons by more energetic growth and elongation of one axis; this is the main axis of the body
and is probably always vertical; its two poles are commonly equal. The Prunoid skeleton is either
a true ellipsoid in the geometrical sense or an "endellipsoidal polyhedron" (i.e., a
polyhedron, all the angles of which lie in an ellipsoidal surface). By further elongation of the
main axis, the ellipsoidal form passes over into the cylindrical, the polar surfaces of the
cylinder being usually rounded, rarely truncated. The rich order Prunoidea (pp. 284-402) contains
numerous modifications of this form of shell which arise on the one hand by the formation of
transverse constrictions, on the other by the apposition of concentric secondary shells. In
respect of the latter, simple and compound Prunoid shells can be distinguished as in the case of
the Sphæroid shells. In the compound Prunoid shells either all the concentric lattice-shells may
be ellipsoidal or the inner may be spherical. More important differences are found in the
transverse annular constrictions, which give the Prunoid skeleton a segmented appearance; in this
respect, three principal forms may be distinguished (p. 288):—(A)
Monoprunida, with unsegmented shell, having no transverse constriction (Pls. 15-17); (B)
Dyoprunida, having a shell with two segments and one (equatorial) transverse constriction
(Pl. 39);
(C) Polyprunida, with three or more parallel transverse constrictions, by means of which
the shell is divided into four or more segments (Pl. 40). In the
same manner as the Prunoidea have arisen from the Sphæroidea among the Spumellaria by greater {lxxxi}development of the vertical main axis, the
ellipsoidal Belonaspida have arisen from the spherical Dorataspida among the Acantharia (p. 859; Pl. 136, figs.
6-9; Pl. 139,
figs. 8, 9). The main axis of the ellipsoid in this case is always occupied by the opposite
equatorial spines of the hydrotomical axis (pp. 719, 860). In the legion Phæodaria a similar prolongation
of the main axis rarely occurs; it is found, however, in Aulatractus (Pl. 111, figs. 6,
7), the lattice-shell of this Aulosphærid being sometimes truly fusiform, sometimes rather
ellipsoidal or even double-conical.
121. The Discoid Skeletons or Lattice-Discs.—The "lattice-discs" or
Discoid skeletons are characteristic of the Spumellarian group Discoidea, and have arisen from the lattice-spheres of the Sphæroidea by a less development of one axis, which is the main axis of the
body, and is probably usually vertical; its two poles are always equal. The Discoid lattice-shell
is either a biconvex lens (with a thin margin), or a plane disc (a shortened cylinder with thick
margin), or some form intermediate between the two. All Discoid shells show a horizontal median
plane or equatorial plane, by which they are divided into two equal halves, an upper and lower;
the margin of the lens itself is originally the equator. The main axis, the shortest of all the
axes of the shell, stands vertically in the centre of the equatorial plane. Among the Phæodaria Discoid shells rarely occur (Aulophacus), as also among the
Acantharia (Hexalaspida).
122. The Larcoid Skeleton or Lentelliptical Lattice-Shell.—The
lentelliptical lattice-shells, which may be shortly designated "Larcoid," are especially
characteristic of the Larcoidea, a large order of Spumellaria (pp. 599-715; Pls. 9, 10, 49, 50). In
addition they recur among the Acantharia, in the small family Hexalaspida
(p. 872, Pl. 139),
and the family Diploconida (p. 881, Pl. 140), which is
derived from it. These lentelliptical lattice-shells are all characterised by the clear
differentiation of three unequal, but isopolar dimensive axes, i.e., the three geometrical
axes, perpendicular to one another, which determine the form of the shell, are of unequal length;
the two poles of each are, however, equal. The geometrical ground-form is, therefore, a triaxial
ellipsoid (§ 34). In the rich order Larcoidea the
lentelliptical lattice-shell shows many variations in its development.
123. The Cyrtoid Skeleton.—Cyrtoid skeletons are those lattice-shells
which possess a vertical main axis with two different poles (Monaxonia allopola); the upper pole
is usually termed the apical, the lower the basal. Such Cyrtoid shells are characteristic of the
great majority of the Nassellaria or Monopylea
(and especially of the Cyrtellaria); they are also found in a large
division of the Phæodaria (the Phæogromia), and
in some Spumellaria. In general the manifold Cyrtoid shells may be divided
into two large groups, those with one and those with several chambers. The monothalamous
Cyrtoid shells are usually ovoid, conical, cap- or helmet-shaped; their {lxxxii}internal cavity is simple, without constrictions or septa. Among
the Nassellaria they occur in the Monocyrtida (Pls. 51-54, 98), where they
have received the name "Cephalis." A form of shell, essentially the same, is found amongst the
Phæodaria in the order Phæogromia, more
especially in the Challengerida (Pl. 99),
Medusettida (Pls. 118-120), and
Tuscarorida (Pl. 100),
many of these latter closely resembling many Monocyrtida. Such monothalamous Cyrtoid shells occur
much more rarely among the Spumellaria (e.g., among the Prunoidea in Lithapium, Lithomespilus, Druppatractus, Pls.
13, 14, &c.).
Polythalamous Cyrtoid shells (Pls. 55-80) occur
exclusively in the Nassellaria, and exhibit in this legion an astonishing
variety of structure; they are distinguished from the monothalamous forms by the development of
internal septa, or of annular incomplete diaphragms, which usually correspond to the external
constrictions; their interior is thus divided into two or more communicating compartments. Among
the polythalamous Cyrtoid shells may be distinguished three principal groups, the Stichocyrtid,
Zygocyrtid, and Polycyrtid. Zygocyrtid shells are characteristic of the Spyroidea (Pls. 84-90), and are
distinguished by a bilobate cephalis (cephalis bilocularis); the median sagittal ring, or a
corresponding constriction, divides the shell into right and left compartments. Polycyrtid shells
(Pl. 96) are
peculiar to the Botryodea, and characterised by a multilobate cephalis
(cephalis multilocularis). Stichocyrtid shells are those in which the primary cephalis remains
simple, and new joints are successively added to its basal pole; such shells occur in the majority
of the Cyrtoidea. Secondary chambers are sometimes added in the other two
groups (Botryodea and Spyroidea). When, as often
happens in these polythalamous Cyrtoid shells, two or three distinct joints follow each other, the
first is called the "cephalis," the second the "thorax," and the third the "abdomen" (Tricyrtida
Pls. 64-75).
124. The Circoid Skeleton.—This is a very important and remarkable
type of skeletal formation, which occurs exclusively in the legion Nassellaria, where it plays a very prominent part; its characteristic element is
the "sagittal ring," a simple, vertical, siliceous ring, which surrounds the central capsule in
its sagittal plane, and is specially differentiated in its basal portion. This "primary sagittal
ring" whose vertical allopolar main axis coincides with that of the Monopylean central capsule
embraced by it, is characteristic of all members of the order Stephoidea
(p. 931, Pls. 81-83, 92-94); here it
forms by itself the skeleton of the Stephanida (Pl. 81); in the
Semantida (Pl. 92)
it is combined with a horizontal basal ring, in the Coronida (Pls. 82, 93) with a
vertical frontal ring and in the Tympanida (Pls. 83, 94) with two
horizontal rings, an upper mitral and a lower basal. In the great majority of these Stephoidea there often develop in definite places characteristic processes or
apophyses, whose branches combine to form a loose tissue or an incomplete lattice-shell. This
becomes complete in the Cyrtellaria, the majority of which retain more or
less {lxxxiii}distinct traces of the sagittal ring.
Hence the skeletons of all Nassellaria may be derived monophyletically
(Hypothesis A, p. 893)
from a simple sagittal ring (Archicircus and Lithocircus, Pl. 81). This
theory, however, encounters the great difficulty that in many Stephoidea
(Cortina, Cortiniscus, &c.) it is combined in a remarkable manner with the basal
tripod of the Plectoidea, whilst in these latter it is entirely wanting
(compare p. 894).
125. The Plectoid Skeleton.—Those forms are distinguished as Plectoid
in which three, four, or more radial siliceous spines proceed from a common point, which lies
excentrically outside the central capsule and at the basal pole of its vertical allopolar main
axis. This peculiar type of skeletal formation only occurs in the legion Nassellaria, and is specially characteristic of the order Plectoidea (p. 898,
Pl. 91). But
since the essential elements of this remarkable skeleton also occur in many other Nassellaria, sometimes combined with the Circoid, sometimes with the Cyrtoid
skeleton, it perhaps has a fundamental significance in this legion; at all events it is possible
to derive monophyletically all the other forms of this legion from it (Hypothesis B, p. 893). The simplest
form of the Plectoid skeleton is a tripod, the three feet of which either lie in a horizontal
plane (Triplagia, Pl. 91,
fig. 2), or correspond to the three edges of a low pyramid (Plagiacantha). A fourth ray is
sometimes added, which stands vertically upon the summit of the pyramid (Plagoniscus,
Plagiocarpa, Pl. 91,
figs. 4, 5). In other Plectoidea three secondary rays are intercalated
between the three primary (Hexaplagida, &c.); seldom the number is greatly increased
(Polyplagida, &c.). The rays are rarely simple, but usually branched; in the Plagonida (Pl. 91, figs. 2-6)
the branches remain free; in the Plectanida (Pl. 91, figs. 7-13)
they are united to form a loose wicker-work. From such a web a perfect Cyrtoid shell may arise.
Several forms of Plagonida may also be readily confounded with the isolated triradiate or
quadriradiate spicula of many Beloid skeletons (Sphærozoum, Lampoxanthium,
&c.).
126. The Spongoid Skeleton.—From the simple lattice-skeleton which the
majority of Radiolaria possess, some of them develop a spongy shell; the trabeculæ of the
lattice-work, situated in one plane in the former, are developed in the latter in different planes
and cross irregularly in all directions; thus arises a kind of wicker-work of more or less spongy
structure, usually with very thin trabeculæ and irregular meshes. Such Spongoid shells are most
common among the Spumellaria, especially in the Sphæroidea (Spongosphærida, Pl. 18) and Discoidea (Spongodiscida, Pls. 41-47), more
rarely in the Prunoidea and Larcoidea.
Lattice-work of similar spongy structure occurs very seldom among the Nassellaria, e.g., in some Plectoidea (Pl. 91) and Cyrtoidea (Spongocyrtis, Spongopyramis, Spongomelissa,
&c., Pl. 56,
fig. 10; Pl. 64,
figs. 5-10, &c.). Among the Phæodaria spongy skeletons are very rare;
they {lxxxiv}are to be seen in some Phæosphæria (Oroplegma, Pl. 107, fig. 1;
Sagoplegma, Pl. 108,
fig. 2; Auloplegma, Pl. 111,
fig. 8). No Spongoid skeletons are known among the Acantharia.
127. The Cannoid Skeleton.—Cannoid or tubular skeletons are those
which are composed of hollow tubes; they occur exclusively in the Phæodaria or Cannopylea. Tubular processes,
nevertheless, occur in some other Radiolaria, as, for example, among the Spumellaria in a portion of the Collosphærida (Siphonosphæra,
Caminosphæra, Pls. 6,
7), and of
the Prunoidea (Pipetta, Cannartus, &c., Pl. 39, figs. 6-10,
&c.), also among the Nassellaria in Theosyringium (Pl. 68, figs. 4-6),
Cannobotrys (Pl. 96,
figs. 3, 4, 8-11, 20-22), &c. In all these cases, however, the tubes are direct processes of
the cavity of the shell, the trabeculæ of the lattice-work being solid. Only in the Cannopylea are the lattice-bars themselves, the radial spines and appendicular
organs, generally tubular (hence the designation "Pansolenia"). The lumen of the thin-walled
siliceous tubes is filled with jelly, and hence the specific gravity of the relatively large
skeleton is considerably diminished. This peculiarity is not found in all Cannopylea; it is wanting in all Sagosphærida and Concharida, as well as in a
part of the Orosphærida and Castanellida; in the latter there are found intermediate stages
between hollow and solid skeletal rods. Very often a fine siliceous thread runs in the axis of the
tubes, which is connected with its wall by lateral branches (Pl. 110, figs. 4,
6; Pl. 115,
figs. 6, 7). More seldom the tubes are divided by horizontal septa into a series of chambers
(Medusettida, Pls. 118-120). The two
families Aulosphærida (Pls. 109-111) and
Cannosphærida (Pl. 112)
are distinguished from all other Phæodaria by the fact that their tubes
are separated by astral septa in the nodal points of the lattice-shell (§§ 112, 134).
128. The Conchoid Skeleton.—By the name "Conchoid skeletons" are
distinguished the bivalved lattice-shells which occur exclusively in the legion Phæodaria; they are quite characteristic of the Phæoconchia or Phæodaria bivalvia, which embrace three
families:—Concharida (Pls. 123-125),
Cœlodendrida (Pls. 121,
122), and
Cœlographida (Pls. 126-128). The two
valves of the lattice-shell of the Concharida are simple, hemispherical, or boat-shaped, whilst in
the Cœlodendrida and Cœlographida tubes grow out from them, which branch and usually
give rise by anastomosis to a second external bivalved shell. In all Phæoconchia the two valves are so disposed about the central capsule that an
open slit remains between them, into which open the apertures of the central capsule; and since
all these Phæodaria conchoidea are Tripylea, with three typical
openings in the central capsule, and since the two lateral accessory openings lie at either side
of the aboral pole, and the unpaired main-opening at the oral pole of the main axis, it follows
that the two valves are to be regarded as dorsal and ventral as in the Brachiopoda (not right and
left as in the Lamellibranchiata). The dorsal and ventral {lxxxv}valves are usually equal, but in a portion of the Concharida they
present constant differences. In this family the two valves are attached to each other by their
free edges, just as in the bivalved Mollusca and Diatoms; and these edges may either be smooth
(Conchasmida, Pl. 123,
figs. 1-6), or dentate (Conchopsida, Pls. 124,
125); the
valvular connection of the latter is sometimes strengthened by a special ligament which unites the
two valves at the aboral pole (Pl. 123,
figs. 8, 9). The form of the valve is sometimes hemispherical, sometimes boat-shaped, with a
sagittal keel.
129. Medullary and Conical Shells.—In all Radiolaria whose skeleton consists of a
double shell or of two concentric lattice-shells united by radial bars, an inner medullary shell
(testa medullaris) and an outer cortical shell (testa corticalis) may be distinguished (see note
A, below). The medullary shell is usually to be regarded as a primary, the cortical as a secondary
structure. Such double shells occur among the Spumellaria in the
Dyosphærida (Pls. 19,
20), as well
as in many Prunoidea (Pls. 39, 40), Discoidea (Pls. 33,
34), and
Larcoidea (Pls. 9,
10); among
the Acantharia only in the family Phractopeltida (Pl. 133); among
the Nassellaria only in very few Cyrtoidea
(e.g., Periarachnium, Pl. 55,
fig. 11), and finally among the Phæodaria in the Cannosphærida (Pl. 112) as well
as in part of the Cœlodendrida (Pl. 121)
and Cœlographida (Pls. 127,
128). In
most cases (if not always?) the cortical shell arises by the growth of radial spines from the
surface of the medullary shell; these become united at equal distances from the centre by
transverse apophyses, the surface of the secondary calymma furnishing the basis for their
secretion (§ 85). Nevertheless, it seems that in many Sphærellaria the formation of the whole cortical shell proceeds simultaneously
(at a definite dictyotic period) like that of the primary medullary shell (see note B). Whilst in
the Phæodaria, Acantharia, and Nassellaria, at most two concentric shells are formed, in many Spumellaria their number increases continuously with additional growth; in many
Sphærellaria it rises to four, eight, or even more, as well as in many
Discoidea (if the concentric, peripherally disposed rings of chambers be
regarded as incomplete flattened shells). In these cases either only the innermost primary
lattice-shell is to be styled "medullary shell," or at most the two innermost (inner and outer
medullary shells), all the others being cortical.
A. The distinction between medullary and cortical shells was originally based in my Monograph
(1862, p. 50) upon the topographical relation of the lattice-shells to the central capsule,
inasmuch as I regarded all intracapsular shells as medullary, all extracapsular as cortical.
Hertwig, however (1879, p. 122), rightly pointed out that this distinction is unpractical,
"because the same lattice-shell in the same species may lie within or without the central
capsule, according to the size of the latter." He proposes, therefore, to restrict the term
medullary shell to the innermost, and to call all the others cortical; a course which seems
justified by the special significance of the primary innermost lattice-shell ("as the point of
origin of the radial spines"). But in most Sphærellaria which form
three or more concentric shells, the two innermost, which lie near together within the {lxxxvi}central capsule, are very different in size and
dictyosis from all the others which lie outside, and are separated by wider interspaces (compare
Pls. 17,
24, 29-32, 40, &c.).
In these cases it appears better to regard the two inner as inner and outer medullary shells,
and all the others as cortical shells. The character of the dictyosis in the intracapsular and
extracapsular shells is often so different that I have made it the basis of separation of
Thecosphæra and Rhodosphæra among the Liosphærida (p. 60),
of Elatommatida and Diplosphærida among the Astrosphærida (p. 208),
&c.
B.—R. Hertwig (1879, L. N. 33, pp. 40, 123) separates
the true (simultaneously formed) "cortical shells" (e.g., of Actinomma,
Cromyomma) from the arachnoid "siliceous networks" (e.g., of Diplosphæra
and Arachnosphæra) which are formed by the successive union of tangential apophyses of
the radial spines. Whether this principle is right in the theory or not, it cannot be carried
out practically. Compare also Pl. 25,
fig. 4.
130. Dictyosis or Lattice Formation of the Skeleton.—In the great
majority of Radiolaria the dictyosis or formation of lattice-work, and especially the formation of
a variously-shaped "lattice-shell," plays such an important part that the whole class has long
been popularly known in Germany by the name "lattice animalcules" ("Gitterthierchen" or
"Gitterlinge") (Protista dictyota). The old name Polycystina also (1838), although
referring only to the Spumellaria and Nassellaria,
is derived from the lattice-work of the siliceous skeleton. The extremely various forms in which
this is manifested furnish the means of distinguishing species. The specific conformation of the
skeletal lattice-work is usually caused by the special disposition of the sarcodictyum (§ 94), whose exoplasmatic threads become silicified or (in the Acantharia) converted into bars of acanthin. In many cases, however, the form of
the lattice is mainly dependent upon the situation and form of the radial spines or of special
processes from them. With respect to their origin, two varieties of lattice may be
distinguished—simultaneous and successive. Simultaneous dictyosis occurs especially
in the simple lattice-shells of the Sphærellaria and Phæodaria, where, at a given moment ("dictyotic moment") the whole
lattice of the shell is excreted on the surface of the calymma. Successive dictyosis, on
the other hand, is found more particularly in the lattice-shells of the Acantharia (and in the concentric cortical shells of many Sphærellaria), which develop from the separate lattice-plates formed by the
apophyses of the radial spines, and hence not at the same moment. The lattice-shells of the Cyrtellaria, which gradually grow out from a sagittal ring or a basal tripod,
arise by successive dictyosis.
131. Dictyosis of the Spumellaria.—Siliceous lattice-structures are
wanting in the first section of the Spumellaria, the Collodaria, but in the second section, Sphærellaria,
they are developed in extraordinary variety of details. In spite of this extreme richness in
different forms, the lattice-shells of the Spumellaria may all be derived
from one and the same primitive ground-form, a simple lattice-sphere with regular hexagonal meshes
(Phormosphæra, p. 61, Pl. 12, figs. 9-11;
Heliosphæra, &c.). {lxxxvii}The siliceous bars which bound these regular and subregular
meshes are at first exceedingly then and filiform; afterwards they become thicker or spread out
laterally, whence the meshes often become round with a hexagonal frame (Pl. 12, fig. 5; Pl.
28, fig. 1).
If the latter vanish, a lattice-shell with simple circular meshes is formed. Very commonly the
regular form of the meshes or pores becomes more or less irregular, polygonal, or roundish. Hence,
in general, four different principal forms of dictyosis may be distinguished among the Spumellaria; viz. (1) regular or subregular hexagonal meshes; (2) regular
or subregular circular meshes; (3) irregular polygonal meshes; (4) irregular
roundish meshes. The three latter forms are to be regarded as secondary, derived from the
primary first form. In those Spumellaria which possess several concentric
lattice-shells enclosed one within another, either these have all the same form of dictyosis, or
the lattice-work of the innermost primary shell is different from that of the outer secondary
shells (Pls. 19,
20);
sometimes these latter also differ more or less among themselves (§ 129).
132. Dictyosis of the Acantharia.—The lattice-structures of the Acantharia differ essentially from those of other Radiolaria in several
particulars. Firstly, they consist not of silica but of acanthin (§ 102);
secondly, they are always secondary formations, usually developed from transverse processes of the
primary centrogenous radial spines; thirdly, their formation is not simultaneous (at the same time
over the same shell), but successive (proceeding from the individual radial spines tangentially
towards the middle of the intervals); fourthly, the configuration of the network is due to the
relative position of the spines and the mode of union of their transverse apophyses. Since they
are at right angles to the spines, and since the branches of the apophyses are at right angles to
them, the original ground-form of their dictyosis is a lattice-work with quadrangular meshes;
these are often quite regular and square (Pl. 130,
figs. 5, 6; Pl. 136,
figs. 2, 9, &c.); more commonly they are rectangular or irregularly quadrangular (Pl. 131, fig. 10;
Pl. 133,
figs. 2, 3, &c.). In the majority of the Acantharia the quadrangular
form of the meshes passes over into an irregularly polygonal or roundish one (Pls. 137, 138). Very
often the primary meshes of the lattice-shells, which immediately surround the radial spines, are
larger and more regular ("aspinal pores"), whilst the numerous secondary meshes between them are
smaller and irregular ("coronal pores"; Pl. 135,
figs. 1-4, &c.).
133. Dictyosis of the Nassellaria.—The siliceous lattice-structures of
the Nassellaria are formed on the whole like those of the Spumellaria, with which they were formerly united under the name "Polycystina."
In this group also there may be distinguished as two main forms the regular and irregular. In the
Nassellaria the regular lattice-structures generally exhibit hexagonal or
circular meshes, whilst the irregular are either polygonal or roundish; the irregular forms are,
however, much more abundant than the {lxxxviii}regular, and a further distinction from the Spumellaria consists in the fact that the primary skeletal elements, from which
the lattice is secondarily developed, exercise a predominant influence upon their form. These
primary elements in the majority of the Nassellaria are to be seen in two
morphologically most important structures:—first, the primary sagittal ring, which
embraces the central capsule in the median plane (§ 124); and secondly, the
basal tripod (§ 125), whose three diverging rays proceed from the
base of the central capsule, whilst commonly a fourth vertical ray supports the dorsal side of
latter (compare Pls. 81-91, p. 892). In the majority
of the Nassellaria these two primary elements appear in combination,
whilst in others only one of them is recognisable. In addition there occur numerous monaxon
lattice-shells in which neither of these elements can be recognised, but a simple ovoid
lattice-shell (cephalis) alone forms the whole skeleton or its primary part (Pl. 51, fig. 13;
Pl. 98, fig.
13). The great difficulty in the morphological interpretation and phylogenetic derivation of the
Nassellarian skeleton lies in the fact that each of these three
elements—the primary sagittal ring, the basal tripod, and the latticed cephalis—may
form the whole skeleton by itself or be combined with one or both of the others (p. 893). Even nearly
related or at all events very similar forms may differ very greatly in this respect. With regard
to the manifold forms of their dictyosis it follows that it is partly dependent upon one of the
two first elements, partly independent. In the Plectellaria (or those
Nassellaria which do not possess a complete lattice-shell) the
lattice-work is usually irregular and arises by union of the ramifications, which proceed either
from the primary sagittal ring (Pls. 81,
82, 92-94) or from the
basal tripod (Pl. 91).
In the Cyrtellaria (or Nassellaria with a
complete lattice-shell, Pls. 51-80), on the
other hand, the lattice-work is sometimes regular, sometimes irregular, being often very different
in the different joints of a segmented shell (Pl. 72); a great
part of it arises independently of the two chief morphological elements, and develops according to
laws similar to those which regulate the dictyosis of the Spumellaria.
134. Dictyosis of the Phæodaria.—The lattice-structures of the Phæodaria, which consist of a silicate of carbon (§ 102),
are on the whole not developed in such variety as those of the other Radiolaria, but exhibit
several essentially different types of structure, not reducible to a common primitive type of
lattice-work. In one portion of this legion there occurs an ordinary simple lattice-work (as in
Spumellaria and Nassellaria), with solid
trabeculæ; of these the Castanellida (Pl. 113)
and Concharida (Pls. 123-125) have
usually regular or subregular, circular meshes, sometimes hexagonally framed; the Orosphærida
(Pls. 106,
107) large
irregular polygonal meshes with thick trabeculæ, the Sagosphærida (Pl. 108) large
triangular meshes with thin filiform trabeculæ. The Challengerida (Pl. 99) are
characterised by a very delicate regular lattice-work, with minute hexagonal pores, like a
Diatomaceous frustule. The Medusettida (Pls. 118-120) {lxxxix}show a peculiar alveolar structure, numerous small
compartments being enclosed between two parallel plates. In the Circoporida (Pls. 114-117) and
Tuscarorida (Pl. 100)
the opaque porcellanous shell has a peculiar cement structure (§ 104), and
the lattice-structure is confined for the most part to characteristic rings of pores at the base
of the hollow tubes, which arise from the shell. The most peculiar lattice-work, however, appears
in the segmented shell of the Aulosphærida (Pls. 109-111) and
Cannosphærida (Pl. 112).
In the former the large meshes of the lattice-work are usually subregular and triangular, in the
latter polygonal; the trabeculæ are hollow cylinders, filled with jelly, and containing usually a
central axial thread. In each nodal point of the lattice, in which three or more tangential tubes
meet, these are separated by stellate or astral septa.
135. Radial Spines of the Skeleton.—The skeleton in the great majority
of Radiolaria is armed with radial spines, which are of great importance in the development of
their general form and of their vital functions. From a morphological point of view the number,
arrangement, and disposition of the spines is usually the determining factor as regards the
general form of the skeleton. Physiologically they discharge distinct functions, as organs of
protection and support; they act also, like the tentacles of the lower animals, as prehensile
organs, since their points, lateral branches, barbed hooks, &c. serve to hold fast nutritive
materials. In general main-spines and accessory spines may be distinguished in most Radiolaria;
the former are of pre-eminent importance in determining the figure of the skeleton; the latter are
merely appendicular organs. The main-spines present such characteristic and important differences
in the various legions of Radiolaria that they must be considered separately.
136. Radial Spines of the Spumellaria.—The radial spines, which
exhibit most manifold variations in the large order Sphærellaria, present
characteristic differences in its four suborders. In the Sphæroidea their
number and disposition serve for the separation into families (p. 59); the
Cubosphærida (Pls. 21-25) always
possess six radial main-spines, which stand opposite to each other in pairs and lie in three
diameters of the shell, which are at right angles to each other and correspond to the axes of the
regular crystallographic system. The Staurosphærida (Pl. 15) have four
spines, which form a regular cross and stand opposite to each other in pairs, in two axes at right
angles. The Stylosphærida (Pls. 13-17) show only
two main-spines, which are opposed to each other in the vertical main axis of the body. Finally,
the Astrosphærida (Pls. 18-20, 26-30) are
characterised by a larger and variable number of radial spines (eight, twelve, twenty or more),
sometimes regularly, sometimes irregularly arranged. Among the other Sphærellaria the Prunoidea (Pls. 13-17, 39, 40) are most
allied to the Stylosphærida with two opposite main-spines; the Discoidea
(Pls. 31-47), on the
other hand, to the Staurosphærida with four crossed spines; there exist, however, Discoidea with two opposite, three marginal, or numerous radial main-spines; it
is {xc}characteristic of this suborder that they all
usually lie in the horizontal median plane of the lenticular shell, arising from its equatorial
margin. The Larcoidea (Pls. 9, 10, 49, 50) show a
great variety in the number and arrangement of their radial main-spines, which in the different
families of this suborder stand in direct causal relation to the various forms of growth of the
shell; usually the primary main-spines lie either in the three different dimensive axes, at right
angles to each other, whose differentiation is characteristic of the lentelliptical Larcoid shell
(§§ 34, 122) or in definite diagonal axes, which cut
the former obliquely. The radial spines of the Spumellaria are
never united in the centre of the body, but arise separately from the surface of the
primary central lattice-shell (medullary shell), more rarely from one of the secondary (cortical)
shells, which enclose it. Their form is originally three-edged (sometimes pyramidal, sometimes
prismatic); the cause of this is to be found in their origin from the nodal points of the
lattice-shell, whose meshes are primitively hexagonal; hence three trabeculæ unite in each nodal
point, and are produced into three edges of the spine. Very commonly, however, the spines are
round (conical or cylindrical), more rarely polygonal. The three edges are often delicately
toothed, not unfrequently spirally twisted around the axis of the spine (Pl. 21, figs. 1,
12).
137. Radial Spines of the Acantharia.—The radial spines of this legion
have a much greater significance than in the other three classes of Radiolaria, since here alone
they are the primary determining factors in the skeletal structure, and grow outwards from the
middle of the central capsule. This centrogenous origin of the radial spines is as characteristic
of the Acantharia as their chemical constitution, which is not siliceous
but acanthinic (§ 102). Furthermore, their form is in most cases so
peculiar that even an isolated Acantharian spine can be generally
distinguished from one belonging to either of the other three legions. In the great majority of
the Acantharia (all Acanthonida and Acanthophracta) twenty radial spines are constantly present, which, disposed
according to a definite geometrical law, make up the skeleton (compare § 110 above and p. 717). The twenty spines are generally
simply apposed to each other in the centre (either by the surfaces or the edges of their pyramidal
base); more rarely they are completely united and form a single star-like piece of acanthin
(Astrolithium). Very rarely (Acanthochiasma) each two opposite spines are united so
that ten diametric bars cross in the middle of the central capsule. Whilst in the great majority
of Acantharia these twenty radial spines are present, the small group
Actinelida is characterised by the possession of an inconstant, often
very large number, sometimes over one hundred. Among these Actinelida are
probably to be found the stem-forms of the whole legion. The variously modified spines of the
Acantharia may be grouped in three main categories: (1) round (cylindrical
or conical); (2) four-edged (prismatic or pyramidal); (3) two-edged (leaf- or sword-shaped). The
latter very commonly bear two {xci}opposite transverse
processes, the former four crossed ones. By ramification and union of these apophyses arise the
lattice-shells of the Acanthophracta (excepting the Sphærocapsida).
138. Radial Spines of the Nassellaria.—The radial spines in this
legion show as great a variety in their form as in the Spumellaria, and,
as in them, are solid, siliceous bars, usually three-edged (prismatic or pyramidal), or round
(cylindrical or conical); more seldom they are polygonal in section. The great majority of the
Nassellaria are, however, distinguished by a triradial structure, three
primary radial bars diverging from the base of the central capsule (usually from the centre of the
porochora); there is usually in addition a fourth apical spine, which passes upwards vertically or
obliquely on the dorsal aspect of the central capsule. These three or four typical radial spines
of the Nassellaria may be derived with great probability from the basal
tripod of the Plectoidea (Plagoniscus, Plectaniscus,
&c., Pl. 91);
and since this tripod is very characteristically combined in Cortina and Cortiniscus
with the primary sagittal ring of the Stephoidea, the three typical rays
may be generally designated "cortinar feet," in contradistinction to the other radial processes of
the Nassellarian skeleton. One of the three descending basal feet ("pes
caudalis," Pls. 91-95, C) is always unpaired, and lies in the vertical median plane (or sagittal
plane), just as does the vertically directed apical spine, which originally forms the dorsal bar
of the sagittal ring, and is produced upwards into the "apical horn," (marked a on the
plates). The other two basal feet are paired, and diverge right and left, forwards and downwards
("pedes pectorales," p.p.). Six-rayed Nassellaria, in which three
secondary (interradial) feet are intercalated between the three primary (perradial) cortinar feet,
are less common than the three-rayed forms. In some groups the number rises still higher, nine,
twelve, or even more secondary feet being intercalated between the three primary. Besides,
accessory radial spines may be developed on different parts of the shell, which have sometimes a
definite relationship to the typical radial spines, sometimes not. Their form and ramification are
very various (Pls. 51-98).
139. Radial Spines of the Phæodaria.—The radial spines of the Phæodaria are very clearly distinguished from those of other Radiolaria by the
fact that they are usually hollow tubes, rarely solid bars. As a rule, the tubes are cylindrical,
often slightly fusiform or conical, their siliceous wall is very thin, and their lumen filled with
jelly; a fine thread of silica usually runs in the axis, and in several families is connected by
fine transverse threads with the wall of the tube (Pl. 110, figs. 4,
6; Pl. 115,
figs. 6, 7). The peculiar family Medusettida is characterised by a very remarkable segmentation of
the hollow spines (Pls. 118-120). Each
tube is divided by a series of septa into chambers, which communicate by a central or excentric
opening in each septum, an arrangement resembling the siphuncle of the chambered Cephalopod
shells. The number and {xcii}arrangement of the radial
tubes in most Phæodaria is indefinite and very variable; only in a few
families is the number constant in each species and genus, and the disposition regular. The
Medusettida (Pls. 118-120) resemble
the Nassellaria, inasmuch as equal radial feet diverge from the base of
their shell, sometimes three in number (Cortinetta, Pl. 117, fig. 9),
sometimes four (Medusetta, Pl. 120,
figs. 1-4), sometimes six (Gazelletta); Gorgonetta is specially distinguished by the
possession of six ascending and six descending spines regularly alternating (Pl. 119). The
Tuscarorida (Pl. 100)
usually have three or four equidistant feet. The Circoporida (Pls. 115-117), on the
other hand, rather approach the Sphæroidea, their spherical or regular
polyhedral shell having a definite number of tubular radial spines, which arise at regular
intervals from their angles; Circoporus has six, Circospathis nine,
Circogonia twelve, and Circorrhegma twenty radial tubes. Very rarely the tubes of
the Phæodaria are angular, usually they are round, more or less
cylindrical, though they are often bifurcated or even ramified, and exhibit a great wealth of the
most delicate appendages; siliceous hairs, bristles, spines, barbed or anchor-like hooks,
spathillæ, brushes, circlets, &c. (compare Pls. 99-128).
140. Main-Spines and Accessory Spines.—As accessory spines (Paracanthæ) we oppose
to the main-spines (Protacanthæ), just described, all those processes which have no determining
influence upon the formation of the skeleton as a whole, but are to be regarded as secondary
constituents of the skeleton, or appendicular organs of inferior significance. They are developed
in the utmost variety, sometimes as hairs or bristles, sometimes as thorns or clubs, either
straight or curved (often zigzag), smooth or barbed; sometimes standing vertically upon the shell,
or directed towards the centre, sometimes obliquely, or rising at a definite angle. In those Spumellaria whose lattice-shell consists of several concentric spheres, the
accessory spines generally arise from the outermost, the main-spines, on the contrary, from the
innermost. In the Nassellaria, multifarious forms of accessory spines are
especially developed in the order Plectellaria. In the Phæodaria they are often furnished with delicate appendages, e.g.,
anchor-hooks, spathillæ, coronets, &c. Among the Acantharia the
accessory spines which arise from the surface of the shell in the Acanthophracta are very characteristic. They are not radially disposed (like
the similar superficial spines of the Spumellaria), but parallel to the
radial main-spines from whose transverse processes they arise. Since in all these Acanthophracta the twenty radial main-spines are opposite to each other in
pairs, all the accessory spines (often several hundred) are parallel to ten different regularly
disposed axes of the lattice-shell (Pls. 134-138).
The skeletons of the Radiolaria, in addition to the general relations which have
been discussed above, present numerous and important special differences in the various larger
and smaller groups. These are indicated in detail in the descriptions of the legions, orders,
and families in the systematic portion of this Report.
{xciii}
BIOGENETICAL SECTION.
A SKETCH OF OUR KNOWLEDGE OF THE DEVELOPMENT OF THE
RADIOLARIA IN THE YEAR 1884.
Chapter V.—ONTOGENY OR INDIVIDUAL DEVELOPMENT.
(§§ 141-152.)
141. Individual Developmental Stages.—The germinal history of the
Radiolaria presents great obstacles to direct observation, and hence is very incompletely known.
The fragmentary observations, however (having been made on Radiolaria of very various groups and
supplemented by comparative anatomical considerations), allow us to draw a general picture of the
essential developmental processes in this great class. It may probably be assumed that in all
Radiolaria, after maturation, the central capsule discharges the function of a sporangium, and its
contents are broken up into numerous flagellate swarm-spores (zoospores). After these flagellate
swarm-spores (resembling Astasia) have emerged from the ruptured central capsule, they
probably pass over into a Heliozoan-stage (Actinophrys) and then after the formation
of a jelly-veil into the condition of Sphærastrum. Afterwards, when a membrane is formed
between the outer jelly-veil and the inner nucleated cell-body, an Actissa-stage arises,
which exhibits in its simplest form the differentiation of the spherical unicellular body into the
central capsule and calymma. Actissa thus represents both ontogenetically and
phylogenetically the primitive condition of the Radiolarian organism, and may thus be regarded as
the point of departure of all other forms.
142. The Astasia-Stage.—The formation of flagellate zoospores in the mature
central capsule is probably to be regarded as the common form of individual development in all
Radiolaria; since the whole contents are utilised in the formation of these swarm-spores, and
since the extracapsulum takes no share in the process and perishes after they are evacuated, the
central capsule may be regarded as a sporangium (see note A, below). The zoospores
of the Radiolaria generally arise in the following way:—the nucleus of the unicellular
organism, sometimes early, sometimes late (and in several different ways, §§ 63-70) breaks up into numerous small nuclei, and each of
these surrounds itself with a small portion of the endoplasm. Very often, perhaps generally, this
endoplasm contains one or several fat-granules and sometimes also a small oblong crystal; from the
protoplasm {xciv}of the small roundish or ovoid cells
protrudes one or more vibratile flagella. The fully developed spores, which commence their
vibrations even within the central capsule, emerge when it ruptures, and swim about freely in the
surrounding water by means of the flagellum. At this stage of its existence the young Radiolarian
represents essentially the simplest form of the Flagellata, such as Astasia or
Euglena; the unicellular body is for the most part ovoid or subcylindrical, sometimes
fusiform or reniform, usually from 0.004 to 0.008 mm. in diameter (Pl. 1, fig.
1c; Pl. 129,
fig. 11). In the anterior part of the flagellate cell, immediately behind the base of the
flagellum, lies a homogeneous, spherical nucleus, whilst in the posterior part are usually several
small fat-granules and often also a small oblong crystal (hence the name "crystal-spore,"
"Krystall-Schwärmer"). The number of vibrating flagella, which are extremely long and fine, seems
to be variable, usually one, sometimes two, occasionally perhaps three, or even four or more (see
note B).
A. The formation of the motile spores in the central capsule was first observed by J. Müller
in Acanthometra (1856, L. N. 10, p. 502), then by A. Schneider in
Thalassicolla (1858, L. N. 13, p. 41), and finally by myself in
Sphærozoum (1859, L. N. 16, p. 141). These older observations were,
however, incomplete, for the origin of the motile corpuscles from the contents of the central
capsule was not observed. The first complete and detailed observations upon the formation of
spores in the Radiolaria were published in 1871 by Cienkowski (L. N. 22, p.
372, Taf. xxix.); they relate to two different Polycyttaria, Collosphæra and
Collozoum. These investigations were supplemented by R. Hertwig on Collozoum and
Thalassicolla (1876, L. N. 26, pp. 28, 43, &c.); on
Collozoum he made the important discovery that the Polycyttaria form two kinds of spores,
one with and the other without crystals, and that the latter are divided into macrospores and
microspores (compare the chapter on "Reproduction," §§ 212-216). Quite recently Karl Brandt has confirmed these observations, and has
extended them to all the genera of Polycyttaria (1881, L. N. 38, p. 393, and
1885, loc. cit.).
B. The number of flagella, projecting from each spore, is very difficult to
determine, owing to their extraordinary length and slenderness. It appeared to me that in the
majority of those Radiolaria whose spores I investigated only a single flagellum could be
demonstrated with certainty, although sometimes two, springing from a common base, seemed to be
present. Compare the chapter on "Reproduction," (§ 215) and the recent
work of Karl Brandt on Sphærozoea
(1885, L. N. 52, pp. 145-174).
143. The Actinophrys-Stage.—The fate of the flagellate zoospores which
emerge from the mature central capsule of the Radiolaria has not hitherto been decided by actual
observation; all attempts to rear the swarming zoospores have been in vain, for they have soon
died. From what we know, however, of the comparative morphology of the Protista, the hypothesis is
fully justified, that between the Astasia-stage of the flagellate swarm-spores, and the
well-known Actissa-stage of the simplest Radiolaria, there lies an intermediate
developmental stage, which may be regarded as being essentially the simplest Heliozoan form,
Actinophrys or Heterophrys. The swarm-spore is very probably converted directly in
to a simple floating Heliozoon by its elongated or ovoid body {xcv}becoming spherical and by fine pseudopodia protruding all round instead
of a single flagellum; the nucleus at the same time assuming a central position.
144. The Sphærastrum-Stage.—The Actinophrys-stage of the young Radiolaria,
which proceeds immediately from the flagellate zoospore, is probably connected with the
Actissa-stage by an intermediate form, which may be regarded as a simple skeletonless
Heliozoon with a jelly-veil; a well-known example of such a form is Sphærastrum (in
the solitary, not the social condition) and Heterophrys. This important intermediate form
has arisen from the simple Actinophrys-stage by the excretion of an external structureless
jelly-veil, such as is formed in many other Protista (e.g., in the encystation of many
Infusoria). The young Radiolarian in this second Heliozoon-stage becomes a simple cell with
pseudopodia radiating on all sides; its body consists of three concentric spheres, the central
nucleus, the protoplasmic body proper, and the surrounding calymma or jelly-veil. When a firm
membrane is developed between the last two spheres this Sphærastrum-stage passes over into
the Actissa.
The gap in our empirical knowledge which still exists between the flagellate
stage (§ 142) and the simplest Radiolarian stage (Actissa, § 145), can be filled hypothetically only by the assumption of several
Heliozoon-stages following one upon another. It is possible also that the
capsule-membrane is not formed between the endoplasm and exoplasm (as here supposed), but that
the membrane was formed first outside the cell and the extracapsulum subsequently secreted
around it.
145. The Actissa-Stage.—The first Spumellarian
genus, Actissa, is not only the simplest form actually observed among the Radiolaria, and
the true prototype of the whole class, but also the simplest form under which the Radiolarian
organisation can be conceived. It is therefore extremely probably that Actissa not only
forms the common stem-form of the whole class in a phylogenetic sense, but is also its common
ontogenetic or germinal form. Probably in all Radiolaria the Sphærastrum-stage develops
immediately into the typical Actissa-stage, by the formation of a firm membrane between the
protoplasmic body of the spherical Heliozoan cell and its jelly-veil. Thus arises the
characteristic central capsule, which is wanting in the nearly related Heliozoa. It is further
probable that all Radiolaria in their early stage will so far conform to the state of things in
Actissa as to have the capsule-membrane of the spherical skeletonless cell perforated
everywhere by fine pores. This structure is retained in all Spumellaria,
whilst in the other three legions those structural relations of the capsule which are
characteristic of each develop from the Actissa-stage.
146. The Ontogeny of the Spumellaria.—In the simplest case the
individual development in the Spumellaria ceases with the
Actissa-stage. In all other genera of this legion diverging forms proceed from this, of
which the different growth of the three dimensive {xcvi}axes on the one hand (§§ 44, 45), and the differentiation of the various parts of the unicellular organism
with the formation of the skeleton on the other, are of pre-eminent significance. Even in the
varying growth of the central capsule in the different dimensions of space in the skeletonless
Colloidea, four different modes may be distinguished, which further, in
the corresponding development of the skeleton, furnish the basis for the origin of the four orders
of Sphærellaria. The most primitive and simplest form of growth, equal
extension in all directions, is found in the spherical central capsule and the concentric
spherical skeletons (Procyttarium, Sphæroidea). When the growth of
the central capsule proceeds more rapidly in the direction of the vertical main axis than in any
other direction, the ellipsoidal or cylindrical central capsule (Actiprunum) arises, and
the vertically elongated skeleton of the Prunoidea, which is derived from
it. When, on the contrary, the growth of the central capsule and lattice-shell is less in the
direction of the vertical main axis than in any other direction, the lenticular or discoid central
capsule (Actidiscus) arises, and the corresponding lenticular shell of the Discoidea. Finally, even quite early in many Spumellaria, the growth of the central capsule and of the corresponding
lattice-shell in the three dimensive axes is different, and hence arise the lentelliptical forms
whose geometrical type is the triaxial ellipsoid or the rhombic octahedron (Actilarcus,
Larcoidea). Thus the origin of the four orders of Sphærellaria is simply explained by a varying growth in the different dimensive
axes. The primary (innermost) lattice-shell is in this legion always simultaneously
developed (suddenly excreted at the moment of lorication from the sarcodictyum). The
secondary lattice-shells, on the other hand, which surround the former concentrically, and
are united with it by radial bars, arise successively from within outwards.
147. The Ontogeny of the Acantharia.—The individual development of the
Acantharia in the simplest case (Actinelius) stops at a point which
differs from the Actissa-stage only in the change of radial axial threads into acanthin
spines. In the small group Actinelida, their number remains variable and
usually indeterminate (Adelacantha), whilst in the great majority of the legion (Acanthonida and Acanthophracta) the number is
constantly twenty, and those spines are regularly arranged according to the Müllerian law in five
parallel circles, each containing four crossed spines (Icosacantha). The simplest form among these
latter is Acanthometron, which may be regarded both ontogenetically and phylogenetically as
the common starting-point of all the Icosacantha. Within this extensive group variations in the
length of the dimensive axes appear, similar to those observed in the Spumellaria. In the Astrolonchida and Sphærophracta the
central capsule remains spherical, extending equally in all directions; and correspondingly the
lattice-shell, which is excreted on the surface of the spherical calymma, remains spherical. In
the Belonaspida (just as in the Prunoidea) {xcvii}this form passes over into an ellipsoid by prolongation of one axis;
on the contrary, in the Hexalaspida (as in the Discoidea) the discoidal
or lenticular form arises by shortening of an axis. Finally, in the Diploconida, and in some
Hexalaspida in which the growth is different in all three dimensive axes (as in the Larcoidea), both the central capsule and the shell assume the lentelliptical
form. The lattice-shell of the Acanthophracta is usually successive in
its development, since from each of the twenty radial spines two or four tangential apophyses
proceed, whose branches subsequently unite and combine to form the lattice-shell. Only in the
peculiar Sphærocapsida can the pavement-like shell arise simultaneously or in a moment of
lorication.
148. The Ontogeny of the Nassellaria.—The individual development of
the Nassellaria in the simplest instance remains stationary at the
skeletonless Nasselid stage (Cystidium, Nassella), which can be immediately derived
from the foregoing Actissa-stage by the disappearance of the pores in the upper (apical)
hemisphere of the central capsule, whilst in the lower (basal) portion they are modified to form a
porochora; the podoconus is developed within the endoplasm upon this latter. Usually the spherical
form of the central capsule passes over into an ovoid or ellipsoidal one, the vertical axis which
passes through the centre of the porochora being elongated. From the skeletonless Nassellida the
other Nassellaria may be derived both ontogenetically and phylogenetically
by the excretion of an extracapsular siliceous skeleton. Unfortunately, the earliest stages in the
formation of this skeleton are unknown, and hence no answer can at present be given to the
important question, in what order the three primary skeletal elements of the Nassellaria (the basal tripod, sagittal ring, and latticed cephalis) appear
(compare §§ 111 and 182). If, for example, in
Cortina and Tripospyris the basal tripod were to appear first in the ontogeny, and
the sagittal ring were developed from this, then the Plectoidea would be
rightly considered to be the oldest forms in the phylogeny of the skeleton-forming Nassellaria; and in the contrary case the Stephoidea
would be so regarded. The relations of growth in the three dimensive axes are very variable in the
Nassellaria; the three most important factors in this respect (partly
separately and partly in combination) are; (1) the development of the basal tripod to a triradial
stauraxon form (the ground-form being a three-sided pyramid); (2) the development of the sagittal
ring in the median plane of the body (the vertical axis having the poles different); (3) the
development of the latticed cephalis outside the central capsule (the poles of the vertical axis
being again different). Since the development both of the skeleton and of the malacoma is
characterised in most Nassellaria by the stronger growth of the vertical
axis and the differentiation of the two poles, the allopolar monaxon ground-form acquires a
predominant significance in this legion (§ 32); the starting point of most
of the further modifications is the basal pole of the vertical main axis. Next to this the
sagittal axis is usually the most important determining factor (its dorsal and ventral poles being
{xcviii}usually different), more rarely the frontal
axis (with equal right and left poles). In the zygothalamous Spyroidea
(as in the Stephoidea) the formation of the shell proceeds from the
sagittal ring, whilst in the polythalamous Cyrtoidea the latticed
cephalis is always the starting point, from which a series of joints (thorax, abdomen, and in the
Stichocyrtida, the numerous post-abdominal joints) successively arise (unipolar growth).
149. The Ontogeny of the Phæodaria.—The individual development of the
Phæodaria in the simplest case stops with the skeletonless condition of
the Phæodinida (Phæodina, Phæocolla), which can be immediately derived from the
foregoing Actissa-stage by the disappearance of the pores in the greater part of the
central capsule, the characteristic astropyle being developed at the basal pole (§ 60). Since this particular form and structure of the spheroidal central capsule
remains the same in all Phæodaria, whilst the formation of their skeleton
follows very different directions, it follows that further common paths of development are
excluded both ontogenetically and phylogenetically. What will be laid down in this respect as
regards the phylogeny of the different groups of Phæodaria (§§ 194-199) holds true also of their ontogeny. The
relations of growth in the three dimensive axes are hence very different in the skeletons of the
various groups of Phæodaria. This difference is best marked in the Phæoconchia, whose bivalved lattice-shells have as their ground-form the
rhomboid pyramid of Ctenophora. In most Phæogromia the monaxon
lattice-shell may develop simultaneously by sudden excretion at a particular moment of lorication;
this is also the case with the polyaxon lattice-shells of the Phæosphæria. In their future growth the development of basal or radial
apophyses is of special importance. In the majority of the Phæodaria these
apophyses are tubes of silicate filled with jelly (often provided with an axial siliceous thread);
thus their development is distinguished by complications which are absent in the case of the other
three legions.
150. Growth.—The growth of the body in the Radiolaria, as in all other organisms,
is the fundamental function of individual development (see note A). All structural relations which
this richest class of the Protista exhibits may be referred to different forms of growth, either
of the unicellular malacoma or of the skeleton which it produces. In general the special
development of the skeleton is dependent upon that of the central capsule, and of the sarcodictyum
on the surface of the calymma; in the further growth, however, the conditions are reversed, and
the condition of the skeleton already formed directly determines the further development of the
central capsule and of the calymma with its sarcodictyum. The four legions of Radiolaria show,
speaking generally, certain characteristic differences in growth, which are due in great part to
the different structure and ground-form of their central capsule. In the two legions of the
Porulosa (Spumellaria and Acantharia), in which
the central capsule is originally spherical and {xcix}the ground-form of the skeleton either polyaxon or isopolar monaxon,
two fundamental and variously combined directions of growth are recognisable; firstly, the
concentric growth (equal increase of volume in all directions), and secondly, multipolar or
diametral growth (hypertrophy of certain parts in the direction of definite pairs of
radii). A different state of things obtains, however, for the most part, in the two legions of the
Osculosa (Nassellaria and Phæodaria), in which the
central capsule possesses a vertical main axis with different poles, and the structure of the
skeleton is determined by this allopolar monaxon ground-form. The two fundamental directions of
growth here combined in the most various ways are, firstly, unipolar growth (starting from
the basal pole of the vertical main axis), and secondly, radial or pyramidal growth
(characterised by the different development of separate parts in the direction of definite radii).
Whilst the growth of the malacoma is dependent on intussusception (as in most organic
structures capable of imbibing), the growth of the skeleton in all Radiolaria takes place
by apposition (see note B).
A. The earliest investigations into the modes of growth in the Radiolaria are due to J.
Müller (L. N. 12, pp. 21-33). More detailed communications I gave myself in
my Monograph (L. N. 16, pp. 150-159). The relations there sketched have now,
in consequence of the examination of the Challenger collection, undergone many important
additions, and in some divisions, important modifications; these are for the most part treated
of in the general account of the separate families.
B. The view here maintained, that the skeleton of all Radiolaria grows only by
apposition, appeared formerly to have certain exceptions. I thought I had shown that in
Cœlodendrum the thin-walled tubes grew not only in length but also in thickness,
with continuous increase in the lumen (L. N. 16, pp. 152, 360). Further K.
Brandt concluded, from the varying size of the median bars in the twin-spicules of
Sphærozoum, that these siliceous structures grow by intussusception (L. N. 38, p. 401). Both suppositions have been proved erroneous and I have come to
the opinion that in all Radiolaria the skeleton grows by apposition.
151. Regeneration.—Whilst the general course of individual development
(perhaps without any exception in the Radiolaria), begins with the formation of zoospores in the
central capsule, there yet occurs in some groups a different form of ontogeny, introduced by
simple division of the unicellular organism, and coming under the term "regeneration" in its wider
sense. This spontaneous division occurs quite commonly in the Polycyttaria (or social Spumellaria), and produces their colonies (compare the chapter on Reproduction,
§ 213). On the
contrary, it has not been observed in the solitary Spumellaria, nor in the
Acantharia and Nassellaria; possibly, however, the
peculiar Acantharian family, Litholophida, has arisen by the division of
Acanthonida (compare p. 734). Among the Phæodaria
division is commonly observed in the order Phæocystina (which have an
incomplete Beloid skeleton or none), and also in the Phæoconchia. In all
these cases the increase by division is nothing else than an ordinary case of cell-division, in
which bisection of the nucleus precedes that of the central capsule. The regeneration by {c}which each of the two daughter-cells develops to a complete
mother-cell depends upon simple growth. Another form of regeneration, different from this, has
been observed in Thalassicolla. If the central capsule be extracted artificially from the
large concentric calymma, the enucleated central capsule produces a new extracapsulum, with
sarcomatrix, pseudopodia, and calymma. This experiment may be repeated several times with the same
result. (Compare A. Schneider, 1867, L. N. 20.)
152. The Formation of Colonies.—The individual development of colonies takes place
in all three families of the Polycyttaria (Collozoida, Sphærozoida, Collosphærida) in the same
simple way, by the repeated division of a single monozootic Spumellarian.
Since these divisions only affect the central capsule and not the extracapsulum, the sister-cells,
which arise by repeated division of the mother, remain enclosed in a common rapidly growing
calymma. Probably in all Polycyttaria the commencement of the formation of colonies immediately
follows the Actissa-stage of the monozootic mother-cell (or takes place in the
Thalassicolla-stage, which arises from the former by the development of alveoles in the
calymma). The simple central nucleus separates (by direct nuclear division) into two halves, and
the central capsule follows this process of bisection, becoming constricted in the middle between
the two daughter nuclei (Pl. 3,
fig. 12). In the further growth of the colony the process of division proceeds in the older, now
multinucleate, central capsules, in which an oil-globule has taken the place of the original
nucleus; then the division of the oil-globules precedes that of the central capsule (Pl. 5, fig. 1).
Another mode of growth of the colonies is the multiplication of the central capsules by
gemmulation, or the formation of the so-called "extracapsular bodies" (Gemmulæ, § 214). The characteristic skeletal structure of the different species appears
at a later stage. Whether ripe central capsules can emerge from the social bond of a
cœnobium, and, having become isolated, establish the formation of a new colony, is very
doubtful. The various forms which the cœnobium assumes in the different species of
Polycyttaria, are due partly to simple growth, partly to the development of large vacuoles in the
calymma.
The form and size of the cœnobia appear in many fully developed
Polycyttaria to exhibit specific differences, which require further investigation; in the young
stage, on the contrary, they are simple spheres or ellipsoids, often cylindrical or
sausage-shaped (Pl. 3,
figs. 1, 4, 6, 11). In some species the cylindrical gelatinous bodies become moniliform, and
separated by transverse constrictions into many segments, each of which encloses a large alveole
(Pl. 3,
fig. 10). The rare ring-shape (Pl. 4,
fig. 1) which I figured in 1862 in the case of Collozoum (L. N. 16,
p. 522, Taf. xxxv. fig. 1), I have recently observed in different species of Polycyttaria; it is
capable of a very simple mechanical explanation, both ends of a sausage-shaped colony having
been accidentally brought into contact by a wave and having united by agglutination. Quite
recently Brandt has given a very complete account of the development, form, and growth of
Polycyttarian colonies in his work on the colonial Radiolaria of the Bay of Naples (1885, L. N.
52, pp. 71-85).
{ci}
Chapter VI.—PHYLOGENY OR GENEALOGICAL DEVELOPMENT.
(§§ 153-200.)
153. Sources of Phylogenetic Knowledge.—For the purpose of
constructing a hypothetical genealogical tree of the Radiolaria, as of all other organisms, three
sources of information are open to us, viz., palæontology, comparative ontogeny, and comparative
anatomy. In the present case, however, these three sources are of very different value; the first
two are at present only very inadequately known and have only been partially investigated, hence
they can only be utilised to a very slight extent. The comparative anatomy of the Radiolaria, on
the other hand, is so completely known, and affords such certain glimpses into the morphological
relations of the related groups, that by its aid we are in a position at all events to lay down
the general features of their phylogeny with some probability, and to lay the foundation of a
natural system.
154. Natural and Artificial Systems.—Although in the classification of
the Radiolaria, as in the case of all other organisms, the natural system must be regarded as the
goal of systematic classification, our phylogenetic knowledge of the Radiolaria is too fragmentary
and inadequate to admit of the systematic arrangement here adopted being regarded as a thoroughly
consistent natural system, that is, as representing the true genealogical tree of the class.
Owing, however, to the extraordinary variety of form of the Radiolaria, and the complicated
relationships of the larger and smaller groups, a synoptical grouping of the different categories
and the erection of a complete, even if to some extent artificial, system, becomes a logical
necessity. Under these circumstances, and regard being had to both these conditions, the following
systematic treatment of the Radiolaria will appear as a compromise between the natural and
artificial systems, like all other zoological and botanical classificatory attempts. On the
one hand, the attempt is made to arrange the larger and smaller groups as nearly as possible
according to their phylogenetic relationships, whilst, on the other hand, the practice of
circumscribing each by a definition as clear and logical as possible has been carried out. Since
these two efforts naturally often come into contact, the insufficiency of many parts of the
arrangement is obvious, hence its hypothetical and provisional character is emphatically
stated.
155. Systematic Categories.—The categories or different orders of divisions have
in the Radiolaria, as in all other organisms, no absolute significance, but only a
relative value. In itself it is quite unimportant whether the whole group be regarded, as
at first, as a family (Ehrenberg, 1847), or as an order (J. Müller, 1858), or as a
class (Haeckel, {cii}1881). These different views
are regulated, on the one hand, by the known extent of the group and by the amount of our
acquaintance with it, and on the other, by comparison with related groups and by reference to
their conventional disposition. When, therefore, the whole class, Radiolaria, is here divided into
two subclasses, four legions, eight orders, eighty-five families, &c., these artificial
categories are drawn up only in the conviction that by this means the easiest survey and most
thorough insight into the system as a whole may be attained; this latter will indeed approach as
far as possible the ideal of a natural system, but must on numerous practical grounds always
remain more or less artificial. Since it is to be expected that with the progress of our
systematic knowledge the rank of the various categories will rise, it is possible that in the
future the arrangement of the group may be somewhat as follows:—Phylum, RADIOLARIA;
Four Classes, Spumellaria, Nassellaria,
Acantharia, Phæodaria; Eight Legions (Nos.
I.-VIII. in the following Table); Twenty Orders (Nos. 1-20 in the Table), &c.
| Four Legions. |
|
Eight Sublegions. |
|
Twenty Orders. |
|
Typical Families. |
I. Legion (or Subclass)
Spumellaria
(Peripylea)
[Porulosa peripylea.] |
|
I. Collodaria
(Spumellaria palliata) |
 |
1. Colloidea, |
|
1a. Thalassicollida. |
| 1b. Collozoida. |
| 2. Beloidea, |
|
2a. Thalassosphærida. |
| 2b. Sphærozoida. |
II. Sphærellaria
(Spumellaria loricata) |
 |
3. Sphæroidea, |
|
3a. Ethmosphærida. |
| 3b. Collosphærida. |
| 4. Prunoidea, |
|
4a. Ellipsida. |
| 4b. Zygartida. |
| 5. Discoidea, |
|
5a. Phacodiscida. |
| 5b. Porodiscida. |
| 6. Larcoidea, |
|
6a. Larnacida. |
| 6b. Pylonida. |
II. Legion (or Subclass)
Acantharia
(Cannopylea).
[Osculosa cannopylea.] |
|
III. Acanthometra
(Acantharia palliata) |
 |
7. Actinelida, |
|
7a. Astrolophida. |
| 7b. Litholophida. |
| 7c. Chiastolida. |
| 8. Acanthonida, |
|
8a. Astrolonchida. |
| 8b. Quadrilonchida. |
| 8c. Amphilonchida. |
IV. Acanthophracta
(Acantharia loricata) |
|
9. Sphærophracta, |
|
9a. Sphærocapsida. |
| 9b. Dorataspida. |
| 9c. Phractopeltida. |
| 10. Prunophracta, |
|
10a. Belonaspida. |
| 10b. Hexalaspida. |
| 10c. Diploconida. |
| {ciii}
III. Legion (or Subclass)
Nassellaria
(Monopylea)
[Osculosa monopylea.]
|
 |
V. Plectellaria
(Nassellaria palliata) |
|
11. Nassoidea, |
|
11. Nassellida. |
| 12. Plectoidea, |
|
12a. Plagonida. |
| 12b. Plectanida. |
| 13. Stephoidea, |
|
13a. Stephanida. |
| 13b. Tympanida. |
VI. Cyrtellaria
(Nassellaria loricata) |
|
14. Spyroidea, |
|
14a. Zygospyrida. |
| 14b. Androspyrida. |
| 15. Botryodea, |
|
15a. Cannobotryida. |
| 15b. Lithobotryida. |
| 15c. Pylobotryida. |
| 16. Cyrtoidea, |
|
16a. Monocyrtida. |
| 16b. Dicyrtida. |
| 16c. Tricyrtida. |
| 16d. Stichocyrtida. |
IV. Legion (or Subclass)
Phæodaria
(Actipylea)
[Porulosa actipylea.] |
|
VII. Phæocystina
(Phæodaria palliata) |
|
17. Phæocystina, |
|
17a. Phæodinida. |
| 17b. Cannorrhaphida. |
| 17c. Aulacanthida. |
| 18. Phæosphæria, |
|
18a. Orosphærida. |
| 18b. Aulosphærida. |
| 18c. Cannosphærida. |
VIII. Phæocoscina
(Phæodaria loricata) |
|
19. Phæogromia, |
|
19a. Challengerida. |
| 19b. Castanellida. |
| 19c. Circoporida. |
| 20. Phæoconchia, |
|
20a. Concharida. |
| 20b. Cœlodendrida. |
| 20c. Cœlographida. |
| Four Legions. |
|
Eight Sublegions. |
|
Twenty Orders. |
|
Typical Families. |
|
I. Legion (or Subclass) (Peripylea)
[Porulosa peripylea.]
|
|
|
I. Collodaria (Spumellaria palliata) |
|
|
|
|
|
|
1. Colloidea, |
|
|
|
|
|
|
|
|
|
|
1a. Thalassicollida. |
|
|
|
|
|
|
|
|
|
|
|
|
1b. Collozoida. |
|
|
|
|
|
|
|
|
|
|
2. Beloidea, |
|
|
|
|
|
|
|
|
|
|
2a. Thalassosphærida. |
|
|
|
|
|
|
|
|
|
|
|
|
2b. Sphærozoida. |
|
|
|
|
|
|
|
|
II. Sphærellaria (Spumellaria loricata) |
|
|
|
|
|
|
3. Sphæroidea, |
|
|
|
|
|
|
|
|
|
|
3a. Ethmosphærida. |
|
|
|
|
|
|
|
|
|
|
|
|
3b. Collosphærida. |
|
|
|
|
|
|
|
|
|
|
4. Prunoidea, |
|
|
|
|
|
|
|
|
|
|
4a. Ellipsida. |
|
|
|
|
|
|
|
|
|
|
|
|
4b. Zygartida. |
|
|
|
|
|
|
|
|
|
|
5. Discoidea, |
|
|
|
|
|
|
|
|
|
|
5a. Phacodiscida. |
|
|
|
|
|
|
|
|
|
|
|
|
5b. Porodiscida. |
|
|
|
|
|
|
|
|
|
|
6. Larcoidea, |
|
|
|
|
|
|
|
|
|
|
6a. Larnacida. |
|
|
|
|
|
|
|
|
|
|
|
|
6b. Pylonida. |
|
|
|
|
|
|
|
II. Legion (or Subclass) Acantharia (Cannopylea).
[Osculosa cannopylea.]
|
|
|
III. Acanthometra (Acantharia palliata) |
|
|
|
|
|
|
7. Actinelida, |
|
|
|
|
|
|
|
|
|
|
7a. Astrolophida. |
|
|
|
|
|
|
|
|
|
|
|
|
7b. Litholophida. |
|
|
|
|
|
|
|
|
|
|
|
|
7c. Chiastolida. |
|
|
|
|
|
|
|
|
|
|
8. Acanthonida, |
|
|
|
|
|
|
|
|
|
|
8a. Astrolonchida. |
|
|
|
|
|
|
|
|
|
|
|
|
8b. Quadrilonchida. |
|
|
|
|
|
|
|
|
|
|
|
|
8c. Amphilonchida. |
|
|
|
|
|
|
|
|
IV. Acanthophracta (Acantharia loricata) |
|
|
|
|
|
|
9. Sphærophracta, |
|
|
|
|
|
|
|
|
|
|
9a. Sphærocapsida. |
|
|
|
|
|
|
|
|
|
|
|
|
9b. Dorataspida. |
|
|
|
|
|
|
|
|
|
|
|
|
9c. Phractopeltida. |
|
|
|
|
|
|
|
|
|
|
10. Prunophracta, |
|
|
|
|
|
|
|
|
|
|
10a. Belonaspida. |
|
|
|
|
|
|
|
|
|
|
|
|
10b. Hexalaspida. |
|
|
|
|
|
|
|
|
|
|
|
|
10c. Diploconida. |
|
|
|
|
|
|
|
III. Legion (or Subclass) Nassellaria (Monopylea)
[Osculosa monopylea.]
|
|
|
V. Plectellaria (Nassellaria palliata) |
|
|
|
|
|
|
11. Nassoidea, |
|
|
|
|
|
|
|
|
|
|
11. Nassellida. |
|
|
|
|
|
|
|
|
|
|
12. Plectoidea, |
|
|
|
|
|
|
|
|
|
|
12a. Plagonida. |
|
|
|
|
|
|
|
|
|
|
|
|
12b. Plectanida. |
|
|
|
|
|
|
|
|
|
|
13. Stephoidea, |
|
|
|
|
|
|
|
|
|
|
13a. Stephanida. |
|
|
|
|
|
|
|
|
|
|
|
|
13b. Tympanida. |
|
|
|
|
|
|
|
|
VI. Cyrtellaria (Nassellaria loricata) |
|
|
|
|
|
|
14. Spyroidea, |
|
|
|
|
|
|
|
|
|
|
14a. Zygospyrida. |
|
|
|
|
|
|
|
|
|
|
|
|
14b. Androspyrida. |
|
|
|
|
|
|
|
|
|
|
15. Botryodea, |
|
|
|
|
|
|
|
|
|
|
15a. Cannobotryida. |
|
|
|
|
|
|
|
|
|
|
|
|
15b. Lithobotryida. |
|
|
|
|
|
|
|
|
|
|
|
|
15c. Pylobotryida. |
|
|
|
|
|
|
|
|
|
|
16. Cyrtoidea, |
|
|
|
|
|
|
|
|
|
|
16a. Monocyrtida. |
|
|
|
|
|
|
|
|
|
|
|
|
16b. Dicyrtida. |
|
|
|
|
|
|
|
|
|
|
|
|
16c. Tricyrtida. |
|
|
|
|
|
|
|
|
|
|
|
|
16d. Stichocyrtida. |
|
|
|
|
|
|
|
IV. Legion (or Subclass) Phæodaria (Actipylea)
[Porulosa actipylea.]
|
|
|
VII. Phæocystina (Phæodaria palliata) |
|
|
|
|
|
|
17. Phæocystina, |
|
|
|
|
|
|
|
|
|
|
17a. Phæodinida. |
|
|
|
|
|
|
|
|
|
|
|
|
17b. Cannorrhaphida. |
|
|
|
|
|
|
|
|
|
|
|
|
17c. Aulacanthida. |
|
|
|
|
|
|
|
|
|
|
18. Phæosphæria, |
|
|
|
|
|
|
|
|
|
|
18a. Orosphærida. |
|
|
|
|
|
|
|
|
|
|
|
|
18b. Aulosphærida. |
|
|
|
|
|
|
|
|
|
|
|
|
18c. Cannosphærida. |
|
|
|
|
|
|
|
|
VIII. Phæocoscina (Phæodaria loricata) |
|
|
|
|
|
|
19. Phæogromia, |
|
|
|
|
|
|
|
|
|
|
19a. Challengerida. |
|
|
|
|
|
|
|
|
|
|
|
|
19b. Castanellida. |
|
|
|
|
|
|
|
|
|
|
|
|
19c. Circoporida. |
|
|
|
|
|
|
|
|
|
|
20. Phæoconchia, |
|
|
|
|
|
|
|
|
|
|
20a. Concharida. |
|
|
|
|
|
|
|
|
|
|
|
|
20b. Cœlodendrida. |
|
|
|
|
|
|
|
|
|
|
|
|
20c. Cœlographida. |
|
|
|
|
|
|
156. Formation of Species.—The totality of similar forms, which we
unite in one species, and which in the earlier dogmatic systems was regarded as a category of
absolute value, possesses only a relative value like all other systematic categories (§ 155). According to the individual views of the systematist and the general
survey which he has attained of the smaller and larger systematic groups, the conception of a
species adopted in his practical work will be wider or narrower. In the present systematic
arrangement a medium extent has been adopted. It is shown that in the Radiolaria, as in all other
extensive groups of organisms, the constancy of the species is very variable in the different
groups. Many families of Radiolaria are very rich in "bad species," i.e., very
variable forms, in which the process of the formation of species is seen in progress; such,
for example, are—among the Spumellaria, the Sphærozoida,
Stylosphærida, Phacodiscida and Pylonida; among the Acantharia, the
Amphilonchida and Phractopeltida; among the Nassellaria, the Stephoidea and Botryodea; and among the Phæodaria, the Aulacanthida, Sagosphærida, Castanellida and Concharida. On the
{civ}other hand, in some families numerous "good
species" may be distinguished, since the intermediate connecting forms are no longer present and
the forms have become relatively constant. As instances of such families may be mentioned,
among the Spumellaria, the Astrosphærida, Cyphinida, Porodiscida and
Tholonida; among the Acantharia the Quadrilonchida and Dorataspida; among
the Nassellaria, the Spyroidea and Cyrtoidea; among the Phæodaria, the Challengerida,
Medusettida, Circoporida and Cœlographida. The more carefully the different groups are
studied, the more numerous the individuals of each species under comparison, the greater becomes
the number of "bad" species among the Radiolaria, and the smaller the number of good ones.
Originally, no doubt, all "species bonæ" were "malæ." There may be observed in the manifold
skeletal forms of the Radiolaria, on the one hand, the utmost accuracy of configuration, and on
the other, the greatest variability, and hence a careful comparative study of them leads to a firm
conviction of the gradual "Transformation of Species," and of the truth of the "Theory of
Descent."
157. Palæontological Development.—The palæontology of the Radiolaria already
offers very considerable material for study; but in consequence of its incompleteness this is of
little value for the study of the phylogeny of the class. By far the larger portion of the fossil
Radiolaria belong to the Tertiary period; only quite recently have numerous well-preserved fossil
Radiolaria been described from the Mesozoic period, and especially from the Jura. Of Palæozoic
Radiolaria (from the coal measures) only slight traces are known. Moreover, the fossil Radiolaria
hitherto known have been found only in very circumscribed and widely separated localities. The
majority of all the species belong to the small island of Barbados. Although our palæontological
acquaintance with the Radiolaria must necessarily be incomplete for this reason, it is still more
so since at least thirty out of the eighty-five families (that is more than a third) could not
possibly leave any fossil remains, either because they possess no skeleton, or because of its
chemical composition.
Of the four legions of the Radiolaria, the Acantharia (on
account of the solubility of their astroid acanthin skeletons) have entirely vanished and have
never been found fossil. Of the Phæodaria, whose silicate skeleton is
not as a rule capable of fossilisation, only one section (Dictyochida) of a single family
(Cannorrhaphida) has been observed fossil. Hence the fossil remains of the Radiolaria belong
almost exclusively to the two legions, Spumellaria and Nassellaria, which were formerly united under the term "Polycystina." Among
these, however, the skeletonless Thalassicollida, Collozoida, and Nassellida could leave no
traces. Hence there only remain fifty-five families of which we might expect to find fossil
siliceous skeletons. Even of these, however, scarcely the half are certainly known in the fossil
condition, whilst of the remainder nothing certain is known; for example, of the large order
Larcoidea (among the Spumellaria) and of the
Stephoidea (among the Nassellaria) with a few
isolated exceptions, no fossils are known. The great majority of fossil Radiolaria belong to the
two Nassellarian orders Cyrtoidea and Spyroidea (two relatively very highly developed groups); next to these follow
the orders {cv}Discoidea and
Sphæroidea among the Spumellaria. From these
palæontological facts it is obvious that our present very incomplete acquaintance with the
fossil Radiolaria is quite insufficient to warrant us in drawing any conclusions from it
regarding the phylogenetic development or palæontological succession of the individual
groups.
158. Origin of the Four Legions.—The agreement of all Radiolaria in
those constant and essential characters of the unicellular body, which distinguish them from all
other Protista (especially the differentiation of the malacoma into a central capsule and
extracapsulum), justifies the conclusion that all members of this class have been developed from a
common undifferentiated stem-form. Only the simplest form of the Spumellaria, a skeletonless spherical cell with concentric spherical nucleus and
calymma, can be regarded as such. The simplest form of the Thalassicollida which is now extant
(Actissa, Procyttarium, p. 12), corresponds so exactly to the
morphological idea of that hypothetical stem-form that it may unhesitatingly be regarded in a
natural system as the common point of origin of the whole class. On the other hand, Actissa
is so closely related to the simple Heliozoa (Actinophrys, Actinosphærium,
Heterophrys, Sphærastrum, &c.) that its origin from this group of Rhizopoda is
exceedingly probable. The three legions Acantharia, Nassellaria, and Phæodaria are to be regarded as three
main diverging branches of the genealogical tree, which have been developed in different
directions and are only connected by their simplest stem-forms (Actinelius,
Nassella, Phæodina) with the stem-form of the Spumellaria,
the primordial Actissa.
159. Phylogeny of the Spumellaria.—The legion Spumellaria or Peripylea is to be regarded as the common
stem-group of the Radiolaria, and its simplest form, Actissa, as the primitive genus or
radical form of the whole class; for it possesses in the simplest and most undifferentiated form
all those characters by which the Radiolaria are distinguished from other Protista; all the other
genera of the class may be derived from it by successive modifications. Considered as a legion the
whole group Spumellaria is undoubtedly monophyletic, for all its members
possess those essential characters by which it is distinctively marked off from the other three
legions, more especially a simple capsule-membrane, which is everywhere evenly perforated by
innumerable small pores; the nucleus lies originally in the centre of the spherical central
capsule. Furthermore, all Spumellaria lack those positive characters which
distinguish the three remaining legions—the centrogenous acanthin skeleton of the Acantharia, the basal porochora and the monaxon podoconus of the Nassellaria, the astropyle and phæodium of the Phæodaria.
160. Origin of the Spumellaria.—The genus Actissa (p. 12, Pl. 1,
fig. 1) presents the Radiolarian type in its simplest and most primitive form—a spherical
central capsule, which encloses in its middle a spherical nucleus, and which is surrounded by a
spherical calymma. The whole unicellular body consists, therefore, of three concentric spheres,
{cvi}and possesses neither skeleton nor alveoles, nor
other differentiated parts. The innumerable fine pseudopodia, which issue from the central capsule
through the evenly distributed pores in its membrane, radiate in all directions through the
calymma and pass out over its surface. Actissa can, therefore, be directly derived
phylogenetically from the simplest skeletonless Heliozoa (Actinophrys, Heterophrys,
Actinosphærium, Sphærastrum). The only essential difference between the two consists
in the development of the central capsule, which in Actissa separates as a distinct
membrane the endoplasm from the exoplasm. This differentiation which we regard is the most
important distinguishing character of the Radiolaria, has been transmitted by inheritance, along
with the formation of flagellate spores in the central capsule, from Actissa, the primitive
parent to all the other Radiolaria.
{cvii}
161. Hypothetical Genealogical Tree of the Spumellaria:—
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Larcoidea
 |
|
Discoidea
 |
|
Streblonida |
|
Phacodiscaria |
|
Tholonida |
|
Coccodiscida |
|
Prunoidea
|
Soreumida |
|
|
| Zygartida |
|
|
|
|
|
|
|
Zonarida |
|
|
|
|
Lithelida |
|
Sphæroidea |
|
|
Cyclodiscaria |
|
|
|
|
|
 |
|
|
Spongodiscida |
|
|
|
|
|
Stylosphærida |
|
Pylodiscida |
|
|
|
Phorticida |
|
|
|
|
|
|
| Panartida |
|
|
|
|
|
|
|
|
|
|
Artiscida |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Phacodiscida |
|
|
Spongodruppida |
|
|
|
|
|
|
|
|
|
|
|
|
|
Staurosphærida |
|
|
|
|
|
Spongel-
lipsida |
|
Pylonida |
|
|
|
|
| |
|
|
|
|
|
|
| Cyphinida |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Porodiscida |
|
|
|
|
|
|
Astrosphærida |
|
|
|
|
|
|
Larnacida |
|
|
|
|
|
|
|
|
Cubosphærida |
Cenodiscida |
|
| Druppulida |
|
|
|
|
|
|
|
|
|
|
|
|
|
Collosphærida |
|
|
Archidiscida |
| |
|
|
Larnacilla
(Trizonium) |
|
|
|
|
|
|
Spongurida |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Ellipsida
(Cenellipsis) |
Larcarida
(Cenolarcus) |
Liosphærida
(Cenosphæra) |
Cenodiscida
(Cenodiscus) |
|
|
|
|
|
[Actiprunum?] |
[Actilarcus?] |
[Procyttarium] |
[Actidiscus?] |
|
|
|
|
| |
|
Cenosphæra (Common stem-form of all Sphærellaria?) |
|
|
|
|
Polycyttaria |
|
|
 |
|
|
Collosphærida |
Collozoida |
|
Sphærozoida |
 |
Beloidea |
|
|
|
|
|
|
|
|
|
|
|
Thalassosphærida |
|
Ethmosphærida |
|
|
|
|
| |
|
Colloidea |
| |
|
Thalassicollida |
| |
|
Actissa |
| |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Larcoidea
|
|
Streblonida |
|
|
Tholonida |
|
Prunoidea
|
Soreumida |
|
| Zygartida |
|
|
|
|
|
|
Zonarida |
|
|
Lithelida |
|
| |
|
|
|
|
|
|
Phorticida |
|
|
| Panartida |
|
|
|
|
|
|
Artiscida |
|
|
|
| |
|
|
|
|
|
|
Spongodruppida |
|
|
| |
|
|
|
|
|
|
Spongel-
lipsida |
|
Pylonida |
| |
|
|
|
| Cyphinida |
|
|
|
|
| |
|
|
|
|
| |
|
|
|
|
|
|
|
|
Larnacida |
| |
|
|
|
| Druppulida |
|
|
|
| |
|
|
|
| |
|
|
Larnacilla
(Trizonium) |
|
|
Spongurida |
| |
|
|
|
| |
|
|
|
Ellipsida
(Cenellipsis) |
Larcarida
(Cenolarcus) |
| |
|
|
[Actiprunum?] |
[Actilarcus?] |
| |
|
|
≈ |
|
≈ |
| |
| |
|
Discoidea
|
|
Phacodiscaria |
|
Coccodiscida |
| |
|
Sphæroidea |
|
|
Cyclodiscaria |
|
|
|
|
Spongodiscida |
|
Stylosphærida |
|
Pylodiscida |
|
|
|
|
|
|
|
Phacodiscida |
|
|
|
|
|
| Staurosphærida |
|
|
|
|
|
|
|
|
|
|
|
|
|
Porodiscida |
| Astrosphærida |
|
|
|
|
|
|
|
Cubosphærida |
Cenodiscida |
|
| |
|
|
|
|
|
Collosphærida |
|
|
Archidiscida |
|
|
|
|
|
| ≈ |
|
|
|
|
|
Liosphærida
(Cenosphæra) |
Cenodiscida
(Cenodiscus) |
Actiprunum
Actilarcus |
|
|
|
[Procyttarium] |
[Actidiscus?] |
|
|
|
| |
| Cenosphæra (Common stem-form of all Sphærellaria?) |
|
|
|
|
Polycyttaria |
|
|
|
|
Collosphærida |
Collozoida |
Sphærozoida |
|
Beloidea |
|
|
|
|
|
|
|
|
|
|
Thalassosphærida |
| Ethmosphærida |
|
|
|
|
|
| |
|
Colloidea |
| |
|
Thalassicollida |
| |
|
Actissa |
{cviii}
162. Collodaria and Sphærellaria.—Whilst in all Spumellaria the malacoma agrees in possessing the characteristic features of the
legion, and thus justifies its derivation monophyletically from the common stem-form
Actissa, the different forms of skeleton, on the other hand, cannot all be referred to the
same fundamental form. More especially the spherical lattice-shell, from which all the
numerous skeletal forms of the Sphærellaria may be derived, cannot have
arisen from the incomplete Beloid skeleton which characterises the Beloidea among the Collodaria. It is probable rather
that the formation of the skeleton has taken place independently in those two groups of Spumellaria. From the skeletonless Colloidea, as the
common stem-group of the Spumellaria, two different main groups have
diverged, on the one hand the Beloidea, whose skeleton consists of
separate spicules scattered in the extracapsulum, and on the other hand, the Sphærellaria, which have formed a simple lattice-sphere around the central
capsule; from this the manifold forms of the remaining Spumellaria may be
derived.
163. Descent of the Sphærellaria.—The extensive order Sphærellaria, which includes all Spumellaria with a
complete lattice-shell, develops an extraordinary variety of skeletal structures; these may,
nevertheless, all be derived without violence from a common stem-form, or simple spherical
lattice-shell, Cenosphæra. The main stem of the order, the extensive suborder Sphæroidea (Pls. 5-30), is derived
immediately from Cenosphæra (p. 61, Pl. 12); three
diverging branches of it being represented by the other three suborders, the Prunoidea (Pls. 16,
17, 39, 40) being
developed by elongation, and the Discoidea (Pls. 31-48) by
shortening of the vertical main axis, whilst the Larcoidea (Pls. 9, 10, 49, 50) have
originated by the modification of the spherical lattice-shell into a lentelliptical or triaxial
ellipsoidal one. Although the monophyletic derivation of all Sphærellaria
from Cenosphæra is exceedingly probable, the possibility of a polyphyletic origin for the
group is by no means excluded. For even in the skeletonless primitive genus of all the Spumellaria, Actissa (as well as in the social Collozoum), there
are found, in addition to the usual spherical types, other species (or subgenera, p. 12) whose central capsule is not spherical but a modification of the sphere; in
Actiprunum ellipsoidal; in Actidiscus lenticular; in Actilarcus
lentelliptical; if such modified forms of Actissa were to develop their lattice-shells
independently, then their form would correspond to that of the central capsule; and such simple
ellipsoidal, discoidal, and lentelliptical lattice-shells might have been the primitive forms of
the Prunoidea, Discoidea and Larcoidea.
164. Genealogical Tree of the Sphæroidea.—Cenosphæra, the simplest form of
the spherical lattice-shell, may be unhesitatingly regarded as the common stem-form of all the
Sphæroidea (pp. 50-284, Pls.
5-30).
Cenosphæra (p. 61, Pl. 12) arose
directly from Actissa simply by the silicification of the spherical exoplasmatic network of
the sarcodictyum around the central capsule, on the surface of the concentric calymma. From this
simple siliceous extracapsular lattice-sphere all other forms of Sphæroidea have arisen, in the main by the manifold combination of two simple
processes, first by the formation of radial spines on the surface of the lattice-sphere, and
second, the addition of concentric spherical lattice-shells. Both processes may be utilised as the
foundation for a systematic treatment of the Sphæroidea (compare pp. 52-58).
If in the Sphæroidea the characteristic number and
disposition of the radial spines be regarded as the most important heritable peculiarity
of the different families, then we have the following natural arrangement:—(1)
Liosphærida, without radial spines; (2) Cubosphærida, with six radial spines (opposite in pairs
in three axes perpendicular to each other); (3) Staurosphærida, with four radial spines (in two
axes crossed at right angles); (4) Stylosphærida, with two opposite radial spines (in the
vertical main axis); and (5) Astrosphærida, with numerous regularly or irregularly distributed
radial spines (eight to twenty or more). If, on the contrary, more stress be laid upon the
number of the concentric lattice-shells, then we have the following artificial
grouping:—(1) Monosphærida, with one simple lattice-sphere; (2) Dyosphærida, with two
concentric lattice-spheres; (3) Triosphærida, with three; (4) Tetrasphærida, with four; (5)
Polysphærida, with numerous (five to twenty or more) concentric lattice-shells; (6)
Spongosphærida, with a spongy spherical shell. In general the former arrangement appears more
natural than the latter, since the number of primary radial spines, which grow out from the
primary lattice-sphere, determines their ground-form from the outset, whatever may be the number
of secondarily added shells. Strictly speaking, according to the view adopted, these Liosphærida
which have several shells, on the outer surface of which there are no radial spines, ought to be
classified according to the number and arrangement of their internal radial connecting beams and
distributed among the other families. The practical application of this correct principle meets,
however, with great difficulties. Also in many cases the phylogenetic relations of the different
Sphæroidea are more complicated than would appear from both these
classificatory principles. In general their phylogeny will quite correspond with their ontogeny,
since from the innermost first formed {cix}lattice-shell (primary medullary shell) a number of radial spines
arises, and upon these the secondary shells are formed from within outwards.
165. Genealogical Tree of the Prunoidea.—The suborder Prunoidea is very closely related to the Sphæroidea,
and is distinguished from it by the elongation of one axis; from the simple lattice-sphere
(Cenosphæra) is developed a latticed ellipsoid (Cenellipsis, Pl. 39, fig. 1).
The development of this vertical isopolar main axis is foreshadowed even among the Sphæroidea, in that family in which two opposite radial spines grow out of the
primary lattice-sphere at the two poles of the vertical main axis (Stylosphærida, Pls. 13, 14). These
latter pass over without any sharp boundary into those forms of Prunoidea
whose ellipsoidal lattice-shell bears two opposite main-spines (Stylatractida, Pls. 15, 16). Other very
intimate relationships between the Sphæroidea and Prunoidea are indicated in certain of the latter by the fact that of the two
concentric lattice-shells the inner (medullary) shell is spherical, the outer (cortical) shell
ellipsoidal (Pl. 39,
figs. 3, 7, 8, 14, 19); often three concentric lattice-shells are present, of which the two inner
are spherical intracapsular medullary shells, whilst the outer is an extracapsular cortical shell,
ellipsoidal or cylindrical in form (Pl. 39,
figs. 4, 12, 17, 18). Owing to the manifold nature of these phylogenetical relations and the
variety of their combinations, the derivation of the individual Prunoidea
from the Sphæroidea is rendered very difficult; in addition to which it
is possible that the simplest Prunoidea (Cenellipsis,
Ellipsidium) have been directly developed from the skeletonless Actiprunum (a form
of Actissa with ellipsoidal central capsule, p. 14) by the excretion
of a simple ellipsoidal lattice-shell on the surface of their calymma.
The phylogeny of the Prunoidea is especially complicated
by the formation of peculiar transverse constrictions, perpendicular to the longitudinal axis.
They are wanting only in the Monoprunida (Ellipsida, Druppulida, and Spongurida); the Dyoprunida
(Artiscida and Cyphinida, Pl. 39,
figs. 9-19) possess only one such constriction (in the equatorial plane); the Polyprunida, on
the other hand, have three, five, or more parallel constrictions (Panartida and Zygartida, Pl.
40). The
chambers, which are separated off by these constrictions, may be regarded as polar sections of
incomplete cortical shells.
166. Genealogical Tree of the Discoidea.—The suborder Discoidea is closely related to the Sphæroidea, but
separated from it by shortening of one axis; from a simple lattice-sphere (Cenosphæra) a
latticed lens or flattened spheroid is developed, whose circular equatorial plane is larger than
any other section (Cenodiscus, Pl. 48,
fig. 1). The formation of this horizontal equatorial plane is perhaps indicated in that family of
Sphæroidea in which four crossed radial spines, lying in one plane, are
developed (Staurosphærida, Pls. 15,
31, 42). The
morphological and phylogenetical relations of the Discoidea to the Sphæroidea are precisely the converse of those of the Prunoidea; in the latter the vertical axis appears longer, in the former
shorter than any {cx}other axis of the body. The Discoidea are probably polyphyletic, having originated from several different
groups of Sphæroidea; at least two essentially different main groups may
be distinguished among them; of these the one is characterised by the formation of a large
extracapsular lenticular cortical shell (Phacodiscaria), whilst in the other this typical "Phacoid
shell" or lattice-lens is wanting (Cyclodiscaria, compare pp. 403-409).
The Phacodiscida (Pls. 31-35) perhaps
constitute the primitive group of the Phacodiscaria, their lenticular or Phacoid cortical shell
being connected by radial bars with one or two concentric spherical medullary shells; they may
have originated directly from the Dyosphærida or Triosphærida by flattening of the spheroidal
cortical shell. From the Phacodiscida the Cenodiscida (if indeed they be not the primitive
stem-form) have been developed by retrogression and loss of those medullary shells. The
Coccodiscida (Pls. 36-38), on the
other hand, have been developed from the Phacodiscida by the addition of concentric rings of
chambers, which may be regarded as incomplete cortical shells, only the equatorial portion of
which is developed. Perhaps the Porodiscida, the primitive group of the Cyclodiscaria, have
arisen in a similar way; they lack, however, the typical Phacoid shell, the concentric rings of
chambers being directly applied to a small spherical medullary shell in the equatorial plane
(Pls. 41-46). If those
rings from the commencement be interrupted by three interradial gaps (gates) the
family Pylodiscida arises (Pl. 38,
figs. 6-20). If, on the contrary, the concentric radially divided chambers of the Porodiscida
become quite irregular and spongy, they pass over into the Spongodiscida (Pls. 46, 47). It is
not, however, impossible that part of the Discoidea (especially the
Cenodiscida) have originated directly from skeletonless Collodaria with
a lenticular central capsule, such as are found in a subgenus of Actissa
(Actidiscus, p. 15).
167. Genealogical Tree of the Larcoidea.—The suborder Larcoidea presents in the structure, composition, and development of its
variously formed lattice-shells much more complicated relations than the other Sphærellaria; it is essentially distinguished from them by the characteristic
ground-form of its lattice-shells, which is a "lentellipsis" or a triaxial ellipsoid (also the
ground-form of the rhombic crystallographic system, the rhombic octahedron). Hence all parts of
the body are regularly disposed with respect to three different dimensive axes; all three axes,
perpendicular one to another, are isopolar but of different lengths; the longest is the vertical
main axis, the mean the horizontal frontal axis, the shortest the horizontal sagittal axis. In the
great majority of the Larcoidea the lentelliptical ground-form is
indicated in the central capsule, even when it is not at once obvious in the skeleton. Since such
lentelliptical central capsules are developed even in Actissa (Actilarcus, p. 16), it is possible that the simplest Larcoidea may
have arisen directly from these by deposition of a simple lentelliptical lattice-shell in the
sarcodictyum, on the surface of the calymma (Cenolarcus, Pl. 50, fig. 7). It
is more probable, however, that these simplest forms (Cenolarcus, Larcarium) have
been developed from the simplest Sphæroidea (Cenosphæra), by the
spherical body growing unequally in the three dimensions of space. It appears especially likely
{cxi}from a study of the concentrically disposed
lattice-shells of some Larcoidea (Coccolarcus, Larcidium,
Pl. 50, fig.
8), in which the inner medullary shell is spherical, the outer cortical shell more or less
elliptical. In the great majority of Larcoidea the latter arises in quite
a peculiar manner, three broad lattice-zones, which are developed in three planes at right angles
to each other, growing out from a small spherical or lentelliptical medullary shell,
Trizonium, Larnacilla (compare pp. 600, 615, 628, &c.).
The trizonal Larnacilla-shell commences by the formation of a transverse
girdle, by the union of two lateral latticed processes, which spring right and left in the
equatorial plane from the poles of the frontal axis of a lentelliptical medullary shell
(Monozonium, p. 633, Pl. 9, fig. 1).
This is followed by a second lateral girdle, which lies in the frontal plane and proceeds from
its lateral poles (Dizonium, p. 634, Pl. 9, figs. 2,
3). Finally the sagittal girdle is formed, lying in the sagittal plane and arising from the
lateral girdle on the two poles of the main axis (Trizonium, p. 637, Pl. 9,
fig. 4). Whilst the gaps between the three zones of this trizonal shell remain open in the
Pylonida, in Larnacilla, the important primitive form of the Larnacida, they are closed
by lattice-work (Pl. 50,
figs. 3-8). From this trizonal Larnacilla-shell the great majority of Larcoid shells may
be derived. Such a system of zones may be repeated (Diplozonaria) or even developed a third time
(Triplozonaria, p. 632). In most Larcoidea the
zones are secondarily connected by lattice-work. In the Tholonida (Pl. 10) each of
the two opposite latticed wings of a zone becomes a closed dome. In the Zonarida (Pl. 50, figs.
9-12) these domes are partially or wholly bisected by constrictions or latticed septa which are
developed in the three dimensive planes. The Lithelida (Pl. 49, figs.
1-7) are characterised by the fact that one of each pair of opposite latticed processes (or half
zones) grows more strongly than the other, and that the larger completely embraces the smaller
so as to form a complicated spiral. Whilst in this case the spiral lies in a plane, in the
Streblonida (Pl. 49,
figs. 8, 9) it becomes turbinoid like a gastropod shell and forms an ascending spiral. Finally,
two small families of Larcoidea are characterised by quite irregular
growth (a very rare occurrence among the Radiolaria); these are the simple-chambered Phorticida
(Pl. 49,
figs. 10, 11) and the many chambered Soreumida (Pl. 49, figs. 12,
13). The phylogenetic relationship of these families of Larcoidea is
probably very complicated and demands closer investigation (compare pp. 599-604).
168. Descent of the Polycyttaria.—The polyzootic or colonial
Radiolaria, which we unite in the group Polycyttaria (sometimes known as "Sphærozoea"), belong
without doubt to the legion Spumellaria, for they possess all the
peculiarities by which these Peripylea are distinguished from the other
legions of the Radiolaria. Only the morphological position of the Polycyttaria in that legion, and
their phylogenetic relation to the monozootic or solitary Spumellaria, can
be variously interpreted. The three families which we distinguish among the Polycyttaria are so
closely related to three different families of the Monocyttaria, that they may be directly derived
from them by the formation of colonies. According to this triphyletic hypothesis the social
skeletonless Collozoida (Pl. 3)
would be descended from the solitary Thalassicollida (Pl. 1), the
polyzootic Sphærozoida with a Beloid skeleton (Pl. 4) from the
monozootic {cxii}Thalassosphærida (Pl. 2), and the
colonial Collosphærida with a Sphæroid skeleton (Pls. 5-8) from the
solitary Ethmosphærida (Pl. 12,
&c.). Many species of monozootic and polyzootic forms in all three groups are so alike that
they can only be distinguished by the fact that the one series are colonial, the others solitary.
On the other hand, there are some reasons which would justify a monophyletic hypothesis for the
Polycyttaria, e.g., the precocious nuclear division; in this case it would be most natural
to hold that the Sphærozoida and Collosphærida have arisen as two diverging branches from the
Collozoida, whilst the latter are nothing else than colonial Thalassicollida.
169. Phylogeny of the Acantharia.—The legion Acantharia or Actipylea is distinguished by its peculiar
acanthin skeleton, which develops centrogenously, as well as by the disposition in groups of the
pores in its central capsule, and its excentric usually precocious nucleus; it is thus so
different from all other Radiolaria as undoubtedly to furnish, phylogenetically considered, an
independent stem (§ 7). This stem is only connected at the root by
Actinelius with the primitive form of the Spumellaria,
Actissa. The stem is monophyletic, since all the forms belonging to it may be derived
without violence from Actinelius as a common primitive form.
170. Origin of the Acantharia.—The genus Actinelius (p. 730, Pl. 129,
fig. 1), which may naturally be regarded as the common primitive form of all Acantharia, possesses a spherical central capsule, which in consequence of the
early division of the nucleus (§ 63), encloses numerous small nuclei; from
its centre arise many simple radial spines of equal size, which penetrate the central capsule. A
large number of radial pseudopodia issue between the spines from the sarcomatrix which surrounds
the capsule. Actinelius may have been directly derived from Actissa, the common
stem-form of all Radiolaria, by the division of the pseudopodia into two groups, myxopodia, which
remained soft, and axopodia, which became firm (§ 95A). As the latter became changed into strong acanthin rods, and touched
each other in the centre, they forced the nucleus from its originally central position and brought
about its early division. Actinelius is also of all Radiolaria the form which, next to
Actissa, most nearly approaches the Heliozoa. If the stiff axial threads of
Actinosphærium be conceived of as partially converted into acanthin spines, and its
nucleated medullary substance as separated from the alveolar cortical layer by a membrane (central
capsule), then Actinelius would be produced.
{cxiii}
171. Hypothetical Genealogical Tree of the Acantharia:—
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Diploconida |
| |
| Phractopeltida |
Hexalaspida |
|
Cenocapsida |
|
Phatnaspida |
|
Lychnaspida |
|
|
|
|
|
Porocapsida |
|
|
|
Coleaspida |
|
|
Ceriaspida |
|
|
|
|
|
|
|
|
|
|
|
Belonaspida |
|
|
|
Phractaspida |
|
Stauraspida |
|
|
|
|
Astrocapsida
Sphærocapsida |
|
|
|
|
|
|
|
|
|
|
Diporaspida
(Dorataspida dipora) |
Tessaraspida
(Dorataspida tetrapora) |
|
|
|
|
|
|
|
|
|
|
|
[Dorataspida] |
|
|
|
|
|
|
Quadrilonchida |
|
| Phractacanthida |
|
|
Stauracanthida |
|
Amphilonchida |
|
|
|
|
|
|
|
|
|
|
Acanthonia |
|
|
|
|
| |
|
Astrolonchida |
| Litholophida |
|
|
Chiastolida |
|
|
Zyganthida |
|
|
|
Acanthonida |
|
Actinastrum |
|
|
Acanthometron |
|
| Astrolophida |
|
|
|
|
Acanthochiasmida |
|
|
|
|
|
|
|
|
|
|
Acanthometron |
|
|
|
|
|
| |
|
Actinelida
Actinelius |
| |
|
Actissa |
| |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Diploconida |
| |
| Phractopeltida |
Hexalaspida |
|
Cenocapsida |
| |
|
|
|
Phatnaspida |
Lychnaspida |
|
| |
|
|
|
|
|
|
|
|
|
Porocapsida |
|
|
|
Coleaspida |
|
|
Ceriaspida |
|
|
|
|
|
|
|
|
|
|
|
Belonaspida |
|
|
| Phractaspida |
|
Stauraspida |
|
|
|
|
Astrocapsida
Sphærocapsida |
|
|
|
|
|
|
|
|
Diporaspida
(Dorataspida dipora) |
Tessaraspida
(Dorataspida tetrapora) |
|
|
|
|
|
|
|
|
|
|
|
[Dorataspida] |
|
|
|
|
|
|
Quadrilonchida |
|
| |
|
|
|
| Phractacanthida |
|
|
Stauracanthida |
|
Amphilonchida |
|
|
|
|
|
|
|
|
|
|
Acanthonia |
|
|
|
|
| |
|
Astrolonchida |
| |
|
Litholophida |
|
Chiastolida |
|
|
|
|
Zyganthida |
|
|
|
Acanthonida |
Actinastrum |
|
Acanthometron |
|
| Astrolophida |
|
|
Acanthochiasmida |
|
|
|
|
|
|
|
|
|
Acanthometron |
|
|
|
|
|
| |
|
Actinelida
Actinelius |
| |
|
Actissa |
{cxiv}
172. Adelacantha and Icosacantha.—The numerous forms of Acantharia, here disposed in twelve families and sixty-five genera, may be
divided phylogenetically into two main groups of very different extent—Adelacantha
and Icosacantha. The more primitive group, Adelacantha, have an indefinite and
variable number of radial spines, which are always quite simple in form and usually irregularly
distributed; this main division includes only the one order Actinelida,
with six genera, among which is Actinelius, the common stem-form of all the Acantharia. The more recent group, Icosacantha, includes all the other Acantharia (fifty-nine genera), and is very markedly distinguished from the
Adelacantha by the fact that the radial spines are always twenty in number, and arranged according
to Müller's law (compare pp. 717-725, and § 110). Since this regular disposition (in five alternating zones each of four
spines) has been retained by inheritance in the whole of the Icosacantha, it is probable that this
large group has been developed monophyletically from a twig of the Adelacantha; Actinastrum
(p. 732) and Chiastolus (p. 738) still
present connecting links between the former and the latter, between Actinelius and
Acanthometron.
173. Acanthonida and Acanthophracta.—The extensive main division
Icosacantha (§ 110), which embraces all Acantharia
with twenty radial spines, disposed according to Müller's law, may be subdivided into two large
groups or orders:—the Acanthonida (p. 740,
Pls. 130-132) and the
Acanthophracta (p. 791, Pls. 133-140). The
latter possess a complete extracapsular lattice-shell, which the former have not. The more recent
Acanthophracta may be derived phylogenetically from the more primitive
Acanthonida simply by the development of this lattice-shell, with which
process are usually (perhaps always) connected certain alterations in the malacoma, e.g.,
degeneration of the myophriscs (§ 96). The most primitive form of all
Icosacantha is the genus Acanthometron (p. 324), in which all the
twenty acanthin spines are of the simplest constitution and of equal dimensions.
174. Differentiation of the Acanthonida.—The order Acanthonida, which embraces all Icosacantha which have no complete
lattice-shell, divides early into three main branches, the three families Astrolonchida,
Quadrilonchida, and Amphilonchida (p. 727, Pls. 130-132). The
first of these constitutes the common stem-group from which the other two as well as the whole
group Acanthophracta have been developed; the common stem-form of all is
Acanthometron (§ 173). All the Astrolonchida (p. 740, Pl. 130)
have twenty radial spines of equal size and similar form. On the other hand, in the Quadrilonchida
(p. 766, Pl. 131)
the four equatorial spines differ from the others in size and sometimes also in form. In the
Amphilonchida (p. 781, Pl. 132) two
opposite equatorial spines (lying in the hydrotomical axis) are much larger than the other
eighteen and of a different shape. Of the three families of the Acanthonida the most important is the primitive group Astrolonchida, for from
this the various stem-forms of the Acanthophracta arise. They are
subdivided according to the formation of the spines into three subfamilies: the Zygacanthida, with
simple spines without apophyses (or transverse processes); the Phractacanthida, with two opposite
apophyses on each radial {cxv}spine, and the
Stauracanthida, with four crossed apophyses on each radial spine. The three genera of the
Zygacanthida represent the stem-forms of the three families, since the radial spines in
Acanthometron (the most primitive form of Acanthonida) are
cylindrical, in Zygacantha two-edged, and in Acanthonia four-edged (p. 741).
175. Capsophracta and Cladophracta.—The extensive order Acanthophracta, which embraces all Acantharia with a
complete lattice-shell, is polyphyletic, its main subdivisions have been developed independently
from different branches of the Acanthonida. The whole order may be
divided directly into two main groups, the Capsophracta and Cladophracta (p. 793), which differ in the structure and
the origin of their lattice-shell. The group (or suborder) Capsophracta
includes only the single family Sphærocapsida (p. 795, Pl. 133, figs.
7-11; Pl. 135,
figs. 6-10); the lattice-shell arises independently of the twenty radial spines, being made up
like a pavement of innumerable small acanthin plates, united by a kind of cement; each plate being
perforated by a fine pore. In addition twenty larger main pores (or groups of four pores each) are
present, corresponding to the twenty radial spines; these are always equal, quadrangular
prismatic, without transverse processes as in Acanthonia. In the Cladophracta, which include the five remaining families of the Acanthophracta, the structure and origin of the lattice-shell are quite
different; the lattice-shell is here made up of the branches of the transverse processes, which
radiate tangentially from the twenty radial spines and are only united secondarily.
176. Ascent of the Dorataspida.—The group Cladophracta, or those Acantharia whose lattice-shell
arises by the union of transverse processes of the twenty radial spines, includes five different
families, whose stem-group is the family Dorataspida, with a simple spherical lattice-shell. This
family itself is, however, diphyletic in origin, being composed of two essentially and originally
different subfamilies—Diporaspida and Tessaraspida (p. 803). The
Diporaspida (p. 808, Pls. 137, 138) have been
developed from the Phractacanthida, and as each radial spine of the latter bears two opposite
apophyses, so the lattice-shell of the former has forty primary aspinal pores (two on the base of
each spine). On the other hand, the Tessaraspida (p. 830, Pls. 135, 136) have been
developed from the Stauracanthida, and as each radial spine of the latter bears four crossed
apophyses, so the lattice-shell of the former has eighty primary aspinal pores (four at the base
of each spine).
177. Descent of the Diporaspida.—Whilst the Tessaraspida (§ 176) have given rise to no new groups which could take rank as independent
families, no less than four separate families of Acantharia have arisen
from the Diporaspida. The Phractopeltida (Pl. 133,
figs. 1-6) are distinguished from all other Acantharia by the possession
of two concentric spherical lattice-shells, and have probably been developed from the {cxvi}Diporaspida in the same way as the Dyosphærida from the
Monosphærida among the Sphæroidea; in that case the smaller inner
lattice-sphere (medullary shell) would be the primary, and the larger outer sphere (cortical
shell) the secondary; this latter shows forty primary aspinal pores like those of the Diporaspida.
The possibility is not excluded, however, that the small inner lattice-sphere of the
Phractopeltida is a secondary product. The three remaining families, which must be regarded as
descendants of the Diporaspida, form together a single phylogenetic series, and are separated from
the primitive group mainly by the fact that the original spherical form of the lattice-shell has
been modified into one distinguished by an elongated equatorial axis (the hydrotomical axis);
hence the Prunophracta (pp. 794-859). The ellipsoidal Belonaspida have arisen directly by hypertrophy of the
two opposite equatorial spines of this hydrotomical axis (p. 859, Pl. 136, figs.
6-9; Pl. 139,
figs. 8, 9; perhaps they have also arisen directly from the Amphilonchida). In the lentelliptical
Hexalaspida (Pl. 139)
all six spines which lie in the hydrotomical meridian plane (two equatorial and four polar) are
very strongly developed, the remaining fourteen being rudimentary. Finally, in the Diploconida the
two conical sheaths of the two opposite hydrotomical equatorial spines are so predominant that
they take the chief part in the formation of the hour-glass-shaped shell.
178. Phylogeny of the Nassellaria.—The legion Nassellaria or Monopylea is so clearly characterised by
the peculiar porochora, which closes the osculum at the oral pole of the monaxon central capsule,
and by the podoconus connected with it, that there can be no doubt that phylogenetically it
represents an independent stem (§ 8). This stem is only connected at its base
by means of Cystidium and Nassella with Actissa and Thalassicolla, the
stem-forms of the Spumellaria. This stem is monophyletic, inasmuch as all
its members may be derived without violence from the skeletonless Nassellida (Nassella,
Cystidium, p. 896,
Pl. 91, fig.
1).
179. Origin of the Nassellaria.—The Nassellida (p. 896), which may
naturally be considered as the common stem-group of the Nassellaria, are
most nearly related among other Radiolaria to the Thalassicollida, and in both these skeletonless
families the simplest forms, Cystidium and Actissa correspond; on the other hand,
those which have arisen from them by the formation of alveoles in the calymma (Nassella and
Thalassicolla) also correspond. The origin of the simplest Nassellida from these primitive
Thalassicollida may be explained by supposing that the numerous (formerly evenly distributed)
pores of the capsule membrane became obliterated in the upper (apical) half of the central
capsule, whilst in the lower (basal) half they became correspondingly more strongly developed;
hence the porochora was formed at the oral pole of the vertical main axis, and a differentiation
of the endoplasm proceeding from this gave rise to the characteristic podoconus. Both these organs
still at present exhibit very various degrees of progressive development.
{cxvii}
180. Hypothetical Genealogical Tree of the Nassellaria.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Cyrtoidea
|
|
Botryodea
|
|
Triradiata |
|
Pylobotryida |
|
Podocampida |
|
Eradiata |
|
|
Multiradiata |
Spyroidea
|
|
Lithocampida |
|
|
Phormocampida |
Androspyrida |
|
Lithobotryida |
|
Podocyrtida |
|
|
Theocyrtida |
|
Phormocyrtida |
|
Tholospyrida |
|
Cannobotryida |
|
Tripocyrtida |
|
Phormospyrida |
|
Sethocyrtida |
|
Anthocyrtida |
|
|
|
|
|
Tripocalpida |
|
|
|
Cyrtocalpida |
|
Phænocalpida |
|
|
Stephoidea
|
|
|
|
|
|
|
Tympanida |
|
|
|
Zygospyrida |
|
|
|
|
(Spyroidea triradiata) |
|
|
Tripocalpida
(Cyrtioidea triradiata monocyrtida) |
|
| Coronida |
|
|
|
|
|
Semantida |
|
|
|
|
|
|
|
|
Cyrtellaria |
|
|
Cortiniscus |
|
| Stephanida |
|
|
|
|
|
|
|
|
|
Cortina |
* |
 |
Cortinida |
|
(Plectellaria) |
|
(Cortina) |
| |
|
Plectaniscus |
|
|
Plagoniscus |
|
|
 |
| |
|
Plectoidea |
|
Tetraplecta |
|
|
Tetraplagia |
|
Plectanida |
| |
|
|
|
Plectophora |
|
|
Plagiacantha |
|
|
| |
|
Plagonida |
|
Triplecta |
|
|
Triplagia |
|
| |
|
Nassoidea
(Nassellida) |
| |
|
Nassella
(Cystidium) |
| |
|
Actissa |
| |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Cyrtoidea
|
Botryodea
 |
Triradiata |
| Pylobotryida |
Podocampida |
|
Eradiata |
Multiradiata |
Spyroidea
|
|
Lithocampida |
Phormocampida |
| |
|
|
|
| Lithobotryida |
|
Podocyrtida |
Androspyrida |
| |
|
|
|
|
|
Theocyrtida |
Phormocyrtida |
Tholospyrida |
| |
|
|
|
|
|
| Cannobotryida |
|
Tripocyrtida |
Phormospyrida |
| |
|
|
|
|
|
|
Sethocyrtida |
Anthocyrtida |
|
| |
|
|
|
|
|
|
|
Tripocalpida |
|
|
| |
|
|
|
|
|
|
Cyrtocalpida |
Phænocalpida |
|
|
|
|
|
|
|
|
|
|
Zygospyrida |
|
|
|
(Spyroidea triradiata) |
|
Tripocalpida
(Cyrtioidea triradiata monocyrtida) |
|
| |
|
|
| |
Stephoidea
 |
|
|
Tympanida |
|
| |
|
Cyrtellaria |
| Coronida |
|
|
Semantida |
|
|
|
|
|
|
|
Cortiniscus |
| Stephanida |
|
|
| |
|
|
| |
|
|
|
Cortina |
* |
—— |
Cortinida |
|
(Plectellaria) |
|
(Cortina) |
| |
| Plectaniscus |
—— |
Plagoniscus |
|
 |
| |
|
Plectoidea |
| Tetraplecta |
—— |
Tetraplagia |
Plectanida |
| |
|
|
| Plectophora |
—— |
Plagiacantha |
|
| |
|
Plagonida |
| Triplecta |
—— |
Triplagia |
| |
|
Nassoidea
(Nassellida) |
| |
|
Nassella
(Cystidium) |
| |
|
Actissa |
{cxviii}
181. Plectellaria and Cyrtellaria.—The extensive legion Nassellaria far surpasses the other three legions in the endless variety of its
skeletal structures, and owing to the complicated relationships of its numerous families presents
no lack of difficult phylogenetic problems. All Nassellaria may be divided
first into two main groups or sublegions, Plectellaria and Cyrtellaria; the latter having a complete lattice-shell, the former not.
Probably the Cyrtellaria have been polyphyletically developed from
several different groups of Plectellaria. These groups are, however,
connected in such manifold ways that a monophyletic origin of all the Nassellarian skeletons from one original element is possible. Such a primitive
element may have been furnished by any one of three different skeletal parts, the sagittal ring,
the basal tripod, and the latticed cephalis (compare pp. 891-895, Bütschli, L. N.
40, 41).
182. Phylogenetic Skeletal Elements of the Nassellaria.—The multiform
skeleton of the Nassellaria may be referred in different ways to one of
the three above-mentioned structural elements. Each of these (p. 891) may by itself
form the skeleton; the sagittal ring in the simplest Stephoidea
(Archicircus, Lithocircus), the basal tripod in the simplest Plectoidea (Triplagia, Plagiacantha), the latticed cephalis in
the simplest Cyrtoidea (Cyrtocalpis, Archicapsa). In the
great majority of the Nassellaria, however, two of these elements, or even
all three, are found combined. In most Cyrtellaria, more especially, both
the sagittal ring and the basal tripod may be recognised in the lattice-shell, though often only
in slight rudiments or scarcely perceptible traces. In the Plectellaria
also (which possess no latticed cephalis) there are individual genera with complete development
both of the sagittal ring and basal tripod; this important combination is especially well
represented in the Cortinida (Cortina, Cortiniscus, Stephanium,
Stephaniscus, Tripocoronis, &c.). The greatest difficulty as regards the
phylogeny of the Nassellaria lies in the fact that the most various
combinations of the three elements are presented by closely related or very similar forms. If, in
spite of this, a monophyletic hypothesis as to the origin of the Nassellaria seems essential all sides of the three possible hypotheses must
receive full consideration and critical comparison (§§ 183-191).
183. Ascent of the Nassellaria from the Plectoidea.—The monophyletic
hypothesis (No. 2, p. 893)
which regards the basal tripod as the common origin of the skeleton of all Nassellaria, starts from the simplest forms of the Plectoidea (Triplagia, Plagoniscus, Triplecta,
Plectaniscus, &c., Pl. 91).
All Plectoidea may be immediately derived as diverging twigs of these, as
well as all triradial and multiradial forms of Cyrtoidea and Spyroidea; for in all these cases the distinctive triradial (or the derived
multiradial) form of skeleton appears directly derivable from the simple basal tripod of the
former. The same is perhaps also true of many Botryodea. {cxix}Furthermore, certain important forms of Stephoidea (Cortina, Cortiniscus, Stephanium,
Stephaniscus, &c.), which have a characteristic combination of the sagittal ring and
basal tripod, may be immediately derived from such forms of Plectoidea as
Plagoniscus cortinaris, Plagiocarpa procortina, Plectaniscus cortiniscus,
&c. On the contrary, those Stephoidea and Cyrtoidea in which the basal tripod is wanting can only be derived from the
Plectoidea by the assumption that this structure has disappeared in
consequence of phylogenetic degeneration. The monophyletic derivation of the Nassellaria from the Plectoidea has more internal
probability than that from the Stephoidea, since it is easier to suppose
that the Cortinida (Cortina, Stephanium, &c.) have been derived from the Plectoidea (Plagoniscus, Plagiocarpa) than the converse. This
view is the basis of the hypothetical tree shown in § 180.
184. Ascent of the Nassellaria from the Stephoidea.—The monophyletic
hypothesis (No. 1, p. 893)
which regards the primary sagittal ring as the common starting point of the skeleton in all Nassellaria, starts from the simplest forms of Stephoidea (Archicircus, Lithocircus, &c., Pl. 81). All Stephoidea and Spyroidea may be immediately derived
from these, as also the majority of the Cyrtoidea and probably of the
Botryodea. Those numerous forms of the last two groups, however, which
possess no trace of a sagittal ring, can only be derived from the former by the supposition that
the latter has completely disappeared in in consequence of gradual phylogenetic degeneration. The
same holds true also of the Plectoidea, although certain forms
(e.g., Plagiocarpa procortina, Pl. 91, fig. 5;
Plectaniscus cortiniscus, Pl. 91,
fig. 9) appear to indicate the commencing formation of the sagittal ring by the concrescence of
two branches, which approach each other from the upper part of the apical rod and the ventral part
of the basal rod. In any case, it is a fact of great phylogenetic significance, that the primary
sagittal ring in the cephalis of the Cyrtoidea shows all conceivable
stages of degeneration (compare Bütschli, L. N. 40, 41, as
well as the general account of and critical comparison of the Nassellaria,
pp. 889-895, &c.).
185. Ascent of the Nassellaria from the Cyrtoidea.—The monophyletic
hypothesis (No. 3, p. 894)
which regards the latticed cephalis as the common point of origin of all the skeletons of the
Nassellaria, starts from the simplest forms of the Cyrtoidea, that is, from the Cyrtocalpida or eradial Monocyrtida
(Archicorida, Archicapsida, Pls. 51,
52, 98). All Cyrtoidea and Botryodea may be regarded as divergent
forms of these monothalamous Cyrtoidea; the polythalamous simply by the
addition of fresh joints at the basal pole, the triradiate and multiradiate by the development of
three or more apophyses. The origin of the sagittal ring (which presents every stage of
development and degeneration in the Cyrtoidea) may be regarded as a
mechanical thickening of the latticed plate in the sagittal circumference of the cephalis. By
stronger {cxx}development of this ring and coincident
sagittal constriction of the cephalis the order Spyroidea may be derived
from the Cyrtoidea. On the other hand, the Plectellaria, which possess no cephalis, and indeed no complete lattice-shell
whatever, may be derived from the Monocyrtida by the assumption of a degeneration of this
structure; the sagittal ring having been preserved in the Stephoidea, and
the tripod of the Tripocalpida in the Plectoidea. Although such a
monophyletic derivation of the Nassellaria from the Cyrtocalpida is
possible, and though here, too, the Cortinida play an important part as connecting links, this
hypothesis has less internal probability than that of the derivation from the Stephoidea (§ 184) or Plectoidea (§ 183).
186. Genealogical Tree of the Plectoidea.—The order Plectoidea includes those Nassellaria whose rudimentary
skeleton does not contain the characteristic sagittal ring of the Stephoidea, but consists of several (at least three) radial spines, which
proceed from a point in the centre of the porochora. The branches of these radial spines remain
free in the Plagonida, whilst in the Plectanida they unite with each other to form a loose
meshwork (not, however, a complete lattice-shell). The number and arrangement of the radial
spines, which serve for generic distinctions, are the same in both families, so that each genus of
the Plectanida has arisen from a corresponding genus of the Plagonida. The simplest Plagonida,
which possess a basal tripod (Triplagia or Plagiacantha with three rays,
Tetraplagia with four rays) are probably to be regarded as forming the common origin of the
whole order. These agree with certain three- and four-rayed skeletal pieces of the Beloidea (Thalassosphærida and Sphærozoida); and also the four and six-rayed
twinned pieces of the latter (spicula bigemina and trigemina) repeat in the same fashion the
skeleton of the former (Plagonidium, Plagonium). This similarity, however, is a mere
analogy and possesses no phylogenetic significance. On the other hand, certain Plagonida
(Plagoniscus, Plagiocarpa), and the corresponding genera of Plectanida
(Plectaniscus, Periplecta) seem to have important phylogenetic relations to certain
Stephoidea (Cortina, Cortiniscus, &c.); the sagittal
ring of the latter having perhaps arisen by the vertical apical spine of the former having been
connected with their horizontal basal rod by two ventral apophyses growing out opposite to each
other (compare pp. 902,
914, Plagiocarpa
procortina, Pl. 91,
fig. 5). In this case the Plectanida would belong to the simplest stem-forms of the Nassellaria.
187. Genealogical Tree of the Stephoidea.—The order Stephoidea includes all those Nassellaria whose
skeleton does not form a complete lattice-shell, but consists of one or more rings, and often of a
loose meshwork which arises by the union of branches of the rings. A vertical sagittal ring
is constantly present, embracing the central capsule in the median sagittal plane, and forming at
its basal pole various processes, the starting point for other skeletal forms. The most important
of these is the tripodal Cortina {cxxi}(p. 950, § 182). The Stephanida are the most archaic family among the Stephoidea (p. 937,
Pl. 81),
perhaps indeed among all the Nassellaria (§ 184);
in them the sagittal ring and its processes alone constitute the skeleton; secondary rings and
meshes are wanting. Two diverging families, the Semantida and Coronida, have been developed from
the Stephanida, and from one of them the family Tympanida has arisen.
The Semantida (p. 953,
Pl. 92)
develop a horizontal basal ring at the oral side of the vertical sagittal ring; the basal meshes
or lattice gates, which remain between the former and the latter, are the important cortinar
pores (one pair jugular, one pair cardinal, p. 954);
they usually appear inherited in the cortinar septum of the Cyrtellaria. In the Coronida (p. 967, Pls. 82, 94) a second
vertical ring (the frontal ring) appears in addition to the sagittal ring; it lies in the
frontal plane at right angles to the latter. Finally the Tympanida (p. 987, Pls. 93, 94) have
probably arisen from the Semantida by the formation of a second horizontal ring (mitral ring)
parallel to the basal and attached to the upper portion of the sagittal ring.
188. Genealogical Tree of the Spyroidea.—The extensive order Spyroidea is of especial interest in connection with the phylogeny of the Nassellaria, since all its members show two well-developed skeletal elements in
combination, the sagittal ring of the Stephoidea and the latticed
cephalis of the Cyrtoidea; the majority possess also the basal tripod of
the Plectoidea (or a radial skeleton derived from it). Hence there is a
possibility of deriving the stem-forms of the Spyroidea from each of
these three groups. The four families of this order exhibit similar relationships to those of the
four families of Cyrtoidea; the common stem-group is the family
Zygospyrida; from this the Tholospyrida have arisen by the development of a galea on the apical
pole, the Phormospyrida by the addition of a thorax on the basal pole. The Androspyrida may be
derived either from the Tholospyrida by the formation of a basal thorax, or from the Phormospyrida
by the development of an apical galea. Some groups, however, such as the peculiar Nephrospyrida
(Pl. 90)
have probably been developed directly from the Stephoidea.
189. Genealogical Tree of the Botryodea.—The peculiar order Botryodea (p. 1103),
which is both difficult to investigate and insufficiently known, presents great phylogenetic
difficulties both as to its ascent and descent. Probably the different genera of this order have
been polyphyletically developed from different groups of Cyrtoidea
(perhaps also to some extent of Spyroidea) by the formation of lobes in
the cephalis. The three families of Botryodea are related to each other
in the same way as are the three first families of the Cyrtoidea. From
the single-jointed Cannobotryida (corresponding to the Monocyrtida), the two-jointed Lithobotryida
(corresponding to the Dicyrtida), may be derived by the development of a basal thorax, and from
the latter the three-jointed Pylobotryida (like the Tricyrtida) by the addition of an abdomen. In
the last two families the forms with an open basal mouth {cxxii}(Botryopylida and Botryocyrtida) are to be regarded as primitive: the
Botryocellida and Botryocampida have arisen by the closure of this mouth with a basal
lattice-plate.
190. Genealogical Tree of the Cyrtoidea.—The multiform and extensive
group Cyrtoidea presents the greatest difficulties to be found in the
phylogeny of the Nassellaria, because their morphological relations are
most complicated, and because similar forms very often appear to be of quite different origin. The
great majority of the Cyrtoidea show more or less clearly a combination
of the three structural elements: sagittal ring, basal tripod, and latticed cephalis (p. 891). There are also,
however, numerous Cyrtoidea, whose skeleton no longer shows any trace of
the sagittal ring. Many of these show as the basis of the skeleton a strong basal tripod with an
apical spine, around which the cephalis has obviously been secondarily developed, e.g., the
remarkable Euscenida (p. 1146,
Pls. 53, 97) and the
interesting Callimitrida (p. 1217,
Pls. 63, 64). These may
have been derived immediately from the Plectoidea without any relation to
the Stephoidea. There are also numerous true Monocyrtida, whose shell
consists of a simple latticed cephalis without a trace of the sagittal ring or basal tripod
(Cyrtocalpida, Pl. 51,
figs. 9-13; Pl. 98,
fig. 13); these may have been developed directly from the skeletonless Nassellida by the formation
of a simple ovoid Gromia-like shell, and may have no relation either to the Stephoidea or Plectoidea. On these grounds, as well as
from the complicated relationships of the many smaller groups of Cyrtoidea, it is probable that the whole order has been developed
polyphyletically from different divisions of the Plectellaria.
191. Systematic Arrangement of the Cyrtoidea.—Although for the reasons just given
no systematic arrangement of the Cyrtoidea can at present, or for a long
time in the future, be regarded as other than artificial, yet some general principles of
classification for this extensive group can be laid down, which may serve as starting points for
some future natural disposition. This is especially true of the relations which in an artificial
system (p. 1129)
were primarily utilised for the distinction of twelve families and twenty-four subfamilies; the
number of segments in the shell, the number of radial apophyses (and parameres), and the
constitution of the basal aperture of the shell.
As regards the number of segments, separated by transverse constrictions,
of which the shell is composed, it is dependent upon the secondary addition of new joints at the
basal pole of the main axis. Hence all many-jointed Cyrtoidea are to be
derived from single-jointed ones, and the four sections thus distinguished (Monocyrtida,
Dicyrtida, Tricyrtida, Stichocyrtida) form a phylogenetic series. Very often, however, the
primary cephalis disappears owing to retrograde metamorphosis; and in such cases the single
joint of the apparent Monocyrtida is formed of the thorax (e.g., {cxxiii}Pls. 52,
54,
&c.). As regards the number of radial apophyses, three sections of Cyrtoidea may be distinguished; the Pilocyrtida with three, the Astrocyrtida
with numerous apophyses, and the Corocyrtida with none (p. 1129). The last two
may in general be regarded as two divergent branches from the first, for the eradiate
Corocyrtida have probably been formed from the triradial Pilocyrtida by entire loss of the
radial apophyses, whilst on the other hand the multiradiate Astrocyrtida have arisen from them
by additions to the primary apophyses (interpolation of interradial between the perradial ones).
As regards the constitution of the shell-aperture, the Cyrtoidea
may be divided into Cyrtaperta and Cyrtoclausa (p. 1129); in general
the Cyrtoclausa (with latticed shell-aperture) have arisen from the Cyrtaperta (with simple open
mouth); in many Monocyrtida the converse may be supposed, the simple basal mouth having been
formed by degeneration of a basal lattice.
192. Phylogeny of the Phæodaria.—The legion Phæodaria or Cannopylea is so clearly marked off from
other Radiolaria by the double membrane of the central capsule and the astropyle at its oral pole,
as well as by the extracapsular phæodium, that it must be regarded phylogenetically as an
independent stem (§ 9). This stem is only connected at its root by
Phæodina with the stem-form of the Spumellaria, Actissa. The
stem itself is monophyletic, inasmuch it its members may be derived without violence from the
skeletonless Phæodinida (Phæodina, Phæocolla). On the other hand, the formation of
the skeleton of the Phæodaria is undoubtedly polyphyletic, different
Phæodinida having independently commenced the formation of a skeleton and having carried it out in
very different ways.
193. Origin of the Phæodaria.—The Phæodinida (p. 1544, Pl. 101), which
may naturally be regarded as the common stem-group of the Phæodaria, have
their nearest relations among other Radiolaria in the Thalassicollida (p. 10); and since this family is to be regarded as the primitive group of all
Radiolaria, they may be directly derived from them phylogenetically. The essential modifications
by which the primitive Phæodinida have arisen from the more archaic Thalassicollida are of three
kinds; (1) the doubling of the membrane of the central capsule; (2) the reduction of the numerous
fine pores in the membrane and the formation of an osculum, and of an astropyle closing it, at the
oral pole of the main axis; (3) the production of an extracapsular phæodium. This last may,
perhaps, be regarded as a unilateral hypertrophy of the voluminous pigment masses which are
deposited in the sarcomatrix of certain Thalassicollida. Of the two genera of Phæodinida hitherto
known, probably Phæodina (Pl. 101,
fig. 2) approaches the original stem of the Phæodaria more nearly than
Phæocolla (Pl. 101,
fig. 1), for the latter exhibits only the large main opening of the central capsule (astropyle),
whilst the former possesses also a pair of accessory openings (parapylæ). The hypothetical
stem-form (Phæometra) presumably had a larger number of small parapylæ (like many
Circoporida and Tuscarorida), and the astropyle was probably but little differentiated from
them.
{cxxiv}
194. Hypothetical Genealogical tree of the Phæodaria:—
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Phæoconchia
|
| Phæosphæria |
|
Cœloplegmida |
|
Phæogromia |
 |
|
|
|
| Aularida |
|
Tuscarorida |
|
|
|
Cœlodrymida |
|
|
Aulonida |
|
|
|
|
|
|
Cœlotholida
Cœlographida |
|
Haeckelinida |
|
|
|
|
|
|
|
|
|
Conchopsida |
|
Cœlodorida |
|
| Aulosphærida |
|
|
|
Cœlodendrida |
Circogonida |
|
|
|
|
|
|
|
|
|
Conchasmida
Concharida |
|
|
|
|
|
|
Sagmarida |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Castanellida |
|
Circoporida |
| Cannosphærida |
|
|
|
|
|
|
|
Oroscenida |
Concharida |
|
|
|
Sagenida
Sagophærida |
|
|
|
Gazellettida |
|
|
|
Oronida
Orosphærida |
|
|
|
|
Pharyngellida |
|
|
|
|
|
|
|
|
|
|
|
|
|
Euphysettida
Medusettida |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Lithogromida |
|
|
|
|
|
Challengerida |
|
Phæodinida |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Phæocystina
|
|
|
|
|
| Aulacanthida |
|
|
|
|
|
|
|
Cannobelida |
Catinulida |
|
|
|
|
|
|
|
|
|
|
Dictyochida |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Phæodinida |
|
Cannorrhaphida |
|
Phæodinida |
|
|
|
|
|
| |
|
Phæodina |
| |
|
(Phæometra) |
| |
|
Actissa |
| |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Phæoconchia
 |
| Phæosphæria |
Cœloplegmida |
|
|
| Aularida |
|
|
|
Cœlodrymida |
|
Aulonida |
|
|
|
|
|
Cœlotholida
Cœlographida |
| |
|
|
|
|
|
|
Conchopsida |
|
Cœlodorida |
| Aulosphærida |
|
|
|
Cœlodendrida |
| |
|
|
|
|
|
Conchasmida
Concharida |
|
|
Sagmarida |
|
|
| |
|
|
|
| |
|
|
| Cannosphærida |
|
|
|
Oroscenida |
Concharida |
|
Sagenida
Sagophærida |
|
|
|
Oronida
Orosphærida |
|
| |
|
|
|
| |
|
|
| |
|
Phæodinida |
| |
|
≈ |
| |
| |
|
Phæogromia |
|
|
|
Tuscarorida |
|
Haeckelinida |
|
|
|
Circogonida |
| |
|
|
| |
|
|
Castanellida |
|
Circoporida |
| |
|
|
|
Gazellettida |
|
|
|
Pharyngellida |
|
|
|
|
|
Euphysettida
Medusettida |
|
|
|
|
|
|
|
Lithogromida |
|
|
Challengerida |
|
|
|
|
| |
|
|
|
Phæocystina
|
|
|
| Aulacanthida |
|
|
|
|
|
Cannobelida |
|
Catinulida |
|
|
|
|
|
|
|
|
Dictyochida |
|
|
|
|
|
|
|
|
|
|
|
|
|
Phæodinida |
≈ |
Phæodinida |
|
|
|
|
|
|
Cannorrhaphida |
Phæodinida |
|
|
|
|
|
| |
|
Phæodina |
| |
|
(Phæometra) |
| |
|
Actissa |
{cxxv}
195. Phæocystina and Phæocoscina.—Whilst the malacoma of all Phæodaria possesses the characteristics of the legion, and hence justifies the
assumption of a monophyletic origin, the skeleton, on the other hand, shows in the different
groups such manifold and fundamental variations that a polyphyletic origin of the latter is
indubitable. Different Phæodinida have commenced the formation of the skeleton independently, and
it has progressed in different directions. In the Phæocystina it remained
incomplete and led to the formation of various Beloid skeletons, whilst the Phæocoscina developed complete lattice-shells. Both of these divisions too are
to be regarded as polyphyletic, since the skeletal forms of the different groups cannot be derived
without violence from a common primitive form.
196. Phæocystina with a Beloid Skeleton.—The order Phæocystina includes all Phæodaria which have no
complete lattice-shell; it contains, firstly, the skeletonless Phæodinida (the common stem-group
of the legion), and secondly, the Phæacanthida, or Phæodaria with a Beloid
skeleton (§ 115). The latter are divisible into several very different
groups (at least two or three) which are probably different in origin. The Aulacanthida (Pls. 102-105) form
radial tubes which perforate the calymma, their proximal end resting upon the surface of the
central capsule, whilst the distal extremity projects freely outwards. The skeleton of the
Cannorrhaphida, on the other hand, is composed of many separate portions which are never radially
arranged but are either placed tangentially to the surface of the calymma or scattered irregularly
in its gelatinous mass. Furthermore, in the three subfamilies of which this family is composed,
the individual skeletal portions are so different that they have probably arisen independently of
each other; in the Cannobelida they form cylindrical tangential tubes (Pl. 101, figs.
3-5), in the Catinulida flat basin or cap-like structures (Pl. 117, fig. 8),
in the Dictyochida hollow rings, from which small pyramids are developed by unilateral formation
of lattice-work (Pl. 101,
figs. 9-14; Pl. 114,
figs. 7-12).
197. Phæosphæria with a Sphæroid Skeleton.—The order Phæosphæria includes those Phæodaria which possess a
spherical (sometimes slightly modified) lattice-shell without the characteristic aperture of the
Phæogromia. They have probably arisen independently of these, though they
may have been derived from the Castanellida by loss of the shell-aperture, which was present
originally. The four families which we have distinguished among the Phæosphæria, are so different in the structure of their lattice-shell that
their phylogenetic connection is doubtful. In the Orosphærida (Pls. 106, 107) and the
Sagosphærida (Pl. 108)
the whole lattice-shell consists of a single piece and is unjointed (without astral septa); in the
former it is very firm and massive, with thick laminated trabeculæ and polygonal meshes; in the
latter it is very delicate and brittle, with filiform trabeculæ and large {cxxvi}triangular meshes. On the other hand, the voluminous shell of the
Aulosphærida (Pls. 109-111), and of
the Cannosphærida (Pl. 112),
is characterised by a very peculiar system of joints; it is composed of numerous separate
cylindrical tubes, which are placed tangentially and united at the nodes by stellate partitions or
astral septa. The Cannosphærida possess further a simple central Cyrtoid shell, connected with the
outer jointed shell by hollow radial trabeculæ. Since many Aulosphærida possess rudiments of such
centripetal trabeculæ it is possible that these latter have been derived from the former by the
loss of the central Cyrtoid shell; the formation of this monaxon shell perhaps indicates descent
from the Phæogromia (Castanellida).
198. Phæogromia with a Cyrtoid Skeleton.—That order of the Phæodaria which we designate Phæogromia, contains many
very different forms, all agreeing in the possession of a Cyrtoid skeleton, or a monaxon
lattice-shell, which has a large aperture at one pole of its vertical main axis (§ 123). This Cyrtoid skeleton is sometimes ovoid or conical, sometimes lentiform
or helmet-shaped, sometimes polyhedral or almost spherical. Although the principle of its
structure is simple and often like that of the Monocyrtida among the Nassellaria, yet the structure of the wall and of the apophyses is so different
in the various groups of the Phæogromia, that the order is probably
polyphyletic, and its Cyrtoid shells have arisen independently of each other. Only in the
Castanellida (Pl. 113)
does the shell-wall usually consist of simple lattice-work; in the Challengerida, on the other
hand (Pl. 99),
it has an extremely fine Diatom-like structure; in the Medusettida (Pls. 118-128) a
peculiar alveolar structure, and in the Circoporida (Pls. 114-117) and
Tuscarorida (Pl. 100)
it possesses a characteristic porcellanous constitution (with tangential spicules in a porous
cement-mass); in the latter of these groups the surface is smooth, in the former peculiarly
tabulate; the two families have also different stem-forms.
199. Phæoconchia with a Conchoid Shell.—The order Phæoconchia (Pls. 121-128) is
separated not only from all other Phæodaria, but also from all other
Radiolaria, by the possession of a bivalved shell resembling that of a Lamellibranch; the two
valves of this Conchoid skeleton are to be interpreted as dorsal and ventral (§ 128). Probably these bivalved shells are independent products, but possibly
they may have been formed by the bisection of a simple spherical lattice-shell; in the former case
the Phæoconchia would be directly descended from the Phæodinida, in the
latter from the Castanellida. The three families which we have distinguished among the Phæoconchia, probably constitute a connected stem, the most primitive group of
which are the Concharida (Pls. 123-125). From
these the Cœlodendrida (Pls. 121,
122) have
next arisen by the formation of a "galea" upon the apex of each valve, and the growth of hollow
tubes from this helmet-like structure. Finally, the Cœlographida {cxxvii}(Pls. 120-128) have been
developed from the Cœlodendrida by the formation of a basal nasal tube (rhinocanna) from
each galea, and the formation of a median or paired frenulum, which connects the opening of the
nasal tube with the apex of the galea. In the Cœlodendrida, as well as in the
Cœlographida, there are two different subfamilies, of which the more primitive
(Cœlodorida, Cœlotholida) have free branches from the hollow radial tubes, whilst
the more recent (Cœlodrymida, Cœloplegmida) form an outer bivalved shell by
anastomosis of the branches of the tubes.
200. The Fundamental Biogenetic Law.—The causal connection between
ontogeny and phylogeny, which finds its most precise statement in the fundamental biogenetic law,
holds in general for the Radiolaria as for all other organisms. In order to furnish direct proof
of this, however, a complete empirical knowledge both of individual and of palæontological
development would be necessary. In both these directions, as has been shown in the foregoing
chapters, our knowledge of the Radiolaria is very incomplete and fragmentary, but still we are
able to convince ourselves indirectly of the validity of the law as applied to Radiolaria by the
aid of comparative anatomy. This is now so fully known to us (§§ 1-140) that we are able not only to draw a complete and satisfactory picture of
their morphology, but also to arrive at most important conclusions regarding the ontogeny and
phylogeny of the individual groups. As regards the formation of the multiform skeleton of the
Radiolaria, most of the ontogenetic series of forms, with which we have become acquainted by
comparative anatomy, are of palingenetic nature; that is, they are primarily due to
inheritance and thus of direct phylogenetic significance. On the other hand, among the ontogenetic
phenomena of the Radiolaria, as far as they have yet been investigated, only very few are
cenogenetic, that is, brought about by adaptive modification and without direct
significance as regards phylogeny.
{cxxviii}
PHYSIOLOGICAL SECTION.
Chapter VII.—VEGETATIVE FUNCTIONS.
(§§ 201-217.)
201. Mechanism of the Functions.—The vital phenomena of the Radiolaria are
dependent upon the mechanical functions of their unicellular body, and like those of all other
organisms, are to be referred to physical and chemical natural laws. All processes which appear in
the life of the Radiolaria are, therefore, ultimately to be explained by the attraction and
repulsion of the smallest particles, which compose the different portions of their unicellular
body; and the sensation of pleasure or the opposite is in its turn the exciting cause of these
elementary movements. Many adaptive arrangements in the Radiolarian organism may produce the
appearance of being the premeditated result of causes working towards an end ("zweckthätig,"
causæ finales), but as opposed to this deceptive appearance it must here be expressly
stated that these may be recognised in accordance with the developmental theory as the necessary
consequence of mechanical causes (causæ efficientes).
Our physiological acquaintance with the Radiolaria has by no means
progressed so far as our morphological, so that the incomplete communications which are
placed here for the sake of completeness must be regarded merely as preliminary fragments, not
as fully elaborated results. Since my recent investigations have been mainly in the direction of
morphology, I can add but little to the physiological conclusions, which I stated at length in
my monograph twenty-four years ago (L. N. 16, pp. 127-165). Recently the
vegetative physiology of the Radiolaria has been much advanced by the recognition of the
symbiosis with the Xanthellæ (§ 205, L. N. 22, 39, 42). In addition Karl Brandt has recently (1885)
published several important contributions to the physiology of the Polycyttaria or Sphaerozoea
(L. N. 52).
202. Distribution of Functions.—The distribution of the functions among the
various parts of the unicellular organism of the Radiolaria corresponds directly to their
anatomical composition, so that physiologically as well as morphologically the central capsule and
the extracapsulum appear as the two coordinated main components. On the one hand the central
capsule with its endoplasm and enclosed nucleus is the central organ of the "cell-soul"
(Zellseele), the unit regulating its animal and vegetative functions, and the special organ of
reproduction and inheritance. The extracapsulum forms, on the other hand, by its calymma
the protective envelope of the central {cxxix}capsule,
the support of the soft pseudopodia and the substratum of the skeleton; the calymma acts also as a
hydrostatic apparatus, whilst the radiating pseudopodia are of the greatest importance both as
organs of nutrition and adaptation, as well as of motion and sensation (§ 15). If, however, the vital functions as a whole be divided in accordance with
the usual convention into the two great groups of vegetative (nutrition and reproduction)
and animal (motion and sensation), then the central capsule would be mainly the organ of
reproduction and sensation, and the extracapsulum the organ of nutrition and motion.
The numerous separate vital phenomena, which by accurate physiological
investigation may be distinguished in the unicellular Radiolarian organism, may be distributed
in the above indicated conventional fashion into a few larger and several smaller groups; it
must always be borne in mind, however, that these overlap in many respects, and that the
division of labour among the different organs in these Protista is somewhat complicated,
notwithstanding the apparent simplicity of their unicellular organization. A general
classification of the groups of functions is difficult, because each individual organ discharges
several different functions. Thus the central capsule is pre-eminently the organ of reproduction
and inheritance, but not less (though less conspicuous) is its importance as the psychical
central organ, the unit regulating the processes of sensation, motion, and also nutrition. In
this last respect it is comparable to the nerve-centres of the Metazoa, whilst the peripheral
nervous system of the latter (including the organs of sense and the muscles) are in the present
instance represented by the pseudopodia, which are at the same time the most important organs of
nutrition and adaptation. In the calymma also in similar fashion several different physiological
functions are united.
203. Metastasis.—The functions of metastasis and nutrition have in all Radiolaria
a purely animal character, so that these Rhizopoda from the physiological standpoint are to be
regarded as truly unicellular animals, or Protozoa ("Urthiere"). Since they do not possess,
like plants, the power of forming synthetically the compounds (protoplasm, carbohydrates, &c.)
necessary for their sustenance, they are compelled to obtain them ready-formed from other
organisms. Like other true animals they evolve carbon dioxide by the partial oxidation of those
products, and hence they successively take up the oxygen necessary to their existence from their
environment.
The question whether the Radiolaria are to be regarded as true animals I
discussed fully from various points of view in 1862, and finally answered in the affirmative (L.
N. 16, pp. 159-165). Afterwards, when in my Generelle Morphologie (1866) I
sought to establish the kingdom Protista, I removed the Radiolaria along with the other
Rhizopoda from the animal kingdom proper and placed them in the kingdom Protista (Bd. i. pp.
215-220; Bd. ii. p. xxix). Compare also my Protistenreich (L. N. 32) and my
Natürliche Schöpfungsgeschichte (vii. Aufl., 1879, p. 364). Both these steps appear fully
justified when considered in the light of our present increased knowledge. From the
physiological standpoint the Radiolaria appear as unicellular animals, for in this
respect the animal character of their metastasis (that proper to an oxidising organism)
furnishes the sole {cxxx}criterion. On the other
hand, from the morphological standpoint, they are to be classed as neutral Protista, for in this
respect their unicellular character is the prominent feature, and distinguishes them from all
true multicellular animals (Metazoa). Compare my Gastræa Theorie (1873, Jena. Zeitschr. für
Naturwiss., Bd. viii. pp. 29, 53).
204. Nutrition.—The nutritive materials which the Radiolaria require for their
sustenance, especially albuminates (plasma) and carbohydrates (starch, &c.), they obtain
partly from foreign organisms which they capture and digest, and partly directly from the
Xanthellæ or Philozoa, the unicellular Algæ, with which they live in symbiosis (§ 205). Zooxanthella intracapsularis, found in the Acantharia (§ 76), is probably of the same significance in
this respect as Zooxanthella extracapsularis of the Spumellaria and
Nassellaria (§ 90); and perhaps the same is true
also of Phæodella extracapsularis (or Zoochlorella phæodaris?) of the Phæodaria (§ 89). The considerable quantity of starch or
amyloid bodies, elaborated by these inquiline symbiontes, as well as their protoplasm and nucleus,
are available, on their death, for the nutrition of the Radiolaria which harbour them. Nutrition
by means of other particles obtained by the pseudopodia from the surrounding medium is by no means
excluded; indeed it may be regarded as certain that numerous Radiolaria (especially such as
contain no symbiotic Algoid cells) are nourished for the most part or exclusively by this means.
Diatoms, Infusoria, Thalamophora (Foraminifera) as well as decaying particles of animal and
vegetable tissues can be seized directly by the pseudopodia and conveyed either to the
sarcodictyum (on the surface of the calymma) or to the sarcomatrix (on the surface of the central
capsule) in order to undergo digestion there. The indigestible constituents (siliceous shells of
Diatoms and Tintinnoidea, calcareous shells of small Monothalamia and Polythalamia, &c.) are
here collected often in large numbers and removed by the streaming of the protoplasm.
The inception and digestion of nutriment, as it usually appears to take place by
the pseudopodia, has already been so fully treated in my Monograph (L. N. 16, pp. 135-140), and since then in my paper on the sarcode body of the
Rhizopoda (L. N. 19, p. 342), that I have nothing of importance to add.
Quite recently Karl Brandt has expressed a doubt as to whether the taking up of formed particles
by the pseudopodia and their aggregation in the calymma be really connected with the process of
nutrition. He is disposed rather to believe that these foreign bodies are usually only
accidentally and mechanically brought into the calymma, and that the nourishment of the
Radiolaria is derived exclusively or pre-eminently from the symbiotic Xanthellæ (L. N. 52, pp. 88-93). I must, however, maintain my former opinion, which I have only
modified insomuch that I now regard the sarcodictyum (on the outer surface of the calymma, § 94) rather than the sarcomatrix (on the outer surface of the central capsule,
§ 92) as the principal seat of true digestion and assimilation. From the
sarcodictyum the dissolved and assimilated nutritive matters may pass by the intracalymmar
pseudopodia (or sarcoplegma, § 93) into the sarcomatrix, and hence may
reach the endoplasm through the openings in the central capsule. To what an extent the
Radiolaria are capable of taking up even large formed bodies into the calymma, is shown by the
{cxxxi}striking instance of Thalassicolla
sanguinolenta, which becomes so deformed by the inception of numerous coccospheres and
coccoliths, that I described it as a special genus under the name Myxobrachia (compare
pp. 23, 30; also L. N. 21, p.
519, Taf. xviii., and L. N. 33, p. 37).
205. Symbiosis.—Very many Radiolaria, but by no means all members of this class,
live in a definite commensal relation with yellow unicellular Algæ of the group Xanthellæ. In the
Acantharia they live within the central capsule (Zooxanthella
intracapsularis, § 76), in the Spumellaria and
Nassellaria, on the other hand, within the calymma but outside the central
capsule (Zooxanthella extracapsularis, § 90); in the Phæodaria a special form of these symbiotic unicellular Algæ appears to inhabit
the phæodium in the extracapsulum, and to compose a considerable portion of the phæodellæ
(Zooxanthella phæodaris, § 90, or better perhaps Zoochlorella
phæodaris, § 89). Undoubtedly this commensal life is in very many cases
of the greatest physiological significance for both the symbiontes, for the animal Radiolarian
cells furnish the inquiline Xanthellæ not only with shelter and protection, but also with carbon
dioxide and other products of decomposition for their nutriment; whilst on the other hand the
vegetable cells of the Xanthellæ yield the Radiolarian host its most important supply of
nutriment, protoplasm and starch, as well as oxygen for respiration. Hence it is not only
theoretically possible, but has been experimentally proved, that Radiolaria which contain numerous
Xanthellæ can exist without extraneous nutriment for a long period in closed vessels of filtered
sea-water, kept exposed to the sunlight; the two symbiontes furnish each other mutually with
nourishment, and are physiologically supplementary to each other by reason of the opposite nature
of their metastasis. This symbiosis is not necessary, however, for the existence of the
Radiolaria; for in many species the number of Xanthellæ is very variable and in many others they
are entirely wanting.
The symbiosis of the Radiolaria and Xanthellæ, or "yellow cells" (§§ 76, 90) was first discovered by Cienkowski in 1871 (L.
N. 22). Ten years later this important and often doubted fact was
established by extended observations and experiments almost simultaneously by Karl Brandt (L. N.
38, 39) and Patrick Geddes (L. N. 42, 43). This commensal life may be compared with that of
the lichens, in which an organism with vegetable metastasis (the Algoid gonidia) and an organism
with animal metastasis (the Fungoid hyphæ) are intimately united for mutual benefit. But the
symbiosis of the Xanthellæ and Radiolaria is not as in the lichens a phenomenon essential for
their development, but has more or less the character of an accidental association. The number
of the inquiline Xanthellæ is so variable even in one and the same species of Radiolaria, that
they do not appear to be exactly essential to its welfare; and in many species they are entirely
wanting. Their significance is questionable in the case of those numerous deep-sea Radiolaria
which live in complete darkness, and in which, therefore, the Xanthellæ, even if present, could
excrete no oxygen on account of the want of light. Nevertheless it is possible that the
phæodellæ of the Phæodaria (usually green, olive, or brown in colour),
which are true cells, represent vegetable symbiontes, {cxxxii}which in the absence of sunlight are able to evolve oxygen by the
aid of the phosphoresence of other abyssal animals. Since the Phæodaria
are, for the most part, dwellers in the deep-sea, and since the voluminous phæodium must be of
great physiological importance, a positive solution of this hypothetical question would be of no
small interest (compare § 89).
206. Respiration.—The respiration of the Radiolaria is animal in nature, since all
Protista of this class, like all other true Rhizopoda, take in oxygen and give off carbon dioxide.
Probably this process goes on continuously and is tolerably active, as may be inferred from the
fact that Radiolaria cannot be kept for long in small vessels of sea-water unless either they
contain numerous Xanthellæ or the water is well aërated. The oxygen is obtained from two sources,
either from the surrounding water or from the enclosed Xanthellæ, which in sunlight evolve
considerable quantities of this gas. Correspondingly, the carbon dioxide which is formed during
the process of oxidation of the Radiolaria is either given up to the surrounding water or to the
inquiline Xanthellæ, which utilise it for their own sustenance (§§ 204, 205).
The significance of the symbiotic Xanthellæ for the respiration of the enclosing
Radiolaria may be shown experimentally in the following way. If two Polycyttarian colonies of
equal size, both of which contain numerous Xanthellæ, be placed in equal quantities of filtered
sea-water in sealed glass tubes, and if one tube be placed in the dark the other in the light,
the colony in the former rapidly perishes, but not that in the latter; the Xanthellæ excrete
only under the influence of sunlight the oxygen necessary for the life of the Radiolarian
(compare Patrick Geddes, L. N. 42, p. 304).
207. Circulation.—In the protoplasm of all Radiolaria, both inside and
outside the central capsule, slow currents may be recognised which fall under the general term
circulation, and have already been compared to the cyclosis in the interior of animal and
vegetable cells, as well as to the sarcode streams in the body of other Rhizopoda. These plasmatic
currents or "plasmorrheumata" probably continue throughout the whole life of the Radiolaria, and
are of fundamental importance for the performance of their vital functions. They depend upon slow
displacements of the molecules of the plasma (plastidules or micellæ) and cause a uniform
distribution of the absorbed nutriment and a certain equalisation of the metastasis. Furthermore
they are of great importance also in the inception of nutriment, the formation of the skeleton,
locomotion, &c. Sometimes the circulation is directly perceptible in the plasma itself; but
usually it is only visible owing to the presence of granules (sarcogranula), which are suspended
in the plasma in larger or smaller numbers. The movements of these granules are usually regarded
as passive, due to the active displacement of the molecules of the plasma. Although the
intracapsular protoplasm is in communication with the extracapsular through the openings in the
capsule membrane, nevertheless the currents exhibit certain differences {cxxxiii}in the two portions of the malacoma. It is sometimes possible,
however, to recognise the direct connection between them and to observe how the granules pass
through the openings in the capsule-membrane.
208. Currents in the Endoplasm.—Intracapsular circulation or a certain
slow flowing of the plasma within the central capsule is probably just as common in the Radiolaria
as without it, but it is not so easy to observe in the former case as in the latter. A more
satisfactory proof of these endoplasmatic currents is furnished by the arrangement of the
protoplasm within the central capsule, since this is (at all events in part) an effect produced by
them. In this respect the two main divisions of the class show characteristic differences. In the
Porulosa (the Spumellaria, § 77, and the Acantharia, § 78) the endoplasm is in general
distinguished by a more or less distinct radial structure, which is to be regarded as the effect
of alternating centripetal and centrifugal radial streams. In the Osculosa, on the other hand,
this radial structure is absent and the intracapsular plasmatic streams converge or diverge
towards the osculum or main-opening in the central capsule which lies at the basal pole of its
main axis, and through which the mass of the endoplasm issues into the calymma. The two legions of
the Osculosa, however, present differences in this respect. In the Nassellaria (§ 79) the endoplasmatic currents appear to
unite in an axial main stream at the apex of the monaxon central capsule, and this apical stream
seems to split into a conical bundle, the individual threads of which pass diverging between the
myophane fibrillæ of the podoconus towards the basis of the central capsule, and issue through the
pores of the porochora. In the Phæodaria (§ 80), on
the other hand, meridional currents of endoplasm are probably present on the inner surface of the
capsule, which flow from the aboral pole of the vertical main axis to its basal pole, and return
in the reverse direction.
209. Currents in the Exoplasm.—Extracapsular circulation, or a distinct flowing of
the plasma outside the central capsule, may be readily observed in all Radiolaria which are
examined alive; this is most readily seen in the astropodia, or those free pseudopodia which
radiate from the sarcodictyum on the surface of the calymma into the surrounding water. The
granular movement is often quite as clear in the sarcodictyum itself, and may be recognised in the
collopodia, which compose the irregular plasmatic network within the calymma. More rarely it is
possible to follow the granular stream thence through the sarcomatrix, and further into the
interior of the central capsule. In general the direction of the extracapsular protoplasmic
streams is radial, and it is frequently possible, even in a single free astropodium, to observe
two streams opposite in direction, the granules on one side of the radial sarcode thread moving
centripetally, those on the other side centrifugally. If the threads branch, and neighbouring ones
{cxxxiv}become united by connecting threads, the
circulation of the granules may proceed quite irregularly in the network thus formed. The rapidity
and character of the extracapsular currents are subject to great variations.
The different forms of extracapsular sarcode currents have been already very
fully described in my Monograph (L. N. 16, pp. 89-126), and in my critical
essay on the sarcode body of the Rhizopoda (L. N. 19, p. 357, Taf.
XXVI.).
210. Secretion.—Under the name secretions, in the strict sense,
all the skeletal formations of the Radiolaria may be included. They may be divided according to
their chemical composition into three different groups: pure silica in the Spumellaria and Nassellaria, a silicate of carbon in the
Phæodaria, and acanthin in the Acantharia (compare
§ 102). It may indeed be assumed that these skeletons arise directly by a
chemical metamorphosis (silicification, acanthinosis, &c.) of the pseudopodia and protoplasmic
network; and this view seems especially justified in the case of the Astroid skeleton of the Acantharia (§ 114), the Spongoid skeleton of the Spumellaria (§ 126), the Plectoid skeleton of the Nassellaria (§ 125), the Cannoid skeleton of the Phæodaria (§ 127), and several other types. On closer
investigation, however, it appears yet more probable that the skeleton does not arise by direct
chemical metamorphosis of the protoplasm, but by secretion from it; for when the dissolved
skeletal material (silica, acanthin) passes from the fluid into the solid state, it does not
appear as imbedded in the plasma, but as deposited from it. However, it must be borne in mind that
a hard line of demarcation can scarcely, if at all, be drawn between these two processes. In the
Acantharia the intracapsular sarcode is the original organ of secretion of
the skeleton; in the other three legions, on the other hand, the extracapsulum performs this
function (§§ 106, 107). In addition to the skeleton,
we may regard as secretions (or excretions) the intracapsular crystals (§ 75) and concretions (§ 75A), and perhaps certain pigment-bodies (§§ 74, 88); and further, the calymma (§ 82) may be considered to
be a gelatinous secretion of the central capsule, and perhaps also the capsule-membrane, in so far
as it represents only a secondary excretory product of the unicellular organism.
211. Adaptation.—The innumerable and very various adaptive phenomena
which we meet with in the morphology of the Radiolaria, and especially in that of their skeleton,
are like other phenomena of the same kind, to be ultimately referred to altered nutritional
relations. These may be caused directly either by the influence of external conditions of
existence (nutrition, light, temperature, &c.), or by the proper activity of the unicellular
organism (use or disuse of its organs, &c.), or, finally, by the combined action of both
causes in the struggle for existence. In very many cases the cause to which the origin of a
particular form of Radiolaria is due may be directly perceived or at least guessed at with
considerable probability; thus, for example, the lattice-shells {cxxxv}may be explained as protective coverings, the radial spines as
defensive weapons, and the anchor-hooks and spathillæ as organs of prehension, which are of
advantage to their possessors in the struggle for existence; the regular arrangement of the radial
spines in the Radiolaria may also be explained on hydrostatic grounds, it being advantageous that
the body should be maintained in a definite position of equilibrium, &c. The well-known laws
of direct or actual adaptation, which we designate cumulative, correlative,
divergent adaptation, &c., here explain a multitude of morphological phenomena. The connection
is less distinct in the case of the laws of indirect or potential adaptation,
although this must play as important a part in the formation of the Radiolaria as in that of other
organisms (compare on this head my Generelle Morphologie, Bd. ii. pp. 202-222).
212. Reproduction.—The most common form of reproduction in the
Radiolaria is the formation of spores in the central capsule, which in this respect is to be
regarded as a sporangium (§ 215). In many Radiolaria (Polycyttaria and
Phæodaria), however, there occurs in addition an increase of the
unicellular organism by simple division (§ 213); upon this the formation of
colonies in the social Radiolaria is dependent (§ 14). Reproduction by
gemmation is much less common, and has hitherto been observed only in the Polycyttaria (§ 214). In this group alone there also occur at certain times two different
forms of swarm-spores which copulate, and thus indicate the commencement of sexual reproduction
(Alternation of Generations, § 216). The general organ of reproduction is
in all cases the central capsule, whilst the extracapsulum never takes an active part in the
process.
213. Cell-Division.—Increase by cell-division among the Radiolaria in the early
stage, before the formation of the skeleton, is widely distributed (perhaps even general?); in the
adults of this class it is rather rare and limited to certain groups. It is most readily observed
in the Polycyttaria; the growth of the colonies in this social group depends mainly (and in many
species exclusively) upon repeated spontaneous division of the central capsule; all the
individuals of each colony (in so far as this has not arisen by the accidental fusion of two or
more colonies) are descendants of a single central capsule, which has arisen from an asexual
swarm-spore (§ 215) or from the copulation of two sexual swarm-spores (§ 216). Whilst the central capsules of the colonies continually increase by
division, their calymma remains a common gelatinous sheath. Among the Spumellaria reproduction by simple cell-division probably occurs also in many
monozootic Collodaria. Among the Acantharia the
peculiar group Litholophida has perhaps arisen by the spontaneous division of Acanthonida (see p. 734). Among the Phæodaria increase by cell-division seems to occur commonly in many groups, as
in the Phæocystina, which have no skeleton (Phæodinida, Pl. 101, {cxxxvi}fig. 2), or only an incomplete Beloid skeleton
(Cannorrhaphida, Pl. 101,
figs. 3, 6, and Aulacanthida, Pl. 104,
figs. 1-3). The Phæosphæria also (Aulosphærida, Cœlacanthida) and
the Phæogromia (Tuscarorida, Challengerida) appear sometimes to divide;
at all events, their central capsule often contains two nuclei. Of special interest is the
spontaneous division of the Phæoconchia, especially the Concharida (Pl.
124, fig.
6). In all monozootic Radiolaria, the nucleus first divides by a median constriction into two
equal halves (usually by the mode of direct division); then the central capsule becomes
constricted in the middle (in the Phæodaria in the vertical main axis),
and each portion of the capsule retains its own nucleus. In the Phæoconchia each half or daughter-cell corresponds to one valve of the shell,
dorsal or ventral, so that probably on subsequent separation each daughter-cell retains one valve
of the mother-cell, and forms a new one for itself by regeneration (as in the Diatoms). In the
polyzootic Radiolaria, which already contain many small nuclei, but usually only a single central
oil-globule in each central capsule, the division of the latter is preceded by that of the
oil-globule. In many Polycyttaria the colony as a whole multiplies by division.
The increase of the central capsule by division was first described in 1862 in my
Monograph (L. N. 16, p. 146); since then R. Hertwig (L. N. 26, p. 24) and K. Brandt (L. N. 52, p. 144) have confirmed
my statement. In the Phæodaria the division of the central capsule
appears always to take place in the main axis; in the bilateral sometimes in the sagittal,
sometimes in the frontal plane. In the Tripylea each daughter-cell seems to retain one parapyle
and half the astropyle (compare the general description of the Phæodaria, Pl. 101,
figs. 1-6, Pl. 104,
figs. 1-3, and also Hertwig, L. N. 33, p. 100, Taf. x. figs. 2, 11).
Regarding the spontaneous division of colonies of the Polycyttaria, see K. Brandt, L. N. 52, p. 142.
214. Cell-Gemmation.—Reproduction by gemmation has hitherto been observed only in
the social Radiolaria, but in them it appears to be widely distributed, and in very young colonies
is perhaps almost universally present. The gemmules or capsular buds (hitherto described as
"extracapsular bodies") are developed on the surface of young central capsules before these had
secreted a membrane. They grow usually in considerable numbers, from the surface of the central
capsule, which is sometimes quite covered with them. Each bud usually contains a raspberry-like
bunch of shining fatty globules, and by means of reagents a few larger or a considerable number of
smaller nuclei may be recognised in them; the naked protoplasmic body of the bud is not enclosed
by any membrane. As soon as the buds have reached a certain size they are constricted off from the
central capsule and separated from it, being distributed in the meshes of the sarcoplegma by the
currents in the exoplasm. Afterwards each bud becomes developed into a complete central capsule by
surrounding itself with a membrane when it has attained a definite size. From the special
relations of the process of nuclear formation, which take place in the multiplication of the {cxxxvii}social central capsules by gemmation and by
cell-division, it would appear that the capsules produced by the former method afterwards produce
anisospores, whilst those in the latter way yield isospores (§ 216).
The gemmules or capsular buds of the Polycyttaria were first accurately described
by Richard Hertwig (L. N. 26, pp. 37-39), under the name "extracapsular
bodies," and their significance rightly indicated; earlier observers had incidentally mentioned
and figured them, but had not seen their origin from the central capsule. Quite recently Karl
Brandt has given a very painstaking account of them in the different Polycyttarian genera (L. N.
52, pp. 179-198). In the Monocyttaria such a formation of buds has not yet
been observed. The basal lobes of the central capsule, which occur in many Nassellaria, are not buds, but simple processes of the capsule, due to its
protrusion through the collar pores of the cortinar septum (§ 55).
215. Sporification.—Asexual reproduction by the formation of movable flagellate
spores has been hitherto observed only in a very small number of genera; but since these belong to
very different groups, and since the comparative morphology of the capsule appears to be similar
throughout as regards the structure and development of its contents, it may be safely assumed that
this kind of reproduction occurs quite generally in the Radiolaria. In all cases it is the
contents of the central capsule, from which the swarm-spores are formed, both nucleus and
endoplasm taking an equal share in the process; in all cases the spores produced are very
numerous, small, ovoid or reniform, and have one or two very long slender flagella at one
extremity (see §§ 141, 142). Since the whole
contents of the mature central capsule are used up in the formation of these flagellate zoospores,
it discharges the function of a sporangium. The division of the simple primary nucleus into
numerous small nuclei, which usually (serotinous Radiolaria) takes place only shortly before
sporification, but sometimes (precocious Radiolaria, § 63) happens very
early, is the commencement of the often repeated process of nuclear division, which terminates
with the production of a very large number of small spore-nuclei. The nucleolus often divides very
peculiarly (§ 69, C). Each spore nucleus becomes surrounded by a portion of
endoplasm and usually receives in addition one or more fatty granules, and sometimes also a small
crystal (hence the "crystal-spores"). The size of the flagellate zoospores which emerge from the
ruptured central capsule and swim freely in the water by means of their flagellum, varies
generally between 0.004 and 0.008 mm. The extracapsulum is not directly concerned in the
sporification, but undergoes degeneration during the process and perishes at its conclusion.
The first complete and detailed observations on the formation of spores in the
Radiolaria were published by Cienkowski in 1871 and related to two genera of Polycyttaria, the
skeletonless Collozoum and the spherical-shelled Collosphæra (L. N. 22, p. 372, Taf. xxix.). These were subsequently continued and supplemented by
R. Hertwig (1876, L. N. 26, pp. 26-42, and L. N. 33, p.
129), and a general summary of these results has been given by Bütschli (L. N. 41, pp. 449-455). {cxxxviii}Recently Karl Brandt has given a very detailed and fully
illustrated account of the sporification of the Polycyttaria (L. N. 52, pp.
145-178). I have also had the opportunity during my sojourn in the Canary Islands (1866), in the
Mediterranean at Corfu (1877), and Portofino (1880), as well as in Ceylon (1881), of observing
the development of flagellate zoospores from the central capsule of individuals of all four
legions: among the Spumellaria in certain Colloidea, Beloidea, Sphæroidea, and Discoidea, among the Acantharia in several Acanthometra and Acanthophracta, among the Nassellaria in individuals
belonging to the Stephoidea, Plectoidea, and
Cyrtoidea, and among the Phæodaria in one
Castanellid. In most zoospores I could distinctly observe only a single long flagellum;
sometimes, however, two or even three appeared to be present, but the determination of their
number is very difficult.
216. Alternation of Generations.—A peculiar form of reproduction, which may be
designated "alternation of generations," appears to occur generally in the Polycyttaria, but has
not yet been observed in the Monocyttaria. All Collozoida, Sphærozoida, and Collosphærida which have hitherto been carefully and
completely examined with respect to their development, are distinguished by the production of two
different kinds of swarm-spores, isospores and anisospores. The Isospores (or monogonous
spores) correspond to the ordinary asexual zoospores of the Monocyttaria (§ 215); they possess a homogeneous, doubly refracting nucleus of uniform
constitution and develop asexually, without copulation. The Anisospores (or amphigonous
spores), on the other hand, are sexually differentiated and possess a heterogeneous, singly
refracting nucleus of twofold constitution; they may therefore be distinguished as female
macrospores and male microspores. The Macrospores (or gynospores, comparable with the
female macrogonidia of many Cryptogams) are larger, less numerous, and possess larger nuclei,
which are less easily stained, and have a fine filiform trabecular network. On the other hand the
Microspores (or androspores, comparable with the male microgonidia) are much smaller
and more numerous, and are distinguished by their smaller nuclei, which have thicker tuberculæ and
become stained more intensely. The gynospores and androspores are developed in the Collozoida and Sphærozoida in the same individual, but
not in the Collosphærida. It is very probable that these two forms of anisospores copulate with
each other after their exit from the central capsule and thus produce a new cell by the simplest
mode of sexual reproduction. But, since the same Polycyttaria, which produce these anisospores, at
other times give rise to ordinary or asexual isospores, it is further possible that these two
forms of reproduction alternate with each other, and that the Polycyttaria thus pass through a
true alternation of generations. This has not yet been observed in the Monocyttaria, and hence
these latter seem to bear to the Polycyttaria a relation similar to that in which the sexless
solitary Flagellata (Astasiea) stand to the sexual social Flagellata (Volvocinea). In the two
analogous cases the sexual differentiation may be regarded as a consequence of the social life in
the gelatinous colonies.
{cxxxix}
The sexual differentiation of the Polycyttaria was first discovered in
1875 by R. Hertwig, and accurately described in the case of Collozoum inerme as occurring
in addition to the formation of the ordinary crystal-spores (L. N. 26, p.
36); compare also the general discussion of Bütschli (L. N. 41, p. 52).
Recently Karl Brandt has demonstrated the formation of both homogeneous isospores
(crystal-spores) and heterogeneous anisospores (macro- and microspores) in seven different
species of Polycyttaria, and has come to the conclusion that in all social Radiolaria there is a
regular alternation between the former and latter generations. Compare his elaborate account of
the colonial Radiolaria of the Gulf of Naples (L. N. 52, pp. 145-178).
217. Inheritance.—Inheritance is to be regarded as the most important
accompaniment to the function of reproduction, and especially in the present case, because the
comparative morphology of the Radiolaria furnishes abundant instances of the action of its
different laws. The laws of conservative inheritance are illustrated by the comparative
anatomy of the larger groups; thus, in the four legions the characteristic peculiarities of the
central capsule are maintained unaltered in consequence of continuous inheritance, although great
varieties appear in the skeleton in each legion. The individual parts of the skeleton furnish by
their development on the one hand and their degeneration on the other, especially in the smaller
groups, examples of progressive inheritance. Thus in the Spumellaria the constant formation of the primary lattice-shell (a central
medullary shell) and its ontogenetic relation to the secondary one, which is developed
concentrically round it, can only be explained phylogenetically by conservative inheritance,
whilst on the other hand the characteristic differentiation of the axes in the various families of
Spumellaria is to be explained by progressive inheritance. In the Acantharia the arrangement of the twenty radial spines (in accordance with
Müller's law, §§ 110, 172) was first acquired by a
group of the most archaic Actinelida (Adelacantha) through hydrostatic
adaptation, and has since been transmitted by inheritance to all the other families of the legion
(Icosacantha). The morphology of the Nassellaria is not less interesting,
because here several different heritable elements (the primary sagittal ring and the basal tripod)
combine in the most manifold ways in the formation of the skeleton (compare §§ 123, 124, 182). The affinities
of the genera in the different families yield an astonishing variety of interesting morphological
phenomena, which can only be explained by progressive inheritance. The same is true also of the
Phæodaria. In this legion the primary inheritance is especially manifested
in the constant and firm structure of the central capsule with its characteristic double wall and
astropyle, whilst the formation of the skeleton in this legion proceeds in different directions by
means of divergent adaptation. The morphology of the Radiolaria thus proves itself a rich source
of materials for the physiological study of adaptation and inheritance.
{cxl}
Chapter VIII.—ANIMAL FUNCTIONS.
(§§ 218-225.)
218. Motion.—In addition to the internal movements which appear
generally in the unicellular Radiolaria and have already been mentioned as plasmatic currents in
treating of the circulation (§§ 207-209), two
different groups of external motor phenomena are to be observed in this class: first, the
contraction of individual parts, which brings about modifications of form (§ 220), and secondly, voluntary or reflex locomotion of the whole body (§
220). These movements are partly due to changes in form of undifferentiated
plasmatic threads or sarcode filaments, partly to the actual contraction of differentiated
filaments which are comparable to muscle fibrillæ, and must therefore be distinguished as
myophanes. In addition to this, endosmose and exosmose probably play an important part in some of
the locomotive phenomena, but nothing is yet certainly known regarding these osmotic processes. We
are at present equally ignorant whether all the movements of the Radiolaria are simply reflex
(direct consequences of irritation) or whether they are in part truly spontaneous.
219. Suspension.—From direct observation of living Radiolaria, as well
as from deductive reasoning, based upon their morphology (and especially their promorphology, §§
17-50), the conclusion appears justified that all
Protista of this class in their normal condition float suspended in the sea-water, either at the
surface or at a definite depth. A necessary condition of this hydrostatic suspension is that the
specific gravity of the Radiolarian organism must be equal to, or but slightly greater than that
of sea-water. The increase in specific gravity brought about by the production of the siliceous
skeleton, is compensated by the lighter fatty globules, and partly perhaps by the calymma,
especially when the latter contains vacuoles or alveoles. The fluid or jelly contained in the
latter appears to be for the most part lighter than sea-water (containing no salt, or only a very
small quantity?). But if the specific gravity of the whole body should be generally (or perhaps
always) slightly greater than that of sea-water, then the organism would be prevented from
sinking, partly by the increased friction, due to the radiating pseudopodia and the radial spines
usually present, and partly perhaps by active (if only feeble) movements of the pseudopodia.
220. Locomotion.—Active locomotion of the whole body, which is very probably to be
regarded as voluntary, occurs in the Radiolaria in three different modes; (1) the vibratile
movement of the flagellate swarm-spores; (2) the swimming of the floating organisms; (3) the slow
creeping of those which rest accidentally upon the bottom. {cxli}The vibratile movement of the swarm-spores is the result of
active sinuous oscillation of the single or multiple flagellum, and is not essentially different
from that of ordinary flagellate Infusoria (see note A). Of the active swimming of mature
Radiolaria, only that form is known which is vertical in direction and causes the sinking and
rising in the sea-water. This is probably, for the most part (perhaps exclusively), due to
increase or diminution in the specific gravity, which is perhaps brought about by the retraction
or protrusion of the pseudopodia; slow, oscillating, sinuous motions of these organs have been
directly observed to take place (though very slowly) in suspended living Radiolaria. The most
important hydrostatic organ is probably the calymma, by the contraction of which the specific
gravity is increased, while it is diminished by its expansion; the contraction is probably brought
about by active contraction of the sarcodictyum, and is connected with exosmosis, while the
expansion is probably due to the elasticity of the calymma and the inception of water by
endosmosis. In the Acanthometra (§ 96) the peculiar
myophriscs appear to be charged with the duty of distending the gelatinous envelope, and thus
diminishing the specific gravity; the latter increases again when the myophriscs are relaxed, and
the calymma contracts by virtue of its own elasticity (see note B). The slow creeping
locomotion exhibited by Radiolaria on a glass slide under the microscope, does not differ from
that of the Thalamophora (Monothalamia and Polythalamia), but can only occur normally when the
animal accidentally comes into contact with a solid surface or sinks to the bottom of the sea.
Whether this actually occurs periodically is not known (see note C). The slow or gliding
locomotion exhibited by creeping Monozoa on a glass slide is due to muscle-like contractions of
bundles of pseudopodia, just as in the case of the social central capsules of Polyzoa, which live
together in the same cœnobium and are able to move within their common calymma sometimes
centrifugally to its surface, sometimes towards the centre where they aggregate into a roundish
mass (see note D).
A. Regarding the movement of the flagella in mature swarm-spores compare L. N. 22, p. 375; L. N. 26, pp. 31, 35; L. N. 41, p. 452, and L. N. 52, p. 170.
B. On the active vertical swimming movements of mature Radiolaria, especially the cause of
sinking and rising, see L. N. 16, p. 134; L. N. 41, p.
443, and L. N. 52, pp. 97-102.
C. On the active horizontal creeping movements of mature Radiolaria on a firm ground, compare
L. N. 12, p. 10, and L. N. 16, pp. 132-134.
D. Regarding the motion of social central capsules within the same
cœnobium and the changes thus brought about in the structure of the calymma, see L. N. 16, pp. 119-127, and L. N. 52, pp. 75-82.
221. Contraction.—Motions, which are due to the contraction of
individual portions and cause changes in volume or form, have been partly already spoken of under
the head of locomotion (§ 220) and are partly connected with other
functions. Examples may be seen in the contraction of the central capsule and of the calymma. A
certain {cxlii}contraction of the central capsule is
probably brought about by the myophanes, which arise by differentiation of the endoplasm and hence
may assume different forms in the four legions. In the Spumellaria, where
numerous radial fibrillæ run from the central nucleus to the capsule membrane (§ 77), the endoplasm is probably driven out evenly through all the pores of the
capsule membrane by their simultaneous contraction, and hence the volume of the capsule is
diminished in all directions. The Acantharia probably behave similarly,
but are different, inasmuch as the number of their contractile radial fibrillæ is less, and
special axial threads (§ 78) are already differentiated. In the Nassellaria it is probable that owing to the contraction of the divergent
myophane fibrillæ in the podoconus the vertical axis of the latter is shortened, the opercular
rods of the porochora are lifted, and the endoplasm driven out of its pores, so that the volume of
the monaxon central capsule is diminished (§ 79). In the Phæodaria the same result is probably brought about by the contraction of the
cortical myophane fibrillæ, which run meridionally along the inside of the capsule membrane from
the apical to the basal pole of the vertical main axis, where they are inserted into the periphery
of the astropyle; since the volume of the capsule is diminished by their contraction (their
spheroidal figure becoming more nearly spherical) the endoplasm will be driven out through the
proboscis of the astropyle. Whilst these contractions of the central capsule are largely due to
differentiated muscle-like threads of endoplasm (myophanes), this appears to be but rarely the
case with the contractions of the extracapsulum (e.g., the myophriscs of the Acanthometra, § 96). Most of the phenomena of contraction
which can be observed in the calymma and pseudopodia depend upon exoplasmatic currents (§ 209).
222. Protection.—Of the utmost importance, both for the physiology and
for the morphology of the Radiolaria are their manifold protective functions, which we now
consider under the heading "protection." From the physiological point of view the consideration of
the exposed situation in which the delicate, free-swimming Radiolarian organism lives, and the
numerous dangers which beset it in the struggle for existence, would lead a priori to the
expectation, that many special protective adaptations would be developed by natural selection. On
the other hand, morphological experience shows us that this latter has been in action for
immeasurable periods, and has gradually produced an abundance of the most remarkable protective
modifications. Examples of these may be found in the formation of the voluminous calymma, as a
gelatinous protective covering for the central capsule, and further, the formation of the capsule
membrane itself, which separates the generative contents of the central capsule from the nutritive
exoplasm. The phosphorescence of the central capsule, too (§ 223), may be
regarded as a useful protective arrangement; as also the radiating of the numerous pseudopodia in
all directions from the surface of the calymma; for they are of great significance to the {cxliii}well-being of the organism, both as sensory organs
and as prehensile organs. By far the most important and most varied means for the actual defence
of the soft body is to be seen in the endless modifications of the skeleton; first, in the
production of the enclosing lattice-shells and projecting radial spines, but especially also in
the very varied structure of the individual parts of the skeleton, and in the special
differentiation of the small appendicular organs which grow out from it (hairs, thorns, spines,
scales, spathillæ, anchors, &c.). Finally "mimicry" possesses a considerable significance
among the different forms of adaptation which are to be observed in this class.
223. Phosphorescence.—Many Radiolarians shine in the dark, and their
phosphorescence presents the same phenomena as that of other luminous marine organisms; it is
increased by mechanical and chemical irritation, or renewed if already extinguished. The light is
sometimes greenish, sometimes yellowish, and appears generally (if not always) to radiate from the
intracapsular fatty spheres (§ 73). Thus these latter unite several
functions, inasmuch as they serve, firstly, as reserve stores of nutriment, secondly, as
hydrostatic apparatus, and thirdly, as luminous organs for the protection of the Radiolaria;
probably the light acts by frightening other animals, for the phosphorescent animals are provided
with spines, nettle-cells, poison glands or other defensive weapons. The production of the light
depends probably, as in other phosphorescent organisms, upon the slow oxidation of the
fat-globules, which combine with active oxygen in the presence of alkalis. Phosphorescence is very
likely widely distributed among the Radiolaria.
The shining of the Radiolaria in the dark has been noticed by the earliest
observers of the class (see L. N. 1, p. 163, L. N. 16, p.
2, and L. N. 52, pp. 136-139). In the winter of 1859 I observed the
production of light in the case of many monozootic and polyzootic Radiolaria, but inadvertently
omitted to record the fact in my Monograph. I made more accurate observations in the winter of
1866 at Lanzerote in the Canary Islands, and convinced myself the the light emanates from the
central capsule, and in particular from the fat-globules contained in it. In most Polycyttaria
(both Collosphærida and Sphærozoida), when
each central capsule contains a large central oil-globule the light radiates from it. In
Collozoum serpentinum (Pl. 3,
figs. 2, 3) each cylindrical central capsule contains a row of luminous spherules like a string
of beads. In Alacorys friderici (Pl. 65,
fig. 1) the four-lobed central capsule contains four shining points. Karl Brandt has recently
made more detailed communication on this point (L. N. 52, p. 137).
224. Sensation.—The general irritability which we ascribe to all organisms, and as
the basis of which we regard the protoplasm, remains at an inferior stage of development in the
Radiolaria. For although they are subject to various stimuli, and certainly possess a power of
discrimination, special sensory organs are not differentiated; the peripheral portions of the
protoplasm, and especially the pseudopodia, rather act both as organs of the different kinds of
sensation and various modes of motion. That different Radiolaria have attained different degrees
of development in this respect may be seen {cxliv}partly by direct observation of the reaction of the living organism
towards various stimuli, and partly by the comparison of the different conditions of existence
under which Radiolarians exist, both in the most various depths of the ocean and in all climatic
zones (see note A). In general the Radiolaria seem to be sensitive to the following stimuli; (1)
pressure (see note B); (2) temperature (see note C); (3) light (see note D); (4) chemical
composition of the sea-water (see note E). The reaction towards these stimuli, corresponding to
the sensation of pleasure or dislike which they call forth, is shown in various forms of motion of
the protoplasm, changes in the currents in it, contraction of the central capsule, changes in the
size, position, and form of the pseudopodia, changes in the volume of the calymma (by the
evocation of water), &c. Among the sensory functions of the Radiolaria must be especially
mentioned their remarkably developed perception of hydrostatic equilibrium (see note F), as well
as their perception of distances, so clearly shown in the production of equal lattice-meshes and
other regularly formed skeletal structures (see note G).
A. I can add but little to the communication which I made twenty-four years ago regarding
sensation in the Radiolaria (L. N. 16, pp. 128-131). The most important
point would be the great difference in irritability which must obtain between the pelagic,
zonarial and abyssal Radiolaria, which may be assumed from a consideration of their very
different conditions of existence as regards pressure, light, warmth, nutrition, &c. It is
natural to suppose that the numerous abyssal Radiolaria, discovered by the Challenger, which
live at great depths (2000 to 4500 fathoms) in complete darkness, in icy cold and under an
enormous pressure, must have quite different sensations of pleasure from their pelagic relatives
which live at the surface of the sea under an equatorial sun. Karl Brandt has recently added
much to our knowledge regarding the special action of different vital conditions upon the
various Polycyttaria and the degrees of their irritability (L. N. 52, pp.
113-132).
B. Regarding the sensation of pressure or sensation of touch of the Radiolaria and the
various degrees of their mechanical irritability, see L. N. 16, p. 129; L.
N. 41, p. 464.
C. Regarding the sensation of warmth or temperature-sense and its dependence upon different
climatic relations, see L. N. 16, p. 129; L. N. 52, pp.
114-129.
D. Regarding the sensation of light, compare L. N. 16, p. 128; L. N. 42, p. 304; L. N. 52, pp. 102-104, 114.
E. Regarding the sense of taste of the Radiolaria or their peculiar sensitiveness towards the
different chemical composition of the water, change in its salinity, presence of organic
impurities, &c., see L. N. 16, p. 130; L. N. 52, pp.
103, 113. This chemical irritability seems to be the most highly developed sense in the
Radiolaria, even more so than their mechanical irritability.
F. The perception of hydrostatic equilibrium among the Radiolaria is immediately visible from
the position which their bodies, floating freely in the water, assume spontaneously, and from
the symmetrical development of the skeleton, which by its gravitation necessitates a definite
position. It may be assumed that the development of the various geometrical ground forms which
correspond to a definite position of equilibrium, is the result of this particular kind of
perception (compare §§ 40-45).
{cxlv}
G. The plastic perception of distance of the pseudopodia is shown by the symmetry
with which the forms composing the regular skeletal structures (e.g., the ordinary
lattice-spheres with regular hexagonal meshes, the radial spines with equidistant branches) are
excreted from the exoplasm. Both this form of sensation and the one first mentioned (note F)
have hitherto received scarcely any attention, but are deserving of a thorough physiological
investigation.
225. The Cell-Soul (Zellseele).—The common central vital principle, commonly
called the "soul," which is considered to be the regulator of all vital functions, appears in the
Radiolaria as in other Protista in its simplest form, as the cell-soul. By the continual activity
of this central "psyche" all vital functions are maintained in unbroken action, and in uniform
correlation. It is also probable that by it the stimulations which the peripheral portions of the
cell receive from the outer world are first transmitted into true sensation, and that, on the
other hand, the volition, which alone calls forth spontaneous movements, proceeds from it. The
central capsule is most likely the sole organ of this cell-soul or central psychic organ, and the
active portion may be either the endoplasm or the nucleus, or both. The central capsule may thus
(apart from its function as a sporangium, § 215) be regarded as a simple
ganglion cell, physiologically comparable to the nervous centre of the higher animals, whilst the
exoplasm (sarcomatrix and pseudopodia) are to be compared to the peripheral nervous system and
sense organs of the latter. The great simplicity of the functions of the cell-soul which appear in
the Radiolaria, and the intimate connection of their different psychic activities, give to these
unicellular Protista a special significance for the comprehension of the monistic elements of a
natural psychology.
Regarding the theory of the cell-soul as the only psychological theory which is
able to explain naturally the true nature of the life of the soul in all organisms as well as in
man, see my address on cell-souls and soul-cells ("Zellseelen und Seelenzellen") in Gesammelte
populäre Vorträge aus dem Gebiete der Entwickelungslehre, Heft 1, p. 143; Bonn, 1878.
{cxlvi}
CHOROLOGICAL SECTION.
Chapter IX.—GEOGRAPHICAL DISTRIBUTION.
(§§ 226-240.)
226. Universal Marine Distribution.—Radiolaria occur in all the seas of the world,
in all climatic zones and at all depths. Probably under normal conditions they always float freely
in the water, whether their usual position be at the surface (pelagic), or at a certain depth
(zonarial), or near to the bottom of the sea (abyssal). This appears both from numerous direct
observations, as well as from conclusions which may be drawn from their organisation (and
especially their promorphology) regarding their floating life (compare §§ 40-50, 219, 220). Hitherto no observation has been recorded, which justifies the
assumption that Radiolaria live anywhere upon the bottom of the sea (on stones, Algæ, or other
firm substances), either sessile or creeping. They perform the latter action, however, when they
fall accidentally upon a firm basis or are accidentally placed upon it, but they seem normally
always to float freely in the water with pseudopodia radiating in all directions. Active
free-swimming movements are only met with in the case of the flagellate zoospores (§ 142). The development of Radiolaria in large masses is very remarkable (see
note A), and in many parts of the ocean is so great that they play an important part in the
economy of marine life, especially as food for other pelagic and abyssal animals (see note B).
Medium salinity of the water seems to be most favourable to their development in masses, although
it is not unknown in seas of high and low salinity (see note C). There are no Radiolaria in fresh
water (see note D).
A. The development of Radiolaria takes place in many parts of the ocean in astonishingly
large masses on the surface, in different strata, and near the bottom. The Collodaria (and especially the Sphærozoida) often cover the surface of the
sea in millions, and form a shining layer, phosphorescent in the dark like the Noctilucæ,
as I observed in 1859 in the Strait of Messina, in 1866 at the Canaries, and in 1881 in the
Indian Ocean. Similar masses of Sphærozoum and Acanthometron were seen by Johannes
Müller on the French and Ligurian coasts (L. N. 12), and John Murray found
another in the Gulf Stream, off the Færöe Islands, from the surface to a depth of 600 fathoms;
considerable masses of large Phæodaria live there also.
B. The alimentary canal of Medusæ, Salpæ, Crustacea, Pteropoda, and many other pelagic
animals is a rich field for the discovery of Radiolaria, and many of the species hereinafter
described are from such sources. Fossil coprolites too (e.g., those from the Jura) often
contain many Polycystina.
C. Some Acantharia (Acanthometra) and Phæodaria (species of Mesocena and Dictyocha) {cxlvii}live in the Baltic; I found their skeletons in
the alimentary canal of Aurelia, Ascidians and Copepods.
D. The so-called "fresh-water Radiolaria," which have been described by Focke,
Greeff, Grenacher and others, are all Heliozoa, without either central capsule or calymma.
227. Local distribution.—As regards their local distribution and its boundaries
the Radiolaria show in general the same relations as other pelagic animals. Since they are only to
a very slight extent, if at all, capable of active horizontal locomotion, the dispersion of the
different species from their point of development (or "centre of creation") is dependent upon
oceanic currents, the play of winds and waves and all the accidental causes which influence the
transport of pelagic animals in general. These passive migrations are here, however, as always, of
the greatest significance, and bring about the wide distribution of individual species in a far
higher degree than any active wanderings could do. Any one who has ever followed a stream of
pelagic animals for hours and seen how millions of creatures closely packed together are in a
short time carried along for miles by such a current, will be in no danger of underestimating the
enormous importance of marine currents in the passive migration of the fauna of the sea. Such
constant currents may, however, be recognised both near the bottom of the sea and at various
depths, as well as at the surface, and are therefore of just as much significance for the abyssal
and zonarial as for the pelagic Radiolaria. It is easy to explain by this means how it is that so
many animals of this class (probably indeed the great majority) have a wide range of distribution.
The number of cosmopolitan species which live in the Pacific, Atlantic and Indian Oceans is
already relatively large. In each of these three great ocean basins, too, many species show a wide
distribution. On the other hand, there are very many species which are hitherto known only from
one locality, and probably many small local faunas exist, characterised by the special development
of particular groups. The observations which we at present possess are too incomplete, and the
rich material of the Challenger is too incompletely worked out, to enable any definite conclusions
to be drawn regarding the local distribution of Radiolaria.
The statements made in the systematic portion of this Report regarding the
distribution of the Challenger Radiolaria are very incomplete. In most cases only one locality
is mentioned, and that is the station (§ 240) in the preparations or
bottom deposit from which I first found the species in question. Afterwards I often found the
same species again in one or more additional stations (not seldom in numerous preparations both
from the Pacific and Atlantic), without the possibility of adding them to the habitat recorded
under the description. The necessary accurate determination and identification of the species
(measuring the different dimensions, counting the pores, &c.), would have occupied too much
time, and the writing of this extensive Report would have lasted not ten but twenty or thirty
years.
228. Horizontal Distribution.—From the extensive collections of the Challenger and
from the other collections which have furnished a welcome supplement to them, it appears {cxlviii}that Radiolaria are distributed throughout all
seas without distinction of zones and physical conditions, even though these latter may be the
cause of differences in their qualitative and quantitative development. In the case of the
Radiolaria as well as of many other classes of animals, the law holds good that the richest
development of forms and the greatest number of species occurs between the tropics, whilst the
frigid zones (both Arctic and Antarctic) exhibit great masses of individuals, but relatively few
genera and species (see note A). In the Challenger collection the greatest abundance of species of
Radiolaria is exhibited by those preparations which were collected at low latitudes in the
immediate neighbourhood of the equator; this is true both of the Atlantic (Stations 346 to 349)
and of the Pacific (Stations 266 to 274); in the former the richest of all is Station 347 (lat. 0°
15′ S.), in the latter Station 271 (lat. 0° 33′ S.) (see note B). From the tropics the
abundance of species seems to diminish regularly towards the poles, and more rapidly in the
northern than in the southern hemisphere; the latter also appears, considered as a whole, to
possess more species than the former. A limit to the life of the Radiolaria towards the poles has
not yet been found; the expeditions towards the North Pole (see note C), like those towards the
South (see note D), have obtained bottom-deposits and ice enclosures which contained Radiolaria;
in some of the most northerly and most southerly positions which were reached the number of
Radiolaria enclosed in the ice was relatively great.
A. The greater abundance of Radiolaria in the tropical seas is probably to be explained by
the more favourable conditions of existence, and in particular the larger quantity of nutritive
material (especially of decayed animals) and not by the higher temperature of the surface, for
at depths of from 2000 to 3000 fathoms where the abyssal Radiolaria live, the temperature is but
little above the freezing point or even below it (compare the bottom temperatures in the list of
Challenger Stations, § 240).
B. Station 271 of the Challenger Expedition, situated almost on the equator in the Mid
Pacific (lat. 0° 33′ S.), exceeds all other parts of the earth, hitherto known, in respect
of its wealth in Radiolaria, and this is true of the pelagic as well as of the zonarial and
abyssal forms. In the Station List the deposit at this point is stated to be "Globigerina ooze";
but after the calcareous matter has been removed by means of acid, the purest Radiolarian ooze
remains, rich in varied and remarkable species. More than one hundred new species have been
described from this Station alone.
C. Regarding the Arctic Radiolaria compare the contributions of Ehrenberg (L. N. 24, pp. 138, 139, 195) and Brady on the English North Polar Expedition, 1875-76
(Ann. and Mag. Nat. Hist., 1878, vol. i. pp. 425, 437).
D. Regarding the Antarctic Radiolaria, compare § 230, note
A, and Ehrenberg, Mikrogeologie (L. N. 6, Taf. xxxv., A.), also L. N. 24, pp. 136-139.
229. Fauna of the Pacific Ocean.—From the splendid discoveries of the Challenger,
and the supplementary observations obtained from other sources, the Pacific seems to be the ocean
basin which is richest both quantitatively and qualitatively in Radiolarian life, {cxlix}excelling both the Indian and Atlantic Oceans in this
respect. It may be assumed with great probability that by far the largest portion of the Pacific
has a depth of between 2000 and 3000 fathoms, and that its bottom is covered either with
Radiolarian ooze (§ 237) or with a red clay (§ 239),
which contains many Spumellaria and Nassellaria,
and has probably been derived for a great part from broken down and metamorphosed Radiolarian ooze
(see note A). Pure Radiolarian ooze was found by the Challenger eastwards in the Central Pacific
(over a wide area between lat. 12° N. and 12° S., Stations 265 to 274), and also westwards in the
latitude of the Philippines, twenty degrees to the east of them (between lat. 5° N. and 15° N.).
The great abundance of Radiolaria present in the neighbourhood of the Philippines and in the Sunda
Sea was already known from other investigations (note B). The red clay also, which covers a great
part of the bottom of the North Pacific, and which was obtained of very constant composition by
the Challenger between lat. 35° N. and 38° N., from Japan to the meridian of Honolulu (from long.
144° E. to 156° W.), is so pre-eminently rich in Radiolaria that it often approaches in
composition the Radiolarian ooze, and has probably been derived from it. The track of the
Challenger through the tropical and northern parts of the Pacific describes nearly three sides of
a rectangle, which includes about half of the enormous Pacific basin, and from this as well as
from other supplementary observations it may with great probability be concluded that by far the
largest part of the bed of the Pacific (at least three-fourths) is covered either with Radiolarian
ooze or with red clay, which contains a larger or smaller amount of the remains of Radiolaria.
With this agrees also the important fact that the numerous preparations of pelagic materials and
collections of pelagic animals, which were collected by the Challenger in the Pacific, almost
always indicate a corresponding amount of Radiolarian life on the surface. This is true in
particular also of the South Pacific, between lat. 33° S. and 40° S. (from long. 133° W. to 73°
W., Stations 287 to 301); the surface of this southern region and the different bathymetrical
zones were rich in new and peculiar species of Radiolaria.
A. Many specimens of bottom-deposits from the Pacific, which are entered in the Challenger
lists either as "red clay" or "Globigerina ooze," contain larger or smaller quantities of
Radiolaria, and the number of different species of Spumellaria and Nassellaria which they contain is often so great that the deposit might have
been almost as appropriately termed "Radiolarian ooze," e.g., Stations 241 to 245, and
270, 271 (compare §§ 236-239).
B. Pacific Radiolarian ooze was first obtained by Lieutenant Brooke (May 11,
1859) between the Philippines and Marianne Islands, from a depth of 3300 fathoms (lat. 18°
3′ N., long. 129° 11′ E.). Ehrenberg, who first described it, found seventy-nine
different species of Polycystina in it, and reported "that their quantity and the number of
different forms increased with the depth" (Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, 1860,
pp. 466, 588, 766).
230. Fauna of the Indian Ocean.—As regards its Radiolarian fauna the Indian Ocean
is the least known of the three great basins. Still the few limited spots, regarding which {cl}investigations are forthcoming, indicate a very rich
development of Radiolarian life. Probably it approaches more nearly the fauna of the Pacific than
that of the Atlantic, both as regards the abundance and the morphological characters of its
species. The researches of the Challenger are very limited and incomplete as regards the Indian
Ocean, for the expedition only just touched upon this great ocean basin (2000 to 3000 fathoms
deep) at its two extremities (westwards at the Cape of Good Hope and eastwards at Tasmania), its
course lying for the most part south of lat. 45° S. and extending beyond lat. 65° S. (from Station
149 to 158, south of lat. 50° S.). It is true that this portion of the South Indian Ocean was
shown to contain Radiolaria everywhere, but these were more plentiful in individuals than in
species. Only from Station 156 to Station 159 (between lat. 62° and 47° S., and long. 95° and 130°
E.) was the bottom, which consisted partly of Diatom ooze and partly of Globigerina ooze, richer
in species (see note A). The gaps left by the Challenger in the investigation of the Indian Ocean,
have, however, been to some extent filled from other sources. As early as 1859 the English
"Cyclops" expedition had shown that the bottom of the Indian Ocean to the east of Zanzibar (lat.
9° 37′ S., long. 61° 33′ W.) is covered with pure Radiolarian ooze (see note B). Also
since the Tertiary rocks of the Nicobar Islands are for the most part of the same composition, and
since a great abundance of Radiolaria has been shown to be present both in the east part of the
ocean, between the Cocos Islands and the Sunda Archipelago (see note C), and in the northern part
or Arabian Sea between Socotra and Ceylon (see note D); it may be assumed with great probability
that the greater part of the basin of the Indian Ocean, like that of the Pacific, is covered
either with Radiolarian ooze or with the characteristic red clay. With this agrees the richness of
the surface of the Indian Ocean in Radiolaria of the most various groups, which has been more
extensively demonstrated.
A. The Radiolarian fauna collected by the Challenger on the voyage from the Cape to
Melbourne, shows in part, namely, from Station 156 to Station 158, very peculiar and
characteristic composition; in particular, the Diatom ooze of Station 157 passes over in great
part into a Radiolarian ooze, mainly composed of Sphærellaria. This is
worthy of a more thorough investigation than I was able, owing to lack of material and time, to
give it.
B. The remarkably pure Radiolarian ooze of Zanzibar, discovered by Ehrenberg in 1859, was the
earliest known recent example of that deposit. It was brought up by Captain Pullen of the
English man-of-war "Cyclops," from a depth of 2200 fathoms, between Zanzibar and the Seychelles,
and "under a magnifying power of 300 diameters, showed at the first glance a mass of almost pure
Polycystina, such as no sample of a deep-sea deposit has hitherto shown. It is very noticeable
that in the whole of this mass of living forms, no calcareous shells are to be seen" (Ehrenberg,
L. N. 24, pp. 148, 149).
C. For the most important material from the Indian Ocean, I am indebted to Captain Heinrich
Rabbe of Bremen, who during many voyages in the Indian Ocean, in his ship "Joseph Haydn," made
numerous collections in different localities with the tow-net and the trawl, and admirably
preserved the rich collections thus made. The greatest abundance of Radiolaria was found in
those {cli}obtained to the east of Madagascar, and
next in those from the neighbourhood of the Cocos Islands. I take this opportunity of expressing
my thanks to Captain Rabbe for the liberality with which he placed all this valuable material at
my disposal.
D. On my voyage from Aden to Bombay, and thence to Ceylon (1881), and especially
on my return journey from Ceylon, between the Maldive Islands and Socotra (1882), I carried on a
number of experiments with a surface net, which yielded a rich fauna of pelagic animals, and
among them many new species of Radiolaria, for observation. On several nights when the smooth
surface of the Indian Ocean, unrippled by any wind, shone with the most lovely phosphorescent
light, I drew up water from the surface with a bucket, and obtained a rich booty. A number of
other new species of Radiolaria from very various parts of the Indian Ocean I obtained from the
alimentary canal of pelagic animals, such as Medusæ, Salpæ, Crustacea, &c. Although the
total number of Radiolaria known to me from the Indian Ocean is much less than from the Atlantic
and Pacific, there are several new genera and numerous species among them, which show that a
careful study of this fauna will be of wide interest.
231. Fauna of the Atlantic Ocean.—The Atlantic Ocean in all parts, of which the
pelagic fauna has been examined, has shown the same constant presence of Radiolaria, and in
certain parts of its abyssal deposits a larger or smaller quantity of different types belonging to
this class; on the whole, however, its Radiolarian fauna is inferior to that of the Pacific, and
probably also to that of the Indian Ocean, both in quantity and quality. Pure Radiolarian ooze,
such as is so extensively found on the floor of the Pacific, and in certain places in that of the
Indian Ocean, has not yet been found in the Atlantic (see § 237). The red
clay, too, of the deep Atlantic does not seem to be so rich in Radiolaria as that of the Pacific;
nevertheless, the number of species peculiar to the Atlantic is very large, and at certain points
the abundance of species as well as of individuals seems to be scarcely less than in the Pacific.
This is especially true of the eastern equatorial zone not far from Sierra Leone, Stations 347 to
352 (see note A); also of the South Atlantic between Buenos Ayres and Tristan da Cunha, Stations
324, 325, 331 to 333 (see note B); and, lastly, in the North Atlantic in the Gulf Stream and near
the Canary Islands (see note C). The fauna of the latter agrees for the most part with that of the
Mediterranean (see note D). In addition to the material collected by the Challenger, other
deep-sea investigations have furnished bottom-deposits from different parts of the ocean, which
have proved very rich in Radiolaria (see note E). Furthermore, since the island of Barbados
consists for the most part of fossil Radiolarian ooze, it is very probable that at certain parts
of the tropical Atlantic true Radiolarian ooze, like that of the Pacific and Indian Oceans, will
eventually be found in depths between 2000 and 3000 fathoms, perhaps over a considerable area.
A. The tropical zone of the eastern Atlantic seems to be especially rich in peculiar
Radiolaria of different species. This is shown by numerous preparations from the surface, and
from various depths (between lat. 3° S. and 11° N., and long. 14° W. to 18° W.), which were made
towards the {clii}end of the cruise. Unfortunately no
bottom-deposits were obtained from the most important stations (except Nos. 346 and 347, depths
2350 and 2250 fathoms) in this region; at these the deposit was a Globigerina ooze containing
numerous different species of Radiolaria.
B. In the South Atlantic, between Buenos Ayres and Tristan da Cunha (between lat. 35° S. and
43° S., long. 8° W. and 57° W.) there appears to be a long stretch covered partly with
Globigerina ooze (Stations 331 to 334), or red clay (Stations 329, 330), partly with blue mud
(Stations 318 to 328), which contains not only large masses of individuals but numerous peculiar
species of Spumellaria and Nassellaria. The
preparations from the surface-takings of this region are also rich in these, as well as in
peculiar Phæodaria.
C. The northern part of the Atlantic appears on the whole to be inferior to the tropical and
southern portions as regards its richness in Radiolaria, and from the western half more
especially, only few species are known. From my researches at Lanzerote in 1866-67, it appears
that the pelagic fauna of the Canary Islands is very rich in them, as is also the Gulf Stream in
the neighbourhood of the Færöe Channel, according to the investigations of John Murray (see his
Report on the "Knight-Errant" Expedition, Proc. Roy. Soc. Edin., vol. xi., 1882).
D. The Radiolaria of the Mediterranean are of special interest, because almost all our
knowledge of these organisms in the living conditions and of their vital functions has been
derived from investigations conducted on its shores. Johannes Müller laid the foundation of this
knowledge by his investigations at Messina, and on the Ligurian and French coasts at Nice,
Cette, and St. Tropez (L. N. 10). The many new Radiolaria which I described
in my Monograph (L. N. 16, 1862), were for the most part taken at Messina,
the place which possesses a richer pelagic fauna than any other, so far as is yet known, in the
Mediterranean. Other new species I found afterwards at Villafranca near Nice, in 1864 (L. N. 19), at Portofino near Genoa (1880), at Corfu (1877), and at other points on
the coast. In Messina also, Richard Hertwig collected the material for his valuable treatise on
the Organisation of the Radiolaria (L. N. 33), after he had previously made
investigations into their histology at Ajaccio in Corsica (L. N. 26).
Lastly, at Naples, Cienkowski (L. N. 22) and Karl Brandt (L. N. 38, 39, 52) carried out their important
investigations into the reproduction and symbiosis of the Radiolaria. With respect to the
character of its Radiolaria, the Mediterranean fauna is to be regarded as a special province of
the North Atlantic.
E. Among the smaller contributions which have been made towards our knowledge of
the Atlantic Radiolarian fauna, the communications of Ehrenberg on the deposits obtained in
sounding for the Atlantic cable, and on the Mexican Gulf Stream near Florida, deserve special
mention (L. N. 24, pp. 138, 139-145).
232. Vertical Distribution.—The most important general result of the discoveries
of the Challenger, as regards the vertical or bathymetrical distribution of the Radiolaria, is the
interesting fact that numerous species of this class are found living at the most various depths
of the sea, and that certain species are limited to particular bathymetrical zones, i.e.,
are adapted to the conditions which obtain there. In this respect three different Radiolarian
faunas may be distinguished, which may be shortly termed "pelagic," "zonarial," and "abyssal." The
pelagic Radiolaria swim at the surface, and when they sink (e.g., in a stormy sea),
only descend to a small depth, probably not more than from {cliii}20 to 30 fathoms (§ 233). The complicated
conditions of existence created by the keen struggle for existence at the surface of the sea, give
rise to the formation of very numerous pelagic species, especially of Porulosa (Spumellaria and Acantharia). The abyssal
Radiolaria are very different from those just mentioned; they live at the bottom of the deep-sea,
not resting upon nor attached to it, but probably floating at a little distance above it, and are
adapted to the conditions of existence which obtain there (§ 235). Here the
Osculosa (Nassellaria and Phæodaria) seem to
predominate. The zonarial Radiolaria live floating at various depths between the pelagic
and abyssal species (§ 234). In their morphological characters they
gradually approach the pelagic forms upwards and the abyssal downwards.
The views which have hitherto been held regarding the bathymetrical or vertical
distribution of the Radiolaria have been entirely altered by the magnificent discoveries of the
Challenger, and especially by the important observations of Sir Wyville Thomson (L. N. 31) and John Murray (L. N. 27). These two distinguished
deep-sea explorers have, as a result of their wide experience, been convinced that Radiolaria
exist at all depths of the ocean, and that there are large numbers of true deep-sea species
which are never found at the surface of the sea nor at slight depths (L. N. 31, vol. i. pp. 236-238; L. N. 27, pp. 523, 525). The
result of my ten years' work upon the Challenger Radiolaria, and the comparative study of more
than a thousand mountings from all depths, has only been to confirm this opinion, and I am
further persuaded that it will some day be possible by the aid of suitable nets (not yet
invented) to distinguish different faunistic zones in the various depths of the sea. In this
connection may be mentioned the specially interesting fact that the species of Radiolaria of one
and the same family present in the different depths characteristic morphological distinctions,
which obviously correspond to their different physiological relations in the struggle for
existence. Owing to those extensive discoveries, the representation which I gave in my Monograph
(1862, L. N. 16, pp. 172-196) of the vertical distribution of the
Radiolaria, and of their life in the greatest depths of the sea, has been entirely changed.
Compare also Bütschli (L. N. 41, p. 466).
233. The Pelagic Fauna.—The surface of the open ocean seems
everywhere, at a certain distance from the coast at least, to be peopled by crowds of living
Radiolaria. In the tropical zone these pelagic crowds consist of many different species, whilst in
the frigid zones, on the other hand, they are made up of many individuals belonging to but few
species. Most of these inhabitants of the surface may be regarded as truly pelagic species, which
either remain always at the surface or descend only very slightly below it. Probably most Porulosa
(both Spumellaria and Acantharia) belong to this
group; whilst but few Osculosa occur in it, and fewer Phæodaria than Nassellaria. In general the pelagic Radiolaria are distinguished from the
abyssal by the more delicate and slender structure of their skeletons; the pores of the
lattice-shells are larger, the intervening trabeculæ thinner; the armature of spines, spathillæ,
anchors, &c., is more various and more highly developed. Numerous forms are to be found among
the pelagic {cliv}Radiolaria which have either an
incomplete skeleton or none at all. When the pelagic forms leave the surface on account of
unfavourable weather, they appear only to sink to slight depths (probably not below 20 or 30
fathoms). Within the limits of the same family the size of the pelagic species seems to be on an
average greater than that of the related abyssal forms.
234. The Zonarial Fauna.—Between the pelagic fauna living at the surface of the
open sea and the abyssal, which floats immediately over the bottom, there appears to be usually a
middle fauna, which inhabits the different bathymetrical zones of the intermediate water, and
hence may be shortly called the "zonarial" fauna. The different species of Radiolaria which
inhabit these different strata in the same vertical column of water present differences
corresponding to those of the plants composing the several zones of vegetation, which succeed each
other at different heights on a mountain; they correspond to the different conditions of existence
which are presented by the different strata of water, and to which they have become adapted in the
struggle for existence. The existence of such bathymetrical zones has been shown by those
important, if not numerous, observations of the Challenger, in which the tow-net was used at
different depths at one and the same Station. In several cases the character of the Radiolarian
fauna at different depths presented characteristic differences.
For the present, and until we are better acquainted with the characters of the
Radiolarian fauna at different depths, we may distinguish provisionally the following five
bathymetrical zones:—(1) The pelagic zone, extending from the surface to a
depth of about 25 fathoms; (2) the pellucid zone, extending from 25 to 150 fathoms, or as
far as the influence of the sunlight makes itself felt; (3) the obscure zone, extending
from 150 to 2000 fathoms, or from the depth at which sunlight disappears to that at which the
influence of the water containing carbonic acid begins and the calcareous organisms vanish; (4)
the siliceous zone, extending from 2000 or 2500 to about 3000 fathoms, in which only
siliceous not calcareous Rhizopoda are found, and in which the peculiar conditions of the lowest
regions have not yet appeared; (5) the abyssal zone, in which the accumulation of the
oceanic deposits, and the influence of the bottom currents, create new conditions of existence.
So far as our isolated and incomplete observations of the zonarial Radiolarian fauna extend, it
appears that the subclass Porulosa (Spumellaria and Acantharia) predominates in the two upper zones, and as the depth increases is
gradually replaced by the subclass Osculosa (Nassellaria and Phæodaria), so that the latter predominates in the two lowest zones. The
obscure zone which lies in the middle is probably the poorest in species. In general, the
morphological characters of the zonarial fauna appear to change gradually upwards into the
delicate form of the pelagic and downwards into the robust constitution of the abyssal; so also
the average size of the individuals (within the limits of the same family) appears to increase
upwards and decrease downwards.
235. The Abyssal Fauna.—The great majority of Radiolaria which have
hitherto been observed, and which are described in the systematic portion of this Report, have
been obtained from the bottom of the deep-sea, and more than half of all the species have been
{clv}derived from the pure Radiolarian ooze, which forms
the bed of the Central Pacific at depths of from 2000 to 4000 fathoms (§ 237). Many of these abyssal forms were brought up with the malacoma uninjured,
and they show, both when mounted immediately in balsam, and when preserved in alcohol, all the
soft parts almost as clearly as fresh preparations of pelagic Radiolaria. These species are to be
regarded as truly abyssal, i.e., as forms which live floating only a little distance above
the bottom of the deep-sea, having become adapted to the peculiar conditions of life which obtain
in the lowest regions of the ocean. Probably the majority of the Phæodaria
belong to these abyssal Radiolaria, as well as a large number of Nassellaria, but on the other hand, only a small number of Acantharia and Spumellaria are found there. A character
common to these abyssal forms, and rarely found in those from the surface or from slight depths,
is found in their small size and their heavy massive skeletons, in which they strikingly resemble
the fossil Radiolaria of Barbados and the Nicobar Islands. The lattice-work of the shell is
coarser, its trabeculæ thicker and its pores smaller than in pelagic species of the same group;
also the apophyses (spines, spathillæ, coronets, &c.), are much less developed than in the
latter. From these true abyssal Radiolaria must be carefully distinguished those species whose
empty skeletons, devoid of all soft parts, occur also in the Radiolarian ooze of the deep-sea, but
are clearly only the sunken remains of dead forms, which have lived at the surface or in some of
the upper zones.
236. Deposits containing Radiolaria.—The richest collection of Radiolaria is found
in the deposits of ooze which form the bed of the ocean. Although the pelagic material skimmed
from the surface of the sea, and the zonarial material taken by sinking the tow-net to various
depths, are always more or less rich in Radiolaria, still the number of species thus obtained is,
on the whole, much less than has hitherto been got merely from deep-sea deposits. Of course the
skeletons found in the mud of the ocean-bed, may belong either to the abyssal species which live
there (§ 235), or to the zonarial (§ 234), or to the
pelagic species (§ 233), for the siliceous skeletons of these latter sink
to the bottom after their death. Almost all these remains found in the deposits belong to the
siliceous "Polycystina" (Spumellaria and Nassellaria); Phæodaria occur but sparingly, and Acantharia are entirely wanting, for their acanthin skeleton readily dissolves.
The abundance of Radiolaria varies greatly according to the composition and origin of the
deposits. In general marine deposits may be divided into two main divisions, terrigenous and
abyssal, or, more shortly, muds and oozes. The terrigenous deposits (or muds) include all
those sediments which are made up for the most part of materials worn away from the coasts of
continents and islands, or brought down into the sea by rivers. Their greatest extent from the
coast is about 200 nautical miles. They contain varying quantities of Radiolaria, but much fewer
than those of the next group. The abyssal deposits (or oozes) usually commence at a
distance of from 100 to 200 nautical miles {clvi}from
the coast. In general they are characterised by great uniformity, corresponding to the constancy
of the conditions under which they are laid down; they may be divided into three categories, the
true Radiolarian ooze (§ 237), Globigerina ooze (§ 238), and red clay (§ 239). Of these three most
important deep-sea formations the first is by far the richest in Radiolaria, although the other
two contain often very many siliceous shells.
The marvellous discoveries of the Challenger have thrown upon the nature of
marine deposits an entirely new light, which justifies most important conclusions regarding the
geographical distribution and geological significance of the Radiolaria. Since Dr. John Murray
and the Abbé Renard will treat fully of these interesting relations in a forthcoming volume of
the Challenger series (Report on the Deep-Sea Deposits), it will be sufficient here to refer to
their preliminary publication already published (Narrative of the Cruise of H.M.S. Challenger,
1885, vol. ii. part ii. pp. 915-926); see also the earlier communications by John Murray (1876,
L. N. 27, pp. 518-537), and by Sir Wyville Thomson (The Atlantic, L. N. 31, vol. i. pp. 206-246). In the Narrative (loc. cit., p. 916) the
following table of marine deposits is given:—
| Terrigenous deposits. |
 |
Shore formations, |
 |
Found in inland seas and along the shores of continents. |
| Blue mud, |
| Green mud and sand, |
| Red mud, |
| |
| Volcanic mud and sand, |
|
Found around oceanic islands and along the shores of
continents. |
| Coral mud and sand, |
| Coralline mud and sand, |
| |
| Abysmal deposits. |
|
Globigerina ooze, |
 |
Found in the abysmal regions of the ocean basins. |
| Pteropod ooze, |
| Diatom ooze, |
| Radiolarian ooze, |
| Red clay, |
237. Radiolarian Ooze.—By Radiolarian ooze, in the strict sense of the term, are
understood those oceanic deposits, the greater part of which (often more than three-quarters) is
composed of the siliceous skeletons of this class. Such pure Radiolarian ooze has only been
found in limited areas of the Pacific and Indian Oceans. It is most conspicuous in the Central
Pacific, between lat. 12° N. and 8° S., long. 148° W. to 152° W., the depth being everywhere
between 2000 and 3000 fathoms (Stations 266 to 268 and 272 to 274). In the deepest of the
Challenger soundings (Station 225, 4475 fathoms) the bottom is composed of pure Radiolarian ooze,
as well as at the next Station in the Western Tropical Pacific (Station 226, 2300 fathoms), the
latitude varying from 12° N. to 15° N., and the longitude from 142° E. to 144° E. In the Indian
Ocean also, pure Radiolarian ooze was found in the year 1859 between Zanzibar and the Seychelles,
this being the first known example of it (§ 230). On the other hand, it has
not yet been found in the bed of the Atlantic; but the Tertiary formations of Barbados (Antilles,
§ 231) like those of the Nicobar Islands (Further India), are to be
regarded as pure Radiolarian {clvii}ooze in the fossil
condition. Mixed Radiolarian ooze is the name given to those deposits in which the
Radiolaria exceed any of the other organic constituents, although they do not make up half the
total mass. To this category belong a large number of the Challenger soundings which are entered
in the Station list either as red clay or Globigerina ooze. Such mixed Radiolarian ooze has been
discovered (A) in the North Pacific in an elongated area of red clay extending from Station 241 to
Station 245 (perhaps even from Station 238 to Station 253), that is, at least, from long. 157° E.
to 175° E., between lat. 35° N. and 37° N.; (B) in the tropical Central Pacific in the Globigerina
ooze of Stations 270 and 271. The ooze from the latter station, situated almost on the equator
(lat. 0° 33′ S., long. 151° 34′ W.), is specially remarkable, for it has yielded more
new species of Spumellaria and Nassellaria than
any other Station, not excluding even the neighbouring Stations 268, 269, and 272. Probably such
mixed Radiolarian ooze is very widely distributed in the depths of the ocean, as, for example, in
the South Pacific (Stations 288, 289, 300, and 302), and in the Southern Ocean (Stations 156 to
159); also in the South Atlantic (Stations 324, 325, 331, 332) and in the tropical Atlantic
(Stations 348 to 352). When carefully purified and decalcified by acids, Radiolarian ooze appears
as a fine shining white powder; in the raw state it is yellowish or reddish, sometimes
reddish-brown or dark brown in colour, according to the quantity of oxides of iron, manganese,
&c., which it contains. Calcareous skeletons (especially the tests of pelagic Foraminifera) do
not occur at all or only in very minute quantities in pure Radiolarian ooze from more than
2000 fathoms, whilst specimens of mixed ooze often contain considerable quantities of
them.
Pure Radiolarian ooze was first described by Dr. John Murray as regards its
peculiar nature and composition under the name "Radiolarian ooze" (1876, L. N. 27, pp. 525, 526); compare also Sir Wyville Thomson (The Atlantic, L. N. 31, vol. i. pp. 231-238), and John Murray (Narr. Chall. Exp., L. N. 53, vol. i. pt. ii. pp. 920-926, pl. N. fig. 2).
The different specimens of pure Radiolarian ooze obtained by the Challenger from the Pacific,
and handed to me for investigation, are from depths of from 2250 fathoms to 4475 fathoms, and
may be divided according to their composition into three different groups:—I. The
Radiolarian ooze of the Western Tropical Pacific, Stations 225 and 226, from depths of 4475 and
2300 fathoms (lat. 11° N. to 15° N., and long. 142° E. to 144° E.). II. The Radiolarian ooze of
the northern half of the Central Pacific, Stations 265 to 269, from depths of 2550 to 2900
fathoms. III. The Radiolarian ooze of the southern half of the Central Pacific, Stations 270 to
274, from depths of 2350 to 2925 fathoms. A fourth group would be constituted by the Radiolarian
ooze from the Philippines, which was brought up by Brooke in 1860 near the Marianne Islands from
3300 fathoms, and described by Ehrenberg (Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, 1860,
p. 765). The Diatom ooze, too, found by the Challenger in the Antarctic regions (Stations 152 to
157) is in some parts so rich in Radiolaria that it passes over into true Radiolarian ooze.
Regarding the Radiolarian ooze from Zanzibar, obtained by Captain Pullen in 1859 from 2200
fathoms (§ 230), we have only the incomplete communications of Ehrenberg
(L. N. 24, p. 147). A more accurate knowledge of these deposits from the
Indian Ocean, and of {clviii}those which we may
with probability expect from the tropical eastern Atlantic, will be sure to increase very widely
our knowledge of the class.
238. Globigerina Ooze.—Next to the Radiolarian ooze proper the Globigerina ooze is
the deposit which is richest in the remains of Radiolaria. Often these are so abundant that it is
doubtful to which category the specimen should be referred (e.g., Stations 270 and 271, see
§ 237). In fact, the two pass without any sharp boundary into each other,
and both present transitions to the Diatom ooze. Next to red clay (§ 239),
Globigerina ooze is the most widely distributed of all sediments, and forms a large part of the
bed of the ocean at depths of 250 to 2900 fathoms (especially between 1000 and 2000 fathoms). It
covers extensive areas at depths below 1800 fathoms, and in still deeper water is replaced by red
clay. It is a fine-grained white, grey, or yellowish powder, which sometimes becomes coloured
rose, red, or brown owing to the admixture of oxides of iron and manganese. True Globigerina ooze
consists for the most part of the accumulated calcareous shells of pelagic Foraminifera,
principally Globigerina and Orbulina, but also Hastigerina,
Pulvinulina, &c. It contains usually from 50 to 80 per cent. of calcium carbonate, the
extreme values being 40 and 95 per cent. After this has been removed by acids, there remains a
residue, which consists partly of the siliceous shells of Radiolaria and Diatoms, and partly of
mineral particles identical with the volcanic elements of the red clay.
Regarding the composition and significance of the Globigerina ooze, see John
Murray (L. N. 27, pp. 523-525, and L. N. 53, vol. i. p.
919). Recently this author has separated from the Globigerina ooze (sensu stricto), the
Pteropod ooze, distinguished from the former by the greater abundance of Pteropod shells
and calcareous shells of larger pelagic organisms which it contains. It is found in moderate
depths (at most 1500 fathoms), and contains fewer Radiolaria.
239. Red Clay.—This is quantitatively the most important of all deep-sea deposits,
covering by far the greatest extent of the three great ocean basins at depths greater than 2200
fathoms. It thus far surpasses in area the other deposits, both Radiolaria and Globigerina oozes,
and commonly forms a still deeper layer beneath them. Probably these three deep-sea deposits
together cover about three-eighths of the whole surface of the earth, that is, about as much as
all the continents together, whilst only two-eighths are covered by the terrigenous deposits. Red
clay is principally composed of silicate of alumina, mixed in various proportions with other
finely granular substances; its usual red colour, which sometimes passes over into grey or brown,
is more especially due to admixture of oxides of iron and manganese. Calcareous matter is usually
entirely wanting, or present only in traces, whilst free silica is found in very variable, often
considerable quantities. The chief mass of the red clay consists of volcanic ashes, pumice,
fragments of lava, &c., whilst a large part of it is generally composed of shells of
Radiolaria or fragments of {clix}them; in many places
the number of well-preserved skeletons contained in the red clay is very considerable, so that it
passes over gradually into the Radiolarian ooze (e.g., in the North Pacific, Stations 238
to 253, see § 237). Hence it may be supposed that a large part of the red
clay consists of decomposed Radiolarian ooze.
The characteristic composition and fundamental significance of the red clay in
the formation of the deep-sea bed were first made known by the discoveries of the Challenger
(compare John Murray, 1876, L. N. 27, p. 527, and Narr. Chall. Exp., L. N.
53, vol. i. pt. ii. pp. 920-926, pl. N; also
Wyville Thomson, The Atlantic, L. N. 31, vol. i. pp. 226-229). The mineral
components of the red clay are for the most part of volcanic origin, due to the decomposition of
pumice, lava, &c. Among the organic remains found in it, the siliceous skeletons of
Radiolaria are by far the most important, and their number is often considerable. A large
portion of the red clay appears to me to consist of broken down Radiolarian shells, in which a
peculiar metamorphism probably has taken place. Sir Wyville Thomson was of opinion that a
considerable proportion of it consisted of the remains of Globigerina ooze, the calcareous
constituents of which had been removed by the carbon dioxide in the deep-sea water (L. N. 31, loc. cit.). Among these remains, however, the siliceous skeletons of
the Radiolaria play a significant and often the most important part. Furthermore, John Murray
has called attention to the fact that in many deep-sea deposits yellow and red insoluble
particles remain, which unmistakably present the form of Radiolarian shells (L. N. 27, p. 513). At Station 303 he found "amorphous clayey matter, rounded yellow
minerals, many Radiolaria-shaped;" at Station 302 there was sediment "consisting almost entirely
of small rounded red mineral particles; many of these had the form of both Foraminifera and
Radiolaria; and it seemed as if some substance had been deposited in and on these organisms."
Similar transitions from well-preserved Radiolarian shells into amorphous mineral particles I
have found in several other specimens of Challenger soundings, and consider them a further
argument for the supposition that the Radiolaria often take an important share in the formation
of the red clay.
240. List of Stations at which Radiolaria were observed on the Challenger
Expedition.—The 168 Stations recorded below, in soundings or surface preparations from
which I found Radiolaria, belong to the most various parts of the sea which the Challenger
traversed during her voyage round the world; they constitute about half of the (364) observing
Stations contained in the official list published in the Narrative of the Cruise (Narr. Chall.
Exp., vol. i. part ii. Appendix ii.).
In addition to the particulars given in the list regarding the geographical
position of the Station, depth, temperature, and composition of the bottom deposit, I have added
the result of my investigations as regards the relative abundance of the Radiolaria in each. The
five letters (A to E) denote the following degrees of frequency:—A, abundant Radiolaria
(AI, pure Radiolarian ooze; AII, mixed
Radiolarian ooze); B, very numerous Radiolaria (but not a predominating quantity); C, many
Radiolaria (medium quantity); D, few Radiolaria; E, very few Radiolaria (as they occur almost
always). In using these symbols regard has been had to abundance of the abyssal as well as of
the zonarial and pelagic forms (§ 232); sometimes also the estimated
number of Radiolaria has been inserted, based upon information given by John Murray in his
Preliminary Report (L. N. 27), and in the Narrative of the Cruise (L. N. 53), as well as by Henry B. Brady in his Report on the {clx}Foraminifera (Zool. Chall. Exp., part xxii., 1884). From Stations 348
to 352 in the Eastern Tropical Atlantic no specimens of the bottom were obtained, but a rich
pelagic Radiolarian fauna was demonstrated by numerous preparations from the surface. The depths
are given in fathoms and the temperature in degrees Fahrenheit. In the column describing the
nature of the bottom the following abbreviations are used:—
rad. oz. = Radiolarian ooze (§ 237).
gl. oz. = Globigerina ooze (§ 238).
r. cl. = red clay (§ 239).
pt. oz. = Pteropod ooze (see p. clviii).
di. oz. = Diatom ooze (see p. clvii). |
bl. m. = blue mud,
gr. m. = green mud,
volc. m. = volcanic mud, |
|
terrigenous deposits
(see p. clvi). |
r. m. = red mud.
|
| rad. oz. = Radiolarian ooze (§ 237). |
| gl. oz. = Globigerina ooze (§ 238). |
| r. cl. = red clay (§ 239). |
| pt. oz. = Pteropod ooze (see p. clviii). |
| di. oz. = Diatom ooze (see p. clvii). |
| bl. m. = blue mud, |
|
terrigenous deposits
(see p. clvi). |
| gr. m. = green mud, |
| volc. m. = volcanic mud, |
| r. m. = red mud. |
 |
Locality 1. |
 |
 |
 |
Relative
Abundance of
Radiolaria. |
Date. |
Latitude and Longitude. |
Nearest Land. |
|
|
|
|
|
|
1873. |
|
|
|
|
|
| 001. |
N. Atl. |
1890 |
36.8 |
gl. oz. |
D few |
Feb. |
15 |
27° |
24′ N., |
16° |
55′ W. |
S. of Tenerife. |
| 002. |
N. " |
1945 |
36.8 |
gl. oz. |
E very few |
F" |
17 |
25° |
52′ N., |
19° |
22′ W. |
S.W. of the Canary Islands. |
| 005. |
N. " |
2740 |
37.0 |
r. cl. |
D few |
F" |
21 |
24° |
20′ N., |
24° |
28′ W. |
S.W. of the Canary Islands. |
| 009. |
N. " |
3150 |
36.8 |
r. cl. |
E very few |
F" |
26 |
23° |
23′ N., |
35° |
11′ W. |
(Ocean). |
| 024. |
Tr. Atl. |
390 |
... |
pt. oz. |
D few |
Mar. |
25 |
18° |
38′ N., |
65° |
5′ W. |
Culebra (Antilles). |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 032. |
N. Atl. |
2250 |
36.7 |
gl. oz. |
E very few |
April |
3 |
31° |
49′ N., |
64° |
55′ W. |
Bermuda. |
| 045. |
N. " |
1240 |
37.2 |
bl. m. |
E very" |
May |
3 |
38° |
34′ N., |
72° |
10′ W. |
S. of New York. |
| 050. |
N. " |
1250 |
38.0 |
bl. m. |
E very" |
F" |
21 |
42° |
8′ N., |
63° |
39′ W. |
S. of Halifax. |
| 064. |
N. " |
2700 |
... |
r. cl. |
D few |
June |
20 |
35° |
35′ N., |
50° |
27′ W. |
(Ocean). |
| 076. |
N. " |
900 |
40.0 |
pt. oz. |
D f" |
July |
3 |
38° |
11′ N., |
27° |
9′ W. |
Azores. |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 098. |
Tr. Atl. |
1750 |
36.7 |
gl. oz. |
C many |
Aug. |
14 |
9° |
21′ N., |
18° |
28′ W. |
W. of Sierra Leone. |
| 106. |
Tr. " |
1850 |
36.6 |
gl. oz. |
C m" |
F" |
25 |
1° |
47′ N., |
24° |
26′ W. |
(Ocean). |
| 108. |
Tr. " |
1900 |
36.8 |
gl. oz. |
C m" |
F" |
27 |
1° |
10′ N., |
28° |
23′ W. |
(Ocean). |
| 111. |
Tr. " |
2475 |
33.7 |
gl. oz. |
C m" |
F" |
31 |
1° |
45′ S., |
30° |
58′ W. |
(Ocean). |
| 120. |
Tr. " |
675 |
... |
r. m. |
D few |
Sept. |
9 |
8° |
37′ S., |
34° |
28′ W. |
Pernambuco. |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 132. |
S. Atl. |
2050 |
35.0 |
gl. oz. |
C many |
Oct. |
10 |
35° |
25′ S., |
23° |
40′ W. |
Tristan da Cunha. |
| 134. |
S. " |
2025 |
36.0 |
gl. oz. |
C m" |
F" |
14 |
36° |
12′ S., |
12° |
16′ W. |
Tristan da Cunha. |
| 137. |
S. " |
2550 |
34.5 |
r. cl. |
D few |
F" |
23 |
35° |
59′ S., |
1° |
34′ E. |
(Ocean). |
| 138. |
S. " |
2650 |
35.1 |
r. cl. |
D f" |
F" |
25 |
36° |
22′ S., |
8° |
12′ E. |
(Ocean). |
| 143. |
S. Ind. |
1900 |
35.6 |
gl. oz. |
E very few |
Dec. |
19 |
36° |
48′ S., |
19° |
24′ E. |
Cape of Good Hope. |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 144. |
S. " |
1570 |
35.8 |
gl. oz. |
E very" |
F" |
24 |
45° |
57′ S., |
34° |
39′ E. |
(Ocean). |
| 145. |
S. " |
140 |
... |
volc. s. |
D few |
F" |
27 |
46° |
43′ S., |
38° |
4′ E. |
Prince Edward Island. |
| 146. |
S. " |
1375 |
35.6 |
gl. oz. |
C many |
F" |
29 |
46° |
46′ S., |
45° |
31′ E. |
(Ocean). |
| 147. |
S. " |
1600 |
34.2 |
di. oz. |
C m" |
F" |
30 |
46° |
16′ S., |
48° |
27′ E. |
W. of the Crozet Islands. |
|
|
|
|
|
|
1874. |
|
|
|
|
|
| 148. |
S. " |
210 |
... |
gravel,
shells |
D few |
Jan. |
3 |
46° |
47′ S., |
51° |
37′ E. |
E. of the Crozet Islands. |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 149H. |
S. " |
127 |
... |
volc. m. |
D f" |
F" |
29 |
48° |
45′ S., |
69° |
14′ E. |
Kerguelen Island. |
| 150. |
S. " |
150 |
35.2 |
gravel |
D f" |
Feb. |
2 |
52° |
4′ S., |
71° |
22′ E. |
N. of Heard Island. |
| 151. |
S. " |
75 |
... |
volc. m. |
D f" |
F" |
7 |
52° |
59′ S., |
73° |
33′ E. |
Heard Island. |
| 152. |
S. " |
1260 |
... |
di. oz. |
C many |
F" |
11 |
60° |
52′ S., |
80° |
20′ E. |
(Ocean). |
| 153. |
S. " |
1675 |
... |
bl. m. |
C m" |
F" |
14 |
65° |
42′ S., |
79° |
49′ E. |
Antarctic Ice. |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 154. |
S. " |
1800 |
... |
bl. m. |
C m" |
F" |
19 |
64° |
37′ S., |
85° |
49′ E. |
Antarctic Ice. |
| 155. |
S. " |
1300 |
... |
bl. m. |
C m" |
F" |
23 |
64° |
18′ S., |
94° |
47′ E. |
Antarctic Ice. |
| 156. |
S. " |
1975 |
... |
di. oz. |
B numerous |
F" |
26 |
62° |
26′ S., |
95° |
44′ E. |
(Ocean). |
| 157. |
S. " |
1950 |
32.1 |
di. oz. |
B num" |
Mar. |
3 |
53° |
55′ S., |
108° |
35′ E. |
(Ocean). |
| 158. |
S. " |
1800 |
33.5 |
gl. oz. |
B num" |
F" |
7 |
50° |
1′ S., |
123° |
4′ E. |
(Ocean). |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 159. |
S. " |
2150 |
34.5 |
gl. oz. |
B num" |
F" |
10 |
47° |
25′ S., |
130° |
22′ E. |
(Ocean). |
| 160. |
S. " |
2600 |
33.9 |
r. cl. |
C many |
F" |
13 |
42° |
42′ S., |
134° |
10′ E. |
(Ocean). |
| 162. |
S. " |
38 |
... |
sand |
E very few |
April |
2 |
39° |
10′ S., |
146° |
37′ E. |
Bass Strait. |
| 163. |
S. Pac. |
2200 |
34.5 |
gr. m. |
E very" |
F" |
4 |
36° |
57′ S., |
150° |
34′ E. |
Port Jackson. |
| 164A. |
S. " |
1200 |
... |
gr. m. |
E very" |
June |
13 |
34° |
9′ S., |
151° |
55′ E. |
W. of Sydney. |
|
Locality 2. |
|
|
|
Relative
Abundance of
Radiolaria. |
|
|
|
|
|
|
| 001. |
N. Atl. |
1890 |
36.8 |
gl. oz. |
D few |
| 002. |
N. " |
1945 |
36.8 |
gl. oz. |
E very few |
| 005. |
N. " |
2740 |
37.0 |
r. cl. |
D few |
| 009. |
N. " |
3150 |
36.8 |
r. cl. |
E very few |
| 024. |
Tr. Atl. |
390 |
... |
pt. oz. |
D few |
| |
|
|
|
|
|
| 032. |
N. Atl. |
2250 |
36.7 |
gl. oz. |
E very few |
| 045. |
N. " |
1240 |
37.2 |
bl. m. |
E very" |
| 050. |
N. " |
1250 |
38.0 |
bl. m. |
E very" |
| 064. |
N. " |
2700 |
... |
r. cl. |
D few |
| 076. |
N. " |
900 |
40.0 |
pt. oz. |
D f" |
| |
|
|
|
|
|
| 098. |
Tr. Atl. |
1750 |
36.7 |
gl. oz. |
C many |
| 106. |
Tr. " |
1850 |
36.6 |
gl. oz. |
C m" |
| 108. |
Tr. " |
1900 |
36.8 |
gl. oz. |
C m" |
| 111. |
Tr. " |
2475 |
33.7 |
gl. oz. |
C m" |
| 120. |
Tr. " |
675 |
... |
r. m. |
D few |
| |
|
|
|
|
|
| 132. |
S. Atl. |
2050 |
35.0 |
gl. oz. |
C many |
| 134. |
S. " |
2025 |
36.0 |
gl. oz. |
C m" |
| 137. |
S. " |
2550 |
34.5 |
r. cl. |
D few |
| 138. |
S. " |
2650 |
35.1 |
r. cl. |
D f" |
| 143. |
S. Ind. |
1900 |
35.6 |
gl. oz. |
E very few |
| |
|
|
|
|
|
| 144. |
S. " |
1570 |
35.8 |
gl. oz. |
E very" |
| 145. |
S. " |
140 |
... |
volc. s. |
D few |
| 146. |
S. " |
1375 |
35.6 |
gl. oz. |
C many |
| 147. |
S. " |
1600 |
34.2 |
di. oz. |
C m" |
|
|
|
|
|
|
| 148. |
S. " |
210 |
... |
gravel,
shells |
D few |
| |
|
|
|
|
|
| 149H. |
S. " |
127 |
... |
volc. m. |
D f" |
| 150. |
S. " |
150 |
35.2 |
gravel |
D f" |
| 151. |
S. " |
75 |
... |
volc. m. |
D f" |
| 152. |
S. " |
1260 |
... |
di. oz. |
C many |
| 153. |
S. " |
1675 |
... |
bl. m. |
C m" |
| |
|
|
|
|
|
| 154. |
S. " |
1800 |
... |
bl. m. |
C m" |
| 155. |
S. " |
1300 |
... |
bl. m. |
C m" |
| 156. |
S. " |
1975 |
... |
di. oz. |
B numerous |
| 157. |
S. " |
1950 |
32.1 |
di. oz. |
B num" |
| 158. |
S. " |
1800 |
33.5 |
gl. oz. |
B num" |
| |
|
|
|
|
|
| 159. |
S. " |
2150 |
34.5 |
gl. oz. |
B num" |
| 160. |
S. " |
2600 |
33.9 |
r. cl. |
C many |
| 162. |
S. " |
38 |
... |
sand |
E very few |
| 163. |
S. Pac. |
2200 |
34.5 |
gr. m. |
E very" |
| 164A. |
S. " |
1200 |
... |
gr. m. |
E very" |
|
Date. |
Latitude and Longitude. |
Nearest Land. |
|
1873. |
|
|
|
|
|
| 001. |
Feb. |
15 |
27° |
24′ N., |
16° |
55′ W. |
S. of Tenerife. |
| 002. |
F" |
17 |
25° |
52′ N., |
19° |
22′ W. |
S.W. of the Canary Islands. |
| 005. |
F" |
21 |
24° |
20′ N., |
24° |
28′ W. |
S.W. of the Canary Islands. |
| 009. |
F" |
26 |
23° |
23′ N., |
35° |
11′ W. |
(Ocean). |
| 024. |
Mar. |
25 |
18° |
38′ N., |
65° |
5′ W. |
Culebra (Antilles). |
| |
|
|
|
|
|
|
|
| 032. |
April |
3 |
31° |
49′ N., |
64° |
55′ W. |
Bermuda. |
| 045. |
May |
3 |
38° |
34′ N., |
72° |
10′ W. |
S. of New York. |
| 050. |
F" |
21 |
42° |
8′ N., |
63° |
39′ W. |
S. of Halifax. |
| 064. |
June |
20 |
35° |
35′ N., |
50° |
27′ W. |
(Ocean). |
| 076. |
July |
3 |
38° |
11′ N., |
27° |
9′ W. |
Azores. |
| |
|
|
|
|
|
|
|
| 098. |
Aug. |
14 |
9° |
21′ N., |
18° |
28′ W. |
W. of Sierra Leone. |
| 106. |
F" |
25 |
1° |
47′ N., |
24° |
26′ W. |
(Ocean). |
| 108. |
F" |
27 |
1° |
10′ N., |
28° |
23′ W. |
(Ocean). |
| 111. |
F" |
31 |
1° |
45′ S., |
30° |
58′ W. |
(Ocean). |
| 120. |
Sept. |
9 |
8° |
37′ S., |
34° |
28′ W. |
Pernambuco. |
| |
|
|
|
|
|
|
|
| 132. |
Oct. |
10 |
35° |
25′ S., |
23° |
40′ W. |
Tristan da Cunha. |
| 134. |
F" |
14 |
36° |
12′ S., |
12° |
16′ W. |
Tristan da Cunha. |
| 137. |
F" |
23 |
35° |
59′ S., |
1° |
34′ E. |
(Ocean). |
| 138. |
F" |
25 |
36° |
22′ S., |
8° |
12′ E. |
(Ocean). |
| 143. |
Dec. |
19 |
36° |
48′ S., |
19° |
24′ E. |
Cape of Good Hope. |
| |
|
|
|
|
|
|
|
| 144. |
F" |
24 |
45° |
57′ S., |
34° |
39′ E. |
(Ocean). |
| 145. |
F" |
27 |
46° |
43′ S., |
38° |
4′ E. |
Prince Edward Island. |
| 146. |
F" |
29 |
46° |
46′ S., |
45° |
31′ E. |
(Ocean). |
| 147. |
F" |
30 |
46° |
16′ S., |
48° |
27′ E. |
W. of the Crozet Islands. |
|
1874. |
|
|
|
|
|
| 148. |
Jan. |
3 |
46° |
47′ S., |
51° |
37′ E. |
E. of the Crozet Islands. |
| |
|
|
|
|
|
|
|
| 149H. |
F" |
29 |
48° |
45′ S., |
69° |
14′ E. |
Kerguelen Island. |
| 150. |
Feb. |
2 |
52° |
4′ S., |
71° |
22′ E. |
N. of Heard Island. |
| 151. |
F" |
7 |
52° |
59′ S., |
73° |
33′ E. |
Heard Island. |
| 152. |
F" |
11 |
60° |
52′ S., |
80° |
20′ E. |
(Ocean). |
| 153. |
F" |
14 |
65° |
42′ S., |
79° |
49′ E. |
Antarctic Ice. |
| |
|
|
|
|
|
|
|
| 154. |
F" |
19 |
64° |
37′ S., |
85° |
49′ E. |
Antarctic Ice. |
| 155. |
F" |
23 |
64° |
18′ S., |
94° |
47′ E. |
Antarctic Ice. |
| 156. |
F" |
26 |
62° |
26′ S., |
95° |
44′ E. |
(Ocean). |
| 157. |
Mar. |
3 |
53° |
55′ S., |
108° |
35′ E. |
(Ocean). |
| 158. |
F" |
7 |
50° |
1′ S., |
123° |
4′ E. |
(Ocean). |
| |
|
|
|
|
|
|
|
| 159. |
F" |
10 |
47° |
25′ S., |
130° |
22′ E. |
(Ocean). |
| 160. |
F" |
13 |
42° |
42′ S., |
134° |
10′ E. |
(Ocean). |
| 162. |
April |
2 |
39° |
10′ S., |
146° |
37′ E. |
Bass Strait. |
| 163. |
F" |
4 |
36° |
57′ S., |
150° |
34′ E. |
Port Jackson. |
| 164A. |
June |
13 |
34° |
9′ S., |
151° |
55′ E. |
W. of Sydney. |
{clxi}
|
Locality 1. |
|
|
|
Relative
Abundance of
Radiolaria. |
Date. |
Latitude and Longitude. |
Nearest Land. |
|
|
|
|
|
|
1874. |
|
|
|
|
|
| 165. |
S. Pac. |
2600 |
34.5 |
r. cl. |
D few |
June |
17 |
34° |
50′ S., |
155° |
28′ E. |
(Ocean). |
| 166. |
S. " |
275 |
50.8 |
gl. oz. |
D f" |
F" |
23 |
38° |
50′ S., |
169° |
20′ E. |
W. of New Zealand. |
| 169. |
S. " |
700 |
40.0 |
bl. m. |
D f" |
July |
10 |
37° |
34′ S., |
179° |
22′ E. |
E. of New Zealand. |
| 175. |
Tr. Pac. |
1350 |
36.0 |
gl. oz. |
E very few |
Aug. |
12 |
19° |
2′ S., |
177° |
10′ E. |
Fiji Islands. |
| 181. |
Tr. " |
2440 |
35.8 |
r. cl. |
E very" |
F" |
25 |
13° |
50′ S., |
151° |
49′ E. |
Louisiades. |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 193. |
Tr. " |
2800 |
38.0 |
bl. m. |
D few |
Sept. |
28 |
5° |
24′ S., |
130° |
37′ E. |
Banda Sea. |
| 195. |
Tr. " |
1425 |
38.0 |
bl. m. |
C many |
Oct. |
3 |
4° |
21′ S., |
129° |
7′ E. |
Banda Sea. |
| 197. |
Tr. " |
1200 |
35.9 |
bl. m. |
D few |
F" |
14 |
0° |
41′ N., |
126° |
37′ E. |
E. of Celebes. |
| 198. |
Tr. " |
2150 |
38.9 |
bl. m. |
C many |
F" |
20 |
2° |
55′ N., |
124° |
58′ E. |
N. of Celebes. |
| 200. |
Tr. " |
250 |
... |
gr. m. |
B numerous |
F" |
23 |
6° |
47′ N., |
122° |
28′ E. |
W. of Mindanao. |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 201. |
Tr. " |
82 |
... |
st. & gra. |
C many |
F" |
26 |
7° |
3′ N., |
121° |
48′ E. |
W. of Mindanao. |
| 202. |
Tr. " |
2550 |
50.5 |
bl. m. |
B numerous |
F" |
27 |
8° |
32′ N., |
121° |
55′ E. |
W. of Mindanao. |
| 205. |
Tr. " |
1050 |
37.0 |
bl. m. |
C many |
Nov. |
13 |
16° |
42′ N., |
119° |
22′ E. |
W. of Luzon. |
|
|
|
|
|
|
1875. |
|
|
|
|
|
| 206. |
Tr. " |
2100 |
36.5 |
bl. m. |
B numerous |
Jan. |
8 |
17° |
54′ N., |
117° |
14′ E. |
W. of Luzon. |
| 211. |
Tr. " |
2225 |
50.5 |
bl. m. |
B num" |
F" |
28 |
8° |
0′ N., |
121° |
42′ E. |
W. of Mindanao. |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 213. |
Tr. " |
2050 |
38.8 |
bl. m. |
C many |
Feb. |
8 |
5° |
47′ N., |
124° |
1′ E. |
S. of Mindanao. |
| 214. |
Tr. " |
500 |
41.8 |
bl. m. |
C m" |
F" |
10 |
4° |
33′ N., |
127° |
6′ E. |
N. of Gilolo. |
| 215. |
Tr. " |
2550 |
35.4 |
r. cl. |
C many |
F" |
12 |
4° |
19′ N., |
130° |
15′ E. |
N. of Gilolo. |
| 216A. |
Tr. " |
2000 |
35.4 |
gl. oz. |
B numerous |
F" |
16 |
2° |
56′ N., |
134° |
11′ E. |
S. of Pelew Islands. |
| 217. |
Tr. " |
2000 |
35.2 |
bl. m. |
C many |
F" |
22 |
0° |
39′ S., |
138° |
55′ E. |
N. of New Guinea. |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 218. |
Tr. " |
1070 |
36.4 |
bl. m. |
C m" |
Mar. |
1 |
2° |
33′ S., |
144° |
4′ E. |
N. of New Guinea. |
| 220. |
Tr. " |
1100 |
36.2 |
gl. oz. |
C m" |
F" |
11 |
0° |
42′ S., |
147° |
0′ E. |
N. of New Guinea. |
| 221. |
Tr. " |
2650 |
35.4 |
r. cl. |
B numerous |
F" |
13 |
0° |
40′ N., |
148° |
41′ E. |
(Ocean). |
| 222. |
Tr. " |
2450 |
35.2 |
r. cl. |
B num" |
F" |
16 |
2° |
15′ N., |
146° |
16′ E. |
(Ocean). |
| 223. |
Tr. " |
2325 |
35.5 |
gl. oz. |
B num" |
F" |
19 |
5° |
31′ N., |
145° |
13′ E. |
Carolines. |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 224. |
Tr. " |
1850 |
35.4 |
gl. oz. |
B num" |
F" |
21 |
7° |
45′ N., |
144° |
20′ E. |
Carolines. |
| 225. |
Tr. " |
4475 |
35.2 |
rad. oz. |
A very many |
F" |
23 |
11° |
24′ N., |
143° |
16′ E. |
Ocean |
|
North-West Pacific,
between Carolines
and Japan. |
| 226. |
Tr. " |
2300 |
35.5 |
rad. oz. |
A very" |
F" |
25 |
14° |
44′ N., |
142° |
13′ E. |
Ocean |
| 230. |
N. Pac. |
2425 |
35.5 |
r. cl. |
C many |
April |
5 |
26° |
29′ N., |
137° |
57′ E. |
Ocean |
| 231. |
N. " |
2250 |
35.2 |
bl. m. |
C m" |
F" |
9 |
31° |
8′ N., |
137° |
8′ E. |
Ocean |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 232. |
N. " |
345 |
41.1 |
gr. m. |
C m" |
May |
12 |
35° |
11′ N., |
139° |
28′ E. |
Ocean |
| 234. |
N. " |
2675 |
35.8 |
bl. m. |
B numerous |
June |
3 |
32° |
31′ N., |
135° |
39′ E. |
S. of Japan. |
| 235. |
N. " |
565 |
38.1 |
gr. m. |
D few |
F" |
4 |
34° |
7′ N., |
138° |
0′ E. |
S. of Japan. |
| 236. |
N. " |
775 |
37.6 |
gr. m. |
C many |
F" |
5 |
34° |
58′ N., |
139° |
29′ E. |
S. of Japan. |
| 237. |
N. " |
1875 |
35.3 |
bl. m. |
C m" |
F" |
17 |
34° |
37′ N., |
140° |
32′ E. |
S. of Japan. |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 238. |
N. " |
3950 |
35.0 |
r. cl. |
B numerous |
F" |
18 |
35° |
18′ N., |
144° |
8′ E. |
Ocean |
 |
North Pacific, between
Japan and San Francisco
(35°-38° N. lat.,
144°-156° W. long.). |
| 239. |
N. " |
3625 |
35.1 |
r. cl. |
B num" |
F" |
19 |
35° |
18′ N., |
147° |
9′ E. |
Ocean |
| 240. |
N. " |
2900 |
34.9 |
r. cl. |
B num" |
F" |
21 |
35° |
20′ N., |
153° |
39′ E. |
Ocean |
| 241. |
N. " |
2300 |
35.1 |
r. cl. |
A very many |
F" |
23 |
35° |
41′ N., |
157° |
42′ E. |
Ocean |
| 242. |
N. " |
2575 |
35.1 |
r. cl. |
AII ver" |
F" |
24 |
35° |
29′ N., |
161° |
52′ E. |
Ocean |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 243. |
N. " |
2800 |
35.0 |
r. cl. |
AII ver" |
F" |
26 |
35° |
24′ N., |
166° |
35′ E. |
Ocean |
| 244. |
N. " |
2900 |
35.3 |
r. cl. |
AII ver" |
F" |
28 |
35° |
22′ N., |
169° |
53′ E. |
Ocean |
| 245. |
N. " |
2775 |
34.9 |
r. cl. |
AII ver" |
F" |
30 |
36° |
23′ N., |
174° |
31′ E. |
Ocean |
| 246. |
N. " |
2050 |
35.1 |
gl. oz. |
B numerous |
July |
2 |
36° |
10′ N., |
178° |
0′ E. |
Ocean |
| 247. |
N. " |
2530 |
35.2 |
r. cl. |
C many |
F" |
3 |
35° |
49′ N., |
179° |
57′ W. |
Ocean |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 248. |
N. " |
2900 |
35.1 |
r. cl. |
C m" |
F" |
5 |
37° |
41′ N., |
177° |
4′ W. |
Ocean |
| 249. |
N. " |
3000 |
35.2 |
r. cl. |
B numerous |
F" |
7 |
37° |
59′ N., |
171° |
48′ W. |
Ocean |
| 250. |
N. " |
3050 |
35.0 |
r. cl. |
B num" |
F" |
9 |
37° |
49′ N., |
166° |
47′ W. |
Ocean |
| 251. |
N. " |
2950 |
35.1 |
r. cl. |
B num" |
F" |
10 |
37° |
37′ N., |
163° |
26′ W. |
Ocean |
| 252. |
N. " |
2740 |
35.3 |
r. cl. |
B num" |
F" |
12 |
37° |
52′ N., |
160° |
17′ W. |
Ocean |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 253. |
N. " |
3125 |
35.1 |
r. cl. |
B num" |
F" |
14 |
38° |
9′ N., |
156° |
25′ W. |
Ocean |
| 254. |
N. " |
3025 |
35.0 |
r. cl. |
C many |
F" |
17 |
35° |
13′ N., |
154° |
43′ W. |
Ocean |
|
North Pacific (35°-23°
N. lat., 154°-156°
W. long.). |
| 255. |
N. " |
2850 |
35.0 |
r. cl. |
C m" |
F" |
19 |
32° |
28′ N., |
154° |
33′ W. |
Ocean |
| 256. |
N. " |
2950 |
35.2 |
r. cl. |
B numerous |
F" |
21 |
30° |
22′ N., |
154° |
56′ W. |
Ocean |
| 257. |
N. " |
2875 |
34.9 |
r. cl. |
C many |
F" |
23 |
27° |
33′ N., |
154° |
55′ W. |
Ocean |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 258. |
N. " |
2775 |
35.2 |
r. cl. |
C m" |
F" |
24 |
26° |
11′ N., |
155° |
12′ W. |
Ocean |
| 259. |
Tr. Pac. |
2225 |
34.9 |
r. cl. |
C m" |
F" |
26 |
23° |
3′ N., |
156° |
6′ W. |
Ocean |
|
Locality 2. |
|
|
|
Relative
Abundance of
Radiolaria. |
|
|
|
|
|
|
| 165. |
S. Pac. |
2600 |
34.5 |
r. cl. |
D few |
| 166. |
S. " |
275 |
50.8 |
gl. oz. |
D f" |
| 169. |
S. " |
700 |
40.0 |
bl. m. |
D f" |
| 175. |
Tr. Pac. |
1350 |
36.0 |
gl. oz. |
E very few |
| 181. |
Tr. " |
2440 |
35.8 |
r. cl. |
E very" |
| |
|
|
|
|
|
| 193. |
Tr. " |
2800 |
38.0 |
bl. m. |
D few |
| 195. |
Tr. " |
1425 |
38.0 |
bl. m. |
C many |
| 197. |
Tr. " |
1200 |
35.9 |
bl. m. |
D few |
| 198. |
Tr. " |
2150 |
38.9 |
bl. m. |
C many |
| 200. |
Tr. " |
250 |
... |
gr. m. |
B numerous |
| |
|
|
|
|
|
| 201. |
Tr. " |
82 |
... |
st. & gra. |
C many |
| 202. |
Tr. " |
2550 |
50.5 |
bl. m. |
B numerous |
| 205. |
Tr. " |
1050 |
37.0 |
bl. m. |
C many |
|
|
|
|
|
|
| 206. |
Tr. " |
2100 |
36.5 |
bl. m. |
B numerous |
| 211. |
Tr. " |
2225 |
50.5 |
bl. m. |
B num" |
| |
|
|
|
|
|
| 213. |
Tr. " |
2050 |
38.8 |
bl. m. |
C many |
| 214. |
Tr. " |
500 |
41.8 |
bl. m. |
C m" |
| 215. |
Tr. " |
2550 |
35.4 |
r. cl. |
C many |
| 216A. |
Tr. " |
2000 |
35.4 |
gl. oz. |
B numerous |
| 217. |
Tr. " |
2000 |
35.2 |
bl. m. |
C many |
| |
|
|
|
|
|
| 218. |
Tr. " |
1070 |
36.4 |
bl. m. |
C m" |
| 220. |
Tr. " |
1100 |
36.2 |
gl. oz. |
C m" |
| 221. |
Tr. " |
2650 |
35.4 |
r. cl. |
B numerous |
| 222. |
Tr. " |
2450 |
35.2 |
r. cl. |
B num" |
| 223. |
Tr. " |
2325 |
35.5 |
gl. oz. |
B num" |
| |
|
|
|
|
|
| 224. |
Tr. " |
1850 |
35.4 |
gl. oz. |
B num" |
| 225. |
Tr. " |
4475 |
35.2 |
rad. oz. |
A very many |
| 226. |
Tr. " |
2300 |
35.5 |
rad. oz. |
A very" |
| 230. |
N. Pac. |
2425 |
35.5 |
r. cl. |
C many |
| 231. |
N. " |
2250 |
35.2 |
bl. m. |
C m" |
| |
|
|
|
|
|
| 232. |
N. " |
345 |
41.1 |
gr. m. |
C m" |
| 234. |
N. " |
2675 |
35.8 |
bl. m. |
B numerous |
| 235. |
N. " |
565 |
38.1 |
gr. m. |
D few |
| 236. |
N. " |
775 |
37.6 |
gr. m. |
C many |
| 237. |
N. " |
1875 |
35.3 |
bl. m. |
C m" |
| |
|
|
|
|
|
| 238. |
N. " |
3950 |
35.0 |
r. cl. |
B numerous |
| 239. |
N. " |
3625 |
35.1 |
r. cl. |
B num" |
| 240. |
N. " |
2900 |
34.9 |
r. cl. |
B num" |
| 241. |
N. " |
2300 |
35.1 |
r. cl. |
A very many |
| 242. |
N. " |
2575 |
35.1 |
r. cl. |
AII ver" |
| |
|
|
|
|
|
| 243. |
N. " |
2800 |
35.0 |
r. cl. |
AII ver" |
| 244. |
N. " |
2900 |
35.3 |
r. cl. |
AII ver" |
| 245. |
N. " |
2775 |
34.9 |
r. cl. |
AII ver" |
| 246. |
N. " |
2050 |
35.1 |
gl. oz. |
B numerous |
| 247. |
N. " |
2530 |
35.2 |
r. cl. |
C many |
| |
|
|
|
|
|
| 248. |
N. " |
2900 |
35.1 |
r. cl. |
C m" |
| 249. |
N. " |
3000 |
35.2 |
r. cl. |
B numerous |
| 250. |
N. " |
3050 |
35.0 |
r. cl. |
B num" |
| 251. |
N. " |
2950 |
35.1 |
r. cl. |
B num" |
| 252. |
N. " |
2740 |
35.3 |
r. cl. |
B num" |
| |
|
|
|
|
|
| 253. |
N. " |
3125 |
35.1 |
r. cl. |
B num" |
| 254. |
N. " |
3025 |
35.0 |
r. cl. |
C many |
| 255. |
N. " |
2850 |
35.0 |
r. cl. |
C m" |
| 256. |
N. " |
2950 |
35.2 |
r. cl. |
B numerous |
| 257. |
N. " |
2875 |
34.9 |
r. cl. |
C many |
| |
|
|
|
|
|
| 258. |
N. " |
2775 |
35.2 |
r. cl. |
C m" |
| 259. |
Tr. Pac. |
2225 |
34.9 |
r. cl. |
C m" |
|
Date. |
Latitude and Longitude. |
Nearest Land. |
|
1874. |
|
|
|
|
|
| 165. |
June |
17 |
34° |
50′ S., |
155° |
28′ E. |
(Ocean). |
| 166. |
F" |
23 |
38° |
50′ S., |
169° |
20′ E. |
W. of New Zealand. |
| 169. |
July |
10 |
37° |
34′ S., |
179° |
22′ E. |
E. of New Zealand. |
| 175. |
Aug. |
12 |
19° |
2′ S., |
177° |
10′ E. |
Fiji Islands. |
| 181. |
F" |
25 |
13° |
50′ S., |
151° |
49′ E. |
Louisiades. |
| |
|
|
|
|
|
|
|
| 193. |
Sept. |
28 |
5° |
24′ S., |
130° |
37′ E. |
Banda Sea. |
| 195. |
Oct. |
3 |
4° |
21′ S., |
129° |
7′ E. |
Banda Sea. |
| 197. |
F" |
14 |
0° |
41′ N., |
126° |
37′ E. |
E. of Celebes. |
| 198. |
F" |
20 |
2° |
55′ N., |
124° |
58′ E. |
N. of Celebes. |
| 200. |
F" |
23 |
6° |
47′ N., |
122° |
28′ E. |
W. of Mindanao. |
| |
|
|
|
|
|
|
|
| 201. |
F" |
26 |
7° |
3′ N., |
121° |
48′ E. |
W. of Mindanao. |
| 202. |
F" |
27 |
8° |
32′ N., |
121° |
55′ E. |
W. of Mindanao. |
| 205. |
Nov. |
13 |
16° |
42′ N., |
119° |
22′ E. |
W. of Luzon. |
|
1875. |
|
|
|
|
|
| 206. |
Jan. |
8 |
17° |
54′ N., |
117° |
14′ E. |
W. of Luzon. |
| 211. |
F" |
28 |
8° |
0′ N., |
121° |
42′ E. |
W. of Mindanao. |
| |
|
|
|
|
|
|
|
| 213. |
Feb. |
8 |
5° |
47′ N., |
124° |
1′ E. |
S. of Mindanao. |
| 214. |
F" |
10 |
4° |
33′ N., |
127° |
6′ E. |
N. of Gilolo. |
| 215. |
F" |
12 |
4° |
19′ N., |
130° |
15′ E. |
N. of Gilolo. |
| 216A. |
F" |
16 |
2° |
56′ N., |
134° |
11′ E. |
S. of Pelew Islands. |
| 217. |
F" |
22 |
0° |
39′ S., |
138° |
55′ E. |
N. of New Guinea. |
| |
|
|
|
|
|
|
|
| 218. |
Mar. |
1 |
2° |
33′ S., |
144° |
4′ E. |
N. of New Guinea. |
| 220. |
F" |
11 |
0° |
42′ S., |
147° |
0′ E. |
N. of New Guinea. |
| 221. |
F" |
13 |
0° |
40′ N., |
148° |
41′ E. |
(Ocean). |
| 222. |
F" |
16 |
2° |
15′ N., |
146° |
16′ E. |
(Ocean). |
| 223. |
F" |
19 |
5° |
31′ N., |
145° |
13′ E. |
Carolines. |
| |
|
|
|
|
|
|
|
| 224. |
F" |
21 |
7° |
45′ N., |
144° |
20′ E. |
Carolines. |
| 225. |
F" |
23 |
11° |
24′ N., |
143° |
16′ E. |
Ocean |
|
North-West Pacific,
between Carolines
and Japan. |
| 226. |
F" |
25 |
14° |
44′ N., |
142° |
13′ E. |
Ocean |
| 230. |
April |
5 |
26° |
29′ N., |
137° |
57′ E. |
Ocean |
| 231. |
F" |
9 |
31° |
8′ N., |
137° |
8′ E. |
Ocean |
| |
|
|
|
|
|
|
|
| 232. |
May |
12 |
35° |
11′ N., |
139° |
28′ E. |
Ocean |
| 234. |
June |
3 |
32° |
31′ N., |
135° |
39′ E. |
S. of Japan. |
| 235. |
F" |
4 |
34° |
7′ N., |
138° |
0′ E. |
S. of Japan. |
| 236. |
F" |
5 |
34° |
58′ N., |
139° |
29′ E. |
S. of Japan. |
| 237. |
F" |
17 |
34° |
37′ N., |
140° |
32′ E. |
S. of Japan. |
| |
|
|
|
|
|
|
|
| 238. |
F" |
18 |
35° |
18′ N., |
144° |
8′ E. |
Ocean |
|
North Pacific, between
Japan and San Francisco
(35°-38° N. lat.,
144°-156° W. long.). |
| 239. |
F" |
19 |
35° |
18′ N., |
147° |
9′ E. |
Ocean |
| 240. |
F" |
21 |
35° |
20′ N., |
153° |
39′ E. |
Ocean |
| 241. |
F" |
23 |
35° |
41′ N., |
157° |
42′ E. |
Ocean |
| 242. |
F" |
24 |
35° |
29′ N., |
161° |
52′ E. |
Ocean |
| |
|
|
|
|
|
|
|
| 243. |
F" |
26 |
35° |
24′ N., |
166° |
35′ E. |
Ocean |
| 244. |
F" |
28 |
35° |
22′ N., |
169° |
53′ E. |
Ocean |
| 245. |
F" |
30 |
36° |
23′ N., |
174° |
31′ E. |
Ocean |
| 246. |
July |
2 |
36° |
10′ N., |
178° |
0′ E. |
Ocean |
| 247. |
F" |
3 |
35° |
49′ N., |
179° |
57′ W. |
Ocean |
| |
|
|
|
|
|
|
|
| 248. |
F" |
5 |
37° |
41′ N., |
177° |
4′ W. |
Ocean |
| 249. |
F" |
7 |
37° |
59′ N., |
171° |
48′ W. |
Ocean |
| 250. |
F" |
9 |
37° |
49′ N., |
166° |
47′ W. |
Ocean |
| 251. |
F" |
10 |
37° |
37′ N., |
163° |
26′ W. |
Ocean |
| 252. |
F" |
12 |
37° |
52′ N., |
160° |
17′ W. |
Ocean |
| |
|
|
|
|
|
|
|
| 253. |
F" |
14 |
38° |
9′ N., |
156° |
25′ W. |
Ocean |
| 254. |
F" |
17 |
35° |
13′ N., |
154° |
43′ W. |
Ocean |
|
North Pacific (35°-23°
N. lat., 154°-156°
W. long.). |
| 255. |
F" |
19 |
32° |
28′ N., |
154° |
33′ W. |
Ocean |
| 256. |
F" |
21 |
30° |
22′ N., |
154° |
56′ W. |
Ocean |
| 257. |
F" |
23 |
27° |
33′ N., |
154° |
55′ W. |
Ocean |
| |
|
|
|
|
|
|
|
| 258. |
F" |
24 |
26° |
11′ N., |
155° |
12′ W. |
Ocean |
| 259. |
F" |
26 |
23° |
3′ N., |
156° |
6′ W. |
Ocean |
{clxii}
|
Locality 1. |
|
|
|
Relative
Abundance of
Radiolaria. |
Date. |
Latitude and Longitude. |
Nearest Land. |
|
|
|
|
|
|
1875. |
|
|
|
|
|
| 261. |
Tr. Pac. |
2050 |
35.2 |
volc. m. |
C m" |
Aug. |
12 |
20° |
18′ N., |
157° |
14′ W. |
Sandwich Islands. |
| 262. |
Tr. " |
2875 |
35.2 |
r. cl. |
C m" |
F" |
20 |
19° |
12′ N., |
154° |
14′ W. |
Sandwich Islands. |
| 263. |
Tr. " |
2650 |
35.1 |
r. cl. |
B numerous |
F" |
21 |
17° |
33′ N., |
153° |
36′ W. |
Ocean |
 |
Tropical Central Pacific,
between Sandwich and
Paumotu (17° N. lat.
to 11° S. lat.). |
| 264. |
Tr. " |
3000 |
35.2 |
r. cl. |
C many |
F" |
23 |
14° |
19′ N., |
152° |
37′ W. |
Ocean |
| 265. |
Tr. " |
2900 |
35.0 |
r. cl. |
A very many |
F" |
25 |
12° |
42′ N., |
152° |
1′ W. |
Ocean |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 266. |
Tr. " |
2750 |
35.1 |
rad. oz. |
A very" |
F" |
26 |
11° |
7′ N., |
152° |
3′ W. |
Ocean |
| 267. |
Tr. " |
2700 |
35.0 |
rad. oz. |
A very" |
F" |
28 |
9° |
28′ N., |
150° |
49′ W. |
Ocean |
| 268. |
Tr. " |
2900 |
34.8 |
rad. oz. |
A very" |
F" |
30 |
7° |
35′ N., |
149° |
49′ W. |
Ocean |
| 269. |
Tr. " |
2550 |
35.2 |
rad. oz. |
A very" |
Sept. |
2 |
5° |
54′ N., |
147° |
2′ W. |
Ocean |
| 270. |
Tr. " |
2925 |
34.6 |
gl. oz. |
A very" |
F" |
4 |
2° |
34′ N., |
149° |
9′ W. |
Ocean |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 271. |
Tr. " |
2425 |
35.0 |
gl. oz. |
A very" |
F" |
6 |
0° |
33′ S., |
151° |
34′ W. |
Ocean |
| 272. |
Tr. " |
2600 |
35.1 |
rad. oz. |
A very" |
F" |
8 |
3° |
48′ S., |
152° |
56′ W. |
Ocean |
| 273. |
Tr. " |
2350 |
34.5 |
rad. oz. |
A very" |
F" |
9 |
5° |
11′ S., |
152° |
56′ W. |
Ocean |
| 274. |
Tr. " |
2750 |
35.1 |
rad. oz. |
A very" |
F" |
11 |
7° |
25′ S., |
152° |
15′ W. |
Ocean |
| 275. |
Tr. " |
2610 |
35.0 |
r. cl. |
B numerous |
F" |
14 |
11° |
20′ S., |
150° |
30′ W. |
Ocean |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 276. |
Tr. " |
2350 |
35.1 |
r. cl. |
C many |
F" |
16 |
13° |
28′ S., |
149° |
30′ W. |
Paumotu. |
| 280. |
Tr. " |
1940 |
35.3 |
gl. oz. |
D few |
Oct. |
4 |
18° |
40′ S., |
149° |
52′ W. |
S. of Tahiti. |
| 281. |
Tr. " |
2385 |
34.9 |
r. cl. |
C many |
F" |
6 |
22° |
21′ S., |
150° |
17′ W. |
Tubuai Islands. |
| 282. |
S. Pac. |
2450 |
35.1 |
r. cl. |
C m" |
F" |
7 |
23° |
46′ S., |
149° |
59′ W. |
Tubuai Islands. |
| 283. |
Tr. " |
2075 |
35.4 |
gl. oz. |
D few |
F" |
9 |
26° |
9′ S., |
145° |
17′ W. |
N. of Oparo Island. |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 284. |
Tr. " |
1985 |
35.1 |
gl. oz. |
C many |
F" |
11 |
28° |
22′ S., |
141° |
22′ W. |
S. of Oparo Island. |
| 285. |
Tr. " |
2375 |
35.0 |
r. cl. |
D few |
F" |
14 |
32° |
36′ S., |
137° |
43′ W. |
Ocean |
|
Open South Pacific
Ocean, between New
Zealand and Valparaiso. |
| 286. |
Tr. " |
2335 |
34.8 |
r. cl. |
D f" |
F" |
16 |
33° |
29′ S., |
133° |
22′ W. |
Ocean |
| 287. |
Tr. " |
2400 |
34.7 |
r. cl. |
D f" |
F" |
19 |
36° |
32′ S., |
132° |
52′ W. |
Ocean |
| 288. |
Tr. " |
2600 |
34.8 |
r. cl. |
B numerous |
F" |
21 |
40° |
3′ S., |
132° |
58′ W. |
Ocean |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 289. |
Tr. " |
2550 |
34.8 |
r. cl. |
B num" |
F" |
23 |
39° |
41′ S., |
131° |
23′ W. |
Ocean |
| 290. |
Tr. " |
2300 |
34.9 |
r. cl. |
C many |
F" |
25 |
39° |
16′ S., |
124° |
7′ W. |
Ocean |
| 291. |
Tr. " |
2250 |
34.6 |
r. cl. |
C m" |
F" |
27 |
39° |
13′ S., |
118° |
49′ W. |
Ocean |
| 292. |
Tr. " |
1600 |
35.2 |
gl. oz. |
C m" |
F" |
29 |
38° |
43′ S., |
112° |
31′ W. |
Ocean |
| 293. |
Tr. " |
2025 |
34.4 |
gl. oz. |
C m" |
Nov. |
1 |
39° |
4′ S., |
105° |
5′ W. |
Ocean |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 294. |
Tr. " |
2270 |
34.6 |
r. cl. |
D few |
F" |
3 |
39° |
22′ S., |
98° |
46′ W. |
Ocean |
| 295. |
Tr. " |
1500 |
35.3 |
gl. oz. |
C many |
F" |
5 |
38° |
7′ S., |
94° |
4′ W. |
Ocean |
| 296. |
Tr. " |
1825 |
35.3 |
gl. oz. |
D few |
F" |
9 |
38° |
6′ S., |
88° |
2′ W. |
Ocean |
| 297. |
Tr. " |
1775 |
35.5 |
gl. oz. |
D f" |
F" |
11 |
37° |
29′ S., |
83° |
7′ W. |
Ocean |
| 298. |
Tr. " |
2225 |
35.6 |
bl. m. |
C many |
F" |
17 |
34° |
7′ S., |
73° |
56′ W. |
W. of Valparaiso. |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 299. |
Tr. " |
2160 |
35.2 |
bl. m. |
C m" |
Dec. |
14 |
33° |
31′ S., |
74° |
43′ W. |
W. of Valparaiso. |
| 300. |
Tr. " |
1375 |
35.5 |
gl. oz. |
B numerous |
F" |
17 |
33° |
42′ S., |
78° |
18′ W. |
N. of Juan Fernandez. |
| 302. |
Tr. " |
1450 |
35.6 |
gl. oz. |
B num" |
F" |
28 |
42° |
43′ S., |
82° |
11′ W. |
(Ocean). |
| 303. |
Tr. " |
1325 |
36.0 |
bl. m. |
D few |
F" |
30 |
45° |
31′ S., |
78° |
9′ W. |
W. of Patagonia. |
| 304. |
Tr. " |
45 |
... |
gr. m. |
E very few |
F" |
31 |
46° |
53′ S., |
75° |
12′ W. |
W. of Patagonia. |
|
|
|
|
|
|
1876. |
|
|
|
|
|
| 318. |
S. Atl. |
2040 |
33.7 |
bl. m. |
C few |
Feb. |
11 |
42° |
32′ S., |
56° |
29′ W. |
(Ocean). |
| 319. |
S. " |
2425 |
32.7 |
bl. m. |
C f" |
F" |
12 |
41° |
54′ S., |
54° |
48′ W. |
(Ocean). |
| 323. |
S. " |
1900 |
33.1 |
bl. m. |
C f" |
F" |
28 |
35° |
39′ S., |
50° |
47′ W. |
W. of Buenos Ayres. |
| 324. |
S. " |
2800 |
32.6 |
bl. m. |
B numerous |
F" |
29 |
36° |
9′ S., |
48° |
22′ W. |
Ocean |
 |
Open South Atlantic
Ocean, between Buenos
Ayres and Tristan
da Cunha (35°-37° S.
lat., 21°-48° W. long.). |
| 325. |
S. " |
2650 |
32.7 |
bl. m. |
B num" |
Mar. |
2 |
36° |
44′ S., |
46° |
16′ W. |
Ocean |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 326. |
S. " |
2775 |
32.7 |
bl. m. |
C many |
F" |
3 |
37° |
3′ S., |
44° |
17′ W. |
Ocean |
| 327. |
S. " |
2900 |
32.8 |
bl. m. |
C m" |
F" |
4 |
36° |
48′ S., |
42° |
45′ W. |
Ocean |
| 328. |
S. " |
2900 |
32.9 |
bl. m. |
B numerous |
F" |
6 |
37° |
38′ S., |
39° |
36′ W. |
Ocean |
| 329. |
S. " |
2675 |
32.3 |
r. cl. |
C many |
F" |
7 |
37° |
31′ S., |
36° |
7′ W. |
Ocean |
| 330. |
S. " |
2440 |
32.7 |
r. cl. |
C m" |
F" |
8 |
37° |
45′ S., |
33° |
0′ W. |
Ocean |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 331. |
S. " |
1715 |
35.4 |
gl. oz. |
B numerous |
F" |
9 |
37° |
47′ S., |
30° |
20′ W. |
Ocean |
| 332. |
S. " |
2200 |
34.0 |
gl. oz. |
B num" |
F" |
10 |
37° |
29′ S., |
27° |
31′ W. |
Ocean |
| 333. |
S. " |
2025 |
35.3 |
gl. oz. |
B num" |
F" |
13 |
35° |
36′ S., |
21° |
12′ W. |
Ocean |
| 334. |
S. " |
1915 |
35.8 |
gl. oz. |
C many |
F" |
14 |
35° |
45′ S., |
18° |
31′ W. |
W. of Tristan da Cunha. |
| 335. |
S. " |
1425 |
37.0 |
pt. oz. |
D few |
F" |
16 |
32° |
24′ S., |
13° |
5′ W. |
N. of Tristan da Cunha. |
|
Locality 2. |
|
|
|
Relative
Abundance of
Radiolaria. |
|
|
|
|
|
|
| 261. |
Tr. Pac. |
2050 |
35.2 |
volc. m. |
C m" |
| 262. |
Tr. " |
2875 |
35.2 |
r. cl. |
C m" |
| 263. |
Tr. " |
2650 |
35.1 |
r. cl. |
B numerous |
| 264. |
Tr. " |
3000 |
35.2 |
r. cl. |
C many |
| 265. |
Tr. " |
2900 |
35.0 |
r. cl. |
A very many |
| |
|
|
|
|
|
| 266. |
Tr. " |
2750 |
35.1 |
rad. oz. |
A very" |
| 267. |
Tr. " |
2700 |
35.0 |
rad. oz. |
A very" |
| 268. |
Tr. " |
2900 |
34.8 |
rad. oz. |
A very" |
| 269. |
Tr. " |
2550 |
35.2 |
rad. oz. |
A very" |
| 270. |
Tr. " |
2925 |
34.6 |
gl. oz. |
A very" |
| |
|
|
|
|
|
| 271. |
Tr. " |
2425 |
35.0 |
gl. oz. |
A very" |
| 272. |
Tr. " |
2600 |
35.1 |
rad. oz. |
A very" |
| 273. |
Tr. " |
2350 |
34.5 |
rad. oz. |
A very" |
| 274. |
Tr. " |
2750 |
35.1 |
rad. oz. |
A very" |
| 275. |
Tr. " |
2610 |
35.0 |
r. cl. |
B numerous |
| |
|
|
|
|
|
| 276. |
Tr. " |
2350 |
35.1 |
r. cl. |
C many |
| 280. |
Tr. " |
1940 |
35.3 |
gl. oz. |
D few |
| 281. |
Tr. " |
2385 |
34.9 |
r. cl. |
C many |
| 282. |
S. Pac. |
2450 |
35.1 |
r. cl. |
C m" |
| 283. |
Tr. " |
2075 |
35.4 |
gl. oz. |
D few |
| |
|
|
|
|
|
| 284. |
Tr. " |
1985 |
35.1 |
gl. oz. |
C many |
| 285. |
Tr. " |
2375 |
35.0 |
r. cl. |
D few |
| 286. |
Tr. " |
2335 |
34.8 |
r. cl. |
D f" |
| 287. |
Tr. " |
2400 |
34.7 |
r. cl. |
D f" |
| 288. |
Tr. " |
2600 |
34.8 |
r. cl. |
B numerous |
| |
|
|
|
|
|
| 289. |
Tr. " |
2550 |
34.8 |
r. cl. |
B num" |
| 290. |
Tr. " |
2300 |
34.9 |
r. cl. |
C many |
| 291. |
Tr. " |
2250 |
34.6 |
r. cl. |
C m" |
| 292. |
Tr. " |
1600 |
35.2 |
gl. oz. |
C m" |
| 293. |
Tr. " |
2025 |
34.4 |
gl. oz. |
C m" |
| |
|
|
|
|
|
| 294. |
Tr. " |
2270 |
34.6 |
r. cl. |
D few |
| 295. |
Tr. " |
1500 |
35.3 |
gl. oz. |
C many |
| 296. |
Tr. " |
1825 |
35.3 |
gl. oz. |
D few |
| 297. |
Tr. " |
1775 |
35.5 |
gl. oz. |
D f" |
| 298. |
Tr. " |
2225 |
35.6 |
bl. m. |
C many |
| |
|
|
|
|
|
| 299. |
Tr. " |
2160 |
35.2 |
bl. m. |
C m" |
| 300. |
Tr. " |
1375 |
35.5 |
gl. oz. |
B numerous |
| 302. |
Tr. " |
1450 |
35.6 |
gl. oz. |
B num" |
| 303. |
Tr. " |
1325 |
36.0 |
bl. m. |
D few |
| 304. |
Tr. " |
45 |
... |
gr. m. |
E very few |
|
|
|
|
|
|
| 318. |
S. Atl. |
2040 |
33.7 |
bl. m. |
C few |
| 319. |
S. " |
2425 |
32.7 |
bl. m. |
C f" |
| 323. |
S. " |
1900 |
33.1 |
bl. m. |
C f" |
| 324. |
S. " |
2800 |
32.6 |
bl. m. |
B numerous |
| 325. |
S. " |
2650 |
32.7 |
bl. m. |
B num" |
| |
|
|
|
|
|
| 326. |
S. " |
2775 |
32.7 |
bl. m. |
C many |
| 327. |
S. " |
2900 |
32.8 |
bl. m. |
C m" |
| 328. |
S. " |
2900 |
32.9 |
bl. m. |
B numerous |
| 329. |
S. " |
2675 |
32.3 |
r. cl. |
C many |
| 330. |
S. " |
2440 |
32.7 |
r. cl. |
C m" |
| |
|
|
|
|
|
| 331. |
S. " |
1715 |
35.4 |
gl. oz. |
B numerous |
| 332. |
S. " |
2200 |
34.0 |
gl. oz. |
B num" |
| 333. |
S. " |
2025 |
35.3 |
gl. oz. |
B num" |
| 334. |
S. " |
1915 |
35.8 |
gl. oz. |
C many |
| 335. |
S. " |
1425 |
37.0 |
pt. oz. |
D few |
|
Date. |
Latitude and Longitude. |
Nearest Land. |
|
1875. |
|
|
|
|
|
| 261. |
Aug. |
12 |
20° |
18′ N., |
157° |
14′ W. |
Sandwich Islands. |
| 262. |
F" |
20 |
19° |
12′ N., |
154° |
14′ W. |
Sandwich Islands. |
| 263. |
F" |
21 |
17° |
33′ N., |
153° |
36′ W. |
Ocean |
|
Tropical Central Pacific,
between Sandwich and
Paumotu (17° N. lat.
to 11° S. lat.). |
| 264. |
F" |
23 |
14° |
19′ N., |
152° |
37′ W. |
Ocean |
| 265. |
F" |
25 |
12° |
42′ N., |
152° |
1′ W. |
Ocean |
| |
|
|
|
|
|
|
|
| 266. |
F" |
26 |
11° |
7′ N., |
152° |
3′ W. |
Ocean |
| 267. |
F" |
28 |
9° |
28′ N., |
150° |
49′ W. |
Ocean |
| 268. |
F" |
30 |
7° |
35′ N., |
149° |
49′ W. |
Ocean |
| 269. |
Sept. |
2 |
5° |
54′ N., |
147° |
2′ W. |
Ocean |
| 270. |
F" |
4 |
2° |
34′ N., |
149° |
9′ W. |
Ocean |
| |
|
|
|
|
|
|
|
| 271. |
F" |
6 |
0° |
33′ S., |
151° |
34′ W. |
Ocean |
| 272. |
F" |
8 |
3° |
48′ S., |
152° |
56′ W. |
Ocean |
| 273. |
F" |
9 |
5° |
11′ S., |
152° |
56′ W. |
Ocean |
| 274. |
F" |
11 |
7° |
25′ S., |
152° |
15′ W. |
Ocean |
| 275. |
F" |
14 |
11° |
20′ S., |
150° |
30′ W. |
Ocean |
| |
|
|
|
|
|
|
|
| 276. |
F" |
16 |
13° |
28′ S., |
149° |
30′ W. |
Paumotu. |
| 280. |
Oct. |
4 |
18° |
40′ S., |
149° |
52′ W. |
S. of Tahiti. |
| 281. |
F" |
6 |
22° |
21′ S., |
150° |
17′ W. |
Tubuai Islands. |
| 282. |
F" |
7 |
23° |
46′ S., |
149° |
59′ W. |
Tubuai Islands. |
| 283. |
F" |
9 |
26° |
9′ S., |
145° |
17′ W. |
N. of Oparo Island. |
| |
|
|
|
|
|
|
|
| 284. |
F" |
11 |
28° |
22′ S., |
141° |
22′ W. |
S. of Oparo Island. |
| 285. |
F" |
14 |
32° |
36′ S., |
137° |
43′ W. |
Ocean |
|
Open South Pacific
Ocean, between New
Zealand and Valparaiso. |
| 286. |
F" |
16 |
33° |
29′ S., |
133° |
22′ W. |
Ocean |
| 287. |
F" |
19 |
36° |
32′ S., |
132° |
52′ W. |
Ocean |
| 288. |
F" |
21 |
40° |
3′ S., |
132° |
58′ W. |
Ocean |
| |
|
|
|
|
|
|
|
| 289. |
F" |
23 |
39° |
41′ S., |
131° |
23′ W. |
Ocean |
| 290. |
F" |
25 |
39° |
16′ S., |
124° |
7′ W. |
Ocean |
| 291. |
F" |
27 |
39° |
13′ S., |
118° |
49′ W. |
Ocean |
| 292. |
F" |
29 |
38° |
43′ S., |
112° |
31′ W. |
Ocean |
| 293. |
Nov. |
1 |
39° |
4′ S., |
105° |
5′ W. |
Ocean |
| |
|
|
|
|
|
|
|
| 294. |
F" |
3 |
39° |
22′ S., |
98° |
46′ W. |
Ocean |
| 295. |
F" |
5 |
38° |
7′ S., |
94° |
4′ W. |
Ocean |
| 296. |
F" |
9 |
38° |
6′ S., |
88° |
2′ W. |
Ocean |
| 297. |
F" |
11 |
37° |
29′ S., |
83° |
7′ W. |
Ocean |
| 298. |
F" |
17 |
34° |
7′ S., |
73° |
56′ W. |
W. of Valparaiso. |
| |
|
|
|
|
|
|
|
| 299. |
Dec. |
14 |
33° |
31′ S., |
74° |
43′ W. |
W. of Valparaiso. |
| 300. |
F" |
17 |
33° |
42′ S., |
78° |
18′ W. |
N. of Juan Fernandez. |
| 302. |
F" |
28 |
42° |
43′ S., |
82° |
11′ W. |
(Ocean). |
| 303. |
F" |
30 |
45° |
31′ S., |
78° |
9′ W. |
W. of Patagonia. |
| 304. |
F" |
31 |
46° |
53′ S., |
75° |
12′ W. |
W. of Patagonia. |
|
1876. |
|
|
|
|
|
| 318. |
Feb. |
11 |
42° |
32′ S., |
56° |
29′ W. |
(Ocean). |
| 319. |
F" |
12 |
41° |
54′ S., |
54° |
48′ W. |
(Ocean). |
| 323. |
F" |
28 |
35° |
39′ S., |
50° |
47′ W. |
W. of Buenos Ayres. |
| 324. |
F" |
29 |
36° |
9′ S., |
48° |
22′ W. |
Ocean |
|
Open South Atlantic
Ocean, between Buenos
Ayres and Tristan
da Cunha (35°-37° S.
lat., 21°-48° W. long.). |
| 325. |
Mar. |
2 |
36° |
44′ S., |
46° |
16′ W. |
Ocean |
| |
|
|
|
|
|
|
|
| 326. |
F" |
3 |
37° |
3′ S., |
44° |
17′ W. |
Ocean |
| 327. |
F" |
4 |
36° |
48′ S., |
42° |
45′ W. |
Ocean |
| 328. |
F" |
6 |
37° |
38′ S., |
39° |
36′ W. |
Ocean |
| 329. |
F" |
7 |
37° |
31′ S., |
36° |
7′ W. |
Ocean |
| 330. |
F" |
8 |
37° |
45′ S., |
33° |
0′ W. |
Ocean |
| |
|
|
|
|
|
|
|
| 331. |
F" |
9 |
37° |
47′ S., |
30° |
20′ W. |
Ocean |
| 332. |
F" |
10 |
37° |
29′ S., |
27° |
31′ W. |
Ocean |
| 333. |
F" |
13 |
35° |
36′ S., |
21° |
12′ W. |
Ocean |
| 334. |
F" |
14 |
35° |
45′ S., |
18° |
31′ W. |
W. of Tristan da Cunha. |
| 335. |
F" |
16 |
32° |
24′ S., |
13° |
5′ W. |
N. of Tristan da Cunha. |
{clxiii}
|
Locality 1. |
|
|
|
Relative
Abundance of
Radiolaria. |
Date. |
Latitude and Longitude. |
Nearest Land. |
|
|
|
|
|
|
1876. |
|
|
|
|
|
| 338. |
Tr. Atl. |
1990 |
36.3 |
gl. oz. |
D f" |
Mar. |
21 |
21° |
15′ S., |
14° |
2′ W. |
(Ocean). |
| 340. |
Tr. " |
1500 |
37.6 |
pt. oz. |
E very few |
F" |
24 |
14° |
33′ S., |
13° |
42′ W. |
Ocean |
|
W. of St. Helena. |
| 341. |
Tr. " |
1475 |
38.2 |
pt. oz. |
E very" |
F" |
25 |
12° |
16′ S., |
13° |
44′ W. |
Ocean |
| 342. |
Tr. " |
1445 |
37.5 |
pt. oz. |
D few |
F" |
26 |
9° |
43′ S., |
13° |
51′ W. |
Ocean |
| 343. |
Tr. " |
425 |
40.3 |
volc. s. |
E very few |
F" |
27 |
8° |
3′ S., |
14° |
27′ W. |
Ascension Island. |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 344. |
Tr. " |
420 |
... |
volc. s. |
E very" |
April |
3 |
7° |
54′ S., |
14° |
28′ W. |
Ascension Island. |
| 345. |
Tr. " |
2010 |
36.8 |
gl. oz. |
D few |
F" |
4 |
5° |
45′ S., |
14° |
25′ W. |
Ocean |
|
Tropical Atlantic,
between Ascension and
Sierra Leone. |
| 346. |
Tr. " |
2350 |
34.0 |
gl. oz. |
C many |
F" |
6 |
2° |
42′ S., |
14° |
41′ W. |
Ocean |
| 347. |
Tr. " |
2250 |
36.2 |
gl. oz. |
B numerous |
F" |
7 |
0° |
15′ S., |
14° |
25′ W. |
Ocean |
| 348. |
Tr. " |
(2450) |
... |
(Pelag.) |
B num" |
F" |
9 |
3° |
10′ N., |
14° |
51′ W. |
Ocean |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 349. |
Tr. " |
... |
... |
(Pelag.) |
B num" |
F" |
10 |
5° |
28′ N., |
14° |
38′ W. |
Ocean |
| 350. |
Tr. " |
... |
... |
(Pelag.) |
B num" |
F" |
11 |
7° |
33′ N., |
15° |
16′ W. |
W. of Sierra Leone. |
| 351. |
Tr. " |
... |
... |
(Pelag.) |
B num" |
F" |
12 |
9° |
9′ N., |
16° |
41′ W. |
W. of Sierra Leone. |
| 352. |
Tr. " |
... |
... |
(Pelag.) |
B num" |
F" |
13 |
10° |
55′ N., |
17° |
46′ W. |
W. of Sierra Leone. |
| 353. |
N. Atl. |
2965 |
37.6 |
r. cl. |
C many |
May |
3 |
26° |
21′ N., |
33° |
37′ W. |
W. of Canary Islands. |
| |
|
|
|
|
|
|
|
|
|
|
|
|
| 354. |
N. " |
1675 |
37.8 |
gl. oz. |
D few |
F" |
6 |
32° |
41′ N., |
36° |
6′ W. |
S. of Azores. |
|
Locality 2. |
|
|
|
Relative
Abundance of
Radiolaria. |
|
|
|
|
|
|
| 338. |
Tr. Atl. |
1990 |
36.3 |
gl. oz. |
D f" |
| 340. |
Tr. " |
1500 |
37.6 |
pt. oz. |
E very few |
| 341. |
Tr. " |
1475 |
38.2 |
pt. oz. |
E very" |
| 342. |
Tr. " |
1445 |
37.5 |
pt. oz. |
D few |
| 343. |
Tr. " |
425 |
40.3 |
volc. s. |
E very few |
| |
|
|
|
|
|
| 344. |
Tr. " |
420 |
... |
volc. s. |
E very" |
| 345. |
Tr. " |
2010 |
36.8 |
gl. oz. |
D few |
| 346. |
Tr. " |
2350 |
34.0 |
gl. oz. |
C many |
| 347. |
Tr. " |
2250 |
36.2 |
gl. oz. |
B numerous |
| 348. |
Tr. " |
(2450) |
... |
(Pelag.) |
B num" |
| |
|
|
|
|
|
| 349. |
Tr. " |
... |
... |
(Pelag.) |
B num" |
| 350. |
Tr. " |
... |
... |
(Pelag.) |
B num" |
| 351. |
Tr. " |
... |
... |
(Pelag.) |
B num" |
| 352. |
Tr. " |
... |
... |
(Pelag.) |
B num" |
| 353. |
N. Atl. |
2965 |
37.6 |
r. cl. |
C many |
| |
|
|
|
|
|
| 354. |
N. " |
1675 |
37.8 |
gl. oz. |
D few |
|
Date. |
Latitude and Longitude. |
Nearest Land. |
|
1876. |
|
|
|
|
|
| 338. |
Mar. |
21 |
21° |
15′ S., |
14° |
2′ W. |
(Ocean). |
| 340. |
F" |
24 |
14° |
33′ S., |
13° |
42′ W. |
Ocean |
|
W. of St. Helena. |
| 341. |
F" |
25 |
12° |
16′ S., |
13° |
44′ W. |
Ocean |
| 342. |
F" |
26 |
9° |
43′ S., |
13° |
51′ W. |
Ocean |
| 343. |
F" |
27 |
8° |
3′ S., |
14° |
27′ W. |
Ascension Island. |
| |
|
|
|
|
|
|
|
| 344. |
April |
3 |
7° |
54′ S., |
14° |
28′ W. |
Ascension Island. |
| 345. |
F" |
4 |
5° |
45′ S., |
14° |
25′ W. |
Ocean |
|
Tropical Atlantic,
between Ascension and
Sierra Leone. |
| 346. |
F" |
6 |
2° |
42′ S., |
14° |
41′ W. |
Ocean |
| 347. |
F" |
7 |
0° |
15′ S., |
14° |
25′ W. |
Ocean |
| 348. |
F" |
9 |
3° |
10′ N., |
14° |
51′ W. |
Ocean |
| |
|
|
|
|
|
|
|
| 349. |
F" |
10 |
5° |
28′ N., |
14° |
38′ W. |
Ocean |
| 350. |
F" |
11 |
7° |
33′ N., |
15° |
16′ W. |
W. of Sierra Leone. |
| 351. |
F" |
12 |
9° |
9′ N., |
16° |
41′ W. |
W. of Sierra Leone. |
| 352. |
F" |
13 |
10° |
55′ N., |
17° |
46′ W. |
W. of Sierra Leone. |
| 353. |
May |
3 |
26° |
21′ N., |
33° |
37′ W. |
W. of Canary Islands. |
| |
|
|
|
|
|
|
|
| 354. |
F" |
6 |
32° |
41′ N., |
36° |
6′ W. |
S. of Azores. |
{clxiv}
CHAPTER X.—GEOGRAPHICAL DISTRIBUTION.
(§§ 241-250.)
241. Historical Distribution.—Radiolaria are found fossil in all the
more important groups of the sedimentary rocks of the earth's crust. Whilst a few years ago their
well-preserved siliceous skeletons were only known in considerable quantity from Cainozoic marls
(§ 242), very many Spumellaria and Nassellaria have recently been found in Mesozoic and a few in Palæozoic strata.
By the aid of improved modern methods of investigation (especially by the preparation of thin
sections of very hard rocks) it has been shown that many hard siliceous minerals, especially
cryptocrystalline quartz, contain numerous well-preserved Radiolaria, and sometimes are mainly
composed of closely compacted masses of such siliceous shells; of this kind are many quartzites of
the Jura (§ 243). These Jurassic quartzes (Switzerland), as well as the
Tertiary marls (Barbados) and clays (Nicobar Islands), are to be regarded as "fossil Radiolarian
ooze" (§ 237). Dense masses of compressed Spumellaria and Nassellaria form the principal part of
these rocks. Isolated or in smaller quantities, fossil Polycystina, belonging to different
families of Spumellaria and Nassellaria, also
occur in in other rocks, and even in some of Palæozoic origin. Since specimens have also been
recently found both in Silurian and Cambrian strata, it may be stated that as regards their
historical distribution, Radiolaria occur in all fossiliferous sedimentary deposits, from the
oldest to those of the present time.
242. Cainozoic Radiolaria.—The great majority of fossil Radiolaria which have
hitherto been described, belong to the Cainozoic or Tertiary period, and in fact, to its middle
portion, the Miocene period. At this period the richest and most important of all the Radiolarian
formations were deposited, such as the pure "Polycystine marl" of Barbados (see note A), also that
of Grotte in Sicily (see note B), and the clay of the Nicobar Islands (see note C). Besides the
above-mentioned deposits, which may be designated "pure" fossil Radiolarian ooze, many deposits
containing these organisms have recently been discovered in widely separated parts of the earth,
partly of the nature of tripoli or marl, partly resembling clay. Among these may be mentioned in
the first place many coasts and islands of the Mediterranean, both on the south coast of Europe
(Sicily, Calabria, Greece), and the north coast of Africa (from Oran to Tripoli). The extensive
layers of tripoli which are found in these Mediterranean Tertiary mountains belong to the upper
Miocene (Tortona stage), and consist partly of marl rich in calcareous matter, and resembling
chalk, partly passing over into plastic clay or "Kieselguhr" (§ 246). The
quantity of Radiolaria contained varies, and is more conspicuous the fewer the calcareous shells
of Foraminifera present. Similar Tertiary Polycystine formations occur in some parts of America
(see note D); probably they have a very wide distribution. In their general morphological
characters, the Tertiary Spumellaria and Nassellaria {clxv}are related to
those forms which are found in the recent Radiolarian ooze of the depths of the Pacific,
especially to the species which are characteristic of the Challenger Stations 225, 226, 265 and
268. Many living genera and families (e.g., most Larcoidea and
Stephoidea) have not yet been found in the Tertiary formations.
A. The famous Polycystine marl of Barbados in the Antilles, which Robert
Schomburgk discovered forty years ago, belongs to the Miocene formation, and is the richest and
best known of all the important Radiolarian deposits (see L. N. 16, pp.
5-8). After Ehrenberg had published in December 1846 the first preliminary communication
regarding its composition out of masses of well-preserved Polycystina, he was able in the
following year to describe no less than 282 species from it; he distributed these in 44 genera
and 7 families (L. N. 4, 1847, p. 54). In the year 1854 Ehrenberg published
figures of 33 species in his Mikrogeologie (L. N. 6, Taf. xxxvi.); but it
was only in 1873 that he published descriptions of 265 species (Monatsber. d. k. preuss. Akad.
d. Wiss. Berlin, Jan. 30, pp. 213-263). Finally there followed in 1875 his Fortsetzung der
Mikrogeologischen Studien, mit specieller Rücksicht auf den Polycystinen-Mergel von Barbados (L.
N. 25). On the thirty plates which accompany this the last work of
Ehrenberg, 282 species are figured and named, of which 54 are Spumellaria (13 Sphæroidea, 8 Prunoidea, 33 Discoidea), and 228 Nassellaria (2 Stephoidea, 38 Spyroidea, and 188 Cyrtoidea). The fourth section of
this memoir contains a survey of the Polycystine formation of Barbados (pp. 106-115), and the
fifth section the special description of a large specimen of rock from Mount Hillaby in Barbados
(see also L. N. 28, p. 117, and L. N. 41, pp. 476-478).
The account given by Ehrenberg of the Polycystina of Barbados is in many respects very
incomplete, and very far from exhausting this rich mine of remarkable forms. This may be readily
seen from the twenty-five plates of figures of Polycystins in the Barbados Chalk Deposit
published by Bury in 1862 (L. N. 17). The number of species here figured
(140 to 142) is about half of those given by Ehrenberg; and there are among them numerous
generic types, some of great interest, which were entirely overlooked by the latter; e.g.
Saturnalis (Sphæroidea), Cannartidium (Prunoidea), Tympanidium (Stephoidea),
Cinclopyramis (Cyrtoidea), &c. Finally, Ehrenberg always
(until 1875) ignored Bury's atlas, which had been published thirteen years ago and was quite
accessible to him. How different were the contents of the two works may easily be seen from the
following abstract.
Comparative View of the Species of Fossil Radiolaria from Barbados made known by the
figures of Bury in 1862 and of Ehrenberg in 1875.
| Legion. |
Order. |
Bury. |
Ehrenberg. |
Total. |
II. Legion
Spumellaria
(Peripylea). |
|
1. Sphæroidea |
16 |
13 |
29 |
| 2. Prunoidea |
10 |
8 |
18 |
| 3. Discoidea |
37 |
33 |
70 |
| |
|
|
|
|
II. Legion
Nassellaria
(Monopylea). |
|
4. Stephoidea |
5 |
2 |
7 |
| 5. Spyroidea |
13 |
38 |
51 |
| 6. Cyrtoidea |
60 |
188 |
248 |
|
Total, |
141 |
282 |
423 |
| Legion. |
Order. |
Bury. |
Ehrenberg. |
Total. |
II. Legion
Spumellaria
(Peripylea). |
1. Sphæroidea |
16 |
13 |
29 |
| 2. Prunoidea |
10 |
8 |
18 |
| 3. Discoidea |
37 |
33 |
70 |
II. Legion
Nassellaria
(Monopylea). |
4. Stephoidea |
5 |
2 |
7 |
| 5. Spyroidea |
13 |
38 |
51 |
| 6. Cyrtoidea |
60 |
188 |
248 |
| Total, |
141 |
282 |
423 |
{clxvi}
In 1882 Bütschli still further increased the number of known Radiolaria from Barbados both by
figures and descriptions (L. N. 40), and gave in particular a very accurate
morphological analysis of 12 new Nassellaria (3 Stephoidea, 3 Spyroidea, and 6 Cyrtoidea; L. N. 40, Taf. xxxii., xxxiii.). The number of
the fossil species collected in the Barbados marl is, however, greater than would appear from
the above-quoted communications. My respected friend, Dr. R. Teuscher, of Jena, has, at my
request, made a large number (about a thousand) of very accurate drawings with the camera lucida
of Polycystina from Barbados (see p. 1760).
From these it appears that the variations in the structure of the shells, with respect to
number, size, and form of the lattice-pores, of the spines, &c., is much greater than would
be supposed from the figures of Ehrenberg and Bury. I have thus come to the conviction that the
number of species from Barbados (using the word "species" in the sense understood by those
authors) is not less than 400 and probably more than 500. Descriptions of some particularly
interesting new species from this series have been included in the systematic account of the
Challenger Radiolaria. A complete critical investigation of the Radiolaria of Barbados, and
especially an accurate comparison of these Cainozoic species with the Mesozoic forms from the
Jura, on the one hand, and with recent types on the other, must be left to the future for its
accomplishment (see § 246).
B. The Cainozoic Polycystine tripoli or marl of the Mediterranean coast, which is probably
always of Miocene origin, forms very extensive mountain ranges both in the south of Europe
(Sicily, Calabria, Greece) and in the north of Africa (from Oran to Tripoli) (§ 246). Hitherto, however, only one locality has been thoroughly investigated,
namely, Grotte in the province of Girgenti in Sicily (L. N. 35). In the
accurate account which was given of it by Stöhr in 1880, 118 species were described, distributed
in 40 genera (L. N. 35; pp. 72-84); of these 118 species 78 are quite new,
25 are identical with previously known fossils, and 29 identical with living forms. Among them
are 73 Spumellaria (28 Sphæroidea, 8 Prunoidea, and 37 Discoidea), but only 40 Nassellaria (1 Stephoidea, 6 Spyroidea, and 33 Cyrtoidea), and 5 Phæodaria (Dictyochida). The other parts of Sicily from which the same upper
Miocene tripoli has been investigated (belonging to the Tortona stage) have proved less rich
than Grotte. The best known of these places is Caltanisetta, since upon three genera discovered
here (Haliomma, Cornutella, Lithocampe) the group Polycystina was founded
by Ehrenberg in 1838 (see L. N. 16, p. 3). Afterwards 31 species were
described from this locality, of which 23 were again found in Grotte. The richest deposit on the
Mediterranean coast, however, appears to be at Oran. A small specimen of the Kieselguhr found
there, which was recently sent to me by Professor Steinman, proved to be pure Radiolarian ooze,
very similar to that now found in the Central Pacific, and contained many hitherto undescribed
species; it is deserving of careful investigation and comparison.
C. Regarding the Tertiary Radiolarian clay of the Nicobar Islands, see § 247 and L. N. 25, pp. 116-120. Its fauna is incompletely
known; probably it is of Miocene or Oligocene origin.
D. Cainozoic tripoli, containing larger or smaller quantities of Radiolaria,
appears to be rather widely distributed in America. Ehrenberg has described such from South
America (polishing-slate from Morro di Mijellones, on the coast between Chili and Bolivia), and
from North America (Richmond and Petersburg in Virginia, Piscataway in Maryland). Similar
deposits are also found in the Bermuda Islands (L. N. 4, 1855-56; L. N. 6, Taf. 18; L. N. 16, pp. 3-9; L. N. 41,
pp. 475-478, and L. N. 25, pp. 2-6).
{clxvii}
243. Mesozoic Radiolaria.—From the Mesozoic or Secondary period numerous
well-preserved Radiolaria have recently been described. They belong for the most part to the
Jurassic formation (see notes A, B, C), whilst the more recent Chalk (see note D) and the older
Trias (see note E) have hitherto yielded but few species. All the main divisions of the Jura, both
the upper (Malm) and the middle (Dogger), and especially the lower (Lias) appear in certain
localities to be very rich in well-preserved shells of fossil Polycystina. Most of these are
aggregated together in coprolites and quartzites (jasper, chert, flint, &c., § 248). The majority are Cyrtoidea, the minority Sphæroidea and Discoidea in almost equal proportions;
a few Beloidea (Sphærozoum) and Phæocystina (Dictyocha) are also found among them. The general morphological
character of these Jurassic Radiolaria is very different from that of the nearly related Tertiary
and living forms. In general, their siliceous shells are firmer and more massive, usually also
somewhat larger, but of simpler structure. The manifold delicate appendages (spines, bristles,
feet, wings, &c.) which are so richly developed in the living Spumellaria and Nassellaria, and are also well shown in
the Tertiary species, are entirely wanting in the majority of the Jurassic Polycystina. The Sphæroidea and Prunoidea are all simple spherical or
ellipsoidal lattice-shells (Monosphærida); concentric lattice-shells (Polysphærida) are entirely
wanting. The Cyrtoidea are, for the most part, devoid of radial processes
or basal feet (Eradiata); triradiate and multiradiate forms, such as are found abundantly in the
recent and Tertiary formations, are very rare. The large number of many-jointed forms
(Stichocyrtida) and of Cyrtoidea with latticed basal opening is very
striking.
A. The most important work on the Jurassic Radiolaria, regarding which but little
was known prior to the year 1885, is the valuable and in some respects very interesting Beiträge
zur Kenntniss der fossilen Radiolarien aus Gesteinen des Jura, by Dr. Rüst of Freiburg i. B.
(1885, Palæontographica, Bd. xxxi. 51 pp. with 12 plates). Unfortunately this important work was
issued only when about half of the present Report was printed off, so that it was no longer
possible to include the 234 species there described in its systematic part. I have therefore
elsewhere given a list of the Jurassic Radiolaria, and at present only make the following
remarks:—Of the 234 species described, the larger half (130) belong to the Nassellaria (Cyrtoidea), the smaller half (102) to
the Spumellaria (38 Sphæroidea, 14 Prunoidea, and 50 Discoidea). In addition, there are
2 Phæodaria depicted, and several spicules which are probably to be
referred to the Beloidea. Among the 130 Cyrtoidea (of which 2 are described as Botryodea),
there are 24 Monocyrtida, 14 Dicyrtida, 22 Tricyrtida, and 70 Stichocyrtida. Just as striking as
the predominant number of the last is the fact that there are only very few triradiate (9) and
multiradiate (4) species found among these 130 Cyrtoidea, as also the
large number of species with latticed basal opening; Stephoidea appear
to be entirely wanting. The rich material of jasper, chert, flint, and coprolites in which Dr.
Rüst found these Radiolaria, is derived for the most part from the Jurassic rocks of Germany
(Hanover, South Bavaria), Tyrol, and Switzerland (compare § 248).
{clxviii}
B. Jurassic Radiolaria from Italy, also found in jasper, which are closely related to the
forms from Germany and Switzerland described by Dr. Rüst, were made known so long ago as 1880 by
Dante Pantanelli in his treatise I Diaspri della Toscana e i loro Fossili (Rome, 1880, 33 pp. 60
figs.). Pantanelli believes, however, that this jasper is for the most part of Eocene origin;
but from his description, and especially from the morphological character of the forms which he
figures, it appears very probable "that these Tuscan jaspers from Galestro, like those of the
Swiss conglomerates, are found in a secondary locality and belong to the Jurassic period" (Rüst,
L. N. 51, p. 3). Unfortunately the figures of Pantanelli are so small and
incomplete that a reliable determination of the species is hardly possible; for example, the
lattice-work is only given in ten of the sixty figures. Among the 32 recorded species 15 are
Spumellaria (6 Sphæroidea and 9 Discoidea) and 17 Nassellaria (4 Stephoidea and 13 Cyrtoidea); many of which seem to
be identical with the forms more accurately described by Dr. Rüst (compare p. 1762).
C. From the Lias of the Alps and more particularly "from the lower Liassic beds of the
Schafberg near Salzburg," Dr. Emil von Dunikowski in 1882 described 18 species of fossil
Radiolaria (L. N. 44, pp. 22-34, Taf. iv.-vi.); most of these are Sphæroidea and Discoidea and appear to have been
more or less altered by petrological changes; their spongy structure is probably secondary.
D. Cretaceous Radiolaria have been hitherto described only in very small numbers; quite
recently Dr. Rüst has found a larger number chiefly in flints from the English chalk, but they
have not yet been published. In 1876 Zittel described 6 very well-preserved species from the
upper chalk of North Germany (L. N. 29, pp. 76-96, Taf. ii.); among them
were 1 Sphæroidea, 1 Discoidea, 1 Dictyocha,
and 3 Cyrtoidea.
E. Triassic Radiolaria have recently been discovered by Dr. Rüst in chert, but
have not yet been described.
244. Palæozoic Radiolaria.—The number of Radiolaria which are known from the
Palæozoic or Primary formations is much less than from either the Mesozoic or Cainozoic periods.
Here, however, the investigations of recent times have yielded important information; a few
species, at all events, of Polycystina (mostly Sphæroidea) are now known
from various Palæozoic formations, and not only from the Permian ("Zechstein") and the
Coal-measures, but also from the older Devonian and Silurian systems. Even in the still older
Cambrian rocks a few fossil Radiolaria have been found. All these Palæozoic Radiolaria are
Polycystina of very simple form and primitive structure, mostly simple Spumellaria (latticed spheres, ellipsoids, lenses, &c.), but partly also
simple Nassellaria.
The important discoveries which have recently been made by Dr. Rüst regarding the
occurrence of Radiolaria in all the Palæozoic formations have not yet been published. From
conversations with this estimable palæontologist I have learned, however, that he has pursued
his fruitful investigation of the Mesozoic quartzites (§ 243), and has
met with no less success in the case of similar Palæozoic structures. Although the number of
species hitherto discovered is relatively small, the important conclusion appears to be
warranted that they extend as far as the Silurian and Cambrian systems. All these very ancient
Spumellaria (Sphæroidea) and Nassellaria (Cyrtoidea) {clxix}exhibit very primitive structural relations. The occurrence of
fossil Polycystina in the Carboniferous formation of England has been incidentally mentioned by
W. J. Sollas:—"In the carboniferous beds of North Wales pseudomorphs of Radiolaria in
calcite occur, along with minute quartz crystals" (Ann. and Mag. Nat. Hist., 1880, ser. 5, vol.
vi. p. 439); and in the siliceous slate-beds of Saxony Rothpletz has shown the existence of a
few Sphæroidea (Zeitschr. d. Deutsch. Geol. Gesellsch., 1800, p.
447).
245. Abundance of Radiolaria in the Various Rocks.—The relative
quantity of well-preserved or at all events recognisable Radiolaria in the different rocks is very
variable. In this respect three different degrees may be distinguished, which may be called
shortly "pure, mixed, and poor" Radiolarian formations. The pure Radiolarian rocks consist
for the greater part (usually much more than half, sometimes even more than three-quarters) of
closely compacted often calcined masses of siliceous Polycystine shells. To this category belong
the pure Miocene Polycystine marls of Barbados (§ 246), the Tertiary
Polycystine clay of the Nicobar Islands (§ 247), and the Polycystine quartz
of the Jura (§ 248). All these pure Radiolarian rocks may be regarded as
fossil Radiolarian ooze (§ 237), and are certainly of deep-sea origin,
having probably been deposited at depths greater than 2000 fathoms. Their palæontological
character also is in favour of this view, for the abyssal Osculosa (§ 235)
are more abundant and richer in species than the pelagic Porulosa (§ 233).
The elevation of this deep-sea layer above the surface of the sea appears to have taken place but
seldom; it has only been observed on a large scale at Barbados and in the Nicobar Islands. The
mixed Radiolarian rocks are much more common; they were probably deposited at much less
depths, or perhaps are not true deep-sea formations at all. The siliceous shells of Polycystina
always constitute less than half (sometimes less than one-tenth) of their mass, and are less
prominent than other siliceous remains (Diatoms), or calcareous remains (Foraminifera), or in some
cases than the mineral constituents (pumice, &c.). To this group belong many of the
above-mentioned Tertiary marls and clays (especially the Mediterranean Tripoli), also many flints,
cherts, and other quartzites from Mesozoic strata (especially from the Jura), and probably also
some palæozoic quartzites. The marine ooze from which they have originated may have been deposited
at very various, even at slight, depths of the ocean. Formations poor in Radiolaria, which
contain only a few species of Spumellaria and Nassellaria mingled with other fossil remains and mineral particles, occur in
all formations and are probably very widely distributed. Further careful examination of thin
sections (especially of coprolites) will yield here a rich harvest of new forms. Both the mixed
and the pure Radiolarian formations may be divided according to their petrographic characters into
three groups, which, however, are connected by intermediate varieties—(1) soft, chalky marl
(§ 246), (2) plastic clay (§ 247), and (3) hard,
flinty quartz (§ 248).
{clxx}
246. Radiolarian Marl.—Those soft, friable rocks, which contain a large quantity
of calcareous matter, but consist for the most part of the shells of Spumellaria and Nassellaria, are called Radiolarian or
Polycystine marl, often more correctly Polycystine tripoli; the best known example of them is the
chalky marl of Barbados in the Antilles (§ 242). The Tertiary mountain
system of this island, which in Mount Hillaby rises to a height of 1147 feet and includes about
15,800 acres, consists almost exclusively of these remarkable masses of rock. Most of it appears
as a soft, earthy, often chalky marl, with a considerable but variable amount of calcareous
matter. Those specimens, the greater half of which is composed of well-preserved siliceous shells
of Polycystina, and which contain little lime, approach the tripoli and "Kieselguhr." Those
specimens, however, which contain the largest amount of calcareous matter resemble common writing
chalk in consistency, and consist for the most part of shells of Foraminifera and their fragments;
of these there are only few species but large numbers of individuals, generally in small fragments
with a fine calcareous powder between them. They may be regarded as fossil Globigerina ooze (§ 238). In a third group of specimens from Barbados the quantity of fragments of
pumice and other volcanic matters predominates; the amount of clay is also very considerable;
these deposits pass over partly into actual clay partly into volcanic tuff. A fourth group
exhibits relations to a coarser often ferruginous material, and although the shells of Polycystina
are less abundant in it, still it may be shown to be composed largely of fragments and
metamorphosed remains of them. The colour of this deposit, which in some places passes over into
sandstone, in others into clay, is usually rather dark, grey, brown, sometimes red and
occasionally black (bituminous). The Radiolarian marls of the first two groups, which sometimes
approach the white chalk, sometimes the Kieselguhr, are grey, or even pure white (see note A). The
same constitution is exhibited by the yellowish or white, very light and friable Polycystine marls
of Sicily, which in Caltanisetta approach the chalk, and in Grotte the Kieselguhr. In Greece
(Ægina, Zante, &c.), on the other hand, they pass over into plastic clay, and the same occurs
in the Baden marl of the Vienna basin. In North Africa, however, on the Mediterranean shores of
which the Radiolarian marl seems to be very widely distributed (from Tripoli to Oran), it
sometimes becomes changed into actual firm polishing slate, sometimes into pulverulent Kieselguhr
or tripoli (Terra tripolitana, see note B). Most of these Radiolarian marls appear to date from
the middle Tertiary (Miocene) period, and to be deep-sea formations.
A. The Polycystine marl of Barbados appears at different parts of the island to
present greater variations in its petrographical and zoographical composition than would appear
from Ehrenberg's description (1875, L. N. 25, pp. 106-116). Through the
kindness of one of my former students, Dr. Dorner, to whom I take this opportunity of expressing
my thanks for the favour, I received a large number of specimens of Barbados rock, taken from
various parts of the island, and they exhibit very great variations in their external
appearance, their chemical composition, and the {clxxi}Radiolaria which they contain. The white specimens resembling
Kieselguhr contained approximately 60 to 70 per cent. by volume of Radiolarian shells, the
yellowish marl 40 to 50 per cent., and the brown and black (bituminous) marl 10 to 20 per cent.
or less. Two analyses of the first, which my friend Dr. W. Weber was good enough to carry out,
yielded different results from those which are given by Ehrenberg on the basis of Rammelsberg's
analyses (L. N. 25, p. 116). The results of both are here given for
comparison.
Ehrenberg-Rammelsberg
(Fragment from Hillaby). |
Weber I.
(Chalk-like Fragment). |
Weber II.
(Tripoli-like Fragment). |
| Silicate of alumina, |
59.47 |
Silica, |
52.2 |
71.3 |
| Alumina and oxide of iron, |
1.95 |
Alumina (with traces of oxide of iron), |
12.3 |
11.2 |
| Calcium carbonate, |
34.31 |
| Water, |
3.67 |
Lime and magnesia, |
31.9 |
14.8 |
|
Carbon dioxide, |
3.2 |
02.7 |
| —— |
—— |
——0 |
| Total, |
99.40 |
Total, |
99.6 |
100.00 |
For further comparison I here add the three different analyses of Miocene
Tripoli-marls from Sicily, given by Stöhr on the authority of Fremy, Schwager, and Mottura
(Tagebl. d. fünfzigsten Versamml. Deutsch. Naturf. u. Aertzte in München, 1877, p. 163).
| Composition. |
Tripoli from
Licata
(Fremy). |
Tripoli from
Grotte
(Schwager). |
Tripoli from
Caltanisetta
(Mottura). |
| Silica, |
30.98 |
58.58 |
68.6 |
| Alumina, |
17.54 |
11.51 |
|
3.6 |
| Oxide of iron, |
0.33 |
1.84 |
| Lime, |
|
38.09 |
|
8.49 |
|
12.1 |
| Magnesia, |
0.41 |
| Water and organic matter, |
|
13.06 |
|
11.26 |
|
15.2 |
| Carbonic acid, |
7.12 |
|
100.00 |
99.21 |
99.5 |
B. The Radiolarian marl of the Mediterranean appears, judging by the accounts
already published, to stretch along a considerable part of the coast in the earlier and middle
Tertiary formations; thus it occurs of similar composition in widely separated localities, in
Sicily, Calabria, Zante, and Greece; in North Africa from Tripoli to Oran and probably much
farther. So long ago as 1854 Ehrenberg, in his Mikrogeologie (L. N. 6) gave a
series of important, even if incomplete, communications regarding the "chalky white calcareous
marl of Caltanisetta" (Taf. xxii.), the "Platten marl of Zante" (Taf. xx.), the "plastic clay of
Ægina" (Taf xix.), and the "polishing slate of Oran" (Taf. xxi.). In 1880 Stöhr showed in his
fundamental description of the Tripoli from {clxxii}Grotte in Sicily (L. N. 35) that its
Radiolarian fauna is much richer than Ehrenberg supposed. The same is the case in the Tripoli of
Caltanisetta, and also in the Baden marl of the Vienna basin. The richest deposit appears to be
the pure Kieselguhr-like Tripoli from Oran; a small specimen, which was recently sent to me by
Professor Steinmann of Freiburg, i. B., contained many hitherto undescribed species, and was at
least as rich as the purest Barbados marl.
247. Radiolarian Clays.—Among the Radiolarian or Polycystine clays we include the
firm, often plastic, formations, which contain a larger proportion of Radiolaria than of other
organic remains. The first of these to be mentioned is the Cainozoic formation of the Nicobar
Islands in Further India, which rises to a height of 2000 feet above the level of the sea, and
consists for the most part of coloured masses of clay of varying constitution; on Car Nicobar
these are mostly grey or reddish, on the Island of Camorta they are partly strongly ferruginous
and red and yellow (e.g. at Frederickshaven), partly white and light, like meerschaum
(e.g. at Mongkata). The latter varieties appear to pass over into pure loose Polycystine
marl like that of Barbados, the former into calcareous sandstone. Although the Polycystine clays
of the Nicobar Islands are as yet only very incompletely known, it may be concluded with great
probability that they are true deep-sea formations and nearly allied to those recent forms of red
clay, which by their abundance in Radiolaria most nearly approach the Radiolarian ooze, such for
example as the red clay of the North Pacific between Japan and the Sandwich Islands (Stations 241
to 245, compare §§ 229 and 239). With this view
agrees also the greater or less quantity of pumice dust and other volcanic products. Probably
Radiolarian clays like those of the Nicobar Islands occur also in other Tertiary rocks; part of
the Barbados marl passes by gradually increasing content of clay into such; and in this case also
the amount of included pumice is often considerable. Many mixed Radiolarian marls of the
Mediterranean (e.g., of Greece and Oran) also appear to pass over at certain points into
Radiolarian clay.
The Radiolarian clays of the Nicobar Islands are unfortunately very incompletely
known both as regards their geological nature and their palæontological composition. The
communications of Rink (Die Nikobaren-Inseln, eine geographische Skizze, Kopenhagen, 1847) and
of Ehrenberg (L. N. 6, p. 160 and L. N. 25, pp. 116 to
120) leave many important questions unanswered. The latter has only figured twenty-three species
in his Mikrogeologie (L. N. 6, Taf. xxxvi.). In his tabular list of names (L.
N. 25, p. 120) he only incompletely records thirty-nine species, although in
1850, immediately after the first examination of the Nicobar clay, he had distinguished "more
than a hundred species, partly new, partly identical with those of Barbados" (L. N. 16, p. 8). I have unfortunately been unable in spite of many efforts, to obtain
for investigation a specimen of Nicobar clay. The only microscopical preparation (from
Ehrenberg's collection), which I was able to examine, contained several hitherto undescribed
species. A thorough systematic examination of these important Radiolarian clays is a pressing
necessity, especially as they seem to be markedly different from those of the Mediterranean
(from Ægina, Zante, &c.).
{clxxiii}
248. Radiolarian Quartzes.—Under the name Radiolarian or Polycystine quartzes are
included those hard, siliceous rocks, which consist for the most part of the closely compacted
shells of Spumellaria and Nassellaria. To these
"cryptocrystalline quartzes," or better, quartzites, belong more especially the pure Radiolarian
formations of the Jura, which have been described as flint, chert, jasper, as well as other
cryptocrystalline quartzites. Most of the rocks of this nature hitherto examined are from Germany
(Hanover, South Bavaria), Hungary, Tyrol, and Switzerland; others are known from Italy (Tuscany).
They occur both in the upper and middle, but especially in the lower Jurassic formation (also in
the lower layers of the Alpine Lias). A small part of them has been examined in their primary
situation (the red jaspers of Allgäu and Tyrol), the greater part, however, only as loose rolled
stones in secondary situations (thus in Switzerland in the breccia of the Rigi, in the
conglomerate of the Uetli-Berg, and in many boulders of the Rhine, the Limmat, the Reuss, and the
Aar). The greatest abundance, however, of Jurassic Radiolaria has been yielded by the silicified
coprolites from the Lias of Hanover. These "Radiolarian coprolites" are roundish or cylindrical
bodies, which may attain the size of a goose-egg; they probably originated from Fish or
Cephalopods, which had fed upon Crustacea, Pteropoda, and similar pelagic organisms, whose
stomachs were already full of Radiolarian skeletons. Next to the coprolites the richest is the red
jasper, whose colour varies from bright to dark red; it constitutes a true "silicified deep-sea
Radiolarian ooze." The "Aptychus beds" also of South Bavaria and Tyrol are very rich, and
have furnished about one-third of all the Radiolaria known from the Jura; most of the species too
are very well preserved (compare § 243).
Regarding the remarkable composition and manifold varieties of the Jurassic
Radiolarian quartz, the very full treatise of Dr. Rüst may be consulted (L. N. 51). The very interesting Radiolarian coprolites, which that author has
discovered in the lower and middle Jura of Hanover, occur in astonishing numbers in the iron
mines at the village of Gross-Ilsede, four and a half miles south of the town of Peine. They
constitute from 2 to 5 per cent. by weight of the Liassic iron ore; of this latter, in the year
1883 alone, not less than two hundred and eighty million kilograms were excavated. It is very
probable that the careful microscopic examination of thin sections of coprolites, as well as of
flints, chert, jasper, and other quartzites, would yield a rich harvest of fossil Radiolaria in
other formations also. In Italy Dante Pantanelli has discovered interesting Polycystine jaspers
in Tuscany (L. N. 36, 45); these also appear to occur in
the Jura (compare § 243, and L. N. 51, pp. 3-10).
249. Fossil Groups.—The preservation of Radiolaria in the fossil state
is, of course, primarily dependent on the composition of their skeleton. Hence the Acantharia, whose acanthin skeleton although firm is readily soluble, are never
found fossil. The same is true of the skeletons of the Phæodaria, which
consist of a silicate of carbon; here, however, a single exception is found in the Dictyochida, a
subfamily of the Cannorrhaphida,
the isolated parts of whose skeletons appear to consist of pure silica, and {clxxiv}are often found fossil. Of the two other legions those families
which possess no skeleton are of course excluded; the Nassellida among the Nassellaria, and the Thalassicollida and Collozoida among the Spumellaria. Thus of the 85 known families there remain scarcely 55 of which the
skeletons may be expected in the fossil state; and of these scarcely half have been actually
observed in this condition. Of the 20 orders of this class enumerated in § 155, the following 9 may be, for palæontological and geological purposes,
completely excluded:—(A) The 4 orders of Acantharia (1, Actinelida; 2, Acanthonida; 3, Sphærophracta; 4, Prunophracta); (B) 3 orders of Phæodaria (5, Phæosphæria; 6, Phæogromia; 7, Phæoconchia); (C) 1 order of Nassellaria (8, Nassoidea); (D) 1 order of Spumellaria (9, Colloidea). From a geological point of
view the following 6 orders, although occasionally found fossil, are of quite subordinate
importance:—(A) Among the Spumellaria (10, Beloidea, and 11, Larcoidea); (B) among the Nassellaria (12, Plectoidea; 13, Stephoidea; 14, Botryodea); (C) among the Phæodaria (15, the Phæocystina). On the other hand the
following 5 orders, which are the main constituents of Radiolarian rocks, are of pre-eminent
geological importance:—(A) Among the Spumellaria (16, Sphæroidea; 17, Prunoidea; 18, Discoidea); (B) among the Nassellaria (19, Spyroidea, and 20, Cyrtoidea). The numerical relation
in which the different families of these orders appear in the Radiolarian formations may be seen
on consulting § 157.
250. Fossil and Recent Species.—The fact that there are many Radiolaria living at
the present day, whose shells are found fossil in Tertiary rocks, is of great phylogenetic and
geological significance. This appeared to be the case even from the older observations upon the
Polycystina of the Barbados marl (see note A), but more recent and extensive observations both
upon these and upon the Miocene Radiolaria of Sicily, have shown that the number of these "living
fossil" forms is much greater than was previously supposed (see note B). Among the Miocene
Radiolaria numerous species, both of Spumellaria (especially Sphæroidea and Discoidea) and of Nassellaria (especially Spyroidea and Cyrtoidea) are not to be distinguished from the corresponding still living
forms (see notes C, D). On the other hand, those genera, which are rich both in species and
individuals (recent as well as fossil), present continuous series of forms which lead gradually
and uninterruptedly from old Tertiary species to others still living, which are specifically
indistinguishable from them. These interesting morphological facts are capable of direct
phylogenetic application, and furnish valuable proofs of the truth of the theory of descent.
A. Ehrenberg, in his list of fossil Polycystina (L. N. 25, pp. 64-85,
1875), records 325 species of which 26 are still living.
{clxxv}
B. Stöhr, in his list of Miocene Radiolaria from Grotte (L. N. 35, p. 84,
1880), records 118 species, of which 29 are still living.
C. Teuscher, who at my request has made a large number of comparative measurements and
drawings, both of fossil and living Radiolaria, comes to the conclusion that numerous Spumellaria and Nassellaria from Barbados are to-day
extant and unchanged in the Radiolarian ooze of the deep Pacific Ocean (compare § 242A, and p. 1760, Note).
D. From the comparative investigations, which I have made during the last ten
years into the recent deep-sea Radiolaria of the Challenger collection and the Miocene
Polycystina of Barbados, it appears that about a quarter of the latter are identical with living
species of the former.
{clxxvi}
BIBLIOGRAPHICAL SECTION.
CHAPTER XI.—LITERATURE AND HISTORY.
251. List of Publications from 1834 to 1884:—
Note.—In the text the references to the following publications are
indicated by the letters L. N.
1. 1834. Meyen, F., Palmellaria (Physematium, Sphærozoum), in
Beiträge zur Zoologie, gesammelt auf einer Reise um die Erde. Nova Acta Acad. Cæs.
Leop.-Carol., vol. xvi., Suppl., p. 160, Taf. xxviii. figs. 1-7.
2. 1838. Ehrenberg, G., Polycystina (Lithocampe, Cornutella,
Haliomma) in Ueber die Bildung der Kreidefelsen und des Kreidemergels durch unsichtbare
Organismen. Abhandl. d. k. Akad. d. Wiss. Berlin, p. 117.
3. 1839. Ehrenberg, G., Ueber noch jetzt lebende Thierarten
der Kreidebildung (Haliomma radians). Abhandl. d. k. Akad. d. Wiss. Berlin, p. 154.
4. 1844-1873. Ehrenberg, G., Vorläufige Mittheilungen über
Beobachtungen von Polycystinen. Monatsber. d. k. preuss. Akad. d. Wiss. Berlin.
Republished with illustrations in the Mikrogeologie (L. N. 6) and in the two
treatises of 1872 (L. N. 24) and 1875 (L. N. 25).
Compare the Monatsberichte of 1844 (pp. 57, 182, 257), of 1846 (p. 382), of 1847 (p. 40),
of 1850 (p. 476), of 1854 (pp. 54, 205, 236), of 1855 (pp. 292, 305), of 1856 (pp. 197, 425), of
1857 (pp. 142, 538), of 1858 (pp. 12, 30), of 1859 (p. 569), of 1860 (pp. 765, 819), of 1861 (p.
222), of 1869 (p. 253), of 1872 (pp. 300-321), of 1873 (pp. 214-263). Only one of these small
papers is of permanent value, The First Systematic Arrangement of the Polycystina in 7 families,
44 genera, and 282 species (Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, 1847, p. 54).
Compare my Monograph (1862, L. N. 16), pp. 3-12, 214-219.
5. 1851. Huxley, Th., Upon Thalassicolla, a new Zoophyte.
Ann. and Mag. Nat. Hist., ser. 2, vol. viii. pp. 433-442, pl. xvi.
6. 1854. Ehrenberg, G., Mikrogeologie. Figures of numerous
Polycystina on 8 plates; Taf. xviii. figs. 110, 111; Taf. xix. figs. 48-56, 60-62; Taf. xx. Nr.
i., figs. 20-25, 42; Taf. xxi. figs. 51-56; Taf. xxii. figs. 20-40; Taf. xxxv. A., Nr. xix. A.
fig. 5; Taf. xxxv. B. figs. 16-23; Taf. xxxvi. figs. 1-33.
7. 1855. Bailey, J. W., Notice of Microscopic Forms of the
Sea of Kamtschatka. Amer. Journ. Sci. and Arts, vol. xxii. p. 1, pl. i.
8. 1855. Müller, Johannes, Ueber Sphærozoum und Thalassicolla.
Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 229.
9. 1855. Müller, Johannes, Ueber die im Hafen von Messina
beobachteten Polycystinen (Haliomma, Eucyrtidium, Dictyospyris, Podocyrtis). Monatsber. d. k.
preuss. Akad. d. Wiss. Berlin, p. 671.
10. 1856. Müller, Johannes, Ueber die Thalassicollen,
Polycystinen und Acanthometren des Mittelmeeres. Monatsber. d. k. preuss. Akad. d. Wiss.
Berlin, p. 474.
11. 1858. Müller, Johannes, Erläuterung einiger bei St. Tropez
am Mittelmeer beobachteter Polycystinen und Acanthometren. Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 154.
12. 1858. Müller, Johannes, Ueber die Thalassicollen,
Polycystinen und Acanthometren des Mittelmeeres, Abhandl. d. k. Akad. d. Wiss. Berlin,
pp. 1-62, Taf. i.-xi. (The fundamental treatise on the Radiolaria.)
{clxxvii}
13. 1858. Schneider, Anton, Ueber zwei neue Thalassicollen
von Messina. Archiv f. Anat. u. Physiol., p. 38, Taf. iii. B, figs. 1-4.
14. 1858. Claparède et Lachmann,
Echinocystida (Plagiacantha et Acanthometra). Études sur les Infusoires et les Rhizopodes, p.
458, pl. xxii. figs. 8, 9; pl. xxiii. figs. 1-6.
15. 1860. Haeckel, Ernst, Ueber neue lebende Radiolarien des
Mittelmeeres. Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, pp. 794, 835.
16. 1862. Haeckel, Ernst, Die Radiolarien (Rhizopoda
radiaria). Eine Monographie. 572 pp. fol. with an Atlas of 35 Copperplates.
17. 1862. Bury, Mrs., Polycystins, figures of remarkable forms
in the Barbados Chalk Deposit. Ed. ii. By M. C. Cooke, 1868. 25 quarto plates, photographed from
drawings by hand, containing many forms overlooked by Ehrenberg from Barbados.
18. 1863 Harting, Paul, Bijdrage tot de Kennis der
mikroskopische Fauna en Flora van de Banda-Zee (Diep-Zee-Polycystinen). Verhandl. d. Kon.
Akad. van. Wetensch. Amsterdam, vol. ix. p. 30, pls. i.-iii.
19. 1865. Haeckel, Ernst, Ueber den Sarcode-Körper der
Rhizopoden (Actinelius, Acanthodesmia, Cyrtidosphæra, &c.). Zeitschr. f. wiss. Zool.,
Bd. xv. p. 342, Taf. xxvi.
20. 1867. Schneider, Anton, Zur Kenntniss des Baues der
Radiolarien (Thalassicolla). Archiv f. Anat. u. Physiol., 1867, p. 509.
21. 1870. Haeckel, Ernst, Beiträge zur Plastiden Theorie
(Myxobrachia; Amylum in den gelben Zellen). Jenaische Zeitschr. für Naturw., Bd. v. p.
519-540, Taf. xviii.
22. 1871. Cienkowski, L., Ueber Schwärmer-Bildung bei
Radiolarien. Archiv f. mikrosk. Anat., Bd. vii. p. 372-381, Taf. xxix.
23. 1872. Wagner, N., Myxobrachia Cienkowskii. Bull. d.
Acad. St. Petersburg, vol. xvii. p. 140.
24. 1872. Ehrenberg, Gottfried, Mikrogeologische Studien über
das kleinste Leben der Meeres-Tiefgründe aller Zonen und dessen geologischen Einfluss.
Abhandl. d. k. Akad. d. Wiss. Berlin, 1872. Mit 12 Tafeln. (The Latin diagnoses of 113
new species here mentioned are given in the Monatsberichte of April 25, 1872, pp.
300-321.)
25. 1875. Ehrenberg, Gottfried, Polycystinen-Mergel von
Barbados (Fortsetzung der Mikrogeologischen Studien). Abhandl. d. k. Akad. d. Wiss.
Berlin, 1875, 168 pag. mit 30 Tafeln. (The Latin diagnoses of 265 species here recorded are
given in Namensverzeichniss der fossilen Polycystinen von Barbados. Monatsber. d. k. preuss.
Akad. d. Wiss. Berlin, Jan. 30, 1873, pp. 213-263.)
26. 1876. Hertwig, Richard, Zur Histologie der Radiolarien.
Untersuchungen über den Bau und die Entwickelung der Sphærozoiden und Thalassicolliden. 91 pp.
with 5 plates.
27. 1876. Murray, John, Challengerida. Preliminary Reports on
Work done on board the Challenger. Proc. Roy. Soc. Lond., vol. xxiv. pp. 471-536, pl.
xxiv.
28. 1876. Zittel, Karl, Palæozoologie, Bd. i. pp. 114-126,
figs. 46-56.
29. 1876. Zittel, Karl, Ueber fossile Radiolarien der oberen
Kreide. Zeitschr. d. deutsch. geol. Gesellsch., Bd. xxviii. pp. 75-96, Taf. ii. (with
figures of six Cretaceous species).
30. 1877. Mivart, St. George, Notes touching recent researches
on the Radiolaria. Journ. Linn. Soc. Lond. (Zool.), vol. xiv. pp. 136-186. (Historical
sketch of previous literature.)
31. 1877. Wyville Thomson, The Voyage of the
Challenger—The Atlantic, vol. i. pp. 231-237, figs. 51-54; vol. ii. pp. 340-343, figs. 58,
59, &c.
32. 1878. Haeckel, Ernst, Das Protistenreich, eine populäre
Uebersicht über das Formengebiet der niedersten Lebewesen, pp. 101-104.
33. 1879. Hertwig, Richard, Der Organismus der Radiolarien.
Jenaische Denkschriften, Bd. ii. Taf. vi.-xvi. pp. 129-277.
34. 1879. Haeckel, Ernst, Ueber die Phæodarien, eine neue
Gruppe kieselschaliger mariner Rhizopoden. Sitzungsb. med.-nat. Gesellsch. Jena, December
12, 1879.
35. 1880. Stöhr, Emil, Die Radiolarien-Fauna der Tripoli von
Grotte (Provinz Girgenti in Sicilien). Palæontographica, Bd. xxvi. pp. 71-124, Taf.
xvii.-xxiii. A preliminary communication regarding this fauna from the tripoli is given in
Tagebl. d. Naturf. Versamml. München, 1877.
{clxxviii}
36. 1880. Pantanelli, Dante, I Diaspri della Toscana e i loro
fossili. Real. Accad. dei Lincei, ser. 3, vol. vii. pp. 13-34, Tab. i. Radiolaria di
Calabria. Atti. Soc. Tosc., p. 59.
37. 1881. Haeckel, Ernst, Prodromus Systematis Radiolarium,
Entwurf eines Radiolarien-Systems auf Grund von Studien der Challenger-Radiolarien. Jenaische
Zeitschr. für Naturw., Bd. xv. pp. 418-472.
38. 1881. Brandt, Karl, Untersuchungen an Radiolarien.
Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, (April 21), pp. 388-404, Taf. i.
39. 1882. Brandt, Karl, Ueber die morphologische und
physiologische Bedeutung des Chlorophylls bei Thieren. I. Artikel. Archiv f. Anat. u.
Physiol., pp. 125-151, Taf. i. II. Artikel. Mittheil. a. d. Zool. Station zu Neapel,
Bd. iv. pp. 193-302, Taf. xix., xx.
40. 1882. Bütschli, Otto, Beiträge zur Kenntniss der
Radiolarien-Skelette, insbesondere der der Cyrtida. Zeitschr. f. wiss. Zool., Bd. xxxvi.
pp. 485-540, Taf. xxxi.-xxxiii.
41. 1882. Bütschli, Otto, Radiolaria. In Bronn's Klassen und
Ordnungen des Thierreichs. Bd. i., Protozoa, pp. 332-478, Taf. xvii.-xxxii.
42. 1882. Geddes, Patrick, Further Researches on Animals
containing Chlorophyll. Nature, pp. 303-305.
43. 1882. Geddes, Patrick, On the Nature and Functions of the
"Yellow Cells" of Radiolarians and Cœlenterates. Proc. Roy. Soc. Edin., p.
377.
44. 1882. Dunikowski, Emil, Die Spongien, Radiolarien und
Foraminiferen der Unter-Liassischen Schichten vom Schafberg bei Salzburg. Denkschr. d. k.
Akad. d. Wiss. Wien, Bd. xlv. pp. 22-34. Taf. iv.-vi.
45. 1882. Pantanelli, Dante, Fauna miocenica di Radiolari del
Appennino settentrional. Boll. Soc. Geol. Ital.
46. 1883. Haeckel, Ernst, Die Ordnungen der Radiolarien
(Acantharia, Spumellaria, Nassellaria, Phæodaria). Sitzungsb. med.-nat. Gesellsch. Jena,
February 16, 1883.
47. 1883. Hertwig, Oscar, Die Symbiose oder das
Genossenschaftsleben im Thierreich. 56. Versamml. Deutscher Naturf. u. Aerzte, Freiburg
i/B.
48. 1883. Rüst, Wilhelm, Ueber das Vorkommen von
Radiolarien-Resten in kryptokrystallinischen Quarzen aus dem Jura und in Koprolithen aus dem
Lias. 56. Versamml. Deutscher Naturf. u. Aerzte, Freiburg i/B.
49. 1884. Car, Lazar, Acanthometra hemicompressa (= Zygacantha
semicompressa). Zool. Anzeiger, p. 94.
50. 1884. Haeckel, Ernst, Ueber die Geometrie der
Radiolarien (Promorphologie). Sitzungsb. med.-nat. Gesellsch. Jena, November 22,
1883.
251 A. Supplementary List of Works Published in 1885:—
51. 1885. D. Rüst, Beiträge zur Kenntniss der fossilen
Radiolarien aus Gesteinen des Jura. 45 pp. 4to, and 20 plates. Palæontographica, Bd.
xxxi. (oder iii. Folge, vii. Band).
52. 1885. Karl Brandt, Die koloniebildenden Radiolarien
(Sphærozoeen) des Golfes von Neapel und der angrenzenden Meeres-Abschnitte. 276 pp. 4to, and 8
plates.
53. 1885. John Murray, Narrative of the Cruise of H.M.S.
Challenger, with a general account of the scientific results of the Expedition. Vol i. First
part, pp. 219-227, pl. A. Second part, pp. 915-926, pl. N. fig. 2.
54. 1885. Ernst Haeckel, System der Acantharien. Sitzungsb.
med.-nat. Gesellsch. Jena, November 13.
Since the printing of this Report began in 1884 and was far advanced in 1885,
it was impossible to include the important works of Rüst and Brandt (L. N. 51, 52) in the descriptive portion, so that they are only
referred to in the Introduction.
251 B. Phaulographic Appendix:—
A list of absolutely worthless literature, which contains either only long
known facts or false statements, and may hence be entirely neglected with advantage. Compare §
252, and also L. N. 26, p. 9.
55. 1865. Wallich, G. C., On the structure and affinities of
Polycystina. Trans. Micr. Soc. Lond., vol. xiii. pp. 57-84. (Compare L. N. 26, p. 9.)
{clxxix}
56. 1879. Wallich, G. C., Observations on the Thalassicollidæ.
Ann. and Mag. Nat. Hist., ser. 4, vol. iii. p. 97.
57. 1866. Stuart, Alexander, Ueber Coscinosphæra ciliosa, eine
neue Radiolarie (= Globigerina echinoides!!). Zeitschr. f. wiss. Zool., Bd. xvi. p. 328,
Taf. xviii. (Compare L. N. 26, p. 9.)
58. 1870. Stuart, Alexander, Neapolitanische Studien.
Göttinger Nachr., p. 99, and Zeitschr. f. wiss. Zool., Bd. xxii. p. 290 ("Blue
Siliceous Crystals" in Collozoum inerme!).
59. 1871. Macdonald, John Denis, Remarks on the Structure of
Polycystina (Astromma Yelvertoni = Euchitonia Mülleri). Ann. and Mag. Nat. Hist., ser. 4,
vol. viii. p. 226.
60. 1871. Doenitz, W., Beobachtungen über
Radiolarien. Archiv f. Anat. u. Physiol., 1871, p. 71, Taf. ii. (Compare L. N. 26, p. 7.)
252. Progress of our Knowledge of the Radiolaria from 1862 to 1885.—The
history of our scientific knowledge of the Radiolaria extends over about half a century (from 1834
to 1885). A historical and critical discussion of the works which appeared within the first
twenty-eight years of this period (from 1834 to 1862) is contained in the historical introduction
to my Monograph (L. N. 16, pp. 1-24); I shall therefore give here only a brief
survey of the investigations published during the last twenty-three years (from 1862 to 1885). The
most important steps in our progress during this period we owe to the following
naturalists:—Cienkowski (1871), Ehrenberg (1872 and 1875), Richard Hertwig (1876 and 1879),
Karl Brandt (1881 and 1885),
Bütschli (1882), and Rüst (1885). To the valuable works of these authors must be added a number of
smaller contributions, which are recorded in the foregoing Bibliography. Some communications from
dilettanti, written with insufficient knowledge of the subject, and hence of no value, are
mentioned for the sake of completeness in the "Phaulographic Appendix" (compare L. N. 55-60, also L. N. 26, p. 9).
The first important advance in our knowledge of the organisation of the Radiolaria, made after
the publication of my Monograph (1862), was the demonstration of the nature of the extracapsular
"yellow cells." In the year 1870 I showed that these yellow cells contain starch (L. N. 21, p. 519). I regarded them, as did all authors up to that time, as integral
parts of the Radiolarian organism, and hence considered this to be multicellular; for no doubt was
possible regarding the true cellular nature of these remarkable, nucleated, yellow globules, which
I had thoroughly studied in 1862. It was first shown by Cienkowski in 1871 that the yellow cells
of the Collodaria remain unchanged even after the death of these
organisms, "that they continue to grow uninterruptedly, and eventually multiply by division" (L.
N. 22, pp. 378-380, Taf. xix. figs. 30-36). Cienkowski concluded from these
important observations that the yellow cells are not integral parts of the Radiolarian body, but
"parasitic structures," independent, unicellular organisms, which live only as parasites in the
body of the Radiolaria (compare § 90).
This important recognition underwent ten years later a further development and complete
establishment by the extensive investigations of Karl Brandt (L. N. 38, 39) {clxxx}and Patrick Geddes (L. N.
42, 43). This arrangement was compared by Brandt to the
remarkable symbiosis of the Algoid gonidia and Fungoid hyphæ in the organisation of the Lichens,
which had been recently discovered, and since he recognised the independent nature of the yellow
cells, as unicellular Algæ, in all divisions of the Radiolaria, he founded for them the genus
Zooxanthella. Geddes named them Philozoon, and showed experimentally that they give
out oxygen under the influence of sunlight (compare § 90). The great
physiological importance of the yellow cells in the metastasis of the Radiolaria, and, when they
are developed in large quantities, in the economy of marine organisms in general, has recently
been insisted upon by Brandt (see § 205 and L. N. 52,
pp. 65-71, 86-94).
The proof that the yellow cells do not belong to the Radiolarian organism itself, but only live
parasitically in it, was a necessary preliminary to the very important step which next took place
in our knowledge of the organisation of the Radiolaria. This step consisted in the demonstration
that the whole body of the Radiolaria, like that of all other Protista, is only a single cell. It
was Richard Hertwig who in two remarkable works (L. N. 26, 33) firmly established this fundamental theorem of the unicellular nature of the
Radiolaria. In his treatise on the histology of the Radiolaria (L. N. 26,
1876) he published complete investigations into the structure and development of the Sphærozoida
and Thalassicollida. Since he made use of the modern methods of histological examination, and
especially of staining fluids, which he was the first to apply to the study of the Radiolaria, he
was able to show that no true cells (apart from the parasitic yellow cells) are to be found in
their bodies, but rather that all their morphological components are to be regarded as
differentiated parts of a single true cell, and in particular that the central capsule includes a
genuine nucleus.
A wider foundation for this important discovery and its applicability to all divisions of this
extensive class, was given by Hertwig in a second work on the organisation of the Radiolaria (L.
N. 33, 1879). Among the numerous discoveries by which this work enriched the
natural history of the Radiolaria must be specially mentioned the recognition of the fundamental
differences exhibited by the main divisions of the class in the structure of their central
capsule. Hertwig first observed that the capsular membrane is double in the Phæodaria but single in the other Radiolaria (§ 56); the
former he named "Tripylea" because he discovered in their capsular
membrane a large, peculiarly constructed main opening and two small accessory openings. The Nassellaria, in which he found a single porous area at the basal pole of the
main axis, with a cone of pseudopodia rising from it, he called on this account "Monopylea"; whilst the other Radiolaria, whose capsular membrane is perforated
on all sides with fine pores, were termed "Peripylea." Besides the central
capsule, Hertwig laid stress upon the significance of the gelatinous envelope as a constant and
important constituent of the body. He also devoted attentive consideration to the morphology of
the skeleton, and on the basis of certain {clxxxi}phylogenetic conclusions which he drew from it, he arrived at an
improved systematic arrangement in which he distinguished six orders:—(1) Thalassicollea, (2) Sphærozoea, (3) Peripylea, (4) Acanthometrea, (5) Monopylea, (6) Tripylea. The numerous isolated
discoveries with which Hertwig enriched the morphology of the Radiolaria, have been already
alluded to in the appropriate paragraphs in the anatomical portion of this Introduction (see L. N.
42, pp. 340, 341).
The new and interesting group, which was thus erected into an order under the name Tripylea, I had already a year previously separated from the other Radiolaria as
"Pansolenia" in my Protistenreich (L. N. 32, p. 102). Since, however,
neither the three capsular openings of the Tripylea nor the skeletal tubes
of the Pansolenia are present in all the families of this extensive order, I substituted in 1879
the more suitable name Phæodaria, which is applicable to all members of
the group (L. N. 34). In the preliminary memoir then published regarding the
Phæodaria, a New Group of Siliceous Marine Rhizopods, I distinguished four orders, ten families,
and thirty-eight genera. The great majority of these new forms (among which were no less than 465
different species) were first discovered by the deep-sea investigations of the Challenger. John
Murray was the first who called attention to the great abundance in the deep sea of these
remarkable Rhizopods, and to the constant presence of their peculiar, dark, extracapsular pigment
body (phæodium); even in 1876 he described a portion of them as Challengerida (L. N. 27, p. 536; L. N. 53, p. 226). The earliest observations on
the Phæodaria were made at Messina in 1859, where I examined five genera
of this remarkable group alive (compare p. 1522
and L. N. 16).
By the discovery that the Phæodaria, although differing in important
respects from the other Radiolaria, still conform to the definition of the class, a new and
extensive series of forms was added to this latter, and by their closer investigation a fresh
source of interesting morphological problems was disclosed. In other groups, however, morphology
was advanced by comparative anatomical studies. In addition to the smaller contributions of
various authors, mentioned in the foregoing bibliography, I may specially refer to the valuable
Beiträge zur Kenntniss der Radiolarien-Skelete, insbesondere der der Cyrtida by O. Bütschli (L. N.
40, 1882). On the basis of careful comparative anatomical studies,
investigations into the skeletal structure of a number of fossil Cyrtoidea and critical application of the recently published researches of
Ehrenberg into the Polycystina of Barbados (L. N. 25), Bütschli attempted to
derive the complicated relations of the Monopylean skeletons phylogenetically from a simple
primitive form,—the primary sagittal ring. Even if this attempt did not actually solve the
very difficult morphological problem in question, still the critical and synthetic mode in which
it was carried out deserves full recognition, and furnishes the proof that the comparative anatomy
of the skeleton in the Radiolaria not less than in the Vertebrata, is a most interesting and
fruitful field of phylogenetic investigation. A {clxxxii}further demonstration of this was furnished by Bütschli in the
general account of the organisation of the Radiolaria which he published in 1882 in Bronn's
Klassen und Ordnungen des Thierreichs (L. N. 41).
In our knowledge of the developmental history of these Protista the last two decades have
witnessed less progress than in their comparative anatomy. The most important advance in this
direction has been the proof that in all the main groups of the class the contents of the central
capsule are used in the formation of swarm-spores. The movements of these zoospores in the central
capsule had indeed been observed by several previous authors in the case of the Spumellaria and Acantharia (L. N. 10, 13, 16; compare also § 142, Note A). The origin of the flagellate spores from the contents of the
central capsule and their peculiar constitution were, however, first described fully by Cienkowski
in 1871 (L. N. 22, p. 372). Soon after this, R. Hertwig discovered that in the
social Radiolaria (Polycyttaria or Sphærozoea) two different forms of zoospores are formed, one
with, the other without crystals, and that the latter are also divided into macrospores and
microspores (compare L. N. 26, and § 142). Recently
this sexual differentiation has been shown by Karl Brandt to exist in all the groups of
Sphærozoea, and its regular interchange with the formation of crystal-spores has been interpreted
as a true "alternation of generations" (compare L. N. 52 and also § 216). The other forms of development also, especially reproduction by
cell-division (§ 213) and gemmation (§ 214), have
been elucidated by the recent investigations of the same author.
The palæontology of the Radiolaria has of late made important and interesting advances. Until
ten years ago fossil remains of this class were known exclusively from the Tertiary period; almost
the only source of our information was to be found in the researches of Ehrenberg, commenced in
1838, continued in his Mikrogeologie in 1854, and concluded in his last work (L. N. 25) published in 1875 (compare L. N. 16, pp. 3-9, 191-193).
In the year 1876 a number of Mesozoic Radiolaria from the chalk were described by Zittel (L. N. 28), and afterwards others from the Jura by Dunikowski (L. N. 44). That fossil Radiolaria occur in Mesozoic formations, especially in the Jura,
as well preserved and as abundantly as in the Tertiary rocks of Barbados, was shown in 1883 by
Rüst (L. N. 48). By the examination of numerous thin sections he discovered
that in all the main divisions of the Jurassic formation (Lias, Dogger, Malm) there are
distributed jaspers, flints, cherts, and other quartzites, which consist largely of the siliceous
shells of Polycystina; the same is true also of many Coprolites found in the Jura. The full
account of these and the descriptions and figures of 234 Jurassic species, distributed in 76
genera, are contained in the Beiträge zur Kenntniss der fossilen Radiolarien aus Gesteinen des
Jura (L. N. 51, 1885). But even in the older rocks, the Trias, the Permian,
and Carboniferous systems, and even as far downwards as the Silurian and Cambrian formations, Rüst
has recently shown the existence of fossil Radiolaria, {clxxxiii}and thus increased the known period of the developmental
history of the class by many millions of years (§ 244).
The great significance of the Radiolaria in geology and palæontology has been brought into new
light not only by these extensive discoveries, but also by the important relations which have been
shown to exist between the Radiolarian rocks and the deep-sea deposits of the present day. In this
direction the wonderful discoveries of the Challenger, and especially the investigation of the
deep-sea deposits by Wyville Thomson (L. N. 31) and John Murray (L. N. 27), have furnished us with new and valuable information (compare §§ 236-239, and §§ 245-250). The Tertiary Polycystine formations of Barbados and the Nicobar Islands,
with which we have been acquainted for the last forty years, as also the Mesozoic Radiolarian
quartzes, which have only recently been made known to us from the Jura, are ascertained to be
fossil representatives of the same deep-sea deposits which now occur in the form of Radiolarian
ooze (§ 237), and to some extent also of Globigerina ooze and red clay (§§
238, 239), on the bottom of the ocean, at depths of
from 2000 to 4500 fathoms.
These investigations into fossil Radiolaria and their comparison with recent deep-sea forms
have a further general significance, inasmuch as the identity of many living and fossil species
from the Tertiary formation has been shown beyond all doubt. In this direction the numerous
measurements and accurate comparisons which I have made during the last ten years of the abyssal
forms in the Challenger collection, and of fossil species from Barbados and Caltanisetta, have
brought to light many important facts. In this I had the able assistance of my friend, Dr.
Reinhold Teuscher (compare § 250, and p. 1760). Further
valuable contributions in this direction are found in the careful observations and comparative
measurements recently published by Emil Stöhr (L. N. 35, 1880), regarding the Radiolarian fauna of
the Tripoli of Grotte in the province of Girgenti, Sicily. From these it appears that the number
of Miocene species which are still extant, is much greater than would appear from the results of
Ehrenberg.
Ehrenberg himself, towards the end of his long and laborious life, collected the results of the
systematic and palæontological researches, which he had begun thirty-seven years previously (L. N.
16, pp. 3-12) into the Polycystina, in two large works (L. N. 24, 25). The first treatise (L. N. 24,
1872) contains the Mikrogeologische Studien über das Kleinste Leben der Meeres-Tiefgründe aller
Zonen und dessen geologischen Einfluss, with a list of 279 Polycystina observed by him from the
deep-sea, as well as figures of 127 species. The second work (L. N. 25, 1875)
contains the Fortsetzung der Mikrogeologischen Studien, mit specieller Rücksicht auf den
Polycystinen-Mergel von Barbados; the list of fossil Polycystina observed by him includes 325
species, of which 26 are still extant; 282 of them are figured on the thirty plates accompanying
the memoir. By means of these numerous figures, as well as by the appended systematic and
chorological tables, Ehrenberg furnished a welcome {clxxxiv}supplement to the numerous communications regarding the
Polycystina, which he had made to the Berlin Academy since 1838, and which he had published in his
Mikrogeologie in 1854. It will always be the merit of this zealous and indefatigable microscopist
that he first called attention to the great wealth of forms existing in this class; he separated
systematically about 500 species, and published drawings of about 400; in addition to which he was
the first to lay stress upon the great chorological and geological importance of the
Radiolaria.
With these systematic and descriptive, chorological and palæontological works, however, which
relate exclusively to the Polycystina, the merits of the famous naturalist of Berlin are exhausted
as regards this class of animals. Of the organisation of the Radiolaria, Gottfried Ehrenberg
remained entirely ignorant up till his death in 1876. All that a number of famous naturalists had
observed during a quarter of a century as to the structure and life-history of the Radiolaria, all
the important discoveries of Huxley (1851), Johannes Müller (1858), Claparède (1858), Cienkowski
(1871), and many others (L. N. 1-22), and all that I had published in my
Monograph (1862) on the basis of three years' study of their anatomy and physiology—all this
Ehrenberg ignored, or rather, he regarded it all as worthless rubbish of science, as a chaos of
devious errors, resting upon incomplete observations and false conclusions. His strange "special
considerations regarding the Polycystina" (L. N. 24, pp. 339-346) and the
general "concluding remarks" (L. N. 25, pp. 146-147) leave no room for doubt
on this point. Ehrenberg indeed doubted to the last whether any observer had seen living
Radiolaria at all (L. N. 25, p. 108).
The invincible obstinacy with which Ehrenberg maintained his preconceived opinion of the high
organisation of the Radiolaria, and entirely ignored the contrary observations of other
naturalists, is explained by the consistency with which he held to the end the "principle peculiar
to himself of the universally equal development of the animal kingdom" (L. N. 16, p. 7). From the complicated arrangement of their siliceous shells he
concluded that the animals inhabiting them must possess a structure correspondingly complex, and
nearly related to that of the Echinodermata (Holothuria). Like all other animals the Radiolaria
must possess systems of organs for locomotion, sensation, nutrition, circulation, and
reproduction. Whilst Ehrenberg originally interpreted the Polycystina as siliceous Infusoria
polygastrica, and regarded them as compound Arcellina, he afterwards classed them sometimes with
the Echinodermata (Holothuria), sometimes with the Bryozoa, sometimes with the Oscillaria (see L.
N. 41, p. 336). Although a decided opponent of the cell-theory he called them
"multicellular animalcules" (Polycystina), interpreting the pores of the siliceous shell as cells.
To-day the opposite term (Monocystina) might be adopted to express their unicellular organisation.
It was a remarkable irony of fate that in the self-same year (1838) in which Schwann of Berlin
made by his foundation of the cell theory the greatest advance in the whole {clxxxv}of Biological Science, that Ehrenberg, all his life the most
zealous opponent of that theory, published his great work on the Infusoria, and at the same time
established the "family of multicellular animalcules or Polycystina" (L. N. 16, p. 4).
The "short systematic survey of the genera of cellular animalcules" given by Ehrenberg in 1875
(L. N. 25, p. 157), is only a new edition, increased by sixteen genera, of his
first systematic arrangement of the Polycystina of 1847 (L. N. 4, p. 53). Since
I have already given a full discussion of this in my Monograph (L. N. 16, pp.
214-219), I need only here remark that a correct understanding of his very inadequate generic
diagnoses is only possible by the aid of his figures. Relying upon these I have retained almost
all Ehrenberg's genera, although entirely new definitions of most of them have been necessary.
The same is true also of the two orders which Ehrenberg distinguished in his class of
"Zellenthierchen." The first order is constituted by his "Netzkörbchen" (Monodictya or Nassellaria) formerly known as "Polycystina solitaria"; they include our Cyrtoidea, the greater part of Hertwig's Monopylea. Ehrenberg's second order is
the "Schaumsternchen" (Polydictya or Spumellaria), previously called
"Polycystina composita"; they include the Peripylea of Hertwig, as well as the Spyridina (our
Spyroidea), which belong properly to the Nassellaria. Although Ehrenberg's statements regarding the organisation of both
these orders were quite erroneous, and his knowledge even of the structure of their shells very
defective, I still thought it advisable to retain his names for the groups, since they constituted
his one successful effort in the systematic treatment of the Radiolaria (compare L. N. 41, p. 336).
The sketch of a systematic arrangement of the Radiolaria (L. N. 37), which
I published in 1881 on the basis of the study of the Challenger Radiolaria, resembles, in respect
of seven orders being distinguished, the new system which R. Hertwig founded in 1879, in
consequence of the variations which he discovered in the structural relations of the central
capsule (L. N. 33, p. 133). It differs, however, inasmuch as his Sphærozoea
(my Polycyttaria) are here divided into two orders, Symbelaria (Collosphærida) and Syncollaria (Sphærozoida). In that
sketch too I separated for the first time the two subclasses Holotrypasta (Porulosa) and
Merotrypasta (Osculosa). The fifteen families established by Hertwig were then raised to
twenty-four. The six hundred and thirty genera, which I then distinguished, are still for the most
part retained, some, however, in a restricted sense, or with amended definitions.
The differential characters of the orders and families of the Radiolaria, given in
the Prodromus in 1881, were amended in a further communication which I gave in 1883 regarding the
orders of the Radiolaria (L. N. 46, p. 17). There I reduced the seven orders
to four, the structural relations of the central capsule being precisely the same in the
Polycyttaria and Collodaria as in the Peripylea.
The survey of the affinities of the class was thus rendered much simpler and clearer, and the
{clxxxvi}hypothetical genealogical tree, which I
then published, has been still further carried out in Chapter VI. of the present Introduction (see
§§ 153-200).
253. General Survey of the Growth of our Systematic Acquaintance with the Radiolaria from
1834 to 1885.
1834. Meyen (L. N. 1) describes 2 genera and
species of Collodaria:—Sphærozoum fuscum and
Physematium atlanticum.
1838. Ehrenberg (L. N. 2) founds the family
Polycystina upon 3 fossil genera (with 6 species):—Lithocampe, Cornutella,
Haliomma.
1847. Ehrenberg (L. N. 4) publishes his
preliminary communications regarding the fossil Polycystina of Barbados and distinguishes 282
species, distributed in 44 genera and 7 families. In the tabular view of the genera he
distinguishes two orders:—I. Solitaria—(1) Halicalyptrina,
(2) Lithochytrina, (3) Eucyrtidina; and II. Composita—(4)
Spyridina, (5) Calodictya, (6) Haliommatina, (7) Lithocyclidina (compare L. N. 16, pp. 214-219).
1851. Huxley (L. N. 5) gives the first accurate
account of living Radiolaria, and describes 2 species of the genus Thalassicolla
(nucleata and punctata); under the latter are included 4 genera of Sphærozoea:—Collozoum, Sphærozoum, Collosphæra,
Siphonosphæra (compare L. N. 16, pp. 12-14).
1854. Ehrenberg (L. N. 6) publishes in his
Mikrogeologie, figures of seventy-two species of fossil Polycystina (without descriptions).
1855. Johannes Müller (L. N. 8, p. 248) describes
the first Acanthometra, and elucidates its affinity to Huxley's Thalassicolla and
Ehrenberg's Polycystina.
1858. Johannes Müller (L. N. 12) establishes the
new group Radiolaria as a special order of the Rhizopoda, and includes in it the Thalassicolla,
Polycystina, and Acanthometra as closely related families. He opposes these radiate Rhizopoda to
the Polythalamia, and describes 50 species observed by him living in the Mediterranean, these he
arranges in 20 genera, of which 10 are new. The figures are contained in eleven plates (see L.
N. 16, pp. 22-24).
1858. Claparède (L. N. 14) describes the first
Plectoidean (Plagiacantha arachnoides) and two species of
Acanthometra, which he had observed living in Norway (see L. N. 16,
p. 18).
1860. Ehrenberg (L. N. 4) gives a short diagnosis
of 22 new genera of Polycystina, based on the investigation of numerous deep-sea species brought
up by Brooke from the depths of the Pacific Ocean. The number of his genera is thus increased to
66 (compare L. N. 16, pp. 10, 11).
1862. Ernst Haeckel (L. N. 16)
embraces in his Monograph of the Radiolaria all the species hitherto known either by figures or
descriptions, and arranges them in 15 families and 113 genera; of which latter 46 are new. The
number of new species observed living amounts to 144. In a "survey of the Radiolarian fauna of
Messina" (p. 565) he records 72 genera and 169 species. Most of these are figured in the
accompanying atlas of thirty-five plates.
{clxxxvii}
1862. Bury (L. N. 17) gives in an atlas of
twenty-five plates, photographed from drawings, the figures of numerous fossil Polycystina of
Barbados (without descriptions), of which many are new species overlooked by Ehrenberg (compare
§ 242, above).
1872. Ehrenberg (L. N. 24) gives a list of names
(without description) of all the Polycystina observed by him from the bottom of the sea, 279
species, of which 127 are figured on twelve plates.
1875. Ehrenberg (L. N. 25) gives a list of names
of all the fossil Polycystina observed by him (from Barbados, the Nicobar Islands and Sicily),
326 species, of which 282 are figured (compare § 242 above). In a new
"Systematic Survey of the Genera" the number of these is given as 63. The 7 families are the
same as given in 1847 (see above), as also the two orders (Nassellaria =
Solitaria, Spumellaria = Composita).
1876. Zittel (L. N. 29) describes the first
fossil Radiolaria from the chalk (6 species) and establishes the new Cyrtoid genus
Dictyomitra.
1876. John Murray (L. N. 27) establishes the new
family Challengerida, and figures 6 new generic types of Phæodaria.
1879. Richard Hertwig (L. N. 33) first describes
the fundamental differences in the structure of the central capsule, and in accordance with them
divides the Radiolaria into six orders:—(1) Thalassicollea, (2) Sphærozoea, (3) Peripylea,
(4) Acanthometrea, (5) Monopylea, (6) Tripylea (p. 133). These are subdivided into 18 families,
and their phylogenetic affinities discussed (p. 137). On the ten plates, several new species
from Messina are figured, among them the types of several new genera (Cystidium,
Cœlacantha, Echinosphæra) (compare § 252).
1879. Ernst Haeckel (L. N. 34) founds the order
Phæodaria as a "new group of marine siliceous Rhizopods," and
distinguishes in it 4 suborders, 10 families and 38 genera.
1880. Emil Stöhr (L. N. 35) describes the Miocene
"Radiolarian fauna of the tripoli from Grotte in Sicily," 118 species, of which 78 are new;
among them is the new genus Ommatodiscus, the type of a new family, Ommatodiscida. The
new species are figured on seven plates.
1880. Dante Pantanelli (L. N. 36) describes 30
species of fossil Polycystina from the jaspers of Tuscany, which he regarded as Eocene, but
which were probably of Jurassic origin (compare § 243, note B,
above).
1881. Ernst Haeckel (L. N. 37) publishes a
"Sketch of a classification of the Radiolaria on the basis of the study of the Challenger
Collection," and distinguishes in his "conspectus ordinum" (p. 421) 2 subclasses and 7 orders,
and in the "prodromus systematis Radiolarium" (pp. 423-472) 24 families with 630 genera, among
which are more than 2000 new species.
1882. Bütschli (L. N. 40) on the basis of studies
of the fossil Monopylea of Barbados, investigates the "mutual relations of the Acanthodesmida,
Zygocyrtida and Cyrtida," and gives a critical revision of the genera of these "Cricoidea;" a
number of new species are described and figured (Tafs. xxxii., xxxiii.), and some new genera of
Stichocyrtida established (Lithostrobus, Lithomitra, &c.).
1882. Dunikowski (L. N. 44) describes 18 new
fossil Polycystina from the lower lias of the Salzburg Alps, among them the types of 3 new
genera (Ellipsoxiphus, Triactinosphæra, and Spongocyrtis).
{clxxxviii}
1883. Ernst Haeckel (L. N. 46) revises the 4
orders and 32 families of Radiolaria, and gives more accurate definitions of them, as well as of
the 2 subclasses (I. Holotrypasta = Acantharia and Spumellaria; II. Merotrypasta = Nassellaria and
Phæodaria).
1885. D. Rüst (L. N. 51) describes
234 new species of fossil Radiolaria from the Jura, and illustrates them by twenty plates. Among
them are 103 Spumellaria, 130 Nassellaria, and 1
Phæodaria; these are contained in 35 genera, of which 20 belong to the
Porulosa, and 15 to the Osculosa.
254. Statistical Synopsis of the Twenty Orders:—
| Legion. |
Sublegion. |
Order. |
 |
 |
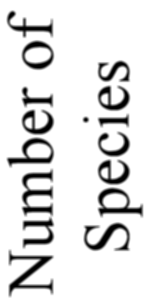 |
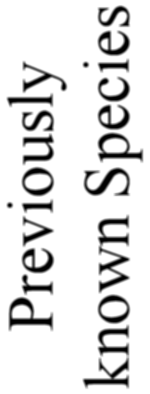 |
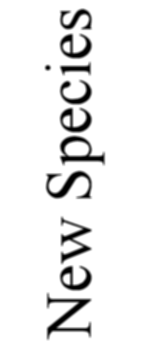 |
 |
 |
 |
Figured on
Plates. |
I. Legion
Spumellaria
(Porulosa peripylea) |
|
I. Collodaria
(Spumellaria palliata) |
|
01. Colloidea |
2 |
6 |
36 |
9 |
27 |
0 |
A |
E |
1,
3 |
| 02. Beloidea |
2 |
8 |
56 |
9 |
47 |
0 |
A |
D |
2,
4 |
II. Sphærellaria
(Spumellaria loricata) |
|
03. Sphæroidea |
6 |
107 |
660 |
105 |
555 |
66 |
A |
B |
|
5-8
11-30 |
| 04. Prunoidea |
7 |
53 |
280 |
35 |
245 |
36 |
B |
B |
|
16,
17
39, 40 |
| 05. Discoidea |
6 |
91 |
503 |
126 |
376 |
102 |
B |
A |
|
31-38
41-48 |
| 06. Larcoidea |
9 |
51 |
260 |
8 |
252 |
0 |
E |
B |
|
9,
10
49, 50 |
|
|
|
|
|
|
|
|
|
|
|
|
II. Legion
Acantharia
(Porulosa actipylea) |
|
III. Acanthometra
(Acantharia palliata) |
|
07. Actinelida |
3 |
6 |
22 |
6 |
16 |
0 |
E |
E |
129
(figs. 1-3) |
| 08. Acanthonida |
3 |
21 |
138 |
50 |
88 |
0 |
A |
C |
130-132 |
IV. Acanthophracta
(Acantharia loricata) |
|
09. Sphærophracta |
3 |
6 |
22 |
6 |
16 |
0 |
E |
E |
133-138 |
| 10. Prunophracta |
3 |
11 |
63 |
5 |
58 |
0 |
D |
B |
139,
140 |
III. Legion
Nassellaria
(Osculosa monopylea) |
|
V. Plectellaria
(Nassellaria palliata) |
|
11. Nassoidea |
1 |
2 |
5 |
1 |
4 |
0 |
E |
E |
91
(fig. 1) |
| 12. Plectoidea |
2 |
17 |
61 |
5 |
56 |
0 |
D |
C |
91
(figs. 2-12) |
| 13. Stephoidea |
4 |
40 |
205 |
14 |
191 |
17 |
C |
B |
|
81,
82
92-94 |
VI. Cyrtellaria
(Nassellaria loricata) |
|
14. Spyroidea |
4 |
45 |
239 |
51 |
188 |
53 |
C |
A |
83-90 |
| 15. Botryodea |
3 |
10 |
55 |
15 |
40 |
10 |
E |
C |
96 |
| 16. Cyrtoidea |
12 |
160 |
1122 |
328 |
794 |
250 |
C |
A |
51-80 |
|
|
|
|
|
|
|
|
|
|
|
|
IV. Legion
Phæodaria
(Osculosa cannopylea) |
|
VII. Phæocystina
(Phæodaria palliata) |
|
17. Phæocystina |
3 |
15 |
112 |
30 |
82 |
24 |
C |
B |
101-105 |
VIII. Phæocoscina
(Phæodaria loricata) |
|
18. Phæosphæria |
4 |
22 |
121 |
5 |
116 |
0 |
C |
A |
106-112 |
| 19. Phæogromia |
5 |
27 |
159 |
5 |
154 |
0 |
C |
A |
|
99,
100
113-120 |
| 20. Phæoconchia |
3 |
20 |
73 |
4 |
69 |
0 |
D |
B |
121-128 |
|
|
Total, |
85 |
739 |
4318 |
810 |
3508 |
558 |
... |
... |
140 |
| Legion. |
Sublegion. |
Order. |
I. Legion
Spumellaria
(Porulosa peripylea) |
|
I. Collodaria
(Spumellaria palliata) |
|
01. Colloidea |
| 02. Beloidea |
II. Sphærellaria
(Spumellaria loricata) |
|
03. Sphæroidea |
| 04. Prunoidea |
| 05. Discoidea |
| 06. Larcoidea |
II. Legion
Acantharia
(Porulosa actipylea) |
|
III. Acanthometra
(Acantharia palliata) |
|
07. Actinelida |
| 08. Acanthonida |
IV. Acanthophracta
(Acantharia loricata) |
|
09. Sphærophracta |
| 10. Prunophracta |
III. Legion
Nassellaria
(Osculosa monopylea) |
|
V. Plectellaria
(Nassellaria palliata) |
|
11. Nassoidea |
| 12. Plectoidea |
| 13. Stephoidea |
VI. Cyrtellaria
(Nassellaria loricata) |
|
14. Spyroidea |
| 15. Botryodea |
| 16. Cyrtoidea |
IV. Legion
Phæodaria
(Osculosa cannopylea) |
|
VII. Phæocystina
(Phæodaria palliata) |
|
17. Phæocystina |
VIII. Phæocoscina
(Phæodaria loricata) |
|
18. Phæosphæria |
| 19. Phæogromia |
| 20. Phæoconchia |
|
|
|
|
|
|
|
|
|
Figured on
Plates. |
| 01. |
2 |
6 |
36 |
9 |
27 |
0 |
A |
E |
1,
3 |
| 02. |
2 |
8 |
56 |
9 |
47 |
0 |
A |
D |
2,
4 |
| 03. |
6 |
107 |
660 |
105 |
555 |
66 |
A |
B |
5-8,
11-30 |
| 04. |
7 |
53 |
280 |
35 |
245 |
36 |
B |
B |
16,
17,
39, 40 |
| 05. |
6 |
91 |
503 |
126 |
376 |
102 |
B |
A |
31-38,
41-48 |
| 06. |
9 |
51 |
260 |
8 |
252 |
0 |
E |
B |
9,
10,
49, 50 |
|
|
|
|
|
|
|
|
|
|
| 07. |
3 |
6 |
22 |
6 |
16 |
0 |
E |
E |
129
(figs.
1-3) |
| 08. |
3 |
21 |
138 |
50 |
88 |
0 |
A |
C |
130-132 |
| 09. |
3 |
6 |
22 |
6 |
16 |
0 |
E |
E |
133-138 |
| 10. |
3 |
11 |
63 |
5 |
58 |
0 |
D |
B |
139,
140 |
| 11. |
1 |
2 |
5 |
1 |
4 |
0 |
E |
E |
91
(fig. 1) |
| 12. |
2 |
17 |
61 |
5 |
56 |
0 |
D |
C |
91
(figs.
2-12) |
| 13. |
4 |
40 |
205 |
14 |
191 |
17 |
C |
B |
81,
82,
92-94 |
| 14. |
4 |
45 |
239 |
51 |
188 |
53 |
C |
A |
83-90 |
| 15. |
3 |
10 |
55 |
15 |
40 |
10 |
E |
C |
96 |
| 16. |
12 |
160 |
1122 |
328 |
794 |
250 |
C |
A |
51-80 |
|
|
|
|
|
|
|
|
|
|
| 17. |
3 |
15 |
112 |
30 |
82 |
24 |
C |
B |
101-105 |
| 18. |
4 |
22 |
121 |
5 |
116 |
0 |
C |
A |
106-112 |
| 19. |
5 |
27 |
159 |
5 |
154 |
0 |
C |
A |
99,
100,
113-120 |
| 20. |
3 |
20 |
73 |
4 |
69 |
0 |
D |
B |
121-128 |
| T. |
85 |
739 |
4318 |
810 |
3508 |
558 |
... |
... |
140 |
Note.—In the tenth and eleventh columns the relative abundance of each
order at or near the surface and near the bottom is approximately indicated by the letters A-E,
which have the following significance:—A, abundant; B, very numerous; C, many (medium
quantity); D, few; E, very few.
{1}
SYSTEMATIC PART.
Class RADIOLARIA.
Radiolaria, Johannes Müller, 1858.
Rhizopoda radiaria, Johannes Müller, 1858.
Polycystina (pro parte), Ehrenberg, 1838.
Echinocystida, Claparède, 1858.
Rhizopoda capsularia, Haeckel, 1861.
Cytophora, Haeckel, 1862.
Definition of the Class:—Rhizopoda with unicellular
body, divided by a porous membrane into an internal or intracapsular part (with nucleus), and an
external or extracapsular part (with calymma); propagating by flagellated spores.
The Radiolaria or Capsulate Rhizopoda, first
constituted by Johannes Müller in the year 1858 as a separate group of the Rhizopoda, form a
peculiar class of the Protista, or unicellular organisms. This class is
exclusively marine, and has in general the characteristic organisation of the Rhizopoda, with the
development of numerous pseudopodia from the surface of the cell; but it differs from all
other Rhizopoda in the possession of a peculiar membrane, dividing the cell-body into two
different parts; the central capsule or the internal part with the nucleus, and the
external part or extracapsulum with the calymma; propagation by flagellated spores
produced in the central capsule; the sarcode or the protoplasm of both parts communicates
by fine pores, piercing the separating membrane, which is called the
capsule-membrane.
The Central Capsule or the inner part of the Radiolarian body is constantly composed of
three essential parts, viz.:—
1. The Central Nucleus (a true cell-nucleus).
2. The Intracapsular Sarcode (endosarc) or surrounding internal protoplasm.
3. The Capsule Membrane or enveloping porous membrane.
{2}
Besides these constant and essential elements, the central capsule contains very commonly (but
not constantly) some other enclosed structures, viz.:—
4. An internal or intracapsular skeleton.
5. Intracapsular vacuoles or alveoli.
6. Fat-granules or oil-globules.
7. Crystals of different composition.
8. Pigment-granules.
The Extracapsulum, or the outer part of the Radiolarian body is also constantly composed
of three essential elements,—
1. The Calymma, or the thick extracapsular jelly-veil, completely enveloping
the whole central capsule.
2. The Matrix, or the maternal tissue of the external protoplasm, enveloping
immediately the capsule-membrane as a thin continuous layer of extracapsular sarcode
(ectosarc).
3. The Pseudopodia, or the very numerous thread-like filaments of
protoplasm, which radiate from the matrix; whilst their inner part is enclosed in the calymma,
their outer part floats freely in the sea-water.
Besides these three constant and essential elements, the extracapsulum contains very commonly
(but not constantly) some other enclosed structures, viz.:—
4. An external or extracapsular skeleton.
5. Extracapsular vacuoles or alveoli.
6. Fat-granules or oil-globules.
7. Pigment-granules or a peculiar large body of dark extracapsular pigment, the
"phæodium."
8. "Xanthellæ" or "zooxanthellæ," peculiar yellow cells, which contain starch and
are unicellular yellow Algæ, living as "Symbiontes" in true Symbiosis with a great many
Radiolaria.
The Nucleus of the Radiolaria is a large true simple cell-nucleus, originally a solid
spherical, roundish or longish body of nuclein. It is placed either in the centre of the capsule
(in most Peripylea) or excentrically (in most other Radiolaria). Originally solid, the nucleus is
commonly differentiated later into an outer dense nuclear-membrane and an inner softer or fluid
content; either with one single nucleolus or with a variable number of nucleoli. Originally always
simple, the nucleus becomes afterwards constantly divided into numerous small nuclei, each of
which, together with a part of the surrounding {3}protoplasm, forms a vibratile-spore or "flagellate-spore." This division in
the Acantharia and in the social (or colonial) Peripylea begins very early, in all other
Radiolaria much later, immediately before propagation.
The Endoplasm or "endosarc," or "intracapsular protoplasm" or "inner sarcode," in all
Radiolaria originally fills that space within the capsule, which is not taken up by the nucleus.
It seems to be employed mainly for the purpose of propagation, becoming divided earlier or later
into numerous small particles, each of which surrounds a small particle of the nucleus and forms
together with it a flagellate-spore. Besides this the endoplasm of the Radiolaria seems to have a
great significance for the nutrition, mainly for the interchange of materials. It becomes very
often vacuolate or alveolate, filled with smaller or larger spherical drops of fluid; it produces
very commonly smaller fat-granules or larger oil-globules, and further pigment-granules of
different colours, more rarely crystals and other peculiar enclosed parts.
The Membrane or "capsule-membrane" is the most typical and characteristic part of the
body of a Radiolarian, sufficient of itself to separate this class from all other Rhizopoda. At
the same time, by its different shape it presents the best means for the systematic distinction of
the four subclasses or "legions" of the class. The membrane is composed of a special organic
matter (probably nearly related to chitin) and combines density with elasticity to a high degree.
Observed with a high power of the microscope its margin (or section) appears commonly
simple-edged, but often in larger forms distinctly double-edged.
The legion Phæodaria is distinguished by a double membrane (the thinner
inner and thicker outer membranes being separated by an interval); in the three other legions it
is simple. The membrane completely separates the intracapsular from the extracapsular body, both
communicating only by certain pores or openings in the membrane. With reference to this important
communication, the whole class can be divided into two subclasses, Holotrypasta and Merotrypasta:
the Holotrypasta contain the Peripylea and Actipylea, in which the
membrane is pierced by innumerable very small pores; the Merotrypasta
consist of the Monopylea and the Cannopylea, in which the membrane exhibits only one large main
opening, distinguished in the former by a peculiar "porous area," in the latter by an "osculum" or
a prolonged tubule.
The Calymma or "jelly-veil" is the most characteristic part of the extracapsular body in
all Radiolaria; in the majority of the class it is the most voluminous part of the whole body,
being much more voluminous than all the other parts taken together. The calymma is a
structureless, clear, and transparent jelly-envelope which always includes the whole central
capsule and often also the whole extracapsular skeleton. Owing to the high degree of its
consistence, this jelly-veil takes a very important part in the formation of the extracapsular
skeleton, furnishing the matrix for the deposition of its tangential parts.
{4}
The Matrix or the "maternal tissue of the pseudopodia" is formed in all Radiolaria by
the thin layer of exoplasm or of extracapsular sarcode, which immediately envelops the central
capsule and is itself enclosed by the calymma. This continuous sarcode-cover of the capsule
communicates by its pores or openings with the endoplasm or the intracapsular sarcode; whilst from
its outer surface arise the pseudopodia. The morphological signification of the matrix is very
small, but the physiological importance is very great, for it seems to be the chief organ of many
vital functions.
The Pseudopodia or the very fine, long, thread-like filaments of exoplasm arise in all
Radiolaria in very great numbers from the surface of the matrix, and exhibit in general the same
characteristic shape as in the other Rhizopoda. Their inner or proximal part is enclosed within
the jelly-veil or calymma, whilst their outer or distal part floats freely in the sea-water. Their
special motions and modifications exhibit considerable variations in different groups, their
tendency to ramify, anastomose, and form networks being in some cases very small, in others very
great. Also the characteristic motion of granules in the pseudopodia is very different. In general
those most important exoplasmatic filaments serve as organs both for the vegetative functions of
nutrition, and for the animal functions of motion and sensation.
The class Radiolaria can be divided according to its varying structure into
four different legions or subclasses, the characters of which are the following:—
I. PERIPYLEA or SPUMELLARIA.
Membrane of the central capsule simple, perforated by innumerable very fine
pores. Fundamental form originally homaxon or spherical. Skeleton wanting or siliceous. No
phæodium in the extracapsular calymma. The Peripylea comprise two orders:—
A. Collodaria (without lattice-shell).
B. Sphærellaria (with lattice-shell).
II. ACTIPYLEA or ACANTHARIA.
Membrane of the central capsule simple, perforated by innumerable fine pores.
Fundamental form originally homaxon or spherical. Skeleton acanthinic (not siliceous). No
phæodium in the extracapsular calymma. The Actipylea consist of two orders:—
A. Acanthometra (without complete lattice-shell).
B. Acanthophracta (with complete lattice-shell).
{5}
III. MONOPYLEA or NASSELLARIA.
Membrane of the central capsule simple, perforated by a porous-area, or by one
single large opening, divided into numerous very fine pores. Fundamental form originally monaxon
or egg-shaped. Skeleton siliceous. No phæodium in the extracapsular calymma. The Monopylea
comprise two orders:—
A. Plectellaria (without complete lattice-shell).
B. Cyrtellaria (with complete lattice-shell).
IV. CANNOPYLEA or PHÆODARIA.
Membrane of the central capsule double, perforated by one simple main-opening,
prolonged into a tubulus, and besides this commonly by one or two (rarely more) small accessory
openings. Fundamental form originally monaxon or egg-shaped. Skeleton siliceous. Constantly a
peculiar dark pigment-body or "phæodium" in the extracapsular calymma. The Cannopylea comprise
two orders:—
A. Phæocystina (without lattice-shell).
B. Phæocoscina (with lattice-shell).
Synopsis of the four Subclasses or Legions of Radiolaria.
| A. HOLOTRYPASTA. |
B. MEROTRYPASTA. |
| Central capsule everywhere perforated by innumerable small
pores. |
Central capsule with one large main-opening (with or without
small accessory openings). |
| Fundamental form originally homaxon (spherical or derived
from a sphere). |
Fundamental form originally monaxon (egg-shaped or perhaps
dipleural). |
| I. |
II. |
III. |
IV. |
| Spumellaria. |
Acantharia. |
Nassellaria. |
Phæodaria. |
| (Peripylea.) |
(Actipylea.) |
(Monopylea.) |
(Cannopylea.) |
| Wall-pores of the capsule equally disposed. |
Wall-pores of the capsule symmetrically disposed. |
Main-opening of the capsule with a porous operculum. |
Main-opening of the capsule with a short tubule. |
| Skeleton siliceous or wanting. |
Skeleton acanthinic (organic). |
Skeleton siliceous (rarely wanting). |
Skeleton siliceous (rarely wanting). |
| Calymma without phæodium. |
Calymma without phæodium. |
Calymma without phæodium. |
Calymma constantly with a phæodium. |
{6}
Legion I. SPUMELLARIA,
vel Peripylea, vel Peripylaria (Pls. 1-50).
Spumellaria (exclusis Spyridinis), Ehrenberg, 1875.
Peripylea (inclusis Thalassicollis et Sphærozois), Hertwig, 1879.
Peripylaria (inclusis Collodariis et Polycyttariis), Haeckel, 1881.
Definition.—Radiolaria with simple membrane of the central capsule, which is
everywhere perforated by innumerable very fine pores. Extracapsulum without phæodium. Skeleton
wanting or siliceous. Fundamental form originally spherical.
The legion Spumellaria vel Peripylea, in the
extent here defined, was constituted by me in 1883 in my paper on Die Ordnungen der Radiolarien.[2] I propose to retain for this legion either the name
Spumellaria of Ehrenberg (1875) or Peripylea of
Hertwig (1879), although both groups have not quite the same extension. We exclude from the Spumellaria the Spyridina (united with them by Ehrenberg) and include the
Collodaria. With the Peripylea of Hertwig we unite his Thalassicollea and Sphærozoea. To avoid any
confusion it would perhaps be better to name this legion "Peripylaria."
The Spumellaria agree with the Acantharia in
the structure of the simple capsule-membrane, which is perforated by numerous small pores (but
devoid of the large main opening, which the Nassellaria and Phæodaria possess), whence we unite both the former as Holotrypasta, both the
latter as Merotrypasta.
The difference between the two legions of Holotrypasta is determined by the skeleton, which in
the Spumellaria is either siliceous or wanting, whilst in the Acantharia
it consists of the peculiar organic substance, acanthin.
The legion Spumellaria is by far the largest and most important of the
four legions of Radiolaria, as well with respect to the number of different forms, as to the
enormous masses of individuals, which we encounter living and fossil. We distinguish in this
legion not less than thirty-two different families, three hundred and sixteen genera, and more
than seventeen hundred species.
The classification of this large group requires for its better comprehension a careful division
into larger and smaller groups. We divide it therefore first of all into two orders, Collodaria and Sphærellaria, as proposed in the paper
mentioned above.[3]
The Collodaria have no perfect latticed skeleton, and comprise two
suborders or sections: in the Colloidea the skeleton is entirely wanting,
in the Beloidea it is represented by a variable number of siliceous
needles or spicules, scattered in the calymma around the central capsule.
{7}
The Sphærellaria differ from the Collodaria
in the possession of a perfect siliceous skeleton, which is originally a latticed spherical shell,
enveloping the central capsule. By modification of this fenestrated sphere arises an enormous mass
of different forms, which we dispose in twenty-eight families, and these in four larger groups,
suborders or sections,—Sphæroidea, Prunoidea, Discoidea, and Larcoidea.
The Sphæroidea, the common ancestral group of the Sphærellaria, possess a skeleton which is either a simple fenestrated sphere,
or composed of two or more concentric latticed spheres, which are united by radial beams; more
rarely it becomes more or less spongy.
The Prunoidea are derived from the Sphæroidea
by prolongation of the latticed sphere in one axis; the skeleton therefore becomes here
ellipsoidal or cylindrical (often with annular transversal constrictions).
The Discoidea on the contrary must be derived from the Sphæroidea by shortening in one axis; here therefore the fenestrated shell
becomes more or less lenticular or iscoidal (often with radial spines or arms in the equatorial
plane, on the circular margin).
The Larcoidea, the fourth section, differ from the three
foregoing sections by the different growth of the shell in three different dimensions of space;
therefore here the fenestrated shell becomes "lentelliptical," or a "triaxial ellipsoid," its
length, breadth, and height being different.
The Skeleton consists in all Spumellaria either of pure
silica or of a peculiar silicate. The siliceous bars and beams constituting it are
invariably solid, as also in the Nassellaria, never hollow, as in
the Phæodaria. Never is the skeleton composed of acanthin, as in all Acantharia. Whilst in the first order of Spumellaria,
Collodaria, the form of the spicula, or the scattered needles, composing
the skeleton, is very simple, never latticed, in the second order, the Sphærellaria, it is constantly latticed or fenestrated, often also spongy.
The geometrical fundamental form of the lattice-shell in the Sphærellaria is originally spherical (homaxon), as preserved in all Sphæroidea; in the Prunoidea and Discoidea it becomes monaxon, with one single axis (prolonged in the former,
shortened in the latter); in the Larcoidea it becomes triaxon, by
different growth in three principal axes, perpendicular one to another. The further development of
radial parts of the skeleton in these three axes is very important for the "promorphology" of the
Radiolaria.
The Malacoma, or the whole soft body of the Spumellaria as
opposed to the skeleton, exhibits some differences of structure in two different groups, which
were separated formerly (1862) as Monocyttaria and Polycyttaria, corresponding to the "Radiolaria
monozoa and polyzoa" of Johannes Müller (1858).
The Monocyttaria (or the Spumellaria solitaria) live isolated as
single cells—like {8}all other Radiolaria—and
are never aggregated in colonies; the calymma includes one single central capsule, and this again
one central nucleus, which does not become divided until full maturity.
The Polycyttaria on the contrary (or the Spumellaria socialia) live
aggregated in large colonies; the calymma includes a variable number of associated central
capsules and each of these commonly one central oil-globule, whilst the original simple nucleus
commonly becomes very early divided into numerous small nuclei.
The Nucleus of the Spumellaria is originally constantly
central, placed quite in the centre of the concentric capsule, and it retains this central
position in all Monocyttaria or solitary Peripylea; whereas in the Polycyttaria—in
consequence of its early division—its place is commonly taken by a central oil-globule.
Whilst the numerous nuclei of the latter are very small, the single nucleus of the former is
comparatively large, extremely large (more than a millimeter in diameter) in some gigantic Collodaria.
The Endoplasm or the intracapsular sarcode exhibits in the greater number of Spumellaria a more or less distinct radial striation. It encloses a great
variety of different parts; vacuoles, oil-globules, pigment-granules, crystals, &c.
The Membrane of the capsule in all Spumellaria is simple (never
double as in the Phæodaria) and everywhere equally perforated by innumerable small pores;
in the thick, double-edged membrane of many large Collodaria these pores
appear (in the optical section of the capsule-wall) as distinct fine radial canals, very densely
and regularly disposed.
The Central Capsule in the Spumellaria is originally a
geometrical sphere, and this simple globular form is preserved in all Sphæroidea, and in the greatest part of Colloidea and
Beloidea. By prolongation of one axis the form becomes ellipsoidal
(or even cylindrical) in the Prunoidea, and in some few forms of Colloidea. By shortening of one axis it becomes lenticular (or even
discoidal) in the Discoidea, and in some few forms of Colloidea.
By unequal growth in three different axes, perpendicular one to another, the capsule becomes
lentelliptical in all Larcoidea. Very rarely the capsule assumes
in the Spumellaria a polyhedral or irregular (sometimes even
amœboid) form, only in a few Colloidea.
The Calymma, or the jelly-veil including the central capsule, is very voluminous in many
Spumellaria of gigantic size, mainly in the large Colloidea, and in all Polycyttaria or social Radiolaria. It includes here a
considerable number of large vacuoles or "alveoli." The calymma never exhibits in this legion the
dark voluminous phæodium, possessed by all Phæodaria.
Xanthellæ or "zooxanthellæ" are numerous in the calymma of most Spumellaria, but by no means constant; they are very variable in number and
size.
The Matrix, placed between the calymma and central capsule, is, in the majority of the
Spumellaria, a rather thick layer of granular exoplasm.
The Pseudopodia arising from it are very numerous, equally disposed over the whole {9}surface, and are in general rather fluid, exhibiting a
considerable tendency to ramify, anastomose, and form networks. The movement of granules is
commonly lively. In the Polycyttaria all capsules of one colony or "cœnobium" are connected
by the dense variable network of anastomosing pseudopodia.
Synopsis of the Orders and Suborders of Spumellaria.
|
I. COLLODARIA.
Skeleton wanting or quite imperfect, not latticed.
|
|
Skeleton entirely wanting, |
1. Colloidea. |
| Skeleton represented by numerous scattered spicules, |
2. Beloidea. |
| |
|
II. SPHÆRELLARIA.
Skeleton a perfect shell of lattice work, or spongy and resembling
wicker-work.
|
|
Lattice-shell spherical or composed of concentric spheres, |
3. Sphæroidea. |
| Lattice-shell ellipsoidal or prolonged in one axis, |
4. Prunoidea. |
| Lattice-shell discoidal or shortened in one axis, |
5. Discoidea. |
| Lattice-shell lentelliptical, with different extent of growth in three
axes, |
6. Larcoidea. |
| I. COLLODARIA. Skeleton wanting or quite imperfect, not latticed. |
|
|
Skeleton entirely wanting, |
|
|
|
|
|
|
1. Colloidea. |
|
|
|
|
|
|
Skeleton represented by numerous scattered spicules, |
|
|
|
|
|
|
2. Beloidea. |
|
|
|
|
| II. SPHÆRELLARIA. Skeleton a perfect shell of lattice work, or spongy and
resembling wicker-work. |
|
|
Lattice-shell spherical or composed of concentric spheres, |
|
|
|
|
|
|
3. Sphæroidea. |
|
|
|
|
|
|
Lattice-shell ellipsoidal or prolonged in one axis, |
|
|
|
|
|
|
4. Prunoidea. |
|
|
|
|
|
|
Lattice-shell discoidal or shortened in one axis, |
|
|
|
|
|
|
5. Discoidea. |
|
|
|
|
|
|
Lattice-shell lentelliptical, with different extent of growth in
three axes, |
|
|
|
|
|
|
6. Larcoidea. |
|
|
|
|
Order I. COLLODARIA, Haeckel, 1881.
Collodaria, Haeckel, Prodromus, 1881, p. 469.
Collida et Sphærozoida, Haeckel, 1862, Monogr. d. Radiol., pp. 246, 522.
Definition.—Spumellaria without latticed shell.
The order Collodaria, the first order of Radiolaria, comprises all
those Spumellaria in which the skeleton is either entirely wanting, or
represented by numerous single, solid, siliceous needles or spicules, loosely scattered in the
calymma around the central capsule. Never in this order is there any trace of the latticed or
fenestrated shell, which characterises the second order, Sphærellaria.
The skeleton exhibits no trace of phylogenetic connection in the two orders.
In my monograph (1862) two families appertaining to this order are described, the Collida (p.
244) and the Sphærozoida (p. 521). Both families
contain forms with and without a skeleton. Of the solitary or monozous Collida the Thalassicollida
are devoid of a skeleton, whilst the Thalassosphærida are provided with a skeleton. Of the social
or polyzous Sphærozoida the Collozoida are without a skeleton, the Rhaphidozoida provided with
one. As the special form in both skeletophorous subfamilies is exactly the same, I prefer now to
associate them in the suborder Beloidea, and to oppose them to the other
two skeletonless subfamilies, which are united under the name of Colloidea.
{10}
Synopsis of the four Families of Collodaria.
|
Suborder I. COLLOIDEA.
Skeleton entirely wanting.
|
|
Solitary cells, living as isolated individuals (Colloidea
monozoa), |
1. Thalassicollida. |
| Associated cells, living in colonies or cœnobia (Colloidea
polyzoa), |
2. Collozoida. |
|
Suborder II. BELOIDEA.
Skeleton composed of numerous needles or spicula,
scattered in the calymma.
|
|
Solitary cells, living as isolated individuals (Beloidea
monozoa), |
3. Thalassosphærida. |
| Associated cells, living in colonies or cœnobia (Beloidea
polyzoa), |
4. Sphærozoida. |
| Skeleton entirely wanting. |
|
|
Solitary cells, living as isolated individuals (Colloidea
monozoa), |
|
|
|
|
|
|
1. Thalassicollida. |
|
|
|
|
|
|
Associated cells, living in colonies or cœnobia
(Colloidea polyzoa), |
|
|
|
|
|
|
2. Collozoida. |
|
|
|
|
| Suborder II. BELOIDEA. Skeleton composed of numerous needles or spicula,
scattered in the calymma. |
|
|
Solitary cells, living as isolated individuals (Beloidea
monozoa), |
|
|
|
|
|
|
3. Thalassosphærida. |
|
|
|
|
|
|
Associated cells, living in colonies or cœnobia
(Beloidea polyzoa), |
|
|
|
|
|
|
4. Sphærozoida. |
|
|
|
|
Suborder I. COLLOIDEA, Haeckel.
Definition.—Spumellaria without skeleton.
The suborder Colloidea comprises all those Spumellaria in which no skeleton is developed. The whole body is therefore
soft—a true malacoma—and is composed only of two essential parts, the central capsule
and the enveloping extracapsulum. The suborder contains only two different families, the solitary
Thalassicollida (or Colloidea monozoa) and the associated Collozoida (or Colloidea polyzoa). Both families are very nearly allied, and
differ only in one single essential character: the solitary life of the former, the social union
of the latter. It seems to be merely in consequence of this difference that the cleavage of the
nucleus commonly takes place very late in the former, very early in the latter.
Therefore the full-grown Thalassicollida (till immediately before propagation) commonly exhibit
one single nucleus in the centre of the capsule, whilst in the Collozoida the capsule is distended
by numerous small nuclei. In these latter the centre of the capsule usually contains one large
oil-globule, whilst in the former oil-globules are either wanting, or scattered in large numbers
in the endoplasm, or disposed in one layer on the inside of the capsule membrane.
In the solitary Thalassicollida each capsule is enclosed in its own peculiar
spherical calymma, whilst in the associated Collozoida all capsules of the colony are united in
one common, very voluminous calymma.
Family I. Thalassicollida, Haeckel,
1862.
Thalassicollida, Haeckel, 1862, Monogr. d. Radiol., p. 246.
Thalassicollida, Haeckel, 1881, Prodromus, p. 469.
Definition.—Colloidea solitaria.
The family Thalassicollida comprises all solitary Spumellaria without a
skeleton. The oldest and best known form of this family is the genus Thalassicolla, as
restricted by {11}Johannes Müller.[4] The most common representative of it, the cosmopolitan
Thalassicolla nucleata, was first described by Huxley in 1851. But as early as 1834 another
large Radiolarian, appertaining either to this or to a nearly allied family, had been described by
Meyen as Physematium atlanticum. A third genus was detected by me in 1859 at Messina and
figured under the name Thalassolampe margarodes.[5] A very accurate histological description of these forms was given
in 1876 by Richard Hertwig.[6] The same author figured
in his Organismus in 1879 a very interesting simpler form under the name Thalassolampe
primordialis (Taf. iii. fig. 5). Some similar forms had already been observed by me, and are
here united with it to form the first genus Actissa.[7]
Actissa is of the highest general interest as the most simple and typical form of all
Radiolaria, and as the common ancestral form, from which all other forms of this large class may
be derived. Its unicellular body exhibits neither the extracapsular alveoli of
Thalassicolla, nor the intracapsular alveoli of Thalassolampe, and shows all
essential characters of the Radiolarian type in its most simple form (Pl. 1, figs. 1 to
1c). The simple cell-body is composed of a spherical central capsule and a concentric,
spherical, enveloping calymma, both separated by a thin membrane which is perforated by
innumerable pores. The capsule includes the endoplasm and in the centre a simple spherical nucleus
with nucleolus; at the time of propagation this latter becomes cleft into numerous small nuclei,
each of which, together with a small piece of the surrounding endoplasm, forms a flagellated
zoospore (fig. 1c). The extracapsulum is formed by the large, structureless, spherical
calymma or concentric jelly-veil enveloping the capsule, and by the thin granular matrix or the
layer of exoplasm which separates the calymma from the membrane. From this matrix or maternal
tissue arise innumerable very long and thin pseudopodia, as simple radiating filaments, the
proximal part of which is included in the calymma, whilst the distal part floats freely in the
sea-water (Pl. 1,
fig. 1).
The other Thalassicollida differ from their common ancestral form, Actissa, mainly by
the higher histological differentiation of the unicellular body. Whilst in Thalassicolla
and Thalassolampe the nucleus remains a single sphere as in Actissa, it becomes
branched or covered with radial blind saccules in Thalassopila and Thalassophysa;
also the intracapsular protoplasm develops here a great variety of peculiar different corpuscles,
as oil-globules, pigment-granules, concentric concretions, crystals, &c. But the most striking
peculiarity by which the other Thalassicollida differ from Actissa is the development of
large vesicular alveoli, either within or without the capsule; the unicellular body reaches by
this inflation the extraordinary size of 5 to 10 mm. or more.
{12}
Synopsis of the Genera of Thalassicollida.
| A. Alveoli neither within nor without the central capsule. |
|
Nucleus spherical (sometimes ellipsoidal), not branched, |
1. Actissa. |
| B. Numerous large alveoli within the central capsule (not
in the calymma). |
|
Nucleus spherical, |
2. Thalassolampe. |
| Nucleus branched or covered with radial sacs, |
3. Thalassopila. |
| C. Numerous large alveoli without the central capsule,
within the jelly-veil or calymma. |
|
Nucleus spherical, |
4. Thalassicolla. |
| Nucleus branched or covered with radial sacs, |
5. Thalassophysa. |
| A. Alveoli neither within nor without the central capsule. |
|
|
Nucleus spherical (sometimes ellipsoidal), not branched, |
|
|
|
|
|
|
1. Actissa. |
|
|
|
|
| B. Numerous large alveoli within the central capsule (not in the
calymma). |
|
|
Nucleus spherical, |
|
|
|
|
|
|
2. Thalassolampe. |
|
|
|
|
|
|
Nucleus branched or covered with radial sacs, |
|
|
|
|
|
|
3. Thalassopila. |
|
|
|
|
| C. Numerous large alveoli without the central capsule, within the jelly-veil
or calymma. |
|
|
Nucleus spherical, |
|
|
|
|
|
|
4. Thalassicolla. |
|
|
|
|
|
|
Nucleus branched, or covered with radial sacs, |
|
|
|
|
|
|
5. Thalassophysa. |
|
|
|
|
Genus 1. Actissa,[8] n. gen.
Definition.—Thalassicollida with simple spherical
nucleus, without any alveoli (either within or outside the central capsule).
The genus Actissa is the most simple and typical form of all Radiolaria, and
may be regarded as the common ancestral form of the whole class. The spherical body is composed of
a simple spherical capsule and a concentric spherical calymma or jelly-envelope. Neither the
former nor the latter contains alveoli. The central capsule possesses a strong membrane perforated
by small pores, and contains in the intracapsular sarcode numerous small pellucid vacuoles, and in
its middle a simple, concentric, spherical nucleus (often with some nucleoli); sometimes also one
or more oil-globules. The extracapsularium forms a soft, voluminous, structureless calymma or
enveloping jelly-sphere, perforated by the numberless, fine pseudopodia, which radiate outwards
from the matrix or the thin granulated sarcode-layer, surrounding the capsule. Often (but not
constantly) xanthellæ or yellow cells are scattered in it. Actissa differs from the
following skeletonless genera in the absence of all alveoli; it has neither intracapsular alveoli
(like Thalassolampe) nor extracapsular alveoli (like Thalassicolla). The first
observed species of this genus is that which I found in 1866 at the Canary Islands, Actissa
prototypus; the second is that which Hertwig accurately described in 1879, from the
Mediterranean (Messina), Actissa primordialis; the third I observed in 1881 at Ceylon,
frequent and sporiparous, Actissa princeps. A fourth species (Actissa radiata)
exhibited a distinct radial segmentation of the capsule-contents. These four species are quite
spherical. Six other species, occurring in different preparations from the Challenger, are
distinguished by modifications of the spherical capsule-form and may represent three different
subgenera (or, perhaps better, genera?)—Actiprunum ellipsoidal, Actidiscus
lenticular, Actilarcus lentelliptical; perhaps these are the ancestral forms of the three
sections: Prunoidea, Discoidea, Larcoidea.
{13}
Subgenus 1. Procyttarium, Haeckel, 1879.
Procyttarium, Haeckel, Natürl. Schöpfungsgeschichte, ed. vii. p. 705.
Definition.—Central capsule spherical.
1. Actissa princeps, n. sp. (Pl. 1,
fig. 1).
Central capsule spherical, colourless or a little reddish, transparent, with a thick
double-edged membrane. Nucleus central, spherical, one-third as broad as the central capsule,
containing a single, central, glossy nucleolus. Protoplasm finely granulated, without
oil-globules, with numerous clear spherical vacuoles of equal size and at equal distances; the
superficial layer of protoplasm (immediately below the membrane) radially striated (fig. 1). In
some older specimens the nucleus was divided into numerous small nuclei (fig. 1a), which by
further division gave the nuclei of the flagellated spores (fig. 1b); each spore with a
very thin lateral flagellum (fig. 1c). Jelly-like calymma twice as broad as the enclosed
capsule, without xanthellæ or yellow cells, pierced by innumerable, very thin and long, undivided
pseudopodia, which arise from the sarcode-matrix on the outside of the membrane (six to eight
times longer than shown in fig. 1).
Dimensions.—Diameter of the central capsule 0.1 to 0.12, of the nucleus 0.03 to
0.04, of the jelly calymma 0.2 to 0.3.
Habitat.—Indian Ocean, Ceylon, Belligemma, Haeckel, 1881; also in a
preparation from Station 271, Central Pacific, surface.
2. Actissa primordialis, Haeckel.
Thalassolampe primordialis, R. Hertwig, 1879, Organismus der Radiolarien, p. 32, Taf.
iii. fig. 5.
Procyttarium primordiale, Haeckel, 1879, Natürl. Schöpf., ed. vii. p. 705,
Taf. xvi. fig. 1.
Central capsule spherical, dim-yellowish, with a thin, simple-edged but compact membrane.
Nucleus large, central (about half as broad), with one or two dark nucleoli; on its side an
excentric oil-globule, nearly of the same size. Protoplasm between nucleus and membrane, in the
younger specimens finely granulated and radially striped; in the older specimens with numerous
hyaline globules (vacuoles). Jelly-envelope or calymma very voluminous, ten to twelve times as
broad as the central capsule, structureless, containing numerous yellow bodies (xanthellæ?),
pierced by very numerous simple pseudopodia.
Dimensions.—Diameter of the central capsule 0.11 to 0.18, of the nucleus 0.04 to
0.09, of the jelly-like calymma 1.2 to 1.5.
Habitat.—Mediterranean, Messina, Hertwig, 1878, surface.
3. Actissa prototypus, n. sp.
Central capsule spherical, red-coloured, with a thick, double-edged membrane. Nucleus central,
spherical, half as large as the radius of the capsule, containing a large number (forty to sixty)
of small {14}dark nucleoli. Protoplasm filled up with
numerous small clear vacuoles, and between them fine red pigment granules; on the inside of the
membrane one layer of dark oil-globules. Jelly-like calymma four times as broad as the enclosed
capsule, with very numerous small xanthellæ.
Dimensions.—Diameter of the capsule 0.2, of the nucleus 0.05, of the calymma
0.8.
Habitat.—Atlantic, Canary Islands (Lanzerote, Haeckel), 1866; also at
Station 348, surface.
4. Actissa radiata, n. sp.
Central capsule spherical, dark, with a thick, double-edged membrane. Nucleus central,
spherical, half as large as the capsule, transparent. Protoplasm divided into numerous cuneiform
radial pieces which are separated by clear intervals, and filled with very fine dark granules
(darker in the distal half). The equatorial optical section exhibits around the circular clear
nucleus a coronal of twenty-five such wedge-shaped pieces (mother-cells of spores?) No
oil-globules in the central capsule. Jelly-like calymma one and a half times as broad as the
enclosed capsule, with numerous small xanthellæ.
Dimensions.—Diameter of the capsule 0.15, of the nucleus 0.07, of the calymma
0.24.
Habitat.—North Pacific, Station 241, surface.
Subgenus 2. Actiprunum, Haeckel, 1882.
Definition.—Central capsule ellipsoidal, with one prolonged axis.
5. Actissa prunoides, n. sp.
Actiprunum prunoideum, Haeckel, 1882, Manuscript.
Central capsule ellipsoidal, colourless, with a thin, simple-edged membrane. Proportion of its
major axis to the minor 4 : 3. Nucleus spherical, its diameter equal to one-third of the
minor axis, in its centre a large, dark, spherical nucleolus. Protoplasm clear, containing
numerous small vacuoles, separated by regular distances, and on the inside of the
capsule-membrane, numerous (forty to fifty) small oil-globules. Calymma (or jelly-veil)
ellipsoidal, with a thin sarcode-stratum on the outside of the capsule; its diameter twice as
large as that of the central capsule.
Dimensions.—Major axis of the capsule 0.16, minor 0.12; diameter of the nucleus
0.04; major axis of the calymma 0.32, minor 0.24.
Habitat.—Central Pacific, Station 274, surface.
6. Actissa ellipsoides, n. sp.
Actiprunum ellipsoides, Haeckel, 1882, Manuscript.
Central capsule ellipsoidal, red-coloured, with a thick, double-edged membrane. Proportion of
its major axis to the minor 5 : 3. Nucleus ellipsoidal, one-third as large as the
capsule, containing eight small dark nucleoli. Protoplasm dusky, filled with dark pink
pigment-granules; in the {15}major axis, on both poles of
the nucleus-axis, two large oil-globules, half as large as the nucleus. Calymma ellipsoidal, with
numerous xanthellæ; its diameter four times as large as that of the capsule.
Dimensions.—Major axis of the capsule 0.2, minor 0.12; major axis of the nucleus
0.07, minor 0.04; major axis of the calymma 0.8, minor 0.5.
Habitat.—Mediterranean, Corfu, 1877, Haeckel, surface.
Subgenus 3. Actidiscus, Haeckel, 1882.
Definition.—Central capsule lenticular, with one shortened axis.
7. Actissa discoides, n. sp.
Actidiscus discoides, Haeckel, 1882, Manuscript.
Central capsule lenticular, red-coloured, about twice as broad as high, with a thick,
double-edged membrane. Nucleus spherical, one-third as broad as the capsule, with one single,
large central nucleolus. Protoplasm dusky, filled with scarlet pigment; granules and small
oil-globules between them. Calymma lenticular, three times as broad as the capsule.
Dimensions.—Major axis of the capsule 0.16, minor 0.08; diameter of the nucleus
0.05; breadth of the calymma 0.5.
Habitat.—North Pacific, Station 236, surface.
8. Actissa lenticularis, n. sp.
Central capsule lenticular, flattened, about three times as broad as high, with a thin,
simple-edged membrane. Nucleus lenticular, one-third as large as the capsule, with ten small dark
nucleoli. Protoplasm transparent, colourless, filled with small vacuoles at regular distances; on
the inside of the membrane in the circular periphery of the lens twenty dark oil-globules. Calymma
lenticular, twice as broad as the capsule.
Dimensions.—Major axis of the capsule 0.15, minor 0.05; breadth of the nucleus
0.05, height 0.02; breadth of the calymma 0.03.
Habitat.—East Pacific, Station 272, surface.
9. Actissa phacoides, n. sp.
Actidiscus phacoides, Haeckel, 1882, Manuscript.
Central capsule lenticular, strongly flattened, about four times as broad as high, with a thin,
simple-edged membrane. Nucleus lenticular, one-fourth as broad as the capsule, with numerous
(twenty or more) small nucleoli. Protoplasm filled with dark pigment-granules; on the inside of
the membrane in the circular periphery of the lens thirty-two dark oil-globules. Calymma
lenticular, three times as broad as the capsule.
{16}
Dimensions.—Major axis of the capsule 0.2, minor 0.05; breadth of the nucleus
0.05, height 0.015; breadth of the calymma 0.6.
Habitat.—Tropical Atlantic, Station 347, surface.
Subgenus 4. Actilarcus, Haeckel.
Definition.—Central capsule lentelliptical, with three different
axes.
10. Actissa larcoides, n. sp.
Central capsule lentelliptical; with three different axes, bearing the proportion
4 : 3 : 2. Nucleus spherical; its diameter equal to the shortest radius of the
capsule. No nucleoli visible. Protoplasm transparent, with small vacuoles; on the inside of the
thin capsule-membrane numerous (fifty to sixty) small oil-globules, regularly disposed. Calymma
lentelliptical, twice as large as the central capsule.
Dimensions.—Major axis or length of the capsule 0.2, middle axis or breadth 0.15,
minor axis or height 0.1; diameter of the nucleus 0.05, of the calymma 0.3-0.4.
Habitat.—Central Pacific, Station 266, surface.
Genus 2. Thalassolampe,[9] Haeckel, 1862,
Monogr. d. Radiol., p. 253.
Definition.—Thalassicollida without extracapsular
alveoles, but with large roundish or globular alveoles within the central capsule, with a simple
spherical, not branched nucleus in the centre.
The genus Thalassolampe is, next to Actissa, the most simple of all
Radiolaria, but differs from it by the large intracapsular alveoles. By these the central capsule
is inflated to an extraordinary size, which in Thalassolampe maxima exceeds that of most
other Radiolaria. From the nearly allied Thalassopila it differs by the simple spherical
nucleus, from Physematium by the absence of spicula. Of the two species of the genus the
first observed Thalassolampe margarodes, 1862, is Mediterranean, the second,
Thalassolampe maxima, 1882, is Indian.
1. Thalassolampe margarodes, Haeckel.
Thalassolampe margarodes, Haeckel, 1862, Monogr. d. Radiol., p. 253, Taf. ii. figs. 4,
5.
Thalassolampe margarodes, R. Hertwig, 1876, Histologie d. Radiol., p. 68,
Taf. iii. figs. 1-5.
Spherical body very soft and limpid, somewhat pearl-like opalescent, yellowish or bluish.
Central capsule with a very thin structureless membrane, its diameter six to eight times as large
as that of the central spherical nucleus. Wall of the vesicular nucleus thick, perforated by fine
{17}pore-canals; on its inside often numerous oval
nucleoli. In the movable protoplasmic network between the large alveoles a considerable number of
large yellowish or orange oil-globules. Extracapsular jelly-envelope very thin, contains small
yellow bodies (zooxanthellæ). (Compare the accurate description of this Mediterranean species in
my monograph and in Hertwig's work.) In the Canary Islands I found very often a large variety of
it, of double and triple the size, distinguished by the delicate orange colour of the
intracapsular oil-globules. This may be distinguished as Thalassolampe aurantiaca.
Dimensions.—Diameter of the whole jelly-sphere 2 to 4 mm., of the central capsule
2 to 3 mm., of its nucleus 0.2 to 0.4 mm.
Habitat.—Mediterranean, Messina, Haeckel, Hertwig; Canary Islands,
Lanzerote, Haeckel; surface.
2. Thalassolampe maxima, n. sp. (Pl. 1,
fig. 2).
Spherical body quite pellucid, like a glass globule, colourless. Central capsule with a
moderately thick, but quite transparent, structureless membrane, its diameter ten to twelve times
as large as that of the central spherical nucleus. Wall of the vesicular nucleus thick, perforated
by fine pore-canals; on its inside numerous small spherical nucleoli. No large oil-globules in the
movable protoplasmic network between the large alveoles. Extracapsular jelly-envelope very thin,
containing no yellow bodies. This differs from the preceding nearly allied species in the want of
the intracapsular oil-globules and of the extracapsular yellow bodies. It possesses the largest
central capsule of all known Radiolaria. I found them living and floating in water taken from the
surface of the Indian Ocean by a bucket.
Dimensions.—Diameter of the whole jelly-body 12 to 15 mm., of the central capsule
10 to 12 mm., of the nucleus 0.8 to 1.2 mm.
Habitat.—Indian Ocean, near the Maldive Islands, Haeckel, 1882,
surface.
Genus 3. Thalassopila,[10] Haeckel, 1881,
Prodromus, p. 469.
Definition.—Thalassicollida without extracapsular alveoles, but with large
roundish or globular alveoles within the central capsule, with a papillate or branched nucleus in
its centre.
The genus Thalassopila has, like Thalassolampe, a voluminous foamy
central capsule, inflated by numerous large alveoles; but it differs in the complicated form of
the nucleus, which is like that of Thalassophysa, and is either branched or occupied by
conical or roundish papillæ.
1. Thalassopila cladococcus, n. sp. (Pl. 1, fig. 3).
Spherical body dark-spotted, with a thin yellowish jelly-envelope. Central capsule with a thick
and firm membrane, perforated by pores; its diameter three times that of the central nucleus,
{18}three-fourths that of the whole jelly-sphere. Nucleus
profusely branched or papillated, its spherical surface covered with numerous (more than a
hundred) finger-shaped obtuse blind sacs, about as long as its radius. Protoplasm of the central
capsule forming a loose network between the large roundish alveoles, in the cortical zone radially
striped and containing one layer of large dark oil-globules. These are regularly distributed on
the inside of the capsule-membrane and separated by intervals, twice as broad as its diameter,
giving to the capsule-surface a spotted appearance. Extracapsular jelly-envelope thin, yellowish,
with very numerous and small xanthellæ.
Dimensions.—Diameter of the whole jelly-sphere 5 mm., of the central capsule 4
mm., of the nucleus 1.3 mm.
Habitat.—Antarctic Ocean, Station 154 (south of Kerguelen),
surface.
Genus 4. Thalassicolla,[11] Huxley, 1851,
Ann. and Mag. Nat. Hist., ser. 2, vol. viii. p. 433.
Definition.—Thalassicollida without intracapsular alveoles, but with large
roundish or globular alveoles within the extracapsular calymma. Nucleus in the centre of the
capsule simple spherical, not branched.
The genus Thalassicolla was proposed by Huxley in 1851, for a certain number
of different voluminous jelly-like Radiolaria, which he had observed living during his voyage in
the "Rattlesnake" through the tropical seas, and of which he gives an excellent
description—the first accurate observations on living Radiolaria. Johannes Müller afterwards
removed from this genus the social genera Sphærozoum and Collosphæra (formerly
Thalassicolla punctata), and retained as type of the genus Thalassicolla nucleata.
In 1862 in my Monograph I added two other species, Thalassicolla pelagica and
Thalassicolla zanclea, and later (1870) Thalassicolla sanguinolenta. Now I think it
better to separate the last two species as a new genus, Thalassophysa, characterised by the
papillate or branched nucleus, and to retain in Thalassicolla only those forms with simple
spherical nucleus. For both genera the extracapsular, voluminous, spherical calymma or
jelly-envelope, with numerous large alveoles, is characteristic. The membrane of the central
capsule in Thalassicolla is now structureless (subgenus Thalassicollarium, with
three species), now characterised by a peculiar structure, prominent ridges on the inside of the
membrane, which form a network with polygonal plates, resembling an epithelium (Pl. 1, fig.
5b; subgenus Thalassicollidium, with four species). Of the seven species here
described, two are cosmopolitan, widely distributed, and common; one is Mediterranean, one
Atlantic, and three Pacific.
Subgenus 1. Thalassicollarium, Haeckel.
Definition.—Membrane of the central capsule structureless, only
perforated by innumerable very small radial pores.
{19}
1. Thalassicolla pellucida, n. sp.
Spherical body very soft, transparent, clear and colourless, without any pigment. Central
capsule soft, hyaline, with a thin, structureless, not areolated membrane. Diameter of the central
capsule about three times that of the nucleus, one-fourth to one-sixth that of the jelly-envelope.
Nucleus delicate, transparent, with one single central nucleolus, about one-third its diameter.
Protoplasm of the central capsule contains only small, pellucid, densely packed globules
(vacuoles?), no oil-globules. Extracapsular body quite transparent, without pigment or
oil-globules, only composed of the large alveoles imbedded in the jelly-cover, and of the fine
protoplasmic network between them. No xanthellæ.
Dimensions.—Diameter of the central capsule 0.8 to 1.2, of the nucleus 0.3 to 0.4,
of the calymma 3 to 6 mm.
Habitat.—Cosmopolitan, Canary Islands, Haeckel; Cape, Australia,
Pacific, Challenger; surface.
2. Thalassicolla spumida, n. sp.
Spherical body nearly transparent, yellowish, without dark pigment. Central capsule pellucid,
with a thick, structureless, not areolated membrane. Diameter of the central capsule about twice
that of the nucleus, one-sixth to one-eighth that of the jelly-cover. Nucleus delicate, somewhat
opaque, with numerous small nucleoli. Protoplasm of the central capsule contains small pellucid
globules (vacuoles?), and immediately under its membrane (on its inside) one single layer of
large, dark, refractive oil-globules. Extracapsular body very voluminous, foamy, with innumerable
alveoles in the jelly, and many xanthellæ between them.
Dimensions.—Diameter of the central capsule 0.6 to 0.8, of the nucleus 0.3 to 0.4,
of the calymma 3 to 5 mm.
Habitat.—Atlantic, Canary Islands, Haeckel; Cape Verde Islands,
Challenger Station 350; Brazil, Rabbe; surface.
3. Thalassicolla zanclea, Haeckel.
Thalassicolla zanclea, Haeckel, 1862, Monogr. d. Radiol., p. 252, Taf. ii.
fig. 3.
Spherical body opaque, transparent only in the periphery, with colourless central capsule, but
with brown or black pigment-powder scattered everywhere through the extracapsular alveolated
jelly-cover. Central capsule soft, transparent, colourless, with a thin structureless, not
areolated membrane. Diameter of the central capsule about one and a half times that of the
nucleus, one-half or one-third that of the jelly-cover. Nucleus delicate, transparent, with a
thin, finely punctated membrane, with one or more nucleoli. Protoplasm of the central capsule
contains only small, pellucid, densely packed globules (vacuoles?), no oil-globules. Extracapsular
body very dark and opaque, with a great mass of brown or blackish-brown pigment between the
alveoles of the jelly-cover. Numerous xanthellæ.
Dimensions.—Diameter of the central capsule 0.1 to 0.12, of the nucleus 0.07 to
0.08, of the calymma 0.2 to 0.4.
Habitat.—Mediterranean, Messina, Haeckel.
{20}
Subgenus 2. Thalassicollidium, Haeckel.
Definition.—Membrane of the central capsule areolated, with small
polygonal plates, resembling an epithelial cell-tissue, spotted by innumerable fine radial
pores.
4. Thalassicolla australis, n. sp.
Spherical body nearly transparent, without dark pigment. Central capsule colourless, somewhat
opaque, with a thick and firm, very elastic membrane, areolated by polygonal, punctated figures
resembling cells. Diameter of the central capsule about three times that of the nucleus, one-third
that of the jelly-cover. Nucleus thin-walled, with many small nucleoli. Protoplasm of the central
capsule finely granulated, containing numerous hyaline globules (vacuoles?) of different size, and
in each of these one single roundish, dark refringing corpuscle, concentrically lamellated like an
amylum grain. Extracapsular body without pigment or oil-globules, only composed of the large
alveoles imbedded in the jelly-cover, and of the fine protoplasmic network between them. No
xanthellæ.
Dimensions.—Diameter of the central capsule 1 to 2, of its nucleus 0.3 to 0.4, of
the nucleoli 0.12 to 0.16, of the hyaline globules in the protoplasm of the capsule 0.02 to 0.05;
calymma, 4 to 6 mm.
Habitat.—South-west Pacific, east coast of Australia, New Zealand,
&c.; Challenger Stations 163, 171; surface.
5. Thalassicolla nucleata, Huxley.
Thalassicolla nucleata, Huxley, 1851, Ann. and Mag. Nat. Hist., ser. 2, vol. viii. p.
435, pl. xvi. fig. 4.
Thalassicolla nucleata, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p.
28.
Thalassicolla coerulea, Schneider, 1858, Archiv f. Anat. u. Physiol., p. 40, Taf. iii.
Bd. i. figs. 5-7.
Thalassicolla nucleata, Haeckel, 1862, Monogr. d. Radiol., p. 249, Taf. iii. figs.
1-5.
Thalassicolla nucleata, R. Hertwig, 1876, Histologie d. Radiol., p. 43, Taf. iii.
figs. 61-5, Taf. iv., v.
Thalassicolla nucleata, R. Hertwig, 1879, Organismus d. Radiol., p.
34.
Spherical body in the central part opaque, black or dark coloured, in the periphery
transparent, whitish, or yellowish. Central capsule rather compact, yellowish, opaque, with a
thick and firm, very elastic membrane, areolated by polygonal, punctated figures resembling cells.
Diameter of the central capsule about twice as large as that of the nucleus, one-half to
one-fourth that of the jelly-cover. Nucleus with a very thick, finely punctated membrane,
containing a viscous fluid (when coagulated finely granular), and sometimes one large, central,
spherical, or ramified nucleolus, sometimes a variable number of smaller roundish nucleoli.
Protoplasm of the central capsule containing many very variable corpuscles, mostly pellucid
(albuminous?) spherules, containing oil-globules, or concentric amyloid concretions, or crystals,
&c. Extracapsular body with dark pigment-powder of variable colour (black, brown, violet,
blue, &c.), densely aggregated around the central capsule, more loosely dissipated between the
alveoles of the outer jelly-cover. Xanthellæ very numerous.
{21}
Dimensions.—Diameter of the central capsule 0.4 to 0.8, of the nucleus 0.02 to
0.05, of the calymma 1 to 5 mm.
Habitat.—Cosmopolitan; common in all warmer seas; Mediterranean,
Atlantic, Indian Ocean, Pacific, mainly between lat. 40° N. and lat. 40° S.; surface.
6. Thalassicolla maculata, n. sp. (Pl. 1, fig. 4).
Spherical body in the central part opaque, black or dark coloured, in the periphery
semi-transparent, spotted. Central capsule compact, yellowish, opaque, with a thick and firm, very
elastic membrane, areolated by polygonal, punctated figures resembling cells. Diameter of the
central capsule about twice that of the nucleus, one-third to one-fifth that of the jelly-cover.
Nucleus thin-walled, with one large spherical nucleolus. Protoplasm of the central capsule
contains innumerable very small, hyaline, spherical vesicles of equal size (or vacuoles?), two to
four times as broad as the separating bridges of protoplasm. Extracapsular body with dark
pigment-powder of black or brown colour, densely accumulated around the central capsule (in the
matrix), loosely scattered between the alveoles of the outer jelly-cover. The latter appears
spotted by numerous large, roundish lumps of protoplasm, scattered between the alveoles. No
xanthellæ.
Dimensions.—Diameter of the central capsule 0.3 to 0.6, of the nucleus 0.2 to 0.3,
of the hyaline globules in the protoplasm of the capsule 0.02 to 0.03; calymma, 2 to 3 mm.
Habitat.—South Pacific, Challenger Station 289.
7. Thalassicolla melacapsa, n. sp. (Pl. 1, fig. 5).
Spherical body in the central part opaque, black or dark coloured, in the periphery
semi-transparent, spotted. Central capsule compact, black, with a thick and firm, very elastic
membrane, areolated by polygonal, punctated figures resembling cells. Diameter of the central
capsule about twice that of the nucleus, one-third or half that of the jelly-cover. Nucleus
thin-walled, transparent, containing very numerous and small nucleoli. Protoplasm of the central
capsule filled with small black pigment-granules, quite intransparent, contains densely packed
hyaline (albuminous?) globules of equal size; every pellucid globule includes a smaller globule
(one-third or one-fourth its diameter), which appears to be composed of aggregated oil-bubbles.
Extracapsular body without pigment, contains between its alveoles in the inner half numerous, dark
refractive oil-globules, in the outer half numerous amœboid lumps of protoplasm,
irregularly scattered. No xanthellæ.
Dimensions.—Diameter of the central capsule 2 to 2.5, of the nucleus 1 to 1.5, of
the hyaline globules in the protoplasm of the capsule 0.03 to 0.04; calymma, 3 to 5 mm.
Habitat.—South-east Pacific (near Valparaiso), Challenger Station 300,
surface.
Genus 5. Thalassophysa,[12] Haeckel, 1881,
Prodromus, p. 470.
Definition.—Thalassicollida without intracapsular alveoles, but with large
roundish or globular alveoles within the extracapsular calymma. Nucleus in the centre of the
capsule papillate or branched.
{22}
The genus Thalassophysa contains those species of Thalassicollida formerly
associated with Thalassicolla, which are distinguished by the complicated, ramose, or
papillate form of the large nucleus. All three species here described are found in the
Mediterranean and the Atlantic. To this genus appertains also that strange form of Radiolaria
which I described in 1870 as Myxobrachia (compare Thalassophysa sanguinolenta).
1. Thalassophysa papillosa, n. sp.
Thalassicolla papillosa, Haeckel, 1867, Manuscript.
Spherical body transparent, colourless, or somewhat yellowish. Central capsule soft,
colourless, with a very thin but firm, elastic, structureless membrane. Diameter of the central
capsule about twice that of the nucleus, one-fourth to one-sixth that of the jelly-envelope.
Nucleus papillated, its spherical surface covered with a great number (50 to 80) of conical or
finger-like protuberances or blind sacs, not longer than half its radius. Protoplasm of the
central capsule filled with very small and numerous spherical vacuoles, without oil-globules.
Extracapsular jelly-body, without dark pigment, oil-globules, and large protoplasmic lumps,
contains between its alveoles very numerous xanthellæ.
Dimensions.—Diameter of the whole jelly sphere 4 to 5 mm., of the central capsule
0.8 to 1 mm., of its nucleus 0.4 to 0.5.
Habitat.—Canary Islands, Lanzerote, common, Haeckel; Cape Verde
Islands, Challenger; surface.
2. Thalassophysa sanguinolenta, Haeckel.
Thalassicolla sanguinolenta, Haeckel, 1870, Jenaische Zeitschr., Bd. v. p. 526, Taf.
18.
Thalassicolla sanguinolenta, Haeckel, 1870, Biolog. Studien, i. p. 113, Taf. iv.
Thalassicolla sanguinolenta, R. Hertwig, 1879, Organismus d. Radiol., p. 37, Taf. iii.
fig. 1.
Myxobrachia rhopalum, Haeckel, 1870, Jenaische Zeitschr., Bd. v. p. 519, Taf. 18 (et
in Biol. Stud., loc. cit.).
Myxobrachia pluteus, Haeckel, 1870, Jenaische Zeitschr., Bd. v. p. 520,
Taf. 18 (et in Biol. Stud., loc. cit.).
Spherical body in the central part opaque, reddish, in the periphery transparent, yellowish.
Central capsule compact, white, red spotted, with a thick elastic membrane, perforated by pores,
but not areolated. Diameter of the central capsule three times that of the nucleus, one-fifth to
one-eighth that of the jelly-envelope. Nucleus papillated, its spherical surface covered with
numerous (80 to 120) conical or finger-like protuberances not longer than one-fourth or one-third
of its radius. On the inside of these blind sacs lie numerous small roundish nucleoli. Protoplasm
of the central capsule in the outer (cortical) zone (on the inside of the membrane) radially
striped, with one layer of very numerous red oil-globules of equal size, producing its
blood-spotted appearance; in the inner (medullary) zone foamy, with numerous small spherical
vacuoles. Extracapsular jelly-body without dark pigment, contains between its alveoles no large
protoplasmic lumps (as in Thalassophysa pelagica), but numerous small oil-globules and
xanthellæ. This species sometimes amasses in its jelly-envelope large accumulations of Coccoliths
and Coccospheres, {23}which are much heavier than the
jelly-body, and produce arm-like protuberances of it; this modified form, often of very regular
and peculiar appearance, I formerly described as Myxobrachia (compare my Biolog. Studien,
loc. cit., and Hertwig, loc. cit., p. 37). Compare also Myxobrachia cienkowski, Wagner, 1872, L. N. 23.
Dimensions.—Diameter of the whole jelly-sphere 5 to 8 mm., of the central capsule
1 to 1.2 mm., of its nucleus 0.3 to 0.4.
Habitat.—Canary Islands, Lanzerote; common, Haeckel; Mediterranean,
Messina, Hertwig; surface.
3. Thalassophysa pelagica, Haeckel.
Thalassicolla pelagica, Haeckel, 1862, Monogr. d. Radiol., p. 247, Taf. i.
Thalassicolla pelagica, R. Hertwig, 1879, Organismus d. Radiol., p. 35,
Taf. iii. fig. 4.
Spherical body in the central part opaque, yellowish, in the periphery semi-transparent,
spotted. Central capsule compact, yellowish-white, with a thick and compact membrane, perforated
by pores, but not areolated. Diameter of the central capsule about twice that of the nucleus,
one-half to one-sixth that of the jelly-envelope. Nucleus papillated, its spherical surface
covered with numerous (20 to 60) conical, roundish, or finger-like protuberances, not longer than
its radius (commonly only one-half or one-third as long). Enclosed in the semi-fluid substance of
the nucleus are very long and thin cylindrical nucleoli snake-like, contorted, and penetrating
into the protuberances of the nucleus. Protoplasm of the central capsule in the outer (cortical)
zone (on the inside of the membrane) radially striped, with one layer of large oil-globules of
different sizes; in the inner (medullary) zone foamy, with numerous small spherical vacuoles,
mostly of equal size. Extracapsular jelly-body without dark pigment, contains between its alveoles
a large number of large roundish or amœboid lumps of protoplasm, and very numerous yellow
cells or xanthellæ (compare the detailed description in my Monograph, and in R. Hertwig's
work).
Dimensions.—Diameter of the whole jelly-sphere 1 to 4 mm., of the central capsule
0.5 to 0.6, of the nucleus 0.2 to 0.3.
Habitat.—Mediterranean—Messina, Corfu, Nizza, Genoa, Haeckel;
Messina, R. Hertwig; surface.
Family II. Collozoida, Haeckel, 1862 (Pl. 3).
Collozoida, Haeckel, 1862, Monogr. d. Radiol., p. 522.
Definition.—Colloidea socialia.
The family Collozoida comprises all associated or colony-building Radiolaria without skeleton.
We unite here all these skeletonless Radiolarian colonies into one single genus Collozoum,
constituted (1862) in my Monograph (p. 522). The oldest known form of it was the Collozoum
inerme, described firstly by Johannes Müller (1856) as Sphærozoum inerme. Two other
species of the genus were figured (1862) in my Monograph (p. 522, Tafn. xxxii., xxxv.). A most
accurate description of its histological structure and {24}development was given in 1876 by Richard Hertwig in his Histologie der
Radiolarien (pp. 12-42, Tafn. i., ii.). A number of other very remarkable forms of
Collozoum have been observed by me during the last few years, and partly figured in Pl. 3.
Collozoum, as the only representative of this family, is sufficiently
distinguished from all other Radiolaria by the definition "Skeletonless Radiolarian
Colonies." These occur floating on the surface of all warmer seas, often in astonishing
masses, and may be easily confounded, owing to their external resemblance, with the jelly-like
egg-masses of certain Mollusca. Collozoum is derived either from Actissa or from
Thalassicolla, simply by multiplication of the unicellular body and by reunion of the
associated capsules in one common calymma or jelly-veil; this is constantly alveolated, as in
Thalassicolla. As in Actissa, the form of the central capsule remains either
spherical, or it becomes ellipsoidal or discoidal, rarely polyhedral or amœboid. In
Collozoum as in all colonial Radiolaria, the original central nucleus commonly undergoes
cleavage very early into numerous small nuclei, whilst its place is usually taken by a central
oil-globule. This peculiarity may serve often (but not constantly) for the distinction of isolated
capsules of Collozoum from Actissa.
Genus 6. Collozoum,[13] Haeckel, 1862,
Monogr. d. Radiol., p. 522.
Definition.—Skeletonless colonies of Radiolaria.
The genus Collozoum, as already mentioned, is the only representative of its
family, and comprises all Radiolaria living associated in colonies, and having no skeleton.
Therefore Collozoum possesses all the peculiarities described above. Although the floating
colonies of this genus occur in enormous masses on the surface of all warmer seas, nevertheless
the number of different species in this genus is not great, and amounts only to thirteen. If this
number increase by further investigations, the subgenera distinguished in the following
description can be advanced to the range of genera; in which case Collodinium (or
Collozoum sensu restricto) will be characterised by the spherical form of its central
capsules, Colloprunum by the ellipsoidal form (Pl. 3, fig. 9),
Collophidium by the cylindrical, very prolonged form (figs. 2, 3), Collodiscus by
the lenticular or discoidal form, and Collodastrum by the indefinite, polyhedral, or
amœboid form (figs. 4, 5).
Subgenus 1. Collodinium, Haeckel.
Definition.—Form of the central capsules spherical or subspherical,
never polyhedral, ellipsoidal, or cylindrical.
{25}
1. Collozoum inerme, Haeckel (Pl. 3,
figs. 10-12).
Collozoum inerme, Haeckel, 1862, Monogr. d. Radiol., p. 522, Taf. xxxv.
Collozoum inerme, Cienkowski, 1871, Archiv. f. mikrosk. Anat., vol. vii. p. 376, Taf.
xxix. figs. 18-36.
Collozoum inerme, R. Hertwig, 1876, Histologie der Radiol., p. 12, Taf. i., ii.
Collozoum inerme, R. Hertwig, 1879, Organismus d. Radiol., p. 31, Taf. iii. fig.
12.
Sphærozoum inerme, J. Müller, 1856, Monatsber. d. k. Akad. d. Wiss. Berlin, p. 478;
Abhandl., p. 54.
Sphærozoum bicellulare, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 54,
Taf. viii. fig. 5.
Thalassicolla punctata, Huxley (pro parte), 1851, Ann. and Mag.
Nat. Hist., ser. 2, vol. viii. p. 433.
Central capsules spherical, with thin, simple-edged membrane, with one single oil-globule in
the centre. (If the capsules multiply by division, the spherical form becomes violin-shaped,
constricted in the middle; and in this condition the number of oil-globules increases; but in the
ordinary mature state the capsule of this species remains spherical, and its oil-globule solitary.
In quite young capsules the oil-globules are wanting; Pl. 3, fig. 12.)
Dimensions.—Diameter of the central capsules 0.04 to 0.16.
Habitat.—Cosmopolitan, common in all warmer seas (Mediterranean,
Atlantic, Indian, and Pacific), surface.
2. Collozoum nostochinum, n. sp.
Central capsules spherical, very large, opaque, distended with red pigment-granules and with
very numerous (two hundred to three hundred) small oil-globules. Membrane thick, double-edged.
Dimensions.—Diameter of the central capsules 0.3 to 0.5.
Habitat.—Indian Ocean, off Socotra, surface, Haeckel.
3. Collozoum volvocinum, n. sp.
Central capsules spherical, very large, opaque, containing a great number (ten to thirty) of
large oil-globules, and between them densely packed masses of dark pigment. Membrane thick,
double-edged. This species differs from Collozoum inerme, mainly by the great size of the
central capsules (three to five times as big as in the former) and the great number of
oil-globules in them.
Dimensions.—Diameter of the central capsules 0.2 to 0.3.
Habitat.—Central Pacific, Station 272, surface.
Subgenus 2. Colloprunum, Haeckel.
Definition.—Form of the central capsules ellipsoidal, with one
prolonged axis.
4. Collozoum ovatum, n. sp.
Colloprunum ovatum, Haeckel, 1882, Manuscript.
Central capsules ovate or ellipsoidal, its longer diameter twice to three times as large as the
shorter. In the centre of every capsule one single oil-globule.
{26}
Dimensions.—Length of the central capsules 0.2 to 0.3, breadth of them 0.1 to
15.
Habitat.—North Pacific, Station 244, surface.
5. Collozoum ellipsoides, n. sp. (Pl. 3, figs. 8,
9).
Colloprunum ellipsoides, Haeckel, 1882, Manuscript.
Central capsules regularly ellipsoidal, very large; their longer diameter once and a half to
twice as large as the shorter. In every capsule fifty to eighty oil-globules.
Dimensions.—Length of the central capsules 0.3 to 4, breadth of them 0.2.
Habitat.—North Atlantic, Færöe Channel (Gulf Stream), surface, John
Murray.
Subgenus 3. Collophidium, Haeckel.
Definition.—Form of the central capsules cylindrical, often
snake-like, contorted, with one axis much prolonged, several times longer than the transverse
axis.
6. Collozoum contortum, n. sp.
Collophidium contortum, Haeckel, 1882, Manuscript.
Central capsules cylindrical, three to four times as long as broad, C- or S-like curved,
transparent, without oil-globules.
Dimensions.—Length of the central capsules 0.2 to 0.3, breadth 0.06 to 0.08.
Habitat.—Tropical Atlantic, Station 347, surface.
7. Collozoum serpentinum, n. sp. (Pl. 3, figs.
1-3).
Collophidium serpentinum, Haeckel, 1882, Manuscript.
Central capsules cylindrical, filiform, much elongated, ten to one hundred times, sometimes two
hundred to four hundred times as long as broad, snake-shaped or worm-shaped, curved and contorted
in the most irregular manner, often spiral or twisted into a large nodule. Numerous oil-vesicles
constantly present, forming one series of globules in the axis of every capsule; distance of the
globules, one from another, and also from the capsule-membrane, about equal to their diameter.
(This interesting and very curious form was very frequently observed living by me in the Canary
Islands, in January 1867; the jelly-colonies were commonly spherical, and contained fifty to two
hundred or more capsules of very different size and form.)
Dimensions.—Length of the central capsules 1 to 10, sometimes 20 to 40 mm.;
average breadth 0.1 mm.
Habitat.—Canary Islands, Lanzerote, Haeckel, surface.
{27}
8. Collozoum vermiforme, n. sp. (Pl. 3,
figs. 6, 7).
Collophidium vermiforme, Haeckel, 1882, Manuscript.
Central capsules cylindrical, much elongated, five to ten times (sometimes twenty to fifty
times) as long as broad, snake-shaped or worm-shaped, very irregularly curved and contorted.
Numerous oil-globules constantly present, forming in the axis of every capsule a double series of
alternating rose-coloured globules. (This species is nearly allied to the preceding; but its
capsules are thicker and shorter, and the oil-vesicles in them are arranged not in a single, but
in a double row.)
Dimensions.—Length of the central capsules 0.6 to 1.2 mm., sometimes 3 to 6 mm.;
breadth 0.12.
Habitat.—Tropical Atlantic, near the west coast of Africa, Station
349, Canary Islands, surface.
Subgenus 4. Collodiscus, Haeckel.
Definition.—Form of the central capsules discoidal or lenticular, with
one shortened axis.
9. Collozoum coeruleum, Haeckel.
Collozoum coeruleum, Haeckel, 1862, Monogr. d. Radiol., p. 523, Taf. xxxii. figs.
6-8.
Collodiscus coeruleus, Haeckel, 1882, Manuscript.
Central capsule lenticular or discoidal, flattened, blue coloured, with one single oil-globule
in the centre. Protoplasm containing numerous crystals and dark blue pigment-granules. Membrane
very thick, double-edged. (Whilst at Messina in 1859 I found this form not constantly discoidal,
in 1867 in the Canary Islands I observed it constantly lenticular.)
Dimensions.—Breadth of the central capsules 0.1 to 0.15, height 0.04 to 0.08.
Habitat.—Mediterranean (Messina), Atlantic (Canary Islands),
surface.
10. Collozoum discoideum, n. sp.
Collodiscus discoideus, Haeckel, 1882, Manuscript.
Central capsule discoidal, flattened, transparent, with a ring of twenty to twenty-five
oil-globules in its circular periphery (on the inside of the thin membrane).
Dimensions.—Breadth of the central capsules 0.2, height 0.05.
Habitat.—South Pacific (Juan Fernandez), Station 300, surface.
Subgenus 5. Collodastrum, Haeckel.
Definition.—Form of the central capsules irregular and indefinite,
variable, commonly polyhedral or polygonal, or amœboid, often with irregular, finger-like
processes.
{28}
11. Collozoum pelagicum, Haeckel.
Collozoum pelagicum, Haeckel, 1862, Monogr. d. Radiol., p. 525, Taf. xxxii. figs. 4,
5.
Sphærozoum pelagicum, Haeckel, 1860, Monatsber. d. k. Akad. d. Wiss.
Berlin, 1860, p. 845.
Central capsules small, quite irregularly formed, roundish-polyhedral or depressed-polygonal,
transparent, without oil-globules. Often many extracapsular oil-vesicles in the common jelly-body
between the central capsules. Membrane very thin and delicate.
Dimensions.—Diameter of the central capsules 0.02 to 0.08.
Habitat.—Mediterranean, Messina, Haeckel; Naples, Brandt; surface.
12. Collozoum stellatum, n. sp.
Collodastrum stellatum, Haeckel, 1882, Manuscript.
Central capsules star-shaped, irregularly radiating, with a great number (eight to twenty or
more) of radial, short, conical, acute processes, very variable in size and number. Membrane thin.
In every capsule several (four to eight) oil-globules.
Dimensions.—Diameter of the central capsules 0.12 to 0.2.
Habitat.—Central Pacific, Station 274, surface.
13. Collozoum amœboides, n. sp. (Pl. 3, figs. 4,
5).
Collodastrum amœboides, Haeckel, 1882, Manuscript.
Central capsules amœbiform, of moderate size, quite irregularly formed, with a variable
number of finger-like, obtuse, irregular prolongations (commonly three to six), very variable in
size and form. Membrane thin. In the centre of every capsule one single oil-globule.
Dimensions.—Diameter of the central capsules 0.04 to 0.08.
Habitat.—Indian Ocean, Ceylon, Haeckel; Madagascar, Rabbe;
surface.
Suborder II. BELOIDEA, Haeckel.
Definition.—Spumellaria with an imperfect
skeleton, composed of numerous solid needles or spicula, scattered irregularly in the calymma.
The suborder Beloidea comprises all those Spumellaria which possess an imperfect or rudimentary skeleton, composed of a
variable number of isolated spicula scattered in the extracapsulum. The suborder contains only two
different families, the solitary Thalassosphærida (or Beloidea monozoa) and the associated
Sphærozoida (or Beloidea polyzoa). Both families are very nearly allied, and differ only in one
single character: the solitary life of the former, the social union of the {29}latter. It seems to be merely a consequence of this difference that the
cleavage of the nucleus commonly takes place very late in the former, very early in the
latter.
Commonly, therefore, the full-grown Thalassosphærida (until immediately before their
propagation) exhibit one single nucleus in the centre of the capsule, whilst in the Sphærozoida
the capsule is distended with numerous small nuclei. In these latter the centre of the capsule
usually contains one large oil-globule, whilst in the former oil-globules are either wanting or
scattered in large numbers in the endoplasm, or disposed in one layer on the inside of the capsule
membrane.
In the solitary Thalassosphærida each capsule is enclosed in its own peculiar
spherical calymma, whilst in the associated Sphærozoida all the capsules of the colony are united
into one common, very voluminous, alveolated calymma.
Family III. Thalassosphærida, Haeckel, 1862,
(Pl. 2).
Thalassosphærida, Monogr. d. Radiol., p. 255.
Definition.—Beloidea solitaria.
The family Thalassosphærida comprises all solitary Spumellaria with an
imperfect skeleton, composed of numerous solid needles or spicula, scattered around the central
capsule in the calymma. The structure of the unicellular soft body is quite the same as in the
Thalassicollida; it differs from these only in the possession of the extracapsular skeleton. All
needles of this skeleton are solid siliceous spicula, never hollow, as in the similar Cannorrhaphida among the Phæodaria. In the special structure and form of the skeleton the
Thalassosphærida agree perfectly with the well-known, colony-building Sphærozoida; they differ
from these only by their hermit-like life and by some peculiarities derived from this solitary
development.
The oldest known form of this family is probably the first Radiolarian, observed in the living
state, described in 1834 by Meyen as Physematium atlanticum (see p. 35). A second form was figured in my Monograph (1862) as Thalassosphæra
bifurca (p. 260, Taf. xii. fig. 1). A third form was there described under the name
Thalassosphæra morum; this remarkable form was first observed by Johannes Müller, and
figured under the name Thalassicolla morum (1858, Abhandl., p. 28, Taf. vii. figs. 1, 2).
The same form was afterwards observed living by myself in the Mediterranean, as well as in the
Atlantic, and in great numbers by the late Sir Wyville Thomson in the Pacific. The latter gave a
good figure of it with some valuable remarks in his excellent work, The Atlantic (1877, vol. i. p.
233, fig. 51). He called this peculiar Rhizopod Calcaromma calcarea, on account of the very
peculiar calcareous bodies "looking in outline like the rowels of spurs," which are
accumulated in great quantity around the central capsule, in the calymma. Further investigations
have convinced me that these peculiar stellate {30}bodies
of carbonate of lime, for which we propose the name "Calcastrella," are not parts of the skeleton
produced by the Radiolarian, but foreign bodies picked up by its extracapsular sarcode (in the
same way as the Coccoliths are picked up by Thalassicolla sanguinolenta = Myxobrachia!).
These Calcastrella occur also in the calymma of some Discoidea and other Radiolaria; they are
either unicellular calcareous Algæ, or foreign bodies of other origin. The Collodarium,
however, described as Thalassicolla morum and Calcaromma calcarea, seems to be a
simple Actissa, which has picked up a number of Calcastrella.
The Challenger collection has yielded a number of other true Thalassosphærida, which partly
agree with Thalassosphæra in the simple structure of the unicellular body (resembling
Actissa), and partly differ from it in the development of alveoles, either within or
without the central capsule (similar to Thalassolampe and Thalassicolla). The solid
siliceous spicula, which occur in great numbers scattered in the calymma, agree perfectly in form
with the spicula of the colony-building Sphærozoida. A characteristic difference between the
social and the solitary Beloidea seems to be determined by the cleavage
of the nucleus, which takes place in the latter very late, in the former very early. Therefore in
the large central capsule of the mature solitary Thalassosphærida, we commonly find one large
nucleus in the centre, and a number of oil-globules around it in the endosarc, or disposed in one
layer on the inside of the capsule-membrane (Pl. 2, figs. 2, 5);
whereas in the much smaller associated capsules of the Sphærozoida one large oil-globule is placed
commonly in the centre, and a great number of small nuclei scattered in the endoplasm (compare
above, p. 24).
Synopsis of the Genera of Thalassosphærida.
| A. Alveoles neither within nor without the central
capsule. |
|
Spicula simple, |
07. Thalassosphæra. |
| Spicula branched, |
08. Thalassoxanthium. |
| B. Numerous large alveoles within the central capsule (not in the
calymma). |
|
Spicula simple, |
09. Physematium. |
| C. Numerous large alveoles within the calymma (not in the
central capsule). |
|
Spicula simple, |
10. Thalassoplancta. |
| Spicula branched, |
11. Lampoxanthium. |
| A. Alveoles neither within nor without the central capsule. |
|
|
Spicula simple, |
|
|
|
|
|
|
7. Thalassosphæra. |
|
|
|
|
|
|
Spicula branched, |
|
|
|
|
|
|
8. Thalassoxanthium. |
|
|
|
|
| B. Numerous large alveoles within the central capsule (not in the
calymma). |
|
|
Spicula simple, |
|
|
|
|
|
|
9. Physematium. |
|
|
|
|
| C. Numerous large alveoles within the calymma (not in the central
capsule). |
|
|
Spicula simple, |
|
|
|
|
|
|
10. Thalassoplancta. |
|
|
|
|
|
|
Spicula branched, |
|
|
|
|
|
|
11. Lampoxanthium. |
|
|
|
|
Genus 7. Thalassosphæra,[14] Haeckel, 1862,
Monogr. d. Radiol., p. 259.
Definition.—Thalassosphærida without alveoles, with simple, unbranched,
needle-shaped spicula in the calymma.
The genus Thalassosphæra was founded by me in 1862 for those solitary Collodaria in which the simple central capsule is surrounded by scattered solid
spicula. {31}I described these two different species, the
new Thalassosphæra bifurca and the Thalassosphæra morum, which J. Müller had
formerly called Thalassicolla morum. This latter form is characterised by peculiar
calcareous bodies "looking in outline like the rowels of spurs, scattered irregularly in
the gelatinous envelope," and was therefore afterwards called "Calcaromma calcarea" by Sir
Wyville Thomson.[15] As already mentioned above, these
calcareous rowels are foreign bodies, picked up by an Actissa (see p. 29). I here confine the genus Thalassosphæra to those solitary Beloidea in which the body exhibits no alveoles, and the siliceous solid
spicula in the calymma are quite simple needles.
Thalassosphæra belonium, n. sp.
Spicula thin cylindrical rods, more or less curved, pointed at both ends, with smooth surface
(similar to the needles of Rhaphidozoum italicum). Central capsule spherical, three times
as large as the central nucleus, without larger oil-globules.
Dimensions.—Diameter of the central capsule 0.1 to 0.12, length of the spicula
0.04 to 0.08.
Habitat.—Central Pacific, Station 272, surface.
Thalassosphæra rhaphidium, n. sp.
Spicula thick cylindrical rods, more or less curved, pointed at both ends, covered with
numerous strong conical thorns, perpendicular to the axis. Central capsule spherical, four times
as broad as the central nucleus, with twenty to thirty large oil-globules on the inside of the
membrane.
Dimensions.—Diameter of the central capsule 0.2, length of the spicula 0.12 to
0.16.
Habitat.—Tropical Atlantic, Station 347, surface.
Genus 8. Thalassoxanthium,[16] Haeckel,
1881, Prodromus, p. 470.
Definition.—Thalassosphærida without alveoles, with numerous branched or compound
spicula in the calymma.
The genus Thalassoxanthium differs from the foregoing Thalassosphæra,
by the ramification of the spicula, and has therefore the same relation to it as Sphærozoum
to Belonozoum. The soft unicellular body is as simple as in Actissa, and exhibits
alveoles neither in the capsule nor in the calymma.
Subgenus 1. Thalassoxanthella, Haeckel.
Definition.—Spicula not geminate, but simply radiate, consisting of
three, four, or more needles or shanks, radiating in different directions from one and the same
point; shanks now simple or needle-like, now furcate or branched.
{32}
1. Thalassoxanthium triactinium, n. sp.
Spicula all (or nearly all) triradiate, composed of three (or sometimes in a few spicula four)
needle-like shanks of equal length, diverging from one common point. Shanks straight or somewhat
curved, smooth, pointed. Central capsule pellucid, twice as broad as its dark nucleus, without
larger oil-globules. Jelly-envelope very thin, with numerous xanthellæ.
Dimensions.—Diameter of the central capsule 0.1, of its nucleus 0.05, length of
the spicule-shanks 0.6 to 0.8.
Habitat.—Central Pacific, Station 266, surface.
2. Thalassoxanthium triradiatum, n. sp.
Spicula all (or nearly all) triradiate, composed of three (or sometimes in a few spicula four)
needle-like shanks of different length, diverging from one common point. Shanks curved or bent,
covered with small conical thorns. Central capsule dark, three times as large as the nucleus, with
numerous large oil-globules. Jelly-envelope thick, without xanthellæ.
Dimensions.—Diameter of the capsule 0.2, of the nucleus 0.07, length of the
spicule-shanks 0.1 to 0.15.
Habitat.—South Pacific, Station 302, surface.
3. Thalassoxanthium medusinum, n. sp. (Pl. 2, fig. 5).
Spicula all (or nearly all) quadriradiate, irregular, composed of four (or sometimes in a few
spicula three) needle-like shanks (mostly of unequal length), diverging from one common point.
Shanks curved, pointed, thorny, covered with small spinules. Central capsule dusky, twice to three
times as large as its dark central nucleus. On the inside of the membrane numerous large
oil-globules. Jelly-envelope thin, with numerous xanthellæ.
Dimensions.—Diameter of the central capsule 0.45 to 0.5, of its nucleus 0.18 to
0.2.
Habitat.—North Pacific, Station 244, surface.
4. Thalassoxanthium hexactinium, n. sp.
Spicula all (or nearly all) hexaradiate, composed of six (or sometimes in a few spicula three)
needle-like shanks, diverging in two opposite hemispheres (three needles upwards, three needles
downwards). Shanks somewhat curved, pointed, smooth. Central capsule yellowish-brown, dark, four
times as broad as its dark central nucleus. Jelly-envelope thick (about twice as broad as the
capsule), with very numerous xanthellæ.
Dimensions.—Diameter of the central capsule 0.3 to 0.4, of its nucleus 0.1 to
0.12.
Habitat.—South Atlantic, Station 325, surface.
{33}
5. Thalassoxanthium cervicorne, n. sp. (Pl. 2, figs. 3,
4).
Spicula all triradiate, trichotomously branched, with three equal shanks, which diverge from
one common point, and are again provided each with three branches on the distal end. These nine
branches are commonly once or twice forked (each fork rarely provided with three ramules). The
ramules are thin, unequal, curved, or bent, and the ramification nearly resembles a stag's horn.
Central capsule transparent, without oil-globules, two to three times as broad as the dark
nucleus, which contains one single long central nucleolus. Calymma thin, scarcely as thick as the
radius of the nucleus.
Dimensions.—Diameter of the central capsule 0.2 to 0.25, of the nucleus 0.08 to
0.1, length of the spicula 0.05 to 0.15.
Habitat.—Central Pacific, Stations 271, 274, surface.
Subgenus 2. Thalassoxanthomma, Haeckel.
Definition.—Spicula all or partly geminate, consisting of one middle
or axial rod, from the two poles of which diverge two, three, or more shanks in different
directions. Shanks or needle-rays sometimes simple, needle-like, sometimes bifurcated or
branched.
6. Thalassoxanthium furcatum, n. sp.
Spicula all (or nearly all) geminate and simply forked, composed of a simple axial rod and two
simple, needle-like shanks on each end of it. Shanks straight, pointed, smooth, somewhat shorter
than the middle rod. Central capsule yellowish, dark, three times as broad as its central dark
nucleus; besides this a single oil-globule, nearly of the same size.
Dimensions.—Diameter of the central capsule 0.1, of its nucleus 0.03; length of
the axial rod of the spicula 0.04, of its shanks 0.03.
Habitat.—Cape Verde Islands.
7. Thalassoxanthium bifurcum, Haeckel.
Thalassosphæra bifurca, Haeckel, 1862, Monogr. d. Radiol., p. 260, Taf. xii. fig.
1.
Sphærozoum bifurcum, Haeckel, 1860, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 845.
Spicula all geminate and double forked, composed of a simple axial rod and two forked branches
on each end; these branches are again forked, so that each spiculum exhibits eight thin distal
ends. All branches are thin, slender and straight. Central capsule red, four times as broad as the
central nucleus, containing between the red pigment-granules numerous, peculiar, violin-shaped
bodies (fat-corpuscles?). Compare fig. 1, loc. cit.
Dimensions.—Diameter of the central capsule 0.08, of the nucleus 0.02, length of
the spicula 0.05 to 0.08.
Habitat.—Mediterranean (Messina), surface, Haeckel.
{34}
8. Thalassoxanthium ovodimare, n. sp.
Spicula all geminate, composed of a simple, very short axial rod and three diverging shanks or
branches on each end of it; the shanks are very thin, straight, or little curved, and eight to ten
times as long as the axial rod. The spicula are quite smooth, as in the similar Sphærozoum
ovodimare (in which, however, the axial rod is much longer). Central capsule transparent,
without oil-globules, twice as broad as the nucleus.
Dimensions.—Diameter of the capsule 0.4, of the nucleus 0.2, length of the spicula
0.1 to 0.2.
Habitat.—Central Pacific, Station 273, surface.
9. Thalassoxanthium punctatum, n. sp.
Spicula all geminate-triradiate, composed of a simple middle rod and of three diverging shanks
on each end of it; the shanks are thorny with small spinules and shorter than the axial rod, very
similar to the common Sphærozoum punctatum. Central capsule dark, with numerous (twenty to
thirty) oil-globules on the inside of the membrane, three times as broad as the nucleus.
Dimensions.—Diameter of the capsule 0.3, of the nucleus 0.1, length of the spicula
0.05 to 0.2.
Habitat.—Central Pacific, Station 274, surface.
10. Thalassoxanthium octoceras, n. sp. (Pl. 2, fig. 6).
Spicula all geminate-quadriradiate, composed of a simple short middle rod and of four diverging
shanks on each end of it; the shanks are quite smooth, irregularly curved or bent, and four to
eight times as long as the middle rod. Central capsule dark, filled with pigment-granules, without
oil-globules, four times as large as the nucleus.
Dimensions.—Diameter of the capsule 0.5, of the nucleus 0.12, length of the
spicula 0.2 to 0.4.
Habitat.—Indian Ocean, Madagascar, Rabbe.
Genus 9. Physematium,[17] Meyen, 1834, Nova
Acta Acad. Nat. Curios., vol. xvi., Suppl., p. 286 (p. 162).
Definition.—Thalassosphærida with large numerous alveoles
within the central capsule (not in the calymma), and with numerous simple, needle-shaped spicula
in the calymma.
The genus Physematium is, together with the colony-forming
Sphærozoum, the first Radiolarian which was observed in the living state, described in 1834
by Meyen. It is most nearly allied to Thalassolampe, and has the same large roundish
alveoles within the central capsule, which reaches therefore an extraordinary size, 5 to 10 mm. It
{35}differs from the latter by the possession of spicula
in the calymma. The peculiar "centripetale Zell-gruppen" on the inside of the capsule-membrane are
probably due to radial cleavages of the endoplasm; as also occurs in other Collodaria.
1. Physematium mülleri, Schneider.
Physematium mülleri, Schneider, 1858, Archiv. f. Anat. u. Physiol., p. 38, Taf. iii.
B, figs. 1-5.
Physematium mülleri, Haeckel, 1862, Monogr. d. Radiol., p. 256, Taf. iii.
figs. 6-9.
Spherical body limpid, somewhat opalescent, sometimes a little yellowish or brownish, rather
soft. Central capsule with a thin, but firm, transparent membrane, its diameter eight to ten times
as large as that of the central spherical nucleus. Membrane of the nucleus thick, porous, on its
inside with some nucleoli. In the protoplasmic network between the large intracapsular alveoles,
numerous large, pale, yellowish, or orange oil-globules. On the inside of the membrane numerous
conical bodies, centripetally directed with the apex towards the centre; every conical body
("kegelförmige centripetale Zellgruppe") composed of three to nine (commonly four or five)
nucleated cells (mother-cells of the spores?); in the axis of the cone there is often a yellowish,
orange, or brown oil-globule. Extracapsular jelly-envelope thin, with short pseudopodia. Xanthellæ
scarce or wanting. Spicula scattered in the jelly numerous, small, simple needles, commonly C- or
S-like curved, smooth.
Dimensions.—Diameter of the whole jelly-sphere 3 to 6 mm., of the central capsule
1 to 5 mm., of the nucleus 0.1 to 0.5, length of the spicula 0.05 to 0.1.
Habitat.—Mediterranean, Messina; Schneider, Haeckel; surface.
2. Physematium atlanticum, Meyen.
Physematium atlanticum, Meyen, 1834, Nova Acta Acad. Nat. Curios., vol.
xvi., Suppl., p. 286 (162), Taf. xxxviii. (xxviii.) figs. 1-3.
Spherical body opalescent, pearl-like, with a violet or purple lustre, very soft. Central
capsule with a strong, semi-transparent membrane, its diameter six to eight times as large as that
of the central spherical nucleus. Membrane of the nucleus very thick, porous, on its inside with
many small nucleoli. In the protoplasmic network between the alveoles very numerous, small, purple
oil-globules. On the inside of the membrane a continuous simple layer of small nuclei, enclosed in
radially striped protoplasm (mother-cells of the spores?). No centripetal conical bodies.
Extracapsular jelly-envelope very thin, with short pseudopodia. No xanthellæ. Spicule scattered in
the jelly numerous, small needles, straight or slightly curved, thorny owing to numerous small
vertical spinules. This species and Thalassolampe maxima (p. 17)
possess the largest central capsule.
Dimensions.—Diameter of the whole jelly-sphere 8 to 12 mm., of the central capsule
5 to 10 mm., of the nucleus 1 to 2 mm., length of the spicula 0.1 to 0.3.
Habitat.—Eastern Atlantic, between Canary Islands and Cape Verde
Islands, Meyen; Lanzerote, Haeckel.
{36}
Genus 10. Thalassoplancta,[18] Haeckel,
1862, Monogr. d. Radiol., p. 261.
Definition.—Thalassosphærida with numerous large alveoles
in the calymma (but not in the central capsule), and with numerous simple, needle-shaped spicula
around the central capsule.
The genus Thalassoplancta was founded by me in 1862 for a Radiolarian with
simple hollow needles in the calymma, which was afterwards recognised as a Phæodarium, belonging
to Cannorrhaphis. We
here retain this name for a true Thalassosphærid, very similar to the latter, but distinguished by
the absence of the phæodium and the solid—not hollow—needle-shaped spicula, which are
scattered in the alveolated calymma. Thalassoplancta can be regarded as the solitary form
of the social Belonozoum.[19]
1. Thalassoplancta longispicula, n. sp.
Spicula long and thin, cylindrical, smooth, more or less bent, pointed at both ends, similar to
those of Thalassoplancta cavispicula. Central capsule thin-walled, without oil-globules,
four times as broad as the nucleus, which encloses one single nucleolus.
Dimensions.—Diameter of the capsule 0.6, of the nucleus 0.15, of the calymma 4
mm.
Habitat.—North Atlantic, Færöe Channel (Gulf Stream), John Murray.
2. Thalassoplancta brevispicula, n. sp. (Pl. 2, fig. 2).
Lampoxanthium brevispiculum, Haeckel, 1882, Atlas.
Spicula short and thick, thorny, irregularly curved, pointed at both ends, very numerous. In
the observed specimen all spicula were aggregated in the outer part of the voluminous calymma,
whilst the inner alveolated part was devoid of them. Central capsule thick walled, with a layer of
large oil-globules on its inner surface, twice as broad as the large nucleus which contains
numerous nucleoli.
Dimensions.—Diameter of the central capsule 0.5, of the nucleus 0.2, of the
calymma 2.5.
Habitat.—South Atlantic, Station 334, surface.
Genus 11. Lampoxanthium,[20] n. gen.
Definition.—Thalassosphærida with numerous large alveoles
in the calymma (but not in the central capsule), and with numerous branched or compound spicula in
the calymma.
{37}
The genus Lampoxanthium differs from the foregoing, Thalassoplancta,
by the composite form of the spicula, which are not simple needles, but radiate or geminate, or
branched in different forms; the former stands therefore in the same relation to the latter as the
social Belonozoum to Sphærozoum. The spicula of some species of Lampoxanthium
are identical with those of some species of Sphærozoum, so that the latter may be derived
from the former by forming colonies. The large central capsule is enveloped by a very voluminous
alveolated calymma, and includes a large central nucleus with numerous nucleoli.
Subgenus 1. Lampoxanthella, Haeckel.
Definition.—Spicula all (or nearly all) of one kind, radiate.
1. Lampoxanthium tetractinium, n. sp.
Spicula all (or nearly all) tetraradiate, with four thorny, straight, pointed shanks, radiating
from one common point. (Intermingled with these are often some few, thorny, triradiate spicula.)
On the inside of the capsule a layer of large oil-globules as in Thalassoplancta, Pl. 2, fig. 2.
Dimensions.—Diameter of the central capsule 0.2, of the nucleus 0.08, of the
calymma 0.8.
Habitat.—South Pacific, Station 288, surface.
Subgenus 2. Lampoxanthomma, Haeckel.
Definition.—Spicula all (or nearly all) of one kind, geminate-radiate,
with a simple middle rod and two to four diverging shanks on each end of it.
2. Lampoxanthium punctatum, n. sp.
Spicula all geminate-triradiate, thorny, of the same form as in the common Sphærozoum
punctatum, of which this species is the large solitary representative. The spicula are
aggregated in a very condensed layer on the surface of the large calymma.
Dimensions.—Diameter of the capsule 0.8, of the nucleus 0.6, of the calymma
2.0.
Habitat.—North Pacific, Station 248, surface.
3. Lampoxanthium octoceras, n. sp.
Spicula all geminate-quadriradiate, with a very short simple middle rod and four very long
divergent shanks on both ends of it; the shanks are smooth, five to ten times as long as the
middle {38}rod, irregularly bent and curved. (Differs
from the similar Thalassoxanthium octoceras, Pl. 2, fig. 6, by
slender, more curved shanks, and by the voluminous calymma, there entirely wanting.)
Dimensions.—Diameter of the capsule 0.5, of the nucleus 0.2, of the calymma
3.0.
Habitat.—South Atlantic, Station 331, surface.
Subgenus 3. Lampoxanthura, Haeckel.
Definition.—Spicula of two or three different kinds, simple, radiate,
and geminate-radiate mixed.
4. Lampoxanthium pandora, n. sp. (Pl. 2, fig. 1).
Spicula mixed, of three different kinds—simple, radiate and geminate-radiate; all three
kinds partly smooth, partly thorny. The simple needles short, thin spindle-shaped, often curved.
The radiate spicula commonly with three or four, rarely five or six, unequal rays, straight or
curved. The radiate-geminate spicula commonly with three, rarely four, shanks on each end, often
different on both ends of the middle rod. The size, number, and form of the irregular spicula are
here quite as variable as in the social Rhaphidozoum pandora, of which it is the solitary
representative. The wall of the large central capsule is very thick, with evident pore-canals,
separated by a clear interval from the coagulated and vacuolated endoplasm, which contains no
oil-globules. Nucleus with numerous nucleoli.
Dimensions.—Diameter of the central capsule 0.5 to 0.6, of the nucleus 0.1 to 0.2,
of the calymma 2 to 4 mm.
Habitat.—North Pacific, Station 244, surface.
Family IV. Sphærozoida, Haeckel (Pl. 4).
Sphærozoida, Haeckel, 1862, Monogr. d. Radiol., p. 521.
Definition.—Beloidea socialia.
The family Sphærozoida comprises all associated or colony-forming
Radiolaria, which are provided with an imperfect skeleton, composed of numerous solid needles or
spicula, scattered around the central capsule in the calymma. The structure and form of this
skeleton is quite the same as in the preceding solitary Thalassosphærida, but on the other hand,
the structure and form of the colonies and of the included numerous central capsules is the same
as in the skeletonless Collozoida.
The oldest well-known form of Sphærozoida is the common cosmopolitan Sphærozoum
punctatum, probably first observed in 1834 by Meyen, and called Sphærozoum fuscum,
afterwards more accurately described by Huxley in 1851.
{39}
Other forms were afterwards described by Müller and by myself in 1862.[21] Further investigations have shown me that some species of this
family are among the most common Radiolaria, and occur in astonishing numbers on the surface of
all warmer seas. But the number of species is comparatively small, and their distinction is very
difficult, as all the different forms are very variable and connected by intermediate
forms—a truly "transformistic" group.
The only character sufficient for the constitution of genera in this transformistic group is
found in the form and composition of the spicula; the very variable form of the jelly-calymma and
the enclosed central capsule being without value for this purpose. But also the form of the
spicula is very variable, and not always constant. In some species the particular form of the
spicula is transmitted by constant heredity, whilst in others it is very inconstant, even in one
and the same individual. (Compare the remarks on variability in the general introduction.)
As the number of various forms is rather great, it seems to be advisable to distinguish the
three following genera.
Synopsis of the Genera of Sphærozoida.
| A. Spicula all of one kind, simple or needle-shaped, |
12. Belonozoum. |
| B. Spicula all of one kind, branched or radiate, or geminate, |
13. Sphærozoum. |
| C. Spicula of two more different kinds, partly simple, partly
branched, |
14. Rhaphidozoum. |
Genus 12. Belonozoum,[22] n. gen.
Definition.—Sphærozoida with simple needle-shaped
spicula, which are neither radiate nor branched.
The genus Belonozoum comprises the Sphærozoida with simple needle-shaped
spicula, and may be regarded as the colonial form of Thalassosphæra or
Thalassoplancta, derived from these solitary Beloidea by
multiplication of the capsules and union in a common calymma.
1. Belonozoum bacillosum, n. sp.
Sphærozoum bacillosum, Haeckel, 1881, Manuscript.
Spicula all simple rods, straight cylindrical, obtuse at both ends, quite smooth. Central
capsule pellucid, with one single central oil-globule.
Dimensions.—Diameter of the central capsule 0.08 to 0.12, length of the spicula
0.05 to 0.08.
Habitat.—Central Pacific, Station 271, surface.
{40}
2. Belonozoum spinulosum, Haeckel.
Sphærozoum spinulosum, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 54,
Taf. viii. fig. 4.
Sphærozoum spinulosum, Haeckel, 1862, Monogr. d. Radiol., p. 527, Taf.
xxxiii. figs. 3, 4.
Spicula all simple rods, straight cylindrical, obtuse on both ends, thorny with numerous small
spines, placed vertically on the rods.
Dimensions.—Diameter of the central capsule 0.08 to 0.1, length of the spicula
0.05 to 0.2.
Habitat.—Mediterranean, Nice, J. Müller; Messina, Haeckel; Naples,
Brandt; surface.
3. Belonozoum italicum, Haeckel.
Sphærozoum italicum, Haeckel, 1862, Monogr. d. Radiol., p. 526, Taf.
xxxiii. figs. 1, 2.
Spicula all simple rods, more or less curved or bent, pointed at both ends, quite smooth.
Central capsule with a variable number (commonly five to twenty) of oil-globules.
Dimensions.—Diameter of the central capsule 0.1 to 0.3, length of the spicula 0.05
to 0.2.
Habitat.—Mediterranean, Nice, Naples, Messina, Haeckel, surface.
4. Belonozoum atlanticum, n. sp.
Sphærozoum atlanticum, Haeckel, 1881, Manuscript.
Spicula all together simple rods, more or less curved or bent, pointed at both ends, thorny
from numerous small spines, placed vertically on the rods.
Dimensions.—Diameter of the central capsule 0.1 to 0.2, length of the spicula 0.07
to 0.15.
Habitat.—Tropical Atlantic, Station 348, surface.
Genus 13. Sphærozoum,[23] Meyen, 1834, Nova
Acta Acad. Nat. Curios., Bd. xvi., Suppl., p. 287 (p. 163).
Definition.—Sphærozoida with branched or radiate spicula
of one kind.
The genus Sphærozoum, with Physematium one of the two oldest
Radiolaria, observed in the living state, was founded 1834 by Meyen for one of the social Beloidea, which was probably the common cosmopolitan Sphærozoum
punctatum, the true type of this genus. Johannes Müller described a number of species, which
were partly skeletonless (Collozoum), partly armed with simple or with compound spicula.
The species with simple spicula we refer here to Belonozoum, the species with two or more
different kinds of spicula to Rhaphidozoum, while we unite in Sphærozoum all species
with one kind of branched or compound spicula. The two following species are incompletely known:—Sphærozoum
orientale, Dana, 1863, Ann. and Mag. Nat. Hist., vol. xii. p. 54. Sphærozoum
sanderi, Dœnitz, 1871, L. N. 60, p. 71.
{41}
Subgenus 1. Sphærozonactis, Haeckel.
Definition.—Spicula radiate, not geminate, consisting of three, four,
or more needles or shanks, radiating in different directions from one common central point.
1. Sphærozoum triactinium, n. sp.
Spicula all (or nearly all) triradiate, composed of three (or sometimes in few spicula four)
needle-like shanks, diverging from one common point. Shanks straight or somewhat curved, smooth,
pointed. Central capsules spherical, with one central oil-vesicle. This species may be regarded as
the social form of Thalassoxanthium triactinium.
Dimensions.—Diameter of the central capsules 0.1 to 0.12, length of the
spicula-shanks 0.05 to 0.1.
Habitat.—South Pacific, Station 295, surface.
2. Sphærozoum medusinum, n. sp.
Spicula all (or nearly all) quadriradiate, composed of four (or sometimes in few spicula three)
needle-like shanks (mostly of unequal length), diverging from one common point. Shanks slightly
curved, pointed, thorny, covered with small spinules. Central capsules ellipsoidal, containing
several (four to eight) oil-vesicles. This species may be regarded as the social form of the
solitary Thalassoxanthium medusinum (Pl. 2, fig. 5).
Dimensions.—Diameter of the central capsules 0.15 to 0.18, length of the
spicula-shanks 0.08 to 0.12.
Habitat.—North Pacific, Station 236, surface.
3. Sphærozoum hamatum, n. sp.
Spicula all (or nearly all) quadriradiate, composed of four (or sometimes in few spicula three)
needle-like shanks, mostly of very different size, diverging from one common point. Shanks strong,
straight, curved, or hook-like; thorny, covered with small spinules on the distal extremity.
Central capsules ellipsoidal, large, containing many (ten to twenty) oil-globules. This large
species is distinguished by the very irregular form and size of the spicula.
Dimensions.—Diameter of the central capsules 0.2 to 0.25, length of the
spicula-shanks 0.12 to 0.18.
Habitat.—Central Pacific, Station 265, surface.
4. Sphærozoum hexactinium, n. sp.
Spicula all (or nearly all) hexaradiate, composed of six (or sometimes in few spicula five or
seven) needle-like shanks, mostly of equal size, diverging from one common point in two opposite
hemispheres (three needles upwards, three needles downwards). Shanks somewhat curved, pointed,
{42}smooth. Central capsules spherical, small, with one
central oil-globule. This species may be regarded as the social form of Thalassoxanthium
hexactinium.
Dimensions.—Diameter of the central capsule 0.06 to 0.08, length of the
spicula-shanks 0.05 to 0.06.
Habitat.—North Atlantic, Færöe Channel (Gulf Stream), John Murray.
Subgenus 2. Sphærozonoceras, Haeckel.
Definition.—Spicula all geminate-radiate, consisting of one middle
rod, which bears an equal and constant number of rays (two, three, or four) at each end.
5. Sphærozoum furcatum, n. sp.
Spicula all (or nearly all) geminate and simply forked, composed of a simple axial rod and two
simple needle-like shanks on each end of it. Shanks straight, pointed, smooth, commonly somewhat
longer than the middle rod.
Dimensions.—Diameter of the central capsules 0.1 to 0.15, length of the axial rod
of the spicula 0.03, of its shanks 0.04 to 0.06.
Habitat.—Tropical zone of the Atlantic, near Ascension Island, Station
344, surface.
6. Sphærozoum furculosum, n. sp.
Spicula all (or nearly all) geminate and simply forked, composed of a simple axial rod and two
simple needle-like shanks on each end of it. Shanks curved or bent, pointed, thorny, with many
small spinules, commonly somewhat shorter than the middle rod.
Dimensions.—Diameter of the central capsules 0.2 to 0.25, length of the axial rod
of the spicula 0.1, of its shanks 0.05 to 0.08.
Habitat.—South Atlantic, near Tristan da Cunha, Station 334,
surface.
7. Sphærozoum ovodimare, Haeckel.
Sphærozoum ovodimare, Haeckel, 1862, Monogr. d. Radiol., p. 527, Taf. xxxiii. figs. 5,
6.
Sphærozoum punctatum, var., Brandt, 1881, Monatsber. d. k. preuss. Akad.
d. Wiss. Berlin, Taf. i. fig. 54.
Spicula all (or nearly all) geminate and triradiate, composed of a long simple axial rod and
three simple needle-like shanks on each end of it. Shanks straight, pointed, smooth, commonly
shorter than the middle rod. (Often few furcate or four-rayed spicula are intermixed, or few
spicula are not smooth, but thorny.)
Dimensions.—Diameter of the central capsules 0.05 to 0.2, length of the middle rod
of the spicula 0.02 to 0.06, of its shanks 0.01 to 0.04.
Habitat.—Mediterranean, Naples, Messina, Haeckel; Atlantic, Canary
Islands, Cape Verde Islands, West Coast of Africa, Stations 351 to 354; surface.
{43}
8. Sphærozoum trigeminum, n. sp.
Spicula all (or nearly all) geminate-triradiate, composed of a short simple axial middle rod
and three simple needle-like shanks on each end of it. Shanks curved or bent, very thin, smooth,
commonly much longer than the middle rod. (Often few quadriradiate or few thorny triradiate
spicules are interspersed among the others.)
Dimensions.—Length of the middle rod of the spicula 0.02 to 0.04, of its shanks
0.03 to 0.09.
Habitat.—North Pacific, Stations 244 to 248, surface.
9. Sphærozoum punctatum, J. Müller.
Sphærozoum punctatum, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 54,
Taf. viii. figs. 1, 2.
Sphærozoum punctatum, Haeckel, 1862, Monogr. d. Radiol., p. 528, Taf. xxxiii. figs.
7-9.
Sphærozoum fuscum, Meyen, 1834, Nova Acta Acad. Nat. Cur., vol. xvi. Taf. xxxviii.
fig. 7.
Thalassicolla punctata, Huxley, 1851, Ann. and Mag. Nat. Hist., ser. 2,
vol. viii. p. 434, pl. xvi. figs. 1, 2, 3.
Spicula all (or nearly all) geminate-triradiate, composed of a long simple axial middle rod and
three simple needle-like shanks on each end of it. Shanks straight, pointed, thorny, with many
small spines, commonly somewhat shorter than the middle rod. (Often few furcate or four-rayed
spicula are intermingled, or some of the spicula are smooth.) This cosmopolitan species is
extremely variable, and produces interesting transitional forms to many other species of the
genus. Compare also the general remarks on the genus, and the chapter on "Transformation" in the
general introduction.
Dimensions.—Length of the middle rod of the spicula 0.02 to 0.06, of its shanks
0.01 to 0.05.
Habitat.—Cosmopolitan, common in nearly all warmer seas,
Mediterranean, Atlantic, Indian Ocean, Pacific; surface.
10. Sphærozoum armatum, n. sp. (Pl. 4,
fig. 9).
Spicula all geminate-triradiate, with a stout and short middle rod and three arborescent shanks
on each end of it. Shanks longer than the middle rod, very stout, straight, pine-shaped, with six
to twelve irregular, spinulated, lateral branches.
Dimensions.—Diameter of the central capsules 0.04 to 0.08, length of the middle
rod of the spicula 0.02 to 0.03, of its shanks 0.05 to 0.08.
Habitat.—North Pacific, Japan, Station 239, surface.
11. Sphærozoum alveolatum, n. sp. (Pl. 4, figs. 2,
3).
Spicula all together geminate-triradiate, with a simple stout middle rod and three arborescent
shanks on each end of it. Shanks more or less curved, slender, pine-shaped, with four to eight
short, thorny lateral branches. In all cœnobia of this remarkable species the central
capsules are enclosed in large thick-walled alveoles (of three times their breadth), and in each
alveole is placed besides {44}the capsule one single very
large spiculum, whilst the others are much smaller (fig. 3). All the alveolated capsules are
placed in one single stratum on the surface of the jelly-like spherical cœnobium,
comparable to the blastoderm-cells of a blastula.
Dimensions.—Diameter of the central capsules 0.08 to 0.1, of the alveoles 0.2 to
0.4, length of the spicula 0.1 to 0.3.
Habitat.—South Pacific (Juan Fernandez), Station 300, surface.
12. Sphærozoum verticillatum, n. sp. (Pl. 4, fig. 7).
Spicula all geminate-triradiate, with a short simple middle rod and three much longer
arborescent shanks on each end of it. Shanks straight, slender, pine-shaped, each in the distal
half with three to four elegant verticils of thorny lateral branches.
Dimensions.—Diameter of the capsules 0.1 to 0.12, middle rod of the spicula 0.03
to 0.05, shanks 0.1 to 0.15.
Habitat.—Indian Ocean, Ceylon, Haeckel; Madagascar, Rabbe;
surface.
13. Sphærozoum octoceras, n. sp.
Spicula all geminate-quadriradiate, with a short simple middle rod and four diverging shanks on
each end of it. Shanks smooth, irregularly curved or bent, three to six times as long as the
middle rod. It may be regarded as the social form of Thalassoxanthium octoceras (Pl. 2, fig. 6).
Dimensions.—Diameter of the capsules 0.12 to 0.16, middle rod of the spicula 0.02,
shanks 0.01.
Habitat.—Australia, south coast, Faber; Station 163, surface.
14. Sphærozoum quadrigeminum, n. sp.
Spicula all geminate-quadriradiate, with a long thick middle rod and four shorter diverging
shanks on each end of it. Shanks straight, thorny.
Dimensions.—Diameter of the capsules 0.06 to 0.08, length of the spicula 0.05 to
0.15.
Habitat.—North Atlantic, Azores, Station 354, surface.
15. Sphærozoum araucaria, n. sp.
Spicula all geminate-quadriradiate, with stout straight middle rod and four longer diverging
shanks on each end of it. Shanks arborescent, with six to twelve thorny lateral branches.
Dimensions.—Diameter of the capsules 0.1 to 0.15, length of the spicula 0.05 to
0.1.
Habitat.—South Atlantic, coast of Brazil, Rabbe; surface.
16. Sphærozoum arborescens, n. sp. (Pl. 4, fig. 8).
Spicula all geminate-quadriradiate, with a stout straight middle rod and four longer diverging
shanks on each end of it. Shanks arborescent, pine-shaped, with four to six verticils of lateral
branches, which again are ramified and thorny.
{45}
Dimensions.—Diameter of the capsules 0.16 to 0.18, length of the spicula 0.1 to
0.2.
Habitat.—South Atlantic (Tristan da Cunha), Station 332, surface.
Subgenus 3. Sphærozonura, Haeckel.
Definition.—Spicula all geminate-radiate, but with a different and
variable number of shanks on each end of the middle rod.
17. Sphærozoum stellatum, n. sp.
Spicula all geminate-radiate, with a strong middle rod and a variable number of shorter
radiating shanks on the two ends of it. Shanks straight, nearly conical, smooth; for the most part
three or four shanks on each end, but sometimes also five or six; very often this number is
unequal on the two ends.
Dimensions.—Diameter of the central capsules 0.1 to 0.2, length of the spicula
0.05 to 0.15.
Habitat.—Central Pacific, Station 270, surface.
18. Sphærozoum geminatum, n. sp. (Pl. 4, fig. 4).
Spicula all geminate-radiate, with a strong middle rod and a variable number of longer radiant
shanks on each end of it. Shanks straight, conical, in the distal half thorny; commonly either
three or four shanks on each end of the middle rod, often also three on one end, four on the other
end; rarely five or six rays on one end.
Dimensions.—Diameter of the capsules 0.15 to 0.2, length of the spicula 0.05 to
0.1.
Habitat.—Indian Ocean, Ceylon, Haeckel; surface.
19. Sphærozoum circumtextum, n. sp.
Spicula all geminate-radiate, with a very variable number of rays (two to six) on each end of
the thin middle rod. All spicula very thin and delicate, smooth, with curved or bent shanks,
densely covering the central capsule like a cobweb. The number of rays on each end is usually
different, generally four or five, often also two or three, rarely six.
Dimensions.—Diameter of the capsule 0.1 to 0.2, length of the spicula 0.04 to
0.12.
Habitat.—Southeast part of the Indian Ocean, Station 160, surface.
20. Sphærozoum variabile, n. sp. (Pl. 4, fig. 5).
Spicula all geminate-radiate, with a short middle rod and a variable number of shanks on each
end of it. Shanks four to eight times as long as the middle rod, curved or bent, in the distal
half thorny; their number is commonly different on the two ends of it, three or five, often also
four or six, rarely two; their size and form very variable.
{46}
Dimensions.—Diameter of the capsules 0.1 to 0.3, length of the spicula 0.1 to
0.2.
Habitat.—North Pacific, Station 248, surface.
Genus 14. Rhaphidozoum,[24] Haeckel, 1862,
Monogr. d. Radiol., p. 529.
Definition.—Sphærozoida with two or more different kinds of spicula; one kind
simple, needle-shaped; the other kinds compound, radiate, or branched.
The genus Rhaphidozoum differs from Sphærozoum by the composition of the skeleton
of two or more different kinds of spicula, and has therefore the same relation to it as the
solitary Lampoxanthura to Lampoxanthella.
In some species nearly all the different forms, which characterise the numerous
species of Beloidea, may be united in one and the same individual.
Subgenus 1. Rhaphidonactis, Haeckel.
Definition.—Spicula of two different kinds; one kind simple,
needle-shaped, the other radiate (composed of three, four, or more shanks, diverging from one
common point).
1. Rhaphidozoum pelagicum, n. sp.
Spicula of two different kinds; one kind simple thin needles, a little curved or bent, the
other kind triradiate, with three thin, curved shanks. Both kinds smooth, without thorns.
Resembles a combination of Belonozoum italicum and Sphærozoum triactinium.
Dimensions.—Diameter of the central capsules 0.1 to 0.12, length of the spicula
0.05 to 0.15.
Habitat.—Central Pacific, Station 267, surface.
2. Rhaphidozoum pacificum, n. sp.
Spicula of two different kinds; one kind simple needles, stout and straight, pointed at both
ends, the other kind triradiate, with three straight and stout shanks. Both kinds thorny.
Dimensions.—Diameter of the central capsule 0.06 to 0.08, length of the spicula
0.05 to 0.1.
Habitat.—Central Pacific, Station 271, surface.
3. Rhaphidozoum acuferum, Haeckel.
Rhaphidozoum acuferum, Haeckel, 1862, Monogr. d. Radiol., p. 529, Taf. xxxii. figs.
9-11.
Sphærozoum acuferum, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 54,
Taf. viii. fig. 3.
Thalassicolla acufera, J. Müller, 1855, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 237.
Spicula of two different kinds, simple needles and quadriradiate; both strong, thorny, covered
with small spinules. Simple needles mostly curved, C-shaped. Four shanks of the quadriradiate
{47}spicula now straight, now curved, commonly of very
different size. (Often one single quadriradiate spiculum is distinguished by its extraordinary
size.) For the detailed description of this species compare my Monograph (loc. cit.).
Dimensions.—Diameter of the central capsules 0.05 to 0.35, length of the simple
needles 0.05 to 0.25, shanks of the quadriradiate spicula 0.05 to 0.15.
Habitat.—Mediterranean, Messina, Naples, Nice.
4. Rhaphidozoum arachnoides, n. sp.
Spicula of two different kinds; one kind simple, needle-like, the other quadriradiate; both
very thin and slender, smooth, without spicules. Simple needles curved, C-shaped. Four shanks of
the quadriradiate spicula also curved, commonly of nearly equal size. The numerous thread-like
spicula of this species are so densely packed around the central capsule, that they extend all
around its surface like the network round a balloon.
Dimensions.—Diameter of the central capsules 0.12 to 0.15, length of the simple
needles 0.1 to 0.12, shanks of the quadriradiate spicula 0.06 to 0.08.
Habitat.—Tropical Atlantic, Station 345, surface.
5. Rhaphidozoum asperum, n. sp.
Spicula of two different kinds; one kind simple, needle-shaped, stout, and straight, the other
kind hexaradiate; its six shanks about half as long as the former, conical. Both kinds very
thorny, covered with short conical spinules.
Dimensions.—Diameter of the capsules 0.06 to 0.08, length of the simple needles
0.05 to 0.07, shanks of the hexaradiate spicula 0.03 to 0.04.
Habitat.—South Pacific, Station 288, surface.
Subgenus 2. Rhaphidoceras, Haeckel.
Definition.—Spicula of two different kinds; one kind simple,
needle-shaped, the other kind geminate-radiate, with rays on both poles of a middle rod.
6. Rhaphidozoum neapolitanum, Haeckel.
Sphærozoum neapolitanum, C. Brandt, 1881, Monatsber. d. k. preuss. Akad.
d. Wiss. Berlin, p. 390, Taf. i. figs. 14, 16-18.
Spicula mixed, of two different kinds; simple needles and geminate-forked. Simple rods, like
those of Belonozoum italicum, more or less curved, pointed at both ends, smooth (sometimes
a little thorny at both ends). Geminate spicula simply forked, like those of Sphærozoum
furcatum, composed of a short, simple, axial rod, and two simple, smooth, straight shanks on
each end of it, commonly somewhat longer than the middle rod. This species, which I have observed
myself in Spezzia in great quantity, is quite as variable as all the other species of the genus,
and has not more claim to specific rights than the others. Commonly the simple needles are much
more numerous {48}than the geminate-forked, but sometimes
the contrary is the case. On their variability compare the general remarks on the genus, and the
chapter on "Transformation" in the general introduction.
Dimensions.—Length of the simple spicula 0.05 to 0.1, of the middle rod of the
forked spicule 0.05 to 0.08, of their shanks 0.01 to 0.03.
Habitat.—Mediterranean, Naples, Spezzia, surface.
7. Rhaphidozoum patagonicum, n. sp.
Spicula mixed, of two different kinds, simple needles and geminate-triradiate. Simple rods,
like those of Belonozoum spinulosum, straight, thorny, pointed at both ends. Geminate
spicula double-triradiate, like those of Sphærozoum punctatum, composed of a simple, short,
axial rod and three simple pointed shanks on each end of it. Shanks straight, thorny, with many
small spinules, commonly somewhat longer than the middle rod. (Often some of the spicula of both
kinds are smooth, not thorny, or not straight, but a little curved, or a few forked or
four-radiated geminate spicula are mingled with the others.)
Dimensions.—Diameter of the central capsules 0.08 to 0.2, length of the simple
spicula 0.1 to 0.15, of the geminate 0.08 to 0.16.
Habitat.—South Pacific, west coast of Patagonia, Station 302,
surface.
8. Rhaphidozoum ascensionis, n. sp.
Spicula mixed, of two different kinds, simple needles and geminate-triradiate; both kinds thin,
smooth, without spinules. Simple needles somewhat curved, C- or S-shaped. Geminate spicula
double-triradiate, composed of a simple, short, straight axial rod and three slender curved shanks
on each end of it. Shanks two to four times longer than the middle rod. (Sometimes few simple
hexaradiate and geminate tetraradiate spicula are mingled.)
Dimensions.—Diameter of the central capsules 0.12 to 0.15, length of the simple
spicula 0.1, of the double-triradiate 0.05 to 0.2.
Habitat.—South Atlantic, off Ascension Island, Station 342,
surface.
9. Rhaphidozoum capense, n. sp.
Spicula of two different kinds; one kind simple, needle-shaped, straight, pointed at both ends,
the other kind geminate-quadriradiate, with a stout short middle rod and four longer bent shanks
on each end of it. Both kinds smooth.
Dimensions.—Diameter of the capsules 0.2 to 0.25, length of the simple needles
0.01 to 0.3, of the geminate spicula 0.05 to 0.15.
Habitat.—Cape of Good Hope (Agulhas), Station 142, surface.
10. Rhaphidozoum australe, n. sp.
Spicula of two different kinds; one kind simple, needle-shaped, curved, thin; the other kind
geminate, with a variable number of shanks on both ends of the shorter middle rod, often {49}different on the two poles of it. The prevalent number of
rays on each end is three or four, often also two or five, rarely six. All spicula smooth, more or
less bent.
Dimensions.—Diameter of the capsules 0.1 to 0.2, length of the spicula 0.05 to
0.15.
Habitat.—South West Pacific, Station 165, surface.
Subgenus 3. Rhaphidonura, Haeckel.
Definition.—Spicula of three different kinds: one kind simple,
needle-shaped; the second kind radiate, with three to six shanks radiating from a common central
point; the third kind geminate-radiate, with rays on both poles of a middle rod.
11. Rhaphidozoum polymorphum, n. sp.
Spicula of three different kinds; simple needles, radiate, and geminate mixed. The simple
needles straight and stout. The radiate spicula commonly with three or six, rarely four or five,
rays. The geminate-radiate spicula prevalent, with three or four, rarely two or five, shanks on
each end of the middle rod. Number very variable. All shanks straight and smooth.
Dimensions.—Diameter of the capsule 0.1 to 0.2, length of the spicula 0.05 to
0.15.
Habitat.—South Pacific, Station 295, surface.
12. Rhaphidozoum pandora, n. sp. (Pl. 4, fig. 6).
Spicula of three different kinds; simple needles, radiate and geminate mixed. The simple
needles thin spindle-shaped, often curved. The radiate spicula commonly with three or four, rarely
five or six, curved rays. The geminate-radiate spicula commonly with three or four, rarely two or
five, shanks on each end, often different on the two ends of the middle rod. Number and form very
variable. All or most of the shanks more or less bent and thorny.
Dimensions.—Diameter of the capsule 0.1 to 0.3, length of the spicula 0.05 to
0.2.
Habitat.—South Atlantic (near Ascension Island), Station 343,
surface.
Order II. SPHÆRELLARIA, Haeckel, 1881.
Sphærellaria, Haeckel, 1881, Prodromus, p. 421.
Sphæridea vel Peripylea, Hertwig, 1879, Organismus der Radiol., p. 133.
Definition.—Spumellaria with latticed or spongy shell.
The order Sphærellaria, the second order of Radiolaria, comprises all
those Spumellaria in which the skeleton is a latticed or fenestrated,
often more or less spongy, siliceous shell. Originally this shell is a simple extracapsular
lattice-sphere, in which the central capsule is included; from this simple ancestral form an
enormous {50}mass of different and often very complicated
forms is derived; this order is by far the largest, and in morphological respects the most
important and most interesting, of all Radiolaria. It contains not less than twenty-eight
different families, three hundred and five genera, and more than sixteen hundred species.
In my Monograph (1862) seven families appertaining to this group are described—the
Ethmosphærida, Cladococcida, Ommatida, Spongurida, Discida, Lithelida, and Collosphærida. The
astonishing increase of this group by the detection of a large series of new and interesting
forms, and particularly of important connecting forms between very different branches of it, now
enables me to give a much better arrangement. I discern now four suborders or sections of Sphærellaria, according to the different geometrical form of the central
capsule and of the latticed shell enveloping it. The first of these, and the common ancestral
group of the whole order, is the Sphæroidea, with spherical capsule; in
the Prunoidea it becomes ellipsoidal or cylindrical by prolongation of
one axis; in the Discoidea lenticular or discoidal by shortening of one
axis; in the Larcoidea lentelliptical, or triaxon-ellipsoid, by different
growth of the capsule in three different "dimensive axes."
Synopsis of the Four Suborders of Sphærellaria.
| Central capsule spherical. |
|
Shell a simple sphere or a system of concentric
spheres, |
1. Sphæroidea. |
| Central capsule ellipsoidal or cylindrical. |
|
Shell a simple ellipsoid or a cylinder with annular transverse
constrictions, |
2. Prunoidea. |
| Central capsule lenticular or discoidal. |
|
Shell a biconvex lens or a flat disk, |
3. Discoidea. |
| Central capsule lentelliptical or triaxon. |
|
Shell a triaxon-ellipsoid, with three different axes, |
4. Larcoidea. |
| Central capsule spherical. |
|
|
Shell a simple sphere or a system of concentric spheres, |
|
|
|
|
|
|
1. Sphæroidea. |
|
|
|
|
| Central capsule ellipsoidal or cylindrical. |
|
|
Shell a simple ellipsoid or a cylinder with annular transverse
constrictions, |
|
|
|
|
|
|
2. Prunoidea. |
|
|
|
|
| Central capsule lenticular or discoidal. |
|
|
Shell a biconvex lens or a flat disk, |
|
|
|
|
|
|
3. Discoidea. |
|
|
|
|
| Central capsule lentelliptical or triaxon. |
|
|
Shell a triaxon-ellipsoid, with three different axes, |
|
|
|
|
|
|
4. Larcoidea. |
|
|
|
|
Suborder I. SPHÆROIDEA, Haeckel.
Sphæroida, Sphæridea, Sphærida, Haeckel, 1878, Protistenreich, p.
103.
Sphæridea, R. Hertwig, 1879, Organismus der Radiol., p. 39.
Definition.—Spumellaria with spherical central capsule
(very rarely somewhat modified, or allomorphous); with spherical fenestrated siliceous shell
(often an endospherical polyhedron, very rarely of more modified, subspherical form or
allomorphous). Growth of the shell in the three dimensive axes equal.
The suborder Sphæroidea, the first and most important of the four of
the Sphærellaria, comprises those Spumellaria in
which the original geometrical {51}spherical form is
quite constantly preserved in the central capsule, and commonly also in the fenestrated shell
enveloping the latter, although in many forms the sphere is more or less modified; very frequently
it is an "endospherical polyhedron," i.e., a polyhedron all the angles (or the nodes of the
network) of which lie upon the surface of a geometrical sphere; more rarely the spherical form is
more or less modified and irregular. In the great majority of Sphæroidea
there is no external indication of the three dimensive axes; but in many forms they are indicated
by the regular position of certain external radial spines or internal radial beams. However, in no
case are those three axes expressed in the form of the shell itself and of the enclosed spherical
central capsule; this is the main character by which the Sphæroidea
differ from the following sections:—Prunoidea, Discoidea, Larcoidea, all three of which arise from
them.
The section Sphæroidea, in the sense here restricted, was founded by
me in my Protistenreich (1878, p. 103) and adopted by Hertwig (1879) in his Organismus der
Radiolarien (p. 39). The different groups appertaining to this large section were characterised
more accurately in my Prodromus (1881, pp. 448-456); there I gave the characters of six
subfamilies with thirty tribes, containing ninety-three genera. Formerly, in my Monograph (1862),
the Sphæroidea were disposed in five different
families:—Ethmosphærida, Cladococcida, Ommatida, Spongosphærida, Collosphærida. At that time
I could not separate them sufficiently from some Acantharia and Phæodaria, which have a similar spherical lattice-shell.
As the number of different genera and species in the Sphæroidea is
much greater than in all other sections of Spumellaria, many forms were
already described by former authors. In the oldest system of Ehrenberg (1847, loc. cit., p.
53) they represent one part of his Haliommatina (with four genera, Haliomma,
Chilomma, Stylosphæra, Spongosphæra). Most species, however, of these genera
are Discoidea. Also in the latest system of Ehrenberg (1875, loc.
cit., p. 157) his Haliommatina are a confused conglomeration of different Spumellaria (Sphæroidea, Discoidea, and Prunoidea).
The section Sphæroidea is the largest division of Sphærellaria, comprising not less than one hundred and seven genera and six
hundred and fifty species. This enormous number (easily to be augmented by further investigations)
requires a careful disposition in different families and subfamilies. For this disposition two
different principles only can be employed: firstly, the number and disposition of the radial
spines; secondly, the number of the concentric latticed spheres, which are connected by
radial beams. I give here the preference to the first principle, whilst in my Prodromus (1881) I
had preferred the second. The question, which of the two principles is more important for the
classification of Sphæroidea, is very difficult to answer; probably in
many cases the former, in many the latter is more important for their phylogeny.
{52}
Regarding the number of the concentric shells which compose the latticed carapace of the Sphæroidea, we can distinguish six families, viz.:—
VIII. Monosphærida (with one single shell).
VIII. Dyosphærida (with two concentric shells).
VIII. Triosphærida (with three concentric shells).
IIIV. Tetrasphærida (with four concentric shells).
IIIV. Polysphærida (with five or more concentric shells).
IIVI. Spongosphærida (with spongy shells).
On the other hand, regarding the number of the radial spines and their regular disposition on
the shell-surface, we can distinguish five families, viz.:—
VIII. Liosphærida (without radial spines).
VIII. Stylosphærida (with two radial spines, opposite in one
axis).
VIII. Staurosphærida (with four radial spines, opposite in pairs in
two axes, perpendicular one to another).
IIIV. Cubosphærida (with six radial spines, opposite in pairs in the
three dimensive axes).
IIIV. Astrosphærida (with numerous—eight, twelve, twenty, or
more—radial spines, often more than a hundred).
All five latter groups contain representatives of all six former groups; therefore we get
together not less than thirty different subfamilies of Sphæroidea,
already enumerated in my Prodromus, 1881, p. 449. I repeat them here to give a better survey of
the system there employed.
| Families and Subfamilies of SPHÆROIDEA. |
Liosphærida
(anacantha). |
Stylosphærida
(diacantha). |
Staurosphærida
(tetracantha). |
Cubosphærida
(hexacantha). |
Astrosphærida
(polyacantha). |
Monosphærida.
(One single shell.) |
Ethmosphærida. |
Xiphostylida. |
Staurostylida. |
Hexastylida. |
Coscinommida. |
Dyosphærida.
(Two concentric shells.) |
Carposphærida. |
Sphærostylida. |
Staurolonchida. |
Hexalonchida. |
Haliommida. |
Triosphærida.
(Three concentric shells.) |
Thecosphærida. |
Amphistylida. |
Stauracontida. |
Hexacontida. |
Actinommida. |
Tetrasphærida.
(Four concentric shells.) |
Cromyosphærida. |
Cromyostylida. |
Staurocromyida. |
Hexacromyida. |
Cromyommida. |
Polysphærida.
(Five or more concentric shells.) |
Caryosphærida. |
Caryostylida. |
Staurocaryida. |
Hexacaryida. |
Arachnommida. |
Spongosphærida.
(Spongy shells.) |
Plegmosphærida. |
Spongostylida. |
Staurodorida. |
Hexadorida. |
Spongiommida. |
| Families and Subfamilies of SPHÆROIDEA. |
Liosphærida
(anacantha). |
Stylosphærida
(diacantha). |
Staurosphærida
(tetracantha). |
Monosphærida.
(One single shell.) |
Ethmosphærida. |
Xiphostylida. |
Staurostylida. |
Dyosphærida.
(Two concentric shells.) |
Carposphærida. |
Sphærostylida. |
Staurolonchida. |
Triosphærida.
(Three concentric shells.) |
Thecosphærida. |
Amphistylida. |
Stauracontida. |
Tetrasphærida.
(Four concentric shells.) |
Cromyosphærida. |
Cromyostylida. |
Staurocromyida. |
Polysphærida.
(Five or more concentric shells.) |
Caryosphærida. |
Caryostylida. |
Staurocaryida. |
Spongosphærida.
(Spongy shells.) |
Plegmosphærida. |
Spongostylida. |
Staurodorida. |
| Families and Subfamilies of SPHÆROIDEA. |
Cubosphærida
(hexacantha). |
Astrosphærida
(polyacantha). |
Monosphærida.
(One single shell.) |
Hexastylida. |
Coscinommida. |
Dyosphærida.
(Two concentric shells.) |
Hexalonchida. |
Haliommida. |
Triosphærida.
(Three concentric shells.) |
Hexacontida. |
Actinommida. |
Tetrasphærida.
(Four concentric shells.) |
Hexacromyida. |
Cromyommida. |
Polysphærida.
(Five or more concentric shells.) |
Hexacaryida. |
Arachnommida. |
Spongosphærida.
(Spongy shells.) |
Hexadorida. |
Spongiommida. |
{53}
The Monosphærida comprise all those Sphæroidea in which the carapace is represented only by one single
lattice-shell. Originally this shell is probably everywhere an extracapsular or "cortical shell,"
which is developed on the outside of the jelly-veil enveloping the central capsule, and serves as
a protective carapace for these soft enclosed parts. But with the progress of growth the central
capsule becomes larger than the including shell, and sends out through its pores club-shaped
prolongations or cæcal-sacs (Pl. 11,
figs. 1, 5; Pl. 19,
figs. 2, 3, 5; Pl. 20,
fig. 1a; Pl. 27,
fig. 3). These protruded sacs may fuse together again outside the shell and form a spherical
bladder, now enveloping the smaller shell; the latter now becomes an intracapsular or "medullary
shell."
As Pliosphærida (or Sphæroidea concentrica) we can oppose to
the simple Monosphærida all other Sphæroidea, the lattice-shell of which
is composed of two or more concentric shells, connected by radial beams. Probably all Pliosphærida
(or at least the greater part of them) arise from the Monosphærida by centrifugal growth; two or
more radial spines are developed from the surface of the simple lattice-sphere, and are united
together by communicating lateral branches, developed at equal distances from the centre; and this
same process may be repeated, two, three, four, or more times. In this way originate the
characteristic systems of concentric spheres, all united by piercing radial beams which arise from
the surface of the innermost sphere (not from its centre). Regarding this mode of growth, we can
distinguish the innermost as "original" or "primary" shell, and all subsequent ones as "apposed"
or "secondary" shells; if the number of concentric shells amount to three or more, commonly both
innermost shells lie within the central capsule and are medullary shells, whilst all others lie
outside it and are therefore cortical shells. This difference can be commonly recognised also in
the isolated shell, without its central capsule; the distance between the cortical and the
medullary shells being commonly much larger than the distance between the two medullary
shells.
The Dyosphærida, or the Sphæroidea with two
concentric shells, are the most numerous among the Pliosphærida. Commonly in this group the inner
or primary shell lies within the central capsule as a true "medullary shell," whilst the outer
lies outside it as a "cortical shell"; therefore the radial beams, connecting both, pierce the
wall of the capsule. But in several forms, mainly in the peculiar group of Diplosphærida, both
concentric shells remain outside the central capsule, and both are therefore "cortical
shells."
The Triosphærida, or the Sphæroidea with
three concentric shells, are also very rich in different forms, though not so numerous by far as
the Dyosphærida. Commonly in the Triosphærida both inner shells lie within the central capsule as
"medullary shells," whilst the third lies outside it as a "cortical shell"; therefore the central
capsule remains intermediate in size between the outer and the middle shell. But in some genera
(e.g., Rhodosphæra) both outer shells are cortical and only the {54}innermost is a medullary shell. In this case the size of
the capsule remains intermediate between the inner and the middle shell.
The Tetrasphærida, or the Sphæroidea with
four concentric shells, are in general not frequent, and not rich in different forms. In most of
the observed species two inner shells are medullary, two outer cortical shells, the former within,
the latter without, the central capsule; and the wall of the capsule, pierced by the connecting
radial beams, lies between the two middle shells. But there are some Tetrasphærida in which all
four shells seem to be external or cortical shells.
The Polysphærida, or the Sphæroidea with five
or more concentric shells, seem of course to offer the greatest possibility for the development of
very different forms; but in reality this group is the poorest and smallest of all; and only one
part of it, the Arachnosphærida, is rather common. In this peculiar division the shell is composed
of five to ten or more, very delicate, cobweb-like concentric shells, which are connected by
radial beams; all are cortical shells, and lie outside the central capsule. Much more rare are
those Polysphærida, in which both innermost shells, as true medullary shells, lie within the
central capsule, all others being outside it. The total number of concentric shells in this group
is commonly between five and ten, rarely more.
The Spongosphærida are distinguished from all other Sphæroidea by the spongy structure of the spherical shell, which is composed
wholly or partially of an irregular spongy framework. The relation of this group to the other
groups of Sphæroidea is probably rather complicated, for in some
Spongosphærida the whole shell is composed of massive spongy reticulation, whilst in others it
contains a spherical central cavity, and in a third group this cavity is filled up by one or two
concentric lattice-shells, connected by radial beams. Many of these Spongosphærida are very
common, and of considerable size.
The Collosphærida form a peculiar separate group of Sphæroidea, distinguished from all others by their social life or aggregation
in colonies (cœnobia). They represent the only group of Sphærellaria in which this association of numerous individual capsules or cells
is realised. The shell is almost constantly simple, without regularly disposed radial spines;
therefore they may be called "social Monosphærida," or better "polyzoic Ethmosphærida." Only in
one small group (Clathrosphærida) the shell, enveloping every central capsule, is double or
surrounded by an external mantle; these may be compared to the Diplosphærida (or better to a part
of the Carposphærida, Liosphæra, p. 76). In most of the Collosphærida
the lattice-shell is more or less irregular in form and structure.
The Lattice Work of the fenestrated shells is in the Sphæroidea
of the greatest variability, and its innumerable modifications serve mainly for the distinction of
species. In general we can distinguish as the most important modifications a regular
network (with equal size, form, and distance of the pores or meshes) and an irregular
network (with {55}differences in the size, form, or
distance of the meshes or pores). In both groups the pores may be either angular or round; so that
there may exist together four different main forms of network—(A) regular lattice with equal
hexagonal pores; (B) regular lattice with equal circular pores; (C) irregular lattice with unequal
polygonal pores; (D) irregular lattice with unequal roundish pores. Besides these modifications,
the pores may be prolonged into tubules which are directed radially towards the outside (rarely
towards the inside) of the sphere. In other cases they are surrounded by elevated or
honeycomb-like frames.
The Radial Spines exhibit in the Sphæroidea the greatest
variety in form, size, disposition, &c., and their numerous modifications serve mainly for the
distinction of genera, their peculiar formation and size also for the distinction of species. In
general we may distinguish as the most important modifications primary and secondary spines. The
primary spines or "main spines" are commonly direct outward prolongations of the internal radial
beams, connecting the concentric shells. The secondary or "by-spines" arise only from the surface
of the lattice-shell, without reference to the internal beams. The by-spines are commonly smaller,
and much more numerous than the main spines. Regarding the form, the radial spines are either
roundish (cylindrical or conical, often also club-shaped, rarely spindle-shaped) or angular
(commonly three-sided, prismatic or pyramidal). The spines are constantly solid, never hollow; the
"internal canals," described by some authors, are only microscopic views of the transparent edges.
In many cases the spines are branched or forked. The most important difference in the variable
shape of the spines is their regular or irregular number and disposition, which afford characters
for the distinction of our five families.
The Three Dimensive Axes—or the three diameters of the sphere, perpendicular one
to another—are in the great majority of the Sphæroidea significant
in the promorphological consideration of the body, and are indicated either by the position of the
external radial spines, or at least of the internal radial beams, connecting the concentric
spheres. Commonly two radial spines are placed opposite in each axis. The most perfect group in
this respect seems to be that of the Cubosphærida, in which the three axes are represented by
three pairs of spines. Next come the Staurosphærida, in which two axes in cross-form are exhibited
by two pairs of spines. The most simple group are the Stylosphærida, in which only one pair of
spines is developed, indicating one single axis. These three families form together a continuous
natural series,—the Sphæroidea with real dimensive axes,—and
exhibit at the same time relations to the three other suborders of Sphærellaria, the Larcoidea, Discoidea, and Prunoidea respectively. At both ends of
this series stand two other families, on one side the Liosphærida, without any radial spines on
the surface of the sphere, on the other side the Astrosphærida, in which the radial spines are
developed in great and variable numbers, at least eight to twelve, commonly twenty to forty, often
more than a hundred or even a thousand.
{56}
The Liosphærida comprise all those Sphæroidea
in which the surface of the shell is smooth, without radial spines (Pls. 12, 20). The
simplest of these are the Ethmosphærida, with one single lattice-shell, enveloping the spherical
central capsule. Cenosphæra, the most simple form of the Ethmosphærida, may be regarded as
the common ancestral form of all Sphæroidea, in an ontogenetical as well
as in a phylogenetical and morphological sense. From this simple lattice sphere all other Sphæroidea can be derived either by radial or by tangential growth. If the
radial beams, arising from the surface of the simple fenestrated sphere, become connected (at
equal distances from the centre) by tangential beams, we get the compound shells of the
"Liosphærida concentrica" (with two, three, four, or more concentric spheres). The radial beams
connecting these exhibit in many Liosphærida the same regular disposition and number as the
external radial spines in the Astrosphærida. Perhaps these forms in a "natural system" would be
better united (e.g., Liosphærida with twelve or twenty internal radial beams, and
Astrosphærida with twelve or twenty external radial spines); but in many cases (mainly for higher
numbers) the certain determination of their number and disposition is very difficult or quite
impossible.
The Cubosphærida (Pls. 21-25) represent
the large and very important family of Sphæroidea, in which all three
dimensive axes are equally distinguished by pairs of spines, corresponding to three axes of a cube
or of a regular octahedron, agreeing therefore also with the three axes of the cubic or regular
crystalline system. In the majority of the Cubosphærida the six radial spines are accurately
opposite each other in pairs in three axes, perpendicular one to another, and commonly they are of
equal size and form; but in some genera the three pairs of spines become differentiated, whilst
both spines of each pair remain equal. Either one pair is larger than the two others (which are
equal), corresponding to the axes of the quadratic crystalline system; or all three pairs are
different (corresponding to the three unequal axes of the rhombic crystalline system); the former
nearer to the Discoidea, the latter to the Larcoidea. We may suppose with some probability, that the Cubosphærida are for
the most part the common ancestral group of those Sphæroidea, in which a
certain number of radial spines or beams is disposed in a regular order; the Staurosphærida may be
derived from them by loss of one pair of spines, the Stylosphærida by loss of two pairs of spines,
and most Astrosphærida by multiplying the radial spines, six to fourteen or more secondary spines
being added to the six primary "dimensive spines." However, in many Astrosphærida (e.g., in
those with eight spines, Centrocubus, Octodendron, &c.) the regular geometrical
disposition of the radial spines seems to follow another mathematical order, quite independent of
the Cubosphærida.
The Staurosphærida (Pl. 15) are
distinguished by the possession of four radial spines, opposite in pairs in two axes,
perpendicular one to another. This rectangular cross determines a certain plane, the "equatorial
plane," and this brings the Staurosphærida near {57}to
the Discoidea, mainly to those which also bear on the periphery of the
circular equatorial plane four crossed spines (such as Staurodisculus,
Stethostaurus, Staurodictya, &c.). But in these cruciform Discoidea the shell and the enclosed central capsule are discoidal or
lenticular, whilst in the Staurosphærida they remain spherical. Commonly the cross is quite
regular, with four right angles and four equal beams; but often also it becomes more or less
irregular. In some genera one pair of equal opposite spines is larger than the other pair. These
forms represent the three different axes of the rhombic crystal system, whilst the common regular
Staurosphærida represent those of the quadratic crystal system. The latter can be derived from the
Cubosphærida (representing the regular crystal system) by reduction of one axis and loss of its
pair of spines. In general the number of species (and particularly of the individuals) is much
smaller in the Staurosphærida than in all other families of Sphæroidea.
The Stylosphærida (Pls. 13-17) can be
derived from the Cubosphærida by reduction of two dimensive axes and loss of two pairs of spines.
Therefore, here one pair of spines only is developed, opposite in one single axis. This
"monaxonial" form brings the Stylosphærida very near to the ellipsoidal Prunoidea (mainly to many two-spined forms of Ellipsida and Druppulida); but
they differ from these by the spherical (not ellipsoidal) form of the central capsule and of the
enclosing lattice-shell. In the greater part of the Stylosphærida both spines are of equal size
and form, accurately opposite in the "main axis." But in many forms both spines become unequal in
size or form, often very different. More rarely they are not accurately opposed, but placed in two
different axes, intersecting at a small variable angle. The small group of Saturnalida presents a
very remarkable and peculiar structure, in which both spines (at equal distances from the centre)
are united by a circular or elliptical ring (Pl. 13, figs. 15,
16; Pl. 16,
figs. 16, 17).
The Astrosphærida are distinguished from the other Sphæroidea by the great and variable number of their external radial spines
(Pls. 11, 18-20, 26-30). Commonly
this number amounts to from twelve to twenty, rarely to only eight to ten, very often to
thirty-two to forty or more; in many species more than one hundred are present. As already
mentioned above, it would be important to distinguish between primary spines (as outer
prolongations of the inner radial beams) and secondary spines (developed from the surface of the
shell), but in many cases this distinction is difficult or impossible. More practical is the
distinction between larger "main spines" and smaller "by-spines." The size and form of the radial
spines is extremely variable. Much more important is their number and disposition. In general we
can here distinguish the following different cases:—(A) radial spines are developed from all
the nodal points of the network on the shell surface; (B) the number of the spines is smaller than
that of the nodal points, but they are irregularly scattered; (C) the radial spines exhibit a
limited number and a certain regular disposition. In this latter case the following modes of
distribution seem to be the most important:—(a) eight spines placed in the four
diagonal axes of the {58}regular cube (Pl. 18, figs. 1-3);
(b) twelve spines (placed in the corner axes of the regular icosahedron); (c)
fourteen spines (six placed in the three dimensive axes of the regular octahedron, eight in the
centres of its eight faces); (d) twenty spines (placed either in the same order as in many
Larcoidea and Acantharia [?], or in the twenty
corners of the regular dodecahedron); (e) thirty-two spines (twelve placed in the twelve
corners of the regular icosahedron, twenty in the centre of its triangular faces). Besides these
most important and quite geometrical modes of disposition there also seem to occur in the
Astrosphærida the following subregular (or symmetrical?) modes: 9, 10, 16, 18, 24, 40, 60, 80. But
it is very difficult to give a correct account of these modes. In every case this manifold and
regular disposition of the radial spines is of the highest interest for the study of general
"Promorphology."
The Central Capsule is in all Sphæroidea (without any
exception) a perfect sphere in the geometrical sense, even in those forms in which the enclosing
lattice-shell is more or less irregular (i.e., many Collosphærida). This is the most
important character, which separates the Sphæroidea from all other Sphærellaria. For in the Prunoidea the capsule is
ellipsoidal, with one prolonged axis; in the Discoidea lenticular, with
one shortened axis; in the Larcoidea lentelliptical, with three different
dimensive axes. The central capsule is originally always enclosed by the lattice-shell; but in
many cases with increasing growth this relation becomes inverted; the capsule sending out many
club-shaped blind sacs through the meshes of the lattice-shell, and these melting together outside
the latter, a new membrane is formed, enclosing a "medullary shell."
The Nucleus of the cell exhibits a very different shape in the solitary and the social
Sphæroidea. In the solitary or monozoic Sphæroidea the centre of the central capsule is occupied by a large spherical
concentric nucleus, with or without nucleoli; also this nucleus is originally always within the
innermost lattice-shell, but with increasing size may overgrow and enclose it. A short time before
the formation of the vibratile spores the central nucleus becomes resolved into many small nuclei.
In the social or polyzoic Sphæroidea—the
Collosphærida—commonly the simple central nucleus very early (a long time before the
formation of the spores) is divided into a great number of small nuclei, whilst the centre of the
capsule becomes filled with a large oil-globule. Therefore we find the same difference between the
solitary and social forms in the Sphæroidea as in the Colloidea. Here also the calymma, or the jelly-mantle, enveloping the central
capsule, is in the social forms very large and voluminous, differentiated into alveoles, whilst in
the solitary forms it is much smaller, without alveoles.
{59}
Synopsis of the Families of Sphæroidea.
| Surface of the spherical shell smooth, rough, or thorny,
but not armed with radial spines. |
 |
A. Liosphærida monozoa. Single cells (each with shell) living
solitary. |
|
Spherical shell commonly quite regular, simple, or composed of two
or more concentric spheres, |
05. Liosphærida. |
| B. Liosphærida polyzoa. Aggregated cells (each with shell) living in
colonies. |
|
Spherical shell commonly more or less irregular, simple (rarely
composed of two concentric spheres), |
06. Collosphærida. |
| Surface of the spherical shell armed with two, four, or six
radial main spines, opposite in pairs in one, two, or three dimensive axes (always
solitary). |
|
Two radial main-spines, opposite in one axis of the
shell |
07. Stylosphærida. |
| Four radial main-spines, opposite in pairs in two
dimensive axes, perpendicular one to another, |
08. Staurosphærida. |
| Six radial main-spines, opposite in pairs in three
dimensive axes (perpendicular one to another), |
09. Cubosphærida. |
| Surface of the spherical shell covered with numerous
(commonly irregularly disposed) radial spines, often also twelve to twenty, more or less
regularly disposed, |
10. Astrosphærida. |
| Surface of the spherical shell smooth, rough, or thorny, but not armed with
radial spines. |
|
|
A. Liosphærida monozoa. Single cells (each with shell) living
solitary. |
|
|
|
|
|
|
Spherical shell commonly quite regular, simple, or composed of two
or more concentric spheres, |
|
|
|
|
|
|
|
|
|
|
5. Liosphærida. |
|
|
|
|
|
|
|
|
B. Liosphærida polyzoa. Aggregated cells (each with shell) living
in colonies. |
|
|
|
|
|
|
Spherical shell commonly more or less irregular, simple (rarely
composed of two concentric spheres), |
|
|
|
|
|
|
|
|
|
|
6. Collosphærida. |
|
|
|
|
|
|
| Surface of the spherical shell armed with two, four, or six radial main
spines, opposite in pairs in one, two, or three dimensive axes (always solitary). |
|
|
Two radial main-spines, opposite in one axis of the shell |
|
|
|
|
|
|
|
|
7. Stylosphærida. |
|
|
|
|
|
|
|
|
Four radial main-spines, opposite in pairs in two dimensive axes,
perpendicular one to another, |
|
|
|
|
|
|
|
|
8. Staurosphærida. |
|
|
|
|
|
|
|
|
Six radial main-spines, opposite in pairs in three dimensive axes
(perpendicular one to another), |
|
|
|
|
|
|
|
|
9. Cubosphærida. |
|
|
|
|
|
|
| Surface of the spherical shell covered with numerous (commonly irregularly
disposed) radial spines, often also twelve to twenty, more or less regularly disposed, |
|
|
|
|
|
|
10. Astrosphærida. |
|
|
|
|
|
|
Family V. Liosphærida, Haeckel, 1881.
Liosphærida, Haeckel, 1881, Prodromus, p. 449.
Definition.—Sphæroidea without radial spines on the
surface of the spherical shell; living solitary (not associated in colonies).
The family Liosphærida comprises all those solitary Sphæroidea in which the surface of the spherical shell is not armed with radial
spines. Nearly the half of this large group is formed by the Ethmosphærida, in which the carapace
is a quite simple, spherical lattice-shell; this subfamily is probably the common ancestral group
from which all other Sphæroidea, or even all Sphærellaria, can be derived in a phylogenetical as well as in a morphological
sense. The central capsule in this first subfamily is constantly enclosed by the fenestrated
shell, and separated from it by the jelly-veil. The shell is therefore an extracapsular or
medullary shell.
To these simple Ethmosphærida all other subfamilies can be opposed as "Liosphærida
concentrica," as their carapace is composed of two or more concentric lattice-shells; two in the
Carposphærida, three in the Thecosphærida, four in the Cromyosphærida, five or more in the
Caryosphærida. In all these four subfamilies the concentric shells are simple (not spongy)
fenestrated shells. In a sixth subfamily, in the Plegmosphærida, the shell is wholly or partially
composed of spongy wicker-work, with or without a latticed medullary shell in the centre.
The internal radial beams, in the "Liosphærida composita" connecting the concentric spheres,
exhibit in their number and disposition similar important differences, such as the external radial
spines in the Astrosphærida. The following eight {60}different cases of regular disposition were observed:—(A) two
opposite beams in one axis; (B) four beams, opposite in pairs in two axes perpendicular one to
another; (C) six beams, opposite in pairs in the three dimensive axes; (D) eight beams, opposite
in pairs in the four diagonals of the regular cube; (E) twelve beams corresponding to the twelve
axes of the regular icosahedron; (F) fourteen beams quite regularly disposed (six corresponding to
the three axes of the regular octahedron, eight to the central points of its faces); (G) twenty
beams (probably corresponding to the twenty corners of a regular dodecahedron); (H) thirty-two
beams, regularly disposed. Rarely the number of the radial beams is intermediate between these
eight cases, and rarely it is higher; then commonly the disposition is irregular. The regularity
of their disposition in the great majority of cases is very remarkable and evident.
Synopsis of the Genera of Liosphærida.
|
I. Subfamily Ethmosphærida.
(Shell one single latticed sphere.)
|
|
Pores of the shell simple, not prolonged into free
tubuli. |
|
Shell cavity simple, |
15. Cenosphæra. |
| Shell cavity with radial beams united in the centre, |
16. Stigmosphæra. |
| Pores prolonged into free conical or cylindrical
tubuli. |
|
Tubuli external, centrifugal, |
17. Ethmosphæra. |
| Tubuli internal, centripetal, |
18. Sethosphæra. |
|
II. Subfamily Carposphærida.
(Two concentric spheres.)
|
|
One shell medullary (intracapsular), the other cortical (extracapsular), |
19. Carposphæra. |
| Both shells cortical (near together), |
20. Liosphæra. |
|
III. Subfamily Thecosphærida.
(Three concentric spheres.)
|
|
Two shells medullary (intracapsular), one shell cortical (extracapsular), |
21. Thecosphæra. |
| One shell medullary (intracapsular), two shells cortical (extracapsular), |
22. Rhodosphæra. |
|
IV. Subfamily Cromyosphærida.
(Four concentric spheres.)
|
|
Two inner medullary shells (intracapsular), and two outer cortical
shells (extracapsular), |
23. Cromyosphæra. |
|
V. Subfamily Caryosphærida.
(Five or more concentric spheres.)
|
|
Two inner medullary shells, and three or more outer cortical
shells, |
24. Caryosphæra. |
|
VI. Subfamily Plegmosphærida.
(Spherical shell wholly or partially of spongy structure.)
|
|
Spongy sphere without latticed medullary shell in the
centre. |
|
Sphere solid, |
25. Styptosphæra. |
| Sphere with a central cavity, |
26. Plegmosphæra. |
| Spongy sphere with one or two latticed medullary shells in
the centre. |
|
One single medullary shell, |
27. Spongoplegma. |
| Two concentric medullary shells, |
28. Spongodictyon. |
| I. Subfamily Ethmosphærida. (Shell one single latticed sphere.) |
|
|
Pores of the shell simple, not prolonged into free tubuli. |
|
|
|
|
|
|
Shell cavity simple, |
|
|
|
|
|
|
|
|
|
|
15. Cenosphæra. |
|
|
|
|
|
|
|
|
|
|
Shell cavity with radial beams united in the centre, |
|
|
|
|
|
|
|
|
|
|
16. Stigmosphæra. |
|
|
|
|
|
|
|
|
Pores prolonged into free conical or cylindrical tubuli. |
|
|
|
|
|
|
Tubuli external, centrifugal, |
|
|
|
|
|
|
|
|
|
|
17. Ethmosphæra. |
|
|
|
|
|
|
|
|
|
|
Tubuli internal, centripetal, |
|
|
|
|
|
|
|
|
|
|
18. Sethosphæra. |
|
|
|
|
|
|
| II. Subfamily Carposphærida. (Two concentric spheres.) |
|
|
One shell medullary (intracapsular), the other cortical
(extracapsular), |
|
|
|
|
|
|
|
|
19. Carposphæra. |
|
|
|
|
|
|
|
|
Both shells cortical (near together), |
|
|
|
|
|
|
|
|
20. Liosphæra. |
|
|
|
|
|
|
| III. Subfamily Thecosphærida. (Three concentric spheres.) |
|
|
Two shells medullary (intracapsular), one shell cortical
(extracapsular), |
|
|
|
|
|
|
|
|
21. Thecosphæra. |
|
|
|
|
|
|
|
|
One shell medullary (intracapsular), two shells cortical
(extracapsular), |
|
|
|
|
|
|
|
|
22. Rhodosphæra. |
|
|
|
|
|
|
| IV. Subfamily Cromyosphærida. (Four concentric spheres.) |
|
|
Two inner medullary shells (intracapsular), and two outer cortical
shells (extracapsular), |
|
|
|
|
|
|
|
|
23. Cromyosphæra. |
|
|
|
|
|
|
| V. Subfamily Caryosphærida. (Five or more concentric spheres.) |
|
|
Two inner medullary shells, and three or more outer cortical
shells, |
|
|
|
|
|
|
|
|
24. Caryosphæra. |
|
|
|
|
|
|
| VI. Subfamily Plegmosphærida. (Spherical shell wholly or partially of spongy
structure.) |
|
|
Spongy sphere without latticed medullary shell in the centre. |
|
|
|
|
|
|
Sphere solid, |
|
|
|
|
|
|
|
|
|
|
25. Styptosphæra. |
|
|
|
|
|
|
|
|
|
|
Sphere with a central cavity, |
|
|
|
|
|
|
|
|
|
|
26. Plegmosphæra. |
|
|
|
|
|
|
|
|
Spongy sphere with one or two latticed medullary shells in the
centre. |
|
|
|
|
|
|
One single medullary shell, |
|
|
|
|
|
|
|
|
|
|
27. Spongoplegma. |
|
|
|
|
|
|
|
|
|
|
Two concentric medullary shells, |
|
|
|
|
|
|
|
|
|
|
28. Spongodictyon. |
|
|
|
|
|
|
{61}
Subfamily Ethmosphærida,[25] Haeckel, 1862, Monogr. d. Radiol., p. 348 (sensu
restricto).
Definition.—Liosphærida with one single
spherical lattice-shell; living solitary, not aggregated in colonies.
Genus 15. Cenosphæra,[26] Ehrenberg, 1854,
Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 237.
Definition.—Liosphærida with one single latticed sphere,
with simple shell-pores (not prolonged into free tubuli) and with simple shell-cavity (without
internal radial beams).
The genus Cenosphæra is the most simple form of all Sphæroidea, and may be regarded as the common ancestral form of this order. The
siliceous latticed shell, in which the central capsule is enclosed, represents a simple regular
sphere, with simple cavity. The pores of the shell-wall are simple, not prolonged into radial
tubuli (as in Ethmosphæra and Sethosphæra). According to the different form of the
pores, the numerous species of this genus can be disposed in four different subgenera. Some
species may be easily confounded with isolated shells of the corresponding social
Collosphæra; but in this latter the spherical shell-form is commonly more or less
irregular, in Cenosphæra quite regular.
Subgenus 1. Phormosphæra, Haeckel, 1881, Prodromus, p. 448.
Definition.—Pores of the shell regular or subregular, hexagonal or
circular, with hexagonal frames or lobes; all nearly of equal size and form.
1. Cenosphæra primordialis, n. sp.
Shell very thin walled, smooth. Pores hexagonal, regular, or subregular; twelve to fifteen on
the half meridian of the shell; bars between them extremely delicate (only visible when three
hundred or four hundred times enlarged). Diameter of the shell nine to ten times that of the
meshes. This species is remarkable for the extreme delicacy of the arachnoidal network of the
simple spherical shell; it may be regarded as the common ancestral form of all Sphæroidea. The shell equals that of Heliosphæra tenuissima (figured in
my Monograph, 1862, pl. ix. fig. 2), but differs from it by the smooth surface and the absence of
all spines or thorns. I observed this species living in the Indian Ocean, near Ceylon, in 1882;
the spherical diameter of the central capsule is about one-third of that of the shell; the
contents of the central capsule are colourless {62}and
transparent, except the central dark globular nucleus. The same shells also occur in some mounted
preparations of surface organisms from the Challenger.
Dimensions.—Diameter of the shell 0.12, of the pores 0.012.
Habitat.—Indian Ocean, Ceylon, Haeckel; Central Pacific, Stations 266,
271, surface.
2. Cenosphæra inermis, Haeckel.
Heliosphæra inermis, Haeckel, 1862, Monogr. d. Radiol., p. 351, Taf. ix.
fig. 1.
Surface of the thin-walled shell smooth. Pores regular, hexagonal, twelve to fifteen times as
broad as the bars, seven to nine on the quadrant.
Dimensions.—Diameter of the shell 0.08 to 0.12, pores 0.012 to 0.015, bars
0.001.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Indian, Pacific,
surface.
3. Cenosphæra hexagonalis, n. sp.
Surface of the thick-walled shell smooth. Pores regular, hexagonal, five to six times as broad
as the bars, six to eight on the quadrant.
Dimensions.—Diameter of the shell 0.1 to 0.15, pores 0.01 to 0.012, bars
0.002.
Habitat.—Central Pacific, Stations 265 to 274, at various depths.
4. Cenosphæra mellifica, n. sp. (Pl. 12,
fig. 9).
Surface of the thick-walled shell smooth. Pores regular, circular, with thin hexagonal frames,
four times as broad as the bars, six to eight on the quadrant.
Dimensions.—Diameter of the shell 0.2, pores 0.12, bars 0.003.
Habitat.—South Pacific, Station 288, surface.
5. Cenosphæra favosa, n. sp. (Pl. 12,
fig. 10).
Surface of the thick-walled shell rough. Pores regular, circular, with thin hexagonal frames,
three times as broad as the bars, ten to twelve on the quadrant.
Dimensions.—Diameter of the shell 0.2, pores 0.09, bars 0.003.
Habitat.—North Atlantic, Færöe Channel (Gulf Stream), John Murray,
surface.
6. Cenosphæra vesparia, n. sp. (Pl. 12,
fig. 11).
Surface of the thick-walled shell smooth. Pores regular, circular, with thick hexagonal
frames, twice as broad as the bars, ten to twelve on the quadrant.
Dimensions.—Diameter of the shell 0.25, pores 0.016, bars 0.008.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms, and
surface.
{63}
7. Cenosphæra bombus, n. sp.
Shell thick walled, rough. Pores regular, circular, with thin hexagonal frames, of the same
breadth as the bars, twenty to twenty-two on the quadrant.
Dimensions.—Diameter of the shell 0.3, pores and bars 0.005.
Habitat.—Tropical Atlantic, Station 347, depth 2250 fathoms.
8. Cenosphæra melecta, n. sp.
Shell thick walled, papillate. Pores regular, circular, double-edged, with thick hexagonal
frames, of the same breadth as the bars; a short conical papilla in the corner of each hexagon;
fourteen to sixteen pores on the quadrant.
Dimensions.—Diameter of the shell 0.2, pores and bars 0.008.
Habitat.—Fossil in Barbados.
9. Cenosphæra anthophora, n. sp.
Shell thick walled, papillate. Pores regular, circular, with an elegant six-lobed frame and a
coronal of six short papillæ; the latter alternating with the six lobes (quite as in Pl. 28, fig.
1b). Pores twice as broad as the bars, ten to twelve on the quadrant.
Dimensions.—Diameter of the shell 0.13, pores 0.01, bars 0.005.
Habitat.—Central Pacific, Station 271, surface.
10. Cenosphæra rosetta, n. sp.
Shell thin walled, smooth. Pores regular, circular, with an elegant six-lobed outer opening,
without papillæ. Pores of the same breadth as the bars, six to eight on the quadrant.
Dimensions.—Diameter of the shell 0.1, pores and bars 0.006.
Habitat.—South Atlantic, Station 323, depth 1900 fathoms.
Subgenus 2. Circosphæra, Haeckel.
Definition.—Pores of the spherical shell regular or subregular,
circular, without hexagonal frames or lobes, all nearly of equal size and form.
11. Cenosphæra porophæna, Ehrenberg.
Cenosphæra porophæna, Ehrenberg, 1858, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 31.
Shell thin walled, smooth. Pores regular, circular, six to eight times as broad as the bars,
five to six on the quadrant.
Dimensions.—Diameter of the shell 0.1, pores 0.012, bars 0.002.
Habitat.—Mediterranean (Crete, depth 1100 fathoms; Corfu,
surface).
{64}
12. Cenosphæra setosa, Ehrenberg.
Cenosphæra setosa, Ehrenberg, 1872, Abhandl. d. k. Akad. d. Wiss. Berlin,
p. 287, Taf. vii. fig. 1.
Shell thin walled, covered with very short and numerous bristles. Pores regular, circular, five
to six times as broad as the bars, six to eight on the quadrant.
Dimensions.—Diameter of the shell 0.17, pores 0.2, bars 0.03.
Habitat.—West Tropical Pacific, 3300 fathoms; Philippine Sea, Station
200, depth 250 fathoms.
13. Cenosphæra plutonis, Ehrenberg.
Cenosphæra plutonis, Ehrenberg, 1854, Mikrogeol., Taf. xxxv. B, B. iv. fig. 20.
Shell thin walled, covered with short conical papillæ. Pores regular, circular, twice as broad
as the bars, eight to nine on the quadrant.
Dimensions.—Diameter of the shell 0.09, pores 0.006, bars 0.003.
Habitat.—North Atlantic, Station 353, depth 2965 fathoms.
14. Cenosphæra proserpinæ, n. sp.
Shell thin walled, smooth. Pores regular, circular, four times as broad as the bars, five to
six on the quadrant.
Dimensions.—Diameter of the shell 0.08, pores 0.008, bars 0.002.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms.
15. Cenosphæra eridani, n. sp.
Shell thin walled, smooth. Pores regular, circular, three times as broad as the bars, eleven to
twelve on the quadrant.
Dimensions.—Diameter of the shell 0.16, pores 0.01, bars 0.003.
Habitat.—South Pacific, Station 295, depth 1500 fathoms.
16. Cenosphæra lethe, n. sp.
Shell thick walled, smooth. Pores regular, circular, double-edged, five times as broad as the
bars, sixteen to eighteen on the quadrant.
Dimensions.—Diameter of the shell 0.25, pores 0.01, bars 0.002.
Habitat.—North Atlantic, Station 64, depth 2700 fathoms.
17. Cenosphæra elysia, n. sp. (Pl. 12,
fig. 8).
Shell thick walled, rough. Pores regular, circular, double-edged, twice as broad as the bars,
twelve to fourteen on the quadrant.
Dimensions.—Diameter of the shell 0.2, pores 0.01, bars 0.005.
Habitat.—Central Pacific, Station 266, depth 2750 fathoms.
{65}
18. Cenosphæra nirwana, n. sp.
Shell thick walled, smooth. Pores regular, circular, twice as broad as the bars, twenty-four to
twenty-five on the quadrant.
Dimensions.—Diameter of the shell 0.25, pores 0.004, bars 0.002.
Habitat.—Indian Ocean, Belligemma, Ceylon, surface; Haeckel.
19. Cenosphæra maxima, n. sp.
Shell thick walled, smooth. Pores regular, circular, twice as broad as the bars, thirty to
thirty-three on the quadrant.
Dimensions.—Diameter of the shell 0.3 to 0.4, pores 0.012, bars 0.006.
Habitat.—West Tropical Pacific, Station 225, depth 4475; also fossil
in Barbados.
20. Cenosphæra compacta, n. sp. (Pl. 12,
fig. 7).
Shell very thick walled, rough (its wall one-fourth to one-third as thick as the radius). Pores
subregular, circular, of the same breadth as the bars, seven to eight on the quadrant.
Dimensions.—Diameter of the shell 0.15, pores and bars 0.012.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms.
Cenosphæra radiata,
Zittel, 1876 (L. N. 29, p. 84, Taf. ii. figs. 7, 8), a fossil Cretaceous
species, is closely allied to Cenosphæra compacta.
21. Cenosphæra crassa, n. sp.
Shell very thick walled, rough (its wall nearly half as thick as the radius). Pores tubular,
double-edged, regular, circular, ten times as broad as the thin united bars, twelve to fourteen on
the quadrant.
Dimensions.—Diameter of the shell 0.14, pores 0.01, bars 0.001.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
22. Cenosphæra solida, n. sp.
Shell very thick walled, covered with innumerable short bristles (its wall one-third as thick
as the radius). Pores regular, circular, four times as broad as the bars, tubular, eight to ten on
the quadrant.
Dimensions.—Diameter of the shell 0.2, pores 0.02, bars 0.005.
Habitat.—Antarctic Ocean, Station 157, depth 1950 fathoms.
Subgenus 3. Cyrtidosphæra, Haeckel, 1862, Monogr. d. Radiol., p. 348.
Definition.—Pores of the spherical shell irregularly polygonal, of
unequal size or form, sometimes roundish with polygonal frames.
{66}
23. Cenosphæra reticulata, Haeckel.
Cyrtidosphæra reticulata, Haeckel, 1862, Monogr. d. Radiol., p. 349, Taf.
xi. fig. 2.
Shell very thin walled, smooth. Pores irregular, polygonal, two to eight times as broad as the
bars, fifteen to twenty on the quadrant (groups of four to eight smaller meshes are scattered on
the surface, and separated by reticular rows of larger meshes).
Dimensions.—Diameter of the shell 0.16, pores 0.004 to 0.016, bars 0.002.
Habitat.—Mediterranean (Messina), surface.
24. Cenosphæra tenerrima, n. sp.
Shell extremely thin walled, smooth, like a cobweb. Pores very irregular and small, polygonal
with thread-like bars, thirty to forty on the quadrant.
Dimensions.—Diameter of the shell 0.25, pores 0.002 to 0.008, bars under
0.001.
Habitat.—Central Pacific, Station 271, surface.
25. Cenosphæra polygonalis, n. sp.
Shell thin walled, smooth, with irregular, polygonal pores, three to four times as broad as the
bars, eight to ten on the quadrant.
Dimensions.—Diameter of the shell 0.2, pores 0.012 to 0.02, bars 0.005.
Habitat.—North Pacific, Station 236, surface.
26. Cenosphæra papillata, n. sp.
Shell thick walled, covered with short conical papillæ. Pores irregular, polygonal, three to
five times as broad as the bars, fourteen to sixteen on the quadrant.
Dimensions.—Diameter of the shell 0.12, pores 0.004 to 0.007, bars 0.0015.
Habitat.—South Atlantic, Station 325, surface.
27. Cenosphæra cristata, n. sp.
Shell thick walled, rough. Pores irregular, roundish, surrounded by polygonal crested frames
two to three times as broad as the bars, eight to twelve on the quadrant.
Dimensions.—Diameter of the shell 0.16, pores 0.006 to 0.01, bars 0.003.
Habitat.—North Pacific, Station 254, surface.
28. Cenosphæra perforata, n. sp. (Pl. 26, fig.
10).
Ceriosphæra perforata, Haeckel, 1881, Prodromus et Atlas, loc.
cit.
Shell thick walled, rough. Pores irregular, roundish, surrounded by high polygonal
funnel-shaped frames, which are solid in the inner half, perforated by numerous very small pores
in the {67}outer half, sieve-shaped. Pores one to three
times as broad as the bars, of very different size, four to six on the quadrant.
Dimensions.—Diameter of the shell 0.15, pores 0.01 to 0.02, bars 0.005 to
0.01.
Habitat.—Tropical Atlantic, Station 347, depth 2250 fathoms.
29. Cenosphæra coronata, n. sp. (Pl. 26,
fig. 11).
Shell thick walled, rough. Pores irregular, roundish, surrounded by high polygonal frames
bearing on their sharp crest a series of small papillæ, so that each pore is surrounded by a
coronal of such spinules. Pores four to eight times as broad as the bars, four to five on the
quadrant.
Dimensions.—Diameter of the shell 0.15, pores 0.01 to 0.03, bars 0.002 to
0.008.
Habitat.—Central Pacific, Station 272, depth 2600 fathoms.
Subgenus 4. Porosphæra, Haeckel.
Definition.—Pores of the spherical shell irregular, roundish, without
polygonal frames, of unequal size or form.
30. Cenosphæra antiqua, Haeckel.
Cenosphæra plutonis, var., Dunikowski, 1882, Denkschr. d. k. Akad. d. Wiss. Wien, Bd.
xlv. p. 25, Taf. iv. figs. 47, 48.
Cenosphæra plutonis, var., Stöhr, 1880, Palæontogr. xxvi. p. 85, Taf. i.
fig. 1.
Shell thin walled, smooth. Pores irregular, roundish, two to eight times as broad as the bars,
ten to fifteen on the quadrant.
Dimensions.—Diameter of the shell 0.15 to 0.2, pores 0.007 to 0.015, bars 0.002 to
0.008.
Habitat.—Fossil in the Jurassic, Cretaceous, and Tertiary formations;
living in the depths of the Atlantic and Pacific; Station 332, depth 2200 fathoms; Station 225,
depth 4475 fathoms, &c.
31. Cenosphæra gigantea, n. sp.
Shell thick walled, smooth. Pores irregular, roundish, two to five times as broad as the bars,
thirty to forty on the quadrant.
Dimensions.—Diameter of the shell 0.4 to 0.5, pores 0.004 to 0.01, bars 0.002.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms; also fossil
in Barbados.
32. Cenosphæra marginata, n. sp.
Shell very thick walled, smooth. Pores irregular, roundish, double-edged, three to eight times
as broad as the bars, six to eight on the quadrant.
{68}
Dimensions.—Diameter of the shell 0.12, pores 0.01 to 0.03, bars 0.004.
Habitat.—Central Pacific, Station 274, depth 2750 fathoms.
33. Cenosphæra aspera, Stöhr.
Cenosphæra aspera, Stöhr, 1880, Palæontogr. Bd. xxvi. p. 85, Taf. i. fig.
2.
Shell thick walled, rough, covered with short conical papillæ. Pores irregular, roundish, of
about the same breadth as the bars, eight to twelve on the quadrant.
Dimensions.—Diameter of the shell 0.17, pores and bars 0.01 to 0.04.
Habitat.—Fossil in Tertiary rocks of Sicily; Grotte, Stöhr.
34. Cenosphæra hirsuta, Ehrenberg.
Cenosphæra hirsuta, Ehrenberg, 1872, Abhandl. d. k. Akad. d. Wiss.,
Berlin, p. 287, Taf. x. fig. 18.
Shell thin walled, rough, covered with innumerable very short bristles. Pores very irregular,
roundish, of about the same breadth as the bars.
Dimensions.—Diameter of the shell 0.11, pores and bars 0.002 to 0.012.
Habitat.—Indian Ocean, Zanzibar, Pullen; depth 2200 fathoms.
Genus 16. Stigmosphæra,[27] Haeckel, n.
gen.
Definition.—Liosphærida with one single latticed sphere,
with simple shell-pores (not prolonged into free tubuli); in the central point of the spherical
shell-cavity are united a number of radial beams, which become forked and inserted into the inner
surface of the shell by their distal ends.
The genus Stigmosphæra differs from Cenosphæra (and from all other
Monosphærida) by internal radial beams, which are united in the centre of the simple spherical
shell; these beams are branched, and the distal ends of the branches inserted on the internal
surface of the shell. I have observed only two, nearly identical specimens of this genus, both
with regular, hexagonal pores and thin bars; the beams were implanted in the corners of the
hexagons. In one specimen the surface was covered with short radial bristles, whilst these in the
other specimen were prolonged into radial spines (like Acanthosphæra). Possibly this
peculiar genus is derived from Carposphæra, by reduction and loss of a central medullary
shell.
Stigmosphæra actinocentra, n. sp.
Shell very thin walled, rough, with regular circular, hexagonally framed pores, six times as
broad as the bars, eight to ten on the quadrant; in the corner of each hexagon a small bristle. In
{69}the central point of the shell are united about
twelve (?) thin and straight radial beams, which are forked, with dichotomous branches; the distal
ends of the branches are inserted in the corners of the hexagons on the inside of the shell.
Dimensions.—Diameter of the shell 0.15, pores 0.02, bars 0.003.
Habitat.—Central Pacific, Station 271, surface.
Genus 17. Ethmosphæra,[28] Haeckel, 1862,
Monogr. d. Radiol., p. 349.
Definition.—Liosphærida with one single latticed sphere,
with simple shell-cavity; with shell-pores which are prolonged on the outside in centrifugal,
conical, or cylindrical tubuli.
The genus Ethmosphæra differs from the simple Cenosphæra, its
ancestral form, by the peculiar formation of the shell-pores; in all observed species of the genus
these are quite regular, of nearly equal size and form; their base in the spherical shell-face is
hexagonal, but on the outside prolonged into centrifugal, external, radial tubuli, which are
either conical or cylindrical (in the latter case both openings of the tubes being equal, in the
former the outer opening being smaller than the inner). The solitary Ethmosphæra
corresponds to the social Siphonosphæra; but in the former the formation of the shell and
of its tubuli is quite regular, in the latter more or less irregular.
Subgenus 1. Ethmosphærella, Haeckel.
Definition.—Tubuli conical, their outer opening smaller than the
inner.
1. Ethmosphæra siphonophora, Haeckel.
Ethmosphæra siphonophora, Haeckel, 1862, Monogr. d. Radiol., p. 350, Taf.
xi. fig. 1.
Tubuli conical, their outer opening half as broad as the inner and three times as broad as
their height. Five to six pores on the quadrant. Diameter of the outer pores one and a half times
as large as their distance from each other.
Dimensions.—Diameter of the shell 0.1, outer pores 0.01, their distance 0.007.
Habitat.—Mediterranean (Messina), surface, Haeckel.
2. Ethmosphæra conosiphonia, n. sp. (Pl. 12, figs. 5,
5a).
Tubuli conical, their outer opening two thirds as broad as the inner, and scarcely broader than
their height. Ten to twelve pores on the quadrant. Diameter of the outer pores twice as large as
their distance from each other.
Dimensions.—Diameter of the shell 0.17, outer pores 0.01, their distance
0.005.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
{70}
3. Ethmosphæra polysiphonia, n. sp. (Pl. 12, fig.
6).
Tubuli conical, their outer opening three-fourths as broad as the inner and three times as
broad as their height. Sixteen to eighteen pores on the quadrant. Diameter of the outer pores
three times as large as their distance apart.
Dimensions.—Diameter of the shell 0.2, outer pores 0.008, their distance
0.003.
Habitat.—West Tropical Pacific, Station 225, depth 4475 fathoms; also
fossil in Barbados and in Sicily.
Subgenus 2. Ethmosphæromma, Haeckel.
Definition.—Tubuli cylindrical, their outer opening about as large as
the inner.
4. Ethmosphæra stenosiphonia, n. sp.
Tubuli cylindrical, short, quite contiguous, so that their diameter is six times as large as
their distance apart, but about equal to their height. Nine to ten pores on the quadrant.
Dimensions.—Diameter of the shell 0.14, outer pores 0.012, their distance
0.002.
Habitat.—Central Pacific, Station 271, surface.
5. Ethmosphæra pachysiphonia, n. sp.
Tubuli cylindrical, twice as long as broad, very thick-walled, and nearly contiguous, so that
their diameter is five times as large as their distance apart. The thickness of their wall is
equal to their lumen. Twelve to fourteen pores on the quadrant.
Dimensions.—Diameter of the shell 0.16, outer pores 0.01, their distance
0.002.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms.
6. Ethmosphæra leptosiphonia, n. sp.
Tubuli cylindrical, short, about as long as broad, very thin-walled and fragile, separated by
wide distances, which are three times as large as their diameter. Six to seven pores on the
quadrant. (Very similar to Siphonosphæra cyathina, Pl. 6, fig. 10, but
quite regular, all tubuli retaining the same size and distance.)
Dimensions.—Diameter of the shell 0.12, outer pores 0.01, their distance 0.03.
Habitat.—North Atlantic, Færöe Channel, John Murray; surface.
7. Ethmosphæra macrosiphonia, n. sp.
Tubuli cylindrical, very elongated, four times as long as broad; their bases separated by
distances which are equal to their breadth. Eight to ten tubuli on the quadrant. (The tubes are
similar to those of Siphonosphæra serpula, Pl. 6, fig. 6, but
quite regular, straight, not curved, all of the same size and at equal distances apart.)
{71}
Dimensions.—Diameter of the shell 0.1, length of the tubes 0.04, breadth 0.01,
basal distance 0.01.
Habitat.—Indian Ocean, Cocos Islands, surface, Rabbe.
Genus 18. Sethosphæra,[29] Hæckel, 1881,
Prodromus, p. 452 (sensu emendato).
Definition.—Liosphærida with one single latticed sphere,
with simple shell-cavity; with shell-pores, which are prolonged on the inside into centripetal,
conical, or cylindrical tubuli.
The genus Sethosphæra differs from its ancestral form, Cenosphæra, by
the production of internal, centripetal, radial tubuli on the inside of the shell (the contrary of
the preceding genus Ethmosphæra). It corresponds therefore to the social
Pharyngosphæra; but in the latter the formation of the shell and its tubes is more or less
irregular, whilst in the former each regular pore is prolonged into a regular tubule.
1. Sethosphæra entosiphonia, n. sp.
Shell with smooth surface and regular circular pores, separated by hexagonal frames, twice as
broad as the bars. Six to eight on the quadrant. Each pore is prolonged on the inside of the shell
in a short conical centripetal tube, twice as long as its diameter.
Dimensions.—Diameter of the shell 0.15, outer pores 0.008, bars 0.004, length of
the tubuli 0.015.
Habitat.—Central Pacific, Station 272, depth 2600 fathoms.
2. Sethosphæra entosolenia, n. sp.
Shell with smooth surface and regular circular pores, without hexagonal frames, of about the
same breadth as the bars. Ten to twelve on the quadrant. Each pore is prolonged on the inside of
the shell into a thin cylindrical centripetal tube, three times as long as its diameter.
Dimensions.—Diameter of the shell 0.2, outer pores and bars 0.006, length of the
tubuli 0.02.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
Subfamily Carposphærida,[30] Haeckel, 1881, Prodromus, p. 449.
Definition.—Liosphærida with two concentric
spherical lattice-shells, which are united by radial beams.
Carposphæra,[31] Haeckel, 1881, Prodromus,
p. 451.
Definition.—Liosphærida with one medullary (intracapsular) and one cortical
(extracapsular) shell, both connected by radial beams piercing the central capsule.
{72}
The genus Carposphæra comprises a large number of double-shelled Sphæroidea, formerly united with Haliomma, but different from this genus
by the absence of radial spines. The shell is composed of two concentric latticed spheres, the
inner of which (or the medullary shell) is situated within the central capsule, the other (or the
cortical shell) outside it. Both shells are connected by radial beams which pierce the wall of the
central capsule. The distance between the shells is at least as large as (commonly much larger
than) the radius of the inner shell, whilst in the following genus, Liosphæra, that
distance is much smaller than this radius.
Subgenus 1. Melittosphæra, Haeckel, 1881, Prodromus, p. 451.
Definition.—Pores of the cortical shell regular, hexagonal (or
circular, with hexagonal frames or lobes), all of nearly equal size and form.
1. Carposphæra capillacea, n. sp.
Cortical shell very delicate, four times as broad as the similarly constructed medullary shell,
with regular hexagonal meshes (twenty to twenty-five on the quadrant) and very thin thread-like
bars. Both shells connected by twenty (?) very thin radial beams. (Similar to Heliosoma
radians, Pl. 28,
fig. 3, 3a, but with smooth surface, without any radial spines.)
Dimensions.—Diameter of the outer shell 0.2, inner 0.05, pores 0.01, bars below
0.001.
Habitat.—Central Pacific, Station 274, surface.
2. Carposphæra cubaxonia, n. sp.
Cortical shell smooth, three times as broad as the medullary shell, with regular hexagonal
pores, four times as broad as the bars. Eight to ten pores on the quadrant. Medullary shell with
regular circular pores, twice as broad as the bars. Both shells connected by six radial beams,
which are three-sided prismatical, opposite in pairs in the three dimensive axes.
Dimensions.—Diameter of the outer shell 0.15, inner 0.05; outer pores 0.01, inner
0.005.
Habitat.—South Pacific, Station 291, surface.
3. Carposphæra infundibulum, Haeckel.
Haliomma infundibuliforme, Stöhr, 1880, Palæontogr. Bd. xxvi. p. 87, Taf.
i. fig. 6.
Cortical shell very thick-walled, two and a half times as broad as the medullary shell, with
rough surface and regular hexagonal, funnel-shaped pores, of about the same breadth as the bars.
Five to six on the quadrant.
Dimensions.—Diameter of the outer shell 0.1, inner 0.04, outer pores and bars
0.01.
Habitat.—North Atlantic, Station 354; fossil in Tertiary rocks
(Barbados and Sicily).
{73}
4. Carposphæra melissa, n. sp.
Cortical shell thick walled, four times as broad as the medullary shell, with regular circular,
hexagonally framed pores, three times as broad as the bars. Eight to ten pores on the quadrant.
Medullary shell with simple circular pores.
Dimensions.—Diameter of the outer shell 0.16, inner 0.04, outer pores 0.012, bars
0.004.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
5. Carposphæra melitomma, n. sp. (Pl. 20, fig.
4).
Melitomma melittosphæra, Haeckel, 1881; Prodromus et Atlas, loc.
cit.
Cortical shell thick walled, with thorny surface, two and a half times as broad as the
medullary shell. Its pores regular, circular, twice as broad as the bars, elegantly six-lobed,
separated by crested hexagonal frames; in each hexagon-corner a short conical papilla (alternating
with a lobe). Eight to ten pores on the quadrant. Medullary shell with small simple circular
pores.
Dimensions.—Diameter of the outer shell 0.17, inner 0.07, outer pores 0.01, bars
0.005, inner pores 0.005.
Habitat.—Central Pacific, Stations 266 to 274, in various depths.
Subgenus 2. Cerasosphæra, Haeckel, 1881, Prodromus, p. 451.
Definition.—Pores of the cortical shell regular, circular, without
hexagonal frames, all of nearly equal size and form.
6. Carposphæra cerasus, n. sp.
Cortical shell thin walled, smooth, twice as broad as the medullary shell; both with regular
circular pores, six times as broad as the bars. Twelve to fifteen pores on the quadrant. Outer
pores twice as large as the inner.
Dimensions.—Diameter of the outer shell 0.24, inner 0.12; outer pores 0.016, inner
0.008.
Habitat.—Central Pacific, Station 271, surface.
7. Carposphæra apiculata, Haeckel.
? Haliomma apiculatum, Ehrenberg, 1872; Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 313.
Cortical shell thin-walled, covered with numerous short conical thorns, three times as broad as
the medullary shell. Pores regular, circular, four times as broad as the bars. Six to eight pores
on the quadrant.
Dimensions.—Diameter of the outer shell 0.15, inner 0.05, outer pores 0.012, bars
0.03.
Habitat.—North Pacific, California, depth 2000 fathoms; Station 254,
depth 3025 fathoms.
{74}
8. Carposphæra entactinia, Haeckel.
Haliomma entactinia, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 74, Taf. xxvi. fig. 4.
Cortical shell thick walled, rough, twice as broad as the medullary shell; both shells with
regular circular pores, twice as broad as the bars. Six to eight pores on the quadrant. Outer
pores half as broad as the inner. Both shells connected by very numerous (twenty-four to
forty-eight or more) radial beams.
Dimensions.—Diameter of the outer shell 0.12, inner 0.06, outer pores 0.008, bars
0.004, inner pores 0.004.
Habitat.—Cosmopolitan; Atlantic and Pacific, in various depths; fossil
in Barbados and Sicily.
9. Carposphæra modesta, Haeckel.
Haliomma modestum, Stöhr, 1880, Palæontogr. Bd. xxvi. p. 86, Taf. i. fig.
5.
Cortical shell thick walled, rough, three times as broad as the medullary shell, with regular
circular pores of the same breadth as the bars. Eight to twelve pores on the quadrant. (Very
common, like the preceding species, and connected with it by numerous intermediate forms.)
Dimensions.—Diameter of the outer shell 0.12 to 0.2, inner 0.04 to 0.07, pores and
bars 0.006 to 0.008.
Habitat.—Atlantic and Pacific, from many Stations and at various
depths; fossil in Barbados and Sicily.
10. Carposphæra belladonna, n. sp.
Cortical shell thick walled, smooth, five times as broad as the medullary shell, with regular
circular pores of the same breadth as the bars. Twenty to twenty-two pores on the quadrant.
Dimensions.—Diameter of the outer shell 0.3, inner 0.06, outer pores and bars
0.004.
Habitat.—North Atlantic, Færöe Channel, John Murray.
11. Carposphæra areca, n. sp.
Cortical shell very thick walled, rough, twice as broad as the medullary shell, with regular
circular pores half as broad as the bars. Eight to ten pores on the quadrant.
Dimensions.—Diameter of the outer shell 0.12, inner 0.06, outer pores 0.03, bars
0.006.
Habitat.—Indian Ocean, Ceylon, Haeckel, surface.
Subgenus 3. Prunosphæra, Haeckel, 1881, Prodromus, p. 451.
Definition.—Pores of the cortical shell irregular polygonal, of
unequal size or dissimilar form.
12. Carposphæra prunulum, n. sp.
Cortical shell thin walled, smooth, four times as broad as the medullary shell, with large
irregular polygonal pores, four to eight times as broad as the bars. Connecting beams between
them numerous.
{75}
Dimensions.—Diameter of the outer shell 0.24, inner 0.06, outer pores 0.008 to
0.016, bars 0.002.
Habitat.—South Atlantic, Station 325, surface.
13. Carposphæra corypha, n. sp.
Cortical shell thin walled, rough, three times as broad as the medullary shell, with irregular
polygonal pores, three to six times as broad as the bars. Connecting beams between the two shells
twenty, regularly disposed.
Dimensions.—Diameter of the outer shell 0.15, inner 0.05, outer pores 0.01 to
0.02, bars 0.003.
Habitat.—South Pacific, Station 300, surface.
14. Carposphæra borassus, n. sp.
Cortical shell thick walled, smooth, three times as broad as the medullary shell, with
irregular polygonal pores, two to four times as broad as the bars. Connecting beams between the
two shells six, opposite by pairs in the three dimensive axes. (Similar to Hexalonche
aristarchi, Pl. 22,
fig. 3, but without external radial spines.)
Dimensions.—Diameter of the outer shell 0.12, inner 0.04, outer pores 0.01 to
0.02, bars 0.005.
Habitat.—Central Pacific, Station 268, surface.
Subgenus 4. Phœnicosphæra, Haeckel.
Definition.—Pores of the cortical shell irregular roundish, of unequal
size or form.
15. Carposphæra nobilis, Haeckel.
Haliomma nobile, Ehrenberg, 1844, Monatsber. d. k. preuss. Akad. d. Wiss.
Berlin, p. 268; Abhandl., 1875, Taf. xxvii. fig. 6.
Cortical shell thin walled, rough, twice as broad as the medullary shell, with irregular
roundish pores, two to four times as broad as the bars.
Dimensions.—Diameter of the outer shell 0.1, inner 0.05, outer pores 0.01 to 0.02,
bars 0.006.
Habitat.—Cosmopolitan; Atlantic, Indian, Pacific, at various depths;
fossil in Jurassic, Cretaceous, and Tertiary rocks.
16. Carposphæra micrococcus, n. sp.
Cortical shell thin walled, rough, seven times as broad as the medullary shell, with irregular
roundish pores, three to six times as broad as the bars.
Dimensions.—Diameter of the outer shell 0.2, inner 0.03, outer pores 0.012 to
0.025, bars 0.004.
Habitat.—South Atlantic, Station 330, surface.
{76}
17. Carposphæra maxima, n. sp.
Cortical shell thin walled, smooth, five times as broad as the medullary shell, with irregular
roundish pores, of about the same breadth as the bars.
Dimensions.—Diameter of the outer shell 0.4, inner 0.08, pores and bars 0.004 to
0.008.
Habitat.—Central Pacific, Station 272, depth 2600 fathoms.
18. Carposphæra nodosa, n. sp. (Pl. 28,
figs. 2, 2a).
Anthomma nodosum, Haeckel, 1879, Atlas, loc. cit.
Cortical shell thick walled, covered with forty to fifty scattered pyramidal nodules, two and a
half times as broad as the medullary shell, connected with it by very numerous thin radial beams.
Outer and inner pores irregular roundish or polygonal, two to three times as broad as the bars.
(This species in consequence of the cortical nodules may represent a peculiar genus, analogous to
Conosphæra, called Anthomma.)
Dimensions.—Diameter of the outer shell 0.13, inner 0.05, inner and outer pores
0.008 to 0.012, bars 0.004.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
Genus 20. Liosphæra,[32] Haeckel, 1881,
Prodromus, p. 449.
Definition.—Liosphærida with two cortical (extracapsular)
shells (without a medullary or intracapsular shell).
The genus Liosphæra agrees with the preceding Carposphæra in the
possession of two concentric latticed spheres; but whilst in the latter genus the inner sphere is
a medullary one (intracapsular), the outer a cortical shell (extracapsular), both connected by
radial beams piercing the capsule-wall, here in Liosphæra the central capsule lies freely
within the inner lattice shell and is not pierced by radial beams. Therefore both shells are here
cortical shells, both separated by a distance, which is constantly much smaller than the radius of
the inner shell; whereas in Carposphæra this distance is at least as large as that radius
(commonly much larger). In Carposphæra the number of pores in both shells is never the
same; in several species of Liosphæra this number is the same, each outer regular hexagonal
pore exactly corresponding to an inner; the six corners of each connected by six short radial
beams.
Subgenus 1. Melitomma, Haeckel.
Definition.—Pores of both shells regular, in each shell all of nearly
equal size and form.
1. Liosphæra hexagonia, n. sp. (Pl. 20,
fig. 3).
Both shells with the same number of pores, exactly corresponding, about ten on the quadrant.
{77}All pores regular, or subregular, hexagonal; the
outer twice as broad as the inner. Outer bars very thin, thread-like; inner bars thick, one-third
as broad as the pores. Surface smooth. Both shells connected by numerous radial beams, their
distance one-third as large as the radius of the inner shell.
Dimensions.—Diameter of the outer shell 0.16, inner 0.12, distance of both 0.02;
outer pores 0.014, inner pores 0.007.
Habitat.—Central Pacific, Station 272, depth 2600 fathoms.
2. Liosphæra rhodococcus, n. sp.
Both shells with the same number of pores, exactly corresponding, about twelve on the quadrant.
All pores regular or subregular; the inner circular, with elegant six-lobed frames, twice as broad
as the bars; the outer hexagonal, with very thin thread-like bars. All corners of the outer and
inner hexagons connected by thin, bristle-shaped radial beams. (Similar to Haliomma
rhodococcus, Pl. 19,
fig. 6; but with smooth surface and regular hexagonal pores of the outer shell.)
Dimensions.—Diameter of the outer shell 0.2, inner 0.16, distance of both 0.02;
outer pores 0.03, inner 0.01.
Habitat.—Central Pacific, Station 266, depth 2750 fathoms.
3. Liosphæra porulosa, n. sp.
Both shells with regular hexagonal pores; their number in the outer shell seven times as great
as in the inner. Pores of the stout inner shell large, three times as broad as the bars, about
eight on the quadrant. From each hexagon-corner arises one bristle-shaped radial beam; their
distal ends are united by threads (three from each), forming the large meshes of the delicate
outer shell. Each of these is divided by very thin threads into seven small circular porules, one
central and six around it.
Dimensions.—Diameter of the outer shell 0.25, inner 0.2, distance of both 0.025;
outer pores 0.04, their porules 0.012, inner pores 0.15.
Habitat.—Indian Ocean, Sunda Strait, Rabbe; surface.
Subgenus 2. Craspedomma, Haeckel.
Definition.—Pores of both shells irregular, in each shell differing
either in form or size.
4. Liosphæra peridromium, n. sp.
Both shells with the same number of large, polygonal, very irregular pores, exactly
corresponding (about eight to ten on the quadrant); both with a very delicate thin framework. From
the thread-like bars of the inner, very large and thin-walled, sphere arise perpendicularly
innumerable short bristles of equal length, which are united at equal distances by tangential
thread-like bars, parallel to the former, composing the outer shell. Each mesh is, therefore,
surrounded by a delicate ballister or rail.
{78}
Dimensions.—Diameter of the outer shell 0.42, inner 0.4, distance of both 0.01;
diameter of the meshes 0.02 to 0.06.
Habitat.—North Pacific, Station 250, surface.
5. Liosphæra polypora, n. sp. (Pl. 20,
fig. 2).
Both shells with small irregular roundish pores, of about the same size as the bars between
them; twenty to thirty on the quadrant. The pores of the outer shell somewhat smaller, therefore
much more numerous than the pores of the inner shell. Distance between the two shells about
one-third as great as the radius of the inner. Both shells connected by numerous thin radial
beams. Surface smooth or a little rough.
Dimensions.—Diameter of the outer shell 0.18, inner 0.14, distance of both 0.02;
pores and bars 0.003 to 0.005.
Habitat.—West Tropical Pacific, Station 225, depth 4475.
Subfamily Thecosphærida,[33] Haeckel, 1881, Prodromus, pp. 449, 452.
Definition.—Liosphærida with three concentric
spherical lattice-shells, which are united by radial beams.
Genus 21. Thecosphæra,[34] Haeckel, 1881,
Prodromus, p. 452.
Definition.—Liosphærida with two medullary
(intracapsular) shells and one cortical (extracapsular) shell.
The genus Thecosphæra comprises a large number of triple-shelled Sphæroidea, formerly united with Actinomma, but different from this
genus in the absence of radial spines. The latticed shell is composed of three concentric spheres,
two of which lie within the central capsule (medullary shells), and one outside (cortical shell).
This latter is connected with the former by radial beams piercing the wall of the central capsule.
From the following Rhodosphæra (with one medullary and two cortical shells)
Carposphæra differs also by the distance of the three shells. In the former the distance
between the two outer shells is much smaller, in the latter much larger, than the distance between
the inner shells.
Subgenus 1. Thecosphærantha, Haeckel.
Definition.—Pores of the cortical shell regular, hexagonal, or
circular, with hexagonal frames or lobes, all of nearly equal size and form.
{79}
1. Thecosphæra triplodictyon, n. sp.
Cortical shell thin walled, smooth, with regular, hexagonal pores, four times as broad as the
bars. Radial proportion of the three spheres = 1 : 2 : 8. Both medullary
shells with regular circular pores, twice as broad as the bars, the inner half as broad as the
outer. All three shells connected by six thin radial beams, opposite in pairs in the three
dimensive axes.
Dimensions.—Diameter of the outer shell 0.2, middle 0.05, inner 0.025; cortical
pores 0.012, bars 0.003.
Habitat.—Central Pacific, Station 271, surface.
2. Thecosphæra phænaxonia, n. sp.
Cortical shell thick walled, rough, with regular, circular, hexagonally framed pores, twice as
broad as the bars. Radial proportion of the three spheres = 1 : 2 : 6. Both
medullary shells with regular hexagonal pores and thin bars. All three shells connected by six
prismatic radial beams, opposite in pairs in the three dimensive axes. (Shell similar to
Hexacontium sceptrum, Pl. 24,
fig. 1, 1a, but without external spines.)
Dimensions.—Diameter of the outer shell 0.12, middle 0.04, inner 0.02; cortical
pores 0.01, bars 0.005.
Habitat.—North Pacific, Station 253, surface.
3. Thecosphæra favosa, n. sp.
Cortical shell thick walled, thorny, with regular, circular, hexagonally framed pores, of the
same breadth as the bars. Radial proportion of the three spheres = 1 : 3 : 10.
Both medullary shells with regular circular pores, connected with the former by twelve short
prismatic, regularly disposed radial beams.
Dimensions.—Diameter of the outer shell 0.2, middle 0.06, inner 0.02; cortical
pores and bars 0.008.
Habitat.—Central Pacific, Station 268, surface; also fossil in
Barbados.
4. Thecosphæra floribunda, n. sp.
Cortical shell thick walled, smooth, with regular, elegantly six-lobed pores, three times as
broad as the bars. Radial proportion of the three spheres = 1 : 2 : 4. Both
medullary shells with simple regular circular pores, connected with the former by six dimensive
radial beams. (Similar to Hexacontium floridum, Pl. 24, fig. 4, but
without external spines.)
Dimensions.—Diameter of the outer shell 0.12, middle 0.06, inner 0.03; cortical
pores 0.01, bars 0.0033.
Habitat.—Central Pacific, Station 266, depth 2750 fathoms.
{80}
Subgenus 2. Thecosphærella, Haeckel.
Definition.—Pores of the cortical shell regular, circular, without
hexagonal frames or lobes, all of nearly equal size and form.
5. Thecosphæra inermis, Haeckel.
Actinomma inerme, Haeckel, 1862, Monogr. d. Radiol., p. 440, Taf. xxiv. fig. 5.
Haliomma inerme, Haeckel, 1860, Monatsber. d. k. preuss. Akad. d. Wiss.
Berlin, p. 815.
Cortical shell thin walled, rough, with regular circular pores, twice as broad as the bars.
Radial proportion of the three spheres and of their circular regular pores =
1 : 2 : 4. All three spheres connected by twelve regularly disposed radial
beams.
Dimensions.—Diameter of the outer shell 0.1, middle 0.05, inner 0.025; cortical
pores 0.006, bars 0.003.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Indian, Pacific, at
many Stations and at various depths.
6. Thecosphæra æquorea, Haeckel.
Haliomma æquorea, Ehrenberg, 1844, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p.
83; Mikrogeol., 1854, Taf. xxii. fig. 35.
Actinomma æquorea, Haeckel, 1862, Monogr. d. Radiol., p. 443.
Cortical shell thick walled, smooth, with regular circular pores of the same breadth as the
bars. Radial proportion of the three spheres and of their regular pores =
1 : 2 : 6 or = 1 : 3 : 9; they are connected by six radial
beams, opposite by pairs in the three dimensive axes.
Dimensions.—Diameter of the outer shell 0.08 to 0.12, middle 0.03 to 0.04, inner
0.09 to 0.12; cortical pores and bars about 0.006.
Habitat.—Mediterranean, Corfu, surface; fossil in Greece and
Sicily.
7. Thecosphæra medusa, Haeckel.
Haliomma medusa, Ehrenberg, 1838, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 130;
Mikrogeol., 1854, Taf. xxii. figs. 33, 34.
Actinomma medusa, Haeckel, 1862, Monogr. d. Radiol., p. 444.
Actinomma medusa, Stöhr, 1880, Palæontogr., Bd. xxvi. p. 90, Taf. ii. fig.
3.
Cortical shell thick walled, rough or thorny, with regular circular pores of the same breadth
as the bars. Radial proportion of the three spheres = 1 : 2 : 4 (or
1 : 2.5 : 6); they are connected by four radial beams, crossed by pairs in two
diameters, perpendicular one to another.
Dimensions.—Diameter of the outer shell 0.08 to 0.12, middle 0.04 to 0.06, inner
0.02 to 0.025; cortical pores and bars in average 0.005.
Habitat.—Fossil in Tertiary rocks of Barbados and the
Mediterranean.
{81}
8. Thecosphæra entactinia, n. sp.
Cortical shell thick walled, smooth, with regular circular pores of the same breadth as the
bars. Radial proportion of the three spheres = 1 : 3 : 12; they are connected
by very numerous (forty to fifty, or more) thin radial beams.
Dimensions.—Diameter of the outer shell 0.24, middle 0.06, inner 0.02; cortical
pores and bars 0.008.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
9. Thecosphæra micropora, n. sp.
Cortical shell thin walled, smooth, with very small and numerous, regular, circular pores, half
as broad as the bars. Radial proportion of the three shells = 1 : 2 : 5; they
are connected by twenty regularly disposed radial beams.
Dimensions.—Diameter of the outer shell 0.2, middle 0.08, inner 0.04; cortical
pores 0.002, bars 0.004.
Habitat.—South Pacific, Station 288, surface.
Subgenus 3. Thecosphærina, Haeckel.
Definition.—Pores of the cortical shell irregular polygonal, of
unequal size or dissimilar form.
10. Thecosphæra capillacea, n. sp.
Cortical shell thin walled, smooth, with irregular polygonal pores, three to six times as broad
as the bars. Both medullary shells with similar, but smaller, pores. Radial proportion of the
three spheres = 1 : 3 : 8; they are connected by very numerous (sixty to
eighty or more) thin radial beams. (Similar to Actinomma capillaceum, Pl. 29, fig. 6, but
without external spines.)
Dimensions.—Diameter of the outer shell 0.2, middle 0.075, inner 0.025; cortical
pores 0.006 to 0.012, bars 0.002.
Habitat.—North Pacific, Station 250, surface.
11. Thecosphæra diplococcus, n. sp.
Cortical shell thick walled, rough, with large irregular polygonal pores, two to three times as
broad as the bars. Both medullary shells with small regular circular pores. Radial proportion of
the three spheres = 1 : 2 : 6; they are connected by twenty (?) stout radial
beams.
Dimensions.—Diameter of the outer shell 0.12, middle 0.04, inner 0.02; cortical
pores 0.008 to 0.012, bars 0.004.
Habitat.—South Atlantic, Station 330, surface.
Subgenus 4. Thecosphæromma, Haeckel.
Definition.—Pores of the cortical shell irregular, roundish, of
unequal size or dissimilar form.
{82}
12. Thecosphæra dodecactis, n. sp.
Cortical shell thin walled, smooth, with large irregular roundish pores, two to eight times as
broad as the bars. Both medullary shells with regular circular pores, twice as broad as the bars.
Radial proportion of the three spheres = 1 : 2 : 5; they are connected by
twelve regularly disposed stout radial beams.
Dimensions.—Diameter of the outer shell 0.2, middle 0.08, inner 0.04; cortical
pores 0.004 to 0.016, bars 0.002.
Habitat.—Central Pacific, Station 263, depth 2650 fathoms.
13. Thecosphæra icosactis, n. sp.
Cortical shell thin walled, with small irregular roundish pores, two to four times as broad as
the bars. Both medullary shells with similar but smaller pores. Radial proportion of the three
spheres = 1 : 3 : 8; they are connected by twenty thin radial beams.
Dimensions.—Diameter of the outer shell 0.32, middle 0.12, inner 0.04; cortical
pores 0.005 to 0.012, bars 0.003.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
14. Thecosphæra maxima, n. sp.
Cortical shell thin walled, with small irregular roundish pores, two to six times as broad as
the bars. Both medullary shells with similar, but smaller, pores. Radial proportion of the three
spheres = 1 : 3 : 9; they are connected by numerous (forty to sixty or more)
thin radial beams.
Dimensions.—Diameter of the outer shell 0.45, middle 0.15, inner 0.05; cortical
pores 0.008 to 0.024, bars 0.004.
Habitat.—Central Pacific, Station 272, depth 2600 fathoms.
Genus 22. Rhodosphæra,[35] Haeckel, 1881,
Prodromus, p. 452.
Definition.—Liosphærida with one medullary
(intracapsular) shell and two cortical (extracapsular) shells.
The genus Rhodosphæra differs from the preceding Thecosphæra in the
same manner in which, among the Dyosphærida, Liosphæra differs from Carposphæra. The
cortical shell is double, composed of two not far distant shells, lying outside the central
capsule. The distance between the shells is much smaller than the radius of the inner shell. This
is connected by radial beams (piercing the central capsule) with the small central medullary
shell.
Subgenus 1. Rhodosphærella, Haeckel.
Definition.—Pores of both cortical shells regular, in each shell all
of nearly equal size and form.
{83}
1. Rhodosphæra hexagonia, n. sp.
Both cortical shells with the same number of regular hexagonal pores; the inner four times as
broad as the bars, and half as broad as the outer pores, which are separated by thread-like bars.
Medullary shell only one-fourth as broad as the inner cortical shell, with regular hexagonal pores
of half the size. (Differs from the similar Liosphæra hexagonia, Pl. 20, fig. 3, by
the possession of a medullary shell.)
Dimensions.—Diameter of the outer shell 0.2, middle 0.16, inner 0.04; outer pores
0.013, middle 0.008, inner 0.004.
Habitat.—Central Pacific, Station 266, depth 2750 fathoms.
2. Rhodosphæra melitomma, n. sp.
Both cortical shells with the same number of regular pores; the inner regular, circular, twice
as broad as the bars, with elegant hexagonal frames and six roundish lobes alternating with the
six radial spines which arise from the hexagon-corners; these short conical spines are connected
at the distal end (at equal distances from the centre) by delicate threads (three from each
spine), which form the delicate external shell. Medullary shell one-third as broad as the inner
cortical shell, with small, simple, regular circular pores. (If in Carposphæra melitomma,
Pl. 20, fig.
4, the distal ends of the spines became united by a cobweb-like outer shell, this species would be
formed.)
Dimensions.—Diameter of the outer shell 0.22, middle 0.18, inner 0.06; outer pores
0.025, middle 0.0125, inner 0.005.
Habitat.—Central Pacific, Station 270, depth 2925 fathoms.
Subgenus 2. Rhodosphæromma, Haeckel.
Definition.—Pores of both cortical shells irregular, in each shell of
unequal size or dissimilar form.
3. Rhodosphæra palliata, n. sp.
Both cortical shells with an unequal number of irregular roundish pores; the outer pores
somewhat smaller and much more numerous than the inner pores; the bars between the latter are
thicker. Medullary shell about one quarter as broad as the inner cortical shell, with regular
circular pores.
Dimensions.—Diameter of the outer shell 0.4, middle 0.36, inner 0.08; outer pores
on an average 0.008, middle 0.012, inner 0.004.
Habitat.—Fossil in Barbados.
4. Rhodosphæra pentaphylla, n. sp.
Both cortical shells with unequal number of irregular roundish pores; the inner pores large,
three to four times as broad as the bars; to each inner pore corresponds a group of five smaller
{84}outer pores, like the five petals of a flower.
Medullary shell half as broad as the inner cortical shell, with regular circular pores.
Dimensions.—Diameter of the outer shell 0.25, middle 0.2, inner 0.1; outer pores
on an average 0.006, middle 0.012, inner 0.004.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
Subfamily Cromyosphærida,[36] Haeckel, 1881, Prodromus, pp. 449, 453.
Definition.—Liosphærida with four concentric
spherical latticed shells, united by radial beams.
Genus 23. Cromyosphæra,[37] Haeckel, 1881,
Prodromus, p. 453.
Definition.—Liosphærida with two intracapsular
(medullary) shells and two extracapsular (cortical) shells; the former united with the latter by
radial beams piercing the wall of the central capsule.
The genus Cromyosphæra is the only known genus of Cromyosphærida, or of such
Sphæroidea, the smooth shell of which is composed of two medullary and
two cortical shells. There may possibly be other Cromyosphærida, in which the shell is composed of
one simple medullary and three cortical shells, or only of four extra-capsular cortical shells;
but such have not as yet been observed. Caryosphæra polysphærica, Bütschli, 1882 (L. N. 41, Taf. xxiii. fig. 12) is probably a Cromyosphæra (fossil in
Barbados).
1. Cromyosphæra quadruplex, n. sp. (Pl. 30, fig.
9).
Radial proportion of the four spheres = 1 : 2 : 4 : 5. Outer
cortical shell smooth, with large regular hexagonal pores, ten times as broad as the bars; inner
cortical shell with irregular polygonal pores, five times as broad as the bars. Both medullary
shells with regular circular pores of the same breadth as the bars.
Dimensions.—Diameter of the four spheres—(A) 0.16, (B) 0.12, (C) 0.06, (D)
0.03.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms.
2. Cromyosphæra rosetta, n. sp.
Radial proportion of the four spheres = 1 : 2 : 8 : 10. Outer
cortical shell smooth, with regular hexagonal pores and very thin bars; inner cortical shell with
the same number of exactly corresponding, regular circular, hexagonally framed pores, twice as
broad as the bars; the corners of the outer and inner hexagons united by radial bristles. Both
medullary shells with regular circular pores, twice as broad as the bars.
Dimensions.—Diameter of the four spheres—(A) 0.2, (B) 0.16, (C) 0.04, (D)
0.02.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
{85}
3. Cromyosphæra bigemina, n. sp.
Radial proportion of the four spheres = 1 : 2 : 7 : 8. Outer
cortical shell smooth, with regular hexagonal pores and very thin bars; inner cortical shell with
the same number of exactly corresponding, regular circular pores. Both medullary shells with
regular circular pores of the same breadth as the bars. (Somewhat similar to Hexacromyon
elegans, Pl. 24,
fig. 9, also with six inner radial beams, but without external radial spines.)
Dimensions.—Diameter of the four spheres—(A) 0.2, (B) 0.17, (C) 0.05, (D)
0.025.
Habitat.—North Pacific, Station 241, depth 2300 fathoms.
4. Cromyosphæra cepa, n. sp.
Radial proportion of the four spheres = 1 : 2 : 4 : 5. All four
shells of the same structure, thick-walled, with regular circular pores, two to four times as
broad as the bars; the size of the pores increases gradually from the inner to the outer shell.
Surface thorny. Distance between the second and third shells twice as great as that between the
others.
Dimensions.—Diameter of the four spheres—(A) 0.125, (B) 0.1, (C) 0.05, (D)
0.025.
Habitat.—Fossil in Barbados.
5. Cromyosphæra scorodonium, n. sp.
Radial proportion of the four spheres = 1 : 2 : 3 : 4. All four
shells of the same structure, thin-walled, with irregular roundish pores, two to four times as
broad as the bars; the size of the pores increasing gradually from the inner to the outer shell.
Surface smooth. Distance between each two shells equal to the diameter of the innermost.
Dimensions.—Diameter of the four spheres—(A) 0.12, (B) 0.09, (C) 0.06, (D)
0.03.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms; also fossil
in Barbados.
6. Cromyosphæra antarctica, n. sp..
Radial proportion of the four spheres = 1 : 2 : 5 : 7. Both
cortical shells with irregular polygonal roundish pores; the outermost with thinner bars and rough
surface, the inner with thicker bars. Both medullary shells with irregular roundish pores.
Dimensions.—Diameter of the four spheres—(A) 0.18, (B) 0.12, (C) 0.05, (D)
0.025.
Habitat.—Antarctic Ocean; in very large number, together with
Rhizosphæra antarctica, in the diatomaceous ooze of Station 157 (3rd March 1874); depth
1950 fathoms.
Subfamily Caryosphærida,[38] Haeckel, 1881, Prodromus, pp. 449, 454.
Definition.—Liosphærida with numerous (five
or more) concentric spherical latticed shells, united by radial beams.
{86}
Genus 24. Caryosphæra,[39] Haeckel, 1881,
Prodromus, p. 454.
Definition.—Liosphærida with two intracapsular
(medullary) shells and three or more extracapsular (cortical) shells; the former united with the
latter by radial beams piercing the wall of the central capsule.
The genus Caryosphæra, the only observed form of this subfamily, comprises
those Liosphærida in which the shell is composed of two medullary and
three or more cortical shells. Such forms (without radial spines) are very rare; I observed only
two species, one with five, the other with six shells. They are derived from Cromyosphæra
by further apposition of outer cortical shells.
1. Caryosphæra pentalepas, n. sp.
Shell composed of five concentric spheres, with radial proportion =
1 : 2 : 8 : 10 : 12. Both medullary shells with regular
circular pores, twice as broad as the bars. First cortical shell with regular, circular,
hexagonally framed pores, three times as broad as the bars; second cortical shell with regular
hexagonal pores, four times as broad as the bars; third (outermost) cortical shell with regular
hexagonal pores and very thin thread-like bars. Surface smooth.
Dimensions.—Diameter of the five shells—(A) 0.02, (B) 0.04, (C) 0.16, (D)
0.2, (E) 0.24.
Habitat.—Central Pacific, Station 274, depth 2750 fathoms.
2. Caryosphæra hexalepas, n. sp.
Shell composed of six concentric spheres, with the radial proportion =
1 : 2 : 4 : 5 : 6 : 8. All six shells with
regular circular pores, two to four times as broad as the bars, with increasing size from the
centrum against the smooth surface.
Dimensions.—Diameter of the six shells—(A) 0.025, (B) 0.05, (C) 0.1, (D)
0.13, (E) 0.16, (F) 0.2.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms; also fossil
in Barbados.
Subfamily Plegmosphærida,[40] Haeckel, 1881, Prodromus, p. 455.
Definition.—Liosphærida with spongy spherical
shell, with or without latticed medullary shell in the centre.
Genus 25. Styptosphæra,[41] Haeckel, 1881,
Prodromus, p. 455.
Definition.—Liosphærida forming a solid sphere of spongy
framework, without enclosed medullary shell, and without central cavity.
{87}
The genus Styptosphæra presents a spherical shell with smooth or rough
surface (without radial spines), the whole mass of which is composed of looser or denser spongy
wicker-work.
1. Styptosphæra spumacea, n. sp.
Spongy framework of the solid sphere loose, with large polygonal meshes of slightly different
size, ten to twenty times as broad as the bars. Structure of the whole spongy sphere the same.
Central capsule filled with crystals. Surface smooth.
Dimensions.—Diameter of the sphere 0.32, of the central capsule 0.26, meshes 0.01
to 0.02, bars 0.001.
Habitat.—North Pacific, Station 236, surface.
2. Styptosphæra spongiacea, n. sp.
Spongy framework in the central part of the solid sphere much more compact than in the
peripheral part, becoming gradually looser towards the rough surface. Meshes in the centre three
to five times, in the periphery fifteen to twenty times as broad as the bent bars.
Dimensions.—Diameter of the sphere 0.45, inner meshes 0.006 to 0.01, outer meshes
0.03 to 0.04, bars 0.002.
Habitat.—Central Pacific, Station 271, surface.
3. Styptosphæra stupacea, n. sp.
Spongy framework of the solid sphere rather compact, everywhere of the same structure, with
roundish, nearly equal meshes, six to eight times as broad as the bars. Surface rough with
prominent thorns.
Dimensions.—Diameter of the sphere 0.22, meshes 0.01 to 0.012, bars 0.0015.
Habitat.—South Pacific, Station 291, surface.
Genus 26. Plegmosphæra, Haeckel,[42] 1881,
Prodromus, p. 455.
Definition.—Liosphærida forming a hollow sphere of spongy framework, without a
medullary shell in the central cavity.
The genus Plegmosphæra develops a large hollow sphere, the wall of which is
composed of looser or denser spongy wicker-work. On the inner as well as on the outer face of the
spongy shell-wall may be present a simple lattice-sphere from which the threads of the framework
arise; but in some species these lattice-plates are quite absent.
{88}
Subgenus 1. Plegmosphærantha, Haeckel.
Definition.—Inside and outside of the spongy shell-wall smooth, closed
by a lattice-plate with polygonal meshes.
1. Plegmosphæra maxima, n. sp.
Radius of the spherical shell-cavity eight to ten times as great as the thickness of the thin
spongy wall. Inside and outside of the wall smooth, closed by a lattice-plate with irregular
polygonal meshes. Only three or four meshes in the thickness of the wall.
Dimensions.—Diameter of the spongy sphere 0.8 to 1.0 mm., of its inner cavity 0.7
to 0.8, meshes 0.01 to 0.02, bars 0.002 to 0.003, central capsule 0.5 to 0.6, nucleus 0.1 to
0.15.
Habitat.—Central Pacific, Station 271, surface.
2. Plegmosphæra coelopila, n. sp.
Radius of the spherical shell-cavity eight to ten times as great as the thickness of the spongy
wall. Inside and outside of the wall closed by a smooth lattice-plate with irregular polygonal
meshes, five to ten times as broad as the bars.
Dimensions.—Diameter of the shell 0.32, of its cavity 0.26, meshes 0.01 to 0.02,
bars 0.002.
Habitat.—North Atlantic, Færöe Channel, Gulf Stream, John Murray.
3. Plegmosphæra pachypila, n. sp.
Radius of the spherical shell-cavity about equal to the thickness of the spongy wall. Inside
and outside of the wall closed by a smooth lattice-plate with irregular polygonal meshes, three to
six times as broad as the bars.
Dimensions.—Diameter of the shell 0.24, of its cavity 0.12.
Habitat.—North Pacific, Station 250, surface.
Subgenus 2. Plegmosphærella, Haeckel.
Definition.—Inside of the spongy shell-wall closed by a smooth
lattice-plate, outside rough, spongy, with prominent thorns.
4. Plegmosphæra entodictyon, n. sp.
Radius of the spherical shell-cavity half as great as the thickness of the spongy wall. Inside
of the wall closed by a smooth lattice-plate, outside rough, spongy.
Dimensions.—Diameter of the shell 0.24, of its cavity 0.08.
Habitat.—South Pacific, Station 300, surface.
{89}
5. Plegmosphæra leptodictyon, n. sp.
Radius of the spongy shell-cavity six times as great as the thickness of the spongy wall.
Inside of the wall closed by a smooth lattice-plate, outside rough spongy.
Dimensions.—Diameter of the shell 0.44, of its cavity 0.36.
Habitat.—Central Pacific, Station 266, surface.
Subgenus 3. Plegmosphæromma, Haeckel.
Definition.—Inside of the spongy shell-wall rough spongy, without
lattice-plate, outside closed by a smooth lattice-plate.
6. Plegmosphæra exodictyon, n. sp. (Pl. 18, fig.
8).
Radius of the spongy shell-cavity only one-fourth as great as the thickness of the spongy
shell-wall. Outside of the wall closed by a smooth lattice-plate, inside rough, spongy.
Dimensions.—Diameter of the shell 0.4, of its cavity 0.08.
Habitat.—South Atlantic, Station 325, surface.
Subgenus 4. Plegmosphærusa, Haeckel.
Definition.—Inside and outside of the spongy shell-wall rough, with
spongy or spiny surface, without lattice-plate.
7. Plegmosphæra leptoplegma, n. sp.
Radius of the spherical shell-cavity half as great as the thickness of the loose spongy
shell-wall. Inside and outside of the wall rough spongy, not closed by a lattice-plate. Meshes ten
to twenty times as broad as the bars.
Dimensions.—Diameter of the shell 0.3, of its cavity 0.088.
Habitat.—North Atlantic, Station 253, surface.
8. Plegmosphæra pachyplegma, n. sp.
Radius of the spherical shell-cavity about equal to the thickness of the dark and dense spongy
shell-wall. Inside and outside of the wall rough spongy, not closed by a lattice-plate. Meshes
three to five times as broad as the bars.
Dimensions.—Diameter of the shell 0.2, of its cavity 0.1.
Habitat.—Central Pacific, Station 270, surface.
Genus 27. Spongoplegma,[43] Haeckel, 1881,
Prodromus, p. 455.
Definition.—Liosphærida forming a sphere of spongy
framework, which encloses in the centre one single latticed medullary shell.
{90}
The genus Spongoplegma may be regarded as a Carposphæra, in which the
simple latticed cortical shell is represented by an irregular spongy framework, immediately
enclosing the simple latticed medullary shell.
1. Spongoplegma antarcticum, n. sp.
Cortical shell with loose spongy framework and rough surface, four to six times as broad as the
enclosed simple medullary shell. Pores of the latter regular circular, twice as broad as the bars.
From its surface arise numerous (forty to fifty or more) short radial beams, which become forked
and compose, by communication of lateral branches, the spongy cortical shell.
Dimensions.—Diameter of the spongy cortical shell 0.2 to 0.3 of the latticed
medullary shell 0.05 to 0.06.
Habitat.—Antarctic Ocean, in large number, together with
Cromyosphæra antarctica; in the Diatom ooze of Station 157 (depth 1950 fathoms).
Genus 28. Spongodictyon,[44] Haeckel, 1862,
Monogr. d. Radiol., p. 459.
Definition.—Liosphærida forming a sphere of spongy
framework, which encloses in the centre a double latticed concentric medullary shell.
The genus Spongodictyon can be regarded as a Thecosphæra, in which
the simple latticed cortical shell is represented by an irregular spongy framework, which
immediately encloses the double latticed medullary shell. Sometimes this latter appears triple,
the inner surface of the spongy cortical shell forming a smooth spherical lattice-plate, separated
by an interval from the double medullary shell.
Subgenus 1. Dictyoplegma, Haeckel, 1862, Monogr. d. Radiol, p. 458.
Definition.—Spongy cortical shell enveloping immediately the double
medullary shell.
1. Spongodictyon spongiosum, Haeckel.
Dictyosoma spongiosum, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 31,
Taf. ii. figs. 9-11.
Dictyoplegma spongiosum, Haeckel, 1862, Monogr. d. Radiol., p. 458.
Spongy framework of the cortical shell loose, with large, polygonal roundish meshes, on an
average as large as the double medullary shell, which is immediately enveloped by it. Both
concentric medullary shells with subregular roundish pores, twice as broad as the bars.
Dimensions.—Diameter of the cortical shell 0.2 to 0.3 or more; of the outer
medullary shell 0.03, inner 0.01.
Habitat.—Mediterranean (French south coast, surface), J. Müller.
{91}
2. Spongodictyon cavernosum, n. sp.
Spongy framework of the cortical shell rather compact in the inner part, which immediately
envelops the double medullary shell; very loose, with large caverns in the outer part, caverns of
the surface larger than the medullary shell. Both medullary shells with regular circular pores,
three times as broad as the bars.
Dimensions.—Diameter of the cortical shell 0.3 to 0.4, outer medullary shell 0.1,
inner 0.03.
Habitat.—Tropical Atlantic, Station 338, surface.
Subgenus 2. Spongodictyoma, Haeckel.
Definition.—Spongy cortical shell on the inner surface with a smooth
lattice-plate (or third medullary shell), which is connected by radial beams with the inner double
medullary shell.
3. Spongodictyon trigonizon, Haeckel.
Spongodictyon trigonizon, Haeckel, 1862, Monogr. d. Radiol., p. 459, Taf. xxvi. figs.
4-6.
Dictyosoma trigonizon, Haeckel, 1860, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 841.
Spongy framework of the cortical shell very loose, with very large, for the most part
triangular meshes, which are two to six times as large as the enclosed double medullary shell.
From the surface of the latter arise numerous radial beams, which are connected by a spherical
lattice-plate, forming the smooth inner surface of the spongy sphere (or a third medullary shell).
The structure of the framework reminds one of the Phæodarium Sagena
(Pl. 108).
Pores of both medullary shells regular circular, twice as broad as the bars.
Dimensions.—Diameter of the cortical shell 0.5 to 1.15, outer medullary 0.05,
inner 0.035.
Habitat.—Mediterranean, Messina, surface.
4. Spongodictyon arcadophoron, n. sp.
Spongy framework of the cortical shell in the inner part very loose, in the outer part more
compact; outer meshes scarcely as large as the inner medullary shell (or only half as large),
inner meshes two to four times as large. From the surface of the double medullary shell arise
numerous radial beams, which are forked at equal distances from the centre; the fork branches are
curved and united together by dichotomous branches, like elegant arcades; and these arcades form
together the large polygonal meshes on the inside of the cortical shell (or a third medullary
shell). Both medullary shells with regular circular pores, of the same breadth as the bars.
Dimensions.—Diameter of the cortical shell 0.2, outer medullary shell 0.04, inner
0.02.
Habitat.—Tropical Atlantic, Station 349, surface.
{92}
Family VI. Collosphærida, J. Müller[45] (Pls. 5-8).
Definition.—Sphæroidea living associated in colonies,
united by an alveolar jelly-body, and connected by the network of anastomosing pseudopodia.
The family Collosphærida comprises all polyzous or social Sphæroidea, and constitutes the only polyzoic group among the Sphærellaria. This group was first constituted by J. Müller as "Radiolaria
polyzoa with shells."[46] Formerly following his
authority, in my Monograph I had separated them from the other Sphæroidea
and united them with the social Collodaria (Sphærozoida).[47] Also R. Hertwig in his Organismus der Radiolarien[48] united them with his Sphærozoea. In my Prodromus[49] I had retained this isolated position. But a further careful
study has convinced me that this isolation is not truly natural, and that the Collosphærida are
only "social Ethmosphærida" which have arisen from this solitary subfamily by adaptation to
colonial life. There are some forms of Collosphærida which are nearly identical with some forms of
Ethmosphærida, only differing from the latter by their association in colonies; and in some forms
of both groups it is quite impossible to decide whether the isolated shells appertain to one or to
the other family.
The isolated shell of the Collosphærida is almost constantly (with few exceptions) a simple
extracapsular lattice-shell, as in the Monosphærida; only the small group of Clathrosphærida (with
the genera Clathrosphæra and Xanthiosphæra) exhibit an exception, the simple
lattice-shell being overgrown by an external mantle or veil of very thin, cobweb-like, irregular
lattice-work (Pl. 8,
figs. 6-11). Therefore these Clathrosphærida bear to the Acrosphærida (or the common simple
Collosphærida) a relation similar to that which Liosphæra (p. 76)
bears to Cenosphæra; both shells are extracapsular "cortical shells" at a very short
distance apart. In the Collosphærida true concentric medullary shells never occur; the central
capsule always lies quite freely in the simple or double cortical shell, separated from it by a
jelly-veil.
Although a well marked difference in the simple lattice-shell of the social Collosphærida and
the solitary Ethmosphærida does not exist, nevertheless in most cases the two shells can be
distinguished by a practiced observer. The simple fenestrated shells of the monozoic Ethmosphærida
are commonly quite regular spheres in a mathematical sense, or regular "endospherical polyhedra";
whereas in the Collosphærida they are commonly more or less irregular, often to an extraordinary
degree (Pls. 5-8). Some species
of Collosphærida, however, also possess quite regular spherical shells. Another difference is
often shown in the lattice-work of the shells, which in the Collosphærida is nearly always very
irregular, and exhibits a peculiar tendency to the {93}production of radial, conical, or cylindrical tubules. These occur as well
on the inside as on the outside of the shell, and the tubules are now more conical, now more
cylindrical; their wall either solid or pierced by pores (Pls. 5-8). The tubules
are commonly very irregular in form, size, and disposition; distinguished, however, by a number of
hereditary peculiarities, which are sufficient for the distinction of genera. Similar tubules
occur also in some genera of solitary Ethmosphærida (Coscinomma, Ethmosphæra,
Conosphæra, &c., Pl. 12);
but the tubules are here much more regular and not so highly developed.
Besides the tubules of the fenestrated shells, in some genera of Collosphærida the surface is
armed with irregular thorns, rarely with more regular radial spines. But these spines obtain
constantly the character of accessory by-spines, and remain short and thin. In this family typical
radial spines never occur in a regular and characteristic disposition, corresponding to dimensive
axes, as is the case in nearly all solitary Sphæroidea, only excepting
the Liosphærida. Commonly these spines or thorns serve as protective arms for the shell-meshes,
surrounding them often in the form of coronels. Often the lattice-plate of the irregular roundish
shell is tubercular, elevated into irregular protuberances, bearing on the top a short spine or
thorn (Pl. 8).
The Central Capsule of the Collosphærida is always a regular sphere, as in all other
Sphæroidea; it is constantly placed within the lattice-shell, and
commonly much smaller than it, separated from it by a thick jelly-veil. A remarkable difference
from the solitary Sphæroidea is shown in the early division of the
nucleus. Commonly the central capsule of the Collosphærida contains in its centre a large
oil-globule, surrounded by very numerous small nuclei. R. Hertwig estimated this difference as so
important, that he separated the social "Sphærozoea" and the solitary "Peripylea" as two different
orders. As already shown above (p. 7, 24), we cannot
support this separation, and are now convinced that this difference in the development of the
spores—just as in the Collodaria—is the consequence of an
adaptation to social life.
The common jelly-body, in which the numerous central capsules and their enveloping shells are
united, exhibits in the Collosphærida quite the same characters as in the other social Radiolaria,
the Collozoida and Sphærozoida. The jelly-body is very voluminous, commonly spherical, often
cylindrical, of considerable size; constantly containing numerous large alveoles. Often each shell
is enclosed in a separate alveole with rather solid wall (Pl. 6, fig. 2).
Sometimes in the dead colonies all shells are united in the central part of the jelly-body, whilst
its peripheral part is composed of a stratum of large alveoles (Pl. 8, fig. 11); at
other times no alveoles are visible (Pl. 7,
fig. 11). In many living colonies I found a very large spherical alveole with thick wall in the
centre of the spherical colony, surrounded by many strata of delicate thin-walled alveoles (Pl. 5, fig. 1). In
this case often the inner younger capsules were naked, {94}without shells, the outer only surrounded by shells. Already in my
Monograph I had described the same peculiar formation.[50]
Synopsis of the Genera of Collosphærida.
| I. Subfamily Acrosphærida. (Lattice-shell simple, without
an external mantle of network.) |
 |
Outside of the shell smooth, without spines or tubuli. |
|
Inside without tubuli. |
|
Inside smooth, |
29. Collosphæra. |
| Inside spiny, |
30. Tribonosphæra. |
| Inside with centripetal tubuli. |
|
Tubuli imperforated, |
31. Pharyngosphæra. |
| Tubuli fenestrated, |
32. Buccinosphæra. |
| Outside of the shell armed with solid spines,
but with hollow tubuli. |
|
Spines irregularly scattered on the surface, |
33. Acrosphæra. |
| Each larger pore with one single spine, |
34. Odontosphæra. |
| Each larger pore with a coronal of spines, |
35. Chœnicosphæra. |
| Outside of the shell with irregular radial tubuli, the wall
of which is solid, not fenestrated. |
|
Tubuli simple, not branched. |
|
Mouth of the tubuli truncated, smooth, |
36. Siphonosphæra. |
| Mouth with one single large tooth, |
37. Mazosphæra. |
| Mouth with a coronal of teeth, |
38. Trypanosphæra. |
| Tubuli irregularly branched, each with two to four or more
openings, |
39. Caminosphæra. |
| Outside of the shell with irregular radial
tubuli, open on both ends, with fenestrated wall. |
|
Mouth of the tubuli truncated, smooth, |
40. Solenosphæra. |
| Mouth with one single large tooth, |
41. Otosphæra. |
| Mouth with a coronal of teeth, |
42. Coronosphæra. |
| II. Subfamily Clathrosphærida. (Lattice-shell
double, with an external mantle of network.) |
|
Surface of the outer shell smooth, |
43. Clathrosphæra. |
| Surface of the outer shell thorny, |
44. Xanthiosphæra. |
| I. Subfamily Acrosphærida. (Lattice-shell simple, without an external mantle
of network.) |
|
|
Outside of the shell smooth, without spines or tubuli. |
|
|
|
|
|
|
Inside without tubuli. |
|
|
|
|
|
|
|
|
|
|
Inside smooth, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
29. Collosphæra. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Inside spiny, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
30. Tribonosphæra. |
|
|
|
|
|
|
|
|
|
|
|
|
Inside with centripetal tubuli. |
|
|
|
|
|
|
|
|
|
|
Tubuli imperforated, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
31. Pharyngosphæra. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Tubuli fenestrated, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
32. Buccinosphæra. |
|
|
|
|
|
|
|
|
|
|
Outside of the shell armed with solid spines, but with hollow
tubuli. |
|
|
|
|
|
|
|
|
Spines irregularly scattered on the surface, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
33. Acrosphæra. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Each larger pore with one single spine, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
34. Odontosphæra. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Each larger pore with a coronal of spines, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
35. Chœnicosphæra. |
|
|
|
|
|
|
|
|
|
|
Outside of the shell with irregular radial tubuli, the wall of
which is solid, not fenestrated. |
|
|
|
|
|
|
Tubuli simple, not branched. |
|
|
|
|
|
|
|
|
|
|
Mouth of the tubuli truncated, smooth, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
36. Siphonosphæra. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Mouth with one single large tooth, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
37. Mazosphæra. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Mouth with a coronal of teeth, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
38. Trypanosphæra. |
|
|
|
|
|
|
|
|
|
|
|
|
Tubuli irregularly branched, each with two to four or more
openings, |
|
|
|
|
|
|
|
|
|
|
|
|
39. Caminosphæra. |
|
|
|
|
|
|
|
|
|
|
Outside of the shell with irregular radial tubuli, open on both
ends, with fenestrated wall. |
|
|
|
|
|
|
|
|
Mouth of the tubuli truncated, smooth, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
40. Solenosphæra. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Mouth with one single large tooth, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
41. Otosphæra. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Mouth with a coronal of teeth, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
42. Coronosphæra. |
|
|
|
|
|
|
|
|
| II. Subfamily Clathrosphærida. (Lattice-shell double, with an external mantle
of network.) |
|
|
|
|
|
|
Surface of the outer shell smooth, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
43. Clathrosphæra. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Surface of the outer shell thorny, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
44. Xanthiosphæra. |
|
|
|
|
|
|
|
|
Subfamily Acrosphærida, Haeckel, 1881, Prodromus, p. 471.
Definition.—Collosphærida with one simple
lattice-shell around every central capsule of the cœnobium.
{95}
Genus 29. Collosphæra,[51] J. Müller,
1855.
Definition.—Collosphærida with simple shells, smooth on
the inside and on the outside, without any spines or tubuli.
The genus Collosphæra is the most simple form of all Collosphærida, and must
be regarded as the common ancestral form of this family. As the lattice-shell is quite a simple
sphere, without any spines, tubules, or other peculiar productions, it agrees perfectly with
Cenosphæra, and represents the social or polyzoid aggregate of this solitary or monozoid
genus. Therefore a certain distinction between the isolated shells of the two genera is often very
difficult or even impossible; but commonly this distinction is possible owing to the circumstance,
that in the majority of the Collosphæræ the shell is more or less irregular roundish or
polyhedral, not quite spherical, as in Cenosphæra. Dermatosphæra, Ehrenberg, is a Collosphæra with small
pores (compare L. N. 16, p. 533).
Subgenus 1. Eucollosphæra, Haeckel.
Definition.—Shell a regular or subregular sphere.
1. Collosphæra primordialis, n. sp.
Shell a regular sphere, with very delicate and regular network of hexagonal meshes. Six to
eight meshes in the half meridian of the shell. Diameter of the meshes ten to fifteen times as
broad as the thin bars between them. Can be regarded as social form of Cenosphæra
primordialis.
Dimensions.—Diameter of the shell 0.1 to 0.12, of the pores 0.008.
Habitat.—Central Pacific, Stations 271 to 274, surface.
2. Collosphæra regularis, n. sp.
Shell a regular sphere, with a perfectly regular network of circular meshes, all of the same
size. Ten to twelve meshes in the half meridian of the shell. Diameter of the meshes the same as
that of the bars between them.
Dimensions.—Diameter of the shell 0.1 to 0.12, of the pores 0.005 to 0.006.
Habitat.—Indian Ocean, Madagascar, surface, Rabbe.
3. Collosphæra globularis, n. sp.
Shell a regular sphere, with subregular network of circular meshes of different sizes; few
large pores between many smaller pores. Ten to twenty meshes in the half meridian of the shell.
Diameter of the meshes from half to twice as broad as that of the bars.
{96}
Dimensions.—Diameter of the shell 0.1 to 0.12, of the pores 0.002 to 0.008,
breadth of the bars 0.004 to 0.008.
Habitat.—Tropical and subtropical zone of both hemispheres, widely
distributed; Canaries, Azores, Cape Verde Islands, Guinea Coast, Brazil Coast, Indian Ocean,
Madagascar, Ceylon, surface.
Subgenus 2. Dyscollosphæra, Haeckel.
Definition.—Shell not a regular sphere, but irregular roundish, in all
degrees of variation between subspherical and polyhedral or quite irregular forms.
4. Collosphæra huxleyi, J. Müller.
Collosphæra huxleyi, J. Müller, 1855, Abhandl. d. k. Akad. d. Wiss. Berlin, pp. 55-59,
Taf. viii. figs. 6-9.
Collosphæra huxleyi, Haeckel, 1862, Monogr. d. Radiol., p. 534, Taf. xxxiv.
Collosphæra huxleyi, Cienkowski, 1871, Archiv f. mikrosk. Anat., Bd. vii. p. 374, Taf.
xxix. figs. 1-6.
Collosphæra ligurina, J. Müller, 1856, Monatsber. d. k. Akad. d. Wiss. Berlin, p.
481.
Thalassicolla punctata, var., Huxley, 1851, Ann. and Mag. Nat. Hist., ser.
2, vol. viii. p. 434, pl. xvi. fig. 6.
Shell subspherical, somewhat irregular, sometimes with more or less superficial impressions,
with irregular network of roundish meshes. Eight to sixteen meshes in the half meridian of the
shell, one to three times as broad as their bars. Very variable, with direct transition-forms to
other species of this genus, especially to Collosphæra globularis, Collosphæra
tuberosa, Collosphæra pyriformis, and Collosphæra polyedra.
Dimensions.—Diameter of the shell 0.1 to 0.16, of the pores 0.004 to 0.012, of the
bridges 0.003 to 0.006.
Habitat.—Cosmopolitan; common in the greater part of the warmer seas,
surface.
5. Collosphæra polygona, n. sp. (Pl. 5,
fig. 13).
Collosphæra huxleyi var., Haeckel, 1862, Monogr. d. Radiol., Taf. xxxiv.
fig. 5.
Shell irregular polygonal, with very delicate, irregular network of polygonal meshes, four to
twelve times as broad as the bars. Ten to twenty pores on the half meridian of the shell.
Dimensions.—Diameter of the shell 0.1 to 0.2, of the pores 0.012 to 0.004, of the
bars 0.001 to 0.002.
Habitat.—Mediterranean, Atlantic, surface; Stations 348 to 354.
6. Collosphæra pyriformis, Haeckel, n. sp.
Shell irregular, rounded, ovate or pear-shaped, with irregular network of rounded or nearly
polygonal meshes. Ten to twenty meshes in the half meridian of the shell, one to three times as
broad as the bars. Commonly one large opening (two to three times as broad as the largest {97}meshes) on the thinner end of the ovate shell
(corresponding to the insertion of a pear-stalk); sometimes two or three such large openings.
Dimensions.—Diameter of the shell 0.1 to 0.15, of the pores 0.008 to 0.016, of the
bridges 0.004 to 0.008.
Habitat.—Tropical zone—Cape Verde Islands, Ceylon; Central
Pacific, Stations 266 to 272, 348 to 352, &c.
7. Collosphæra polyedra, n. sp.
Trisolenia zanguebarica, Ehrenberg, 1872, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 301, Taf. x. fig. 11.
Shell irregular, polyhedral, with even or somewhat vaulted sides, and obtuse ridges between
them. Network more or less irregular, with small rounded meshes, quite as broad or twice as broad
as their bars. Besides these small pores constantly some large round openings (commonly three to
six), situated on the corners of the polyhedral shell, four to six times as large as the pores.
Often an acute tooth on the edge of each large opening. Transition-form between Collosphæra
and Solenosphæra or Odontosphæra.
Dimensions.—Diameter of the shell 0.1 to 0.15, of the pores 0.004 to 0.008, of
their bridges 0.004, of the large openings 0.24 to 0.032.
Habitat.—Tropical zone of the Pacific and the Indian Ocean; Stations
266 to 272, surface.
8. Collosphæra tuberosa, n. sp.
Collosphæra huxleyi, var., Haeckel, 1862, Monogr. d. Radiol., Taf. xxxiv.
figs. 3, 9.
Shell very irregular, between subspherical and polyhedral in form, but always with irregular
impressions, boils or bosses, and between these different rounded prominent tubercles and ridges.
Network irregular, strong, with rounded, subcircular or nearly polygonal meshes. Ten to thirty
pores in the half meridian of the shell. Diameter of the meshes half to four times as broad as
that of the thick bars.
Dimensions.—Diameter of the shell very variable in the same cœnobium, 0.05
to 0.2, of the pores 0.002 to 0.008, breadth of the bridges 0.004 to 0.006.
Habitat.—Cosmopolitan, common in all warmer seas, surface.
9. Collosphæra irregularis, n. sp.
Collosphæra huxleyi, var., Haeckel, 1862, Monogr. d. Radiol., Taf. xxxiv. fig. 8.
Shell quite irregular, knotty or bulbous, with irregular impressions, and prominent knobs or
bulbs between them. Network thin, fragile, quite irregular, with polygonal meshes of most unequal
size and form. Five to twenty pores in the half meridian of the shell. Diameter of the meshes two
to ten times as broad as that of the thin bars.
Dimensions.—Diameter of the shell very variable in the same cœnobium, 0.04
to 0.24, of the pores 0.005 to 0.05, of the bridges 0.002 to 0.004.
Habitat.—Mediterranean, Atlantic, not common; Stations 348, 352,
&c., surface.
{98}
Genus 30. Tribonosphæra,[52] Haeckel, 1881,
Prodromus, p. 471.
Definition.—Collosphærida with simple shells, on the
inside with radial centripetal beams.
The genus Tribonosphæra differs from Collosphæra by a very peculiar
and rare character, the development of centripetal radial sticks on the internal face of the
shell; these beams are not united in the centrum, but finish freely in a certain distance from
it.
1. Tribonosphæra centripetalis, n. sp. (Pl. 5, fig. 12).
Shell roundish or subspherical, with numerous small circular or roundish pores, about twice as
broad as the bars. Twenty to thirty pores on the half meridian of the shell. Outside of the shell
smooth, inside a variable number (ten to twenty) of thin, radial, centripetal sticks or spines,
one-third or one-half as long as the radius of the shell. (In the central capsule many very large
crystals, resting after the destruction of the capsule.)
Dimensions.—Diameter of the shell 0.1 to 0.12, of the pores 0.003 to 0.005, of the
bridges 0.001 to 0.002; length of the inner centripetal sticks 0.02 to 0.03.
Habitat.—Central Pacific, Station 271, surface.
Genus 31. Pharyngosphæra,[53] n. gen.
Definition.—Collosphærida with simple shells, having on
the inside radial centripetal tubes, the walls of which are solid.
The genus Pharyngosphæra differs from Collosphæra by the development
of radial tubules on the inside of the shell, which are directed centripetally towards its centre.
The wall of the tubule is solid, not latticed as in the following genus.
1. Pharyngosphæra stomodæa, n. sp. (Pl. 5, fig. 10).
Shell irregular polyhedral, with ten to fifteen polygonal faces and rounded edges. Pores very
small, circular, irregularly scattered, smaller than the bars. Twelve to fifteen pores on the half
meridian of the shell. On the inside of every shell-face one short, nearly cylindrical,
centripetal tubule, twice as long as broad, and about one-third as long as the shell radius. Outer
umbilical mouth of the tubules somewhat wider than the inner truncated mouth.
Dimensions.—Diameter of the shell 0.11 to 0.12, of the pores 0.003 to 0.005, of
the bars 0.01 to 0.02; length of the inner tubuli 0.02, breadth of them 0.01.
Habitat.—South Pacific, Station 288, depth 2600 fathoms.
{99}
Genus 32. Buccinosphæra,[54] n. gen.
Definition.—Collosphærida with simple shells, having on
the inside radial centripetal tubes, the walls of which are fenestrated.
The genus Buccinosphæra exhibits, on the inner surface of the shell, radial
centripetal tubules similar to those of the foregoing Pharyngosphæra; but the walls of
these tubes are here latticed, not solid; they represent therefore true invaginations of the whole
shell-wall.
1. Buccinosphæra invaginata, n. sp. (Pl. 5, fig. 11).
Shell irregular roundish or nearly polyhedral, with a variable number of umbilical depressions,
which are prolonged on the inside into cylindrical or somewhat conical, centripetal, fenestrated
tubes, about one-third as long as the shell radius. Inner mouth of the tubes narrower, scarcely
half as broad as the outer mouth, about equal to one-fourth the shell radius, truncated. Pores of
the tubes and of the shell small, roundish, irregular in size and distribution, about as broad as
the bars. Twenty-five to thirty pores in the half meridian of the shell. In all observed specimens
the spherical central capsule (half as broad as the shell) contained a large number of
crystals.
Dimensions.—Diameter of the shell 0.1 to 0.12, of the pores 0.001 to 0.003, of the
bars 0.002 to 0.003; length of the tubuli 0.02; outer mouth 0.026, inner mouth 0.013; crystals in
the central capsule 0.002 to 0.004, sometimes 0.008.
Habitat.—Philippine Islands (Samboangan), Station 213, surface.
2. Buccinosphæra tubaria, n. sp.
Shell irregular polyhedral with rounded edges, with a variable number of umbilical depressions,
which are prolonged on the inside into large, nearly cylindrical, centripetal, fenestrated tubes,
half as long as the shell radius. In the middle the tubes are somewhat constricted and narrower.
Inner mouth of the tubes dilated, nearly as broad as the outer mouth, about equal to one-half the
shell-radius, truncated. Pores of the tubes and of the shell large, roundish polygonal, irregular
in size and distribution, three to four times as broad as the bars. Fifteen to twenty pores in the
half meridian of the shell.
Dimensions.—Diameter of the shell 0.12 to 0.14, of the pores 0.008 to 0.012, of the bars 0.002 to 0.004;
length of the tubuli 0.03; outer mouth 0.04, inner mouth 0.03.
Habitat.—North coast of New Guinea, Station 217, surface.
Genus 33. Acrosphæra,[55] Haeckel, 1881,
Prodromus, p. 471.
Definition.—Collosphærida with simple shells, the outer
surface of which is covered with radial, irregularly scattered spines.
{100}
The genus Acrosphæra differs from its ancestral genus Collosphæra by
the development of spines on the outer surface of the shell. These are either short, straight,
radial spines, or oblique and often curved; their base is often inflated; they are irregularly
scattered on the whole surface between the pores.
1. Acrosphæra erinacea, n. sp.
Shell a regular sphere, everywhere covered with small, very numerous, straight radial spines,
regularly scattered between the pores. In the half meridian of the shell ten to twelve circular
pores, all of the same form and size, double as broad as the bars. Spines bristle-shaped, very
thin, solid, about as long as the diameter of the pores.
Dimensions.—Diameter of the shell 0.1 to 0.12, of the pores 0.008 to 0.012; length
of the spines 0.01.
Habitat.—Tropical zone of the Atlantic, coast of Brazil, Rabbe,
surface.
2. Acrosphæra echinoides, n. sp. (Pl 8,
fig. 1).
Shell a regular sphere, covered with numerous, straight, radial spines, irregularly scattered
over the whole surface. In the half meridian of the shell twenty to thirty irregular roundish
pores of variable size, one to four times as broad as the bars. Spines conical, strong, quite
radial, at the top of small conical elevations, which are perforated by from three to six
pores.
Dimensions.—Diameter of the shell 0.12 to 0.15, of the pores 0.002 to 0.008;
length of the spines 0.015, of their basal zones 0.01.
Habitat.—South-east corner of the Pacific, Valparaiso, Station 298,
surface.
3. Acrosphæra setosa, Haeckel.
Polysolenia setosa, Ehrenberg, 1872, Abhandl. d. k. Akad. d. Wiss. Berlin,
p. 299, Taf. viii. fig. 10.
Shell a regular sphere, covered with numerous bristle-shaped radial spines, irregularly
scattered between the pores. In the half meridian of the shell two to four very large circular
pores (equal to one-third the radius), and between them numerous very small, point-like pores.
Dimensions.—Diameter of the shell 0.05 to 0.08, of the large pores 0.01, of the
small 0.001; length of the spines 0.01 to 0.02.
Habitat.—West Tropical Pacific, Philippine Sea, Station 206, depth
2100 fathoms.
4. Acrosphæra spinosa, Haeckel.
Collosphæra spinosa, Haeckel, 1862, Monogr. d. Radiol., p. 536, Taf. xxxiv. figs. 12,
13.
Collosphæra spinosa, Cienkowsky, 1871, Archiv f. mikrosk. Anat., vii. p.
374, Taf. xxix. figs. 7-17.
Shell a regular or subregular sphere, covered with numerous, obliquely standing spines,
irregularly scattered over the surface. In the half meridian of the shell fifteen to twenty
irregular {101}roundish pores of very different form and
size, one to four times as broad as their bars. Spines conical, irregularly diverging and curved,
their hollow base perforated by several pores, not longer than the diameter of the largest
pores.
Dimensions.—Diameter of the shell 0.1 to 0.2, of the pores 0.001 to 0.04; length
of the spines 0.01 to 0.02.
Habitat.—Mediterranean, Messina; Canary Islands, Haeckel.
5. Acrosphæra collina, n. sp. (Pl. 8,
fig. 2).
Shell quite irregular, polyhedrical, hilly, with a variable number (eight to sixteen) of large
conical hill-like prominences; every cone or hill about as high as broad, perforated by the same
pores as the shell, on its top bearing a larger irregular roundish pore, and on its edge one
single bristle-like spine, not larger than the diameter of this pore, obliquely inserted. In the
half meridian of the shell twenty to thirty irregular roundish pores of very different size, one
to six times as broad as the bars. A very characteristic species, closely resembling the following
Odontosphæra.
Dimensions.—Diameter of the shell 0.15 to 0.25, of the pores 0.005 to 0.02; length
of the spines 0.01 to 0.02, height of the hills from which they rise 0.03 to 0.04.
Habitat.—North coast of New Guinea, Station 218, surface.
6. Acrosphæra inflata, n. sp. (Pl. 5,
fig. 7).
Mazosphæra inflata, Haeckel, 1879, Atlas, loc. cit.
Shell more or less irregular, polyhedral, hilly, with a variable number (six to twelve) of
large pyramidal, hill-like prominences; every hill about as high as broad, on the top a strong
conical, radial, or obliquely inserted spine, inflated, with three to six very large polygonal
meshes, much larger than the other pores between the hills, which are also polygonal, two to six
times as broad as the bars. Ten to fifteen pores on the half meridian.
Dimensions.—Diameter of the shell 0.1 to 0.14, of the largest pores 0.05, of the
smallest 0.005; length of the spines 0.02 to 0.03.
Habitat.—North Atlantic, Station 64, surface.
Genus 34. Odontosphæra,[56] n. gen.
Definition.—Collosphærida with simple shells, the outside
of which bears single scattered spines, one single spine on the margin of each larger pore.
The genus Odontosphæra is distinguished from the foregoing Acrosphæra
by the peculiar disposition of the spines, which are not scattered on the outside of the shell
between the pores, but so disposed that each larger pore is protected by one single spine,
obliquely placed over it.
{102}
1. Odontosphæra monodon, n. sp. (Pl. 5,
fig. 5).
Shell spherical or subspherical, with very small and numerous circular pores, much smaller than
the bars. Twelve to fifteen pores on the half meridian of the shell. Between them a variable
number of larger roundish apertures (mostly twelve to sixteen) irregularly scattered, one-fourth
to one-fifth as broad as the shell radius. On the margin of every larger aperture a single (rarely
two or three) sharp conical tooth, about as long as the diameter of the aperture, and obliquely
laid over them.
Dimensions.—Diameter of the shell 0.1 to 0.13, of the pores 0.001 to 0.003, of the
bars 0.01 to 0.02, of the larger apertures 0.01 to 0.02.
Habitat.—Sunda Archipelago, Station 192, surface.
2. Odontosphæra cyrtodon, n. sp. (Pl. 5, fig. 6).
Shell spherical or subspherical, with numerous roundish pores of very irregular size and
distribution, mostly larger than the bars. Ten to twelve pores on the half meridian of the shell.
Between them a variable number (mostly six to nine) of large roundish pores, about half as broad
as the shell radius, armed on one side of the margin with one single large tooth, about as long as
the diameter of the aperture, hook-like, curved, and obliquely laid over them.
Dimensions.—Diameter of the shell 0.12 to 0.14, of the pores 0.01 to 0.02, of the
bars 0.003 to 0.006, of the larger apertures 0.03 to 0.04.
Habitat.—Indian Ocean, near the Cocos Islands, Rabbe, surface.
Genus 35. Chœnicosphæra,[57] n.
gen.
Definition.—Collosphærida with simple shells, armed on
the outside with radial spines, forming elegant coronals around the larger pores.
The genus Chœnicosphæra is characterised by the peculiar disposition
of its radial spines, which form protective coronals around the larger pores, or even around all
pores of the shell.
Subgenus 1. Chœnicosphærula.
Definition.—A coronal of spines around every pore of the shell.
1. Chœnicosphæra murrayana, n. sp. (Pl. 8, fig. 4).
Shell spherical, with large circular or roundish pores of unequal size, two to four times as
broad as the bars. Ten to twelve pores in the half meridian of the shell. Margin of every pore
{103}with a coronal of six to nine short and acute
spines, not longer than the half diameter of the pore. No spines between the pores.
Dimensions.—Diameter of the shell 0.16 to 0.2, of the pores 0.02 to 0.03; length
of the spines 0.008 to 0.012.
Habitat.—Færöe Channel (Gulf Stream), common. Expedition of H.M.S.
"Triton," John Murray.
2. Chœnicosphæra flosculenta, n. sp.
Shell spherical, with large circular or roundish pores of different size, three to six times as
broad as the bars. Six to eight pores in the half meridian of the shell. Margin of every pore
somewhat prominent, with a coronal of ten to twenty parallel acute spines of different length, the
largest somewhat longer than the diameter of the pore. No spines between the pores.
Dimensions.—Diameter of the shell 0.12 to 0.15, of the pores 0.02 to 0.04; length
of the spines 0.02 to 0.05.
Habitat.—Central Pacific, Station 272, depth 2600 fathoms.
Subgenus 2. Chœnicosphærium.
Definition.—A coronal of spines only around the larger pores, not
around the smaller.
3. Chœnicosphæra nassiterna, n. sp. (Pl. 8, fig. 3).
Shell spherical, with circular or roundish pores of very different size. The smaller pores very
numerous, without coronal of spines, roundish, about as broad as the bridges. Twenty to thirty
pores in the half meridian of the shell. Between them, irregularly scattered, a small number
(eight to twelve) of very large circular pores, one-third to one-half as broad as the radius of
the shell, armed with a coronal of six to nine parallel, straight, acute spines, about half as
long as the radius of the shell.
Dimensions.—Diameter of the shell 0.1 to 0.13, of the smaller pores 0.002 to
0.004, of the larger armed pores 0.02 to 0.03; length of the spines 0.03 to 0.04.
Habitat.—Philippine Islands, Mindanao, Station 213, depth 2050
fathoms.
4. Chœnicosphæra flammabunda, n. sp. (Pl. 8, fig. 5).
Shell spherical, with circular or roundish pores of very different size. The smaller pores very
numerous and unequal, very irregularly scattered, hardly one-fourth to one-half as broad as the
bridges between them. Twelve to twenty-four pores in the half meridian of the shell. Between them,
irregularly scattered, a variable number (ten to twenty) of very large circular pores, about
one-fourth as broad as the radius of the shell. The margin of these large pores is armed with a
very irregular coronal of four to twelve unequal, curved acute spines, partly simple, partly
branched like {104}a deer-horn, one-fourth to one-half
as long as the radius of the shell. Some other small spines irregularly scattered over the
shell.
Dimensions.—Diameter of the shell 0.12 to 0.16, of the smaller pores 0.001 to
0.004, of the larger armed pores 0.016 to 0.024; length of the spines 0.02 to 0.04.
Habitat.—Central area of the Tropical Pacific, Stations 266 to 272,
depth 2425 to 2925 fathoms.
Genus 36. Siphonosphæra,[58] J. Müller,
1858, Abhandl. d. k. Akad. d. Wiss.
Definition.—Collosphærida with simple shells, the pores
of which are prolonged into external simple radial tubuli with solid wall; outer mouth of the
tubuli truncated, smooth.
The genus Siphonosphæra is, next to Collosphæra, the most common of
all Collosphærida, and rich in different species; all agreeing in the tubular prolongation of the
pores, and corresponding therefore to Ethmosphæra among the simple Liosphærida.
Subgenus 1. Holosiphonia, Haeckel.
Definition.—All the pores or apertures of the shell prolonged into
tubules.
1. Siphonosphæra pansiphonia, n. sp.
Shell a regular sphere, everywhere occupied by short, regular cylindrical tubes, all of the
same size and form. Length of the tubules equal to their breadth and to the intervals between
them. Ten to twelve tubules in the half meridian of the shell. This most regular species is nearly
allied to Collosphæra regularis, and may be derived from it by tubular prolongation of all
the regular pores.
Dimensions.—Diameter of the shell 0.1 to 0.12, length and breadth of the tubules
0.005 to 0.006.
Habitat.—Indian Ocean, Sunda Strait, Rabbe, surface.
2. Siphonosphæra marginata, n. sp.
Shell a regular or subregular sphere, occupied by numerous short cylindrical tubules of
different sizes. Six to eight tubules in the half meridian of the shell. Diameter of the tubules
about equal to their distance apart, but two to four times as large as their length.
Dimensions.—Diameter of the shell 0.1 to 14; length of the tubules 0.004 to 0.006,
breadth of the tubules and the intervals 0.01 to 0.02.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
{105}
3. Siphonosphæra tubulosa, J. Müller (Pl. 6, fig. 4).
Siphonosphæra tubulosa, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p.
59.
Siphonosphæra tubulosa, Haeckel, 1862, Monogr. d. Radiol., p. 532.
Collosphæra tubulosa, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p.
59.
Thalassicolla punctata, var., Huxley, 1851, Ann. and Mag. Nat. Hist., ser.
2, vol. viii. p. 435, pl. xvi. fig. 5.
Shell subspherical or roundish, somewhat irregular, occupied by a small number (five to ten) of
short cylindrical tubules, irregularly scattered at great distances; intervals between the tubules
very large, two to four times as broad as the length of the tubules, which is equal to their
breadth and about one-fifth or one-sixth of the shell diameter. Only two or three tubules in the
half meridian of the shell.
Dimensions.—Diameter of the shell 0.12 to 0.15, length and breadth of the tubules
0.02 to 0.03.
Habitat.—Equatorial zone of the Pacific, Stations 225, 265, 268,
surface.
4. Siphonosphæra cyathina, n. sp. (Pl. 6, fig. 10).
Shell a regular sphere, everywhere occupied by short cup-like tubules of somewhat different
size. In the half meridian of the shell about ten to twelve tubules, nearly cylindrical, but
constricted in the middle. Inner and outer aperture of the tubule of the same size; their diameter
equal to their length and distance.
Dimensions.—Diameter of the shell 0.1 to 0.12, length and breadth of the tubules
0.005 to 0.01.
Habitat.—West coast of Tropical Africa, Stations 348 to 352,
surface.
5. Siphonosphæra patinaria, n. sp. (Pl. 6, figs. 7,
8).
Shell a regular or subregular sphere, occupied by numerous broad cup-like tubules of very
different sizes. In the half meridian of the shell about six to eight tubules, very flat, circular
or subcircular, much constricted in the middle. Diameter of the inner aperture larger than that of
the middle stricture, smaller than that of the outer aperture of the tubule; outer diameter two to
four times as great as their length.
Dimensions.—Diameter of the shell 0.1 to 0.15, of the pores 0.01 to 0.04; length
of the tubules 0.01, breadth 0.02 to 0.04.
Habitat.—Tropical Atlantic, Station 348, depth 2450 fathoms.
6. Siphonosphæra infundibulum, n. sp.
Shell subspherical or irregular rounded, occupied by a small number (six to twelve) of large
funnel-like tubules of very different size, scattered irregularly at great distances. Intervals
between the tubules larger than their length, which surpasses the radius of the shell. Outer
opening of the funnels three to four times as broad as the inner.
Dimensions.—Diameter of the shell 0.15; length of the tubules 0.05 to 0.09.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
{106}
7. Siphonosphæra conifera, n. sp. (Pl. 6, fig. 9).
Shell subspherical, everywhere occupied by irregular conical tubules, the length of which
nearly reaches the shell radius. Inner aperture of the cones two to three times as large as the
outer. Four to five tubules in the half meridian of the shell. Distances between the bases of the
cones small and irregular.
Dimensions.—Diameter of the shell 0.06 to 0.08, length of the tubules 0.03 to
0.04; inner aperture of the cones 0.01 to 0.02, outer aperture 0.008 to 0.01.
Habitat.—Indian Ocean, Cocos Islands, Rabbe, surface.
8. Siphonosphæra fragilis, n. sp.
Shell quite irregular roundish or nearly ovate, very thin and fragile, everywhere occupied by
irregular, short, and broad cylindrical or conical tubes. Six to ten tubes in the half meridian of
the shell. Diameter of the tubules about one-eighth that of the shell, three to four times as
large as the length of the tubules, and the distance apart of their bases.
Dimensions.—Diameter of the shell 0.16 to 0.24, of the tubules 0.02 to 0.03;
length of the tubules 0.006 to 0.008, distance of them 0.005 to 0.009.
Habitat.—East coast of Australia, Sydney, Faber; Station 165,
surface.
Subgenus 2. Merosiphonia, Haeckel.
Definition.—Only part of the shell apertures prolonged in tubules, the
others simple.
9. Siphonosphæra socialis, n. sp. (Pl. 6, figs. 1,
2).
Shell a regular or subregular sphere, bearing only a small number (one to four, commonly two to
three) of short and broad cylindrical tubules, irregularly scattered. Between them many small
circular or subcircular pores of different sizes, double as broad as their bars. Eight to ten
pores in the half meridian of the shell. Tubules three to six times as broad as the pores, about
as long as broad, now quite cylindrical, now somewhat dilated at the outer opening. (Although the
shells and cells of this common species are among the smallest, their colonies are among the
largest, often containing more than one hundred social individuals, often enclosed in
alveoles.)
Dimensions.—Diameter of the shell 0.04 to 0.05, of the pores 0.002 to 0.004, of
the tubules 0.015 to 0.02; length of them about the same.
Habitat.—Tropical and subtropical part of the Eastern Atlantic, Cape
Verde Islands, Canaries, very common, Haeckel; Stations 338 to 353, surface.
10. Siphonosphæra polysiphonia, n. sp.
Shell a regular or subregular sphere, bearing twelve to sixteen circular pores in its half
meridian. Nearly one half the pores simple, very small; the other half prolonged into short
cylindrical tubules, {107}half as long as broad, two to
four times as broad as the pores and their intervals. This species is nearly related to the
foregoing, which it represents in the western tropical part of the Atlantic, but differs
constantly in the double size of the shell and the much larger number of the tubules.
Dimensions.—Diameter of the shell 0.06 to 0.09, of the pores 0.002 to 0.004, of
the tubules 0.01 to 0.012; their length 0.006 to 0.008, surface.
Habitat.—Tropical and subtropical part of the Western Atlantic, coast
of Brazil, &c., Rabbe.
11. Siphonosphæra macrosiphonia, n. sp.
Shell a regular sphere, with numerous very small pores of equal size and distribution. Twelve
to sixteen pores in its half meridian. Bars (between the pores) three to four times as broad as
their diameter. Only a small number (two to four) of very long cylindrical tubes, irregularly
scattered, nearly as long or somewhat longer than the shell diameter; now quite straight, now
somewhat curved. Diameter of the cylinders four to six times as large as that of the pores.
Dimensions.—Diameter of the shell 0.1 to 0.12, of the pores 0.002 to 0.004, of the
tubules 0.015 to 0.018; length of the tubules 0.08 to 0.16.
Habitat.—Central Tropical Pacific, Station 266 to 272, surface.
12. Siphonosphæra serpula, n. sp. (Pl. 6, fig. 6).
Shell a regular sphere, with numerous very small pores of equal size and distribution. Eight to
ten pores in its half meridian; bars between them three to four times as broad as their diameter.
Only a small number (six to eight) of very long and snake-like, contorted, cylindrical tubes,
irregularly scattered. The shape of the latter is very much like that of the calcareous tubes of
some species of Serpula; they are nearly as long as, or longer than, the shell diameter,
and four to six times as broad as the pores.
Dimensions.—Diameter of the shell 0.12 to 0.14, of the pores 0.003 to 0.005, of
the tubules 0.02 to 0.022; length of the tubules 0.1 to 0.2.
Habitat.—North Eastern Pacific, Station 252 to 262, Sandwich Islands,
Haltermann.
13. Siphonosphæra chonophora, n. sp. (Pl. 6, fig. 5).
Shell a regular or subregular sphere, with numerous very small pores of equal size and
distribution, ten to twelve in its half meridian. Bars between the pores four to six times their
diameter. Only a small number (two to six) of very large funnel-like tubules, irregularly
scattered. The inner half of these tubules is a short cylindrical tube, of the same thickness as
the shell, three to four times as broad as the pores; the outer half is an irregular funnel,
suddenly expanded, with siliceous walls of the utmost tenuity and fragility, often irregularly
folded and contorted, like a decayed flower-calyx, often half as large as the shell.
Dimensions.—Diameter of the shell 0.1 to 0.12, of the pores 0.003 to 0.005, inner
half of the tubules 0.015 to 0.02, outer funnel-like half 0.05 to 0.08.
Habitat.—South Pacific, Stations 285 to 295, surface.
{108}
14. Siphonosphæra pipetta, n. sp. (Pl. 6, fig. 3).
Shell more or less irregularly roundish or subspherical, occupied in part by very small pores,
in part by very large cylindrical tubules, inflated in the middle. Number, form, and size of the
tubes very irregular; commonly there are five to ten, half as long or as long as the shell radius;
their inner and outer aperture about half as broad as their inflated middle part; three to nine
times as broad as the pores. A very irregular and variable species.
Dimensions.—Diameter of the shell 0.1 to 0.15, of the pores 0.001 to 0.005, of the
tubules 0.015 to 0.03; length of the tubules 0.03 to 0.08.
Habitat.—North Pacific, Stations 242 to 253, surface.
Genus 37. Mazosphæra,[59] Ehrenberg,
1860.
Mazosphæra, Ehrenberg, 1860, Monatsber. d. k. preuss. Akad. d. Wiss.
Berlin, p. 833.
Definition.—Collosphærida with simple shells, the pores
of which are prolonged into external simple radial tubuli with solid wall; outer mouth of each
tubulus armed with a single tooth.
The genus Mazosphæra is intermediate between Siphonosphæra and
Odontosphæra, agreeing with the former in the tubular prolongation of the pores, with the
latter in the possession of a single large protective tooth on the outer opening.
1. Mazosphæra hippotis, n. sp. (Pl. 5,
fig. 8).
Shell spherical, with circular pores of irregular size and distribution, scarcely half as broad
as the bars; fifteen to twenty on the half meridian of the shell. Between them, irregularly
distributed, a variable number (ten to fifteen) of short cylindrical radial tubules, about twice
as long as broad, and half as long as the shell radius. Mouth of the tubuli obliquely truncated,
having on one side a strong acute tooth.
Dimensions.—Diameter of the shell 0.11 to 0.12, of the pores 0.002 to 0.004, of
the bridges 0.006 to 0.009; length of the tubuli 0.03, breadth of them 0.01 to 0.015.
Habitat.—North Pacific, Station 253, depth 3125 fathoms.
2. Mazosphæra lagotis, n. sp. (Pl. 5,
fig. 9).
Shell spherical, with circular pores of irregular size and distribution, about as broad as the
bars; twelve to sixteen on the half meridian. Between them, irregularly distributed, a variable
number (eight to twelve) of long, cylindrical, curved tubules, three to six times as long as
broad, and about as long as the shell radius; their external mouth lateral, obliquely truncated,
ovate, having on one side a strong conical tooth.
{109}
Dimensions.—Diameter of the shell 0.1 to 0.12, of the pores 0.002 to 0.01; length
of the tubuli 0.05 to 0.07.
Habitat.—Central Pacific, Station 266, depth 2750 fathoms.
3. Mazosphæra lævis, Ehrenberg.
Mazosphæra lævis, Ehrenberg, 1872, Abhandl. d. k. Akad. d. Wiss. Berlin,
p. 297, Taf. vii. fig. 7.
Shell spherical, with very small pores, scarcely one-fourth as broad as the bars. Fifteen to
twenty pores on the half meridian of the shell. Between them, irregularly distributed, a variable
number (fifteen to twenty) of short conical tubules, about as long as broad, only one-fifth to
one-sixth as long as the shell radius. Mouth of the tubuli truncated, with an obtuse short tooth
on one side.
Dimensions.—Diameter of the shell 0.08 to 0.09, of the pores 0.001 to 0.002, of
the bridges 0.005 to 0.008; length of the tubuli 0.01 to 0.02, breadth of them the same.
Habitat.—Philippine Islands (depth 3300 fathoms), Ehrenberg; Station
206, depth 2100 fathoms; Station 225, depth 4575 fathoms.
4. Mazosphæra apicata, Ehrenberg.
Mazosphæra apicata, Ehrenberg, 1872, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 316.
Shell spherical, without small pores, only with a variable number (ten to twenty) of short
conical tubules, twice as long as broad, and half as long as the shell radius. Mouths of the
tubuli obliquely truncated, with a strong acute tooth on one side. (This species differs from the
two preceding by the want of the small pores between the tubules.)
Dimensions.—Diameter of the shell 0.08 to 0.1, of the tubules 0.01.
Habitat.—Philippine Islands (depth 3300 fathoms), Ehrenberg; north
coast of New Guinea, depth 2000 fathoms; Station 217.
Genus 38. Trypanosphæra,[60] n. gen.
Definition.—Collosphærida with simple shells, the pores
of which are prolonged into external simple radial tubuli with solid walls; outer mouth of each
tubulus armed with a coronal of spines.
The genus Trypanosphæra is intermediate between Siphonosphæra and
Chœnicosphæra agreeing with the former in the tubular prolongation of the pores,
with the latter in the possession of a coronal of teeth on their outer opening.
Subgenus 1. Trypanosphærula, Haeckel.
Definition.—All pores of the shell prolonged into short coronated
tubules.
{110}
1. Trypanosphæra trepanata, n. sp. (Pl. 5, fig. 4).
Shell regular spherical, with regular circular pores of nearly equal size, at unequal
distances, one to four times as broad as the bars. Eight to ten pores on the half meridian. All
the pores prolonged into short cylindrical tubuli about as long as broad, armed on the external
mouth with an elegant coronal of twenty to thirty straight bristle-shaped, parallel teeth.
Dimensions.—Diameter of the shell 0.12 to 0.14, of the pores 0.015 to 0.02; length
of the tubuli 0.02.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
2. Trypanosphæra dentata, n. sp.
Shell regular spherical, with regular circular pores of equal size, but at very different
distances. Only three to four pores on the half meridian. All the pores prolonged into irregular
curved, cylindrical tubuli, about as long as the shell radius, with a coronal of ten to twelve
short conical teeth on the distal end.
Dimensions.—Diameter of the shell 0.08 to 0.09; length of the tubuli 0.04, breadth
0.02.
Habitat.—Central Pacific, Station 274, depth 2750 fathoms.
Subgenus 2. Trypanosphærium, Haeckel.
Definition.—Only part of the shell-pores prolonged into coronated
tubules.
3. Trypanosphæra coronata, n. sp. (Pl. 5, fig. 3).
Shell regular spherical, with irregular roundish pores of very different sizes. On the half
meridian four to six large and twelve to sixteen very small pores. About half of the large pores
prolonged into short cylindrical tubuli, the outer mouth of each being armed with an elegant
coronal of ten to twenty thin irregular teeth.
Dimensions.—Diameter of the shell 0.1 to 0.12, of the large pores 0.01 to 0.02, of
the small pores 0.001; length of the tubuli 0.012.
Habitat.—North Pacific, Station 241, depth 2300 fathoms.
4. Trypanosphæra terebrata, n. sp.
Shell irregular roundish, with unequal, small, roundish pores. Sixteen to twenty pores on the
half meridian. Six to eight larger pores are prolonged into curved cylindrical tubuli, about as
long as the shell radius, with a coronal of ten to twelve strong conical straight teeth on the
distal end.
Dimensions.—Diameter of the shell 0.15, of the pores 0.001 to 0.003; length of the
tubuli 0.08, breadth 0.02.
Habitat.—West Tropical Pacific, Station 225, depth 4575 fathoms.
{111}
5. Trypanosphæra transformata, n. sp. (Pl. 5, figs. 1,
2).
Shell quite irregular, of very variable, roundish, or polyhedral form, with small irregular
roundish pores, two to four times as broad as the bars. Ten to thirty on the half meridian. The
different form of the shell depends upon the variable number of tubuli, which arise at irregular
distances from the shell; commonly three to four, often also five to six, more rarely one or two.
The tubuli are now more conical, now more cylindrical, about as long as the shell radius, at other
times scarcely one-half or one-third as long, with a coronal of ten to twenty more or less curved
teeth on the narrower distal mouth. All the different forms are to be found in one and the same
colony, as shown in fig. 1. This cœnobium, which I observed living in Ceylon, exhibited the
same peculiar formation as I figured in Collosphæra huxleyi in my Monograph 1862 (Taf.
xxxiv. fig. 1). In the centre of the jelly-sphere lies a large globular alveole, surrounded by
numerous small, young central capsules without shell; whilst in the surface lies one layer of
older capsules, enclosed in shells. Some of the younger capsules exhibit self-division.
Dimensions.—Diameter of the shells 0.08 to 0.12, pores 0.002 to 0.006; length and
breadth of the tubuli 0.03 to 0.05.
Habitat.—Indian Ocean, Belligemma, Ceylon, surface.
Genus 39. Caminosphæra,[61] n. gen.
Definition.—Collosphærida with simple shells, the pores
of which are prolonged into external branched radial tubuli with solid wall.
The genus Caminosphæra differs from Siphonosphæra (and from all other
Collosphærida) in the ramification of the tubuli, which arise from the pores; the walls of the
tubuli are solid, not fenestrated.
1. Caminosphæra furcata, n. sp.
Shell spherical or subspherical, with a variable number (four to eight) of short cylindrical
tubes, irregularly scattered, about as long as the radius of the shell. Every tube forked, with
two cylindrical branches of the same size as the simple basal part of the tube. Mouth of the
branches truncated, not dilated. Pores of the shell between the tubes very small, all of the same
size, half as broad as their bars. Fifteen to twenty pores in the half meridian of the shell.
Dimensions.—Diameter of the shell 0.1 to 0.12, of the pores 0.001 to 0.002; length
of the tubules 0.05 to 0.06, breadth of them 0.012 to 0.015.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
2. Caminosphæra elongata, n. sp.
Shell spherical, with a large number (twelve to twenty) of long cylindrical tubes, irregularly
formed and scattered, somewhat longer than the diameter of the shell. Every tube forked at {112}the distal end, with two or three short irregular
branches of unequal size and form; branches much shorter than the undivided basal part of the
tube. Mouth of the branches narrowed, truncated. Pores of the shell between the tubes about half
as broad, irregularly roundish or polygonal, two to three times as broad as their bars. Ten to
twelve pores in the half meridian of the shell.
Dimensions.—Diameter of the shell 0.05 to 0.07, of the pores 0.006 to 0.009;
length of the tubules 0.06 to 0.09, breadth of them 0.015 to 0.02.
Habitat.—Tropical Central Pacific, Station 271, depth 2425
fathoms.
3. Caminosphæra dichotoma, n. sp. (Pl. 7, fig. 2).
Shell spherical, with a variable number (ten to fifteen) of cylindrical tubes, irregularly
scattered, about as long as the radius of the shell. Every tube furcated, with two cylindrical
branches of the same size as the simple basal part of the tube. Mouth of the branches dilated,
funnel-like, twice as broad as the tube; the edges irregularly dentated or lacerated. Pores
between the tubes small, one-third to one-sixth as broad as these, half as broad as their bars.
Ten to twelve pores in the half meridian of the shell.
Dimensions.—Diameter of the shell 0.12 to 0.14, of the pores 0.003 to 0.005;
length of the tubules 0.06 to 0.08, breadth of them 0.02 to 0.03.
Habitat.—Southern Pacific, Station 295, depth 1500 fathoms.
4. Caminosphæra dendrophora, n. sp. (Pl. 7, fig. 1).
Shell spherical, with a variable number (eight to twelve) of long cylindrical tubes,
irregularly branched and scattered, nearly as long as the diameter of the shell. Every tube with
two to six (commonly three to four) branches of different sizes. Mouth of the branches dilated,
funnel-like; the edges irregularly dentated or lacerated. Pores between the tubes half as broad as
these, irregularly roundish, twice as broad as their bars. Ten to twelve pores in the half
meridian of the shell.
Dimensions.—Diameter of the shell 0.12 to 0.15, of the pores 0.006 to 0.01; length
of the tubules 0.1 to 0.13, breadth of them 0.02 to 0.025.
Habitat.—Central Pacific, Station 266, depth 2750 fathoms.
Genus 40. Solenosphæra,[62] n. gen.
Definition.—Collosphærida with simple shells, the pores
of which are prolonged into external simple radial tubuli with fenestrated wall; outer mouth of
the tubuli truncated, smooth.
The genus Solenosphæra differs from Siphonosphæra in the fenestration
of the external radial tubes. A large number of shells, appertaining to this genus, were already
described by Ehrenberg, and disposed in five different genera corresponding to the {113}different numbers of the tubuli:—Disolenia
with two tubes, Trisolenia with three tubes, Tetrasolenia with four tubes,
Pentasolenia with five tubes, Polysolenia with six or more tubes. All these five
genera are without value, as those different numbers of tubes occur frequently intermingled in the
individual cells of one and the same colony, wherever the form and structure of the tubes is
inherited with sufficient constancy to determine the species.
Subgenus 1. Solenosphactra, Haeckel.
Definition.—Tubuli of the shell cylindrical or nearly cylindrical, the
outer and inner apertures nearly of the same size.
1. Solenosphæra variabilis, Haeckel.
Tetrasolenia quadrata, Ehrenberg, 1872, Abhandl. d. k. Akad. d. Wiss.
Berlin, Taf. x. fig. 20.
Shell quite irregular roundish or polyhedral, with roundish pores of different size. Ten to
fifteen pores in the half meridian of the shell, two to three times as broad as the bars. Porous
tubuli of the shell in variable number (in one and the same colony), three to nine, mostly four to
six; cylindrical or subcylindrical or somewhat conical, two to three times as broad as long, not
longer than the half radius of the shell. Inner aperture of the tubuli commonly as broad as the
half radius of the shell (or somewhat smaller), about as large as the truncated outer aperture.
This species is closely related to Collosphæra polyedra (p. 97), and
may be derived from it by a short tube-like prolongation of the larger apertures.
Dimensions.—Diameter of the shell 0.1 to 0.16, of the pores 0.005 to 0.015; length
of the tubules 0.02 to 0.03, breadth of them 0.04 to 0.06.
Habitat.—Central area of the Tropical Pacific, Stations 270, 271, 272,
depths 2425 to 2925 fathoms.
2. Solenosphæra pandora, n. sp. (Pl. 7,
figs. 10, 11).
Shell irregular roundish or subglobular, with roundish pores of different sizes, mostly
somewhat broader than the bars. About twelve to sixteen pores on the half meridian of the shell.
Porous tubuli of the shell of variable number (in one and the same colony), one to six, mostly
three to four; cylindrical or nearly cylindrical, somewhat longer than broad, not longer than the
radius of the shell. Inner aperture of the tubuli commonly as broad as the half radius of the
shell, and a little smaller than the truncated outer aperture.
Dimensions.—Diameter of the shell 0.07 to 0.1, of the pores 0.003 to 0.006; length
of the tubuli 0.03 to 0.05, breadth of them 0.02 to 0.03.
Habitat.—Central area of the Tropical Pacific, Stations 266 to 274,
depths 2350 to 2925 fathoms.
{114}
3. Solenosphæra megalactis, Haeckel.
Trisolenia megalactis, Ehrenberg, 1872, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 301, Taf. viii. fig. 19.
Shell irregularly polyhedrical, with very small roundish pores, scarcely half as broad as the
bars. Only eight to ten pores on the half meridian of the shell. Porous tubuli of the shell of
variable number (in one and the same colony), two to five, mostly three or four; cylindrical,
about as long as the radius of the shell. Inner aperture of the tubuli commonly as broad as the
half radius of the shell, and quite as broad as the truncated outer aperture.
Dimensions.—Diameter of the shell 0.07 to 0.09, of the pores 0.002 to 0.004, of
the bars 0.005 to 0.009; length of the tubuli 0.03 to 0.04, breadth of them 0.02.
Habitat.—Pacific; California, Philippine Sea, Ehrenberg; Stations 256
to 285, depths 310 to 3000 fathoms.
4. Solenosphæra serpentina, n. sp. (Pl. 7, fig. 7).
Shell nearly spherical, with very small circular pores, scarcely one-third or one-fourth as
broad as the bars. Only five to seven pores in the half meridian of the shell. Porous tubuli of
the shell of variable number (in one and the same colony), two to nine, mostly seven or eight;
cylindrical, somewhat curved or contorted, once and a half or twice as long as the diameter of the
shell, with few very small and widely scattered pores. Inner and outer aperture of the tubuli have
the same diameter, about one-fifth or one-fourth that of the shell. (This species is closely
allied to Siphonosphæra serpula, but is distinguished from it by the long tortuous tubuli
and the small scarce pores.)
Dimensions.—Diameter of the shell 0.08 to 0.1, of the pores 0.001 to 0.002, of the
bars 0.004 to 0.008; length of the tubuli 0.12 to 0.18, breadth of them 0.02 to 0.025.
Habitat.—North-eastern Pacific, between Sandwich Islands and
California, Haltermann, surface.
Subgenus 2. Solenosphenia, Haeckel.
Definition.—Tubuli of the shell more or less conical, the inner
aperture much larger than the outer aperture.
5. Solenosphæra venosa, Haeckel.
Tetrasolenia venosa, Ehrenberg, 1872, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 301, Taf. vii. fig. 22.
Shell irregular polyhedral or roundish, with a delicate network of large irregular polyhedral
meshes, five to ten times as broad as the thin bars. Eight to twelve meshes on the half meridian
of the shell. Fenestrated tubuli of the shell of variable number (in one and the same colony), one
to five, commonly three or four, shaped like a short truncated cone, about half as long as broad
on its base, shorter than the radius of the shell. Inner aperture of the cone nearly as broad as
the half radius of the shell, about twice as broad as the truncated outer aperture.
{115}
Dimensions.—Diameter of the shell 0.07 to 0.12, of the pores 0.008 to 0.016, of
the bars 0.001; length of the tubuli 0.02 to 0.03, inner aperture 0.03 to 0.04, outer aperture
0.02 to 0.03.
Habitat.—Indian Ocean, Sunda Strait, Rabbe.
6. Solenosphæra ascensionis, n. sp. (Pl. 7, fig. 9).
Shell somewhat irregular, subspherical, with polygonal pores of different size. Twelve to
fifteen pores in the half meridian of the shell, two to eight times as broad as their bars. Porous
tubuli of the shell of variable number (in one and the same colony), three to nine, mostly five to
seven; conical or nearly cylindrical, irregular, about as long as broad at their base. Inner
aperture of the tubuli two to four times as broad as the broadest pores, and double as broad as
the truncated circular outer aperture.
Dimensions.—Diameter of the shell 0.1 to 0.12, of the pores 0.004 to 0.018, of the
bars 0.002, length of the tubuli 0.04, inner aperture 0.04, outer 0.02.
Habitat.—South Atlantic, near Ascension Island, Station 343,
surface.
Subgenus 3. Solenosphyra, Haeckel.
Definition.—Tubuli of the shell funnel-like, the outer aperture much
larger than the inner.
7. Solenosphæra cornucopia, n. sp. (Pl. 7, fig. 8).
Shell spherical or subspherical, with roundish pores of different size. Ten to twelve pores in
the half meridian of the shell, two to three times as broad as the bars. Porous tubuli of the
shell of variable number (in one and the same colony), four to eight, mostly five to seven,
funnel-like, about as long as the diameter of the shell. Inner aperture of the tubuli commonly
two-thirds or three-fourths as broad as the radius of the shell (or somewhat smaller), only
one-half or two-thirds as broad as the dilated and truncated outer aperture.
Dimensions.—Diameter of the shell 0.07 to 0.09, of the pores 0.006 to 0.018;
length of the tubuli 0.06 to 0.08, diameter of the inner aperture 0.04 to 0.05, of the outer 0.06
to 0.08.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
8. Solenosphæra amalthea, n. sp.
Shell irregular roundish or spherical, with small circular pores of different size. Fifteen to
twenty pores in the half meridian of the shell, but still not as broad as the bars. Porous tubuli
of the shell of variable number (in one and the same colony), three to six, commonly four or five,
funnel-like, about as long as the radius of the shell. Inner aperture of the tubuli about half as
broad as the radius of the shell, only one-half or one-third as broad as the truncated outer
aperture. (This species is intermediate between the preceding and Siphonosphæra chonophora,
Pl. 6, fig.
5.)
{116}
Dimensions.—Diameter of the shell 0.09 to 0.11, of the pores 0.002 to 0.004, of
the bars 0.003 to 0.006; length of the tubuli 0.05 to 0.06; diameter of the inner aperture 0.02 to
0.03, of the outer aperture 0.05 to 0.07.
Habitat.—Western part of the South Atlantic, Station 325, surface.
Genus 41. Otosphæra,[63] n. gen.
Definition.—Collosphærida with simple shells, the pores
of which are prolonged into external simple radial tubuli with fenestrated walls; outer mouth of
the tubuli armed with a single tooth.
The genus Otosphæra differs from Solenosphæra by the single tooth on
the external mouth of the tubuli, from Mazosphæra by the fenestration of the walls of the
tubuli.
1. Otosphæra polymorpha, n. sp. (Pl. 7,
fig. 6).
Shell quite irregular, polyhedral or roundish, very variable in size and form, with numerous
very small pores, much smaller than the bars. Twenty to thirty pores in the half meridian of the
shell. Porous tubuli of the shell commonly in variable number (one to four), but sometimes
constant in number (one, two, three, or four) in the one and same colony. Tubuli irregular
conical, commonly about as long as the radius of the shell; their outer aperture obliquely
truncated, on one side prolonged into one large, prominent, bill-like, curved, acute tooth.
Dimensions.—Diameter of the shell 0.12 to 0.15, of the pores 0.001 to 0.002;
length of the tubuli 0.06 to 0.08, inner aperture 0.03, outer aperture 0.02.
Habitat.—Indian Ocean, Madagascar, Rabbe, surface.
2. Otosphæra auriculata, n. sp. (Pl. 7,
fig. 5).
Shell quite irregular, of extremely variable form, now inclining to roundish, now to
polyhedral, with very numerous small pores, irregularly formed and distributed. Twelve to
twenty-four pores in the half meridian of the shell, of very different size, for the most part
larger than the bars. Porous tubuli of the shell of variable number (in one and the same colony),
one to five, mostly three or four, of conical form, irregularly formed and scattered, commonly
about half as long as the radius of the shell. Outer aperture of the tubuli obliquely truncated,
with one large prominent, often curved, acute tooth.
Dimensions.—Diameter of the shell 0.1 to 0.2, of the pores 0.003 to 0.005; length
of the tubuli 0.01 to 0.05, inner aperture 0.04, outer aperture 0.03.
Habitat.—Central Tropical Pacific, Stations 268 to 272, depths 2425 to
2925 fathoms.
{117}
Genus 42. Coronosphæra,[64] n. gen.
Definition.—Collosphærida with simple shells, the pores
of which are prolonged into external simple radial tubuli with fenestrated walls; outer mouth of
the tubuli armed with a coronal of spines.
The genus Coronosphæra differs from Solenosphæra by the coronated
mouth of the tubuli, from Trypanosphæra by the fenestration of the walls of the tubuli.
1. Coronosphæra diadema, n. sp. (Pl. 7,
fig. 3).
Shell spherical or subspherical, with a variable number (fifteen to twenty) of short,
coronal-like tubules, irregularly scattered, about half as long as the radius of the shell. Outer
aperture of the tubuli irregularly dentated, a little dilated, and not much broader than the inner
aperture, one-half or one-third as broad as the shell radius. Pores of the shell and of the tubuli
circular or roundish, very irregularly scattered, mostly one-half or one-third as broad as the
bars.
Dimensions.—Diameter of the shell 0.11 to 14, of the pores 0.002 to 0.004, of the
bars 0.006 to 0.012; length of the tubuli 0.03, inner aperture 0.02 to 0.03, outer aperture 0.03
to 0.04.
Habitat.—Central Pacific, Stations 268 to 270, depths 2550 to 2925
fathoms.
2. Coronosphæra calycina, n. sp. (Pl. 7, fig. 4).
Shell spherical or subspherical, with a variable number (eight to twelve) of large, funnel-like
tubules, irregularly scattered, about as long as the radius of the shell. Outer aperture of the
tubuli irregularly dentated, much dilated, somewhat broader than the shell radius, three to four
times as broad as the inner circular aperture. Pores of the shell and of the tubuli circular or
roundish, of very different size, one to three times as broad as the bars.
Dimensions.—Diameter of the shell 0.1 to 0.3, of the pores 0.003 to 0.01, of the
bars 0.002 to 0.004; length of the tubuli 0.1, inner aperture 0.02 to 0.03, outer aperture 0.06 to
0.18.
Habitat.—Central Pacific, Stations 271, 272, depths 2425 and 2600
fathoms respectively.
3. Coronosphæra convolvulus, n. sp.
Shell irregular roundish, with a variable number (five to ten) of long, curved tubules, about
as long as the shell diameter. The inner half of the tubuli is narrow, cylindrical; the outer half
funnel-like dilated, similar to the flower of Convolvulus. The outer aperture is elegantly
dentated, five to six times as broad as the inner aperture. Pores of the shell and of the tubuli
very irregular roundish, about as broad as the bars.
Dimensions.—Diameter of the shell 0.08 to 0.09, of the pores and bars 0.004 to
0.008; length of the tubuli 0.07 to 0.1, inner aperture 0.01, outer aperture 0.05.
Habitat.—Tropical Atlantic, Station 347, surface.
{118}
Subfamily Clathrosphærida, Haeckel, 1881, Prodromus, p. 472.
Definition.—Collosphærida with a double
lattice-shell around every central capsule of the cœnobium; both concentric shells
connected by irregular or subradial beams, commonly solid or lamellar staffs, rarely hollow
tubes.
Genus 43. Clathrosphæra,[65] Haeckel, 1881,
Prodromus, p. 472.
Definition.—Collosphærida with a double lattice-shell
around every central capsule of the cœnobium; surface of the outer shell smooth.
The genus Clathrosphæra (with smooth surface) and the following
Xanthiosphæra (with spiny surface) form together the small subfamily, Clathrosphærida,
different from the other Collosphærida by the double lattice-shell. From the surface of the inner
primary shell arise either solid spines or hollow tubes, which unite by the anastomosis of
irregular branches and so form the outer secondary shell, often very incomplete and irregular. All
Clathrosphærida seem to inhabit great depths.
Subgenus 1. Clathrosphærula, Haeckel.
Definition.—The connecting staffs between both shells are hollow tubes
(derived from Siphonosphæra).
1. Clathrosphæra circumtexta, n. sp. (Pl. 8, fig. 6).
Inner shell spherical, with irregular roundish large meshes, now broader now smaller than their
bars. Eight to ten meshes in the half meridian of the shell. All these meshes are prolonged into
short cylindrical hollow tubes, about as long as broad, somewhat constricted in the middle. From
the margins of the outer openings of these tubes proceed very numerous and delicate siliceous
filaments, which all lie on the same spherical face, branch, anastomose, and twine over the
openings and the intervals between them, forming a very thin, arachnoid spherical outer shell. The
meshes of this are quite irregular polygonal, of very different size and form. The radius of the
inner shell bears to that of the outer a ratio = 5 : 6.
Dimensions.—Diameter of the inner shell 0.11 to 0.13, of the outer 0.13 to 0.16;
meshes of the inner shell 0.005 to 0.02, of the outer 0.005 to 0.04.
Habitat.—North Pacific, Stations 238 to 253, depths 2050 to 3950
fathoms.
Subgenus 2. Clathrosphærium, Haeckel.
Definition.—The connecting staffs between the two shells are solid
rods or lamellar spines (derived from Acrosphæra).
{119}
2. Clathrosphæra arachnoides, n. sp. (Pl. 8, fig. 7).
Inner shell spherical, with irregular roundish meshes, two or three times as broad as the bars.
Ten to twelve meshes in the half meridian of the shell. From its surface arise numerous conical
radial spines (with base often fenestrated), which at equal distances from the surface send out
lateral branches. All these branches lie on a spherical face, and form by communications the
irregular, very delicate, arachnoid network of the outer shell, quite unlike that of the inner,
with large polygonal meshes of very different size. Eight to sixteen meshes in the half meridian
of the shell. Surface of the outer shell nearly spherical, somewhat uneven, like a spider's web.
The radius of the inner shell bears to that of the outer a ratio = 3 : 4.
Dimensions.—Diameter of the inner shell 0.12 to 0.14, of the outer 0.15 to 0.18;
pores of the inner shell 0.003 to 0.02, of the outer 0.01 to 0.04.
Habitat.—Central area of the Tropical Pacific, Station 268, depth 2900
fathoms.
3. Clathrosphæra lamellosa, n. sp. (Pl. 8, fig. 8).
Inner shell spherical or subspherical, with irregular roundish meshes, about half as broad as
the bars. Twelve to sixteen meshes in the half meridian of the shell. From its surface arise
numerous oblique irregular staffs or broad and thin lamellæ, which branch quite irregularly, and
by communications of the branches form the thin outer shell. This is quite irregular roundish or
subspherical, very unlike the inner, with large polygonal meshes of different size, six to twelve
in the half meridian of the shell. Bridges between the meshes very variable, now very thin
filamentous, now very broad lamellar. Outer surface very uneven or tuberculated, but not spinous.
The radius of the inner shell bears to that of the outer a ratio = 5 : 6.
Dimensions.—Diameter of the inner shell 0.1 to 0.13, of the outer 0.12 to 0.18;
pores of the inner shell 0.003 to 0.009, of the outer 0.01 to 0.04.
Habitat.—Central area of the Tropical Pacific, Stations 270 to 274,
depths 2350 to 2925 fathoms.
Genus 44. Xanthiosphæra,[66] Haeckel, 1881,
Prodromus, p. 472.
Definition.—Collosphærida with a double lattice-shell
around every central capsule of the cœnobium; surface of the outer shell thorny or
spiny.
The genus Xanthiosphæra differs from the foregoing Clathrosphæra by
spines or thorns arising from the surface of the outer shell, commonly very irregular.
1. Xanthiosphæra capillacea, n. sp.
Inner shell spherical, with irregular polygonal meshes, three to five times as broad as their
narrow bars. Six to eight meshes in the half meridian of the shell. From its surface arise at the
nodes of the network numerous thin radial spines, which, at equal distances from the surface,
{120}send out lateral branches. All these branches lie
on a spherical face, and form by communications the irregular delicate network of the outer shell,
very like that of the inner, with large polygonal meshes, six to eight meshes in the half meridian
of the shell. Surface of the outer shell covered with numerous straight spines, prolongations of
the inner spines, but only half as long as these. The radius of the inner shell bears to that of
the outer a ratio = 3 : 5.
Dimensions.—Diameter of the inner shell 0.1 to 0.12, of the outer 0.15 to 0.19;
pores of the inner shell 0.02 to 0.04 to 0.06, of the outer 0.04 to 0.06 to 0.08; length of the
outer spines 0.01 to 0.02.
Habitat.—Central area of the Tropical Pacific, Station 263, depth 2650
fathoms.
2. Xanthiosphæra erinacea, n. sp. (Pl. 8, fig. 9).
Inner shell spherical, with irregular roundish meshes, one-half to two times as broad as the
bars. Fifteen to twenty meshes in the half meridian of the shell. From its surface arise numerous
thin radial spines, which at equal distances from the surface send out lateral branches. All these
branches lie on the face of a sphere, and form by communications the irregular delicate network of
the outer shell, very unlike that of the inner, with large polygonal meshes, twelve to twenty-four
in the half meridian of the shell. Surface of the outer shell covered with numerous straight
spines, prolongations of the inner spines, and of the same length. The radius of the inner shell
bears to that of the outer a ratio = 3 : 4.
Dimensions.—Diameter of the inner shell 0.1 to 0.12, of the outer 0.13 to 0.16;
pores of the inner shell 0.002 to 0.008, of the outer 0.01 to 0.03; length of the outer spines
0.02 to 0.03.
Habitat.—Central area of the Tropical Pacific, Stations 270, 272,
depth 2925 and 2600 fathoms respectively.
3. Xanthiosphæra lappacea, n. sp. (Pl. 8, figs. 10,
11).
Inner shell spherical or subspherical, with very small roundish pores, quite irregularly
scattered, one-fourth to three-fourth as broad as their bars. Ten to twenty pores in the half
meridian of the shell. From its surface arise in an extremely irregular and variable manner
numerous oblique spines, often curved, often lamellar, and perforated by pores, sometimes hollow,
fenestrated cones. At different distances from the surface these spines send out lateral curved
branches, which by communications form the delicate and very irregular network of the outer shell.
This network is often incomplete and very unlike that of the inner shell, with large polygonal
meshes, six to eighteen in the half meridian of the shell. Surface of the outer shell covered with
numerous small, curved, and oblique spines, prolongations of the inner spines, but scarcely
one-third to one-half as long as these. The radius of the inner shell bears to that of the outer a
ratio = 3 : 4.
Dimensions.—Diameter of the inner shell 0.08 to 0.12, of the outer 0.11 to 0.15;
pores of the inner shell 0.001 to 0.009, of the outer 0.01 to 0.04; length of the outer spines
0.005 to 0.009.
Habitat.—Central area of the Tropical Pacific, Stations 263 to 274,
depths 2350 to 3000 fathoms.
{121}
Family VII. Stylosphærida, Haeckel (Pls. 13-17).
Stylosphærida, Haeckel, 1881, Prodromus, p. 449.
Definition.—Sphæroidea with two radial spines on the
surface of the spherical shell, opposite in one axis; living solitary (not associated in
colonies).
The family Stylosphærida comprises a large number of very common Sphæroidea, and is distinguished from all others by the possession of two
radial spines which are placed in one axis of the spherical shell.[67] By the expression of this "main axis" as a solid rod they form
the transition to the Prunoidea, in which the whole shell is more or less
transformed according to this "monaxial growth." But in these latter the shell, as well as the
central capsule, becomes ellipsoidal, prolonged in one axis, whilst in the former they remain
spherical. However, the distinction of both nearly allied groups is sometimes difficult.
The most simple Stylosphærida are the Xiphostylida, with one single spherical
lattice-shell. To this ancestral group all other subfamilies can be opposed as "Stylosphærida
concentrica," as their carapace is composed of two or more concentric lattice-shells: two in the
Sphærostylida, three in the Amphistylida, four in the Cromyostylida, five or more in the
Caryostylida. In all these four subfamilies the concentric shells are simple (not spongy)
fenestrated spheres. In a sixth subfamily, in the Spongostylida, the shell is wholly or partially
composed of a spongy irregular wicker-work, with or without a medullary shell in the centre.
Both the radial spines in all Stylosphærida are opposed normally in one axis; but in many
species besides the normal form occur individual abnormalities, in which the two spines are not
accurately opposed in this main axis, but placed in two different axes, intersecting at a smaller
or larger angle. In the majority of the Stylosphærida both opposite spines have the same size and
form; but in some genera they are more or less different, often in a very striking degree. The
same differences occur in the nearly allied groups of Prunoidea, in the
Ellipsida and Druppulida.
The distal ends of both spines are commonly free; but in the small group of Saturnalida
(Saturnalis with one single shell, Saturnulus with two concentric shells,
Saturninus with three concentric shells) the distal ends of both spines are united, at
equal distances from the centre, by a circular or elliptical ring. This remarkable peculiarity
occurs in no other group of Sphæroidea, and consequently brings the
Saturnalida into close relation with the Discoidea.
{122}
Synopsis of the Genera of Stylosphærida.
|
I. Subfamily Xiphostylida.
(Spherical shell simple.)
|
|
Polar spines free, without connecting ring on the distal
ends. |
|
Both spines equal, |
45. Xiphosphæra. |
| Spines different in size or form, |
46. Xiphostylus. |
| Both polar spines united by a circular or elliptical
ring, |
47. Saturnalis. |
|
II. Subfamily Sphærostylida.
(Two concentric spheres.)
|
|
Polar spines free. |
|
Both spines equal, |
48. Stylosphæra. |
| Spines different in size or form, |
49. Sphærostylus. |
| Both polar spines united by a circular or elliptical
ring, |
50. Saturnulus. |
|
III. Subfamily Amphistylida.
(Three concentric spheres.)
|
|
Polar spines free. |
|
Both spines equal, |
51. Amphisphæra. |
| Spines different in size or form, |
52. Amphistylus. |
| Both polar spines united by a circular or elliptical
ring, |
53. Saturninus. |
|
IV. Subfamily Cromyostylida.
(Four concentric spheres.)
|
|
Polar spines free. |
|
Both spines equal, |
54. Stylocromyum. |
| Spines different, |
55. Cromyostylus. |
|
V. Subfamily Caryostylida.
(Five or more concentric spheres.)
|
|
Polar spines free. |
|
Both spines equal, |
56. Caryostylus. |
|
VI. Subfamily Spongostylida.
(Spherical shell partially or wholly of a spongy structure.)
|
|
Shell a solid spongy sphere without central medullary
shell, |
57. Spongolonchis. |
| In the centre of the spongy sphere one or two medullary
shells. |
|
One central medullary shell, |
58. Spongostylus. |
| Two concentric medullary shells, |
59. Spongostylidium. |
| I. Subfamily Xiphostylida. (Spherical shell simple.) |
|
|
Polar spines free, without connecting ring on the distal
ends. |
|
|
|
|
|
|
Both spines equal, |
|
|
|
|
|
|
|
|
|
|
45. Xiphosphæra. |
|
|
|
|
|
|
|
|
|
|
Spines different in size or form, |
|
|
|
|
|
|
|
|
|
|
46. Xiphostylus. |
|
|
|
|
|
|
|
|
Both polar spines united by a circular or elliptical ring, |
|
|
|
|
|
|
|
|
47. Saturnalis. |
|
|
|
|
|
|
| II. Subfamily Sphærostylida. (Two concentric spheres.) |
|
|
Polar spines free. |
|
|
|
|
|
|
Both spines equal, |
|
|
|
|
|
|
|
|
|
|
48. Stylosphæra. |
|
|
|
|
|
|
|
|
|
|
Spines different in size or form, |
|
|
|
|
|
|
|
|
|
|
49. Sphærostylus. |
|
|
|
|
|
|
|
|
Both polar spines united by a circular or elliptical ring, |
|
|
|
|
|
|
|
|
50. Saturnulus. |
|
|
|
|
|
|
| III. Subfamily Amphistylida. (Three concentric spheres.) |
|
|
Polar spines free. |
|
|
|
|
|
|
Both spines equal, |
|
|
|
|
|
|
|
|
|
|
51. Amphisphæra. |
|
|
|
|
|
|
|
|
|
|
Spines different in size or form, |
|
|
|
|
|
|
|
|
|
|
52. Amphistylus. |
|
|
|
|
|
|
|
|
Both polar spines united by a circular or elliptical ring, |
|
|
|
|
|
|
|
|
53. Saturninus. |
|
|
|
|
|
|
| IV. Subfamily Cromyostylida. (Four concentric spheres.) |
|
|
Polar spines free. |
|
|
|
|
|
|
Both spines equal, |
|
|
|
|
|
|
|
|
|
|
54. Stylocromyum. |
|
|
|
|
|
|
|
|
|
|
Spines different, |
|
|
|
|
|
|
|
|
|
|
55. Cromyostylus. |
|
|
|
|
|
|
| V. Subfamily Caryostylida. (Five or more concentric spheres.) |
|
|
Polar spines free. |
|
|
|
|
|
|
Both spines equal, |
|
|
|
|
|
|
|
|
|
|
56. Caryostylus. |
|
|
|
|
|
|
| VI. Subfamily Spongostylida. (Spherical shell partially or wholly of a spongy
structure.) |
|
|
Shell a solid spongy sphere without central medullary shell, |
|
|
|
|
|
|
|
|
57. Spongolonchis. |
|
|
|
|
|
|
|
|
In the centre of the spongy sphere one or two medullary
shells. |
|
|
|
|
|
|
One central medullary shell, |
|
|
|
|
|
|
|
|
|
|
58. Spongostylus. |
|
|
|
|
|
|
|
|
|
|
Two concentric medullary shells, |
|
|
|
|
|
|
|
|
|
|
59. Spongostylidium. |
|
|
|
|
|
|
Subfamily Xiphostylida, Haeckel, 1881, Prodromus, pp. 449, 450.
Definition.—Stylosphærida with one simple
spherical lattice-shell.
Genus 45. Xiphosphæra,[68] Haeckel, 1881,
Prodromus, p. 450.
Definition.—Stylosphærida with one single lattice-sphere
and two free spines of equal size and form.
The genus Xiphosphæra is the most simple form of all Stylosphærida, and may
be regarded as the common ancestral form of this family. On the surface of a simple {123}spherical lattice-shell, enclosing the central capsule,
arise two equal, free, radial spines, opposite to each other on the poles of one axis.
Subgenus 1. Xiphosphærantha, Haeckel.
Definition.—Pores of the spherical shell regular, of nearly equal size
and form; surface smooth or a little rough, without spines or thorns (other than the two polar
spines).
1. Xiphosphæra planeta, n. sp.
Pores regular, hexagonal, eight to nine times as broad as the thin bars. Ten to twelve pores on
the half equator. Shell very thin walled; surface smooth. Polar spines three-sided pyramidal,
about as long as the axis of the sphere, as broad at the base as one pore.
Dimensions.—Diameter of the sphere 0.12 to 0.13, pores 0.016 to 0.018, bars 0.002;
length of the polar spines 0.1 to 0.15, basal thickness 0.02.
Habitat.—Pacific, central area, surface; Stations 271 to 274, depths
2425 to 2750 fathoms.
2. Xiphosphæra gæa, n. sp. (Pl. 14,
fig. 5).
Pores regular, circular, with prominent hexagonal crests between them. On the half equator ten
to twelve pores, of the same breadth as the crested bars. Shell thin walled; surface smooth. Polar
spines three-sided prismatic, about twice as long as the axis of the sphere, twice as broad at the
base as one pore.
Dimensions.—Diameter of the sphere 0.07 to 0.09, pores and bars 0.005; length of
the polar spines 0.15 to 0.2, basal thickness 0.01.
Habitat.—Pacific, central area, Station 274, depth 2750 fathoms.
3. Xiphosphæra venus, n. sp. (Pl. 14,
fig. 2).
Pores regular, circular, with prominent hexagonal frames. On the half equator fifteen to
eighteen pores, of the same breadth as the bars. Shell very thick walled; surface smooth,
honeycomb-like. Polar spines conical, smooth, about as long as the axis of the shell, twice as
broad at the base as one pore.
Dimensions.—Diameter of the sphere 0.12 to 0.13, pores and bars 0.005; thickness
of the shell wall 0.013; length of the polar spines 0.12 to 0.15, basal breadth 0.01.
Habitat.—Pacific, central area, Station 272, depth 2600 fathoms.
4. Xiphosphæra luna, n. sp.
Pores regular, circular, hexagonally lobed or rosette-shaped, three times as broad as the bars.
Ten to twelve pores on the half equator. Shell thick walled; surface smooth. Polar spines
three-sided pyramidal, one to two times as long as the axis of the shell, as broad at the base as
one pore {124}(very similar to Xiphostylus
phasianus, Pl. 13,
fig. 9, but different in the equal size and similar form of the two large polar spines).
Dimensions.—Diameter of the sphere 0.12, pores 0.015, bars 0.005; length of the
polar spines 0.1 to 0.2, basal breadth 0.02.
Habitat.—Indian Ocean, Cocos Islands, surface, Rabbe.
5. Xiphosphæra hebe, n. sp.
Pores regular, circular, three times as broad as the bars. On the half equator sixteen to
twenty pores. Shell thick walled; surface smooth. Polar spines conical or nearly cylindrical,
about as long as the axis of the sphere, as broad at the base as two pores.
Dimensions.—Diameter of the sphere 0.1 to 0.13, pores 0.006, bars 0.002; polar
spines 0.1 to 0.15 long, 0.01 thick.
Habitat.—Pacific, central area, Stations 265 to 268, depths 2700 to
2900 fathoms.
6. Xiphosphæra maxima, n. sp.
Pores regular, circular, twice as broad as the bars, funnel-shaped. Twenty to thirty pores on
the half equator. Shell very thick walled; surface smooth. Polar spines three-sided pyramidal,
about as long as the radius of the sphere, as broad at the base as two pores.
Dimensions.—Diameter of the sphere 0.22 to 0.35, pores 0.008 to 0.01, bars 0.005;
polar spines 0.1 to 0.15 long, 0.02 thick.
Habitat.—Equatorial Atlantic, Station 347, depth 2250 fathoms.
7. Xiphosphæra euphrosyne, n. sp.
Pores regular, circular, about as broad as the bars, double contoured. Eight to ten on the half
equator. Shell thin walled; surface smooth. Polar spines conical, about as long as the radius of
the sphere, as broad at the base as one pore.
Dimensions.—Diameter of the sphere 0.12 to 0.15, pores and bars 0.02; polar spines
0.06 to 0.09 long, 0.02 thick.
Habitat.—South Atlantic, Station 323, depth 1900 fathoms.
Subgenus 2. Xiphosphærella, Haeckel.
Definition.—Pores of the spherical shell regular, of nearly equal size
and form; surface thorny or spiny, covered with regularly distributed papillæ or thorns (in
addition to the two large polar spines).
8. Xiphosphæra pallas, n. sp. (Pl. 14,
fig. 4).
Pores regular, circular, separated by hexagonal elevated frames, the sharp crest of which is
elegantly denticulated; in each corner of the hexagons (between three pores) is a short radial
spine, {125}about as long as one pore. On the half
equator sixteen to twenty pores, of the same breadth as the bars. Shell thick walled; whole
surface spiny. Polar spines cylindrical, at the apex conical, about as long as the axis of the
sphere, three to four times as broad as one pore.
Dimensions.—Diameter of the sphere 0.1, pores and bars 0.005; length of the polar
spines 0.07 to 0.11, thickness 0.015 to 0.02.
Habitat.—Western Tropical Pacific, Station 225, depth 4475
fathoms.
9. Xiphosphæra flora, n. sp.
Pores regular, circular, with hexagonal frames, twice as broad as the bars. Ten to twelve pores
on the half equator. Shell thin walled, with spiny surface; in each corner of the hexagons is one
bristle-like radial spine twice as long as one pore. Polar spines three-sided prismatic, at the
apex pyramidal, nearly twice as long as the axis of the sphere, as broad at the base as two pores
(similar to Ellipsoxiphus palliatus, Pl. 14, fig.
7).
Dimensions.—Diameter of the sphere 0.15, pores 0.01, bars 0.005; length of the
polar spines 0.2 to 0.25, breadth 0.02.
Habitat.—Tropical Atlantic, Station 342, depth 1445 fathoms.
10. Xiphosphæra juno, n. sp.
Pores regular, circular, as broad as the bars, funnel-shaped. Fifteen to twenty pores on the
half equator. Shell thick walled, covered with bristle-like spines, about twice as long as one
pore. Polar spines conical, thick, about as long as the axis of the sphere, twice as broad at the
base as one pore.
Dimensions.—Diameter of the sphere 0.12, pores and bars 0.01; length of the polar
spines 0.14, basal breadth 0.02.
Habitat.—Fossil in the Barbados rocks; living in the greatest depth of
the Tropical Pacific, Station 225, depth 4475.
11. Xiphosphæra gigantea, n. sp.
Pores regular, circular, two to three times as broad as the bars; twenty-eight to thirty-two on
the half equator. Shell thick walled, covered with short conical thorns. Polar spines three-sided
pyramidal, about as long as the radius of the sphere, as broad at the base as three pores.
Dimensions.—Diameter of the sphere 0.25 to 0.3, pores 0.01, bars 0.004; polar
spines 0.1 to 0.15 long, 0.03 broad.
Habitat.—Fossil in the Tertiary rocks of Barbados and Sicily
(Cattanisetta).
Subgenus 3. Xiphosphærissa, Haeckel.
Definition.—Pores of the spherical shell irregular, of different size
or form; surface smooth or a little rough, without spines or thorns (other than the polar
spines).
{126}
12. Xiphosphæra ceres, n. sp.
Pores irregular, roundish, of different sizes, two to four times as broad as the bars. Sixteen
to twenty pores on the half equator. Shell thin walled, with smooth surface. Polar spines conical,
about as long as the axis of the sphere, very thick at the base.
Dimensions.—Diameter of the sphere 0.15 to 0.2, pores 0.004 to 0.008, bars 0.002;
polar spines 0.18 to 0.24 long, at the base 0.02 thick.
Habitat.—North Atlantic, Station 353, surface.
13. Xiphosphæra clavigera, n. sp.
Pores irregular, roundish, double contoured, of very unequal size, two to seven times as broad
as the bars; ten to twelve on the half equator. Shell thick walled; surface a little rough. Polar
spines club-shaped, with prominent edges, about half as long as the axis of the sphere; thinner at
both ends than in the middle. (Differs from Ellipsoxiphus claviger, Pl. 14, fig. 3, in
the spherical shell and shorter spines.)
Dimensions.—Diameter of the sphere 0.2, pores 0.005 to 0.02, bars 0.003; polar
spines 0.06 long, 0.02 broad.
Habitat.—Pacific, central area, Station 274, depth 2750 fathoms.
Subgenus 4. Xiphosphæromma, Haeckel.
Definition.—Pores of the spherical shell irregular, of different size
or form; surface thorny or spiny (besides the two large polar spines).
14. Xiphosphæra vesta n. sp. (Pl. 14,
fig. 6).
Pores irregular, roundish, three to five times as broad as the bars; fourteen to sixteen on the
half equator. Scattered on the surface of the thick-walled shell are from twenty to thirty strong
three-sided pyramidal spines of unequal size, the largest twice as long as the largest pores.
Polar spines very strong, nearly three-sided prismatic, with curved edges, nearly as long as the
axis of the sphere and twice as broad as the largest pores.
Dimensions.—Diameter of the sphere 0.17, pores 0.01 to 0.02, bars 0.004; length of
the polar spines 0.13, thickness 0.02 to 0.03.
Habitat.—Pacific, central area. Station 266, depth 2750 fathoms,
15. Xiphosphæra astræa, n. sp.
Pores irregular, roundish, one to two times as broad as the bars; ten to twelve on the half
equator. Surface of the thick-walled shell covered with numerous short conical thorns. Polar
spines cylindro-conical, one and a half to two times as long as the axis of the sphere.
{127}
Dimensions.—Diameter of the sphere 0.18, pores 0.01 to 0.015, bars 0.008; length
of the polar spines 0.25 to 0.3, thickness 0.02.
Habitat.—Indian Ocean, surface; Ceylon, Haeckel.
Genus 46. Xiphostylus,[69] Haeckel, 1881,
Prodromus, p. 450.
Definition.—Stylosphærida with one single lattice-sphere
and two free spines of different size or form.
The genus Xiphostylus differs from the foregoing Xiphosphæra in the
unequal size or form of both polar spines, which become more or less differentiated.
Subgenus 1. Xiphostylantha, Haeckel.
Definition.—Pores of the spherical shell regular, of nearly equal size
and form; surface smooth or a little rough, without spines or thorns.
1. Xiphostylus alcedo, n. sp. (Pl. 13,
fig. 4).
Pores regular, circular, with elevated hexagonal frames, twice as broad as the bars. Eight to
ten pores on the half equator. Surface smooth. Polar spines three-sided pyramidal, as broad at the
base as one hexagon; the major spine four to five times as long as the minor, which is about equal
to the radius of the sphere.
Dimensions.—Diameter of the sphere 0.12, pores 0.012, bars 0.006; length of the
major polar spine 0.16 to 0.2, of the minor 0.04 to 0.06, basal breadth 0.02.
Habitat.—Western Tropical Pacific, Station 225, depth 4475.
2. Xiphostylus phasianus, n. sp. (Pl. 13, fig.
9).
Pores regular, circular, twice as broad as the bars. Eight to ten pores on the half equator.
Outer opening of each pore elegantly lobed, with eight indentations. Surface a little rough. Polar
spines very unequal; major spine sword-like, sharply edged, about as long as the diameter of the
sphere; minor spine scarcely half so long, pommel-shaped, with nine (?) wing-like edges.
Dimensions.—Diameter of the sphere 0.13, inner circular opening of the pores 0.01,
outer eight-lobed opening 0.015, bars 0.005; length of the major polar spine 0.14, of the minor
0.06, breadth 0.03.
Habitat.—Australian Sea, Station 162, surface.
3. Xiphostylus motacilla, n. sp.
Pores regular, circular, three times as broad as the bars; sixteen to twenty on the half
equator. Surface smooth. Polar spines compressed, two-edged, at the base three to four times as
broad as {128}one pore; the major spine somewhat longer
than the diameter of the shell, the minor scarcely one-third or one-half as long.
Dimensions.—Diameter of the sphere 0.14, pores 0.006, bars 0.002; length of the
major spine 0.16 to 0.18, of the minor 0.05 to 0.07, basal breadth 0.02.
Habitat.—Indian Ocean, Zanzibar, 2200 fathoms, Pullen.
4. Xiphostylus gallus, n. sp.
Pores regular, circular, five times as broad as the bars. Twelve to sixteen pores on the half
equator. Surface smooth. Polar spines very unequal; the major conical spine one and a half to
three times as long as the diameter of the sphere; the minor pommel-shaped, scarcely one-third as
long (length of both spines very variable).
Dimensions.—Diameter of the sphere 0.13, pores 0.01, bars 0.002; length of the
major spine 0.2 to 0.4, of the minor 0.05 to 0.08.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
5. Xiphostylus alauda, n. sp. (Pl. 14,
fig. 15).
Lithomespilus alauda, Haeckel, 1881, Prodrom. et Atlas, loc.
cit.
Pores subregular, circular, three to four times as broad as the bars; fifteen to eighteen on
the half equator. Surface a little rough. Polar spines irregularly conical or pyramidal, scarcely
as long as the radius of the sphere; one spine simple, the other composed of a bunch of four or
five spines united at the base.
Dimensions.—Diameter of the sphere 0.11, pores 0.01, bars 0.003; length of the
polar spines 0.03 to 0.05, basal breadth 0.02.
Habitat.—Pacific, central area, Station 272, depth 2600 fathoms.
6. Xiphostylus anhinga, Haeckel.
Rhabdolithis pipa, Bury, 1862, Polycystins of Barbados, pl. iii. fig.
4.
Pores subregular, circular, about the same breadth as the bars; eight to ten on the half
equator. Surface smooth or a little rough. Polar spines cylindrical, very irregularly curved like
S or contorted, the major three to six times as long as the diameter of the sphere, the minor
scarcely one-fourth as long as the former, at the end truncated.
Dimensions.—Diameter of the sphere 0.07, pores and bars 0.005; length of the major
polar spine 0.2 to 0.4, of the minor 0.06 to 0.09, basal breadth 0.01.
Habitat.—Fossil in the Barbados rocks.
Subgenus 2. Xiphostyletta, Haeckel.
Definition.—Pores of the spherical shell regular, of nearly equal size
and form; surface thorny or spiny (other than the two large polar spines).
{129}
7. Xiphostylus cuculus, n. sp.
Pores regular, circular, hexagonally framed, three times as broad as the bars; ten to twelve on
the half equator. Surface thorny, between every three pores a short conical thorn. Polar spines
three-sided prismatic, the major somewhat longer than the diameter of the sphere, the minor
scarcely one-third as long, pommel-shaped.
Dimensions.—Diameter of the sphere 0.17, pores 0.012, bars 0.004; length of the
major polar spine 0.2, of the minor 0.05, basal breadth 0.015.
Habitat.—South Atlantic, surface; Station 335, depth 1425 fathoms.
8. Xiphostylus trochilus, n. sp. (Pl. 13, fig.
10).
Pores regular, circular, four times as broad as the bars; eight to nine on the half equator.
Polar spines cylindrical, the major somewhat longer than the axis of the sphere, the minor
shorter, surrounded by a group of from four to eight shorter conical spines. Surface of the
opposite hemisphere smooth, without by-spines.
Dimensions.—Diameter of the sphere 0.07 to 0.08, pores 0.01, bars 0.0025.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
9. Xiphostylus picus, n. sp. (Pl. 14,
fig. 13).
Lithomespilus picus, Haeckel, 1881, Prodrom. et Atlas.
Pores regular, circular, twice as broad as the bars; sixteen to eighteen on the half equator.
Polar spines cylindrical, conical at the apex, the major once and a half to twice as long as the
diameter of the shell, the minor scarcely half so long; around the latter a group of twelve to
twenty shorter conical spines, irregularly scattered. Surface of the other hemisphere smooth.
Dimensions.—Diameter of the sphere 0.13, pores 0.006, bars 0.003; length of the
major polar spine 0.2 to 0.24, of the minor 0.08 to 0.09, basal breadth 0.02.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms.
Subgenus 3. Xiphostylissa, Haeckel.
Definition.—Pores of the spherical shell irregular, of unequal size or
form; surface smooth or a little rough, without thorns.
10. Xiphostylus trogon, n. sp. (Pl. 14,
fig. 12).
Lithomespilus trogon, Haeckel, 1881, Prodrom. et Atlas.
Pores irregular, roundish or subcircular, two to three times as broad as the bars; ten to
twelve on the half equator. Surface smooth. Major polar spine three-sided prismatic, once and a
half to twice as long as the axis of the sphere; minor spine quite rudimentary, scarcely longer
than broad, but surrounded by a group of from three to six similar short spines.
{130}
Dimensions.—Diameter of the sphere 0.1, pores 0.005 to 0.015, bars 0.005 to 0.008;
length of the major spine 0.15 to 0.18, of the minor 0.01 to 0.02, basal breadth 0.02.
Habitat.—Western Tropical Pacific, Station 225, depth 4475
fathoms.
11. Xiphostylus falco, n. sp. (Pl. 13,
fig. 14).
Pores irregular, roundish, two to five times as broad as the bars; sixteen to eighteen on the
half equator. Surface smooth. Polar spines cylindrical, very stout, nearly half as thick as the
radius of the shell; major spine two to four times as long as the diameter of the shell; minor
spine obliquely inserted, scarcely longer than the diameter, divided at the end into two short,
hook-shaped, curved branches.
Dimensions.—Diameter of the sphere 0.08, pores 0.002 to 0.005, bars 0.001; breadth
of the spines 0.02, length of the major spine 0.15 to 0.2, of the minor 0.09.
Habitat.—South Pacific, Station 302, depth 1450 fathoms.
12. Xiphostylus alca, n. sp. (Pl. 13,
fig. 13).
Pores irregular, roundish, two to six times as broad as the bars; six to eight on the half
equator. Each pore with three to six lobes, composed of three to six confluent smaller pores.
Surface smooth. Major spine conical, curved, somewhat longer than the axis of the sphere; minor
spine somewhat shorter, pommel-like, edged.
Dimensions.—Diameter of the sphere 0.07, pores 0.01 to 0.02, bars 0.003; length of
the major spine 0.08, of the minor 0.06, basal thickness 0.02.
Habitat.—Indian Ocean, Sunda Strait, Rabbe, surface.
13. Xiphostylus edolius, n. sp. (Pl. 13,
fig. 5).
Pores irregular, roundish, composed of two to six smaller confluent pores. On the half equator
six to eight large pores, and twenty to thirty small pores; bars between the smaller very thin.
Surface a little rough. Major polar spine conical, S-shaped, about twice as long as the axis of
the shell; minor spine pommel-shaped, edged, scarcely as long as its radius.
Dimensions.—Diameter of the sphere 0.12, large pores 0.01 to 0.03, small pores
0.004 to 0.008, bars 0.001 to 0.004; length of the major spine 0.2, of the minor 0.05, basal
breadth 0.02.
Habitat.—Central Pacific, Station 273, surface.
Subgenus 4. Xiphostylomma, Haeckel.
Definition.—Pores of the spherical shell irregular, of different size
or form; surface thorny or spiny.
{131}
14. Xiphostylus emberiza, n. sp. (Pl. 13, fig.
11).
Pores irregular, roundish, one to four times as broad as the bars; six to eight on the half
equator. Polar spines very unequal; major cylindrical, twice as long as the axis of the sphere;
minor scarcely half as long, obliquely inserted, like a bird's head, surrounded by a group of ten
to twenty smaller conical spines. Opposite hemisphere smooth.
Dimensions.—Diameter of the sphere 0.005, pores 0.002 to 0.008, bars 0.002; length
of the major spine 0.09, of the minor 0.05, basal breadth 0.01.
Habitat.—South Atlantic, Station 332, surface.
15. Xiphostylus ardea, n. sp.
Pores irregular, roundish, one to three times as broad as the bars; twelve to sixteen on the
half equator. Whole surface spiny. Major polar spine three-sided pyramidal, somewhat longer than
the diameter of the sphere; minor scarcely so long as its half radius, pommel-like, edged.
Dimensions.—Diameter of the sphere 0.12, pores 0.003 to 0.01, bars 0.003; length
of the major polar spine 0.15, of the minor 0.03, basal breadth 0.02.
Habitat.—North Atlantic, Station 64, surface.
Genus 47. Saturnalis,[70] Haeckel, 1881,
Prodromus, p. 450.
Definition.—Stylosphærida with one single lattice-sphere
and two equal opposite spines, connected at the distal end by a circular or elliptical ring.
The genus Saturnalis (with simple lattice-sphere) and the two similar genera
Saturnulus (with two concentric spheres) and Saturninus (with three spheres) form
together the small peculiar group of Saturnalida, distinguished by a remarkable circular or
elliptical ring, connecting the distal ends of the two equal opposite polar spines. This ring
indicates a certain equatorial plane, and therefore brings these Sphæroidea into relation with the Discoidea.
Subgenus 1. Saturnalina, Haeckel.
Definition.—Ring smooth, without spines or thorns.
1. Saturnalis circularis, n. sp.
Pores of the spherical shell regular, circular, hexagonally framed, twice as broad as the bars.
Ten to twelve pores on the half equator. Ring circular, smooth, its diameter three times as great
as that of the sphere.
{132}
Dimensions.—Diameter of the sphere 0.07, pores 0.005, bars 0.0025; diameter of the
circular ring 0.2, thickness of the axial beams and the ring 0.01.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
2. Saturnalis annularis, n. sp. (Pl. 13,
fig. 16).
Pores of the spherical shell regular, circular, with elevated hexagonal frames, of the same
breadth as the bars. Sixteen to twenty pores on the half equator. Ring elliptical, smooth,
somewhat constricted at the poles of the axis, its diameter three times as great as that of the
sphere.
Dimensions.—Diameter of the sphere 0.09, pores and bars 0.005, major axis of the
elliptical ring 0.27 to 0.3, minor
axis 0.19 to 0.2; thickness of the ring and of the axial beams 0.01.
Habitat.—Pacific, central area, Stations 270 to 274, surface.
3. Saturnalis cyclus, n. sp.
Lithocircus mesocena, Bury, 1862, Polycystins of Barbados, pl. iii. fig.
1.
Pores of the spherical shell regular, circular, without hexagonal frames, twice as broad as the
bars. Eight to ten pores on the half equator. Ring circular, smooth, its diameter four times as
great as that of the sphere.
Dimensions.—Diameter of the sphere 0.07, pores 0.006, bars 0.003; diameter of the
circular ring 0.28, thickness of the ring and both axial beams 0.01.
Habitat.—Fossil in the Barbados rocks.
4. Saturnalis circoides, n. sp. (Pl. 13,
fig. 12).
Pores of the spherical shell irregular, roundish, often somewhat lobed, one to three times as
broad as the bars; fifteen to twenty on the half equator. Ring circular, smooth, with four
prominent edges, its diameter twice as great as that of the sphere. (The figured specimen is a
young or not fully developed one; afterwards I found in the same locality other specimens with
quite perfect rings, similar to the edged ring of Saturnulus annulus, Pl. 16, fig.
17.)
Dimensions.—Diameter of the sphere 0.09 to 0.1, pores 0.003 to 0.01, bars 0.004;
diameter of the circular ring 0.2 to 0.24, thickness of the ring and the polar beams 0.01.
Habitat.—Indian Ocean; fossil in the Nicobar rocks; living at great
depths near Zanzibar, 2200 fathoms, Pullen.
Subgenus 2. Saturnalium, Haeckel.
Definition.—Ring armed on the periphery with numerous spines or
thorns.
5. Saturnalis trochoides, n. sp.
Haliomma species, Bury, 1862, Polycystins of Barbados, pl. xx. fig. 2.
Pores of the spherical shell subregular, circular, twice as broad as the bars. Twelve to
sixteen pores on the half equator. Ring circular, armed with ten to twelve strong conical,
irregular spines, its diameter twice as great as that of the sphere.
{133}
Dimensions.—Diameter of the sphere 0.08, pores 0.006, bars 0.003; diameter of the
circular ring 0.16; length of the radial spines 0.02 to 0.04; thickness of the ring and the axial
beams 0.01.
Habitat.—Fossil in the Barbados rocks.
6. Saturnalis rotula, n. sp. (Pl. 13,
fig. 15).
Pores of the spherical shell regular, circular, twice as broad as the bars; sixteen to twenty
on the half equator. Ring circular, armed with fifteen to twenty strong, conical, irregular
spines, partly simple, partly divided into two or three irregular branches; diameter of the ring
two and a half times as great as that of the sphere.
Dimensions.—Diameter of the sphere 0.08, pores 0.004, bars 0.002; diameter of the
circular ring 0.2, length of its spines 0.02 to 0.03; thickness of the ring and the radial beams
0.01.
Habitat.—North Pacific, Station 244, surface.
Subfamily Sphærostylida, Haeckel, 1881, Prodromus, pp. 449, 451.
Definition.—Stylosphærida with two
concentric, spherical lattice-shells.
Genus 48. Stylosphæra,[71] Ehrenberg, 1847,
Monatsber. d. Berlin Akad., p. 54.
Definition.—Stylosphærida with two concentric
lattice-spheres and two free spines of equal size and similar form.
The genus Stylosphæra, the most simple form of the Sphærostylida, can be
derived either from Xiphosphæra by duplication of the spherical shell, or from
Carposphæra by development of two opposite polar spines. The inner or medullary shell is
enclosed in the central capsule, whilst the outer or cortical shell lies outside it; the two are
connected by two or more radial beams, piercing the wall of the capsule.
Subgenus 1. Stylosphærantha, Haeckel.
Definition.—Pores of the cortical shell regular, of nearly equal size
and similar form; surface smooth or a little rough, without spines or thorns.
1. Stylosphæra musa, n. sp.
Radial proportion of the two concentric spheres = 3 : 1. Cortical shell thin walled,
smooth, with regular, hexagonal pores, three times as broad as the thin bars; twelve on the half
equator. Polar spines three-sided pyramidal, as long as the axis of the cortical shell, one-tenth
as broad at the base.
Dimensions.—Diameter of the outer shell 0.2, pores 0.01, bars 0.003; diameter of
the inner shell 0.06; length of the polar spines 0.2, basal breadth 0.02.
Habitat.—Tropical Atlantic, Station 347, depth 2250 fathoms.
{134}
2. Stylosphæra urania, n. sp.
Radial proportion of the two shells = 4 : 1. Cortical shell thin walled, smooth;
pores regular, circular, hexagonally framed, twice as broad as the bars; ten on the half equator.
Polar spines conical, as long as the radius of the outer shell.
Dimensions.—Diameter of the cortical shell 0.24, pores 0.012, bars 0.006;
medullary shell 0.06; length of the polar spines 0.12, basal breadth 0.024.
Habitat.—South Pacific, Station 285, depth 2375 fathoms.
3. Stylosphæra calliope, n. sp. (Pl. 16,
fig. 6).
Radial proportion of the two shells = 3 : 1. Cortical shell thick walled, smooth;
pores regular, circular, three times as broad as the bars. Each pore on its outer opening with
eight regular lobules, flower-like. Nine to ten pores on the half equator. Polar spines
three-sided pyramidal, with three strong prominent edges, about as long as the axis, as broad as
one pore. (Sometimes, as in the figured specimen, one spine is smaller than the other; this
variety, otherwise identical, may be called Sphærostylus calliope.)
Dimensions.—Diameter of the outer shell 0.12, pores 0.015, bars 0.005; inner shell
0.04; length of the polar spine 0.08 to 0.12, breadth 0.02.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
4. Stylosphæra clio, n. sp. (Pl. 16,
fig. 7).
Radial proportion of the two shells = 2 : 1. Cortical shell thick walled, smooth;
pores regular, circular, three times as broad as the bars; fourteen to sixteen on the half
equator. Polar spines three-sided pyramidal, very robust, with thick prismatic edges, about as
long as the axis of the cortical shell, one-third as broad at the base. (Sometimes, as in the
figured specimen, one spine is greater than the other; this form may be called Sphærostylus
clio.)
Dimensions.—Diameter of the outer shell 0.12, pores 0.01, bars 0.003; inner shell
0.06; length of the polar spines 0.08 to 0.12, basal breadth 0.03 to 0.04.
Habitat.—Pacific, central area; Station 272, depth 2600 fathoms.
5. Stylosphæra polyhymnia, n. sp.
Radial proportion of the two spheres = 3 : 1. Cortical shell very thin walled,
smooth, with regular, circular pores, three times as broad as the bars; sixteen to twenty on the
half equator. Polar spines cylindrical, pointed, once and a half to twice as long as the axis of
the outer sphere, scarcely broader than one pore. The two spheres are connected only by the two
opposite beams.
Dimensions.—Diameter of the outer shell 0.12 to 0.16, pores 0.006 to 0.009, bars
0.002 to 0.003; inner shell 0.04 to 0.05; length of the polar spines 0.18 to 0.22, breadth
0.01.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Indian, Pacific,
surface.
{135}
6. Stylosphæra dixyphos, Haeckel.
Haliomma dixyphos, Ehrenberg, 1854, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p.
83; Mikrogeol., Taf. xxii. fig. 31.
Haliomma dixyphos, Haeckel, 1862, Monogr. d. Radiol. p. 433.
Radial proportion of the two spheres = 2 : 1. Cortical shell thin walled, smooth,
with regular, circular pores, twice as broad as the bars; ten to twelve on the half equator. Polar
spines about as long as the axis of the outer shell, three-sided pyramidal, at the base twice as
broad as one pore. (The two spheres connected by four beams, two opposite in the main axis, two
opposite in the equatorial axis.)
Dimensions.—Diameter of the outer shell 0.1, pores 0.01, bars 0.05; inner shell
0.05; length of the polar spines 0.08 to 0.1, basal breadth 0.02.
Habitat.—South Atlantic, Station 332, surface; fossil in Tertiary
rocks of Sicily.
Subgenus 2. Stylosphærella, Haeckel.
Definition.—Pores of the cortical shell regular, of nearly equal size
and similar form; surface thorny or spiny.
7. Stylosphæra setosa, Ehrenberg, 1872.
Stylosphæra setosa, Ehrenberg, 1872, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 320; Abhandl. d. k. Akad. d. Wiss. Berlin, Taf. viii. fig. 15.
Radial proportion of the two shells = 2 : 1. Cortical shell thin walled, spiny;
pores regular, hexagonal, four times as broad as the bars. Six to eight pores on the half equator.
Polar spines conical, thin, scarcely as long as the radius of the cortical shell.
Dimensions.—Diameter of the cortical shell 0.1, pores 0.002, bars 0.005; medullary
shell 0.05; length of the polar spines 0.04, basal breadth 0.01.
Habitat.—Philippine Sea, depth 3300 fathoms, Ehrenberg; Station 206,
depth 2100 fathoms.
8. Stylosphæra euterpe, n. sp.
Radial proportion of the two shells = 3 : 1. Cortical shell thin walled, spiny; pores
regular, circular, with hexagonal frames, twice as broad as the bars; eight to ten on the half
equator. Polar spines conical, as thick as one pore at the base, about as long as the axis of the
cortical shell.
Dimensions.—Diameter of the cortical shell 0.12, pores 0.012, bars 0.006;
medullary shell 0.04; length of the polar spines 0.1, basal breadth 0.012.
Habitat.—South Pacific, Station 302, depth 1450 fathoms.
9. Stylosphæra melpomene, n. sp. (Pl. 16, fig.
1).
Radial proportion of the two shells = 3 : 1. Cortical shell thin walled, spiny, with
regular, circular pores, four times as broad as the bars; eight to ten on the half equator. Polar
spines three-sided prismatic, pointed, as broad as one pore, only one-third as long as the axis of
the sphere (the two shells connected by four thin beams, two opposite in the main axis, two in the
equatorial axis).
{136}
Dimensions.—Diameter of the outer shell 0.12, pores 0.012, bars 0.003; inner shell
0.04; length of the polar spines 0.04, thickness 0.013.
Habitat.—Indian Ocean, Cocos Islands, Rabbe.
10. Stylosphæra hispida, Ehrenberg, 1854.
Stylosphæra hispida, Ehrenberg, 1854, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin,
p. 246; Mikrogeol, Taf. xxxvi. fig. 26.
Haliomma hispidum, Haeckel, 1862, Monogr. d. Radiol., p. 433.
Radial proportion of the two spheres = 3 : 1. Cortical shell thick walled, spiny,
with regular, circular pores of the same breadth as the bars; ten to fifteen on the half equator.
Polar spines three-sided prismatic, pointed, about as long as the axis of the outer sphere, nearly
as broad at the base as the inner sphere. (Compare Sphærostylus hispidus; also Ehrenberg,
Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, 1874, p. 259.)
Dimensions.—Diameter of the outer shell 0.1 to 0.12, pores and bars 0.004; inner
shell 0.04; length of the polar spines 0.1 to 0.15, basal breadth 0.03.
Habitat.—Fossil in the Tertiary rocks of Sicily, Barbados, Nicobars,
&c.
11. Stylosphæra liostylus, Ehrenberg, 1875.
Stylosphæra liostylus, Ehrenberg, Abhandl. d. k. Akad. d. Wiss. Berlin, p.
84, Taf. xxv. fig. 3.
Radial proportion of the two spheres = 3 : 1. Cortical shell thick walled, thorny,
with regular, circular pores, three times as broad as the bars; eight to ten on the half equator.
Polar spines conical, once and a half to twice as long as the axis of the outer sphere, half as
broad at the base as its radius. (This species, common in the Barbados rocks, is different from
Sphærostylus liostylus, loc. cit., fig. 2, which Ehrenberg believed identical.)
Dimensions.—Diameter of the outer shell 0.1, pores 0.01, bars 0.0035; inner shell
0.03; length of the polar spines 0.14 to 0.18, basal breadth 0.02.
Habitat.—Fossil in the Barbados rocks; living in the depths of the
North Atlantic, Gulf Stream, Florida.
Subgenus 3. Stylosphærissa, Haeckel.
Definition.—Pores of the cortical shell irregular, of different size
or form; surface smooth or a little rough, without thorns or spines.
12. Stylosphæra nana, n. sp. (Pl. 16,
figs. 12, 13).
Radial proportion of the two spheres = 2 : 1. Cortical shell thick walled, somewhat
irregular, smooth, with irregular, roundish pores, one to three times as broad as the bars; eight
to ten on the half equator. Polar spines three-sided pyramidal, scarcely as long as the axis of
the outer sphere, and nearly as broad at the base as its radius. (A very variable and irregular
form.)
Dimensions.—Diameter of the outer shell 0.07 to 0.09, pores 0.003 to 0.009, bars
0.003; inner shell 0.03 to 0.04; length of the polar spines 0.04 to 0.07, basal breadth 0.03.
Habitat.—North Pacific, Stations 241 to 253, surface.
{137}
13. Stylosphæra jugata, n. sp.
Radial proportion of the two shells = 2 : 1. Cortical shell thick walled, smooth,
with irregular, roundish, double-contoured pores, confluent in groups of two to six. On the half
equator six to nine groups and fifteen to twenty pores; bars between them of very variable
breadth. Polar spines very strong, three-sided pyramidal, twice as long as the axis of the outer
sphere, half as broad at the base as its radius. (Nearly allied to Lithatractus jugatus,
Pl. 16, fig.
2, but differs in the truly spherical form of both shells and the double length of the polar
spines.)
Dimensions.—Diameter of the outer shell 0.15, pores 0.005 to 0.02; inner shell 0.07; length of the
polar spines 0.25 to 0.3, basal breadth 0.03.
Habitat.—Western Tropical Pacific, Station 224, depth 1850
fathoms.
14. Stylosphæra terpsichore, n. sp.
Radial proportion of the two shells = 3 : 1 or 4 : 1. Cortical shell thick
walled, smooth, with irregular, roundish pores, one to three times as broad as the bars; fifteen
to twenty-five on the half equator. Polar spines conical, about as long as the axis of the outer
sphere, as broad at the base as the inner shell.
Dimensions.—Diameter of the outer shell 0.15 to 0.2, pores 0.005 to 0.02, bars
0.004 to 0.008; inner shell 0.05; length of the polar spines 0.15 to 0.25, basal breadth 0.05.
Habitat.—Western Indian Ocean, Zanzibar, depth 2200 fathoms,
Pullen.
Subgenus 4. Stylosphæromma, Haeckel.
Definition.—Pores of the cortical shell irregular, of different size
or form; surface spiny or thorny.
15. Stylosphæra thalia, n. sp.
Radial proportion of the two shells = 2 : 1. Cortical shell thin walled, thorny, with
irregular, roundish pores, two to four times as broad as the bars; eight to twelve on the half
equator. Polar spines conical, one to one and a half times as long as the axis of the outer
sphere, one-fourth to one-sixth as thick at the base. (Resembles Sphærostylus ophidium, Pl.
16, fig. 14,
but differs in the straight regular conical polar spines, both of equal length and similar
form.)
Dimensions.—Diameter of the outer shell 0.12, pores 0.01 to 0.02, bars 0.005;
inner shell 0.06; polar spines 0.1 to 0.16 long, 0.03 broad.
Habitat.—South Pacific, Station 302, depth 1450 fathoms.
16. Stylosphæra erato, n. sp.
Radial proportion of the two shells = 3 : 1. Cortical shell thick walled, thorny,
with irregular, roundish pores, two to five times as broad as the bars; fourteen to eighteen on
the half equator. {138}Polar spines three-sided
pyramidal, about as long as the axis of the outer sphere, one-fourth as broad as its radius.
(Similar to Xiphosphæra vesta, Pl. 14,
fig. 6.)
Dimensions.—Diameter of the outer shell 0.15, pores 0.008 to 0.015, bars 0.003;
inner shell 0.05; polar spines 0.12 long, 0.02 broad.
Habitat.—South Atlantic, Station 335, depth 1425 fathoms.
Genus 49. Sphærostylus,[72] Haeckel, 1881,
Prodromus, p. 451.
Definition.—Stylosphærida with two concentric
lattice-spheres and two free spines, of different size or form.
The genus Sphærostylus differs from Stylosphæra in the different size
or form of the two polar spines, and therefore has the same relation to it that Xiphostylus
bears to Xiphosphæra.
Subgenus 1. Sphærostylantha, Haeckel.
Definition.—Pores of the cortical shell regular, of nearly equal size
and similar form; surface smooth or a little rough, without thorns.
1. Sphærostylus liostylus, Haeckel.
Stylosphæra liostylus, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss.
Berlin, Taf. xxv. fig. 2.
Cortical shell thin walled, with rough surface, three times as broad as the medullary shell.
Pores of the cortical shell regular, circular, twice as broad as the bars; ten to twelve on the
half equator. Polar spines cylindrical, as broad as one pore, with conical apex; the minor spine
about as long as the axis of the outer shell, the major three to four times as long.
Dimensions.—Diameter of the outer sphere 0.12, pores 0.12, bars 0.006; diameter of
the inner sphere 0.04; length of the major polar spine 0.3 to 0.4, of the minor 0.1 to 0.15,
breadth 0.012.
Habitat.—Fossil in the Barbados rocks.
2. Sphærostylus flexuosus, Haeckel.
Stylosphæra flexuosa, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss.
Berlin, Taf. xxv. fig. 5.
Cortical shell thick walled, with rough surface, three times as broad as the medullary shell.
Pores of the cortical shell regular circular, of the same breadth as the bars; eight to ten on the
half equator. Polar spines cylindrical, S-like curved, irregular; the minor scarcely as long as
the axis of the outer sphere, the major two to three times as long. (In the figure of Ehrenberg
the spines are broken off; I have found them myself constantly irregular and of unequal length,
sometimes with conical apex.)
{139}
Dimensions.—Diameter of the outer sphere 0.08, pores and bars 0.006; inner sphere
0.03; length of the major polar spine 0.15 to 0.25, of the minor 0.07 to 0.09, breadth 0.02.
Habitat.—Fossil in the Barbados rocks.
3. Sphærostylus clio, n. sp.
Cortical shell thick walled, with smooth surface, twice as broad as the medullary shell; its
network has regular, circular pores, three times as broad as the bars; sixteen to twenty on the
half equator. Polar spines very strong, three-sided pyramidal; the major nearly twice as long as
the axis of the outer sphere, the minor scarcely as long as its radius. (Nearly related to
Stylosphæra clio, Pl. 16,
fig. 7, but differs in the slender form and unequal length of the polar spines.)
Dimensions.—Diameter of the outer sphere 0.14, pores 0.01, bars 0.03; inner sphere
0.07; length of the major spine 0.25, of the minor 0.06, basal breadth 0.02.
Habitat.—Pacific, central area, Station 274, depth 2750 fathoms.
4. Sphærostylus hippocampus, n. sp. (Pl. 16, figs. 10,
11).
Cortical shell thick walled, with smooth surface and regular network; the pores circular,
hexagonally-lobed, three times as broad as the bars; ten to twelve on the half equator. Medullary
shell half as large, with very small circular pores, eight to ten on the half equator. Major polar
spine scarcely as long as the axis of the outer shell, curved like a horn; minor spine scarcely
half as long, pommel-like, edged. (Sometimes, by prolongation of the main axis, the spherical
shells become ellipsoidal and thus the species is transformed into Druppatractus
hippocampus.)
Dimensions.—Diameter of the outer sphere 0.08, pores 0.009, bars 0.003; inner
sphere 0.04; length of the major spine 0.07, of the minor 0.03, breadth 0.02.
Habitat.—Pacific, central area, Station 270, depth 2925 fathoms.
Subgenus 2. Sphærostyletta, Haeckel.
Definition.—Pores of the cortical shell regular, of nearly equal size
and form; surface spiny or thorny.
5. Sphærostylus diadema, n. sp.
Cortical shell thick walled, spiny, twice as broad as the medullary shell, with regular,
circular pores, twice as broad as the bars; eight to ten on the half equator. Major polar spine
about as long as the axis of the outer sphere, three-sided pyramidal; minor spine scarcely half as
long, pommel-shaped, edged. (Similar in general form to Xiphatractus glyptodon, Pl. 17, figs. 9,
10; but differs in the simple medullary shell, the spherical form of both shells, and the simple
circular regular pores.)
{140}
Dimensions.—Diameter of the outer sphere 0.12, pores 0.014, bars 0.007; inner
sphere 0.06; length of the major spine 0.13, of the minor 0.05, greatest breadth 0.04.
Habitat.—Southern Pacific, surface, Station 289.
Subgenus 3. Sphærostylissa, Haeckel.
Definition.—Pores of the cortical shell irregular, of different size
or form; surface smooth or a little rough.
6. Sphærostylus cottus, n. sp.
Cortical shell thick walled, smooth, about twice as broad as the medullary shell, with
irregular, roundish pores, scarcely larger than the bars; fifteen to twenty on the half equator.
Polar spines conical, the major once and a half to twice as long as the axis of the outer sphere,
the minor scarcely as long as its radius.
Dimensions.—Diameter of the outer sphere 0.14, pores and bars 0.008 to 0.012;
inner sphere 0.065; length of the major spine 0.2 to 0.3, of the minor 0.05 to 0.07, basal breadth
0.03.
Habitat.—Northern Atlantic, Færöe Channel, surface, John Murray.
7. Sphærostylus trigla, n. sp.
Cortical shell thin walled, smooth, three times as broad as the medullary shell, with
irregular, roundish pores, two to three times as broad as the bars; ten to twelve on the half
equator. Polar spines very unequal; the major three-sided pyramidal, one and a half times as long
as the axis of the outer sphere; the minor scarcely as long as its radius, edged, pommel-like.
Dimensions.—Diameter of the outer sphere 0.12, pores 0.01 to 0.015, bars 0.005;
inner sphere 0.04; length of the major spine 0.2, of the minor 0.05, breadth 0.03.
Habitat.—Northern Pacific, Station 241, depth 2300 fathoms.
Subgenus 4. Sphærostylomma, Haeckel.
Definition.—Pores of the cortical shell irregular, of different size
or form; surface spiny or thorny.
8. Sphærostylus ophidium, n. sp. (Pl. 16, figs. 14,
15).
Stylosphæra ophidium, Haeckel, 1878, Atlas, loc. cit.
Cortical shell thin walled, thorny, twice as broad as the medullary shell, with irregular,
roundish pores; eight to ten on the half equator. Polar spines conical, more or less curved or
S-shaped; the minor scarcely as long as the axis of the outer sphere, the major two to three times
as long.
{141}
Dimensions.—Diameter of the outer sphere 0.11, pores 0.01 to 0.017, bars 0.003 to
0.007; diameter of the inner sphere 0.06, pores 0.005 to 0.008, bars 0.002 to 0.004; length of the major
polar spine 0.25, of the minor 0.1, basal breadth 0.03.
Habitat.—Indian Ocean, Madagascar, Rabbe, surface.
Genus 50. Saturnulus,[73] Haeckel, 1881,
Prodromus, p. 451.
Definition.—Stylosphærida with two concentric
lattice-spheres and two equal opposite spines, the distal ends of which are connected by a
circular or elliptical ring.
The genus Saturnulus differs from the similar Saturnalis by the
duplication of the spherical lattice-shell; the inner lies within, the outer without the central
capsule.
1. Saturnulus circulus, n. sp.
Cortical shell smooth, twice as broad as the medullary shell, with regular, circular,
hexagonally framed pores, of the same breadth as the bars; fourteen to sixteen on the half
equator. Ring circular, smooth, without edges, its diameter three times as great as that of the
sphere.
Dimensions.—Diameter of the outer sphere 0.08, pores and bars 0.004; of the inner
sphere 0.04, of the ring 0.24.
Habitat.—Central Pacific, Station 272, surface.
2. Saturnulus annulus, n. sp.
Cortical shell smooth, three times as broad as the medullary shell, with regular, circular,
hexagonally framed pores, of the same breadth as the bars; eighteen to twenty on the half equator.
Ring elliptical, smooth, without edges, somewhat constricted at the poles of the minor axis, its
major diameter four times as great as that of the sphere. Differs from Saturnalis
annularis, Pl. 13,
fig. 16, mainly in the possession of a medullary shell.
Dimensions.—Diameter of the outer sphere 0.1, pores and bars 0.005; inner sphere
0.033; major axis of the ring 0.4, minor 0.3.
Habitat.—North Pacific, Station 244, surface.
3. Saturnulus ellipticus, n. sp. (Pl. 16, fig.
16).
Cortical shell smooth, three times as broad as the medullary shell, with regular, circular
pores, three times as broad as the bars; sixteen to eighteen on the half equator. Ring elliptical,
smooth, without edges, its major diameter three times as great as that of the sphere.
Dimensions.—Diameter of the outer sphere 0.09, pores 0.006, bars 0.002; inner
sphere 0.03; major axis of the elliptical ring 0.28, minor 0.24; thickness of the ring and the
axial beams 0.008.
Habitat.—South Pacific, Station 300, surface.
{142}
4. Saturnulus planetes, n. sp. (Pl. 16,
fig. 17).
Cortical shell smooth, twice as broad as the medullary shell, with regular, circular pores, of
the same breadth as the bars; sixteen to eighteen on the half equator. Ring elliptical, smooth,
with strong prominent edges, constricted at the poles of the minor axis, its major diameter three
times as great as that of the outer sphere.
Dimensions.—Diameter of the outer sphere 0.08, pores and bars 0.005; inner sphere
0.035; major axis of the ring 0.25, minor axis 0.2; thickness of the ring and the axial beams
0.012.
Habitat.—Indian Ocean, Sunda Strait, Rabbe; Station 200, surface.
Subfamily Amphistylida, Haeckel, 1881, Prodromus, pp. 449, 452.
Definition.—Stylosphærida with three
concentric spherical lattice-shells.
Genus 51. Amphisphæra, Haeckel,[74] 1881,
Prodromus, p. 452.
Definition.—Stylosphærida with three concentric
lattice-spheres and two free spines of equal size and similar form.
The genus Amphisphæra differs from its probable ancestral form,
Stylosphæra, in the triple spherical lattice-shell. Commonly two of these lie within the
central capsule (medullary shell), whilst the third lies outside it (cortical shell). But
sometimes this order is inverted, the cortical shell being double, the medullary shell simple; and
perhaps these forms may better represent a peculiar genus, Amphisphæridium.
Subgenus 1. Amphisphærantha, Haeckel.
Definition.—Pores of the cortical shell regular, of nearly equal size
and similar form; surface smooth or a little rough, without spines or thorns.
1. Amphisphæra neptunus, n. sp.
Radial proportion of the three concentric spheres = 4 : 2 : 1. Cortical
shell thick walled, smooth, with regular, circular, hexagonally framed pores, of the same breadth
as the bars; twelve to fifteen on the half equator. Polar spines three-sided pyramidal, with
strong prominent edges, about as long as the radius of the outer shell, half as broad at the base.
(Similar to Stylatractus neptunus, Pl. 17,
fig. 6, but differs in the purely spherical form of the three concentric shells and the regular
form of the network and of the polar spines.)
Dimensions.—Diameter of the outer shell 0.16, middle shell 0.08, inner shell 0.04;
pores and bars of the cortical shell 0.008; length of the polar spines 0.08, basal breadth
0.04.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
{143}
2. Amphisphæra uranus, n. sp.
Radial proportion of the three spheres = 4 : 2 : 1. Cortical shell thick
walled, smooth, with regular, circular pores, three times as broad as the bars; ten to twelve on
the half equator. Polar spines three-sided pyramidal, about as long as the diameter of the outer
shell, twice as broad at the base as one pore.
Dimensions.—Diameter of the outer shell 0.12, middle 0.06, inner 0.03; pores of
the cortical shell 0.012, bars 0.004; length of the polar spines 0.1, basal breadth 0.024.
Habitat.—Western Indian Ocean, Zanzibar, 2200 fathoms, Pullen.
3. Amphisphæra jupiter, n. sp.
Radial proportion of the three spheres = 10 : 3 : 2. Cortical shell thick
walled, smooth, with regular, circular pores, twice as broad as the bars; twenty to twenty-five on
the half equator. Polar spines cylindro-conical, nearly as long as the axis of the outer shell,
twice as broad at the base as one pore.
Dimensions.—Diameter of the outer shell 0.2, middle 0.06, inner 0.04; pores of the
cortical shell 0.01, bars 0.005; length of the polar spines 0.15 to 0.18, breadth 0.02.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
Subgenus 2. Amphisphærella, Haeckel.
Definition.—Pores of the cortical shell regular, of nearly equal size
and similar form; surface thorny or spiny.
4. Amphisphæra apollo, n. sp.
Radial proportion of the three spheres = 3 : 1.5 : 1. Cortical shell thick
walled, spiny, with regular, circular, hexagonally framed pores, of the same breadth as the bars;
twenty to twenty-two on the half equator. In each hexagon-corner (between three pores) a
bristle-like radial spine. Polar spines cylindrical, with conical apex; one to one and a half
times as long as the axis of the outer shell, as broad as three pores at the base. (Resembles
closely Xiphosphæra pallas, Pl. 14,
fig. 4, but differs in the presence of two medullary shells and the absence of the fine
denticulations on the hexagonal crests.)
Dimensions.—Diameter of the outer shell 0.12, middle 0.06, inner 0.04; pores and
bars of the cortical shell 0.006; length of the polar spines 0.15, breadth 0.02.
Habitat.—Western Tropical Pacific, Station 224, depth 1850
fathoms.
5. Amphisphæra mercurius, n. sp.
Radial proportion of the three spheres = 3 : 2 : 1. Cortical shell thin
walled, spiny, with regular, circular pores, twice as broad as the bars; fifteen to eighteen on
the half equator; between {144}them short bristle-like
radial spines. Polar spines three-sided pyramidal, about as long as the radius of the outer shell,
one-third as broad at the base.
Dimensions.—Diameter of the outer shell 0.15, middle 0.09, inner 0.05; pores of
the cortical shell 0.01, bars 0.005; length of the polar spines 0.09, basal breadth 0.03.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
Subgenus 3. Amphisphærissa, Haeckel.
Definition.—Pores of the cortical shell irregular, of different size
or form; surface smooth or a little rough, without spines or thorns.
6. Amphisphæra cronos, n. sp. (Pl. 17,
fig. 5).
Radial proportion of the three spheres = 3 : 2 : 1. Cortical shell thin
walled, smooth, with irregular, roundish pores, two to four times as broad as the bars; eight to
ten on the half equator. Outer medullary shell similar, but with pores of half the size, connected
with the cortical shell by numerous radial beams; inner medullary shell with very small pores.
Polar spines three-sided pyramidal, as long as the axis of the inner medullary shell, half as
broad at the base.
Dimensions.—Diameter of the outer shell 0.12, middle 0.08, inner 0.04; pores of
the cortical shell 0.01 to 0.02, bars 0.05; length of the polar spines 0.04, basal breadth
0.02.
Habitat.—South Atlantic, Station 330, surface.
7. Amphisphæra pluto, n. sp. (Pl. 17,
figs. 7, 8).
Radial proportion of the three spheres about = 4 : 2 : 1 (or
11 : 7 : 3). Cortical shell thick walled, smooth, with very irregular,
roundish pores, two to four times as broad as the bars; eight to ten on the half equator; often
two to four pores confluent. Margin of their outer aperture double. Polar spines conical, double
contoured, as long as the radius of the outer shell, one-third as broad at the base.
Dimensions.—Diameter of the outer shell 0.11, middle shell 0.07, inner shell 0.03;
pores of the cortical shell 0.01 to 0.02, bars 0.06, length of the polar spines 0.06, basal
breadth 0.02.
Habitat.—Central Pacific, Station 268, surface.
Subgenus 4. Amphisphæromma, Haeckel.
Definition.—Pores of the cortical shell irregular, of different size
or form; surface spiny or thorny.
8. Amphisphæra mars, n. sp.
Radial proportion of the three spheres = 10 : 3 : 2. Cortical shell thin
walled, thorny, with irregular, roundish pores, one to three times as broad as the bars; sixteen
to twenty on the {145}half equator. Irregularly
scattered between them short conical thorns. Polar spines conical, about as long as the axis of
the outer shell, as broad at the base as the inner shell.
Dimensions.—Diameter of the outer shell 0.2, middle shell 0.06, inner shell 0.04;
pores of the cortical shell 0.005 to 0.02, bars 0.006; length of the polar spines 0.17, basal
breadth 0.04.
Habitat.—North Atlantic, Gulf Stream, near Florida, depth 1500
fathoms, Schaffner.
Genus 52. Amphistylus,[75] Haeckel, 1881,
Prodromus, p. 452.
Definition.—Stylosphærida with three concentric
lattice-spheres and two free spines of different size or form.
The genus Amphistylus differs from its ancestral form, Amphisphæra,
by the differentiation of both polar spines, and exhibits therefore the same relation to it that
Sphærostylus bears to Stylosphæra.
1. Amphistylus clio, n. sp.
Radial proportion of the three spheres = 4 : 2 : 1. Cortical shell thick
walled, smooth; its pores regular, circular, three times as broad as the bars; eighteen to twenty
on the half equator. Polar spines three-sided pyramidal, very stout, as broad at the base as the
inner medullary shell; major spine somewhat longer than the diameter of the cortical shell; minor
spine scarcely half as long. (Similar to Stylosphæra clio, Pl. 16, fig. 7, but
different in the double medullary shell and the different length of the polar spines.)
Dimensions.—Diameter of the outer sphere 0.15, middle 0.08, inner 0.04; pores of
the outer shell 0.01, bars 0.003; length of the major spine 0.17, minor 0.07, basal breadth
0.04.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms.
2. Amphistylus hippocampus, n. sp.
Radial proportion of the three spheres = 3 : 2 : 1. Cortical shell thick
walled, smooth; its pores regular, circular, hexagonally lobed, three times as broad as the bars;
twelve to fifteen on the half equator. Major polar spine three-sided pyramidal, about as long as
the axis of cortical shell; minor spine pommel-like, edged, scarcely one-third as long. (Similar
to Sphærostylus hippocampus, Pl. 16,
figs. 10, 11, but differs in the larger size, the double medullary shell, and the straight, not
curved, major spine.)
Dimensions.—Diameter of the outer sphere 0.11, middle 0.07, inner 0.035; pores of
the outer shell 0.011, bars 0.004; length of the major spine 0.12, of the minor 0.04, breadth
0.03.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
3. Amphistylus glyptodon, n. sp.
Radial proportion of the three spheres = 6 : 3 : 1. Cortical shell thick
walled, spiny, with irregular, roundish pores, two to four times as broad as the bars; ten to
twelve on the half equator. {146}Inner aperture of each
pore closed by a thin fenestrated lamella with five to seven small pores. Major polar spine
three-sided pyramidal, longer than the diameter of the outer sphere; minor polar spine scarcely as
long as its radius, pommel-shaped. (Similar to Xiphatractus glyptodon, Pl. 17, figs. 9,
10, but differs in the spherical, not ellipsoidal, form of the three shells and the size of the
polar spines.)
Dimensions.—Diameter of the outer sphere 0.12, middle 0.06, inner 0.02; pores and
spines of the outer shell 0.01 to 0.02, bars and porules 0.006; length of the major polar spine
0.15, of the minor 0.05, breadth 0.04.
Habitat.—South Pacific, Station 285, depth 2375 fathoms.
Genus 53. Saturninus,[76] n. gen.
Definition.—Stylosphærida with three concentric
lattice-spheres and two equal opposite spines, connected at the distal end by a circular or
elliptical ring.
The genus Saturninus differs from the similar Saturnulus by the
triplication of the spherical lattice-shell; the inner shell is enclosed in the central capsule,
whilst both the others lie outside it.
1. Saturninus triplex, n. sp.
Radial proportion of the three spheres = 4 : 3 : 1. Inner cortical shell
with regular, circular pores, of the same breadth as the bars, sixteen to eighteen on the half
equator; outer cortical shell connected with the inner by numerous bristle-like radial spines,
network very delicate, cobweb-shaped, with irregular polygonal meshes. Ring elliptical, two and a
half times as broad as the outer shell.
Dimensions.—Diameter of the inner sphere 0.03, middle 0.09, outer 0.12; pores and
bars of the middle shell 0.004; major axis of the elliptical ring 0.3, minor 0.25; thickness of
the ring and the axial beams 0.008.
Habitat.—Central Pacific, Station 272, surface.
Subfamily Cromyostylida, Haeckel, 1881, Prodromus, pp. 449, 453.
Definition.—Stylosphærida with four
concentric spherical lattice-shells (two medullary and two cortical).
Genus 54. Stylocromyum,[77] Haeckel, 1881,
Prodromus, p. 453.
Definition.—Stylosphærida with four concentric
lattice-spheres and two free spines of equal size and similar form.
{147}
The genus Stylocromyum differs from its probable ancestral form,
Amphisphæra, by the duplication of the cortical shell; two opposite radial beams, piercing
the wall of the central capsule, connect it with the double medullary shell, and are prolonged
outside into two equal spines.
1. Stylocromyum amphiconus, n. sp.
Surface of the shell smooth. Radial proportion of the four spheres =
1 : 2 : 8 : 9. Two medullary shells inside the central capsule, two
cortical shells outside it. Interval between the second and third shells the greatest. Pores of
all the shells regular, circular, two to four times as broad as the bars. Both polar spines equal,
conical, about as long as the axis of the outermost sphere, as broad at the base as the
innermost.
Dimensions.—Diameter of the four spheres—(A) inner medullary shell 0.03, (B)
outer medullary shell 0.06, (C) inner cortical shell 0.25, (D) outer cortical shell 0.28; length
of the polar spines 0.3, basal breadth 0.03.
Habitat.—Central Pacific, Station 273, depth 2350 fathoms.
2. Stylocromyum amphipyramis, n. sp.
Surface of the shell smooth. Radial proportion of the four spheres =
2 : 3 : 6 : 8. Both medullary shells with regular, circular pores,
twice as broad as the bars. Both cortical shells with irregular, roundish, much larger pores,
three to six times as broad as the bars. Both polar spines equal or nearly equal, three-sided
pyramidal, about as long as the axis of the third shell.
Dimensions.—Diameter of the four spheres—(A) 0.04, (B) 0.06, (C) 0.12, (D)
0.16; length of the polar spines 0.11, basal breadth 0.02.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms.
Genus 55. Cromyostylus,[78] Haeckel, 1881,
Prodromus, p. 453.
Definition.—Stylosphærida with four concentric
lattice-spheres and two free spines of different size or form.
The genus Cromyostylus differs from its ancestral form, Stylocromyum,
in the differentiation of the two unequal polar spines.
1. Cromyostylus gladius, n. sp.
Surface of the shell smooth. Radial proportion of the four spheres =
1 : 3 : 10 : 12. Both medullary shells with regular, circular,
simple pores. Inner cortical shell with regular, circular, hexagonally framed pores, twice as
broad as the bars. From each hexagon-corner arises a {148}bristle-shaped radial spine, which at the distal end gives off three
thread-like branches; by communication of these threads (at equal distances from the centre) the
delicate outer medullary shell is formed. The polar spines very different; major spine six-sided
pyramidal, longer than the diameter of the shell; minor pommel-shaped, shorter than the radius
(similar to Xiphatractus glyptodon, Pl. 17,
figs. 9, 10, but different in the double spherical cortical shell).
Dimensions.—Diameter of the four spheres—(A) 0.02, (B) 0.06, (C) 0.2, (D)
0.24; length of the major spine 0.3, of the minor 0.1.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
Subfamily Caryostylida, Haeckel, 1881, Prodromus, pp. 449, 454.
Definition.—Stylosphærida with five or more
concentric, spherical lattice-shells.
Genus 56. Caryostylus, Haeckel, 1881, Prodromus, p. 454.
Definition.—Stylosphærida with five to six or more
concentric lattice-shells and two free opposite spines of equal size and similar form.
The genus Caryostylus differs from its ancestral form, Stylocromyum,
by the multiplication of the concentric spheres, the number of which amounts to five or six or
more. I have only observed one single species of this genus. Some similar forms which in my
Prodromus (1881, p. 454) were annexed to it, and disposed in three nearly allied genera
(Caryoxiphus, Caryodoras, Caryolonche), have now been proved to belong to
other groups, mainly ellipsoidal Druppulida.
1. Caryostylus hexalepas, n. sp.
Surface of the spherical shell smooth. Radial proportion of the component six concentric shells
= 1 : 2 : 7 : 9 : 12 : 15. Both medullary shells
connected only by six radial beams, opposite in pairs in the three dimensive axes. Between second
and third shell numerous (twenty regularly disposed?) radial beams. Four cortical shells connected
by very numerous (sixty to eighty or more?) short radial beams. Pores of all six shells regular,
circular, the size increasing towards the surface, two to three times as broad as the bars. Two
opposite polar spines very large, of equal size, three times as long as the shell radius,
cylindrical, club-shaped at the thicker distal end. (The whole shell structure is similar to Pl.
15, fig. 2,
but the shells are spherical, not ellipsoidal.)
Dimensions.—Diameter of the six spheres—(A) 0.02, (B) 0.04, (C) 0.15, (D)
0.18, (E) 0.24, (F) 0.3; length of the spines 0.5.
Habitat.—West Tropical Pacific, Station 225, depth 4475 fathoms.
Subfamily Spongostylida, Haeckel, 1881, Prodromus, pp. 449, 455.
Definition.—Stylosphærida with spherical
spongy shell (with or without enclosed latticed medullary shells).
{149}
Genus 57. Spongolonchis,[79] Haeckel, 1881,
Prodromus, p. 455.
Definition.—Stylosphærida with a solid sphere of spongy
framework, and with two opposite free radial spines.
The genus Spongolonchis differs from its probable ancestral form, Styptosphæra,
by the development of two opposite radial spines situated in one axis.
1. Spongolonchis compacta, n. sp.
Spongy framework of the spherical shell very compact, with small meshes, three to four times as
broad as the bars. Surface rough, but not spiny. Two polar spines pyramidal, only as long as the
radius of the shell, one-third as broad at the base.
Dimensions.—Diameter of the shell 0.2; length of the spines 0.1, basal breadth
0.03.
Habitat.—Central Pacific, Station 274, depth 2750 fathoms.
2. Spongolonchis laxa, n. sp.
Spongy framework loose, with large meshes, ten to twelve times as broad as the bars. Surface
spiny. Two polar spines three-sided prismatic, longer than the diameter of the shell (broken off
in the observed specimen).
Dimensions.—Diameter of the shell 0.5; length of the spine 0.6 and more, breadth
0.02.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms.
Genus 58. Spongostylus,[80] Haeckel, 1881,
Prodromus, p. 455.
Definition.—Stylosphærida with spongy spherical cortical
shell, enclosing in the centre a simple latticed medullary shell, and with two opposite free
radial spines.
The genus Spongostylus may probably be derived from Spongoplegma by
development of two opposite radial spines in one axis.
1. Spongostylus hastatus, n. sp.
Spongy cortical shell with compact framework, twice as broad as the enclosed medullary shell,
the pores of which are regular, circular, twice as broad as the bars. Two polar spines, three
times as long as the radius of the shell, in the basal two-thirds cylindrical, in the distal third
compressed, two-edged, spear-shaped.
Dimensions.—Diameter of the cortical shell 0.1, medullary shell 0.05; length of
the spines 0.15.
Habitat.—South Atlantic, Station 335, depth 1425 fathoms.
{150}
2. Spongostylus gladiatus, Haeckel.
Stylosphæra holosphæra, Ehrenberg, 1872, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 299, Taf. viii. fig. 14.
Spongy cortical shell with rather compact framework, four times as broad as the enclosed
medullary shell, the pores of which are irregular, roundish, three to four times as broad as the
bars. Two polar spines sword-shaped, two-edged, as long as the shell diameter.
Dimensions.—Diameter of the cortical shell 0.24, medullary shell 0.06; length of
the spines 0.25.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms; Philippine
Sea, depth 3300 fathoms.
3. Spongostylus serratus, n. sp.
Spongy cortical shell with lax framework, ten to twelve times as broad as the enclosed
medullary shell, the pores of which are irregular, roundish, once and a half to twice as broad as
the bars. Two polar spines longer than the shell diameter (broken off in the observed specimen),
three-sided prismatical, with three straight serrated edges.
Dimensions.—Diameter of the cortical shell 0.5, medullary shell 0.04; length of
the spines 0.6 or more.
Habitat.—South Pacific, Station 285, depth 2375 fathoms.
Genus 59. Spongostylidium,[81] Haeckel,
1881, Prodromus, p. 455.
Definition.—Stylosphærida with spongy spherical cortical
shell, enclosing two concentric spherical latticed medullary shells, and with two opposite, free
radial spines.
The genus Spongostylidium differs from Spongostylus by duplication of
the latticed medullary shell, and therefore bears the same relation to it as Spongodictyon
to Spongoplegma.
1. Spongostylidium streptacanthum, n. sp.
Both medullary shells spherical, with small, regular, circular pores, twice as broad as the
bars (outer twice as broad as the inner). Spongy cortical shell enclosing it with dense framework,
four times as broad as the outer medullary shell. Two polar spines very large, four times as long
as the diameter of the outer shell; as broad as the inner medullary shell, with three dentated,
spirally contorted edges. (Very similar to the common Spongosphæra streptacantha and to
Hexadoridium streptacanthum, but with only two opposite spines in one axis.)
Dimensions.—Diameter of the cortical shell 0.16, of the outer medullary shell
0.04, inner 0.02; length of the spines 0.7 and more, breadth 0.02.
Habitat.—Indian Ocean, Cocos Islands, Rabbe.
{151}
Family VIII. Staurosphærida, Haeckel (Pl. 15).
Staurosphærida, Haeckel, 1881, Prodromus, p. 449.
Definition.—Sphæroidea with four radial spines on the
surface of the spherical shell, forming a regular cross, being opposite in pairs in two axes
perpendicular to one another; living solitary (not associated in colonies).
The family Staurosphærida is distinguished from the other Sphæroidea by the possession of four radial spines, which are opposite in pairs
in two perpendicularly crossed axes. By these "two main axes" an equatorial plane is determined,
which approximates them to the Discoidea. But in the latter the shells as
well as the central capsule become more or less flattened, lenticular, or discoidal, whilst in the
former they remain spherical. However, some forms of both groups are very similar, and inspection
from different sides (and mainly from the margin of the equatorial plane) is required to determine
the spherical (not compressed) shell-form of the Staurosphærida. As a rule the species of this
family are much rarer, and much less numerous, than those of all the other Sphæroidea.
The most simple Staurosphærida are the Staurostylida, with one single spherical lattice-shell.
To this ancestral group all other subfamilies can be opposed as "Staurosphærida concentrica,"
since their carapace is composed of two or more concentric lattice-shells; two in the
Staurolonchida, three in the Stauracontida, four in the Staurocromyida, five or more in the
Staurocaryida. In all these four subfamilies the concentric shells are simple (not spongy)
fenestrated spheres. In a sixth subfamily, in the Staurodorida, the shell is wholly or partially
composed of irregular spongy wickerwork, with or without a medullary shell in the centre.
The Four Radial Spines in all Staurosphærida are normally opposed in pairs in two axes
perpendicular one to another, and therefore together form a rectangular cross. But in many species
besides this normal form individual abnormalities occur, in which the four spines in the
equatorial plane are not quite accurately opposed, so that the four angles between them are not
right angles, but more or less unequal. More rarely also their position in the equatorial plane is
not accurately retained, so that they are placed in two, three, or four different meridian planes,
intersecting at very small variable angles.
In the greater part of Staurosphærida all four radial spines are quite equal, and
of the same size and form. But in some genera there takes place a more or less considerable
differentiation of the four spines, commonly in pairs, so that the two opposite spines of each
pair are equal, but the pairs different (Staurostylus, Staurolonchidium). More
rarely also both spines of one pair become unequal, whilst those of the other pair remain equal
(Stauroxiphos). Very rarely all four spines assume a different size or form.
{152}
Synopsis of the Genera of Staurosphærida.
|
I. Subfamily Staurostylida.
(Shell one single lattice-sphere.)
|
|
All four spines equal, |
60. Staurosphæra. |
| Four spines different in pairs, |
61. Staurostylus. |
| One spine larger than the three others, |
62. Stylostaurus. |
|
II. Subfamily Staurolonchida.
(Shell with two concentric lattice-spheres.)
|
|
All four spines equal, |
|
Simple, |
63. Staurolonche. |
| Branched, |
64. Staurancistra. |
| Four spines different in pairs, |
65. Staurolonchidium. |
| One spine larger than the three others, |
66. Stauroxiphos. |
|
III. Subfamily Stauracontida.
(Shell with three concentric spheres.)
|
|
All four spines equal, simple, |
67. Stauracontium. |
|
IV. Subfamily Staurocromyida.
(Shell with four concentric spheres.)
|
|
All four spines equal, |
|
Simple, |
68. Staurocromyum. |
| Branched, |
69. Cromyostaurus. |
|
V. Subfamily Staurocaryida.
(Shell with five or more concentric spheres.)
|
|
All four spines equal, |
70. Staurocaryum. |
|
VI. Subfamily Staurodorida.
(Shell a spongy sphere.)
|
|
Solid spongy sphere without medullary shell, |
71. Staurodoras. |
| I. Subfamily Staurostylida. (Shell one single lattice-sphere.) |
|
|
All four spines equal, |
|
|
|
|
|
|
|
|
60. Staurosphæra. |
|
|
|
|
|
|
|
|
Four spines different in pairs, |
|
|
|
|
|
|
|
|
61. Staurostylus. |
|
|
|
|
|
|
|
|
One spine larger than the three others, |
|
|
|
|
|
|
|
|
62. Stylostaurus. |
|
|
|
|
|
|
| II. Subfamily Staurolonchida. (Shell with two concentric
lattice-spheres.) |
|
|
All four spines equal, |
|
|
|
|
|
|
Simple, |
|
|
|
|
|
|
|
|
|
|
63. Staurolonche. |
|
|
|
|
|
|
|
|
|
|
Branched, |
|
|
|
|
|
|
|
|
|
|
64. Staurancistra. |
|
|
|
|
|
|
|
|
Four spines different in pairs, |
|
|
|
|
|
|
|
|
65. Staurolonchidium. |
|
|
|
|
|
|
|
|
One spine larger than the three others, |
|
|
|
|
|
|
|
|
66. Stauroxiphos. |
|
|
|
|
|
|
| III. Subfamily Stauracontida. (Shell with three concentric spheres.) |
|
|
All four spines equal, simple, |
|
|
|
|
|
|
|
|
67. Stauracontium. |
|
|
|
|
|
|
| IV. Subfamily Staurocromyida. (Shell with four concentric spheres.) |
|
|
All four spines equal, |
|
|
|
|
|
|
Simple, |
|
|
|
|
|
|
|
|
|
|
68. Staurocromyum. |
|
|
|
|
|
|
|
|
|
|
Branched, |
|
|
|
|
|
|
|
|
|
|
69. Cromyostaurus. |
|
|
|
|
|
|
| V. Subfamily Staurocaryida. (Shell with five or more concentric spheres.) |
|
|
All four spines equal, |
|
|
|
|
|
|
|
|
70. Staurocaryum. |
|
|
|
|
|
|
| VI. Subfamily Staurodorida. (Shell a spongy sphere.) |
|
|
Solid spongy sphere without medullary shell, |
|
|
|
|
|
|
|
|
71. Staurodoras. |
|
|
|
|
|
|
Subfamily Staurostylida,[82] Haeckel, Prodromus, 1881, pp. 449, 450.
Definition.—Staurosphærida with one single
spherical lattice-shell.
Genus 60. Staurosphæra,[83] Haeckel, 1881,
Prodromus, p. 450.
Definition.—Staurosphærida with a single lattice-sphere
and four crossed equal spines.
The genus Staurosphæra may be regarded as the common ancestral form of this
subfamily, since it represents their most simple and primitive form. From the surface of the
simple lattice-sphere, enclosing the central capsule, arise four equal, simple, radial spines,
opposite in pairs in two diameters, perpendicular one to another. Staurosphæra may be
derived phylogenetically either from Cenosphæra by production of the four spines, or from
Hexastylus by reduction of two opposite spines.
{153}
Subgenus 1. Staurosphærantha, Haeckel.
Definition.—Pores regular, all of nearly equal size and similar form;
surface smooth.
1. Staurosphæra cruciata, n. sp.
Shell thin walled, smooth, with regular, hexagonal pores, four times as broad as the bars; ten
to twelve on the quadrant. Four crossed radial spines three-sided pyramidal, as long as the
diameter of the shell, as broad at the base as one pore (very similar to Hexastylus
phænaxonius, Pl. 21,
fig. 3, but with only four spines).
Dimensions.—Diameter of the shell 0.12, of the pores 0.008, bars 0.002.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
2. Staurosphæra christiana, n. sp.
Shell thick walled, smooth, with regular, circular, hexagonally framed pores, three times as
broad as the bars; six to eight on the quadrant. Four crossed spines six-sided pyramidal, as long
as the radius, as broad as one mesh.
Dimensions.—Diameter of the shell 0.13, pores 0.012, bars 0.004.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
3. Staurosphæra johannis, n. sp.
Shell thick walled, smooth, with regular, circular pores, four times as broad as the bars; five
to six on the quadrant. Four spines six-sided pyramidal, half as long as the radius, as broad as
one mesh.
Dimensions.—Diameter of the shell 0.15, pores 0.02, bars 0.005.
Habitat.—North Pacific, Station 241, depth 2300 fathoms.
4. Staurosphæra pauli, n. sp.
Shell very thick walled, smooth, with regular, circular, double-edged pores, four times as
broad as the bars; eight to ten on the quadrant. Four spines conical, as long as the radius, as
broad as one mesh.
Dimensions.—Diameter of the shell 0.15, pores 0.016, bars 0.004.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms.
5. Staurosphæra petri, n. sp.
Shell thin walled, smooth, with regular, circular pores, three times as broad as the bars;
twenty to twenty-two on the quadrant. Four spines conical, half as long as the radius, as broad as
one mesh.
Dimensions.—Diameter of the shell 0.22, pores 0.006, bars 0.002.
Habitat.—South Atlantic, Station 325, depth 2650 fathoms; also fossil
in Barbados.
{154}
6. Staurosphæra jacobi, n. sp.
Shell thick walled, smooth, with regular, circular pores, twice as broad as the bars; eleven to
twelve on the quadrant. Four spines cylindrical, three times as long as the radius, three times as
broad as one mesh.
Dimensions.—Diameter of the shell 0.1, pores 0.006, bars 0.003.
Habitat.—North Atlantic, Station 353, depth 2965 fathoms.
7. Staurosphæra simonis, n. sp.
Haliomma with four spines, Bury, 1862, Polycystins of Barbados, pl. iv.
fig. 4.
Shell thick walled, smooth, with regular, circular pores, of the same breadth as the bars;
eight to ten on the quadrant. Four spines cylindrical, twice as long as the radius, five times as
broad as one mesh.
Dimensions.—Diameter of the sphere 0.1, pores and bars 0.003.
Habitat.—Fossil in Barbados.
Subgenus 2. Staurosphærella, Haeckel.
Definition.—Pores regular, all of nearly equal size and similar form;
surface covered with by-spines or accessory thorns.
8. Staurosphæra philippi, n. sp. (Pl. 15, fig.
6).
Shell thin walled, covered with bristle-shaped by-spines, as long as the radius. Pores regular,
circular, twice as broad as the bars; six to eight on the quadrant. Four main spines cylindrical,
five to ten times as long as the radius, as broad as one mesh.
Dimensions.—Diameter of the shell 0.08, pores 0.006, bars 0.003.
Habitat.—Central Pacific, Station 272, surface.
9. Staurosphæra andreæ, n. sp.
Shell thick walled, with regular, circular, hexagonally framed pores, three times as broad as
the bars; nine to ten on the quadrant. From each hexagon-corner arises a bristle-shaped by-spine,
half as long as the radius. Four main spines three-sided pyramidal, with spirally contorted edges,
as long as the radius (very similar to Hexastylus solonis, Pl. 21, fig. 11,
but with only four spines).
Dimensions.—Diameter of the shell 0.16, pores 0.01, bars 0.003.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms.
{155}
10. Staurosphæra thomæ.
Shell thick walled, with regular, circular pores, twice as broad as the bars; twelve to
fourteen on the quadrant; surface covered with short conical by-spines. Four main spines conical,
twice as long as the radius, twice as broad as one mesh.
Dimensions.—Diameter of the sphere 0.2, pores 0.012, bars 0.006.
Habitat.—South Atlantic, Station 332, depth, 2200 fathoms.
Subgenus 3. Staurosphærissa, Haeckel.
Definition.—Pores irregular, of different size or form; surface
smooth.
11. Staurosphæra judæ, n. sp.
Shell thin walled, smooth, with irregular, polygonal pores, twice to four times as broad as the
bars; six to ten on the quadrant. Four main spines three-sided pyramidal, as long as the
radius.
Dimensions.—Diameter of the sphere 0.15, pores 0.006 to 0.012, bars 0.003.
Habitat.—Central Pacific, Station 273, depth, 2350 fathoms.
12. Staurosphæra crassa, Dunikowski.
Staurosphæra crassa, 1882, Denkschr. d. k. Akad. d. Wiss. Wien, Bd. xlv.
p. 27, Taf. v. figs. 52-55.
Shell thick walled, smooth, with irregular, roundish pores, scarcely broader than the bars;
eight to ten on the quadrant. Four spines three-sided pyramidal, nearly as long as the shell
diameter.
Dimensions.—Diameter of the sphere 0.19, pores and bars 0.015.
Habitat.—Fossil in the Alpine Lias (Schafberg near Salzburg,
Dunikowski).
13. Staurosphæra apostolorum, Haeckel.
? Cenosphæra megapora, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 66,
Taf. iii. fig. 1.
? Cenosphæra micropora, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 66, Taf. iii. fig. 2.
Shell thin walled, smooth, with large, irregular, roundish pores, twice to six times as broad
as the bars; four to six on the quadrant. Four spines conical, very stout, about as long as the
shell diameter, often more or less irregularly disposed.
Dimensions.—Diameter of the sphere 0.12 to 0.2, pores 0.01 to 0.03, bars
0.005.
Habitat.—Fossil in Barbados.
{156}
Subgenus 4. Staurosphæromma, Haeckel.
Definition.—Pores irregular, of different size or form; surface
covered with by-spines or accessory thorns.
14. Staurosphæra bartholomæi, n. sp.
Shell thin walled, with irregular, polygonal pores, three times as broad as the bars; six to
eight on the quadrant; surface covered with short bristle-shaped by-spines. Four main spines
three-sided pyramidal, twice as long as the radius.
Dimensions.—Diameter of the shell 0.14, pores 0.008 to 0.016, bars 0.003 to
0.005.
Habitat.—South Pacific, Station 285, depth 2375 fathoms.
15. Staurosphæra thaddæi, n. sp.
Shell thick walled, with irregular, roundish, polygonally framed pores, twice to three times as
broad as the bars; five to seven on the quadrant; surface covered with bristle-shaped spines, half
as long as the radius. Four main spines pyramidal, as long as the radius.
Dimensions.—Diameter of the shell 0.12, pores 006 to 0.012, bars 0.004.
Habitat.—Central Pacific, Station 266, depth 2750 fathoms; also fossil
in Barbados.
16. Staurosphæra matthæi, n. sp.
Shell thick walled, with irregular, roundish pores, twice to five times as broad as the bars;
ten to twelve on the quadrant; surface covered with short conical thorns or by-spines. Four main
spines conical, as long as the radius.
Dimensions.—Diameter of the shell 0.15, pores 0.004 to 0.01, bars 0.002.
Habitat.—North Pacific, Station 253, depth 3125 fathoms.
Genus 61. Staurostylus,[84] Haeckel, 1881,
Prodromus, p. 450.
Definition.—Staurosphærida with one single lattice-sphere
and four crossed spines which are arranged in opposite pairs, one pair opposite being larger than
the other.
The genus Staurostylus has been developed from Staurosphæra by
differentiation of the four crossed spines in pairs; two opposite spines growing much more
strongly than the other two.
1. Staurostylus græcus, n. sp.
Shell thick walled, smooth, with regular, circular, hexagonally framed pores, twice as broad as
the bars; six to eight on the quadrant. Spines three-sided prismatic, pointed, as broad as {157}one mesh; two opposite larger spines as long as the
diameter of the shell, two smaller only as long as the radius.
Dimensions.—Diameter of the shell 0.15, pores 0.01, bars 0.005; length of the
major spines 0.16, minor 0.08.
Habitat.—Central Pacific, Station 274, depth 2750 fathoms.
2. Staurostylus latinus, n. sp.
Shell thin walled, smooth, with regular, circular, double-edged pores, three times as broad as
the bars; twelve to fourteen on the quadrant. Spines conical, twice as broad at the base as one
mesh; two opposite larger spines as long as the radius of the shell, two smaller only one-third as
long.
Dimensions.—Diameter of the shell 0.24, pores 0.012, bars 0.004; length of the
major spines 0.12, minor 0.04.
Habitat.—South Pacific, Station 295, depth 1500 fathoms.
3. Staurostylus germanicus, n. sp.
Shell thick walled, covered with bristle-shaped by-spines, half as long as the radius. Pores
regular, circular, hexagonally framed, twice as broad as the bars; eight to ten on the quadrant.
From each hexagon-corner arises one short by-spine. Four main spines three-sided pyramidal, as
broad at the base as one mesh; two opposite larger spines one and a half times as long as the
radius, two smaller two-thirds as long.
Dimensions.—Diameter of the shell 0.16, pores 0.008, bars 0.004; length of the
major spines 0.12, minor 0.08.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
Genus 62. Stylostaurus,[85] 1881,
Prodromus, p. 450.
Definition.—Staurosphærida with one single lattice-sphere
and four crossed spines, one of which is much larger than the other three.
The genus Stylostaurus differs from the preceding in the extraordinary
development of one of the four spines, which is much longer than the other three; these may be
equal or different.
1. Stylostaurus caudatus, n. sp. (Pl. 13, fig.
7).
Shell thick walled, smooth, with regular, circular, hexagonally framed pores, twice as broad as
the bars; five to six on the quadrant. Spines three-sided pyramidal, as broad at the base as {158}one mesh; one of the spines longer than the shell
diameter, the opposite spine nearly as long as the shell radius; both lateral spines scarcely
one-third as long.
Dimensions.—Diameter of the shell 0.1, pores 0.01, bars 0.005; length of the major
spine 0.14, of the opposite 0.04, of both lateral spines 0.012.
Habitat.—Central Pacific, Station 266, depth 2750 fathoms.
2. Stylostaurus gladiatus, n. sp.
Shell thick walled, smooth, with regular, circular pores, three times as broad as the bars;
nine to ten on the quadrant. Spines three-sided prismatic, as broad at the base as one mesh; one
of the spines longer than the shell diameter, and much larger than the other three, which are
nearly equal (half as long as the radius).
Dimensions.—Diameter of the shell 0.2, pores 0.015, bars 0.005; length of the
major spine 0.25, of the three others 0.005.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
Subfamily Staurolonchida,[86] Haeckel, 1881, Prodromus, pp. 449, 451.
Definition.—Staurosphærida with two
concentric spherical lattice-shells.
Genus 63. Staurolonche,[87] Haeckel, 1881,
Prodromus, p. 451.
Definition.—Staurosphærida with two concentric
lattice-spheres and four crossed, equal, simple spines.
The genus Staurolonche may be derived either from Staurosphæra by the
duplication of the lattice-sphere, or from Carposphæra by the production of four crossed
radial spines, lying in one meridional plane, or from Hexalonche by the reduction of two
opposite spines.
Subgenus 1. Staurolonchantha, Haeckel.
Definition.—Pores of the cortical shell regular, and of nearly equal
size and similar form; surface smooth.
1. Staurolonche hexagona, n. sp.
Haliomma hexagonum, Ehrenberg, 1854, Mikrogeol., Taf. xxxvb., Bd. iv. fig. 17.
Haliomma hexagonum, Haeckel, 1862, Monogr. d. Radiol., p. 434.
Cortical shell thin walled, smooth, three times as broad as the medullary shell, with regular,
hexagonal pores, four times as broad as the bars; seven to eight on the quadrant. Four spines
three-sided pyramidal, somewhat longer than the radius, as broad at the base as one mesh.
{159}
Dimensions.—Diameter of the outer shell 0.12, inner 0.04; cortical pores 0.012,
bars 0.003; length of the spines 0.08.
Habitat.—North Atlantic, Station 64, depth 2700 fathoms.
2. Staurolonche spinozæ, n. sp.
Cortical shell thick walled, smooth, twice as broad as the medullary shell, with regular,
circular, hexagonally framed pores, three times as broad as the bars; five to six on the quadrant.
Four spines three-sided pyramidal, half as long as the radius, as broad at the base as one
mesh.
Dimensions.—Diameter of the outer shell 0.16, inner 0.08; cortical pores 0.012,
bars 0.004; length of the spines 0.04.
Habitat.—Central Pacific, Station 271, surface.
3. Staurolonche aperta, Haeckel.
Haliomma apertum, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss. Berlin,
p. 74, Taf. xxviii. fig. 5.
Cortical shell thin walled, smooth, three times as broad as the medullary shell; pores regular,
circular, five times as broad as the bars; four to five on the quadrant. Four spines conical, as
long as the radius, as broad as one mesh.
Dimensions.—Diameter of the outer shell 0.1, inner 0.03; cortical pores 0.015,
bars 0.003; length of the spines 0.05.
Habitat.—Fossil in Barbados.
4. Staurolonche brunonis, n. sp.
Cortical shell thick walled, smooth, twice as broad as the medullary shell, with regular,
circular pores, three times as broad as the bars; eight to ten on the quadrant. Four spines
cylindro-conical, longer than the diameter, twice as broad as one mesh.
Dimensions.—Diameter of the outer shell 0.08, inner 0.04; cortical pores 0.006,
bars 0.002; length of the spines 0.12.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
5. Staurolonche pertusa, n. sp. (Pl. 15,
figs. 5, 5a).
Cortical shell thin walled, smooth, three times as broad as the medullary shell, with regular,
circular pores, three times as broad as the bars; eight to ten on the quadrant. Pores of the
medullary shell only one-third as large, also circular. Four spines three-sided prismatic, three
times as long as the radius; each of their three thin wings perforated by a single row of small
pores.
Dimensions.—Diameter of the outer shell 0.1, inner 0.033; cortical pores 0.01,
bars 0.003; length of the spines 0.15.
Habitat.—North Atlantic, Station 353, surface.
{160}
Subgenus 2. Staurolonchella, Haeckel.
Definition.—Pores of the cortical shell regular, and of nearly equal
size and similar form; surface covered with by-spines or thorns.
6. Staurolonche straussii, n. sp.
Cortical shell thin walled, four times as broad as the medullary shell, and covered with
numerous bristle-shaped by-spines, half as long as the radius. Pores regular, circular,
hexagonally framed, twice as broad as the bars; six to eight on the radius. On each hexagonal
frame twelve by-spines (six at the corners, six in the middle between them). Four main spines
six-sided pyramidal, about as long as the radius, twice as broad as one mesh.
Dimensions.—Diameter of the outer shell 0.16, inner 0.04; cortical pores 0.016,
bars 0.008; length of the spines 0.1.
Habitat.—West Tropical Pacific, Station 225, depth 4475 fathoms.
7. Staurolonche feuerbachii, n. sp.
Cortical shell thick walled, five times as broad as the medullary shell, and covered with short
conical by-spines. Pores regular, circular, twice as broad as the bars; twenty to twenty-two on
the quadrant. Four main spines three-sided prismatic, with pyramidal apex, two to three times as
long as the radius, twice as broad as one mesh.
Dimensions.—Diameter of the outer shell 0.27, of the inner 0.055; cortical pores
0.008, bars 0.004; length of the spines 0.3 to 0.4.
Habitat.—Fossil in Barbados.
8. Staurolonche moleschottii, n. sp.
Cortical shell thick walled, three times as broad as the medullary shell, and covered with
short conical by-spines. Pores regular, circular, of the same breadth as the bars; fourteen to
sixteen on the quadrant. Four main spines conical, as long as the radius, three times as broad at
the base as one mesh.
Dimensions.—Diameter of the outer shell 0.15, inner 0.05; cortical pores and bars
0.004; length of the spines 0.08.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
Subgenus 3. Staurolonchissa, Haeckel.
Definition.—Pores of the cortical shell irregular, of different size
or form; surface smooth.
9. Staurolonche holbachii, n. sp.
Cortical shell thin walled, with smooth surface, three times as broad as the medullary shell.
Pores irregular, polygonal, twice to four times as broad as the bars. Four main spines three-sided
prismatic, longer than the shell diameter.
{161}
Dimensions.—Diameter of the outer shell 0.12, inner 0.04; cortical pores 0.006 to
0.012, bars 0.003; length of the spines 0.15.
Habitat.—South Pacific, Station 295, depth 1500 fathoms.
10. Staurolonche gassendii, n. sp.
Cortical shell thick walled, with smooth surface, four times as broad as the medullary shell.
Pores irregular, roundish, three to five times as broad as the bars. Four main spines conical, as
long as the radius.
Dimensions.—Diameter of the outer shell 0.2, inner 0.05; cortical pores 0.01 to
0.02, bars 0.004; length of the spines 0.1.
Habitat.—North Atlantic, Station 353, depth 2965 fathoms.
Subgenus 4. Staurolonchura, Haeckel.
Definition.—Pores of the cortical shell irregular, of different size
or form; surface covered with by-spines or thorns.
11. Staurolonche epicurii, n. sp.
Cortical shell thick walled, five times as broad as the medullary shell, and covered with
numerous bristle-shaped by-spines (half as long as the radius). Pores irregular, roundish,
polygonally framed, twice to three times as broad as the bars. Four main spines pyramidal, as long
as the radius.
Dimensions.—Diameter of the outer shell 0.22, inner 0.044; cortical pores 0.02 to
0.04, bars 0.012; length of the spines 0.1.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
12. Staurolonche lucretii, n. sp.
Cortical shell thick-walled, three times as broad as the medullary shell, and covered with
numerous short, conical thorns. Pores irregular, roundish, twice to four times as broad as the
bars. Four main spines conical, half as long as the radius.
Dimensions.—Diameter of the outer shell 0.15, inner 0.05; cortical pores 0.015 to
0.03, bars 0.008; length of the spines 0.04.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms.
Genus 64. Staurancistra,[88] Haeckel, 1881,
Prodromus, p. 451.
Definition.—Staurosphærida with two concentric
lattice-spheres and four crossed, equal, branched spines.
{162}
The genus Staurancistra differs from its ancestral form,
Staurolonche, in the ramification of the four crossed spines.
1. Staurancistra quadricuspis, n. sp.
Cortical shell thin walled, with rough, thorny surface, and irregular, roundish pores, twice to
four times as broad as the bars; six to eight on the quadrant. Medullary shell one-third as large,
with regular, circular pores; connected with the cortical shell by four crossed radial beams.
These are prolonged outside into four strong three-sided prismatic spines, nearly as long as the
shell diameter, each having three curved branches below the distal end. (Similar to Hexancistra
quadricuspis, Pl. 22,
fig. 11, but with only four spines.)
Dimensions.—Diameter of the cortical shell 0.16, of the medullary shell 0.05;
pores of the former 0.06 to 0.012, bars 0.003; length of the spines 0.14, breadth 0.012.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
Genus 65. Staurolonchidium,[89] n.
gen.
Definition.—Staurosphærida with two concentric
lattice-spheres and four crossed simple spines which are disposed in two different pairs, two
opposite being larger than the other two.
The genus Staurolonchidium has arisen from Staurolonche by the
stronger growth of the two opposite spines, the other two remaining stationary; both spines of
each pair equal.
1. Staurolonchidium artioscelides, n. sp.
Cortical shell thin walled, with smooth surface, three times as broad as the medullary shell.
Pores regular, circular, hexagonally framed, twice as broad as the bars; eight to ten on the
quadrant. Four radial spines three-sided pyramidal, as broad as one mesh; two opposite larger
spines three times as long as the two smaller, which are about equal to the radius.
Dimensions.—Diameter of the cortical shell 0.16, medullary shell 0.05; pores of
the former 0.012, bars 0.006; length of the major spines 0.24, minor 0.08.
Habitat.—Central Pacific, Station 265, surface.
2. Staurolonchidium perspicuum, Haeckel.
Haliomma perspicuum, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 74, Taf. xxix. fig. 1.
Cortical shell thin walled, covered with small conical by-spines, five times as broad as the
medullary shell. Pores regular, circular, eight times as broad as the bars; three to four on the
quadrant. Four radial spines three-sided pyramidal, scarcely one-third as broad as one mesh; two
opposite larger spines four times as long as the two smaller, which are about equal to half the
radius.
{163}
Dimensions.—Diameter of the cortical shell 0.12, medullary shell 0.025; pores of
the former 0.03, bars 0.004; length of the major spines 0.12, minor 0.03.
Habitat.—Fossil in Barbados.
Genus 66. Stauroxiphos,[90] n. gen.
Definition.—Staurosphærida with two concentric
lattice-spheres and four crossed simple spines, one of which is much larger than the other
three.
The genus Stauroxiphos differs from its ancestral form, Staurolonche,
in the greater development of one single spine, and exhibits therefore the same relation to it
that Stylostaurus bears to Staurosphæra.
1. Stauroxiphos gladius, n. sp. (Pl. 15,
fig. 7).
Cortical shell thick walled, smooth, three times as broad as the medullary shell. Pores of the
outer shell regular, circular, regularly six-lobed on the outer opening, four times as broad as
the bars; about six on the quadrant. Pores of the inner shell only one-third as large, simple,
circular. Three of the four radial spines of nearly equal size, pommel-shaped, with three
prominent, dentated wings, somewhat shorter than the shell radius and about half as broad as long;
the fourth spine much larger, sword-like, about three times as long as the shell radius.
Dimensions.—Diameter of the outer shell 0.12, of the inner 0.04; pores of the
former 0.01, bars 0.0025; pores of the latter 0.003, bars 0.001; length of the major spine 0.18,
of the three minor 0.04.
Habitat.—Central Pacific, Station 266, depth 2750 fathoms.
Subfamily Stauracontida,[91] Haeckel, 1881, Prodromus, p. 52.
Definition.—Staurosphærida with three
concentric spherical lattice-shells.
Genus 67. Stauracontium,[92] Haeckel, 1881,
Prodromus, p. 452.
Definition.—Staurosphærida with three concentric
lattice-spheres and four crossed, equal, simple spines.
The genus Stauracontium differs from its probable ancestral form,
Staurolonche, in the duplication of the cortical shell.
Subgenus 1. Stauracontarium, Haeckel.
Definition.—Pores of the cortical shell regular, all of nearly equal
size and similar form; surface smooth.
{164}
1. Stauracontium cruciferum, n. sp.
Cortical shell thin walled, smooth, with regular, hexagonal pores, four times as broad as the
bars; eight to ten on the quadrant. Radial proportion of the three spheres =
1 : 3 : 9. Connecting radial beams between them six (opposite in pairs in the
three dimensive axes), but only four of them are prolonged outside into four stout three-sided
prismatic spines, lying in one equatorial plane, as long as the shell diameter.
Dimensions.—Diameter of the outer shell 0.22, middle 0.07, inner 0.025; cortical
pores 0.012, bars 0.003; length of the spines 0.2.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
2. Stauracontium tetracanthum, Haeckel.
Haliomma tetracanthum, Ehrenberg, 1872, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 295, Taf. x. figs. 7, 8.
Cortical shell thin walled, smooth, with regular, circular pores, three times as broad as the
bars; five to six on the quadrant. Radial proportion of the three spheres =
1 : 3 : 12. Radial spines three-sided pyramidal, about as long as the shell
diameter.
Dimensions.—Diameter of the outer shell 0.12, middle 0.03, inner 0.01; cortical
pores 0.01, bars 0.003; length of the spines 0.1.
Habitat.—Indian Ocean, Zanzibar, depth 2200 fathoms, Pullen.
3. Stauracontium tetracontium, Haeckel.
Actinomma tetracanthum, Stöhr, 1880, Palæontogr. 26, p. 91, Taf. ii. fig.
6.
Cortical shell thin walled, rough, with regular, circular pores, of the same breadth as the
bars; six to seven on the quadrant. Radial proportion of the three spheres =
1 : 2.5 : 8. Radial spines three-sided pyramidal, nearly as long as the shell
diameter.
Dimensions.—Diameter of the outer shell 0.1, middle 0.03, inner 0.013; cortical
pores and bars 0.006; length of the spines 0.08.
Habitat.—Fossil in Tertiary rocks of Sicily; Grotte, Caltanisetta.
Subgenus 2. Stauracontellium, Haeckel.
Definition.—Pores of the cortical shell regular, and of nearly equal
size and similar form; surface covered with numerous small thorns or by-spines.
4. Stauracontium daturæforme, Haeckel.
Actinomma daturæforme, Stöhr, 1880, Palæontogr. 26, p. 93, Taf. ii. fig.
13.
Cortical shell thick walled, covered with short thorns, and with regular, hexagonal pores, five
times as broad as the bars; six to seven on the quadrant. Radial proportion of the three spheres =
1 : 3 : 3.5. Radial spines six-sided pyramidal, about half as long as the
radius and one-fourth as broad.
{165}
Dimensions.—Diameter of the outer shell 0.15, middle 0.13, inner 0.04; cortical
pores 0.017, bars 0.003; length of the spines 0.03.
Habitat.—Fossil in Tertiary rocks of Sicily; Grotte, Stöhr.
5. Stauracontium sparganium, n. sp.
Cortical shell thick walled, covered with very numerous, short, conical spines, and with
regular, circular pores, five times as broad as the bars; sixteen to eighteen on the quadrant.
Radial proportion of the three spheres = 2 : 3 : 9. Radial main spines
three-sided prismatic, as long as the radius or longer.
Dimensions.—Diameter of the outer shell 0.275, middle 0.09, inner 0.06; cortical
pores 0.01, bars 0.002; length of the spines 0.15.
Habitat.—Fossil in Barbados.
Subgenus 3. Stauracontidium, Haeckel.
Definition.—Pores irregular, of different size or form; surface
smooth.
6. Stauracontium antarcticum, n. sp.
Cortical shell thin walled, smooth. Pores irregular, roundish, two to four times as broad as
the bars. Radial proportion of the three spheres = 1 : 2 : 8. Radial spines
conical, as long as the radius.
Dimensions.—Diameter of the outer shell 0.2, middle 0.05, inner 0.025; cortical
pores 0.006 to 0.012, bars 0.003; length of the spines 0.1.
Habitat.—Antarctic Ocean, Station 157, depth 1950 fathoms.
Subgenus 4. Stauracontonium, Haeckel.
Definition.—Pores irregular, of different size or form; surface
covered with thorns or by-spines.
7. Stauracontium setosum, n. sp.
Cortical shell thin walled, covered with thin bristle-shaped by-spines, half as long as the
radius. Pores irregular, polygonal, three to five times as broad as the bars. Proportion of the
three spheres = 1 : 2 : 6. Radial main spines pyramidal, nearly as long as the
shell diameter.
Dimensions.—Diameter of the outer shell 0.15, middle 0.05, inner 0.025; cortical
pores 0.01 to 0.015, bars 0.003; length of the spines 0.12.
Habitat.—Central Pacific, Station 266, depth 2750 fathoms.
8. Stauracontium papillosum, n. sp.
Cortical shell thick walled, covered with short conical papillæ or by-spines. Pores irregular,
roundish, two to four times as broad as the bars. Proportion of the three spheres =
1 : 3 : 12. Radial main spines conical, as long as the radius.
{166}
Dimensions.—Diameter of the outer shell 0.24, middle 0.06, inner 0.02; cortical
pores 0.007 to 0.015, bars 0.004; length of the spines 0.12.
Habitat.—Central Pacific, Station 274, depth 2750 fathoms.
Subfamily Staurocromyida,[93] Haeckel, 1881, Prodromus, pp. 449, 453.
Definition.—Staurosphærida with four
concentric spherical lattice-shells.
Genus 68. Staurocromyum,[94] Haeckel, 1881,
Prodromus, p. 453.
Definition.—Staurosphærida with four concentric
lattice-spheres and four crossed, equal, simple spines.
The genus Staurocromyum has arisen probably from Stauracontium by
duplication of the cortical shell, two concentric shells lying within, two others outside the
central capsule.
1. Staurocromyum quadruplex, n. sp.
Radial proportion of the four spheres = 1 : 2 : 8 : 10. Both
medullary shells with very small, regular, circular pores. Inner cortical shell with regular,
circular, hexagonally-framed pores, twice as broad as the bars; from each hexagon-corner arises a
small by-spine, and these, connected by tangential branches at equal distances from the centre,
form the delicate outer cortical shell, with spiny surface. Four main spines three-sided
pyramidal, as long as the shell radius.
Dimensions.—Diameter of the four shells—(A) 0.25, (B) 0.2, (C) 0.05, (D)
0.025; length of the spines 0.12.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
2. Staurocromyum quadrispinum, n. sp.
Radial proportion of the four spheres = 1 : 3 : 9 : 12. Both
medullary shells with small, regular, circular pores. Both cortical shells with irregular,
roundish pores. Surface covered with short, conical by-spines. Four main spines cylindro-conical,
somewhat longer than the shell diameter.
Dimensions.—Diameter of the four shells—(A) 0.24, (B) 0.18, (C) 0.06, (D)
0.02.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms.
Genus 69. Cromyostaurus,[95] Haeckel, 1881,
Prodromus, p. 453.
Definition.—Staurosphærida with four concentric
lattice-spheres and four crossed, equal, branched spines.
{167}
The genus Cromyostaurus differs from the preceding Staurocromyum, its
ancestral form, in the ramification of the four crossed spines.
1. Cromyostaurus verticillatus, n. sp.
Radial proportion of the four spheres = 1 : 3 : 11 : 13. Both
medullary shells with small, regular, circular pores; inner cortical shell with regular, hexagonal
pores; from the hexagon-corners arise small, radial by-spines, which at equal distances from the
centre send out forked tangential branches, three from each spine, and by communication of these
form the outer, delicate, cortical shell. Four main spines nearly as long as the shell diameter,
three-sided prismatic, with four to six verticils of ramified lateral branches, each verticil
composed of three forked branches, which ramify again.
Dimensions.—Diameter of the four shells—(A) 0.26, (B) 0.22, (C) 0.06, (D)
0.02; length of the spines 0.24.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
Subfamily Staurocaryida,[96] Haeckel, 1881, Prodromus, pp. 449, 454.
Definition.—Staurosphærida with five or more
concentric spherical lattice-shells.
Genus 70. Staurocaryum,[97] Haeckel, 1881,
Prodromus, p. 454.
Definition.—Staurosphærida with five or more concentric
lattice-spheres and four crossed, equal spines.
The genus Staurocaryum has arisen from the preceding Staurocromyum by
the further multiplication of the concentric spheres; in the only observed form there are six, at
nearly equal distances apart.
1. Staurocaryum arborescens, n. sp. (Pl. 15, fig.
8).
Shell composed of six concentric latticed spheres, at nearly equal distances apart, and with
somewhat regular, circular pores, the size of which gradually increases from the first to the
sixth shell. The surface of the outermost shell is densely covered with numerous arborescent
by-spines, which bifurcate from three to four times, and are three-sided pyramidal at the base,
and twice as long as the distance between each two shells. The six shells are connected only by
four crossed, conical, radial beams, which increase in diameter from the centre, and are prolonged
outside into very stout cylindrical, radial spines, irregularly covered with small thorns and
forked ramules, and nearly as long as the shell diameter. Only a single specimen was observed.
{168}
Dimensions.—Diameter of the whole shell 0.22; distance between each two shells
0.02; length of the by-spines 0.05, of the main spines 0.2.
Habitat.—Indian Ocean, Cocos Islands, surface, Rabbe.
Subfamily Staurodorida,[98] Haeckel, 1881, Prodromus, pp. 449, 455.
Definition.—Staurosphærida with spongy,
spherical shell (with or without enclosed concentric lattice-shells).
Genus 71. Staurodoras,[99] Haeckel, 1881,
Prodromus, p. 455.
Definition.—Staurosphærida with solid, spongy, spherical
shell and four crossed simple spines.
The genus Staurodoras may be developed from Styptosphæra by the
production of four crossed spines on the simple, spongy, spherical shell, which is composed of
looser or denser irregular wicker-work, without enclosed medullary shell.
1. Staurodoras spongosphæra, n. sp.
Four crossed spines, two to three times as long as the diameter of the spongy sphere,
three-sided prismatic, with three dentated and spirally contorted edges. (Form of Spongosphæra
streptacantha, but without medullary shell and with four equal spines, crossed regularly at
right angles.)
Dimensions.—Diameter of the sphere 0.25; length of the spines 0.4 to 0.7.
Habitat.—Central Pacific, Station 271, surface.
2. Staurodoras mojsisovicsi, Dunikowski.
Staurodoras mojsisovicsi, Dunikowski, 1882, Denkschr. d. k. Akad. d. Wiss.
Wien, Bd. xlv. p. 28, Taf. v. fig. 56.
Four crossed spines, one and a half times as long as the diameter of the spongy sphere,
three-sided pyramidal, with three smooth edges. (What Dunikowski describes as "inner canals" of
the spines are their edges.)
Dimensions.—Diameter of the sphere 0.14; length of the spines 0.18 to 0.2. basal
breadth 0.04.
Habitat.—Fossil in the Alpine Lias (Schafberg near Salzburg).
3. Staurodoras liassica, Dunikowski.
Staurodoras liassica, Dunikowski, 1882, Denkschr. d. k. Akad. d. Wiss.
Wien, Bd. xlv. p. 28, Taf. v. fig. 57.
Four crossed spines, shorter than the diameter of the spongy sphere, conical or pyramidal
(?).
Dimensions.—Diameter of the sphere 0.19; length of the spine 0.13, basal breadth
0.045.
Habitat.—Fossil in the Alpine Lias (Schafberg near Salzburg).
{169}
4. Staurodoras wandae, Dunikowski.
Staurodoras wandae, Dunikowski, 1882, Denkschr. d. k. Akad. d. Wiss. Wien,
Bd. xlv. p. 28, Taf. v. fig. 58.
Four crossed spines shorter than the radius of the spongy sphere, conical. (May be the young
form of the preceding species.)
Dimensions.—Diameter of the sphere 0.14; length of the spines 0.06, basal breadth
0.05.
Habitat.—Fossil in the Alpine Lias (Schafberg near Salzburg).
Family IX. Cubosphærida, Haeckel (Pls. 21-25).
Cubosphærida, Haeckel, 1881, Prodromus, p. 449.
Definition.—Sphæroidea with six radial spines on the
surface of the spherical shell, opposite in pairs in the three dimensive axes, perpendicular one
to another; living solitary (not associated in colonies).
The family Cubosphærida is distinguished from the other Sphæroidea by the possession of six radial spines, which are opposite in pairs
in three different axes, one perpendicular to the other two. These three axes are the typical
"dimensive axes," which are more or less differentiated in the Larcoidea.
But in these latter the shell itself and the enclosed central capsule become affected by the
unequal growth in the three axes, whilst in the former the capsule constantly, and commonly also
the shell, remains spherical. Sometimes the shell assumes the form of a regular octahedron, from
the six corners of which arise the six radial spines, indicating its three axes.
The most simple Cubosphærida are the Hexastylida, with one single, spherical lattice-shell. To
this ancestral group all other subfamilies can be opposed as "Cubosphærida concentrica," as their
carapace is composed of two or more concentric lattice-shells—two in the Hexalonchida, three
in the Hexacontida, four in the Hexacromyida, five or more in the Hexacaryida. In all these four
subfamilies the concentric shells are simple (not spongy), fenestrated spheres. In a sixth
subfamily, in the Hexadorida, the shell is wholly or partially composed of irregular, spongy
wicker-work or loose reticulations, with or without a medullary shell in the centre.
The Six Radial Spines of the Cubosphærida are normally opposite in pairs in the three
dimensive axes, each of which is perpendicular to the other two. But in many species besides this
normal form occur individual abnormalities, in which the six spines are not quite accurately
opposed, but more or less divergent; and often also the three dimensive planes (determined each by
two axes) are not quite regular, but more or less uneven. More rarely the six spines appear
disposed in quite an irregular manner.
In the greater part of the Cubosphærida all six spines are quite equal, of the same size and
form. But in some genera a more or less considerable differentiation takes place, so {170}that two pairs or all three pairs of spines become
different; very rarely, also both spines of one pair become unequal (probably only an individual
abnormality). Those variations correspond to the differences between the crystalline systems. The
common Cubosphærida, with three equal spine-pairs, correspond to the regular or cubic system, with
three equal axes. The rarer forms (Hexastylarium, Hexaloncharium,
Hexacontarium) exhibit two equal pairs and one different pair; they correspond to the
quadratic system, with three perpendicular axes, two of which are equal, the third unequal. Still
more rare are those forms (Hexastylidium, Hexalonchidium), in which all three pairs
of spines are different, corresponding to the three unequal axes of the rhombic crystalline
system.
Synopsis of the Genera of Cubosphærida.
|
I. Subfamily Hexastylida.
(Shell one simple latticed sphere.)
|
|
All six radial spines simple, of equal size, |
72. Hexastylus. |
| Six spines of different sizes, all six simple. |
|
Two pairs equal, one pair different, |
73. Hexastylarium. |
| All three pairs different, |
74. Hexastylidium. |
|
II. Subfamily Hexalonchida.
(Shell composed of two concentric latticed spheres.)
|
|
All six radial spines of equal size. |
|
Spines simple, |
75. Hexalonche. |
| Spines branched, |
76. Hexancistra. |
| Six spines simple, in pairs of different sizes. |
|
Two pairs equal, one pair different, |
77. Hexaloncharium. |
| All three pairs different, |
78. Hexalonchidium. |
|
III. Subfamily Hexacontida.
(Shell composed of three spheres.)
|
|
All six radial spines of equal size. |
|
Spines simple, |
79. Hexacontium. |
| Spines branched, |
80. Hexadendron. |
| Six spines simple, of different sizes. |
|
Two pairs equal, one pair different, |
81. Hexacontarium. |
|
IV. Subfamily Hexacromyida.
(Four concentric spheres.)
|
|
All six radial spines of equal size. |
|
Spines simple, no branched, |
82. Hexacromyum. |
|
V. Subfamily Hexacaryida.
(Five or more spheres.)
|
|
All six radial spines of equal size. |
|
Spines simple, |
83. Cubosphæra. |
| Spines branched, |
84. Hexacaryum. |
|
VI. Subfamily Hexadorida.
(Shell a spongy sphere, with or without an enclosed central medullary
shell.)
|
|
All six radial spines of equal size, simple (not
branched). |
|
Without latticed medullary shell, |
85. Cubaxonium. |
| With one single medullary shell, |
86. Hexadoras. |
| With two medullary shells, |
87. Hexadoridium. |
| I. Subfamily Hexastylida. (Shell one simple latticed sphere.) |
|
|
All six radial spines simple, of equal size, |
|
|
|
|
|
|
|
|
72. Hexastylus. |
|
|
|
|
|
|
|
|
Six spines of different sizes, all six simple. |
|
|
|
|
|
|
Two pairs equal, one pair different, |
|
|
|
|
|
|
|
|
|
|
73. Hexastylarium. |
|
|
|
|
|
|
|
|
|
|
All three pairs different, |
|
|
|
|
|
|
|
|
|
|
74. Hexastylidium. |
|
|
|
|
|
|
| II. Subfamily Hexalonchida. (Shell composed of two concentric latticed
spheres.) |
|
|
All six radial spines of equal size. |
|
|
|
|
|
|
Spines simple, |
|
|
|
|
|
|
|
|
|
|
75. Hexalonche. |
|
|
|
|
|
|
|
|
|
|
Spines branched, |
|
|
|
|
|
|
|
|
|
|
76. Hexancistra. |
|
|
|
|
|
|
|
|
Six spines simple, in pairs of different sizes. |
|
|
|
|
|
|
Two pairs equal, one pair different, |
|
|
|
|
|
|
|
|
|
|
77. Hexaloncharium. |
|
|
|
|
|
|
|
|
|
|
All three pairs different, |
|
|
|
|
|
|
|
|
|
|
78. Hexalonchidium. |
|
|
|
|
|
|
| III. Subfamily Hexacontida. (Shell composed of three spheres.) |
|
|
All six radial spines of equal size. |
|
|
|
|
|
|
Spines simple, |
|
|
|
|
|
|
|
|
|
|
79. Hexacontium. |
|
|
|
|
|
|
|
|
|
|
Spines branched, |
|
|
|
|
|
|
|
|
|
|
80. Hexadendron. |
|
|
|
|
|
|
|
|
Six spines simple, of different sizes. |
|
|
|
|
|
|
Two pairs equal, one pair different, |
|
|
|
|
|
|
|
|
|
|
81. Hexacontarium. |
|
|
|
|
|
|
| IV. Subfamily Hexacromyida. (Four concentric spheres.) |
|
|
All six radial spines of equal size. |
|
|
|
|
|
|
Spines simple, no branched, |
|
|
|
|
|
|
|
|
|
|
82. Hexacromyum. |
|
|
|
|
|
|
| V. Subfamily Hexacaryida. (Five or more spheres.) |
|
|
All six radial spines of equal size. |
|
|
|
|
|
|
Spines simple, |
|
|
|
|
|
|
|
|
|
|
83. Cubosphæra. |
|
|
|
|
|
|
|
|
|
|
Spines branched, |
|
|
|
|
|
|
|
|
|
|
84. Hexacaryum. |
|
|
|
|
|
|
| VI. Subfamily Hexadorida. (Shell a spongy sphere, with or without an enclosed
central medullary shell.) |
|
|
All six radial spines of equal size, simple (not branched). |
|
|
|
|
|
|
Without latticed medullary shell, |
|
|
|
|
|
|
|
|
|
|
85. Cubaxonium. |
|
|
|
|
|
|
|
|
|
|
With one single medullary shell, |
|
|
|
|
|
|
|
|
|
|
86. Hexadoras. |
|
|
|
|
|
|
|
|
|
|
With two medullary shells, |
|
|
|
|
|
|
|
|
|
|
87. Hexadoridium. |
|
|
|
|
|
|
{171}
Subfamily Hexastylida,[100] Haeckel, 1881, Prodromus, pp. 449, 450.
Definition.—Cubosphærida with one single
spherical lattice-shell.
Genus 72. Hexastylus,[101] Haeckel, 1881,
Prodromus, p. 450.
Definition.—Cubosphærida with one simple lattice-sphere
and six simple spines of equal size.
The genus Hexalonche is the most simple form of all Cubosphærida, and may be
regarded as the common ancestral form of this family. It can be derived phylogenetically from
Cenosphæra, by development of six radial spines on the surface of the simple spherical
lattice-shell. These six simple spines are of equal size and opposite in pairs in the three
dimensive axes, corresponding to the three equal axes of a cubic crystal.
Subgenus 1. Hexastylanthus, Haeckel.
Definition.—Pores regular or subregular, of nearly equal size and
form; surface of the cortical shell smooth, without radial by-spines (other than the six main
spines).
1. Hexastylus phænaxonius, n. sp. (Pl. 21, fig.
3).
Shell thin walled, with smooth surface. Pores subregular, hexagonal, five to six times as broad
as the bars; nine to ten on the radius. Six spines triangular pyramidal, as long as the radius of
the shell, as broad at the base as one pore.
Dimensions.—Diameter of the shell 0.13, pores 0.008 to 0.01, bars 0.0015; length
of the spines 0.07, basal breadth 0.008 to 0.012.
Habitat.—Central Pacific, Station 272, depth 2600 fathoms.
2. Hexastylus sapientum, n. sp.
Shell thin walled, with smooth surface. Pores regular, hexagonal, eight to ten times as broad
as the bars; six to seven on the radius. Six spines bristle-shaped, longer than the diameter of
the shell. (Lattice-work and spines similar to those of Heliosphæra actinota, Monogr. d.
Radiol., Taf. ix. fig. 3.)
Dimensions.—Diameter of the shell 0.15, pores 0.016, bars 0.002; length of the
spines 0.2, breadth 0.002.
Habitat.—North Atlantic, Station 354, surface.
{172}
3. Hexastylus thaletis, n. sp. (Pl. 21,
fig. 4).
Shell thin walled, with smooth surface. Pores subregular, circular, hexagonally framed,
somewhat funnel-shaped, of the same breadth as the bars; eight to nine on the radius. Six spines
triangular pyramidal, with prominent edges, as long as the radius, three to four times as broad at
the base as one pore.
Dimensions.—Diameter of the shell 0.1, pores and bars 0.005; length of the spines
0.05, basal breadth 0.02.
Habitat.—Tropical Pacific, Station 225, depth 4575 fathoms.
4. Hexastylus favosus, n. sp.
Shell thick walled, with smooth surface. Pores regular, circular, hexagonally framed, deep
funnel-shaped, of the same breadth as the bars; six to seven on the radius. Six spines triangular
pyramidal, as long as the diameter, twice as broad at the base as one pore.
Dimensions.—Diameter of the shell 0.12, pores and bars 0.01; length of the spines
0.12, basal breadth 0.02.
Habitat.—South Atlantic, Station 320, surface.
5. Hexastylus longissimus, n. sp.
Shell thick walled, with smooth surface. Pores regular, circular, hexagonally framed, of the
same breadth as the bars; five to six on the radius. Six spines triangular prismatic, extremely
elongated, ten to twenty times as long as the diameter of the shell, twice as broad as one
pore.
Dimensions.—Diameter of the shell 0.08, pores and bars 0.008; length of the spines
1.0 to 1.5, breadth 0.015.
Habitat.—Tropical Atlantic, Station 347, surface.
6. Hexastylus minimus, n. sp. (Pl. 21,
fig. 5).
Shell thin walled, with smooth surface. Pores subregular, circular, twice as broad as the bars;
five to six on the radius. Six spines triangular pyramidal, scarcely as long as the radius, at the
base half as broad as long.
Dimensions.—Diameter of the shell 0.05, pores 0.004, bars 0.002; length of the
spines 0.02, basal breadth 0.01.
Habitat.—Tropical Pacific, Station 225, depth 4575 fathoms.
7. Hexastylus biantis, n. sp.
? Haliphormis
hexacantha, Ehrenberg, 1872, L. N. 24, Taf. x. fig. 6.
Shell thin walled, smooth. Pores regular, circular, four times as broad as the bars; eight to
nine on the radius. Six spines conical, as long as the radius, at the base as broad as one
pore.
{173}
Dimensions.—Diameter of the shell 0.12, pores 0.012, bars 0.003; length of the
spines 0.06, basal breadth 0.012.
Habitat.—Indian Ocean, Madagascar, surface, Rabbe.
8. Hexastylus pittaci, n. sp.
Shell thin walled, smooth. Pores regular, circular, ten to twelve times as broad as the bars;
six to seven on the radius. Six spines triangular pyramidal, as long as the radius, as broad as
one pore.
Dimensions.—Diameter of the shell 0.12, pores 0.02, bars 0.002; length of the
spines 0.06, basal breadth 0.02.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
9. Hexastylus maximus, n. sp.
Shell thick walled, smooth. Pores regular, circular, twice as broad as the bars; twelve to
sixteen on the radius. Six spines six-sided pyramidal, half as long as the radius, three times as
broad as one pore.
Dimensions.—Diameter of the shell 0.3, pores 0.01, bars 0.005; length of the
spines 0.08, basal breadth 0.03.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
10. Hexastylus periandri, n. sp.
Shell thick walled, smooth. Pores regular, circular, double-edged, four times as broad as the
bars; six to seven on the radius. Six spines conical, as long as the radius, twice as broad as one
pore.
Dimensions.—Diameter of the shell 0.2, pores 0.012, bars 0.003; length of the
spines 0.1, basal breadth 0.025.
Habitat.—Tropical Atlantic, Station 335, depth 1425 fathoms.
Subgenus 2. Hexastylettus, Haeckel.
Definition.—Pores regular or subregular, of nearly equal size and
form; surface of the spherical shell spiny, covered with numerous conical or bristle-shaped radial
by-spines.
11. Hexastylus solonis, n. sp. (Pl. 21,
fig. 11).
Shell thick walled, covered with numerous bristle-shaped radial spines (one-third to one-half
as long as the six main spines, arising from the lattice-knots). Pores regular, circular, enclosed
by prominent, hexagonal frames, four to six times as broad as the bars; seven to eight on the
radius. {174}Six main spines triangular pyramidal, with
spirally contorted prominent edges, as long as the radius, as broad at the base as one pore.
Dimensions.—Diameter of the shell 0.13, pores 0.01, bars 0.002; length of the
spines 0.06 to 0.8, basal breadth 0.01.
Habitat.—Central Pacific, Station 266, depth 2750 fathoms.
12. Hexastylus cochleatus, n. sp. (Pl. 21, fig.
1).
Shell thick walled, covered with numerous short, conical spines (about as large as one pore).
Pores regular, circular, three times as broad as the bars; eight to nine on the radius. Six main
spines triangular pyramidal, with prominent, spirally-twisted edges, as long as the diameter of
the shell, and twice as broad at the base as one pore.
Dimensions.—Diameter of the shell 0.17, pores 0.015, bars 0.005; length of the
spines 0.16, breadth 0.03.
Habitat.—Mediterranean, Corfu, Haeckel, surface.
13. Hexastylus setosus, Haeckel.
Acanthosphæra setosa, Ehrenberg, 1872, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 285, Taf. ix. fig. 11.
Shell thin walled, covered with numerous short, bristle-shaped spines (not larger than one
pore). Pores regular, circular, four times as broad as the bars; thirteen to sixteen on the
radius. Six main spines three-sided pyramidal, scarcely one-third as long as the radius. (In the
figure of Ehrenberg, loc. cit., only four spines are in regular, crossed disposition, two
others opposite in oblique direction; this is either an individual abnormality, or an error of
drawing; the same species occurs with six spines exactly regularly disposed in the three dimensive
axes.)
Dimensions.—Diameter of the shell 0.15; pores 0.008, bars 0.002; length of the
spines 0.02 to 0.03, basal breadth 0.005.
Habitat.—Indian Ocean, Zanzibar, Pullen; Madagascar, Rabbe.
14. Hexastylus chilonis, n. sp.
Shell thin walled, covered with numerous bristle-shaped spines (as long as the radius). Pores
regular, circular, twice as broad as the bars; eight to nine on the radius. Six main spines
triangular pyramidal, with straight edges, as long as the diameter of the shell, as broad as two
pores.
Dimensions.—Diameter of the shell 0.2, pores 0.02, bars 0.01; length of the spines
0.2, basal breadth 0.05.
Habitat.—South Atlantic, Station 325, surface.
15. Hexastylus cleobuli, n. sp.
Shell thick walled, covered with numerous short, conical spines (twice as large as one pore).
Pores regular, circular, of the same breadth as the bars; twelve to thirteen on the radius. Six
main spines conical, as long as the radius, three times as broad as one pore.
{175}
Dimensions.—Diameter of the shell 0.24, pores and bars 0.005; length of the spines
0.12, basal breadth 0.016.
Habitat.—South Pacific, Station 300, depth 1375 fathoms.
Subgenus 3. Hexastylissus, Haeckel.
Definition.—Pores irregular, of unequal size or form; surface of the
spherical shell smooth, without radial by-spines (other than the six main spines).
16. Hexastylus triaxonius, n. sp. (Pl. 21, fig.
2).
Shell thin walled, with smooth surface. Pores irregular, polygonal, three to six times as broad
as the bars; four to six on the radius. Six spines triangular pyramidal, as long as the diameter
of the shell, as broad at the base as one pore.
Dimensions.—Diameter of the shell 0.05, pores 0.004 to 0.008, bars 0.0015; length
of the spines 0.04, basal breadth 0.006.
Habitat.—North Pacific, Station 253, depth 3125 fathoms.
17. Hexastylus dimensivus, n. sp. (Pl. 21, fig.
6).
Shell thin walled, with smooth surface. Pores irregular, polygonal, four to eight times as
broad as the bars; eleven to thirteen on the radius. Six spines hexagonal pyramidal, as long as
the radius, about three times as broad at the base as one pore.
Dimensions.—Diameter of the shell 0.13, pores 0.006 to 0.009, bars 0.0012; length
of the spines 0.06, basal breadth 0.025.
Habitat.—North Pacific, Station 256, depth 2950 fathoms.
18. Hexastylus brevispinus, n. sp.
Shell thin walled, with smooth surface. Pores irregular, roundish, two to four times as broad
as the bars; six to eight on the radius. Six spines triangular pyramidal, half as long as the
radius, twice as broad as one pore.
Dimensions.—Diameter of the shell 0.2, pores 0.008 to 0.016, bars 0.004; length of
the spines 0.05, basal breadth 0.025.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
19. Hexastylus longispinus, n. sp.
Shell thick walled, smooth. Pores irregular, roundish, two to three times as broad as the bars;
three to four on the radius. Six spines triangular prismatical, two to three times as long as the
diameter of the shell.
{176}
Dimensions.—Diameter of the shell 0.08, pores 0.008 to 0.012, bars 0.004; length
of the spines 0.2, breadth 0.01.
Habitat.—North Atlantic, Station 353, surface.
20. Hexastylus marginatus, n. sp. (Pl. 21, fig.
10).
Shell thick walled, rough. Pores irregular, roundish, somewhat funnel-shaped double-edged, two
to three times as broad as the bars; twelve to fourteen on the radius. Six spines three-sided
pyramidal, somewhat longer than the radius, three times as broad as one pore.
Dimensions.—Diameter of the shell 0.15, pores 0.007 to 0.01, bars 0.004; length of
the spines 0.1, basal breadth 0.025.
Habitat.—South Pacific, Station 295, depth 1500 fathoms.
21. Hexastylus conifer, n. sp.
Shell thick walled, rough. Pores irregular, roundish, scarcely broader than the bars; fifteen
to sixteen on the radius. Six spines conical, as long as the radius, five to seven times as broad
as one pore.
Dimensions.—Diameter of the shell 0.18, pores and bars 0.004 to 0.006; length of
the spines 0.1, basal breadth 0.03.
Habitat.—Indian Ocean, Sunda Strait, Rabbe, surface.
Subgenus 4. Hexastylurus, Haeckel.
Definition.—Pores irregular, of unequal size or form; surface of the
spherical shell spiny, covered with numerous conical or bristle-shaped by-spines.
22. Hexastylus dictyotus, n. sp. (Pl. 21, figs. 8,
9).
Shell thin walled, spiny; short spines conical, smaller than the pore-breadth. Pores irregular,
polygonal, five to seven times as broad as the bars; four to six on the radius. Six spines
triangular pyramidal, longer than the radius, about as broad as one smaller pore.
Dimensions.—Diameter of the shell 0.09, pores 0.01 to 0.015, bars 0.002; length of
the spines 0.06, basal breadth 0.01.
Habitat.—Central Pacific, Station 272, depth 2600 fathoms.
23. Hexastylus hirsutus, n. sp.
Shell thin walled, densely covered with bristle-shaped, radial spines, half as long as the six
main spines. Pores irregular, polygonal, three to four times as broad as the bars; eight to ten on
the radius. Six spines triangular pyramidal, as long as the radius, twice as broad as one
pore.
{177}
Dimensions.—Diameter of the shell 0.14, pores 0.005 to 0.008, bars 0.002; length
of the spines 0.06, basal breadth 0.01.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
24. Hexastylus contortus, n. sp. (Pl. 21, fig.
12).
Shell thick walled, covered with bristle-shaped, radial spines, half as long as the radius.
Pores irregular, roundish, two to three times as broad as the bars; seven to eight on the radius.
Six spines triangular prismatic, in the distal half spirally twisted like a cork-screw (fig. 12);
longer than the diameter of the shell, about as broad as one pore.
Dimensions.—Diameter of the shell 0.12, pores 0.006 to 0.008, bars 0.003; length
of the spines 0.15, breadth 0.015.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms.
25. Hexastylus spiralis, n. sp. (Pl. 21,
fig. 7).
Shell thick walled, covered with short conical spines. Pores irregular, roundish, two to three
times as broad as the bars; five to six on the radius. Six spines triangular prismatic, with three
thin, spirally contorted edges, two to three times as long as the diameter of the shell, about as
broad as one large pore.
Dimensions.—Diameter of the shell 0.16, pores 0.01 to 0.02, bars 0.006; length of
the spines 0.3 to 0.5, basal breadth 0.02.
Habitat.—Tropical Western Pacific, Station 225, depth 4475
fathoms.
Genus 73. Hexastylarium,[102] n.
gen.
Definition.—Cubosphærida with one simple lattice-sphere
and six simple spines of different sizes; one opposite pair larger than the other two.
The genus Hexastylarium differs from its probable ancestral form,
Hexastylus, by the unequal growth of the six simple spines; two opposite spines of one pair
being more strongly developed than the four others, which are equal. They correspond therefore to
the three axes of a quadratic crystal.
1. Hexastylarium heteraxonium, n. sp.
Cortical shell thin walled, smooth, with regular, hexagonal pores, three times as broad as the
bars; eight to ten on the radius. Six spines three-sided pyramidal, at the base as broad as one
pore. Two opposite major spines longer than the shell diameter; four others scarcely as long as
{178}the radius. (Similar to Hexastylus
phænaxonius, Pl. 21,
fig. 3, but differing in the unequal length of the spines.)
Dimensions.—Diameter of the shell 0.15, pores 0.12, bars 0.004; length of the two
major spines 0.2, of the four minor 0.07.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms.
2. Hexastylarium quadratum, n. sp.
Shell very delicate, with smooth surface, and irregular, polygonal pores, separated by very
thin bars. The form of the shell is not, as commonly, a sphere, but a geometrical square
octahedron, one axis (with two opposite major spines) being nearly twice as long as the other two
dimensive axes; four spines, opposite by pairs in the latter, are only half as long. The eight
sides of the octahedral shell are even, equilateral-triangular. Spines angular, thin.
Dimensions.—Diameter of the shell in the major axis 0.18, in the minor 0.1; length
of the major spines 0.24, minor 0.12.
Habitat.—South Pacific, Station 295, depth 1500 fathoms.
3. Hexastylarium elongatum, n. sp.
Shell thick walled, with spiny surface, and with irregular, roundish pores, two to four times
as broad as the bars; eight to ten on the radius. Two opposite major spines, three to four times
as long as the shell diameter, whilst the four other spines are very short, scarcely as long as
the radius. All six spines at the base three-sided pyramidal, the two longer being
cylindrical.
Dimensions.—Diameter of the shell 0.1; length of the major spines 0.3 to 0.4, of
the minor 0.04, basal breadth 0.02.
Habitat.—North Pacific, Station 244, surface.
Genus 74. Hexastylidium,[103] Haeckel,
1881, Prodromus, p. 450.
Definition.—Cubosphærida with one simple lattice-sphere
and six simple spines in pairs different; the two opposite spines of each pair equal, the three
pairs unequal.
The genus Hexastylidium differs from its probable ancestral form,
Hexastylus, by the unequal growth of the six simple spines; the two spines of each pair
reaching the same dimensions, whilst the three pairs are different. They correspond therefore to
the three axes of a rhombic crystal.
1. Hexastylidium rhomboides, n. sp.
Shell thin walled, smooth, with irregular, polygonal pores and very thin bars. Its form is not,
as commonly, a sphere, but a rhombic octahedron. The radial proportion of the three unequal {179}dimensive axes = 1 : 2 : 4. The
radial proportion of the three pairs of spines = 1 : 3 : 8. Spines thin
cylindrical, at the base angular.
Dimensions.—Diameter of the major shell axis 0.2, middle 0.1, minor 0.05; length
of the major spines 0.3, middle 0.12, minor 0.04.
Habitat.—Indian Ocean, Madagascar, Rabbe, surface.
2. Hexastylidium spirale, n. sp.
Shell thick walled, spherical, thorny, with irregular, roundish pores, three to five times as
broad as the bars; six spines very stout, prismatic (as broad as one large pore), with three
ring-like, spirally convoluted edges. Two opposite major spines of extraordinary length, ten to
twelve times as long as the shell diameter, the two middle opposite spines being about as long as
the latter, the two minor scarcely one-third as long. (Similar to Hexastylus spiralis, Pl.
21, fig. 7,
but distinguished by the very unequal length of the spines.)
Dimensions.—Diameter of the shell 0.12; length of the major spines 1 to 1.5 mm.,
middle 0.15, minor 0.04.
Habitat.—Central Pacific, Station 266, surface.
Subfamily Hexalonchida,[104] Haeckel, 1881, Prodromus, pp. 449, 451.
Definition.—Cubosphærida with two concentric
spherical lattice-shells.
Genus 75. Hexalonche,[105] Haeckel, 1881,
Prodromus, p. 451.
Definition.—Cubosphærida with two concentric
lattice-spheres and six simple spines of equal size.
The genus Hexalonche is the most simple form, and probably the common
ancestral form, of all Hexalonchida, or those Cubosphærida which possess two concentric latticed
spheres, connected by six radial beams. Commonly one shell is intracapsular (medullary shell) and
the other extracapsular (cortical shell); but sometimes also both shells are extracapsular, and
these forms may perhaps be better separated as a peculiar genus Hexadilemma. In
Hexalonche all six simple spines are of equal size, and opposite by pairs in three equal
dimensive axes, corresponding to the three equal axes of a tesseral crystal. It can be derived
from Hexastylus by duplication of the lattice-shell.
Subgenus 1. Hexalonchara, Haeckel.
Definition.—Pores of the cortical shell regular or subregular, of
nearly equal size and similar form; surface smooth, without radial by-spines (other than the six
main spines).
{180}
1. Hexalonche phænaxonia, n. sp.
Cortical shell thin walled, smooth; its pores regular, hexagonal, six to eight times as broad
as the bars; eight to ten to twelve on the radius. Medullary shell one-third as broad, with
regular, hexagonal pores of half size. Six spines triangular pyramidal, as long as the radius of
the shell, at the base as broad as one pore. (Differs from Hexastylus phænaxonius, Pl. 21, fig. 3,
only in the medullary shell and the six inner radial beams, connecting it with the cortical
shell.)
Dimensions.—Diameter of the outer shell 0.15, pores 0.01 to 0.015, bars 0.015 to
0.02; inner shell 0.05; length of the spines 0.08, basal breadth 0.01.
Habitat.—Central Pacific, Stations 266 to 271, depth 2425 to 2925
fathoms.
2. Hexalonche rosetta, n. sp. (Pl. 25,
figs. 3, 3a, 3b).
Cortical shell thick walled, smooth, two and a half time as broad as the medullary shell. Pores
of the latter (fig. 3a) regular circular; eight to ten on the half meridian, about as broad
as the bars between them. Pores of the outer shell regular, hexagonal, remarkable for a very
peculiar form and arrangement. In the transverse section of the shell (fig. 3b) they appear
as narrow, hexagonal prismatic canals, twice as high as broad, and four to five times as broad as
the thin elevated bars between them. Every seven meshes form together a larger, regular hexagon
(six pores surrounding one central pore). The periphery of these larger, rosette-like hexagons
projects more strongly from the surface than the walls between the smaller hexagons. On the half
meridian of the shell may be counted six to seven larger and eighteen to twenty smaller hexagons.
The six radial beams between the two shells are thin, three-sided prismatic, not broader than the
bars of the network, the prominent prolongations of which form six very strong spines of peculiar
club-like shape (fig. 3), as long as the radius of the outer shell. The three wings of the club
are lower in the inner, higher in the outer half; the broadest part of the spine (at the base and
in the outer third) is as broad as a hexagonal rosette (equal to three meshes of the outer shell);
its outer apex is pyramidal.
Dimensions.—Diameter of the outer shell 0.13, pores 0.008 to 0.01, bars 0.02;
inner shell 0.05; length of the spines 0.07, distal breadth 0.03.
Habitat.—Tropical West Pacific, Station 225, depth 4475 fathoms.
3. Hexalonche favosa, n. sp.
Cortical shell thick walled, smooth, four times as broad as the medullary shell. Pores of the
former regular circular, hexagonally framed, deep funnel-shaped, of the same breadth as the bars;
six to eight on the radius. Six spines triangular-pyramidal, as long as the radius, at the base
twice as broad as one pore. (Differs from the similar Hexastylus favosus mainly in the
possession of a medullary shell.)
Dimensions.—Diameter of the outer shell 0.16, pores and bars 0.012; inner shell
0.04; length of the spines 0.08, basal breadth 0.025.
Habitat.—Equatorial Atlantic, Station 348, depth 2450 fathoms; also
fossil in Barbados.
{181}
4. Hexalonche octahedra, n. sp. (Pl. 22,
figs. 8, 8a).
Cortical shell a regular octahedron, thin walled, with twelve more or less rounded edges
(between the bases of the spines), and with smooth surface, three times as broad as the spherical
medullary shell. Pores of the former regular circular (three times as large as those of the
latter), four times as broad as the bars; five to seven on the radius. Six spines three-sided
prismatic, with thickened base and cuspidated end, somewhat longer than the radius, and once to
twice as broad as one pore.
Dimensions.—Diameter of the outer shell 0.11, pores 0.01, bars 0.0025; inner shell
0.04; length of the spines 0.08, bars 0.01.
Habitat.—Central Pacific, Station 272, depth 2600 fathoms.
5. Hexalonche conicornis, n. sp. (Pl. 22, fig.
2).
Cortical shell thick walled, smooth, three times as broad as the medullary shell. Pores of both
spheres regular circular, three to four times as broad as the bars; those of the thick walled
outer shell six to seven on the radius, twice as large as those of the thin walled inner shell;
six radial beams between the two spheres, very thin, cylindrical; six spines short, conical,
scarcely as long as the radius of the outer shell, at the base twice as broad as one pore.
Dimensions.—Diameter of the outer shell 0.12 to 0.14, pores 0.01, bars 0.003;
inner shell 0.04 to 0.05; length of the spines 0.04 to 0.06, basal breadth 0.02.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
6. Hexalonche curvicornis, n. sp. (Pl. 25, fig.
4).
Cortical shell thick walled, smooth, not much larger than the medullary shell (=
4 : 3). Pores of both spheres regular circular, nearly of the same size, three times as
broad as the bars between them; seven to nine on the radius. Six spines three-sided prismatic,
inside and outside of the exterior shell of equal thickness, twice as broad as one pore, longer
than the diameter of the outer shell, and in a singular manner curved like an ox horn; the three
edges of each horn somewhat spirally twisted.
Dimensions.—Diameter of the outer shell 0.16, pores 0.01, bars 0.003; inner shell
0.12; length of the spines 0.2, breadth 0.02.
Habitat.—Central Pacific, Station 266, depth 2750 fathoms.
7. Hexalonche brevicornis, n. sp.
Cortical shell thin walled, smooth, three times as broad as the medullary shell. Pores of the
former regular circular, double-edged, six to eight times as broad as the thin bars, five to six
on the radius. Six spines triangular pyramidal, as broad as one pore and only twice as long.
Dimensions.—Diameter of the outer shell 0.14, pores 0.02, bars 0.003; inner shell
0.045; length of the spines 0.05, basal breadth 0.02.
Habitat.—North Pacific, Station 253, depth 3125 fathoms.
{182}
8. Hexalonche grandis, n. sp.
Cortical shell thick walled, smooth, four times as broad as the medullary shell. Pores of the
outer shell regular circular, four times as broad as the bars; thirteen to fifteen on the radius.
Six spines conical, scarcely half as long as the radius, at the base as broad as one pore.
Dimensions.—Diameter of the outer shell 0.2 to 0.32, pores 0.01 to 0.02, bars
0.003 to 0.005; inner shell 0.06 to 0.08; length of the spines 0.06, breadth 0.02.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
Subgenus 2. Hexalonchetta, Haeckel.
Definition.—Pores of the cortical shell regular or subregular, of
nearly equal size and similar form; surface covered with numerous conical or bristle-shaped radial
by-spines.
9. Hexalonche amphisiphon, n. sp. (Pl. 25, figs. 2,
2a, 2b).
Cortical shell thin walled, armed with very numerous bristle-shaped, radial by-spines, a
quarter to a half as long as the six main spines. Pores regular hexagonal, twelve to fourteen on
the radius, with very thin bars, prolonged on the outer as well as the inner surface into a short
truncated conical tube (Pl. 25,
fig. 2b). Medullary shell octahedral, with irregular polygonal meshes and very thin bars
between them (fig. 2a), connected with the outer (six to eight times larger) shell by six
very thin radial beams. These are prolonged outside into six strong pyramidal spines, nearly as
long as the diameter of the outer shell, with sharp straight edges, at the base twice as broad as
one pore.
Dimensions.—Diameter of the outer shell 0.15, pores 0.01, bars 0.001; inner shell
0.02; length of the spines 0.12, basal breadth 0.02.
Habitat.—Central Pacific, Station 271, surface.
10. Hexalonche anaximandri, n. sp. (Pl. 22, fig.
5).
Cortical shell thin walled, covered with short conical spines (smaller than the pores), three
times as broad as the medullary shell. Pores of the outer shell subregular hexagonal, four times
as broad as the bars; five to seven on the radius. Inner shell with regular hexagonal pores of
half the size, connected with the outer by six strong, three-sided prismatic beams, which are
prolonged outside into six very stout pyramidal spines, with three prominent edges, longer than
the radius and twice as broad as one pore.
Dimensions.—Diameter of the outer shell 0.12, pores 0.01, bars 0.0025; inner shell
0.04; length of the spines 0.07, basal breadth 0.02.
Habitat.—Central Pacific, Station 272, depth 2600 fathoms.
{183}
11. Hexalonche octocolpa, n. sp. (Pl. 22, figs. 6,
6a).
Cortical shell thin walled, nearly octahedral, with eight hemispherical or bosom-shaped
vaultings, corresponding to the eight faces of a regular octahedron, the three axes of which are
indicated by the six spines. Surface covered with short bristle-shaped spines. Pores regular
hexagonal, ten to fifteen times as broad as the thin bars; seven to nine on the radius. Medullary
shell (fig. 6a) spherical, with regular circular pores, one-third as broad as the cortical
shell, and connected with it by six strong triangular radial beams, which are prolonged outside
into pyramidal spines, one-third as long as the shell diameter.
Dimensions.—Diameter of the outer shell 0.2, pores 0.03, bars 0.002; inner shell
0.06; length of the spines 0.06, breadth 0.02.
Habitat.—Central Pacific, Station 274, depth 2750 fathoms.
12. Hexalonche cristata, n. sp.
Cortical shell thick walled, four times as broad as the medullary shell, and covered with short
conical spines arising from the nodal points of elevated crests which form regular hexagonal
frames around the funnel-shaped circular pores. These are two to three times as broad as the
crested bars; eight to ten on the radius. Six spines three-sided pyramidal, with strong prominent
edges, about as long as the radius of the shell.
Dimensions.—Diameter of the outer shell 0.16 to 0.2, pores 0.01 to 0.015, bars
0.003 to 0.005; inner shell 0.04 to 0.05; length of the spines 0.08 to 0.12, basal breadth 0.01 to
0.03.
Habitat.—Central Pacific, Station 266 to 274, depth 2350 to 2925
fathoms.
13. Hexalonche serrata, n. sp.
Cortical shell thick walled, four times as broad as the medullary shell, with spiny surface.
Pores regular circular, twice as broad as the bars, five to seven on the radius, funnel-shaped,
separated by hexagonal frames, the sharp crests of which are serrated; at the nodal-points longer
bristle-shaped by-spines. Six main spines triangular prismatic, longer than the diameter of the
shell, twice as broad as one pore.
Dimensions.—Diameter of the outer shell 0.15, pores 0.01, bars 0.005; inner shell
0.04; length of the spines 0.2, breadth 0.02.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
14. Hexalonche anaximenis, n. sp. (Pl. 25, fig.
5).
Cortical shell thin walled, twice as broad as the medullary shell, and covered with short
conical by-spines. Pores regular circular, twice as broad as the bars; five to six on the radius.
Circular pores of the medullary shell one-third as broad. Radial main spines conical, nearly as
long as the radius of the shell.
Dimensions.—Diameter of the outer shell 0.1, pores 0.01, bars 0.005; inner shell
0.05; length of the spines 0.04, basal breadth 0.015.
Habitat.—Central Pacific, Station 272, depth 2600 fathoms.
{184}
15. Hexalonche aspera, n. sp.
Cortical shell thin walled, three times as broad as the medullary shell, and covered with short
conical spines. Pores regular circular, four to six times as broad as the bars; eight to ten on
the radius. Radial spines conical, about as long as the diameter of the shell.
Dimensions.—Diameter of the outer shell 0.12, pores 0.08, bars 0.0015; inner shell
0.04; length of the spines 0.1, basal breadth 0.02.
Habitat.—South Pacific, Station 300, depth 1375 fathoms.
16. Hexalonche castanella, n. sp.
Cortical shell thick walled, five times as broad as the medullary shell, and covered with
numerous short conical spines. Pores regular circular, twice as broad as the bars; ten to twelve
on the radius. Radial spines triangular pyramidal, about as long as the radius of the shell.
Dimensions.—Diameter of the outer shell 0.22, pores 0.01, bars 0.005; inner shell
0.045; length of the spines 0.12, basal breadth 0.02.
Habitat.—North Atlantic, Station 353, depth 3125 fathoms.
Subgenus 3. Hexalonchilla, Haeckel.
Definition.—Pores of the cortical shell irregular, of unequal size or
form; surface smooth, without radial by-spines (other than the six main spines).
17. Hexalonche hexacantha, Haeckel.
Haliomma hexacanthum, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 35,
Taf. iv. fig. 5.
Haliomma hexacanthum, Haeckel, 1862, Monogr. d. Radiol., p. 430.
Cortical shell thin walled, smooth, with irregular polygonal pores (commonly hexagonal or
pentagonal), two to three times as broad as the bars; eight to ten on the radius. Medullary shell
one quarter as broad, connected with the former by six thin radial beams which are prolonged
outside into six triangular pyramidal spines (not quadrangular, as Müller describes), longer than
the radius of the shell.
Dimensions.—Diameter of the outer shell 0.2, pores 0.01 to 0.015, bars 0.005;
inner shell 0.05; length of the spines 0.15, basal breadth 0.015.
Habitat.—Mediterranean (Messina, Nice, Genoa).
18. Hexalonche geometrica, n. sp.
Cortical shell thick walled, smooth, with very peculiar geometrical formation of its network,
the pores of which are of very different size and form, but highly regular disposition. Each of
the six {185}triangular spines is surrounded at the base
by three small roundish pores (between the three wings of its base), and further by a coronal of
six very large pores, three of which are pentagonal, and the other three (alternating) heptagonal.
The six basal coronals are separated by irregular smaller pores. Medullary shell thin walled, with
regular hexagonal pores (three on the radius) and thin bars, connected with the outer shell by six
thin triangular prismatic radial beams, which are prolonged outside into short pyramidal spines
(half as long as the radius).
Dimensions.—Diameter of the outer shell 0.2, thirty-six larger pores of the
coronals 0.03 to 0.04, smaller pores between them 0.01 to 0.02, bars 0.005; inner shell 0.05 (with
pores of 0.008); length of the spines 0.05, basal breadth 0.03.
Habitat.—South Atlantic, off Buenos Ayres, Station 323, depth 1900
fathoms.
19. Hexalonche pythagoræa, n. sp. (Pl. 22, fig.
1).
Cortical shell thick walled, smooth, three times as broad as the medullary shell. Pores
irregular roundish, twice to eight times as broad as the bars, of very different size; usually in
the space between every three spines are three or six larger pores, separated by numerous smaller
pores (often the disposition of the larger pores is much more regular than in the figured
specimen). Medullary shell with regular circular pores, connected with the outer by six very thin
radial beams, which are prolonged outside into six short triangular pyramidal spines about as long
and broad as the half radius.
Dimensions.—Diameter of the outer shell 0.12, pores 0.008 to 0.03, bars 0.004;
inner shell 0.04; length of the spines 0.04, basal breadth 0.03.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
20. Hexalonche aristarchi, n. sp. (Pl. 22, fig.
3).
Cortical shell thin walled, smooth, four times as broad as the medullary shell. Pores irregular
polygonal, of very variable size and form, twice to six times as broad as the bars; six to ten on
the radius. Medullary shell with regular hexagonal pores, connected with the outer by six very
thin radial beams, which are prolonged outside into six triangular pyramidal spines, nearly as
long as the radius.
Dimensions.—Diameter of the outer shell 0.1, pores 0.005 to 0.02, bars 0.003;
inner shell 0.025; length of the spines 0.05, basal breadth 0.01.
Habitat.—North Pacific, Station 241, depth 2300 fathoms.
21. Hexalonche ekphantæa, n. sp.
Cortical shell thin walled, smooth, four times as broad as the medullary shell. Pores irregular
roundish, of very variable size and form, twice to six times as broad as the bars, ten to fifteen
on the radius. Six spines cylindro-conical, longer than the diameter of the shell.
Dimensions.—Diameter of the outer shell 0.2, pores 0.004 to 0.012, bars 0.002;
inner shell 0.05; length of the spine 0.3, basal breadth 0.01.
Habitat.—Indian Ocean, Sunda Strait, surface, Rabbe.
{186}
Subgenus 4. Hexalonchusa, Haeckel.
Definition.—Pores of the cortical shell irregular, of different size
or form; surface covered with numerous conical or bristle-shaped, radial by-spines.
22. Hexalonche philosophica, n. sp. (Pl. 22, fig.
4).
Cortical shell thin walled, covered with short conical spines, three times as broad as the
medullary shell. Pores irregular polygonal, or more roundish, twice to six times as broad as the
bars; six to eight on the radius. Inner shell of the same structure, pores three times smaller,
connected with the outer by six very thin radial beams, which are prolonged outside into six
strong, triangular pyramidal spines, as long as the radius. (Similar to Hexalonche
anaximandri, Pl. 22,
fig. 5, but different in the irregular network and the shorter by-spines.)
Dimensions.—Diameter of the outer shell 0.11, pores 0.005 to 0.015, bars 0.0025;
inner shell 0.04; length of the spines 0.06, breadth 0.02.
Habitat.—North Atlantic, Færöe Channel, John Murray; Iceland, Krabbe,
surface.
23. Hexalonche seleuci, n. sp.
Cortical shell thick walled, covered with numerous short, bristle-shaped spines. Pores of very
different size and form, and of a peculiar, subregular disposition, similar to those of
Hexalonche geometrica (though in this case more regular). Each of the six triangular main
spines (which reach nearly the length of the radius) is surrounded at the base by three small
roundish pores, and these are further supplemented by a coronal of six very large polygonal pores;
the six coronals are separated by irregular rows of smaller pores. Inner shell equal to one-third
of the outer. Six main spines pyramidal, as long as the radius.
Dimensions.—Diameter of the outer shell 0.16, larger pores 0.03 to 0.04, smaller
pores 0.01 to 0.02, bars 0.05 to 0.01; inner shell 0.05; length of the spines 0.08, basal breadth
0.03.
Habitat.—North Atlantic, Station 354, surface.
24. Hexalonche sexaculeata, Haeckel.
Haliomma sexaculeatum, Stöhr, 1880, Palæontogr., vol. xxvi. p. 87, Taf. i.
fig. 8.
Cortical shell thick walled, twice as broad as the medullary shell, and covered with short
conical spines. Pores irregular polygonal (mostly hexagonal), twice to three times as broad as the
bars; six to eight on the radius. Six spines triangular-pyramidal (not quadrangular), somewhat
longer than the radius of the shell.
Dimensions.—Diameter of the outer shell 0.11, pores 0.06 to 0.08, bars 0.03; inner
shell 0.06; length of the spines 0.07, basal breadth 0.015.
Habitat.—Fossil in Tertiary rocks of Sicily, Grotte, Caltanisetta.
{187}
25. Hexalonche heracliti, n. sp. (Pl. 22, fig.
7).
Cortical shell thick walled, covered with bunches of small spines, each bunch at the nodule
point between every three pores composed of four to eight conical spinules. Pores irregular
roundish, twice to four times as broad as the bars; four to six on the radius. Inner shell equal
to one-third of the outer, connected with it by six thin radial beams, which are prolonged outside
into six strong triangular spines about as long as the radius.
Dimensions.—Diameter of the outer shell 0.15, pores 0.015 to 0.03, bars 0.08;
inner shell 0.05; length of the spines 0.06, basal breadth 0.02.
Habitat.—Indian Ocean, Madagascar, Rabbe, surface.
26. Hexalonche xenophanis, n. sp.
Cortical shell thin walled, covered with numerous short conical spines, four times as broad as
the medullary shell. Pores irregular circular, twice to eight times as broad as the bars; five to
seven on the radius. Six spines conical or more cylindrical, as long as the diameter of the shell
or longer.
Dimensions.—Diameter of the outer shell 0.16 to 0.24, pores 0.01 to 0.04, bars
0.005; inner shell 0.04 to 0.06; length of the spines 0.2 to 0.3, breadth 0.02.
Habitat.—Central Pacific, Stations 266 to 272, surface.
27. Hexalonche setosa, n. sp.
Cortical shell thin walled, covered with numerous bristle-shaped spines, half as long as the
six main spines. Pores irregular roundish, twice to five times as broad as the bars; ten to twelve
on the radius. Inner shell equal to one-fifth of the outer. Six spines conical, as long as the
radius.
Dimensions.—Diameter of the outer shell 0.25, pores 0.008 to 0.02, bars 0.004;
inner shell 0.05; length of the spines 0.12, basal breadth 0.03.
Habitat.—South Pacific, Station 291, surface.
28. Hexalonche hystricina, n. sp. (Pl. 25, fig.
6).
Cortical shell thick walled, covered with numerous bristle-shaped spines, half as long as the
six main spines. Pores irregular roundish, twice to four times as broad as the bars; five to seven
on the radius. Medullary shell with very small circular pores, one-fourth of the cortical shell,
connected with it by six thin prismatic radial beams, which are prolonged outside into six strong,
short, three-sided pyramidal, cuspidated spines, only half as long as the radius of the outer
shell.
Dimensions.—Diameter of the outer shell 0.2, pores 0.015 to 0.03, bars 0.008;
inner shell 0.05; length of the spines 0.05, basal breadth 0.02.
Habitat.—Indian Ocean, Sunda Strait, surface, Rabbe.
{188}
Genus 76. Hexancistra,[106] Haeckel,
1881, Prodromus, p. 451.
Definition.—Cubosphærida with two concentric
lattice-spheres and six branched spines of equal size.
The genus Hexancistra differs from its ancestral form, Hexalonche, in
the ramification of the six radial spines. These are very different in the two subgenera; in
Hexancora each spine bears only three simple lateral branches, while in Hexapitys
there are three rows of verticillate lateral branches on each spine.
Subgenus 1. Hexancora, Haeckel.
Definition.—Each radial spine with three simple lateral branches only
(one branch from each edge of the triangular spine).
1. Hexancistra tricuspis, n. sp. (Pl. 22, fig.
9).
Cortical shell thin walled, covered with short conical by-spines, three times as broad as the
medullary shell; its pores regular circular, three times as broad as the bars; ten to twelve on
the radius. Pores of the medullary shell half as large, also regular circular. The two shells
connected by six thin prismatic radial beams, which are prolonged outside into six very stout main
spines, three-sided prismatic, as long as the shell diameter, with three thin wing-like edges.
Each edge at the distal end prolonged into a strong curved lateral branch.
Dimensions.—Diameter of the outer shell 0.13, inner 0.4; cortical pores 0.01, bars
0.003; medullary pores 0.005, bars 0.003; length of the six spines 0.13, breadth 0.02.
Habitat.—Central Pacific, Station 266, depth 2750 fathoms.
2. Hexancistra ancorata, n. sp.
Cortical shell thick walled, smooth, twice as broad as the medullary shell; its pores regular
circular, twice as broad as the bars; six to eight on the radius. Six radial spines, three-sided
prismatic, as long as the shell radius, with three recurved lateral branches at the distal end
like the three teeth of an anchor.
Dimensions.—Diameter of the outer shell 0.1, inner 0.05; cortical pores 0.006,
bars 0.003; length of the six spines 0.05, breadth 0.02.
Habitat.—South Pacific, Station 296, depth 1825 fathoms.
3. Hexancistra triserrata, n. sp. (Pl. 22, fig.
10).
Cortical shell thin walled, thorny, twice to three times as broad as the medullary shell; the
two shells connected by six strong triangular beams. Inner shell spherical, with very small,
regular, circular {189}pores; ten to twelve on the half
meridian. Bars as broad as the pores. Outer shell regularly octahedral, with eight triangular
perfectly regular even faces, separated by eight prominent edges; the meshes circular, regular,
about twelve to fourteen on the half meridian, two to three times as broad as the bars between
them. Six strong radial spines, about as long as the diameter of the outer shell, three-sided
prismatic, cuspidate; their three edges prominent, serrate, spirally twisted. Each spine bears
about its middle three strong flattened lateral branches, nearly perpendicular to it, not
serrated, and slightly curved.
Dimensions.—Diameter of the outer shell 0.13, inner 0.045; pores of the former
0.01, of the latter 0.004; length of the spine 0.12, breadth 0.014.
Habitat.—Central Pacific, Station 274, depth 2750 fathoms.
4. Hexancistra quadricuspis, n. sp. (Pl. 22, fig.
11).
Cortical shell thin walled, covered with short bristle-shaped by-spines, three and a half times
as broad as the medullary shell. Pores of the latter regular circular, small; pores of the former
three to six times as large, very irregular, roundish, double-edged. Six radial spines,
three-sided prismatic, about as long as the shell diameter, with three wing-like, slightly twisted
edges, which are prolonged towards the distal end into three curved horn-shaped branches.
Dimensions.—Diameter of the outer shell 0.17, inner 0.05; cortical pores 0.01 to
0.03, bars 0.005; medullary pores 0.003, bars 0.001; length of the spines 0.15, breadth 0.013.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
Subgenus 2. Hexapitys, Haeckel, 1881, Prodromus, p. 451.
Definition.—Each radial spine with three rows of verticillate lateral
branches (a row arising from each edge of the spine).
5. Hexancistra mirabilis, n. sp. (Pl. 23, fig.
3).
Hexapitys mirabilis, Haeckel, 1881, Prodromus, p. 451.
Cortical shell very thin walled, three times as broad as the medullary shell. Inner shell
spherical, with very delicate, subregular hexagonal meshes; seven to eight on the half diameter.
Outer shell octahedral, with irregular polygonal meshes of very different size; on the surface
numerous thin accessory radial spines, equal in length to its radius. Six main spines, extremely
long and stout, many times longer than the diameter of the outer shell, nearly as broad as the
radius of the inner shell, three-sided prismatic, with sharp, prominent, spirally twisted edges;
on every edge a great number of thin lateral branches, arranged perpendicularly to it, as long as
the diameter of the outer shell, and pinnated by ten to twenty pairs of delicate secondary
spinules, biserial and perpendicular to the primary branches. (In the figured specimen the
spherical central capsule, between both shells, was well preserved; its nucleus nearly filled the
medullary shell. The thick jelly-veil around it was radially striped and octahedral.)
{190}
Dimensions.—Diameter of the outer shell 0.13, of the inner 0.05; length of the
spines 0.5 to 0.8 or more, breadth 0.02.
Habitat.—Central Pacific, Station 271, surface.
Genus 77. Hexaloncharium,[107] n.
gen.
Definition.—Cubosphærida with two concentric
lattice-spheres and six simple spines of different sizes; one opposite pair larger than the two
others.
The genus Hexaloncharium exhibits the same relation to its ancestral form,
Hexalonche, that Hexastylarium bears to Hexastylus. Two opposite spines of
one pair are larger than the four others, and correspond to the three axes of a quadratic
crystal.
1. Hexaloncharium octahedrum, n. sp.
Cortical shell smooth, three to four times as broad as the spherical medullary shell, each
having regular circular pores, twice to three times as broad as the bars. Form of the outer shell
not a sphere, but a regular octahedron, with eight congruent, equilateral triangular even faces.
Two opposite spines twice as long as the shell diameter, whilst the four others are scarcely equal
to it. Basal breadth of all six the same (three times as large as one pore); form, three-sided
prismatic, with cuspidate distal end. (Similar to Hexalonche octahedra, Pl. 22, fig. 8, but
distinct in the unequal length of the spines.)
Dimensions.—Diameter of the outer shell 0.16, inner 0.045; length of the major
spine 0.3, minor 0.12, basal breadth 0.03.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
2. Hexaloncharium philosophicum, n. sp.
Cortical shell spherical, covered with short conical by-spines, twice as broad as the medullary
shell; both with regular circular pores three to four times as broad as the bars. Two major spines
cylindrical, with conical apex, three times as long as the four others, which are conical and
about as long as the shell radius; basal breadth of all six the same (equal to one pore). Similar
to Hexalonche anaximenis (Pl. 25,
fig. 5), but distinct in the enormous prolongation of two opposite spines.
Dimensions.—Diameter of the outer shell 0.12, inner 0.06; length of the two major
spines 0.2, of the four minor 0.06, basal breadth 0.02.
Habitat.—Central Pacific, Station 267, depth 2700 fathoms.
{191}
3. Hexaloncharium hystricinum, n. sp.
Cortical shell spherical, three times as broad as the medullary shell, and densely covered with
oblique bristle-shaped by-spines. Pores irregular roundish. Two opposite major spines three-sided
prismatic, longer than the shell diameter; four minor spines pyramidal, scarcely half as long as
the shell radius. (Somewhat similar to Hexalonche hystricina, but distinct in the
prolongation of two major spines.)
Dimensions.—Diameter of the outer shell 0.15, inner 0.05; length of the two major
spines 0.2, four minor 0.03, basal breadth 0.02.
Habitat.—South Pacific, Station 288, surface.
Genus 78. Hexalonchidium,[108] Haeckel,
1881, Prodromus, p. 451.
Definition.—Cubosphærida with two concentric
lattice-spheres and six simple spines of different sizes in pairs; the two opposite spines of each
pair equal, the three pairs unequal.
The genus Hexalonchidium exhibits the same relation to Hexalonche
that Hexastylidium bears to Hexastylus; the growth of the three spine-pairs is
different, whilst both spines of each pair are equal; they correspond therefore to the three axes
of a rhombic crystal.
1. Hexalonchidium axonometrum, n. sp.
Cortical shell thin walled, covered with short bristle-shaped by-spines, twice as broad as the
medullary shell; both with regular hexagonal meshes, twice to three times as broad as the bars
(inner meshes half as broad as the outer). All three spine-pairs three-sided prismatic, of very
different length but of equal breadth (equal to three pores). Major spine-pair twice as long as
the shell diameter; middle pair about equal to the latter, minor scarcely half as long. (Similar
to Hexalonche anaximandri, Pl. 22,
fig. 5, but distinct in the different length of the spines.)
Dimensions.—Diameter of the outer shell 0.12, inner 0.06; length of the major
spines 0.25, middle 0.1, minor 0.04.
Habitat.—Indian Ocean, Madagascar, surface, Rabbe.
Subfamily Hexacontida,[109] Haeckel, 1881, Prodromus, pp. 449, 452.
Definition.—Cubosphærida with three
concentric, spherical, or octahedral lattice-shells.
{192}
Genus 79. Hexacontium, Haeckel,[110]
1881, Prodromus, p. 452.
Definition.—Shell with three concentric lattice-spheres and six simple spines of
equal size.
The genus Hexacontium, the ancestral form of the Hexacontida, is probably
derived from Hexalonche by duplication of the medullary shell. As in the latter, all six
spines are of equal size, opposite in pairs in the three dimensive axes, and correspond therefore
to the three equal axes of a tesseral crystal.
Subgenus 1. Hexacontanna, Haeckel.
Definition.—Pores of the cortical shell regular or subregular, of
nearly equal size and similar form; surface smooth, without radial spines or papillæ (other than
the six main spines).
1. Hexacontium phænaxonium, n. sp.
Cortical shell thin walled, smooth; its pores regular hexagonal, six to nine times as broad as
the bars; twelve to sixteen on the radius. Radial proportion of the three spheres =
1 : 2 : 4. Both medullary shells of the same structure as the cortical shell,
only with smaller pores. The three spheres connected by six thin radial beams, which are prolonged
on the outside into six strong triangular pyramidal spines, as long as the radius of the cortical
shell, and, at the base, as broad as one of its pores. (Differs from Hexastylus phænaxonius
and from Hexalonche phænaxonia, Pl. 21,
fig. 3, in the larger size and the triple shell.)
Dimensions.—Diameter of the outer sphere 0.2, middle 0.1, inner 0.05; cortical
pores 0.008, bars 0.0012; length of the spines 0.1.
Habitat.—Central Pacific, Station 270, surface.
2. Hexacontium axotrias, n. sp. (Pl. 24,
fig. 3).
Cortical shell thin walled, smooth; its pores regular circular, five to six times as broad as
the bars; eleven to thirteen on the radius. Radial proportion of the three spheres =
1 : 3 : 8. Outer medullary shell with hexagonal frames around the regular
circular pores (five to six on the radius). Inner medullary shell with simple small circular pores
(three on the radius). The two outer shells connected by six triangular prismatic beams, which are
prolonged outside to the length of the cortical radius or more.
Dimensions.—Diameter of the outer sphere 0.2, middle 0.07, inner 0.025; cortical
pores 0.016, bars 0.003; length of the spines 0.1 to 0.015, basal breadth 0.02.
Habitat.—North Pacific, Station 253, surface.
3. Hexacontium hexactis, Haeckel.
Actinomma hexactis, Stöhr, 1880, Palæontogr., vol. xxvi. p. 91, Taf. ii.
fig. 7.
Cortical shell thick walled, smooth, or a little rough. Pores regular circular, of the same
breadth as the bars; five to seven on the radius. Radial proportion of the three spheres {193}= 1 : 3 : 6. Both medullary shells of
the same structure, but with smaller pores. Six spines triangular pyramidal, nearly as long as the
diameter of the outer shell, three times as broad at the base as one pore.
Dimensions.—Diameter of the outer sphere 0.1, middle 0.05, inner 0.016; cortical
pores and bars 0.008; length of the spines 0.08, basal breadth 0.025.
Habitat.—Fossil in Tertiary rocks of Sicily, Grotte, Stöhr.
4. Hexacontium lævigatum, n. sp. (Pl. 24, fig.
6).
Cortical shell thick walled, quite smooth. Pores regular circular, with double margins, eight
to ten on the radius, of the same breadth as the smooth bars. Radial proportion of the three
spheres = 1 : 2 : 6. All three spheres connected by six very thin radial
beams, which are prolonged outside into six short, stout, triangular, pyramidal spines, half as
long as the radius of the outer shell.
Dimensions.—Diameter of the outer shell 0.13, middle 0.04, inner 0.02; cortical
pores and bars 0.008; length of the spines 0.04, basal breadth 0.02.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
5. Hexacontium triplosphærium, n. sp.
Cortical shell thick walled, smooth. Pores regular circular, three times as broad as the bars;
ten to twelve on the radius. Radial proportion of the three spheres =
1 : 3 : 10. Six spines conical, about as long as the radius of the cortical
shell.
Dimensions.—Diameter of the outer shell 0.16, middle 0.05, inner 0.016; cortical
pores 0.012, bars 0.004; length of the spines 0.07, basal breadth 0.02.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
6. Hexacontium octahedrum, n. sp.
Cortical shell a regular octahedron, thin walled, with twelve more or less rounded edges
(between the spine-bases) and with smooth surface; its pores regular circular, five to seven on
the radius, four times as broad as the bars. Radial proportion of the three shells =
1 : 3 : 9. Both medullary shells spherical, with very small circular pores.
Six inner bars very thin; six outer spines (their prolongations) triangular pyramidal, as long as
the radius of the outer shell. (Differs from Hexalonche octahedra, Pl. 22, fig. 8,
almost solely in the duplication of the medullary shell.)
Dimensions.—Diameter of the outer shell 0.12, middle 0.04, inner 0.013.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
7. Hexacontium circumtextum, n. sp. (Pl. 25, figs. 7,
7a).
Cortical shell double, enclosing a simple medullary shell. Radial proportion of the three
spheres = 3 : 10 : 12. Inner cortical shell very thick walled, with regular
circular, double-edged pores, four times as broad as the bars; seven to nine on the radius. From
each nodal-point between {194}every six pores arises a
small, bristle-shaped, radial spine, as long as the diameter of one pore. The distal ends of all
these spines are connected by very delicate tangential threads, and consequently form by their
attachment an outer cortical shell, with regular hexagonal meshes and smooth surface. Six main
spines short, three-sided prismatic, scarcely as long as half the radius of the outer shell.
Dimensions.—Diameter of the outer shell 0.12, middle 0.1, inner 0.03; pores of the
middle shell 0.004, bars 0.001; length of the spines 0.025, basal breadth 0.01.
Habitat.—Central Pacific, Station 274, depth 2750 fathoms.
Subgenus 2. Hexacontella, Haeckel.
Definition.—Pores of the cortical shell regular or subregular, of
nearly equal size and similar form; surface covered with numerous conical or bristle-shaped,
radial spines (other than the six main spines).
8. Hexacontium hexagonale, n. sp.
Cortical shell thin walled, bristly, with very delicate network; its pores regular hexagonal,
twelve to sixteen on the radius, ten to twelve times as broad as the thin thread-like bars. At
each nodal-point of the network (between every three meshes) arises a bristle-shaped, radial
spine, as long as the diameter of one mesh. Six main spines hexagonal, as long as the radius of
the outer shell, as broad at its base as one mesh. Radial proportion of the three spheres =
1 : 2 : 4. Pores of both medullary shells also regular hexagonal, but much
smaller, in the middle shell eleven to twelve, in the inner, six to seven on the half
meridian.
Dimensions.—Diameter of the outer shell 0.12, of the middle 0.06, of the inner
0.03; meshes of the outer shell 0.008; length of the six spines 0.05, basal thickness 0.008.
Habitat.—Indian Ocean, Ceylon, surface, Haeckel.
9. Hexacontium favosum, n. sp. (Pl. 24,
figs. 2, 2a).
Cortical shell very thick walled, spiny, with regular, honeycomb-like network; its meshes
funnel-shaped, with circular inner, hexagonal outer aperture, twice as broad as the bars; five to
seven on the radius. Between every three meshes (at each corner of the hexagon) arises a short
radial thorn, not so long as the thickness of the shell-wall. Six radial spines very short and
stout, three-sided pyramidal, scarcely half so long as the radius of the outer shell. Radial
proportion of the three spheres = 1 : 2 : 5. Pores of both medullary shells
regular, circular, about as broad as the bars, six to eight on the half meridian.
Dimensions.—Diameter of the outer shell 0.1 to 0.12, of the middle 0.04 to 0.05,
of the inner 0.02; meshes of the outer shell 0.008; length of the six spines 0.2 to 0.3, basal
breadth 0.015.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms.
10. Hexacontium sceptrum, n. sp. (Pl. 24, figs. 1,
1a).
Cortical shell thick walled, spiny, with regular, honeycomb-like network; its meshes
funnel-shaped, with circular inner, hexagonal outer aperture, three to four times as broad as the
bars; {195}five to six on the radius. Between every
three meshes arises a short radial thorn, as long as the thickness of the shell-wall. Six radial
spines sceptre-shaped, six-sided, somewhat constricted towards their middle part, strong, as long
as the radius of the outer shell, as broad as one of its meshes. Radial proportion of the three
spheres = 1 : 2 : 6. Pores of the two inner shells regular hexagonal, with
thin bars, six to eight on the half meridian (fig. 1a).
Dimensions.—Diameter of the outer shell 0.13, of the middle 0.04, of the inner
0.02; pores of the outer shell 0.01, of the middle 0.008, of the inner 0.004; length of the six
spines 0.06, basal breadth 0.015.
Habitat.—Equatorial Atlantic, Station 347, surface.
11. Hexacontium prionacanthum, n. sp. (Pl. 24, figs. 7,
7a).
Cortical shell thick walled, thorny; its pores regular circular, with elevated hexagonal frames
six to nine on the radius, twice as broad as the crest-shaped bars. At each nodal-point of the
hexagon arises one short conical papilla or thorn. Radial proportion of the three spheres =
1 : 3 : 10. Pores of both medullary shells much smaller, regular circular. Six
main spines three-sided prismatic, longer than the radius of the outer shell, as broad as one of
its meshes; their three edges serrated, with ten to twelve teeth.
Dimensions.—Diameter of the outer shell 0.15, middle 0.045, inner 0.015; cortical
pores 0.01, bars 0.005; length of the spines 0.1, breadth 0.012.
Habitat.—West Tropical Pacific, Station 225, depth 4475 fathoms.
12. Hexacontium clavigerum, n. sp. (Pl. 23, fig.
5).
Cortical shell very thick walled, thorny; its pores regular circular, hexagonally framed, three
times as broad as the bars, nine to twelve on the radius; between them short conical papillæ or
thorns arising at the nodal-points. Radial proportion of the three spheres =
1 : 2 : 5. Outer medullary shell with regular circular, much smaller pores;
connected with the cortical shell by numerous (twenty to thirty) thin radial beams. Six main
spines very stout, club-shaped, scarcely as long as the radius of the outer shell, three times as
broad as one of its pores, three-sided, with three to four teeth on each edge.
Dimensions.—Diameter of the outer shell 0.2, of the middle 0.08, of the inner
0.04; cortical pores 0.012, bars 0.004; length of the six spines 0.08, breadth 0.03.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
13. Hexacontium floridum, n. sp. (Pl. 24, fig.
4).
Cortical shell papillose, thick walled. Pores regular, six-lobed, four times as broad as the
bars, five to seven on the radius; each pore with six (sometimes also five or seven) concave
indentations or lobules; between the pores at the nodal-points arise short conical papillæ or
thorns. Radial proportion of the three spheres = 1 : 2 : 3. Radial main spines
pommel-shaped, three-sided prismatic, longer than the radius of the outer shell, two to three
times as broad as one pore.
{196}
Dimensions.—Diameter of the outer shell 0.12, middle 0.06, inner 0.03; cortical
pores 0.012, bars 0.003; length of the six spines 0.08, breadth 0.03.
Habitat.—Central Pacific, Station 266, depth 2750 fathoms.
14. Hexacontium hexaconicum, n. sp.
Cortical shell spiny, thick walled. Pores regular circular, of the same breadth as the bars,
ten to twelve on the radius; between them long bristle-shaped by-spines, half as long as the six
main spines. Radial proportion of the three spheres = 1 : 3 : 9. Six main
spines conical, as long as the radius of the outer shell, four times as broad at the base as one
pore.
Dimensions.—Diameter of the outer shell 0.18, middle 0.06, inner 0.02; cortical
pores and bars 0.005; length of the spines 0.1, basal breadth 0.02.
Habitat.—Antarctic Sea, Station 157, depth 1950 fathoms.
15. Hexacontium asteracanthion, Haeckel.
Haliomma asteracanthion, Haeckel, 1860, Monatsber. d. k. preuss. Akad. d. Wiss.
Berlin, p. 816.
Actinomma asteracanthion, Haeckel, 1862, Monogr. d. Radiol., p. 441, Taf.
xxiii. figs. 5, 6.
Cortical shell thin walled, spiny; its pores circular, without hexagonal frame (as a rule very
regular, but in other specimens more or less irregular), often very variable in size, three to six
times as broad as the bars, commonly seven to eight on the radius. Radial proportion of the three
spheres = 1 : 2 : 4, or sometimes 1 : 3 : 8. Pores of the
two inner shells also circular, but much smaller; those of the middle shell about half as broad,
those of the inner shell one-fourth to one-sixth as broad as the pores of the outer shell. Six
radial spines strong, three-sided prismatic, about as long as (or somewhat longer than) the radius
of the outer shell, as broad as one of its large meshes. Accessory spines very numerous and very
thin, bristle-like, usually half as long as the six main spines. (For the variability of this
common species compare my Monograph, p. 442.)
Dimensions.—Diameter of the outer shell 0.1 to 0.12, of the middle 0.04 to 0.06,
of the inner 0.02 to 0.03; cortical pores 0.005 to 0.01; length of the six spines 0.06 to
0.08.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific, surface.
Subgenus 3. Hexacontosa, Haeckel.
Definition.—Pores of the cortical shell irregular, of different size
or form; surface smooth, without radial by-spines or papillæ (other than the six main spines).
16. Hexacontium axophænum, n. sp.
Cortical shell thin walled, smooth. Pores irregular polygonal, three to nine times as broad as
the bars; seven to ten on the radius. Radial proportion of the three spheres =
1 : 3 : 8. Both {197}medullary
shells with smaller, irregular roundish pores. Six spines three-sided pyramidal, longer than the
radius of the outer shell, one to two times as broad as one larger pore.
Dimensions.—Diameter of the outer shell 0.12, middle 0.045, inner 0.015; cortical
pores 0.006 to 0.018, bars 0.002; length of the spines 0.06 to 0.12, basal breadth 0.02 to
0.03.
Habitat.—Central Pacific, Stations 266 to 272, at various depths.
17. Hexacontium polygonale, n. sp.
Cortical shell thick walled, smooth. Pores irregular roundish, with polygonal frames, two to
four times as broad as the bars; eight to twelve on the radius. Radial proportion of the three
spheres = 1 : 3 : 10. Six spines pyramidal, nearly as long as the diameter of
the outer shell, with six to nine prominent edges (along the corner number of the polygonal
meshes).
Dimensions.—Diameter of the outer shell 0.2, middle 0.06, inner 0.02; cortical
pores 0.008 to 0.016, bars 0.004; length of the spines 0.16 to 0.18, basal breadth 0.02.
Habitat.—South Pacific, Station 295, depth 1500 fathoms.
18. Hexacontium antarcticum, n. sp.
Cortical shell thin walled, smooth, with irregular roundish pores, eight to ten on the radius,
two to four times as broad as the bars. Radial proportion of the three spheres =
1 : 3 : 9. Both medullary shells with smaller roundish irregular pores. Six
spines conical, pyramidal at their origin, about as long as the radius, as broad as one larger
mesh.
Dimensions.—Diameter of the outer shell 0.18, middle 0.06, inner 0.02; cortical
pores 0.006 to 0.012, bars 0.003; length of the spines 0.1, basal breadth 0.12.
Habitat.—Antarctic Ocean, Station 157, depth 1950 fathoms.
Subgenus 4. Hexacontura, Haeckel.
Definition.—Pores of the cortical shell irregular, of different size
or form; surface covered with numerous conical papillæ or bristle-shaped, radial by-spines (other
than the six main spines).
19. Hexacontium papillosum, n. sp. (Pl. 24, fig.
5).
Cortical shell thick walled, papillose. Pores irregular roundish, often somewhat lobed, five to
six on the radius, two to four times as broad as the bars. Radial proportion of the three spheres
= 1 : 2.5 : 5. Both medullary shells with subregular circular pores (inner
much smaller). Six radial spines short and stout, pommel or club shaped, with three prominent
wings, about as long as the shell radius, and as broad as the medullary shell.
Dimensions.—Diameter of the outer shell 0.1, middle 0.05, inner 0.02, cortical
pores 0.006 to 0.012, bars 0.003; length of the spines 0.05, bars 0.02.
Habitat.—Central Pacific, Stations 266 to 272, at various depths.
{198}
20. Hexacontium gladiatum, n. sp. (Pl. 25, fig.
8).
Cortical shell very thick walled, spiny, with irregular network; its meshes roundish, two to
five times as broad as the bars, of very different size, ten to fifteen on the half meridian. Bars
between them very strong, three-sided prismatic, armed with a great number of small thorns, and
with large spines at the nodal-points between every three meshes. Radial proportion of the three
spheres = 1 : 3 : 12. Six strong radial main spines three-sided prismatic,
with three prominent, somewhat contorted wings, acute, broader than the diameter of the inner
shell and as long as the diameter of the outer shell. These six main spines are situated in the
same three dimensive axes as the six thin radial beams connecting the two medullary shells. But
the six radial beams which connect the middle with the outer shell alternate with the former and
lie in three other dimensive axes,—a very rare and remarkable disposition.
Dimensions.—Diameter of the outer shell 0.2, middle 0.05, inner 0.016; cortical
pores 0.01 to 0.03, bars 0.006; length of the six spines 0.2, breadth 0.02 to 0.03.
Habitat.—Tropical Atlantic, Station 342, depth 1445 fathoms.
21. Hexacontium setosum, n. sp.
Cortical shell thin walled, with numerous bristle-shaped, simple, radial by-spines of variable
length. Pores irregular roundish, five to seven on the radius, two to eight times as broad as the
bars. Radial proportion of the three spheres = 1 : 3 : 9. Main spines
three-sided pyramidal, as long as the radius.
Dimensions.—Diameter of the outer shell 0.1 to 0.015, middle 0.035 to 0.05, inner
0.013 to 0.016; cortical pores 0.01 to 0.03, bars 0.003 to 0.004; length of the six spines 0.05 to
0.08, basal breadth 0.012 to 0.015.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific, surface; also
fossil in Barbados.
22. Hexacontium furcatum, n. sp.
Cortical shell thin walled, covered with numerous thin, bristle-shaped by-spines, which are
forked and nearly as long as the main spines. Pores irregular roundish, eight to ten on the
radius, two to eight times as broad as the bars. Radial proportion of the three spheres =
1 : 2.5 : 10. Main spines triangular pyramidal, shorter then the radius.
Dimensions.—Diameter of the outer shell 0.18, middle 0.045, inner 0.018; cortical
pores 0.005 to 0.015, bars 0.002; length of the six spines 0.07, basal breadth 0.015.
Habitat.—North Atlantic, Station 354, surface.
23. Hexacontium drymodes, Haeckel.
Actinomma drymodes, Haeckel, 1862, Monogr. d. Radiol., p. 442, Taf. xxiv.
fig. 9.
Cortical shell thin walled, covered with numerous thin, bristle-shaped spines, which are double
forked and half as long as the main spines. Pores irregular roundish, eight to ten on the radius,
{199}two to eight times as broad as the bars. Radial
proportion of the three spheres = 1 : 2 : 4. Main spines three-sided
prismatic, with prominent, often somewhat contorted edges, at the distal end cuspidated; longer
than the radius of the outer shell.
Dimensions.—Diameter of the outer shell 0.15, middle 0.07, inner 0.035; cortical
pores 0.01 to 0.03, bars 0.04; length of the six spines 0.11, basal breadth 0.03.
Habitat.—Mediterranean (Messina), surface; Canary Islands (Lanzerote);
Haeckel.
24. Hexacontium periplectum, n. sp.
Cortical shell double, enclosing a simple medullary shell. Radial proportion of the three
shells = 1 : 4 : 5. Inner cortical shell thick walled, with irregular roundish
pores, two to eight times as broad as the bars; five to seven on the radius. Numerous radial
spines, arising from it, are connected below their distal ends (at equal distances from the
centre) by delicate branched threads, and so form an outer, irregular, thin, cortical shell, with
spiny surface. Six main spines three-sided pyramidal, about as long as the radius.
Dimensions.—Diameter of the outer shell 0.22, middle 0.2, inner 0.045; cortical
pores (of both outer shells) 0.01 to 0.03, bars 0.001 to 0.005; length of the spines 0.1, basal
breadth 0.02.
Habitat.—North Pacific, Station 241, surface.
Genus 80. Hexadendron,[111] Haeckel,
1881, Prodromus, p. 452.
Definition.—Cubosphærida with three concentric
lattice-spheres and six branched spines of equal size.
The genus Hexadendron differs from Hexacontium in the ramification of
the six dimensive spines, and from the similar Hexancistra in the duplication of the
medullary shell. As in the latter instance, each spine can bear either three simple lateral
branches or three rows of pinnate lateral branches.
1. Hexadendron quadricuspis, n. sp.
All three shells spherical, with radial proportion = 1 : 2 : 6. Pores of
both medullary shells regular circular, twice as broad as the bars. Pores of the cortical shell
irregular roundish, four to six times as broad as the bars; surface a little thorny. Six radial
spines prismatic, with three prominent wing-like edges, which are prolonged below the distal end
in three curved lateral branches. (Differs from Hexalonche quadricuspis, Pl. 22, fig. 11,
mainly in the double medullary shell.)
Dimensions.—Diameter of the outer shell 0.15, middle 0.05, inner 0.025; length of
the spines 0.12, breadth 0.01.
Habitat.—Central Pacific, Station 272, depth 2600 fathoms.
{200}
2. Hexadendron bipinnatum, n. sp. (Pl. 23, fig.
1).
All three shells regular octahedral, with very delicate network of irregular polygonal meshes,
and very thin, thread-like bars between them; their radial proportion =
1 : 2.5 : 7.5. Surface of the cortical shell covered with numerous
bristle-shaped by-spines, as long as the radius. Six main spines very large, three-sided
prismatic, with three rows of pinnate, lateral branches, on the three wing-like, spirally twisted
edges (similar to those of Hexancistra mirabilis, p. 189, Pl. 23, fig.
3).
Dimensions.—Diameter of the outer shell 0.12, middle 0.04, inner 0.016.
Habitat.—Central Pacific, Station 272, surface.
Genus 81. Hexacontarium,[112] n.
gen.
Definition.—Cubosphærida with three concentric
lattice-spheres and six simple spines of different size; one opposite pair larger than the two
others.
The genus Hexacontarium exhibits to its ancestral form Hexacontium
the same relation that Hexaloncharium bears to Hexalonche; the former is developed
from the latter by duplication of the medullary shell. As two opposite spines of one pair are
larger than the four others, they correspond to the three axes of a quadratic crystal.
1. Hexacontarium dentatum, n. sp.
Cortical shell with regular circular, hexagonally framed pores, twice as broad as the bars,
with smooth surface. Radial proportion of the three spheres = 1 : 2 : 5. Six
radial spines three-sided prismatic, half as broad as the inner medullary shell, with three
dentated edges. Two opposite major spines longer than the shell diameter; four other minor spines
only half as long as the radius. (Similar to Hexacontium clavigerum, Pl. 23, fig. 5, but
distinct by the prolongation of the spines of one axis.)
Dimensions.—Diameter of the outer shell 0.15, middle 0.06, inner 0.03; length of
the major spines 0.2, minor 0.04.
Habitat.—Central Pacific, Station 266, depth 2750 fathoms.
2. Hexacontarium clavatum, n. sp.
Cortical shell covered with short conical by-spines and irregular roundish pores, three to four
times as broad as the bars. Radial proportion of the three spheres =
1 : 3 : 8. Six radial spines three-sided prismatic, at the distal end
club-shaped; two major spines three times as long as the four others, which are equal to the shell
radius.
Dimensions.—Diameter of the outer shell 0.16, middle 0.06, inner 0.02.
Habitat.—Central Pacific, Station 272, depth 2600 fathoms.
{201}
Subfamily Hexacromyida,[113] Haeckel, 1881, Prodromus, pp. 449, 453.
Definition.—Cubosphærida with four concentric spherical
lattice-shells.
Genus 82. Hexacromyum,[114] Haeckel,
1881, Prodromus, p. 453.
Definition.—Cubosphærida with four concentric
lattice-spheres and six simple spines of equal size.
The genus Hexacromyum possesses four concentric, spherical, or octahedral
lattice-shells; two inner medullary shells within the central capsule, two outer cortical outside
it. The four spheres are connected by six radial beams, which are prolonged outside into simple
spines of equal size, opposite in pairs in the three dimensive axes. This genus can be derived
from Hexacontium by duplication of the cortical shell.
1. Hexacromyum elegans, n. sp. (Pl. 24,
fig. 9).
Shell composed of four concentric shells, with radial proportion =
1 : 2.5 : 7.5 : 10. First (innermost) shell with very small circular
pores, second shell with larger circular pores. Third shell (inner cortical shell) with large,
subregular, circular, hexagonally framed pores (eight to nine on the radius), twice as broad as
the bars; from the elevated nodal-points of the hexagonal frames (between every three pores) arise
thin bristle-shaped radial beams, which are united at the distal end by vaulted branches forming
the delicate fourth shell. Surface smooth. Radial spines three-sided pyramidal, as long as the
radius, as broad at the base as the innermost shell.
Dimensions.—Diameter of the four shells—(A) 0.02, (B) 0.05, (C) 0.15, (D)
0.2; length of the six radial spines 0.1, basal breadth 0.02.
Habitat.—Central Pacific, Station 271, surface.
2. Hexacromyum quadrigatum, n. sp.
Shell composed of four concentric shells, with radial proportion =
1 : 3 : 8 : 10. Structure of all four shells the same, with regular,
circular pores, twice to three times as broad as the bars; size of the pores gradually increasing
from the innermost to the outermost shell; surface smooth. Radial spines three-sided pyramidal, as
long as the radius, half as broad at the base as the innermost shell.
Dimensions.—Diameter of the four shells—(A) 0.025, (B) 0.08, (C) 0.20, (D)
0.25; length of the spines 0.12, basal breadth 0.012.
Habitat.—North Pacific, Station 253, depth 3125 fathoms.
{202}
3. Hexacromyum arachnoides, n. sp.
Hexacromidium arachnoides, Haeckel, 1881, Prodromus, p. 453.
Shell composed of four concentric shells, with radial proportion =
1 : 2 : 3 : 4. Innermost shell with regular, hexagonal meshes; the
three other shells with irregular, polygonal meshes; bars between the large meshes in all four
shells very thin, cobweb-like. Surface covered with thin bristle-shaped by-spines, as long as the
radius. Six radial main spines three-sided prismatic, longer than the diameter of the whole
shell.
Dimensions.—Diameter of the four shells—(A) 0.08, (B) 0.16, (C) 0.24, (D)
0.32; length of the spines 0.4, breadth 0.01.
Habitat.—South Pacific, Station 300, depth 1375 fathoms.
4. Hexacromyum octahedrum, n. sp. (Pl. 23, fig.
2).
Shell composed of four concentric shells which are not spherical (as in the three preceding
species), but regular octahedral. Radial proportion =
1 : 2.5 : 6 : 9. Network in all four shells delicate, with irregular
polygonal meshes and thin bars; the thickness of the bars and size of the meshes increasing from
the innermost to the outermost shell. Six radial spines three-sided prismatic, increasing slowly
in breadth towards the distal end, much longer than the shell diameter.
Dimensions.—Diameter of the four shells—(A) 0.02, (B) 0.05, (C) 0.12, (D)
0.18; length of the radial spines 0.2 to 0.3 and more, breadth 0.01.
Habitat.—Central Pacific, Station 263, depth 2650 fathoms.
Subfamily Hexacaryida,[115] Haeckel, 1881, Prodromus, p. 454.
Definition.—Cubosphærida with five or more
concentric lattice-shells.
Genus 83. Cubosphæra,[116] n. gen.
Definition.—Cubosphærida with five to six or more
concentric lattice-shells and six simple spines of equal size.
The genus Cubosphæra is developed from the preceding Hexacromyum by
further addition of the concentric lattice-shells, their number amounting to five, six, or more.
The innermost two of these are medullary shells, the others being cortical shells. All are
connected by six radial beams, prolonged outside into six simple spines of equal size; these lie
opposite in pairs in three dimensive axes, corresponding to the three axes of a cube.
{203}
1. Cubosphæra cubaxonia, n. sp. (Pl. 24,
fig. 8).
Hexacromyon cubaxonium, Haeckel, 1881, Prodrom. et Atlas.
Shell composed of five concentric spheres, with the radial proportion =
1 : 3 : 8 : 10 : 13. The two medullary shells with small
regular, circular pores of the same breadth as the bars. Inner cortical shell (third shell) with
large regular, circular pores, four times as broad as the bars, hexagonally framed. From each
hexagon-corner arises a thin bristle-shaped radial by-spine, which at a fixed distance from the
centre is united with the middle cortical shell (fourth shell), which has very small circular
pores. The beginning of the fifth shell (outermost) is indicated by six small reticula, produced
by the six main spines at equal distances from the centre. All five shells are united by six
prismatic radial beams, ending outside in pyramidal furrowed spines.
Dimensions.—Diameter of the five shells—(A) 0.2, (B) 0.15, (C) 0.12, (D)
0.045, (E) 0.015.
Habitat.—Central Pacific, Station 272, depth 2600 fathoms.
2. Cubosphæra concentrica, n. sp.
Shell composed of six concentric spheres, with the radial proportion =
1 : 2 : 6 : 7.5 : 9 : 11. The two medullary
shells with regular, circular pores, twice as broad as the bars. The four cortical shells of the
same structure, with irregular, roundish pores, three to four times as broad as the bars. The size
of these pores and the breadth of their bars gradually increase towards the smooth surface. All
six shells are connected by six thin three-sided prismatic radial beams, which are prolonged
outside in six stronger spines, angular pyramidal, with smooth edges, as long as the shell
radius.
Dimensions.—Diameter of the six shells—(A) 0.02 (B) 0.04, (C) 0.12, (D) 0.15, (E) 0.18, (F) 0.22.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
Genus 84. Hexacaryum,[117] Haeckel, 1881,
Prodromus, p. 454.
Definition.—Cubosphærida with five to six or more
concentric lattice-shells and six branched spines of equal size.
The genus Hexacaryum is distinguished from the foregoing Cubosphæra
by ramification of the six radial spines, and therefore exhibits the same relation to it that
Hexancistra bears to Hexastylus, &c.
1. Hexacaryum arborescens, n. sp. (Pl. 23, figs. 4,
4a).
Shell composed of five, six, or more concentric shells, which are united by six very large
radial spines. The two medullary shells spherical, inner with regular, circular, outer with
irregular polygonal pores. All cortical shells (third and following) not spherical, but regular
octahedral, with irregular polygonal meshes and thin bars. Radial spines prismatic, with three
wing-like, spirally twisted edges, which at equal distances send out thin forked lateral branches
(six on each {204}verticil); by further ramification and
communication of these branches the triangular net-plates arise, filling out the sides of the
octahedral cortical shells. Diameter of all shells little different. The outer free parts of the
six spines are arborescent, twice to three times as long as the enclosed inner parts, and bear six
to eight verticils of free lateral branches, decreasing in size towards the distal end (similar to
Arachnosphæra).
Dimensions.—Diameter of the six shells—(A) 0.015, (B) 0.04, (C) 0.1, (D)
0.16, (E) 0.22, (F) &c.; average distance of the concentric octahedra = 0.06; length of the
radial spines 0.3 to 0.4 and more, breadth 0.02.
Habitat.—Central Pacific, Station 274, surface.
Subfamily Hexadorida,[118] Haeckel, 1881, Prodromus, pp. 449, 455.
Definition.—Cubosphærida with spongy
spherical or octahedral shell (with or without enclosed concentrical lattice-shells).
Genus 85. Cubaxonium,[119] n. gen.
Definition.—Cubosphærida with solid spongy spherical or
octahedral shell, without latticed medullary shell in the centre, and with six simple radial
spines of equal size.
The genus Cubosphæra may be regarded as a form of Styptosphæra, which
develops six radial spines, opposite in pairs in the three dimensive axes. The solid spongy
framework of the shell assumes the outer form either of a sphere, or of a regular octahedron.
1. Cubaxonium spongiosum, n. sp.
Spongy shell spherical, composed of a very dense spongy framework of nearly uniform structure;
the meshes three to four times as broad as the bars; surface almost smooth. Six spines
cylindrical, twice to three times as long as the diameter of the spongy sphere, about as broad as
one half mesh.
Dimensions.—Diameter of the sphere 0.2, meshes 0.006 to 0.008, bars 0.002; length
of the spines 0.4 to 0.5, breadth 0.004.
Habitat.—South Pacific, Station 288, surface.
2. Cubaxonium octahedrum, n. sp.
Spongy shell octahedral, composed of a loose spongy framework of nearly uniform structure; the
meshes ten to twelve times as broad as the bars; surface thorny. Six spines three-sided {205}pyramidal, longer than the diameter of the octahedron,
arising from its six corners, as broad at the base as one mesh.
Dimensions.—Diameter of the shell 0.15, meshes 0.01 to 0.012, bars 0.001; length
of the spines 0.2, basal breadth 0.01.
Habitat.—South Pacific, Station 300, surface.
Genus 86. Hexadoras,[120] Haeckel, 1881,
Prodromus, p. 455.
Definition.—Cubosphærida with spongy spherical shell and
one simple latticed medullary shell in its centre, having six simple spines of equal size.
The genus Hexadoras exhibits the same structure of the spongy shell as
Spongoplegma, but differs from it in the production of six dimensive spines. In the centre
lies one simple, latticed, medullary shell, which is either spherical or octahedral. In the latter
case the six spines arise from the six corners of the octahedron.
1. Hexadoras axophæna, n. sp.
Medullary shell spherical, with regular hexagonal meshes, twice as broad as the bars. Cortical
shell entirely enclosing it, with loose irregular framework and thorny surface. Diameter of the
outer shell three times as large as that of the inner. Six radial spines arising from the
medullary shell, four to five times as long as the radius of the cortical shell, three-sided
prismatic, with straight dentated edges.
Dimensions.—Diameter of the outer shell 0.15, inner 0.05; length of the spines 0.3
to 0.4.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms.
2. Hexadoras lychnosphæra, n. sp.
Medullary shell spherical, with regular, circular, hexagonally framed pores, three times as
broad as the bars. Cortical shell enveloping it, with loose irregular framework and spiny surface.
Diameter of the outer shell eight times as large as that of the inner. Six radial spines arising
from the inner shell, three-sided prismatic, with dentated, spirally contorted edges, seven times
as long as the diameter of the medullary shell, with three lateral branches at the distal end
(similar to Lychnosphæra, Pl. 11,
fig. 1).
Dimensions.—Diameter of the outer shell 0.5, inner 0.06; total length of the
spines 0.4, breadth 0.01.
Habitat.—Central Pacific, Station 270, depth 2925 fathoms.
3. Hexadoras octahedrum, n. sp.
Medullary shell regular octahedral, with irregular polygonal meshes, five times as broad as the
bars. Cortical shell enveloping it, with dense spongy framework, also octahedral with rough
surface. {206}Diameter of the outer shell five times as
large as that of the inner. Six radial spines very long, arising from the six corners of the inner
and piercing the spongy mass of the outer shell, considerably exceeding it at the free distal end,
three-sided prismatic, with elegantly denticulate edges.
Dimensions.—Diameter of the outer shell 0.2, inner 0.04; total length of the
spines 0.3 or more, breadth 0.02.
Habitat.—Central Pacific, Station 266, depth 2750 fathoms.
Genus 87. Hexadoridium,[121] Haeckel,
1881, Prodromus, p. 455.
Definition.—Cubosphærida with spongy
spherical shell and two concentric latticed medullary shells in its centre, having six simple
spines of equal size.
The genus Hexadoridium differs from Hexadoras in the duplication of
the medullary shell, and exhibits therefore the same relation to it that Spongodictyon
bears to Spongoplegma. In the only known species the six spines are opposite, arranged
quite regularly in pairs in the three dimensive axes, and consequently represent the three axes of
a regular crystal or cube.
1. Hexadoridium streptacanthum, n. sp. (Pl. 25, figs. 1,
1a).
Both medullary shells spherical, with small regular, circular pores, twice as broad as the
bars; outer twice as broad as the inner. Spongy cortical shell enclosing it with dense framework,
five times as broad as the outer medullary shell, regular octahedral. Six radial spines, arising
from the latter, are thinned at the inner end, three to five times as long as the diameter of the
cortical shell, and nearly as broad as the inner medullary shell, with three dentated and spirally
contorted edges. (Very similar to the common Spongosphæra streptacantha, with irregular and
variable number and dispositions of spines; possibly its ancestral form?).
Dimensions.—Diameter of the cortical shell 0.2, of the outer medullary shell 0.04,
inner 0.02; length of the spines 1 mm. and more, breadth 0.02.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
Family X. Astrosphærida (Pls. 11, 18-20, 26-30).
Astrosphærida, Haeckel, 1881, Prodromus, p. 449.
Definition.—Sphæroidea with numerous (eight to twelve or
more, commonly between twenty and sixty) radial spines on the surface of the spherical shell;
living solitary (not associated in colonies).
The family Astrosphærida, the largest and most varied of all Sphæroidea, is distinguished from the other members of this group by the
possession of numerous {207}radial spines, which are
either regularly or irregularly disposed on the surface of the spherical shell. The extreme
variability and richness of form in this family is mainly due to the different size, shape, and
disposition of these radial spines.
The simplest Astrosphærida are the Coscinommida, with a single spherical or polyhedral
lattice-shell. To this ancestral group all other subfamilies can be opposed as "Astrosphærida
composita," since their skeleton is composed of two or more concentric lattice-shells: two in the
Haliommida, three in the Actinommida, four in the Cromyommida, five or more in the Caryommida. In
these four subfamilies the concentric shells are all simple (not spongy) fenestrated spheres or
endospherical polyhedra. In the sixth subfamily, the Spongiommida, the shell is wholly or
partially composed of spongy irregular wicker-work, with or without a medullary shell in the
centre.
The Number of the Radial Spines in the Astrosphærida is extremely variable, and ranges
from eight to forty or more; in many cases more than one hundred. Often each nodal-point of the
network develops on the shell surface one spine. Still more frequently the number of the spines is
less than that of the nodal-points. In all concentric Astrosphærida, having two or more
concentrical shells, we can distinguish "primary spines," as outer prolongations of the inner
radial beams connecting the shells, and "secondary spines," developed only on the outer surface of
the shell. Naturally the former are of much greater importance than the latter. But we can also
often distinguish among the latter larger "main spines" and smaller "by-spines," the latter
commonly much more numerous than the former.
The Disposition of the Radial Spines, either regular or irregular, is a subject of great
morphological interest, and remains to be exhausted by further observations. The following cases
of regular disposition have been observed by me—(A) eight spines, opposite in pairs in four
axes corresponding to the four diagonal axes of a cube; (B) nine spines, regularly disposed at
equal distances (?) (not opposed in pairs); (C) ten spines, disposed at equal distances (?); (D)
twelve spines, regularly disposed, corresponding to the twelve corners of the regular icosahedron;
(E) fourteen spines, quite regularly disposed (six corresponding to the three axes of a regular
octahedron, eight to the central points of its eight faces); (F) sixteen spines, regularly
disposed (?); (G) twenty spines (very common!), either disposed in the same manner (after the law
of Johannes Müller) as in the Acantharia (?), or corresponding to the
twenty corners of the regular or pentagonal dodecahedron, or disposed in the same manner as in
many Larcoidea (Tholonida, &c., to be described afterwards); (H)
twenty-four spines, regularly disposed (?); (I) thirty-two spines, quite regularly disposed
(twenty corresponding to the twenty corners of the regular dodecahedron, twelve to the central
points of its twelve faces); (K) forty spines, nearly regularly (or quite symmetrically?)
disposed. If the number of the spines amounts to more than forty, it is as a rule impossible to
determine their regular disposition in a satisfactory manner.
{208}
Synopsis of the Genera of Astrosphærida.
|
I. Subfamily Coscinommida.
(One single latticed shell.)
|
|
Spines all simple, not branched and not tubulous. |
|
All spines of the same kind, |
088. Acanthosphæra. |
| Larger main spines and smaller by-spines, |
089. Heliosphæra. |
| Spines hollow conical tubes with porous walls, |
090. Conosphæra. |
| Between simple spines the pores prolonged in hollow
tubes, |
091. Coscinomma. |
| Spines branched or forked. |
|
Spines with lateral branches, |
092. Cladococcus. |
| Spines forked or dichotomous, |
093. Elaphococcus. |
|
II. Subfamily Haliommida.
(Two concentric latticed shells.)
|
 |
A. Elatommida, one medullary and one cortical shell. |
|
All spines equal, simple, |
094. Haliomma. |
| All spines simple, of two different kinds, |
095. Heliosoma. |
| Spines branched, |
096. Elatomma. |
| B. Diplosphærida, both shells cortical. |
|
No by-spines, |
097. Leptosphæra. |
| Inner by-spines, |
098. Diplosphæra. |
| Outer by-spines, |
099. Drymosphæra. |
| Outer and inner by-spines, |
100. Astrosphæra. |
|
III. Subfamily Actinommida.
(Three concentric latticed shells.)
|
|
Radial spines not branched. |
|
All spines equal, |
101. Actinomma. |
| Larger main spines and smaller by-spines, |
102. Echinomma. |
| Radial spines branched or forked, |
103. Pityomma. |
|
IV. Subfamily Cromyommida.
(Three concentric latticed shells.)
|
|
Radial spines not branched. |
|
All spines equal, |
104. Cromyomma. |
| Larger main spines and smaller by-spines, |
105. Cromyechinus. |
| Radial spines branched or forked, |
106. Cromyodrymus. |
|
V. Subfamily Caryommida.
(Numerous, five to ten or more, concentric latticed shells.)
|
|
Two medullary shells and three or more cortical shells
(lattice work ordinary), |
107. Caryomma. |
| No medullary shells (lattice work arachnoidal); three-sided
prismatic spines, with verticils of three forked branches. (Arachnosphærida.) |
|
Triangular meshes, simple bars, without diagonal bars, |
108. Arachnopila. |
| Triangular meshes, simple bars; diagonal bars between the shells, |
109. Arachnopegma. |
| Polygonal meshes, bars branched, |
110. Arachnosphæra. |
| I. Subfamily Coscinommida. (One single latticed shell.) |
|
|
Spines all simple, not branched and not tubulous. |
|
|
|
|
|
|
All spines of the same kind, |
|
|
|
|
|
|
|
|
|
|
88. Acanthosphæra. |
|
|
|
|
|
|
|
|
|
|
Larger main spines and smaller by-spines, |
|
|
|
|
|
|
|
|
|
|
89. Heliosphæra. |
|
|
|
|
|
|
|
|
Spines hollow conical tubes with porous walls, |
|
|
|
|
|
|
|
|
90. Conosphæra. |
|
|
|
|
|
|
|
|
Between simple spines the pores prolonged in hollow tubes, |
|
|
|
|
|
|
|
|
91. Coscinomma. |
|
|
|
|
|
|
|
|
Spines branched or forked. |
|
|
|
|
|
|
Spines with lateral branches, |
|
|
|
|
|
|
|
|
|
|
92. Cladococcus. |
|
|
|
|
|
|
|
|
|
|
Spines forked or dichotomous, |
|
|
|
|
|
|
|
|
|
|
93. Elaphococcus. |
|
|
|
|
|
|
| II. Subfamily Haliommida. (Two concentric latticed shells.) |
|
|
A. Elatommida, one medullary and one cortical shell. |
|
|
|
|
|
|
All spines equal, simple, |
|
|
|
|
|
|
|
|
|
|
94. Haliomma. |
|
|
|
|
|
|
|
|
|
|
All spines simple, of two different kinds, |
|
|
|
|
|
|
|
|
|
|
95. Heliosoma. |
|
|
|
|
|
|
|
|
|
|
Spines branched, |
|
|
|
|
|
|
|
|
|
|
96. Elatomma. |
|
|
|
|
|
|
|
|
B. Diplosphærida, both shells cortical. |
|
|
|
|
|
|
No by-spines, |
|
|
|
|
|
|
|
|
|
|
97. Leptosphæra. |
|
|
|
|
|
|
|
|
|
|
Inner by-spines, |
|
|
|
|
|
|
|
|
|
|
98. Diplosphæra. |
|
|
|
|
|
|
|
|
|
|
Outer by-spines, |
|
|
|
|
|
|
|
|
|
|
99. Drymosphæra. |
|
|
|
|
|
|
|
|
|
|
Outer and inner by-spines, |
|
|
|
|
|
|
|
|
|
|
100. Astrosphæra. |
|
|
|
|
|
|
| III. Subfamily Actinommida. (Three concentric latticed shells.) |
|
|
Radial spines not branched. |
|
|
|
|
|
|
All spines equal, |
|
|
|
|
|
|
|
|
|
|
101. Actinomma. |
|
|
|
|
|
|
|
|
|
|
Larger main spines and smaller by-spines, |
|
|
|
|
|
|
|
|
|
|
102. Echinomma. |
|
|
|
|
|
|
|
|
Radial spines branched or forked, |
|
|
|
|
|
|
|
|
103. Pityomma. |
|
|
|
|
|
|
| IV. Subfamily Cromyommida. (Three concentric latticed shells.) |
|
|
Radial spines not branched. |
|
|
|
|
|
|
All spines equal, |
|
|
|
|
|
|
|
|
|
|
104. Cromyomma. |
|
|
|
|
|
|
|
|
|
|
Larger main spines and smaller by-spines, |
|
|
|
|
|
|
|
|
|
|
105. Cromyechinus. |
|
|
|
|
|
|
|
|
Radial spines branched or forked, |
|
|
|
|
|
|
|
|
106. Cromyodrymus. |
|
|
|
|
|
|
| V. Subfamily Caryommida. (Numerous, five to ten or more, concentric latticed
shells.) |
|
|
Two medullary shells and three or more cortical shells (lattice
work ordinary), |
|
|
|
|
|
|
|
|
107. Caryomma. |
|
|
|
|
|
|
|
|
No medullary shells (lattice work arachnoidal); three-sided
prismatic spines, with verticils of three forked branches. (Arachnosphærida.) |
|
|
|
|
|
|
Triangular meshes, simple bars, without diagonal bars, |
|
|
|
|
|
|
|
|
|
|
108. Arachnopila. |
|
|
|
|
|
|
|
|
|
|
Triangular meshes, simple bars; diagonal bars between the
shells, |
|
|
|
|
|
|
|
|
|
|
109. Arachnopegma. |
|
|
|
|
|
|
|
|
|
|
Polygonal meshes, bars branched, |
|
|
|
|
|
|
|
|
|
|
110. Arachnosphæra. |
|
|
|
|
|
|
{209}
|
VI. Subfamily Spongiommida.
(Spherical shell whole or partly spongy, with or without enclosed
latticed medullary shells in the centre.)
|
 |
I. Tribe Spongodrymida, without latticed medullary
shell. |
|
Spongy sphere solid. |
|
Spines simple, |
111. Spongiomma. |
| Spines branched, |
112. Spongodrymus. |
| Spongy sphere hollow. |
|
Spines simple, |
113. Spongechinus. |
| Spines branched, |
114. Spongothammus. |
| II. Tribe Rhizoplegmida, with one single latticed medullary
shell. |
|
Medullary shell spherical. |
|
Framework arising from the medullary shell, |
115. Spongopila. |
| Framework separate from the medullary shell. |
|
No medullary by-spines, |
116. Rhizoplegma. |
| Medullary by-spines, |
117. Lychnosphæra. |
| Medullary shell a simple cube. |
|
Framework arising immediately from the medullary
shell, |
118. Centrocubus. |
| Framework separate from the medullary shell, |
119. Octodendron. |
| III. Tribe Rhizosphærida, with two concentric
latticed medullary shells. |
|
Framework arising from the medullary shell, |
120. Spongosphæra. |
| Framework separate from the medullary shell, |
121. Rhizosphæra. |
| VI. Subfamily Spongiommida. (Spherical shell whole or partly spongy, with or
without enclosed latticed medullary shells in the centre.) |
|
|
I. Tribe Spongodrymida, without latticed medullary shell. |
|
|
|
|
|
|
Spongy sphere solid. |
|
|
|
|
|
|
|
|
|
|
Spines simple, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
111. Spongiomma. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Spines branched, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
112. Spongodrymus. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Spongy sphere hollow. |
|
|
|
|
|
|
|
|
|
|
Spines simple, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
113. Spongechinus. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Spines branched, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
114. Spongothammus. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Medullary shell spherical. |
|
|
|
|
|
|
|
|
|
|
Framework arising from the medullary shell, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
115. Spongopila. |
|
|
|
|
|
|
|
|
|
|
|
|
II. Tribe Rhizoplegmida, with one single latticed medullary
shell. |
|
|
|
|
|
|
|
|
Framework separate from the medullary shell. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
No medullary by-spines, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
116. Rhizoplegma. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Medullary by-spines, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
117. Lychnosphæra. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Medullary shell a simple cube. |
|
|
|
|
|
|
|
|
|
|
Framework arising immediately from the medullary shell, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
118. Centrocubus. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Framework separate from the medullary shell, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
119. Octodendron. |
|
|
|
|
|
|
|
|
|
|
|
|
III. Tribe Rhizosphærida, with two concentric latticed medullary
shells. |
|
|
|
|
|
|
|
|
Framework arising from the medullary shell, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
120. Spongosphæra. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Framework separate from the medullary shell, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
121. Rhizosphæra. |
|
|
|
|
|
|
|
|
|
|
Subfamily Coscinommida, Haeckel.
Heliosphærida, Haeckel, 1881, Prodromus, pp. 449, 450.
Definition.—Astrosphærida with one single
spherical lattice-shell.
Genus 88. Acanthosphæra,[122] Ehrenberg,
1858, Monatsber. d. k. preuss. Akad. d.
Definition.—Astrosphærida with one simple lattice-sphere,
covered with simple radial spines of the same kind.
The genus Acanthosphæra exhibits the most simple form of all
Astrosphærida; a simple spherical lattice-shell, the surface of which is covered by radial
spines of one and the same kind. The number of the latter is very variable, often twelve to
twenty, regularly disposed; in other cases forty to sixty or more; and sometimes at each
nodal-point of the network a spine is developed.
{210}
Subgenus 1. Rhaphidococcus, Haeckel, 1862, Monogr. d. Radiol., p. 365 (sensu
emendato).
Definition.—Pores of the spherical shell regular or subregular, all of
nearly equal size and similar form. Radial spines arising from all the nodal-points of the
network.
1. Acanthosphæra tenuissima, Haeckel.
Heliosphæra tenuissima, Haeckel, 1862, Monogr. d. Radiol., p. 351, Taf.
ix. fig. 2.
Shell extremely thin walled, eight to ten times as broad as one pore. Meshes or pores regular,
hexagonal, with thread-like bars; five to seven on the radius. At each nodal-point (between every
three meshes) arises a bristle-shaped radial spine, as long as the diameter of one pore.
Dimensions.—Diameter of the shell 0.2 to 0.25, of the meshes or pores 0.025 to
0.03, bars below 0.0001; length of the spines 0.03.
Habitat.—Mediterranean (Messina), North Atlantic (Canary Islands).
2. Acanthosphæra tenuis, n. sp.
Shell very thin walled, about twenty times as broad as one pore. Meshes subregular, hexagonal,
with thread-like bars; twelve to fourteen on the radius. At each nodal-point arises a
bristle-shaped radial spine, about as long as the radius.
Dimensions.—Diameter of the shell 0.3 to 0.35, pores 0.015 to 0.018, bars below
0.001; length of the spines 0.12 to 0.16.
Habitat.—Central Pacific, Stations 266 to 274, depth 2350 to 2925
fathoms.
3. Acanthosphæra macropora, n. sp.
Shell thin walled, five to six times as broad as one mesh. Pores regular, circular, hexagonally
framed, ten to twelve times as broad as the bars. Radial spines bristle-shaped, as long as the
diameter of one pore, arising from all the nodal-points.
Dimensions.—Diameter of the shell 0.07 to 0.08, pores 0.012, bars 0.001; length of
the spines 0.012.
Habitat.—Central Pacific, Station 271, surface.
4. Acanthosphæra micropora, n. sp.
Shell thick walled, forty to fifty times as broad as one pore. Pores regular, circular,
hexagonally framed, half as broad as the bars. Radial spines bristle-shaped, as long as the radius
of the shell, arising from all the nodal-points.
Dimensions.—Diameter of the shell 0.22, pores 0.003, bars 0.006; length of the
spines 0.1.
Habitat.—South Pacific, Station 288, surface.
{211}
5. Acanthosphæra dentata, Haeckel.
Cladococcus dentatus, Haeckel, 1862, Monogr. d. Radiol., p. 367, Taf.
xiii. fig. 10.
Shell thin walled, eight to ten times as broad as one pore. Pores regular, circular,
hexagonally framed, three times as broad as the bars. Radial spines arising from all the
nodal-points of the network, three-sided prismatic, with dentated or serrated edges, longer than
the shell diameter.
Dimensions.—Diameter of the shell 0.08, pores 0.01, bars 0.003; length of the
spines 0.1, breadth 0.003.
Habitat.—Mediterranean (Messina), surface.
6. Acanthosphæra acufera, Haeckel.
Rhaphidococcus acufer, Haeckel, 1862, Monogr. d. Radiol., p. 366, Taf. xiv. fig.
1.
Cladococcus acufer, Haeckel, 1860, Monatsber. d. k. preuss. Akad. d. Wiss.
Berlin, p. 800.
Shell thin walled, eight to ten times as broad as one mesh. Pores regular, circular, three
times as broad as the bars. Radial spines bristle-shaped, arising with thicker conical bases from
all the nodal-points, about as long as the diameter of the shell.
Dimensions.—Diameter of the shell 0.08, pores 0.01, bars 0.003; length of the
spines 0.08.
Habitat.—Mediterranean (Messina); North Atlantic, Station 354,
surface.
7. Acanthosphæra castanea, n. sp. (Pl. 26, fig.
3).
Shell thick walled, thirty times as broad as one mesh. Pores regular, circular, nearly of the
same breadth as the bars. Radial spines bristle-shaped, arising with thicker conical bases from
all the nodal-points, scarcely half as long as the radius of the shell.
Dimensions.—Diameter of the shell 0.15, pores and bars 0.005; length of the spines
0.03.
Habitat.—North Pacific, Station 244, surface.
8. Acanthosphæra flosculenta, n. sp.
Shell thick walled, thirteen times as broad as one mesh. Pores regular, circular, three times
as broad as the bars, with an elegant six-lobed frame (Pl. 28, fig.
1b). In the intervals between the six lobes of each mesh arise six conical radial spines
(half as long as the radius), six around each pore. (Differs from the similar Haliomma
flosculentum, Pl. 28,
fig. 1, in the absence of an enclosed medullary shell and the stronger development of the
spines.)
Dimensions.—Diameter of the shell 0.13, pores 0.01, bars 0.003; length of the
spines 0.03.
Habitat.—Indian Ocean, Cocos Islands, surface, Rabbe.
Subgenus 2. Rhaphidocapsa, Haeckel.
Definition.—Pores of the spherical shell regular or subregular, all of
nearly equal size and similar form. Radial spines scattered at some distance apart, not at all the
nodal-points.
{212}
9. Acanthosphæra insignis, Haeckel.
Heliosphæra insignis, R. Hertwig, 1879, Organismus der Radiol., p. 40,
Taf. v. fig. 7.
Shell thin walled, about ten times as broad as one mesh. Pores regular, hexagonal, ten to
twelve times as broad as the bars. Radial spines about one hundred and twenty, arising from
certain nodal-points of the network, being as long as the diameter of the sphere, three-sided
prismatic, with three thin denticulated edges.
Dimensions.—Diameter of the shell 0.2, pores 0.02, bars 0.002; length of the
spines 0.18, breadth 0.003.
Habitat.—Cosmopolitan; Mediterranean (Messina), Atlantic (Stations
325, 347), Indian (Ceylon), Pacific (Stations 270 to 274), surface.
10. Acanthosphæra fortispina, n. sp.
Shell thin walled, about six times as broad as one mesh. Pores subregular, hexagonal, with
thread-like bars; three to four on the radius. Radial spines about twenty, three-sided pyramidal,
as long as the diameter, and one-third as broad at the base as one pore.
Dimensions.—Diameter of the shell 0.06 to 0.09, pores 0.01 to 0.014, bars below
0.001; length of the spines 0.07 to 0.08, basal breadth 0.03 to 0.04.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
11. Acanthosphæra mucronata, n. sp. (Pl. 26, fig.
7).
Shell thick walled, fifteen times as broad as one pore. Pores regular, circular, hexagonally
framed, funnel-shaped, three times as broad as the bars. Radial spines twenty to thirty,
dagger-shaped or spindle-shaped, angular, twice as broad in the middle as one pore, about as long
as the radius.
Dimensions.—Diameter of the shell 0.11, pores 0.0075, bars 0.0025; length of the
spines 0.06, breadth 0.015.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
12. Acanthosphæra clavata, n. sp. (Pl. 26, fig.
8).
Shell thick walled, ten to twelve times as broad as one mesh. Pores subregular, circular, three
times as broad as the bars. Radial spines twenty, club-shaped, angular, with prominent edges,
twice as broad at the distal end as one pore, about as long as the radius.
Dimensions.—Diameter of the shell 0.11, pores 0.01, bars 0.003; length of the
spines 0.06, breadth 0.02.
Habitat.—Western Tropical Pacific, Station 225, depth 4475
fathoms.
13. Acanthosphæra marginata, n. sp.
Shell thick walled, twelve times as broad as one mesh. Pores regular, circular double-edged,
four times as broad as the bars. Radial spines conical, fourteen in number, about as long as the
{213}radius, and as broad at the base as one mesh. Six
spines correspond to the six corners of a regular octahedron, eight to the centre of the eight
faces.
Dimensions.—Diameter of the shell 0.1, pores 0.008, bars 0.002; radial spines
0.06, basal breadth 0.01.
Habitat.—Central Pacific, Station 274, surface.
14. Acanthosphæra florida, n. sp.
Shell thick walled, fifteen times as broad as one mesh. Pores regular, circular, hexagonally
lobed, separated by prominent funnel-shaped crests of half the breadth. Twenty conical radial
spines, half as long as the radius, as broad at the base as one funnel.
Dimensions.—Diameter of the shell 0.15, pores 0.01, bars 0.005; length of the
spines 0.04, basal breadth 0.02.
Habitat.—South Atlantic, Station 325, depth 2650 fathoms.
15. Acanthosphæra enneacantha, n. sp.
Shell thin walled, fourteen times as broad as one mesh. Pores regular, circular, twice as broad
as the bars. Nine radial spines, regularly disposed, as long as the shell diameter, three-sided
prismatic, with pyramidal apex, as broad as one mesh.
Dimensions.—Diameter of the shell 0.14, pores 0.01, bars 0.005; length of the
spines 0.15, breadth 0.01.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms.
16. Acanthosphæra octahedralis, n. sp.
Shell thick walled, octahedral, fourteen times as broad as one mesh. Pores regular, circular,
three times as broad as the bars. Fourteen radial spines, regularly disposed, conical, as long as
the radius of the shell, twice as broad at the base as one mesh. Six spines correspond to the six
corners of a regular octahedron, eight to the central points of its eight faces.
Dimensions.—Diameter of the shell 0.14, pores 0.01, bars 0.003; length of the
spines 0.08, basal breadth 0.02.
Habitat.—North Pacific, Station 253, depth 3125 fathoms.
17. Acanthosphæra compacta, n. sp.
Shell thick walled, about sixty times as broad as one mesh. Pores regular, circular, twice as
broad as the bars. Thirty to forty radial spines, three-sided pyramidal, scarcely half as long as
the radius, five to six times as broad at the base as one pore.
Dimensions.—Diameter of the shell 0.24, pores 0.004, bars 0.002; length of the
radial spines 0.05, basal breadth 0.02.
Habitat.—Fossil in Barbados.
{214}
Subgenus 3. Raphidodrymus, Haeckel.
Definition.—Pores of the spherical shell irregular, of different size
or form. Radial spines arising from all the nodal-points of the network.
18. Acanthosphæra capillaris, n. sp.
Shell thin walled, with irregular polygonal meshes, twelve to twenty times as broad as the
bars; eight to ten on the radius. Radial spines bristle-shaped, arising from all the nodal-points
of the network, about as long as the diameter of the largest meshes.
Dimensions.—Diameter of the shell 0.15 to 0.2, pores 0.012 to 0.02, bars 0.001;
length of the spines 0.02.
Habitat.—Central Pacific, Stations 260 to 274, surface.
19. Acanthosphæra arctica, n. sp.
Shell thin walled, with irregular roundish, polygonally framed meshes, three to four times as
broad as the bars. Radial spines arising from all the nodal-points of the network, pyramidal at
the base; in the distal half bristle-shaped, as long as the radius.
Dimensions.—Diameter of the shell 0.12, pores 0.006 to 0.008, bars 0.002; length
of the spines 0.07.
Habitat.—Arctic Ocean, Greenland (in the stomach of Periphylla
hyacinthina).
20. Acanthosphæra antarctica, n. sp.
Shell thick walled, with irregular, roundish pores, about as broad as the bars. Radial spines
arising from all nodal-points of the network, conical at the base, half as long as the radius.
Dimensions.—Diameter of the shell 0.15, pores and bars 0.005 to 0.008; length of
the spines 0.04.
Habitat.—Antarctic Ocean, Station 157, depth 1950 fathoms.
Subgenus 4. Rhaphidosphæra, Haeckel, 1881, Prodromus, p. 450.
Definition.—Pores of the spherical shell irregular, of different size
or form. Radial spines scattered at intervals, not at all the nodal-points.
21. Acanthosphæra echinoides, Haeckel.
Cyrtidosphæra echinoides, Haeckel, 1865, Zeitschr. f. wiss. Zool., xv. p.
367, Taf. xxvi. fig. 5.
Shell thin walled, with irregular polygonal or more roundish pores of very different size.
Forty to fifty very large meshes, separated by rows of much smaller meshes. Radial spines forty to
sixty, half as long as the shell radius, bristle-shaped, with conical bases.
{215}
Dimensions.—Diameter of the shell 0.13, larger pores 0.03, smaller 0.003; length
of the spines 0.03.
Habitat.—Mediterranean (Nice), surface.
22. Acanthosphæra longispina, n. sp.
Shell thin walled, with irregular polygonal meshes, four to six times as broad as the bars; six
to eight on the radius. Thirty to forty radial spines, three-sided prismatic, twice as broad as
the bars, twice to three times as long as the diameter of the shell.
Dimensions.—Diameter of the shell 0.12 to 0.15, pores 0.012 to 0.02, bars 0.003;
length of the radial spines 0.2 to 0.4, breadth 0.006.
Habitat.—North Pacific, Station 253, surface.
23. Acanthosphæra brevispina, n. sp.
Shell thick walled, with irregular polygonal meshes, twice to four times as broad as the bars;
twelve to sixteen on the radius. Sixty to eighty radial spines, pyramidal, half as long as the
radius of the shell, one-fourth as broad at the base.
Dimensions.—Diameter of the shell 0.2 to 0.22, pores 0.006 to 0.012, bars 0.003;
length of the spines 0.05, basal breadth 0.02.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
24. Acanthosphæra acanthica, Haeckel.
Cenosphæra acanthica, Stöhr, 1880, Palæontogr., vol. xxvi. p. 86, Taf. i.
fig. 3.
Shell thick walled, with irregular, roundish pores, twice to three times as broad as the bars;
seven to nine on the radius. Ten to twenty radial spines pyramidal, shorter than the radius, twice
as broad at the base as one mesh.
Dimensions.—Diameter of the shell 0.12 to 0.16, pores 0.006 to 0.009, bars 0.003;
length of the spines 0.04 to 0.06, basal breadth 0.01 to 0.015.
Habitat.—Fossil in Tertiary rocks of Sicily and Barbados.
25. Acanthosphæra haliphormis, Ehrenberg.
Acanthosphæra haliphormis, Ehrenberg, 1861, Abhandl. d. k. Akad. d. Wiss.
Berlin, 1872, Taf. ii. fig. 1.
Shell thick walled, with irregular, roundish pores, twice to four times as broad as the bars;
four to five on the radius. Twelve to twenty radial spines pyramidal, longer than the radius,
scarcely as broad as one mesh at the base.
Dimensions.—Diameter of the shell 0.11, pores 0.01 to 0.02, bars 0.005; length of
the spines 0.06 to 0.08, basal breadth 0.01.
Habitat.—Arctic Ocean, near Greenland, depth 1000 fathoms.
{216}
26. Acanthosphæra angulata, n. sp. (Pl. 26, fig.
4).
Shell thin walled; its pores irregular, roundish, with angular, double-edged margin, two to
four times as broad as the bars; six to eight on the radius. Twenty to thirty radial spines
pyramidal, angular, with prominent edges, shorter than the radius, as broad at the base as one
small mesh.
Dimensions.—Diameter of the shell 0.2, pores 0.02 to 0.04, bars 0.01; length of
the spines 0.06, basal breadth 0.02.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
27. Acanthosphæra conifera, n. sp.
Shell thick walled, with irregular, roundish pores, twice to five times as broad as the bars;
ten to twelve on the radius. Twenty radial spines conical, regularly disposed, half as long as the
radius, as broad at the base as one of the largest meshes.
Dimensions.—Diameter of the shell 0.2, pores 0.008 to 0.02, bars 0.004; length of
the spines 0.05, basal breadth 0.02.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
28. Acanthosphæra maxima, n. sp.
Shell thick walled, with irregular, roundish pores, twice to eight times as broad as the bars;
twelve to twenty on the radius. Radial spines very numerous (two to three hundred), short,
conical, scarcely as long as the diameter of the largest meshes, and one-third as broad.
Dimensions.—Diameter of the shell 0.3 to 0.4, pores 0.008 to 0.03, bars 0.004;
length of the radial spines 0.03, basal breadth 0.01.
Habitat.—Tropical Atlantic, Station 348, depth 2450 fathoms.
29. Acanthosphæra simplex, Haeckel.
Rhaphidococcus simplex, Haeckel, Monogr. d. Radiol., 1862, p. 366, figs. 5, 6.Taf. xiii.
Cladococcus simplex, Haeckel, 1860, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 800.
Shell thick walled, with irregular, roundish pores, three to six times as broad as the bars;
eight to nine on the radius. Forty to sixty radial spines, about as long as the diameter of the
shell, three-sided prismatic, not straight, but more or less bent.
Dimensions.—Diameter of the shell 0.12, pores 0.005 to 0.012, bars 0.0015 to 0.02;
length of the spines 0.12, breadth 0.003.
Habitat.—Mediterranean (Messina), surface, Haeckel.
30. Acanthosphæra gibbosa, n. sp.
Shell thin walled, rugged or tuberculate, covered by about twenty hill-shaped tubercles or
protuberances with flat valleys between them. Network very delicate, with thread-like bars and
{217}irregular, polygonal pores; twenty to thirty on the
radius. Radial spines very numerous, bristle-shaped, twice to three times as long as the diameter
of the pores.
Dimensions.—Diameter of the shell 0.3, pores 0.01 to 0.02; length of the spines
0.02 to 0.05.
Habitat.—Central Pacific, Station 274, surface.
31. Acanthosphæra reticulata, n. sp. (Pl. 26, fig.
5).
Rhaphidosphæra reticulata, Haeckel, 1881, Prodromus.
Shell thick walled, with irregular, roundish pores, twice to four times as broad as the bars;
six to eight on the radius. Surface of the bars covered with a peculiar delicate network of very
fine crests. Twenty to forty radial spines, angular, pyramidal, scarcely one-third as long as the
radius of the shell, as broad at the base as the bars.
Dimensions.—Diameter of the shell 0.22, pores 0.02 to 0.04, bars 0.01; length of
the spines 0.04, basal breadth 0.01.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
Genus 89. Heliosphæra,[123] Haeckel,
1862, Monogr. d. Radiol., p. 350 (sensu emendato).
Definition.—Astrosphærida with one simple lattice-sphere,
covered with simple radial spines of two different kinds: larger main spines and smaller
by-spines.
The genus Heliosphæra (in the mended definition here employed) differs from
the foregoing Acanthosphæra in the possession of two different kinds of radial spines:
larger main spines scattered on the surface or disposed regularly in limited numbers (twelve to
twenty, sometimes forty to fifty or more), and smaller by-spines in much larger numbers, arising
from all the nodal-points of the network (or sometimes also from its bars).
Subgenus 1. Heliosphærella, Haeckel.
Definition.—Pores of the shell regular or subregular, all of nearly
equal size and similar form.
1. Heliosphæra hexagonaria, n. sp. (Pl. 26, fig.
2).
Shell very thin walled, about twenty times as broad as one pore. Meshes or pores subregular,
hexagonal, with thread-like bars; fifteen to seventeen on the radius. Radial spines at the
nodal-points of the network; about forty main spines three-sided pyramidal, half as broad at the
base as one pore, and twice as long as the bristle-shaped by-spines, which are very numerous, and
as long as the diameter of one pore.
{218}
Dimensions.—Diameter of the shell 0.25 to 0.3, of the meshes or pores 0.012 to
0.015, bars below 0.001; length of the main spines 0.03, basal breadth 0.007.
Habitat.—Central Pacific, Stations 272 to 274, depth 2350 to 2750
fathoms.
2. Heliosphæra actinota, Haeckel.
Heliosphæra actinota, Haeckel, 1862, Monogr. d. Radiol., p. 352, Taf. ix.
fig. 3.
Shell very thin walled, about ten times as broad as one mesh. Pores regular, hexagonal, with
thread-like bars; six to eight on the radius. Radial spines at the nodal-points of the network,
bristle-shaped, scarcely broader than the bars; about twenty main spines as long as the diameter
of the shell, and numerous by-spines, only one-third to one-half as long as the former.
Dimensions.—Diameter of the shell 0.2 to 0.25, of the meshes 0.02 to 0.03, bars
below 0.001; length of the main spines 0.2 to 0.3.
Habitat.—Mediterranean (Messina), Atlantic (Canaries, Azores),
surface.
3. Heliosphæra echinoides, Haeckel.
Heliosphæra echinoides, Haeckel, 1862, Monogr. d. Radiol., p. 352, Taf.
ix. fig. 4.
Shell thin walled, about six times as broad as one mesh. Pores regular, hexagonal, eight times
as broad as the bars. Radial spines arising, not from the nodal-points of the network, but from
the midst of the bars (very rare disposition!); twenty main spines regularly disposed, as long as
the radius, four times as long as the numerous by-spines; all spines bristle-shaped, of the same
breadth as the bars.
Dimensions.—Diameter of the shell 0.09, pores 0.015, bars 0.002; length of the
main spines 0.04.
Habitat.—Mediterranean (Messina), surface, Haeckel.
4. Heliosphæra elegans, Haeckel.
Heliosphæra elegans, Haeckel, 1862, Monogr. d. Radiol., p. 353, Taf. ix.
fig. 5.
Shell very thin walled, about ten times as broad as one mesh. Pores regular, hexagonal, with
thread-like bars, which are crossed by tangential bars, so that each side of a hexagon exhibits a
regular rectangular cross (exactly the same as in Pl. 19, fig. 5).
All radial spines bristle-shaped, as thin as the bars, and arising from the nodal-points; twenty
main spines as long as the radius, numerous by-spines scarcely one-sixth as long.
Dimensions.—Diameter of the shell 0.26, pores 0.026, bars below 0.001; length of
the main spines 0.13.
Habitat.—Mediterranean (Messina), surface, Haeckel.
5. Heliosphæra pectinata, n. sp. (Pl. 26, fig.
9).
Acanthosphæra pectinata, Haeckel, 1881, Atlas.
Shell thick walled, combed, about fourteen times as broad as one mesh. Pores subregular,
circular, with elevated hexagonal frames, three times as broad as the bars. Radial spines very
{219}numerous and stout; thirty to forty main spines,
three-sided pyramidal, nearly as long as the radius, as broad as one mesh; by-spines small,
conical, everywhere scattered at the nodal-points of the network and on the high combs of the
bars.
Dimensions.—Diameter of the shell 0.16, pores 0.012, bars 0.004; length of the
main spines 0.07, basal breadth 0.016.
Habitat.—West Tropical Pacific, Station 225, depth 4475 fathoms.
6. Heliosphæra coronata, n. sp. (Pl. 26,
figs. 6, 6a).
Acanthosphæra coronata, Haeckel, 1881, Atlas.
Shell thick walled, about ten times as broad as one mesh. Pores regular, circular, five times
as broad as the bars; each pore surrounded by a regular coronal of six short, conical by-spines
(fig. 6a); twenty to thirty main spines, also conical, half as long as the radius, as broad
as one pore.
Dimensions.—Diameter of the shell 0.15, pores 0.015, bars 0.003; length of the
main spines 0.04, basal breadth 0.015.
Habitat.—Central Pacific, Station 272, depth 2600 fathoms.
7. Heliosphæra floribunda, n. sp.
Shell thick walled about ten times as broad as one mesh. Pores regular, six-lobed, twice as
broad as the bars; each pore surrounded by six small conical by-spines (as in Pl. 28, figs. 1,
1b); twenty main spines regularly disposed cylindro-conical, as long as the diameter of the
shell or longer.
Dimensions.—Diameter of the shell 0.15, pores 0.015, bars 0.008; length of the
main spines 0.16, breadth 0.008.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
8. Heliosphæra cristata, n. sp.
Shell thick walled, about twelve times as broad as one mesh. Pores subregular, circular, twice
as broad as the bars; each pore surrounded by an elegant coronal of ten to twenty small, conical
by-spines; twenty main spines regularly disposed, conical, only one-third as long as the radius,
as broad at the base as one mesh.
Dimensions.—Diameter of the shell 0.25, pores 0.02, bars 0.01; length of the main
spines 0.04, basal breadth 0.02.
Habitat.—South Pacific, Station 285, depth 2375 fathoms.
9. Heliosphæra castanella, n. sp.
Shell thick walled, about twenty times as broad as one mesh. Pores regular, circular, of the
same breadth as the bars. Whole surface densely covered with innumerable bristle-shaped by-spines,
half as long as the radius; fifty to eighty main spines, conical, nearly as long as the diameter,
{220}twice as broad at the base as one pore. (Very
similar to some species of Castanella, Pl. 113,
but without the osculum characteristic of this Phæodarian; may be easily confounded with it.)
Dimensions.—Diameter of the shell 0.3, pores 0.015, bars 0.015; length of the main
spines 0.25, basal breadth 0.03.
Habitat.—North Pacific (Japan), Station 234, surface.
Subgenus 2. Heliosphæromma, Haeckel.
Definition.—Pores of the shell irregular, of different size or
form.
10. Heliosphæra polygonaria, n. sp.
Shell very thin walled, with thread-like bars and irregular, polygonal pores (having four to
eight angles, commonly five to seven); eight to ten on the radius. Radial spines at all the
nodal-points of the network, bristle-shaped; forty to sixty main spines, as long as the radius,
twice as thick as the numerous by-spines, which are not larger than one mesh.
Dimensions.—Diameter of the shell 0.15 to 0.2, pores 0.012 to 0.02, bars 0.001;
length of the main spines 0.08 to 0.1.
Habitat.—Central Pacific, Stations 266 to 272, surface.
11. Heliosphæra heteracantha, n. sp.
Shell thin walled, with irregular, polygonal pores, twice to four times as broad as the bars;
six to eight on the radius. Twenty radial main spines, three-sided pyramidal, nearly as long as
the diameter of the shell, as broad as a larger mesh; innumerable bristle-shaped by-spines
variously distributed on the bars and at the nodal-points of the net; half as long as the
radius.
Dimensions.—Diameter of the shell 0.12, pores 0.006 to 0.012, bars 0.003; length
of the main spines 0.1, basal breadth 0.012.
Habitat.—Equatorial Atlantic, Station 347, surface.
12. Heliosphæra hyperionis, n. sp.
Shell thick walled. Pores irregular, roundish, with polygonal frames, three to six times as
broad as the bars; twelve to fourteen on the radius. Thirty to forty main spines, angular,
pyramidal, scarcely as long as the radius and twice as long as the numerous bristle-shaped
by-spines.
Dimensions.—Diameter of the shell 0.24, pores 0.006 to 0.012, bars 0.002; length
of the main spines 0.1, basal breadth 0.012.
Habitat.—North Pacific, Station 256, surface.
13. Heliosphæra elector, n. sp.
Shell thick walled, with irregular, roundish pores, twice to three times as broad as the bars;
eight to ten on the radius. Twenty main spines, three-sided pyramidal, somewhat longer than the
radius and four times as long as the short bristle-shaped by-spines.
{221}
Dimensions.—Diameter of the shell 0.12, pores 0.005 to 0.01, bars 0.003; length of
the main spines 0.08, basal breadth 0.01.
Habitat.—South Atlantic, Station 325, surface.
14. Heliosphæra solaris, n. sp.
Shell thick walled, with irregular, roundish pores, about the same breadth as the bars; six to
eight on the radius. Fifty to eighty main spines, conical, as long as the radius; by-spines very
numerous, also conical, but only as large as one pore.
Dimensions.—Diameter of the shell 0.15, pores and bars 0.006 to 0.008; length of
the main spines 0.08, basal breadth 0.012.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
Genus 90. Conosphæra,[124] Haeckel, 1881,
Prodromus, p. 451.
Definition.—Astrosphærida with one simple latticed
sphere, covered with radial spines having the form of hollow cones with porous walls.
The genus Conosphæra differs from Acanthosphæra in the peculiar
formation of the radial spines, which are not simple solid sticks, but hollow cones with porous
walls, as immediate elevations of the hollow sphere.
1. Conosphæra platyconus, n. sp.
Pores of the shell regular, circular, twice as broad as the bars; ten to twelve on the radius.
Conical spines about sixty, regular, broader than they are high, with six to nine pores in the
wall.
Dimensions.—Diameter of the shell 0.16, pores 0.008, bars 0.004; length of the
spines 0.012, basal breadth 0.024.
Habitat.—Central Pacific, Station 272, depth 2600 fathoms.
2. Conosphæra orthoconus, n. sp. (Pl. 12, fig.
2).
Pores of the shell irregular, roundish, twice to three times as broad as the bars; fourteen to
sixteen on the radius. Conical spines about forty, regular, higher than they are broad, as long as
the radius, with sixteen to twenty pores in the wall.
Dimensions.—Diameter of the shell 0.2, pores 0.008 to 0.012, bars 0.004; length of
the spines 0.05, basal breadth 0.03.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
{222}
3. Conosphæra plagioconus, n. sp. (Pl. 12, fig.
4).
Pores of the shell irregular, polygonal, twice to five times as broad as the bars; twenty to
twenty-four on the radius. Conical spines about eighty, irregularly formed and scattered, with
oblique (not radial) axes; about as high as broad, with eight to twelve pores in the wall.
Dimensions.—Diameter of the shell 0.25, pores 0.005 to 0.015, bars 0.003; length
of the spines 0.02, basal breadth 0.02.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms.
Genus 91. Coscinomma,[125] n. gen.
Definition.—Astrosphærida with one simple lattice-sphere,
covered with simple radial spines; the pores between them prolonged into hollow, conical, or
cylindrical tubuli.
The genus Coscinomma exhibits among the Astrosphærida the same peculiar
formation that distinguishes Ethmosphæra and Sethosphæra among the Liosphærida; each
pore of the simple shell is prolonged into a short conical or cylindrical tubulus, as a rule
either on the outside or on the inside of the shell, but sometimes on both sides.
Subgenus 1. Coscinommarium, Haeckel.
Definition.—Pores prolonged into short tubes both on the inside as
well as the outside of the shell.
1. Coscinomma amphisiphon, Haeckel (Pl. 26, figs. 1,
1a, 1b).
Pores regular, circular, hexagonally framed, twice as broad as the bars, prolonged on the
inside as well as on the outside of the shell into a short truncated conical tube; fifteen to
eighteen pores on the radius. In each hexagon-corner arises a bristle-shaped radial spine, half as
long as the radius.
Dimensions.—Diameter of the shell 0.22, pores 0.01, bars 0.005; length of the
spines 0.1.
Habitat.—Central Pacific, Station 271, surface.
Subgenus 2. Coscinommidium, Haeckel.
Definition.—Pores prolonged into external tubes on the outside of the
shell.
2. Coscinomma ectosiphon, n. sp.
Pores regular, circular, hexagonally framed, twice as broad as the bars, prolonged on the
outside of the shell into a short truncated conical tube; ten to twelve pores on the radius. In
each {223}hexagon-corner arises a short bristle-shaped
spine, twice as long as the tube, one-third as long as the radius. (Very similar to Ethmosphæra
conulosa, Pl. 12,
fig. 5, but differs in the possession of radial spines.)
Dimensions.—Diameter of the shell 0.2, pores 0.012, bars 0.006; length of the
spines 0.04, of the tubes 0.02.
Habitat.—North Pacific, Station 253, surface.
3. Coscinomma macrosiphon, n. sp.
Pores regular, circular, without hexagonal frames, of the same breadth as the bars, prolonged
on the outside of the shell into a long cylindrical tube, half as long as the radius (eight to
nine pores on the radius); between them bristle-shaped, at the base conical, radial spines of
double length.
Dimensions.—Diameter of the shell 0.16, pores and bars 0.008; length of the spines
0.08, of the tubes 0.04.
Habitat.—Central Pacific, Station 274, surface.
Subgenus 3. Coscinommonium, Haeckel.
Definition.—Pores prolonged into internal tubes on the inside of the
shell.
4. Coscinomma endosiphon, n. sp.
Pores regular, circular, hexagonally framed, twice as broad as the bars (fourteen to sixteen on
the radius), prolonged on the inside of the shell into a short truncated cylindrical tube. In each
hexagon-corner arises a thin, bristle-shaped, radial spine with pyramidal base, half as long as
the radius, twice as long as the tube.
Dimensions.—Diameter of the shell 0.25, pores 0.012, bars 0.006; length of the
spines 0.066, of the tubes 0.03.
Habitat.—South Pacific, Station 288, surface.
Genus 92. Cladococcus,[126] J. Müller,
1856, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 485.
Definition.—Astrosphærida with one simple lattice-sphere,
covered with branched radial spines (the stem of the spine never forked).
The genus Cladococcus, together with the following Elaphococcus, is
distinguished from the other Coscinommida by the ramification of the radial spines
covering the surface of the simple hollow lattice-sphere. In Cladococcus each spine sends
out three or more lateral branches, which are either simple or again ramified; but the stem of the
spine itself is not forked, as in Elaphococcus.
{224}
Subgenus 1. Cladococcalis, Haeckel.
Definition.—Branches of the spines simple, not ramified. Pores
regular, all of nearly equal size and similar form.
1. Cladococcus arborescens, J. Müller.
Cladococcus arborescens, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 31, Taf. i. fig. 2.
Pores of the spherical shell regular, hexagonal, three times as broad as the bars; three to
four on the radius. Ten to twenty spines, three-sided prismatic, two to three times as long as the
shell diameter; towards the distal end each spine with three branches (one lateral simple straight
branch on each edge of the spine).
Dimensions.—Diameter of the shell 0.1, pores 0.012, bars 0.004; length of the
spines 0.2 to 0.3, breadth 0.02.
Habitat.—Mediterranean (Nice); North Atlantic, Canary Islands,
surface.
2. Cladococcus spinifer, Haeckel.
Cladococcus spinifer, Haeckel, 1862, Monogr. d. Radiol., p. 368, Taf.
xiii. fig. 9.
Pores regular, circular, hexagonally framed, three times as broad as the bars; five to six on
the radius. Radial spines, arising from all the nodal-points of the network, three-sided, longer
than the shell diameter, with six to twelve simple verticillate branches (two to four branches on
each edge).
Dimensions.—Diameter of the shell 0.08, pores 0.01, bars 0.003; length of the
spines 0.1, breadth 0.03.
Habitat.—Mediterranean (Messina), surface.
3. Cladococcus penicillus, n. sp.
Pores subregular, hexagonal, twice as broad as the bars; eight to ten on the radius. Sixty to
eighty radial spines, three-sided prismatic, pencil-shaped, longer than the shell diameter; each
at the distal end with a brush or pencil composed of nine to twenty-one short, simple, curved
branches (three to seven on each edge).
Dimensions.—Diameter of the shell 0.1, pores 0.006, bars 0.003; length of the
spines 0.12 to 0.16, breadth 0.006.
Habitat.—Central Pacific, Station 271, surface.
Subgenus 2.
Cladococcinus, Haeckel.
Definition.—Branches of the spines simple, not ramified. Pores
irregular, of different size or form.
{225}
4. Cladococcus antarcticus, n. sp.
Pores irregular, polygonal, twice to four times as broad as the bars; five to six on the
radius. Forty to fifty radial spines, angular, curved, of variable size and form, with three to
nine irregular, simple, blunt curved branches.
Dimensions.—Diameter of the shell 0.14, pores 0.01 to 0.02, bars 0.05; length of
the spines 0.1 to 0.18, breadth 0.007.
Habitat.—Antarctic Ocean, Station 157, depth 1950 fathoms.
5. Cladococcus japonicus, n. sp.
Pores irregular, roundish, of the same breadth as the bars; six to eight on the radius. Twenty
to thirty radial spines, angular, longer than the shell diameter, with thirty to forty simple
branches, decreasing in size from the middle part of the spine to the distal end (ten to thirteen
branches on each edge).
Dimensions.—Diameter of the shell 0.12, pores and bars 0.007; length of the spines
0.15 to 0.2, breadth 0.004.
Habitat.—North Pacific (Japan), Station 240, surface.
6. Cladococcus quadricuspis, n. sp.
Pores irregular, roundish, twice to eight times as broad as the bars; eight to ten on the
radius. About twenty radial spines, three-sided prismatic, as long as the shell diameter; in the
proximal half simple, with smooth edges; in the distal half with three diverging simple branches,
half the length of the spine, with dentated edges.
Dimensions.—Diameter of the shell 0.15, pores 0.004 to 0.016, bars 0.002; length
of the spines 0.17, breadth 0.01.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms.
Subgenus 3. Cladococcodes, Haeckel.
Definition.—Branches of the spines again ramified. Pores regular, of
nearly equal size and similar form.
7. Cladococcus scoparius, n. sp. (Pl. 27, fig.
2).
Pores regular, circular, three times as broad as the bars; five to six on the radius. Twenty
radial spines, broom-shaped, three times as long as the shell diameter; in the proximal half
simple, in the distal half branched, with three to nine branches, which are again ramified; spines
and their branches with three smooth edges, not dentated.
Dimensions.—Diameter of the shell 0.055, pores 0.0075, bars 0.0025; length of the
spines 0.15, basal thickness 0.007.
Habitat.—Central Pacific, Station 271, surface.
{226}
8. Cladococcus viminalis, Haeckel.
Cladococcus viminalis, Haeckel, 1862, Monogr. d. Radiol., p. 369, Taf.
xiv. figs. 2, 3.
Pores regular, circular, hexagonally framed, four times as broad as the bars; five to six on
the radius. Radial spines, arising from all the nodal-points of the network, twice as long as the
shell diameter; in the basal half simple, in the distal half with three to six long, thin, curved
branches, which are partly forked; three edges of the spines dentated.
Dimensions.—Diameter of the shell 0.08, pores 0.01, bars 0.003; length of the
radial spines 0.16, basal breadth 0.003.
Habitat.—Mediterranean (Messina).
9. Cladococcus bifurcus, Haeckel.
Cladococcus bifurcus, Haeckel, 1862, Monogr. d. Radiol., p. 368, Taf.
xiii. figs. 7, 8.
Pores regular, circular, hexagonally framed, four times as broad as the bars; five to seven on
the radius. Radial spines arising from all the nodal-points, three-sided prismatic, with dentated
edges, longer than the shell diameter. On each spine six to nine branches, which are for the most
part forked, the distal branches only being simple. (May be regarded as a further developmental
stage of Cladococcus spinifer and Cladococcus viminalis.)
Dimensions.—Diameter of the shell 0.08, pores 0.01, bars 0.0025; length of the
spines 0.1, basal breadth 0.003.
Habitat.—Mediterranean (Messina), Canary Islands, Azores, surface.
10. Cladococcus pinetum, n. sp. (Pl. 27,
fig. 1).
Pores regular, circular, polygonally framed, about the same breadth as the bars: two to three
on the radius. About twenty radial spines, very large, three to four times as long as the shell
diameter, branched like a pine tree, with straight, stout, three-sided prismatic stem; three
prominent edges dentated. On each edge five to seven lateral branches, the distal ends of which
are simple, the proximal again ramified, with numerous ramules. The figured specimen is a young
one, with branches but little developed.
Dimensions.—Diameter of the shell 0.06, pores and bars 0.008; length of the spines
0.2 to 0.25 breadth 0.01.
Habitat.—Central Pacific, Station 266 to 274, surface.
Subgenus 4. Cladococcurus, Haeckel.
Definition.—Branches of the spines again ramified. Pores irregular, of
different size and form.
11. Cladococcus abietinus, n. sp. (Pl. 27, fig.
3).
Pores irregular, roundish, twice to four times as broad as the bars; six to ten on the radius.
About twenty radial spines, two to three times as long as the shell diameter, branched like a pine
{227}tree, with straight, stout, three-sided pyramidal
stem. From the dentated edges arise numerous verticillate branches, the proximal ends of which are
thickly ramified. (Differs from the preceding and similar species mainly in the large size and
irregular lattice-work of the shell.)
Dimensions.—Diameter of the shell 0.12 to 0.14, pores 0.006 to 0.012, bars 0.003;
length of the spines 0.25 to 0.4, breadth 0.012.
Habitat.—North Pacific, Station 256, surface.
12. Cladococcus tricladus, n. sp.
Pores irregular, polygonal, twice to eight times as broad as the bars; eight to ten on the
radius. About twenty radial spines, three-sided prismatic, twice as long as the shell diameter; in
the proximal half simple, with smooth edges; in the distal half with three diverging curved
branches, which bifurcate two to three times or ramify irregularly. (Closely related to the
simpler Cladococcus quadricuspis.)
Dimensions.—Diameter of the shell 0.16, pores 0.04 to 0.016, bars 0.002; length of
the spines 0.3, breadth 0.012.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
13. Cladococcus stalactites, n. sp. (Pl. 27, fig.
4).
Pores irregular, roundish, once to five times as broad as the bars; eight to ten on the radius.
About twenty radial spines, very stout, longer than the shell diameter, with three wing-like,
prominent smooth edges. At the middle, or in the distal half, each spine bears a verticil of three
strong, irregularly formed ramified branches.
Dimensions.—Diameter of the shell 0.14, pores 0.003 to 0.015, bars 0.003; length
of the spines 0.16 to 0.2, breadth 0.02.
Habitat.—West Tropical Pacific, Station 220, depth 1100 fathoms.
14. Cladococcus dendrites, n. sp. (Pl. 27, fig.
5).
Pores irregular, roundish, with denticulate margin, twice to four times as broad as the bars;
eight to twelve on the radius. Fifty to eighty radial spines, three-sided prismatic, with
elegantly denticulated edges; in the proximal two-thirds simple, in the distal third with a bunch
of ten to twenty short simple spines (three to seven on each edge).
Dimensions.—Diameter of the shell 0.16 to 0.2, pores 0.006 to 0.012, bars 0.003;
length of the spines 0.2 to 0.3, breadth 0.01.
Habitat.—Antarctic Ocean, Station 157, depth 1950 fathoms.
Genus 93. Elaphococcus,[127] Haeckel,
1881, Prodromus, p. 450.
Definition.—Astrosphærida with one simple lattice-sphere,
covered with forked or dichotomously branched radial spines (the stem of the spine always
forked).
{228}
The genus Elaphococcus differs from the preceding Cladococcus in the
mode of ramification of the radial spines. These are forked; and the forked branches are either
simple, again forked, or dichotomously ramified.
Subgenus 1. Elaphococcinus, Haeckel.
Definition.—Pores of the shell regular, of nearly equal size and
similar form.
1. Elaphococcus furcatus, n. sp.
Pores of the spherical shell regular, hexagonal, four times as broad as the bars; four to five
on the radius. Radial spines, arising from all the nodal-points of the network, cylindrical, as
long as the radius, simply forked at the distal end; both branches half as long as the undivided
part.
Dimensions.—Diameter of the shell 0.08, pores 0.01, bars 0.0025; length of the
spines 0.04, breadth 0.004.
Habitat.—Tropical Atlantic, Station 348, surface.
2. Elaphococcus cervicornis, Haeckel.
Cladococcus cervicornis, Haeckel, 1862, Monogr. d. Radiol., p. 370, Taf.
xiv. figs. 4-6.
Pores regular, hexagonal, ten to twenty times as broad as the bars; five to seven on the
radius. Fifty to ninety radial spines (or more), arising not only from the nodal-points of the
network but also from the bars between them. Each spine is cylindrical, longer than the shell
diameter, and repeatedly forked (three to six times), having, therefore, numerous (thirty to sixty
or more) curved branches. The distal ends of all branches fall in one spherical face.
Dimensions.—Diameter of the shell 0.07 to 0.09, pores 0.01 to 0.015, bars 0.001 to
0.015; length of the spines 0.1 to 0.15, breadth 0.005.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Indian, Pacific,
surface.
3. Elaphococcus elaphoceras, n. sp.
Pores regular, circular, hexagonally framed, three to four times as broad as the bars; six to
eight on the radius. At each nodal-point of the hexagon arises a short bristle-shaped, simple
by-spine. In addition, there arise from the bars thirty to sixty large main spines, longer than
the shell diameter, repeatedly forked in the same way as in the preceding species.
Dimensions.—Diameter of the shell 0.1, pores 0.008, bars 0.002; length of the
spines 0.12, breadth 0.002.
Habitat.—Central Pacific, Station 272. depth 2600 fathoms.
4. Elaphococcus umbellifer, n. sp.
Pores regular, circular, three times as broad as the bars; ten to twelve on the radius. Twenty
to forty straight cylindrical spines, as long as the shell diameter, having at the distal end a
regular {229}umbel composed of nine to twelve curved
branches of equal length, which are again ramified and resemble the inflorescence of an
umbelliferous plant, the distal ends of all ramules falling in a spherical face.
Dimensions.—Diameter of the shell 0.12, pores 0.006, bars 0.002; length of the
spines 0.14, breadth 0.002.
Habitat.—South Atlantic, Station 325, surface.
Subgenus 2. Elaphococculus, Haeckel.
Definition.—Pores of the shell irregular, of different size or
form.
5. Elaphococcus dichotomus, n. sp.
Pores irregular, polygonal, twice to four times as broad as the bars; six to eight on the
radius. Thirty to sixty radial spines cylindrical, curved, as long as the shell diameter, simply
forked at the distal end; both branches one-third as long as the undivided part.
Dimensions.—Diameter of the shell 0.15, pores 0.007 to 0.015, bars 0.004; length
of the spines 0.2, breadth 0.004.
Habitat.—Arctic Ocean, Greenland, surface (Koch).
6. Elaphococcus umbellatus, n. sp.
Pores irregular, roundish, or polygonal, twice to five times as broad as the bars; six to eight
on the radius. Forty to sixty radial spines cylindrical, curved, as long as the shell radius,
having at the distal end an irregular umbel, composed of six to twelve short branches, which are
irregularly ramified or forked. (Differs from the regular Elaphococcus umbellifer mainly in
the irregularity.)
Dimensions.—Diameter of the shell 0.14, pores 0.007 to 0.015, bars 0.003; length
of the spines 0.12, breadth 0.01.
Habitat.—South-east Pacific (Juan Fernandez), Station 299,
surface.
7. Elaphococcus drymodes, n. sp.
Pores irregular, roundish, little broader than the bars; ten to twelve on the radius. Eighty to
one hundred and twenty (or more) radial spines, cylindrical, three to four times as long as the
shell diameter, irregularly forked or repeatedly dichotomous (each spine with forty to sixty
forked branches); the distal ends of all branches fall in a spherical plane. (Differs from the
regular Elaphococcus cervicornis mainly in the irregularity.)
Dimensions.—Diameter of the shell 0.1, pores and bars 0.003 to 0.008; length of
the spines 0.3 to 0.4, breadth 0.01.
Habitat.—Arctic Ocean, Iceland (Steenstrup).
{230}
Subfamily Haliommida,[128] Haeckel.
Diplosphærida, Haeckel, 1881, Prodromus, pp. 449, 451.
Definition.—Astrosphærida with two concentric
spherical lattice-shells, united by radial beams.
Genus 94. Haliomma,[129] Ehrenberg, 1838,
Abhandl. d. k. Akad. d. Wiss. Berlin, p. 128.
Definition.—Astrosphærida with one medullary
(intracapsular) and one cortical (extracapsular) shell, which are connected by radial beams,
piercing the central capsule. Shell surface covered with simple radial spines of the same
kind.
The genus Haliomma, one of the oldest known Radiolarian genera, contained in
the catalogue of its discoverer, Ehrenberg, a large number of very different Sphærellaria, belonging to at least sixteen different genera. We limit here the
conception of the genus to those Haliommida which bear simple radial spines of one kind on the
surface of the cortical shell (the latter being separated from the medullary shell by the central
capsule).
Subgenus 1. Haliommantha, Haeckel.
Definition.—Pores of the cortical shell regular, of nearly equal size
and similar form; spines on the whole surface (commonly one spine at each nodal-point).
1. Haliomma hexagonium, n. sp.
Cortical shell four times as broad as the medullary shell, both having very thin thread-like
bars, and regular, hexagonal pores (eighteen to twenty on the radius of the outer, five to six on
the radius of the inner shell). The two shells connected by twenty thin thread-like radial beams.
At each nodal-point of the outer shell arises a bristle-shaped radial spine, half as long as the
radius. (Similar to Heliosoma radians, Pl. 28, fig. 3, but
with all the spines equal.)
Dimensions.—Diameter of the outer shell 0.2, inner 0.05; pores of the outer 0.015,
of the inner 0.01; length of the spines 0.05.
Habitat.—Equatorial Atlantic, Station 347, surface.
2. Haliomma arachnium, n. sp.
Cortical shell three times as broad as the medullary shell; pores of the former regular,
hexagonal, with very thin thread-like bars (fourteen to sixteen on the radius); pores of the
latter regular, {231}circular, three times as broad as
the bars (four to five on the radius). The two shells connected by about forty radial beams. At
each nodal-point of the outer shell arises one bristle-shaped radial spine, twice as long as the
diameter of one hexagonal mesh.
Dimensions.—Diameter of the outer shell 0.15, inner 0.05; pores of the outer
0.012, of the inner 0.006; length of the spines 0.025.
Habitat.—Central Pacific, Station 266, surface.
3. Haliomma favosum, n. sp.
Cortical shell thick walled, three times as broad as the medullary shell, with regular,
circular, hexagonally framed pores, twice as broad as the bars; eight to ten on the radius.
Medullary shell with simple, circular pores, of the same breadth as the bars. At each nodal-point
of the outer shell arises a short triangular spine three times as long as one pore.
Dimensions.—Diameter of the outer shell 0.12, inner 0.04; pores of the former
0.01, of the latter 0.005; bars 0.005; length of the spines 0.015.
Habitat.—Central Pacific, Station 271 to 274, surface.
4. Haliomma regulare, n. sp.
Cortical shell thin walled, nearly twice as broad as the medullary shell, and connected with it
by forty to sixty thin radial beams. Both shells of the same perfectly regular structure, with an
identical number of regular, circular pores, which are hexagonally framed, four times as broad as
the bars, nine to eleven on the radius. The outer pores are twice as broad as the inner, exactly
corresponding pores. Between every three pores of the outer surface (in each corner of the
cortical hexagon) arises one short three-sided pyramidal spine, twice as long as the diameter of
one pore.
Dimensions.—Diameter of the outer shell 0.2, of the inner 0.11; pores of the
former 0.018, of the latter 0.009; length of the spines 0.04.
Habitat.—South Atlantic (Tristan da Cunha), Station 332, depth 2200
fathoms.
5. Haliomma melitomma, n. sp. (Pl. 20,
fig. 4).
Melitomma formosum, Haeckel, 1879, Atlas, loc. cit.
Cortical shell thick walled, two and a half times as broad as the medullary shell, with regular
pores of very elegant structure, twice as broad as the bars; eight to ten on the radius. The inner
opening of each pore is simple, circular, the outer regular, six-lobed; corresponding to the six
lobes are six short conical spines, which arise from the six corners of the regular hexagonal
frames separating the pores. Pores of the medullary shell simple, circular, of the same breadth as
the bars. The two shells connected only by six radial beams (in three dimensive axes).
Dimensions.—Diameter of the outer shell 0.17, of the inner 0.07; pores of the
former 0.012, of the latter 0.004; length of the spines 0.005 to 0.01.
Habitat.—Indian Ocean, Zanzibar, Pullen, depth 2200 fathoms.
{232}
6. Haliomma lirianthus, n. sp. (Pl. 28,
figs. 1, 1a, 1b).
Cortical shell thick walled, three times as broad as the medullary shell, with regular,
rosette-shaped pores, twice as broad as the bars; eight to ten on the radius. The regular
structure of the elegant pores is the same as in the preceding species, but without prominent
crested frames (fig. 1b). Also the medullary shell (fig. 1a) is different, much
thinner and smaller, with simple, circular pores, which are three times as broad as the bars. The
two shells are connected by numerous (twenty?) radial beams.
Dimensions.—Diameter of the outer shell 0.15, of the inner 0.05; pores of the
former 0.01, of the latter 0.005; length of the spines 0.005 to 0.02.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
7. Haliomma castanea, Haeckel.
Haliomma castanea, Haeckel, 1862, Monogr. d. Radiol., p. 428, Taf. xxiv.
fig. 4.
Cortical shell thick walled, three times as broad as the medullary shell; pores of both
regular, circular, twice as broad as the bars (seven to eight on the radius of the outer, four to
five on the radius of the inner shell), the two connected by six to twelve (?) strong radial
beams. Radial spines bristle-shaped, with conical bases, twice as long as the diameter of the
cortical pores (one spine at the nodal-point between every three pores).
Dimensions.—Diameter of the outer shell 0.1 to 0.15, inner 0.03 to 0.05; pores of
the former 0.005, of the latter 0.003; length of the spines 0.005 to 0.01.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Indian, Pacific,
surface.
8. Haliomma horridum, Stöhr.
Haliomma horridum, Stöhr, 1880, Palæontogr., vol. xxvi. p. 87, Taf. i.
fig. 10.
Cortical shell thick walled, two and a half times as broad as the medullary shell. Pores
regular, circular, of the same breadth as the bars; eight to ten on the radius. Radial spines
conical, stout, nearly half as long as the radius. (Differs from the closely allied Haliomma
castanea in the smaller pores and larger spines.)
Dimensions.—Diameter of the outer shell 0.14, of the inner 0.06; pores of the
former 0.004, of the latter 0.02.
Habitat.—Fossil in Tertiary rocks of Barbados and Sicily; living in
the Atlantic, Station 9, depth 3150 fathoms, and Station 353, depth 2965 fathoms.
9. Haliomma datura, n. sp.
Cortical shell thin walled, only one and a half times as broad as the medullary shell, both
having regular, circular pores, four to six times as broad as the bars (five to six on the radius
of the outer, three to four on the radius of the inner shell), the two connected by numerous
(forty to sixty?) thin, short, radial beams. Radial spines conical, twice as long as the diameter
of one cortical pore, arising from all the nodal-points between them.
{233}
Dimensions.—Diameter of the outer shell 0.08 to 0.12, inner 0.06 to 0.08; pores of
the former 0.015 to 0.02, of the latter 0.007 to 0.012; length of the spines 0.02.
Habitat.—Central Pacific, Stations 268 to 274, surface.
Subgenus 2. Haliommetta, Haeckel.
Definition.—Pores of the cortical shell regular, of nearly equal size
and similar form; the spines not covering the entire surface, but scattered at intervals (their
number smaller than that of the nodal-points of the network).
10. Haliomma circumtextum, n. sp. (Pl. 28, figs. 7,
7a).
Cortical shell very delicate, with thin thread-like bars, and regular, hexagonal pores, little
larger than the thick-walled medullary shell (= 7 : 6). Pores of the latter regular,
circular, double-edged, with hexagonal frames, of the same breadth as the bars; from all the
hexagon-corners arise thin bristle-shaped, radial spines, twice as long as the diameter of the
pores, becoming connected at equal distances from the centre by tangential threads, regularly
disposed, forming the cortical shell. Twelve strong, three-sided pyramidal, radial spines, as
broad at the base as one mesh, and about half as long as the radius of the shell.
Dimensions.—Diameter of the outer shell 0.14, of the inner 0.12; pores of the
former 0.012, of the latter 0.005; length of the radial spines 0.04, basal breadth 0.012.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
11. Haliomma duodecinum, n. sp.
Cortical shell thick walled, three times as broad as the medullary shell. Both shells with
regular, circular pores, twice as broad as the bars; eight to ten on the radius of the outer, five
to six on the radius of the inner shell. Twelve conical, regularly disposed radial spines, as long
as the radius, and as broad at the base as one mesh.
Dimensions.—Diameter of the outer shell 0.12, of the inner 0.04; pores of the
former 0.005, of the latter 0.002; length of the spines 0.05, basal breadth 0.005.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms.
12. Haliomma megaporum, Ehrenberg.
Haliomma megaporum, Ehrenberg, 1872, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 313.
Cortical shell thin walled, three times as broad as the medullary shell; the pores regular,
circular, three to four on the radius, eight times as broad as the bars, quite as broad as the
medullary shell, the pores of which are much smaller, twice as broad as the bars. Eight radial
spines regularly disposed, conical, as long as the radius. (As the diagnosis of Ehrenberg is very
{234}incomplete, and no figure is given with it, it
remains doubtful whether his Mediterranean species be identical with my Atlantic variety.)
Dimensions.—Diameter of the outer shell 0.12, of the inner 0.04; pores of the
former 0.04, of the latter 0.008; length of the spines 0.06.
Habitat.—Mediterranean (Grecian shore); North Atlantic, Station 354,
surface.
13. Haliomma oculatum, Ehrenberg.
Haliomma oculatum, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss. Berlin,
p. 74, Taf. xxviii. figs. 2, 3.
Cortical shell thick walled, four times as broad as the medullary shell; its pores regular,
circular, four times as broad as the bars, six to eight on the radius. Pores of the medullary
shell regular, hexagonal, with very thin bars, three to four on the radius. Nine radial spines,
regularly disposed, conical, as long as the radius, as broad at the base as one cortical pore. (In
the specimen figured by Ehrenberg, only two spines were preserved, seven being accidentally broken
off.)
Dimensions.—Diameter of the outer shell 0.16, of the inner 0.04; pores of the
former 0.01, of the latter 0.005.
Habitat.—Fossil in Barbados.
14. Haliomma enneaxiphos, n. sp.
Cortical shell thick walled, four times as broad as the dark medullary shell, with regular,
circular, hexagonally framed pores, twice as broad as the bars; eight to ten on the radius. Nine
radial spines regularly disposed, three sided pyramidal, as long as the radius, as broad at the
base as one mesh.
Dimensions.—Diameter of the outer shell 0.11, of the inner 0.03; cortical pores
0.008, bars 0.004; length of the radial spines 0.05, basal breadth 0.012.
Habitat.—Central Pacific, Station 272, surface.
15. Haliomma tenuispinum, J. Müller.
Haliomma tenuispinum, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 39,
Taf. iv. fig. 9.
Haliomma tenuispinum, Haeckel, 1862, Monogr. d. Radiol., p. 431.
Cortical shell thin walled, three times as broad as the medullary shell. Both shells with very
fine, thread-like bars, and regular, hexagonal pores; five to seven on the radius. Twenty radial
spines, very thin, bristle-shaped, as long as the radius.
Dimensions.—Diameter of the outer shell 0.12, of the inner 0.04; pores of the
former 0.016, of the latter 0.006.
Habitat.—Mediterranean (Nice); North Atlantic, Station 353,
surface.
{235}
16. Haliomma longispinum, J. Müller.
Haliomma longispinum, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 39,
Taf iv. fig. 8.
Haliomma longispinum, Haeckel, 1862, Monogr. d. Radiol., p. 431.
Cortical shell thick walled, three times as broad as the dark medullary shell. Pores regular,
circular, hexagonally framed, twice as broad as the bars; four to six on the radius. Twenty radial
spines, very long, regularly disposed, four to five times as long as the diameter of the shell,
three-sided prismatic, with elegantly dentated edges, as broad as one cortical mesh.
Dimensions.—Diameter of the outer shell 0.08, of the inner 0.03; pores of the
former 0.01, bars 0.005; length of the spines 0.3 to 0.4.
Habitat.—Mediterranean (Nice); Central Pacific, Station 272,
surface.
17. Haliomma capense, n. sp.
Cortical shell thick walled, twice as broad as the medullary shell, both with regular, circular
pores, twice as broad as the bars; eight to ten on the radius of the outer, four to six on the
radius of the inner shell. Forty to sixty radial spines, conical, as long as the radius, as broad
as one cortical pore.
Dimensions.—Diameter of the outer shell 0.11, of the inner 0.05; pores of the
former 0.01, bars 0.005; length of the spines 0.006, basal breadth 0.012.
Habitat.—Cape of Good Hope, Station 142, surface.
18. Haliomma denticulatum, n. sp.
Cortical shell thick walled, three times as broad as the dark medullary shell, with regular,
circular, double-edged pores, of the same breadth as the bars; eight to ten on the radius. Thirty
to fifty radial spines, three-sided pyramidal, half as long as the radius, with three denticulated
edges. (Very similar to Actinomma denticulatum, Pl. 29, fig. 3, but
with simple medullary shell.)
Dimensions.—Diameter of the outer shell 0.12, inner 0.04; pores and bars of the
former 0.008; length of the spines 0.04, basal breadth 0.01.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
19. Haliomma grande, n. sp.
Cortical shell thick walled, five times as broad as the dark medullary shell, with small,
regular, circular pores, half as broad as the bars; twenty to twenty-four on the radius. One
hundred to one hundred and twenty conical radial spines, five times as long as broad, only half as
long as the radius.
Dimensions.—Diameter of the outer shell 0.4, inner 0.08; pores of the outer 0.006,
bars 0.012; length of the spines 0.1, basal breadth 0.02.
Habitat.—Equatorial Atlantic, Station 348, depth 2450 fathoms.
{236}
Subgenus 3. Haliommilla, Haeckel.
Definition.—Pores of the cortical shell irregular, of different size
and form; spines covering the entire surface (commonly one spine at each nodal-point).
20. Haliomma capillaceum, Haeckel.
Haliomma capillaceum, Haeckel, 1862, Monogr. d. Radiol., p. 426, Taf.
xxiii. fig. 2.
Cortical shell very thin walled, seven to eight times as broad as the medullary shell, both
with irregular, polygonal pores, and very thin thread-like bars; outer pores twice to three times
as broad as the inner. Radial spines very numerous, straight, bristle-shaped, about as long as the
diameter of the medullary shell.
Dimensions.—Diameter of the outer shell 0.2, inner 0.025 to 0.03; outer pores 0.02
to 0.04, inner 0.008 to 0.016, bars 0.001; length of the spines 0.02 to 0.03.
Habitat.—Mediterranean (Messina), Atlantic (Canary Islands),
surface.
21. Haliomma erinaceum, Haeckel.
Haliomma erinaceum, Haeckel, 1862, Monogr. d. Radiol., p. 427, Taf. xxiii.
figs. 3, 4.
Cortical shell thin walled, seven to eight times as broad as the medullary shell, both with
irregular, polygonal pores, two to ten times as broad as the bars, outer pores much larger than
the inner. Radial spines very numerous, bristle-shaped, as long as the diameter of the medullary
shell, the majority or all being either curved or obliquely depressed, the greater part neither
straight nor radial.
Dimensions.—Diameter of the outer shell 0.2, inner 0.025 to 0.03; outer pores
0.005 to 0.03, inner 0.008 to 0.016, bars 0.002 to 0.004; length of the spines 0.03.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific, surface.
22. Haliomma tenellum, Haeckel.
Haliomma tenellum, Haeckel, 1862, Monogr. d. Radiol., p. 428.
Haliomma spinuloso affine, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 40, Taf. iv. fig. 7.
Cortical shell thin walled, three times as broad as the medullary shell, with irregular,
roundish pores, and very thin bars. Pores of the inner shell regular, circular. Radial spines very
numerous, bristle-shaped, straight, as long as the radius of the outer shell.
Dimensions.—Diameter of the outer shell 0.15, inner 0.05; outer pores 0.008 to
0.016, inner 0.005; length of the spines 0.07.
Habitat.—Mediterranean (Nice); Atlantic, Stations 349 to 354,
surface.
{237}
23. Haliomma spinulosum, J. Müller.
Haliomma spinulosum, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 39, Taf. iv. fig. 6.
Cortical shell thin walled, twice as broad as the medullary shell, with irregular, polygonal
pores, and very thin bars. Pores of the inner shell subregular, hexagonal, ten to twelve times as
broad as the bars. Radial spines very numerous, bristle-shaped, one-sixth to one-fourth as long as
the radius of the outer shell.
Dimensions.—Diameter of the outer shell 0.16, inner 0.08; outer pores 0.01 to
0.03, inner 0.02; length of the spines 0.02 to 0.03.
Habitat.—Mediterranean (Nice); Central Pacific, Station 266,
surface.
24. Haliomma rhodococcus, n. sp. (Pl. 19, fig.
6).
Sethosphæra rhodococcus, Haeckel, 1879, Atlas, pl. xix. fig. 6.
Cortical shell very thin walled, little larger than the thick walled medullary shell (=
10 : 9). Pores of the outer shell very irregular, roundish, twice to six times as broad
as the bars, about twenty on the radius. Pores of the inner shell twice as broad as the bars, six
to eight on the radius, very regular, circular on the inner, six-lobed on the outer opening,
separated by prominent hexagonal crests; in the hexagon-corners arise short conical radial spines,
which at equal distances from the centre are united by the outer shell, but are prominent over its
surface. (A very peculiar form; the two shells may better be regarded as inner and outer cortical
shell.)
Dimensions.—Diameter of the outer shell 0.2, inner 0.18; outer pores 0.04 to
0.012, inner 0.01; length of the spines 0.01.
Habitat.—Tropical Atlantic, Station 338, depth 1990 fathoms; also
fossil in Barbados.
25. Haliomma boreale, n. sp.
Cortical shell thick walled, four times as broad as the medullary shell, with irregular,
roundish pores, twice to four times as broad as the bars; eight to ten on the radius. Pores of the
inner shell regular, circular, twice as broad as the bars, four to six on the radius. Radial
spines very numerous, conical, nearly as long as the diameter of the shell.
Dimensions.—Diameter of the outer shell 0.24, inner 0.06; outer pores 0.008 to
0.02, bars 0.005, inner pores 0.006, bars 0.003; length of the spines 0.2.
Habitat.—Arctic Ocean, Greenland ("Alert" Expedition).
Subgenus 4. Haliommura, Haeckel.
Definition.—Pores of the cortical shell irregular, of different size
and form; spines not covering the entire surface, but scattered at intervals (their number smaller
than that of the nodal-points in the network).
{238}
26. Haliomma macrodoras, n. sp. (Pl. 28,
figs. 6, 6a).
Cortical shell thin walled, twice as broad as the medullary shell, with irregular, polygonal
meshes, three to six times as broad as the bars. Inner shell with regular, hexagonal meshes, ten
times as broad as the bars. Twenty radial spines, very long, stout, three-sided prismatic, two to
four times as long as the diameter of the shell, as broad as one medullary mesh, with three
wing-like, denticulated edges. At the base of each spine three supporting curved beams.
Dimensions.—Diameter of the outer shell 0.14 to 0.16, of the inner 0.07 to 0.08;
outer pores 0.01 to 0.02, bars 0.003; inner pores 0.01, bars 0.001; length of the spines 0.3 to
0.6, breadth 0.01.
Habitat.—Central Pacific, Station 271, surface.
27. Haliomma antarcticum, n. sp.
Cortical shell very thin walled, three times as broad as the medullary shell, with irregular,
polygonal pores, and very thin thread-like bars. Inner shell with regular, hexagonal meshes, six
times as broad as the bars. About forty radial spines, angular, pyramidal, half as long as the
radius of the shell, as broad at the base as the largest mesh.
Dimensions.—Diameter of the outer shell 0.2, of the inner 0.07; outer pores 0.006
to 0.015, inner 0.008; length of the spines 0.05, basal breadth 0.012.
Habitat.—Antarctic Ocean, Station 154, surface.
28. Haliomma wyvillei, Haeckel.
Haliomma wyvillei, Haeckel, 1878, Protistenreich, p. 44, fig. 31.
Haliomma species, Wyville Thomson, 1877, Atlantic, vol. i. p. 236, fig.
54.
Cortical shell thin walled, three times as broad as the medullary shell. Both shells with
irregular, large, polygonal pores, and very thin bars. From the inner shell arise very numerous
(eighty to one hundred and twenty or more) radial spines, which pierce the outer shell, and are
outside it, as long as the radius of the inner shell, straight, bristle-shaped, and as thick as
the bars.
Dimensions.—Diameter of the outer shell 0.18, of the inner 0.06; pores 0.01 to
0.02, bars 0.001 to 0.002; length of the free spines 0.06.
Habitat.—Central Pacific, Stations 266 to 274, surface.
29. Haliomma beroes, Ehrenberg.
Haliomma beroes, Ehrenberg, 1854, Mikrogeol., Taf. xxxvB.
B. iv. fig. 19.
Haliomma beroes, Haeckel, 1862, Monogr. d. Radiol., p. 434.
Cortical shell thin walled, three times as broad as the medullary shell, with irregular,
roundish pores, twice to four times as broad as the bars. Inner shell with regular, circular
pores, twice as broad as the bars. Both shells connected by four (or six ?) radial beams,
perpendicularly crossed. Thirty to forty radial spines, conical, thin, shorter than the
radius.
Dimensions.—Diameter of the outer shell 0.11, inner 0.04; outer pores 0.007 to
0.012, inner 0.005; length of the spines 0.03, basal breadth 0.005.
Habitat.—Atlantic, depth 2000 fathoms.
{239}
30. Haliomma compactum, n. sp. (Pl. 28,
figs. 5, 5a).
Cortical shell very thick walled, four times as broad as the thin walled medullary shell; pores
of the former irregular, roundish, with high polygonal frames of very different size, twice to
four times as broad as the bars. Inner shell with simple, small, polygonal pores, connected with
the outer shell by eight (?) regularly disposed thin radial beams, opposite in pairs, prolonged
outside into strong conical spines, longer than the radius.
Dimensions.—Diameter of the outer shell 0.08, inner 0.02; outer pores 0.005 to
0.01, inner 0.005; length of the spines 0.06, basal breadth 0.012.
Habitat.—Central Pacific, Stations 265 to 272, depth 2425 to 2925
fathoms.
31. Haliomma permagnum, n. sp.
Cortical shell thick walled, six times as broad as the medullary shell, with irregular,
roundish, double-edged pores, three to six times as broad as the bars. Inner shell with regular,
circular pores, twice as broad as the bars. One hundred and twenty to one hundred and fifty radial
spines, conical, only one-fourth as long as the radius.
Dimensions.—Diameter of the outer shell 0.42, inner 0.07; outer pores 0.012 to
0.025, bars 0.004; inner pores 0.008, bars 0.004; length of the spines 0.05, basal breadth
0.03.
Habitat.—Central Pacific, Station 268, surface.
32. Haliomma patagonicum, n. sp.
Cortical shell thick walled, four times as broad as the medullary shell, with irregular,
roundish pores, twice to three times as broad as the bars. Inner shell also with irregular,
roundish pores, of half the size. Forty to fifty radial spines, cylindro-conical, about as long as
the shell diameter.
Dimensions.—Diameter of the outer shell 0.24, inner 0.06; outer pores 0.012 to
0.02, bars 0.006; inner pores 0.005 to 0.01, bars 0.004; length of the spines 0.3, breadth
0.02.
Habitat.—South-east Pacific, west coast of Patagonia, Station 302,
surface.
33. Haliomma clavatum, n. sp.
Cortical shell thick walled, three times as broad as the medullary shell, with irregular,
roundish pores little larger than the bars; eight to twelve on the radius. Inner shell with
similar but smaller pores. About twenty radial spines, club-shaped, as long as the radius,
three-sided, with prominent edges, twice as broad at the distal end as at the base. (Similar to
Acanthosphæra clavata, Pl. 26,
fig. 8, but differs from it in the possession of a medullary shell.)
Dimensions.—Diameter of the outer shell 0.2, inner 0.06; outer pores and bars
0.008 to 0.02, inner 0.006 to 0.01; length of the spines 0.1, distal breadth 0.03.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms.
{240}
Genus 95. Heliosoma,[130] Haeckel, 1881,
Prodromus, p. 452.
Definition.—Astrosphærida with one medullary
(intracapsular) and one cortical (extracapsular) shell, the two being connected by radial beams
piercing the central capsule. Shell surface covered with simple radial spines of two different
kinds; larger main spines and smaller by-spines.
The genus Heliosoma differs from the preceding Haliomma in the
possession of two different kinds of radial spines, and exhibits therefore the same relation to it
that Heliosphæra bears to Acanthosphæra among the Coscinommida. The smaller
by-spines are much more numerous than the larger main spines.
Subgenus 1. Heliosomantha, Haeckel.
Definition.—Pores of the cortical shell regular, of nearly equal size
and similar form.
1. Heliosoma radians, n. sp. (Pl. 28,
figs. 3, 3a).
Cortical shell very thin walled, with thread-like bars and regular, hexagonal meshes; twenty to
twenty-two on the radius. Medullary shell only one-fifth as large, of the same structure. The two
shells connected by twenty very thin radial beams, which are prolonged outside into twenty stout,
three-sided pyramidal main spines, as long as the diameter of the inner shell. Between these,
arising from the surface, numerous bristle-shaped by-spines.
Dimensions.—Diameter of the outer shell 0.25, inner 0.05, pores 0.012; length of
the main spines 0.05, basal breadth 0.012.
Habitat.—Central Pacific, Stations 266 to 274, surface.
2. Heliosoma elegans, n. sp.
Cortical shell thick walled, with regular, circular, hexagonally framed pores, twice as broad
as the bars; ten to twelve on the radius. Medullary shell half as large, with simple, regular,
circular pores, twice as broad as the bars. The two shells connected by twenty very thin radial
beams, which are prolonged outside into twenty slender, three-sided pyramidal main spines as long
as the radius. In each corner of the surface hexagons a bristle-shaped by-spine one-fourth as
long.
Dimensions.—Diameter of the outer shell 0.12, inner 0.06; outer pores 0.006, bars
0.003; length of the main spines 0.05, basal breadth 0.01.
Habitat.—Central Pacific, Station 271, surface.
3. Heliosoma echinaster, Haeckel.
Haliomma echinaster, Haeckel, 1862, Monogr. d. Radiol., p. 429, Taf. xxiv.
figs. 1-3.
Cortical shell thin walled, with regular, circular, hexagonally framed pores, twice as broad as
the bars; ten to twelve on the radius. Medullary shell one-fourth as large, with few irregular,
large, {241}polygonal pores and very thin thread-like
bars, connected with the former by nine similar thin radial beams, which are prolonged outside
into nine pyramidal main spines, as long as the radius. From each corner of the surface hexagon
arises a bristle-shaped by-spine, half as long as the main spine.
Dimensions.—Diameter of the outer shell 0.16, inner 0.04; outer pores 0.008, bars
0.004; length of the main spines 0.08, basal breadth 0.01.
Habitat.—Mediterranean (Messina), surface.
4. Heliosoma duodecilla, n. sp.
Cortical shell thick walled, with regular, circular pores, of the same breadth as the bars;
sixteen to eighteen on the radius. Medullary shell one-third as large, of the same structure.
Twelve radial main spines, regularly disposed, cylindro-conical, longer than the shell diameter,
as broad as two pores. Whole surface covered with short conical by-spines.
Dimensions.—Diameter of the outer shell 0.15, inner 0.05; outer pores and bars
0.007; length of the main spines 0.2, breadth 0.02.
Habitat.—South Atlantic, Station 333, surface.
Subgenus 2. Heliosomura, Haeckel.
Definition.—Pores of the cortical shell irregular, of different size
and form.
5. Heliosoma hastatum, n. sp. (Pl. 28,
fig. 4).
Cortical shell very thin walled, with large, irregular, polygonal meshes, three to six times as
broad as the bars; four to six on the radius. Medullary shell one-third as large, with small,
regular, circular pores. The two shells connected by twelve regularly disposed radial beams, which
are prolonged outside into twelve very stout three-sided pyramidal main spines, somewhat longer
than the radius, spear-shaped; each of the three wings in the middle part with one tooth.
Scattered on the surface numerous by-spines of the same form, half as long and only one-fourth as
broad.
Dimensions.—Diameter of the outer shell 0.1, inner 0.033; outer pores 0.006 to 0.013, inner pores 0.002, bars
0.002; length of the main spines 0.06, breadth 0.015.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
6. Heliosoma indicum, n. sp.
Cortical shell thick walled, with irregular, roundish pores, twice to five times as broad as
the bars. Medullary shell of the same structure, only one-fourth as large. Thirty to forty conical
main spines, as long as the radius; between them numerous bristle-shaped by-spines, only half as
long.
Dimensions.—Diameter of the outer shell 0.2, inner 0.05; outer pores 0.004 to
0.01, bars 0.002; length of the main
spines 0.1, breadth 0.01.
Habitat.—Indian Ocean, between Aden and Socotora, surface,
Haeckel.
{242}
Genus 96. Elatomma,[131] n. gen.
Definition.—Astrosphærida with one medullary
(intracapsular) and one cortical (extracapsular) shell, the two being connected by radial beams
piercing the central capsule. Shell surface covered with branched radial spines.
The genus Elatomma differs from Haliomma in the ramification of the
radial spines, and exhibits therefore the same relation to it that Cladococcus, among the
Coscinommida, bears to Acanthosphæra.
Subgenus 1. Elatommella, Haeckel.
Definition.—Pores of the cortical shell regular, of nearly equal size
and similar form.
1. Elatomma pinetum, n. sp.
Cortical shell thick walled, with regular, circular, hexagonally framed pores, three times as
broad as the bars. From its surface arise twenty to thirty large, three-sided prismatic, radial
spines as long as the shell diameter, branched like a pine tree; on each edge of the spine five to
six ramified branches, decreasing in size towards the distal end. Similar to Cladococcus
pinetum (Pl. 27,
figs. 1, 3), but differing in the possession of a medullary shell (one-third as large as the
outer), with regular, circular pores, as broad as the bars.
Dimensions.—Diameter of the outer shell 0.2, inner 0.07; cortical pores 0.012,
bars 0.004; length of the spines 0.2 to 0.24.
Habitat.—Central Pacific, Stations 266 to 272, surface.
2. Elatomma scoparium, n. sp.
Cortical shell thick walled, with regular, circular pores, twice as broad as the bars,
connected with the medullary shell by twenty thin radial beams, which are prolonged outside into
twenty large cylindrical radial spines somewhat longer than the radius; in the proximal half
simple, in the distal half with six to nine dichotomous, irregular branches. Similar to
Cladococcus scoparius (Pl. 27,
fig. 2), but much larger, and with a large medullary shell (one-third as broad as the outer), and
regular, circular pores.
Dimensions.—Diameter of the outer shell 0.12, inner 0.04; cortical pores 0.01,
bars 0.005; length of the spines 0.08.
Habitat.—North Pacific, Station 240, surface.
Subgenus 2. Elatommura, Haeckel.
Definition.—Pores of the cortical shell irregular, of different size
or form.
{243}
3. Elatomma juniperinum, n. sp. (Pl. 28,
fig. 8).
Cortical shell thin walled, with regular, polygonal, or roundish pores, twice to four times as
broad as the bars, connected with the small medullary shell by about twenty thin radial beams.
Inner shell only one-eighth as broad as the outer, with few irregular, polygonal meshes. Surface
covered with very numerous (one hundred to one hundred and fifty or more) branched conical radial
spines, one-third as long as the shell radius, with six to twelve short lateral branches.
Dimensions.—Diameter of the outer shell 0.2, inner 0.025; cortical pores 0.08 to
0.016, bars 0.004; length of the spines 0.04.
Habitat.—Central Pacific, Station 271, surface.
4. Elatomma penicillus, n. sp.
Cortical shell thin walled, with irregular, polygonal pores, twice to four times as broad as
the bars, connected with the medullary shell by twenty (or more?) very thin beams. These are
prolonged outside into twenty straight, three-sided prismatic, radial spines as long as the
radius, with a brush-like bunch of six to nine short, irregularly ramified branches at the end.
Medullary shell very delicate (half as broad as the outer), with regular, hexagonal meshes and
thread-like bars.
Dimensions.—Diameter of the outer shell 0.15, inner 0.08; length of the spines
0.08.
Habitat.—West coast of Norway, Bergen, Haeckel.
5. Elatomma irregulare, n. sp.
Cortical shell thin walled, with irregular, polygonal, or roundish meshes, twice to six times
as broad as the bars; the medullary shell of the same structure, half as broad, with smaller
irregular pores. Forty to sixty curved, three-sided, radial spines, as long as the shell, with a
bunch of very irregular, curved, and partly ramified branches at the distal end.
Dimensions.—Diameter of the outer shell 0.2, inner 0.1; length of the spines 0.2,
breadth 0.01.
Habitat.—North Atlantic, Gulf Stream, Færöe Channel, surface, John
Murray.
Genus 97. Leptosphæra,[132] n. gen.
Definition.—Astrosphærida with two extracapsular cortical
shells without by-spines, connected by long prismatic radial spines.
The genus Leptosphæra, together with the three following genera, forms the
very peculiar and typical small group of Sphæroidea which we call the
Diplosphærida (with four genera and twenty-four species); their shell is composed of two
concentric spheres as in the three foregoing genera; but whilst in these the inner shell is an
intracapsular medullary shell and the outer an extracapsular cortical shell, in the Diplosphærida
{244}both shells are extracapsular or cortical shells,
therefore the inner shell of the latter corresponds to the outer of the former. The inner
spherical shell of all Diplosphærida is composed of very delicate beams and large pores, which are
either regular hexagonal or irregular polygonal (never roundish). From its surface arise a
variable number (twenty to thirty) of stout long radial spines, which are invariably longer than
the shell diameter (often two to three times as long or more), and of three-sided prismatic form,
the three edges either smooth or serrate, often with three rows of lateral branches (commonly
three to five branches in each row); the latter are invariably of the same form, concavely curved
towards the spine, and decrease in size towards the distal end. From the three edges of each main
spine in all Diplosphærida, at equal distances from the centre, arise six very thin, thread-like
lateral branches (a pair from each edge) and connect the spine in a tangential direction with all
neighbouring spines. In this manner the polyhedral outer shell is formed, the meshes of which
therefore are always very large and triangular. Sometimes each of these primary triangular meshes
becomes filled up with a secondary network, either of regular quadrangular or of irregular
polygonal secondary meshes. Besides the constant twenty to thirty large main spines, in the
majority of Diplosphærida bristle-shaped radial by-spines arise, either from the inner shell
(Diplosphæra) or from the outer (Drymosphæra), or from both (Astrosphæra).
They are absent only in Leptosphæra. Commonly the by-spines are simple, rarely forked or
branched. The central capsule in the Diplosphærida is usually enclosed in the inner shell; often
it completely fills up the latter, or drives out a cæcal protuberance through each mesh; but these
processes rarely unite outside. The average size of the Diplosphærida, which are all pelagic
organisms, is much larger than that of the other Haliommida.
Subgenus 1. Leptosphærella, Haeckel.
Definition.—Radial spines simple, without lateral branches.
1. Leptosphæra hexagonalis, n. sp. (Pl. 19, fig.
2).
Inner shell with regular, hexagonal meshes, and very thin, thread-like bars; outer shell twice
as broad, with simple triangular meshes. Radial spines with three smooth edges. (Fig. 2 represents
the central capsule with numerous club-shaped saccules, prominent externally through the meshes;
in the centre a large simple spherical nucleus, one-third as broad as the capsule. The skeleton of
this species is identical with that of Diplosphæra hexagonalis, Pl. 19, fig. 3, but
has no by-spines.)
Dimensions.—Diameter of the outer shell 0.3, of the inner 0.15.
Habitat.—Cosmopolitan; Mediterranean (Corfu), North Atlantic (Canary
Islands), Tropical Pacific, surface.
{245}
2. Leptosphæra serrata, n. sp.
Inner shell with regular, hexagonal meshes, five to six times as broad as the bars; outer shell
three times as broad, with simple triangular meshes. Radial spines with three serrated edges (of
the same form as the main spines of Drymosphæra dendrophora, Pl. 20, fig.
1).
Dimensions.—Diameter of the outer shell 0.45, of the inner 0.15.
Habitat.—Central Pacific, Station 271, surface.
3. Leptosphæra polygonalis, n. sp.
Inner shell with irregular, polygonal meshes and very thin thread-like bars; outer shell twice
as broad, with simple triangular meshes. Radial spines with three smooth edges. (Resembles
Drymosphæra polygonalis, Pl. 19,
fig. 1, but has no by-spines.)
Dimensions.—Diameter of the outer shell 0.35, inner 0.175.
Habitat.—North Pacific, Station 256, surface.
Subgenus 2. Leptosphæromma, Haeckel.
Definition.—Radial spines with three rows of lateral branches (one row
on each edge).
4. Leptosphæra ciliata, n. sp.
Inner shell with regular, hexagonal meshes and very thin thread-like bars; outer shell three
times as broad, with simple triangular meshes. Radial spines with smooth edges and three rows of
simple, smooth, curved, lateral branches (six branches on each edge), similar to those of
Diplosphæra gracilis.
Dimensions.—Diameter of the outer shell 0.6, inner 0.2.
Habitat.—South Atlantic, Station 325, surface.
5. Leptosphæra spinosa, Haeckel.
Diplosphæra spinosa, R. Hertwig, 1879, Organismus d. Radiol., p. 40, Taf.
v. fig. 2.
Inner shell with regular, hexagonal meshes, five times as broad as the bars; outer shell three
times as broad, with simple triangular meshes. Radial spines with three densely serrated edges,
and with three rows of serrated, simple, curved, lateral branches (three branches on each edge).
Differs from the preceding in the thicker bars and the elegantly denticulated spines and
branches.
Dimensions.—Diameter of the outer shell 0.66, inner 0.22.
Habitat.—Mediterranean (Messina), R. Hertwig.
6. Leptosphæra stellata, n. sp.
Inner shell with regular, hexagonal meshes, six times as broad as the bars, each bar crossed by
a transverse tangential rod, so that each mesh represents an elegant six-rayed star (as {246}in Astrosphæra stellata, Pl. 19, fig. 5, but
without radial by-spines); outer shell twice as broad, with simple triangular meshes. Radial
spines with serrated edges and three rows of simple lateral branches (four branches on each
edge).
Dimensions.—Diameter of the outer shell 0.44, inner 0.22.
Habitat.—Tropical Atlantic (Ascension Island), Station 343,
surface.
7. Leptosphæra reticulum, n. sp.
Inner shell with irregular, polygonal meshes and very thin thread-like bars; outer shell four
times as broad, also with irregular, polygonal meshes, the sides of the triangular main meshes
being connected by irregular lateral ramules, forming an extremely delicate reticulum.
Dimensions.—Diameter of the outer shell 0.64, inner 0.16.
Habitat.—South Pacific, Station 291, surface.
Genus 98. Diplosphæra,[133] Haeckel,
1860, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 804.
Definition.—Astrosphærida with two extracapsular cortical
shells, connected by long, prismatic, radial spines; inner shell with thin radial by-spines.
The genus Diplosphæra differs from its ancestral form Leptosphæra in
the development of radial by-spines on the surface of the inner shell, the outer shell being
smooth.
Subgenus 1. Diplosphærella, Haeckel.
Definition.—Radial main spines simple, without lateral branches.
1. Diplosphæra hexagonalis, n. sp. (Pl. 19, fig.
3).
Inner shell with regular, hexagonal meshes and very thin thread-like bars; at each nodal-point
one bristle-shaped by-spine; outer shell twice as broad, with simple triangular meshes. Radial
spines with three smooth edges. (The radially striped central capsule, enclosed in the inner
shell, envelops a large central nucleus one-third its size, fig. 3.)
Dimensions.—Diameter of the outer shell 0.36, inner 0.18.
Habitat.—Central Pacific, Stations 265 to 274, surface.
2. Diplosphæra ornata, n. sp.
Inner shell with regular, hexagonal meshes, four times as broad as the bars; outer shell three
times as broad, with simple triangular meshes. Radial main spines as well as the bars of both
{247}shells very elegantly denticulated; radial
by-spines (very numerous in the surface of the inner shell) smooth.
Dimensions.—Diameter of the outer shell 0.7, inner 0.24.
Habitat.—Indian Ocean, Madagascar, surface, Rabbe.
3. Diplosphæra dictyota, n. sp.
Inner shell with regular, hexagonal meshes, six times as broad as the bars; at each nodal-point
one radial by-spine; outer shell two and a half times as broad, with very delicate square meshes,
separated by thread-like bars which arise from the sides of the triangular main meshes. Radial
main spines with three serrated edges. (Differs from the similar Diplosphæra gracilis in
the simple main spines, with delicate dentition of the edges.)
Dimensions.—Diameter of the outer shell 0.5, inner 0.2.
Habitat.—North Atlantic, Canary Islands, surface.
4. Diplosphæra polygonalis, n. sp.
Inner shell with irregular, polygonal meshes and very thin thread-like bars, covered with very
numerous bristle-shaped by-spines; outer shell twice as broad, with simple triangular meshes.
Radial main spines with three smooth edges.
Dimensions.—Diameter of the outer shell 0.44, inner 0.22.
Habitat.—South Pacific, Station 288, surface.
Subgenus 2. Diplosphæromma, Haeckel.
Definition.—Radial main spines with three rows of lateral branches
(one row on each edge).
5. Diplosphæra gracilis, Haeckel.
Diplosphæra gracilis, Haeckel, 1862, Monogr. d. Radiol., p. 354, Taf. x.
fig. 1.
Inner shell with regular, hexagonal meshes and very thin bars; at each nodal-point one simple
bristle-shaped radial by-spine; outer shell twice as broad, with very delicate square meshes,
separated by thread-like bars which arise from the sides of the triangular main meshes. Radial
spines with smooth edges and with three rows of simple curved lateral branches (four to six
branches on each edge).
Dimensions.—Diameter of the outer shell 0.54, inner 0.27.
Habitat.—Mediterranean (Messina).
6. Diplosphæra denticulata, n. sp.
Inner shell with regular, hexagonal meshes, four times as broad as the bars; at each
nodal-point is a simple bristle-shaped by-spine; outer shell three times as broad, with simple
triangular meshes. {248}Radial spines very strong, with
dentated edges and with three rows of lateral branches (six dentated curved branches on each
edge).
Dimensions.—Diameter of the outer shell 0.4, inner 0.13.
Habitat.—North Atlantic, Azores, surface.
7. Diplosphæra reticulata, n. sp.
Inner shell with regular, hexagonal meshes, five times as broad as the bars, which are densely
covered with very numerous, simple bristle-shaped by-spines; outer shell four times as broad, with
very numerous small irregular polygonal meshes, forming a very delicate network, and filling out
the large triangular main meshes. Radial spines with three serrated edges and three rows of short
curved branches (four to five branches on each edge).
Dimensions.—Diameter of the outer shell 0.6, inner 0.15.
Habitat.—North Atlantic, Gulf Stream, Færöe Channel, surface, John
Murray.
8. Diplosphæra triglochin, n. sp.
Inner shell with irregular, polygonal meshes and very thin thread-like bars; outer shell three
times as broad, with similar irregular polygonal meshes, filling out the large triangular main
meshes. Radial spines with three serrated edges, each of which bears one single large lateral
branch, concavely curved towards the distal end. By-spines of the inner shell very numerous.
Dimensions.—Diameter of the outer shell 0.5, inner 0.17.
Habitat.—Central Pacific, Station 264, surface.
Genus 99. Drymosphæra,[134] Haeckel,
1881, Prodromus, p. 452.
Definition.—Astrosphærida with two extracapsular cortical
shells, connected by long prismatic, radial spines; outer shell with thin radial by-spines.
The genus Drymosphæra differs from its ancestral form Leptosphæra in
the development of radial by-spines on the surface of the outer shell, the inner shell being
smooth.
Subgenus 1. Drymosphærella, Haeckel.
Definition.—Radial by-spines simple, not branched (main spines
simple).
1. Drymosphæra hexagonalis, n. sp.
Inner shell with regular, hexagonal meshes and very thin thread-like bars; outer shell twice as
broad, with simple triangular meshes and thin thread-like bars, which bear very numerous, {249}bristle-shaped radial by-spines. Radial main spines with
three smooth edges. (Differs from the first species of the other three genera of Diplosphærida in
the presence of by-spines on the outer, and their absence on the inner shell.)
Dimensions.—Diameter of the outer shell 0.32, inner 0.16.
Habitat.—Central Pacific, Station 274, surface.
2. Drymosphæra polygonalis, n. sp. (Pl. 19, fig.
1).
Diplosphæra polygonalis, Haeckel, 1879, Atlas.
Inner shell with irregular, polygonal meshes and very thin thread-like bars; outer shell one
and a half times as broad, with simple triangular meshes and thicker bars, which bear numerous
bristle-shaped radial by-spines (in the figure the majority of these are broken off). Radial main
spines with three smooth edges. (Similar to Leptosphæra polygonalis and Diplosphæra
polygonalis, but differs from both in the presence of by-spines on the outer, and their
absence on the inner shell.)
Dimensions.—Diameter of the outer shell 0.33, inner 0.22.
Habitat.—North Pacific, Station 236 south of Japan, surface.
Subgenus 2. Drymosphæromma, Haeckel.
Definition.—Radial by-spines forked or branched (main spines
simple).
3. Drymosphæra furcata, n. sp.
Inner shell with irregular, polygonal meshes and thin bars, the former five times as broad as
the latter; outer shell two and a half times as broad, with simple triangular meshes and smooth
bars, bearing numerous long smooth bristle-shaped by-spines, which in the distal half are forked.
Radial main spines with three smooth edges.
Dimensions.—Diameter of the outer shell 0.4, inner 0.16.
Habitat.—Indian Ocean, Ceylon, surface, Haeckel.
4. Drymosphæra cladophora, n. sp.
Inner shell with irregular, polygonal meshes, three to six times as broad as the smooth bars;
outer shell twice as broad, with simple triangular meshes and very thin smooth bars, bearing
numerous curved, irregularly branched, smooth by-spines. Radial main spines with three serrated
edges.
Dimensions.—Diameter of the outer shell 0.4, inner 0.2.
Habitat.—Central Pacific, Station 276, surface.
5. Drymosphæra dendrophora, n. sp. (Pl. 20, figs. 1,
1a, 1b).
Inner shell with irregular, polygonal meshes and very thin thread-like bars; outer shell one
and a half times as broad, with simple triangular meshes and thicker bars, which bear a forest of
{250}very numerous, repeatedly dichotomous or
irregularly branched, curved by-spines, longer than the diameter of the outer shell. Radial main
spines with three dentated edges. All parts of the skeleton, the net bars as well as the radial
beams and spines, are very elegantly denticulated (fig. 1b). The central capsule (fig.
1a) completely distends the inner shell and forces out protuberances through all its pores;
in its centre lies a nucleus one-third its size.
Dimensions.—Diameter of the outer shell 0.25, inner 0.16.
Habitat.—Central Pacific, Station 271, surface.
Genus 100. Astrosphæra,[135] n. gen.
Definition.—Astrosphærida with two extracapsular cortical
shells, connected by long, prismatic, radial spines; inner and outer shell with thin radial
by-spines.
The genus Astrosphæra differs from its ancestral form, Leptosphæra,
in the development of radial by-spines on the surface of both shells.
Subgenus 1. Astrosphærella, Haeckel.
Definition.—Radial main spines simple without lateral branches.
1. Astrosphæra hexagonalis, n. sp. (Pl. 19, fig.
4).
Inner shell with regular, hexagonal meshes and very thin bars, having a bristle-shaped, short
radial by-spine in each hexagon-corner; outer shell twice as broad, with simple triangular meshes
and thicker bars, bearing one row of simple bristle-shaped, curved, radial by-spines. Radial main
spines with three smooth edges.
Dimensions.—Diameter of the outer shell 0.32, inner 0.16.
Habitat.—South Pacific (West Patagonia), Station 302, surface.
2. Astrosphæra splendens, n. sp.
Inner shell with regular, hexagonal meshes, four times as broad as the bars, and covered with
numerous curved, long, bristle-shaped by-spines; outer shell three times as broad, with simple
triangular meshes and thin bars, bearing a row of very long, curved, bristle-shaped by-spines.
Radial main spines with three dentated edges. All parts of the skeleton elegantly denticulated, as
in Drymosphæra dendrophora (Pl. 20,
fig. 1).
Dimensions.—Diameter of outer shell 0.7, inner 0.24.
Habitat.—Tropical Atlantic, Station 347, surface.
{251}
Subgenus 2. Astrosphæromma.
Definition.—Radial main spines with three rows of lateral branches
(one row on each edge).
3. Astrosphæra sideræa, n. sp.
Inner shell with regular, hexagonal meshes, six times as broad as the bars, and covered with
numerous short bristle-shaped by-spines; outer shell four times as broad, with simple triangular
meshes, and rows of long bristle-shaped by-spines arising from the bars. Radial main spines with
three serrated edges and three rows of lateral branches (four branches on each edge).
Dimensions.—Diameter of the outer shell 0.6, inner 0.15.
Habitat.—Central Pacific, Station 266, surface.
4. Astrosphæra stellata, n. sp. (Pl. 19,
fig. 5).
Diplosphæra stellata, Haeckel, 1881, Prodrom. et Atlas.
Inner shell with regular, hexagonal, six-rayed meshes, each bar being crossed by a transverse
tangential rod, at each nodal-point a long bristle-shaped by-spine; outer shell three times as
broad, with simple triangular meshes and denticulated thin bars, bearing a row of bristle-shaped
radial by-spines. Radial main spines in the proximal half with three serrated edges, in the distal
half with three rows of curved lateral branches (five branches on each edge).
Dimensions.—Diameter of the outer shell 0.6, inner 0.2.
Habitat.—Central Pacific, Station 274, surface.
Subfamily Actinommida,[136] Haeckel, 1862, Monogr. d. Radiol., p. 440 (sensu
emendato).
Definition.—Astrosphærida with three
concentric, spherical, lattice-shells, united by radial beams.
Genus 101. Actinomma,[137] Haeckel, 1862,
Monogr. d. Radiol., p. 440.
Definition.—Astrosphærida with three concentric
lattice-spheres and numerous simple radial spines of one kind.
The genus Actinomma is here restricted to those Sphæroidea which combine the possession of three concentric lattice-shells with
numerous radial spines on the surface; the spines are all simple, being of one and the same kind.
Commonly two of the three shells are intracapsular medullary shells, connected by radial beams
(piercing {252}the central capsule) with the outer,
extracapsular, cortical shell. But in some species only one medullary shell is enclosed in the
central capsule, whilst both other shells lie outside it. In such case the distance between these
two cortical shells is much smaller than their distance from the simple internal medullary shell.
These forms correspond more to aculeate Rhodosphæræ, whilst the others resemble aculeate
Thecosphæræ.
Subgenus 1. Actinommantha, Haeckel.
Definition.—Pores of the cortical shell regular, of nearly equal size
and similar form; spines on the entire surface (commonly one spine at each nodal-point).
1. Actinomma hexagonium, n. sp.
Cortical shell, as well as both medullary shells, very thin walled, with regular, hexagonal
pores and thread-like bars between them. Pores of the outer shell twice as broad as those of the
middle, and three times as broad as those of the inner shells. Radial proportion of the three
spheres = 1 : 3 : 9; about twenty thin radial beams between them. At each
nodal-point of the surface arises one bristle-shaped radial spine, half as long as the radius.
Dimensions.—Diameter of the outer shell 0.22, middle 0.07, inner 0.025; meshes of
the cortical shell 0.01; length of the spines 0.05.
Habitat.—Central Pacific, Stations 270 to 274, surface.
2. Actinomma facetum, n. sp.
Cortical shell thick walled, with regular, circular, hexagonally framed pores, three times as
broad as the bars. Pores of both medullary shells regular, circular. Radial proportion of the
three spheres = 1 : 2 : 4; about forty thin radial beams between them. At each
nodal-point of the surface arises one short, three-sided pyramidal, radial spine, about one-third
as long as the radius.
Dimensions.—Diameter of the outer shell 0.16, middle 0.08, inner 0.04; cortical
pores 0.012, bars 0.004; length of the spines 0.03, basal breadth 0.015.
Habitat.—North Pacific, Station 253, depth 3125 fathoms.
3. Actinomma anthomma, n. sp.
Cortical shell thick walled, with regular, six-lobed pores, twice as broad as the bars. At each
nodal-point of the surface is one short conical radial spine, as long as the diameter of the
pores; one corresponding to each lobe, and there is therefore around each pore a regular corona of
six spines, as in Haliomma lirianthus (Pl. 28, fig.
1b). Both medullary shells with simple, circular, regular pores. Radial proportion of the
three spheres = 1 : 2.5 : 7; radial beams between them only six, opposite in
pairs in the three dimensive axes.
{253}
Dimensions.—Diameter of the outer shell 0.2, middle 0.075, inner 0.003; cortical
pores 0.01, bars 0.005; length of the spines 0.01.
Habitat.—Central Pacific, Station 271, surface.
4. Actinomma castanomma, n. sp.
Cortical shell as well as both medullary shells thick walled, with regular, circular pores,
twice as broad as the bars; between them at each nodal-point one bristle-shaped radial spine,
one-third as long as the radius, with conical base. Radial proportion of the three spheres =
2 : 3 : 8; only six radial beams between them (opposed in pairs in the three
dimensive axes).
Dimensions.—Diameter of the outer shell 0.15, middle 0.06, inner 0.04; cortical
pores 0.008, bars 0.004; length of the spines 0.025.
Habitat.—South Atlantic, Station 333, surface.
5. Actinomma entactinia, Stöhr.
Actinomma entactinia, Stöhr, 1880, Palæontogr., vol. xxvi. p. 93, Taf. ii.
fig. 12.
Cortical shell as well as both medullary shells thick walled, with regular, circular pores, of
the same breadth as the bars. Radial proportion of the three spheres =
1 : 3 : 8; radial beams between them very numerous (thirty to fifty or more?).
Entire surface covered with short thick conical spines, only one-fifth as long as the radius.
Dimensions.—Diameter of the outer shell 0.16, middle 0.06, inner 0.02; cortical
pores and bars 0.007; length of the spines 0.015.
Habitat.—Fossil in the Tertiary rocks of Sicily, Grotte,
Caltanisetta.
Subgenus 2. Actinommetta, Haeckel.
Definition.—Pores of the outer shell regular, of nearly equal size and
similar form; spines not over the entire surface, but scattered at intervals (their number smaller
than that of the nodal-points).
6. Actinomma japonicum, n. sp.
Cortical shell thick walled, with regular, circular, hexagonally framed pores, three times as
broad as the bars. Both medullary shells with simple, circular, regular pores, of the same breadth
as the bars. Radial proportion of the three spheres = 2 : 5 : 9; radial beams
between them twenty, prolonged outside into twenty symmetrically disposed, three-sided pyramidal
spines, as long as the radius.
Dimensions.—Diameter of the outer shell 0.18, middle 0.1, inner 0.04; cortical pores 0.01, bars 0.003; length of
the spines 0.1, basal breadth 0.012.
Habitat.—North Pacific, Station 240, east of Japan, surface.
{254}
7. Actinomma denticulatum, n. sp. (Pl. 29, fig.
3).
Cortical shell thick walled, with regular, circular, double-edged pores, twice as broad as the
bars. Forty to fifty radial spines, three-sided pyramidal, half as long as the radius, with three
elegantly denticulated edges. Radial proportion of the three spheres =
1 : 2 : 5. (Differs from Haliomma denticulatum only in the double
medullary shell and larger size.)
Dimensions.—Diameter of the outer shell 0.14, middle 0.06, inner 0.03; cortical
pores 0.01, bars 0.005; length of the spines 0.04, basal breadth 0.01.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
8. Actinomma trinacrium, Haeckel.
Actinomma trinacrium, Haeckel, 1862, Monogr. d. Radiol., p. 441, Taf. xxiv. figs.
6-8.
Haliomma trinacrium, Haeckel, 1860, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 815.
Cortical shell as well as both medullary shells thin walled, with subregular, circular pores,
twice as broad as the bars. Radial proportion of the three spheres =
1 : 3 : 9; radial beams between them twenty, prolonged outside into strong,
three-sided pyramidal spines, as long as the radius; between them, on the surface, numerous
similar spines. Sometimes the latter remain smaller, the pores more irregular, and then this
species corresponds to Echinomma trinacrium.
Dimensions.—Diameter of the outer shell 0.09, middle 0.03, inner 0.01; cortical
pores 0.008, bars 0.004; length of the spines 0.04, basal breadth 0.01.
Habitat.—Mediterranean (Messina, Corfu, Haeckel), surface.
9. Actinomma pachyderma, n. sp. (Pl. 29,
figs. 4, 5).
Cortical shell very thick walled, with regular, circular, double-edged pores, twice as broad as
the bars. Both medullary shells with simple, small, circular pores, of the same breadth as the
bars. Radial proportion of the three spheres = 1 : 2 : 4; radial spines about
twenty, conical, shorter than the radius, prolonged inside into twenty very thin connecting beams
(fig. 4).
Dimensions.—Diameter of the outer shell 0.08, middle 0.04, inner 0.02; cortical
pores 0.01; bars 0.005; length of the spines 0.03, basal breadth 0.01.
Habitat.—South Pacific, Station 297, depth 1775 fathoms.
Subgenus 3. Actinommilla, Haeckel.
Definition.—Pores of the cortical shell irregular, of different size
and form; spines over the entire surface (commonly one spine at each nodal-point).
10. Actinomma spinigerum, Stöhr.
Actinomma spinigerum, Stöhr, 1880, Palæontogr., vol. xxvi. p. 92, Taf. ii.
fig. 10.
Cortical shell thick walled, with irregular, roundish, or subcircular pores, twice to four
times as broad as the bars. Radial proportion of the three spheres =
1 : 3 : 8; between them eight (?) {255}connecting radial beams. Entire surface densely covered with small
conical spines, not larger than the pores.
Dimensions.—Diameter of the outer shell 0.17, middle 0.07, inner 0.023; cortical
pores 0.006 to 0.012, bars 0.003; length of the spines 0.01.
Habitat.—Fossil in Tertiary rocks of Sicily, Grotte, Stöhr.
11. Actinomma hirsutum, n. sp.
Cortical shell thick walled, with irregular, roundish pores, about the same breadth as the
bars. Radial proportion of the three spheres = 1 : 2 : 8; between them
numerous (twenty to thirty or more) cylindrical connecting beams. Entire surface densely covered
with innumerable small bristle-shaped spines, as long as the diameter of the inner shell.
Dimensions.—Diameter of the outer shell 0.25, middle 0.06, inner 0.03; cortical
pores and bars 0.004 to 0.008; length of the spines 0.03.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms.
Subgenus 4. Actinommura, Haeckel.
Definition.—Pores of the cortical shell irregular, of different size
and form; spines not over the entire surface, but scattered at intervals (their number smaller
than that of the nodal-points).
12. Actinomma capillaceum, n. sp. (Pl. 29, fig.
6).
Cortical shell very thin walled, with irregular, polygonal meshes (sixteen to eighteen on the
radius), three to six times as broad as the bars. Both medullary shells with smaller pores of the
same structure. Radial proportion of the three spheres = 1 : 2.5 : 7; radial
connecting beams between them very thin and numerous (one hundred and twenty to one hundred and
fifty or more), each prolonged outside into a short three-sided pyramidal spine, as long as the
radius of the inner shell. (Similar to Haliomma capillaceum.)
Dimensions.—Diameter of the outer shell 0.27, middle 0.1, inner 0.04; cortical
pores 0.01 to 0.02, bars 0.003; length of the spines 0.02, basal breadth 0.005.
Habitat.—Central Pacific, Stations 266 to 274, surface.
13. Actinomma arcadophorum, n. sp. (Pl. 29, figs. 7,
8).
Cortical shell very thin walled, with irregular, polygonal meshes, ten to twenty times as broad
as the bars; twelve to sixteen on the radius. Both medullary shells with similar delicate network.
Radial proportion of the three spheres = 1 : 2 : 6; numerous thin radial beams
(forty to eighty or more) connect both medullary shells and alternate with other beams, which
arise from arcade-shaped {256}protuberances of the outer
medullary shell, and connect it with the cortical shell. Outer prolongations of these form the
bristle-shaped spines of the surface.
Dimensions.—Diameter of the outer shell 0.22, middle 0.09, inner 0.04; cortical
pores 0.01 to 0.02, bars 0.001 to 0.002; length of the radial spines 0.2.
Habitat.—Central Pacific, Station 266, surface.
14. Actinomma schwageri, Stöhr.
Actinomma schwageri, Stöhr, 1880, Palæontogr., vol. xxvi. p. 92, Taf. ii.
fig. 9a, b.
Cortical shell thick walled, with irregular, large, polygonal meshes, five to twelve times as
broad as the bars; six to eight on the radius. Both medullary shells with very small, circular,
regular pores. Radial proportion of the three spheres = 1 : 2 : 9; connecting
radial beams between them nine, prolonged outside into nine short stout, three-sided pyramidal
spines, as long as the diameter of the inner shell.
Dimensions.—Diameter of the outer shell 0.17, middle 0.04, inner 0.02; cortical
pores 0.01 to 0.03, bars 0.006 and less; length of the spines 0.02, basal breadth 0.01.
Habitat.—Fossil in Tertiary rocks of Sicily, Grotte, Stöhr.
15. Actinomma dodecomma, n. sp.
Cortical shell thin walled, with irregular, roundish pores, three to six times as broad as the
bars. Both medullary shells with regular, circular pores, twice as broad as the bars. Radial
proportion of the three shells = 1 : 2.5 : 7; connecting radial beams between
them twelve, regularly disposed, prolonged outside into twelve strong, three-sided pyramidal
spines, as long as the radius.
Dimensions.—Diameter of the outer shell 0.14, middle 0.05, inner 0.02; cortical
pores 0.008 to 0.018, bars 0.003; length of the spines 0.08, breadth 0.02.
Habitat.—Tropical Atlantic, Station 348, depth 2450 fathoms.
16. Actinomma pachycapsa, n. sp.
Cortical shell very thick walled, with irregular, roundish pores, twice to four times as broad
as the bars; twelve to sixteen on the radius. Radial proportion of the three spheres =
1 : 3 : 12. Both medullary shells of similar irregular structure, connected
with the cortical shell by twelve regularly disposed radial beams, which are prolonged outside
into twelve short stout conical spines, as long as the diameter of the middle shell.
Dimensions.—Diameter of the outer shell 0.25, middle 0.06, inner 0.02; cortical
pores 0.006 to 0.012, bars 0.003; length of the spines 0.05, breadth 0.02.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
17. Actinomma giganteum, n. sp.
Cortical shell thick walled, little larger than the outer medullary shell, whilst the inner is
only one-tenth as large. Pores of all three shells irregular, roundish, or subcircular, twice to
three times {257}as broad as the bars, on an average
twice as large in the outer and middle as in the inner shell. Sixty to eighty short conical radial
spines on the surface, about as long as the diameter of the inner shell, and one-fourth as broad
at the base. Radial proportion of the three spheres = 1 : 9 : 10.
Dimensions.—Diameter of the outer shell 0.4 to 0.5, middle 0.35 to 0.45, inner
0.05 to 0.06.
Habitat.—Fossil in Barbados.
Genus 102. Echinomma,[138] Haeckel, 1881,
Prodromus, p. 453.
Definition.—Astrosphærida with three concentric
lattice-spheres and numerous simple radial spines of two different kinds; larger main spines and
smaller by-spines.
The genus Echinomma, differs from its ancestral form, Actinomma, in
the differentiation of the radial spines. Whilst a large number of small by-spines cover the
entire surface, a smaller number of large main spines are scattered over it, or limited to certain
regularly distributed points. It represents Heliosoma among the Haliommida.
Subgenus 1. Echinommetta, Haeckel.
Definition.—Pores of the cortical shell regular, of nearly equal size
and similar form.
1. Echinomma echinidium, n. sp.
Cortical shell thin walled, with regular, hexagonal pores, four times as broad as the bars; ten
to twelve on the radius. Both medullary shells with regular, circular pores, twice as broad as the
bars. Radial proportion of the three spheres = 1 : 2 : 5. Surface covered with
short bristle-shaped by-spines (one in every hexagon-corner); twenty main spines regularly
disposed, three-sided pyramidal, as long as the diameter of the inner shell.
Dimensions.—Diameter of the outer shell 0.1, middle 0.045, inner 0.02; cortical
pores 0.008, bars 0.002; length of the main spines 0.02, breadth 0.01.
Habitat.—South Pacific, Station 295, surface.
2. Echinomma cidaris, n. sp.
Cortical shell thin walled, with regular, circular, hexagonally framed pores, twice as broad as
the bars; six to eight on the radius. Both medullary shells with simple, circular pores. Radial
proportion of the three spheres = 1 : 2 : 6. Surface covered with short
bristle-shaped by-spines, half as long as the forty to fifty main spines, which are three-sided
pyramidal, and reach the radius of the shell.
Dimensions.—Diameter of the outer shell 0.18, middle 0.06, inner 0.03; cortical
pores 0.01, bars 0.005; length of the main spines 0.08, basal breadth 0.015.
Habitat.—Central Pacific, Station 272, surface.
{258}
3. Echinomma aculeatum, Haeckel.
Actinomma aculeatum, Stöhr, 1880 Palæontogr., vol. xxvi. p. 92, Taf. ii.
fig. 11.
Cortical shell thick walled with regular circular pores, of the same breadth as the bars; eight
to ten on the radius. Radial proportion of the three spheres = 1 : 2 : 8.
Surface covered with short conical by-spines about twenty strong main spines, three-sided
pyramidal, one-third as long as the shell diameter.
Dimensions.—Diameter of the outer shell 0.16, middle 0.04, inner 0.02; cortical
pores and bars 0.003; length of the main spines 0.05, basal breadth 0.013.
Habitat.—Fossil in the Tertiary rocks of Sicily and Barbados; living
in the Atlantic, Station 332, depth 2200 fathoms.
4. Echinomma diadema, n. sp.
Cortical shell thick walled with regular circular pores, twice as broad as the bars; sixteen to
eighteen on the radius. Radial proportion of the three spheres = 1 : 3 : 11.
Surface covered with short bristle-shaped by-spines one third as long as the twelve stout conical
main spines, which reach in length the radius, and lie opposite in pairs in six axes.
Dimensions.—Diameter of the outer shell 0.22, middle 0.06, inner 0.02; cortical
pores 0.006, bars 0.003 length of the main spines 0.12, basal breadth 0.02.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
Subgenus 2. Echinommura, Haeckel.
Definition.—Pores of the cortical shell irregular, of different size
or form.
5. Echinomma sphærechinus, n. sp. (Pl. 29, fig.
2).
Actinomma sphærechinus, Haeckel, 1879, Atlas (pl. xxix. fig. 2).
Cortical shell thin walled, with irregular, roundish pores, twice to five times as broad as the
bars; five to seven on the radius. Both medullary shells with regular, circular pores, twice as
broad as the bars. Radial proportion of the three spheres = 1 : 2 : 4. Surface
covered with thirty to fifty pyramidal main spines, as long as the diameter of the inner shell,
and with numerous small conical by-spines of half the length.
Dimensions.—Diameter of the outer shell 0.11, middle 0.05, inner 0.025; cortical
pores 0.01 to 0.02, bars 0.004; length of the main spines 0.03, basal breadth 0.005.
Habitat.—North Atlantic, Station 353, surface.
6. Echinomma trinacrium, Haeckel.
Actinomma trinacrium, Haeckel, 1862, Monogr. d. Radiol., p. 441, Taf.
xxiv. figs. 6-8.
Cortical shell thin walled, with irregular, roundish pores, twice to three times as broad as
the bars. Both medullary shells with regular, circular pores. Radial proportion of the three
spheres {259}= 1 : 3 : 9. On the
surface about twenty three-sided pyramidal main spines, as long as the radius, and numerous (forty
to sixty) by-spines of half the length. (Compare with this species Actinomma trinacrium,
with which it is connected by transitional forms.)
Dimensions.—Diameter of the outer shell 0.09, middle 0.03, inner 0.01; cortical
pores 0.008 to 0.012, bars 0.004; length of the spines 0.02 to 0.05, basal breadth 0.01.
Habitat.—Mediterranean (Messina).
7. Echinomma toxopneustes, n. sp. (Pl. 29, fig.
1).
Cortical shell thin walled, with large, irregular roundish, polygonally framed pores, twice to
four times as broad as the crested bars. Both medullary shells of similar structure. Radial
proportion of the three spheres = 1 : 2.5 : 6. Numerous (thirty to fifty or
more) thin radial beams connecting them, prolonged outside into strong three-sided pyramidal
spines, shorter than the radius; each of the three wings with two teeth. Between these, numerous
smaller angular by-spines of one-quarter to one-half the length.
Dimensions.—Diameter of the outer shell 0.12, middle 0.05, inner 0.02; cortical
pores 0.01 to 0.03, bars 0.008; length of the main spines 0.05, basal breadth 0.01.
Habitat.—South-east Pacific (South of Juan Fernandez), Station 300,
surface.
Genus 103. Pityomma,[139] Haeckel, 1881,
Prodromus, p. 453.
Definition.—Astrosphærida with three concentric
lattice-shells and numerous branched radial spines.
The genus Pityomma differs from its ancestral form, Actinomma, in the
ramification of its radial spines, and exhibits therefore the same relation to it that
Elatomma, among the Haliommida, bears to Haliomma.
1. Pityomma scoparium, n. sp.
Cortical shell thick walled, connected with both concentric medullary shells by twenty thin
radial beams, which are prolonged outside into twenty large cylindrical radial spines; these are
somewhat shorter than the shell radius, simple in the proximal inner half, irregularly branched in
the outer half. All three spheres with regular, circular pores, twice to three times as broad as
the bars; radial proportion = 1 : 2 : 6. (Similar to Cladococcus
scoparius, Pl. 27,
fig. 2, and Elatomma scoparium, but differs from both in the double medullary shell.)
Dimensions.—Diameter of the outer shell 0.15, middle 0.05, inner 0.025; length of
the spines 0.1.
Habitat.—Central Pacific, Station 273, surface.
{260}
2. Pityomma piniferum, n. sp.
Cortical shell thin walled, with irregular, roundish pores, connected with both medullary
shells by twenty stout radial beams, which are prolonged outside into twenty large three-sided
prismatic spines. These are longer than the shell diameter and have three dentate edges, being
branched like a pine tree (with four to six ramified branches on each edge, decreasing in size
towards the distal end); similar to those of Cladococcus abietinus (Pl. 27, fig. 3).
Radial proportion of the three spheres = 1 : 2 : 8. Both medullary shells with
regular, circular pores, twice as broad as the bars.
Dimensions.—Diameter of the outer shell 0.2, middle 0.05, inner 0.025; length of the spines 0.25.
Habitat.—South Pacific, Station 299 (Juan Fernandez), surface.
3. Pityomma drymodes, n. sp. (Pl. 29,
fig. 9).
Cortical shell thin walled, connected with both medullary shells by very numerous (one hundred
to two hundred or more) very thin radial beams, which are prolonged outside into thin branched
radial spines, scarcely half as long as the shell radius, each spine bearing six to nine simple
branches. Outer and middle shell uneven, with hill-shaped protuberances and valleys between them;
the connecting radial beams, which arise from the top of the protuberances of the middle shell,
are inserted into the deepest part of the valleys of the outer shell. Radial proportion of the
three spheres = 1 : 3 : 8. Outer medullary shell with small, irregular,
roundish, or polygonal pores, not much broader than the bars; innermost shell with polygonal pores
and very thin bars.
Dimensions.—Diameter of the outer shell 0.28, middle 0.1, inner 0.033; length of the spines 0.07, breadth
0.004.
Habitat.—Central Pacific, Station 271, surface.
Subfamily Cromyommida,[140] Haeckel, 1881, Prodromus, pp. 449, 453.
Definition.—Astrosphærida with four
concentric spherical lattice-shells.
Genus 104. Cromyomma,[141] Haeckel, 1881,
Prodromus, p. 454.
Definition.—Astrosphærida with four concentric
lattice-spheres and numerous simple radial spines of one kind.
The genus Cromyomma is the common ancestral form of the Cromyommida, or of
those Astrosphærida in which the shell is composed of four concentric spheres, connected by radial
beams. Usually two of these are intracapsular or medullary {261}shells, two extracapsular or cortical shells; the distance between the
former and the latter is greater than the distance between either the two inner or the two outer
shells. In some cases, however (e.g., Cromyomma zonaster), the distance between all
four shells is equal.
Subgenus 1. Cromyommetta, Haeckel.
Definition.—Pores of the outer cortical shell regular, of nearly equal
size and similar form.
1. Cromyomma villosum, n. sp. (Pl. 30,
fig. 2).
Radial proportion of the four spheres = 1 : 2 : 4 : 5. Outer
cortical shell thick walled, densely covered with innumerable bristle-shaped radial spines, half
as long as the radius. Pores regular, circular, double-edged, of the same breadth as the bars; ten
to twelve on the radius. The thick outer shell is so dark, that the outlines only of the other
three shells can be seen.
Dimensions.—Diameter of the four shells—(A) outer cortical shell 0.2, (B)
inner cortical shell 0.16, (C) outer medullary shell 0.08, (D) inner medullary shell 0.04; pores
and bars of the outer shell 0.008; length of the spines 0.04.
Habitat.—Central Pacific, Station 267, depth 2700 fathoms.
2. Cromyomma zonaster, Haeckel.
Acanthosphæra zonaster, Ehrenberg, 1872, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 285, Taf. ii. fig. 2.
Radial proportion of the four spheres = 2 : 3 : 4 : 5. Outer
cortical shell thick walled, densely covered with numerous bristle-shaped radial spines, as long
as the radius. Pores regular, circular, twice as broad as the bars; eight to ten on the radius.
The thick outer shell is so dark, that the outlines only of the other three shells can be
seen.
Dimensions.—Diameter of the four shells—(A) 0.15, (B) 0.12, (C) 0.09, (D)
0.06; pores of the outer shell 0.01, bars 0.005; length of the spines 0.08.
Habitat.—Arctic Ocean, Greenland (depth 1000 fathoms), "Alert"
Expedition.
3. Cromyomma perplexum, Stöhr.
Cromyomma perplexum, Stöhr, 1880, Palæontogr., vol. xxvi. p. 95, Taf. iii.
fig. 5.
Radial proportion of the four spheres = 1 : 2 : 3 : 4. Outer
cortical shell thick walled, densely covered with short conical spines, as long as the diameter of
the pores. These are regular, circular, three times as broad as the bars; six to eight on the
radius. The thick outer shell is so dark, that the outlines only of the three other shells can be
seen.
Dimensions.—Diameter of the four shells—(A) 0.17, (B) 0.13, (C) 0.087, (D) 0.043; pores of the
outer shell 0.015, bars 0.005; length of the spines 0.02.
Habitat.—Fossil in the Tertiary rocks of Sicily (Grotte) Stöhr.
{262}
Subgenus 2. Cromyommura, Haeckel.
Definition.—Pores of the outer cortical shell irregular, of different
size and form.
4. Cromyomma perspicuum, n. sp. (Pl. 30,
fig. 8).
Cromyosphæra perspicua, Haeckel, 1879, Atlas (pl. xxx. fig. 8).
Radial proportion of the four shells = 1 : 2.5 : 6 : 9. Outer
cortical shell very thin walled, with thread-like bars, and irregular, polygonal pores; inner
cortical shell with similar pores, but with thicker bars. Both medullary shells with regular,
circular pores (the outer three times as broad as the inner). Numerous thin radial beams connect
the latter with the former; other very thin and numerous beams connect the two cortical shells,
and are prolonged outside into short bristle-shaped spines (often longer than in the figure).
Dimensions.—Diameter of the four shells—(A) 0.18, (B) 0.12, (C) 0.05, (D)
0.02; pores of the outer shell 0.01 to 0.02; length of the spines 0.01 to 0.03.
Habitat.—Central Pacific, Station 274, surface.
5. Cromyomma quadruplex, Haeckel.
Cromyomma quadruplex, Haeckel, 1862, Monogr. d. Radiol., p. 446.
Haliomma quadruplex, Ehrenberg, 1854, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 243.
Radial proportion of the four spheres = 1 : 3 : 7 : 10. Outer
cortical shell thin walled, irregularly covered with numerous angular spines, as long as the
radius. Pores of all four shells irregular, roundish, increasing in size from the first to the
fourth.
Dimensions.—Diameter of the four shells—(A) 0.15, (B) 0.1, (C) 0.045, (D)
0.015; pores of the outer shell 0.01 to 0.02; length of the spines 0.07.
Habitat.—North Atlantic, Station 353, depth 2965 fathoms.
6. Cromyomma macroporum, Stöhr.
Cromyomma macroporum, Stöhr, 1880, Palæontogr., vol. xxvi. p. 95, Taf.
iii. fig. 4.
Radial proportion of the four spheres = 1 : 3 : 6 : 9. Outer
cortical shell thick walled, densely covered with short conical spines, as long as the diameter of
the pores; the latter are irregular, roundish, or polygonal, larger and smaller alternating.
Dimensions.—Diameter of the four shells—(A) 0.1, (B) 0.066, (C) 0.033, (D)
0.011; pores of the outer shell 0.033 to 0.016, bars 0.003; length of the spines 0.01.
Habitat.—Fossil in the Tertiary rocks of Sicily, Grotte, Stöhr.
7. Cromyomma circumtextum, n. sp. (Pl. 30, fig.
4).
Radial proportion of the four spheres = 1 : 2 : 5 : 7. Outer
cortical shell very delicate and thin walled, with thread-like bars and large, irregular,
polygonal meshes (larger than the innermost shell); {263}inner cortical shell also with irregular, polygonal pores, three to nine
times as broad as the thick bars. Both medullary shells with subregular, circular pores (the outer
three times as broad as the inner). Radial spines twelve to twenty, three-sided prismatic, stout,
piercing the three outer shells, inserted on the innermost; their outer part pyramidal, half as
long as the radius.
Dimensions.—Diameter of the four shells—(A) 0.22, (B) 0.15, (C) 0.07, (D)
0.03; meshes of the outer shell 0.02 to 0.04; length of the outer spines 0.05.
Habitat.—Central Pacific, Station 266, depth 2750 fathoms.
8. Cromyomma mucronatum, n. sp. (Pl. 30,
figs. 5, 5a).
Radial proportion of the three spheres = 1 : 2 : 3 : 6. Outer
cortical shell thin walled, with large, irregular, roundish pores and crested three-sided bars;
inner cortical shell with regular, circular, hexagonally formed pores (four times as broad as the
bars). Both medullary shells with simple, small, regular, circular pores (fig. 5a). Twelve
radial spines three-sided prismatic, dagger-shaped, with three teeth, as long as the diameter of
the innermost shell.
Dimensions.—Diameter of the four shells—(A) 0.24, (B) 0.12, (C) 0.08, (D)
0.04; meshes of the outer shell 0.02 to 0.04; length of the spines 0.04.
Habitat.—Central Pacific, Station 274, depth 2750 fathoms.
Genus 105. Cromyechinus,[142] Haeckel,
1881, Prodromus, p. 454.
Definition.—Astrosphærida with four concentric
lattice-spheres and numerous simple spines of two different kinds; larger main spines and smaller
by-spines.
The genus Cromyechinus differs from Cromyomma in the same way as
Echinomma from Actinomma; the radial spines being differentiated into two different
kinds; a larger number of small by-spines, and a smaller number of large main spines.
1. Cromyechinus icosacanthus, n. sp. (Pl. 30, fig.
1).
Radial proportion of the three spheres = 1 : 2 : 6 : 8. Outer
cortical shell very delicate, with very small, regular, circular pores, covered with very numerous
bristle-shaped by-spines, half as long as the radius; inner cortical shell with very large,
irregular, polygonal pores, three to eight times as broad as the bars. Both medullary shells with
regular, circular pores, connected with the former by twenty regularly disposed, very thin bars;
these are prolonged outside into twenty very stout, three-sided prismatic main spines, as long as
the radius, which arise from the inner cortical shell, and at the distal end are cuspidate.
Dimensions.—Diameter of the four shells—(A) 0.16, (B) 0.13, (C) 0.04, (D)
0.02; pores of the outer cortical shell 0.003, of the inner 0.03, bars 0.003; length of the spines
0.08.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
{264}
2. Cromyechinus dodecacanthus, n. sp. (Pl. 30, figs. 3,
3a).
Radial proportion of the four spheres = 1 : 3 : 10 : 12. Outer
cortical shell very delicate, with very small, regular, circular pores, covered with numerous
short bristle-shaped by-spines, scarcely one-fourth as long as the radius; inner cortical shell
with very large, irregular, polygonal pores, twice to six times as broad as the bars. Both
medullary shells with very small, regular, circular pores, connected with the former by twelve
regularly disposed, thin radial beams; these are prolonged outside into twelve strong, three-sided
pyramidal main spines, one-third as long as the shell diameter. (Differs from the foregoing only
in the number and form of the radial spines.)
Dimensions.—Diameter of the four shells—(A) 0.12, (B) 0.1, (C) 0.03, (D)
0.01; pores of the outer cortical shell 0.002, of the inner 0.02, bars 0.002; length of the spines
0.04, basal breadth 0.01.
Habitat.—South Atlantic, Station 325, surface.
3. Cromyechinus polyacanthus, n. sp.
Radial proportion of the four spheres = 1 : 2 : 8 : 10. Outer
cortical shell of the same structure as the inner, with irregular, roundish pores, twice to six
times as broad as the bars. Both medullary shells with small, regular, circular pores, twice as
broad as the bars. Connecting radial beams forty to sixty, thin, cylindrical, prolonged outside
into forty to sixty strong conical main spines, about as long as the radius. Between these,
numerous thin bristle-shaped by-spines, nearly of the same length.
Dimensions.—Diameter of the four shells—(A) 0.25, (B) 0.2, (C) 0.05, (D)
0.025; pores of both cortical shells 0.01 to 0.03; length of the spines 0.12.
Habitat.—North Pacific, Station 237, surface.
Genus 106. Cromyodrymus,[143] Haeckel,
1881, Prodromus, p. 454.
Definition.—Astrosphærida with four concentric
lattice-spheres and numerous branched radial spines.
The genus Cromyodrymus differs from its ancestral form, Cromyomma, in
the ramification of the radial spines, and exhibits therefore the same relation to it that
Pityomma among the Actinommida bears to Actinomma.
1. Cromyodrymus quadricuspis, n. sp. (Pl. 30, figs. 7,
7a).
Radial proportion of the four shells = 1 : 3 : 6 : 12. All four
shells with regular, circular pores, about twice as broad as the bars. Radial proportion of the
pores in the four shells = 2 : 4 : 2 : 1. Radial spines fifty to
sixty, three-sided prismatic, half as long as the radius, with three recurved teeth, each spine in
this way bearing four points.
{265}
Dimensions.—Diameter of the four shells—(A) 0.16, (B) 0.08, (C) 0.04, (D)
0.013; length of the spines 0.03 to 0.04, breadth 0.007.
Habitat.—Central Pacific, Station 272, depth 2600 fathoms.
2. Cromyodrymus abietinus, n. sp. (Pl. 30, fig.
6).
Radial proportion of the four shells = 1 : 2 : 5 : 11. All four
shells with irregular, roundish pores, twice to four times as broad as the bars. Radial proportion
of the pores in the four shells = 9 : 3 : 2 : 1. Radial spines
eighty to one hundred and twenty, about as long as the radius, branched like a pine tree, with six
to twelve ramified branches.
Dimensions.—Diameter of the four shells—(A) 0.22, (B) 0.1, (C) 0.04, (D)
0.02; length of the spines 0.12, breadth 0.006.
Habitat.—South-east Pacific (Juan Fernandez), Station 299,
surface.
Subfamily Caryommida,[144] Haeckel.
Arachnosphærida, Haeckel, 1862, Monogr. d. Radiol., p. 354;
Prodromus, 1881, p. 454.
Definition.—Astrosphærida with five or more
concentric spherical lattice-shells.
Genus 107. Caryomma,[145] n. gen.
Definition.—Arachnosphærida with five to six or more
concentric spherical lattice-shells; two inner (intracapsular) medullary and three or more outer
(extracapsular) cortical shells, composed of ordinary lattice-work; distance between the former
and the latter greater than between any other two shells.
The genus Caryomma may be regarded as a Cromyomma, in which the
number of the cortical shells is increased to three, four, or more. These lie outside the central
capsule, whilst two medullary shells lie within it, and are connected with the former by numerous
radial spines piercing the wall of the capsule. The ordinary lattice-work is not arachnoidal, as
in the three following genera.
1. Caryomma regulare, n. sp.
Radial proportion of the five shells = 1 : 2 : 6 : 8 : 10.
All five shells with regular, circular pores, twice to three times as broad as the bars, and
gradually increasing in size from the innermost to the outermost shell. Radial beams connecting
them twenty, prolonged on the surface into twenty stout, three-sided pyramidal, radial spines,
half as long as the radius, regularly disposed.
Dimensions.—Diameter of the five shells—(A) 0.25, (B) 0.2, (C) 0.15, (D)
0.05, (E) 0.025.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
{266}
2. Caryomma irregulare, n. sp.
Radial proportion of the six shells = 1 : 2.5 : 8 : 10.5 : 13 : 15.
Both inner (medullary) shells with regular, circular pores, the other four (cortical) shells with
irregular, roundish pores, gradually increasing in size from the innermost to the outermost shell.
Radial spines sixty to eighty, pyramidal, irregularly disposed.
Dimensions.—Diameter of the six shells—(A) 0.3, (B) 0.26, (C) 0.21, (D)
0.16, (E) 0.05, (F) 0.02.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms.
Genus 108. Arachnopila,[146] n. gen.
Definition.—Astrosphærida with five to ten or more
cortical, concentric, polyhedral, or spherical lattice-shells, composed of a very thin cobweb-like
network; innermost shell with hexagonal (regular) or polygonal (irregular) meshes; other shells
with simple triangular meshes, without diagonal threads between them.
The genus Arachnopila, together with the two following genera, forms the
peculiar small group of large Arachnosphærida, separated from the true Caryommida by the totally
different structure and disposition of the numerous concentric shells; the former exhibits a
similar relation to the latter that the Diplosphærida bears to the Elatommida among the
dispherical Haliommida. The concentric shells (five to ten or more) lie outside the central
capsule, and are composed of very delicate, cobweb-like threads. From the innermost shell arise
numerous, three-sided prismatic, very long spines, from which at equal regular distances arise
lateral branches (three pairs from each spine, and one pair from each corner). In
Arachnopila these threads pass directly from one spine to the other, and form simple,
large, triangular meshes between them. The concentric shells are not connected by interwoven
diagonal threads.
1. Arachnopila hexagonella, n. sp.
Innermost shell with regular hexagonal pores; its diameter twice as long as the equal distance
between every two concentric shells. Radial spines twenty to forty, each with twenty to
twenty-four verticils.
Dimensions.—Diameter of the innermost shell 0.1; distance between the concentric
shells 0.05.
Habitat.—Central Pacific, Station 271, surface.
2. Arachnopila polygonella, n. sp.
Innermost shell with irregular, polygonal pores; its diameter fully as long as the equal
distance between every two concentric shells. Radial spines sixty to eighty, each with twelve to
sixteen verticils.
Dimensions.—Diameter of the innermost shell 0.04; distance between the concentric
shells 0.04.
Habitat.—Central Pacific, Station 266, surface.
{267}
Genus 109. Arachnopegma,[147] Haeckel,
1881, Prodromus, p. 454.
Definition.—Astrosphærida with five to ten or more
cortical, concentric, polyhedral, or spherical lattice-shells, composed of a very thin,
cobweb-like network; innermost shell with hexagonal or polygonal meshes; other shells with simple
triangular meshes, connected to one another by diagonal threads between them.
The genus Arachnopegma differs from its ancestral form Arachnopila,
in the possession of peculiar diagonal threads, which connect the verticils or nodal-points of
every two neighbouring radial spines in two different neighbouring concentric shells. In this
case, therefore, not only do triangular meshes lie in the spherical faces of the concentric
spheres, but also between them, in numerous oblique diagonal planes; a very rare and remarkable
structure, and forming a transition to spongy shells.
1. Arachnopegma verticillatum, n. sp.
Innermost shell with regular, hexagonal meshes; its diameter is the same as the equal distance
between each of the two shells. Threads of the network smooth. Radial spines twenty to forty, each
with fifteen to twenty verticils.
Dimensions.—Diameter of the innermost shell 0.04, distance between the concentric
shells 0.03 to 0.04.
Habitat.—Central Pacific, Station 266, surface.
2. Arachnopegma longispinum, n. sp.
Innermost shell with regular, hexagonal meshes; its diameter half as large as the equal
distance between every two shells. Threads of the network dentated or with small knots. Radial
spines fifty to sixty, each with twenty to twenty-five verticils.
Dimensions.—Diameter of the innermost shell 0.02; distance between the concentric
shells 0.04 to 0.05.
Habitat.—Central Pacific, Station 272, surface.
3. Arachnopegma increscens, n. sp.
Innermost shell with regular, hexagonal meshes; its diameter quite as large as the distance
between it and the second shell; only half as large as the distance between the fourth and fifth
shells. Forty to fifty radial spines, each with fifteen to twenty verticils.
Dimensions.—Diameter of the innermost shell (A) 0.025; distance between the
following shells—A, B = 0.025, B, C = 0.03, C, D = 0.04, D, E = 0.048, E, F = 0.056, F, G =
0.064, &c.
Habitat.—Central Pacific, Station 274, surface.
{268}
Genus 110. Arachnosphæra,[148] Haeckel,
1862, Monogr. d. Radiol., p. 355.
Definition.—Astrosphærida with five to ten or more
cortical, concentric, polyhedral, or spherical lattice-shells, composed of a very thin cobweb-like
network; innermost shell with hexagonal (regular) or polygonal (irregular) meshes; other shells
with larger, irregular, polygonal meshes.
The genus Arachnosphæra (accurately described by me in 1862, loc.
cit.) differs from the two preceding genera in the irregular form of the large meshes in all
shells (except often the innermost). This is caused by the ramification of the thin threads, which
on each shell connect the neighbouring spines. In Arachnopila and Arachnopegma the
threads pass directly and undivided from each spine to the neighbouring spine (three pairs from
the three edges), and therefore all meshes are triangular. In Arachnosphæra they become
polygonal by irregular ramification of the threads. There are here no diagonal threads.
Subgenus 1. Arachnosphærella, Haeckel.
Definition.—Pores of the innermost shell regular, hexagonal.
1. Arachnosphæra oligacantha, Haeckel.
Arachnosphæra oligacantha, Haeckel, 1862, Monogr. d. Radiol., p. 356, Taf.
x. fig. 2, Taf. xi. fig. 3.
Innermost shell with regular, hexagonal meshes; its diameter three times as long as the equal
distances between every two concentric shells. Fifteen to twenty radial spines scattered at wide
intervals, each with six to eight verticils.
Dimensions.—Diameter of the innermost shell 0.12; distance between the concentric
shells 0.04 to 0.05.
Habitat.—Mediterranean (Messina), surface, Haeckel.
2. Arachnosphæra myriacantha, Haeckel.
Arachnosphæra myriacantha, Haeckel, 1862, Monogr. d. Radiol., p. 357, Taf.
x. fig. 3, Taf. xi. fig. 4.
Innermost shell with regular, hexagonal meshes; its diameter twice as long as the equal
distances between every two concentric shells. At each nodal-point occurs one radial spine with
six to eight verticils (spines in all one hundred to one hundred and twenty or more).
Dimensions.—Diameter of the innermost shell 0.1; distance between the concentric
shells 0.04 to 0.05.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Indian, Pacific,
surface.
{269}
3. Arachnosphæra dolichacantha, n. sp.
Innermost shell with regular, hexagonal meshes, very small; its diameter only as long as the
equal distances between every two concentric shells. At each nodal-point occurs one radial spine
with twelve to sixteen verticils (altogether thirty to forty spines).
Dimensions.—Diameter of the innermost shell 0.05; distance between the concentric
shells 0.04 to 0.05.
Habitat.—Central Pacific, Station 265 to 274, surface.
4. Arachnosphæra increscens, n. sp.
Innermost shell with regular, hexagonal meshes; its diameter three times as long as the
distance between it and the second shell, quite as long as the distance between the fifth and
sixth shells; the distances between the concentric shells gradually increasing from the centre.
Radial spines about fifty to sixty, each with six to eight verticils.
Dimensions.—Diameter of the innermost shell (A) 0.75; distances between the
following shells—A, B = 0.025, B, C = 0.037, C, D = 0.05, D, E = 0.062, E, F = 0.075,
&c.
Habitat.—Central Pacific, Station 271, surface.
Subgenus 2. Arachnosphæromma, Haeckel.
Definition.—Pores of the innermost shell irregular, polygonal.
5. Arachnosphæra tenuissima, n. sp.
Innermost shell with irregular, polygonal meshes; its diameter twice as long as the equal
distance between every two concentric shells. Forty to fifty radial spines, each with twelve to
sixteen verticils.
Dimensions.—Diameter of the innermost shell 0.1, distance between the concentric
shells 0.04 to 0.05.
Habitat.—South Atlantic, Station 332, surface.
6. Arachnosphæra velaris, n. sp.
Innermost shell with irregular, polygonal meshes; its diameter twice as long as the distance
between it and the second shell, quite as long as the distance between the third and fourth
shells; the distance between the concentric shells gradually increasing from the centre. Radial
spines twenty to thirty, each with ten to twelve verticils.
Dimensions.—Diameter of the innermost shell (A) 0.5; distances of the following
shells—A, B = 0.025, B, C = 0.037, C, D = 0.05, D, E = 0.062, E, F = 0.075.
Habitat.—Tropical Atlantic, Station 347, surface.
{270}
Subfamily Spongiommida,[149] Haeckel.
Definition.—Astrosphærida with spongy
spherical or polyhedral shell (with or without enclosed concentric lattice-shells).
Genus 111. Spongiomma,[150] n. gen.
Definition.—Astrosphærida with solid spongy sphere, with
numerous simple radial spines, but without latticed medullary shells.
The genus Spongiomma differs from its ancestral form, Styptosphæra,
in the development of simple radial spines on the surface of the solid sphere, the entire mass of
which is composed of an irregular, spongy wicker-work.
Subgenus 1. Spongiommella, Haeckel.
Definition.—Radial spines on the surface of the spongy sphere all of
the same shape.
1. Spongiomma radiatum, n. sp.
Spongy framework of the solid sphere of the same structure throughout, with small, irregular,
polyhedral meshes and very thin thread-like bars. From the surface arise very numerous (one
hundred and twenty to one hundred and sixty or more) straight, bristle-shaped radial spines, as
long as the radius of the sphere (counting from the middle part of it).
Dimensions.—Diameter of the spongy sphere 0.2; length of the radial spines
0.1.
Habitat.—Central Pacific, Station 272, surface.
2. Spongiomma denticulatum, n. sp.
Spongy framework of the solid sphere of the same structure throughout, with small, almost equal
meshes, four to eight times as broad as the thin, elegantly denticulated bars; from the surface
arise very numerous (two hundred to three hundred or more) curved, radial spines, as long as the
radius, and of the same form as the bars.
Dimensions.—Diameter of the sphere 0.25; length of the spines 0.12.
Habitat.—Central Pacific, Station 274, surface.
3. Spongiomma spathillatum, n. sp.
Spongy framework in the central part of the sphere much denser and darker, and with smaller
meshes than in the peripheral part in which are very thin bars. Entire surface covered with {271}innumerable short, bristle-shaped radial spines, only
one-eighth as long as the radius, of the same elegant form as in Octodendron spathillatum (Pl. 18, figs. 2,
4); each spine developed in a zig-zag fashion, with very small beards, with a delicate spathillum
(or coronal of beard spines) at the distal end.
Dimensions.—Diameter of the spheres 0.16; length of the spines 0.01.
Habitat.—South Pacific, Station 295, surface.
4. Spongiomma clavatum, n. sp.
Spongy framework in the central part of the sphere much denser and darker than in the
peripheral part. On the surface are sixty to eighty stout, club-shaped radial spines, as long as
the radius of the sphere, in the proximal half three-sided prismatic, with three dentated edges;
they begin at the middle of the radius (where the denser inner framework changes into the looser
outer) and are very thin at first but increase slowly in thickness towards the truncated distal
end. (Similar to Centrocubus rhopalophorus, Pl. 18, fig. 1, but
without the cubical medullary shell.)
Dimensions.—Diameter of the sphere 0.4; length of the spines 0.2.
Habitat.—North Pacific, Station 241, surface.
Subgenus 2. Spongiommura, Haeckel.
Definition.—Radial spines on the surface of the spongy sphere of two
different kinds; large main spines and small by-spines.
5. Spongiomma helioides, n. sp.
Spongy framework of the sphere everywhere of the same structure, with almost equal meshes, ten
to twelve times as broad as the bars. Sixteen to twenty radial main spines, longer than the shell
diameter, three-sided prismatic, with three serrated edges, beginning about the middle of the
radius and increasing in thickness to the truncated distal end. Between them occur numerous thin,
bent, bristle-shaped by-spines. (Very similar to Spongosphæra helioides, Monogr. d.
Radiol., Taf. xii. figs. 11-13, but without medullary shells.)
Dimensions.—Diameter of the sphere 0.3; length of the main spines 0.4, of the
by-spines 0.01.
Habitat.—Tropical Atlantic, Station 352, surface.
6. Spongiomma multiaculeum, Haeckel.
Spongechinus multiaculeatus, Dunikowski, 1882, Denkschr. d. k. Akad. d.
Wiss. Wien, Bd. xlv. p. 29, Taf. v. figs. 60-63.
Spongy framework of the sphere very compact, with small meshes, scarcely broader than the bars;
four to eight large main spines, three-sided pyramidal, longer than the shell radius; numerous
(thirty to forty) thin by-spines, scarcely half as long.
{272}
Dimensions.—Diameter of the sphere 0.16; length of the main spines 0.11, of the
by-spines 0.03.
Habitat.—Fossil in the Alpine Jura, Schafberg near Salzburg
(Dunikowski).
7. Spongiomma asteroides, n. sp.
Spongy framework in the central part of the sphere much denser and darker than in the
peripheral part. Sixty to eighty cylindro-conical main spines, as long as the radius, between them
are numerous straight, bristle-shaped by-spines half that length.
Dimensions.—Diameter of the sphere 0.28; length of the main spines 0.16, of the
by-spines 0.08.
Habitat.—South Atlantic, Station 325, surface.
Genus 112. Spongodrymus,[151] Haeckel,
1881, Prodromus, p. 456.
Definition.—Astrosphærida with solid spongy sphere,
without latticed medullary shell, with numerous branched radial spines.
The genus Spongodrymus differs from the preceding Spongiomma in the
ramification of the numerous radial spines, covering the surface of the solid spongy sphere.
1. Spongodrymus elaphococcus, n. sp. (Pl. 18, fig.
9).
Spongy framework of the solid sphere equal throughout, on the surface arising in the form of
very numerous (sixty to eighty or more) spongy cones with rather rhomboidal meshes. Each cone is
prolonged into a very thin, irregularly curved radial spine, which is twice as long as the shell
diameter, simple in the proximal half, branched like a tree in the distal half; each arborescent
spine has from sixteen to thirty-two terminal branches, which fall in one spherical face. The
branches of neighbouring spines are partly connected by anastomoses, so that they begin to form an
outer spherical shell (cortical shell) with a looser spongy framework.
Dimensions.—Diameter of the whole spherical skeleton (sphere formed by the distal
ends of the dichotomous branches) 0.9, of the inner solid spongy sphere 0.02.
Habitat.—Tropical Atlantic, Station 349, surface.
2. Spongodrymus abietinus, n. sp.
Spongy framework denser and darker in the central part of the sphere than in the peripheral
part, with rather coarse meshes and thick bars. From the surface arise very numerous (sixty to
eighty) stout, radial branches, as long as the shell radius, branched like a pine tree (with six
to twelve ramified branches). Similar in structure to Cromyodrymus abietinus (Pl. 30, fig. 6),
but with a quite irregular spongy texture in the central sphere.
Dimensions.—Diameter of the sphere 0.3; length of the spines 0.015.
Habitat.—Central Pacific, Station 272, surface.
{273}
Genus 113. Spongechinus,[152] Haeckel,
1881, Prodromus, p. 456.
Definition.—Astrosphærida with hollow, spongy sphere,
without latticed medullary shell in the central cavity, and with numerous simple radial
spines.
The genus Spongechinus differs from its ancestral form, Plegmosphæra,
in the development of numerous radial spines on the surface of the spongy sphere, within which is
enclosed a large spherical central cavity.
1. Spongechinus setosus, n. sp.
Spongy sphere three times as broad as its inner cavity, with a very delicate, equal framework.
Entire surface covered with short, straight, bristle-shaped radial spines, about half as long as
the radius.
Dimensions.—Diameter of the sphere 0.2, of its inner cavity 0.07; length of the
spines 0.05.
Habitat.—North Atlantic, Azores, surface.
2. Spongechinus serrulatus, n. sp.
Spongy sphere twice as broad as its inner cavity, with a delicate, equal framework. Entire
surface covered with short, curved, radial spines, which are elegantly denticulated, and as long
as the shell radius.
Dimensions.—Diameter of the sphere 0.3, of its cavity 0.15; length of the spines
0.15.
Habitat.—Central Pacific, Station 274, surface.
3. Spongechinus cavus, n. sp.
Spongy sphere only one-fourth broader than the large inner cavity, its spongy wall being only
one-fourth as thick as the radius, composed of three to four strata of irregular, small meshes.
Surface covered with short, bristle-shaped, curved spines, one-third as long as the radius.
Dimensions.—Diameter of the sphere 0.5, of its inner cavity 0.4; length of the
spines 0.08.
Habitat.—North Atlantic, Station 354, surface.
Genus 114. Spongothamnus,[153] n.
gen.
Definition.—Astrosphærida with hollow, spongy sphere,
without latticed medullary shell in the central cavity, and with numerous branched radial
spines.
The genus Spongothamnus differs from the preceding Spongechinus in
the ramification of the numerous radial spines, covering the surface of the hollow spongy sphere;
{274}it therefore exhibits the same relation to the
latter that Spongodrymus bears to Spongiomma.
1. Spongothamnus furcatus, n. sp.
Spongy sphere twice as broad as its inner cavity, with very delicate bars, and irregular, dense
framework. From the surface arise one hundred and fifty to one hundred and eighty thin, forked,
bristle-shaped spines, half as long as the radius; both fork branches one-third as long as the
basal or simple part.
Dimensions.—Diameter of the sphere 0.3, of its inner cavity 0.15; length of the
spines 0.08.
Habitat.—North Pacific, Station 241, surface.
2. Spongothamnus scoparius, n. sp.
Spongy sphere four times as broad as its inner cavity, with thick bars and rather loose
framework. From the surface arise sixty to eighty broom-shaped radial spines, as long as the
radius, each in the basal half simple, in the distal half with six to twelve irregularly ramified
branches (similar to the spines of Cromyodrymus abietinus, Pl. 30, fig.
6).
Dimensions.—Diameter of the sphere 0.4, of the inner cavity 0.1; length of the
spines 0.2.
Habitat.—Central Pacific, Station 271, surface.
Genus 115. Spongopila,[154] Haeckel,
1881, Prodromus, p. 456.
Definition.—Astrosphærida with a single, spherical,
latticed medullary shell, immediately enveloped by the spongy framework of the cortical shell; on
the surface of the latter are numerous radial spines.
The genus Spongopila is a Spongoplegma with radial spines. On the
other hand it may be derived either from Elaphococcus, by communication of the branched
spines, or from Arachnosphæra, by development of spongy branches between the concentric
spheres.
1. Spongopila dichotoma, n. sp.
Medullary shell with regular, hexagonal meshes, six times as broad as the bars. From each
nodal-point (between every three meshes) arises a bristle-shaped radial spine, which is
dichotomously branched. By communication of the neighbouring branches the loose spongy framework
of the spherical cortical shell is formed, which is four times as broad as the medullary shell. On
the surface occur very numerous bristle-shaped radial spines, as long as the diameter of the
medullary shell. (May be derived from Elaphococcus.)
Dimensions.—Diameter of the spongy cortical shell 0.3, of the medullary shell
0.08.
Habitat.—Tropical Atlantic, Station 347, surface.
{275}
2. Spongopila verticillata, n. sp.
Medullary shell with regular, hexagonal meshes, four times as broad as the bars. From its
surface arise forty to sixty, three-sided prismatic radial spines, bearing eight to ten verticils
of lateral branches, each verticil with six forked branches (two from each edge). By irregular
ramification of these branches, and communication in all directions, the loose spongy framework of
the cortical shell originates, which is six times as broad as the medullary shell. The free distal
ends of the spines are as long as the shell radius. (May be derived from
Arachnosphæra.)
Dimensions.—Diameter of the spongy cortical shell 0.6, of the medullary shell
0.1.
Habitat.—Tropical Pacific, Station 200, surface.
Genus 116. Rhizoplegma,[155] Haeckel,
1881, Prodromus, p. 456.
Definition.—Astrosphærida with a single, spherical,
latticed medullary shell, which is not armed with by-spines, but connected by stout radial main
spines with the spongy cortical shell.
The genus Rhizoplegma is immediately allied to the foregoing
Spongopila, but differs from it in the large interval separating both shells. In this
interval lies the wall of the central capsule, which is only pierced by the radial spines
connecting both shells.
Subgenus 1. Rhizoplegmarium, Haeckel.
Definition.—No free lateral branches of the radial spines between the
two shells.
1. Rhizoplegma polyacanthum, n. sp.
Spongy cortical shell with a very fine dense framework and nearly smooth surface (without
superficial by-spines). Its inner cavity is four times as broad as the medullary shell, which
exhibits regular, hexagonal meshes. Radial spines sixty to eighty, three-sided prismatic, with
three smooth edges, without lateral branches between both shells; their free outer part as long as
the shell radius.
Dimensions.—Diameter of the spongy cortical shell 0.3, of its inner cavity 0.2, of
the medullary shell 0.05.
Habitat.—Central Pacific, Station 274, surface.
2. Rhizoplegma spirale, n. sp.
Spongy cortical shell with a very delicate loose framework and bristly surface. Its inner
cavity three times as broad as the medullary shell, which exhibits regular, hexagonal meshes.
{276}Radial spines thirty to forty, prismatic, with
three smooth, spirally contorted edges, without lateral branches between the two shells; their
free outer part half as long as the shell radius.
Dimensions.—Diameter of the spongy cortical shell 0.2, of its inner cavity 0.12,
of the medullary shell 0.04.
Habitat.—Central Pacific, Station 265, surface.
3. Rhizoplegma trigonacantha, n. sp.
Spongy cortical shell with a rather loose framework and coarse bars, with nearly smooth surface
(without by-spines). Its inner cavity twice as broad as the medullary shell, which exhibits
irregular, roundish pores. Radial spines forty to sixty, prismatic, with three smooth straight
edges, without free branches between the two shells; their outer pyramidal part only one-third of
the shell radius. (Very similar to the common Rhizosphæra trigonacantha, but with simple
medullary shell.)
Dimensions.—Diameter of the spongy shell 0.2, of its inner cavity 0.16, of the
medullary shell 0.08.
Habitat.—North Atlantic, Station 353, surface.
Subgenus 2. Rhizoplegmidium, Haeckel.
Definition.—Between the two shells free lateral branches arise from
the three edges of the radial spines.
4. Rhizoplegma radicatum, n. sp. (Pl. 15, figs. 9,
9a).
Spongy cortical shell with a very loose framework; on the surface are innumerable thin, forked,
or repeatedly dichotomous by-spines. Its inner cavity three times as broad as the medullary shell,
which exhibits regular, hexagonal meshes (fig. 9a). Radial spines twelve, prismatic, with
straight dentated edges, their outer pointed part as long as the shell radius. Each spine between
the two shells has a verticil of three forked lateral branches (fig. 9a). The central
capsule completely distends the medullary shell, and forces out through each mesh a hernia-shaped
process (fig. 9).
Dimensions.—Diameter of the spongy shell 0.5, of its inner cavity 0.25, of the
medullary shell 0.08.
Habitat.—Central Pacific, Station 271, surface.
5. Rhizoplegma lychnosphæra, n. sp. (Pl. 11, fig.
5).
Lychnosphæra rhizoplegma, Haeckel, 1879, Atlas (pl. xi. fig. 5).
Spongy cortical shell with a very loose framework, composed of long thin beams as in
Lychnosphæra regina (Pl. 11,
figs. 1-4). Surface covered with short bristles. Its inner cavity six times as broad as the
medullary shell, which exhibits regular, hexagonal meshes. Radial spines twelve, prismatic, with
three smooth edges; their outer pointed part half as long as the shell radius. Each spine has
three verticils of three forked branches; the first verticil is free {277}between the two shells, while the two following verticils, by
communication of their ramules, form the irregular framework. Central capsule with many herniæ,
forced out through the meshes of the medullary shell (fig. 5).
Dimensions.—Diameter of the spongy shell 0.7, of its inner cavity 0.45, of the
medullary shell 0.07.
Habitat.—South Pacific, Station 284, surface.
Genus 117. Lychnosphæra,[156] Haeckel,
1881, Prodromus, p. 453.
Definition.—Astrosphærida with a single, spherical,
latticed medullary shell, which is armed with free radial by-spines, and connected by stout radial
main spines with the spongy cortical shell.
The genus Lychnosphæra, known only by one single, large, and very remarkable
species, mainly differs from the foregoing in the development of free radial by-spines on the
surface of the medullary shell, and in the free interval between it and the cortical shell; but
beyond this the loose spongy framework of the latter exhibits a very remarkable structure, figured
in Pl. 11.
1. Lychnosphæra regina (Pl. 11,
figs. 1-4).
Medullary shell (fig. 3) with regular, circular, hexagonally framed pores, twice as broad as
the bars; from each hexagon-corner arises a radial, bristle-shaped by-spine, as long as the
diameter. Twelve radial main spines each as broad as one of the meshes, three-sided prismatic, six
to eight times as long as the medullary shell. From their three leaf-shaped (often somewhat
denticulated or spirally contorted) edges arise four verticils of lateral branches, each composed
of three forked, thin branches. The forked branches of the first verticil end free between the two
shells (figs. 2, 3), while the two following verticils are ramified, and, by anastomosis of their
branches, compose the loose spongy framework of the cortical shell. On the surface of the latter
arise numerous radial (zig-zag-shaped) by-spines. The fourth verticil is terminal, with three
shorter, thicker, dentated, simple branches, which constitute, together with the distal apex of
the spine itself, a bunch of four terminal spines. The large central capsule completely distends
the medullary shell, and forces out by its pores numerous club-shaped herniæ (fig. 1).
Dimensions.—Diameter of the spongy cortical shell 0.6, of its inner cavity 0.4, of
the central capsule 0.22, of the medullary shell 0.06; length of the radial spines 0.4, breadth
0.01.
Habitat.—Central Pacific, Station 271, surface.
Genus 118. Centrocubus,[157] n. gen.
Definition.—Astrosphærida with a single, cubical
medullary shell, immediately surrounded by the spongy framework of the cortical shell; from the
eight corners {278}of the central cube arise eight
primary radial spines, and often others from the framework between them.
The genus Centrocubus and the following closely allied Octodendron
may represent a peculiar small group of Spongiommida, remarkable for the regular, cubical form of
the medullary shell, which is composed of twelve thin rods, corresponding to the twelve edges of a
mathematical cube; from the eight corners invariably arise eight primary radial spines, the
branches of which form the spongy cortical shell.
1. Centrocubus octostylus, n. sp.
Radial spines eight, arising from the eight corners of the cubiform, regular, medullary shell,
gradually increasing in thickness towards the club-shaped distal end, which is five to six times
as broad as the central end. From the three denticulate edges of each spine arise six to eight
lateral branches, which ramify irregularly and form by their anastomosis the spongy framework
which is of nearly similar structure throughout, and with large loose meshes. The free distal part
of each spine is half as long as the enclosed part.
Dimensions.—Diameter of the sphere 0.6, of the central cube 0.02; length of the
spines 0.15, distal thickness 0.02.
Habitat.—Central Pacific, Station 271, surface.
2. Centrocubus cladostylus, n. sp. (Pl. 18, fig.
1).
Radial spines thirty-two, club-shaped, at the distal end eight to ten times as broad as at the
basal end. Eight primary spines arise from the eight corners of the regular, cubiform medullary
shell, and from these, in the form of lateral branches, twenty-four secondary spines arise with
concavely curved bases (three from the three denticulate edges of each spine, at nearly equal
distances from the centre). The free distal end of each of the thirty-two spines is of the same
shape, about half as long as the radius of the spongy sphere; framework much looser in the outer
than in the inner part.
Dimensions.—Diameter of the sphere 0.8, of the central cube 0.02; length of the
spines 0.2, distal thickness 0.02.
Habitat.—North Pacific, Station 256, surface.
3. Centrocubus polystylus, n. sp.
Radial spines sixty to eighty, club-shaped, four to six times as broad at the distal as at the
basal end. Eight primary spines arise from the eight corners of the regular, cubiform medullary
shell, the remainder either springing as lateral branches from the three denticulate corners of
the former, or arising within the spongy framework, which is much denser and darker in the central
than in the peripheral part. The free distal end of each spine is one-third as long as the
radius.
{279}
Dimensions.—Diameter of the sphere 0.9, of the central cube 0.02; length of the
spines 0.15, distal thickness 0.02.
Habitat.—South Pacific, Station 295, surface.
Genus 119. Octodendron,[158] n. gen.
Definition.—Astrosphærida with a single, cubical
medullary shell, and eight primary radial spines arising from its eight corners; these are
connected at equal distances by a latticed, spherical, cortical shell, from which the spongy
framework directly springs; often from the latter secondary radial spines arise.
The genus Octodendron has the same regular, cubical medullary shell as the
preceding Centrocubus, but differs from it in the wide interval separating the medullary
shell from the inner spherical face of the spongy cortical shell, the two being connected only by
eight radial beams, arising from the eight corners of the cube.
Subgenus 1. Octodendridium, Haeckel.
Definition.—Only eight primary radial spines, arising from the eight
corners of the central cube.
1. Octodendron cubocentron, n. sp. (Pl. 18, fig.
3).
Radial spines eight, club-shaped, with three denticulate, straight edges, twice as long as the
diameter of the central cavity of the cubical spongy shell; the inner face of the latter (or the
"inner cortical shell") exhibits the form of a large cube, the spongy sides of which are parallel
with the simple square sides of the central cube. The thin eight radial beams, connecting the
corresponding corners of both cubes, are not thicker than the edge-bars of the central cube,
whilst their outer prolongations are much thicker, with four to six verticils of lateral branches.
Surface of the thin walled, loose, spongy shell armed with short simple thorns.
Dimensions.—Diameter of the spongy shell 0.2, of its inner cavity 0.1, of the
central cube 0.02; length of the spines 0.2 (from the centre 0.3).
Habitat.—Central Pacific, Station 266, surface.
2. Octodendron spirale, n. sp.
Radial spines eight, three-sided prismatic, three times as long as the diameter of the inner
shell-cavity; their three edges denticulated and spirally contorted around the spine axis.
Thickness of the loose spongy shell-wall equal to the radius of the inner cavity, three times as
large as the central cube. Surface covered, with numerous thin, bristle-shaped, bent spinules.
{280}
Dimensions.—Diameter of the spongy shell 0.24, of its inner cavity 0.12, of the
central cube 0.02; length of the spines 0.4, breadth 0.01.
Habitat.—North Pacific, Station 254, surface.
3. Octodendron pinetum, n. sp.
Radial spines eight, three-sided prismatic, three to four times as long as the diameter of the
inner shell-cavity, with three spirally contorted edges. From each spine arise at equal distances
(equal to the half radius of the cavity) fifteen to twenty verticils of branches which increase in
size from the base of the spine. Each verticil is composed of six forked branches (two from each
spine-edge); the inferior are richly branched, and form by their connection the loose network of
the spongy cortical shell, the surface of which is covered with numerous bent threads. Each spine
bears a resemblance to a pine tree.
Dimensions.—Diameter of the spongy shell 0.3, of its inner cavity 0.15, of the
central cube 0.02; length of the spines 0.5 to 0.6, breadth 0.01.
Habitat.—Central Pacific, Station 272, surface.
4. Octodendron araucaria, n. sp.
Radial spines eight, three-sided prismatic, with three spirally contorted edges, eight to ten
times as long as the diameter of the inner shell-cavity (the free distal portion twice to three
times as long). From each spine arise at equal distances thirty to thirty-five verticils of
branches, which decrease in size towards the distal end. Each verticil is composed of six forked
branches (two from each spine-edge); the inferior are richly ramified and form by their connection
the loose spongy framework; the distal branches bear on the free end elegant spathillæ (as in the
following species). Surface covered with innumerable straight bristles, as long as the cavity
radius, and ending with a spathilla. Diameter of the central capsule about equal to the radius of
the spongy sphere, its membrane is double-edged.
Dimensions.—Diameter of the spongy shell 1.4 mm., of its inner cavity 0.14, of the
central cube 0.02, of the central capsule 0.5 to 0.6; length of the spines 0.9 to 1.2, breadth
0.02.
Habitat.—South Pacific, Station 288, surface.
5. Octodendron spathillatum, n. sp. (Pl. 18, figs. 2,
4).
Radial spines eight, three-sided prismatic, with contorted edges, five to eight times as long
as the diameter of the shell-cavity (the free part twice as long). From each spine arise ten to
twelve verticils of lateral branches; each verticil composed of three forked branches. The
proximal larger branches ramify richly, and form by their connection the loose spongy framework of
the spherical shell; the distal smaller branches are simple or bifurcated, and the ramules are
provided with an elegant spathillum at the end (fig. 4). Entire surface of the spongy sphere
covered with innumerable bristle-shaped radial spines (half as long as the cavity radius),
zig-zag, bent, with beards, and with a spathillum at the end.
{281}
Dimensions.—Diameter of the spongy sphere 1 mm., of its central cavity 0.15, of
the central cube 0.02; length of the spines 1 to 1.2 mm., breadth 0.008.
Habitat.—Central Pacific, Station 271, surface.
Subgenus 2. Octodendronium, Haeckel.
Definition.—Radial spines thirty-two or more, eight primary (arising
from the eight corners of the central cube) and twenty-four or more secondary (between them).
6. Octodendron verticillatum, n. sp.
Radial spines thirty-two, with three denticulate straight edges, six to eight times as long as
the diameter of the shell-cavity; the distal parts half free. Eight primary spines arise from the
eight corners of the central cube, twenty-four secondary from the edges of these (a verticil of
every three from each primary spine). Distal free parts of all thirty-two spines equal. Each spine
with eight to ten verticils of forked lateral branches, without spathillæ. Surface of the spongy
sphere covered with short simple bristles.
Dimensions.—Diameter of the spongy shell 0.3, of its cavity 0.12, of the central
cube 0.02; length of the spines 0.7 to 1 mm., breadth 0.02.
Habitat.—South Pacific, Station 291, surface.
7. Octodendron contortum, n. sp.
Radial spines thirty-two, disposed in a similar manner to those of the former species (eight
primary and twenty-four secondary); also the spongy shell of the same shape. The difference arises
in the form of the spines, the three edges of which are much broader and spirally contorted around
the axis; and thus the corresponding branches of the verticil do not lie in the same
meridian-plane, but alternate one with another.
Dimensions.—Diameter of the spongy shell 0.4, of its cavity 0.1, of the central
cube 0.02; length of the spines 0.8 to 1.2, breadth 0.04.
Habitat.—Central Pacific, Station 274, surface.
8. Octodendron arboretum, n. sp.
Radial spines sixty to ninety, three-sided prismatic, with spirally contorted and denticulate
edges, six to eight times as long as the diameter of the inner shell-cavity. Eight primary arise
from the eight corners of the central cube, twenty-four others from their three edges (as in the
two former species); the remaining thirty to sixty spines seem to arise between the former and
immediately from the dense spongy framework, which is twice as thick as the diameter of the inner
shell-cavity. The numerous verticils of the free distal part are of equal shape in all the spines,
composed of three forked branches in the terminal, and of more ramified branches in the inferior
parts. Entire surface of the spongy shell covered with simple radial bristles, without
spathillæ.
{282}
Dimensions.—Diameter of the spongy shell 0.75, of its central cavity 0.15, of the
central cube 0.02; length of the spines 0.9 to 1.2, breadth 0.02.
Habitat.—Tropical Pacific, Station 225, surface.
Genus 120. Spongosphæra,[159] Ehrenberg,
1847, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 54.
Definition.—Astrosphærida with two concentric latticed
medullary shells, connected by radial beams; the outer is immediately enveloped by the spongy
framework, and bears numerous radial spines.
The genus Spongosphæra (in the definition here restricted) differs from
Spongopila in the double medullary shell, which in the latter is simple; it exhibits to the
latter the same relation that Spongodictyon among the Liosphærida bears to
Spongoplegma. The outer medullary shell of Spongosphæra is immediately enveloped by
the spongy wicker-work, which everywhere pierces the wall of the central capsule.
1. Spongosphæra streptacantha, Haeckel.
Spongosphæra streptacantha, Haeckel, 1862, Monogr. d. Radiol., p. 455,
Taf. xxvi. figs. 1-3.
Spongy shell of polyhedric, irregular outline, the framework being prolonged sheath-like into
the eight to twelve radial spines, which are quite irregularly distributed, very large,
three-sided prismatic, with three serrated, spirally contorted edges; their length is twice to
four times as great as the diameter of the spongy body; they arise with thinner bases from the
outer medullary shell, which is three times as broad as the inner, both having roundish pores,
twice to four times as broad as the bars; surface without radial by-spines.
Dimensions.—Diameter of the spongy shell 0.2 to 0.6, outer medullary shell 0.04 to
0.06, inner 0.012 to 0.016.
Habitat.—Cosmopolitan; common in all warmer seas, surface.
2. Spongosphæra polyacantha, J. Müller.
Spongosphæra polyacantha, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 32, Taf. iv. figs. 1-4.
Spongy shell spherical, with ten to twenty large radial spines, which arise with thinner bases
from the medullary shell, and are prominent on the surface at different lengths; they are
three-sided prismatic (not four-sided), with three leaf-shaped straight edges. Whilst the spongy
framework is much looser than in the preceding species, both medullary shells have nearly the same
shape.
Dimensions.—Diameter of the spongy shell 0.2 to 0.5, outer medullary shell 0.04 to
0.06, inner 0.012 to 0.016.
Habitat.—Mediterranean (Nice); Atlantic, Station 353, surface.
{283}
Subgenus Spongosphæromma, Haeckel.
Definition.—Radial spines on the surface of the spongy shell of two
different kinds; large piercing main spines and small superficial by-spines.
3. Spongosphæra helioides, Haeckel.
Spongosphæra helioides, Haeckel, 1862, Monogr. d. Radiol., p.456, Taf.
xii. figs. 11-13.
Spongy shell spherical, with numerous curved, bristle-shaped by-spines on the surface, as long
as the radius. Main spines ten to twenty, irregularly disposed, prismatic, with three dentated,
spirally contorted edges, broader toward the distal end. Outer medullary shell three times as
broad as the inner, with polygonal meshes (of the same size as those in the spongy framework) and
fine bars.
Dimensions.—Diameter of the spongy shell 0.2, outer medullary shell 0.02, inner
0.007.
Habitat.—Mediterranean (Messina).
4. Spongosphæra quadricuspis, n. sp.
Spongy shell spherical, with numerous curved, bristle-shaped by-spines on the surface, half as
long as the radius. Main spines twenty to thirty, irregularly disposed, prismatic, with three
dentated, spirally contorted edges, and at the distal end four strong pyramidal divergent teeth
(three as terminations of the edges, the fourth as end of the spine axis). Outer medullary shell
twice as broad as the inner, both having circular, regular pores, twice to three times as broad as
the bars.
Dimensions.—Diameter of the spongy shell 0.3, outer medullary shell 0.04, inner
0.013.
Habitat.—Central Pacific, Station 272, surface.
Genus 121. Rhizosphæra,[160] Haeckel,
1860, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 840.
Definition.—Astrosphærida with two concentric latticed
medullary shells, connected by radial beams; from the outer arise numerous radial spines, which at
equal distances are connected by a latticed spherical cortical shell, surrounded by a spongy
framework.
The genus Rhizosphæra exhibits the same relation to Spongosphæra that
Rhizoplegma bears to Spongopila; but in the latter the latticed medullary shell is
simple, in the two former double. The wall of the central capsule is pierced only by the radial
spines connecting the medullary and the cortical shells.
1. Rhizosphæra trigonacantha, Haeckel.
Rhizosphæra trigonacantha, Haeckel, 1862, Monogr. d. Radiol., p. 452, Taf.
xxv. figs. 1-7.
Central cavity of the spongy cortical shell twice as broad as the diameter of the outer
medullary shell. Bars of all three shells of the same breadth as the thirty to fifty (or more)
radial spines, {284}which are three-sided prismatic;
their outer free distal end only as long as the diameter of the inner medullary shell.
Dimensions.—Diameter of the spongy shell 0.25, of its inner cavity 0.2, outer
medullary shell 0.1, inner 0.05.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Indian, Pacific,
surface from many Stations.
2. Rhizosphæra serrata, n. sp. (Pl. 18,
figs. 5-7).
Central cavity of the spongy shell five times as broad as the diameter of the outer medullary
shell. Bars of all three shells of the same breadth as the forty to sixty (or more) thin radial
beams between them. These are three-sided prismatic, with denticulate edges, scarcely half as
broad as their outer prolongations, which are half as long as the shell radius, and possess three
spirally contorted serrated edges. (The figured specimen is a young one; in the older specimens
the spongy framework of the cortical shell is much more developed.)
Dimensions.—Diameter of the spongy shell 0.3, of its central cavity 0.22, outer
medullary shell 0.06, inner 0.02.
Habitat.—Central Pacific, Stations 270 to 274, surface.
3. Rhizosphæra leptomita, Haeckel.
Rhizosphæra leptomita, Haeckel, 1862, Monogr. d. Radiol., p. 453, Taf.
xxv. figs. 8-10.
Central cavity of the spongy cortical shell twice as broad as the diameter of the outer
medullary shell; bars of both very thin, only one-third as broad as the bars of the inner
medullary shell. Radial spines thirty to fifty (or more), curved, three-sided prismatic; inside
the spongy shell as thin as their bars, outside three times as broad.
Dimensions.—Diameter of the spongy shell 0.27, of its inner cavity 0.2, outer
medullary shell 0.1, inner 0.05.
Habitat.—Mediterranean (Messina); Atlantic, Stations 348 to 354,
surface.
Suborder PRUNOIDEA, Haeckel, 1883 (Pls. 13-17, 39, 40).
Definition.—Spumellaria with an ellipsoidal or cylindrical
central capsule, prolonged into one axis (sometimes articulate by annular transverse strictures);
with an ellipsoidal or cylindrical, fenestrated siliceous shell (often articulate by annular
strictures), invariably prolonged into one axis. Fundamental form monaxon, usually with the poles
of the prolonged dimensive main axis equal.
The suborder Prunoidea comprises those Spumellaria in which the fenestrated spherical shell appears prolonged into one
axis. The geometric fundamental form of the shell, which in the Sphæroidea was a sphere, in this case therefore becomes an ellipsoid, and
whilst in the former all axes originally have the same value (Homaxonia), {285}here one main axis is constantly larger than all other axes (Monaxonia).
Usually both poles of this main axis are equal (Haplopola); but in some genera both poles become
different (Diplopola).
In the Sphæroidea all planes going through the centre of the shell are
circular, whereas in the Prunoidea only those planes are circular which
are perpendicular to the main axis; all other planes going through the centre are elliptical; the
largest of these are the meridian planes, in which is situated the main axis. Commonly all
meridian planes are equal, as no transverse axes (or cross axes) are differentiated.
In my Monograph (1862) only very few forms of Prunoidea, such as
Didymocyrtis and Spongurus, are described, and the greater part of them are
distributed under different genera of Sphæroidea, such as Haliomma
and Actinomma. In my Prodromus (1881) I separated them as the family Zygartida (p. 462). But it seems now much more convenient to restrict this term to a
particular family and to give a wider extension to the whole suborder under the name Prunoidea (called after the characteristic ellipsoidal form of a plum, or
Prunus, with its stone).
The suborder Prunoidea comprises seven different families, of which
the Ellipsida constitutes the simplest and the probable common ancestral group. In all Ellipsida
the fenestrated shell is simple, and never composed of concentric or twin shells. In their
primitive genus Cenellipsis, the whole shell is geometrically nothing more than a simple
ellipsoid (Pl. 39,
figs. 1, 2). By development of radial spines it passes into Ellipsidium, by development of
spongy framework into Spongellipsis. In the greater part of this family large spines are
developed on both poles of the main axis of the shell, but sometimes instead of these solid
spines, two opposite hollow fenestrated tubes are developed (Pipettella, Pl. 39, fig.
6).
The second family of Prunoidea, the Druppulida, is much richer in
different forms than the simple Ellipsida. In this case the ellipsoidal shell is composed of two
or three (rarely more) concentric shells. Constantly one or two of these fenestrated shells are
enclosed in the central capsule, and may therefore be called "medullary shells"; and one or two
(rarely more) lie outside the central capsule, "cortical shells." The inner medullary shells (one
or two) are either spherical or ellipsoidal; the outer cortical shells (one or two, rarely more)
are always ellipsoidal. All concentric shells are connected by radial beams. In the simplest form
of the subfamily, Druppula (Pl. 39,
fig. 3), one medullary shell is connected with one cortical shell. By duplication of the medullary
shell arises Prunulum (Pl. 39,
fig. 4), and by duplication or multiplication of the cortical shell Cromyodruppa (Pl. 15, figs. 1-4)
is formed. In by far the greater portion of this subfamily large spines are developed on both
poles of the main axis of the cortical shell (Pls. 16, 17), but
sometimes also instead of these solid spines two opposite hollow fenestrated tubes are developed
(Pipetta, Pl. 39,
figs. 7, 8).
{286}
A third family of Prunoidea, closely allied to the two preceding
families, is the Spongurida, in which we include all Prunoidea with an
ellipsoidal or cylindrical, unjointed shell, in which the lattice-work of the cortical shell is
transformed into an irregular, siliceous framework. In the simplest form, Spongellipsis,
the simple lattice-shell of Cenellipsis is substituted by an external spongy envelope. In
other cases (Spongurus and allied genera) the whole cavity of this external spongy shell is
distended with a fine spongy framework. The subfamily of Spongodruppida is distinguished by the
possession of a simple or double latticed medullary shell; this lies in the midst of the central
capsule, and is connected by radial beams (perforating its membrane) with the enveloping spongy
cortical shell. The surface of the latter may bear either radial spines, or two opposite strong
polar spines, at the poles of the main axis (Pl. 17, fig.
12).
Closely allied to the Ellipsida and Druppulida are two other families of the Prunoidea, the Artiscida and Cyphinida, which differ from the former by a
circular constriction in the equatorial plane of the ellipsoidal shell; and in this way assume a
characteristic twin form, like a figure of eight. In the Artiscida the shell is simple (as in the
Ellipsida), whereas in the Cyphinida it is composed of two or more concentric shells (as in the
Druppulida). The simplest form of the Artiscida is Artiscus (Pl. 39, fig. 9),
differing from Cenellipsis in the ring-shaped, equatorial constriction. In other Artiscida
polar appendages are developed on both poles of the main axis, either in the form of solid, strong
spines (Stylartus), or hollow fenestrated tubes (Cannartus, Pl. 39, fig.
10).
The family Cyphinida differs from the Druppulida in the equatorial constriction of the shell,
and from the Artiscida in the presence of two or more concentric shells. One or two of these
concentric fenestrated shells are enclosed in the central capsule (and therefore may be called
"medullary shells"); the others (one or two, rarely more) lie outside of the central capsule
(therefore "cortical shells"). The internal "medullary shells" are always spherical or somewhat
lenticular, compressed from both sides; the external "cortical shells" have constantly a ring-like
constriction in the equatorial plane, and "twin-shells" are therefore like a figure of eight. The
simplest form of this subfamily is Cyphanta, composed of a simple medullary shell and a
simple cortical shell, the two being connected in the equatorial plane by radial beams. In
Cyphonium (Pl. 39,
fig. 12) the medullary shell is doubled, and in Cypassis (Pl. 39, fig. 13)
the cortical shell likewise. On both poles of the main axis strong spines are often developed
(Cyphinus, Pl. 39,
fig. 14), or hollow fenestrated tubes (Cannartidium, Pl. 39, figs.
16-19).
The equatorial constriction of the ellipsoidal shell, which characterises the Artiscida and
Cyphinida, is repeated or multiplied in the two following families, in the Panartida and
Zygartida; in the former we find three ring-like strictures, in the latter five or more (lying in
parallel transverse planes); therefore the fenestrated shell is composed in the one instance of
four chambers, in the other of six or {287}more; all the
chambers form a single series and have a common main axis. All constrictions lie in planes
parallel to the equatorial plane of the original ellipsoid; in the centre of the latter constantly
lies a double "medullary shell," composed of two concentric, either spherical or lenticular,
compressed shells. In all Panartida we call the two inner chambers (on both sides of the
equatorial constriction) "proximal chambers," the two outer chambers (on the poles of the main
axis) "distal chambers." The four-chambered cortical shell of the Panartida is either simple (in
Panartus, Pl. 40,
figs. 1-4) or double, with an external mantle (as in Peripanartus, Pl. 40, figs. 5-7).
The simplest form of the subfamily is Panartus (loc. cit.). In this case also on
both poles of the main axis may be developed solid spines, or hollow fenestrated tubes
(Panarium, Pl. 40,
fig. 9).
The seventh and last family of the Prunoidea, the Zygartida is most
nearly allied to the Panartida, and appears as a further developmental step from that family.
Whilst in the Panartida the cortical shell is constantly four-chambered, with three parallel
ring-like constrictions, in the Zygartida it is always prolonged and composed of six or more
chambers, separated by five or more ring-shaped constrictions, in the middle of which is the
equatorial stricture. In the centre of the latter (as also in the Panartida) always lies the
double medullary shell, composed of two concentric, spherical, or lenticular shells. The number of
the chambers of the cortical shells is commonly six or eight (with five to seven ring strictures),
but it often mounts to ten and sometimes to twenty (with nineteen strictures), as in some species
of Zygartus (Pl. 40,
fig. 13). All the chambers lie in one series, one behind another, with a common main axis. The
cortical shell is usually simple (in Ommatocampe, Pl. 40, fig. 10),
sometimes double (in Desmocampe, Pl. 40,
fig. 12), rarely triple (in Zygocampe, Pl. 40, fig. 13).
In all three cases hollow fenestrated tubes may be developed on the poles of the main axis.
The morphological references and the phylogenetic affinities of all Prunoidea are so complex, that they seem to represent a quite natural group;
all forms of it may be derived from the common ancestral form Cenellipsis. But a far more
difficult question is the manner in which its pedigree may be constructed. The oldest family is
probably the simplest, namely, Ellipsida. From this the Druppulida may be derived by production of
medullary shells, the Artiscida by equatorial constriction. The Cyphinida can be produced either
from the Druppulida by equatorial constriction or from the Artiscida by development of medullary
shells. The Panartida appear as further developmental steps of the Cyphinida, by duplication of
the chamber number; and the Zygartida as further productions of the Panartida, by increasing the
number of the chambers.
The seven subfamilies of the Prunoidea can be arranged in two sections
according to the presence or absence of medullary shells. The Ellipsida, Spongellipsida, and
Artiscida possess a simple cortical shell, without a medullary shell; they represent the {288}section Cenoprunida. All other families possess medullary
shells, and so represent the section Coccoprunida.
Another character, which can be employed in the arrangement of the seven subfamilies in some
larger groups, is the presence or absence of ring-like constrictions, by which the cortical shell
is divided into chambers. I. The Monoprunida comprise all forms without any constriction, of which
the Ellipsida are without a medullary shell, the Druppulida with one or two medullary shells, and
the Spongurida with a spongy cortical shell. II. The Dyoprunida contain all forms with a cortical
twin shell, or with two chambers separated by one equatorial constriction, of which the Artiscida
are without a medullary shell and the Cyphinida have one or two medullary shells. III. The
Polyprunida comprise all forms with several (three or more) constrictions, which separate four or
more chambers, of which the Panartida have three constrictions and four chambers, and the
Zygartida five or more constrictions and six or more chambers.
The Central Capsule of the Prunoidea is originally ellipsoidal
(monaxial), and preserves this form in the greater part of the genera. In some groups, where the
axis of the ellipsoid is much prolonged, it passes over to the cylindrical form (with
hemispherical vaultings on both poles), as in Spongurus and Spongocore, in many
Panartida and Zygartida. Very often the ellipsoidal or cylindrical capsule gets annular transverse
constrictions, corresponding to those of the enveloping cortical shell (one single, equatorial
stricture in the Artiscida and Cyphinida, three strictures in the Panartida, five or more in the
Zygartida). In the Cenoprunida (Ellipsida and Artiscida, also in Spongellipsis) the central
capsule lies freely in the cavity of the cortical shell, separated from its inner surface by the
jelly-envelope; in all other groups it contains a part of the skeleton, the medullary shell and
the beams which connect it with the enveloping cortical shell.
Synopsis of the Families of Prunoidea.
|
A. MONOPRUNIDA.
Shell without transverse stricture
|
|
a. Shell simple, latticed (not spongy), without enclosed
internal shells, |
1. Ellipsida. |
| b. Shell composed of two or more concentric latticed shells (not
spongy), |
2. Druppulida. |
| c. Shell partially or wholly composed of an irregular spongy
framework, |
3. Spongurida. |
|
B. DYOPRUNIDA.
Shell bilocular, divided by an equatorial stricture into two
communicating hemi-ellipsoidal shells.
|
|
d. Shell simple, without enclosed internal shells, |
4. Artiscida. |
| e. Shell composed of two or more concentric shells, |
5. Cyphinida. |
|
C. POLYPRUNIDA.
Shell multilocular, divided by three or more parallel transverse
strictures into four or more serial cameræ.
|
|
f. Shell with three parallel strictures and therefore four
cameræ, |
6. Panartida. |
| g. Shell with five or more parallel strictures and therefore six
or more cameræ, |
7. Zygartida. |
| A. MONOPRUNIDA. Shell without transverse stricture |
|
|
a. Shell simple, latticed (not spongy), without enclosed
internal shells, |
|
|
|
|
|
|
1. Ellipsida. |
|
|
|
|
|
|
b. Shell composed of two or more concentric latticed shells
(not spongy), |
|
|
|
|
|
|
2. Druppulida. |
|
|
|
|
|
|
c. Shell partially or wholly composed of an irregular
spongy framework, |
|
|
|
|
|
|
3. Spongurida. |
|
|
|
|
| B. DYOPRUNIDA. Shell bilocular, divided by an equatorial stricture into two
communicating hemi-ellipsoidal shells. |
|
|
d. Shell simple, without enclosed internal shells, |
|
|
|
|
|
|
4. Artiscida. |
|
|
|
|
|
|
e. Shell composed of two or more concentric shells, |
|
|
|
|
|
|
5. Cyphinida. |
|
|
|
|
| C. POLYPRUNIDA. Shell multilocular, divided by three or more parallel
transverse strictures into four or more serial cameræ. |
|
|
f. Shell with three parallel strictures and therefore four
cameræ, |
|
|
|
|
|
|
6. Panartida. |
|
|
|
|
|
|
g. Shell with five or more parallel strictures and
therefore six or more cameræ, |
|
|
|
|
|
|
7. Zygartida. |
|
|
|
|
{289}
Family XI. Ellipsida, Haeckel, 1882 (Pls. 13, 14, 39).
Definition.—Prunoidea with simple ellipsoidal shell,
without equatorial stricture (without enclosed medullary shell); network a simple lattice lamella,
not spongy. Central capsule ellipsoidal or cylindrical, without annular equatorial
constriction.
The family Ellipsida comprises the simplest forms of Prunoidea, and probably represents the ancestral forms of this whole suborder.
The fenestrated shell, which encloses the ellipsoidal central capsule, is a perfectly simple
"cortical shell" of the same form, without enclosed "medullary shell." Its form is commonly a
regular monaxial ellipsoid; sometimes a little modified by unequal growth of the two poles of the
main axis. Two opposite large spines are often developed at these poles, or it may be that instead
of these, two hollow fenestrated tubes are present.
The ellipsoidal fenestrated shell exhibits in the regular Ellipsida all the characters of a
geometric ellipsoid; one main axis surpasses in length all other possible axes. All sections going
through this main axis are "meridian sections," with elliptical periphery; all sections
perpendicular to the main axis are "transverse sections," with circular periphery. The largest of
these is the equatorial section, which divides the main axis into halves. The diameter of this
equatorial plane is the "minor axis" of the ellipsoid.
The proportion of the two axes of the ellipsoidal shell, of the major vertical or main axis and
the minor horizontal or equatorial axis, is commonly between 6 : 5 and 3 : 2.
In the former case it approaches the spherical shell, from which it is derived; in the latter case
it becomes almost fusiform or cylindrical. The network of silex, constituting the shell, is
constantly a simple latticed lamella, never composed of concentric shells (as in Druppulida) or
spongy (as in Spongurida). The network is often very regular and elegant, in other cases
irregular.
The simplest genus among the Ellipsida, and probably the common ancestral form of the whole
subfamily, is the genus Cenellipsis, possessing a simple ellipsoidal shell without any
appendages. It is derived from Cenosphæra (the simplest spherical shell) by the
prolongation of one axis. Cenellipsis passes over into Ellipsidium by the production
of radial spines on the surface (corresponding to Heliosphæra). Axellipsis is a
peculiar genus differing from Cenellipsis in an axial rod, which corresponds to the minor
or equatorial axis. In all other genera of the subfamily both poles of the main axis are
distinguished by peculiar polar prolongations, either hollow fenestrated tubes (as in
Pipettella) or strong solid spines. Both polar spines are of equal size and similar form in
Ellipsoxiphus, unequal in Ellipsostylus. From the latter is derived
Lithapium, by reduction and loss of one spine (so that only one remains);
Lithomespilus, by production of a bunch of several spines at one pole. In the three latter
genera both poles of the main axis are unequal, in all others equal.
{290}
The central capsule of the Ellipsida is in all cases ellipsoidal, and occupies the largest part
of the shell, being separated from its inner surface by a thinner or thicker jelly-mantle.
Synopsis of the Genera of Ellipsida.
| Ellipsoidal shell without polar appendages (neither
solid spines nor hollow tubes at the poles of the axis). |
|
Surface without radial spines. |
|
Shell cavity simple, without an axial rod, |
122. Cenellipsis. |
| Shell cavity with a transverse axial rod, |
123. Axellipsis. |
| Surface covered with radial spines, |
124. Ellipsidium. |
| Ellipsoidal shell with polar appendages (either solid
spines or hollow fenestrated tubes) at the poles of the main axis. |
 |
Solid spines of similar shape, at both poles of the main
axis. |
|
Shell cavity simple, without axial rods, |
125. Ellipsoxiphus. |
| Shell cavity with a cross of axial rods, |
126. Axoprunum. |
| Solid spines of different shape, at both poles of the main
axis. |
|
Two polar spines of different shape, |
127. Ellipsostylus. |
| A bunch of spines at one pole only, |
128. Lithomespilus |
| Only a single spine at one one pole, |
129. Lithapium. |
| Two hollow fenestrated tubes opposite, at the poles of the
main axis, |
130. Pipettella. |
| Ellipsoidal shell without polar appendages (neither solid spines nor hollow
tubes at the poles of the axis). |
|
|
Surface without radial spines. |
|
|
|
|
|
|
Shell cavity simple, without an axial rod, |
|
|
|
|
|
|
|
|
|
|
122. Cenellipsis. |
|
|
|
|
|
|
|
|
|
|
Shell cavity with a transverse axial rod, |
|
|
|
|
|
|
|
|
|
|
123. Axellipsis. |
|
|
|
|
|
|
|
|
Surface covered with radial spines, |
|
|
|
|
|
|
|
|
124. Ellipsidium. |
|
|
|
|
|
|
| Ellipsoidal shell with polar appendages (either solid spines or hollow
fenestrated tubes) at the poles of the main axis. |
|
|
Solid spines of similar shape, at both poles of the main
axis. |
|
|
|
|
|
|
Shell cavity simple, without axial rods, |
|
|
|
|
|
|
|
|
|
|
125. Ellipsoxiphus. |
|
|
|
|
|
|
|
|
|
|
Shell cavity with a cross of axial rods, |
|
|
|
|
|
|
|
|
|
|
126. Axoprunum. |
|
|
|
|
|
|
|
|
Solid spines of different shape, at both poles of the main
axis. |
|
|
|
|
|
|
Two polar spines of different shape, |
|
|
|
|
|
|
|
|
|
|
127. Ellipsostylus. |
|
|
|
|
|
|
|
|
|
|
A bunch of spines at one pole only, |
|
|
|
|
|
|
|
|
|
|
128. Lithomespilus |
|
|
|
|
|
|
|
|
|
|
Only a single spine at one one pole, |
|
|
|
|
|
|
|
|
|
|
129. Lithapium. |
|
|
|
|
|
|
|
|
Two hollow fenestrated tubes opposite, at the poles of the main
axis, |
|
|
|
|
|
|
|
|
130. Pipettella. |
|
|
|
|
|
|
Genus 122. Cenellipsis,[161] n. gen.
Definition.—Ellipsida with simple ellipsoidal shell,
without radial spines and without polar tubes.
The genus Cenellipsis is the simplest and most primitive form, not only
among the Ellipsida, but also among the Prunoidea, and it may therefore
be regarded as the common ancestral form of the whole family. It corresponds to Cenosphæra
among the Sphæroidea, to Cenodiscus among the Discoidea, to Cenolarcus among the Larcoidea.
Probably it is derived from Cenosphæra by prolongation of one axis.
Subgenus 1. Cenellipsium, Haeckel.
Definition.—Network of the shell regular, with meshes of equal size
and similar form.
{291}
1. Cenellipsis primitiva, n. sp.
Proportion of the longer axis of the ellipsoid to the shorter = 3 : 2. Network of the
thin wall very delicate and regular, with hexagonal pores. All pores of nearly the same size and
form, ten to twelve times as broad as the bars; nine to twelve on the half equator of the shell.
Surface smooth.
Dimensions.—Longer axis of the ellipsoid 0.09 to 0.11, shorter axis 0.06 to 0.08;
pores 0.01, bars between them 0.001.
Habitat.—Pacific, central area, Station 272, surface.
2. Cenellipsis faceta, n. sp. (Pl. 39,
fig. 1).
Proportion of the longer axis of the ellipsoid to the shorter = 4 : 3. Network of the
thin wall delicate and regular. All the pores of the same size and form, circular, with a
hexagonal frame, twice to three times as broad as the bars; eight to ten on the half equator.
Surface smooth or slightly spiny.
Dimensions.—Longer axis of the ellipsoid 0.12 to 0.13, shorter axis 0.09 to 0.1;
pores 0.013, bars 0.005.
Habitat.—Pacific, central area. Station 274, surface.
3. Cenellipsis ehrenbergii, Haeckel.
Haliomma (?) cenosphæra, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 74, Taf xxvi. fig. 5.
Proportion of the longer axis to the shorter = 2 : 1. Network of the thin wall
regular or subregular. All pores circular, nearly of the same size, without hexagonal frame, three
to four times as broad as the bars; eight to nine on the half equator. Surface covered with small
thorns.
Dimensions.—Longer axis of the ellipsoid 0.16 to 0.2, shorter axis 0.08 to 0.1; pores 0.1, bars 0.003.
Habitat.—Fossil in the Tertiary rocks of Barbados (Ehrenberg,
Haeckel).
4. Cenellipsis circopora, n. sp.
Proportion of the longer axis to the shorter = 5 : 3. Network of the thin wall
delicate and regular. All pores circular, nearly of the same size, without hexagonal frame, twice
as broad as the bars; fifteen to twenty on the half equator. Surface quite smooth.
Dimensions.—Longer axis of the ellipsoid 0.15 to 0.2, shorter axis 0.09 to 0.12;
pores 0.08, bars 0.4.
Habitat.—North Pacific, Station 236, surface.
5. Cenellipsis micropora, n. sp.
Proportion of the longer axis to the shorter = 5 : 4. Network of the thick wall quite
regular. All pores circular, without hexagonal frame, very small, scarcely half as broad as the
bars; six to eight on the half equator. Surface quite smooth.
{292}
Dimensions.—Longer axis of the ellipsoid 0.1, shorter axis 0.08; pores 0.004, bars
0.01.
Habitat.—South Pacific, Station 289, surface.
Subgenus 2. Cenellipsula, Haeckel.
Definition.—Network of the shell irregular, with meshes of different
size or form.
6. Cenellipsis heteropora, n. sp.
Proportion of the longer axis to the shorter = 3 : 2 or = 5 : 3. Network of
the thin wall very irregular, with polygonal meshes of different size and form (for the most part
pentagonal or hexagonal, but also many tetragonal or heptagonal). The largest pores four to six
times as broad as the smallest; ten to twenty pores on the half equator; bars between them thin.
Surface smooth or somewhat spiny.
Dimensions.—Longer axis of the ellipsoid 0.18 to 0.22, shorter axis 0.1 to 0.12;
pores 0.004 to 0.008, bars 0.001.
Habitat.—Indian Ocean, near the Cocos Islands, surface, Rabbe.
7. Cenellipsis maxima, n. sp.
Proportion of the longer axis to the shorter = 3 : 2. Network of the thin wall very
irregular, with polygonal meshes of very different size and form, twice to eight times as broad as
the bars; fifty to sixty on the half equator. Surface smooth.
Dimensions.—Longer axis of the ellipsoid 0.36, shorter axis 0.25; pores 0.004 to
0.015, bars 0.002.
Habitat.—Equatorial Atlantic, Station 347, surface.
8. Cenellipsis ovulum, n. sp.
Proportion of the longer axis to the shorter = 3 : 2. Network of the thin wall very
irregular, with roundish or longish pores of very different size and form. Pores flat, with even
margins, thirty to forty on the half equator, twice to six times as broad as the bars. Surface
smooth. (This species being observed alive, showed a transparent central capsule of ellipsoidal
form, two-thirds as large as the shell.)
Dimensions.—Longer axis of the ellipsoid 0.3, shorter axis 0.2; pores 0.04 to
0.012, bars 0.002.
Habitat.—Indian Ocean, Ceylon, Haeckel; Pacific, central area, Station
266, surface.
9. Cenellipsis infundibulum, n. sp. (Pl. 39, fig.
2).
Proportion of the longer axis to the shorter = 3 : 2. Network of the thick wall
irregular, with roundish pores of different size and form. Pores funnel-like, with prominent,
irregular, spiny {293}crests between them, their outer
aperture three to four times as broad as the inner; twelve to fifteen pores on the half equator.
Surface prickly.
Dimensions.—Longer axis of the ellipsoid 0.14, shorter axis 0.11; outer aperture
of the pores 0.01 to 0.012, inner aperture 0.003 to 0.004.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
10. Cenellipsis oblonga, n. sp.
Proportion of the longer axis to the shorter = 2 : 1. Network of the thick wall
irregular, with roundish pores of different size and form. Pores with prominent conical edges,
about as broad as the bars; twenty to twenty-five on the half equator. Sometimes each pore is
prolonged into a short conical tubulus.
Dimensions.—Longer axis of the ellipsoid 0.22, shorter axis 0.12; pores and bars
0.008 to 0.012.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms.
Genus 123. Axellipsis,[162] n. gen.
Definition.—Ellipsida with simple ellipsoidal shell,
without radial spines or polar tubes, but with an inner transverse axial rod, which corresponds to
the shorter axis of the ellipsoid.
The genus Axellipsis differs from the simple Cenellipsis in a very
peculiar character, namely, the presence of an inner siliceous bar marking the minor axis of the
ellipsoidal shell. If in some forms of Druppula (e.g., Pl. 39, fig. 3) we
remove the central medullary shell and prolong both beams (connecting it with the cortical shell)
till they unite in the centre, we shall arrive at Axellipsis. Therefore Axellipsis
may be derived in the same way from Druppula as Axoprunum is from
Lithatractus, by phyletic loss of the medullary shell.
1. Axellipsis perforata, n. sp.
Proportion of the major axis of the ellipsoid to the minor = 3 : 2. Pores of the
shell regular, circular, twice as broad as the bars; ten to twelve on the half equator. Surface
smooth.
Dimensions.—Longer axis of the shell 0.12, shorter axis 0.08; pores 0.006, bars
0.003.
Habitat.—Central area of the Pacific, Station 266, depth 2750
fathoms.
2. Axellipsis lobata, n. sp.
Proportion of the major axis to the minor = 5 : 4. Meshes of the shell irregular,
roundish, lobed, with three to six indentations (each mesh formed by the confluence of three to
six, commonly {294}four or five, smaller pores); eight
to nine meshes on the half equator, these being twice to four times as broad as the bars.
Dimensions.—Longer axis of the shell 0.1, shorter axis 0.08; pores 0.007 to 0.01,
bars 0.003.
Habitat.—Tropical zone of the Western Pacific, Station 225,
surface.
Genus 124. Ellipsidium,[163] n. gen.
Definition.—Ellipsida with simple ellipsoidal shell, with
numerous radial spines on the surface, without polar spines or tubes.
The genus Ellipsidium differs from Cenellipsis solely in the
development of numerous radial spines on the surface, and exhibits the same phylogenetic relation
to it that Heliosphæra bears to Cenosphæra.
1. Ellipsidium pandanidium, n. sp.
Proportion of the longer axis of the ellipsoid to the shorter = 5 : 4. Shell thick
walled, with regular and hexagonal meshes, twice as broad as the bars; eighteen to twenty on the
half equator. In each hexagon-corner (between three pores) arises a short, three-sided pyramidal
spine, half as long as the equatorial radius, and as thick at the base as a single mesh.
Dimensions.—Major axis of the ellipsoid 0.15, minor 0.12; pores 0.006, bars 0.003;
length of the radial spines 0.03, basal breadth 0.006.
Habitat.—Fossil in the Tertiary rocks of Barbados (Haeckel).
2. Ellipsidium datura, n. sp.
Proportion of the longer axis of the ellipsoid to the shorter = 4 : 3. Shell thick
walled, with regular, circular meshes, twice as broad as the thick bars between them eight to ten
meshes on the half equator. Outer surface of the shell thorny, covered with short, conical, radial
spines, which are regularly distributed (one spine between every three meshes), and about as long
as the diameter of the meshes. (The shell is similar to the outer shell of Haliomma
castanea, Haeckel, Monogr. d. Radiol., Taf. xxiv. fig. 4.)
Dimensions.—Major axis of the ellipsoid 0.12, minor axis 0.09; meshes 0.012, bars
0.006.
Habitat.—Western part of the Tropical Pacific, Station 225, depth 4475
fathoms.
3. Ellipsidium artocarpus, n. sp.
Proportion of the longer axis to the shorter = 3 : 2. Shell thick walled, with
regular, circular meshes, separated by deep furrows, which represent a regular, hexagonal
framework; on the half equator twelve to fifteen meshes, scarcely broader than the broad bars
between them. Outer {295}surface spiny; between every
three meshes arises a strong radial spine, twice to three times as long as the diameter of the
meshes; the base of the spine is like a three-sided pyramid.
Dimensions.—Major axis of the ellipsoid 0.12, minor axis 0.08; meshes 0.006, bars
0.005.
Habitat.—Central area of the Pacific, Station 268, depth 2900
fathoms.
4. Ellipsidium opuntia, n. sp.
Proportion of the longer axis to the shorter = 5 : 4. Shell thin walled, with
irregular, roundish meshes of different size and form, about twice to three times as broad as the
irregular, thin bars between them; ten to fifteen meshes on the half equator. Between the meshes
arise numerous thin, bristle-like, radial spines, about as long as the shorter radius of the
shell. The number of the meshes may be three to four times as great as the number of the
spines.
Dimensions.—Major axis of the ellipsoid 0.15, minor axis 0.12; pores 0.006 to
0.01, bars 0.003 to 0.004.
Habitat.—Southern Pacific, Station 284, surface.
5. Ellipsidium echinidium, n. sp.
Proportion of the longer axis to the shorter = 4 : 3. Shell thick walled, with
irregular, roundish pores of different size and form, about as large or somewhat smaller than the
broad bars; twelve to sixteen pores on the half equator. On the surface, irregularly scattered,
twenty to thirty strong, three-sided pyramidal, radial spines, one-fourth to one-half as long as
the main axis.
Dimensions.—Major axis of the ellipsoid 0.16, minor 0.12; pores and bars 0.002 to
0.008; length of the radial spines 0.04 to 0.08, basal breadth 0.01.
Habitat.—Equatorial Atlantic, Station 347, depth 2250 fathoms.
Genus 125. Ellipsoxiphus,[164]
Dunikowski, 1882, Denkschr. d. k. Akad. d. Wiss. Wien, vol. xlv. p. 25.
Definition.—Ellipsida with simple ellipsoidal shell, the
main axis of which is prolonged at both poles into two strong opposite spines of equal size and
similar form.
The genus Ellipsoxiphus was established by Dunikowski (in 1882, loc.
cit.) for those simple amphistylous fenestrated shells, formerly united with
Xiphosphæra, in which the mathematical form of the shell itself is not a true sphere, but
an ellipsoid. It may therefore be derived from Xiphosphæra by prolongation of the axis in
which lie both polar spines; but it may also be derived from Cenellipsis by the production
of two equal spines at the poles of the main axis.
{296}
Subgenus 1. Ellipsoxiphetta, Haeckel.
Definition.—Network of the shell regular, with pores of equal size and
similar form.
1. Ellipsoxiphus elegans, n. sp. (Pl. 14, fig.
7).
Proportion of the major axis of the ellipsoid to the minor = 5 : 4. Shell thick
walled, with regular, circular meshes, separated by a regular, hexagonal elevated framework. From
each hexagon-corner arises (between every three meshes) a short thin spine, about as long as the
diameter of a mesh; twelve to fourteen meshes on the half equator, of the same breadth as the
bars. Polar spines three-sided prismatic, cuspidate, about as long as the axis of the shell. (This
elegant and not uncommon species is remarkable for its variations. By unequal development of both
spines it passes over to Ellipsostylus.) In the figured variety (Pl. 14, fig. 7) the
ends of the surface spines are connected by a very delicate network, so as to form an outer veil.
This may further be separated as a peculiar genus Ellipsoxiphium palliatum.
Dimensions.—Longer axis 0.08 to 0.13, shorter axis 0.06 to 0.11; pores and bars
0.008; length of the polar spines 0.06 to 0.12, basal breadth 0.01.
Habitat.—Pacific, central area, Stations 270 to 274, from 2350 to 2925
fathoms.
2. Ellipsoxiphus flosculus, n. sp.
Proportion of the major axis to the minor = 6 : 5. Shell very thick walled, with
regular, hexagonal meshes, eight to nine on the half equator, three to four times as broad as the
bars. The inner contour of the meshes is circular, the outer six-lobed (similar to Xiphostylus
phasianus, Pl. 13,
fig. 9). Polar spines three-sided pyramidal, about as long as the major axis of the shell; their
basal thickness equals one mesh.
Dimensions.—Longer axis 0.12, shorter axis 0.1; pores 0.012 to 0.015, bars 0.004;
length of the polar spines 0.12 to 0.15, basal breadth 0.015.
Habitat.—Fossil in the Tertiary rocks of Barbados (Teuscher).
3. Ellipsoxiphus fragilis, n. sp.
Proportion of the major axis to the minor = 4 : 3. Shell thin walled, with regular,
circular meshes, twice as broad as the bars; ten to twelve on the half equator. Surface smooth.
Polar spines cylindrical, at the apex conical, half as long as the major axis.
Dimensions.—Longer axis 0.16, shorter axis 0.12; pores 0.008, bars 0.004; length
of the polar spines 0.09, basal breadth 0.008.
Habitat.—South Atlantic, Station 325, surface.
Subgenus 2. Ellipsoxiphilla, Haeckel.
Definition.—Network of the shell irregular, with pores of different
size or form.
{297}
4. Ellipsoxiphus claviger, n. sp. (Pl. 14, fig.
3).
Xiphosphæra clavigera, Haeckel, 1881, Prodromus et Atlas.
Proportion of the major axis to the minor = 5 : 4. Shell thick walled, with irregular
network; meshes roundish or circular, double contoured, of unequal size, twice to four times as
broad as the bars; eight to ten on the half equator. Polar spines club-shaped, with prominent
edges, about as long as the minor axis; thicker in their middle part than at both ends (differs
from Xiphosphæra clavigera by the ellipsoidal form of the shell, and the double length of
the spines).
Dimensions.—Longer axis 0.15. shorter axis 0.12; pores 0.007 to 0.015, bars 0.004;
length of the polar spines 0.12, thickness in the middle part 0.02.
Habitat.—Central area of the Pacific, Station 272, depth 2600
fathoms.
5. Ellipsoxiphus suessi, Dunikowski.
Ellipsoxiphus suessi, Dunikowski, 1882, Denkschr. d. k. Akad. d. Wiss.
Wien, Bd. xlv. p. 26, Taf. v. fig. 50.
Proportion of the major axis to the minor = 7 : 5. Shell thick walled, with
irregular, roundish meshes, six to eight on the half equator, twice to three times as broad as the
bars. Polar spines nearly cylindrical, blunt, as long as the major axis.
Dimensions.—Longer axis 0.14; shorter axis 0.1; pores 0.018 to 0.023, bars 0.01;
length of the polar spines 0.12; thickness of them 0.035.
Habitat.—Fossil in the Alpine Lias, Schafberg near Salzburg,
Dunikowski.
6. Ellipsoxiphus parvoforaminus, Dunikowski.
Ellipsoxiphus parvoforaminus, Dunikowski, 1882, Denkschr. d. k. Akad. d.
Wiss. Wien, Bd. xlv. p. 26, Taf. v. fig. 51.
Proportion of the major axis to the minor = 7 : 6. Shell thick walled, with
irregular, roundish or oval meshes, ten to twelve on the half equator, scarcely as broad as the
bars. Polar spines conical, shorter than the minor axis.
Dimensions.—Longer axis 0.14, shorter axis 0.12; pores 0.016 to 0.01, bars 0.01 to
0.02; length of the polar spines 0.1, basal thickness 0.045.
Habitat.—Fossil in the Alpine Lias, Schafberg near Salzburg,
Dunikowski.
7. Ellipsoxiphus bipolaris, n. sp. (Pl. 14, fig.
11).
Xiphosphæra bipolaris, 1881, Prodromus et Atlas.
Proportion of the major axis to the minor = 8 : 7. Shell thick walled, with
irregular, roundish meshes, twice to three times as broad as the bars; sixteen to twenty on the
half equator. Polar spines short and thick, three-sided pyramidal, scarcely half as long as the
shorter radius of the shell; surrounding the base of every spine is a circle of eight to nine
shorter spines.
Dimensions.—Longer axis 0.08, shorter axis 0.07; pores 0.003 to 0.007, bars 0.002;
length of the polar spines 0.02 to 0.03, basal thickness 0.02.
Habitat.—Tropical part of the Western Pacific, Station 225, depth 4475
fathoms.
{298}
8. Ellipsoxiphus atractus, n. sp. (Pl. 14, fig.
1).
Proportion of the major axis to the minor = 3 : 2. Shell spindle-shaped, very thick
walled, with roundish, very irregular meshes, twice to five times as broad as the bars; eight to
twelve on the half equator. The meshes are partly simple, oblong, partly lobed or composed of two
to four (commonly three) confluent meshes. Surface smooth. Polar spines very short and thick,
shorter than the minor radius of the shell, three-sided pyramidal, with prominent, somewhat
contorted edges.
Dimensions.—Longer axis 0.15, shorter axis 0.1; meshes 0.007 to 0.02, bars 0.004;
length of the polar spines 0.04, basal thickness 0.03.
Habitat.—Central area of the Pacific, Station 268, depth 2900
fathoms.
Genus 126. Axoprunum,[165] n. gen.
Definition.—Ellipsida with simple ellipsoidal shell, the
main axis of which is prolonged at both poles into two opposite spines of equal size and similar
form. Within the cavity of the shell four radial rods arise from its inner surface, two in the
main axis, two in the smallest axis, perpendicular to the former; their free inner ends are at the
same distance from the centre.
The genus Axoprunum possesses precisely the same shell as the foregoing
Ellipsoxiphus, but differs from it in a very remarkable peculiarity. The two polar spines
are centripetally prolonged into two internal beams, and perpendicular to these are two other,
opposite, transverse beams, marking the minor axis of the ellipsoid. The free inner ends of all
four radial rods bear little thickened knobs, and are at the same distance from the centre. It
therefore appears as though a central, spherical, medullary shell had been lost, and this gives a
strong support to the important hypothesis, that in many Sphærellaria,
where the medullary shell is absent, it may have been lost by phylogenetic reduction or retrograde
metamorphosis. In this case Axoprunum (and Ellipsoxiphus) would arise from
Lithatractus.
1. Axoprunum stauraxonium, n. sp. (Pl. 48, fig.
4).
Shell ellipsoidal, one and one-third times as long as broad, with smooth surface. Network
regular, with circular meshes four times as broad as the bars. Two polar spines three-sided
pyramidal, half as long as the shell, as thick at the base as a single mesh. Four inner radial
beams (lying, two in the major and two in the minor axis of the ellipsoid) very thin, at the
central free ends knob-like, thickened. The distance between two opposite beams equals one-third
of the minor axis, and indicates probably the diameter of the lost spherical medullary shell.
(Three perfect and complete specimens of the same size and shape were observed.)
{299}
Dimensions.—Major axis of the ellipsoid 0.14, minor axis 0.11; pores 0.012, bars
0.003; length of the polar spines 0.07, basal breadth 0.012. (Diameter of the lost medullary shell
0.03?)
Habitat.—South Atlantic, Station 338, depth 1990 fathoms.
Genus 127. Ellipsostylus,[166] n.
gen.
Definition.—Ellipsida with simple ellipsoidal shell, the
main axis of which is prolonged at both poles into two opposite spines of different size or
form.
The genus Ellipsostylus differs from Ellipsoxiphus only in the
unequal size or different form of the polar spines, which in the former are equal. This difference
does not seem important at first, but in the further development it produces very singular and
strange forms. Theoretically it is always important, because the fundamental haplopolar form of
the monaxon body becomes diplopolar by this differentiation.
Subgenus 1. Ellipsostyletta, Haeckel.
Definition.—Network of the shell regular, with pores of equal size and
similar form.
1. Ellipsostylus ornithoides, n. sp.
Proportion of the major axis of the ellipsoid to the minor = 4 : 3. Shell thin
walled, with regular, circular meshes and hexagonal framework between them, like that of
Xiphostylus favosus (Pl. 13,
fig. 4). Circular pores about as broad as the bars; eight to nine on the half equator. Surface
smooth. Polar spines three-sided pyramidal, as broad at the base as a single hexagon; the longer
spine as long as the major axis, the shorter half as long.
Dimensions.—Longer axis of the ellipsoidal shell 0.16, shorter axis 0.12; pores
and bars 0.006; length of the longer polar spine 0.16, of the shorter 0.08.
Habitat.—North Pacific, Station 241, depth 2300 fathoms.
2. Ellipsostylus avicularis, n. sp.
Proportion of the major axis to the minor = 3 : 2. Shell thick walled, with regular,
rosette-shaped meshes, three times as broad as the bars; ten to twelve on the half equator. Every
mesh on the outer margin with five to six rounded lobes, as in Xiphostylus phasianus (Pl.
13, fig. 9).
Surface smooth. Polar spines sharp edged, thick; the longer straight and equal to the longer axis,
the shorter half as long, shaped like a bird's head.
Dimensions.—Longer axis 0.18, shorter axis 0.12; pores 0.015, bars 0.005.
Habitat.—South Pacific, Station 300, depth 1375 fathoms.
{300}
3. Ellipsostylus psittacus, n. sp. (Pl. 13, fig.
6).
Proportion of the major axis to the minor = 4 : 3. Shell thin walled, with regular or
subregular rosette-shaped meshes, twice as broad as the bars; eight to nine on the half equator;
each mesh with three to four rounded lobes. Surface smooth. Polar spines sharp edged, very
unequal; longer spine about twice as long as the major axis, somewhat curved; shorter spine
scarcely longer than the radius, shaped like a bird's head.
Dimensions.—Longer axis 0.08, shorter axis 0.06; pores 0.01, bars 0.005; length of
the polar spines—longer 0.15, shorter 0.05.
Habitat.—Central area of the Pacific, Station 266, depth 2750
fathoms.
4. Ellipsostylus aquila, n. sp. (Pl. 13,
fig. 1).
Proportion of the major axis to the minor = 7 : 6. Shell thick walled, with
subregular, circular meshes, five to six times as broad as the bars; ten to twelve on the half
equator. Surface smooth. Polar spines sharp edged (six-sided?), very unequal; longer spine
straight, about equal to the minor axis; shorter spine obliquely inserted, scarcely half as long,
shaped like a bird's head.
Dimensions.—Longer axis 0.15, shorter axis 0.13; pores 0.02, bars 0.004; length of
the polar spines—longer 0.1, shorter 0.05.
Habitat.—Central area of the Pacific, Station 272, depth 2600
fathoms.
5. Ellipsostylus columba, n. sp. (Pl. 13, fig.
3).
Proportion of the major axis to the minor = 4 : 3. Shell egg-shaped, thin walled,
with regular, circular meshes, three times as broad as the bars; ten to twelve on the half
equator. Surface somewhat thorny. Polar spines nearly of equal length (equal to the major axis),
but of very different form; one straight, pyramidal, and obliquely inserted, the other like a
bird's head.
Dimensions.—Longer axis 0.08, shorter 0.06; pores 0.003, bars 0.01; length of the
spines 0.09.
Habitat.—South Atlantic, Station 332, surface.
6. Ellipsostylus ciconia, n. sp. (Pl. 13, fig.
8).
Proportion of the major axis to the minor = 7 : 5. Shell egg-shaped, thick walled;
the meshes very small, regular, circular, three times as broad as the bars; sixteen to twenty on
the half equator. Surface uneven. Polar spines nearly cylindrical, pointed; the smaller obliquely
inserted, equal to the major axis; the larger more than twice as long and thick, furrowed at its
base. (Compare with this and the allied species Rhabdolithis pipa, Ehrenberg, 1875,
Abhandl. d. k. Akad. d. Wiss. Berlin, Taf. i. fig. 27.)
Dimensions.—Longer axis 0.07, shorter 0.05; pores 0.003, bars 0.001; length of the
polar spines—longer 0.2, shorter 0.08.
Habitat.—North Pacific, Station 253, depth 3125 fathoms.
{301}
Subgenus 2. Ellipsostylissa, Haeckel.
Definition.—Network of the shell irregular, with pores of different
size or form.
7. Ellipsostylus megadictya, Haeckel.
Stylosphæra megadictya, Ehrenberg, 1872, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 299, Taf. viii. fig. 13.
Proportion of the major axis to the minor = 5 : 4. Shell thin walled, with irregular,
roundish, large meshes, four to five times as broad as the bars; only four to five on the half
equator. Surface smooth. Polar spines straight, thin, angular; the shorter equal to the minor
axis, the longer four times as large.
Dimensions.—Longer axis 0.05, shorter axis 0.04; meshes 0.01, bars 0.002; length
of the polar spines—longer 0.12, shorter 0.03.
Habitat.—Philippine Sea, 3300 fathoms, Ehrenberg.
8. Ellipsostylus gallinula, n. sp.
Proportion of the major axis to the minor = 3 : 2. Shell thick walled, with
irregular, roundish meshes, twice to four times as broad as the bars; eight to ten on the half
equator. Surface thorny. Polar spines conical, straight; the longer twice as long as the major
axis, the minor scarcely half as long.
Dimensions.—Longer axis 0.15, shorter 0.12; pores 0.01 to 0.02, bars 0.002 to
0.004.
Habitat.—North Pacific, Station 253, surface.
9. Ellipsostylus hirundo, n. sp. (Pl. 13, fig.
2).
Proportion of the major axis to the minor = 4 : 3. Shell thick walled, with
irregular, roundish meshes, three to five times as broad as the bars; eight to ten on the half
equator. The inner aperture of every mesh is fenestrated by a delicate lamella of silex,
perforated by six to eight very small circular pores. Polar spines sharp edged, more or less
curved, the shorter equal to the minor axis, the longer twice as long.
Dimensions.—Longer axis 0.16, shorter 0.12; pores 0.01 to 0.02, bars 0.003 to
0.006; length of the polar spines—longer 0.24, shorter 0.12.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms; the
same form also fossil in the rocks of Barbados.
Genus 128. Lithomespilus,[167] Haeckel,
1881, Prodromus, p. 450.
Definition.—Ellipsida with simple ellipsoidal or oviform
shell, the main axis of which bears at one pole a single spine, at the other a bunch of several
spines.
{302}
The genus Lithomespilus differs from the closely allied Ellipsoxiphus
in the further differentiation of both poles of the main axis. One pole exhibits only a single
polar spine, the other pole a group of several spines, peculiarly grouped together. It differs
from the similar Sphæromespilus (Pl. 14,
figs. 12, 13) in the ellipsoidal form of the shell.
1. Lithomespilus phloginus, n. sp. (Pl. 14, fig.
16).
Proportion of the major axis of the ellipsoid to the minor = 4 : 3. Shell thick
walled, with circular pores of different size, the breadth of which equals that of the bars;
twelve to fifteen pores on the half equator. Surface smooth, with the exception of a circumpolar
region covered with numerous thick spines of unequal size, which surround the large single polar
spine and are curved like a bow against its axis. This larger polar spine is straight, and equals
in length the major axis of the shell. The other and opposite polar spine is scarcely one-fourth
as long, perfectly simple, very stout, and of a three-sided pyramidal shape.
Dimensions.—Longer axis of the ellipsoidal shell 0.08, shorter axis 0.06; pores
0.002 to 0.006, bars 0.003 to 0.005; length of the polar spines—longer 0.08, shorter
0.02.
Habitat.—Indian Ocean, near Madagascar, depth 1200 fathoms, Smith.
2. Lithomespilus phlogoides, n. sp. (Pl. 14, fig.
17).
Proportion of the major axis to the minor = 5 : 4. Shell very thick walled, with
roundish, double-contoured pores of different size and form, twice to three times as broad as the
bars; sixteen to eighteen pores on the half equator. Surface smooth, with the exception of a
circumpolar region covered with numerous thin spines of unequal size, which surround the large
single polar spine and are curved like a bow against its axis. This larger polar spine is
straight, and equals half the size of the major axis of the shell; the other polar spine
(sometimes double, as in the figured specimen) is much shorter, being scarcely one-fourth as
long.
Dimensions.—Longer axis of the shell 0.08, shorter axis 0.065; pores 0.003 to
0.006, bars 0.002 to 0.004; length of the polar spines—longer 0.04, shorter 0.01.
Habitat.—Central area of the Pacific, Stations 270 to 272, depth 2425
to 2925 fathoms.
3. Lithomespilus flammeus, n. sp.
Stylosphæra species, Bury, 1862, Polycystins of Barbados, pl. xi. fig.
3.
Proportion of the major axis to the minor = 4 : 3. Shell thick walled, with
irregular, roundish pores, about as broad as the bars; ten to twelve pores on the half equator.
Surface spiny. Length of the conical straight spines increasing towards the poles; each polar
spine surrounded by a circumpolar group of larger, somewhat curved spines. The larger polar spine
equals in length the major axis; the smaller is scarcely half as long. (The figure, given by Bury,
is not quite exact; there the spines are situated in the pores, instead of between them. In my
specimen the polar spines were not branched.)
{303}
Dimensions.—Longer axis of the shell 0.12, shorter axis 0.09; pores and bars 0.004
to 0.006; length of the polar spines—longer 0.12, shorter 0.06.
Habitat.—Fossil in Barbados.
4. Lithomespilus flammabundus, n. sp. (Pl. 14, fig.
14).
Proportion of the major axis to the minor = 4 : 3. Shell thin walled, with irregular,
roundish pores, partly simple, partly composed of three to six confluent pores; only six to eight
pores on the half equator, twice to four times as broad as the bars. Surface spiny. Length of the
conical irregular spines increasing towards the poles; each polar spine surrounded by a
flame-shaped, circumpolar area of longer spines; all large spines (also the polar spines) curved
or contorted at one pole and much stronger and more numerous than at the other; length variable,
often equal to the longer axis.
Dimensions.—Longer axis of the shell 0.12, shorter axis 0.09; pores 0.005 to
0.015, bars 0.003 to 0.005; length of the polar spines 0.1 to 0.15.
Habitat.—Western part of the Tropical Atlantic, Station 347, depth
2250 fathoms.
Genus 129. Lithapium,[168] n. gen.
Definition.—Ellipsida with simple ellipsoidal or
pear-shaped shell; with a single spine only situated at one pole of the main axis.
The genus Lithapium represents a peculiar modification of
Ellipsoxiphus; one of the two opposite polar spines disappears by reduction, and in this
way only a single spine remains, at one pole of the main axis. For this reason the shell assumes a
characteristic pear-shape, and may easily be confounded with some similar Monocyrtida
(Halicapsa).
1. Lithapium pyriforme, n. sp. (Pl. 14,
fig. 9).
Proportion of the longer axis to the shorter = 6 : 5. Shell thin walled, with
regular, circular pores, four times as broad as the bars; six to eight on the half equator.
Surface a little thorny. The single polar spines three-sided pyramidal, as broad at the base as
one mesh, about as long as the radius of the shell. (In the specimen figured, there was on the
opposite pole a little rudiment of the other lost polar spine; it is missing in other
specimens.)
Dimensions.—Major axis of the ellipsoidal shell 0.12, minor axis 0.1; pores 0.02,
bars 0.005; length of the single polar spine 0.05, basal thickness 0.02.
Habitat.—Central area of the Pacific, Station 266, depth 2750
fathoms.
2. Lithapium halicapsa, n. sp. (Pl. 14,
fig. 8).
Proportion of the longer axis to the shorter = 6 : 5. Shell thin walled, with
irregular, lobed meshes, six to eight on the half equator, twice to five times as broad as the
bars; each mesh {304}composed of two to five confluent
roundish pores. Surface a little thorny. The single polar spine pyramidal, as broad at the base as
one mesh, one-third as long as the axis.
Dimensions.—Major axis 0.12, minor axis 0.1; pores 0.01 to 0.02, bars 0.002 to
0.006; length of the single polar spine 0.03, basal thickness 0.02.
Habitat.—Central area of the Pacific, Station 270, depth 2925
fathoms.
3. Lithapium monocyrtis, n. sp. (Pl. 14,
fig. 10).
Proportion of the longer axis to the shorter = 7 : 6. Shell thick walled, with
irregular, lobed meshes, four to six on the half equator, three to six times as broad as the bars;
each mesh composed of three to six confluent roundish pores. Surface quite smooth, without thorns.
The single polar spine pyramidal, as broad at the base as the largest mesh, half as long as the
radius. (This species is closely allied to the foregoing, but differs in the smooth surface and
the larger meshes.)
Dimensions.—Major axis 0.15, minor axis 0.13; pores 0.01 to 0.03, bars 0.003 to
0.01; length of the single spine 0.04, basal thickness 0.03.
Habitat.—Central area of the Pacific, Station 268, depth 2900
fathoms.
Genus 130. Pipettella,[169] n. gen.
Definition.—Ellipsida with simple ellipsoidal shell, the
main axis of which is prolonged at the pole into two opposite hollow fenestrated tubes of equal
size and similar form.
The genus Pipettella is distinguished from the other Ellipsida by two hollow
perforated tubes, which are directed in the longer axis of the ellipsoidal shell and arise from
opposite poles of this axis. It may be derived from Cenellipsis by prolongation of both
poles of the main axis. As the same peculiar production of two opposite latticed tubuli at the
poles of the main axis obtains in nearly all families of Prunoidea
(Pipetta, Cannartus, Panarium, Zygartus, &c.), it may possess a
peculiar value in this group.
1. Pipettella fusiformis, n. sp.
Shell spindle-shaped, thin walled, the two opposite tubes being conical and not longer than the
short transverse axis of the ellipsoidal shell; no sharp demarcation between them. The two axes of
the ellipsoid bear the proportion of 3 : 2. Network of the shell and of the tubes equal,
delicate, regular, hexagonal, with circular apertures of equal size; fourteen to sixteen on the
half equator of the shell. Bars very thin. The shell of this species is similar to that of
Cannartiscus amphiconiscus (Pl. 39,
fig. 19), but possesses no equatorial stricture and no medullary shell.
Dimensions.—Longer axis of the ellipsoid 0.18, shorter axis 0.12; length of the
tubes 0.1, their basal breadth 0.05; pores of the network 0.01, bars 0.001.
Habitat.—Northern Pacific, Station 244, depth 2900 fathoms.
{305}
2. Pipettella tubulosa, n. sp.
Shell ellipsoidal, thin walled, distinctly separated from the two opposite tubes, which are
cylindrical, longer than the main axis of the ellipsoid, and one-sixth as broad as the shorter
axis. The longer axis of the ellipsoid bears to the shorter the proportion of 5 : 4.
Network of the shell and of the tubes equal, regular, with very small circular pores, about as
broad as the bars; sixteen to eighteen pores on the half equator of the shell.
Dimensions.—Longer axis of the ellipsoid 0.15, shorter axis 0.12; length of the
tubes 0.16 to 0.2, breadth of them 0.02; pores of the network 0.003, bars 0.003.
Habitat.—Western Tropical Pacific, Station 225, depth 4475
fathoms.
3. Pipettella elongata, n. sp.
Shell ellipsoidal, thick walled, distinctly separated from both opposite tubes, which are
cylindrical, much prolonged, twice to three times as long as the main axis of the ellipsoid, and
one-fifth as broad as the shorter axis. Both axes of the ellipsoid bear the proportion of
3 : 2. Network of the shell and of the tubes irregular with small rounded pores of
different size, separated by broader bars (often twice to three times as broad), four to six pores
on the half equator of the shell. (This species somewhat recalls Solenosphæra serpentina,
Pl. 7, fig.
7; but the tubes are straight, not contorted.)
Dimensions.—Longer axis of the ellipsoid 0.2, shorter axis 0.14; length of the
tubes 0.4 to 0.5, breadth 0.03; pores of the network 0.001 to 0.002, bars 0.003 to 0.004.
Habitat.—Central area of the Pacific, Station 271, depth 2425
fathoms.
4. Pipettella prismatica, n. sp. (Pl. 39, fig.
6).
Shell ellipsoidal, thick walled, distinctly marked off from the two opposite tubes, which are
longer than its main axis and as broad as one-fifth of it; they are nearly four-sided prismatic,
with four strong ribs or edges; these are directed parallel to the main axis, in two meridian
planes, perpendicular to one another. The wall of the shell is thickened in the equatorial plane,
so as to form a slight stricture on the inside, separating its two halves. Both axes of the
ellipsoid bear the proportion of 7 : 6. Network regular, with circular pores, somewhat
broader than the bars. The meshes of the shell (fourteen to sixteen on the half equator) are twice
as great as those of the tubes, which are arranged in two longitudinal rows between every two ribs
(there being eight longitudinal rows on the whole tube). This species is very remarkable for the
rudimentary internal equatorial stricture of the shell (transition to the genus Cannartus,
Pl. 39, fig.
10), and by the four edges of the tubes, which indicate two of the dimensive axes, the third being
represented by the main axis.
Dimensions.—Longer axis of the ellipsoid 0.14, shorter axis 0.12; length of the
tubes 0.15 to 0.16, breadth 0.03; pores of the shell 0.008 to 0.01, pores of the tubes 0.004.
Habitat.—Western Tropical Pacific, Station 225, depth 4475
fathoms.
{306}
Family XII. Druppulida, Haeckel, 1882 (Pls.
15, 16, 17, 39).
Definition.—Prunoidea with ellipsoidal, latticed (not
spongy) shell, composed of two or more concentric shells; a simple or double cortical shell
enclosing one or two internal concentric shells (medullary shells), without equatorial stricture.
Central capsule ellipsoidal or cylindrical, without annular equatorial constriction.
The family Druppulida differs from the Ellipsida only in the
possession of a simple or double medullary shell, which is enclosed in the centre of the central
capsule, and connected with the ellipsoidal cortical shell (lying outside it) by radial beams,
perforating the membrane of the capsule. The Druppulida exhibit therefore the same relation to the
Ellipsida that the Dyosphærida among the Sphæroidea bear to the
Monosphærida. The cortical shell may be simple or multiple. The whole fenestrated shell is thus
composed of a variable number (two at least) of concentric shells, which are connected by radial
beams.
The Medullary Shell—enclosed in the centre of the central capsule—is either
simple or double, and composed of two small concentric shells. Their form is either spherical, or
ellipsoidal, or lenticular. If the medullary shell be ellipsoidal, the main axis of the ellipsoid
is the same as in the cortical shell. Sometimes the inner medullary shell is spherical, the outer
ellipsoidal or lenticular. If the medullary shell be lenticular (arising from both poles of the
main axis) its vertical axis is also identical with that of the cortical shell (Pl. 39, fig.
5).
The Radial Beams, which connect the medullary and cortical shell, are either developed
in all possible directions (Pls. 16,
17), or
limited to the equatorial plane, more rarely to the meridional plane. Sometimes the connection is
produced only by two opposite beams which lie in the minor or equatorial axis (Pl. 39, figs. 3, 7,
8); more rarely in the major or meridional axis (Pl. 17, figs. 7,
8).
The Cortical Shell is constantly ellipsoidal, rarely with modifications, similar to
those which appear in the simple shell of some Ellipsida. As a rule it is simple, rarely composed
of two or more (sometimes six or more) concentric ellipsoidal shells (in Cromyodruppa and
Cromyocarpus). The outer surface is commonly smooth, more rarely covered with radial spines
(in Druppocarpus, Prunocarpus, &c.). In the majority peculiar polar appendages
are developed at both poles of the main axis, these being prolongations of them, either in the
form of hollow fenestrated tubes (Pipetta, Pipettaria, Pl. 39, figs. 7,
8), or solid strong spines. The two polar spines are either equal in size and similar in form (as
in Lithatractus, Stylatractus) or unequal (as in Druppatractus,
Xiphatractus, Pls. 16,
17).
The most primitive of all Druppulida is Druppula, with simple medullary shell and simple
cortical shell (Pl. 39,
fig. 3); Prunulum differs from it only in the possession {307}of a double medullary shell (Pl. 39, fig. 4).
From these two genera all other forms of the subfamily may be derived.
The Central Capsule of the Druppulida is constantly ellipsoidal, larger than the
concentric enclosed medullary shells, smaller than the surrounding cortical shell; it is separated
from the inner surface of the latter by a thinner or thicker jelly-mantle.
Synopsis of the Genera of the Druppulida.
| Cortical shell without polar appendages (neither solid
spines nor hollow tubes at the poles of the main axis). |
|
Cortical shell simple, ellipsoidal. |
|
Medullary shell simple. |
|
Surface smooth, |
131. Druppula. |
| Surface spiny, |
132. Druppocarpus. |
| Medullary shell double. |
|
Surface smooth, |
133. Prunulum. |
| Surface spiny, |
134. Prunocarpus. |
| Cortical shell composed of two or more
concentric shells. |
|
Medullary shell double. |
|
Surface smooth, |
135. Cromyodruppa. |
| Surface spiny, |
136. Cromyocarpus. |
| Two opposite solid spines, arising from the poles of
the main axis. |
|
Cortical shell simple, ellipsoidal. |
|
Medullary shell simple. |
|
Both polar spines equal, |
137. Lithatractus. |
| Both polar spines unequal, |
138. Druppatractus. |
| Medullary shell double. |
|
Both polar spines equal, |
139. Stylatractus. |
| Both polar spines unequal, |
140. Xiphatractus. |
| Cortical shell composed of two or more concentric shells;
medullary shell double; both polar spines equal, |
141. Cromyatractus. |
| Two opposite hollow fenestrated tubes, arising from the
poles of the main axis. |
|
Cortical shell ellipsoidal. |
|
Medullary shell simple, |
142. Pipetta. |
| Medullary shell double, |
143. Pipettaria. |
| Cortical shell without polar appendages (neither solid spines nor hollow tubes
at the poles of the main axis). |
|
|
Cortical shell simple, ellipsoidal. |
|
|
|
|
|
|
Medullary shell simple. |
|
|
|
|
|
|
|
|
|
|
Surface smooth, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
131. Druppula. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Surface spiny, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
132. Druppocarpus. |
|
|
|
|
|
|
|
|
|
|
|
|
Medullary shell double. |
|
|
|
|
|
|
|
|
|
|
Surface smooth, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
133. Prunulum. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Surface spiny, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
134. Prunocarpus. |
|
|
|
|
|
|
|
|
|
|
Cortical shell composed of two or more concentric shells. |
|
|
|
|
|
|
Medullary shell double. |
|
|
|
|
|
|
|
|
|
|
Surface smooth, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
135. Cromyodruppa. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Surface spiny, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
136. Cromyocarpus. |
|
|
|
|
|
|
|
|
| Two opposite solid spines, arising from the poles of the main axis. |
|
|
Cortical shell simple, ellipsoidal. |
|
|
|
|
|
|
Medullary shell simple. |
|
|
|
|
|
|
|
|
|
|
Both polar spines equal, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
137. Lithatractus. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Both polar spines unequal, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
138. Druppatractus. |
|
|
|
|
|
|
|
|
|
|
|
|
Medullary shell double. |
|
|
|
|
|
|
|
|
|
|
Both polar spines equal, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
139. Stylatractus. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Both polar spines unequal, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
140. Xiphatractus. |
|
|
|
|
|
|
|
|
|
|
Cortical shell composed of two or more concentric shells;
medullary shell double; both polar spines equal, |
|
|
|
|
|
|
|
|
|
|
141. Cromyatractus. |
|
|
|
|
|
|
|
|
| Two opposite hollow fenestrated tubes, arising from the poles of the main
axis. |
|
|
Cortical shell simple, ellipsoidal. |
|
|
|
|
|
|
|
|
Medullary shell simple, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
142. Pipetta. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Medullary shell double, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
143. Pipettaria. |
|
|
|
|
|
|
|
|
Genus 131. Druppula,[170] n. gen.
Definition.—Druppulida with simple ellipsoidal, cortical
shell, and simple medullary shell, without spines or polar tubes.
The genus Druppula, as the simplest form of the Druppulida, may be regarded
as the common ancestral form of this subfamily. It may be derived phylogenetically either {308}from Carposphæra by prolongation of one axis, or
from Cenellipsis by duplication of the fenestrated shell. The outer (or cortical) shell is
always more or less ellipsoidal; the inner (or medullary) shell also is sometimes ellipsoidal,
sometimes spherical. Both shells are concentric, connected by a variable number of radial beams.
Compare Haliomma oblongum, Harting,
1863, L. N. 18, p. 15, pl. 2, fig. 42.
Subgenus 1. Druppuletta, Haeckel.
Definition.—Pores of the cortical shell regular, hexagonal, or
circular.
1. Druppula drupa, n. sp.
Cortical shell thin walled, with smooth surface and regular network; both its axes bear the
proportion of 4 : 3. Pores regular, hexagonal, three times as broad as the bars; twelve
to fifteen on the half equator. Medullary shell spherical, about one-third as broad as the
cortical shell.
Dimensions.—Major axis of the ellipsoidal cortical shell 0.1 to 0.12, minor axis
0.08 to 0.09; pores 0.06, bars 0.02; diameter of the medullary shell 0.03.
Habitat.—Western Tropical Pacific, Station 225, depth 4475 fathoms;
also fossil in Barbados.
2. Druppula pandanus, n. sp. (Pl. 39,
fig. 3).
Cortical shell thin walled, somewhat rough, with regular network; both its axes bear the
proportion of 3 : 2. Pores subregular, circular, hexagonally framed, one and a half
times as broad as the elevated bars; ten to twelve on the half equator. Medullary shell spherical,
one-fourth as broad as the cortical shell.
Dimensions.—Major axis of the cortical shell 0.16 to 0.17, minor axis 0.11 to
0.12; pores 0.01, bars 0.007; diameter of the medullary shell 0.035.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
3. Druppula cocos, n. sp.
Cortical shell thick walled, somewhat rough, with regular network; proportion of both axes =
6 : 5. Pores regular, circular, hexagonally framed, quite as broad as the elevated bars;
fourteen to sixteen on the half equator. Medullary shell ellipsoidal (with longitudinal main
axis), nearly half as large as the cortical shell.
Dimensions.—Major axis of the cortical shell 0.07 to 0.08, minor axis 0.6 to 0.65;
pores and bars 0.003 to 0.004; diameter of the medullary shell 0.03 and 0.035.
Habitat.—Western Tropical Pacific, Station 222, surface, Indian Ocean,
Ceylon, Haeckel.
4. Druppula phœnix, n. sp.
Cortical shell thick walled, smooth, with regular network; proportion of both axes =
5 : 4. Pores regular, circular, hexagonally lobulate (in the same manner as in
Stauroxiphus gladius, {309}Pl. 15, fig. 7),
three times as broad as the bars; ten to twelve on the half equator. Medullary shell ellipsoidal
(with longitudinal main axis), one-third as broad as the cortical shell.
Dimensions.—Major axis of the cortical shell 0.1, minor axis 0.08; pores 0.009, bars 0.003; axis of the
medullary shell 0.03 and 0.025.
Habitat.—Mediterranean, Smyrna, Haeckel, surface.
5. Druppula areca, n. sp.
Cortical shell thin walled, with smooth surface and regular network; both its axes bear the
proportion of 4 : 3. Pores regular, circular, four times as broad as the bars; fifteen
to eighteen on the half equator. Medullary shell spherical, half as broad as the cortical
shell.
Dimensions.—Major axis of the ellipsoidal cortical shell 0.12 to 0.14, minor axis
0.1 to 0.11; pores 0.008, bars 0.002; diameter of the medullary shell 0.05 to 0.06.
Habitat.—Indian Ocean, off Bombay, Haeckel.
6. Druppula ovata, Haeckel.
Haliomma ovatum, Ehrenberg, 1854, Mikrogeol., pl. xx. I. fig. 20.
Haliomma ovatum, Haeckel, 1862, Monogr. d. Radiol., p. 432.
Cortical shell thin walled, with smooth surface and regular network; proportion of both axes =
3 : 2. Pores regular, circular, three times as broad as the bars; nine to ten on the
half equator. Medullary shell spherical, one-third as broad as the cortical shell.
Dimensions.—Major axis 0.08 to 0.1, minor axis 0.05 to 0.06; pores 0.003, bars
0.001; diameter of the medullary shell 0.02.
Habitat.—Fossil in the Tertiary rocks of the Mediterranean coast,
Greece (Zante), Ehrenberg; Sicily (Caltanisetta), Haeckel.
7. Druppula caryota, n. sp.
Cortical shell thick walled, with rough surface and regular network; proportion of both axes =
5 : 4. Pores regular, circular, twice as broad as the bars; sixteen to twenty on the
half equator. Medullary shell ellipsoidal, one-third as large as the cortical shell.
Dimensions.—Major axis 0.2 to 0.22, minor 0.16 to 0.18, pores 0.008, bars 0.004;
axes of the medullary shell 0.07 and 0.055.
Habitat.—Pacific, central area, Stations 266 to 268, depth 2700 to
2900 fathoms.
Subgenus 2. Druppulissa, Haeckel.
Definition.—Pores of the cortical shell irregular, of different form
or size, usually subcircular or roundish, sometimes lobed or compound.
{310}
8. Druppula nucula, n. sp.
Cortical shell thin walled, smooth, with irregular network; its two axes bear the proportion
3 : 2. Pores subcircular or irregular, roundish, twice to four times as broad as the
bars; fifteen to twenty on the half equator. Medullary shell spherical, about one-third as broad
as the cortical shell.
Dimensions.—Major axis of the cortical shell 0.15 to 0.2, minor axis 0.1 to 0.14;
pores 0.006 to 0.012, bars 0.03; diameter of the medullary shell 0.04.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific, surface.
9. Druppula elliptica, Haeckel.
Haliomma ellipticum, Stöhr, 1880, Palæontogr., loc. cit., p. 88,
Taf. i. fig. 11.
Cortical shell thin walled, rough, or thorny, with irregular network; proportion of the axes =
3 : 2. Pores subcircular or irregular, roundish, about as broad as the bars; ten to
twelve on the half equator. Medullary shell half as broad as the cortical shell, irregularly polyhedral (with
crooked beams in its interior).
Dimensions.—Major axis 0.15, minor 0.06; pores and bars 0.006; diameter of the
medullary shell 0.03.
Habitat.—Fossil in the Tertiary rocks of Sicily: Grotte (Stöhr),
Caltanisetta (Haeckel).
10. Druppula prunum, n. sp.
Cortical shell thick walled, smooth, with irregular network; proportion of the axes =
5 : 4. Pores subcircular or irregular, roundish, three to four times as broad as the
bars; seven to eight on the half equator; every pore is closed at the bottom by a thin membrane,
perforated by four to six irregular pores (like Stylatractus sethoporus, Pl. 17, fig. 3).
Medullary shell spherical, one-third as broad as the cortical shell.
Dimensions.—Major axis 0.2, minor axis 0.16; pores 0.03, bars 0.01; small enclosed
porules 0.01; diameter of the medullary shell 0.05.
Habitat.—Mediterranean, surface, Portofino near Genoa, Haeckel.
11. Druppula oliva, n. sp.
Cortical shell thick walled, rough, with irregular network; proportion of the axes =
5 : 4. Pores irregular, roundish, three to four times as broad as the bars; lobed or
composed of several conjugated porules (as in Lithapium halicapsa, Pl. 14, fig. 8);
five to six large pores on the half equator. Medullary shell ellipsoidal, about one-third as broad
as the cortical shell.
Dimensions.—Major axis of the cortical shell 0.15, minor 0.12; pores 0.02 to 0.03,
bars 0.007; diameter of the medullary shell 0.04.
Habitat.—Mediterranean (Corfu), Canary Islands (Lanzerote),
Haeckel.
{311}
Genus 132. Druppocarpus,[171] n.
gen.
Definition.—Druppulida with simple ellipsoidal cortical
shell and simple medullary shell, with numerous radial spines, without polar tubes.
The genus Druppocarpus differs from Druppula only in the radial
spines, which arise between the pores of the cortical shell, and therefore exhibits the same
relation to it that Ellipsidium bears to Cenellipsis, or Prunocarpus to
Prunulum.
Subgenus 1. Druppocarpetta, Haeckel.
Definition.—Network of the cortical shell regular, with meshes of
equal size and similar form.
1. Druppocarpus ananassa, n. sp.
Cortical shell thin walled, with regular, circular, hexagonally framed pores, of about the same
breadth as the elevated bars; fourteen to sixteen on the half equator. From each hexagon-corner
(between every three pores) arises a short thin radial spine, about as long as two pores,
three-sided pyramidal at the base. Proportion of both cortical axes = 4 : 3. Medullary
shell spherical, half as broad as the cortical shell. (Greatly resembles Druppula pandanus,
Pl. 39, fig.
3, but differs in the larger medullary shell and the spines on the surface.)
Dimensions.—Major axis of the ellipsoidal cortical shell 0.16, minor 0.12; pores
and bars 0.006; length of the radial spines 0.02; diameter of the medullary shell 0.06.
Habitat.—South Pacific, Station 289, surface.
2. Druppocarpus castanea, n. sp.
Cortical shell thin walled, with regular circular pores (without hexagonal frames), of about
the same breadth as the bars; twelve to fourteen on the half equator. Between every three meshes
arise a short conical spine with bristle-like apex, twice to four times as long as one pore.
Proportion of both cortical axes = 3 : 2. Medullary shell spherical, one-third as broad
as the cortical. (Resembles very closely the spherical Haliomma castanea, Haeckel, figured
in my Monograph. pl. xxiv. fig. 4.)
Dimensions.—Major axis 0.15, minor axis 0.1; pores and bars 0.005; length of the
radial spines 0.01 to 0.02; medullary shell 0.035.
Habitat.—North Atlantic, surface; Canary Islands, Lanzerote,
Haeckel.
Subgenus 2. Druppocarpissa, Haeckel.
Definition.—Network of the cortical shell irregular, with meshes of
different size or form.
{312}
3. Druppocarpus chamaerops, n. sp.
Cortical shell thin walled, with irregular, roundish, or subcircular pores of very different
size, twice to six times as broad as the thin bars; eight to twelve on the half equator. Between
them arise numerous thin, bristle-like radial spines, about half as long as the equatorial axis,
and equal to the diameter of the spherical medullary shell; pores of the latter subregular,
circular, very small. (Resembles Prunocarpus artocarpus, Pl. 39, fig. 5, but
differs in the simple spherical medullary shell.)
Dimensions.—Major axis 0.12, minor 0.09; meshes 0.005 to 0.02, bars 0.003; length
of the radial spines 0.05; diameter of the medullary shell 0.04.
Habitat.—Mediterranean, in the Strait of Gibraltar, Algesiras,
Haeckel, surface.
4. Druppocarpus borassus, n. sp.
Cortical shell thick walled, with irregular, roundish, or subcircular pores, three to five
times as broad as the bars; twelve to fifteen on the half equator. Irregularly scattered on the
surface about twenty to thirty short conical spines; their length equals their basal breadth and
the diameter of the largest pores. Medullary shell ellipsoidal, half as large as the
cortical.
Dimensions.—Major axis 0.1, minor 0.08; pores 0.006 to 0.01, bars 0.002; length
and thickness of the radial spines 0.01; axes of the medullary shell 0.05 and 0.04.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
5. Druppocarpus corypha, n. sp.
Cortical shell thick walled, with irregular, funnel-like, roundish pores, scarcely as broad as
the bars; sixteen to twenty on the half equator. Irregularly scattered on the surface about
fifteen to twenty three-sided pyramidal radial spines, half as long as the equatorial axis, and as
the diameter of the medullary shell.
Dimensions.—Major axis 0.17, minor 0.14; pores and bars 0.003 to 0.009; length of
the radial spines 0.08; diameter of the medullary shell 0.09.
Habitat.—Fossil in the Tertiary rocks of the Nicobars, Haeckel.
Genus 133. Prunulum,[172] n. gen.
Definition.—Druppulida with simple ellipsoidal cortical
shell and double medullary shell; without spines or polar tubes.
The genus Prunulum differs from Druppula in the double (not simple)
medullary shell, which is sometimes spherical, sometimes ellipsoidal; it may be derived either
from Druppula by duplication of the medullary shell, or from Thecosphæra by
prolongation of one axis.
{313}
Subgenus 1. Prunuletta, Haeckel.
Definition.—Network of the cortical shell regular, with meshes of
equal size and similar form.
1. Prunulum frugulum, n. sp.
Cortical shell thin walled, smooth, with regular, hexagonal pores, three times as broad as the
bars; ten to twelve on the half equator. Proportion of the major axis of the ellipsoid to the
minor = 4 : 3. Both medullary shells spherical. (Differs from Druppula drupa
almost entirely in the double medullary shell.)
Dimensions.—Major axis of the ellipsoidal cortical shell 0.12, minor 0.09; pores
0.006, bars 0.002; diameter of both medullary shells 0.05 and 0.03.
Habitat.—Pacific, central area, Station 266, depth 2750 fathoms.
2. Prunulum cerasum, n. sp.
Cortical shell thick walled, smooth, with circular, hexagonally framed pores, of the same
breadth as the bars; sixteen to eighteen on the half equator. Proportion of the two axes of the
ellipsoid = 5 : 4. Both medullary shells spherical. (The cortical shell resembles that
of Pipetta tuba, Pl. 39,
fig. 7, without the polar tubes.)
Dimensions.—Major axis 0.15, minor 0.12; pores and bars 0.007; diameter of the
medullary shells 0.08 and 0.04.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
3. Prunulum amygdalum, n. sp.
Cortical shell thick walled, smooth, with regular, circular, hexagonally lobulated pores, three
times as broad as the bars; twelve to fourteen on the half equator (of the same form as in
Druppula phœnix and in Stauroxiphos gladius, Pl. 15, fig. 7).
Proportion of the two axes = 6 : 5. Inner medullary shell spherical, outer
ellipsoidal.
Dimensions.—Major axis 0.12, minor 0.1; pores 0.009, bars 0.003; diameter of the
medullary shells 0.07 and 0.04.
Habitat.—Mediterranean (Corfu), surface.
4. Prunulum coccymelium (Pl. 39,
fig. 4).
Cortical shell thin walled, slightly rough, with regular, circular pores, twice as broad as the
bars; twelve to fifteen on the half equator. Proportion of both axes = 4 : 3. Both
medullary shells spherical.
Dimensions.—Major axis 0.12, minor 0.09; pores 0.006, bars 0.003; diameter of the
medullary shells 0.06 and 0.03.
Habitat.—Pacific, central area, Station 273, depth 2350 fathoms.
{314}
5. Prunulum crenatum, Haeckel.
? Haliomma crenatum, Ehrenberg, 1854, Mikrogeol., Taf. xxii. fig. 36.
Actinomma crenatum, Stöhr, 1880, Palæontogr., vol. xxvi. p. 94, Taf. iii. fig. 3.
Caryolithis crenata, Ehrenberg, 1847, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 43.
Cortical shell thick walled, slightly rough or thorny, with small, regular, circular pores, of
the same breadth as the bars; sixteen to eighteen on the half equator. Proportion of the two axes
= 4 : 3. Both medullary shells ellipsoidal. (The figure of Stöhr represents exactly the
fossil form, as I have observed it myself in the Caltanisetta rocks, whilst the figure of
Ehrenberg is inaccurate and doubtful. The same form, somewhat variable in size and in the number
of the pores, I have also observed in the Pacific ooze.)
Dimensions.—Major axis 0.14 to 0.17, minor 0.1 to 0.13; pores and bars 0.006; main
axes of the medullary shells 0.09 and 0.03.
Habitat.—Fossil in the Tertiary rocks of Sicily (Grotte and
Caltanisetta); living in the Central Pacific, Station 268, depth 2900 fathoms.
6. Prunulum triplex, Haeckel.
Haliomma triplex, Ehrenberg, 1854, Microgeol., Taf. xxxvB. fig. Biv., q.
Actinomma triplex, Haeckel, 1862, Monogr. d. Radiol., p. 444.
Cortical shell thin walled, covered with numerous very thin, short, bristle-like spines. Pores
small, regular, circular, of the same breadth as the bars; fourteen to sixteen on the half
equator. Proportion of the two axes = 3 : 2. Both medullary shells ellipsoidal. The
description of Ehrenberg—as is very often the case—is quite incongruent with his
figure. From a combination of both I give here the diagnosis of a deep-sea species, which is
possibly identical with it. The velvet-like covering of very short and thin bristles is peculiarly
characteristic of this species.
Dimensions.—Major axis 0.1, minor 0.065; pores and bars 0.004; main axes of the medullary
shells 0.04 and 0.02.
Habitat.—North Atlantic, 1800 fathoms, Ehrenberg; Station 353, depth
2965 fathoms.
Subgenus 2. Prunulissa, Haeckel.
Definition.—Network of the cortical shell irregular, with meshes of
different form or size (usually roundish, but sometimes lobed or compound).
7. Prunulum persicum, n. sp.
Cortical shell thick walled, with smooth surface and irregular, roundish, double-contoured
pores, twice to four times as broad as the bars; eight to ten on the half equator. Some of the
pores are simple, often subcircular, others lobed, i.e., composed of from two to three confluent
pores as in Amphisphæra pluto (Pl. 17,
fig. 7). Proportion of the two axes = 3 : 2. Both medullary shells ellipsoidal (or the
inner spherical).
{315}
Dimensions.—Major axis 0.12, minor 0.08; pores 0.06 to 0.012, bars 0.003; main
axes of the medullary shells 0.05 and 0.025.
Habitat.—Indian Ocean, between Socotra and Ceylon, surface,
Haeckel.
8. Prunulum fenestratum, Haeckel.
Actinomma fenestratum, Stöhr, 1880, Palæontogr., vol. xxvi. p. 94, Taf.
iii. fig. 2.
Cortical shell thick walled, rough, with irregular, roundish pores, once to three times as
broad as the bars; twelve to fourteen on the half equator. Proportion of the two axes =
9 : 8. Both medullary shells ellipsoidal. (The pores in Stöhr's description are by
mistake called "regular"; in the figure they are very irregular, as also in the fossil specimens
observed by myself. Between the cortical pores arise very short irregular thorns.)
Dimensions.—Major axis 0.12 to 0.15, minor 0.1 to 0.12; pores 0.003 to 0.01, bars
0.003; main axis of the outer medullary shell 0.07 to 0.09, of the inner 0.03 to 0.04.
Habitat.—Fossil in the Tertiary rocks of Sicily; Grotte, Stöhr,
Caltanisetta, Haeckel.
9. Prunulum pyrenium, n. sp.
Cortical shell very thick walled, smooth, with large irregular, roundish pores, three to four
times as broad as the bars; six to eight on the half equator. The bottom of each funnel-like pore
is closed by a thin siliceous membrane, perforated by three to five irregular roundish pores.
Proportion of the two axes = 4 : 3. Both medullary shells ellipsoidal. (Closely
resembles Stylatractus sethoporus, Pl. 17,
figs. 2, 3, but is devoid of the polar spines.)
Dimensions.—Major axis 0.16, minor 0.12; pores 0.015 to 0.02, bars 0.005; porules
0.01; main axes of the medullary shells 0.07 and 0.03.
Habitat.—Western Tropical Pacific, Station 225, depth 4475
fathoms.
Genus 134. Prunocarpus,[173] n. gen.
Definition.—Druppulida with simple ellipsoidal cortical
shell and double medullary shell, with numerous radial spines, but without polar tubes.
The genus Prunocarpus differs from Prunulum only in the radial spines
of the cortical shell, and exhibits therefore the same relation to it that Druppocarpus
bears to Druppula. While in the latter the medullary shell is simple, in the former it is
double.
Subgenus 1. Prunocarpetta, Haeckel.
Definition.—Network of the cortical shell regular, with meshes of
equal size and similar form.
{316}
1. Prunocarpus datura, n. sp.
Cortical shell thick walled, with regular, circular pores, twice as broad as the bars; ten to
twelve on the half equator. Between every three meshes arises a short conical spine, twice to
three times as long as one pore. Both medullary shells spherical. (Differs mainly from
Ellipsidium datura and from Druppocarpus castanea in the double medullary shell. The
outer network resembles Haliomma castanea, figured 1862 in my Monograph, Taf. xxiv. fig.
4.)
Dimensions.—Major axis of the ellipsoid 0.16, minor 0.12; pores 0.012, bars 0.006;
length of the radial spines 0.03; diameter of the medullary shells 0.06 and 0.04.
Habitat.—North Atlantic, Færöe Channel, John Murray, surface.
2. Prunocarpus sparganium, n. sp.
Cortical shell thick walled, with very small, numerous, regular, circular pores, of the same
size as the bars; forty to fifty on the half equator. Between them over the entire surface occur
small conical spines. Irregularly scattered over the surface ten to twenty larger conical spines,
three to six times as thick at the base as one pore, one-fourth to one-half as long as the main
axis. Both medullary shells ellipsoidal.
Dimensions.—Major axis of the cortical shell 0.16, of the outer medullary shell
0.11, of the inner 0.06; equatorial axis of the first shell 0.12, of the second 0.08, of the third
0.04; pores and bars on an average 0.005; length of the spines 0.05 to 0.1, basal breadth 0.02 to
0.03.
Habitat.—Fossil in the Barbados deposits (Haeckel).
Subgenus 2. Prunocarpilla, Haeckel.
Definition.—Network of the cortical shell irregular, with meshes of
different size or form.
3. Prunocarpus artocarpium, n. sp. (Pl. 39, fig.
5).
Cortical shell thin walled with irregular, roundish pores of very different sizes, twice to
nine times as broad as the thin bars; ten to fifteen on the half equator. Between them arise
numerous bristle-shaped, radial spines, with conical base, on an average one-fourth to two-thirds
as long as the equatorial axis. The outer medullary shell, with irregular, roundish pores,
presents a transverse ellipsoid, its main axis lying in the equatorial axis of the cortical shell,
whilst its equatorial axis corresponds to the main axis of the latter. Inner medullary shell very
small, spherical, with very small pores.
Dimensions.—Major axis of the cortical shell 0.14, of the outer medullary shell
0.05; minor axis of the former 0.1, of the latter 0.035; diameter of the inner medullary shell
0.014; pores of the cortical shell 0.005 to 0.02, bars 0.002; length of the radial spines 0.02 to
0.06.
Habitat.—Indian Ocean, surface; Ceylon, Belligemma, Haeckel.
{317}
4. Prunocarpus melocactus, n. sp.
Cortical shell thick walled, with irregular, roundish pores, twice to five times as broad as
the bars; eighteen to twenty-four on the half equator. Between them arise over the entire surface
small conical thorns, not longer than the largest pores. Irregularly scattered over the surface
twenty to thirty strong, conical, radial spines, about half as long as the main axis, as broad at
the base as a large pore. Both medullary shells ellipsoidal, their main axis identical with that
of the cortical shell.
Dimensions.—Major axis of the outer shell 0.18, of the middle 0.09, of the inner
0.05; minor axis of the first 0.14, of the second 0.07, of the third 0.04; pores of the cortical
shell 0.006 to 0.02, bars 0.004; length of the spines 0.1, basal breadth 0.01.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms; also
fossil in the Tertiary rocks of Sicily; Caltanisetta, Haeckel.
Genus 135. Cromyodruppa,[174] n.
gen.
Definition.—Druppulida with four or more concentric
shells (two medullary and two or more cortical shells), without spines or polar tubes.
The genus Cromyodruppa is characterised by the multiplication of the
concentric fenestrated shell, which is composed of two medullary shells (enclosed in the central
capsule) and two or more cortical shells (outside it). The former may be either spherical or
ellipsoidal. The latter are always ellipsoidal, and in this it differs from Cromyosphæra.
Probably Cromyodruppa has arisen from Prunulum by secondary apposition of more
cortical envelopes.
Subgenus 1. Cromyodruppium, Haeckel.
Definition.—Shell composed of four concentric shells, two medullary
and two cortical.
1. Cromyodruppa cepa, n. sp.
Shell composed of two ellipsoidal, cortical, and two spherical medullary shells. Proportion of
the main axes of the four shells = 1 : 2 : 4 : 5. Network of all
four shells nearly of the same form, subregular, with circular pores of almost equal size in every
shell. The absolute size of the pores increases from the innermost to the outermost shell, but the
breadth of the bars does not increase in a similar degree. The bars of the outermost shell are
only twice as broad as those of the innermost; but the pores are three to four times larger.
Surface of all four shells smooth. (The shell greatly resembles that of Cromyatractus
tetraphractus, Pl. 15,
fig. 2, but is devoid of polar spines; it differs also in the spherical form of both medullary
shells and their relative size.)
{318}
Dimensions.—Main axes of the four shells—(A) innermost 0.035, (B) second
0.08, (C) third 0.15, (D) outermost 0.2; their equatorial axes—(A) 0.03, (B) 0.08, (C) 0.12,
(D) 0.16; pores 0.004 to 0.016, bars 0.002 to 0.004.
Habitat.—Western Tropical Pacific, Station 198, depth 2150
fathoms.
Subgenus 2. Caryodruppula, Haeckel.
Definition.—Shell composed of five or more concentric shells (two
medullary and three or more cortical).
2. Cromyodruppa mango, n. sp.
Shell composed of six ellipsoidal, concentric shells, two medullary and four cortical.
Proportion of their main axes = 2 : 3 : 8 : 11 : 14 : 18.
Network of the two medullary shells and of the innermost cortical shell subregular, with
subcircular pores, about the same breadth as the bars. Network of the three outer cortical shells
more lax, with larger, irregular, roundish pores, twice to six times as broad as the bars. The
size of the pores and bars increases gradually from the innermost to the outermost shell. Surface
smooth.
Dimensions.—Main axis of the six shells—(A) innermost 0.04, (B) 0.06, (C)
0.16, (D) 0.22, (E) 0.28, (F) 0.35; their equatorial axes—(A) innermost 0.03, (B) 0.05, (C)
0.12, (D) 0.16, (E) 0.2, (F) 0.25; pores 0.003 to 0.02, bars 0.003 to 0.005.
Habitat.—Indian Ocean, surface; Matura, Ceylon, Haeckel.
Genus 136. Cromyocarpus,[175] n.
gen.
Definition.—Druppulida with four or more concentric
shells (two medullary and two or more cortical shells), with numerous radial spines, but without
polar tubes.
The genus Cromyocarpus differs from Cromyodruppa only in the
development of numerous large radial spines which start from the outer surface of the shell. It
exhibits therefore the same relation to the latter that Prunocarpus bears to
Prunulum, or Druppocarpus to Druppula. It differs from both in the
multiplication of the cortical shell.
1. Cromyocarpus quadrifarius, n. sp.
Shell composed of four concentric shells, two ellipsoidal cortical shells, and two spherical
medullary shells. Proportion of their main axes = 10 : 8 : 3 : 2.
Pores of the two cortical shells irregular, roundish, twice to four times as broad as the bars.
Pores of the two medullary shells subregular, circular, about the same size as the bars. Outer
surface covered with numerous conical radial spines, about half as long as the main axis, half as
broad at the base as the innermost shell.
{319}
Dimensions.—Major axis of the outer cortical shell 0.2, of the inner 0.15;
equatorial axis of the former 0.16, of the latter 0.12; diameter of the outer medullary shell
0.06, of the inner 0.04; pores of the cortical shells 0.01 to 0.02, of the medullary shells 0.004
to 0.008, bars 0.003 to 0.006; length of the radial spines 0.1, basal breadth 0.02.
Habitat.—Indian Antarctic Ocean, Station 157, depth 1950 fathoms.
Genus 137. Lithatractus,[176] n.
gen.
Definition.—Druppulida with simple ellipsoidal cortical
shell and simple medullary shell, with two large opposite polar spines in the main axis of equal
size and similar form.
The genus Lithatractus, rich in common and widely distributed species,
begins the series of those Druppulida which are characterised by peculiar polar spines at both
poles of the main axis. It repeats the formation of Stylosphæra and Ellipsostylus,
and differs from the former in the ellipsoidal form of the cortical shell, from the latter in the
possession of a medullary shell. Formerly all these forms were united in the one genus
Stylosphæra (see above, p. 121).
Subgenus 1. Lithatractara, Haeckel.
Definition.—Network of the outer shell regular or subregular, with
meshes of nearly equal size and similar form; surface smooth, without thorns or papillæ.
1. Lithatractus hexagonalis, n. sp.
Outer shell thin walled, smooth, without thorns or papillæ, with regular delicate network; the
meshes hexagonal, three to four times as broad as the thin bars; ten to twelve on the half
equator. Proportion of the major axis of the ellipsoid to the minor = 4 : 3. Minor axis
three times as large as that of the inner spherical shell; pores of the latter, small, circular.
Two spines three-sided pyramidal, as long as the radius of the outer shell, each as broad at its
base as one mesh.
Dimensions.—Longer axis of the ellipsoidal cortical shell 0.16, shorter axis 0.12;
pores 0.011, bars 0.003; diameter of the spherical medullary shell 0.04; length of the polar
spines 0.07; basal breadth 0.01.
Habitat.—North Pacific, Station 253, depth 3125 fathoms.
2. Lithatractus fragilis, n. sp. (Pl. 16, fig.
3).
Stylosphæra fragilis, Haeckel, 1881, Prodromus et Atlas, pl. xvi. fig.
3.
Outer shell thin walled, smooth, without thorns or papillæ, with regular network; the meshes
circular, of equal size, eight to ten times as broad as the thin bars; six to eight on the half
equator. {320}Proportion of the major axis of the
ellipsoid to the minor = 5 : 4. Minor axis once and one-third as broad as that of the
ellipsoidal inner shell, the pores of which are also circular, but of half the size. Two spines
strong, three-sided pyramidal, acute, as broad as a large mesh, as long as the equatorial axis.
The stout inner prolongations of these form the only connection between the two shells.
Dimensions.—Longer axis of the ellipsoidal cortical shell 0.12, shorter axis 0.1;
pores 0.02, bars 0.002; longer axes of the ellipsoidal medullary shell 0.09, shorter axis 0.07;
pores 0.01, bars 0.002; length of the polar spines 0.1, basal breadth 0.002.
Habitat.—Central area of the Pacific, Station 272, depth 2600
fathoms.
3. Lithatractus leptostylus, n. sp.
Outer shell thin walled, smooth, with regular, circular pores of equal size, three to four
times as broad as the thin bars; ten to twelve on the half equator. Proportion of the major axis
to the minor = 7 : 6. Minor axis three times as large as that of the inner spherical
shell; pores of the latter half as large. Polar spines cylindrical, blunt, longer than the major
axis, scarcely half as broad as one larger pore.
Dimensions.—Longer axis of the cortical shell 0.14, shorter axis 0.12; pores 0.015
to 0.02, bars 0.004; diameter of the medullary shell 0.04; length of the polar spines 0.15 to 0.2,
its thickness 0.01.
Habitat.—Central area of the Pacific, Stations 270 to 272, depth 2425
to 2925 fathoms.
4. Lithatractus pachystylus, n. sp.
Outer shell thick walled, smooth, with regular, circular pores of equal size; twelve to fifteen
on the half equator. Each pore is deep, funnel-shaped, its outer aperture double the size of the
inner, its breadth about three times that of the high bars. Proportion of the major axis to the
minor = 6 : 5. Major axis double as long as the diameter of the spherical medullary
shell. Polar spines very thick and short, tetrahedral, one-fourth as long and broad as the major
axis.
Dimensions.—Longer axis of the cortical shell 0.17, shorter axis 0.14; pores 0.01,
bars 0.003; diameter of the medullary shell 0.08; length and thickness of the polar spines
0.04.
Habitat.—Central area of the Pacific, Station 271, depth 2425
fathoms.
5. Lithatractus convallaria, n. sp.
Outer shell thick walled, smooth, with elegant regular network; the meshes circular, six-lobed,
rosette-like (of the same form as Stauroxiphos gladius, Pl. 15, fig. 7),
twice to three times as broad as the bars; six to eight on the half equator. Proportion of the
major axis of the ellipsoid to the minor = 4 : 3. Minor axis twice as long as the
diameter of the inner spherical shell, which has regular, simple, circular pores of half the size.
Polar spines short and thick, conical, only one-fourth to one-sixth as long as the minor axis, and
quite as thick.
Dimensions.—Longer axis of the cortical shell 0.16, shorter axis 0.12; pores
0.016, bars 0.006; diameter of the medullary shell 0.06; length of the polar spines 0.02 to 0.03,
basal thickness the same.
Habitat.—Central area of the Pacific, Station 268, depth 2900
fathoms.
{321}
Subgenus 2. Lithatractylis, Haeckel.
Definition.—Network of the outer shell regular, with meshes of equal
size and similar form; surface thorny or papillose, covered with small conical spines or
tubercles.
6. Lithatractus echiniscus, n. sp.
Outer shell thorny, thin walled, with regular network; meshes circular, with hexagonal frame,
four to five times as broad as the thin bars; eighteen to twenty on the half equator. From every
corner between the three meshes, where three hexagons unite, starts one short, straight,
triangular thorn (as in Ellipsoxiphus elegans, Pl. 14, fig. 7).
Proportion of the major axis of the ellipsoid to the minor = 4 : 3. Minor axis twice the
diameter of the inner spherical shell, the pores of which are half as broad, circular. Polar
spines three-sided pyramidal, about as long as the minor axis, as broad at the base as one
hexagonal frame.
Dimensions.—Longer axis of the cortical shell 0.08, shorter axis = 0.06; pores
0.004, bars 0.001; length of the polar spines 0.05, basal thickness 0.005; diameter of the inner
shell 0.03.
Habitat.—South Atlantic, off Tristan da Cunha, Station 334,
surface.
7. Lithatractus carduelis, n. sp.
Outer shell thin walled, thorny, with regular network; meshes circular, simple, four to six
times as broad as the thin bars; ten to twelve on the half equator. Between every three meshes is
one short conical spine. Proportion of the major axis to the minor = 6 : 5. Minor axis
equals three times the diameter of the inner spherical shell. Polar spines conical, thick, half as
long as the major axis, thicker at the base than one pore. (This species differs from
Stylatractus carduus only by the simple medullary shell.)
Dimensions.—Longer axis of the ellipsoid 0.12 to 0.16, shorter axis 0.1 to 0.14;
pores 0.02 to 0.03, bars 0.005; length of the polar spines 0.05 to 0.10, basal breadth 0.04;
diameter of the medullary shell 0.03 to 0.04.
Habitat.—Central area of the Pacific, Stations 268 to 274, depth 2350
to 2900 fathoms; also fossil in the rocks of Barbados.
8. Lithatractus cirsium, n. sp.
Outer shell thin walled, thorny, with regular network; pores circular, simple, small, two to
three times as broad as the thin bars; eighteen to twenty-four on the half equator. Proportion of
the major axis to the minor = 4 : 3. Inner shell ellipsoidal, half as large as the
outer. Polar spines cylindrical, blunt, thin, very variable in length (one-fourth to three-fourths
of the major axis, not thicker than a mesh).
Dimensions.—Longer axis of the ellipsoid 0.07 to 0.09, shorter axis 0.05 to 0.07;
pores 0.005 to 0.006, bars 0.002; length of the polar spines 0.02 to 0.06, basal breadth 0.004;
length of the inner shell 0.04, breadth 0.03.
Habitat.—North Pacific, Station 244, surface.
{322}
9. Lithatractus rosetta, n. sp.
Outer shell thick walled, thorny, with regular network; meshes circular, funnel-shaped, its
outer aperture elegant, rosette-like, with eight to ten incisions (like Stylosphæra
calliope, Pl. 16,
fig. 6); eight to ten meshes on the half equator, three to four times as broad as the bars.
Proportion of the longer axis to the shorter = 5 : 4. Shorter axis equals three times
the diameter of the inner spherical shell. Polar spines three-sided pyramidal, about half as long
as the shorter axis, as broad at the base as a mesh.
Dimensions.—Longer axis of the cortical shell 0.15, shorter 0.12; pores 0.02, bars
0.005; length of the polar spines 0.05, basal breadth 0.02; diameter of the inner shell 0.04.
Habitat.—South-eastern part of the Pacific, Station 302, depth 1450
fathoms.
Subgenus 3. Lithatractona, Haeckel.
Definition.—Network of the outer shell irregular, with meshes of
unequal size or dissimilar form; surface smooth, without thorns or papillæ.
10. Lithatractus conifer, n. sp.
Outer shell thin walled, smooth, with irregular, roundish pores, two to four times as broad as
the thin bars; fifteen to twenty on the half equator. Margin of the pores simple. Proportion of
the major axis to the minor = 3 : 2. Minor axis twice as large as the diameter of the
inner spherical shell, the pores of which are also irregular, roundish, but of half the size.
Polar spines conical, somewhat shorter than the main axis, on the base two to three times as thick
as a large pore.
Dimensions.—Longer axis of the outer shell 0.15, shorter axis 0.1; pores 0.002 to
0.004, bars 0.001; diameter of the inner shell 0.05; length of the polar spines 0.12, basal
breadth 0.012.
Habitat.—South Atlantic, Station 325, surface.
11. Lithatractus lobatus, n. sp.
Outer shell thick walled, smooth, with irregular, roundish pores, four to eight times as broad
as the bars; six to eight on the half equator. Margin of the pores lobed, very irregular, bluntly
dentate, by five to twenty slight incisions. Proportion of the major axis to the minor very
variable, between 3 : 2 and 9 : 8. Diameter of the inner shell also variable,
between one-third and one-half of the outer; pores of the former scarcely half the size of the
latter, simple, roundish, or circular. The inner shell is at some parts quite spherical, at other
parts more or less ellipsoidal. Polar spines conical, very variable in size and form, sometimes in
the basal half triangular; they are sometimes somewhat longer than the main axis, at other times
considerably shorter; their basal breadth is occasionally the same as that of the largest pores,
sometimes, however, scarcely half as large. This deep-sea species is very common in the central
area of the Tropical Pacific (Stations 266 to 272), and occurs also fossil in the Barbados
deposits. It is interesting from its great variability, and sometimes constitutes a transitional
form to Stylosphæra.
{323}
Dimensions.—Longer axis of the outer shell 0.1 to 0.15, shorter axis 0.05 to 0.12;
pores 0.018 to 0.024, bars 0.002 to 0.004; diameter of the inner shell 0.04 to 0.08; length of the
polar spines 0.08 to 0.2, basal thickness 0.01 to 0.02.
Habitat.—Central area of the Pacific, common; Stations 266 to 272,
depth 2425 to 2900 fathoms; also fossil in Barbados.
12. Lithatractus jugatus, n. sp. (Pl. 16, fig.
2).
Stylosphæra jugata, Haeckel, 1881, Prodromus et Atlas (pl. xvi. fig.
2).
Outer shell thick walled, smooth, or somewhat reticulated, with a peculiar, irregular, double
network. The pores are roundish, of very different sizes, with double margin of the outer
aperture, and so irregularly distributed in polygonal groups that every group contains two to six
pores immediately touching each other; the groups are separated by broader bars. On the half
equator of the shell are about six to eight groups and twelve to eighteen pores. Proportion of the
longer axis of the ellipsoidal shell to the shorter = 6 : 5 or 5 : 4. Inner
ellipsoidal shell about half the size; its pores are not easy to make out. Polar spines
three-sided pyramidal, with prominent edges, about half as long as the major axis, as broad at the
base as a group of pores. (Nearly allied to Stylosphæra jugata, p. 137.)
Dimensions.—Longer axis of the cortical shell 0.12 to 0.16, shorter axis 0.1 to
0.13; pores 0.01 to 0.02; length of the polar spines 0.06 to 0.08, basal breadth 0.03 to 0.4.
Habitat.—Central area of the Pacific, Stations 266 to 272, depth 2425
to 2900 fathoms.
Subgenus 4. Lithatractium, Haeckel.
Definition.—Network of the outer shell irregular, with meshes of
unequal size or dissimilar form; surface thorny or papillose, covered with small conical spines or
tubercles.
13. Lithatractus conostylus, n. sp.
Outer shell thorny, thin walled, with irregular network; pores roundish, of unequal size, twice
to four times as broad as the thin bars; sixteen to twenty on the half equator. Proportion of the
longer axis to the shorter = 5 : 4. Shorter axis twice the diameter of the inner
spherical shell. Polar spines conical, very stout, longer than the main axis, its base equals
one-third of the minor axis.
Dimensions.—Major axis of the cortical shell 0.1, shorter axis 0.08; pores 0.004
to 0.002, bars 0.001; diameter of the medullary shell 0.04; polar spines—length 0.12,
thickness 0.03.
Habitat.—Central area of the Pacific, Station 271, depth 2425
fathoms.
14. Lithatractus gamoporus, n. sp.
Outer shell thorny, thick walled, with irregular network; pores roundish, of unequal size, so
irregularly distributed in polygonal groups that in every group two to six pores (commonly three
to four) are near together; the groups are separated by broader bars. On the half equator {324}six to eight groups and eighteen to twenty-four pores.
Proportion of the longer axis to the shorter = 6 : 5. Shorter axis equals twice the
diameter of the inner ellipsoidal shell. Polar spines three-sided pyramidal, about as long as the
equatorial axis, as broad at the base as a group of pores.
Dimensions.—Major axis of the cortical shell 0.12, minor axis 0.1; pores 0.004 to
0.012, bars 0.003; length of the polar spines 0.1, basal breadth 0.03.
Habitat.—Central area of the Pacific, Stations 271 to 274, depth 2350
to 2750 fathoms.
Genus 138. Druppatractus,[177] n.
gen.
Definition.—Druppulida with simple ellipsoidal cortical
shell and simple medullary shell, with two large opposite polar spines in the main axis of
different size or dissimilar form.
The genus Druppatractus differs from its near relation Lithatractus
in the differentiation of the two polar spines, which are different in size or form, often to a
very considerable degree. It has therefore the same relation to the latter that
Ellipsoxiphus bears to Ellipsostylus.
Subgenus 1. Druppatractara, Haeckel.
Definition.—Network of the cortical shell regular or subregular, with
meshes of nearly equal size and similar form; surface smooth, without thorns or papillæ.
1. Druppatractus ichthydium, n. sp.
Cortical shell thin walled, with smooth surface and regular network. Proportion of the major
axis to the minor = 3 : 2. Pores circular, hexagonally framed, twice as broad as the
bars; seven to eight on the half equator (as in Xiphostylus alcedo, Pl. 13, fig. 4).
Medullary shell spherical, one-third as broad as the cortical shell. Polar spines straight,
three-sided pyramidal, as broad at the base as one mesh; the longer equals the main axis of the
cortical shell, the shorter only its half.
Dimensions.—Major axis of the ellipsoidal cortical shell 0.18, minor axis 0.12;
pores 0.01, bars 0.005; diameter of the medullary shell 0.04; length of the major polar spine 0.16
to 0.2, of the minor 0.01 to 0.12, basal breadth 0.01.
Habitat.—Mediterranean (Corfu), surface, Haeckel.
2. Druppatractus
hippocampus, n. sp. (Pl. 16,
figs. 10, 11).
Stylosphæra hippocampus, Haeckel, 1881, Atlas, pl. xvi. figs. 10, 11.
Cortical shell thick walled, with smooth surface and regular network. Proportion of both axes =
7 : 6. Pores circular, hexagonally lobed, three times as broad as the bars; nine to ten
on the half {325}equator. Medullary shell ellipsoidal,
nearly half as large as the cortical. Larger polar spine horn-like curved, as long as the main
axis, smaller scarcely half as long, pommel-shaped.
Dimensions.—Major axis 0.09, minor 0.075; pores 0.01, bars 0.003; axes of the
medullary shell 0.04 and 0.03; length of the larger polar spine 0.06 to 0.09, of the smaller 0.04,
basal breadth 0.02.
Habitat.—Pacific, central area, Stations 270 to 272, surface.
3. Druppatractus belone, n. sp.
Cortical shell thin walled, with smooth surface and regular network. Proportion of both axes =
4 : 3. Pores circular, twice as broad as the bars; ten to twelve on the half equator.
Medullary shell spherical, one-third as broad as the cortical shell. Polar spines strong,
straight, cylindrical, with conical apex, as broad at the base as two pores; the larger once and a
half to twice as long as the main axis, the smaller only one-third to one-fourth as long.
Dimensions.—Major axis 0.1 to 0.12, minor 0.08 to 0.09; pores 0.008, bars 0.004;
diameter of the medullary shell 0.03; length of the larger polar spine 0.15 to 0.2, of the minor
0.04 to 0.05, basal thickness of them 0.02.
Habitat.—Cosmopolitan; Atlantic, Indian, and Pacific, surface.
4. Druppatractus testudo, Haeckel.
Stylosphæra testudo, Ehrenberg, 1872, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 299, Taf. viii. fig. 16.
Cortical shell thick walled, spindle-shaped, with smooth surface and regular network.
Proportion of the two axes = 3 : 2. Pores circular, five times as broad as the bars;
only six to seven on the half equator. Medullary shell ellipsoidal, about one-third as large as
the cortical shell. Polar spines strong, short, three-sided pyramidal; the larger as long as half
the transverse axis, the smaller only one-fourth as long.
Dimensions.—Major axis 0.12, minor 0.8; pores 0.015, bars 0.003; axes of the
medullary shell 0.04 and 0.03; length of the larger polar spine 0.04, of the smaller 0.02, basal
thickness 0.01.
Habitat.—Pacific, Philippine and Californian Sea (Ehrenberg), Stations
244, 266, 289, &c., depth 2550 to 2900 fathoms.
Subgenus 2. Druppatractylis, Haeckel.
Definition.—Network of the cortical shell regular or subregular, with
meshes of nearly equal size and similar form; surface thorny or papillose, covered with small
conical spines or tubercles.
5. Druppatractus accipenser, n. sp.
Cortical shell thick walled, thorny, with regular network. Proportion of the two axes =
6 : 5. Pores circular, each with a six-lobed outer opening, funnel-shaped, twice as
broad as the bars; nine {326}to ten on the half equator.
Medullary shell spherical, one-third as broad as the cortical shell. Polar spines very strong,
three-sided pyramidal, as broad at the base as three pores; the larger about as long as the main
axis, the smaller only one-third as long.
Dimensions.—Major axis 0.12, minor 0.1; pores 0.01, bars 0.005; medullary shell
0.035; length of the larger polar spine 0.11, of the smaller 0.04, basal breadth 0.03.
Habitat.—Indian Ocean, Madagascar, surface, Rabbe.
6. Druppatractus ostracion, n. sp. (Pl. 16, figs. 8,
9).
Cortical shell thick walled, thorny, with regular network. Proportion of the two axes =
4 : 3. Pores circular, hexagonally framed, three to four times as broad as the crested
bars; from every corner of the hexagon (between three meshes) arises a short papilla; the bottom
of every funnel-like mesh is closed by a very thin plate with three regular, circular pores
(occasionally four), sometimes confluent, at other times separate. Medullary shell (fig. 9) half
as large as the cortical, ellipsoidal and papillate, with regular, circular pores. Polar spines
very strong, three-sided prismatic, often somewhat irregular, with short apex; the larger once to
twice as long as the main axis, the smaller scarcely half as long.
Dimensions.—Major axis 0.16, minor 0.12; pores 0.02 (porules at their base 0.01),
bars 0.006; axes of the medullary shell 0.07 and 0.06, its pores 0.01, bars 0.003; length of the
larger polar spine 0.15 to 0.3, of the smaller 0.07 to 0.09, basal thickness 0.03.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms.
7. Druppatractus coronatus, Haeckel.
Stylosphæra coronata, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss. Berlin,
p. 84, Taf. xxv. fig. 4.
Cortical shell thick walled, thorny, with regular network. Proportion of the two axes =
5 : 4. Pores circular, simple, twice as broad as the bars; eight to ten on the half
equator. Medullary shell circular, one-third as broad as the cortical shell. Polar spines furrowed
and angular; the longer and thinner pyramidal about as long as the main axis, the shorter and
thicker scarcely half as long, thickened towards the short conical apex.
Dimensions.—Major axis 0.08 to 0.1, minor 0.06 to 0.08; pores 0.01, bars 0.005;
diameter of the medullary shell 0.03; length of the large polar spine 0.08 to 0.12, of the shorter
0.04 to 0.05.
Habitat.—Fossil in the Tertiary rocks of Barbados; also living in the
depths of the Pacific, central area, Stations 265 to 268, depth 2700 to 2900 fathoms.
Subgenus 3. Druppatractona, Haeckel.
Definition.—Network of the cortical shell irregular, with meshes of
unequal size or dissimilar form; surface smooth, without thorns or papillæ.
{327}
8. Druppatractus lævis, Haeckel.
Stylosphæra lævis, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss. Berlin,
p. 84, Taf. xxv. fig. 6.
Cortical shell thin walled, smooth, with irregular network. Proportion of the two axes =
3 : 2. Pores roundish or subcircular, of different size, one-half to twice as broad as
the bars; eight to ten on the half equator. Medullary shell ellipsoidal, one-third as large as the
cortical shell. Polar spines conical, the larger and thinner nearly as long as the main axis, the
shorter and thicker scarcely one-third as long.
Dimensions.—Major axis 0.07 to 0.1, minor 0.05 to 0.07; pores and bars 0.005 to
0.01; axes of the medullary shell 0.03 and 0.02; length of the major polar spine 0.06 to 0.09, of
the shorter 0.03 to 0.04.
Habitat.—Cosmopolitan; Atlantic, Indian, Pacific, surface.
9. Druppatractus xiphias, n. sp.
Cortical shell thick walled, smooth, with irregular network. Proportion of the two axes =
6 : 5. Pores funnel-shaped, composed of two to four confluent smaller porules, twice to
four times as broad as the bars; ten to twelve on the half equator. Medullary shell spherical,
half as broad. Polar spines three-sided pyramidal, the larger about as long as the main axis, the
shorter one-third to one-half as long.
Dimensions.—Major axis 0.12, minor 0.1; pores 0.012 to 0.018, bars 0.004; diameter
of the medullary shell 0.05; length of the major polar spine 0.1, of the minor 0.03 to 0.05.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
Subgenus 4. Druppatractium, Haeckel.
Definition.—Network of the cortical shell irregular, with meshes of
unequal size or dissimilar form; surface thorny or papillose, covered with small spines or
tubercles.
10. Druppatractus diodon, n. sp.
Cortical shell thin walled, thorny, with irregular network. Proportion of the two axes =
5 : 4. Pores simple, irregular, roundish, twice to four times as broad as the bars;
eight to ten on the half equator. Medullary shell spherical, half as broad. Polar spines conical,
more or less curved; the major longer than the main axis, the minor scarcely half as long.
(Resembles Sphærostylus ophidium, Pl. 16,
figs. 14, 15, but differs from it in the prolongation of the main axis.)
Dimensions.—Major axis 0.15, minor 0.12; pores 0.01 to 0.02, bars 0.005; diameter
of the medullary shell 0.06; length of the larger polar spine 0.12 to 0.18, of the shorter 0.06 to
0.08, basal thickness 0.003.
Habitat.—Pacific, central area, Station 274, surface.
{328}
11. Druppatractus pisciculus, n. sp.
Cortical shell thick walled, spiny, with irregular network. Proportion of the two axes =
3 : 2. Pores irregular, roundish, lobed, or composed of three to five confluent porules,
twice to six times as broad as the bars; six to eight on the half equator. (Similar to
Ellipsoxiphus atractus, Pl. 14,
fig. 1.) Medullary shell ellipsoidal, nearly half as large as the cortical shell. Polar spines
three-sided pyramidal, the major longer than the main axis, the minor scarcely one-third to
one-fourth as long.
Dimensions.—Major axis 0.18, minor 0.12; pores 0.01 to 0.03, bars 0.004; axes of
the medullary shell 0.08 and 0.05; length of the major polar spine 0.2, of the minor 0.05, basal
breadth 0.02.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
Genus 139. Stylatractus,[178] n.
gen.
Definition.—Druppulida with simple ellipsoidal cortical
shell and double medullary shell, in the main axis with two large opposite polar spines of equal
size and similar form.
The genus Stylatractus differs from Lithatractus in the double
medullary shell, from Amphisphæra in the ellipsoidal form of the cortical shell. As in
these two genera, both opposite polar spines have the same shape and size.
Subgenus 1. Stylatractara, Haeckel.
Definition.—Network of the cortical shell regular or subregular, with
meshes of nearly equal size and similar form; surface smooth, without thorns or papillæ.
1. Stylatractus neptunus, n. sp. (Pl. 17, fig.
6).
Amphistylus neptunus, Haeckel, 1878, Atlas, pl. xvii. fig. 6.
Cortical shell thick walled, smooth, with subregular, circular, polygonally framed pores, quite
as broad as the bars; ten to twelve on the half equator. Polar spines three-sided pyramidal, about
as long as the half main axis, three times as broad at the base as the pores. (Much resembles
Amphisphæra neptunus, p. 142, but differs from it by the
prolongation of the main axis, which equals one and a half or one and a fourth the equatorial
axis, and by the somewhat irregular formation of the cortical hexagonal network. Sometimes also,
as in the figured specimen, the length of both polar spines is somewhat different.)
Dimensions.—Major axis of the cortical shell 0.15, minor axis 0.12; pores and bars
0.007; main axes of both ellipsoidal medullary shells 0.08 and 0.05; length of the polar spines
0.08 to 0.1, basal breadth 0.02.
Habitat.—Pacific, central area, Stations 266 to 268, depth 2700 to
2900 fathoms.
{329}
2. Stylatractus fusiformis, n. sp.
Cortical shell thick walled, smooth, with regular, simple, circular pores, quite as broad as
the bars; fourteen to sixteen on the half equator. Polar spines three-sided pyramidal, half as
long as the main axis, as broad at the base as the inner medullary shell. (Resembles very nearly
Xiphatractus armadillo, Pl. 17,
fig. 11, but differs in the regular form and equal length of the polar spines.)
Dimensions.—Major axis 0.17, minor axis 0.13; pores and bars 0.007; main axes of
both ellipsoidal medullary shells 0.09 and 0.05; length of the polar spines 0.08, basal breadth
0.04.
Habitat.—Cosmopolitan; Atlantic, Indian, Pacific, at various
depths.
3. Stylatractus compactus, n. sp. (Pl. 17, fig.
4).
Cortical shell thick walled, smooth, with subregular, circular, double-contoured pores, smaller
than the bars; sixteen to eighteen on the half equator. The thickness of the shell-wall equals the
radius of the inner medullary shell. Polar spines short, three-sided pyramidal; their length and
basal thickness variable, but commonly equal to the diameter of the inner medullary shell.
Dimensions.—Major axis 0.16, minor axis 0.13; pores 0.007, bars 0.01; main axes of
both ellipsoidal medullary shells 0.09 and 0.04; length and basal breadth of the polar spines 0.04
to 0.05.
Habitat.—Pacific, central area, Stations 270 to 272, depth 2425 to
2925 fathoms.
Subgenus 2. Stylatractylis, Haeckel.
Definition.—Network of the outer shell regular, with meshes of equal
size and similar form; surface thorny or papillose, covered with small spinules or tubercles.
4. Stylatractus giganteus, n. sp. (Pl. 17, fig.
1).
Amphistylus giganteus, Haeckel, 1879, Atlas (pl. xvii. fig. 1).
Cortical shell papillose, very thick walled, with regular network; pores circular, with double
margin, about twice as broad as the bars; ten to twelve on the half equator. The cortical shell is
connected with the outer medullary shell by numerous strong beams, and the inner prolongations of
both polar spines are much stronger. The circular pores of the outer medullary shell are three
times as large as those of the inner, and equal to those of the cortical shell, but the bars are
much thinner. Polar spines very strong, three-sided pyramidal, with spirally contorted edges, as
long as the main axis, as broad at the base as the inner medullary shell.
Dimensions.—Major axis of the cortical shell 0.3, minor axis 0.22; pores 0.02,
bars 0.01; main axes of the ellipsoidal medullary shells 0.14 and 0.07; length of the polar spines
0.3, basal thickness 0.06.
Habitat.—Western Tropical Pacific, Station 225, depth 4475
fathoms.
{330}
5. Stylatractus carduus, Haeckel.
Stylosphæra carduus, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 84, Taf. xxv. fig. 7.
Cortical shell thin walled, spiny, with regular, circular pores, five times as broad as the
bars; ten to twelve on the half equator. Polar spines conical, half as long as the main axis, at
the base broader than the pores. (The cortical shell is ellipsoidal as well as both medullary
shells; the figure of Ehrenberg, as is generally the case, is more correct than his description.
This interesting species occurs in the Barbados rocks, with double as well as with simple
medullary shell, and may in the latter case be distinguished as Lithatractus
carduelis.)
Dimensions.—Major axis of the cortical shell 0.12 to 0.16, minor 0.1 to 0.14;
pores 0.01 to 0.02, bars 0.002 to 0.005; main axes of the ellipsoidal medullary shells 0.05 and
0.03; length of the polar spines 0.06, basal breadth 0.02.
Habitat.—Fossil in the Tertiary rocks of Barbados.
Subgenus 3. Stylatractona, Haeckel.
Definition.—Network of the outer shell irregular, with meshes of
different size or form; surface smooth, without thorns or papillæ.
6. Stylatractus variabilis, n. sp.
Cortical shell thin walled, smooth, with irregular, roundish pores, twice to six times as broad
as the bars; twelve to sixteen on the half equator. Polar spines conical, about half as long as
the main axis, and as broad at the base as the inner medullary shell. (The size and form of the
pores and of the polar spines in this species are very variable, so that the proportions given are
to be understood as averages.)
Dimensions.—Major axis 0.13, minor axis 0.11; pores 0.004 to 0.012, bars 0.002;
main axes of the ellipsoidal medullary shells 0.07 and 0.04; length of the polar spines 0.06,
basal breadth 0.03.
Habitat.—Pacific, central area. Stations 272 to 274, depth 2350 to
2750 fathoms.
7. Stylatractus sethoporus, n. sp. (Pl. 17, figs. 2,
3).
Cortical shell thick walled, smooth, with large irregular, roundish pores, twice to four times
as broad as the bars; seven to eight on the half equator. At the bottom of each pore a thin
lamella of silex, perforated by four to six irregular, roundish, double-contoured porules.
Medullary shells resemble those of Stylatractus giganteus. Polar spines three-sided
pyramidal; their length and basal thickness scarcely equal the diameter of the inner medullary
shell.
Dimensions.—Major axis 0.15, minor axis 0.13; pores 0.02, bars 0.005, porules
0.01; length of the polar spines 0.04.
Habitat.—Pacific, central area, Station 272, depth 2600 fathoms.
{331}
Subgenus 4. Stylatractium, Haeckel.
Definition.—Network of the outer shell irregular, with meshes of
different size and form; surface thorny or papillose, covered with small spinules or
tubercles.
8. Stylatractus papillosus, n. sp.
Cortical shell thin walled, with papillose surface, and irregular, simple, roundish pores, ten
to sixteen on the half equator, twice to three times as broad as the bars. Conical papillæ of the
surface irregularly scattered. Polar spines conical, about as long as the main axis, at the base
half as broad as the inner medullary shell.
Dimensions.—Major axis 0.12, minor 0.1; pores 0.006 to 0.01, bars 0.003; main axes
of the medullary shells 0.09 and 0.06; length of the polar spines 0.13, basal breadth 0.04.
Habitat.—Indian Ocean, surface (between Socotra and Ceylon),
Haeckel.
9. Stylatractus disetanius, n. sp.
Cortical shell thin walled, with spiny surface, and irregular, roundish pores, eight to ten on
the half equator, twice to three times as broad as the bars. Every pore is divided by thinner bars
into four to six small roundish porules. Polar spines three-sided pyramidal, nearly as long as the
main axis, as broad at the base as the inner medullary shell. (Resembles Xiphatractus
glyptodon, Pl. 17,
figs. 9, 10, but differs from it by the irregular network and the equal size of both polar
spines.)
Dimensions.—Major axis 0.14, minor 0.1; pores 0.02, bars 0.008, porules 0.007;
main axes of the two medullary shells 0.08 and 0.05; length of the polar spines 0.13, basal
breadth 0.04.
Habitat.—South Pacific, near New Zealand, Station 169, surface.
Genus 140. Xiphatractus,[179] n.
gen.
Definition.—Druppulida with a simple ellipsoidal cortical
shell and double medullary shell, in the main axis with two large opposite polar spines of
different size or form.
The genus Xiphatractus differs from the nearly related Stylatractus
in the differentiation of two polar spines (in the same manner as Druppatractus differs
from Lithatractus). From the spherical Amphistylus it differs in the ellipsoidal
form of the cortical shell.
Subgenus 1. Xiphatractara, Haeckel.
Definition.—Network of the cortical shell regular or subregular, with
meshes of nearly equal size and similar form; surface smooth, without thorns or papillæ.
{332}
1. Xiphatractus armadillo, n. sp. (Pl. 17, fig.
11).
Cortical shell thick walled, smooth, with regular, circular pores, about as broad as the bars;
fifteen to sixteen on the half equator. Polar spines three-sided pyramidal, with thick, prominent
edges, of irregular form; the longer about as long as the main axis, the shorter scarcely
one-third as long; their basal breadth equals the diameter of the inner medullary shell.
Dimensions.—Major axis of the cortical shell 0.13, minor axis 0.1, thickness of
its wall 0.01; pores and bars 0.005; main axes of the two medullary shells 0.07 and 0.03; length
of the major polar spine 0.12, of the minor 0.04, basal thickness 0.04.
Habitat.—Pacific, central area, Station 272, depth 2600 fathoms.
2. Xiphatractus euphractus, n. sp.
Cortical shell thick walled, smooth, with regular, circular pores, twice as broad as the bars;
eight to nine on the half equator. Each pore has six to nine excisions (commonly eight), elegantly
lobed, flower-like (as in Xiphostylus phasianus, Pl. 13, fig. 9).
Polar spines strong, angulate; the major pyramidal, longer than the main axis, the shorter
elegantly pommel-like, scarcely one-third as long.
Dimensions.—Major axis 0.15, minor 0.12; pores 0.014, bars 0.007; main axes of the
two medullary shells 0.09 and 0.05; length of the larger polar spine 0.16, of the shorter 0.05,
greatest thickness 0.03.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
Subgenus 2. Xiphatractylis, Haeckel.
Definition.—Network of the cortical shell regular or subregular, with
meshes of nearly equal size and similar form; surface thorny or papillose.
3. Xiphatractus spinulosus, Haeckel.
Stylosphæra spinulosa, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 84, Taf. xxv. fig. 8.
Cortical shell thin walled, with spiny surface, and large, regular, circular pores, four times
as broad as the bars; seven to eight on the half equator. Spines between the pores of the same
size, conical. Polar spines also conical, but much larger; the major about as long as the main
axis, the minor scarcely half as long; their basal thickness equals the largest pores.
Dimensions.—Major axis 0.1, minor 0.08; pores 0.012, bars 0.003; diameter of the
spherical medullary shells 0.06 and 0.03; length of the major polar spine 0.09, of the minor 0.04,
basal thickness 0.02.
Habitat.—Fossil in the Tertiary rocks of Barbados.
{333}
4. Xiphatractus sulcatus, Haeckel.
Stylosphæra sulcata, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 84, Taf. xxiv, fig. 6.
Cortical shell thick walled, with thorny surface, and regular, circular pores, quite as broad
as the bars; nine to ten on the half equator. Polar spines cylindrical, with longitudinal basal
furrows and conical apex, three times as broad as the pores; the larger about as long as the main
axis, the shorter only one-half or two-thirds as long.
Dimensions.—Major axis 0.01, minor 0.08; pores and bars 0.007; main axes of the
two medullary shells 0.08 and 0.04; length of the major polar spine 0.1, of the minor 0.06,
breadth of them 0.02.
Habitat.—Fossil in the Barbados deposits.
5. Xiphatractus dasypus, n. sp.
Cortical shell thick walled, with spiny surface, and regular, elegant network; pores circular,
hexagonally framed, about as broad as the bars; from every corner of the network (between three
pores) arises a short bristle-like spine. Polar spines three-sided prismatic, as broad as one
hexagon; the major once to twice as long as the main axis, the minor only one-half to one-fourth
as long. (The cortical shell much resembles Ellipsoxiphus elegans, Pl. 14, fig.
7.)
Dimensions.—Major axis 0.15, minor 0.13; pores and bars 0.01; main axes of the two
medullary shells 0.1 and 0.06; length of the major polar spine 0.1 to 0.3, of the minor 0.05 to
0.1, thickness 0.01.
Habitat.—Tropical Western Pacific, Station 222, surface.
Subgenus 3. Xiphatractona, Haeckel.
Definition.—Network of the cortical shell irregular, with meshes of
different size or form; surface smooth, without thorns or papillæ.
6. Xiphatractus chlamydophorus, n. sp.
Cortical shell thin walled, smooth, with irregular, roundish pores, twice to four times as
broad as the bars; eight to twelve on the half equator. Polar spines angular, irregularly curved
or contorted; the larger once to twice as long as the main axis, the shorter and thicker
pommel-shaped, only one-third to one-half as long; their basal breadth equals two pores.
Dimensions.—Major axis 0.14, minor 0.1; pores 0.006 to 0.012, bars 0.003; main
axis of the two ellipsoidal medullary shells 0.09 and 0.04; length of the major polar spine 0.1 to
0.2, of the minor 0.05 to 0.08, breadth 0.02.
Habitat.—Cosmopolitan; Atlantic, Pacific, many Stations, surface.
{334}
Subgenus 4. Xiphatractium, Haeckel.
Definition.—Network of the cortical shell irregular, with meshes of
unequal size or dissimilar form; surface thorny or papillose.
7. Xiphatractus radiosus, Haeckel.
Stylosphæra radiosa, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 84, Taf. xxiv. fig. 5.
Cortical shell very thick walled, with thorny surface, and irregular, roundish pores, once to
three times as broad as the bars; nine and ten on the half equator. Polar spines conical, on the
base about as broad as the largest pores; the larger as long as the main axis, the smaller
scarcely one-fourth as long. (The radial striation, figured by Ehrenberg and applied to the name
of this species, is produced by the contours of the funnel-shaped pores in the thick walls seen in
optical section; the ellipsoidal cortical shell is double.)
Dimensions.—Major axis 0.13, minor 0.09; pores 0.003 to 0.01, bars 0.003; main
axes of the two medullary shells 0.05 and 0.03; length of the major polar spine 0.12, of the minor
0.03, basal breadth 0.01.
Habitat.—Fossil in the Tertiary rocks of Barbados.
8. Xiphatractus glyptodon, n. sp. (Pl. 17, figs. 9,
10).
Cortical shell thick walled, covered with radial spines of the size of the pores; on the half
equator eight to ten irregular or subregular roundish pores, twice to four times as broad as the
bars. In the bottom of each pore a thin lamella of silex, perforated by four to six smaller
roundish pores. Both medullary shells (fig. 10) spherical, with smaller, regular, circular pores.
Larger polar spine three-sided pyramidal, in length equals the main axis; shorter spine only half
as long, but twice as thick, of elegant pommel-form (fig. 9), with nine prominent edges.
Dimensions.—Major axis 0.12, minor, 0.1; pores and spines 0.01 to 0.02, bars and
porules 0.005; diameter of the two medullary shells 0.05 and 0.02; length of the major polar spine
0.1, of the minor 0.06, greatest thickness 0.04.
Habitat.—Pacific, central area, Station 263, surface.
Genus 141. Cromyatractus,[180] n.
gen.
Definition.—Druppulida with four or more concentric
shells (two medullary shells and two or more cortical shells), in the main axis with two large
opposite polar spines of equal size and similar form.
The genus Cromyatractus is very nearly allied to all the foregoing genera,
which bear two equal spines opposite to one another on the poles of the main axis; it differs
{335}from them in the multiplication of the cortical
shell, and may be considered as the most highly developed form of this amphistylous series, which
proceeds from Ellipsoxiphus to Lithatractus and Stylatractus.
Subgenus 1. Cromyatractium, Haeckel.
Definition.—Shell composed of two medullary shells and two cortical
shells.
1. Cromyatractus tetraphractus, n. sp. (Pl. 15, fig.
2).
Stylocromium tetraphractum, Haeckel, 1879, Atlas (pl. xv. fig. 2).
Proportion of the main axes of the four concentric shells about =
1 : 3 : 5 : 7. Network of all four shells nearly of the same
structure, subregular, with circular pores of equal size (in one and the same shell). The absolute
size of the pores increases gradually from the innermost to the outermost shell. The bars between
the pores are smaller and quite smooth. Surface of the outermost shell smooth. Polar spines
cylindrical, with conical apex, of variable length, nearly as broad as the innermost shell.
Dimensions.—Main axes of the four shells—(A) inner medullary shell 0.03, (B)
outer medullary shell 0.08, (C) inner cortical shell 0.15, (D) outer cortical shell 0.2;
equatorial axes of them—(A) 0.025, (B) 0.06, (C) 0.12, (D) 0.15; pores of (A) 0.004, (B)
0.007, (C) 0.01, (D) 0.013, bars 0.002 to 0.004; length of the polar spines 0.1 to 0.3 (and more),
breadth 0.015.
Habitat.—Antarctic (Indian) Ocean, Station 157, depth 1950
fathoms.
2. Cromyatractus tetralepas, n. sp.
Proportion of the main axes of the four concentric shells about =
1 : 2 : 4 : 6. Network of the two medullary shells regular, with
small circular pores, little larger than the bars. Network of the two cortical shells irregular,
with much larger polygonal pores, three to nine times as broad as the thin bars. (Somewhat similar
to Cromyatractus ceparius, Pl. 15,
fig. 4.) Surface of the outermost shell smooth or a little thorny. Polar spines cylindrical, very
large, twice to three times as long as the main axis of the outermost shell, about as broad as the
innermost shell, with conical apex; smooth or a little thorny.
Dimensions.—Main axes of the four shells—(A) 0.03, (B) 0.05, (C) 0.13, (D)
0.18; equatorial axes of them—(A) 0.2, (B) 0.45, (C) 0.11, (D) 0.14; pores of the two
medullary shells 0.004 and 0.006, of both cortical shells 0.01 to 0.03, bars 0.003.
Habitat.—Indian Ocean, Zanzibar, Pullen, 2200 fathoms.
3. Cromyatractus tetracelyphus, n. sp. (Pl. 15, figs. 1,
1a).
Proportion of the main axes of the four concentric shells about =
1 : 3 : 10 : 11. Network of the two spherical medullary shells (fig.
1a) regular, with small circular pores, twice as broad as {336}the bars. Network of the inner cortical shell regular with circular,
hexagonally-framed pores, twice as broad as the bars. From every corner of the hexagons (between
three pores) arises a very thin, hair-like, short radial spinule. These spinules communicate with
one another by tangential branches (at equal distances from the inner cortical shell), and form
thereby an outer, delicate cortical network, with large polygonal meshes and very thin bars. Polar
spines very strong, pyramidal, with prominent edges, as long as the main radius of the cortical
shell, and half as broad at the base.
Dimensions.—Main axes of the four shells—(A) 0.02, (B) 0.06, (C) 0.2, (D)
0.22; equatorial axes of them—(A) 0.02, (B) 0.06, (C) 0.13, (D) 0.15; pores of the four
shells—(A) 0.002, (B) 0.004, (C) 0.008, (D) 0.02; bars 0.001 to 0.003; length of the polar
spines 0.1, basal breadth 0.05.
Habitat.—South Atlantic, Station 335, depth 1425 fathoms.
Subgenus 2. Caryatractus, Haeckel.
Definition.—Shell composed of two medullary shells and three or more
cortical shells.
4. Cromyatractus cepicius, n. sp. (Pl. 15, fig.
3).
Caryostylus cepicius, Haeckel, 1879, Atlas (pl. xv. fig. 3).
Caryodoras cepicius, Haeckel, 1881, Prodromus, p. 454.
Proportion of the main axes of the five concentric shells about =
1 : 2 : 5 : 7 : 8. Network of both spherical medullary
shells regular, with small circular pores, about as broad as the bars. Network of the three
ellipsoidal cortical shells irregular, with large polygonal meshes, five to ten times as broad as
the thin bars. Surface of the outermost shell smooth. Polar spines cylindrical, thorny, about as
thick as the innermost shell and as long as the outermost shell, against the apex with a
spindle-like intumescence, formed by delicate spongy framework (fig. 3).
Dimensions.—Main axes of the five concentric shells—(A) 0.03, (B) 0.05, (C)
0.16, (D) 0.2, (E) 0.25; equatorial axes of them—(A) 0.03, (B) 0.05, (C) 0.12, (D) 0.16, (E)
0.2; pores of the two medullary shells 0.002 to 0.004, of the three cortical shells 0.02 to 0.04,
bars 0.002; length of the polar spines 0.3, breadth 0.03.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
5. Cromyatractus ceparius, n. sp. (Pl. 15, fig.
4).
Caryostylus ceparius, Haeckel, 1881, Prodromus et Atlas (pl. xv. fig.
4).
Proportion of the main axes of the six concentric shells about =
1 : 2 : 7 : 10 : 13 : 18. Network of the three
inner shells regular, with small, circular pores, which are about as broad as the bars and in the
third shell hexagonally framed. Network of the three outer shells irregular, with large polygonal
meshes, six to twelve times as broad as the bars. Surface smooth. Polar spines strong,
spindle-shaped, thorny, outside of the sixth shell about as long as inside of it; broader than
{337}the innermost shell; the thorns of their outer free
part (arising at equal distances) represent perhaps the beginnings of three to four further
shells.
Dimensions.—Main axes of the six concentric shells—(A) 0.02, (B) 0.05, (C)
0.14, (D) 0.2, (E) 0.26, (F) 0.37; equatorial axes—(A) 0.02, (B) 0.05, (C) 0.09, (D) 0.15,
(E) 0.21, (F) 0.3; pores of the three inner shells 0.003 to 0.006, of the three outer shells 0.02
to 0.04, bars 0.003 to 0.005; length of the polar spines, from the base of the innermost shell to
the apex, 0.35, greatest breadth 0.03.
Habitat.—South Pacific, Station 285, depth 2375 fathoms.
Genus 142. Pipetta,[181] n. gen.
Definition.—Druppulida with simple ellipsoidal cortical
shell and simple medullary shell, with two hollow fenestrated tubes opposite on both poles of the
main axis.
The genus Pipetta differs from Pipettella (p. 304) in the possession of an inner (medullary) shell; it exhibits the same
tubular prolongations of the cortical shell at both poles of the main axis.
1. Pipetta fusus, n. sp. (Pl. 39,
figs. 8, 8a).
Cortical shell spindle-shaped, the middle ellipsoidal part gently passing over on both poles
into the conical tubes, which attain about the same length. Network regular, with circular,
hexagonally-framed pores, twice as broad as the bars, sixteen to eighteen on the half equator. In
the middle part of the shell the pores arise from their hexagonal bases in the form of short
conical funnels; on both tubes they are much smaller and simple, without frame. Medullary shell
(fig. 8a) spherical, scarcely one-third as broad as the cortical, with regular, circular
pores, twice as broad as the bars, eight to nine on the half equator. The two shells are connected
only by two opposite beams, lying in the equatorial axis and ramified at the distal insertion
(fig. 8a).
Dimensions.—Middle ellipsoidal part of the cortical shell 0.13 to 0.15; polar
tubes 0.15 long, 0.06 broad at the base; pores of the former 0.01, bars 0.005; pores of the tubes
0.06, bars 0.003. Medullary shell 0.04, pores 0.004, bars 0.002.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
2. Pipetta tuba, n. sp. (Pl. 39,
fig. 7).
Cortical shell nearly spherical in the middle part, which is sharply separated at both poles
from the long, nearly cylindrical tubes; these are longer than the main axis, at the distal end
open (always broken off). Network regular, with circular, hexagonally-framed pores, of the same
breadth as the bars, eighteen to twenty on the half equator. The pores of the polar tubes are of
the same shape, only much smaller, and arranged in sixteen to eighteen longitudinal rows.
Medullary shell {338}spherical, about one-third as broad
as the cortical, with simple circular pores. The connection between the two shells is only made by
two simple opposite beams, lying in the equatorial axis.
Dimensions.—Middle spheroidal part of the cortical shell 0.14 to 0.16; polar tubes
0.15 to 0.2 or longer, 0.03 broad; pores and bars of the former 0.007, of the latter 0.003.
Medullary shell 0.04, pores 0.006, bars 0.003.
Habitat.—Western Tropical Pacific, Station 225, depth 4475
fathoms.
3. Pipetta salpinx, n. sp.
Cortical shell ellipsoidal, sharply separated from the cylindrical polar tubes, which reach
about the same length (or more). Network regular, with circular pores, three times as broad as the
bars, without hexagonal frames, fourteen to fifteen on the half equator. Pores of the polar tubes
smaller, arranged in eight to ten longitudinal rows. Medullary shell spherical, one-fourth as
broad as the cortical, connected with it by a circle of four to six radial beams, lying in the
equatorial plane. Differs from Pipettella prismatica (Pl. 39, fig. 6) in
the possession of a medullary shell and the absence of the tube edges.
Dimensions.—Main axis of the ellipsoid 0.14, equatorial axis 0.12, pores 0.01,
bars 0.003; length of the tubes 0.15 or more, breadth 0.03. Medullary shell 0.03, pores 0.002,
bars 0.001.
Habitat.—Pacific, central area, Stations 265 to 268, depth 2700 to
2900 fathoms.
4. Pipetta conus, n. sp.
Cortical shell spindle-shaped, the middle ellipsoidal part gently passing over at both poles
into the conical tubes, which attain only half its length. Network irregular, with roundish or
subcircular pores, twice to three times as broad as the bars, twelve to sixteen on the half
equator. Medullary shell spherical, one-fourth as broad as the cortical. (Differs from the
foregoing species in the short conical tubes and the irregular reticulation.)
Dimensions.—Middle part of the cortical shell 0.16 long, 0.13 broad; tubes 0.09
long, 0.05 on the base broad; pores 0.005 to 0.01, bars 0.003. Medullary shell 0.04.
Habitat.—North Pacific, Station 241, depth 2300 fathoms.
Genus 143. Pipettaria,[182] n. gen.
Definition.—Druppulida with simple ellipsoidal cortical
shell and double medullary shell, with two hollow fenestrated tubes opposite on both poles of the
main axis.
The genus Pipettaria differs from the foregoing Pipetta only in the
duplication of the medullary shell; as in this, the tubular prolongations of the main axis of the
cortical shell may be either conical (with closed apex) or cylindrical (with apical opening?).
{339}
1. Pipettaria fusaria, n. sp.
Cortical shell spindle-shaped, the middle ellipsoidal part gently passing over on both poles
into the conical tubes, which attain about half its length. Pores regular, circular, twice as
broad as the bars, sixteen to eighteen on the half equator; pores of the tubes smaller. Both
medullary shells spheroidal, compressed. (The appearance of the cortical shell resembles that of
Cannartiscus amphiconus, Pl. 39,
fig. 19, but without the equatorial constriction.)
Dimensions.—Main axis of the ellipsoid 0.15, equatorial axis 0.13; length of the
polar tubes 0.08, basal breadth 0.05; pores of the former 0.008, bars 0.004; diameter of the
medullary shells 0.04 and 0.02.
Habitat.—South Pacific, Station 300, depth 1375 fathoms.
2. Pipettaria tubaria, n. sp. (Pl. 39,
fig. 15).
Cannartidium tubarium, Haeckel, 1882, Atlas (pl. xxxix. fig. 15).
Cortical shell ellipsoidal, on both poles distinctly separated from the short conical tubes,
the length and breadth of which equal the outer medullary shell. In the equatorial plane arises a
circle of four to six short conical protuberances, similar to the polar tubes. Pores subregular,
circular, or roundish, scarcely broader than the bars, sixteen to twenty on the half equator. Both
medullary shells spheroidal, somewhat compressed in the direction of the two poles (as in fig.
18a).
Dimensions.—Main axis of the ellipsoid 0.12, equatorial axis 0.09; pores 0.005,
bars 0.004; size of the equatorial protuberances and of the polar tubes 0.02.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms.
Family XIII. Spongurida, Haeckel (Pl. 48, figs. 6,
7).
Spongurida, Haeckel, 1862, Monogr. d. Radiol., p. 447 (sensu
emendato).
Definition.—Prunoidea with spongy ellipsoidal or
cylindrical shell, composed wholly or partially of a spongy framework, without equatorial
stricture, with or without an enclosed medullary shell.
The family Spongurida comprises, in the sense here restricted, all
those Prunoidea in which the ellipsoidal or cylindrical shell is composed
wholly or partially of an irregular siliceous framework, not of simple lattice-work. It contains
two subfamilies, differing in the absence or presence of a latticed medullary shell in the middle
of the central capsule; in the Spongellipsida it is absent, in the Spongodruppida present; the
former are most nearly related to the Ellipsida, the latter to the Druppulida, the difference
consisting only in the spongy structure of the cortical shell.
In my Monograph (1862, p. 447) the family Spongurida had a much wider extent, comprising also a
number of Sphæroidea and Discoidea, agreeing in
the spongy {340}structure of the shell. Here we restrict
the definition to those spongy Sphærellaria in which the central capsule
and the enclosing spongy cortical shell are ellipsoidal or cylindrical, therefore each transverse
section is a circle, and each meridional section an ellipse, as in all Prunoidea.
The Cortical Shell in all Spongurida is composed of a delicate framework of irregularly
branched and interwoven siliceous threads; commonly this spongy structure is rather dense or
compact, but sometimes also very loose. In the simplest form, Spongellipsis, the spongy
cortical shell contains a large cavity, in which lies freely the central capsule. In
Spongurus this cavity is completely distended by a spongy framework. In this case the solid
spongy shell becomes often prolonged, and its original ellipsoidal form passes over into a
cylindrical one (as in many Zygartida). Sometimes (particularly in Spongocore, Pl. 48, fig. 6) the
cylinder becomes three-jointed by two more or less distinct annular constrictions.
Spongocore is distinguished by an outer veil, a thin lattice-lamella, which envelops the
spongy shell and is connected with it by radial beams.
The Medullary Shell, absent in the Spongellipsida, is constant in the second subfamily
Spongodruppida. It is either a simple latticed shell (Spongodruppa) or double, composed of
two concentric latticed shells (Spongoliva); its form is either spherical or ellipsoidal.
It lies in the middle of the central capsule, and is connected by radial beams (perforating the
membrane of the latter) with the enveloping spongy cortical shell.
In many Spongurida the surface of the shell is armed with radial spines, and in some genera
(Spongoprunum, Spongatractus, &c., Pl. 17, fig. 12),
on both poles of the main axis, are developed two strong opposite polar spines, as in many other
Prunoidea.
The Central Capsule of the Spongurida is either ellipsoidal or cylindrical. Only in
Spongellipsis it lies freely in the internal cavity of the spongy shell. In all other
genera it is perforated by a part of the skeleton; in the Spongodruppida it contains the simple or
double medullary shell, and the radial beams which perforate its membrane and connect the latter
with the external spongy cortical shell. In Spongurus and the allied genera
(Spongocore, Spongoprunum) the whole central capsule is filled with a spongy
framework which also envelops its surface.
{341}
Synopsis of the Genera of Spongurida.
| I. Subfamily Spongellipsida. (Ellipsoidal shell
composed only of a spongy framework without a latticed medullary shell in the centre. |
|
Spongy shell with internal cavity. |
|
No polar spines, |
144. Spongellipsis. |
| Spongy shell solid, without internal cavity. |
|
Without lattice mantle. No polar spines, |
145. Spongurus. |
| With lattice mantle. No polar spines, |
146. Spongocore. |
| Without lattice mantle. Two opposite spines on the poles of the
axis, |
147. Spongoprunum |
| II. Subfamily Spongodruppida. (Ellipsoidal shell
composed of an outer spongy cortical shell and an inner latticed medullary shell.) |
|
Medullary shell simple. |
|
No polar spines, |
148. Spongodruppa. |
| Two opposite spines on the poles of the axis, |
149. Spongatractus. |
| Medullary shell double. |
|
No polar spines, |
150. Spongoliva. |
| Two opposite spines on the poles of the axis, |
151. Spongoxiphus. |
| I. Subfamily Spongellipsida. (Ellipsoidal shell composed only of a spongy
framework without a latticed medullary shell in the centre. |
|
|
Spongy shell with internal cavity. |
|
|
|
|
|
|
No polar spines, |
|
|
|
|
|
|
|
|
|
|
144. Spongellipsis. |
|
|
|
|
|
|
|
|
Spongy shell solid, without internal cavity. |
|
|
|
|
|
|
Without lattice mantle. No polar spines, |
|
|
|
|
|
|
|
|
|
|
145. Spongurus. |
|
|
|
|
|
|
|
|
|
|
With lattice mantle. No polar spines, |
|
|
|
|
|
|
|
|
|
|
146. Spongocore. |
|
|
|
|
|
|
|
|
|
|
Without lattice mantle. Two opposite spines on the poles of the
axis, |
|
|
|
|
|
|
|
|
|
|
147. Spongoprunum |
|
|
|
|
|
|
| II. Subfamily Spongodruppida. (Ellipsoidal shell composed of an outer spongy
cortical shell and an inner latticed medullary shell.) |
|
|
Medullary shell simple. |
|
|
|
|
|
|
No polar spines, |
|
|
|
|
|
|
|
|
|
|
148. Spongodruppa. |
|
|
|
|
|
|
|
|
|
|
Two opposite spine on the poles of the axis, |
|
|
|
|
|
|
|
|
|
|
149. Spongatractus. |
|
|
|
|
|
|
|
|
Medullary shell double. |
|
|
|
|
|
|
No polar spines, |
|
|
|
|
|
|
|
|
|
|
150. Spongoliva. |
|
|
|
|
|
|
|
|
|
|
Two opposite spine on the poles of the axis, |
|
|
|
|
|
|
|
|
|
|
151. Spongoxiphus. |
|
|
|
|
|
|
Subfamily 1. Spongellipsida, Haeckel.
Definition.—Spongurida with a spongy
ellipsoidal or cylindrical shell, without an internal latticed medullary shell.
Genus 144. Spongellipsis,[183] n.
gen.
Definition.—Spongurida with an ellipsoidal or cylindrical
spongy shell, containing an internal cavity, without a latticed medullary shell. Polar spines
absent.
The genus Spongellipsis embraces those very simple Spongurida in which the
ellipsoidal central capsule is enclosed in a spongy cortical shell of the same form. It
corresponds, therefore, to Plegmosphæra among the Sphæroidea, to
Plegmodiscus among the Discoidea, and to Spongolarcus among
the Larcoidea. In some species the ellipsoidal form is prolonged and
passes into a cylindrical one.
Subgenus 1. Spongellipsarium, Haeckel.
Definition.—Surface of the shell smooth or rough, without radial
spines.
{342}
1. Spongellipsis lævis, n. sp.
Shell ellipsoidal, with smooth surface, its spongy wall scarcely one-tenth as thick as the
minor axis of the inner cavity. Spongy framework very compact, with small meshes, three to six
times as broad as the bars. Proportion of the major axis to the minor = 3 : 2.
Dimensions.—Major axis (or length) of the shell 0.36, minor axis (or breadth)
0.24.
Habitat.—Pacific, central area, Station 265, surface.
2. Spongellipsis aspera, n. sp.
Shell ellipsoidal, with rough surface, its spongy wall nearly half as thick as the minor axis
of the inner cavity. Spongy framework very loose, with large meshes, ten to twenty times as broad
as the bars. Proportion of the major axis to the minor = 4 : 3.
Dimensions.—Length of the shell 0.24, breadth 0.18.
Habitat.—North Pacific, Station 253, surface.
3. Spongellipsis aplysina, n. sp.
Shell nearly cylindrical, with rough surface, its spongy wall about one-fourth as thick as the
minor axis of the internal cavity. Spongy framework loose, with large meshes, four to eight times
as broad as the bars. Proportion of both axes = 6 : 1. (Similar to a spongy cylinder of
Aplysina.)
Dimensions.—Length of the shell 0.3, breadth 0.05.
Habitat.—North Atlantic, Station 64, surface.
Subgenus 2. Spongellipsidium, Haeckel.
Definition.—Surface of the shell covered with radial spines.
4. Spongellipsis setosa, n. sp.
Shell ellipsoidal, covered with numerous (sixty to eighty) thin, bristle-shaped, radial spines,
about half as long as the major axis. Spongy framework loose, with large meshes, ten to twenty
times as broad as the bars. Minor axis of the inner cavity twice as long as the thickness of the
spongy wall. Proportion of both axes = 5 : 3.
Dimensions.—Length of the shell 0.2, breadth 0.12.
Habitat.—South Atlantic, Station 325, surface.
5. Spongellipsis spinosa, n. sp.
Shell nearly cylindrical, covered with numerous thorns and thirty to forty larger conical
radial spines, somewhat longer than the major axis. Spongy framework compact, with small meshes,
four {343}to eight times as broad as the bars. Minor
axis of the inner cavity about six times as large as the thickness of the spongy wall. Proportion
of both axes = 4 : 1.
Dimensions.—Length of the shell 0.24, breadth 0.06.
Habitat.—South Pacific, Station 300, surface.
Genus 145. Spongurus,[184] Haeckel, 1862,
Monogr. d. Radiol., p. 465.
Definition.—Spongurida with ellipsoidal or cylindrical
(sometimes three-jointed) shell, of solid spongy framework, without internal cavity and without
latticed medullary shell. Polar spines and outer lattice-mantle absent.
The genus Spongurus was founded by me in 1862 for the common cosmopolitan
Spongurus cylindricus, a massive spongy cylinder with radial spines. I enlarge here the
conception of the genus, in receiving also ellipsoidal, massive spongy Prunoidea, with or without radial spines. Sometimes the cylindrical shell is
more or less distinctly three-jointed, with two annular strictures, as also in the following and
nearly allied genus.
Subgenus 1. Spongurantha, Haeckel.
Definition.—Spongy framework everywhere of the same structure; surface
smooth or rough, but without radial spines.
1. Spongurus stuparius, n. sp.
Shell ellipsoidal, one and a half times as long as broad, with nearly smooth surface. Spongy
framework everywhere of equal structure, with small meshes, four to six times as broad as the
bars.
Dimensions.—Length of the shell (or major axis) 0.2, breadth of it (or minor axis)
0.14.
Habitat.—South Pacific, Station 295, depth 1500 fathoms.
2. Spongurus stypticus, n. sp.
Shell ellipsoidal, twice as long as broad, with thorny surface. Spongy framework everywhere of
equal structure, with large meshes, ten to twenty times as broad as the bars.
Dimensions.—Length of the shell 0.3, breadth 0.15.
Habitat.—Pacific, central area, Station 274, depth 2750 fathoms.
3. Spongurus phalanga, Haeckel.
Spongurus cylindricus, Stöhr, 1880, Palæontogr., vol. xxvi. p. 119, Taf.
vii. fig. 3.
Shell nearly cylindrical, five times as long as broad, with thorny surface. Spongy framework
compact, everywhere of equal structure, with small meshes, scarcely broader than the bars.
{344}
Dimensions.—Length of the shell 0.24, breadth 0.05.
Habitat.—Pacific, central area, Station 268, depth 2600 fathoms;
fossil in the Tertiary rocks of Sicily, Stöhr.
Subgenus 2. Spongurella, Haeckel.
Definition.—Spongy framework everywhere of the same structure; surface
armed with radial spines.
4. Spongurus asper, Haeckel.
Haliomma asperum, Joh. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 40, Taf.
ii. fig. 2.
Haliomma asperum, Haeckel, 1862, Monogr. d. Radiol., p. 431.
Shell ellipsoidal, one and a third times as long as broad, with thorny surface and twenty
symmetrically disposed, thin, bristle-shaped, radial spines, about as long as the shell. Spongy
framework everywhere of the same structure, very compact, with small meshes, three to four times
as broad as the bars.
Dimensions.—Length of the shell 0.11, breadth 0.08.
Habitat.—Mediterranean, Ligurian coast (J. Müller), Portofino
(Haeckel).
5. Spongurus cylindricus, Haeckel.
Spongurus cylindricus, Haeckel, 1862, Monogr. d. Radiol., p. 465, Taf.
xxvii. fig. 1.
Shell cylindrical, four to five times as long as broad, with nearly smooth surface, and twenty
to thirty thin, bristle-shaped, radial spines, about half as long as the shell. Spongy framework
everywhere of the same structure, very compact, with small meshes, scarcely broader than the
bars.
Dimensions.—Length of the shell 0.2, breadth 0.04 to 0.05.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, and Pacific,
surface.
6. Spongurus tricolus, n. sp.
Shell nearly cylindrical, with two slight annular transverse strictures; its middle part is
somewhat broader. Surface thorny, with numerous (forty to sixty or more) short, thin, radial
spines, not longer than the breadth of the shell. Spongy framework everywhere of the same
structure, compact, with small meshes, twice to three times as broad as the bars.
Dimensions.—Length of the shell 0.24, breadth 0.04 to 0.06.
Habitat.—Pacific, central area, Station 272, 2600 fathoms.
Subgenus 3. Sponguroma, Haeckel.
Definition.—Spongy framework in the inner part of the shell very
compact, in the outer part very loose; surface armed with radial spines.
{345}
7. Spongurus radians, n. sp.
Shell ellipsoidal, one and a half times as long as broad, with thorny surface, and eighty to
one hundred and twenty (or more) thin, bristle-shaped, radial spines, somewhat longer than the
shell. Spongy framework in the inner part very compact, in the outer part very loose, with a
gradual transition between the two parts. Meshes in the central part not broader than the bars, in
the superficial part ten to twenty times as broad.
Dimensions.—Length of the shell 0.24, breadth 0.16.
Habitat.—Mediterranean, Portofino, near Genoa, Haeckel, surface.
8. Spongurus spongechinus, n. sp.
Shell ellipsoidal, one and a third times as long as broad, with thorny surface, and thirty to
forty thick, conical radial spines, about half as long as the shell. Spongy framework in the inner
part of the shell compact, in the outer loose, with a gradual transition between the two parts.
Meshes in the central part smaller than the bars, in the superficial part four to eight times as
broad.
Dimensions.—Length of the shell 0.18, breadth 0.14.
Habitat.—North Atlantic, Station 353, surface.
Genus 146. Spongocore,[185] n. gen.
Definition.—Spongurida with ellipsoidal or cylindrical
(sometimes three-jointed) shell of solid spongy framework, without internal cavity and without
latticed medullary shell. Polar spines absent. An outer lattice-mantle is connected with the
spongy shell by radial beams.
The genus Spongocore contains some widely distributed Spongurida, which are
distinguished from the nearly related Spongurus by the development of a peculiar veil or
mantle of delicate lattice-work, which envelops either the whole shell or only the middle part of
it, and is connected with it by numerous radial beams. The distance of the simple fine
lattice-lamella from the spongy shell is everywhere the same. Often the cylindrical shell is
three-jointed, with two annular strictures, as also in the foregoing Spongurus. It is
derived from the latter by development of the veil connecting the points of the radial spines.
Subgenus 1. Spongocorina, Haeckel.
Definition.—Shell without distinct annular strictures, not evidently
three-jointed.
{346}
1. Spongocore velata, n. sp.
Shell ellipsoidal, one and a half times as long as broad, without annular strictures. Spongy
framework loose, with rather large meshes, four to six times as broad as the bars. The whole
spongy shell enveloped by a delicate veil with smooth surface, connected with it by numerous thin
radial beams. Breadth of the spongy ellipsoid (or minor axis) six times as large as its distance
from the veil.
Dimensions.—Length of the whole shell (with veil) 0.3, breadth 0.2; distance of
the veil from the spongy ellipsoid 0.035.
Habitat.—South Atlantic, Station 325, surface.
2. Spongocore diplocylindrica, n. sp.
Shell cylindrical, three times as long as broad, without annular strictures. Spongy framework
compact, with small meshes, twice to three times as broad as the bars. The whole surface of the
spongy cylinder, with exception of both rounded polar faces, enveloped by a delicate cylindrical
veil with smooth surface, connected with it by numerous radial beams. Diameter of the spongy
cylinder twice as large as its distance from the veil.
Dimensions.—Length of the shell 0.3, breadth (with veil) 0.1; distance of the veil
0.025.
Habitat.—South Pacific, Station 302, surface.
Subgenus 2. Spongocorisca, Haeckel.
Definition.—Shell distinctly three-jointed, with two transverse
annular strictures.
3. Spongocore chrysalis, n. sp.
Spongy shell cylindrical, without the veil six times as long as broad, three-jointed, with two
annular strictures; all three joints of the same length. The whole spongy shell enveloped by a
thin veil with thorny surface, connected with it by numerous radial beams. Breadth of the spongy
cylinder three times as large as its distance from the veil.
Dimensions.—Length of the shell (with veil) 0.4, breadth 0.1; distance of the veil
from the spongy shell 0.02.
Habitat.—South Atlantic (off Patagonia), Station 318, surface.
4. Spongocore cincta, n. sp.
Spongy shell cylindrical, without the veil four times as long as broad, three-jointed, with two
annular strictures; all three joints of the same length. Only the middle joint enveloped by a thin
veil with smooth surface, connected with it by radial beams. Both terminal joints with long,
{347}bristle-shaped, radial spines. Breadth of the
spongy cylinder twice as large as its distance from the veil.
Dimensions.—Length of the shell 0.24, breadth (with veil) 0.12; distance of the
veil 0.03.
Habitat.—Indian Ocean, Madagascar, Rabbe, surface.
5. Spongocore pupula, n. sp.
Spongy shell cylindrical, without the veil six times as long as broad, three-jointed, with two
annular strictures; the middle joint half as long as either terminal joint. Only the middle joint
enveloped by a thin veil with spiny surface, connected with it by twenty to thirty radial beams,
which are prolonged on the outside into short radial spines. Breadth of the spongy cylinder about
equal to its distance from the veil.
Dimensions.—Length of the shell 0.2, breadth (with veil) 0.1; distance of the veil
0.033.
Habitat.—North Pacific, Station 244, surface.
6. Spongocore puella, n. sp. (Pl. 48,
fig. 6).
Spongy shell cylindrical, without the veil five times as long as broad, three-jointed, with two
annular strictures; the middle joint twice as long as either terminal joint. Only the middle joint
enveloped by a thin veil with delicate lattice-work and smooth surface; both terminal joints armed
with numerous thin, bristle-shaped, radial spines. Breadth of the spongy cylinder twice as large
as the distance from the veil.
Dimensions.—Length of the shell 0.25 to 0.3, breadth (with veil) 0.1 to 0.12;
distance of the veil 0.028 to 0.032.
Habitat.—South Pacific, Stations 295 to 304, surface.
Genus 147. Spongoprunum,[186] n.
gen.
Definition.—Spongurida with ellipsoidal or cylindrical
shell of solid spongy framework, without internal cavity and without latticed medullary shell. On
the poles of the axis occur two opposite strong spines.
The genus Spongoprunum differs from the most nearly allied Spongurus
by the possession of two strong solid spines, lying in the axis of the ellipsoidal or cylindrical
massive spongy shell, on its two opposite poles. Spongoprunum bears therefore the same
relation to Spongurus that Ellipsoxiphus does to Cenellipsis.
1. Spongoprunum amphilonche, n. sp. (Pl. 48, fig.
7).
Shell ellipsoidal, two and a third times as long as broad, with nearly smooth surface. Spongy
framework very compact, in the whole solid shell of the same structure, with very small meshes,
{348}scarcely broader than the bars. Two polar spines
conical, very strong, about half as long as the main axis of the shell and three times as long as
broad.
Dimensions.—Length of the shell (without spines) 0.2, breadth 0.08.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms; fossil in
the rocks of Barbados.
2. Spongoprunum atractus, n. sp.
Shell spindle-shaped, twice as long as broad, with thorny surface. Spongy framework loose, in
the whole shell of the same structure, with large meshes, eight to twelve times as broad as the
bars. Two polar spines angular, pyramidal, very strong, about one-third as long as the shell, and
twice as long as broad.
Dimensions.—Length of the shell 0.2, breadth 0.11.
Habitat.—Western Tropical Pacific, Station 225, depth 4475
fathoms.
3. Spongoprunum amphicylindrus, n. sp.
Shell nearly cylindrical, four times as long as broad, with rough surface. Spongy framework
compact, in the whole shell of equal structure, with small meshes, twice to three times as broad
as the bars. Two polar spines cylindrical, very large, longer than the shell, and about one-fourth
as thick as its diameter.
Dimensions.—Length of the shell 0.2, breadth 0.05.
Habitat.—South Atlantic, Station 333, surface.
Subfamily Spongodruppida, Haeckel.
Definition.—Spongurida with latticed
medullary shell, enclosed by a spongy cortical shell.
Genus 148. Spongodruppa,[187] n.
gen.
Definition.—Spongurida with spongy ellipsoidal cortical
shell, enclosing a simple, spherical or ellipsoidal, latticed medullary shell. Polar spines
absent.
The genus Spongodruppa opens the series of Spongodruppida, or of those
Spongurida in which the ellipsoidal spongy cortical shell encloses a simple or double, latticed,
medullary shell. In Spongodruppa, the simplest and probably the ancestral form of this
subfamily, the medullary shell is simple and the polar spines absent. It may be derived from
Druppula by a spongy thickening of the simple latticed cortical shell.
{349}
Subgenus 1. Spongodruppula, Haeckel.
Definition.—Surface of the shell smooth or rough, without radial
spines.
1. Spongodruppa terebintha, n. sp.
Cortical shell one and a third times as long as broad, with smooth surface, three times as
broad as the spherical medullary shell. Spongy framework very compact, with small meshes of the
same breadth as the bars. Thickness of the spongy wall equal to the radius of the medullary
shell.
Dimensions.—Length of the cortical shell (or major axis of the ellipsoid) 0.16,
breadth (or minor axis) 0.12; thickness of its wall 0.02; medullary shell 0.04.
Habitat.—Western Tropical Pacific, Station 225, depth 4475
fathoms.
2. Spongodruppa pistacia, n. sp.
Cortical shell one and a half times as long as broad, with thorny surface, nearly five times as
broad as the spherical medullary shell. Spongy framework compact, its thickness equal to the
diameter of the medullary shell.
Dimensions.—Length of the cortical shell 0.2, breadth 0.14; thickness of its wall
0.03; medullary shell 0.03.
Habitat.—Fossil in the rocks of Barbados.
3. Spongodruppa lentisca, n. sp.
Cortical shell twice as long as broad, with rough surface, four times as large as the
ellipsoidal medullary shell. Spongy framework loose, with large meshes, its thickness equal to the
length of the medullary shell.
Dimensions.—Length of the cortical shell 0.25, breadth 0.13; thickness of its wall
0.06; length of the medullary shell 0.06, breadth 0.04.
Habitat.—Pacific, central area, Station 266, surface.
Subgenus 2. Spongodruppium, Haeckel.
Definition.—Surface of the shell armed with radial spines.
4. Spongodruppa frangula, n. sp.
Cortical shell one and a third times as long as broad, three times as broad as the spherical
medullary shell. Spongy framework loose, with large meshes, its thickness half as large as the
diameter of the medullary shell. Surface covered with numerous (forty to fifty) thin,
bristle-shaped, radial spines, about as long as the cortical shell.
{350}
Dimensions.—Length of the cortical shell 0.24, breadth 0.18; thickness of its wall
0.035; diameter of the medullary shell 0.06.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
5. Spongodruppa elliptica, Haeckel.
Acanthosphæra elliptica, Ehrenberg, 1872, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 285, Taf. vii. fig. 4.
Cortical shell nearly twice as long as broad, four times as broad as the ellipsoidal medullary
shell. Spongy framework compact, with small meshes. Surface covered with very numerous short
radial spines, about as long as the medullary shell.
Dimensions.—Length of the cortical shell 0.2, breadth 0.11; length of the
medullary shell 0.04, breadth 0.03.
Habitat.—Philippine Sea, Ehrenberg, Station 206, depth 2100
fathoms.
6. Spongodruppa polyacantha, Haeckel.
Haliomma polyacanthum, J. Müller, 1858, Abhandl., p. 36, Taf. i. figs. 10, 11.
Haliomma polyacanthum, Haeckel, 1862, Monogr. d. Radiol., p. 430.
Cortical shell one and a third times as long as broad, twice as broad as the ellipsoidal
medullary shell. Spongy framework loose on the surface, with large regular meshes. Surface covered
with fifteen to twenty thick, conical spines, about as long as the radius of the shell.
Dimensions.—Length of the cortical shell 0.12, of the medullary shell 0.09;
diameter of the medullary shell 0.05 to 0.06.
Habitat.—Mediterranean (south coast of France); Atlantic (Canary
Islands), Station 354, surface.
Genus 149. Spongatractus,[188] n.
gen.
Definition.—Spongurida with spongy ellipsoidal cortical
shell, enclosing a simple, spherical or ellipsoidal, latticed medullary shell. On the poles of the
axis occur two opposite strong spines.
The genus Spongatractus differs from Spongodruppa by development of
two strong spines in the axis of the shell, on its two opposite poles; therefore it bears the same
relation to the latter that Lithatractus does to Druppula, and can be derived from
Lithatractus by a spongy thickening of the cortical shell.
1. Spongatractus pachystylus, Haeckel.
Spongosphæra pachystyla, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 82, Taf. xxvi. fig. 3.
Cortical shell one and a half times as long as broad, with rough surface. Spongy framework
compact, with small meshes, about as thick as the spherical medullary shell. Polar spines very
{351}stout, conical, slightly sulcated, about as long as
the breadth of the cortical shell, as broad at the base as the medullary shell.
Dimensions.—Length of the cortical shell 0.2, breadth 0.13; thickness of the
spongy wall 0.045; diameter of the medullary shell 0.04.
Habitat.—Fossil in the rocks of Barbados, Ehrenberg; living in the
Equatorial Atlantic, Station 348, depth (2450) fathoms.
2. Spongatractus fusiformis, n. sp.
Cortical shell one and a third times as long as broad, with nearly smooth surface. Spongy
framework very compact, with very small meshes, about as thick as the radius of the spherical
medullary shell. Polar spines very strong, three-sided pyramidal, about as long as the breadth of
the cortical shell, as broad at the base as the medullary shell.
Dimensions.—Length of the cortical shell 0.16 to 0.18, breadth 0.12 to 0.14;
thickness of the spongy wall 0.02; diameter of the medullary shell 0.04.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
3. Spongatractus streptacanthus, n. sp.
Cortical shell twice as long as broad, with thorny surface. Spongy framework loose, with large
meshes, about as thick as the ellipsoidal medullary shell. Polar spines very long, three-sided
prismatic, with three spirally contorted edges, much longer than the cortical shell, half as broad
as the medullary shell.
Dimensions.—Length of the cortical shell 0.25, breadth 0.13; thickness of the
spongy wall 0.03; diameter of the medullary shell 0.03 to 0.04.
Habitat.—North Atlantic, off Canary Islands, Haeckel.
Genus 150. Spongoliva,[189] n. gen.
Definition.—Spongurida with spongy ellipsoidal cortical
shell, enclosing a double, spherical or ellipsoidal, latticed medullary shell. Polar spines
absent.
The genus Spongoliva differs from the nearest Spongodruppa by
duplication of the medullary shell; it bears therefore the same relation to this that
Prunulum does to Druppula, and can be regarded as a Prunulum, in which the
simple latticed cortical shell is replaced by a spongy framework.
Subgenus 1. Spongolivetta, Haeckel.
Definition.—Surface of the shell smooth or rough, without radial
spines.
{352}
1. Spongoliva cerasina, n. sp.
Cortical shell one and a fourth times as long as broad, with smooth surface, four times as
broad as the outer spherical medullary shell. Spongy framework very compact, with very small
meshes, its thickness equal to the diameter of the outer medullary shell.
Dimensions.—Length of the cortical shell 0.2, breadth 0.16; thickness of its wall
0.04; diameter of the outer medullary shell 0.04, of the inner 0.014.
Habitat.—Fossil in the rocks of Barbados.
2. Spongoliva prunulina, n. sp.
Cortical shell one and a third times as long as broad, with smooth surface, three times as
large as the outer ellipsoidal medullary shell. Spongy framework very compact, with small meshes,
its thickness equal to the diameter of the inner medullary shell.
Dimensions.—Length of the cortical shell 0.24, breadth 0.18; thickness of its wall
0.03; size of the outer medullary shell 0.08 to 0.06, of the inner 0.03.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms.
3. Spongoliva persicina, n. sp.
Cortical shell one and a half times as long as broad, with rough surface, five to six times as
large as the ellipsoidal outer medullary shell. Spongy framework loose, with large meshes, its
thickness about equal to the outer medullary shell.
Dimensions.—Length of the cortical shell 0.26, breadth 0.18; thickness of its wall
0.04; size of the outer medullary shell 0.045 to 0.035, of the inner 0.015.
Habitat.—Equatorial Atlantic, Station 348, depth (2450) fathoms.
4. Spongoliva amygdalina, n. sp.
Cortical shell twice as long as broad, with thorny surface, ten times as long as the small
spherical outer medullary shell. Spongy framework loose, with large meshes, its thickness one and
a half times as large as the outer medullary shell.
Dimensions.—Length of the cortical shell 0.3, breadth 0.16; thickness of its wall
0.045; size of the outer medullary shell 0.03, of the inner 0.015.
Habitat.—Fossil in the Tertiary rocks of Sicily (Caltanisetta).
Subgenus 2. Spongolivina, Haeckel.
Definition.—Surface of the shell armed with radial spines.
5. Spongoliva opuntina, n. sp.
Cortical shell one and a third times as long as broad, armed with numerous thin,
bristle-shaped, radial spines, nearly as long as the shell. Spongy framework loose, with large
meshes, its thickness nearly equal to the diameter of the outer spherical medullary shell.
{353}
Dimensions.—Length of the cortical shell 0.18, breadth 0.14; thickness of its wall
0.035; diameter of the outer medullary shell 0.04, of the inner 0.02.
Habitat.—South Atlantic, Station 325, surface.
6. Spongoliva daturina, n. sp.
Cortical shell one and a half times as long as broad, armed with numerous, thick and short,
conical radial spines, about as long as the outer medullary shell. Spongy framework compact, with
small meshes, its thickness equal to half the length of the outer ellipsoidal medullary shell.
Dimensions.—Length of the cortical shell 0.24, breadth 0.16; thickness of its wall
0.025; diameter of the outer medullary shell 0.045 to 0.035, of the inner 0.025.
Habitat.—Pacific, central area, Station 272, surface.
Genus 151. Spongoxiphus,[190] n.
gen.
Definition.—Spongurida with spongy ellipsoidal cortical
shell, enclosing a double, spherical or ellipsoidal, latticed medullary shell. On the poles of the
axis occur two opposite strong spines.
The genus Spongoxiphus differs from Spongoliva by the development of
two strong spines in the axis of the shell, on its two opposite poles. From the similar
Spongatractus it differs in the duplication of the medullary shell. It may also be regarded
as a Stylatractus, in which the simple latticed cortical shell is replaced by an irregular
spongy framework.
1. Spongoxiphus sphærococcus, n. sp.
Cortical shell one and a half times as long as broad, with thorny surface. Spongy framework
compact, with small meshes, its thickness equal to the radius of the outer medullary shell. Both
medullary shells spherical, the outer three times as broad as the inner. Polar spines very stout,
conical, slightly sulcated, about half as long as the cortical shell, as broad at the base as the
outer medullary shell. (Differs from Spongatractus pachystylus almost only in the double
medullary shell.)
Dimensions.—Length of the cortical shell 0.18, breadth 0.12; thickness of the
spongy wall 0.025; diameter of the outer medullary shell 0.05, of the inner 0.016.
Habitat.—Fossil in the rocks of Barbados (Haeckel); also living in the
Pacific, central area, Station 265, depth 2900 fathoms.
{354}
2. Spongoxiphus prunococcus, n. sp. (Pl. 17, figs. 12,
13).
Spongostylus prunococcus, Haeckel, 1881, Prodromus, p. 455, et Atlas, pl.
xvii. figs. 12, 13.
Cortical shell one and a third times as long as broad, with smooth surface. Spongy framework
very compact, with very small meshes, its thickness about equals the breadth of the outer
medullary shell. Both medullary shells ellipsoidal, the outer three times as large as the inner.
Polar spines very stout, three-sided pyramidal, about half as long as the cortical shell, on the
base about as broad as the outer medullary shell. (Differs from the preceding in the ellipsoidal
form of both medullary shells, and in the finer structure of the spongy framework.)
Dimensions.—Length of the cortical shell 0.18 to 0.2, breadth 0.14 to 0.16;
thickness of the spongy wall 0.035 to 0.04; length of the outer medullary shell 0.04 to 0.06,
breadth 0.03 to 0.04.
Habitat.—Pacific, central area, Stations 265 to 268, depths 2700 to
2900 fathoms.
Family XIV. Artiscida, Haeckel (Pl. 39, figs. 9,
10; Pl. 48,
fig. 5).
Artiscida, Haeckel, 1881, Prodromus, p. 462.
Definition.—Prunoidea with an ellipsoidal twin-shell
divided by an equatorial stricture into two communicating hemiellipsoidal or hemispherical
chambers, without enclosed medullary shell. Central capsule ellipsoidal, with or without
equatorial stricture.
The family Artiscida has a simple fenestrated outer shell, like that
of the Ellipsida, but differs from these in the presence of an equatorial constriction, by which
it assumes a characteristic twin form, somewhat similar to a violin (Pl. 39, figs. 9,
10). From the Cyphinida, which have the same form of the cortical shell, the Artiscida differ in
the absence of the medullary shell. It is possible that the Artiscida are descended from the
Cyphinida (by loss of the medullary shell), but it is more probable that they arise from the
Ellipsida by an annular constriction in the equatorial plane (perhaps the formation of the shell
originally took place while the central capsule was undergoing division). Both halves of the twin
shell are always of the same size and form. Its outer surface is either smooth or covered with
radial spines (Artiscus, Pl. 39,
fig. 9). Sometimes on the opposite poles of the main axis are developed solid spines
(Stylartus, Pl. 48,
fig. 5) or hollow fenestrated tubes (Cannartus, Pl. 39, fig.
10).
The Central Capsule is either simply ellipsoidal, or has also an equatorial
constriction, which divides it into two equal halves. It is constantly smaller than the
surrounding shell, and separated from its inner surface by a thicker or thinner jelly-mantle, the
calymma.
Synopsis of the Genera of Artiscida.
| On the poles of the main axis neither solid spines nor hollow tubes, |
152. Artiscus. |
| On the poles of the main axis two solid spines (or bunches of spines), |
153. Stylartus. |
| On the poles of the main axis two hollow fenestrated tubes, |
154. Cannartus. |
| On the poles of the main axis neither solid spines nor hollow tubes, |
|
|
152. Artiscus. |
|
|
| On the poles of the main axis two solid spines (or bunches of spines), |
|
|
153. Stylartus. |
|
|
| On the poles of the main axis two hollow fenestrated tubes, |
|
|
154. Cannartus. |
|
|
{355}
Genus 152. Artiscus,[191] Haeckel, 1881,
Prodromus, p. 462.
Definition.—Artiscida without peculiar polar appendages
(without strong solid spines or hollow fenestrated tubes on the poles of the main axis).
The genus Artiscus represents the most simple and primitive form of the
Artiscida, and may therefore be regarded as the ancestral form of this subfamily. It resembles an
ordinary double loaf, composed of two equal, nearly ellipsoidal fenestrated shells separated by
the stricture in the equatorial plane. Artiscus can be derived from Cenellipsis
simply by this equatorial constriction. But it can also be derived from Cyphanta by loss of
the medullary shell.
Subgenus 1. Artiscium, Haeckel.
Definition.—Surface of the shell smooth or rough, without radial rods
or spines.
1. Artiscus paniscus, n. sp.
Pores of the shell regular, hexagonal, four times as broad as the bars; ten to twelve pores on
the half meridian, eight to nine on the half equator. Surface smooth. (The network of this species
is quite the same as that of the proximal cameræ of Panartus diploconus, Pl. 40, fig.
1.)
Dimensions.—Main axis of the shell 0.12, equatorial axis 0.07; meshes 0.013, bars
0.003.
Habitat.—Pacific, central area, Station 271, depth 2425.
2. Artiscus facetus, n. sp.
Pores of the shell regular, circular, with prominent hexagonal frame, twice as broad as the
bars; twenty-two to twenty-four pores on the half meridian, fourteen to fifteen on the half
equator. Surface a little thorny. (The shell of this species exhibits nearly the same formation as
that of Cyphonium virgineum, Pl. 39,
fig. 12, but without enclosed medullary shells.)
Dimensions.—Main axis of the shell 0.15, equatorial axis 0.09; meshes 0.01, bars
0.005.
Habitat.—Pacific, central area, Station 266, depth 2750 fathoms.
3. Artiscus paniculus, n. sp.
Pores of the shell regular, circular, without hexagonal frame, as broad as the bars; fourteen
to sixteen pores on the half meridian, ten to eleven on the half equator. Surface quite
smooth.
Dimensions.—Main axis of the shell 0.13, equatorial axis 0.06; meshes 0.01, bars
0.01.
Habitat.—Fossil in the Tertiary rocks of Barbados (Haeckel).
{356}
4. Artiscus panarius, n. sp.
Pores of the shell irregular, roundish, twice to four times as broad as the bars; sixteen to
eighteen pores on the half meridian, twelve to thirteen on the half equator. Surface smooth.
Dimensions.—Main axis of the shell 0.11, equatorial axis 0.06; meshes 0.005 to
0.02, bars 0.001 to 0.004.
Habitat.—Western Tropical Pacific, Station 224, depth 1850
fathoms.
Subgenus 2. Artidium, Haeckel, 1881, Prodromus, p. 462.
Definition.—Surface of the shell covered with radial rods or
spines.
5. Artiscus elegans, n. sp.
Pores of the shell regular, circular, with hexagonal frames, twice as broad as the bars;
fourteen pores on the half meridian, eight to nine on the half equator. From every corner of the
hexagonal frames (between every three pores) starts a thin, three-sided pyramidal spine, twice as
large as a pore.
Dimensions.—Main axis 0.13, equatorial axis 0.08; meshes 0.007, bars 0.004; spines
0.015 long.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
6. Artiscus nodosus, n. sp. (Pl. 39,
fig. 9).
Pores of the shell subregular, circular, without hexagonal frame, three times as broad as the
bars; sixteen to eighteen on the half meridian, ten to twelve on the half equator. Irregularly
scattered on the whole surface a variable number (twenty-five to thirty in all) of stout short
radial spines or rather blunt rods; the length and thickness of these is the same, and equals the
size of two to three meshes; its form resembles a truncated six-sided pyramid.
Dimensions.—Main axis 0.11, equatorial axis 0.08; meshes 0.01, bars 0.003; length
and thickness of the radial sticks 0.02.
Habitat.—Pacific, central area, Station 272, depth 2600 fathoms.
7. Artiscus hystrix, n. sp.
Pores of the shell irregular, roundish, of very unequal size and dissimilar form, twice to
eight times as broad as the bars; ten to fifteen on the half meridian, seven to nine on the half
equator. Irregularly scattered on the whole surface a large number of thin conical spines, about
as large as the meshes, partly directed radially, partly obliquely. (The shell of this species
resembles very much that of Cyphonium ceratospyris (p. 366) =
Didymocyrtis ceratospyris, Monogr. d. Radiol., 1862, Taf. xxii. fig. 14, but is without the
enclosed inner shells.)
{357}
Dimensions.—Main axis 0.12, equatorial axis 0.07; meshes 0.004 to 0.02, bars 0.002
to 0.005; spines 0.01 to 0.02.
Habitat.—Atlantic, Eastern Tropical part, Station 346, surface.
Genus 153. Stylartus,[192] Haeckel, 1881,
Prodromus, p. 462.
Definition.—Artiscida with two strong, solid, polar
spines, or two bunches of polar spines, opposite on the two poles of the main axis.
The genus Stylartus differs from Artiscus by the production of two
large opposite spines in the main axis, starting from both poles of it; sometimes every spine is
surrounded by a group of smaller radial spines. The genus is nearly allied to Ellipsoxiphus
(p. 295), and differs from it only in the equatorial stricture of the
ellipsoidal shell. But it may also be derived from the similar Cyphinus (Pl. 39, fig. 14) by
loss of the medullary shell.
Subgenus 1. Stylartella, Haeckel.
Definition.—On each pole of the main axis only one single large
spine.
1. Stylartus bipolaris, n. sp. (Pl. 48,
fig. 5).
Shell thick walled, rough; both its chambers nearly spherical, with irregular, roundish pores,
twice to four times as broad as the bars; eight to ten on the half equator of each chamber. Polar
spines very strong, conical, as long as the greatest breadth.
Dimensions.—Length of the shell (without spines) 0.18, greatest breadth 0.13;
length of the polar spines 0.13, basal breadth 0.03.
Habitat.—Western Tropical Pacific, Station 224, depth 1850
fathoms.
2. Stylartus bicuspis, n. sp.
Shell thin walled, smooth, with regular, circular pores, twice as broad as the bars; twelve to
fourteen on the half equator of each chamber. Polar spines very stout, straight, three-sided
pyramidal, half as long as the shell.
Dimensions.—Length of the shell 0.16, greatest breadth 0.12; length of the polar
spines 0.08, basal thickness 0.025.
Habitat.—Pacific, central area. Station 266, depth 2750 fathoms.
Subgenus 2. Stylartura, Haeckel.
Definition.—On each pole of the main axis a bunch of several
spines.
{358}
3. Stylartus palatus, n. sp.
Shell thick walled, thorny, with irregular, roundish pores, twice to three times as broad as
the bars; fifteen to eighteen on the half equator of each chamber. On each pole of the main axis a
large conical spine, surrounded by a group of ten to fifteen smaller spines, one-third to
two-thirds as long as the breadth of the deep equatorial stricture.
Dimensions.—Length of the shell 0.14, greatest breadth 0.09; length of the polar
spines 0.03 to 0.06, basal breadth 0.005 to 0.015.
Habitat.—Indian Ocean, Madagascar, Rabbe, surface.
4. Stylartus penicillus, n. sp.
Shell thin walled, smooth, with irregular, roundish pores, three to four times as broad as the
bars; ten to twelve on the half equator of each chamber. On each pole of the main axis a
brush-like bunch of twenty to thirty thin conical radial spines, half as long as the shell.
Dimensions.—Length of the shell 0.15, greatest breadth 0.1; length of the polar
spines 0.08, basal breadth 0.005.
Habitat.—Equatorial Atlantic, Station 347, surface.
Genus 154. Cannartus,[193] Haeckel, 1881,
Prodromus, p. 462.
Definition.—Artiscida with two hollow polar tubes
(fenestrated cylindrical or conical tubuli, opposite on the two poles of the main axis).
The genus Cannartus differs from Artiscus by the production of two
opposite hollow tubes on both poles of the main axis. The cavity of the conical or cylindrical
fenestrated tubes communicates freely with that of the shell; the network in both is the same. The
distal end of the tubes is nearly always broken off, sometimes closed, with a conical apex.
Cannartus can be derived either from Pipettella by a transverse equatorial
constriction, or from Cannartiscus by the loss of the medullary shell, or from
Artiscus by the production of the polar tubes.
1. Cannartus violina, n. sp. (Pl. 39,
fig. 10).
Pores of the shell subregular, circular, twice to three times as broad as the bars; eighteen to
twenty on the half meridian, fourteen to sixteen on the half equator. Shell-wall in the "tropical
zone" of both halves thickened. Polar tubes nearly cylindrical, about as long as the main axis,
distal ends broken off.
Dimensions.—Main axis 0.14, equatorial axis 0.09; meshes 0.008, bars 0.003; length
of the polar tubes 0.12, breadth 0.02 to 0.03.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
{359}
2. Cannartus bitubulus, n. sp.
Pores of the shell regular, circular, hexagonally framed, of the same breadth as the bars;
fourteen to sixteen on the half meridian, ten to twelve on the half equator. Polar tubes
cylindrical, longer than the main axis, sulcated, distal ends broken off. (Similar to Pipetta
tuba, Pl. 39,
fig. 7, but with equatorial stricture and without medullary shell.)
Dimensions.—Main axis 0.16, equatorial axis 0.12; meshes and bars 0.006; length of
the polar tubes 0.2, breadth 0.025.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
3. Cannartus biscottus, n. sp.
Pores of the shell irregular, roundish, twice to three times as broad as the bars; ten to
twelve on the half meridian, six to eight on the half equator. Polar tubes conical, shorter than
the main axis, with closed apex. (Similar to Cannartidium bicinctum, Pl. 39, fig. 18,
but without enclosed medullary shell.)
Dimensions.—Main axis 0.11, equatorial axis 0.07; meshes 0.005 to 0.01, bars
0.003; length of the polar tubes 0.08, breadth on the base 0.03.
Habitat.—Equatorial Atlantic, Station 348, depth 2450 fathoms.
Family XV. Cyphinida, Haeckel (Pl. 39, figs.
11-19).
Cyphinida, Haeckel, 1881, Prodromus, p. 462.
Definition.—Prunoidea with ellipsoidal twin-shell,
divided by an equatorial stricture into two communicating hemiellipsoidal or hemispherical
chambers; this external twin-shell (cortical shell) is either simple or double, and encloses one
or more internal concentric shells (medullary shells). Central capsule ellipsoidal, commonly with
an equatorial constriction.
The family Cyphinida have the same characteristic twin-form of the
cortical shell as the Artiscida, but differ from them in the presence of a simple or double
internal medullary shell, connected with the cortical shell by radial beams. The fenestrated
shell is therefore composed of two or more concentric shells as in the Druppulida, but differs
from these in the ring-like equatorial constriction.
The Medullary Shell, in the middle of the central capsule, is either single or double,
composed of two concentric shells. As in the Druppulida, the form of the medullary shells is
either spherical (Pl. 39,
fig. 12a) or lenticular, compressed from both poles of the main axis (Pl. 39, fig.
18a); sometimes the inner medullary shell is spherical, the outer lenticular.
The Radial Beams, which connect the medullary shell with the equatorial constriction of
the cortical shell, lie either all in the equatorial plane (Pl. 39, fig. 11) or
near it on both sides (Pl. 39,
figs. 12a, 18a).
{360}
The Cortical Shell is commonly simple (Pl. 39, figs. 12,
18), sometimes composed of two concentric shells (Pl. 39, fig. 13),
rarely of three. As in the Artiscida, also in the Cyphinida, from both poles of the main axis
solid spines or hollow fenestrated tubes are often developed (Pl. 39, figs. 14,
16-18).
The Central Capsule of the Cyphinida (Pl. 39, fig. 13) is
of the same form as in the Artiscida, generally ellipsoidal, but with an equatorial ring-like
stricture, which divides it into two equal halves. It encloses the simple or double medullary
shell, and is perforated by the radial beams starting from this. From the inner surface of the
surrounding cortical shell it is separated by a thicker or thinner jelly-layer, the calymma.
(Compare also Taf. xxii. fig. 14 of my Monograph, 1862.)
Synopsis of the Genera of Cyphinida.
| Cortical shell without peculiar spines or hollow
fenestrated tubes on both poles of the main axis. |
|
Cortical twin-shell simple |
|
Medullary shell simple, |
155. Cyphanta. |
| Medullary shell double, |
156. Cyphonium. |
| Cortical twin-shell double or triple; medullary shell
double. |
|
Cortical shell double, |
157. Cypassis. |
| Cortical shell triple, |
158. Cyphocolpus. |
| Cortical shell simple, with peculiar spines or hollow
fenestrated tubes on both poles of the main axis. |
|
Two opposite polar spines (or bunches of spines). |
|
Medullary shell simple, |
159. Cyphinus. |
| Medullary shell double, |
160. Cyphinidium. |
| Two opposite hollow fenestrated polar tubes. |
|
Medullary shell simple, |
161. Cannartiscus. |
| Medullary shell double, |
162. Cannartidium. |
| Cortical shell without peculiar spines or hollow fenestrated tubes on both
poles of the main axis. |
|
|
Cortical twin-shell simple |
|
|
|
|
|
|
Medullary shell simple, |
|
|
|
|
|
|
|
|
|
|
155. Cyphanta. |
|
|
|
|
|
|
|
|
|
|
Medullary shell double, |
|
|
|
|
|
|
|
|
|
|
156. Cyphonium. |
|
|
|
|
|
|
|
|
Cortical twin-shell double or triple; medullary shell double. |
|
|
|
|
|
|
Cortical shell double, |
|
|
|
|
|
|
|
|
|
|
157. Cypassis. |
|
|
|
|
|
|
|
|
|
|
Cortical shell triple, |
|
|
|
|
|
|
|
|
|
|
158. Cyphocolpus. |
|
|
|
|
|
|
| Cortical shell simple, with peculiar spines or hollow fenestrated tubes on
both poles of the main axis. |
|
|
Two opposite polar spines (or bunches of spines). |
|
|
|
|
|
|
Medullary shell simple, |
|
|
|
|
|
|
|
|
|
|
159. Cyphinus. |
|
|
|
|
|
|
|
|
|
|
Medullary shell double, |
|
|
|
|
|
|
|
|
|
|
160. Cyphinidium. |
|
|
|
|
|
|
|
|
Two opposite hollow fenestrated polar tubes. |
|
|
|
|
|
|
Medullary shell simple, |
|
|
|
|
|
|
|
|
|
|
161. Cannartiscus. |
|
|
|
|
|
|
|
|
|
|
Medullary shell double, |
|
|
|
|
|
|
|
|
|
|
162. Cannartidium. |
|
|
|
|
|
|
Genus 155. Cyphanta,[194] n. gen.
Definition.—Cyphinida with simple cortical shell and
simple medullary shell, without polar spines or tubes.
The genus Cyphanta is the most simple of all Cyphinida, and can be regarded
as the common ancestral form of this family. It may be derived phylogenetically from
Druppula by a ring-like constriction in the equatorial plane of the ellipsoidal cortical
shell, or from Artiscus by secondary formation of a central (spherical or ellipsoidal)
medullary shell.
Subgenus 1. Cyphantella, Haeckel.
Definition.—Surface of the cortical shell smooth, without spines or
thorns.
{361}
1. Cyphanta colpodes, n. sp.
Cortical shell thin walled, with smooth surface; its pores regular, hexagonal, three to four
times as broad as the bars; ten to twelve on the half meridian, seven to eight on the half
equator. Medullary shell spherical, its diameter one-third of the equatorial axis of the cortical
shell. (The cortical shell of this species is nearly identical with the central half of the
cortical shell of Panartus diploconus, Pl. 40, fig.
1.)
Dimensions.—Main axis of the cortical shell 0.12, equatorial axis 0.07; meshes
0.013; bars 0.003; diameter of the medullary shell 0.025.
Habitat.—South Pacific, Station 285, depth 2375 fathoms.
2. Cyphanta circopora, n. sp.
Cortical shell thick walled, with smooth surface; pores subregular, circular, twice as broad as
the bars; eighteen to twenty on the half meridian, twelve to fourteen on the half equator.
Medullary shell spherical, its diameter one-half of the equatorial axis of the cortical shell.
(The shell of this species is very similar to that of Cannartiscus amphiconiscus, Pl. 39, fig. 19,
but without the polar tubes of that species.)
Dimensions.—Main axis of the cortical shell 0.13, equatorial axis 0.08; meshes
0.007, bars 0.003; diameter of the medullary shell 0.04.
Habitat.—Pacific, central area. Station 266, depth 2750 fathoms.
3. Cyphanta lævis, Haeckel.
Ommatospyris lævis, Ehrenberg, 1872, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 318.
Cortical shell thin walled, with smooth surface; pores of it irregular, roundish, of very
different size (some very large in the tropical circles of both hemispheres). Pores twice to six
times as broad as the bars; ten to twelve on the half meridian, six to eight on the half equator.
Medullary shell spheroidal, compressed, its main axis somewhat shorter than its equatorial axis,
which attains half the length of that of the cortical shell. (This species may be perhaps
identical with Ommatospyris lævis of Ehrenberg, the diagnosis of which is insufficient and
figure not given.)
Dimensions.—Main axis of the cortical shell 0.1, equatorial axis 0.06; meshes
0.005 to 0.02, bars 0.002 to 0.04; diameter of the medullary shell 0.03.
Habitat.—Philippine Sea (Ehrenberg), Station 213, depths 2650 and 3300
fathoms.
4. Cyphanta arachnoides, n. sp.
Cortical shell very delicate and thin walled, cobweb-like, with smooth surface; pores
irregular, polygonal (mostly pentagonal or hexagonal), eight to ten times as broad as the
thread-like bars; ten to twelve on the half meridian, six to eight on the half equator. Medullary
shell spheroidal, compressed, its main axis shorter than its equatorial axis, which is about
one-third that of the cortical shell. (This species is very similar to the middle part of the
shell of Ommatocampe profundissima, Ehrenberg, 1872, Abhandl. d. k. Akad. d. Wiss. Berlin,
Taf. viii. fig. 6.)
{362}
Dimensions.—Main axis of the cortical shell 0.11, equatorial axis 0.06; meshes
0.01 to 0.02, bars 0.002; diameter of the medullary shell 0.02.
Habitat.—Atlantic, Canary Islands (Lanzerote), surface.
Subgenus 2. Cyphantissa, Haeckel.
Definition.—Surface of the cortical shell spiny, everywhere scattered
with numerous thorns or spines.
5. Cyphanta hispida, n. sp.
Cortical shell thick walled, with spiny surface; pores regular, circular, with hexagonal frame,
twice as broad as the bars; twelve to fourteen on the half meridian, eight to ten on the half
equator. From the corners of the hexagonal frames (between every three pores) arise short,
straight, conical radial spines, somewhat longer than the breadth of the pores. Medullary shell
subspherical, its diameter about one-third of the equatorial axis of the cortical shell. (The
appearance of the cortical shell is the same as that of the middle part of Peripanartus
amphiconus, Pl. 40,
fig. 5.)
Dimensions.—Main axis of the cortical shell 0.12, equatorial axis 0.07; meshes
0.01, bars 0.005; length of the spines 0.012; diameter of the medullary shell 0.02.
Habitat.—Pacific, central area, Station 274, depth 2750 fathoms.
6. Cyphanta hystrix, n. sp.
Cortical shell thin walled, with thorny surface; pores irregular, roundish, of very unequal
size, twice to six times as broad as the bars; twelve to eighteen on the half meridian, eight to
thirteen on the half equator. Between the pores irregularly scattered, bristle-like, thin spines
about the same size, partly straight, partly oblique, rising from the surface. Medullary shell
lenticular, compressed, its equatorial axis one and a half times the length of the main axis, and
about half that of the cortical shell.
Dimensions.—Main axis of the cortical shell 0.15, equatorial axis 0.09; meshes
0.004 to 0.02, bars 0.003 to 0.006; length of the spines 0.02; diameter of the medullary shell
0.04.
Habitat.—South Atlantic, Station 318, surface.
Genus 156. Cyphonium,[195] n. gen.
Definition.—Cyphinida with simple cortical shell and
double medullary shell, without polar spines or tubes.
The genus Cyphonium contains a number of very common species, among which
are the earliest known forms of this family, partly described by Ehrenberg as Ommatospyris {363}(which genus contains also a number of other Prunoidea), partly by me (1862) as Didymocyrtis. Both names are
inadequate, as allusions to quite different families of Nassellaria, but may be retained as
significations of subgeneric divisions. Cyphonium differs from Cyphanta by the
double medullary shell, which is either spherical or lenticular.
Subgenus 1. Ommatospyris, Ehrenberg (partim).
Definition.—Surface of the cortical shell smooth, without thorns or
spines.
1. Cyphonium coscinoides, n. sp.
Ommatospyris coscinoides, Haeckel, 1881, Prodromus, p. 462.
Cortical shell thin walled, with smooth surface; pores regular, hexagonal, three to four times
as broad as the bars; five to six on the half meridian, eight to nine on the half equator of each
chamber. Medullary shells both spherical. (The cortical shell of this species is nearly the same
as that of Cyphanta colpodes, and as the middle part of the cortical shell of Panartus
diploconus, Pl. 40,
fig. 1.)
Dimensions.—Main axis of the cortical shell 0.11, equatorial axis 0.06; greatest
breadth of the chambers 0.08; pores 0.013, bars 0.003.
Habitat.—North Pacific, Station 244, surface.
2. Cyphonium virgineum, n. sp. (Pl. 39,
figs. 12, 12a).
Ommatospyris virginea, Haeckel, 1881, Prodromus et Atlas (pl. xxxix. fig.
12a).
Cortical shell thick walled, with smooth surface (sometimes a little rough with very small
thorns); pores subregular, circular, with hexagonal elevated frames, about as broad as the bars;
ten to twelve on the half meridian, sixteen to eighteen on the half equator of each chamber.
Medullary shells (fig. 12a) both spherical. (Sometimes, as in the figured specimen, the
shell is somewhat irregular, an individual abnormality.)
Dimensions.—Main axis of the cortical shell 0.13 to 0.15; equatorial axis of the
structure 0.08 to 0.09, of each chamber 0.1 to 0.12, pores and bars 0.005 to 0.007.
Habitat.—Pacific, western tropical part, Station 225, depth 4475
fathoms.
3. Cyphonium ethmarium, n. sp.
Ommatospyris ethmaria, Haeckel, 1881, Prodromus, p. 462.
Cortical shell thin walled, with quite smooth surface; pores subregular, circular (without
hexagonal frame), twice as broad as the bars; nine to ten on the half meridian of each chamber,
sixteen to eighteen on its half equator. Medullary shells both spherical. (This species resembles
the proximal internal chambers of Peripanartus atractus, Pl. 40, fig.
7.)
Dimensions.—Main axis 0.13, equatorial axis 0.07; greatest breadth 0.09; pores
0.006, bars 0.003.
Habitat.—Equatorial Atlantic, Station 347, surface.
{364}
4. Cyphonium trinacrium, n. sp.
Cortical shell thick walled, with rough surface; pores subregular, circular (without hexagonal
frame), not broader than the bars; five to six on the half meridian of each chamber, nine to ten
on its half equator. Medullary shells both compressed, lenticular. This species resembles
Cypassis entomocora vel Ommatocampe trinacria, Stöhr, 1880, loc. cit., p. 90,
Taf. ii. fig. 1, but has not its external mantle. It may be the ancestral form of it (both in an
ontogenetic and phylogenetic sense).
Dimensions.—Main axis 0.11, equatorial axis 0.07; greatest breadth 0.08; pores
0.005, bars 0.005.
Habitat.—Fossil in Tertiary rocks of Sicily, Caltanisetta, Haeckel
(Grotte, Stöhr?).
5. Cyphonium diattus, n. sp.
Cortical shell thin walled, with quite smooth surface; pores irregular, polygonal, mostly
pentagonal or hexagonal, three to six times as broad as the bars; six to seven on the half
meridian of the chamber, ten to twelve on its half equator. Medullary shells both spherical.
(Resembles Cyphonium profundum, Ehrenberg, 1872, loc. cit., Taf. x. fig. 5, but does
not possess the spines of the surface.)
Dimensions.—Main axis 0.13, equatorial axis 0.06; greatest breadth 0.08; pores
0.005 to 0.012, bars 0.002 to 0.004.
Habitat.—Indian Ocean, western part (Zanzibar), Pullen, depth 2200
fathoms.
6. Cyphonium mammarium, n. sp.
Cortical shell thick walled, with smooth surface (sometimes a little rough); pores irregular,
roundish, twice to three times as broad as the bars; eleven to twelve on the half meridian of each
chamber, sixteen to nineteen on its half equator. Internal medullary shell spherical, external
lenticular compressed, sometimes both spherical or both compressed. (Resembles the internal
cortical twin-shell of Cyphocolpus virginis, Pl. 40, fig.
11.)
Dimensions.—Main axis 0.14, equatorial axis 0.08, greatest breadth 0.09; pores
0.004 to 0.006, bars 0.002.
Habitat.—South Pacific, Station 285, depth 2375 fathoms.
Subgenus 2. Ommatocyrtis, Haeckel.
Definition.—Surface of the cortical shell thorny or spiny.
7. Cyphonium hexagonium, n. sp.
Didymocyrtis hexagonia, Haeckel, 1881, Prodromus.
Cortical shell thin walled, with spiny surface; pores regular or subregular, hexagonal, twice
to three times as broad as the bars; five to six on the half meridian, nine to ten on the half
equator {365}of each chamber. Radial spines between them
conical, about as long as the pores. Medullary shells both spherical.
Dimensions.—Main axis 0.14, equatorial axis 0.08; greatest breadth of each chamber
0.1; pores 0.012, bars 0.004.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
8. Cyphonium facettarium, n. sp.
Didymocyrtis facettaria Haeckel, 1881, Prodromus.
Cortical shell thick walled, with spiny surface; pores subregular, circular, with elevated
hexagonal frames, twice as broad as the bars; eight to nine on the half meridian, fourteen to
sixteen on the half equator of each chamber. In the corners of the hexagons (between every three
meshes) arise radial spines, about twice as long as the pores. Both medullary shells spheroidal
compressed. (This species is nearly identical with the internal cortical twin-shell of
Peripanartus amphiconiscus, Pl. 40,
fig. 5.)
Dimensions.—Main axis 0.12, equatorial axis 0.07; greatest breadth of both
chambers 0.09; pores 0.01, bars 0.005, spines 0.02.
Habitat.—Western Tropical Pacific, Station 225, depth 4475
fathoms.
9. Cyphonium cribellum, n. sp. (Pl. 39,
fig. 11).
Cortical shell thick walled, with spiny surface; pores regular, circular, about as broad as the
bars; seven to eight on the half meridian of each chamber, thirteen to fifteen on its half
equator. Radial spines between them conical, twice as long as the pores. Medullary shells both
lenticular.
Dimensions.—Main axis 0.11, equatorial axis 0.06; greatest breadth 0.08; pores and
bars 0.004.
Habitat.—North Atlantic, Station 353, surface.
10. Cyphonium profundum, Haeckel.
Ommatospyris profunda, Ehrenberg, 1872, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 297, Taf. viii. fig. 5, Taf. x. fig. 5.
Cortical shell thin walled, with thorny surface; pores irregular, polygonal, three to six times
as broad as the thin bars; five to six on the half meridian of each chamber, ten to twelve on its
half equator. Thorns of the surface short. Medullary shells both spherical.
Dimensions.—Main axis 0.1, equatorial axis 0.06; greatest breadth 0.07; pores
0.005 to 0.012, bars 0.002, spines 0.005.
Habitat.—Pacific, tropical zone; Philippine Sea, depth 3300 fathoms,
Ehrenberg; Stations 266 to 274, depth 2350 to 2925 fathoms.
{366}
11. Cyphonium ceratospyris, Haeckel.
Didymocyrtis ceratospyris, Haeckel, 1862, Monogr. d. Radiol., p. 445, pl. xxii. figs.
14-16.
Haliomma didymocyrtis, Haeckel, 1860, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 816.
Cortical shell thin walled, with spiny surface; pores irregular, roundish, three to nine times
as broad as the bars; five to six on the half meridian of each chamber, eight to nine on its half
equator. Spines of the surface partly very short, partly as long as the largest pores, either
radially or obliquely inserted. Medullary shells both spherical. (Compare the detailed description
and figures also of the soft body in my Monograph, loc. cit.)
Dimensions.—Main axis of the cortical shell 0.12, equatorial axis (in the
stricture) 0.06; greatest breadth 0.08; pores 0.03 to 0.2, bars 0.002 to 0.006, spines 0.05 to
0.015.
Habitat.—Mediterranean (Messina), Canary Islands (Lanzerote),
Haeckel.
Genus 157. Cypassis,[196] n. gen.
Definition.—Cyphinida with double cortical shell and
double medullary shell, without polar spines or tubes.
The genus Cypassis differs from Cyphonium by duplication of the
cortical twin-shell; from the outer surface of the simple twin-shell arise numerous radial spines,
which become connected by anastomosing transverse branches, and in this manner form an outer
envelope or mantle. Cypassis may also be regarded as a Cromyodruppa, the double
cortical shell of which is constricted in the equatorial plane.
Subgenus 1. Didymospyris, Haeckel (1881).
Definition.—Surface of the shell smooth, without thorns or spines.
1. Cypassis palliata, n. sp.
Inner cortical shell thin walled, with regular, circular, hexagonally-framed pores, twice as
broad as the bars; seven to eight on the half meridian of each chamber, ten to twelve on its half
equator. Outer cortical shell very thin, with smooth surface, and very small, irregular, roundish
pores, twice to five times smaller than those of the inner shell. Distance between the two
cortical shells equals the diameter of the outer medullary shell, which, like the inner, is
spherical. (The network of this species resembles that of Peripanartus amphiconiscus, Pl.
40, fig.
5.)
Dimensions.—Main axis of the external cortical shell 0.18, of the internal 0.12;
greatest breadth (in the equator of each chamber) of the former 0.13, of the latter 0.09; pores of
the outer 0.002 to 0.005, of the inner shell 0.01, bars 0.002 to 0.005.
Habitat.—South Pacific, Station 288, surface.
{367}
2. Cypassis eucolpos, n. sp.
Inner cortical shell thick walled, with regular, circular pores, twice as broad as the bars
(without hexagonal frames); nine to ten on the half meridian of each chamber, fifteen to seventeen
on its half equator. Outer cortical shell very thin, with smooth surface, and very small,
irregular, roundish pores, twice to three times smaller than those of the inner shell. Distance
between the two cortical shells about one and a half times the diameter of the inner medullary
shell, which, like the outer, is spherical. (The inner cortical shell resembles that of
Cannartiscus amphiconiscus, Pl. 39,
fig. 19, but without polar tubes.)
Dimensions.—Main axis of the external cortical shell 0.2, of the internal 0.15;
greatest breadth of the former 0.16, of the latter 0.12; pores of the outer 0.002, of the inner
0.006, bars 0.003.
Habitat.—South Atlantic, Station 319, surface.
Subgenus 2. Didymocyrtis, Haeckel (1862).
Definition.—Surface of the cortical shell thorny or spiny.
3. Cypassis entomocora, Haeckel.
Astromma entomocora, Ehrenberg, 1847, Mikrogeol., Taf. xxii. fig. 32.
Haliomma didymum, Ehrenberg, 1844, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p.
83.
? Haliomma amphisiphon, Ehrenberg, 1844, Monatsber. d. k. Akad. d. Wiss. Berlin, p.
267.
Ommatospyris entomocora, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss. Berlin, pp.
66, 74.
? Ommatocampe trinacria, Stöhr, 1880, Palæontogr., vol. xxvi. p. 90, Taf. ii. fig.
1.
Didymospyris entomocora, Haeckel, 1881, Prodromus.
Didymocyrtis entomocora, Haeckel, 1862, Monogr. d. Radiol., p. 445.
Inner cortical shell thick walled, with regular, circular pores, twice as broad as the bars
(without hexagonal frames); eight to nine on the half meridian of each chamber, ten to eleven on
its half equator. Outer cortical shell thin walled, with regular, circular pores like those of the
inner. Distance between the two cortical shells equals about twice the diameter of one pore (or
the short axis of the lenticular double medullary shell). Surface covered with short conical
spines (in the figure of Ehrenberg broken off).
Dimensions.—Main axis of the external cortical shell 0.2, of the internal 0.15;
greatest breadth of the former 0.13, of the latter 0.1; pores of the outer 0.005 to 0.01, of the
inner 0.01, bars 0.004; length of the surface spines 0.005 to 0.01.
Habitat.—Fossil in the Tertiary rocks of Barbados, Bermuda, and Sicily
(Caltanisetta); also living in the greatest depth of the Pacific, Stations 268, 225, depth 2900 to
4475.
4. Cypassis puella, n. sp. (Pl. 39,
fig. 13).
Didymospyris colpodes, Haeckel, 1881, Prodromus et Atlas (pl. xxxix., fig.
13).
Internal cortical shell thin walled, with irregular, circular pores of very different size,
once to four times as broad as the bars; fourteen to fifteen on the half meridian of each chamber,
eighteen {368}to twenty on its half equator. Outer
cortical shell thin walled, with irregular, circular pores, only one-third to one-half as large as
those of the inner. Distance between the two shells equals the largest diameter of the double
lenticular medullary shell. Surface of both cortical shells covered with short conical spines (not
longer than the largest pores).
Dimensions.—Main axis of the external cortical shell 0.2, of the internal 0.15, of
the central capsule 0.1; greatest breadth of the first 0.14, of the second 0.1, of the third 0.07;
pores of the outer shell 0.001 to 0.005, of the inner 0.002 to 0.01, bars 0.002 to 0.003; length
of the surface spines 0.005 to 0.01.
Habitat.—Pacific, central area; Stations 266 to 274, surface;
Atlantic, Canary Islands (Haeckel), Station 354, surface.
5. Cypassis halicora, n. sp.
Internal cortical shell thin walled, with irregular, roundish pores, once to three times as
broad as the bars; ten to eleven on half meridian of each chamber, fourteen to sixteen on its half
equator. Outer cortical shell thin walled, with very delicate network, spindle-like, with conical
prolongations at both poles. Distance between the two shells larger than the greatest diameter of
the double lenticular medullary shell. Surface of both cortical shells covered with innumerable
small thorns. (Resembles closely Cyphocolpus virginis, Pl. 40, fig. 11,
but without the third shell.)
Dimensions.—Main axis of the external cortical shell 0.21, of the internal 0.14,
greatest breadth of the former 0.13, of the later 0.09; pores 0.003 to 0.009; bars of the outer
shell 0.001, of the inner 0.004.
Habitat.—Indian Ocean, Ceylon (Haeckel), surface.
Genus 158. Cyphocolpus,[197] n. gen.
Definition.—Cyphinida with triple cortical shell and
double medullary shell, without polar spines or tubes.
The genus Cyphocolpus differs from both foregoing genera by the increased
number of the cortical twin-shells. Whilst these are simple in Cyphonium, double in
Cypassis, they are triple in Cyphocolpus, composed of three concentric envelopes.
The three genera named represent a phylogenetic series, which is repeated in the ontogenetic
development of Cyphocolpus.
1. Cyphocolpus didymus, n. sp.
Inner cortical shell with circular, subregular pores, twice as broad as the bars; five to six
on the half meridian of each chamber, eight to ten on its half equator. Middle cortical shell also
with subregular, circular pores of the same size. Outer cortical shell with smooth surface, with
more irregular, roundish pores of very different size. The distance between every two shells
equals the diameter of the inner medullary shell, which, like the outer, is spheroidal.
{369}
Dimensions.—Main axis of the outer cortical shell 0.3, of the middle 0.22, of the
inner 0.15; greatest breadth of the inner cortical shell 0.1, its pores 0.01, bars 0.005.
Habitat.—Indian Ocean, Madagascar, Rabbe, surface.
2. Cyphocolpus virginis, n. sp. (Pl. 40,
fig. 11).
Zygartus virginis, Haeckel, 1881, Prodromus et Atlas (pl. xl. fig.
11).
Inner cortical shell with irregular, roundish pores, twice to three times as broad as the bars;
eleven to twelve on the half meridian of each chamber, sixteen to eighteen on its half equator.
Middle cortical shell with very delicate network, its irregular, polygonal pores of the same size
as those of the inner, but separated by extremely thin bars. Outer cortical shell with spiny
surface, and with more irregular, roundish pores of different size. The distance between each two
shells is variable, and attains more than the diameter of the outer medullary shell, which, like
the inner, is spheroidal, slightly compressed.
Dimensions.—Main axis of the outer cortical shell 0.26, of the middle 0.22, of the
inner 0.14; greatest breadth of the inner cortical shell 0.09, its pores 0.004 to 0.006, bars
0.002.
Habitat.—Pacific, central area, Station 271, surface.
Genus 159. Cyphinus,[198] Haeckel, 1881,
Prodromus, p. 463.
Definition.—Cyphinida with simple cortical shell and
simple medullary shell, with two opposite polar spines (or bunches of polar spines) on the poles
of the main axis.
The genus Cyphinus differs from Cyphanta by the development of two
opposite spines (or bunches of spines) on both poles of the main axis. It simulates therefore the
bipolar formation of Stylartus, and differs from this in the possession of a medullary
shell.
Subgenus 1. Cyphinoma, Haeckel.
Definition.—On both poles of the main axis only one single large
spine.
1. Cyphinus amphacanthus, n. sp.
Cortical shell with smooth surface, and circular, regular pores, twice as broad as the bars;
five to six pores on the half meridian of each chamber, ten to twelve on its half equator. On both
poles of the main axis, one single, very strong, conical spine, about half as long as this axis,
and half as broad at the base as the spherical medullary shell.
Dimensions.—Main axis of the cortical shell (without spines) 0.2, greatest breadth
(in the equator of each chamber) 0.1; pores 0.006, bars 0.003; length of the polar spines 0.1,
basal thickness of them 0.02.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
{370}
2. Cyphinus dixiphus, n. sp.
Cortical shell with rough surface, and irregular, roundish pores, twice to four times as broad
as the bars; eight to nine pores on the half meridian of each chamber, twelve to fourteen on its
half equator. On both poles of the main axis, one single, strong, conical spine, with prominent
edges at the base, and nearly as broad as the spherical medullary shell, nearly as long as the
main axis.
Dimensions.—Main axis (without spines) 0.18, greatest breadth 0.13; pores 0.003 to
0.008, bars 0.002; length of the polar spines 0.16, basal thickness 0.03.
Habitat.—South Pacific, Station 295, depth 1500 fathoms.
Subgenus 2. Cyphinura, Haeckel.
Definition.—On both poles of the main axis a bunch or circle of
several spines.
3. Cyphinus amphilophus, n. sp. (Pl. 39,
fig. 14).
Cyphinidium amphilophus, Haeckel, 1882, Atlas (pl. xxxix. fig. 14).
Cortical shell thick walled, with rough surface, and subregular, circular pores of different
size, twice to five times as broad as the bars; nine to ten pores on the half meridian of each
chamber, twelve to fourteen on its half equator. On both poles of the main axis a bunch of six to
eight strong conical spines, the largest of which are about as long as the greatest breadth of the
shell; their basal part is furrowed, and nearly half as thick as the spherical medullary
shell.
Dimensions.—Main axis of the cortical shell (without spines) 0.18 to 0.2, greatest
breadth 0.1 to 0.12; pores 0.004 to 0.01, bars 0.002; length of the polar spines 0.04 to 0.12,
basal thickness 0.02 to 0.04.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
4. Cyphinus penicillatus, Haeckel.
Ommatospyris penicillata, Ehrenberg, 1872, Monatsber. d. k. preuss. Akad.
d. Wiss. Berlin, p. 318; Abhandl. d. k. Akad. d. Wiss. Berlin, Taf. viii. fig. 4.
Cortical shell thin walled, with rough surface, and irregular, polygonal pores, six to eight
times as broad as the bars; five to six pores on the half meridian of each chamber, nine to ten on
its half equator. Around both poles of the main axis a polar circle of eight to ten divergent
radial spines, scarcely thicker than the bars, and about as long as the diameter of the spherical
medullary shell. (In the figure of Ehrenberg the network is too thin and the spines too
short.)
Dimensions.—Main axis (without spines) 0.1, greatest breadth 0.09; pores 0.008 to
0.014, bars 0.001 to 0.002; length of the spines 0.02 to 0.04.
Habitat.—Pacific, Philippine Sea, 3300 fathoms, Ehrenberg; Station
206, depth 2100 fathoms.
{371}
Genus 160. Cyphinidium,[199] n. gen.
Definition.—Cyphinida with simple cortical shell and
double medullary shell, with two opposite polar spines (or bunches of polar spines) on the poles
of the main axis.
The genus Cyphinidium differs from Cyphinus only in the duplication
of the medullary shell, which is composed of two concentric spheres or somewhat compressed
lenticular spheroids. It exhibits therefore the same relation to Cyphinus that
Stylatractus in the Druppulida bears to Lithatractus. Possibly the two former genera
are derived from the two latter by an annular constriction in the equatorial plane.
Subgenus 1. Cyphinidoma, Haeckel.
Definition.—On both poles of the cortical shell one single polar spine
or a bunch of polar spines.
1. Cyphinidium amphistylium, n. sp.
Cortical shell thick walled, with smooth surface, and subregular, circular pores, twice to
three times as broad as the bars; eight to ten on the half meridian, sixteen to eighteen on the
half equator of each chamber. Both medullary shells spherical. The cortical shell resembles very
much that of Cannartiscus amphiconiscus (Pl. 39, fig. 19),
but exhibits instead of the hollow polar tubes two strong solid polar spines of conical form, half
as long as the main axis, and as broad at the base as the inner medullary shell.
Dimensions.—Main axis of the cortical shell (without polar spines) 0.14, greatest
breadth 0.1; pores 0.008, bars 0.004; length of the polar spines 0.08, basal thickness 0.02.
Habitat.—North Pacific, off Japan, Station 241, depth 2300
fathoms.
2. Cyphinidium apicatum, Haeckel.
Ommatospyris apicata, Ehrenberg, 1872, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 317.
Cortical shell thin walled, with thorny surface, and irregular, polygonal pores, three to five
times as broad as the thin bars; eight to nine on the half meridian, thirteen to fifteen on the
half equator of each chamber. On both poles a bunch of four to eight strong conical spines, the
central of which (in the main axis) is much larger than the others, and twice as long as the
diameter of the outer spheroidal medullary shell; size and number of the polar spines is very
variable (commonly five to six).
{372}
Dimensions.—Main axis (without spines) 0.12, greatest breadth 0.08; pores 0.005 to
0.01, bars 0.002; length of the
polar spines 0.02 to 0.06, basal thickness 0.01.
Habitat.—Northern Pacific, Californian Sea, depth 2500 fathoms,
Ehrenberg; Station 253, depth 3125 fathoms.
Subgenus 2. Cyphinidura, Haeckel.
Definition.—On both poles of the cortical shell, a circle of divergent
polar spines.
3. Cyphinidium coronatum, n. sp.
Cortical shell with thorny surface, and irregular, roundish pores, twice to four times as broad
as the bars; seven to eight on the half meridian, eleven to thirteen on the half equator of each
chamber. Around both poles of the main axis occurs a circle of ten to twelve strong, radially
divergent spines, about as long as the diameter of the outer medullary shell, which, like the
inner, is spheroidal. (This species is identical with the younger developmental stage of
Panicium coronatum, Pl. 40,
fig. 4, before the formation of two distal chambers.)
Dimensions.—Main axis 0.12, greatest breadth 0.1; pores 0.008 to 0.016, bars
0.004; length of the polar spines 0.02, thickness 0.005.
Habitat.—Pacific, central area, Stations 270 to 274, depth 2350 to
2925 fathoms.
Genus 161. Cannartiscus,[200] n.
gen.
Definition.—Cyphinida with simple cortical shell and
simple medullary shell, with two hollow fenestrated polar tubes, opposite on both poles of the
main axis.
The genus Cannartiscus differs from Cyphanta in the development of
two hollow fenestrated tubes, opposite on both poles of the main axis. It simulates therefore the
characteristic formation of Cannartus, and differs from this Artiscid in the possession of
a medullary shell.
1. Cannartiscus amphiconiscus, n. sp. (Pl. 39, fig.
19).
Cannartidium amphiconiscus, Haeckel, 1882, Atlas (pl. xxxix. fig. 19).
Cortical shell thick walled, smooth, with regular, circular pores, twice as broad as the bars;
nine to ten on the half meridian, sixteen to eighteen on the half equator of each chamber. Polar
tubes conical, about as long as one single chamber, on the base somewhat broader than the simple
spherical medullary shell. Pores of the tubes only one-third as great as those of the
chambers.
Dimensions.—Main axis of the cortical shell (without tubes) 0.14, greatest breadth
0.1; {373}pores 0.008, bars 0.004. Length of the polar
tubes 0.07, basal breadth 0.04; pores 0.003, bars 0.002.
Habitat.—North Pacific, Station 253, depth 3125 fathoms.
2. Cannartiscus amphicylindrus, n. sp.
Cortical shell thick walled, rough, with subregular, circular pores, twice to four times as
broad as the bars; six to seven on the half meridian, ten to twelve on the half equator of each
chamber. Polar tubes cylindrical, on the distal end open (broken off?) nearly as long as the main
axis, somewhat narrower than the spherical medullary shell. Pores of the tubes much smaller than
those of the chambers.
Dimensions.—Main axis (without tubes) 0.17, greatest breadth 0.12; pores 0.006 to
0.012, bars 0.003. Length of the polar tubes 0.15, breadth of them 0.03; pores 0.003, bars
0.002.
Habitat.—Pacific, central area, Station 268, 2900 fathoms; the same
form occurs fossil in the rocks of Barbados.
Genus 162. Cannartidium,[201] n.
gen.
Definition.—Cyphinida with simple cortical shell and
double medullary shell, with two hollow fenestrated polar tubes, opposite on both poles of the
main axis.
The genus Cannartidium differs from Cannartiscus and Cannartus
in the duplication of the medullary shell, which is composed of two concentric spheres or
compressed lenticular spheroids. The three genera named form therefore one morphological series,
with identical cortical shell, and only differing in the absence or presence of a simple or double
medullary shell.
Subgenus 1. Cannartidella, Haeckel.
Definition.—Surface of the cortical shell smooth or a little rough,
but without spines or fenestrated protuberances.
1. Cannartidium amphiconicum, n. sp.
Cortical shell thick walled, smooth with regular circular pores twice as broad as the bars;
eight to nine on the half meridian, fifteen to sixteen on the half equator of each chamber. Polar
tubes conical, with smaller pores, tapering towards the closed apex, about as long as one single
chamber, at the base one-third as broad as the equatorial constriction. Both concentric medullary
shells spherical. (Nearly identical with Cannartiscus amphiconiscus, Pl. 39, fig. 19,
but differs in the possession of the double medullary shell.)
{374}
Dimensions.—Main axis of the cortical shell (without tubes) 0.15, greatest breadth
0.11; pores 0.009, bars 0.004; length of the polar tubes 0.08, basal breadth 0.04.
Habitat.—Pacific, western tropical part, Station 225, depth 4475
fathoms.
2. Cannartidium amphicanna, Haeckel.
"Curious twin Polycystin," Bury, 1862, Polycystins of Barbados, pl. xx. fig.
4.
Cortical shell thick walled, rough, with regular, circular pores, not broader than the bars;
six to seven on the half meridian, ten to twelve on the half equator of each chamber. Polar tubes
cylindrical, at the distal end open (broken off?), nearly as long as the main axis, about
one-third as broad as the equatorial constriction. Both concentric medullary shells spherical.
Dimensions.—Main axis 0.14, greatest breadth 0.11; pores and bars 0.005; length of
the polar tubes 0.13, breadth of them 0.03.
Habitat.—Fossil in the Barbados deposits.
3. Cannartidium amphisiphon, Haeckel.
? Haliomma amphisiphon, Ehrenberg, 1844, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 267.
Cortical shell thick walled, rough, with irregular, roundish pores, twice to five times as
broad as the bars; five to six on the half meridian, nine to ten on the half equator of each
chamber. Polar tubes conical, with smaller pores, about as long as one single chamber, half as
broad at the base as the equatorial constriction. (This deep-sea form is probably identical with
that fossil species which Ehrenberg, in 1844, described as Haliomma amphisiphon, and which
he afterwards, in 1875, erroneously identified with his Astromma entomocora (= Cypassis
entomocora). But the figure given of the latter is quite different from the given diagnosis of
the former.)
Dimensions.—Main axis of the cortical shell (without tubes) 0.13, greatest breadth
0.1; pores 0.006 to 0.12, bars 0.003; length of the polar tubes 0.07, basal breadth of them
0.04.
Habitat.—Tropical Atlantic, Antilles, Station 24, depth 390 fathoms;
fossil in the Tertiary rocks of Bermuda.
4. Cannartidium bicinctum, n. sp. (Pl. 39, fig.
18).
Cortical shell thick walled, with a thickened ring-shaped protuberance in the greatest breadth
of both chambers. Pores subregular, circular, twice to three times as broad as the bars; seven to
eight on the half meridian, thirteen to fifteen on the half equator of each chamber. Polar tubes
conical, with smaller pores, somewhat shorter than the greatest breath, nearly as thick at the
base as the outer spheroidal medullary shell (fig. 18a).
Dimensions.—Main axis (without tubes) 0.12, greatest breadth 0.08; pores 0.003 to
0.006, bars 0.002; length of the polar tubes 0.08, basal thickness 0.03.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
{375}
Subgenus 2. Cannartidissa, Haeckel.
Definition.—Surface of the cortical shell with conical fenestrated
protuberances in the greatest breadth of both chambers.
5. Cannartidium mammiferum, n. sp. (Pl. 39, fig.
16).
Cortical shell-thin walled, having a circle of six to eight conical protuberances in the
greatest breadth of both chambers, each of which exhibits a central apical pore, surrounded by a
circle of six to eight oblique larger pores. Between the protuberances occur nine to ten pores on
the half meridian of each chamber, circular, subregular, twice to three times as broad as the
bars. Polar tubes cylindrical, conical at the closed end, with smaller pores, about as long as the
greatest breadth of the chambers, as broad as the spherical outer medullary shell.
Dimensions.—Main axis (without tubes) 0.13, greatest breadth (including the
protuberances) 0.1; pores 0.005 to 0.008, bars 0.003; length of the polar tubes 0.09, basal
thickness 0.02.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
6. Cannartidium mastophorum, n. sp. (Pl. 39, fig.
17).
Cortical shell thin walled, of the same form and structure as in the foregoing species, differs
from this mainly in the form of the polar tubes, which are not cylindrical but conical, tapering
gradually from the base towards the closed apex. Besides this, the conical protuberances (six in
the greatest periphery of each chamber) are more regular and acute, with smaller and more numerous
pores. Both medullary shells are here also spherical.
Dimensions.—Main axis (without tubes) 0.14, greatest breadth (including the
protuberances) 0.13; pores 0.006 to 0.009, bars 0.004; length of the polar tubes 0.07, basal
thickness 0.03.
Habitat.—Pacific, central area, Station 266, depth 2750 fathoms.
Family XVI. Panartida, Haeckel (Pl. 40, figs.
1-9).
Definition.—Prunoidea with a four-jointed cortical shell,
the external shell being divided by three parallel transverse constrictions into four chambers, in
the centre enclosing two internal concentric shells (medullary shells). Central capsule
cylindrical, commonly four-jointed (with three transverse annular constrictions).
The family Panartida (Pl. 40, figs. 1-9)
is characterised by its four-jointed cortical shell, constantly composed of four chambers, lying
one behind another in the elongated main axis. They are separated by three ring-like constrictions
lying in three parallel transverse planes; the middle of these is the equatorial plane. In the
centre of this latter lies constantly a double medullary shell.
The Panartida must be derived from the Cyphinida by further increase in the longitudinal or
main axis, and by repetition of the equatorial constriction in two transverse planes parallel to
it and at an equal distance from it.
{376}
The Medullary Shell is always double in this family, composed of two small concentric
shells lying in the middle of the central capsule. They are either spherical or lenticular,
compressed towards both poles of the main axis; sometimes the inner is spherical, the outer
lenticular. From the equatorial circumference of the latter starts a number of radial beams, which
perforate the central capsule, and are inserted into the equatorial constriction of the cortical
shell. Commonly all radial beams lie in the equatorial plane, sometimes also a part of them on
both sides of it.
The Cortical Shell is constantly composed of four completely latticed chambers, which
are only separated by the three annular constrictions. In many species all four chambers have the
same form and size (Pl. 40,
fig. 3), but in the greater number of species both proximal (or central) chambers are different in
size, shape, or structure from both distal (or polar) chambers. The cortical shell is either
simple or double, composed of two concentric four-jointed shells; rarely it is triple or multiple,
composed of three or more shells fitting one inside the other. Very often the outer cortical shell
is incomplete, and only developed around the two proximal chambers of the complete inner cortical
shell, both distal chambers of the latter remaining simple.
On both poles of the main axis often are developed solid spines (Panicium, Pl. 40, fig. 4) or
hollow fenestrated tubes (Panarium, Pl. 40,
fig. 9), as in the foregoing families. Besides this, the surface of the cortical shell may be
armed with spines.
The Central Capsule of the Panartida is always cylindrical, on both poles hemispherical;
commonly it is more or less distinctly four-jointed, with three ring-like constrictions
corresponding to those of the enclosing cortical shell. From the inner surface of the latter it is
separated by a thinner or thicker jelly-mantle.
Synopsis of the Genera of Panartida.
| On both poles of the main axis neither peculiar
polar spines nor fenestrated tubes. |
|
Cortical shell simple, |
163. Panartus. |
| Cortical shell double or triple, |
164. Peripanartus. |
| On both poles of the main axis a large apical spine or
a group of polar spines. |
|
Cortical shell simple, |
165. Panicium. |
| Cortical shell double or triple, |
166. Peripanicium. |
| On both poles of the main axis a conical or cylindrical
fenestrated tube. |
|
Cortical shell simple, |
167. Panarium. |
| Cortical shell double or triple, |
168. Peripanarium. |
| On both poles of the main axis neither peculiar polar spines nor fenestrated
tubes. |
|
|
Cortical shell simple, |
|
|
|
|
|
|
163. Panartus. |
|
|
|
|
|
|
Cortical shell double or triple, |
|
|
|
|
|
|
164. Peripanartus. |
|
|
|
|
| On both poles of the main axis a large apical spine or a group of polar
spines. |
|
|
Cortical shell simple, |
|
|
|
|
|
|
165. Panicium. |
|
|
|
|
|
|
Cortical shell double or triple, |
|
|
|
|
|
|
166. Peripanicium. |
|
|
|
|
| On both poles of the main axis a conical or cylindrical fenestrated tube. |
|
|
Cortical shell simple, |
|
|
|
|
|
|
167. Panarium. |
|
|
|
|
|
|
Cortical shell double or triple, |
|
|
|
|
|
|
168. Peripanarium. |
|
|
|
|
Genus 163. Panartus,[202] n. gen.
Definition.—Panartida with simple cortical shell and
double medullary shell, without polar spines or tubes on both poles of the main axis.
{377}
The genus Panartus represents the common ancestral form of all Panartida, as
all other genera of this subfamily must be derived from it and are only further developmental
stages, from an ontogenetic as well as from a phylogenetic point of view. The cortical shell of
Panartus is constantly composed of four fenestrated chambers jointed to one another in the
main axis; both proximal chambers are separated from one another by the equatorial ring-like
constriction, in the centre of which lies the double medullary shell; both distal chambers are
separated from the former by two other annular constrictions (in planes parallel to the equatorial
plane). All four chambers may exhibit the same (kidney-shaped) form and structure (in the
subgenera Panartella and Panartoma); or the proximal chamber may differ more or less
in shape and size from the distal (in the subgenera Panartissa and Panartura). The
outer surface of the cortical shell is sometimes smooth (as in Panartella and
Panartissa), at other times spiny or thorny (as in Panartoma and Panartura).
The double medullary shell is sometimes spherical, or commonly compressed at both poles and
spheroidal or lenticular.
Subgenus 1. Panartella, Haeckel.
Definition.—Surface of the cortical shell smooth, without spines or
thorns. All its four chambers exhibit nearly the same form and structure.
1. Panartus tetraplus, n. sp.
All four chambers of the cortical shell of the same form, size, and structure, kidney-shaped,
twice as broad as long. Pores rather regular, hexagonal, all nearly of the same size and form,
four times as broad as the thin bars; five to six pores on the half meridian, nine to ten on the
half equator of each chamber. Surface of the cortical shell smooth, its main axis three times as
long as its equatorial axis (in the median constriction). Both concentric medullary shells
spherical; the equatorial axis of the outer half as long as that of the cortical shell. (All four
chambers of this species have the same appearance as the two proximal chambers of Panartus
diploconus, Pl. 40,
fig. 1.)
Dimensions.—Main axis of the cortical shell 0.21, equatorial axis 0.07; breadth of
every chamber 0.09; meshes 0.013, bars 0.03.
Habitat.—Central area of the Pacific, Stations 270 to 274, depth 2350
to 2925 fathoms.
2. Panartus tetracolus, n. sp.
All four chambers of the cortical shell of the same form, size, and structure, kidney-shaped,
twice as broad as long. Pores regular, circular, with hexagonal frame, twice as broad as the bars;
six to seven pores on the half meridian, ten to twelve on the half equator of each chamber.
Surface of the cortical shell smooth; its main axis three times as long as the equatorial axis.
Both concentric medullary shells spheroidal, somewhat compressed at both poles; the equatorial
axis {378}of the outer almost equals half that of the
cortical shell. (All four chambers of this species exhibit the structure of the two proximal
chambers of Peripanartus amphiconus, Pl. 40, fig.
5.)
Dimensions.—Main axis of the cortical shell 0.24, equatorial axis 0.08; breadth of
every chamber 0.01; meshes 0.01, bars 0.005.
Habitat.—Equatorial Atlantic, Station 348, depth 2450 fathoms.
3. Panartus tetrameres, n. sp.
All four chambers of the cortical shell of the same form, size, and structure, kidney-shaped,
twice as broad as long. Pores rather regular, circular, without hexagonal frame, about as broad as
the bars; five to six pores on the half meridian, ten to twelve on the half equator of each
chamber. Surface of the cortical shell smooth, its main axis three times as long as the equatorial
axis. Both concentric medullary shells spheroidal, compressed; the outer half as broad as the
constriction. (This species is very much like Ommatocampe increscens, Stöhr, 1880, loc.
cit., Taf. ii. fig. 2, and may be considered as the ancestral form of it.)
Dimensions.—Main axis of the cortical shell 0.2, equatorial axis 0.065; breadth of
every chamber 0.08; meshes and bars 0.005.
Habitat.—Fossil in the Tertiary rocks of Sicily, Caltanisetta,
Haeckel.
4. Panartus tetraphalangus, n. sp.
All four chambers of the cortical shell of the same form, size, and structure, kidney-shaped,
twice as broad as long. Pores irregular, polygonal (mostly pentagonal or hexagonal), four to eight
times as broad as the thin bars; four to seven on the half meridian, eight to fifteen on the half
equator of each chamber. Surface of the cortical shell smooth, its main axis three times as long
as its equatorial axis. Both concentric medullary shells compressed, the outer, half as broad as
the constriction.
Dimensions.—Main axis of the cortical shell 0.26, equatorial axis 0.09; breadth of
every chamber 0.12; meshes 0.003 to 0.02, bars 0.001 to 0.003.
Habitat.—Fossil in the rocks of Barbados, Haeckel.
5. Panartus tetrathalamus, n. sp. (Pl. 40, fig.
3).
All four chambers of the cortical shell of the same form, size, and structure, kidney-shaped,
twice as broad as long. Pores irregular, roundish, twice to three times as broad as the bars; a
circle of larger pores at the base of both distal chambers; five to six pores on the half
meridian, ten to twelve on the half equator of each chamber. Surface of the cortical shell smooth;
its main axis two and a half times as long as the equatorial axis. Both medullary shells
spheroidal, compressed, the outer one-third as broad as the constriction. (In the specimen figured
spines begin to arise from both distal chambers; this forms a transition to Panartus
quadrijugus.)
Dimensions.—Main axis of the cortical shell 0.18, equatorial axis 0.07; breadth of
every chamber 0.09; meshes 0.003 to 0.01, bars 0.002 to 0.004.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific, in various
depths.
{379}
Subgenus 2. Panartissa.
Definition.—Surface of the cortical shell smooth, without spines or
thorns. Both its proximal chambers differ in form and structure from the two distal chambers.
6. Panartus diploconus, n. sp. (Pl. 40,
fig. 1).
Both proximal chambers nearly ellipsoidal, one and a half times as broad as long, with regular
hexagonal pores, four times as broad as the bars; five to six pores on the half meridian, nine to
ten on the half equator of each chamber. Both distal chambers conical, with a circle of ten to
twelve large square pores at the base; the other pores very small and numerous, roundish, sixteen
to twenty in the height of each cone, and the same number on the half periphery of its base. Both
concentric medullary shells compressed, the outer, half as broad as the constriction of the
cortical shell, the surface of which is quite smooth.
Dimensions.—Total length of the shell (or main axis) 0.34; greatest breadth of
each chamber 0.09; pores of the proximal chambers 0.013, bars 0.003; pores and bars of the distal
chambers 0.003; large basal square pores 0.02.
Habitat.—Pacific, central area, Station 272, depth 2600 fathoms.
7. Panartus amphiconus, n. sp.
Both proximal chambers nearly kidney-shaped, one and a half times as broad as long, with
regular, circular pores, with hexagonal frames, twice as broad as the bars; six to seven pores on
the half meridian, ten to twelve on the half equator of each chamber. Both distal chambers
conical, with a circle of ten to twelve very large pores at the base; the other pores very small,
roundish, ten to fifteen in the height of each cone, twenty to thirty on the half basal periphery.
Both concentric medullary shells compressed, the outer one-third as broad as the constriction of
the cortical shell, whose surface is smooth. (This species resembles Peripanartus
amphiconus, Pl. 40,
fig. 5, but is without the surface spines and the outer envelope.)
Dimensions.—Total length of the shell 0.28; greatest breadth of the proximal
chambers 0.1, of the distal chambers 0.14; pores of the former 0.008, of the latter 0.004; bars of
both 0.003 to 0.005.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
8. Panartus fusiformis, n. sp.
Both proximal chambers nearly ellipsoidal, one and a half times as long as broad, with
irregular, roundish pores, twice to three times as broad as the bars; eight to ten pores on the
half meridian, twelve to fifteen on the half equator of each chamber. Both distal chambers
hemiellipsoidal or egg-shaped, somewhat higher and narrower than the proximal chambers, with a
circle of larger square pores at their base; the other pores roundish and very irregular. Both
medullary shells spheroidal, one-third as broad as the constriction of the cortical shell, whose
surface is quite smooth. (This {380}species is similar
to Peripanartus atractus, Pl. 40,
fig. 7, but is without the superficial spines and the equatorial girdle-like envelope of the
latter.)
Dimensions.—Total length of the shell 0.25 to 0.3; greatest breadth (in the
equator of the proximal chambers) 0.08 to 0.1; pores of the proximal chambers 0.008 to 0.012, pores of
the distal chambers half their size, bars 0.002 to 0.006.
Habitat.—North Pacific, Station 237, surface.
9. Panartus profundissimus, Haeckel.
Ommatocampe profundissima, Ehrenberg, 1872, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 297, Taf. viii. fig. 6.
Both proximal chambers nearly ellipsoidal, with irregular, polygonal pores, six to eight times
as broad as the bars; four to five pores on the half meridian, eight to nine on the half equator
of each chamber. Both distal chambers hemispherical, with a very delicate irregular network.
Medullary shells spherical; surface of the shell smooth.
Dimensions.—Total length of the shell 0.15; greatest breadth in the equator of the
proximal chambers 0.08; pores of the proximal chambers 0.015, bars 0.002.
Habitat.—Philippine Sea, depth 3300 fathoms, Ehrenberg; Station 213,
depth 2050 fathoms.
Subgenus 3. Panartoma, Haeckel.
Definition.—Surface of the cortical shell thorny, covered with
scattered spines. All its four chambers exhibit the same form and structure.
10. Panartus quadriceps, n. sp.
All four chambers of the cortical shell of the same form, size, and structure, kidney-shaped,
twice as broad as long. Pores rather regular, circular, with hexagonal frames, twice to three
times as broad as the bars; six to seven pores on the half meridian, ten to twelve on the half
equator of each chamber. Surface spiny; from the corners of the hexagonal frames (between every
three pores) arise short radial spines. Both medullary shells spherical, the outer one-third as
broad as the constriction of the cortical shell. (All the four chambers of this species have the
same form as the two proximal chambers of Peripanartus amphiconus, Pl. 40, fig.
5.)
Dimensions.—Main axis of the cortical shell 0.24, equatorial axis 0.08; meshes
0.01, bars 0.003 to 0.005; length of the spines 0.01 to 0.02.
Habitat.—Pacific, central area, Station 274, depth 2750 fathoms.
11. Panartus quadrijugus, n. sp.
All four chambers of the cortical shell of the same form and size, kidney-shaped, twice as
broad as long. Pores irregular, roundish, twice to three times as broad as the bars; a circle of
larger pores at the base of both distal chambers; six to eight pores on the half meridian, twelve
to {381}fourteen on the half equator of each chamber.
Surface spiny, everywhere covered with small irregular thorns. (This species differs from the
smooth Panartus tetrathalamus, Pl. 40,
fig. 3, almost entirely by the thorny surface.)
Dimensions.—Main axis 0.2, equatorial axis 0.08; breadth of every chamber 0.1;
meshes 0.005 to 0.01, bars 0.003, spines 0.01 to 0.02.
Habitat.—Cosmopolitan; on the surface of the Atlantic, Indian, and
Pacific Oceans.
12. Panartus quadrigeminus, n. sp.
All four chambers of the cortical shell nearly of the same size and form, kidney-shaped, twice
as broad as long. Pores irregular, polygonal (mostly pentagonal or hexagonal), six to eight times
as broad as the thin bars; five to seven on the half meridian, twelve to sixteen on the half
equator of each chamber. Surface of the cortical shell spiny, covered with numerous irregularly
scattered, often oblique, bristle-like thorns.
Dimensions.—Main axis 0.27, equatorial axis 0.1; breadth of each chamber 0.13;
meshes 0.01 to 0.02, bars 0.001 to 0.005.
Habitat.—Cosmopolitan; surface of the Atlantic and Pacific, many
Stations.
Subgenus 4. Panartura, Haeckel.
Definition.—Surface of the cortical shell thorny, covered with
scattered spines. Both its proximal chambers differ in form or structure from the two distal
chambers.
13. Panartus spinosus, n. sp.
Both proximal chambers nearly ellipsoidal, with regular, circular pores enclosed by hexagonal
frames. Both distal chambers conical, with a circle of ten to twelve large square pores at the
base; the other pores small, roundish. The cortical shell of this species is quite the same as
that of Panartus amphiconus (Pl. 40,
fig. 5), but differs in the absence of the external envelope (or the second cortical shell), and
is covered with short, conical spines on the whole surface.
Dimensions.—Total length of the cortical shell 0.25, greatest breadth (on the base
of the distal chambers) 0.12; meshes of the proximal chambers 0.008, of the distal chambers 0.004;
bars 0.002 to 0.04, spines 0.01 to 0.02.
Habitat.—North Pacific, Station 240, surface.
14. Panartus setosus, Haeckel.
Ommatocampe setosa, Ehrenberg, 1872, Abhandl. d. k. Akad. d. Wiss. Berlin,
p. 297, Taf. viii. fig. 7.
Both proximal chambers kidney-shaped, broader than the two hemispherical distal chambers.
Meshes of the cortical shell irregular, polygonal, or subregular hexagonal; at the base of each
{382}distal chamber a circle of large square meshes. The
breadth of the pores is eight to twelve times that of the bars between them. The whole surface is
covered with small bristle-like spines.
Dimensions.—Total length of the cortical shell 0.14, greatest breadth (in the
equator of each proximal chamber) 0.07; meshes 0.008 to 0.015, bars 0.002, spines 0.01.
Habitat.—Pacific (Philippine and Californian Sea) 3300 and 2300
fathoms, Ehrenberg; Station 200, depth 250 fathoms; Station 253, depth 3125 fathoms.
15. Panartus pluteus, n. sp. (Pl. 40,
fig. 2).
Both proximal chambers kidney-shaped, with irregular, roundish pores, twice to four times as
broad as the bars; their surface everywhere covered with short conical thorns. From both polar
circles arise ten to twelve radial rods, which bear a fenestrated cap, and form thus either the
beginning of a second, external, cortical shell, or (if remaining thus) two imperfect distal
chambers. The spherical segment, which forms their surface, is concentric with the proximal
chambers, is perforated by the same irregular, roundish pores, and covered with numerous
bristle-like spines.
Dimensions.—Main axis of the cortical shell 0.17, equatorial axis (in the
constriction) 0.06; distance between the proximal and distal chambers 0.03; pores 0.003 to 0.01,
bars 0.003 to 0.005.
Habitat.—Pacific, central area, Station 272, depth 2600 fathoms.
Genus 164. Peripanartus,[203] n.
gen.
Definition.—Panartida with double cortical shell and
double medullary shell, without polar spines or tubes on both poles of the main axis.
The genus Peripanartus differs from Panartus only in the development
of an outer reticulated envelop, which mantle-like surrounds the shell and represents a second or
external cortical shell. This mantle or veil envelops either the whole internal cortical shell or
only both proximal chambers. Peripanartus exhibits the same relation to Panartus as
in the foregoing ancestral family Cypassis bears to Cyphonium.
Subgenus 1. Peripanartula, Haeckel.
Definition.—Surface of the outer cortical shell smooth, without spines
or thorns.
1. Peripanartus palliatus, n. sp.
Internal cortical shell with four kidney-shaped chambers of equal size and similar form,
identical with the cortical shell of Panartus tetrathalamus and Panartus quadrijugus
(Pl. 40,
fig. 3), with irregular, roundish pores, twice to three times as broad as the bars. It differs
from these two species {383}only by the external
cortical shell enveloping the whole internal one, in the same manner as in Peripanartus
cylindrus (Pl. 40,
fig. 6). The two shells are connected by numerous radial rods. The surface of the outer cortical
shell is quite smooth, its irregular, roundish pores about half as large as those of the inner
cortical shell.
Dimensions.—Main axis of the external cortical shell 0.27, of the internal 0.2;
greatest breadth of the former (in the equator of each chamber) 0.13, of the latter 0.09; pores of
the external cortical shell 0.002 to 0.004, of the internal 0.004 to 0.008; bars of the former
0.002, of the latter 0.003.
Habitat.—Pacific, central area, Stations 271, 272, depth 2425 to 2600
fathoms.
2. Peripanartus lævigatus, n. sp.
Internal cortical shell with four unequal chambers, both proximal kidney-shaped (with
subregular polygonal pores, twice to three times as broad as the bars), both distal nearly
hemispherical, with subregular, roundish pores of half that size. External cortical shell envelops
only the proximal chambers, and appears as the direct continuation of the internal shell of the
distal chambers, with the same small roundish pores. The outer surface is quite smooth. This
species has nearly the same structure as Peripanicium amphicorona (Pl. 40, fig. 8),
but differs from it by the absence of the polar spine-circles, and by the perfect smoothness of
the surface.
Dimensions.—Main axis of the shell 0.25, greatest breadth (in the equator of the
proximal chambers) 0.15; equatorial stricture of the external shell 0.10, of the internal 0.07;
pores of the internal shell of the proximal chambers 0.013, of the external 0.006; bars 0.003.
Habitat.—Pacific, central area, Station 274, depth 2750 fathoms.
3. Peripanartus amphiconus, n. sp. (Pl. 40, fig.
5).
Inner cortical shell with four very unequal chambers; both proximal chambers kidney-shaped
(with regular, circular, hexagonally framed pores, twice as broad as the bars); both distal
chambers conical, with a circle of large square pores at the base, the other pores very small,
roundish. Outer cortical shell very thin, quite smooth, with irregular, roundish pores, envelops
in younger specimens only the distal chambers. But the radial spines on the surface of the
internal cortical shell of the proximal chambers indicate that these will also be afterwards
enclosed by the growing of the external shell. This species exhibits the progressive (ontogenetic
as well as phylogenetic) development of Panartus amphiconus.
Dimensions.—Main axis of the outer cortical shell 0.3, its greatest breadth (at
the prominent distal ends of the radial rods between the large square pores) 0.15; pores 0.003 to
0.001, bars 0.005; main axis of the inner cortical shell 0.26, its greatest breadth 0.13; pores of
its distal chambers 0.003, of its proximal chambers 0.01; large square pores 0.03; bars 0.004.
Habitat.—Pacific, central area, Station 270, depth 2925 fathoms.
{384}
Subgenus 2. Peripanartium, Haeckel.
Definition.—Surface of the outer cortical shell covered with spines or
thorns.
4. Peripanartus atractus, n. sp. (Pl. 40, fig.
7).
Inner cortical shell with four very unequal chambers; both proximal chambers spheroidal (with
subregular, roundish pores, twice to three times as broad as the bars), both distal chambers
nearly conical, with a circle of eight to ten very large square pores at the base; the other pores
very small, roundish. Outer cortical shell envelops only the proximal chambers and the basal half
of the distal chambers, appearing as the direct continuation of the microporous covering of their
apical half. The whole external cortical shell is inflated in the equatorial zone, and hence
assumes an irregular, spindle-like appearance. Its whole surface is rough with innumerable very
short thorns.
Dimensions.—Main axis 0.28, equatorial axis 0.17; pores of the external cortical
shell 0.002 to 0.004, bars 0.002; main axis of the proximal chambers of the internal cortical
shell 0.07, equatorial axis 0.09; pores 0.007, bars 0.004.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
5. Peripanartus cylindrus, n. sp. (Pl. 40, fig.
6).
Internal cortical shell with four unequal chambers; both proximal chambers kidney-shaped (with
regular, circular, hexagonally framed pores of the same breadth as the bars), both distal chambers
nearly hemispherical, with a circle of eight to ten very large square pores at their base; the
other pores very small, roundish. External cortical shell envelops the whole internal like a
perfect hollow cylinder, which is closed at both poles by a hemispherical cap. The whole surface
of this cylindrical mantle is perforated by innumerable very small roundish pores, and covered
with small bristle-like spines.
Dimensions.—Main axis of the cylinder 0.3, equatorial axis 0.12; pores and bars
0.006; spines of its surface 0.012 to 0.15; main axis of the internal cortical shell 0.2;
equatorial axis (in the constriction) 0.06; pores and bars of its proximal chambers 0.007, of its
distal chambers 0.003; large square pores at their base 0.02.
Habitat.—Pacific, central area, Stations 270 to 274, depth 2350 to
2925 fathoms.
Genus 165. Panicium,[204] n. gen.
Definition.—Panartida with simple cortical shell and
double medullary shell, with two opposite apical spines on both poles of the main axis, or with a
group of polar spines.
The genus Panicium comprises those forms of Panartus which develop a
peculiar armature on both poles of the cortical shell. This may be only a single strong spine
{385}on each pole, lying in the main axis (subgenus
Panicidium); or a bunch or a circle of polar spines diverging radially (subgenus
Panartidium); in the latter case the polar spines can either form a bunch on the pole
itself, or a crown of thorns around it.
Subgenus 1. Panicidium, Haeckel.
Definition.—On both poles of the main axis only a single large
spine.
1. Panicium amphacanthum, n. sp.
All four chambers of the cortical shell nearly of the same size and form, kidney-shaped, with
subregular, circular pores, twice as broad as the bars; five to six pores on the half meridian,
ten to eleven on the half equator of each chamber. Surface covered with small spines. On both
poles of the main axis is a very strong, conical, apical spine, half as long as the main axis, on
the base one-fourth as broad as the equatorial axis.
Dimensions.—Main axis (without polar spines) 0.22, equatorial axis 0.06; pores
0.01, bars 0.005; length of the polar spines 0.12, basal thickness 0.015.
Habitat.—Antarctic, Station 152, depth 1260 fathoms.
2. Panicium amphistylus, n. sp.
Both proximal chambers kidney-shaped, both distal chambers nearly hemispherical, somewhat
smaller. Pores irregular, roundish, twice to three times as broad as the bars; four to six on the
half meridian, eight to ten on the half equator of each chamber; a circle of larger square pores
on the base of each distal chamber. Surface thorny. On both poles of the main axis one strong,
pyramidal, apical spine, one-third as long as the main axis, on the base one-fourth as broad as
the equatorial constriction. (Similar to Panarium tubularium, Pl. 40, fig. 9, but
without tubes.)
Dimensions.—Main axis (without polar spines) 0.2, equatorial axis 0.05; pores
0.005 to 0.01, bars 0.003; length of the polar spines 0.07, basal thickness 0.012.
Habitat.—Antarctic, Station 156, depth 1975 fathoms.
Subgenus 2. Panartidium, Haeckel.
Definition.—On both poles of the main axis a bunch or a circle of
aggregated spines.
3. Panicium scoparium, n. sp.
Both proximal chambers kidney-shaped, both distal chambers nearly hemispherical, somewhat
smaller. Pores irregular, polygonal, twice to five times as broad as the bars; six to eight on the
half meridian, twelve to fourteen on the half equator of each chamber. Surface spiny, covered with
thin, {386}bristle-like spines. On both poles of the
main axis a bunch of six to eight aggregated larger conical spines, about as high as a single
chamber.
Dimensions.—Main axis (without polar spines) 0.2, equatorial axis 0.06; pores
0.005 to 0.015, bars 0.003; length of the polar spines 0.05, basal thickness 0.005.
Habitat.—Pacific, central area, Stations 270 to 274, depth 2350 to
2925 fathoms.
4. Panicium coronatum, n. sp. (Pl. 40,
fig. 4).
Both proximal chambers kidney-shaped, covered with short stout spines. Pores irregular,
roundish, twice to four times as broad as the bars; seven to eight on the half meridian, eleven to
thirteen on the half equator of each chamber. Both distal chambers cap-like, separated from the
former by a circle of ten to twelve very large square pores; the other pores very small, roundish.
The circumpolar area is smooth, surrounded by a circle of ten to twelve very stout, conical,
radial spines, which arise from the distal ends of the bars separating the large square
meshes.
Dimensions.—Main axis (without spines) 0.22, equatorial axis 0.08; greatest
breadth in the equator of the chambers 0.1; pores of the proximal chambers 0.007 to 0.015, bars
0.004; square pores of the distal chambers 0.02 to 0.03, small pores 0.005, bars 0.004; length of
the radial spines of the polar circles 0.07, basal thickness 0.007.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
Genus 166. Peripanicium,[205] n.
gen.
Definition.—Panartida with double cortical shell and
double medullary shell, on both poles of the main axis with two opposite apical spines, or with a
bunch or circle of polar spines.
The genus Peripanicium differs from Panicium only in the development
of an outer reticulated envelope, which mantle-like surrounds the shell, and represents a second
or external cortical shell. As in Panicium, its ancestral form, so also in
Peripanicium, there can be distinguished two subgenera:—Peripanicea, where
only a single large spine arises from each pole (lying in the main axis), and Peripanicula,
where a variable number of spines is to be found, either diverging bunch-like from the pole
itself, or surrounding it as a crown-like polar circle.
Subgenus 1. Peripanicea, Haeckel.
Definition.—On both poles of the main axis only a single large
spine.
1. Peripanicium amphixiphus, n. sp.
Internal cortical shell with four unequal chambers, with thorny surface. Both proximal chambers
kidney-shaped (with subregular hexagonal pores, three times as broad as the bars). {387}Both distal chambers nearly hemispherical (with a circle
of ten to twelve very large square pores at their base, the other pores very small). From both
poles arises a single, strong, conical spine, half as long as the main axis. The internal cortical
shell is very much like that of Panarium tubularium (Pl. 40, fig. 9),
but differs by the solid polar spines (instead of the hollow tubules) and by the formation of an
external, cylindrical, very thin, cortical shell, which envelops the whole internal, and is
connected with it by numerous beams. The surface of the external shell is quite smooth, and
perforated by innumerable very small roundish pores.
Dimensions.—Main axis of the external cortical shell 0.25, of the internal 0.19;
transverse axis of the former 0.13, of the latter 0.08; pores of the proximal chambers of the
internal shell 0.01, bars 0.003; pores of external cortical shell 0.002, bars 0.001; length of
both polar spines 0.12, basal thickness of them 0.02.
Habitat.—South Atlantic, near Tristan da Cunha, Station 333, depth
2025 fathoms.
Subgenus 2. Peripanicula, Haeckel.
Definition.—On both poles of the main axis a bunch or a circle of
aggregated polar spines.
2. Peripanicium amphicorona, n. sp. (Pl. 40, fig.
8).
Peripanartus amphicorona, Haeckel, 1881, Prodromus et Atlas (pl. xl. fig.
8).
Internal cortical shell with four very unequal chambers. Both proximal chambers kidney-shaped,
with subregular, polygonal pores, three times as broad as the bars. Both distal chambers double
cone-shaped; the proximal cone of these formed by a circle of ten to twelve radial beams, which
separate the same number of large square meshes; the distal cone with five to six circles of very
small roundish, irregular pores. The continuation of the thin porous lamella of this distal cone
forms the external cortical shell, whose surface is a little rough with innumerable very small
thorns. On both poles is a large circular opening (nearly as broad as the equatorial constriction
of the inner shell), surrounded by a delicate crown of thorns. The spines of this crown equal the
diameter of the polar opening; they are very thin and bristle-like, connected by a few bridges and
diverge outwards.
Dimensions.—Main axis (without the crowns of polar thorns) 0.26, greatest breadth
(in the equator of the proximal chambers) 0.15; equatorial constriction of the external shell
0.12, of the internal 0.07; pores of the proximal chambers of the internal cortical shell 0.01, of
the external 0.005, bars 0.003; diameter of the circular polar opening 0.05; length of the polar
spines 0.05.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
3. Peripanicium coronarium, n. sp.
Internal cortical shell with four unequal chambers. Both proximal chambers kidney-shaped, with
irregular, roundish pores, twice to four times as broad as the bars. Both distal chambers
cap-like, with a basal circle of ten to twelve large square meshes, the other pores very small.
External {388}cortical shell very thin, with smooth
surface, and very small, irregular, roundish pores; it envelops the whole internal shell at a
constant distance, which equals the breadth of the square meshes. From the distal ends of the
radial beams between the square meshes arise on every polar circle ten to twelve strong spines, as
direct prolongations of those beams. These form two regular, polar crowns of thorns. The inner
part of the thorns (between both shells) has only one-third to one-fourth the length of the outer
free part. (This species represents a further development of Panicium coronatum, Pl. 40, fig. 4, by
secondary formation of an external mantle, like that of Peripanartus atractus, Pl. 40, fig.
7.)
Dimensions.—Main axis 0.27, greatest breadth 0.15; pores of the internal proximal
chambers 0.008 to 0.016, bars 0.004; square meshes of the distal chambers 0.03; pores of the outer
cortical shell 0.003; length of the spines of the polar circles 0.1 to 0.15.
Habitat.—Pacific, central area, Station 274, depth 2750 fathoms.
Genus 167. Panarium,[206] Haeckel, 1881,
Prodromus, p. 463.
Definition.—Panartida with simple cortical shell and
double medullary shell, with two hollow fenestrated tubes, opposite on the poles of the main
axis.
The genus Panarium differs from Panartus by two hollow latticed
tubes, which start from both poles of the main axis and lie in it. It repeats therefore in this
family the same peculiar and remarkable formation, which we find in Pipettella among the
Ellipsida, in Pipetta among the Druppulida, in Cannartidium among the Cyphinida, in
Cannartus among the Artiscida, &c.
Subgenus 1. Panarelium, Haeckel.
Definition.—Surface of the shell smooth, without spines or thorns.
1. Panarium facettarium, n. sp.
Surface of the cortical shell smooth. All the four chambers nearly of the same size and form,
kidney-shaped, about twice as broad as long. Pores of these subregular, circular, with hexagonal
frames, about as broad as the bars; nine to ten on the half meridian, twelve to fourteen on the
half equator of each chamber. Polar tubuli nearly cylindrical, longer than half the main axis of
the cortical shell, about one-fourth as broad as the equatorial constriction. Pores of the tubuli
of the same shape as those of the chambers, but only half as large. (This species is like
Pipetta tuba, Pl. 39,
fig. 7, but distinguished by three parallel transverse constrictions.)
Dimensions.—Total length of the shell (without tubuli) 0.26; breadth of each
chamber 0.12; pores 0.01, bars 0.01; length of the tubuli 0.15, breadth of them 0.03.
Habitat.—Pacific central area, Station 268, depth 2900 fathoms.
{389}
2. Panarium pipettarium, n. sp.
Surface of the cortical shell smooth. Both proximal chambers nearly kidney-shaped; both distal
chambers somewhat smaller, hemispherical. Pores of all four chambers subregular, circular, without
hexagonal frames, twice as broad as the bars; six to seven pores on the half meridian, ten to
eleven on the half equator of each chamber. Polar tubuli slender, conical, nearly half as long as
the main axis of the shell, one-third as broad at the base as the equatorial constriction. Pores
of the tubuli of the same shape as those of the chambers, but only half as large. (This species
resembles somewhat Cannartiscus amphiconiscus, Pl. 39, fig. 19,
but with double the number of chambers.)
Dimensions.—Total length of the shell (without tubuli) 0.25, greatest breadth 0.1;
pores 0.008, bars 0.004; length of the polar tubuli 0.11, basal breadth 0.03.
Habitat.—Pacific, western tropical part, Station 225, depth 4475
fathoms.
Subgenus 2. Panaromium, Haeckel.
Definition.—Surface of the shell spiny, covered with short thorns.
3. Panarium annularium, n. sp.
Surface of the cortical shell thorny, everywhere covered with small bristle-like spines. All
four chambers nearly of the same size and form, kidney-shaped. Their pores subregular, circular,
three to four times as broad as the bars; seven to eight on the half meridian, ten to eleven on
the half equator of each chamber. Polar tubuli conical, nearly half as long as the main axis, half
as broad at the base as the equatorial constriction. Pores of the tubuli like those of the
chambers, but only half as broad.
Dimensions.—Total length of the shell (without tubuli) 0.16; breadth of each
chamber 0.07; pores 0.006 to 0.008, bars 0.002; length of the polar tubuli 0.07, basal breadth
0.03.
Habitat.—Northern Pacific, Station 253, depth 3125 fathoms.
4. Panarium artophorum, n. sp.
Surface of the shell thorny, with scattered small spines. Both proximal chambers nearly
kidney-shaped; both distal hemispherical, somewhat smaller. Pores of all chambers irregular,
roundish, little broader than the bars; four to five on the half meridian, six to eight on the
half equator of each chamber. Polar tubuli cylindrical, longer than the half main axis, only
one-fourth as broad the equatorial constriction. Pores of the tubuli only one-third to one-fourth
as broad as those of the chambers.
Dimensions.—Total length of the shell (without tubuli) 0.22, greatest breadth (in
the equator of the proximal chambers) 0.09; pores and bars 0.008 to 0.012; length of the polar
tubuli 0.14, breadth of them 0.02.
Habitat.—Southern Pacific, Station 289, depth 2550 fathoms.
{390}
5. Panarium tubularium, n. sp. (Pl. 40,
fig. 9).
Surface of the cortical shell thorny, covered with small spines. Both proximal chambers nearly
kidney-shaped, with irregular, polygonal pores, three to four times as broad as the bars; five to
six pores on the half meridian, nine to ten on the half equator of each chamber. Both distal
chambers nearly hemispherical, with a circle of ten to twelve large square pores at their base;
the other pores much smaller, irregular, roundish. Polar tubuli prismatic, with prominent edges,
nearly half as long as the main axis, only one-fourth as broad as the equatorial constriction.
Pores of the tubuli very small, in longitudinal series between the edges.
Dimensions.—Total length of the shell (without tubuli) 0.18 to 0.2, greatest
breadth (in the equator of the proximal chambers) 0.07 to 0.09; pores 0.003 to 0.01, bars 0.002 to
0.004; length of the polar tubuli 0.06 to 0.09, breadth of them 0.015 to 0.02.
Habitat.—Pacific, central area, Stations 270 to 272, depth 2425 to
2925 fathoms.
Genus 168. Peripanarium,[207] n.
gen.
Definition.—Panartida with double cortical shell and
double medullary shell, with two hollow fenestrated tubes, opposite on the poles of the main
axis.
The genus Peripanarium differs from Panarium only in the development
of an outer reticulated envelop, which mantle-like surrounds the shell and represents a second or
external cortical shell. It bears therefore to Panarium the same relation as
Peripanartus to Panartus, &c.
1. Peripanarium cenoconicum, n. sp.
Internal cortical shell with four kidney-shaped chambers of nearly the same form and size;
their pores irregular, roundish, twice to four times as broad as the bars. External cortical
shell cylindrical, hemispherical at both poles, with irregular, very small, roundish pores, only
one-third to one-half as large as those of the internal. From both poles of the internal cortical
shell arises a hollow conical fenestrated tube, which perforates the external. The outer free
part of this slender cone equals in length the inner part or the distance between the two cortical
shells, which is about equal to the length of one chamber. External surface covered with small
thorns. (This species has the same appearance as Desmartus larvalis, Pl. 40, fig. 12,
but has only four chambers instead of six.)
Dimensions.—Main axis (without cones) 0.23, greatest breadth 0.11; pores of the
inner cortical shell 0.004 to 0.009, of the outer 0.002 to 0.003; bars of the former 0.002, of the
latter 0.001; total length of the polar cones 0.05, basal thickness of them 0.03.
Habitat.—West Tropical Pacific, Station 225, depth 4475 fathoms.
{391}
2. Peripanarium cenocylindricum, n. sp.
Internal cortical shell with four unequal chambers, the two proximal kidney-shaped, the two
distal conical, and somewhat smaller; pores irregular, polygonal-roundish, twice to five times as
broad as the bars. External cortical shell nearly spindle-shaped, in the equatorial zone inflated,
conical, tapering towards both poles, its pores very small, roundish, its surface covered with
small thorns. The shell of this species resembles very much Peripanartus atractus (Pl. 40, fig. 7),
but differs by the hollow cylindrical polar tubes, which arise from both poles of the inner
cortical shell and attain more than half the length of the main axis. They are open at their ends
(broken away), about as broad as the outer medullary shell, and perforated by irregular, roundish
pores.
Dimensions.—Main axis (without polar tubes) 0.28, greatest breadth (in the
equator) 0.16; pores of the inner cortical shell 0.004 to 0.01, of the outer 0.002 to 0.005; bars
of the former 0.004, of the latter 0.002; total length of the polar tubes 0.16, breadth of them
0.03.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
Family XVII. Zygartida, Haeckel (Pl. 40, figs.
10-13).
Zygartida, Haeckel, 1881, Prodromus, p.462 (sensu
restricto).
Definition.—Prunoidea with annulated cortical shell, the
external shell being divided by five or more parallel transverse constrictions into six or more
chambers, enclosing in the centre two internal concentric shells (medullary shells). Central
capsule cylindrical, commonly annulated (corresponding to the transverse annular constrictions of
the cortical shell).
The family Zygartida, the seventh and last of the Prunoidea, comprises, in the sense here restricted, all those Prunoidea in which the cortical shell is annulated and composed of six to
twenty or more (at least six) cameræ, lying one behind another in the elongated main axis, and
separated by five or more annular constrictions. When I constituted this family in 1881 (loc.
cit.), I had given to it a much wider extent, embracing all those Prunoidea which exhibit annular constrictions of the cortical shell; as the
number of these, in consequence of further researches, is much increased, it seems now more
convenient to restrict the family to the extent here given.
No doubt the Zygartida must be derived from the Panartida by progressive growth of the cortical
shell in the main axis and accompanying increase of the number of its chambers. Whilst this number
in the Panartida is constantly restricted to four, in the Zygartida it amounts to ten, twenty, or
more, and is in the lowest case six (Pl. 40,
fig. 10). The maximum number is variable in the different species, but of course also different in
the various degrees of individual development. Each Zygartid is at the beginning of its growth a
Cyphinid, later a Panartid. The number of the annular constrictions separating the chambers and
lying in parallel transverse planes is at least {392}five, but may amount to nineteen, to twenty-one or more; they lie
constantly in pairs on both sides of the equatorial constriction (Pl. 40, figs. 12,
13).
The cortical shell may either remain simple (Ommatocampe, Pl. 40, fig. 10),
or become double (Desmocampe, Pl. 40,
fig. 12), or sometimes triple (Zygocampe, (Pl. 40, fig. 13).
In the latter cases the outer (secondary and tertiary) cortical shells are commonly incomplete,
and only developed around the proximal chambers of the complete first (primary) cortical shell,
its distal chambers remaining simple.
The Medullary Shell is constantly double, as in the Panartida; its form is either
spherical or lenticular, compressed in the direction of the main axis. It is always connected
with the equatorial constriction of the cortical shell by a number of radial beams, lying either
in the equatorial plane or on each side of it (Pl. 40, figs.
10-13).
The Central Capsule of the Zygartida is constantly cylindrical; its increasing growth on
both poles of the axis corresponds to that of the including cortical shell. Commonly (but not
always) its cylindrical surface is annulated, with five or more transverse strictures,
corresponding to those of the cortical shell. From the inner surface of the latter it is separated
by a jelly-mantle, the calymma.
Synopsis of the Genera of Zygartida.
III. Ommacampida.
1. Cortical shell simple.
(Medullary shell double.) |
|
Without polar tubes, |
169. Ommatocampe. |
| With two hollow fenestrated tubes, on the poles of the axis, |
170. Ommatartus. |
III. Desmocampida.
2. Cortical shell double.
(Medullary shell double.) |
|
Without polar tubes, |
171. Desmocampe. |
| With two hollow fenestrated tubes, on the poles of the axis, |
172. Desmartus. |
III. Zygocampida.
3. Cortical shell triple.
(Medullary shell double.) |
|
Without polar tubes, |
173. Zygocampe. |
| With two hollow fenestrated tubes, on the poles of the axis, |
174. Zygartus. |
| I. Ommacampida. 1. Cortical shell simple. (Medullary shell double.) |
|
|
Without polar tubes, |
|
|
|
|
|
|
169. Ommatocampe. |
|
|
|
|
|
|
With two hollow fenestrated tubes on the poles of the axis, |
|
|
|
|
|
|
170. Ommatartus. |
|
|
|
|
| II. Desmocampida. 2. Cortical shell double. (Medullary shell double.) |
|
|
Without polar tubes, |
|
|
|
|
|
|
171. Desmocampe. |
|
|
|
|
|
|
With two hollow fenestrated tubes on the poles of the axis, |
|
|
|
|
|
|
172. Desmartus. |
|
|
|
|
| III. Zygocampida. 3. Cortical shell triple. (Medullary shell double.) |
|
|
Without polar tubes, |
|
|
|
|
|
|
173. Zygocampe. |
|
|
|
|
|
|
With two hollow fenestrated tubes on the poles of the axis, |
|
|
|
|
|
|
174. Zygartus. |
|
|
|
|
Genus 169. Ommatocampe,[208] Ehrenberg,
1860, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 832.
Definition.—Zygartida with simple cortical shell and
double medullary shell, without polar tubes.
The genus Ommatocampe was founded by Ehrenberg in 1860 for one of his
"Haliommatina," with the following diagnosis:—"Shell rod-like, long, articulate, with
nucleus, without spines, with four or more joints." The species figured by him, Ommatocampe
{393}polyarthra, exhibits five pairs of chambers on
both sides of the meridian plane of the cortical shell, the centre of which includes a double
medullary shell. It represents the most simple form of all Zygartida, and may be derived
phylogenetically from Cyphonium simply by multiplication of the chambers of the cortical
shell, growing on both poles of the main axis.
Subgenus 1. Ommatocampium.
Definition.—Surface of the cortical shell smooth or rough, without
thorns or spines, also without polar spines.
1. Ommatocampe polyarthra, Ehrenberg.
Ommatocampe polyarthra, Ehrenberg, 1872, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 279, Taf. vi. fig. 9.
Cortical shell with smooth surface, composed of six to ten chambers of the same size and form.
Every chamber kidney-shaped, about twice as broad as long, with three transverse rows of circular,
subregular pores, twice to three times as broad as the bars. Both medullary shells spherical.
Dimensions.—Length of the six-chambered cortical shell 0.15; greatest breadth of
each chamber 0.04; pores 0.005 to 0.007, bars 0.002 to 0.003.
Habitat.—Californian Sea, depth 2600 fathoms, Ehrenberg; Pacific,
central area, Station 268, depth 2900 fathoms; fossil in the Tertiary rocks of Barbados,
Haeckel.
2. Ommatocampe increscens, Stöhr.
Ommatocampe increscens, Stöhr, 1880, Palæontogr., vol. xxvi. p. 90, Taf.
ii. fig. 2, a, b.
Cortical shell with rough surface, composed of six to ten chambers of nearly the same size and
form; the breadth of the chambers a little increasing towards both poles. Every chamber
kidney-shaped, the proximal chambers twice as broad as long, with four or five transverse rows of
pores; the distal chambers three to four times as broad as long, with two or three transverse rows
of pores. Form of the pores irregular, roundish, bars between them in the distal part smaller, in
the proximal part larger than the pores. Both medullary shells compressed lenticular.
Dimensions.—Length of the six-chambered cortical shell 0.22; greatest breadth of
the chambers 0.09 to 0.1; pores and bars 0.003 to 0.006.
Habitat.—Fossil in the Tertiary rocks of Sicily, Grotte, Stöhr.
3. Ommatocampe annulata, n. sp.
Cortical shell with smooth surface, composed of six to twelve chambers of nearly the same size
and form. Each chamber kidney-shaped, twice as broad as long, with four to five transverse rows of
{394}irregular, roundish pores, once to five times as
broad as the bars. On the base of each distal chamber (beginning in the second or third pair) a
circle of twelve to sixteen larger square pores. Both medullary shells compressed lenticular.
(Resembles the inner cortical shell of Desmartus larvalis, Pl. 40, fig. 12,
but is without the polar tubes and the outer envelope of this species.)
Dimensions.—Length of the six-chambered cortical shell 0.23; greatest breadth of
each chamber 0.07; pores 0.002 to 0.01, bars 0.002.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific, in various
depths.
Subgenus 2. Ommatocampula, Haeckel.
Definition.—Surface of the cortical shell thorny or spiny, but without
peculiar polar spines and without regular coronals of spines.
4. Ommatocampe erucæformis, n. sp.
Cortical shell with spiny surface, composed of six to twelve chambers of nearly the same size
and form, the distal chambers a little smaller. The form and structure of the cortical shell is
nearly the same as in the foregoing species, only the pores are somewhat larger, and the whole
surface is covered with bristle-like radial spines, about half as long as the breadth of one
chamber. Both medullary shells compressed and lenticular.
Dimensions.—Length of the six-chambered cortical shell 0.24; greatest breadth of
each chamber 0.08; pores 0.005 to 0.012, bars 0.002.
Habitat.—North Atlantic, Canary Islands, Station 353, depth 2965
fathoms.
5. Ommatocampe nereis, n. sp. (Pl. 40,
fig. 10).
Cortical shell with spiny surface, composed of six (or more) chambers of different size and
structure. Both proximal chambers kidney-shaped, with subregular, circular, hexagonally framed
pores, twice as broad as the bars. All following chambers cap-like, with much smaller, irregular,
roundish pores, at the base of every chamber a circle of ten to twelve large square pores. Spines
of the surface short, irregularly scattered. Both medullary shells spherical. (All the observed
specimens possessed only six chambers.)
Dimensions.—Length of the six-chambered cortical shell 0.25; greatest breadth of
each chamber 0.08; pores of both proximal chambers 0.01, bars 0.005; basal pores of the other
chambers 0.02; pores of their distal caps 0.003 to 0.006, bars 0.003.
Habitat.—Pacific, central area, Stations 271 to 274, depth 2350 to
2750 fathoms.
Subgenus 3. Ommatocorona, Haeckel.
Definition.—Surface of the cortical shell spiny, on every chamber a
regular circle or coronal of radial spines.
{395}
6. Ommatocampe chætopodum, n. sp.
Cortical shell with spiny surface, composed of six (or more) kidney-shaped chambers of
different size and form. Both proximal chambers kidney-shaped, with subregular, circular,
hexagonally framed pores, about as broad as the bars. All the following chambers hemispherical,
with irregular, roundish pores; at the base of every chamber a circle of ten to twelve larger
square pores. The beams between these latter are prolonged into free radial spines, twice as thick
as the bars. Therefore every chamber is surrounded by a circle of radial spines, like those of
Panicium coronatum (Pl. 40,
fig. 4). Both medullary shells lenticular.
Dimensions.—Length of the six-chambered cortical shell 0.3, greatest breadth 0.08,
pores and bars of both proximal chambers 0.01; pores of the other chambers 0.002 to 0.008; square
pores 0.02, bars 0.005; length of the coronal spines 0.03, thickness 0.01.
Habitat.—Indian Ocean, Madagascar, Rabbe.
Subgenus 4. Ommatacantha, Haeckel.
Definition.—Surface of the shell smooth or spiny, on the poles of the
main axis occur two strong opposite polar spines.
7. Ommatocampe amphilonche, n. sp.
Cortical shell composed of six kidney-shaped chambers of nearly the same size and structure;
every chamber twice as broad as long, with four to five transverse rows of irregular, roundish
pores, once to four times as broad as the bars. Both medullary shells lenticular. Surface of the
cortical shell covered with bristle-like spines. On both poles of the main axis one larger,
strong, conical spine, about half as long as this axis, and on the base as broad as the inner
medullary shell. (Resembles on the whole Desmartus larvalis, Pl. 40, fig. 12,
but without external mantle, and with two solid polar spines instead of the polar tubes.)
Dimensions.—Length of the six-chambered cortical shell 0.24; greatest breadth of
each chamber 0.08; pores 0.002 to 0.08, bars 0.002; length of the polar spines 0.12, basal
thickness 0.02.
Habitat.—Pacific, central area, Station 266, depth 2750 fathoms.
Genus 170. Ommatartus,[209] Haeckel,
1881, Prodromus, p. 463.
Definition.—Zygartida with simple cortical shell and
double medullary shell, with two hollow fenestrated tubes, opposite on both poles of the main
axis.
The genus Ommatartus differs from Ommatocampe by the development of
two hollow fenestrated tubes on both poles of the main axis, and bears therefore the same relation
to it as Cannartidium to Cyphonium. The former two genera differ from the two latter
by the augmentation of the chambers of the cortical shell.
{396}
1. Ommatartus amphicanna, n. sp.
Cortical shell with spiny surface, composed of six chambers of equal size and kidney-shaped;
every chamber with four to five transverse rows of irregular, roundish pores, twice to three times
as broad as the bars. Both medullary shells lenticular. Polar tubes conical, about as long as one
chamber, with smaller pores. Both medullary shells spheroidal, more or less compressed. (Resembles
Desmartus larvalis, Pl. 40,
fig. 12, but is without the external shell.)
Dimensions.—Length of the six-chambered cortical shell 0.24; greatest breadth of
each chamber 0.07; pores 0.008 to 0.012, bars 0.004; length of the polar tubes 0.04 to 0.05, basal
thickness 0.02.
Habitat.—Pacific, central area, Stations 270 to 274, depth 2350 to
2925 fathoms.
2. Ommatartus amphisiphon, n. sp.
Cortical shell with spiny surface, composed of six or eight chambers of different size and
form. Both proximal chambers kidney-shaped, each with five to six transverse rows of irregular,
polygonal pores, three to four times as broad as the bars. The other (four to six) chambers
somewhat smaller, more hemispherical, with smaller, irregular pores, but on the base of each a
circle of ten to twelve larger square pores. Polar tubuli prismatic, about half as long as the
main axis, with prominent edges and longitudinal rows of smaller pores between them. Both
medullary shells lenticular. (Resembles closely Panarium tubularium, Pl. 40, fig. 9, but
with six to eight chambers instead of four, and with longer tubuli.)
Dimensions.—Length of the six-chambered cortical shell 0.26; greatest breadth of
the proximal chambers 0.09, of the distal chambers 0.07; pores of the former 0.01, of the latter
0.004; large square pores 0.013, bars 0.003; length of the polar tubes 0.1 to 0.12, thickness
0.02.
Habitat.—South Pacific, Station 297, depth 1775 fathoms.
3. Ommatartus amphobolus, n. sp.
Cortical shell with spiny surface, composed of six chambers of different size and form. Both
proximal chambers kidney-shaped, with four to five transverse rows of subregular, circular pores,
twice as broad as the bars. Both middle chambers cap-like, with a basal circle of ten to twelve
larger square pores, and small roundish pores on the cap. Both distal chambers smaller, conical,
with very small roundish pores. Polar tubuli conical, nearly half as long as the main axis, also
with very small pores. Both medullary shells lenticular.
Dimensions.—Length of the six-chambered shell 0.28, greatest breadth 0.08; pores
of the proximal chambers 0.01, bars 0.005; pores of the distal chambers and the polar tubes 0.002
to 0.004, bars 0.002; length of the polar tubes 0.13, basal thickness 0.03.
Habitat.—North Pacific, Station 253, depth 3125 fathoms.
Genus 171. Desmocampe,[210] n. gen.
Definition.—Zygartida with double cortical shell and
double medullary shell, without polar tubes.
{397}
The genus Desmocampe differs from Ommatocampe in the duplication of
the jointed cortical shell; the radial spines, which start from the surface of the inner cortical
shell, are connected one with another by transverse communicating branches which form an outer
envelop around it; but this reticulated mantle is commonly not quite perfect and more or less
irregular.
1. Desmocampe catenula, n. sp.
Inner cortical shell with six to eight chambers of the same size and form. Every chamber
kidney-shaped, with three to four transverse rows of circular, subregular pores, twice as broad as
the bars. Outer cortical shell cylindrical, hemispherical at both poles, with smooth surface and
irregular, roundish pores of very different size. Distance between the two cortical shells equals
the diameter of the outer medullary shell, which, like the inner, is spherical. (Resembles
Ommatocampe polyarthra, Ehrenberg, 1872, loc. cit., Taf. vi, fig. 9, but differs in the
external mantle.)
Dimensions.—Length of the six-chambered inner cortical shell 0.16, of the outer
0.21; greatest breadth of each chamber of the former 0.05, of the latter 0.09; pores of the inner
0.006, bars 0.003; pores of the outer 0.002 to 0.008, bars 0.002.
Habitat.—North Pacific, Station 237, off Japan, surface.
2. Desmocampe tænioides, n. sp.
Inner cortical shell with six to ten chambers of nearly the same size and form; the distal
chambers somewhat smaller. Every chamber kidney-shaped, with four to five transverse rows of
irregular roundish pores, twice to three times as broad as the bars. Outer cortical shell
cylindrical, hemispherical at both poles, with spiny surface; its pores like those of the inner,
but the bars between them much thinner. Distance between the two cortical shells equals the
diameter of the outer medullary shell. Both medullary shells lenticular. (Resembles Desmartus
larvalis, Pl. 40,
fig. 12, but is without polar tubes.)
Dimensions.—Length of the six-chambered inner cortical shell 0.25, of the outer
0.3; greatest breadth of the former 0.07, of the latter 0.11; pores 0.005 to 0.012; bars of the
inner shell 0.004, of the outer 0.001.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
3. Desmocampe aphrodite, n. sp.
Inner cortical shell with six to ten chambers of different size and structure. Both proximal
chambers kidney-shaped, with five to six transverse rows of subregular, circular,
hexagonally-framed pores, twice as broad as the bars. All following chambers cap-like, with much
smaller, irregular, roundish pores, on the base of each a circle of ten to twelve large square
pores. Outer cortical shell cylindrical, on both poles hemispherical, with spiny surface and very
delicate network of small polygonal pores. Both medullary shells spherical. (The inner cortical
shell of this species resembles that of Ommatocampe nereis, Pl. 40, fig. 10;
the outer that of Cyphocolpus virginis, Pl. 40, fig.
11.)
{398}
Dimensions.—Length of the six-chambered internal cortical shell 0.25, of the outer
0.3; greatest breadth of the former 0.08, of the latter 0.12; pores of the proximal chambers of
the inner shell 0.01, bars 0.005; pores of the distal chambers 0.003 to 0.005, square pores 0.02,
bars 0.003; pores of the external cortical shell 0.002 to 0.006, bars 0.001.
Habitat.—North Atlantic, Station 353, depth 2965 fathoms.
4. Desmocampe atractus, n. sp.
Inner cortical shell with six chambers of very different size and structure. Both proximal
chambers kidney-shaped, with seven to eight transverse rows of subregular, roundish pores, twice
to three times as broad as the bars. Both middle chambers cap-like, on the base with a circle of
eight to ten very large square pores, on the distal cap with small irregular, roundish pores. Both
distal chambers conical, also with small irregular, roundish pores. Outer cortical shell
spindle-shaped, inflated in the equatorial zone, tapering conically towards both poles, with very
delicate network of small roundish pores and thin bars. Surface covered with innumerable very
small spines. Both medullary shells lenticular. (Resembles very much Peripanartus atractus,
Pl. 40, fig.
7, but differs in the number of the chambers and their proportion to the outer mantle, which
envelops spindle-like all six chambers.)
Dimensions.—Length of the six-chambered internal cortical shell 0.27, of the outer
0.32; greatest breadth of the former 0.09, of the latter 0.15; pores of the inner cortical shell
(on an average)—proximal chambers 0.008, middle chambers 0.02, distal chambers 0.004, bars
0.004; pores of the outer cortical shell 0.002 to 0.004, bars 0.002.
Habitat.—North Pacific, Station 241, depth 2300 fathoms.
Genus 172. Desmartus,[211] n. gen.
Definition.—Zygartida with double cortical shell and
double medullary shell, with two hollow fenestrated tubes, opposite on both poles of the main
axis.
The genus Desmartus differs from Desmocampe by the development of two
hollow fenestrated tubes, opposite on both poles of the main axis, and bears therefore the same
relation to it as Ommatartus to Ommatocampe. Both the former genera can be produced
from the two latter by duplication of the cortical shell.
1. Desmartus larvalis, n. sp. (Pl. 40,
fig. 12).
Zygartus larvalis, Haeckel, 1881, Prodromus et Atlas (pl. xl, fig.
12).
Inner cortical shell composed of six kidney-shaped chambers of nearly the same size and
structure; every chamber twice as broad as long, with four to five transverse rows of irregular,
roundish pores, twice to five times as broad as the bars; the basal pores of the distal chambers
{399}somewhat larger and more square. Outer cortical
shell cylindrical, hemispherical at both poles, with irregular, polygonal pores, on an average
twice as large as those of the inner, but the bars between them much thinner. Outer surface spiny.
Both medullary shells lenticular. Polar tubes conical, a little longer than one internal chamber,
as broad at the base as the inner medullary shell. Sometimes the tubes exhibit prominent edges (as
in the lower spine of fig. 12); the pores of these are very small, and roundish.
Dimensions.—Length of the six-chambered internal cortical shell 0.23, of the
external 0.3; greatest breadth of the former 0.07, of the latter 0.11; pores of the inner shell
0.004 to 0.01, of the outer 0.01 to 0.02; bars of the former 0.002, of the latter 0.001; length of
the polar tubes 0.05, basal thickness 0.02.
Habitat.—North Pacific, Station 241, depth 2300 fathoms.
2. Desmartus tubulatus, n. sp.
Inner cortical shell composed of six to ten kidney-shaped chambers, tapering in size towards
both poles, every chamber with six to seven transverse rows of irregular, roundish pores, twice to
six times broader than the bars. Outer cortical shell spindle-shaped, in the equatorial zone
inflated, tapering towards both poles, its network similar to the inner, only more delicate. Polar
tubes conical, twice as long as an inner chamber, as broad at the base as the outer medullary
shell. Both medullary shells lenticular.
Dimensions.—Length of the six-chambered internal cortical shell 0.25, of the
external 0.32; greatest breadth of the former 0.08, of the latter 0.12; pores of the inner shell
0.002 to 0.012, of the outer 0.003 to 0.01; bars of the former 0.002, of the latter 0.001; length
of the polar tubes 0.07, basal thickness 0.03.
Habitat.—North Pacific, Station 256, depth 2950 fathoms.
Genus 173. Zygocampe,[212] n. gen.
Definition.—Zygartida with triple (or multiple) cortical
shell and double medullary shell, without polar tubes.
The genus Zygocampe differs from Desmocampe and Ommatocampe by
the multiplication of the cortical shell, which is composed of three or more concentric envelopes.
The three mentioned genera form therefore a phylogenetic series, produced by the concentric
increase on the outside of the jointed cortical shell. Commonly the second cortical shell is not
as complete as the first (or innermost), and the third (or outermost) is yet more incomplete.
Rarely the number of the concentric cortical shells surpasses three.
1. Zygocampe pupula, n. sp.
Inner cortical shell with six to twelve chambers of nearly the same size and form. Every
chamber kidney-shaped, with four to five transverse rows of circular, subregular pores, twice as
{400}broad as the bars. Middle cortical shell
cylindrical, hemispherical at both poles, with subregular circular pores somewhat smaller than
those of the inner; the bars also thinner. Outer cortical shell of the same form as the middle,
but with very delicate network, and quite irregular, roundish pores and very thin bars. Surface
quite smooth. Both medullary shells lenticular. (May be regarded in a phylogenetic as well as an
ontogenetic sense, as the further developmental form of Desmocampe catenula and
Ommatocampe polyarthra.)
Dimensions.—Length of the six-chambered inner cortical shell 0.17, of the middle
0.22, of the outer 0.27; greatest breadth of the first 0.05, of the second 0.09, of the third
0.12; pores of the inner cortical shell 0.005, of the middle 0.004, of the outer 0.003 to 0.012;
bars of the first 0.003, of the second 0.002, of the third 0.001.
Habitat.—Pacific, central area, Station 266, depth 2750 fathoms.
2. Zygocampe corasium, n. sp.
Inner cortical shell with six to eight chambers of different size and form. Both proximal
chambers kidney-shaped, with five to six transverse rows of subregular, circular, hexagonally
framed pores, twice as broad as the bars. All following chambers cap-like, the distal somewhat
smaller; their pores much smaller, irregular, roundish; only at the base of each chamber a circle
of ten to twelve large square pores. Middle cortical shell cylindrical, in the equatorial zone a
little constricted, hemispherical at both poles, with irregular delicate network of roundish,
polygonal meshes. Outer cortical shell of the same form as the middle, but with a very delicate
and quite irregular network of polygonal meshes. Commonly this outer mantle is incomplete, and
sometimes interwoven in a spongy manner with the middle (or also with the inner). Surface covered
with many irregular, thin, bristle-like spines. Both medullary shells lenticular. (May be
considered in a phylogenetic and ontogenetic sense as a further developmental stage of
Desmocampe aphrodite and Ommatocampe nereis, Pl. 40, fig.
10.)
Dimensions.—Length of the six-chambered inner cortical shell 0.24, of the middle
0.3, of the outer 0.36; greatest breadth of the first 0.08, of the second 0.12, of the third 0.16,
pores of the first (on an average) 0.005 to 0.02, of the second 0.003 to 0.015, of the third 0.01
to 0.05; bars corresponding 0.005 or 0.003 or 0.001.
Habitat.—North Pacific, Station 253, depth 3125 fathoms.
3. Zygocampe chrysalidium, n. sp. (Pl. 40, fig.
13).
Inner cortical shell with six to eighteen chambers of different size and form. Both proximal
chambers kidney-shaped, with six to seven transverse rows of subregular, circular pores, three to
four times as broad as the bars. All following chambers with more irregular pores, with a circle
of ten to twelve larger square pores at the base. Middle cortical shell with smaller, irregular,
roundish pores. Outer cortical shell with larger polygonal, quite irregular pores. Both outer
shells appear commonly incomplete or somewhat irregularly developed, and sometimes in a spongy
manner interwoven with one another. Surface covered with irregular, bristle-like spines. Both
medullary shells spherical. The breadth of the chambers decreases gradually towards both poles, so
that the whole shell assumes a spindle form. Sometimes on both poles is developed a short conical
{401}polar tube (in fig. 13 only on the upper pole), and
so this species is transformed into Zygartus chrysalis.
Dimensions.—Length of the six-chambered inner cortical shell 0.25, of the middle
0.3, of the outer 0.35; greatest breadth of the first 0.07, of the second 0.11, of the third 0.14;
pores (on an average) 0.01 (0.002 to 0.02), bars 0.002 to 0.005.
Habitat.—Pacific, central area, Stations 272 to 274, depth 2350 to
2750 fathoms.
Genus 174. Zygartus,[213] Haeckel, 1881,
Prodromus, p. 463.
Definition.—Zygartida with triple (or multiple) cortical
shell and double medullary shell, with two hollow fenestrated tubes, opposite on both poles of the
main axis.
The genus Zygartus differs from Zygocampe by the development of two hollow
fenestrated tubes, on the two opposite poles of the main axis. It exhibits therefore to the latter
the same relation that Desmartus does to Desmocampe and Ommatartus to
Ommatocampe; it differs from these by the multiplication of the cortical shell, which is
composed of at least three concentric envelopes.
1. Zygartus doliolum, n. sp.
Inner cortical shell composed of six (or more) kidney-shaped chambers, all nearly of the same
size and structure, every chamber with four to five transverse rows of irregular, roundish pores,
twice to four times as broad as the bars; the basal pores scarcely larger than the apical pores.
Middle cortical shell nearly of the same structure as the inner, only the pores larger and the
bars thinner. Outer cortical shell cylindrical, hemispherical at both poles, its network very
delicate, with large irregular, polygonal pores, and very thin bars between them; its surface
covered with thin bristle-like spines. Both medullary shells lenticular. Polar tube cylindrical,
with conical apex, and with very small pores; its length equals the breadth of two internal
chambers, its breadth that of the inner medullary shell. (This species appears to be a further
developed form of Ommatocampe annulata and Desmartus larvalis, Pl. 40, fig.
12.)
Dimensions.—Length of the six-chambered inner cortical shell 0.25, of the middle
0.3, of the outer 0.35; greatest breadth of the first 0.07, of the second 0.11, of the third 0.15;
pores of the inner shell 0.005 to 0.01, of the middle 0.003 to 0.007, of the outer 0.008 to 0.02;
bars of the first 0.002, of the second and third 0.001; length of the polar tubes 0.15, basal
thickness 0.02.
Habitat.—Pacific, central area, Station 270, depth 2925 fathoms.
2. Zygartus chrysalis, n. sp. (Pl. 40,
fig. 13).
Inner cortical shell composed of six to twenty (commonly eight to twelve) chambers of different
size and form. Both proximal chambers nearly hemispherical, with spiny surface and subregular,
{402}circular pores, three to four times as broad as the
bars. All following chambers cap-like, tapering towards both poles, with more irregular, roundish
pores, with a circle of ten to twelve larger square pores at the base. Middle cortical shell with
smaller roundish pores. Outer cortical shell with larger polygonal, quite irregular pores. Both
medullary shells lenticular. Polar tubes conical or cylindrical with conical apex, of very
variable length, sometimes not longer than one internal chamber, at other times twice to four
times as long (in the figured specimen not fully developed, as also a part of the shells). Differs
from Zygocampe chrysalis only by the production of polar tubes.
Dimensions.—Length of the six-chambered inner cortical shell 0.25, of the middle
0.3, of the outer 0.35; greatest breadth of the first 0.07, of the second 0.11, of the third 0.14;
other measures the same as in Zygocampe chrysalis; length of the tubes 0.05 to 0.12 or
more.
Habitat.—Pacific, central area, Stations 270 to 274, depth 2350 to
2925 fathoms.
Suborder V. DISCOIDEA, Haeckel (Pls. 31-38, 41-48).
Discida vel Discoidea, Haeckel, 1862, Monogr. d. Radiol., pp. 56, 476.
Discoida, Discoidea, Discida, Haeckel, 1878, Protistenreich, p. 103.
Definition.—Spumellaria with discoidal or lenticular
central capsule (often with radial prolongations, rarely allomorphic); with discoidal or
lenticular fenestrated siliceous shell (often with radial spines or fenestrated arms, rarely
allomorphic). Growth reduced or diminished in the direction of one dimensive axis.
The section Discoidea comprises those Spumellaria in which the fenestrated shell is more or less discoidal or
lenticular, flattened or compressed in the direction of one axis. The geometric fundamental form
of the latticed shell, which in the Sphæroidea is a sphere, here becomes
a flat disk, like a medal, or a biconvex lens, sometimes also a biconcave lens. The Discoidea can be derived from the Sphæroidea by
shortening of one axis. This shortened vertical axis is the main axis of the disk; both its poles
are constantly equal. Perpendicular to this axis is the equatorial plane of the disk by which it
becomes divided into equal halves. In the simplest forms of Discoidea all
axes of this horizontal equatorial plane (all "equatorial axes" or "cross axes") are equal; in the
most of the genera and species these cross axes are different, so that rays of stronger growth
("perradii") alternate with rays of weaker growth ("interradii"). The number of these cross axes
distinguishable is commonly two to four, rarely more. In the direction of these are developed
either radial marginal spines or spongy arms.
The order Discoidea was founded in my Monograph (1862, p. 476) as the
family "Discida" (Radiolaria with flat discoidal or biconvex lenticular shell), comprising the
{403}"Calodictya and Lithocyclidina" of
Ehrenberg and a great part of his "Haliommatina." As three different subfamilies of that
family I separated the Coccodiscida (with five genera), the Trematodiscida (with seven genera),
and the Discospirida (with two genera; Monogr. d. Radiol., p. 485). A fourth group of Discoidea was constituted by the Spongodiscida (with eight genera, including
the Spongocyclida), which at that time I united with the Spongurida, because of their
spongy structure (loc. cit., p. 452).
As the number of fossil Discoidea found in the Tertiary rocks of
Barbados and of the Mediterranean shores (Sicily and Greece) is comparatively very large, we find
even in the first system of Polycystina of Ehrenberg (1847), not less than twelve genera
distinguished, viz., six Calodictya, two Haliommatina, and four
Lithocyclidina (Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, 1847, p. 53). The whole
number of Radiolarian genera distinguished in that first system was forty-four. The diagnoses of
them given by Ehrenberg were as usual very insufficient. The characters of the three families
given by him were the following:—Calodictya—"Testarum intus spongiosarum et
nucleo destitutarum orbes; Haliommatina—Testæ subglobosæ nucleus radiatus;
Lithocyclidina—Testarum disci in media parte nucleati margine celluloso." In the
latest work of Ehrenberg (1875, p. 157) the same system was repeated, but some new genera added;
and thirty-eight different species, appertaining to the Discoidea, were
figured in the same work (Abhandl. d. k. Akad. d. Wiss. Berlin, 1875, Tafs. xx.-xxx.).
Richard Hertwig, 1879, in his excellent work, Der Organismus der Radiolarien (pp. 57-68), gave
a detailed description of the skeleton of some Discoidea, and arrived at
the conclusion that this whole family had a spirally constructed skeleton, and should therefore be
derived from the Lithelida. But this conclusion is certainly erroneous, and in my opinion the
whole explanation of that spiral structure, and of its signification in the development of Discoidea, is the weakest part of that otherwise very important work.
In my Prodromus (1881, p. 456) I gave a provisional system of the Discida or Discoidea from the immense quantity of new
material collected by the Challenger, and could distinguish not less than eighty-four genera. This
number is from subsequent research only augmented by seven, so that in the following pages
ninety-one genera with five hundred and one species are described. In the Prodromus I had disposed
them in four different families, which number is now increased to six. These six families can be
again disposed in two main groups or sections, the Phacodiscaria and the
Cyclodiscaria, each section with three families.
The Phacodiscaria are characterised by the possession of a typical
"phacoid shell," and contain the three families Cenodiscida, Phacodiscida, and Coccodiscida. On
the other hand, the Cyclodiscaria are distinguished by the absence of
such a "phacoid shell," and contain the three families Porodiscida, Pylodiscida, and
Spongodiscida. Both sections exhibit an analogous development.
{404}
The Cenodiscida (Pl. 48, figs. 1-3)
open the series of the Discoidea as their simplest forms; a discoidal or
lenticular simple lattice-shell encloses a central capsule of the same form, and is separated from
it by the calymma or jelly-veil. The common ancestral form of this family is Cenodiscus,
without radial marginal spines; it can be derived from Cenosphæra in the most simple way,
by flattening in one axis. If on the equatorial margin of the lens a peculiar solid girdle be
developed, we obtain Zonodiscus; in all other genera of the Cenodiscida radial spines are
developed on the margin. As the simple lenticular cortical shell of Cenodiscus, in which
the central capsule is enclosed, is most characteristic not only of this family, but also of the
two following families, we call it the phacoid shell (that is, a lenticular extracapsular or
cortical lattice-shell).
The Phacodiscida (Pls. 31-35), the second
family, have the same extracapsular "phacoid shell" as the Cenodiscida, but differ from these by
the possession of one or two intracapsular concentric medullary shells, which are connected with
the former by radial beams, perforating the lenticular central capsule. The radial beams are
commonly numerous, and arranged in two opposite bunches around the shortened main axis. But often
also besides these occur other longer radial beams, situated in the equatorial plane; the number
of these is commonly four, and they form a regular rectangular cross, lying opposite in pairs in
two equatorial diameters, perpendicular one to another. In the simplest genera of this family (the
Sethodiscida) the equatorial margin of the phacoid shell is simple or surrounded by a solid
smooth girdle; in all other genera are developed on the margin solid radial spines lying in the
equatorial plane, either regularly disposed in a somewhat constant number (two to eight,
Heliosestrida), or irregularly disposed, in a larger and more variable number (ten to
twenty or more, Heliodiscida).
The Coccodiscida (Pls. 36-38) form a
third family of the Discoidea, directly associated with the Phacodiscida;
both have the same characteristic extracapsular "phacoid shell," which is connected by radial
beams with a simple or double, intracapsular medullary shell. But whilst in the foregoing family
the equatorial margin of the phacoid shell is simple or only armed with radial spines, in the
Coccodiscida it is surrounded by peculiar concentric chambered girdles, or rings, which resemble
those of the following family, the Porodiscida. Each of these "chambered girdles" is composed of a
circular ring in the equatorial plane, a variable number of radial beams dividing it into
incomplete chambers, and two porous cover-plates or "sieve-plates," covering the upper and lower
face of the disk. These sieve-plates may be regarded as incomplete lenticular cortical shells,
which are only developed in the peripheral part of the disk, whilst their central part is
represented by the only complete cortical shell, the "phacoid shell." The number of these
concentric chamber-girdles amounts to from one to ten or more. The margin of the disk is either
simple (Lithocyclida) or armed with radial spines (Stylocyclida), or provided with two to five
chambered radial arms (Astracturida); the structure of the arms is the same as that of the
girdles.
{405}
The Porodiscida (Pls. 41-47), the
largest family of all Discoidea, begins the series of Cyclodiscaria, or those Discoidea in which there is no
"phacoid shell," but a small simple central chamber surrounded by a number of small latticed
chambers of nearly the same size and form. In the Porodiscida these chambers are arranged in
complete circular concentric rings or spiral convolutions; in the small family of Pylodiscida the
central chamber is surrounded by three radial arm-chambers separated by three open spaces; in the
third family of Cyclodiscaria, the Spongodiscida, all the chambers are
arranged more or less irregularly, and the whole disk becomes spongy; also the surface of the disk
is spongy, whilst in both former families it is covered by two regular even porous plates or
"sieve-plates." Probably all Cyclodiscaria can be derived
phylogenetically from Archidiscus, a very small and simple lenticular disk, which is
composed of a small spherical latticed central chamber and of a single concentric chambered ring
or girdle; the margin of this ring is connected with the central chamber by a variable number of
radial beams. This Archidiscus can be derived either from Saturnalis (Pl. 13, fig. 16.) by the development of lattice-work
between the equatorial ring and both polar faces of the concentric central chamber, or from
Sethodiscus (Pl. 33,
figs. 1-3) by flattening of the lenticular shell, so that the enclosed inner medullary shell (the
central chamber) meets the outer phacoid shell at both poles.
The Porodiscida are commonly flat or biconvex (rarely biconcave) disks, the central chamber of
which is surrounded not by a single, but by a variable number (commonly three to six) of
concentric chambered girdles or rings; they arise from Archidiscus by apposition of new
concentric chambered rings around the first ring, all lying in the equatorial plane. Afterwards
the disk often becomes thickened by apposition of concentric chamber-rings on both flat sides
also, so that two to four or more layers are stratified one over the other. The circular
concentric rings often become interrupted, or spirally convoluted (wholly or partially); also the
chambers sometimes become irregularly crowded. But in all cases both surfaces of the disk (upper
and lower) continue to be porous plates or sieve-plates, at least in the centre, but they never
become spongy.
The margin of the disk exhibits in the Porodiscida a great variety of different forms, serving
for distinction of subfamilies and of genera. In the Trematodiscida the margin remains quite
simple, as in the Archidiscida, or is only surrounded by a hyaline equatorial girdle. In the
Ommatodiscida it is distinguished by one or two peculiar oscula, surrounded by a corona of spines.
The Stylodictyida are distinguished by a number of solid radial spines, and the Euchitonida by a
number of chambered, or spongy, radial arms, arising from the margin of the disk and lying in the
equatorial plane. The variety of these radial marginal appendages is in the Porodiscida much
greater than in the Coccodiscida.
The Pylodiscida (Pl. 48, figs.
12-20) represent a new, small, but very remarkable family of Discoidea,
all triradial, and distinguished by the peculiar formation of {406}large open spaces in the latticed discoidal shell, which reappear in a
similar shape among the Larcoidea in the Pylonida (Tetrapyle,
&c.). We get the best understanding of this peculiar formation if we return to
Archidiscus, probably the common ancestral form of all Cyclodiscaria, of the Porodiscida as well as of the Pylodiscida and
Spongodiscida. In some species of Archidiscus (Archidiscus hexoniscus,
Archidiscus pyloniscus, &c.) the small lenticular shell is composed of a spherical
latticed central chamber and of a concentric equatorial girdle composed of six such chambers,
either all six equal, or alternately larger and smaller. This latter form is nearly identical with
Triodiscus, and if we imagine the lattice-work of only three ring-chambers complete, whilst
that of the three alternating chambers is reduced to the marginal ring, we get Triopyle, by
loss of this ring Triolene (a disk composed of four simple lattice-chambers, lying in one
plane, three radial around one central spherule). The genera mentioned form together the subfamily
of Triopylida. In the second subfamily, Hexapylida, the same formation is doubled; here three
double arm-chambers are separated by three double spaces (two in each radius). Also here the three
distal spaces may be either quite open (Pylolena), or half closed by the marginal girdle
(Hexapyle), or quite loosely latticed (Pylodiscus). If the margin of this latter
form become surrounded by a perfect chambered equatorial girdle, we get Discozonium, and if
this acquire a peculiar marginal ostium (surrounded by a corona of spines) we arrive at
Discopyle. These two latter genera form the third subfamily, the Discopylida. The eight
genera of Pylodiscida represent therefore a continuous phylogenetic series.
The Spongodiscida are the sixth and last family of the Discoidea, differing from the five other families in the irregular, spongy
structure of the discoidal skeleton; both surfaces of the flat disk (upper and lower) are here
principally covered with a rough, spongy framework, whilst in the five other families they are
covered by the flat and smooth porous plates or sieve-plates. Nevertheless there is no sharp
boundary between the Spongodiscida and the closely allied Porodiscida. In these latter also the
discoidal shell becomes often more or less spongy (mainly in the peripheral part, e.g., in
Myelastrum, Pl. 47);
but at least the central part of the disk here remains constantly covered by sieve-plates. The
massive skeleton of the Spongodiscida is either of perfectly irregular structure, only composed of
innumerable fine branched siliceous threads, interwoven in all possible directions; or only the
outer part of the disk is composed of such spongy framework, whilst the central part is more or
less distinctly composed of concentric chambered rings, as in the Porodiscida. These latter forms
indeed exhibit an immediate transition to this family, and were formerly (in 1862) separated by me
as Spongocyclida. Also the polymorphous shape of the disk margin in the Spongodiscida is quite
analogous to that of the Porodiscida. Whilst in the first subfamily, the Spongophacida, the margin
is quite simple; in the second, the Spongotrochida, it is armed with solid radial spines; and in
the third, the Spongobrachida, it is provided with two, three, {407}or four spongy, radial arms—the former as well as the latter lying
in the equatorial plane of the disk, either regularly or irregularly disposed.
The Equatorial Margin of the lenticular disk exhibits in all six families of Discoidea similar characters, mainly serving for the distinction of subfamilies
and genera. In the most primitive genera of all six families the margin is simple, without radial
prolongations (spines or arms); it is quite simple in Cenodiscus, Sethodiscus,
Phacodiscus, Lithocyclia, Coccodiscus, Archidiscus, Porodiscus,
Pylodiscus, and Spongodiscus. In some genera the simple margin of the lenticular
disk is bordered and surrounded by a thin, hyaline, equatorial girdle of silex, either quite solid
or slightly porous (Zonodiscus, Periphæna, Perizona, Perichlamydium,
and Spongophacus).
A quite peculiar and remarkable character of few genera is the development of one or two
oscula, larger marginal openings, which are surrounded by a corona of spines, and probable are fit
for the issue a peculiar bunch of pseudopodia or of a "sarcode flagellum." Such oscula occur only
in two families of Cyclodiscaria; in the Porodiscida and Pylodiscida; in
the former Ommatodiscus, in the latter Discopyle (Pl. 48, figs. 19,
20) is distinguished by a single marginal osculum; besides this, in the former occurs
Stomatodiscus, with two such oscula, opposite on the poles of one axis (Pl. 48, fig. 8).
These oscula may be compared with the similar polar formations in some Ellipsida
(Lithomespilus) and in many Cyrtoidea; but they do not prove a nearer affinity with the
latter, and are only analogous, not homologous.
Radial Spines occur on the margin in the equatorial plane of the Discoidea in the greatest variety of number, form, size, and disposition. If
the number be low (between two and eight) they are commonly regularly disposed; if the number be
larger (ten to twenty or more) their disposition becomes commonly more or less irregular. The
regular disposition is of great promorphological importance, as indicating the axes in which the
growth is preponderant, and introduces other peculiar radial formations. Regarding these axes we
can generally distinguish two groups, Artiacantha with a paired number (two, four, eight),
and Perissacantha with odd numbers (usually three). The section of Artiacantha could
be divided into three following groups:—A. Stylodiscida, with two radial spines only, lying
opposite on both poles of one equatorial axis (the "first cross axis")—Stylodiscus,
Sethostylus, Stylocyclia, Xiphodictya, Spongolonche (Pl. 31, figs. 9-12;
Pl. 38, fig.
1; Pl. 42,
figs. 10-12, &c.); B. Staurodiscida, with four radial spines, lying opposite in pairs on the
poles of two crossed equatorial axes, perpendicular to one another (first and second cross
axes)—Crucidiscus, Sethostaurus, Staurocyclia, Staurodictya,
Spongostaurus (Pl. 31,
figs. 1-8; Pl. 37,
figs. 1-4; Pl. 42,
figs. 1-6; Pl. 48,
fig. 2, &c.); C. Octostylida, with eight radial spines, opposite in pairs in four axes, which
are crossed at angles of 45°—Heliosestrum, Astrosestrum, &c. (Pl. 32, figs. 4, 5;
Pl. 34,
figs. 3, 6); in this latter case sometimes the radial symmetry is the same as in many Medusæ, four
larger (perradial) {408}spines alternating with four
smaller (interradial), indicating radii of first and second order. The section of
Perissacantha is much smaller, and commonly represented only by triradial forms, with three
spines at equal distances (120°)—Triactis, Tripocyclia, Tripodictya,
Spongotripus, &c. (Pl. 33,
fig. 6; Pl. 37,
fig. 5; Pl. 42,
figs. 7-9).
Radial Arms on the margin of the disk appear in similar variety of number, form, and
disposition as the radial spines; but the number is here commonly limited to from two to four,
rarely five to six. The arms are absent in the families Cenodiscida and Phacodiscida; in the four
other families they return under similar forms. These arms are direct prolongations of the disk,
and exhibit the same structure, so that they may be regarded both as centrifugal productions of
certain radii, and also inversely as peripheral parts of a disk, the interjacent radii of which
are reduced. The regular disposition and shape of the arms, an important character for the
distinction of genera and species, is repeated in a quite analogous manner in the four above
mentioned families, so that we can distinguish the following groups—A. Amphibrachida, with
two radial arms, opposite on the poles of one equatorial axis (the first cross
axis)—Diplactura, Amphibrachium, Spongobrachium (Pl. 38, figs. 3-5;
Pl. 44,
figs. 6-11); B. Triobrachida, with three radial arms; the most important group (with all
Pylodiscida); either all three arms are equal and disposed at equal distances
(Trigonactura, Dictyastrum, Rhopalodictyum, Pl. 38, figs. 6-9;
Pl. 43,
figs. 5, 13, 16; Pl. 48,
figs. 12-19), or a single odd arm differs in size and position, and is often larger than both the
opposite paired arms (Rhopalastrum, Euchitonia, Pl. 43, figs. 6,
10, 15, &c.); C. Tetrabrachida, with four radial arms, opposite in pairs in two crossed axes,
commonly perpendicular one to another, Stauractura, Stauralastrum,
Spongaster, &c. (Pls. 46,
47).
The arms are commonly simple, undivided, but sometimes also forked or branched (Pl. 43, figs. 15,
16; Pl. 47).
Their basal parts are either free, separately inserted into the margin of the circular central
disk, or they are connected by a "patagium," a peculiar connecticulum, like a web-membrane, which
is composed of a chambered, commonly more or less spongy framework, different in texture from the
lattice-work of the arms (Pl. 38,
figs. 8, 9; Pl. 43,
figs. 9-16; Pl. 46).
Sometimes the patagium overgrows the whole shell. A peculiar modification of it appears in
Stephanastrum (Pl. 44,
fig. 1), where only the distal parts of the arms are connected by the ring-shaped patagium, whilst
the basal parts are free; therefore open gates rest between them, like those of the Pylodiscida
(Pl. 48,
figs. 12-20).
The Central Capsule of the Discoidea is constantly discoidal,
more or less lenticular; in some cases more biconvex, with vaulted faces and thin margin; in
others more medal-shaped, with flat faces and thick margin. In the Cenodiscida alone the capsule
lies freely inside the simple phacoid shell, and is separated from it by the jelly-veil. In the
other five families the capsule encloses the central parts of the skeleton, and is enveloped by
the superficial parts of it, whilst its membrane is perforated by radial beams {409}connecting the latter with the former. In the
Phacodiscida and Coccodiscida the capsule encloses the simple or double medullary shell, but is
itself enclosed by the cortical phacoid shell. In all Cyclodiscaria (in
the Porodiscida, Pylodiscida, and Spongodiscida) the capsule fills out the greatest part of the
chambered or spongy skeleton, and is only protected by the superficial parts of it, in the
Porodiscida and Pylodiscida by the covering sieve-plates, in the Spongodiscida by the spongy
cortical substance of the shell. The growth of the capsule corresponds to that of the including
shell, gradually increasing on the margin in the equatorial plane. Whilst in the greater number of
Discoidea its form continues circular, in many forms provided with radial
arms it enters into the arms and assumes their form. The protoplasm of the capsule is commonly
coloured by brown or red pigment, and often contains many oil-globules. The nucleus is originally
enclosed by the medullary shell or the central chamber, and with increasing size enters into the
surrounding parts; in the Cyclodiscaria it often fills out the internal
concentric rings. The extracapsular jelly or the calymma is commonly thick, and envelops the
greater part or the whole body.
Synopsis of the Families of the Discoidea.
| I. Section Phacodiscaria.
Discoidea with external phacoid shell (or lenticular latticed cortical shell). |
|
Phacoid shell simple, without enclosed medullary
shell, |
1. Cenodiscida. |
| Phacoid shell with simple or double enclosed medullary
shell. |
|
Margin without chambered girdles, |
2. Phacodiscida. |
| Margin surrounded by by chambered girdles. |
3. Coccodiscida. |
| II. Section Cyclodiscaria.
Discoidea without external phacoid shell (no lenticular latticed cortical shell). |
|
Surface of the shell covered by convex or even porous
sieve-plates (not spongy). |
|
Concentric rings around the central chamber complete without open
spaces), |
4. Porodiscida. |
| Concentric rings around the central chamber interrupted by three open
spaces, |
5. Pylodiscida. |
| Surface of the shell spongy, not covered by peculiar porous
sieve-plates, |
6. Spongodiscida. |
| I. Section Phacodiscaria. Discoidea with external
phacoid shell (or lenticular latticed cortical shell). |
|
|
Phacoid shell simple, without enclosed medullary shell, |
|
|
|
|
|
|
|
|
1. Cenodiscida. |
|
|
|
|
|
|
|
|
Phacoid shell with simple or double enclosed medullary shell. |
|
|
|
|
|
|
Margin without chambered girdles, |
|
|
|
|
|
|
|
|
|
|
2. Phacodiscida. |
|
|
|
|
|
|
|
|
|
|
Margin surrounded by by chambered girdles. |
|
|
|
|
|
|
|
|
|
|
3. Coccodiscida. |
|
|
|
|
|
|
| II. Section Cyclodiscaria. Discoidea without external
phacoid shell (no lenticular latticed cortical shell). |
|
|
Surface of the shell covered by convex or even porous sieve-plates
(not spongy). |
|
|
|
|
|
|
Concentric rings around the central chamber complete without open
spaces), |
|
|
|
|
|
|
|
|
|
|
4. Porodiscida. |
|
|
|
|
|
|
|
|
|
|
Concentric rings around the central chamber interrupted by three
open spaces, |
|
|
|
|
|
|
|
|
|
|
5. Pylodiscida. |
|
|
|
|
|
|
|
|
Surface of the shell spongy, not covered by peculiar porous
sieve-plates, |
|
|
|
|
|
|
|
|
6. Spongodiscida. |
|
|
|
|
|
|
Family XVIII. Cenodiscida, n. fam. (Pl. 31, fig. 11;
Pl. 48,
figs. 1-3).
Definition.—Discoidea with simple extracapsular phacoid
shell (or lenticular latticed cortical shell), without medullary shell and without chambered
equatorial girdles.
The new family Cenodiscida opens the long series Discoidea, as their most simple and primitive form. The circular lenticular
central capsule is enclosed by a {410}simple latticed
shell of the same form, only separated from it by a thinner or thicker jelly-veil. The lenticular
or discoidal fenestrated shell is therefore an extracapsular or "cortical shell," without an
enclosed medullary shell.
The few genera of the Cenodiscida differ only in the shape of the equatorial margin of the
lenticular disk. In the first subfamily, Zonodiscida, the margin is either quite simple
(Cenodiscus) or surrounded by a smooth, solid equatorial girdle (Zonodiscus). In the
second subfamily, Trochodiscida, the margin is armed with solid radial spines, lying in the
equatorial plane. According to the number and disposition of these marginal spines, we distinguish
Stylodiscus (with two spines, opposite in one equatorial axis), Crucidiscus (with
four spines, opposite in pairs in two equatorial axes, perpendicular one to another),
Theodiscus (with three marginal spines), and Trochodiscus (with numerous, commonly
twenty to thirty, irregularly disposed spines). The spines are constantly simple, not branched;
sometimes more conical or cylindrical, at other times more angular or pyramidal.
The two convex faces of the lenticular shell are constantly of similar shape, commonly smooth,
sometimes more or less thorny, or armed with bristle-shaped radial spines. The pores are commonly
more or less regular, circular, and disposed in series, which are occasionally more radial, at
other times more concentric. If the wall of the hollow lens be rather thick, the difference in the
shape of the central and peripheral pores is often striking. The central pores perforating the
thick wall perpendicularly are short cylindrical tubes; the marginal pores perforating it in an
oblique direction are longer conical tubes. The bars between the central pores are often somewhat
smaller.
The Central Capsule of the Cenodiscida is in all cases a perfect, circular, biconvex
lens, the equatorial diameter of which is commonly between two-thirds and three-fourths of the
enclosing lattice-shell. The interval between the two is filled up by the jelly-veil, or the
hyaline "calymma," which is perforated by the numerous pseudopodia that pass through the
shell-pores.
As the Cenodiscida possess the most simple shell-form of all Discoidea, we may regard Cenodiscus as the common ancestral form of this
large section, in the same manner as Cenosphæra is the ancestral form of Sphæroidea, Cenellipsis of the Prunoidea, Cenolarcus of the Larcoidea.
But it is also possible that a part of Cenodiscida (or all?) arises from the Phacodiscida by
reduction and loss of the medullary shell. For in some cases we find arising from the inside of
the shell centripetal radial beams, which end at a certain equal distance from the hollow centre
(Pl. 31,
fig. 11; Pl. 38,
fig. 2). Cenodiscus itself can be derived either from Cenosphæra by compression of the
spheroidal shell in one axis, or from Sethodiscus by loss of the intracapsular medullary
shell, or from Actidiscus (the lenticular Actissa) by formation of a cortical shell around
the lenticular central capsule.
{411}
Synopsis of the Genera of the Cenodiscida.
|
I. Subfamily Zonodiscida.
Margin of the disk without radial spines.
|
|
Margin quite simple, without equatorial girdle, |
175. Cenodiscus. |
| Margin surrounded by a solid siliceous equatorial girdle, |
176. Zonodiscus. |
|
II. Subfamily Trochodiscida.
Margin of the disk armed with radial spines (lying in the equatorial
plane).
|
|
Two spines opposite in one equatorial axis, |
177. Stylodiscus. |
| Three spines on the margin of the disk, |
178. Theodiscus. |
| Four spines opposite in pairs in two perpendicularly crossed equatorial
axes, |
179. Crucidiscus. |
| Ten to twenty or more radial spines (variable in number and commonly
irregular in disposition), |
180. Trochodiscus. |
| I. Subfamily Zonodiscida. Margin of the disk without radial spines. |
|
|
Margin quite simple, without equatorial girdle, |
|
|
|
|
|
|
175. Cenodiscus. |
|
|
|
|
|
|
Margin surrounded by a solid siliceous equatorial girdle, |
|
|
|
|
|
|
176. Zonodiscus. |
|
|
|
|
| II. Subfamily Trochodiscida. Margin of the disk armed with radial spines
(lying in the equatorial plane). |
|
|
Two spines opposite in one equatorial axis, |
|
|
|
|
|
|
177. Stylodiscus. |
|
|
|
|
|
|
Three spines on the margin of the disk, |
|
|
|
|
|
|
178. Theodiscus. |
|
|
|
|
|
|
Four spines opposite in pairs in two perpendicularly crossed
equatorial axes, |
|
|
|
|
|
|
179. Crucidiscus. |
|
|
|
|
|
|
Ten to twenty or more radial spines (variable in number and
commonly irregular in disposition), |
|
|
|
|
|
|
180. Trochodiscus. |
|
|
|
|
Subfamily 1. Zonodiscida, Haeckel.
Definition.—Cenodiscida without radial spines
on the margin of the disk.
Genus 175. Cenodiscus,[214] n. gen.
Definition.—Cenodiscida with simple margin of the
circular disk, without surrounding equatorial girdle and without radial spines.
The genus Cenodiscus is the most simple and primitive form of all Discoidea, and represents possibly the common ancestral form of this order. The
latticed shell is a simple biconvex lens, and encloses a smaller central capsule of the same form,
separated from it by the jelly-veil. Cenodiscus can be derived phylogenetically either from
Cenosphæra by lenticular flattening of a simple latticed sphere, or directly from
Actissa by formation of a lenticular fenestrated shell around the lentiform central
capsule. Possibly also some forms of Cenodiscus can be derived from Sethodiscus by
reduction and loss of the medullary shell.
1. Cenodiscus phacoides, n. sp. (Pl. 48,
figs. 1, 1a).
Disk with smooth surface, without radial ribs or spines. Margin of the lenticular biconvex disk
thin, simple. Pores regular, circular; fifteen to sixteen on the radius of the disk. (Very similar
to Sethodiscus phacoides, but without medullary shell.)
Dimensions.—Diameter of the disk 0.2, of the pores 0.005.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
{412}
2. Cenodiscus rotula, n. sp.
Disk with smooth surface, without medial ribs or spines. Margin of the disk blunt, very thick,
rounded; both faces little convex. Pores regular, circular; thirteen to fourteen on the radius of
the disk. (Similar to Phacodiscus rotula, Pl. 35, fig. 7, but
without medullary shell.)
Dimensions.—Diameter of the disk 0.16, of the pores 0.006.
Habitat.—North Pacific, Station 241, depth 2300 fathoms.
3. Cenodiscus lenticula, n. sp.
Disk with thorny surface, scattered with small, conical spines. Margin of the lenticular
biconvex disk thin. Pores irregular, roundish; ten to eleven on the radius of the disk.
Dimensions.—Diameter of the disk 0.15, of the pores 0.008.
Habitat.—South Pacific, Station 295, depth 1500 fathoms.
Genus 176. Zonodiscus,[215] n. gen.
Definition.—Cenodiscida with surrounding solid equatorial
girdle on the margin of the lenticular disk, without radial spines.
The genus Zonodiscus differs from the preceding Cenodiscus only in
the development of a solid siliceous girdle around the keen margin of the lenticular disk. This
form can also be derived from Periphæna or Perizona by reduction and loss of the
medullary shell. The same girdle formation returns not only in both these Phacodiscida (Pl. 32, fig. 7; Pl.
33, fig. 4),
but also in the Porodiscid Perichlamydium.
1. Zonodiscus saturnalis, n. sp.
Disk with smooth surface, without radial spines. Pores regular, circular, fifteen to sixteen on
the radius of the disk, in its distal half arranged in fifty to sixty radial series, which are
separated by prominent radial crests or ribs. The crests are prolonged into the proximal half of
the thin solid equatorial girdle, which is one-third as broad as the radius of the disk.
Dimensions.—Diameter of the disk 0.25, of the pores 0.05.
Habitat.—South Atlantic, Station 335, depth 1425 fathoms.
Subfamily 2. Trochodiscida, Haeckel.
Definition.—Cenodiscida with radial spines on
the margin of the disk, disposed in the equatorial plane.
Genus 177. Stylodiscus,[216] n. gen.
Definition.—Cenodiscida with two radial spines on the
margin of the disk, opposite in one equatorial axis.
{413}
The genus Stylodiscus opens the series of the Trochodiscida or of those
Cenodiscida in which the thin margin of the hollow lenticular disk is armed with a number of solid
radial spines, situated in its equatorial plane. Stylodiscus is at the same time the most
simple form of the Stylodiscida, or of the numerous Discoidea (belonging
to different families) in which the disk bears only two spines, opposite in one equatorial axis.
Hitherto only two species of this genus have been observed, but they seem to represent two
different subgenera.
Subgenus 1. Stylentodiscus, Haeckel.
Definition.—Internal cavity of the shell with centripetal axial
rods.
1. Stylodiscus endostylus, n. sp. (Pl. 31, fig.
11).
Sethostylus endostylus, Haeckel, 1881, Prodromus et Atlas (pl. xxxi, fig.
11).
Disk with smooth surface and dentated margin. Pores irregular, roundish; ten to twelve on the
radius of the disk. Marginal teeth conical, short, irregular. Both opposite polar spines
cylindrical, longer than the diameter of the disk, and as broad as one larger pore. On the inside
of the hollow disk both spines are prolonged into two strong centripetal axial rods, which do not
reach the centre. Also a number of smaller centripetal axial rods surrounds the central cavity, so
that an original medullary shell (Sethostylus) seems to have been lost (comp. above, p. 410).
Dimensions.—Diameter of the disk 0.25, of the pores 0.005 to 0.02; length of the
polar spines 0.3 and more, thickness 0.012.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms.
Subgenus 2. Stylexodiscus, Haeckel.
Definition.—Internal cavity of the shell without axial rods.
2. Stylodiscus amphistylus, n. sp.
Disk with smooth surface and smooth margin. Pores regular, circular; eight to nine on the
radius of the disk. Both opposite polar spines pyramidal, sulcated, about as long as the radius of
the disk, and three times as long as broad at the base. Inner cavity of the disk simple, without
centripetal rods.
Dimensions.—Diameter of the disk 0.15, of the pores 0.007; length of the polar
spines 0.08, basal breadth 0.03.
Habitat.—Pacific, central area, Station 274, depth 2750 fathoms.
{414}
Genus 178. Theodiscus,[217] n. gen.
Definition.—Cenodiscida with three radial spines on the
margin of the disk, placed in its equatorial plane.
The genus Theodiscus is the most simple form of those very numerous Discoidea in which three rays are developed on the margin of the disk; a symbol
of the Christian dogma of the Trinity. Commonly the three angles between the three spines are
equal, more rarely one angle is larger than the two others which are equal. The shell of some
species is nearly spherical (formerly separated by me as Theosphæra), whilst in others it
is a flattened biconvex lens.
Subgenus 1. Theodiscoma, Haeckel.
Definition.—Angles between the three radial spines equal (triangle
equilateral).
1. Theodiscus divinus, n. sp.
Disk nearly spherical, with smooth surface. Pores regular, circular, hexagonally framed; seven
to eight on the radius. Three angles between the spines equal. Spines prismatic, straight, twice
as broad as one pore, eight to eleven times as long as the shell diameter.
Dimensions.—Diameter of the disk 0.1, of the pores 0.006; length of the spines 0.8
to 1.1, breadth 0.01.
Habitat.—Central Pacific, Station 265, surface.
2. Theodiscus christianus, n. sp.
Disk nearly spherical, with smooth surface. Pores regular, circular, prolonged into short
cylindrical, prominent tubuli, half as high as broad; eight to nine on the radius. Three angles
between the spines equal. Spines prismatic, straight, twice as broad as one pore, twelve to
fifteen times as long as the shell diameter.
Dimensions.—Diameter of the disk 0.08, of the pores 0.005; length of the spines
1.1 to 1.2, breadth 0.01.
Habitat.—Central Pacific, Station 271, surface.
3. Theodiscus trinitatis, n. sp.
Disk a rather flattened, biconvex lens, about twice as broad as thick. Pores regular, circular,
not prolonged into tubuli; five to six on the radius. Three angles between the spines equal.
Spines pyramidal, three times as broad as one pore, about twice as long as the shell diameter.
{415}
Dimensions.—Diameter of the disk 0.06, of the pores 0.007; length of the spine
0.12, basal breadth 0.02.
Habitat.—Central Pacific, Station 274, depth 2750 fathoms.
Subgenus 2. Theodiscura, Haeckel.
Definition.—Angles between the three radial spines unequal, two paired
angles equal, larger or smaller than the odd angle (triangle isosceles).
4. Theodiscus vanitatis, n. sp.
Disk nearly spherical, with smooth surface. Pores irregular, roundish; eight to ten on the
radius. Three angles between the spines unequal; one odd angle larger than both others. Spines
pyramidal, of unequal size; one odd spine larger, both others smaller than the shell diameter.
Dimensions.—Diameter of the disk 0.12, pores 0.004 to 0.006; length of the odd
spine 0.2, of the paired spines 0.1.
Habitat.—South Atlantic, Station 335, depth 1425 fathoms.
5. Theodiscus nirvana, n. sp.
Disk a flat biconvex lens, about twice as broad as thick. Pores regular, circular; ten to
twelve on the radius. Three angles between the spines unequal; one odd angle smaller than both
others. Spines prismatic, very long and thin, twice as broad as one pore, ten to twelve times as
long as the shell diameter.
Dimensions.—Diameter of the disk 0.1, pores 0.005; length of the spines 1 to 1.2
or more, breadth 0.01.
Habitat.—Indian Ocean, Ceylon, surface, Haeckel.
Genus 179. Crucidiscus,[218] n. gen.
Definition.—Cenodiscida with four radial spines on the
margin of the disk, crossed in the equatorial plane.
The genus Crucidiscus is the most simple form of the Staurodiscida, or of
the numerous Discoidea (belonging to different families) in which the
margin of the disk bears four radial spines, lying in the equatorial plane, and crossed at right
angles. Whilst commonly the internal shell-cavity of Crucidiscus is quite simple, in one
case it bears four centripetal axial rods, as inner prolongations of the outer radial
cross-spines, perhaps indications of a lost medullary shell (comp. p. 410).
{416}
Subgenus 1. Staurentodiscus, Haeckel.
Definition.—Internal cavity of the shell with centripetal axial
rods.
1. Crucidiscus endostaurus, n. sp. (Pl. 48, fig.
2).
Disk with smooth surface and smooth simple margin. Pores regular, circular; thirteen to
fourteen on the radius of the disk. Four crossed spines conical, strong, longer than the radius of
the disk, on the inside prolonged into four thinner centripetal axial rods, which do not reach the
centre. In the middle part of the disk also some other short axial rods arise from the inside, not
reaching the centre (as in Stylodiscus endostylus, Pl. 31, fig.
11).
Dimensions.—Diameter of the disk 0.16, of the pores 0.004; length of the spines
0.2, breadth 0.014.
Habitat.—North Pacific, Station 253, depth 3125 fathoms.
Subgenus 2. Staurexodiscus, Haeckel.
Definition.—Internal cavity of the shell without centripetal axial
rods.
2. Crucidiscus cuspidatus, n. sp.
Disk with smooth surface and simple smooth margin. Pores regular, circular; seven to eight on
the radius of the disk. Four crossed spines pyramidal, with prominent edges, somewhat shorter than
the radius of the disk, twice as broad at their thick base as one pore. No internal axial rods in
the shell cavity.
Dimensions.—Diameter of the disk 0.12, of the pores 0.005; length of the spines
0.05, basal breadth 0.025.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
3. Crucidiscus echinatus, n. sp.
Disk with thorny surface and thorny margin. Pores regular, circular, six to seven on the
radius. Four crossed spines pyramidal (four sided?), about as long as the radius of the disk, as
broad at their base as one pore. No internal axial rods. Spines of the surface short, conical.
Dimensions.—Diameter of the disk 0.14, of the pores 0.008; length of the spines
0.08, basal breadth 0.02.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
4. Crucidiscus cruciatus, n. sp.
Disk with smooth surface and broad dentated margin. Pores regular, circular; eleven to twelve
on the radius. Four crossed spines pyramidal, sulcated, about as long as the radius of the disk,
as {417}broad at their base as the radiated margin,
which bears fifty to sixty triangular pointed teeth of irregular size. (Similar to Sethostaurus
cruciatus, Pl. 31,
fig. 5, but without internal axial rods and medullary shell.)
Dimensions.—Diameter of the disk 0.2, of the pores 0.007; length of the spines
0.1, basal breadth 0.02.
Habitat.—Pacific, central area, Station 274, depth 2750 fathoms.
Genus 180. Trochodiscus,[219] n.
gen.
Definition.—Cenodiscida with numerous (ten to twenty or
more) radial spines on the margin of the disk, situated in its equatorial plane.
The genus Trochodiscus bears on the thin margin of the latticed disk a
variable number of radial spines (commonly ten to twenty, sometimes fifty to eighty or more).
Their size and disposition are commonly more or less irregular. For the most part all the marginal
spines lie in the equatorial plane; but sometimes part of them are crowded into two or four
parallel girdles. Several species of this genus are very similar to some species of
Heliodiscus, and only differ in the absence of the medullary shell, perhaps in consequence
of the phylogenetic loss of it; if this be the case, the former are to be derived from the latter
(compare p. 410).
Subgenus 1. Trochodisculus, Haeckel.
Definition.—Bases of the marginal spines free, not connected by a
solid equatorial girdle.
1. Trochodiscus cenophacus, n. sp.
Disk with smooth surface. Pores regular, circular; eleven to twelve on the radius. Marginal
spines sixteen to twenty, conical, of irregular size and disposition, about three times as long as
broad, and as long as the radius of the disk.
Dimensions.—Diameter of the disk 0.15, of the pores 0.006; length of the spines
0.06 to 0.09, basal breadth 0.02 to 0.03.
Habitat.—Central Pacific, Station 266, depth 2750 fathoms.
2. Trochodiscus medusinus, n. sp.
Disk with smooth surface. Pores regular, circular; nine to ten on the radius. Marginal spines
ten to twelve, equilateral triangular, deeply sulcated, about half as long and broad as the radius
of the disk. (Very similar to Heliosestrum medusinum, Pl. 34, fig. 6, but
without enclosed medullary shell. Compare p. 410.)
Dimensions.—Diameter of the disk 0.2, of the pores 0.01; length of the spines
0.05, basal breadth 0.05.
Habitat.—North Pacific, Station 241, depth 2300 fathoms.
{418}
3. Trochodiscus odontotrochus, n. sp.
Disk with smooth surface, in the distal part radially sulcated. Pores irregular, roundish;
twelve to fourteen on the radius. Marginal spines very short and numerous (thirty to forty),
conical, scarcely one-fourth as long as the radius of the disk, which resembles a wheel with
marginal teeth.
Dimensions.—Diameter of the disk 0.18, of the pores 0.004; length of the spines
0.02, basal breadth 0.01.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
4. Trochodiscus solaris, n. sp.
Disk with smooth surface. Pores subregular, circular; eight to nine on the radius. Marginal
spines very numerous (sixty to eighty), in two to four girdles densely crowded together, the
largest as long as the diameter of the disk, bent and conical. (Similar to Heliodiscus solaster, Pl. 34, fig. 4, but
without medullary shell.)
Dimensions.—Diameter of the disk 0.15, of the pores 0.008; length of the spines
0.05 to 0.15, basal breadth 0.01.
Habitat.—South Pacific, Station 300, depth 1375 fathoms.
5. Trochodiscus echiniscus, n. sp.
Disk with spiny surface. Pores irregular, roundish; ten to eleven on the radius. Marginal
spines conical, very numerous (forty to fifty), irregular in size and disposition, the largest
half as long as the radius of the disk. (Similar to Heliodiscus echiniscus, Pl. 34, fig. 5, but
without medullary shell.)
Dimensions.—Diameter of the disk 0.18, of the pores 0.005 to 0.015; length of the
spines 0.02 to 0.04, basal breadth 0.01.
Habitat.—North Atlantic, Færöe Channel, John Murray.
Subgenus 2. Pristodiscus, Haeckel.
Definition.—Bases of the marginal spines connected by a solid
siliceous equatorial girdle.
6. Trochodiscus stellaris, n. sp. (Pl. 48, fig.
3).
Disk with smooth surface. Pores subregular, circular; seven to eight on the radius. Marginal
spines twelve to sixteen, triangular, of subregular size and disposition, about half as long as
the radius of the disk, connected at their broad base by a solid equatorial girdle of half the
breadth; girdle and spines radially striped.
Dimensions.—Diameter of the disk 0.25, of the pores 0.015; length of the spines
0.04 to 0.06, basal breadth 0.02 to 0.04.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
{419}
7. Trochodiscus cingillum, n. sp.
Disk with smooth surface, in the distal part radially sulcated. Pores regular, circular; twelve
to thirteen on the radius. Marginal spines twenty to twenty-four, triangular, of equal size and at
regular distances, only one-fourth as long as the radius of the disk, connected at their broad
base by a solid equatorial girdle of the double breadth. (Similar to Heliodiscus cingillum,
Pl. 33, fig.
7, but without medullary shell.)
Dimensions.—Diameter of the disk 0.22, of the pores 0.004; length of the spines
0.02, basal breadth 0.02
Habitat.—South Pacific, Station 285, depth 2375 fathoms.
Family XIX. Phacodiscida, Haeckel (Pls. 31-35).
Phacodiscida, Haeckel, 1881, Prodromus, p. 456.
Definition.—Discoidea with simple extracapsular phacoid
shell (or lenticular latticed cortical shell), connected by radial beams with an intracapsular,
simple or double, concentric medullary shell, without chambered equatorial girdles.
The family Phacodiscida comprises a large number of splendid forms
(about a hundred species), which agree with the preceding Cenodiscida in the possession of the
characteristic extracapsular "phacoid shell," but differ from them in having one or two
intracapsular "medullary shells"; these concentric spherical medullary shells are connected with
the lenticular cortical shell or phacoid shell by means of radial beams perforating the central
capsule. The Phacodiscida bear therefore the same relation to the Cenodiscida that the Disphærida
and Trisphærida do to the Monosphærida.
Formerly several species belonging to the family were described by Ehrenberg and Johannes
Müller, but not distinguished from the Sphæroidea, genus Haliomma (e.g., Haliomma
sol et Haliomma humboldtii of the former, Haliomma amphidiscus of the latter).
For these oldest known species I constituted in 1862 my genus Heliodiscus (Monogr. d.
Radiol., p. 436). Some other genera were afterwards (1875) figured by Ehrenberg as
Periphæna and Chilomma. The rich material of the Challenger revealed this family as
very polymorphic and widely distributed, so that in my Prodromus (1881, p. 457) I could enumerate
eighteen different genera of Phacodiscida. This number is here reduced to fifteen, uniting several
of them into one genus as "subgenera."
The Medullary Shell of the Phacodiscida, or the intracapsular latticed shell, is either
simple and spherical, or double, composed of two concentric spheres, which are united by a
variable number of radial beams. We could distinguish therefore as two subfamilies the
Carpodiscida (with simple medullary shell) and the Thecodiscida (with double concentric medullary
shell); the former corresponding to the Carposphærida (or Dyosphæria), the latter to the
Thecosphærida (or Triosphæria). But as this difference seems not to be so important as the
different shape of the disk margin, we prefer this latter as a character {420}for the separation of subfamilies. The form of the medullary shells is
commonly quite spherical (as also in the above mentioned Sphæroidea);
sometimes it is a little compressed in the same direction as the enclosing lenticular phacoid
shell. The diameter of the latter is commonly three to four times as large as the diameter of the
medullary shell; if this be double (in the Thecodiscida) then the diameter of the outer medullary
shell is commonly three to four times as large as that of the inner. The radial beams connecting
the two seem to be very variable in number and disposition (compare Pl. 31, fig. 8; Pl.
32, figs. 3,
4a, 7, 8a; Pl. 33,
figs. 2, 3; Pl. 35,
figs. 4, 8, 9, &c.).
The Radial Beams, which connect the medullary shell with the cortical or "phacoid
shell," and which pierce the membrane of the lenticular central capsule, are commonly aggregated
into two polar bunches around the shortened axis of the disk (compare Pl. 31, fig. 8; Pl.
32, figs. 3,
8; Pl. 33,
figs. 2, 3, &c.). Their number seems to be usually between ten and thirty. More rarely
piercing radial beams lie also in the equatorial plane, and then commonly as inner prolongations
of the outer marginal spines; so we find two opposite in one axis, in Heliostylus (Pl. 34, figs. 1,
2), or four opposite in pairs in two crossed axes, in Phacostaurus (Pl. 31, figs. 1, 2,
7). Often the thickness of the beams increases from the centre towards the periphery.
The Phacoid Shell, or the lenticular extracapsular cortical shell, exhibits in the
Phacodiscida quite the same general character as in the Cenodiscida, above described (p. 410). Commonly the equatorial diameter of the lens is twice to three times as
large as the vertical diameter or the shortened "main axis." The convexity of both opposite faces
is either quite even to the sharp margin (Pl. 31,
fig. 2; Pl. 33,
figs. 2, 5), or the central part of the lens is more strongly vaulted, and often the margin is
thickened or truncated (Pl. 31,
fig. 10; Pl. 35,
fig. 7). The surface of the lens is commonly smooth, but sometimes also covered with
bristle-shaped radial spines (Pl. 34,
figs. 3, 5; Pl. 32,
figs. 2, 3); rarely these spines are prolonged and branched (Pl. 35, figs. 3,
5). The pores of the phacoid shell are usually quite regular, circular, and regularly arranged,
either in more concentric or in more radial rows; the latter are sometimes separated by radial
crests arising towards the margin (Pl. 35,
fig. 6). If the wall of the phacoid shell be much thickened, the pores in its central part are
shorter and cylindrical, in its marginal part longer and conical (Pl. 31, fig. 7; Pl.
32, fig.
1).
The Margin of the Lens of the Phacodiscida is very polymorphic, and serves mainly for
the separation of genera. In the first subfamily, Sethodiscida, it is either quite simple (Pl. 35, figs. 6-8)
or surrounded by a thin solid equatorial girdle, the basal part of which is often radially striped
(Pl. 32,
figs. 7, 8). In the second subfamily, Heliosestrida, we find on the margin a small number of
radial spines in the equatorial plane regularly disposed, either two opposite in one axis (Pl. 31, figs. 9-12)
or four opposite in pairs in two crossed axes (Pl. 31, figs. 1-8),
or eight opposite in pairs in four axes, crossed at angles of 45° (Pl. 34, figs. 3,
6); in the latter case we can sometimes distinguish {421}(as in many Medusæ) four larger perradial spines alternating with four
smaller interradial spines. Often in one and the same species occur abnormalities in number and
disposition of the radial spines, three or five spines instead of four, or also seven or nine
spines instead of eight; often both halves of the disk become asymmetrical. If the number of the
marginal spines exceed eight to ten, they commonly become very variable in size and irregular in
disposition; these variations characterise the third subfamily, Heliodiscida. Commonly also here
all spines lie in the equatorial plane; but sometimes they become crowded in several parallel
circles on both sides of the margin (Pl. 32,
fig. 1). The form of the marginal spines is commonly conical or flattened triangular, often also
pyramidal or deeply furrowed (Pl. 31,
figs. 6-9). Very rarely the spines are fenestrated (Pl. 35, fig. 1);
only in one genus (Heliodrymus) they are all or partly branched (Pl. 33, fig. 9; Pl.
35, figs. 3,
5).
The peculiar development of the phacoid shell has been already described by J. Müller (compare
my Monograph, 1862, pp. 156, 438).
The Central Capsule of the Phacodiscida is everywhere circular, lenticular, envelops the
medullary shell, and is enclosed by the phacoid shell, perforated by the radial beams, which
connect the latter to the former.
Synopsis of the Genera of the Phacodiscida.
|
III. Subfamily Sethodiscida.
Margin of the disk without radial spines.
|
|
Margin simple, without equatorial girdle. |
|
Medullary shell simple, |
181. Sethodiscus. |
| Medullary shell double, |
182. Phacodiscus. |
| Margin surrounded by a hyaline equatorial girdle. |
|
Medullary shell simple, |
183. Periphæna. |
| Medullary shell double, |
184. Perizona. |
|
III. Subfamily Heliosestrida.
Margin of the disk with two to eight solid radial spines, usually quite
regularly disposed. (Number usually constant.)
|
|
Two radial spines (opposite in one axis). |
|
Medullary shell simple, |
185. Sethostylus. |
| Medullary shell double, |
186. Phacostylus. |
| Three radial spines. |
|
Medullary shell simple, |
187. Triactiscus. |
| Four radial spines (in cross form). |
|
Medullary shell simple, |
188. Sethostaurus. |
| Medullary shell double, |
189. Phacostaurus. |
| Six radial spines. |
|
Medullary shell simple, |
190. Distriactis. |
| Eight radial spines. |
|
Medullary shell simple, |
191. Heliosestrum. |
| Medullary shell double, |
192. Astrosestrum. |
|
III. Subfamily Heliodiscida.
Margin of the disk with numerous (ten to twenty or more) radial spines,
usually irregularly disposed. (Number variable.)
|
|
Radial spines all simple, not branched. |
|
Medullary shell simple, |
193. Heliodiscus. |
| Medullary shell double, |
194. Astrophacus. |
| Radial spines all or partly |
|
Medullary shell simple, |
195. Heliodrymus. |
| I. Subfamily Sethodiscida. Margin of the disk without radial spines. |
|
|
Margin simple, without equatorial girdle. |
|
|
|
|
|
|
Medullary shell simple, |
|
|
|
|
|
|
|
|
|
|
181. Sethodiscus. |
|
|
|
|
|
|
|
|
|
|
Medullary shell double, |
|
|
|
|
|
|
|
|
|
|
182. Phacodiscus. |
|
|
|
|
|
|
|
|
Margin surrounded by a hyaline equatorial girdle. |
|
|
|
|
|
|
Medullary shell simple, |
|
|
|
|
|
|
|
|
|
|
183. Periphæna. |
|
|
|
|
|
|
|
|
|
|
Medullary shell double, |
|
|
|
|
|
|
|
|
|
|
184. Perizona. |
|
|
|
|
|
|
| II. Subfamily Heliosestrida. Margin of the disk with two to eight solid radial
spines, usually quite regularly disposed. (Number usually constant.) |
|
|
Two radial spines (opposite in one axis). |
|
|
|
|
|
|
Medullary shell simple, |
|
|
|
|
|
|
|
|
|
|
185. Sethostylus. |
|
|
|
|
|
|
|
|
|
|
Medullary shell double, |
|
|
|
|
|
|
|
|
|
|
186. Phacostylus. |
|
|
|
|
|
|
|
|
Three radial spines. Medullary shell simple, |
|
|
|
|
|
|
|
|
187. Triactiscus. |
|
|
|
|
|
|
|
|
Four radial spines (in cross form). |
|
|
|
|
|
|
Medullary shell simple, |
|
|
|
|
|
|
|
|
|
|
188. Sethostaurus. |
|
|
|
|
|
|
|
|
|
|
Medullary shell double, |
|
|
|
|
|
|
|
|
|
|
189. Phacostaurus. |
|
|
|
|
|
|
|
|
Six radial spines. Medullary shell simple, |
|
|
|
|
|
|
|
|
190. Distriactis. |
|
|
|
|
|
|
|
|
Eight radial spines. |
|
|
|
|
|
|
Medullary shell simple, |
|
|
|
|
|
|
|
|
|
|
191. Heliosestrum. |
|
|
|
|
|
|
|
|
|
|
Medullary shell double, |
|
|
|
|
|
|
|
|
|
|
192. Astrosestrum. |
|
|
|
|
|
|
| III. Subfamily Heliodiscida. Margin of the disk with numerous (ten to twenty
or more) radial spines, usually irregularly disposed. (Number variable.) |
|
|
Radial spines all simple, not branched. |
|
|
|
|
|
|
Medullary shell simple, |
|
|
|
|
|
|
|
|
|
|
193. Heliodiscus. |
|
|
|
|
|
|
|
|
|
|
Medullary shell double, |
|
|
|
|
|
|
|
|
|
|
194. Astrophacus. |
|
|
|
|
|
|
|
|
Radial spines all or partly branched. |
|
|
|
|
|
|
Medullary shell simple, |
|
|
|
|
|
|
|
|
|
|
195. Heliodrymus. |
|
|
|
|
|
|
{422}
Subfamily 1. Sethodiscida, Haeckel, 1881, Prodromus, p. 457.
Definition.—Phacodiscida without radial
spines on the margin of the disk.
Genus 181. Sethodiscus,[220] Haeckel,
1881, Prodromus, p. 457.
Definition.—Phacodiscida with simple medullary shell and
simple margin of the circular disk, without surrounding equatorial girdle and without radial
spines.
The genus Sethodiscus is the most simple and primitive form of all
Phacodiscida, and may be regarded as the common ancestral form of this family. The simple
spherical medullary shell is connected by a variable number of radial beams with the lenticular or
discoidal cortical shell (or "phacoid shell"). The margin of this latter is quite simple,
circular, without solid equatorial girdle or radial spines. From the nearly allied genus
Carposphæra of the Sphæroidea, its probable ancestral form,
Sethodiscus can be derived simply by lenticular compression of the spheroidal cortical
shell.
Subgenus 1. Sethodiscinus, Haeckel.
Definition.—Surface of the disk smooth, without radial ribs or
spines.
1. Sethodiscus phacoides, n. sp.
Disk with smooth surface, three times as broad as the medullary shell. Pores regular, circular;
fourteen to fifteen on the radius of the disk. (Very similar to Periphæna cincta, Pl. 33, fig. 4, but
without the girdle of the margin.)
Dimensions.—Diameter of the disk 0.2, of the medullary shell 0.07, of the pores
0.005.
Habitat.—Pacific, central area, Stations 270 to 274, in various
depths.
2. Sethodiscus macroporus, n. sp.
Disk with smooth surface, twice as broad as the medullary shell. Pores regular, circular, very
large; five to six on the radius of the disk. (Remarkable for the extraordinary size of the pores,
which reaches half the radius of the medullary shell.)
Dimensions.—Diameter of the disk 0.1, of the medullary shell 0.05, of the pores
0.012.
Habitat.—North Atlantic, Gulf Stream, Færöe Channel, John Murray.
3. Sethodiscus microporus, n. sp.
Disk with smooth surface, four times as broad as the medullary shell. Pores regular, circular,
very small; twenty-two to twenty-four on the radius of the disk. (The small pores are scarcely
half as broad as the thick bars between them.)
{423}
Dimensions.—Diameter of the disk 0.25, of the medullary shell 0.06, of the pores
0.002.
Habitat.—South Pacific, Station 288, surface.
4. Sethodiscus lenticula, n. sp. (Pl. 33, figs. 1,
2).
Disk with smooth surface, four times as broad as the medullary shell. Pores irregular,
polygonal; eight to nine on the radius of the disk. (The pores of the medullary shell, fig. 2, are
also irregular, polygonal, or roundish.)
Dimensions.—Diameter of the disk 0.17, of the medullary shell 0.04, of the pores
0.01.
Habitat.—Pacific, central area, Station 274, depth 2750 fathoms.
5. Sethodiscus macrococcus, n. sp. (Pl. 33, fig.
3).
Disk with smooth surface, two and a half times as broad as the medullary shell. Pores
irregular, polygonal; eleven to twelve on the radius of the disk. (The pores of the medullary
shell, fig. 3, are regular, circular, with elevated hexagonal frames between them. The figured
specimen is a young one, both halves of the biconvex disk being not yet united in the equatorial
plane.)
Dimensions.—Diameter of the disk 0.16, of the medullary shell 0.065, of the pores
0.01.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
6. Sethodiscus micrococcus, n. sp.
Disk with smooth surface, five times as broad as the medullary shell. Pores irregular,
roundish, very small; twenty-six to twenty-eight on the radius of the disk. (The pores of the
small medullary shell are regularly circular, of the same size as those of the disk.)
Dimensions.—Diameter of the disk 0.2, of the medullary shell 0.04, of the pores
0.002.
Habitat.—South Atlantic, Station 335, depth 1425 fathoms.
Subgenus 2. Sethodisculus, Haeckel.
Definition.—Surface of the disk with elevated radial ribs or
spines.
7. Sethodiscus radiatus, Haeckel.
Haliomma radians, Ehrenberg, 1854, Mikrogeol., Taf. xix. fig. 50.
Haliomma radiatum, Ehrenberg, 1854, Mikrogeol., Taf. xxi. fig. 54.
Disk with radiated surface, four times as broad as the medullary shell. Pores regularly
circular; nine to ten on the radius of the disk, arranged in radial series (about forty), which
are separated by smooth crests, not prominent on the smooth margin.
Dimensions.—Diameter of the disk 0.12, of the medullary shell 0.03, of the pores
0.003.
Habitat.—Fossil in Tertiary deposits of the Mediterranean, Greece,
Sicily, Oran, &c.
{424}
8. Sethodiscus echinatus, Haeckel.
Haliomma echinatum, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss. Berlin,
p. 74, Taf. xxvii. fig. 2.
Disk with radiated thorny surface, three times as broad as the medullary shell. Pores regular,
circular; eleven to twelve on the radius of the disk, arranged in radial series (about sixty),
which are separated by thorny crests, prominent a little on the dentated margin.
Dimensions.—Diameter of the disk 0.18, of the medullary shell 0.06, of the pores
0.01.
Habitat.—Fossil in the rocks of Barbados.
9. Sethodiscus cristatus, n. sp.
Disk with radiated surface, four times as broad as the medullary shell. Pores regularly
circular; sixteen to seventeen on the radius of the disk, arranged in radial series (about
eighty), which are separated in the distal half by smooth elevated crests, strongly prominent on
the dentated margin. (Very similar to Phacodiscus cristatus, Pl. 35, fig. 6, but
with simple medullary shell.)
Dimensions.—Diameter of the disk 0.2, of the medullary shell 0.05, of the pores
0.005.
Habitat.—South Pacific, Station 285, depth 2375 fathoms.
Genus 182. Phacodiscus,[221] Haeckel,
1881, Prodromus, p. 457.
Definition.—Phacodiscida with double medullary shell and
simple margin of the circular disk, without surrounding equatorial girdle and without radial
spines.
The genus Phacodiscus differs from the ancestral genus Sethodiscus
only in the duplication of the medullary shell, and has therefore the same relation to it that
Thecosphæra in the Sphæroidea exhibits to Carposphæra. Both
former discoidal genera differ from the two latter spheroidal by the lenticular flattening of the
cortical shell.
Subgenus 1. Phacodiscinus, Haeckel.
Definition.—Surface of the disk smooth, without radial ribs or
spines.
1. Phacodiscus rotula, n. sp. (Pl. 35,
fig. 7).
Disk with smooth surface, four and a half times as broad as the outer and fourteen times as
broad as the inner medullary shell. Pores regularly circular; sixteen to eighteen on the radius of
the disk. Margin of the lens very thick, truncated, nearly as broad as the outer medullary
shell.
Dimensions.—Diameter of the disk 0.21, of the outer medullary shell 0.045, of the
inner 0.015; pores 0.008.
Habitat.—North Pacific, Station 224, depth 1850 fathoms.
{425}
2. Phacodiscus lentiformis, n. sp. (Pl. 35, fig.
8).
Disk with smooth surface, three and a half times as broad as the outer, and ten times as broad
as the inner medullary shell. Pores regularly circular; twelve to thirteen on the radius of the
disk. Margin of the disk sharp, as in a biconvex lens.
Dimensions.—Diameter of the disk 0.25, of the outer medullary shell 0.07, of the
inner 0.025; pores 0.01.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms; fossil in
Barbados.
3. Phacodiscus grandis, n. sp.
Disk with thorny surface, five times as broad as the outer, and fifteen times as broad as the
inner medullary shell. Pores regularly circular; twenty-two to twenty-four on the radius of the
disk. Margin of the disk thick, rounded.
Dimensions.—Diameter of the disk 0.45, of the outer medullary shell 0.09, of the
inner 0.03; pores 0.01.
Habitat.—Western Tropical Pacific, Station 225, depth 4475
fathoms.
Subgenus 2. Phacodisculus, Haeckel.
Definition.—Surface of the disk with elevated radial ribs or
spines.
4. Phacodiscus echiniscus, n. sp.
Disk with spiny surface, four times as broad as the outer, and ten times as broad as the inner
medullary shell. Pores irregularly roundish; fourteen to fifteen on the radius of the disk. Margin
of the disk sharp, as in a biconvex lens.
Dimensions.—Diameter of the disk 0.3, of the outer medullary shell 0.75, of the
inner 0.03; pores 0.007.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
5. Phacodiscus clypeus, n. sp. (Pl. 35,
figs. 6, 9).
Disk with radiated surface, four times as broad as the outer, and twelve times as broad as the
inner medullary shell. Pores regularly circular; sixteen to eighteen on the radius of the disk; in
the distal half disposed in sixty to seventy radial series, which are separated by prominent
crests; the sharp margin therefore a little jagged.
Dimensions.—Diameter of the disk 0.2, of the outer medullary shell 0.05, of the
inner 0.017; pores 0.006.
Habitat.—Pacific, central area, Station 266, depth 2750 fathoms.
{426}
Genus 183. Periphæna,[222] Ehrenberg,
1873 Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 246.
Definition.—Phacodiscida with simple medullary shell,
without radial spines, but with a solid equatorial girdle around the margin of the lenticular
disk.
The genus Periphæna, founded by Ehrenberg in 1873 for the fossil
Periphæna decora of Barbados, differs from its ancestral form Sethodiscus in the
development of a very thin siliceous solid girdle around the margin of the lenticular disk; this
girdle lies in the equatorial plane of the shell, and reappears in similar form in
Perichlamydium among the Porodiscida, in Spongophacus among the Spongodiscida, and
in Zonodiscus among the Cenodiscida.
1. Periphæna cincta, n. sp. (Pl. 33,
fig. 4).
Disk with smooth surface, four times as broad as the medullary shell. Pores regularly circular;
fourteen to sixteen on the radius of the disk. Girdle of the margin about half as broad as the
radius of the medullary shell, in the distal half structureless, in the proximal half with seventy
to eighty short radial ribs.
Dimensions.—Diameter of the disk 0.2, of the medullary shell 0.05, of the pores
0.005.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
2. Periphæna decora, Ehrenberg.
Periphæna decora, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss. Berlin,
p. 80, Taf. xxviii. fig. 6.
Disk with smooth surface, three times as broad as the medullary shell. Pores regular, circular;
twenty to twenty-two on the radius of the disk, disposed in radial series. Girdle of the margin
nearly as broad as the radius of the medullary shell, in the distal half structureless, in the
proximal half with eighty to ninety short radial ribs.
Dimensions.—Diameter of the disk 0.25, of the medullary shell 0.08, of the pores
0.06.
Habitat.—Fossil in Barbados.
3. Periphæna statoblastus, n. sp.
Disk with smooth surface, five times as broad as the medullary shell. Pores regularly circular;
twenty-four to twenty-six on the radius of the disk, disposed in radial series, those in the
distal half of the disk being separated by piercing radial beams. Girdle of the margin twice as
broad as the radius of the medullary shell, in the whole breadth with one hundred and twenty to
one hundred and thirty piercing radial ribs, the prolongations of the beams.
Dimensions.—Diameter of the disk 0.3, of the medullary shell 0.06, of the pores
0.04.
Habitat.—South Pacific, Station 300, depth 1375 fathoms.
{427}
Genus 184. Perizona,[223] Haeckel, 1881,
Prodromus, p. 457.
Definition.—Phacodiscida with double medullary shell,
without radial spines, but with a solid equatorial girdle around the margin of the lenticular
disk.
The genus Perizona differs from its ancestral form Phacodiscus in the
development of a thin solid siliceous girdle, lying in the equatorial plane around the margin of
the disk. The two genera bear the same relation to each other that Periphæna bears to
Sethodiscus. But in the two latter genera the spherical medullary shell is simple, in the
two former double.
1. Perizona scutella, n. sp. (Pl. 32,
fig. 7).
Disk with smooth surface, in the distal third radiated, four times as broad as the outer, and
ten times is broad as the inner medullary shell. Pores subregular, circular; thirteen to fourteen
on the radius of the disk, in the marginal part separated by eighty to ninety prominent radial
crests, which are prolonged into the proximal half of the solid girdle. Breadth of the girdle
equal to that of the inner medullary shell.
Dimensions.—Diameter of the disk 0.25, of the outer medullary shell 0.06, of the
inner 0.025; pores 0.007.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
2. Perizona pterygota, n. sp. (Pl. 32,
figs. 8, 8a).
Disk with smooth surface, six times as broad as the outer, and fifteen times as broad as the
inner medullary shell. Pores regularly circular; eighteen to twenty on the radius of the disk.
Margin much thickened and truncated, abruptly separated in the equatorial circumference from the
peripheral sharp margin of the solid girdle; breadth of the girdle equal to that of the outer
medullary shell.
Dimensions.—Diameter of the disk 0.25, of the outer medullary shell 0.04, of the
inner 0.015; pores 0.006.
Habitat.—Pacific, central area, Station 266, depth 2750 fathoms.
Subfamily 2. Heliosestrida, Haeckel.
Definition.—Phacodiscida with a constant
number of radial spines on the margin of the disk (two, three, four, six, or eight), which are
commonly regularly disposed (sometimes more or less irregularly).
{428}
Genus 185. Sethostylus,[224] Haeckel,
1881, Prodromus, p. 457.
Definition.—Phacodiscida with simple medullary shell and
with two radial spines on the margin of the disk, opposite in the equatorial axis.
The genus Sethostylus opens the series of the Heliosestrida or of those
Phacodiscida in which a constant number of radial spines (two to eight) is more or less regularly
disposed on the margin of the disk. All these marginal spines lie in the equatorial plane of the
lens, and have in the same species a rather constant number and similar size, a certain form and
disposition. Sometimes they incline more or less to irregular variations. In Sethostylus
there are only two spines, opposite in the equatorial diameter of the lens; it corresponds to
Stylocyclia among the Coccodiscida, and to Xiphodictya among the Porodiscida.
Subgenus 1. Sethostylium, Haeckel.
Definition.—Margin of the disk simple, smooth, without equatorial
girdle and without a corona of marginal spines.
1. Sethostylus distyliscus, n. sp. (Pl. 31, fig.
9).
Disk with smooth surface, three times as broad as the medullary shell. Pores regular, circular;
nine to ten on the radius of the disk, in the outer rows cylindrical. Margin of the disk simple,
smooth, thin, without peculiar equatorial girdle, radially striped by the prominent beams of the
peripheral series of pores. Both marginal spines of equal size, pyramidal, sulcated, about as long
as the radius of the disk and as broad at the base as the medullary shell.
Dimensions.—Diameter of the disk 0.13 to 0.16, of the medullary shell 0.04 to
0.05; length of both opposite spines 0.06 to 0.08, basal breadth 0.03 to 0.04.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms.
2. Sethostylus dicylindrus, n. sp. (Pl. 31, fig.
10).
Disk with smooth surface, four times as broad as the medullary shell. Pores regular, circular;
sixteen to eighteen on the radius of the disk. Margin of the disk simple, smooth, thick, without
peculiar equatorial girdle. Both marginal spines cylindrical, about as long as the diameter of the
disk, about twice as broad as a single pore. (Walls of the disk in the central part twice to three
times as thick as in the peripheral part.)
Dimensions.—Diameter of the disk 0.2, of the medullary shell 0.05; length of both
spines 0.2 to 0.3, breadth 0.008.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
{429}
Subgenus 2. Heliostylus, Haeckel, 1881, Prodromus, p. 457.
Definition.—Margin of the disk with a solid equatorial girdle or a
corona of radial spines.
3. Sethostylus dentatus, n. sp. (Pl. 34,
fig. 1).
Heliostylus dentatus, Haeckel, 1881, Prodromus et Atlas (pl. xxxiv. fig.
1).
Disk with smooth surface, six times as broad as the medullary shell. Pores regular, circular;
eighteen to twenty on the radius of the disk. Margin of the disk with a broad solid equatorial
girdle, which bears fifty to sixty strong conical teeth on the periphery; teeth about as long as
the diameter of the medullary shell. Both opposite marginal spines cylindro-conical, about as long
as the diameter of the disk, and as broad as the girdle.
Dimensions.—Diameter of the disk 0.3, of the medullary shell 0.05; length of both
main spines 0.2 to 0.3, breadth 0.02.
Habitat.—Pacific, central area, Station 272, depth 2600 fathoms.
4. Sethostylus serratus, n. sp. (Pl. 34,
fig. 2).
Heliostylus serratus, Haeckel, 1881, Prodromus et Atlas (pl. xxxiv. fig.
2).
Disk with smooth surface, four times as broad as the medullary shell. Pores regular, circular;
fourteen to sixteen on the radius of the disk. Margin with a broad solid equatorial girdle, which
bears forty to fifty strong conical teeth on the periphery; teeth about as long as the radius of
the medullary shell. Both opposite marginal spines spindle-shaped, about as long as the diameter
of the disk, and as broad as the girdle.
Dimensions.—Diameter of the disk 0.3, of the medullary shell 0.075; length of both
main spines 0.2 to 0.3, breadth 0.03.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
5. Sethostylus hastatus, n. sp.
Disk with smooth surface, two and a half times as broad as the medullary shell. Pores
irregular, roundish; ten to twelve on the radius of the disk. Margin with a broad solid equatorial
girdle, which bears on the periphery six conical teeth, three on each half between the two main
spines, which are cylindrical, longer than the diameter of the disk, and half as broad as the
girdle. (This species can be derived from Heliosestrum, two opposite spines of the eight
marginal spines being much stronger developed than the other six.)
Dimensions.—Diameter of the disk 0.15, of the medullary shell 0.06; length of both
main spines 0.2, breadth 0.01.
Habitat.—North Atlantic, Station 354, surface.
{430}
6. Sethostylus spicatus, n. sp.
Disk with spiny surface, four times as broad as the medullary shell. Pores irregular roundish,
with many unequal conical spines between them. Margin with an equatorial girdle, composed of three
to four concentric series of conical radial spines, about as long as the radius of the medullary
shell. Both main spines very large, cylindrical, longer than the diameter of the disk, and as
broad as the radius of the medullary shell.
Dimensions.—Diameter of the disk 0.2, of the medullary shell 0.05; length of the
main spines 0.3 or more, breadth 0.025.
Habitat.—Fossil in the rocks of Barbados, Haeckel; living in the
depths of the Equatorial Atlantic, Station 348, depth (2450) fathoms.
Genus 186. Phacostylus,[225] Haeckel,
1881, Prodromus, p. 457.
Definition.—Phacodiscida with double medullary shell and
with two radial spines on the margin of the disk, opposite in the equatorial axis.
The genus Phacostylus differs from the foregoing Sethostylus by the
duplication of the medullary shell, and bears the same relation to it as Phacodiscus does
to Sethodiscus, or as Amphicyclia in the Coccodiscida does to
Stylocyclia.
Subgenus 1. Phacostylium, Haeckel.
Definition.—Margin of the disk simple, smooth, without equatorial
girdle, and without a corona of marginal spines.
1. Phacostylus amphistylus, n. sp. (Pl. 31, fig.
12).
Disk with smooth surface, four times as broad as the outer, and twelve times as broad as the
inner medullary shell. Pores regular, circular; twelve to fourteen on the radius of the disk.
Margin of the disk simple, smooth, without spines and equatorial girdle. Both polar spines
conical, longer than the diameter of the disk, furrowed at the base, and twice as broad as the
inner medullary shell.
Dimensions.—Diameter of the disk 0.16 to 0.18, of the outer medullary shell 0.045,
of the inner 0.015; length of the polar spines 0.2, basal breadth 0.03.
Habitat.—Pacific, central area, Station 272, depth 2600 fathoms.
2. Phacostylus amphixiphus, n. sp.
Disk with smooth surface, three times as broad as the outer, and eight times as broad as the
inner medullary shell. Pores irregular, roundish; ten to eleven on the radius. Margin of the disk
{431}simple, smooth, without spines and equatorial
girdle. Both polar spines sword-shaped triangular, two-edged, about as long as the radius of the
disk, and as broad at the base as the inner medullary shell.
Dimensions.—Diameter of the disk 0.15, of the outer medullary shell 0.05, of the
inner 0.02; length of the polar spines 0.08, basal breadth 0.02.
Habitat.—Indian Ocean, Madagascar, Rabbe, surface.
Subgenus 2. Astrostylus, Haeckel.
Definition.—Margin of the disk with a solid equatorial girdle or a
corona of radial spines.
3. Phacostylus amphipyramis, n. sp.
Disk with spiny surface, four and a half times as broad as the outer, and fourteen times as
broad as the inner medullary shell. Pores irregular, roundish; eight to ten on the radius. Margin
of the disk with a corona of irregular, radial spines. Both opposite polar spines pyramidal, twice
as long as broad, and nearly as long as the radius of the disk.
Dimensions.—Diameter of the disk 0.22, of the outer medullary shell 0.05, of the
inner 0.016; length of the polar spines 0.1, basal breadth 0.05.
Habitat.—Western Tropical Pacific, Station 225, depth 4475
fathoms.
4. Phacostylus caudatus, n. sp. (Pl. 32,
fig. 6).
Astrosestrum caudatum, Haeckel, 1881, Prodromus et Atlas (pl. xxxii. fig.
6).
Disk with smooth surface, two and a half times as broad as the outer, and six times as broad as
the inner medullary shell. Pores regular, circular; six to seven on the radius of the disk. Margin
with a solid equatorial girdle, and irregularly bordered with eight to ten conical spines; two
opposite of these are much longer than the others. (This species can be derived from
Astrosestrum, two opposite marginal spines being much more strongly developed than the six
to eight others.)
Dimensions.—Diameter of disk 0.12, of the outer medullary shell 0.05, of the inner
0.02; length of the polar spines 0.1 to 0.25, basal breadth 0.025.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
5. Phacostylus maximus, n. sp.
Disk with smooth surface, five times as broad as the outer, and ten times as broad as the inner
medullary shell. Pores regular, circular; twenty to twenty-two on the radius. Margin with a solid
equatorial girdle, bearing on the periphery one hundred to one hundred and twenty plain teeth, and
two very large polar spines, which are cylindrical, longer than the diameter of the disk, and as
broad at the furrowed base as the radius of the outer medullary shell. (Similar to Sethostylus
dentatus, Pl. 34,
fig. 1, but much larger, and with a double medullary shell.) {432}Dimensions.—Diameter of the disk 0.4, of the outer medullary
shell 0.08, of the inner 0.04; length of the polar spines 0.5, basal breadth 0.04.
Habitat.—Fossil in the rocks of Barbados.
Genus 187. Triactiscus,[226] Haeckel,
1881, Prodromus, p. 457.
Definition.—Phacodiscida with simple medullary shell, and
with three radial spines on the margin of the disk, placed in the equatorial plane.
The genus Triactiscus exhibits on the margin of the lenticular phacoid shell
three radial solid spines, the distance between which is sometimes equal, at other times unequal.
Either all the three spines are of equal size and similar form, or one odd arm is smaller than the
two paired arms. The triradial form of this genus is repeated in Tripodocyclia among the
Coccodiscida, and in Tripodictya among the Porodiscida. The medullary shell of
Triactiscus is simple.
1. Triactiscus tripyramis, n. sp. (Pl. 33, fig.
6).
Disk with smooth surface and smooth margin, three times as broad as the medullary shell. Pores
regular, circular; twelve to thirteen on the radius of the disk. Three radial spines of nearly
equal size and equidistant, or one odd spine a little smaller, and the opposite angle (between the
paired spines) also smaller. Spines pyramidal, with broad prominent edges, one to one and a half
times as long as the radius of the disk, as broad at the base as the radius of the medullary
shell.
Dimensions.—Diameter of the disk 0.15; of the medullary shell 0.05, pores
0.006.
Habitat.—Pacific, central area, Station 272, depth 2600 fathoms.
2. Triactiscus tricuspis, n. sp. (Pl. 33, fig.
5).
Disk with smooth surface and smooth margin, five times as broad as the medullary shell. Pores
irregular, roundish; ten to eleven on the radius of the disk. Three radial spines of unequal size
and at unequal distances, very short, conical, not longer than the radius of the medullary
shell.
Dimensions.—Diameter of the disk 0.15, of the medullary shell 0.03; pores 0.004 to
0.008.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
3. Triactiscus tripodiscus, Haeckel.
Haliomma triactis, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss. Berlin,
p. 236, Taf. xxviii. fig, 4.
Disk with thorny surface and spiny margin, three times as broad as the medullary shell. Pores
regular, circular; eight to nine on the radius of the disk. Three radial spines of different {433}size and at unequal distances, one odd spine as long as
the radius, both paired spines as long as the diameter of the disk; the odd angle between the
latter is smaller. Spines pyramidal, with broad edges.
Dimensions.—Diameter of the disk 0.16, of the medullary shell 0.05, pores
0.008.
Habitat.—Fossil in the rocks of Barbados.
Genus 188. Sethostaurus,[227] Haeckel,
1881, Prodromus, p. 457.
Definition.—Phacodiscida with simple medullary shell and
with four radial spines on the margin of the disk, crossed in the equatorial plane.
The genus Sethostaurus exhibits four marginal spines, which form commonly a
more or less regular cross in the equatorial plane. Sometimes the size and disposition of the four
spines become more or less different, and also the angles between them vary; the regular
rectangular cross passes over into a bilateral or irregular form. The medullary shell is simple.
The same cross-form of the disk is seen in Staurocyclia among the Coccodiscida, and in
Staurodictya among the Porodiscida.
Subgenus 1. Sethostaurium, Haeckel.
Definition.—Margin of the disk without a solid equatorial girdle or a
corona of spines.
1. Sethostaurus orthostaurus, n. sp. (Pl. 31, figs. 1,
2).
Disk with smooth surface, three times as broad as the medullary shell. Pores regular, circular;
thirteen to fourteen on the radius of the disk. Margin simple, smooth, without girdle. Four
crossed spines equal, pommel-shaped, angular, contracted at the base, scarcely as long as the
diameter of the medullary shell.
Dimensions.—Diameter of the disk 0.16, of the medullary shell 0.05; length of the
crossed spines 0.04, breadth 0.02.
Habitat.—Pacific, central area, Station 266, depth 2750 fathoms.
2. Sethostaurus conostaurus, n. sp. (Pl. 34, figs. 7,
8).
Disk with spiny surface, three times as broad as the medullary shell. Pores irregular,
roundish; seven to eight on the radius. Margin simple, smooth, without girdle. Four crossed
spines subregular, equal, conical, about as long as the diameter and as broad at the base as the
radius of the medullary shell. (Fig. 7 exhibits the normal form, fig. 8 an abnormality with five
spines.)
Dimensions.—Diameter of the disk 0.2, of the medullary shell 0.06; length of the
crossed spines 0.05, basal breadth 0.03.
Habitat.—South Atlantic, Station 333, surface.
{434}
3. Sethostaurus rhombostaurus, n. sp. (Pl. 31, fig.
4).
Disk with smooth surface, two and a half times as broad as the medullary shell. Pores regular,
circular; ten to eleven on the radius. Margin simple, smooth, without girdle. Two opposite spines
longer than the diameter of the disk, four times as long as the two others, which only equal its
radius. Spines sword-shaped, angular.
Dimensions.—Diameter of the disk 0.14, of the medullary shell 0.06; length of the
major spines 0.2, of the minor 0.05, breadth 0.025.
Habitat.—North Atlantic, Station 353, depth 2965 fathoms.
4. Sethostaurus recurvatus, n. sp. (Pl. 31, fig.
3).
Disk with smooth surface, three times as broad as the medullary shell. Pores irregular,
roundish; eight to nine on the radius. Margin simple, smooth, without girdle. Four spines
cylindrical, irregularly curved, of different sizes; one single very large, much longer than the
three others; two opposite lateral spines recurved, hook-shaped.
Dimensions.—Diameter of the disk 0.15, of the medullary shell 0.05; length of the
major spine 0.32, of the opposite spine 0.16, of both lateral spines 0.08.
Habitat.—South Pacific, Station 295, depth 1500 fathoms.
Subgenus 2. Heliostaurus, Haeckel 1881, Prodromus, p. 457.
Definition.—Margin of the disk with a solid equatorial girdle or a
corona of spines.
5. Sethostaurus cruciatus, n. sp. (Pl. 31, fig.
5).
Heliostaurus cruciatus, Haeckel, 1881, Prodromus et Atlas (pl. xxxi. fig.
5).
Disk with smooth surface, three times as broad as the medullary shell. Pores regular, circular;
eleven to twelve on the radius. Margin with a solid, radially striped girdle, which bears on the
periphery forty to fifty triangular pointed teeth of unequal length. Four crossed spines of equal
size and similar form, pyramidal, sulcated, about as long as the radius of the disk, as broad at
the base as the girdle.
Dimensions.—Diameter of the disk 0.2, of the medullary shell 0.06; length of the
crossed spines 0.1, basal breadth 0.02.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
6. Sethostaurus coronatus, n. sp.
Disk with spiny surface, twice as broad as the medullary shell. Pores irregular, roundish; nine
to ten on the radius. Margin of the disk with a solid broad girdle, bearing on the periphery a
corona of thirty to forty flat tongue-shaped teeth of different length. Four crossed spines {435}prismatic with prominent edges, about as long as the
diameter of the disk, and as broad as the girdle.
Dimensions.—Diameter of the disk 0.16, of the medullary shell 0.08, length of the
crossed spines 0.18, breadth 0.02.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
7. Sethostaurus gigas, n. sp.
Disk with smooth surface, four times as broad as the medullary shell. Pores regular, circular;
twenty to twenty-two on the radius. Margin of the disk with a narrow solid girdle, bearing on the
periphery one hundred to one hundred and twenty slender flat pointed teeth. Four crossed spines
equal, conical, about as long as the radius of the disk, as broad at their furrowed base as the
radius of the medullary shell.
Dimensions.—Diameter of the disk 0.4, of the medullary shell 0.1; length of the
crossed spines 0.5, basal breadth 0.05.
Habitat.—Western Tropical Pacific, Station 225, depth 4475
fathoms.
Genus 189. Phacostaurus,[228] Haeckel,
1881, Prodromus, p. 457.
Definition.—Phacodiscida with double medullary shell and
with four radial spines on the margin of the disk, crossed in the equatorial plane.
The genus Phacostaurus differs from the foregoing Sethostaurus by the
duplication of the medullary shell, and bears to it the same relation as Phacodiscus does
to Sethodiscus. Also in this genus the cross of the shell is commonly regular, rectangular,
sometimes more or less irregular.
Subgenus 1. Phacostaurium, Haeckel.
Definition.—Margin of the disk without a solid equatorial girdle or a
corona of spines.
1. Phacostaurus oceanidum, n. sp. (Pl. 31, fig.
6).
Disk with smooth surface, three times as broad as the outer and eight times as broad as the
inner medullary shell. Pores regular, circular; eleven to twelve on the diameter of the disk.
Margin smooth. Four crossed spines pyramidal, deeply sulcate, about as long as the diameter of the
outer medullary shell, and two-thirds as broad at the base.
Dimensions.—Diameter of the disk 0.15, of the outer medullary shell 0.05, of the
inner 0.02; length of the spines 0.05, basal breadth 0.03.
Habitat.—Central Pacific, Station 266, depth 2750 fathoms.
{436}
Subgenus 2. Astrostaurus, Haeckel, 1881, Prodromus, p. 457.
Definition.—Margin of the disk with a solid equatorial girdle, or a
corona of spines.
2. Phacostaurus quadrigatus, n. sp.
Disk with smooth surface, four times as broad as the outer and ten times as broad as the inner
medullary shell. Pores irregular, roundish; fifteen to sixteen on the diameter of the disk. Margin
with a broad solid equatorial girdle, from which arise four crossed spines, conical, as long as
the radius of the disk.
Dimensions.—Diameter of the disk 0.2, of the outer medullary shell 0.05, of the
inner 0.02; length of the spines 0.1, basal breadth 0.03.
Habitat.—South Pacific, Station 300, depth 1375 fathoms.
3. Phacostaurus magnificus, n. sp. (Pl. 31, figs. 7,
8).
Astrostaurus magnificus, Haeckel, 1881, Prodromus et Atlas (pl. xxxi.
figs.7, 8).
Disk with smooth surface, three times as broad as the outer and nine times as broad as the
inner medullary shell. Pores regular, circular; thirteen to fourteen on the diameter of the disk.
Margin with a corona of fifty to sixty conical spines of unequal length. Four spines of the cross
very large pyramidal, deeply sulcated, longer than the radius of the disk, and as broad at the
base as the radius of the medullary shell.
Dimensions.—Diameter of the disk 0.22, of the outer medullary shell 0.07, of the
inner 0.025; length of the four spines 0.16, basal breadth 0.04.
Habitat.—Indian Ocean, Madagascar, Rabbe, surface.
Genus 190. Distriactis,[229] n. gen.
Definition.—Phacodiscida with simple medullary shell and
with six radial spines on the margin of the disk placed in the equatorial plane.
The genus Distriactis exhibits on the margin of the lenticular phacoid shell
six radial solid spines. Their distance is commonly equal, rarely unequal. Either all six spines
are of equal size and similar form, or three larger (perradial) alternating with three smaller
(interradial), so that Distriactis appears to be derived from Triactiscus by
development of three secondary between three primary spines.
1. Distriactis liriantha, n. sp.
Disk smooth, four times as broad as the medullary shell, with regular, circular pores (about
ten on the radius). Six marginal spines of equal size and at equal distances, triangular, sulcate,
half as long as the radius. (Similar to Heliosestrum medusinum, Pl. 34, fig.
6.)
{437}
Dimensions.—Diameter of the disk 0.2, of the medullary shell 0.05; length of the
spines 0.05, basal breadth 0.04.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms.
2. Distriactis corallantha, n. sp.
Disk thorny, three times as broad as the medullary shell, with regular, circular pores, about
eight on the radius. Six marginal spines of equal size and equidistant, conical, nearly as long as
the diameter of the disk, half as broad at the base as the medullary shell.
Dimensions.—Diameter of the disk 0.12, of the medullary shell 0.04; length of the
spines 0.1, breadth 0.02.
Habitat.—South Pacific, Station 300, surface.
3. Distriactis alterna, n. sp.
Disk smooth, five times as broad as the medullary shell, about twelve pores on the radius. Six
marginal spines pyramidal, at equal distances, but unequal alternating size; three larger
(perradial) spines as long as the shell radius, three smaller between them (interradial) half as
long and broad.
Dimensions.—Diameter of the disk 0.15, of the medullary shell 0.03; length of the
three major spines 0.08, of the three minor 0.05.
Habitat.—North Atlantic, Station 353, surface.
4. Distriactis palmantha, n. sp.
Disk rough, four times as broad as the medullary shell, about ten pores on the radius. Six
marginal spines conical, at equal distances, but unequal alternating size; three larger
(perradial) spines as long as the shell diameter, three smaller between them (interradial) as long
as the radius.
Dimensions.—Diameter of the disk 0.2, of the medullary shell 0.05; length of the
three major spines 0.2, of the three minor 0.1,
Habitat.—South Atlantic, Station 325, surface.
5. Distriactis amphithecta, n. sp.
Disk thorny, three times as broad as the medullary shell. Six marginal spines conical, of very
different size; one single (anterior) much larger than the five others, the opposite (posterior)
much smaller. The other four spines are paired, the anterior pair larger than the posterior.
Dimensions.—Diameter of the disk 0.15, of the medullary shell 0.05; length of the
longest spine 0.2, of the shortest 0.04, of the anterior pair 0.1, of the posterior 0.07.
Habitat.—Indian Ocean, Cocos Islands, surface, Rabbe.
{438}
Genus 191. Heliosestrum,[230] Haeckel,
1881, Prodromus, p. 457.
Definition.—Phacodiscida with simple medullary shell and
with eight radial spines on the margin of the disk (more or less regularly disposed, sometimes
seven or nine).
The genus Heliosestrum (with simple medullary shell) and the following
Astrosestrum (with double medullary shell) contain those Phacodiscida in which the margin
of the lenticular disk bears eight radial spines, commonly more or less regularly disposed in the
equatorial plane, so that the equal angles between them amount to 45°. There are, however, many
exceptions to this regular eight-rayed form, either the angles between the eight spines becoming
unequal or the number of the spines amounting to seven or nine (sometimes also six or ten) in one
and the same species. But the regular form is as prevalent, and also in the abnormal forms
indicated by the position of the spines, that we separate the genus Heliosestrum from
Heliodiscus. Both genera are rich in common species, and in some of the former four
perradial (larger) spines alternate regularly with four interradial (smaller) spines.
Subgenus 1. Heliosestantha, Haeckel.
Definition.—Surface of the disk smooth, without radial spines. Bases
of the marginal spines free, not connected by an equatorial girdle.
1. Heliosestrum medusinum, n. sp. (Pl. 34, fig.
6).
Heliodiscus medusinus, Haeckel, 1881, Atlas (pl. xxxiv. fig. 6).
Disk with smooth surface, four times as broad as the medullary shell. Pores regular, circular;
about ten on the radius of the disk. Eight marginal spines regularly distributed (sometimes seven
or nine, more or less irregular), angular, nearly pyramidal (with equilateral triangular outline),
about as long and broad as the diameter of the medullary shell, without connecting equatorial
girdle. From the broad base of each spine run eight to ten deep furrows convergent to its
apex.
Dimensions.—Diameter of the disk 0.2, of the medullary shell 0.05; length of the
radial spines 0.05, basal breadth 0.05.
Habitat.—North Pacific, Station 266, surface.
2. Heliosestrum octastrum, n. sp.
Disk with smooth surface, three times as broad as the medullary shell. Pores irregular,
roundish; eight to nine on the radius. Eight marginal spines regularly distributed (sometimes
{439}seven or nine, more or less irregular), conical,
about as long as the diameter of the disk, without a connecting equatorial girdle.
Dimensions.—Diameter of the disk 0.15, of the medullary shell 0.05; length of the
radial spines 0.16, basal breadth 0.01.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
3. Heliosestrum solarium, Haeckel.
Haliomma sol (partim), Ehrenberg, 1875 (non 1844), Abhandl.
d. k. Akad. d. Wiss. Berlin, p. 74, Taf. xxviii. fig. 1.
Surface of the disk smooth; its diameter six times as large as that of the medullary shell.
Pores regular, circular; nine to ten on the radius. Eight marginal spines regularly distributed
(sometimes seven or nine, more or less irregular), conical, about as long as the diameter of the
medullary shell, without a connecting equatorial girdle.
Dimensions.—Diameter of the disk 0.18, of the medullary shell 0.03; length of the
radial spines 0.03, basal breadth 0.01.
Habitat.—Fossil in the Tertiary rocks of Barbados.
4. Heliosestrum liriope, n. sp.
Surface of the disk smooth; its diameter four times as large as that of the medullary shell.
Pores subregular, circular; seven to eight on the radius. Eight marginal spines regularly
distributed, conical, compressed, alternating longer and shorter; the longer equal to the diameter
of the disk, the shorter to the radius.
Dimensions.—Diameter of the disk 0.16, of the medullary shell 0.4; length of the
major radial spines 0.15, of the minor 0.09.
Habitat.—North Pacific, Station 236, surface.
5. Heliosestrum quadrigeminum, n. sp.
Surface of the disk smooth; its diameter four times as large as that of the medullary shell.
Pores irregular, roundish; six to seven on the radius. Eight marginal spines triangular, flat,
regularly disposed, alternating longer and shorter, the major as long as the radius of the shell,
and half as broad at the base, with three to four deep furrows on both sides, without a connecting
equatorial girdle.
Dimensions.—Diameter of the disk 0.16, of the medullary shell 0.04; length of the
larger radial spines 0.08, of the smaller 0.04, basal breadth 0.04.
Habitat.—Pacific, central area, Station 266, depth 2750 fathoms.
6. Heliosestrum contiguum, Haeckel.
Haliomma contiguum, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss. Berlin,
p. 74, Taf. xxvii. fig. 5.
Disk with smooth surface, four times as broad as the medullary shell. Pores irregular,
roundish, touching, with very thin bars; nine to ten on the radius. Eight marginal spines (often
{440}seven or nine) conical, half as long as the radius,
twice as long as broad, without a connecting equatorial girdle.
Dimensions.—Diameter of the disk 0.17, of the medullary shell 0.04; length of the
radial spines 0.04, basal breadth 0.02.
Habitat.—Fossil in Barbados.
7. Heliosestrum irregulare, n. sp.
Disk with smooth surface, four times as broad as the medullary shell. Pores irregular,
roundish; nine to eleven on the radius. Eight conical radial spines of irregular size and
distribution, often seven or nine, about as long as the radius of the disk (in some cases longer,
in others shorter). No connecting equatorial girdle.
Dimensions.—Diameter of the disk 0.2, of the medullary shell 0.05; length of the
radial spines 0.1, basal breadth 0.02.
Habitat.—Central Pacific, Stations 270 to 272, depth 2425 to 2925
fathoms.
Subgenus 2. Heliosestilla, Haeckel.
Definition.—Surface of the disk armed with radial spines. Bases of the
marginal spines free, without a connecting equatorial girdle.
8. Heliosestrum octonum, n. sp. (Pl. 34,
fig. 3).
Disk with spiny surface, three times as broad as the medullary shell. Pores circular, of very
different sizes; eight to ten on the radius. Eight conical marginal spines of equal size and
equidistant, as long as the radius of the disk, and one-third as broad at the base as the
medullary shell. Numerous spines on the surface bristle-shaped, on the margin half as long as the
eight main spines, in the central part shorter.
Dimensions.—Diameter of the disk 0.18, of the medullary shell 0.06; length of the
marginal spines 0.1, basal breadth 0.02.
Habitat.—Central Pacific, Station 271, surface.
9. Heliosestrum ægineta, n. sp.
Disk with spiny surface, four times as broad as the medullary shell. Pores irregular,
roundish, of different sizes; twelve to fourteen on the radius. Eight pyramidal marginal spines
regularly disposed, four larger (perradial) alternating with four smaller (interradial); the
former as long as the radius of the disk, the latter half as long.
Dimensions.—Diameter of the disk 0.2, of the medullary shell 0.05; length of the
marginal spines 0.05 to 0.1, basal breadth 0.03.
Habitat.—South Atlantic, Station 330, surface.
{441}
Subgenus 3. Heliosestomma, Haeckel.
Definition.—Surface of the disk smooth, without radial spines. Bases
of the marginal spines connected by a solid equatorial girdle.
10. Heliosestrum octangulum, n. sp.
Disk with smooth surface, octagonal, twice as broad as the medullary shell. Pores regular,
circular; nine to ten on the radius. Equatorial girdle narrow, radially striped, connecting the
points of the eight short, regularly disposed, marginal spines in such a manner that the whole
shell forms a regular octagon with rectilinear sides.
Dimensions.—Diameter of the disk 0.11 to 0.12, of the medullary shell 0.05 to
0.06; length of the sides of the octagon 0.06.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
11. Heliosestrum octogonium, n. sp.
Disk with smooth surface, octagonal, four times as broad as the medullary shell. Pores
irregular, roundish; eight to nine on the radius. Equatorial girdle broad, smooth, connecting the
points of the eight short, triangular, flat marginal spines in such a manner that the whole shell
forms a subregular octagon with concave, nearly equal sides. (Sometimes seven or nine spines
instead of eight are developed.)
Dimensions.—Diameter of the disk 0.16 to 0.18, of the medullary shell 0.04 to
0.05; length of the radial spines 0.02 to 0.04, basal breadth 0.03.
Habitat.—Central Pacific, Station 266, depth 2750 fathoms.
12. Heliosestrum craspedotum, n. sp.
Haliomma humboldti, var., Bury, 1862, Polycystins of Barbados, pl. viii.
fig. 4 (below).
Disk with smooth surface, three times as broad as the medullary shell. Pores subregular,
circular; ten to twelve on the radius. Eight marginal spines (sometimes seven or nine) triangular,
deeply sulcated, half as long and one-third as broad as the radius of the disk, connected by a
narrow, radially striped equatorial girdle.
Dimensions.—Diameter of the disk 0.18, of the medullary shell 0.06; length of the
radial spines 0.05, basal breadth 0.03.
Habitat.—Central Pacific, Stations 271 to 274, at various depths, also
fossil in Barbados.
Genus 192. Astrosestrum,[231] Haeckel,
1881, Prodromus, p. 457.
Definition.—Phacodiscida with double medullary shell and
with eight radial spines on the margin of the disk (more or less regularly disposed, sometimes
seven or nine).
{442}
The genus Astrosestrum differs from the foregoing Heliosestrum by the
duplication of the medullary shell. The eight marginal spines in the majority of individuals are
regularly formed and disposed, of equal size and equidistant. But there are frequent exceptions to
this rule, either the angles between the eight spines being more or less different, or the number
amounting to seven or nine, instead of eight. Here also in some species four larger (perradial)
spines alternate regularly with four smaller (interradial spines), after the same law of symmetry,
which is common in the Medusæ.
Subgenus 1. Astrosestantha, Haeckel.
Definition.—Surface of the disk smooth, without radial spines. Bases
of the marginal spines free, not connected by an equatorial girdle.
1. Astrosestrum ephyra, n. sp. (Pl. 32,
figs. 4, 4a).
Disk with smooth surface, three times as broad as the outer and nine times as broad as the
inner medullary shell. Pores subregular, circular; seven to eight on the radius of the disk. Eight
marginal spines (sometimes seven or nine) more or less irregularly disposed, of variable size,
commonly as long as the radius of the disk, twice as long as broad, pyramidal, sulcate, without a
connecting equatorial girdle.
Dimensions.—Diameter of the disk 0.12, of outer medullary shell 0.04, of the inner
0.014; length of the marginal spines 0.05 to 0.07, basal breadth 0.03.
Habitat.—Central Pacific, Stations 270 to 274, depths 2350 to 2925
fathoms.
2. Astrosestrum nauphanta, n. sp. (Pl. 32, fig.
5).
Disk with smooth surface, two and a half times as broad as the outer and five times as broad as
the inner medullary shell. Pores regular, circular; eight to nine on the radius of the disk. Eight
marginal spines (often seven or nine) more or less regularly disposed, of equal size, half as long
as the radius of the disk, and quite as broad at the base, compressed triangular, sulcate, without
a connecting equatorial girdle.
Dimensions.—Diameter of the disk 0.15, of the outer medullary shell 0.06, of the
inner 0.03; length of the marginal spines 0.04, basal breadth 0.04.
Habitat.—Pacific, central area, Stations 266 to 268, depth 2700 to
2900 fathoms.
3. Astrosestrum octacanthum, Haeckel.
Haliomma octacanthum, Ehrenberg, 1872, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 295, Taf. viii. fig. 11.
Disk with smooth surface, twice as broad as the outer and six times as broad as the inner
medullary shell. Pores regular, circular; six to seven on the radius of the disk. Eight marginal
spines {443}regularly disposed, of variable size, the
longest as long as the radius, pyramidal, not broader at the base than one pore, without a
connecting equatorial girdle.
Dimensions.—Diameter of the disk 0.12, of the outer medullary shell 0.06, inner
0.02; length of the marginal spines 0.03 to 0.06, basal breadth 0.01.
Habitat.—Western Pacific, Philippine Sea, 3300 fathoms, Ehrenberg;
Station 225, depth 4475 fathoms.
4. Astrosestrum acraspedum, n. sp.
Disk with smooth surface, three times as broad as the outer and six times as broad as the inner
medullary shell. Pores regular, circular; nine to ten on the radius of the disk. Eight marginal
spines regularly disposed, of alternating size; four major (perradial) spines as long as the
diameter of the disk, four minor (interradial) half as long; spines cylindrical, not broader at
the base than one pore, without a connecting equatorial girdle.
Dimensions.—Diameter of the disk 0.2, of the outer medullary shell 0.07, of the
inner 0.035; length of the larger spines 0.2, of the smaller 0.1, basal breadth 0.01.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
Subgenus 2. Astrosestilla, Haeckel.
Definition.—Surface of the disk covered with radial spines. Bases of
the marginal spines free, not connected by an equatorial girdle.
5. Astrosestrum acanthastrum, n. sp.
Disk with spiny surface, three times as broad as the outer and nine times as broad as the inner
medullary shell. Pores subregular, circular; ten to eleven on the radius. Eight marginal spines
(sometimes seven or nine) regularly disposed, triangular, about half as long as the radius of the
disk, and one-third as broad at the base, without a connecting equatorial girdle.
Dimensions.—Diameter of the disk 0.18, of the outer medullary shell 0.06, of the
inner 0.02; length of the radial spines 0.05, basal breadth 0.03.
Habitat.—North Atlantic, Gulf Stream, Færöe Channel, surface, John
Murray.
6. Astrosestrum echinastrum, n. sp.
Disk with spiny surface, twice as broad as the outer and four times as broad as the inner
medullary shell. Pores irregular, roundish; eight to nine on the radius. Eight marginal spines
more or less regularly disposed, pyramidal, nearly as long as the radius of the disk, and not
broader at the base than one large pore, without a connecting equatorial girdle.
Dimensions.—Diameter of the disk 0.15, of the outer medullary shell 0.07, of the
inner 0.04; length of the radial spines 0.07, basal breadth 0.01.
Habitat.—Fossil in the Tertiary rocks of Sicily, Caltanisetta,
Teuscher.
{444}
Subgenus 3. Astrosestomma, Haeckel.
Definition.—Surface of the disk smooth, without radial spines. Bases
of the marginal spines connected by a solid equatorial girdle.
7. Astrosestrum pelagia, n. sp.
Disk with smooth surface, three times as broad as the outer and eight times as broad as the
inner medullary shell. Pores regular, circular; eight to nine on the radius. Eight marginal spines
conical, about as long as the radius of the disk, of nearly equal length, one-third as broad at
the base, connected by a radially striped equatorial girdle.
Dimensions.—Diameter of the disk 0.18, of the outer medullary shell 0.06, of the
inner 0.023; length of the radial spines 0.09, basal breadth 0.03.
Habitat.—Indian Ocean, Ceylon, Haeckel, surface.
8. Astrosestrum floscula, n. sp.
Disk with smooth surface, two and a half times as broad as the outer and five times as broad as
the inner medullary shell. Pores irregular, roundish; ten to eleven on the radius. Eight marginal
spines triangular, plain, scarcely half as long as the radius of the disk, nearly as broad at the
base, connected by an even equatorial girdle.
Dimensions.—Diameter of the disk 0.2, of the outer medullary shell 0.08, of the
inner 0.04; length of the radial spines 0.09, basal breadth 0.07.
Habitat.—South Atlantic, Station 325, surface.
Subfamily 3. Heliodiscida, Haeckel, 1881, Prodromus, p. 457.
Definition.—Phacodiscida with a variable
number of radial spines on the margin of the disk (ten to twenty or more), which are commonly more
or less irregular (sometimes regularly formed and disposed).
Genus 193. Heliodiscus,[232] Haeckel,
1862, Monogr. d. Radiol., p. 436.
Definition.—Phacodiscida with simple medullary shell and
with numerous (ten to twenty or more) simple radial spines on the margin of the disk (commonly
with a variable number and an irregular disposition of the undivided spines).
The genus Heliodiscus, the most common and polymorphic of all Phacodiscida, was founded by me in 1862 as the first known type of this family
(loc. cit.). I use {445}here the diagnosis of
this genus in a restricted sense, including only the species, in which the number of marginal
spines amounts to ten, twenty, or more (sometimes fifty to eighty, rarely more than one hundred).
The number of spines is in the different species variable, and their disposition commonly more or
less irregular, whilst in the preceding genera (with two, three, four, six, or eight marginal
spines) their distance and form are commonly regular.
Subgenus 1. Heliodiscetta, Haeckel.
Definition.—Surface of the disk smooth, without radial spines. Bases
of the marginal spines free, without a connecting equatorial girdle.
1. Heliodiscus asteriscus, n. sp. (Pl. 33, fig.
8).
Disk with smooth surface, three times as broad as the medullary shell. Pores regular, circular;
ten to twelve on the radius of the disk. Marginal spines fifteen to twenty, conical, often double
contoured, of variable size and disposition, the largest as long as the radius of the disk, as
broad at the base as one pore.
Dimensions.—Diameter of the disk 0.15, of the medullary shell 0.05; length of the
radial spines 0.04 to 0.07, basal breadth 0.01.
Habitat.—Cosmopolitan; Mediterranean (Corfu), Indian Ocean
(Madagascar), Atlantic and Pacific, many Stations, surface and various depths.
2. Heliodiscus trochiscus, n. sp. (Pl. 34, figs. 10,
13).
Disk with smooth surface, four times as broad as the medullary shell. Pores regular, circular;
fourteen to sixteen on the radius. Marginal spines twelve to sixteen, conical, of variable size
and disposition, the largest as long as the diameter of the medullary shell, twice as broad as one
pore.
Dimensions.—Diameter of the disk 0.2, of the medullary shell 0.05; length of the
radial spines 0.01 to 0.025, basal breadth 0.005.
Habitat.—Central Pacific, Stations 272 to 274, surface.
3. Heliodiscus trigonodon, n. sp.
Disk with smooth surface, four times as broad as the medullary shell. Pores regular, circular;
eighteen to twenty on the radius. Marginal spines sixteen to twenty, equilateral triangular, flat,
smooth, about as long and broad as the radius of the medullary shell.
Dimensions.—Diameter of the disk 0.2, of the medullary shell 0.05; length of the
radial spines 0.03, basal breadth 0.03.
Habitat.—Central Pacific, Stations 266 to 274, depths 2350 to 2925
fathoms.
{446}
4. Heliodiscus glyphodon, n. sp. (Pl. 35, fig.
2).
Heliosestrum glyphodon, Haeckel, 1881, Prodromus et Atlas (pl. xxxv. fig.
2).
Disk with smooth surface, four times as broad as the medullary shell. Pores subregular,
circular; seven to eight on the radius. Marginal spines ten to twelve, equilateral triangular,
flat, deeply furrowed, twice as long as broad, and as long as the radius of the disk.
Dimensions.—Diameter of the disk 0.14, of the medullary shell 0.035; length of the
radial spines 0.07, basal breadth 0.035.
Habitat.—North Pacific, Station 253, depth 3125 fathoms.
5. Heliodiscus helianthus, Haeckel.
Haliomma helianthus, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 74, Taf. xxvii. fig. 1.
Disk with smooth surface, five times as broad as the medullary shell. Pores regular, oblong,
disposed regularly in sixty to eighty radial series; fourteen to sixteen on the radius. Marginal
spines sixty to eighty, conical, about as long as the diameter of the medullary shell, twice as
broad as one pore.
Dimensions.—Diameter of the disk 0.2, of the medullary shell 0.04; length of the
radial spines 0.04, basal breadth 0.005.
Habitat.—Fossil in Barbados.
6. Heliodiscus sol, Haeckel.
Haliomma sol, Ehrenberg, 1854, Mikrogeol., Taf. xix. fig. 52.
Heliodiscus sol, Haeckel, 1862, Monogr. d. Radiol., p. 438.
Disk with smooth surface, four times as broad as the medullary shell. Pores irregular,
roundish; eight to ten on the radius. Marginal spines twenty to thirty, conical, the largest as
long as the radius of the medullary shell, their bases widely distant. (The species from Barbados
figured by Ehrenberg, 1875, as Haliomma sol, appertains to Heliosestrum solare.)
Dimensions.—Diameter of the disk 0.2, of the medullary shell 0.05; length of the
radial spines 0.02 to 0.025, basal breadth 0.01.
Habitat.—Fossil in Tertiary rocks of Greece (Ægina), Ehrenberg.
7 Heliodiscus siculus, Stöhr.
Heliodiscus siculus, Stöhr, 1880, Palæontogr., vol. xxvi. p. 89, Taf. i.
fig. 14.
Disk with smooth surface, three times as broad as the medullary shell. Pores irregular
roundish; seven to eight on the radius. Marginal spines forty to fifty, conical, very irregular,
the largest as long as the radius of the medullary shell, their bases coming in contact.
Dimensions.—Diameter of the disk 0.13 to 0.15, of the medullary shell 0.04 to
0.05; length of the radial spines 0.02 to 0.03, basal breadth 0.002 to 0.006.
Habitat.—Fossil in Tertiary rocks of Sicily, Grotte (Stöhr),
Caltanisetta (Teuscher).
{447}
8. Heliodiscus polymorphus, n. sp. (Pl. 34, figs. 11,
12).
Disk with smooth surface, four times as broad as the medullary shell. Pores irregular,
roundish; ten to twelve on the radius. Marginal spines ten to fifteen, pyramidal, angular, very
irregular and variable in size and distribution, the largest nearly as long as the diameter of the
disk, as broad at the base as the radius of the medullary shell.
Dimensions.—Diameter of the disk 0.15 to 0.2, of the medullary shell 0.04 to 0.05;
length of the radial spines 0.06 to 0.18, basal breadth 0.02 to 0.03.
Habitat.—Central Pacific, Stations 265 to 268, depths 2700 to 2900
fathoms.
9. Heliodiscus solaster, n. sp. (Pl. 34,
fig. 4).
Disk with smooth surface, three times as broad as the medullary shell. Pores subregular,
roundish; nine to ten on the radius. Marginal spines fifty to sixty, cylindro-conical, flexuose,
very variable in size, the largest nearly as long as the diameter of the disk. The spines lie not
only in the equatorial plane (as usual), but also in two to four crowded girdles on both sides of
it.
Dimensions.—Diameter of the disk 0.15, of the medullary shell 0.05; length of the
radial spines 0.05 to 0.15, basal breadth 0.008.
Habitat.—Indian Ocean, Madagascar, Rabbe, surface.
Subgenus 2. Heliodiscilla, Haeckel.
Definition.—Surface of the disk covered with radial spines. Bases of
the marginal spines free, without a connecting equatorial girdle.
10. Heliodiscus phacodiscus, Haeckel.
Heliodiscus phacodiscus, Haeckel, 1862, Monogr. d. Radiol., p. 437, Taf. xvii. figs.
5-7.
Haliomma phacodiscus, Haeckel, 1860, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 815.
Disk with spiny surface, three times as broad as the medullary shell. Pores regular, circular;
six to eight on the radius. Marginal spines twelve to sixteen, conical, as long as the radius of
the disk, and one-third as broad as the diameter of the medullary shell.
Dimensions.—Diameter of the disk 0.12 to 0.16, of the medullary shell 0.04 to
0.05; length of the marginal spines 0.06 to 0.08, basal breadth 0.01 to 0.02.
Habitat.—Mediterranean (Messina), Canary Islands (Lanzerote).
11. Heliodiscus amphidiscus, Haeckel.
Heliodiscus amphidiscus, Haeckel, 1862, Monogr. d. Radiol., p. 437.
Haliomma amphidiscus, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 154, Taf. ii. figs. 3-7.
Disk with spiny surface, three times as broad as the medullary shell. Pores regular, circular;
eight to ten on the radius. Marginal spines twelve to fifteen, bristle-shaped, not larger than the
surface spines, about as long as the radius of the medullary shell.
{448}
Dimensions.—Diameter of the disk 0.16, of the medullary shell 0.05; length of the
marginal spines 0.03, basal breadth 0.002.
Habitat.—Mediterranean (south shore of France), J. Müller.
12. Heliodiscus echiniscus, n. sp. (Pl. 34, fig.
5).
Disk with spiny surface, three times as broad as the medullary shell. Pores irregular, roundish
or polygonal; eight to ten on the radius. Marginal spines thirty to fifty, pyramidal, angular, of
very variable size, number, and disposition, gradually passing into the surface-spines; the
largest nearly as long as the diameter of the medullary shell.
Dimensions.—Diameter of the disk 0.14, of the medullary shell 0.045; length of the
radial spines 0.02 to 0.04, basal breadth 0.006.
Habitat.—Central Pacific, Station 272, depth 2600 fathoms.
13. Heliodiscus pertusus, n. sp. (Pl. 35, fig.
1).
Heliosestrum pertusum, Haeckel, 1881, Prodromus et Atlas (pl. xxxv. fig.
1).
Disk with spiny surface, three times as broad as the medullary shell. Pores irregular,
circular, hexagonally framed; eight to ten on the radius. Marginal spines ten to thirty, very
variable in size and disposition; commonly eight to twelve larger spines, which are pyramidal,
about as long as the radius of the disk and perforated by two to four irregular, longish pores.
Between these fenestrated large spines are commonly ten to twenty smaller conical spines,
gradually passing into those of the surface.
Dimensions.—Diameter of the disk 0.13 to 0.16, of the medullary shell 0.04 to
0.05; length of the marginal spines 0.04 to 0.08, basal breadth 0.01 to 0.015.
Habitat.—North Pacific, Stations 241 to 244, depths 2300 to 2900
fathoms.
Subgenus 3. Heliodiscomma, Haeckel.
Definition.—Surface of the disk smooth, without radial spines. Bases
of the marginal spines connected by a solid equatorial girdle.
14. Heliodiscus cingillum, n. sp. (Pl. 33, fig.
7).
Disk with smooth surface, five times as broad as the medullary shell. Pores regular, circular;
twelve to fourteen on the radius. Equatorial girdle about as broad as the medullary shell, in the
proximal half radially striped, on the margin with twenty to twenty-four short, flat, triangular
spines, which are shorter than the breadth of the girdle.
Dimensions.—Diameter of the disk 0.25, of the medullary shell 0.05; length of the
marginal spines 0.02, basal breadth 0.02.
Habitat.—Central Pacific, Station 274, depth 2750 fathoms.
{449}
15. Heliodiscus humboldti, Haeckel.
Heliodiscus humboldti, Haeckel, 1862, Monogr. d. Radiol., p. 438.
Haliomma humboldti, Ehrenberg, 1854, Mikrogeol., Taf. xxxvi. fig. 27; Abhandl. d. k.
Akad. d. Wiss. Berlin, 1875, Taf. xxvii. fig. 3.
Haliomma humboldti, Bury, 1862, Polycystins of Barbados, pl. viii. fig. 3
(at left).
Disk with smooth surface, four times as broad as the medullary shell. Pores regular, circular;
eighteen to twenty on the radius. Equatorial girdle half as broad as the medullary shell, in the
proximal half radially striped, on the margin with sixteen to twenty short, flat, triangular
teeth, which are longer than the breadth of the girdle.
Dimensions.—Diameter of the disk 0.12 to 0.2, of the medullary shell 0.03 to 0.05;
length of the marginal spines 0.02 to 0.04, basal breadth 0.02.
Habitat.—Fossil in Barbados (common and very variable).
16. Heliodiscus marginatus, n. sp. (Pl. 34, fig.
9).
Disk with smooth surface, three times as broad as the medullary shell. Pores regular, circular;
nine to ten on the radius. Equatorial girdle one-fourth to one-half as broad as the medullary
shell, in the whole breadth radially striped, on the margin with twelve to eighteen very short and
broad, triangular, marginal spines.
Dimensions.—Diameter of the disk 0.15 to 0.2, of the medullary shell 0.05 to 0.06;
length of the marginal spines 0.01 to 0.02, basal breadth 0.02.
Habitat.—Central Pacific, Stations 265 to 268, depth 2900 fathoms.
17. Heliodiscus sulcatus, n. sp.
Disk with smooth surface, four times as broad as the medullary shell. Pores subregular,
circular; twelve to fifteen on the radius. Equatorial girdle as broad as the medullary shell, in
the whole breadth radially striped on the margin with ten to fifteen triangular, deeply sulcated
teeth, about as long and broad as the medullary shell. (Similar to Heliodiscus glyphodon,
Pl. 35, fig.
2, but with broad sulcated girdle and shorter, more numerous spines.)
Dimensions.—Diameter of the disk 0.15 to 0.2, of the medullary shell 0.04 to 0.05;
length of the radial spines 0.05 to 0.06, basal breadth 0.04 to 0.05.
Habitat.—Atlantic and Pacific, tropical part, many Stations,
surface.
18. Heliodiscus umbonatus, Haeckel.
Haliomma umbonatum, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss. Berlin,
p. 74, Taf. xxvii. fig. 4.
Disk with smooth surface, three times as broad as the medullary shell. Pores irregular,
roundish; eight to ten on the radius. Equatorial girdle half as broad as the medullary shell,
hyaline, not radially striped, on the margin with ten to twenty triangular, irregular, smooth
teeth, very variable in size and disposition.
{450}
Dimensions.—Diameter of the disk 0.15 to 0.2, of the medullary shell 0.05 to 0.06;
length of the marginal spines 0.02 to 0.08, basal breadth 0.01 to 0.03.
Habitat.—Cosmopolitan; Atlantic, Pacific, in various depths; also
fossil in Barbados and Sicily.
Subgenus 4. Heliodiscura, Haeckel.
Definition.—Surface of the disk covered with radial spines. Bases of
the marginal spines connected by a solid equatorial girdle.
19. Heliodiscus apollinis, n. sp.
Disk with spiny or bristly surface, three times as broad as the medullary shell. Pores regular,
circular; eleven to twelve on the radius. Equatorial girdle narrow, on the margin with sixteen to
twenty broad, flat, triangular teeth, which are half as long and one-fourth as broad as the
medullary shell. (Very similar to Astrophacus apollinis, Pl. 32, fig. 2, but
with simple medullary shell.)
Dimensions.—Diameter of the disk 0.18, of the medullary shell 0.06; length of the
marginal spines 0.03, basal breadth 0.015.
Habitat.—Mediterranean (Corfu), Haeckel, surface.
20. Heliodiscus zoroaster, n. sp.
Disk with spiny surface, four times as broad as the medullary shell. Pores subregular,
circular; fourteen to sixteen on the radius. Equatorial girdle broad, radially striped, on the
margin with ten to twelve pyramidal, deeply sulcated radial spines, which are nearly as long as
the radius of the disk, and one-fourth as broad at the base.
Dimensions.—Diameter of the disk 0.24, of the medullary shell 0.06; length of the
marginal spines 0.11, basal breadth 0.03.
Habitat.—Indian Ocean, between Aden and Ceylon, Haeckel, surface.
Genus 194. Heliodrymus,[233] Haeckel,
1881, Prodromus, p. 457.
Definition.—Phacodiscida with simple medullary shell and
with numerous (ten to twenty or more) branched radial spines on the margin of the disk (commonly
with a variable number and an irregular disposition of the ramified spines).
The genus Heliodrymus differs from the nearly allied Heliodiscus by
the ramification of the marginal spines, a character hitherto observed in no other genus of
Phacodiscida. The branching is more or less irregular, either a simple bifurcation or a repeated
fissure; the spines and their branches are commonly more or less flexuose. {451}We can distinguish two subgenera: in Heliocladus the surface of
the disk is smooth, in Heliodendrum covered with bristle-shaped radial spines, which are
either simple or also branched, sometimes longer than the thick marginal spines.
Subgenus 1. Heliocladus, Haeckel, 1881, Prodromus, p. 457.
Definition.—Surface of the disk smooth, without radial spines.
1. Heliodrymus dendrocyclus, n. sp. (Pl. 33, fig.
9).
Heliocladus dendrocyclus, Haeckel, 1881, Prodromus et Atlas (pl. xxxiii.
fig. 9).
Disk with smooth surface, three times as broad as the medullary shell. Pores regular, circular,
hexagonally framed; eight to nine on the radius. Marginal spines sixteen to twenty, cylindrical,
very strong, flexuose, irregularly branched, nearly as long as the diameter of the disk. Between
these main spines, each of which bears two to six irregular branches, are scattered on the margin
numerous smaller simple spines.
Dimensions.—Diameter of the disk 0.16, of the medullary shell 0.05; length of the
main spines 0.1 to 0.14, breadth 0.01 to 0.02.
Habitat.—Central Pacific, Station 271, surface.
2. Heliodrymus furcatus, n. sp.
Disk with smooth surface, four times as broad as the medullary shell. Pores irregular,
roundish; ten to twelve on the radius. Marginal spines twenty to twenty-five, cylindrical,
flexuose, forked, about as long as the radius of the disk; fork-branches irregular, of unequal
size. Some smaller simple spines are scattered between the forked ones.
Dimensions.—Diameter of the disk 0.15, of the medullary shell 0.04; length of the
radial spines 0.07 to 0.09, breadth 0.01.
Habitat.—Central Pacific, Station 265, surface.
3. Heliodrymus grottensis, Haeckel.
Heliodiscus grottensis, Stöhr, 1880, Palæontogr., vol. xxvi. p. 89, Taf.
i. fig. 13.
Disk with smooth surface, two and a half times as broad as the medullary shell. Pores
irregular, roundish; eight to nine on the radius. Marginal spines twenty to thirty, conical, very
irregular in form, size, and disposition; the smaller simple, the larger irregularly branched and
half as long as the radius of the disk.
Dimensions.—Diameter of the disk 0.17, of the medullary shell 0.07; length of the
marginal spines 0.02 to 0.04, basal breadth 0.01 to 0.02.
Habitat.—Fossil in Tertiary rocks of Sicily, Grotte, Stöhr.
{452}
Subgenus 2. Heliodendrum, Haeckel.
Definition.—Surface of the disk armed with simple or branched radial
spines.
4. Heliodrymus setosus, n. sp.
Disk with spiny surface, four times as broad as the medullary shell. Pores regular, circular;
twelve to thirteen on the radius. Marginal spines ten to twelve, cylindrical, irregularly
branched, each with two to eight flexuose branches of different sizes; the largest as long as the
diameter of the disk. Spines of the surface bristle-shaped, half as long, not branched.
Dimensions.—Diameter of the disk 0.2, of the medullary shell 0.05; length of the
marginal spines 0.12 to 0.18, breadth 0.025.
Habitat.—North Pacific, Station 254, surface.
5. Heliodrymus ramosus, n. sp. (Pl. 35,
figs. 3, 4).
Disk with spiny surface, three times as broad as the medullary shell. Pores regular, circular,
hexagonally framed; eleven to twelve on the radius. Marginal spines sixteen to twenty,
cylindrical, about as long as the radius, irregularly forked or branched, with unequal flexuose
branches. Spines of the surface nearly as long, bristle-shaped, also irregularly branched.
Dimensions.—Diameter of the disk 0.15, of the medullary shell 0.05; length of the
spines 0.06 to 0.08, basal breadth 0.01 to 0.015.
Habitat.—South Pacific, Station 288, surface.
6. Heliodrymus viminalis, n. sp. (Pl. 35, fig.
5).
Disk with spiny surface, two and a half times as broad as the medullary shell. Pores irregular,
roundish; ten to twelve on the radius. Marginal spines fifteen to twenty, cylindro-conical,
strong, partly simple, partly forked, about as long as the diameter of the disk. Spines of the
surface very numerous, bristle-shaped, longer than the marginal spines, and more branched.
Dimensions.—Diameter of the disk 0.15, of the medullary shell 0.06; length of the
marginal spines 0.11 to 0.14, basal breadth 0.01 to 0.015; length of the surface spines 0.2 to
0.03.
Habitat.—Central Pacific, Station 271, surface.
Genus 195. Astrophacus,[234] Haeckel,
1881, Prodromus, p. 457.
Definition.—Phacodiscida with double medullary shell and
with numerous (ten to twenty or more) simple radial spines on the margin of the disk (commonly
with a variable number and an irregular disposition of the undivided spines).
The genus Astrophacus differs from the similar Heliodiscus in the
duplication of the medullary shell. The number and disposition of the radial spines of the margin
{453}(commonly between ten and twenty) is also here
variable in one and the same species. The greater number of observed species of Astrophacus
resemble in a very remarkable manner the corresponding species of Heliodiscus, and differ
only in the double medullary shell.
Subgenus 1. Astrophacetta, Haeckel.
Definition.—Surface of the disk smooth, without radial spines. Bases
of the marginal spines free, without a connecting equatorial girdle.
1. Astrophacus asteriscus, n. sp.
Disk with smooth surface, three times as broad as the outer and eight times as broad as the
inner medullary shell. Pores regular, circular; twelve to fourteen on the radius. Marginal spines
fifteen to twenty, of variable size and disposition; the largest as long as the radius of the
disk, as broad at the base as one pore. (Very similar to Heliodiscus asteriscus, Pl. 33, fig. 8, but
differing in the double medullary shell.)
Dimensions.—Diameter of the disk 0.2, of the outer medullary shell 0.07, of the
inner 0.025; length of the marginal spines 0.05 to 0.1, basal breadth 0.01.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms.
2. Astrophacus trochiscus, n. sp. (Pl. 34, fig.
14).
Disk with smooth surface, three times as broad as the outer and seven times as broad as the
inner medullary shell. Pores irregular, roundish; twelve to thirteen on the radius. Marginal
spines sixteen to twenty, conical, of irregular variable size and disposition; the largest as long
as the inner medullary shell. (Differs from Heliodiscus trochiscus in the double medullary
shell.)
Dimensions.—Diameter of the disk 0.22, of the outer medullary shell 0.08, of the
inner 0.03.
Habitat.—North Atlantic, Station 354, surface.
3. Astrophacus solaris, n. sp. (Pl. 32,
fig. 1).
Disk with smooth surface, three times as broad as the outer and seven times as broad as the
inner medullary shell. Pores subregular, roundish; twelve to fourteen on the radius. Marginal
spines one hundred to one hundred and twenty, conical, flexuose, of irregular size and form; the
largest one-third as long as the diameter of the disk. The spines lie not only in the equatorial
plane (as is usual) but also in two to four crowded girdles on both sides of it. (Very similar to
Heliodiscus solaster, Pl. 34,
fig. 4, but of double the size, with double the number of spines and with a double medullary
shell.)
Dimensions.—Diameter of the disk 0.3, of the outer medullary shell 0.11, of the
inner 0.045; length of the spines 0.03 to 0.1, basal breadth 0.01 to 0.02.
Habitat.—Indian Ocean, Sunda Strait, Rabbe, surface.
{454}
Subgenus 2. Astrophacilla, Haeckel.
Definition.—Surface of the disk covered with radial spines. Bases of
the marginal spines free, without a connecting equatorial girdle.
4. Astrophacus phacodiscus, n. sp. (Pl. 32, fig.
3).
Disk with spiny surface, two and a half times as broad as the outer and seven times as broad as
the inner medullary shell. Pores subregular, circular; ten to twelve on the radius. Marginal
spines twelve to sixteen, conical, stout, nearly as long as the radius of the disk, and as broad
at the base as the inner medullary shell. The numerous bristle-shaped spines of the surface are
scarcely half as long. (Similar to Heliodiscus phacodiscus, Haeckel, Monogr. d. Radiol.,
Taf. xvii. figs. 5-7, but differing in the double medullary shell.)
Dimensions.—Diameter of the disk 0.18, of the outer medullary shell 0.07, of the
inner 0.025; length of the marginal spines 0.08 to 0.09, basal breadth 0.03.
Habitat.—South Pacific, Station 300, surface.
Subgenus 3. Astrophacomma, Haeckel.
Definition.—Surface of the disk smooth, without radial spines. Bases
of the marginal spines connected by a solid equatorial girdle. (Perhaps = Chilomma(?)
Ehrenberg, 1847, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 54.)
5. Astrophacus cingillum, n. sp.
Haliomma humboldti, var., Bury, 1862, Polycystins of Barbados, pl. viii.
fig. 3 (at right).
Disk with smooth surface, three times as broad as the outer and eight times as broad as the
inner medullary shell. Pores regular, circular; eleven to thirteen on the radius. Equatorial
girdle about as broad as the inner medullary shell, in the proximal half radially striped, on the
margin with eighteen to twenty-four short, flat, triangular spines, about as long as the breadth
of the girdle. (Similar to Heliodiscus cingillum, but with double medullary shell.)
Dimensions.—Diameter of the disk 0.2, of the outer medullary shell 0.07, of the
inner 0.025; length of the spines 0.02, basal breadth 0.02.
Habitat.—Western Tropical Pacific, Station 225, depth 4475 fathoms;
also fossil in Barbados.
6. Astrophacus saturnus, Haeckel.
? Chilomma saturnus, Ehrenberg, 1861, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin,
p. 297; 1872, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 286, Taf. ii. fig. 5.
? Chilomma saturnus, Haeckel, 1862, Monogr. d. Radiol., p. 447.
Disk with smooth surface, two and a half times as broad as the outer, six times as broad as the
inner medullary shell. Pores large, irregular, roundish; five to seven on the radius (?).
Equatorial girdle very broad, radially striped, nearly as broad as the outer medullary shell,
perforated by {455}twenty to thirty (or more?) radial
spines. (The position of this species, and the identity of Chilomma with
Astrophacomma, remains doubtful, as the imperfect figure given by Ehrenberg of Chilomma
saturnus, the only species of the genus, is in contradiction with his vague description, as is
very often the case.)
Dimensions.—Diameter of the disk 0.12 (with girdle 0.22) of the outer medullary
shell 0.05, of the inner 0.02.
Habitat.—Arctic Ocean (Greenland, depth 1000 fathoms), Ehrenberg.
Subgenus 4. Astrophacura, Haeckel.
Definition.—Surface of the disk covered with radial spines. Bases of
the marginal spines connected by a solid equatorial girdle.
7. Astrophacus apollinis, n. sp. (Pl. 32, fig.
2).
Disk with spiny surface, three times as broad as the outer, eight times as broad as the inner
medullary shell. Pores regular, circular; eleven to twelve on the radius of the disk. Equatorial
girdle narrow, smooth, on the margin with twelve to sixteen broad, flat, triangular spines, of the
same length as the numerous bristle-shaped spines of the surface, which reach half the radius of
the disk. (Very similar to Heliodiscus apollinis, but differing in the double medullary
shell.)
Dimensions.—Diameter of the disk 0.24, of the outer medullary shell 0.08, of the
inner 0.03; length of the radial spines 0.06, basal breadth 0.03.
Habitat.—Western Tropical Pacific, Station 225, depth 4475
fathoms.
Family XX. Coccodiscida, Haeckel (Pls. 36-38).
Coccodiscida, Haeckel, 1862, Monogr. d. Radiol., p. 485.
Coccodiscida, Haeckel, 1881, Prodromus, p. 458.
Lithocyclidina, Ehrenberg, 1847, Monatsber. d. k. preuss. Akad.
d. Wiss. Berlin, p. 214 (partim).
Definition.—Discoidea with extracapsular phacoid shell
(or lenticular latticed cortical shell), connected by radial beams with an intracapsular, simple
or double, concentric medullary shell, and surrounded by one or more concentric chambered
equatorial girdles on the margin.
The family Coccodiscida was founded by me in 1862 for those Discoidea which agree with the Phacodiscida in the formation of the lenticular
"phacoid shell" (including a simple or double medullary shell), but differ from them in the
development of peculiar concentric chambered rings or girdles around the equatorial margin of the
disk, similar to those of the Porodiscida.
The Coccodiscida represent a polymorphic family, in which we here distinguish sixteen genera
with fifty-seven species; it comprises the greater part of those {456}Discoidea which Ehrenberg united in his group
Lithocyclidina (1875, represented by four genera and eight species); several of these, however,
appertain to quite different families, as his Astromma entomocora, Lithocyclia
amphitrites, &c. His knowledge of the structure was very imperfect. The peculiar
differentiation of the genera and species exhibits the greatest analogy to that of the following
family, Porodiscida, though the structure of the central disk in both families is quite
different.
The Phacoid Shell, or the circular, lenticular cortical shell exhibits in the
Coccodiscida quite the same structure and composition as in the Phacodiscida, described above (p.
420), so that there can be no doubt as to the phylogenetic origin of the
former from the latter. Quite in the same way in both families, the lenticular "phacoid shell" is
connected by numerous, short, radial beams with the intracapsular, simple or double, medullary
shell; and also here these beams are commonly disposed in two groups around the poles of the
shortened main axis of the lens, so that their distal ends are implanted in both circumpolar areas
(Pl. 36,
figs. 2-6; Pl. 37,
figs. 3, 7; Pl. 38,
figs. 2, 7). The medullary shell is commonly simple, spherical, sometimes a little lenticularly
compressed; more rarely it is double, composed of two concentric lattice-shells, which are
connected by radial beams; in this case either both concentric medullary shells are spherical, or
the inner is spherical, and the outer lenticular, very rarely the inner is lenticular also. In
average size and structure they agree perfectly with those of the Phacodiscida.
The Chamber Girdles or "chambered rings" around the equatorial margin of the disk, which
constitute the only difference between the Coccodiscida and the Phacodiscida, seem to exhibit a
considerable degree of difference of structure in the numerous species of this family; but I
regret that I cannot explain them here satisfactorily. The study of these structures is extremely
difficult because of the thickness and darkness of the massive opaque shells; to get a perfect
knowledge of them, it is indispensable to compare slides made in different directions (horizontal
slides through the equatorial and parallel planes, vertical slides through radial and parallel
planes, oblique slides in different directions). But this requires a long time and a most careful
study of the slides, which are very difficult to get in satisfactory condition. Therefore the
following remarks can have only a provisional value.
In all Coccodiscida we can distinguish on the equatorial chamber-girdle of the lens-margin
(even on superficial inspection) three different elements of structure, viz.:—(A) concentric
circular rings in the equatorial plane; (B) numerous radial beams piercing the former and dividing
them into imperfect chambers; (C) porous plates or sieve-plates on both convex faces of the disk.
The probable morphological significance of these three elements is the following:—Each ring
or girdle corresponds to an outer lenticular cortical shell, which is only developed on the
marginal part, whilst its central part is represented by the phacoid shell. Therefore the radial
beams (separating the imperfect chambers) are the same as in the concentric Polysphærida, and the
{457}sieve-plates of the surface are the porous walls of
the cortical shell itself. The correctness of this explanation seems to be proved by such forms as
figured in Pl. 38,
figs. 2, 4, where the whole surface of the phacoid shell is covered by a concentric chamber-work,
as a central continuation of the marginal concentric rings. If we imagine a system of perfect
concentric lenticular phacoid shells, compressed strongly from both poles of the shortened main
axis, we get the same figure.
Rarely one single girdle only is developed on the equatorial margin of the lenticular disk (Pl.
37, figs. 2,
3, 5). Commonly the number of concentric girdles amounts to three to six, often to ten to twelve
or more. Some of these largest Coccodiscida reach a considerable size. Commonly all girdles are of
the same breadth, which is about equal to the radius or to the diameter of the inner medullary
shell. Rarely the first (or innermost) girdle differs by its greater breadth from the succeeding
ones (Pl. 36,
fig. 8).
Only in few Coccodiscida the girdle-building remains restricted to the equatorial planes, so
that all the chambers lie in it. Commonly on both sides of this plane become developed several
layers, and often the number of these (three to six or more) increases towards the periphery; in
other cases not their number, but their height increases. Therefore very often the margin of the
discoidal shell is much thickened, as thick as the centre of the lenticular phacoid shell (or even
more); whilst between the latter and the former (on the proximal girdles) the disk is considerably
thinner (Pl. 36,
figs. 2, 4; Pl. 37,
figs. 7, 8; Pl. 38,
figs. 2, 4). The stratified layers communicate by large openings between their chambers. The
radial beams are commonly more or less regular and piercing, but also frequently irregular and
interrupted; often their number increases towards the margin by intercalation of new beams.
The Pores of the sieve-plates, which cover both sides of the chambered disk, appear on
the margin of the phacoid shell as direct continuations of the pores of the latter, and sometimes
they are so regularly disposed that one single circular pore is situated on the surface of each
chamber (Pl. 36,
fig. 7; Pl. 37,
fig. 1). But commonly the pores are of variable size and number, two to three on each chamber, and
often quite irregularly scattered.
The Margin of the chambered disk exhibits many differences, which afford characters for
the distinction of genera. In the first subfamily, the Lithocyclida, the margin is quite simple
without radial appendages. In the second subfamily, the Stylocyclida, it is armed with solid
radial spines lying in the equatorial plane, and often regularly disposed in the same manner as in
the Phacodiscida (compare above, p. 421). In the third subfamily, the
Astracturida, the margin bears two or more (commonly three or four) chambered arms, also situated
in the plane of the disk, and of the same structure as the circular chambered girdles (Pl. 38). In some
cases even the whole system of chamber-girdles is represented only by the radial arms, which are
inserted immediately on the margin of the phacoid shell. We may regard therefore these formations
as imperfect chambered disks, which are developed only in the direction of certain rays {458}(perradii), and reduced in the direction of the
alternating rays (interradii). In some Astracturida the chambered arms exhibit a structure
different from the more irregular chamber-work between them, so that we can distinguish the
latter, connecting the arms like a web-membrane, as a peculiar "patagium" (as in many Porodiscida)
(Pl. 38,
figs. 8, 9). Often the distal ends of the chambered arms are armed with a radial spine (Pl. 38, figs. 5, 6,
9).
The Central Capsule of the Coccodiscida is originally always of the same form and shape
as in their ancestral group, the Phacodiscida; a circular lenticular disk, which envelops the
simple or double medullary shell and is enclosed by the cortical phacoid shell. But whilst in the
Phacodiscida the phacoid shell envelops the central capsule perfectly, in the Coccodiscida it
envelops only the capsule from the two flat sides (by the upper and lower sieve-plates); the
marginal part of the lenticular capsule overgrows the margin of the phacoid shell by peripheral
extension, and fills out the chambered cavity of the concentric girdles. In the Lithocyclida and
the Stylocyclida, where there are no chambered arms, the central capsule remains a simple circular
lens or disk; in the Astracturida, where chambered radial arms surround the margin of the circular
central disk, the capsule enters also into these arms and fills out the greatest part of their
chambered cavities.
Synopsis of the Genera of Coccodiscida.
|
I. Subfamily Lithocyclida.
Margin of the disk simple, circular, without radial appendages.
|
|
Neither radial spines nor chambered arms on the circular
margin. |
|
Medullary shell simple, |
196. Lithocyclia. |
| Medullary shell double, |
197. Coccodiscus. |
|
II. Subfamily Stylocyclida.
Margin of the disk armed with solid radial spines.
|
|
Two opposite spines. |
|
Medullary shell simple, |
198. Stylocyclia. |
| Medullary shell double, |
199. Amphicyclia. |
| Three radial spines. |
|
Medullary shell simple, |
200. Trigonocyclia. |
| Four crossed spines. |
|
Medullary shell simple, |
201. Staurocyclia. |
| Five to ten or more radial spines. |
|
Medullary shell simple, |
202. Astrocyclia. |
| Medullary shell double, |
203. Coccocyclia. |
|
III. Subfamily Astracturida.
Margin of the disk with two to five or more (commonly three or four) hollow
radial chambered arms (with or without a connecting patagium). (Medullary shell commonly
simple.)
|
|
Two arms, opposite in one axis. |
|
Without patagium, |
204. Diplactura. |
| With patagium, |
205. Amphiactura. |
| Three radial arms (at equal distances). |
|
Without patagium, |
206. Trigonactura. |
| With patagium, |
207. Hymenactura. |
| Four arms (in two crossed diameters). |
|
Without patagium, |
208. Astractura. |
| With patagium, |
209. Stauractura. |
| Five radial arms (at variable distances). |
|
Without patagium, |
210. Pentactura. |
| With patagium, |
211. Echinactura. |
| I. Subfamily Lithocyclida. Margin of the disk simple, circular, without radial
appendages. |
|
|
Neither radial spines nor chambered arms on the circular
margin. |
|
|
|
|
|
|
Medullary shell simple, |
|
|
|
|
|
|
|
|
|
|
196. Lithocyclia. |
|
|
|
|
|
|
|
|
|
|
Medullary shell double, |
|
|
|
|
|
|
|
|
|
|
197. Coccodiscus. |
|
|
|
|
|
|
| II. Subfamily Stylocyclida. Margin of the disk armed with solid radial
spines. |
|
|
Two opposite spines. |
|
|
|
|
|
|
Medullary shell simple, |
|
|
|
|
|
|
|
|
|
|
198. Stylocyclia. |
|
|
|
|
|
|
|
|
|
|
Medullary shell double, |
|
|
|
|
|
|
|
|
|
|
199. Amphicyclia. |
|
|
|
|
|
|
|
|
Three radial spines. |
|
|
|
|
|
|
Medullary shell simple, |
|
|
|
|
|
|
|
|
|
|
200. Trigonocyclia. |
|
|
|
|
|
|
|
|
Four crossed spines. |
|
|
|
|
|
|
Medullary shell simple, |
|
|
|
|
|
|
|
|
|
|
201. Staurocyclia. |
|
|
|
|
|
|
|
|
Five to ten or more radial spines. |
|
|
|
|
|
|
Medullary shell simple, |
|
|
|
|
|
|
|
|
|
|
202. Astrocyclia. |
|
|
|
|
|
|
|
|
|
|
Medullary shell double, |
|
|
|
|
|
|
|
|
|
|
203. Coccocyclia. |
|
|
|
|
|
|
| III. Subfamily Astracturida. Margin of the disk with two to five or more
(commonly three or four) hollow radial chambered arms (with or without a connecting patagium).
(Medullary shell commonly simple.) |
|
|
Two arms, opposite in one axis. |
|
|
|
|
|
|
Without patagium, |
|
|
|
|
|
|
|
|
|
|
204. Diplactura. |
|
|
|
|
|
|
|
|
|
|
With patagium, |
|
|
|
|
|
|
|
|
|
|
205. Amphiactura. |
|
|
|
|
|
|
|
|
Three radial arms (at equal distances). |
|
|
|
|
|
|
Without patagium, |
|
|
|
|
|
|
|
|
|
|
206. Trigonactura. |
|
|
|
|
|
|
|
|
|
|
With patagium, |
|
|
|
|
|
|
|
|
|
|
207. Hymenactura. |
|
|
|
|
|
|
|
|
Four arms (in two crossed diameters). |
|
|
|
|
|
|
Without patagium, |
|
|
|
|
|
|
|
|
|
|
208. Astractura. |
|
|
|
|
|
|
|
|
|
|
With patagium, |
|
|
|
|
|
|
|
|
|
|
209. Stauractura. |
|
|
|
|
|
|
|
|
Five radial arms (at variable distances). |
|
|
|
|
|
|
Without patagium, |
|
|
|
|
|
|
|
|
|
|
210. Pentactura. |
|
|
|
|
|
|
|
|
|
|
With patagium, |
|
|
|
|
|
|
|
|
|
|
211. Echinactura. |
|
|
|
|
|
|
{459}
Subfamily 1. Lithocyclida, Haeckel, 1881, Prodromus, p. 458.
Definition.—Coccodiscida with simple circular
disk, without any radial appendages of the margin (either solid radial spines or chambered
arms).
Genus 196. Lithocyclia,[235] Ehrenberg,
1847, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 54.
Definition.—Coccodiscida with simple circular margin of
the disk, without radial appendages. Medullary shell simple.
The genus Lithocyclia is the most simple form of Coccodiscida, and
represents the common ancestral form of this family, from which all other genera of it can be
derived. The lenticular, biconvex disk is quite simple, composed of a variable number of
concentric, circular, chambered rings, which are pierced by radial beams, and which surround the
circular lenticular cortical shell or "phacoid shell." The latter contains a simple spherical
medullary shell in its centre, and is connected with it by radial beams. The margin of the disk is
circular, quite simple, without radial spines or chambered arms.
1. Lithocyclia cingulata, n. sp.
Phacoid shell (or lenticular porous cortical shell) three times as broad as the spherical
enclosed medullary shell, surrounded by one single chambered girdle or ring (with about forty
chambers of equal size, separated by radial beams). Margin of the disk circular, smooth. Pores of
the convex covering plates regular, circular; eight on the radius of the phacoid shell, two on the
breadth of the ring.
Dimensions.—Diameter of the disk (with one ring) 0.13, of the phacoid shell 0.1,
of the medullary shell 0.033.
Habitat.—Western Tropical Pacific, Station 225, depth 4475
fathoms.
2. Lithocyclia lenticula, n. sp. (Pl. 36, figs. 3,
4).
Phacoid shell two and a half times as broad as the enclosed medullary shell, surrounded by
three chambered girdles of equal size, which are divided by piercing radial beams each into about
fifty chambers. Margin of the disk thorny. Pores irregular, roundish; seven on the radius of the
phacoid shell, two on the breadth of each girdle.
Dimensions.—Diameter of the disk (with three girdles) 0.2, of the phacoid shell
0.11, of the medullary shell 0.045.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
{460}
3. Lithocyclia ocellus, Ehrenberg.
Lithocyclia ocellus, Ehrenberg, 1854, Mikrogeol., Taf. xxxvi. fig. 30;
Abhandl. d. k. Akad. d. Wiss. Berlin, 1875, Taf. xxix. fig. 3.
Phacoid shell three times as broad as the medullary shell, surrounded by numerous (seven to
eleven) chambered rings, which are divided by piercing radial beams each into sixty to ninety
chambers. Margin of the disk smooth. Pores regular, circular; nine on the radius of the phacoid
shell, one single pore on each chamber.
Dimensions.—Diameter of the disk (with eleven rings) 0.22, of the phacoid shell
0.1, of the medullary shell 0.035.
Habitat.—Fossil in the rocks of Barbados.
4. Lithocyclia monococcus, n. sp.
Stephanopyxis dubiosa (?), Bury, 1862, Polycystins of Barbados, pl. xiii.
figs. 1, 2.
Phacoid shell four times as broad as the medullary shell, surrounded by numerous (five to
eight) chambered rings, which are divided by piercing radial beams each into fifty to seventy
chambers. Margin of the disk thickened, thorny. Pores regular, circular; seven on the radius of
the phacoid shell, one single pore on each chamber.
Dimensions.—Diameter of the disk (with eight rings) 0.18, of the phacoid shell
0.12, of the medullary shell 0.03.
Habitat.—Pacific, central area, Station 267, depth 2700 fathoms; also
fossil in the rocks of Barbados.
5. Lithocyclia heteropora, n. sp.
Phacoid shell two and a third times as broad as the medullary shell, surrounded by five to nine
chambered rings, which are divided by piercing radial beams each into fifty to seventy chambers.
Margin of the disk smooth. Pores very different in the inner and outer part of the surface; in the
phacoid shell larger, regular, circular, eight on its radius, in the chambered periphery very
small and irregular, somewhat spongy.
Dimensions.—Diameter of the disk (with nine rings) 0.2, of the phacoid shell 0.13,
of the medullary shell 0.055.
Habitat.—Pacific, central area, Station 263, depth 2650 fathoms.
Genus 197. Coccodiscus,[236] Haeckel,
1862, Monogr. d. Radiol., p. 485.
Definition.—Coccodiscida with simple circular margin of
the disk, without radial appendages. Medullary shell double.
The genus Coccodiscus has quite the same form and structure as the preceding
Lithocyclia, and differs from it only in the double medullary shell, composed of two
concentric latticed spheres; sometimes the inner medullary shell is spherical, the outer {461}lenticular; the latter is connected with the lenticular
phacoid shell (or cortical shell) by radial beams.
1. Coccodiscus lamarckii, n. sp. (Pl. 36, fig.
1).
Phacoid shell (or lenticular porous cortical shell) very thick walled (as thick as one ring),
three times as broad as the outer and seven times as broad as the inner medullary shell,
surrounded by two to three chambered rings of equal breadth, each of which is divided by piercing
radial beams into thirty-six to forty square chambers. Margin of the disk circular, smooth. Pores
regular, circular, of equal size; ten on the radius of the phacoid shell, two on the breadth of
each chamber.
Dimensions.—Diameter of the disk (with two rings) 0.2, of the phacoid shell 0.14,
outer medullary shell 0.05, inner 0.02.
Habitat.—Western Tropical Pacific, Station 220, depth 1100
fathoms.
2. Coccodiscus darwinii, Haeckel.
Coccodiscus darwinii, Haeckel, 1862, Monogr. d. Radiol., p. 486, Taf.
xxviii. figs. 11, 12.
Phacoid shell three times as broad as the outer and nine times as broad as the inner medullary
shell, surrounded by five to eight chambered rings of equal breadth, each of which is divided by
forty piercing radial beams into forty square chambers of equal size. Margin of the disk smooth,
circular. Pores irregular, roundish, of unequal size; eleven on the radius of the phacoid shell,
one to two on each chamber.
Dimensions.—Diameter of the disk (with eight rings) 0.32, of the phacoid shell
0.11, outer medullary shell 0.036, inner 0.012.
Habitat.—Mediterranean, Messina, surface.
3. Coccodiscus goethei,
n. sp. (Pl. 36,
fig. 2).
Phacoid shell two and a half times as broad as the outer and five times as broad as the inner
medullary shell, surrounded by three to seven chambered rings of equal breadth, each of which is
divided by piercing radial beams into sixty to eighty chambers. Margin of the disk thorny. Pores
regular, circular; eight on the radius of the phacoid shell, a single one on the breadth of each
chamber.
Dimensions.—Diameter of the disk (with seven rings) 0.25, of the phacoid shell
0.1, of the medullary shell 0.04, inner 0.02.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
Subfamily 2. Stylocyclida, Haeckel, 1881, Prodromus, p. 458.
Definition.—Coccodiscida with solid radial
spines on the margin of the circular disk, situated in its equatorial plane (without chambered
arms).
{462}
Genus 198. Stylocyclia,[237] Ehrenberg,
1847, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 54.
Definition.—Coccodiscida with two opposite solid radial
spines on the margin of the circular disk. Medullary shell simple.
The genus Stylocyclia opens the series of the Stylocyclida or of those
Coccodiscida in which the margin of the chambered disk is armed with solid radial spines, situated
in its equatorial plane, but without chambered arms. Stylocyclia is the most simple form of
this subfamily, and bears only two marginal spines, opposite in one equatorial axis of the disk.
The medullary shell is simple. This genus corresponds to Xiphodictya in the family
Porodiscida. The genus was previously known only by one single species described by Ehrenberg.
1. Stylocyclia dimidiata, Ehrenberg.
Stylocyclia dimidiata, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 84, Taf. xxix. fig. 4.
Phacoid shell three times as broad as the medullary shell, surrounded by five to eight
chambered rings of equal breadth, divided by eighty to ninety piercing radial beams into square
chambers. Pores subregular, circular; six to seven on the radius of the phacoid shell, a single
one on each chamber. Both opposite marginal spines strong, club-shaped, their thickness decreasing
from the margin towards the centre of the disk. (The figure of Ehrenberg is very incomplete.)
Dimensions.—Diameter of the disk (with eight rings) 0.25, of the phacoid shell
0.11, of the medullary shell 0.035.
Habitat.—Fossil in the Tertiary rocks of Barbados; living in the
depths of the Equatorial Atlantic, Station 348, depth (2450) fathoms.
2. Stylocyclia prionacantha, n. sp. (Pl. 37, fig.
6).
Phacoid shell two and a half times as broad as the medullary shell, surrounded by five to six
chambered rings, which are divided by fifty to sixty piercing radial beams into square chambers.
Pores in the thick-walled phacoid shell regularly circular, increasing in size from the centre;
eight to nine on its radius. Pores on the surface of the chambered girdle smaller, very irregular,
two to three on each ring. Both marginal spines longer than the diameter of the disk, with broad
serrated edges.
Dimensions.—Diameter of the disk (with five rings) 0.23, of the phacoid shell
0.12, of the medullary 0.05.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
{463}
3. Stylocyclia excavata, n. sp. (Pl. 37,
fig. 8).
Phacoid shell four times as broad as the medullary shell, surrounded by four chambered rings,
which are divided by twenty to thirty radial beams into broad chambers. The height of the rings
increases strongly from the centre, so that the fourth ring is two and a half times as high as the
first. Pores irregular, roundish; five to six on the radius of the phacoid shell, one to two on
the breadth of each ring. Margin of the disk smooth. Both marginal spines thin and long,
cylindrical, arising from the medullary shell, longer than the diameter of the disk.
Dimensions.—Diameter of the disk (with four rings) 0.25, of the phacoid shell 0.1,
of the medullary shell 0.025.
Habitat.—Fossil in the rocks of Barbados. Indian Ocean, Cocos Islands, Rabbe.
4. Stylocyclia amphacantha, n. sp.
Phacoid shell twice as broad as the medullary shell, surrounded by five chambered rings,
divided by piercing beams each into forty to fifty chambers. Pores regular, circular; eight to
nine on the radius of the phacoid shell, one on each chamber. Margin of the disk thorny. Both
marginal spines strong, conical, as long as its radius.
Dimensions.—Diameter of the disk (with five rings) 0.24, of the phacoid shell 0.1,
of the medullary shell 0.05.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms.
Genus 199. Amphicyclia,[238] Haeckel,
1881, Prodromus, p. 458.
Definition.—Coccodiscida with two opposite solid radial
spines on the margin of the circular disk. Medullary shell double.
The genus Amphicyclia has the same form and structure as Stylocyclia,
and differs from it only in the double concentric medullary shell. It bears therefore to the
latter the same relation that Coccodiscus does to Lithocyclia.
1. Amphicyclia chronometra, n. sp. (Pl. 38, fig.
1).
Phacoid shell three times as broad as the outer and nine times as broad as the inner medullary
shell, surrounded by two broad chambered rings, which are divided into irregular chambers by fifty
to sixty radial beams of different distance. Margin of the disk thorny, lacerated. Pores
irregular, roundish; twelve to sixteen on the radius of the phacoid shell, two to three on the
breadth of each ring. The two opposite marginal spines strong, prismatic, with prominent edges,
about as long as the diameter of the disk.
Dimensions.—Diameter of the disk (with two rings) 0.24, of the phacoid shell 0.15,
outer medullary shell 0.05, inner 0.017.
Habitat.—Pacific, central area, Stations 265 to 268, depths 2700 to
2900 fathoms.
{464}
2. Amphicyclia amphistyla, n. sp. (Pl. 37, fig.
7).
Stylocyclia amphistyla, Haeckel, 1879, MS. et Atlas (pl. xxxvii. fig.
7).
Phacoid shell thin walled, two and a half times as broad as the outer and seven times as broad
as the inner medullary shell, divided by eighty to ninety radial beams into irregular chambers,
which are stratified in four to five floors. Pores regular, circular; eight to nine on the radius
of the phacoid shell, two on the breadth of each chamber. Both marginal spines cylindrical.
Dimensions.—Diameter of the disk (with seven rings) 0.27, of the phacoid shell
0.1, outer medullary shell 0.04, inner 0.014.
Habitat.—North Atlantic, Canary Islands, Station 354, surface.
3. Amphicyclia pachydiscus, n. sp. (Pl. 38, fig.
2).
Phacoid shell very thick, spongy, twice as broad as the outer and five times as broad as the
inner medullary shell, divided by fifty to seventy radial beams into subregular chambers, which
are stratified in five to six floors. Pores irregular, roundish; ten to twelve on the radius of
the phacoid shell, two to three on the breadth of each chamber. The two opposite marginal spines
quadrangular prismatic, very long, as broad as the radius of the inner medullary shell. Fig. 2
exhibits a vertical section (slide) through the centre.
Dimensions.—Diameter of the disk (with five rings) 0.22, of the phacoid shell 0.1,
outer medullary shell 0.05, inner 0.02.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms; fossil in
Barbados.
Genus 200. Trigonocyclia,[239] Haeckel,
1881, Prodromus, p. 458.
Definition.—Coccodiscida with three solid radial spines
on the margin of the circular disk. Medullary shell simple.
The genus Trigonocyclia (only known by one single species, once observed)
bears on the margin of the chambered disk three solid radial spines, at equal distances one from
another. It corresponds to the genus Tripodictya amongst the Porodiscida.
1. Trigonocyclia triangularis, n. sp. (Pl. 37, fig.
5).
Phacoid shell three times as broad as the medullary shell, connected with it by six equidistant
radial beams, three of which are prolonged into marginal spines. In the equatorial plane only one
single chambered ring, divided by forty-four radial beams into narrow chambers. Pores large,
irregular, roundish; five on the radius of the phacoid shell, two on each chamber. Surface and
margin of the disk spiny. Three strong conical marginal spines, divergent at equal angles, as long
as the radius of the disk.
Dimensions.—Diameter of the disk 0.16, of the phacoid shell 0.12, of the medullary
shell 0.04.
Habitat.—Indian Ocean, Zanzibar, depth 2200 fathoms, Pullen.
{465}
Genus 201. Staurocyclia,[240] Haeckel,
1881, Prodromus, p. 458.
Definition.—Coccodiscida with four solid radial spines on
the margin of the disk, crossed in two equatorial diameters perpendicular one to another.
Medullary shell simple.
The genus Staurocyclia is characterised by four radial spines on the margin
of the disk, which are opposite in pairs and situated in two equatorial diameters, commonly
perpendicular one to another. They form therefore a rectangular cross (rarely more or less
irregular). The genus corresponds exactly to Staurodictya in the family Porodiscida. The
medullary shell is simple.
1. Staurocyclia cruciata, n. sp. (Pl. 37, fig.
1).
Phacoid shell two and a half times as broad as the medullary shell, surrounded by six to eight
regular rings, which are divided by fifty to seventy piercing beams into square chambers. Pores
regular, circular; six on the radius of the phacoid shell, one single pore on each chamber. Four
crossed radial spines, arising from the medullary shell by thin bases, prolonged over the dentated
margin as four strong quadrangular swords, as long as the radius of the disk.
Dimensions.—Diameter of the disk (with eight rings) 0.23, of the phacoid shell
0.09, of the medullary shell 0.035.
Habitat—Pacific, central area, Station 267, depth 2700 fathoms.
2. Staurocyclia serrata, n. sp.
? Haliomma sp., Bury, 1862, Polycystins of Barbados, pl. xxii. fig. 2.
Phacoid shell twice as broad as the medullary shell, surrounded by three to four spongy rings,
which are divided by thirty to forty radial beams into irregular chambers. Pores in the phacoid
shell regular, circular, five on its radius; in the chambered spongy girdle much smaller and
irregular. Margin dentated, with four crossed, very large, quadrangular spines, as long as the
radius of the disk, with wing-shaped, serrated edges.
Dimensions.—Diameter of the disk (with four rings) 0.2, of the phacoid shell 0.1,
of the medullary shell 0.05.
Habitat.—Fossil in the rocks of Barbados.
3. Staurocyclia phacostaurus, n. sp. (Pl. 37, figs. 2,
3).
Phacostaurus pyramidalis, Haeckel, 1879, MS. (as a separate genus).
Phacoid shell two and a quarter times as broad as the medullary shell, connected with it by
twelve radial beams (four equatorial and eight divergent) and surrounded by one single ring, which
is divided by forty radial beams into regular chambers. Pores regular, honeycomb-like in the {466}phacoid shell, seven on its radius; smaller on the
equatorial ring, three on its breadth. Four marginal spines pyramidal four-sided, as long and as
broad at the base as the ring.
Dimensions.—Diameter of the disk (with one ring) 0.14, of the phacoid shell 0.11,
of the medullary shell 0.048.
Habitat.—South Atlantic, Station 335, depth 1425 fathoms.
4. Staurocyclia magniducis, n. sp. (Pl. 37, fig.
4).
Coccostaurus magniducis, Haeckel, 1881, MS. et Atlas (pl. xxxvii. fig.
4).
Phacoid shell twice as broad as the medullary shell, connected with it by numerous radial beams
and surrounded by eight chambered rings, which are divided by one hundred to one hundred and
twenty piercing radial beams into small chambers. Pores subregular, circular; ten on the radius of
the phacoid shell, two on the breadth of each chamber. Margin of the disk armed with numerous
bristle-shaped radial spines, as long as the breadth of the chambered girdle. Four very large
crossed spines, nearly as long as the diameter of the disk, quadrangular, with four dentated
edges; at the club-shaped distal end thorny, as broad as the medullary shell and three times as
broad as at the narrow base. I name this splendid species in the honour of H.R.H. the Grand Duke
of Saxe-Weimar, Carl Alexander, the magnanimous protector of arts and sciences, the rector
magnificentissimus of the University of Jena.
Dimensions.—Diameter of the disk (with eight rings) 0.27, of the phacoid shell
0.11, of the medullary shell 0.05; length of the four crossed club-spines 0.2, basal breadth 0.02,
distal breadth 0.06.
Habitat.—Indian Ocean, Ceylon, Belligemma, surface (Haeckel).
Genus 202. Astrocyclia,[241] Haeckel,
1881, Prodromus, p. 458.
Definition.—Coccodiscida with numerous (five or more,
commonly thirty to sixty) solid radial spines on the margin of the circular disk. Medullary shell
simple.
The genus Astrocyclia exhibits on the margin of the circular chambered disk
a large but variable number of solid radial spines, commonly between thirty and sixty. They are
the external prolongations of the inner piercing radial beams, which divide the concentric rings
of the disk into chambers. All the spines lie in the equatorial plane of the disk. The genus
corresponds to Stylodictya in the family Porodiscida.
1. Astrocyclia solaster, n. sp. (Pl. 36,
fig. 7).
Phacoid shell two and a half times as broad as the medullary shell, surrounded by four to six
regular rings of equal breadth, which are divided by thirty to forty piercing radial beams into
broad {467}chambers. Pores regular, circular; seven on
the radius of the phacoid shell, one single pore on each chamber (the size increasing from the
centre). Margin with thirty to forty angular, broad, dentate spines, the prolongations of the
inner radial beams.
Dimensions.—Diameter of the disk (with six rings) 0.22, of the phacoid shell 0.1,
of the medullary shell 0.04.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
2. Astrocyclia stella, Haeckel.
Lithocyclia stella, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss. Berlin,
p. 78, Taf. xxix. fig. 2.
Phacoid shell three times as broad as the medullary shell, surrounded by four to eight rings of
increasing breadth from the centre, the outer of which are more or less spongy. Each ring is
divided by forty to fifty piercing radial beams into chambers of increasing size. Pores regular,
circular; a single one on each chamber, ten on the radius of the phacoid shell. On the margin are
scattered ten to twenty thick and short spines, four-sided pyramidal, about as long and as broad
at the base as the diameter of the medullary shell.
Dimensions.—Diameter of the disk (with eight rings) 0.25, of the phacoid shell
0.09, of the medullary shell 0.03.
Habitat.—Fossil in the rocks of Barbados.
3. Astrocyclia rotula, n. sp.
Phacoid shell four times as broad as the medullary shell, surrounded by one single ring, which
is divided by forty radial beams into equal chambers. Pores regular, circular; two on the breadth
of the ring, six on the radius of the phacoid shell. Margin with forty short, conical spines, the
prolongations of the inner radial beams, twice as long as the breadth of the ring.
Dimensions.—Diameter of the disk 0.13, of the phacoid shell 0.1, of the medullary
shell 0.025.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
4. Astrocyclia arachnia, n. sp.
Phacoid shell twice as broad as the medullary shell, surrounded by four to six rings of equal
breadth, which are divided by twelve to twenty radial beams into broad chambers. Pores regular,
circular; two on the breadth of each ring, eight on the radius of the phacoid shell. Margin with
twelve to twenty very long and thin, bristle-shaped, radial spines, the prolongations of the inner
beams, longer than the diameter of the disk. (Resembles Stylodictya arachnia, but differs
in the possession of a phacoid shell.)
Dimensions.—Diameter of the disk (with six rings) 0.22, of the phacoid shell 0.09,
of the medullary shell 0.045.
Habitat.—North Atlantic, Canary Islands.
{468}
5. Astrocyclia heterocycla, n. sp. (Pl. 36, fig. 8).
Phacoid shell two and a half times as broad as the medullary shell, surrounded by three rings of
unequal breadth, the first ring as broad as the second and third together. They are divided by
fifty to sixty piercing radial beams into chambers, which are square in the second and third
rings, and half as large as in the first. The radial beams are alternately thicker and thinner,
prolonged into short marginal spines. Pores subregular, circular; eight on the radius of the
phacoid shell, two on the breadth of the first ring, a single one on each chamber of the second
and third rings.
Dimensions.—Diameter of the disk (with three rings) 0.2, of the phacoid shell
0.12, of the medullary shell 0.05.
Habitat.—Equatorial Atlantic, Station 348, depth (2450) fathoms; also
fossil in Barbados.
Genus 203. Coccocyclia,[242] Haeckel,
1881, Prodromus, p. 458.
Definition.—Coccodiscida with numerous (five or more)
solid radial spines on the margin of the circular disk. Medullary shell double.
The genus Coccocyclia has the same form and structure as the foregoing
Astrocyclia, and differs from it only in the double concentric medullary shell. It bears
therefore to the latter the same relation as Coccodiscus to Lithocyclia.
1. Coccocyclia liriantha, n. sp.
Phacoid shell three times as broad as the outer and ten times as broad as the inner medullary
shell, connected with both by six equidistant piercing radial beams. The margin of the phacoid
shell is surrounded by one or two equatorial rings, each as broad as the outer medullary shell,
and divided by thirty to forty radial beams into narrow chambers. Pores subregular, circular,
about nine to ten on the radius of the phacoid shell, three to four on the breadth of each ring.
Margin of the disk ciliated, with six regularly disposed radial spines as prolongations of the
inner six piercing beams. Each spine cylindrical, delicately crenulated, about as broad as the
inner medullary shell, and once to twice as long as the diameter of the whole disk, elegantly
undulated, with a conical terminal point.
Dimensions.—Diameter of the disk (with two rings) 0.32, of the phacoid shell 0.2,
of the outer medullary shell 0.06, of the inner 0.02.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms.
2. Coccocyclia
heliantha, n. sp. (Pl. 36,
figs. 5, 6).
Phacoid shell two and a half times as broad as the outer and six times as broad as the inner
medullary shell, surrounded by six to eight rings of nearly equal breadth, the outer somewhat
smaller. They are divided by fifty to sixty piercing radial beams into irregular chambers. Pores
{469}irregular, roundish; ten on the radius of the
phacoid shell, a single one on each chamber of the equatorial girdle. Margin of the disk with
numerous strong conical radial spines of different breadths, arranged in several circles, the
strongest in the equatorial plane, about as broad as one ring and twice as long. Surface
thorny.
Dimensions.—Diameter of the disk (with seven rings) 0.3, of the phacoid shell
0.12, of the outer medullary shell 0.05, of the inner 0.02.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms; also
fossil in Barbados.
Subfamily 3. Astracturida, Haeckel, 1881, Prodromus, p. 458.
Definition.—Coccodiscida with two or more
(commonly three or four) chambered radial arms on the margin of the disk, situated in its
equatorial plane (with or without a connecting patagium between the arms).
Genus 204. Diplactura,[243] Haeckel,
1881, Prodromus, p. 458.
Definition.—Coccodiscida with two opposite chambered arms
on the margin of the circular disk, without a connecting patagium.
The genus Diplactura is the simplest form of the Astracturida, or of those
Coccodiscida in which the margin of the disk bears chambered arms, situated in its equatorial
plane. This subfamily corresponds to the Euchitonida among the Porodiscida, to the Spongobrachida
among the Spongodiscida. In Diplactura there are only two free arms, opposite in one
equatorial diameter (corresponding to Amphibrachium among the former, to Spongolena among the
latter).
Subgenus 1. Diplacturium, Haeckel.
Definition.—Arms blunt at the distal end, without radial spines.
1. Diplactura diplobrachia, n. sp.
Phacoid shell twice as broad as the medullary shell, with six pores on its radius, without a
perfect chambered ring around it. Arms club-shaped, one and a half times as long as the diameter
of the phacoid shell, at the distal end rounded, blunt, as broad as the latter, at the base only
half as broad.
Dimensions.—Diameter of the phacoid shell 0.08, of the medullary shell 0.04;
length of the arms 0.12, basal breadth 0.04, distal breadth 0.08.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms.
{470}
Subgenus 2. Diplactinium, Haeckel.
Definition.—Arms on the distal end armed with a radial terminal
spine.
2. Diplactura diploconus, n. sp. (Pl. 38, fig.
5).
Amphiactura diploconus, Haeckel, 1877, MS. et Atlas (pl. xxxviii. fig.
5).
Phacoid shell three times as broad as the medullary shell, with eight pores on its radius,
surrounded by a single chambered ring. Arms club-shaped, twice as long as the diameter of the
phacoid shell, at the distal end as broad as the latter, at the base only half as broad. Both
poles of the common axis of the arms are armed with a strong spindle-shaped terminal spine.
Dimensions.—Diameter of the phacoid shell 0.09, of the medullary shell 0.03;
length of the arms (without terminal spines) 0.17, basal breadth 0.05, distal breadth 0.09.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
Genus 205. Amphiactura,[244] Haeckel,
1881, Prodromus, p. 458.
Definition.—Coccodiscida with two opposite chambered arms
on the margin of the circular disk, connected by a spongy patagium.
The genus Amphiactura differs from the foregoing Diplactura in the
development of a patagium between the arms, and therefore bears the same relation to it as
Amphymenium in the Porodiscida does to Amphibrachium, or Spongobrachium in
the Spongodiscida does to Spongolene. In this and in the following Discoidea provided with a patagium, this connecting web constantly exhibits a
different texture of its framework, which is sometimes more regularly chambered, at other times
more irregularly spongy.
1. Amphiactura amphibrachia, n. sp. (Pl. 38, figs. 3,
4).
Phacoid shell three times as broad as the medullary shell, with eight pores on its radius. Arms
nearly equilateral triangular, twice as long as the diameter of the phacoid shell, at the
truncated distal end as broad as the latter, at the base only one-third as broad. Patagium a
circular lenticular disk, enveloping only the basal third of the arms, with three to four
concentric circular rings, divided into chambers by about forty radial beams, which are prolonged
beyond the margin of the patagium into radial spines. The vertical section (fig. 4) shows that the
chambers of each arm (eleven to twelve transverse rows in the radius) are disposed in two
layers.
Dimensions.—Diameter of the phacoid shell 0.09, of the medullary shell 0.03;
length of the arms 0.18, basal breadth 0.03, distal breadth 0.09.
Habitat.—Pacific, central area, Station 263, depth 2650 fathoms.
{471}
Genus 206. Trigonactura,[245] Haeckel,
1881, Prodromus, p. 459.
Definition.—Coccodiscida with three chambered arms on the
margin of the circular or triangular disk, without a connecting patagium.
The genus Trigonactura exhibits three radial arms, which in all known
species are separated by three equal angles. The terminal points of the arm-axes are the corners
of an equilateral triangle. It corresponds therefore to Dictyastrum among the
Porodiscida.
Subgenus 1. Trigonacturium, Haeckel.
Definition.—Distal end of the arms blunt or truncated, without a
terminal spine.
1. Trigonactura pythagoræ, Haeckel.
Astromma pythagoræ, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss. Berlin,
p. 66, Taf. xxx. fig. 2.
Phacoid shell circular, twice as broad as the medullary shell, with five pores on its radius,
without a completely surrounding chambered girdle. Arms nearly square, at the truncated distal end
as broad as long, and scarcely broader than at the base, two-thirds as long as the diameter of the
central disk.
Dimensions.—Diameter of the phacoid shell 0.1, of the medullary shell 0.05; length
of the arms 0.07, distal breadth 0.07.
Habitat.—Cosmopolitan; Atlantic, Indian, Pacific, in various depths;
also fossil in Tertiary rocks of Barbados and Nicobar.
2. Trigonactura rhopalastrella, n. sp.
Stephanastrum sp., Bury, 1862, Polycystins of Barbados, pl. xv. fig.
7.
Phacoid shell twice as broad as the medullary shell, with five pores on its radius, without a
perfect chambered ring around it. Arms club-shaped, at the blunt distal end rounded, as long as
the diameter of the phacoid shell; their basal breadth is one-fourth, their distal breadth
one-half of its length.
Dimensions.—Diameter of the phacoid shell 0.1, of the medullary shell 0.05; length
of the arms 0.1, basal breadth 0.025, distal breadth 0.05.
Habitat.—Pacific, central area, Stations 270 to 274, depths 2400 to
2800 fathoms; also fossil in Tertiary rocks of Barbados.
{472}
3. Trigonactura trigonobrachia, n. sp.
Phacoid shell two and a half times as broad as the medullary shell, with six pores on its
radius, without a perfect chambered girdle. Arms nearly equilateral triangular, twice as long as
the diameter of the phacoid shell, at the truncated distal end as broad as its diameter and three
times as broad as at the narrow base. (Resembles Hymenactura hexagona, but without a
patagium.)
Dimensions.—Diameter of the phacoid shell 0.11, of the medullary shell 0.045;
length of the arms 0.2, basal breadth 0.035, distal breadth 0.1.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
4. Trigonactura lanceolata, n. sp.
Phacoid shell three times as broad as the medullary shell, with eight pores on its radius,
surrounded by one perfect chambered ring. Arms lanceolate, three times as long as the diameter of
the phacoid shell, and four times as long as broad in the middle part, at both ends very narrow,
blunt.
Dimensions.—Diameter of the phacoid shell 0.09, of the medullary shell 0.03;
length of the arms 0.25, breadth in the midst 0.06.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
Subgenus 2. Trigonactinium, Haeckel.
Definition.—Distal end of the arms armed with a radial spine.
5. Trigonactura triacantha, n. sp. (Pl. 38, figs. 6,
7).
Phacoid shell two and a half times as broad as the medullary shell, with eight pores on its
radius, surrounded by one perfect chambered ring. Arms nearly lanceolate, in the middle part one
and a half times as broad as at both ends, twice as long as the phacoid shell, at the distal end
with a strong pyramidal terminal spine. Through the spongy framework of each arm shine sixteen to
eighteen transverse bars (as septa of the joints) and six to eight longitudinal piercing
beams.
Dimensions.—Diameter of the phacoid shell 0.09, of the medullary shell 0.038;
length of the arms 0.2, greatest breadth 0.08.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms.
6. Trigonactura trigonodiscus, n. sp.
? Astromma sp., Bury, 1862, Polycystins of Barbados, Taf. xv. fig. 2.
Phacoid shell triangular, twice as broad as the spherical medullary shell, with six to seven
pores on its radius, surrounded by one chambered ring. Arms equilateral triangular, as long as
{473}the diameter of the central disk and at the base
half as broad, slowly decreasing in breadth towards the distal end, which is armed with a strong
pyramidal spine.
Dimensions.—Diameter of the phacoid shell 0.1, of the medullary shell 0.05; length
of the arms 0.12, basal breadth 0.07.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms; also
fossil in Barbados.
7. Trigonactura trixiphos, n. sp.
Phacoid shell circular, twice as broad as the medullary shell, without a completely surrounding
chambered ring, with four pores on its radius. Arms club-shaped, about as long as the diameter of
the central disk, at the base half as broad as long, at the rounded distal end broader, and armed
with a strong pyramidal terminal spine, nearly as long as the arm itself.
Dimensions.—Diameter of the phacoid shell 0.08, of the medullary shell 0.04;
length of the arms (without terminal spine) 0.08, basal breadth 0.04, distal breadth 0.06.
Habitat.—Fossil in the rocks of Barbados.
Genus 207. Hymenactura,[246] Haeckel,
1881, Prodromus, p. 459.
Definition.—Coccodiscida with three chambered arms on the
margin of the circular or triangular disk, connected by a spongy patagium.
The genus Hymenactura differs from the foregoing Trigonactura in the
spongy patagium between the arms, and bears therefore to it the same relation that among the
Porodiscida Hymeniastrum does to the simpler Dictyastrum. The oldest known species
of this genus is Hymenactura pythagoræ, described by Ehrenberg as Hymeniastrum
pythagoræ, but differing from this in the structure of the central disk.
Subgenus 1. Hymenacturium, Haeckel.
Definition.—Distal end of the arms blunt or truncated, without
terminal spines.
1. Hymenactura archimedis, n. sp. (Pl. 38, fig.
8).
Phacoid shell three times as broad as the medullary shell, with eight pores on its radius. Arms
nearly trapezoidal, somewhat longer than the diameter of the central disk, at the truncated distal
end nearly as broad, at the base only half as broad. In each arm eleven to twelve transverse rows
of square chambers, each of which exhibits on the surface one large pore. Patagium enveloping the
basal half of the arms, with four to five convex rows of chambers, forming together a circular
concentric disk.
Dimensions.—Diameter of the phacoid shell 0.11, of the medullary shell 0.035;
length of the arms 0.12, basal breadth 0.05, distal breadth 0.1.
Habitat.—Western Tropical Pacific, Station 225, depth 4475
fathoms.
{474}
2. Hymenactura pythagoræ, Haeckel.
Hymeniastrum pythagoræ, Ehrenberg, 1854, Mikrogeol., Taf. xxxvi. fig. 31;
Abhandl. d. k. Akad. d. Wiss. Berlin, 1875, p. 76, Taf. xxx. fig. 5.
Phacoid shell three times as broad as the medullary shell, with six pores on its radius. Arms
nearly trapezoidal, somewhat longer than the diameter of the central disk, at the truncated distal
end nearly as broad, at the base two-thirds as broad. In each arm eight to nine transverse rows of
square chambers, each chamber with one pore on the surface. Patagium enveloping the basal half of
the arms, with four to five rectilinear parallel rows of chambers, forming together a regular
triangle. Differs from the preceding species mainly in the quite different structure of the
patagium, from the true Hymeniastrum pythagoræ in the phacoid shell of the central disk,
which encloses only one simple medullary shell.
Dimensions.—Diameter of the phacoid shell 0.11, of the medullary shell 0.04;
length of the arms 0.13, basal breadth 0.07, distal breadth 0.12.
Habitat.—Fossil in the rocks of Barbados.
3. Hymenactura trigona, n. sp.
Astromma sp., Bury, 1862, Polycystins of Barbados, pl. xv. fig. 1.
Phacoid shell twice as broad as the medullary shell, with four to five pores on its radius.
Arms slender, lanceolate, three times as long as the diameter of the phacoid shell and five times
as long as broad in the middle part; distal ends blunt. Patagium enveloping only the basal half of
the arms, forming a regular triangle with concave sides.
Dimensions.—Diameter of the phacoid shell 0.08, of the medullary shell 0.035;
length of the arms 0.2, greatest breadth 0.04.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms; fossil in
the rocks of Barbados.
4. Hymenactura hexagona.
Hymeniastrum sp., Bury, 1862, Polycystins of Barbados, pl. xv. fig. 3.
Phacoid shell twice as broad as the medullary shell, with four to five pores on its radius.
Arms nearly equilateral triangular, three times as long as the diameter of the phacoid shell, at
the truncated distal end nearly three-fourths as broad as long, at the narrow base only one-fourth
as broad. Patagium complete, enveloping the arms completely, so that the whole body represents a
regular hexagonal disk.
Dimensions.—Diameter of the phacoid shell 0.08, of the medullary shell 0.04;
length of the arms 0.18, basal breadth 0.04, distal breadth 0.16.
Habitat.—Fossil in the rocks of Barbados and Nicobar.
{475}
Subgenus 2. Hymenactinium, Haeckel.
Definition.—Distal ends of the arms armed with terminal spines.
5. Hymenactura ptolemæi, n. sp.
Astromma sp., Bury, 1862, Polycystins of Barbados, pl. xv. figs. 5, 6.
Phacoid shell twice as broad as the medullary shell, with six to seven pores on its radius.
Arms nearly square, about as large as the phacoid shell, at the truncated distal end little
broader than at the base, and armed with a strong pyramidal terminal spine. Patagium incomplete,
enveloping the basal half of the arms.
Dimensions.—Diameter of the phacoid shell 0.1, of the medullary shell 0.05; length
and greatest breadth of the arms 0.08.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms;
fossil in the rocks of Barbados.
6. Hymenactura copernici, n. sp. (Pl. 38, fig.
9).
Phacoid shell three times as broad as the medullary shell, with six pores on its radius. Arms
lanceolate, nearly twice as long as the phacoid shell, in the middle part twice as broad as the
medullary shell, with a strong conical terminal spine at the distal end. In each arm about ten
transverse rows of chambers. Patagium enveloping the basal half of the arms, with four convex
parallel rows of chambers.
Dimensions.—Diameter of the phacoid shell 0.1, of the medullary shell 0.035;
length of the arms 0.17, greatest breadth 0.07.
Habitat.—Pacific, central area, Station 266, depth 2750 fathoms.
Genus 208. Astractura,[247] Haeckel,
1881, Prodromus, p. 459.
Definition.—Coccodiscida with four chambered arms on the
margin of the circular or quadrangular disk, crossed in two equatorial diameters, without a
connecting patagium.
The genus Astractura has the form of a regular cross, four radial arms being
opposite in two equatorial diameters perpendicular one to another. In the Porodiscida the same
form is repeated by Stauralastrum, in the Spongodiscida by Spongasteriscus. The
oldest known species of the genus is Astromma aristotelis of Ehrenberg, in which genus this
author confounded triradial and four-radial forms.
{476}
Subgenus 1. Astracturium, Haeckel.
Definition.—Distal end of the arms blunt or truncated, without
terminal spines.
1. Astractura ordinata, n. sp.
? Astromma sp., Bury, 1862, Polycystins of Barbados, pl. xiv. fig. 3.
Phacoid shell three times as broad as the medullary shell, with six pores on its radius,
without a completely developed chambered ring. Arms trapezoidal, somewhat longer than the radius
of the disk, at the truncated distal end as broad as long, at the base one-third smaller.
Dimensions.—Diameter of the phacoid shell 0.08, of the medullary shell 0.03;
length of the arms 0.05, distal breadth 0.05, basal breadth 0.035.
Habitat.—Tropical Atlantic, Station 348, depth 2450 fathoms; also
fossil in Barbados.
2. Astractura clavigera, n. sp.
Astromma aristotelis, Bury, 1862, Polycystins of Barbados, pl. iv. fig.
2.
Phacoid shell twice as broad as the medullary shell, with ten pores on its radius, surrounded
by one perfect chambered ring. Arms club-shaped, about as long as the radius of the central disk,
at the rounded distal end two-thirds, at the base one-third as broad as long.
Dimensions.—Diameter of the phacoid shell 0.1, of the medullary shell 0.05; length
of the arms 0.06, basal breadth 0.02, distal breadth 0.04.
Habitat.—Pacific, central area, Stations 270 to 274, depths 2350 to
2925 fathoms; fossil in the rocks of Barbados.
Subgenus 2. Astractinium, Haeckel.
Definition.—Distal end of the arms furnished with a radial spine.
3. Astractura aristotelis, Haeckel.
Astromma aristotelis, Ehrenberg, 1856, Microgeol., Taf. xxxvi. fig. 32; Abhandl. d. k.
Akad. d. Wiss. Berlin, 1875, p. 66, Taf. xxx. fig. 4.
Astromma aristotelis, Haeckel, 1862, Monogr. d. Radiol., p. 489.
? Astromma aristotelis, Bury, 1862, Polycystins of Barbados, pl. xiv. fig.
4.
Phacoid shell three times as broad as the medullary shell, with six to eight pores on its
radius, surrounded by one perfect chambered ring. Arms nearly trapezoidal, about as long as the
diameter of the central disk, at the base half as broad, at the truncated distal end nearly as
broad as long, and armed with a large pyramidal terminal spine. Each arm with about eight
transverse and longitudinal rows of chambers.
{477}
Dimensions.—Diameter of the phacoid shell 0.11, of the medullary shell 0.04;
length of the arms 0.1, basal breadth 0.05, distal breadth 0.09.
Habitat.—Cosmopolitan; Atlantic, Indian, Pacific, at various depths;
fossil in the Tertiary rocks of Barbados and Nicobar.
4. Astractura democriti, n. sp.
Astromma aristotelis, var., Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss. Berlin, p.
66, Taf, xxx. fig. 3.
? Astromma aristotelis, var., Bury, 1862, Polycystins of Barbados, pl.
xiv. fig. 4.
Phacoid shell twice as broad as the medullary shell, with five to seven pores on its radius,
without perfect chambered ring. Arms about twice as long as broad, longer than the diameter of the
central disk, at the truncated distal end a little broader than at the base, and armed with a
strong, pyramidal, terminal spine.
Dimensions.—Diameter of the phacoid shell 0.1, of the medullary shell 0.05; length
of the arms 0.15, basal breadth 0.04, distal breadth 0.08.
Habitat.—Pacific, central area, Station 266, depth 2750 fathoms;
fossil in Barbados.
5. Astractura hippocratis, n. sp.
Phacoid shell three times as broad as the medullary shell, with seven pores on its radius,
surrounded by two to three perfect chambered rings. Arms nearly square, scarcely as long and
nearly as broad as the diameter of the central disk, at the rounded distal end armed with a very
strong, pyramidal, terminal spine, longer than the arm itself, and at the base as broad as the
medullary shell.
Dimensions.—Diameter of the phacoid shell 0.09, of the medullary shell 0.03;
length of the arms 0.08, breadth 0.1.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms.
Genus 209. Stauractura,[248] Haeckel,
1881, Prodromus, p. 459.
Definition.—Coccodiscida with four chambered arms on the
margin of the circular or quadrangular disk, crossed in two equatorial diameters, connected by a
spongy patagium.
The genus Stauractura differs from the foregoing in the patagium between the
arms, and bears therefore the same relation to it as Histiastrum in the Porodiscida does to
Stauralastrum. All known species of this genus form a regular square, if we connect the end
points of the arm axes by lines.
Subgenus 1. Stauracturium, Haeckel.
Definition.—Distal end of the arms blunt or truncated, without
terminal spines.
{478}
1. Stauractura octogona, n. sp.
Phacoid shell twice as broad as the medullary shell, with eight pores in its radius. Arms
nearly square, little larger than the phacoid shell, with broad truncated distal ends. The corners
of the latter are so connected by the complete patagium, that the whole shell forms a regular
octagon.
Dimensions.—Diameter of the phacoid shell 0.1, of the medullary shell 0.05; length
of the arms 0.12, distal breadth 0.11.
Habitat.—Pacific, central area, Station 266, depth 2750 fathoms.
2. Stauractura tetragona, n. sp.
Phacoid shell three times as broad as the medullary shell, with eleven pores on its radius.
Arms club-shaped, nearly as long as the diameter of the phacoid shell, and one and a half times as
long as broad at the distal part; at the distal end rounded, blunt, without terminal spine. The
terminal points of the arms are so connected by a thin, complete patagium, that the whole shell
becomes a regular square.
Dimensions.—Diameter of the phacoid shell 0.13, of the medullary shell 0.045;
length of the arms 0.12, greatest breadth 0.08.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms.
Subgenus 2. Stauractinium, Haeckel.
Definition.—Distal end of the arms furnished with a radial spine.
3. Stauractura medusina, n. sp.
Phacoid shell two and a half times as broad as the medullary shell, with nine pores on its
radius. Arms club-shaped, one and a half times as long as the diameter of the phacoid shell, and
in the outer third nearly as broad as the latter, at the base scarcely one-third as broad; their
truncated distal end armed with a strong pyramidal terminal spine. Patagium incomplete, enveloping
only the basal half of the arms, with three rectilinear parallel rows of chambers, forming a
square.
Dimensions.—Diameter of the phacoid shell 0.1, of the medullary shell 0.04; length
of the arms 0.14, greatest breadth 0.08.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
4. Stauractura quadrata, n. sp.
Phacoid shell twice as broad as the medullary shell, with seven pores on the radius. Arms
nearly square, about as large as the phacoid shell, with a strong pyramidal terminal spine at the
distal end. Patagium complete, between every two arms triangular, with rectilinear parallel {479}rows of chambers; it connects the end points of the arm
radius in such a manner that the whole shell becomes a regular square.
Dimensions.—Diameter of the phacoid shell 0.09, of the medullary shell 0.045;
length of the arms 0.1, greatest breadth 0.08.
Habitat.—South Pacific, Station 295, depth 1500 fathoms.
Genus 210. Pentactura,[249] Haeckel,
1881, Prodromus, p. 459.
Definition.—Coccodiscida with five chambered arms on the
margin of the circular or pentagonal disk, without a connecting patagium.
The genus Pentactura possesses five free radial arms, and resembles
Pentalastrum among the Porodiscida. The distance of the five arms seems to be sometimes
equal, at other times different in one and the same species.
1. Pentactura pentactis, Haeckel.
Astromma pentactis, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss. Berlin,
p. 66, Taf. xxx. fig. 1.
Phacoid shell two and a half times as broad as the medullary shell, with eight pores on its
radius. Arms regularly or irregularly disposed, nearly square, about half as large as the phacoid
shell, at the truncated distal end little broader than at the base. In the specimen figured, and
very imperfectly described by Ehrenberg, (loc. cit.), the arms were asymmetrically
disposed, two opposite in one axis of the disk, two others on one side of this axis, the fifth
opposite to these on the other side. Another specimen which I found in the Barbados rocks had five
arms of equal size, regularly disposed, at equal intervals.
Dimensions.—Diameter of the phacoid shell 0.1, of the medullary shell 0.04; length
and breadth of the arms 0.05 to 0.06.
Habitat.—Fossil in the rocks of Barbados.
2. Pentactura astropecten, n. sp.
Phacoid shell three times as broad as the medullary shell, with ten pores on its radius. Arms
regularly disposed, club-shaped, nearly twice as long as the diameter of the phacoid shell, and
equal to it in breadth at the rounded distal end. The latter is armed with a short conical spine,
and is twice as broad as the base.
Dimensions.—Diameter of the phacoid shell 0.09, of the medullary shell 0.03;
length of the arms 0.17, basal breadth 0.05, distal breadth 0.08.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms.
{480}
Genus 211. Echinactura,[250] n. gen.
Definition.—Coccodiscida with five chambered arms on the
margin of the circular or pentagonal disk, connected by a spongy patagium.
The genus Echinactura differs from the foregoing Pentactura in the
patagium between the arms, and bears therefore the same relation to it as Pentinastrum in
the Porodiscida does to Pentalastrum.
1. Echinactura culcita, n. sp.
Phacoid shell pentagonal, three times as broad as the medullary shell, with nine pores on its
radius. Arms egg-shaped, about as long as the diameter of the phacoid shell, and in the distal
part half as broad. The blunt ends of the arms are so connected by the complete patagium that the
whole shell becomes a regular pentagonal disk.
Dimensions.—Diameter of the phacoid shell 0.1, of the medullary shell 0.035;
length of the arms 0.11, basal breadth 0.04, distal breadth 0.06.
Habitat.—Pacific, central area, Station 266, depth 2750 fathoms.
2. Echinactura asteriscus, n. sp.
Phacoid shell twice as broad as the medullary shell, with seven pores on its radius. Arms
lanceolate, twice as long as the diameter of the phacoid shell, in the middle part nearly as broad
as the latter, at the distal end with a strong conical terminal spine. Patagium incomplete,
enveloping only the basal half of the arms.
Dimensions.—Diameter of the phacoid shell 0.09, of the medullary shell 0.045;
length of the arms 0.2, greatest breadth 0.08.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
3. Echinactura goniaster, n. sp.
Phacoid shell three times as broad as the medullary shell, with nine pores on its radius. Arms
club-shaped, one and a half times as long as the diameter of the phacoid shell, in the distal part
nearly as broad as the latter, in the basal part one-third as broad; at the end a strong pyramidal
spine. Patagium nearly complete, with concave dentated margin between each pair of arms, therefore
the pentagonal disk of the whole shell with concave sides.
Dimensions.—Diameter of the phacoid shell 0.12, of the medullary shell 0.04;
length of the arms (without terminal spine) 0.18, basal breadth 0.04, distal breadth 0.1.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms.
{481}
Family XXI. Porodiscida, Haeckel (Pls. 41-47).
Porodiscida, Haeckel, 1881, Prodromus, p. 459.
Trematodiscida et Discospirida, Haeckel, 1862, Monogr. d. Radiol.,
pp. 485, 491, 513.
Calodictya, Ehrenberg, 1847, Monatsber. d. k. preuss. Akad. d. Wiss.
Berlin, p. 53 (partim).
Definition.—Discoidea without phacoid shell, with flat
discoidal shell, in which a simple spherical central chamber is surrounded by concentric chambered
rings (each ring divided by radial beams into imperfect chambers). Surface of the disk on the two
flat sides covered by a porous sieve-plate.
The family Porodiscida is by far the largest and richest in different
and common forms among all Discoidea; already in my Monograph (1862) nine
genera and twenty-eight species have been enumerated. Their number is here increased to more than
thirty genera and two hundred species. Many of these species appertain to the most common and
widely distributed Spumellaria, both living and fossil. But the study of
their structure is not easy, and requires (as in the foregoing Coccodiscida) not only careful
examination of the facial views of the disk, but also of the marginal view and of slides and
sections through different planes.
In my Monograph (1862, pp. 485, 491, 513) I had constituted for these Discoidea two different families, the Trematodiscida and Discospirida; but the
comparative study of a far greater number of different types in the Challenger collection has
since convinced me that those two families are but little different, and united by transitional
forms within one and the same genus, so that they must be united as Porodiscida. Of the group,
which Ehrenberg formerly had called "Calodictya," many genera appertain to the Porodiscida, whilst
many others are true Spongodiscida.
The Porodiscida represent the first and the most important family of the Cyclodiscaria, or of those Discoidea which are devoid
of the peculiar extracapsular lenticular "phacoid shell," characteristic of the three preceding
families (united therefore as Phacodiscaria). Probably all Cyclodiscaria can be derived from Archidiscus, from a morphological as
well as a phylogenetic point of view. Archidiscus seems to be the common ancestral form not
only of the Porodiscida, but also of the nearly allied Pylodiscida and Spongodiscida. This
important Archidiscus (Pl. 48,
figs. 9-11) is a small lenticular circular disk, in which a simple latticed spherical central
chamber is surrounded by one single concentric ring, connected with it by a variable number of
radial beams in the equatorial plane. From this typical Archidiscus, as from their
"architype," all other Cyclodiscaria may be derived; the Porodiscida by
regular apposition of new concentric chambered rings on the margin, the Spongodiscida by irregular
apposition of a spongy framework, the Pylodiscida by a peculiar interrupted, concentric, triradial
growth, three radial arm-chambers alternating with three open gates or holes, so that already the
first chambered ring is not complete.
{482}
Archidiscus (Pl. 48,
figs. 9-11) is not only the common phylogenetic ancestral form of all Cyclodiscaria, but also the common ontogenetic original form of all
Porodiscida, or at least of the greater part of them. The numerous species of Archidiscus,
which are distinguished in the sequel, are at the same time the embryonic forms of different
Porodiscida, corresponding to the "biogenetic main law of development." The small shell of
Archidiscus is sometimes completely lenticular, circular, at other times more or less
polygonal; commonly on the biconvex centre much thicker than on the margin, but sometimes also of
nearly equal thickness (like a medal or a short cylinder). The latticed central chamber of it is
probably in the majority of species spherical, but in some more or less compressed, lenticular;
the number of small pores on its surface is probably commonly between ten and twenty (four to
eight usually being visible on each hemisphere). The number of radial beams, which connect it with
the equatorial ring, varies commonly from four to eight; but sometimes only two or three are to be
found, in other cases nine to ten or more. The regular disposition of these beams (in certain
equatorial axes of the disk) is probably of great importance, as determining the later development
of characteristic radial appendages of the margin in the more highly developed Porodiscida. The
equatorial ring itself, forming the margin of the lenticular disk, is either a simple solid ring
or a broader latticed girdle; in the latter case it merges slowly into the opposite sieve-plates
of the two flat disk sides, or the porous "cover-plates," covering its parallel or convex
surfaces. These latter can be regarded as direct peripheral continuations of the polar regions of
the spherical central chamber. The ring-chambers, surrounding the latter in a single circle, are
commonly of nearly the same breadth, but often also of different irregular size. Their number
varies between two and ten or more, but commonly between four and eight; each ring-chamber is
covered on the upper and lower side by the sieve-plate, bounded on the inner (proximal) side by
the wall of the central chamber, on the outer (distal) side by the marginal ring, on both lateral
sides by the contiguous neighbouring ring-chambers.
The important question as to the phylogenetic origin of Archidiscus can be answered in a
twofold way. The most simple form of Archidiscus (Archidiscus dioniscus) can be
derived immediately from the Stylosphærida, Saturnalis (Pl. 13, fig. 16),
only by the development of lattice-work between the equatorial ring and the two polar faces of the
concentric central chamber (on the surface of the biconvex jelly-mantle). But on the other hand
Archidiscus may also be derived from the simplest Phacodiscida, Sethodiscus (Pl. 33, figs. 1-3),
by the stronger compression of the biconvex lenticular shell, so that the enclosed medullary shell
on the two poles runs together with the lenticular phacoid shell, of which only the peripheral
part remains free, and thus forms the chambered ring; this latter explanation seems the more
natural in many cases, as often in the Porodiscida the central chamber is enclosed in one or two
concentric spherical or lenticular lattice-shells.
{483}
The second subfamily of the Porodiscida are the Trematodiscida, which are derived from the
preceding Archidiscida by concentric growth in the equatorial plane. In the same way in which the
simple chambered ring of Archidiscus is connected by radial beams with the central chamber,
so in Porodiscus a variable number of concentric rings is connected with that first ring.
The number of these concentric rings varies between two and ten or more, but commonly amounts to
between three and five. The radial beams connecting them are either piercing or interrupted;
their number increases gradually from the centre towards the periphery. The chambers between them
are sometimes more regular, at other times more irregular in size and form. Their upper and lower
wall is formed by the two covering "sieve-plates," or the porous cover-plates, which are continued
from the central disk to the margin. If these two sieve-plates continue being parallel, the disk
becomes medal-shaped or a short cylinder; if the two plates become more or less concavely vaulted
one against the other, the disk becomes biconvex lenticular, the middle part thicker than the
marginal part. Rarely the contrary is the case, the margin thicker than the centre, and then the
disk biconcave.
In my Monograph I had separated as two different subfamilies the true Trematodiscida (with
circular concentric rings) from the Discospirida (with spirally convoluted rings). But the
enormous mass of specimens, which I afterwards examined in the Challenger collection, has
convinced me that this separation was not natural. For in one and the same genus of most nearly
allied forms we find on one hand quite regular concentric circular forms (Trematodiscus),
on the other hand spirally convoluted forms (Discospira), and connecting between them such
forms as are in the central part concentric, in the marginal part spiral
(Perispira)—or conversely, these in the centre spiral, on the margin concentric
(Centrospira)—and frequently also more or less irregular forms with interrupted rings
(Atactodiscus); therefore, all those genera (Prodromus, 1881, Nos. 448-452) have only the
value of subgenera of Porodiscus. But a distinct genus is Perichlamydium, in which
the two sieve-plates run on the margin of the lens and form a broad hyaline porous or solid
girdle. More important is the distinction of the Ommatodiscida, in which the margin of the disk
exhibits one larger osculum, armed with a corona of spines (Ommatodiscus), or two oscula,
opposite on the poles of one axis (Stomatodiscus). Whilst in many Porodiscida all chambers
of the concentric rings lie in one and the same (equatorial) plane, in many others with further
growth they become stratified in floors, and the whole disk is therefore composed of two to four
or more parallel disks, each with a system of concentric chambered rings or girdles, quite as in
the majority of the Coccodiscida (p. 457). Often the central part of the
lenticular disk becomes thickened by apposition of such floors or strata, whilst the marginal part
remains simple, with one single stratum. The communication between the chambers of the different
strata seems to be the same as in the similar Coccodiscida.
Also the margin of the disk exhibits in the Porodiscida the same characteristic {484}differences as in the foregoing family. In the subfamily
of Stylodictyida it bears a certain number of solid radial spines, often regularly disposed (as in
the Stylocyclida). In the subfamily of Euchitonida the margin is distinguished by the possession
of two to six or more (commonly three or four) chambered arms, also situated in the plane of the
disk, and of the same structure (sometimes more or less irregular, spongy). These arms are very
variable in size, form, and structure, exhibit the same peculiarities as in the Astracturida, and
are sometimes free, at other times connected by a "patagium" or an interbrachial spongy skeleton
of different structure, like a web membrane (compare above, p. 458). In
some genera the arms become forked or branched on the distal end. Sometimes their distal end
bears a terminal radial spine.
The Central Capsule of the Porodiscida assumes generally the form of the including
shell, with or without arms, but is constantly somewhat smaller, as it remains enclosed by the
sieve-plates of the disk surface. Often the capsule is filled with many coloured oil-globules,
disposed regularly in the chamber rows. The nucleus of it is enclosed by the central chamber, and
in many cases by this and the innermost concentric rings.
Synopsis of the Genera of the Porodiscida.
I. Section of the Porodiscida—Archidiscaria.
Central chamber of the disk surrounded only by one single chambered ring. |
|
1. Subfamily Archidiscida.
Only one single chambered girdle surrounds the central chamber; margin smooth
or spiny.
|
|
Margin smooth, without spines, |
212. Archidiscus. |
| Margin armed with radial spines, |
213. Axodiscus. |
II. Section of the Porodiscida—Astrodiscaria.
Central chamber of the disk surrounded by two or more (commonly three to six) concentric
chambered rings or radiated girdles. |
|
2. Subfamily Trematodiscida.
Margin of the disk quite simple, without radial appendages (spines or
chambered arms), without peculiar oscula.
|
|
Margin simple, without an equatorial girdle, |
214. Porodiscus. |
| Margin with a thin porous equatorial girdle, |
215. Perichlamydium. |
|
3. Subfamily Ommatodiscida.
Margin of the disk without chambered arms but distinguished by one or two
large oscula, or wide openings armed with a crown of spines.
|
|
Disk with a single marginal osculum, |
216. Ommatodiscus. |
| Disk with two opposite marginal oscula, |
217. Stomatodiscus. |
|
4. Subfamily Stylodictyida.
Margin of the disk without peculiar oscular openings and without chambered
arms, but armed with solid radial spines.
|
|
Radial spines of the margin two, three, or
four, usually quite regularly disposed. |
|
Two opposite spines, |
218. Xiphodictya. |
| Three equidistant spines, |
219. Tripodictya. |
| Four crossed spines, |
220. Staurodictya. |
| Radial spines of the margin five or more,
commonly irregularly disposed (generally ten to twelve). |
|
Margin simple, without an equatorial girdle, |
221. Stylodictya. |
| Margin with a thin porous equatorial girdle, |
222. Stylochlamydium. |
|
5. Subfamily Euchitonida.
Margin of the disk with radial chambered arms (or hollow chambered
appendages), on the distal end of the arms with or without solid radial spines.
|
 |
Two arms, opposite in one main axis. |
|
Two arms simple, undivided. |
|
Without a patagium, |
223. Amphibrachium. |
| With a patagium, |
224. Amphymenium. |
| Two arms forked (or one simple, other forked). |
|
Without a patagium, |
225. Amphirrhopalum. |
| With a patagium, |
226. Amphicraspedum. |
| Three arms (either regular, equal, or bilateral, with
unequal arms). |
|
Three arms simple, undivided. |
|
Without a patagium. |
|
Regular, |
227. Dictyastrum. |
| Bilateral, |
228. Rhopalastrum. |
| With a patagium. |
|
Regular, |
229. Hymeniastrum. |
| Bilateral, |
230. Euchitonia. |
| Three arms forked. |
|
Either regular or bilateral. |
|
Without a patagium, |
231. Chitonastrum. |
| With a patagium, |
232. Trigonastrum. |
| Four arms (either regular, with four equal crossed arms, or
bilateral, with paired arms). |
|
Four arms simple, undivided. |
|
Without a patagium. |
|
Regular, |
233. Stauralastrum. |
| Bilateral, |
234. Hagiastrum. |
| With a patagium. |
|
Regular, |
235. Histiastrum. |
| Bilateral, |
236. Tessarastrum. |
| With a terminal patagial girdle, |
237. Stephanastrum |
| Four arms forked. |
|
Without a patagium. |
|
Regular, |
238. Dicranastrum. |
| Bilateral, |
239. Myelastrum. |
| Five arms (equal or unequal). |
|
Five arms simple, undivided. |
|
Without a patagium, |
240. Pentalastrum. |
| With a patagium, |
241. Pentinastrum. |
| Five arms forked. |
|
Without a patagium, |
242. Pentophiastrum. |
| Six arms (equal or unequal). |
|
Six arms simple, undivided. |
|
Without a patagium, |
243. Hexalastrum. |
| With a patagium, |
244. Hexinastrum. |
I. Section of the Porodiscida—Archidiscaria.
Central chamber of the disk surrounded only by one single chambered ring. |
| 1. Subfamily Archidiscida. Only one single chambered girdle surrounds the
central chamber; margin smooth or spiny. |
|
|
|
|
|
|
Margin smooth, without spines, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
212. Archidiscus. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Margin armed with radial spines, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
213. Axodiscus. |
|
|
|
|
|
|
|
|
|
|
II. Section of the Porodiscida—Astrodiscaria.
Central chamber of the disk surrounded by two or more (commonly three to six) concentric
chambered rings or radiated girdles. |
| 2. Subfamily Trematodiscida. Margin of the disk quite simple, without radial
appendages (spines or chambered arms), without peculiar oscula. |
|
|
|
|
|
|
Margin simple, without an equatorial girdle, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
214. Porodiscus. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Margin with a thin porous equatorial girdle, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
215. Perichlamydium. |
|
|
|
|
|
|
|
|
|
|
| 3. Subfamily Ommatodiscida. Margin of the disk without chambered arms but
distinguished by one or two large oscula, or wide openings armed with a crown of spines. |
|
|
|
|
|
|
Disk with a single marginal osculum, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
216. Ommatodiscus. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Disk with two opposite marginal oscula, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
217. Stomatodiscus. |
|
|
|
|
|
|
|
|
|
|
| 4. Subfamily Stylodictyida. Margin of the disk without peculiar oscular
openings and without chambered arms, but armed with solid radial spines. |
|
|
Radial spines of the margin two, three, or four, usually quite
regularly disposed. |
|
|
|
|
|
|
|
|
Two opposite spines, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
218. Xiphodictya. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Three equidistant spines, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
219. Tripodictya. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Four crossed spines, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
220. Staurodictya. |
|
|
|
|
|
|
|
|
|
|
|
|
Radial spines of the margin five or more, commonly irregularly
disposed (generally ten to twelve). |
|
|
|
|
|
|
|
|
Margin simple, without an equatorial girdle, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
221. Stylodictya. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Margin with a thin porous equatorial girdle, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
222. Stylochlamydium. |
|
|
|
|
|
|
|
|
|
|
| 5. Subfamily Euchitonida. Margin of the disk with radial chambered arms (or
hollow chambered appendages), on the distal end of the arms with or without solid radial
spines. |
|
|
Two arms, opposite in one main axis. |
|
|
|
|
|
|
Two arms simple, undivided. |
|
|
|
|
|
|
|
|
|
|
Without a patagium, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
223. Amphibrachium. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
With a patagium, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
224. Amphymenium. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Two arms forked (or one simple, other forked). |
|
|
|
|
|
|
|
|
|
|
Without a patagium, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
225. Amphirrhopalum. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
With a patagium, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
226. Amphicraspedum. |
|
|
|
|
|
|
|
|
|
|
|
|
Three arms (either regular, equal, or bilateral, with unequal
arms). |
|
|
|
|
|
|
Three arms simple, undivided. |
|
|
|
|
|
|
|
|
|
|
Without a patagium. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Regular, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
227. Dictyastrum. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Bilateral, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
228. Rhopalastrum. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
With a patagium. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Regular, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
229. Hymeniastrum. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Bilateral, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
230. Euchitonia. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Three arms forked. |
|
|
|
|
|
|
|
|
|
|
Either regular or bilateral. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Without a patagium, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
231. Chitonastrum. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
With a patagium, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
232. Trigonastrum. |
|
|
|
|
|
|
|
|
|
|
|
|
Four arms (either regular, with four equal crossed arms, or
bilateral, with paired arms). |
|
|
|
|
|
|
Four arms simple, undivided. |
|
|
|
|
|
|
|
|
|
|
Without a patagium. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Regular, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
233. Stauralastrum. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Bilateral, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
234. Hagiastrum. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
With a patagium. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Regular, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
235. Histiastrum. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Bilateral, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
236. Tessarastrum. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
With a terminal patagial girdle, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
237. Stephanastrum |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Four arms forked. |
|
|
|
|
|
|
|
|
|
|
Without a patagium. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Regular, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
238. Dicranastrum. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Bilateral, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
239. Myelastrum. |
|
|
|
|
|
|
|
|
|
|
|
|
Five arms (equal or unequal). |
|
|
|
|
|
|
Five arms simple, undivided. simple, |
|
|
|
|
|
|
|
|
|
|
Without a patagium, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
240. Pentalastrum. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
With a patagium, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
241. Pentinastrum. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Five arms forked. |
|
|
|
|
|
|
|
|
|
|
Without a patagium, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
242. Pentophiastrum. |
|
|
|
|
|
|
|
|
|
|
|
|
Six arms (equal or unequal). |
|
|
|
|
|
|
Six arms simple, undivided. |
|
|
|
|
|
|
|
|
|
|
Without a patagium, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
243. Hexalastrum. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
With a patagium, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
244. Hexinastrum. |
|
|
|
|
|
|
|
|
|
|
Subfamily 1. Archidiscida, Haeckel.
Definition.—Porodiscida with a simple
spherical or lenticular latticed central chamber, surrounded by a single concentric latticed ring,
which is divided by radial beams into two to six or more radial chambers.
{486}
Genus 212. Archidiscus,[251] n. gen.
Definition.—Porodiscida with a simple central chamber,
surrounded by a single concentric ring, which is divided by radial beams into two to six or more
radial chambers, without radial spines on the margin.
The genus Archidiscus begins the long and polymorphous series of the Cyclodiscaria or of those Discoidea which do not
possess the peculiar "phacoid shell" characteristic of the three preceding families, united as
"Phacodiscaria." As already mentioned above, both these groups are
probably of independent origin, derived from the Sphæroidea in different
ways (compare pp. 402, 405, &c.). Among all
Cyclodiscaria Archidiscus is the most simple, and probably the
common ancestral form, from which the other genera may be derived.
Subgenus 1. Dioniscus, Haeckel.
Definition.—Ring with two chambers, separated by two radial beams.
1. Archidiscus dioniscus, n. sp.
Ring circular, connected with the central chamber by two radial beams, opposite in one axis,
therefore two equal semicircular ring chambers. (This primitive form has an interesting reference
to Saturnalis, Pl. 13,
fig. 16, and differs from it only in the lattice-work covering both faces of the lenticular disk,
the margin of which forms the ring.)
Dimensions.—Diameter of the ring 0.05, of the central chamber 0.016.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
2. Archidiscus dithalamus, n. sp.
Ring roundish, connected with the central chamber by two radial beams, not opposite in one
axis; both semicircular ring chambers more or less unequal, one of them larger than the other, and
sometimes much more prominent. (If this prominence increase, we can regard it as the beginning of
spiral convolutions, Discospira.)
Dimensions.—Diameter of the ring 0.06, of the central chamber 0.014.
Habitat.—Central Pacific, Station 266, depth 2750 fathoms.
Subgenus 2. Trioniscus, Haeckel.
Definition.—Ring with three chambers, separated by three radial
beams.
{487}
3. Archidiscus trioniscus, n. sp.
Ring triangular, roundish, equilateral, connected with the central chamber by three radial
beams at equal distances (120°); therefore three equal ring chambers. (Resembles the central part
of the disk of Tripodictya trigonaria, Pl. 42, fig. 8, and
may be the ancestral form of it.)
Dimensions.—Diameter of the ring 0.04, of the central chamber 0.014.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms.
4. Archidiscus trithalamus, n. sp.
Ring irregular, roundish, connected with the central chamber by three radial beams at unequal
distances; therefore three ring chambers of different size. (If these differences be important,
they introduce a spiral convolution in the further development of the Porodiscus arising
from it.)
Dimensions.—Diameter of the ring 0.06, of the central chamber 0.016.
Habitat.—South Pacific, Station 288, surface.
Subgenus 3. Tetroniscus, Haeckel.
Definition.—Ring with four chambers, separated by four radial
beams.
5. Archidiscus stauroniscus, n. sp. (Pl. 48, figs. 9,
9a).
Ring regular, square, connected with the central chamber by four radial beams at equal
distances, opposite in pairs in two axes perpendicular one to another; therefore four equal ring
chambers (or congruent quadrants of the ring). Resembles the central part of the disk of
Staurodictya medusa, &c. (Pl. 42,
figs. 1-3); also of Staurodictya gracilis, Ehrenberg, 1875 (Abhandl. d. k. Akad. d. Wiss.
Berlin, Taf. xxiii. fig. 3).
Dimensions.—Diameter of the ring 0.05, of the central chamber 0.016.
Habitat.—North Pacific, Station 253, depth 3125 fathoms.
6. Archidiscus tetroniscus, n. sp.
Ring elliptical, of rhomboidal fundamental form, connected with the central chamber by four
radial beams, halving the sides of the rhombus and opposite in pairs in two axes which are not
perpendicular one to another; therefore four ring chambers in pairs different, two opposite equal
and larger than the two others.
Dimensions.—Diameter of the ring 0.05 to 0.07, of the central chamber 0.016.
Habitat.—Central Pacific, Station 263, depth 2650 fathoms.
7. Archidiscus tetrathalamus, n. sp.
Ring irregular, quadrangular, connected with the central chamber by four radial beams of
increasing unequal length; therefore all four ring chambers of different size, gradually
increasing {488}in the following quadrants of the ring.
Important as an ancestral type of such spiral and semi-spiral forms as Staurodictya
cruciata (Pl. 42,
figs. 4, 12, &c.) and Stylodictya clavata, Stylodictya stellata, &c., of
Ehrenberg, 1875 (Abhandl. d. k. Akad. d. Wiss. Berlin, Taf. xxiii. figs. 2, 7, 8, 9).
Dimensions.—Diameter of the ring 0.05 to 0.08, of the central chamber 0.06.
Habitat.—South Pacific, Station 295, depth 1500 fathoms.
Subgenus 4. Pentoniscus, Haeckel.
Definition.—Ring with five chambers, separated by five radial
beams.
8. Archidiscus pentoniscus, n. sp.
Ring pentagonal or nearly circular, regular, connected with the central chamber by five radial
beams of equal length and at equal distances (72°); therefore all five chambers of the ring of
equal size and similar form. (Resembles the central part of the disk of Pentinastrum
asteriscus, Pl. 44,
fig. 2.)
Dimensions.—Diameter of the ring 0.05, of the central chamber 0.015.
Habitat.—North Pacific, Station 244, surface.
Subgenus 5. Hexoniscus, Haeckel.
Definition.—Ring with six chambers, separated by six radial beams.
9. Archidiscus hexoniscus, n. sp. (Pl. 48, figs. 10,
10a).
Ring regular, hexagonal, or nearly circular, connected with the central chamber by six radial
beams of equal length and at equal distances (60°); therefore all six chambers of the same size
and form. (Resembles the central part of the disk of Hexinastrum geryonidum, Pl. 44, fig.
4.)
Dimensions.—Diameter of the ring 0.06, of the central chamber 0.018.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
10. Archidiscus pyloniscus, n. sp. (Pl. 48, figs. 11,
11a).
Ring triangular, connected with the central chamber by six radial beams at alternating
distances; therefore three larger chambers (of looser network) alternate with three smaller
chambers (of denser network); pores of the former twice to three times as large as those of the
latter. This species is of peculiar importance, as an immediate transitional form to the
Pylodiscida. If these three larger ring chambers lose their few lattice-beams and so became open
gates, we get Triolene or Triopyle, the original forms of the Pylodiscida.
Dimensions.—Diameter of the ring 0.05, of the central chamber 0.015.
Habitat.—Central Pacific, Station 266, depth 2750 fathoms.
{489}
11. Archidiscus hexathalamus, n. sp.
Ring irregular, roundish, or hexagonal, connected with the central chamber by six radial beams
of unequal increasing length; therefore all six ring chambers of gradually increasing size
(beginning a spiral convolution, original form of some Discospira).
Dimensions.—Diameter of the ring 0.05 to 0.07, of the central chamber 0.014.
Habitat.—South Pacific, Station 300, depth 1375 fathoms.
Subgenus 6. Circoniscus, Haeckel.
Definition.—Ring with seven or more chambers, separated by seven or
more radial beams.
12. Archidiscus octoniscus, n. sp.
Ring circular or regular octagonal, connected with the central chamber by eight equidistant
radial beams; therefore eight ring chambers of equal size. (Compare the central part of the disk
of Porodiscus quadrigatus, Pl. 41,
fig. 3.)
Dimensions.—Diameter of the ring 0.04, of the central chamber 0.014.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
13. Archidiscus polythalamus, n. sp.
Ring circular, connected with the central chamber by nine to ten or more radial beams at nearly
equal distances. Ring chambers nine to ten, more or less equal, sometimes also eleven to twelve,
more different. (This species is very variable, and may perhaps be divided into a number of
different "transformistic species.")
Dimensions.—Diameter of the ring 0.04 to 0.06, of the central chamber 0.015.
Habitat.—Central Pacific, Stations 265 to 274, depths 2350 to 2925
fathoms.
Genus 213. Axodiscus,[252] n. sp.
Definition.—Porodiscida with a simple central chamber,
surrounded by one single concentric ring, which is divided by radial beams into two to six or more
radial chambers; margin of the disk armed with radial spines.
The genus Axodiscus differs from the preceding Archidiscus only in
the shape of the margin of the small lenticular disk, which is armed with a variable number of
radial spines, indicating certain axes or radii of the shell. If these marginal spines at certain
equal distances from the margin branch and their distal ends become united by these branches
forming a concentric second ring, the genus passes into Porodiscus. The different number
and disposition of the marginal spines are probably very important, {490}as determining the later development of two to four or more radii in the
different genera of Porodiscida.
1. Axodiscus stylophorus, n. sp.
Ring circular, with two equal semicircular chambers, connected with the central chamber by two
opposite radial beams, which are prolonged outside into two strong conical spines.
Dimensions.—Diameter of the ring 0.05, of the central chamber 0.016.
Habitat.—Central Pacific, Station 266, depth 2750 fathoms.
2. Axodiscus triradiatus, n. sp.
Ring regular, triangular, equilateral, connected with the central chamber by three equidistant
radial beams, which are prolonged outside into three short conical spines. (Differs from
Archidiscus trioniscus in the possession of marginal spines.)
Dimensions.—Diameter of the ring 0.04, of the central chamber 0.013.
Habitat.—Central Pacific, Station 267, depth 2700 fathoms.
3. Axodiscus staurophorus, n. sp.
Ring regular, square, connected with the central chamber by four radial beams, which lie
opposite in pairs in two perpendicularly crossed axes, and are prolonged outside into four
delicate cylindrical spines.
Dimensions.—Diameter of the ring 0.045, of the central chamber 0.015.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
4. Axodiscus hexagonus, n. sp.
Ring regular, hexagonal, connected with the central chamber by six radial beams at equal
distances, which are prolonged outside into six strong pyramidal spines.
Dimensions.—Diameter of the ring 0.06, of the central chamber 0.018.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
5. Axodiscus trigonus, n. sp.
Ring regular, triangular, equilateral, connected with the central chamber by six radial beams
of alternating length and distance; therefore three smaller (perradial) chambers (with denser and
darker network) alternating with three larger (interradial) chambers (with looser and finer
network). On the margin three strong conical radial spines, arising in the radius of the larger
chambers. (Differs from Archidiscus pyloniscus, Pl. 48, fig. 11,
mainly in the strong angular spines on the three corners of the triangular shell.)
Dimensions.—Diameter of the ring 0.05, of the central chamber 0.014.
Habitat.—South Pacific, Station 296, depth 1825 fathoms.
{491}
6. Axodiscus octogonus, n. sp.
Ring circular or nearly octagonal, connected with the central chamber by eight radial beams at
nearly equal distances, which are prolonged outside into eight short conical spines. (Differs from
Archidiscus octoniscus in the marginal prolongation of the eight beams.)
Dimensions.—Diameter of the ring 0.04, of the central chamber 0.013.
Habitat.—Central Pacific, Station 274, depth 2750 fathoms.
7. Axodiscus spinosus, n. sp.
Ring circular, connected with the central chamber by ten to twelve radial beams at nearly equal
distances, which are prolonged outside into short conical spines of variable length. (May be
regarded as an aculeate variety of Archidiscus polythalamus.)
Dimensions.—Diameter of the ring 0.04 to 0.05, of the central chamber 0.014.
Habitat.—Central Pacific, Stations 265 to 274, depths 2350 to 2950
fathoms.
Subfamily 2. Trematodiscida, Haeckel, 1862, Monogr. d. Radiol., p. 491
(sensu emendato et restricto).
Definition.—Porodiscida without radial
appendages of the disk (solid spines or chambered arms on the margin), and without peculiar oscula
on the margin of the disk, which is composed of two to four or more concentric rings.
Genus 214. Porodiscus,[253] Haeckel,
1881, Prodromus, p. 459.
Definition.—Porodiscida with simple circular disk,
composed of several rings (without radial appendages or peculiar oscula on the margin of the
disk).
The genus Porodiscus is, next to its ancestral form, Archidiscus, the
simplest and most primitive form of the Porodiscida, from which all other genera of this family
can be derived. The disk is quite simple, without any marginal appendages, composed of a variable
number of rings, commonly of circular form, sometimes more or less polygonal, elliptical, or
irregular. In my Monograph (1862, pp. 491, 513) I had separated the species, here united in
Porodiscus, into two different genera: Trematodiscus with concentric rings, and
Discospira with spiral rings. But the extended study of these very common forms in a great
number of specimens in the Challenger collection has convinced me that the separation of those two
genera cannot be maintained. In one and the same locality, where one single characteristic
disk-form is very common, we find intermingled quite regular disks with only concentric, circular
rings (Trematodiscus), and other disks with one single perfect spiral ring
(Discospira); and between {492}these a smaller
number of specimens, in which the rings of the disk are partly concentric, partly spiral; either
the rings of the central part of the disk are concentric, the outer spiral (Perispira), or
the proportion is inverse (Centrospira); and sometimes the whole disposition of the
concentric and spiral rings is irregular, and the rings often interrupted (Atactodiscus).
Therefore it appears more natural to give to all these different forms only the value of subgenera
of Porodiscus, as I have already proposed in my Prodromus (1881, p. 459). Even the numerous
species of Porodiscus (mainly characterised by the equal or different breadth of the rings,
and by the number, form, and size of the connecting radial beams and of the superficial pores) are
for the most part very variable and hard to distinguish, as all those characters are not constant.
Porodiscus is a quite "transformistic genus."
Subgenus 1. Trematodiscus, Haeckel, 1860, Monatsber. d. k. preuss. Akad. d. Wiss.
Berlin, p. 841.
Definition.—All rings of the disk concentric (commonly circular,
rarely a little elliptical or polygonal).
1. Porodiscus orbiculatus, Haeckel.
Trematodiscus orbiculatus, Haeckel, 1862, Monogr. d. Radiol., p. 492, Taf. xxix. fig.
1.
Trematodiscus orbiculatus, Stöhr, 1880, Palæontogr., vol. xxvi. p.
108.
All rings of the disk circular, concentric, of equal breadth, connected by numerous alternating
radial beams. Chambers differing little in size, about as large as the central chamber. Pores
regular, circular, two to two and a half on the breadth of each ring.
Dimensions.—Diameter of the disk (with nine rings) 0.18; breadth of each ring
0.01; pores 0.003.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Indian, Pacific,
surface; also fossil in Tertiary rocks of Barbados, Sicily, and Nicobar.
2. Porodiscus concentricus, Haeckel.
Trematodiscus concentricus, Haeckel, 1862, Monogr. d. Radiol., p. 493.
Trematodiscus concentricus, Stöhr, 1880, Palæontogr., vol. xxvi. p. 108.
Flustrella concentrica, Ehrenberg, 1838, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 132;
Ibid., 1875, p. 72, Taf. xxii. fig. 13.
? Flustrella concentrica, Ehrenberg, 1854, Mikrogeol., Taf. xix. fig. 61,
Taf. xx. fig. 42, Taf. xxi. fig. 51, Taf. xxxvi. fig. 29.
All rings of the disk circular, concentric, of equal breadth, connected by numerous piercing
radial beams. Chambers different in size, increasing from the centre towards the periphery.
Pores regular, circular, one and half to two on the breadth of each ring.
Dimensions.—Diameter of the disk (with eight rings) 0.16; breadth of each ring
0.01; pores 0.003.
Habitat.—Fossil in many Tertiary rocks—Barbados, Sicily, Greece, &c.
Porodiscus haeckelii =
Trematodiscus haeckelii, Bütschli, 1882, L. N. 41, Taf. xxiv. figs.
5a, 5b.
{493}
3. Porodiscus flustrella, n. sp. (Pl. 41, fig.
1).
Trematodiscus flustrella, Haeckel, 1866, MS. Canar. Ins.
All rings of the disk circular, concentric, of equal breadth, connected by numerous piercing
radial beams. Chambers different in size, increasing from the centre towards the periphery. Pores
very irregular, polygonal, or roundish, one to three on the breadth of each ring.
Dimensions.—Diameter of the disk (with six rings) 0.12; breadth of each ring 0.01;
pores 0.002 to 0.006.
Habitat.—Cosmopolitan; Atlantic, Pacific, &c., many Stations,
surface.
4. Porodiscus sorites, Haeckel.
Trematodiscus sorites, Haeckel, 1862, Monogr. d. Radiol., p. 492, Taf.
xxix. fig. 2.
All rings of the disk circular, concentric, of equal breadth, connected by numerous alternating
radial beams. Chambers different in size, increasing from the centre. Pores regular, circular, one
to one and a half on the breadth of each ring.
Dimensions.—Diameter of the disk (with six rings) 0.12; breadth of each ring 0.01;
pores 0.006.
Habitat.—Mediterranean (Messina), surface.
5. Porodiscus macroporus, Haeckel.
Trematodiscus macroporus, Haeckel, 1879, MS.
Flustrella macropora, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 160.
All rings of the disk circular, concentric, of equal breadth, connected by numerous piercing
radial beams. Chambers of different breadth, increasing from the centre. Pores regular, very
large, nearly square, one single pore on the breadth of each ring.
Dimensions.—Diameter of the disk (with five rings) 0.11; breadth of each ring
0.01; pores 0.005.
Habitat.—Fossil in the Tertiary rocks of Barbados.
6. Porodiscus microporus, Haeckel.
Trematodiscus microporus, Stöhr, 1880, Palæontogr., vol xxvi. p. 108, Taf.
iv. fig. 17.
All rings of the disk concentric, either circular or a little elliptical; the innermost rings
of the same breadth as the central chamber, the third ring much broader. Radial beams between them
alternating; chambers of different size. Pores very small, subregular, everywhere of the same
size, four to five pores on the breadth of each ring.
Dimensions.—Diameter of the disk (with three rings) 0.15; breadth of the inner
rings 0.02, of the outer 0.03; pores 0.0016.
Habitat.—Fossil in Tertiary rocks of Sicily, Grotte, Stöhr.
{494}
7. Porodiscus ellipticus, Haeckel.
Trematodiscus ellipticus, Stöhr, 1880, Palæontogr., vol. xxvi. p. 108,
Taf. iv. fig. 16.
All rings of the disk concentric, either circular or a little elliptical, connected by eight
piercing radial beams; central chamber elliptical, of the same breadth as the first ring, broader
than the following rings, the breadth of which decreases towards the periphery. Pores irregular,
in the inner rings twice as broad as in the outer (third) ring, four to five on the breadth of
each ring.
Dimensions.—Diameter of the disk (with three rings) 0.18; breadth of the inner
rings 0.03, of the outer 0.02; pores 0.003 to 0.006.
Habitat.—Fossil in Tertiary rocks of Sicily, Grotte, Stöhr.
8. Porodiscus heterocyclus, Haeckel.
Trematodiscus heterocyclus, Haeckel, 1862, Monogr. d. Radiol., p. 493, Taf. xxix. fig.
3.
? Flustrella cyclia, Harting, 1863, Fauna Banda-Zee, p. 11, pl. i. fig. 19.
Trematodiscus heterocyclus, Stöhr, 1880, Palæontogr., vol. xxvi. p.
108.
All rings of the disk concentric, circular, connected by numerous radial beams, which are
partly piercing, partly alternating. The breadth of the rings increases gradually from the centre
towards the periphery, corresponding also to the size of the pores; two to three circular pores on
the breadth of each ring.
Dimensions.—Diameter of the disk (with six rings) 0.17; breadth of the second ring
0.007, of the sixth 0.02; pores 0.002 to 0.006.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific, surface; also
fossil in Tertiary rocks of Barbados and the Mediterranean.
9. Porodiscus quadrigatus, n. sp. (Pl. 41, fig.
3).
All rings of the disk concentric, of nearly equal breadth, connected by four interradial beams,
perpendicular one to another. The first ring (surrounding the central chamber) with eight
chambers, the second only with four (alternating with the four radial beams). Size of the four
chambers of each ring increases much towards the periphery. Pores regular, circular, about two on
the breadth of each ring.
Dimensions.—Diameter of the disk (with six rings) 0.18; breadth of each ring 0.05;
pores 0.004.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
Subgenus 2. Perispira, Haeckel, 1881, Prodromus, p. 459.
Definition.—The inner rings of the disk concentric (commonly
circular); the outer rings spirally convoluted.
{495}
10. Porodiscus perispira, n. sp. (Pl. 41, fig.
2).
Perispira perforata, Haeckel, 1881, Prodromus.
Inner rings of the disk (two or three) concentric, outer rings (three or four) spirally
convoluted, spiral line simple. All rings nearly of the same breadth, connected by alternating
irregular radial beams. Pores irregular, two to three on the breadth of each ring.
Dimensions.—Diameter of the disk (with six rings) 0.17; breadth of each ring
0.016; pores 0.003 to 0.005.
Habitat.—Pacific, central area, Stations 272 to 274, surface.
11. Porodiscus radiatus, n. sp.
Perispira radiata, Haeckel, 1881, Prodromus.
Inner rings of the disk (four or five) concentric, outer rings (three or four) spirally
convoluted, spiral line simple. All rings connected by piercing radial beams (eight in the inner
half, sixteen in the outer half). Breadth of the rings and of the pores increasing from the centre
towards the periphery, three to four pores on the breadth of each ring.
Dimensions.—Diameter of the disk (with eight rings) 0.18; breadth of the second
ring 0.006, of the eighth ring 0.02; pores 0.002 to 0.006.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
Subgenus 3. Centrospira, Haeckel, 1881, Prodromus, p. 459.
Definition.—The inner rings of the disk spirally convoluted, the outer
rings concentric (commonly circular).
12. Porodiscus centrospira, n. sp. (Pl. 41, fig.
6).
Centrospira perispongidium, Haeckel, 1881, Prodromus.
Inner rings of the disk (two or three) spirally convoluted (with simple or double spiral line),
outer rings (three or four) concentric, subcircular. All rings nearly of the same breadth,
connected by alternating radial beams. Pores subregular, two to three on the breadth of each ring.
Lattice-work in the periphery of the disk a little spongy (as in Pl. 41, fig.
11).
Dimensions.—Diameter of the disk (with five rings) 0.15; breadth of each ring
0.015; pores 0.003.
Habitat.—Pacific, central area, Station 267, depth 2700 fathoms.
Subgenus 4. Discospira, Haeckel, 1862, Monogr. d. Radiol., p. 513.
Definition.—All rings of the disk spirally convoluted, forming parts
of a simple or double spiral turning.
{496}
13. Porodiscus helicoides, Haeckel.
Discospira helicoides, Haeckel, 1862, Monogr. d. Radiol., p. 514, Taf. xxix. fig.
7.
Trematodiscus helicoides, Haeckel, 1860, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 841.
All rings of the disk spirally convoluted around the central chamber; spiral line regular,
simple. All rings nearly of the same breadth, connected by numerous alternating radial beams.
Chambers little different in size, little longer than broad. Pores of equal size, regular, two on
the breadth of each ring.
Dimensions.—Diameter of the disk (with ten rings) 0.2; breadth of each ring 0.01;
pores 0.0025.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific; also fossil
in the Tertiary rocks of the Mediterranean, Sicily, Oran, &c.
14. Porodiscus spiralis, Haeckel.
Flustrella spiralis, Ehrenberg, 1840, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin,
p. 210; Mikrogeol., 1854, Taf. xix. fig. 62.
Discospira spiralis, Haeckel, 1862, Monogr. d. Radiol., p. 513.
All rings of the disk spirally convoluted around the central chamber; spiral line regular,
simple. All rings nearly of the same breadth, connected by numerous (twelve to sixteen) piercing
radial beams. Chambers increasing in size from the centre towards the periphery. Pores irregular,
of different size, one and a half to two on the breadth of each ring.
Dimensions.—Diameter of the disk (with eight rings) 0.16; breadth of each ring
0.01; pores 0.004.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Indian, Pacific; also
fossil in the Tertiary rocks of Barbados, Sicily, Greece, &c.
15. Porodiscus operculina, Haeckel.
Discospira operculina, Haeckel, 1862, Monogr. d. Radiol., p. 514, Taf.
xxix. fig. 8.
All rings of the disk spirally convoluted around the central chamber, spiral line regular,
simple. Breadth of the rings and length of the chambers increasing in size from the centre towards
the periphery. Most part of chambers twice as long as broad. Pores irregular, of very different
sizes, two to three on the breadth of each ring.
Dimensions.—Diameter of the disk (with six rings) 0.16; breadth of the rings 0.012
to 0.016; pores 0.002 to 0.006.
Habitat.—Mediterranean (Messina), surface.
16. Porodiscus bilix, Haeckel.
Discospira bilix, Stöhr, 1880, Palæontogr., vol. xxvi. p. 113, Taf. vi.
fig. 3.
All rings of the disk spirally convoluted around the large elliptical central chamber. Spiral
convolutions of the inner rings simple, of the outer rings double; the latter twice as broad as
the {497}former. Pores regular, circular, in the inner
rings one pore on the breadth, in the outer rings two to three pores. Radial beams piercing,
numerous, on the margin prominent.
Dimensions.—Diameter of the disk (with six rings) 0.14; breadth of the inner rings
0.007, of the outer 0.013; pores 0.0017.
Habitat.—Fossil in the Tertiary rocks of Sicily, Grotte, Stöhr.
17. Porodiscus bispiralis, Haeckel.
Stylodictya bispiralis, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 160. Taf. xxiv. fig. 1.
All rings of the disk spirally convoluted, in a perfect double spiral, with increasing breadth
of the rings; the fourth ring twice as broad as the first. Pores regular, circular; in the inner
rings one pore on the breadth, in the outer rings two to three pores. Radial beams piercing,
numerous, prominent on the margin.
Dimensions.—Diameter of the disk (with four rings) 0.1; breadth of the inner rings
0.006, of the outer 0.012; pores 0.0015.
Habitat.—Fossil in the rocks of Barbados; living in the depths of the
Central Pacific, Station 268, depth 2900 fathoms.
18. Porodiscus duplex, Haeckel.
Discospira duplex, Stöhr, 1880, Palæontogr., vol. xxvi. p. 114, Taf. vi.
fig. 4.
All rings of the disk spirally convoluted, in a perfect double spiral, with increasing breadth
of the broad rings. Pores regular, circular; in the inner rings two to three, in the outer four to
five on the breadth. Radial beams interrupted. Margin of the rings thorny.
Dimensions.—Diameter of the disk (with three rings) 0.15; breadth of the rings
0.02 to 0.04; pores 0.007.
Habitat.—Fossil in Tertiary rocks of Sicily, Grotte, Stöhr.
19. Porodiscus semispiralis, n. sp. (Pl. 41, fig.
4).
All rings of the disk spirally convoluted, divided by four radial, perpendicularly crossed and
zigzag shaped beams into four quadrants; the quarter rings of each quadrant halving the rings of
each adjacent quarter. Half spiral line often irregular or partly interrupted. Breadth of all
rings nearly equal. Pores irregular, roundish, two on the breadth of each ring. Margin of the disk
dentated.
Dimensions.—Diameter of the disk (with nine rings) 0.22; breadth of each ring
0.012; pores 0.002 to 0.006.
Habitat.—Pacific, central area, Station 272, depth 2600 fathoms.
Subgenus 5. Atactodiscus, Haeckel, 1881, Prodromus, p. 459.
Definition.—Rings of the disk more or less irregular, partly
concentric, partly spirally convoluted, often interrupted.
{498}
20. Porodiscus deformis, Haeckel.
Discospira deformis, Stöhr, 1880, Palæontogr., vol. xxvi. p. 114, Taf. vi.
fig. 6.
Rings of the disk irregular, partly concentric, partly spiral, often interrupted, increasing in
breadth from the centre. Radial beams not piercing; pores regular, circular, two to six on the
breadth of the different rings. (Very variable and irregular, sometimes more spiral, at other
times more concentric, but always with equal pores.)
Dimensions.—Diameter of the disk (with four rings) 0.18; breadth of the first ring
0.014, of the second 0.03, of the fourth 0.06; pores 0.005.
Habitat.—Fossil in the Tertiary rocks of Barbados and Sicily; living
in the Central Pacific, Stations 266 to 268, depths 2700 to 2900 fathoms.
21. Porodiscus irregularis, n. sp. (Pl. 41, fig.
7).
Atactodiscus irregularis, Haeckel, 1881, Prodromus, p. 459.
Perispongidium irregulare, Haeckel, 1878, MS. et Atlas (pl. xli. fig.
7).
Rings of the disk irregular, partly concentric, partly spiral, often interrupted, with nearly
equal breadth. Radial beams not piercing; pores irregular, roundish, two to four on the breadth of
each ring; network in the periphery of the disk spongy. (Very variable and irregular, sometimes
more spiral, at other times more concentric; disk in the peripheral part often more or less
spongy. Differs from the preceding by the equal breadth of the rings and the different size of the
pores.)
Dimensions.—Diameter of the disk (with six rings) 0.15; breadth of each ring
0.011; pores 0.002 to 0.004.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific, surface; also
fossil in Barbados.
Genus 215. Perichlamydium,[254]
Ehrenberg, 1847, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 54.
Definition.—Porodiscida with a simple circular disk
(without radial spines and chambered arms), surrounded on the margin by a thin porous (but not
chambered) equatorial girdle.
The genus Perichlamydium differs from Porodiscus only in the
development of a thin, porous, equatorial girdle, which surrounds the circular margin of the
chambered disk. This girdle lies in the equatorial plane of the lenticular disk, and represents a
very delicate siliceous plate, perforated by numerous small pores. Sometimes the proximal part of
the girdle is ribbed by thin radial beams, the distal prolongations of the radial rods of the
central disk. If these ribs reach the margin of the girdle and are prominent over it,
Perichlamydium passes over into Stylochlamydium.
{499}
1. Perichlamydium praetextum, Ehrenberg.
Perichlamydium praetextum, Ehrenberg, 1847, Monatsber. d. k. preuss. Akad. d. Wiss.
Berlin, p. 43; Mikrogeol., 1854, Taf. xxii. fig. 21 (non 20).
Perichlamydium praetextum, Haeckel, 1862, Monogr. d. Radiol., p. 495.
All rings of the disk (three to four) concentric, circular, of equal breadth, with interrupted
(not piercing) radial beams. Equatorial girdle without radial beams, nearly as broad as the disk;
its circular pores of the same size as those of the disk; about two pores on the breadth of each
ring.
Dimensions.—Diameter of the disk (without the girdle, with four rings) 0.11;
breadth of each ring 0.012; breadth of the girdle 0.06 to 0.1; pores 0.004.
Habitat.—Cosmopolitan; Atlantic, Indian, Pacific, surface; also fossil
in the Tertiary rocks of Barbados and Sicily.
2. Perichlamydium saturnus, n. sp. (Pl. 41, fig.
5).
All rings of the disk (three to four) concentric, circular (sometimes partly concentric,
circular, partly spiral, or irregular), with interrupted (not piercing) radial beams. Equatorial
girdle without radial beams, about half as broad as the disk; its circular pores very small,
scarcely half as large as those of the disk; about three pores on the breadth of each ring. (Very
variable in the ring-form, differs from the preceding in the small pores of the girdle.)
Dimensions.—Diameter of the disk (without the girdle, with three rings) 0.11;
breadth of each ring 0.02, pores 0.004, breadth of the girdle 0.05, pores 0.002.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms.
3. Perichlamydium accrescens, Haeckel.
Discospira accrescens, Stöhr, 1880, Palæontogr., vol. xxvi. p. 114, Taf.
vi. fig. 5.
All rings of the disk (six to seven) not concentric, convoluted in a simple spiral, of nearly
equal breadth, with interrupted (not piercing) radial beams. Equatorial girdle in the proximal
part with numerous radial beams, which do not reach its margin; its pores half as large as those
of the disk, where one to two pores arise on the breadth of each ring. (The girdle becomes twice
to three times as broad as in the figure of Stöhr.)
Dimensions.—Diameter of the disk (with seven rings, without the girdle) 0.13;
breadth of each ring 0.007 to 0.01; pores 0.0036; breadth of the girdle 0.05, pores of it
0.0017.
Habitat.—Fossil in Tertiary rocks of Sicily, Grotte (Stöhr),
Caltanisetta (Haeckel); living in the Central Pacific, Station 266, depth 2750 fathoms.
4. Perichlamydium spirale, Ehrenberg.
Perichlamydium spirale, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 80, Taf. xxii. fig. 12.
All rings of the disk (three to four) not concentric, convoluted in a simple spiral, of
increasing breadth from the centre; connected by numerous piercing radial beams. Equatorial girdle
about {500}half as broad as the disk, without radial
beams; its pores of the same size as those of the disk, regular, circular; three pores on the
breadth of the first ring, six pores of the fourth ring.
Dimensions.—Diameter of the disk (with four rings, without the girdle) 0.12;
breadth of the first ring 0.01, of the fourth 0.02; breadth of the girdle 0.05; pores 0.003.
Habitat.—Fossil in the rocks of Barbados.
Subfamily 3. Ommatodiscida, Stöhr, 1880, Palæontographica, vol. xxvi.
p. 115.
Definition.—Porodiscida without radial
appendages of the concentrically annulated disk, but distinguished by one single or two opposite
large marginal oscula, or wide openings on the margin of the disk, armed with a coronet of
spines.
Genus 216. Ommatodiscus,[255] Stöhr,
1880, Palæontographica, vol. xxvi. p. 115.
Definition.—Porodiscida without chambered arms and radial spines on the margin of
the circular or elliptical disk, but with one large marginal osculum or opening surrounded by a
coronet of spines.
The genus Ommatodiscus, together with the following Stomatodiscus,
form the peculiar small subfamily of Ommatodiscida, founded by Stöhr in 1880 (loc. cit.).
These remarkable Porodiscida, very nearly allied to Porodiscus, are distinguished by one or
two large openings on the margin of the disk, and these "marginal oscula" are constantly armed
with a coronet of spines (comparable to the osculum coronatum of Sycon in the
Calcispongiæ). Probably in the living Ommatodiscida the osculum is the door from which a
"sarcode-flagellum" issues (comp. above, p. 407). Perhaps this osculum is
comparable to the peculiar coronet of spines which is developed on one pole of the shell axis in
some Ellipsida (Lithomespilus, Lithapium). The internal structure of the disk is
commonly more or less obscure, as the lenticular disk is much thickened, sometimes nearly
ellipsoidal. It is possible that the Ommatodiscida are more nearly related to the Lithelida than
to the Porodiscida; but there is no indication of an internal trizonal medullary shell. Also the
apparent resemblance to the Cyrtida is of no morphological value; both groups are of quite
independent phylogenetic origin.
Subgenus 1. Ommatodiscinus, Haeckel.
Definition.—Disk circular.
1. Ommatodiscus decipiens, Stöhr.
Ommatodiscus decipiens, Stöhr, 1880, Palæontogr., vol. xxvi. p. 115, Taf.
vi. figs. 8, 8a.
Disk circular, with two chambered rings around the spherical central chamber, of equal breadth.
Chambers twice as high as broad. Pores very small, one-third as broad as the bars {501}between them, two on the breadth of each ring. Osculum of
the same breadth as the central chamber, surrounded by numerous very short teeth.
Dimensions.—Diameter of the disk 0.12; breadth of the central chamber and of each
ring 0.03; pores 0.0015.
Habitat.—Fossil in Tertiary rocks of Sicily, Grotte, Stöhr.
2. Ommatodiscus stöhrii, n. sp.
Disk circular with three chambered rings around the spherical central chamber, of equal
breadth. Chambers broader than high. Pores of the same breadth as the bars between them, four on
the breadth of each ring. Osculum of the same breadth as the central chamber, surrounded by a
corona of ten to twenty thin, bristle-shaped teeth, as long as its diameter.
Dimensions.—Diameter of the disk 0.2; breadth of the central chamber and of each
ring 0.03; pores 0.004.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
3. Ommatodiscus circularis, n. sp.
Disk circular, with four circular chambered rings around the spherical central chamber, of
increasing breadth; the fourth ring twice as broad as the second. Chambers about as broad as high.
Pores large, twice as broad as the bars, one to two on the breadth of each ring. Osculum twice as
broad as the central chamber, surrounded by a coronal of strong conical teeth, twice as long as
its diameter.
Dimensions.—Diameter of the disk 0.18; breadth of the central chamber and inner
rings 0.015, of the outer rings 0.03; pores 0.01.
Habitat.—South Pacific, Station 295, depth 1500 fathoms.
Subgenus 2. Ommatodisculus, Haeckel.
Definition.—Disk elliptical.
4. Ommatodiscus haeckelii, Stöhr.
Ommatodiscus haeckelii, Stöhr, 1880, Palæontogr., vol. xxvi. p. 115, Taf.
vi. figs. 7, 7a.
Disk elliptical (6 : 7), with four chambered rings around the elliptical central
chamber, of equal breadth. Chambers about as high as broad. Pores small, half as broad as the
bars, two on the breadth of each ring. Osculum three times as broad as the central chamber,
surrounded by a crown of strong conical teeth.
Dimensions.—Length of the disk 0.18, breadth 0.16; breadth of each ring and of the
central chamber 0.02; pores 0.003.
Habitat.—Fossil in Tertiary rocks of Sicily, Grotte (Stöhr),
Caltanisetta (Haeckel).
{502}
5. Ommatodiscus lævigatus, Stöhr.
Ommatodiscus lævigatus, Stöhr, 1880, Palæontogr., vol. xxvi. p. 116, Taf.
vi. figs. 9, 9a.
Disk elliptical (3 : 4), with three chambered rings around the circular central
chamber, the third ring half as broad as the second. Chambers twice as high as broad. Pores very
small, one-third as broad as the bars. Osculum twice as broad as the central chamber, armed with a
crown of short conical teeth.
Dimensions.—Length of the disk 0.15, breadth 0.11; breadth of inner rings 0.02, of
the outer 0.01; pores 0.0017.
Habitat.—Fossil in Tertiary rocks of Sicily, Grotte, Stöhr.
6. Ommatodiscus fragilis, Stöhr.
Ommatodiscus fragilis, Stöhr, 1880, Palæontogr., vol. xxvi. p. 116, Taf.
vi. figs. 10, 10a.
Disk elliptical (4 : 5), with five chambered rings around the elliptical central
chamber, the fifth ring twice as broad as each of the others. Chambers about as high as broad.
Pores very small, one-fifth as broad as the bars. Osculum three times as broad as the central
chamber, surrounded by a coronet of short teeth.
Dimensions.—Length of the disk 0.17, breadth 0.13; breadth of the inner rings
0.01, of the outer 0.02; pores 0.001.
Habitat.—Fossil in Tertiary rocks of Sicily and Barbados; living in
depths of the Tropical Atlantic and Pacific, Station 353, depth 2965 fathoms, Station 265, depth
2900 fathoms, &c.
Genus 217. Stomatodiscus,[256] n.
gen.
Definition.—Porodiscida without chambered arms and radial
spines on the margin of the circular or elliptical disk, but with two large, opposite, marginal
oscula, or openings surrounded by a coronet of spines.
The genus Stomatodiscus has the same characteristic structure as the
foregoing Ommatodiscus, and differs from it only in the duplication of the large marginal
openings. Whilst in the latter there is only one such marginal osculum, here we find on the margin
of the disk two oscula, opposite on the poles of one equatorial axis.
1. Stomatodiscus amphistomus, n. sp.
Disk circular, with three concentric rings of equal breadth around the central chamber. Pores
irregular, roundish, about two on the breadth of each ring. Surface of the lenticular shell spiny.
On two opposite points of the margin a large osculum, three to four times as broad as the central
chamber, armed with a coronet of strong pyramidal spines of different length, the longest equal to
the radius of the disk.
{503}
Dimensions.—Diameter of the disk (with three rings) 0.12; breadth of each ring
0.016; pores 0.004.
Habitat.—South Pacific, Station 302, depth 1450 fathoms.
2. Stomatodiscus osculatus, n. sp. (Pl. 48, fig.
8).
Disk elliptical, nearly twice as long as broad, with three concentric rings around the
elliptical central chamber, one piercing radial beam in the main axis, the other beams
interrupted. Surface of the shell with small scattered thorns. Pores very irregular, roundish,
partly aggregated in groups of four to eight smaller porules. On both poles of the main axis a
large elliptical marginal osculum, about as large as the central chamber, armed with a coronet of
short conical spines.
Dimensions.—Length of the disk (with three rings) 0.18, breadth 0.1; pores 0.001
to 0.006.
Habitat.—Western Tropical Pacific, Station 225, depth 4475
fathoms.
Subfamily 4. Stylodictyida, Haeckel, 1881, Prodromus, p. 459.
Definition.—Porodiscida with solid radial
spines on the margin of the concentrically annulated disk, situated in the equatorial plane of the
disk (without chambered arms and marginal oscula).
Genus 218. Xiphodictya,[257] Haeckel,
1881, Prodromus, p. 460.
Definition.—Porodiscida with two opposite, solid, radial
spines on the margin of the circular or elliptical disk.
The genus Xiphodictya opens the series of the Stylodictyida or of those
Porodiscida in which the margin of the chambered disk is armed with solid radial spines, all
situated in the equatorial plane of the disk. Xiphodictya exhibits the minimum number of
spines, two being opposite on the poles of one equatorial axis of the disk. It repeats, therefore,
in this family the same amphistylic formation as Sethostylus in the Phacodiscida and
Stylocyclia in the Coccodiscida.
Subgenus 1. Xiphodictyon, Haeckel.
Definition.—All rings of the disk concentric, circular.
1. Xiphodictya amphibelonia, n. sp. (Pl. 42, fig.
10).
All rings of the disk concentric, circular, of equal breadth. Pores irregular, roundish, one
and a half to two on the breadth of each ring. Margin of the disk thorny, of the same thickness as
{504}the central part of the medal-shaped or cylindrical
disk. Two opposite radial spines very long and thin, cylindrical, twice to three times as long as
the diameter of the disk, only half as thick as the breadth of one ring.
Dimensions.—Diameter of the disk (with five rings) 0.17; breadth of each ring
0.014; pores 0.003 to 0.012.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
2. Xiphodictya amphirrhopalia, n. sp. (Pl. 42, fig.
11).
All rings of the disk concentric, circular, with increasing breadth from the centre; the fifth
ring twice as broad as the second. Central chamber very large. Margin of the lenticular disk
thorny, much thinner than the central part. Pores irregular, roundish, two to three on the breadth
of each ring. Two opposite radial spines club-shaped, about as long as the diameter of the disk,
in the outer spindle-shaped part three to four times as broad as at the base.
Dimensions.—Diameter of the disk (with six rings) 0.17; breadth of the second ring
0.008, of the fifth ring 0.016.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms; also
fossil in the Tertiary rocks of Sicily (Caltanisetta).
Subgenus 2. Xiphospira, Haeckel.
Definition.—All rings of the disk or a part of them not concentric,
spirally convoluted; sometimes irregular or interrupted.
3. Xiphodictya staurospira, n. sp. (Pl. 42, fig.
12).
All rings of the disk not concentric, half spiral. Four radial beams, crossed perpendicularly
and zigzag-shaped, divide each ring into four quadrants; the quarter-ring of each quadrant halves
the two adjacent rings. All rings of equal breadth. Pores irregular, roundish, two on the breadth
of each ring. Two opposite radial spines pyramidal, somewhat shorter than the diameter of the
disk.
Dimensions.—Diameter of the disk (with six rings) 0.16; breadth of each ring:
0.014; pores 0.003 to 0.007.
Habitat.—South Pacific, Station 295, depth 1500 fathoms.
4. Xiphodictya heliospira, n. sp.
All rings of the disk not concentric, convoluted in a simple, regular, spiral line; all nearly
of equal breadth. Pores subregular, circular, two on the breadth of each ring. Two opposite radial
spines conical, about as long as the radius of the disk. Margin of the disk thorny. (Differs from
Stylodictya heliospira, Pl. 41,
fig. 8, mainly by the two strong, opposite, radial spines.)
{505}
Dimensions.—Diameter of the disk (with six rings) 0.14; breadth of each ring 0.01
to 0.012; pores 0.002 to 0.006.
Habitat.—Pacific, central area, Station 272, depth 2600 fathoms.
Genus 219. Tripodictya,[258] Haeckel,
1881, Prodromus, p. 460.
Definition.—Porodiscida with three solid, equidistant,
radial spines on the margin of the circular or triangular disk.
The genus Tripodictya exhibits three radial spines on the margin of the
disk, divergent at equal angles; rarely in some specimens the angles differ more or less. Perhaps
this genus bears a near relation to the Dictyastrida (or to the Euchitonida with three chambered
arms—Dictyastrum, Euchitonia, &c.).
1. Tripodictya trigonaria, n. sp. (Pl. 42, fig.
8).
All rings of the disk concentric, triangular, with three equal convex sides, all of the same
breadth; first and second rings with three simple chambers, third and fourth rings with six
chambers, fifth ring with twelve chambers. Central chamber also equilateral triangular, from its
three corners arise three piercing perradial beams; from the second ring arise three interradial
beams, alternate with the latter; from the fourth ring between these and the former arise six
adradial beams. Pores subregular, two on the breadth of each ring. Three marginal spines
pyramidal, as long as the radius of the disk and as broad as one ring.
Dimensions.—Diameter of the disk (with five rings) 0.11; breadth of each ring
0.011; pores 0.004.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms.
2. Tripodictya triacantha, n. sp. (Pl. 42, fig.
7).
All rings of the disk concentric, circular, of the some breadth, connected by very numerous
irregular radial beams, the number of which increases towards the periphery. Pores irregular,
roundish, two on the breadth of each ring. Three marginal spines spindle-shaped, as long as the
radius of the disk, and in the middle part as broad as one ring.
Dimensions.—Diameter of the disk (with eight rings) 0.2; breadth of each ring
0.012; pores 0.004.
Habitat.—North Pacific, Station 253, depth 3125 fathoms.
3. Tripodictya tribelonia, n. sp. (Pl. 42, fig.
9).
All rings of the disk, or a part of them, not concentric, spirally convoluted, of equal
breadth, connected by numerous irregular, interrupted radial beams, the number of which increases
from the centre. (In one marginal view of the disk, Pl. 42, fig. 9, the
disk seemed to be composed of seven {506}parallel
chambered plates in the central part, and five similar plates in the peripheral part.) Pores
irregular, roundish, three on the breadth of each ring. Three marginal spines long and thin,
cylindrical (as in Staurodictya cruciata, Pl. 42, fig, 4),
longer than the diameter of the disk, and half as thick as one ring.
Dimensions.—Diameter of the disk (with nine rings) 0.18; breadth of each ring
0.01; pores 0.002 to 0.004.
Habitat.—Pacific, central area, Station 263, depth 2650 fathoms.
Genus 220. Staurodictya,[259] Haeckel,
1881, Prodromus, p. 460.
Definition.—Porodiscida with four solid radial spines
(commonly crossed at right angles) on the margin of the circular or quadrangular disk.
The genus Staurodictya is characterised by four marginal spines, lying
opposite in pairs in two crossed equatorial diameters of the disk, perpendicular one to another.
Sometimes the regular rectangular position of the spines becomes more or less irregular. Commonly
the marginal spines are the extreme prolongations of four internal crossed radial beams, which are
either rectilinear or zigzag-shaped. In the latter case the concentric disposition of the circular
rings becomes more or less spiral. Perhaps Staurodictya has a near relation to the
Stauralastrida (or to the Euchitonida with four crossed chambered arms—Stauralastrum,
Histiastrum, &c.).
Subgenus 1. Staurodictyon, Haeckel.
Definition.—All rings of the disk concentric (commonly circular,
sometimes with four incisions, produced by two crossed constrictions, or nearly square).
1. Staurodictya medusa, n. sp. (Pl. 42,
fig. 3).
All rings of the disk concentric, of nearly equal breadth, divided into four quadrants by two
perradial constrictions (perpendicular one to another). Pores irregular, roundish, two to three on
the breadth of each ring. Four marginal spines strong, pyramidal, nearly as long as the radius of
the disk. Margin between them denticulated.
Dimensions.—Diameter of the disk (with four rings) 0.13; breadth of each ring
0.015; pores 0.002 to 0.01.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
2. Staurodictya ciliata, n. sp. (Pl. 42,
fig. 2).
All rings of the disk concentric, of nearly equal breadth, circular or roundish, without
perradial constrictions. Pores subregular, circular, three to four on the breadth of each ring.
Four marginal {507}spines short, pyramidal, about as
long as the breadth of one ring. Margin between them ciliated, with thin radial bristles.
Dimensions.—Diameter of the disk (with four rings) 0.13; breadth of each ring
0.015; pores 0.0025.
Habitat.—South Pacific, Station 295, depth 1500 fathoms.
3. Staurodictya elegans, n. sp. (Pl. 42,
fig. 1).
All rings of the disk concentric, circular, or roundish, of increasing breadth towards the
margin; the fifth ring twice as broad as the first. Pores irregular, roundish, two to three on the
breadth of each ring. Four marginal spines very large, conical, with a thin pedicle at the base,
cancellated by ten to twelve deep furrows, about as long as the radius of the disk and three times
as long as broad at the base (above the pedicle). Margin between them ciliated, with numerous
short radial spines.
Dimensions.—Diameter of the disk (with six rings) 0.14; breadth of the inner rings
0.006, of the outer 0.012; pores 0.002 to 0.008.
Habitat.—Pacific, central area, Station 272, depth 2600 fathoms.
4. Staurodictya quadrispina, Haeckel.
Stylodictya quadrispina, Haeckel, 1862, Monogr. d. Radiol., p. 496, Taf.
xxix. fig. 4.
All rings of the disk concentric, circular, of increasing breadth towards the margin; the
fourth ring twice as broad as the second. Pores irregular, roundish, two to three on the breadth
of each ring. Four marginal spines short and thin, conical, about as long as the breadth of one
ring. Margin between them smooth.
Dimensions.—Diameter of the disk (with six rings) 0.12; breadth of the inner rings
0.006, of the outer rings 0.012; pores 0.001 to 0.006.
Habitat.—Mediterranean, Atlantic (Canary Islands), surface.
Subgenus 2. Staurospira, Haeckel.
Definition.—All rings of the disk or a part of them not concentric,
spirally convoluted; spiral line simple or double, sometimes half or irregular, interrupted.
5. Staurodictya cruciata, n. sp. (Pl. 42, figs. 4,
5).
All rings of the disk nearly of the same breadth, not concentric, half-spiral, interrupted by
four zigzag-shaped radial beams crossed in two diameters perpendicular one to another. The quarter
ring of each quadrant halving both neighbouring quarters. Pores irregular, roundish, two on the
breadth of each ring. Pores of the outermost (eighth) ring much smaller than the others. Four
marginal spines cylindrical or nearly spindle-shaped, thick, about as long as the radius of the
disk. Margin between them smooth.
Dimensions.—Diameter of the disk (with eight rings) 0.2; breadth of each ring
0.011; pores 0.004.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms.
{508}
6. Staurodictya splendens, Haeckel.
Stylodictya splendens, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 84, Taf. xxiii. fig. 9.
All rings of the disk nearly of the same breadth, not concentric, half-spiral, interrupted by
four zigzag-shaped perradial beams, crossed in two diameters. The quarter ring of each quadrant
halving both neighbouring quarters. Pores regular, circular, only one single on the breadth of
each ring. Four marginal spines conical or spear-shaped, about half as long as the radius of the
disk. Margin between them ciliated, with short bristle-shaped radial spines.
Dimensions.—Diameter of the disk (with eight rings) 0.2; breadth of each ring
0.012; pores 0.004.
Habitat.—Fossil in the rocks of Barbados.
7. Staurodictya grandis, n. sp.
All rings of the disk (twelve to sixteen) of the same breadth, not concentric, irregular,
partly spiral, interrupted by irregular turnings, and by ramified radial beams, which divide each
ring into numerous square chambers. Pores subregular, circular, only one single on the breadth of
each ring (and on each chamber). Four marginal spines short and stout, conical, twice as long as
broad at the base, four to five times as long as the breadth of one ring. Margin between them
dentated.
Dimensions.—Diameter of the disk (with sixteen rings) 0.3; breadth of each ring
0.01; pores 0.005.
Habitat.—Pacific, central area, Stations 265 to 268, depth 2900
fathoms; also fossil in the Tertiary rocks of Barbados.
8. Staurodictya ocellata, Haeckel.
Stylodictya
ocellata, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 84, Taf. xxiii. fig.
7.
All rings of the disk irregular, not concentric, half-spiral, interrupted by four
zigzag-shaped, crossed, radial beams; the quarter ring of each quadrant halving both neighbouring
quarters. Breadth of the rings increasing towards the periphery; the fourth ring twice as broad as
the second. Pores irregular, with increasing size from the centre, three on the breadth of each
ring. Four marginal spines thick and long, cylindrical; margin between them smooth.
Dimensions.—Diameter of the disk (with four rings) 0.1; breadth of the inner rings
0.01, of the outer 0.02; pores 0.002 to 0.006.
Habitat.—Fossil in the rocks of Barbados.
Stylodictya haeckelii,
Zittel, 1876, L. N. 29, p. 85, Taf. ii. fig. 9, is a fossil Cretaceous
species, related to Stylodictya multispina.
{509}
Genus 221. Stylodictya,[260] Ehrenberg,
1847, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 54.
Definition.—Porodiscida with numerous (five or more,
commonly eight to twelve) solid radial spines, regularly or irregularly disposed on the margin of
the circular or polygonal disk; margin simple, without a porous equatorial girdle.
The genus Stylodictya comprises the majority of this subfamily, in which the
number of the marginal spines exceeds four. Commonly we find eight to twelve spines, more or less
regularly disposed (four perradial and four interradial, or four perradial and eight adradial);
but often also the number and disposition become irregular (sometimes very large). In my Monograph
(1862 pp. 495, 515) I had separated the concentric disks with closed circular rings (as true
Stylodictya, s. str.) from the spiral disks with convoluted rings (Stylospira). But
I retain these two groups here only as two subgenera, as intermediate forms between them are very
common, and often a part of the disk concentric, a part spiral (compare above, p. 492).
Subgenus 1. Stylodictyon, Haeckel, 1862, Monogr. d. Radiol., p. 495.
Definition.—All rings of the disk concentric, commonly circular
(rarely a little elliptical or polygonal).
1. Stylodictya gracilis, Ehrenberg.
Stylodictya gracilis, Ehrenberg, 1854, Mikrogeol., Taf. xxxvi. fig. 28.
Stylodictya gracilis, Ehrenberg, 1873, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin,
p. 257; Abhandl. d. k. Akad. d. Wiss. Berlin, 1875, Taf. xxiii. fig. 3.
Stylodictya gracilis, Haeckel, 1862, Monogr. d. Radiol., p. 499.
All rings of the disk concentric, circular, of equal breadth (the first ring sometimes, but not
constantly, four-lobed). Pores regular, circular, small, three on the breadth of each ring. Four
perradial beams (crossed in two perpendicular diameters) beginning from the circular central
chamber, four interradial beams from the first or second ring (sometimes others between them).
Beams prolonged into eight to twelve (or more) marginal spines, bristle-shaped, as long as the
radius of the shell.
Dimensions.—Diameter of the disk (with four rings) 0.12; breadth of each ring
0.013; pores 0.0025.
Habitat.—Fossil in Tertiary rocks of Barbados and Nicobar; living in
the depths of the Pacific and Atlantic.
{510}
2. Stylodictya multispina, Haeckel.
Stylodictya multispina, Haeckel, 1862, Monogr. d. Radiol., p. 496, Taf. xxix. fig.
5.
Stylodictya forbesii, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 160, Taf. xxiii. fig. 6.
All rings of the disk concentric, circular, of equal breadth. Pores regular, circular, two and
a half to three on the breadth of each ring. Eight to twelve radial beams beginning from the
central chamber, others from the inner rings. Commonly from the third or fourth ring arise
twenty-four to thirty (sometimes forty or more) piercing beams, which are prolonged at the margin
into bristle-shaped radial spines, as long as the breadth of two to four rings.
Dimensions.—Diameter of the disk (with seven rings) 0.2; breadth of each ring
0.013; pores 0.004.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Indian, Pacific,
surface and various depths.
3. Stylodictya hastata, Ehrenberg.
Stylodictya hastata, Ehrenberg, 1873, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin,
p. 257; Abhandl. d. k. Akad. d. Wiss. Berlin, 1875, Taf. xxiii. fig. 5.
Stylodictya gracilis, Bury, 1862, Polycystins of Barbados, pl. ii. fig.
1.
All rings of the disk concentric, circular, of equal breadth. Pores regular, circular, very
small, two on the breadth of each ring. Eight to twelve marginal spines very large, spear-shaped,
sulcated, pyramidal, nearly as long as the diameter of the disk, with a thin pedicle at the base,
above this as thick as the breadth of one ring.
Dimensions.—Diameter of the disk (with five rings) 0.13; breadth of each ring
0.012; pores 0.003.
Habitat.—Fossil in the rocks of Barbados.
4. Stylodictya stellata, Bailey.
Stylodictya stellata, Bailey, 1856, Amer. Journ., vol. xxii. p. 6, pl. i. fig. 20.
Stylodictya stellata, Haeckel, 1862, Monogr. d. Radiol., p. 499.
All rings of the disk concentric, circular, of equal breadth. Pores regular, circular, very
small, two on the breadth of each ring. Marginal spines fourteen (probably variable in number,
twelve to sixteen), very thick and short, conical; their length seems to be equal to their basal
breadth and to the breadth of one ring. Related to Stylochlamydium?
Dimensions.—Diameter of the disk (with five rings) 0.11; breadth of each ring
0.01; pores 0.003.
Habitat.—North Pacific, depths of the Kamtschatka Sea, Bailey; Station
241, depth 2300 fathoms.
5. Stylodictya arachnia, Haeckel.
Stylodictya arachnia, Haeckel, 1862, Monogr. d. Radiol., p. 497.
Stylocyclia arachnia, J. Müller, 1856, p. 492; Abhandl. d. k. Akad. d.
Wiss. Berlin, 1858, p. 41, Taf. i. figs. 8, 9.
All rings of the disk concentric, circular, or polygonal; their breadth increases from the
centre towards the periphery, so that the fourth or fifth ring is twice as broad as the second.
Pores {511}subregular, circular, two on the breadth of
each ring. Radial beams partly piercing. Marginal spines eight to sixteen (commonly twelve),
bristle-shaped, very thin, once to three times as long as the diameter of the disk. (On the
numerous varieties of this common species compare my Monograph, 1862, p. 498.)
Dimensions.—Diameter of the disk (with ten rings) 0.22; breadth of the inner rings
0.004 to 0.008, of the outer 0.012 to 0.015; pores 0.003 to 0.005.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Indian, Pacific,
surface.
6. Stylodictya solmaris, n. sp.
All rings of the disk concentric, polygonal, with eight to sixteen rounded corners; their
breadth increases from the centre; eighth ring twice as broad as the second. Pores subregular
circular, large, only one pore on the breadth of each ring. Twenty to thirty marginal spines,
bristle-shaped, undulating, about as long as (or longer than) the diameter of the disk.
Dimensions.—Diameter of the disk (with eight rings) 0.2; breadth of the inner
rings 0.005, of the outer 0.012; pores 0.004 to 0.008.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
7. Stylodictya octogonia, n. sp.
All rings of the disk concentric, increasing in breadth from the centre. The outer rings (five
to eight) regular, octogonal, twice as broad as the circular inner rings. Pores subregular,
circular, two to three on the breadth of each ring. Eight piercing perradial spines (alternating
with the eight corners of the octogonal rings) bristle-shaped, longer than the diameter of the
disk.
Dimensions.—Diameter of the disk (with eight rings) 0.24; breadth of the inner
rings 0.006, of the outer 0.014; pores 0.005 to 0.01.
Habitat.—North Atlantic, Station 353, depth 2965 fathoms.
Subgenus 2. Stylodictula, Haeckel.
Definition.—Rings of the disk partly concentric, completely annular;
partly spiral or interrupted, often irregular.
8. Stylodictya perispira, n. sp.
Inner rings of the disk (two to four) concentric, circular, or roundish, outer rings forming a
simple spiral, breadth a little increasing from the centre. Pores subregular, circular, about two
on the breadth of each ring. Marginal spines bristle-shaped, ten to fifteen, about as long as the
diameter of the disk.
Dimensions.—Diameter of the disk (with eight rings) 0.2; breadth of each ring
0.013; pores 0.004.
Habitat.—Pacific, central area, Station 272, depth 2600 fathoms.
{512}
9. Stylodictya centrospira, n. sp. (Pl. 41, fig.
9).
Inner rings of the disk (three or four) convoluted in a simple or double spiral, often
interrupted or irregular; outer rings (two to three) concentric, circular, or roundish. Breadth of
the rings variable, irregular. Pores irregular, of very different sizes, in the outer concentric
part twice to four times as large as in the inner spiral part. Radial beams partly interrupted,
partly piercing. Marginal spines fifty to eighty, very variable in size and number, commonly
fifteen to twenty strong conical spines, twice to three times as long as the ring-breadth, and
numerous (thirty to sixty) smaller spines. Very variable.
Dimensions.—Diameter of the disk (with seven rings) 0.2; breadth of the rings 0.01
to 0.02; pores 0.002 to 0.01.
Habitat.—Pacific, central area, Stations 263 to 274, depth 2350 to
2925 fathoms.
10. Stylodictya setigera, Ehrenberg.
Stylodictya setigera, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 84, Taf. xxiii. fig. 4.
Inner rings of the disk (two to three) convoluted in a simple or double spiral, outer rings
(two to three) concentric, circular, or roundish. Breadth of the rings nearly equal. Pores
regular, circular, three on the breadth of each ring. Marginal spines twenty to forty,
bristle-shaped, about twice as long as the ring-breadth. (The specimen figured by Ehrenberg was a
young one; in older specimens I found the inner spiral disk surrounded by two to three concentric
circular rings.)
Dimensions.—Diameter of the disk (with six rings) 0.16; breadth of the rings 0.12;
pores 0.003.
Habitat.—Fossil in the rocks of Barbados; also living in the depths of
the Central Pacific, Station 266, depth 2750 fathoms.
Subgenus 3. Stylospira, Haeckel, 1862, Monogr. d. Radiol., p. 515.
Definition.—All rings of the disk convoluted in a simple or double
spiral (sometimes also in quartered half-spirals).
11. Stylodictya heliospira, n. sp. (Pl. 41, fig.
8).
Stylospira heliospira, Haeckel, 1879, MS.
All rings of the disk convoluted in a simple regular spiral, increasing in breadth from the
centre towards the periphery; the sixth ring twice as broad as the second. Pores irregular,
roundish, two on the breadth of each ring. Marginal spines numerous, thirty to forty,
bristle-shaped, nearly half as long as the radius of the disk.
Dimensions.—Diameter of the disk (with seven rings) 0.16; breadth of the inner
rings 0.008, of the outer 0.016; pores 0.002 to 0.006.
Habitat.—Mediterranean, Portofino near Genoa, surface; also fossil in
the Tertiary rocks of Sicily; Caltanisetta, Haeckel.
{513}
12. Stylodictya hertwigii, Haeckel.
Stylospira arachnia, R. Hertwig, 1879, Organismus d. Radiol., p. 59, Taf.
vi. fig. 8.
All rings of the disk convoluted in a simple regular spiral, with increasing breadth from the
centre; the fifth ring twice as broad as the second. Pores regular, circular, two on the breadth
of each ring. Twelve piercing radial beams and some others interrupted, prolonged into twelve to
twenty radial marginal spines, bristle-shaped, about as long as the diameter of the disk.
Dimensions.—Diameter of the disk (with five rings) 0.15; breadth of the second
ring 0.01, of the fifth 0.02; pores 0.006.
Habitat.—Mediterranean (Messina), R. Hertwig.
13. Stylodictya
dujardinii, Haeckel.
Stylospira dujardinii, Haeckel, 1862, Monogr. d. Radiol., p. 515, Taf.
xxix. figs. 9, 10.
All rings of the disk convoluted in a simple regular spiral, of nearly equal breadth. Pores
regular, circular, two on the breadth of each ring. Very numerous (twenty to thirty or more)
piercing radial beams, prolonged into bristle-shaped marginal spines, about as long as the
diameter of the disk.
Dimensions.—Diameter of the disk (with six rings) 0.12; breadth of each ring 0.01;
pores 0.004.
Habitat.—Mediterranean (Messina), Haeckel.
14. Stylodictya echinastrum, Ehrenberg.
Stylodictya echinastrum, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 84, Taf. xxiii. fig. 1.
All rings of the disk convoluted in a double spiral, of nearly equal breadth. Pores irregular,
roundish, two to three on the breadth of each ring. Marginal spines numerous, twenty to thirty, of
very different size, the largest conical, strong, about as long as the diameter of the disk, and
on the base as broad as one ring.
Dimensions.—Diameter of the disk (with five rings) 0.12; breadth of each ring
0.01; pores 0.002 to 0.006.
Habitat.—Fossil in the rocks of Barbados.
15. Stylodictya clavata, Ehrenberg.
Stylodictya clavata, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 84, Taf. xxiii. fig. 2.
All rings of the disk convoluted in a half spiral, of nearly equal breadth; each ring by four
zigzag beams (crossed in two perpendicular diameters) divided into four equal quarters; the spiral
line of each quarter ring halving both neighbouring quarters. Pores regular, circular, two on the
breadth of each ring. Eight marginal spines short, conical, with thin pedicle; four perradial (as
{514}prolongations of the four internal beams)
alternating with four interradial spines arising from the margin of the disk.
Dimensions.—Diameter of the disk (with five rings) 0.13; breadth of each ring
0.014; pores 0.003.
Habitat.—Fossil in the rocks of Barbados.
Genus 222. Stylochlamydium,[261] Haeckel,
1881, Prodromus, p. 460.
Definition.—Porodiscida with numerous (five or more,
commonly eight to twelve) solid radial spines, regularly or irregularly disposed on the margin of
the circular or polygonal disk; margin of the disk surrounded by a thin, porous (but not
chambered), equatorial girdle.
The genus Stylochlamydium is intermediate between Perichlamydium
(with which it was formerly united) and Stylodictya. It deals with the former in the
peculiar equatorial girdle, with the latter in the radial spines of the disk margin, which pierce
the girdle. To both these genera it shows slow transitions, and can hardly be subjected to a sharp
definition.
Subgenus 1. Stylochlamys, Haeckel.
Definition.—All rings of the disk concentric, circular (or somewhat
polygonal).
1. Stylochlamydium asteriscus, n. sp. (Pl. 41, fig.
10).
Perichlamydium asteriscus, Haeckel, 1879, MS.
All rings of the disk concentric, circular, or polygonal, with increasing breadth from the
centre; the fifth ring twice as broad as the second. Pores irregular, roundish, three to four on
the breadth of each ring; in the rings two to six times as large as in the equatorial girdle,
which is half as broad or two-thirds as broad as the radius of the disk. Twelve bristle-shaped
radial spines are connected by the girdle near to the points; four crossed spines arising from the
central chamber; two others between these in each quadrant arising from the first ring.
Dimensions.—Diameter of the disk (with five rings) 0.15, with the girdle 0.25;
breadth of the second ring 0.01, of the fifth ring 0.02; pores 0.001 to 0.005.
Habitat.—Pacific, central area, Station 272, depth 2600 fathoms.
2. Stylochlamydium limbatum, Haeckel.
Perichlamydium limbatum, Ehrenberg, 1847, Monatsber. d. k. preuss. Akad. d. Wiss.
Berlin, p. 43; Mikrogeol., 1854, Taf. xxii. fig. 20.
Perichlamydium limbatum, Haeckel, 1862, Monogr. d. Radiol., p. 494.
All rings of the disk concentric, circular, of equal breadth. Pores regular, circular, two on
the breadth of each ring; twice to three times as large as the fine pores of the equatorial
girdle, which is {515}about half as broad as the radius
of the disk. Twelve (or eleven) bristle-shaped radial spines are connected by the girdle near to
the points, irregularly disposed.
Dimensions.—Diameter of the disk (with five rings) 0.12, with the girdle 0.2,
breadth of each ring 0.01; pores 0.001 to 0.003.
Habitat.—Fossil in the Tertiary rocks of Sicily, Caltanisetta,
Grotte.
3. Stylochlamydium venustum, Haeckel.
Perichlamydium venustum, Bailey, 1856, Amer. Journ., vol. xxii. p. 5, pl. i. figs. 16,
17.
Perichlamydium venustum, Haeckel, 1862, Monogr. d. Radiol., p. 495.
All rings of the disk concentric, circular, of equal breadth. Pores regular, circular,
everywhere of nearly equal size, three on the breadth of each ring, a little smaller in the
equatorial girdle, which is about as broad as the radius of the disk. Twenty to twenty-four
bristle-shaped radial spines, irregularly disposed, proceed with their free points over the
margin.
Dimensions.—Diameter of the disk (with four rings) 0.1, with the girdle 0.3;
breadth of each ring 0.011; pores 0.002.
Habitat.—North Pacific, Kamtschatka, Bailey.
4. Stylochlamydium æquale, Haeckel.
Perichlamydium æquale, Stöhr, 1880, Palæontogr., vol. xxvi. p. 109, pl. v.
fig. 2.
All rings of the disk concentric, circular, with increasing breadth from the centre; the sixth
ring twice as broad as the second. Pores regular, circular, everywhere of equal size; on the
breadth of the inner rings one, of the outer two, of the girdle three pores. Girdle only
one-fourth as broad as the radius of the disk. About twenty bristle-shaped radial spines,
irregularly disposed, are connected by the girdle near to the points.
Dimensions.—Diameter of the disk (with six rings) 0.11, with the girdle 0.17;
breadth of the second ring 0.006, of the sixth 0.013; pores 0.006.
Habitat.—Fossil in Tertiary rocks of Sicily, Grotte, Stöhr.
Subgenus 2. Stylochlamyum, Haeckel.
Definition.—Rings of the disk all (or in part) not concentric,
spirally convoluted or irregular.
5. Stylochlamydium perispirale, Haeckel.
Perichlamydium limbatum, var. Stöhr, 1880, Palæontogr., vol. xxvi. p. 109,
Taf. v. fig. 1.
Inner rings of the disk concentric, circular, outer rings convoluted spirally, all rings of
equal breadth. Pores regular, circular, two on the breadth of each ring, twice as large as in the
equatorial {516}girdle, which is about half as broad as
the radius of the disk. Twelve to sixteen bristle-shaped radial spines, irregularly disposed, are
connected by the girdle near to the points.
Dimensions.—Diameter of the disk (with six rings) 0.12, with the girdle 0.2;
breadth of each ring 0.011; pores in the central disk 0.004, in the girdle 0.002.
Habitat.—Fossil in Tertiary rocks of Sicily, Grotte, Stöhr.
6. Stylochlamydium spongiosum, Haeckel.
Perichlamydium spongiosum, Stöhr, 1880, Palæontogr., vol. xxvi, p. 109,
Taf. v. fig. 3.
Rings of the disk partly concentric, partly spiral, more or less irregular and often
interrupted, with increasing breadth from the centre. Central part of the disk more or less spongy
and obscure. Equatorial girdle half as broad as the radius of the chambered disk, with smaller
pores than the latter, pierced by twenty to thirty thin, bristle-shaped radial beams, which
proceed over the margin of the disk.
Dimensions.—Diameter of the disk (with ten rings) 0.2, with the girdle 0.3;
breadth of the rings 0.005 to 0.015; pores 0.001 to 0.005.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms; also
fossil in the Tertiary rocks of Barbados and Sicily.
Subfamily 5. Euchitonida, Haeckel.
Definition.—Porodiscida with two or more
(commonly three or four) radial chambered or spongy arms on the margin of the concentrically
annulated disk, situated in its equatorial plane (with or without a connecting patagium between
the arms).
Genus 223. Amphibrachium,[262] Haeckel,
1881, Prodromus, p. 460.
Definition.—Porodiscida with two simple, undivided,
chambered arms, opposite in one axis, without a patagium.
The genus Amphibrachium opens the long series of the Euchitonida, or of
those Porodiscida which bear on the margin of the circular central disk a certain number of
chambered arms, composed of a series of chambers which are separated by transverse septa. The
first group or tribe of this subfamily is formed by the Amphibrachida, in which the disk bears
only two arms opposite on the poles of one axis. The simplest form of these is
Amphibrachium, in which both arms are simple, equal, and without a patagium or spongy
connecticulum.
{517}
Subgenus 1. Amphibrachella, Haeckel.
Definition.—Both arms equal, of the same form and size, blunt at the
distal end, without a terminal spine.
1. Amphibrachium sponguroides, n. sp.
Both opposite arms of the same form and size, nearly cylindrical, three times as long as broad,
with six to eight transverse septa or joints, at the distal end rounded, blunt, without a terminal
spine.
Dimensions.—Radius of each arm 0.22, breadth 0.065.
Habitat.—Antarctic Ocean, Station 154, surface.
2. Amphibrachium lanceolatum, n. sp.
Both arms equal, lanceolate, in the middle part three times as broad as at the two ends, two
and a half times as long as broad, with eight to nine transverse septa, at the distal end blunt,
without a terminal spine.
Dimensions.—Radius of each arm 0.3, greatest breadth 0.09.
Habitat.—Pacific, central area, Station 267, depth 2700 fathoms.
3. Amphibrachium dilatatum, n. sp. (Pl. 44, fig.
6).
Both arms equal, trapezoidal, somewhat broader than long, on the convex distal end three times
as broad as on the narrow base, with five to six transverse septa, without a terminal spine.
Central disk large, somewhat irregular, with three to four rings, twice as broad as the base of
the arms.
Dimensions.—Radius of each arm 0.32, basal breadth 0.15, terminal breadth
0.44.
Habitat.—South Atlantic, off Patagonia, Station 319, surface.
Subgenus 2. Amphibrachidium, Haeckel.
Definition.—Both arms equal, of the same size and form, with terminal
spines.
4. Amphibrachium amphilonche, n. sp.
Both arms equal, lanceolate, in the middle part four times as broad as at the two ends, twice
as long as broad; on the distal end of each arm is a long conical spine.
Dimensions.—Radius of each arm (without terminal spine) 0.2, breadth 0.08.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
{518}
5. Amphibrachium capitatum, n. sp.
Both arms equal, club-shaped, three times as long as broad, in the outer distal half thickened,
three times as broad as at the narrow base; on the distal end of each arm a strong, angular,
terminal spine. (The form of the arms like that of Stephanastrum capitatum, Pl. 44, fig.
1.)
Dimensions.—Radius of each arm 0.22, basal breadth 0.02, terminal breadth
0.06.
Habitat.—Equatorial Atlantic, Station 347, surface.
6. Amphibrachium armatum, n. sp.
Both arms equal, four times as long as broad, in the thickened distal part twice as broad as at
the base, thorny, with twenty to thirty larger spines on the distal end, and a very large
pyramidal spine in the longitudinal axis. (The form of the arms like that of the odd arm in
Euchitonia carcinus, Pl. 43,
fig. 10.)
Dimensions.—Radius of each arm 0.24, basal breadth 0.03, distal breadth 0.06.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
Subgenus 3. Amphibrachoma, Haeckel.
Definition.—Both arms of different size or form, without terminal
spines.
7. Amphibrachium indicum, n. sp.
Both arms club-shaped, but very different in size and form; larger arm three times as long and
twice as broad as the smaller arm; the larger with nine joints slowly increasing in size, the
smaller with four joints, rapidly increasing; the terminal joint three times as broad as the
basal. Distal end blunt, rounded, without spines.
Dimensions.—Radius of the larger arm 0.24, of the smaller 0.08; distal breadth of
the former 0.06, of the latter 0.03; basal breadth 0.015.
Habitat.—Indian Ocean, Ceylon, Haeckel, surface.
Subgenus 4. Amphibrachura, Haeckel.
Definition.—Both arms of different size or form, with terminal
spines.
8. Amphibrachium clavula, n. sp.
Both arms different in size and form; larger arm club-shaped, four times as long as broad, at
the distal end three times as broad as at the base, and twice as long as the smaller arm, which
resembles a stalked knob, with thin basal peduncle and spherical distal part. Ends of the two arms
{519}thorny (with numerous smaller, and three to five
larger spines); one very large conical terminal spine on each pole of the main axis.
Dimensions.—Radius of the larger arm 0.3, of the smaller 0.15; distal breadth of
the former 0.06, of the latter 0.04; basal breath 0.02.
Habitat.—South Atlantic, Station 333, surface.
Genus 224. Amphymenium,[263] Haeckel,
1881, Prodromus, p. 460.
Definition.—Porodiscida with two simple, undivided,
chambered arms, opposite in one axis, connected by a patagium.
The genus Amphymenium differs from the preceding Amphibrachium, its
ancestral form, by development of a patagium or connecticulum between both arms. This forms a
latticed or more spongy envelop, which surrounds either the middle part of the shell, or the whole
shell with exception of the distal ends of both arms. If the envelop become very spongy, the shell
may be confounded with the cylindrical Ellipside Spongocore (nearly allied to
Spongurus); possibly also Ommatogramma of Ehrenberg belongs to this genus.
Subgenus 1. Ommatogramma, Ehrenberg (?).
Definition.—Both opposite arms of the same size and form, blunt,
without terminal spines.
1. Amphymenium pupula, n. sp. (Pl. 44,
fig. 8).
Both arms equal, twice as long as broad, three-jointed; the terminal joint egg-shaped, as large
as both other joints together; distal end rounded, blunt. Patagium nearly complete, enveloping the
arms with exception of the distal end. Perimeter nearly spindle-shaped.
Dimensions.—Radius of each arm 0.17, greatest breadth 0.06; transverse breadth of
the patagium 0.13.
Habitat.—Pacific, central area, Station 273, depth 2350 fathoms.
2. Amphymenium naviculare, Haeckel.
? Ommatogramma navicularis, Ehrenberg, 1872, Monatsber. d. k. preuss.
Akad. d. Wiss. Berlin, p. 317; Abhandl. d. k. Akad. d. Wiss. Berlin, Taf. vi. fig. 7.
Both arms equal, three times as long as broad, spongy, not jointed; distal end a little
club-shaped, blunt. Patagium nearly complete, enveloping the arms with exception of the distal
end. Perimeter nearly lanceolate. The imperfect diagnosis and figure of Ehrenberg make it doubtful
{520}whether this species belongs to the Porodiscida
(Amphymenium) or to the Spongodiscida (Spongobrachium) or perhaps to the Spongurida
(Spongocore).
Dimensions.—Radius of each arm 0.1, greatest breadth 0.03; transverse breadth of
the patagium 0.05.
Habitat.—North Pacific, Californian Sea, depth 2600 fathoms,
Ehrenberg.
3. Amphymenium zygartus, n. sp. (Pl. 44,
fig. 7).
Both arms equal, four times as long as broad, with seven to eight joints, slowly decreasing in
size towards the blunt end. Patagium incomplete, protecting only the middle part of the shell on
both sides; on each side two parallel lattice-plates, connected by transverse radial beams,
perpendicular to the surface. Perimeter nearly rectilinear. (Resembles much certain forms of
Zygartus, Pl. 40,
but is a true Discoid, no Prunoid.) Compare also Pl. 45, fig. 8.
Dimensions.—Radius of each arm 0.2, greatest breadth 0.05; transverse breadth of
the patagium 0.11.
Habitat.—Pacific, central area, Station 271, surface.
4. Amphymenium monstrosum, n. sp. (Pl. 44, fig.
11).
Both arms equal, little longer than broad, with six to seven convex joints. The axis of both
arms is not common and straight, as in all other species of this genus, but broken, therefore the
incomplete patagium, which envelops only two to three joints of the arms, is on one side convex,
on the other side concave; it is formed by a simple lattice-plate, connected with the arms by
numerous radial beams. This anomalous form, seen only once, may perhaps be a monstrosity of
Euchitonia.
Dimensions.—Radius of each arm 0.12, greatest breadth 0.06; transverse breadth of
the patagium 0.13.
Habitat.—South Pacific, Station 290, surface.
Subgenus 2. Ommathymenium, Haeckel.
Definition.—Both opposite arms of the same size and form, armed at the
distal end with terminal spines.
5. Amphymenium amphistylium, n. sp. (Pl. 44, fig.
9).
Both arms equal, three times as long as broad, thickened towards the truncated distal end, and
armed with a strong pyramidal terminal spine. Each arm with seven joints, separated by convex,
transverse septa, and halved by a radial beam lying in the longitudinal axis. Patagium incomplete,
cylindrical, enveloping only the middle part of the shell.
Dimensions.—Radius of each arm 0.18, greatest breadth 0.06; transverse breadth of
the patagium 0.08.
Habitat.—North Atlantic, Station 354, surface.
{521}
6. Amphymenium fusiforme, n. sp.
Both arms equal, lanceolate, three times as long as broad in the width, with seven to eight
joints. Distal end pointed, armed with a strong conical terminal spine. Patagium complete,
enveloping the whole shell with exception of the terminal spines. Whole form spindle-shaped.
Dimensions.—Radius of each arm 0.2, greatest breadth 0.07; transverse breadth of
the patagium 0.15.
Habitat.—South Atlantic, Station 330, surface.
Genus 225. Amphirrhopalum,[264] Haeckel,
1881, Prodromus, p. 460.
Definition.—Porodiscida with two chambered arms, opposite
in one axis, without a patagium; one arm or both forked at the distal end.
The genus Amphirrhopalum differs from Amphibrachium, its ancestral
form, by bifurcation of the distal ends of the arms, which may affect either both arms, or only
one of them.
Subgenus 1. Amphirrhopalium, Haeckel.
Definition.—Both opposite arms of similar size and form, with blunt
branches, without terminal spines.
1. Amphirrhopalum ximorphum, n. sp.
Both arms equal, in the proximal half simple, in the distal half forked, with six to seven
transverse septa; distal end of each branch blunt, without terminal spine, somewhat broader than
the base of the whole arm. Axis of the branches concavely curved. (Resembles Amphicraspedum
maclaganium, Pl. 45,
fig. 11, but wants the patagium.)
Dimensions.—Radius of the arms 0.18, basal breadth 0.065; terminal breadth of each
branch 0.075.
Habitat.—North Pacific, off Japan, Station 240, surface.
2. Amphirrhopalum amphidicranum, n. sp.
Both arms equal, in the proximal half simple, in the distal half forked, with irregular septa;
distal end of each arm blunt, without a terminal spine, smaller than the basal breadth of the arm.
Axis of the branches straight. (Resembles Dicranastrum furcatum, Pl. 47, fig. 2, but
without lateral arms.)
Dimensions.—Radius of each arm 0.15, basal breadth 0.03; terminal breadth of each
branch 0.02.
Habitat.—Pacific, central area, Station 263, depth 2650 fathoms.
{522}
Subgenus 2. Amphirrhopella, Haeckel.
Definition.—Both opposite arms of similar size and form, with terminal
spines.
3. Amphirrhopalum bigeminum, n. sp.
Both arms equal, in the proximal larger half simple, in the distal smaller half forked; each
branch triangular, with a strong conical terminal spine. Axis of the branches straight. (Resembles
Dicranastrum cornutum, Pl. 45,
fig. 2, but without lateral arms.)
Dimensions.—Radius of the arms (without spines) 0.15, basal breadth 0.03; breadth
of the bifurcation 0.08.
Habitat.—South Pacific, Station 284, surface.
4. Amphirrhopalum echinatum, n. sp. (Pl. 45, fig.
10).
Both arms equal, in the proximal smaller half simple, nearly square, in the distal larger half
forked; the branches thorny, armed at the end with numerous spines, one larger on the terminal
pole of the concavely curved arm-axis.
Dimensions.—Radius of the arms (without spines) 0.15, basal breadth 0.05; breadth
of branches 0.03.
Habitat.—Pacific, central area, Station 266, depth 2750 fathoms.
Subgenus 3. Amphirrhopoma, Haeckel.
Definition.—Both opposite arms of different size or form, without
terminal spines.
5. Amphirrhopalum ypsilon, n. sp.
Both arms very different. Larger arm simple, egg-shaped, twice as long as broad; smaller arm in
the basal half simple, nearly square, in the distal half forked; both branches egg-shaped, blunt.
(Resembles Amphicraspedum wyvilleanum, Pl. 45, fig. 12,
but wants the patagium.)
Dimensions.—Radius of the larger simple arm 0.18, breadth 0.09; radius of the
smaller forked arm 0.15, breadth of its branches 0.05.
Habitat.—Indian Ocean, Madagascar, Rabbe, surface.
Genus 226. Amphicraspedum,[265] Haeckel,
1881, Prodromus, p. 460.
Definition.—Porodiscida with two chambered arms, opposite
in one axis, connected by a lateral patagium; one arm or both forked at the distal end.
The genus Amphicraspedum exhibits the same bifurcation of the arms as does
Amphirrhopalum, but differs from this ancestral form in the development of a patagium, an
external connecticulum between the arms, which envelops the disk totally or partially.
{523}
Subgenus 1. Amphicraspedon, Haeckel.
Definition.—Both arms of equal size and form, without terminal spines
of the branches.
1. Amphicraspedum maclaganium, n. sp. (Pl. 45, fig.
11).
Both arms equal, in the proximal half simple, in the distal half forked, with six to seven
transverse septa; distal end of each branch rounded, blunt, somewhat broader than the base of the
whole arm. Divergent axes of both branches concavely curved. Patagium incomplete, with elliptical
perimeter, enveloping only the middle part of the shell. I call this interesting species in honour
of Miss Nellie Maclagan, the learned translator of several zoological papers from German into
English.
Dimensions.—Radius of each arm 0.25, basal breadth 0.07; distal breadth of each
branch 0.08; equatorial breadth of the patagium 0.25.
Habitat.—North Atlantic, off Halifax, Station 50, surface.
Subgenus 2. Amphicraspedina, Haeckel.
Definition.—Both arms of different size or form, without terminal
spines on the branches.
2. Amphicraspedum wyvilleanum, n. sp. (Pl. 45, fig.
12).
Both arms different. Larger arm simple, egg-shaped, with eleven convex joints, one and a half
times as long as broad; smaller arm in the basal half simple, triangular, with six cap-like
joints, in the distal half forked; both branches egg-shaped, with five joints and blunt ends.
Patagium nearly complete with four to five concave chamber-rows. Called in honour of Sir C.
Wyville Thomson.
Dimensions.—Radius of the larger simple arm 0.18, breadth 0.08; radius of the
smaller forked arm 0.16; breadth of the branches 0.05; transverse breadth of the patagium 0.2.
Habitat.—South Atlantic, Station 333, surface.
Subgenus 3. Amphicraspedula, Haeckel.
Definition.—Both arms of different size or form, with terminal spines
of the branches.
3. Amphicraspedum murrayanum, n. sp. (Pl. 44, fig.
10).
Amphymenium murrayanum, Haeckel, 1879, MS. et Atlas (pl. xliv. fig.
10).
Both arms different in size; the larger one and a half times as long and broad as the smaller.
Both arms triangular, forked at the broader distal end, with two very strong, conical, divergent,
{524}straight terminal spines. Patagium broad,
incomplete, with circular perimeter. Called in honour of my friend Dr. John Murray.
Dimensions.—Radius of the larger arm (including the spines) 0.24, of the smaller
0.16; distance of the terminal points of the former 0.18, of the latter 0.09; diameter of the
patagium 0.2.
Habitat.—North Atlantic, Færöe Channel, Gulf Stream, surface, John
Murray.
Genus 227. Dictyastrum,[266] Ehrenberg,
1860, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 830.
Definition.—Porodiscida with three simple, undivided,
chambered arms, without a patagium; triangular shell regular, with three equal arms and three
equal angles.
The genus Dictyastrum is the simplest form of the Trigonastrida, or of the
Porodiscida, in which the margin of the central disk is furnished with three chambered arms. In
Dictyastrum these are quite simple and regular, without a patagium, separated by equal
angles, so that the whole shell represents a regular, equilateral triangle, if we connect the
distal points of the arms by lines. The genus Dictyastrum, founded by Ehrenberg in 1860,
differs from his Rhopalodictyum—after his own diagnosis—only by an
insignificant difference in the form of the simple arms, which is scarcely a specific character. I
therefore apply this name here in the above amended sense, seeing that the only figured species of
Ehrenberg (Dictyastrum angulatum) occurs in two different, but externally very similar
forms: one of these is a true Porodiscid (Dictyastrum) with two porous covering-plates and
concentric rings; the other is a true Spongodiscid (Rhopalodictyum) with quite spongy,
irregular network, and is probably identical with the Rhopalodictyum truncatum of
Ehrenberg.
Subgenus 1. Dictyastrella, Haeckel.
Definition.—Arms with blunt ends, without terminal spines.
1. Dictyastrum angulatum, Ehrenberg.
Dictyastrum angulatum, Ehrenberg, 1872, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 289, Taf. viii. fig. 18.
Arms nearly square with straight edges, towards the truncated end a little broader, about the
same diameter as the triangular central disk. The figure of Ehrenberg seems to represent a
Spongodiscid (Rhopalodictyum angulatum), but in the same locality (Philippine Sea) occurs
also a true Dictyastrum of quite the same form, but with three to four concentric rings of
the central disk, and with jointed arms.
{525}
Dimensions.—Radius of each arm (length from the centre to the distal end) 0.13;
breadth of the truncated end 0.1.
Habitat.—Tropical Pacific, Philippine Sea, Station 200, depth 250
fathoms.
2. Dictyastrum bandaicum, Haeckel.
Rhopalastrum bandaicum, Harting, 1863, Mikr. Fauna Banda-Zee, p. 16, Taf.
iii. fig. 45.
Arms nearly square, with convex edges, in the middle a little broader than at both ends, about
half the diameter of the central disk. Differs from the nearly allied preceding species by the
half size of the arms and the convex edges.
Dimensions.—Radius of each arm 0.12, its greatest breadth 0.07.
Habitat.—Tropical Pacific, Banda Sea, Harting.
3. Dictyastrum hexagonum, n. sp. (Pl. 43, fig.
7).
Rhopalastrum hexagonum, Haeckel, 1880, Atlas (pl. xliii. fig. 1).
Arms nearly triangular, one and a third times as broad at the distal end as long, and three
times as broad as at the base. Central disk about the same diameter. In each arm six simple broad
chambers. If we connect the six corners of the truncated distal ends by straight lines, we get a
regular hexagon.
Dimensions.—Radius of each arm 0.17, basal breadth 0.06, terminal breadth
0.17.
Habitat.—Equatorial Atlantic, Station 347, surface.
4. Dictyastrum trirrhopalum, n. sp.
Arms club-shaped, five times as long as broad at the base, at the thickened end three times as
broad as at the base. Diameter of the central disk equals half the length of the arms. (Similar to
Rhopalastrum malleus, Pl. 43,
fig. 1, but with three equal angles and much smaller disk.)
Dimensions.—Radius of each arm 0.25, basal breadth 0.04, distal breadth 0.12.
Habitat.—Pacific, central area, Station 273, surface.
Subgenus 2. Dictyastromma, Haeckel.
Definition.—Arms on the distal end provided with terminal spines.
5. Dictyastrum trispinosum, n. sp. (Pl. 43, fig.
5).
Rhopalastrum trispinosum, Haeckel, 1881, Prodromus et Atlas (pl. xliii.
fig. 5).
Arms trapezoid, at the rounded distal end twice as broad as at the base, with a strong and
short, conical, terminal spine. Diameter of the circular central disk about equal to the length
and the greatest breadth of the arms.
Dimensions.—Radius of each arm 0.15, basal breadth 0.06, distal breadth 0.11.
Habitat.—Equatorial Atlantic, Station 347, surface.
{526}
6. Dictyastrum triactis, Ehrenberg.
Dictyastrum triactis, Ehrenberg, 1872, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 306.
Arms rectilinear, four times as long as broad, with parallel edges, pointed at the distal end,
with a short terminal spine. Diameter of the circular central disk equal to double the breadth of
the arms.
Dimensions.—Radius of each arm 0.2, breadth 0.04.
Habitat.—Pacific, Philippine Sea, Station 206, depth 2100 fathoms.
7. Dictyastrum aculeatum, n. sp.
Arms lanceolate, three times as long as broad, twice as broad in the middle as at either end,
with thorny surface and numerous conical terminal spines, one very large in the radius. Central
disk triangular, about as broad as the arms. (Resembles Rhopalastrum arcticum, Pl. 43, fig. 6, but
differs by the equal angles and the triangular disk.)
Dimensions.—Radius of each arm 0.2, breadth 0.06.
Habitat.—Western Tropical Pacific, Station 225, depth 4475
fathoms.
Genus 228. Rhopalastrum,[267] Ehrenberg,
1847, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 54.
Definition.—Porodiscida with three simple undivided,
chambered arms, without a patagium; triangular shell bilateral, one odd arm opposite to the odd
angle between two paired arms.
The genus Rhopalastrum, founded by Ehrenberg (1847) with a very insufficient
diagnosis, is here retained for those Trigonastrida that agree in the generic characters with the
only species figured by him, viz., Rhopalastrum lagenosum (compare my Monograph, 1862, p.
500). It comprises, therefore, such Euchitonida as agree with the preceding Dictyastrum in
the simple form of the three arms and the absence of a patagium, but differ from it in the
different size of the three angles, and often also in the divergent form and size of the three
arms; one odd arm is opposite to the odd angle between the two paired arms.
Subgenus 1. Rhopalastrella, Haeckel.
Definition.—Arms with blunt ends, without terminal spines.
1. Rhopalastrum truncatum, Haeckel.
Rhopalastrum truncatum, Haeckel, 1862, Monogr. d. Radiol., p. 500, Taf.
xxix. fig. 6.
Distance of both paired arms about half as large as their distance from the odd arm. All three
arms nearly of the same form and size, very short and broad; their breadth nearly equals that of
{527}the roundish central disk, whilst their length
reaches only one-fourth of it. End of the arms convex rounded, without spines.
Dimensions.—Radius of each arm 0.1, breadth 0.15.
Habitat.—Mediterranean (Messina), Atlantic (Canary Islands), surface,
Haeckel.
2. Rhopalastrum pistillum, Stöhr.
Rhopalastrum pistillum, Stöhr, 1880, Palæontogr., vol. xxvi. p. 110, Taf.
v. fig. 4.
Distance between the paired arms about two-thirds as large as their distance from the odd arm.
All three arms nearly of the same form and size, about three times as long as the diameter of the
central disk, at the base one-third as broad as at the convex rounded end, without spines. Stöhr
has only observed a fragment with one arm; some perfect specimens, which I found in the
Caltanisetta-rock, exhibited nearly the same form as Rhopalastrum malleus (Pl. 43, fig. 1),
but differ from this by the smaller disk, the broader arms, and the smaller angle between the
paired arms.
Dimensions.—Radius of all three arms 0.2; basal breadth of each arm 0.045,
terminal breadth 0.13.
Habitat.—Fossil in Tertiary rocks of Sicily, Grotte (Stöhr),
Caltanisetta (Haeckel).
3. Rhopalastrum malleus, n. sp. (Pl. 43,
fig. 1).
Distance between the paired arms one and a third times as large as their distance from the odd
arm. All three arms nearly of the same form and size, hammer-shaped, three times as broad at the
truncated distal end as at the base. Central disk broader than the arms.
Dimensions.—Radius of each arm 0.25, basal breadth 0.05, distal breadth 0.15.
Habitat.—South Atlantic, Station 325, surface.
4. Rhopalastrum lagenosum, Ehrenberg.
Rhopalastrum lagenosum, Ehrenberg, 1847, Monatsber. d. k. preuss. Akad. d. Wiss.
Berlin, p. 43; Mikrogeol., 1854, Taf. xxii. fig. 22.
Rhopalastrum lagenosum, Haeckel, 1862, Monogr. d. Radiol., p. 501.
Flustrella bilobata, Ehrenberg, 1844, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin,
p. 81.
Haliomma lagena, Ehrenberg, 1840, Monatsber. d. k. preuss. Akad. d. Wiss.
Berlin, p. 200.
Distance between the paired arms one and a half times as great as their distance from the odd
arm. All three arms nearly of the same form and size, about one and a half times as long as the
diameter of the central disk, at the base half as broad as at the convex rounded end, without
spines. Ehrenberg has only observed a fragment with two paired arms; some perfect specimens with
three arms, observed by me, differed from the nearly allied Rhopalastrum pistillum (from
the same locality) by the larger disk, the broader arms, and the larger unpaired angle.
Dimensions.—Radius of all three arms 0.2, basal breadth 0.05, terminal breadth
0.09.
Habitat.—Fossil in Tertiary rocks of Sicily, Caltanisetta.
{528}
5. Rhopalastrum ypsilinum, n. sp. (Pl. 43, fig.
2).
Distance between the paired arms half as large as their distance from the odd arm. All three
arms at the concavely curved distal end three times as broad as at the narrow base. Odd arm twice
as long and broad as the paired arms. Central disk smaller than the latter. No terminal
spines.
Dimensions.—Radius of the odd arm 0.32, of the paired arms 0.2; terminal breadth
of the former 0.2, of the latter 0.12.
Habitat.—Indian Ocean, Cocos Islands, Rabbe, surface.
6. Rhopalastrum clavatum, n. sp.
Distance between the paired arms half as large as their distance from the odd arm. All three
arms club-shaped, at the thickened, nearly spherical, distal end three times as broad as at the
narrow base. Odd arm nearly twice as long and broad as the paired arms. Central disk equal to the
distal knob of the latter. No terminal spines.
Dimensions.—Radius of the odd arm 0.3, of the paired arms 0.18; distal breadth of
the former 0.16, of the latter 0.08.
Habitat.—South Atlantic, Station 332, surface.
7. Rhopalastrum irregulare, n. sp. (Pl. 43, fig.
8).
Distance between all three arm-points different. All three arms cylindrical, nearly of the same
length, but of different form, irregularly curved, about four times as long as broad, with blunt
ends.
Dimensions.—Length of each arm about 0.2, breadth 0.05.
Habitat.—Antarctic Sea, Station 157, depth 1950 fathoms.
Subgenus 2. Rhopalastromma, Haeckel.
Definition.—Arms on the distal end provided with one or more terminal
spines.
8. Rhopalastrum martellum, n. sp.
Distance between all three arm-points nearly the same; but the odd arm is one and a half times
as large as both paired arms, and is perpendicular to the common axis of the latter, therefore the
shell has the form of a hammer. Each arm is twice as broad at the distal end as at the base, and
armed with a conical terminal spine; the latter is vertical in the odd arm, horizontal in the
paired arms.
Dimensions.—Radius of the odd arm 0.3, of the paired arms 0.15; distal breadth of
the former 0.12, of the latter 0.06.
Habitat.—North Atlantic, Station 354, surface.
{529}
9. Rhopalastrum triceros, n. sp. (Pl. 43, fig.
4).
Distance between the paired arms about two-thirds as large as their distance from the odd arm.
All three arms club-shaped, three times as broad at the thickened distal part as at the base, and
armed with one single, conical, terminal spine. Odd arm of the same breadth, but twice as long as
the paired arms.
Dimensions.—Radius of the odd arm 0.35, of the paired arms 0.2; basal breadth
0.04, distal breadth 0.12.
Habitat.—Pacific, central area, Station 274, surface.
10. Rhopalastrum hexaceros, n. sp. (Pl. 43, fig.
3).
Distance between the paired arms equals four-fifths of their distance from the odd arm. All
three arms nearly of the same size, about square, a little broader at the truncated distal end,
which is armed at both corners with a strong, conical, radial spine.
Dimensions.—Radius of each arm 0.2, basal breadth 0.1, distal breadth 0.12.
Habitat.—Indian Ocean, Ceylon, Belligemma, Haeckel, surface.
11. Rhopalastrum arcticum, n. sp. (Pl. 43, fig.
6).
Distance between the paired arms half as large as their distance from the odd arm, which is a
little larger. All three arms of the same form, lanceolate, twice to three times as long as broad,
twice as broad in the middle as at either end. Each arm with twelve to fourteen transverse septa,
at the distal end with a bunch of conical spines, and one single, very large, pyramidal, terminal
spine.
Dimensions.—Radius of each arm (without spine) 0.17, greatest breadth of it 0.05
to 0.06.
Habitat.—Arctic Ocean, lat. 83° 19' N., North Polar expedition of the
"Alert."
Genus 229. Hymeniastrum,[268] Ehrenberg,
1847, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 54.
Definition.—Porodiscida with three simple, undivided,
chambered arms, connected by a patagium; triangular shell regular, with three equal arms and three
equal angles.
The genus Hymeniastrum was founded by Ehrenberg (1847) with a very
incomplete diagnosis, and hitherto known only by one single species, figured by him as
Hymeniastrum pythagoræ (Mikrogeol., 1854, Taf. xxxvi. fig. 31). This form occurs in two
different states, externally quite identical; in one state the central disk (as figured, loc.
cit.), is a simple lens or hollow disk, containing a medullary shell or "central chamber"; in
the other state the central disk is composed of two concentric rings surrounding the "central
chamber." We retain here the name Hymeniastrum for this latter state, {530}expressed in the diagnosis given above, and call the former state (the
Coccodiscid) Hymenactura (compare above, p. 473). One practical advantage, obtained in this
way, is that all genera of Discoidea ending with "-astrum" belong
to the Porodiscida. Hymeniastrum differs from Dictyastrum by the possession of a
patagium, and from Euchitonia by the equal size of the angles and the arms.
Subgenus 1. Hymenastrella, Haeckel.
Definition.—Arms with blunt ends, without terminal spines.
1. Hymeniastrum leydigii, Haeckel.
Euchitonia leydigii, Haeckel, 1862, Monogr. d. Radiol., p. 510, Taf. xxxi. figs. 4,
5.
Hymeniastrum leydigii, Haeckel, 1881, Prodromus, p. 460.
Histiastrum trinacrium, Haeckel, 1860, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 843.
Arms twice as long as broad, two-thirds as broad at the base as at the blunt, nearly truncated
distal end. Patagium nearly complete, perfectly filling out the interbrachial spaces, with six to
seven regular, concave chamber-rows; only the truncate terminal faces of the arms free.
Dimensions.—Radius of the arms 0.15 to 0.18, breadth on their base 0.04 to 0.05,
on their broadest distal end 0.06 to 0.07.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific, surface. Very
common and variable.
2. Hymeniastrum köllikeri, Haeckel.
Euchitonia köllikeri, Haeckel, 1862, Monogr. d. Radiol., p. 511, Taf. xxxi. figs. 6,
7.
Hymeniastrum köllikeri, Haeckel, 1881, Prodromus, p. 460.
Arms nearly lanceolate, twice as long as broad, half as broad at the base as at the broadest
middle part, pointed at the distal end. Patagium complete, enveloping the whole triangular disk,
with irregular chamber-rows.
Dimensions.—Radius of the arms 0.2, breadth on their base 0.04, on the broadest
part 0.08.
Habitat.—Mediterranean (Messina), surface.
3. Hymeniastrum gümbelii, Haeckel.
Stylactis gümbelii, Stöhr, 1880, Palæontogr., vol. xxvi. p. 112, Taf. vi.
fig. 1.
Arms twice as long as broad, nearly twice as broad in the circular distal half as in the square
proximal half. Patagium incomplete, circular, with eight to nine convex chamber-rows, enveloping
only the proximal square halves of the arms; the lenticular distal halves remain free.
Dimensions.—Radius of the arms 0.14; breadth at the base 0.04, at the distal
lenticular part 0.07.
Habitat.—Fossil in the Tertiary rocks of Sicily, Grotte, Stöhr.
{531}
4. Hymeniastrum euclidis, n. sp. (Pl. 43, fig.
13).
Euchitonia euclidis, Haeckel, 1881, Prodromus, p. 460 et Atlas (pl. xliii.
fig. 13).
Arms one and a half times as long as broad, club-shaped, three times as broad in the oval
distal part as in the narrow square proximal part. Patagium complete, enveloping the whole
triangular disk, and also the convex ends of the arms, with eight to nine convex chamber-rows.
Dimensions.—Radius of the arms 0.18; breadth at the base 0.025, at the broadest
distal part 0.07.
Habitat.—Pacific, central area, Stations 270 to 274, surface.
5. Hymeniastrum pythagoræ, Ehrenberg.
Hymeniastrum pythagoræ, Ehrenberg, 1854 (partim), Mikrogeol., Taf.
xxxvi. fig. 31.
Arms nearly as broad as long, two-thirds as broad at the base as at the truncated distal end.
Patagium incomplete, enveloping only the basal half of the arms, with four to five rectilinear
parallel chamber-rows. (This form has the greatest resemblance to the figure given by Ehrenberg,
loc. cit., but differs by the central disk, which is composed of two concentric rings
surrounding the small central chamber; compare above Hymenactura pythagoræ, p. 474.)
Dimensions.—Radius of the arms 0.18; breadth at the base 0.08, at the truncated
end 0.12.
Habitat.—Equatorial Atlantic, Station 347, depth 2250 fathoms.
6. Hymeniastrum archimedis, n. sp.
Arms nearly triangular, at the truncated, slightly convex end three times as broad as at the
narrow base and one and a third times as broad as long; each arm with six simple chambers.
Patagium incomplete, enveloping only the basal half of the arms with three to four convex
chamber-rows. (Differs from Rhopalastrum hexagonum, Pl. 43, fig. 7,
only by the patagium.)
Dimensions.—Radius of the arms 0.2; breadth at the base 0.07, at the truncated end
0.2.
Habitat.—Pacific, central area, Station 274, surface.
Subgenus 2. Hymenastromma, Haeckel.
Definition.—Arms at the distal end provided with radial spines.
7. Hymeniastrum ternarium, Haeckel.
Histiastrum ternarium, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 76, Taf. xxiv. fig. 2.
Arms two and a half times as long as broad, three times as broad at the truncated end as at the
narrow base, with a strong, conical, radial spine at the end. Patagium incomplete, enveloping
only the basal half of the arms, with three to four convex chamber-rows.
Dimensions.—Radius of the arms (without terminal spine) 0.02; breadth at the base
0.025, at the broadest terminal part 0.08.
Habitat.—Fossil in the rocks of Barbados.
{532}
8. Hymeniastrum trigonarium, n. sp.
Arms one and a half times as long as broad, a little broader at the rounded end than at the
base, with three strong conical radial spines, one larger (in the radius of each arm) and two
smaller on both sides of this. Patagium complete, with four to five rectilinear parallel
chamber-rows, enveloping the whole arms (with exception of the terminal spines) and forming a
perfect equilateral triangle.
Dimensions.—Radius of the arms (without terminal spines) 0.18; breadth at the base
0.1, at the distal part 0.12.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
Genus 230. Euchitonia,[269]
Ehrenberg, 1860, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 831 (sensu
emendato).
Definition.—Porodiscida with three simple, undivided,
chambered arms, connected by a patagium; triangular shell bilateral, one odd arm opposite to the
odd angle between the two paired arms.
The genus Euchitonia, quite insufficiently characterised by Ehrenberg, was
founded by him (1860) for one single species, afterwards (1872) described and figured as
Euchitonia furcata. Retaining this species correctly as the type of this genus, I give to
it here the above diagnosis. In my Monograph (1862, p. 503) I described seven Mediterranean
species of Euchitonia. Three of these have in common the characters according to the
present diagnosis: Euchitonia mülleri, Euchitonia virchowii, Euchitonia
beckmanni; two others appertain (on account of the regular, not bilateral form) to
Hymeniastrum, and two others (on account of the forked, not simple arms) to
Trigonastrum. Afterwards (1880) three true fossil species of Euchitonia were
described by Stöhr (Euchitonia cruciata, Euchitonia zittelii, and Euchitonia
acuta). Some species of this genus are cosmopolitan, and appertain to the most common and
everywhere represented Discoidea.
Subgenus 1. Stylactis, Ehrenberg, 1872 (loc. cit.).
Definition.—Arms with blunt ends, without terminal spines.
1. Euchitonia furcata, Ehrenberg.
Euchitonia furcata, Ehrenberg, 1872, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 308; Abhandl. d. k. Akad. d. Wiss. Berlin, Taf. vi. iii. fig. 6.
Distance between the paired arms about half as large as their distance from the odd arm. All
three arms nearly of the same size and form, about twice as long as broad, one and a half times at
the {533}blunt convex end as broad as at the base.
Patagium incomplete, with concave chamber-rows and irregular network, only enveloping the basal
half of the arms.
Dimensions.—Radius of all three arms 0.15, basal breadth of each arm 0.04,
terminal breadth 0.06.
Habitat.—North Pacific, Californian Sea, depth 2600 fathoms,
Ehrenberg.
2. Euchitonia mülleri, Haeckel.
Euchitonia mülleri, Haeckel, 1862, Monogr. d. Radiol., p. 508, Taf. xxx. figs.
5-10.
Euchitonia mülleri, Stöhr, 1880, Palæontogr., vol. xxvi. p. 110, Taf. v.
fig. 5.
Distance between the paired arms about two-thirds as large as their distance from the odd arm,
which is somewhat larger. Length of the arms equals two and a half times the breadth of the blunt
convex end or five times the breadth of the base. Patagium with concave chamber-rows, nearly
complete, enveloping the arms with exception of the terminal face. (This common species is very
variable; compare my Monograph.)
Dimensions.—Radius of all three arms 0.16 to 0.22, basal breadth 0.03 to 0.05,
terminal breadth 0.06 to 0.08.
Habitat.—Cosmopolitan; one of the most common Discoidea in all seas, on the surface as well as at different depths; also
fossil in the Tertiary rocks of Barbados and Sicily.
3. Euchitonia triangulum, Haeckel.
Stylactis triangulum, Ehrenberg, 1872, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin,
p. 320; Abhandl. d. k. Akad. d. Wiss. Berlin, Taf. viii. fig. 9.
Stylactis triangulum, Stöhr, 1880, Palæontogr., vol. xxvi. p. 113, Taf.
vi. fig. 2.
Distance between the paired arms about two-thirds as large as their distance from the odd arm.
All three arms nearly of the same size and form, about one and a half times as long as broad,
nearly as broad at the base as at the blunt rounded end. Patagium incomplete, with convex
chamber-rows, enveloping about two-thirds of the arms.
Dimensions.—Radius of all three arms 0.15, basal breadth 0.05, terminal breadth
0.06.
Habitat.—Pacific and Atlantic, Stations 253, 272, 354, surface; also
fossil in the Tertiary rocks of Barbados and Sicily.
4. Euchitonia cruciata, Stöhr.
Euchitonia cruciata, Stöhr, 1880, Palæontogr., vol. xxvi. p. 111, Taf. v.
fig. 7.
Distance between the paired arms about one and a half times as large as their distance from the
odd arm, which is a little larger. Length of the arm nearly equals twice the breadth, which is the
same at the base and at the rounded blunt ends. Patagium incomplete, with concave chamber-rows,
enveloping only the base of the arms, and forms between them three other smaller arms; therefore
the whole shell forms six angles with six alternating arms.
Dimensions.—Radius of all three arms 0.15, basal breadth 0.05.
Habitat.—Fossil in Tertiary rocks of Sicily, Grotte, Stöhr.
{534}
5. Euchitonia lanceolata, n. sp. (Pl. 43, fig.
9).
Distance between the paired arms about one and a third times as large as their distance from
the odd arm. All three arms nearly of the same size and form, three times as long as broad,
lanceolate, much broader in the middle part than at both ends; distal end cuspidate, but not
spiny. Patagium incomplete, with convex chamber-rows, envelops about two-thirds of the arms.
Dimensions.—Radius of all three arms 0.18, greatest breadth (in the width)
0.06.
Habitat.—North Atlantic, Station 353, depth 2965 fathoms.
6. Euchitonia zittelii, Haeckel.
Stylactis zittelii, Stöhr, 1880, Palæontogr., vol. xxvi. p. 112, Taf. v.
fig. 8.
Distance between the paired arms about one-fifth as large as their distance from the odd arm.
All three arms nearly of the same size and form, in the proximal half thinner and nearly square,
in the distal half thicker and circular; the latter half twice to three times as broad as the
former. Patagium incomplete, with convex chamber-rows, envelops only the proximal square half of
the arms.
Dimensions.—Radius of all three arms 0.15, basal breadth 0.03, terminal breadth
0.08.
Habitat.—Fossil in Tertiary rocks of Sicily, Grotte, Stöhr.
7. Euchitonia stöhrii, n. sp. (Pl. 43,
fig. 12).
Distance between the paired arms about four-fifths as large as their distance from the odd arm,
which is one and a half times as long as the former. Arms about three times as long as broad,
twice as broad in the distal half as in the proximal half, with rounded blunt ends. Patagium
complete, with concave chamber-rows, enveloping the whole shell, also the ends of the arms.
Dimensions.—Radius of the paired arms 0.2, of the odd arm 0.25; basal breadth
0.03, distal breadth 0.06.
Habitat.—Fossil in the rocks of Barbados and of Nicobar Islands,
Haeckel. Indian Ocean, Cocos Islands (Rabbe).
Also in the Central Pacific, Station 267.
8. Euchitonia beckmannii, Haeckel.
Euchitonia beckmannii, Haeckel, 1862, Monogr. d. Radiol., p. 505, Taf.
xxxi. fig. 1.
Distance between the paired arms scarcely half as large as their distance from the odd arm,
which is somewhat larger. Each arm with six simple broad chambers (without radial septa), the
terminal chambers semilunar, convex, blunt, four times as broad as the first (basal) chamber.
Patagium incomplete, with convex chamber-rows, enveloping the arms with exception of the broad
blunt terminal face.
Dimensions.—Radius of the paired arms 0.15, of the odd arm 0.18; basal breadth
0.02 to 0.03, terminal breadth 0.1 to 0.15.
Habitat.—Mediterranean (Messina), Haeckel, surface.
{535}
9. Euchitonia virchowii, Haeckel.
Euchitonia virchowii, Haeckel, 1862, Monogr. d. Radiol., p. 503, Taf. xxx. figs.
1-4.
Histiastrum fasciatum, Haeckel, 1860, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 842.
Distance between the paired arms about half as large as their distance from the odd arm, which
is somewhat larger. Each arm with six broad chambers, bisected by a radial septum; the terminal
chamber convex, blunt, twice as broad as the basal chamber. Patagium incomplete, with concave
chamber-rows, enveloping the arms with exception of the broad blunt terminal face.
Dimensions.—Radius of the paired arms 0.15, of the odd arm 0.16; basal breadth
0.05, terminal breadth 0.1 to 0.12.
Habitat.—Mediterranean (Messina), Atlantic (Canary Islands).
Subgenus 2. Pteractis, Ehrenberg, 1872 (loc. cit.).
Definition.—Arms provided with radial spines at the distal end.
10. Euchitonia elegans, Haeckel.
Pteractis elegans, Ehrenberg, 1872, Abhandl. d. k. Akad. d. Wiss. Berlin,
p. 299, Taf. viii. fig. 3.
Distance between the paired arms half as large as their distance from the odd arm. This latter
is straight, while both the former are concavely curved towards the middle line. Arms five times
as long as broad, at the distal end pointed and armed with a short conical terminal spine.
Patagium nearly complete, enveloping four-fifths of the arms, with four to five concave
chamber-rows.
Dimensions.—Radius of the arms 0.2, breadth of them 0.03.
Habitat.—Tropical Pacific, Philippine Sea, depth 3300 fathoms
(Ehrenberg).
11. Euchitonia carcinus, n. sp. (Pl. 43,
fig. 10).
Distance between the paired arms scarcely one-fourth as great as their distance from the odd
arm. This latter is straight, twice as long as the former, which are concavely curved towards the
middle line. The odd arm is three times as broad at the distal end as at the narrow base. The end
of each arm is furnished with a strong triangular radial spine and a group of smaller spines.
Patagium incomplete, with two to three concave chamber-rows, enveloping only the basal half of the
arms.
Dimensions.—Radius of the paired arms 0.15, breadth 0.03; radius of the odd arm
0.3, breadth on its base 0.02, on its distal end 0.07.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
{536}
12. Euchitonia acuta, Stöhr.
Euchitonia acuta, Stöhr, 1880, Palæontogr., vol. xxvi. p. 111, Taf. v.
fig. 6.
Distance between the paired arms two-thirds as great as their distance from the odd arm. All
three arms nearly of the same size and form, two and a half times as long as broad, nearly
lanceolate, at their broadest part one and a half times as broad as at their base. Patagium
complete, enveloping the whole triangular disk, with five to six concave chamber-rows. In the
figure of Stöhr the ends of the arms are simply pointed, while I find in the same fossil form a
short terminal conical spine.
Dimensions.—Radius of all three arms 0.14; breadth at the base 0.04, at the
broadest part 0.06.
Habitat.—Fossil in Tertiary rocks of Sicily, Grotte (Stöhr),
Caltanisetta (Haeckel).
13. Euchitonia ypsiloides, Haeckel.
Histiastrum ypsiloides, Haeckel, 1860, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 843.
Distance between the paired arms two-thirds as great as their distance from the odd arm, which
is somewhat larger. Length of the arms equals five times the breadth of the narrow base, which is
half that of the distal end; this latter is armed with three to five short conical spines.
Patagium complete, with six to seven concave chamber-rows, enveloping the whole arms with the
exception of the terminal spines. (Differs from Euchitonia mülleri almost solely by the
possession of terminal spines.)
Dimensions.—Radius of the arms 0.18 to 0.2; breadth at the base 0.04, at the
distal end 0.08.
Habitat.—Mediterranean (Messina), Atlantic (Canary Islands),
surface.
14. Euchitonia echinata, n. sp. (Pl. 43,
fig. 11).
Distance between paired arms three-fourths as great as their distance from the odd arm, which
is somewhat larger. Arms one and a half times as long as broad, somewhat constricted in the middle
part, armed at the rounded ends with numerous (thirty to forty) strong, conical spines. Patagium
complete, with four or five rectilinear parallel chamber-rows, enveloping the whole of the arms
with the exception of the spiny ends.
Dimensions.—Radius of the arms (without spines) 0.2, breadth 0.06 to 0.09.
Habitat.—North Pacific, Station 253, depth 3125 fathoms.
Genus 231. Chitonastrum,[270] Haeckel,
1881, Prodromus, p. 460.
Definition.—Porodiscida with three forked, chambered
arms, without a patagium. (Arms and angles between them either equal or unequal.)
The genus Chitonastrum differs from its ancestral form, Dictyastrum,
by the bifurcation of the distal ends of the arms. The few species of this genus are partly {537}regular (like Dictyastrum), partly bilateral (like
Rhopalastrum). If the number of species increases much, these two subgenera may be
separated into two genera: Chitonastrella corresponding to the former,
Chitonastromma to the latter.
Subgenus 1. Chitonastrella, Haeckel.
Definition.—All three arms of the same size and form, equidistant;
fundamental form of the shell therefore an equilateral triangle.
1. Chitonastrum triglochin, n. sp.
All three arms equal and equidistant. Each arm has the form of an isosceles triangle, twice as
high as broad; the truncated apex of the triangle is inserted into the large central disk, whilst
its distal base (four times as broad) is divided by a deep incision (half as long as the arm).
Each arm with ten to twelve joints, simple in its basal half, double in its distal half. Axes of
the six branches straight. (Resembles Trigonastrum regulare, Pl. 43, fig. 16,
but differs in the absence of a patagium.)
Dimensions.—Radius of each arm 0.24, greatest breadth 0.11, basal breadth
0.03.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
2. Chitonastrum bathybium, n. sp.
All three arms equal and equidistant, in the basal two-thirds simple, rectilinear, three times
as long as broad, in the distal third forked, both branches equal, straight, blunt, half as broad
as the basal part.
Dimensions.—Radius of each arm 0.18, basal breadth 0.04, breadth of the branches
0.02.
Habitat.—Western Tropical Pacific, Station 225, depth 4475
fathoms.
Subgenus 2. Chitonastromma, Haeckel.
Definition.—One odd arm different in size or form from the two other
arms, which are paired; distance between them different; fundamental form of the shell therefore a
bilateral, isosceles triangle.
3. Chitonastrum jugatum, n. sp. (Pl. 43,
fig. 14).
Dictyastrum jugatum, Haeckel, 1881, Prodromus et Atlas (pl. xliii. fig.
14).
Arms very different; odd arm club-shaped, twice as long as broad, at the blunt distal end twice
as broad as at the base; its axis is perpendicular to the common axis of both paired arms, which
are only two-thirds as long, not so broad, and in the distal half divided into two branches; the
anterior branch is straight, nearly horizontal, the posterior shorter and curved backwards.
{538}
Dimensions.—Radius of the odd arm 0.2, of the paired arms 0.15; distal breadth of
the former 0.08, basal breadth 0.04; breadth of the paired arms 0.04.
Habitat.—South Atlantic, Station 325, surface.
4. Chitonastrum dicranoides,
n. sp.
All three arms in the basal half simple, nearly square, in the distal half forked; branches
straight, blunt. Odd arm twice as large as the paired arms; angle between the latter larger than
the angles between them and the odd arm. (The form of the arms resembles Dicranastrum
furcatum, Pl. 47,
fig. 2.)
Dimensions.—Radius of the odd arm 0.24, of the paired arms 0.12; basal breadth
0.06.
Habitat.—North Atlantic, Station 353, surface.
5. Chitonastrum lyra, n. sp. (Pl. 43,
fig. 15).
Dictyastrum lyra, Haeckel, 1881, Prodromus et Atlas (pl. xliii. fig.
15).
All three arms forked and nearly of the same size, but different in form and position. The
distance between the branches of the two paired arms is only one-fourth of the distance between
them and the odd arm. Each arm in the basal two-thirds is simple, with eleven to twelve transverse
septa, in the distal third forked, each branch with four to five transverse septa. The branches of
each arm are curved convexly one to another, ending obtusely. The axis of the simple proximal part
is straight in the odd arm, in the paired arms curved concavely towards the middle line. In the
figured specimen, which I observed living in Portofino (in September 1880), the central chamber of
the central disk and the first surrounding ring were filled with the nucleus of the cell; both
external rings were filled (like all chambers of the arms) with pink oil-globules of the red
central capsule. From the mantle, enveloping the shell, radiated innumerable fine pseudopodia
(much too short in the figure), and between the two paired arms a long "sarcode-flagellum."
Dimensions.—Radius of each arm 0.16; greatest breadth of the odd arm 0.04; basal
breadth of the paired arms 0.02; distance of both branches of each arm 0.08.
Habitat.—Mediterranean, Portofino, near Genoa, Haeckel.
Genus 232. Trigonastrum,[271] n.
gen.
Definition.—Porodiscida with three forked, chambered
arms, connected by a patagium. (Arms and angles between them either equal or unequal.)
The genus Trigonastrum differs from the preceding Chitonastrum, its
ancestral form, in the development of a patagium between the arms. It bears therefore to the
latter the same relation that Euchitonia does to Rhopalastrum.
{539}
Subgenus 1. Trigonastrella, Haeckel.
Definition.—All three arms of the same size and form, equidistant;
fundamental form of the shell therefore a regular, equilateral triangle.
1. Trigonastrum regulare, n. sp. (Pl. 43, fig.
16).
Chitonastrum regulare, Haeckel, 1881, Prodromus et Atlas (pl. xliii. fig.
16).
All three arms equal and equidistant. Each arm has the form of an isosceles triangle, twice as
high as broad, the truncated apex of which is inserted into the large circular central disk,
whilst its distal base (four times as broad) is divided by a deep incision (half as long as the
arm). Each arm with ten to twelve joints. Patagium between the arms nearly complete, spongy.
(Differs from Chitonastrum triglochin mainly in the possession of a patagium.)
Dimensions.—Radius of each arm 0.24, greatest breadth 0.11, basal breadth 0.03;
length of the sides of the regular triangle 0.45.
Habitat.—Pacific, central area, Station 274, surface.
Subgenus 2. Trigonastromma, Haeckel.
Definition.—One odd arm different in form or size from the other two
arms, which are paired; distance between them different; fundamental form of the shell therefore
an isosceles triangle.
2. Trigonastrum krohnii, Haeckel.
Euchitonia krohnii, Haeckel, 1862, Monogr. d. Radiol., p. 507.
All three arms different; distance between the two paired arms smaller than their distance from
the odd arm. Odd arm with six joints. Each paired arm with seven joints, increasing in breadth
towards the distal end. Odd arm and one paired arm forked at the end, the other paired arm simple.
Patagium nearly complete. (The asymmetry in this form may perhaps be an individual anomaly, as
also in Myelastrum anomalum, Pl. 47,
fig. 9.)
Dimensions.—Radius of the arms about 0.13 to 0.16, breadth 0.05 to 0.08.
Habitat.—North Atlantic, Funchal, Madeira, Krohn, surface.
3. Trigonastrum gegenbauri, Haeckel.
Euchitonia gegenbauri, Haeckel, 1862, Monogr. d. Radiol., p. 506, Taf.
xxxi. figs. 2, 3.
Arms different; distance between the paired arms larger than their distance from the odd arm,
which is one-third shorter. Odd arm egg-shaped, simple, with seven joints, undivided. Both paired
arms equal, with ten joints, in the distal third forked. Patagium nearly complete.
Dimensions.—Radius of the odd arm 0.14, greatest breadth 0.08; radius of the
paired arms 0.2.
Habitat.—Mediterranean (Messina), Haeckel, surface.
{540}
Genus 233. Stauralastrum,[272] n.
gen.
Definition.—Porodiscida with four simple, undivided,
chambered arms, without a patagium; quadrangular shell a regular cross, with four equal arms
placed at right angles.
The genus Stauralastrum is the most simple form of the Tessarastrida, or of
those Porodiscida in which the margin of the central disk is armed with four chambered arms. In
Stauralastrum these four arms are quite simple and equal, without a patagium, separated by
four right angles, so that the whole shell represents a regular rectangular cross. If we connect
the distal points of the arms by lines, we get a complete square. (In my Prodromus, 1881, the
species of this genus were united with Hagiastrum, which genus I now retain for the simple
bilateral Tessarastrida.)
Subgenus 1. Stauralastrella, Haeckel.
Definition.—Ends of the arms blunt, without terminal spines.
1. Stauralastrum cruciforme, n. sp. (Pl. 45, fig.
6).
Arms very thin, nearly linear, four to five times as long as broad, of equal breadth at the
base and at the truncated distal end; their breadth equals one-third of the radius of the central
disk. Edges of the arms parallel.
Dimensions.—Radius of each arm 0.8, breadth 0.016.
Habitat.—South Pacific, Station 293, surface.
2. Stauralastrum lanceolatum, n. sp.
Arms lanceolate, three times as long as broad, in their middle part three times as broad as at
both ends; their greatest breadth nearly equals the diameter of the central disk. (The arms have
the same form as in Euchitonia lanceolata, Pl. 43, fig. 9.)
Edges of the arms convex.
Dimensions.—Radius of each arm 0.3, greatest breadth (in the middle part)
0.08.
Habitat.—Pacific, central area, Station 273, depth 2350 fathoms.
3. Stauralastrum ordo, n. sp.
Arms trapezoid, about as long as broad, twice as broad at their truncated distal end as at the
base; their basal breadth equals the radius of the central disk, which exhibits two to three
rings. (The arms have nearly the same form as those in Hagiastrum mosis, Pl. 45, fig. 3.)
Edges of the arms rectilinear, divergent towards the ends.
Dimensions.—Radius of each arm 0.12, basal breadth 0.04, distal breadth 0.08.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms.
{541}
4. Stauralastrum clavigerum, n. sp.
Arms twice as long as broad, in their distal half lenticular, nearly circular, twice as broad
as in their square proximal half; their distal breadth equals the diameter of the central disk,
which exhibits three to four rings. Edges of the arms concave.
Dimensions.—Radius of each arm 0.2, basal breadth 0.04, distal breadth 0.08.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
5. Stauralastrum dilatatum, n. sp.
Arms of nearly equal length and breadth, at their convex distal end three times as broad as at
their narrow base; their distal breadth three times as large as the radius of the central disk,
which exhibits three to four rings. (Resembles Histiastrum quadrigatum, Pl. 46, fig. 3, but
has no patagium.) Edges of the arms concave.
Dimensions.—Radius of each arm 0.15, basal breadth 0.04, terminal breadth
0.12.
Habitat.—South Pacific, Station 300, depth 1375 fathoms.
Subgenus 2. Stauralastromma, Haeckel.
Definition.—Ends of the arms with one or more terminal spines.
6. Stauralastrum rhopalophorum, n. sp. (Pl. 45, fig.
1).
Hagiastrum rhopalophorum, Haeckel, 1881, Prodromus, p. 460.
Arms cylindrical, eight times as long as broad at their base, at their distal end club-shaped,
three times as broad as at their base; their distal breadth twice as large as the diameter of the
central disk, which exhibits two to three rings. Surface thorny, with larger spines towards the
end, and one radial, very strong, angular terminal spine. Edges of the arms parallel.
Dimensions.—Radius of each arm (without terminal spine) 0.32, basal breadth 0.03,
terminal breadth 0.1.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms.
7. Stauralastrum antiquum, n. sp.
Rhopalastrum sp. Bury, 1862, Polycystins of Barbados, pl. xiv. fig. 5.
Arms six times as long as broad at their base, in their distal half nearly spherical, three
times as broad as in their cylindrical basal half; their distal breadth nearly equals the diameter
of the central disk which exhibits three to four rings. On the end of each arm one strong,
angular, terminal spine. (Differs from the preceding species by larger central disk and stouter
arms, also by less developed spines.) Edges of the arms parallel.
Dimensions.—Radius of each arm 0.25, basal breadth 0.04, terminal breadth
0.12.
Habitat.—Fossil in the Barbados rocks; and living in the depths of the
Central Pacific, Station 266, depth 2750 fathoms.
{542}
8. Stauralastrum staurolonche, n. sp.
Arms four times as long as broad at their base, gradually increasing towards their truncated
end, which is one and a half times as broad as their base; their distal breadth equals the radius
of the central disk, which exhibits four to five rings. At the end of each arm is a very strong
conical terminal spine. (Resembles Histiastrum quaternarium, Abhandl. k. Akad. Wiss.
Berlin, 1875, Taf. xxiv. fig. 3, but has no patagium.) Edges of the arms rectilinear,
divergent.
Dimensions.—Radius of each arm 0.25, basal breadth 0.045, distal breadth 0.07.
Habitat.—Fossil in the Barbados rocks; and living in the depth of the
Equatorial Atlantic, Station 348, depth (2450) fathoms.
9. Stauralastrum horridum, n. sp.
Arms three times as long as broad at their base, gradually increasing towards their rounded
end, which is twice as broad as their base, their distal breadth equals the diameter of the
central disk, which exhibits four to five rings. Surface thorny, at the distal end of each arm is
a group of twenty to twenty-five smaller and five to six larger, straight, conical spines. Edges
of the arms rectilinear, divergent.
Dimensions.—Radius of each arm 0.15, basal breadth 0.05, distal breadth 0.1.
Habitat.—Pacific, central area, Station 270, depth 2925 fathoms.
Genus 234. Hagiastrum,[273] Haeckel,
1881, Prodromus, p. 460.
Definition.—Porodiscida with four simple, undivided,
chambered arms, without a patagium; quadrangular shell bilateral, two opposite arms of the main
axis (or principal arms) different from the two others (or lateral arms).
The genus Hagiastrum, as here defined, was formerly united by me with the
foregoing Stauralastrum, but differs from it by the bilateral or symmetrical form. Whilst
in the latter all four arms and the four angles between them are equal, they are here
differentiated into pairs.
Subgenus 1. Hagiastrella, Haeckel.
Definition.—Both longitudinal arms of equal size and form.
1. Hagiastrum buddhae, n. sp. (Pl. 45,
fig. 5).
Cross rectangular. Both longitudinal arms of equal size, twice as long as the transverse arms;
all arms smooth, club-shaped, twice as broad at their globose distal part as at their base, each
with three large conical terminal spines.
{543}
Dimensions.—Radius of the principal arms 0.4, of the lateral arms 0.2; basal
breadth 0.06, distal breadth 0.12.
Habitat.—Indian Ocean, Belligemma, Ceylon, surface, Haeckel.
2. Hagiastrum bramae, n. sp.
Cross rectangular. Both longitudinal arms of equal size, one and a half times as long as the
transverse arms; all arms thorny, club-shaped, at their pear-shaped distal part three times as
broad as at their base, provided with numerous conical spines, one larger terminal spine at their
distal point. (Resembles Stauralastrum rhopalophorum, Pl. 45, fig. 1, but
is distinguished by the different size of the arm-pairs, and by the stronger spines.)
Dimensions.—Radius of the principal arms 0.3, of the lateral arms 0.2; basal
breadth 0.04, distal breadth 0.12.
Habitat.—Indian Ocean, off Maldive Islands, surface, Haeckel.
Subgenus 2. Hagiastromma, Haeckel.
Definition.—The two longitudinal arms different in size or form.
3. Hagiastrum mosis, n. sp. (Pl. 45,
fig. 3).
Cross rectangular. All four arms nearly isosceles, triangular, at their narrow base half as
broad as at their truncated, concavely fluted, distal end. The posterior principal arm with twelve
to thirteen joints, twice as long as the anterior arm, which has six to seven joints and is one
and a half times as long as the two lateral arms (with four to five joints).
Dimensions.—Radius of the posterior arm 0.3, of the anterior 0.15, of the lateral
arms 0.1; basal breadth 0.05, terminal breadth 0.1.
Habitat.—Mediterranean (Smyrna), surface, Haeckel.
4. Hagiastrum mohammedis, n. sp.
Cross with unequal angles, the anterior little smaller than the posterior. All four arms
club-shaped, thorny, three times as broad at their globose distal end as at their narrow base, and
furnished with ten to twelve conical spines. Posterior principal arm twice as long as the
anterior, and four times as long as the rudimentary lateral arms.
Dimensions.—Radius of the posterior arm 0.4, of the anterior 0.2, of the lateral
arms 0.1; basal breadth 0.02 to 0.03, distal breadth 0.06 to 0.1.
Habitat.—Philippine Sea, Samboangan, Station 200, surface.
5. Hagiastrum christi, n. sp.
Cross with unequal angles, the anterior somewhat smaller than the posterior. All four arms of
similar form, lanceolate, in their middle twice as broad as at either obtuse end. The posterior
{544}principal arm with twelve joints, one and a half
times as long as the anterior (with nine joints) and twice as long as the two lateral arms (each
with six joints). The form and structure of the arms in this species are nearly the same as in
Tessarastrum
straussi (Pl. 45,
fig. 8); but the arms are broader in the middle, and are not connected by a patagium.
Dimensions.—Radius of the principal posterior arm 0.2, of the anterior 0.15, of
each lateral arm 0.1; greatest breadth (in the width) 0.01, basal breadth 0.03.
Habitat.—North Atlantic, Færöe Channel, Gulf Stream, surface, John
Murray.
Genus 235. Histiastrum,[274] Ehrenberg,
1847, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 54.
Definition.—Porodiscida with four simple, undivided,
chambered arms, connected by a patagium; square shell a regular cross, with four equal arms and
four right angles between them.
The genus Histiastrum, quite insufficiently characterised by Ehrenberg
(1847), was afterwards (1875) illustrated by the figures of two different fossil species. One of
these, Histiastrum ternarium, with three arms, belongs to Hymeniastrum; the other,
Histiastrum quaternarium, is here retained as the true, typical representative species of
the genus. It differs from its ancestral form Stauralastrum, by the possession of a
patagium, from Tessarastrum by the regular square form of the shell.
Subgenus 1. Histiastrella, Haeckel.
Definition.—Distal ends of the arms blunt, without terminal
spines.
1. Histiastrum quadrigatum, n. sp. (Pl. 46, fig.
3).
Arms at their distal end nearly as broad as long, and four times as broad as at their narrow
base; their lateral edges concave, their terminal edge convex, without spines. Each arm is divided
by seven to eight convex transverse septa into eight to nine simple, broad chambers. Central disk
with three to four rings, about as broad as the fifth chamber. Patagium complete, connecting all
the lateral edges of the arms.
Dimensions.—Radius of each arm 0.15, basal breadth 0.03, terminal breadth
0.12.
Habitat.—Equatorial Atlantic, Station 347, surface.
2. Histiastrum excisum, n. sp.
Arms four times as long as broad at their base, and twice as broad at their rounded blunt
distal end as at their base; their lateral edges rectilinear, divergent. Central disk with three
to four rings, {545}somewhat broader than their distal
end. Patagium incomplete, connecting only the basal half of the arms, with three to four concave
chamber-rows, on the margin concave. (May be regarded as Euchitonia mülleri, with four
arms.)
Dimensions.—Radius of each arm 0.2, basal breadth 0.05, distal breadth 0.1.
Habitat.—Atlantic, surface, Canary Islands.
3. Histiastrum velatum, n. sp. (Pl. 46,
fig. 4).
Dictyastrum velatum, Haeckel, 1879, Atlas (pl. xlvi. fig. 4).
Arms pear-shaped, rapidly increasing from their narrow base, nearly circular, little longer
than broad; each with eight to nine transverse chamber-rows; their lateral edges at their base
concave, at their end circular. Central disk with three to four rings, somewhat smaller than one
arm. Patagium complete, with six to seven radial beams, filling out perfectly the intervals
between the arms. A peculiar girdle of finer network and equal breadth surrounds the whole
equatorial periphery of the disk, and gives it the appearance of a square with rounded
corners.
Dimensions.—Radius of each arm 0.2, basal breadth 0.03, distal breadth 0.12.
Habitat.—South Atlantic, Station 330, surface.
Subgenus 2. Histiastromma, Haeckel.
Definition.—Distal ends of the arms spiny, furnished with one or more
terminal spines.
4. Histiastrum quaternarium, Ehrenberg.
Histiastrum quaternarium, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 74, Taf. xxiv. figs. 3, 4.
Arms six times as long as broad at their base, with rectilinear, little divergent edges; at
their truncated distal end a little broader, with one single, very strong, conical, terminal
spine. Central disk with four to five rings, somewhat broader than the arm. Patagium incomplete,
enveloping only the basal half of the arms.
Dimensions.—Radius of each arm 0.18, basal breadth 0.03, distal breadth 0.05.
Habitat.—Fossil in the rocks of Barbados.
5. Histiastrum gladiatum, n. sp.
Astromma sp., Bury, 1862, Polycystins of Barbados, pl. v. fig. 1.
Arms triangular, eight times as long as broad at their base; at their distal end three times as
broad as at their base, with rectilinear, divergent edges; their truncated end with a large
conical, terminal spine. Central disk with four to five rings, broader than the arms. Patagium
incomplete, enveloping only the basal half of the arms.
{546}
Dimensions.—Radius of each arm 0.12, basal breadth 0.013, distal breadth 0.04.
Habitat.—Fossil in the rocks of Barbados; and living in the depth of
the Central Pacific, Station 268, depth 2900 fathoms.
6. Histiastrum boseanum, n. sp. (Pl. 46,
fig. 1).
Arms linear, twelve times as long as broad at their base (at their distal end twice as broad as
at their base), with rectilinear, parallel edges. The club-shaped end thickened, dentate, with two
lateral rows of strong teeth in the equatorial plane, and with one very stout, angular, terminal
spine. Central disk with two to three rings, broader than the arms. Patagium incomplete, with four
to five concave chamber-rows, enveloping only the basal half of the arms. I call this splendid
species in honour of Dr. Graf Bose, the great friend of nature and patron of the University of
Jena.
Dimensions.—Radius of each arm 0.25, basal breadth 0.02, distal breadth 0.05.
Habitat.—Indian Ocean, Madagascar, Rabbe.
7. Histiastrum coronatum, n. sp.
Stephanastrum sp., Bury, 1862, Polycystins of Barbados, pl. iv. fig.
1.
Arms linear, eight times as long as broad, with rectilinear parallel edges; the thickened,
nearly spherical, distal end three times as broad, with five strong conical spines, one middle
(perradial) larger and two smaller on each side of it. Central disk with two to three rings, of
the same breadth as the terminal knot of the arms. Patagium incomplete, square, enveloping the
arms, with the exception of the knot.
Dimensions.—Radius of each arm 0.16, basal breadth 0.02, terminal breadth
0.06.
Habitat.—Fossil in the rocks of Barbados.
8. Histiastrum circulare, n. sp.
Stephanastrum sp., Bury, 1862, Polycystins of Barbados, pl. xxiii. fig.
1.
Arms linear in their inner half, egg-shaped and three times as broad in their outer half, with
ten to twelve strong terminal spines, the middle (perradial) larger, in all three times as long as
broad. Central disk with three to four rings, broader than their distal knobs. Patagium nearly
complete, circular, enveloping the whole arms, with exception of the outermost end.
Dimensions.—Radius of each arm 0.22, basal breadth 0.02, terminal breadth
0.06.
Habitat.—Fossil in the rocks of Barbados; and living in the depths of
the Central Pacific, Station 268, depth 2900 fathoms.
9. Histiastrum pentadiscus, n. sp. (Pl. 46, fig.
2).
Arms in their inner half linear, twice as long as broad, in their outer half circular, three
times as broad, with the same structure as the central disk, exhibiting three concentric rings
around one {547}central chamber. Patagium complete,
spongy, with radiating beams, enveloping the whole disk, with the exception of the outermost end
of the arms, which is armed with twelve to sixteen strong conical spines, the middle (perradial)
spine much larger.
Dimensions.—Radius of each arm (without terminal spine) 0.18, basal breadth 0.025,
terminal breadth 0.08.
Habitat.—South Atlantic, Station 333, surface.
Genus 236. Tessarastrum,[275] n.
gen.
Definition.—Porodiscida with four simple, undivided,
chambered arms, connected by a patagium; quadrangular shell bilateral, two opposite arms of the
main axis (or principal arms) different from the two others (or lateral arms).
The genus Tessarastrum, formerly united by me with Histiastrum,
differs from the latter in its bilateral or symmetrical form, and bears therefore the same
relation to it that Hagiastrum does to Stauralastrum.
Subgenus 1. Tessarastrella, Haeckel.
Definition.—Both longitudinal arms of equal size and form.
1. Tessarastrum straussii, n. sp. (Pl. 45, fig.
8).
Histiastrum straussii, Haeckel, 1881, Prodromus et Atlas (pl. xlv. fig.
8).
Cross not rectangular. Both principal arms of equal size and form, four times as long as broad,
and twice as long as the broader lateral arms; the former with ten to eleven, the latter with five
to six joints, separated by convex transverse septa. Distal ends of the arms blunt. Axes of the
smaller arms not perpendicular to that of the larger arms; therefore the anterior angles between
them smaller than the posterior angles. Patagium between the arms incomplete. I call this
remarkable species after the great German philosopher David Strauss.
Dimensions.—Radius of the principal arms 0.24, of the lateral arms 0.12; greatest
breadth (in the middle) of the former 0.05, of the latter 0.06.
Habitat.—North Pacific, Hyalonema-ground, March 5, 1875.
2. Tessarastrum spinozæ, n. sp.
Cross rectangular. Both principal arms of equal size and form, ten times as long as broad, and
twice as long as the lateral arms, which are only five times as long as broad. All arms linear, at
their distal end club-shaped, and armed with twenty to thirty very strong angular spines. Patagium
incomplete, enveloping only the basal half of the arms. (Resembles Histiastrum boseanum,
{548}Pl. 46, fig. 1, but
differs in the broader arms and the unequal size of both pairs.) I call this species after the
great monistic philosopher Benedictus Spinoza.
Dimensions.—Radius of the principal arms 0.3, of the lateral arms 0.16; basal
breadth 0.025, distal breadth 0.05.
Habitat.—South Atlantic, Station 333, surface.
3. Tessarastrum brunonis, n. sp. (Pl. 45, fig.
9).
Histiastrum brunonis, Haeckel, 1881, Prodromus et Atlas (pl. xlv. fig.
9).
Cross rectangular. Both principal arms of equal size and form, three times as long as broad,
each with ten joints, three times as long as the lateral arms, which are nearly square, with four
joints. All arms rounded, at their truncated end little broader than at their base. No spines.
Patagium complete, envelops the whole shell, and is composed of two parallel lattice-lamellæ on
each side of the flat disk, which are connected by very fine perpendicular bars. This is shown
clearly in fig. 9, Pl. 45,
where the disk is seen from the edge. I call this species after the great Italian philosopher
Giordano Bruno.
Dimensions.—Radius of the principal arms 0.22, of the lateral arms 0.12; basal
breadth 0.04, distal breadth 0.05.
Habitat.—South Pacific, Station 285, depth 2375 fathoms.
Subgenus 2. Tessarastromma,
Haeckel.
Definition.—The two principal arms of different size or form.
4. Tessarastrum democriti, n. sp. (Pl. 45, fig.
7).
Histiastrum democriti, Haeckel, 1881, Prodromus et Atlas (pl. xlv. fig.
7).
Cross not rectangular; the two anterior angles smaller than the two posterior. All four arms
club-shaped, twice as broad at their rounded obtuse distal end as at their base, of unequal
length. Posterior principal arm one and a fourth times as long as the posterior, and one and
two-thirds as long as the lateral arms. Patagium incomplete, enveloping only the basal half of the
arms. I call this species after the great Greek philosopher Democritus.
Dimensions.—Radius of the posterior arm 0.3, of the anterior 0.25, of each lateral
arm 0.22; basal breadth 0.05, distal breadth 0.1.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
Genus 237. Stephanastrum,[276] Ehrenberg,
1847, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 54.
Definition.—Porodiscida with four simple, undivided,
chambered arms, connected on the distal ends by a spongy, square or rhomboidal, patagial girdle
(or a patagium with four large, interbrachial openings). Shell either regular or bilateral (with
equal or unequal arms).
{549}
The genus Stephanastrum, founded (1847) by Ehrenberg for the very peculiar
Stephanastrum rhombus, differs from the nearly allied foregoing genera in the imperfect
development of the peculiar patagium, connecting only the distal ends of the four arms, while it
is absent at their base. Two new species, different from Stephanastrum rhombus by the
regular square form, were found in the Challenger collection.
Subgenus 1. Stephanastrella, Haeckel.
Definition.—All four arms of the cross have the same size.
1. Stephanastrum quadratum, n. sp. (Pl. 46, fig.
5).
All four arms of the same size, six times as long as broad at their base, ending with a strong,
short, four-sided pyramidal spine. In the outer half of each arm are two opposite lateral spongy
wings, which form an equilateral triangle, and from union of the bases of the four triangles
arises the peculiar patagium, which forms a square with four large interbrachial openings.
Dimensions.—Radius of each arm 0.25, basal breadth 0.035; length of the sides of
the square patagium 0.3.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
2. Stephanastrum capitatum, n. sp. (Pl. 44, fig.
1).
All four arms of the same size, five times as long as broad at their base, at their distal end
with a spongy, nearly spherical capitulum of twice their breadth, provided with a very strong,
angular, pyramidal, terminal spine (half as long as the arm). All four arms connected by a square
patagium, arising immediately below the capitula, and perforated by four large interbrachial
openings.
Dimensions.—Radius of each arm (without the terminal spine) 0.25, basal breadth
0.05; length of the sides of the square patagium 0.3.
Habitat.—Pacific, central area. Station 265, depth 2900 fathoms.
Subgenus 2. Stephanastromma, Haeckel.
Definition.—Two opposite arms of the cross larger than the two
others.
3. Stephanastrum rhombus, Ehrenberg.
Stephanastrum rhombus, Ehrenberg, 1854, Mikrogeol., Taf. xxxvi. fig. 33;
Abhandl. d. k. Akad. d. Wiss. Berlin, 1875, Taf. xxv. fig. 1.
Two arms of the longitudinal axis one and a third times as long as two arms of the transverse
axis. All four arms linear, about eight times as long as broad, at their distal end somewhat {550}thickened, club-shaped, with a pyramidal terminal spine.
The ends of the arms are connected by a riband-shaped, straight, spongy patagium of the same
breadth as the arms. Between the rhomboidal patagium and the arms remain four large rectangular
triangles as interbrachial openings.
Dimensions.—Radius of the longer arms 0.2, of the shorter 0.15; basal breadth
0.02; length of the sides of the rhombic patagium 0.25.
Habitat.—Fossil in the rocks of Barbados.
Genus 238. Dicranastrum,[277] Haeckel,
1881, Prodromus, p. 460.
Definition.—Porodiscida with four forked, spongy, or
chambered arms, without a patagium; shell regular (not bilateral), with four equal arms crossed at
right angles.
The genus Dicranastrum comprises a number of very remarkable, hitherto
unknown, Euchitonida, which are rather common in the Pacific (mainly on the surface), and
characterised by the bifurcation of the cross-arms of the regular square shell. It bears therefore
to its probable ancestral form, Stauralastrum, the same relation that in the triradiate
Euchitonida Chitonastrum does to Dictyastrum. The arms are commonly of very delicate
structure, more or less spongy.
Subgenus 1. Dicranaster, Haeckel.
Definition.—Each cross-arm with two simple branches.
1. Dicranastrum furcatum, n. sp. (Pl. 47, fig.
2).
Arms simply forked, three times as long as broad at their base, with eight blunt ends of the
fork-branches. The simple proximal half of each arm about the same size as each branch of the
dichotomous distal part, twice as long as broad. Edges of the arms ragged.
Dimensions.—Radius of each arm 0.38, basal breadth 0.12; breadth of the forked
part 0.3.
Habitat.—Pacific, central area, Station 271, surface.
2. Dicranastrum dichotomum, n. sp.
Arms simply forked, four times as long as broad at their base; each arm with two blunt
branches. The simple proximal part of each arm is three times as long and twice as broad as each
branch of the dichotomous distal part. Ends of the arms blunt, truncated.
Dimensions.—Radius of each arm 0.35, basal breadth 0.08; breadth of the forked
part 0.2.
Habitat.—South Pacific, Station 281, surface.
{551}
3. Dicranastrum cornutum, n. sp. (Pl. 45, fig.
2).
Hagiastrum cornutum, Haeckel, 1879, Atlas (pl. xlv. fig. 2).
Four arms simply forked, four times as long as broad; each arm with two triangular diverging
branches ending in strong conical spines. The simple basal part of each arm about of the same
length as each branch of the distal part, twice as long as broad. Distance between the terminal
spines of each arm nearly as great as its length.
Dimensions.—Radius of the arm 0.24, basal breadth 0.05; breadth across the
bifurcation 0.12.
Habitat.—South Atlantic, Station 325, surface.
4. Dicranastrum antilope, n. sp.
Four arms simply forked, three times as long as broad; each arm with two lanceolate diverging
branches, ending in strong angular spines. The simple basal part of each arm is twice as long as
each branch of the distal part. Distance of the two terminal spines of each arm scarcely half as
great as its length.
Dimensions.—Radius of the arm 0.32, basal breadth 0.11; breadth of the branches
0.07.
Habitat.—Tropical Atlantic (off Ascension), Station 342, surface.
Subgenus 2. Tricranastrum, Haeckel, 1881, Prodromus, p. 460.
Definition.—Each cross arm with three terminal branches, one middle
(perradial) and two lateral (adradial) branches.
5. Dicranastrum wyvillei, n. sp. (Pl. 47, fig.
3).
Tricranastrum wyvillei, Haeckel, 1879, Natürl. Schöpfungsgesch., p. 705,
Taf. xvi. fig. 5.
Arms trifid, one and a half times as long as broad; each arm with three blunt terminal branches
of equal size. The simple basal part of each arm twice as long as the trifid distal part. (The
central capsule depicted in fig. 3, Pl. 47,
has the same form as the skeleton, and is only a little smaller.)
Dimensions.—Radius of the arm 0.4, basal breadth 0.12, greatest breadth (in the
distal part) 0.24.
Habitat.—Pacific, central area, Station 271, surface.
6. Dicranastrum tricuspis, n. sp.
Arms trifid, twice as long as broad; each arm with three pointed terminal branches, ending in
strong conical spines, the middle branch somewhat larger than the other two. The simple basal part
of each arm three times as long as the trifid distal part.
Dimensions.—Radius of the arm 0.3, basal breadth 0.06, greatest breadth (in their
distal part) 0.15.
Habitat.—North Pacific, Station 244, surface.
{552}
7. Dicranastrum trifarium, n. sp.
Arms trifid, three times as long as broad at their base; each arm with three pointed terminal
branches, ending in strong sulcate spines; the middle branch twice as large as the two others. The
simple basal part of each arm two and a half times as long as the trifid distal part.
Dimensions.—Radius of the arm 0.35, basal breadth 0.03, greatest breadth (in the
distal part) 0.05.
Habitat.—South Pacific, Station 298, surface.
Subgenus 3. Tetracranastrum, Haeckel.
Definition.—Each cross-arm with four terminal branches, the two
fork-branches being again bifurcated.
8. Dicranastrum bifurcatum, n. sp. (Pl. 47, figs. 1,
1a).
Arms doubly forked or quadripartite, six times as long as broad at their base; each arm in its
proximal half simple, three times as long as broad; in its distal half doubly forked; the
secondary branches with blunt, roundish ends, nearly as large as the primary branches. Central
disk (fig. 1a) with three concentric rings around the central chamber; from its periphery
radiate thin radial beams in the spongy framework of the delicate arms. The central capsule has
the same form as the skeleton, and is only a little smaller.
Dimensions.—Radius of the arm 0.45, basal breadth 0.08; breadth of the terminal
branches 0.03.
Habitat.—North Pacific, east of Japan, Station 241, surface.
Genus 239. Myelastrum,[278] Haeckel,
1881, Prodromus, p. 460.
Definition.—Porodiscida with four forked, spongy, or
chambered arms, without a patagium; shell bilateral, with paired different arms; two equal
anterior arms of different shape from the two equal posterior arms.
The genus Myelastrum differs from the foregoing Dicranastrum, the
ancestral form, by the twofold differentiation of the four arms. Whilst in the latter all four
arms are equal, separated by equal angles, here the two anterior arms are constantly different
from the two posterior. The lateral angles between the two arm-pairs are equal, while the
posterior and the anterior angle (between the two arms of each pair) are more or less different.
The shell assumes, therefore, a very characteristic bilateral form, similar to the "quadricorn
cross" of the grey central substance in the transverse section of the human medulla spinalis.
Though the spongy shell is commonly a most delicate and {553}thin disk it nevertheless reaches unusual dimensions, its diameter in
some species being more than a millimetre.
Subgenus 1. Myelastrella, Haeckel.
Definition.—Posterior arms simple, undivided; anterior arms lobated or
cleft, with one or more incisions at the distal end.
1. Myelastrum medullare, n. sp. (Pl. 47,
fig. 13).
Anterior arms bifid, nearly square, with a shallow incision at their broad truncated end.
Posterior arms somewhat smaller, nearly triangular, with simple blunt ends. Sagittal constriction
three-fourths as large as the transverse one. Surface smooth.
Dimensions.—Radius of the anterior arms 0.36, of the posterior 0.3; longitudinal
constriction 0.24, transverse 0.36.
Habitat.—Pacific, central area, Station 271, surface.
2. Myelastrum spinale, n. sp.
Anterior arms bifid, twice as long as broad, with a deep incision at their truncated end.
Posterior arms slender, half as large, with simple blunt ends. Sagittal constriction one and a
half times as large as the transverse. Surface spiny.
Dimensions.—Radius of the anterior arms 0.4, of the posterior 0.2; longitudinal
constriction 0.3, transverse 0.2.
Habitat.—North Pacific, Station 248, surface.
3. Myelastrum heteropterum, n. sp. (Pl. 47, fig.
8).
Anterior arms trifid, about as long as broad, with two incisions at their broad truncated end.
Posterior arms about half as large, simple, with rounded blunt ends. Sagittal constriction
two-thirds as large as the transverse. Surface bristly.
Dimensions.—Radius of the anterior arms 0.65, of the posterior 0.035; longitudinal
constriction 0.4, transverse 0.6.
Habitat.—South Pacific, Station 291, surface.
Subgenus 2. Myelastromma, Haeckel.
Definition.—All four arms (posterior as well as anterior) lobated or
cleft.
4. Myelastrum octocorne, n. sp. (Pl. 47,
fig. 12).
All four arms with a deep incision in their distal half; anterior arms broader, but shorter
than the posterior; in the anterior arms the two lobes are of the same size, in the posterior arms
the {554}median lobe is longer than the lateral lobe.
Sagittal constriction little smaller than the transverse. Surface of the disk rough.
Dimensions.—Radius of the anterior arms 0.45, of the posterior 0.62; longitudinal
constriction 0.25, transverse 0.3.
Habitat.—Pacific, central area, Station 271, surface.
5. Myelastrum farfalla, n. sp. (Pl. 47,
fig. 10).
All four arms with a shallow incision at their distal end. Anterior arms broader, but shorter
than the posterior; in each arm the anterior lobe is shorter than the posterior. Sagittal
constriction smaller than the transverse. Surface smooth.
Dimensions.—Radius of the anterior arms 0.4, of the posterior 0.6; longitudinal
constriction 0.5, transversal stricture 0.6.
Habitat.—Pacific, central area, Station 274, surface.
6. Myelastrum papilio, n. sp. (Pl. 47,
fig. 6).
All four arms with a shallow incision at their distal end. Anterior arms triangular, of the
same breadth, but of the double length of the square posterior arms. Sagittal and transverse
constrictions of the same length. Surface ciliated. (Resembles a butterfly.)
Dimensions.—Radius of the anterior arms 0.7, of the posterior 0.4; longitudinal
and transverse constrictions 0.4.
Habitat.—North Pacific, near Japan, Station 241, surface.
7. Myelastrum decaceros, n. sp. (Pl. 47,
fig. 7).
Anterior arms bifid, with one shallow incision at their distal end. Posterior arms of the same
length, but much broader, trifid, with two incisions (one larger and one smaller). Sagittal and
transverse constrictions of the same length. Surface spiny.
Dimensions.—Radius of all four arms 0.4; longitudinal and transverse constrictions
0.35.
Habitat.—Pacific, central area, Station 272, surface.
8. Myelastrum dodecaceros, n. sp. (Pl. 47, figs. 11,
11a).
Anterior arms trifid, with two shallow incisions at their distal end. Posterior arms about
one-third larger, also trifid, with one deeper anterior and one shallower posterior incision.
Sagittal constriction much longer than the transverse. Surface ciliated, covered with numerous
thin and long radial bristles (fig. 11a).
Dimensions.—Radius of the anterior arms 0.5, of the posterior 0.7; longitudinal
constriction 0.5, transverse 0.35.
Habitat.—Pacific, central area, Station 270, surface.
{555}
9. Myelastrum ciliatum, n. sp.
Anterior arms trifid, with two shallow incisions. Posterior arms somewhat smaller, also trifid,
with two deeper incisions. Sagittal constriction a little larger than the transverse. Margin of
the disk ciliated, with radial bristle-shaped spines, as prolongations of the inner radial beams,
arising from the central disk.
Dimensions.—Radius of the anterior arms 0.6, of the posterior 0.5; longitudinal
constriction 0.4, transverse 0.35.
Habitat.—South Pacific, Station 288, surface.
10. Myelastrum lobatum, n. sp.
Anterior arms somewhat broader but shorter than the posterior arms. Each arm four-lobed, with
three terminal shallow incisions of nearly equal size. Sagittal constriction smaller than the
transverse. Surface of the disk bristly.
Dimensions.—Radius of the anterior arms 0.4, of the posterior 0.5; longitudinal
constriction 0.3, transverse 0.35.
Habitat.—North Pacific, Station 253, surface.
11. Myelastrum rotula, n. sp.
Anterior arms little broader than the posterior, but of the same length. Each arm four-lobed,
with three terminal deep incisions of equal size. Sagittal and transverse constrictions equal. The
whole disk nearly circular, resembles a wheel with sixteen spokes. Surface smooth.
Dimensions.—Radius of all four arms 0.5; longitudinal and transverse constrictions
0.3.
Habitat.—Pacific, central area, Station 274, surface.
12. Myelastrum giganteum, n. sp.
Anterior arms somewhat broader and shorter than the posterior. Each arm four-lobed, with three
shallow terminal incisions, the middle incision twice as deep as the two laterals. Sagittal
constriction a little larger than the transverse. Margin ciliated, with radial bristle-shaped
spines as prolongations of the inner radial beams, proceeding from the central disk (as in
Myelastrum dodecaceros, Pl. 47,
figs. 11, 11a).
Dimensions.—Radius of the anterior arms 0.6, of the posterior 0.8; longitudinal
constriction 0.6, transverse 0.5.
Habitat.—Pacific, central area, Station 271, surface.
{556}
13. Myelastrum anomalum, n. sp. (Pl. 47,
fig. 9).
All four arms of different size and form; anterior arms broader, posterior longer; one anterior
arm trifid, the three other arms bifid; length of the branches unequal; all four angles between
the arms unequal. (This anomalous form, seen only once, may be an individual abnormality.)
Dimensions.—Radius of the arms 0.4 to 0.7; constrictions 0.3.
Habitat.—North Pacific, Station 237, surface.
Genus 240. Pentalastrum,[279] Haeckel,
1881, Prodromus, p. 461.
Definition.—Porodiscida with five simple, undivided,
chambered arms, without a patagium.
The genus Pentalastrum opens the small series of Euchitonida, in which the
shell is not provided with three or four arms, as usual, but with five. All forms of this little
group are rare. Some species resemble in their external form and in the articulation of their arms
certain forms of Asterida. In Pentalastrum, the most simple genus, the five arms are
simple, not forked, and without a patagium. It can be derived from Dictyastrum or
Stauralastrum by increase of the number of arms.
Subgenus 1. Pentalastrella, Haeckel.
Definition.—All five arms equal, with equal angles between them. Shell
a regular pentagon.
1. Pentalastrum asteracanthion, n. sp.
All five arms equal, club-shaped, at their thickened obtuse end three times as broad as at
their base, twice as long as broad. Angles between the arms equal.
Dimensions.—Radius of each arm 0.2, basal breadth 0.03, distal breadth 0.08.
Habitat.—Pacific, central area, Station 272, depth 2600 fathoms.
2. Pentalastrum astropecten, n. sp.
All five arms equal, with five to six distinct, simple joints, the basal joint two-thirds as
broad as the terminal joint, which bears a strong conical spine. Angles between the arms equal.
(Resembles Pentinastrum asteriscus, Pl. 44,
fig. 2, but has no patagium.)
Dimensions.—Radius of each arm 0.14, basal breadth 0.024, distal breadth
0.036.
Habitat.—Pacific, central area, Station 263, depth 2650 fathoms.
{557}
Subgenus 2. Pentalastromma, Haeckel.
Definition.—Arms of different sizes, one odd arm larger than the two
others; the opposite odd angle generally different from the four other angles.
3. Pentalastrum ophidiaster, n. sp. (Pl. 44, fig.
3).
Arms nearly triangular, at their obtuse truncated distal end twice as broad as at their base.
Four arms equal, with five joints each; the fifth arm twice as long, with seven joints. Angles
between the arms nearly equal; the odd angle a little larger.
Dimensions.—Radius of the larger odd arm 0.25, of the four smaller arms 0.15;
basal breadth 0.035, distal breadth 0.07.
Habitat.—Pacific, central area, Station 274, surface.
4. Pentalastrum cometa, n. sp.
Arms nearly cylindrical, at their obtuse truncated distal end one and a half times as broad as
at their base. Posterior odd arm very large, with eleven joints, about three times as long as the
two lateral arms (with five joints each) and four times as long as the two anterior arms (with
three joints each). Angles between the paired arms different; the two lateral angles smaller than
the two posterior, and these smaller than the odd anterior angle.
Dimensions.—Radius of the odd posterior arm 0.5, of the lateral arms 0.25, of the
anterior arms 0.18; basal breadth 0.05, distal breadth 0.08.
Habitat.—South Pacific, Station 288, surface.
Genus 241. Pentinastrum,[280] Haeckel,
1881, Prodromus, p. 461.
Definition.—Porodiscida with five simple, undivided,
chambered arms, connected by a patagium.
The genus Pentinastrum differs from the foregoing Pentalastrum only
in the development of a patagium or connecticulum between the arms, and bears therefore the same
relation to it that Histiastrum
does to Stauralastrum, or Hymeniastrum to Dictyastrum.
1. Pentinastrum asteriscus , n. sp. (Pl. 44, fig.
2).
All arms equal, twice as long as broad, at their base two-thirds as broad as at their truncated
distal end, which bears a strong, pyramidal, terminal spine. Each arm is divided by five
transverse septa into six joints or chambers, and each of these by a radial beam into a pair of
chambers. The five radial beams arise from the innermost chamber of the central disk, and end in
the five terminal {558}spines. The diameter of the
central disk is larger than the length of the arms. The angles between the arms are equal and
filled up by an incomplete patagium, so that the whole disk forms a regular pentagon with five
concave sides.
Dimensions.—Radius of each arm (without terminal spine) 0.14; breadth at their
base 0.02, at their terminal joint 0.03; radius of the central disk 0.06.
Habitat.—Pacific, central area, Station 266, depth 2750 fathoms.
2. Pentinastrum goniaster, n. sp.
? Stephanastrum sp., Bury, 1862, Polycystins of Barbados, pl. xx. fig.
1.
All five arms equal, four times as long as broad, club-shaped, at their globose distal end
twice as broad as at their base, and armed with a strong conical terminal spine. Diameter of the
central disk equals only one-third of the length of the arms. The articulation of the spongy arms
is somewhat obscure. Patagium complete, totally fills up the interbrachial spaces, so that the
whole disk forms a regular pentagon with five rectilinear sides, except that the terminal spines
project at the corners.
Dimensions.—Radius of each arm 0.18, basal breadth 0.02, distal breadth 0.04;
radius of the central disk 0.05.
Habitat.—South Pacific, Station 295, depth 1500 fathoms; also fossil
in the rocks of Barbados.
Genus 242. Pentophiastrum,[281] n.
gen.
Definition.—Porodiscida with five forked chambered arms,
without a patagium.
The genus Pentophiastrum differs from the two preceding genera by the
bifurcation of the five arms, and can be derived either from Pentalastrum by the
ramification of the distal ends of the arms, or from the similar Myelastrum by the increase
in the number of arms.
1. Pentophiastrum dicranastrum, n. sp.
All five arms equal, with equal angles between them. Each arm in the basal half simple, in the
distal half forked; both branches of it equal, with obtuse ends. (This regular species resembles
Dicranastrum furcatum, Pl. 47,
fig. 2, but with five rays instead of four; also the form of the arms is more slender and the
edges smooth.)
Dimensions.—Radius of each arm 0.25, breadth 0.05.
Habitat.—Equatorial Atlantic, Station 347, depth 2250 fathoms.
Subgenus Pentophiastromma, Haeckel.
Definition.—Arms of different size, one odd arm opposite to the angle
between both arm-pairs.
{559}
2. Pentophiastrum caudatum, n. sp. (Pl. 47, fig.
5).
Arms in pairs different; four arms in the basal half simple, in the distal half forked; the
fifth (posterior) odd arm simple, undivided, cylindrical; the anterior pair a little smaller than
the posterior; the neighboring branches of the two pairs on each side larger than the two others.
Axes of the arms and their branches straight.
Dimensions.—Radius of the arms about 0.5, breadth 0.12.
Habitat.—North Atlantic, Station 353, depth 2965 fathoms.
3. Pentophiastrum forcipatum, n. sp. (Pl. 47, fig.
4).
Arms in pairs different, all in the basal two thirds simple, in the distal third forked. Only
in the posterior (odd) arm both branches are equal, in the four others unequal. The common axis of
the posterior lateral pair is horizontal, perpendicular to the median line; the axes of the
anterior pair are pincer-like, concavely curved towards the median line or principal axis.
Dimensions.—Radius of the arms about 0.5, breadth 0.14.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
Genus 243. Hexalastrum,[282] Haeckel,
1881, Prodromus, p. 461.
Definition.—Porodiscida with six simple chambered arms,
without a patagium.
The genus Hexalastrum, together with the following Hexinastrum,
encloses those Euchitonida in which the number of the chambered arms surrounding the
central disk amounts to six. This is the highest number of these articulated marginal appendages
which is reached in any Discoidea. Formerly (1881) in my Prodromus, p.
459, I supposed that the same number was reached also by one Coccodiscid, and called this genus
Hexactura. Afterwards I was convinced that this form was also a Hexalastrum.
1. Hexalastrum palmanthum, n. sp.
All six arms equal, with equal angles between them. Each arm club-shaped, three times as long
as broad, twice as broad at the thickened distal end as at the base, without a terminal spine.
Dimensions.—Radius of each arm 0.2, basal breadth 0.02, distal breadth 0.06.
Habitat.—Pacific, central area, Station 274, depth 2750 fathoms.
2. Hexalastrum crinanthum, n. sp.
All six arms equal, with equal angles between them. Each arm club-shaped, four times as long as
broad, at the egg-shaped distal end three times as broad as in the linear basal part, provided
{560}with numerous short conical spines and one longer
terminal spine. (Resembles Stauralastrum rhopalophorum, Pl. 45, fig. 1, but
with six rays instead of four.)
Dimensions.—Radius of each arm 0.3, basal breadth 0.03, distal breadth 0.08.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
Subgenus Hexalastromma, Haeckel.
Definition.—Arms more or less different in size or form; shell
bilateral.
3. Hexalastrum orchidaceum, n. sp. (Pl. 44, fig.
5).
Hexactura orchidacea, Haeckel, 1881, Prodromus, p. 459.
Arms different in length, so that two unequal opposite odd arms determine the main axis, and
the four other arms lie on both sides of this as two different pairs. The proportion of their
relative length is the following:—anterior lateral arms five, anterior odd arm six;
posterior lateral arms seven, posterior odd arm eight. Each arm is club-shaped, two to three times
as long as broad, and divided into six to eight joints by five to seven transverse septa; its
distal end is armed with a terminal spine and twice as broad as its base.
Dimensions.—Radius of the posterior odd arm 0.4, of the anterior odd arm 0.3; of
the posterior lateral pair 0.35, of the anterior lateral pair 0.25; basal breadth 0.08, distal
breadth 0.16.
Habitat.—Pacific, central area, Station 266, depth 2750 fathoms.
Genus 244. Hexinastrum,[283] Haeckel,
1881, Prodromus, p. 461.
Definition.—Porodiscida with six simple, undivided,
chambered arms, connected by a patagium.
The genus Hexinastrum differs from its ancestral form Hexalastrum by
the development of a patagium between the arms. The only observed species is regular.
1. Hexinastrum geryonidum, n. sp. (Pl. 44, fig.
4).
Hexalastrum geryonidum, Haeckel, 1879, Atlas (pl. xliv. fig. 4).
Disk quite regular with six radii; all six arms of the same size and form, at their broad,
convexly rounded, smooth end five times as broad as at their narrow base, and little longer than
broad. Each arm is divided by eight transverse septa into nine simple joints or chambers of the
same height; the breadth of the distal chambers increases rapidly. The regular, hexagonal, central
disk exhibits four concentric rings around the central chamber. Patagium between the arms
incomplete, with concavely fluted edge.
Dimensions.—Radius of each arm 0.15, basal breadth 0.016, distal breadth 0.08;
radius of the central disk 0.04.
Habitat.—Pacific, central area, Station 272, depth 2600 fathoms.
{561}
Family XXII. Pylodiscida, n. fam. (Pl. 48, figs.
12-20).
Definition.—Discoidea without phacoid shell, with flat
discoidal shell, in which a simple spherical central chamber is surrounded by one or two
concentric triradial girdles; each girdle with three gates, separated by three simple
arm-chambers. Surface of the disk with three open or latticed gates on each flat side.
The family Pylodiscida represents a new small but interesting group of
Discoidea, which exhibits rather complex affinities to different groups
of Sphærellaria. In my Prodromus (1881, p. 464) I had enumerated only two
genera of this family, Triopyle and Hexapyle, and had united them with
Tetrapyle and allied genera in the family Pylonida. Indeed, the resemblance of skeletal
structure in the two groups is very great. The most simple forms of both groups exhibit a simple
spherical latticed central chamber, which is surrounded by few latticed chambers of similar size
and form, separated by open gates. But in the Pylonida these chambers are opposite in pairs, and
form together a complete lattice-girdle around the central chamber, whereas in the Pylodiscida the
chambers are not opposite in pairs in one axis, and form therefore only latticed half girdles,
which arise from the central chamber like radial arms, and may perhaps better be called
"arm-chambers"; their number is constantly three. The free open spaces between these three
arm-chambers form three gates, comparable to the two or four gates of Amphipyle,
Tetrapyle, &c., and become afterwards closed by lattice-work in a similar way in both
groups. A more important difference between them is indicated by the further mode of growth. The
Pylonida build new girdles in all three dimensive planes (alternating in the transverse, lateral
and sagittal planes); their geometric fundamental form is therefore the "lentellipsis" or the
"triaxial ellipsoid." The Pylodiscida, however, grow only at the periphery of the discoidal shell
in one single plane (the equatorial plane); their fundamental form is therefore the biconvex lens
or the flat disk (a shortened cylinder). This important difference is my deciding motive, in
separating the latter from the former and in regarding the Pylodiscida as true Discoidea, the more so as they can easily be derived from Archidiscus,
the fundamental and ancestral form of the Porodiscida.
One single form of Archidiscus seems to be of peculiar importance in this relation,
viz., Archidiscus hexoniscus
(Pl. 48,
fig. 10). In this species the simple central chamber is surrounded by a latticed ring or girdle,
composed of six equal chambers of the same size and form, all lying in the same plane with the
central chamber. In a nearly allied species, viz., Archidiscus pyloniscus, the six
ring-chambers are different, three smaller (with denser network) alternating with three larger (of
looser network); if we imagine the network of the latter reduced to a marginal bar we get
Triopyle, and if also this bar disappear by reduction we get Triolena, the most
simple form of the Pylodiscida. {562}Even Archidiscus
pyloniscus may possibly represent the same form among the Porodiscida as Triodiscus
lenticula among the Pylodiscida; this important form indicates clearly the close affinity of
the two families.
If we take the latter, nearly identical form as the common starting point of both families of
Cyclodiscaria, then probably Triopyle and Triolene must be regarded as retrograde
forms, derived from Triodiscus in the one case, from Archidiscus in the other, by
reduction of three interradial arm-chambers, whilst three perradial only remain. But it is also
possible that the most simple form, Triolene, originated independently from some
Cenosphæra, three simple radial chambers, like the latticed central chamber, being derived
from the latter by apposition in three equidistant radii, whilst three other radii between them
remained free. In this case the other genera of Pylodiscida are derived from their ancestral form
Triolene.
Adopting this latter view, we find that all eight genera of Pylodiscida, here distinguished,
may be regarded as following members of a continuous series. If the three simple arm-chambers of
Triolene, surrounding the equal central chamber, become united at their distal ends by a
concentric equatorial ring, then originates Triopyle; and this graduates into
Triodiscus by fenestration of the three open gates between the three latticed arms. Whilst
these three genera form together the subfamily Triopylida, a second family, Hexapylida, is
composed of three other analogous genera, in which the same process of development becomes
repeated.
Pylolena, the most simple form of Hexapylida, arises from Triodiscus by the
development of three new arm-chambers (of the second order) which are apposed at the distal end of
the three primary arm-chambers (of the first order) in the same radius. If the distal ends of
these three secondary arm-chambers become united by a concentric latticed ring or girdle, we get
Hexapyle (with six open gates, two in each radius), and if its six gates become afterwards
closed by loose lattice-work, we arrive at Pylodiscus (a repetition of
Triodiscus).
A third subfamily, Discopylida, is formed by the building of a chambered equatorial girdle
around the margin of Pylodiscus. This girdle has quite the same structure as the similar
chambered rings or girdles of the Porodiscida and Coccodiscida. Between the two sieve-plates of
the disk surface is enclosed a variable number (twelve to twenty-four or more) of chambers,
imperfectly separated by radial beams, which connect the margin of the Pylodiscus-shell
with an outer peripheral concentric ring. In Discozonium this marginal ring is perfect,
whilst in Discopyle it is interrupted by a peculiar large opening, a "marginal osculum"
surrounded by a corona of spines, quite the same remarkable formation which we encountered in
Ommatodiscus among the Porodiscida.
All Pylodiscida are therefore triradial (with three perradial arms and three interradial gates
between them), and many of them have a great resemblance to certain triradial Porodiscida and
Spongodiscida, perhaps not only a morphological resemblance, but also {563}a true phylogenetic relation. But it is remarkable that we do not find
further forms of development in this family, by multiplication either of the arm-chambers (further
growth in the three perradii) or of the concentric chambered rings (in the periphery of the disk
margin).
The central capsule of the Pylodiscida is constantly flat, discoidal, and enclosed between the
two sieve-plates of the surface. Its form is either circular or triangular.
Synopsis of the Genera of the Pylodiscida.
|
I. Subfamily Triopylida.
Three gates between three simple arm-chambers.
|
|
Three gates open, without a barring equatorial girdle, |
245. Triolena. |
| Three gates barred by a latticed equatorial girdle. |
|
Gate-faces simple, |
246. Triopyle. |
| Gate-faces latticed, |
247. Triodiscus. |
|
II. Subfamily Hexapylida.
Six gates between three double arm-chambers (three inner and three outer); no
chambered marginal girdle.
|
|
Three outer gates open, without a barring equatorial
girdle, |
248. Pylolena. |
| Three outer gates barred by a latticed (second) equatorial
girdle. |
|
Both faces of the outer gates simple, |
249. Hexapyle. |
| Both faces of the outer gates latticed, |
250. Pylodiscus. |
|
III. Subfamily Discopylida.
Six gates between three double arm-chambers.
|
|
In the equatorial plane on the margin of the
Pylodiscus-shell is a chambered equatorial girdle. |
|
No peculiar osculum on the margin of the disc, |
251. Discozonium. |
| One peculiar osculum (with a corona of spines) on the margin of the
disc, |
252. Discopyle. |
| I. Subfamily Triopylida. Three gates between three simple arm-chambers. |
|
|
Three gates open, without a barring equatorial girdle, |
|
|
|
|
|
|
|
|
245. Triolena. |
|
|
|
|
|
|
|
|
Three gates barred by a latticed equatorial girdle. |
|
|
|
|
|
|
Gate-faces simple, |
|
|
|
|
|
|
|
|
|
|
246. Triopyle. |
|
|
|
|
|
|
|
|
|
|
Gate-faces latticed, |
|
|
|
|
|
|
|
|
|
|
247. Triodiscus. |
|
|
|
|
|
|
| II. Subfamily Hexapylida. Six gates between three double arm-chambers (three
inner and three outer); no chambered marginal girdle. |
|
|
Three outer gates open, without a barring equatorial girdle, |
|
|
|
|
|
|
|
|
248. Pylolena. |
|
|
|
|
|
|
|
|
Three outer gates barred by a latticed (second) equatorial
girdle. |
|
|
|
|
|
|
Both faces of the outer gates simple, |
|
|
|
|
|
|
|
|
|
|
249. Hexapyle. |
|
|
|
|
|
|
|
|
|
|
Both faces of the outer gates latticed, |
|
|
|
|
|
|
|
|
|
|
250. Pylodiscus. |
|
|
|
|
|
|
| III. Subfamily Discopylida. Six gates between three double arm-chambers. |
|
|
In the equatorial plane on the margin of the Pylodiscus-shell is a
chambered equatorial girdle. |
|
|
|
|
|
|
No peculiar osculum on the margin of the disc, |
|
|
|
|
|
|
|
|
|
|
251. Discozonium. |
|
|
|
|
|
|
|
|
|
|
One peculiar osculum (with a corona of spines) on the margin of
the disc, |
|
|
|
|
|
|
|
|
|
|
252. Discopyle. |
|
|
|
|
|
|
Subfamily 1. Triopylida, Haeckel.
Definition.—Pylodiscida with a simple,
spherical or lenticular, central chamber, surrounded by three simple arm-chambers, which are
separated by three notches or gates.
Genus 245. Triolena,[284] n. gen.
Definition.—Pylodiscida with a simple, spherical or
lenticular, central chamber, surrounded by three simple arm-chambers. Notches between the three
arms open.
The genus Triolena is the most simple form of all Pylodiscida, and must be
regarded as their common ancestral form, from an ontogenetic as well as a phylogenetic point of
view. The small shell is composed of a simple, spherical or lenticular, latticed, central chamber,
and of three simple, surrounding equal arms, which are also simple latticed chambers, lie in the
equatorial plane, and are separated by three equal angles or open gates.
{564}
1. Triolena primordialis, n. sp. (Pl. 48, fig.
12).
Arm-chambers trapezoid, nearly square, of the same size as the circular, lenticular,
primordial, central chamber. Surface of the disk smooth.
Dimensions.—Diameter of the shell 0.045, of the central chamber 0.015, of each arm
0.015.
Habitat.—Central Pacific, Station 272, depth 2600 fathoms.
2. Triolena tribelone, n. sp.
Arm-chambers lanceolate, of the same breadth as and twice the length of the triangular, central
chamber; at the pointed end of each arm is a conical terminal spine (in the equatorial plane).
Surface of the disk thorny.
Dimensions.—Diameter of the shell 0.05, of the central chamber 0.015; length of
the arms 0.02, breadth 0.016.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
3. Triolena trispinosa, n. sp.
Arm-chambers ovate, in the basal half nearly as broad as the hexagonal central chamber, at the
pointed distal end with a strong conical radial spine of double the length. Surface rough.
Dimensions.—Diameter of the shell 0.055, of the central chamber 0.02.
Habitat.—Western Tropical Pacific, Station 225, depth 4475
fathoms.
4. Triolena hexabelone, n. sp.
Arm-chambers nearly triangular, at the base half as broad as the hexagonal central chamber, at
the truncate distal end one and a half times as broad, and armed with two radial conical spines
(in the equatorial plane). Surface of the disk smooth.
Dimensions.—Diameter of the shell 0.06, of the central chamber 0.02.
Habitat.—South Pacific, Station 295, depth 1500 fathoms.
5. Triolena trigonalis, n. sp.
Arm-chambers nearly triangular, at the base half as broad as the circular central chamber, at
the concave lunulate distal end twice as broad, and armed with four conical radial spines (two on
each side of the equatorial plane). Surface thorny.
Dimensions.—Diameter of the shell 0.05, of the central chamber 0.015.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
{565}
Genus 246. Triopyle,[285] Haeckel, 1881,
Prodromus, p. 464.
Definition.—Pylodiscida with a simple, spherical or
lenticular, central chamber, surrounded by three simple arm-chambers. Notches between the three
arms transformed into gates by a connecting equatorial girdle.
The genus Triopyle differs from the preceding Triolene in the
development of a simple ring or latticed equatorial girdle, which connects the distal ends of the
three arm-chambers, and transforms the open notches between them into three gates. The ring may be
circular, triangular, or hexagonal.
1. Triopyle circulus, n. sp.
Disk circular, three times as broad as the hexagonal central chamber. Three arm-chambers
trapezoidal, at the convex distal end as broad, at the base half as broad as the three circular
gates between them. Surface smooth. No marginal spines on the girdle.
Dimensions.—Diameter of the disk 0.045, of the gates 0.015.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
2. Triopyle hexagona, n. sp. (Pl. 48,
fig. 13).
Disk hexagonal, three times as broad as the circular central chamber. Three arm-chambers
trapezoidal, at the truncated distal end as broad, at the base half as broad as the three
triangular gates between them. Surface smooth. No marginal spines.
Dimensions.—Diameter of the disk 0.05, of the gates 0.02.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
3. Triopyle trigona, n. sp.
Disk triangular, four times as broad as the circular central chamber. Three arm-chambers nearly
triangular, at the narrow base half as broad, at the distal end twice as broad as the circular or
roundish gates. Surface smooth. On the margin (in the equatorial plane) three strong pyramidal
spines (at the end of the arms).
Dimensions.—Diameter of the disk 0.04, of the gates 0.015.
Habitat.—Central Pacific, Station 274, depth 2750 fathoms.
4. Triopyle cordigera, n. sp.
Disk hexagonal, four times as broad as the hexagonal central chamber. Three arm-chambers
triangular, at the base half as broad as at the truncated distal end, about the same size as the
{566}three heart-shaped gates. On the six corners of the
margin (which forms a regular hexagon) are six pyramidal radial spines, as prolongations of the
arm-edges.
Dimensions.—Diameter of the disk 0.05, of the gates 0.02.
Habitat.—Indian Ocean, Ceylon, surface, Haeckel.
5. Triopyle renigera, n. sp.
Disk hexagonal, five times as broad as the circular central chamber. Three arm-chambers
trapezoidal, at the base one-third, at the distal end two-thirds as broad as the three
kidney-shaped gates. On the six corners of the margin (which forms an irregular hexagon) six
conical radial spines, as prolongations of the arm-edges.
Dimensions.—Diameter of the disk 0.06, of the gates 0.02.
Habitat.—South Pacific, Station 295, depth 1500 fathoms.
6. Triopyle spinigera, n. sp.
Disk roundish, triangular, four times as broad as the triangular central chamber, which is
armed with three radial spines between the arms. Arm-chambers club-shaped, at the narrow base
one-fourth, at the distal end half as broad as the square gates. On the margin twelve large
conical spines, two opposite on each face of the distal end of each arm-edge. Three smaller radial
spines on the three corners of the girdle (in the same interradial meridian planes as the three
spines of the central chamber). Compare Triodiscus spinosus.
Dimensions.—Diameter of the disk 0.05, of the gates 0.02.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
Genus 247. Triodiscus,[286] n. sp.
Definition.—Pylodiscida with a simple, spherical or
lenticular, central chamber, surrounded by three simple arm-chambers. Notches between the three
arms closed by lattice-work and by an equatorial girdle.
The genus Triodiscus differs from the preceding Triopyle in the
development of loose lattice-work on both sides of the discoidal shell. This network closes the
gates and transforms the whole shell into a fenestrated lens. The singular species of
Triodiscus correspond to certain species of Triopyle.
1. Triodiscus lenticula, n. sp.
Disk circular, lenticular, three times as broad as the central chamber. Three arm-chambers
trapezoidal, of the same size and form as the three gates between them, which are closed by a
loose {567}delicate network (differs from Triopyle
circulus by the production of the two convex latticed plates, which envelop the whole lens).
Margin smooth.
Dimensions.—Diameter of the disk 0.045, of the gates 0.015.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
2. Triodiscus trigonus, n. sp.
Disk triangular, four times as broad as the central chamber. Three arm-chambers at the base
half as broad, at the distal end twice as broad as the roundish gates. Surface smooth. On the
three corners of the margin (in the arm-radius) three strong spines. (Differs from Triopyle
trigona only in the loose framework closing the gates.)
Dimensions.—Diameter of the disk 0.04, of the gates 0.015.
Habitat.—Central Pacific, Station 272, depth 2600 fathoms.
3. Triodiscus spinosus, n. sp. (Pl. 48,
fig. 14).
Disk subcircular, four times as broad as the triangular central chamber. Three arm-chambers
club-shaped, at the base one-third, at the distal end two-thirds as broad as the semicircular
gates. Surface thorny. On the margin fifteen larger radial spines, three on the corners of the
disk (in the radius of the gates), twelve on the two faces of the arms ends (two opposite on the
edge of each end.) (Differs from Triopyle spinigera mainly by the delicate hexagonal
network closing the gates.)
Dimensions.—Diameter of the disk 0.05, of the gates 0.02.
Habitat.—North Atlantic, Canary Islands (Lanzerote, Haeckel).
Subfamily 2. Hexapylida, Haeckel.
Definition.—Pylodiscida with triopyle-shaped
medullary shell, surrounded by three distal arm-chambers, which are separated by three open
notches or gates.
Genus 248. Pylolena,[287] n. gen.
Definition.—Pylodiscida with triopyle-shaped medullary
shell, surrounded by three distal arm-chambers. Notches between the three arms open.
The genus Pylolena opens the series of the Hexapylida, or of those
Pylodiscida in which the centre of the shell is formed by a tri-radiated medullary shell like
Triopyle. In the equatorial plane of this triopyle-shaped disk are developed on its margin
three distal arm chambers, as prolongations of the three arms of Triopyle, but much larger.
In Pylolene the three angles or notches between the distal arms remain open, repeating the
form of Triolene.
{568}
1. Pylolena inermis, n. sp.
Arms club-shaped, at the base half as broad, at the rounded distal end as broad as the
triopyle-shaped medullary shell. Surface and margin smooth.
Dimensions.—Diameter of the cortical shell 0.15, of the medullary shell 0.05.
Habitat.—Equatorial Atlantic, Station 347, surface; Central Pacific,
Station 272, surface.
2. Pylolena armata, n. sp. (Pl. 48,
fig. 15).
Arms trapezoid, at the base half as broad, at the distal end twice as broad as the
triopyle-shaped medullary shell. Surface and margin thorny. Twelve strong conical spines at the
distal end of the three arms, two opposite on both faces of the arm-edges.
Dimensions.—Diameter of the cortical shell 0.2, of the medullary shell 0.06.
Habitat.—South Atlantic, Station 325, surface.
Genus 249. Hexapyle,[288] Haeckel, 1881,
Prodromus, p. 464.
Definition.—Pylodiscida with triopyle-shaped medullary
shell, surrounded by three distal arm-chambers. Notches between the three arms transformed into
gates by a connecting equatorial girdle.
The genus Hexapyle differs from the preceding Pylolene in the
development of an equatorial ring or latticed girdle, which connects the free extremities of the
three distal ends and transforms the open notches between them into gates. It simulates therefore
the formation of Triopyle, from which it differs by duplication of the arm-joints and of
the gates (in each radius occur one proximal and one distal gate).
1. Hexapyle triangula, n. sp.
Cortical shell triangular, with three rounded corners, three times as broad as the triangular
triopyle-shaped medullary shell. Surface smooth or rough, but not spiny. Three arms two-thirds as
broad as the three egg-shaped gates of each side.
Dimensions.—Diameter of the cortical shell (or length of one side of the triangle)
0.15, of the medullary shell 0.05; breadth of the gates 0.06, of the bridges between them
0.04.
Habitat.—Pacific, central area, Station 272, depth 2600 fathoms.
2. Hexapyle sexangula, n. sp.
Cortical shell hexagonal, with six equal sides, four times as broad as the triangular
triopyle-shaped medullary shell. Surface rough, but not spiny. Three arms (on their smallest part)
half as broad as the three triangular gates (on their broadest part).
{569}
Dimensions.—Diameter of the cortical shell 0.16, of the medullary shell 0.04;
breadth of the gates 0.06, of the bridges 0.03.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
3. Hexapyle circularis, n. sp.
Cortical shell circular, four times as broad as the triangular, Triopyle-shaped medullary
shell. Surface smooth. Three arms half as broad as the three kidney-shaped gates on each side of
the shell.
Dimensions.—Diameter of the cortical shell 0.2, of the medullary shell 0.05;
breadth of the gates 0.08, of the bridges between them 0.04.
Habitat.—Pacific, central area, Station 266, depth 2750 fathoms.
4. Hexapyle triacantha, n. sp.
Cortical shell triangular, thorny, with three pointed corners, prolonged into three strong
conical radial spines; its diameter four times as great as that of the medullary shell. Three arms
of the same breadth as the three egg-shaped gates of each side.
Dimensions.—Diameter of the cortical shell (or length of one side of the triangle,
without spines) 0.2, of the medullary shell 0.05; breadth of the gates and of the bridges between
them 0.06.
Habitat.—Indian Ocean, surface; Madagascar, Rabbe.
5. Hexapyle hexacantha, n. sp.
Cortical shell hexagonal, thorny, three times as broad as the medullary shell. Six stronger
conical radial spines on the six corners at equal distances, lying in the equatorial plane of the
disk. Three half girdles half as broad as the three triangular gates of each side.
Dimensions.—Diameter of the cortical shell 0.15, of the medullary shell 0.05;
breadth of the gates 0.05, of the bridges 0.025.
Habitat.—North Pacific, Station 253, depth 3125 fathoms.
6. Hexapyle dodecantha, n. sp. (Pl. 48,
fig. 16).
Cortical shell triangular, thorny, with rounded corners, three and a half times as broad as the
medullary shell. Twelve stronger radial spines, six opposite in pairs on each side of the
discoidal shell, as prolongations of the lateral edges of the triangular gates, which are about
the same breadth as the half girdles between them.
Dimensions.—Diameter of the cortical shell 0.21, of the medullary shell 0.06;
breadth of the gates and of the bridges 0.06.
Habitat.—Pacific, central area, Station 270, surface.
{570}
7. Hexapyle polyacantha, n. sp.
Cortical shell circular, thorny, with a circle of numerous conical radial spines in the
periphery; its diameter nearly three times as great as that of the medullary shell. Three
half-girdles half as broad as the three kidney-shaped gates on each side of the discoidal
shell.
Dimensions.—Diameter of the cortical shell 0.11, of the medullary shell 0.04;
breadth of the gates 0.04, of the bridges between them 0.02.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
Genus 250. Pylodiscus,[289] n. gen.
Definition.—Pylodiscida with Triopyle-shaped
medullary shell, surrounded by three distal arm-chambers. Notches between the three arms closed by
lattice-work and by an equatorial girdle.
The genus Pylodiscus differs from Hexapyle in the development of two
convex plates of lattice-work, which close the six open gates and transform the disk into a
biconvex fenestrated lens. Pylodiscus simulates therefore the characteristic form of
Triodiscus; but whilst in the latter we find only three arm-joints and three simple gates,
their number is doubled in the former.
1. Pylodiscus triangularis, n. sp. (Pl. 48, fig.
17).
Cortical shell triangular, with rounded corners, three times as broad as the triangular,
Triopyle-shaped medullary shell. Three arms about as broad as the three egg-shaped gates
between them. (Differs from Hexapyle triangula only by the delicate network of both
triangular convex covering plates, which close the open gates.) Six larger and many smaller spines
on the margin of the disk.
Dimensions.—Diameter of the disk 0.15, of the gates 0.05.
Habitat.—Pacific, central area, Station 270, depth 2925 fathoms.
2. Pylodiscus sexangularis, n. sp.
Cortical shell hexangular, equilateral, four times as broad as the circular,
Triopyle-shaped medullary shell. Surface spiny. Three arms nearly of the same form and size
as the three triangular gates between them. (Similar to Hexapyle sexangularis, but
differing in the irregular loose network closing the gates.)
Dimensions.—Diameter of the disk 0.18, of the gates 0.06.
Habitat.—North Pacific, south of Japan, Station 237, surface
{571}
3. Pylodiscus cardiopylus, n. sp.
Cortical shell hexangular, equilateral, five times as broad as the triangular,
Triopyle-shaped medullary shell. Surface smooth. Three arms triangular, at the distal end
as broad as the three heart-shaped gates. On the margin six strong pyramidal spines in the
equatorial plane (adradial, as prolongations of the lateral arm-edges).
Dimensions.—Diameter of the disk 0.15, of the gates 0.05.
Habitat.—Indian Ocean, Sunda Strait, Rabbe.
4. Pylodiscus nephropylus, n. sp.
Cortical shell circular, three times as broad as the circular, Triopyle-shaped medullary
shell. Surface spiny. Three arms trapezoidal, half as broad as the three kidney-shaped gates. On
the margin twelve stronger, conical, radial spines, opposite in pairs on both sides of the disk,
disposed in six pairs at the distal end of the arms.
Dimensions.—Diameter of the disk 0.2, of the medullary shell 0.06.
Habitat.—Central Pacific, Station 268, depth 2900 fathoms.
Subfamily 3. Discopylida, Haeckel.
Definition.—Pylodiscida with
Triopyle-shaped medullary shell and Pylodiscus-shaped cortical shell, which is
surrounded by a marginal chambered equatorial girdle.
Genus 251. Discozonium,[290] n. gen.
Definition.—Pylodiscida with Triopyle-shaped
medullary shell and Pylodiscus-shaped cortical shell, which is surrounded by an equatorial
chambered girdle. No peculiar osculum on the margin of the disk.
The genus Discozonium and the following Discopyle make up together
the small group of the Discopylida, or those Pylodiscida in which a discoidal shell like
Pylodiscus is surrounded by a marginal equatorial girdle; this girdle is divided into
twelve to twenty-four or more chambers by radial beams, which are the external prolongations of
the radial marginal spines of Pylodiscus. The latter genus bears therefore to
Discozonium the same relation as Sethodiscus in the other Discoidea does to Lithocyclia, or Phacodiscus to
Coccodiscus.
1. Discozonium cyclonium, n. sp.
Disk circular, lenticular, with smooth margin, three times as broad as the triangular,
Triopyle-shaped medullary shell. Three gates of the cortical shell kidney-shaped, twice as
broad as the three {572}arms. Chambered equatorial
girdle with twenty to twenty-four subregular chambers (similar to Discopyle osculata, Pl.
48, fig. 19,
but without any marginal osculum).
Dimensions.—Diameter of the disk 0.2, of the cortical shell 0.14, of the medullary
shell 0.07.
Habitat.—Central Pacific, Station 265, depth 2900 fathoms.
2. Discozonium trigonium, n. sp.
Disk triangular, with thorny margin, four times as broad as the triangular,
Triopyle-shaped medullary shell. Three gates of the cortical shell egg-shaped, scarcely as
broad as the three arms. Chambered equatorial girdle with twenty-four to thirty irregular
chambers.
Dimensions.—Diameter of the disk 0.24, of the cortical shell 0.2, of the medullary
shell 0.06.
Habitat.—Central Pacific, Station 274, depth 2750 fathoms.
3. Discozonium hexagonium, n. sp. (Pl. 48, fig.
18).
Disk hexagonal, with spiny margin, four times as broad as the triangular,
Triopyle-shaped medullary shell. Three gates of the cortical shell kidney-shaped, one and a
half times as broad as the arms. Chambered equatorial girdle with twelve large regular chambers;
the radial beams between them are prolonged into twelve strong pyramidal marginal spines (three
perradial on the ends of the arms, three interradial on the radii of the gates, six adradial
between the former and latter).
Dimensions.—Diameter of the disk 0.2, of the cortical shell 0.12, of the medullary
shell 0.05.
Habitat.—Central Pacific, Station 271, depth 2425 fathoms.
Genus 252. Discopyle,[291] n. gen.
Definition.—Pylodiscida with Triopyle-shaped
medullary shell and Pylodiscus-shaped cortical shell, which is surrounded by an equatorial
chambered girdle. One peculiar osculum, surrounded by a corona of spines, on the margin of the
disk.
The genus Discopyle differs from the preceding genus Discozonium in
the development of a peculiar marginal osculum, and bears therefore to it the same relation as, in
the Porodiscida, Ommatodiscus does to Porodiscus (compare above, p. 500). This peculiar osculum is here also surrounded by a corona of spines, and
serves probably for the exit or outlet of a bunch of pseudopodia or a "sarcode-flagellum." Only
two species of Discopyle have been observed, which represent perhaps better two different
genera; in one species the disk is circular, in the other elliptical. In this latter the osculum
lies on one pole of the main axis.
{573}
1. Discopyle osculata, n. sp. (Pl. 48,
fig. 19).
Disk circular with spiny margin, three times as broad as the triangular, Triopyle-shaped
medullary shell. Three gates of the cortical shell kidney-shaped, on the inside with an
interradial spine, twice as broad as the three pentagonal arms. Chambered equatorial girdle with
twenty-four subregular chambers, in the radius of one odd gate with a large marginal osculum,
which is as broad as the medullary shell, and surrounded by a dense corona of twenty to thirty
strong conical spines.
Dimensions.—Diameter of the disk 0.15, of the cortical shell 0.1, of the medullary
shell 0.05, of the marginal osculum 0.06.
Habitat.—Central Pacific, Station 272, depth 2600 fathoms.
2. Discopyle elliptica, n. sp. (Pl. 48,
fig. 20).
Disk elliptical, four-fifths as broad as long, with spiny margin, three times as broad as the
triangular, Triopyle-shaped medullary shell. Three gates of the cortical shell roundish, on
the inside with an interradial spine, little broader than the quadrangular arms. Chambered
equatorial girdle with twenty to thirty irregular chambers, on one pole of the main axis with a
large marginal osculum, which is one-third as broad as the length of the main axis, and armed with
a corona of twenty to thirty short conical spines. The osculum does not correspond to a certain
radius.
Dimensions.—Diameter of the disk 0.15, of the cortical shell 0.08, of the
medullary shell 0.04, of the marginal osculum 0.05.
Habitat.—Central Pacific, Station 267, depth 2700 fathoms.
Family XXIII. Spongodiscida, Haeckel (Pl. 41, fig.
11).
Spongodiscida et Spongocyclida, Haeckel, 1862, Monogr. d. Radiol.,
pp. 452, 460, 469.
Spongodiscida, Haeckel, 1881, Prodromus, p. 461.
Calodictya, Ehrenberg, 1847, Monatsber. d. k. preuss. Akad. d. Wiss.
Berlin, p. 53 (partim).
Definition.—Discoidea without a phacoid shell, with a
flat discoidal shell, in which a simple spherical central chamber is surrounded by an irregular
spongy framework (sometimes with concentric rings around the central chamber). Surface of the disk
quite spongy, without porous sieve-plates.
The family Spongodiscida is the sixth and last family of the Discoidea, and bears to the other families of this group the same relation as
the family Spongosphærida does to the other Sphæroidea, or the Spongurida
to the other Prunoidea. Its characteristic structure consists in the
irregular spongy framework of the disk, and mainly in the rough, irregular shape of its spongy
surface, which is never covered with porous plates (neither phacoid shell nor corresponding
sieve-plates), as in all other Discoidea. Of course a little spongy
structure occurs also in many Porodiscida {574}and even
in some Coccodiscida (principally in the peripheral part of the disk or its chambered arms); but
both flat (or convex) surfaces of the disk (at least in the central part) remain here constantly
as simple lattice-plates, whilst in all Spongodiscida the whole surface of the disk is spongy.
When I constituted the family Spongodiscida in my Monograph (1862, pp. 452, 460) I had
separated from them the Spongocyclida, exhibiting in the central part of the disk a more or less
distinct concentric arrangement of the spongy chambers, whilst in the former the delicate spongy
framework is quite irregular, composed of branched siliceous threads, connected and interwoven in
all directions. But in all Spongocyclida the whole surface of the spongy disk is quite as
irregularly rough and deprived of smooth sieve-plates as in all true Spongodiscida, and the more
or less concentric structure of the central part of the disk in the former (very variable and
often scarcely able to be recognised) seems not sufficient to separate both groups; even the
single genera cannot be sufficiently separated by this character. I now therefore give up entirely
the group of Spongocyclida (as already done in my Prodromus, 1881). Nevertheless the concentric
annular structure in the dark central part of some Spongodiscida is very interesting as transition
to the Porodiscida; it indicates already that the former are derived from the latter. Even the
single genera in both families are corresponding.
In the new system of "Polycystina," which Ehrenberg gave, 1875 (Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 157), are enumerated under the Calodictya four genera "with spongy disk," viz.,
Spongodiscus, Rhopalodictyum, Dictyocoryne, Spongaster. Indeed these
four genera, which I here retain, are true Spongodiscida, and must be separated from the other
Calodictya, the greater part of which are Porodiscida. The number of species of true
Spongodiscida now amounts to sixty-seven, which I dispose in thirteen genera.
The whole family may be divided into three subfamilies. The first of these are the
Spongophacida (corresponding to the Trematodiscida among the Porodiscida), in which the circular
margin of the spongy disk bears no radial appendages; either the margin is quite simple, spongy
(Spongodiscus), or surrounded by a hyaline, solid, or porous equatorial girdle
(Spongophacus). The disk is either more lenticular (biconvex) or more flat discoidal (a
shortened cylinder), rarely a little biconcave (thicker at the margin than in the centre). The
spongy framework of the solid disk is either quite irregular (Spongodisculus), or in the
central part with concentric circular rings (Spongocyclia), or in the central part spirally
convoluted (Spongospira).
The second subfamily, Spongotrochida, corresponds to the Stylodictyida (among the Porodiscida), and is distinguished by solid radial spines on the margin of the
disk, disposed in the equatorial plane either irregularly or regularly (after the same order as in
the other families of Discoidea).
The third subfamily, Spongobrachida, correspond perfectly to the Euchitonida {575}(among the Porodiscida), bearing on the disk-margin two,
three, or four spongy arms, commonly disposed regularly in the equatorial plane. Here also occurs
the peculiar formation of a "patagium," or of an interbrachial spongy framework different from
that of the arms, which connects the arms like a web-membrane in the equatorial plane.
The spongy framework exhibits in all these Spongodiscida no remarkable differences, being
everywhere composed of fine branched solid siliceous threads, interwoven in all directions, with
irregular meshes of very different size.
The central capsule of all Spongodiscida is filled up with the same spongy framework which
covers also both its sides. It grows according to the enveloping skeleton, but remains constantly
smaller. The form of the central capsule is circular (lenticular or discoidal) in the
Spongophacida and Spongotrochida, whilst in the Spongobrachida it enters into the radial spongy
arms, developed from the margin of the spongy disk.
Synopsis of the Genera of the Spongodiscida.
|
III. Subfamily Spongophacida.
Spongy disk without radial appendages.
|
|
Spongy disk with simple margin (without peculiar equatorial
girdle), |
253. Spongodiscus. |
| Spongy disk with a peculiar (solid or porous) equatorial
girdle, |
254. Spongophacus. |
|
III. Subfamily Spongotrochida.
Spongy disk with solid radial spines on the margin (in the equatorial
plane).
|
|
Few (two, three, or four) radial spines regularly
disposed. |
|
Two opposite spines, |
255. Spongolonche. |
| Three marginal spines, |
256. Spongotripus. |
| Four crossed spines, |
257. Spongostaurus. |
| Numerous (five to ten or more) radial spines, often
irregularly disposed. |
|
Spines only on the margin (equatorial), |
258. Stylotrochus. |
| Spines on both sides of the disk, |
259. Spongotrochus. |
|
III. Subfamily Spongobrachida.
Spongy disk with spongy radial arms on the margin (in the equatorial
plane).
|
|
Two arms, opposite in one axis. |
|
Without a patagium, |
260. Spongolena. |
| With a patagium, |
261. Spongobrachium. |
| Three arms on the margin. |
|
Without a patagium, |
262. Rhopalodictyum. |
| With a patagium, |
263. Dictyocoryne. |
| Four arms in cross form. |
|
Without a patagium, |
264. Spongasteriscus. |
| With a patagium, |
265. Spongaster. |
| I. Subfamily Spongophacida. Spongy disk without radial appendages. |
|
|
Spongy disk with simple margin (without peculiar equatorial
girdle), |
|
|
|
|
|
|
|
|
253. Spongodiscus. |
|
|
|
|
|
|
|
|
Spongy disk with a peculiar (solid or porous) equatorial
girdle, |
|
|
|
|
|
|
|
|
254. Spongophacus. |
|
|
|
|
|
|
| II. Subfamily Spongotrochida. Spongy disk with solid radial spines on the
margin (in the equatorial plane). |
|
|
Few (two, three, or four) radial spines regularly disposed. |
|
|
|
|
|
|
Two opposite spines, |
|
|
|
|
|
|
|
|
|
|
255. Spongolonche. |
|
|
|
|
|
|
|
|
|
|
Three marginal spines, |
|
|
|
|
|
|
|
|
|
|
256. Spongotripus. |
|
|
|
|
|
|
|
|
|
|
Four crossed spines, |
|
|
|
|
|
|
|
|
|
|
257. Spongostaurus. |
|
|
|
|
|
|
|
|
Numerous (five to ten or more) radial spines, often irregularly
disposed. |
|
|
|
|
|
|
Spines only on the margin (equatorial), |
|
|
|
|
|
|
|
|
|
|
258. Stylotrochus. |
|
|
|
|
|
|
|
|
|
|
Spines on both sides of the disk, |
|
|
|
|
|
|
|
|
|
|
259. Spongotrochus. |
|
|
|
|
|
|
| III. Subfamily Spongobrachida. Spongy disk with spongy radial arms on the
margin (in the equatorial plane). |
|
|
Two arms, opposite in one axis. |
|
|
|
|
|
|
Without a patagium, |
|
|
|
|
|
|
|
|
|
|
260. Spongolena. |
|
|
|
|
|
|
|
|
|
|
With a patagium, |
|
|
|
|
|
|
|
|
|
|
261. Spongobrachium. |
|
|
|
|
|
|
|
|
Three arms on the margin. |
|
|
|
|
|
|
Without a patagium, |
|
|
|
|
|
|
|
|
|
|
262. Rhopalodictyum. |
|
|
|
|
|
|
|
|
|
|
With a patagium, |
|
|
|
|
|
|
|
|
|
|
263. Dictyocoryne. |
|
|
|
|
|
|
|
|
Four arms in cross form. |
|
|
|
|
|
|
Without a patagium, |
|
|
|
|
|
|
|
|
|
|
264. Spongasteriscus. |
|
|
|
|
|
|
|
|
|
|
With a patagium, |
|
|
|
|
|
|
|
|
|
|
265. Spongaster. |
|
|
|
|
|
|
Subfamily 1. Spongophacida, Haeckel, 1881, Prodromus, p. 461.
Definition.—Spongodiscida with a simple
circular disk, without radial appendages on the margin (neither solid spines nor chambered
arms).
{576}
Genus 253. Spongodiscus,[292] Ehrenberg,
1854, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 237.
Definition.—Spongodiscida with a simple circular disk,
without radial appendages and without an equatorial girdle on the margin.
The genus Spongodiscus represents the most simple and primitive form of the
Spongodiscida, or of those Discoidea in which the central disk is more or
less spongy, composed of an irregular fine framework. In my Monograph (1862, pp. 452, 460, 469) I
had separated the true Spongodiscus (first described by Ehrenberg, loc. cit.) and
the Spongocyclia; the former being characterised by the irregular spongy framework of the
whole disk, whilst in the latter this framework includes in the central part some concentric
circular rings (approaching Porodiscus). In the same way afterwards Stöhr (1880, loc.
cit.) separated the genus Spongospira as spongy disks, which include in the central
part some spiral convolutions. But as these differences are rather inconstant and not sharply
discernible, I think it now better to regard these three forms as subgenera of
Spongodiscus. All three have the common simple circular disk, without any marginal
appendages.
Subgenus 1. Spongodisculus, Haeckel.
Definition.—Spongy framework of the disk quite irregular, without
concentric rings or spiral convolutions.
1. Spongodiscus mediterraneus, Haeckel.
Spongodiscus mediterraneus, Haeckel, 1862, Monogr. d. Radiol., p. 461,
Taf. xii. figs. 14, 15.
Spongy disk plain on both sides, with quite an irregular framework, without concentric rings
and without radial piercing beams. Texture everywhere uniform; meshes eight to ten times as broad
as the bars.
Dimensions.—Diameter of the disk 0.08 to 0.24, of the meshes 0.005 to 0.008.
Habitat.—Mediterranean (Messina), Haeckel; also fossil in the Tertiary
rocks of Sicily (Grotte), Stöhr.
2. Spongodiscus radiatus, n. sp.
Spongy disk plain on both sides, with quite irregular framework, without concentric rings, but
with numerous piercing radial beams which are not prolonged into marginal spines. Texture
everywhere uniform; meshes once and a half to twice as broad as the bars.
Dimensions.—Diameter of the disk 0.12 to 0.15, of the meshes 0.002 to 0.003.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
{577}
3. Spongodiscus resurgens, Ehrenberg.
Spongodiscus resurgens, Ehrenberg, 1854, Mikrogeol., Taf. xxxvB. B. iv., fig. 16.
Spongodiscus resurgens, Stöhr, 1880, Palæontogr., xxvi. p. 117, Taf. vi.
fig. 11.
Spongy disk lenticular, biconvex, in the darker centre much thicker than towards the thin
periphery, with an irregular framework, without concentric rings, but with numerous piercing
radial beams. Texture equal; meshes three to four times as broad as the bars.
Dimensions.—Diameter of the disk 0.1 to 0.3, of the meshes 0.003 to 0.006.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific, surface; also
fossil in the Tertiary rocks of Sicily and Barbados.
4. Spongodiscus favus, Ehrenberg.
Spongodiscus favus, Ehrenberg, 1861, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 301.
Spongy disk lenticular, biconvex, in the darker centre much thicker than towards the periphery,
with an irregular framework, without concentric rings. Texture different, in the outer half with
numerous radial beams and loose network, the meshes of which are three to four times as large as
in the darker and denser framework of the centre.
Dimensions.—Diameter of the disk 0.2, of the outer meshes 0.008 to 0.012, of the inner meshes 0.002 to 0.003.
Habitat.—North Atlantic, Greenland, Færöe Channel (John Murray),
surface.
5. Spongodiscus biconcavus, n. sp.
Spongy disk biconcave, in the peripheral, ring-like, thickened part twice as thick as in the
hollowed central part, with an irregular framework, without concentric rings. Texture different,
in the outer half looser than in the inner, darker part; meshes of the outer part five to six
times, of the inner twice to three times, as broad as the bars.
Dimensions.—Diameter of the disk 0.25, of the outer meshes 0.01 to 0.012, of the
inner 0.004 to 0.006.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms.
Subgenus 2. Spongocyclia, Haeckel, 1862, Monogr. d. Radiol., p. 469.
Definition.—Spongy framework of the disk in the inner part with
concentric, circular rings, in the outer part irregular.
{578}
6. Spongodiscus cycloides, Haeckel.
Spongodiscus cycloides, Haeckel, 1860, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin,
p. 843.
Spongocyclia cycloides, Haeckel, 1862, Monogr. d. Radiol., p. 469, Taf.
xxviii. fig. 1.
Spongy disk on both sides plain, in the central part with five to ten concentric, circular
rings, in the peripheral part quite irregularly and densely spongy. Meshes twice to four times as
broad as the bars.
Dimensions.—Diameter of the disk 0.1 to 0.2, of the meshes 0.003 to 0.006.
Habitat.—Mediterranean (Messina), North Atlantic (Canary Islands).
7. Spongodiscus spongocyclia, Haeckel.
Spongocyclia triangularis, Stöhr, 1880, Palæontogr., vol. xxvi. p. 119,
Taf. vii. fig. 5.
Spongy disk lenticular, in the thicker central part with eleven to twelve circular, concentric
rings, in the thinner, peripheral zone irregularly spongy. Meshes twice to four times as broad as
the bars. (The triangular form in the specimen figured by Stöhr is accidental, produced by the
broken margin.)
Dimensions.—Diameter of the disk 0.2 to 0.3, of the meshes 0.006 to 0.008.
Habitat.—Fossil in Tertiary rocks of Barbados (Haeckel) and Sicily
(Stöhr).
Subgenus 3. Spongospira, Stöhr, 1880, Palæontogr., vol xxvi. p. 120.
Definition.—Spongy framework of the disk in the inner part with spiral
convolutions, in the outer part irregular.
8. Spongodiscus florealis, Haeckel.
Spongospira florealis, Stöhr, 1880, Palæontogr., vol. xxvi. p. 120, Taf.
vii. fig. 6.
Spongy disk lenticular, in the thicker central part with five to six spiral convolutions, in
the outer peripheral zone irregularly spongy. No radial beams piercing the framework. Meshes three
to four times as broad as the bars.
Dimensions.—Diameter of the disk 0.26, of the meshes 0.006 to 0.008.
Habitat.—Fossil in Tertiary rocks of Sicily (Grotte), Stöhr.
9. Spongodiscus spiralis, n. sp.
Spongospira spiralis, Haeckel, 1881, Prodromus.
Spongy disk on both sides plain, with twelve to sixteen spiral convolutions in the central
part, with irregular, spongy framework in the outer part, pierced by numerous interrupted radial
beams. Meshes five to six times as broad as the bars.
Dimensions.—Diameter of the disk 0.2 to 0.3, of the meshes 0.01 to 0.012.
Habitat.—Antarctic Ocean, Station 157, depth 1950 fathoms.
{579}
Genus 254. Spongophacus,[293] Haeckel,
1881, Prodromus, p. 461.
Definition.—Spongodiscida with a simple circular disk,
without radial appendages, on the margin surrounded by a thin porous or solid equatorial
girdle.
The genus Spongophacus, represented hitherto only by a single but
interesting species, differs from Spongodiscus by the peculiar girdle which surrounds the
margin of the disk in the equatorial plane. It simulates the same formation as
Perichlamydium in the Porodiscida, and resembles also Periphæna, &c., in the
Phacodiscida.
1. Spongophacus periphæna, n. sp.
Spongy disk lenticular, with an irregular, dense framework, in the centre darker than in the
peripheral part. Margin of the disk very thin, surrounded by a broad, circular girdle, lying in
the equatorial plane, about as broad as the half radius of the spongy disk. The inner part of the
girdle is perforated by numerous irregular, small pores, which pass over gradually into the spongy
meshes; the outer part is quite homogeneous, solid, transparent, with an extremely thin
margin.
Dimensions.—Diameter of the disk 0.2; breadth of the girdle 0.05.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
Subfamily 2. Spongotrochida, Haeckel, 1881, Prodromus, p. 461.
Definition.—Spongodiscida with a circular
disk, the margin of which is armed with solid radial spines, situated in the equatorial plane
(rarely also on both sides of the disk with radial spines).
Genus 255. Spongolonche,[294] Haeckel,
1881, Prodromus, p. 461.
Definition.—Spongodiscida with two solid marginal spines,
opposite in one equatorial diameter of the disk.
The genus Spongolonche opens the series of the Spongotrochida, or of those
Spongodiscida in which the margin of the disk is armed with solid radial spines, situated in the
equatorial plane. Spongolonche possesses only two such spines, opposite in one equatorial
diameter of the disk; it corresponds therefore to Stylocyclia among the Coccodiscida, to
Xiphodictya among the Porodiscida.
{580}
1. Spongolonche conostyla, n. sp..
Spongy disk circular, with irregular framework, without concentric rings. Both opposite radial
spines conical, about as long as the radius of the disk, and four times as long as broad at the
base. Margin of the disk nearly smooth.
Dimensions.—Diameter of the disk 0.16; length of the radial spines 0.18, basal
thickness 0.045.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
2. Spongolonche amphistyla, n. sp.
Spongy disk circular, with four to six concentric rings in the inner part, with quite an
irregular framework in the outer part. Both opposite radial spines cylindrical, twice to three
times as long as the diameter of the disk, at the base about as broad as two meshes of the
framework. Margin of the disk ciliated.
Dimensions.—Diameter of the disk 0.2; length of the radial spines 0.4 to 0.6,
basal thickness 0.01.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
Genus 256. Spongotripus,[295] Haeckel,
1881, Prodromus, p. 461.
Definition.—Spongodiscida with three solid radial spines
on the margin of the circular or triangular disk.
The genus Spongotripus is characterised by three marginal spines, which are
commonly regularly disposed, more rarely in a bilateral or an irregular manner. It corresponds to
Trigonocyclia among the Coccodiscida, to Tripodictya among the Porodiscida.
Subgenus 1. Spongotripodiscus, Haeckel.
Definition.—Radial spines of equal size and distance; triangle
regular.
1. Spongotripus regularis, n. sp.
Spongy disk circular; three radial spines on its margin of equal size and equidistant, strong,
conical, about as long as the diameter of the disk, and five times as long as broad at the
base.
Dimensions.—Diameter of the disk 0.15; length of the spines 0.16, basal breadth
0.03.
Habitat.—Pacific, central area, Station 272, surface.
{581}
2. Spongotripus neumayri, Haeckel.
Spongechinus neumayri, Dunikowski, 1882, Denskchr. d. k. Akad. d. Wiss.
Wien, xlv. p. 28, Taf. v. fig. 59.
Spongy disk circular, nearly spherical; three radial spines on its margin of equal size and
equidistant, pyramidal, nearly as long as the diameter of the disk, and three times as long as
broad at the base.
Dimensions.—Diameter of the disk 0.15 to 0.2; length of the spines 0.11, basal
breadth 0.04.
Habitat.—Fossil in the Jura of the Alps and in Tertiary rocks of
Barbados.
3. Spongotripus strepsiceros, n. sp.
Spongy disk circular; three radial spines on its margin of equal size and equidistant, angular,
twice as long as the diameter of the disk, spirally twisted like the horns of Antilope
strepsiceros.
Dimensions.—Diameter of the disk 0.15; length of the spines 0.3, basal breadth
0.02.
Habitat.—Pacific, central area, Station 272, depth 2600 fathoms.
Subgenus 2. Spongotripodium, Haeckel.
Definition.—Radial spines of different size or at different distances;
triangle either isosceles or irregular.
4. Spongotripus ypsilon, n. sp.
Spongy disk triangular, isosceles, with convex sides; three spines angular, of different size
and at different distances; the odd spine straight and twice as long as the two paired spines,
which are more approximated and curved concavely one to the other, like  .
.
Dimensions.—Diameter of the disk 0.12; length of the odd spine 0.2, of the paired
spines 0.1, basal breadth 0.04.
Habitat.—Pacific, central area, Station 274, depth 2750 fathoms.
5. Spongotripus irregularis, n. sp.
Spongy disk circular; three spines conical, irregularly curved, all three of different size,
and at irregular distances; once to three times as long as the diameter of the disk; margin of the
disk thorny.
Dimensions.—Diameter of the disk 0.1; length of the spines 0.1 to 0.3, basal
breadth 0.05.
Habitat.—South Pacific, Station 300, depth 1375 fathoms.
{582}
Genus 257. Spongostaurus,[296] Haeckel,
1881, Prodromus, p. 461.
Definition.—Spongodiscida with four solid radial spines
on the margin of the circular or square disk, commonly crossed in two equatorial diameters
perpendicular one to another.
The genus Staurodictya exhibits four marginal spines in cross form. Commonly
the cross is regular and rectangular, the four spines being opposite in pairs in two perpendicular
diameters; sometimes more or less irregular. The genus repeats Staurodictya among the
Porodiscida, Staurocyclia among the Coccodiscida.
1. Spongostaurus cruciatus, n. sp.
Spongy disk circular; four radial spines on its margin opposite in two crossed equatorial
diameters, perpendicular one to another, conical, about as long as the diameter of the disk.
Dimensions.—Diameter of the disk 0.16; length of the spines 0.18, basal breadth
0.04.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
2. Spongostaurus serratus, n. sp.
Spongy disk circular; four crossed radial spines very large, twice to three times as long as
the diameter of the central disk, serrated on both edges, with two rows of strong conical
perpendicular teeth (similar to the saw of Pristis antiquorum).
Dimensions.—Diameter of the disk 0.1; length of the spines 0.2 to 0.3, breadth
0.02.
Habitat.—Indian Ocean, Madagascar, Rabbe, surface.
3. Spongostaurus hastatus, n. sp.
Spongy disk square, with concave sides; four crossed radial spines cylindrical, on the broader
distal end spear-shaped or nearly lanceolate, one and a half times as long as the diameter of the
disk.
Dimensions.—Diameter of the disk 0.13; length of the spines 0.2, basal breadth
0.01, distal breadth 0.03.
Habitat.—Tropical Atlantic, Station 347, surface.
4. Spongostaurus quadratus, n. sp.
Spongy disk square, with rectilinear sides; four crossed radial spines pyramidal, arising from
the corners of the square, about as long as its half diagonal.
Dimensions.—Diameter of the disk 0.16; length of the spines 0.08, basal breadth
0.02.
Habitat.—Pacific, central area, Station 271, surface.
{583}
Genus 258. Stylotrochus,[297] Haeckel,
1862, Monogr. d. Radiol., p. 463.
Definition.—Spongodiscida with numerous solid radial
spines on the margin of the disk (five to ten or more), all situated in the equatorial plane.
The genus Stylotrochus comprises those Spongodiscida in which the margin of
the circular disk bears numerous radial spines. All these spines lie in the same equatorial plane,
whilst in the following genus they are disposed over the whole surface of the disk.
Stylotrochus corresponds to Astrocyclia among the Coccodiscida, to
Stylodictya among the Porodiscida. The spongy framework of the disk is either quite
irregular (Stylotrochiscus), or includes in the middle part some concentric circular rings
(Stylospongia).
Subgenus 1. Stylotrochiscus, Haeckel.
Definition.—Spongy framework of the whole disk irregular, without
concentric circular rings or spiral convolutions.
1. Stylotrochus arachnius, Haeckel.
Spongotrochus arachnius, Haeckel, 1862, Monogr. d. Radiol., p. 464.
Spongy framework of the whole disk irregular. Eight to twelve marginal spines very long and
thin, bristle-shaped, twice to four times as long as the diameter of the disk. (Very similar to
the common Stylodictya arachnia, but without concentric circular rings and sieve-plates,
with quite irregular network of fine bars.)
Dimensions.—Diameter of the disk 0.12 to 0.15; length of the radial spines 0.2 to
0.6, basal breadth 0.001.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Indian, Pacific,
surface; also fossil in Tertiary rocks of Barbados and the Mediterranean.
2. Stylotrochus craticulatus, Haeckel.
Spongotrochus craticulatus, Stöhr, 1880, Palæontogr., vol. xxvi. p. 118, Taf. vi. fig.
12.
? Spongodiscus aculeatus, Ehrenberg, 1854, Monatsber. d. k. preuss. Akad.
d. Wiss. Berlin, p. 246.
Spongy framework of the whole disk irregular. Sixteen to twenty short marginal spines (twice to
four times as long as the diameter of one mesh of the framework), free prolongations of internal
radial beams which arise from the darker centre of the disk. (The interruption of the disk-margin
on one point of its circumference, figured by Stöhr as osculum or "Mündungs-Oeffnung," is probably
an accidental abnormality; I did not find it in other specimens.)
{584}
Dimensions.—Diameter of the disk 0.2 to 0.25; length of the radial spines 0.005 to
0.018, basal breadth 0.001 to 0.003.
Habitat.—Pacific, central area, Stations 266 to 268, surface; fossil
in Barbados and Sicily.
3. Stylotrochus helianthus, n. sp.
Spongy framework of the whole disk irregular. Thirty to fifty very large, conical radial
spines, about as long as the diameter of the disk, and at the base two to four times as broad as
one mesh of the framework (without internal prolongations).
Dimensions.—Diameter of the disk 0.15 to 0.2; length of the radial spines 0.16 to
0.24, basal breadth 0.008 to 0.016.
Habitat.—Pacific, central area, Station 272, surface.
4. Stylotrochus rhabdostylus, Haeckel.
Spongosphæra rhabdostyla, Ehrenberg, 1875, Abhandl. d. k. Akad. d. Wiss. Berlin,
p. 256, Taf. xxvi. figs. 1, 2.
Spongy framework of the whole disk irregular. Four very large marginal primary spines crossed
in two equatorial diameters, perpendicular one to another, and between them numerous (twelve to
twenty or more) smaller secondary spines. The latter arise from the margin, whilst the former
pierce the disk and are nearly united in its centre. All the spines are cylindrical, the smaller
as broad as one mesh, the larger three to six times as broad.
Dimensions.—Diameter of the disk 0.2; length of the four main spines 0.15 to 0.3,
breadth 0.01 to 0.02; length of the accessory spines 0.05 to 0.1, breadth 0.005.
Habitat.—Fossil in the rocks of Barbados.
5. Stylotrochus heteracanthus, Haeckel.
Spongotrochus heteracanthus, Haeckel, 1862, Monogr. d. Radiol., p.
464.
Spongy framework of the whole disk irregular. Ten very long needle-shaped marginal spines
(symmetrically distributed), and between them numerous very fine, shorter, accessory,
bristle-shaped spines.
Dimensions.—Diameter of the disk 0.16; length of the main spines 0.16, basal
breadth 0.002; length of the secondary spines 0.03.
Habitat.—Mediterranean (Messina), surface.
Subgenus 2. Stylospongia, Haeckel, 1862, Monogr. d. Radiol., p. 473.
Definition.—Spongy framework of the disk in the inner part with
concentric rings or spiral convolutions, in the outer part quite irregular.
{585}
6. Stylotrochus huxleyi, Haeckel.
Stylospongia huxleyi, Haeckel, 1862, Monogr. d. Radiol., p. 473, Taf.
xxviii. fig. 7.
Spongy framework of the disk in the inner part with five concentric rings, in the outer part
quite irregular. Ten marginal spines, conical at the base, about as long as the radius of the
disk, without inner piercing prolongations.
Dimensions.—Diameter of the disk 0.12; length of the radial spines 0.06, basal
breadth 0.003.
Habitat.—Mediterranean (Messina), Haeckel.
7. Stylotrochus geddesii, n. sp. (Pl. 41, fig.
11).
Stylospongidium geddesii, Haeckel, 1881, Atlas (pl. xli. fig. 11).
Spongy framework of the disk in the inner part with four to eight concentric rings (or
partially spiral convolutions), in the outer part quite irregular. Thirty to fifty pyramidal
marginal spines of variable size, one-fourth to one-half as long as the radius of the disk, outer
prolongations of inner piercing radial beams, which arise from various concentric rings. I call
this interesting species, which is intermediate between Stylodictya and
Stylotrochus, in honour of the morphologist Mr. Patrick Geddes of Edinburgh.
Dimensions.—Diameter of the disk 0.15 to 0.25; length of the radial spines 0.03 to
0.06, basal breadth 0.004 to 0.01.
Habitat.—Pacific, central area, Stations 270 to 274, in 2350 to 2925
fathoms.
Genus 259. Spongotrochus,[298] Haeckel,
1860, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 844.
Definition.—Spongodiscida with numerous solid radial
spines (five to ten or more), which are scattered over the whole surface and the margin of the
disk, or regularly disposed on both sides of it.
The genus Spongotrochus differs from the foregoing and nearly allied genus
by the distribution of the numerous radial spines. These are not confined to the margin of the
disk, but also scattered on its whole surface, and sometimes symmetrically disposed on both its
sides in a regular manner. Also in this genus the spongy framework is sometimes quite irregular
(Spongotrochiscus), at other times in the middle part with enclosed concentric rings
(Stylospongidium).
Subgenus 1. Spongotrochiscus, Haeckel, 1862, Monogr. d. Radiol., p. 463.
Definition.—Spongy framework of the whole disk irregular, without
concentric rings or spiral convolutions.
{586}
1. Spongotrochus brevispinus, Haeckel.
Spongotrochus brevispinus, Haeckel, 1862, Monogr. d. Radiol., p. 462, Taf.
xxvii. figs. 4, 5.
Spongy framework of the whole disk irregular, everywhere equal. The whole surface of the disk
covered with numerous short, needle-shaped, radial spines, about as long as the thickness of the
disk, which is one-fifth of its diameter.
Dimensions.—Diameter of the disk 0.18; length of the radial spines 0.03 to 0.04,
basal breadth 0.002.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific, surface.
2. Spongotrochus multispinus, n. sp.
Spongy framework of the whole disk irregular, in the centre darker. The whole surface of the
disk covered with numerous conical radial spines, about as long as the radius of the disk.
Dimensions.—Diameter of the disk 0.22; length of the radial spines 0.1, basal
breadth 0.008.
Habitat.—Pacific, central area, Stations 271 to 274, surface.
3. Spongotrochus longispinus, Haeckel.
Spongotrochus longispinus, Haeckel, 1862, Monogr. d. Radiol., p. 463, Taf
xxvii. figs. 2, 3.
Spongy framework of the whole disk irregular, everywhere equal. Surface thorny. Twenty long,
needle-shaped, radial spines, about as long as the diameter of the disk, symmetrically distributed
in pairs on both flat sides of the disk, so that the opposite pairs lie in five equidistant,
meridian planes (compare the figures).
Dimensions.—Diameter of the disk 0.2; length of the twenty radial spines 0.2,
basal breadth 0.001.
Habitat.—Mediterranean (Messina), surface.
Subgenus 2. Stylospongidium, Haeckel, 1881, Prodromus, p. 460.
Definition.—Spongy framework of the disk in the inner part with
concentric rings or spiral convolutions, in the outer part quite irregular.
4. Spongotrochus scutella, n. sp.
Spongotrochus
ehrenbergii, Bütschli, 1882, L. N. 41, Taf. xxvi. figs. 1a,
1b.
Spongy framework of the disk in the inner part with four to six concentric rings, in the outer
part quite irregular. The whole surface of the disk covered with bristle-shaped radial spines,
about half as long as the radius of the disk.
Dimensions.—Diameter of the disk 0.24; length of the radial spines 0.06, basal
breadth 0.02.
Habitat.—Antarctic Ocean, Station 157, depth 1950 fathoms.
{587}
5. Spongotrochus parma, n. sp.
Spongy framework of the disk in the inner part with five to eight spiral convolutions (or
partly concentric rings), in the outer part quite irregular. Thirty to forty long, bristle-shaped,
radial spines, about as long as the diameter of the disk, disposed on both flat sides of the disk,
but not on the margin.
Dimensions.—Diameter of the disk 0.16; length of the radial spines 0.18, basal
breadth 0.003.
Habitat.—South Atlantic, Station 325, surface.
Subfamily 3. Spongobrachida, Haeckel, 1881, Prodromus, p. 461.
Definition.—Spongodiscida with two or more
(commonly three or four) spongy radial arms on the margin of the disk, situated in its equatorial
plane (with or without a connecting patagium between the arms).
Genus 260. Spongolena,[299] n. gen.
Definition.—Spongodiscida with two opposite spongy arms
on the margin of the disk, without a connecting patagium.
The genus Spongolena opens the series of the Spongobrachida, or of the
Spongodiscida provided with radial spongy arms on the margin of the disk. Spongolena is the
most simple form of this subfamily, and bears only two simple opposite arms, without a connecting
patagium. It corresponds to Amphibrachium (Porodiscida) and to Diplactura
(Coccodiscida). Also there is no patagium. Spongolena may easily be confounded with
Spongurus (compare my Prodromus, 1881, p. 461); but in the true Spongurus (an
ellipsoid) the transverse section is circular, in Spongolena elliptical.
1. Spongolena rhopalura, n. sp.
Arms club-shaped, three times as long as broad, at the rounded distal end twice as broad as at
the base, and twice as long as the diameter of the circular central disk. Surface nearly
smooth.
Dimensions.—Radius of the arms (or the distance from the centre to the distal
arm-end) 0.16, distal breadth 0.05, basal breadth 0.03.
Habitat.—Pacific, central area, Station 273, depth 2350 fathoms.
2. Spongolena spongura, n. sp.
Arms nearly cylindrical, twice as long as broad, and a little longer than the diameter of the
elliptical central disk. Surface thorny, some longer bristle-shaped spines on the distal end of
the {588}arms. (Very similar to the ellipsoid
Spongurus cylindricus, Monogr. d. Radiol., p. 465, Taf. xxvii. fig. 1, but differs in the
compressed lenticular (not ellipsoidal) form of the central disk; the transverse section of the
arm is elliptical, not circular.)
Dimensions.—Radius of the arms 0.1, breadth 0.04.
Habitat.—Pacific, central area, Station 270, depth 2925 fathoms.
3. Spongolena cypselura, n. sp.
Arms nearly triangular, not longer than broad, about half as large as the elliptical central
disk, at the broader distal end with two very large, widely divergent lateral spines, and between
them several smaller, like the tail of a swallow. Surface thorny.
Dimensions.—Radius of the arms 0.2, distal breadth (without spines) 0.15, basal
breadth 0.07.
Habitat.—Pacific, central area, Station 272, depth 2600 fathoms.
Genus 261. Spongobrachium,[300] Haeckel,
1881, Prodromus, p. 461.
Definition.—Spongodiscida with two opposite spongy arms
on the margin of the disk, connected by a spongy patagium of different texture.
The genus Spongobrachium differs from the foregoing only in the loose spongy
patagium, which envelops both opposite spongy arms. It corresponds to Amphymenium among the
Porodiscida and to Amphiactura among the Coccodiscida.
1. Spongobrachium ellipticum, Haeckel.
Spongocyclia elliptica, Haeckel, 1862, Monogr. d. Radiol., p. 470, Taf. xxviii. fig.
2.
Spongodiscus ellipticus, Haeckel, 1860, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 844.
Arms nearly square, scarcely as long and broad as the radius of the circular central disk, at
the broader distal end truncated. Patagium complete, enveloping the whole disk with the arms, and
forming a larger elliptical disk of looser framework. (In my Monograph, 1862, loc. cit., I
had not distinguished the opposite darker arms, opposite in the longer axis of the elliptical
disk, from the enveloping looser framework of the patagium. In larger specimens of the Challenger
collection this distinction is very evident.)
Dimensions.—Radius of the arms 0.12, breadth 0.05; major axis of the elliptical
patagium 0.24, minor 0.16.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific, surface.
2. Spongobrachium lanceolatum, n. sp.
Arms club-shaped, twice as long as broad, at the distal end pointed, five times as long as the
radius of the circular central disk. Patagium complete, enveloping the whole disk with the arms,
{589}and forming a larger lanceolate disk of looser
framework. (Similar in form to Amphymenium pupula, Pl. 44, fig. 8, but
with an irregular spongy framework and pointed ends.)
Dimensions.—Radius of the arms 0.16, breadth 0.05; major axis of the lanceolate
patagium 0.32, minor axis 0.12.
Habitat.—Pacific, central area, Station 270, surface.
Genus 262. Rhopalodictyum,[301]
Ehrenberg, 1860, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 830.
Definition.—Spongodiscida with three spongy arms on the
margin of the circular or triangular disk, without a connecting patagium.
The genus Rhopalodictyum comprises those very common forms of Spongodiscida
in which the margin of the disk is provided with three simple, free, spongy arms. It corresponds
to Dictyastrum and Rhopalastrum among the Porodiscida, to Trigonactura among
the Coccodiscida. The typical species of this genus is Rhopalodictyum abyssorum of
Ehrenberg, the only species figured by him. His diagnosis of the genus was very insufficient, and
agreed with that of his Dictyastrum (compare above the improved diagnosis of this genus, p.
526, and my Monograph, p. 466).
Subgenus 1. Rhopalodictya, Haeckel.
Definition.—Triangular shell regular, with three arms of equal size
and equidistant.
1. Rhopalodictyum abyssorum, Ehrenberg.
Rhopalodictyum abyssorum, Ehrenberg, 1872, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 299, Taf. viii. fig. 17.
Arms of equal size and equidistant, club-shaped, about as long as the diameter of the
triangular central disk, and a little longer than the breadth of the pear-shaped rounded distal
end. Surface rough.
Dimensions.—Radius of the arms 0.11, basal breadth 0.03, distal breadth 0.06.
Habitat.—Cosmopolitan; Atlantic, Indian, Pacific; tropical zone,
surface.
2. Rhopalodictyum truncatum, Ehrenberg.
Rhopalodictyum truncatum, Ehrenberg, 1861, Monatsber. d. k. preuss. Akad. d. Wiss.
Berlin, p. 301.
? Dictyastrum angulatum, Ehrenberg, 1872, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 289, Taf. viii. fig. 18.
Arms of equal size and equidistant, nearly square, with straight edges, towards the truncated
end a little broader, about of the same diameter as the central triangular disk. This species is
indicated {590}by Ehrenberg only by the short diagnosis,
"Radiis stellæ tribus apice truncatis." It is probably identical with his figure of Dictyastrum
angulatum (loc. cit.). This latter name I have retained for the similar Porodiscid
(above, p. 526), mainly because the genera Dictyastrum and
Rhopalodictyum, according to the insufficient diagnosis of Ehrenberg, seem to be identical.
(Compare my Monograph, 1862, p. 466.)
Dimensions.—Radius of each arm 0.13, basal breadth 0.09, distal breadth 0.1.
Habitat.—Tropical Pacific, Philippine Sea, Station 200, depth 250
fathoms; Atlantic (Mexican Gulf Stream).
3. Rhopalodictyum subacutum, Ehrenberg.
Rhopalodictyum subacutum, Ehrenberg, 1861, Monatsber. d. k. preuss. Akad.
d. Wiss. Berlin, p. 301.
Arms of equal size and equidistant, club-shaped, three times as long as the diameter of the
central disk, which equals the breadth of the thickened distal end; the latter is armed with a
strong pyramidal terminal spine. (The diagnosis of Ehrenberg is "Radiis stellæ tribus, apice
cuneatis subacutis.")
Dimensions.—Radius of the arms 0.2, basal breadth 0.04, distal breadth 0.07.
Habitat.—North Atlantic, surface and various depths; Greenland;
Mexican Gulf Stream; Færöe Channel (John Murray).
4. Rhopalodictyum bifidum, n. sp.
Arms of equal size and equidistant, in the distal half forked, twice as long as the diameter of
the central disk; both fork branches half as broad as the simple basal part, truncated at the
distal end.
Dimensions.—Radius of the arms 0.16, basal breadth 0.08, distal breadth 0.04.
Habitat.—North Pacific, Station 237, surface.
Subgenus 2. Triactinosphæra, Dunikowski, 1882, Denkschr. d. k. Akad. d.
Definition.—Triangular shell bilateral or irregular, with three arms
of different size or distance.
5. Rhopalodictyum zittelii, Haeckel.
Triactinosphæra zittelii, Dunikowski, 1882, Denkschr. d. k. Akad. d. Wiss,
Wien, vol. 45, ii. p. 192.
Arms of different size and at unequal distances, one odd arm being a little shorter than the
two paired arms; the odd angle between the latter is larger than the paired angles between them
and the {591}former. Arms club-shaped, their basal
semi-cylindrical, half as broad as the spherical distal half, which bears a strong, conical,
terminal spine. The discoverer of this remarkable, very old, Liassic species, Dunikowski, supposes
that it is not a true Discoid, from the absence of a central disk; in my opinion the central disk
(nearly spherical) has the same relation to the arms as in many other Discoidea, their equatorial plane is the same.
Dimensions.—Radius of the odd arm 0.18, of the paired arms 0.24; basal breadth
0.08, distal breadth 0.16.
Habitat.—Fossil in the Alpine Lias, Schafberg, near Salzburg,
Dunikowski.
6. Rhopalodictyum elongatum, n. sp.
Arms of different size and at unequal distances, one odd arm being twice as long as both paired
arms; the odd angle between the latter much larger than both equal paired angles. Arms nearly
cylindrical, little flattened, three to six times as long as broad, with rounded blunt distal
end.
Dimensions.—Radius of the arms 0.15 to 0.3, breadth 0.05.
Habitat.—Pacific, central area, Station 274, surface.
7. Rhopalodictyum curvatum, n. sp.
Arms of different size and at unequal distances, irregular, more or less curved, nearly
cylindrical, five to ten times as long as broad, with rounded blunt distal end. (There were
observed only two specimens of this remarkable irregular species; in one specimen all three arms
were simple, in the other one odd arm forked. The length of the arms and the size of the angles
between them seems to be very different and variable.)
Dimensions.—Radius of the arms 0.3 to 0.6 to 1.15, breadth 0.06 to 0.12.
Habitat.—Pacific, central area, Station 271, surface.
Genus 263. Dictyocoryne,[302] Ehrenberg,
1860, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 830.
Definition.—Spongodiscida with three spongy arms on the
margin of the circular or triangular disk, connected by a spongy patagium of different
texture.
The genus Dictyocoryne differs from the foregoing Rhopalodictyum only
in the patagium, connecting the three spongy arms, and bears to it the same relation as in the
Porodiscida Euchitonia does to Rhopalastrum. The typical species of this genus is
Dictyocoryne profunda, the only species figured by Ehrenberg. After his {592}insufficient diagnosis Dictyocoryne was identical
with his Spongaster (compare my Monograph, p. 467); but as the type of this latter genus
(Spongaster tetras) figured by him, 1872, had four crossed arms, we retain this genus here
separate.
Subgenus 1. Dictyocorynula, Haeckel.
Definition.—Triangular shell regular, with three arms of equal size
and equal distance.
1. Dictyocoryne profunda, Ehrenberg.
Dictyocoryne profunda, Ehrenberg, 1872, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 307, Taf. vii. fig. 23.
Arms of equal size and equidistant, club-shaped, in the outer circular half three times as
broad as in the inner cylindrical half, and much larger than the small triangular central disk.
Patagium complete, forming an equilateral triangle with rounded corners.
Dimensions.—Radius of the arms 0.14, basal breadth 0.025, distal breadth 0.07.
Habitat.—Pacific, Philippine Sea, depth 3300 fathoms, Ehrenberg;
Station 198, depth 2150 fathoms; Station 274, depth 2750 fathoms.
2. Dictyocoryne tetradiscus, n. sp.
Arms of equal size and equidistant, club-shaped, their outer circular half is quite as large as
the circular central disk, and is connected with it by the thin cylindrical inner half, which is
scarcely one-fourth as broad. Patagium complete, forming an equilateral triangle with rounded
corners and convex sides.
Dimensions.—Radius of the arms 0.16, basal breadth 0.02, distal breadth 0.08.
Habitat.—Pacific, central area, Stations 271 to 274, in 2350 to 2750
fathoms.
3. Dictyocoryne euchitonia, Haeckel.
Dictyocoryne euchitonia, Haeckel, 1862, Monogr. d. Radiol., p. 468.
Arms of equal size and equidistant, lanceolate, twice as long as broad in the distal part, and
three times as long as the small circular central disk. Patagium complete, forming an equilateral
triangle with pointed corners. (Very similar to Euchitonia köllikeri, Monograph, 1862, p.
511, Taf. xxxi. fig. 6, but quite spongy, not concentric.)
Dimensions.—Radius of the arms 0.15, basal breadth 0.03, distal breadth 0.06.
Habitat.—Mediterranean (Messina), Haeckel.
{593}
4. Dictyocoryne trigona, n. sp.
Arms of equal size and equidistant, lanceolate, three times as long as broad in the middle
part, and as the diameter of the triangular central disk; their distal end armed with a strong
conical radial spine. Patagium complete, forming an equilateral triangle with pointed corners.
Dimensions.—Radius of the arms 0.2, basal breadth 0.02, distal breadth 0.06.
Habitat.—North Atlantic, Canary Islands, surface.
Subgenus 2. Dictyocorynium, Haeckel.
Definition.—Triangular shell bilateral or irregular, with three arms
of different sizes or at different distances.
5. Dictyocoryne charybdaea, Haeckel.
Spongocyclia charybdaea, Haeckel, 1862, Monogr. d. Radiol., p. 472, Taf. xxviii. figs.
5, 6.
Spongodiscus charybdaeus, Haeckel, 1860, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 844.
Arms at different distances, nearly equilateral triangular, scarcely half as long as the radius
of the large circular central disk. Both paired arms touching at their bases, separated by a great
distance from the opposite odd arm. Patagium complete, nearly pentagonal. (The illustration in my
Monograph, in the coloured plate xxviii., is better than my description, as I had not exactly
separated the arms from the patagium.)
Dimensions.—Radius of the arms 0.18, basal breadth 0.08.
Habitat.—Mediterranean (Messina), surface.
6. Dictyocoryne pentagona, Stöhr.
Dictyocoryne pentagona, Stöhr, 1880, Palæontogr., vol. xxvi. p. 118, Taf.
vii. fig. 2.
Arms at different distances, club-shaped, little longer than broad at their rounded distal end.
Both paired arms smaller and more approximate than the opposite odd arm. Patagium complete, very
large, enveloping the whole shell, and forming a pentagon with rectilinear base and transverse
constriction, and with five rounded corners.
Dimensions.—Radius of the arms 0.1 to 0.15, middle breadth 0.04.
Habitat.—Fossil in the Tertiary rocks of Sicily, Grotte, Stöhr.
7. Dictyocoryne agrigentina, Stöhr.
Dictyocoryne agrigentina, Stöhr, 1880, Palæontogr., vol. xxvi. p. 118,
Taf. vii. fig. 1.
Arms at different distances, club-shaped, about twice as long as broad, and smaller than the
large triangular central disk. Both paired arms smaller and more approximate than the opposite odd
{594}arm. Patagium incomplete, enveloping only the basal
half of the arms as a circular spongy disk of loose framework.
Dimensions.—Radius of the arms 0.18, basal breadth 0.05, distal breadth 0.08.
Habitat.—Fossil in the Tertiary rocks of Sicily, Grotte, Stöhr.
8. Dictyocoryne echinata, n. sp.
Arms unequal and at different distances, club-shaped, in the distal half twice as broad as in
the basal half, three times as long as the small triangular central disk, at their distal end
armed with one larger and several smaller conical spines. Both paired arms with touching bases,
and only half as large as the doubly remote odd arm. Patagium incomplete, enveloping only the
basal half of the arms, and forming a circular disk of looser framework. Surface thorny.
Dimensions.—Radius of the arms 0.15 to 0.2, basal breadth 0.03, distal breadth
0.06.
Habitat.—South Atlantic, Station 325, surface.
Genus 264. Spongasteriscus,[303] Haeckel,
1862, Monogr. d. Radiol., p. 474 (sensu restricto).
Definition.—Spongodiscida with four spongy arms on the
margin of the circular or quadrangular disk, crossed in two equatorial diameters, without a
connecting patagium.
The genus Spongasteriscus (in the restricted definition here stated)
exhibits on the margin of the disk four spongy arms, which form either a regular or a bilateral
cross. It corresponds to Stauralastrum and Myelastrum among the Porodiscida, to
Astractura among the Coccodiscida.
Subgenus 1. Spongasteriscinus, Haeckel.
Definition.—Cross formed by the four arms regular, rectangular, with
equal-sized and equidistant arms.
1. Spongasteriscus ovatus, n. sp.
Arms at equal distances, forming a regular, rectangular cross, egg-shaped, with a broader
rounded distal end, one and a third times as long as broad, and three times as long as the radius
of the central disk; in the latter three to four concentric rings.
Dimensions.—Radius of the arms 0.1, greatest breadth 0.06.
Habitat.—Western Tropical Pacific, Station 225.
{595}
2. Spongasteriscus clavatus, n. sp.
Arms at equal distances, forming a regular, rectangular cross, club-shaped, about as long as
the diameter of the central disk, and at their rounded distal end one and a third times as long as
broad, at their narrow base only one third as broad. In the centre five to six concentric
rings.
Dimensions.—Radius of the arms 0.13, distal breadth 0.06, basal breadth 0.02.
Habitat.—Pacific, central area. Station 271, surface.
3. Spongasteriscus mucronatus, n. sp.
Arms at equal distances, forming a regular, rectangular cross, club-shaped, three times as long
as the radius of the central disk, in the distal half nearly circular, three to four times as
broad as at the narrow base. The rounded distal end armed with a strong pyramidal spine. In the
centre no concentric rings. (Similar to Stauralastrum rhopalophorum, Pl. 45, fig. 1, but
quite spongy.)
Dimensions.—Radius of the arms 0.12, basal breadth 0.03, distal breadth 0.02.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms.
4. Spongasteriscus furcatus, n. sp.
Arms at equal distances, forming a regular, rectangular cross, in the distal third forked; both
fork branches half as long as the basal undivided part of the arm, which is twice as long as
broad. Distal ends of the eight branches blunt, rounded. In the central disk no concentric
rings.
Dimensions.—Radius of the arms 0.12, basal breadth 0.03; distal breadth of the
branches 0.02.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
5. Spongasteriscus armatus, n. sp.
Arms at equal distances, forming a rectangular, regular cross, in the distal half forked; both
fork branches of the same length as the basal undivided part of the arm, which is nearly square.
Distal ends of the eight branches armed with a strong pyramidal spine. In the central disk no
concentric rings. (Similar to Dicranastrum cornutum, Pl. 45, fig. 2, but
quite spongy.)
Dimensions.—Radius of the arms 0.18, basal breadth 0.03, distal breadth 0.015.
Habitat.—South Pacific, Station 295, depth 1500 fathoms.
Subgenus 2. Spongasterisculus, Haeckel.
Definition.—Cross formed by the four arms, bilateral or irregular,
with the arms at different distances.
{596}
6. Spongasteriscus quadricornis, Haeckel.
Spongasteriscus quadricornis, Haeckel, 1862, Monogr. d. Radiol., p. 474, Taf. xxviii.
figs. 8-10.
Spongodiscus quadricornis, Haeckel, 1860, Monatsber. d. k. preuss. Akad.
d. Wiss. Berlin, p. 844.
Arms at different distances, forming a bilateral or irregular cross, grouped in two opposite
pairs; their form equilateral triangular; their length smaller than the radius of the large
circular central disk, which exhibits in the interior eight to sixteen concentric rings.
Dimensions.—Radius of the arms 0.2, of the central disk 0.13; basal breadth of the
arms 0.08.
Habitat.—Mediterranean (Messina), Haeckel, surface.
7. Spongasteriscus tetraceros, Haeckel.
Spongasteriscus tetraceros, Haeckel, 1862, Monogr. d. Radiol., p. 475.
Arms at different distances, forming a bilateral or irregular cross, grouped in two opposite
pairs; their form isosceles triangular; their length larger than the radius of the large
elliptical central disk, which exhibits in the interior six to twelve concentric rings.
Dimensions.—Radius of the arms 0.16, of the central disk 0.1; basal breadth of the
arms 0.06.
Habitat.—Mediterranean (Messina), North Atlantic (Canary Islands),
surface.
8. Spongasteriscus myelastrum, n. sp.
Arms at different distances, forming a bilateral or irregular cross, grouped in two opposite
pairs; the arms of one pair broader and shorter than the arms of the other pair. Each arm in its
basal half simple, in the distal half forked; ends of the fork branches blunt. In the central disk
no concentric rings. (Similar to Myelastrum octocorne, Pl. 47, fig. 12,
but quite spongy.)
Dimensions.—Radius of the arms 0.2, basal breadth 0.05, distal breadth 0.02.
Habitat.—North Pacific, Station 236, surface.
Genus 265. Spongaster,[304] Ehrenberg,
1860, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 833.
Definition.—Spongodiscida with four spongy arms on the
margin of the circular or quadrangular disk, connected by a spongy patagium of different
texture.
The genus Spongaster differs from the foregoing Spongasteriscus in
the patagium connecting the spongy arms, and bears therefore to it the same relation as, in the
Porodiscida, Histiastrum does to Stauralastrum, or, in the Coccodiscida,
Stauractura does to Astractura. The typical specimen, figured by Ehrenberg
(Spongaster tetras), exhibits a regular, square disk, as also some other species. In a
certain number of other species (formerly united by me with Spongocyclia) the quadrangular
disk is bilateral.
{597}
Subgenus 1. Spongastrella, Haeckel.
Definition.—Cross formed by the four arms regular, rectangular, with
the arms of equal size and equidistant.
1. Spongaster tetras, Ehrenberg.
Spongaster tetras, Ehrenberg, 1872, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 299, Taf.
vi. (iii.) fig. 8.
Dictyocoryne tetras, Haeckel, 1862, Monogr. d. Radiol., p. 469.
Arms at equal distances, forming a rectangular, regular Myelastrum, papiliocross, club-shaped,
about twice as long as the diameter of the square central disk and eight times as long as broad at
their base. Distal ends rounded, perfectly enclosed by the complete patagium, which forms a
regular square, with slightly concave sides.
Dimensions.—Radius of the arms 0.12, distal breadth 0.03, basal breadth 0.015;
length of the square side 0.2.
Habitat.—Cosmopolitan; Atlantic, Indian, Pacific, surface and in
various depths.
2. Spongaster quadratus, n. sp.
Arms at equal distances, forming a regular, rectangular cross, club-shaped, about four times as
long as the diameter of the central circular disk, and five times as long as broad at the base.
Basal third of the arms square; distal two thirds triangular, three times as broad, with a
truncated distal end. Patagium complete, perfectly enveloping the arms, and bordered by an
elegant, radially striated, broad edge, forming a regular square. (Similar to Histiastrum
quadratum, Pl. 46,
fig. 4, but quite spongy.)
Dimensions.—Radius of the arms 0.15, distal breadth 0.06, basal breadth 0.02;
length of the square side 0.25.
Habitat.—Pacific, central area, Stations 270 to 274, depths 2350 to
2925 fathoms.
3. Spongaster cruciatus, n. sp.
Arms at equal distances, forming a rectangular, regular cross, lanceolate, three times as long
as broad and as the diameter of the central circular disk. Ends of the arms provided with a short
conical spine. Patagium incomplete, enveloping only the basal half of the arms, forming a regular
square with concave sides.
Dimensions.—Radius of the arms 0.18, greatest breadth 0.05; length of the square
side 0.2.
Habitat.—South Pacific, Station 288, surface.
{598}
4. Spongaster pentacyclus, n. sp.
Arms at equal distances, forming a rectangular, regular cross, of the same size and form as the
circular central disk, so that the dark interior part of the shell is composed of five equal
circular disks, situated in a quincuncial manner. The clearer complete patagium, enveloping the
whole cross perfectly, forms a regular square with rounded edges.
Dimensions.—Radius of the arms 0.2; diameter of each of the five circular disks
0.01; length of the square side 0.35.
Habitat.—West Indies, Cuba, surface (Thomson).
Subgenus 2. Spongastromma, Haeckel.
Definition.—Cross formed by the four arms bilateral or irregular, with
the arms at different distances.
5. Spongaster orthogonus, Haeckel.
Spongocyclia orthogona, Haeckel, 1862, Monogr. d. Radiol., p. 471, Taf. xxviii. fig.
3.
Spongodiscus orthogonus, Haeckel, 1860, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 844.
Arms at different distances, forming a bilateral cross, grouped in two opposite pairs of equal
size and similar form. Arms club-shaped, about as long as the diameter of the central circular
disk, enveloped perfectly by the complete patagium, which forms a regular rectangle, the longer
side of which is one and a half times as long as the shorter side. (The arms in my figure are not
distinctly enough marked.)
Dimensions.—Radius of the arms 0.08, breadth 0.02; length of the larger side of
the rectangle 0.15, of the smaller 0.1.
Habitat.—Mediterranean (Messina), surface.
6. Spongaster scyllaeus, Haeckel.
Spongocyclia scyllaea, Haeckel, 1862, Monogr. d. Radiol., p. 471, Taf. xxviii. fig.
4.
Spongodiscus scyllaeus, Haeckel, 1860, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 844.
Arms at different distances, forming a bilateral cross, grouped in two opposite pairs of
different size and form, one pair smaller and less divergent than the other. Arms club-shaped,
little longer than the radius of the central circular disk, enveloped perfectly by the complete
patagium, which forms a trapezium; the convergent longer sides of the latter are one and a half
times as long as the larger, and twice as long as the smaller parallel side. (The arms are in my
figure, loc. cit., not distinctly enough marked.)
Dimensions.—Radius of the arms 0.12 to 0.14, breadth 0.02; length of both
convergent sides 0.24, of the larger parallel side 0.18, of the smaller 0.12.
Habitat.—Pacific, central area, Station 272, surface; Mediterranean
(Messina).
{599}
Suborder VI. LARCOIDEA, Haeckel, 1883 (Pls. 9, 10, 49, 50).
Definition.—Spumellaria with lentelliptical central
capsule (rarely somewhat modified or allomorphic), with a lentelliptical fenestrated siliceous
shell (often modified or allomorphic, and sometimes quite irregular). Growth different in the
three unequal dimensive axes, perpendicular one to another. The typical Lentellipsis is
characterised by three elliptical dimensive planes of different sizes, perpendicular one to
another.
The section Larcoidea, the fourth and last of the Sphærellaria, comprises all those forms of this group in which the fenestrated
shell originally is lentelliptical, characterised by different growth in three different axes,
perpendicular one to another, all three equal on both poles. The geometrical fundamental form of
the shell is therefore a lentellipsis or a triaxial ellipsoid; and this typical form is preserved
completely in the majority of Larcoidea in the pure geometrical form of
the central capsule.
The three dimensive axes, which determine the typical form of Larcoidea, are commonly differentiated in such a way, that the first, the
longitudinal or principal axis, is the longest; both its poles, oral and aboral (or anterior and
posterior) are equal. The second, the lateral or transverse axis, is commonly less than the first,
greater than the third axis; both its poles are the equal lateral poles (right and left not
differentiated). The third dimensive axis, the equatorial or sagittal axis, is commonly the
shortest; both its equal poles are the sagittal poles (dorsal and ventral poles not different).
The relative size of the three dimensive axes in the human body exhibits similar relations.
The three dimensive planes of the Larcoidea, the sagittal, lateral,
and transverse planes, are elliptical, all three of different sizes. The first plane, the median
or sagittal plane, is commonly as regards size between the two others; its major axis is the
principal, its minor the sagittal axis; it separates the right half of the body from the left. The
second plane or lateral plane, is commonly larger than the two others; its major axis the
principal, its minor the transverse axis; it separates the dorsal half of the body from the
ventral. The third plane, the equatorial or zonal plane, is commonly less than the two others; its
major axis the lateral, its minor the sagittal axis; it separates the two principal halves of the
body, the oral and aboral halves.
In my Monograph (1862) only very few forms of Larcoidea are described,
Tetrapyle and Lithelius (the latter representing a peculiar family, Lithelida). In
my Prodromus (1881, pp. 463, 464) I disposed all observed forms of Larcoidea in two different families, the Pylonida and Lithelida. The rich
materials of the Challenger collection have since offered an astonishing number of new and
interesting forms of this section, so that I can enumerate here fifty-one genera and two hundred
and sixty-five species. I dispose them here in four subsections and nine families. Three of these
have regular lentelliptical shells, which are not articulate, and {600}without annular constrictions (Larcarida, Larnacida, Pylonida); these
form the subsection Pylolarcida. Two other families (Tholonida and Zonarida) are distinguished by
annular constrictions, which divide the regular lentelliptical shell into a number of dome-shaped
chambers or cupolas; we call these Thololarcida. A third group, Spirolarcida, comprises the Larcoidea with spiral growth; the two families of Lithelida and Streblemida.
Finally a fourth group the Sorolarcida is formed by the Larcoidea with
irregular shells, also two families, the Phorticida and Soreumida.
The first family of Larcoidea, the Larcarida, contains the most simple
forms, beginning with Cenolarcus, a quite simple lentelliptical latticed shell. In
Coccolarcus we find already two concentric shells, connected by radial beams, an inner
medullary and an outer cortical shell. In Spongolarcus the lentelliptical shell becomes
spongy.
The second family, Larnacida, is very similar to the Larcarida, and seems to diverge only by
the different mode of connection between the two concentric lentelliptical shells. But in truth
this slight difference is of great morphological importance, as it depends on a quite different
and peculiar mode of growth. In the foregoing Larcarida (Coccolarcus, &c.), the
concentric shells originate in the same manner as in the concentric Prunoidea and Sphæroidea, by radial beams, which arise
from the surface of the inner (medullary) shell and become connected by a network to form the
outer (cortical) shell. Here, in the Larnacida, a quite similar shell originates in a quite
different way, first arrived at in the Pylonida (Trizonium). Both concentric shells become
here connected by peculiar lattice girdles, which are developed in the perimeter of the three
elliptical dimensive planes. Firstly, on both sides of a simple, spherical, or lentelliptical
central chamber, arise two lateral wings (on the poles of the transverse axis), and build around
the former a transverse girdle. This is crossed by a larger lateral girdle, the minor axis of
which is the major of the former, and perpendicular to both girdles is yet developed a third, the
sagittal girdle. If the open fissures or "gates" between these three girdles become closed by
network, we obtain Larnacilla, the probable ancestral form of all Larnacida.
Whilst in Larnacilla and Larnacidium this typical trizonal lentelliptical shell
constitutes by itself alone the whole skeleton, in the other Larnacida it becomes overgrown by
outer envelops, and so becomes enclosed in the interior of the central capsule as a
"Larnacilla-shaped medullary shell." If the enclosing external envelops be simply latticed,
we get the subfamily Larnacalpida; if they be spongy, we get the Larnacospongida.
The third family, Pylonida, is the most important of all Larcoidea, as
not only the largest and most interesting number of species belong to it, but also many other
genera (far the greater part of all Larcoidea) may be derived from it.
The peculiar character of the Pylonida is determined by the imperfect fenestration of the
lentelliptical shell growing in the three dimensive axes in a quite different manner. Each
elliptical dimensive plane becomes circumscribed by an elliptical latticed girdle (or fenestrated
{601}ring), and between these three girdles
(perpendicular one to another) remain wide open fissures of the shell or "gates" (Pylæ).
The beginning of the shell-building is the same as in Larnacilla, the most simple form of
Larnacida. From a quite simple medullary shell, a spherical, subspherical, ellipsoidal, or
lentelliptical central chamber, arise two latticed wings, opposite on the poles of the transverse
axis (Monozonium). Both wings are short and wide hollow fenestrated tubes, the axes of
which are parallel to the principal axis. Therefore they form together with the central chamber an
elliptical transverse girdle. This first girdle becomes crossed by a second lateral girdle; from
both poles of the transverse axis arise latticed wings, which unite on the poles of the principal
axis, therefore the minor axis of this second larger ring is the major axis of the first smaller
ring (Dizonium). Between the two crossed rings remain four wide open gates. Now follows the
development of a third sagittal girdle, arising from both poles of the principal axis and
overgrowing the four gates. But as this third girdle is larger than the second, four other larger
gates arise between the two (in planes perpendicular to the former four gates). Now we have the
characteristic and most important trizonal shell (Trizonium), composed of three elliptical
lattice-girdles of different size, perpendicular one to another, and enclosing a simple central
chamber. If the four gates of this Trizonium become closed by lattice-work, it passes over
into Larnacilla, the most important ancestral form of the Larnacida.
This most significant "trizonal shell," either incompletely latticed in Trizonium (with
four open gates), or completely latticed by fenestration of the four gates, in Larnacilla,
is to be found in far the greater part of all Larcoidea, representing the
medullary shell, which is overgrown by an outer cortical shell. In many Larcoidea, in which this "Larnacilla-shell" is absent, it is perhaps
lost by phylogenetic reduction, or retrograde metamorphosis.
The same process of triple girdle-building, by which the typical Trizonium-shell or
Larnacilla-shell is produced (Haplozonaria), is repeated once or twice in the larger forms
of Pylonida. The first system of three girdles (perpendicular one to another) becomes overgrown by
a second system of the same formulation in the Diplozonaria, and this becomes overgrown by a third
system in the Triplozonaria; in the highest genus of this group, Pylozonium, we find not
less than nine girdles (three systems, each of three girdles). Till now only one genus of the
whole polymorphous family was well known, Tetrapyle (with five girdles, three of the
medullary, two of the cortical shell). If the gates between the girdles remain open, all these
forms must be regarded as Pylonida; if the gates afterwards become closed by a network, they pass
over into other families.
The fourth family of the Larcoidea is the Tholonida, distinguished by the polythalamous
shell being composed of a certain number of roundish or hemispherical chambers (domes or cupolas),
which surround a primordial central chamber in quite regular disposition, lying opposite in pairs
on the poles of the three dimensive axes. {602}If we
imagine that each "wing" (or open half-girdle) of the Pylonida becomes closed by a lattice-work,
and so transformed into a hemispherical or roundish cupola, we obtain the characteristic shell of
the Tholonida. Indeed every girdle of the former corresponds to a pair of opposite domes of the
latter. The axis of each pair of domes is one of the three dimensive axes.
The primordial chamber of the Tholonida (or the central chamber, around which all cupolas are
regularly disposed) is either a simple lentelliptical lattice-shell, like Cenolarcus, or it
is a trizonal shell (with an enclosed concentric medullary shell), like Larnacilla. As in
both cases the building and the disposition of the cupolas around it are quite the same, we can
suppose that the whole family of Tholonida may have been derived originally from Larnacilla
(or Trizonium), and that the Cenotholida (with a simple central chamber) are sprung from
the Coccotholida (with a Larnacilla-shaped central chamber) by reduction and loss of the
original medullary shell.
The family Tholonida can be divided into three subfamilies according to the disposition of the
cupola-pairs in one, two, or three dimensive axes. In the Cubotholida lie two cupolas on the poles
of the transverse axis of the central chamber (corresponding to Amphipyle); in the
Staurotholida we find four cupolas crosswise disposed, on the poles of the transverse and
principal axes (corresponding to Tetrapyle); in the Cubotholida are at least six cupolas,
on the poles of all three dimensive axes (corresponding to Tholonium). In all three cases
the number of cupolas may be augmented by the secondary apposition of other chambers or domes in
the same disposition. Sometimes also the whole cortical shell becomes enclosed by an external veil
or mantle of delicate network. The lentelliptical (or often nearly cubical) central chamber
becomes often reduced, so that its sides are incompletely latticed or widely opened; in some
Cubotholida only the twelve edges of the eight cornered cubical central chamber remain; its six
sides are quite open and only over-vaulted by the six hemispherical cupolas. From the opposite
points of the latter (in the deep annular constrictions between them) often arise radial spines,
and these lie commonly in diagonal planes, separating the dome-pairs.
A similar dome-building or a composition of the polythalamous shell by pairs of cupolas we find
also in the next (fifth) family, the Zonarida. But here the true cause of the peculiar
dome-structure is quite different, not an apposition of new chambers, but the constriction of a
cortical shell-like Larnacalpis by two or more constrictions. These constrictions lie in
dimensive planes (or in planes parallel to these), and therefore the cupolas are (all or partly)
in diagonal planes, a condition quite opposite to that found in the Tholonida. One of the annular
constrictions is constantly in the sagittal plane (separating the right and left halves of the
shell). The number of the constrictions in the few genera is two, three, and four, and therefore
the number of the cupolas four, six, or eight. As this cortical shell constantly encloses a
trizonal medullary shell (or Larnacilla-shell), we cannot doubt that the Zonarida must be
derived from the Larnacida.
{603}
Whilst in all the foregoing five families of Larcoidea the shell-form
is regular and their geometrical fundamental form is a lentellipsis (or a triaxial ellipsoid, with
three unequal isopolar dimensive axes), in the four remaining families of this suborder the shell
becomes bilateral or irregular (with the poles of the axes unequal). In two of these families
(Lithelida and Streblemida) the growth of the shell becomes spiral, in the last two families
(Soreumida and Phorticida) quite irregular. But as in all four families we encounter the typical
trizonal medullary shell (or Larnacilla-shell), we are convinced that they must be derived
(wholly or partially) from the Larnacida.
The Lithelida (the sixth family) are Larcoidea with spiral growth and
bilateral form (like Nautilus); therefore the spiral line lies in one plane and this spiral
plane divides the whole shell into two symmetrical halves (right and left). The axis of the spiral
(around which the shell winds) is a straight line, one of the three dimensive axes. In the greater
part of Lithelida (in the Larcospirida) the primordial of central chamber of the polythalamous
shell is a trizonal medullary shell or Larnacilla-shell, and the growth of the first spiral
turning begins as the development of the first (transverse) cortical girdle of Amphipyle;
but as one wing (or lateral half) of this girdle grows more rapidly than the other, it overgrows
the latter and begins the spiral winding; if the other wing follow and overgrow the first, the
spiral becomes double. Each of the three dimensive girdles (of the Pylonida) may begin the spiral
winding. There can be no doubt that all these Lithelida (the Larcospirida) must be derived from
the Pylonida, by unequal growth of the two halves of one girdle. Perhaps from these may also
derived the other part of this family, the Spiremida (Spirema and Lithelius); in
these the primordial chamber of the spiral shell is simple, and may be derived by reduction of the
original Larnacilla-shell. But it is also possible that the Spiremida proceed directly from
the Larcarida, and that their ancestors did not possess a Larnacilla-shell.
The Streblemida (the seventh family) are Larcoidea with spiral growth
and asymmetrical form of the polythalamous shell (like Helix or Turrilites);
therefore the spiral line is twisted like a winding stair, and the spiral face is curved and
divides the shell into two unequal halves. The Streblemida have the same likeness and relation to
the turbinoid Foraminifera (Rotalia, Globigerina, &c.) as the Lithelida to the
nautiloid Foraminifera (Polystomella, Nummulina, &c.). As in these calcareous
Rhizopods also the peculiar growth of the siliceous Streblemida begins from a primordial chamber
to which a variable number of roundish chambers (of increasing size) is apposed. But the building
of these chambers and of their septa is by no means so regular and complete as in the greater
number of turbinoid Foraminifera. As in a part of this family the primordial chamber is
Larnacilla-shell, these also may be derived from the Larnacida, but the other part (with
simple central chamber) is perhaps produced directly from the Larcarida.
The eighth family, Soreumida, is perhaps derived from the Streblemida by the loss of the spiral
growth. The polythalamous shell is similar to the latter, but the chambers are {604}aggregated without any order, like the Acervulinida among
the Foraminifera. In some cases also here the primordial chamber is a trizonal
Larnacilla-shell, in other cases it is a simple, subspherical or lentelliptical shell.
The last family, the Phorticida, is formed of irregular Larcoidea, in
which a lentelliptical trizonal Larnacilla-shell (as an inner medullary shell) is enveloped
by an irregular, latticed, or spongy cortical shell. They can be regarded as abnormalities or
irregular deformities of Larnacida or Pylonida.
The central capsule of the Larcoidea is originally lentelliptical and
preserves this form, the "triaxial ellipsoid," in the greater number of genera. In some groups it
follows the prevalent growth of the shell in the direction of one of the three dimensive axes, and
becomes prolonged in this way. In many chambered forms (particularly Tholonida and Zonarida) the
growing central capsule gets constricted, corresponding to the constrictions of the shell. In the
Soreumida and Phorticida its form often becomes irregular. But in general for the greater number
of Larcoidea the lentelliptical form of their central capsule is quite
characteristic.
Synopsis of the Families of Larcoidea.
| Larcoidea with a regular or
symmetrical shell, the growth of which is determined by the three dimensive axes. (Both poles
of each axis are equal.) |
|
Cortical shell completely latticed, without external gates
(or interzonal fissures), without annular constrictions and domes. |
|
Medullary shell absent or simple (spherical or lentelliptical), |
1. Larcarida. |
| Medullary shell trizonal or Larnacilla-shaped (composed of three
dimensive girdles), |
2. Larnacida. |
| Cortical shell incompletely latticed, with two to four or
more symmetrically disposed gates or fissures remaining between latticed dimensive
girdles, |
3. Pylonida. |
| Cortical shell completely latticed, without external gates
(or interzonal fissures), with two or more annular constrictions, which separate three to six
or more dome-shaped protuberances. |
|
Constrictions of the cortical shell in diagonal planes; domes in
dimensive axes, |
4. Tholonida. |
| Constrictions of the cortical shell in dimensive planes; domes in
diagonal axes, |
5. Zonarida. |
| Larcoidea with a symmetrical
or irregular shell, either with spiral growth or with quite irregular growth. (Both poles of
one axis are different.) |
|
Cortical shell with spiral growth. |
|
Spiral cortical shell bilateral (with plane spiral), |
6. Lithelida. |
| Spiral cortical shell asymmetrical (with ascending spiral), |
7. Streblemida. |
| Cortical shell with quite irregular growth. |
|
Cortical shell simple, with one single chamber, |
8. Phorticida. |
| Cortical shell composed of a number of heaped up or aggregated
chambers, |
9. Soreumida. |
| Larcoidea with a regular or symmetrical shell, the
growth of which is determined by the three dimensive axes. (Both poles of each axis are
equal.) |
|
|
Cortical shell completely latticed, without external gates (or
interzonal fissures), without annular constrictions and domes. |
|
|
|
|
|
|
Medullary shell absent or simple (spherical or
lentelliptical), |
|
|
|
|
|
|
|
|
|
|
1. Larcarida. |
|
|
|
|
|
|
|
|
|
|
Medullary shell trizonal or Larnacilla-shaped (composed of
three dimensive girdles), |
|
|
|
|
|
|
|
|
|
|
2. Larnacida. |
|
|
|
|
|
|
|
|
Cortical shell incompletely latticed, with two to four or more
symmetrically disposed gates or fissures remaining between latticed dimensive girdles, |
|
|
|
|
|
|
|
|
3. Pylonida. |
|
|
|
|
|
|
|
|
Cortical shell completely latticed, without external gates (or
interzonal fissures), with two or more annular constrictions, which separate three to six or
more dome-shaped protuberances. |
|
|
|
|
|
|
Constrictions of the cortical shell in diagonal planes; domes in
dimensive axes, |
|
|
|
|
|
|
|
|
|
|
4. Tholonida. |
|
|
|
|
|
|
|
|
|
|
Constrictions of the cortical shell in dimensive planes; domes in
diagonal axes, |
|
|
|
|
|
|
|
|
|
|
5. Zonarida. |
|
|
|
|
|
|
| Larcoidea with a symmetrical or irregular shell,
either with spiral growth or with quite irregular growth. (Both poles of one axis are
different.) |
|
|
Cortical shell with spiral growth. |
|
|
|
|
|
|
Spiral cortical shell bilateral (with plane spiral), |
|
|
|
|
|
|
|
|
|
|
6. Lithelida. |
|
|
|
|
|
|
|
|
|
|
Spiral cortical shell asymmetrical (with ascending spiral), |
|
|
|
|
|
|
|
|
|
|
7. Streblemida. |
|
|
|
|
|
|
|
|
Cortical shell with quite irregular growth. |
|
|
|
|
|
|
Cortical shell simple, with one single chamber, |
|
|
|
|
|
|
|
|
|
|
8. Phorticida. |
|
|
|
|
|
|
|
|
|
|
Cortical shell composed of a number of heaped up or aggregated
chambers, |
|
|
|
|
|
|
|
|
|
|
9. Soreumida. |
|
|
|
|
|
|
{605}
Family XXIV. Larcarida, Haeckel, 1883 (Pl. 50, figs. 1,
2).
Definition.—Larcoidea with a regular, completely
latticed, lentelliptical cortical shell, without open gates and annular constrictions; medullary
shell absent or simple (not trizonal), connected with the cortical shell by radial beams.
The family Larcarida opens the long series of Larcoidea as the most simple group of this suborder. It commences with
Cenolarcus, a quite simple lentelliptical latticed shell, which is characterised by three
unequal isopolar dimensive axes, perpendicular one to another. The major of these three axes is
the longitudinal or principal, the middle is the lateral or transverse, and the minor is the
equatorial or sagittal axis (as in the human body). Among the three dimensive planes, which are
determined by pairs of these axes, the lateral plane is the largest (halved by the crossed
principal and lateral axes). The intermediate is the sagittal plane or median plane (halved by the
crossed principal and sagittal axes). The smallest is the equatorial plane or transverse plane
(halved by the crossed lateral and sagittal axes). Therefore the shell has all the characters of
the true Lentellipsis or of the "triaxial ellipsoid," and its axes agree with the three
axes of the "rhombic crystalline system."
In the three subfamilies of Larcarida this lentelliptical shell assumes a different shape: in
the Cenolarcida it remains simple, in the Spongolarcida it becomes spongy (sometimes quite filled
out with a spongy framework), in the Coccolarcida it is composed of two or more concentric
lentelliptical shells (at least an inner medullary and one outer cortical shell). These shells are
simply connected by radial beams, and not, as in the Larnacida, by latticed wings (or half
girdles).
The network of the Larcarida shell is sometimes regular, commonly irregular (as in the greater
number of Larcoidea). The surface of the shell is sometimes smooth or
thorny, at other times covered with radial spines. These are often symmetrically disposed, either
on the poles of the dimensive axes or in crossed diagonal planes.
The central capsule is a true "lentellipsis" in a geometrical sense; it is halved by three
elliptical dimensive planes of different sizes, perpendicular one to another. In the Cenolarcida
the central capsule lies freely inside the simple (cortical) shell, only separated from it by the
jelly-mantle. In the Coccolarcida it contains the medullary shell, and is enclosed by the simple
or double cortical shell, perforated by the radial beams connecting the two shells. The spongy
shell of the Spongolarcida exhibits a different relation to the central capsule: in
Spongolarcus the latter lies freely in the internal cavity of the spongy shell; in
Stypolarcus, where this cavity is quite filled with a spongy network, the central capsule
also contains a part of it.
The morphological and phylogenetic relations of the Larcarida to the other families of Spumellaria admit of a different explanation. As this family contains the {606}most simple forms of all Larcoidea, we can regard the Cenolarcus as the common ancestral form of
this group, having originated from Actilarcus (or the lentelliptical
Actissa) by the building of a simple lentelliptical lattice-shell. But it is also possible
that a part of the Larcarida (or all?) descend from Larnacida by reduction or loss of the original
Larnacilla-shell (compare Cenolarcus triaxonius, p. 607).
Synopsis of the Genera of the Larcarida.
|
III. Subfamily Cenolarcida.
Shell simple, latticed (lentelliptical cortical shell).
|
|
Without radial spines, |
266. Cenolarcus. |
| With radial spines, |
267. Larcarium. |
|
III. Subfamily Coccolarcida.
Shell composed of two or more concentric latticed shells (inner medullary and
outer cortical).
|
|
Without radial spines, |
268. Coccolarcus. |
| With radial spines, |
269. Larcidium. |
|
III. Subfamily Spongolarcida.
Shell spongy, partly or whole composed of a spongy framework.
|
|
With an internal cavity, |
270. Spongolarcus. |
| Without an internal cavity, |
271. Stypolarcus. |
| I. Subfamily Cenolarcida. Shell simple, latticed (lentelliptical cortical
shell). |
|
|
Without radial spines, |
|
|
|
|
|
|
266. Cenolarcus. |
|
|
|
|
|
|
With radial spines, |
|
|
|
|
|
|
267. Larcarium. |
|
|
|
|
| II. Subfamily Coccolarcida. Shell composed of two or more concentric latticed
shells (inner medullary and outer cortical). |
|
|
Without radial spines, |
|
|
|
|
|
|
268. Coccolarcus. |
|
|
|
|
|
|
With radial spines, |
|
|
|
|
|
|
269. Larcidium. |
|
|
|
|
| III. Subfamily Spongolarcida. Shell spongy, partly or whole composed of a
spongy framework. |
|
|
With an internal cavity, |
|
|
|
|
|
|
270. Spongolarcus. |
|
|
|
|
|
|
Without an internal cavity, |
|
|
|
|
|
|
271. Stypolarcus. |
|
|
|
|
Subfamily 1. Cenolarcida, Haeckel.
Definition.—Larcarida with a simple,
lentelliptical latticed shell (cortical shell without a medullary shell).
Genus 266. Cenolarcus,[305] n. gen.
Definition.—Larcarida with a simple, lentelliptical
latticed shell, without a medullary shell, without radial spines.
The genus Cenolarcus begins the group of Larcoidea
as the most simple form of this suborder. It corresponds to Cenosphæra among the Sphæroidea, to Cenodiscus among the Discoidea,
to Cenellipsis among the Prunoidea. The simple latticed shell is
distinguished from that of the three other genera by its typical lentelliptical form, a triaxial
ellipsoid with three dimensive axes of unequal length. Probably Cenolarcus is the original
ancestral form of the Larcoidea, derived from Actilarcus (the lentelliptical Actissa)
by the formation of a simple fenestrated shell around the lentelliptical central capsule. But
possibly also some species of Cenolarcus may be derived from Coccolarcus or
Larnacilla by reduction and loss of the medullary shell (compare Cenolarcus
triaxonius, n. sp.).
{607}
1. Cenolarcus primordialis, n. sp. (Pl. 50, figs. 7,
7a, 7b).
Network of the shell regular, with circular, hexagonally framed pores, twice as broad as the
elevated bars; about ten pores on the half meridian, eight on the half equator. Surface a little
rough. Proportion of the three dimensive axes = 2 : 2.5 : 3.
Dimensions.—Principal axis (or length) 0.12, transverse axis (or breadth) 0.1,
sagittal axis (or thickness) 0.08; pores 0.01, bars 0.005.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
2. Cenolarcus dimensivus, n. sp.
Network of the shell regular, with circular pores (without hexagonal frames), three times as
broad as the thin bars; about twelve pores on the half meridian, nine on the half equator. Surface
thorny. Proportion of the three dimensive axes = 3 : 4 : 5.
Dimensions.—Principal axis 0.15, transverse axis 0.12, sagittal axis 0.09; pores
0.012, bars 0.004.
Habitat.—North Pacific, Station 244, surface.
3. Cenolarcus triaxonius, n. sp.
Network of the shell regular, with circular, hexagonally framed pores, four times as broad as
the thin bars; about twelve pores on the half meridian, nine on the equator. Surface smooth. From
the inner surface of the shell arise six very thin radial beams, opposite in pairs in the three
dimensive axes; all six beams end freely in a little knob, at an equal distance from the centre;
therefore this remarkable species seems to have lost a medullary shell (descending from
Coccolarcus or Larnacilla?). Proportion of the three dimensive axes =
2 : 2.5 : 3.
Dimensions.—Principal axis 0.13, transverse axis 0.11, sagittal axis 0.09; pores
0.012, bars 0.03.
Habitat.—Tropical Atlantic, Station 338, depth 1990 fathoms.
4. Cenolarcus lentellipticus, n. sp.
Network of the shell regular, with circular pores of the same breadth as the thick bars; about
eighteen pores on the half meridian, fourteen on the half equator. Surface smooth. Proportion of
the three dimensive axes = 2 : 3 : 4.
Dimensions.—Principal axis 0.16, transverse axis 0.12, sagittal axis 0.08; pores
and bars 0.004.
Habitat.—Western Tropical Pacific, Station 224, surface.
5. Cenolarcus minimus, n. sp.
Network of the shell subregular, with very small circular pores of the same breadth as the
bars; only four pores on the half meridian, three on the half equator. Surface smooth. Proportion
of the three dimensive axes = 3 : 4 : 5.
{608}
Dimensions.—Principal axis 0.05, transverse 0.04, sagittal axis 0.03; pores and
bars 0.006.
Habitat.—Pacific, central area, Station 266, depth 2750 fathoms.
Genus 267. Larcarium,[306] n. gen.
Definition.—Larcarida with a simple, lentelliptical
latticed shell, without a medullary shell; surface covered with radial spines.
The genus Larcarium differs from Cenolarcus only in the possession of
radial spines on the surface of the simple fenestrated lentelliptical shell. These spines are
commonly disposed symmetrically, opposite in pairs, either on the poles of the three dimensive
axes, or on the poles of certain diagonal axes. Larcarium differs from the similar genera
Larcidium, Larnacidium, and Larnacantha by the absence of any medullary
shell.
1. Larcarium amphistylum, n. sp.
Shell thorny, with two large conical spines, opposite on both poles of the principal axis,
somewhat longer than it. Network of the shell regular, with circular, hexagonally framed pores,
twice as broad as the bars; about twelve pores on the half meridian, ten on the half equator.
Proportion of the three dimensive axes = 3 : 4 : 5.
Dimensions.—Principal axis (or length) 0.15, transverse axis (or breadth) 0.12, sagittal axis (or thickness) 0.09;
pores 0.01, bars 0.005.
Habitat.—Pacific, central area, Station 274, depth 2750 fathoms.
2. Larcarium staurostylum, n. sp.
Shell smooth, with four short three-sided pyramidal spines of equal length, opposite in pairs
on the poles of the principal and lateral axes. Network of the shell regular, with circular pores
of the same breadth as the bars; about eight pores on the half meridian, six on the half equator.
Proportion of the three dimensive axes = 2 : 2.5 : 3.
Dimensions.—Length of the shell 0.12, breadth 0.1, thickness 0.08; pores and bars
0.006.
Habitat.—South Pacific, Station 300, depth 1375 fathoms.
3. Larcarium hexastylum, n. sp.
Shell smooth, with six short conical spines of equal length (= the shortest axis of the shell),
opposite in pairs on the poles of the three dimensive axes. Network of the shell subregular, with
{609}circular pores three times as broad as the bars;
about eleven pores on the half meridian, nine on the half equator. Proportion of the three
dimensive axes = 3 : 4 : 5.
Dimensions.—Length of the shell 0.1, breadth 0.08, thickness 0.06, pores 0.006;
bars 0.002.
Habitat.—North Pacific, Station 244, surface.
4. Larcarium axostylum, n. sp.
Shell thorny, with six strong conical radial spines, opposite in pairs on the poles of the
three dimensive axes. All three pairs are of different sizes, the length of each spine
corresponding nearly to the size of the shell-axis, of which it is the prolongation. Network of
the shell subregular, with circular pores twice as broad as the bars; ten on the half meridian,
eight on the half equator. Proportion of the three dimensive axes =
2 : 3 : 4.
Dimensions.—Length of the shell 0.12, breadth 0.09, thickness 0.06; pores 0.01,
bars 0.005.
Habitat.—Western Tropical Pacific, Station 225, depth 4475
fathoms.
5. Larcarium octostylum, n. sp.
Shell thorny, with eight thin cylindrical radial spines, opposite in pairs in two crossed
diagonal planes. Network of the shell irregular, with roundish pores, twice to four times as broad
as the bars; nine to eleven on the half meridian, five to seven on the half equator. Proportion of
the three dimensive axes = 3 : 4 : 5.
Dimensions.—Length of the shell 0.11, breadth 0.09, thickness 0.07; pores 0.006 to
0.012, bars 0.003.
Habitat.—Pacific, central area, Station 267, surface.
6. Larcarium polystylum, n. sp.
Shell thorny, with numerous (twenty to thirty) stronger conical radial spines, about as long as
the shortest axis of the shell. Network of the shell irregular, with roundish pores, twice to four
times as broad as the bars; seven to eight on the half meridian, five to six on the half equator.
Proportion of the three dimensive axes = 1 : 2 : 3.
Dimensions.—Length of the shell 0.09, breadth 0.06, thickness 0.03; pores 0.006 to
0.012, bars 0.003.
Habitat.—South Atlantic, Station 323, surface.
7. Larcarium chætostylum, n. sp.
Shell bristly, with very numerous (sixty to eighty or more) very thin, bristle-like, radial
spines, somewhat longer than the longest axis of the shell. Network irregular, with very small
roundish pores, about the same size as the bars; sixteen to eighteen on the half meridian,
thirteen to fifteen on the half equator. Proportion of the three dimensive axes =
1 : 1.5 : 2.
{610}
Dimensions.—Length of the shell (without spines) 0.13, breadth 0.1, thickness
0.07; pores and bars 0.004 to 0.006.
Habitat.—Pacific, central area, Station 273, surface.
Subfamily 2. Coccolarcida, Haeckel.
Definition.—Larcarida with encased
lentelliptical shell, composed of two or more concentric lentelliptical latticed shells, which are
united by radial beams (at least one inner medullary shell and one outer cortical shell).
Genus 268. Coccolarcus,[307] n. gen.
Definition.—Larcarida with two concentric latticed
shells, an outer lentelliptical cortical shell, and an inner (spherical or lentelliptical)
medullary shell, both connected by radial beams. Surface without radial spines.
The genus Coccolarcus differs from Cenolarcus by the possession of an
internal medullary shell. This is quite simple, either spherical or lentelliptical, and connected
with the outer cortical shell by a number of radial beams. In the similar Larnacilla this
connection is effected by four internal latticed lamellæ (the half lateral wings of the transverse
girdle); therefore we find here four internal gates (on the poles of the principal axis), absent
in Coccolarcus.
1. Coccolarcus lentellipsis, n. sp.
Cortical shell with smooth surface and regular network; pores circular, twice as broad as the
bars; about thirteen on the half meridian, eleven on the half equator. Proportion of the three
dimensive axes = 3 : 4 : 5. Medullary shell spherical, half as broad as the
transverse radius.
Dimensions.—Principal axis (or length) of the cortical shell 0.15, transverse axis
(or breadth) 0.12, sagittal axis (or thickness) 0.09; pores 0.01, bars 0.005; diameter of the
medullary shell 0.03.
Habitat.—Pacific, central area, Station 272, depth 2600 fathoms.
2. Coccolarcus platellipsis, n. sp.
Cortical shell with thorny surface and irregular network; pores roundish, twice to three times
as broad as the bars; sixteen to eighteen on the half meridian, ten to twelve on the half equator.
Proportion of the three dimensive axes = 1 : 2 : 3. Medullary shell
lentelliptical, one-third as large as the cortical shell.
{611}
Dimensions.—Length of the cortical shell 0.17, breadth 0.11, thickness 0.06; pores
0.008 to 0.012, bars 0.004; medullary shell 0.03 to 0.06.
Habitat.—Pacific, central area, Station 274, depth 2750 fathoms.
Genus 269. Larcidium,[308] n. gen.
Definition.—Larcarida with two concentric latticed
shells, an outer lentelliptical cortical shell, and an inner (spherical or lentelliptical)
medullary shell, both connected by radial beams. Surface covered with radial spines.
The genus Larcidium differs from the foregoing Coccolarcus only in
the possession of radial spines, and bears to it the same relation that Larcarium does to
Cenolarcus. The spines are commonly symmetrically disposed, opposite in pairs in the
dimensive axes, sometimes also in diagonal axes.
1. Larcidium dissacanthum, n. sp.
Cortical shell with thorny surface and regular network; pores circular, hexagonally framed,
three times as broad as the bars; about thirteen on the half meridian, eleven on the half equator.
Proportion of the three dimensive axes = 2 : 3 : 5. Medullary shell spherical,
one-fourth as broad as the cortical, connected with it by two thin beams, lying in the principal
axis, and prolonged on its poles into two strong conical spines, somewhat longer than the greatest
axis.
Dimensions.—Length of the cortical shell (or principal axis) 0.15, breadth 0.09,
thickness 0.07; pores 0.015, bars 0.005; medullary shell 0.025.
Habitat.—North Pacific, Station 253, depth 3125 fathoms.
2. Larcidium hexacanthum, n. sp.
Cortical shell with smooth surface and regular network; pores circular, twice as broad as the
bars; about eleven on the half meridian, nine on the half equator. Proportion of the three axes =
2 : 2.5 : 3. Medullary shell spherical, one-third as broad as the cortical. On
the surface six strong, three-sided pyramidal spines, all about as long as the breadth of the
cortical shell.
Dimensions.—Length of the cortical shell 0.12, breadth 0.1, thickness 0.08; pores
0.01, bars 0.005; medullary shell 0.035.
Habitat.—Pacific, central area. Station 266, surface.
3. Larcidium axacanthum, n. sp.
Cortical shell with rough surface and irregular network; pores roundish, twice to four times as
broad as the bars; about fifteen to nineteen on the half meridian, twelve to fourteen on the half
{612}equator. Proportion of the three axes =
2 : 3 : 4. Medullary shell lentelliptical, of the same form and structure as
the cortical, but only one-third as large, connected with it by six thin radial beams, lying in
pairs in the three dimensive axes; on the outside they are prolonged into six strong conical
radial spines, which are in pairs of different size (as in Larcarium axostylum); the length
of each spine nearly equals the axis of the cortical shell, of which it is the prolongation.
Dimensions.—Length of the cortical shell (and the principal spines) 0.18, breadth
of it (and length of the lateral spines) 0.135, thickness of it (and length of the sagittal
spines) 0.09; pores 0.005 to 0.013, bars 0.003.
Habitat.—Pacific, central area, Station 271, surface.
4. Larcidium octacanthum, n. sp.
Cortical shell thorny, with irregular network; pores roundish, once to four times as broad as
the bars; about thirteen to fifteen on the half meridian, eleven to thirteen on the half equator.
Proportion of the three axes = 1 : 2.5 : 4. Medullary shell lentelliptical,
one-fifth as large as the cortical, connected with it by eight radial beams, which are situated in
two crossed diagonal planes (opposite in pairs), and are prolonged on the surface into eight long
and thin cylindrical radial spines similar to Tetrapyle octacantha.
Dimensions.—Length of the cortical shell 0.16, breadth 0.11, thickness 0.045;
pores 0.003 to 0.012, bars 0.003; medullary shell 0.02 to 0.03.
Habitat.—Pacific, central area, Station 263, depth 2650 fathoms.
5. Larcidium dodecanthum, n. sp. (Pl. 50, figs. 8,
8a).
Cortical shell rough, with irregular network; pores roundish, twice to four times as broad as
the bars; about twelve to fourteen on the half meridian, eight to ten on the half equator.
Proportion of the three axes = 2 : 3 : 4. Medullary shell lentelliptical,
one-third as large as the cortical, connected with it by twelve thin radial beams, which are
prolonged outside into twelve strong conical radial spines, about half as long as the breadth of
the shell.
Dimensions.—Length of the cortical shell 0.14, breadth 0.1, thickness 0.07; pores
0.008 to 0.015, bars 0.004; medullary shell 0.04 to 0.05.
Habitat.—South Pacific, Station 288, surface.
6. Larcidium polyacanthum, n. sp.
Cortical shell spiny, with irregular network; pores roundish, once to twice as broad as the
bars; about ten to twelve on the half meridian, six to eight on the half equator. Proportion of
the three axes = 1 : 2 : 3. Medullary shell lentelliptical, half as large as
the cortical shell, connected with it by numerous (twenty to twenty-five) radial beams, which are
prolonged outside into three-sided pyramidal spines, about as long as the breadth of the
shell.
Dimensions.—Length of the cortical shell 0.18, breadth 0.12, thickness 0.06; pores
0.01 to 0.015, bars 0.008; medullary shell 0.03 to 0.09.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
{613}
Subfamily 3. Spongolarcida, Haeckel.
Definition.—Larcarida with spongy
lentelliptical shell (with or without enclosed medullary shell).
Genus 270. Spongolarcus,[309] n.
gen.
Definition.—Larcarida with lentelliptical spongy shell,
containing a central cavity of the same form, without medullary shell (without radial spines).
The genus Spongolarcus differs from Cenolarcus (its probable
ancestral form) only in the development of spongy framework forming the wall of the hollow
lentelliptical shell. It corresponds, therefore, to Plegmosphæra among the Sphæroidea, and to Spongellipsis among the Prunoidea. From these two similar spongy Sphærellaria
it differs in its characteristic lentelliptical form, with three dimensive axes of unequal
length.
1. Spongolarcus lentellipsis, n. sp.
Spongy network of the shell very loose, its meshes fifteen to twenty times as broad as the
bars. Surface of the shell nearly smooth; diameter of its internal cavity twice as large as the
thickness of its wall. Proportion of the three dimensive axes = 2 : 3 : 4.
Dimensions.—Length 0.16, breadth 0.12, height 0.08; thickness of the spongy wall
0.05.
Habitat.—Pacific, central area, Station 274, surface.
2. Spongolarcus triaxonius, n. sp.
Spongy network of the shell rather loose, its meshes twelve to sixteen times as broad as the
bars. Surface of the shell rough; diameter of its internal cavity about eight times as large as
the thickness of its wall. Proportion of the three dimensive axes =
2 : 2.5 : 3.
Dimensions.—Length 0.25, breadth 0.2, height 0.16; thickness of the spongy wall
0.025.
Habitat.—North Pacific, Station 253, surface.
3. Spongolarcus dimensivus, n. sp.
Spongy network of the shell dense, its meshes four to eight times as broad as the bars. Surface
of the shell thorny; diameter of its internal cavity about fifteen times as large as the thickness
of its wall. Proportion of the three dimensive axes = 3 : 4 : 5.
Dimensions.—Length 0.2, breadth 0.16, height 0.12; thickness of the spongy wall
0.01.
Habitat.—Western Tropical Pacific, Station 225, depth 4475
fathoms.
{614}
4. Spongolarcus amphicentria, Haeckel.
? Amphicentria salpa, Ehrenberg, 1861, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin,
p. 296; Abhandl. d. k. Akad. d. Wiss. Berlin, 1872, Taf. ii. fig. 18.
? Spongurus salpa, Haeckel, 1862, Monogr. d. Radiol., p. 466.
Spongy network of the shell compact, its meshes about the same breadth as the bars. Surface of
the shell spiny, with some larger spines around the poles of the axis; diameter of the internal
cavity about six times as large as the thickness of its wall. Proportion of the three dimensive
axes = 1 : 2 : 3. (Perhaps this Spongolarcus is identical with
Amphicentria salpa, very imperfectly described and figured by Ehrenberg, loc.
cit.?)
Dimensions.—Length 0.14, breadth 0.09, height 0.05; thickness of the spongy wall
0.015.
Habitat.—North Atlantic; off Greenland, 1000 fathoms, Ehrenberg;
Station 64, depth (2700) fathoms.
Genus 271. Stypolarcus,[310] n. gen.
Definition.—Larcarida with lentelliptical spongy shell,
composed of compact spongy framework, without central cavity and medullary shell (without radial
spines).
The genus Stypolarcus differs from Spongolarcus in the absence of any
central cavity. This is quite filled up by spongy framework, which forms the whole mass of the
lentelliptical body. Stypolarcus bears therefore the same relation to Spongolarcus
that Styptosphæra does to Plegmosphæra.
1. Stypolarcus spongiosus, n. sp.
Lentelliptical shell composed in the whole mass of loose, spongy framework of similar texture,
with irregular meshes, about ten to twenty times as broad as the thin bars. Surface rough, without
radial spines. Proportion of the three axes = 3 : 4 : 5.
Dimensions.—Length 0.2, breadth 0.16, height 0.12.
Habitat.—Antarctic Ocean, Station 157, depth 1950 fathoms.
Family XXV. Larnacida, Haeckel, 1883 (Pl. 50, figs.
3-8).
Definition.—Larcoidea with a regular, completely
latticed, lentelliptical cortical shell, without open gates and annular constrictions; either this
cortical shell or the enclosed medullary shell is trizonal, composed of three elliptical,
latticed, dimensive girdles of different sizes, perpendicular one to another.
{615}
The family Larnacida immediately follows the Larcarida as the next
simple group of all Larcoidea; some genera of both groups (such as
Larnacalpis and Coccolarcus, or Larnacantha and Larcidium) may easily
be confounded from their being so much alike. In both the lentelliptical shell is composed of two
concentric shells, an inner (medullary) and an outer (cortical) shell. But the connection between
these shells and the construction of the inner shell is quite different in the two groups. Whilst
in the Larcarida the medullary shell is connected with the cortical shell simply by radial beams,
here in the Larnacida this connection is effected by two latticed lamellæ, which are the lateral
wings of a transverse girdle. Therefore we encounter here for the first time that peculiar mode of
growth which characterises the greater part of the Larcoidea, but
particularly the Pylonida. But whilst in the Pylonida between the three crossed lattice-girdles,
remain four open gates, here in the Larnacida the gates become closed by lattice-work; the
lentelliptical cortical shell becomes perfect.
The most simple genus of Larnacida, and no doubt the common ancestral form of this whole
family, is Larnacilla (Pl. 50,
figs. 1, 1a, 1b). The most important shell of this typical genus is composed of a
simple lentelliptical medullary shell and of three elliptical latticed girdles surrounding it,
perpendicular one to another. These three "dimensive girdles" lie in the perimeter of the three
dimensive planes, the minor (and first) in the equatorial plane, the second (and major) in the
lateral plane, the third (and intermediate) in the sagittal plane. Therefore we have before us the
same "trizonal shell" as in the important genus Trizonium among the Pylonida. But whilst in
Trizonium, as in all Pylonida, the four gates between the girdles remain open, here in
Larnacilla they become perfectly closed by lattice-work.
The formation of the typical "Larnacilla-shell" begins with a simple, spherical or
lentelliptical lattice-shell, from both sides of which arise two latticed "lateral wings" opposite
on the poles of the transverse axis. These two wings are comparable to the lateral chambers of
Tholartus (among the Tholonida), but differ by two large openings. Each wing is a short
cylindrical tube with latticed wall, open at both ends; the axis of the tube (going through the
centre of the open ends) is parallel to the principal axis of the whole shell (and of the central
chamber). Therefore both wings form together a transverse ring, the middle of which encloses the
central chamber. The distal parts of both wings grow towards the poles of the principal axis; if
they became united here, the second (lateral) girdle would be complete. Between it and the first
girdle four open gates remain ("Tetrapyle"); but these become overgrown by the third or
sagittal girdle, and at last the gates between this and the two other girdles become closed by
lattice-work. This perfect fenestration of the trizonal cortical shell, and the complete closing
of the gates between the girdles by network, is the only difference between Trizonium and
Larnacilla.
In Larnacilla and in the nearly allied Larnacidium (only differing by radial
spines {616}on the surface) the "trizonal shell" is an
external or "cortical shell," enclosing the central capsule, the interior of which only contains
the simple central chamber and the jointed proximal parts of both lateral wings. In the other
genera of Larnacida (by proceeding growth) this trizonal Larnacilla-shell becomes enclosed
by the growing central capsule and is now only a "medullary shell," whilst on the outside of the
central capsule in the same manner is developed an outer cortical shell (Larnacalpis,
Larnacantha); and perhaps the same process may be repeated. But sometimes also this
cortical shell becomes doubled by a simple envelop of network (Larnacoma). In the
Larnacospongida the cortical shell is composed of a spongy framework (corresponding to the
Spongolarcida in the foregoing family).
The lattice-work of the Larnacida is commonly irregular (as in most other Larcoidea), and its pores have little signification for the different species.
On the outer surface often arise radial spines, symmetrically disposed either in dimensive planes
or in diagonal planes.
The central capsule is constantly a true lentellipsis or a "triaxial ellipsoid," characterised
by three halving, elliptical dimensive planes, perpendicular one to another. It bears a different
relation to the skeleton in the two subfamilies of Larnacida. In the first subfamily, the
Larnacillida (Larnacilla, Larnacidium), the central capsule encloses only the simple
spherical or lentelliptical central chamber ("simple medullary shell"), and is enveloped by the
trizonal cortical shell. In the other subfamily, the Larnacalpida (Larnacalpis,
Larnacospongus, &c.), this trizonal "Larnacilla-shell" becomes enclosed by the
overgrowing central capsule, which now becomes enveloped by an external, latticed or spongy,
lentelliptical "cortical shell."
Synopsis of the Genera of Larnacida.
|
II. Subfamily Larnacillida.
Medullary shell simple, spherical or subspherical. Cortical shell
lentelliptical, trizonal; between them four gates.
|
|
Shell without radial spines, |
272. Larnacilla. |
| Shell with radial spines, |
273. Larnacidium. |
|
II. Subfamily Larnacalpida.
Medullary shell Larnacilla-shaped, double; the inner spherical or
subspherical, the outer lentelliptical and trizonal.
|
|
Cortical shell simple or double, but not spongy. |
|
Cortical shell simple. |
|
Without radial spines, |
274. Larnacalpis. |
| With radial spines, |
275. Larnacantha. |
| Cortical shell double, without radial spines, |
276. Larnacoma. |
| Cortical shell simple or double, wholly or partly
spongy. |
|
Shell without radial spines, |
277. Larnacospongus. |
| Shell with radial spines, |
278. Larnacostupa. |
| I. Subfamily Larnacillida. Medullary shell simple, spherical or subspherical.
Cortical shell lentelliptical, trizonal; between them four gates. |
|
|
|
|
Shell without radial spines, |
|
|
|
|
|
|
|
|
|
|
|
|
272. Larnacilla. |
|
|
|
|
|
|
|
|
|
|
|
|
Shell with radial spines, |
|
|
|
|
|
|
|
|
|
|
|
|
273. Larnacidium. |
|
|
|
|
|
|
|
|
| II. Subfamily Larnacalpida. Medullary shell Larnacilla-shaped, double;
the inner spherical or subspherical, the outer lentelliptical and trizonal. |
|
|
Cortical shell simple or double, but not spongy. |
|
|
|
|
|
|
Cortical shell simple. |
|
|
|
|
|
|
|
|
|
|
Without radial spines, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
274. Larnacalpis. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
With radial spines, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
275. Larnacantha. |
|
|
|
|
|
|
|
|
|
|
|
|
Cortical shell double, without radial spines, |
|
|
|
|
|
|
|
|
|
|
|
|
276. Larnacoma. |
|
|
|
|
|
|
|
|
|
|
Cortical shell simple or double, wholly or partly spongy. |
|
|
|
|
|
|
Shell without radial spines, |
|
|
|
|
|
|
|
|
|
|
|
|
277. Larnacospongus. |
|
|
|
|
|
|
|
|
|
|
|
|
Shell with radial spines, |
|
|
|
|
|
|
|
|
|
|
|
|
278. Larnacostupa. |
|
|
|
|
|
|
|
|
{617}
Subfamily 1. Larnacillida, Haeckel.
Definition.—Larnacida with a simple,
spherical or lentelliptical, medullary shell, connected by the lateral wings of a latticed
transverse girdle with the simple lentelliptical trizonal cortical shell; the central capsule
encloses the former and is enveloped by the latter.
Genus 272. Larnacilla,[311]
n. gen.
Definition.—Larnacida with a simple lentelliptical
cortical shell, connected by the lateral wings of a latticed transverse girdle with the simple,
spherical or lentelliptical, medullary shell. Surface without radial spines.
The genus Larnacilla represents the most simple form of Larnacida, and at
the same time the most important common ancestral form, from which the greater number of Larcoidea may be derived, viz., all those genera which possess the
characteristic "Larnacilla-shaped medullary shell." This typical form of medullary shell
may be derived from the genus Trizonium among the Pylonida by the closing of the four open
gates of this genus. The free opening of these four gates becomes overgrown and closed by
lattice-work, developed from the free edges of the three crossed girdles, and thus finally all
three girdles are united in the form of a simple lentelliptical shell (Pl. 50, figs. 1,
1a, 1b). Seen from the sagittal poles (or from the poles of the shortest axis, fig.
1), the shell exhibits on both sides of the small spherical medullary shell the two lateral wings
of the transverse girdle from the face; seen from the lateral poles (or from the poles of the
transverse axis, fig. 1a), one of these wings appears in the optical section as an oblong
ring, which seemingly encloses the concentric medullary shell, and on both sides is grown together
with the sagittal girdle; seen from the principal poles (or from the poles of the longitudinal
axis, fig. 1b), both wings exhibit their elliptical opening (at the right and left from the
central medullary shell). The two concentric shells are only connected by the two lateral
tube-like wings of the transverse girdle; the lateral and the sagittal girdles have no connection
with the medullary shell. The latter is sometimes spherical, at other times lentelliptical.
1. Larnacilla typus, n. sp. (Pl. 50,
fig. 1, 1a, 1b).
Cortical shell with smooth surface and with subregular network; pores twice as broad as the
bars; about twelve pores on the half meridian, ten on the half equator. Proportion of the three
dimensive axes = 2 : 3 : 4. Internal four gates (between transverse and
lateral girdles) roundish-triangular, little broader than high. Medullary shell spherical,
scarcely one-third as broad as the lentelliptical cortical shell.
{618}
Dimensions.—Length of the cortical shell (or principal axis) 0.13, breadth of it
(or transverse axis) 0.1, height of it (or sagittal axis) 0.07; pores 0.006, bars 0.003; medullary
shell (diameter) 0.03.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
2. Larnacilla prometor, n. sp.
Cortical shell with rough surface and regular network; pores three times as broad as the bars;
about ten on the half meridian, eight on the half equator. Proportion of the three axes =
1 : 2 : 3. Internal four gates (between transverse and lateral girdles)
kidney-shaped, twice as broad as high. Medullary shell lentelliptical, of the same form as the
external cortical shell, but only one third as large.
Dimensions.—Length of the cortical shell 0.15, breadth 0.1, height 0.05, pores
0.015, bars 0.005; medullary shell 0.02 to 0.05.
Habitat.—Indian Ocean, Zanzibar, Pullen, depth 2200 fathoms.
3. Larnacilla subglobosa, n. sp.
Cortical shell nearly spherical, with thorny surface and irregular network; pores roundish,
twice to four times as broad as the bars; twelve to sixteen in the half circumference. Proportion
of the three axes very little different = 1.3 : 1.4 : 1.5. Internal four gates
elliptical, one and a half times as broad as high. Medullary shell spherical, one-fourth as broad
as the cortical shell.
Dimensions.—Length of the cortical shell 0.15, breadth 0.14, height 0.13; pores
0.006 to 0.012, bars 0.003; medullary shell 0.035.
Habitat.—Pacific, central area. Station 266, depth 2750 fathoms.
4. Larnacilla medullaris, n. sp.
Cortical shell lentelliptical, with smooth surface and irregular network; pores roundish, very
small, scarcely as broad as the bars; about eight to nine on the half meridian, six to seven on
the half equator. Proportion of the three axes = 3 : 4 : 6. Internal four
gates elliptical. Medullary shell spherical, scarcely one-third as broad as the cortical shell.
(This small species may be only the medullary shell of some other Larcoid, the cortical shell of
which is not yet formed.)
Dimensions.—Length of the cortical shell 0.06, breadth 0.04, height 0.03; pores
and bars about 0.004; medullary shell 0.013.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms.
Genus 273. Larnacidium,[312] n. gen.
Definition.—Larnacida with a simple lentelliptical
cortical shell, connected by the lateral wings of a latticed transverse girdle with the simple,
spherical or lentelliptical, medullary shell. Surface armed with radial spines.
{619}
The genus Larnacidium has the same shell-formation as the foregoing
Larnacilla, and differs from it only in the possession of radial spines on the surface,
which in all known species exhibit a symmetrical disposition. From the nearly allied genus
Larcidium it differs in the characteristic mode of connection between the two shells, owing
to the different kind of growth. In Larcidium this connection is effected only by radial
beams, whereas in Larnacidium (as in all Larnacida) by two lateral latticed tubes, the
wings of the primary transverse girdle.
1. Larnacidium staurobelonium, n. sp.
Cortical shell smooth, with four strong conical, radial spines in the lateral plane, opposite
in pairs, two on the poles of the principal and two on the poles of the transverse axis. Pores
subregular, circular, twice as broad as the bars; about twelve on the half meridian. Proportion of
the three axes = 2 : 3 : 4. Medullary shell spherical, one-third as broad as
the cortical shell.
Dimensions.—Length of the cortical shell 0.12, breadth 0.09, height 0.06; pores
0.008, bars 0.04; medullary shell 0.03.
Habitat.—North Pacific Station 244, depth 2900 fathoms.
2. Larnacidium hexabelonium, n. sp.
Cortical shell thorny, with six strong, three-sided pyramidal, radial spines, lying opposite in
pairs on the poles of the three dimensive axes. Pores subregular, circular, three times as broad
as the bars; about fourteen on the half meridian. Proportion of the three axes =
2 : 2.5 : 3. Medullary shell spherical, one-third as broad as the cortical
shell.
Dimensions.—Length of the cortical shell 0.14, breadth 0.11, height 0.08; pores
0.01, bars 0.003; medullary shell 0.04.
Habitat.—South Pacific, Station 295, depth 1500 fathoms.
3. Larnacidium polybelonium, n. sp.
Cortical shell very spiny, with numerous (twenty to thirty or more) larger thin radial spines,
about as long as the shell. Pores irregular, twice to five times as broad as the bars; about
sixteen on the half meridian. Proportion of the three axes = 2 : 2.5 : 3.
Medullary shell lentelliptical, half as large as the cortical shell.
Dimensions.—Length of the cortical shell 0.11, breadth 0.09, height 0.07; pores
0.004 to 0.01, bars 0.002; length of the medullary shell 0.06, breadth 0.05, height 0.04.
Habitat.—Western Tropical Pacific, Station 225. depth 4475
fathoms.
Subfamily 2. Larnacalpida, Haeckel.
Definition.—Larnacida with a double,
trizonal, Larnacilla-shaped medullary shell, enclosed in the central capsule, and enveloped
by a simple or double, latticed or spongy, lentelliptical, cortical shell.
{620}
Genus 274. Larnacalpis,[313] n. gen.
Definition.—Larnacida with a simple lentelliptical
cortical shell, without radial spines. Medullary shell double, Larnacilla-shaped.
The genus Larnacalpis represents the most simple form of the sub-family
Larnacalpida, and is very important as the common original form of all those Larcoidea in which a double Larnacilla-shaped medullary shell is
surrounded by a simple, perfectly closed, latticed, lentelliptical cortical shell. Therefore the
same typical, trizonal, lentelliptical shell, which in Larnacilla represents the external
envelop (or cortical shell) of the central capsule, here in Larnacalpis becomes enclosed as
an internal nucleus (or medullary shell) in the interior of the central capsule, and this latter
becomes overgrown by a new lentelliptical cortical shell. The connection between the two shells of
Larnacalpis is either effected by a number of radial beams (e.g., in Larnacalpis
triaxonia by six beams situated in the three dimensive axes), or by two lateral, latticed,
tube-like wings, which are repetitions of the smaller lateral wings connecting its external shell
with the internal medullary shell (as in Larnacalpis lentellipsis). The latter species may
be regarded as a Pylonium with a completely latticed shell.
1. Larnacalpis lentellipsis, n. sp. (Pl. 50, figs. 2,
2a, 2b).
Cortical shell with thorny surface and irregular network; pores roundish, twice to four times
as broad as the bars; about sixteen on the half meridian, twelve on the half equator. Proportion
of the three dimensive axes = 2 : 3 : 4. Medullary shell one-third as large as
the cortical, with four elliptical internal gates, connected with it by two opposite beams in the
principal axis and by two latticed wings in the transverse axis; therefore between the two shells
are four large kidney-shaped gates, halved by the polar beams (as in Octopyle).
Dimensions.—Length of the cortical shell (or principal axis) 0.14, breadth (or
transverse axis) 0.11, height (or sagittal axis) 0.07; pores 0.01 to 0.02, bars 0.005; length of
the medullary shell 0.05, breadth 0.04, height 0.03.
Habitat.—Pacific, central area, Station 273, depth 2600 fathoms.
2. Larnacalpis phacodiscus, n. sp.
Cortical shell with thorny surface and regular network; pores circular, twice as broad as the
bars; about ten on the half meridian, eight on the half equator. Proportion of the three axes =
2 : 2.5 : 3. Medullary shell half as large as the cortical, with four
kidney-shaped internal gates.
Dimensions.—Length of the cortical shell 0.13, breadth 0.11, height 0.09; pores
0.012, bars 0.006; length of the medullary shell 0.06, breadth 0.045, height 0.03.
Habitat.—North Pacific, Station 253, depth 3125 fathoms.
{621}
3. Larnacalpis macrococcus, n. sp.
Cortical shell with spiny surface and regular network; pores circular, small, of the same
breadth as the bars; about twenty-two on the half meridian, nineteen on the half equator.
Proportion of the three axes = 2 : 3 : 4. Medullary shell two-thirds as large
as the cortical, with four wide internal semicircular gates.
Dimensions.—Length of the cortical shell 0.1, breadth 0.075, height 0.05; pores
and bars 0.003; length of the medullary shell 0.066, breadth 0.05, height 0.032.
Habitat.—Western Tropical Pacific, Station 224, depth 1850
fathoms.
4. Larnacalpis subsphærica, n. sp.
Cortical shell with rough surface and irregular network; roundish pores twice to five times as
broad as the bars; about twenty-four on the half meridian, twenty-one on the half equator.
Proportion of the three axes = 1.2 : 1.3 : 1.4. Medullary shell half as large
as the cortical, with four elliptical internal gates.
Dimensions.—Length of the cortical shell 0.14, breadth 0.13, height 0.12; pores
0.004 to 0.01, bars 0.002; length of the medullary shell 0.08, breadth 0.07, height 0.06.
Habitat.—Pacific, central area, Station 266, depth 2750 fathoms.
5. Larnacalpis triaxonia, n. sp. (Pl. 50, fig.
3).
Cortical shell with smooth surface and peculiar network, composed of four meridian rows of
larger pores (five large elliptical pores on each half meridian, the largest in the equator) and
numerous small irregular pores between them. Proportion of the three axes =
2 : 3 : 4. Medullary shell with four semicircular internal gates, about
one-fourth as large as the cortical, connected with it by six thin radial beams, opposite in pairs
in the three dimensive axes.
Dimensions.—Length of the cortical shell 0.14, breadth 0.1, height 0.07; large
pores 0.03, small pores 0.002 to 0.01, bars 0.002 to 0.01; length of the medullary shell 0.04,
breadth 0.03, height 0.02.
Habitat.—Pacific, central area, Station 263, depth 2650 fathoms.
Genus 275. Larnacantha,[314] n. gen.
Definition.—Larnacida with a simple lentelliptical
cortical shell, armed with symmetrically disposed radial spines. Medullary shell double,
Larnacilla-shaped.
The genus Larnacantha has the same characteristic shell-formation as the
foregoing Larnacalpis, and differs from it only in the possession of radial spines, which
are symmetrically distributed on the surface in a definite order. Commonly these spines are
external prolongations of the internal radial beams, which connect the double
Larnacilla-shaped medullary shell with the simple lentelliptical cortical shell.
{622}
1. Larnacantha dissacantha, n. sp.
Cortical shell smooth, with two large cylindrical spines, opposite on the poles of the
principal axis, and somewhat longer than it. Pores regular, circular, three times as broad as the
bars; about eleven on the half meridian. Proportion of the three axes =
2 : 3 : 4. Medullary shell one-third as large as the cortical shell.
Dimensions.—Length of the cortical shell (principal axis) 0.13, breadth
(transverse axis) 0.1, height (sagittal axis) 0.07; pores 0.009, bars 0.003; length of the
Larnacilla-shaped medullary shell 0.045.
Habitat.—North Pacific, Station 256, depth 2950 fathoms.
2. Larnacantha stauracantha, n. sp.
Cortical shell smooth, with four large conical spines in the lateral plane, two larger opposite
on the poles of the principal, two smaller on those of the transverse axis. Pores regular,
circular, twice as broad as the bars; about seventeen on the half meridian. Proportion of the
three axes = 3 : 3.5 : 4. Medullary shell one-third as large as the cortical
shell.
Dimensions.—Length of the cortical shell 0.15, breadth 0.13, height 0.11; pores
0.008, bars 0.004; length of the medullary shell 0.05.
Habitat.—Pacific, central area, Station 274, surface.
3. Larnacantha quadricornis, n. sp.
Cortical shell spiny, with four strong, horn-like curved spines in the lateral plane, opposite
in pairs on the poles of the crossed diagonal axes. Pores irregular, roundish, twice to four times
as broad as the bars; about fourteen on the half meridian. Proportion of the three axes =
2 : 3 : 4. Medullary shell one-third as large as the cortical shell.
Dimensions.—Length of the cortical shell 0.14, breadth 0.11, height 0.07; pores
0.006 to 0.012, bars 0.003; length of the medullary shell 0.05.
Habitat.—Pacific, central area, Station 270, surface.
4. Larnacantha hexacantha, n. sp. (Pl. 50, fig.
4).
Cortical shell thorny, with six strong conical radial spines in the lateral plane, two opposite
on the poles of the principal axis, four others opposite in pairs on the poles of the two crossed
diagonal axes. Pores with peculiar distribution; twelve large elliptical pores (nearly of the size
of the medullary shell) symmetrically disposed in four crossed meridians (between the sagittal and
the lateral meridians), separated by bands of smaller irregular pores. Proportion of the three
axes = 3 : 4 : 5. Medullary shell hexagonal, one-third as large as the
cortical shell.
Dimensions.—Length of the cortical shell 0.14, breadth 0.11, height 0.08; large
pores 0.04, small pores 0.003 to 0.01, bars 0.004; length of the medullary shell 0.05.
Habitat.—Pacific, central area, Station 263, depth 2650 fathoms.
{623}
5. Larnacantha bicruciata, n. sp. (Pl. 50, fig.
5).
Cortical shell thorny, in the lateral plane with eight strong conical radial spines,
alternating with eight smaller spines; four of the eight stronger spines opposite on the poles of
the principal and transverse axes (in the figure 5, by mistake, not represented large enough),
four others between those, opposite on the poles of the two crossed diagonal axes. Pores with a
peculiar disposition; on both flat sides of the lentellipsis an elliptical ring of eight large
elliptical pores (alternating with the eight stronger radial spines), separated by bands of
smaller irregular pores. Proportion of the three axes = 2 : 3 : 4. Medullary
shell nearly half as long as the cortical shell.
Dimensions.—Length of the cortical shell 0.16, breadth 0.13, height 0.08; large
pores 0.03, small pores 0.003 to 0.01, bars 0.006; length of the medullary shell 0.07.
Habitat.—Indian Ocean, surface, Madagascar (Rabbe).
6. Larnacantha octacantha, n. sp.
Cortical shell thorny, with eight long and thin, cylindrical, radial spines, lying opposite in
pairs in two crossed diagonal planes. Pores irregular, roundish, twice to five times as broad as
the bars; about sixteen on the half meridian. Proportion of the three axes =
1 : 1⅓ : 2. Medullary shell scarcely one-fourth as long as the cortical
shell. (This species resembles closely the common Tetrapyle octacantha, from which it seems
to be developed by a complete over-growing of the four gates, which become closed by a network
connecting the free edges of the transverse and lateral girdles.)
Dimensions.—Length of the cortical shell 0.18, breadth 0.13, height 0.1; pores
0.005 to 0.015, bars 0.003; length of the medullary shell 0.04.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific, surface.
7. Larnacantha cladacantha, n. sp.
Cortical shell very spiny, with eight longer ramified spines, lying opposite in pairs in two
crossed diagonal planes; each spine with two to six irregular, lateral branches. Pores irregular,
roundish, twice to three times as broad as the bars; about twelve on the half meridian. Proportion
of the three axes = 5 : 6 : 7. Medullary shell nearly half as long as the
cortical shell. (Differs from the foregoing by the branching spines end the larger medullary
shell.)
Dimensions.—Length of the cortical shell 0.15, breadth 0.13, height 0.11; pores
0.01 to 0.015, bars 0.005; length of the medullary shell 0.07.
Habitat.—Indian Ocean, surface, Cocos Islands (Rabbe).
8. Larnacantha prismatica, n. sp. (Pl. 50, fig.
6).
Cortical shell smooth, four-sided prismatic, with eight short, parallel, three-sided pyramidal
spines; these lie opposite in pairs in four parallel longitudinal lines, as prolongations of the
four {624}lateral edges of the prism, and arise from its
eight corners. Pores regular, circular, three times as broad as the bars; about fourteen on the
half meridian. Proportion of the three axes = 2 : 3 : 4. Medullary shell half
as long as the cortical shell.
Dimensions.—Length of the cortical shell 0.11, breadth 0.07, height 0.05; pores
0.006, bars 0.002; length of the medullary shell 0.06.
Habitat.—Pacific, central area, Station 263, depth 2650 fathoms.
9. Larnacantha decacantha, n. sp.
Cortical shell thorny, with ten short and stout, conical, radial spines, two opposite on the
poles of the principal axis (as prolongations of inner axial beams), eight others opposite in
pairs in two crossed diagonal planes. Pores irregular, roundish, twice to four times as broad as
the bars; about sixteen on the half meridian. Proportion of the three axes =
3 : 3.75 : 4. Medullary shell about one-third as long as the cortical
shell.
Dimensions.—Length of the cortical shell 0.16, breadth 0.15, height 0.12; pores
0.008 to 0.016, bars 0.004; length of the medullary shell 0.06.
Habitat.—North Atlantic, Station 354, surface.
10. Larnacantha dodecantha, n. sp.
Cortical shell nearly smooth, but with twelve strong conical radial spines; four in the lateral
plane opposite in pairs (two on the poles of the principal, and two on the poles of the transverse
axis); eight others opposite in pairs in two crossed diagonal planes. Pores with a peculiar
disposition: twelve large elliptical pores in two crossed meridian planes (alternating with the
twelve spines), separated by bands of irregular small pores. Proportion of the three axes =
1 : 1.5 : 2. Medullary shell hexagonal, one-third as long as the cortical
shell.
Dimensions.—Length of the cortical shell 0.15, breadth 0.11, height 0.08; large
pores 0.03, small pores 0.005 to 0.01, bars 0.003; length of the medullary shell 0.05.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms.
11. Larnacantha drymacantha, n. sp.
Cortical shell very spiny, on the whole surface covered with a forest of numerous (thirty to
fifty or more) large branched spines, about the length of the shell; each spine with three to nine
lateral branches, simple or forked (very similar to Cromyodrymus abietinus, Pl. 30, fig. 6).
Pores very irregular, roundish. Proportion of the three axes = 2 : 2.5 : 3.
Medullary shell half as long as the cortical shell.
Dimensions.—Length of the cortical shell 0.16, breadth 0.13, height 0.1; pores
0.005 to 0.015, bars 0.03; length of the medullary shell 0.08.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
{625}
Genus 276. Larnacoma,[315] n. gen.
Definition.—Larnacida with double lentelliptical cortical
shell, without radial spines. Medullary shell double, Larnacilla-shaped.
The genus Larnacoma has originated from the nearly allied Larnacalpis
by duplication of the cortical shell. Whilst in both genera the connection between the
Larnacilla-shaped medullary shell and the primary cortical shell is the same, many short
radial beams arise from the surface of the latter in Larnacoma, which at constant equal
distances from it unite by a network forming the secondary or outer cortical shell. It differs
from the similar Druppulida (Cromyodruppa) by the sagittal flattening of the lentelliptical
shell and the Larnacilla-form of the double medullary shell.
1. Larnacoma lentellipticum, n. sp.
Shell with smooth surface and elliptical perimeter, one and a third times as long as broad. All
four shells lentelliptical. Distance between the two cortical shells about twice as great as the
distance of the inner cortical and outer medullary shell. Network of both outer shells irregular,
with large roundish pores, twice to six times as broad as the bars.
Dimensions.—Length (or principal axis) of the first (innermost) shell 0.03, second
0.08, third 0.16, fourth (outermost) 0.27, breadth (or transverse axis) corresponding—(A)
0.02, (B) 0.05, (C) 0.11, (D) 0.2.
Habitat.—South Atlantic, Station 323, depth 1900 fathoms.
2. Larnacoma quadruplex, n. sp.
Shell with thorny surface and elliptical perimeter, one and a fifth times as long as broad. All
four shells lentelliptical. Distance between the two cortical shells somewhat smaller than the
distance between the inner cortical and outer medullary shell. Network of both outer shells
irregular, with large roundish pores, twice to ten times as broad as the bars.
Dimensions.—Length of the first shell 0.02, second 0.06, third 0.15, fourth 0.24;
breadth corresponding—(A) 0.016, (B) 0.04, (C) 0.11, (D) 0.2.
Habitat.—South Atlantic, Station 335, depth 1425 fathoms.
3. Larnacoma hexagonium, n. sp.
Shell with thorny surface and hexagonal perimeter, as long as broad. All four shells hexagonal,
connected by six piercing radial beams (two in the principal axis, four others in two crossed
diagonals). Distance between the two cortical shells somewhat greater than the distance between
the inner cortical and outer medullary shell. Network of both
outer shells subregular, with small circular pores, twice as broad as the bars.
{626}
Dimensions.—Length of the first shell 0.02, second 0.05, third 0.09, fourth 0.16;
breadth corresponding—(A) 0.015, (B) 0.035, (C) 0.065, (D) 0.12.
Habitat.—South Atlantic, west of Tristan da Cunha, Station 332, depth
2200 fathoms.
Genus 277. Larnacospongus,[316] n.
gen.
Definition.—Larnacida with spongy lentelliptical cortical
shell, without radial spines. Medullary shell double, Larnacilla-shaped.
The genus Larnacospongus differs from the nearly allied genera
Larnacalpis and Larnacoma by the spongy texture of the lentelliptical cortical shell
whilst the enclosed medullary shell in both genera is the same trizonal Larnacilla-shell.
Therefore Larnacospongus (and the following nearly related Larnacostupa) can be
derived directly by development of a spongy envelop either from Larnacilla and
Larnacalpis, or from Trizonium and Amphipyle. But some species of these
spongy genera appear to be derived rather from Tetrapyle or Pylonium, perhaps also
from Cubotholus. Their phylogenetic origin may be explained in different ways.
1. Larnacospongus larnacillifer, n. sp.
Cortical shell lentelliptical, one and a half times as long as broad, with rough surface and
rather loose spongy framework, directly enclosing a trizonal Larnacilla-shell of the same
form, but of only one-third its size.
Dimensions.—Length of the whole shell 0.17, breadth 0.12; length of the medullary
shell 0.06, breadth 0.04.
Habitat.—South Atlantic, east coast of Patagonia, Station 319,
surface.
2. Larnacospongus tetrapylifer, n. sp.
Cortical shell lentelliptical, one and a third times as long as broad, with thorny surface;
composed of an outer envelop of loose spongy framework and an inner lattice-shell with four
kidney-shaped gates, like Tetrapyle; the latter encloses a trizonal medullary shell of
one-fourth its size.
Dimensions.—Length of the whole shell 0.22, breadth 0.16; length of the medullary
shell 0.045, breadth 0.035.
Habitat.—South Atlantic, west of Tristan da Cunha, Station 332,
surface.
{627}
Genus 278. Larnacostupa,[317] n.
gen.
Definition.—Larnacida with spongy lentelliptical cortical
shell, with radial spines on the surface. Medullary shell double, Larnacilla-shaped.
The genus Larnacostupa differs from the preceding Larnacospongus only in the
possession of radial spines, covering either the whole surface irregularly or disposed in a
certain symmetrical order.
1. Larnacostupa octacantha, n. sp.
Cortical shell lentelliptical, with thorny surface, and rather dense, irregular, spongy
framework, which arises from an inner latticed cortical shell, like that of Tetrapyle or
Pylonium. This latter is twice as large as the enclosed Larnacilla-shell. Eight long
and thin, cylindrical, radial spines, opposite in pairs in two diagonal planes. (Seems to be the
common Tetrapyle octacantha, enveloped by an outer spongy framework mantle.)
Dimensions.—Length of the whole shell (without spines) 0.22, breadth 0.16; length
of the medullary shell 0.06, breadth 0.04.
Habitat.—Indian Ocean, Madagascar, Rabbe, surface.
2. Larnacostupa spinosa, n. sp.
Cortical shell lentelliptical, about one and a half times as long as broad, with very lax and
irregular spongy framework, arising from a nearly quadrangular lattice-shell like that of
Tetrapyle; this latter encloses a Larnacilla-shell of half its size. Whole surface
covered with thin bristle-like radial spines, of about the length of the shell.
Dimensions.—Length of the whole shell (without spines) 0.18, breadth 0.12; length
of the medullary shell 0.05, breadth 0.035.
Habitat.—Antarctic Ocean, off Kerguelen, Station 150, surface.
3. Larnacostupa dendrophora, n. sp.
Cortical shell nearly spherical, scarcely longer than broad, with lax, irregular spongy
framework, arising from a lentelliptical trizonal medullary shell (like Larnacilla). Whole
surface covered with thin arborescent radial spines, about half as long as the shell, each spine
with three to six irregular branches.
Dimensions.—Length of the whole shell (without spines) 0.15, breadth 0.13; length
of the medullary shell 0.07, breadth 0.04.
Habitat.—Antarctic Ocean, Station 154, surface.
{628}
Family XXVI. Pylonida, Haeckel, 1881 (Pl. 9).
Pylonida, Haeckel, 1881, Prodromus, p. 463.
Definition.—Larcoidea with regular, incompletely latticed
cortical shell, distinguished by two to four or more symmetrically disposed gates or large
fissures remaining between one to three latticed dimensive girdles (perpendicular one to another).
One, two, or three concentric systems of such girdles (each system with three girdles) may be
developed.
The family Pylonida is the most important and interesting among all
the Larcoidea, not only because it is much richer in different and
peculiar forms than the other families of this section, but also because it has direct and very
complex relations to all the other families of Larcoidea. It is even
possible that the Pylonida represent the original ancestral group of the whole section, and that
the apparently simpler group of the Larcarida must be derived from the former by retrogressive
metamorphosis.
Till the year 1881 the family Pylonida, which here now exhibits ten genera with eighty-six
species, was only represented by one single species, accurately described and extensively
illustrated by Johannes Müller in 1858, the well known and widely distributed cosmopolitan
Tetrapyle octacantha (Abhandl. d. k. Akad. d. Wiss. Berlin, p. 33, Taf. iii.). A slight
modification of it was afterwards described by Ehrenberg as Schizomma quadrilobum (Abhandl.
d. k. Akad. d. Wiss. Berlin, 1872, Taf. ii. fig. 12). A more accurate description of it, with a
good explanation of its characteristic growth, was given in 1879 by Richard Hertwig in his
Organismus der Radiolarien (pp. 52-54, Taf. iv. figs. 7, 8; Taf. vi. figs. 2, 5). In my Prodromus
(1881, p. 463) I constituted for a large number of allied species, detected in the Challenger
collection, the special family Pylonida, and distinguished among it twelve different genera.
However, I think it now better to restrict the definition of the family as given in the above
definition, and to remove from it a number of genera formerly with it united, as the genera
Triopyle and Hexapyle, appertaining to the Discoidea.
The characteristic type of all true Pylonida is clearly demonstrated by their peculiar mode of
growth, the consequence of which is the imperfect lattice-work of the fenestrated larcoid shell.
This remarkable growth is effected by the development of elliptical latticed girdles (or rings),
which enclose a quite simple, spherical, subspherical, or lentelliptical primordial shell. The
girdles lie in three different planes, perpendicular to one another, and are of different sizes;
each girdle being somewhat larger than the foregoing and somewhat smaller than the following
girdle. Between these latticed girdles remain on the surface of the shell large openings or
"gates," which are not closed by network, and it is just the symmetrical disposition and form of
these open "gates," separated and enclosed by the fenestrated girdles, which give to the Pylonida
their characteristic appearance.
{629}
To understand clearly this peculiar constitution of the Pylonida-shell by a system of
alternating girdles, developing one after the other it is indispensable to pay careful attention
to the three different elliptical dimensive planes, which characterise all Larcoidea, and to the three different dimensive axes, which bisect those
planes. The girdle which first develops around simple primordial shell or central chamber is the
transverse girdle, lying in the equatorial plane; then comes, secondly, the lateral girdle, lying
in the lateral plane; and thirdly follows the sagittal girdle, lying in the sagittal or median
plane. The three simplest genera of the Pylonida—Monozonium, Dizonium,
Trizonium—represent these three different stages, with one, two, or three girdles.
These three genera constitute the first subfamily, Haplozonaria (with one single system of
girdles); all three girdles lie in the surface of a simple lentelliptical cortical shell.
From this first subfamily the other two subfamilies of Pylonida must be derived, by repetition
of the same characteristic process of growth. In the Diplozonaria a second system of girdles has
been developed, constituting a second (outer) cortical shell of lentelliptical form, concentric
with the first. Also in this second system the transverse girdle is first developed, secondly the
lateral girdle, thirdly the sagittal girdle. The three genera Amphipyle, Tetrapyle
(with Octopyle), and Pylonium represent these three different stages of growth.
Commonly the growth of the Pylonida stops with the completion of the second system but
sometimes the same process is once repeated and a third system of girdles is formed, constituting
a third lentelliptical shell; in this case also the succession of the three latticed girdles is
the same; firstly the (third) transverse girdle is formed, secondly the (third) lateral girdle,
and thirdly the (third) sagittal girdle. Each of these three girdles of the third system encloses
concentrically the corresponding girdles of the second and first system. The three corresponding
genera of this third subfamily (Triplozonaria) are Amphipylonium, Tetrapylonium, and
Pylozonium. But in general this highest number of girdles (nine) is very seldom reached;
commonly the growth of the Pylonida stops with five girdles (Tetrapyle and
Octopyle). More than nine girdles I have never observed, though there remains the
possibility of the apposition of a fourth system owing to the peculiar imperfect character of the
growth itself.
The central or primordial chamber of the shell, with which in all Pylonida the shell-building
commences, is a quite simple, very small fenestrated shell. Commonly one sees on the surface only
five to ten small pores (three to four on the diameter). Its form seems to be sometimes spherical,
sometimes elongated, ellipsoidal or probably lentelliptical. It may be originally a small
Cenolarcus. This simple central chamber, the true "medullary shell" of the small
Haplozonaria, is quite different from the medullary shell of the larger Diplozonaria, and
particularly of the well-known Tetrapyle. The former observers, J. Müller as well as R.
Hertwig, have described in these forms also the medullary shell as a simple spherical or oblong
body. But a careful {630}comparison of many hundred
specimens of them and of their dimensions, has convinced me that this was an error, and that the
small spherical or elliptical medullary shell of Tetrapyle and the other Diplozonaria
possesses already the same complex structure, composed of a system of three girdles, as
Trizonium and Larnacilla. Whilst in the Haplozonaria probably the simple central
chamber only represents the medullary shell (enclosed in the central capsule), and the first
system of girdles (complete in Trizonium) the external cortical shell, with the progressive
growth this latter becomes enclosed in the central capsule and so constitutes the "trizonal
medullary shell" of the Diplozonaria and Triplozonaria.
A very difficult matter is the mode of connection between the cortical and medullary shell. In
most of the Pylonida it seems that the first or transverse girdle (in each system) is produced by
the formation of two lateral wings or chambers (one on each side of the medullary shell), so that
each wing (or half girdle) represents a short and wide, nearly cylindrical tube, the axis of which
(with free openings on both poles) is parallel to the principal axis of the medullary shell. In
this case (probably the ordinary one) both principal faces of the medullary shell itself (dorsal
and ventral face) constitute the middle part of the first girdle whilst its lateral parts are
formed by the wings (comparable to the lateral chambers of Amphitholus).
In the second case (probably a much rarer one) there is a free ring-shaped space between the
medullary shell and the first (transverse) girdle, and both are connected by a small number of
very short and small radial beams (R. Hertwig, loc. cit., p. 52, line 19 to 21 from above).
This mode of connection would be the same as is common between the concentric shells of the Sphæroidea and Prunoidea. The distinction between
these two different modes of connection is often very difficult.
The second or lateral girdle is commonly not in direct connection with the medullary shell, or
only by some scattered radial beams (mainly in the principal axis). This lateral guide arises by
prolongation of both wings of the transverse girdle in the lateral plane, so that from both sides
(right and left) they become united on the poles of the principal axis. The minor axis of the
elliptical lateral ring (thus formed) is therefore the major axis of the foregoing (transverse)
elliptical ring; the major axes of both are perpendicular one to another. The major axis of the
lateral ring is the principal (or longitudinal) axis of the whole body.
The third or sagittal girdle becomes developed from the second almost in the same manner as the
second from the first. On both poles of the principal axis two latticed wings arise from the
lateral girdle, growing further in the direction of an elliptical ring, which represents the
perimeter of the sagittal plane or median plane. These wings are already mentioned by J. Müller as
"prominent roofs, protecting the gates of the Tetrapyle-shell." If these roofs grow towards
the equatorial plane of the shell and become united in pairs on the poles of the sagittal axis,
the third girdle becomes {631}complete. R. Hertwig
supposes that the minor axis of this sagittal girdle is constantly at the same time the major axis
of the lateral girdle, but this is not always the case. Very often the size of both these girdles
is nearly the same, or one is not much larger than the other. In this case the principal axis of
the body is the major axis of the second as well as of the third girdle.
The characteristic "gates" of the Pylonida, or the large wide openings in their cortical shell,
remaining between the crossed latticed girdles, are in general roundish, sometimes nearly
circular, commonly more elliptical, kidney-shaped or semilunar, their special form varying much
according to the different form of the girdles. The narrowest part of each girdle, or its
"isthmus," in the case of the halves of the transverse girdle is commonly at their origin from the
medullary shell, in the case of the halves of the lateral girdle at the poles of the principal
axis, and in the case of the halves of the sagittal girdle at the poles of the sagittal axis. The
number of the gates is quite constant in the different genera. If only one girdle (the transverse)
be developed, we find only two large gates, between the two wings on the poles of the principal
axis (in Monozonium, Amphipyle, Amphipylonium). In all other cases there are
four gates (determining the original name "Tetrapyle"), as well if only two or if all three
girdles be completed. If two girdles be complete (in Dizonium, Tetrapyle,
Tetrapylonium) the four gates lie opposite in pairs on the sagittal faces (two anterior and
two posterior gates), and are limited by the transverse and lateral girdles. If all three girdles
be complete (in Trizonium, Pylonium, Pylozonium) the four gates lie opposite
in pairs on the lateral faces (two dorsal and two ventral gates), and are limited by the sagittal
and the transverse girdles. If we turn the shell through an angle of 90°, we have the same aspect
as in the former group. A sagittal septum sometimes becomes developed, beginning with two polar
beams, rising from the poles of the principal axis. If these polar beams become branched and
connected with the middle part of the lateral girdle, we get a latticed vertical septum, which
divides the four gates of Tetrapyle into eight gates, Octopyle.
The lattice-work of the Pylonida is commonly very variable and irregular, with roundish meshes
of very unequal size, therefore without value in the determination of the species. Commonly the
outside of the shell is thorny, and often distinguished by larger radial spines, symmetrically
disposed. We can separate these into two groups; "dimensive" spines, lying in one of the three
dimensive axes (principal, transverse, or sagittal), and "diagonal" spines, lying crossed in pairs
in diagonal axes. Among these latter eight diagonal wing-spines, which arise from the lateral
edges of the four gates, are particularly remarkable; they are not only characteristic of
Tetrapyle octacantha, but also of a large number of other Pylonida, and form the
starting-point for many specific forms.
The shell of the Pylonida is characterised by extraordinary variability and great {632}inclination to individual abnormalities, formation of
varieties and transitions into other families, hence derived, as Larnacida, Tholonida, Lithelida,
&c.
The central capsule in all Pylonida, in which I could observe it, was a true lentellipsis (or a
"triaxial ellipsoid" in the geometrical sense, with the three unequal isopolar axes of the
"rhombic octahedron"). In the living Pylonida it is commonly coloured pink or scarlet. During
growth its dimensions are probably more or less changed, and perhaps the axes alternate. Regarding
the relation of the central capsule to the skeleton, we can distinguish two different groups,
quite as in the nearly allied Larnacida. In the Haplozonaria (as also in the Larnacillida) the
central capsule encloses only the central chamber and is enveloped by the first system of girdles,
whereas in the Diplozonaria and Triplozonaria (as in the Larnacalpida) that "trizonal shell"
becomes enclosed (as the "medullary shell") in the central capsule, which is now enveloped by the
second system of girdles as the "cortical shell."
Synopsis of the Genera of Pylonida.
|
III. Subfamily Haplozonaria.
One system of girdles. Medullary shell simple, spherical or lentelliptical;
cortical shell simple, with one, two, or three girdles.
|
|
Cortical shell only with one latticed (transverse)
girdle, |
279. Monozonium. |
| Cortical shell with two perfect girdles (transverse and
lateral), |
280. Dizonium. |
| Cortical shell with three perfect girdles (transverse,
lateral, and sagittal |
281. Trizonium. |
|
III. Subfamily Diplozonaria.
Two systems of concentric girdles. Medullary shell trizonal, with three
perfect girdles; cortical shell simple, with one, two or three girdles.
|
|
Cortical shell only with one perfect (transverse)
girdle, |
282. Amphipyle. |
| Cortical shell with two perfect girdles (the transverse and
lateral). |
|
Four gates simple, |
283. Tetrapyle. |
| Four gates bisected by a sagittal septum, |
284. Octopyle. |
| Cortical shell with three perfect girdles (transverse,
lateral, and sagittal), |
285. Pylonium. |
|
III. Subfamily Triplozonaria.
Three systems of concentric girdles. Medullary shell trizonal, with three
perfect girdles, quite as the inner cortical shell; outer cortical shell with one, two, or
three girdles.
|
|
Outer cortical shell (third system) only with one perfect
(transverse) girdle, |
286. Amphipylonium. |
| Outer cortical shell with two perfect girdles (transverse
and lateral), |
287. Tetrapylonium. |
| Outer cortical shell with three perfect girdles
(transverse, lateral, and sagittal), |
288. Pylozonium. |
| I. Subfamily Haplozonaria. One system of girdles. Medullary shell simple,
spherical or lentelliptical; cortical shell simple, with one, two, or three girdles. |
|
|
Cortical shell only with one latticed (transverse) girdle, |
|
|
|
|
|
|
|
|
279. Monozonium. |
|
|
|
|
|
|
|
|
Cortical shell with two perfect girdles (transverse and
lateral), |
|
|
|
|
|
|
|
|
280. Dizonium. |
|
|
|
|
|
|
|
|
Cortical shell with three perfect girdles (transverse, lateral,
and sagittal |
|
|
|
|
|
|
|
|
281. Trizonium. |
|
|
|
|
|
|
| II. Subfamily Diplozonaria. Two systems of concentric girdles. Medullary shell
trizonal, with three perfect girdles; cortical shell simple, with one, two or three
girdles. |
|
|
Cortical shell only with one perfect (transverse) girdle, |
|
|
|
|
|
|
|
|
282. Amphipyle. |
|
|
|
|
|
|
|
|
Cortical shell with two perfect girdles (the transverse and
lateral). |
|
|
|
|
|
|
Four gates simple, |
|
|
|
|
|
|
|
|
|
|
283. Tetrapyle. |
|
|
|
|
|
|
|
|
|
|
Four gates bisected by a sagittal septum, |
|
|
|
|
|
|
|
|
|
|
284. Octopyle. |
|
|
|
|
|
|
|
|
Cortical shell with three perfect girdles (transverse, lateral,
and sagittal), |
|
|
|
|
|
|
|
|
285. Pylonium. |
|
|
|
|
|
|
| III. Subfamily Triplozonaria. Three systems of concentric girdles. Medullary
shell trizonal, with three perfect girdles, quite as the inner cortical shell; outer cortical
shell with one, two, or three girdles. |
|
|
Outer cortical shell (third system) only with one perfect
(transverse) girdle, |
|
|
|
|
|
|
|
|
286. Amphipylonium. |
|
|
|
|
|
|
|
|
Outer cortical shell with two perfect girdles (transverse and
lateral), |
|
|
|
|
|
|
|
|
287. Tetrapylonium. |
|
|
|
|
|
|
|
|
Outer cortical shell with three perfect girdles (transverse,
lateral, and sagittal), |
|
|
|
|
|
|
|
|
288. Pylozonium. |
|
|
|
|
|
|
Subfamily 1. Haplozonaria, Haeckel.
Definition.—Pylonida with one single system
of fenestrated girdles (with one, two, or three girdles, lying in one lentelliptical face).
{633}
Genus 279. Monozonium,[318] n. gen.
Definition.—Pylonida with simple, spherical or
subspherical, central chamber, surrounded by one single (transverse) latticed girdle.
The genus Monozonium is the most simple and primitive of all Pylonida, and
may be regarded as their common ancestral form; it may probably be derived phylogenetically either
directly from Cenosphæra or from Lentellipsis, either by apposition of two imperfect
lateral chambers, or by surrounding it with an equatorial latticed girdle. This transverse girdle
is composed of three parts, the central chamber and two lateral wings, which represent two short
hollow latticed tubes, the axes of which are parallel to the principal axis. On both principal
sides (on the anterior and posterior faces) there are two large open gates as in Amphipyle.
If we imagine the openings of the tube-shaped lateral wings closed by lattice-work,
Monozonium becomes transformed into Tholartus, the most simple form of Tholonida.
Probably in all Pylonida the ontogeny of the shell begins with the formation of a
Monozonium.
Subgenus 1. Monozonaris, Haeckel.
Definition.—Shell smooth or rough, without radial spines or
thorns.
1. Monozonium primordiale, n. sp.
Central chamber of the shell spherical, smooth, with three to four pores on the half equator.
Both wings of the girdle of the same breadth, but of twice the length of the central chamber, with
three to four longitudinal rows of pores. No radial spines.
Dimensions.—Diameter of the central chamber 0.02; breadth of the wings 0.02,
length 0.04; pores and bars 0.003.
Habitat.—Pacific, central area, Station 271, surface.
2. Monozonium alatum, n. sp. (Pl. 9,
fig. 1).
Central chamber of the shell lentelliptical, smooth, one and a half times as long as broad,
with three to four pores on the half equator. Both wings of the girdle twice as broad, and
somewhat longer than the central chamber, with five to six longitudinal rows of pores. No radial
spines.
Dimensions.—Length of the central chamber 0.03, breadth 0.02; breadth of the wings
0.04, length 0.05; pores and bars 0.004.
Habitat.—North Pacific, Station 241, surface.
Monozonium hartingii =
Haliomma amphiaspis, Harting, 1863 (L. N. 18, p. 15, pl. 2, fig.
43).
{634}
Subgenus 2. Monozonitis, Haeckel.
Definition.—Shell with radial spines or thorns, symmetrically
disposed.
3. Monozonium pleurostylum, n. sp.
Central chamber spherical, smooth, with four to five pores on the half equator. Both wings of
the girdle of the same breadth, but somewhat longer than the central chamber, with four to five
longitudinal rows of pores. On the poles of the lateral axis two opposite strong conical spines
(one in the middle of each wing).
Dimensions.—Diameter of the central chamber 0.025; breadth of the wings 0.025,
length 0.03; pores and bars 0.005.
Habitat.—Pacific, central area, Station 265, surface.
4. Monozonium amphistylum, n. sp.
Central chamber lentelliptical, one and a half times as long as broad, with three to four pores
on the half equator. Breadth and length of each wing somewhat greater than that of the central
chamber. On the latter two strong conical spines, opposite on the poles of the principal or
longitudinal axis.
Dimensions.—Length of the central chamber 0.03, breadth 0.02; breadth of the wings
0.025, length 0.04; pores and bars 0.004.
Habitat.—South Atlantic, Station 332, surface.
5. Monozonium staurostylum, n. sp.
Central chamber lentelliptical, one and a third times as long as broad, with four to five pores
on the half equator. Both wings of the girdle have the same breadth, but one and a half times the
length of the central chamber. Four conical radial spines, two opposite on the poles of the
lateral, two on the poles of the principal axis.
Dimensions.—Length of the central chamber 0.028, breadth 0.021; breadth of the
wings 0.02, length 0.04.
Habitat.—South Pacific, Station 295, surface.
Genus 280. Dizonium,[319] n. gen.
Definition.—Pylonida with simple, spherical or
subspherical, central chamber, surrounded by two crossed latticed girdles, one smaller (primary)
transverse and one larger (secondary) lateral girdle.
The genus Dizonium differs from the preceding Monozonium in the
possession of two crossed elliptical lattice-girdles. The smaller girdle, immediately surrounding
the {635}central chamber, is the transverse girdle (the
single girdle of Monozonium). On the poles of its major axis (the transverse axis) it is
connected with the larger girdle, the elliptical perimeter of which circumscribes the lateral
plane. The minor axis of this latter is the major axis of the former. Between the two crossed
girdles remain four larger openings or gates, quite as in Tetrapyle. But whilst in
Tetrapyle the enclosed medullary shell is a trizonal Larnacilla-shell, here in
Dizonium it is a simple spherical or lentelliptical chamber.
Subgenus 1. Dizonaris, Haeckel.
Definition.—Shell smooth or rough, without radial spines or
thorns.
1. Dizonium circulare, n. sp.
Central chamber of the shell spherical, smooth, with four to five pores on the half equator.
Lateral girdle circular, three times as broad as the former. Four gates semilunar, twice as broad
as high. No radial spines.
Dimensions.—Diameter of the central chamber 0.02, of the lateral girdle 0.06;
height of the gates 0.02, breadth 0.04.
Habitat.—Pacific, central area, Station 274, surface.
2. Dizonium ellipticum, n. sp.
Central chamber of the shell elliptical, smooth, with three to four pores on the half equator.
Lateral girdle elliptical, three times as long and broad as the central chamber. Four gates
kidney-shaped, twice as broad as high. No radial spines.
Dimensions.—Length of the central chamber 0.03, breadth 0.02; length of the
lateral girdle 0.09, breadth 0.06; height of the gates 0.02, breadth 0.038.
Habitat.—South Pacific, Station 288, surface.
3. Dizonium transversum, n. sp.
Central chamber of the shell spherical, rough, with five to six pores on the half equator.
Lateral girdle transverse-elliptical, so that its longer axis corresponds to the lateral axis of
the transverse girdle, and equals four times the diameter of the central chamber. Four gates
kidney-shaped, four times as broad as high. No radial spines.
Dimensions.—Diameter of the central chamber 0.02; length of the lateral girdle
0.05, breadth 0.08; height of the gates 0.01, breadth 0.04.
Habitat.—Pacific, central area, Station 274, depth 2750 fathoms.
{636}
Subgenus 2. Dizonitis, Haeckel.
Definition.—Shell with radial spines or thorns, symmetrically
disposed.
4. Dizonium pleuracanthum, n. sp. (Pl. 9, fig. 2).
Central chamber of the shell nearly spherical, smooth. Lateral girdle elliptical, one and a
third times as long as broad, three times as long as the central chamber. Four gates
transverse-elliptical, one and a half times as broad as high. Two conical spines, opposite on the
poles of the lateral axis.
Dimensions.—Diameter of the central chamber 0.03; length of the lateral girdle
0.1, breadth 0.075; height of the gates 0.026, breadth 0.04.
Habitat.—Western Tropical Pacific, Station 224, depth 1850
fathoms.
5. Dizonium amphacanthum, n. sp.
Central chamber elliptical. Lateral girdle elliptical, twice as long as broad. Four gates
heart-shaped, about as high as broad. Two conical spines, opposite on the poles of the principal
axis.
Dimensions.—Length of the central chamber 0.03, breadth 0.02; length of the
lateral girdle 0.1, breadth 0.05; height and breadth of the gates 0.03.
Habitat.—Pacific, central area, Station 270, depth 2925 fathoms.
6. Dizonium stauracanthum, n. sp. (Pl. 9, fig. 3).
Central chamber spherical. Lateral girdle elliptical, one and a third times as long as broad.
Four gates elliptical, one and a half times as broad as high, halved by an axial beam (as in
Octopyle). Four conical radial spines opposite in pairs, two on the poles of the principal,
two on the poles of the transverse axis.
Dimensions.—Diameter of the central chamber 0.025; length of the lateral girdle
0.09, breadth 0.06; height of the gates 0.024, breadth 0.036.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
7. Dizonium octacanthum, n. sp.
Central chamber spherical. Lateral girdle elliptical, one and a half times as long as broad.
Four gates kidney-shaped, twice as broad as high. Eight radial spines, opposite in pairs in two
crossed diagonal planes.
Dimensions.—Diameter of the central chamber 0.02; length of the lateral girdle
0.075, breadth 0.05; height of the gates 0.02, breadth 0.04.
Habitat.—Pacific, central area, Station 272, surface.
{637}
Genus 281. Trizonium,[320] n. gen.
Definition.—Pylonida with simple, spherical or
subspherical, central chamber, surrounded by three latticed girdles, one smaller (primary)
transverse, one larger (secondary) lateral, and one (tertiary) sagittal girdle.
The genus Trizonium represents the most highly developed form of the
Haplozonaria, with three complete elliptical latticed girdles, crossed at right angles and lying
in the perimeter of the three dimensive planes. Commonly the first (transverse) girdle is the
smallest, immediately connected with the spherical or lentelliptical central chamber. The minor
axis of the second (lateral) girdle is identical with the major axis of the first girdle. The
third (sagittal) girdle is either larger than both others, or intermediate between them. If the
four gates between the girdles become afterwards closed by lattice-work, the "trizonal shell" of
Trizonium passes over into the typical Larnacilla, the ancestral form of a great
many Larcoidea.
Subgenus 1. Trizonaris, Haeckel.
Definition.—Shell smooth or rough, without radial spines or
thorns.
1. Trizonium tricinctum, n. sp. (Pl. 9,
fig. 4).
Central chamber of the shell elliptical. Lateral girdle elliptical, one and a half times as
long as broad. Four gates kidney-shaped, one and a half times as broad as high. Ten to eleven
pores on the half equator, fourteen to sixteen on the half meridian of the lentelliptical shell.
No radial spines on the surface.
Dimensions.—Length of the central chamber 0.024, breadth 0.016; length of the
lateral girdle 0.075, breadth 0.05; height of the four gates 0.026, breadth 0.04.
Habitat.—Pacific, central area, Station 272, depth 2600 fathoms.
2. Trizonium constrictum, n. sp.
Central chamber elliptical. Lateral girdle violin-shaped, on both sides in the middle
constricted, twice as long as broad. Four gates transverse-elliptical, twice as broad as high.
Eight to nine pores on the half equator, twelve to thirteen on the half meridian. No radial
spines.
Dimensions.—Length of the central chamber 0.03, breadth 0.02; length of the
lateral girdle 0.09, breadth (in the middle) 0.045; height of the gates 0.022, breadth 0.044.
Habitat.—Pacific, central area, Station 263, surface.
{638}
3. Trizonium hexagonium, n. sp.
Central chamber spherical. Lateral girdle hexagonal, with parallel sides, twice as long as
broad. Four gates triangular, twice as broad as high. Eleven to twelve pores on the half equator,
sixteen to eighteen on the half meridian. No radial spines.
Dimensions.—Diameter of the central chamber 0.025; length of the lateral girdle
0.08, breadth 0.04; height of the gates 0.018, breadth 0.036.
Habitat.—Pacific, central area, Station 266, depth 2750 fathoms.
4. Trizonium octogonium, n. sp.
Central chamber spherical. Lateral girdle octagonal; two lateral sides of the octagon twice as
long as the two polar sides and the four diagonal sides. Four gates hexagonal, one and a half
times as broad as high. Nine to ten pores on the half equator, twelve to fourteen on the half
meridian. No radial spines.
Dimensions.—Diameter of the central chamber 0.02; length of the lateral girdle
0.01, breadth 0.07; height of the gates 0.04, breadth 0.06.
Habitat.—Indian Ocean, surface; Ceylon, Haeckel.
Subgenus 2. Trizonitis, Haeckel.
Definition.—Shell with radial spines or thorns, symmetrically
disposed.
5. Trizonium pleurobelonium, n. sp.
Central chamber spherical. Lateral girdle elliptical, one and a third times as long as broad.
Four gates nearly circular. Ten to eleven pores on the half equator, fifteen to sixteen on the
half meridian. Two opposite conical spines on the poles of the lateral axis.
Dimensions.—Diameter of the central chamber 0.02; length of the lateral girdle
0.08, breadth 0.06; height and breadth of the gates 0.025.
Habitat.—South Atlantic, off Buenos Ayres, Station 323, depth 1900
fathoms.
6. Trizonium amphibelonium, n. sp.
? Echinosphæra datura, R. Hertwig (partim), 1879, Organismus d. Radiol.,
p. 54, Taf. iv. figs. 8, 8a.
Central chamber lentelliptical. Lateral girdle lanceolate, nearly one and a half times as long
as broad. Four gates subtriangular. Eight to nine pores on the half equator, ten to twelve on the
half meridian. Two opposite thin and long spines on the poles of the principal axis, numerous
shorter radial spines on the whole surface. Possibly this species is identical with one of the
three different Larcoidea which R. Hertwig has described as
Echinosphæra datura. His fig. 8, Taf. iv., would be the aspect from the sagittal
girdle.
{639}
Dimensions.—Length of the central chamber 0.04, breadth 0.025; length of the
lateral girdle 0.11, breadth 0.08; height of the gates 0.03, breadth 0.04.
Habitat.—Mediterranean (Genoa), Haeckel, (Messina), R. Hertwig,
surface; Tropical Atlantic, Station 348, surface.
7. Trizonium staurobelonium, n. sp.
Central chamber spherical, with two opposite polar beams. Lateral girdle elliptical, one and a
half times as long as broad. Four gates transverse-elliptical, half as high as broad. Ten to
eleven pores on the half equator, fourteen to fifteen on the half meridian. Four conical radial
spines, two on the poles of the lateral, two on the poles of the principal axis.
Dimensions.—Diameter of the central chamber 0.02; length of the lateral girdle
0.075, breadth 0.05; height of the gates 0.014, breadth 0.028.
Habitat.—North Pacific, Station 241, depth 2300 fathoms.
8. Trizonium hexabelonium, n. sp.
Central chamber elliptical. Lateral girdle hexagonal, one and a half times as long as broad.
Four gates rhombic, two-thirds as high as broad. Eight to nine pores on the half equator, twelve
to thirteen on the half meridian. Six radial spines in the lateral plane, opposite in pairs on the
six corners of the lateral girdle (two principal and four diagonal).
Dimensions.—Length of the central chamber 0.024, breadth 0.018; length of the
lateral girdle 0.06, breadth 0.04; height of the gates 0.02, breadth 0.03.
Habitat.—Pacific, central area, Station 263, depth 2650 fathoms.
9. Trizonium octobelonium, n. sp.
Central chamber elliptical. Lateral girdle hexagonal, one and one-third times as long as broad.
Four gates kidney-shaped or nearly rhombic, twice as broad as high. Eleven to twelve pores on the
half equator, sixteen to seventeen on the half meridian. Eight radial spines, opposite in pairs in
two crossed diagonal planes. (An intermediate stage between Dizonium octacanthum and
Amphipyle octostyle.)
Dimensions.—Length of the central chamber 0.03, breadth 0.024; length of the
lateral girdle 0.09, breadth 0.07; height of the gates 0.025, breadth 0.05.
Habitat.—Indian Ocean, Madagascar, Rabbe, surface.
10. Trizonium decabelonium, n. sp.
Central chamber elliptical. Lateral girdle hexagonal, one and a half times as long as broad.
Four gates rhombic, twice as broad as high. Twelve to thirteen pores on the half equator,
seventeen to eighteen on the half meridian. Ten radial spines, two opposite on the two poles of
the principal axis, eight others opposite in pairs in two crossed diagonal planes.
{640}
Dimensions.—Length of the central chamber 0.04, breadth 0.03; length of the
lateral girdle 0.12, breadth 0.08; height of the gates 0.03, breadth 0.06.
Habitat.—North Atlantic, Station 353, surface.
11. Trizonium dodecabelos, n. sp.
Central chamber spherical. Lateral girdle octagonal, nearly twice an long as broad. Four gates
hexagonal, one and a half times as broad as high. Nine to ten pores on the half equator, fifteen
to sixteen on the half meridian. Twelve radial spines, four in the lateral plane, in pairs on both
sides of the poles of the principal axis, eight others opposite in pairs in two crossed diagonal
axes.
Dimensions.—Diameter of the central chamber 0.025; length of the lateral girdle
0.1, breadth 0.06; height of the gates 0.033, breadth 0.05.
Habitat.—Pacific, central area, Station 272, surface.
Subfamily 2. Diplozonaria, Haeckel.
Definition.—Pylonida with two concentric
systems of fenestrated girdles, lying in two concentric lentelliptical faces (every one system
with one to three girdles, lying in one lentelliptical face).
Genus 282. Amphipyle,[321] Haeckel, 1881,
Prodromus, p. 463.
Definition.—Pylonida with trizonal lentelliptical
medullary shell, surrounded by one single (transverse) latticed cortical girdle.
The genus Amphipyle opens the large series of Diplozonaria, comprising all
Pylonida, the shell of which is composed of two concentric systems of latticed girdles; the first
system constituting the characteristic "trizonal medullary shell" or "Larnacilla-shell;"
the second system composed of one to three girdles of the second order. The first system lies
inside, the second outside the central capsule. In Amphipyle only the first (transverse)
girdle of the second system becomes developed, and therefore on both poles of the principal axis
are two large open gates. Amphipyle repeats the two-winged form of Monozonium; but
whilst the medullary shell in this latter is a simple central chamber, it is here a trizonal
Larnacilla-shell.
Subgenus 1. Amphipylissa, Haeckel.
Definition.—Cortical shell smooth or thorny, but without large,
symmetrically disposed spines.
{641}
1. Amphipyle aceros, n. sp.
Cortical shell quite smooth, without any spines or thorns. Both lateral wings (or opposite half
girdles) semilunar, with convex lateral crest, three times as long as broad; twice as broad as the
trizonal lentelliptical medullary shell.
Dimensions.—Length of the medullary shell 0.05, breadth 0.03; length of each
lateral wing (or principal dimension of the latticed girdle) 0.18, breadth of it (or transverse
dimension of each girdle-tube) 0.06.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
2. Amphipyle stenoptera, n. sp.
Cortical shell smooth, without spines. Both lateral wings of the same breadth as the
lentelliptical medullary shell, but five times as long as broad, prolonged on both ends into
cylindrical latticed tubes, with rectilinear lateral crest.
Dimensions.—Length of the medullary shell 0.06, breadth 0.04; length of each
cylindrical lateral wing 0.2, breadth 0.04.
Habitat.—South Atlantic, off Ascension Island, Station 343,
surface.
3. Amphipyle platyptera, n. sp.
Cortical shell thorny, but without larger spines. Both lateral wings nearly triangular, very
broad and short, four times as broad as the medullary shell between them, and only twice as long
as broad, with rectilinear lateral crest.
Dimensions.—Length of the medullary shell 0.05, breadth 0.25; length of each
lateral wing 0.18, breadth 0.09.
Habitat.—Indian Ocean, Ceylon, Haeckel, surface.
Subgenus 2. Amphipylura, Haeckel.
Definition.—Cortical shell armed with large, symmetrically disposed
spines.
4. Amphipyle amphiceros, n. sp.
Cortical shell with two conical spines on the poles of the longitudinal axis (of the
lentelliptical medullary shell), without other large spines, but with small thorns on the surface;
two to three longitudinal rows of irregular, roundish pores on each half wing of the cortical
girdle.
Dimensions.—Length of the medullary shell (or principal axis) 0.06, breadth (or
transverse axis) 0.04; length of each lateral wing 0.2, breadth of it 0.05.
Habitat.—Western Tropical Pacific, Station 222, surface.
{642}
5. Amphipyle tetraceros, n. sp. (Pl. 9,
fig. 5).
Cortical shell with smooth surface, and with four strong conical spines in the lateral plane,
opposite in pairs on the ends of the concave lateral crests (one pair on the end-points of the
lateral line of each wing); four to five longitudinal rows of irregular, roundish pores on each
half wing of the cortical girdle. Surface of the medullary shell spiny.
Dimensions.—Length of the medullary shell 0.08, breadth of the same 0.05; length
of each lateral wing 0.16, breadth 0.04.
Habitat.—Pacific, central area, Station 271, surface.
6. Amphipyle stauroceros, n. sp.
Cortical shell with four strong conical spines, two on the poles of the longitudinal axis, two
opposite on the poles of the transverse axis (perpendicular to the former). Three to four
longitudinal rows of irregular, roundish pores on each half wing of the cortical girdle, about six
pores in the longest row. Lateral crest convex.
Dimensions.—Length of the medullary shell 0.05, breadth 0.03; length of each
lateral wing 0.15, breadth 0.08.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
7. Amphipyle amphiptera, n. sp. (Pl. 9,
fig. 7).
Cortical shell with six strong conical spines, lying in the lateral plane in three parallel
longitudinal lines; two on the poles of the longitudinal axis, two others on the poles of the wing
axes, which are parallel to the former. Two to three longitudinal rows of irregular, roundish
pores on each half wing of the cortical girdle, about eight pores in the longest row. Lateral
crest convex, thorny.
Dimensions.—Length of the medullary shell 0.06, breadth 0.045; length of each
lateral wing 0.09, breadth 0.035.
Habitat.—Western Tropical Pacific, Station 224, surface.
8. Amphipyle hexaceros, n. sp.
Cortical shell thorny, with six strong conical spines lying in the lateral plane; two on the
poles of the lateral axis (in the central point of each wing), two on the end-points of the
triangular crest of each wing (on both poles of the longitudinal wing-axis).
Dimensions.—Length of the medullary shell 0.06, breadth 0.04; length of each
lateral wing 0.13, breadth 0.07.
Habitat.—South Pacific, Station 291, surface.
{643}
9. Amphipyle octoceros, n. sp.
Cortical shell thorny, with eight long radial spines, lying in two crossed diagonal planes,
opposite in pairs. These eight horns are the prolongations of the anterior and posterior edges of
each wing, on both sides of its lateral plane; they are of great importance, as appearing (by
heredity) in many other Pylonida (e.g., in Tetrapyle octacantha). Lateral crest of
each wing without spines, slightly convex.
Dimensions.—Length of the medullary shell 0.07, breadth 0.05; length of each
lateral wing 0.15, breadth 0.06.
Habitat.—Pacific, central area, Stations 270 to 274, surface.
10. Amphipyle cladoceros, n. sp.
Cortical shell spiny, with eight strong ramified radial spines, lying in the same two crossed
diagonal planes as in the preceding species. It differs from this in the ramification of the eight
horns, which bear six to nine irregular lateral branches, partly simple, partly bifurcate. Lateral
crest of each wing spiny, convex.
Dimensions.—Length of the medullary shell 0.06, breadth 0.045; length of each
lateral wing (without spines) 0.2, breadth 0.09.
Habitat.—Indian Ocean, surface, Madagascar, Rabbe.
11. Amphipyle decaceros, n. sp.
Cortical shell spiny, with ten strong conical spines; eight lying in two crossed diagonal
planes, in the same disposition (opposite in pairs) as in both preceding species; two others
opposite on the poles of the principal axis, arising from both poles of the medullary shell
(columella-beams). Spiny crest of the wings convex, semilunar.
Dimensions.—Length of the medullary shell 0.04, breadth 0.03; length of each
lateral wing 0.12, breadth 0.04.
Habitat.—North Atlantic, Station 354, surface.
12. Amphipyle dodecaceros, n. sp.
Cortical shell thorny, with twelve large cylindrical spines; eight lying in two crossed
diagonal planes, in the same disposition (opposite in pairs) as in the three preceding species;
four others crossed at right angles in the lateral plane, two opposite on the poles of the
principal, two on the poles of the lateral axis (the latter shorter). Both lateral wings nearly
rhombic.
Dimensions.—Length of the medullary shell 0.06, breadth 0.04; length of the
lateral wings 0.15, breadth 0.05.
Habitat.—South Atlantic, coast of Brazil, Rabbe, surface.
{644}
13. Amphipyle callizona, n. sp. (Pl. 9,
fig. 6).
Cortical shell smooth, with sixteen strong and short conical spines, lying opposite in pairs in
two crossed diagonal planes; each lateral wing four-sided prismatic, its lateral face concave,
both ends truncated, and each end provided with four divergent spines. Length of the wings
somewhat greater, but breadth smaller, than that of the medullary shell.
Dimensions.—Length of the medullary shell 0.07, breadth 0.05; length of the
lateral wings 0.09, breadth 0.03.
Habitat.—Western Tropical Pacific, Station 225, depth 4475
fathoms.
Genus 283. Tetrapyle,[322] J. Müller,
1858, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 154.
Definition.—Pylonida with trizonal lentelliptical
medullary shell, surrounded by two crossed latticed cortical girdles, one smaller (primary)
transverse, and one larger (secondary) lateral girdle. Four gates between the two cortical girdles
simple, without a sagittal septum.
The genus Tetrapyle, till 1881 the only known genus of the whole family, was
founded by Johannes Müller in 1858, and clearly illustrated by the Mediterranean (and common
cosmopolitan) Tetrapyle octacantha, hitherto the best known type of this family. Afterwards
(1860, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 832) Ehrenberg founded the genus
Schizomma for a nearly allied form, which exhibits only slight specific differences
(compare my Monograph, 1862, p. 434). Some good remarks on the structure of this typical genus and
its relations to other Pylonida are to be found in Richard Hertwig's Organismus, &c., 1879, p.
52, but the true trizonal structure of the medullary shell in this genus was not recognised by
him, so that his description agrees more with Dizonium. We confine here the genus
Tetrapyle to those Pylonida for which Tetrapyle octacantha of J. Müller remains the
determining type; the cortical shell is composed only of two perfect lattice-girdles (the
transverse and lateral), between which four wide gates remain open. This structure is similar to
that of Dizonium; but whilst here the medullary shell is a simple central chamber, in
Tetrapyle it is a complete trizonal or Larnacilla-shell.
Subgenus 1. Tetrapylissa, Haeckel.
Definition.—Cortical shell smooth or thorny, but without large,
symmetrically disposed spines.
{645}
1. Tetrapyle circularis, n. sp. (Pl. 9,
fig. 8).
Cortical shell rough, without radial spines. Lateral girdle circular, therefore the
longitudinal axis is equal to the transverse. Four gates kidney-shaped, twice as broad as high. In
each half wing of the transverse girdle six to seven longitudinal rows of irregular, roundish
pores.
Dimensions.—Medullary shell 0.05 long, 0.04 broad; cortical shell 0.15 long and
broad; gates 0.04 high, 0.08 broad.
Habitat.—Pacific, central area, Station 263, surface.
2. Tetrapyle circopyle, n. sp.
Cortical shell smooth, without radial spines. Lateral girdle elliptical, one and a half times
as long as broad. Four gates nearly circular. On each half wing of the transverse girdle four to
five rows of subregular, circular pores. (Similar to Tetrapyle pleuracantha, Pl. 9, fig. 9, but
without lateral spines.)
Dimensions.—Medullary shell 0.04 long, 0.03 broad; cortical shell 0.16 long, 0.11
broad; gates 0.05 high and broad.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
3. Tetrapyle quadriloba, Haeckel.
Tetrapyle quadriloba, Haeckel, 1862, Monogr. d. Radiol., p. 436.
Schizomma quadrilobum, Ehrenberg, 1860, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 815; Abhandl. d. k. Akad. d. Wiss. Berlin, 1872, Taf. x. figs. 12-14.
Cortical shell thorny, without regular, radial spines. Lateral girdle elliptical, one and a
half times as long as broad. Four gates nearly circular. On each half wing of the transverse
girdle three to four rows of irregular, roundish pores.
Dimensions.—Medullary shell 0.06 long, 0.04 broad; cortical shell 0.16 long, 0.1
broad; diameter of the gates 0.05.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Indian Ocean, Pacific,
surface.
4. Tetrapyle nephropyle, n. sp.
Cortical shell thorny, without radial spines. Lateral girdle elliptical, one and a third times
as long as broad. Four gates kidney-shaped, nearly twice as broad as high, with two prominent
polar spines on the poles of the longitudinal axis of the medullary shell. On each half wing of
the transverse girdle three to four longitudinal rows of large roundish pores.
Dimensions.—Medullary shell 0.06 long, 0.04 broad; cortical shell 0.24 long, 0.18
broad; gates 0.05 high, 0.08 broad.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
{646}
5. Tetrapyle cardiopyle, n. sp.
Cortical shell rough, without radial spines. Lateral girdle with hexagonal contour, nearly
cylindrical in the middle part, conical at both ends, twice as long as broad. Four gates
heart-shaped, about as high as broad. On each half wing of the transverse girdle three to four
longitudinal rows of subregular, polygonal pores.
Dimensions.—Medullary shell 0.04 long, 0.02 broad; cortical shell 0.14 long, 0.07
broad; gates 0.05 high and broad.
Habitat.—Eastern Tropical Atlantic, Station 348, depth (2450)
fathoms.
Subgenus 2. Tetrapylura, Haeckel.
Definition.—Cortical shell armed with large, symmetrically disposed
spines.
6. Tetrapyle fusiformis, n. sp.
Cortical shell rough, with two strong conical spines on the poles of the longitudinal axis.
Lateral girdle spindle-shaped, one and a half times as long as broad. Four gates kidney-shaped,
twice as broad as high. On each half wing of the transverse girdle four to five longitudinal rows
of irregular, roundish pores.
Dimensions.—Medullary shell 0.07 long, 0.05 broad; cortical shell 0.18 long, 0.12
broad; gates 0.04 high, 0.08 broad.
Habitat.—Western Tropical Atlantic, Mexican Gulf Stream, depth 1500
fathoms, Schaffner.
7. Tetrapyle pleuracantha, n. sp. (Pl. 9, fig. 9).
Cortical shell rough, with two strong conical spines on the poles of the transverse axis.
Lateral girdle elliptical, one and half times as long as broad. Four gates subcircular or nearly
hexagonal, about as high as broad. On each half wing of the transverse girdle three to four
longitudinal rows of very irregular pores.
Dimensions.—Medullary shell 0.045 long, 0.035 broad; cortical shell 0.16 long,
0.11 broad; gates 0.045 high and broad.
Habitat.—Western Tropical Pacific, off New Guinea, Stations 222 to
224, depths 1850 to 2450 fathoms.
8. Tetrapyle transversa, n. sp.
Cortical shell thorny, with two strong conical lateral spines on the poles of the transverse
axis. Lateral girdle elliptical, only four-fifths as long as broad. Four gates elliptical, twice
as broad as high, with two prominent spines on the poles of the longitudinal axis of the medullary
shell. On each half wing of the transverse girdle five to six longitudinal rows of subregular,
roundish pores. (Similar to Octopyle transversa, but without sagittal septum and with
stronger lateral spines.)
{647}
Dimensions.—Medullary shell 0.05 long, 0.035 broad; cortical shell 0.12 long, 0.15
broad; gates 0.03 high, 0.06 broad.
Habitat.—Southern Pacific, Station 290, surface.
9. Tetrapyle cruciata, n. sp.
Cortical shell smooth, with four strong radial spines, two on the poles of the longitudinal
axis, two on the poles of the transverse axis. Lateral girdle nearly circular, quite as long as
broad. Four gates kidney-shaped, twice as broad as high. On each half wing of the transverse
girdle five to six rows of subregular, circular pores.
Dimensions.—Medullary shell 0.06 long, 0.05 broad; cortical shell 0.16 long and
broad; gates 0.05 high, 0.09 broad.
Habitat.—South Atlantic, Station 330, surface.
10. Tetrapyle staurophora, n. sp.
Cortical shell thorny, with four strong radial spines, two on the poles of the longitudinal
axis, two on the poles of the transverse axis. Lateral girdle elliptical, one and a half times as
long as broad. Four gates nearly hexagonal, quite as high as broad. On each half wing of the
transverse girdle three to four longitudinal rows of irregular, roundish pores.
Dimensions.—Medullary shell 0.06 long, 0.04 broad; cortical shell 0.18 long, 0.12
broad; gates 0.08 high and broad.
Habitat.—Western Tropical Atlantic, coast of Brazil, Rabbe,
surface.
11. Tetrapyle quadricornis, n. sp.
Cortical shell thorny, with four strong horn-like curved spines, lying in the lateral plane and
converging in pairs towards the poles of the transverse axis. Lateral girdle elliptical, one and a
third times as long as broad. Four gates kidney-shaped, twice as broad as high. On each half wing
of the transverse girdle four to five rows of irregular, roundish pores. (May be only the young
form of Pylonium quadricorne, Pl. 9,
fig. 14.)
Dimensions.—Medullary shell 0.05 long, 0.04 broad; cortical shell 0.16 long, 0.12
broad; gates 0.03 high, 0.07 broad.
Habitat.—Pacific, central area, Stations 270 to 274, surface.
12. Tetrapyle tetracantha, n. sp.
Cortical shell thorny, with four strong radial spines lying in the lateral plane and in pairs
in its two crossed diagonal axes. Lateral girdle elliptical, constricted in the middle, nearly
twice as long as broad. Four gates kidney-shaped, one and a half times as broad as high. On each
half wing of the transverse girdle three to four longitudinal rows of large, irregular, roundish
pores. {648}(Similar to Tetrapylonium
quadrangulare, Pl. 49,
fig. 15, but without a sagittal girdle and with stronger lateral spines.)
Dimensions.—Medullary shell 0.06 long, 0.04 broad; cortical shell 0.18 long, 0.1
broad; gates 0.007 high, 0.01 broad.
Habitat.—North Atlantic, Canary Islands, Station 353, surface.
13. Tetrapyle quadrigata, n. sp.
Cortical shell thorny, with four brush-like groups of radial spines on the four diagonal
corners of the lateral plane, opposite in pairs in diagonal axes. Lateral girdle nearly square,
somewhat broader than long. Four gates kidney-shaped, twice as broad as high. On each half wing of
the transverse girdle five to six longitudinal rows of irregular, roundish pores.
Dimensions.—Medullary shell 0.06 long, 0.04 broad; cortical shell 0.14 long, 0.15
broad; gates 0.03 high, 0.07 broad.
Habitat.—Pacific, central area, Station 270, surface.
14. Tetrapyle octacantha, J. Müller.
Tetrapyle octacantha, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 33,
Taf. ii. figs. 1-6.
Tetrapyle octacantha, Haeckel, 1862, Monogr. d. Radiol., p. 435.
Tetrapyle octacantha, R. Hertwig, 1879, Organismus d. Radiol., p. 52, Taf.
iv. fig. 7, Taf. vi. figs. 2, 5, 5a.
Cortical shell thorny, with eight long and thin, cylindrical radial spines, lying in two
crossed diagonal planes, opposite in pairs. These eight characteristic diagonal spines (or
"angular spines") are the same as in Amphipyle octoceros and many other Pylonida, and arise
as prolongations of the proximal edges of the four gates (or of the lateral wings on the eight
points, where they are intersected by the edges of the lateral ring). This cosmopolitan, widely
distributed and very variable species was a long time the only known species of all the Pylonida,
and very accurately first described (1858) by Johannes Müller, afterwards (1879) by R. Hertwig.
But in the descriptions of these authors also some different species (such as the following) may
be confounded with the true typical Tetrapyle octacantha. The four gates of this species
are transverse-elliptical or almost triangular, nearly twice as broad as high.
Dimensions.—Length of the medullary shell 0.04, breadth 0.03; length of the
cortical shell 0.18, breadth 0.13; height of the gates 0.05, breadth 0.08.
Habitat.—Cosmopolitan; very common in all warmer seas, Mediterranean,
Atlantic, Indian, Pacific, surface.
15. Tetrapyle cladacantha, n. sp.
Cortical shell thorny, with eight strong ramified radial spines, lying opposite in pairs in two
crossed diagonal planes, as in the preceding species. It differs from this by the peculiar
ramification of the eight spines, which bear two to eight simple or furcate lateral branches
arising under {649}right angles (already commencing in a
transitional variety of Tetrapyle octacantha, J. Müller, loc. cit., Taf. ii. figs.
5, 6). Four gates transverse-elliptical, one and a half times as broad as high.
Dimensions.—Medullary shell 0.05 long, 0.04 broad; cortical shell 0.16 long, 0.13
broad; gates 0.05 high, 0.08 broad.
Habitat.—Mediterranean (Corfu), Haeckel, surface.
16. Tetrapyle pluteus, Haeckel.
Tetrapyle octacantha, var., J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss.
Berlin, Taf. iii. figs. 7-12.
Cortical shell thorny, with eight strong radial spines, lying opposite in pairs, as in the two
foregoing species. On the distal edge each of the four triangular gates is protected by a
prominent roof of lattice-work (or "pluteus"), which arises from the distal part of the lateral
girdle and connects the two angular diagonal spines of each gate. Therefore the shell assumes the
characteristic form very well represented by J. Müller in his fig. 11 (seen from the lateral side)
and fig. 7 (seen from the pole of the principal axis).
Dimensions.—Medullary shell 0.04 long, 0.03 broad; cortical shell 0.15 long, 0.11
broad; gates 0.045 high, 0.07 broad.
Habitat.—Mediterranean, Atlantic, Stations 348 to 353, &c.,
surface.
17. Tetrapyle turrita, n. sp. (Pl. 9,
fig. 10).
Cortical shell smooth, but with ten strong conical spines, two on the poles of the principal
axis, eight others symmetrically distributed on both sides of the distal edge of the four roundish
gates, and directed towards the transverse axis. Lateral girdle lanceolate, twice as long as
broad.
Dimensions.—Medullary shell 0.05 long, 0.04 broad; cortical shell 0.2 long, 0.1
broad; gates 0.05 high, 0.07 broad.
Habitat.—Pacific, central area, Station 274, depth 2750 fathoms.
18. Tetrapyle dodecaceros, n. sp.
Cortical shell thorny, with twelve stronger conical spines, eight diagonal spines in two
crossed planes (as in Tetrapyle octacantha), and four others in the lateral plane, in pairs
on both sides of a deep sagittal constriction of the quadrangular lateral girdle. Four gates
kidney-shaped, twice as broad as high. On each half wing of the transverse girdle five to six
longitudinal rows of irregular, roundish pores.
Dimensions.—Medullary shell 0.06 long, 0.04 broad; cortical shell 0.18 long, 0.14
broad; gates 0.04 high, 0.08 broad.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
{650}
Genus 284. Octopyle,[323] Haeckel, 1881,
Prodromus, p. 464.
Definition.—Pylonida with trizonal lentelliptical
medullary shell, surrounded by two crossed, latticed, cortical girdles; one smaller (primary)
transverse, and one larger (secondary) lateral girdle. Four gates between the two cortical girdles
divided by a sagittal septum into eight gates.
The genus Octopyle comprises those Pylonida which are distinguished from the
nearly allied Tetrapyle by the development of a sagittal septum, dividing the four gates of
the latter into eight separate gates. The septum begins with the formation of two axial rods or
columellæ, which afterwards become branched; the branches communicating one with another and with
the middle parts of the lateral girdle (on the poles of the principal axis), there is formed a
latticed septum in the sagittal plane, which separates more or less incompletely the right and
left halves of the shell. The four gates of Tetrapyle become halved by this septum, and
their number doubled.
Subgenus 1. Octopylissa, Haeckel.
Definition.—Cortical shell smooth or thorny, but without larger
symmetrically disposed spines.
1. Octopyle ovulina, n. sp.
Cortical shell lentelliptical, smooth, without thorns. Lateral girdle elliptical, one and a
third times as long as broad. Transverse girdle narrow, with two pores on the isthmus (or on the
narrowest part of each quadrant). Sagittal septum as long as the elliptical medullary shell. Eight
gates egg-shaped.
Dimensions.—Length of the medullary shell 0.05, breadth 0.035; length of the
cortical shell 0.16, breadth 0.12.
Habitat.—Pacific, central area, Station 274, surface.
2. Octopyle quadrata, n. sp.
Cortical shell thorny, nearly square, of equal length and breadth. Lateral girdle quadrangular,
with rounded edges, of equal length and breadth. Transverse girdle narrow, with three pores on the
isthmus. Sagittal septum of the same length as the subspherical medullary shell. Eight gates
nearly circular.
Dimensions.—Diameter of the medullary shell 0.04, of the cortical shell 0.14.
Habitat.—Northern Pacific, Station 244, depth 2900 fathoms.
{651}
3. Octopyle subglobosa, n. sp.
Cortical shell nearly spherical, thorny, of equal length and breadth. Lateral girdle broad,
nearly circular. Transverse girdle broad, with four pores on the isthmus. Sagittal septum shorter
than the subspherical medullary shell. Eight gates egg-shaped.
Dimensions.—Diameter of the medullary shell 0.05, of the cortical shell 0.13.
Habitat.—Southern Pacific, Station 300, depth 1375 fathoms.
4. Octopyle transversaria, n. sp.
Cortical shell thorny, transverse-elliptical. Lateral girdle very broad, one and a third times
as broad as long. Transverse girdle also very broad, with five pores on the isthmus. Sagittal
septum shorter than the elliptical medullary shell. Eight gates small, roundish.
Dimensions.—Length of the medullary shell 0.05, breadth 0.04; length of the
cortical shell 0.11, breadth 0.15.
Habitat.—Western Tropical Pacific, Station 224, depth 1850
fathoms.
Subgenus 2. Octopylura, Haeckel.
Definition.—Cortical shell armed with large, symmetrically disposed
spines.
5. Octopyle amphistyle, n. sp.
Cortical shell smooth, nearly spindle-shaped, about twice as long as broad. Transverse girdle
broad, with four large pores on the isthmus. Lateral girdle lanceolate. Sagittal septa about as
long as the medullary shell, prolonged at both poles of the principal axis into two opposite,
strong, angular spines. Eight gates obliquely quadrangular.
Dimensions.—Length of the medullary shell 0.07, breadth 0.05; length of the
cortical shell 0.2, breadth 0.12.
Habitat.—Pacific, central area, Station 272, depth 2600 fathoms.
6. Octopyle staurostyle, n. sp.
Cortical shell nearly rhombic, smooth, one and a half times as long as broad, with four strong,
conical, radial spines, two on the poles of the principal axis (as prolongations of the sagittal
septa), two others on the poles of the transverse axis. Transverse girdle narrow, with two pores
on the isthmus. Sagittal septa longer than the medullary shell. Eight gates triangular.
Dimensions.—Length of the medullary shell 0.04, breadth 0.03; length of the
cortical shell 0.18, breadth 0.12.
Habitat.—Tropical Atlantic, Station 347, depth 2250 fathoms.
{652}
7. Octopyle tetrastyle, n. sp.
Cortical shell lentelliptical, with rough surface and four angular radial spines in the lateral
plane, opposite in pairs in two crossed diagonals. Lateral girdle nearly rectangular (each angle
with one spine), one and a third times as long as broad. Sagittal septum longer than the hexagonal
medullary shell. Eight gates egg-shaped.
Dimensions.—Length of the medullary shell 0.05, breadth 0.03; length of the
cortical shell 0.16, breadth 0.12.
Habitat.—North Atlantic, Station 354, surface.
8. Octopyle tetraptera, n. sp.
Cortical shell quadrangular, with thorny surface and four latticed wing-like prolongations on
the corners of the quadrangle, opposite in pairs in two crossed diagonals, each wing supported by
a strong radial spine. Lateral girdle quadrangular, one and a half times as long as broad. On the
isthmus of the transverse girdle four pores. Sagittal septum longer than the hexagonal medullary
shell. Eight gates triangular.
Dimensions.—Length of the medullary shell 0.07, breadth 0.04; length of the
cortical shell 0.24, breadth 0.16.
Habitat.—Pacific, central area, Station 271, surface.
9. Octopyle stenozona, n. sp. (Pl. 9,
fig. 11).
Cortical shell quadrangular, with thorny surface, and four conical spines in the lateral plane,
on the points, where both cortical girdles are crossed. Lateral girdle very narrow, nearly square,
with rounded corners, of equal length and breadth. On the isthmus of the narrow transverse girdle
only one pore. Sagittal septum of the same length as the subspherical medullary shell. Eight gates
irregular roundish or nearly circular.
Dimensions.—Diameter of the medullary shell 0.04, of the cortical shell 0.14.
Habitat.—Tropical Atlantic, Station 338, depth 1990 fathoms.
10. Octopyle euryzona, n. sp.
Cortical shell nearly quadrangular, very thorny, with four bunches of stronger radial spines on
the four corners. Lateral girdle broad, with rounded edges of the quadrangle, little longer than
broad. On the isthmus of the broad transverse girdle four pores. Sagittal septum of the same
length as the elliptical medullary shell, with many radial beams. Eight gates nearly square.
Dimensions.—Length of the medullary shell 0.05, breadth 0.035; length of the
cortical shell 0.16, breadth 0.14.
Habitat.—Southern Atlantic, Station 332, depth 2200 fathoms.
{653}
11. Octopyle hexastyle, n. sp.
Cortical shell nearly rectangular, thorny, with six strong radial spines in the lateral plane,
two opposite on the poles of the principal axis (prolongations of the principal beam at the
sagittal septum), four others on the truncate corners of the quadrangular lateral girdle, which is
one and a half times as long as broad. On the isthmus of the broad transverse girdle three pores.
Sagittal septum longer than the hexagonal medullary shell. Eight egg-shaped gates.
Dimensions.—Length of the medullary shell 0.05, breadth 0.04; length of the
cortical shell 0.18, breadth 0.12.
Habitat.—Northern Atlantic, surface; Canary Islands, Station 9, depth
3150 fathoms.
12. Octopyle sexangulata, n. sp. (Pl. 9, fig. 12).
Cortical shell thorny, hexagonal, with six strong conical radial spines on the six corners of
the lateral girdle, lying in the lateral plane, two opposite on the poles of the transverse axis,
four others in pairs on both sides of the annular sagittal constriction, which is only twice as
long as the elliptical medullary shell. Transverse girdle broad, with five pores on the isthmus.
Eight gates roundish, smaller than the medullary shell.
Dimensions.—Length of the medullary shell 0.045, breadth 0.035; length of the
cortical shell 0.11, breadth 0.13.
Habitat.—Pacific, central area, Station 270, surface.
13. Octopyle octostyle, n. sp.
Cortical shell elliptical, thorny, with eight long cylindrical radial spines, lying in two
crossed diagonal planes, opposite in pairs, quite as in the nearly related Tetrapyle
octacantha. It differs from this species by the perfect, latticed, vertical sagittal septum,
which divides every gate into two triangular halves. Lateral girdle elliptical, one and a third
times as long as broad. Transverse girdle on the isthmus with three pores. Septum longer than the
elliptical medullary shell.
Dimensions.—Length of the medullary shell 0.06, breadth 0.045; length of the
cortical shell 0.16, breadth 0.12.
Habitat.—South Pacific, Station 288, surface.
14. Octopyle obtecta, n. sp.
Cortical shell quadrangular, thorny, with eight strong diagonal spines, lying in two crossed
planes, and arising from the proximal edges of the gates in the same way as in the foregoing
species. It differs from this by four prominent latticed roofs, which arise from the distal edge
of the gates and connect the diagonal spines of one gate. Octopyle obtecta bears therefore
the same relation to Octopyle octostyle that Tetrapyle pluteus does to Tetrapyle
octacantha.
Dimensions.—Length of the medullary shell 0.055, breadth 0.04; length of the
cortical shell 0.16, breadth 0.12.
Habitat.—Indian Ocean, surface, Madagascar, Rabbe.
{654}
15. Octopyle decastyle, n. sp. (Pl. 9,
fig. 13).
Cortical shell thorny, nearly quadrangular, with ten stronger radial spines, two opposite on
the poles of the principal axis (as prolongations of the axial beams), eight others opposite in
pairs in two diagonal planes (as in the foregoing species). Lateral girdle nearly square, with
slightly convex lateral faces, slightly concave principal faces. Transverse girdle broad, with
four pores on the isthmus. Sagittal septum of the same length as the hexagonal medullary shell.
Eight gates elliptical or irregular roundish.
Dimensions.—Length of the medullary shell 0.05, breadth 0.032; diameter of the cortical shell 0.15.
Habitat.—Western Tropical Pacific, Station 224, depth 1850
fathoms.
Genus 285. Pylonium,[324] Haeckel, 1881,
Prodromus, p. 464.
Definition.—Pylonida with trizonal lentelliptical
medullary shell surrounded by three crossed latticed cortical girdles, one smaller (primary)
transverse, one larger (secondary) lateral, and one (tertiary) sagittal girdle.
The genus Pylonium is the most highly developed among the Diplozonaria, with
six complete latticed girdles, three on the medullary shell, three on the cortical shell, the
latter corresponding to the former. In each of the two systems the transverse girdle is smaller
than the lateral; the sagittal girdle may be smaller or larger than the lateral girdle.
Pylonium repeats the typical form of Trizonium; but whilst in this latter the
medullary centre of the shell is a simple spherical chamber, here in Pylonium it is a true
trizonal or Larnacilla-shaped medullary shell.
Subgenus 1. Pylonissa, Haeckel.
Definition.—Cortical shell smooth or thorny, but without large,
symmetrically disposed spines.
1. Pylonium circozonium, n. sp.
Cortical shell thorny, without radial spines; all its three girdles nearly of the same size,
subcircular, very narrow, only with one to two rows of pores. Four gates subcircular.
Dimensions.—Principal axis 0.12, transverse axis 0.11; diameter of the
subspherical medullary shell 0.05.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
{655}
2. Pylonium hexazonium, n. sp.
Cortical shell smooth, without symmetrical radial spines. Transverse girdle elliptical, one and
a half times as long as broad. Lateral and sagittal girdles nearly circular. Four gates
egg-shaped.
Dimensions.—Principal axis 0.16, transverse axis 0.15; diameter of the
subspherical medullary shell 0.04.
Habitat.—South Pacific, Station 295, depth 1500 fathoms.
3. Pylonium nephropylium, n. sp.
Cortical shell thorny, without radial spines. All its three girdles elliptical, one and a third
times as long as broad. Four gates kidney-shaped, nearly twice as broad as high.
Dimensions.—Principal axis 0.17, transverse axis 0.13; length of the
lentelliptical medullary shell 0.04, breadth 0.03.
Habitat.—Pacific, central area, Station 273, depth 2350 fathoms.
Subgenus 2. Pylonura, Haeckel.
Definition.—Cortical shell armed with large, symmetrically disposed
spines.
4. Pylonium quadricorne, n. sp. (Pl. 9,
fig. 14).
Cortical shell thorny, with four strong horn-like curved spines in the lateral plane. All its
three girdles elliptical, one and a third times as long as broad. Four gates kidney-shaped, twice
as broad as high. (This species is a further developmental stage of Tetrapyle quadricornis,
with perfect sagittal girdle.)
Dimensions.—Principal axis 0.16, transverse axis 0.12; length of the medullary
shell 0.05, breadth 0.04.
Habitat.—Pacific, central area, Stations 270 to 274, surface; Indian
Ocean, Madagascar, Rabbe, surface.
5. Pylonium octacanthum, n. sp.
Cortical shell thorny, with eight long and thin cylindrical radial spines, lying in two crossed
diagonal planes, opposite in pairs, and arising from the eight points where the transverse girdle
is crossed by the lateral girdle. All three girdles elliptical, one and a half times as long as
broad; four gates triangular. (This species is a further developmental stage of Tetrapyle
octacantha, with perfect sagittal girdle.)
Dimensions.—Principal axis 0.18, transverse axis 0.13; length of the medullary
shell 0.03 to 0.04.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific, surface.
{656}
6. Pylonium stenozonium, n. sp.
Cortical shell thorny, with eight short conical radial spines, lying in two crossed diagonal
planes (as in the foregoing species). All three girdles nearly of the same size, subcircular, very
narrow, only with one to two rows of pores. Four gates kidney-shaped.
Dimensions.—Principal axis 0.15, transverse axis 0.14; diameter of the
quadrangular medullary shell 0.04.
Habitat.—Equatorial Atlantic, Station 347, surface.
Subfamily 3. Triplozonaria, Haeckel.
Definition.—Pylonida with three concentric
systems of fenestrated girdles, lying in three concentric lentelliptical faces (every system with
one to three girdles lying in one lentelliptical face).
Genus 286. Amphipylonium,[325] Haeckel,
1881, Prodromus, p. 463.
Definition.—Pylonida with trizonal lentelliptical
medullary shell, surrounded by a double latticed cortical shell; inner cortical shell
Pylonium-shaped, with three perfect crossed girdles; outer cortical shell only represented
by a single (transverse) girdle.
The genus Amphipylonium opens the series of Triplozonaria, or of those
Pylonida in which the shell is composed of three concentric systems of latticed girdles. The first
(and innermost) system represents the complete trizonal medullary shell, which is probably a
lentelliptical Larnacilla-shell. The first and intermediate system is formed of a complete
trizonal cortical shell of the same form, but much larger (like Pylonium). The third (and
outermost) system is represented by one to three latticed girdles, corresponding to the former and
forming an outer or second cortical shell. In Amphipylonium (as the most simple form of the
Triplozonaria) there is only developed the first (transverse) girdle of the third system. It
repeats therefore the form of Amphipyle, the cortical shell of which is here double
(Prodromus, 1881, p. 463).
1. Amphipylonium semilunare, n. sp.
Inner cortical shell lentelliptical, one and a half times as long as broad, with smooth surface
and four semilunar gates. Transverse girdle of the outer cortical shell with two smooth semilunar
wings, with convex, smooth lateral crests; each wing as broad as the transverse girdle of the
inner cortical shell and twice as long as its lateral girdle.
Dimensions.—Length of the lentelliptical medullary shell 0.04, breadth 0.03;
length of the inner cortical shell 0.15, breadth 0.1; length of each lateral wing of the outer
cortical shell 0.3, breadth 0.1.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
{657}
2. Amphipylonium spinosissimum, n. sp.
Inner cortical shell lentelliptical, one and a third times as long as broad, with thorny
surface and four kidney-shaped gates. Transverse girdle of the outer cortical shell very spiny,
with two semilunar wings, with convex thorny lateral crests; each wing one and a half times as
broad as the transverse girdle of the inner cortical shell, and one and a half times as long as
its lateral girdle.
Dimensions.—Length of the lentelliptical medullary shell 0.05, breadth 0.03;
length of the inner cortical shell 0.16, breadth 0.12; length of each lateral wing of the
outer cortical shell 0.24, breadth 0.18.
Habitat.—Western Tropical Pacific, Station 225, depth 4475
fathoms.
3. Amphipylonium tetraceros, n. sp.
Inner cortical shell hexagonal, one and a half times as long as broad, with smooth surface and
four transverse elliptical gates. Transverse girdle of the outer cortical shell smooth, with
rectilinear smooth lateral crests, which are prolonged at both ends into strong conical spines
(therefore four spines in the lateral plane); each wing of the same breadth as the transverse
girdle of the inner cortical shell, and twice as long as its lateral girdle.
Dimensions.—Length of the hexagonal medullary shell 0.03, breadth 0.02; length of the inner cortical shell 0.12,
breadth 0.08; length of each lateral wing of the outer cortical shell 0.24, breadth 0.08.
Habitat.—Northern Pacific, Station 253, depth 3125 fathoms.
4. Amphipylonium octoceros, n. sp.
Inner cortical shell quadrangular, one and a third times as long as broad, with thorny surface
and four kidney-shaped gates. Transverse girdle of the outer cortical shell thorny, constricted at
the equator, with concave thorny lateral crests, and with eight strong radial spines, opposite in
pairs and lying in two crossed diagonal planes; each wing narrower than the transverse girdle of
the inner cortical shell and twice as long as its lateral girdle.
Dimensions.—Length of the subspherical medullary shell 0.04, breadth 0.035; length
of the inner cortical shell 0.14, breadth 0.11; length of each lateral wing of the outer cortical
shell 0.26, breadth 0.09.
Habitat.—Southern Pacific, Station 295, depth 1500 fathoms.
Genus 267. Tetrapylonium,[326] Haeckel,
1881, Prodromus, p. 464.
Definition.—Pylonida with trizonal lentelliptical
medullary shell, surrounded by a double latticed cortical shell; inner cortical shell
Pylonium-shaped, with three perfect crossed girdles; outer cortical shell represented by
two crossed girdles, one (smaller) transverse and one (larger) lateral girdle.
{658}
The genus Tetrapylonium represents a further developmental stage of the
foregoing Amphipylonium; whilst in this latter the outer cortical shell (or the third
system of girdles) is formed only by a transverse girdle, here this is crossed by a lateral
girdle. Tetrapylonium repeats therefore the typical form of Tetrapyle, but with
doubled cortical shell (Prodromus, 1881, p. 464).
1. Tetrapylonium pantellipticum, n. sp.
Outer cortical shell elliptical, one and a third times as long as broad, with smooth surface
and four elliptical gates. Inner cortical shell elliptical, with smooth surface and four
elliptical gates which repeat the form of the four outer gates, but are of half the size.
Transverse girdle broad, with five pores on the isthmus.
Dimensions.—Length (or principal axis) of the first shell (medullary shell) 0.04,
breadth (or transverse axis) 0.03; length of the second (or inner cortical) shell 0.12, breadth
0.09; length of the third (or outer cortical) shell 0.18, breadth 0.14.
Habitat.—Pacific, central area, Station 274, depth 2750 fathoms.
2. Tetrapylonium reniforme, n. sp.
Outer cortical shell elliptical, one and a half times as long as broad, with thorny surface and
four large kidney-shaped gates. Inner cortical shell of the same form, but one-third smaller, also
with four kidney-shaped gates. Transverse girdle small, but with two pores on the isthmus.
Dimensions.—Length of the first (innermost) shell 0.03, breadth 0.02; length of
the second (middle) shell 0.14, breadth 0.08; length of the third (outermost) shell 0.2, breadth
0.11.
Habitat.—Indian Ocean, Zanzibar, Pullen, depth 2200 fathoms.
3. Tetrapylonium quadrangulare, n. sp. (Pl. 9, fig. 15).
Outer cortical shell nearly quadrangular, one and a third times as long as broad, with four
rounded corners, from which arise in the lateral plane four three-sided pyramidal radial spines,
opposite in pairs in two crossed diagonal axes. Surface thorny; four gates elliptical or nearly
quadrangular. Inner cortical shell more elliptical, by one half smaller, with broader transverse
girdle and four kidney-shaped gates. Medullary shell nearly spherical, of half the size.
Dimensions.—Length of the first shell 0.045, breadth 0.035; length of the second
shell 0.1, breadth 0.07; length of the third shell 0.16, breadth 0.11.
Habitat.—South Atlantic, off Tristan da Cunha, Station 332, depth 2200
fathoms.
4. Tetrapylonium octacanthum, n. sp.
Outer cortical shell elliptical, one and a third times as long as broad, with thorny surface,
and eight long and thin radial spines arising from the corners of the four triangular gates, and
lying {659}opposite in pairs in two crossed diagonal
planes. Inner cortical shell of the same shape, but two-thirds smaller, also thorny. Transverse
girdle broad, with four pores on the isthmus. (This species appears to be the common Tetrapyle
octacantha, with doubled cortical shell.)
Dimensions.—Length of the first shell 0.04, breadth 0.03; length of the second
shell 0.16, breadth 0.12; length of the third shell 0.24, breadth 0.18.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
5. Tetrapylonium armatum, n. sp.
Outer cortical shell nearly quadrangular, one and a fourth times as long as broad, with very
spiny surface. Between numerous smaller (simple or branched) thorns arise twelve larger radial
spines, symmetrically distributed, eight wing-spines crossed in two diagonal planes (as in
Tetrapylonium octacanthum) and four corner spines on the four rounded corners of the
lateral plane (as in Tetrapylonium quadrangulare). Therefore this species combines the
armature of both foregoing species. Four gates kidney-shaped. Inner cortical shell elliptical,
smooth, three-fourths smaller.
Dimensions.—Length of the first shell 0.045, breadth 0.035; length of the second
shell 0.18, breadth 0.15; length of the third shell 0.25, breadth 0.2.
Habitat.—Pacific, central area, Station 266, depth 2750 fathoms.
Genus 288. Pylozonium,[327] n. gen.
Definition.—Pylonida with trizonal lentelliptical
medullary shell, surrounded by double latticed cortical shell; inner cortical shell as well as the
outer Pylonium-shaped, each with three perfect crossed girdles (transverse, lateral, and
sagittal).
The genus Pylozonium is the most highly developed form of the Pylonida, as
all three systems of concentric girdles here become fully developed, each system with three
perfect girdles. In this genus only among all the Pylonida we find nine complete latticed girdles,
and in each of the three dimensive planes three concentric elliptical girdles. The first system
forms the medullary shell, the second system the inner cortical shell, and the third system the
outer cortical shell.
1. Pylozonium novemcinctum, n. sp.
Outer cortical shell lentelliptical, thorny, one and a third times as long as broad, without
radial spines, with four kidney-shaped gates. Inner cortical shell of the same shape, but
one-third smaller, four times as large as the lentelliptical medullary shell.
Dimensions.—Length of the medullary shell 0.04, breadth 0.03; length of the inner
cortical shell 0.16, breadth 0.12; length of the outer cortical shell 0.24, breadth 0.18.
Habitat.—North Pacific, Station 253, depth 3125 fathoms.
{660}
2. Pylozonium octacanthum, n. sp. (Pl. 9, fig. 16).
Outer cortical shell lentelliptical, one and a third times as long as broad, with thorny
surface and eight long and thin radial spines arising from the corners of the four elliptical
gates, and lying opposite in pairs in two crossed diagonal planes. Inner cortical shell of the
same shape but one-fourth smaller, about twice as large as the lentelliptical medullary shell.
Dimensions.—Length of the medullary shell 0.065, breadth 0.045; length of the
inner cortical shell 0.18, breadth 0.12; length of the outer cortical shell 0.24, breadth
0.18.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
Family XXVII. Tholonida, n. fam. (Pl. 10).
Definition.—Larcoidea with regular, completely latticed
cortical shell, which is composed of two to six or more hemispherical or cap-shaped domes (vaulted
chambers or cupolas). The domes lie opposite in pairs on the poles of the three dimensive axes,
are separated by annular constrictions, and surround a simple or Larnacilla-shaped central
chamber.
The family Tholonida represents a peculiar and very remarkable group
of the Larcoidea, distinguished from the other groups of this suborder by
the characteristic form of the shell, composed of a variable number of hemispherical domes or
cupolas. The middle and original part of the shell is constantly formed of an elliptical or
subspherical central chamber, which often, but not constantly, contains a small medullary shell.
An even number (two, four, six, or more) of domes is attached to the poles of the three dimensive
axes of the central chamber; according as only one, or two, or all three axes develop cupolas, we
distinguish in this family
three different subfamilies (the Amphitholida, Staurotholida, and Cubotholida). The Amphitholida
(or Tholonida monaxonia) form cupolas only on the two poles of one single axis, and this axis
corresponds to the minor (or transverse) axis of the central chamber, we find here therefore
constantly at least two lateral cupolas (Pl. 10,
figs. 1-7). The Staurotholida (or Tholonida diaxonia) form cupolas on the poles of two axes
perpendicular one to another; these two axes are the major (principal) and the minor (transverse)
axis of the central chamber, we find here therefore constantly at least four cupolas crossed in
pairs (Pl. 10,
figs. 8-11). The Cubotholida (or Tholonida triaxonia) form cupolas on the poles of all three
dimensive axes (perpendicular one to another); corresponding to the principal, transverse, and
sagittal axes of the lentelliptical central chamber; therefore we find here constantly at least
six cupolas, attached in pairs on the six sides of the central chamber (Pl. 10, figs.
12-17).
The number of genera and species in the family Tholonida is rather
large, but the number of individuals is much smaller than in most of the other Spumellaria, and {661}particularly
than in the nearly allied Pylonida; the greater number of the species are very rare. Besides this
they seem to be very variable and inclined to produce numerous abnormalities. Very often
intermediate forms of transition are to be found between the Tholonida and other Larcoidea, particularly the Pylonida and Lithelida. In many species the
thick-walled shell is very opaque, and offers great difficulties to the clear study; in the
greater number the structure of the shell cannot be understood completely without rolling the
shell to the different sides; and the distinction between the Staurotholida and Cubotholida is
often very difficult.
The primordial chamber of the Tholonida, or the central chamber in which its growth begins, is
either a simple lentelliptical shell (without enclosed medullary shell), like Cenolarcus,
or it is a trizonal shell, like Larnacilla, and contains a small concentric medullary
shell. But this important inner shell of the central chamber offers peculiar difficulties for
study. In many cases (probably in the greater number) there is no doubt the same characteristic
trizonal medullary shell, which we found in the Larnacida and Pylonida, and this is our principal
argument, if we regard the Tholonida as Larcoidea, which are most nearly
allied to both these families, and in which the fenestrated open cortical girdles of the Pylonida
are replaced by fenestrated closed cupolas or domes; the characteristic "gates" or large fissures
in the cortical shell of the former are therefore here perfectly closed by network. The Tholonida
agree in this point with the Larnacida, but are distinguished from them by the prominent vaultings
of the hemispherical cupolas or domes, which give them a peculiar appearance. Each pair of domes
(opposite on both poles of one dimensive axis) corresponds to one single girdle of the
Pylonida.
Regarding the absence or presence of a medullary shell in the central chamber, we may divide
the Tholonida into two groups, Cenotholida (without medullary shell) and Coccotholida (with
medullary shell). The absence of the medullary shell in many Tholonida may be either primary
(original) or secondary (by reduction and loss of it). Probably in the larger proportion (if not
always?) its absence is the consequence of reduction and loss, and in this case the Cenotholida
must be derived phylogenetically from the Coccotholida, but possibly often (or always?) the
contrary may also be the case. At present we cannot find certain arguments for one opinion or the
other.
The connection between the medullary (internal) and the cortical (external) shell of the
central chamber in the Coccotholida is effected by two lateral wings of a latticed transverse
girdle; on both sides of the latter remain the four internal open "gates" of the Pylonida and
Larnacida. Very often the limiting edges of these four gates are prolonged into eight external
radial spines which lie opposite in pairs in two crossed diagonal planes, and correspond to the
eight portal-spines of Tetrapyle octacantha. Besides these, we often find four other beams
opposite in pairs in the two crossed axes (principal and lateral). Very often also these radial
beams (between medullary and {662}cortical shells) are
prolonged on the outside into prominent free radial spines. But other spines may also arise from
the surface. Very remarkable is the presence of twenty symmetrically disposed radial spines in
some forms of Amphitholida (Amphitholus acanthometra and Amphitholonium acanthonium,
&c.). It recalls the twenty radial spines of the Acanthonida, though
the laws of distribution in the two similar cases are quite different.
The cortical shell of the Tholonida remains either simple, or it becomes doubled by formation
of an outer veil or envelope. This outer cortical shell or "veil" may either repeat completely the
typical form of the inner with its cupolas, or it may form an ellipsoidal envelope without
dome-shaped partitions. Its network is either like that of the inner, or it is very delicate and
irregular, spider-web like. The connection between the two cortical shells is effected by a
variable number of radial beams, often prolonged on the outside into radial spines.
The growth of the Tholonida by apposition of new chambers is very characteristic, constantly
pairs of chambers originating at both opposite poles of the three dimensive axes, firstly on the
transverse axis, secondly on the principal axis, thirdly on the sagittal axis. It is possible that
by repeated apposition of new chambers the Tholonida are transformed into Discoidea, but commonly the number of domes is restricted, two in the
Amphitholida, four in the Staurotholida, six in the Coccotholida. If new chambers in an irregular
manner or in a spiral order be opposed to those first formed pairs of chambers, the Tholonida may
be transformed into Soreumida, Streblemida, or Lithelida. The phylogenetic connection between
these families is probably very complex and very worthy of further research.
The network of the shell in the Tholonida is sometimes regular, with circular (often
hexagonally framed) pores of equal size, sometimes irregular, with roundish pores of unequal size.
Commonly the shell is thick and compact; in those genera in which the cortical shell is doubled,
the inner is commonly compact, the outer a very delicate veil.
The central capsule of the Tholonida is originally always enclosed by the central chamber, and
like this of lentelliptical form, a triaxial ellipsoid. With the apposition of cupolas the growing
central capsule may form dome-shaped protuberances which enter into the former. So we may find in
the Amphitholida a central capsule with three joints (separated by two annular parallel
constrictions), in the Staurotholida a cross-shaped capsule (with four caps around the central
lentellipsis), in the Cubotholida a clustered capsule with six caps, surrounding six sides of the
cubical central mass. In the Coccotholida (with medullary shell in the central chamber) the latter
encloses in the centre the nucleus of the cell. The calymma, or the jelly-veil between the central
capsule and the enveloping shell, probably always exhibits annular constrictions, corresponding to
those which separate the cupolas of the shell. All these anatomical peculiarities in the Tholonida
require a further accurate study.
{663}
Synopsis of the Genera of Tholonida.
|
III. Subfamily Amphitholida.
Cupolas developed only in the direction of one single axis (commonly two
cupolas). (Tholonida unidimensiva.)
|
|
Central chamber simple, without medullary shell. |
|
Two simple cupolas (without veil), |
289. Tholartus. |
| Two double cupolas (with veil), |
290. Tholodes. |
| Central chamber Larnacilla-shaped, with enclosed
medullary shell. |
|
Two simple cupolas (without veil), |
291. Amphitholus. |
| Two double cupolas (with veil), |
292. Amphitholonium. |
|
III. Subfamily Staurotholida.
Cupolas developed in the direction of two axes perpendicular one to another
(commonly four cupolas). (Tholonida bidimensiva.)
|
|
Central chamber simple, without medullary shell. |
|
Four simple cupolas (without veil), |
293. Tholostaurus. |
| Four double cupolas (with veil), |
294. Tholoma. |
| Central chamber Larnacilla-shaped, with enclosed
medullary shell. |
|
Four simple cupolas (without veil), |
295. Staurotholus. |
| Four double cupolas (with veil), |
296. Staurotholonium. |
|
III. Subfamily Cubotholida.
Cupolas developed in the direction of three axes perpendicular one to another
(commonly six cupolas). (Tholonida tridimensiva.)
|
|
Central chamber simple, without medullary shell. |
|
Six simple cupolas (without veil), |
297. Tholocubus. |
| Six double cupolas (with veil), |
298. Tholonium. |
| Central chamber Larnacilla-shaped, with enclosed
medullary shell. |
|
Six simple cupolas (without veil, |
299. Cubotholus. |
| Six double cupolas (with veil), |
300. Cubotholonium. |
| I. Subfamily Amphitholida. Cupolas developed only in the direction of one
single axis (commonly two cupolas). (Tholonida unidimensiva.) |
|
|
Central chamber simple, without medullary shell. |
|
|
|
|
|
|
Two simple cupolas (without veil), |
|
|
|
|
|
|
|
|
|
|
289. Tholartus. |
|
|
|
|
|
|
|
|
|
|
Two double cupolas (with veil), |
|
|
|
|
|
|
|
|
|
|
290. Tholodes. |
|
|
|
|
|
|
|
|
Central chamber Larnacilla-shaped, with enclosed medullary
shell. |
|
|
|
|
|
|
Two simple cupolas (without veil), |
|
|
|
|
|
|
|
|
|
|
291. Amphitholus. |
|
|
|
|
|
|
|
|
|
|
Two double cupolas (with veil), |
|
|
|
|
|
|
|
|
|
|
292. Amphitholonium. |
|
|
|
|
|
|
| II. Subfamily Staurotholida. Cupolas developed in the direction of two axes
perpendicular one to another (commonly four cupolas). (Tholonida bidimensiva.) |
|
|
Central chamber simple, without medullary shell. |
|
|
|
|
|
|
Four simple cupolas (without veil), |
|
|
|
|
|
|
|
|
|
|
293. Tholostaurus. |
|
|
|
|
|
|
|
|
|
|
Four double cupolas (with veil), |
|
|
|
|
|
|
|
|
|
|
294. Tholoma. |
|
|
|
|
|
|
|
|
Central chamber Larnacilla-shaped, with enclosed medullary
shell. |
|
|
|
|
|
|
Four simple cupolas (without veil), |
|
|
|
|
|
|
|
|
|
|
295. Staurotholus. |
|
|
|
|
|
|
|
|
|
|
Four double cupolas (with veil), |
|
|
|
|
|
|
|
|
|
|
296. Staurotholonium. |
|
|
|
|
|
|
| III. Subfamily Cubotholida. Cupolas developed in the direction of three axes
perpendicular one to another (commonly six cupolas). (Tholonida tridimensiva.) |
|
|
Central chamber simple, without medullary shell. |
|
|
|
|
|
|
Six simple cupolas (without veil), |
|
|
|
|
|
|
|
|
|
|
297. Tholocubus. |
|
|
|
|
|
|
|
|
|
|
Six double cupolas (with veil), |
|
|
|
|
|
|
|
|
|
|
298. Tholonium. |
|
|
|
|
|
|
|
|
Central chamber Larnacilla-shaped, with enclosed medullary
shell. |
|
|
|
|
|
|
Six simple cupolas (without veil, |
|
|
|
|
|
|
|
|
|
|
299. Cubotholus. |
|
|
|
|
|
|
|
|
|
|
Six double cupolas (with veil), |
|
|
|
|
|
|
|
|
|
|
300. Cubotholonium. |
|
|
|
|
|
|
Subfamily 1. Amphitholida, Haeckel.
Definition.—Tholonida with monaxial growth;
cupolas opposite on the poles of one axis. (Shell commonly three-chambered, with two domes on both
sides of the central chamber.)
Genus 289. Tholartus,[328] n. gen.
Definition.—Tholonida with simple cortical shell (without
external veil); with two hemispherical cupolas, opposite on the poles of one axis; central chamber
between them simple (without medullary shell).
{664}
The genus Tholartus (Pl. 10,
fig. 1) represents the most simple form of all Tholonida, a three-jointed simple cortical shell
without medullary shell; two hemispherical cupolas or domes are attached on both lateral sides of
a simple triaxial ellipsoidal central chamber, corresponding to the poles of its lateral or
transverse axis. Tholartus may be regarded as the common ancestral form of all Tholonida,
and may be derived either from the Pylonida Monozonium (by complete lattice-locking of the
lateral wings) or from the Ellipsida Cenellipsis (by development of two lateral
protuberances on the poles of the transverse axis). But it is also possible that the absence of
the medullary shell is produced by reduction, and in this case Tholartus may be descended
from Amphitholus.
Subgenus 1. Tholartella, Haeckel.
Definition.—Surface of the shell smooth or rough, without radial
spines.
1. Tholartus tricolus, n. sp. (Pl. 10,
fig. 1).
Central chamber one and a half times as high and as broad as both cupolas. Surface of the shell
rough. Pores regular, circular, without hexagonal frames, three times as broad as the bars; about
sixteen on the half meridian of the central chamber (or its vertical diameter).
Dimensions.—Length of the shell (major axis of the central chamber, vertical) 0.1,
breadth of the shell (major axis of the whole three-chambered shell, horizontal) 0.13; pores 0.01,
bars 0.003.
Habitat.—Pacific, central area, Station 270, depth 2925 fathoms.
2. Tholartus paniscus, n. sp.
Central chamber of the same breadth, but of the double height of both cupolas. Surface of the
shell rough. Pores regular, circular, with hexagonal frames, of the same breadth as the bars;
eight to ten on the half meridian of the central chamber.
Dimensions.—Length of the shell (major axis of the central chamber, vertical)
0.12, breadth of the shell (major axis of the whole three-chambered shell, horizontal) 0.15; pores
and bars 0.008.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
3. Tholartus isocolus, n. sp.
Central chamber of the same breadth and height as both cupolas. Surface of the shell smooth.
Pores irregular, roundish, once to three times as broad as the bars; ten to fifteen on the half
meridian of the central chamber.
Dimensions.—Length of the shell 0.11, breadth 0.14; pores 0.004 to 0.01, bars
0.003.
Habitat.—Pacific, central area, Station 270, depth 2925 fathoms.
{665}
Subgenus 2. Tholartissa, Haeckel.
Definition.—Surface of the shell with radial spines.
4. Tholartus tripanis, n. sp.
Central chamber of the same size as both cupolas. Surface of the shell thorny, everywhere
covered with short conical radial spines, about as long as the cupolas. Pores regular or
subregular, circular, twice as broad as the bars; eight to ten on the half meridian.
Dimensions.—Length of the shell 0.1, breadth 0.12; pores 0.01, bars 0.005.
Habitat.—South Pacific, Station 295, surface.
5. Tholartus sagitta, n. sp.
Central chamber twice as large as both cupolas. Surface of the shell spiny, with eight to
twelve regularly (?) distributed radial spines, radiating from the two constrictions between the
three chambers. Spines needle-shaped, very thin and long. Pores subregular or irregular, roundish
or circular, about three times as broad as the bars; twelve to sixteen on the half meridian.
Dimensions.—Length of the shell 0.11, breadth 0.15; pores 0.01, bars 0.003.
Habitat.—North Pacific, Station 244, surface.
Genus 290. Tholodes,[329] n. gen.
Definition.—Tholonida with double cortical shell (with
external veil), with two hemispherical cupolas, opposite on the poles of one axis; central chamber
between them simple (without medullary shell).
The genus Tholodes (Pl. 10,
fig. 2) differs from Tholartus,
its probable ancestral form, only in the duplication of the shell; the outer shell has exactly the
same three-jointed form as the inner; both are connected by eight radial beams, lying in two
diagonal planes and corresponding to the eight frontal spines of Tetrapyle octacantha.
Possibly Tholodes may also be descended from Amphitholonium by loss of the medullary
shell. I have observed only one single specimen of this genus.
1. Tholodes cupula, n. sp. (Pl. 10,
fig. 2).
Outer shell of the same form and structure as the inner shell. Central chamber larger than both
cupolas. Surface a little rough, without radial spines. Distance of both shells equals the height
of the outer cupolas. Pores of both shells regular, circular, three times as broad as the {666}bars; six to seven in the semicircle of one inner cupola,
ten to twelve in the semicircle of one outer cupola. Both shells are connected by eight radial
beams, crossed in two diagonal planes.
Dimensions.—Major axis of the outer shell 0.1, minor 0.065; major axis of the
inner shell 0.05, minor 0.04; pores 0.006, bars 0.002.
Habitat.—Western Tropical Pacific, Station 225, depth 4475
fathoms.
Genus 291. Amphitholus,[330] n. gen.
Definition.—Tholonida with simple cortical shell (without
external veil), with two hemispherical cupolas, opposite on the poles of one axis; central chamber
(between them) Larnacilla-shaped, with medullary shell.
The genus Amphitholus is the most simple form of all Coccotholida (or all
Tholonida provided with medullary shell). Its simple cortical shell exhibits, like
Tholartus, two lateral hemispheric cupolas, attached on both sides of a lentelliptical central chamber
(on the poles of its transverse axis). The central chamber has the form of a triaxial ellipsoid,
and contains, like Larnacilla, a small medullary shell, connected with it by the wings of a
latticed transverse girdle. Amphitholus may be derived either from Tholartus by
secondary production of a cortical shell, or (more probably) from Larnacilla by apposition
of two lateral dome-shaped protuberances. In the latter case it may be regarded as an
Amphipyle, the lateral open wings of which are closed by lattice-work.
Subgenus 1. Amphitholissa, Haeckel.
Definition.—Surface of the shell smooth or rough, without radial
spines.
1. Amphitholus artiscus, n. sp. (Pl. 10,
fig. 3).
Central chamber of the same height as both cupolas. Surface of the cortical shell rough. Pores
regular, circular, with prominent hexagonal frames, twice as broad as the bars; eight to ten in
the basal semicircle of one cupola.
Dimensions.—Major axis of the shell 0.16, minor axis 0.1; pores 0.008, bars
0.004.
Habitat.—Pacific, central area, Station 272, depth 2600 fathoms.
2. Amphitholus artidium, n. sp.
Central chamber one-fourth higher than both cupolas. Surface of the cortical shell smooth.
Pores irregular, roundish, once to three times as broad as the bars. (Very similar to
Tholartus tricolus, Pl. 10,
fig. 1, but different by the medullary shell.)
Dimensions.—Major axis of the shell 0.15, minor axis 0.12; pores 0.004 to 0.01,
bars 0.003.
Habitat.—North Pacific, Station 241, depth 2300 fathoms.
{667}
Subgenus 2. Amphitholura, Haeckel.
Definition.—Surface of the shell with radial spines or thorns.
3. Amphitholus octacanthus, n. sp.
Central chamber about twice as high and broad as both cupolas. From the surface arise,
symmetrically distributed, eight stout conical radial spines, lying in two diagonal planes, as
prolongations of the eight wing-beams, which connect the Tetrapyle-shaped medullary shell
with the two ring-like constrictions of the cortical shell. Pores of the latter regular, circular,
twice as broad as the bars; ten to twelve in the basal semicircle of one cupola.
Dimensions.—Major axis of the shell 0.15, minor 0.12; pores 0.01, bars 0.005.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
4. Amphitholus dodecanthus, n. sp.
Central chamber a little larger than both cupolas. From the surface arise twelve strong conical
radial spines, four on the poles of the major and the minor axis of the shell, lying in the
lateral plane; eight others lying in two diagonal planes, as prolongations of the eight
wing-beams, which connect the Tetrapyle-shaped medullary shell with the two ring-like
constrictions of the cortical shell. Pores of the latter irregular, roundish, twice to four times
as broad as the bars; sixteen to eighteen in the basal semicircle of one cupola.
Dimensions.—Major axis of the shell 0.15, minor 0.11; pores 0.004 to 0.008, bars
0.002.
Habitat.—North Pacific, Station 256, surface.
5. Amphitholus acanthometra, n. sp. (Pl. 10, figs. 5,
6).
Central chamber about twice as broad and as high as both cupolas. On the surface quite
symmetrically distributed twenty long and strong cylindrical radial spines; eight in the sagittal
plane or in the meridian plane of the central chamber (four opposite in pairs in the principal and
the sagittal axes, four others in the middle between the principal and the sagittal spines); four
in the horizontal transverse plane, on both sides of the poles of the major or lateral axis; and
eight in diagonal planes (corresponding to the eight wing-spines of Tetrapyle octacantha).
In the centre of the central chamber is a distinct trizonal medullary shell like that of
Tetrapyle, with two vertical columella beams. Pores of the cortical shell subregular,
circular, with elevated hexagonal frames, about four times as broad as the bars; five to seven in
the semicircle of one cupola. (This remarkable species differs from Tholartus vicenus in
the possession of a medullary shell, from Amphitholonium acanthometra by the simple
cortical shell. Compare these species).
Dimensions.—Major axis of the shell 0.14 to 0.16, minor axis 0.10 to 0.12; pores
0.015, bars 0.004; major axis of the medullary shell 0.04 to 0.05, minor (horizontal) axis 0.02 to
0.03; length of the twenty radial spines 0.18 to 0.24, basal breadth 0.01.
Habitat.—South Pacific, Station 300, depth 1375 fathoms.
{668}
6. Amphitholus armatus, n. sp.
Central chamber twice as broad, but of the same height, as the flat vaulted cupolas. Surface
armed with thirty to forty very large three-sided prismatic spines, longer than the major axis of
the shell. Pores irregular, roundish, twice to five times as broad as the bars; eight to nine on
the basal semicircle of one cupola.
Dimensions.—Major axis of the shell 0.15, minor axis 0.1; pores 0.006 to 0.015,
bars 0.003; length of the spines 0.2 to 0.25, breadth 0.01.
Habitat.—North Atlantic, Station 353, surface.
7. Amphitholus polyacanthus, n. sp.
Central chamber of the same breadth and height as both cupolas. Surface armed with very
numerous (sixty to eighty) strong conical spines, about as long as the minor axis of the shell.
Pores subregular, circular, twice as broad as the bars; sixteen to eighteen in the basal
semicircle of one cupola.
Dimensions.—Major axis of the shell 0.13, minor 0.08; pores 0.006, bars 0.003;
length of the radial spines 0.09, breadth 0.006.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
8. Amphitholus panicium, n. sp. (Pl. 10,
fig. 4).
Central chamber ellipsoidal, nearly twice as broad, but of the same height, as both cupolas.
Whole surface thorny; the largest radial thorns about as long as the medullary shell. Pores
irregular, roundish, twice to four times as broad as the bars. On the base of each cupola a circle
of ten to twelve larger square pores, separated by radial beams, which are prolonged into stouter
free spines. This remarkable formation of the cupolas, very rare in this family, recalls the
characteristic formation of the distal chambers of the Panartida and Zygartida (e.g., Pl.
40, figs. 4,
8, &c.), where it is very common.
Dimensions.—Major axis of the shell 0.15, minor 0.1; pores 0.005 to 0.012, bars
0.003; large square pores 0.02; axes of the medullary shell 0.03 and 0.02.
Habitat.—Indian Ocean, Madagascar, Rabbe, surface.
Genus 292. Amphitholonium,[331] n.
gen.
Definition.—Tholonida with double cortical shell (with
external veil), with two hemispherical cupolas, opposite on the poles of one axis; central chamber
(between them) Larnacilla-shaped, with medullary shell.
The genus Amphitholonium differs from the nearly allied Amphitholus
(probably its ancestral form) only in the duplication of the cortical shell; the outer has the
{669}same three-jointed form as the inner, and is
connected with it by a variable number of radial beams. From the similar Tholodes it
differs in the possession of a medullary shell.
1. Amphitholonium tricolonium, n. sp. (Pl. 10, fig.
7).
Outer cortical shell smooth, of the same three-jointed form as the inner, at an equal distance
from it throughout the whole circumference; central chamber higher-vaulted than both cupolas.
Network of the outer shell delicate, with subregular, circular pores. Pores of the thick-walled
inner shell subregular, circular, with hexagonal frames, three times as broad as the bars; eight
to ten in the basal semicircle of one cupola.
Dimensions.—Major axis of the outer cortical shell 0.2, minor axis 0.15; major
axis of the inner cortical shell 0.16, minor axis 0.11; pores 0.01, bars 0.0035.
Habitat.—South Pacific, Station 302, depth 1450 fathoms.
2. Amphitholonium octostylium, n. sp.
Outer cortical shell lentelliptical,
smooth, not articulated, at a varying distance from the three-jointed inner shell; central chamber
of the latter twice as high as both hemispherical cupolas. Network of the outer shell very thin,
cob-web like. Pores of the inner shell subregular, roundish, twice as broad as the bars; six to
seven in the semicircle of one cupola. Eight thin and long, bristle-shaped, radial spines,
opposite in pairs in two crossed diagonal planes.
Dimensions.—Major axis of the outer cortical shell 0.16, minor 0.12; major axis of
the inner cortical shell 0.12, minor 0.09; pores 0.01, bars 0.005.
Habitat.—Pacific, central area, Station 273, depth 2350 fathoms.
3. Amphitholonium acanthonium, n. sp.
Outer cortical shell of the same three-jointed form as the inner, at a uniform distance from
it; central chamber higher vaulted than both cupolas. Network of the outer shell irregular,
delicate, of the inner regular, strong, with circular, hexagonally framed pores, three times as
broad as the bars; eight to nine in the semicircle of one cupola. On the surface, quite
symmetrically disposed, twenty long and strong, cylindrical, radial spines; eight in the sagittal
plane, four in the transverse plane; eight in two diagonal planes between the former and the
latter. The remarkable geometric disposition of the twenty spines is in this species quite the
same as in Amphitholus acanthometra. It differs from this nearly allied species in the
double cortical shell.
Dimensions.—Major axis of the outer cortical shell 0.18, minor 0.14; major axis of
the inner cortical shell 0.14, minor 0.1; pores 0.012, bars 0.004.
Habitat.—South Pacific, Station 295, surface.
{670}
Subfamily 2. Staurotholida, Haeckel.
Definition.—Tholonida with diaxial growth;
cupolas crossed in pairs, opposite on the poles of two axes, perpendicular one to another. (Shell
commonly five-chambered, with four domes, cross-wise surrounding the central chamber.)
Genus 293. Tholostaurus,[332] n.
gen.
Definition.—Tholonida with simple cortical shell (without
external veil), composed of four hemispherical cupolas in cross-form, opposite in pairs on the
poles of two axes perpendicular one to another; central chamber simple (without medullary
shell).
The genus Tholostaurus is the most simple form of the Staurotholida, or the
Tholonida with four crossed hemispherical cupolas, lying on the poles of two axes perpendicular
one to another. The central chamber communicates by four wide openings with the four domes, and
contains no medullary shell. Tholostaurus may originate either from Tholartus by
apposition of two opposite cupolas between the first pair, or from Staurotholus by loss of
the medullary shell.
Subgenus 1. Tholostaurantha, Haeckel.
Definition.—Surface of the shell smooth or rough, without radial
spines.
1. Tholostaurus quadrigatus, n. sp.
All four cupolas nearly of the same size and form, subregular. Surface smooth, without radial
spines. Pores subregular, circular, four times as broad as the bars; eight to ten pores in the
basal semicircle of one cupola.
Dimensions.—Diameter of the shell 0.12; pores 0.008, bars 0.002.
Habitat.—Pacific, central area, Station 272, surface.
2. Tholostaurus cruciformis, n. sp.
Two opposite cupolas larger than the two others. Surface rough, without radial spines. Pores
regular, circular, hexagonally framed, twice as broad as the bars; twelve to fourteen in the basal
semicircle of one cupola.
Dimensions.—Major axis of the shell 0.16, minor axis 0.12; pores 0.006, bars
0.003.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms.
{671}
Subgenus 2. Tholostauroma, Haeckel.
Definition.—Surface of the shell with radial spines or thorns.
3. Tholostaurus tetrabelonis, n. sp.
All four cupolas nearly of the same size and form, subregular. Pores regular, circular,
hexagonally framed, twice as broad as the bars; twelve to fourteen in the semicircle of one
cupola. Four long and stout conical radial spines, two vertical on the poles of the principal
axis, two horizontal on the poles of the transverse axis.
Dimensions.—Major axis of the shell 0.15, minor 0.13; pores 0.006, bars 0.003.
Habitat.—South Pacific, Station 288, surface.
4. Tholostaurus octobelonis, n. sp.
Two opposite cupolas larger than the two others. Pores subregular, circular, three times as
broad as the bars; ten to twelve in the semicircle of one cupola. Eight long and thin,
needle-shaped, radial spines, in pairs crossed in two diagonal planes.
Dimensions.—Major axis of the shell 0.16, minor 0.12; pores 0.01, bars 0.003.
Habitat.—South Pacific, Station 291, surface.
5. Tholostaurus dodecabelos, n. sp.
Two opposite cupolas larger than the two others. Pores irregular, roundish, twice to three
times as broad as the bars; six to eight in the semicircle of one cupola. Twelve thin and long
radial spines, four on the poles of the two crossed axes (principal and lateral), eight crossed in
two diagonal planes.
Dimensions.—Major axis of the shell 0.14, minor 0.11; pores 0.006 to 0.01, bars
0.003.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
6. Tholostaurus polybelonis, n. sp.
Two opposite cupolas larger than the two others. Pores circular, hexagonally framed, of the
same breadth as the bars; ten to twelve in the semicircle of one cupola. Numerous (twenty to
thirty or more) thin, bristle-like, radial spines, about as long as the radius of the shell.
Dimensions.—Major axis of the shell 0.15, minor axis 0.12; pores and bars
0.007.
Habitat.—Indian Ocean, Zanzibar, Pullen, depth 2200 fathoms.
Genus 294. Tholoma,[333] n. gen.
Definition.—Tholonida with double cortical shell (with
external veil), composed of four hemispherical cupolas in cross-form, opposite in pairs on the
poles of two axes perpendicular one to another; central chamber simple (without medullary
shell).
{672}
The genus Tholoma (Pl. 10,
figs. 10, 13) differs from the preceding Tholostaurus (its probable ancestral form) only in
the duplication of the shell. The outer (secondary) shell has the same cross-form as the inner
(primary) shell. In the two observed species the growth seems to be different, in the first
species all four cupolas of each cross being of the same size, form, and age; in the second two
opposite cupolas, larger and apparently older than the other two. Tholoma is possibly the
offspring of Staurotholonium, from which it may have been produced by loss of the medullary
shell.
Subgenus 1. Tholomantha, Haeckel.
Definition.—Surface of the shell smooth, without radial spines.
1. Tholoma quadrigeminum, n. sp. (Pl. 10, fig.
10).
All four chambers in each cortical shell nearly equal, subregular. Surface smooth, without
radial spines. Structure of the network in both shells similar. Pores regular, circular, twice as
broad as the bars; six to eight in the basal semicircle of one inner, ten to twelve in the
semicircle of one outer cupola.
Dimensions.—Diameter of the outer shell 0.14, of the inner 0.09; pores 0.006, bars
0.003.
Habitat.—South Pacific, Station 302, surface.
Subgenus 2. Tholomura, Haeckel.
Definition.—Surface of the shell with radial spines.
2. Tholoma metallasson,[334] n. sp. (Pl.
10, fig.
13).
Two opposite chambers in each cortical shell larger than the two others; the larger chambers of
the inner shell corresponding to the smaller chambers of the outer shell. Network in both shells
of the same structure, regular, with circular, hexagonally framed pores of the same breadth as the
bars; eight to ten in the semicircle of an inner, fourteen to sixteen in the semicircle of an
outer cupola. Numerous (twenty to thirty or more) thin, cylindrical, bristle-shaped, long radial
spines, symmetrically disposed.
Dimensions.—Major axis of the outer shell 0.26, minor axis 0.2; major axis of the
inner shell 0.16, minor axis 0.13; pores and bars 0.006.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms.
Genus 295. Staurotholus,[335] n.
gen.
Definition.—Tholonida with simple cortical shell (without
external veil), composed of four hemispherical cupolas in cross-form, opposite in pairs on the
poles of {673}two axes perpendicular one to another;
central chamber Larnacilla-shaped (with enclosed medullary shell).
The genus Staurotholus differs from Tholostaurus in the possession of
a medullary shell in the central chamber, and may be derived from this genus by its production.
But it may also be derived from Amphitholus by apposition of two secondary opposite cupolas
between the two primary cupolas. The symmetrical position of an increasing number of radial spines
in the different species is remarkable (resembling Tholostaurus as well as
Amphitholus).
Subgenus 1. Staurotholissa, Haeckel.
Definition.—Surface of the shell smooth or rough, without radial
spines.
1. Staurotholus quadratus, n. sp.
Surface of the cortical shell smooth. All four cupolas nearly of the same size and form;
therefore principal and lateral axes equal. Pores regular, circular, with hexagonal frames, twice
as broad as the bars; eight to ten on the basal semicircle of one cupola. Medullary shell
square.
Dimensions.—Diameter of the cortical shell 0.15; pores 0.01, bars 0.005.
Habitat.—South Atlantic, Station 323, depth 1900 fathoms.
2. Staurotholus cruciatus, n. sp.
Surface of the cortical shell rough. The two principal cupolas somewhat larger than the two
lateral cupolas; therefore the longitudinal axis longer than the transverse. Pores irregular,
roundish, twice to three times as broad as the bars; twelve to sixteen in the semicircle of one
cupola.
Dimensions.—Major axis of the shell 0.16, minor 0.13; pores 0.005 to 0.009, bars
0.003.
Habitat.—Pacific, central area, Station 266, depth 2750 fathoms.
Subgenus 2. Staurotholura, Haeckel.
Definition.—Surface of the shell with radial spines.
3. Staurotholus tetrastylus, n. sp. (Pl. 10, fig.
8).
The two principal cupolas smaller than the two lateral cupolas. Pores subregular, circular,
three times as broad as the bars; six to eight in the semicircle of one cupola. Medullary shell
elliptical. On the surface four long cylindrical radial spines; two in the principal and two in
the lateral axis.
Dimensions.—Major axis of the cortical shell 0.15, minor 0.12; pores 0.012, bars
0.004.
Habitat.—South Pacific, Station 166, surface.
{674}
4. Staurotholus octostylus, n. sp.
The two lateral cupolas larger than the two principal cupolas. Pores subregular, circular,
twice as broad as the bars; ten to twelve in the semicircle of one cupola. Medullary shell
hexagonal, connected with each ring-like constriction by eight strong cylindrical radial spines
(longer than the whole shell), lying in two crossed meridian planes, and corresponding to the
eight diagonal wing-spines of Tetrapyle octacantha.
Dimensions.—Major axis of the cortical shell 0.12, minor 0.1; pores 0.008, bars
0.004.
Habitat.—Pacific, central area, Station 274, depth 2750 fathoms.
5. Staurotholus decastylus, n. sp.
The two principal cupolas twice as broad and high as the two lateral cupolas. Pores subregular,
circular, four times as broad as the bars; six to eight in the semicircle of one cupola. Ten long
and thin radial spines with thickened conical bases, about as long as the major axis of the shell,
by inner prolongations connected with the lentelliptical medullary shell; two polar spines in the
principal axis, eight wing-spines in two crossed diagonal planes.
Dimensions.—Major axis 0.12, minor 0.1; pores 0.012, bars 0.003.
Habitat.—Western Tropical Pacific; Station 224, depth 1850
fathoms.
6. Staurotholus dodecastylus, n. sp. (Pl. 10, fig.
9).
The two lateral cupolas larger than the two principal cupolas. Pores subregular, circular,
three times as broad as the bars; five to seven in the semicircle of one cupola. Twelve strong
cylindrical radial spines, four on the poles of the two larger axes (two principal and two
lateral), eight wing-spines in two crossed diagonal planes. Medullary shell hexagonal.
Dimensions.—Major (lateral) axis 0.11, minor (principal) 0.09; pores 0.009, bars
0.003.
Habitat.—Tropical Atlantic, Station 348, surface.
7. Staurotholus polystylus, n. sp.
The two principal cupolas larger than the two lateral cupolas. Pores irregular, roundish, twice
to three times as broad as the bars; ten to twelve in the semicircle of one cupola. On the surface
numerous thin bristle-shaped spines (twenty to thirty or more).
Dimensions.—Major axis 0.14, minor 0.11; pores 0.006 to 0.01, bars 0.003.
Habitat.—North Pacific, Station 231, surface.
Genus 296. Staurotholonium,[336] n.
gen.
Definition.—Tholonida with double cortical shell (with
external veil), composed of four hemispherical cupolas in cross-form, opposite in pairs on the
poles of two axes perpendicular one to another; central chamber Larnacilla-shaped (with
medullary shell).
{675}
The genus Staurotholonium differs from the nearly allied Staurotholus
(its probable ancestral form) only in the duplication of the cortical shell. The outer shell
commonly repeats the cross-form of the inner, with four corresponding cupolas; but sometimes the
four cupolas of the outer shell alternate in size and form with those of the inner, or the outer
shell forms a simple lenticular envelope around the inner. From the similar Tholoma,
Staurotholonium differs in the possession of a medullary shell in the central chamber.
Subgenus 1. Staurotholodes, Haeckel.
Definition.—Surface of the outer cortical shell smooth or rough,
without radial spines.
1. Staurotholonium biquadratum, n. sp.
Outer cortical shell smooth, of the same regular crucial form as the inner, equidistant from it
everywhere. Form and structure of both cortical shells nearly the same, but the outer about twice
as large as the inner; both connected only by eight diagonal beams. All four cupolas of each
cortical shell regular, of the same size. Pores regular, circular, twice as broad as the bars;
six to eight in the basal semicircle of each cupola. Medullary shell square. (Similar to
Staurotholus quadratus, but differs by the double cortical shell.)
Dimensions.—Diameter of the outer cortical shell 0.16, of the inner 0.08, of the
medullary shell 0.04; pores of the inner cortical shell 0.006, bars 0.003.
Habitat.—South Pacific, Station 295, depth 1500 fathoms.
2. Staurotholonium bicruciatum, n. sp.
Outer cortical shell rough, twice as large as the inner, of the same form and structure, at an
equal distance from it. In both shells the principal cupolas are larger than the lateral. Pores
irregular, roundish, in the outer shell four times, in the inner twice as large as the bars; eight
to ten in the basal semicircle of one cupola. (Similar to Staurotholus cruciatus, but
differs mainly in the double cortical shell.) Medullary shell lentelliptical.
Dimensions.—Major (longitudinal) axis of the outer cortical shell 0.016, of the
inner 0.08; minor (transverse) axis of the former 0.14, of the latter 0.07; medullary shell 0.03
to 0.04.
Habitat.—Pacific, central area, Station 267, depth 2700 fathoms.
3. Staurotholonium alternatum, n. sp.
Outer cortical shell smooth, very different from the inner; in the outer the two principal
cupolas are larger than the two lateral; in the inner inversely smaller. Pores subregular,
circular, in the outer shell four times, in the inner shell twice as broad as the bars; ten to
twelve pores in the basal semicircle of one cupola.
{676}
Dimensions.—Major axis (length) of the outer cortical shell 0.15, minor (breadth)
0.13; pores 0.012, bars 0.003; major axis (length) of the inner cortical shell 0.11, minor axis
(breadth) 0.09; pores 0.006, bars 0.003.
Habitat.—Indian Ocean, surface, Madagascar, Rabbe.
4. Staurotholonium lenticulare, n. sp.
Outer cortical shell smooth, lenticular, with circular circumference, with very delicate
irregular network and small roundish pores. Its distance from the inner much greater in the four
diagonal points than in the four polar points. Inner cortical shell regular, cross-like, with four
cupolas of similar size and form. Pores subregular, circular, twice as broad as the bars; eight to
ten in the basal semicircle of one cupola. Medullary shell lenticular.
Dimensions.—Diameter of the outer cortical shell 0.16, of the inner 0.12; pores of
the inner 0.008, bars 0.004.
Habitat.—North Atlantic, Station 353, surface.
Subgenus 2. Staurotholoma, Haeckel.
Definition.—Surface of the outer cortical shell with radial spines or
thorns.
5. Staurotholonium octodoratium, n. sp.
Outer cortical shell of the same form and structure as the inner, but twice as large, both
principal domes somewhat larger than both lateral. Pores subregular, circular, of the same breadth
as the bars; five to six in the semicircle of one inner, eight to ten of one outer cupola. Eight
thin and long bristle-like spines, opposite in pairs in two crossed diagonal planes.
Dimensions.—Major axis of the outer cortical shell 0.15, minor 0.12; major axis of
the inner cortical shell 0.07, minor 0.06; pores and bars 0.005.
Habitat.—North Pacific, Station 231, surface.
6. Staurotholonium octodoronium, n. sp. (Pl. 10, fig.
11).
Outer cortical shell of the same form as the inner, both little distant; network of the outer
irregular and delicate. Pores of the inner regular, circular, twice as broad as the bars; seven to
nine in the basal semicircle of one cupola. Both lateral domes larger than the principal. Eight
long and thin, cylindrical radial spines opposite in pairs in two crossed diagonal planes.
Dimensions.—Major axis of the outer cortical shell 0.2, minor 0.18; major axis of
the inner cortical shell 0.16, minor 0.14; pores 0.008, bars 0.004.
Habitat.—South Pacific, Station 288, surface.
{677}
Subfamily 3. Cubotholida, Haeckel.
Definition.—Tholonida with triaxial growth;
cupolas lying in pairs on the six sides of a cubical central chamber, opposite at the poles of
three axes perpendicular one to another. (Shell commonly seven-chambered, with six domes
surrounding the central chamber.)
Genus 297. Tholocubus,[337] n. gen.
Definition.—Tholonida with simple cortical shell (without
external veil), composed of six hemispherical cupolas, opposite in pairs on the poles of three
axes perpendicular one to another, covering six sides of the simple cuboidal central chamber
(without medullary shell).
The genus Tholocubus is the most simple form of the Cubotholida, or of the
Tholonida with domes situated in three axes perpendicular one to another; six hemispherical
cupolas lying on the six sides of a cuboidal central chamber; this latter contains no medullary
shell. Tholocubus may be derived phylogenetically either from Tholostaurus by
apposition of two opposite domes on the flat sides of the cross-shell, or from Cubotholus
by loss of the medullary shell.
Subgenus 1. Tholocubulus, Haeckel.
Definition.—Surface of the shell smooth or rough, without radial
spines.
1. Tholocubus tessellatus, n. sp. (Pl. 10, fig.
12).
Surface of the shell smooth, without radial spines. Pores regular, circular, hexagonally
framed, twice as broad as the bars; eight to twelve pores on the semicircle of one cupola.
Principal cupolas smaller than the lateral, larger than the sagittal cupolas.
Dimensions.—Diameter of the shell 0.15; pores 0.01, bars 0.005.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
2. Tholocubus tesserarius, n. sp.
Surface of the shell rough, without radial spines. Pores irregular, roundish, once to three
times as broad as the bars; twelve to sixteen pores in the semicircle of one cupola. All six
cupolas nearly of the same size.
Dimensions.—Diameter of the shell 0.16; pores 0.004 to 0.008, bars 0.003.
Habitat.—North Atlantic, Station 353, depth 2965 fathoms.
{678}
Subgenus 2. Tholocubitus, Haeckel.
Definition.—Surface of the shell with radial spines or thorns.
3. Tholocubus tesseralis, n. sp. (Pl. 10, fig.
16).
Surface of the shell with numerous (eight to sixteen) thin and long, bristle-shaped radial
spines (the greater part broken off in the figured specimen); pores subregular, circular, three to
four times as broad as the bars; ten to twelve in the semicircle of one cupola.
Dimensions.—Major axis of the shell 0.16, minor axis 0.14; pores 0.01, bars
0.0027.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
Genus 298. Tholonium,[338] n. gen.
Definition.—Tholonida with double cortical shell (with
external veil), composed of six hemispherical cupolas, opposite in pairs on the poles of three
axes perpendicular one to another, covering six sides of the simple cuboidal central chamber
(without medullary shell).
The genus Tholonium (Pl. 10,
fig. 17) differs from the preceding Tholocubus only in the duplication of the cortical
shell. The outer (secondary) shell forms either a simple (spheroidal or ellipsoidal) thin veil
around the inner (primary) shell, or both shells are of the same form, with six corresponding
cupolas. Possibly Tholonium is the offspring of Cubotholonium, having originated by
loss of the medullary shell.
Subgenus 1. Tholonetta, Haeckel.
Definition.—Surface of the shell smooth or rough, without radial
spines or thorns.
1. Tholonium bicubicum, n. sp.
Outer shell with six hemispherical dome-shaped protuberances, corresponding in those of the
inner shell. Both shells connected by numerous radial beams. Surface of the outer shell smooth;
its network nearly of the same shape as that of the inner, with subregular, circular pores, twice
as broad as the bars; ten to fifteen pores on the semicircle of one cupola.
Dimensions.—Diameter of the outer shell 0.14, of the inner 0.12; pores of the
latter 0.006, bars 0.003.
Habitat.—South Atlantic, Station 325, depth 2650 fathoms.
{679}
2. Tholonium ellipticum, n. sp.
Outer shell ellipsoidal, without dome-shaped protuberances, with smooth surface, without radial
spines; network delicate, with subregular, circular pores. Inner shell with regular, circular,
hexagonally framed pores, twice as broad as the bars; ten to twelve pores on the semicircle of one
cupola.
Dimensions.—Major axis of the outer shell 0.16, minor axis 0.14; major axis of the
inner shell 0.14, minor axis 0.12; pores 0.008, bars 0.004.
Habitat.—Pacific central area, Station 267, depth 2700 fathoms.
3. Tholonium sphæricum, n. sp.
Outer shell spherical, without dome-shaped protuberances, with smooth surface, without radial
spines; network very delicate, with very small subregular, circular pores. Inner shell with
regular, circular pores of the same breadth as the bars; fourteen to sixteen on the basal
semicircle of one cupola.
Dimensions.—Diameter of the spherical outer shell 0.15, inner shell 0.12; pores
and bars of the inner shell 0.005.
Habitat.—Indian Ocean, Zanzibar, Pullen, depth 2200 fathoms.
Subgenus 2. Tholonilla, Haeckel.
Definition.—Surface of the outer shell covered with radial spines or
thorns.
4. Tholonium hexonium, n. sp. (Pl. 10,
fig. 17).
Outer shell ellipsoidal, without dome-shaped protuberances, covered with numerous (thirty to
fifty) thin and long, bristle-shaped radial spines; network very delicate, irregular, with
roundish pores. Inner shell with six marked hemispherical domes of somewhat different sizes; pores
regular, circular, with prominent hexagonal frames, twice as broad as the bars; ten to twelve on
the basal semicircle of one cupola.
Dimensions.—Major axis of the outer shell 0.15, minor axis 0.14; major axis of the
inner shell 0.13, minor axis 0.12; pores 0.008, bars 0.004.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
5. Tholonium sphæronium, n. sp.
Outer shell very thin walled, spherical, without dome-shaped protuberances, with smooth
surface, excepting eight large radial spines, rising from the eight corners of the inner cubical
central chamber. Pores of the outer shell very small, subcircular. Inner shell very thick walled,
with six marked hemispherical domes; pores subregular, circular, with prominent hexagonal frames,
twice as broad as the bars; six to eight on the semicircle of one cupola.
{680}
Dimensions.—Diameter of the spherical outer shell 0.15, inner shell 0.11; pores of
the inner 0.01, bars 0.005.
Habitat.—Pacific, central area, Station 274, depth 2750 fathoms.
Genus 299. Cubotholus,[339] n. gen.
Definition.—Tholonida with simple cortical shell (without
external veil), composed of six hemispherical cupolas, opposite in pairs on the poles of three
axes perpendicular one to another, covering six sides of the cuboidal Larnacilla-shaped
central chamber (with medullary shell).
The genus Cubotholus differs from Tholocubus in the possession of a
medullary shell in the central chamber, and may be derived from this genus by its production; but
it may also be derived from Staurotholus by apposition of two opposite domes on the flat
sides of the cross-shell. Sometimes all six domes are of the same size and form, but commonly
different in pairs.
Subgenus 1. Cubotholissa, Haeckel.
Definition.—Surface of the shell smooth or rough, without radial
spines.
1. Cubotholus regularis, n. sp. (Pl. 10,
fig. 14).
On the six sides of the cubical central chamber six hemispherical cupolas of the same size and
form. Surface smooth. Pores regular, circular, twice as broad as the bars; eight in the basal
semicircle of each cupola. Medullary shell apparently spherical (?), connected with the eight
corners of the central chamber by eight radial beams, regularly disposed. (This species is
remarkable for the perfect symmetry of the shell, the six sides of which appear to be quite
similar. It differs from the similar Tholocubus regularis in the possession of a medullary
shell.)
Dimensions.—Diameter of the cortical shell (equal in all three dimensive axes)
0.15; pores 0.01, bars 0.005; diameter of the medullary shell 0.04.
Habitat.—Pacific, central area, Station 273, depth 2350 fathoms.
2. Cubotholus quadraticus, n. sp.
Surface of the shell smooth. Both principal cupolas (on the poles of the longitudinal axis)
larger than the four other cupolas, which have the same size and lie cross-wise in the equatorial
plane. (Therefore two of the three fundamental axes equal, the third larger.) Pores subregular,
circular, three times as broad as the bars; ten to twelve in the semicircle of each
cupola-basis.
Dimensions.—Major axis 0.16, minor axis 0.12; pores 0.012, bars 0.004.
Habitat.—Indian Ocean, Zanzibar, Pullen; depth 2200 fathoms.
{681}
3. Cubotholus rhombicus, n. sp.
Surface of the shell smooth. Both principal cupolas (on the poles of the longitudinal axis)
larger than the two lateral (on the poles of the transverse axis), and these larger than the two
sagittal cupolas (on the poles of the sagittal axis). Therefore all three fundamental axes
unequal. Pores irregular, roundish, twice to four times as broad as the bars; eight to twelve in
the basal semicircle of each cupola. Medullary shell lentelliptical.
Dimensions.—Major (principal) axis 0.16, middle (lateral) axis 0.14, minor
(sagittal) axis 0.12; pores 0.006 to 0.012, bars 0.003.
Habitat.—South Atlantic, Station 335, depth 1425 fathoms.
Subgenus 2. Cubotholura, Haeckel.
Definition.—Surface of the shell with radial spines or thorns.
4. Cubotholus octoceras, n. sp.
All six cupolas in pairs of different sizes. Both principal domes larger than the lateral
domes, and these larger than the sagittal domes. Pores subregular, circular, twice as broad as the
bars; eight to ten in the semicircle of each cupola. From the surface arise (at the intersecting
points of every three cupolas) eight strong conical radial spines, about as long as the
shell-axis; they lie in two diagonal planes, and are the external free prolongations of eight
inner beams (homologous with the eight wing-spines of Tetrapyle octacantha), which connect
the lentelliptical medullary shell with the eight corners of the cuboidal central chamber.
Dimensions.—Major axis of the cortical shell 0.15, middle 0.13, minor 0.11; pores
0.008, bars 0.004; diameters of the medullary shell corresponding to 0.05, 0.04, 0.03.
Habitat.—Western Tropical Pacific, Station 224, depth 1850
fathoms.
Genus 300. Cubotholonium,[340] n.
gen.
Definition.—Tholonida with double (or sometimes triple)
cortical shell (with external veil), composed of six hemispherical cupolas, opposite in pairs on
the poles of three axes perpendicular one to another, covering six sides of the cuboidal
Larnacilla-shaped central chamber (with medullary shell).
The genus Cubotholonium differs from the nearly allied Cubotholus
(its probable ancestral form) only in duplication of the cortical shell. I have observed only two
species of this rare form, both rather different. In the first species the outer cortical shell
forms a simple spherical thin veil around the inner, the six cupolas of which are nearly of the
same form and size. In the second species each of the domes of the {682}inner cortical shell is protected by an outer larger cupola, and besides
this the whole shell is enveloped by a thin ellipsoidal veil (Pl. 10, fig. 15).
Therefore this species may be the representative of a peculiar genus, the most highly developed of
all Tholonida—Tholothauma.
1. Cubotholonium sphæroides, n. sp.
Outer cortical shell (or veil) spherical, with smooth surface; network very delicate, with very
thin bars and very small irregular, roundish pores. Inner cortical shell simple, composed of six
hemispherical cupolas, surrounding the six sides of the cubical central chamber, which encloses a
spherical medullary shell (one-third as large as itself). Network of the inner cortical shell
regular, with circular pores of the same breadth as the bars, twelve to fourteen in the basal
semicircle of one cupola.
Dimensions.—Diameter of the spherical outer shell 0.2, of the inner cortical shell
0.15; pores and bars of the latter 0.006.
Habitat.—Indian Ocean, Zanzibar, Pullen, depth 2200 fathoms.
2. Cubotholonium ellipsoides, n. sp. (Pl. 10, fig.
15).
Tholothauma ellipsoides, Haeckel, 1883, MS.
Outer cortical shell (or veil) ellipsoidal, with very thin irregular network and thorny
surface. Inner cortical shell double, with six double, flatly vaulted cupolas, surrounding the six
sides of the Larnacilla-shaped central chamber; the double domes of each shell are in
opposite pairs somewhat larger than the alternating pairs. Pores subregular, circular, about the
same breadth as the bars; eight to twelve in the basal semicircle of one cupola. Central chamber
with ellipsoidal medullary shell. Radial spines short, very numerous.
Dimensions.—Major axis of the outer cortical shell 0.28, minor 0.24; major axis of
the inner cortical shell 0.16, minor axis 0.14; pores and bars 0.006; medullary shell 0.03.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
Family XXVIII. Zonarida, n. fam. (Pl. 50, figs.
9-12).
Definition.—Larcoidea with regular, completely latticed
cortical shell, distinguished by two to four or more annular constrictions, which lie (all or
partly) in the dimensive planes (sagittal, transverse, or lateral), and by which four to eight or
more vaulted cupolas or dome-like chambers become separated. In the centre of this chambered
cortical shell lies constantly a trizonal or Larnacilla-shaped medullary shell.
The family Zonarida comprises a small number of peculiar Larcoidea, resembling the Tholonida in the composition of the polythalamous
cortical shell by a number of cupolas or dome-shaped protuberances. But the disposition and origin
of these latter {683}are quite different. Whilst in the
Tholonida the axes of the domes are dimensive axes, and these are separated by annular
constrictions lying in diagonal planes, in the Zonarida we find the contrary; the axes of the
domes are here diagonal axes, and these are separated by annular constrictions lying in dimensive
planes. However, this definition agrees absolutely only in the four-chambered Zonarium and
in the eight-chambered Zonidium, whilst in the six chambered Zoniscus only four
domes are disposed according to this law, two others, however, in the same manner as in the
Tholonida. Therefore this genus is intermediate between both families.
The Cortical Shell of the Zonarida is in all cases completely latticed and of regular
lentelliptical fundamental form, as in the nearly allied Larnacida and Tholonida. The three
dimensive axes are constantly of different sizes, each with two equal poles; commonly (as in the
human body) the principal or longitudinal axis is the longest, the sagittal (or dorso-ventral)
axis the shortest; the transverse (or lateral) axis being intermediate between them. Of the three
dimensive planes the lateral plane is the largest (determined by the principal and transverse
axes); the smallest is the equatorial plane (crossed by the transverse and sagittal axes); the
sagittal plane (determined by the sagittal and principal axes) being intermediate between
them.
The annular constrictions of the cortical shell which produce the dome-shaped protuberances are
different in number in the three known genera—two, three, or four. To each constriction
often (but not always) corresponds an internal latticed septum, which connects the cortical with
the medullary shell. The number of the cupolas is always double the number of the annular
constrictions by which they are separated, therefore four, six, or eight.
In all known Zonarida the sagittal septum is quite constant, but derived from the original
axial rod, which lies in the principal axis. By ramification of this axial beam and reticular
connection with the sagittal girdle arises the sagittal septum, which we found first in
Octopyle, halving the four gates of Tetrapyle. Whilst this sagittal septum (between
right and left halves of the body) is common to all three known genera of this family, the number
and shape of the other annular constrictions are different. In Zonarium (Pl. 50, fig. 9) we
find only one transverse constriction (in the equatorial plane), in Zoniscus (figs. 10, 11)
two parallel transverse constrictions (parallel to the equatorial plane, on both sides of it).
Zonidium (fig. 12) is a combination of both foregoing genera; it has three parallel
transverse constrictions (one in the equatorial plane, and one on each side of it).
The Latticed Domes (cupolas or chambers) of the cortical shell exhibit correspondingly a
different number and disposition in the three known genera. In Zonarium are found only four
crossed chambers, separated by the sagittal and transverse septa; the axes of the four crossed
domes are diagonal axes, whilst in the similar Staurotholus they are dimensive axes
(principal and transverse axes). In Zonidium we find eight domes, each cupola of
Zonarium being halved by a diagonal septum. Zoniscus is intermediate {684}between the two foregoing genera, and has six cupolas,
three on each side of the sagittal septum.
The inner communication of the cupolas or chambers is more or less free, the lattice-work of
the separating septa between them commonly remaining more or less imperfect, or represented only
by some isolated beams or meshes. The outer network of the cupolas is commonly irregular (as in
the majority of Larcoidea), but sometimes distinguished by a small number
of regularly disposed larger apertures (similar to the "gates" of the Pylonida). From the surface
radial spines often arise in characteristic number and symmetrical disposition, commonly as
prolongations of the septal axes or of the constricted edges.
The Medullary Shell in all Zonarida is a true trizonal or Larnacilla-shaped
lattice-shell (compare above, p. 600); its perimeter (or the first lateral
girdle) is sometimes more elliptical, at other times more hexagonal; the hexagon is amphithect;
both its lateral sides are often concave and commonly longer than the four other sides.
The Central Capsule in all Zonarida is in a strict geometrical sense a true lentellipsis
(compare above, p. 599); its principal axis is commonly one and a third to
one and a half times as great as the transverse axis, and twice to three times as great as the
sagittal axis. The lentelliptical central capsule encloses the trizonal medullary shell, whilst it
is externally enveloped by the chambered cortical shell.
Synopsis of the Genera of Zonarida.
| Two annular constrictions and four cupolas, |
301. Zonarium. |
| Three annular constrictions and six cupolas, |
302. Zoniscus. |
| Four annular constrictions and eight cupolas, |
303. Zonidium. |
Genus 301. Zonarium,[341] n. gen.
Definition.—Zonarida with four dome-shaped chambers of
the cortical shell, separated by two annular constrictions (one sagittal and one transverse).
The genus Zonarium is the most simple form of the Zonarida, and differs from
the nearly allied Larnacalpis by two ring-like constrictions, which are crossed at right
angles, one in the sagittal (or median) plane, and one in the transverse (or equatorial) plane. By
these two annular constrictions four egg-shaped or kidney-shaped chambers become imperfectly
separated, which correspond to the quadrants of the lateral plane. The first cause of the marked
constrictions may be the formation of the latticed sagittal septum, which is found first in
Octopyle, as halving the four gates of {685}Tetrapyle. Between this septum and the constricted narrow
transverse girdle the cortical shell grows out in the form of four vaulted cupolas; every two
opposite domes are congruent, two neighbouring are symmetrically equal.
1. Zonarium quadrigatum, n. sp.
Cortical shell quadrangular, one and a half times as long as broad, with four rounded corners.
Surface thorny, with numerous short radial spines. Sagittal constriction twice as long as the
hexagonal medullary shell. Four cupolas kidney-shaped.
Dimensions.—Length of the cortical shell 0.16, breadth 0.11; length of the
medullary shell 0.06, breadth 0.04.
Habitat.—Pacific, central area, Station 272, depth 2600 fathoms.
2. Zonarium quadrispinum, n. sp.
Cortical shell quadrangular, nearly rectangular, about twice as long as broad, with four
corners, from which arise four strong, three-sided pyramidal, radial spines (crossed in two
diagonals of the lateral plane). Surface thorny, only smooth in the transverse constriction.
Sagittal constriction three times as long as the lentelliptical medullary shell. Four cupolas
elliptical or nearly quadrangular.
Dimensions.—Length of the cortical shell 0.22, breadth 0.1; length of the
medullary shell 0.07, breadth 0.05.
Habitat.—Pacific, central area, Station 274, depth 2750 fathoms.
3. Zonarium octangulum, n. sp. (Pl. 50,
fig. 9).
Cortical shell octangular, one and a fourth times as long as broad, with eight strong conical
spines on the eight corners; these are separated by four deep constrictions on the poles of the
principal and transverse axes, and by four truncated planes on the poles of the crossed diagonal
axes between the former. Sagittal constriction twice as long as the hexagonal medullary shell.
Four cupolas kidney-shaped or nearly pentagonal.
Dimensions.—Length of the cortical shell 0.15, breadth 0.12; length of the
medullary shell 0.07, breadth 0.04.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
4. Zonarium tetratholium, n. sp.
Cortical shell quadrangular, with four rounded corners. Surface thorny, with sixteen stronger
radial spines; eight of these lie in the lateral plane, in the same symmetrical disposition as in
the foregoing species; eight others lie on both sides of the lateral plane, opposite in pairs in
two crossed diagonal planes, in the same symmetrical disposition as in Tetrapyle
octacantha. Sagittal {686}constriction three times
as long as the lentelliptical medullary shell. Four cupolas obliquely elliptical.
Dimensions.—Length of the cortical shell 0.16, breadth 0.12; length of the
medullary shell 0.04, breadth 0.03.
Habitat.—North Pacific, Station 241, depth 2300 fathoms.
Genus 302. Zoniscus,[342] n. gen.
Definition.—Zonarida with six dome-shaped chambers of the
cortical shell, separated by three annular constrictions (one sagittal and two transverse,
parallel to the equatorial plane).
The genus Zoniscus differs from Zonarium as well as from
Zonidium by the development of the transverse girdle, which is not constricted, but on the
contrary prominently vaulted in the equatorial plane. Therefore both wings of the transverse
girdle form here two opposite lateral or "equatorial cupolas," as in Amphitholus. These are
separated from four other domes (the "corner cupolas") by two transverse annular constrictions,
which correspond to the free edges of the original transverse girdle. The corner domes of each
pair are separated from each other by the sagittal septum.
1. Zoniscus rectangulus, n. sp.
Cortical shell nearly rectangular, with rounded corners, nearly one and a half times as long as
broad. Surface thorny, without larger radial spines. Sagittal constriction scarcely half as long
as the hexagonal medullary shell. Both equatorial cupolas (or wings of the transverse girdle)
scarcely half as large as the four corner cupolas.
Dimensions.—Length of the cortical shell 0.15, breadth 0.11; length of the
medullary shell 0.07, breadth 0.045.
Habitat.—North Pacific, Station 253, depth 3125 fathoms.
2. Zoniscus hexathalamus, n. sp.
Cortical shell nearly elliptical, one and a third times as long as broad. Surface nearly
smooth, without radial spines. Sagittal constriction three times as long as the lentelliptical
medullary shell. Both equatorial cupolas about as large as the four corner cupolas.
Dimensions.—Length of the cortical shell 0.12, breadth 0.09; length of the
medullary shell 0.035, breadth 0.025.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
{687}
3. Zoniscus tetracanthus, n. sp. (Pl. 50, fig.
10).
Cortical shell nearly quadrangular, one and a half times as long as broad, with four prominent
corners, from which arise in the lateral plane four strong, three-sided pyramidal radial spines
(opposite in pairs in two crossed diagonal axes). Surface thorny, with exception of the concave
lateral sides. Sagittal constriction two and a half times as long as the hexagonal medullary
shell. Both equatorial cupolas scarcely half as large as the four corner cupolas.
Dimensions.—Length of the cortical shell 0.18, breadth 0.12; length of the
medullary shell 0.07, breadth 0.04.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
4. Zoniscus octacanthus, n. sp.
Cortical shell nearly four-sided, prismatic, one and a third times as long as broad, with spiny
surface. Eight longer thin radial spines opposite in pairs in two crossed diagonal planes (as in
Tetrapyle octacantha). Sagittal constriction two and a half times as long as the hexagonal
medullary shell. Both equatorial cupolas somewhat larger than the four corner cupolas.
Dimensions.—Length of the cortical shell 0.16, breadth 0.12; length of the
medullary shell 0.055, breadth 0.04.
Habitat.—Pacific, central area, Station 263, depth 2650 fathoms.
5. Zoniscus hexatholius, n. sp. (Pl. 50,
fig. 11).
Cortical shell nearly rectangular, with four rounded corners and deep sagittal constriction,
one and a fourth times as long as broad with thorny surface. Twelve longer edged radial spines;
eight opposite in pairs in two crossed diagonal planes (as in the foregoing species), four others
in the lateral plane, opposite in pairs on both sides of the sagittal constriction, which is
scarcely twice as long as the hexagonal, in the equatorial plane constricted medullary shell. Both
equatorial cupolas nearly of the same size as the four corner cupolas.
Dimensions.—Length of the cortical shell 0.16, breadth 0.13; length of the
medullary shell 0.07, breadth 0.04.
Habitat.—South Pacific, Station 300, depth 1375 fathoms.
Genus 303. Zonidium,[343] n. gen.
Definition.—Zonarida with eight dome-shaped chambers of
the cortical shell, separated by four annular constrictions (one sagittal, one transverse, and two
others parallel to the latter).
The genus Zonidium must be regarded as a Zonarium, in which the four
cupolas (in the quadrants of the lateral plane) are halved by diagonal beams or latticed septa,
{688}lying opposite in pairs in two crossed diagonal
planes. Therefore the number of the septa (four) and the domes (eight) is here doubled. Among the
eight cupolas we distinguish four median (on both sides of the sagittal plane) and four lateral
(on both sides of the equatorial plane); both groups are of different size and form. Probably
Zonidium is derived from Zonarium by lattice-connection between the eight diagonal
wing-spines, which in both species of this genus are present, the same as in Tetrapyle
octacantha.
1. Zonidium octostylium, n. sp.
Cortical shell nearly quadrangular, with rounded corners and thorny surface. Eight long and
thin radial spines on both sides of the lateral plane opposite in pairs and lying in two crossed
diagonal planes (as in Tetrapyle octacantha). Sagittal constriction three times as long as
the lentelliptical medullary shell. Four medial cupolas (on both sides of the sagittal plane)
somewhat larger than the four lateral cupolas (on both sides of the equatorial plane).
Dimensions.—Length of the cortical shell 0.15, breadth 0.12; length of the
medullary shell 0.05, breadth 0.03.
Habitat.—Indian Ocean, Madagascar, Rabbe, surface.
2. Zonidium octotholium, n. sp. (Pl. 50,
fig. 12).
Cortical shell nearly octangular, with spiny surface; twenty long and stout radial spines
between numerous smaller spines; eight wing-spines opposite in pairs in two crossed diagonal
planes (as in the preceding species); twelve other strong spines in the lateral plane (four longer
opposite in pairs on the poles of the principal and transverse axes, eight others smaller,
alternating between these and the diagonal spines). Sagittal constriction twice as long as the
hexagonal medullary shell. Four median cupolas somewhat smaller than the four lateral cupolas.
Dimensions.—Length of the cortical shell 0.18, breadth 0.15; length of the
medullary shell 0.07, breadth 0.05.
Habitat.—Indian Ocean, Zanzibar, Pullen, depth 2200 fathoms.
Family XXIX. Lithelida, Haeckel (Pl. 49, figs.
1-7).
Lithelida, Haeckel, 1862, Monogr. d. Radiol., p. 515.
Definition.—Larcoidea with symmetrical spiral shell,
divided by the spiral plane into two symmetrical halves; all windings of the spiral lie in this
plane. Primordial chamber either simple or Larnacilla-shaped.
The family Lithelida comprises all those Larcoidea in which the growth of the latticed shell is spirally winding in one
plane, Nautilus-like. They agree in the spiral growth with the following family. But in the
Streblonida the spiral is screw-shaped, {689}ascending
(like Helix). Therefore in these latter the geometrical fundamental form of the shell is
asymmetrical or "dysdipleural," whereas in the Lithelida bilateral-symmetrical or "eudipleural."
The lentelliptical or nearly spherical shell may be divided by a median section into two
symmetrical halves; the right half is the mirror image of the left half.
When in 1862 I founded the family Lithelida in my Monograph (p. 515), I knew only one genus,
Lithelius, with two species. The rich material of the Challenger collection contains a
great number of similar spirally constructed Larcoidea, so that at the
present time we may distinguish at least six genera. These belong to two different subfamilies,
which may possibly be afterwards better separated as families. The first subfamily, Spiremida,
possess a simple, spherical or subspherical, medullary shell; the second subfamily, Larcospirida,
possess a trizonal or Larnacilla-shaped medullary shell. No doubt these latter must be
derived from Pylonida, as we observe all stages of development starting from a simple
Trizonium; but perhaps also the Spiremida have the same origin, their simple, spherical or
subspherical, medullary shell being derived from a trizonal or Larnacilla-shaped medullary
shell by reduction.
The general appearance in both subfamilies of the Lithelida is quite the same, and it requires
a careful study of the medullary shell to distinguish certainly the Spiremida from the
Larcospirida. This distinction is often not easy, particularly in the larger forms; the shell is
often very opaque and difficult to understand. Only in one position, if the spiral axis be
parallel to the axis of the eye of the observer, and the spiral plane be therefore fully seen in
the optical plane of the microscope, the spiral line (or the axial section of the latticed spiral
lamella) is distinctly observed; in all other positions the figure of the spiral is more or less
indistinct, and the whole microscopical image often quite intricate and confused. The sufficient
study of this family requires therefore the contemplation of the shell from different sides, and
is the more difficult, as the variability of the Lithelida—as of the Pylonida—is
extraordinarily great.
The description which I gave of Lithelius (1862) in my Monograph is in some points
erroneous, and was afterwards (1879) corrected by R. Hertwig, who explained particularly the near
relation of it to Tetrapyle. Indeed the intermediate forms between the Lithelida and the
Pylonida are so numerous and so evident in all stages of development, that the derivation of the
former (at least of the Larcospirida) from the latter is quite clear. The analogy between the
structure of the Lithelida and the calcareous (foraminiferous) Alveolinida is not so complete as I
supposed it to be in my Monograph (1862); particularly the formation of the small chambers between
the turnings of the spiral lamella is much more complete in the Alveolinida than in the
Lithelida.
The cortical shell of all Lithelida has the same geometrical fundamental form as
Nautilus or as the nautiloid Polythalamia (Polystomella, Nummulites,
&c.); therefore the shell is dipleural, being divided by the median plane into two symmetrical
lateral halves. {690}Since the spiral line lies in the
median plane, we will call it the spiral plane; it separates the right half from the left. The
axis of the body, around which the spiral turns (without touching it), is the spiral axis. The
latticed part of the cortical shell, which turns around them, is the spiral lamella. Only in one
genus of our family, viz., Tholospira, are the spiral axis, the lateral axis, the spiral
plane, and the sagittal plane quite as in Nautilus. In all other genera this disposition is
different or is uncertain. This depends on the different part of the cortical shell, from which
the spiral growth begins. In this respect we can distinguish four different modes.
In the Larcospirida (or the Lithelida with Larnacilla-shaped medullary shell) the spiral
growth exhibits four quite different forms. It begins here with Larcospira, in which
already the first cortical girdle of the Diplozonaria determines the spiral growth; one wing of
this girdle, the transverse girdle of Amphipyle, grows more swiftly than the other,
overgrows it, and thus turns around the principal axis. In Pylospira the first or
transverse girdle is already perfectly formed (as in Amphipyle), and the spiral growth is
introduced by the second or lateral girdle of Tetrapyle; one wing of it (the right or the
left) grows more swiftly than the other, overgrows it, and thus turns around the sagittal axis. In
Tholospira also the second girdle is complete, and the spiral growth begins from the third
or sagittal girdle. One of its wings grows more swiftly than the other, overgrows it, and thus
turns around the transverse axis. Consequently we see that each of the three dimensive planes of
the lentelliptical Larcoid-body may be the spiral plane: in Larcospira the transverse
plane, in Pylospira the lateral plane, in Tholospira the sagittal plane.
Correspondingly the spiral axis in the first genus is the principal, in the second the sagittal,
in the third the transverse axis of the central Larnacilla-shell. Therefore in these three
genera the spiral plane is the plane of the latticed girdle, which determines the spiral growth,
one of both its wings overgrowing the other.
In each of the three above mentioned genera the spiral may be simple or double; it remains
simple if only one of both wings of the turning girdle overgrow the other, and this latter remain
a simple half girdle (or tube-like wing). Whereas the spiral becomes double if the second wing of
the girdle afterwards follow the example of the first wing and now turn around it in the same
direction. As this happens in all three genera, we can subdivide them into six subgenera.
A quite peculiar form of spiral growth is produced in Spironium, in which the direction
of growth in both lateral wings of the transverse girdle is inverse from the beginning. The left
wing grows against the posterior, the right wing against the anterior pole of the principal axis,
turning around it in crossed, eight-like spirals. The whole shell afterwards assumes a
lentelliptical form.
Commonly between the embracing spiral turnings or convolutions a great number of radial beams
is developed, irregularly disposed and often branching; they support the {691}thin spiral lamellæ and give to the whole shell more solidity. Often
these beams form imperfect radial septa, by which the spiral cavity of the turnings is divided
into a variable number of chambers. But these chambers never become so regular and perfect as in
the analogous nautiloid Polythalamia.
In many Lithelida the growth of the shell reaches a certain limit, concluding with the
formation of a superficial latticed lamella of lentelliptical or nearly spherical form. In many
other forms of the family this seems not to be the case; but these may possibly be younger forms,
afterwards reaching the same limit.
The network of the shell in the Lithelida is commonly quite irregular, and so variable that its
special conformation has usually no value in the determination of the species. The surface of the
shell is often covered with radial spines, which are sometimes arborescent.
The central capsule seems always to preserve the same lentelliptical form (or triaxial
ellipsoid) as in all other Larcoidea. With the increase of growth it
encloses successively a larger part of the spiral cortical shell, but on the outside is constantly
protected by the last turnings of the spiral, or by the lattice-lamella of the surface.
Synopsis of the Genera of Lithelida.
|
II. Subfamily Spiremida.
Central medullary shell simple, spherical or lentelliptical.
|
|
Surface of the cortical shell smooth or thorny, withou radial
spines, |
304. Spirema. |
| Surface of the cortical shell covered with numerous simple or branched
radial spines, |
305. Lithelius. |
|
II. Subfamily Larcospirida.
Central medullary shell double, trizonal or Larnacilla-shaped.
|
|
The transverse girdle turns around the principal axis, |
306. Larcospira. |
| The lateral girdle turns around the sagittal axis, |
307. Pylospira. |
| The sagittal girdle turns around the transverse axis, |
308. Tholospira. |
| Both wings of the transverse girdle turn around the principal axis in
an opposite diagonal direction, |
309. Spironium. |
| I. Subfamily Spiremida. Central medullary shell simple, spherical or
lentelliptical. |
|
|
Surface of the cortical shell smooth or thorny, withou radial
spines, |
|
|
|
|
|
|
304. Spirema. |
|
|
|
|
|
|
Surface of the cortical shell covered with numerous simple or
branched radial spines, |
|
|
|
|
|
|
305. Lithelius. |
|
|
|
|
| II. Subfamily Larcospirida. Central medullary shell double, trizonal or
Larnacilla-shaped. |
|
|
The transverse girdle turns around the principal axis, |
|
|
|
|
|
|
306. Larcospira. |
|
|
|
|
|
|
The lateral girdle turns around the sagittal axis, |
|
|
|
|
|
|
307. Pylospira. |
|
|
|
|
|
|
The sagittal girdle turns around the transverse axis, |
|
|
|
|
|
|
308. Tholospira. |
|
|
|
|
|
|
Both wings of the transverse girdle turn around the principal axis
in an opposite diagonal direction, |
|
|
|
|
|
|
309. Spironium. |
|
|
|
|
Subfamily 1. Spiremida, Haeckel, 1881, Prodromus, p. 464.
Definition.—Lithelida with simple, spherical
or subspherical, medullary shell.
Genus 304. Spirema,[344] Haeckel, 1881,
Prodromus, p. 464.
Definition.—Lithelida with simple, spherical or
subspherical, medullary shell, and lentelliptical or subspherical, spirally constructed cortical
shell; surface smooth or thorny, without radial spines.
{692}
The genus Spirema begins the series of the Spiremida, or of those Lithelida
in which the medullary shell presents a simple latticed sphere or ellipsoid, never composed of a
double, trizonal or Larnacilla-shaped shell. In the present state of our knowledge we
cannot say whether this simple medullary shell be a primary formation, or effected by secondary
means, by reduction of a double Larnacilla-shaped medullary shell, which is constantly
found in the Larcospirida. The species of this genus (as of all Lithelida) are difficult to
distinguish, are transformistic, and incline very much to variations and abnormalities. The spiral
may be simple or double.
Subgenus 1. Spiremarium, Haeckel.
Definition.—Spiral convolutions of the cortical shell simple.
1. Spirema lentellipsis, n. sp.
Cortical shell lentelliptical, with smooth surface; proportion of its three dimensive axes =
4 : 5 : 6. In the median plane are visible four perfect turnings of the simple
spiral, the breadth of which gradually increases towards the third convolution, finally
decreasing; the broadest (third) turning three times as broad as the simple spherical medullary
shell.
Dimensions.—Length of the lentelliptical cortical shell 0.18, breadth 0.15, height
0.12; diameter of the spherical medullary shell 0.02.
Habitat.—North Atlantic, Station 353, surface.
2. Spirema melonia, n. sp. (Pl. 49,
fig. 1).
Cortical shell nearly spherical, with smooth surface; proportion of its three dimensive axes =
1.4 : 1.5 : 1.6. In the median plane are visible three perfect turnings of the
simple spiral, all of the same breadth as the simple spherical medullary shell; the breadth of
each convolution somewhat greater at the poles of the principal than at the poles of the sagittal
axis.
Dimensions.—Length of the shell 0.16, breadth 0.15, height 0.14; medullary shell
0.018.
Habitat.—Pacific, central area, Station 271, surface.
3. Spirema flustrella, Haeckel.
Flustrella haliomma, Ehrenberg (1861), Abhandl. d. k. Akad. d. Wiss.
Berlin, 1872, p. 293, Taf. ii. fig. 6.
Cortical shell egg-shaped, with thorny surface; proportion of its three axes =
4 : 5 : 6. In the median plane are visible three perfect turnings of the
simple spiral, the first and second of about the same breadth as the simple spherical medullary
shell, the third suddenly increasing, and finally three to four times as broad. Network of the
surface irregular, with roundish pores.
Dimensions.—Length of the shell 0.2, breadth 0.17, height 0.14; medullary shell
0.013.
Habitat.—North Atlantic, Greenland, 1000 fathoms, Ehrenberg; Færöe
Channel, John Murray.
{693}
Subgenus 2. Spiremidium, Haeckel.
Definition.—Spiral convolutions of the cortical shell double.
4. Spirema diplospira, n. sp.
Cortical shell lentelliptical, with smooth surface; proportion of its three dimensive axes =
6 : 7 : 8. In the median plane are visible three perfect turnings of a double
spiral, the breadth of which gradually increases; the broadest (third) convolution three times as
broad as the simple lentelliptical medullary shell.
Dimensions.—Length of the shell 0.24, breadth 0.21, height 0.18; medullary shell
0.02.
Habitat.—Tropical Pacific, Philippines, Station 200, depth 250
fathoms.
5. Spirema subglobosum, n. sp.
Cortical shell nearly spherical, with thorny surface; proportion of its three axes =
2 : 2.1 : 2.2. In the median plane are visible two perfect turnings of a
double spiral, the breadth of which in the second convolution is four times as great as that of
the first convolution and the simple spherical medullary shell.
Dimensions.—Length of the shell 0.22, breadth 0.21, height 0.2; medullary shell
0.02.
Habitat.—Tropical Atlantic, off Sierra Leone, Station 348, depth
(2450) fathoms.
Genus 305. Lithelius,[345] Haeckel, 1862,
Monogr. d. Radiol., p. 519.
Definition.—Lithelida with simple, spherical or
subspherical, medullary shell, and lentelliptical or subspherical, spirally constructed cortical
shell; surface covered with numerous, simple or branched, radial spines.
The genus Lithelius, founded by me in 1862, and represented by two
Mediterranean species, was at that time the only known form of this family, which now contains six
genera and twenty-seven species. It differs from the foregoing Spirema in the possession of
numerous radial spines on the surface. These may be either simple or branched. The spiral may be
simple or double, and according to this latter modification we distinguish two different
subgenera.
Subgenus 1. Lithospira, Haeckel.
Definition.—Spiral convolutions of the cortical shell simple.
1. Lithelius spiralis, Haeckel.
Lithelius spiralis, Haeckel, 1862, Monogr. d. Radiol., p. 519, Taf. xxvii.
figs. 6, 7.
Cortical shell lentelliptical, one and a third times as long as broad, covered with very
numerous (one hundred to one hundred and fifty or more) simple, bristle-shaped radial spines,
about as long {694}as the shell. Spiral turnings simple,
all nearly of the same breadth and scarcely broader than the simple spherical medullary shell.
Dimensions.—Length of the cortical shell (with six spiral convolutions) 0.15,
breadth 0.13; diameter of the medullary shell 0.012.
Habitat.—Mediterranean, Messina, Haeckel, surface; Atlantic, Stations
348 to 353, surface.
2. Lithelius primordialis, R. Hertwig.
Lithelius primordialis, R. Hertwig, 1879, Organism. d. Radiol., p. 54,
Taf. vi. figs. 4, 4a.
Cortical shell subspherical, covered with numerous simple, bristle-shaped radial spines, longer
than the shell. Spiral turnings simple, with gradually increasing breadth, so that the beginning
of the third spiral is twice as broad as the first and as the simple spherical medullary
shell.
Dimensions.—Diameter of the cortical shell (with two spiral convolutions) 0.12;
diameter of the medullary shell 0.02.
Habitat.—Mediterranean, Messina, R. Hertwig, surface.
3. Lithelius alveolina, Haeckel, 1862.
Lithelius alveolina, Haeckel, 1862, Monogr. d. Radiol., p. 520, Taf.
xxvii. figs. 8, 9.
Cortical shell spherical, covered with simple, very numerous (two hundred to three hundred or
more), short, bristle-shaped radial spines, scarcely half as long as the radius of the shell.
Spiral turnings simple, with gradually increasing breadth, so that the beginning of the third
spiral is three times as broad as the first and as the simple spherical medullary shell.
Dimensions.—Diameter of the cortical shell (with four spiral convolutions) 0.2;
diameter of the medullary shell 0.01.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific, on many
Stations, surface.
4. Lithelius capreolus, n. sp.
Cortical shell lentelliptical, one and a fourth times as long as broad, covered with numerous
(eighty to one hundred and twenty or more) short, branched radial spines, about half as long as
the radius of the shell; each spine once or twice dichotomous, with curved divergent branches.
Spiral turnings simple, with gradually increasing breadth, so that the beginning of the third
spiral is twice as broad as the first, and as the simple spherical medullary shell.
Dimensions.—Length of the cortical shell (with four spiral convolutions) 0.18,
breadth 0.14; diameter of the medullary shell 0.015.
Habitat.—Pacific, central area, Station 274, surface.
Subgenus 2. Drymospira, Haeckel, 1881, Prodromus, p. 464.
Definition.—Spiral convolutions of the cortical shell double.
{695}
5. Lithelius solaris, n. sp. (Pl. 49,
fig. 2).
Cortical shell spherical, covered with simple, very numerous (two hundred to three hundred)
bristle-shaped radial spines, longer than the diameter of the shell. Spiral turnings double, both
of the same breadth, gradually increasing with the growth of the shell and several times
surpassing the diameter of the simple spherical medullary shell. (Pl. 49, fig. 2,
exhibits only the first convolutions in the centre of the shell.)
Dimensions.—Diameter of the cortical shell (with four spiral convolutions) 0.18;
diameter of the medullary shell 0.02.
Habitat.—Pacific, central area, Stations 266 to 272, surface and in
various depths.
6. Lithelius arborescens, n. sp.
Cortical shell lentelliptical, one and a half times as long as broad, covered with numerous
(fifty to eighty or more) branched radial spines, about as long as the greatest diameter of the
shell; each spine with two to four lateral branches, which are again branched or dichotomous.
Spiral turnings double, both of little different breadth, which increases considerably with the
growth of the shell, so that the third turn is four times as broad as the simple spherical
medullary shell.
Dimensions.—Length of the cortical shell (with three spiral convolutions) 0.18,
breadth 0.12; diameter of the medullary shell 0.01.
Habitat.—North Atlantic, Færöe Channel, surface, John Murray.
Subfamily 2. Larcospirida, Haeckel.
Definition.—Lithelida with double, trizonal,
or Larnacilla-shaped medullary shell.
Genus 306. Larcospira,[346] n. gen.
Definition.—Lithelida with double, trizonal, or
Larnacilla-shaped medullary shell; cortical shell subspherical or lentelliptical,
constructed of a simple or double spiral of the transverse girdle (or primary cortical girdle);
the spiral lamella revolving around the principal axis.
The genus Larcospira begins the interesting series of the Larcospirida, or
of those Lithelida in which the medullary shell is formed by a trizonal or
Larnacilla-shaped lattice-shell, and the cortical shell by spiral turnings of one of the
three girdles, which compose the cortical shell of the Pylonida. In Larcospira, as the
oldest and most simple form of Larcospirida, the spiral is formed by the transverse girdle, or the
first girdle of the Diplozonaria, the only cortical girdle of Amphipyle. If in this genus
one of both wings of the transverse girdle grow stronger than the other and overgrow the latter,
turning {696}around the principal axis, we reach the
characteristic form of Larcospirema, the first subgenus of Larcospira; but if
afterwards the second wing follow the example of the first, and overgrow it from the other side,
we reach the typical form of the second subgenus, Larcospironium. In this latter subgenus
the spiral becomes double, whilst in the former it remains simple.
Subgenus 1. Larcospirema, Haeckel.
Definition.—Spiral convolutions or turns of the cortical shell simple;
only one single wing of the transverse girdle turning around the principal axis.
1. Larcospira lentelliptica, n. sp.
Cortical shell with smooth surface, four times as long as the lentelliptical medullary shell.
Perimeter of the lateral plane elliptical, one and a half times as long as broad, without
constrictions. One lateral wing of the transverse girdle is more strongly developed and turns
around the other in two to three simple spiral turns.
Dimensions.—Length of the cortical shell 0.2, breadth 0.16; length of the
medullary shell 0.05, breadth 0.04.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms.
2. Larcospira quadrangula, n. sp. (Pl. 49, fig.
3).
Cortical shell with thorny surface, three times as long as the lentelliptical medullary shell.
Perimeter of the lateral plane nearly quadrangular, with four rounded corners (on the poles of two
crossed diagonal axes), one and a third times as long as broad, with one sagittal constriction at
the poles of the principal axis. One lateral wing of the transverse girdle turns around the other
in one and a half to two simple spiral turns.
Dimensions.—Length of the cortical shell 0.18, breadth 0.14; length of the
medullary shell 0.06, breadth 0.04.
Habitat.—Pacific, central area, Station 274, depth 2750 fathoms.
3. Larcospira sexangula, n. sp.
Cortical shell with spiny surface, four times as long as the hexagonal medullary shell.
Perimeter of the lateral plane nearly hexagonal, one and a half times as long as broad, with six
rounded corners (two on the poles of the principal axis, four on the poles of two crossed diagonal
axes), with three slight ring-like constrictions. One lateral wing of the transverse girdle turns
around the other in two to two and a half simple spiral turns.
Dimensions.—Length of the cortical shell 0.24, breadth 0.16; length of the
medullary shell 0.06, breadth 0.04.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
{697}
Subgenus 2. Larcospironium, Haeckel.
Definition.—Spiral convolutions or turns of the cortical shell double;
both wings of the transverse girdle turning around the principal axis.
4. Larcospira oliva, n. sp.
Cortical shell with smooth surface, six times as long as the lentelliptical medullary shell.
Perimeter of the lateral plane elliptical, one and a fourth times as long as broad, without
constrictions. Both lateral wings of the transverse girdle turn round one another and form one and
a half to two double spiral turns.
Dimensions.—Length of the cortical shell 0.25, breadth 0.2; length of the
medullary shell 0.04, breadth 0.035.
Habitat.—Antarctic Ocean, Station 157, depth 1950 fathoms.
Genus 307. Pylospira,[347] n. gen.
Definition.—Lithelida with double, trizonal or
Larnacilla-shaped medullary shell; cortical shell subspherical or lentelliptical,
constructed of a single or double spiral of the lateral girdle (or second cortical girdle); the
spiral lamella revolving round the sagittal axis.
The genus Pylospira follows after Larcospira as the second genus of
Larcospirida; but in this latter the spiral of the cortical shell is formed by the transverse
girdle (or the first lattice-girdle of the Diplozonaria), whilst in Pylospira it is
produced by the lateral girdle, or the second lattice-girdle of that group. Therefore
Pylospira may be derived phylogenetically from Tetrapyle in the same manner as
Larcospira from Amphipyle. Whilst in this latter the first cause of the spiral
turning, the unequal growth of both girdle-wings, proceeds from the transverse girdle, in
Pylospira it proceeds from the lateral girdle. One of its wings overgrows the other,
turning around the sagittal axis. If the second wing do not become developed, the spiral remains
simple and represents the subgenus Pylospirema; but if afterwards the second wing follow
the example of the first and overgrow it from the other side, we reach the typical form of the
second subgenus, Pylospironium, with a double spiral.
Subgenus 1. Pylospirema, Haeckel.
Definition.—Spiral convolutions or turns of the cortical shell simple,
only one single wing of the lateral girdle turning around the sagittal axis.
{698}
1. Pylospira tetrapyle, n. sp.
Cortical shell with thorny surface, five times as long as the lentelliptical medullary shell.
Perimeter of the lateral plane elliptical, one and a half times as long as broad. Four internal
gates (as in Tetrapyle, between the complete lateral wings of the transverse girdle)
kidney-shaped. One principal wing of the lateral girdle turns around the other in one and a half
to two simple spiral turns.
Dimensions.—Length of the cortical shell 0.21, breadth 0.15; length of the
medullary shell 0.04, breadth 0.03.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
2. Pylospira octopyle, n. sp. (Pl. 49,
fig. 4).
Cortical shell with thorny surface, three times as long as the lentelliptical medullary shell.
Perimeter of the lateral plane elliptical or nearly hexagonal, with four rounded corners (on the
poles of two crossed diagonal axes), one and a third times as long as broad. Eight internal
egg-shaped gates (as in Octopyle), between the complete lateral wings of the transverse
girdle, and two axial beams in the principal axis. One single wing of the lateral girdle turns
around the other in two to three simple spiral turns.
Dimensions.—Length of the cortical shell 0.18, breadth 0.14; length of the
medullary shell 0.06, breadth 0.035.
Habitat.—North Pacific, Station 253, depth 3125 fathoms.
Subgenus 2. Pylospironium, Haeckel.
Definition.—Spiral convolutions or turns of the cortical shell double;
both wings of the lateral girdle turning around the sagittal axis.
3. Pylospira cymbium, n. sp.
Cortical shell with smooth surface, seven times as long as the lentelliptical medullary shell.
Perimeter of the lateral plane elliptical, one and a third times as long as broad. Four internal
kidney-shaped gates between the complete lateral wings of the transverse girdle (as in
Tetrapyle). Both principal wings of the lateral girdle turn round one another in two to two
and a half double spiral turns.
Dimensions.—Length of the cortical shell 0.22, breadth 0.17; length of the
medullary shell 0.03, breadth 0.025.
Habitat.—Indian Ocean, Madagascar, Rabbe, surface.
{699}
Genus 308. Tholospira,[348] n. gen.
Definition.—Lithelida with double, trizonal or
Larnacilla-shaped medullary shell; cortical shell subspherical or lentelliptical
constructed of a simple or double spiral of the sagittal girdle (or third cortical girdle); the
spiral lamella revolving around the transverse axis.
The genus Tholospira represents the third genus of Larcospirida. Whilst the
spiral growth of the cortical shell is produced in Larcospira by the first (transverse)
girdle of the Diplozonaria, in Pylospira by the second (lateral) girdle, in
Tholospira it is effected by the third or sagittal girdle, which we found complete in
Pylonium. If in this genus one wing of the sagittal girdle overgrow the other remaining one
and turn around the transverse axis, we get Tholospirema, the first subgenus of our genus,
with simple spiral; but if afterwards the second wing follow the example of the first, and
overgrow it from the other side, we get Tholospironium, with double spiral.
Subgenus 1. Tholospirema, Haeckel.
Definition.—Spiral convolutions or turns of the cortical shell simple,
only one single wing of the sagittal girdle turning around the transverse axis.
1. Tholospira nautiloides, n. sp.
Cortical shell with smooth surface, five times as long as the lentelliptical medullary shell.
Perimeter of the lateral plane elliptical, one and a half times as long as broad. Four internal
kidney-shaped gates (between the lateral wings of the transverse girdle, as in Tetrapyle).
One wing of the sagittal girdle turns around the other in two to two and a half simple spiral
turns.
Dimensions.—Length of the cortical shell 0.24, breadth 0.18; length of the
medullary shell 0.05, breadth 0.04.
Habitat.—Indian Ocean, between Ceylon and Socotra, Haeckel,
surface.
2. Tholospira spinosa, n. sp.
Cortical shell covered with numerous (sixty to eighty or more) simple, bristle-like radial
spines, about the length of the shell. Perimeter of the lateral plane nearly quadrangular, with
four rounded corners (on the poles of two crossed diagonal axes), one and a half times as long as
broad. Eight internal egg-shaped gates (between the lateral wings of the transverse girdle and two
axial {700}beams in the principal axes, as in
Octopyle). One wing of the sagittal girdle turns around the other in two to three simple
spiral turns.
Dimensions.—Length of the cortical shell 0.27, breadth 0.18; length of the
medullary shell 0.04, breadth 0.03.
Habitat.—South Pacific, Station 288, surface.
3. Tholospira dendrophora, n. sp. (Pl. 49, fig.
6).
Cortical shell covered with numerous (forty to fifty or more) branched radial spines, about
half as long as the shell; each spine with two to six dichotomous branches. Perimeter of the
lateral plane elliptical, one and a third times as long as broad. Eight internal egg-shaped gates,
as in the foregoing species. One single wing of the sagittal girdle turns around the other in
three to four spiral turns.
Dimensions.—Length of the cortical shell 0.22, breadth 0.17; length of the
medullary shell 0.05, breadth 0.04.
Habitat.—Pacific, central area, Station 272, depth 2600 fathoms.
Subgenus 2. Tholospironium, Haeckel.
Definition.—Spiral convolutions of the cortical shell double; both
wings of the sagittal girdle turning around the transverse axis.
4. Tholospira hystrix, n. sp.
Cortical shell covered with numerous (sixty to eighty or more) simple conical spines, about
half as long as the shell. Perimeter of the lateral plane hexagonal, one and a half times as long
as broad. Four internal kidney-shaped gates, as in Tetrapyle. Both wings of the sagittal
girdle turn round one another in two to two and a half double spiral turns.
Dimensions.—Length of the cortical shell 0.21, breadth 0.14; length of the
medullary shell 0.04, breadth 0.03.
Habitat.—North Atlantic, Station 354, surface.
5. Tholospira cervicornis, n. sp. (Pl. 49, fig.
5).
Cortical shell covered with numerous (forty to fifty or more) branched radial spines; each
spine antler-shaped, about as long as the medullary Larnacilla-shell, with eight to twelve
dichotomous branches (similar to Cromyodrymus abietinus, Pl. 30, fig. 6).
Perimeter of the lateral plane elliptical, one and a third times as long as broad. Four internal
kidney-shaped gates, as in Tetrapyle. Both wings of the sagittal girdle turn round one
another in one and a half to two double spiral turns.
Dimensions.—Length of the cortical shell 0.2, breadth 0.15; length of the
medullary shell 0.03, breadth 0.025.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
{701}
Genus 309. Spironium,[349] n. gen.
Definition.—Lithelida with double, trizonal or
Larnacilla-shaped medullary shell; cortical shell subspherical or lentelliptical,
constructed of two crossed spirals, which arise from both lateral wings of one girdle (commonly
the lateral girdle) and revolve in an opposite diagonal direction around the principal axis.
The genus Spironium differs in a very remarkable manner from all foregoing
Lithelida, and is distinguished by a quite peculiar mode of growth. It is most nearly allied to
Larcopyle, and may, like this, be derived from Amphipyle (or rather from
Larnacilla, beginning to transform into Amphipyle). But whilst in Larcopyle
one of the two wings of the lateral girdle overgrows the other in the direction of the transverse
axis (turning around the principal axis), here in Spironium both lateral wings begin at the
same time to grow out from the lateral sides of the Larnacilla-shaped medullary shell; the
most remarkable thing is, however, that the direction of growth in the wings is diverse from the
beginning: the left wing grows downwards and turns around the lower (aboral) pole of the principal
axis, the right wing grows upwards and turns around the upper (oral) pole of the same axis. Thus
both wings of the lateral girdle are crossed in diagonal axes, and with increasing growth one
overgrows the other in the direction of these diagonals, so as to resemble the figure 8 in shape.
The open gates remaining between the turnings of the girdle become afterwards closed on the
surface by irregular lattice-work, and so the whole cortical shell assumes finally a spherical,
ellipsoidal, or lentelliptical form. Its surface sometimes becomes covered with simple or branched
radial spines. In the interior the eight characteristic egg-shaped gates of Octopyle are
commonly (or constantly?) visible, two strong radial beams in the principal axis arising from the
poles of the lentelliptical medullary shell.
Subgenus 1. Spironetta, Haeckel.
Definition.—Surface of the shell smooth or rough, but not with radial
spines.
1. Spironium octonium, n. sp. (Pl. 49,
fig. 7).
Cortical shell lentelliptical, its breadth (or transverse axis) surpassing considerably the
length (or the principal axis). Surface thorny and somewhat hump-backed. The spiral wings of the
transverse girdle have about the same breadth as the eight internal gates between them and the
axial beams.
Dimensions.—Length of the cortical shell (or principal axis) 0.15, breadth (or
transverse axis) 0.2; length of the medullary shell 0.05, breadth 0.04.
Habitat.—South Atlantic, west of Tristan da Cunha, Station 332, depth
2200 fathoms.
{702}
2. Spironium diagonale, n. sp.
Cortical shell nearly spherical, four times as great as the subspherical medullary shell.
Surface rough. The spiral wings of the transverse girdle about half as broad as the eight internal
gates between them and the axial beams.
Dimensions.—Diameter of the cortical shell 0.16, of the medullary shell 0.04.
Habitat.—Indian Ocean, Zanzibar, Pullen, depth 2200 fathoms.
Subgenus 2. Spironilla, Haeckel.
Definition.—Surface of the shell covered with simple or branched
radial spines.
3. Spironium spinosum, n. sp.
Cortical shell subspherical, five times as great as the subspherical medullary shell. Surface
covered with numerous (sixty to eighty or more) simple, bristle-like radial spines, longer than
the shell. The spiral wings of the transverse girdle of about the same breadth as the eight
internal gates between them and the axial beams.
Dimensions.—Diameter of the cortical shell 0.2, of the medullary shell 0.04.
Habitat.—Pacific, central area, Station 274, surface.
4. Spironium arbustum, n. sp.
Cortical shell lentelliptical, its breadth surpassing its length considerably. Surface covered
with numerous (forty to sixty or more) thin radial spines, about as long as the greatest diameter
of the shell; each spine with two to six lateral branches, which are either simple or again
branched (similar to Cromyodrymus abietinus, Pl. 30, fig. 6).
The spiral wings of the transverse girdle only half as broad as the eight internal gates between
them and the strong beams of the principal axis.
Dimensions.—Length of the cortical shell 0.12, breadth 0.15; length of the
hexagonal medullary shell 0.05, breadth 0.04.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
Family XXX. Streblonida, n. fam. (Pl. 49, figs. 8,
9).
Definition.—Larcoidea with asymmetrical, spiral,
polythalamous shell, composed of a variable number of roundish chambers, which form together an
ascending spiral; both halves of the shell unequal. Primordial chamber either simple or
Larnacilla-shaped.
The family Streblonida comprises those Larcoidea in which a number of chambers is arranged in an ascending spiral,
round a simple or trizonal primordial chamber, like winding stairs. They show the same spiral
structure as in the foregoing {703}family, but whilst in
the Lithelida the spiral line lies in one plane (as in Nautilus), in the Streblonida it
ascends like a screw (as in Helix). Therefore the former have the same relation, regarding
the spiral structure, to the nautiloid Polythalamia as the latter to the turbinoid Foraminifera.
Indeed the single forms of Streblemida repeat in their special structure the characteristic genera
of Turbinoida, such as Globigerina, Rosalina, Pulvinulina,
Hastigerina, &c. As in these calcareous turbinoid Foraminifera, so also in the
analogous siliceous Streblonida the distinction of species is very difficult and open to many
objections.
The number of species in this family is very small; all are rare and for the most part very
opaque and difficult to understand, so that the following distinction of a dozen species can have
only a provisional value. To get a complete idea of their peculiar structure, the shell must be
turned and observed from different sides, and thus their full study requires yet much time and
work. There are to be found evident transitional forms between them and the Lithelida on the one
hand and the Soreumida on the other. Besides this, most species of Streblonida seem to have more
inclination to individual varieties and abnormalities than the majority of the other
Radiolaria.
The general form of the whole shell is in the Streblonida sometimes more egg-shaped or even
subspherical, at other times more top-like or conical, sometimes nearly discoidal. The height of
the shell (or the vertical axis of the ascending spiral) is occasionally larger, at other times
smaller than the breadth (or the greatest horizontal diameter, perpendicular to the height). Some
very flat forms seem to approach the Lithelida. With regard to the internal screw-formation, the
shell of all Streblonida is asymmetrical.
The number of the aggregated incomplete chambers is commonly between ten and twenty, but
ascends sometimes to thirty, forty, or more. Sometimes the size of all the chambers is nearly the
same, sometimes they increase gradually, occasionally also very rapidly. The primordial chamber
(or the first and oldest) seems to be commonly the smallest, and inversely, the last and youngest
chamber, the largest. But sometimes (in Streblopyle) also the contrary may be the case. The
form of the single chambers is very variable, from the spherical or hemispherical through all
transitions leading to irregular roundish or longish forms. The network is commonly irregular,
with small roundish pores of different sizes, but sometimes also regular, circular. The surface of
the shell is commonly smooth or rough, rarely covered with radial spines. In most species the
reticulation and particularly the separation of the chambers is more or less incomplete.
As in the Lithelida, so also in the Streblonida we can distinguish two subfamilies. In the
Streblacanthida (Streblonia, Streblacantha) the primordial chamber is a simple,
spherical, subspherical, or lentelliptical latticed shell. In the Streblopylida
(Streblopyle) the primordial chamber is trizonal or Larnacilla-shaped, as in the
greater number of Larcoidea, composed of three elliptical latticed
girdles of unequal size, perpendicular one to another, and surrounding a simple central chamber.
As in the Lithelida, so also {704}here we cannot
certainly say whether the former have originated from the latter by reduction of the
Larnacilla-shell, or whether both groups be of different origin. The latter is perhaps more
probable. This family as well as the foregoing requires a much more careful study than I could
give to it.
Synopsis of the Genera of Streblonida.
|
I. Subfamily Streblacanthida.
Primordial chamber simple, spherical or lentelliptical.
|
|
Shell without radial spines, |
310. Streblonia. |
| Shell with radial spines, |
311. Streblacantha. |
|
II. Subfamily Streblopylida.
Primordial chamber trizonal or Larnacilla-shaped
|
|
Shell without radial spines, |
312. Streblopyle. |
| I. Subfamily Streblacanthida. Primordial chamber simple, spherical or
lentelliptical. |
|
|
Shell without radial spines, |
|
|
|
|
|
|
310. Streblonia. |
|
|
|
|
|
|
Shell with radial spines, |
|
|
|
|
|
|
311. Streblacantha. |
|
|
|
|
| II. Subfamily Streblopylida. Primordial chamber trizonal or
Larnacilla-shaped |
|
|
Shell without radial spines, |
|
|
|
|
|
|
312. Streblopyle. |
|
|
|
|
Genus 310. Streblonia,[350] n. gen.
Definition.—Streblonida with simple, spherical,
subspherical, or lentelliptical primordial chamber, beginning the screw-like series of spirally
ascending chambers. Surface smooth or thorny, without radial spines.
The genus Streblonia contains those Streblonida in which a variable number
of roundish, subspherical, or longish chambers form a screw-like aggregate, beginning with a quite
simple primordial chamber. The special order of the complex spiral offers interesting resemblances
to different genera of the calcareous Foraminifera, from which I have taken the corresponding name
of the species. The whole form of the shell is sometimes more egg-shaped or subspherical, at other
times more top-like or conical, occasionally very flat. Its surface is smooth or rough, but not
covered with radial spines.
1. Streblonia globigerina, n. sp.
Shell subspherical, thick walled, clustered, with eight to ten nearly spherical chambers, of
rapidly increasing size, the tenth chamber about twelve times as broad as the first. Breadth of
the shell nearly equal to the height. Pores subregular, circular, hexagonally formed, of about the
same breadth as the bars; about sixteen on the breadth of the tenth chamber. (Resembles very much
the common Globigerina.)
Dimensions.—Breadth of the shell 0.18, height 0.16.
Habitat.—North Pacific, Station 253, depth 3125 fathoms.
2. Streblonia uvigerina, n. sp.
Shell nearly egg-shaped, clustered, with eight to eleven subspherical chambers of gradually
increasing size, the tenth chamber about six times as broad as the first. Breadth of the shell
about {705}half its height. Pores of the shell
subregular, circular, about twice as broad as the bars; about twenty on the breadth of the tenth
chamber. (Resembles some species of Uvigerina.)
Dimensions.—Breadth of the shell 0.11, height 0.23.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
3. Streblonia polymorphina, n. sp.
Shell egg-shaped, thin walled, with twelve to fourteen roundish chambers of rapidly increasing
size, the tenth chamber about seven times as broad as the first. Breadth of the shell about
two-thirds of the height. Pores irregular roundish, twice as broad as the bars; about twenty on
the tenth chamber. (Resembles certain forms of Polymorphina.)
Dimensions.—Breadth of shell 0.22, height 0.14.
Habitat.—Pacific, central area, Station 266, depth 2750 fathoms.
4. Streblonia bulimina, n. sp.
Shell nearly egg-shaped, thick walled, clustered, with fourteen to eighteen egg-shaped chambers
of rapidly increasing size, the tenth chamber about eight times as broad as the first. Breadth of
the shell about two-thirds of the height. Pores irregular, roundish, half as broad as the bars;
about twelve on the breadth of the tenth chamber. (Resembles closely Bulimina.)
Dimensions.—Breadth of the shell 0.17, height 0.24.
Habitat.—Pacific, central area, Station 268, depth 2900 fathoms.
5. Streblonia rosalina, n. sp.
Shell top-shaped or flatly conical, with twelve to sixteen chambers of gradually increasing
size, the tenth chamber about four times as broad as the first. Breadth of the shell twice as
large as the height. Pores subregular, circular, very small, of the same breadth as the bars;
about twelve on the breadth of the tenth chamber. (Resembles some forms of Rosalina.)
Dimensions.—Breadth of the shell 0.27, height 0.13.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
6. Streblonia planorbulina, n. sp.
Shell very flatly conical, nearly lenticular or discoidal, with twenty to twenty-five chambers
of nearly equal size, the tenth chamber a little broader than the first. Breadth of the shell
exceeds five to six times the height. Pores subregular, circular, very small, half as broad as the
bars; about eight on the breadth of the tenth chamber. (Resembles closely
Planorbulina.)
Dimensions.—Breadth of the shell 0.3 to 0.4, height 0.06 to 0.07.
Habitat.—Indian Ocean, Zanzibar, Pullen, depth 2200 fathoms.
{706}
7. Streblonia pulvinulina, n. sp.
Shell top-shaped or flatly conical, with thirty to forty chambers of slowly increasing size,
the tenth chamber about three times as broad as the first. Breadth of the shell exceeds three
times the height. Pores circular, subregular, very small, about one-third as broad as the bars;
about thirty in the breadth of the tenth chamber. (Resembles closely Pulvinulina.)
Dimensions.—Breadth of the shell 0.25, height 0.08.
Habitat.—Pacific, central area, Station 263, depth 2650 fathoms.
Genus 311. Streblacantha,[351] n.
gen.
Definition.—Streblonida with simple, spherical,
subspherical, or lentelliptical primordial chamber, beginning the screw-like series of spirally
ascending chambers. Surface covered with radial spines.
The genus Streblacantha differs from the nearly allied Streblonia
only in the covering of radial spines, and bears therefore the same relation to it as
Hastigerina has to Globigerina amongst the similar calcareous Polythalamia.
1. Streblacantha siderolina, n. sp. (Pl. 49, figs. 8,
8a).
Streblonia siderolina, Haeckel, 1883, MS.
Shell flatly conical, with fourteen to sixteen nearly hemispherical chambers of gradually
increasing size, the tenth chamber about six times as broad as the first. Breadth of the shell
nearly equal to the height. Pores subregular, circular, hexagonally framed, twice as broad as the
bars; about nine pores on the breadth of the tenth chamber. Surface covered with numerous short
conical radial spines, one-fourth to one-sixth as long as the diameter of the shell.
Dimensions.—Breadth of the shell 0.15, height 0.17.
Habitat.—South Pacific, Station 298, depth 2225 fathoms.
2. Streblacantha calcarina, n. sp.
Shell conical, with sixteen to eighteen roundish chambers of gradually increasing size, the
tenth chamber about three times as broad as the first. Breadth of the shell about one and a half
times the height. Pores irregular, roundish. Surface covered with numerous strong conical radial
spines, about half as long as the diameter of the shell.
Dimensions.—Breadth of the shell 0.24, height 0.17.
Habitat.—South Pacific, Station 285, depth 2375 fathoms.
{707}
3. Streblacantha hastigerina, n. sp.
Shell nearly spherical, clustered, with nine to eleven nearly spherical chambers of rapidly
increasing size, the tenth chamber about six times as broad as the first. Breadth of the shell
nearly equal to the height. Pores subregular, circular, of about the same breadth as the bars.
Surface bristly, covered with numerous very thin and long, needle-shaped radial spines, longer
than the diameter of the shell. (Resembles closely Hastigerina.)
Dimensions.—Breadth of the shell 0.18, height 0.15.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
Genus 312. Streblopyle,[352] n. gen.
Definition.—Streblonida with trizonal lentelliptical
medullary shell (composed like Larnacilla of three elliptical dimensive girdles surrounding
one simple central primordial chamber). From this begins a screw-like series of spirally ascending
chambers. Surface smooth or thorny, without radial spines.
The genus Streblopyle presents externally the same appearance and contour as
Streblonia, and is composed like this of a variable number of chambers, ascending
screw-like around the axis of the spiral shell. The first or primordial chamber, however, in which
the growth begins, is in Streblonia a simple spherical shell, but in Streblopyle a
trizonal shell or Larnacilla-shell (compare above, p. 600). The
chambers are very incompletely separated, and comparatively much larger, their number much smaller
than in Streblonia. The structure in the species of this genus is difficult to
understand.
1. Streblopyle helicina, n. sp. (Pl. 49,
fig. 9).
Shell helicoid, one and a third times as high as broad, with eight to twelve incomplete
semizonal chambers, ascending spirally from the lateral half girdle of the lentelliptical
medullary shell, octopyle-shaped, and enveloping it in three to four spiral turnings. The height
of the whole cortical shell equals nearly five times the height of the trizonal medullary shell.
Pores irregular, roundish. Surface of the shell rough or nearly smooth. (This species seems to be
nearly allied to Spironium octonium.)
Dimensions.—Breadth of the spiral cortical shell 0.18, height 0.24; breadth of the
medullary shell 0.04, height 0.05.
Habitat.—Pacific, central area, Station 266, depth 2750 fathoms.
2. Streblopyle spirulina, n. sp.
Shell egg-shaped or nearly spherical, about as high as broad, with eight to nine semizonal
chambers, ascending spirally from the subspherical trizonal medullary shell, and enveloping it in
{708}four to five turnings. The sixth chamber twice as
broad as the trizonal medullary shell. Pores irregular, roundish. Surface of the shell
thorny.
Dimensions.—Breadth of the spiral cortical shell 0.27, height 0.25; breadth of the
medullary shell 0.05, height 0.06.
Habitat.—Pacific, central area, Station 265, depth 2900 fathoms.
Family XXXI. Phorticida, Haeckel (Pl. 49, figs. 10,
11).
Phorticida, Haeckel, 1881, Prodromus, p. 464.
Definition.—Larcoidea with quite irregular monothalamous
shell, representing irregular modifications of an original lentelliptical latticed shell; the
irregular cortical shell encloses a regular or subregular, lentelliptical or trizonal medullary
shell.
The family Phorticida comprises a small number of Larcoidea in which a
subregular, trizonal, lentelliptical medullary shell is enclosed by an irregular simple or spongy
cortical shell. The lattice-work of the latter is sometimes simple and complete, at other times
incomplete, with open gates (as in the Pylonida), sometimes also spongy. Its form is always more
or less irregular, roundish, often dimply or tuberous; different from most other Larcoidea.
The medullary shell is constantly a regular or subregular Larnacilla-shell, composed of
three elliptical latticed girdles of different sizes, perpendicular one to another. This leaves no
doubt that the Phorticida are true Larcoidea. The connection of it with
the cortical shell is rarely effected by radial beams, commonly by two opposite latticed wings,
which are identical with the lateral halves of the transverse girdle in the Pylonida diplozonaria
(Amphipyle, Tetrapyle). Often also between this transverse and a second (lateral)
girdle there remain large open gates, so that the affinity between these Phorticida and the
Pylonida cannot be doubted. In other cases these gates become closed, so that they more nearly
approach the Larnacida. From both families they differ by the irregularity of the papillate or
tuberous cortical shell. The network is more or less irregular, its surface often thorny, but
never covered with symmetrically disposed radial spines.
The central capsule is lentelliptical, encloses the medullary shell, and is enveloped by the
cortical shell, as in the nearly allied Pylonida and Larnacida, of which the Phorticida may be
regarded as irregular aberrant forms.
Synopsis of the Genera of Phorticida.
| Cortical shell simply latticed, |
313. Phorticium. |
| Cortical shell spongy, |
314. Spongophortis. |
{709}
Genus 313. Phorticium,[353] Haeckel,
1881, Prodromus, p. 464.
Definition.—Phorticida with irregular cortical shell of
simple lattice-work, enclosing a lentelliptical Larnacilla-shaped medullary shell.
The genus Phorticium comprises all Phorticida in which the irregular
cortical shell is formed by simple lattice-work, not by spongy framework. We can divide this genus
into two subgenera: in Phortopyle (as in the Pylonida) the lattice-work of the
cortical shell exhibits large openings or gates; in Phortolarcus these gates are perfectly
closed by network; the former may be regarded as abnormal or irregular Pylonida, the latter as
modifications of Larnacida.
Subgenus 1. Phortopyle, Haeckel.
Definition.—Lattice-work of the irregular cortical shell incomplete,
with large openings or gates.
1. Phorticium pylonium, n. sp. (Pl. 49,
fig. 10).
Cortical shell irregular, roundish, about three times as large as the enclosed lentelliptical,
regular, Larnacilla-shell, connected with it by some radial beams and irregularly latticed
girdles; between these remain four to eight large open gates of irregular roundish form and size;
and these gates are the same as in Tetrapyle and Octopyle. This very variable
species may be regarded as a monstrosity of those genera of Pylonida; it is very common, but all
individuals are more or less unequal; some specimens approach to some common species of
Tetrapyle. The surface of the shell is more or less spiny.
Dimensions.—Diameter of the irregular cortical shell 0.12 to 0.18; length of the
lentelliptical medullary shell 0.05 to 0.06, breadth 0.035 to 0.045.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific, &c.,
common, surface and in various depths.
2. Phorticium spironium, n. sp.
Cortical shell irregular, roundish, tuberous, about four times as large as the enclosed
subregular Larnacilla-shell, connected with it by some irregular radial beams, and by
opposite, spirally begining, irregularly latticed girdles, comparable to those of
Spironium; between them remain six to twelve large open gates of irregular size and form.
Surface rough. The resemblance to some forms of Spironium makes it probable that this
species is a deformity or monstrosity of that genus.
Dimensions.—Diameter of the irregular cortical shell 0.12 to 0.2, of the
lentelliptical medullary shell 0.03 to 0.06.
Habitat.—Pacific, central area, Stations 270 to 274, surface, and in
various depths.
{710}
Subgenus 2. Phortolarcus, Haeckel.
Definition.—Lattice-work of the irregular cortical shell complete,
without large openings or gates.
3. Phorticium deforme, n. sp.
Cortical shell irregular, roundish or longish, three times as large as the enclosed subregular,
lentelliptical Larnacilla-shell, connected with it by two opposite latticed wings (the
halves of the transverse girdle of Tetrapyle). Network of the cortical shell irregular,
dense, perfectly closed, without larger openings or gates. Surface thorny. (May be regarded as a
monstrous form of Larnacalpis.)
Dimensions.—Diameter of the irregular cortical shell 0.15 to 0.18, of the
medullary shell 0.04 to 0.06.
Habitat.—Pacific, central area, Station 272, depth 2600 fathoms.
4. Phorticium abnorme, n. sp.
Cortical shell irregular, roundish, tuberous, with five to ten quite irregular or nearly
hemispherical protuberances, which resemble the cupolas of Zonarida. The regular lentelliptical
Larnacilla-shell is one-third to one-fourth as large as the enclosing cortical shell, and
is connected with it by some irregular radial beams. Lattice-work completely closed, without
gates. Surface spiny. (May be regarded as an anomalous form of Zonidium; as in the other
species of this variable genus, the individuals are very unequal.)
Dimensions.—Diameter of the irregular cortical shell 0.12 to 0.2, of the medullary
shell 0.03 to 0.05.
Habitat.—Atlantic and Pacific, tropical zone, surface, and at various
depths.
Genus 314. Spongophortis,[354] Haeckel,
1881, Prodromus, p. 464.
Definition.—Phorticida with irregular cortical shell of
spongy framework, enclosing a lentelliptical Larnacilla-shaped medullary shell.
The genus Spongophortis differs from Phorticium in the spongy
framework of the cortical shell. This encloses the inner Larnacilla-shaped medullary shell
either directly, or both shells are separated by a hollow interval, and connected either by radial
beams or by latticed lamellæ. Perhaps both these subgenera might be better separated as
genera.
{711}
Subgenus 1. Stypophorticium, Haeckel.
Definition.—Spongy cortical shell immediately enclosing the
lentelliptical medullary shell, without hollow interval.
1. Spongophortis spongiosa, n. sp.
Cortical shell irregular, roundish, rough or tuberous, composed of loose spongy framework,
which immediately envelops the lentelliptical central Larnacilla-shell; the diameter of the
former becomes about five to six times as large as that of the latter.
Dimensions.—Diameter of the spongy cortical shell 0.15 to 0.25, of the trizonal
medullary shell 0.03 to 0.04.
Habitat.—Pacific, central area, Station 274, surface.
Subgenus 2. Spongophorticium, Haeckel.
Definition.—Spongy cortical shell separated by a hollow interval from
the lentelliptical medullary shell.
2. Spongophortis radiosa, n. sp.
Cortical shell irregular, roundish, four to five times as large as the enclosed lentelliptical
Larnacilla-shell, with which it is connected by ten to twenty irregularly disposed radial
beams. Spongy framework compact, about as thick as the medullary shell. Surface covered with
numerous short, bristle-shaped, radial spines.
Dimensions.—Diameter of the spongy cortical shell 0.15 to 0.2, of the trizonal
medullary shell 0.035 to 0.045.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
3. Spongophortis larnacilla, n. sp. (Pl. 49, figs.
11a-11d).
Cortical shell irregular, roundish, tuberous, three to four times as large as the enclosed
lentelliptical Larnacilla-shell, connected with it by two opposite latticed wings (the
halves of the transverse girdle of Tetrapyle). Spongy framework compact, about half as
thick as the medullary shell. Surface rough. (May be regarded as an abnormal Tetrapyle or
Larnacalpis, with an irregular spongy cortical shell.)
Dimensions.—Diameter of the spongy cortical shell 0.16 to 0.2, of the trizonal
medullary shell 0.04 to 0.06.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
{712}
Family XXXII. Soreumida, Haeckel (Pl. 49, figs. 12,
13).
Soreumida, Haeckel, 1881, Prodromus, p. 464.
Definition.—Larcoidea with quite irregular polythalamous
shell, composed of a variable number of chambers, aggregated without any definite order.
Primordial chamber either simple or Larnacilla-shaped.
The family Soreumida contains a small number of Larcoidea, different from most other Sphærellaria in
the complete irregularity of the polythalamous shell, which is composed of a variable number of
roundish chambers or subspherical latticed shells, aggregated in the form of an irregular heap. We
can distinguish in this family only two genera, with very different structure of the central
medullary shell or the first chamber beginning the growth; and these correspond to the two
subfamilies of the nearly allied Streblonida (p. 704). In Soreuma
(as in Streblonia) the first or primordial chamber, from which the growth begins, is like
the others, a simple spherical or irregular roundish lattice-shell. In Sorolarcus, however
(as in Streblopyle), the first or primordial chamber is a trizonal or
Larnacilla-shell. It is not improbable that the former originated phylogenetically from
Streblonia, the latter from Streblopyle, by loss of the original spiral order of
growth. But it is also possible that these groups have no nearer relation. Among the calcareous
Foraminifera a very similar form is represented by Acervulina and its allies.
The general form of the whole shell in the Soreumida is sometimes more egg-shaped or
lentelliptical, at other times even subspherical, occasionally quite irregular, tuberous, or
clustered. The number of the aggregated chambers is very variable, in Sorolarcus between
ten and thirty, in Soreuma ascending to fifty to eighty, sometimes from one hundred to one
hundred and fifty and more. Their size is sometimes nearly equal, at other times very different,
their form commonly very irregular, roundish, but sometimes also subspherical or egg-shaped. The
network of the shell is also commonly irregular, with roundish pores of different sizes. The
surface is usually smooth or rough, rarely covered with radial spines.
The central capsule is not known, as I observed only a few skeletons of this family.
Synopsis of the Genera of Soreumida.
| Primordial chamber of the shell simple, subspherical or
roundish, |
315. Soreuma. |
| Primordial chamber of the shell trizonal or
Larnacilla-shaped, |
316. Sorolarcus. |
Genus 315. Soreuma,[355] Haeckel, 1881,
Prodromus, p. 464.
Definition.—Soreumida with numerous chambers, aggregated
without any regularity around one simple, spherical or subspherical, central chamber.
{713}
The genus Soreuma contains those Soreumida in which no trace of any regular
structure is found, but all the chambers of the irregular shell are without any order, aggregated
around a simple spherical or subspherical central chamber or medullary shell. Soreuma may
have originated either from Sorolarcus by loss of the central Larnacilla-shell or
from Cenolarcus by irregular apposition of new chambers around the lentelliptical central
chamber or simple Larcoid-shell. Some species seem to exhibit a transition to Sorolarcus.
Owing to the absolute irregularity of the polythalamous shell Soreuma resembles
Acervulina among the Foraminifera.
Subgenus 1. Soreumium, Haeckel.
Definition.—Shell without radial spines.
1. Soreuma irregulare, n. sp. (Pl. 49,
fig. 12).
Shell irregular, clustered, or tuberous, composed of a large number (one hundred and twenty to
one hundred and fifty or more) of irregular, roundish chambers of very different sizes, the
largest four to five times as broad as the smallest. Network very delicate, with very small
roundish pores, to five times as broad as the bars. Surface thorny.
Dimensions.—Diameter of the shell 0.3, of the single chambers 0.02 to 0.08.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
2. Soreuma acinosum, n. sp.
Shell irregularly lentelliptical, with different growth in the three dimensions, composed of a
large number (forty to sixty or more) of irregular, roundish chambers of very different sizes, the
largest six to eight times as broad as the smallest. Pores subregular, circular, twice as broad as
the bars. Surface smooth.
Dimensions.—Length of the shell 0.21, breadth 0.17, height 0.13; diameter of the
largest chambers 0.03, of the smallest 0.004.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
3. Soreuma subglobosum, n. sp.
Shell nearly spherical, composed of a variable number (twelve to fifteen or more) of
irregularly aggregated subspherical chambers of nearly equal size. Pores subregular, circular,
twice as broad as the bars. Surface thorny.
Dimensions.—Diameter of the shell 0.25, of the largest chambers 0.04, of the
smallest 0.005.
Habitat.—Pacific, central area, Station 266, depth 2750 fathoms.
{714}
4. Soreuma acervulina, n. sp.
Shell quite irregular, cloddy, or tuberous, composed of twenty to thirty (or more) roundish
chambers of almost uniform size, the largest twice to three times as broad as the smallest. Pores
irregular, roundish. Surface smooth.
Dimensions.—Diameter of the shell 0.18 to 0.24, of the largest chambers 0.06, of
the smallest 0.02.
Habitat.—South Pacific, Station 295, depth 1500 fathoms.
Subgenus 2. Soreumidium, Haeckel.
Definition.—Shell with radial spines.
5. Soreuma spinosum, n. sp.
Shell quite irregular, cloddy, or tuberous, composed of thirty to forty subspherical chambers
of nearly the same size. Pores subregular, circular, twice as broad as the bars; on the equator of
each chamber six to eight pores. Surface thorny, covered with irregularly scattered conical radial
spines, about as long as the diameter of the chambers.
Dimensions.—Diameter of the shell 0.17 to 0.25, of the chambers 0.04.
Habitat.—North Pacific, Station 241, depth 2300 fathoms.
6. Soreuma setosum, n. sp.
Shell nearly spherical, composed of sixty to seventy (or more) irregular, roundish chambers of
very different sizes, the largest five to six times as broad as the smallest. Pores irregular,
roundish. Surface bristly, covered with very numerous, long and thin, bristle-shaped radial
spines, about as long as the diameter of the shell.
Dimensions.—Diameter of the shell 0.28, of the chambers 0.005 to 0.03.
Habitat.—Pacific, central area, Station 271, depth 2425 fathoms.
Genus 316. Sorolarcus,[356] n. gen.
Definition.—Soreumida with numerous chambers, aggregated
irregularly around a trizonal medullary shell or Larnacilla-shell.
The genus Sorolarcus comprises those Soreumida in which the heap of
irregularly aggregated chambers encloses a central trizonal medullary shell, by which they
demonstrate clearly their descent from Larnacida or Pylonida. The lentelliptical medullary {715}shell exhibits quite the same characteristic structure as
that of Larnacilla, being composed of three elliptical latticed girdles, perpendicular one
to another. In some species also the beginning of a second system of girdles is clearly indicated,
so that there can be no doubt as to their derivation from Amphipyle or
Tetrapyle.
Subgenus 1. Sorolarcium, Haeckel.
Definition.—Shell without radial spines.
1. Sorolarcus larnacillifer, n. sp. (Pl. 49, fig.
13).
Shell irregular, clustered, or tuberous, composed of twenty to thirty irregular, roundish
chambers of very different size, the largest four to eight times as broad as the smallest,
aggregated without order around a central, lentelliptical, Larnacilla-shaped medullary
shell. Pores irregular, roundish, twice to four times as broad as the bars. Surface smooth or a
little spiny.
Dimensions.—Diameter of the whole shell 0.18, of the central
Larnacilla-shell 0.05.
Habitat.—Pacific, central area, Station 266, depth 2750 fathoms.
2. Sorolarcus tetrapylifer, n. sp.
Shell irregularly roundish, clustered, composed of ten to twelve irregular rather long chambers
of almost uniform size, the largest twice as broad as the smallest, aggregated without order
around a central shell of the structure of Tetrapyle, which encloses an inner trizonal
Larnacilla-shell of half the size. Pores irregular, roundish, twice to four times as broad
as the bars. Surface spiny.
Dimensions.—Diameter of the whole shell 0.25, of the outer (Tetrapyle-like)
medullary shell 0.12, of the inner (Larnacilla-like) shell 0.06.
Habitat.—Pacific, central area, Station 272, depth 2600 fathoms.
Subgenus 2. Sorolarcidium, Haeckel.
Definition.—Shell with radial spines.
3. Sorolarcus terminalis, n. sp.
Shell nearly spherical, composed of fifteen to eighteen irregularly aggregated roundish
chambers of nearly equal size; in the centre a lentelliptical Larnacilla-shell. Surface
covered with numerous thin, bristle-like radial spines, somewhat longer than the shell.
Dimensions.—Diameter of the whole shell 0.21, of the central
Larnacilla-shell 0.05.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
{716}
Legion II. ACANTHARIA,
vel Actipylea, vel Acanthometrea (Pls. 129-140).
Acantharia, Haeckel, 1881.
Actipylea, Haeckel, 1882.
Acanthometrea, Hertwig, 1879.
Panacantha, Haeckel, 1878.
Definition.—Radiolaria with simple membrane bounding the central capsule, which is
everywhere perforated by innumerable fine pores (disposed either equally or symmetrically).
Extracapsulum without phæodium. Skeleton centrogenous (its growth proceeding from the centre),
acanthinic (organic, not siliceous). Fundamental form originally spherical.
The legion Acantharia vel Actipylea, to the
extent here defined, was constituted by me, 1878, in my Protistenreich (p. 102) under the name
"Panacantha." A more accurate definition of this group was given in 1879
by Hertwig under the name Acanthometrea. Both names were replaced by me,
1881, in my Prodromus (pp. 421, 465) by the more convenient name Acantharia. This legion comprises all those Radiolaria which were first
described by Johannes Müller, 1858, as Acanthometrae, and also an important part of his
Haliomma. In my Monograph (1862, pp. 371-424) I disposed them in three families,
Acanthometrida, Diploconida, and Dorataspida.
Although the number of genera and species in this legion is much increased by the rich
collection of the Challenger, we can divide all Acantharia into two
different orders: Acanthometra (without complete lattice-shell) and Acanthophracta (provided with a complete lattice-shell).
The Acantharia agree with the Spumellaria in
the structure of the simple capsule-membrane, which is perforated by numerous small pores (but
constantly devoid of the large main opening, which the Nassellaria and
Phæodaria possess, being hence united as "Merotrypasta"). We can therefore
unite both former legions as "Holotrypasta" (compare above, pp. 5, 6); but in many Acantharia (if not in all?) the numerous
small pores of the capsule-membrane exhibit a certain peculiar arrangement not observed in the
Spumellaria; therefore the latter can be regarded as true "Peripylea" in
opposition to the former as "Actipylea."
The peculiar main character of all Actipylea or Acantharia is determined by the chemical constitution of their skeleton, which
is not silex, but a peculiar organic substance, called by me in 1862 "acanthin" (Monogr. d.
Radiol., pp. 30, 32). In all other Radiolaria the skeleton is composed of silex or of a silicate.
But besides this {717}chemical difference, an important
morphological character of the skeleton also separates the Acantharia from
all other Radiolaria: in the latter the skeleton is never centrogenous or arising from the centre
of the capsule; in strict opposition to this general fact the skeleton of all Acantharia is centrogenous, composed of radial spines, which arise from the
central point of the capsule and pierce its membrane. These characteristic "radial spines of
acanthin," arising from the centre, are never hollow (as formerly was supposed), but constantly
solid. Their form is extremely variable, and most important for the distinction of genera and
species; but more interesting from a general point of view is their peculiar arrangement or
disposition.
The regular disposition of twenty radial spines has general value almost for all Acantharia, with the exception only of the small group of Actinelida. In this latter group the number of radial spines is either more or
less than twenty, and their disposition is either quite irregular or follows a peculiar rule. The
number of individuals of these Actinelida, compared with that of the
other Acantharia, may be scarcely 1 per cent., whilst the latter have more
than 99 per cent.; the number of observed species is in the former about 5 per cent., in the
latter about 95 per cent. Nevertheless the small group of Actinelida is
very important, being probably the ancestral group from which all other Acantharia have been phylogenetically derived. These other Acantharia, with twenty regularly disposed radial spines, represent the two
large groups of Acanthonida and Acanthophracta.
For short and clear distinction of these two groups of Acantharia, we will
call the Actinelida (with irregular number and disposition of radial
spines) Adelacantha, in opposition to the Icosacantha (Acanthonida and
Acanthophracta), which all possess twenty regularly disposed radial
spines.
Johannes Müller, the great zoologist, to whom we are indebted for the first detection and
accurate knowledge of the Acanthometra, already recognised the regularity
in the peculiar disposition of their twenty radial spines (Abhandl. d. k. Akad. d. Wiss. Berlin,
1858, pp. 12, 37). In honour of my great master I have called this regular disposition the
"Müllerian law of spine disposition," and have given a full explanation of it in my Monograph
(1862, pp. 40-45, 371, 372). With regard to its general value for all Icosacantha (Acanthonida and Acanthophracta), we might also call
this promorphological Müllerian law "the Icosacanthan law."
In 1862 I had already given the following precise definition of this "Icosacanthan law"
(loc. cit., p. 40):—"Between two poles of a spineless axis are regularly disposed
five parallel zones, each with four radial spines; the four spines of each zone are equidistant
one from another, and also equidistant from each pole; and the four spines of each zone are so
alternating with those of each neighbouring zone, that all twenty spines together lie in four
meridian planes, which intersect one another at an angle of 45°." For the clear conception of
this remarkable Müllerian law, and for the complete understanding of its high value for the
complicated {718}morphology of all Icosacantha, it is
the most profitable way to retain constantly in mind for comparison the figure of a terrestrial
globe with its axis and zones. The axis of the globe is the spineless axis of all Icosacantha,
around which all twenty spines are symmetrically disposed; it is perpendicular to the bisecting
equatorial plane, in which lies the middle of the five parallel zones; therefore the four spines,
crossed perpendicularly in this equatorial plane, are called the equatorial spines (c1 to
c4 in the figures of Pls. 131-140); often,
and mainly in the family Quadrilonchida (Pl. 131),
these four equatorial spines are much larger or of a peculiar form, different from that of the
sixteen other spines. Each pair of the four equatorial spines lies in one equatorial axis, and
this latter is perpendicular to the crossing axis, in which lies the other pair of opposite
spines. We may regard these two equatorial diameters, perpendicular one to another and to the
spineless axis, as the two perradial axes or primary axes. Correspondingly the two meridian
planes, which are determined by one perradial axis and the spineless axis, may be called the two
primary or perradial meridian planes.
The globe is divided by the equatorial plane into two equal halves, the northern and the
southern hemisphere. In each hemisphere there are disposed quite symmetrically eight radial
spines, the distal ends of which fall in two parallel circles, a larger tropical circle (nearer to
the equator) and a smaller polar circle (nearer to the pole of the spineless axis). Therefore we
call the four spines of the former the "tropical spines" and the four spines of the latter the
"polar spines." The angle between the former and the equatorial plane is about 30°, the angle
between the latter and that plane about 60°.
The eight polar spines (four northern and four southern) lie in the same two meridian planes as
the four equatorial spines. Therefore in each of these two perradial planes lie six radial spines,
opposite in pairs; two equatorial and four polar spines. Commonly all eight polar spines are of
the same size and form; and often they are also equal to the eight tropical spines; but in some
cases (e.g., in some species of Quadrilonchida) they are much smaller than the twelve other
spines, and sometimes even rudimentary. In all figures of the Pls. 131-140 (and also
in my Monograph, 1862, Taf. xv.-xxii.) the polar spines of the northern circle are marked by the
characters a1 to a4, the polar spines of the southern circle by the characters
e1 to e4. In the first perradial meridian plane lie a1 and a3,
e1 and e3, in the second a2 and a4, e2 and e4.
The eight tropical spines lie between the eight polar and the four equatorial spines, four in
each hemisphere; their distal points fall in two parallel circles, which correspond exactly to the
two tropics of the globe. Therefore the four northern tropical spines may be called "canceral
spines" (as their ends fall in the Tropic of Cancer) and the four southern correspondingly
"capricornal spines" (as their points lie {719}in the
Tropic of the Capricorn). In the figures of the Pls. 131-140 (as well
as in my Monograph, 1862, Taf. xv.-xxii.) the four northern or canceral spines are marked by the
characters b1 to b4, and the four southern or capricornal spines by the characters
d1 to d4. Also the eight tropical spines lie (crossed in pairs) in two meridian
planes; they do not lie, however, in those perradial planes, in which are placed the twelve other
spines; but in two different meridian planes, crossing the former at angles of 45°; we call these
the "secondary" or "interradial" meridian planes. Each of these planes is determined by the
spineless axis and by two crossed interradial or secondary axes; in each of the latter lie two
opposite tropical spines. In the first interradial meridian plane lie b1 and b3,
d1 and d3, in the second b2 and b4, d2 and d4.
It is a most interesting and important fact, that in all Icosacantha (Acanthonida and Acanthophracta) this regular
disposition of the twenty spines (in five parallel zones and four meridian planes) becomes
constantly preserved by heredity, whilst the form and size of the different spines are extremely
varied by adaptation.
Only in a minority of the Icosacantha are all twenty spines perfectly equal or nearly equal in
size and form; and then it is often very difficult to distinguish the different zones in their
disposition. But in far the greater part the size or the form of the twenty spines becomes
different in different zones; and then we can commonly distinguish easily the five different
zones. Firstly, in all Quadrilonchida and Dorataspida, the four equatorial are distinguished from
the sixteen other spines either by form or by size, and often in a very remarkable degree. As soon
as these four principal spines are recognised, it is easy to determine also the sixteen others;
for the eight polar spines lie in the same two (perradial) meridian planes as the former, whilst
the eight tropical spines lie in two different (interradial) meridian planes, intersecting the two
former at angles of 45°. Commonly, therefore, this distinction is rather easy.
In the majority of the Icosacantha all four equatorial spines are exactly of the same form and
size. But in four families the two opposite spines of one equatorial axis are much larger, or of
another form, than those of the crossing axis. This is the case in the Amphilonchida, Belonaspida,
Hexalaspida, and Diploconida. Therefore we here call the major equatorial axis (with larger
spines) the "hydrotomical axis," and the minor axis (with smaller spines) the "geotomical axis."
Correspondingly, the meridian plane, in which the two larger equatorial spines are placed
(c1, c3) and the appertaining four polar spines (a1, a3, e1,
e3) may be called the "hydrotomical plane"; in the remarkable family of Hexalaspida (Pl. 139) all six
spines of this hydrotomical plane are much larger than the other fourteen. Perpendicular to this
plane is the second perradial meridian plane, which we call the "geotomical plane"; in it lie the
two smaller equatorial spines (c2, c4) and the corresponding four polar spines (a2,
a4, e2, e4). In some Hexalaspida (Hexonaspis and Hexacolpus)
the six spines of the hydrotomical plane become so preponderant that {720}the other fourteen spines appear rudimentary; and in some of them the two
equatorial spines of the hydrotomical plane are much larger than the four polar spines of the same
plane. This curious relation reaches its maximum in the Diploconida (Pl. 140).
The different development of the two equatorial axes (of the larger hydrotomical and the
smaller geotomical axis) is the first and most important cause of the peculiar forms, which are
produced in the four cited families. We derive these terms also from the metaphor of the
terrestrial globe. The hydrotomical plane is that meridian plane of the globe which intersects
almost only the water-hemisphere (the island of Ferro in the Atlantic, the island of Pandora in
the Pacific). Perpendicular to this is the geotomical plane, the meridian of which intersects
great land-masses in both hemispheres (Bombay in India, Athabasca in Canada). Both poles of the
smaller geotomical axis are everywhere equal (the East Indian and the Western American). However,
both poles of the larger hydrotomical axis (the eastern Atlantic and the western Pacific) are in
some genera very different, e.g., in Amphibelone among the Amphilonchida, and in
Zygostaurus among the Quadrilonchida. In this case we call the anterior (commonly more
developed) pole of the hydrotomical axis the frontal pole, the opposite posterior (commonly
smaller) the caudal pole (Pl. 131,
figs. 7, 8; Pl. 132,
figs. 9, 10). On both sides of these (right and left) lie symmetrically the two equal poles of the
geotomical lateral axis.
The promorphology of the Acantharia demonstrates that the geometrical
fundamental form in those groups is different. In the majority of the Acantharia, where the two equatorial axes are equal, that form is a double
square-pyramid or a "quadrate octahedron"; the four equal equatorial spines indicate the two
diagonals of the square, which is the common base of the united regular four-sided pyramids; their
common axis is the spineless axis of the body; the ends of the polar spines fall on the edges of
the pyramids, while the ends of the tropical spines fall on the halving lines of their faces.
However, in those Acantharia in which the two equatorial axes become
different, the square double pyramid becomes changed into a rhombic double pyramid; the common
base of the united pyramids is thus a rhombus; the hydrotomical axis is the larger, the geotomical
axis the smaller diagonal of the rhombus.
Opposed to the Icosacantha, under the name "Adelacantha," is the small group of Actinelida, in
which the number and disposition of the radial spines is variable, not determined by the Müllerian
law. Probably this group is the common ancestral stock, from which the Icosacantha have been
derived by gradual development of their peculiar disposition. Probably the oldest and most
primitive form of all Acantharia is Actinelius, in which a variable
and undetermined (often very large) number of radial spines is united in one common central point,
and therefore forms a needle-sphere. Whilst here all spines (often more than a hundred) are of
equal size and form, in the nearly allied Astrolophus large and small spines are
intermingled. Both genera together form the small ancestral family of Astrolophida. In the strange
family of Litholophida the radial spines do not {721}radiate within a spherical space (equally disposed in all directions),
but within a quadrant or even an octant, forming a conical brush or pencil.
One very remarkable form of Actinelida is Actinastrum, forming
the transition from these Adelacantha to the common regular Icosacantha. In the two observed
species of Actinastrum we find thirty-two radial spines, twenty of which are disposed after
the Müllerian law, as in the Icosacantha. The other twelve are four interradial equatorial spines
(lying in the two secondary meridian planes) and eight perradial tropical spines (lying in the two
primary meridian planes). Therefore here in each primary meridian plane are placed ten spines (two
equatorial, four tropical, and four polar spines), whereas in each secondary meridian plane are
placed six spines (two equatorial and four tropical). But here also all thirty-two spines are so
regularly placed that their free distal ends fall into five parallel zones, four in each polar
zone, eight in each tropical zone, and eight in the equatorial zone.
The Central Junction of the radial spines in the Acantharia
becomes effected in four different ways:—(1) by simple apposition of the pyramidal central
ends or bases; (2) by a basal leaf-cross, or by broad wings, four on each spine, supported one
upon the other; (3) by a central concrescence of the meeting bases of all the twenty spines,
growing perfectly together; and (4) by a concrescence in pairs of every two opposite spines. The
most common and probably the original mode of junction is the first—by pyramidal apposition;
the spines at the central base are pointed in the form of a pyramid, and the triangular faces of
the neighbouring pyramids are simply placed upon one another. Often the small basal pyramids are
imperfectly separated from the spines by an annular constriction. Commonly the basal pyramids of
the four equatorial spines are six-sided, those of the sixteen other spines five-sided.
The second mode of junction, by a basal leaf-cross, is developed from the first and appears as
a strengthening or a mechanical elaboration of it. Immediately above the basal pyramid arise from
its radial edges four thin and broad triangular leaves or wings, and the meeting edges of the
neighbouring wings are in apposition one with the other, so that between the bases of every three
or four neighbouring spines a hollow pyramidal space remains open. The apex of such a pyramidal
space is directed towards the centre of the body, but separated from it by the small basal
pyramid; its open base is directed outwards. The twenty-two hollow pyramidal spaces are disposed
regularly in four different groups:—(A) Four equatorial spaces, four-sided, each limited by
two equatorial and two tropical spines (one canceral and one capricornal); (B) eight perizonal
spaces (four northern and four southern), four-sided, each limited by one equatorial, two
tropical, and one polar spine; (C) eight peripolar spaces (four northern and four southern),
three-sided, each limited by one tropical and two polar spines; (D) two polar spaces (one northern
and one southern), four-sided, each limited by four neighbouring polar spines.
{722}
The third mode of junction, by central concrescence of all twenty spines, was formerly regarded
by me as an important peculiarity, sufficient for the separation of subfamilies and genera
(Monogr. d. Radiol., 1862, pp. 399, 401; Prodromus, 1881, p. 466). But I found afterwards that in
many species where the twenty spines commonly remain separated, accidentally they grow perfectly
together and form one single piece of acanthin—a starrulet with twenty rays. Therefore I now
think it is more natural to divide those species only into different subgenera.
A fourth and a very different mode of junction, quite sufficient for the distinction of
different families, is the concrescence in pairs of every two opposite spines, lying in one
diameter (in Acanthochiasma and Chiastolus). Here we obtain a number of "diametral
spines" (each composed of two originally opposed radial spines) and all these diametral spines are
crossed loosely near the central point of the body without any solid and permanent apposition
(Chiastolida). However, in some species of this peculiar family the central part of the diametral
spines is twisted like a screw or spirally convoluted (Pl. 129, figs. 2,
3).
The Form of the Radial Spines in the Acantharia is extremely
varied, and constitutes the main characters for the distinction of nearly four hundred species.
But all these different forms may be reduced phylogenetically to three different fundamental
forms:—(a) the cylindrical (with circular transverse section), (b) the
two-edged (with elliptical or lanceolate transverse section), and (c) the four-edged (with
square transverse section). No doubt the first (a) is the original primitive form, from
which the two others are secondarily derived. Triangular spines never occur in the Acantharia, whilst, however, they are common in the Sphærellaria. The first and original form, the cylindrical spine, is either a
true cylinder of equal thickness in its whole length, or it is more or less conical. Rarely the
spine is in the distal half spindle-shaped, and thicker than in the basal half. The second form,
the two-edged spine, is more or less compressed from two opposite sides; its two edges are either
more blunt, rounded, or more acute, sharp; its transverse section in the former case is
elliptical, in the latter case lanceolate or rhomboidal. Sometimes the two edges are broader and
in the form of two thin opposite wings. The two-edged spines may be occasionally shorter,
triangular or lanceolate, at other times longer sword-shaped or linear. The third form, the
four-edged spine, has constantly a square transverse section; the sides of this square are either
even or concave; in the latter case the four edges are broadened and wing-like, but in the former
case not. The quadrangular spines are either prismatic (of equal breadth throughout their whole
length) or pyramidal (becoming gradually thinner towards the distal apex).
The Apex of the Radial Spines, or their free distal end, is in the majority of Acantharia simple, conical. In the minority it is either truncated or bifid, or
four-sided pyramidal, often with two, rarely with four prominent parallel teeth. In some forms
{723}the bifid spines are so deeply cleft that they
become forked. Much more interesting and more varied than these different forms of the distal end
are those of the apophyses of the radial spines.
The Apophyses of the Radial Spines, or their "lateral transverse processes," are of the
greatest importance for the morphological development of the whole subclass. Only in sixteen among
the sixty-five genera of Acantharia are the apophyses perfectly wanting;
in the other genera they determine in the first place their general character. In the Acanthometra the apophyses remain perfectly free, whilst in the Acanthophracta their meeting ends or branches compose the latticed shell. All
differences in form and shape of the apophyses can be reduced to only two primary modes; either
the spine bears two opposite or four crossed apophyses; correspondingly all Acantharia apophysaria
may be divided into two different main groups, the Zygapophysica (with two opposite lateral
processes) and the Staurapophysica (with four crossed lateral processes opposite in pairs). Both
groups have probably no direct phylogenetic connection, but seem to be derived independently from
different stocks, and produce different families. The Zygapophysica are probably derived from
Astrolonchida with two-edged spines (Zygacantha), and from this group arise the
Diporaspida, the ancestral group of the majority of Acanthophracta. On
the other hand the Staurapophysica are probably derived from Astrolonchida with four-edged spines
(Acanthonia), and from this group arise the Tessaraspida. The apophyses of the Acanthonida are partly simple, partly branched or even latticed; the apophyses
of the Acanthophracta are never simple, constantly branched and commonly
latticed.
The Malacoma (or the whole soft body of the Acantharia as
opposed to the skeleton) exhibits some peculiarities which distinguish them from the other
Radiolaria, as well in the structure of the central capsule and its nucleus as in that of the
enveloping extracapsular body and the pseudopodia.
The Central Capsule is constantly spherical in the far greater number of the Acantharia, viz., in the following six families:—Astrolophida,
Chiastolida, Astrolonchida, Dorataspida, Sphærocapsida, and Phractopeltida. Among these six
families the Astrolonchida and Dorataspida are far greater and far richer in different forms than
all the other families. The central capsule becomes ellipsoidal or cylindrical, prolonged in one
axis, in the three families, Amphilonchida, Belonapsida, and Diploconida; it becomes discoidal or
lenticular, by the shortening of one axis, in two families, viz., in the Quadrilonchida and
Hexalaspida. Finally, the peculiar family Litholophida is distinguished by the conical form of its
central capsule.
The Membrane of the central capsule in all Acantharia is simple,
commonly thin, sometimes very delicate; in some species it seems to be developed late, just
immediately before the formation of the spores; but in no species is it completely missing. The
membrane is constantly pierced by innumerable fine pores, for the emission of the {724}pseudopodia; but in many species (and probably more or
less in all Acantharia) there is recognisable a certain regularity in the
disposition of the numerous pseudopodia and of the pores by which they radiate from the capsule.
Sometimes these pores are disposed in a regular network of ramified lines, whilst the meshes of
this network are devoid of pores; in other cases they form regular tufts or bushes between the
radial spines. Probably in no Acantharia are the pores of the capsule
membrane so numerous and so equally distributed throughout as in the Spumellaria; we may therefore call the former Actipylea
(in opposition to the latter, as Peripylea).
The Nucleus of the Acantharia is constantly excentric, whilst it
is originally constantly central in the Spumellaria. This excentric
position is a necessary consequence of the centrogenous development of the radial spines. Probably
connected with this peculiarity is the other, that the nucleus assumes a peculiar, complicated
structure, and that in the greater number of Acantharia it becomes cleft
very early, and that this cleavage is effected by a peculiar kind of gemmation, first detected and
very accurately described by R. Hertwig (compare his Organismus d. Radiol., 1879, pp. 10-24).
However, in the young Acantharia the nucleus is constantly simple, and in
a certain number of species its cleavage takes place late (as in the greater number of Spumellaria).
The Endoplasm, or the intracapsular sarcode, exhibits in the greater number of Acantharia a more or less distinct radial arrangement; but this is often
concealed by the different enclosed products of the endoplasm—oil-globules, vacuoles, red or
different coloured pigment-granules, crystals, &c. Often it encloses a variable number of
"yellow cells" (becoming green by mineral acids) to be considered as symbiotic xanthellæ.
The Calymma or the jelly-veil, including the central capsule, in the Acantharia is more or less voluminous, and commonly envelops the skeleton
perfectly. In its surface is sometimes developed a peculiar network of "supporting fibres." A very
peculiar product are the remarkable "Myophrisca" of the Acanthometra,
which are wanting in the Acanthophracta; they were first detected by
Johannes Müller, and figured as "Cilien-Kränze," afterwards explained by Hertwig as "contractile
Fäden," similar to muscular fibrillæ (compare below).
The Matrix, placed between the calymma and central capsule, in the majority of the Acantharia is a rather thin layer of granular exoplasm.
The Pseudopodia arising from it are not so numerous as in the Spumellaria, and not so equally disposed over the whole surface. Also their
tendency to ramify, anastomose, and form networks seems to be much less developed. Commonly they
are simple or little ramified. In many cases (and perhaps everywhere) there may be distinguished
two different kinds of pseudopodia:—(1) Axopodia, or permanent pseudopodia (with axial
filaments?), piercing the wall of the central capsule, and arising from the central mass of
endoplasm; and (2) Collopodia, or variable pseudopodia (without axial filaments), arising outside
the capsule from the matrix of extracapsular sarcode or from the {725}exoplasm on the surface of the calymma. These and other differentiations
seem to indicate that the pseudopodia in the Acantharia are more highly
developed than in the Spumellaria, and justify the denomination of the
former as "Actipylea."
Synopsis of the Orders and Suborders of Acantharia.
|
II. ACANTHOMETRA.
Skeleton composed only of acanthinic radial spines not forming a complete
lattice-shell.
|
|
Radial spines in variable and indefinite number, disposed
irregularly, |
1. Actinelida. |
| Radial spines constantly twenty, disposed regularly after the Müllerian
law of Icosacantha, |
2. Acanthonida. |
|
II. ACANTHOPHRACTA.
Skeleton composed of twenty acanthinic radial spines (disposed after the
Müllerian law) and of a spherical or variously shaped complete lattice-shell.
|
|
Radial spines all twenty of equal size; shell and central capsule
spherical, |
3. Sphærophracta. |
| Radial spines of different sizes; shell and central capsule
ellipsoidal, discoidal, or heteromorphous, |
4. Prunophracta. |
| I. ACANTHOMETRA. Skeleton composed only of acanthinic radial spines not
forming a complete lattice-shell. |
|
|
Radial spines in variable and indefinite number, disposed
irregularly, |
|
|
|
|
|
|
1. Actinelida. |
|
|
|
|
|
|
Radial spines constantly twenty, disposed regularly after the
Müllerian law of Icosacantha, |
|
|
|
|
|
|
2. Acanthonida. |
|
|
|
|
| II. ACANTHOPHRACTA. Skeleton composed of twenty acanthinic radial spines
(disposed after the Müllerian law) and of a spherical or variously shaped complete
lattice-shell. |
|
|
Radial spines all twenty of equal size; shell and central capsule
spherical, |
|
|
|
|
|
|
3. Sphærophracta. |
|
|
|
|
|
|
Radial spines of different sizes; shell and central capsule
ellipsoidal, discoidal, or heteromorphous, |
|
|
|
|
|
|
4. Prunophracta. |
|
|
|
|
Order III. ACANTHOMETRA, Johannes Müller, 1855.
Acanthometra, J. Müller, 1855, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin.
Acanthometrida, Haeckel, 1862, Monogr. d. Radiol., p. 371.
Acanthometrea, R. Hertwig, 1879, Organismus d. Radiol., p. 133.
Acanthonida et Litholophida, Haeckel, 1881, Prodromus, pp. 465, 469.
Definition.—Acantharia without complete latticed
shell.
The order Acanthometra, the third order of Radiolaria, comprises all
those Acantharia in which the acanthinic skeleton is only composed of
radial spines arising from one common central point, but never forms a complete latticed shell. By
the absence of such a latticed or fenestrated shell the Acanthometra
differ principally from the nearly allied Acanthophracta, the second
order of Acantharia, which constantly possess such a complete shell.
Johannes Müller, who first detected and described the Acanthometra (in
1855, loc. cit.), defined them as follows:—"Radiolaria without shell, with siliceous
radial spines" (1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 46). He described and figured
eighteen species of them, disposed in four genera (Acanthometra with fifteen species, and
Zygacantha, Lithophyllium, Lithoptera, each with a single species). Among
those eighteen species, however, were two "Acanthometræ cataphractæ," appertaining to the
following order, the Acanthophracta.
In my Monograph (1862, p. 371) all true Acanthometra were united into
a single family, Acanthometrida, with the following definition:—"Skeleton composed of a
number of radial spines, piercing the central capsule and united in its centre, without {726}latticed shell." In the majority of them I observed that
the skeleton did not consist of silex, but of a very peculiar organic substance, which I called
"acanthin." At that time I divided the family Acanthometrida into four subfamilies:—(1)
Acanthostaurida, (2) Astrolithida, (3) Litholophida, (4) Acanthochiasmida. The two former now
represent the suborder Acanthonida, the two latter the suborder Actinelida. The number of genera which I distinguished in my Monograph amounted
to nine, the number of species to fifty. By the rich collections of the Challenger this number is
so much increased that we can here describe twenty-seven genera and one hundred and sixty
species.
Richard Hertwig in his work on the Organismus der Radiolarien (1879, pp. 6-25) adopted my
family Acanthometrida, and gave a very accurate description of its anatomical structure. He
confirmed my observations that the radial spines of this family are never hollow, but solid, and
that their chemical substance is not silex, but the organic matter "acanthin." He found that the
simple nucleus of the Acanthometrida is commonly very early cleft, and that the peculiar brushes
of filaments on the calymma, described by Johannes Müller and by me as "Gallert-cilien," are
peculiar "contractile filaments," comparable to the "muscle-fibrillæ" of some Infusoria, or the
"Myophan-filaments" (Myophrisca).
The order Acanthometra is here divided into two different suborders of
very unequal extent and value, the Actinelida and Acanthonida. The first may be regarded as the common ancestral stock, not only
of the second, but of all Acantharia. In the small group of Actinelida the number of radial spines is variable and commonly indefinite,
often very large (more than a hundred); they are therefore Adelacantha. The second suborder, the
Acanthonida, comprise by far the greatest part of the order, and possess
constantly twenty radial spines, regularly disposed after the Müllerian law; they are therefore
(like all Acanthophracta) Icosacantha (compare above, p. 717).
The Actinelida possess constantly simple radial spines, without any
apophyses; their form is commonly very simple and primitive. This suborder comprises three small
but very different families, the Astrolophida, Litholophida, and Chiastolida. The first family,
the Astrolophida, is the original ancestral group. A large and variable, commonly indefinite
number of radial spines is here united in the centre of the spherical central capsule and
radiating within a spherical space. In the second family, the Litholophida, a small and variable
number of radial spines (between ten and twenty) is united in the apex of a conical central
capsule and radiating within the quadrant or octant of a spherical space. In the third family, the
Chiastolida, a variable number of radial spines is grown together by pairs, in such a manner that
every two opposite spines (placed originally in one axis of the spherical central capsule) forms a
single "diametral spine"; all these diametral spines are not united in the centre of the central
capsule but only crossed loosely near the centre.
{727}
The Acanthonida, the second suborder of Acanthometra, embraces by far the greatest number in this order, viz., all
those forms in which twenty radial spines are regularly disposed after the Müllerian
law—Icosacantha (compare above, p. 717). The radial spines of this
suborder are either simple or provided with transverse processes (either two opposite or four
crossed apophyses). They are commonly united in the middle of the central capsule by their opposed
basal ends, forming small pyramids; the meeting triangular faces of the neighbouring pyramids
being propped one upon another. Above these small basal pyramids often arises a basal leaf-cross
formed by four broad triangular leaves or wings with straight edges; the meeting thin edges of the
neighbouring spines serve for strengthening the basal junction and form hollow pyramidal spaces or
compartments, filled with the contents of the central capsule (compare p. 721). The suborder Acanthonida comprises three
different families, the Astrolonchida, Quadrilonchida, and Amphilonchida. The first family, the
Astrolonchida, comprises by far the greater number of the Acanthonida;
those genera in which all twenty spines are perfectly equal or nearly equal in size and form. In
the second family, the Quadrilonchida, the four equatorial spines are much larger (and often also
of another form) than the sixteen other spines (often also the eight tropical larger than the
eight polar spines). The third family, the Amphilonchida, is distinguished by the preponderating
development of only two opposite equatorial spines, which are much larger (and often also of
another form) than the eighteen other spines.
Synopsis of the Suborders and Families of Acanthometra.
|
Suborder I. ACTINELIDA.
Number of the radial spines variable, either more or less than twenty,
commonly disposed irregularly and not according to the Müllerian law.
|
|
Radial spines very numerous (thirty to a hundred or more), radiating
from a common centre within a spherical space, |
1. Astrolophida. |
| Radial spines between ten and twenty, radiating from one common point
within a sphere-quadrant, |
2. Litholophida. |
| Radial spines of variable number; every two opposite spines grown
together in the centre; therefore numerous diametral spines are crossed freely in the
centre, |
3. Chiastolida. |
|
Suborder II. ACANTHONIDA.
Number of the radial spines constantly twenty, disposed regularly according
to the Müllerian law.
|
|
All twenty radial spines nearly equal, and of the same size and
form, |
4. Astrolonchida. |
| Four equatorial spines much larger than (and often also of different
form from) the sixteen other spines, |
5. Quadrilonchida. |
| Two opposite equatorial spines (or principal spines) much larger than
(and often also of different form from) the eighteen other spines, |
6. Amphilonchida. |
| Suborder I. ACTINELIDA. Number of the radial spines variable, either more or
less than twenty, commonly disposed irregularly and not according to the Müllerian law. |
|
|
Radial spines very numerous (thirty to a hundred or more),
radiating from a common centre within a spherical space, |
|
|
|
|
|
|
1. Astrolophida. |
|
|
|
|
|
|
Radial spines between ten and twenty, radiating from one common
point within a sphere-quadrant, |
|
|
|
|
|
|
2. Litholophida. |
|
|
|
|
|
|
Radial spines of variable number; every two opposite spines grown
together in the centre; therefore numerous diametral spines are crossed freely in the
centre, |
|
|
|
|
|
|
3. Chiastolida. |
|
|
|
|
| Suborder II. ACANTHONIDA. Number of the radial spines constantly twenty,
disposed regularly according to the Müllerian law. |
|
|
All twenty radial spines nearly equal, and of the same size and
form, |
|
|
|
|
|
|
4. Astrolonchida. |
|
|
|
|
|
|
Four equatorial spines much larger than (and often also of
different form from) the sixteen other spines, |
|
|
|
|
|
|
5. Quadrilonchida. |
|
|
|
|
|
|
Two opposite equatorial spines (or principal spines) much larger
than (and often also of different form from) the eighteen other spines, |
|
|
|
|
|
|
6. Amphilonchida. |
|
|
|
|
{728}
Suborder I. ACTINELIDA, Haeckel, 1882.
Definition.—Acanthometra with a variable
number of radial spines, which are commonly irregularly disposed, not according to the
Icosacantha.
Family XXXIII. Astrolophida, Haeckel.
Astrolophida, Haeckel, 1881, Prodromus, p. 469.
Definition.—Acantharia with a variable number of simple
radial spines, radiating within a spherical space from one common central point which is the
centre of the spherical central capsule. No lattice shell.
The family Astrolophida comprises the simplest and the most primitive
forms among all Acantharia, and may therefore be regarded as the common
ancestral stock of this whole legion or subclass of Radiolaria. The acanthinic skeleton is
composed of a variable number of quite simple radial spines, which are united in the centre of the
spherical central capsule and radiate, piercing its walls and the surrounding jelly-veil, within a
spherical space.
The first observed form of this family is the ancestral genus Actinelius, two different
species of which I detected in 1864 in the northern Mediterranean, at Villafranca, near Nice
(compare Zeitschr. f. wiss. Zool., 1865, Bd. xv. p. 364, Taf. xxvi. fig. 4). Three other species
of the same genus were afterwards found by me in the Challenger collections. Whilst in this
Actinelius all radial spines are of the same size, a new nearly allied genus,
Astrolophus (with two species), differs from it by the different size of the radial spines,
a small number of very large spines being intermingled with a very large number of small spines.
In these two genera, Actinelius and Astrolophus (the true "Astrolophida" sensu
strictiori), the number of the radial spines is quite indeterminable and their arrangement
quite irregular and variable.
A third remarkable genus, Actinastrum, differs from these two genera in the definite
number and regular order of thirty-two radial spines, and may therefore perhaps better represent
a peculiar family, Actinastrida. In this genus (of which two species were observed) the thirty-two
radial spines are disposed in such a regular manner that they lie in four meridian planes, and
that their distal ends fall into five parallel zones. These five zones and these four planes are
the same as we find in all Icosacantha (compare above, p. 717). Also the
constant twenty spines of these latter are present in Actinastrum; but their number is here
enlarged by twelve other spines missing in the Icosacantha; four of these are secondary or
interradial equatorial spines, lying opposite in pairs between the four primary or perradial
equatorial spines; and eight are perradial tropical spines, lying between the eight interradial
tropical spines. Therefore the {729}distal ends of the
thirty-two radial spines are disposed regularly in five parallel zones, and while two zones (the
two polar) contain only the points of every four spines, three zones (the single equatorial and
the two tropical) contain the points of every eight spines. The four meridian planes are in
Actinastrum the same as in the Icosacantha, crossed in the spineless axis at angles of 45°.
But in the Icosacantha each of the two perradial meridian planes contains six radial spines (two
equatorial and four polar), each of the two interradial meridian planes only four tropical spines.
Whereas in Actinastrum each of the two primary or perradial meridian planes contains ten
spines (two equatorial, four tropical, and four polar), each of the two secondary or interradial
meridian planes six spines (two equatorial and four tropical). We find therefore altogether
thirty-two radial spines in three orders; eight equatorial, sixteen tropical, and eight polar
spines.
Only one other genus of Radiolaria exhibits the same characteristic disposition of thirty-two
radial spines as Actinastrum, and this is Chiastolus; but here the two opposite
spines of each pair are grown together and form one diametral spine; and the sixteen diametral
spines are crossed in the centre of the capsule. In Actinastrum, as in Astrolophus
and Actinelius, the central ends or bases of all the spines are pyramidal, and the
triangular faces of the neighbouring spines rest one upon another (as in the greater number of
Acanthonida). The form of the radial spines in all Astrolophida is quite
simple, without lateral processes or apophyses; chiefly cylindrical, more rarely compressed,
two-edged or quadrangular.
The central capsule in all Astrolophida is spherical, and in the younger specimens contains a
single large concentric and lobed nucleus, but in the older specimens a large number of small
nuclei. The surrounding jelly-veil or calymma seems commonly to envelop the spines perfectly. The
piercing pseudopodia radiate everywhere between the spines, and are very numerous and thin. The
circulating granules in them are sometimes red (Actinelius purpureus).
Synopsis of the Genera of Astrolophida.
| Radial spines of indefinite number and of irregular
disposition. |
|
Spines of equal size, |
317. Actinelius. |
| Spines of unequal size, |
318. Astrolophus. |
| Radial spines thirty-two, disposed regularly in five
parallel zones, |
319. Actinastrum. |
| Radial spines of indefinite number and of irregular disposition. |
|
|
Spines of equal size, |
|
|
|
|
|
|
317. Actinelius. |
|
|
|
|
|
|
Spines of unequal size, |
|
|
|
|
|
|
318. Astrolophus. |
|
|
|
|
| Radial spines thirty-two, disposed regularly in five parallel zones, |
|
|
|
|
319. Actinastrum. |
|
|
|
|
Genus 317. Actinelius,[357] Haeckel,
1865, Zeitschr. f. wiss. Zool., Bd. xv. p. 364.
Definition.—Astrolophida with a variable and undetermined
number of simple radial spines, all of equal size, united in the centre of the spherical central
capsule.
{730}
The genus Actinelius comprises the most simple and primitive forms among all
Acantharia, and may be regarded as the common ancestral stock of this
whole legion. The spherical central capsule is pierced by numerous simple radial spines of equal
size, the pyramidal bases of which are supported one upon another with their triangular faces in
the centre of the capsule. The number and position of the spines are quite indefinite and
variable. We may derive Actinelius either from Actissa (Colloidea) by development of acanthinic radial spines, or directly from
Actinosphærium (Heliozoa) by formation of a central capsule.
Subgenus 1. Actinelarium, Haeckel.
Definition.—Radial spines cylindrical, conical, or spindle-shaped,
their transverse section circular.
1. Actinelius primordialis, n. sp. (Pl. 129, fig.
1).
Spines sixty to eighty or more, cylindrical, at the distal end thickened, spindle-shaped. Apex
simple. Base a small slender pyramid. Central capsule yellow. Granules of the sarcode
colourless.
Dimensions.—Length of the spines 0.3 to 0.4, breadth in the distal part 0.02, in
the basal part 0.008.
Habitat.—Central Pacific, Stations 265 to 274, surface.
2. Actinelius purpureus, Haeckel.
Actinelius purpureus, Haeckel, 1865, Zeitschr. f. wiss. Zool., Bd. xv. p.
364, Taf. xxvi. fig. 4.
Spines thirty to forty or more, cylindrical, very thin, a little thinner towards both ends.
Apex simple. Base a small sulcate pyramid. Central capsule opaque, purple. Granules of the sarcode
also purple.
Dimensions.—Length of the spines 0.2 to 0.3, breadth 0.002.
Habitat.—Mediterranean (Nice), Haeckel.
Subgenus 2. Actinelidium, Haeckel.
Definition.—Radial spines compressed, two-edged; their transverse
section elliptical or lanceolate.
3. Actinelius protogenes, n. sp.
Spines fifty to sixty, compressed, two-edged, gradually broadened towards the truncated distal
end. Basal or proximal end thin, pyramidal. The spines of this species are similar to those of
{731}Actinastrum pentazonium (p. 733) and of Chiastolus amphicopium (Pl. 129, fig. 3),
but much more numerous, smaller, and not regularly disposed. These latter two Actinelida must be
separated on account of the regular disposition of the thirty-two spines.
Dimensions.—Length of the spines 0.2, basal breadth 0.008, distal breadth
0.02.
Habitat.—South Pacific, Station 165, surface.
Subgenus 3. Actinelonium, Haeckel.
Definition.—Radial spines quadrangular, prismatic, or pyramidal, their
transverse section square.
4. Actinelius pallidus, Haeckel.
Actinelius pallidus, Haeckel, 1865, Zeitschr. f. wiss. Zool., Bd. xv. p.
364.
Spines eighty to one hundred and twenty or more, quadrangular, prismatic, of equal breadth
throughout their whole length. Apex simple, truncate or pyramidal. Base a four-sided slender
pyramid. Central capsule pale yellowish. Granules of the sarcode colourless.
Dimensions.—Length of the spines 0.2 to 0.3, breadth 0.005.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific.
5. Actinelius polyacanthus, n. sp.
Spines two hundred to three hundred or more, quadrangular, pyramidal, gradually thinned towards
the simple apex. Base a small three-sided pyramid. Central capsule opaque.
Dimensions.—Length of the spines 0.12 to 0.18, basal breadth 0.012.
Habitat.—South Pacific, Station 291, surface.
Genus 318. Astrolophus,[358] Haeckel,
1881, Prodromus, p. 469.
Definition.—Astrolophida with a variable and undetermined
number of simple radial spines of different sizes (large and small spines intermingled), which are
united in the centre of the spherical central capsule.
The genus Astrolophus differs from the nearly allied ancestral genus
Actinelius only in the unequal size of the numerous radial spines. In both observed species
very numerous small spines are intermingled with a small number of large spines, and between them
numerous spines of medium size. The small spines fill up the hollow spaces between the basal parts
of the large spines.
{732}
1. Astrolophus stellaris, n. sp.
Radial spines from one hundred to two hundred, of very different sizes, but of similar form;
about sixteen to twenty very large spines, forty to fifty of medium size, and one hundred to one
hundred and twenty much smaller. All spines cylindrical in the greater part of their length, with
simple apex, gradually thickened towards the central part, conical, without edges. The base itself
is a slender pyramid with four to eight edges.
Dimensions.—Length of the largest spines 0.3 to 0.4, of the majority 0.1 to 0.2,
of the smallest 0.05 to 0.1.
Habitat.—South Pacific, Station 288, surface.
2. Astrolophus solaris, n. sp. (Pl. 132,
figs. 12a, 12b).
Radial spines from two hundred to three hundred, of very different sizes, but of similar form;
about twenty to thirty very large spines, sixty to eighty of medium size, and one hundred and
twenty to one hundred and fifty much smaller. All spines cylindrical in the greater part of their
length, with simple apex, gradually thickened and four-edged towards the central base. The base
itself is a slender pyramid with four to eight edges; partly the faces, partly the edges of these
basal pyramids rest one upon another, the points of the larger spines meeting in the centre.
Dimensions.—Length of the largest spines 0.4 to 0.5, of the majority 0.2 to 0.3,
of the smallest 0.1 to 0.16; basal thickness of the largest spines 0.015.
Habitat.—South-east Pacific (off Juan Fernandez), Station 296,
surface.
Genus 319. Actinastrum,[359] n. gen.
Definition.—Astrolophida with thirty-two simple radial
spines, regularly disposed within four meridian planes in such an order that their distal ends
fall into five parallel zones. Central ends of the thirty-two spines supported one upon another in
the centre of the spherical central capsule.
The genus Actinastrum differs from the two preceding genera in the definite
number and order of the thirty-two radial spines, which are disposed in a very remarkable manner.
Twenty radial spines are disposed after the Müllerian law of Icosacantha (compare above, p. 717). The remaining twelve spines are four equatorial spines lying in the two
secondary meridian planes, and eight tropical spines lying in the two primary meridian planes. We
have therefore together eight equatorial, sixteen tropical, and eight polar spines (compare above,
p. 729).
{733}
1. Actinastrum legitimum, n. sp.
All thirty-two radial spines of equal size and similar form, cylindrical, conical at the distal
end, at the central base pyramidal. Central capsule pellucid, colourless.
Dimensions.—Length of the radial spines 0.3, breadth 0.004.
Habitat.—South Pacific, Station 288, surface.
2. Actinastrum pentazonium, n. sp.
All thirty-two radial spines of equal size and similar form, compressed, two-edged, gradually
becoming broader and thinner from the pyramidal central base towards the truncated distal end.
Central capsule dark, opaque. (Compare the similar Chiastolus amphicopium, Pl. 129, fig.
3.)
Dimensions.—Length of the radial spines 0.2, breadth at the base 0.005, at the
distal end 0.02.
Habitat.—South Pacific (west coast of Patagonia), Station 302,
surface.
Family XXXIV. Litholophida, Haeckel.
Litholophida, Haeckel, 1862, Monogr. d. Radiol., p. 401.
Definition.—Acantharia with a variable number of simple
radial spines radiating within a conical space (or within the quadrant of a sphere) from one
common central point, which is the apex of the conical central capsule. No lattice-shell.
The family Litholophida, represented only by a single genus,
Litholophus, differs from all other Acantharia in the remarkable
fact that the common point, from which the radial spines arise, is not the geometrical central
point of the whole body, but is quite excentric in position, the apex of the conical or pyramidal
central capsule. Therefore the spines form together a kind of brush or broom.
When I founded the family Litholophida in my Monograph (1862, p. 401) I knew only a single
species, Litholophus rhipidium, observed very frequently in Messina. Another species,
Litholophus ligurinus, was afterwards (1864) found by me at Nice. Six other species were
detected in the preparations of the Challenger, some of them very frequent. All these eight
species of Litholophus are very nearly allied, and exhibit only slight differences in the
form and number of the radial spines; their mode of excentric connection and the structure of the
peculiar soft body is everywhere the same.
The radial spines in all observed Litholophida possess the form of the genus Acanthonia,
i.e., they are quite simple, four-sided prismatic or quadrangular, with square transverse
section; their four edges are sometimes smooth, at other times elegantly denticulate, commonly
more or less prominent or wing-shaped. In the greater number of species they are very long and of
nearly equal breadth, prismatic; in some species they are more pyramidal, thinned towards the
distal end; the latter {734}is commonly truncated or
broken off, sometimes pyramidal. The central end is everywhere thinned, more or less pyramidal,
and the neighbouring spines are propped one upon another by the triangular faces of their small
basal pyramids. A slight pressure is sufficient to destroy their connection.
The number and disposition of the radial spines seem to be variable and irregular, but require
further researches. In four of the observed eight species I found constantly ten spines, in two
other species from ten to twenty (commonly twelve or sixteen), and in two species twenty or more.
A certain order or disposition of the spines within the conical space in which they radiate could
nowhere be ascertained.
When I first observed Litholophus, I supposed that it might only be a
mutilated or altered form of an Acanthonia. Afterwards, observing many specimens with ten
spines, I was led to the suggestion that they were produced by self-division of an
Acanthonia, and that the number of the spines in each half of the body might be afterwards
doubled. But this suggestion seems to be refuted by the fact that in no other genus of the
numerous Acantharia is self-division observed, and that many hundreds of
Litholophus which I observed exhibit quite constantly only a single form of radial spines,
that of Acanthonia—simple quadrangular spines without any apophyses.
Genus 320. Litholophus,[360] Haeckel,
1862, Monogr. d. Radiol., p. 401.
Definition.—Litholophida with a variable number of
quadrangular diverging radial spines, united with pyramidal bases in the apex of the conical
central capsule.
The genus Litholophus, the only one of this family, exhibits the peculiarities just
described, but might more nearly be defined as a typical "genus" by the quadrangular form of the
radial spines, identical with those of Acanthonia.
The central capsule of Litholophus is constantly conical or pyramidal, commonly opaque,
of a dark brownish or reddish colour; it contains many small nuclei. It envelops the basal half of
all radial spines in such a manner that their basal parts are united in its apex, and their distal
parts pierce the rounded base of the conical capsule (Pl. 129, fig.
2).
The calymma or the jelly envelope of the central capsule is only developed at its
base, where the spines radiate; at the conical mantle of the capsule it is very thin. The spines
seem to be perfectly enclosed in the calymma and connected with it by the same contractile
retinacula or "myophrisca" which we observe in the Acanthonida. The
pseudopodia arise only from the rounded base of the conical capsule, and radiate between the
spines, piercing the calymma, diverging within the conical space occupied by the fascicle of
spines.
{735}
Subgenus 1. Litholopharium, Haeckel.
Definition.—Ten radial spines.
1. Litholophus decimalis, n. sp.
Ten radial spines, four-sided prismatic, with prominent smooth edges, of equal breadth
throughout their whole length.
Dimensions.—Length of the spines 0.2 to 0.3, breadth 0.006.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Indian, Pacific,
surface.
2. Litholophus pyramidalis, n. sp.
Ten radial spines, four-sided pyramidal, with prominent smooth edges, gradually thickened from
the small pyramidal base towards the truncated distal end.
Dimensions.—Length of the spines 0.3 to 0.4, breadth in the basal part 0.002, in
the middle part 0.006, in the distal part 0.012 to 0.02.
Habitat.—Central Pacific, Station 266, surface.
3. Litholophus decapristis, n. sp. (Pl. 129, fig.
2).
Ten radial spines, four-sided prismatic, with prominent, elegantly denticulated edges, of equal
breadth in their whole length.
Dimensions.—Length of the spines 0.2 to 0.4, breadth 0.008.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific, surface.
4. Litholophus decastylus, n. sp.
Ten radial spines, four-winged pyramidal, with broad and thin, elegantly denticulated edges,
gradually thickened from the small pyramidal base towards the truncated distal end.
Dimensions.—Length of the spines 0.2 to 0.3, breadth in the basal part 0.002, in
the middle part 0.005, in the distal part 0.015.
Habitat.—South Atlantic, Station 332, surface.
Subgenus 2. Litholophidium, Haeckel.
Definition.—Number of the radial spines variable, between ten and
twenty, commonly twelve to sixteen.
{736}
5. Litholophus ligurinus, Haeckel.
Litholophus ligurinus, Haeckel, 1865, Zeitschr. f. wiss. Zool., Bd. xv. p.
366.
Spines of variable number, from eleven to twenty, commonly twelve to sixteen, four-sided
prismatic, with smooth thin edges, of equal breadth in their whole length or a little thinner
towards the proximal end.
Dimensions.—Length of the spines 0.3, breadth 0.005.
Habitat.—Mediterranean (Nice); Central Pacific, Station 274,
surface.
6. Litholophus rhipidium, Haeckel.
Litholophus rhipidium, Haeckel, 1862, Monogr. d. Radiol., p. 402, Taf.
xix. fig. 6.
Spines of variable number, from eleven to twenty, commonly twelve to sixteen, four-sided
prismatic, with distantly denticulated edges, of equal breadth in their whole length or a little
thinner towards both ends.
Dimensions.—Length of the spines 0.3, breadth 0.006.
Habitat.—Mediterranean (Messina); North Atlantic, Station 352,
surface.
Subgenus 3. Litholophonium, Haeckel.
Definition.—Number of the radial spines twenty (or more?).
7. Litholophus fasciculus, n. sp.
Spines constantly (?) twenty, four-sided prismatic, with smooth prominent edges, nearly of
equal breadth throughout their whole length.
Dimensions.—Length of the spines 0.3 to 0.5, breadth 0.008.
Habitat.—South Pacific, Station 291, surface.
8. Litholophus penicillus, n. sp.
Spines constantly twenty (or more?), four-sided prismatic, with distantly denticulated edges,
gradually thickened from the small pyramidal base to the middle part, of equal breadth in the
distal half.
Dimensions.—Length of the spines 0.3 to 0.4, breadth 0.005 to 0.007.
Habitat.—North Pacific, Station 252, surface.
Family XXXV. Chiastolida, Haeckel.
Acanthochiasmida, Haeckel, 1862, Monogr. d. Radiol., p. 402.
Definition.—Acantharia with a variable number of simple
radial spines, which are grown together in pairs (two opposite spines of each pair representing
together a single diametral spine). Diametral spines crossed loosely in the centre of the
spherical or irregular roundish central capsule. No lattice-shell.
{737}
The family Chiastolida (or Acanthochiasmida) differs from all other
Acantharia in the peculiar mode of the radial spines; these grown together
in pairs in the centre of the body, so that every two spines opposite in one axis of the body form
together one single diametral spine. All diametral spines are loosely crossed in the middle of the
central capsule, or connected by a peculiar screw-like winding, but not united firmly.
I established the family Acanthochiasmida in my Monograph (1862, p. 402) upon the single genus
Acanthochiasma (with three species), in which only ten diametral spines are constantly
found; I derived these from the twenty radial spines of the common Acanthometra, supposing that every two opposite spines of the latter (lying in
one axis) were grown together in the centre, whilst the intimate connection of the twenty radial
spines in the common centre was dissolved. This opinion was afterwards confirmed by Richard
Hertwig, who observed Acanthochiasma intact in the living state. Although two species of
this genus are cosmopolitan and very common, the number of species is very small; I could add to
those three older known forms only a single new species.
Another genus of this family, Chiastolus, was observed by me only in a single specimen
but it is extremely interesting. It has sixteen diametral spines, disposed quite regularly after
the same law of the thirty-two spines of Actinastrum which we described above (compare
above, p. 729). Therefore we cannot doubt that the former is derived from
the latter in the same way, every two opposite radial spines (of one axis) being grown together to
form a single diametral spine. As we place Acanthometron (with twenty spines) and
Actinastrum (with thirty-two spines) in two different families, it would perhaps be more
convenient to separate also Acanthochiasma and Chiastolus as representatives of two
different families—Acanthochiasmida (with ten diametral spines) and Chiastolida (with
sixteen diametral spines).
As we derive Acanthochiasma from Acanthometron by concrescence in pairs of the
twenty radial spines, the Müllerian law of Icosacantha must be employed also to the ten diametral
spines of the former, therefore two of them are equatorial, four tropical, and four polar spines.
In the same way we may employ the new law of disposition found in the thirty-two radial spines of
Actinastrum equally to the sixteen diametral spines of Chiastolus, which we derive
from the former, four of them are equatorial, eight tropical, and four polar spines (compare
above, p. 732).
The Central Capsule of the Chiastolida is spherical, and exhibits in general the same
shape as in the Acanthonida, and specially in the Astrolonchida. Of
course every diametral spine pierces the capsule twice, at two points diametrically opposed. In
some species of Acanthochiasma the central capsule is formed very late, so that it seems
often to be absent. A very accurate description of the capsule and its nucleus, as well as of the
calymma and the pseudopodia, is given by Richard Hertwig in his Organismus der Radiolarien (1879,
pp. 10-18). The pseudopodia are very numerous, and sometimes bear reddish granules.
{738}
Synopsis of the Genera of Chiastolida.
|
I. Subfamily Chiastolidina.
Sixteen diametral spines, derived by concrescence of thirty-two radial
spines,
|
321. Chiastolus. |
|
II. Subfamily Acanthochiasmida.
Ten diametral spines, derived by concrescence of twenty radial spines,
|
322. Acanthochiasma. |
Genus 321. Chiastolus,[361] n. gen.
Definition.—Chiastolida with sixteen diametral spines,
derived from thirty-two radial spines opposite and grown together in pairs.
The genus Chiastolus, hitherto known only by a single observed specimen,
comprises the Chiastolida with sixteen diametral spines, which are loosely crossed in the centre
of the body. Four of these spines are equatorial, four polar, and eight tropical. We derive
Chiastolus from Actinastrum by concrescence of every two opposite spines in one axis
of the body (compare above, pp. 729, 732).
1. Chiastolus amphicopium, n. sp. (Pl. 129, figs. 3,
3a, 3b).
Spines in the central part cylindrical, spirally convoluted in a very peculiar manner,
broadened towards both ends, strongly compressed, two-edged; the broadest parts are the two
truncated distal ends, five to seven times as broad as the thinnest central part, which is placed
between two spindle-shaped intumescences. These cochleary central parts of the sixteen spines seem
to be resting one upon another. Each spine (composed of two opposite equal radial spines) has
nearly the form of a double oar. The single observed specimen (preserved in glycerine) exhibited a
most regular disposition of the thirty-two spines (grown together in pairs in the centre). The
diameter of the dark non-transparent spherical central capsule equalled one-fifth to one-fourth of
the total length of the double spines. When the soft parts of the body were destroyed by sulphuric
acid, the sixteen single spines were suddenly dispersed.
Dimensions.—Length of the sixteen double spines 0.5, distal breadth (of the
truncated ends) 0.05, central breadth 0.01; diameter of the central capsule 0.12.
Habitat.—South-east Pacific (near Juan Fernandez), Station 297,
surface.
Genus 322. Acanthochiasma,[362] Krohn,
1860, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 810.
Definition.—Chiastolida with ten diametral spines,
derived from twenty radial spines opposite and grown together in pairs.
{739}
The genus Acanthochiasma with a small number of common species, comprises
the Chiastolida with ten diametral spines, which are loosely crossed in the centre of the body. I
could distinguish only four species, two of which are cosmopolitan and very widely distributed. In
all four species the diametral spines are quite simple, cylindrical; only in one species
distinguished by a spiral winding or torsion in the middle part, where they are crossed one to
another. We derive Acanthochiasma from Acanthometron by concrescence of every two
spines opposite in one axis of the body.
1. Acanthochiasma krohnii, Haeckel.
Acanthochiasma krohnii, Haeckel, 1862, Monogr. d. Radiol., p.403, Taf. xix. fig.
7.
Acanthochiasma krohnii, R. Hertwig, 1879, Organismus d. Radiol., Taf. ii.
fig. 6.
Spines needle-shaped, cylindrical, very thin and long, of equal breadth in their whole length,
distinguished by a high degree of elasticity. Central capsule colourless or yellowish-white,
transparent. Granules of the sarcode colourless.
Dimensions.—Length of the spines 0.5 to 1.0, breadth 0.001 to 0.002.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Indian, Pacific, very
common.
2. Acanthochiasma rubescens, Krohn.
Acanthochiasma rubescens, Haeckel, 1862, Monogr. d. Radiol., p. 403.
Spines cylindrical, of equal breadth in their whole length, not very elastic, pointed at the
two ends. Central capsule intransparent, reddish, with violin-shaped concretions. Granules of the
sarcode red coloured.
Dimensions.—Length of the spines 0.2 to 0.6, breadth 0.004 to 0.006.
Habitat.—North Atlantic, Madeira, Krohn; Lanzerote, Haeckel.
3. Acanthochiasma fusiforme, Haeckel.
Acanthochiasma fusiforme, Haeckel, 1862, Monogr. d. Radiol., p. 404, Taf.
xix. fig. 8.
Spines spindle-shaped, from the thicker central part thinned towards the two thin conical ends,
perfectly straight and smooth, rigid, inelastic. Central capsule non-transparent, brown.
Dimensions.—Length of the spines 0.3 to 0.5, breadth in the central part 0.006 to
0.009.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific.
4. Acanthochiasma spirale, n. sp.
Spines spindle-shaped, tapering from the thicker central part towards the two thin conical
ends, rigid, inelastic; their central part is spirally convoluted in a very peculiar cochlea-like
manner, as in Chiastolus amphicopium (Pl. 129, figs.
3a, 3b). The ten spines are propped one upon another by the central screw. Central
capsule dark, opaque.
Dimensions.—Length of the spines 0.2 to 0.3, breadth of the central spiral part
0.01.
Habitat.—Central Pacific, Station 266, surface.
{740}
Suborder II. ACANTHONIDA, Haeckel (Pls. 130-132).
Acanthonida, Haeckel, 1881, Prodromus, p. 465.
Definition.—Acanthometra with twenty radial
spines, disposed according to the Müllerian or Icosacanthan law in five zones each of four
spines.
Family XXXVI. Astrolonchida, Haeckel (Pl. 130).
Astrolonchida, Haeckel, 1881, Prodromus, p. 465.
Definition.—Acanthometra with twenty radial spines of
nearly equal size and similar form, disposed according to the law of the Icosacantha. No
lattice-shell.
The family Astrolonchida, the first and oldest of the Acanthonida, is no doubt the ancestral stock not only of this suborder but also
all Acanthophracta, i.e., of all Icosacantha or all Acantharia in which twenty radial spines are regularly disposed according to the
Müllerian law, forming five zones each of four alternating spines (compare above, p. 717). The Astrolonchida differ from the Acanthophracta in the absence of a complete lattice-shell, from the other two
families of Acanthonida (the Quadrilonchida and Amphilonchida) in the
equal size and similar form of all the spines. Probably this equality is nowhere quite perfect,
since in all Icosacantha the central bases of the twenty spines exhibit originally certain slight
differences of form and junction, effected by the regular disposition itself. But setting aside
this slight difference, only recognisable by means of a very accurate investigation of the central
junction (and in thinner spines often not at all recognisable), the twenty spines of the
Astrolonchida appear perfectly equal. Therefore the four equatorial spines are not distinguished
from the sixteen other spines, as is constantly the case in the two following families.
The number of genera (eleven) and of species (seventy-six) in the Astrolonchida is far larger
than in the five other families of Acanthometra, and requires a
distinction into three different subfamilies. (A) In the Zygacanthida the form of the radial
spines is quite simple, without apophyses or transverse processes; (B) in the Phractacanthida each
spine bears two opposite apophyses (rarely two longitudinal rows of these opposite apophyses); (C)
in the Stauracanthida each spine bears a cross of four apophyses, opposite in pairs (rarely four
longitudinal crossed rows of apophyses, opposite in pairs). The Phractacanthida and Stauracanthida
appear as two divergent branches of the pedigree, derived independently from the common ancestral
stock of Zygacanthida.
In the Zygacanthida, constantly devoid of apophyses, we can distinguish only three genera,
characterised by the different fundamental form of the radial spines; these are:—(1)
Acanthometron, with cylindrical or conical spines (without edges); (2) Zygacantha,
with compressed and two-edged spines; (3) Acanthonia, with four-edged, prismatic or
pyramidal spines. The transverse section of the spines is in the first case {741}circular, in the second elliptical or lanceolate, in the third square.
All the different forms of spines, which we find in the numerous Acantharia, may be reduced to these three forms, and among these the second and
third are derived from the first.
The development of apophyses or of lateral transverse processes (wanting in the Zygacanthida)
is of the greatest value for the further differentiation of the Acantharia. For from the Phractacanthida (with two opposite apophyses on each
spine) we must derive the Phrastaspida, the common ancestral stock of the Diporaspida (and
therefore also the Belonaspida, Hexalaspida, Diploconida, and Phractopeltida). On the other hand
the Stauracanthida (with four crossed apophyses on each spine) have produced the Stauraspida, or
the ancestral group of the Tessaraspida and Sphærocapsida. From all these Acanthophracta, possessing a complete lattice-shell, the Astrolonchida differ
in the absence of such a complete shell. Also in the few cases in which the apophyses become
latticed (Doracantha among the Phractacanthida, and Phatnacantha among the
Stauracanthida), the lattice-plates of the neighbouring spines never meet with their edges, as is
the case in all Acanthophracta. But in a phylogenetic as well as in an
ontogenetic sense the former are the ancestral stock of the latter.
The Central Capsule in the Astrolonchida is commonly spherical, sometimes with twenty
roundish elevations or conical papillæ, extending radially to the basal half of the radial spines.
The calymma is voluminous, and forms around the radial spines conical or cylindrical
"jelly-sheaths," which are connected with the spines by coronas of Myophrisca (or of the bodies
formerly called "Gallert-cilien," afterwards recognised as "contractile Filamente").
Synopsis of the Genera of Astrolonchida.
|
I. Subfamily Zygacanthida.
Twenty radial spines simple, without apophyses or lateral transverse
processes.
|
|
Spines cylindrical, with circular transverse section, |
323. Acanthometron. |
| Spines compressed, two-edged or lamellar, with elliptical
or rhomboidal transverse section, |
324. Zygacantha. |
| Spines quadrangular (prismatic or pyramidal), with four
edges, with square transverse section, |
325. Acanthonia. |
|
II. Subfamily Phractacanthida.
Twenty radial spines provided each with two opposite apophyses (or two
longitudinal rows of apophyses).
|
|
Two apophyses opposite on each spine. |
|
Apophyses simple, |
326. Lithophyllium. |
| Apophyses branched, |
327. Phractacantha. |
| Apophyses latticed, |
328. Doracantha. |
| Two opposite longitudinal rows of apophyses (four to eight
or more apophyses on each spine, opposite in pairs), |
329. Astrolonche. |
|
III. Subfamily Stauracanthida.
Twenty radial spines provided each with four crossed apophyses (or four
crossed longitudinal rows of apophyses).
|
|
Four apophyses (in cross form) opposite in pairs on each
spine. |
|
Apophyses simple, |
330. Xiphacantha. |
| Apophyses branched, |
331. Stauracantha. |
| Apophyses latticed, |
332. Phatnacantha. |
| Four longitudinal rows of apophyses, opposite in pairs in
cross form on each spine, |
333. Pristacantha. |
| I. Subfamily Zygacanthida. Twenty radial spines simple, without apophyses or
lateral transverse processes. |
|
|
Spines cylindrical, with circular transverse section, |
|
|
|
|
|
|
|
|
323. Acanthometron. |
|
|
|
|
|
|
|
|
Spines compressed, two-edged or lamellar, with elliptical or
rhomboidal transverse section, |
|
|
|
|
|
|
|
|
324. Zygacantha. |
|
|
|
|
|
|
|
|
Spines quadrangular (prismatic or pyramidal), with four edges,
with square transverse section, |
|
|
|
|
|
|
|
|
325. Acanthonia. |
|
|
|
|
|
|
| II. Subfamily Phractacanthida. Twenty radial spines provided each with two
opposite apophyses (or two longitudinal rows of apophyses). |
|
|
Two apophyses opposite on each spine. |
|
|
|
|
|
|
Apophyses simple, |
|
|
|
|
|
|
|
|
|
|
326. Lithophyllium. |
|
|
|
|
|
|
|
|
|
|
Apophyses branched, |
|
|
|
|
|
|
|
|
|
|
327. Phractacantha. |
|
|
|
|
|
|
|
|
|
|
Apophyses latticed, |
|
|
|
|
|
|
|
|
|
|
328. Doracantha. |
|
|
|
|
|
|
|
|
Two opposite longitudinal rows of apophyses (four to eight or more
apophyses on each spine, opposite in pairs), |
|
|
|
|
|
|
|
|
329. Astrolonche. |
|
|
|
|
|
|
| III. Subfamily Stauracanthida. Twenty radial spines provided each with four
crossed apophyses (or four crossed longitudinal rows of apophyses). |
|
|
Four apophyses (in cross form) opposite in pairs on each
spine. |
|
|
|
|
|
|
Apophyses simple, |
|
|
|
|
|
|
|
|
|
|
330. Xiphacantha. |
|
|
|
|
|
|
|
|
|
|
Apophyses branched, |
|
|
|
|
|
|
|
|
|
|
331. Stauracantha. |
|
|
|
|
|
|
|
|
|
|
Apophyses latticed, |
|
|
|
|
|
|
|
|
|
|
332. Phatnacantha. |
|
|
|
|
|
|
|
|
Four longitudinal rows of apophyses, opposite in pairs in cross
form on each spine, |
|
|
|
|
|
|
|
|
333. Pristacantha. |
|
|
|
|
|
|
{742}
Subfamily 1. Zygacanthida, Haeckel.
Definition.—Astrolonchida with twenty simple
radial spines, without apophyses or lateral transverse processes.
Genus 323. Acanthometron,[363] J. Müller,
1855, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 229.
Definition.—Astrolonchida with simple cylindrical or
needle-shaped radial spines, without edges and without apophyses; their transverse section is
circular.
The genus Acanthometron, with the restricted definition here given, is the
most simple form of all Acanthonida, and may be regarded as the common
ancestral form not only of this suborder but also of all Acanthophracta,
in general of all Icosacantha, or all Acantharia in which twenty radial
spines are regularly disposed after the Müllerian law (p. 717). In the
wider sense, given originally to Acanthometra by Johannes Müller, its
discoverer, this genus comprised all Acantharia constituting here our
order "Acanthometra" (Radiolaria without lattice-shell, with radial
spines united in the centre). In my Monograph (1862, p. 375) I restricted this genus to those
"Acanthometrida" in which twenty simple spines of equal size (and without apophyses) are supported
one upon another in the centre, and I separated as Astrolithium those forms in which they
are grown together in the centre. But this difference now appears not so important, and I restrict
here the genus Acanthometron (not Acanthometra) to those most
simple forms in which the simple radial spines are cylindrical or conical, without edges.
Subgenus 1. Acanthometrella, Haeckel.
Definition.—Spines at the central base without leaf-cross, united by
the opposed triangular faces of their pyramidal bases, resting one upon another.
1. Acanthometron
elasticum, Haeckel.
Acanthometra elastica, Haeckel, 1862, Monogr. d. Radiol., p. 376, Taf. xv. fig. 1,
Taf. xviii. fig. 1.
Acanthometra elastica, R. Hertwig, 1879, Organismus d. Radiol., Taf. i.
figs. 2, 2a, 2b.
Spines cylindrical, very thin and long, needle-shaped, at the central base four-sided
pyramidal, without leaf-cross. Distal apex conical. The spines are very elastic, of nearly equal
thickness in their whole length. Central capsule quite pellucid, colourless, with a variable
number of yellow pigment-bodies (xanthellæ?).
Dimensions.—Length of the spines 0.3 to 0.6, breadth 0.001 to 0.002.
Habitat.—Cosmopolitan, very common in all warmer seas; Mediterranean,
Atlantic, Indian, Pacific, surface.
{743}
2. Acanthometron cylindricum, n. sp. (Pl. 130, fig.
2).
Spines cylindrical, thick and long, at the central base thickened with a pear-shaped knob, and
with very small central fulcral pyramid, without leaf-cross. Distal apex rounded or truncated.
Central capsule opaque, filled with red pigment-bodies.
Dimensions.—Length of the spines 0.4 to 0.8, breadth 0.01 to 0.015.
Habitat.—Central Pacific, Stations 266 to 274, surface.
3. Acanthometron fuscum, J. Müller.
Acanthometra fusca, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 47, Taf.
xi. fig. 4.
Acanthometra fusca, Haeckel, 1862, Monogr. d. Radiol, p. 377.
Spines very thin and long, in the proximal half cylindrical, in the distal half conical,
gradually thinned towards the simple conical apex. Central base a small four-sided pyramid,
without leaf-cross. Central capsule opaque, filled with brown pigment-bodies.
Dimensions.—Length of the spines 0.2 to 0.4, breadth 0.002 to 0.004.
Habitat.—Mediterranean (Cette, Portofino, Messina).
4. Acanthometron bulbosum, Haeckel.
Acanthometra bulbosa, Haeckel, 1862, Monogr. d. Radiol., p. 377, Taf. xv.
fig. 2, Taf. xviii fig. 2.
Spines very thin and long, cylindrical, with simple needle-shaped apex; in the basal part
conical, towards the centre much thickened, with a short four-sided fulcral pyramid, without
leaf-cross. Central capsule opaque, filled with brown pigment-bodies.
Dimensions.—Length of the spines 0.3 to 0.5, breadth in the middle and outer part
0.001, in the basal part 0.008 to 0.012.
Habitat.—Mediterranean; Atlantic, Station 354, surface.
5. Acanthometron dolichoscion, Haeckel (Pl. 129, figs.
6-8).
Acanthometra dolichoscia, Haeckel, 1862, Monogr. d. Radiol., p. 377, Taf.
xviii. figs. 3a, 3b.
Spines very long and thin, cylindrical, in the thickened outer half about twice as thick as in
the thin inner half, at the simple apex compressed, two-edged. Central base little thickened, with
a small four-sided pyramid, without leaf-cross. Central capsule transparent, whitish or
yellowish.
Dimensions.—Length of the spines 0.6 to 0.8, breadth in the proximal part 0.004,
in the distal part 0.008.
Habitat.—Mediterranean (Messina, Portofino), surface.
6. Acanthometron conicum, n. sp.
Spines short and thick, conical, gradually thinner from the thick conical base towards the
simple apex. Central part of the base with a large fulcral pyramid, but without leaf-cross.
Central capsule transparent, colourless (?).
Dimensions.—Length of the spines 0.08 to 0.12, basal breadth 0.02.
Habitat.—North Pacific, Station 244, surface.
{744}
7. Acanthometron pellucidum, J. Müller.
Acanthometra pellucida, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 47,
Taf. xi. figs. 1-3.
Acanthometra pellucida, Haeckel, 1862, Monogr. d. Radiol., p. 378.
Spines cylindrical, very thin and long, needle-shaped, nearly of equal thickness in their whole
length. Central base with a small fulcral pyramid, without leaf-cross. Distal apex bifid or
bifurcate, with two thin parallel teeth. Central capsule pellucid, colourless, with yellow
pigment-bodies. (Differs from Acanthometron elasticum mainly in the bifid apex.)
Dimensions.—Length of the spine 0.1 to 0.3, breadth 0.002 to 0.004.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific, surface.
8. Acanthometron wageneri, Haeckel.
Acanthometra wageneri, Haeckel, 1862, Monogr. d. Radiol., p. 378.
Spines cylindrical, in the thickened outer half about twice as broad as in the thin inner half.
Central base a little thickened, with large fulcral pyramid, but without leaf-cross. Distal apex
bifid, with two divergent, often denticulated teeth. Central capsule pellucid, with yellow
pigment-bodies. (Differs from Acanthometron dolichoscion mainly in the bifid apex.)
Dimensions.—Length of the spines 0.2 to 0.5, breadth in the inner part 0.003, in
the outer 0.006.
Habitat.—Adriatic Sea; Triest, Wagner; Corfu, Haeckel, surface.
Subgenus 2. Phyllostaurus, Haeckel, 1862, Monogr. d. Radiol., p. 381.
Definition.—Spines at the central base with a broad leaf-cross,
composed of four prominent triangular lamellæ; the meeting edges of the neighbouring lamellæ are
propped one upon another in such a manner that there are formed twenty-two hollow pyramidal spaces
or compartments (compare p. 721).
9. Acanthometron siculum, Haeckel.
Acanthometra sicula, Haeckel, 1862, Monogr. d. Radiol., p. 382, Taf. xvii.
figs. 1, 2; Taf. xviii. fig. 8.
Spines elongate, conical, tapering gradually from the thick base towards the simple distal
apex. Conical circular base supported by a basal leaf-cross of double the breadth. Central capsule
yellowish-brown, opaque.
Dimensions.—Length of the spines 0.3 to 0.4, basal breadth 0.008 to 0.012,
leaf-cross 0.024.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific, surface.
{745}
10. Acanthometron catervatum, Haeckel.
Acanthometra brevispina, Haeckel, 1862, Monogr. d. Radiol., p. 382, Taf.
xv. fig. 5, Taf. xviii. fig. 9.
Spines cylindrical, nearly of equal breadth throughout their whole length. Apex either simple,
conical, or bifid. Base with a large leaf-cross, four to six times as broad as the spine itself.
Central capsule transparent, yellow. The Atlantic specimens have much longer spines than those
figured from the Mediterranean, but are otherwise not different. Therefore I have changed the
inconvenient name brevispinum into catervatum.
Dimensions.—Length of the spines 0.1 to 0.3, breadth 0.005 to 0.008; basal
leaf-cross 0.02 to 0.032.
Habitat.—Mediterranean (Messina); North Atlantic, Station 354, Gulf
Stream (Færöe Channel) in great abundance, John Murray, surface.
Subgenus 3. Astrolithium, Haeckel, 1860, Monatsber. d. k. preuss. Akad. d. Wiss.
Berlin, p. 810.
Definition.—Spines in the basal part grown perfectly together, so that
the whole skeleton forms a single piece of acanthin; a star with twenty equal rays.
11. Acanthometron bulbiferum, n. sp. (Pl. 130, fig.
1).
Astrolithium bulbiferum, Haeckel, 1881, Prodromus, p. 466.
Spines needle-shaped, cylindrical, very thin and long, with simple apex; suddenly thickened and
forming a broad regular cone at the central base; all twenty conical bulbs have their broad bases
grown together and forming a central icosahedron of acanthin; from its twenty faces arise the
circular bases of the cones in regular disposition.
Dimensions.—Length of the spines 0.2 to 0.5, breadth 0.001 to 0.003; height of the
basal cones 0.06, basal breadth of them 0.02.
Habitat.—Central Pacific, Stations 265 to 274, surface.
12. Acanthometron bifidum, Haeckel.
Astrolithium bifidum, Haeckel, 1862, Monogr. d. Radiol., p. 400, Taf. xx.
fig. 5.
Spines needle-shaped, cylindrical, of equal breadth in their whole length. Distal apex bifid,
with two parallel straight teeth. Central bases of all twenty spines grown perfectly together and
forming a central sphere of acanthin. Central capsule brown opaque.
Dimensions.—Length of the spines 0.2, breadth 0.003.
Habitat.—Mediterranean (Messina), Haeckel, surface.
{746}
Genus 324. Zygacantha,[364] J. Müller,
1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 51.
Definition.—Astrolonchida with simple, compressed, and
two-edged radial spines, without apophyses; their transverse section is elliptical or
rhomboidal.
The genus Zygacantha comprised in the original definition of J. Müller only
a single species, Zygacantha furcata, distinguished from the other Acanthometra by forked spines with two long parallel teeth. It seems now
advisable to unite in this genus all those Astrolonchida in which the simple spines are two-edged,
compressed, or leaf-shaped. The term Zygacantha may be conceived as the general expression
of the important fact, that in all Icosacantha the twenty spines are opposite in pairs.
Subgenus 1. Zygacantharium, Haeckel.
Definition.—Spines at the central base without leaf-cross and without
hollow pyramidal compartments, united by the opposed triangular faces of their pyramidal bases,
resting one upon another.
1. Zygacantha lanceolata, Haeckel.
Acanthometra lanceolata, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 48,
Taf. xi. fig. 12.
Acanthometra lanceolata, Haeckel, 1862, Monogr. d. Radiol., p. 380.
Spines lanceolate, from the broader middle part equally thinned towards the two ends. Apex
simple. Base pyramidal, without leaf-cross. Each flat lamellar spine exhibits an elevated middle
rib (like a lanceolate leaf), and is therefore compressed quadrangular.
Dimensions.—Length of the spines 0.1 to 0.15, greatest breadth (in the width) 0.03
to 0.04.
Habitat.—Mediterranean (Saint Tropez, French shore), J. Müller; North
Atlantic (Canary Islands), Haeckel, surface.
2. Zygacantha costata, n. sp.
Spines compressed, two-edged, linear, of nearly equal breadth in their whole length. Apex
truncate. Base pyramidal, without leaf-cross. Each flat lamellar spine exhibits an elevated middle
rib, which in the distal half is cleft into two divergent rods ending in the corners of the
truncated apex. (Similar to Zygacantha dicopa, but with broader free spines, which are not
grown together in the centre.)
Dimensions.—Length of the spines 0.1 to 0.15, breadth 0.02.
Habitat.—Central Pacific, Stations 265 to 274, surface.
{747}
3. Zygacantha compressa, Haeckel.
Acanthometra compressa, Haeckel, 1862, Monogr. d. Radiol., p. 378, Taf.
xviii. figs. 4a, 4b.
Spines in the proximal half compressed, broad lanceolate, four to six times as broad as in the
thin cylindrical distal half. Apex simple or short bifid. Base pyramidal, thickened, without
leaf-cross. No middle rib. Central capsule opaque, yellowish or reddish-brown.
Dimensions.—Length of the spines 0.3 to 0.5, breadth of the inner half 0.015 to
0.02, of the outer half 0.002 to 0.003.
Habitat.—Mediterranean (Messina); Atlantic (Canary Islands), Station
352, surface.
4. Zygacantha furcata, J. Müller.
Zygacantha furcata, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 51, Taf.
ix. fig. 6.
Acanthometra furcata, J. Müller, 1856, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin,
p. 499.
Acanthometra furcata, Haeckel, 1862, Monogr. d. Radiol., p. 380.
Spines compressed, fork-shaped, divided by two thickened knots into three sections of nearly
equal length; middle section broader than the proximal, but smaller than the distal section, which
is cleft by a deep fissure into two long parallel straight branches. Base pyramidal, without
leaf-cross. Central capsule purple, with yellow bodies.
Dimensions.—Length of the spines 0.1 to 0.15, basal breadth 0.003 to 0.005, distal
breadth 0.02 to 0.03.
Habitat.—Mediterranean (Cette). J. Müller, surface.
Subgenus 2. Zygacanthidium, Haeckel.
Definition.—Spines at the central base with a cross of four prominent
leaves; the meeting edges of the neighbouring lamellæ so rest one upon another that twenty-two
hollow pyramidal compartments are formed (compare p. 721).
5. Zygacantha dichotoma, Haeckel.
Zygacantha dichotoma, Haeckel, 1862, Monogr. d. Radiol., p. 381.
Acanthometra dichotoma, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 50, Taf. ix. fig. 5.
Spines compressed, pincer-shaped, cleft nearly throughout their whole length into two thin
parallel straight rods or fork branches, which are united only in their middle by a narrow bridge,
and at their central base by the pyramidal small central apex; above this pyramid each rod is
divided into two broad triangular leaves, forming a large basal leaf-cross. Central capsule
purple, with yellow bodies.
Dimensions.—Length of the spines 0.1 to 0.15, breadth 0.01 to 0.02.
Habitat.—Mediterranean (Nice), J. Müller, surface.
{748}
6. Zygacantha complanata, n. sp.
Spines compressed, two-edged, linear, of equal breadth throughout their whole length. Apex
bifid. Base thickened, with a large leaf-cross. (Similar to Amphilonche complanata, but
different in the equal size of all twenty spines, which are somewhat broader.)
Dimensions.—Length of the spines 0.12 to 0.18, breadth 0.002.
Habitat.—North Pacific, Station 244, surface.
7. Zygacantha semicompressa, Haeckel.
Acanthometra hemicompressa, Car, 1884, Zool. Anzeiger, p. 94, with
woodcut.
Spines in the proximal half compressed, two-edged, linear, three to four times as broad as in
the needle-shaped cylindrical distal half. Apex simple. Base pyramidal, with a small leaf-cross.
Central capsule transparent.
Dimensions.—Length of the spines 0.1 to 0.2, breadth in the proximal half 0.02, in
the distal half 0.006.
Habitat.—Mediterranean (Genoa), Haeckel; Adriatic Sea (Trieste), Car, surface.
8. Zygacantha foliacea, n. sp.
Spines lanceolate, tapering from the broader middle part towards the two ends, with two
dentated or serrated edges and a prominent middle rib. Apex simple. Base with a small leaf-cross.
The spines like the leaves of Agave americana.
Dimensions.—Length of the spines 0.15 to 0.2, middle breadth 0.02 to 0.03.
Habitat.—Indian Ocean, near the Island of Socotra, Haeckel.
Subgenus 3. Zygacanthonium, Haeckel.
Definition.—Spines in the centre grown perfectly together and forming
a single star-shaped piece of acanthin.
9. Zygacantha dicopa, Haeckel.
Astrolithium dicopum, Haeckel, 1862, Monogr. d. Radiol., p. 400, Taf. xx.
figs. 3, 4.
Spines compressed, two-edged, linear, of nearly equal breadth throughout their whole length,
with a prominent middle rib, which in the distal third is cleft into two divergent teeth, ending
in the two corners of the broad, obliquely truncated apex. All twenty spines with their central
bases grown together and forming one single piece of acanthin—a star with twenty rays.
Dimensions.—Length of the spines 0.12 to 0.2, breadth 0.01 to 0.02.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific, many
Stations, surface.
{749}
Genus 325. Acanthonia,[365] Haeckel,
1881, Prodromus, p. 465.
Definition.—Astrolonchida with simple, four-edged,
prismatic or pyramidal radial spines, without apophyses; their transverse section is square.
The genus Acanthonia comprises all those Astrolonchida (formerly united with
Acanthometron) in which the simple spines either in their whole length or in the greatest
part of it are four-edged, with square transverse section. They are sometimes more prismatic (with
equal breadth), at other times more pyramidal (with decreasing breadth towards the distal apex).
If Acanthometron be the common simple ancestral form of the Acanthonida, then the two-edged
Zygacantha, and the four-edged Acanthonia may be regarded as two divergent main
lines arising from it; the former leading to the Phractacanthida and Diporaspida, the latter
leading to the Stauracanthida and Tessaraspida.
Subgenus 1. Acanthonarium, Haeckel.
Definition.—Spines at the central base, without leaf-cross and without
hollow pyramidal compartments, united by the triangular faces of their pyramidal bases, resting
one upon another.
1. Acanthonia tetracopa, Haeckel (Pl. 129, figs.
9-11).
Acanthometra tetracopa, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 47,
Taf. vii. figs. 3 to 5, Taf. xi. fig. 5.
Acanthometra tetracopa, Haeckel, 1862, Monogr. d. Radiol., p. 379, Taf.
xviii. fig. 5.
Spines quadrangular prismatic, with prominent lamellar edges, of equal breadth in their whole
length. Base pyramidal, without leaf-cross. Apex truncated, or with four short teeth (sometimes
only two teeth). Central capsule opaque, brown or yellow.
Dimensions.—Length of the spines 0.15 to 0.2, breadth 0.01 to 0.012.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Indian, Pacific, many
Stations, surface.
2. Acanthonia prismatica, n. sp.
Spines quadrangular prismatic, without prominent edges, with four plane lateral faces, of equal
breadth throughout their whole length. Base pyramidal, without leaf-cross. Apex truncate, with
square apical face.
Dimensions.—Length of the spines 0.1 to 0.16, breadth 0.005 to 0.008.
Habitat.—Antarctic Ocean, Station 154, surface.
{750}
3. Acanthonia denticulata, n. sp.
Spines quadrangular prismatic, with prominent elegantly denticulated edges, of equal breadth
throughout their whole length. Both ends pyramidal, without leaf-cross. (Similar to Amphilonche
denticulata, but with the twenty spines all equal, and with spherical central capsule.)
Dimensions.—Length of the spines 0.12 to 0.16, breadth 0.008.
Habitat.—Central Pacific, Stations 266 to 274, surface.
4. Acanthonia mülleri, Haeckel.
Acanthometra mülleri, Haeckel, 1862, Monogr. d. Radiol., p. 379, Taf. xv.
fig. 3, Taf. xviii. fig. 6.
Spines quadrangular, nearly prismatic, but tapering gradually from the pyramidal base towards
the distal bifid apex, which bears two thin, parallel, or little divergent teeth; four edges
elegantly denticulated; base without leaf-cross. Central capsule yellowish or reddish.
Dimensions.—Length of the spines 0.12 to 0.18, basal breadth 0.005.
Habitat.—Mediterranean (Messina); Atlantic, Stations 348 to 352,
surface.
5. Acanthonia fragilis, Haeckel.
Acanthometra fragilis, Haeckel, 1862, Monogr. d. Radiol., p. 380, Taf. xv.
fig. 4, Taf. xviii. fig. 7.
Spines quadrangular, nearly prismatic, but gradually thickened from the pyramidal base towards
the distal truncated end; four edges regularly denticulated; base without leaf-cross. Central
capsule opaque.
Dimensions.—Length of the spines 0.5 and more, basal breadth 0.002 to 0.003,
distal breadth 0.008 to 0.016 or more.
Habitat.—Mediterranean (Messina); North Atlantic, Station 354,
surface.
6. Acanthonia convexa, n. sp.
Spines quadrangular, with four broad lamellar, convex, prominent edges, which from the broader
middle part are thinned towards the two emarginated ends; each end with a small quadrangular
pyramid, without leaf-cross. All the twenty spines of this species exhibit nearly the same form,
which is seen in the caudal or posterior (geotomical) spine of Amphilonche anomala (Monogr.
d. Radiol., Taf. xviii. fig. 23, b).
Dimensions.—Length of the spines 0.12 to 0.16, breadth in the middle part 0.02 to
0.025.
Habitat.—North Pacific, Station 244, surface.
7. Acanthonia concava, n. sp.
Spines quadrangular, with four broad lamellar, concave, prominent edges, which from the smaller
middle part are broadened towards the two emarginated ends; each end with a small {751}quadrangular pyramid, without leaf-cross. All the twenty
spines of this species exhibit nearly the same form, which is seen in the frontal or anterior
(hydrotomical) spine of Amphilonche anomala (Monogr. d. Radiol., Taf. xviii. fig. 23,
a).
Dimensions.—Length of the spines 0.15, breadth in the middle part 0.012, on both
ends 0.025.
Habitat.—South Pacific, Station 288, surface.
8. Acanthonia quadrangula, n. sp.
Spines quadrangular prismatic, with smooth prominent straight edges, but of very different
breadth in both halves; the inner or proximal half (inside the central capsule) twice to four
times as broad as the outer or distal half (outside the central capsule); both ends suddenly
separated by a constriction, in which is inserted the membrane of the capsule. The latter is
pellucid, with a number of yellow bodies. Apex of the spines simple, base pyramidal, without
leaf-cross.
Dimensions.—Length of the spines 0.2 to 0.4, breadth of the basal half 0.01, of
the distal half 0.002 to 0.004.
Habitat.—Atlantic, Canary Islands, Azores, Station 354, surface.
Subgenus 2. Acanthonidium, Haeckel.
Definition.—Spines at the central base with a broad leaf-cross,
composed of four prominent triangular lamellæ; the meeting edges of these crossed lamellæ between
every three or four neighbouring spines so rest one upon another that twenty-two hollow pyramidal
compartments are formed (compare p. 721).
9. Acanthonia echinoides, Haeckel.
Acanthometra echinoides, Claparède et Lachmann, 1858, Études sur les Infusoires et les
Rhizopodes, &c., p. 459, pl. xxiii. figs. 1-5.
Acanthometra echinoides, Haeckel, 1862, Monogr. d. Radiol., p. 383.
Spines quadrangular prismatic without prominent edges, with four plane lateral faces, of equal
breadth throughout their whole length. Central base three to four times as broad, with large
leaf-cross. Apex truncated or bifid (sometimes with four short teeth).
Dimensions.—Length of the spines 0.3 to 0.5, breadth 0.004 to 0.008; leaf-cross
0.02 to 0.03.
Habitat.—North Atlantic, west coast of Norway (Claparède et Lachmann);
Færöe Channel (Gulf Stream), John Murray, surface.
10. Acanthonia claparedei, Haeckel.
Acanthometra claparedei, Haeckel, 1862, Monogr. d. Radiol., p. 383, Taf.
xviii. fig. 12.
Spines quadrangular prismatic, with four prominent lamellar edges, of equal breadth throughout
their whole length. Apex bifid. Central base twice as broad, with large leaf-cross. Central
capsule opaque, reddish-brown.
Dimensions.—Length of the spines 0.6, breadth 0.016; leaf-cross 0.032.
Habitat.—Mediterranean (Messina); Atlantic, Station 347, surface.
{752}
11. Acanthonia cuspidata, Haeckel.
Acanthometra cuspidata, Haeckel, 1862, Monogr. d. Radiol., p. 383, Taf.
xviii. fig. 11.
Spines quadrangular pyramidal, with four prominent lamellar edges, tapering gradually from the
broad base towards the simple distal apex. Basal leaf-cross as broad, with a small central
pyramid.
Dimensions.—Length of the spines 0.5, basal breadth 0.012; leaf-cross 0.025.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific, surface.
12. Acanthonia quadrifolia, Haeckel.
Acanthometra quadrifolia, Haeckel, 1862, Monogr. d. Radiol., p. 382, Taf.
xviii. fig. 10.
Spines in the basal part quadrangular pyramidal, with four prominent lamellar edges and a large
basal leaf-cross; in the distal part three to six times as long, cylindrical, of equal breadth.
Apex simple, conical, or bifid. Central capsule yellow or whitish. This common species differs
from the nearly allied Acanthometron catervatum mainly by the strong development of the
large basal leaves or wings.
Dimensions.—Length of the spines 0.3 to 0.5, breadth in the distal half 0.002, in
the basal part 0.02.
Habitat.—Mediterranean; North Atlantic, Stations 252 to 254; Færöe
Channel, Gulf Stream, in enormous numbers, John Murray, surface and at various depths.
13. Acanthonia diplopyramis, n. sp.
Spines formed like a quadrangular double pyramid or an irregular octahedron; the basal
leaf-cross being extremely developed, with four very large and thin lamellar leaves; the basal
halves of the twenty double pyramids are united by the meeting edges of those leaves, while their
distal halves are free, with simple apices. Therefore the four triangular leaves of each spine are
equally thinned from the middle towards the two ends.
Dimensions.—Length of the spines 0.1 to 0.2, greatest breadth 0.04 to 0.08.
Habitat.—Central Pacific, Stations 266 to 274, surface.
14. Acanthonia multispina, Haeckel.
Acanthometra multispina, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 47,
Taf. vii. figs. 6-9.
Acanthometra multispina, Haeckel, 1862, Monogr. d. Radiol., p. 384.
Spines quadrangular, with four lamellar prominent edges, in the proximal half nearly prismatic,
in the distal half pyramidal; both halves separated by a prominent short tooth on each edge. Here
in the middle part the breadth (including the four teeth) is equal to the basal leaf-cross.
Central capsule opaque, brown.
Dimensions.—Length of the spines 0.2 to 0.3, greatest breadth 0.01 to 0.02.
Habitat.—Mediterranean (Messina); Tropical Atlantic, Station 348.
{753}
15. Acanthonia serrulata, n. sp.
Spines quadrangular pyramidal, with simple apex; the large basal leaf-cross nearly half as long
as the prolonged distal part. The four prominent triangular edges of the latter are very thin and
broad lamellæ, finely dentate or serrate.
Dimensions.—Length of the spines 0.2 to 0.3, greatest breadth 0.05 to 0.06.
Habitat.—Central Pacific, Station 272, surface.
Subgenus 3. Acantholithium, Haeckel.
Definition.—Spines in the basal part grown together, so that the whole
skeleton is not composed of twenty separated pieces, but represents a single piece of
acanthin—a star with twenty equal rays.
16. Acanthonia stellata, n. sp.
Spines quadrangular pyramidal, with simple distal apex, in the basal part grown perfectly
together, so that the whole skeleton forms a single piece of acanthin—a starlet with twenty
equal rays. The free pyramidal part of each spine is twice to four times as long as the basal
part.
Dimensions.—Length of the spines 0.1 to 0.15, greatest breadth (on the surface of
the central solid sphere) 0.02 to 0.04.
Habitat.—Central Pacific, Station 271, surface.
Subfamily 2. Phractacanthida, Haeckel.
Definition.—Astrolonchida with twenty radial
spines, each of which bears two opposite apophyses or lateral transverse processes; sometimes two
longitudinal rows of opposite apophyses.
Genus 326. Lithophyllium,[366] J. Müller,
1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 52.
Definition.—Astrolonchida with two simple, not branched,
opposite apophyses on each radial spine.
The genus Lithophyllium was founded by J. Müller for a single species
(Lithophyllium foliosum), which we also here retain as the type of the genus. It is the
first observed Astrolonchid, which bears two opposite lateral apophyses on each spine, and may
therefore be regarded as the ancestral form of the subfamily Phractacanthida. The two opposite
apophyses are here simple, whilst in the other genera of the subfamily they are branched or
multiplied.
{754}
1. Lithophyllium cruciatum, Haeckel.
Acanthometra cruciata, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 49,
Taf xi. fig. 11.
Xiphacantha cruciata, Haeckel, 1862, Monogr. d. Radiol., p. 385, Taf.
xviii. fig. 13.
Spines cylindrical, very thin, crossed perpendicularly in the distal third by a thin transverse
beam; both lateral rods of the cross have the same length as the distal end. Apex simple. Base
pyramidal, without leaf-cross.
Dimensions.—Length of the spines 0.05 to 0.15, breadth 0.001 to 0.002.
Habitat.—Mediterranean, Atlantic, Stations 352 to 354, &c.,
surface.
2. Lithophyllium gladiatum, n. sp. (Pl. 130, fig.
3).
Spines compressed, sword-shaped, two-edged, crossed perpendicularly in the middle part by a
broad, somewhat curved transverse beam; both lateral rods of the cross are triangular compressed,
and have the same length as the proximal end. Apex simple. Base pyramidal, without leaf-cross.
Dimensions.—Length of the spines 0.2 to 0.25, breadth 0.01.
Habitat.—North Pacific, Station 241, surface.
3. Lithophyllium condylatum, n. sp.
Spines compressed quadrangular, tapering from the broader middle towards the two ends; from the
middle part arise two opposite perpendicular apophyses, which bear on the rounded end a thickened
condyle. Apex simple. Base pyramidal, with a small leaf-cross.
Dimensions.—Length of the spines 0.1 to 0.12, breadth in the middle part 0.02.
Habitat.—Central Pacific, Station 266 to 272, surface.
4. Lithophyllium foliosum, J. Müller.
Lithophyllium foliosum, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 52,
Taf. xi. figs. 6-10.
Xiphacantha foliosa, Haeckel, 1862, Monogr. d. Radiol., p. 385.
Spines lanceolate, tapering from the broader middle towards the two ends. From the middle part
or from the outer third arise two opposite triangular apophyses, which are not perpendicular to
the axis of the spine, but form an acute angle with its distal part. Therefore each spine
represents a broad leaf with a middle rib and with three lobes or truncated teeth. The distal apex
is distinguished by a violet colour. Base pyramidal, without leaf-cross. Central capsule
yellow.
Dimensions.—Length of the spines 0.05 to 0.1, breadth 0.02 to 0.03.
Habitat.—Mediterranean, French shore, Saint Tropez, J. Müller.
{755}
Genus 327. Phractacantha,[367] Haeckel,
1881, Prodromus, p. 465.
Definition.—Astrolonchida with two branched, but not
latticed, opposite apophyses on each radial spine.
The genus Phractacantha differs from its ancestral form, the preceding
Lithophyllium by the ramification of the apophyses, which are either forked or bear lateral
branches. If the prolonged fork-branches of the neighbouring spines meet and form a lattice-shell,
this genus passes over into Phractaspis, the common ancestral form of the Diporaspida.
1. Phractacantha bifurca, n. sp.
Spines cylindrical, thin, of equal breadth throughout their whole length. Apex simple. Base
pyramidal, without leaf-cross. From the outer third arise two opposite, thin, forked apophyses;
ends of the fork branches acute.
Dimensions.—Length of the spines 0.1, breadth 0.006.
Habitat.—Central Pacific, Station 274, surface.
2. Phractacantha bipennis, n. sp.
Spines compressed, two-edged, tapering from the broader middle towards the two ends. Apex
simple. Base pyramidal, without leaf-cross. From the middle arise two opposite broad, forked
apophyses; ends of the fork-branches broad and obtuse.
Dimensions.—Length of the spines 0.15, breadth in the middle 0.012.
Habitat.—Central Pacific, Station 266, surface.
Genus 328. Doracantha,[368] Haeckel,
1881, Prodromus, p. 465.
Definition.—Astrolonchida with two latticed or
fenestrated apophyses on each radial spine.
The genus Doracantha arises from the foregoing Phractacantha by union
of the fork-branches of the apophyses on each spine. By this concrescence is formed a polygonal or
roundish plate with two pores, pierced by the radial spine between them. Doracantha may be
regarded as a Dorataspis, in which the twenty small fenestrated plates have not become
united.
{756}
1. Doracantha dorataspis, n. sp.
Spines compressed, two-edged, tapering from the broader middle towards the two ends. Apex
simple. Base pyramidal, without leaf-cross. From the middle part of each spine arise two opposite
forked apophyses; the neighbouring fork-branches are recurved and united in the tangential plane;
so that each spine bears a plate or shield with two elliptical pores; the margin of the roundish
plate bears a variable number of short teeth.
Dimensions.—Length of the spines 0.2, breadth 0.02.
Habitat.—Central Pacific, Station 271, surface.
Genus 329. Astrolonche,[369] Haeckel,
1881, Prodromus, p. 465.
Definition.—Astrolonchida with numerous simple apophyses
(four to eight or more on each radial spine), which are arranged in two opposite longitudinal rows
(rarely in six such rows, opposite in three parallel planes).
The genus Astrolonche differs from the preceding Phractacanthida in the
multiplication of the simple apophyses. Commonly these are opposite in pairs in two longitudinal
rows (on the two edges of the compressed spine, at least two on each side). But sometimes (in the
subgenus Astrolonchidium) there are six instead of two longitudinal rows of teeth, and
these are placed in three parallel planes.
Subgenus 1. Astroloncharium, Haeckel.
Definition.—Each spine with two longitudinal rows of apophyses,
opposite in one meridian plane.
1. Astrolonche bicruciata, n. sp.
Spines compressed, two-edged, almost of equal breadth throughout their whole length, with
simple apex and small leaf-cross at the base. From the two edges of the middle part of each spine
arise two pairs of opposite apophyses, which are compressed and a little curved (convex on the
distal, concave on the proximal margin). All four apophyses are of the same size and form; the
radial distance of each pair much smaller than their common distance from the centre.
Dimensions.—Length of the spines 0.25, breadth 0.02.
Habitat.—Central Pacific, Station 270, surface.
2. Astrolonche mucronata, Haeckel.
Acanthometra mucronata, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 49,
Taf. x. fig. 9.
Aspidomma mucronatum, Haeckel, 1862, Monogr. d. Radiol., p. 424.
Spines conical, little compressed, with simple apex and small leaf-cross at the base. From the
basal half of each spine arise two pairs of opposite apophyses, which exhibit a very different
form. {757}The upper or distal apophyses (nearly in the
middle of the spine) are flat, leaf-shaped, broadened in the periphery, often lobed, and sometimes
branched or even fenestrated. (Transition to Phractaspis and Dorataspis, or to
Phractopelta?) The lower or proximal apophyses are thick, simple, all slightly curved; they
are equidistant from the former and from the centre.
Dimensions.—Length of the spines 0.2 to 0.3, greatest breadth 0.01.
Habitat.—Mediterranean (Cette on the French shore), surface, J.
Müller.
3. Astrolonche pectinata, Haeckel.
Acanthometra pectinata, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 48,
Taf. x. figs. 1, 2.
Xiphacantha pectinata, Haeckel, 1862, Monogr. d. Radiol., p. 386.
Spines compressed quadrangular, with short, simple or bifid apex, pyramidal on the base,
without leaf-cross. From the two broader edges of the proximal half arise two longitudinal rows of
opposite apophyses; three to four slender teeth in each row.
Dimensions.—Length of the spines 0.2, breadth 0.08.
Habitat.—Mediterranean, Cette (French shore), surface, J. Müller.
4. Astrolonche pinnata, n. sp.
Spines in the distal half compressed, linear, two-edged, with bifid apex; in the proximal half
three times as broad lanceolate, at the base pyramidal, without leaf-cross. From the two broader
edges of the proximal half arise two longitudinal rows of opposite apophyses; four to six broad
triangular teeth in each row.
Dimensions.—Length of the spines 0.3 to 0.4, breadth in the outer half 0.012, in
the inner half 0.03 to 0.04.
Habitat.—South Atlantic, Station 332, surface.
Subgenus 2. Astrolonchidium, Haeckel.
Definition.—Each spine with three parallel double rows of opposite
apophyses.
5. Astrolonche serrata, Haeckel.
Xiphacantha serrata, Haeckel, 1862, Monogr. d. Radiol., p. 386, Taf. xvii. fig. 3,
Taf. xviii. figs. 14a, 14b.
Xiphacantha serrata, R. Hertwig, 1879, Organismus d. Radiol., p. 11, Taf. ii. fig.
4.
Acanthometra serrata, Haeckel, 1860, Monogr. d. Radiol., p. 807.
Spines in the distal half thin, nearly cylindrical or a little compressed, thinned towards the
short simple or bifid apex; in the proximal half three to six times as broad, four-winged, with
six longitudinal rows of opposite apophyses. From the edges of the two broader (lateral) wings
arise three {758}to four pairs of strong conical teeth;
from the edges of the two smaller (sagittal) wings arise also three to four pairs of teeth, but
very short and broad, triangular; each of these teeth bears on both its (lateral) sides two
opposite slender conical teeth, which are parallel to the large conical teeth of the lateral
wings. Therefore all teeth (eighteen to twenty-four) are placed opposite in pairs in three
parallel planes. Base of the spines pyramidal, with a small leaf-cross. The central capsule of
this large and very remarkable species commonly entirely includes the apophyses of the spines,
and is opaque, whitish.
Dimensions.—Length of the spines 0.2 to 0.3, breadth of the distal half 0.002 to
0.004, of the proximal half 0.01 to 0.02.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific, surface.
Subfamily 3. Stauracanthida, Haeckel.
Definition.—Astrolonchida with a cross of
four free apophyses (or four crossed longitudinal rows of apophyses) on each radial spine.
Genus 330. Xiphacantha,[370] Haeckel,
1862, Monogr. d. Radiol., p. 384.
Definition.—Astrolonchida with four simple apophyses on
each radial spine, opposite in pairs in the form of a cross.
The genus Xiphacantha was founded by me in 1862 for all those
Acanthometrida which bear simple or branched apophyses on their twenty equal spines. I
restrict here the genus to those Astrolonchida which bear on each spine a cross of four simple,
not branched, apophyses. These are either conical teeth or broad wings, sometimes extremely thin
leaves. Xiphacantha may be regarded as the ancestral form not only of the subfamily
Stauracanthida, but also of the Tessaraspida, derived from the latter.
Subgenus 1. Xiphacanthonia, Haeckel.
Definition.—Apophyses of the radial spines small, formed like a tooth
or a hook, not wing-shaped. Edges of the spines commonly narrow, little prominent.
1. Xiphacantha quadridentata, Haeckel.
Xiphacantha quadridentata, Haeckel, 1862, Monogr. d. Radiol., p. 387, Taf. xviii.
figs. 15a, 15b.
Acanthometra quadridentata, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 48, Taf. x. fig. 3.
Spines slender, four-sided prismatic, gradually thinner towards the simple pyramidal apex. Base
with large wing-cross. Four apophyses about in the middle of each spine, conical, straight,
smooth, about as long as the basal breadth of the spine. Central capsule opaque,
reddish-brown.
{759}
Dimensions.—Length of the spines 0.2 to 0.3, breadth in the middle part 0.012, on
the base 0.02; length of the apophyses 0.02 to 0.03.
Habitat.—Mediterranean, Atlantic, Stations 348, 354, surface.
2. Xiphacantha crucifera, n. sp.
Spines slender, four-sided prismatic, thin, of nearly equal breadth throughout the whole
length. Base with small wing-cross. Four apophyses much nearer the proximal than the distal end,
thin, straight smooth, three to six times as long as the basal breadth of the spine. Central
capsule pellucid, with yellow bodies.
Dimensions.—Length of the spines 0.4 to 0.5, breadth 0.002 to 0.004; length of the
apophyses 0.01 to 0.02.
Habitat.—Central Pacific, Station 266, surface.
3. Xiphacantha spinulosa, Haeckel.
Xiphacantha spinulosa, Haeckel, 1862, Monogr. d. Radiol., p. 388, Taf. xvii. fig.
4.
Acanthometra spinulosa, Haeckel, 1860, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 807.
Spines stout, four-sided prismatic, of nearly equal breadth throughout the whole length, on the
distal apex truncated, two-edged. Base with large wing-cross. Four apophyses about in the middle
of each spine, conical, straight, spinulated, twice to four times as long as the basal breadth of
the spine. Central capsule opaque, yellow.
Dimensions.—Length of the spines 0.27, breadth 0.014; length of the apophyses
0.05.
Habitat.—Mediterranean (Messina), surface.
4. Xiphacantha emarginata, n. sp.
Spines stout, four-sided, in the proximal half prismatic, in the distal half pyramidal, with
emarginated apex. Base with very large wing-cross. Four apophyses about in the middle of each
spine, compressed, straight, two-edged, with emarginated ends, about twice as long as the basal
breadth of the spine. Central capsule small, pellucid.
Dimensions.—Length of the spines 0.2 to 0.3, breadth 0.02 to 0.03; length of the
apophyses 0.04 to 0.06.
Habitat.—South Pacific, Station 291, surface.
5. Xiphacantha falcata, n. sp.
Spines slender, four-sided prismatic, gradually thickened from both ends towards the middle
part, with simple apex. Base with very small wing-cross. Four apophyses about in the middle part
of each spine, falcated, compressed, recurved, twice to three times as long as the greatest
breadth of the spine.
{760}
Dimensions.—Length of the spines 0.3 to 0.4, greatest breadth 0.015; length of the
apophyses 0.03 to 0.05.
Habitat.—Central Pacific, Station 270, surface.
6. Xiphacantha ancorata, n. sp.
Spines slender, four-sided prismatic, gradually thickened from the narrow base towards the
short, simple, pyramidal apex. Base with large wing-cross. Four apophyses on the distal end,
immediately below the pyramidal apex, falcated, strongly recurved, like an anchor with four strong
teeth.
Dimensions.—Length of the spines 0.4 to 0.5, basal breadth 0.005, distal breadth
0.015; length of the apophyses 0.04.
Habitat.—Indian Ocean, Madagascar, Rabbe.
Subgenus 2. Xiphacanthidium, Haeckel.
Definition.—Apophyses of the radial spines broad, compressed,
wing-shaped, formed like a large thin plate. Edges of the spines commonly broad, strongly
prominent.
7. Xiphacantha stauroptera, n. sp.
Spines four-winged, from the middle part little thinner towards both ends. Apex simple
pyramidal. Base with small wing-cross. Four apophyses wing-shaped, isosceles triangular or
truncated quadrangular; their base is about half as broad as their length, and occupies in the
basal half of each spine about one-sixth to one-eighth of its length.
Dimensions.—Length of the spines 0.1 to 0.15; greatest breadth (diagonal of the
cross of the apophyses) 0.006 to 0.009.
Habitat.—South Atlantic, Station 332, surface.
8. Xiphacantha trigonoptera, n. sp.
Spines four-winged, tapering rapidly from the broad middle part towards both ends. Apex simple
pyramidal. Base with a small wing-cross. Four apophyses equilateral triangular; their base
occupies the middle of each spine, extending to about one-fifth of its length.
Dimensions.—Length of the spines 0.15 to 0.2, greatest breadth (diagonal of the
cross of the apophyses) 0.06 to 0.08.
Habitat.—Central Pacific, Station 272, surface.
9. Xiphacantha macroptera, n. sp.
Spines four-winged, from the broad middle part gradually thinner towards both ends. Apex simple
pyramidal. Base with small wing-cross. Four apophyses wing-shaped, very long and broad, {761}triangular; their base occupies about the middle third of
each spine; the proximal side of each wing is the shortest, truncated or concave.
Dimensions.—Length of the spines 0.4 to 0.5, greatest breadth (diagonal of the
middle leaf-cross) 0.08 to 0.12.
Habitat.—North Pacific, Stations 253 to 256, surface.
10. Xiphacantha platyptera, n. sp.
Spines four-winged, nearly prismatic in the basal and distal parts. Apex truncated. Base with
small wing-cross. Four apophyses wing-shaped, very broad and thin, extremely delicate, of
irregular quadrangular or nearly rhomboidal form; their base occupies the middle half of the
length of the spines (second and third quarter). Very variable in form. Often the edges of the
wing-apophyses of the neighbouring spines seem to meet.
Dimensions.—Length of the spines 0.2 to 0.3, greatest breadth (diagonal of the
leaf-cross) 0.06 to 0.08.
Habitat.—Pacific, widely distributed, Stations 239, 253, 272, 288,
&c., surface.
11. Xiphacantha ciliata, n. sp. (Pl. 129,
figs. 4, 5).
Spines four-winged, prismatic in the basal and the distal third. Apex pyramidal, short. Base
with large leaf-cross. Four apophyses wing-shaped, nearly semicircular, with dentated edges and
rough spinulate or porous faces; their base occupies the middle third of the length of each
spine.
Dimensions.—Length of the spines 0.2 to 0.3, greatest breadth (diagonal of the
apophysial cross) 0.08 to 0.12.
Habitat.—Tropical Atlantic, Stations 342 to 352, surface.
12. Xiphacantha alata, Haeckel.
Xiphacantha alata, Haeckel, 1862, Monogr. d. Radiol., p. 388.
Acanthometra alata, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin,
p. 48, Taf. ix. figs. 1-3.
Spines four-winged, prismatic in the basal quarter, more slender in the distal half. Apex
simple pyramidal. Base with small wing-cross. Four apophyses wing-shaped, nearly semicircular,
with denticulated edges (and often also with spinulate faces); their base occupies the second
quarter of the length of each spine.
Dimensions.—Length of the spines 0.3 to 0.4, greatest breadth (diagonal of the
apophysial cross) 0.05 to 0.06.
Habitat.—Mediterranean (Nice), Müller, (Portofino), Haeckel.
Genus 331. Stauracantha,[371] Haeckel,
1881, Prodromus, p. 465.
Definition.—Astrolonchida with four branched (but not
latticed) apophyses on each radial spine, opposite in pairs in the form of a cross.
{762}
The genus Stauracantha differs from its ancestral form, Xiphacantha,
in the ramification of the four crossed apophyses on each spine. These bear either lateral
branches, which are parallel to the cross axes of the radial spine itself (subgenus
Stauracanthonium), or they are forked, with divergent branches not parallel to those cross
axes (subgenus Stauracanthidium). Both subgenera may perhaps be better separated as genera.
They form the transition to the Stauraspida.
Subgenus 1. Stauracanthonium, Haeckel.
Definition.—Apophyses of the radial spines not forked, but crossed by
perpendicular branches, which are placed in tangential planes and parallel to the cross axes of
the quadrangular spine itself.
1. Stauracantha orthostaura, n. sp. (Pl. 130, fig.
5).
Spines four-sided prismatic, with simple apex and small leaf-cross at the base; with four thin
slender conical apophyses in the proximal third. Each apophysis represents a regular rectangular
cross, being intersected in its middle by one perpendicular rod of its own length.
Dimensions.—Length of the spines 0.3, breadth 0.02; distance of the apophyses from
the apex 0.15 to 0.2.
Habitat.—South Pacific, Station 169, surface.
2. Stauracantha tetrastaura, n. sp.
Spine four-winged prismatic, with four broad prominent edges, pyramidal apex, and large basal
leaf-cross, with four broad compressed apophyses about in the middle. Each apophysis represents a
vertical lamella (placed in a meridian) with convex distal and concave proximal edge, and is
crossed in its apical part by one perpendicular short rod.
Dimensions.—Length of the spines 0.5, breadth 0.03; distance of the apophyses from
the centre 0.2.
Habitat.—Central Pacific, Station 272, surface.
3. Stauracantha diplostaura, n. sp.
Spines four-sided prismatic, with truncated apex and broad basal leaf-cross; in the proximal
half with four slender conical apophyses. Each apophysis is crossed in its apical half at right
angles by two short parallel transverse rods.
Dimensions.—Length of the spines 0.32, breadth 0.012; distance of the apophyses
from the centre 0.11.
Habitat.—South Pacific, Station 291, surface.
{763}
4. Stauracantha scalaris, n. sp.
Spines four-winged prismatic, with four broad prominent edges, truncated apex, and small basal
leaf-cross; in the proximal third with four compressed lamellar apophyses. Each apophysis is
pinnate with opposite pinnulæ, or crossed at right angles by three to six parallel transverse
rods.
Dimensions.—Length of the spines 0.24, breadth 0.016; central distance of the
apophyses 0.12.
Habitat.—Indian Ocean (Madagascar), Rabbe, surface.
5. Stauracantha johannis, n. sp. (Pl.
132, fig.
11).
Spines four-sided pyramidal, with simple apex and broad basal leaf-cross, with four large
conical apophyses about in their middle. Each apophysis bears two irregular rows of alternating
perpendicular lateral branches, the length of which decreases from the base towards the apex of
the apophysis.
Dimensions.—Length of the spines 0.2, middle breadth 0.02; central distance of the
apophyses 0.1.
Habitat.—South-east Pacific (off Juan Fernandez), Station 300,
surface.
6. Stauracantha murrayana, Haeckel.
Xiphacantha species, Wyville Thomson, 1877, The Atlantic, vol. i. p. 235, fig. 53.
Xiphacantha murrayana, Haeckel, 1878, Protistenreich, p. 45, fig. 33.
Spines quadrangular prismatic, with pyramidal apex and small basal leaf-cross, with four
regular conical apophyses in the proximal part. Each apophysis is crossed at right angles by two
parallel transverse rods; the smaller apical rod is simple; the larger basal rod is crossed again
on each side by a secondary perpendicular branch, which therefore is parallel to the apophysis
itself.
Dimensions.—Length of the spines 0.3, breadth 0.01, distance of the apophyses
0.08.
Habitat.—South Atlantic, Station 332, surface.
7. Stauracantha pinnulata, n. sp.
Spines four-winged pyramidal, with prominent edges, simple apex and large basal leaf-cross, in
the middle with doubly pinnate triangular apophyses. Each apophysis is crossed at right angles by
three to four transverse parallel rods, which bear again perpendicular secondary branches; the
outline of the doubly pinnate apophysis is an isosceles triangle.
Dimensions.—Length of the spines 0.3, middle breadth 0.02; distance of the
apophyses 0.12.
Habitat.—North Pacific, Station 244, surface.
{764}
Subgenus 2. Stauracanthidium, Haeckel.
Definition.—Apophyses of the radial spines forked, each with two
divergent terminal branches, which are not parallel to the cross axes of the quadrangular
spine.
8. Stauracantha stauraspis, Haeckel.
Dorataspis polyancistra, juvenis, Haeckel, 1862, Monogr. d. Radiol., p.
418, Taf. xxi. fig. 7.
Spines slender, quadrangular prismatic, with simple apex and pyramidal base (without basal
leaf-cross); with four slender apophyses, which are simply forked in the outer third. Each
apophysis with two recurved fork branches.
Dimensions.—Length of the spines 0.12, breadth 0.004; distance of the apophyses
0.06.
Habitat.—Mediterranean (Messina), surface.
9. Stauracantha bifurca, n. sp.
Spines stout, four-winged prismatic, with pyramidal apex and pyramidal base (without
leaf-cross); in their middle with four compressed, little curved apophyses (convex on the outer,
concave on the inner edge), which are simply forked at the end. Each apophysis with two short
straight fork branches.
Dimensions.—Length of the spines 0.2, breadth 0.006; distance of the apophyses
0.08.
Habitat.—Central Pacific, Station 266, surface.
10. Stauracantha quadrifurca, n. sp. (Pl. 130, fig.
4).
Spines slender, four-sided prismatic, with simple apex and pyramidal base (without leaf-cross);
with four slender apophyses, which are doubly forked about in their middle. Each apophysis with
four short and thin terminal branches.
Dimensions.—Length of the spines 0.2, breadth 0.002; distance of the apophyses
from the centre 0.08.
Habitat.—South Atlantic, Stations 325 to 330, surface.
Genus 332. Phatnacantha,[372] Haeckel,
1881, Prodromus, p. 465.
Definition.—Astrolonchida with four apophyses on each
radial spine, opposite in pairs in cross form, and forming a lattice-plate by communicating
branches.
The genus Phatnacantha has been derived from the preceding
Stauracantha by concrescence of the branches of the apophyses. Therefore each spine bears a
lattice-plate or a fenestrated shield. If the growing plates of the neighbouring spines {765}meet together, then the characteristic lattice-shell of
the Tessaraspida is
perfect. Either each plate bears four crossed pores (like Tessaraspis), or a larger number of pores (four
aspinal and four to eight or more coronal) like Icosaspis.
1. Phatnacantha tessaraspis, n. sp.
Spines quadrangular prismatic, with simple pyramidal apex and small basal leaf-cross. Each
spine bears in the basal half a square plate, which is perforated by four square pores; margin of
the plate with twelve short straight teeth.
Dimensions.—Length of the spines 0.12, breadth 0.005.
Habitat.—Central Pacific, Station 265, surface.
2. Phatnacantha icosaspis, n. sp. (Pl. 130, fig.
6).
Spines quadrangular with prominent edges; prismatic in the basal half, with a broad basal
leaf-cross, pyramidal in the distal half, with a simple or truncate apex. Each spine bears in the
middle part a square plate, which is perforated by eight to sixteen or more square pores (four
aspinal and four to eight or more coronal); margin of the plate with twenty-four to forty-eight or
more short straight irregular compressed teeth.
Dimensions.—Length of the spines 0.18, breadth 0.008.
Habitat.—Central Pacific, Station 273, surface.
Genus 333. Pristacantha,[373] n.
gen.
Definition.—Astrolonchida with numerous simple apophyses
(eight to sixteen or more on each radial spine) which are arranged in four longitudinal rows
opposite in pairs in the form of a cross.
The genus Pristacantha differs from all other Stauracanthida in the
multiplication of the crossed apophyses (at least eight on each spine), and exhibits therefore to
them the same relation as Astrolonche exhibits to the other Phractacanthida. The remarkable
Astrolonchidium serratum appears intermediate between both groups.
1. Pristacantha octodon, n. sp. (Pl. 130,
fig. 9).
Spines four-sided prismatic, a little broader in the middle part than at either end, with thin
prominent edges. Apex truncate or pyramidal. Base pyramidal, with a small leaf-cross. From the
four edges arise in the basal half (between first and second third of the length) eight slender,
conical, or triangular apophyses (two on each edge).
Dimensions.—Length of the spines 0.4 to 0.6, breadth 0.02 to 0.04.
Habitat.—Central Pacific, Station 271, surface.
{766}
2. Pristacantha dodecodon, n. sp. (Pl. 130, fig.
8).
Spines four-winged, of lanceolate outline, tapering from the broader middle towards the two
ends. Apex pyramidal. Base with a large leaf-cross. From the four wings arise in the basal part
(between first and second third of the length) twelve triangular apophyses (three from each
wing).
Dimensions.—Length of the spines 0.3, breadth in the middle 0.03.
Habitat.—North Pacific, Station 244, surface.
3. Pristacantha polyodon, n. sp. (Pl. 130, fig.
7).
Spines in the distal half four-sided prismatic, thin, with pyramidal apex; in the basal half
much broader, four-winged, with a large leaf-cross on the base. From the four wings of the basal
half arises a variable number of slender teeth or triangular apophyses, commonly sixteen to
twenty-four, often irregular (four to six from each wing).
Dimensions.—Length of the spines 0.3 to 0.4, breadth in the distal part 0.007 to
0.01, in the basal part 0.02 to 0.03.
Habitat.—South Pacific (off New Zealand), Station 169, surface.
Family XXXVII. Quadrilonchida, Haeckel (Pl.
131).
Acanthostaurida, Haeckel, 1881, Prodromus, p. 466.
Definition.—Acantharia with twenty radial spines of very
unequal size, disposed according to the law of the Icosacantha; four equatorial spines much larger
than the sixteen others. No lattice-shell.
The family Quadrilonchida differs from the foregoing ancestral family,
the Astrolonchida, in the unequal development of the twenty radial spines. The four equatorial
spines are constantly much larger, and often also of another form and shape, than the sixteen
other spines; often also among these the eight tropical spines are larger and of another form than
the eight polar spines. Therefore the five parallel girdles or zones of every four spines, which
in the Astrolonchida are equal, are here distinctly unequal. The whole body is flattened and
compressed in the direction of the spineless axis, so that the equatorial plane is larger than any
other plane, laid through the centre. In consequence of this flattening the central capsule is
also commonly compressed and flattened, lenticular or discoidal, rarely spherical. In the
Astrolonchida the "promorph" or the "geometrical fundamental form" is constantly a square double
pyramid, the axes of which are of equal length. In the Quadrilonchida it becomes a square double
pyramid, the two equatorial axes of which (or the diagonals of the square) are constantly longer
than all other axes.
In the simpler forms of Quadrilonchida are found only two different kinds of spines, the four
larger equatorial spines being of the same size and form, and the sixteen smaller {767}spines also not differing from one another
(Acanthostaurus, &c.). But in the majority of this family there are three different
kinds: four larger equatorial spines, eight tropical spines of middle size, and eight smaller
polar spines (Belonostaurus, &c.); the latter become sometimes rudimentary, so that
only twelve spines are developed (four larger equatorial and eight smaller tropical spines). In
this case the development of the flat discoidal body is much stronger in the equatorial plane than
in all other planes. These discoidal or lenticular Quadrilonchida exhibit a relation to the
spherical Astrolonchida similar to that which the Discoidea exhibit to
the Sphæroidea among the Sphærellaria.
A further morphological differentation takes place in the remarkable genera
Lonchostaurus and Zygostaurus. In the former (Pl. 131, figs.
4-6) the four larger equatorial spines becomes differentiated in pairs, so that the opposite equal
spines of one pair, c1, c3 (in the longitudinal or hydrotomical axis), are larger,
and often also of another form, than the opposite equal spines of the other pair, c2,
c4 (in the transverse or geotomical axis). The most peculiar form is the rather common
Zygostaurus (Pl. 131,
figs. 7, 8). Here the two opposite spines of one equatorial axis (of the longitudinal axis) become
very different, so that the anterior or frontal spine (c1) is very unequal to the posterior
or caudal spine (c3), whereas the two opposite spines, of the other equatorial axis (of the
transverse axis) remain equal (c2, c4). Therefore the fundamental forms become here
"amphithect," as in the Ctenophora.
The numerous Quadrilonchida may be disposed in two different subfamilies: in the
Acanthostaurida all twenty radial spines are simple, without apophyses; in the Lithopterida all
twenty spines (or only one part of them) bear two opposite apophyses (or lateral transverse
processes). The former correspond to the Zygacanthida, the latter to the Phractacanthida among the
Astrolonchida. The two opposite apophyses are simple in Quadrilonche (Pl. 133, fig. 1).
In Xiphoptera they are provided on their distal side with lateral branches which are
parallel to the spine itself. In Lithoptera the spines bear two to four parallel pairs of
transverse apophyses, and these are crossed by perpendicular branches, parallel to the spine
itself, so that there arise fenestrated wings or latticed plates, comparable to the sails of a
wind-mill. The lattice-work of these plates lies in the same meridian plane with the radial spine
itself, and is therefore not comparable to the fenestrated apophyses of Doracantha, of
Phatnacantha, and of the Dorataspida; in these the lattice-plates lie in tangential planes,
perpendicular to the radial spine.
The apophyses of the Lithopterida may be developed either on all twenty spines equally, or only
on twelve spines (four equatorial and eight tropical, whilst the eight polar spines are simple,
Pl. 131,
fig. 10), or only on the four equatorial spines (whilst the sixteen others are simple, Pl. 131, fig.
9).
The Central Capsule of the Quadrilonchida is rarely spherical, commonly more or {768}less compressed from both poles of the spineless axis,
lenticular or discoidal, sometimes square. It is enveloped by a voluminous calymma constantly
bearing coronals of "Myophrisca" (compare p. 724).
Synopsis of the Genera of Quadrilonchida.
|
II. Subfamily Acanthostaurida.
All twenty spines simple, without lateral apophyses (sometimes forked, but
neither branched nor latticed).
|
|
Four equatorial spines of equal size and form. |
|
Eight tropical and eight polar spines nearly equal, |
334. Acanthostaurus. |
| Eight tropical and eight polar spines very different, |
335. Belonostaurus. |
| Four equatorial spines of very different size or form (the
two lateral constantly equal). |
|
Two principal spines of equal size and form, |
336. Lonchostaurus. |
| Two principal spines (frontal and caudal) very different, |
337. Zyostaurus. |
|
II. Subfamily Lithopterida.
Either all twenty spines or a part of them provided with two opposite lateral
branches or apophyses.
|
|
Apophyses simple, neither branched nor latticed, |
338. Quadrilonche. |
| Apophyses branched or pinnate, but not latticed, |
339. Xiphoptera. |
| Apophyses latticed, with fenestrated network, |
340. Lithoptera. |
| I. Subfamily Acanthostaurida. All twenty spines simple, without lateral
apophyses (sometimes forked, but neither branched nor latticed). |
|
|
Four equatorial spines of equal size and form. |
|
|
|
|
|
|
Eight tropical and eight polar spines nearly equal, |
|
|
|
|
|
|
|
|
|
|
|
|
34. Acanthostaurus. |
|
|
|
|
|
|
|
|
|
|
|
|
Eight tropical and eight polar spines very different, |
|
|
|
|
|
|
|
|
|
|
|
|
35. Belonostaurus. |
|
|
|
|
|
|
|
|
|
|
Four equatorial spines of very different size or form (the two
lateral constantly equal). |
|
|
|
|
|
|
Two principal spines of equal size and form, |
|
|
|
|
|
|
|
|
|
|
|
|
336. Lonchostaurus. |
|
|
|
|
|
|
|
|
|
|
|
|
Two principal spines (frontal and caudal) very different, |
|
|
|
|
|
|
|
|
|
|
|
|
337. Zyostaurus. |
|
|
|
|
|
|
|
|
| II. Subfamily Lithopterida. Either all twenty spines or a part of them
provided with two opposite lateral branches or apophyses. |
|
|
Apophyses simple, neither branched nor latticed, |
|
|
|
|
|
|
|
|
|
|
338. Quadrilonche. |
|
|
|
|
|
|
|
|
|
|
Apophyses branched or pinnate, but not latticed, |
|
|
|
|
|
|
|
|
|
|
339. Xiphoptera. |
|
|
|
|
|
|
|
|
|
|
Apophyses latticed, with fenestrated network, |
|
|
|
|
|
|
|
|
|
|
340. Lithoptera. |
|
|
|
|
|
|
|
|
Subfamily 1. Acasthostaurida, Haeckel, 1881, Prodromus, p. 466.
Definition.—Quadrilonchida with simple radial
spines, without apophyses.
Genus 334. Acanthostaurus,[374] Haeckel,
1862, Monogr. d. Radiol., p. 395.
Definition.—Quadrilonchida with four equatorial spines of
equal size and form, which are much larger than the sixteen other spines. Eight tropical and eight
polar spines nearly equal. No apophyses.
The genus Acanthostaurus is the most simple and primitive form of the
Quadrilonchida, and the common ancestral genus of this family; it is at the same time its most
common and widely distributed form. Some species appear in astonishing numbers in different seas.
It has been derived from Acanthometron by stronger development of the four equatorial
spines, which are all of equal size and much larger than the sixteen others.
{769}
Subgenus 1. Acostaurus, Haeckel.
Definition.—All twenty spines separated, in the centre united only by
the triangular faces or the meeting leaf-shaped edges of their pyramidal bases.
1. Acanthostaurus aequatorialis, n. sp.
Spines cylindrical, of nearly equal breadth throughout their whole length. Apex simple conical.
Base with a large leaf-cross. Four equatorial spines of the same form as the sixteen others, but
much longer and about three times as broad.
Dimensions.—Length of the four major spines 0.2 to 0.25, breadth 0.008; length of
the sixteen minor spines 0.01 to 0.15, breadth 0.003.
Habitat.—Equatorial Pacific, Station 271, surface.
2. Acanthostaurus bipennis, n. sp.
Spines linear, sword-shaped, strongly compressed, two edged; of nearly equal breadth in their
whole length. Apex bifid. Base with a small leaf-cross. Four equatorial spines of the same form as
the sixteen others, but twice as long and as broad.
Dimensions.—Length of the four major spines 0.4 to 0.5, of the sixteen minor 0.2
to 0.3; breadth of the former 0.02, of the latter 0.01.
Habitat.—North Pacific, Station 244, surface.
3. Acanthostaurus conacanthus, n. sp.
Spines conical, short, gradually tapering from the thick base towards the simple apex. Base
with large leaf-cross. Four equatorial spines more cylindrical, twice to three times as long and
as broad as the sixteen others.
Dimensions.—Length of the four major spines 0.15 to 0.2, of the sixteen minor 0.05
to 0.08; basal breadth of the former 0.03, of the latter 0.012.
Habitat.—South Atlantic, Station 332, surface.
4. Acanthostaurus purpurascens, Haeckel.
Acanthostaurus purpurascens, Haeckel, 1862, Monogr. d. Radiol., p. 395, Taf. xix.
figs. 1, 2.
Acanthostaurus purpurascens, R. Hertwig, 1879, Organism. d. Radiol., Taf. i. figs. 8,
9, Taf. iii. figs. 13, 15.
Acanthometra purpurascens, Haeckel, 1860, Monatsber. d. k. preuss. Akad.
d. Wiss. Berlin, p. 809.
Spines nearly cylindrical, with four blunt (often scarcely visible) edges, tapering slightly
from the central to the distal end. Apex bifid, with two short parallel teeth. Base with a broad
leaf-cross. {770}Four equatorial spines one and a half
times to twice as long and broad as the sixteen others. Central capsule cruciate, with four arms
(enveloping the basal part of the four large spines); filled up with yellow bodies and purple
granules. Calymma large, with a network of purple granules. The four main spines are constantly
much larger than the sixteen others, but in variable proportion. The eight polar spines are
sometimes rudimentary.
Dimensions.—Length of the four major spines 0.3 to 0.4, of the sixteen minor 0.15
to 0.3; breadth of the former 0.012, of the later 0.008 or less.
Habitat.—Mediterranean (Messina); Atlantic, Stations 348 to 354,
surface.
5. Acanthostaurus pallidus, Haeckel.
Acanthostaurus pallidus, Haeckel, 1862, Monogr. d. Radiol., p. 396.
Acanthometra pallida, Claparède et Lachmann, 1858, Études sur les
Infusoires, &c., p. 461, Taf. xxiv. fig. 6.
Spines four-sided prismatic, with four blunt edges, of nearly equal breadth throughout their
whole length. Apex bifid. Base with a small leaf-cross. Four equatorial spines much larger than
the sixteen others. Central capsule spherical, transparent, containing some yellow bodies.
Dimensions.—Length of the four main spines 0.12 to 0.2, of the sixteen others 0.04
to 0.08; breadth of the former 0.01, of the latter 0.004.
Habitat.—North Atlantic, west coast of Norway, Claparède; Færöe
Channel, John Murray.
6. Acanthostaurus forceps, Haeckel.
Acanthostaurus forceps, Haeckel, 1862, Monogr. d. Radiol., p. 396, Taf. xix. figs. 3,
4.
Acanthometra forceps, Haeckel, 1860, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 809.
Spines pincer-shaped, linear, compressed, cleft nearly throughout their whole length into two
parallel thin, straight branches, which are united only at the pyramidal base and by means of a
small bridge in the middle part. Four main spines twice as long and as broad as the sixteen
others. Central capsule square, filled up with red pigment, and containing yellow bodies. The
diagonals of the square are the two equatorial axes.
Dimensions.—Length of the four main spines 0.1, of the sixteen others 0.05;
breadth of the former 0.01, of the latter 0.005.
Habitat.—Mediterranean (Messina); North Atlantic, Canary Islands,
Station 354, surface.
7. Acanthostaurus hastatus, Haeckel.
Acanthostaurus hastatus, Haeckel, 1862, Monogr. d. Radiol., p. 397, Taf. xix. fig.
5.
Acanthometra hastata, Haeckel, 1860, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 809.
Spines of very different size and form; four equatorial spines cylindrical in the basal half,
spear-shaped or lanceolate in the distal half, with thin edges, little longer, but five to eight
times as {771}broad as the sixteen smaller spines, which
are cylindrical, of equal breadth throughout their whole length, with bifid apex. Base of all
twenty spines pyramidal, without leaf-cross. Central capsule yellow, spherical, or lenticular.
Dimensions.—Length of the four major spines 0.066, breadth in the middle 0.005, on
the apex 0.008; length of the sixteen smaller spines 0.054, breadth 0.001.
Habitat.—Mediterranean (Messina), surface.
Subgenus 2. Staurolithium, Haeckel, 1862, Monogr. d. Radiol., p. 401.
Definition.—All twenty spines in the centre melted and grown together,
forming one single, star-like piece of acanthin.
8. Acanthostaurus cruciatus, Haeckel.
Staurolithium cruciatum, Haeckel, 1862, Monogr. d. Radiol., p. 401, Taf. xx. fig.
6.
Astrolithium cruciatum, Haeckel, 1860, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 811.
Spines cylindrical, of equal breadth throughout their whole length. Apex simple conical. Four
equatorial spines twice as long and broad as the sixteen other spines. All twenty spines in the
centre melted and grown together, forming a single piece of acanthin. Central capsule spherical,
red-brown, opaque.
Dimensions.—Length of the four equatorial spines 0.12, breadth 0.006; length of
the sixteen smaller spines 0.06, breadth 0.003.
Habitat.—Mediterranean (Messina), surface.
Genus 335. Belonostaurus,[375] n.
gen.
Definition.—Quadrilonchida with four equatorial spines of
equal size and form, which are much larger than the sixteen other spines. Eight tropical and eight
polar spines very different. No apophyses.
The genus Belonostaurus differs from the preceding Acanthostaurus in
the unequal size and form of the eight tropical and the eight polar spines; the latter are much
smaller than the former and often quite rudimentary, so that the skeleton appears composed only of
twelve spines, four larger equatorial and eight smaller tropical spines. The central bases of the
polar spines are constantly preserved. The central capsule is a flattened square disk.
1. Belonostaurus quadratus, n. sp.
Four equatorial spines little compressed, almost cylindrical in the basal half, lanceolate,
broader, with simple apex in the distal half. Eight tropical spines a little shorter, but only
half as broad, {772}sword-shaped, two-edged, of equal
breadth, with simple apex. Eight polar spines very short, conical, also with simple apex. Base
without leaf-cross, a simple pyramid.
Dimensions.—Length of the four equatorial spines 0.18, of the eight tropical
spines 0.14, of the eight polar spines 0.04.
Habitat.—Central Pacific, Station 271, surface.
2. Belonostaurus bicuspis, n. sp.
Four equatorial spines compressed, almost lanceolate in the basal half, bifid, with two large,
bent, little divergent horns in the distal half. Eight tropical spines nearly of the same form,
but only half as large, with much smaller horns. Eight polar spines rudimentary, very short,
conical, with simple apex. Base with a large leaf-cross.
Dimensions.—Length of the four equatorial spines 0.2, of the eight tropical spines
0.1, of the eight polar spines 0.02.
Habitat.—Central Pacific, Station 263, surface.
Genus 336. Lonchostaurus,[376] Haeckel,
1862, Monogr. d. Radiol., p. 397 (sensu emendato).
Definition.—Quadrilonchida with four equatorial spines of
unequal size and form; the principal spines (in the longitudinal axis) larger than the lateral
spines (in the transverse axis); both opposite spines of each pair equal. Sixteen other spines
much smaller (often the eight tropical larger than the eight rudimentary polar spines). No
apophyses.
The genus Lonchostaurus differs from the two preceding genera in the unequal
size and shape of both equatorial pairs of spines, whilst the two opposite spines of each pair are
equal. Therefore the equatorial plane is here a rhombus, not a square, as in the preceding genera.
We encounter here for the first time the differentiation of the two equatorial axes, the
longitudinal (or hydrotomical) and the transverse (or geotomical) axis. The sixteen other spines
are constantly smaller (often also the eight polar more or less rudimentary).
1. Lonchostaurus rhomboides, n. sp. (Pl. 131, fig.
6).
Four equatorial spines cylindrical, twice to three times as broad as the sixteen smaller
spines, conical at the distal apex; the two opposite spines of the longitudinal axis twice as long
as those of the transverse axis. Sixteen smaller spines conical at the base, with simple
bristle-shaped apex. The surface of the rhombical calymma, including the spines, was covered in
the figured specimen with small plates like those of Sphærocapsa.
Dimensions.—Length of the cross (longitudinal axis) 0.24, breadth (transverse
axis) 0.12.
Habitat.—South Pacific, Station 302, depth 1450 fathoms.
{773}
2. Lonchostaurus lanceolatus, n. sp.
Four equatorial spines compressed, lanceolate, tapering equally from the middle towards the two
ends; the two opposite spines of the longitudinal axis one and a half times as long as those of
the transverse axis. Sixteen smaller spines compressed, linear, two-edged, half as long and only
one-fourth as broad as the two longitudinal spines.
Dimensions.—Length of the equatorial cross 0.3, breadth 0.2.
Habitat.—Central Pacific, Station 272, surface.
3. Lonchostaurus bifidus, n. sp.
Four equatorial spines compressed, nearly rectangular, forked, with two long and thin, bent,
little divergent horns; the two opposite spines of the longitudinal axis about one-third longer
and broader than those of the transverse axis. Sixteen smaller spines of the same form, also
forked, but the eight tropical only half as large, and the eight polar spines scarcely one-fourth
as large as the two longitudinal spines.
Dimensions.—Length of the equatorial cross 0.36, breadth 0.26.
Habitat.—South-west Pacific, Station 166, surface.
4. Lonchostaurus bifurcus, n. sp. (Pl. 131, fig.
4).
Four equatorial spines nearly rhomboidal, about one and a half times as large as the eight
tropical and three times as large as the eight polar spines; the two opposite spines of the
longitudinal axis only with two large divergent straight horns; the two spines of the transverse
axis somewhat shorter, with four short horns (the two outer horns longer than the two inner).
Eight tropical spines doubly forked, each with four thin bristle-shaped horns. Eight polar spines
very short, each with two thin bristle-shaped, much divergent horns.
Dimensions.—Length of the equatorial cross 0.34, breadth 0.28.
Habitat.—North-west Pacific, Station 236, surface.
5. Lonchostaurus rhombicus, n. sp.
Four equatorial spines nearly rhomboidal, tapering from the broader middle towards the two
ends. Leaf-cross of their base very large. The two opposite spines of the longitudinal axis about
one-third larger than the two spines of the transverse axis. Eight tropical spines of the same
form, but only half as large. Eight polar spines rudimentary, very short, conical.
Dimensions.—Length of the equatorial cross 0.4, breadth 0.3.
Habitat.—North Pacific, Station 253, surface.
6. Lonchostaurus crystallinus, n. sp. (Pl. 131, fig.
5).
Four equatorial spines lanceolate or rhomboidal, with broad leaf-cross at the base, of a
peculiar crystalline structure and a lamellated surface, both opposite spines of the longitudinal
axis twice as {774}large as the two spines of the
transverse axis. Eight tropical spines of similar form, but only half as large, and with a very
large, extremely prominent, lamellated leaf-cross. Eight polar spines quite rudimentary, scarcely
prominent.
Dimensions.—Length 0.36, breadth 0.2.
Habitat.—Central Pacific, Station 271, surface.
Genus 337. Zygostaurus,[377] n. gen.
Definition.—Quadrilonchida with four equatorial spines of
unequal size and form; both lateral spines (in the transverse axis) equal; but both principal
spines (in the longitudinal axis) very different, the frontal larger than the caudal. Sixteen
other spines much smaller (often the eight tropical larger than the eight rudimentary polar
spines). No apophyses.
The genus Zygostaurus differs from all other Staurolonchida in the peculiar
differentiation of the four equatorial spines; the two opposite spines of the hydrotomical or
longitudinal axis being very different in size and form (the frontal spine forked, the caudal
spine simple); whilst the two opposite spines of the geotomical or lateral axis (perpendicular to
the former) are equal, forked, but different in shape from the former. Therefore the geometrical
fundamental form of the body in this remarkable genus becomes "amphithect" or "bilateral" in the
widest signification of this term (comp. my General Morphology, vol. i. pp. 480, 482). Of the
three different dimensive axes one (the longitudinal) exhibits unequal poles, whilst the two other
(the sagittal and lateral) axes exhibit equal poles.
1. Zygostaurus amphithectus, n. sp. (Pl. 131, fig.
7).
Frontal spine (c1) little different from the two lateral spines; each with two divergent
curved horns, which are equal and of about the same length as the simple broad basal part. Caudal
spine (c3) simple, spindle-shaped, about as long as the frontal spine. Tropical spines
little smaller than the three former, and of the same symmetrical forked form. Eight polar spines
much smaller, but also forked, with thin bristle-shaped branches.
Dimensions.—Length of the cross (longitudinal axis) 0.5, breadth (lateral axis)
0.4.
Habitat.—Central Pacific, Station 272, surface.
2. Zygostaurus longicornis, n. sp.
Frontal spine (c1) little different from the two lateral spines, each with two divergent
curved horns, which are equal and of about the same length as the simple broad basal part. Caudal
{775}spine (c3) simple, lanceolate, shorter than
the frontal spine. Eight tropical spines in the basal part smaller than the three former, but with
much longer fork-branches, which are prolonged into very thin and long bristles. Eight polar
spines rudimentary, very short.
Dimensions.—Length of the equatorial cross 0.4, breadth 0.3.
Habitat.—South Pacific, Station 295, surface.
3. Zygostaurus cornutus, n. sp.
Frontal spine (c1) little different from the two lateral spines, each with two divergent
straight horns, which are shorter than the basal part. Caudal spine (c3) simple,
triangular, shorter than the three former. Eight tropical spines of equal size and form, doubly
forked, each with four thin, bristle-shaped, little divergent teeth. Eight polar spines short,
simply forked.
Dimensions.—Length of the equatorial cross 0.5, breadth 0.4.
Habitat.—Indian Ocean (Madagascar), Rabbe, surface.
4. Zygostaurus caudatus, n. sp.
Frontal spine (c1) very different from the others, pincer-shaped, with two long, nearly
parallel slightly bent horns. Lateral spines (c2 and c4) only half as long, with two
short, nearly parallel horns. Caudal spine (c3) very long and stout, spindle-shaped, about
twice as long as the frontal spine. Eight tropical spines of equal size and form, symmetrical,
with two long and thin, divergent horns. Eight polar spines short and stout, with two divergent
horns.
Dimensions.—Length of the equatorial cross 0.8, breadth 0.3.
Habitat.—Central Pacific, Station 265, surface.
5. Zygostaurus frontalis, n. sp.
Frontal spine (c1) very different from the others, with two very long, divergent, stout
branches. Lateral spines (c2 and c4) only half as long, each with two stout, nearly
parallel horns of equal length. Caudal spine (c3) simple, sword-like, shorter than the
three former. Eight tropical spines of equal size and form, symmetrical, of the same shape as the
two lateral, but only half as large. Eight polar spines very small, rudimentary, each with two
short teeth.
Dimensions.—Length of the equatorial cross 0.7, breadth 0.5.
Habitat.—Central Pacific, Station 274, surface.
6. Zygostaurus sagittalis, n. sp. (Pl. 131, fig.
8).
Frontal spine (c1) very different from the others, with two equal, strongly divergent,
bent horns. Lateral spines (c2 and c4) little smaller, but with two very unequal
horns (the anterior shorter than the posterior). Caudal spine (c3) simple, sword-like or
triangular, two-edged, longer {776}than the frontal
spine. Eight tropical spines different, the four anterior (b1, b4, d1,
d4) smaller, with longer horns; the four posterior (b2, b3, d2,
d3) broader, with shorter horns; the anterior horn of each tropical spine is longer than
the posterior. Eight polar spines rudimentary, simple, very short.
Dimensions.—Length of the equatorial spine-cross 0.6, breadth 0.4.
Habitat.—Central Pacific, Station 271, surface.
Subfamily 2. Lithopterida, Haeckel.
Definition.—Quadrilonchida with two opposite
transverse apophyses either on all twenty spines or only on a part of them.
Genus 338. Quadrilonche,[378] n.
gen.
Definition.—Quadrilonchida with two simple opposite
apophyses either on each radial spine or only on a part of the twenty spines.
The genus Quadrilonche is the ancestral form of the Lithopterida, or of
those Quadrilonchida which bear two opposite apophyses or transverse processes. In
Quadrilonche these apophyses are simple, whilst they are branched in Xiphoptera and
fenestrated in Lithoptera. Each of these three genera may be divided into three subgenera;
in the first only the four equatorial spines are provided with apophyses, in the second twelve
spines (four equatorial and eight tropical), in the third subgenus all twenty spines.
Subgenus 1. Quadriloncharium, Haeckel.
Definition.—Four large equatorial spines provided with transverse
apophyses; sixteen other smaller spines simple, without apophyses.
1. Quadrilonche tetrastaura, n. sp.
Four equatorial spines very large, compressed, two-edged, each crossed in the distal third by
two opposite simple apophyses. Sixteen other spines also compressed, linear, somewhat shorter than
the former and only half as broad, without apophyses.
Dimensions.—Length of the four major spines 0.12, breadth 0.02; length of the
sixteen minor spines 0.08, breadth 0.01.
Habitat.—Central Pacific, Station 266, surface.
{777}
2. Quadrilonche platystaura, n. sp. (Pl. 131, fig.
2).
Four equatorial spines lanceolate, compressed, with two opposite triangular simple apophyses in
the broadest middle part. Sixteen other spines also lanceolate, much smaller, of about half the
length, but only of one-fourth the breadth, without apophyses.
Dimensions.—Length of the four major spines 0.2, breadth 0.018; length of the
sixteen minor spines 0.1, breadth 0.004.
Habitat.—North Pacific, Station 235, surface.
Subgenus 2. Quadrilonchidium, Haeckel.
Definition.—All twenty spines provided with transverse apophyses.
3. Quadrilonche mesostaura, n. sp. (Pl. 131, fig.
1).
Four equatorial spines almost of the some form and length as the sixteen others, but of double
or triple the breadth; each spine in its middle third with two opposite broad triangular
apophyses, in the basal third rectangular, compressed, in the distal third isosceles triangular,
with simple apex.
Dimensions.—Length of the four major spines 0.16, breadth 0.01 to 0.015; length of
the sixteen minor spines 0.12, breadth 0.005.
Habitat.—Tropical Atlantic, Station 345, surface.
4. Quadrilonche telostaura, n. sp.
Four equatorial spines one and a half times as long and three times as broad as the sixteen
others; all twenty spines cylindrical, of equal breadth throughout their whole length, with simple
conical apex; each spine crossed in the distal third by two opposite, simple, conical
apophyses.
Dimensions.—Length of the four major spines 0.3, breadth 0.012; length of the
sixteen minor spines 0.2, breadth 0.004.
Habitat.—South Atlantic, Station 325, surface.
Genus 339. Xiphoptera,[379] Haeckel,
1881, Prodromus, p. 466.
Definition.—Quadrilonchida with two opposite branched
(but not latticed) apophyses, either on each radial spine or only on a part of the twenty
spines.
The genus Xiphoptera differs from the preceding ancestral genus
Quadrilonche in the ramification of the apophyses, which bear perpendicular branches on
their distal side. These are therefore parallel to the spine itself. If the branches become united
by transverse beams then we get Lithoptera.
{778}
1. Xiphoptera tessaractena, n. sp.
Four equatorial spines in the outer third crossed by two opposite transverse apophyses, each of
which bears on its distal side two to three branches, perpendicular to the apophysis and parallel
to the spine itself. Sixteen other spines much smaller, in the outer third crossed by two simple
opposite transverse apophyses.
Dimensions.—Length of the four major spines 0.18, of the sixteen minor 0.05 to
0.1.
Habitat.—North Pacific, Station 238, surface.
2. Xiphoptera dodecactena, n. sp. (Pl. 131, fig.
3).
Four equatorial spines one and a half times to twice as long and broad as the eight tropical
spines. These twelve spines have the same form and are crossed in their distal third by two large
opposite transverse apophyses, each of which bears on its distal side two to four branches,
perpendicular to the apophysis and parallel to the spine. Eight polar spines much smaller than the
twelve others, simple, without apophyses. The central capsule of this species exhibited a conical
protuberance around the base of each individual spine.
Dimensions.—Length of the twelve larger spines 0.1 to 0.2, of the eight smaller
0.02 to 0.06.
Habitat.—Central Pacific, Station 272, surface.
3. Xiphoptera icosactena, n. sp.
Four equatorial spines about twice as long and four times as broad as the sixteen other spines.
All twenty spines crossed in the outer third by two large opposite transverse apophyses, each of
which bears on its distal side two to four branches perpendicular to the apophysis and parallel to
the spine itself.
Dimensions.—Length of the four equatorial spines 0.26, of the sixteen smaller 0.11
to 0.14.
Habitat.—South Pacific, Station 288, surface.
Genus 340. Lithoptera,[380] J. Müller,
1858, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 155.
Definition.—Quadrilonchida with two opposite branched and
latticed apophyses, either on each radial spine or only on a part of the twenty spines.
The genus Lithoptera, founded by Johannes Müller in 1858 for the first
observed Mediterranean species, Lithoptera fenestrata, differs from all other
Quadrilonchida in the fenestrated form of the apophyses, which he compared to the sails of a
windmill. This peculiar fenestration is effected by two to four parallel pairs of opposite
apophyses, which are crossed by perpendicular branches, parallel to the spine itself. {779}Therefore the wings or lattice-plates have quadrangular
meshes and lie in one meridian plane of the spine, not in a tangential plane (as in the
Acanthophracta). Commonly the lateral ends of the four broad equatorial wings are so
crossed that one lateral corner of each wing lies on the upper, the other corner on the under side
of both its neighbours; but sometimes the meeting corners have grown together.
Subgenus 1. Lithopteranna, Haeckel.
Definition.—Four equatorial spines with transverse apophyses; sixteen
others (eight tropical and eight polar spines) simple, without apophyses.
1. Lithoptera tetraptera, n. sp. (Pl. 131, fig.
9).
Four equatorial spines spindle-shaped, with latticed apophyses, each crossed by two transverse
beams which are connected at equal distances by four rods parallel to the spine (therefore each
wing with four square meshes in a single row). Lateral corners of the neighbouring wings not
meeting. Sixteen smaller spines (eight tropical and eight polar) simple, conical, without
apophyses.
Dimensions.—Diagonal of the square body 0.24; length of the inner square 0.16.
Habitat.—Tropical Atlantic, Station 348, surface.
2. Lithoptera mülleri, Haeckel.
Lithoptera mülleri, Haeckel, 1862, Monogr. d. Radiol., p. 398, Taf. xx.
figs. 1, 2.
Four equatorial spines four-edged, with large latticed apophyses, each crossed by three
transverse beams, which are connected at irregular distances by eight to twelve rods parallel to
the spine (therefore each wing with two rows of rectangular meshes). The wings are placed not
perfectly in the equatorial plane, but a little obliquely, so that each wing lies with one lateral
corner on the upper, with the other corner on the under side of its neighbours. Sixteen smaller
spines simple, thin, cylindrical, without apophyses.
Dimensions.—Diagonal of the square body 0.3; length of the inner square 0.17.
Habitat.—Mediterranean (Messina), surface.
3. Lithoptera lamarckii, n. sp.
Four equatorial spines four-edged, with large latticed apophyses; each crossed by three
transverse beams, which are connected at short distances by twelve to sixteen rods parallel to the
spine (therefore each wing with two rows of hexagonal or nearly elliptical meshes). Wings placed
as in Lithoptera mülleri. Sixteen smaller spines simple, thin, prismatic, without
apophyses.
Dimensions.—Diagonal of the square body 0.35; length of the inner square 0.2.
Habitat.—Central Pacific, Station 271, surface.
{780}
4. Lithoptera tetragona, n. sp.
Four equatorial spines compressed, four-edged, with large latticed apophyses, each crossed by
four transverse beams, which are connected at regular distances by eight to twelve rods parallel
to the spine. (Therefore each wing with three rows of square meshes.) Wings placed in the
equatorial plane and grown together by their meeting lateral ends, forming a square equatorial
girdle of lattice-work. Sixteen smaller spines simple, thin, prismatic, without apophyses.
Dimensions.—Diameter of the square body 0.4; length of the inner square 0.22.
Habitat.—North Pacific, Station 244, surface.
Subgenus 2. Lithopterella, Haeckel.
Definition.—Twelve spines (four equatorial and eight tropical) with
transverse apophyses; the eight polar spines simple, without apophyses.
5. Lithoptera quadrata, n. sp. (Pl. 131,
fig. 10).
Twelve spines with transverse apophyses; eight (polar) spines simple, small, without apophyses.
Four equatorial spines very large and stout, compressed, each crossed by four transverse beams,
which are connected by eight to ten rods parallel to the spine (therefore each wing with three
rows of irregular rectangular meshes). Wings placed in the equatorial plane and grown together by
their meeting lateral ends, forming a square equatorial girdle of lattice-work. Eight tropical
spines thin, crossed by a long and thin transverse beam, which bears on its distal side eight to
ten rods parallel to the spine.
Dimensions.—Diagonal of the square body 0.35; length of the inner square 0.18.
Habitat.—South Atlantic, Station 326, surface.
6. Lithoptera dodecaptera, n. sp.
Twelve spines with transverse apophyses; eight (polar) spines simple, small, without apophyses.
Four equatorial spines very large, quadrangular; each crossed by three transverse beams, which are
connected by ten to twelve rods parallel to the spine (therefore each wing with two rows of square
meshes). Wings placed as in Lithoptera mülleri. Eight tropical spines much thinner, crossed
each by two transverse beams, which are connected by six to eight rods parallel to the spine
(therefore each wing with one row of rectangular meshes).
Dimensions.—Diagonal of the square body 0.4; length of the inner square 0.25.
Habitat.—Central Pacific, Station 271, surface.
Subgenus 3. Lithopteromma, Haeckel.
Definition.—All twenty spines with transverse apophyses.
{781}
7. Lithoptera darwinii, Haeckel.
Lithoptera darwinii, Haeckel, 1879, Natürl. Schöpfungsgesch., Aufl. vii.
p. 706, Taf. xvi. fig. 12.
All twenty spines with transverse apophyses; these are quite simple in the eight small
cross-shaped polar spines; also simple, but bearing some perpendicular rods, in the eight thin
tropical spines. Four equatorial spines very large and stout, each crossed by four transverse
beams, which are connected by eight to ten rods parallel to the spine (therefore each wing with
three rows of irregular rectangular meshes). Wings placed in the equatorial plane and grown partly
together by their meeting lateral ends.
Dimensions.—Diagonal of the square body 0.4; length of the inner square 0.2.
Habitat.—South-east Pacific, west coast of Patagonia, Station 302,
surface.
8. Lithoptera icosaptera, n. sp.
All twenty spines with transverse apophyses, bearing one to three rows of rectangular meshes;
eight polar spines small, each with a single row; eight tropical spines long and thin, each with
two parallel rows. Four equatorial spines much larger and thicker, each with three parallel
transverse rows of meshes. All twenty wings free, not grown together by their lateral ends.
Dimensions.—Diagonal of the square body 0.48; length of the inner square 0.24.
Habitat.—Indian Ocean, Belligemma, Ceylon (Haeckel), surface.
9. Lithoptera fenestrata, J. Müller.
Lithoptera fenestrata, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 154, Taf. xi. fig. 13.
All twenty spines with transverse apophyses, bearing one to three rows of irregular,
rectangular meshes; eight polar and eight tropical spines small, each with a single row of meshes
(between two parallel transverse beams). Four equatorial spines little larger, each with two rows
(between three parallel transverse beams). Central capsule filled up with green pigment.
Dimensions.—Diagonal of the square body 0.3; length of the inner square 0.2.
Habitat.—Mediterranean (Saint Tropez, Nice).
Family XXXVIII. Amphilonchida, Haeckel (Pl.
132).
Acantholonchida (Amphilonchida et Amphilithida), Haeckel, 1881,
Prodromus, p. 466.
Definition.—Acantharia with twenty radial spines of very
unequal size, disposed according to the law of the Icosacantha; two opposite equatorial spines (in
the longitudinal axis) much larger than the eighteen others. No lattice-shell.
The family Amphilonchida is distinguished from all other Acanthometra by the excessive development of two opposite equatorial spines,
which are much larger {782}than the eighteen other
spines. In my Monograph (1862, p. 389) I had united all these Acanthonida in a single genus
Amphilonche (with ten species). Some species of it are very common and widely distributed;
but in general the number of different forms in this family is much smaller than in the two
foregoing families.
The two principal spines, which in all Amphilonchida are much larger than the eighteen other
spines, characterise the "hydrotomical axis" or the larger equatorial axis. The two other
equatorial spines or the "geotomical spines" are much smaller, and commonly of the same size as
the eight tropical and the eight polar spines. In the genera Amphilonche and
Amphibelone these eighteen smaller spines are rather equally developed; sometimes they are
very small or quite rudimentary, so that the skeleton seems to be represented only by the two very
long principal spines (Pl. 132,
fig. 4). The genus Acantholonche is distinguished by the unequal size of the eight tropical
and the eight polar spines, the latter being more or less rudimentary. However, the central bases
of all twenty spines, by which they are united in the centre, are constantly present. The genus
Amphibelone is distinguished by the unequal size and form of the two principal spines, one
of them the "caudal spine," being larger (and often of another form) than the opposite "frontal
spine."
The form of the radial spines is in the Amphilonchida far less varied and complicated than in
the other Acanthonida (the Astrolonchida and Quadrilonchida). Apophyses or lateral transverse
processes are never developed. The three main forms of spines are the same as in the other
Acanthonida; they are (1) either cylindrical or conical (like Acanthometron), or (2)
compressed or two-edged (like Zygacantha), or (3) quadrangular or four-edged (like
Acanthonia). Often the spines are angular in the inner or proximal, roundish in the outer
or distal part. The distal apex is commonly simple, conical or pyramidal. The central base is
commonly also pyramidal, as in the majority of the Acanthonida; and the triangular faces of the
neighbouring bases are simply propped one upon another. More rarely a basal leaf-cross is
developed above the basal pyramid. Very rarely the central bases of the united spines grow
together in the centre, so that the whole skeleton forms a single piece of acanthin.
The Central Capsule is rarely spherical, commonly prolonged in the direction of the
hydrotomical axis; ellipsoidal or cylindrical, sometimes also four-sided prismatic; it commonly
envelops the greater part of the two principal spines; its structure and contents are the same as
in the other Acanthonida.
Synopsis of the Genera of Amphilonchida.
| Eighteen smaller spines of nearly equal size and
similar form. |
|
Two opposite large principal spines (frontal and caudal) equal, |
341. Amphilonche. |
| Caudal spine larger than the frontal spine, |
342. Amphibelone. |
| Eight tropical and eight polar spines of different
sizes (the latter often rudimentary), |
343. Acantholonche. |
| Eighteen smaller spines of nearly equal size and similar form. |
|
|
Two opposite large principal spines (frontal and caudal)
equal, |
|
|
|
|
|
|
341. Amphilonche. |
|
|
|
|
|
|
Caudal spine larger than the frontal spine, |
|
|
|
|
|
|
342. Amphibelone. |
|
|
|
|
| Eight tropical and eight polar spines of different sizes (the latter often
rudimentary), |
|
|
|
|
343. Acantholonche. |
|
|
|
|
{783}
Genus 341. Amphilonche,[381] Haeckel,
1862, Monogr. d. Radiol., p. 389.
Definition.—Amphilonchida with two equal principal spines
(frontal and caudal spines not different); the eighteen smaller spines nearly equal.
The genus Amphilonche represents the original and at the same time the most
common form of Amphilonchida; the two opposite principal spines are of equal size and form, much
larger (and often also of another form) than the eighteen smaller spines; these latter exhibit no
marked differences in size and form.
Subgenus 1. Amphiloncharium, Haeckel.
Definition.—Spines in the centre united by triangular faces of their
pyramidal bases, propped one upon another. No basal leaf-cross.
1. Amphilonche belonoides, Haeckel.
Amphilonche belonoides, Haeckel, 1862, Monogr. d. Radiol., p. 392, Taf. xvi. fig. 6,
Taf. xviii. fig. 21.
Acanthometra belonoides, Haeckel, 1860, Monatsber. d. k. Akad. d. Wiss.
Berlin, p. 808.
Two principal spines cylindrical, of equal breadth throughout their whole length, with simple
conical apex; base a small pyramid, without leaf-cross. Eighteen smaller spines conical, much
shorter, with simple bristle-shaped apex. Central capsule spindle-shaped or cylindrical,
yellow.
Dimensions.—Length of the two major spines 0.4 to 0.8, of the eighteen minor 0.05
to 0.2.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific, surface.
2. Amphilonche lanceolata, n. sp. (Pl. 132, fig.
1).
Two principal spines compressed, lanceolate, two-edged, gradually tapering from the broader
middle towards the two ends; apex simple; base a large pyramid, with broad leaf-cross. Eighteen
smaller spines about half as long, pyramidal, with simple conical or bristle-shaped apex. Central
capsule lentelliptical, pink, opaque.
Dimensions.—Length of the two major spines 0.2, of the eighteen minor 0.1.
Habitat.—Tropical Atlantic, Station 343, surface.
3. Amphilonche diodon, n. sp. (Pl. 132,
fig. 3).
Two principal spines compressed, lanceolate, two-edged, tapering from the broader middle
towards the two ends; apex simple; base a small pyramid, without leaf-cross. Eighteen smaller
spines {784}sword-shaped, only two-thirds or one-half as
long, and one-third or one-fourth as broad, each with two opposite teeth in the distal part.
Central capsule ellipsoidal.
Dimensions.—Length of the two major spines 0.13, breadth 0.02; length of the
eighteen minor spines 0.1.
Habitat.—North Atlantic, Station 352, surface.
4. Amphilonche tenuis, Haeckel.
Amphilonche tenuis, Haeckel, 1862, Monogr. d. Radiol., p. 389, Taf. xvi. fig. 1, Taf.
xviii. fig. 16.
Acanthometra tenuis, Haeckel, 1860, Monatsber. d. k. Akad. d. Wiss.
Berlin, p. 807.
Two principal spines quadrangular prismatic, with smooth edges, slightly or not at all
prominent, of equal breadth throughout their whole length; apex simple or bifid; base a small
pyramid, without leaf-cross. Eighteen smaller spines of the same form, but only one-fourth or
one-third as long. Central capsule spherical or ellipsoidal, colourless.
Dimensions.—Length of the two major spines 0.2 to 0.3, of the eighteen minor
spines 0.05 to 0.1.
Habitat.—Mediterranean (Messina); Atlantic, Station 354, Canary
Islands, surface.
5. Amphilonche denticulata, Haeckel.
Amphilonche denticulata, Haeckel, 1862, Monogr. d. Radiol., p. 390, Taf.
xvi. fig. 2, Taf.
xviii. fig. 17.
Two principal spines quadrangular prismatic, with prominent, elegantly denticulated edges, of
equal breadth throughout their whole length; apex simple or truncated; base a small pyramid,
without leaf-cross. Eighteen smaller spines of the same form, but only one-fourth or one-third as
long. Central capsule ellipsoidal, opaque, yellowish-brown.
Dimensions.—Length of the two major spines 0.3 to 0.4, of the eighteen minor 0.1;
breadth of the former 0.009, of the latter 0.006.
Habitat.—Mediterranean (Messina), Haeckel, surface.
6. Amphilonche heteracantha, Haeckel.
Amphilonche heteracantha, Haeckel, 1862, Monogr. d. Radiol., p. 293, Taf. xvi. fig.
7.
Acanthometra heteracantha, Haeckel, 1860, Monatsber. d. k. Akad. d. Wiss.
Berlin, p. 808.
Two principal spines quadrangular prismatic, very large, with four broad, prominent, lamellar
edges, of equal breadth throughout their whole length. Apex as well as the base a four-sided
pyramid. Eighteen smaller spines very thin, conical, with bristle-shaped apex; base a small
pyramid, without leaf-cross. Central capsule cylindrical or violin-shaped, opaque yellow.
Dimensions.—Length of the two major spines 0.2 to 0.3, breadth 0.015 to 0.02;
length of the eighteen minor spines 0.1 to 0.15, basal breadth 0.004 to 0.008.
Habitat.—Mediterranean (Messina); Central Pacific, Station 271,
surface.
{785}
7. Amphilonche elongata, Haeckel.
Amphilonche elongata, Haeckel, 1862, Monogr. d. Radiol., p. 392, Taf. xviii. figs.
22a, 22b.
Acanthometra elongata, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss.
Berlin, p. 48, Taf. vii. fig. 13.
Two principal spines quadrangular prismatic in the proximal half, cylindrical or spindle-shaped
in the distal half, with simple apex; base a small pyramid, without leaf-cross. Eighteen smaller
spines very thin, bristle-shaped, conical on the base, with small pyramid. Central capsule
spindle-shaped, transparent, yellow.
Dimensions.—Length of the two major spines 0.3 to 0.5, breadth 0.01 to 0.03;
length of the eighteen smaller spines 0.05 to 0.3.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Indian, Pacific,
surface.
Subgenus 2. Amphilonchidium, Haeckel.
Definition.—Spines at the central base with a broad leaf-cross,
composed of four prominent triangular lamellæ; between the meeting edges of the latter twenty-two
pyramidal compartments or basal funnels.
8. Amphilonche ovata, Haeckel.
Amphilonche ovata, Haeckel, 1862, Monogr. d. Radiol., p. 390.
Acanthometra ovata, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin,
p. 47, Taf. vii. fig. 10, Taf. ix. fig. 4.
Two principal spines cylindrical, of equal breadth throughout their whole length, with simple
conical or bifid apex; base with a large leaf-cross of double the breadth. Eighteen smaller spines
of the same form, but thinner and only half as long. Central capsule ellipsoidal, brown,
opaque.
Dimensions.—Length of the two major spines 0.3 to 0.4, breadth 0.008 to 0.012;
length of the eighteen minor spines 0.1 to 0.2.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific, surface.
9. Amphilonche conica, n. sp. (Pl. 132,
fig. 6).
Two principal spines conical, elongate, with simple apex; base thickened, with large
leaf-cross. Eighteen smaller spines also conical, of about the same basal breadth, but only
one-fourth to one-half as long. Central capsule ellipsoidal, pellucid.
Dimensions.—Length of the two major spines 0.15 to 0.2, basal breadth 0.03 to
0.04; length of the eighteen minor spines 0.05 to 0.1.
Habitat.—Central Pacific, Stations 265 to 274, surface.
{786}
10. Amphilonche lancetta, n. sp.
Two principal spines compressed, lanceolate, two edged, gradually tapering from the broader
middle towards the two ends; apex simple; base of double the breadth, with a large leaf-cross.
Eighteen smaller spines of the same form, but shorter and only half as broad. Central capsule
lentelliptical, pellucid.
Dimensions.—Length of the two major spines 0.2 to 0.3, breadth in the middle part
0.02 to 0.03; length of the eighteen smaller spines 0.1 to 0.2.
Habitat.—South Pacific, Station 288, surface.
11. Amphilonche complanata, Haeckel.
Amphilonche complanata, Haeckel, 1862, Monogr. d. Radiol., p. 390, Taf. xvi. fig. 3,
Taf. xviii. fig. 18, a, b.
Acanthometra complanata, Haeckel, 1860, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 808.
Two principal spines compressed, two-edged, linear, of equal breadth throughout their whole
length; apex emarginate or bifid; base with a large leaf-cross of double the breadth. Eighteen
smaller spines of the same form, but only one-fourth to one-half as large. Central capsule
ellipsoidal, yellow, pellucid.
Dimensions.—Length of the two major spines 0.15 to 0.25, breadth 0.004 to 0.006;
length of the eighteen minor spines 0.05 to 0.1.
Habitat.—Mediterranean (Messina), surface.
12. Amphilonche messanensis, Haeckel.
Amphilonche messanensis, Haeckel, 1862, Monogr. d. Radiol., p. 391, Taf. xvi. fig. 4,
Taf. xviii. fig. 19.
Acanthometra messanensis, Haeckel, 1860, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 808.
Two principal spines quadrangular prismatic, often a little compressed from two sides, of
nearly equal breadth throughout their whole length; apex either truncate or emarginate, with two
opposite teeth; base with a large leaf-cross of double the breadth. Eighteen smaller spines of
similar form or more compressed, much shorter. Central capsule spherical or ellipsoidal, yellow,
transparent.
Dimensions.—Length of the two major spines 0.12 to 0.18, breadth 0.016; length of
the minor spines 0.05 to 0.09.
Habitat.—Mediterranean (Messina, Corfu), Haeckel, surface.
13. Amphilonche hydrotomica, n. sp. (Pl. 132, fig.
2).
Two principal spines quadrangular prismatic, with four broad prominent lamellar edges or wings,
of equal breadth throughout their whole length; apex pyramidal; base with a large leaf-cross.
Eighteen smaller spines cylindrical or bristle-shaped, only half as long and very thin. Central
capsule cylindrical or spindle-shaped, very long, opaque.
{787}
Dimensions.—Length of the two major spines 0.1 to 0.2, breadth 0.02 to 0.03;
length of the eighteen minor spines 0.05 to 0.1.
Habitat.—Central Pacific, Stations 270 to 274, surface.
14. Amphilonche violina, n. sp. (Pl. 132,
fig. 5).
Two principal spines quadrangular prismatic, nearly violin-shaped, with four very broad,
prominent, lamellar wings, which are constricted in the middle part, and broadened towards the two
ends; apex truncate pyramidal; base with a large leaf-cross. Eighteen smaller spines much shorter,
of equal breadth at the base, assuming the form of a quadrangular pyramid, thin prismatic in the
distal half. Central capsule spindle-shaped, opaque.
Dimensions.—Length of the two major spines 0.15 to 0.18, breadth 0.02 to 0.025;
length of the eighteen minor spines 0.04 to 0.08.
Habitat.—Central Pacific, Station 271, surface.
15. Amphilonche tetraptera, Haeckel.
Amphilonche tetraptera, Haeckel, 1862, Monogr. d. Radiol., p. 391, Taf. xvi. fig. 5,
Taf. xviii. fig. 20.
Acanthometra tetraptera, Haeckel, 1860, Monatsber. d. k. preuss. Akad. d.
Wiss. Berlin, p. 808.
Two principal spines four-sided pyramidal, with four broad lamellar prominent edges; apex
simple or bifid; base with a large leaf-cross. Eighteen smaller spines of similar form, but only
half as large. Central capsule spherical, opaque.
Dimensions.—Length of the two major spines 0.2, breadth on the base 0.025; length
of the eighteen minor spines 0.1.
Habitat.—Mediterranean (Messina); North Atlantic, Canary Islands,
Stations 352 to 354, surface.
Subgenus 3. Amphilithium, Haeckel, 1881, Prodromus, p. 466.
Definition.—Spines in the basal part grown together, so that the whole
skeleton represents a single piece of acanthin; a star with two larger and eighteen smaller
rays.
16. Amphilonche concreta, n. sp. (Pl. 132, figs. 4,
4a).
Two principal spines cylindrical, very long, of equal breadth throughout their whole length,
with simple conical apex. Eighteen smaller spines short, conical or bristle-shaped, scarcely
one-fourth or one-tenth as long, often quite rudimentary. All twenty spines perfectly grown
together in the centre, forming a single piece of acanthin (derived from Amphilonche
belonoides by central concrescence; often the sutures of the concreted bases are visible, fig.
4a).
Dimensions.—Length of the two major spines 0.1 to 0.4, breadth 0.005 to 0.015;
length of the eighteen minor spines 0.005 to 0.15.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific, surface.
{788}
17. Amphilonche acufera, n. sp.
Two principal spines thick, four-sided prismatic in the basal half, cylindrical or
spindle-shaped in the distal half, with simple conical apex. Eighteen smaller spines shorter,
thin, bristle-shaped or conical on the base. All twenty spines in the centre perfectly grown
together, forming a single piece of acanthin. (Derived from Amphilonche elongata by central
concrescence.)
Dimensions.—Length of the two major spines 0.3 to 0.5, breadth 0.01 to 0.03;
length of the eighteen minor spines 0.08 to 0.2.
Habitat.—Central Pacific, Stations 266 to 274, surface.
Genus 342. Amphibelone,[382] Haeckel,
1862, Monogr. d. Radiol., p. 392.
Definition.—Amphilonchida with two unequal principal
spines (the frontal spine very different from the caudal spine); the eighteen smaller spines
nearly equal.
The genus Amphibelone exhibits among the Amphilonchida the same remarkable
differentiation of the two principal or longitudinal spines, as Zygostaurus among the
Quadrilonchida; the frontal spine differs commonly from the caudal spine not only in its size, but
also in its peculiar form; commonly one pole of the longitudinal axis is much more strongly
developed than the other. The eighteen smaller spines are nearly equal.
Subgenus 1. Amphibelonium, Haeckel.
Definition.—All twenty spines separate, but in contact in the centre
and resting one against another by the triangular sides of their pyramidal bases, without a
prominent basal leaf-cross.
1. Amphibelone aciculata, n. sp.
Two principal spines thick, without edges and wings, the frontal spine short, spindle-shaped,
the caudal three to six times as long, cylindrical, both of equal breadth, with simple conical
apex and simple pyramidal base, without leaf-cross. Eighteen smaller spines very thin,
bristle-shaped, of the same length as the frontal spine. Central capsule cylindrical or
spindle-shaped, enveloping the two principal spines nearly throughout their whole length.
Dimensions.—Length of the frontal spine 0.2 to 0.4, of the caudal spine 1.0 to
2.0; breadth of both 0.01; length of the eighteen smaller spines 0.1 to 0.3.
Habitat.—South Atlantic, Station 325, surface.
{789}
2. Amphibelone cultellata, n. sp. (Pl. 132, fig.
10).
Two principal spines broad, two-edged, knife-shaped; two opposite in the equatorial plane, thin
and broad wings or lamellæ are developed, and these enclose the proximal part of both spines,
whilst their distal part is free, cylindrical, with conical apex: the two wings of the longer
caudal spine are lanceolate, twice to four times as broad as the two wings of the shorter frontal
spine; their base is a simple pyramid without leaf-cross. Eighteen smaller spines conical, with
bristle-shaped prolongation, one-fourth to one-half as long as the frontal spine. Central capsule
green, semitransparent.
Dimensions.—Length of the frontal spine 0.1 to 0.2, breadth 0.02 to 0.03; length
of the caudal spine 0.2 to 0.4, breadth 0.04 to 0.08; length of the eighteen smaller spines 0.05
to 0.15.
Habitat.—Indian Ocean, Maldive Islands (Haeckel), surface.
3. Amphibelone pyramidata, n. sp. (Pl. 132, fig.
9).
Two principal spines four-winged, with four crossed, very thin and broad wings, which are
somewhat broader in the convex middle part than at either end; both ends of each spine four-sided
pyramidal, with four concave edges and a very small terminal pyramid; base without leaf-cross. The
longer caudal spine is twice as broad at the distal end as at its proximal end, and three times as
broad as the distal end of the shorter frontal spine. Eighteen smaller spines bristle-shaped, with
four-sided pyramidal bases. Central capsule pyramidal, enveloping the two principal spines
throughout their whole length; the base of the slender quadrangular pyramid is on the caudal, the
apex on the frontal pole of the longitudinal axis.
Dimensions.—Length of the frontal spine 0.2 to 0.25, distal breadth 0.01; length
of the caudal spine 0.3 to 0.4, distal breadth 0.03; length of the eighteen smaller spines 0.04 to
0.08.
Habitat.—Cape of Good Hope, Station 143, depth 1900 fathoms.
4. Amphibelone anomala, Haeckel.
Amphilonche anomala, Haeckel, 1862, Monogr. d. Radiol., p. 394, Taf. xvi. fig. 8, Taf.
xviii. figs. 23a, 23b.
Acanthometra anomala, Haeckel, 1860, Monatsber. d. k. Akad. d. Wiss.
Berlin, p. 808.
Two principal spines four-winged, nearly prismatic, with four crossed, very broad and thick
wings; both ends of each spine four-sided pyramidal, with four concave edges and a very small
terminal pyramid; base without leaf-cross. The edges of the four wings are concave on the shorter
frontal, convex on the longer caudal spine. Eighteen smaller spines only one-fourth to one-half as
long, linear, two-edged, with two parallel teeth on the distal end. Central capsule four-sided
prismatic, olive-green, enveloping almost completely the two principal spines.
Dimensions.—Length of the frontal spine 0.12, middle breadth 0.016; length of the
caudal spine 0.14, middle breadth 0.024; length of the eighteen smaller spines 0.04 to 0.06.
Habitat.—Mediterranean (Messina), surface.
{790}
Subgenus 2. Amphibelithium, Haeckel.
Definition.—All twenty spines grown together in the centre, forming a
single star-shaped piece of acanthin.
5. Amphibelone clavaria, n. sp.
Two principal spines roundish, without edges and wings; the smaller frontal spine elongate
conical, the larger caudal spine two to four times as long, cylindrical, in the distal part
spindle-shaped or club-shaped, with conical apex. Central base simple pyramidal, without
leaf-cross. Eighteen smaller spines much shorter, bristle-shaped. Central capsule very long,
club-shaped. All twenty spines perfectly grown together in the centre.
Dimensions.—Length of the frontal spine 0.12, of the caudal spine 0.4 to 0.8;
frontal breadth 0.01, caudal breadth 0.03; length of the eighteen smaller spines 0.05.
Habitat.—Central Pacific, Station 272, surface.
Genus 343. Acantholonche,[383] Haeckel,
1881, Prodromus, p. 466.
Definition.—Amphilonchida with two equal principal spines
(frontal and caudal spines not different). The eighteen smaller spines are very unequal, ten of
them (eight tropical and two transverse equatorial spines) much larger than the rudimentary eight
polar spines.
The genus Acantholonche differs from its ancestral genus Amphilonche
in the different shape of the eight tropical and the eight polar spines; these latter are much
smaller than the former, which are almost equal to the two transverse equatorial spines. The two
principal spines are equal, but in size and shape very different from the others.
1. Acantholonche amphipolaris, n. sp. (Pl. 132, fig.
7).
Two principal spines stout, quadrangular prismatic in the basal half, cylindrical or
spindle-shaped in the distal half, with simple conical apex; base a small pyramid without
leaf-cross. Two transverse and eight tropical spines, about half as long as the former, very thin,
bristle-shaped, conical at the basal part. Eight polar spines very small, scarcely one-eighth or
one-fourth as long as the latter, short pyramidal or conical, often quite rudimentary. Central
capsule cylindrical, enveloping the basal half of the two principal spines.
Dimensions.—Length of the two principal spines 0.4 to 0.5, of the ten smaller
spines 0.2 to 0.3, of the eight rudimentary polar spines 0.01 to 0.06; breadth of the two large
spines 0.02 to 0.03.
Habitat.—Central Pacific, Stations 266 to 274, surface.
{791}
2. Acantholonche peripolaris, n. sp. (Pl. 132, fig.
8).
Two principal spines quadrangular prismatic, with four broad prominent lamellar wings, of
increasing breadth towards the pyramidal distal apex. Both ends of each spine four-sided
pyramidal, base without leaf-cross. Two transverse and eight tropical spines about two-thirds as
long as the former, four-sided pyramidal in the basal half, conical in the distal half, often
curved. Eight polar spines very small, about one-fourth as long as the latter, short conical or
pyramidal. Central capsule four-sided prismatic, enveloping both principal spines.
Dimensions.—Length of the two principal spines 0.2, of the ten smaller spines
0.12, of the eight rudimentary polar spines 0.04.
Habitat.—Central Pacific, Station 274, surface.
Order IV. ACANTHOPHRACTA, Richard Hertwig, 1879.
Acanthometræ cataphractæ, Johannes Müller, 1858, Abhandl. d. k.
Akad. d. Wiss. Berlin, pp. 12, 22, 49.
Dorataspida et Diploconida, Haeckel, 1862, Monogr. d. Radiol.,
pp. 404, 412.
Acanthophractida, Richard Hertwig, 1879, Organismus d. Radiol.,
pp. 25, 137.
Dorataspida, Diploconida, et Sphærocapsida, Haeckel, 1881,
Prodromus, p. 467.
Definition.—Acantharia with complete latticed shell.
The order Acanthophracta, the fourth order of Radiolaria, comprises
all those Acantharia in which the acanthinic skeleton is a complete
latticed or fenestrated shell, supported by radial spines arising from one common central point.
By the possession of such a complete shell the Acanthophracta differ from
their ancestral group, the nearly allied Acanthometra, which represent
the older and simpler, first order of Acantharia. All Acanthophracta are Icosacantha (like the Acanthonida,
their ancestral group), and possess twenty radial spines disposed according to the Müllerian law
(compare above, p. 717).
Johannes Müller, who first observed five representatives of this order, called a part of them
"Acanthometræ cataphractæ," and united these with the true Acanthometra (Acanthometra costata and Acanthometra cataphracta;
Abhandl. d. k. Akad. d. Wiss. Berlin, 1858, pp. 12, 49). Another part was united by him with the
true Haliomma (Haliomma echinoides, Haliomma hystrix, Haliomma
tabulatum; Abhandl. d. k. Akad. d. Wiss. Berlin, 1858, pp. 36, 37). He supposed that these
latter formed the immediate transition from the true Acanthometra to the
true Haliomma, and that their skeleton was siliceous.
{792}
In my Monograph (1862, p. 412) I founded a separate subfamily, Dorataspida, for the
"Acanthometræ cataphractæ," which I considered as the first subfamily of the "Ommatida."
That subfamily contained at that time only two genera, Dorataspis (with seven species) and
Haliommatidium (with five species). A third genus, Aspidomma (with two species), was
united by me with the Haliommatida (because of its double shell). For a fourth genus
(Diploconus) with a single species I founded the peculiar family of Diploconida. Therefore
the whole number of Acanthophracta described in my Monograph amounted
only to four genera and fifteen species. Now the rich collections of the Challenger have added
such a great number of new forms, that we may distinguish here thirty-eight genera and two hundred
and twelve species.
Richard Hertwig in his excellent work (Der Organismus der Radiolarien, 1879, p. 25) separated
his "Acanthophractida" perfectly from the "Ommatida" (or the siliceous Sphæroidea), and united them with the "Acanthometrida" in his order
"Acanthometrea." But he separated them also from the nearly allied Diploconida, following my
former arrangement. He distinctly noted that the skeleton in all these Acanthophractida (as well
as in the Acanthometrida) consists not of silex but of the organic substance "acanthin."
The astonishing number of new and interesting forms of Acanthophracta
which I have found in the rich collection of the Challenger enables me to distinguish now in this
suborder six different families, two of which are perfectly new (the Sphærocapsida and the
Hexalaspida). But the four other families also are so much enlarged that their interesting
morphology appears in quite a new and clear light. Far the largest and most important of these six
families is that of the true Dorataspida, which embraces seventeen genera and one hundred and
eight species (more than the other five families together). From this largest and oldest ancestral
family four other families have afterwards arisen, whilst a single family, the Sphærocapsida,
seems to possess no direct phylogenetic connection with the five other families.
The peculiar and quite new family of Sphærocapsida (Pl. 133, figs.
7-11; Pl. 135,
figs. 6-10) differs from all other Acanthophracta in the singular
structure of the spherical acanthinic shell, composed of innumerable small plates or aglets, each
of which is pierced by a very small porule. This peculiar pavemented shell (enclosing the central
capsule and separated from it by the jelly-like calymma) seems to be produced on the surface of
the spherical calymma, immediately by secretion of the pseudopodia, and independently from the
twenty radial spines, united in the centre of the sphere. On the twenty points, where the spines
perforate the shell, there are originally eighty larger pores (four around each piercing spine);
but there is no certain indication that the shell is produced by the meeting apophyses of the
twenty spines, as is the case in the five other families of Acanthophracta. Therefore perhaps it is {793}more natural to unite these latter into another suborder as Cladophracta, and to separate them from the Sphærocapsida, which may be called
Capsophractæ.
The Dorataspida (Pls. 134-138), the
common ancestral stock of the Cladophracta, in the definition here
restricted embraces all those Acanthophracta in which the spherical
lattice-shell is simple and composed of the meeting branches of twenty radial spines united in its
centre. As already pointed out above, this family is probably diphyletic, and embraces two
subfamilies which have been derived originally from two different forms of Acanthonida—the Diporaspida (with two opposite apophyses on each spine)
derived from the Phractacanthida, and the Tessaraspida (with four crossed apophyses on each spine)
derived from the Stauracanthida; in the former we find originally forty apophyses, in the latter
eighty apophyses, by the meeting branches of which the spherical lattice-shell originates. The
four following families of Acanthophracta have probably been derived from
the Diporaspida.
The Phractopeltida (Pl. 133,
figs. 1-6) differ from all other Acanthophracta in the possession of a
double lattice-shell, composed of two concentric spheres which are united by the twenty radial
spines meeting in the centre. As all Phractopeltida possess originally only two apophyses on each
radial spine, they must be derived from the Diporaspida (Orophaspis), and bear to them the
same relation as the Dyosphærida do to the Monosphærida. As the spherical central capsule of the
Phractopeltida is enclosed between both shells, smaller than the outer, larger than the inner
shell, the latter may be called "medullary shell," the former "cortical shell." This family
represents among the Acanthophracta only the "Diplophracta," whilst all
others are "Haplophracta."
The three families here characterised may be called together "Sphærophracta," as their central capsule and the enveloping shell are
constantly spherical (or the shell sometimes an "endospherical polyhedron"). On the contrary the
following three families of Acanthophracta may be united as "Prunophracta," as their central capsule and shell are never spherical, but
either ellipsoidal or lenticular or of another form. The common ancestral stock of this suborder
are the Belonaspida, in which the form of the central capsule and the enclosing lattice-shell is
ellipsoidal; they are derived from the Dorataspida (and probably all from the subfamily
Diporaspida) by the prolongation of two opposite radial spines which are larger than the eighteen
others; they are the two equatorial spines of the "hydrotomical axis" (compare above, p. 719, and Pl. 136,
figs. 6-9).
The Hexalaspida (Pl. 139)
represent a new and very remarkable family, distinguished from all other Acanthophracta by the preponderating development of six stout radial spines,
which are much larger than the fourteen others. These six principal spines lie in one meridian
plane of the shell (in the "hydrotomical plane," p. 720), and are the two
opposite equatorial spines and the four appertaining polar spines of the same plane. {794}As the fourteen smaller spines develop their apophyses at
smaller distances from the centre, the shell assumes a peculiar lenticular or discoidal form, and
the margin of this disk bears the six larger spines. Moreover the enclosed small central capsule
is lenticular. The Hexalaspida may be derived immediately from the Belonaspida.
The Diploconida (Pl. 140)
form the last and the most modified family of all Acanthophracta. The
remarkable shell exhibits the strange form of a double cone, bearing in its axis two very large
opposite spines; these are the two equatorial spines of the "hydrotomical axis" (p. 719). The double-conical or nearly cylindrical shell is composed of three
different parts or segments; the small middle part is the true lattice-shell of the Hexalaspida
and Belonaspida, and bears the eighteen smaller (often quite rudimentary) radial spines. The two
other parts (opposite on both poles of its hydrotomical axis) are the conical or cylindrical,
solid, basal sheaths of the two large equatorial spines, enveloping their major part. In
consequence of this peculiar metamorphosis of the shell the Diploconida represent the last and the
most aberrant group of all Acantharia.
Synopsis of the Suborders and Families of Acanthophracta.
|
Suborder I. SPHÆROPHRACTA.
Twenty radial spines of equal size. Shell spherical (or an endospherical
polyhedron).
|
|
Shell spherical, simple, pierced by twenty or eighty aspinal pores and
composed of a pavement of innumerable very small plates or aglets, each pierced by one
porule, |
1. Sphærocapsida. |
| Shell spherical, simple, composed of the meeting branches of two or
four apophyses of the twenty radial spines, |
2. Dorataspida. |
| Shell spherical, double, composed of two concentric lattice-spheres,
which are connected by the twenty radial spines and composed of the meeting branches of their
apophyses, |
3. Phractopeltida. |
|
Suborder II. PRUNOPHRACTA.
Twenty radial spines of unequal size; two or six hydrotomical spines much
larger than the eighteen or fourteen others. Shell not spherical.
|
|
Shell ellipsoidal, with prolonged hydrotomical axis, the two spines of
which are larger than the eighteen others, |
4. Belonaspida. |
| Shell lenticular or discoidal, with six larger spines placed in the
hydrotomical plane (fourteen other spines much smaller), |
5. Hexalaspida. |
| Shell diploconical or nearly cylindrical, with two opposite large
funnels, the sheaths of the enlarged two spines of the hydrotomical axis (eighteen other
spines much smaller or rudimentary), |
6. Diploconida. |
| Suborder I. SPHÆROPHRACTA. Twenty radial spines of equal size. Shell spherical
(or an endospherical polyhedron). |
|
|
Shell spherical, simple, pierced by twenty or eighty aspinal pores
and composed of a pavement of innumerable very small plates or aglets, each pierced by one
porule, |
|
|
|
|
|
|
1. Sphærocapsida. |
|
|
|
|
|
|
Shell spherical, simple, composed of the meeting branches of two
or four apophyses of the twenty radial spines, |
|
|
|
|
|
|
2. Dorataspida. |
|
|
|
|
|
|
Shell spherical, double, composed of two concentric
lattice-spheres, which are connected by the twenty radial spines and composed of the meeting
branches of their apophyses, |
|
|
|
|
|
|
3. Phractopeltida. |
|
|
|
|
| Suborder II. PRUNOPHRACTA. Twenty radial spines of unequal size; two or six
hydrotomical spines much larger than the eighteen or fourteen others. Shell not
spherical. |
|
|
Shell ellipsoidal, with prolonged hydrotomical axis, the two
spines of which are larger than the eighteen others, |
|
|
|
|
|
|
4. Belonaspida. |
|
|
|
|
|
|
Shell lenticular or discoidal, with six larger spines placed in
the hydrotomical plane (fourteen other spines much smaller), |
|
|
|
|
|
|
5. Hexalaspida. |
|
|
|
|
|
|
Shell diploconical or nearly cylindrical, with two opposite large
funnels, the sheaths of the enlarged two spines of the hydrotomical axis (eighteen other
spines much smaller or rudimentary), |
|
|
|
|
|
|
6. Diploconida. |
|
|
|
|
{795}
Suborder I. SPHÆROPHRACTA, Haeckel.
Definition.—Shell spherical, with twenty radial beams of equal
size.
Family XXXIX. Sphærocapsida, Haeckel (Pl. 133, figs.
7-11; Pl. 135,
figs. 6-10).
Sphærocapsida, Haeckel, 1881, Prodromus, p. 469.
Definition.—Acantharia with simple spherical porous shell,
composed of innumerable very small plates, each of which is pierced by one radial porule. Twenty
radial spines of equal size meeting in the centre of the shell and disposed according to the
Müllerian law of the Icosacantha, sometimes short and enclosed in the shell, at other times long
and piercing it (rarely rudimentary or quite absent). Shell pierced therefore either by twenty
larger perspinal pores or by eighty smaller aspinal pores. Central capsule spherical, enclosed in
the porous shell.
The family Sphærocapsida, founded by me in 1881 for the single genus
Sphærocapsa, represents a very peculiar and remarkable group of the Acanthophracta, very different from the five other families of this suborder,
and probably derived, independently of them, directly from the Acanthonida. Whilst the lattice-shell of the five other families is composed of
the meeting branches of lateral apophyses of the twenty spines, and its meshes are all or partly
the intervals between these apophyses, in the Sphærocapsida the spherical shell has quite another
structure, and is composed of innumerable small plates (each with one pore) which are secreted on
the surface of the spherical calymma, independently of the twenty radial spines, which do not
possess true apophyses.
In all Sphærocapsida the structure of the spherical shell is quite peculiar and different from
that of all other Radiolaria. It is composed everywhere of innumerable very small plates or
aglets, which are connected irregularly like paving-stones, and form a single continuous layer or
pavement on the surface of the spherical calymma (Pl. 133, fig. 11,
a; Pl. 135,
figs. 8, 10). The small plates or paving-stones, which we will call "aglets," are connected at
their meeting edges by a kind of cement, and form together with it a continuous thick capsule of
acanthin. The form of the aglets is commonly more or less irregular, roundish or polygonal,
sometimes longish (Pl. 133,
fig. 11, a), more rarely it becomes rather regular, hexagonal, square, or roundish (Pl. 135, fig. 8).
Usually all aglets of one and the same individual are of nearly equal size, between 0.01 and 0.02
in diameter, rarely less or more. The outer face of the aglets is more or less concave, so that
the elevated meeting edges of the neighbouring aglets commonly form together a prominent network
of crests (Pl. 135,
figs. 8, 10); rarely the meeting edges partly cover one another like squamules (Pl. 133, fig. 11,
a). {796}Each small plate or aglet is pierced in
its centre by a single radial canalicule or porule. The dimpled surface, so produced, resembles
somewhat the dimpled plates of Ceriaspis, &c. Different from these innumerable very
small dimples of the surface are the twenty larger "spinal dimples," or the concave larger plates,
which are originally pierced by the twenty radial spines. Before we describe these, we must
examine the spines themselves.
The twenty radial spines of all observed Sphærocapsida (sixteen species) agree perfectly with
those of the genus Acanthonia (p. 749), and especially with
Acanthonia tetracopa, Acanthonia denticulata, &c. All twenty spines, regularly
disposed according to the Müllerian law of the Icosacantha, are of equal size, constantly
four-edged prismatic, of equal breadth throughout their whole length. The prominent four edges are
parallel, sometimes smooth, at other times elegantly denticulated. The central bases of the twenty
spines are pyramidal, without leaf-cross, and propped one upon another with their triangular
faces, as in the majority of the Acanthonida.
The relation of the twenty radial spines to the spherical shell exhibits in the five genera
described very peculiar and important differences. In the first described genus, in
Sphærocapsa, the spines are exactly as long as the shell-radius, and therefore are not
prominent over the surface of the shell, with which they are firmly connected; the truncated
distal end of the spine lies therefore here in the surface of the shell itself, and is connected
with it by its four edges, between which four open aspinal pores remain, as in Tessaraspis,
&c. (Pl. 135,
figs. 6-10). In the next allied genus, Astrocapsa (Pl. 133, figs. 9,
10), the spines are longer than the shell-radius, and therefore more or less prominent over its
surface; the piercing part of each spine is also surrounded by four aspinal pores. In the two
following genera, Porocapsa and Cannocapsa (Pl. 133, figs. 7,
8), the radial spines are shorter than the shell-radius and therefore quite hidden and withdrawn
inside the shell, which they do not reach. But in the ideal prolongation of each spine the shell
is pierced by a single large opening, the "perspinal pore" or "perspinal hole," composed of the
four united aspinal pores. Whilst in Porocapsa the perspinal pores are simple, they are
prolonged in Cannocapsa into cylindrical tubes, open at both ends. The twenty perspinal
holes of these Porocapsida are therefore derived by confluence of the eighty original aspinal
pores of the Astrocapsida and preserve the same regular disposition, according to the Müllerian
law of the Icosacantha. Finally, the same law as is valid also in the last genus is found in
Cenocapsa; here the radial spines have completely disappeared, and the whole skeleton is a
simple sphere, but of the same structure, and with the same twenty perspinal pores as in
Porocapsa. It is very interesting that this spineless Cenocapsa among the Acantharia exhibits the same shell (a simple hollow sphere) as a last reduced
form, which Cenosphæra among the Sphærellaria produces as a
primitive ancestral form of numerous genera.
{797}
The Central Capsule of the Sphærocapsida is spherical, constantly smaller than the
enclosing concentric shell, and separated from it by the calymma. Its structure seems to be the
same as in the Acanthonida, and specially in the Astrolonchida. The
pseudopodia (not yet observed) are probably protruded only through the twenty perspinal holes or
the eighty aspinal pores.
Synopsis of the Genera of Sphærocapsida.
|
III. Subfamily Astrocapsida.
Radial spines connected with the porous shell, as long or longer than its
radius. Eighty aspinal pores.
|
|
Spines as long as the radius, without external prolongation, |
344. Sphærocapsa. |
| Spines longer than the radius, with external prolongation, |
345. Astrocapsa. |
|
III. Subfamily Porocapsida.
Radial spines not connected with the porous shell, shorter than its radius.
Twenty perspinal pores.
|
|
Perspinal holes of the shell simple, without external
prolongation, |
346. Porocapsa. |
| Perspinal holes of the shell prolonged into radial centrifugal
tubuli, |
347. Cannocapsa. |
|
III. Subfamily Cenocapsida.
Radial spines disappeared.
|
|
Twenty perspinal holes of the shell simple, without tubular
prolongation, |
348. Cenocapsa. |
| I. Subfamily Astrocapsida. Radial spines connected with the porous shell, as
long or longer than its radius. Eighty aspinal pores. |
|
|
Spines as long as the radius, without external prolongation, |
|
|
|
|
|
|
344. Sphærocapsa. |
|
|
|
|
|
|
Spines longer than the radius, with external prolongation, |
|
|
|
|
|
|
345. Astrocapsa. |
|
|
|
|
| II. Subfamily Porocapsida. Radial spines not connected with the porous shell,
shorter than its radius. Twenty perspinal pores. |
|
|
Perspinal holes of the shell simple, without external
prolongation, |
|
|
|
|
|
|
346. Porocapsa. |
|
|
|
|
|
|
Perspinal holes of the shell prolonged into radial centrifugal
tubuli, |
|
|
|
|
|
|
347. Cannocapsa. |
|
|
|
|
| III. Subfamily Cenocapsida. Radial spines disappeared. |
|
|
Twenty perspinal holes of the shell simple, without tubular
prolongation, |
|
|
|
|
|
|
348. Cenocapsa. |
|
|
|
|
Subfamily 1. Astrocapsida, Haeckel.
Definition.—Radial spines connected with the porous shell, as long as
or longer than its radius. Therefore the shell pierced by eighty aspinal pores (four around each
spine).
Genus 344. Sphærocapsa,[384] Haeckel,
1881, Prodromus, p. 469.
Definition.—Sphærocapsida with twenty radial spines as
long as the radius of the shell, without external prolongation; therefore their distal ends
inserted in the perspinal holes, each of which is composed of four aspinal pores.
The genus Sphærocapsa is the most common form of the Sphærocapsida, and
comprises those species in which the radial spines are as long as the radius of the shell, and
therefore are connected with the margin of its aspinal holes, but not prolonged beyond its
surface.
{798}
1. Sphærocapsa cruciata, n. sp. (Pl. 135,
figs. 6, 7).
Aspinal holes nearly circular, with flat radially striated margin. Four aspinal pores of each
hole rounded equilateral triangular. Porules of the shell simple, without ring and dimple. Four
edges of the spines smooth.
Dimensions.—Diameter of the shell 0.4 to 0.5, of the central capsule 0.3 to
0.4.
Habitat.—North Atlantic, Færöe Channel (Gulf Stream), 1880, John
Murray, surface.
2. Sphærocapsa dentata, n. sp. (Pl. 135,
fig. 9).
Aspinal holes four-lobed, with broad concave margin denticulated on the periphery. Four aspinal
pores of each hole pear-shaped, oblong, elevated in the centre. Porules of the shell simple,
without ring and dimple. Four edges of the spines denticulate.
Dimensions.—Diameter of the shell 0.3 to 0.4, of the central capsule 0.2 to
0.3.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
3. Sphærocapsa quadrata, n. sp. (Pl. 135,
fig. 8).
Aspinal holes square, with concave umbilicus in the centre, surrounded by a convex denticulated
margin. Four aspinal pores of each hole nearly square. Shell-porules with a polygonal elevated
smooth ring, in the bottom of a shallow dimple. Four edges of the spines smooth.
Dimensions.—Diameter of the shell 0.22, of the central capsule 0.17.
Habitat.—Indian Ocean, south of Australia, Station 159, surface.
4. Sphærocapsa pavimentata, n. sp. (Pl. 135, fig.
10).
Aspinal holes four-lobed, with broad concave, irregularly crenated and figured margin. Four
aspinal pores of each hole violin-shaped. Porules of the shell surrounded by an irregularly oblong
ring with thick elevated, elegantly crenated margin. Four edges of the spines smooth.
Dimensions.—Diameter of the shell 0.36, of the central capsule 0.3.
Habitat.—South-east Pacific (off Valparaiso), Station 298,
surface.
Genus 345. Astrocapsa,[385] n. gen.
Definition.—Sphærocapsida with twenty radial spines
longer than the radius of the shell, piercing its perspinal holes, with free external
prolongation; therefore with four aspinal pores around each spine.
The genus Astrocapsa differs from the preceding Sphærocapsa in the
external prolongation of the radial spines piercing the perspinal holes; it assumes therefore the
common shape of the Dorataspida more than the other Sphærocapsida do.
{799}
1. Astrocapsa tritonis, n. sp.
Aspinal holes circular, with smooth convex margin and four circular aspinal pores. Porules of
the shell simple, without ring and dimple. Four edges of the spines smooth, their outer free part
about as long as the diameter of the shell.
Dimensions.—Diameter of the shell 0.25; outer length of the spines 0.3.
Habitat.—North Atlantic, Færöe Channel, Gulf Stream (expedition of
H.M.S "Triton," August, 1882), John Murray, surface.
2. Astrocapsa stellata, n. sp. (Pl. 133,
fig. 10).
Aspinal holes cruciform, with high crenated margin. Four aspinal pores of each hole egg-shaped.
Porules of the shell simple, without ring and dimple. Four edges of the spines denticulate; their
outer free part twice to three times as long as the diameter of the shell.
Dimensions.—Diameter of the shell 0.36; outer length of the spines 0.5 to 0.8.
Habitat.—Antarctic Ocean (near Kerguelen), Station 152, surface.
3. Astrocapsa quadrifida, n. sp.
Aspinal holes four-lobed, with four prominent teeth between the four roundish aspinal pores.
Porules of the shell surrounded by a high polygonal smooth ring. Four edges of the spines
denticulate; their outer free part about as long as the radius of the shell.
Dimensions.—Diameter of the shell 0.28; outer length of the spines 0.15.
Habitat.—North Pacific, Station 241, surface.
4. Astrocapsa coronata, n. sp. (Pl. 133,
fig. 9).
Aspinal holes circular, with a coronet of numerous thin parallel teeth. Porules of the shell in
dimples on irregular polygonal small plates, with coronated ring. Four edges of the spines
denticulate; their outer free part about twice as long as the diameter of the shell.
Dimensions.—Diameter of the shell 0.4 to 0.5 ; outer length of the spines 1.0 to
1.2.
Habitat.—North Atlantic, Færöe Channel, Gulf Stream (expedition of the
"Knight Errant," 1880), John Murray, surface and at depths varying from 10 to 200 fathoms.
Subfamily 2. Porocapsida, Haeckel.
Definition.—Radial spines not connected with the porous shell, shorter
than its radius; therefore the shell pierced by twenty perspinal pores (each one in the ideal
radial prolongation of one spine).
{800}
Genus 346. Porocapsa,[386] n. gen.
Definition.—Sphærocapsida with twenty radial spines
shorter than the radius of the shell; therefore their distal ends not connected with the twenty
perspinal holes, which are simple, not prolonged into radial tubes.
The genus Porocapsa and the following Cannocapsa form together the
small sub-family of Porocapsida, distinguished by the peculiar reduction or retrograde development
of the twenty radial spines; all these twenty are present and disposed according to the Müllerian
law of the Icosacantha, but they are shorter than the radius of the shell and therefore do not
reach it. In the ideal prolongation of the spines the shell is pierced by twenty simple
quadrangular or circular perspinal holes.
1. Porocapsa murrayana, n. sp. (Pl. 133,
fig. 7).
Perspinal holes cruciform, with smooth thickened margin or with four short teeth between the
four lobes. Porules of the shell simple, without ring and dimple. Four edges of the spines smooth.
(Differs from Sphærocapsa cruciata, Pl. 135,
figs. 6, 7, living in the same locality, mainly in the reduction of the radial spines, which do
not reach the shell.)
Dimensions.—Diameter of the shell 0.28; length of the spines 0.2.
Habitat.—North Atlantic, Færöe Channel, Gulf Stream (expedition of
H.M.S. "Triton," August 1882), John Murray, surface and in depths from 40 to 640 fathoms.
2. Porocapsa tetrodon, n. sp.
Perspinal holes cruciform, with four triangular prominent teeth between the four lobes of the
cross. Porules of the shell with an elevated polygonal ring, in the bottom of a dimple. Four edges
of the spines elegantly denticulate.
Dimensions.—Diameter of the shell 0.36; length of the spines 0.12.
Habitat.—North Pacific, Station 253, surface.
3. Porocapsa octodon, n. sp.
Perspinal holes square, with four larger prominent teeth on the sides of the square, and four
smaller teeth on its corners. Porules of the shell surrounded by an elevated ring with elegantly
crenated irregular margin. Four edges of the spines smooth.
Dimensions.—Diameter of the shell 0.48; length of the spines 0.18.
Habitat.—Arctic Ocean (Greenland), in the contents of the stomach of
the Peromedusa, Periphylla hyacinthina.
{801}
4. Porocapsa coronodon, n. sp.
Perspinal holes circular, with ciliated margin, which forms a crown of sixteen to twenty-four
thin parallel teeth. Poruli of the shell surrounded by an elevated ring with high crenated margin.
Four edges of the spines denticulate.
Dimensions.—Diameter of the shell 0.55; length of the spines 0.15.
Habitat.—Antarctic Ocean, Station 154, surface.
Genus 347. Cannocapsa,[387] n. gen.
Definition.—Sphærocapsida with twenty radial spines
shorter than the radius of the shell; therefore their distal ends not connected with the twenty
perspinal holes, which are prolonged outside into radial tubes (each one in the radial ideal
prolongation of one inner spine).
The genus Cannocapsa, exhibits the same peculiar reduction of the radial
spines as the foregoing Porocapsa; the spines are also here shorter than the shell-radius
and do not therefore reach the perspinal holes of the shell. But whilst these latter are simple in
Porocapsa, they are prolonged into radial tubules in Cannocapsa; the outer surface
bears therefore twenty such cylindrical tubules, separated by a short distance from the inner
enclosed spines, but disposed quite regularly according to the law of Icosacantha.
1. Cannocapsa osculata, n. sp.
Perspinal holes prolonged into short cylindrical tubuli, the length of which about equals their
diameter. Both ends of the tubuli with smooth thickened margin. Poruli of the shell simple,
without ring and dimple. Four edges of the spines smooth; their length equals about four-fifths of
the shell-radius.
Dimensions.—Diameter of the shell 0.24; length of the spines 0.09, of the tubuli
0.01.
Habitat.—North Atlantic, Færöe Channel (Gulf Stream), surface, John
Murray.
2. Cannocapsa stethoscopium, n. sp. (Pl. 133, fig.
8).
Perspinal holes prolonged into cylindrical tubuli, half as long as the shell radius. Both ends
of the tubuli with smooth thin trumpet-shaped margins. Poruli of the shell simple, without ring
and dimple. Four edges of the spines smooth; their length about equals three-fourths of the
shell-radius.
Dimensions.—Diameter of the shell 0.2; length of the spines 0.08, of the tubuli
0.05.
Habitat.—South Atlantic (west of Tristan da Cunha), Station 333,
surface.
3. Cannocapsa tubulosa, n. sp.
Perspinal holes prolonged into cylindrical tubuli, which are as long as or longer then the
shell-radius. Both ends of the tubuli with smooth thickened margin. Poruli of the shell simple,
{802}without ring and dimple. Four edges of the spines
smooth; their length scarcely equals half the shell-radius.
Dimensions.—Diameter of the shell 0.15; length of the spines 0.03, of the tubuli
0.08.
Habitat.—South Atlantic (east coast of Patagonia), Station 318,
surface.
Subfamily 3. Cenocapsida, Haeckel.
Definition.—Radial spines completely reduced and absent; shell cavity
therefore simple; shell pierced by twenty perspinal pores (each placed in the direction of one
radial spine which has disappeared).
Genus 348. Cenocapsa,[388] n. gen.
Definition.—Sphærocapsida without radial spines, with
simple cavity of the spherical shell, which is pierced by twenty perspinal holes (each one placed
in the radial direction of one spine which has disappeared).
The genus Cenocapsa comprises only a single species, but is very remarkable
in that it is the most reduced form among all Sphærocapsida. The twenty radial spines of the
Icosacantha have perfectly disappeared by complete retrograde metamorphosis, and the only evidence
of their former existence (in the ancestral genus Porocapsa) are the twenty perspinal holes
remaining in the shell. Cenocapsa is the only form of Acantharia
which possesses no radial spines.
1. Cenocapsa nirvana, n. sp. (133,
fig. 11, 11a, 11c).
Perspinal holes four-lobed, cruciform, with four short triangular teeth between the four lobes
of the cross. Poruli of the shell in the bottom of an elliptical dimple surrounded by an elevated
ring.
Dimensions.—Diameter of the shell 0.2 to 0.3, of the aspinal holes 0.02.
Habitat.—North Pacific, Station 248, surface.
Family XL. Dorataspida, Haeckel (Pls.
134-138).
Dorataspida, Haeckel, 1862, Monogr. d. Radiol., p. 412.
Definition.—Acantharia with simple spherical
lattice-shell, composed of the branched apophyses of twenty equal radial spines meeting in its
centre and disposed according to the Müllerian law of Icosacantha. Central capsule spherical,
enclosed in the fenestrated shell.
The family Dorataspida is the most important family of the Acanthophracta, or of those Acantharia in which the
radial spines are connected by a complete extracapsular lattice-shell. The Dorataspida represent
probably the ancestral {803}stock of this whole order,
with the exception of the Sphærocapsida. The four following families of the order may be easily
derived from the Dorataspida. The number of genera (seventeen) and of species (one hundred and
eight) is in this family greater than in the other five families together. When I constituted that
family in my Monograph 1862, it comprised only one genus, Dorataspis, with seven species.
The nearly allied genus Haliommatidium (Phatnaspis) belongs to the Belonaspida.
The Dorataspida differ from the other Acanthophracta in the simple
spherical lattice-shell, which is composed of the meeting apophyses of the twenty radial spines.
In three other families of the suborder the shell is not spherical, but ellipsoidal (Belonaspida),
discoidal (Hexalaspida), or diploconical (Diploconida). In the Phractopeltida the spherical shell
is double, composed of two concentric lattice-spheres. In the Sphærocapsida the simple spherical
shell is not composed of the apophyses of the spines, but of innumerable small plates.
The family Dorataspida may be divided into two very different subfamilies, which are probably
derived, independently of one another, from two different subfamilies of the Astrolonchida. The
first subfamily, Diporaspida, exhibits on each radial spine two opposite apophyses, like its
ancestral group, the Phractacanthida (p. 753); whereas the second
subfamily, Tessaraspida, possesses on each radial spine four crossed apophyses (opposite in
pairs), like its ancestral group, the Stauracanthida (p. 758). Therefore
the composition of the spherical shell, produced by the meeting branches of the tangential
apophyses, is essentially different in the two subfamilies: in the Diporaspida each radial spine
is surrounded by two opposite primary aspinal meshes, in the Tessaraspida by four crossed primary
aspinal meshes.
Another principle of division may be established for the whole family by the different mode of
composition of the shell, and regarding this important difference we may distinguish also two
different subfamilies as Cladophracta and Peltophracta. In the first and simpler subfamily, the Cladophracta, the shell is composed totally (or sometimes partially) of the
meeting branches of the apophyses of the neighbouring spines; but in each single spine (or in the
most part of them) the branches of the apophyses are not united, and form no lattice-plate (Pl. 137, figs. 1
to 8). Whereas in the Peltophracta the shell is composed constantly of
twenty perforated plates, as in each single spine the branches of its apophyses are united and
form a fenestrated shield with two or four (and sometimes numerous) pores (Pls. 135, 136, 138).
In the Diporaspida as well as in the Tessaraspida we find numerous representatives of the two
groups of the Cladophracta and of the Peltophracta; therefore the whole family of Dorataspida may be divided into
four different tribes. The Diporaspida (with two opposite apophyses on each spine) are partly
Cladophracta (the Phractaspida, Pl. 137, figs.
1-4), partly Peltophracta (the Ceriaspida, Pl. 138). On the
other hand the Tessaraspida (with four crossed apophyses on each {804}spine) are also partly Cladophracta (the
Stauraspida, Pl. 137,
figs. 5-8), partly Peltophracta (the Lychnaspida, Pls. 135-136). The
differences and relations of these tribes are placed synoptically in the following table:—
| Synopsis of the four tribes of Dorataspida. |
A. Diporaspida.
Two opposite apophyses on each radial spine. Two primary aspinal meshes. |
B. Tessaraspida.
Four crossed apophyses on each radial spine. Four primary spinal meshes. |
|
a. Cladophracta
All twenty spines (or a part of them) without lattice-plates.
|
1. Tribe Phractaspida. |
3. Tribe Stauraspida. |
|
b. Peltophracta
All twenty spines with lattice-plates.
|
2. Tribe Ceriaspida. |
4. Tribe Lychnaspida. |
All Dorataspida are true Icosacantha, and the twenty spines, composing the spherical shell, are
equally developed, regularly disposed according to the Müllerian law, and of equal size; also the
distance of their plates from the common centre is equal. Nevertheless they are never of perfectly
the same form; in consequence of their peculiar disposition in five zones (each with four spines)
certain slight differences are effected, so that with accurate knowledge of the peculiar
shell-composition it is generally not difficult to distinguish the spines of the equatorial, the
two tropical, and the two polar zones.
Already the central bases, by which the twenty spines are united in the centre of the sphere,
exhibit certain differences in the five zones. Commonly these bases are small pyramids, all
meeting with their apex in the centre, and the triangular faces of the neighbouring pyramids are
supported one upon another. The four equatorial pyramids are commonly six-sided, the other sixteen
five-sided; but sometimes there are eight six-sided and twelve five-sided basal pyramids; two
opposite polar spines on each pole having a six-sided base (like the four equatorial), the other
two polar spines on each pole having a five-sided base (like the eight tropical). Rarely the
central bases are perfectly grown together, forming a single spherical central piece of
acanthin.
The three different fundamental forms of radial spines, which are found in all Acantharia, the cylindrical, the two-edged, and the four-edged (spines with
circular, with elliptical, and with square transverse section respectively) occur also in the
different groups of Dorataspida; but commonly the two-edged or compressed form is prevalent in the
Diporaspida, the four-edged or quadrangular form in the Tessaraspida. In the majority of species
the spines are thickened in the shell-face, where the apophyses arise, and thinner towards the two
ends. Usually the outer or distal part of the spine (outside the shell) is longer than the inner
or proximal part (inside {805}the shell). The distal
apex is commonly simple, conical or pyramidal, rarely bifid or truncate. The edges of the spines
are commonly smooth, rarely denticulate or serrate.
The apophyses, or the lateral transverse processes of the radial spines, in the Dorataspida
assume the greatest variety and complexity in form, size, mode of ramification, and in composition
of the shell. An expert and practised observer may determine easily the range of each spine,
whether it be an equatorial (c), or a tropical (b, d), or a polar spine
(a, e, Pls. 133-138). The two
opposite apophyses of the Diporaspida, as well as the four crossed apophyses of the Tessaraspida,
lie constantly in certain meridian planes of the spine, which have a legitimate signification for
each of the five zones. The comparative morphology of this regular disposition of the apophyses
and the regular meeting of their branches is of the greatest interest, and necessary for the
complete understanding of the complicated structure of these wonderful shells.
The pores or meshes of the spherical shell, offering the most varied forms, may generally be
divided into two different groups, into sutural and parmal meshes. The sutural pores are bordered
by the meeting branches of the apophyses of two, three, or four neighbouring spines, and therefore
also by the sutures in which they meet. The parmal pores on the other hand are bordered only by
the united branches of the apophyses of a single spine and pierce the shield or lattice-plate
formed by them. Therefore the shell-meshes of the Cladophracta are all
sutural pores (Pl. 137,
figs. 1-8; rarely and only in a part of the spines parmal pores also: Zonaspis,
Dodecaspis); whereas the shell-meshes of the Peltophracta,
piercing the shields or lattice-plates of all twenty spines, are always partly sutural, partly
parmal pores (Pls. 135,
136, 138). The
parmal pores again may be divided into two different groups—aspinal and coronal pores.
Aspinal pores ("ad spinam") are those which lie immediately on the sides of the radial spine and
are bordered by the primary branches of its apophyses; therefore constantly only two in the
Diporaspida, four in the Tessaraspida. Coronal pores on the contrary are those which lie in the
periphery of the lattice-plates, surrounding in a circle or crown the aspinal pores and not
touching the spine itself. In Dorataspis, Ceriaspis, Tessaraspis,
Lychnaspis, &c., all parmal meshes are only aspinal pores (Pl. 135, figs.
2-5; Pl. 136);
whilst in Coscinaspis, Acontaspis, Icosaspis, Hylaspis, &c., one
part of the parmal pores is aspinal, one part coronal (Pl. 136). The
number, form, and size of the coronal pores is very variable and often very large (sometimes more
than a hundred in one plate).
The Cladophracta exhibit a comparatively simple shell-formation;
either all twenty spines or at least a part of them not forming lattice-plates. The most primitive
form among these is Phractaspis (Pl. 137,
figs. 1, 2). The forty apophyses of its twenty spines are simply forked, and their eighty
fork-branches united by forty sutures, enclosing twenty-two sutural meshes: two square polar
meshes (between the four polar spines on the poles of the spineless axis, a a a a and e
e e e); eight triangular {806}circumpolar meshes
(each between two polar and one tropical spine, a b a and e d e); eight quadrangular
tropical meshes (each between one polar, one equatorial, and two tropical spines, a b c b
and e d c d); and four rhomboidal equatorial meshes (between two tropical and two
equatorial spines, c b c d). If the fork-branches be again forked (Phractaspidium,
Pl. 137,
fig. 3), the number of the sutures and sutural meshes is doubled, and the same is the case in
Stauraspis, the most simple form of the Tessaraspida.
A peculiar small group, and an interesting transition for the Cladophracta to the Peltophracta, is presented by the
Zonaspida among the Tessaraspida (Zonaspis and Dodecaspis). Here only one part of
the radial spines bears lattice-plates, the other part not. In Zonaspis the four equatorial
spines bear lattice-plates, the sixteen other only free branches of the apophyses. In
Dodecaspis twelve spines are provided with lattice-plates (four equatorial and eight polar
spines), whilst the eight other (tropical) spines are devoid of them.
The Peltophracta exhibit a great variety in the form and composition
of their twenty lattice-plates or fenestrated shields. In the most simple case (a part of
Dorataspis and Diporaspis) the shell is composed of four (equatorial) hexagonal
plates, and sixteen pentagonal plates (four tropical and four polar); in this case the four polar
plates meet on each pole in one common point. More commonly, however, the shell seems to be
composed of eight hexagonal plates (four equatorial and the four polar spines of the hydrotomical
plane) and twelve pentagonal plates (eight tropical and the four polar spines of the geotomical
plane); in this case only two (hexagonal) polar plates meet on each pole in a suture which
separates the two other (pentagonal) polar plates (Pl. 138, fig. 4).
In the majority of the Dorataspida the composition of the shell is much more complicated and often
very difficult to understand. Often the surface of the plates is covered with a network of
elevated crests, by which concave blind dimples are separated (Ceriaspis,
Hystrichaspis, Pl. 138);
and sometimes these dimples become pierced by coronal pores (Pl. 138, fig. 11,
&c.).
Peculiar by-spines or "accessory spines" cover the outer surface of the shell in a great part
of Dorataspida, and commonly these most characteristic by-spines are not placed radially, but
parallel to the radial spine, from the lattice-plate of which they arise (Pl. 135, figs. 1,
5; Pl. 137,
figs. 4-8; Monogr. d. Radiol., 1862, Taf. xxi. figs. 8, 9). They are commonly placed perpendicular
to the sutural condyles, or the branch-ends of the apophyses; so that close to each suture arises
a pair of divergent by-spines, belonging to the meeting apophyses of the two neighbouring spines,
which meet in the suture (Pl. 137,
fig. 4). Rarely these thin, bristle-shaped by-spines are quite simple and straight, commonly they
undulate or are zigzag and often armed with recurved hooks. Sometimes they are also forked or
arborescent (Pl. 138,
fig. 7).
The Central Capsule of the Dorataspida is constantly spherical and about one-third
smaller than the enclosing shell, from which it is separated by the spherical calymma. {807}The membrane of the central capsule is commonly rather
thin, and pierced by the twenty radial spines, meeting in the centre of the capsule. Between these
lie innumerable small pores for the radiating pseudopodia; however, in many cases (and perhaps
constantly) these pores exhibit a certain regular disposition. In many species the central capsule
encloses Xanthellæ or symbiotic yellow unicellular Algæ. The nucleus becomes cleft very early in
the majority of Dorataspida.
Synopsis of the Genera of Dorataspida.
|
II. Subfamily Diporaspida.
Each radial spine with two opposite primary apophyses; therefore the whole
shell with forty primary apophyses.
|
 |
I. Tribe Phractaspida. Twenty radial spines
without lattice-plates; no normal pores. |
|
No by-spines, |
349. Phractaspis. |
| With by-spines, |
350. Pleuraspis. |
| II. Tribe Ceriaspida. Twenty radial spines all with
lattice-plates produced by union of the branched apophyses of each plate. |
|
Forty parmal pores (two in each plate)—no coronal
pores in the plates. |
|
Plates not dimpled, without network of crests. |
|
No by-spines, |
351. Dorataspis. |
| With by-spines, |
352. Diporaspis. |
| No by-spines, but free apophyses, |
353. Orophaspis. |
| Plates dimpled, with a network of crests. |
|
No by-spines, |
354. Ceriaspis. |
| With by-spines, |
355. Hystrichaspis. |
| Eighty to two hundred or more parmal pores (in
each plate two aspinal and two to ten or more coronal pores). |
|
No by-spines, |
356. Coscinaspis. |
| With by-spines, |
357. Acontaspis. |
|
II. Subfamily Tessaraspida.
Each radial spine with four crossed primary apophyses; therefore the whole
shell with eighty primary apophyses.
|
 |
III. Tribe Stauraspida. Twenty radial spines all or partly
without lattice-plates. |
|
All twenty spines without lattice-plates. |
|
No by-spines, |
358. Stauraspis. |
| With by-spines, |
359. Echinaspis. |
| Four plates with, sixteen without, lattice-plates. |
|
With by-spines, |
360. Zonaspis. |
| Twelve plates with, eight without, lattice-plates. |
|
With by-spines, |
361. Dodecaspis. |
| IV. Tribe Lychnaspida. Twenty radial spines all with
lattice-plates (produced by union of the branched apophyses of each plate). |
|
Eighty parmal pores (four on each
plate)—no coronal pores in the plates. |
|
No by-spines, |
362. Tessaraspis. |
| With by-spines, |
363. Lychnaspis. |
| One hundred and sixty to three hundred or more
parmal pores (in each plate four aspinal and four to twelve or more coronal pores). |
|
No by-spines, |
364. Icosaspis. |
| With by-spines, |
365. Hylaspis. |
| I. Subfamily Diporaspida. Each radial spine with two opposite primary
apophyses; therefore the whole shell with forty primary apophyses. |
|
|
I. Tribe Phractaspida. Twenty radial spines without
lattice-plates; no normal pores. |
|
|
|
|
|
|
|
|
|
|
No by-spines, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
349. Phractaspis. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
With by-spines, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
350. Pleuraspis. |
|
|
|
|
|
|
|
|
|
|
|
|
II. Tribe Ceriaspida. Twenty radial spines all with lattice-plates
produced by union of the branched apophyses of each plate. |
|
|
|
|
|
|
Forty parmal pores (two in each plate)—no coronal pores in
the plates. |
|
|
|
|
|
|
|
|
|
|
Plates not dimpled, without network of crests. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
No by-spines, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
351. Dorataspis. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
With by-spines, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
352. Diporaspis. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
No by-spines, but free apophyses, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
353. Orophaspis. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Plates dimpled, with a network of crests. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
No by-spines, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
354. Ceriaspis. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
With by-spines, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
355. Hystrichaspis. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Eighty to two hundred or more parmal pores (in each plate two
aspinal and two to ten or more coronal pores). |
|
|
|
|
|
|
|
|
|
|
|
|
No by-spines, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
356. Coscinaspis. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
With by-spines, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
357. Acontaspis. |
|
|
|
|
|
|
|
|
|
|
| II. Subfamily Tessaraspida. Each radial spine with four crossed primary
apophyses; therefore the whole shell with eighty primary apophyses. |
|
|
III. Tribe Stauraspida. Twenty radial spines all or partly without
lattice-plates. |
|
|
|
|
|
|
All twenty spines without lattice-plates. |
|
|
|
|
|
|
|
|
|
|
|
|
No by-spines, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
358. Stauraspis. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
With by-spines, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
359. Echinaspis. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Four plates with, sixteen without, lattice-plates. |
|
|
|
|
|
|
|
|
|
|
|
|
With by-spines, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
360. Zonaspis. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Twelve plates with, eight without, lattice-plates. |
|
|
|
|
|
|
|
|
|
|
|
|
With by-spines, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
361. Dodecaspis. |
|
|
|
|
|
|
|
|
|
|
|
|
IV. Tribe Lychnaspida Twenty radial spines all with lattice-plates
(produced by union of the branched apophyses of each plate). |
|
|
|
|
|
|
Eighty parmal pores (four on each plate)—no coronal pores in
the plates. |
|
|
|
|
|
|
|
|
|
|
|
|
No by-spines, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
362. Tessaraspis. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
With by-spines, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
363. Lychnaspis. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
One hundred and sixty to three hundred or more parmal pores (in
each plate four aspinal and four to twelve or more coronal pores). |
|
|
|
|
|
|
|
|
|
|
|
|
No by-spines, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
364. Icosaspis. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
With by-spines, |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
365. Hylaspis. |
|
|
|
|
|
|
|
|
|
|
{808}
Subfamily 1. Diporaspida, Haeckel.
Definition.—Dorataspida with twenty radial
spines, each of which bears two opposite apophyses. The spherical shell is composed either of the
meeting branches of these apophyses (Phractaspida), or of twenty perforated plates, produced by
concrescence of their branches (Ceriaspida).
A. Tribe I. Phractaspida, Haeckel.
Definition.—Dorataspida without perforated
plates; the spherical shell is composed only of the meeting branches of the two opposite
apophyses, which arise from each radial spine. Therefore the meshes of the shell are all
sutural.
Genus 349. Phractaspis,[389] Haeckel,
1881, Prodromus, p. 467.
Definition.—Dorataspida without perforated plates; shell
composed only of the meeting branches of the two opposite apophyses, which arise from each radial
spine. Condyles of the branch-ends without by-spines.
The genus Phractaspis is the most simple and primitive form among all
Dorataspida, and may be regarded as the common ancestral form either of this whole family, or at
least of its first subfamily, the Diporaspida. In all members of this subfamily the spherical
shell is composed of twenty radial spines, each of which bears two opposite apophyses; but the
mode of composition is different in the two tribes of the subfamily, in the Phractaspida and
Ceriaspida. In the simpler tribe, the Phractaspida, the shell is composed only of the meeting
branches of the apophyses of neighbouring spines; there are no peculiar perforated plates or
shields. In the Ceriaspida, however, both apophyses of each single spine form a perforated plate
or shield by union of their branches, and the shell is formed of the meeting edges of these
shields. Of course the Ceriaspida must be derived from the simpler Phractaspida.
Phractaspis, as the common ancestral form of both, exhibits a very simple structure of the
shell (Pl. 137,
figs. 1, 2). Commonly, if the fork-branches of each apophysis be not again branched, the shell
possesses only twenty-two large meshes and forty sutures. More rarely their number increases, the
fork-branches of the apophyses being again branched (Phractaspidium, Pl. 137, fig.
3).
Subgenus 1. Phractasparium, Haeckel.
Definition.—Shell with twenty-two meshes, and forty sutures, each
spine with only four branches, its two apophyses being simply forked.
{809}
1. Phractaspis prototypus, n. sp. (Pl. 137, fig.
2).
Radial spines cylindrical, thin, pointed; the outer and inner halves of nearly equal size. Each
spine bears in its middle part two opposite apophyses, which are simply forked; the four condyles
of each spine (or the thickened ends of the fork-branches) are united with the meeting condyles of
the neighbouring spines by sutures. Therefore the network of the spherical shell is composed of
twenty-two large meshes: two square polar meshes on each pole (a a a a and e e e e);
eight triangular circumpolar meshes (each between two polar and one tropical spine, a b a
and e d e); eight tropical rhomboidal meshes (each between one polar, one equatorial, and
two tropical spines: a b c b and e d c d); and four rhomboidal equatorial meshes
(each between two tropical and two equatorial spines c b c d).
Dimensions.—Diameter of the shell 0.1, of the meshes 0.03 to 0.04; breadth of the
spines and bars 0.005.
Habitat.—Cosmopolitan; Mediterranean (Corfu), Atlantic (Stations 348,
354), Indian Ocean (Ceylon), Pacific (Stations 253, 265, 274), &c., surface.
2. Phractaspis complanata, n. sp. (Pl. 137, fig.
1).
Radial spines leaf-shaped, strongly compressed, two-edged, pointed; their outer part longer
than the inner. Each spine with two opposite apophyses which are simply forked, therefore with
four condyles. The network with forty sutures and twenty-two large meshes, as in the foregoing
species. The broad faces of the eight polar spines lie in two meridian planes, of the four
equatorial spines in the equatorial plane, of the eight tropical spines in two planes parallel to
the latter. The planes of the leaf-shaped fork-branches lie in the spherical face.
Dimensions.—Diameter of the shell 0.1, of the meshes 0.02 to 0.03; breadth of the
fork-branches 0.012.
Habitat.—South Atlantic, Station 343, surface.
3. Phractaspis condylophora, n. sp.
Radial spines quadrangular, thin; their outer pyramidal part shorter than the inner. Each spine
with four branches, its two opposite apophyses being simply forked. The eighty condyles (or
sutural ends of the branches) much thickened, twice to four times as broad as the branches
themselves. Network with forty sutures and twenty-two large meshes, as in both foregoing
species.
Dimensions.—Diameter of the shell 0.12, of the meshes 0.03 to 0.04; breadth of the
condyles 0.01.
Habitat.—South Pacific, Station 288, surface.
4. Phractaspis bipennis, Haeckel.
Dorataspis bipennis, Haeckel, 1862, Monogr. d. Radiol., p. 413, Taf. xxi. figs. 1,
2.
Phractasplenium bipenne, Haeckel, 1882, Manuscript.
Radial spines very thin, quadrangular; their outer pyramidal part shorter than the inner. Each
spine with four bent branches, its two opposite apophyses being simply forked. Eighty {810}condyles, very thin, pointed. In the specimens of this
remarkable species, which I first observed in Messina, two opposite equatorial spines had quite
free apophyses, not connected with the neighbouring spines; therefore the thin lattice-work of the
shell exhibited only thirty-six sutures and twenty meshes (two meshes with six sutures, six meshes
with four sutures, and twelve meshes with three sutures). In similar specimens, which I afterwards
observed in the Canary Islands, all four equatorial spines were connected in the same manner with
the neighbouring spines; therefore they possessed forty sutures and twenty-two meshes, like
Phractaspis prototypus (Pl. 137,
fig. 2). Perhaps the Mediterranean species represents a peculiar genus, Phractasplenium
bipenne.
Dimensions.—Diameter of the shell 0.1, of the meshes 0.03 to 0.05; breadth of the
spines 0.002.
Habitat.—Mediterranean (Messina), Canary Islands (Lanzerote),
surface.
Subgenus 2. Phractaspidium, Haeckel.
Definition.—Shell with forty to eighty or more meshes, and eighty to
one hundred or more sutures. Each spine with six to eight or more branches, its two apophyses
being doubly forked or more ramified.
5. Phractaspis constricta, n. sp. (Pl. 137, fig.
3).
Radial spines strongly compressed, two-edged, pointed; their outer half twice constricted and
somewhat longer than the inner half. Each spine with two opposite forked apophyses, the branches
of which are again forked; therefore eight condyles on each spine. The network of the spherical
shell with eighty sutures and sixty-two meshes (twenty-two large primary meshes and forty smaller
secondary meshes, the latter between the distal fork-branches).
Dimensions.—Diameter of the shell 0.11, of the large meshes 0.04 to 0.05, of the
small meshes 0.01; breadth of the spines 0.01.
Habitat.—South Atlantic, Station 348, depth 2450 fathoms.
6. Phractaspis cataphracta, Haeckel.
Acanthometra cataphracta, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p.
49, Taf. x. figs. 7, 8.
Dorataspis cataphracta, Haeckel, 1862, Monogr. d. Radiol., p. 415.
Radial spines thin, quadrangular; their outer pyramidal part shorter than the inner. Each spine
with six to eight condyles, the fork-branches of their two opposite apophyses being (all or
partly) again forked. The network with sixty to eighty sutures and meshes: sometimes as regular as
in the preceding species, at other times more or less irregular.
Dimensions.—Diameter of the shell 0.1, of the meshes 0.02 to 0.04; breadth of the
bars 0.004 to 0.008.
Habitat.—Mediterranean (Cette), Müller (Messina), Haeckel; North
Atlantic, Station 353, surface.
{811}
Genus 350. Pleuraspis,[390] Haeckel,
1881, Prodromus, p. 467.
Definition.—Dorataspida without perforated plates; shell
composed only of the meeting branches of the two opposite apophyses, which arise from each radial
spine. Condyles of the branch ends bearing by-spines.
The genus Pleuraspis has quite the same structure of the shell as the
foregoing Phractaspis, and differs from it only in the development of external by-spines;
commonly each condyle of the branch end of the apophyses bears one zigzag by-spine, which is
directed parallel to the radial main-spine from which the apophyses arise. Therefore each suture
of the shell is armed with two divergent by-spines (Pl. 137, fig.
4).
Subgenus 1. Pleurasparium, Haeckel.
Definition.—Shell regularly developed with twenty-two meshes and forty
sutures (sometimes twenty to twenty-four meshes and thirty-six to forty-eight sutures); each spine
commonly with four branches, its two apophyses being simply forked.
1. Pleuraspis horrida, n. sp. (Pl. 137,
fig. 4).
Radial spines roundish, somewhat compressed, very thick, conical, pointed at both ends, outer
part nearly twice as long as the inner. Two apophyses of each spine simply forked, with short and
broad branches and thickened condyles. Forty sutures, very broad. Twenty-two large meshes three
to four times as broad as the bars. By-spines much developed, as long as the radius, zigzag, with
alternating recurved hooks.
Dimensions.—Diameter of the shell 0.15, of the meshes 0.04 to 0.06; breadth of the
bars 0.01 to 0.015.
Habitat.—Tropical Pacific (east of Philippines), Station 215,
surface.
2. Pleuraspis amphithecta, n. sp.
Radial spines two-edged, leaf-shaped, strongly compressed, pointed at both ends; outer half
longer than the inner. Two apophyses of each spine simply forked, with short and very broad
branches; condyles not thickened. Forty sutures, broad. Twenty-two large meshes twice to three
times as broad as the bars. By-spines zigzag, half as long as the radius. (Resembles
Phractaspis complanata, Pl. 137,
fig. 1.)
Dimensions.—Diameter of the shell 0.12, of the meshes 0.02 to 0.03; bars
0.012.
Habitat.—Central Pacific, Station 274, surface.
{812}
3. Pleuraspis costata, Haeckel.
Acanthometra costata, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 49,
Taf. ii. fig. 1, Taf. x. figs. 4-6.
Dorataspis costata, Haeckel, 1862, Monogr. d. Radiol., p. 414, Taf. xxiii.
fig. 1.
Radial spines roundish, conical, pointed at both ends; outer and inner halves of nearly equal
length. Two apophyses of each spine simply forked, with broad branches and thickened condyles.
Forty sutures, broad. Twenty-two large meshes, four to five times as broad as the bars. By-spines
straight, denticulate, half as long as the radius. (This common species is rather variable in size
and details.)
Dimensions.—Diameter of the shell 0.1 to 0.13, meshes 0.03 to 0.06, bars 0.005 to
0.01.
Habitat.—Cosmopolitan; Mediterranean, Atlantic, Pacific, surface.
4. Pleuraspis pyramidalis, n. sp.
Radial spines quadrangular, their outer part pyramidal, very thick, about half as long as the
prismatic inner part. Two apophyses of each spine simply forked, with very short and broad
branches. Condyles much thickened. Forty sutures, very broad. Twenty-two meshes small, the largest
twice as broad as the bars; the smaller only half as broad. By-spines short, straight,
denticulate, half as long as the radius.
Dimensions.—Diameter of the shell 0.14, meshes 0.005 to 0.03.
Habitat.—Western Tropical Pacific, Station 222, surface.
Subgenus 2. Pleuraspidium, Haeckel.
Definition.—Shell with forty to eighty or more meshes and eighty to
one hundred or more sutures; each spine with six to eight or more branches, its two apophyses
being forked twice or oftener.
5. Pleuraspis ramosa, n. sp.
Radial spines roundish, stout, somewhat compressed; their inner and outer half nearly of equal
length. Two apophyses of each spine doubly forked or more irregularly branched; therefore commonly
eight (sometimes six, seven, nine, or ten to twelve) thickened condyles on each spine. Sutures
eighty to one hundred, broad. Irregular meshes sixty to eighty; three to five times as broad as
the bars. By-spines zigzag, ramified, half as long as the radius.
Dimensions.—Diameter of the shell 0.15, of the meshes 0.007 to 0.014, breadth of
the bars 0.003.
Habitat.—Central Pacific, Station 272, surface.
{813}
B. Tribe II. Ceriaspida, Haeckel.
Definition.—Dorataspida, the spherical shell
of which is composed of twenty perforated plates, produced by union of the branches of the two
opposite apophyses, which arise from each radial spine. Therefore the meshes of the shell are
partly sutural, partly parmal.
Genus 351. Dorataspis,[391] Haeckel,
1860, Monatsber. d. k. preuss. Akad. d. Wiss. Berlin, p. 811.
Definition.—Dorataspida with twenty plates, which are
perforated by forty aspinal pores (two pores in each plate). Surface of the shell without combs,
dimples, and by-spines.
The genus Dorataspis opens the long series of the Ceriaspida, or of those
Dorataspida in which the shell is composed of twenty plates, each of which is perforated by two
primary aspinal pores. This tribe has arisen from the simpler Phractaspida by reunion of the
branches of the apophyses in each single spine. If in Phractaspis, their common ancestral
form (Pl. 137,
figs. 1, 2), the neighbouring fork-branches of both opposite apophyses in each radial spine become
recurved and united, they will produce a shield, which is perforated by two pores and between them
by the spine itself. These two "primary aspinal pores" are characteristic of all Ceriaspida; among
these Dorataspis itself is the most simple form. Its shell exhibits therefore constantly
forty aspinal pores, and besides these a variable number of "sutural pores" (in the sutures
between the twenty plates). If in each suture there be only a single pore, we get on the whole
fifty-two or fifty-four pores; if in each suture there be two or three pores, that number becomes
doubled or tripled.
Subgenus 1. Doratasparium, Haeckel.
Definition.—Shell with fifty-two sutures; four polar plates on each
pole of the main axis meeting in one point (in the pole itself); therefore all eight polar plates
pentagonal and of equal size. Spherical shell therefore composed of four (equatorial) hexagonal
plates and of sixteen pentagonal (eight tropical and eight polar plates).
1. Dorataspis fusigera, n. sp. (Pl. 138,
fig. 2).
Shell with fifty-two sutures and fifty-two sutural meshes, with four hexagonal and sixteen
pentagonal plates. Both aspinal meshes of each plate elliptical, twice to four times as broad as
the {814}sutural meshes. Radial spines in the outer half
fusiform, somewhat longer than the inner cylindrical half.
Dimensions.—Diameter of the shell 0.08 to 0.09, of the parmal meshes 0.1 to 0.2,
of the sutural meshes 0.004 to 0.01.
Habitat.—Central Pacific, Station 271, surface.
2. Dorataspis macracantha, n. sp.
Shell with fifty-two sutures and fifty-two sutural meshes; with four hexagonal and sixteen
pentagonal plates. Both aspinal meshes of each plate circular, small, twice as broad as the small
sutural meshes. Radial spines quadrangular, prismatic, their outer half twice to three times as
long as the inner half.
Dimensions.—Diameter of the shell 0.12, of the parmal pores 0.008, of the sutural
meshes 0.004.
Habitat.—South Pacific, Station 288, surface.
3. Dorataspis macropora, n. sp.
Shell with fifty-two sutures and one hundred to one hundred and fifty sutural meshes, with four
hexagonal and sixteen pentagonal plates. Both aspinal meshes nearly circular, very large, five to
ten times as broad as the small circular sutural meshes; the number of the latter is in this
species multiplied, in each suture being two to three (commonly three) small pores. Radial spines
in the outer half leaf-shaped, compressed, nearly lanceolate, longer than in the inner cylindrical
half.
Dimensions.—Diameter of the shell 0.16, of the parmal pores 0.03 to 0.04, of the
sutural pores 0.004 to 0.006.
Habitat.—Indian Ocean (Madagascar), Rabbe, surface.
Subgenus 2. Dorataspidium, Haeckel.
Definition.—Shell with fifty-four sutures; four polar plates on each
pole of the main axis different in pairs; two major hexagonal meeting in a polar suture (the
"hydrotomical suture"); two minor pentagonal, not meeting together (separated by that hydrotomical
suture). Spherical shell therefore composed of eight hexagonal plates (four equatorial and four
polar) and by twelve pentagonal plates (eight tropical and four polar plates).
4. Dorataspis loricata, Haeckel.
Dorataspis loricata, Haeckel, 1862, Monogr. d. Radiol., p. 415, Taf. xxi.
figs. 3-6.
Shell with fifty-four sutures and fifty-four sutural meshes, with eight hexagonal and twelve
pentagonal plates. Both aspinal meshes of each plate kidney-shaped or roundish, nearly of the
{815}same size as the sutural meshes. Radial spines
cylindrical; their outer half on the pointed distal end two-edged and longer than the inner
half.
Dimensions.—Diameter of the shell 0.14, of its meshes 0.008 to 0.016.
Habitat.—Mediterranean (Messina), North Atlantic (Canary Islands),
Azores, surface.
5. Dorataspis typica, n. sp. (Pl. 138,
figs. 4, 4a).
Shell with fifty-four crest-like sutures and fifty-four circular sutural meshes, with eight
hexagonal and twelve pentagonal plates. Both aspinal meshes of each plate elliptical, twice to
three times as broad as the sutural meshes. Aspinal meshes of the four equatorial plates twice as
broad as those of the sixteen other plates. The meshes are surrounded by elevated and denticulated
crests (incipient spine-sheaths). Radial spines compressed, more or less two-edged; outer pointed
half somewhat larger than the inner half. (In this typical species the composition of the shell
prevailing in the majority of Ceriaspida is very clear.)
Dimensions.—Diameter of the shell 0.15 to 0.18, of the aspinal pores 0.02 to 0.03,
of the sutural pores 0.01.
Habitat.—Central Pacific, Stations 270 to 274, surface and in various
depths.
6. Dorataspis micropora, n. sp. (Pl. 138,
fig. 3).
Shell with fifty-four sutures and fifty-four very small sutural meshes, with eight hexagonal
and twelve pentagonal plates. Both aspinal meshes of each plate elliptical, four to six times as
broad as the sutural meshes. Radial spines in the outer half conical, somewhat shorter than in the
inner cylindrical half.
Dimensions.—Diameter of the shell 0.11, of the aspinal pores 0.01 to 0.02, of the
sutural pores 0.003.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
7. Dorataspis gladiata, n. sp.
Shell with fifty-four sutures and one hundred to one hundred and twenty sutural meshes, with
eight hexagonal and twelve pentagonal plates. Both aspinal meshes elliptical, three times as broad
as the small sutural meshes; commonly two meshes in each suture. Radial spines sword-shaped,
two-edged, tapering from the shell towards the two ends. (Resembles Belonaspis datura, Pl.
139, fig.
9, but differs in the spherical shell, the equal size of the four equatorial spines, and the
absence of by-spines.)
Dimensions.—Diameter of the shell 0.12, of the aspinal pores 0.012, of the sutural
pores 0.004.
Habitat.—Indian Ocean (Madagascar), Rabbe, surface.
8. Dorataspis polypora, n. sp.
Shell with fifty-four sutures and one hundred and sixty to two hundred sutural meshes, with
eight hexagonal and twelve pentagonal plates. Commonly three meshes (sometimes four) on {816}each suture. Both aspinal meshes kidney-shaped, four
times as broad as the small sutural meshes. Radial spines thick, in the inner longer part
cylindrical, in the outer shorter part conical. (Differs from all other species in the
multiplication of the sutural pores.)
Dimensions.—Diameter of the shell 0.2, of the aspinal pores 0.016, of the sutural
pores 0.004.
Habitat.—Equatorial Atlantic, Station 347, surface.
Genus 352. Diporaspis,[392] n. gen.
Definition.—Dorataspida with twenty plates, which are
perforated by forty aspinal pores (two pores in each plate). Surface of the shell without combs
and dimples, but armed with numerous by-spines.
The genus Diporaspis has the same characteristic structure of the shell as
the typical Dorataspis, and differs from it only in the development of numerous by-spines
on the surface. The number of the sutures between the twenty plates is sometimes fifty-two, at
other times fifty-four, and in each suture we find occasionally a single pore, at other times two
or three such pores.
Subgenus 1. Diporasparium.
Definition.—Shell with fifty-two sutures, four polar plates on each
pole of the main axis meeting in one common point; therefore all eight polar plates pentagonal and
of equal size. Shell therefore composed of four (equatorial) hexagonal plates, and of sixteen
pentagonal (eight tropical and eight polar) plates.
1. Diporaspis nephropora, n. sp. (Pl. 134, fig.
15).
Shell with fifty-two sutures and fifty-two sutural pores, with four hexagonal and sixteen
pentagonal plates. Both aspinal pores of each plate kidney-shaped, about twice as broad as the
circular sutural pores. Radial spines thin, cylindrical, longer than the radius. By-spines forked,
only one-third as long as the radius.
Dimensions.—Diameter of the shell 0.13, parmal pores 0.03, sutural pores 0.07.
Habitat.—North Pacific, Station 244, surface.
2. Diporaspis circopora, n. sp.
Shell with fifty-two sutures and one hundred to one hundred and fifty sutural pores, with four
hexagonal and sixteen pentagonal plates. Both aspinal pores of each plate circular, very large,
six to eight times as broad as the small circular sutural pores (in each suture two to three
pores). {817}Radial spines strongly compressed,
two-edged; outer half shorter than the inner. By-spines undulate, half as long as the radius.
Dimensions.—Diameter of the shell 0.12, of the parmal pores 0.02 to 0.03, of the
sutural pores 0.003 to 0.004.
Habitat.—South Pacific, Station 287, surface.
Subgenus 2. Diporaspidium, Haeckel.
Definition.—Shell with fifty-four sutures, four polar plates on each
pole of the main axis different in pairs: two major hexagonal meeting in a polar ("geotomical")
suture, two minor pentagonal, not meeting together (separated by that suture). Shell therefore
composed of eight hexagonal plates (four equatorial and four polar) and of twelve hexagonal plates
(eight tropical and four polar).
3. Diporaspis zygopora, n. sp.
Shell with fifty-four sutures and fifty-four circular sutural pores: with eight hexagonal and
twelve pentagonal plates. Both aspinal pores of each plate elliptical, three times as broad as the
sutural pores. Radial spines compressed, two-edged; outer half shorter than the inner. By-spines
very numerous, simple, one-third as long as the radius, forming coronels or elegant circles around
the pores (a small coronel around each sutural pore, a large one around each couple of aspinal
pores).
This typical species is nearly allied to Dorataspis typica (Pl. 138, fig. 4),
and may be derived from it by development of the coronels of by-spines.
Dimensions.—Diameter of the shell 0.16, aspinal pores 0.03, sutural pores
0.01.
Habitat.—Central Pacific, Station 271, surface
Genus 353. Orophaspis,[393] Haeckel,
1881, Prodromus, p. 468.
Definition.—Dorataspida with twenty plates, which are
perforated by forty aspinal pores (two pores in each plate). Surface of the shell without combs,
dimples, and by-spines. Each radial spine bears outside of the shell two opposite free apophyses,
which are either simple or branched.
The genus Orophaspis differs not only from its ancestral form,
Dorataspis, but from all other Dorataspida in the development of peculiar free apophyses on
the radial spines, outside the shell. These apophyses, two being opposite on each spine, appear as
a repetition of the primary apophyses of Phractaspis; they are either simple or branched,
and sometimes the branches are united together, forming an outer free shield with two or four
pores. These outer plates represent the beginning of a second outer shell and form the transition
to Phractopelta, the ancestral form of the Phractopeltida.
{818}
Subgenus 1. Orophasparium, Haeckel.
Definition.—Free apophyses of the radial spines simple, not
branched.
1. Orophaspis astrolonche, n. sp.
Parmal pores of the shell circular, twice as large as the sutural pores and as the breadth of
the bars. Radial spines very long, three to six times as long as the diameter of the shell,
compressed, two-edged; each with two simple, opposite, triangular apophyses; their distance from
the shell equal to its diameter.
Dimensions.—Diameter of the shell 0.05, parmal pores 0.004, sutural pores
0.002.
Habitat.—North Pacific, Station 239, surface.
2. Orophaspis gladiata, n. sp.
Parmal pores of the shell circular, of the same size as the sutural pores and the bars. Radial
spines thick, spindle-shaped, scarcely longer than the diameter of the shell, each with two simple
opposite conical apophyses; their distance from the shell about equal to its radius.
Dimensions.—Diameter of the shell 0.06, pores 0.003.
Habitat.—Central Pacific, Station 265, surface.
Subgenus 2. Orophaspidium, Haeckel.
Definition.—Free apophyses of the radial spines branched, their
branches free (not anastomosing).
3. Orophaspis furcata, n. sp. (Pl. 133,
fig. 6).
Parmal pores of the shell roundish or elliptical, three times as broad as the sutural pores and
the bars. Radial spines very long, compressed, each with two opposite apophyses, which are simply
forked; their distance from the shell somewhat greater than its diameter.
Dimensions.—Diameter of the shell 0.06 to 0.08, parmal pores 0.006, sutural pores
0.002.
Habitat.—Central Pacific, Station 274, surface.
4. Orophaspis ramosa, n. sp.
Parmal pores of the shell circular, of the same size as the sutural pores and the bars. Radial
spines compressed, very long, each with two opposite apophyses, which are more or less irregularly
branched (commonly between eight and twelve thin branches on each spine); their distance from the
shell smaller than its diameter.
Dimensions.—Diameter of the shell 0.08, pores 0.006.
Habitat.—North-West Pacific, Station 235, surface.
{819}
Subgenus 3. Stegaspis, Haeckel, 1881, Prodromus, p. 468.
Definition.—Free apophyses of the radial spines branched, and by
reunion of their anastomosing branches forming perforated shields.
5. Orophaspis diporaspis, n. sp.
Parmal pores of the shell circular, of the same size as the sutural pores and the bars. Radial
spines sword-shaped, two-edged, very broad; each spine bears an elliptical free shield with two
longish pores (opposite on the two flat sides of the sword). Distance of the twenty free two-pored
shields from the shell about equal to its diameter.
Dimensions.—Diameter of the shell 0.04, of the pores 0.004.
Habitat.—South Pacific, Station 295, surface.
6. Orophaspis tessaraspis, n. sp.
Parmal pores of the shell roundish, somewhat larger than the sutural pores and the bars. Radial
spines very long, compressed; each spine bears a roundish free shield with four irregular pores
disposed in a cruciform manner, the two larger pores being opposite on the flat sides of the
spine, the two smaller being opposite but on its two edges. Distance of the twenty free four-pored
shields from the shell about equal to its radius.
Dimensions.—Diameter of the shell 0.05, of the pores 0.003.
Habitat.—North Pacific, Station 239, surface.
Genus 354. Ceriaspis,[394] Haeckel, 1881,
Prodromus, p. 468.
Definition.—Dorataspida with twenty plates, which are
perforated by forty aspinal pores (two pores in each plate). Surface of the shell with numerous
dimples, separated by a network of elevated combs. No by-spines.
The genus Ceriaspis has the same structure of the shell, as its ancestral
form Dorataspis, differing from it only in the development of high combs or crests, which
form on the surface of the shell a peculiar elevated network. The dimples or funnel-shaped pits
between these combs are either all or partly perforated by the pores of the shell. Both the
aspinal pores of each plate are usually placed in one common dimple, whilst each sutural pore is
placed in its peculiar smaller dimple.
Subgenus 1. Ceriasparium, Haeckel.
Definition.—Shell-surface with seventy to one hundred or more small
funnel-shaped dimples, each of which opens on the bottom by one pore or by a couple of {820}apertures; twenty larger dimples in the centre of the
plates (each with a couple of aspinal pores) and fifty to one hundred or more smaller dimples,
each of which contains one sutural pore. No blind dimples between the perforated dimples.
1. Ceriaspis lacunosa, n. sp.
Shell spherical with seventy-two funnel-shaped dimples, each of which is perforated on the
bottom by one or two apertures; twenty larger dimples in the centre of the plates, each with two
elliptical aspinal pores, and fifty-two smaller sutural dimples between them, each with one
circular pore of half the size. No blind dimples. Radial spines quadrangular, stout; their outer
part shorter than the inner.
Dimensions.—Diameter of the shell 0.1, of the forty parmal pores 0.01, of the
fifty-two sutural pores 0.005.
Habitat.—South Atlantic, Station 330, surface.
2. Ceriaspis scrobiculata, n. sp.
Shell spherical, with seventy-four funnel-shaped dimples, each of which is perforated on the
bottom by one or two apertures; twenty larger dimples in the centre of the plates, each with two
kidney-shaped large pores, and fifty-four smaller dimples on the sutures, each with one circular
pore of one-fourth of the size of the reniform pores. No blind dimples. Radial spines cylindrical,
the outer part longer than the inner.
Dimensions.—Diameter of the shell 0.12, of the forty parmal pores 0.016, of the
fifty-four sutural pores 0.004.
Habitat.—Central Pacific, Station 270, surface.
Subgenus 2. Ceriaspidium, Haeckel.
Definition.—Shell-surface with funnel-shaped dimples (commonly one
hundred and seventy-six or one hundred and eighty-two), which on the bottom are partly closed,
partly perforated by one aperture (or by a pair of pores). The blind dimples are placed on the
corners of the twenty plates, and are therefore either one hundred and four or one hundred and
eight; if there be no polar suture, the blind dimples are one hundred and four (twenty-four on the
four hexagonal equatorial plates, forty on the eight pentagonal tropical plates, and forty on the
eight pentagonal polar plates); if, however, there be a polar suture on both main poles, the
number of blind dimples is one hundred and eight (twenty-four on the four hexagonal equatorial and
twenty-four on the four hexagonal polar plates, two opposite on each pole; forty on the eight
pentagonal tropical plates and twenty on the four pentagonal polar plates, two opposite on each
pole). Between the blind dimples there are usually seventy-two to seventy-four perforated {821}dimples, twenty larger parmal dimples (enclosing a radial
spine and a couple of aspinal pores) and fifty-two to fifty-four sutural dimples (sometimes one
hundred or more), each of which encloses one sutural pore.
3. Ceriaspis inermis, n. sp. (Pl. 138,
fig. 5).
Shell spherical, with one hundred and seventy-six funnel-shaped dimples, one hundred and four
of which are blind and seventy-two perforated; of the latter, each of the fifty-two smaller
contains a single sutural pore, each of the twenty larger a couple of aspinal pores. The
elliptical or kidney-shaped aspinal pores are about twice as broad as the circular sutural pores.
Radial spines thin, compressed, two-edged; their outer prolongation very short, rudimentary,
scarcely higher than the axis of the surrounding funnel, and projecting but slightly from its
aperture.
Dimensions.—Diameter of the shell 0.15, of the forty parmal pores 0.016, of the
fifty-two sutural pores 0.008.
Habitat.—South Pacific, Station 289, surface.
4. Ceriaspis favosa, n. sp. (Pl. 138,
fig. 6).
Shell polyhedral (icosahedral), with one hundred and eighty-two funnel-shaped dimples, one
hundred and eight of which are blind and seventy-four perforated; of the latter, each of the
fifty-four smaller contains a single sutural pore, each of the twenty larger a couple of aspinal
pores. All pores roundish, nearly of the same size. Radial spines thin, compressed, two-edged;
their outer part somewhat longer than the inner.
Dimensions.—Diameter of the shell 0.12 to 0.13, of the pores 0.01 to 0.015.
Habitat.—Central Pacific, Station 274, surface.
5. Ceriaspis icosahedra, n. sp.
Shell polyhedral (icosahedral), with one hundred and eighty-two funnel-shaped dimples, one
hundred and eight of which are blind and seventy-four perforated; of the latter, each of the
fifty-four smaller contains one small sutural pore, each of the twenty larger a couple of aspinal
pores. The kidney-shaped aspinal pores are very large, four to six times as broad as the small
circular sutural pores. Radial spines strong, quadrangular, their outer part from two to three
times as long as the inner part.
Dimensions.—Diameter of the shell 0.16, of the parmal pores 0.015 to 0.02, of the
sutural pores 0.003 to 0.004.
Habitat.—Tropical Atlantic, Station 349, surface.
6. Ceriaspis cicatricosa, n. sp.
Shell spherical, with two hundred and fifty to three hundred (or more) small funnel-shaped
dimples, the majority of which are blind, the minority perforated by pores; of the latter forty
{822}are aspinal pores (two at the base of each spine),
the others sutural pores. All pores nearly of the same size. As the shell of this species is very
dark and thick-walled, it was impossible to obtain more information with regard to the pores.
Radial spines very stout; the outer conical part shorter than the inner cylindrical part.
Dimensions.—Diameter of the shell 0.2, of the pores 0.006 to 0.008.
Habitat.—North Pacific, Station 236, surface.
Genus 355. Hystrichaspis,[395] n.
gen.
Definition.—Dorataspida with twenty plates, which are
perforated by forty aspinal pores (two pores in each plate). Surface of the shell with numerous
dimples, separated by a network of elevated combs. Numerous by-spines.
The genus Hystrichaspis has the same structure with regard to the shell, as
its ancestral form Ceriaspis, and differs from the latter only in possessing numerous
by-spines. Moreover, in the former the dimples of the shell-surface are either all perforated by
pores, or some of them are blind.
Subgenus 1. Hystrichasparium, Haeckel.
Definition.—Shell with seventy to one hundred or more funnel-shaped
dimples, each of which opens on the bottom by one aperture or by a couple of pores; twenty larger
dimples in the centre of the plates (each with a couple of pores, sometimes also three couples)
and fifty to one hundred or more smaller dimples, each of which contains one sutural pore. No
blind dimples between the perforated dimples.
1. Hystrichaspis pectinata, n. sp. (Pl. 138, fig.
8).
Shell with numerous (one hundred to two hundred?) funnel-shaped dimples, each of which is
pierced at the bottom by one or two pores; twenty larger dimples in the centre of the plates (each
with two aspinal pores), and fifty to one hundred and fifty or more smaller dimples (each with one
sutural pore). No blind dimples. Crests between the dimples armed with a continuous series of
simple by-spines. Radial main spines stout, in the inner half cylindrical, in the outer conical,
more or less compressed.
Dimensions.—Diameter of the shell 0.13, of the pores 0.004 to 0.006.
Habitat.—Tropical Atlantic, Station 352, surface.
2. Hystrichaspis furcata, n. sp. (Pl. 138, fig.
9).
Shell with numerous (one hundred to two hundred?) funnel-shaped dimples, each of which is
pierced at the bottom by one or two pores; twenty larger dimples in the centre of the plates (each
{823}with two large aspinal pores), and fifty to one
hundred and fifty or more smaller dimples (each with one small sutural pore). No blind dimples.
Crests between the dimples armed with forked by-spines. Radial main spines stout, leaf-shaped,
tapering towards both ends.
Dimensions.—Diameter of the shell 0.12, parmal pores 0.01, sutural pores
0.005.
Habitat.—Central Pacific, Station 276, surface.
3. Hystrichaspis cristata, n. sp. (Pl. 138, fig.
11).
Siphonasphis cristata, Haeckel, 1882, Manuscript.
Shell with numerous (one hundred to two hundred?) funnel-shaped dimples, each of which is
pierced at the bottom by one or more pores. Twenty larger dimples in the centre of the plates are
pierced by the radial main-spines; among these fourteen contain each a couple of aspinal pores;
six others are much larger, and contain each six larger pores; these six plates are two opposite
equatorial plates and four polar plates, placed in the same meridian plane (the "hydrotomical
plane"); in each of these six "hydrotomical dimples" two pores are placed opposite to one another
on the two edges of the leaf-shaped spine, four others being opposite in pairs on both flat sides
of it. By this peculiar structure this species connects the true Hystrichaspis with
Hexalaspis and Diploconus; however the twenty spines are of equal length, and the
shell continues to be spherical. The twenty radial main-spines are leaf-like and compressed. The
crests between the dimples are dentated by a series of small by-spines.
Dimensions.—Diameter of the shell 0.15, of the aspinal pores 0.01, of the sutural
pores 0.005.
Habitat.—North Pacific, Station 240, surface.
Subgenus 2. Hystrichaspidium, Haeckel.
Definition.—Shell-surface with numerous funnel-shaped dimples
(commonly one hundred and seventy-six to one hundred and eighty-two), which on the bottom are
partly closed, partly perforated by one aperture (or by a pair of pores). The blind dimples are
situated on the corners of the twenty plates; their number is commonly one hundred and four or one
hundred and eight, sometimes more. The perforated dimples, alternating with the former, are
usually seventy-two to seventy-four, sometimes more; twenty larger parmal dimples (each with a
couple of aspinal pores, sometimes also with three such couples) and fifty-two to fifty-four
sutural dimples, sometimes one hundred or more (each with one sutural pore). (Compare the
definition of Ceriaspidium, p. 820.)
4. Hystrichaspis dorsata, n. sp. (Pl. 138, fig.
10).
Shell with one hundred and seventy-six funnel-shaped dimples, one hundred and four of which are
blind and seventy-two perforated; of the latter, each of the fifty-two smaller contains a single
{824}sutural pore, each of the twenty larger a couple of
aspinal pores; the elliptical aspinal pores are twice as broad as the circular sutural pores. The
crests between the dimples are armed with simple scattered short by-spines; usually in the nodal
points, where three combs meet, there exist three or two divergent by-spines. The twenty radial
main-spines are thin and long, compressed.
Dimensions.—Diameter of the shell 0.22, of the aspinal pores 0.02, of the sutural
pores 0.01.
Habitat.—Central Pacific, Station 271, surface.
5. Hystrichaspis armata, n. sp.
Shell with one hundred and seventy-six funnel-shaped dimples, one hundred and four of which are
blind and seventy-two perforated; of the latter, each of the fifty-two smaller contains a single
sutural pore, each of the twenty larger a couple of aspinal pores. Crests between the dimples
comb-like, armed with a series of simple by-spines. Twenty main-spines very stout, in the inner
part cylindrical, in the outer shorter part conical.
Dimensions.—Diameter of the shell 0.25, of the aspinal pores 0.02, of the sutural
pores 0.01.
Habitat.—South-East Pacific, Station 300, surface.
6. Hystrichaspis sulcata, n. sp.
Shell with one hundred and eighty-two funnel-shaped dimples, one hundred and eight of which are
blind and seventy-four perforated; of the latter, each of the fifty-four smaller contains a single
sutural pore, each of the twenty larger a couple of aspinal pores. All pores nearly of the same
size, very small. Crests between the dimples very high, sulcated, on the free edge serrate with
short very numerous, simple by-spines. Radial main-spines very stout and short, conical.
Dimensions.—Diameter of the shell 0.2, of the pores 0.003.
Habitat.—South Atlantic, Station 325, surface.
7. Hystrichaspis foveolata, n. sp.
Shell with very numerous (three hundred to four hundred?) small and deep funnel-shaped dimples,
the majority of which are blind, the minority perforated; among the latter there are forty larger
aspinal pores; the other smaller (scarcely half as large) pores are sutural. Crests between the
dimples densely armed with simple short by-spines. Radial main-spines stout, leaf-shaped.
Dimensions.—Diameter of the shell 0.12, of the aspinal pores 0.01, of the sutural
pores 0.004.
Habitat.—Tropical Atlantic, Station 338, surface.
8. Hystrichaspis divaricata, n. sp.
Shell with numerous (two hundred to three hundred?) funnel-shaped dimples, the majority of
which are blind, the minority perforated; among the latter there are forty larger aspinal pores.
{825}Crests between the dimples armed with long forked
by-spines with divergent fork-branches. Radial spines cylindrical, very long and thick, longer
than the diameter of the shell.
Dimensions.—Diameter of the shell 0.14, aspinal pores 0.08, sutural pores
0.03.
Habitat.—Central Pacific, Station 266, surface.
9. Hystrichaspis fruticata, n. sp. (Pl. 138, fig,
7).
Shell with numerous (one hundred and fifty to two hundred?) funnel-shaped dimples, the majority
of which are blind, the minority perforated; forty aspinal pores elliptical, of the same size as
the circular sutural pores. Crests between the dimples bearing elegant arborescent by-spines.
Twenty radial main-spines long and thin, cylindrical or a little compressed.
Dimensions.—Diameter of the shell 0.17, pores 0.01.
Habitat.—Tropical Pacific, Station 275, surface.
10. Hystrichaspis serrata, n. sp.
Shell with numerous (two hundred to three hundred?) funnel-shaped dimples, the majority of
which are blind, the minority perforated; forty aspinal pores of the same size as the sutural
pores. Crests between the dimples covered with denticulated by-spines. Twenty radial main-spines
compressed, two-edged, with serrated edges, about as long as the diameter of the shell.
Dimensions.—Diameter of the shell 0.21, pores 0.012.
Habitat.—North Pacific, Station 238, surface.
Genus 356. Coscinaspis,[396] n. gen.
Definition.—Doratispida with twenty plates, which are
perforated by eighty to two hundred or more parmal pores (two aspinal and two to ten or more
coronal pores in each plate). Surface without by-spines.
The genus Coscinaspis, together with the following nearly allied genus
Acontaspis, may be separated from the other Ceriaspida as a peculiar tribe,
Coscinaspida. This tribe is characterised by the larger number of the parmal pores. Whilst
in all other Ceriaspida this number is constantly forty (only two pores in each plate), here it
may be from eighty to two hundred or more; in each shield the two primary "aspinal pores" are
surrounded by a circle of two to ten or more (commonly eight to twelve) "coronal pores." The
number of sutural pores in this group is also usually larger.
{826}
Subgenus 1. Coscinasparium, Haeckel.
Definition.—Plates of the shell smooth, without crests, not
dimply.
1. Coscinaspis peripora, n. sp. (Pl. 138,
fig. 1).
Shell thin walled, with smooth surface, without crests and dimples, perforated by pores of very
different sizes. Forty aspinal pores, roundish or elliptical, of about the same size as the fifty
or sixty violin-shaped sutural pores, and three to four times as broad as the small roundish
coronal pores, which are irregularly formed and distributed, four to eight occurring on each plate
(altogether one hundred to one hundred and twenty). Radial spines roundish, somewhat compressed;
inner and outer half nearly of the same length.
Dimensions.—Diameter of the shell 0.12, aspinal pores 0.01 to 0.012, coronal pores
0.003.
Habitat.—North-West Pacific, Station 235, surface.
2. Coscinaspis stigmopora, n. sp.
Shell very thin walled, with smooth surface, without crests and dimples, perforated by pores of
very different sizes. Forty aspinal pores, roundish, of about the same size as the fifty or sixty
violin-shaped sutural pores, their diameter being about one-fifth of that of the plates. Coronal
pores very numerous (two hundred to four hundred) and very small, irregularly scattered, ten to
twenty occurring in each plate. Sutures strongly denticulated. Radial spines very thin and short,
cylindrical.
Dimensions.—Diameter of the shell 0.12 to 0.15, aspinal and sutural pores 0.01,
coronal pores 0.001 to 0.003.
Habitat.—North Pacific, Station 257, surface.
3. Coscinaspis rhacopora, n. sp.
Shell very thin walled, with smooth surface, without crests and dimples, perforated by very
irregular pores of very different sizes and forms; commonly more or less lobed or sinuate. Forty
aspinal pores and fifty to eighty sutural pores, larger than the numerous (one hundred to two
hundred) irregularly scattered coronal pores. Sutures very sinuate. Radial spines cylindrical,
thin and long.
Dimensions.—Diameter of the shell 0.15 to 0.18, aspinal and sutural pores 0.01 to 0.02,
coronal pores 0.001 to 0.005.
Habitat.—Central Pacific, Station 274.
4. Coscinaspis coscinopora, n. sp.
Shell thin walled, with smooth surface, without crests and dimples, perforated by very numerous
pores of circular form, but of different sizes. Forty aspinal pores and one hundred to two hundred
{827}sutural pores much larger than the coronal pores,
which are very small and very numerous (fifty to sixty on each plate). Sutures sinuate. Radial
spines compressed, outer and inner half nearly of equal length.
Dimensions.—Diameter of the shell 0.3, aspinal and sutural pores 0.01 to 0.015,
coronal pores 0.001 to 0.002.
Habitat.—Tropical Atlantic, Station 338, surface.
5. Coscinaspsis polypora, n. sp. (Pl. 136, fig.
8).
Shell very thin walled, with smooth surface, without crests and dimples, the two aspinal pores
of each plate narrow, lanceolate, ten to twelve times as long as broad and half as long as the
radius of each plate. Coronal pores irregular, polygonal, very numerous (two hundred to three
hundred on each plate), commonly arranged more or less regularly in ten to twelve series parallel
to the longitudinal diameter of the aspinal spines (ten to twenty pores in each series). Sutural
pores irregular, polygonal, very numerous. Radial spines very thin and long, cylindrical.
Dimensions.—Diameter of the shell 0.24; length of the aspinal pores 0.04, breadth
0.004; coronal and sutural pores 0.002 to 0.01.
Habitat.—Tropical Atlantic, Station 349, surface.
6. Coscinaspis orthopora, n. sp.
Shell very thin walled, with smooth surface, without crest and dimples. All pores of nearly
equal size and form; quadrangular, mostly rectangular; one hundred to two hundred, separated by
two systems of parallel bars, perpendicular one to another, occur in each plate. Sutural pores
mostly triangular. Radial spines very thin and long, cylindrical, somewhat compressed. (Similar to
those of Phatnaspis lacunaria,
Pl. 136,
fig. 9, but spherical, not ellipsoidal.)
Dimensions.—Diameter of the shell 0.2, pores 0.008 to 0.012.
Habitat.—Central Pacific, Station 272, surface.
7. Coscinaspis parmipora, n. sp. (Pl. 137, fig.
9).
Craniaspis parmipora, Haeckel, 1866, Manuscript.
Dorataspis parmipora, Haeckel, 1881, Prodromus, Atlas.
Shell thin walled, with smooth surface, without crests and dimples. There are no sutural pores,
since all twenty plates are connected by perfect sinuate sutures (therefore this excellent species
may perhaps better represent a peculiar genus, called by me in 1866 Craniaspis). All pores
are parmal pores; each plate with two elliptical aspinal pores, which are twice to five times
broader than the numerous, roundish irregularly scattered coronal pores (thirty to fifty occurring
on each plate). The radial spines are quite internal, that is, not prolonged on the outside of the
shell; in this respect they resemble those of Sphærocapsa.
Dimensions.—Diameter of the shell 0.16, aspinal pores 0.01, coronal pores 0.002 to
0.005.
Habitat.—North Atlantic, Canary Islands (Lanzerote), surface.
{828}
8. Coscinaspis isopora, n. sp. (Pl. 134,
figs. 13, 14).
Shell thick walled, with smooth surface, without crests and dimples. All pores of the shell
nearly of the same size and of similar form, about as broad as the bars between them, and about
two hundred in number, viz., fifty to sixty (regularly fifty-two or fifty-four) sutural pores and
one hundred and forty to one hundred and fifty parmal pores (forty aspinal kidney-shaped pores,
and one hundred to one hundred and ten coronal circular pores: six in the angles of each
equatorial plate, and five in the angles of each tropical and polar plate; if the disposition be
quite regular, one hundred and four or one hundred and eight). Radial spines short, rudimentary,
conical.
Dimensions.—Diameter of the shell 0.2, of the pores and bars 0.012.
Habitat.—Tropical Pacific, Station 218 (off New Guinea), surface.
Subgenus 2. Coscinaspidium, Haeckel.
Definition.—Surface of the shell dimply, with a network of prominent
crests.
9. Coscinaspis ceriopora, n. sp.
Shell thick-walled, with numerous (one hundred and sixty to two hundred?) funnel-shaped
dimples, which are separated by prominent crests; on the bottom of each dimple there is a simple
or double circular pore. If this species be quite regularly developed, it closely resembles the
preceding, differing from it mainly in the prominent combs of the surface. It resembles also
Ceriaspis favosa, Pl. 138,
fig. 6; but whilst in this latter the majority of the dimples are blind, here they are all
perforated. The twenty aspinal dimples (in the centre of each plate) present at the bottom a
couple of pores, all other dimples a single pore. Among the latter there are fifty to sixty
sutural pores and one hundred to one hundred and ten coronal pores, viz., six in each equatorial
plate, and five in each of the other plates; but the number is not quite constant. Radial spines
strong, in the inner part cylindrical, in the outer conical.
Dimensions.—Diameter of the shell 0.15, of the pores and bars 0.01.
Habitat.—Tropical Pacific, Station 215, surface.
Genus 357. Acontaspis,[397] Haeckel,
1881, Prodromus, p. 468.
Definition.—Dorataspida with twenty plates, which are
perforated by eighty to two hundred or more parmal pores (in each plate two aspinal and two to ten
or more coronal pores). Surface covered with by-spines.
The genus Acontaspis has the same characteristic structure of the shell as
Ceriaspis, differing from it only in the presence of numerous by-spines. Each plate is
perforated by four to sixteen or more (commonly ten to twelve) parmal pores, the two central of
which are primary "aspinal pores," all the others being secondary "coronal pores."
{829}
Subgenus 1. Acontasparium, Haeckel.
Definition.—Plates of the shell not dimply, without prominent
crests.
1. Acontaspis lanceolata, n. sp.
Shell thin walled, even, without crests and dimples between them, perforated by about three
hundred pores of different sizes: forty aspinal pores elliptical, about as large as the irregular
(fifty to sixty) sutural pores, and two to four times as broad as the small circular coronal pores
(eight to twelve being on each plate, altogether about two hundred). Between the pores numerous
short conical by-spines. Radial main spines lanceolate, about as long as the radius. (Similar to
Coscinaspis peripora, Pl. 138,
fig. 1, but with broad lanceolate spines and numerous short by-spines.)
Dimensions.—Diameter of the shell 0.15, aspinal and sutural pores 0.012 to 0.015,
coronal pores 0.004 to 0.008.
Habitat.—South Atlantic, Station 325, surface.
Subgenus 2. Acontaspidium, Haeckel.
Definition.—Surface of the shell dimply, with a network of prominent
crests.
2. Acontaspis hastata, n. sp. (Pl. 134,
fig. 16).
Shell thick walled, with numerous (one hundred and seventy to one hundred and ninety) dimples,
which are separated by an elevated network of prominent crests. In the centre of each plate a
larger dimple with a couple of aspinal pores, surrounded by a corona of ten or twelve smaller
dimples, each of which contains a single pore (a coronal and sutural alternately). All pores
circular or roundish, nearly of the same size. If the shell be quite regularly developed, there
are fifty to sixty sutural pores and one hundred to one hundred and ten coronal pores (six in each
equatorial plate, five in each of the other plates). The knobs of the meeting crests are conical,
and bear each a simple short by-spine. Radial main spines compressed, at the distal end
spear-shaped, with a rhomboidal plate below the apex, about as long as the radius.
Dimensions.—Diameter of the shell 0.18, of the pores and bars 0.01.
Habitat.—Western Tropical Pacific (off the Philippines), Station
215.
3. Acontaspis furcata, n. sp.
Shell thick walled, dimply, with a network of crests, of the same composition as in the
foregoing species, with one hundred and seventy to one hundred and ninety dimples (twenty aspinal,
one hundred to one hundred and ten coronal and fifty to sixty sutural dimples). All dimples and
pores nearly of the same size. Each node of the crested network bears a stout by-spine, which is
forked {830}on the base, with two divergent straight
branches (similar to those of Hystrichaspis furcata, Pl. 138, fig. 9).
Radial main spines two-edged, sword-like, nearly as long as the diameter of the shell.
Dimensions.—Diameter of the shell 0.13, of the pores 0.008.
Habitat.—Central Pacific, Station 272, surface.
4. Acontaspis capillata, n. sp.
Shell very dark and thick walled, non-transparent, with very numerous (three hundred to four
hundred or more?) deep funnel-shaped dimples, each of which is perforated by a small circular pore
(forty aspinal, two hundred to three hundred coronal, and fifty to one hundred sutural pores?).
The high crests between the dimples bear very numerous simple by-spines, nearly half as long as
the radius, so that the shell appears covered with hairs. Radial main spines very long and thin,
cylindrical, longer than the diameter of the shell.
Dimensions.—Diameter of the shell 0.2, of the pores 0.012.
Habitat.—North Pacific, Station 244, surface.
Subfamily 2. Tessaraspida, Haeckel.
Definition.—Dorataspida with twenty radial
spines, each of which bears four crossed apophyses (opposite in pairs). The spherical shell is
composed either of the meeting branches of these apophyses (Stauraspida), or of twenty perforated
plates, produced by concrescence of their branches (Lychnaspida).
A. Tribe II. Stauraspida, Haeckel, 1881, Prodromus, p.
467.
Definition.—Dorataspida with spherical shell,
which is composed either of the meeting branches of the four crossed apophyses only, or exhibits
four to twelve perforated plates which are produced by the crossed apophyses of four to twelve
radial spines (but never of all twenty spines). Each plate bears four crossed pores.
Genus 358. Stauraspis,[398] Haeckel,
1881, Prodromus, p. 467.
Definition.—Dorataspida without perforated plates; shell
composed only of the meeting branches of the four crossed apophyses, which arise (opposite in
pairs) from each radial spine. Condyles of the branch-ends without by-spines.
The genus Stauraspis is the most simple and primitive form among all
Tessaraspida, or that subfamily of Dorataspida, in which the shell is composed of twenty {831}radial spines, each of which bears four crossed
apophyses. The subfamily may be divided into two different tribes, the Stauraspida and
Lychnaspida. In the Stauraspida either all twenty spines, or a part of them, bear no perforated
plates, and the shell is composed wholly or partially of the meeting branches of their apophyses.
In the Lychnaspida, however, the four apophyses of each single spine form, by reunion of their
recurved branches, a plate or shield with four crossed aspinal pores. The Lychnaspida represent
therefore a more developed stage in the shell-formation than the simpler Stauraspida.
Stauraspis, as the common ancestral form of both, may be derived phylogenetically from
Xiphacantha or Stauracantha, which differ only by the apophyses or branches of the
apophyses not meeting. These branches (originally eight on each spine) are either simple or again
branched.
Subgenus 1. Staurasparium, Haeckel.
Definition.—Apophyses of the spines simple, not branched; therefore
each spine with four sutural condyles.
1. Stauraspis cruciata, n. sp. (Pl. 134,
fig. 5).
Radial spines thin, quadrangular, prismatic; outer and inner half nearly of equal length.
Central bases pyramidal, with wing-like edges. Four apophyses of each spine simple, not branched,
with thin condyles. Large meshes of the shell ten to twenty times as broad as the bars. This and
the following species greatly resemble the simplest forms of Phractaspis (Pl. 137, figs. 1,
2); they differ from these, however, by the equal size and distance of the four branches of each
spine, which thus form a rectangular cross.
Dimensions.—Diameter of the shell 0.1; breadth of the spines and bars 0.002.
Habitat.—Central Pacific, Station 268, surface.
2. Stauraspis xiphacantha, n. sp.
Radial spines stout, cylindrical in the inner half, conical in the shorter outer half. Four
apophyses of each spine simple, not branched, broad, with thick condyles. Meshes of the shell six
to eight times as broad as the bars.
Dimensions.—Diameter of the shell 0.12; breadth of the spines and bars 0.008 to
0.01.
Habitat.—South Pacific, Station 290, surface.
Subgenus 2. Stauraspidium, Haeckel.
Definition.—Apophyses of the spines branched; therefore each spine
with eight to twenty or more sutural condyles.
{832}
3. Stauraspis furcata, n. sp.
Radial spines thin, quadrangular, prismatic; outer and inner halves nearly of equal length. The
four apophyses of each spine simply forked (or partly with bifid fork-branches); each spine with
eight to twelve sutural condyles. Meshes of the shell ten to twenty times as broad as the
bars.
Dimensions.—Diameter of the shell 0.15; breadth of the spines and bars 0.003.
Habitat.—Central Pacific, Station 266, surface.
4. Stauraspis stauracantha, n. sp. (Pl. 137, figs. 5,
6).
Radial spines thin, in the inner longer half cylindrical, in the outer half conical with
thickened base. Four apophyses of each spine doubly forked or dichotomously (more or less
irregularly) branched; each spine with sixteen to twenty-four sutural condyles. Meshes of the
shell of very different sizes and forms; the largest ten to fifteen, the smallest two to three
times as broad as the bars.
Dimensions.—Diameter of the shell 0.14; breadth of of the spines 0.002 to 0.01, of
the bars 0.002.
Habitat.—Tropical Atlantic, Station 343, surface.
Genus 359. Echinaspis,[399] Haeckel,
1881, Prodromus, p. 467.
Definition—Dorataspida without perforated plates; shell
composed only of the meeting branches of the four crossed apophyses, which arise (opposite in
pairs) from each radial spine. Condyles of the branch-ends bearing by-spines.
The genus Echinaspis exhibits the same structure of the shell as its
ancestral form Stauraspis, and differs from it only in the development of by-spines on the
sutural condyli.
1. Echinaspis dichotoma, n. sp.
Radial spines cylindrical, thin, outer half longer than the inner. Four apophyses of each spine
simply forked (or partly with bifid fork-branches); therefore each spine usually possesses eight
(sometimes ten to twelve) sutural condyles. Meshes of the shell ten to twelve times as broad as
the bars. Each condyle bears a zigzag by-spine, half as long as the radius.
Dimensions.—Diameter of the shell 0.12, of the spines 0.004 to 0.006.
Habitat.—North Pacific, Station 253, surface.
{833}
2. Echinaspis diadema, n. sp.
Radial spines thin and long, quadrangular, prismatic; outer half two to three times as long as
the inner. Four apophyses of each spine simply forked (or partly with bifid fork-branches); each
spine with eight to twelve sutural condyles. Meshes of the shell fifteen to twenty-five times as
broad as the bars. Each condyle bears a simple bristle-shaped by-spine, nearly as long as the
radius.
Dimensions.—Diameter of the shell 0.11; breadth of the spines and bars 0.002.
Habitat.—South Atlantic, Station 333, surface.
3. Echinaspis echinoides, n. sp. (Pl. 137, figs.
7,8).
Radial spines roundish, in the outer half conical, and two to three times as broad as in the
inner half. Four apophyses of each spine doubly forked or dichotomously (more or less irregularly)
branched; each spine with sixteen to twenty-four sutural condyles. Meshes of the shell four to
eight times as broad as the bars. Each condyle bears a zigzag by-spine, one-third as long as the
radius.
Dimensions.—Diameter of the shell 0.15; breadth of the spines 0.004 to 0.01, of
the bars 0.002.
Habitat.—Central Pacific, Station 266, surface.
Genus 360. Zonaspis,[400] n. gen.
Definition.—Dorataspida with four plates, each of which
is perforated by four crossed aspinal pores; these four plates are formed by the united branches
of the other spines (eight polar and eight tropical) form no lattice-plates by union. Each condyle
bears a by-spine.
The genus Zonaspis differs from all other Dorataspida in the peculiar
formation of the four equatorial spines, which form by union of the recurved branches of their
apophyses four perforated plates (each with four crossed pores), whilst the branches of the
sixteen other plates do not unite to form plates, but simply meet the branches of the neighboring
spines.
1. Zonaspis fragilis, n. sp.
Radial spines very thin and long, cylindrical. Four meshes of each equatorial plate pentagonal,
ten to twelve times as broad as the bars. By-spines zig-zag, nearly as long as the radius.
Dimensions.—Diameter of the shell 0.16, of the parmal pores 0.018.
Habitat.—South Atlantic, Station 332, surface.
{834}
2. Zonaspis cingulata, n. sp. (Pl. 134,
figs. 3, 4).
Radial spines cylindrical in the inner half, with thickened pyramidal bases (fig. 3), conical
in the outer half; both halves of equal length. The four meshes of each equatorial plate
egg-shaped, four to six times as broad as the bars. By-spines zigzag, half as long as the
radius.
Dimensions.—Diameter of the shell 0.15, of the parmal meshes 0.015.
Habitat.—Tropical Pacific (east coast off Philippines), Station 275,
surface.
3. Zonaspis æquatorialis, n. sp. (Pl. 135, fig.
5).
Radial spines stout, cylindrical in the shorter inner half, conical in the longer outer half.
The four meshes of each equatorial plate circular, only twice as broad as the bars. By-spines
zigzag, nearly as long as the radius.
Dimensions.—Diameter of the shell 0.11, of the parmal meshes 0.008.
Habitat.—Equatorial Atlantic, Station 347, surface.
Genus 361. Dodecaspis,[401] n. gen.
Definition.—Dorataspida with twelve plates, each of which
is perforated by four crossed aspinal pores; these twelve plates are formed by the united branches
of the apophyses of four equatorial and eight polar spines. The branches of the apophyses of the
eight tropical spines form no lattice-plates by union. Each condyle bears a by-spine.
The genus Dodecaspis differs from all other Dorataspida in the peculiar
composition of its shell; twelve spines (four equatorial and eight polar spines) form by union of
the branches of their apophyses twelve plates, each of which exhibits four crossed pores, whilst
the eight other (tropical) spines form no perforated plates, but simply unite with the
neighbouring spines by meeting branches. Some irregularity is often to be found in this genus;
instead of four pores in each plate there may be two closed, the other two which are opposite
being open. Several times I observed a peculiar "hemihedral" variety: only six spines of one
meridian plane (the two equatorial and the four polar spines of the "hydrotomical" plane)
exhibited complete shields, whilst the branches of the other fourteen plates remained open. This
"hemihedral" form may perhaps represent a peculiar genus, Hemiaspis.
1. Dodecaspis tricinata, n. sp. (Pl. 134,
fig. 1).
Radial spines thin and long, cylindrical. The four meshes of each equatorial and each polar
plate of equal size, pentagonal, about eight times as broad as the bars. By-spines zigzag, nearly
as long as the radius. In this species the hemihedral variety, Hemiaspis, often occurs.
Dimensions.—Diameter of the shell 0.15, of the parmal pores 0.016.
Habitat.—South-east Pacific (west coast of Patagonia), Station 302,
surface.
{835}
2. Dodecaspis trizonia, n. sp.
Radial spines thin, cylindrical in the longer inner half, conical in the shorter outer half.
The four meshes of each equatorial and each polar plate of equal size, roundish or nearly
circular, five to six times as broad as the bars. By-spines straight, denticulated, scarcely half
as long as the radius.
Dimensions.—Diameter of the shell 0.12, of the parmal pores 0.012.
Habitat.—South-west Pacific (east coast of New Zealand), Station 169,
surface.
B. Tribe II. Lychnaspida, Haeckel, 1881, Prodromus, p.
467.
Definition.—Dorataspida with twenty
perforated plates or fenestrated shields (each plate at least with four pores), produced by union
of the branches of the four crossed apophyses, which arise, opposite in pairs, from each radial
spine. The spherical shell is composed of the twenty plates united by sutures (rarely by
concrescence).
Genus 362. Tessaraspis,[402] Haeckel,
1881, Prodromus, p. 468.
Definition.—Dorataspida with twenty plates, which are
perforated by eighty aspinal pores (four crossed pores in each plate). Surface smooth, without
by-spines.
The genus Tessaraspis introduces the series of Lychnaspida, which comprise
all those Dorataspida in which the shell is composed of twenty plates, each of which is perforated
by four primary aspinal pores. In Tessaraspis and Lychnaspis each plate exhibits
only these four primary pores, whilst in Icosaspis and Hylaspis they become
surrounded by a circle of secondary or coronal pores. If in Stauraspis, the common
ancestral form of the Tessaraspida, the four crossed apophyses of each single radial spine became
recurved and united together, we should have the typical plate of Tessaraspis, in which the
piercing radial spine is surrounded by four crossed pores of equal size. The number of sutural
pores, between the neighbouring plates, is variable; usually each plate is surrounded by a circle
of eight to twelve sutural pores. The sutures between the meeting condyles of the apophyses
usually remain open; but in some species they become obliterated (subgenus
Tessaraspidium).
Subgenus 1. Tessarasparium, Haeckel.
Definition.—Condyles of the neighbouring plates connected by permanent
open sutures; therefore the whole shell is composed of twenty separated pieces of acanthin.
{836}
1. Tessaraspis arachnoides, n. sp. (Pl. 136, fig.
1).
Parmal pores pentagonal, ten to twenty times as broad as the thin thread-like bars, on an
average of about the same size as the irregular sutural meshes; the majority of the latter are
either triangular or hexagonal. Radial spines very thin and long, cylindrical, their outer part
two to four times as long as the inner. As the insertion of the spines is on the highest point of
the plates, the shell becomes polyhedral (dodecahedral?).
Dimensions.—Diameter of the shell 0.15 to 0.17, of the parmal meshes 0.02 to
0.025, sutural meshes 0.01 to 0.03, bars 0.002.
Habitat.—Equatorial Atlantic, Station 347, surface.
2. Tessaraspis pentagonalis, n. sp.
Parmal meshes pentagonal, three to four times as broad as the thick bars, and on an average
smaller than the irregular polygonal sutural meshes. Radial spines stout, quadrangular; their
outer pyramidal part shorter than the inner prismatic part.
Dimensions.—Diameter of the shell 0.12, of the parmal pores 0.01 to 0.012, bars
0.003.
Habitat.—Central Pacific, Station 269, surface.
3. Tessaraspis tetragonalis, n. sp.
Parmal meshes tetragonal, or nearly square, six to eight times as broad as the thin bars, and
on an average larger than the irregular polygonal sutural meshes. Radial spines thin,
quadrangular, prismatic, their outer part longer than their inner.
Dimensions.—Diameter of the shell 0.16, of the parmal pores 0.012 to 0.016, bars
0.002.
Habitat.—South Pacific, Station 288, surface.
4. Tessaraspis hexagonalis, n. sp.
Parmal meshes hexagonal, ten to twelve times as broad as the thin bars, and on an average
larger than the polygonal sutural meshes (the two proximal sides of each hexagonal parmal mesh two
to three times as long as the four distal sides). Radial spines cylindrical, thin; their outer
part longer than the inner.
Dimensions.—Diameter of the shell 0.15, of the parmal pores 0.01 to 0.015, bars
0.001.
Habitat.—North Pacific, Station 254, surface.
5. Tessaraspis trigonalis, n. sp.
Parmal meshes triangular, three to four times as broad as the thick bars, and on an average
smaller than the irregular sutural meshes (in each plate all four parmal meshes of the same size,
formed like an isosceles triangle, the distal base of which is somewhat curved, and convex towards
{837}the periphery, whilst the two proximal sides are
straight). Radial spines thick, rounded; their outer conical part shorter than the inner
cylindrical part.
Dimensions.—Diameter of the shell 0.1, of the parmal pores 0.01, of the sutural
pores 0.01 to 0.02.
Habitat.—Tropical Atlantic (near Ascension), Station 345, depth 2010
fathoms.
6. Tessaraspis circularis, n. sp.
Parmal meshes circular, all eighty of equal size, quite regular, five to six times as broad as
the thick bars, on an average larger than the roundish irregular sutural meshes. Radial spines
quadrangular, prismatic, the outer part longer than the inner.
Dimensions.—Diameter of the shell 0.18, parmal pores 0.015 to 0.018, bars
0.003.
Habitat.—South-east Pacific, Station 300, surface.
7. Tessaraspis micropora, n. sp.
Parmal meshes circular, very small, all eighty of the same size, regular, scarcely as broad as
the separating thick bars, and much smaller than the large irregular sutural meshes. Radial spines
rounded, their outer conical part about as long as the inner cylindrical part.
Dimensions.—Diameter of the shell 0.08, of the parmal pores 0.003, of the sutural
pores 0.005 to 0.015, bars 0.004.
Habitat.—Western Tropical Pacific, Station 225, surface.
8. Tessaraspis diodon, Haeckel.
Dorataspis diodon, Haeckel, 1862, Monogr. d. Radiol., p. 417, Taf. xxii.
figs. 1-5.
Parmal meshes roundish, for the most part nearly circular, three to four times as broad as the
bars, and on an average smaller than the roundish sutural meshes. Radial spines in the inner part
cylindrical; the outer part very short (only one-third or one-fourth of the radius), divided
completely into two parallel conical pointed teeth by a deep incision.
Dimensions.—Diameter of the shell 0.12, of the parmal pores 0.01 to 0.012, of the
sutural pores 0.01 to 0.03, bars 0.004.
Habitat.—Mediterranean (Messina), surface.
9. Tessaraspis quadriforis, n. sp.
Parmal meshes irregular, roundish, of nearly equal size, four to six times as broad as the
bars, and on an average of the same size as the roundish sutural meshes. Radial spines
quadrangular, prismatic in the inner part, which is somewhat longer than the conical outer
part.
Dimensions.—Diameter of the shell 0.16, of the pores 0.012 to 0.018, bars
0.003.
Habitat.—North Atlantic, Canary and Cape Verde Islands, Station 353,
surface.
{838}
10. Tessaraspis irregularis, n. sp.
Parmal meshes irregular, roundish, of unequal size, three to six times as broad as the bars,
and on an average larger than the irregular sutural meshes. Radial spines cylindrical, the outer
part longer than the inner part.
Dimensions.—Diameter of the shell 0.13, parmal pores 0.006 to 0.012, sutural pores
0.005 to 0.01; bars 0.002.
Habitat.—Central Pacific, Station 270, surface.
Subgenus 2. Tessaraspidium, Haeckel.
Definition.—Condyles of the neighbouring plates grown together and
sutures obliterated; therefore the whole shell forms a single piece of acanthin.
11. Tessaraspis quadrata, n. sp.
Tessaraspidium quadratum, Haeckel, 1882, Manuscript.
Parmal meshes square, four times as broad as the thin bars, on an average of the same size as
the polygonal sutural meshes. Radial spines stout, tetrapterous, prismatic, in the outer part
longer than in the inner. Sutures of the shell completely obliterated, therefore the whole shell
forms one piece.
Dimensions.—Diameter of the shell 0.16, of the parmal pores 0.012, sutural pores
0.01 to 0.015.
Habitat.—South Atlantic, Station 332, depth 2200 fathoms.
12. Tessaraspis rotunda, n. sp.
Parmal meshes circular, of equal size, twice as broad as the thick bars, on an average smaller
than the roundish sutural meshes. Radial spines cylindrical in the inner part, which is somewhat
longer than the outer conical part. Sutures of the shell completely obliterated, therefore the
whole shell forms one piece.
Dimensions.—Diameter of the shell 0.12, parmal pores 0.008, sutural pores 0.005 to
0.015.
Habitat.—North Atlantic, Station 354, surface.
13. Tessaraspis concreta, n. sp. (Pl. 136 fig.
5).
Parmal meshes irregular, polygonal or roundish, six to eight times as broad as the narrow and
high bars, much larger than the irregular sutural meshes. Radial spines leaf-shaped, compressed,
two-edged, pointed at both ends; inner and outer part nearly of equal size. Shell very thick
walled; meshes therefore funnel-shaped; sutures completely obliterated, therefore the whole shell
forms a single piece.
Dimensions.—Diameter of the shell 0.09, parmal pores 0.02, sutural pores 0.01.
Habitat.—Central Pacific, Station 274, depth 2750 fathoms.
{839}
Genus 363. Lychnaspis,[403] Haeckel,
1862, Prodromus, p. 468.
Definition.—Dorataspida with twenty plates, which are
perforated by eighty aspinal pores (four crossed pores in each plate). Surface covered with
numerous by-spines.
The genus Lychnaspis, the largest and most common of all Dorataspida,
exhibits the same structure of the shell as its ancestral form Tessaraspis, and differs
from it only in the development of by-spines on the sutural condyles. Many species of this genus
are very widely distributed, and appear in large numbers, and some of them are amongst the most
graceful and elegant of the Radiolaria.
Subgenus 1. Lychnasparium, Haeckel.
Definition.—Condyles of the neighbouring plates connected by permanent
open sutures; therefore the whole shell is composed of twenty separate pieces of acanthin.
1. Lychnaspis giltschii, n. sp. (Pl. 135,
fig. 3).
Parmal meshes pentagonal, about ten or twelve times as broad as the thin bars, on an average of
the same size as the irregular polygonal sutural meshes. By-spines (two hundred to three hundred)
very delicate, half as long as the radius, barbed, and zigzag. Radial main-spines very thin and
long, straight, cylindrical; their outer part longer than the inner part.
Dimensions.—Diameter of the shell 0.2, of the parmal pores 0.02, sutural pores
0.01 to 0.03, bars 0.002.
Habitat.—Tropical Atlantic, Stations 338 to 348, surface.
2. Lychnaspis capillaris, n. sp.
Parmal meshes pentagonal, twenty to thirty times as broad as the very thin thread-like bars, on
an average of the same size as the irregular polygonal sutural meshes. By-spines (two hundred to
two hundred and fifty) very delicate, zigzag, with very small denticles, one-third as long as the
radius. Radial main-spines very thin and long, cylindrical, more or less undulated.
Dimensions.—Diameter of the shell 0.25, parmal pores 0.025, sutural pores 0.02 to
0.04, bars 0.001.
Habitat.—North Pacific, Station 250, surface.
3. Lychnaspis maxima, n. sp.
Parmal meshes pentagonal, six to eight times as broad as the thick bars, for the most part
larger than the irregular sutural meshes. By-spines (four hundred to five hundred) nearly as {840}long as the radius, zigzag, with short denticles. Radial
spines very long and stout, quadrangularly-prismatic, with four smooth edges.
Dimensions.—Diameter of the shell 0.3, parmal pores 0.03, sutural pores 0.01 to
0.025, bars 0.004.
Habitat.—Central Pacific, Stations 271 to 274, surface.
4. Lychnaspis serrata, n. sp.
Parmal meshes pentagonal, three to four times as broad as the thick bars, smaller than the
irregular sutural meshes. By-spines (two hundred to three hundred) scarcely one-third as long as
the radius, zigzag, with strong denticles. Radial spines very long, stout,
quadrangularly-prismatic, with four serrated edges.
Dimensions.—Diameter of the shell 0.22, parmal pores 0.015, sutural pores 0.012 to
0.03, bars 0.004.
Habitat.—South Atlantic, Station 330, surface.
5. Lychnaspis wagenschieberi, n. sp.
Parmal meshes tetragonal, nearly rhombic, three to four times as broad as the thick bars,
smaller than the large irregular sutural meshes. By-spines (about two hundred) large, as long as
the radius, with long recurved denticles, very zigzag. Radial main-spines very long, four-sided
prismatic. This species differs by the form and size of the spines, and by the square form of the
parmal pores from the similar Lychnaspis polyancistra, of which the late excellent engraver
Wagenschieber, of Berlin, has given such a beautiful figure in my monograph (Taf. xxi. fig.
8).
Dimensions.—Diameter of the shell 0.16, parmal pores 0.014, sutural pores 0.015 to
0.02, bars 0.004.
Habitat.—Central Pacific, Station 266, surface.
6. Lychnaspis polyancistra, Haeckel.
Dorataspis polyancistra, Haeckel, 1862, Monogr. d. Radiol., p. 418, Taf.
xxi. figs. 7-9.
Parmal meshes roundish, tetragonal, or nearly square, two to three times as broad as the thick
bars, smaller than the irregular sutural meshes. By-spines (about two hundred) thin, half as long
as the radius, zigzag, with blunt denticles. Radial main-spines stout, in the inner half
cylindrical, in the outer half (of the same length) conical, pointed.
Dimensions.—Diameter of the shell 0.12, parmal pores 0.012, sutural pores 0.01 to
0.04, bars 0.004.
Habitat.—Mediterranean (Messina), surface.
{841}
7. Lychnaspis rottenburgii, n. sp. (Pl. 135, fig.
4).
Parmal meshes roundish, about twice as broad as the thick bars, and for the most part smaller
than the irregular sutural meshes. By-spines (about two hundred to two hundred and fifty) thin,
zigzag, half as long as the radius. Radial main-spines very strong, cylindrical in the inner half,
in the outer half much thicker and conical, pointed. I call this species in honour of my learned
friend, the great patron of zoological studies, Mr. Paul Rottenburg, of Glasgow.
Dimensions.—Diameter of the shell 0.15, parmal pores 0.012, sutural pores 0.01 to
0.03, bars 0.006.
Habitat.—Central Pacific, Station 270, depth 2925 fathoms.
8. Lychnaspis undulata, n. sp. (Pl. 135,
fig. 2).
Parmal meshes circular, twice as broad as the thick bars, on an average of the same size as the
irregular sutural meshes. By-spines (one hundred and fifty to two hundred) very thin, barbed and
zigzag, as long as the radius. Radial main-spines cylindrical; their outer pointed part longer
than the inner part.
Dimensions.—Diameter of the shell 0.1, parmal pores 0.01, sutural pores 0.01, bars
0.005.
Habitat.—Tropical Atlantic, Station 343, surface.
9. Lychnaspis longissima, n. sp. (Pl. 134, fig.
6).
Parmal meshes circular, very small, of the same breadth as the thick bars, much smaller than
the irregular sutural meshes. By-spines (one hundred to one hundred and ten) very long and thin,
zigzag, twice as long as the diameter of the shell. Radial main-spines very long and strong,
cylindrical (at the base quadrangular, pyramidal), four to six times as long as the diameter of
the shell, and one fourth as thick as its radius.
Dimensions.—Diameter of the shell 0.08, parmal pores 0.002, sutural pores 0.01 to
0.015, bars 0.003; length of the radial spines 0.3 to 0.5.
Habitat.—Tropical Pacific (Philippines), Stations 200 to 215,
surface.
10. Lychnaspis minima, n. sp. (Pl. 134,
figs. 2, 7, 8).
Parmal meshes circular, very small, only half as broad as the thick bars, much smaller than the
irregular sutural meshes. By-spines (one hundred to one hundred and ten) half as long as the
radius, zigzag. Radial main-spines thick, in the inner part cylindrical, in the outer shorter part
conical, of very variable size.
Dimensions.—Diameter of the shell 0.05, parmal pores 0.002, sutural pores 0.006 to
0.012, bars 0.004.
Habitat.—Antarctic Ocean, Station 154, surface.
{842}
Subgenus 2. Lychnaspidium, Haeckel.
Definition.—Condyles of the neighbouring plates grown together and
sutures obliterated, therefore the whole shell forms a single piece of acanthin.
11. Lychnaspis echinoides, Haeckel.
Haliomma echinoides, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p.36, Taf.
v. figs. 3, 4.
Haliommatidium echinoides, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p.
22.
Haliommatidium echinoides, Haeckel, 1862, Monogr. d. Radiol., p. 422.
Parmal meshes pentagonal or somewhat roundish, four times as broad as the bars, and of about
the same size as the polygonal meshes. By-spines (about two hundred) short, zigzag. Radial spines
thin; their outer conical part shorter than the inner cylindrical part. Sutures perfectly
obliterated, but recognisable by the characteristic pair of divergent by-spines. (Some recent
observations on this species, made during 1880 in Portofino, have convinced me that the
interpretation of it given in my Monograph, 1862, loc. cit., was quite correct.)
Haliomma ligurinum, J. Müller (=
Haliommatidium ligurinum, Haeckel, L. N. 16, p. 423) seems to be
closely allied to the preceding.
Dimensions.—Diameter of the shell 0.12 to 0.14, parmal pores 0.015, sutural pores
0.01 to 0.02, bars 0.004.
Habitat.—Mediterranean, Nice, Saint Tropez (J. Müller); Portofino near
Genoa (Haeckel).
12. Lychnaspis haliommidium, n. sp.
Lychnaspidium haliommidium, Haeckel, 1882, Manuscript.
Parmal meshes circular, twice as broad as the bars, smaller than the irregular sutural meshes.
By-spines (about two hundred) barbed and zigzag, as long as the radius. Radial main-spines
four-sided; their outer pyramidal part shorter than the inner prismatic part. Sutures perfectly
obliterated.
Dimensions.—Diameter of the shell 0.1, parmal pores 0.01, sutural meshes 0.015 to
0.02.
Habitat.—South Atlantic, Station 335, depth 1425 fathoms.
13. Lychnaspis rabbeana, n. sp.
Parmal meshes circular, very small, of the same breadth as the bars, and much smaller than the
irregular sutural meshes. By-spines (about one hundred) very long and thin, zigzag, about as long
as the diameter of the shell. Radial main-spines cylindrical, thick, twice to three times as long
as the diameter of the shell. Sutures perfectly obliterated, with thickened condyles. Named in
honour of Captain Henrik Rabbe (of Bremen), to whom I am indebted for many new Indian and Atlantic
Radiolaria.
Dimensions.—Diameter of the shell 0.086, parmal pores 0.002, sutural pores 0.012,
bars 0.002.
Habitat.—Indian Ocean (Madagascar), Rabbe, surface.
{843}
14. Lychnaspis cataplasta, n. sp.
Parmal pores very small, circular, half as broad as the bars, and much smaller than the
irregular sutural pores. By-spines zigzag, as long as the diameter of the shell. Radial
main-spines very thin and long, needle-shaped, cylindrical, five to six times as long as the
diameter of the shell. Sutures perfectly obliterated. (This stunted species is one of the smallest
of the Dorataspida.)
Dimensions.—Diameter of the shell 0.05, parmal pores 0.0015, sutural pores 0.01,
bars 0.002.
Habitat.—Antarctic Ocean (off Kerguelen Island), Station 149,
surface.
Genus 364. Icosaspis,[404] Haeckel, 1881,
Prodromus, p. 468.
Definition.—Dorataspida with twenty plates, which are
perforated by one hundred and sixty to three hundred or more parmal pores (in each plate four
crossed aspinal pores, and around them four to twelve or more coronal pores). Surface without
by-spines.
The genus Icosaspis and the closely allied Hylaspis differ from all
other Tessaraspida in the increased number of the parmal pores. Whilst this number in all other
genera is eighty (only four crossed pores in each plate), here it amounts to one hundred and sixty
to three hundred or more (sometimes more than a thousand); in each shield four primary, crossed
"aspinal pores" being surrounded by a circle of four to twelve or more "coronal pores." The number
of sutural pores in these two genera is also increased.
Subgenus 1. Icosasparium, Haeckel.
Definition.—Condyles of the neighboring plates connected by permanent
open sutures; therefore the whole shell composed of twenty separated pieces of acanthin.
1. Icosaspis tabulata, n. sp. (Pl. 136,
fig. 2).
Parmal meshes all of nearly equal size and form, square, four times as broad as the bars,
little larger than the triangular or polygonal sutural meshes. In each plate fifty to seventy
(regularly sixty-four) quadrangular pores, viz., four primary square aspinal meshes, forming
together a regular square surrounded by two to three coronas of rectangular (not quite regular)
coronal meshes (six to eight in each transverse row). Radial spines tetrapterous, prismatic, with
four thin and broad wings, from which arise the crossed bars between the four primary pores. Outer
part of the spines longer than the inner. Commonly the condyles of the plates are only contiguous;
sometimes they grow together, and this form approaches Icosaspis tetragonopa.
Dimensions.—Diameter of the shell 0.25 to 0.3, of the pores 0.02, bars 0.005.
Habitat.—North Pacific, Station 244, depth 2900 fathoms.
{844}
2. Icosaspis elegans, n. sp. (Pl. 136,
fig. 4; Pl. 134,
fig. 9).
Tessaraspis elegans, Haeckel, 1882, Manuscript et Atlas.
Parmal meshes of very different size and form; in the centre of each plate a cross of four
primary, pear-shaped "aspinal pores" (the largest of all); between them four secondary, little
smaller, crossed, egg-shaped "angular pores"; around this rosette of eight larger meshes an inner
complete circle of sixteen to twenty polygonal coronal pores, and an outer incomplete circle of
thirty to forty very small marginal pores. The latter are smaller than the irregular sutural
meshes, which are constricted in the middle, about forty to sixty around each plate. Radial spines
thin, cylindrical, or a little compressed; their outer part longer than the inner. In this elegant
and very common species the condyles usually remain separated by sutures; but sometimes the latter
become obliterated, and the whole shell then forms a single piece, Icosaspidium
elegans.
Dimensions.—Diameter of the shell 0.2 to 0.3, commonly 0.25; larger parmal pores
0.02, smaller 0.001 to 0.003;
sutural pores 0.005 to 0.015; bars 0.003.
Habitat.—Tropical and Subtropical Atlantic, Canary Islands to
Ascension Island, Stations 340 to 354, surface.
3. Icosaspis cruciata, n. sp. (Pl. 134,
fig. 10).
Parmal meshes very different; in the centre of each plate a cross of four primary, nearly
oblong, rectangular "aspinal pores"; between these four secondary, triangular, egg-shaped "angular
pores" (the largest of all), and around this rosette a single circle of twelve to twenty-four
small "coronal pores." The latter are of about the same size as the irregular sutural meshes, of
which there are twenty to thirty around each plate. Radial spines thin, cylindrical, or a little
compressed; their outer part longer than the inner.
Dimensions.—Diameter of the shell 0.2 to 0.3, commonly 0.25; larger parmal pores
0.025, smaller 0.005 to 0.01; bars 0.005.
Habitat.—Tropical and Subtropical Pacific, Sandwich to Marquesas
Islands, Stations 256 to 274, surface.
4. Icosaspis ornata, n. sp.
Parmal meshes very different; in the centre of each plate a cross of four primary octagonal
aspinal pores (the largest of all); between them four secondary, rhombic angular pores, and around
this rosette a circle of twelve to sixteen smaller, polyhedral coronal pores, which are however
larger than the irregular sutural pores (surrounding each plate to the number of twenty to
thirty). Radial spines thin, quadrangular, prismatic; the outer part longer than the inner.
Dimensions.—Diameter of the shell 0.25; larger parmal pores 0.03, smaller 0.01;
sutural pores 0.004 to 0.008; bars 0.006.
Habitat.—South Pacific, Station 295, depth 1500 fathoms.
{845}
5. Icosaspis spectabilis, n. sp.
Parmal meshes very different; in the centre of each plate four very large, pentagonal aspinal
pores, and around these two to three circles of smaller polygonal coronal pores, which are very
numerous, and not larger than the small sutural pores. Radial spines quadrangular, prismatic,
stout, very long; the outer part two to three times as long as the inner.
Dimensions.—Diameter of the shell 0.4, of the larger aspinal meshes 0.03, of the
outer meshes 0.002 to 0.02, bars 0.005.
Habitat.—South Atlantic, Station 333, surface.
6. Icosaspis multiforis, n. sp.
Parmal meshes very numerous, more than one hundred in each plate; in the centre four larger
pear-shaped pores, and around these four to five circles of smaller pores, gradually smaller
towards the margin of the plate; the sutural meshes also very small and numerous (more than fifty
around each plate), so that the number of all the pores together amounts to two thousand or even
more. Radial spines thin, cylindrical, very long.
Dimensions.—Diameter of the shell 0.32; larger aspinal pores 0.02, smaller 0.002
to 0.01; bars 0.003.
Habitat.—Indian Ocean (Madagascar), Rabbe, surface.
Subgenus 2. Icosaspidium, Haeckel.
Definition.—Condyles of the neighbouring plates grown together, and
sutures obliterated; therefore the whole shell forms a single piece of acanthin.
7. Icosaspis tetragonopa, Haeckel.
Haliommatidium tetragonopum, Haeckel, 1862, Monogr. d. Radiol., p.421,
Taf. xxii. fig. 13.
Parmal meshes all of nearly equal size and form, square, three times as broad as the bars,
little larger than the sutural meshes. In each plate commonly sixteen equal square meshes, viz.,
four primary aspinal and twelve secondary, surrounding the former as a square corona. Radial
spines tetrapterous, stout; the outer pyramidal half somewhat longer than the inner. This species
differs from the similar Icosaspis tabulata (Pl. 136, fig. 2)
in the concrescence of the sutures, the smaller number of pores, and the form of the stouter
spines. The figure in my Monograph, drawn from a broken fragment, is not quite correct.
Dimensions.—Diameter of the shell 0.18, pores 0.009, bars 0.003.
Habitat.—Mediterranean (Messina, Corfu), surface.
8. Icosaspis icosahedra, n. sp.
Parmal meshes of different size and form; in the centre of each plate a cross of four
pentagonal, primary aspinal pores, surrounded by a complete corona of twelve to sixteen polygonal
coronal {846}pores and an incomplete corona of thirty to
forty very small marginal pores; the latter are not to be distinguished from the pores of the
obliterated sutures. Radial spines quadrangular, the outer pyramidal part shorter than the inner
prismatic part. Condyles grown together. As the plates are quite even, the shell becomes
icosahedral.
Dimensions.—Diameter of the shell 0.16, pores 0.002 to 0.02, bars 0.005.
Habitat.—North Pacific, Station 241, surface.
9. Icosaspis icosastaura, n. sp. (Pl. 136, fig.
3).
Tessaraspis icosastaura, Haeckel, 1882, Manuscript et Atlas.
Parmal plates of different size and form; in the centre of each plate a cross of four larger
primary, pyriform aspinal pores; between these four smaller roundish angular pores, and around
this rosette a circle of ten to twenty (commonly sixteen) coronal pores, little larger than the
very small sutural pores. Radial spines very thin and long, cylindrical or bristle-shaped.
Condyles grown together; no suture visible.
Dimensions.—Diameter of the shell 0.14; larger pores of the cross 0.01, smaller
pores 0.002 to 0.008; bars 0.002 to 0.004.
Habitat.—Central Pacific, Station 268, surface.
Genus 365. Hylaspis,[405] n. gen.
Definition.—Dorataspida with twenty plates, which are
perforated by one hundred and sixty to three hundred or more parmal pores (in each plate four
crossed aspinal pores, and around them four to twelve or more coronal pores). Surface covered with
numerous by-spines.
The genus Hylaspis exhibits the same structure of the shell as the nearly
allied ancestral genus Icosaspis, and differs from it only in the development of by-spines.
Some species of these two genera exhibit the highest degree of complication in the structure of
the shell seen among the Dorataspida.
1. Hylaspis serrulata, n. sp. (Pl. 135,
fig. 1).
Parmal meshes four hundred to five hundred; in the centre of each plate a cross of four very
large pentagonal or roundish aspinal pores, and around this a circle of sixteen to twenty much
smaller irregular, polygonal, coronal pores; the latter of about the same size as the sutural
pores. On each condyle one thin zigzag-shaped by-spine, nearly as long as the radius. Twenty
radial spines very long, quadrangular, prismatic; on the inside thinner and smooth, on the outside
thickened, and armed with four rows of recurved teeth, serrated.
Dimensions.—Diameter of the shell 0.18, aspinal spines 0.02, other pores 0.002 to
0.01, bars 0.003.
Habitat.—South Atlantic, Station 326, surface.
{847}
2. Hylaspis coronata, n. sp.
Parmal meshes five hundred to six hundred; in the centre of each plate a cross of four long
rectangular aspinal pores, between these four larger egg-shaped angular pores; around this rosette
a circle of sixteen to twenty much smaller, irregular, roundish, coronal pores; the latter about
of the same size as the sutural pores. On each condyle one bearded by-spine about one-third or
one-fourth as long as the radius. Twenty radial spines, very long, smooth, quadrangular,
prismatic.
Dimensions.—Diameter of the shell 0.25, aspinal pores 0.02, other pores 0.005 to
0.01, bars 0.004.
Habitat.—Central Pacific, Station 271, surface.
3. Hylaspis barbata, n. sp.
Parmal meshes twelve hundred to sixteen hundred; in the centre of each plate a cross of four
large, somewhat oblong, octahedral aspinal pores, between these four rhombic smaller angular
pores; around this rosette an inner circle of twelve to sixteen larger and an outer circle of
fifty to sixty very small coronal pores; the latter smaller than the sutural pores. On each
condyle one bristle-shaped zigzag by-spine, with recurved thin hooks, half as long as the radius.
Twenty radial spines, very long, cylindrical, smooth.
Dimensions.—Diameter of the shell 0.25 to 0.3, aspinal pores 0.022, other pores
0.002 to 0.015, bars 0.002.
Habitat.—South Pacific, Station 295, surface.
Family XLI. Phractopeltida, Haeckel (Pl. 133, figs.
1-6).
Phractopeltida, Haeckel, 1881, Prodromus, p. 468.
Definition.—Acantharia with double spherical
lattice-shell, composed of the branched apophyses of twenty radial spines meeting in its centre,
and disposed according to the Müllerian law of Icosacantha. Central capsule spherical, enclosing
the inner and surrounded by the outer concentric shell.
The family Phractopeltida differs from all other Acantharia in the development of a double spherical shell, composed of two
concentric lattice-spheres, which are united by twenty radial spines meeting in the common centre.
We could therefore oppose the Phractopeltida as Diplophracta to all other Acanthophracta as Haplophracta (with simple shell). The former exhibit a
relation to the latter, similar to that exhibited by the Dyosphærida to the simple Monosphærida
among the Sphæroidea.
In my Monograph (1862, p. 423) I described only one genus appertaining to this family,
Aspidomma. I founded it upon the singular Phractopelta, described by {848}J. Müller as Haliomma hystrix. A second species of
Aspidomma, the Acanthometra mucronata of J. Müller, was probably an
Astrolonche. At that time I placed Aspidomma among the Haliommatida, led by the
erroneous opinion that it might represent a transition-form between Dorataspis and
Haliomma. But I afterwards gave up this view, as I was convinced that there is no true
phylogenetic connection between the acanthinic Dorataspida (Actipylea) and the siliceous
Haliommatida (Peripylea). Therefore in my Prodromus (1881, p. 468) I placed
Aspidomma among the Dorataspida and changed its name to Phractopelta, to avoid
further confusion with the unrelated Ommatida (Sphæroidea). It formed
there, with three nearly related genera, the "subfamily Phractopeltida," which we now advance to
the higher rank of a separate family. (By a typographical mistake the words are printed in the
Prodromus Phractopelma and Phractopelmida, &c., instead of Phractopelta and
Phractopeltida, &c.). The detection of other new species appertaining to this family, and a
closer anatomical investigation of them, has now led to the distinction of five different genera,
characterised by other differences than were employed in 1881 in the provisional system of the
"Prodromus."
The two concentric spherical lattice-shells of the Phractopeltida, connected by radial beams,
correspond perfectly to those of the double-shelled Dyosphærida (Haliomma,
Diplosphæra, &c.), and in both cases we may call the smaller inner the "medullary
shell," and the larger outer the "cortical shell." There is no doubt that the double-shelled
Phractopeltida must be derived phylogenetically from the simple-shelled Dorataspida (just as we
derive the double Dyosphærida from the simple Monosphærida). But it is not yet possible to decide
positively which of the two shells is the first formed. Probably the small inner or medullary
shell of the Phractopeltida is the first formed, and corresponds to the simple spherical
lattice-shell of the Dorataspida; and the larger outer or cortical shell of the former is a later
new formation, absent in the latter family. This opinion seems to be confirmed by the genus
Orophaspis, the only form among the Dorataspida, in which the radial spines outside the
shell bear free latticed apophyses. If these twenty apophyses grow further and meet one another,
the second or outer shell of Phractopelta may be formed. But some objections may be raised
to this opinion from the peculiar structure and the very small size of the inner shell; and there
is some possibility that this latter is a secondary later product inside of the primary cortical
shell. The probably phylogenetic series which reveals the origin of the Phractopeltida is the
following:—Acanthometron, Zygacantha, Lithophyllium,
Phractacantha, Doracantha, Dorataspis, Orophaspis,
Phractopelta.
The twenty radial spines exhibit in all Phractopeltida the same characteristic position and
relation as in all other Icosacantha, and are constantly arranged according to the Müllerian law
in four meridian planes, their distal ends falling into five parallel zones. Their distinction in
the majority of the Phractopeltida is not difficult, {849}since the spines of the different zones bear apophyses of different
shapes. Sometimes the four equatorial spines are stouter than the sixteen other spines, and often
the eight tropical spines are somewhat different in form from the eight polar and from the four
equatorial spines. The length of all twenty spines is commonly equal. Their form is usually more
or less compressed, two-edged (as in Zygacantha), more rarely cylindrical (as in
Acanthometron), or somewhat quadrangular (but not truly prismatic); therefore the
transverse section of the spines is commonly elliptical or lanceolate, rarely circular or rhombic,
never square; this seems to indicate their origin from Zygacantha. As in all Acantharia, the spines consist of acanthin, not of silex. Their central ends are
either perfectly grown together, and form a single star of acanthin, or the triangular faces of
their small pyramidal bases are supported one upon another, without true concrescence.
The apophyses of the radial spines, by which the two concentric spherical shells are formed,
seem to be constantly four on each spine, two being opposite in each shell. The proximal pair of
opposite apophyses, forming the inner or medullary shell, is constantly much smaller than the
distal pair composing the outer or cortical shell (Pl. 133, fig. 5).
The proximal pair corresponds probably to the two primary apophyses of the Diporaspida
(Phractaspis, Dorataspis, &c.), whilst the distal pair corresponds to the free
apophyses of Orophaspis (Pl. 133,
fig. 6). Therefore the Phractopeltida may be derived phylogenetically from the Diporaspida (not
from the Tessaraspida). In the common ancestral genus of this family, Phractopelta, the
free part of the radial spines (outside the outer shell) is quite simple, without free apophyses;
in all other genera of the family that free part of the spines (either in all twenty spines or
only in some of them) bears a third pair of lateral apophyses; these may be either simple or
branched or even latticed; but the outer apophyses (of the third rank) remain constantly free, and
a third lattice-shell is never formed by union of their edges (Pl. 133, figs.
2-4).
The inner lattice-shell of the Phractopeltida, or their "medullary shell," is constantly very
small (commonly 0.03 to 0.05 mm. in diameter, rarely more). Its structure is difficult to make
out; in the unbroken shell it is concealed by the dense network of the outer shell; in the broken
shell it is commonly destroyed. Usually the pores of the inner shell are very small, circular or
subcircular, scarcely as broad as the small separating bars. In the majority of Phractopeltida
their number seems to be about forty, being probably the forty primary aspinal pores of the
Diporaspida; in some species this number seems to be exceeded, so that perhaps some sutural pores
may exist between the aspinal pores; but commonly the twenty plates composing the inner shell
(each with two aspinal pores) seems to grow together perfectly by their meeting edges, so that
there are no sutural pores between them. Evident sutures were not recognisable in the inner shell
of any Phractopeltida.
{850}
The outer lattice-shell of the Phractopeltida, or their "cortical shell," is at least twice as
broad, commonly about three times as broad, as the enclosed inner shell; it is much more varied in
composition than the latter. Like the greater part of the Dorataspida we may distinguish here in
the lattice-work two kinds of pores—parmal pores and sutural pores. The parmal pores are
produced by the union of the meeting branches of the apophyses of each single spine, and are
therefore visible on each isolated spine; whilst the sutural pores are formed by the meeting
branches of the apophyses of neighbouring spines. The distinction of the parmal and the sutural
pores, easy in most Dorataspida, is difficult in most Phractopeltida, because the sutures between
the meeting branches are usually very early obliterated. However, the place of the obliterated
suture is often indicated by the thickened condyles of the apophyses on both sides of the suture.
Commonly also the form of the sutural pores is much more irregular than that of the parmal pores;
the former are more or less constricted in the middle by the intumescence of the sutural condyles,
whilst the latter are more roundish, elliptical, kidney-shaped, or square. The number of the pores
in the outer shell in the typical normal form of Phractopeltida seems to be the same as in the
most species of Dorataspis, Diporaspis, &c., between ninety and one hundred,
viz., forty parmal pores and from fifty to sixty sutural pores. However, in many species this
number is increased. Since in all Phractopeltida, each of the twenty plates is composed only of
the meeting branches of two opposite apophyses, we find originally in each plate only two primary
parmal pores or "aspinal pores." But in some species there occur four, six, or more pores in each
plate; in this case two of them only are aspinal pores, all the others being "coronal pores."
Moreover, in those species which exhibit on the base of each spine in the outer shell four crossed
pores (Pl. 133,
fig. 2), there are not four equivalent aspinal pores (as in the Tessaraspida), but the two
opposite are primary or aspinal pores and the other two (different from them in size and form)
coronal pores. However, the number of coronal pores in the Phractopeltida is never so large as in
many Dorataspida, and the same holds good also for the increasing number of the irregular sutural
pores. In none of the species observed does the total number of the pores in the outer shell reach
two hundred.
The original mode of development of the apophyses composing the outer shell seems to be
imitated by the free apophyses of the third order, which are developed from the radial spines
outside the outer shell in all Phractopeltida, with the single exception of the simple ancestral
genus Phractopelta. These apophyses of the third rank are also originally constantly two,
opposite to one another (after the type of Lithophyllium, Dorataspis, &c.).
Commonly they do not remain simple, but become branched, and by communication of the neighbouring
branches small lattice-plates arise. Originally each of these free lattice-plates has only two
parmal pores, but the number of the parmal pores increases afterwards, so that we may distinguish
two (primary) {851}aspinal pores, and two, four, or more
(secondary) coronal pores. In the majority of species the two opposite apophyses are first crossed
at right angles by a transverse beam, and the two parallel transverse beams are again crossed by
perpendicular tertiary branches (again parallel to the apophyses). In this case the network of the
free lattice-plates becomes more or less rectangular. But in other species the ramification of the
apophyses assumes more the form of bifurcation or of irregular branching. As already said, the
neighbouring free lattice-plates of this third order never meet, and therefore a complete third
shell is never formed.
The different genera of Phractopeltida exhibit very remarkable differences in the development
of free apophyses (or lattice-plates of the third order). Whilst in the numerous species of the
ancestral genus Phractopelta all twenty spines remain simple, without such apophyses, only
in a single observed species (representing the genus Pantopelta) were all twenty spines
protected by them. In the three other genera only one part of the spines bears free apophyses, but
not the other part. The most frequent form is Dorypelta (Pl. 133, fig. 2);
here eight spines are simple (four equatorial and four polar spines of the hydrotomical plane),
whilst twelve spines bear apophyses (eight tropical and four polar spines of the geotomical
plane). In Octopelta the eight tropical spines only bear apophyses, whilst the twelve other
(four equatorial and eight polar) are simple. In Stauropelta finally the four equatorial
spines only are simple, whilst the sixteen other bear free apophyses (eight tropical and eight
polar spines).
The Central Capsule of the Phractopeltida is constantly spherical, and enclosed between
the two concentric spherical shells; it is therefore larger than the inner and smaller than the
outer shell. Its wall is pierced by the twenty radial beams connecting the two shells. The shape
of the central capsule and of the enveloping calymma is the same as in the other Acanthophracta and specially in the Dorataspida.
Synopsis of the Genera of Phractopeltida.
| All twenty spines of the same form, |
|
All twenty spines without apophyses in the free external part, |
366. Phractopelta. |
| All twenty spines with apophyses in the free part, |
367. Pantopelta. |
| Twenty radial spines, partly without, partly with
apophyses in the free external part, |
|
Eight tropical spines with apophyses, twelve others (eight polar and
four equatorial) simple, |
368. Octopelta. |
| Twelve radial spines (eight tropical and four polar) with apophyses,
eight others (four polar and four equatorial) simple, |
369. Dorypelta. |
| Sixteen radial spines (eight tropical and eight polar) with apophyses,
four equatorial, simple, |
370. Stauropelta. |
| All twenty spines of the same form, |
|
|
All twenty spines without apophyses in the free external
part, |
|
|
|
|
|
|
366. Phractopelta. |
|
|
|
|
|
|
All twenty spines with apophyses in the free part, |
|
|
|
|
|
|
367. Pantopelta. |
|
|
|
|
| Twenty radial spines, partly without, partly with apophyses in the free
external part, |
|
|
Eight tropical spines with apophyses, twelve others (eight polar
and four equatorial) simple, |
|
|
|
|
|
|
368. Octopelta. |
|
|
|
|
|
|
Twelve radial spines (eight tropical and four polar) with
apophyses, eight others (four polar and four equatorial) simple, |
|
|
|
|
|
|
369. Dorypelta. |
|
|
|
|
|
|
Sixteen radial spines (eight tropical and eight polar) with
apophyses, four equatorial, simple, |
|
|
|
|
|
|
370. Stauropelta. |
|
|
|
|
{852}
Genus 366. Phractopelta,[406] Haeckel,
1881, Prodromus, p. 468.
Definition.—Phractopeltida with twenty simple radial
spines, bearing no free apophyses
outside the outer shell.
The genus Phractopelta is the most simple form among the Phractopeltida, and
may be regarded as the common ancestral form of this family. All twenty spines are of nearly equal
form and size, and bear no free apophyses on their free part, outside the two concentric shells.
Phractopelta may be derived from Orophaspis by further development of the free
apophyses, which by union of their branches form a second outer shell around the primary shell of
that Dorataspid.
Subgenus 1. Phractopeltaris, Haeckel.
Definition.—Outer shell composed of twenty separated plates, the
sutures of their meeting condyles not being grown together.
1. Phractopelta dorataspis, n. sp. (Pl. 133, fig.
1).
Outer shell composed of twenty plates, the meeting condyles of which are separated by permanent
sutures. Each plate commonly with two elliptical aspinal pores, which are two to three times as
broad as the irregular sutural pores. Radial spines (in the outer free part) compressed, linear,
twice as long as the diameter of the shell.
Dimensions.—Diameter of the outer shell 0.11, of the inner 0.05.
Habitat.—North Pacific (off Japan), Station 239, surface.
2. Phractopelta dyadopora, n. sp.
Outer shell composed of twenty plates, the meeting condyles of which are separated by permanent
sutures. Each plate commonly with two kidney-shaped aspinal pores, which are three to four times
as broad as the irregular sutural pores. Radial spines conical, about as long as the radius of the
shell.
Dimensions.—Diameter of the outer shell 0.08, of the inner 0.04.
Habitat.—North Pacific, Station 256, surface.
3. Phractopelta diporaspis, n. sp.
Outer shell composed of twenty plates, the meeting condyles of which are separated by permanent
sutures. Each plate commonly with two quadrangular aspinal pores, which are four to five times
{853}as broad as the irregular sutural pores. Radial
spines compressed, sword-shaped, about as long as the diameter of the shell.
Dimensions.—Diameter of the outer shell 0.09, of the inner 0.04.
Habitat.—South Atlantic, Station 332, surface.
4. Phractopelta tessaraspis, n. sp.
Outer shell composed of twenty plates, the meeting condyles of which are separated by permanent
sutures. Each plate commonly with four crossed, quadrangular, aspinal pores, the two opposite of
which are much larger than the two others. Sutural pores small, roundish. Radial spines
compressed, linear, about twice as long as the diameter of the shell.
Dimensions.—Diameter of the outer shell 0.084, of the inner 0.032.
Habitat.—North Pacific (off Japan), Station 238, surface.
5. Phractopelta tetradopora, n. sp.
Outer shell composed of twenty plates, the meeting condyles of which are separated by permanent
sutures. Each plate commonly with four crossed quadrangular (or nearly circular) aspinal pores,
all of nearly the same size. Sutural pores polygonal or roundish. Radial spines cylindrical, two
to three times as long as the diameter of the shell.
Dimensions.—Diameter of the outer shell 0.11, of the inner 0.045.
Habitat.—North Pacific, Station 253, surface.
6. Phractopelta hexadopora, n. sp.
Outer shell composed of twenty plates, the meeting condyles of which are separated by permanent
sutures. Each plate commonly with six aspinal pores, the two opposite of which are much larger
than the four others. Sutural pores small, roundish. Radial spines compressed, two-edged, larger
than the diameter of the shell.
Dimensions.—Diameter of the outer shell 0.1, of the inner 0.04.
Habitat.—Central Pacific, Station 272, surface.
7. Phractopelta octadopora, n. sp.
Outer shell composed of twenty plates, the meeting condyles of which are separated by permanent
sutures. Each plate commonly with eight aspinal pores, the four crossed being larger than the four
others alternating with them. Sutural pores irregular. Radial spines conical, shorter than the
diameter of the shell.
Dimensions.—Diameter of the outer shell 0.12, of the inner 0.04.
Habitat.—Indian Ocean (Cocos Islands, surface), Rabbe.
{854}
Subgenus 2. Phractopeltidium, Haeckel.
Definition.—Outer shell composed of twenty united plates, the sutures
of their meeting condyles being grown together.
8. Phractopelta aspidomma, n. sp.
Outer shell composed of twenty united plates, the meeting condyles of which are grown together.
On the base of each radial spine (where its outer free part arises from the surface of the outer
shell), two large elliptical aspinal pores, two to four times as large as the other roundish
pores. Radial spines compressed, sword-shaped, about as long as the radius of the shell.
Dimensions.—Diameter of the outer shell 0.1, of the inner 0.05.
Habitat.—Equatorial Atlantic, Station 347, surface.
9. Phractopelta haliomma, n. sp.
Outer shell composed of twenty united plates, the meeting condyles of which are grown together.
On the base of each radial spine two large kidney-shaped aspinal pores, little larger than the
other irregular pores. Radial spines compressed, linear, two-edged, much longer than the diameter
of the shell.
Dimensions.—Diameter of the outer shell 0.09, of the inner 0.03.
Habitat.—South Pacific, Station 295, surface.
10. Phractopelta tessaromma, Haeckel.
Outer shell composed of twenty united plates, the meeting condyles of which are grown together.
On the base of each radial spine four crossed egg-shaped aspinal pores, two opposite of which are
much larger than the two others. Sutural pores irregular. Radial spines cylindrical or little
compressed, thin, longer than the diameter of the shell.
Dimensions.—Diameter of the outer shell 0.1, of the inner 0.04.
Habitat.—Central Pacific, Station 266, surface.
11. Phractopelta hystrix, Haeckel.
Haliomma hystrix, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 37, Taf.
v. figs. 1, 2.
Aspidomma hystrix, Haeckel, 1862, Monogr. d. Radiol., p. 424.
Outer shell composed of twenty united plates, the meeting condyles of which are grown together.
On the base of each radial spine four crossed circular aspinal spines of equal size. Sutural pores
roundish, of about the same size. Radial spines conical, about as long as the radius of the
shell.
Dimensions.—Diameter of the outer shell 0.07, of the inner 0.024.
Habitat.—Mediterranean (Nice), J. Müller, surface.
{855}
Genus 367. Pantopelta,[407] n. gen.
Definition.—Phractopeltida with twenty radial spines,
each of which is protected by two free external apophyses.
The genus Pantopelta differs from all other Phractopeltida in the
development of free protecting apophyses on all twenty radial spines. Only a single specimen of
this rare form was seen, and in this all twenty spines exhibited no marked differences in form and
size, but were more or less irregularly developed.
1. Pantopelta icosaspis, n. sp. (Pl. 133,
fig. 4).
Apophyses of all twenty radial spines with forked anastomosing branches, forming a more or less
irregular cup-like fenestrated shield, pierced by a variable number of pores. Distance of the
apophyses from the outer shell equal to half its radius. Pores of the outer shell irregular,
roundish, four to six times as broad as those of the inner shell. Radial spines compressed,
sword-shaped, longer than the radius of the shell.
Dimensions.—Diameter of the outer shell 0.08, of the inner 0.03.
Habitat.—Antarctic Ocean, Station 157, depth 1950 fathoms.
Genus 368. Octopelta,[408] n. gen.
Definition.—Phractopeltida with twelve simple spines
(four equatorial and eight polar), and with eight tropical spines protected by external free
apophyses.
The genus Octopelta differs from the other Phractopeltida in the possession
of eight shields or pairs of free apophyses outside the outer lattice-shell; the spines bearing
these apophyses are the eight tropical spines. The twelve other spines (eight polar and four
equatorial) are quite simple, without apophyses.
1. Octopelta cultella, n. sp.
Apophyses of the eight tropical spines simple, compressed, knife-shaped, about as long as their
distance from the outer shell. On the base of each spine (in the network of the outer shell) two
orthogonal aspinal pores, two to three times as long as the other polygonal pores.
Dimensions.—Diameter of the outer shell 0.08, of the inner 0.03.
Habitat.—Tropical Atlantic, Station 338, surface.
{856}
2. Octopelta furcella, n. sp.
Apophyses of the eight tropical spines forked, each with two parallel simple fork-branches,
about as long as their distance from the outer shell. On the base of each spine (in the outer
shell) two kidney-shaped aspinal pores, about twice as broad as the other irregular pores.
Dimensions.—Diameter of the outer shell 0.1, of the inner 0.04.
Habitat.—South Atlantic (off Tristan d'Acunha), Station 332,
surface.
3. Octopelta scutella, n. sp. (Pl. 133,
fig. 5).
Apophyses of the eight tropical spines crossed by a transverse beam, which is again crossed by
two perpendicular branches; by union of these branches each tropical spine forms a square shield
with four crossed pores and twelve to sixteen marginal spikes. On the base of each spine (in the
surface of the outer shell) four crossed aspinal pores which correspond to those of the free
shields; these are somewhat larger than the other pores.
Dimensions.—Diameter of the outer shell 0.12, of the inner 0.06.
Habitat.—Tropical Atlantic, Station 348, surface.
Genus 369. Dorypelta,[409] Haeckel,
Prodromus, p. 369.
Definition.—Phractopeltida with eight simple spines (four
equatorial and four hydrotomical polar spines) and with twelve spines protected by external free
apophyses (eight tropical and four geotomical polar spines).
The genus Dorypelta, the most common of all Phractopeltida which are
protected by free apophyses, exhibits a very peculiar differentiation of its twenty radial spines.
There are constantly eight simple spines and twelve spines with apophyses. The eight simple spines
are the four equatorial spines and four polar spines placed in the hydrotomical median plane. The
four other polar spines (placed in the geotomical meridian plane) and the eight tropical spines
are protected by two opposite apophyses, which are now simple, now branched or shield-shaped. A
similar differentiation occurs in no other Acanthophractida.
Subgenus 1. Dorypeltarium, Haeckel.
Definition.—Free apophyses of the radial spines simple, not
branched.
{857}
1. Dorypelta stauroptera, n. sp.
Apophyses of the long compressed radial spines simple, conical, about as long as their distance
from the shell. At the base of each spine (in the outer shell) two elliptical aspinal pores, two
to three times as large as the other pores.
Dimensions.—Diameter of the outer shell 0.1, of the inner 0.044.
Habitat.—North Pacific, Station 238, surface.
2. Dorypelta gladiata, n. sp.
Apophyses of the long cylindrical radial spines simple, triangular, about half as long as their
distance from the shell. At the base of each spine (in the outer shell) four crossed aspinal
pores, the two opposite of which are much larger than the other two pores.
Dimensions.—Diameter of the outer shell 0.09, of the inner 0.04.
Habitat.—Central Pacific, Station 266, surface.
Subgenus 2. Dorypeltidium, Haeckel.
Definition.—Free apophyses of the radial spines branched, with free
(not anastomosing) branches.
3. Dorypelta furcata, n. sp.
Apophyses of the long compressed spines simply forked, each with two simple parallel
fork-branches. At the base of each spine two elliptical aspinal pores, much larger than the other
roundish pores.
Dimensions.—Diameter of the outer shell 0.08, of the inner 0.04.
Habitat.—Central Pacific, Station 274, surface.
4. Dorypelta tetrodon, n. sp.
Apophyses of the long compressed spines crossed by a transverse beam, which bears four simple
parallel fork-branches or spikes (perpendicular to the beam), two larger medial and two smaller
lateral. At the base of each spine four crossed aspinal pores of nearly equal size, little
different from the other pores.
Dimensions.—Diameter of the outer shell 0.09, of the inner 0.035.
Habitat.—South Pacific, Station 284, surface.
5. Dorypelta ramosa, n. sp.
Apophyses of the long cylindrical spines crossed by a transverse beam, which bears four to six
irregularly branched spikes or fork-branches, the medial of which are larger than the lateral. At
{858}the base of each spine four crossed aspinal pores,
two opposite of which are much larger than the two other pores.
Dimensions.—Diameter of the outer shell 0.09, of the inner 0.04.
Habitat.—South Pacific, Station 302, surface.
Subgenus 3. Dorypeltonium, Haeckel.
Definition.—Free apophyses of the radial spines branched, and forming
perforated shields by union of their anastomosing branches.
6. Dorypelta lithoptera, n. sp.
Apophyses of the long compressed radial spines with anastomosing branches; each spine bearing
two free separated shields (each with two roundish aspinal pores). At the base of each spine (in
the network of the outer shell) two elliptical aspinal pores, larger than the other pores.
Dimensions.—Diameter of the outer shell 0.09, of the inner 0.03.
Habitat.—North Pacific, Station 239, surface.
7. Dorypelta tessaraspis, n. sp. (Pl. 133, fig.
2).
Apophyses of the long cylindrical spines with anastomosing branches; each spine bearing a
single free shield with four crossed aspinal pores of equal size (on the margin of the shield
twelve to sixteen spikes). At the base of each spine (in the outer shell) four crossed aspinal
pores, about as large as the other roundish pores.
Dimensions.—Diameter of the outer shell 0.11, of the inner 0.05.
Habitat.—Central Pacific, Station 263, surface.
8. Dorypelta dodecaspis, n. sp.
Apophyses of the long two-edged spines with anastomosing branches; each spine bearing a single
free shield with six to eight parmal pores (four crossed aspinal pores alternating with four outer
coronal pores), on the margin of each shield twelve to sixteen spikes. At the base of each spine
(in the outer shell) four crossed aspinal pores, about as large as the other irregular pores.
Dimensions.—Diameter of the outer shell 0.1, of the inner 0.04.
Habitat.—North Pacific (off Japan), Station 235, surface.
Genus 370. Stauropelta,[410] Haeckel,
1881, Prodromus, p. 468.
Definition.—Phractopeltida with four simple equatorial
spines and sixteen spines protected by external free apophyses (eight tropical and eight polar
spines).
{859}
The genus Stauropelta is distinguished from the other Phractopeltida by the
possession of sixteen pairs of free apophyses (on the eight tropical and the eight polar spines);
only the other four equatorial spines remain simple, without apophyses, and form a simple cross in
the equatorial plane.
1. Stauropelta cruciata, n. sp. (Pl. 133,
fig. 3).
Apophyses of the long polar and tropical spines crossed by a transverse beam, which is again
crossed by two perpendicular branches parallel to the apophyses; branches with free ends, not
united by concrescence. At the base of each spine (in the outer shell) two large elliptical
aspinal pores, larger than the other irregular pores. Four equatorial spines simple, without
apophyses, of the same size as the other sixteen spines.
Dimensions.—Diameter of the outer shell 0.12, of the inner 0.05.
Habitat.—Indian Ocean (Madagascar), Rabbe, surface.
2. Stauropelta stauropora, n. sp.
Apophyses of the long polar and tropical spines crossed by a transverse beam, which is again
crossed by two perpendicular branches parallel to the apophyses; these branches are united by
concrescence, and form a square shield with four crossed pores, the centre of which is pierced by
the spine. At the base of each spine (in the outer shell) four crossed aspinal pores,
corresponding to those of the shield, of about the same size as the other roundish pores. Four
equatorial spines, simple, without apophyses, somewhat larger than the other sixteen spines.
Dimensions.—Diameter of the outer shell 0.1, of the inner 0.04.
Habitat.—Indian Ocean, Sunda-Archipelago, Singapore, Trebing,
surface.
Suborder II. PRUNOPHRACTA.
Definition.—Shell ellipsoidal, lenticular or diploconical, with radial
beams of different size.
Family XLII. Belonaspida, n. fam. (Pl. 136, figs.
6-9; Pl. 139,
figs. 8, 9).
Definition.—Acantharia with a simple ellipsoidal
lattice-shell, composed of the branched apophyses of twenty radial spines meeting in its centre
and disposed according to the Müllerian law of Icosacantha. Two opposite equatorial spines larger
than the two others. Central capsule ellipsoidal, enclosed in the fenestrated shell.
The family Belonaspida, formerly united by me with the Dorataspida (in
the Prodromus, 1881, p. 468), differs from it in the ellipsoidal form of the lattice-shell and
{860}the enclosed central capsule. In the spherical
Dorataspida, their ancestral family, all twenty radial spines are of equal size, whilst here in
the Belonaspida two opposite spines are larger than the eighteen others. These two larger or
principal spines are both equatorial spines, placed in the longitudinal or major axis of the
ellipsoid, or the "hydrotomical axis" (compare above, p. 719). The two
other equatorial spines are constantly smaller, and lie in the transverse or minor axis of the
ellipsoid, or the "geotomical axis." The geometrical proportion of these two determining axes of
the ellipsoidal shell is very variable (even in one and the same species), commonly
4 : 3 or 3 : 2, rarely 2 : 1, often only 5 : 4 or
6 : 5. All meridian planes, passing through the principal spines (or the longitudinal
axis of the shell) are elliptical, and of equal size. All transverse planes, perpendicular to that
axis, are circular; the largest of these circular parallel planes is the geotomical plane, which
passes through the smaller equatorial spines and the spineless axis.
In the spherical Dorataspida the internal length of the radial spines (or the distance between
the shell and the centre) is equal in all twenty spines. In the ellipsoidal Belonaspida this
internal length is different, and commonly exhibits four different degrees; in the two principal
spines it is of first rank, in the eight tropical spines of second rank, in the four
(hydrotomical) polar spines of third rank, and in the six spines (four geotomical polar spines and
two transverse equatorial spines) of fourth rank. These differences of the internal length become
more important the more the hydrotomical axis is prolonged. Regarding all other qualities (in
form, disposition, and mode of junction at the centre) the ellipsoidal Belonaspida do not differ
from their ancestral group, the spherical Dorataspida (compare above, p. 802). In both families the pyramidal central bases of the twenty spines are
commonly supported one upon another with their triangular neighbouring faces; but sometimes also
here (particularly in Phatnaspis) the central bases are perfectly grown together. In this
case also the sutures of the meeting branches of the apophyses are obliterated, whilst usually
they remain open. Such forms, with spines and plates perfectly grown together, form a single piece
of acanthin, and were formerly separated by me as a peculiar genus Haliommatidium (Monogr.
d. Radiol., 1862, p. 419). But as I now find this concrescence to be an accidental and inconstant
peculiarity of some species, it no longer seems to me to be of generic importance.
The apophyses of the twenty radial spines, the branches of which compose the ellipsoidal
lattice-shell, exhibit in all Belonaspida the same appearance as in the Diporaspida (and
especially the Ceriaspida) among the spherical Dorataspida. Everywhere each radial spine gives off
only two opposite primary apophyses, the fork-branches of which unite together and form a plate or
shield with two aspinal pores (as in Dorataspis). Commonly the shell exhibits only these
forty parmal pores, the other meshes between them being sutural pores. Only in one genus,
Phatnaspis (Pl. 136,
{861}figs. 6-9), is this number multiplied; the two
opposite primary apophyses are here crossed by numerous perpendicular lateral branches, and these
are again united by secondary perpendicular ramules which are parallel to the apophyses
themselves; therefore each plate here forms a rhombic shield pierced by very numerous (often more
than one hundred) small quadrangular or circular pores. Only two of these numerous parmal pores
are the primary aspinal pores (sometimes, as in Pl. 136, fig. 8,
distinguished by their larger size); all others are secondary or coronal pores. Therefore
Phatnaspis repeats the characteristic formation of Coscinaspis, from which it
differs in its ellipsoidal shell.
Moreover, the four other genera of the ellipsoidal Belonaspida correspond perfectly to certain
genera of the spherical Dorataspida, and may be derived from these by hypertrophy or stronger
development of two opposite equatorial spines. In Thoracaspis and Belonaspis (Pl. 139, figs. 8,
9) the surface of the shell-plates is without crests and dimples (in the former without, in the
latter with accessory by-spines); they thus correspond to Dorataspis and Diporaspis
among the Dorataspida. In two other genera the surface of the shell-plates bears a network of
elevated crests, separating funnel-shaped dimples, in Dictyaspis without, in
Coleaspis with by-spines; these correspond to Ceriaspis and Hystrichaspis
among the Dorataspida. The small by-spines, covering the surface, exhibit the same forms as in the
Dorataspida; but they are usually smaller, and less developed in the Belonaspida.
The twenty radial spines are commonly more or less compressed or two-edged, sometimes very
flat, broad, and triangular (Pl. 139,
figs. 8, 9). In some species (mainly of Phatnaspis) they are very long, thin, and
needle-shaped (Monogr. d. Radiol., 1862, Taf. xxii. figs. 10-12). Rarely they are cylindrical,
four-edged, or prismatic. The Belonaspida testify by this and other peculiarities to their near
relation to the Diporaspida and Phractacanthida and their older origin from Zygacantha.
The Central Capsule is in the Belonaspida ellipsoidal, and more or less prolonged in the
hydrotomical or longitudinal axis of the shell. It is constantly smaller than the enveloping
ellipsoidal shell, and separated from it by the calymma. Its structure and the shape of its
nucleus are the same as in the nearly allied Dorataspida.
Synopsis of the Genera of Belonaspida.
|
II. Subfamily Coleaspida.
Forty parmal pores (two aspinal pores on each plate, no coronal pores).
|
|
Plates not dimpled, without prominent crests. |
|
No by-spines, |
371. Thoracaspis. |
| With by-spines, |
372. Belonaspis. |
| Plates dimpled, with a network of prominent crests, |
|
No by-spines, |
373. Dictyaspis. |
| With by-spines, |
374. Coleaspis. |
|
II. Subfamily Phatnaspida.
Eighty to two thousand or more parmal pores (two aspinal pores on each plate,
surrounded by two to one hundred or more coronal pores).
|
|
No by-spines (plates not dimpled), |
375. Phatnaspis. |
| I. Subfamily Coleaspida. Forty parmal pores (two aspinal pores on each plate,
no coronal pores). |
|
|
Plates not dimpled, without prominent crests. |
|
|
|
|
|
|
No by-spines, |
|
|
|
|
|
|
|
|
|
|
371. Thoracaspis. |
|
|
|
|
|
|
|
|
|
|
With by-spines, |
|
|
|
|
|
|
|
|
|
|
372. Belonaspis. |
|
|
|
|
|
|
|
|
Plates dimpled, with a network of prominent crests, |
|
|
|
|
|
|
No by-spines, |
|
|
|
|
|
|
|
|
|
|
373. Dictyaspis. |
|
|
|
|
|
|
|
|
|
|
With by-spines, |
|
|
|
|
|
|
|
|
|
|
374. Coleaspis. |
|
|
|
|
|
|
| II. Subfamily Phatnaspida. Eighty to two thousand or more parmal pores (two
aspinal pores on each plate, surrounded by two to one hundred or more coronal pores). |
|
|
|
|
No by-spines (plates not dimpled), |
|
|
|
|
|
|
|
|
|
|
375. Phatnaspis. |
|
|
|
|
|
|
{862}
Genus 371. Thoracaspis,[411] Haeckel,
1862, Prodromus, p. 468.
Definition.—Belonaspida with forty parmal pores (two on
each plate), without dimples and crests, also without by-spines, therefore with smooth
surface.
The genus Thoracaspis is the simplest form among the Belonaspida, and
repeats in this family the nearly allied genus Dorataspis among the Dorataspida. It differs
from this genus only in the characteristic prolongation of one equatorial axis, which effects an
ellipsoidal transformation of the spherical central capsule and the enclosing lattice-shell.
1. Thoracaspis ellipsoides, n. sp.
Parmal pores large, elliptical, five or six times as large as the small circular sutural pores.
Each plate surrounded by five or six sutural pores (a single one on each side). Spines compressed
triangular, sword-like, two-edged; inner and outer half of nearly equal length.
Dimensions.—Length of the ellipsoidal shell (or major axis) 0.15, breadth (or
minor axis) 0.12; length of the spines 0.07, basal breadth 0.016.
Habitat.—North Pacific, Station 253, surface.
2. Thoracaspis nephropora, n. sp.
Parmal pores large, kidney-shaped, three to four times as large as the roundish small sutural
pores. Each plate surrounded by five or six sutural pores (a single one on each side). Spines in
the inner longer part cylindrical, in the outer shorter part conical, not compressed.
Dimensions.—Length of the shell 0.2, breadth 0.15; length of the spines 0.06,
basal breadth 0.018.
Habitat.—Central Pacific, Station 268, surface.
3. Thoracaspis circopora, n. sp.
Parmal pores circular, of the same size as the circular sutural pores. Each plate surrounded by
five or six sutural pores (a single one on each side). Spines compressed, thin, linear, two-edged;
outer part one and a half times as long as the inner.
Dimensions.—Length of the shell 0.17, breadth 0.13; length of the spines 0.12,
basal breadth 0.003.
Habitat.—South Pacific, Station 288, surface.
4. Thoracaspis bipennis, n. sp. (Pl. 139,
fig. 8).
Parmal pores elliptical, about twice as large as the circular sutural pores, which are of
rather variable size. Each plate surrounded by eight to twelve sutural pores (commonly two on each
side). {863}Spines compressed, triangular, gradually
tapering towards both ends; outer half a little longer, and much broader than the inner.
Dimensions.—Length of the shell 0.14, breadth 0.12; length of the spines 0.08,
basal breadth 0.025.
Habitat.—Central Pacific, Station 274, surface.
Genus 372. Belonaspis,[412] Haeckel,
1862, Prodromus, p. 468.
Definition.—Belonaspida with forty parmal pores (two on
each plate), without dimples and crests, but with numerous by-spines on the surface.
The genus Belonaspis differs from its ancestral genus Thoracaspis
only in the possession of numerous superficial by-spines, and bears therefore to it the same
relation as Diporaspis does to Dorataspis. The two latter differ from the two former
in the spherical form of the central capsule and the enclosing shell, which here becomes
ellipsoidal.
1. Belonaspis pandanus, n. sp.
Parmal pores elliptical, three or four times as large as the circular sutural pores. Each plate
surrounded by five or six sutural pores (a single one on each side). Spines compressed,
triangular; outer part half as long as the inner, and twice as long as the numerous, simple,
bristle-shaped by-spines.
Dimensions.—Length of the ellipsoidal shell (or major axis) 0.12, breadth (or
minor axis) 0.1; length of the spines 0.03, basal breadth 0.02.
Habitat.—South Pacific, Station 295, surface.
2. Belonaspis furcata, n. sp.
Parmal pores elliptical, twice as broad as the circular sutural pores. Each plate surrounded by
five or six sutural pores (a single one on each side). Spines compressed, linear, very thin; outer
part longer than the inner. By-spines very numerous, half as long as the radius, furcate, with
divergent fork-branches.
Dimensions.—Length of the shell 0.1, breadth 0.08; length of the spines 0.12,
breadth 0.003.
Habitat.—Indian Ocean (Cocos Islands), Rabbe, surface.
3. Belonaspis datura, n. sp. (Pl. 139,
fig. 9).
Parmal pores elliptical, three or four times as large as the circular sutural pores. Each plate
surrounded by ten or twelve sutural pores (two on each side). Spines triangular, two-edged, about
{864}as long as the radius of the shell; gradually
tapering from its surface towards both ends. By-spines very short and numerous, simple.
Dimensions.—Length of the shell 0.13, breadth 0.11; length of the spines 0.08,
basal breadth 0.025.
Habitat.—Central Pacific, Station 271, surface.
4. Belonaspis lanceolata, n. sp.
Parmal pores elliptical, six or eight times as large as the small circular sutural pores. Each
plate surrounded by ten to twelve sutural pores (two on each side). Spines lanceolate, flat, in
the distal part needle-shaped, about as long as the diameter of the shell. By-spines very
numerous, zigzag, half as long as the shell-radius.
Dimensions.—Length of the shell 0.15 breadth 0.13; length of the spines 0.14,
basal breadth 0.03; length of the by-spines 0.04.
Habitat.—North Pacific, Station 235, surface.
5. Belonaspis conifera, n. sp.
Parmal pores kidney-shaped, three or four times as large as the small circular pores. Each
plate surrounded by ten to twelve sutural pores (two on each side). Spines very thick, half as
long in the outer conical part as in the inner cylindrical part. By-spines very short,
conical.
Dimensions.—Length of the shell 0.2, breadth 0.16; length of the spines 0.05,
basal breadth 0.03.
Habitat.—Central Pacific, Station 267, surface.
6. Belonaspis multiforis, n. sp.
Parmal pores circular, of the same size as the circular sutural pores. Each plate surrounded by
fifteen to eighteen sutural pores (three on each side). Spines compressed, about as long as the
radius. By-spines very numerous, zigzag, half as long as the radius.
Dimensions.—Length of the shell 0.22, breadth 0.18; length of the spines 0.12,
basal breadth 0.012; length of the by-spines 0.06.
Habitat.—Equatorial Atlantic, Station 347, surface.
Genus 373. Dictyaspis,[413] n. gen.
Definition.—Belonaspida with forty parmal pores (two on
each plate), without by-spines, but with a network of prominent crests on the dimply surface.
The genus Dictyaspis repeats among the Belonaspida the characteristic
structure of Ceriaspis (among the Dorataspida), by the development of prominent crests
forming a {865}network with dimples on the outer surface
of the shields. Either all the dimples or only a part of them are pierced by a pore. The forty
aspinal pores are united in pairs in twenty larger dimples. The shell is usually very thick-walled
and non-transparent.
1. Dictyaspis solidissima, Haeckel.
Dorataspis solidissima, Haeckel, 1862, Monogr. d. Radiol., p. 416, Taf.
xxii. figs. 6-9.
Surface of the shell with seventy to eighty funnel-shaped dimples, each of which is perforated
at the base by one or two small meshes; twenty larger dimples in the centre of the plates (each
with two elliptical aspinal pores), and fifty to sixty smaller sutural dimples between them (each
with a single circular pore). All pores of nearly the same size. No blind dimples. Outer conical
part of the thick radial spines of the same length as the inner cylindrical part. Radius of the
shell four times as large as the thickness of its wall.
Dimensions.—Length of the shell 0.12, breadth 0.1; length of the spines 0.06,
basal breadth 0.012.
Habitat.—Mediterranean (Messina), surface.
2. Dictyaspis favosa, n. sp.
Surface of the shell with one hundred and seventy to one hundred and ninety deep funnel-shaped
dimples, seventy to eighty of which are perforated, the others blind; among the former each of the
fifty to sixty smaller contains a single sutural pore, each of the twenty larger a couple of
aspinal pores. All pores of nearly the same size. Outer part of the compressed two-edged spines
triangular, half as long as the inner part. Radius of the shell six times as large as the
thickness of its wall. (Differs from Ceriaspis favosa, Pl. 138, fig. 6,
mainly in the ellipsoidal form of the shell and the different size and form of the spines.)
Dimensions.—Length of the shell 0.16, breadth 0.13; length of the spines 0.04,
basal breadth 0.025.
Habitat.—Central Pacific, Station 265, surface.
3. Dictyaspis compacta, n. sp.
Surface of the shell with one hundred and seventy to one hundred and ninety deep funnel-shaped
dimples, seventy to eighty of which are perforated, the others blind; among the former each of
fifty to sixty each contain a single sutural pore, twenty each a couple of aspinal pores. All
dimples nearly of the same size, very deep. Outer part of the compressed sword-like spines
somewhat longer than the inner part. Radius of the dark shell only twice as large as the thickness
of its compact wall.
Dimensions.—Length of the shell 0.18, breadth 0.15; length of the spines 0.12,
basal breadth 0.02.
Habitat.—Central Pacific, Station 274, surface.
{866}
Genus 374. Coleaspis,[414] Haeckel, 1881,
Prodromus, p. 468.
Definition.—Belonaspida with forty parmal pores (two on
each plate), with numerous by-spines and with a network of prominent crests on the dimpled
surface.
The genus Coleaspis differs from its ancestral genus Dictyaspis in
the development of numerous by-spines, and bears therefore among the ellipsoidal Belonaspida the
same relation to it as Hystrichaspis does to Coscinaspis among the spherical
Dorataspida. If the four equatorial spines in Coleaspis become different in pairs, it
passes over into Hexalaspis. The shell is usually very dark and thick-walled.
1. Coleaspis coronata, n. sp.
All twenty spines of nearly equal size and form, about as long as the radius of the shell,
compressed, sword-like, two-edged; both equatorial spines of the hydrotomical axis little longer
than the eighteen others. Crests between the dimples of the shell-surface dentated, forming around
the basal half of each spine a cylindrical sheath with a dentated, crown-like mouth, the teeth of
which are prolonged into simple by-spines. (Similar to Hexaconus coronatus, Pl. 139, fig. 5,
but differing in the nearly equal size of the spines and their sheaths, and in the longer
by-spines.)
Dimensions.—Major axis of the shell 0.2, minor 0.17; length of the spines 0.12,
basal breadth 0.02.
Habitat.—Central Pacific, Station 266, surface.
2. Coleaspis vaginata, n. sp. (Pl. 140,
fig. 13).
All twenty spines of nearly equal size and form, nearly as long as the diameter of the shell,
conical; both equatorial spines of the hydrotomical axis about one-third longer than the eighteen
others. Crests between the dimples of the shell-surface serrated, forming around the basal half of
each spine a conical truncate sheath with crested surface and serrated mouth, the teeth of which
are prolonged into short by-spines. (Similar to Hexaconus vaginatus, Pl. 139, fig. 7,
but differing in the nearly equal size of all the spines and in the double thickness of the
shell-wall.)
Dimensions.—Major axis of the shell 0.12, minor 0.1, length of the spines 0.08,
basal breadth 0.01.
Habitat.—North Pacific, Station 253, surface.
3. Coleaspis obscura, n. sp.
All twenty spines of nearly equal size and form, little longer than their large cylindrical
sheaths, which are nearly as long as the radius of the shell, and armed on the truncated mouth
with strong triangular by-spines, both equatorial spines of the hydrotomical axis one and a half
times as long as the eighteen others. Shell very dark and thick walled, quite opaque.
{867}
Dimensions.—Major axis of the shell 0.1, minor 0.08; length of the spines 0.06, of
their sheaths 0.04.
Habitat.—Arctic Ocean, Greenland, Koch, surface.
4. Coleaspis
occulta, n. sp.
All twenty spines of nearly equal size and form, quite concealed in their long cylindrical
sheaths, which are somewhat longer than the radius of the shell, and armed on the truncated mouth
with acute simple teeth; each sheath deeply sulcated, apparently resulting from the concrescence
of numerous parallel leaf-shaped by-spines. Small shell very dark and thick-walled, quite
opaque.
Dimensions.—Major axis of the shell 0.08, minor 0.06; length of the spines and
their sheaths 0.05 to 0.06.
Habitat.—Antarctic Ocean (off Kerguelen), Station 159, surface.
Subgenus 2. Coleaspidium, Haeckel.
Definition.—Both equatorial spines of the hydrotomical axis much
larger and of peculiar form, different from the eighteen other spines.
5. Coleaspis amphilonche, n. sp.
Both equatorial spines of the hydrotomical axis much longer than the eighteen others, and of
very different form; one and a half times as long as the diameter of the shell, prismatic, with
six prominent edges, pyramidal at the distal point. The eighteen other spines triangular,
compressed, two-edged, scarcely as long as the radius of the thick-walled shell. Pores of the
shell irregular polygonal, separated by ciliated crests, which bear simple by-spines (one-third as
long as the radius).
Dimensions.—Major axis of the shell 0.15, minor 0.12; length of the two larger
spines 0.22, of the eighteen smaller 0.06 to 0.08; basal breadth of the former 0.03, of the latter
0.008.
Habitat.—South Atlantic, Station 341, surface.
6. Coleaspis hydrotomica, n. sp. (Pl. 140, fig.
14).
Both equatorial spines of the hydrotomical axis much larger than the eighteen others, and of
very different form; about as long as the diameter of the shell, in the basal half four-sided
pyramidal, with four thick prominent edges, in the middle part constricted, in the distal half
lanceolate, two-edged. Both equatorial spines of the geotomical axis of similar form, but much
smaller, scarcely one-third as long. The sixteen other spines sword-like, thin, two-edged, about
as long as the radius of the thick-walled shell. Pores of the shell irregular roundish, separated
by high dentated crests, which bears zigzag by-spines (half as long as the radius).
Dimensions.—Major axis of the shell 0.12, minor axis 0.1; length of the two larger
spines 0.11, of the eighteen others 0.04 to 0.06; basal breadth of the former 0.04, of the latter
0.01.
Habitat.—Cape of Good Hope, Station 143, surface.
{868}
Genus 375. Phatnaspis,[415] Haeckel,
1881, Prodromus, p. 468
Definition.—Belonaspida with eighty to two thousand or
more parmal pores (four to one hundred or more on each plate), without by-spines on the
surface.
The genus Phatnaspis corresponds to the spherical Coscinaspis among
the Dorataspida, and differs from the other Belonaspida in the great number of the parmal pores;
whilst the four other preceding genera exhibit only two opposite aspinal pores in the centre of
each plate, in this there are constantly numerous coronal pores in addition to these, and the
plates always possess the characteristic form of a wainscotted or panelled work, with quadrangular
meshes. The number of these parmal pores amounts in each plate to from ten to twenty, often one
hundred and twenty or more; therefore the number of parmal pores in the whole shell amounts to two
thousand or more. Sometimes the pores are circular, but in this case too they are surrounded by
quadrangular frames. The quadrangles are sometimes quite regular squares, sometimes more or less
irregular. The thin and fragile bars between the quadrangular pores form in each plate two
peculiar systems of parallel crests, which cross at right angles. Commonly the parallel crests of
one system (parallel to the major diameter of the compressed radial spines) are equidistant, and
pierce from one edge of the plate to the opposite, whilst the parallel crests of the other system
(parallel to the minor diameter of the spines) are interrupted and at different distances (Pl. 136, fig. 9);
but in other species both crossed systems are quite regular. In each plate there is one primary
diagonal rib (often stronger than the parallel crests) which connects the two opposite corners of
the rhomboidal plate. We can distinguish in this genus three different subgenera: A. in
Phatnasparium the primary diagonal rib arises from both flat sides of the compressed
sword-like, radial spines; B. in Phatnasplenium from both sharp edges of them; C. in
Phatnaspidium two crossed diagonal ribs arise from four edges of the spines (combination of
A and B). Therefore in the first subgenus (A) two primary aspinal pores are placed opposite on the
sharp edges of the spines, but in the second (B) inversely on their flat sides; in the third (C)
there are apparently four primary aspinal pores, which are probably derived from B or A by
division of the two pores.
Subgenus 1. Phatnasparium, Haeckel.
Definition.—In the centre of each rhomboidal plate two primary aspinal
pores, opposite on both edges of the compressed spines, from both flat sides of which the primary
diagonal crest arises.
{869}
1. Phatnaspis lacunaria, n. sp. (Pl. 136,
fig. 9).
Parmal pores irregular quadrangular, of unequal size and form, ten to twelve on each side of
the primary diagonal rib, which arises from both flat sides of the two-edged spines; two primary
aspinal pores opposite on both edges of the latter. Spines much compressed, sword-like, their
outer half shorter than the inner.
Dimensions.—Length of the shell 0.21 to 0.23, breadth 0.18 to 0.2; basal breadth
of the spines 0.01.
Habitat.—Central Pacific, Station 268, surface.
2. Phatnaspis ensiformis, n. sp.
Parmal pores irregular quadrangular, of unequal size and form, six to eight on each side of the
primary diagonal rib, which arises from both flat sides of the two-edged spines; two primary
aspinal pores opposite on the two edges of the latter. Spines very broad, strongly compressed,
sword-like, their outer half larger than the inner.
Dimensions.—Length of the shell 0.16, breadth 0.11; basal breadth of the spines
0.02.
Habitat.—Tropical Atlantic, Station 348, surface.
3. Phatnaspis loculata, n. sp.
Parmal pores regular quadrangular, of nearly equal size and form, twelve to sixteen on each
side of the primary diagonal rib, which arises from both flat sides of the two-edged spines; two
primary aspinal pores on the two edges of the latter. Spines linear, compressed, very long and
thin, their outer half three to four times as long as the inner.
Dimensions.—Length of the shell 0.24, breadth 0.18; breadth of the spines
0.004.
Habitat.—North Atlantic, Station 354, surface.
4. Phatnaspis fenestrata, Haeckel.
Haliommatidium fenestratum, Haeckel, 1862, Monogr. d. Radiol., p. 421.
Parmal pores regular quadrangular, of nearly equal size and form, six to eight on each side of
the primary diagonal rib, which arises from both flat sides of the two-edged spines; two primary
aspinal pores on the two edges of the latter. Spines linear, little compressed, or nearly
needle-shaped, very long and thin, their outer half five to ten times as long as the inner.
Dimensions.—Length of the shell 0.11 to 0.12, breadth 0.07 to 0.08; breadth of the
spines 0.002.
Habitat.—Mediterranean (Messina), surface.
5. Phatnaspis cristata, n. sp. (Pl. 136,
fig. 6).
Parmal pores circular, of very different size, separated by high square crests, ten to twelve
on each side of the high, comb-like, primary diagonal rib, which arises from both flat sides of
the {870}twoedged spines; two primary aspinal pores on
the two edges of the latter. Each circular pore is surrounded by a square frame. Spines
sword-like, strongly compressed, their outer part longer than the inner.
Dimensions.—Length of the shell 0.2 to 0.22, breadth 0.16 to 0.18; basal breadth
of the spines 0.01.
Habitat.—North Pacific, Station 254, surface.
6. Phatnaspis coscinoides, n. sp.
Parmal pores circular, regular, all of nearly equal size, twelve to sixteen on each side of the
primary diagonal rib, which arises from both flat sides of the two-edged spines; two primary pores
on the two edges of the latter. Spines linear, little compressed, their outer part much longer
than the inner.
Dimensions.—Length of the shell 0.18, breadth 0.15; breadth of the spines
0.006.
Habitat.—South Atlantic, Station 325, surface.
Subgenus 2. Phatnasplenium, Haeckel.
Definition.—In the centre of each rhomboidal plate two primary aspinal
pores, opposite on the two flat sides of the compressed spines, from the two edges of which the
primary diagonal crest arises.
7. Phatnaspis orthopora, n. sp.
Parmal pores irregular quadrangular, of unequal size and form, four to six on each side of the
primary diagonal rib, which arises from both edges of the compressed spines; two larger primary
aspinal pores opposite on the two flat sides of the latter. Spines very thin and long, linear,
their outer half three to four times as long as the inner.
Dimensions.—Length of the shell 0.08, breadth 0.06; breadth of the spines
0.003.
Habitat.—North Atlantic, Station 353, surface.
8. Phatnaspis polypora, n. sp.
Parmal pores irregular polygonal, of very unequal size and form, sixteen to twenty on each side
of the primary diagonal rib, which arises from both edges of the compressed spines; two very
large, lanceolate, primary aspinal pores opposite on the two flat sides of the latter. Spines very
thin, linear, on the outside of the shell rudimentary. (This remarkable species somewhat resembles
Coscinaspis polypora, Pl. 136,
fig. 8.)
Dimensions.—Length of the shell 0.32, breadth 0.24; breadth of the spines
0.002.
Habitat.—South Atlantic, Station 332, surface.
{871}
9. Phatnaspis quadratura, n. sp.
Parmal pores regular, square, all of nearly equal size and form, ten to twelve on each side of
the primary diagonal rib, which arises from both edges of the compressed spines; two primary
aspinal pores opposite on the two flat sides of the latter, not different from the other pores.
Spines sword-like, their outer part about as long as the inner.
Dimensions.—Length of the shell 0.2, breadth 0.16; basal breadth of the spines
0.004.
Habitat.—Central Pacific, Station 274, surface.
10. Phatnaspis tabulata, Haeckel.
Haliomma tabulatum, J. Müller, 1858, Abhandl. d. k. Akad. d. Wiss. Berlin, p. 37, Taf.
v. figs. 5-8.
Haliomma tabulatum, Haeckel, 1862, Monogr. d. Radiol., p. 429.
Parmal pores regular, circular, with square frames, all of nearly equal size, eight to ten on
each side of the primary diagonal rib, which arises from both edges of the compressed spines; two
primary aspinal pores opposite on the flat sides of the latter. Spines sword-like, short, scarcely
half as long as the radius of the shell, their central ends are thickened and cause by their union
the deceptive appearance of an enclosed "medullary shell."
Dimensions.—Length of the shell 0.12 to 0.15, breadth 0.1 to 0.12; breadth of the
spines 0.006.
Habitat.—Mediterranean; Nice, Müller; Portofino, near Genoa, Haeckel,
surface.
Subgenus 3. Phatnaspidium, Haeckel.
Definition.—In the centre of each rhomboidal plate four primary
aspinal spines, forming a regular cross, the centre of which receives the cylindrical or
four-sided prismatic spine.
11. Phatnaspis haliommidium, n. sp. (Pl. 136, fig.
7).
Parmal pores irregular quadrangular, of unequal size and form, eight to ten on each side of the
crossed diagonal ribs, which arise at right angles from the four edges of the prismatic spines;
four primary aspinal pores not different from the others. Outer part of the strong spines scarcely
longer than the inner. (Resembling Icosaspis tabulata, Pl. 136, fig. 2,
which, however, is distinguished by the spherical shell and the larger pores.)
Dimensions.—Length of the shell 0.22, breadth 0.17; breadth of the spines
0.016.
Habitat.—North Pacific, Station 244, surface.
12. Phatnaspis mülleri, n. sp.
Haliommatidium mülleri, Haeckel, 1862, Monogr. A. Radiol, p. 419, pl.
xiii. figs. 10-12.
Parmal pores regular, square, all of nearly equal size and form, twelve to sixteen on each side
of the crossed diagonal ribs, which arise at right angles from the needle-shaped spines; the four
{872}primary aspinal pores not different from the
others. Spines very thin and long, cylindrical; four to six times longer in the outer than in the
inner part.
Dimensions.—Length of the shell 0.24, breadth 0.16; breadth of the spines
0.002.
Habitat.—Mediterranean (Messina), surface.
Family XLIII. Hexalaspida, n. fam. (Pl. 139).
Definition.—Acantharia with a simple discoidal or
lenticular lattice-shell, composed of the branched apophyses of twenty radial spines meeting in
the centre and disposed according to the Müllerian law of Icosacantha. Six larger spines in the
hydrotomical plane, prominent on the margin of the circular or elliptical biconvex lens. Fourteen
other spines much smaller or rudimentary. Central capsule biconvex lenticular, enclosed in the
fenestrated shell.
The family Hexalaspida represents a new small, but very interesting
group of Acanthophracta, which differs from all others in the lentelliptical or triaxial form of
the lenticular lattice-shell, the margin of which bears six larger spines placed in the
hydrotomical plane (compare above, p. 719). They may therefore be
characterised shortly as "Acanthophracta lentelliptica," with three different dimensive
axes and six larger marginal spines. A closer comparison with the other Acantharia leaves no doubt that the Hexalaspida must be derived from the
Belonaspida by stronger development of six radial spines placed in the hydrotomical plane, namely,
two equatorial and four associated polar spines; whilst the six spines of the geotomical plane
(perpendicular to the former) are much smaller; the eight tropical spines are intermediate in size
between the former and the latter.
The geometrical fundamental form of the Hexalaspida (of the central capsule as well as of the
enclosing shell) is therefore lentelliptical, with three different dimensive axes, and they
exhibit among the Acantharia a relation to the spherical Dorataspida and
the ellipsoidal Belonaspida similar to that which the lentelliptical Larcoidea exhibit to the spherical Sphæroidea and the
ellipsoidal Prunoidea among the Sphærellaria
(compare above, p. 599). The largest of the three dimensive axes (which are
perpendicular to one another) is here the hydrotomical axis, the shortest, on the contrary, the
geotomical axis; the intermediate in size being the spineless axis. The development of the whole
body is strongest in the hydrotomical meridian plane, in which the six principal spines are
placed; it is weakest in the geotomical plane, in which the six smallest spines are placed; the
eight tropical spines are intermediate in size between the others. This peculiar development is
illustrated by the figures of Pl. 139,
where the four equatorial spines are everywhere marked by c, the eight tropical spines by
b and d, the eight polar spines by a and e.
{873}
Rarely the six hydrotomical or principal spines are of equal size, and thus the margin of the
shell may be quite circular or regularly hexagonal (Pl. 139, figs.
1-3); the two equatorial spines of the hydrotomical plane are usually larger than its four polar
spines, and thus the margin of the shell becomes more or less elliptical (Pl. 139, figs.
4-7). Usually (almost constantly) these six larger spines are more or less compressed, triangular,
often very broad and flat; their two edges lie in the hydrotomical plane. Their two flat surfaces
are often furrowed, with longitudinal ribs or crests converging towards the simple apex of the
spines.
The fourteen smaller spines are regularly disposed according to the Müllerian law of
Icosacantha on both convex sides of the lenticular shell; they are not only smaller than the six
principal spines, but often also of different form, much thinner and shorter, sometimes
needle-shaped. In the genera Hexonaspis and Hexacolpus (Pl. 139, figs. 1,
2) only their inner part (inside of the shell) is developed, whilst their outer part is quite
rudimentary and not prominent on the surface. Therefore these genera appear to possess only six
marginal spines externally.
The Lenticular Shell itself offers in the Hexalaspida great difficulties in the way of
accurate study, as its wall is constantly very thick and dark, often quite opaque and
non-transparent. However, prolonged accurate researches have convinced me that its structure is
essentially the same as in the Belonaspida and especially in the genera Dictyaspis and
Coleaspis. As in these latter the twenty plates of the shell bear high crests or combs on
the outer surface, and by these funnel-shaped dimples are separated. The network of these crests
is more or less regular (Pl. 139,
figs. 1-7). Around the base of each radial spine the shell is usually elevated in the form of a
conical or cylindrical sheath; the crests are prolonged into the sheaths as longitudinal ribs,
parallel to the spine or convergent towards its apex. Whilst in Hexalaspis and
Hexonaspis these basal sheaths are not at all or but little prominent (Pl. 139, fig. 2;
Pl. 140,
fig. 16), in Hexaconus and Hexacolpus they envelop the basal half (or even more) of
the spines, and very often the circular or elliptical free distal edge of the sheath is elegantly
denticulated or serrated (Pl. 139,
figs. 1, 3-7; pl. 140, figs. 9-16).
The Pores of the shell exhibit in the Hexalaspida the same shape as in the majority of
the Belonaspida. Each spine bears only two broad opposite apophyses, the fork-branches of which
unite to form a polygonal shield with two pores. The number of parmal pores is constantly (?)
forty, as each plate possesses only two primary aspinal pores; there are no secondary or coronal
pores. The numerous (between fifty and one hundred, rarely more) smaller pores between the forty
parmal pores are probably always sutural pores; however, their number and position is very
difficult to determine, on account of the high protecting crests; the majority of the
funnel-shaped dimples between the latter seem to be blind, not perforated. Sometimes all the
dimples, except the twenty spinal ones, seem to be blind and the sutural pores appear {874}to have disappeared completely, so that there remain only
twenty aspinal pores (Pl. 139,
fig. 4).
The internal cavity of the shell in all Hexalaspida is very small, on account of the thickness
of the massive wall; the latter is often greater than the diameter of the cavity. Setting aside
this disproportion, the space of the cavity is further much reduced by the internal parts of the
thick radial spines, which are united in the centre by their pyramidal bases (Pl. 140, fig. 15).
Usually the twenty bases seem to remain free (supported one upon another by means of their
triangular faces); but sometimes they seem to grow together perfectly and to form a single central
star of acanthin. In some species, too, the sutures of the meeting branches of the apophyses of
neighbouring spines seem to grow together, so that the whole shell exhibits a single piece of
acanthin.
The Central Capsule of the Hexalaspida is therefore very small and seems to fill up the
greatest part of the shell-cavity. Its form is constantly more or less lenticular, sometimes
lentelliptical. On account of the opacity of the shell I could not make out its shape more
closely.
Synopsis of the Genera of Hexalaspida.
| All twenty spines externally developed, prominent
on the surface of the lenticular shell. |
|
Sheaths of the spines not prominent, |
376. Hexalaspis. |
| Sheaths of the spines prominent, |
377. Hexaconus. |
| Only six spines (the hydrotomical spines) externally
developed and prominent on the margin of the disk; the other fourteen spines not
prominent. |
|
Sheaths of the spines not prominent, |
378. Hexonaspis. |
| Sheaths of the spines prominent, |
379. Hexacolpus. |
| All twenty spines externally developed, prominent on the surface of the
lenticular shell. |
|
|
Sheaths of the spines not prominent, |
|
|
|
|
|
|
376. Hexalaspis. |
|
|
|
|
|
|
Sheaths of the spines prominent, |
|
|
|
|
|
|
377. Hexaconus. |
|
|
|
|
| Only six spines (the hydrotomical spines) externally developed and prominent
on the margin of the disk; the other fourteen spines not prominent. |
|
|
Sheaths of the spines not prominent, |
|
|
|
|
|
|
378. Hexonaspis. |
|
|
|
|
|
|
Sheaths of the spines prominent, |
|
|
|
|
|
|
379. Hexacolpus. |
|
|
|
|
Genus 376. Hexalaspis,[416] n. gen.
Definition.—Hexalaspida with twenty prominent radial
spines, which are not surrounded by prominent sheaths; the six hydrotomical spines much larger
than the fourteen others.
The genus Hexalaspis is the simplest form among the Hexalaspida, and may be
derived directly from Dictyaspis among the Belonaspida, by stronger development of the six
hydrotomical spines. As in the following genus Hexaconus all twenty spines are prominent
externally.
Subgenus 1. Hexalasparium, Haeckel.
Definition.—Six hydrotomical spines of equal size.
{875}
1. Hexalaspis heliodiscus, n. sp. (Pl. 139, fig.
2).
All six hydrotomical spines of nearly equal size (or sometimes the two equatorial a little
larger than the four polar spines), isosceles triangular, compressed, smooth; about as long as the
radius of the shell, and half as broad at the base. The fourteen other spines very small, also
triangular and compressed, but little prominent on the two convex sides of the lenticular
shell.
Dimensions.—Diameter of the shell 0.11; length of the six hydrotomical spines
0.05, basal breadth 0.03.
Habitat.—Central Pacific, Station 271 to 274, surface.
2. Hexalaspis stellata, n. sp.
All six hydrotomical spines of nearly equal size, lanceolate compressed, with two longitudinal
furrows on each flat side, about as long as the diameter of the shell, and one-fourth as broad at
the base. The fourteen other spines very thin, also compressed, half as long and only one-fourth
as broad as the six large spines.
Dimensions.—Diameter of the shell 0.14; length of the six hydrotomical spines
0.13, basal breadth 0.035.
Habitat.—South Pacific, Station 284, surface.
Subgenus 2. Hexalaspidium, Haeckel.
Definition.—Six hydrotomical spines of unequal size, two opposite
(equatorial) much larger than the four other (polar) spines.
3. Hexalaspis sexalata, n. sp.
Six hydrotomical spines of unequal size; the two equatorial spines about as long as the
shell-radius and twice as long as the four polar spines, which are isosceles triangular. The
fourteen other spines are only half as long and one-fourth as broad as the latter, little
prominent. (Resembles Hexonaspis hastata, Pl. 140, fig. 16,
which is distinguishable by the furrows on the six spines and by the total absence of the fourteen
external rudimentary spines.)
Dimensions.—Diameter of the shell 0.15; length of the two equatorial spines 0.08,
of the four polar spines 0.04, of the fourteen other spines 0.02.
Habitat.—North Pacific, Station 240, surface.
4. Hexalaspis hexalastrum, n. sp.
Six hydrotomical spines of unequal size; the two equatorial spines somewhat longer than the
diameter of the shell and three times as long as the four polar spines, all six triangular,
smooth, of the same basal breadth (equal to half the radius). The fourteen other spines very thin,
conical at the base, nearly as long as the radius.
{876}
Dimensions.—Diameter of the shell 0.21; length of the two equatorial spines 0.24,
of the eighteen others 0.08 to 0.1; basal breadth of the six larger spines 0.05, of the fourteen
others 0.01.
Habitat.—Western Tropical Pacific, Station 224, surface.
5. Hexalaspis hexaglypha, n. sp.
Six hydrotomical spines of unequal size; the two equatorial very large, six-sided prismatic,
twice as long as the diameter of the shell and four times as long as the four pyramidal polar
spines; each of these six spines with six deep furrows between the six prominent edges. The
fourteen other spines very thin, compressed, two-edged, about as long as the radius of the
shell.
Dimensions.—Diameter of the shell 0.11; length of the two equatorial spines 0.2,
of the eighteen others 0.04 to 0.06; basal breadth of the six larger spines 0.03, of the fourteen
others 0.01.
Habitat.—Tropical Atlantic, Station 352, surface.
Genus 377. Hexaconus,[417] n. gen.
Definition.—Hexalaspida with twenty prominent radial
spines, which are surrounded at the base by prominent sheaths; the six hydrotomical spines much
larger than the fourteen others.
The genus Hexaconus differs from the preceding Hexalaspis in the
development of conical or cylindrical sheaths surrounding the basal parts of the radial spines;
these sheaths are developed sometimes around all twenty spines, sometimes only around the six
larger hydrotomical spines.
Subgenus 1. Hexaconarium, Haeckel.
Definition.—Six hydrotomical spines of equal size.
1. Hexaconus ciliatus, n. sp. (Pl. 139,
fig. 3).
All six hydrotomical spines of nearly equal size, pyramidal, with six prominent edges, somewhat
longer than the radius of the shell. Sheaths sulcated, finely ciliated at the mouth, twice as
broad as long and only one-fourth as long as the spines. The fourteen smaller spines very thin,
bristle-shaped, shorter than the radius of the shell.
Dimensions.—Diameter of the shell 0.16; length of the six hydrotomical spines
0.11; basal breadth 0.03.
Habitat.—Central Pacific, Station 270, surface.
{877}
2. Hexaconus coronatus, n. sp. (Pl. 139,
fig. 5).
All six hydrotomical spines of equal size, conical, somewhat compressed, with two prominent
edges, scarcely as long as the radius of the shell. Sheaths crested, strongly dentated on the
mouth, three times as broad as long and only one-sixth as long as the spines. Fourteen smaller
spines about half as large as the six principal spines, of the same form, but without coronated
sheaths.
Dimensions.—Diameter of the shell 0.2; length of the six hydrotomical spines 0.1;
basal breadth 0.02.
Habitat.—North Pacific, Station 253, surface.
3. Hexaconus velatus, n. sp. (Pl. 139,
fig. 6).
All six hydrotomical spines of equal size, conical, somewhat compressed, shorter than the
radius of the shell. Sheaths very large, truncated conical, enveloping the spines almost entirely,
with crested wall, only half as broad at the constricted mouth as at the base. The fourteen
smaller spines about half as large as the six principal spines, of the same form, but without
large sheaths.
Dimensions.—Diameter of the shell 0.12; length of the six hydrotomical spines
0.05; basal breadth 0.01.
Habitat.—North Atlantic, Station 354, surface.
Subgenus 2. Hexaconidium, Haeckel.
Definition.—Six hydrotomical spines of unequal size, two opposite
(equatorial) much larger than the four other (polar) spines.
4. Hexaconus serratus, n. sp. (Pl. 139,
fig. 4).
Six hydrotomical spines of unequal size, the two equatorial being as long as the radius of the
shell and one and a half times as large as the four polar; all of the same form, triangular,
compressed, with six prominent edges. Sheaths cylindrical, half as long as the spines, with
prominent crests, and with strong serrated teeth at the distal mouth. The fourteen smaller spines
scarcely one-fourth or one-sixth as large as the six principal spines, without prominent
sheaths.
Dimensions.—Diameter of the shell 0.2; length of the equatorial spines 0.1, basal
breadth 0.05; length and breadth of the sheaths 0.05.
Habitat.—South Pacific, Station 288, surface.
5. Hexaconus vaginatus, n. sp. (Pl. 139,
fig. 7).
Six hydrotomical spines of unequal size; the two equatorial very stout, pyramidal, six-edged,
nearly as long as the diameter of the shell, and on the base three to four times as broad as the
eighteen other spines, which are much thinner, all nearly of equal length, and two-edged. All
{878}twenty spines are provided with prominent basal
sheaths, which are truncate conical, sulcate, and dentate on the narrowed distal mouth. The
sheaths of the six hydrotomical spines are twice to three times as large as those of the fourteen
smaller spines.
Dimensions.—Diameter of the shell 0.15; length of the spines 0.12; basal breadth
of the equatorial spines 0.04, of the other spines 0.02; length of the hydrotomical sheaths 0.05,
of the other sheaths 0.02.
Habitat.—Central Pacific, Station 274, surface.
6. Hexaconus echinatus, n. sp. (Pl. 140,
fig. 12).
Six hydrotomical spines of unequal size; the two equatorial, and their sheaths twice as large
as the four polar spines. These six spines are six-edged, pyramidal, and their basal half
enveloped by very large conical sheaths which are sulcate, and twice as broad on the dentate
distal mouth as on the narrower base. The other fourteen spines are very thin, two-edged, half as
long, with low sheaths. Approaches some forms of Diploconus.
Dimensions.—Diameter of the shell 0.15; length of the equatorial spines 0.13, of
the polar spines 0.08.
Habitat.—Indian Ocean, Belligemma, Ceylon, Haeckel, surface.
Genus 378. Hexonaspis,[418] n. gen.
Definition.—Hexalaspida with six prominent radial spines
(in the hydrotomical plane) which are not surrounded by prominent sheaths; the fourteen other
spines quite rudimentary, not prominent.
The genus Hexonaspis and the following Hexacolpus differ from the two
preceding genera in the rudimentary shape of the fourteen reduced and stunted smaller spines;
these are only developed inside the shell, and are not prominent outside over its surface.
Subgenus 1. Hexonasparium, Haeckel.
Definition.—Six hydrotomical spines of equal size.
1. Hexonaspis heliosestrum, n. sp.
All six hydrotomical spines of nearly equal size, isosceles triangular, compressed, smooth,
about as long as the diameter of the shell and three times as long as broad at the base. This
species is very similar to Hexalaspis heliodiscus (Pl. 139, fig. 2),
but differs in the larger size of the six marginal spines and in the complete external absence of
the fourteen smaller spines.
{879}
Dimensions.—Diameter of the shell 0.13; length of the six marginal spines 0.12,
basal breadth 0.04.
Habitat.—Tropical Atlantic, Station 348, surface.
2. Hexonaspis hexapleura, n. sp. (Pl. 140, fig.
15).
All six hydrotomical spines of nearly equal size, triangular, compressed, six-edged, about as
long as the radius of the shell and twice as long as broad on the base. Two strong prominent ribs
on each flat side of the spines. Shell very thick-walled, with a very small cavity.
Dimensions.—Diameter of the shell 0.11; length of the six marginal spines 0.06,
basal breadth 0.03.
Habitat.—South Atlantic, Station 332, surface.
3. Hexonaspis hexagona, n. sp.
All six hydrotomical spines of equal size, very short, triangular, only half as long as broad
at the base. The whole shell accordingly forms a regular hexagon, the six corners of which are
formed by the distal points of the rudimentary spines, the six sides by their straight lateral
edges. Shell very dark.
Dimensions.—Diameter of the shell 0.2; length of the six marginal spines 0.03,
basal breadth 0.06.
Habitat.—South Atlantic (east of Patagonia), Station 318, surface.
Subgenus 2. Hexonaspidium, Haeckel.
Definition.—Six hydrotomical spines of unequal size, two opposite
(equatorial) much larger than the four other (polar) spines.
4. Hexonaspis hastata, n. sp. (Pl. 140,
fig. 16).
Six hydrotomical spines of unequal size; two larger equatorial spines with six prominent wings,
about as long as the radius of the shell and half as broad at the base; the four polar spines
quite as broad, but only half as long, nearly equilateral triangular, with two shallow furrows on
each flat side at the broader base. The fourteen smaller spines not visible on the surface, quite
rudimentary. Crests of the surface elegantly denticulated.
Dimensions.—Diameter of the shell 0.14; length of the two equatorial spines 0.08,
of the four polar spines 0.04; basal breadth 0.04.
Habitat.—Central Pacific, Station 271, surface.
{880}
Genus 379. Hexacolpus,[419] n. gen.
Definition.—Hexalaspida with six prominent radial spines
(in the hydrotomical plane), which are surrounded by prominent sheaths at the base; the fourteen
other spines quite rudimentary, not prominent.
The genus Hexacolpus differs from the preceding Hexonaspis in the
development of conical or cylindrical sheaths around the basal parts of the radial spines. It
bears to the latter the same relation as Hexaconus exhibits to Hexalaspis.
Subgenus 1. Hexacolparium, Haeckel.
Definition.—Six hydrotomical spines all nearly of equal size.
1. Hexacolpus nivalis, n. sp. (Pl. 139,
fig. 1).
Hexalaspis nivalis, Haeckel, 1882, Manuscript et Atlas.
All six hydrotomical spines of nearly equal size and equidistant, thin, lanceolate, compressed,
somewhat shorter than the diameter of the hexagonal shell. Sheaths nearly prismatic, conical in
the distal third, each with twelve prominent parallel crests, which are separated by twelve deep
furrows; their contracted distal opening or mouth denticulate. The sheaths are as long as the
radius of the shell, and envelop two-thirds of the spines. (Resembles certain forms of
snow-crystals.)
Dimensions.—Diameter of the shell 0.15, thickness of its wall 0.08; length of the
spines 0.1, of the sheath 0.007; breadth of the latter 0.04.
Habitat.—West Tropical Pacific, Station 225, surface.
2. Hexacolpus conifer, n. sp.
All six hydrotomical spines of equal size and equidistant, triangular, compressed, about as
long as the radius of the circular shell. Sheaths conical, sulcate, half as broad on the serrate
distal end as on the base. The sheaths envelop the basal half of the spines.
Dimensions.—Diameter of the shell 0.18; length of the spines 0.1, of the sheaths
0.06; basal breadth of the latter 0.07, distal breadth 0.04.
Habitat.—North Pacific, Station 236, surface.
Subgenus 2. Hexacolpidium, Haeckel.
Definition.—Six hydrotomical spines of unequal size, two opposite
(equatorial) much larger than the four other (polar) spines.
{881}
3. Hexacolpus trypanon, n. sp. (Pl. 140,
fig. 11).
Six hydrotomical spines of unequal size, two equatorial about as long as the diameter of the
shell and nearly twice as large as the four polar; all of the same form, quadrangular prismatic,
with prominent edges and pointed distal ends. Sheaths six-sided prismatic, with strong prominent
edges and thinner parallel ribs between them; their distal mouth with six strong denticulated
teeth. The sheaths envelop two-thirds of the spines, and are as broad as the radius of the
shell.
Dimensions.—Diameter of the shell 0.13; length of the equatorial spines 0.12, of
their sheaths 0.08; breadth of the latter 0.06.
Habitat.—South Pacific, Station 288, surface.
4. Hexacolpus dodecodus, n. sp.
Six hydrotomical spines of unequal size, two equatorial about as long as the radius of the
shell and twice as large as the four polar; all of the same form, triangular, compressed,
sulcated. Sheaths prismatic, with twelve prominent, parallel edges, and twelve strong serrated
triangular teeth on the mouth. The sheaths envelop the basal half of the spines. This species
greatly resembles Hexaconus serratus (Pl. 139, fig. 4),
but differs in the larger size of the equatorial spines and the complete reduction of the fourteen
smaller spines.
Dimensions.—Diameter of the shell 0.2; length of the equatorial spines 0.1, of
their sheaths 0.06; breadth of the latter 0.08.
Habitat.—North Pacific, Station 248, surface.
5. Hexacolpus infundibulum, n. sp. (Pl. 140, fig.
10).
Six hydrotomical spines of unequal size, two equatorial about twice as long as the diameter of
the shell and as the four polar spines; all six spines of the same form, quadrangular prismatic,
pointed at the distal pyramidal end. Sheaths conical, two to three times as broad at the
denticulate distal mouth as at the narrow base, sulcate; the mantle of the cone concavely vaulted.
The large sheaths envelop two-thirds or three-fourths of the spines.
Dimensions.—Diameter of the shell 0.1 to 0.13; length of the equatorial spines
0.15 to 0.2, of their sheaths 0.1 to 0.15; basal breadth of the latter 0.04, distal breadth
0.12.
Habitat.—Central Pacific, Station 272, surface.
Family XLIV. Diploconida, Haeckel (Pl. 140).
Diploconida, Haeckel, 1862, Monogr. d. Radiol., p.404.
Definition.—Acantharia with simple diploconical shell,
composed of two very large equatorial spines which are opposite in the hydrotomical axis, are
surrounded by conical or cylindrical, often compressed sheaths, and arise from a small central
{882}lattice-shell. Eighteen other spines (disposed
according to the Müllerian law of Icosacantha) much smaller, often rudimentary. Central capsule
ellipsoidal or diploconical.
The family Diploconida, founded by me in 1862 for a single
Mediterranean species (Diploconus fasces), appears to be the most aberrant and strange form
among the Acantharia. As I had met with only a single specimen, very dark
and intransparent in its central part, my observations on its structure were imperfect and the
explanation of it partly erroneous (compare my Monograph, pp. 46, 404, Taf. xx. figs. 7, 8).
However, I regarded Diploconus as the representative of quite a peculiar family, derived
from the Acanthometrida, and I correctly compared the large opposite radial spines of one
equatorial axis with the corresponding parts in Amphilonche.
Afterwards Richard Hertwig observed some specimens of Diploconus fasces in the same
locality (Messina), and gave an accurate description of its central capsule, including numerous
small nuclei (1879, Organismus d. Radiol., p. 28, Taf. ii. fig. 3). He found also that the
peculiar diploconical skeleton is not composed of silex, but of acanthin. In the explanation of
the shell-structure he adopted my opinion.
In the rich collections of the Challenger I detected ten different forms of Diploconida, all
very rare, and for the most part represented only by single specimens. A twelfth species was found
by me in the collection of Captain Rabbe from the Indian Ocean. By the study of these new forms,
and particularly by their comparison with the most nearly allied Hexalaspida and Belonaspida, it
was possible for me to correct some errors in my former description and to give a much more
correct description and natural explanation of this very peculiar and strange family of Radiolaria
(compare Pl. 140).
The most characteristic and the most voluminous part of the acanthinic skeleton in all
Diploconida appears as the diploconical or nearly cylindrical solid "mantle" giving them their
name and odd appearance (Pl. 140).
Usually this mantle is broader on its two opposite terminal openings than in its more or less
constricted middle part. This latter is now more spherical or ellipsoidal, now more lenticular,
and usually separated from the two cones by two slight transverse strictures. On the surface of
this middle part twelve to eighteen radial spines, which in Diplocolpus are rudimentary or
absent, are visible in Diploconus. The longitudinal axis of this shell is constantly
occupied by a very large pair of opposite stout prismatic or cylindrical principal spines, which
are united in the centre and usually more or less prominent with their distal apex over the two
openings of the double cone.
In my first communication on the Diploconus (1862, loc. cit.) I correctly
compared these two large spines in the prolonged main axis of the shell to the principal
equatorial spines of Amphilonche (or to the "hydrotomical spines," c1, c3);
but my explanation of the two peculiar cones enveloping them was erroneous. I supposed at that
time that they were formed by the eight flattened and leaf-shaped curved {883}tropical spines, so that around each principal spine the four
neighbouring tropical spines (two of the northern and two of the southern hemisphere) had grow
together by their edges and formed the peculiar conical sheath. I can now say that this opinion
(afterwards adopted also by Hertwig) was quite erroneous, the two conical or funnel-shaped sheaths
being the enlarged basal sheaths of the two hydrotomical spines, which we have already seen in the
Hexalaspida (Pl. 139).
But whilst in these latter all six principal spines of the hydrotomical meridian plane are
hypertrophied (two equatorial and four polar spines), in the nearly allied Diploconida only the
two opposite equatorial spines are developed, whilst all other eighteen spines are more or less
atrophied or quite rudimentary. In Diploconus the latter are more or less evident, whilst
in Diplocolpus they disappear externally.
The true lattice-shell of the Acanthophracta (constantly composed of
the meeting apophyses of twenty radial spines) is therefore represented in the Diploconida by the
small roundish middle part of the whole shell, which is usually much smaller than the two opposite
cones, and separated from them by the two slight transverse strictures. Usually this small but
most important middle part of the shell is very dark and opaque, on account of its very thick wall
and small pores; but in some species it is clear enough to ascertain that the structure of this
lattice-shell is the same as in the lenticular Hexalaspida, there being a network of thick crests
on the outer surface and small pores in the dimples between them. Indeed, in many (and probably in
all) Diploconida the forty aspinal pores are present which we found in all Hexalaspida,
Belonaspida, and Diporaspida, so that these four families of Acanthophracta represent one continuous phylogenetical series;
Phractaspis among the Diporaspida is at the beginning, and Diplocolpus among the
Diploconida at the end of this remarkably transformed morphological series.
The twenty radial spines in all Diploconida are probably united very firmly (or even perfectly
grown together) in the centre of the small thick-walled lattice-shell, the inner space of which is
extremely reduced. Probably, too, the sutures between the meeting apophyses of the thick radial
spines are often (or even constantly) obliterated by concrescence, so that the whole shell forms a
single piece of acanthin. But I regret that I cannot ascertain these and other points in the
structure of the shell, as the small number of specimens observed did not permit an anatomical
examination to be made. I have no doubt, however, that the structure of the whole of the middle
main part of the shell is quite the same as in the lenticular shell of the thick-walled
Hexalaspida, and that in both families each of the twenty radial spines bears originally only two
opposite apophyses.
The characteristic mantle of the double cone of the Diploconida, or the basal sheath of their
two large, perfectly developed principal spines is usually much larger than the shell itself, and
more or less compressed from both poles of the shortened geotomical axis. Therefore the transverse
section of the two cones is usually elliptical, more {884}rarely circular. Their widest part is generally the distal opening; more
rarely this is a little constricted. The thin transparent lamella of acanthin, representing the
mantle of the double cone, is commonly ribbed or furrowed by longitudinal, parallel or divergent
crests, and elegantly denticulated on the edge of the distal opening.
The two conical or cylindrical halves of the mantle are connected with the two enclosed
principal spines not only at the base, where they arise from the small central lattice-shell, but
also throughout a certain part of their length, by means of two, four, or six wings or leaves,
which lie opposite and in pairs in the meridian planes of those spines. These meridian wings are
more or less triangular (with broader concave outer bases), and connected by their axial edge with
the spine and by their peripheral edge with the mantle. They separate two, four, or six conical
spaces or pyramidal compartments in each cone. But these aspinal compartments and the separating
septa are not new productions of the Diploconida, but are inherited from their ancestral family,
the Hexalaspida (compare above, p. 873).
The eighteen smaller spines in Diploconus are either of nearly equal size or more or
less differentiated. The eight tropical spines are often much larger than the eight polar spines.
The two geotomical spines, (or the two opposite equatorial spines of the shortened geotomical
axis) are often quite rudimentary. In Diplocolpus the external part (outside the shell) is
in all eighteen smaller spines rudimentary or atrophied.
The Central Capsule, as shown by Hertwig, contains numerous small nuclei, and is divided
into three parts by the above named two transverse strictures; the smaller central part (in the
original lenticular lattice-shell) and the two opposite larger parts, filling up the greater part
of the two conical or cylindrical sheaths, and more or less adopting their form. Corresponding to
the shell itself the enclosed capsule is often more or less flattened, being compressed at both
poles of the geotomical axis. The pseudopodia seem to proceed only from the two large polar
apertures of the sheaths, and form therefore two opposite conical tufts or bunches.
Synopsis of the Genera of Diploconida.
| All twenty spines more or less developed (sometimes eight of them
rudimentary), |
380. Diploconus. |
| Only the two hydrotomical spines developed (all the eighteen
others rudimentary), |
381. Diplocolpus. |
Genus 380. Diploconus,[420] Haeckel,
1862, Monogr. d. Radiol., p. 404.
Definition.—Diploconida with two very large spines
(opposite in the hydrotomical axis) and ten to eighteen other much smaller spines externally
visible.
{885}
The genus Diploconus must be derived from Hexacolpus (among the
Hexalaspida) by the stronger development of the two hydrotomical spines and their large
sheaths; all the other eighteen spines are much smaller, and usually devoid of prominent sheaths.
Sometimes the eight tropical spines are rudimentary.
Subgenus 1. Diploconulus, Haeckel.
Definition.—Mantle of the double cone not compressed; its transverse
section therefore circular.
1. Diploconus amalla, n. sp. (Pl. 140,
fig. 1).
Mantle of the double cone thick walled, not compressed, its transverse section circular; its
contour little convex; its surface nearly smooth; its distal margin regularly denticulated.
Diameter of its mouth twice as large as the equatorial diameter of the shell and one-third as long
as its total length. The two large spines prismatic, one-fourth longer than their conical sheath.
The eighteen smaller spines very thin, about half as long as the former, a little curved.
Dimensions.—Length of the shell 0.3; equatorial breadth 0.05, polar breadth
0.1.
Habitat.—Central Pacific, station 271, surface.
2. Diploconus cyathiscus, n. sp. (Pl. 140, fig.
3).
Mantle of the double cone thin walled, not compressed; its transverse section circular; its
contour strongly convex; its surface with six stronger and many smaller ribs; its margin with
numerous straight and long, parallel denticles. Diameter of its mouth half as long as the whole
shell and one and a half times as long as its equatorial diameter. The two principal spines
one-fourth longer than their conical sheath. The eighteen smaller spines about half as long, thin,
straight. Both geotomical spines thick and short.
Dimensions.—Length of the shell 0.2; equatorial breadth 0.07, polar breadth
0.1.
Habitat.—West Tropical Pacific, Station 225, surface.
3. Diploconus cylindrus, n. sp.
Mantle of the double cone thick walled, dark, cylindrical, not compressed; its transverse
section circular; its contours straight, parallel; its surface with strong parallel straight
longitudinal ribs; its margin irregularly dentated. Diameter of its mouth one-fifth as long as the
whole shell and two-thirds as long as the diameter of the equatorial intumescence. The two
principal spines prismatic, nearly twice as long as their cylindrical sheath. The smaller spines
about half as long, thin, conical, straight. (Resembles the medial part of Hexacolpus
trypanon, Pl. 140,
fig. 11.)
Dimensions.—Length of the shell 0.25; equatorial breadth 0.08, polar breadth
0.05.
Habitat.—North Pacific, Station 241, surface.
{886}
Subgenus 2. Diploconium, Haeckel.
Definition.—Mantle of the double cone compressed from both poles of
the shortened geotomical axis; its transverse section therefore elliptical.
4. Diploconus fasces, Haeckel.
Diploconus fasces, Haeckel, 1862, Monogr. d. Radiol., p. 405, Taf. xx. figs. 7, 8.
Diploconus fasces, R. Hertwig, 1879, Organismus d. Radiol., p. 28, Taf.
ii. fig. 3.
Mantle of the double cone compressed, with elliptical transverse section; its contour straight;
its surface with numerous longitudinal furrows; its margin finely denticulated. Diameter of its
mouth two-fifths as long as the whole shell and three times as broad as its equatorial diameter.
The two main spines one-third longer than their conical sheath, four-edged. The eighteen smaller
spines thick and short, cylindrical or a little compressed.
Dimensions.—Length of the shell 0.18; equatorial breadth 0.025, polar breadth
0.07.
Habitat.—Mediterranean (Messina), surface.
5. Diploconus cotyliscus, n. sp. (Pl. 140, fig.
4).
Mantle of the double cone compressed, thin walled, with elliptical transverse section; its
contour convex; surface and margin nearly smooth. Diameter of its mouth half as long as the whole
shell and twice as long as its equatorial diameter. The two main spines scarcely longer than their
hemispherical sheath. The smaller spines conical, straight, nearly of the same length.
Dimensions.—Length of the shell 0.3; equatorial breadth 0.08, polar breadth
0.15.
Habitat.—Tropical Atlantic, Station 347, surface,
6. Diploconus saturnus, Haeckel.
Diploconus saturnus, Haeckel, 1879, Natürl. Schöpfungsgesch., Aufl. vii.
p. 706, Taf. xvi. fig. 11.
Mantle of the double cone compressed, thick walled, with elliptical transverse section; its
contour convex; its surface with numerous deep and irregular longitudinal furrows and marginal
incisions. Diameter of its mouth one-third as long as the whole shell and twice as broad as its
equatorial diameter. The two hydrotomical spines short, scarcely longer than their sheath. The
eighteen other spines thin, cylindrical, nearly of the same length, curved. (Resembles a
sheaf.)
Dimensions.—Length of the shell 0.24; equatorial breadth 0.04, polar breadth
0.07.
Habitat.—South Pacific, Station 300, surface.
7. Diploconus hexaphyllus, n. sp. (Pl. 140, fig.
2).
Mantle of the double cone compressed, thick walled, with elliptical transverse section; its
contour straight; surface with six stronger and numerous smaller straight ribs and six marginal
{887}denticulate incisions. Diameter of its mouth
one-third as long as the whole shell and somewhat smaller than the equatorial diameter.
Hydrotomical spines about twice as long as their sheath, pyramidal, with emarginate point and six
wings. Geotomical spines short and strong, pyramidal. Sixteen other spines thin, straight,
conical.
Dimensions.—Length of the shell 0.3; equatorial breadth 0.11, polar breadth
0.1.
Habitat.—Central Pacific, Station 266, surface.
Genus 381. Diplocolpus,[421] n. gen.
Definition.—Diploconida with only two developed, very
large spines (opposite in the hydrotomical axis); all the other eighteen spines quite rudimentary
or externally atrophied.
The genus Diplocolpus is the last and the most modified genus among the
Acantharia. The eighteen smaller spines of Diploconus are here
quite rudimentary or have even perfectly disappeared by atrophy, so that the shell seems to
consist only of the large hydrotomical spines and their enveloping sheaths.
Subgenus 1. Diplocolpulus, Haeckel.
Definition.—Mantle of the double cone not compressed; its transverse
section therefore circular.
1. Diplocolpus costatus, n. sp. (Pl. 140,
fig. 7).
Mantle of the double cone thin walled, not compressed; its transverse section circular or
nearly hexagonal; its contour little convex; surface with six thick longitudinal equidistant ribs
and many thinner ribs between them; margin denticulate. Transverse diameter of the shell one-third
as long as the longitudinal. The two hydrotomical spines thick and short, little prominent over
the polar mouth. No rudiments of other spines visible externally.
Dimensions.—Length of the shell 0.15; equatorial and polar breadth 0.05.
Habitat.—Indian Ocean (Madagascar), Rabbe, surface.
2. Diplocolpus cristatus, n. sp. (Pl. 140, fig.
6).
Mantle of the double cone thick walled, not compressed; its transverse section circular or
hexagonal; its contour nearly straight; surface with six thick prominent longitudinal equidistant
ribs; margin irregularly dentated. Transverse diameter of the shell half as long as the
longitudinal. {888}The two hydrotomical spines very
thick and short, pyramidal, little prominent. Short rudiments of the eighteen other spines
present, bristle-shaped.
Dimensions.—Length of the shell 0.1, breadth 0.05.
Habitat.—South Atlantic, Station 335, surface.
Subgenus 2. Diplocolpium, Haeckel.
Definition.—Mantle of the double cone compressed from both poles of
the shortened geotomical axis; its transverse section therefore elliptical.
3. Diplocolpus serratus, n. sp. (Pl. 140,
fig. 5).
Mantle of the double cone thick walled, compressed, with elliptical transverse section; its
surface with numerous delicate ribs; contour little convex; margin regularly serrate. Hydrotomical
spines short, pyramidal, with six serrate wings. Transverse diameter of the shell nearly twice as
great at the polar mouth as at the equator and half as long as the whole shell.
Dimensions.—Length of the shell 0.15; breadth on the equator 0.035, on the poles
0.07.
Habitat.—Central Pacific, Station 274, surface.
4. Diplocolpus dentatus, n. sp. (Pl. 140,
fig. 9).
Mantle of the double cone thin walled, compressed, with elliptical transverse section; its
contour straight; surface with six deeper and numerous shallower furrows; margin denticulated,
with six triangular prominent larger teeth. Hydrotomical spines very large prismatic, pyramidal at
the ends. Transverse diameter of the shell a little larger at the polar mouth than at the equator
and half as long as the whole shell.
Dimensions.—Length of the shell 0.2; breadth on the equator 0.08, on the poles
0.1.
Habitat.—North Pacific, Station 244, surface.
5. Diplocolpus sulcatus, n. sp. (Pl. 140,
fig. 8).
Mantle of the double cone thin walled, compressed, with elliptical transverse section; its
contour straight; surface with twelve to sixteen deep longitudinal furrows, which are separated by
thin, undulating, double edged ribs; margin little dentated. Hydrotomical spines short, pyramidal,
with six dentate wings. Transverse diameter of the shell a little larger at the polar mouth than
at the equator, about one-third of the whole length of the shell.
Dimensions.—Length of the shell 0.17; breadth on the equator 0.05, on the poles
0.06.
Habitat.—South Pacific, Station 291, surface.
Notes.