
CAMBRIDGE BIOLOGICAL SERIES.
General Editor:—Arthur E. Shipley, M.A., F.R.S.
FELLOW AND TUTOR OF CHRIST’s COLLEGE, CAMBRIDGE.
GRASSES.
CAMBRIDGE UNIVERSITY PRESS WAREHOUSE,
C. F. CLAY, Manager.
London: FETTER LANE, E.C.
Edinburgh: 100, PRINCES STREET.
ALSO
London: H. K. LEWIS, 136, GOWER STREET, W.C.
Leipzig: F. A. BROCKHAUS.
New York: G. P. PUTNAM’S SONS.
Bombay and Calcutta: MACMILLAN & CO. Ltd.
[All Rights reserved.]
A HANDBOOK FOR USE IN THE FIELD
AND LABORATORY.
BY
H. MARSHALL WARD, Sc.D., F.R.S.
LATE PROFESSOR OF BOTANY IN THE UNIVERSITY OF CAMBRIDGE.
CAMBRIDGE:
AT THE UNIVERSITY PRESS.
1908
First Edition 1901
Reprinted 1908
THE following pages have been written in the hope that they may be used in the field and in the laboratory with specimens of our ordinary grasses in the hand. Most of the exercises involved demand exact study by means of a good hand-lens, a mode of investigation far too much neglected in modern teaching. The book is not intended to be a complete manual of grasses, but to be an account of our common native species, so arranged that the student may learn how to closely observe and deal with the distinctive characters of these remarkable plants when such problems as the botanical analysis of a meadow or pasture, of hay, of weeds, or of “seed” grasses are presented, as well as when investigating questions of more abstract scientific nature.
I have not hesitated, however, to introduce general statements on the biology and physiological peculiarities of grasses where such may serve the purpose of interesting the reader in the wider botanical bearings of the subject, though several reasons may be urged against extending this part of the theme in a book intended to be portable, and of direct practical use to students in the field.
I have pleasure in expressing my thanks to Mr R. H. Biffen for carefully testing the classification of “seeds" on pp. 135-174, and to him and to Mr Shipley for kindly looking over the proofs; also to Mr Lewton-Brain, who has tested the classification of leaf-sections put forward on pp. 72-82, and prepared the drawings for Figs. 21-28.
That errors are entirely absent from such a work as this is perhaps too much to expect: I hope they are few, and that readers will oblige me with any correctionsvi they may find necessary or advantageous for the better working of the tables.
The list of the chief authorities referred to, which students who desire to proceed further with the study of grasses should consult, is given at the end.
I have pleasure in acknowledging my indebtedness to the following works for illustrations which are inserted by permission of the several publishers:—Stebler’s Forage Plants (published by Nutt & Co.), Nobbe’s Handbuch der Samenkunde (Wiegandt, Hempel and Parey, Berlin), Harz’s Landwirthschaftliche Samenkunde (Paul Parey, Berlin), Strasburger and Noll’s Text-Book of Botany (Macmillan & Co.), Figuier’s Vegetable World (Cassell & Co.), Lubbock’s Flowers, Fruits and Seeds (Macmillan & Co.), Kerner’s Natural History of Plants (Blackie & Son), and Oliver’s First Book of Indian Botany (Macmillan & Co.).
It is impossible to avoid the question of variation in work of this kind, and students will without doubt come across instances—especially in such genera as Agropyrum, Festuca, Agrostis and Bromus—of small variations which show how impossible it is to fit the facts of living organisms into the rigid frames of classification. It may possibly be urged that this invalidates all attempts at such classifications: the same argument applies to all our systems, though it is perhaps less disastrous to the best Natural Systems which attempt to take in large groups of facts, than to artificial systems selected for special purposes. Perhaps something useful may be learned by showing more clearly where and how grasses vary, and I hope that the application to them of these preliminary tests may elucidate more facts as we proceed.
H. M. W.
Cambridge, April, 1901.
| CHAPTER I. | |
| PAGE | |
| The Vegetative Organs | 1 |
| CHAPTER II. | |
| The Vegetative Organs (continued) | 17 |
| CHAPTER III. | |
Grasses Classified according to their Vegetative Characters | 39 |
| CHAPTER IV. | |
| Anatomy and Histology | 62 |
| CHAPTER V. | |
Grasses Classified according to the Anatomical Characters of the Leaf | 72 |
| viiiCHAPTER VI. | |
| Grasses in Flower | 83 |
| CHAPTER VII. | |
Grasses Grouped according to their Flowers and Inflorescences | 99 |
| CHAPTER VIII. | |
| The Fruit and Seed | 119 |
| CHAPTER IX. | |
| Classification of Grasses by the “Seeds" (Grains) | 135 |
| Bibliography | 175 |
| Index, Glossary and List of Synonyms | 177 |
That grasses are interesting and important plants is a fact recognised by botanists all the world over, yet it would appear that people in general can hardly have appreciated either their interest or their importance seeing how few popular works have been published concerning their structure and properties.
Apart from their almost universal distribution, and quite apart from the fascinating interest attaching to those extraordinary tropical giants, the Bamboos, West Indian Sugar-cane, the huge Reed-grasses of Africa, the Pampas-grasses of South America; and from the utilitarian value of the cereals—Maize, Rice, Wheat and other corn, &c.—everyone must be struck by the significance of the enormous tracts of land covered by grasses in all parts of the world, the Prairies of North America and the Savannahs of the South, the Steppes of Russia and Siberia, and the extensive tracts of meadow and pasture-land in Europe being but a few examples.
Although in the actual number of species the Grass family is by no means the largest in the vegetable kingdom, for there are far more Composites or Orchids, the curious sign of success in the struggle for existence comes out in grasses in that the number of individuals far transcends those of any other group, and that they have taken possession of all parts of the earth’s surface. Some species are cosmopolitan—e.g. our common Reed, Arundo Phragmites; while others—e.g. several of our native species of Festuca and Poa—are equally common in both hemispheres. On the whole the Tropics afford most species and fewest individuals, and the temperate regions most individuals.
Considering their multifarious uses as fodder and food, for brewing, weaving, building and a thousand other purposes, it is perhaps not too much to say that if every other species of plant were displaced by grasses of all kinds—as many indeed gradually are—man would still be able to supply his chief needs from them.
The profound significance of the grass-carpet of the earth, however, comes out most clearly when we realise the enormous amounts of energy daily stored up in the countless myriads of green blades as they fix their carbon. By decomposing the carbon-dioxide of the air in their chlorophyll apparatus by the action of the radiant energy of the sun, they build up starches and sugars and other plant-substances, which are then consumed and turned into flesh by our cattle and sheep and other herbivorous animals, and so furnish us with food. The whole theory of agriculture turns on this pivot, and the by no means3 small modicum of truth in such sayings as “All flesh is grass,” and that the man who can make two blades of grass grow where one grew before deserves well of his country, obtains a larger significance when it is realised that the only real gain of wealth is that represented by the storage of energy from without which comes to us by the action of green leaves waving in the sunshine.
The true Grasses, comprising the Natural Order Graminaceæ—also written Gramineæ—are often popularly confounded with other herbs which possess narrow green ribbon-like leaves, or even with plants of very different aspects—e.g. Cotton-grass (Eriophorum) and other Sedges, and the names Rib-grass (Plantago), Knot-grass (Polygonum), Scorpion-grass (Myosotis) and Sea-grass (Zostera), as well as the general usage of the word grass to signify all kinds of leguminous and other hay-plants in agriculture, point to the wider use of the word in former times. This has been explained by the use of the words gaers, gres, gyrs, and grass in the old herbals to indicate any kind of small herbage.
In view of the importance of our British grasses in agriculture, I have here put together some results of observation and reading in the hope that they may aid students in recognising easily our ordinary agricultural and wild grasses. During several years of work in the fields, principally directed at first to the study of the parasitic fungi on grasses, and subsequently to that of the importance of grasses in forestry and agriculture, and to the variations they exhibit, the need of some guide to the identification of a grass at any time of the year,4 whether in flower or not, forced itself on the attention, and although a botanist naturally turns to a good Flora when he has the grass in flower, as the best and quickest way of ascertaining the species, it soon became evident that much may be done by the study of the leaves and vegetative parts of most grasses. Indeed some are recognisable at a glance by certain characters well known to continental observers: in the case of others the matter is more difficult, and perhaps with a few it is impossible to be certain of the species from such characters only.
Nevertheless, while the best means for the determination of species are always in the floral characters so well worked up in the Floras of Hooker, Bentham and others, there is unquestionably much value in the characters of the vegetative organs also, as the works of Jessen, Lund, Stebler, Vesque and others abroad, and Sinclair, Parnell, Sowerby and others in this country attest.
Almost the only plants confounded with true grasses by the ordinary observer are the sedges and a few rushes. Apart from the very different floral structures, there are two or three easily discoverable marks for distinguishing all our grasses from other plants (Fig. 1). The first is their leaves are arranged in two rows, alternately, up the stems; and the second that their stems are circular or flattened in section, or if of some other shape they are never triangular and solid1 (Figs. 6 and 7). Moreover the leaves are always of some elongated shape, and without 5leaf-stalks2, but pass below into a sheath, which runs some way down the stem and is nearly always perceptibly split 6(Figs. 8-13). Further, the stems themselves are usually terete, and distinctly hollow except at the swollen nodes, and only branch low down at the surface of the ground or below it3.
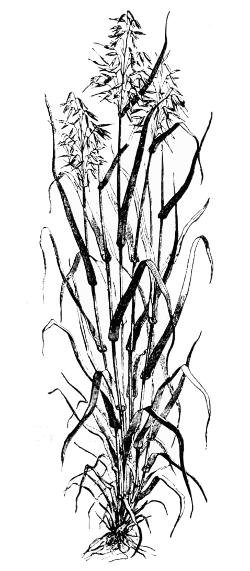
Fig. 1. A plant of Oat (Avena), an example of a typical grass, showing tufted habit and loose paniculate inflorescence (reduced). Figuier.
All our native grasses are herbaceous, and none of
them attain very large dimensions. In the following lists
I term those small which average about 6-18 inches in
the height of the tufts, whereas those over 3 feet high
may be termed large, the tufts being regarded as in
flower. The sizes cannot be given very accurately, and
starved specimens are frequently found dwarfed, but in
most cases these averages are not far wrong for the
species freely growing as ordinarily met with, and in
some cases are useful. I have omitted the rare species
throughout, and in the annexed lists have added the
popular names.
Large Grasses.
(Over 3 feet.)
Milium effusum (Millet-grass).
Digraphis arundinacea (Reed-grass).
Aira cæspitosa (Tufted Hair-grass).
Arrhenatherum avenaceum (False Oat).
Elymus arenarius (Lyme-grass).
Bromus asper (Hairy Brome).
B. giganteus (Tall Brome).
Festuca elatior (Meadow Fescue).
F. sylvatica (Reed Fescue).
Glyceria aquatica (Reed Sweet-grass).
G. fluitans (Floating Sweet-grass).
Arundo Phragmites (Common Reed).
Medium Grasses.
(1-3 feet.)
Phleum pratense (Timothy).
Avena pratensis (Perennial Oat-grass).
Anthoxanthum odoratum (Sweet Vernal).
Alopecurus agrestis (Slender Foxtail).
A. pratensis (Meadow Foxtail).
Agrostis alba (Fiorin).
Psamma arenaria (Sea Mat-grass).
Avena flavescens (Yellow Oat-grass).
Holcus lanatus (Yorkshire Fog).
Hordeum sylvaticum (Wood Barley).
H. pratense (Meadow Barley).
Agropyrum repens (Couch-grass).
A. caninum (Fibrous Twitch).
Lolium italicum (Italian Rye-grass).
Brachypodium sylvaticum (Wood False-Brome).
B. pinnatum (Heath False-Brome).
Bromus erectus (Upright Brome).
B. sterilis (Barren Brome).
B. arvensis (Field Brome).
Festuca ovina (var. rubra, &c.). (Sheep’s Fescue).
F. elatior (var. pratensis). Meadow Fescue.
Dactylis glomerata (Cock’s-foot).
Cynosurus cristatus (Crested Dog’s-tail).
Poa pratensis (Meadow-grass).
P. trivialis (Rough stalked Meadow-grass).
P. nemoralis (Wood Poa).
Molinia cærulea (Flying Bent).
Melica nutans (Mountain Melick).
M. uniflora (Wood Melick).
Small Grasses.
(6-18 inches.)
Phleum arenarium (Sand Cat’s-tail).
Alopecurus geniculatus (Marsh Foxtail).
Agrostis canina (Brown Bent).
Aira flexuosa (Wavy Hair-grass).
8 Aira canescens (Grey Hair-grass).
A. præcox (Early Hair-grass).
A. caryophyllea (Silvery Hair-grass).
Nardus stricta (Moor Mat-grass).
Hordeum murinum (Wall Barley).
H. maritimum (Sea Barley).
Lolium perenne (Rye-grass).
L. temulentum (Darnel).
Brœmus arvensis (var. mollis). (Field Brome).
Festuca ovina (Sheep’s Fescue).
F. Myuros (Rat’s-tail Fescue).
Briza media (Quaking-grass).
Poa maritima (Sea Poa).
P. annua (Annual Meadow-grass).
P. compressa (Flattened Meadow-grass).
P. alpina (Alpine Poa).
P. bulbosa (Bulbous Poa).
Triodia decumbens (Heath-grass).
Kœleria cristata (Crested Kœleria).
The roots of our grasses are almost always thin and
fibrous and are adventitious from the nodes, frequently
forming radiating crowns round the base and easily pulled
up, and usually broken in the process; but in the case
of a few moor grasses—especially Nardus (Fig. 2) and
Molinia—the roots are so tough and thick (stringy) as to
resist breakage very efficiently. In stoloniferous grasses a
similar difficulty of removal may be caused in a slighter
degree by the underground stems. In a few cases, e.g.
Alopecurus bulbosus (Fig. 3), Poa bulbosa, Phleum pratense
and P. Bœhmeri, Arrhenatherum avenaceum, and to a
slighter extent in Poa alpina and one or two others, the
lowermost internodes and sheaths of the stems may be
swollen and stored with food-materials, and a sort of tuber
or bulb results; this is especially apt to occur in dry sandy
9
10soils. In old lawns, pastures, &c., the roots of Poa annua
and others may have nodules on them due to the presence
of certain small Nematode worms, Heterodera.
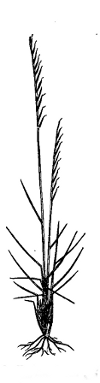 |
 |
Fig. 2. Nardus stricta. Plant showing tufted habit, and simple spikate inflorescence, with pointed spikelets all turned towards one side (secund) on the rachis (reduced). Note also the bristle-like (setaceous) leaves at length reflexed. Parnell. |
Fig. 3. Alopecurus geniculatus, var. bulbosus. Plant (reduced) showing habit, bulbous shoots and cylindrical spike-like inflorescences (Foxtail type). Notice the inflated sheaths, and the “kneed" lower parts of the ascending stems. Parnell. |
Grasses are annual, biennial, or perennial, and it is
often of importance to know which. The point may usually
be determined by examining the shoots. If all the shoots
have flowering stems in them, and are evidently of the
current year, the grass is an annual; but if any shoots
have leaves only, it is either biennial or perennial: to
determine which is not always easy, but in perennial
grasses there will generally be evident remains of older
leaf-bases and shoots, and if there are distinct underground
stolons or creeping rhizomes as well the point
may be considered decided, and the grass is perennial, as
is the case with most of our important species. If all the
shoots are barren, the grass is a biennial in its first year
of growth: if all have flowering stems in them, but show
traces of old leaf-bases of the previous year, then the grass
is a biennial in its second year. The proof of biennial
character is not always easy, however, and a few grasses
may be either annual or biennial, or biennial or perennial,
according to conditions—e.g. species of Hordeum, Bromus,
&c. In the following lists I have given the duration of
the principal grasses, where the character is especially
important.
Annuals.
Phleum arenarium.
Aira præcox.
A. caryophyllea.
Hordeum murinum.
H. maritimum.Lolium temulentum.
Festuca Myurus.
Briza minor.
Poa rigida.
P. annua.Annuals
which may become biennial or perennial.
Alopecurus geniculatus.
Hordeum pratense.
Lolium perenne.
L. italicum (may be perennial).
Bromus asper (may be perennial).
B. sterilis.
B. arvensis (may be perennial).Perennials.
Holcus lanatus.
H. mollis.
Nardus.
Hordeum sylvaticum.
Agropyrum.
Brachypodium.
Bromus erectus.
B. giganteus.
Festuca ovina.
F. elatior.
F. sylvatica.
Dactylis.
Cynosurus cristatus.
Briza media.
Milium.
Anthoxanthum.
Digraphis.
Phleum pratense.
Alopecurus pratensis.
Agrostis alba.
A. canina.Psamma.
Aira cæspitosa.
A. flexuosa.
A. canescens.
Avena pratensis.
A. flavescens.
Arrhenatherum.
Glyceria aquatica.
G. fluitans.
Poa maritima.
P. compressa.
P. pratensis.
P. trivialis.
P. nemoralis.
P. alpina.
P. bulbosa.
Molinia.
Melica.
Triodia.
Kœleria.
Arundo.
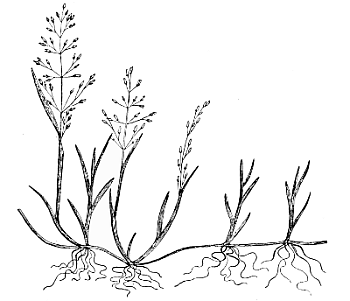
Fig. 4. Catabrosa aquatica. Plant showing the creeping habit, rooting nodes, and paniculate inflorescence (reduced). Parnell.
The rhizome of a perennial grass is continued sympodially
by means of buds branching from the lowermost
joints of the flowering shoots, and some importance is
attached to the mode of spreading of these lateral sprout12ing
shoots. The buds always arise in the axils of the lower
leaf-sheaths—i.e. they are intra-vaginal. If they remain
intra-vaginal during further growth, the shoots are forced
upwards and only tufts (Fig. 2) are formed, except in so far
as such shoots may fall prostrate on the surface of the
ground later, and throw out roots from their nodes, and
so act as runners or offsets, or put out a few roots &c.
as they ascend through the soil. But in many cases the
buds soon burst through the leaf-sheaths, and develope
as extra-vaginal shoots, and may then run horizontally
as underground stolons. Only creeping grasses of these
latter kinds can rapidly cover large areas4: the grasses
13
with intra-vaginal shoots only can only make tufts or
"tussocks." Several peculiarities in the habits of grasses
depend on these facts. The following are the most
important creeping, or stoloniferous species, contrasted
with the much more common tufted and the far rarer
grasses with runners above ground (Fig. 4). Some of
these (Elymus, Psamma, &c.) are of great importance as
sand-binders.
With intra-vaginal branches only.
Lolium—slightly stoloniferous.
Festuca elatior—slightly stoloniferous.
Avena flavescens—slightly stoloniferous.
Phleum pratense—no stolons, but may be bulbous.
Dactylis—no stolons.
Festuca ovina—no stolons.
Poa alpina—no stolons.
Cynosurus—no stolons.With extra-vaginal shoots.
Arrhenatherum—short stolons, sometimes bulbous.
Holcus lanatus—creeping.
Alopecurus pratensis—long stolons.
Anthoxanthum—slightly stoloniferous.
Agrostis alba (var. stolonifera)—long stolons and runners.
Digraphis—long stolons.
Poa pratensis—long stolons.
P. trivialis—runners only.
Festuca heterophylla, Lam.—a variety of F. ovina with slight stolons.
F. rubra (Linn.)—a variety of F. ovina with long stolons.
Bromus erectus—no stolons.
B. inermis—long stolons.Creeping below ground and truly stoloniferous.
Agropyrum.
Elymus.
Psamma.
Poa pratensis.
P. compressa.
Agrostis alba (var. stolonifera).
Alopecurus pratensis.
Brachypodium (slightly).Bromus erectus (slightly).
Festuca ovina (var. rubra, Linn.).
F. elatior (slightly).
Briza (slightly).
Glyceria.
Poa maritima.
Melica.
Arundo.Tufted Grasses.
Milium.
Agrostis alba (on downs, &c.).
Aira cæspitosa.
A. flexuosa.
A. canescens.
A. præcox.
A. caryophyllea.
Avena pratensis (slightly creeping).
Arrhenatherum.
Nardus (Fig. 2).
Hordeum sylvaticum.
Lolium.
Bromus.
Festuca ovina (except some varieties).F. sylvatica.
F. Myurus.
Dactylis.
Cynosurus.
Poa rigida.
P. annua.
P. trivialis.
P. nemoralis.
P. alpina.
P. bulbosa.
Molinia.
Triodia.
Kœleria.Creeping above ground (with runners).
Holcus lanatus.
Alopecurus geniculatus.
Agrostis alba (var. stolonifera).
Hordeum pratense (slightly).
H. murinum (slightly).
Catabrosa (Fig. 4).
Cynodon (Fig. 5).
Hackel has pointed out that a distinction must be drawn
between the true nodes of the culm, and the swellings15
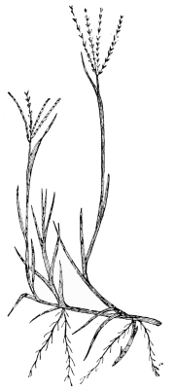 Fig. 5. Cynodon Dactylon. Plant (reduced) showing creeping and stoloniferous habit, and peculiar inflorescence of digitate
spikes. Parnell.often found at the base of the sheaths themselves over
these: the latter are often conspicuous when the
former are inconspicuous—e.g.
most species of Agrostis, Avena,
Festuca, &c.
Fig. 5. Cynodon Dactylon. Plant (reduced) showing creeping and stoloniferous habit, and peculiar inflorescence of digitate
spikes. Parnell.often found at the base of the sheaths themselves over
these: the latter are often conspicuous when the
former are inconspicuous—e.g.
most species of Agrostis, Avena,
Festuca, &c.
The nodes are of importance in the description of a few species only—e.g. they are usually dark coloured in certain Poas such as P. compressa and P. nemoralis; they are sharply bent in Alopecurus geniculatus, and may be so in other species if “layed" by wind, rank growth, &c.
A point of considerable classificatory value is the shape of the transverse section of the shoot, which is correlated with the mode of folding up of the young leaf-blades.
In most grasses the blades are convolute—i.e. rolled up like the paper of a cigarette, one edge over the other—and the section of the shoot is round (Fig. 7). In some cases, however, the leaves are conduplicate—i.e. each half of the lamina is folded flat on the other, the upper sides being turned face to face inwards, with the mid-rib as the hinge—and in this case the shoots are more or less compressed (Fig. 6).
In these latter cases the transverse section may be
elliptical—e.g. Poa pratensis and P. alpina, Briza, &c.,
or more flattened and linear-oblong—e.g. Glyceria fluitans—with
the flattened sides straight, or the section is
oval but pointed more or less at each end owing to projecting
keels and leaf-edges, and the form is naviculate—e.g.
Glyceria aquatica, Dactylis (Fig. 6)—or, the sides being
less flattened, more or less rhomboidal as in Poa trivialis.
In Melica the leaves are convolute and the shoot-section
quadrangular.
Flat, and usually sharp-edged shoots.
Dactylis glomerata (Fig. 6).
Poa trivialis, P. annua, P. pratensis, P. compressa, P. maritima,
and P. alpina.
Glyceria aquatica and G. fluitans.
Avena pubescens.
Lolium perenne.
 |
 |
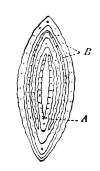
Fig. 6. Dactylis glomerata. Transverse section of a leaf-shoot (× 5). A, conduplicate leaf-blade. B, sheath. Stebler. |
Fig. 7. Digraphis arundinacea. Transverse section of a leaf-shoot (× 5). A, sheath. B, convolute leaves. Compare Fig. 14. Stebler. |
The leaves of all our grasses consist of the blade, which passes directly into the sheath, without any petiole or leaf-stalk (Fig. 1).
The sheath is usually obviously split, and so rolled
round the internode that one edge overlaps the other,
but in the following grasses the sheath is either quite
entire, or only slit a short way down, the two edges being
fused as it were for the greater part of its length.
Sheath more or less entire.
Glyceria aquatica and G. fluitans.
Melica uniflora and M. nutans.
Dactylis glomerata.
Poa trivialis (Fig. 8), P. pratensis, P. alpina.
Sesleria cærulea.
Bromus (all the species).
Briza media and B. minor.
In some cases—e.g. Arrhenatherum, Bromus asper, and Holcus lanatus—the sheath is marked with a more or less18 prominent ridge down its back, due to the continuation of the keel of the leaf. The sheath may also be glabrous or hairy, and grooved or not.
A few grasses are so apt to develope characteristic
colours in their sheaths, especially below, that they may
often be recognised in winter by this peculiarity.
Sheaths coloured.
Lolium—all red.
Holcus—red with purple veins.
Festuca elatior—red.
Cynosurus—yellow.
Alopecurus pratensis, and
A. agrestis—violet-brown, &c.
Festuca ovina, var. rubra—red.
 |
 |
 |
Fig. 8. Poa trivialis. A, base of blade. B, ligule. C, sheath. D, culm (× about 3). |
Fig. 9. Alopecurus pratensis. A, base of blade. B, ligule. C, sheath. Slightly magnified. |
Fig. 10. Avena flavescens. Lettering as before (× 2). Note the split sheath, the hairs and ridges. Stebler. |
At the junction of the blade with the sheath there is in most cases a delicate membranous upgrowth of the former, more or less appressed to the stem, and called the Ligule (Figs. 8-13). Its use is probably to facilitate the shedding19 of water which has run down the leaf; and so lessen the danger of rotting between the sheath and stem: possibly the shelves and ears commonly met with at the base of the lamina (Fig. 12) aid in the same process. This ligule may be long or short, acute or obtuse, toothed or entire, or it may be reduced to a mere line, or tuft of hairs, or even be obsolete, and is of considerable value in classification—e.g. the ligule is obsolete or wanting in Melica, Festuca ovina, F. Myurus, F. elatior, Kœleria and Panicum.
It is represented by a tuft of hairs in Molinia, Triodia and Arundo.
 |
 |
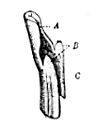 |
Fig. 11. Lolium perenne. A, base of lamina, B, ligule. C, sheath (× 3). Note the low ribs, and absence of hairs (glabrous). |
Fig. 12. Festuca elatior, var. pratensis. A, base of lamina. B, the extremely short ligule, with pointed ears. C, sheath (× 3). |
Fig. 13. Festuca ovina. A, base
of lamina. B,
ligular ears. C,
sheath (× about
4). Stebler. |
Our other ordinary grasses have a more or less well-developed membranous ligule (Fig. 8).
The leaf-blade is long or short, broad or narrow, but always of some elongated form such as linear, linear-lanceolate or linear-acuminate, or subulate, setaceous, &c., varying as to the degree of acuteness of the apex, and the tapering of the base.
In the following native grasses the form of the lamina affords a useful character.
The base tapers to the sheath below—i.e. the leaf is more or less linear-lanceolate—in Molinia, Brachypodium, Melica, Milium, Kœleria, and the very rare Hierochloe; less distinctly so in Bromus asper and species of Hordeum. The base is rounded in Arundo. In the following cases the leaves are setaceous, due to the very narrow blade remaining permanently folded or inrolled at its edges, and usually being thickened and hardened also (Figs. 13 and 18). The habitat of these moor-and heath-grasses suggests that these are no doubt adaptations to prevent excessive evaporation by the exposure of too large a surface—e.g. various species of Aira, Festuca ovina, F. Myurus and allies, Nardus, and several other species; whereas, conversely, the thin flat leaves of shade-grasses facilitate exposure to light and transpiration. In Avena pratensis and Agrostis canina some of the leaves are involute and subulate, and the thickened leaves of Poa maritima also are turned up at the edges, and are U-shaped in cross-section.
As we shall see later the degree of inrolling of many grass leaves varies with circumstances.
In most others the blades are either flat (Figs. 8-12), or more or less conduplicate on the mid-rib. The latter case occurs, for example, in grasses with flattened shoots, especially at the lower part of the blade—e.g. Lolium perenne, Dactylis, Glyceria, and some species of Poa, and the cross-section of the leaf below, just before it enters the sheath, is V-shaped. In Glyceria the leaf-bases may show yellow or brownish triangles.
Further characters of the leaves are derived from their texture, apex, margins, mid-ribs and venation, hairiness, and especially the presence and characters of the longitudinal ridges which run along the upper or lower surface in many cases.
The venation is parallel from base to apex in nearly all our grasses, but such is not always the case—e.g. in the exotic Panicum plicatum the mid-rib, which enters the leaf with several vascular bundles, gives off strong and weak veins below, which first diverge and then run in arches which converge upwards: this leaf is also remarkable in being plaited (plicate) in vernation. In Arundo Donax also the veins, though approximately parallel, do not all run to the apex of the tapering leaf; the outer ones end above in the margins and are shorter than the mid-rib.
As regards texture, the leaves of most grasses are thin and herbaceous; but in some they are dry and harsh to the touch. They are thin and dry in Agropyrum caninum, Hordeum pratense, H. murinum, Avena pratensis, &c., very hard and leathery (coriaceous) in Psamma, Nardus, species of Festuca, Aira, Agropyrum junceum, Elymus, &c. In aquatic grasses like Glyceria, the leaf is almost spongy owing to the large air-chambers developed in the tissues. These are easily visible with a lens.
The apex is in most cases slender and tapering—acuminate; but in some it is merely brought to a point (acute) as in Catabrosa, Glyceria and several species of Poa and Avena, &c., usually flat, but somewhat hooded or curved up in some Poas. In cases where the leaves are setaceous or subulate, the apex is like a thin tapering22 bristle, and even flatter leaves may be so inrolled at the tips as to have the apex prolonged into a sharp needle-like pungent or spinescent point—e.g. Hordeum pratense, Avena pratensis to a slight extent, and pronounced in Elymus, &c. In Sesleria the apex is rounded with a short, sharp, prickle-like median projection (mucronate).
The passage of blade into sheath has already been described, but the base of the blade may have its margins projecting as horizontal shelves, like a Byron collar, round the sides of the throat of the sheath, sometimes tinged with yellow or pink—e.g. Lolium, Holcus, Bromus inermis, Hordeum; the ends of these may project as auricles or ears—e.g. Festuca elatior, Elymus, Agropyrum, Anthoxanthum, Bromus asper, Hordeum, &c. In Festuca ovina the ears are short, stiff, and erect (Fig. 13).
The margin may be perfectly even, as in most grasses, or it is more or less scabrid or scaberulous, as in Aira cæspitosa, Poa maritima, Festuca elatior, Avena pratensis, Agrostis, Milium, Phleum, Briza, the minute teeth (serrulæ) pointing up or down.
The surface may be bright green, or glaucous, harsh, hairy or glabrous, and is not uncommonly also scabrid, like a file or emery-paper, and sometimes only when rubbed in one direction up or down, owing to the minute teeth being directed all one way. These teeth are developed on the ridges.
All our ordinary grass leaves are parallel-veined, and the vascular strands (the veins) can usually be seen on holding the leaf up to the light. In most cases the tissue is raised over the veins, as ridges or “ribs," and according23 to the height of these ridges the thinner parts between look like deep or shallow furrows (cf. Figs. 8-16 and Chapter IV.). If the leaf is held up to the light the ridges appear dark in proportion to their opacity—i.e. height or thickness—and the furrows light in proportion to the thinness of the tissues there. If the contrast is very great, as in Aira cæspitosa (Fig. 23), the furrows seem like transparent sharp lines, and when, as in Poa, which is practically devoid of ridges, the difference of thickness is small they appear merely as fine striæ. These characters must be determined on the fresh leaves, however, because the contraction in drying draws the ridges closer together and tends to obliterate the lines.
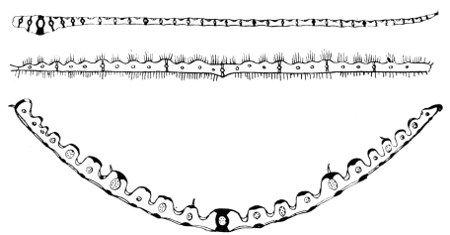
Fig. 14. Digraphis arundinacea. Transverse section of mid-rib and half the leaf (× about 6).
Fig. 15. Holcus lanatus. Transverse section of leaf-blade (× 10).
Fig. 16. Cynosurus cristatus. Transverse section of the leaf-blade (× 20). Stebler.
The ridges are almost always evident—Catabrosa, Poa, and Avena furnishing the chief exceptions—and are nearly invariably on the upper surface: they are below in Melica,24 however; and their relative numbers, heights and breadths, section—acute, rounded, or flattened—furnish valuable characters; as also does the coexistence or absence of hairs, asperities, &c.

Fig. 17. Transverse section of the leaf of Festuca elatior, var. pratensis (× 12).
Fig. 18. Ditto of the leaf of F. ovina (× 15).
Fig. 19. Ditto of the leaf of F. ovina, var. rubra (× 35).
Fig. 20. Festuca ovina, var. rubra. Transverse section of the blade of an upper leaf (× 35). Stebler.
A very interesting anatomical adaptation is met with in the leaves of many grasses which grow in dry situations (xerophytes) such as on sandy sea-shores, exposed mountains and so forth. When the air is moist, in wet weather or in the dews, and the sun’s rays not too powerful, the leaf is spread out with its upper surface flat or nearly so, but when the scorching sun and dry air or winds prevail, the leaves fold or roll up, with the upper sides apposed or overlapping inside the hollow cylinder thus made.
In such leaves some of the upper epidermal cells,
either next the mid-rib (Sesleria &c.) or between the other
ribs (Festuca &c.) are large and very thin-walled, full of
sap when distended, and so placed that as they lose water
by evaporation they contract, and so draw together
the two halves of the lamina (Sesleria) or each ribbed
segment (Festuca), thus causing the infolding or inrolling
(see Chapter IV). Not only from the structure
and actions of these motor-cells, but also from the fact that
the stomata are on the upper surfaces and thus protected,
and that the lower surfaces which alone are exposed to
the drought are defended by hard and impenetrable
tissues, we must look upon these as adaptations to the
xerophytic conditions.
Leaves prominently ridged.
Elymus.
Psamma.
Aira cæspitosa.
Lolium.
Cynosurus (Fig. 16).
Agrostis.Alopecurus.
Glyceria fluitans.
Kœleria.
Festuca elatior.
Festuca Myurus (var. sciuroides).
Melica has ridges on the lower surface.
Ridges are less prominent in Phleum pratense, Briza, Agropyrum, Triodia, Arrhenatherum avenaceum.
Leaves practically devoid of ridges.
Poa—all common species.
Glyceria aquatica.
Catabrosa aquatica.
Avena pratensis.
In some grasses the tissue over the mid-rib is considerably
raised and strengthened on the dorsal side of the
blade as a “keel."
Keel more or less prominent.
Arrhenatherum (sheath keeled).
Poa (all except P. maritima).
Dactylis.
Bromus.
Bromus asper (sheath keeled, often a white line).
Holcus lanatus (slight and decurrent) (Fig. 15).
Digraphis (Fig. 14).
Glyceria.
Most grasses are glabrous, but there are a number in
which hairs are nearly always a prominent feature. It
must be remarked, however, that with grasses, as with
other plants, the character of pubescence is apt to vary
with the situation. In general it may be stated that a
hairy grass tends to become more glabrous in a moist
situation, and more pubescent in a dry one, but the rule
is by no means absolute. In some cases,—e.g. Avena
pubescens, A. flavescens, Agropyrum, the hairs are almost
entirely confined to the crests of the ridges (Figs. 10, 15).
The following is a list of hairy grasses.
Hairy Grasses.
Holcus (Fig. 15).
Molinia cærulea.
Brachypodium sylvaticum.
Agropyrum (variable).
Bromus asper.
B. mollis.Hordeum.
Anthoxanthum.
Avena flavescens (Fig. 10).
A. pubescens.
Triodia.
Kœleria.To a less extent. Festuca sciuroides (on ribs). Melica.
Grasses as a rule are devoid of strong scents5 or tastes, but Anthoxanthum has a faint but distinct sweet odour, especially as it dries—it is one of the grasses which give the scent to new-mown hay—and a bitter flavour, and Milium, Hierochloe and Holcus are also more or less bitter. Spartina stricta emits a strong unpleasant odour.
The habitat of grasses is of great importance as an aid to determination. No one would expect to find a sea-shore grass growing in a beech-forest, or an aquatic grass on a dry chalk-down; but they are even more true to their habitats than this, and I append the following lists of habitats of British grasses as of use in determining them, though it is not pretended that the limits are absolute.
In the following list “pasture-grass" (P) means useful
for grazing, and "meadow-grass" (M) one that is especially
valuable for mowing—i.e. for hay. A “weed" (W) is used
in its agricultural sense for a grass not useful and not
wanted on cultivated land, though often found there.
Meadow- and Pasture-grasses.
(P and M) Dactylis glomerata (fields, &c.).
(P and M) Poa trivialis (meadow and pasture).
(W) Bromus arvensis (cultivated and waste places, meadow and pasture).
(W) B. sterilis (ruderal).
(P and M) Poa pratensis (meadow and pasture).
(W) Briza media (meadow and pasture).
(P) Avena pratensis (meadow and pasture, especially hilly).
(P) A. pubescens (var.)—dry.
28(P and M) Lolium perenne (meadow, pasture and waste places).
(P and M) L. italicum (valuable culture grass).
(P) Cynosurus cristatus (downs).
(M and P) Festuca elatior (meadow and moist pasture, banks and river-sides).
(W) Agrostis alba and A. canina (pasture and waste places, wet or dry).
(P and M) Alopecurus pratensis (meadow and pasture).
(W) A. geniculatus (moist meadows and marshes).
(P and M) Phleum pratense (meadow and pasture).
(P) Arrhenatherum avenaceum (meadow, hedges and copse).
(P and M) Anthoxanthum odoratum (fields generally).
(W) Hordeum pratense (moist meadow and pasture).
(W) Holcus lanatus and H. mollis (meadow, pasture and waste).
(P and M) Avena flavescens (dry meadow and pasture).
(W) Avena fatua (corn-weed).
(P) Festuca ovina (light limestone pastures and chalk downs).Shade-grasses.
Found in woods, copses, &c., under shade.
Melica uniflora (woods, &c.).
Bromus asper (hedges, thickets, and edges of woods).
B. giganteus (hedges and woods).
Aira cæspitosa (moist shade and damp hedges).
Poa nemoralis (woods, shady places and damp mountain rocks).
Milium effusum (moist woods, &c.).
Agropyrum caninum (woods and shady places).
Hordeum sylvaticum (woods and copse).
Brachypodium sylvaticum (woods, hedges and thickets).
Arrhenatherum avenaceum (meadows, hedges and copse).
Festuca sylvatica (mountain woods).Aquatic and Semi-aquatic Grasses.
Found in wet ditches, ponds, and on marshes, river-banks, &c.
Glyceria fluitans (wet ditches and slow waters).
G. aquatica (wet ditches and shallow waters).
29Alopecurus geniculatus (moist meadow and marsh lands).
Digraphis arundinacea (river-banks, marshes).
Arundo Phragmites (wet ditches, marshes and shallow waters).
Molinia cærulea (wet heaths and moors, woods and waste places).
Triodia decumbens, Agrostis alba, Catabrosa and Calamagrostis.Moor-and Heath-grasses.
Downs and dry hill-pastures.
Nardus stricta (moors, heaths and hilly pastures).
Aira flexuosa (heaths and hill pastures).
Molinia cærulea (wet heathy moors, woods and waste places).
Kœleria cristata (dry pasture).
Triodia decumbens (dry heathy and hilly pastures).
Festuca ovina (hilly pastures—especially dry and open—rarer in moist situations).
Agrostis vulgaris and A. canina.Maritime or Seaside Grasses.
Poa maritima (maritime).
P. distans (sandy pastures and wastes near sea).
Elymus arenarius (coasts).
Psamma arenaria (coasts).
Poa bulbosa (waste places in S.E. of England).
Agropyrum junceum (coasts).
Hordeum maritimum (S. and E. coast).
Phleum arenarium (coasts).Ruderal or Vagabond Grasses.
Waste places, walls, road-sides and dry sandy situations.
Molinia cærulea (wet, heathy moors, woods and waste places).
Festuca Myurus (waste places, walls, road-sides).
F. ovina (hilly pastures and especially dry, rarely moist situations).
Aira caryophyllea (sandy and hilly pastures).
30Aira præcox (sandy and hilly pastures).
Poa distans (sandy wastes near the sea).
P. compressa (dry, barren, waste ground).
P. annua (cultivated and waste lands and fields).
Agropyrum repens (fields and waste places).
Hordeum murinum (waste places and road-sides).
Holcus lanatus (meadow, pasture, and waste lands).
H. mollis (same—rarer).
Alopecurus agrestis (waste lands and roads in S. of England).
Lolium perenne (meadows, pastures and waste places).
L. temulentum (fields and waste places, not common).
Bromus sterilis (on way-sides, &c.).
B. arvensis (cultivated and waste meadows and pastures).
Poa rigida (dry, rocky places).
It is also often useful to know whether a grass is rare or local, especially for the purpose we have in view, and I have therefore drawn up the following list of rare, local or introduced foreign grasses either not noticed at all, or only referred to incidentally in this work.
In many cases these introduced foreign grasses have sprung up from seeds brought over in cargoes of hay, wool, and other products and packing materials, which in part accounts for their occurrence only near certain sea-ports, manufacturing towns and so forth. Such plants are frequently termed ballast plants. Foreign plants are also introduced in seed, as mixtures or impurities, and frequently escape from corn-fields &c.
Leersia oryzoides (ditches of Hants., Sussex and Surrey).
Panicum sanguinale (S. England).
P. verticillatum (fields in S. and E.).
P. glaucum (rarely introduced).
Hierochloe borealis (Thurso only).
Phleum alpinum (Highlands only).
31P. Bœhmeri (Eastern counties, rare).
P. asperumxEas"terncou"nties"
Phalaris canariensis (rare weed).
Alopecurus alpinus (Highlands).
Mibora verna (Anglesea and Channel Islands).
Lagurus ovatus (Suffolk coasts).
Polypogon monspeliensis (rare, in S. England near sea).
P. littoralis (salt marshes S. England).
Agrostis setacea (dry heaths of S. Wales).
A. Spica-venti (sandy fields of E. counties).
Gastridium lendigerum (fields and waste places in S. Wales and Norfolk).
Calamagrostis Epigeios (moist glades &c. in Scotland).
C. lanceolata (moist shades, scattered in England).
C. stricta (bogs, &c., very rare).
Cynodon Dactylon (waste and cultivated lands near sea in Scotland).
Spartina stricta (salt marshes S. and E. coast).
Lepturus incurvatus (scattered on shores).
Bromus maximus (Jersey).
B. madritensis (roads and waste, Scotland and Tipperary).
B. inermis (introduced from Hungary).
Lolium italicum (introduced from Lombardy).
Festuca uniglumis (Irish and S.E. coast).
Poa procumbens (waste ground near sea).
P. loliacea (sandy sea-shores).
P. laxa (Ben Nevis, &c.).
P. alpina (Highlands and N.).
Catabrosa aquatica (shallow pools and ditches, scattered).
Finally, a few words may be said on a subject still in its infancy—that of Indicator-plants. In many cases certain plants are found so confined to certain classes of soil, that foresters and agriculturists have claimed to be able to infer from their presence the presence or absence of certain chemical or other constituents of soils: on the contrary we find other plants so universally distributed32 without reference to the quality of the soil, that they are not indicative. The latter are often termed ruderal or vagabonds (see p. 29). Without attempting too rigid a classification of Grasses in this connection—which would be premature in this early state of our knowledge—the following remarks are at least generally true.
A few grasses are Indicators of chalk and limestone—e.g. Briza media, Kœleria cristata, and the exotic species Stipa pennata and Melica ciliata.
The following are said to indicate a sufficiency of potassium salts,
In moister soils.
Digraphis arundinacea.
Phleum pratense.
Avena pubescens.Arundo Phragmites.
Molinia cærulea.
Glyceria fluitans.In drier soils.
Anthoxanthum odoratum.
Alopecurus pratensis.
Agrostis alba.
Holcus lanatus.
Arrhenatherum.
Kœleria cristata.
Briza media.Dactylis glomerata.
Cynosurus cristatus.
Poa pratensis.
P. trivialis.
P. compressa.
Festuca elatior.
Lolium perenne.
Grasses like Bromus arvensis indicate the existence of clay in the soil.
While the following are indicative of sand,In drier soils.
Aira caryophyllea.
A. præcox.
A. canescens.Festuca ovina.
Bromus sterilis.
And only if the sandy soil is moist and of better quality, owing to a certain proportion of humus, the following,
Anthoxanthum odoratum.
Agrostis alba.
Dactylis glomerata.Arrhenatherum avenaceum.
Avena pubescens.
Poa pratensis.
That the soil contains considerable quantities of common salt—sodium chloride—may be inferred if the following grasses occur,
Psamma arenaria.
Elymus arenarius.Hordeum maritimum.
Agropyrum junceum, &c.
The existence of much humus is indicated by such shade grasses as
Melica uniflora.
M. nutans.
Milium effusum.Bromus giganteus.
B. asper.
Brachypodium sylvaticum.
Whereas soils known as “sour," though containing much vegetable remains, may be suspected if the following grasses abound on them,
Aira cæspitosa.
Nardus stricta.Alopecurus geniculatus.
Molinia cærulea;
especially if sedges and rushes coexist with them.
When cuttings are made in forests, such grasses as the following are very apt to appear, and may do harm to young plants,
Festuca ovina and varieties.
Agrostis alba.Holcus mollis.
Aira flexuosa, &c.
The grasses more especially indicative of particular classes of forest-soils are chiefly the wood-species (see34 p. 28), and need not be further specified. In gaps, borders, and copses—half-shade—we find several common grasses—e.g.
Anthoxanthum odoratum.
Agrostis alba.
Aira flexuosa.
Holcus lanatus.
Arrhenatherum avenaceum.Triodia decumbens.
Dactylis glomerata.
Festuca rubra.
Brachypodium pinnatum.
Hordeum sylvaticum.
Whereas
Poa nemoralis,
Festuca sylvatica,
Agropyrum caninum,
Melica,Milium,
Bromus asper,
B. giganteus,
Brachypodium sylvaticum,
are more likely to be met with in the deep shade inside the forest.
On the other hand there are vagabond grasses which seem to show no signs of preference for one soil over another—e.g. Poa annua—though in some cases these ruderal plants indicate the presence of rotting substances, on ash-heaps and rubbish of various kinds.
With reference to the above, however, the student must not forget that very complex relations are concerned in changes of soil, shade, moisture, elevation, &c. and that although experienced observers can draw conclusions of some value from the presence of numerous species and individuals on a given soil, no one must conclude too readily that a soil is so and so, from observing solely that a particular kind of grass will grow there.
An excellent example of what may be done by applying such knowledge as exists of the habits of grasses, is afforded by the historic case of the planting up of shifting35 sand-dunes with species like Psamma arenaria, Elymus arenarius, Agropyrum junceum, &c. (together with sand-binding species of sedges) and so not only fixing the sand, but preparing it for gradual afforestation with bushes and eventually trees, and so saving enormous tracts of land and sums of money, as has been done on the West coasts of France.
Moreover, the action of ruderal plants—including grasses—is to completely alter the nature of the poor soil and gradually fit it for other plants. Coverings of grass greatly affect the actions of heat and sunshine on the surface soil, and modify the effects of radiation and evaporation, to say nothing of the penetrating and other effects of the roots.
Rhizomes and stolons break up stiff soils; and every engineer and forester knows how useful certain grasses are in keeping the surface-soil from being washed down by heavy rains on steep hill-sides or embankments.
On the other hand, luxuriant growths of tall grasses may do harm to young plants, by their action as weeds and especially as shade-plants; though foresters can employ them in the latter capacity, under restrictions, to shelter young trees from the sun. Again, too much dry grass near a forest offers dangers from fire; and it is a well known fact that certain injurious animals, e.g. mice and other vermin, are favoured by a covering of grass.
Graminaceæ are for the most part chalk-fleeing plants, in spite of the fact that certain species can grow in very thin layers of soil on chalk downs. They must be regarded as requiring moderate supplies of humus as a36 rule, and even sand-loving grasses are not real exceptions.
The physiognomy of the grasses has always been regarded as a striking one, and Humboldt classed it as one of his 19 types of vegetation. As is well known they are sociable plants, often covering enormous areas—prairies, alps, steppes, &c.—with a few species, alone or densely scattered throughout a mixed herbage. They also represent characteristically the sun-plants, the erect leaves exposing their surfaces obliquely to the solar rays, and being often folded and nearly always narrow.
The dead remains of these sociable grasses are an important factor in protecting the soil against drought and in facilitating humification, as well as in covering up plants during long winters or dry seasons, keeping the ground warmer and moister, and generally lessening the effect of extremes.
Many Graminaceæ are pronounced xerophytes, the epidermis often being developed as a water-storing tissue, while the erect leaves roll themselves in intense light, the stomata being situated accordingly. The halophytic strand-plants Psamma arenaria, Elymus arenarius, Agropyrum junceum, and other Dune-species, as well as species of Aira, Festuca, Anthoxanthum, Stipa, Lygeum, Aristida, &c. are examples. The heath-grasses—e.g. Festuca ovina, Nardus stricta, Molinia cærulea—also come under this category.
Many of the strand-plants (halophytes) Agropyrum, Psamma, Elymus, are covered with waxy bloom, and have long rhizomes which bind the sand and form new soil, a37 property largely taken advantage of in certain forest operations.
Other grasses, particularly annual species, show their adaptation to xerophytic habits by forming bulbous store-houses at the base of the culms—e.g. Phleum arenarium.
Some Graminaceæ are hydrophytes, such as Arundo, Glyceria, &c., with large intercellular spaces in their tissues; while many species—e.g. Aira cæspitosa, Agrostis canina, Molinia cærulea—grow on wet moor-lands, forming perennial tufts, with or without creeping rhizomes.
The mesophyte grasses are especially characteristic of what may be termed carpets—a lawn is a good example on a small scale, though of course we must remember that here the struggle for existence has been artificially interfered with more or less. Such carpets consist of the densely interwoven rootlets and rhizomes forming sod, and contain much humus from the accumulated débris of former years. These grass-carpets may be composed of nearly pure growths of a few species, or of very many different grasses and other herbage. They are common in Arctic regions, on Alps, and in temperate climates generally, where we know them as meadows, hay-fields, pasture and lawns.
The Bamboos in the wider sense have a physiognomy of their own, e.g. in India, and may drive out most other plants and form dense undergrowths or jungle of interlaced stems and leaves and thorny shoots. Similar growths occur on the Andes and elsewhere in South America. In some parts of India and tropical Asia the taller bamboos form aggregates comparable to dense forests, and such forests are common on the banks of several large tropical38 rivers. Most of these Bamboos are xerophytes. Bamboos are neither confined to the tropics, nor to warmer regions, however, for species are known from distinctly cool regions—e.g. South America—or even from near the snow line—e.g. Chili, the Himalayas, Japan, &c., and the number of species known as hardy is increasing annually, as is evident on examining our larger English gardens.
The permanence and character of extensive grasslands, especially prairies, savannahs, and steppes, are much affected by the periodical firing they are exposed to in the dry season, and large tracts of country in various parts of the world would doubtless bear forests or other vegetation if not thus fired, while in other cases the herbage would be differently constituted were firing discontinued.
The following chapter embodies an attempt to classify our British grasses solely for purposes of identification when not in flower. It is not claimed that the arrangement is the best possible, nor that it is complete, and I need hardly say that corrections will be gratefully received.
I. Sheaths entire except where those of lower leaves are burst by branches, &c.
A. Aquatics with the sheaths reticulated, owing to large air-cavities. Leaves equitant, linear acute, often floating.
Glyceria fluitans (Br.). Floating sweet grass. Somewhat coarse, but useful pasture in water-meadows and fens. Sweet-tasting.
Section of sheathed leaves linear oblong; sheath striate or furrowed, keeled; leaf ribbed; ligule broad acute. Leaf-base with a yellow triangle. Smooth.
Glyceria aquatica (Sm.). Reed sweet grass. Especially given to growing in the water-courses and on banks instead of spreading in the water-meadows, &c. Sweet-tasting.
Section of sheathed leaves broadly naviculate; sheath smooth, no keel; leaf not ribbed, thick and inflated with 40 large air-cavities; ligule short. Leaf-base with a brown triangle. Margins and keel rather rough.
These two species of Glyceria are distinguished by their shoot-sections and the ridges of the leaves of G. fluitans: they often occur in the same ditch.
They cannot readily be confused with others on account of their aquatic habit, and the characters given. The only other aquatic or semi-aquatic species are forms of Catabrosa, Digraphis, Arundo, Alopecurus geniculatus, Molinia cærulea and the rare Calamagrostis.
The ligule and flat shoots with closed sheaths alone suffice to distinguish it from the round and split sheathed Arundo Phragmites; and the round shoots of Digraphis, its split sheath and firm leaves, suffice to distinguish it.
Molinia also has a tuft of hairs instead of a ligule, and a split sheath, and its habit is different.
Alopecurus geniculatus, with its “kneed" shoots, has a totally different habit from Glyceria, and its very high ridges and want of visible air-chambers complete the diagnosis.
Catabrosa is a small creeping aquatic with very flaccid leaves, quite glabrous and soft. Also sweet-tasting.
B. Not aquatic, and devoid of visible air-chambers in leaf or sheath. Often perennial, i.e. having stolons or other branches with no rudiments of flowers in them, and with relics of old leaf-bases.
(α) Sections of sheathed leaves acute: either two-edged or four-edged.
(1) Section of sheathed leaves quadrangular. Blades of leaf thin and dry, sparsely hairy. Sheath quite entire. Woods and shady places.
Melica uniflora, L. (Wood Melick). Lamina slightly tapered below, convolute. Ligule obsolete, with a stiff subulate process on the sheath opposite the blade-insertion. Ridges below, but not above.
Melica nutans, L. (Mountain Melick). Ligule longer, and without the awl-shaped peg. Only in Scotland and W. of England.
Both are shade grasses of no agricultural value.
M. uniflora, with its quadrangular shoots and anti-ligular peg, cannot be confounded with any other grass.
(2) Sections of sheathed leaves more or less acutely two-edged, owing to the keels of the compressed equitant leaves.
(i) Shoots broad and fan-like, much compressed, with old brown leaf-sheaths below, sometimes burst by the intra-vaginal branches: leaf ridgeless, with prominent keel. No underground stolons.
Dactylis glomerata, L. (Cock’s-foot). An early and quick-growing pasture-grass, which forms much aftermath. Grows on all soils. Often coarse. Coarse tussocks, and harsh, with broad thick succulent bluish-green leaves.
Section of sheathed leaves acutely naviculate. Prominent obtuse ligule, torn above. Lamina long, rough, acute, with white lines if held up, and serrulate edges. No flanking lines6. No stolons (Fig. 6).
There is a cultivated variety of Dactylis with broad opaque white stripes down the leaves: these are totally different from the translucent white stripes seen on holding the wild form, or Aira cæspitosa, up to the light. Another cultivated “ribbon-grass"—Digraphis—has round shoots, split sheaths, and a different habit, and the same applies to its wild form.
Probably the only serious chances of confusion with Dactylis are between it and Poa pratensis, which also has flattened shoots and closed sheath; but in the latter the section of the shoot is elliptical—not naviculate,—the keel is far less prominent, and the ligule 42 shorter. Moreover P. pratensis is a creeping stoloniferous grass, less harsh, and with less pointed leaves.
The distance to which the sheath is torn may be from 1/8 to 1/2 down. Leaves tend to remain conduplicate. Margins serrulate with teeth extremely short and directed forwards.
(ii) Shoots compressed but narrow: the section almost
rhomboid with rounded edges.
Poa trivialis, L. (Rough-stalked Meadow-grass). Conspicuous in deep rich pastures and orchards, preferring slight shade and rich soil. Valuable pasture and hay grass.
Rootstock shortly creeping, branches extra-vaginal and above ground, shoots rough. Blade narrow, harsh, with an acute point, thin, shining below, ridgeless, with flanking lines and keel. Ligule acute, and short or long (Fig. 8).
Sesleria cærulea, Ard. (Blue Moor-grass), of our northern limestone hills, has narrow, flat, glaucous blue, stiff, mucronate leaves, with scabrid apex. Ligule ciliate.
Poa trivialis is most likely to be confounded with other Poas, especially P. annua and P. pratensis, since they both have thin leaves and flat shoots; but P. annua has a split sheath, less acute and duller leaves, is annual, and less harsh, and the shoot-section is flatter at the sides and rounder at the ends.
Poa pratensis, L. is larger and more stoloniferous, with both extra- and intra-vaginal branches, culms erect and smooth, sheaths smooth, and the shoot-sections elliptical—not cornered or rhomboidal—and with darker green and larger, thicker, 7-veined, more glossy, and less harsh leaves, with shorter, blunter ligule.
Poa compressa, L. also presents difficulties, but the sheath is split, and the ligule is shorter than in P. trivialis, the leaves thicker, and the shoot-sections more linear-oblong or elliptical.
(β) Sections of sheathed leaves rounded, circular or oval, there being no prominent keels.
(1) Section of sheathed leaves circular or nearly so, the shoots being only slightly compressed.
✲ Perennial.
Bromus inermis (Awnless Brome).
Sections circular, the leaves being convolute, base shelving. Glabrous sheaths and leaves. Stoloniferous. Ligule short, truncate, and finely toothed. A forage grass of the Hungarian steppes. Now being grown in this country, but of doubtful value here.
Bromus erectus, Huds. (Upright Brome). A weed.
Sections oval and rounded, but leaves equitant. Radical leaves remain folded and almost subulate, hairy edges. No stolons. Fields, &c. It is a weed on dry lands, and of little or no value.
Bromus asper, Murr. (Hairy Brome). In thickets, &c.: a weed, and useless. Leaves green, long, flat, hanging, and eared. Sheath with scattered deflexed hairs. Lamina tapering at the base. Keel a white line, ridges inconspicuous: distance between veins 2-3 times breadth of latter. Ligule very short, toothed.
B. giganteus, L. (Tall Brome), also comes here. It is less common and glabrous. Woods, &c., a useless weed.
✲✲ Annual or biennial.
Bromus mollis (B. arvensis, var. mollis, L.), Field Brome. A too abundant and useless weed in water-meadows and hay-fields. Softly downy. Blades very thin and not eared: dry.
Bromus sterilis, L. (Barren Brome). A useless weed. Rough and downy, but less so than the last. Moist waysides, &c.
The Bromes are extremely variable and difficult to determine by the leaves. The annual species are apt to be biennial or (B. sterilis) perennial; and some vary much as regards hairiness—e.g. B. mollis is connected by a series of semi-glabrous forms to varieties quite smooth, all grouped by Bentham under B. arvensis.
Bromus asper, being auriculate and a shade-species, runs some risk of confusion with Hordeum sylvaticum, but Hordeum has a split sheath and in B. asper the translucent interspace between the ridges is 2-3 times as broad as in Hordeum sylvaticum.
The other species of Bromus are not eared, and their entire sheaths at once distinguish them from Hordeum.
Bromus giganteus has leaves glabrous and very like Festuca elatior. The red split sheaths of the latter, its sharp ears and prominent ridges afford the best distinctions; and B. giganteus has broader leaves and more evident serrulation or descending bristles at the basal margins.
(2) Section of sheathed leaves elliptical, owing to the shoots being compressed. Sheaths often only slightly split above. No hair on surface of leaves or sheaths.
✲ Margins of leaves smooth and even. Blades without ridges, a keel and flanking lines, acute, base rounded. Ligule of lower leaves very short.
Poa pratensis, L. (Smooth-stalked Meadow-grass). An early and valuable dry pasture-grass, but though deep-rooted, it yields thin hay: its chief value is for "bottom grass" and in lawn mixtures, &c. Leaves stiff and pointed. Extra-vaginal rooting underground stolons, and intra-vaginal branches. Shoots smooth. Keel slight: seven principal veins and smaller ones between. Leaves blunter and broader than in P. trivialis.
Poa alpina, L. (Alpine Poa). On mountains in the north. No stolons. 4-5 veins on each side of the median one.
Poa pratensis presents similar difficulties to P. trivialis: for diagnoses see p. 42. It is distinguished from P. nemoralis by its closed sheath, thicker, blunter and harder leaves, linear-elliptical shoot-sections, and light coloured nodes, as well as by its habit. All other Poas have shallow and poorly developed roots.
P. fertilis is a form very like P. nemoralis, with rougher leaves and longer ligule, introduced into cultivation.
✲✲ Margins of leaves scaberulous with descending hairs. Very low flat ridges. Sheath smooth.
Briza media, L. (Quaking Grass). A weed in meadows, indicating poor soil—e.g. moorlands and chalk—but eaten by sheep. Tufted and slightly creeping perennial. Ligules very short, entire.
Briza minor, L. (Lesser Quaking-grass). Annual. Leaves broader and shorter, and ligules longer. In the south and rarer.
II. Sheaths split, at least some distance down.
A. Glabrous—i.e. with no obvious hairs7.
(a) Grasses with setaceous or bristle-like leaves;—i.e. the lamina of the lower leaves remains permanently folded instead of opening out flat.
(1) Ligule obsolete, auricled at the junction of blade and sheath.
Festuca ovina (Sheep’s Fescue). Densely tufted perennial. Leaves hard, glabrous and often glaucous, with 5-7 ridges if forcibly unrolled, ears short, stiff and erect. Branches in permanent sheaths. Chiefly useful as pastures 46 on downs and dry chalk-soils. Several varieties are recognised by agriculturists, as hard, red, various-leafed, fine-leafed Fescue, &c. (see Figs. 13 and 18).
Festuca Myurus, L. (Rat’s-tail Fescue). Annual, longer auricles, and hair on the ribbed inrolled surface. A roadside weed.
Festuca ovina presents difficulties with its varieties and with F. Myurus, L. (var. sciuroides, Roth.).
The chief varieties of F. ovina are Hard Fescue (F. duriuscula, L.), taller and with some of the upper leaves flat, and found in moister and rich soils: Red Fescue (F. sabulicola, Duf. or F. rubra, L.) more or less creeping and with red sheaths to the lower leaves, on poor stony land—F. heterophylla is a form of this on chalky soils, with flat leaves above: and F. tenuifolia a very wiry form on sheep-lands. They all pass into one another, however, and cannot be distinguished by the leaves (see Figs. 18-20).
F. Myurus (var. sciuroides) is ruderal and annual, and has longer hairs on the ridges of the folded leaves. It has no agricultural value.
(2) Ligule membranous, not auricled.
(α) Bristle-like (setaceous) leaves, very hard and stiff, and more or less solid.
Nardus stricta, L. (Moor Mat-grass). Roots very tough and stringy: ligule small, but thick and blunt. Leaves channelled: upper erect, lower horizontal. Sheath smooth. Moors and sandy heaths: useless (Figs. 2 and 26).
Aira flexuosa, L. (Wavy Hair-grass). Roots fibrous. Leaves short, filiform, terete, solid—the channel hardly discernible. Ligule short, obtuse. Heaths, &c. Of little use, even for sheep (Fig. 28).
(β) Leaves bristle-like, but distinctly due to inrolling of edges.
Aira caryophyllea, L. (Silvery Hair-grass), is scabrid. A weed, with very slight foliage.
A. præcox, L. (Early Hair-grass). Greener and more glabrous. Habit more rigid.
A. canescens, L. (Grey Hair-grass). Glaucous or purplish; rare, on S.E. coasts.
(γ) Leaves narrow and more or less involute, and subulate upwards, but easily unrolled, and apt to become flatter as they age.
Avena pratensis, L. (Perennial Oat). Leaves rather thin, dry, harsh, ridgeless, with flanking lines and a keel8; glaucous, glabrous, but edges scabrous. Usually involute, but may open out. Ligule long ovate-acute. Dry pastures, especially on calcareous soil, and of little value.
Poa maritima, Huds. (Sea-grass). Leaves narrow, rather short, and U-shaped in section. Involute: ridgeless, with flanking lines, but no keel; soft and rather thick. Ligule rather long, obtuse and decurrent. Useless agriculturally.
For difficulties with other species of Avena and Poa see pp. 44, 54 and 60.
(b) Grasses with the leaves expanded, more or less flat.
(1) Blades conspicuously ridged—i.e. the surface is raised in prominent longitudinal ridges with furrows between.
(i) Leaves rigid and hard, sharp pointed. Sheath and outer leaf-surface usually glabrous.
Aira cæspitosa, L. (Tufted Hair-grass). Forms large tufts. A coarse weed forming bad tussocks in wet meadows and pastures: useless for fodder. Leaves flat. Ligule long, acute. Ridges equal, high and sharp, and scabrid, 48 with 5-6 white lines between, if viewed by transmitted light. Wet meadows.
A. cæspitosa cannot easily be mistaken for any other species. Alopecurus geniculatus is also a moisture-loving grass with strongly ridged leaves, but the interspaces are far less translucent and the whole habit is different.
All the other species of Aira have involute and setaceous leaves, and even A. cæspitosa is apt to roll in its leaves in mountain varieties, but they are easily flattened out, and show the ridges.
Psamma arenaria, Beauv. (Sea Mat-grass). This is one of the most valuable “sand-binders," its long matted rhizomes holding loose sand together. It is a sea-shore grass, of no use for fodder. It was formerly much used for mats and thatching. Leaves concave, long, narrow, erect, scabrid and glaucous above, and polished below: pungent. Ridges rounded, alternately high and low. Sheath long. Ligule very long and bifid.
Elymus arenarius, L. (Sand Lyme-grass). Like Psamma, this is a “sand-binder" and of no use for fodder. Leaves concave, and eared at the base of the blade: ears pointed and tend to cross in front. Ligule very short and obtuse. Ridges flattened above, not scabrid. Apex of blade rolled, forming a hard spine.
Psamma cannot easily be mistaken for the much less common Elymus, as it is not eared, and the ridges and ligule are very different.
(ii) Leaves not specially rigid and hard, and often thin; glabrous, or shining below. Ridges less evident.
✲ Ligule very short or obsolete; blade firm but not hard, glabrous or nearly so, and shining below. Sheath often coloured red or yellow at the base.
† Sections of sheathed leaves narrow, oblong, owing to compression of shoots. Sheath nearly entire.
Lolium perenne, L. (Perennial Rye-grass). Very valuable pasture-grass, especially on clay. Less successful as hay. Deep rooted tufts. Glossy dark green. Ligule short (Fig. 11). Sheath red or purplish below. Blade conduplicate and keeled, often rounded, collared or eared at the base; with rounded ridges and rough above, shining below. When the ears are well developed their points often cross one over the other in front of the sheath.
L. italicum, Braun. (Italian Rye-grass), is an earlier and better variety for hay and sewage farms. Shoot more rounded in section, and has less marked veins on the more rolled leaf.
L. temulentum, L. (Common Darnel), is annual and a weed of corn-fields. Foliage usually rougher.
Lolium perenne presents some difficulties in relation to such forms as L. italicum, species of Agrostis and Festuca, Alopecurus pratensis, Cynosurus and Agropyrum.
Owing to the leaves not being always strictly conduplicate in the first year, the flat shoots may not sharply mark it off from L. italicum. Its somewhat looser, almost stoloniferous tufts, and darker green foliage, less polished below and usually narrower and harder, have then to be taken into account.
The ridges of Lolium are often like those of Festuca pratensis; and the shining lower surface and rather firm leaves and red sheaths, present other points of confusion. The smooth basal margins of Lolium, absence of white translucent lines when held up, and the different ligule and ears afford distinctions—the ligule of Festuca being a mere line, and the ears pointed and projecting, whereas they may be mere lateral ledges in Lolium.
Cynosurus has the ligule and ears very like those of Lolium, the ears being mere ledges; but the former has yellow sheaths, firmer and thicker leaves with more evident ridges, and the old plants usually have the characteristic crested spikes remaining. Cynosurus, moreover, has the sheath split only a short way down.
With regard to Agrostis, there is no colour in the sheath, the ligule is longer and pointed, and the leaves drier and thinner than in Lolium, and harsher on both surfaces. Agrostis has also no ears.
Alopecurus pratensis has much broader and flatter ridges than Lolium and a longer ligule, and its sheaths are dark-brown or black—not red; but A. agrestis has very similar ridges to Lolium and may easily be confounded at first.
Agropyrum is sometimes nearly glabrous, and may then be confused with Lolium by beginners: its low ridges, curled and pointed ears, obsolete ligule, and thinner, drier, harsher blade, as well as the stolons, distinguish it.
Lolium temulentum and Hordeum murinum occasionally cause difficulty, but the latter is always more or less hairy, its blades thinner and drier, and the ridges less raised.
†† Sections rounded—elliptical or nearly circular. Sheath distinctly split, at least above.
Cynosurus cristatus, L. (Crested Dog’s-tail). Useful as pasture on dry soils, but only moderately so as hay. Blade narrow, slightly eared or collared below, tapered above; firmer than Lolium. Sheath only split a short way down. Yellow or yellowish-white at the base. Leaves conduplicate or convolute, short and narrow, the ligule short: minute ears at base. Usually easily recognised by the withered culms and persistent pectinate spikes (Fig. 15).
Festuca elatior, L. (Meadow Fescue). A valuable meadow and pasture grass, though somewhat coarse. Several varieties are known. Best on heavy soils. Deep rooted. Blade flat and broad, conduplicate, sharp-eared at the base, and there rough at the margin: lower surface polished. Rich green. Mid-rib flat above, numerous ribs with white lines between if held up and examined with a lens. Ligule obsolete (Figs. 12 and 17).
Arundo Phragmites, L. (Common Reed). A large aquatic, reed-like creeping grass, with broad leaves (3/4 to 1 in.), flat, rather rigid, acuminate, glaucous below, hispid at edges. Sheath smooth, striate, bearded at mouth. Ligule a mere fringe of hair. (Cf. Digraphis, p. 54.)
Cynosurus is not very liable to confusion; but it has resemblances to Lolium (see p. 49) and to species of Agrostis. The leaves of Cynosurus are firmer, thicker, less dry, and with a shining undersurface, and the sheath is only split above, and yellow below; whereas Agrostis has relatively thin and dry leaves, rough surfaces and margin, distinct ridges, and converging margins as the blade nears the sheath.
Festuca elatior is easily confused with the glabrous Bromes. For B. giganteus see p. 43.
Bromus erectus is distinguished by the entire sheath, usually hairy, the want of auricles, and the conduplicate—not convolute—leaves.
Agrostis has thinner, duller, and drier leaves, and no red sheath.
Alopecurus pratensis has more depressed, flatter and broader ridges than Festuca, and a longer ligule, and lacks the pointed ears.
✲✲ Ligule whitish, membranous, long, or at least well developed. Sheaths not coloured or brown. Leaves thin and rough, at least at the base. Ridges not very prominent, but numerous and distinct.
Agrostis stolonifera, L. (Fiorin). Stolons, with numerous short offsets bursting through the leaf-sheaths. Blade flat, rough, tapering, with rounded ridges, and convolute in bud: there are no auricles, but the blade may narrow, and form ledges, as it runs into the sheath. Sheaths nearly smooth. Ligule long and pointed, and often toothed at the margins. The leaves vary in breadth.
This and A. vulgaris, With. with shorter ligules, and, possibly, A. canina, L. with finer leaves, are varieties of52 A. alba, L. Only the variety A. stolonifera is of moderate value for pasture, especially on poor soils, as it lasts late into autumn: the others are weeds, like couch-grass.
Agrostis is full of difficulties for the beginner. The weed-forms often spring up after wheat has been cut, and count as “twitch," like Agropyrum.
All the ordinary forms—A. stolonifera, A. vulgaris, and A. canina—may be included in A. alba (Linn.). On dry hills a close tufted grass, with setaceous leaves, and in rich soils creeping and luxuriant with broad leaves. It is one of the few grasses that thrive in wet soils.
The chief points in the flat-leafed forms are the thin, dry leaves, rough on both sides and on the margins, with distinct raised ridges, and the base of the leaf narrowing suddenly into its insertion with the sheath, with no auricle, but with a long membranous ligule. The sheath not coloured, and the blade convolute.
Again, A. stolonifera has a long, serrated, acute ligule, while A. vulgaris has a much shorter, entire and truncate one, and narrower leaves.
Agropyrum is the grass most likely to lead to confusion. Its ears, lower ridges, very short or obsolete ligule, and pubescence (sometimes glabrous) distinguish it.
Cynosurus sometimes gives trouble (see p. 50) with Alopecurus pratensis: the sheaths, ligule and flattened ridges should suffice for distinction.
Alopecurus geniculatus is even more like Agrostis, but its ridges are more prominent and sharp, and its aquatic habit and bent "knees" distinguish it.
Alopecurus agrestis, in dry corn-fields, has a thickened ligule, sometimes coloured, and is annual or biennial, but otherwise very like Agrostis.
Alopecurus pratensis, L. (Meadow Foxtail). Large grass with stolons; very early, and much prized as pasture and hay, but soon dies out on light poor soils. Especially good for stiff soils. Sheaths long, ridged, brown or nearly black at the base as they age. Ligule distinct and obtuse,53 entire. Leaves numerous. Blades long, dark green, succulent and scabrous: ridges numerous and flat above, but distinct (Fig. 9).
A. agrestis, L. in S. England has shorter leaves, and ridges not flattened; it is a troublesome pest of arable land, but does not usually invade pasture.
A. geniculatus, L. is semi-aquatic, and like the last. It is easily recognised by its sharply bent “knees," and is of little value (Fig. 3).
Alopecurus shows resemblances to Lolium (see p. 49), Festuca (see p. 50), and Agrostis (see p. 52). If well grown its ridged sheath and leaves, the former brown or black at the base, aid in distinguishing it.
(2) Blades either devoid of ridges or with very inconspicuous ones.
✲ No trace of ridges, and the mid-ribs not prominent, but the leaves show median lines flanked by finer ones when held up. Blades thin and narrow. Somewhat keeled.
(i) Shoots compressed.
Poa compressa, L. (Flat-stemmed Meadow-grass). Leaves rather short, more or less glabrous or glaucous, and V-shaped at the base; shoots compressed, and naviculate in section. Ligule short and thin. Sheath tends to be closed below. A creeping perennial on commons and waste lands, and of little or no value.
Poa annua, L. (Annual Meadow-grass). Small annual. Compressed shoots, limp. Leaves linear, pale, sub-acute, thin, often wavy, flat, flaccid, bright green; dull or slightly shining and V-shaped in section below. Ligule long, pointed, whitish and clasping the shoot. It is a harmless weed, and since it puts out shoots all the year round, furnishes a certain amount of pasturage.
(ii) Shoots terete or nearly so.
Poa nemoralis, L. (Wood Poa). Leaves and sheaths smooth. Blade bright green, thin, often glaucous, linear-narrow, flaccid, acute. Ligule almost obsolete. Section of shoots round. Of little value.
Poa bulbosa, L. (Bulbous Meadow-grass). Stems bulbous at the base. Ligule long and acute. Leaves very narrow and tapering. Sections of shoot round. Coasts of S. and E.
All the Poas, except the aquatic ones (Glyceria) and P. maritima, have glabrous ribless blades with the median lines, and slight keel.
P. pratensis, P. alpina and P. trivialis (Fig. 8) have entire sheaths (as have Glyceria fluitans, G. aquatica and P. maritima), but the others have them split some way down.
The leaves of P. pratensis and P. compressa are firmer than the thin leaves of P. annua, P. trivialis and P. nemoralis.
Sheaths flattened in P. pratensis, P. compressa, P. annua, and P. trivialis; but rounded in P. nemoralis. Glyceria aquatica and G. fluitans have netted sheaths.
Poa annua is annual, and P. bulbosa has the bulbous base. P. maritima has involute leaves and no keel, and the rare P. alpina has short rigid keeled mucronate leaves, with tip often inflexed and thickened scabrid edges.
The leaves of Avena are apt to appear similar to those of the Poas at first sight, but the former are hairy, and ridged, dry as well as thin, and the peculiar median lines of Poa are wanting. Poa bulbosa has drier leaves than usual, but its leaves are devoid of ridges.
✲✲ Ridges can be detected, but are slight and not distinct. Margins scabrid, at least at the base.
† Leaves firm, flat, linear, acuminate, not narrowed below. Glabrous. Ligule membranous.
Digraphis arundinacea, Trin. (Reed-grass). Sheathed leaves round in section; blades convolute, tapering above, 55 flat, firm, long and broad (1 in.) below. Mid-rib and veins numerous, and prominent below. Stoloniferous: branches extra-vaginal, often with deep red basal scales. Sheaths with much overlapping membranous margins, with a collar-like ledge above. Ligule long and somewhat acute. Wet ditches, &c., of no value (Figs. 7, 14).
For distinction between Digraphis and Dactylis see p. 41. Poa pratensis is at once distinguished by its flattened shoots, more rounded leaf apex and shorter ligule. Arundo Phragmites is easily distinguished by the ligule (see p. 51), and the other aquatic grasses are quite different (see p. 39).
Phleum pratense, L. (Timothy-grass). No stolons, but bulbous on dry ground. Early, and a heavy cropping hay grass: also excellent pasture; branches intra-vaginal, but burst the glabrous sheaths. Old sheaths fibrous. Leaves short, convolute, with scabrid margins owing to deflexed teeth: ridges obsolete above, no keel; broader and greyer green than Alopecurus. Ligule short on radical leaves, thin. Pastures. Perennial.
The smooth ligule, deflexed marginal teeth, and no keel distinguish it from Arrhenatherum.
Phleum arenarium, L. (Sand Phleum). Shoot annual, with no bulbs. Leaves broad, flat and glabrous, but rough at the edges, with descending teeth. Ridges low and flat. Sheaths smooth: leaves conduplicate. Ligule long. Sandy coasts, &c. A weed.
Phleum asperum, Jacq. and P. Bœhmeri, Schrad. are rare ruderal plants, and P. alpinum, L. is confined to the Scotch Highlands. P. arenarium is sharply distinct by its conduplicate leaves and habitat.
Alopecurus pratensis has narrower and less grey-green leaves than Phleum pratense, its ligule is shorter and blunter, its sheath 56more grooved and dark below, and the ridges more distinct and flat. It is relatively well rooted and is stoloniferous.
Arrhenatherum avenaceum, Beauv. (False Oat-grass). Loose tufts with short stolons, or bulbous below. Leaves few, narrow, thin, dry, rough, with very low flat ribs, convolute in bud, and practically glabrous. Sheath smooth. Ligule truncate, hairy on its outer surface. Bitter, and commonly undervalued by agriculturists, but useful in mixed pasture, and yields bulky, coarse hay.
There are often a few sparse isolated hairs on the low ribs. The base enters the sheath with slight and sometimes pinkish ledges. Ridges hardly observable. Traces of roughness if rubbed downwards. White lines, about 5 each side on holding up to the light. The not very long leaves taper slightly below.
Arrhenatherum is liable to confusion with Holcus, Anthoxanthum, Molinia and Avena, but it is typically glabrous, whereas the others are hairy.
From Holcus it is easily distinguished by the sheaths, ligule and soft hairs of that genus.
Anthoxanthum differs in its habit, ears, scent, sheath and ligule.
Molinia differs in habit, ligule, sheath, and tough stringy roots, and the shape of the leaves.
Avena pratensis differs in its narrow leaves, less prominent ridges, and ligule; A. flavescens in its much broader and coarser hairy leaves, and the ligule; and A. pubescens in pubescence and flat-shoots and ligule.
†† Leaves very thin, blade tapering below. Keel prominent, but no ridges above. Ligule long and torn.
Milium effusum, L. (Spreading Millet-grass). Tufted perennial. Leaves linear-lanceolate, scabrid above. Sheath smooth. Bitter tasting. May be slightly hairy. Woods. It is much liked by birds, but is of no value in agriculture.
B. Leaves or sheaths, or both, distinctly hairy.
(α) Leaf-blades eared at the base.
(a) Ears sharply pointed. Leaves convolute, and sections of shoots round. Ligule short and inconspicuous.
Agropyrum repens, Beauv. (Couch-grass). A troublesome weed of arable land and gardens, &c., owing to the extraordinary vitality of its underground stolons. The young shoots are readily eaten by stock. Perennial, and extensively stoloniferous; bright or glaucous green. Blade thin, dry, rough edged, hairy and rough above, glabrous or hairy below. The short ligule fringed. Ears long, or sometimes short, pointed; often obliquely crossing in front of the sheath. Ridges inconspicuous. Hairs may be absent from the sheath, and nearly so from the blade.
The sea-shore varieties are stiffer and more glaucous, the leaves more ribbed, involute and pointed—e.g. A. junceum, Beauv.
Agropyrum caninum, Beauv. (Bearded Wheat-grass). Tufted weed, not creeping, in woods, &c. Blade thinner and rougher beneath, but very variable. Properties similar to those of A. repens.
Agropyrum may be confused with Lolium (see p. 49) and Agrostis (see p. 51), but hardly with any other grass, and with these only because it is liable to be glabrous or nearly so on poor soil.
Elymus has much more pronounced ridges than the sea-shore varieties of Agropyrum.
(b) Base of blade with inconspicuous rounded ears. Leaves convolute. Ligule conspicuous.
Anthoxanthum odoratum, L. (Sweet Vernal-grass). Compact tuft. Common in pastures and hay, but it only58 forms a small proportion of the crop. One of the earliest grasses, and the principal one, which gives the scent to new-mown hay: a perfume has been extracted from it. Its value as fodder is probably overrated. Bitter tasted. Leaves more or less hairy at margins, especially at throat of sheath, flat, and slightly ridged. Sheath furrowed, often pubescent. Ligule long and blunt, with ciliate margins. Sweet scented when dried. The most shallow rooted of all meadow-grasses. Leaves often short and few.
Anthoxanthum is sometimes confused with Arrhenatherum (see p. 5) and Molinia; the latter differs in its ligule—a tuft of hair—its stringy roots, tapering leaf-base, less obvious ridges, and smooth sheaths, &c.
Anthoxanthum is deep green and often very luxurious in rich wet soils—e.g. in Devonshire.
(c) Ears as mere collar-like ledges where the blade joins the sheath. Sheath usually pubescent or hispid with reflexed hairs. Ridges inconspicuous. Ligule very short.
✲ Perennial, with firmer leaves.
Hordeum sylvaticum, Huds. (Wood Barley). Leaves flat, thin but firm, rather broad, scaberulous. Sheath hispid, with reflexed hairs. Blade not tapering below. Translucent spaces between the veins as broad as the latter. Ligule short and blunt. Shady places. Useless.
Hordeum pratense, Huds. (Meadow Barley). Tufted, or bulbous below. Leaves narrower, flat, tending to roll up, scabrid above and hairy beneath. Sheath narrow, hairy. Moist meadows, and of some use as pasture in the young state.
✲✲ Annual, with thin dry leaves.
Hordeum murinum, L. (Wall Barley). Coarse tufts; leaves small, narrow, hairy or scabrid. Sheaths sparsely hispid, or very downy, inflated. Roads, &c. A useless weed.
H. maritimum, With. is a sea-side form, smaller and more glaucous. Sheaths hairy.
(β) Leaf-blades not eared at the base.
✲ Sheaths of radical leaves veined with red-purple.
Holcus lanatus, L. (Yorkshire Fog). A useless weed, but very common in pasture and hay; forming tussocks, greyish-green, softly hairy (tomentose). Blades with roundish ridges. Ligule short and obtuse. Sheath somewhat keeled, with trace of collar ledge. It is said to have a bitter taste (Fig. 15).
Ligule pilose. Tufted hairs along the broad rounded ridges, and on the lower surface and prominent keel.
The much rarer H. mollis, L. is not so long-haired, except on the nodes, and is more creeping and slender in habit. It is a “twitch."
The Hordeums present several points of difficulty to beginners. The differences between the species are given above. H. maritimum has narrower and thicker leaves than the rest.
Bromes are most likely to be confounded with Hordeums, but they have entire sheaths and no ears (see p. 43).
For distinctions between H. murinum and Lolium see p. 49. H. sylvaticum and Bromus asper (p. 44).
✲✲ No conspicuously red-veined sheaths.
† Ligule absent, or a tuft of hairs.
Molinia cærulea, Mœnch. (Purple Molinia). Tussocks, with tough stringy roots. Leaves narrowed below, and tapering above to a long point, ridges obsolete; very thin60 and dry but fairly stiff, and hairy above, especially at the base. Ligule absent, or a tuft of hairs. Sheaths smooth. Moors. Useless as forage, but used locally for brooms.
Molinia is not easily confounded with any others but Anthoxanthum (see p. 57), Arrhenatherum (see p. 56) or Brachypodium.
Brachypodium sylvaticum is distinguished by habitat, its broad leaves, membranous ligule, fibrous roots, &c.
Kœleria cristata, Pers. (Crested Kœleria). Very short, perennial in dry pastures, pubescent, pale green. Leaves narrow, tapering below, soon involute, ciliated. Ridges prominent, alternately high and low. Ligule obsolete, or a mere jagged yellowish line. Useless.
Triodia decumbens, Beauv. (Decumbent Heath-grass). Low perennial. Leaves narrow, obtuse, slightly ridged, tough, at length involute, with long, soft hairs, especially below and on the edges. Sheath grooved, hairy, especially at the throat. Ligule a tuft of hairs. Section of shoot flat; leaves conduplicate. Of no known use as fodder.
The rare grasses Panicum glabrum, Gaud., P. viride, L. and P. Crus-galli, L. introduced in the S.E. counties also come here.
†† Ligule membranous.
Avena flavescens, L. (Yellow Oat-grass). Loose tufted perennial, pale green, with rounded shoots bursting the sheaths. Leaves flat, slender, soft, fine-ribbed and hairy, especially on the low ridges above. Sheath hairy, especially below, not keeled. Ligule short, obtuse, often truncate, ciliate. A valuable pasture and meadow-grass, also in water-meadows. Its roots are abundant, and it will grow well in calcareous soils (see Fig. 10).
Avena pubescens, Huds. (Downy Oat-grass). A variety of A. pratensis (see p. 47), but less densely tufted, and the leaves flat and pubescent, and especially the sheaths very pubescent. Ligule ovate-acute. Shoots flat. Dry districts, and a weed.
Avena flavescens is not easily confounded with any other grass if well grown. All the Poas otherwise like it are glabrous, and without the ridges. The same applies to A. pubescens.
Arrhenatherum is also glabrous, its leaves narrower, its ridges much flatter and broader, and its ligule is hairy outside (see p. 56).
Brachypodium sylvaticum, Beauv. (Wood False-brome). Rather slender, perennial. Leaves flat and devoid of ridges; long, very thin and dry, limp, slightly tapering below, hirsute. Sheath round, hairy. Ligule fairly long, obtuse, toothed. Copses, &c. Useless.
Brachypodium pinnatum, L. (Heath False-brome), is a species growing in the open, with narrow, firm, rigid, erect leaves, hardly hairy; with distinct ridges, and tending to roll up. Ligule fringed with hair. Open heaths. Useless.
The only grasses likely to be confounded here are the Bromes, and they have entire sheaths.
The principal anatomical features observed in the leaves of grasses—apart from finer histological details into which it is not my purpose to enter—concern the characters of the epidermis and distribution of the stomata and hairs, the arrangement of the chlorophyll-tissue, that of the mechanical tissue (sclerenchyma) and the vascular bundles to which the venation and ribbing of the leaves are due, and the presence or absence of those peculiar thin-walled cells (motor-cells) which bring about the infolding or inrolling of the lamina (see p. 25) as they lose water, and, finally, the presence or absence of conspicuous lacunæ or air-spaces so characteristic of aquatic species. Several observers have occupied themselves with these matters, and the researches of Schwendener, Duval Jouve, Pfitzer, Pée-Laby, and others have rendered it possible to group most of our grasses according to the microscopic characters of the leaves, somewhat as I have done in Chapter V.
Reference has been made to the rolling and folding of leaves, due to the thin-walled cells on the upper surfaces capable of varying in turgescence (motor-cells). These are specially adapted epidermal cells found on the upper surfaces only. In the leaves of Poa compressa, P. annua (Fig. 21), P. nemoralis, P. alpina, Catabrosa, Sesleria, &c., a row of these motor-cells, easily distinguished by their large size, thin walls and clear contents, is found on each side of the mid-rib; as they dry the leaf folds its two halves together (conduplicate), and on the re-absorption of water they flatten the two halves out again. In Dactylis these flanking rows coalesce into one over the mid-rib. In other leaves, e.g. Avena pratensis, Festuca elatior (Figs. 17, 22), Melica, Elymus (Fig. 25), &c., there are in addition to these two flanking rows, other sets of motor-cells between the other ribs, and their combined action causes the halves of the lamina to inroll, usually one-half inside the other—convolute.

Fig. 21. Transverse section of left-half of leaf of Poa annua (× about 50) showing keel below, and two flanking lines of motor-cells (slightly shaded) above the median vascular bundle of the mid-rib. Hence the leaf folds. The half lamina has six smaller vascular bundles, only the stronger one girdered. Ridges practically obsolete and subtending bands of sclerenchyma slight: hence the leaf-surfaces are parallel.
It is easy to observe leaves of such grasses as Festuca pratensis (Fig. 22), Aira cæspitosa (Fig. 23), &c., which64 are wide open in the dewy mornings in summer, close up as the air gets dry and hot; and any such leaf may be seen to roll up after plucking and can be reopened by moistening it.

Fig. 22. Transverse section of left-half of leaf of Festuca elatior, var. pratensis (× about 50). The ridges are well marked and flattened above. The vascular bundles of two orders are girdered below, but only slightly above. There is no keel. There are well marked motor-cells—not shown in the figure—in each groove.
The epidermis of grasses has been closely investigated by Grob, but unfortunately his results concern very few of our native species. The principal elements are ordinary elongated cells, with plane or sinuous walls, various kinds of short cells intercalated between the ends of these, several forms of papillæ, hairs, &c. and stomata.
The epidermis over the parenchyma of Digraphis arundinacea consists of rectangular cells with plane walls.
Series or bands of long cells only may alternate with other series where short cells intervene between the long ones—e.g. Nardus.
Nardus has some of the bands devoid of stomata, but abounding in short cells, whereas others (above) have stomata throughout.
In Nardus stricta, Glyceria fluitans, Sesleria, &c., there are two kinds of short cells, some siliceous, others cutinized only.
Nardus has closely appressed small 2-celled hairs bent65 at right-angles, and some epidermal and parenchyma cells—especially below the stomata—have solid masses of silica filling the lumina.

Fig. 23. Part of transverse section of leaf of Aira cæspitosa (× about 30). Ridges very high and acute, each tipped with sclerenchyma, and containing an isolated vascular bundle—sometimes one or more small ones also. Motor-cells well developed at the base of each groove. The bundles are not girdered, but numerous bands of sclerenchyma almost join into a continuous band below. The leaf rolls inwards.
Short cells occur in Holcus lanatus, Hierochloe borealis and Dactylis glomerata interspersed between plane-walled cells. They may be silicified and vary in shape—square, saddle-shaped, elliptical, irregular, &c.; or they may be replaced here and there by asperities—e.g. Elymus—or in rarer cases by stomata. Grob has attempted the classification of their distribution in different grasses, but the subject is too complex for treatment here.
The epidermis of many grasses is studded with short two-celled hairs bent sharply at right-angles; so that the pointed or blunt, hollow or solid, apical portion is appressed to the surface. Grob says that these are absent from the Hordeæ, whereas 90% of the Panicoideæ and many species of all other groups have them. Examples of the sharply pointed form occur in Nardus, of blunt ones in Cynodon &c.
In Nardus they occur on the leaf surface both66 between and above the veins, but in Hierochloe &c. they are confined to the margins.
The following grasses have no hairs of either type:
| Agrostis vulgaris, Calamagrostis lanceolata, Avena pratensis, Arrhenatherum avenaceum, |
Dactylis glomerata, Briza media, Arundo Phragmites, Glyceria fluitans. |
The sharp, hard prickle-hairs which give the pronounced roughness to many leaves of grasses are longer than the foregoing, and stand off more from the leaf. They occur both on the surface and at the margins, and may be isolated—e.g. Avena pratensis,—or mixed with the short cells—Aira canescens, Elymus arenarius. They are very abundant on Kœleria cristata.
Leersia oryzoides has asperities at the margin of the leaf with their points directed upwards on the upper part of the leaf, downwards on the basal parts, and the direction of such minute marginal asperities often affords a useful distinctive character—e.g. Phleum, Arrhenatherum. The marginal asperities in Nardus are siliceous.
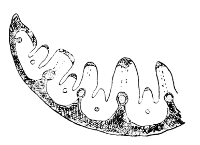
Fig. 24. Transverse section of part of leaf of Agropyrum junceum (× about 40) partly inrolled; showing unequal ridges. The principal vascular bundles are girdered below, the sclerenchyma joining into a strong continuous sheath. Each ridge is tipped with sclerenchyma, and each groove has motor-cells—not shown in the figure—below.
Bristles—i.e. long, sharp, stiff hairs—are not very common. They occur on Nardus, Anthoxanthum Puelii, Panicum, Cynodon.
Papillæ occur on the leaves of Glyceria, Nardus, Leersia, &c.
Poa pratensis has soft hairs on the upper epidermis.
The stomata of Sesleria cærulea are depressed and six-celled, two guard-cells being overgrown by four accessory cells, but in most grasses they are of the ordinary type with two elongated guard-cells only.
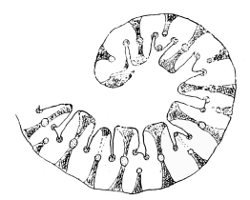
Fig. 25. Transverse section of part of leaf of Elymus arenarius, partly inrolled (× about 30), showing ridges of unequal height, of which the higher are flat above. Vascular bundles girdered, the stronger above and below. Motor-cells in each groove cause the inrolling of the lamina by their contraction.
As regards the vascular bundles constituting the venation, they are as is well known parallel from base to apex in our common grasses, with linear leaves, and are usually of four orders as regards strength. Those of the first (e.g. mid-rib) and second orders have conspicuous vessels, but those of the third and fourth orders may be practically devoid of vessels, though xylem and phloem68 elements are always present. Contrary to the general assumption, there are frequent though minute transverse bundles joining the parallel veins.
The rule is that one vascular bundle runs up each mid-rib or ridge, but exceptions occur—e.g. in Arundo several bundles run up the mid-rib, and in Aira cæspitosa (Fig. 23) and others even the strong ribs may have two or three bundles.
Each vascular bundle has its own sclerenchyma sheath, and very often the stronger veins are accentuated owing to the vascular bundle having a girder-like band of sclerenchyma running conjointly with its sheath and joining the latter above and below—or below only—to the epidermis (Figs. 24 and 25). In many cases these lower girders spread out laterally below—fan-shaped in section—and nearly join the neighbouring girders.
In other cases the strands of sclerenchymatous supporting tissue do not join the bundles, but run parallel to them, above or below, as separate strands just beneath the epidermis.
Finally, these strands may separate from the bundles, and fuse below into a continuous layer under the epidermis; this occurs especially in leaves of xerophytes where the cuticle is well developed—e.g. in varieties of Festuca ovina (Fig. 18), Aira flexuosa (Fig. 28).
The distribution of the strands of isolated sclerenchyma affords good characters. While there are none in Mibora, we find one large strand at the ridge of the keel and one at each margin, in addition to smaller ones subtending each vascular bundle, in Avena pubescens, Sesleria, Poa69 annua (Fig. 21), P. bulbosa, P. compressa and Dactylis glomerata. In Festuca ovina, F. rubra, F. heterophylla (Figs. 18, 27) there are groups more or less pronounced at the keel and margins, or even a continuous band below, but none above the bundles.
 |
 |
Fig. 26. Transverse section of leaf of Nardus stricta (× about 50). The upper surface is represented by the four grooves and five ridges, each of the former with traces of motor-cells at its base. The deep shaded portions are sclerenchyma, strong girders of which join the vascular bundle of each ridge to the lower surface. This type is obviously derived from that in Fig. 19, and may be regarded as a permanently rolled leaf. |
Fig. 27. Transverse section of
leaf of Festuca ovina, var.
duriuscula (× about 50), the
type of a permanently folded
leaf. Seven ridges and six
intervening grooves are seen:
each of the latter with traces
of motor-cells below. In each
ridge is an isolated vascular
bundle, and a narrow sclerenchyma
band below. |
Many grasses have an isolated band above and below each primary bundle only—e.g. Panicum, Cynodon—or above and below each of the other bundles as well—e.g. Spartina, Arundo, Polypogon, Agrostis alba, Aira cæspitosa (Fig. 23), Holcus lanatus, Glyceria aquatica, G. fluitans, Digraphis, Elymus (Fig. 25), Agropyrum70 (Fig. 24), Brachypodium, Nardus (Fig. 26). In Psamma arenaria the lower bands join into a continuous layer.
In the following there is a band like a girder above and below each bundle, and contiguous with it, joining it to the epidermis above and below—Leersia, Phleum pratense, Calamagrostis Epigeios, Bromus erectus, &c.
Güntz points out that xerophilous grasses are apt to have upright, narrow (Figs. 26-28), grooved or folded leaves, with strong cuticle, and marked motor-cells when the leaves open. It is in grasses of this kind, especially such as inhabit dry sandy districts, that the subulate, solid or grooved leaves shown in Figures 18, 19 occur—e.g. Festuca ovina and its varieties, Aira flexuosa, Nardus stricta, &c. The epidermal cell-walls are sinuous, the stomata protected—e.g. on the flanks of ribs and in grooves—and waxy or hairy coverings occur. Colourless water-storing cells are apt to occur between or around the vascular bundles, and the chlorophyll-tissues tend to be dense and well protected inside the leaf: strongly developed bast-sclerenchyma is also frequent (Fig. 18).
In shade-grasses, on the other hand, and in hygrophilous species, the leaves are as a rule flat, with thin epidermal cell-walls, which have plane sides, free stomata, and no wax &c. Water-storing tissue (apart from tropical species) is sparse or absent, and the chlorophyll-tissues have well aerated lacunar spaces. Bast-sclerenchyma is in these cases feebly developed.
In the following chapter I have brought together some of the principal anatomical features, in such form that the characters can be employed in checking other determina71tions of grass leaves. The results, which are based on the elaborate investigations of Duval Jouve, Schroeter, Pée-Laby and Grob, as well as on my own observations, are not complete in all respects, and much more should be done to extend the theme, but the account given will serve to show the student how such results may be employed. It is as yet impossible to decide how far these characters are constant—they are known to be fairly so in many cases—but several grasses cannot yet be distinguished by them alone.
Fig. 28. Transverse section of subulate leaf of Aira flexuosa (× about 50), the upper surface represented by a mere ridge with two flanking grooves each with but traces of motor-cells below. One large vascular bundle and four much smaller ones are seen. There are no girders, but slender bands of sclerenchyma at the lower surface nearly join into a continuous sub-epidermal sheath. This type is the extreme form of that in Fig. 26.
It should also be added that some grasses develope two types of leaves (heterophylly), solid or subulate below, flat or slightly inrolled above—e.g. Festuca heterophylla—and the following arrangement is intended to apply to the vegetative lower leaves and not to those on the upper parts of the flowering specimen. Moreover the sections should be cut from the basal third of the lamina, and not from the tip of the leaf.
I. The chlorophyll-tissue, on transverse sections, is arranged in rings round the vascular bundles. There are motor-cells between the ribs, and the stomata are sunk and occur on both faces.
Cynodon Dactylon. The larger lateral nerves have as a rule three smaller ones between each pair, hardly projecting as ribs. Chlorophyll chiefly in a ring round the vascular bundle. Long hairs on lower surface, a few papillæ above. Motor-cells in each shallow furrow. Short cells occur between the long epidermal cells over the bast-region.
The Panicums also come here, and differ according to the disposition of the sclerenchyma sheaths around the bundles.
II. The chlorophyll-tissue is between the vascular bundles, and not confined to rings surrounding them.
A. Conspicuous lacunæ between the vascular bundles. Stomata on both faces. Motor-cells occur.
Lacunæ large and rectangular. Motor-cells confined to a flanking line on each side of the mid-rib.
Glyceria aquatica. Leaves folded and in section V-shaped, hardly keeled, with sclerenchyma at apex. Motor-cells each side of the mid-rib only. The large square or rectangular lacunæ bounded by stellate cells. Papillæ on epidermal cells. Vascular bundles midway between upper and lower surfaces.
Glyceria fluitans. Section V-shaped and keeled, the roof of each polygonal lacuna arched, hence the “ribs" on the upper surface are between the vascular bundles. The latter lie nearer the lower epidermis. The epidermis has papillæ.
Catabrosa aquatica and Hierochloe also come here, the former with small lacunæ, the latter with larger ones chiefly towards the upper surface of the leaf.
Digraphis is also apt to have a few air cavities near the mid-rib.
B. Lacunæ none, or inconspicuous, the chlorophyll-tissue filling up between the ribs.
(a) Upper and lower leaf-surfaces parallel, or nearly so, and much alike, the ridges being very low or obsolete. Stomata equal or nearly so on both surfaces.
(1) Motor-cells absent; vascular bundles feeble and very few.
Mibora verna. The small leaves are flat, or nearly so, and have three isolated and very feebly developed bundles, devoid of girders or sclerenchyma bands.
(2) Motor-cells present, vascular bundles of various orders, with sclerenchyma bands or girders.
✲ Leaf keeled, and folded—not inrolled. Motor-cells confined to the neighbourhood of the mid-rib. No hairs.
† Motor-cells conspicuous and conjoined into a band above the mid-rib.
Dactylis glomerata. Keel pronounced, with one large vascular bundle and a sclerenchyma band occupying its crest. Motor-cells forming one conjoint band along the upper course of the mid-rib only. Stomata on both faces, but no hairs or thick cuticle. Ribs low, and all bundles have feeble girders. A little sclerenchyma at the margins. A few pale cells in the chlorophyll-tissue.
†† Motor-cells inconspicuous and in two flanking lines, one on each side of the mid-rib.
Poa trivialis. Keel with sclerenchyma at its apex, and a small band of the same at the margins. Vascular bundles of three orders, isolated, without girders, but with a small band of sclerenchyma above and below. Ridges obsolete. Short hook-asperities above. No thickened cuticle.
Other species of Poa also come here: I cannot distinguish them by the leaf anatomy; but P. annua, P. compressa, P. nemoralis and
P. pratensis are devoid of the hooked asperities; P. nemoralis has a thicker lamina than the rest, and girders to the secondary bundles. P. annua agrees in the latter point.
✲✲ Leaf not keeled: rolling up. Motor-cells distributed between the ridges.
† Hairs none or rare, or at most a few asperities.
≡ Veins numerous, 30-40 on each half lamina. Motor-cells very large.
⊙ All vascular bundles with girders above and below.
Digraphis arundinacea. No keel. Marginal sclerenchyma conspicuous. A few asperities below. Leaf thin, and all the bundles joined to the epidermis above and below by girders (Fig. 14). Stomata on both surfaces, fairly large: epidermal cells with plane walls. There may be a few irregular air cavities, especially near the mid-rib.
⊙⊙ Only the principal bundles girdered.
Arundo Phragmites. Ridges very numerous and low. No keel. Marginal sclerenchyma strong. Vascular bundles with sheaths of large colourless cells, a few of the strongest girdered below, but most have only sclerenchyma bands above and below. Motor-cells particularly large, between all the bundles. There are no conspicuous lacunæ. Hairs very rare. Epidermal cells small, with sinuous walls: all the cell-walls contain silica. Stomata on both faces, sunk, small and more difficult to see than in Digraphis, where the epidermal cells are plane walled, or nearly so.
Arundo Donax is very like A. Phragmites, but has larger bundles each with a horse-shoe shaped sclerenchymatous mass below, and larger lacunæ.
≡ ≡ Veins not more than 10-20 in each half lamina.
⊙ More or less conspicuously hairy. The smaller bundles isolated and devoid of girders.
Bromus sterilis. Girders to the stronger bundles only. Stiff hairs above and below. Motor-cells poorly developed between each pair of low ridges. No pronounced cuticle. A faint sclerenchyma-band at margin, and at apex of low rounded keel. Stomata on both faces.
Bromus arvensis. Similar to B. sterilis, with stiff hairs commoner below. Harsh in cutting.
B. giganteus shows no hairs, but I cannot distinguish the Bromes generally by the leaf anatomy.
Anthoxanthum odoratum. No keel, ridges obsolete, the stronger bundles only with girders. Motor-cells conspicuous between all the ribs. Marginal sclerenchyma, and that above and below the bundles, poorly developed. A few coarse hairs both above and below, and stomata on both faces. Leaf thin and narrow.
Hordeum murinum. Few girdered bundles, and sclerenchyma at margins poor. Hairs sparse and coarse.
Bromus asper, Brachypodium sylvaticum and Lagurus also come here.
In all these grasses the epidermal cells are chiefly long, rectangular or slightly hexagonal, with thin and plane walls.
⊙⊙ Hairs none or very rare on the sections.
Phleum pratense. Low rounded ribs with motor-cells between. The larger vascular bundles girdered. Stomata about equal on both faces. No hairs. No keel. Marginal sclerenchyma scanty.
Arrhenatherum avenaceum. Very rare hairs above: a few blunt asperities here and there. No keel. Ridges low. Girders to the primary bundles, but not very strong: marginal and other sclerenchyma faint, as is also the cuticle. Stomata on both faces. Motor-cells fairly developed between the ridges.
Briza media. No keel, and mere traces of marginal sclerenchyma. Ribs practically obsolete, but well developed motor-cells in furrows. Principal bundles girdered. Stomata on both sides. No hairs or thickened cuticle.
Avena fatua, Molinia and Leersia also come here.
(b) Upper and lower leaf-surfaces dissimilar, or at least not parallel, owing to the conspicuous ridges and grooves above.
(1) No stomata below.
✲ Leaves flat or nearly so, or at least exhibit a conspicuous concave upper surface.
⊙ Motor-cells between each pair of ribs: sclerenchyma not forming a continuous layer below.
≡ Ridges at least 5-6 times as high as the leaf-thickness between.
Aira cæspitosa. Ridges high, 7-10 times as high as the breadth of leaf between, triangular, each with 1-3 vascular bundles devoid of girders, with an upper isolated band of sclerenchyma at the acute tip, and another below the principal bundle. Also small bands below each group of motor-cells. Small conical asperities on the ridges and below. No mid-rib. Stomata on flanks of ridges only, and few motor-cells between (Fig. 23).
Each vascular bundle has a sheath, but is isolated. Sclerenchyma at tips of the ridges dense: smaller bands below: strong at margins. Lower cuticle strong. Leaf rolls up.
The flat upper leaves of Festuca rubra (Fig. 20) and F. heterophylla are somewhat similar in type. They have stiff hairs on the ridges.
≡ ≡ Ridges not more than 2-3 times as high as the tissue between; each furrow with motor-cells, and each vascular bundle joined to epidermis above and below by a sclerenchyma girder.
Brachypodium pinnatum. Smooth. Ridges rounded. Hairs rare. The strong sclerenchyma girders below almost continuous laterally. Epidermal cells with sinuous thick walls, and a few tooth-hairs.
Note the differences from B. sylvaticum, p. 76.
Melica nutans, M. uniflora, and Calamagrostis Epigeios also come here.
⊙⊙ Motor-cells confined to the innermost 2-4 furrows. Sclerenchyma in a continuous band just inside the thick cuticle below.
Festuca duriuscula. The ridges are only about half to one-third as high again as the thickness between, and the motor-cells in four series at the base of the three innermost ridges. Each ridge has only one isolated sheathed bundle, without girders. Stomata on the flanks of the ridges, and few in number. The sclerenchyma forms a thick band just inside the strong cuticle below. The leaf is conduplicate, not convolute.
This applies particularly to the more open leaves: the subulate leaves belong to the next type (see Fig. 27).
Aira canescens and Spartina stricta also come here.
Psamma arenaria. Inrolled. Smooth below and devoid of keel, with sub-epidermal band of sclerenchyma, and similar tissue at the margin. Ridges of three sizes, the largest twice or three times as high as the leaf-tissue between is thick, all rounded above, and very hairy. Stomata above only. Motor-cells in each sinus not large. Vascular bundles isolated, without girders or bands of sclerenchyma.
Elymus is very like Psamma, but has a few stomata below and the sub-epidermal sclerenchyma is not continuous (see Fig. 25).
✲✲ Leaves (subulate) not opening out, the upper surface represented by a groove or a few ridges above the angular or ovate solid section.
⊙ Section pentagonal or angular-ovate: sclerenchyma below in a continuous band.
Aira flexuosa. Upper surface a depression, with one ridge flanked by two grooves at its base, the depression extending about one-fifth through the whole thickness of the nearly solid leaf. Vascular bundles about 3-5, isolated, sheathed. Sclerenchyma band extending all round the lower surface just inside the thick cuticle. Stomata very few, flanking the ridge; motor-cells in the furrows, poorly developed (Fig. 28).
⊙⊙ Section elliptical or angular-ovate; sclerenchyma not always in a continuous band below.
Festuca ovina. Upper surface a deep fold, with three ridges and 2-4 grooves at its base. Vascular bundles several, with girders. Motor-cells in four series, in the80 grooves. The lower girders may not fuse laterally into a continuous band of sclerenchyma below (Fig. 18).
The folded lower leaves of F. rubra and F. heterophylla come here also. For the flatter leaves of F. duriuscula see p. 78 and compare Fig. 27.
The epidermal cells in this series have sinuous thickened walls, and here and there small tooth-like hairs.
Nardus also comes here (see Fig. 26).
(2) There are stomata below, but fewer than on the upper surface. Motor-cells usually conspicuous between the ridges.
✲ Stronger bundles with girders of sclerenchyma joining them to the epidermis, at least below.
⊙ Hairs sparse or none.
Cynosurus cristatus. Mid-rib obsolete, except the strong vascular bundle. Ridges low and rounded, with 2-4 flanking stomata, and well developed motor-cells in furrows. Secondary vascular bundles with strong girders below, the smaller bundles sheathed only and isolated. Each ridge with slight sclerenchyma above. A few stiff short hairs above, and the leaves are convolute. Ridges about twice the height of the leaf-thickness between (Fig. 16).
Agropyrum repens. Mid-rib and margin with strong sclerenchyma-groups: ridges unequal, low and rounded and each vascular bundle girdered. A few pointed hairs above, and motor-cells in all the grooves. A slight keel, stomata on both surfaces.
Agropyrum caninum. All the bundles have girders. Slight keel. Marginal sclerenchyma. Few, very short, hard, hooked asperities above and below. Ridges low,81 and motor-cells poorly developed between. Few stomata on lower surface. Very like A. repens, but the principal ridges are more prominent below and those nearer the mid-rib have asperities.
A. junceum resembles Psamma, but the ridges are much lower, and there are a few stomata on the under surface (Fig. 24).
⊙⊙ Leaf obviously hairy.
† Hairs more especially above.
Avena flavescens is very similar to Cynosurus, but is evidently hairy, and A. pratensis also comes here.
†† Hairs abundant on both surfaces.
Holcus lanatus. Very hairy above and below, and at the margins. Slight keel with sclerenchyma band: sclerenchyma at margin slight. Ridges rounded, about twice as high as thickness between. Stomata more abundant above. Cuticle very thin and leaf soft. All bundles except the mid-rib with girders. Motor-cells fairly well developed between the ridges (Fig. 15).
Kœleria cristata. Very hairy on both surfaces. Ridges irregular, the largest flat and high, the others rounded or triangular. Vascular bundles isolated, and the sclerenchyma reduced to a few cells in a single layer beneath the epidermis at the apex of each ridge and below the bundle. Motor-cells well developed in each furrow. Stomata more numerous above.
✲✲ No girders to the vascular bundles.
Lolium perenne. Ridges numerous and unequal. Vascular bundles sheathed and isolated—i.e. devoid of82 girders: small patches of sclerenchyma at the apex of each stronger ridge, and on the opposite side below only. No hairs.
Lolium temulentum is similar but is more apt to be convolute, whereas L. perenne is more folded.
Alopecurus pratensis. Leaf thin and somewhat like Phleum, but the ridges somewhat higher and more rounded, and only the principal bundles girdered below. Stomata on both faces.
Festuca elatior, Bromus giganteus and most species of Agrostis come near Lolium. See Figs. 17, 22.
When the flowering shoot of a grass pushes up into the light and air from the enveloping leaves, it forms a more or less branched collection of flowers known as the Inflorescence, and in all our grasses this inflorescence consists of a principal stalk, haulm or culm, on which shorter stalks—branched or not—are arranged. The mode of branching is usually such that the youngest branches are nearest the top, and the oldest nearest the bottom. It is evident at once, on comparing the Moor Mat-grass (Nardus), Vernal-grass (Anthoxanthum), Cock’s-foot (Dactylis), Meadow-grass (Poa) that considerable differences exist as to the extent of this primary branching of the inflorescence.
In Nardus (Fig. 2) we find a number of long cylindrical-tapering bud-like structures each seated on one side of the principal stem, and one over the other: in the Vernal-grass and Cock’s-foot we find tufts of such bud-like structures closely crowded round the upper end of the84 principal stalk, the whole forming an elongated tuft of tufts: in the Poa we find a number of radiating, slender, long branches springing from the principal stalk, and each of these ramifies again, and yet again, until each of the ultimate hair-like branches bears one of the bud-like structures. See also Catabrosa (Fig. 4).

Fig. 29. A spikelet of Festuca elatior, var. pratensis, from which the glumes and one palea (the outer) have been removed to show the flower in situ (× 12). The two lodicules are in front: the inner palea behind. Strasburger.
The first thing for the student to apprehend is the nature of the bud-like structures referred to.
Each of these is in itself a small tuft or bud of leaf85like organs or scales arranged on a short twig (Rachis, Rachilla), as it were, and is called a Spikelet, and the true flowers of the grass are contained in the angles between the scales—the scales being popularly known as "chaff": technically as Glumes and Paleæ.
In order to understand the structure of a spikelet the student should carefully dissect a large one, such as that of an Oat (Fig. 1). Proceeding from outside, he will find two large scales, like two boats, fixed below to the stalk (rachis) one just below the other, and shutting together as if hinged. These are called the glumes—the inner and outer glume respectively—and they enclose the rest of the spikelet.
Fig. 30. Diagram of a spikelet of a grass as it would appear if the internodes between each set of organs were elongated. g1 lower and g2 upper glume. P lower and p upper palea of the second oldest flower F2. f a barren flower represented only by the axis and paleæ. Above it a single palea and the termination of the axis (a) of the spikelet.
Inside them the axis or stalk (rachis) is continued for a short distance only and on its sides are hinged two86 other pairs of more or less boat-shaped scales, smaller and more delicate than the glumes, and known as the pales (paleæ), while a third pair of still smaller pales is fixed to the end of the axis. In each case one smaller and more delicate inner palea is hinged just inside its more obvious outer palea. In the closed condition of the spikelet each of the three pairs of pales is shut together, and pressed close to the axis, and the pair of glumes shut in the whole.
On opening each of the lower pairs of pales we find a flower inside; but the terminal pair usually contain only the barren end of the axis. Hence the latter is barren and the former are fertile.
Each fertile flower is found on careful dissection to consist of a small swollen Ovary, or young grain, covered with silky hairs and with a couple of delicate plumes (the Stigma) at its apex, and three long and slender Stamens; while the magnifying glass will show two tiny scales at the base—the Lodicules. All our ordinary grasses have their flowers thus constructed—a pair of lodicules, three dangling stamens and an ovary with a feathery two-plumed stigma: each such flower is also enclosed in its pair of pales, and the several pairs of pales of each spikelet, with their contents, are enclosed in the pair of glumes (Figs. 29-32).
Returning now to the inflorescence. It is clear that we have to distinguish between the entire branched total Inflorescence, and the Spikelets or partial inflorescences of which it is composed. In Botany it is agreed to call any inflorescence consisting of a stalk or axis on which87 the flowers are arranged without stalks—i.e. sitting directly on it—and so that the youngest are above and the older below, a Spike, and each spikelet is a little spike.
 |
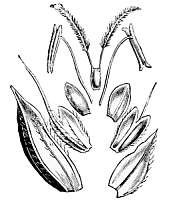 |
Fig. 31. Diagram of a spikelet of Wheat dissected (× about 5) showing—from below upwards—the two glumes, two paleæ, two lodicules, three stamens, and the ovary of the typical grass. Oliver. |
Fig. 32. Diagram of a spikelet of
Anthoxanthum dissected (× about
8), and showing—from below
upwards—two outer and two
(awned) inner glumes, two paleæ,
two stamens, and the ovary.
There are no lodicules. Oliver. |
On looking at the total inflorescence of the Nardus we see that we have a number of spikelets seated on the sides of an axis: this is then a spike of spikelets, or, shortly, a Spike9 (Fig. 5). Mibora and Lepturus afford other examples. In Panicum, Cynodon (Fig. 2) and Spartina we have groups of such spikes.
The Poa inflorescence is, however, different. It consists of a loose branched system of spikelets. Botanists term such a loosely branching system, where each branch ends in a flower, a panicle: here then we have a panicle of spikelets, or, shortly, a Panicle. Aira, Agrostis, Calamagrostis, Avena, Catabrosa (Fig. 4) and many others afford further examples.
In Dactylis we have a condition of affairs between the two extremes given: the inflorescence is not so close a spike as Nardus, and not so open a panicle as Poa—it is rather a spike-like panicle, partaking of the nature of both. A special type of this (Foxtail) occurring in certain grasses—e.g. Phleum, Alopecurus, Phalaris and Lagurus,—is so characteristic as to be worth noting (Fig. 3).
There is also another aspect of these inflorescences which is not without interest as showing how diagnostic characters may be obtained from purely external features, easily observed in the field. We have seen that in Nardus the spikelets are arranged on one side only of the rachis, or main axis, so that about three quarters of the circumference of the latter is bare; whereas in Lolium—with which Agropyrum and Brachypodium agree in this respect—the spikelets are on opposite sides, leaving the intervening two quarters, i.e. half its surface, of the circumference of the axis naked.
In Cynosurus and the simpler forms of Dactylis, we find the spikelets crowded round about three quarters of the surface of the rachis, leaving the fourth quarter naked; and, finally, in Phleum, Alopecurus, Hordeum, and Anthoxanthum the spikelets cover the entire surface.
In the first (Nardus) and third examples (Cynosurus, Dactylis) where the spikelets are turned to one side, the inflorescence is said to be secund.
The next point to notice is that not every grass has so many as two fertile flowers and one barren one in its spikelet, as the Oat has. A spikelet may have one (Phleum), two (Aira) or three (Avena) or more (Poa) fertile flowers, and no barren ones or several, the number of flowers being counted by the number of pairs of pales found inside the pair of glumes. Moreover every flower is not necessarily fertile (e.g. Arrhenatherum, Holcus) and several grasses have one or more flowers in each spikelet containing stamens only—male flowers—while others may have ovaries only—female flowers. In some exotics the male and female flowers are on different parts of the same plant (Maize) or even on different plants (Gynerium), an arrangement not met with in our grasses. Accordingly, it is of importance in determining a grass to discover how many flowers the spikelet contains, and whether any are male only, or barren, &c., as well as to make out the character of its inflorescence.
In the following lists I have brought together some of the chief points with illustrative examples.
Spikelets with only one perfect flower
(without rudimentary ones).
Psamma.
Milium.
Nardus.
Alopecurus.Phleum.
Hordeum.
Agrostis.
And species of the rare grasses Calamagrostis, Mibora,90 Lepturus, Spartina, Cynodon, Gastridium, Lagurus, Polypogon, Leersia.
Spikelets with one perfect flower and one or more male or rudimentary ones.
Digraphis.
Anthoxanthum.Holcus.
Arrhenatherum.
And the rare genera Hierochloe and Panicum.
Spikelets with at least two perfect flowers.
Molinia.
Arundo.
Avena.
Aira (some species).
Bromus.
Cynosurus.
Dactylis.Triodia.
Sesleria.
Kœleria.
Melica (one species).
Briza.
Poa trivialis.
Catabrosa.Spikelets with at least three perfect flowers and usually more.
Elymus.
Agropyrum.
Brachypodium.
Lolium.Festuca.
Poa.
Glyceria.Inflorescence, a spike of single spikelets.
Agropyrum.
Brachypodium.Lolium.
Nardus.
And varieties of Festuca Myurus and F. loliacea, &c.
Inflorescence, a spike of pairs or tufts of three or more spikelets.
Elymus (pairs).
Hordeum (threes).Cynosurus (clusters). Inflorescence, a cylindrical closely tufted spike-like panicle.
Phleum.
Alopecurus.Psamma.
Anthoxanthum.Inflorescence, a compact more or less tufted panicle.
Molinia.
Kœleria.
Triodia.
Dactylis.Aira præcox.
A. canescens.
Sesleria.
And rare grasses like Polypogon, Gastridium, &c.
Inflorescence, a loose plume-like or branched panicle.
Avena.
Bromus.
Arrhenatherum.
Catabrosa.
Aira (except A. præcox and
A. canescens).
Arundo.
Digraphis.Milium.
Agrostis.
Melica.
Holcus.
Poa (most of the species).
Glyceria.
Briza.
Festuca (except F. Myurus).
And the rare Hierochloe.
The Glumes are always present in our grasses, and rudimentary only in the rare grass Leersia; but Lolium and Nardus have only one glume to the spikelet, and Hierochloe, Digraphis and Anthoxanthum (Fig. 32) have four or six. Our other grasses have two, but often unequal in size.
In shape they are usually boat-like, pointed or obtuse (Briza) and frequently with a distinct keel (Anthoxanthum, Digraphis, Phleum, &c.) or with ridges, green lines (veins) and other characteristic markings (e.g. Digraphis). The tip may be extended into a stiff long point or awn (Agropyrum, Phleum, Nardus) and the keel, ribs, and awn may have hairs or serrulæ on them. The rule is, however, that the glumes are not awned. In texture the glumes may be herbaceous and green-brown or purple92 (e.g. Melica) or membranous or stiff, or scarious (i.e. browned, as if scorched) at the edges. In Hordeum some of the glumes are so narrow and pointed as to resemble stiff awns. In Catabrosa the glumes are truncate, as if bitten off at the top.
The Paleæ are also often more or less boat-shaped, or flat ovate or oblong scales, usually more delicate than the glumes and frequently pointed, or (especially the outer pale) awned at the tip: in some cases, however, the awn springs from the middle or base of the back of the pale, and the latter may be bifid at its apex. The pale has usually a distinct middle nerve. The inner pale is commonly the smaller and more delicate of the two, and is sometimes difficult to see.
Conspicuously awned Grasses.
Avena.
Arrhenatherum (Fig. 33).
Hordeum.
Bromus.Agropyrum caninum.
Lolium temulentum.
Brachypodium sylvaticum.
Festuca Myurus.And a few rare grasses like Panicum, Polypogon, Lagurus.
Grasses with no true awns.
Kœleria.
Milium.
Digraphis.
Elymus.
Festuca (except F. Myurus
and F. uniglumis).
Briza.Poa.
Glyceria.
Catabrosa.
Molinia.
Melica.
Psamma.
Agrostis alba.And a few rare forms like Leersia, Hierochloe, &c.
Grasses with inconspicuous awns, or mere points, to some of the Glumes or Pales.
Arundo.
Phleum.
Alopecurus.
Holcus.
Nardus.
Agropyrum repens.Lolium perenne.
Brachypodium pinnatum.
Dactylis.
Cynosurus.
Sesleria.
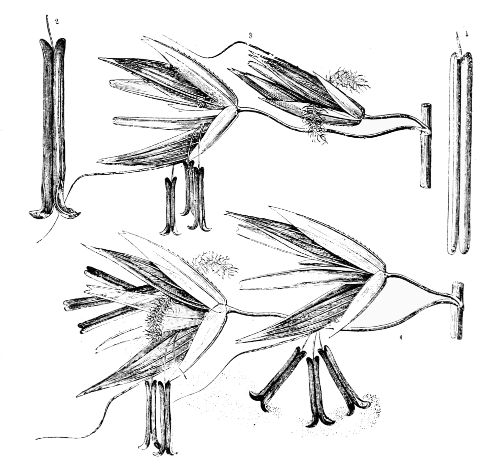
Fig. 33. Arrhenatherum. 1 unopened and 2 open anther (× 12). 3, spikelets open and exposing the stamens and stigmas; 4, the pollen escaping and being dusted on to the stigmas (× about 5). Kerner.
As regards the flower proper, all our British grasses except Anthoxanthum (which has two only) have three stamens; but many exotic grasses have six stamens, and a few have a large number—even 40. The stamens have slender filaments and large versatile anthers, which dangle from between the paleæ when the flowers are mature, scattering their clouds of fine pollen in the wind (Fig. 33).
All our ordinary grasses except Nardus—where there
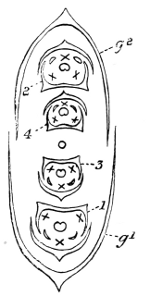 Fig. 34. Diagram of a
spikelet of a grass.
The two glumes—g1
outer, g2 inner—embrace
four flowers, of
which 1 is the lowermost
and 4 the uppermost.
is a simple straight hairy style—have
two spreading feathery stigmatic
plumes, which stand out right and
left from between the paleæ when
the pollen is flying about on the
wind. (Fig. 33.)
Fig. 34. Diagram of a
spikelet of a grass.
The two glumes—g1
outer, g2 inner—embrace
four flowers, of
which 1 is the lowermost
and 4 the uppermost.
is a simple straight hairy style—have
two spreading feathery stigmatic
plumes, which stand out right and
left from between the paleæ when
the pollen is flying about on the
wind. (Fig. 33.)
Much interesting speculation has been expended in attempting to explain the morphological or theoretical significance of the parts of the spikelet of a grass. If we project the various organs on a flat surface in the form of a plan, keeping their relative positions intact, we obtain a diagram such as that shown in Fig. 34.
The comparison of numerous cases, and the study of the development of the parts on the microscopic growing point dissected out from young buds, have suggested that the inner and outer glumes are bracts, or covering leaves,95 at the base of the true spikelet. In like manner the two paleæ are bracteoles which subtend the flower proper. On this assumption they can be compared with the corresponding structures in other plants; whereas any attempt to compare the paleæ or glumes with the sepals and petals of ordinary flowers breaks down.
A curious interest attaches to the awns so often found on the backs of paleæ, and especially to those where the (sub-terminal) awn springs from just below the bifid apex (e.g. Avena, &c.).
Hackel showed by comparison with a rolled leaf attached to its sheath and ligule (e.g. Psamma) that such an awn as that of Bromus Alopecurus attached to its palea stands in the relation of a leaf to its sheath, the part of the palea above the insertion corresponding to the ligule, the awn itself to the lamina, and the palea below its insertion to the sheath. This view is rendered the more probable by the anatomy of the awn and by the observations of Schmid, who has shown that the awns of cereals contain chlorophyll-tissue and a vascular bundle, and have stomata, and his experiments led him to conclude that in the young condition they transpire and assimilate, and probably even contribute to the nutrition of the ripening grain. When dry and mature the awns subserve biological functions of quite another kind, and as we shall see are of importance in the distribution and sowing of the grains. (Fig. 42.)
Returning to the floral diagram, we see that the two lodicules, the three stamens and the ovary still remain to96 be explained. Much discussion has been held regarding the lodicules. Functionally they are said to aid in the divarication of the paleæ when the period of anthesis arrives, and the stamens and stigmatic lobes are to be exposed, by swelling and driving the valve-like paleæ apart. Morphologically they have been explained as representing the rudimentary perianth, here reduced to two minute scales, but in some exotic grasses (Bambusa, Stipa, &c.) three lodicules, or even more, are present. (Figs. 35, 36.) On the other hand they may be, and probably are, scales of the nature of minute bracteoles and of no significance to the flower itself.
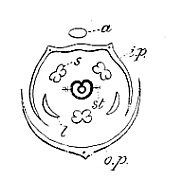 |
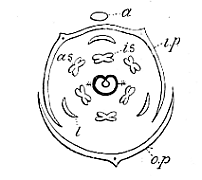 |
Fig. 35. Floral diagram of ordinary grass. Each pair of paleæ—i.p. inner and o.p. outer palea—encloses three stamens (s), two lodicules (l) and the ovary. st, stigmatic plumes. a, axis. |
Fig. 36. Floral diagram of a
Bamboo, showing six stamens,
three inner (i.s.) and three
outer (a.s.), and three lodicules
(l) in addition to the
ovary. i.p. inner and o.p.
outer palea. a, axis. |
If this is so the flower of the grass is perfectly naked,97 and consists in the typical case of three stamens and one carpel. The development of the ovary lends no support to the view that there are two carpels: the stigmatic plumes are not separate styles. Nor does the fact that some grasses have six stamens lend support to the idea that the flower is derived from the trimerous type so common in Monocotyledons: other numbers may occur—e.g. as few as two (Anthoxanthum, Fig. 32) or even one only (Uniola, Cinna, &c.) or as many as 20 or 40 in certain other exotic grasses.
Even when three stigmatic plumes are developed, as in some Bamboos, close investigation does not confirm the view that the ovary consists of more than one carpel.
We must therefore regard the flower of the grass as typically composed of one carpel and three stamens, with no perianth whatever. It is subtended by one or more bracteoles (the lodicules), and enclosed in a pair of bracteoles one higher than the other (the paleæ). The glumes are bracts of the partial inflorescence—the spikelet.
That there are some departures from this type in detail does not invalidate the importance of the fact that most grasses conform to it.
I now pass to the consideration of a grouping of our ordinary grasses according to their floral arrangements.
The student should distinctly understand that the following notes are intended to serve as an introduction to the floral characters of our grasses, and not to replace98 the study of the Flora. I have dealt with this section of the subject less in detail, because our best Floras give so much information that it seemed undesirable to do more than attempt to guide the reader in the recognition of the genera and principal species by means of external features easily observed by anyone with a little care. The detailed and critical examination of species, varieties and rare forms should always be done with reference to a complete Flora.
I. Grasses with only one perfect flower in the spikelet.
(1) Inflorescence spikate10.
A. Inflorescence a spike of simple spikelets.
Nardus stricta, L. A tough wiry tufted moor-grass, with setaceous leaves, secund spikelets with a single rudimentary glume, and a stiff simple hairy style. (Fig. 2.)
The much rarer Mibora verna about three inches or so high, and Lepturus, both with flat leaves, also come here.
Certain superficial resemblances in the habit or inflorescence in Festuca Myurus and rare forms like Spartina, may occasionally cause hesitation until the spikelets are examined.
B. Spike with sessile or sub-sessile tufts of three or more spikelets.
(a) A spike of tufted awned spikelets, in triplets at each notch of the rachis, and one or two of each triplet barren.
Hordeum.
No other genus of our grasses is like Hordeum. The purely superficial resemblances in the inflorescences of Polypogon, Lagurus, and Cynosurus echinatus—all extremely rare species—disappear at once on examination.
In Bromus erectus the equally superficial resemblance is due to the stiff awns: the spikelet has six to twelve flowers and is stalked.
It should also be noted that Hordeum sylvaticum occasionally has a rudimentary second flower in the lateral spikelets (see note p. 105).
(α) A shade-grass with the central spikelet only imperfect; staminate, or rudimentary, or entirely wanting.
H. sylvaticum, Huds.
(β) The central spikelet is the perfect one, the two lateral barren. Growing in open land.
(i) A perennial meadow-grass. All the glumes scabrid and bristle-like.
H. pratense, Huds.
(ii) Annuals with some of the glumes at least, lanceolate or broad below.
✲ Ruderal plant, with cylindrical spikes, long awns; glumes of the central flower dilated below.
H. murinum, With.
✲✲ Maritime plant, more or less glaucous, with short ovoid spikes: glumes of the central flower bristle-like.
H. maritimum, With.
(b) Spike cylindrical, of sessile or nearly sessile awned spikelets, densely crowded round the axis, the whole resembling a fox’s brush or cat’s tail.
Species of Lagurus, Polypogon, Phalaris (not truly awned), Panicum (with bristles between the spikelets), and Gastridium are other British grasses approaching this type of inflorescence: they are all rare or very local.
Sesleria has an ovoid spike, but the spikelets are two-flowered and not truly awned.
Kœleria may present resemblances, but the spikelets are very different in detail (see p. 109).
(i) Awns inserted into the back of the single palea, and hair-like. Glumes connate below, keeled. Only one palea.
Alopecurus.
✲ Annual corn-weed, with a long and slender spike, pointed above. Glumes almost glabrous, and connate to the middle.
A. agrestis, L.
✲✲ Perennials, with shorter and stouter spikes, rounded above. Glumes connate at the base only, and obviously hairy on the keel.
† Procumbent and kneed at the nodes below. Spike 1-2 inches long. In marshy places.
A. geniculatus, L.
†† A meadow-grass, with erect stems, and spikes 2-3 inches or more and stouter.
A. pratensis, L.
The rare A. alpinus, Sm. with short ovoid spikes, about one inch long, only occurs in the Highlands.
(ii) Awns, when present, merely the stiff, pointed termination of the keel. Glumes free below. Paleæ two.
✲ A glaucous shore plant with long creeping stolons (sand-binder). Inflorescence harsh, 5—6 inches long. Glumes tapering, simply acute.
Psamma arenaria, Beauv.
Elymus, a much rarer sand-binder of similar habit, may lead to confusion until the 3—4 flowered spikelets and different arrangement are observed. (see p. 108.)
Agropyrum repens (var. junceum) is similar in habit and station, but its spikes and spikelets are very different (see p. 107).
Phleum arenarium is much smaller (see below).
✲✲ Erect. Inflorescence rarely longer than three inches. Spikelets flat: glumes keeled, the keel suddenly produced into a sharp stiff awn or mucronate point. Paleæ two.
Phleum.
† Tall perennial meadow-grass. Awn bristle-like, almost as long as the nearly glabrous glume: spike long, cylindrical.
P. pratense, L.
†† Small, compact annual shore plant, with the glumes acute only and the keel ciliate above. Inflorescence not more than 1—1-1/4 inch long.
P. arenarium, L.
The rare P. Bœhmeri, Schrad. has the glumes merely tapering to a sharp point; and the rare P. alpinum, L. has a much shorter spike and glumes ciliate on the keels.
The rare P. asperum, Jacq. has broad, shortly mucronate glumes and a longer and more slender spike.
(2) Inflorescence a panicle—i.e. tufts or spreading stalked groups of spikelets are arranged on the main axis.
A. Inflorescence compact and irregular; a spike of tufts (spike-like panicle). Glumes four, the inner pair awned: paleæ minute. Stamens two only.
Anthoxanthum odoratum, L.
The four glumes and two stamens distinguish this grass at once. Other grasses with occasionally tuft-like inflorescences—e.g. species of Agrostis, Gastridium, Aira, Dactylis, Cynosurus, Poa, Triodia, Kœleria—are distinguished at once by having three stamens (Bromus occasionally has but two), only two glumes, several flowered spikelets, &c.
B. Inflorescence a distinctly branched panicle, more or less loose and spreading.
(a) Tall reed-like perennials, growing in water or in marshes, with plume-like inflorescences, and silky hairs at the base of the paleæ. Glumes with a keel and point, but not awned.
(i) Spikelets purplish: outer palea with a slender dorsal awn: basal hairs longer than the paleæ. Leaves narrow. Not common.
Calamagrostis Epigeios, Roth.
(ii) Spikelets greenish. No awns: basal hairs much shorter than the paleæ. Leaves broad. Common.
Digraphis arundinacea, Trin.
A variety of Digraphis with white stripes in the leaves is grown in gardens. Other aquatic reed-like grasses are Arundo and Glyceria aquatica: both have several flowers in the spikelet.
The rare Calamagrostis lanceolata, Roth., C. stricta, Nutt. and C. strigosa, Hartm. also come here.
(b) Slender grasses, not reed-like, with delicate loosely spreading panicles of small spikelets.
(i) A tall, slender shade-grass, in woods. Paleæ very smooth and glistening. Spikelets few, distant and turgid, awnless.
Milium effusum, L.
Melica uniflora, also a wood-grass, has the spikelets two-flowered, though the upper one may be a mere rudiment, and much fewer, on a flexuous nodding axis. The much rarer M. nutans has two perfect flowers in the spikelet (see p. 105).
(ii) Grasses of open situations, with numerous small pointed spikelets. Inner palea minute or absent.
Agrostis.
The genus most likely to give trouble here is Aira, which though normally with two flowers in the spikelet, occasionally has but one. As regards the common species of similar habit, &c., Aira has bristle-like leaves and Agrostis flat ones.
Again, Agrostis alba has no protruding awn, as in the spikelets of Aira. For Aira cæspitosa and other details, see p. 117.
✲ Awnless, or at most a short bristle not equalling the palea. Leaves flat and short.
A. alba, L.
An exceedingly variable plant. The variety A. stolonifera has a less spreading panicle, and broader leaves with a long serrated and pointed ligule: the variety A. vulgaris has a spreading loose panicle, narrower leaves, and a short truncated ligule. There is, however, much difficulty in distinguishing the intermediate forms on mountain-pastures, &c.
✲✲ Distinctly awned. Leaves narrower, the lower ones inrolled and almost setaceous.
A. canina, L.
The rarer A. setacea, Curt. with subulate leaves and A. Spicaenti, L. with long awns also come here.
The lax spreading type of panicle characteristic of Aira and Agrostis, &c., described above, occurs in species of Avena, Bromus, Briza, Poa and Catabrosa, but the basal silky hairs and twisted awns (Avena), long sub-terminal bristle-awns (Bromus), habit 105 (Catabrosa), and the presence of two, three, or more flowers in the considerably larger spikelets of all, easily distinguish them.
The rare grasses Leersia, with two glumes and no paleæ, and Gastridium, with curiously bullate bases to the glumes, also come into this group with one-flowered spikelets, but their characters must be studied with the flora. The same remark applies to species of Panicum, Cynodon, Spartina and other aliens, occasionally met with.
II. Grasses with at least two flowers—of which one may be imperfect—in the spikelet.
A. Only one perfect flower, the other staminate only or altogether rudimentary. Inflorescence a loose or racemous panicle.
In Hordeum sylvaticum the two lateral spikelets occasionally conform to this heading (see p. 100), and the same is said to be the case sometimes in Aira (see p. 104). The rudiments are extremely minute, however, and hardly suffice to justify the removal of these grasses to this group.
In one or two species of Aira the panicle may be somewhat contracted and tuft-like.
The very rare Hierochloe has one perfect flower, and two lower staminate ones in each spikelet.
(a) A shade-grass found in woods. Awnless. One flower perfect, the other (upper) reduced to a small stalked knob. Inflorescence racemous.
Melica uniflora, L.
The much rarer M. nutans has two perfect flowers and a similar rudiment (see note, p. 104).
(b) Grasses of open situations, spikelets with awns. The reduced flower staminate.
✲ Upper flower perfect, lower staminate only. Tall oat-like meadow-grass, with a bent and twisted dorsal awn to the outer palea of the lower flower: silky hairs at the base of paleæ.
Arrhenatherum avenaceum, Beauv.
The grasses most like this are species of Avena and Aira. The former have two or more perfect flowers, and the only broad-leafed Aira—A. cæspitosa, see p. 117—is easily distinguished by its leaves and its very small spikelets and short simple awns.
✲✲ Upper flower staminate: lower perfect. Small hairy grasses, with red-veined basal leaf-sheaths and short simple awns.
Holcus.
† Erect, evenly hairy, glumes blunt, awn not protruding. Common.
H. lanatus, L.
†† More or less procumbent, hairs chiefly at the nodes. Glumes pointed. Awns simple and exserted. Rarer.
H. mollis, L.
B. Each spikelet with at least two perfect flowers, often more.
(1) Inflorescence spikate, the main axis bearing sessile or sub-sessile spikelets, each containing three or more flowers.
(a) Spike simple11, axis stout and notched, each notch having one spikelet closely sessile in it.
(i) Spikelets distichous, the flat side of each—i.e. the edges of the glumes—being next the axis (rachis).
Agropyrum.
✲ A weed with creeping stolons, and no awns or mere points to the glumes.
A. repens, Beauv.
The variety A. junceum found as a sand-binder on sea-coasts is glaucous, stiffer, with sharply pointed leaves, and blunt glumes. For other sand-binders see note, p. 102.
✲✲ Not creeping. Awns long and prominent.
A. caninum, Beauv.
(ii) The rounded backs of the glumes are next the rachis.
✲ Spikelets flat and closely sessile in the notches of the rachis.
Lolium.
† Awnless or nearly so. Perennial.
L. perenne, L.
†† With conspicuous awns. Annual, not common.
L. temulentum, L.
There are several cultivated varieties of L. perenne: L. temulentum is notoriously poisonous (see note, p. 168). The lowermost glume of each spikelet is often alone developed or conspicuous, and looks like a bract in the axis of which the spikelet sits.
✲✲ Spikelets elongated and hardly flattened, and not quite sessile, especially the lower: rachis scarcely notched, the spikelets with their sides (edges of glumes) next the axis.
Brachypodium.
† A shade-grass with long, conspicuous awns to the more or less drooping spikelets. Common.
B. sylvaticum, Beauv.
†† Growing in the open. Spikelets stouter, stiffer and more erect, with short awns. Not common.
B. pinnatum, L.
Brachypodium may easily be confounded with Bromus, but the spikelets are nearly sessile: their shape and the absence of conspicuous notches distinguish this genus from Agropyrum. Lolium 108 has a conspicuously notched rachis and the spikelets arranged in the other plane.
Poa loliacea, Huds., an uncommon sea-shore weed, may also be placed here; as also Festuca elatior, var. loliacea, Curt. and some forms of Bromus arvensis, var. mollis, L.
No other British grasses resemble Brachypodium: any superficial likeness remarked in species of Hordeum, Festuca, &c. disappears at once on examination.
(b) Spike compound—i.e. with clusters of two or more sessile or sub-sessile spikelets arranged along the rachis.
(i) Spike elongated, fertile spikelets with 3-5 flowers.
✲ Pasture-grass with wiry rachis, on which the spikelets are secund and sessile in clusters: in each cluster a comb-like group of barren glumes subtends one of fertile spikelets.
Cynosurus cristatus, L.
The rare C. echinatus has the pectinate groups of barren glumes even more prominent.
✲✲ Stout glaucous sand-binder with pairs of spikelets sessile in the notches of the rachis, and all fertile. Spike cylindrical. Not common.
Elymus arenarius, L.
For other sand-binders see p. 102. The only grass likely to be confounded is Agropyrum, in which the spikelets are not paired. Lolium, Brachypodium, &c. are easily distinguished.
(ii) Spike short and ovoid: spikelets sub-sessile and imbricate, in clusters. Bluish.
Sesleria cærulea, Ard.
Not easily mistaken for any other grass. As some of the spikelets are shortly stalked, the inflorescence is strictly paniculate, but the fact is not obvious. The glume-like bract at the base of the spike, and the general appearance suggest resem109blances to certain moor-sedges at first sight. The only other grasses with similarly shaped spikes are species of Alopecurus, Phleum, and the rare Lagurus, Polypogon, Phalaris, Panicum, all with very different spikelets and easily distinguished.
(2) Inflorescence a panicle or raceme—i.e. the spikelets on evident stalks, simple or branched, from the main axis.
It does not follow that every spikelet is distinctly stalked, and cases occur where the stalks are very short and stiff: when this happens to the stalks arising from the main axis, and the latter is elongated, the type of the spike is closely approached, and the inflorescence resembles that of Brachypodium, Lolium, &c. In some depauperated varieties of Poa, Festuca, &c., an actual spike results (see note, p. 111): the number of flowers in the spikelet is important.
(a) Panicle short, contracted and tuft-like, owing to the shortness and stiffness of most of the stalks and their tendency to remain erect, at least until anthesis. Glumes keeled and ending in a point.
(i) Panicle with a few branches, at first erect, ending in tuft-like secund clusters. Spikelets harsh, with 3-5 flowers. Coarse meadow-grass.
Dactylis glomerata, L.
Not easily confounded with any other grass if attention is paid to the folded coarse leaves, the tufted perennial habit, and the harsh inflorescence, the glumes on the spikelets being stiffly hairy on the keel.
(ii) Panicle contracted, more or less ovoid or cylindroid, but most of the spikelets stalked, and not aggregated into dense clusters, spikelets silvery, containing 2-3 flowers.
Kœleria cristata, Pers.
Any resemblances to forms of Aira are at once nullified by the absence of true awns in Kœleria. Festuca and Poa have more numerous flowers in the spikelet. Anthoxanthum has only one flower, with two stamens, in the spikelets. Phleum and Alopecurus present resemblances, but see pp. 101 and 102.
(b) Panicle elongated and lax—i.e. all or most of the spikelets on distinct slender stalks, longer than themselves.
(i) Panicle close: the spikelets on distinct and even long stalks, but the latter erect and keeping the spikelets near the main axis.
In some cases the stalks from the main axis bear only one spikelet each and the type of inflorescence is that of a raceme; usually, however, one or more of the lowermost stalks branch and disclose the panicle.
✲ Panicle with few simple branches, racemous, each spikelet large and containing about four flowers, palea three-toothed at the apex. A small heath-grass, with hairs at the ligule.
Triodia decumbens, Beauv.
The inflorescence may have some resemblance to Melica (see p. 105), but the four-flowered spikelet, ligule, habitat and stature distinguish it at once.
✲✲ Panicle with divided branches, which are more or less erect.
† Spikelets awnless.
⊙ Spikelets small, purplish, with two or at most three florets; the upper on a stalk. Coarse moor-grass.
Molinia cærulea, Mœnch.
⊙⊙ Spikelets with six or more florets.
‡ Tall aquatic grass with long, lax, narrow panicle and spikelets, somewhat nodding, and with 8-20 flowers in each.
Glyceria fluitans, Br.
The only similar grass is Glyceria aquatica, Sm. which differs in its more reed-like habit, open panicle, and fewer flowers.
‡‡ Small land-plants not more than a foot high, with short, stiff, somewhat crowded panicles, and spikelets containing 6-8 flowers.
≡ Maritime perennial with creeping stolons, and about a foot high: panicle somewhat open below.
Poa maritima, Huds.
≡ ≡ Inland plant with secund panicle, about six inches high.
P. rigida, L.
The rare P. procumbens, Curt. and P. bulbosa, L. also come here, as well as starved forms of P. compressa, L. Some forms of Festuca elatior, var. pratensis, Huds. growing in dry situations may give trouble, and will have to be examined with the flora. See also note, p. 109.
†† Spikelets awned.
⊙ Awns bristle-like and terminal on the paleæ: flowers about six in each spikelet. Leaves setaceous. Panicle secund.
≡ Awns long and bristle-like. An annual weed.
Festuca Myurus, L.
≡ ≡ Awns very short, or merely the drawn out points of the paleæ. Perennial.
F. ovina.
The rare F. uniglumis, Soland., and some other varieties may also come here.
Some forms of F. ovina are viviparous (see p. 134), and several varieties have to be distinguished.
⊙⊙ Awns hair-like and dorsal on the paleæ. Flowers two in each spikelet. Annual, six inches or less in height.
Aira præcox, L.
The rare A. canescens, L. also comes here: its awn is jointed and with a minute tuft of hairs about the middle.
(ii) Panicle lax and open, the fine hair-like branches spreading widely during florescence, or even pendent or drooping.
✲ Spikelets awnless.
† A small aquatic grass with prostrate habit and two-flowered spikelets with broad truncate glumes and paleæ.
Catabrosa aquatica, Beauv. (Fig. 4).
The two-flowered spikelets distinguish it at once from Glyceria aquatica, to say nothing of its softer and smoother texture and small stature. Poa trivialis may have two flowers, but it is an erect meadow-grass, with keeled and pointed glumes and paleæ. Aira and Agrostis are awned, or differ entirely in habit.
†† Spikelets with at least three or four, but usually more flowers.
⊙ A perennial field-grass with few large, compressed, bluntly triangular or ovate spikelets, dangling at the end of capillary branchlets; with membranous, loosely imbricated, concave and inflated paleæ and glumes, and 6-8 flowers.
Briza media, L.
The much rarer B. minor, L. is an annual and smaller.
⊙⊙ Spikelets small and numerous, more or less elongated and pointed, not dangling: glumes and paleæ not inflated.
Poa and Festuca (see p. 116) are difficult genera for the beginner; several of the species vary considerably in detail. Generally speaking the spikelets of Festuca are hard, harsh and sharply pointed, or with short or evident awns; in Poa they are softer, and with blunter points, and never awned. The most obviously awned species of Festuca have more or less setaceous leaves and contracted inflorescences (see p. 111). Some of the mountain species of Poa are extremely variable in small characters.
≡ Reed-like aquatic, with obtuse glumes rounded dorsally; the large richly branched panicle bearing numerous 5-10 flowered spikelets. Sheath entire or nearly so. No web to the paleæ.
Glyceria aquatica, Sm.
For other reed-grasses, see note, p. 103. Glyceria fluitans has a more contracted panicle and usually more flowers in the spikelet &c. (see p. 111).
≡ ≡ Meadow and pasture grasses &c., usually small. Glumes more or less keeled and acute. Sheaths split.
Poa.
‡ Spikelets very small and containing 2-3 flowers only. Stem slender, terete and rough; ligule long and acute, palea with prominent lateral nerves.
P. trivialis, L.
The flowers are fewer than in any other Poa, and may be only two. P. nemoralis, with 3-5 flowers, may also occasionally be found with two only: it differs from others in its more acute glumes, smooth stem and short ligule.
P. pratensis differs in its creeping stolons, short ligule and more numerous 4-5 flowers, and in the indistinct nerves of the palea.
‡‡ Spikelets 4-6 flowered.
≡ Annuals, about six inches high, with relatively large and numerous spikelets for so small a panicle. No “web" at the base of the paleæ.
P. annua, L.
This is the small grass so common as a weed on roadsides and on lawns, flowering even in winter. For other annual species of Poa—P. rigida, P. loliacea and P. procumbens—see note p. 108 and p. 111.
≡ ≡ Perennials.
Δ With creeping stolons.
x Stems and leaf-sheaths flattened. Panicle somewhat close and secund, some spikelets being nearly sessile.
P. compressa, L.
xx Stems and leaf-sheaths terete. Panicle spreading widely.
P. pratensis, L.
ΔΔ Without creeping stolons. Stems and panicle slender, round. A shade-plant.
P. nemoralis, L.
The rarer species of Poa are P. bulbosa with the stems swollen below; P. alpina, also slightly bulbous and often viviparous; P. laxa an allied Highland species; and P. distans a maritime form allied to P. maritima (see p. 111).
✲✲ Spikelets with awns or with sharp terminal points (awn-points) to the glumes or paleæ.
In some species and varieties of Festuca awns are not evident (see p. 116): the hard paleæ simply end in acute or acuminate points, but these are sharp and in most cases much more so than in any Poa. When not so evident, the student will probably think the specimen is a Poa and the flora must be consulted.
The difficulty is most likely to occur with varieties of F. ovina and F. elatior: in the former the leaves are narrower, setaceous and stiffer than in any Poa. The ribbing of the leaf, the ligule, and other characters of the vegetative organs (see pp. 45 and 50) will help in these doubtful cases.
† Awns terminal, or sub-terminal; often very short or nearly obsolete and the paleæ merely drawn out at the tips to a hard sharp awn-point.
⊙ Awns long and sub-terminal, inserted between the teeth of the cleft apex of the paleæ. Sheaths entire. Panicle usually nodding.
Bromus.
With the exception of Festuca Myurus and one or two other rare Fescues with setaceous leaves, Hordeum and Brachypodium are the only other genera with awns much resembling the Bromes. The former has a totally different inflorescence, and in the latter the spikelets are practically sessile (see p. 107).
≡ Spikelets short and fat, and relatively heavy. Paleæ broad and distinctly nerved. Awns fine, about as long as the paleæ.
B. arvensis, L.
There are several varieties, of which B. secalinus with a looser panicle, and B. mollis with a more compact panicle and very downy, are the chief.
≡ ≡ Spikelets lanceolate and with conspicuous awns. Nerves on the paleæ obsolete.
x Panicle conspicuously loose and drooping and awns long, paleæ narrow and elongated.
z Sheaths with long often reflexed hairs. A shade-grass over three feet high.
B. asper, Mull.
zz Sheaths downy. A weed of open lands and hedges, two feet or less.
B. sterilis, L.
B. giganteus is rarer than B. asper and has smaller spikelets and more slender awns. B. maximus and B. madritensis are rare.
⊙⊙ Awns, if present, merely the points of the paleæ, very short or obsolete. Sheaths split: ligule short.
Festuca.
≡ Leaves flattened. Panicle somewhat close. Often tall grasses.
z Meadow-grass, with 5-6 to a dozen or more flowers in the spikelet.
F. elatior, L.
zz Shade-grass, with 4-5 flowers only in the spikelet.
F. sylvatica, Vill.
≡ ≡ Leaves subulate or setaceous, at least below. Mountain pasture grass, usually small.
F. ovina.
Concerning difficulties between Poa and the awnless forms of Festuca, see p. 114. F. elatior and F. sylvatica are practically awnless, the awn-points being merely the acuminate tips of the paleæ.
The long-awned species of Festuca have compact stiff panicles and narrow or setaceous leaves (see p. 111).
Regarding varieties of F. ovina, see p. 112.
†† Awns, twisted below and bent above, inserted into the middle of the back of the bifid palea or below it.
⊙ Awns long and conspicuously projecting from the spikelet, which is gaping and contains 3-5 flowers. Leaves flat.
Avena.
≡ Tall annual corn-weed, with large (18-20 mm.) heavy, pendent, hairy spikelets on the long slender stalks of the lax open panicle. Leaves glabrous.
A. fatua, L.
This is the so-called Wild Oat, and the type of this group.
≡ ≡ Tufted perennials with spikelets more or less erect on stiffer stalks, the panicle therefore less open.
z Panicle nearly simple; spikelets silvery or reddish, 12-15 mm. long.
A. pratensis, L.
zz Panicle branched but not very open; spikelets glistening yellow and only 5-6 mm. long.
A. flavescens, L.
There is no other genus closely resembling Avena. The superficial likeness of some Bromes disappears at once on examination. The spikelets of Aira are much smaller, and the leaves quite different (see below and p. 47).
⊙⊙ Awns fine and hair-like and not conspicuously protruding from the spikelets; the latter small, 2-5 mm. Flowers two in each spikelet.
Aira.
≡ Coarse and tall tufted grass with flattened, harsh, and conspicuously ribbed leaves: the very short awns hardly protruding.
A. cæspitosa, L.
No other grass can be confused with this if the very high ridges of the leaves are observed (see p. 47).
≡ ≡ Small grasses with setaceous or very narrow inrolled leaves. Awns slightly protruding.
x Palea distinctly bifid at the apex: awn nearly twice its length. A very small grass about six inches high: leaves subulate, fine and short.
A. caryophyllea, L.
xx Paleæ almost imperceptibly bidentate: awn not much longer than palea. About 12-18 inches high: leaves inrolled.
A. flexuosa, L.
Other species of Aira, with more condensed inflorescences, are dealt with on p. 112. Agrostis is distinguished by the leaves and one-flowered spikelets (see p. 104). Poa and Catabrosa have no awns.
The stigma of an ordinary grass consists of two divaricating plume-like structures composed of thin-walled cells. When the paleæ open these stigmatic plumes protrude, one on either side, and readily catch pollen shed from the dangling stamens and carried by the wind, and since the pollen of the same flower is usually shed at a time when the stigmas of many neighbouring plants are mature, there is every opportunity for cross-fertilisation. (Fig. 33.)
In some cases, however, e.g. Anthoxanthum, Alopecurus,
the flowers are proterogynous, the stigmatic plumes being
ready for pollination some time before the pollen is shed
from the anthers of the same flower; whereas in most of
our grasses the pollen begins to scatter before the stigmas
 Fig. 37. Pollen-grains adherent to
the papillæ of the stigma, on
which they germinate, sending
the pollen-tubes down between
the cells. Kerner. × about 100.
are ready (protandrous). Among exotic grasses, many are
diœcious or monœcious—i.e. the flowers contain stamens
only or ovary only, on each plant, or on different inflorescences
of the same plant respectively—and even in our
own Holcus and Arrhenatherum this state of affairs is
partially represented, since one flower of the spikelet is
male only.
Fig. 37. Pollen-grains adherent to
the papillæ of the stigma, on
which they germinate, sending
the pollen-tubes down between
the cells. Kerner. × about 100.
are ready (protandrous). Among exotic grasses, many are
diœcious or monœcious—i.e. the flowers contain stamens
only or ovary only, on each plant, or on different inflorescences
of the same plant respectively—and even in our
own Holcus and Arrhenatherum this state of affairs is
partially represented, since one flower of the spikelet is
male only.
In some grasses, e.g. Rye, however, it appears im120probable that cross-fertilisation ever occurs, since the paleæ do not open, and the pollen falls on to the stigma direct; and in Leersia and the foreign Amphicarpum the spikelets are completely cleistogamous, those of the latter being on stalks close to the ground which push the flowers into the soil, where pollination and fructification are accomplished.
Hybrid grasses are by
no means uncommon. To
say nothing of the numerous
cross-bred Wheats and Barleys,
artificial hybrids have
been raised between Wheat
and Rye. In the Maize
an astonishing number of
selected cross-breeds have
been obtained, and, among
 Fig. 38. Longitudinal median
section of the caryopsis of a
grass—e.g. Lolium (× about
35). p, pericarp; t, attachment
to axis; m, position of
micropyle; E, endosperm; A,
its aleurone layer; l, folded
leaves of plumule; r, radicle;
r´´, secondary roots; RC, root-cap;
s, scutellum. The dark
line e represents the surface
where the face of the scutellum
is applied to the endosperm
and where absorption
of the latter takes place.others, certain forms in
which the seeds have a
violet outer layer and a
sugary endosperm, are found
to transmit these characters to the resulting seed of a
variety which would normally have produced seed with
white outer layers and starchy endosperm, if the pollen of
the former is used on the stigma of the latter. Such
direct influences of the pollen are termed Xenia.
Fig. 38. Longitudinal median
section of the caryopsis of a
grass—e.g. Lolium (× about
35). p, pericarp; t, attachment
to axis; m, position of
micropyle; E, endosperm; A,
its aleurone layer; l, folded
leaves of plumule; r, radicle;
r´´, secondary roots; RC, root-cap;
s, scutellum. The dark
line e represents the surface
where the face of the scutellum
is applied to the endosperm
and where absorption
of the latter takes place.others, certain forms in
which the seeds have a
violet outer layer and a
sugary endosperm, are found
to transmit these characters to the resulting seed of a
variety which would normally have produced seed with
white outer layers and starchy endosperm, if the pollen of
the former is used on the stigma of the latter. Such
direct influences of the pollen are termed Xenia.
When the pollen-grain, having adhered to the hairy stigma, has begun to germinate, the resulting pollen-tube creeps down between the cells of the stigma, and hands over its enclosed nuclei to the embryo-sac, where fertilisation of the egg-cell is accomplished, by the fusion of one of the pollen nuclei with the nucleus of the egg-cell. As the resulting embryo developes, the sac becomes filled with endosperm-cells charged with starch-grains or sugar, and in the ripe seed the embryo is always found affixed laterally and below to this endosperm—a point of distinction from Sedges, where the embryo is buried in the endosperm.
The ripe seed fills the ovary, and its outer walls usually fuse with those of the carpel, forming the well-known Caryopsis or “grain."
If such a “grain" is carefully examined, three chief parts are visible in addition to the embryo. (Fig. 38.) Firstly, we find on the outside the fused seed-and fruit122coats, differing in the number of layers and in the microscopic characters of the cells, some of which characters can be employed in diagnoses. (Fig. 39.)
Secondly, the great mass of the “grain" internally is composed of delicate cells filled in most cases with starch-grains, the sizes, shapes and arrangement of which can also be employed for diagnoses—e.g. the compound grains of the Aveneæ and Festuceæ are different from the simple polyhedral or rounded grains of the Andropogoneæ and Maydeæ, and some races of Maize have sugar and soluble starch instead of grains of the latter.
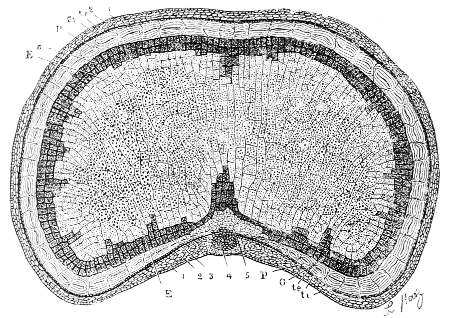
Fig. 39. Transverse section (highly magnified) through a grain of
Brachypodium pinnatum taken about half-way up. 1, epidermis;
2, pericarp; 3, remains of the true seed-coat; 4, vascular bundle;
5, remains of nucellus; P, epidermis of nucellus; G, aleurone
layer: remarkable in being several cells thick; E, endosperm.
Harz.
Thirdly, the outermost layer or layers of cells of the endosperm are filled with proteids, and are known as the Aleurone layer. (Fig. 38, A.)
The embryo consists of the folded embryonic leaves in
bud (plumule) above, which will grow up on germination
as the shoot or “spear," the short primary root (radicle)
below, with in many cases two or more secondary rootlets
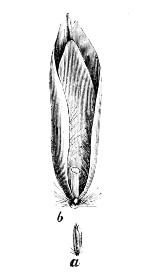 Fig. 40. Triodia decumbens.
a, “seed," nat.
size; b, ditto, × 6.
Note the basal hair-tuft
and rachilla, and
the ciliate, toothed
outer palea with a
short awn-point, enclosing
the inner more
delicate palea. Between
the two lies the
caryopsis. Nobbe.already showing in its tissues, and
from the common “collar" uniting
these a more or less prominent
shield-shaped organ (scutellum)
standing out laterally in contact
with the endosperm, the dissolved
contents of which it absorbs on
germination. (Fig. 38, s.)
Fig. 40. Triodia decumbens.
a, “seed," nat.
size; b, ditto, × 6.
Note the basal hair-tuft
and rachilla, and
the ciliate, toothed
outer palea with a
short awn-point, enclosing
the inner more
delicate palea. Between
the two lies the
caryopsis. Nobbe.already showing in its tissues, and
from the common “collar" uniting
these a more or less prominent
shield-shaped organ (scutellum)
standing out laterally in contact
with the endosperm, the dissolved
contents of which it absorbs on
germination. (Fig. 38, s.)
Although typical grasses form a caryopsis as described, exceptions occur. In the exotic Sporobolus, Eleusine, Crypsis and Heleochloa the fruit becomes truly dehiscent, the seed being loose in the fruit, and the latter opens and allows it to fall out; and in many Bamboos the seed is loose in the achene, while in a few cases—e.g. Melocanna—the fruit is fleshy and may be as big as a walnut.
Returning to the typical grasses. When the fruit ripens in the spikelet, several events may happen.
In most of our grasses the caryopsis comes away trapped between its two paleæ, and the latter bring away with them the small piece of the axis of the spikelet on which they stand: this bit of axis—the rachilla—often affords valuable characters in diagnosis. (Fig. 41.) It is the pair of paleæ enclosing their caryopsis which goes by the name of “seed" in most of our grasses. (Fig. 40.)
In a few cases, however, e.g. Panicum, the spikelet comes away as a whole, so that here the “seed" consists of the glumes, enclosing one, two or more pairs of paleæ with their contained caryopses.
 Fig. 41. Diagram of a spikelet
of a grass—e.g. Festuca—comprising
six matured
flowers and their paleæ,
embraced by the two
glumes (g1g2) at the moment
of disarticulation as
the fruits ripen. The small
piece of axis (a) left attached
to each segment is the
rachilla R. At p2 the tip of
the inner palea is visible
protruding from the outer
one p1: in the rest it is still
enclosed in the latter.Even among our native
grasses, however, cases occur
where the separation takes
place below some of the
glumes, and so the “seed," as
met with in samples, consists
of glumes as well as enclosed
paleæ and caryopsis—e.g. Anthoxanthum,
Alopecurus, &c.—and
some care is necessary in
examining grass “seed" in these circumstances (see p. 134).
Fig. 41. Diagram of a spikelet
of a grass—e.g. Festuca—comprising
six matured
flowers and their paleæ,
embraced by the two
glumes (g1g2) at the moment
of disarticulation as
the fruits ripen. The small
piece of axis (a) left attached
to each segment is the
rachilla R. At p2 the tip of
the inner palea is visible
protruding from the outer
one p1: in the rest it is still
enclosed in the latter.Even among our native
grasses, however, cases occur
where the separation takes
place below some of the
glumes, and so the “seed," as
met with in samples, consists
of glumes as well as enclosed
paleæ and caryopsis—e.g. Anthoxanthum,
Alopecurus, &c.—and
some care is necessary in
examining grass “seed" in these circumstances (see p. 134).
Interesting biological adaptations are met with in the distribution of grass “seeds." The very small and light125 fruits of Agrostis easily fall and are scattered by the wind, but in many cases the glumes (Holcus) or paleæ (Briza) are expanded and serve as “wings" offering extensive surfaces to the wind. In Arundo, Calamagrostis, Aira, &c., fine silky hairs attached to the rachilla serve a similar function, reminding us of the coma of true seeds and the pappus of Composites. In Hordeum jubatum of the prairies, the axis breaks up and the disarticulated portions with their attached tufts of fruits are blown away by the wind, and something similar occurs in our own H. murinum to a less extent. In the exotic Spinifex whole heads of fruits are thus detached and blown over the sands as “tumble weeds.”
In Stipa pennata we have an example of perhaps the most complex of all such adaptations: the exceedingly long awn terminating the palea is plumose at the upper end and twisted below, and the hard sharp rachilla at the base of the fruit is furnished with short, stiff hairs directed upwards. The plumed awn serves as a wind surface, the whole fruit flying like an arrow through the air. The stiff hairs below serve to fix the lower end between particles of soil, and by their alternate drying and wetting, the warping of these and of the twisting and untwisting awn drives the sharp base into the soil. (Fig. 42.) Similar mechanisms exist in Avena and others.
These bristles and awns also subserve dissemination in other ways, especially by clinging to the wool and fur of sheep and other animals, and cases occur where the twisting awns and reflexed hairs on the hard pointed fruit-base drive the latter into the bodies of sheep with fatal effects—e.g. Stipa capillata in Russia, S. spartea in 126 America, Aristida hygrometrica in New Zealand, Heteropogon contortus in New Caledonia.
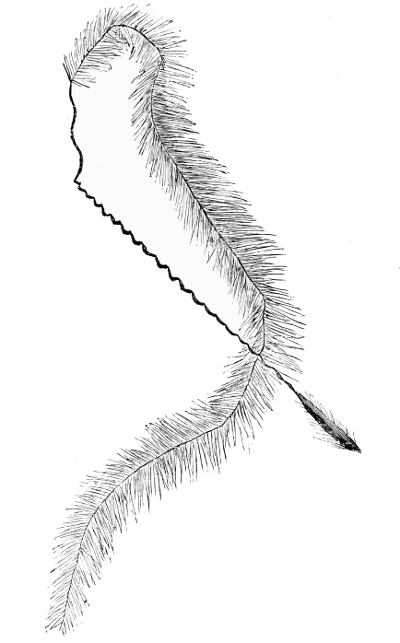
Fig. 42. Awned fruit of Stipa. The reflexed stiff hairs and hard point favour penetration into the soil. The long twisted awn performs hygroscopic movements, and its terminal plume offers surface to the wind. Lubbock.
America, Aristida hygrometrica in New Zealand, Heteropogon contortus in New Caledonia.
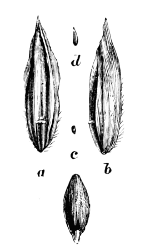 Fig. 43. Poa annua.
d, the “seed," nat.
size; a and b, ditto,
× about 7; c, caryopsis,
natural size
(above) and × about
7. Note the ribs
and silky keel. No
"web." Nobbe. Cf.
Figs. 54-56.
The driving action of even small
reflexed asperities on awns is well
illustrated by the fruits of Hordeum,
which are often made by children to
creep up the sleeve.
Fig. 43. Poa annua.
d, the “seed," nat.
size; a and b, ditto,
× about 7; c, caryopsis,
natural size
(above) and × about
7. Note the ribs
and silky keel. No
"web." Nobbe. Cf.
Figs. 54-56.
The driving action of even small
reflexed asperities on awns is well
illustrated by the fruits of Hordeum,
which are often made by children to
creep up the sleeve.
When we come to examine the external features of the “seeds" of grasses—usually the caryopsis enclosed in one or more paleæ, but sometimes in glumes as well—the following diagnostic characters are of importance.
The size varies from lengths of 2 mm. or less (Poa, Aira) (Figs. 43, 44) to 15-20 mm. or more (Arrhenatherum) (Fig. 47) and distinction must be made between the various characters of the caryopsis devoid of its coverings (paleæ, glumes) and such as really belong to the latter.
The caryopsis proper may be short and stout and devoid of a groove—"Millet-seed" type (Fig. 45)—but is oftener elongated, like a grain of Wheat, and then has the characteristic longitudinal groove on the face opposite that where the embryo is situated (Fig. 46). Details of shape—e.g. cylindroid, fusiform, flattened, &c.—are also of diagnostic value.
In many cases the “seed," consisting of the caryopsis closely compressed between the paleæ, is boat-shaped, e.g.128 Lolium perenne, Festuca elatior. If this “boat" is long and narrow it may be termed barge-shaped, e.g. Brachypodium sylvaticum, &c. (Figs. 71, 72): if short, broad, and open or shallow, the term coracle-shaped seems to apply, e.g. Bromus arvensis (Fig. 73).
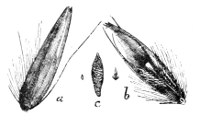 |
 |
|
Fig. 44. Aira cæspitosa. The minute “seed" (nat. size) is seen to the right, and the caryopsis (nat. size) to the left of c, the caryopsis, × 8. a and b, the "seed," × 10. The basal awn is about as long as the palea, and the rachilla is very hairy. Nobbe. |
Fig. 45. Phleum pratense. Millet-seed type. a, “seed," nat. size; b, ditto, × about 7; d, caryopsis, nat. size; c, ditto, × 7. The rounded caryopsis in b is only loosely covered by one palea. Nobbe. |
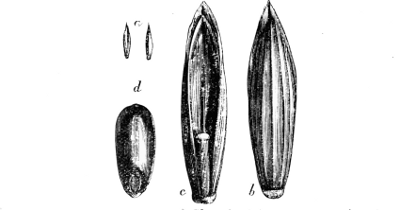
Fig. 46. Boat-shaped “seed" of Glyceria fluitans. a, nat. size; b and c, × about 6. d, caryopsis, × about 10. Note the 5-7 strong nerves, and the slender cylindrical rachilla. Cf. Figs. 57 and 58. Nobbe.
 |
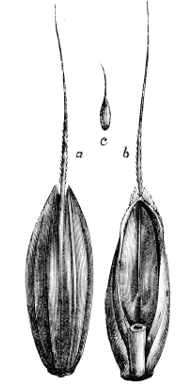 |
|
Fig. 47. Arrhenatherum avenaceum. The “double seed” (a and b) consists of two pairs of paleæ with their enclosures. The lower pair (γ) are devoid of fruit, and the outer palea has a long twisted and kneed basal awn: the upper pair (β) enclose the fruit, and the palea has a short straight awn. Note the stiff basal hairs. a, nat. size; b, × about 6; c, caryopsis, nat. size; d, ditto, × 6. Nobbe. |
Fig. 48. Lolium temulentum. Type of boat-shaped “seed" with sub-terminal awn arising from between two teeth. c, "seed," nat. size; a and b, ditto, × 6. Note the large smooth rachilla. Nobbe. Cf. Fig. 57. |
Further important distinctive characters are obtained
from the absence or presence of awns, and the peculiarities—length,
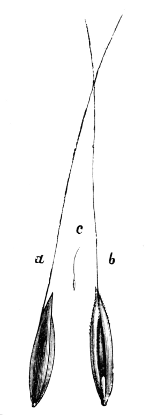 Fig. 49. Agrostis Spicaventi.
c, “seed," nat.
size; a and b, ditto,
× about 9. The long
slender awn is inserted
below the bifid tip
of the palea. Nobbe.
stiffness, &c.—of the latter. The awns may be130
straight, hooked, or kneed (i.e. sharply bent); twisted or
not; smooth or serrulate. They may be inserted at the
base or near the middle of the back of the investing
palea, or glume, or near its apex, as indicated by the
terms basal, dorsal, sub-terminal (see Figs. 47-50).
Fig. 49. Agrostis Spicaventi.
c, “seed," nat.
size; a and b, ditto,
× about 9. The long
slender awn is inserted
below the bifid tip
of the palea. Nobbe.
stiffness, &c.—of the latter. The awns may be130
straight, hooked, or kneed (i.e. sharply bent); twisted or
not; smooth or serrulate. They may be inserted at the
base or near the middle of the back of the investing
palea, or glume, or near its apex, as indicated by the
terms basal, dorsal, sub-terminal (see Figs. 47-50).
Some difficulty arises in connection with terminal awns. In some cases there is a true awn—i.e. a distinct bristle or hair—at the apex of the palea, and apparently continuing its substance without interruption; but in most instances close examination shows that this awn arises from between two minute teeth, and is really inserted at the back of the slight depression between them—e.g. Lolium temulentum (Fig. 48), Brachypodium pinnatum (Fig. 77), &c.
In another class of cases the awn appears to be really the prolongation of the palea—e.g. Nardus (Fig. 81), Festuca Myurus (Fig. 80), &c.—and when it runs out into a distinct bristle we may speak of a terminal awn without staying to discuss whether or no it is really terminal in development.
In Arundo, Cynosurus and some Fescues, where the palea tapers off into a stiff long point, I have not spoken of it as an awn, but have described the131 palea as tapering into a sharp point (awn-point). It must be admitted that the distinction is somewhat artificial, but it has its advantages in practice.
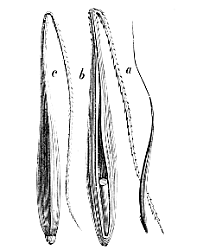 |
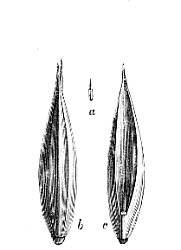 |
|
Fig. 50. Bromus sterilis. a, “seed," nat. size; b and c, ditto, × about 2. Nobbe. Cf. Figs. 80 and 81. |
Fig. 51. Festuca sylvatica. a, "seed," nat. size; b and c, ditto, × 8. Nobbe. Cf. Fig. 78. |
The rachilla (Fig. 41), when present, often affords good characters, and in a few cases is relied on for the distinction of “seeds" otherwise much alike—e.g. Lolium perenne and Festuca elatior; and similarly with the presence or absence of hairs (Digraphis, Arundo, &c.) or “web" (Poa) at the base of the “seed." (Figs. 55, 69.)
On germination the primary root of the embryo usually emerges and at once plunges into the soil, but soon ceases to grow, and the secondary roots (and subsequently adventitious rootlets from the lower internodes) soon give the peculiar fibrous character so well known in grass-roots.
The plumule either pushes out from the same end of the caryopsis as the root (e.g. Wheat, Rye) or drives its way between the coats to the opposite end (e.g. Barley) and appears as a pointed cylinder of rolled leaves (the "spear") the outermost of which is sheath only, no lamina being developed. As Darwin showed, the pointed apex of such a plumule is hardened, facilitating the piercing of the soil above, and when the blade attains the open air it performs spiral movements during growth, indicating that similar attempts to rock from side to side have aided the plumule in forcing its way through the soil. It has also been shown that the apices of roots and plumules are sensitive to differences of temperature, of light-intensity, moisture and contact, all of which aid the seedling in establishing its position in the soil and in liberating the "spear." For particulars the student should read Darwin’s Power of Movement in Plants.
With regard to the established seedlings of grasses, many interesting details of structure are to be found in them. I have not sufficient material to draw up a diagnostic arrangement of grass-seedlings, but it is evident that such could be done. It may be useful to illustrate this by the following tabular view of the characters of the larger seedlings of our common cereals, in part adapted from Vesque and Percival; though I find that some variations may occur, especially in the development of the auricles.
Seedlings.
I. First expanded aerial leaves broad, bright green, with 18-24 ribs.
Barley.
The blade tends to twist to the left: auricles, when well developed, long, pointed, and claw-like, embracing the glabrous sheath in front. Ligule long, membranous, pointed and irregularly toothed. The plumule emerges at the upper end of the grain, and the embryo has 5-6 secondary roots.
II. First aerial leaves narrower, with not more than 11-13 ribs.
A. Ligule long, rounded and toothed.
Wheat.
The blade tends to twist to the left: auricles filiform and embrace the densely pubescent sheath. Leaves green. Embryo with three roots.
B. Ligule short and toothed. Leaves with a reddish cast.
(a) Blade and sheath softly hairy, the latter with scattered long hairs, the former tending to twist to the left.
Rye.
The first leaf-sheath purple. Auricle-claws more triangular and smaller than those of wheat, and the accompanying bristles are fewer and shorter. Embryonic roots four.
(b) Blade and sheath glabrous or merely ciliate or silky, the former tending to twist to the right.
Oat.
Auricles filiform. The plumule emerges above. The embryo has three roots.
A curious phenomenon is observed in some grasses growing in high latitudes, or mountainous regions, or in134 moist situations. The flowers, or even entire spikelets, grow out into minute leafy buds, with rudimentary roots at the base, and fall off like the bulbils of other monocotyledons, taking root directly in the damp soil. The phenomenon must be looked upon as a case of apogamy, since the development of sexual organs is entirely passed over; the parts which would normally have become ovary and stamens being transformed into leaves. In some species or varieties—e.g. Poa alpina, Festuca ovina—this viviparous condition may coexist with normal flowers and spikelets; in others—e.g. Poa laxa, var. stricta—only the viviparous state occurs.
In the following arrangement the student should note that the terms “Seed" and “Fruit" are used in the ordinary sense of the farmer and seedsman: by the former is meant the “seed" as it comes in samples into the market, when the true fruit or grain (Caryopsis) is almost invariably invested by adherent “chaff"—i.e. paleæ or glumes or both. When the word Caryopsis is employed, I mean it strictly in the botanical sense explained above. In Hordeum, for example, we never see the true fruit, the grain consisting of the caryopsis with paleæ so closely adherent to it, that we are apt to take them as part of the grain itself. The true seed, in the strict botanical sense, is never seen as a naturally separate organ in our native grasses; and, as already explained, only very few exotic grasses ever shed it—e.g. Sporobolus.
I. “Seed" rounded (Millet-seed12 type). Caryopsis ovoid or sub-globose, devoid of distinct groove, and distending the awnless paleæ, or falling out free.
A. Glumes cuspidate, “fruit" yellowish.
Phleum pratense.
The student should familiarise himself with the “seed" of Phleum pratense, as a type, and an important grass easily obtained pure, but sometimes with minute round seeds of weeds intermixed.
Phleum pratense, L. (Fig. 45).
Palea 2·3 mm. long, five-ribbed, with a short point, delicate but hard, smooth, with a silvery lustre. Inner pale two-nerved. Closely investing the yellow-brown
caryopsis, which easily falls out and is ovoid-acute, about 2 mm. long by 1 broad, and finely punctate.
The much rarer P. arenarium and the very rare P. Bœhmeri and P. asperum also come here.
Phleum is not easily confounded with any other of our grasses, and Briza and Glyceria are almost the only other common grasses of about the same size of which the caryopsis often falls free from the palea ("naked fruits"). Glyceria is longer and corn-shaped, and Briza usually larger. Anthoxanthum and Phalaris are easily distinguished. Agrostis is smaller and “corn-shaped." Certain species of Panicum present resemblances, but the enveloping paleæ, &c. are very different. Melica also occurs as “naked fruits,” but is rarely seen, and its colour and brilliant lustre distinguish it.
B. Glumes not cuspidate. “Fruit" not yellow.
(a) “Fruit" white, owing to the closely investing palea.
Milium effusum.
A common grass, but not often met with in “seed" grasses. It is eagerly eaten by birds.
Milium effusum, L. (Fig. 52).
Palea about 3 mm. long, with
 Fig. 52. Milium effusum.
"Seed," a,
nat. size; b and c,
× 7. Nobbe.
few nerves, closely investing the
caryopsis: the seed is ellipsoid-acute,
slightly compressed, and 2·2-2·7
mm. long. The inner pale becomes
hard and shines like porcelain.
Fig. 52. Milium effusum.
"Seed," a,
nat. size; b and c,
× 7. Nobbe.
few nerves, closely investing the
caryopsis: the seed is ellipsoid-acute,
slightly compressed, and 2·2-2·7
mm. long. The inner pale becomes
hard and shines like porcelain.
(b) “Fruit" dark-brown shining; slightly flattened and grooved.
Melica.
Melica is rarely met with as “seed." When it is, it has to be distinguished from the other “Millet-seed" types which readily fall as naked fruits—see Phleum, Milium, &c. Agrostis, Glyceria and other corn-shaped “seeds" are easily distinguished.
Melica nutans, L. (Fig. 53).
"Seed" coracle-shaped. Palea broad, parchment-like, elliptic convex, 5-6 mm. long, 5-7 nerved and keeled, awnless, loose, purplish. Caryopsis ellipsoid-acute, 2-3 mm. long, and easily separating, wrinkled, dark shining brown as if lacquered.

Fig. 53. Coracle-shaped “seed" of Melica nutans, showing the broad, ribbed and keeled palea (c), and small rachilla (d). a, the “seed," nat. size; c and d, ditto, × 8; b, the caryopsis, nat. size; e and f, ditto, × 10. Nobbe.
M. uniflora, Retz. is commoner and very similar, but neither is often met with in “seed" grass, except as impurities among Fescues.
The rare Panicum Crus-galli and allies, and the rice-like Leersia oryzoides as well as Cynodon Dactylon, come here.
Panicums may occur in grass “seed" from America: Burchard describes them in detail.
II. “Seed" long (Corn and Barley type). Caryopsis oblong or flattened, usually trapped between the boat-shaped paleæ: if otherwise, with an awn.
A. “Seed" awnless. There may be a short sharp point to the palea (mucronate), but no prominent hair-or bristle-awn exceeding half the length of the palea.
(a) A “web" or tuft of hairs at the base or on the rachilla.
(1) Hairs of basal tuft silky and erect.
✲ Pencil of hairs as long as palea or longer.
Calamagrostis lanceolata, Roth.
Palea 3 mm. long, thin, two-toothed and with a short bristle at the apex.
Other species of Calamagrostis are awned. None occurs as an ordinary impurity in “seed."
Arundo Phragmites, L.
Palea narrow and long, 10-11 mm., delicate, entire, tapering to an acuminate point, violet, three-nerved, smooth. Caryopsis about 2 mm. A pencil of long silky hairs on the rachilla.
The long acuminate point is almost an awn.
Calamagrostis also has long basal hairs: both are useless grasses agriculturally. For Glyceria see note, p. 146. Avena, Aira and Psamma are easily distinguished.
✲✲ Pencil of hairs short.
† Palea mucronate, 11-12 mm. long: caryopsis 4·5 mm.
Psamma arenaria.
Digraphis differs in the stout caryopsis, smaller size, double hair-tuft. Arundo has a long pointed palea and long silky basal hairs and is larger.
Psamma arenaria, Beauv.
Palea 11·5 mm. ovate-lanceolate, papery, 4-5 nerved, as rolled round the fruit about 1·3 mm. diameter, yellow, and with a small tuft of stiff fine hairs at the base. Mere trace of awn, sub-terminal. Fruit 4-5 mm. long, obovate, pale-brown.
A shore-grass, not often seen as “seed": more valuable as a sand-binder than as fodder, though the young shoots are grazed.
†† Palea acuminate, and only about 4 mm. long: caryopsis 1·4 mm.
Digraphis arundinacea.
A Fen-grass, but coarse and not in use except the young growth, and for thatching.
Digraphis arundinacea, Trin.
Paleæ ovate-lanceolate, nerveless and awnless, but silky with double basal hair-tuft, and polished at the base; smooth, hard and shining, and closely investing the caryopsis which has no groove. Dark grey in colour. The glumes are without awns or wings, and are left behind.
The allied Phalaris canariensis has wing-like keels to the glumes.
Triodia also comes into this group, with short basal hairs; but its broad ciliate palea, 6 mm. long, has a short mucronate point between two teeth (Fig. 40).
(2) Hairs at the base forming a fluffy “web." Paleæ thin, 2-3 mm. long.
Poa pratensis.
P. trivialis.
P. compressa.
P. nemoralis.
The “seeds" of Poa proper are nearly all small—not more than 3-4 mm., more or less lanceolate, with tough, keeled glumes, and when “webbed" tend to adhere together as if stuck with cobweb. The keeled glumes give them an angular appearance—triangular in section—and make them tend to lie on the side. Rachilla evident. Nerves of paleæ distinct.
Poa pratensis, L. (Fig. 55).
"Seed “ 2-3 mm. long, brownish: caryopsis about 1-1·5 mm. Outer palea acute, indistinctly 3-5 nerved, edges and keel, silky; margins overlapping the hyaline inner palea.
The “web" looks like hyphæ of a mould.
Caryopsis ellipsoid-acute, with traces of the stigma. It falls easily. No distinct groove, section somewhat triangular. Rachilla truncate.
 |
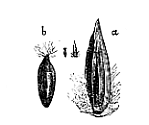 |
 |
Fig. 54. Poa nemoralis. c, the minute “seed" (to the right) and caryopsis (to the left) nat. size. a, the "seed," × 8; b, the caryopsis, × 8. Traces of a basal “web," not shown here, occasionally occur. Nobbe. |
Fig. 55. Poa pratensis. Type of a “webbed seed." a, “seed," × about 7; b, caryopsis (inverted), × 7. Between are the same, nat. size. Note the nerves on the palea and the conspicuous "web" at the base a. Nobbe. |
Fig. 56. Poa trivialis. a, “seed," nat. size and × 8; b, caryopsis, nat. size and × 7. Note the conspicuous "web." Nobbe. Cf. Fig. 43. |
Poa compressa, L.
Palea obtuse, nearly glabrous and nerveless, or faintly141 3-5 nerved and with a faint web. It is close to P. pratensis.
Poa trivialis, L. (Fig. 56).
"Seed" 2-2·5 mm., and with a bluish or bronzed cast. Caryopsis 1-1·5 mm.
Palea acute, distinctly five-ribbed, glabrous, the margin scarcely overlapping. Caryopsis grooved, blunt and more tightly held in the palea.
P. trivialis differs from P. pratensis chiefly in the distinct ribs and smoother palea, grooved caryopsis, and bluer hue.
P. nemoralis, L., hardly differs from P. trivialis, but the paleæ are sub-acute, nerves obsolete, and scarcely hyaline at the margins. The “seed" is shorter and more acute, and the colour less pronounced, and with hardly a trace of hairs (Fig. 54).
When the basal hairs are absent this should go with P. annua, &c. (see p. 146).
It is practically impossible to distinguish these species by the "seed." Care is necessary to distinguish Kœleria, which is somewhat larger in size, keeled, compressed and nerved similarly, but is more pointed and curved and paler yellowish white in colour. Hard and devoid of web or hairs. Aira is distinguished by the awn: Agrostis by the very different paleæ and caryopsis.
The other Poas are devoid of web, though they may have hairs below, and require very careful examination. The whole group is excessively difficult to deal with in “seed," and a special study of it is needed, since several species are important, and it forms a type.
(b) No web or pencil of hairs below the “seed."
(1) Apex of palea rounded, blunt, notched, or at most bluntly pointed, but with no trace of awn-or bristle-tip.
✲ Palea inflated, round-backed and somewhat winged, membranous.
Briza media.
B. minor.
Briza media, L.
Paleæ blunt, 2·5-3 mm., without lateral projections, markedly convex—almost conduplicate; nerves several. Caryopsis ovoid, 1·5-2·5 mm., dark brown.
Sometimes called a good grass, but meagre and only found in poor meadows in this country.
It should perhaps go with the “Millet-seed" type and should be compared with Phleum, Melica, Panicum, &c. “Seed" seldom met with.
B. minor is smaller.
✲✲ Paleæ not inflated or winged.
† Paleæ broad, flat and truncate, prominently three-ribbed. Caryopsis 2 mm. long.
Catabrosa aquatica, Beauv.
Cattle like it, but it only grows in ditches &c. in water-meadows: distinction from Glyceria aquatica easy if the paleæ are examined.
†† Paleæ acute or sub-acute, boat-shaped.
⊙ Paleæ and “seed" at least 6-8 mm. long.
≡ Rachilla flattened or angular, and tapering below.
Lolium perenne.
Occasionally awned, and then less easily distinguished from L. temulentum. One of our most valuable grasses. It forms a distinct type of boat-shaped “seeds."
Lolium perenne, L. (Fig. 57).
Palea distinctly five-ribbed, glabrous, oblong-lanceolate, obtuse or sub-acute, awnless, about 7 mm. long. Margins143 membranous. Inner palea nearly as long, delicate, ciliate.
Rachilla short, flattened and angular, tapering below (see Festuca elatior).
Caryopsis about 3·5 mm. long, obovate, yellowish brown.
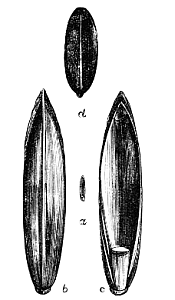 |
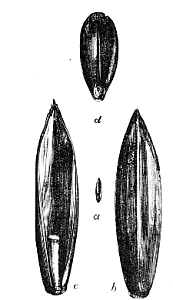 |
|
Fig. 57. Lolium perenne with typical boat-shaped “seed." a, natural size; b and c, × about 8. d, caryopsis, × 8. The rachilla, in c, is broader upwards and flatter than in Fig. 58. Nobbe. |
Fig. 58. Festuca elatior, var. pratensis. Lettering and sizes as in Fig. 57: compare the more cylindrical and slender rachilla. Nobbe. |
Although Bromus mollis, Holcus lanatus, &c. occur as impurities, they are easily distinguished: the real difficulties are with Festuca elatior and F. pratensis.
≡ ≡ Rachilla cylindrical.
Festuca elatior.
This is an important grass, and should be thoroughly mastered as a type of the boat-shaped “seed."
This, with Lolium, Agropyrum, other Fescues, and even some Bromes and Poas, will give trouble until the student thoroughly masters the importance of the minute characters of size, nervature, of the rachilla, awn, palea, &c.
Festuca elatior, L. (var. pratensis) (Fig. 58).
Palea ovate-lanceolate, five-nerved, slightly scabrid, with a slight membranous margin and cilia, rounded back, and no awn. 6-7·5 mm. long.
Caryopsis oblong-obovate, somewhat flattened, hardly grooved and adhering to the paleæ, about 3-3·5 mm. long.
Rachilla cylindrical, smooth.
The other varieties of F. elatior cannot be distinguished by the "seed" alone: in my samples F. elatior proper is longer than F. pratensis, and both have occasionally a trace of awn. Var. arundinacea has a more acuminate, stiff point. Festuca pratensis has its palea as a rule somewhat more acute than Lolium perenne, and the flatter tapering rachilla of the latter is more closely appressed to the palea. The caryopsis of Festuca also tends to adhere to the paleæ.
⊙⊙ Paleæ and “seed" at most 3-3·5 mm. long.
≡ Palea devoid of hairs or keel, notched or blunt, and with no trace of awn.
Δ Palea hyaline, nerves indistinct. Tips notched or blunt.
Agrostis alba.
There is occasionally a very short basal hair (awn).
Agrostis and Aira will present difficulties to the beginner, not only on account of their small sizes, but also from the variability as regards awns, basal hair-tuft, &c. None are of value, and most of them are weeds.
Agrostis alba, L. (var. stolonifera).
Palea enclosing the fruit 1·8-2 mm. long, white, delicate, membranous, with a blunt notched apex and three (or five) nerves: occasionally there is a fine kneed basal awn, not projecting.
The glumes may remain attached: the outer has a serrulate keel, and often a violet hue. No awn. About 2-3 mm.
Caryopsis about 1·1 mm. long, oblong, yellow, corn-shaped, furrowed, shining through.
A. vulgaris, L., is a variety with slightly smaller fruits, no awn, and usually three nerves to the palea, but they cannot be distinguished with certainty by the "seed." Rachilla obsolete.
The colour of the glumes may vary considerably and is sometimes yellow.
The absence of awn, rachilla, and basal hairs distinguishes Agrostis from Aira.
ΔΔ Palea membranous, ribbed, tips scarious.
Glyceria aquatica.
G. fluitans.
Glyceria aquatica, Sm.
Palea strongly 5-7 ribbed, obtuse and scarious, about 4 mm. long. Green with a purple cast. Caryopsis dark-brown, about 2 mm. long.
Inner palea punctate. Rachilla slender and cylindrical.
Glyceria fluitans, Br., is very similar, but longer (6 mm.) and more slender; the palea scabrid with truncate or ragged tips, and the inner palea not punctate. Yellow. (Fig. 46.)
Glyceria aquatica and G. fluitans are sweet grasses of value in water-meadows only. The “seed" is not often met with.
Not only with the Poas, but also with Catabrosa are there difficulties in determination. Molinia and Kœleria also present difficulties with Glyceria: the former has fewer and feebler nerves. Digraphis and Arundo are easily distinguished by the basal hairs.
≡ ≡ Palea acute and somewhat compressed and keeled, and with hairs on the ribs. Tips nerved.
Poa annua.
P. maritima.
P. distans.
P. rigida.
P. loliacea.
P. bulbosa.
P. alpina.Their “seeds" are all small, about 2-3·5 mm. only, and angular, brown, and present many difficulties (see note, p. 141).
Poa annua, L. (Fig. 43).
Palea 3·5 and caryopsis 2 mm. long, so that the “seed" is much larger than P. trivialis, and there is no web. Minutely silky-hairy on the keel, and the ribs are strong for a Poa.
Poa alpina, L., has the palea five-ribbed with stiffish hairs below, but no web. Bronzy green-violet. “Seed" 3-3·5 mm. long. Caryopsis 1·5-2 mm., and slightly grooved. It is rare in England, and is interesting as it becomes viviparous in Alpine situations.
(2) Apex of palea distinctly pointed, acuminate or mucronate, but not giving rise to a true, long, bristle-like awn.
✲ Palea acuminate—i.e. taper pointed.
† Not compressed or obviously keeled.
Festuca ovina.
F. sylvatica.
Cynosurus cristatus.It is doubtful whether these should not be regarded as awned: if so they come near Nardus—see p. 130.
Festuca ovina, L. (Fig. 59).
Palea rounded on the back, narrow, terete-lanceolate and five-nerved, tip scaberulous and drawn out to a stiff scabrid point half as long as the palea, or less. About 3-4 mm. long without the tip-point, 4-5·5 mm. with it. Rachilla obliquely truncate and concave at its apex.

Fig. 59. Festuca ovina. a, spikelet, × about 3-1/2. b, “seed," nat. size and × about 7. c, caryopsis, nat. size and × 7.
Caryopsis oblong, somewhat flattened, dark-brown.
The numerous varieties of F. ovina cannot be distinguished by the “seed": the allied varieties F. rubra, F. duriuscula, and F. heterophylla, &c. are sometimes more distinctly awned (see p. 172). But difficulties will be found with the whole group, which needs revision. This group comprises the grasses so valued on downs and dry hill-pastures for sheep-feeding.
Festuca sylvatica, Vill., not a common plant, is also with difficulty separable (Fig. 51).
Cynosurus cristatus, L. (Fig. 60).
Palea canary yellow to light-brown, lanceolate with
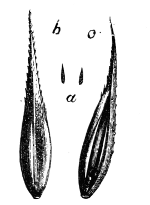 Fig. 60. Cynosurus
cristatus. a, “seed"
and caryopsis, nat.
size; b and c, “seed"
× about 8. Note the
scabrid and slightly
curved awn-point.
Nobbe. Cf. Fig. 64.
rounded back and the mid-rib prominent
as a slight keel and margins infolded,
about 4 mm. long (varies from
3-4·5 × 0·5 to 1 mm. broad), scabrid
above, dotted below and acute to
acuminate with a scabrid awn-point,
often curved. Caryopsis 2-2·3 mm.,
somewhat flattened. Rachilla short
and smooth, dilated above.
Fig. 60. Cynosurus
cristatus. a, “seed"
and caryopsis, nat.
size; b and c, “seed"
× about 8. Note the
scabrid and slightly
curved awn-point.
Nobbe. Cf. Fig. 64.
rounded back and the mid-rib prominent
as a slight keel and margins infolded,
about 4 mm. long (varies from
3-4·5 × 0·5 to 1 mm. broad), scabrid
above, dotted below and acute to
acuminate with a scabrid awn-point,
often curved. Caryopsis 2-2·3 mm.,
somewhat flattened. Rachilla short
and smooth, dilated above.
A valuable and easily recognised grass. Molinia, Holcus and similar impurities are easily detected: some Fescues are much more like it; e.g. F. tenuifolia (see p. 46) in which the apex tapers suddenly to a point. Dactylis should also be compared.
†† Paleæ compressed or distinctly keeled.
Molinia cærulea.
Kœleria cristata.Unimportant, except as impurities among other grass “seeds." Molinia is said to occur as such in samples of Cynosurus, Dactylis, Poa trivialis, Fescues, &c., but it should give little difficulty except in comparison with Glyceria (see p. 146).
Molinia cærulea (Mœnch.) (Fig. 61).
"Seed" 3-4 mm. or more, leather coloured or with a bluish tinge. Paleæ divaricating, the lower keeled and compressed, and tapering to a sharp point. Rachilla long, oblique and prominent. Smooth tapering, palea 5·5, fruit 2 mm. long.
Kœleria cristata, Pers. (Fig. 62).
Palea 6 mm., keeled, entire, no awn but stiffly tapering, ribbed, no hairs. Rachilla large. Fruit 4·5 mm.
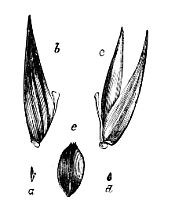 |
 |
|
Fig. 61. Molinia cærulea. a, "seed," nat. size; b and c, ditto, × about 8; d, the caryopsis, nat. size; e, ditto, × about 7. Note the compressed, keeled and glabrous palea, and the long slender bone-shaped rachilla. Nobbe. |
Fig. 62. Kœleria cristata. a, the “seed" (to the left) and caryopsis (to the right), nat. size; b, “seed," × about 7. The palea is compressed and keeled, and, like the large rachilla, glabrous. Nobbe. |
✲✲ Palea not taper pointed, but mucronate, or two-toothed, with a sub-terminal point or spine.
Elymus arenarius.
Agropyrum repens.
Dactylis glomerata.
Sesleria cærulea.
Elymus arenarius, L. (Fig. 63).
Fruiting palea lanceolate, 13-15 × 3 mm., tough and stiffly hairy or velvety: 5-7 ribs. Rachilla stout, hairy. Fruit hairy above, 5-11 mm. × 2·5 × 1·2, somewhat grooved.150 No keel. Apex of inner palea bifid, outer mucronate. Fruit shelled.
Easily distinguished from Digraphis arundinacea, which has a basal hair-tuft and very different caryopsis; less easily from Agropyrum and Lolium, except in the velvety surface.
 |
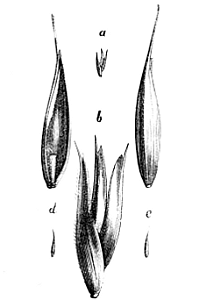 |
|
Fig. 63. Elymus arenarius. a, “seed," nat. size; b and c, the same, × 3. Note the stiff, velvety, bifid and mucronate outer palea, and the large hairy rachilla. Nobbe. |
Fig. 64. Dactylis glomerata. a, spikelet with glumes removed, nat. size; b, ditto, × about 6; c and d, “seed," nat. size (below) and × about 6. The stiff and slightly curved awn-point is sub-terminal and arises from between two teeth. Rachilla dilated above. Nobbe. Cf. Fig. 60. |
Agropyrum repens may have a mucronate awn-point or not. The small palea 10-12, the caryopsis 5-6 mm. (see p. 169 for description).
This troublesome weed (Couch-grass) presents similar difficulties with Lolium perenne that Agropyrum caninum does with Lolium temulentum—see p. 168. It is moreover extremely variable.
Dactylis glomerata, L. (Fig. 64).
Stiff awn-point just below the two teeth of apex of pale. Rachilla dilated above. Paleæ 5-6 mm. long exclusive of point, compressed and inflated, ribbed and ciliate-bristly on keel, the tip incurved. Caryopsis yellow-brown, ellipsoid, 2 mm.
An important grass, said to be adulterated with Lolium, Fescues, Holcus, Molinia, Bromus, &c., most of which should be readily detected. The curved tip is an important distinctive character. Cynosurus and Festuca arundinacea, F. ovina and F. rubra present resemblances. The seed is larger than that of Poa.
F. ovina and F. rubra are smaller, rounded dorsally, and have no keel or cilia: the nerves are less prominent and the rachilla smaller.
Sesleria cærulea, Ard.
Palea distinctly toothed at the apex, making it almost three-lobed and very characteristic; with awn-point; five ribs, serrulate. Fruit grey-yellow, obovate.
It is a mountain-and moor-grass of little value.
B. “Seed" awned—i.e. the investing palea bears, or tapers into, a distinct bristle or hair at least as long as itself or nearly so.
(a) Awn not terminal.
(1) Awn stout, either obviously twisted and bent or “kneed."
(i) “Seed" consisting of the glumes as well as paleæ investing the caryopsis13.
⊙ Awn bent but not twisted; glumes free below, ribbed, and with stiff short hairs on the keel.
≡ Awn sub-terminal, shorter than the palea, hooked.
Holcus lanatus.
"Yorkshire Fog," of little use or importance, except that it is frequently found as an impurity of other hairy grasses—e.g. Alopecurus.
It cannot easily be confounded with any other grass: Anthoxanthum and Alopecurus, Arrhenatherum, &c. present superficial resemblances only.
These glumed hairy “seeds" are uncommon and form an easily recognised type.
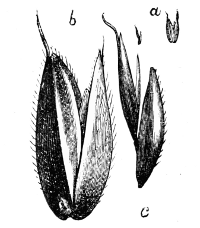
Fig. 65. Holcus lanatus. a, “seed"—i.e. complete spikelet—and ditto devoid of glumes, nat. size; b, spikelet, and c the same devoid of glumes, × 7. The “seed" is here composed of the keeled glumes enclosing two pairs of paleæ and their flowers (c): the upper of these is barren and has a hooked sub-terminal awn to its outer palea. The lower awnless one is fertile. Nobbe.
Holcus lanatus, L. (Fig. 65).
The “seed" consists of the complete spikelet, separated below the compressed and acute, keeled glumes; these have hairs on the keel, and completely enclose the two flowers and their paleæ. Palea ribless, white to grey, shining, obtuse, that of the upper (male) flower with a sub-terminal short hook-like awn. Total length about 4-5 mm. Caryopsis oblong-ovate, grooved. In its palea about 2-3 mm. long and with a few hairs at the base.
≡ ≡ Awn dorsal, kneed, longer than the palea.
Holcus mollis, L. (Fig. 66).
Palea 2·5 mm., fruit 3·1 mm. long; glumes with stiff hairs or serrulæ on keel.
The seeds of Holcus often fall from the glumes, but may bring them away.
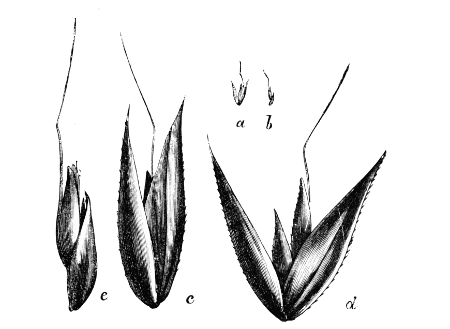
Fig. 66. Holcus mollis. Type of “double seed,” which really consists of the entire spikelet (a, nat. size; c and d, × 8) comprised of the two glumes enclosing two pairs of paleæ and their enclosures (b, nat. size; e, × about 8). The lower pair of paleæ are devoid of awns and enclose the caryopsis: the upper pair have stamens only, and the outer palea has a dorsal kneed awn, not twisted or hooked. Nobbe.
Holcus is of little use. The two-flowered spikelets and peculiar awns sufficiently distinguish it among the smaller forms. It is said to occur as an adulterant in “seed" of Alopecurus.
⊙⊙ Awn bent and twisted, basal or nearly so; glumes very hairy.
Δ Glumes golden brown: one awn straight, one “kneed."
Anthoxanthum odoratum.
The grass which gives the scent to new-mown hay. The “seed" is often impure, containing a continental species A. Puelii and other hairy forms.
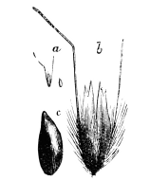
Fig. 67. Anthoxanthum odoratum. a, “seed" and caryopsis, nat. size; b, the “seed," and c, caryopsis, × about 7. The “seed" consists of the inner hairy glumes, each with a dorsal awn—one kneed—enclosing the paleæ and caryopsis. The outer pair of unawned glumes has been removed. Nobbe.
Anthoxanthum odoratum, L. (Fig. 67).
The one-flowered spikelet has four hairy glumes, the outer pair of which are unequal and awnless: the “seed" consists of the inner pair of golden-brown hairy and dorsally awned glumes, covering the thin, membranous, shining, smooth, blunt paleæ and the caryopsis, to which the inner palea adheres. Total length, without awns, about 3-4 mm. Palea 1·6 mm. Caryopsis 1·5 mm., brown, shiny, and easily separating. One awn is short, stiff and straight and inserted above the middle of its glume;155 the other longer and sharply bent, inserted about the middle.
Hierochloe, a rare northern grass, also comes here.
ΔΔ Glumes connate below, hairy on the keels.
Alopecurus pratensis.
A. geniculatus.
A. agrestis.Alopecurus is very characteristic, and should be carefully examined as a type. A. pratensis is a valuable grass and said to be often adulterated with the undesirable A. agrestis, Holcus, &c. Arrhenatherum and Avena are easily distinguished by the basal hairs, rachilla, &c.
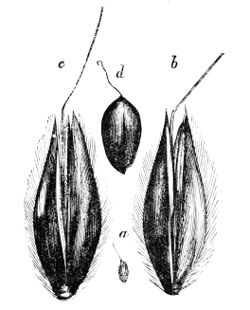
Fig. 68. Alopecurus pratensis. a, “seed," nat. size, and b, c, ditto, × 8; d, caryopsis, × 7. The “seed" here consists of the two glumes, connate below and hairy on the keel and ribs, including the caryopsis enveloped in a single palea with a dorsal awn. Note remains of stigma in d. Nobbe.
Alopecurus pratensis, L. (Fig. 68).
Inner palea absent. Outer palea 5 mm., caryopsis 3 mm. long, often with remains of the stigma.
The greyish-brown spikelet of Alopecurus easily falls as a whole, included in the glumes; the palea is loose and delicate.
Glumes connate below, silky-hairy on keel and ribs all the way up. The palea has a bent dorsal awn, about 5 mm. long.
Alopecurus geniculatus, L.
Palea 2 mm., fruit 1·3 mm. long.
The glumes hairy on ribs and keel as in A. pratensis: glumes more obtuse.
Alopecurus agrestis, L.
Palea 6 mm., fruit 3 mm. long.
Glumes only ciliate, on ribs and keel, except at the base where the hairs are longer.
Keel slightly winged.
Best distinguished from A. pratensis by the glumes, which are smoother, broader above the middle, harder and with a few basal hairs.
The species A. fulvus, A. bulbosus, and A. alpinus are rare.
(ii) “Seed" consisting of the paleæ investing the caryopsis, freed from the glumes.
✲ Palea at least 14 or 15 mm. long: caryopsis 5-7 mm. [Oat type.]
Avena fatua.
A. pratensis.
Avena fatua, L.
Palea parchment-like, lanceolate-terete, acuminate and bifid at the apex, rounded on the back and with yellow 157 hairs on the lower part. 15 × 3 mm. Awn brown, twisted and bent, arising from the middle, about 30 mm. long. Caryopsis 7 mm. long, hairy at the top and distinctly grooved. Rachilla with fine bristles.
Avena pratensis, L.
Palea very similar to A. fatua, apex 2-4-toothed or bifid, rachilla with stiffer hairs. Caryopsis about 5 mm. long.
Avena pubescens, Huds. is a variety of A. pratensis found on dry calcareous soils. The caryopsis is very narrow, and the rachilla long and feathered with hairs. "Seed" shorter than A. pratensis, 10-13 mm., and darker in colour.
A. fatua is an excellent type of the grasses with dorsal twisted and bent awns, and easily examined on account of its size. The student will find difficulties with other species of Avena and Arrhenatherum.
Harz says the stiff hairs of Avena bring about the formation of intestinal concretions (phytobezoars) and are therefore dangerous to domestic animals.
✲✲ Palea not more than 7-10 mm. long.
† Caryopsis 4-5 mm. long.
Arrhenatherum avenaceum, Beauv. (Fig. 47).
Paleæ papery, ribbed. Awn 13-15 mm., twisted below; paleæ, with a tuft of hairs below and a ciliate keel, investing the fruit: the whole being cylindrical, 8-10 mm. × 1·5.
Caryopsis 4-5 mm. × 1·2, fusiform, pubescent. Rachilla hairy.
The distinctive difference between Avena and Arrhenatherum is in the spikelets as a whole. The latter has the lower flower of each spikelet male only. Samples contain the “double seeds,” and the awn of the upper fertile portion is short (see Fig. 69).
†† Caryopsis not more than 3 mm. long. A tuft of hairs on rachilla and base.
Avena flavescens.
Aira flexuosa, distinguished by its nearly basal awn, harder texture and darker colour of paleæ, and Dactylis—awnless and with curved long drawn-out apex—are found as impurities in foreign "seed."
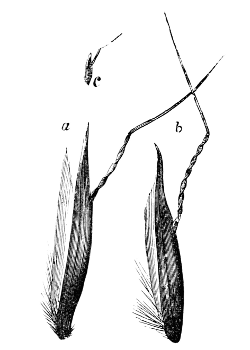
Fig. 69. Avena flavescens. c, “seed," nat. size; a and b, ditto, × about 7. The dorsal twisted and kneed awn is very characteristic. Note also the hairy rachilla. The palea is bifid above—not visible in the lateral view. Nobbe.
Avena flavescens, L. (Fig. 69).
Yellow. Palea about 5 mm. long, five-ribbed, bifid at the apex into two long slender teeth, closely investing the brownish caryopsis, and with a sub-dorsal awn 10 mm. long with little or no twist, and hairy at the base. Rachilla flattened and with white hairs. Caryopsis not 159 much grooved, fusiform, 2-3 mm. long, glumes unequal, somewhat keeled and rough.
(2) Awn dorsal or basal, fine and hair-like, and little or not at all twisted or kneed. “Seed" small.
✲ A pencil of silky hairs on base or rachilla. Palea bifid at the tip.
† Basal hairs longer than palea, and obscuring the awn.
Calamagrostis Epigeios.
Of little importance. Digraphis has no awn.
Calamagrostis Epigeios, Roth.
Palea about 3 mm. long with toothed apex and very thin caryopsis 1 mm. A conspicuous tuft of fine silky hairs, longer than the palea at the base and on the rachilla.
Awn slender, dorsal, about as long as the palea.
C. stricta, Nutt. and C. strigosa, Hartm. are rare.
†† Basal hairs shorter than palea.
⊙ Awn brown, bearded in the middle, thickened and white above.
Aira canescens.
The student will find considerable difficulties in the various species of Aira and Agrostis, owing to their small size and variability as regards awn and basal hair-tufts. All these are weeds, but some occur as adulterants.
Aira canescens, L.
"Seed" (apart from the awn, it is very like Agrostis) 1·5-2 mm., with a very thin, smooth, keeled, two-nerved160 palea, bifid at its apex, through which the yellow caryopsis shines.
Awn slender, basal, dark-brown, with oblique colourless hairs above the middle and with a slight swelling above.
The palea has a few hairs on its mid-rib below.
Caryopsis about 1 mm. long.
This grass is somewhat rare in England. The jointed basal awn, with a minute hair-tuft above the middle, is unique, and readily distinguishes it when mixed with Agrostis.
⊙⊙ Awn not bearded or thickened.
≡ Palea bifid at apex.
Aira caryophyllea.
A. præcox.
Aira caryophyllea, L.
"Seed" 1·4-1·6 mm., comma-shaped and dark-brown.
Palea acute, nerveless, inrolled at edges and with a dorsal, kneed, very fine, dark twisted awn twice as long as the palea: the latter with two-toothed apex and a basal hair-tuft.
A common impurity in commercial seed.
Aira præcox, L. is very similar, and cannot be distinguished with certainty by the “seed," but has a shorter awn and no basal hairs. Apex of palea two-toothed.
≡ ≡ Palea jagged or toothed, but not bifid.
Aira flexuosa.
A. cæspitosa.
Aira flexuosa, L. (Fig. 70).
Palea 5 mm. long, four-toothed at the apex, and caryopsis 2·6 mm. The fine basal awn waved or kneed, about twice as long as the palea. Fruit with a groove.
Rachilla hairy. Whole “seed" browner than Avena flavescens (see p. 158).
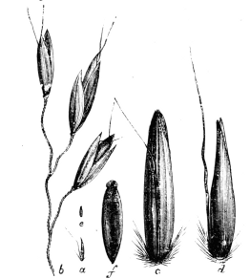
Fig. 70. Aira flexuosa. a, “seed," nat. size; c and d, the same, × about 7; b, portion of fruiting panicle, showing the flexuous rachis; e, the caryopsis, nat. size; f, ditto, × about 7. Note the basal hairs and twisted awn. Nobbe. Cf. Fig. 44.
An almost worthless grass, on sandy soils, but important as an impurity in Avena flavescens (see note, p. 158).
Aira cæspitosa, L. (Fig. 44).
The palea, about 2·5 mm. long, toothed above, has a fine straight hair-like basal awn not longer than itself. Caryopsis 1·3 mm., grooved. Rachilla distinctly hairy.
Easily distinguished when mixed with Poa nemoralis, &c.—e.g. by the awn, hairs on rachilla, &c. It requires scrutiny in regard to Agrostis, which also has no awn or basal hairs.
✲✲ No basal tuft, or merely a few short microscopic hairs below. Paleæ not bifid above.
Agrostis canina.
A. Spica-venti.
A. setacea.
Agrostis canina, L., is somewhat larger than A. alba. Palea (only one present, sometimes minute) punctate, 2 mm., with a fine dorsal awn a little longer than the palea. Caryopsis 1·2 mm. Rachilla obsolete.
The awn is usually absent from A. alba and its varieties; and when present is so small as to be almost negligible.
A. canina is easily distinguished from Poa by the rachilla in the latter, and absence of awn. Aira has basal hairs, &c.
Agrostis setacea, Curt., has the awn basal and just projecting beyond the pointed glumes, and the palea very minute, with a few silky hairs below.
Agrostis Spica-venti, L. (Fig. 49).
Palea 2·4 mm. long, with infolded edges and punctate; a minute rachilla at its base, and a slender awn, 8-9 mm. or more long, inserted below the bifid tip. Caryopsis 1·5 mm. A few silky hairs at the base of the palea.
(b) Awn terminal or slightly sub-terminal, and straight, or at least not twisted or “kneed."
(1) Paleæ so closely adherent to the fruit that the terminal awn appears to come from the latter [Barley type]. The three florets generally coherent.
✲ Awn at least twice the length of the grain.
Hordeum sylvaticum.
H. murinum.
Hordeum sylvaticum, Huds.
Palea scabrid, ribbed above, awn at least twice as long as the very narrow grain.
Hordeum murinum, L.
Lower palea 9-10 mm. long, five-ribbed, lanceolate and wrapped round and adhering to the fruit (grain), upper palea also ribbed. The grain about 7-8 mm. × 2 × 1·2, hardly grooved. Awn about 30 mm. long, serrulate. Rachilla slightly serrulate.
✲✲ Awn not twice the length of the grain.
Hordeum pratense.
H. maritimum.
Hordeum pratense, Huds.
Spikelet with a reddish tinge; awn almost smooth, less than 20 mm. long: not twice the length of the smooth and obscurely nerved grain.
Hordeum maritimum, With., similar to H. pratense, but softer and the awn somewhat longer.
The principal features of the Hordeum-grain are the closely adherent paleæ and angular fruit, the stiff awn appearing to come off from it as in Barley. The rachilla remains.
They are all weeds of no use in agriculture, though H. pratense is not uncommon in good pastures.
(2) Palea investing the caryopsis, often closely, but not fused with it, and its tip and awn quite free.
✲ Awn not strictly terminal but sub-terminal, or arising from between two teeth or in a sinus at the apex of the palea.
† Caryopsis thin, flattened and usually 6-10 mm. long, and the paleæ hairy.
≡ Palea inrolled, terete-lanceolate-acuminate, or linear-lanceolate (barge-shaped). Apical teeth minute and pointed.
Δ Palea scaberulous or hairy, awn from half to about its own length.
Bromus erectus.
B. asper.
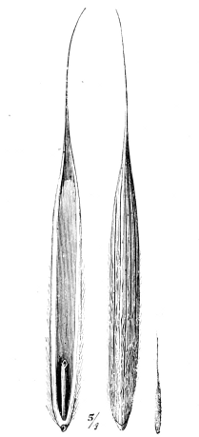 |
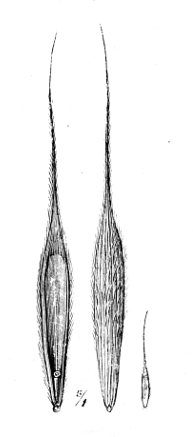 |
|
Fig. 71. Bromus asper, a barge-shaped "seed" with terminal awn, nat. size and × about 5. Cf. Fig. 50. Stebler. |
Fig. 72. Brachypodium sylvaticum, a barge-shaped “seed" with terminal awn, natural size and × about 5. Cf. Fig. 77. Stebler. |
Bromus erectus, Huds.
Palea with 5-7 scaberulous ribs, but not hairy, flat above, incurved below, so that the inner palea is clearly165 visible. About 10-14 mm. long and narrowing somewhat suddenly into the smooth stiff awn, shorter than itself. Indications of a tooth at each side of the base of the awn. Caryopsis somewhat flattened, thin and long, barge-shaped, 8-10 × 1·5-2 mm., pointed at both ends, brown, hairy at the tip with the remains of the stigma. “Seed" often purplish. Rachilla long and slender.
Must be carefully distinguished from Brachypodium, which has the palea less inrolled and a stouter rachilla.
Bromus giganteus has a shorter and flatter “seed" and longer awn.
Bromus asper, Murr. (Fig. 71).
Palea often reddish, 5-7 ribbed and usually roughly hairy, mid-rib scaberulous, about 15 mm. long, linear-lanceolate (barge-shaped), the tip more distinctly two-toothed and tapering more gradually into the smooth awn, about its own length. Rachilla distinct. Otherwise resembling B. erectus.
B. giganteus has a shorter “seed" and longer awn.
ΔΔ Palea scarcely hairy: awn up to twice its length.
Bromus sterilis.
B. giganteus.
Bromus sterilis, L. (Fig. 50).
Palea nearly smooth, somewhat inrolled, terete-lanceolate-acuminate, tough, red-brown, 12-15 mm. long, seven-ribbed, with a long serrulate awn (20-30 mm. or more) practically terminal. Ribs serrate. Caryopsis 9-10 mm. long, flattened, thin. Rachilla distinct, smooth, much wrapped in, widens upwards and is somewhat flattened.
The very slender and long “seed" and caryopsis distinguish this from all but B. erectus, which has a shorter awn. Perhaps the awn is truly terminal: if so it should go with Festuca Myurus, &c. (see p. 171). The length varies greatly.
Bromus giganteus, L.
Palea 7 mm. long and inrolled, the base and rachilla bristly, indistinctly 5-7 nerved, the tip bifid: sub-terminal awn serrulate, 12-15 mm. long.
Caryopsis 4·5 mm. long, flattened, thin, glabrous.
The caryopsis is shorter and more ovoid than in other Bromes.
≡ ≡ Palea expanded above (coracle-shaped), awn from sinus between two blunt or triangular teeth.
Bromus arvensis.
Bromus arvensis, var. mollis, L. (Fig. 73).
Palea oblong or obovate, somewhat flattened above (coracle-shaped), distinctly 5-7 ribbed, 8-9 mm. long, not hairy, though the ribs may be ciliate. Awn smooth, arising from the depression between two teeth or rounded projections. Caryopsis flattened and thin, 6 × 1·5 mm.; apt to fall loose from the paleæ.
In the variety B. secalinus the awn is usually shorter than the elliptical palea, and originates in the sinus of a notch (Fig. 74): in B. mollis from between two teeth, and is as long as the obovate palea.
Bromus maximus and B. madritensis are both rare.
The broader nerved paleæ of B. mollis and shorter, fatter caryopsis, as well as the longer awn distinguish it from B. secalinus and B. arvensis. In the other Bromes the palea is more inrolled and the “seed" more slender as a rule. The rachilla is usually bent above into a slight kink.
The Bromes are an extremely difficult group. The awn is inserted between the two teeth of the palea or just below. The palea smooth or ciliate on the nerves or roughly hairy. The fruit is 167 flattened in the antero-posterior plane, and usually shines through the closely investing palea. A rachilla is visible. In Brachypodium the caryopsis is less flattened, and the apex tapers into the awn without teeth. In Lolium and Agropyrum the caryopsis is still more wheat-like and grooved.
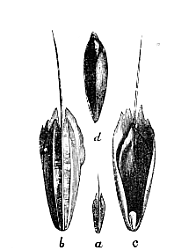 |
 |
|
Fig. 73. Bromus arvensis, var. mollis. a, “seed," nat. size; b and c, the same, × 3. d, caryopsis, × about 3. The awn originates between two teeth of the strongly ribbed palea. Coracle-shaped type. Nobbe. |
Fig. 74. Bromus arvensis, var. secalinus. a, spikelet, b and c, “seed," nat. size; d and e, the same, × 6. Nobbe. Cf. Fig. 73. |
All our Bromes are weeds, but some of them are important adulterants, requiring careful examination.
B. inermis is cultivated, and is a valuable grass. Awn short or wanting. Paleæ strongly 5-7 ribbed, inner with a marked central rib, closely adhering to caryopsis.
B. Schraderi is also said to be valuable.
†† Caryopsis not thin and flattened, corn-like, 3·5-5 mm. Palea smooth, inflated, boat-like.
Lolium temulentum.
A weed of cultivation, said to be poisonous to cattle, a possible explanation of which may be found in a fungus recently discovered as a very constant inhabitant of the tissues under the seed coats.
Lolium temulentum, L. (Fig. 48).
Palea ovate, 6-7·5 × 2·5 mm., papyraceous, inflated, smooth, the tip dry and emarginate with a slender sub-terminal awn its own length or longer. It tends to adhere to the fruit. Fruit 3·5-5 mm. × 2·5 × 1·5, somewhat depressed. Rachilla large, cylindrical and smooth.
L. perenne has no awn, or the merest trace of one. “Seed" 10-12 mm. long. Boat-shaped. Palea yellow, three-nerved, margins papery. Rachilla flat, see p. 143.
✲✲ Awn terminal, and with no evident teeth at its base.
≡ Palea not much inrolled, ovate-lanceolate to linear-lanceolate, and therefore boat-or barge-shaped.
Δ Very hairy.
Brachypodium sylvaticum, Beauv. (Fig. 72).
Palea ribbed and hirsute much like that of Bromus asper, but straw-coloured, shorter (10-12 mm.), linear-lanceolate, more boat-shaped, and tapering without teeth into the longer (10-13 mm.) hairy awn. Caryopsis 7-8 mm., less flattened than in Bromus, with a shallow groove. Rachilla smooth.
B. pinnatum (p. 171) has a much shorter awn, and is nearly glabrous.
These grasses are weeds, but are said to occur frequently as adulterants.
ΔΔ Glabrous or nearly so.
++ Palea five-nerved, pubescent.
Agropyrum caninum.
Agropyrum repens is also often awned and may be described here (see also p. 150).
 |
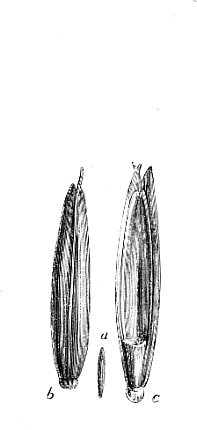 |
|
Fig. 75. Agropyrum caninum. Type of boat-shaped “seed" with sub-terminal awn. a, nat. size; b, × about 9, but with awn cut short. Rachilla thicker above and hairy; palea not toothed. Nobbe. Cf. Fig. 48. |
Fig. 76. Agropyrum repens. Awned variety. The awn is very short and sub-terminal: the rachilla smooth. a, the "seed," nat. size; b and c, ditto, × about 4-1/2. Boat-shaped "seed" with rounded back. Nobbe. Cf. Fig. 57. |
Agropyrum repens, Beauv. (Fig. 76).
Palea tough, 10-12 mm. long, ovate-lanceolate, 3-5 ribbed and smooth; with a short (2-3 mm.) or long170 (8 mm.) terminal awn or a mere tooth. Rounded back. Fruit grey-brown, ciliate above, 5-6 × 1·4 × 0·7-0·9 mm., depressed, hardly grooved, adhering to the palea. Rachilla smooth or not, and narrowed below.
A noxious weed (see p. 150) and hence should be carefully excluded from “seeds."
A. caninum, Beauv., has a longer (10 mm. or more) and rougher awn, and is smaller (about 8 mm.), but otherwise very similar. Palea smooth except at the five-nerved apex. Rachilla thickened above, and hairy (Fig. 75).
Elymus differs from Agropyrum in its harder velvety palea, no awn, and its club-like velvety rachilla. It is also larger altogether, as a rule.
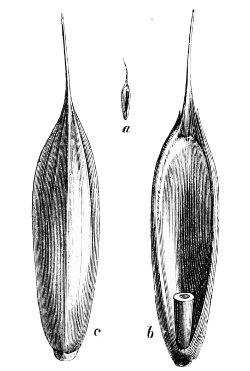
Fig. 77. Brachypodium pinnatum. Type of boat-shaped “seed" with awn. a, nat. size; b and c, × 7. Nobbe. Cf. Fig. 72.
++++ Palea seven-ribbed, smooth.
Brachypodium pinnatum.
Brachypodium pinnatum, L., is similar, but the palea more open and boat-like and narrows more suddenly above into the smooth awn which is shorter than itself (Fig. 77). Length 8-9 mm. The caryopsis is, however, much more slender than in Agropyrum.
For distinction from B. sylvaticum, see above. The inner palea is ciliate and more delicate than in Agropyrum.
≡ ≡ Paleæ rolled round the “seed" which is terete and tapering gradually into a stiff awn.
Δ Awn longer than the palea.
Festuca Myurus.
See also Festuca ovina, Cynosurus, &c., p. 147, and Arundo.
Festuca Myurus, L. (Fig. 80).
"Seed" 5-7 mm. without the long (10 mm.) awn, tapering and slender (like Nardus), grey or brown. Finely mamillate upwards. Rachilla small.
ΔΔ Awn shorter than palea.
Festuca ovina (var. rubra).
Nardus stricta.
Aira flexuosa, Molinia and other moor-grasses occur with these narrow-leafed Fescues, but are easily distinguished: Molinia by having no awn and being shorter and stouter, and Aira flexuosa by the dorsal awn and basal hairs.
Festuca rubra, L. (Fig. 78).
"Seed" pale brown with rosy tinge, 3-5 mm. long without the awn, smooth, or slightly hairy upwards. Caryopsis 2-3·5 mm., compressed, with a shallow broad172 flat furrow on the ventral face and a median long “hilum." Rachilla cylindrical, smooth, dilated above.
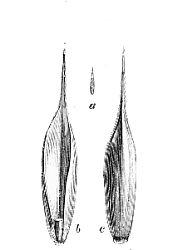 |
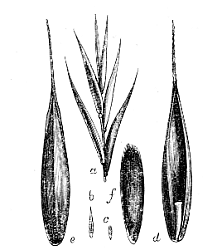 |
|
Fig. 78. Festuca ovina, var. rubra. a, “seed," nat. size; b and c, ditto, × about 7. Nobbe. Cf. Fig. 59. |
Fig. 79. Festuca ovina, var. heterophylla. a, spikelet, × about 3-1/2; b, “seed," and c, caryopsis, nat. size; d and e, “seed," and f, caryopsis, × 7. Nobbe. |
The variety F. heterophylla has narrower and longer "seeds," and a hispid rachilla. Palea 5-6 mm.: caryopsis 3·5-4·5 mm. (Fig. 79).
Perhaps Bromus sterilis should also come here (see p. 165).
With regard to these Fescues the student may note that F. ovina, var. tenuifolia, has the smallest (4-5 mm.) and most ovoid “seeds," usually golden brown in colour. F. rubra is darker and larger (5-6 mm.), as is also F. duriuscula (6-7 mm.): the latter is also 173 narrower, with toothed palea, tapering suddenly at the base and more awned. F. heterophylla is much like F. duriuscula, but the palea less toothed, and it tapers gradually at the base. The whole group is very difficult, and needs attention.
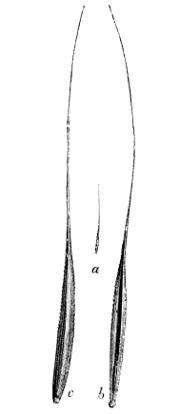 |
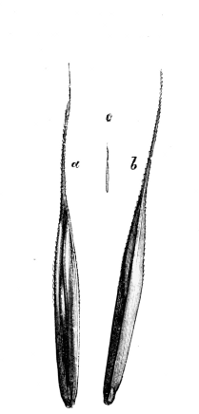 |
|
Fig. 80. Festuca Myurus. a, "seed," nat. size; b and c, ditto, × about 6. Compare Fig. 50. Nobbe. |
Fig. 81. Nardus stricta. Type of cylindrical “seed" with a terminal stiff awn, here serrulate; c, “seed," nat. size; a and b, ditto, × 8. Compare Fig. 50. Nobbe. |
Nardus stricta, L. (Fig. 81).
Outer palea tough and fused, 10-12 mm. long, narrow, lanceolate and tapering gradually into the stiff, serrulate,174 terminal awn shorter than itself. Inner palea minutely toothed at the apex and without awn, loosely enclosing the red-brownish yellow to bluish grey fruit which has no groove. “Seed" 4-6 mm. × 0·6 × 0·5 (without the awn), more or less triangular in section and serrulate on angles above.
Bentham and Hooker, British Flora, London, 1896.
Bonnier and Layens, Nouvelle Flore de France, Paris, 1887.
Bruns, Der Grasembryo in Flora, 1892, vol. LXXVI.
Burchard, Die Unkrautsamen der Klee-und Grasarten, Berlin, 1900.
Darwin, Power of Movement in Plants, London, 1880.
Duval Jouve, in Mémoires de l’Acad. des Sciences de Montpellier, 1871, vol. VII.
Fream, Elements of Agriculture, London, 1892.
Grob, in Bibliotheca Botanica, H. 36, 1896.
Guérin, Recherches sur le développement du Tégument &c. des Graminées in Ann. d. Sc. Nat. 1899.
Güntz, Unters. ü. d. anatomische Structur d. Grasblätter, Leipzig, 1886.
Hackel, in Engler’s Natürliche Pflanzenfamilien, II. Th. 2 Abth. 1887.
Harz, Landwirthschaftliche Samenkunde, Berlin, 1885.
Holzner, Beitr. zur Kenntn. d. Gerste, Munich, 1888.
Index Kewensis, London, 1893-95.
Jackson, A Glossary of Botanic Terms, London, 1900.
Kienitz-Gerloff, Botanik für Landwirte, Berlin, 1886.
Nobbe, Handbuch der Samenkunde, Berlin, 1876.
Parnell, British Grasses, London, 1845.
Pée-Laby, in Annales des Sc. Naturelles, 1898, vol. VIII.
Percival, Agricultural Botany, London, 1900.
Pfitzer, in Pringsh. Jahrb. f. wiss. Bot. B. VII.
Schmid, in Bot. Centralbl. 1898, B. LXXVI.
Schwarz, Forstliche Botanik, Berlin, 1892.
Schwendener, in Sitzungsber. d. Akad. Berlin, 1889 and 1890.
Settegast, Die landwirthschaftl. Sämereien u. d. Samenbau, Leipzig, 1892.
Sinclair, Hortus gramineus Woburnensis, London, 1824.
Sowerby, The Grasses of Great Britain, London, 1861.
Stebler and Schroeter, The Best Forage Plants, London, 1889.
Sutton, Permanent and Temporary Pastures, London, 1886.
Trimen, Article Grasses, in Encyclopædia Britannica, 9th ed.
Van Tieghem, Morphol. de l’embryon &c. chez les Graminées in Ann. des Sc. Nat. 1897.
Vesque, Traité de Botanique Agricole &c. 1885.
Warming, Lehrb. d. Oekologischen Pflanzengeogr. Berlin, 1896.
Wittmack, Gras-und Kleesamen, Berlin, 1873.
[1] Some foreign grasses (Andropogon, Panicum, &c.) have solid stems, and in Psamma and some others the lower parts may be solid.
[2] Leaf-stalks occur in tropical Bamboos.
[3] Tropical Bamboos branch in the upper parts and are woody. Dinochloa and Olyra are climbing grasses.
[4] Except, of course, in cases of virgin ground rapidly occupied by the seedlings.
[5] The most marked exceptions are the lemon-scented grasses (especially Andropogon) of India and Ceylon.
[6] The pale flanking lines seen in many grasses on each side of the mid-rib are the series of motor-cells referred to on p. 25.
[7] They may have short microscopic asperities, but there are no distinct long hairs.
[8] Very like a Poa when opened out, but the leaves are scabrid at the sheaths.
[9] Strictly speaking a spike is an axis bearing sessile flowers—not sessile spikelets: in Grasses, however, the conventional abbreviated term is sanctioned by long usage. The same applies to the panicle, &c.
[12] This term does not necessarily imply any botanical relationship with the true Millets (Panicum), but merely that the caryopsis is short and broad as in these grasses.
[13] In cases where the “seed" has fallen from the glumes the sample will usually show some of the latter lying loose.
Figures in bold type (25) refer to pages whereon the species or subject is more particularly dealt with; figures in italics (25) refer to pages containing illustrations.
CAMBRIDGE: PRINTED BY JOHN CLAY, M.A. AT THE UNIVERSITY PRESS.
CAMBRIDGE BIOLOGICAL SERIES
The Elements of Botany. By Francis Darwin, Sc.D., M.B., F.R.S., Fellow of Christ’s College. Second edition. Crown 8vo. With 94 illustrations. 4s. 6d.
Journal of Education. A noteworthy addition to our botanical literature.
Practical Physiology of Plants. By Francis Darwin, Sc.D., F.R.S., and E. Hamilton Acton, M.A. Third edition. Crown 8vo. With 45 illustrations. 4s. 6d.
Nature. The authors are much to be congratulated on their work, which fills a serious gap in the botanical literature of this country.
Morphology and Anthropology. By W. L. H. Duckworth, M.A., M.D., Fellow and Lecturer of Jesus College, University Lecturer in Physical Anthropology. Demy 8vo. With 333 illustrations. 15s. net.
Athenæum. Mr Duckworth has managed to produce in his “Morphology and Anthropology" just such a text-book as students have long been asking for.... It is no easy task to have undertaken such a work and the author is to be congratulated on the success which has attended his efforts. The volume can be confidently recommended to all whose studies lead them in this direction.
Lectures on the History of Physiology during the Sixteenth, Seventeenth and Eighteenth Centuries. By Sir M. Foster, K.C.B., M.D., D.C.L. Demy 8vo. With a frontispiece. 9s.
Nature. There is no more fascinating chapter in the history of science than that which deals with physiology, but a concise and at the same time compendious account of the early history of the subject has never before been presented to the English reader. Physiologists therefore owe a debt of gratitude to Sir Michael Foster for supplying a want which was widely felt.... No higher praise can be given to the book than to say that it is worthy of the reputation of its author.
The Soluble Ferments and Fermentation. By J. Reynolds Green, Sc.D., F.R.S., Professor of Botany to the Pharmaceutical Society of Great Britain. Second edition. Demy 8vo. 12s.
Nature. It is not necessary to recommend the perusal of the book to all interested in the subject since it is indispensable to them, and we will merely conclude by congratulating the Cambridge University Press on having added to their admirable series of Natural Science Manuals an eminently successful work on so important and difficult a theme, and the author on having written a treatise cleverly conceived, industriously and ably worked out, and on the whole, well written.
Conditions of Life in the Sea. A short account of Quantitative Marine Biological Research. By James Johnstone, Fisheries Laboratory, University of Liverpool. Demy 8vo. With a chart and 31 illustrations. 9s. net.
The Natural History of some Common Animals. By Oswald H. Latter, M.A., Senior Science Master at Charterhouse. Crown 8vo. With 54 illustrations. 5s. net.
Nature. An excellent book, written by a man who is equally in his element whether he writes as an outdoor naturalist or as a laboratory student. This combination is by no means a common one, and it is just the combination that is wanted for a book of this kind.... Altogether the book is an admirable one.
Athenæum. A book that may be judiciously placed in the hands of any boy who evinces a reasonable interest in the animal life around him.
The Classification of Flowering Plants. By Alfred Barton Rendle, M.A. (Cantab.), D.Sc. (Lond.), F.L.S., Keeper of the Department of Botany, British Museum. Vol. I. Gymnosperms and Monocotyledons. Demy 8vo. With 187 illustrations. 10s. 6d. net.
Gardener’s Chronicle. Numerous illustrations and an excellent index add to the value of the work. We heartily congratulate the author on the partial accomplishment of a difficult and laborious task. The part before us does but whet our appetite for what is to follow.
Athenæum. The first instalment of a text book which will well represent the state of our knowledge in the early years of the century. In the present volume the Gymnosperms and the Monocotyledons alone are dealt with; but they are treated with such excellent co-ordination of detail and such clear-headed sense of proportion, that we eagerly await the publication of the next instalment.
The Origin and Influence of the Thorough-bred Horse. By W. Ridgeway, Sc.D., F.B.A., Disney Professor of Archæology and Fellow of Gonville and Caius College. Demy 8vo. With 143 illustrations. 12s. 6d. net.
Westminster Gazette. There has never been a more learned contribution to equine literature than Professor Ridgeway’s comprehensive and exhaustive book.
Spectator. It would be difficult for Professor Ridgeway to write a book which did not contain at least one wholly novel thesis, and the present work is no exception to his practice. It is also an encyclopaedia of information on the history of the Equidae, collected from every source, from post-Pleiocene deposits to modern sporting newspapers. No detail escapes the author’s industry, and ... the result is a monument of sound learning, unique of its kind.
Manual of Practical Morbid Anatomy, being a Handbook for the Post-mortem Room. By H. D. Rolleston, M.A., M.D., F.R.C.P., and A. A. Kanthack, M.D., M.R.C.P. Crown 8vo. 6s.
Fossil Plants: a text-book for students of Botany and Geology. By A. C. Seward, M.A., F.R.S., Professor of Botany in the University of Cambridge. In 3 vols. Demy 8vo. Vol. I. with a frontispiece and 111 illustrations, 10s. net. Vol. II. with a frontispiece and 265 illustrations. 15s. net.
[Vol. III in the Press.
Revue Scientifique. Nous ne pouvons entrer dans le détail; mais il est évident que M. Seward, praticien distingué lui-même, est très au courant des travaux des autres, il les cite et utilise abondamment; et ceci est fait pour inspirer confiance. Au total, son œuvre est appuyée sur des bases solides, et elle restera sans doute longtemps le bréviaire, le manuel de ceux qui veulent, non pas seulement s’initier à la paléobotanique, mais retrouver les renseignements qui sont épars dans des centaines de monographies qu’on a souvent peine à se procurer. Le livre de M. Seward fait partie des Cambridge Natural Science Manuals, et il est digne de cette collection, qui est elle-même digne du foyer scientifique universellement réputé, où il a vu le jour.
Zoology. An Elementary Text-Book. By A. E. Shipley, Sc.D., F.R.S., and E. W. MacBride, M.A. (Cantab.), D.Sc. (London). Second edition. Demy 8vo. With 349 illustrations. 10s. 6d. net.
Pall Mall Gazette. Precisely the sort of book which, if it came into a thoughtful boy’s hands, would turn him from a smatterer into a student.... One of the most instructive and attractive books that could be put into the hands of a young naturalist.
Trees: A Handbook of Forest Botany for the Woodlands and the Laboratory. By H. Marshall Ward, Sc.D., F.R.S. Vol. I. Buds and Twigs. Vol. II. Leaves. Vol. III. Flowers and Inflorescences. Vol. IV. Fruits. Vol. V. Form and Habit, with an Appendix on Seedlings. Crown 8vo. With numerous illustrations. 4s. 6d. net each. Price for the set of five volumes, 20s. net.
Nature. The clear and simple way in which the author treats the subject is sure to inspire many with interest and enthusiasm for the study of forest botany.... The work will be found indispensable to those students who wish to make an expert study of forest botany. At the same time it is expressed in language so clear and devoid of technicalities that the amateur who wishes to know something about our trees and shrubs will find this one of the most useful guides to which he can turn.... The work is a many sided one, acting not only as a guide to the naturalist in the field, but also as a laboratory handbook, where the use of the lens and microscope may be employed to amplify the study of objects already observed in their natural habitats. Botanists generally, and especially forest botanists will welcome the appearance of this book as supplying a decided want, and filling a distinct gap in our literature of forest botany.
Grasses: a Handbook for use in the Field and Laboratory. By H. Marshall Ward, Sc.D., F.R.S. Crown 8vo. With 81 figures. 6s.
Field. The work is essentially suited to the requirements of those desirous of studying the grasses commonly grown in this country, and it can fairly be said that it furnishes an amount of information seldom obtained in more pretentious volumes.
P. T. O.
A Treatise on the British Freshwater Algæ. By G. S. West, M.A., A.R.C.S., F.L.S., Lecturer in Botany in the University of Birmingham. Demy 8vo. With a frontispiece and 166 illustrations. 10s. 6d. net.
Nature. Its aim is stated as “to give the student a concise account of the structure, habits and life-histories of Freshwater Algæ, and also to enable him to place within the prescribed limits of a genus any Algæ he may find in the freshwater of the British Islands.” To do this within the limits of an octavo volume of less than 400 pages, in which are numerous illustrations, is a task possible of accomplishment only by one very familiar with the subject and skilled in concise expression; but that it has been successfully done will, we think, be the verdict after testing the book thoroughly.... Prof. West’s treatment of his subject is instructive and stimulating.
A Manual and Dictionary of the Flowering Plants and Ferns. By J. C. Willis, M.A., Sc.D., Director of the Royal Botanic Gardens, Ceylon. Third edition. Crown 8vo. 10s. 6d.
Field. Taking this handy volume and a local flora, the traveller or student may do an enormous amount of practical field work without any other botanical literature whatever.... The result is a work that ought to be included in every library of botany and horticulture or agriculture, and it is certainly one that the nomadic botanist cannot afford to leave at home.... We have used the original edition of this work since its publication, and have found it to be one of the most useful and comprehensive works on plants ever produced.
Athenæum. The whole is well abreast of modern research, and a thoroughly business-like volume, lucid though compact.
Agriculture in the Tropics. An elementary Treatise. By J. C. Willis, M.A., Sc.D. Demy 8vo. With 25 plates. 7s. 6d. net.
Palæontology—Invertebrate. By Henry Woods, M.A., F.G.S., University Lecturer in Palæozoology. Fourth edition. Crown 8vo. With 151 illustrations. 6s.
Outlines of Vertebrate Palæontology for students of Zoology. By Arthur Smith Woodward, M.A., F.R.S., Keeper of the Department of Geology in the British Museum. Demy 8vo. With 228 illustrations. 14s.
Athenæum. The author is to be congratulated on having produced a work of exceptional value, dealing with a difficult subject in a thoroughly sound manner.
CAMBRIDGE UNIVERSITY PRESS
London: FETTER LANE, E.C.
C. F. CLAY, Manager
Edinburgh: 100, Princes Street
ALSO
London: H. K. Lewis, 136, Gower Street, W.C.