
The subject of Physiological Chemistry, or Biochemistry, is enlarging its borders to such an extent at the present time that no single text-book upon the subject, without being cumbrous, can adequately deal with it as a whole, so as to give both a general and a detailed account of its present position. It is, moreover, difficult, in the case of the larger text-books, to keep abreast of so rapidly growing a science by means of new editions, and such volumes are therefore issued when much of their contents has become obsolete.
For this reason, an attempt is being made to place this branch of science in a more accessible position by issuing a series of monographs upon the various chapters of the subject, each independent of and yet dependent upon the others, so that from time to time, as new material and the demand therefor necessitate, a new edition of each monograph can be issued without reissuing the whole series. In this way, both the expenses of publication and the expense to the purchaser will be diminished, and by a moderate outlay it will be possible to obtain a full account of any particular subject as nearly current as possible.
The editors of these monographs have kept two objects in view: firstly, that each author should be himself working at the subject with which he deals; and, secondly, that a Bibliography, as complete as possible, should be included, in order to avoid cross references, which are apt to be wrongly cited, and in order that each monograph may yield full and independent information of the work which has been done upon the subject.
It has been decided as a general scheme that the volumes first issued shall deal with the pure chemistry of physiological products and with certain general aspects of the subject. Subsequent monographs will be devoted to such questions as the chemistry of special tissues and particular aspects of metabolism. So the series, if continued, will proceed from physiological chemistry to what may be now more properly termed chemical physiology. This will depend upon the success which the first series achieves, and upon the divisions of the subject which may be of interest at the time.
THE NATURE OF ENZYME ACTION. By W. M. Bayliss, D.Sc., F.R.S. Third Edition. 5s. net.
THE CHEMICAL CONSTITUTION OF THE PROTEINS. By R. H. A. Plimmer, D.Sc. Part I.—Analysis. Second Edition, Revised and Enlarged. 5s. 6d. net. Part II.—Synthesis, etc. Second Edition, Revised and Enlarged. 3s. 6d. net.
THE GENERAL CHARACTERS OF THE PROTEINS. By S. B. Schryver, Ph.D., D.Sc. 2s. 6d. net.
THE VEGETABLE PROTEINS. By Thomas B. Osborne, Ph.D. 3s. 6d. net.
THE SIMPLE CARBOHYDRATES AND THE GLUCOSIDES. By E. Frankland Armstrong, D.Sc., Ph.D. Second Edition, Revised and Enlarged. 5s. net.
THE FATS. By J. B. Leathes, F.R.S., M.A., M.B., F.R.C.S. 4s. net.
ALCOHOLIC FERMENTATION. By A. Harden, Ph.D., D.Sc., F.R.S. Second Edition. 4s. net.
THE PHYSIOLOGY OF PROTEIN METABOLISM. By E. P. Cathcart, M.D., D.Sc. 4s. 6d. net.
SOIL CONDITIONS AND PLANT GROWTH. By E. J. Russell, D.Sc. 5s. net.
OXIDATIONS AND REDUCTIONS IN THE ANIMAL BODY. By H. D. Dakin, D.Sc., F.I.C. 4s. net.
THE SIMPLER NATURAL BASES. By G. Barger, M.A., D.Sc. 6s. net.
NUCLEIC ACIDS. THEIR CHEMICAL PROPERTIES AND PHYSIOLOGICAL CONDUCT. By Walter Jones, Ph.D. 3s. 6d. net.
THE DEVELOPMENT AND PRESENT POSITION OF BIOLOGICAL CHEMISTRY. By F. Gowland Hopkins, M.A., M.B., D.Sc., F.R.S.
THE POLYSACCHARIDES. By Arthur R. Ling, F.I.C.
COLLOIDS. By W. B. Hardy, M.A., F.R.S.
RESPIRATORY EXCHANGE IN ANIMALS. By A. Krogh, Ph.D.
PROTAMINES AND HISTONES. By A. Kossel, Ph.D.
LECITHIN AND ALLIED SUBSTANCES. By H. Maclean, M.D., D.Sc.
THE ORNAMENTAL PLANT PIGMENTS. By A. G. Perkin, F.R.S.
CHLOROPHYLL AND HAEMOGLOBIN. By.H. J. Page, B.Sc.
ORGANIC COMPOUNDS OF ARSENIC AND ANTIMONY. By Gilbert T. Morgan, D.Sc., F.I.C.
LONGMANS, GREEN AND CO.,
LONDON, NEW YORK, BOMBAY, CALCUTTA, AND MADRAS.

The following chapters are based on courses of lectures delivered at the London University and the Royal Institution during 1909–1910. In them an account is given of the work done on alcoholic fermentation since Buchner's epoch-making discovery of zymase, only in so far as it appears to throw light on the nature of that phenomenon. Many interesting subjects, therefore, have perforce been left untouched, among them the problem of the formation of zymase in the cell, and the vexed question of the relation of alcoholic fermentation to the metabolic processes of the higher plants and animals.
My thanks are due to the Council of the Royal Society, and to the Publishers of the "Journal of Physiology" for permission to make use of blocks (Figs. 2, 4 and 7) which have appeared in their publications.
In the New Edition no change has been made in the scope of the work. The rapid progress of the subject has, however, rendered necessary many additions to the text and a considerable increase in the bibliography.
May,1914.
| CHAPTER | PAGE | |
|---|---|---|
| I. | Historical Introduction | 1 |
| II. | Zymase and its Properties | 18 |
| III. | The Function of Phosphates in Alcoholic Fermentation | 41 |
| IV. | The Co-Enzyme of Yeast-Juice | 59 |
| V. | Action of Some Inhibiting and Accelerating Agents on the Enzymes of Yeast-Juice | 70 |
| VI. | Carboxylase | 81 |
| VII. | The By-Products of Alcoholic Fermentation | 85 |
| VIII. | The Chemical Changes involved in Fermentation | 96 |
| IX. | The Mechanism of Fermentation | 119 |
| Bibliography | 136 | |
| Index | 155 | |
The problem of alcoholic fermentation, of the origin and nature of that mysterious and apparently spontaneous change which converted the insipid juice of the grape into stimulating wine, seems to have exerted a fascination over the minds of natural philosophers from the very earliest times. No date can be assigned to the first observation of the phenomena of the process. History finds man in the possession of alcoholic liquors, and in the earliest chemical writings we find fermentation, as a familiar natural process, invoked to explain and illustrate the changes with which the science of those early days was concerned. Throughout the period of alchemy fermentation plays an important part; it is, in fact, scarcely too much to say that the language of the alchemists and many of their ideas were founded on the phenomena of fermentation. The subtle change in properties permeating the whole mass of material, the frothing of the fermenting liquid, rendering evident the vigour of the action, seemed to them the very emblems of the mysterious process by which the long sought for philosopher's stone was to convert the baser metals into gold. As chemical science emerged from the mists of alchemy, definite ideas about the nature of alcoholic fermentation and of putrefaction began to be formed. Fermentation was distinguished from other chemical changes in which gases were evolved, such as the action of acids on alkali carbonates (Sylvius de le Boë, 1659); the gas evolved was examined and termed gas vinorum, and was distinguished from the alcohol with which it had at first been confused (van Helmont, 1648); afterwards it was found that like the gas from potashes it was soluble in water (Wren, 1664). The gaseous product of fermentation and putrefaction was identified by MacBride, in 1764, with the fixed air of Black, whilst Cavendish in 1766 showed that fixed air alone was evolved in alcoholic fermentation and that a mixture of this with inflammable air was produced by putrefaction. In the meantime it had been recognised that only sweet liquors could be fermented ("Ubi notandum, nihil fermentare quod non sit dulce," Becher, 1682), and finally Cavendish [p002] [1776] determined the proportion of fixed air obtainable from sugar by fermentation and found it to be 57 per cent. It gradually became recognised that fermentation might yield either spirituous or acid liquors, whilst putrefaction was thought to be an action of the same kind as fermentation, differing mainly in the character of the products (Becher).
As regards the nature of the process very confused ideas at first prevailed, but in the time of the phlogistic chemists a definite theory of fermentation was proposed, first by Willis (1659) and afterwards by Stahl [1697], the fundamental idea of which survived the overthrow of the phlogistic system by Lavoisier and formed the foundation of the views of Liebig. To explain the spontaneous origin of fermentation and its propagation from one liquid to another, they supposed that the process consisted in a violent internal motion of the particles of the fermenting substance, set up by an aqueous liquid, whereby the combination of the essential constituents of this material was loosened and new particles formed, some of which were thrust out of the liquid (the carbon dioxide) and others retained in it (the alcohol).
Stahl specifically states that a body in such a state of internal disquietude can very readily communicate the disturbance to another, which is itself at rest but is capable of undergoing a similar change, so that a putrefying or fermenting liquid can set another liquid in putrefaction or fermentation.
Taking account of the gradual accumulation of fact and theory we find at the time of Lavoisier, from which the modern aspect of the problem dates, that Stahl's theoretical views were generally accepted. Alcoholic fermentation was known to require the presence of sugar and was thought to lead to the production of carbon dioxide, acetic acid, and alcohol.
The composition of organic compounds was at that time not understood, and it was Lavoisier who established the fact that they consisted of carbon, hydrogen, and oxygen, and who made systematic analyses of the substances concerned in fermentation (1784–1789). Lavoisier [1789] applied the results of these analyses to the study of alcoholic fermentation, and by employing the principle which he regarded as the foundation of experimental chemistry, "that there is the same quantity of matter before and after the operation," he drew up an equation between the quantities of carbon, hydrogen, and oxygen in the original sugar and in the resulting substances, alcohol, carbon dioxide, and acetic acid, showing that the products contained the whole matter of the sugar, and thus for the first time giving a clear view of the chemical [p003] change which occurs in fermentation. The conclusion to which he came was, we now know, very nearly accurate, but the research must be regarded as one of those remarkable instances in which the genius of the investigator triumphs over experimental deficiencies, for the analytical numbers employed contained grave errors, and it was only by a fortunate compensation of these that a result so near the truth was attained.
Lavoisier's equation or balance sheet was as follows:—
| Carbon. | Hydrogen. | Oxygen. | |
|---|---|---|---|
| 95·9 pounds of sugar (cane sugar) consist of | 26·8 | 7·7 | 61·4 |
| These yield:— | |||
| 57·7 pounds of alcohol containing | 16·7 | 9·6 | 31·4 |
| 35·3 pounds of carbon dioxide containing | 9·9 | — | 25·4 |
| 2·5 pounds of acetic acid containing | 0·6 | 0·2 | 1·7 |
| ――― | ――― | ――― | |
| Total contained in products | 27·2 | 9·8 | 58·5 |
The true composition of the sugar used was carbon 40·4, hydrogen 6·1, oxygen 49·4.
Lavoisier expressed no view as to the agency by which fermentation was brought about, but came to a very definite and characteristic conclusion as to the chemical nature of the change. The sugar, which he regarded in harmony with his general views as an oxide, was split into two parts, one of which was oxidised at the expense of the other to form carbonic acid, whilst the other was deoxygenised in favour of the former to produce the combustible substance alcohol, "so that if it were possible to recombine these two substances, alcohol and carbonic acid, sugar would result".
From this point commences the modern study of the problem. Provided by the genius of Lavoisier with the assurance that the hitherto mysterious process of fermentation was to be ranked along with familiar chemical changes, and that it proceeded in harmony with the same quantitative laws as these simpler reactions, chemists were stimulated in their desire to penetrate further into the mysteries of the phenomenon, and the importance and interest of the problem attracted many workers.
So important indeed did the matter appear to Lavoisier's countrymen that in the year 8 of the French Republic (1800) a prize—consisting of a gold medal, the value of which, expressed in terms of the newly introduced metric system, was that of one kilogram of gold—was offered by the Institute for the best answer to the question: "What are the characteristics by which animal and vegetable substances which act as ferments can be distinguished from those which they are capable of fermenting?" [p004]
This valuable prize was again offered in 1802 but was never awarded, as the fund from which it was to be drawn was sequestrated from the Institute in 1804. The first response to this stimulating offer was an important memoir by citizen Thenard [1803], which provided many of the facts upon which Liebig subsequently based his views. Thenard combats the prevailing idea, first expressed by Fabroni (1787–1799), that fermentation is caused by the action of gluten derived from grain on starch and sugar, but is himself uncertain as to the actual nature of the ferment. He points out that all fermenting liquids deposit a material resembling brewer's yeast, and he shows that this contains nitrogen, much of which is evolved as ammonia on distillation. His most important result is, however, that when yeast is used to ferment pure sugar, it undergoes a gradual change and is finally left as a white mass, much reduced in weight, which contains no nitrogen and is without action on sugar. Thenard, moreover, it is interesting to note, differs from Lavoisier, inasmuch as he ascribes the origin of some of the carbonic acid to the carbon of the ferment, an opinion which was still held in various degrees by many investigators (see Seguin, quoted by Thenard).
Thenard's memoir was followed by a communication of fundamental importance from Gay-Lussac [1810]. A process for preserving food had been introduced by Appert, which consisted in placing the material in bottles, closing these very carefully and exposing them to the temperature of boiling water for some time. Gay-Lussac was struck by the fact that when such a bottle was opened fermentation or putrefaction set in rapidly. Analysis of the air left in such a sealed bottle showed that all the oxygen had been absorbed, and these facts led to the view that fermentation was set up by the action of oxygen on the fermentable material. Experiment appeared to confirm this in the most striking way. A bottle of preserved grape-juice was opened over mercury and part of its contents passed through the mercury into a bell-jar containing air, the remainder into a similar vessel free from air. In the presence of air fermentation set in at once, in the absence of air no fermentation whatever occurred. This connection between fermentation and the presence of air was established by numerous experiments and appeared incontestable. Fermentation, it was found, could be checked by boiling even after the addition of oxygen, and hence food could be preserved in free contact with the air, provided only that it was raised to the temperature of boiling water at short intervals of time. Gay-Lussac's opinion was that the ferment was formed by the action of the oxygen on the [p005] liquid, and that the product of this action was altered by heat and rendered incapable of producing fermentation, as was also brewer's yeast, which, however, he regarded, on account of its insolubility, as different from the soluble ferment which initiated the change in the limpid grape-juice. Colin, on the other hand [1825], recognised that alcoholic fermentation by whatever substance it was started, resulted in the formation of an insoluble deposit more active than the original substance, and he suggested that this deposit might possibly in every case be of the same nature.
So far no suspicion appears to have arisen in the minds of those who had occupied themselves with the study of fermentation that this change differed in any essential manner from many other reactions familiar to chemists. The origin and properties of the ferment were indeed remarkable and involved in obscurity, but the uncertainty regarding this substance was no greater than that surrounding many, if not all, compounds of animal and vegetable origin. Although, however, the purely chemical view as to the nature of yeast was generally recognised and adopted, isolated observations were not wanting which tended to show that yeast might be something more than a mere chemical reagent. As early as 1680 in letters to the Royal Society Leeuwenhoek described the microscopic appearance of yeast of various origins as that of small, round, or oval particles, but no further progress seems to have been made in this direction for nearly a century and a half, when we find that Desmazières [1826] examined the film formed on beer, figured the elongated cells of which it was composed, and described it under the name of Mycoderma Cerevisiæ. He, however, regarded it rather as of animal than of vegetable origin, and does not appear to have connected the presence of these cells with the process of fermentation.
Upon this long period during which yeast was regarded merely as a chemical compound there followed, as has so frequently occurred in similar cases, a sudden outburst of discovery. No less than three observers hit almost simultaneously upon the secret of fermentation and declared that yeast was a living organism.
First among these in strict order of time was Cagniard-Latour [1838], who made a number of communications to the Academy and to the Société Philomatique in 1835–6, the contents of which were collected in a paper presented to the Academy of Sciences on 12 June, 1837, and published in 1838. The observations upon which this memoir was based were almost exclusively microscopical. Yeast was recognised as consisting of spherical particles, which were capable of [p006] reproduction by budding but incapable of motion, and it was therefore regarded as a living organism probably belonging to the vegetable kingdom. Alcoholic fermentation was observed to depend on the presence of living yeast cells, and was attributed to some effect of their vegetative life (quelque effet de leur végétation). It was also noticed that yeast was not deprived of its fermenting power by exposure to the temperature of solid carbonic acid, a sample of which was supplied to Cagniard-Latour by Thilorier, who had only recently prepared it for the first time.
Theodor Schwann [1837], whose researches were quite independent of those of Cagniard-Latour, approached the problem from an entirely different point of view. During the year 1836 Franz Schulze [1836] published a research on the subject of spontaneous generation, in which he proved that when a solution containing animal or vegetable matter was boiled, no putrefaction set in provided that all air which was allowed to have access to the liquid was previously passed through strong sulphuric acid. Schwann performed a very similar experiment by which he showed that this same result, the absence of putrefaction, was attained by heating all air which came into contact with the boiled liquid. Wishing to show that other processes in which air took part were not affected by the air being heated, he made experiments with fermenting liquids and found, contrary to his expectation, that a liquid capable of undergoing vinous fermentation and containing yeast did not undergo this change after it had been boiled, provided that, as in the case of his previous experiments, only air which had been heated was allowed to come into contact with it.
Schwann's experiments on the prevention of putrefaction were unexceptionable and quite decisive. The analogous experiments dealing with alcoholic fermentation were not quite so satisfactory. Yeast was added to a solution of cane sugar, the flask containing the mixture placed in boiling water for ten minutes, and then inverted over mercury. About one-third of the liquid was then displaced by air and the flasks corked and kept inverted at air temperature. In two flasks the air introduced was ordinary atmospheric air, and in these flasks fermentation set in after about four to six weeks. Into the other two flasks air which had been heated was led, and in these no fermentation occurred. As described, the experiment is quite satisfactory, but Schwann found on repetition that the results were irregular. Sometimes all the flasks showed fermentation, sometimes none of them. This was correctly ascribed to the experimental difficulties, but none [p007] the less served as a point of attack for hostile and damaging criticism at the hands of Berzelius (p. 8).
The origin of putrefaction was definitely attributed by Schwann to the presence of living germs in the air, and the similarity of the result obtained with yeast suggested the idea that alcoholic fermentation was also brought about by a living organism, a conception which was at once confirmed by a microscopical examination of a fermenting liquid. The phenomena observed under the microscope were similar to those noted by Cagniard-Latour, and in accordance with these observations alcoholic fermentation was attributed to the development of a living organism, the fermentative function of which was found to be destroyed by potassium arsenite and not by extract of Nux vomica, so that the organism was regarded rather as of vegetable than of animal nature. This plant received the name of "Zuckerpilz" or sugar fungus (which has been perpetuated in the generic term Saccharomyces). Alcoholic fermentation was explained as "the decomposition brought about by this sugar fungus removing from the sugar and a nitrogenous substance the materials necessary for its growth and nourishment, whilst the remaining elements of these compounds, which were not taken up by the plant, combined chiefly to form alcohol".
Kützing's memoir, the third of the trio [1837], also dates from 1837, and his opinions, like those of Cagniard-Latour, are founded on microscopical observations. He recognises yeast as a vegetable organism and accurately describes its appearance. Alcoholic fermentation depends on the formation of yeast, which is produced when the necessary elements and the proper conditions are present and then propagates itself. The action on the liquid thus increases and the constituents not required to form the organism combine to form unorganised substances, the carbonic acid and alcohol. "It is obvious," says Kützing, in a passage which roused the sarcasm of Berzelius, "that chemists must now strike yeast off the roll of chemical compounds, since it is not a compound but an organised body, an organism."
These three papers, which were published almost simultaneously, were received at first with incredulity. Berzelius, at that time the arbiter and dictator of the chemical world, reviewed them all in his "Jahresbericht" for 1839 [1839] with impartial scorn. The microscopical evidence was denied all value, and yeast was no more to be regarded as an organism than was a precipitate of alumina. Schwann's experiment (p. 6) was criticised on the ground that the fermenting power of the added yeast had been only partially destroyed in the [p008] flasks in which fermentation ensued, completely in those which remained unchanged, the admission of heated or unheated air being indifferent, a criticism to some extent justified by Schwann's statement, already quoted, of the uncertain result of the experiment.
Berzelius himself regarded fermentation as being brought about by the yeast by virtue of that catalytic force, which he had supposed to intervene in so many reactions, both between substances of mineral and of animal and vegetable origin [1836], and which enabled "bodies, by their mere presence, and not by their affinity, to arouse affinities ordinarily quiescent at the temperature of the experiment, so that the elements of a compound body arrange themselves in some different way, by which a greater degree of electro-chemical neutralisation is attained".
To the scorn of Berzelius was soon added the sarcasm of Wöhler and Liebig [1839, 1839]. Stimulated in part by the publications of the three authors already mentioned, and in part by the report of Turpin [1838], who at the request of the Academy of Sciences had satisfied himself by observation of the accuracy of Cagniard-Latour's conclusions, Wöhler prepared an elaborate skit on the subject, which he sent to Liebig, to whom it appealed so strongly that he added some touches of his own and published it in the "Annalen," following immediately upon a translation of Turpin's paper. Yeast was here described with a considerable degree of anatomical realism as consisting of eggs which developed into minute animals, shaped like a distilling apparatus, by which the sugar was taken in as food and digested into carbonic acid and alcohol, which were separately excreted, the whole process being easily followed under the miscroscope.
Close upon this pleasantry followed a serious and important communication from Liebig [1839], in which the nature of fermentation, putrefaction, and decay was exhaustively discussed. Liebig did not admit that these phenomena were caused by living organisms, nor did he attribute them like Berzelius to the catalytic action of a substance which itself survived the reaction unchanged. As regards alcoholic fermentation, Liebig's chief arguments may be briefly summarised. As the result of alcoholic fermentation, the whole of the carbon of the sugar reappears in the alcohol and carbon dioxide formed. This change is brought about by a body termed the ferment, which is formed as the result of a change set up by the access of air to plant juices containing sugar, and which contains all the nitrogen of the nitrogenous constituents of the juice. This ferment is a substance remarkably susceptible of change, which undergoes an uninterrupted and progressive metamorphosis, of [p009] the nature of putrefaction or decay, and produces the fermentation of the sugar as a consequence of the transformation which it is itself undergoing.
The decomposition of the sugar is therefore due to a condition of instability transferred to it from the unstable and changing ferment, and only continues so long as the decomposition of the ferment proceeds. This communication of instability from one substance undergoing chemical change to another is the basis of Liebig's conception, and is illustrated by a number of chemical analogies, one of which will suffice to explain his meaning. Platinum is itself incapable of decomposing nitric acid and dissolving in it; silver, on the other hand, possesses this power. When platinum is alloyed with silver, the whole mass dissolves in nitric acid, the power possessed by the silver being transferred to the platinum. In like manner the condition of active decomposition of the ferment is transferred to the sugar, which by itself is quite stable. The central idea is that of Stahl (p. 2) which was thus reintroduced into scientific thought.
In a pure sugar solution the decomposition of the ferment soon comes to an end and fermentation then ceases. In beer wort or vegetable juices, on the other hand, more ferment is continually formed in the manner already described from the nitrogenous constituents of the juice, and hence the sugar is completely fermented away and unexhausted ferment left behind. Liebig's views were reiterated in his celebrated "Chemische Briefe," and became the generally accepted doctrine of chemists. There seems little doubt that both Berzelius and Liebig in their scornful rejection of the results of Cagniard-Latour, Schwann and Kützing, were influenced, perhaps almost unconsciously, by a desire to avoid seeing an important chemical change relegated to the domain of that vital force from beneath the sway of which a large part of organic chemistry had just been rescued by Wöhler's brilliant synthetical production of urea and by the less recognised synthesis of alcohol by Hennell (see on this point Ahrens [1902]). A strong body of evidence, however, gradually accumulated in favour of the vegetable nature of yeast, so that it may be said that by 1848 a powerful minority adhered to the views of Cagniard-Latour, Schwann, and Kützing [see Schrohe, 1904, p. 218, and compare Buchner, 1904]. Among these must be included Berzelius [1848], who had so forcibly repudiated the idea only ten years before, whereas Liebig in the 1851 edition of his letters does not mention the fact that yeast is a living organism (Letter XV).
The recognition of the vegetable nature of yeast, however, by no [p010] means disproved Liebig's view of the nature of the change by which sugar was converted into carbon dioxide and alcohol, as was carefully pointed out by Schlossberger [1844] in a research on the nature of yeast, carried out in Liebig's laboratory but without decisive results.
Mitscherlich was also convinced of the vegetable character of yeast, and showed [1841] that when yeast was placed in a glass tube closed by parchment and plunged into sugar solution, the sugar entered the glass tube and was there fermented, but was not fermented outside the tube. He regarded this as a proof that fermentation only occurred at the surface of the yeast cells, and explained the process by contact action in the sense of the catalytic action of Berzelius, rather than by Liebig's transference of molecular instability. Similar results were obtained with an animal membrane by Helmholtz [1843], who also expressed his conviction that yeast was a vegetable organism.
In 1854 Schröder and von Dusch [1854, 1859, 1861] strongly reinforced the evidence in favour of this view by succeeding in preventing the putrefaction and fermentation of many boiled organic liquids by the simple process of filtering all air which had access to them through cotton-wool. These experiments, which were continued until 1861, led to the conclusion that the spontaneous alcoholic fermentation of liquids was due to living germs carried by the air, and that when the air was passed through the cotton-wool these germs were held back.
At the middle of the nineteenth century opinions with regard to alcoholic fermentation, notwithstanding all that had been done, were still divided. On the one hand Liebig's theory of fermentation was widely held and taught. Gerhardt, for example, as late as 1856 in the article on fermentation in his treatise on organic chemistry [1856], gives entire support to Liebig's views, and his treatment of the matter affords an interesting glimpse of the arguments which were then held to be decisive. The grounds on which he rejects the conclusions of Schwann and the other investigators who shared the belief in the vegetable nature of yeast are that, although in some cases animal and vegetable matter and infusions can be preserved from change by the methods described by these authors, in others they cannot, a striking case being that of milk, which even after being boiled becomes sour even in filtered air, and this without showing any trace of living organisms. The action of heat, sulphuric acid, and filtration on the air is to remove, or destroy, not living organisms but particles of decomposing matter, that is to say, ferments which would add their activity to that of the oxygen of the air. Moreover, many ferments, as for example diastase, act without [p011] producing any insoluble deposit whatever which can be regarded as an organism.
"Evidemment," he concludes, "la théorie de M. Liebig explique seule tous les phénomènes de la manière la plus complète et la plus logique; c'est à elle que tous les bons esprits ne peuvent manquer de se rallier."
On the other hand it was held by many to have been shown that Liebig's view of the origin of yeast by the action of the air on a vegetable infusion was erroneous, and that fermentation only arose when the air transferred to the liquid an active agent which could be removed from it by sulphuric acid (Schulze), by heat (Schwann), and by cotton-wool (Schröder and von Dusch). Accompanying alcoholic fermentation there was a development of a living organism, the yeast, and fermentation was believed, without any very strict proof, to be a phenomenon due to the life and vegetation of this organism. This doctrine seems indeed [Schrohe, 1904] to have been widely taught in Germany from 1840–56, and to have established itself in the practice of the fermentation industries.
In 1857 commenced the classical researches of Pasteur which finally decided the question as to the origin and functions of yeast and led him to the conclusion that "alcoholic fermentation is an act correlated with the life and organisation of the yeast cells, not with the death or putrefaction of the cells, any more than it is a phenomenon of contact, in which case the transformation of sugar would be accomplished in presence of the ferment without yielding up to it or taking from it anything" [1860]. It is impossible here to enter in detail into Pasteur's experiments on this subject, or indeed to do more than indicate the general lines of his investigation. His starting-point was the lactic acid fermentation.
The organism to which this change was due had hitherto escaped detection, and as we have seen the spontaneous lactic fermentation of milk was one of the phenomena adduced by Gerhardt (p. 10) in favour of Liebig's views. Pasteur [1857] discovered the lactic acid producing organism and convinced himself that it was in fact a living organism and the active cause of the production of lactic acid. One of the chief buttresses of Liebig's theory was thus removed, and Pasteur next proceeded to apply the same method and reasoning to alcoholic fermentation. Liebig's theory of the origin of yeast by the action of the oxygen of the air on the nitrogenous matter of the fermentable liquid was conclusively and strikingly disproved by the brilliant device of producing a crop of yeast in a liquid medium containing only comparatively [p012] simple substances of known composition—sugar, ammonium tartrate and mineral phosphate. Here there was obviously present in the original medium no matter which could be put into a state of putrefaction by contact with oxygen and extend its instability to the sugar. Any such material must first be formed by the vital processes of the yeast. In the next place Pasteur showed by careful analyses and estimations that, whenever fermentation occurred, growth and multiplication of yeast accompanied the phenomenon. The sugar, he proved, was not completely decomposed into carbon dioxide and alcohol, as had been assumed by Liebig (p. 8). A balance-sheet of materials and products was constructed which showed that the alcohol and carbon dioxide formed amounted only to about 95 per cent. of the invert sugar fermented, the difference being made up by glycerol, succinic acid, cellulose, and other substances [1860, p. 347]. In every case of fermentation, even when a paste of yeast was added to a solution of pure cane sugar in water, the yeast was found by quantitative measurements to have taken something from the sugar. This "something" was indeterminate in character, but, including the whole of the extractives which had passed from the yeast cells into the surrounding liquid, it amounted to as much as 1·63 per cent. of the weight of the sugar fermented [1860, p. 344].
Pasteur was therefore led to consider fermentation as a physiological process accompanying the life of the yeast. His conclusions were couched in unmistakable words: "The chemical act of fermentation is essentially a phenomenon correlative with a vital act, commencing and ceasing with the latter. I am of opinion that alcoholic fermentation never occurs without simultaneous organisation, development, multiplication of cells, or the continued life of cells already formed. The results expressed in this memoir seem to me to be completely opposed to the opinions of Liebig and Berzelius. If I am asked in what consists the chemical act whereby the sugar is decomposed and what is its real cause, I reply that I am completely ignorant of it.
"Ought we to say that the yeast feeds on sugar and excretes alcohol and carbonic acid? Or should we rather maintain that yeast in its development produces some substance of the nature of a pepsin, which acts upon the sugar and then disappears, for no such substance is found in fermented liquids? I have nothing to reply to these hypotheses. I neither admit them nor reject them, and wish only to restrain myself from going beyond the facts. And the facts tell me simply that all true fermentations are correlative with physiological phenomena."
Liebig felt to the full the weight of Pasteur's criticisms; his reply [p013] was long delayed [1870], and, according to his biographer, Volhard [1909], caused him much anxiety. In it he admits the vegetable nature of yeast, but does not regard Pasteur's conclusion as in any way a solution of the problem of the nature of alcoholic fermentation. Pasteur's "physiological act" is for Liebig the very phenomenon which requires explanation, and which he still maintains can be explained by his original theory of communicated instability. On some of Pasteur's results, notably the very important one of the cultivation of yeast in a synthetic medium, he casts grave doubt, whilst he explains the production of glycerol and succinic acid as due to independent reactions. The phenomenon of fermentation is still for him one which accompanies the decomposition of the constituents of the cell, rather than their building up by vegetative growth. "When the fungus ceases to grow, the bond which holds together the constituents of the cell contents is relaxed, and it is the motion which is thus set up in them which is the means by which the yeast cells are enabled to bring about a displacement or decomposition of the elements of sugar or other organic molecules." Pasteur replied in a brief and unanswerable note [1872]. All his attention was concentrated on the one question of the production of yeast in a synthetic medium, which he recognised as fundamental. The validity of this experiment he emphatically reaffirmed, and finally undertook, from materials supplied by Liebig himself, to produce as much yeast as could be reasonably desired. This challenge was never taken up, and this communication formed the last word of the controversy. Pasteur had at this time firmly established his thesis, no fermentation without life, both for alcoholic fermentation and for those other fermentations which are produced by bacteria, and had put upon a sound and permanent basis the conclusions drawn by Schulze, Cagniard-Latour, Schwann, and Kützing from their early experiments. It became generally recognised that putrefaction and other fermentative changes were due to specific organisms, which produced them in the exercise of their vital functions.
Pasteur subsequently [1875] came to the conclusion that fermentation was the result of life without oxygen, the cells being able, in the absence of free oxygen, to avail themselves of the energy liberated by the decomposition of substances containing combined oxygen. This view, which did not involve any alteration of Pasteur's original thesis but was an attempt to explain the physiological origin and function of fermentation, gave rise to a prolonged controversy, which cannot be further discussed in these pages. [p014]
Nevertheless, Liebig's desire to penetrate more deeply into the nature of the process of fermentation remained in many minds, and numerous endeavours were made to obtain further insight into the problem. In spite of an entire lack of direct experimental proof, the conception that alcoholic fermentation was due to the chemical action of some substance elaborated by the cell and not directly to the vital processes of the cell as a whole found strenuous supporters even among those who were convinced of the vegetable character of yeast. As early as 1833 diastase, discovered still earlier by Kirchhoff and Dubrunfaut, had been extracted by means of water from germinating barley and precipitated by alcohol as a white powder, the solution of which was capable of converting starch into sugar, but lost this power when heated [Payen and Persoz, 1833]. Basing his ideas in part upon the behaviour of this substance, Moritz Traube [1858] enunciated in the clearest possible manner the theory that all fermentations produced by living organisms are caused by ferments, which are definite chemical substances produced in the cells of the organism. He regarded these substances as being closely related to the proteins and considered that their function was to transfer the oxygen and hydrogen of water to different parts of the molecule of the fermentable substance and thus bring about that apparent intramolecular oxidation and reduction which is so characteristic of fermentative change and had arrested the attention of Lavoisier and, long after him, of Liebig.
Traube's main thesis, that fermentation is caused by definite ferments or enzymes, attracted much attention, and received fresh support from the separation of invertase in 1860 from an extract of yeast by Berthelot, and from the advocacy and authority of this great countryman of Pasteur, who definitely expressed his opinion that insoluble ferments existed which could not be separated from the tissues of the organism, and further, that the organism could not itself be regarded as the ferment, but only as the producer of the ferment [1857, 1860]. Hoppe-Seyler [1876] also supported the enzyme theory of fermentation, but differed in some respects from Traube as to the exact function of the ferment [see Traube, 1877; Hoppe-Seyler, 1877].
Direct experimental evidence was, however, still wanting, and Pasteur's reiterated assertion [1875] that all fermentation phenomena were manifestations of the life of the organism remained uncontroverted by experience.
Numerous and repeated direct experimental attacks had been made [p015] from time to time upon the problem of the existence of a fermentation enzyme, but all had yielded negative or unreliable results.
As early as 1846 a bold attempt had been made by Lüdersdorff [1846] to ascertain whether fermentation was or was not bound up with the life of the yeast by grinding yeast and examining the ground mass. A single gram of yeast was thoroughly ground, the process lasting for an hour, and the product was tested with sugar solution. Not a single bubble of gas was evolved. A similar result was obtained in a repetition of the experiment by Schmidt in Liebig's laboratory [1847], the grinding being continued in this case for six hours, but the natural conclusion that living yeast was essential for fermentation was not accepted, on the ground that during the lengthy process of trituration in contact with air the yeast had become altered and now no longer possessed the power of producing alcoholic fermentation, but instead had acquired that of changing sugar into lactic acid [see Gerhardt, 1856, p. 545].
Similar experiments made in 1871 by Marie von Manasseïn [1872, 1897], in which yeast was ground for six to fifteen hours with powdered rock crystal, yielded products which fermented sugar, but they contained unbroken yeast cells, so that the results obtained could not be considered decisive [Buchner and Rapp, 1898, 1], although Frau von Manasseïn herself drew from them and from others in which sugar solution was treated with heated yeast, but not under aseptic conditions, the conclusion that living yeast cells were not necessary for fermentation.
Quite unsuccessful were also the attempts made to accomplish the separation of fermentation from the living cell by Adolf Mayer [1879, p. 66], and, as we learn from Roux, by Pasteur himself, grinding, freezing, and plasmolysing the cells, having in his hands proved alike in vain. Extraction by glycerol or water, a method by which many enzymes can be obtained in solution, gave no better results [Nägeli and Loew, 1878], and the enzyme theory of alcoholic fermentation appeared quite unjustified by experiment.
Having convinced himself of this, Nägeli [1879] suggested a new explanation of the facts based on molecular-physical grounds. According to this view, which unites in itself some of the conceptions of Liebig, Pasteur, and Traube, fermentation is the transference of a state of motion from the molecules, atomic groups, and atoms of the compounds constituting the living plasma of the cell to the fermentable material, whereby the equilibrium existing in the molecules of the latter is disturbed and decomposition ensues [1879, p. 29]. [p016]
This somewhat complex idea, whilst including, as did Liebig's theory, Stahl's fundamental conception of a transmission of a state of motion, satisfies Pasteur's contention that fermentation cannot occur without life, and at the same time explains the specific action of different organisms by differences in the constitution of their cell contents. The really essential part of Nägeli's theory consisted in the limitation of the power of transference of molecular motion to the living plasma, by which the failure of all attempts to separate the power of fermentation from the living cell was explained. This was the special phenomenon which required explanation; to account for this the theory was devised, and when this was experimentally disproved, the theory lost all significance.
For nearly twenty years no further progress was made, and then in 1897 the question which had aroused so much discussion and conjecture, and had given rise to so much experimental work, was finally answered by Eduard Buchner, who succeeded in preparing from yeast a liquid which, in the complete absence of cells, was capable of effecting the resolution of sugar into carbon dioxide and alcohol [1897, 1].
In the light of this discovery the contribution to the truth made by each of the great protagonists in the prolonged discussion on the problem of alcoholic fermentation can be discerned with some degree of clearness. Liebig's main contention that fermentation was essentially a chemical act was correct, although his explanation of the nature of this act was inaccurate. Pasteur, in so far as he considered the act of fermentation as indissolubly connected with the life of the organism, was shown to be in error, but the function of the organism has only been restricted by a single stage, the active enzyme of alcoholic fermentation has so far only been observed as the product of the living cell. Nearest of all to the truth was Traube, who in 1858 enunciated the theorem, which was only proved for alcoholic fermentation in 1897, that all fermentations produced by living organisms are due to ferments secreted by the cells.
Buchner's discovery of zymase has introduced a new experimental method by means of which the problem of alcoholic fermentation can be attacked, and the result has been that since 1897 a considerable amount of information has been gained with regard to the nature and conditions of action of the enzymes of the yeast cell. It has been found that the machinery of fermentation is much more complex than had been surmised. The enzyme zymase, which is essential for fermentation, cannot of itself bring about the alcoholic fermentation of sugar, but is dependent on the presence of a second substance, termed, for [p017] want of a more reasonable name, the co-enzyme. The chemical nature and function of this mysterious coadjutor are still unknown, but as it withstands the temperature of boiling water and is dialysable, it is probably more simple in constitution than the enzyme. This, however, is not all; for the decomposition of sugar a phosphate is also indispensable. It appears that in yeast-juice, and therefore also most probably in the yeast cell, the phosphorus present takes an active part in fermentation and goes through a remarkable cycle of changes. The breakdown of sugar into alcohol and carbon dioxide is accompanied by the formation of a complex hexosephosphate, and the phosphate is split off from this compound and thus again rendered available for action by means of a special enzyme, termed hexosephosphatase. In addition to this complex of ferments, the cell also possesses special enzymes by which the zymase and the co-enzyme can be destroyed, and, further, at least one substance, known as an anti-enzyme, which directly checks this destructive action. It seems probable, moreover, that the decomposition of the sugar molecule takes place in stages, although much doubt yet exists as to the nature of these.
At the present moment the subject remains one of the most interesting in the whole field of biological chemistry, the limited degree of insight which has already been gained into the marvellous complexity of the cell lending additional zest to the attempt to penetrate the darkness which shrouds the still hidden mysteries.
The history of Buchner's discovery is of great interest [Gruber, 1908; Hahn, 1908]. As early as 1893 Hans and Eduard Buchner found that the cells of even the smallest micro-organism could be broken by being ground with sand [Buchner, E. and H., and Hahn, 1903, p. 20], and in 1896 the same process was applied by these two investigators to yeast, with the object of obtaining a preparation for therapeutic purposes. Difficulties arose in the separation of the cell contents from the ground-up mixture of cell membranes, unbroken cells, and sand, but these were overcome by carrying out the suggestion of Martin Hahn (at that time assistant to Hans Buchner) that kieselguhr should be added and the liquid squeezed out by means of a hydraulic press [Buchner, E. and H., and Hahn, 1903, p. 58]. The yeast-juice thus obtained was, in the first instance, employed for animal experiments, but underwent change very rapidly. The ordinary antiseptics were found to be unsuitable, and hence sugar was added as a preservative, and it was the marked action of the juice upon this added cane sugar that drew Eduard Buchner's attention to the fact that fermentation was proceeding in the absence of yeast-cells.
As in the case of so many discoveries, the new phenomenon was brought to light, apparently by chance, as the result of an investigation directed to quite other ends, but fortunately fell under the eye of an observer possessed of the genius which enabled him to realise its importance and give to it the true interpretation.
In his first papers [1897, 1, 2; 1898], Buchner established the following facts: (1) yeast-juice free from cells is capable of producing the alcoholic fermentation of glucose, fructose, cane sugar, and maltose; (2) the fermenting power of the juice is neither destroyed by the addition of chloroform, benzene, or sodium arsenite [Hans Buchner, 1897], by filtration through a Berkefeld filter, by evaporation to dryness at 30° to 35°, nor by precipitation with alcohol; (3) the fermenting power is completely destroyed when the liquid is heated to 50°. [p019]
From these facts he drew the conclusion "that the production of alcoholic fermentation does not require so complicated an apparatus as the yeast cell, and that the fermentative power of yeast-juice is due to the presence of a dissolved substance". To this active substance he gave the name of zymase.
Buchner's discovery was not received without some hesitation. A number of investigators prepared yeast-juice, but failed to obtain an active product [Will, 1897; Delbrück, 1897; Martin and Chapman, 1898; Reynolds Green, 1897; Lintner, 1899]. A more accurate knowledge of the necessary conditions and of the properties of yeast-juice, however, led to more successful results [Will, 1898; Reynolds Green, 1898; Lange, 1898], and it was soon established that, given suitable yeast, an active preparation could be readily procured by Buchner's method. Criticism was then directed to the effect of the admitted presence of a certain number of micro-organisms in yeast-juice [Stavenhagen, 1897], but Buchner [Buchner and Rapp, 1897] was able to show by experiments in the presence of antiseptics and with juice filtered through a Chamberland candle that the fermentation was not due to living organisms of any kind.
The most weighty criticism of Buchner's conclusion consisted in an attempt to show that the properties of yeast-juice might be due to the presence, suspended in it, of fragments of living protoplasm, which, although severed from their original surroundings in the cell, might retain for some time the power of producing alcoholic fermentation. This, it will be seen, was an endeavour to extend Nägeli's theory to include in it the newly discovered fact.
In favour of this view were adduced the similarity between the effects of many antiseptics on living yeast and on the juice, the ephemeral nature of the fermenting agent present in the juice, the effect of dilution with water, and the phenomenon of autofermentation which is exhibited by the juice in the absence of added sugar [Abeles, 1898; v. Kupffer, 1897; v. Voit, 1897; Wehmer, 1898; Neumeister, 1897; Macfadyen, Morris, and Rowland, 1900; Bokorny, 1906; Fischer, 1903; Beijerinck, 1897, 1900; Wroblewski, 1899, 1901].
A brief general description of the actual properties of yeast-juice and of the phenomena of fermentation by its means is sufficient to show the great improbability of this view.
The juice prepared by Buchner's method forms a somewhat viscous opalescent brownish-yellow liquid, which is usually faintly acid in reaction [compare Ahrens, 1900] and almost optically inactive. It has a specific gravity of 1·03 to 1·06, contains 8·5 to 14 per cent. [p020] of dissolved solids, and leaves an ash amounting to 1·4 to 2 per cent. About 0·7 to 1·7 per cent. of nitrogen is present, nearly all in the form of protein, which coagulates to a thick white mass when the juice is heated.
A powerful digestive enzyme of the type of trypsin is also present, so that when the juice is preserved its albumin undergoes digestion at a rate which depends on the temperature [Hahn, 1898; Geret and Hahn, 1898, 1, 2; 1900; Buchner, E. and H., and Hahn, 1903, pp. 287–340], and is converted into a mixture of bases and amino-acids. After about six days at 37°, or 10 to 14 days at the ordinary temperature, the digestion is so complete that no coagulation occurs when the juice is boiled. As this proteoclastic enzyme, like the alcoholic enzyme, cannot be extracted from the living cells, it is termed yeast endotrypsin or endotryptase. Fresh yeast-juice produces a slow fermentation of sugar, which lasts for forty-eight to ninety-six hours at 25° to 30°, about a week at the ordinary temperature, and then ceases, owing, not to exhaustion of the sugar, but to the disappearance of the fermenting agent. When the juice is preserved or incubated in the absence of a fermentable sugar this disappearance occurs considerably sooner, so that even after standing for a single day at room temperature, or two days at 0°, no fermentation may occur when sugar is added. The reason for this behaviour has not been definitely ascertained. As will be seen later on (p. 64) the phenomenon is a complex one, but the disappearance of the enzyme was originally ascribed by Buchner to the digestive action upon it of the endotrypsin of the juice [1897, 2], and no better explanation has yet been found. Confirmation of this view is afforded by the fact that the addition of a tryptic enzyme of animal origin greatly hastens the disappearance of the alcoholic enzyme [Buchner, E. and H., and Hahn, 1903, p. 126], and that some substances which hinder the tryptic action favour fermentation [Harden, 1903]. The amount of fermentation produced is almost unaffected by the presence of such antiseptics as chloroform or toluene, although some others, such as arsenites and fluorides, decrease it when added in comparatively high concentrations, and it is only slightly diminished by dilution with three or four volumes of sugar solution, somewhat more considerably by dilution with water. When it is filtered through a Chamberland filter the first portions of the filtrate are capable of bringing about fermentation, but the fermenting power diminishes in the succeeding portions and finally disappears. The juice can be spun in a centrifugal machine without being in any way altered, and no separation into more or less active layers takes place under these conditions. [p021]
The amorphous powder obtained by drying the precipitate produced when the juice is added to a mixture of alcohol and ether is also capable of producing fermentation, and the process of precipitation may be repeated without seriously diminishing the fermenting power of the product.
These facts clearly show that the various phenomena adduced by the supporters of the theory of protoplasmic fragments are quite consistent with the presence of a dissolved enzyme as the active agent of the juice, and at the same time that the properties demanded of the living fragments of protoplasm to which fermentation is ascribed are such as cannot be reconciled with our knowledge of living matter. If living protoplasm is the cause of alcoholic fermentation by yeast-juice, a new conception of life will be necessary; the properties of the postulated fragments of protoplasm must be so different from those which the protoplasm of the living cell possesses as to deprive the theory of all real value [Buchner, 1900, 2; Buchner, E. and H., and Hahn, 1903, p. 33].
Further and very convincing evidence against the protoplasm theory is afforded by the behaviour of yeast towards various desiccating agents. When yeast is dried at the ordinary temperature it retains its vitality for a considerable period. If, however, the dried yeast be heated for six hours at 100° it loses the power of growth and reproduction but still retains that of fermenting sugar, and when ground with sand, kieselguhr and 10 per cent. glycerol solution yields an active juice [Buchner, 1897, 2; 1900, 1]. Preparations (known as zymin) obtained by treating yeast with a mixture of alcohol and ether [Albert, 1900, 1901, 1], or with acetone and ether [Albert, Buchner, and Rapp, 1902], show precisely similar properties (p. 38). The proof in this case has been carried a step further, for the active juice obtained by grinding such acetone-yeast, when precipitated with alcohol and ether, yields an amorphous powder, still capable of fermenting sugar.
Buchner's process for the preparation of active yeast-juice is characterised by extreme simplicity. The yeast employed, which should be fresh brewery yeast, is washed two or three times by being suspended in a large amount of water and allowed to settle in deep vessels. It is then collected on a filter cloth, wrapped in a press cloth, and submitted to a pressure of about 50 kilos, per sq. cm. for five minutes. The resulting friable mass contains about 70 per cent. of water and is free from adhering wort. The washed yeast is then [p022] mixed with an equal weight of silver sand and 0·2 to 0·3 parts of kieselguhr, care being taken that this is free from acid. The correct amount of kieselguhr to be added can only be ascertained by experience, and varies with different samples of yeast. The dry powder thus obtained is brought in portions of 300 to 400 grams into a large porcelain mortar and ground by hand by means of a porcelain pestle fastened to a long iron rod which passes through a ring fixed in the wall (Fig. 1). The mortar used by Buchner has a diameter of 40 cm. and the pestle and rod together weigh 8 kilos.
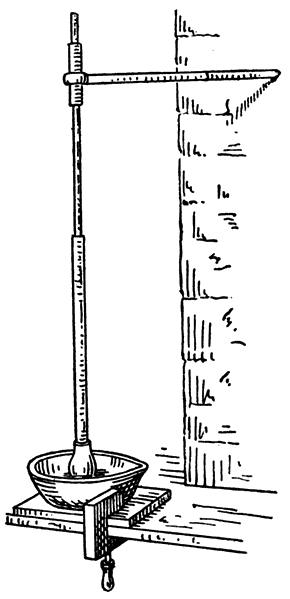
As the grinding proceeds the light-coloured powder gradually darkens and becomes brown, and the mass becomes moist and adheres to the pestle, until finally, after two to three minutes' grinding, it takes the consistency of dough, at which stage the process is stopped. The mass is next enveloped in a press cloth and submitted to a pressure of 90 kilos, per sq. cm. in a hydraulic hand press, the pressure being very gradually raised in order to avoid rupture of the cloth. The cloth required for 1000 grams of yeast measures 60 by 75 cm. and is previously soaked in water and then submitted to a pressure of 50 kilos, per sq. cm., retaining about 35 to 40 c.c. of water.
The juice runs from the press on to a folded filter paper, to remove kieselguhr and yeast cells, and passes into a vessel standing in ice water.
The yield of juice obtained by Buchner in an operation of this kind from 1 kilo. of yeast amounts to 320 to 460 c.c. It may be increased by re-grinding the press cake and again submitting it to pressure, and then amounts on the average to 450 to 500 c.c.
Since the cell membranes constitute about 20 per cent. of the weight of the dry yeast, this yield corresponds to more than 60 per cent. of the total cell contents of the yeast. It has been computed by Will [quoted by Buchner, E. and H., and Hahn, 1903, p. 66] that [p023] only about 20 per cent. of the cells are left unaltered by one grinding and pressing, and only 4 per cent. after a repetition of the process, at least 57 per cent. of the cells being actually ruptured by the double process, and the remainder to some extent altered. It seems probable from these figures that a certain amount of the juice may be derived from the unbroken cells, and Will expressly states that many unbroken cells have lost their vacuoles.
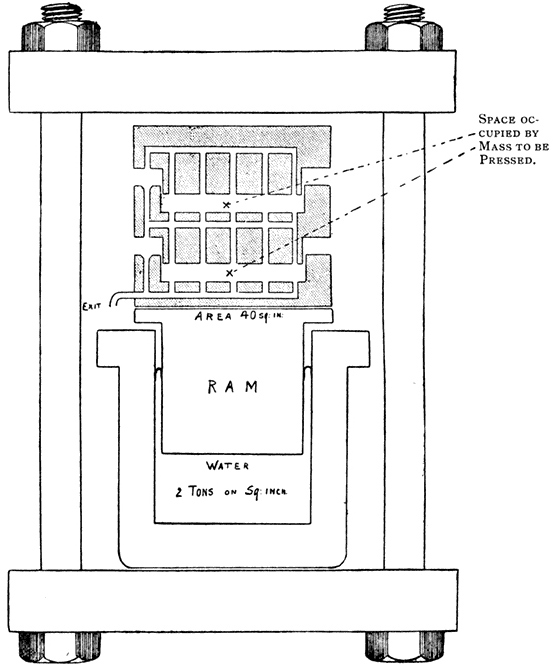
If the yeast be submitted to a process of regeneration, which consists in exposure to a well-aerated solution of sugar and mineral salts until fermentation is complete, the juice subsequently obtained [p024] is more active than that yielded by the original yeast [Albert, 1899, 1].
A modified method of grinding yeast was introduced by Macfadyen, Morris, and Rowland [1900], who placed a mixture of yeast and sand in a jacketed and cooled vessel, in which a spindle carrying brass flanges was rapidly rotated [Rowland, 1901]. One kilo. of yeast ground in this way for 3·5 hours yielded 350 c.c. of juice.
This grinding process was at first adopted by Harden and Young in their experiments but was afterwards abandoned in favour of Buchner's hand-grinding process, as it was found liable to yield juices of low fermenting power, probably on account of inefficient cooling during the grinding process. A slight modification of Buchner's process has, however, been introduced, the hand-ground mass being mixed with a further quantity of kieselguhr until a nearly dry powder is formed, and the mass packed between two layers of chain cloth in steel filter plates and pressed out in a hydraulic press at about 2 tons to the square inch (300 kilos. per sq. cm.). The press and plates are shown in section in Fig. 2. It has also been found convenient to remove yeast cells and kieselguhr from the freshly pressed juice by centrifugalisation instead of by filtration through paper, and to wash the yeast before grinding by means of a filter-press.
Working with English top yeasts Harden and Young have found the yield of juice extremely variable, the general rule being that the amount of juice obtainable from freshly skimmed yeast is smaller than that yielded by the same yeast after standing for a day or two after being skimmed. The yield for 1000 grams of pressed brewer's yeast varies from 150 to 375 c.c., and is on the average about 250 c.c.
Very fresh yeast occasionally presents the peculiar phenomenon that scarcely any juice can be expressed from the ground mass, although the latter does not differ in appearance or consistency from a mass which gives a good yield.
A valuable addition to the methods of obtaining an active solution of zymase was made in 1911 by Lebedeff [1911, 2; 1912, 2; see also 1911, 3, 7, and 1912, 1]. This investigator had been in the habit of grinding dried yeast with water for preparing samples of yeast-juice of uniform character and observed that when the dried yeast was digested with sugar solution and the mixture heated, coagulation [p025] took place throughout the whole liquid, the proteins of the yeast having passed out of the cells. Further examination revealed the interesting fact that dried yeast readily yielded an active extract when macerated in water for some time. The quality of the resulting "maceration extract" depends on a considerable number of factors, the chief of which are: (1) the temperature of drying of the yeast; (2) the temperature of maceration; (3) the duration of maceration; and (4) the nature of the yeast, as well as, of course, the amount of water added in maceration.
In general the yeast should be dried at 25°–30° and then macerated with 3 parts of water for 2 hours at 35°.
The temperature of maceration may as a rule be varied, without detriment to the product provided that the time of maceration is also suitably altered; thus with dried Munich yeast, maceration for 4·5 hours at 25° is about as effective as 2 hours at 35°, whereas treatment for a shorter time at 25° or a longer time at 35° produces in general a less efficacious extract. Yeast dried at a lower temperature than 25° tends to yield an extract poor in co-enzyme (p. 59) and hence of low fermenting power, this being especially marked at air temperature.
The subsequent treatment of the yeast during maceration may, however, be of great influence in such cases. Thus a yeast dried at 15° gave by maceration at 25° for 4·5 hours a weak extract (yielding with excess of sugar 0·33g. CO2), whereas when macerated at 35° for 2 hours it yielded a normal extract (1·36g. CO2).
The nature of the yeast is of paramount importance. Thus while Munich (bottom) yeast usually gives a good result, a top yeast from a Paris brewery was found to yield extracts containing neither zymase nor its co-enzyme in whatever way the preparation was conducted. The existence of such yeasts is of great interest, and it was probably due to the unfortunate selection of such a yeast for his experiments that Pasteur was unable to prepare active fermenting extracts and therefore failed to anticipate Buchner by more than 30 years (see p. 15). The English top yeasts as a rule give poor results [see Dixon and Atkins, 1913] and sometimes yield totally inactive maceration extract. It is not understood why the enzyme passes out of the cell during the process of maceration and the whole method gives rise to a number of extremely interesting problems.
Method.—A suitable yeast is washed by decantation, filtered through a cloth, lightly pressed by means of a hand press, and then passed through a sieve of 5mm. mesh, spread out in a layer 1–1·5cm. thick and left at 25°–35° for two days. Fifty grams of the dried yeast is [p026] thoroughly and carefully mixed with 150 c.c. of water in a basin by means of a spatula and the whole digested for two hours at 35°. The mass often froths considerably. It is then filtered through ordinary folded filter paper, preferably in two portions, and collected in a vessel cooled by ice. The separation may also be effected by centrifuging or pressing out the mass, and the maceration may be conveniently conducted in a flask immersed in the water of a thermostat. It is not advisable to macerate more than 50 grams in one operation. Under these conditions 25–30 c.c. of extract are obtained after 20 minutes' filtration, 70–80 c.c. in twelve hours. Dried Munich yeast can be bought from Messrs. Schroder of Munich and serves as a convenient source of the extract.[1]
This extract closely resembles in properties the juice obtained by grinding the same yeast, but it is usually more active and contains more inorganic phosphate (see p. 46).
Attempts to prepare active extracts from undried yeast in an analogous manner have so far not been very successful. Thus Rinckleben [1911] found that plasmolysis by glycerol (8 per cent.) or sodium phosphate (5 per cent.) sometimes yielded an active juice and sometimes a juice which contained enzyme but no co-enzyme, but more often an inactive juice incapable of activation (p. 64) [see also Kayser, 1911].
Giglioli [1911] by the addition of chloroform also obtained an active liquid. It appears in fact as though almost any method of plasmolysing the yeast cell may yield a certain proportion of zymase in the exudate.
An ingenious process has been devised by Dixon and Atkins [1913] who applied the method of freezing in liquid air which they had found efficacious for obtaining the sap from various plant organs. They thus succeeded in obtaining from yeast, derived from Guinness' brewery in Dublin, liquids capable of fermenting sugar and of about the same efficacy as the maceration extracts prepared by Lebedeff's method from the same yeast. The results were, however, in both cases very low, the maximum total production of CO2 by 25 c.c. of liquid from excess of sugar being 32·5 c.c. (air temperature) or about 0·06g. Munich yeast on the other hand yields, either by maceration or grinding, a liquid giving as much as 1·5–2g. of CO2 per 25 c.c., whilst [p027] English yeast-juice prepared by grinding often gives as much as 0·5–0·7g. of CO2.
No direct comparison with the juice prepared by grinding was made by Dixon and Atkins, but it may be concluded from their results that the best method of obtaining an active preparation from the top yeasts used in this country is that of grinding. Maceration, freezing and plasmolysis alike yield poor results. With Munich yeast on the other hand the maceration process yields excellent results, whilst the liquid air process has not so far been tried.
In order to estimate the amount of carbon dioxide evolved in a given time and the total amount evolved by the action of yeast-juice on sugar, Buchner adopted an extremely simple method, which consisted in carrying out the fermentation in an Erlenmeyer flask provided with a small wash-bottle, which contained sulphuric acid and was closed by a Bunsen valve, and ascertaining the loss of weight during the experiment. Corrections are necessary for the carbon dioxide present in the original juice and retained in the liquid at the close of the experiment and for that present in the air space of the apparatus, but it was found that for most purposes these could be neglected. In cases in which greater accuracy was desired, the carbon dioxide was displaced by air before the weighings were made. A typical experiment of this kind, without displacement of carbon dioxide, is the following:—
| March 22, 1899, Berlin bottom yeast V. 20 c.c. juice + 8 grams cane sugar + 0·2 c.c. toluene as antiseptic at 16°. Grams of carbon dioxide after | ||||
|---|---|---|---|---|
| 24 | 48 | 72 | 96 hours. | |
| 0·40 | 0·64 | 0·99 | 1·11 | |
The total weight of carbon dioxide evolved under these conditions is termed the fermenting power of the juice (Buchner).
A more accurate method [Macfadyen, Morris, and Rowland, 1900] consists in passing the carbon dioxide into caustic soda solution and estimating it by titration. The yeast-juice, sugar, and antiseptic are placed in an Erlenmeyer flask provided with a straight glass tube, through which air can be passed over the surface of the liquid, and a conducting tube leading into a second flask which contains 50 c.c. of 10 per cent. caustic soda solution and is connected with the air by a guard tube containing soda lime. The juice can be freed from carbon dioxide by agitation in a current of air before the flask is connected to [p028] that containing the caustic soda solution, and at the end of the period of incubation air is passed through the apparatus, the liquid being boiled out if great accuracy is required. The absorption flask is then disconnected and the amount of absorbed carbon dioxide estimated by titration. This is carried out by making up the contents of the flask to 200 c.c., taking out an aliquot portion, rendering this exactly neutral to phenophthalein by the addition first of normal and finally of decinormal acid, adding methyl orange and titrating with decinormal acid to exact neutrality. Each c.c. of decinormal acid used in this last titration represents 0·0044 gram of carbon dioxide in the quantity of solution titrated.
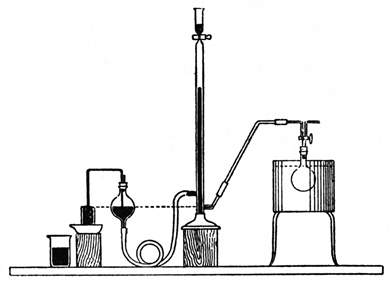
These methods are only suitable for observations at considerable intervals of time. For the continuous observation of the course of fermentation Harden, Thompson and Young [1910] connect the fermentation flask with a Schiff's azotometer filled with mercury and measure the volume of gas evolved, the liquid having been previously saturated with carbon dioxide (Fig. 3). The level of the mercury in the reservoir is kept constant by a syphon overflow, as shown in the figure, or, according to a modification introduced by S. G. Paine, by a specially constructed bottle provided with two tubulures near the bottom. This ensures that no change in the pressure in the flask occurs, and the volume of gas observed is reduced to normal pressure by means of a table. Before making a reading it is necessary to shake the fermenting mixture thoroughly, as the albuminous liquid very readily becomes greatly supersaturated with carbon dioxide, so much so in fact that very little gas is evolved in the intervals between the shakings. The exact procedure in making an observation consists in shaking the flask [p029] thoroughly, replacing in the thermostat, allowing to remain for one minute, and then reading the level of the mercury in the azotometer. After the required time, say five minutes, has elapsed from the time at which the flask was first shaken, it is again removed from the bath, shaken as before, replaced, allowed to remain for one minute and the reading then taken. In this way readings can be conveniently made at intervals of three or five minutes or even less, and much more detailed information obtained about the course of the reaction than is possible by means of observations made at intervals of several hours.
Another form of volumetric apparatus, designed by Walton [1904], has been used by Lebedeff [1909].
An apparatus on a different principle has been designed by Slator [1906] for use with living yeast, but is equally applicable to yeast-juice, and a very similar form has been more recently employed by Iwanoff [1909, 2]. In this apparatus the change of pressure produced by the evolution of carbon dioxide is measured at constant volume, and comparative rates of evolution can be obtained with considerable accuracy, although the method has the disadvantage that the absolute volume of gas evolved is not measured. The apparatus consists of a bottle or flask connected with a mercury manometer. The fermenting mixture is placed in the bottle along with glass beads to facilitate agitation, the pressure is reduced to a small amount by the water-pump, and the rise of pressure is then observed at intervals, this being proportional to the volume of gas produced. As in the preceding case, the liquid must be well shaken before a reading is made.
Yeast-juice brings about a slow fermentation of those sugars which are fermented by the yeast from which it is prepared as well as of dextrin, and of starch and glycogen, which are not fermented by living yeast.
Both in rate of fermentation and in the total fermentation produced, yeast-juice stands far behind the equivalent amount of living yeast. Taking 25 c.c. of yeast-juice to be equivalent to at least 36 grams of pressed yeast containing 70 per cent. of moisture, it is found that whereas the yeast-juice (from English top yeast) gives with glucose a maximum rate of fermentation of about 3 c.c. in five minutes, the living yeast ferments the sugar at the rate of about 126 c.c. in the same time, or [p030] about forty times as quickly. The total carbon dioxide obtainable from the yeast-juice, moreover, corresponds to the fermentation of only 2 to 3 grams of sugar, whilst the living yeast will readily ferment a much larger quantity, although the exact limit in this respect has not been accurately determined. The reasons for this great difference in behaviour will be discussed later on, after the various factors concerned in fermentation have been considered (p. 123).
In all cases of fermentation by yeast-juice and zymin, the relative amounts of carbon dioxide and alcohol produced are substantially in the ratio of the molecular weights of the compounds, that is as 44: 46, so that for 1 part of carbon dioxide 1·04 of alcohol are formed. This has been shown for the juice and zymin from bottom yeasts by Buchner [Buchner, E. and H., and Hahn, 1903, pp. 210, 211], who obtained the ratios 1·01, 0·98, 1·01, and 0·99 from experiments in which from 8 to 15 grams of alcohol were produced. Similar numbers, 0·90, 1·12, 0·95, 0·91 and 0·92, have been obtained for the juice from top yeasts by Harden and Young [1904], who worked with much smaller quantities. The variable results obtained with juice from top yeast by Macfadyen, Morris and Rowland [1900], have not been confirmed.
The construction of a balance-sheet between the sugar fermented and the products formed is of special interest in the case of alcoholic fermentation by yeast-juice, because, there being no cell growth as in the case of living yeast, an opportunity appears to be afforded of ascertaining whether the whole of the sugar is converted into alcohol and carbon dioxide, or whether some fraction of the sugar passes into any of the well-known subsidiary products of alcoholic fermentation by yeast, such as glycerol, fusel oil, or succinic acid. Unfortunately the question cannot be settled in this way. When the loss of sugar during the fermentation is estimated directly, it is usually found to be considerably greater than the sum of the alcohol and carbon dioxide produced from it. This fact was first observed by Macfadyen, Morris and Rowland [1900], and was then confirmed by Buchner [Buchner, E. and H., and Hahn, 1903, p. 212], in one instance, the excess of sugar lost over products being in this case about 15 per cent. of the total sugar which had disappeared. The matter was then more thoroughly investigated by Harden and Young [1904]. [p031]
The conditions under which the experiment must be carried out are not very favourable to the attainment of extreme accuracy. Yeast-juice contains glycogen and a diastatic enzyme which converts this into dextrins and finally into sugar. This process goes on throughout fermentation, tending to increase the sugar present and to make the apparent loss of sugar less than the sum of the products. In spite of this it was found that a certain amount of sugar invariably disappeared without being accounted for as alcohol or carbon dioxide, and this whether the fermentation lasted sixty or a hundred and eight hours, and independently of the dilution of the juice. This disappearing sugar amounted in some cases to 44 per cent. of the total loss of sugar, and on the average of twenty-five experiments was 38 per cent. Further information was sought by converting all the sugar-yielding constituents of the juice into sugar by hydrolysis before and after the fermentation. This process revealed the fact that when the glucose equivalent of the juice before and after fermentation was determined after hydrolysis with three times normal acid for three hours (and a correction made for the loss of reducing power experienced by glucose itself when submitted to this treatment), the difference was almost exactly equal to the alcohol and carbon dioxide produced. In other words, accompanying fermentation, a change proceeds by which sugar is converted into a less reducing substance, reconvertible into sugar by hydrolysis with acids. Similar results were subsequently obtained by Buchner and Meisenheimer [1906], who employed 1·5 normal acid and observed a small nett loss of sugar. Still more recently Lebedeff [1909, 1910, see also 1913, 2] has carried out similar estimations with the same result. It is doubtful whether the experiments which have so far been made on this point are sufficiently accurate to decide with certainty whether or not the loss of sugar is exactly equal to the sum of the carbon dioxide and alcohol produced. It has been shown by Buchner and Meisenheimer [1906] that glycerol is a constant product of alcoholic fermentation by yeast-juice (p. 95), and no other source for this than the sugar has yet been found, so that it is not improbable that a small amount of sugar is converted into non-carbohydrate substances other than carbon dioxide and alcohol.
It has also been shown [Harden and Young, 1913] that the deficit of sugar is not due to the formation of hexosephosphate (p. 47), which has a lower reduction than glucose, and that the solution from which the sugar (either glucose or fructose) has disappeared actually contains some substance of relatively high dextrorotation and of low reducing power. [p032]
However this may be, it may be considered as established that during alcoholic fermentation sugar is converted by an enzyme into some compound of less reducing power, which again yields sugar on hydrolysis with acids. The exact nature of this substance has not been ascertained, but it appears likely that the process is a synthetical one resulting in the formation of some polysaccharide, possibly intermediate between the hexoses and glycogen.
A similar phenomenon has been observed with living yeast by Euler and Johansson [1912, 1], and Euler and Berggren [1912], whose interpretation of the observation is discussed later on (p. 57).
Yeast-juice and zymin ferment all the sugars which are fermented by the yeast from which they are prepared, and, in addition, a number of colloidal substances which cannot pass through the membrane of the living yeast cell, but which are hydrolysed by enzymes in the juice and thus converted into simpler sugars capable of fermentation [Buchner and Rapp, 1898, 3; 1899, 2]. Of the simple sugars which have been examined, glucose, fructose, and mannose are freely fermented, l-arabinose not at all, whilst the case of galactose is doubtful. Galactose is, however, fermented by juice prepared from a yeast which has been "trained" to ferment galactose [Harden and Norris, 1910]. As regards both the rate of fermentation and the total amount of carbon dioxide evolved from glucose and fructose by the action of a definite amount of yeast-juice, Buchner and Rapp obtained practically identical numbers. Harden and Young [1909], using juice from top yeast, found that fructose was slightly more rapidly fermented and gave a somewhat larger total than glucose, whilst mannose was initially fermented at almost the same rate as glucose, but gave a decidedly lower total, the following being the average result:—
| Sugar. | Relative Rates. |
Relative Totals. |
|---|---|---|
| Glucose | 1 | 1 |
| Fructose | 1·29 | 1·15 |
| Mannose | 1·04 | 0·67 |
Among the disaccharides, cane sugar and maltose are freely fermented, and the juice can be shown like living yeast to contain invertase and maltase. The extent of fermentation does not differ materially from that attained with glucose. Lactose is not fermented.
Of the higher sugars raffinose is fermented by juice from bottom yeast, but more slowly than cane sugar or maltose. No experiments seem to have been made with juice from top yeast. [p033]
As regards the fermentation of the higher carbohydrates, very little experimental work has been carried out. Buchner and Rapp found that the fermentation of starch paste was doubtful, but that soluble starch and commercial dextrin were fermented with some freedom. No special study has been made of the diastatic enzymes which bring about the hydrolysis of these substances.
The fermentation of glycogen by yeast-juice is of considerable interest, since it is known that the characteristic reserve carbohydrate of the yeast cell is glycogen [see Harden and Young, 1902, where the literature is cited], and moreover that in living yeast the intracellular fermentation of glycogen proceeds readily, whereas glycogen added to a solution in which yeast is suspended is not affected. Yeast-juice contains a diastatic enzyme which hydrolyses glycogen to a reducing and fermentable sugar, so that in a juice poor in zymase to which glycogen has been added, the amount of sugar is found to increase, the hydrolysis of the glycogen proceeding more quickly than the fermentation of the resulting sugar [Harden and Young, 1904], but the course of this enzymic hydrolysis of glycogen by yeast-juice has not yet been studied. As a rule, it is found both with juices from top and bottom yeast that the evolution of carbon dioxide from glycogen proceeds less rapidly and reaches a lower total than from an equivalent amount of glucose.
Since nearly all samples of yeast contain glycogen, yeast-juice and also zymin usually contain this substance as well as the products of its hydrolysis. These provide a source of sugar which enters into alcoholic fermentation, so that a slow spontaneous production of carbon dioxide and alcohol proceeds when yeast-juice is preserved without any addition of sugar. The extent of this autofermentation varies considerably, as might be expected, with the nature of the yeast employed or the preparation of the material, but is generally confined within the limits of 0·06 to 0·5 gram of carbon dioxide for 25 c.c. of juice.
In juice from bottom yeast it amounts to about 5 to 10 per cent. of the total fermentation obtainable with glucose [Buchner, 1900, 2], whereas in juice from top yeasts, which gives a smaller total fermentation with glucose, it may occasionally equal, or even exceed, the glucose fermentation, and frequently amounts to 30 to 50 per cent. of it. It is therefore generally advisable in studying the effect of yeast-juice on any particular substance to ascertain the extent of autofermentation by means of a parallel experiment.
The maceration extract of Lebedeff (p. 24) is usually, but not invariably [Oppenheimer, 1914, 2], free from glycogen, which is hydrolysed [p034] and fermented during the processes of drying and macerating, and therefore as a rule shows no appreciable autofermentation.
The kinetics of fermentation by zymase will be considered later on (p. 120), but the effect on the total fermentation of different concentrations of sugar, this substance being present throughout in considerable excess, may be advantageously discussed at this stage. The subject has been investigated by Buchner [Buchner, E. and H., and Hahn, 1903, pp. 150–8; Buchner and Rapp, 1897] using cane sugar, and he has found both for yeast-juice and for dried yeast-juice dissolved in water that (a) the total amount of fermentation increases with the concentration of the sugar; (b) the initial rate of fermentation decreases with the concentration of the sugar. The following are the results of a typical experiment, 20 c.c. of yeast-juice being employed in presence of toluene at 22°:—
| Cane Sugar. | CO2 in grams after | |||
|---|---|---|---|---|
| Weight. | Per cent. | 6 hours. | 24 hours. | 96 hours. |
| 2·2 | 10 | 0·17 | 0·50 | 0·55 |
| 3·52 | 15 | 0·14 | 0·53 | 0·64 |
| 5 | 20 | 0·13 | 0·54 | 0·73 |
| 6·66 | 25 | 0·13 | 0·52 | 0·80 |
| 8·56 | 30 | 0·12 | 0·46 | 0·81 |
| 10·76 | 35 | 0·12 | 0·40 | 0·82 |
| 13·33 | 40 | 0·11 | 0·36 | 0·82 |
The results as to the total fermentations in experiments of this kind are liable to be vitiated by the circumstance that when a low initial concentration of sugar is employed, the supply of sugar may be so greatly exhausted before the close of the experiment as to cause a marked diminution in the rate of fermentation and hence an unduly low total. Even allowing, however, for any effect of this kind, the foregoing table clearly shows the increase in total fermentation and the decrease in initial rate accompanying the increase of sugar concentration from 10 to 40 per cent. Working with a greater range of concentrations (3·3–53·3 grm. per 100 c.c.) Lebedeff has obtained similar results with maceration extract [1911, 4], but has found that the total amount fermented diminishes after a certain optimum concentration (about 33·3 grm. per 100 c.c.) is reached.
A practical conclusion from these experiments is that a high [p035] concentration of sugar tends to preserve the enzyme in an active state for a longer time. Simultaneously it prevents the development of bacteria and yeast cells.
This subject, which is of considerable importance with reference to the question of the protoplasmic or enzymic nature of the active agent in yeast-juice, has been examined in some detail by Buchner [Buchner, E. and H., and Hahn, 1903, pp. 158–65] and by Meisenheimer [1903] for juices from bottom yeast, by Harden and Young [1904] for those from top yeast, and by Lebedeff [1911, 4] for maceration extract, the results obtained being in substantial agreement.
Dilution of yeast-juice with sugar solution, so that the concentration of the sugar remains constant, produces a small progressive diminution in the total fermentation, which only becomes marked when more than 2 volumes are added, and this independently of the actual concentration of the sugar. Dilution with water produces a somewhat more decided diminution, which, however, does not exceed 50 per cent. of the total for the addition of 3 volumes of water. The effect on maceration extract is somewhat greater but of the same kind. The autofermentation of juice from top yeast is scarcely affected by dilution with 4 volumes of water.
| Nature of Juice. |
Per cent. of Sugar Employed by Weight. |
Volumes of Sugar Solution Added. |
Volumes of Water Added. |
Total Fermentation in g. of CO2. |
|
|---|---|---|---|---|---|
| Bottom Yeast |
1 | 29 | 0 | — | 0·99 |
| 1 | — | 1·13 | |||
| 2 | — | 0·92 | |||
| 4 | — | 0·79 | |||
| 2 | 9 | 0 | — | 0·43 | |
| 1 | — | 0·60 | |||
| 2 | — | 0·53 | |||
| 4 | — | 0·41 | |||
| 3 | 9 | — | 0 | 0·46 | |
| — | 1 | 0·32 | |||
| — | 2 | 0·33 | |||
| — | 3 | 0·36 | |||
| Top Yeast |
1 | 0 (Auto- ferment- ation) |
— | 0 | 0·29 |
| — | 2 | 0·29 | |||
| — | 3 | 0·28 | |||
| 2 | 29 | 0 | — | 0·31 | |
| 1 | — | 0·34 | |||
| 2 | — | 0·31 | |||
| 4 | — | 0·35 | |||
| 6 | — | 0·28 | |||
| 3 | 7·4 | — | 0 | 0·44 | |
| — | 1 | 0·35 | |||
| — | 2 | 0·30 | |||
| — | 3 | 0·28 | |||
On the whole, therefore, yeast-juice may be said to be only slightly affected by dilution even with pure water, and the effect of the latter can in no way be regarded as comparable with the poisonous effect which it exerts on living protoplasm, as suggested by Macfadyen, Morris, and Rowland [1900].
Buchner has paid special attention to the effect of antiseptics on the course of fermentation by yeast-juice [Buchner and Rapp, 1897; 1898, 2, 3; 1899, 1; Buchner and Antoni, 1905, 1; Buchner and Hoffmann, 1907; Buchner, E. and H., and Hahn, 1903, pp. 169–205; see also Albert, 1899, 2; Gromoff and Grigorieff, 1904; Duchaček, 1909] in order (1) to obtain evidence as to the possibility of the active agent in yeast-juice consisting of fragments of protoplasm and not of a soluble enzyme, and (2) also to provide a safe method of avoiding contamination, by the growth of bacteria or yeasts, of the liquids used which were often kept at 25° for several days. The results of these experiments are briefly summarised in the following table, in which the effect of each substance on the total fermentation produced is noted:—
| Substance. | Effect on Total Fermentation. |
|
|---|---|---|
| Concentrated solution of glycerol | Slight diminution | |
| Concentrated solution of sugar | Slight increase | |
| Toluene (to saturation or excess) | Less than 10 per cent. diminution | |
| Chloroform | 0·5 per cent. | Slight increase |
| 0·8 per cent. (saturation) | No change | |
| Large excess (17 per cent.) | 64 per cent. diminution | |
| Chloral hydrate | 0·7 per cent. | Increase up to 27 per cent. |
| 3·5–5·4 per cent. | Completely destroyed | |
| Phenol | 0·1 per cent. | No change |
| 0·5 " | 40 per cent. diminution | |
| 1·2 " | Completely destroyed | |
| Thymol | 1 " | Slight diminution |
| 5 " | Marked " | |
| Benzoic acid | 0·1 " | 7 per cent. diminution |
| 0·25 " | 26 " | |
| Salicylic acid | 0·1 " | 10 " |
| 0·27 " | 35 " | |
| Formaldehyde | 0·12 " | 20 " |
| 0·24 " | 30–60 " | |
| Acetone | 6 " | 20 " |
| 14 " | 80 " | |
| Alcohol | 6 " | 0–20 " |
| 14 " | 75 " | |
| Sodium fluoride | 0·5 " | 90 " |
| 2 " | Almost completely destroyed | |
| Ammonium fluoride | 0·55 per cent. | Completely destroyed |
| Sodium azoimide, NaN3, | 0·36 per cent. | Slight diminution |
| 0·71 " | Marked " | |
| Quinine hydrochloride | 1 " | Slight increase |
| Ozone | 10·4–34·8 mgs. per 20 c.c. | Marked diminution |
| Hydrocyanic acid | 1·2 per cent. | Completely destroyed |
The general result of these experiments is to show that quantities of antiseptics which are sufficient to inhibit the characteristic action of living cells have only a slight effect on the fermentative activity of yeast-juice. A large excess of the antiseptic in many cases produces a very decided diminution or total destruction of the fermenting power, and accompanying this a precipitation of the constituents of the juice. The decided increase of activity produced by small quantities of chloral hydrate, and to a less marked extent by chloroform and a few other substances, is of considerable interest. It is ascribed by Duchaček to a selective action on the proteoclastic enzyme, but without satisfactory evidence.
Hydrocyanic acid, even in dilute solution, completely suspends the fermenting power of the juice, without, however, producing any permanent change in the fermenting complex, as is shown by the fact that when the hydrocyanic acid is removed by a current of air, the juice regains its fermenting power. In this respect hydrocyanic acid behaves precisely as with many other enzymes and with colloidal platinum [Bredig, 1901]. Sodium arsenite is a pronounced protoplasmic poison, which rapidly destroys the power of growth and reproduction in living cells, and was therefore applied to yeast-juice to differentiate between protoplasmic and enzymic action. It was, however, found that the action of this substance was complicated by some unknown factor and very irregular results were obtained [Buchner, E. and H., and Hahn, 1903, pp. 193 ff.]. These phenomena appear to be of the same order as those produced by the addition of arsenates to yeast-juice [Harden and Young, 1906, 3], and will be discussed along with the latter (p. 77).
A considerable number of preparations have been obtained in the dry state which retain some proportion of the fermenting power of yeast or yeast-juice.
Starting with yeast-juice, it is possible to arrive at this result either by evaporation or precipitation. When the juice is very rapidly evaporated to a syrup at 20° to 25° and then further dried at 35°, either in the air or in a vacuum and finally exposed over sulphuric acid in a vacuum desiccator, a dry brittle mass is obtained which is soluble in water and retains practically the whole of the fermenting power of the juice. The success of the preparation depends on the nature of the yeast from which the juice is derived, Berlin yeasts V and S yielding much less satisfactory results than Munich yeast. The powder when [p038] thoroughly dry is found to retain its properties almost unimpaired for at least a year, and can be heated to 85° for eight hours without undergoing any serious loss of fermenting power [Buchner and Rapp, 1898, 4; 1901; Buchner, E. and H., and Hahn, 1903, pp. 132–9].
Active powders can also be obtained by precipitating yeast-juice with alcohol, alcohol and ether, or acetone. The preparation is best effected by bringing the juice into 10 volumes of acetone, centrifuging at once and as rapidly as possible, washing, first with acetone and then with ether, and finally drying over sulphuric acid. The white powder thus obtained is not completely soluble in water but is almost entirely dissolved by aqueous glycerol (2·5 to 20 per cent.), forming a solution which has practically the same fermenting power as the original juice. The precipitation can be repeated without any serious loss of fermenting power. Prolonged contact of the precipitate with the supernatant liquid, especially when alcohol or alcohol and ether are used, causes a rapid loss of the characteristic property [Albert and Buchner, 1900, 1, 2; Buchner, E. and H., and Hahn, 1903, pp. 228–246; Buchner and Duchaček, 1909].
Dry preparations capable of fermenting sugar can also be readily obtained from yeast without any preliminary rupture of the cells. Heat alone (yielding a product known as hefanol) or treatment with dehydrating agents may be used for this purpose, and a brief allusion has already been made (p. 21) to the different varieties of permanent yeast (Dauerhefe) obtainable in these ways. The most important of these products are the dried Munich yeast (Lebedeff, see p. 25), and the material known as zymin, which is now made under patent rights for medicinal purposes by Schroder of Munich. The latter has proved of value in the investigation of the production of zymase in the yeast cell [Buchner and Spitta, 1902], and of many other problems concerned with alcoholic fermentation. In order to prepare it 500 grams of finely divided pressed brewer's yeast, containing about 70 per cent. of water, are brought into 3 litres of acetone, stirred for ten minutes, and filtered and drained at the pump. The mass is then well mixed with 1 litre of acetone for two minutes and again filtered and drained. The residue is roughly powdered, well kneaded with 250 c.c. of ether for three minutes, filtered, drained, and spread on filter paper or porous plates. After standing for an hour in the air it is dried at 45° for twenty-four hours. About 150 grams of an almost white powder containing only 5·5 to 6·5 per cent. of water are obtained. This is quite incapable of growth or reproduction but produces a very considerable amount of alcoholic fermentation, far greater indeed than a corresponding [p039] quantity of yeast-juice. Two grams of the powder corresponding to 6 grams of yeast and about 3·5 to 4 c.c. of yeast-juice, are capable of fermenting about 2 grams of sugar, whereas the 4 c.c. of yeast-juice would on the average only ferment from one-quarter to one-sixth of this amount of sugar. The rate produced by this amount of zymin is about one-eighth of that given by the corresponding amount of living yeast [Albert, 1900; Albert, Buchner, and Rapp, 1902]. The proteoclastic ferment is still present in zymin, which undergoes autolysis in presence of water in a similar manner to yeast-juice [Albert, 1901, 2].
As already mentioned an active juice can be prepared by grinding acetone-yeast with water, sand, and kieselguhr, and this process presents the advantage that samples of yeast-juice of approximately constant composition can be prepared at intervals from successive portions of a uniform supply of acetone-yeast.
Preparations of acetone-yeast, made from yeast freed from glycogen by exposure in a thin layer to the air for three or four hours at 35° to 45°, or eight hours at the ordinary temperature [Buchner and Mitscherlich, 1904], show practically no autofermentation and may be used analytically for the estimation of fermentable sugars.
All the foregoing preparations exhibit the same general properties as yeast-juice, as regards their behaviour towards the various sugars, antiseptics, etc.
When zymin is mixed with sugar solution without being previously ground, it exhibits a peculiarity which is of some practical interest. The time which elapses before the normal rate of fermentation is attained and the total fermentation obtainable vary with the amount of sugar solution added, the time increasing and the total diminishing as the quantity of this increases. This phenomenon appears to have been noticed by Trommsdorff [1902], and a single experiment of Buchner shows the influence of the same conditions [Buchner, E. and H., and Hahn, 1903, p. 265, Nos. 700–1]. Harden and Young have found that when 2 grams of zymin are mixed with varying quantities of 10 per cent. sugar solution the following results are obtained:—
| Volumes of Sugar Solution |
Total Gas Evolved in | ||||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 22·5 hours. |
|
| 5 c.c. | 15·7 | 31·6 | 44·8 | 56·5 | 233·3 |
| 10 | 2·2 | 10·5 | 23 | 31·8 | 202·3 |
| 20 | 0·9 | 2·4 | 13·6 | 23·7 | 125·5 |
| 40 | 1·4 | 1·7 | 2·3 | 2·9 | 56·3 |
This behaviour appears to be due to the removal of soluble matter essential for fermentation from the cell, which is discussed later on. It follows that when zymin is being tested for fermenting power, a uniform method should be adopted, and all comparative tests should be made with the same volumes of added sugar solution. Ground zymin appears to begin to ferment somewhat more slowly than unground (2 grm. to 12·4 c.c. of sugar solution in each case), but eventually produces the same total volume of gas [Buchner and Antoni, 1905, 1].
In the course of some preliminary experiments (commenced by the late Allan Macfadyen, but subsequently abandoned) on the production of anti-ferments by the injection of yeast-juice into animals, the serum of the treated animals was tested for the presence of such antibodies both for the alcoholic and proteoclastic enzymes of yeast-juice, and it was then observed that the serum of normal and of treated animals alike greatly diminished the autolysis of yeast-juice.
As the explanation of the comparatively rapid disappearance of the fermenting power from yeast-juice had been sought, as already mentioned (p. 20), in the hydrolytic action of the tryptic enzyme which always accompanies zymase, the experiment was made of carrying out the fermentation in the presence of serum, with the result that about 60 to 80 per cent. more sugar was fermented than in the absence of the serum [Harden, 1903].
This fact was the starting-point of a series of attempts to obtain a similar effect by different means, in the course of which a boiled and filtered solution of autolysed yeast-juice was used, in the hope that the products formed by the action of the tryptic enzyme on the proteins of the juice would, in accordance with the general rule, prove to be an effective inhibitant of that enzyme. This solution was, in fact, found to produce a very marked increase in the total fermentation effected by yeast-juice, the addition of a volume of boiled juice equal to that of the yeast-juice doubling the amount of carbon dioxide evolved [Harden and Young, 1905, 1]. This effect was found to be common to the filtrates from boiled fresh yeast-juice and from boiled autolysed yeast-juice, and was ultimately traced in the main, not to the antitryptic effect which had been surmised, but to two independent factors, either of which was capable in some degree of bringing about the observed result.
Boiled yeast-juice was indeed found to possess a decided anti-autolytic effect, as determined by a comparison of the amounts of nitrogen rendered non-precipitable by tannic acid in yeast-juice alone [p042] and in a mixture of yeast-juice and boiled juice on preservation [Harden, 1905]. The anti-autolytic effect, however, appeared to vary independently of the effect on the fermentation, and the conclusion was drawn, as stated above, that the increase in the alcoholic fermentation was not directly dependent on the decrease in the action of the proteoclastic enzyme but was due to some independent cause. The property possessed by boiled yeast-juice of diminishing the autolysis of yeast-juice has now been carefully examined by Buchner and Haehn [1910, 2] and ascribed by them to a soluble antiprotease (p. 65).
The two factors to which the increase in fermentation produced by the addition of boiled juice were ultimately traced were (1) the presence of phosphates in the liquid, and (2) the existence in boiled fresh yeast-juice of a co-ferment or co-enzyme, the presence of which is indispensable for fermentation [Harden and Young, 1905, 1, 2].
The former of these factors will be here discussed and the co-enzyme will form the subject of the following chapter.
The general fact that sodium phosphate increases the total fermentation produced by a given volume of yeast juice was observed on several occasions by Wroblewski [1901] and also by Buchner [Buchner, E. and H., and Hahn, 1903, pp. 141–2], who ascribed the action of this salt to its alkalinity, comparing it in this respect with potassium carbonate and remarking that the increase in both cases took place chiefly in the first twenty hours of fermentation. The increased amount of fermentation following the addition of boiled yeast-juice was also noted by Buchner and Rapp [1899, 2, No. 265, p. 2093] in a single experiment.
Observations made at intervals of a few minutes instead of twenty hours have, however, revealed the fact that phosphates play a part of fundamental importance in alcoholic fermentation and that their presence is absolutely essential for the production of the phenomenon.
When a suitable quantity[2] of a soluble phosphate is added to a fermenting mixture of glucose, fructose, or mannose with yeast-juice, the rate of fermentation rapidly rises, sometimes increasing as much as twenty-fold, continues at this high value for a certain period and then falls again to a value approximately equal to, but generally [p043] somewhat higher than, that which it originally had. Careful experiments have shown that during this period of enhanced fermentation the amounts of carbon dioxide and alcohol produced exceed those which would have been formed in the absence of added phosphate by a quantity exactly equivalent to the phosphate added in the ratio CO2 or C2H6O:R′2HPO4 [Harden and Young, 1906, 1].
This result is of fundamental importance, and the evidence on which it rests deserves some consideration. Quantitative experiments on this subject require certain preliminary precautions. The acid phosphates are too acid to permit of any extended fermentation and the phosphates of the formula R′2HPO4 absorb a considerable volume of carbon dioxide with production of a bicarbonate, according to the reaction:—
The method which has been adopted, therefore, is to employ either a secondary phosphate saturated with carbon dioxide at the temperature of the experiment, or a mixture of five molecular proportions of the secondary phosphate with one molecular proportion of a primary phosphate, in which the amount of bicarbonate formed is negligible. In the former case it is necessary to ascertain whether any of the carbon dioxide evolved is derived from the bicarbonate by the action of acid originally present or produced in the yeast-juice or by a disturbance of the original equilibrium owing to the chemical change which occurs. This is done by acidifying duplicate samples with hydrochloric acid before and after the fermentation and measuring the gas evolved in each case. Any necessary correction can then be made. The calculation of the extra amount of carbon dioxide evolved from yeast-juice containing sugar when a phosphate is added involves an estimation of the amount which would have been evolved in the absence of added phosphate, and this is a matter of some difficulty. Since the final steady rate of fermentation attained is often slightly different from the initial rate, the practice has been adopted of ascertaining this final rate and then calculating the total evolution corresponding to it for the whole period from the time of the addition of the phosphate to the end of the observations. This amount deducted from the observed total leaves the extra amount of carbon dioxide formed, and it is this quantity which is equivalent to the phosphate added. Alcohol is simultaneously produced in the normal ratio. The justification for this method of calculation will be found later (p. 54).
The following table, containing the results of experiments with [p044] glucose, fructose, and mannose, indicates very clearly the nature of the method of calculation and also of the agreement between observation and theory.
Three quantities of 25 c.c. of yeast-juice + 5 c.c. of a solution containing 1 gram of the sugar to be examined (a large excess) were incubated with toluene at 25° for one hour, in order to remove all free phosphate, and to each were then added 5 c.c. of a solution of sodium phosphate corresponding to 0·1632 gram of Mg2P2O7 and equivalent to 32·6 c.c. of carbon dioxide at N.T.P. The rates of fermentation were then observed until they had passed through the period of acceleration and had fallen and attained a steady value, the gases being measured moist at 19·3° and 760·15 mm.
| Glucose. | Mannose. | Fructose. | |
|---|---|---|---|
| Maximum rate attained, c.cs. per five minutes | 9·6 | 7 | 11·3 |
| Final rate of fermentation | 1·1 | 0·96 | 1·08 |
| Total carbon dioxide produced by fermentation in fifty-five minutes after addition of phosphate | 49·7 | 47·8 | 47·6 |
| Correction for evolution in absence of phosphate in fifty-five minutes | 12·1 | 10·6 | 11·9 |
| Extra carbon dioxide equivalent to phosphate | 37·6 | 37·2 | 35·7 |
| Extra carbon dioxide equivalent to phosphate at N.T.P. | 34·4 | 34 | 32·6 |
These numbers agree well with the value calculated from the phosphate added, viz. 32·6 [Harden and Young, 1909].
Another experiment is illustrated graphically in Fig. 4, in which the volume of carbon dioxide evolved is plotted against time. The determination was in this case made by adding 25 c.c. of an aqueous solution containing 5 grams of glucose to one quantity of 25 c.c. of yeast-juice (curve A) and 5 c.c. of 0·3 molar solution of the mixed primary and secondary sodium phosphates, and 20 c.c. of a solution containing 5 grams of glucose to a second equal quantity of yeast-juice (curve B). Curve A shows the normal course of fermentation of yeast-juice with glucose. There is a slight preliminary acceleration during the first twenty minutes, due to free phosphate in the juice, and the rate then becomes steady at about 1·4 c.c. in five minutes. During this preliminary acceleration 10 c.c. of extra carbon dioxide are evolved, this number being obtained graphically by continuing the line of steady rate back to the axis of zero time. Curve B shows the effect of the added phosphate. The rate rises to about 9·5 c.c. in five minutes, i.e. to more than six times the normal rate, and then gradually falls until after an hour it is again steady and almost exactly equal to 1·4 c.c. per five minutes. Continuing the line of steady rate back to the axis of zero [p045] time it is found that the extra amount of carbon dioxide is 48 c.c. Subtracting from this the 10 c.c. shown in curve A as due to the juice alone, a difference of 38 c.c. is obtained due to the added phosphate. The amount calculated from the phosphate added in this case is, at atmospheric temperature and pressure, 38·9 c.c.
After the expiration of seventy minutes from the commencement of the experiment, a second addition is made of an equal amount of phosphate. The whole phenomenon then recurs, as shown in curve C, the maximum rate being slightly lower than before, about 6 c.c. per five minutes, and the rate again becoming finally steady at 1·4 c.c. as before. The extra amount of carbon dioxide evolved in this second period obtained graphically as in the former case, is 107–68 = 39 c.c.
It may be noted that in this case the observations after each addition last fifty to seventy minutes, so that an error of 0·1 c.c. per five minutes in the estimated final rate would make an error of 1 to 1·4 c.c. in the extra amount of carbon dioxide, i.e. about 3 to 4 per cent. of the total, and this is approximately the limit of accuracy of the method. [p046] The results are more precise when the yeast-juice employed is an active one, since, when the fermenting power of the juice is low, the initial period of accelerated fermentation is unduly prolonged and the calculation of the extra amount of carbon dioxide is rendered uncertain.
Zymin (p. 38) yields precisely similar results to yeast-juice, but in this case the rate of fermentation is not so largely increased. This has the effect that the extra amount of carbon dioxide cannot be quite so accurately estimated for zymin, because a slight error in the determination of the final rate of fermentation has a greater influence on the result. The equivalence between the extra amount of carbon dioxide evolved and the phosphate added is, however, unmistakable, as is shown by the following results of an experiment with zymin, in which 6 grams of zymin (Schroder) + 3 grams of fructose (Schering) + 25 c.c. of water were incubated at 25° in presence of toluene until a steady rate had been attained. Five c.c. of a solution of sodium phosphate equivalent to 32·2 c.c. carbon dioxide at N.T.P. were then added.
| Maximum rate attained, c.c. per five minutes | 14·1 |
| Final rate of fermentation | 6·2 |
| Total evolved by fermentation in eighty minutes after addition of phosphate | 131 |
| Correction for evolution in absence of phosphate in eighty minutes | 99·2 |
| Extra carbon dioxide at 16° and 767·1 mm | 31·8 |
| Extra carbon dioxide at N.T.P | 29·8 |
Considering the small proportional rise in rate and the long period of accelerated fermentation, the agreement between the volume observed, 29·8 c.c., and that calculated from the phosphate, 32·2, is quite satisfactory [Harden and Young, 1910, 1.] Precisely the same relations hold for maceration extract, but in this case it must be remembered that a large amount of free phosphate is present in the extract, as much as 0·3129 grm. Mg2P2O7 being obtained from 20 c.c. in one preparation, so that the original extract had the concentration of a 0·14 molar solution of sodium phosphate. It is in fact not improbable that the delay in the onset of fermentation sometimes observed with maceration extract (see Lebedeff, 1912, 2; Neuberg and Rosenthal, 1913) may be due to the presence of phosphate in so great an excess of the amount which can be rapidly esterified by the enzymes that the rate of fermentation is at first greatly lowered (see p. 71). When this phosphate is removed by incubation with glucose or fructose, the subsequent addition of phosphate produces the characteristic action and the extra carbon dioxide evolved is, as with other yeast preparations, equivalent to the phosphate added. An actual estimation carried out in this way gave 35 c.c. of CO2 for an addition of phosphate equivalent to 32·9 c.c. [Harden and Young, 1912]. [p047]
Within the limits imposed by the experimental conditions, then, the fact is well established that the addition of a soluble phosphate to a fermenting mixture of a hexose with yeast-juice, maceration extract, dried yeast, or zymin causes the production of an equivalent amount of carbon dioxide and alcohol.
This fact indicates that a definite chemical reaction occurs in which sugar and phosphate are concerned, and this conclusion is confirmed when the fate of the added phosphate is investigated. If an experiment, such as one of those described above, be interrupted as soon as the rate of fermentation has again become normal, and the liquid be boiled and filtered, it is found that nearly the whole of the phosphorus present passes into the filtrate, but that only a small proportion of this exists as mineral phosphate, whilst the remainder, including that added in the form of a soluble phosphate, is no longer precipitable by magnesium citrate mixture [Harden and Young, 1905, 2].
A similar observation was made at a later date by Iwanoff [1907], who had previously observed [1905] that living yeast, like many other vegetable organisms, converted mineral phosphates into organic derivatives. Iwanoff employed zymin and hefanol (p. 38) instead of yeast-juice, and found that phosphates were thereby rendered non-precipitable by uranium acetate solution, but did not observe the accelerated fermentation caused by their addition.
The foregoing conclusions have been strikingly confirmed by experiments with maceration extract carried out by Euler and Johansson [1913], in which both the carbon dioxide evolved and the phosphate rendered non-precipitable by magnesia were determined at intervals. When dried yeast is employed as the fermenting agent, the amount of phosphate esterified in the earlier stages is greater than would be expected, but ultimately becomes exactly equivalent to the carbon dioxide evolved.
The formation and properties of the compound produced from phosphates in the manner just described have been investigated by Harden and Young [1905, 2; 1908, 1; 1909; 1911, 2], Young [1909; 1911], Iwanoff [1907; 1909, 1], Lebedeff [1909; 1910; 1911, 5, 6; 1912, 3; 1913, 1]; and Euler [1912, 1; Euler and Fodor, 1911; Euler and Kullberg, 1911, 3; Euler and Ohlsén, 1911; 1912; Euler and Johansson, 1912, 4; Euler and Bäckström, 1912], but its exact constitution cannot as yet be regarded as definitely known. [p048]
Phosphates undergo this characteristic change when the sugar undergoing fermentation is glucose, mannose, or fructose, and it may be said at once that no distinction can be established between the products formed from these various hexoses; they all appear to be identical. The compound produced is, as already mentioned, not precipitated by ammoniacal magnesium citrate mixture, nor by uranium acetate solution. It can, however, be precipitated by copper acetate (Iwanoff) and by lead acetate (Young). The preparation of the pure lead salt from the liquid obtained by fermenting a sugar with yeast-juice or zymin in presence of phosphate is commenced by boiling and filtering the liquid. Magnesium nitrate solution and a small quantity of caustic soda solution are then added to precipitate any free phosphate, and the liquid well stirred and allowed to stand over night. To the neutralised filtrate lead acetate is then added together with sufficient caustic soda solution to maintain the reaction neutral to litmus, until no further precipitate is formed. The liquid is then filtered or, better, centrifugalised, and the precipitate repeatedly washed with water until a portion of the clear filtrate gives no reduction when boiled with Fehling's solution. It is essential that this washing should be thorough as evidence has recently been obtained of the formation under certain conditions of a hexosephosphate, the lead salt of which is not so sparingly soluble as that of the hexosediphosphate [Harden and Robison, 1914]. The lead precipitate is then suspended in water, decomposed by a current of sulphuretted hydrogen, the clear filtrate freed from sulphuretted hydrogen by a current of air, and finally neutralised with caustic soda. The removal of phosphate and conversion into lead salt are repeated twice, and the resulting lead salt is then found to be free from nitrogen and to have a composition represented by the formula C6H10O4(PO4Pb)2. Lebedeff carries out the preparation in a somewhat different manner. The fermentation is effected by means of air-dried yeast (150 grams to 1 litre of water, 210 grams cane-sugar and 105 grams of a mixture of 2 parts Na2HPO4 and 1 part NaH2PO4) and the liquid (about 700 c.c.) after boiling and filtering, is treated with an equal volume of acetone. About 300 c.c. of a thick liquid is precipitated and this is redissolved in water and precipitated by an equal volume of acetone two or three times. The final liquid is then precipitated with warm lead acetate solution and filtered and washed with dilute lead acetate solution until the filtrate is clear and no longer reduces Fehling's solution after removal of the lead [1910]. Euler and Fodor [1911] on the other hand precipitate the free phosphate with magnesia mixture and then add acetone, dissolve the syrup thus precipitated in water and add copper [p049] acetate solution. A blue copper salt is precipitated which is thoroughly washed with water and used for the preparation of solutions of the acid. A solution of the free acid can readily be prepared by the action of sulphuretted hydrogen on the lead salt suspended in water. It forms a strongly acid liquid, which requires exactly two equivalents of base for each atom of phosphorus present to render it neutral to phenolphthalein. It decomposes when evaporated, leaving a charred mass containing free phosphoric acid. The acid is slightly optically active, and has [aD] = + 3·4°. A number of amorphous salts have been prepared by precipitation from a solution of the sodium salt, and of these the silver, barium, and calcium salts have been analysed with results agreeing with the general formula C6H10O4(PO4R′2)2. The magnesium, calcium, barium, and manganese salts, which are only sparingly soluble, are all precipitated when their solutions are boiled but re-dissolve on cooling, and this property can be utilised for their purification. The alkali salts have only been obtained as viscid residues.
A difference of opinion exists as to the molecular weight and constitution of this substance. Iwanoff [1909, 1] regards it as a triosephosphoric acid, C3H5O2(PO4H2), basing this view on the preparation of an osazone which melted at 142°, but when recrystallised from benzene gave a product melting at 127°–8°, which had the same appearance, melting-point, and nitrogen content as the triosazone formed by the action of phenylhydrazine on the oxidation products of glycerol. Neither Lebedeff [1909] nor Young could obtain Iwanoff's osazone, and all attempts to reduce the acid with formation of glycerol either by sodium amalgam or hydriodic acid were unsuccessful (Young). There is therefore practically no serious experimental evidence in favour of Iwanoff's view.
On the other hand, Harden and Young regard the acid as a diphosphoric ester of a hexose. This view is based on the fact that when the acid is boiled with water, or an acid, free phosphoric acid is produced along with a levo-rotatory solution containing fructose and possibly a small proportion of some other sugar or sugars. (Euler and Fodor however did not obtain a hexose in this way [1911].) The acid itself only reduces Fehling's solution after some hours in the cold, rapidly when boiled, whereas when its solution is first boiled, and then treated with Fehling's solution in the cold, the products of decomposition bring about reduction in a few minutes. The reduction brought about when the acid is boiled with Fehling's solution is considerably less (33 per cent.) than that produced by an equivalent amount of glucose. The behaviour of the compound towards phenylhydrazine is also in complete agreement [p050] with this view. Lebedeff found [1909, 1910] that the acid or its salts heated with phenylhydrazine in presence of acetic acid gave an insoluble compound which was ultimately found to be the phenylhydrazine salt of hexosemonophosphoric acid osazone
[Lebedeff, 1910; 1911, 6; Young, 1911]. After recrystallisation from alcohol this compound forms yellow needles, melting at 151°–152°. It is decomposed by caustic soda yielding a sodium salt
and on boiling with caustic soda decomposes giving a hexosazone (free from phosphorus) which is probably glucosazone, and in addition glyoxalosazone, probably as the result of a secondary decomposition. Towards acids it is remarkably stable yielding with hydrochloric acid a hexosonephosphoric ester from which the original osazone can be regenerated (Lebedeff). Lebedeff at first [1910] argued from the formation of this osazone that the original hexosephosphate contained only one phosphoric acid group per molecule of hexose. It was however shown by Young [1911] and subsequently confirmed by Lebedeff [1911, 6] that one molecule of phosphoric acid is split off during the formation of the osazone, even in neutral solution. Moreover it has been found that in the cold hexosediphosphoric acid reacts with 3 molecules of phenylhydrazine forming the diphenylhydrazine salt of hexosediphosphoric acid phenylhydrazone
This compound crystallises out when 1 volume of alcohol is added to a solution of 3 molecules of phenylhydrazine in one of the acid and forms colourless needles melting at 115°–117°. p-Bromophenylhydrazine yields an analogous compound melting at 127°–128°.
Precisely the same products are given with phenylhydrazine by the hexosephosphoric acid prepared from glucose, mannose, and fructose, proving that all these sugars yield the same hexosediphosphoric acid, a point of fundamental importance.
Direct measurements of the molecular weight of the acid by the freezing-point method, combined with the determination of the degree of dissociation by the rate of cane-sugar inversion, are indecisive, but indicate that the acid has a molecular weight considerably higher than that required for a triosephosphoric acid.
A similar uncertainty attaches to the determination of the molecular weight from the freezing-point depression and conductivity of the acid potassium salt [Euler and Fodor, 1911]. Euler however concludes [p051] that both a hexosediphosphoric acid and a triosemonophosphoric acid are formed, but has not prepared any derivatives of the latter.
As regards the constitution of the hexosephosphoric ester several suggestions have been made by Young, but no decisive evidence at present exists. The identity of the products from glucose, mannose, and fructose may be explained by regarding the acid as a derivative of the enolic form common to these three sugars (p. 97), or by supposing that portions of two sugar molecules may be concerned in its production. The formation and composition of the hydrazone and osazone are of great importance as they indicate that in all probability one of the phosphoric acid residues is united with the carbon atom adjacent to the carbonyl group of the hexose. They moreover render it certain that the original phosphoric ester is a hexosediphosphoric ester and not a triosemonophosphoric ester.
Hexosediphosphoric acid has not as yet been discovered in the animal body. The action of a number of enzymes upon it has been examined [Euler, 1912, 2; Euler and Funke, 1912; Harding, 1912; Plimmer, 1913] with the following results.
The lipase of castor oil seeds, a glycerol extract of the intestinal mucous membrane of the rabbit and pig, and an aqueous extract of bran have a slow hydrolytic action, whereas pepsin and trypsin are without effect. Feeding experiments with rabbits and dogs indicate that the ester is capable of hydrolysis in the animal body, a large proportion of the phosphorus being excreted as inorganic phosphate. The ester is also decomposed by Bacillus coli communis.
It is remarkable that the hexosephosphate is not fermented nor hydrolysed by living yeast, a fact observed by Iwanoff, Harden and Young, and Euler, although, according to the experiments of Paine [1911], the yeast cell is at all events partially permeable to the sodium salt.
An equation can readily be constructed for the reaction in which hexosephosphate is formed, the data available being the formula of the product and the relation between the phosphate added and the carbon dioxide and alcohol produced:—
According to this, two molecules of sugar are concerned in the change, the carbon dioxide and alcohol being equal in weight to one [p052] half of the sugar used, and the hexosephosphate and water representing the other half.
Additional confirmation of this equation is afforded by the determination of the ratio between sugar used and carbon dioxide evolved when a known weight of sugar together with an excess of phosphate is added to yeast-juice at 25°. The phenomena then observed are precisely similar to those which occur when a phosphate is added to a fermenting mixture of yeast-juice and excess of sugar as described above. The rate of fermentation rapidly rises and then gradually falls until a rate is attained approximately equal to that of the autofermentation of the juice in presence of phosphate. At this point it is found that the extra amount of carbon dioxide evolved, beyond that which would have been given off in the absence of added sugar, bears the ratio expressed in equation (1) to the sugar added [Harden and Young, 1910, 2]. The results of four estimations made in this way were (a) 0·2 grams of glucose gave 26·5 and 27·9 c.c. of carbon dioxide at N.T.P.; (b) 0·2 grams of fructose gave 27·9 and 28·9 c.c. The carbon dioxide calculated from the sugar added in each of the four cases is 26·96 c.c.
It has also been shown by Euler and Johansson [1913] that in the fermentation of a mixture of equivalent amounts of phosphate and glucose, the whole of the glucose had disappeared when the whole of the phosphate had become esterified.
According to equation (1) the free phosphate present is used up in the reaction, and the question at once arises whether there is any source from which a constant supply of free phosphate can be elaborated in the juice, or whether some other change occurs which results in the formation of carbon dioxide and alcohol in the absence of free phosphate. The experimental evidence points in the direction of the former of these alternatives, but the question is a very difficult one to decide with absolute certainty.
When a mixture of a phosphate with yeast-juice and sugar is examined at intervals and the amount of free phosphate estimated, the following stages are observed:—
1. During the initial period of accelerated fermentation following the addition of the phosphate, the concentration of free phosphate rapidly diminishes.
2. At the close of this period, the amount of free phosphate [p053] present is very low, and, as long as active fermentation continues, no marked increase in it occurs.
3. As alcoholic fermentation slackens and finally ceases, a marked and rapid rise in the amount of free phosphate occurs at the expense of the hexosephosphate, which steadily diminishes in amount, and this change is brought about by an enzyme in the juice and ceases if the liquid be boiled.
This last reaction may be represented by the equation
In the light of this equation, together with equation No. 1, given above, all these facts can be simply and easily understood.
The rapid diminution in the amount of free phosphate during stage 1 corresponds with the occurrence of reaction (1). During the whole period of fermentation the enzymic hydrolysis of the hexosephosphate is proceeding according to equation (2). Up to the end of stage 2 the phosphate thus produced enters into reaction, according to equation (1), with the sugar which is present in excess and is thus reconverted into hexosephosphate, so that as long as alcoholic fermentation is proceeding freely, no accumulation of free phosphate can occur.
As soon as alcoholic fermentation ceases, however, it is no longer possible for the phosphate to pass back into hexosephosphate, and hence it accumulates in the free state.
A similar hydrolysis of hexosephosphate and accumulation of phosphate occur when a solution of hexosephosphate is treated with yeast-juice which has been deprived of the power of fermentation by dialysis, or with zymin freed from co-enzyme by washing (p. 63).
The actual rate of fermentation observed in any particular case in presence of excess of sugar, enzyme, and co-enzyme must on this view depend on the supply of phosphate which is available.
In presence of an adequate amount of phosphate, as well as of sugar, the highest rate attained represents the maximum velocity at which reaction (1) can proceed in that sample of yeast-juice or zymin, and this high rate is characteristic of the initial period of accelerated fermentation which follows the addition of a suitable quantity of phosphate. By the simple expedient of renewing the supply of phosphate as rapidly as it is converted into hexosephosphate, this high rate can be maintained for a considerable time [Harden and Young, 1908, 1]. In this way, for example, an average rate of evolution of carbon dioxide of 15 c.c. in five minutes was maintained for an hour and a [p054] quarter, whereas the normal rate in the absence of added phosphate was 3 c.c.
As soon as all the free phosphate has entered into the reaction, however, the supply of phosphate depends in the main on the rate at which the resulting hexosephosphate is decomposed, and the rate of fermentation now attained is conditioned by the rate at which reaction (2) proceeds, and this evidently depends on the existing concentration of the hydrolytic enzyme, which may be provisionally termed hexosephosphatase.
The rates attained during the initial period of rapid fermentation and the subsequent period of slow fermentation are thus seen to represent the velocities of two entirely different chemical reactions.
These considerations also explain why it is the extra carbon dioxide evolved during the initial period, and not the total carbon dioxide, which is equivalent to the added phosphate. As the production of phosphate is proceeding throughout the whole period at a rate which is equivalent to the normal rate of fermentation, it is obviously necessary to deduct the corresponding amount of carbon dioxide from the total evolved in order to ascertain the amount equivalent to the added phosphate.
An explanation is also afforded of the fact that a considerable increase in the concentration of hexosephosphate does not materially increase the normal rate of fermentation. This is probably due to the circumstance that, in accordance with the general behaviour of enzymes in presence of excess of the fermentable substance, the hexosephosphatase hydrolyses approximately equal amounts of hexosephosphate in equal times whatever the concentration of the latter may be, above a certain limit.
According to the experiments of Euler and Johansson [1913] the hydrolytic activity of the hexosephosphatase is greatly diminished by the presence of toluene.
The addition of a phosphate to yeast-juice not only produces the effect already described, but also enables a given volume of yeast-juice to effect a larger total fermentation, even after allowance is made for the carbon dioxide equivalent to the quantity of phosphate added. The increase in the case of ordinary yeast-juice has been found to amount to from 10 to 150 per cent. of the original total fermentation [p055] produced by the juice in the absence of added phosphate. The numbers contained in columns 1 and 2 of the table on p. 56 illustrate this effect, the ratio of the total in the presence of phosphate to that obtained in its absence being given, as well as that of the total in presence of phosphate less the equivalent of the phosphate added, to the original fermentation. The cause of this increase in the total fermentation is probably to be sought mainly in a protective action of the excess of hexosephosphate on the various enzymes, for, as has been stated above, the rate of fermentation after the termination of the initial period, is practically the same as in the absence of added phosphate (see p. 43).
Now it follows from equation (1) (p. 51) that in the total absence of phosphate no fermentation should occur, and the experimental realisation of this result would afford very strong evidence in favour of this interpretation of the phenomenon.
Hitherto, however, it has not been found possible to free the materials employed completely from phosphorus compounds which yield phosphates by enzymic hydrolysis during the experiment, but it has been found that when the phosphate contents are reduced to as low a limit as possible, the amount of sugar fermented becomes correspondingly small, and, further, that in these circumstances the addition of a small amount of phosphate or hexosephosphate produces a relatively large increase in the fermenting power of the enzyme.
When the total phosphorus present is thus largely reduced, the increase produced by the addition of a small amount of phosphate may amount to as much as eighty-eight times the original, in addition to the quantity equivalent to the phosphate, whilst the actual total evolved, including this equivalent, may be as much as twenty times the original fermentation. This result must be regarded as strong evidence in favour of the view that phosphates are indispensable for alcoholic fermentation.
The results indicated above were experimentally obtained in three different ways and are exhibited in the following table. In the first place (cols. 3 and 4), advantage was taken of the fact that the residues obtained by filtering yeast-juice through a Martin gelatin filter (p. 59) are sometimes found to be almost free from mineral phosphates, whilst they still contain a small amount of co-enzyme. The experiment then consists in comparing the fermentation produced by such a residue poor in phosphate with that observed when a small amount of phosphate is added. The second method (col. 5) consisted in carrying out two parallel fermentations by means of a residue rendered inactive by filtration [p056] and a solution of co-enzyme free from phosphate and hexosephosphate (p. 67) [Harden and Young, 1910, 2].
The third method (col. 6) consisted in washing zymin with water, to remove soluble phosphates, and then adding to it a solution of co-enzyme containing only a small amount of phosphate, and ascertaining the effect of the addition of a small known amount of hexosephosphate upon the fermentation produced by this mixture [Harden and Young, 1911, 1].
| 1 c.c. |
2 c.c. |
3 c.c. |
4 c.c. |
5 c.c |
6 c.c. |
|
|---|---|---|---|---|---|---|
| Gas evolved in absence of added phosphate | 369 | 220 | 1·4 | 1·2 | 20·3 | 1·5 |
| In the presence of added phosphate | 629 | 629 | 25·8 | 26·8 | 92·3 | 132·7 |
| Increase due to phosphate | 260 | 409 | 24·4 | 25·6 | 72·0 | 131·2 |
| Carbonic acid equivalent to phosphate | 63 | 61 | 16·9 | 16·8 | 16·8 | — |
| Increase after initial period | 197 | 348 | 7·5 | 8·8 | 55·2 | — |
| Ratio of totals | 1·7 | 2·9 | 18·4 | 21·3 | 4·5 | 88 |
| Ratio of increase after initial period to original fermentation | 0·5 | 1·6 | 5·3 | 7·3 | 2·7 | — |
The sugar which, according to equation (2) accompanies the phosphate formed by the enzymic hydrolysis of hexosephosphate is under ordinary circumstances fermented by the alcoholic enzyme of the juice and thus escapes detection.
When, however, a solution of a hexosephosphate is exposed to the action of either yeast-juice or zymin, entirely or partially freed from co-enzyme, this sugar, being no longer fermented, accumulates and can be examined. It has thus been found [Harden and Young, 1910, 2] that a sugar is in fact produced in this way which can be fermented by living yeast and exhibits the reactions of fructose, although the presence of other hexoses is not excluded. The products of the enzymic hydrolysis of the hexosephosphates therefore appear to be the same as, or similar to, those formed by the action of acids [Young, 1909].
A further consequence of these facts is that a hexosephosphate will yield carbon dioxide and alcohol when it is added to yeast-juice or zymin, and this has also been found to be the case [Harden and Young, 1910, 2; Iwanoff, 1909, 1]. [p057]
On this subject little is yet known, but a number of extremely interesting results, the interpretation of which is still doubtful, have been obtained by Euler and his colleagues. Euler has obtained a yeast [Yeast H of the St. Erik's brewery in Stockholm] which differs from Munich yeast in several respects. A maceration extract prepared from the yeast dried at 40° in a vacuum produces no effect on a glucose solution containing phosphate. If, however, the glucose solution be previously partially fermented with living yeast and then boiled and filtered, the addition of the extract prepared from Yeast H brings about the esterification of phosphoric acid without any accompanying evolution of carbon dioxide [Euler and Ohlsén, 1911, 1912].
Euler interprets this as follows: (a) Glucose itself is not directly esterified, but must first undergo some preliminary change, which is brought about by the action of living yeast. No proof of the existence of a new modification of glucose in this solution has however been advanced, other than its behaviour to extract of Yeast H, so that Euler's conclusion cannot be unreservedly accepted. It is moreover possible and even more probable that some thermostable catalytic substance (perhaps a co-enzyme) passes from the yeast into the glucose solution and enables the yeast extract to attack the glucose and phosphoric acid. A very small degree of esterification was also produced when an extract having no action on glucose and phosphate was added to glucose which had been treated with 2 per cent. caustic soda for forty hours, but the nature of the compound formed was not ascertained [Euler and Johansson, 1912, 4]. (b) The esterification of phosphoric acid without the evolution of carbon dioxide implies that the enzyme by which this process is effected is distinct from that which causes the actual decomposition of the sugar. Euler goes further than this and regards the enzyme as a purely synthetic one, giving it the name of hexosephosphatese to distinguish it from the hexosephosphatase which hydrolyses the hexosephosphate.
The evidence on which this conclusion is based cannot be regarded as satisfactory, inasmuch as it consists in the observation that in presence of sugar yeast extract does not hydrolyse the phosphoric ester. This, however, could not be expected since hydrolysis and synthesis under these conditions would ultimately proceed at equal rates.
In any case the adoption of this nomenclature is inconsistent with the conception of an enzyme as a catalyst and is therefore inadvisable until the reaction has been much more thoroughly studied. [p058]
It may further be pointed out that no proof has yet been advanced that the phosphoric ester produced without evolution of carbon dioxide is identical with hexosediphosphoric acid produced with evolution of carbon dioxide. It is by no means improbable that it represents some intermediate stage in the production of the latter (see p. 117).
Euler's other results on this subject may be briefly summarised as follows:—
(1) In presence of excess of sugar the esterification of the phosphoric acid proceeds by a monomolecular reaction and is most rapid in faintly alkaline reaction [Euler and Kullberg, 1911, 3].
(2) When yeast extract has been heated for 30 minutes to 40° it effects the esterification of phosphoric acid at a much greater rate than the unheated extract (2–10 times). Heating at 50° for 30 minutes however completely inactivates the extract. The cause of the activation is as yet unknown. The temperature coefficient for the unheated extract (17·5°–30°) is 1·4–1·5 for 10° rise of temperature [Euler and Ohlsén, 1911].
(3) Yeasts which in the dried state all produce rapid esterification of phosphoric acid, yield extracts of very unequal powers in this respect [Euler, 1912, 1].
In the previous chapter reference was made to the fact that the addition of boiled yeast-juice greatly increases the amounts of carbon dioxide and alcohol formed from sugar by the action of a given volume of yeast-juice.
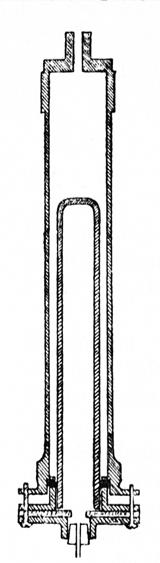
When the boiled juice is dialysed the substance or substances to which this effect is due pass into the dialysate, the residue being quite inactive. In order to ascertain the effect on the process of alcoholic fermentation of the complete removal of these unknown substances from yeast-juice itself, dialysis experiments were instituted with fresh yeast-juice, capable of bringing about an active production of carbon dioxide and alcohol from sugar. It was already known from the experiments of Buchner and Rapp [1898, 1] that dialysis in parchment paper for seventeen hours at 0° against water or physiological salt solution only produced a diminution of about 20 per cent. in the total amount of fermentation obtainable, and in view of the less permanent character of the juice from top yeasts a more rapid method of dialysis was sought. This was found in the process of filtration under pressure through a film of gelatin, supported in the pores of a Chamberland filter candle, which had been introduced by Martin [1896].
In this way it was found possible to divide the juice into a residue and a filtrate, each of which was itself incapable of setting up the alcoholic fermentation of glucose, whereas, when they were reunited, the mixture produced almost as active a fermentation as the original juice [Harden and Young, 1905, 1; 1906, 2].
The apparatus employed for this purpose consists of a brass tube provided with a flange in which the gelatinised candle is held by a compressed india-rubber ring, and is shown in section in Fig. 5. Two such apparatus are used, each capable of holding about 70 c.c. of the liquid to be filtered. The tubes, after being filled with the yeast-juice, are connected by means of a screw joint with a cylinder of compressed air and the filtration carried out under a pressure of 50 atmospheres, [p060] the arrangement employed being shown in Fig. 6. In the earlier experiments 25 to 50 c.c. of yeast-juice were placed in each tube and the filtration continued until no more liquid passed through. The residue was then washed several times in situ by adding water and forcing it through the candle. The time occupied in this process varied from six to twelve hours with different preparations of yeast-juice. The candle was then removed from the brass casing and the thick, brown-coloured residue scraped off, dissolved in water, and at once examined. It was subsequently found to be possible to dry this residue in vacuo over sulphuric acid without seriously altering the fermenting power, and this led to a slight modification of the method, which is now conducted as follows. Two quantities of 50 c.c. each of yeast-juice are filtered, without washing, and the residues spread on watch-glasses and dried in vacuo. Two fresh quantities of 50 c.c. are then filtered through the same candles and the residues also dried. The 200 c.c. of juice treated in this way give a dry residue of 17 to 24 grams. The residue is then dissolved in 100 c.c. of water and filtered in quantities of 50 c.c. through two fresh gelatinised candles and the residue again dried. A considerable diminution in weight occurs, partly owing to incomplete removal from the candle and brass casing, and the final residue only amounts to about 9 to 12 grams. Occasionally it is necessary to repeat the processes of dissolving in water, filtering, and drying, but a considerable loss both of material and fermenting power attends each such operation.
The sticky residue dries up very rapidly in vacuo to a brittle, scaly mass, which is converted by grinding into a light yellow powder.
The filtrate was invariably found to be quite devoid of fermenting power, none of the enzyme passing through the gelatin.
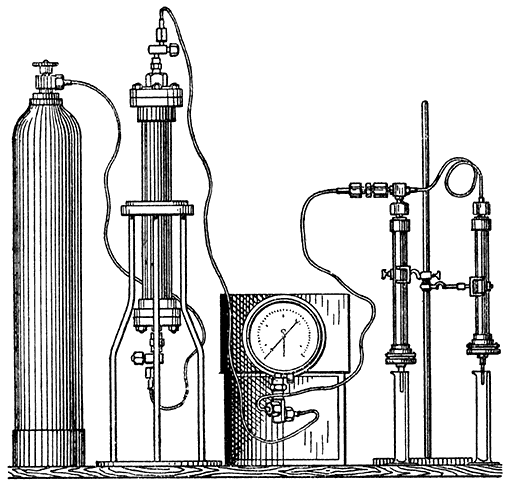
Properties of the Filtered and Washed Residue.—The residue prepared as described above consists mainly of the protein, glycogen, and dextrins of the yeast-juice, and is almost free from mineral phosphates, but contains a certain amount of combined phosphorus. It also contains the enzymes of the juice, including the proteoclastic enzyme, and the hexosephosphatase (p. 54). Its solution in water is usually quite inactive to glucose or fructose, but in some cases produces a small and evanescent fermentation. When the original filtrate or a corresponding [p061] quantity of the filtrate from boiled fresh yeast-juice is added, the mixture ferments glucose or fructose quite readily. The following table shows the quantitative relations observed, the sugar being in all cases present in excess:—
| No. | Material. | Volume. | Fil- trate added. c.c. |
Boiled Juice added. c.c. |
Water added. c.c. |
CO2 evolved. |
|---|---|---|---|---|---|---|
| 1 | Undried and unwashed residue | 15 c.c. | 0 | 0 | 15 | 0 g. |
| 15 " | 15 | 0 | 0 | 0·035 " | ||
| 2 | 15 " | 0 | 0 | 15 | 0·024 " | |
| 15 " | 0 | 15 | 0 | 0·282 " | ||
| 3 | Undried and washed residue | 25 " | 0 | 0 | 0 | 0·4 c.c. |
| 25 " | 0 | 25 | 0 | 268 " | ||
| 4 | 20 " | 0 | 0 | 0 | 8·3 " | |
| 20 " | 20 | 0 | 0 | 90·3 " | ||
| 5 | Washed and dried residue | 1 gram in 15 c.c. | 0 | 0 | 0 | 0 " |
| 0 | 12 | 0 | 108 " | |||
| 6 | 1 gram in 25 c.c. | 0 | 0 | 0 | 0 " | |
| 0 | 25 | 0 | 364 " |
These experiments lead to the conclusion that the fermentation of glucose and fructose by yeast-juice is dependent upon the presence, not only of the enzyme, but also of another substance which is dialysable and thermostable.
Precisely similar results were subsequently obtained by Buchner and Antoni [1905, 2] by the dialysis of yeast-juice. One hundred c.c. of juice were dialysed for twenty-four hours at 0° against 1300 c.c. of distilled water, and the dialysate was then evaporated at 40° to 50° to 20 c.c. The fermenting power of 20 c.c. of the dialysed juice was then determined with the following additions:—
It was shown in the previous chapter that phosphates are essential to fermentation, and hence it becomes necessary to inquire whether the effect of dialysis is simply to remove these. Experiment shows that this is not the case. Soluble phosphates do not confer the power of producing fermentation on the inactive residue obtained by filtration. Moreover, when yeast-juice is digested for some time before being boiled, it is found, as will be subsequently described, that the boiled autolysed juice is quite incapable of setting up fermentation in the inactive residue, although free phosphates are abundantly present [Harden and Young, 1906, 2].
The filtration residue is never obtained quite free from combined phosphorus, but the production from this of the phosphate necessary for fermentation to proceed, may be so slow as to render the test for co-enzyme uncertain, owing to the absence of sufficient phosphate. When a filtration residue is being tested it is therefore necessary to secure the presence of sufficient phosphate to enable the characteristic reaction to proceed, and at the same time to avoid adding phosphate in too great concentration, as this may, in the presence of only small amounts of enzyme or co-enzyme, inhibit the fermentation (p. 71). The proof that a filtration residue or dialysed juice is quite free from co-enzyme is therefore a somewhat complicated matter, and not only involves the experimental demonstration that the material will not ferment sugar, but also that this power is not imparted to it by the addition of a small concentration of phosphate. As it has been found (p. 73) that the fermentation of fructose is less affected than that of glucose by the presence of excess of phosphate, the practical method of examining a filtration residue for co-enzyme is to test its action on a solution of fructose (1) alone and (2) in presence of a small concentration of phosphate. If the residue produces no action [p063] in either case, but produces fermentation when a solution of co-enzyme is added in the presence of the same concentration of phosphate as was previously employed, it may be concluded that this sample was free from co-enzyme but contained enzyme; such an experiment also affords a definite proof that the co-enzyme does not consist of phosphate.
This dialysable, thermostable substance, without which alcoholic fermentation cannot proceed, has been provisionally termed the co-ferment or co-enzyme of alcoholic fermentation. This expression was first introduced by Bertrand [1897], to denote substances of this kind, and he applied it in two instances—to the calcium salt which he considered was necessary for the action of pectase on pecten substances, and to the manganese which he supposed to be essential for the activity of laccase. Without inquiring whether these substances are precisely comparable in function with that contained in yeast-juice, the term may be very well applied to signify the substance of unknown constitution without the co-operation of which the thermolabile enzyme of yeast-juice is unable to set up the process of alcoholic fermentation. The active agent of yeast-juice consisting of both enzyme and co-enzyme may be conveniently spoken of as the fermenting complex, and this term will occasionally be employed in the sequel.
The co-enzyme is present alike in the filtrates from fresh yeast-juice and from boiled yeast-juice, and is also contained in the liquids obtained by boiling yeast with water and by washing zymin or dried yeast with water.
Practically the only chemical property of the co-enzyme, other than that of rendering possible the process of alcoholic fermentation, which has so far been observed, is that it is capable of being decomposed, probably by hydrolysis, by a variety of reagents, prominent among which is yeast-juice. This was observed by Harden and Young in the course of their attempts to prepare a completely inactive residue by filtration. In many cases a residue was obtained which still possessed a very limited power of fermentation, only a small amount of carbon dioxide being formed and the action ceasing entirely after the expiration of a short period; on the subsequent addition of boiled juice, however, a very considerable evolution of carbon dioxide was produced. This was interpreted to mean that the residue in question contained an ample supply of enzyme but only a small proportion of co-enzyme, and that the latter was rapidly destroyed, so that the fermentation soon ceased. The boiled juice then added provided a further proportion of co-enzyme by the aid of which the surplus enzyme was [p064] enabled to carry on the fermentation. This view was confirmed by adding to a solution of a completely inactive filtration residue and glucose successive small quantities of boiled juice and observing the volumes of carbon dioxide evolved after each such addition. Thus in one case successive additions of volumes of 3 c.c. of boiled juice produced evolutions of 8·2, 6, and 6 c.c. of carbon dioxide. In another case two successive additions of 15 c.c. of boiled juice produced evolutions of 54 and 41·2 c.c. On the other hand, the enzyme itself also gradually disappears from yeast-juice when the latter is incubated either alone or with sugar (p. 20).
The cessation of fermentation in any particular mixture of enzyme and co-enzyme may, therefore, be due to the disappearance of either of these factors from the liquid. If the amount of co-enzyme present be relatively small it is the first to disappear, and fermentation can then only be renewed by the addition of a further quantity, whilst the addition of more enzyme produces no effect. If, on the other hand, the amount of co-enzyme be relatively large, the inverse is true; the enzyme is the first to disappear, and fermentation can only be renewed by the addition of more enzyme, a further quantity of co-enzyme producing no effect. It has, moreover, been found that the co-enzyme, like the enzyme, disappears more rapidly in the absence of glucose than in its presence, incubation at 25° for two days being as a rule sufficient to remove all the co-enzyme from yeast-juice from top yeasts in the absence of sugar, whilst in the presence of fermentable sugar co-enzyme may still be detected at the end of four days.
In all the experiments carried out by Harden and Young with juice from English top yeast it was found that when a mixture of the juice with glucose was incubated until fermentation had ceased, the further addition of co-enzyme in the form of boiled juice did not cause any renewal of the action; in other words, the whole of the enzyme had disappeared.
On the other hand, Buchner and Klatte [1908], working with juice and zymin prepared from bottom yeast, observed the extremely interesting fact that after the cessation of fermentation the addition of an equal volume of boiled juice caused a renewed decomposition of sugar, and that the processes of incubation until no further evolution of gas occurred and re-excitation of fermentation by the boiled juice could be repeated as many as six times. Thus in one experiment the duration of the fermentation was extended from three to a total of twenty-four days, and the total gas evolved from 0·73 gram to 2·19 grams. The phenomenon has been found to be common to yeast from Munich and [p065] from Berlin as well as to zymin and maceration extract, and it was further observed that the boiled juice from one yeast could regenerate the juice from another, although the quantitative relations were different.
In these samples of yeast-juice, therefore, there is present a natural condition of affairs precisely similar to that obtaining in the artificial mixtures of inactive filtration residue and co-enzyme solution made by Harden and Young. The balance of quantities is such that the co-enzyme disappears before the enzyme, leaving a certain amount of enzyme capable of exercising its usual function as soon as sufficient co-enzyme is added. This establishes an interesting point of contrast with the juice prepared from top yeast in England, in which the enzyme does not outlast the co-enzyme [Harden and Young, 1907]. The difference may be due to some variation in the relative proportions of enzyme and co-enzyme or of the enzymes to which the disappearance of each of these is presumptively due, or to a combination of these two causes. It was, however, found, even in the juice from bottom yeast, that incubation for three days at 22° without the addition of sugar caused the disappearance of the enzyme as well as of the co-enzyme, and left a residue alike incapable of being regenerated by the addition of co-enzyme or of restoring the power of producing fermentation to an inactive mixture containing enzyme and sugar.
If the fermenting power of the juice is to be preserved by repeated regeneration for a long period, it is absolutely necessary to add the co-enzyme solution each time as soon as fermentation has ceased, since the enzyme in the absence of this addition rapidly disappears, even in the presence of sugar.
This result is probably to be explained, at all events in the main, by the presence in the co-enzyme solution of the antiprotease to which reference has already been made [Buchner and Haehn, 1910, 2]. This agent, the constitution of which is still unknown, protects proteins in general from the action of digestive enzymes, and on the assumption that the alcoholic enzyme of yeast-juice belongs to the class of proteins, may be supposed to lessen the rate at which this enzyme is destroyed by the endotryptase of the juice. This antiprotease is, like the co-enzyme (p. 68), destroyed by lipase but is more stable than the co-enzyme towards hydrolytic agents, and can be obtained free from co-enzyme by boiling the solution for some hours alone or by heating with dilute sulphuric acid. Such a solution possesses no regenerative power, but still retains its power of protecting proteins against digestion and of preserving the fermenting power of yeast-juice. [p066]
It must, however, be remembered that the addition of a phosphate alone may greatly prolong the period of fermentation of yeast-juice (p. 55), and sugar is well known to exert a similar action. It appears, therefore, that the existence of the enzyme is prolonged not only by the presence of the antiprotease but also by that of sugar and hexosephosphate, into which phosphate passes in presence of sugar. Similar effects are exerted on the co-enzyme by sugar and probably also by hexosephosphate.
The fermenting complex, therefore, in the presence of these substances, either separately or together, falls off more slowly in activity and is present for a longer time, and for both of these reasons produces an increased amount of fermentation. It seems probable also that the hexosephosphatase is similarly affected, so that the supply of free phosphate is at the same time better maintained, and the rate of fermentation for this reason decreases more slowly than would otherwise be the case.
It is in this way that an explanation may be found of the remarkable increase in total fermentation, which is produced by the addition to yeast-juice and sugar of boiled yeast-juice, containing free phosphate (which passes into hexosephosphate) as well as co-enzyme, of boiled autolysed yeast-juice, containing free phosphate but no co-enzyme, or of phosphate solution alone.
In no case is the original rate of fermentation greatly increased after the initial acceleration has disappeared, but in every case the total fermentation is considerably augmented, and this is no doubt mainly to be attributed, as just explained, to the diminished rate of decomposition of the fermenting complex and probably of the hexosephosphatase.
Although both enzyme and co-enzyme are completely precipitated from yeast-juice, as already described (p. 38), by 10 volumes of acetone, the co-enzyme is less easily precipitated than the enzyme, and a certain degree of separation can therefore be attained by fractional precipitation [Buchner and Duchaček, 1909]. The enzyme cannot, however, be completely freed from co-enzyme in this manner, and the process is attended by a very considerable loss of enzyme. This is probably due to the fact that only small quantities of acetone can be added (1·5 to 3 volumes), in order to avoid precipitation of co-enzyme, and that the precipitates thus formed contain a large proportion of water, a condition which appears to be fatal to the preservation of the enzyme.
It is, however, not quite certain whether it is the zymase or the hexosephosphatase which is destroyed in these cases, as no attempt [p067] was made to distinguish between them. In any case the precipitates obtained by fractional treatment with acetone, even when reunited, produce a much smaller fermentation than the original juice or the powder prepared by bringing it into 10 volumes of acetone.
Attempts to isolate the co-enzyme from boiled yeast-juice have also been hitherto unsuccessful. It has, however, been found possible to remove a considerable amount of material from the solution without affecting the co-enzyme. When 1 volume of alcohol is added to boiled yeast-juice, a bulky precipitate, consisting largely of carbohydrates, is produced, and the filtrate from this is found to contain the co-enzyme and can be freed from alcohol by evaporation. Further precipitation with alcohol has not led to useful results.
When a solution which has been treated in this way is precipitated with lead acetate and kept neutral to litmus, the free phosphate and hexosephosphate are thrown down and the co-enzyme remains in solution. The filtrate can be freed from lead by means of sulphuretted hydrogen and neutralised, and then forms a solution of co-enzyme free from phosphate and hexosephosphate but still containing combined phosphorus. More complete purification than this has not yet been accomplished. Occasionally the precipitate of lead salts retains some of the co-enzyme, apparently by adsorption, but usually the greater part remains in the solution (Harden and Young).
The co-enzyme is partially removed from yeast-juice by means of a colloidal solution of ferric hydroxide (Resenscheck). A precipitate is thus obtained which contains phosphorus and resembles boiled yeast-juice in its regenerative action on yeast-juice rendered inactive by fermentation. It has not, however, so far been found possible to isolate any definite compound from this precipitate. There are also indications that when yeast-juice, either fresh or boiled, is electrolysed, the co-enzyme tends to accumulate at the cathode [Resenscheck, 1908, 1, 2].
Buchner and Klatte [1908] made use of yeast-juice rendered free from co-enzyme by incubation with sugar solution to examine the nature of the agent by which the co-enzyme is destroyed. This agent is certainly an enzyme, since boiled yeast-juice can be preserved with unimpaired powers for a considerable length of time, and suspicion fell naturally, in the first instance, on the endotryptase of the yeast cell. Direct experiment showed, however, that yeast-juice, which, when fresh, rapidly destroyed the co-enzyme of boiled juice, lost this power on preservation, but retained its proteoclastic properties without diminution, so that the tryptic enzyme could not be the one concerned. The direct action of commercial trypsin on boiled yeast-juice also yielded [p068] a negative result, although this cannot strictly be regarded as an indication of the effect of the specific proteoclastic enzymes of yeast-juice. On the other hand, it was found that when boiled juice was treated for some time with an emulsion containing the lipase of castor oil seeds, the co-enzyme was completely destroyed. This is a result of great importance, inasmuch as it probably indicates that the co-enzyme is chemically allied to the class of substances hydrolysable by lipase, i.e. to the fats and other esters.
Further, observations by Buchner and Haehn [1909] have shown that digestion with potassium carbonate solution containing 2·5 grams per 100 c.c. also brings about the destruction of the co-enzyme, and that this is also slowly accomplished by the repeated boiling of the juice. The co-enzyme is also destroyed both by acid and alkaline hydrolysis, and when the solution is evaporated to dryness and the residue ignited.
Beyond this general indication nothing is known of the chemical nature of the co-enzyme. The intimate relation of phosphoric acid to the process of fermentation renders it not impossible that the co-enzyme may contain this group, but there is no definite evidence for such a belief. Purely negative results have been obtained with all the substances of known composition which have yet been tested, among these being soluble phosphates, hexosephosphates and a number of oxidisable and reducible substances, such as quinol, p-phenylenediamine, methylene blue, peptone beef broth, etc. (Harden and Young; Harden and Norris [1914]; see also Euler and Bäckström [1912]), glycero-phosphates (Buchner and Klatte).
The precise function of the co-enzyme is even more obscure than its chemical nature. The system of reacting substances consisting of fermentable material, enzyme and co-enzyme, bears, however, an obvious superficial resemblance to many of the systems required for the accomplishment of chemical changes in the animal or vegetable organism. Such a triad of substances is, for example, requisite for the process by which the red blood corpuscles of an animal are broken up by the serum of a different animal into the blood of which the red corpuscles of the first animal have been injected. This effect is only produced when two substances are present, the amboceptor or immune body and the complement. The analogy may be carried to a further stage since the amboceptor is, like the co-enzyme, more thermostable than the complement, which therefore corresponds with the enzyme. Immune serum can, in fact, be freed from complement by being heated at 57–60° for half an hour, whereas the amboceptor is unaffected by this treatment. On the other hand, the complement and amboceptor do not [p069] appear to act like enzymes but rather like ordinary chemical reagents, remaining in combination even after the blood corpuscle has been broken up, whereas the enzyme and co-enzyme of yeast-juice are again liberated when the reaction between sugar and phosphate has been completed.
One of the most interesting and at the same time most difficult problems concerning enzyme action in general is the nature of the inhibiting or accelerating effect produced by many substances upon the rate or total result of the chemical process set up in presence of the enzyme. Inhibition, it is usually supposed, involves either the decomposition of the enzyme, in which case it is irreversible, its removal from the sphere of action by some change in its mode of solution, or the formation of an inactive or less active compound between the enzyme and the inhibiting agent. This compound it may sometimes be possible to decompose, with the result that the activity of the enzyme is restored. A striking example of this, to which allusion has already been made, is the effect of hydrocyanic acid on alcoholic fermentation (p. 37).
Acceleration of enzyme action can in some cases be ascribed to the fact that the accelerating substance possesses an assignable chemical function in the reaction, so that an increase in the concentration of this substance causes an increase in the rate of the reaction. As we have seen in Chapter III, this is the explanation of the accelerating effect of phosphates on fermentation by yeast-juice. In many other cases, however, no such chemical function can be traced, as, for example, in the effect of neutral salts on the hydrolytic action of invertase, or the effect of the addition of the co-enzyme to zymase, and it is necessary to fall back on some assumption, such as that the accelerating agent acts by increasing the effective concentration of the enzyme or by combining either with the enzyme or the substrate, forming a compound which undergoes the reaction more readily.
The interest in the following examples of inhibition and acceleration of fermentation by yeast-juice lies not only in their relation to these general problems but also, and perhaps chiefly, in their bearing on the specific problem of the nature and mode of action of the various agents concerned in the production of alcohol and carbon dioxide from sugar in the yeast-cell. [p071]
Prominent among these instances of inhibition and acceleration are the phenomena attendant on the addition of excess of phosphate to yeast-juice.
When a phosphate is added to a fermenting mixture of a sugar and yeast-juice, the effect varies with the concentration of the phosphate and the sugar and with the particular specimen of yeast-juice employed. With low concentrations of phosphate in presence of excess of glucose the acceleration produced is so transient that no accurate measurements of rate can be made. As soon as the amount of phosphate added is sufficiently large, it is found that the rate of evolution of carbon dioxide very rapidly increases from five to ten times, and then quickly falls approximately to its original value.
As the concentration of phosphate is still further increased, it is first observed that the maximum velocity, which is still attained almost immediately after the addition of the phosphate, is maintained for a certain period before the fall commences, and then, as the increase in concentration of phosphate proceeds, that the maximum is only gradually attained after the addition, the period required for this increasing with the concentration of the phosphate. Moreover, with still higher concentrations, the maximum rate attained is less than that reached with lower concentrations, and further, the rate falls off more slowly. The concentration of phosphate which produces the highest rate, which may be termed the optimum concentration, varies very considerably with different specimens of yeast-juice [Harden and Young, 1908, 1].
All these points are illustrated by the accompanying curves (Fig. 7) which show the rate of evolution per five minutes plotted against the time for four solutions in which the initial concentrations of phosphate were (A) 0·033, (B) 0·067, (C) 0·1, and (D) 0·133 molar, the volumes of 0·3 molar phosphate being 5, 10, 15, and 20 c.c. in each case added to 25 c.c. of yeast-juice, and made up to 45 c.c, each solution containing 4·5 grams of glucose. The time of addition is taken as zero, the rate before addition being constant, as shown in the curves.
It will be observed that 5 and 10 c.c. (A and B) give the same maximum, whilst 15 c.c. (C) produce a much lower maximum, and 20 c.c. (D) a still lower one, the rate at which the velocity diminishes after the attainment of the maximum being correspondingly slow in these last two cases. By calculating the amount of phosphate which has disappeared as such from the amount of carbon dioxide evolved, [p072] it is found that the maximum does not occur at the same concentration of free phosphate in each case.
These results suggest that the phosphate is capable of forming two or more different unstable associations with the fermenting complex. One of these, formed with low concentrations of the phosphate, has the composition most favourable for the decomposition of sugar, whilst the others, formed with higher concentrations of phosphate, contain more of the latter, probably associated in such a way with the fermenting complex as to render the latter partially or wholly incapable of effecting the decomposition of the sugar molecule. As the fermentation proceeds slowly in the presence of excess of phosphate, the concentration of the latter is reduced by conversion into hexosephosphate, and a re-distribution of phosphate occurs, resulting in the gradual change of the less active into the more active association of phosphate with fermenting complex, and a consequent rise in the rate of fermentation.
In those cases in which the maximum rate corresponding to the optimum concentration of phosphate is never attained, some secondary cause may be supposed to intervene, such as a permanent change in a portion of the fermenting complex, accumulation of the products of the reaction, etc.
It is also possible as suggested by Buchner for the analogous case of arsenite (p. 78) that the addition of increasing amounts of phosphate causes a progressive but reversible change in the mode of dispersion [p073] of the colloidal enzyme, and that this has the secondary effect of altering the rate of fermentation. No decisive evidence is as yet available upon the subject.
The results obtained by Euler and Johansson [1913] to which reference has already been made indicate that in presence of a moderate excess of phosphate esterification is more rapid than production of carbon dioxide. No explanation of this phenomenon has yet been given, but it might obviously be due either to the production of some phosphorus compound which subsequently takes part in the production both of hexosediphosphate and of carbon dioxide, or, less probably, to the entire independence of the two changes—esterification of phosphate and production of carbon dioxide—which might then be differently affected by the presence of excess of phosphate and therefore take place at different rates.
Although, as has been pointed out (p. 42), glucose, mannose, and fructose all react with phosphate in a similar manner in presence of yeast-juice, there are nevertheless certain quantitative differences between the behaviour of glucose and mannose on the one hand, and fructose on the other, which appear to be of considerable importance. Fructose differs from the other two fermentable hexoses in two particulars: (1) the optimum concentration of phosphate is much greater; (2) the maximum rate of fermentation attainable is much higher [Harden and Young, 1908, 2; 1909].
These points are clearly illustrated by the following results, which all refer to 10 c.c. of yeast-juice, and show that the optimum concentration of phosphate for the fermentation of fructose is from 1·5 to 10 times that of glucose, and that the maximum rate of fermentation for fructose in presence of phosphate is 2 to 6 times that of glucose.
| Sugar in Grams. | Total Volume. | Optimum Volume of 0·6 Molar Phosphate in c.c. | Maximum Rate in Cubic Centimetres of CO2 per Five minutes. | ||
|---|---|---|---|---|---|
| Glucose. | Fructose. | Glucose. | Fructose. | ||
| 2 | 35 | 2 | 5 | 7·5 | 32·2 |
| 4 | 50 | 1 | 10 | 5·4 | 28·4 |
| 1·6 | 23 | 2 | 5 | 8 | 17 |
| 1 | 25 | 1·75 | 5 | 5·2 | 25·9 |
| 2 | 25 | 5 | 7·5 | 16·2 | 31·2 |
| 2 | 20 | 2 | 3·5 | 7·9 | 22·6 |
| 2 | 22·5 | 0·75 | 2 | 3·4 | 22·2 |
It is interesting to note that the two high rates, 32·2 and 31·2 c.c. per five minutes, are equal to about half the rate obtainable with an amount of living yeast corresponding to 10 c.c. of yeast-juice, assuming that about 16 to 20 grams of yeast are required to yield this volume of juice, and that this amount of yeast would give about 56 to 70 c.c. of carbon dioxide per five minutes at 25°, which has been found experimentally to be about the rate obtainable with the top yeast employed for these experiments.
When the maximum rate of fermentation of glucose or mannose by yeast-juice in presence of phosphate is greatly lowered by the addition of a large excess of phosphate, the addition of a relatively small amount of fructose (as little as 2·5 per cent. of the weight of the glucose) causes rapid fermentation to occur. This induced activity is not due solely to the selective fermentation of the added fructose, since the amount of gas evolved may be greatly in excess of that obtainable from the quantity added.
Another way of expressing the same thing is to say that the optimum concentration of phosphate (p. 71) is greatly raised when 2·5 per cent. of fructose is added to glucose, and that consequently the rate of fermentation rises. The effect is extremely striking, since a mixture of glucose and yeast-juice fermenting in the presence of a large excess of phosphate at the rate of less than 1 c.c. of carbon dioxide in five minutes may be made to ferment at six to eight times this rate by the addition of only 0·05 gram of fructose (2·5 per cent. of the glucose present), and to continue until the total gas evolved is at least five to six times as great as that obtainable from the added fructose, the concentration of the phosphate being the whole time at such a height as would limit the fermentation of glucose alone to its original value.
The effect is not produced when the concentration of the phosphate is so high that the rate of fermentation of fructose is itself greatly lowered.
This remarkable inductive effect is specific to fructose and is not produced when glucose is added to mannose or fructose, or by mannose when added to glucose or fructose, under the proper conditions of concentration of phosphate in each case.
This interesting property of fructose, taken in connection with the [p075] facts that this sugar in presence of phosphate is much more rapidly fermented than glucose or mannose, and that the optimum concentration of phosphate for fructose is much higher than for glucose or mannose, appears to indicate that fructose when added to yeast-juice does not merely act as a substance to be fermented, but in addition, bears some specific relation to the fermenting complex.
All the phenomena observed are, indeed, consistent with the supposition that fructose actually forms a permanent part of the fermenting complex, and that, when the concentration of this sugar in the yeast-juice is increased, a greater quantity of the complex is formed. As the result of this increase in the concentration of the active catalytic agent, the yeast-juice would be capable of bringing about the reaction with sugar in presence of phosphate at a higher rate, and at the same time the optimum concentration of phosphate would become greater, exactly as is observed. The question whether, as suggested above, fructose actually forms part of the fermenting complex, and the further questions, whether, if so, it is an essential constituent, or whether it can be replaced by glucose or mannose with formation of a less active complex, remain at present undecided, and cannot profitably be more fully discussed until further information is available.
It must, moreover, be remembered that different samples of yeast-juice vary to a considerable extent in their relative behaviour to glucose and fructose, so that the phenomena under discussion may be expected to vary with the nature and past history of the yeast employed.
The close analogy which exists between the chemical functions of phosphorus and arsenic lends some interest to the examination of the action of sodium arsenate upon a mixture of yeast-juice and sugar, and experiments reveal the fact that arsenates produce a very considerable acceleration in the rate of fermentation of such a mixture [Harden and Young, 1906, 3; 1911, 1]. The phenomena observed, however, differ markedly from those which accompany the action of phosphate.
The acceleration produced is of the same order of magnitude as that obtained with phosphate, but it is maintained without alteration for a considerable period, so that there is no equivalence between the amount of arsenate added and the extra amount of fermentation effected. Further, no organic arsenic compound corresponding in composition with the hexosephosphates appears to be formed.
Increase of concentration of arsenate produces a rapid inhibition of [p076] fermentation, probably due to some secondary effect on the fermenting complex, possibly to be interpreted as the formation of compounds incapable of combining with sugar and hence unable to carry on the process of fermentation. An optimum concentration of arsenate therefore exists just as of phosphate, at which the maximum rate is observed, and this optimum concentration and the corresponding rate vary with different samples of juice and are less for glucose than for fructose. The rate of fermentation by zymin is relatively less increased than that by yeast-juice.
Owing to the fact that the rate is permanently maintained the addition of a suitable amount of arsenate increases the total fermentation produced to a much greater extent than phosphate.
The nature of these effects may be gathered from the result of a few typical experiments. In one case the rate of fermentation of glucose by yeast-juice was raised by the presence of 0·03 molar arsenate from 2 to 23 c.c. per five minutes, and the total evolved in ninety-five minutes from 51 to 459 c.c. The accelerating effect on 20 c.c. of juice, of as little as 0·005 c.c. of 0·3 molar arsenate, containing 0·11 mgrm. of arsenic, can be distinctly observed, but the maximum effect is usually produced by about 1 to 3 c.c., the concentration being therefore 0·015 to 0·045 molar. Greater concentrations than this produce a less degree of acceleration accompanied by a shorter duration of fermentation, as shown by the following numbers which refer to 20 c.c. of yeast-juice in a total volume of 40 c.c. containing 10 per cent. of glucose:—
| C.cs. of 0·3 Molar Arsenate in 40 c.c. | Molar Concentration of Arsenate. | Maximum Rate of Fermentation. |
|---|---|---|
| 0 | 0 | 3·5 |
| 0·005 | 0·0000375 | 6·3 |
| 0·01 | 0·000075 | 8 |
| 0·02 | 0·00015 | 14·2 |
| 0·04 | 0·0003 | 19·9 |
| 0·1 | 0·00075 | 29·7 |
| 0·2 | 0·0015 | 35 |
| 0·5 | 0·00375 | 34·9 |
| 1·0 | 0·0075 | 29·5 |
| 2·0 | 0·015 | 23·2 |
| 5·0 | 0·0375 | 14·5 |
| 10·0 | 0·075 | 8·7 |
| 15·0 | 0·1125 | 5·3 |
| 20 | 0·15 | 3·2 |
The contrast between glucose and fructose in their relations to [p077] arsenate are well exhibited in the following table, in which the rates of fermentation produced by arsenate in presence of excess of glucose and fructose respectively are given:—
| Concentration of Arsenate. | Rate. | |
|---|---|---|
| Glucose. | Fructose. | |
| 0·0075 molar | 12·1 | 26·6 |
| 0·0225 (opt. for glucose) | 13·4 | — |
| 0·0525 (opt. for fructose) | — | 45·8 |
| 0·1125 | 5·1 | 39 |
Here the optimum concentration for fructose is more than twice that for glucose, whilst the maximum rate of fermentation obtainable with fructose is between three and four times the maximum given by glucose.
Effects somewhat similar to those produced by arsenates were observed by Buchner [Buchner and Rapp, 1897; 1898, 1, 2, 3; 1899, 2; Buchner, E. and H., and Hahn, 1903, pp. 184–205] when potassium arsenite was added to yeast-juice. This substance, the action of which on yeast had been adduced by Schwann as a proof of the vegetable nature of this organism, was employed by Buchner on account of its poisonous effect on vegetable cells as an antiseptic and as a means of testing for the protoplasmic nature of the agent present in yeast-juice. Its effect on the fermentation was, however, found to be irregular, and at the same time it did not act as an efficient antiseptic in the concentrations which could be employed. Even 2 per cent. of arsenious oxide, added as the potassium salt, had in many cases a decided effect in diminishing the total fermentation obtained with cane sugar, and this effect increased with the concentration. A number of irregularities were also observed which cannot here be discussed. It was further found that in some cases 2 per cent. of arsenious oxide inhibited the fermentation of glucose but not of saccharose, or of a mixture of glucose and fructose, whilst its effect on fructose alone was of an intermediate character.
The important observation was also made by Buchner that the addition of a suitable quantity of arsenite as a rule caused a greatly increased fermentation during the first sixteen hours even in experiments in which the total fermentation was diminished. By examining the effect of arsenite on fermentation in a similar manner to that of arsenate, Harden and Young [1911, 1] have found that a close analogy exists [p078] between the effects and modes of action of these substances, but that arsenite produces a much smaller acceleration than arsenate. An optimum concentration of arsenite exists, just as in the case of arsenate, which produces a maximum rate of fermentation. Further increase in concentration leads to inhibition, and in no case is there any indication of the production of an exactly equivalent amount of fermentation as in the case of phosphate. In various experiments with dialysed, evaporated, and diluted yeast-juice in which 2 per cent. of arsenious oxide was found by Buchner to inhibit fermentation, it is probable that, owing to the small amount of fermenting complex left, this amount of arsenious oxide was considerably in excess of the optimum concentration, although Buchner ascribes the effect to the removal of some of the protective colloids of the juice, owing to the prolonged treatment to which it had been subjected.
The extent of the action of arsenite appears from the following results. In one case a rate of 1·7 c.c. was increased to 7 c.c. by 0·06 molar arsenite. In another experiment it was found that the optimum concentration was 0·04 molar arsenite, the addition of which increased the rate three-fold. As in the case of arsenate the optimum concentration and the corresponding maximum rate of fermentation are considerably greater for fructose than for glucose. The relative rates produced by the addition of equivalent amounts of arsenate and arsenite (1 c.c. of 0·3 molar solution in each case to 20 c.c. of yeast-juice) were 27·5 and 3·1, the original rate of the juice being 1·7. In general the optimum concentration of arsenite is considerably greater than that of arsenate.
The inhibiting effects of higher concentrations of arsenite and arsenate also present close analogies, but this most interesting aspect of the question has not yet been sufficiently examined to repay detailed discussion. Buchner [Buchner, E. and H., and Hahn, 1903, pp. 199–205] has suggested that the inhibition is due primarily to some change in the colloidal condition of the enzyme and has shown that certain colloidal substances appear to protect it, as does also sugar. The possibility is also present that inactive combinations of some sort are formed between the fermenting complex and the inhibiting agent, in the manner suggested to account for the inhibiting effect of excess of phosphate (p. 72). It seems most probable that the effect is a complex one, in which many factors participate.
In explanation of the remarkable accelerating action of arsenates and arsenites two obvious possibilities present themselves. In the [p079] first place the arsenic compound may actually replace phosphate in the reaction characteristic of alcoholic fermentation, the resulting arsenic analogue of the hexosephosphate being so unstable that it undergoes immediate hydrolysis, and is therefore only present in extremely small concentration at any period of the fermentation and cannot be isolated. In the second place it is possible that the arsenic compound may accelerate the action of the hexosephosphatase of the juice, and thus by increasing the rate of circulation of the phosphate produce the permanent rise of rate. With this effect may possibly be associated a direct acceleration of the action of the fermenting complex.
The experimental decision between these alternative explanations is rendered possible by the use of a mixture of enzyme and co-enzyme free from phosphate and hexosephosphate. As has already been described (p. 55) a mixture of boiled yeast-juice, which has been treated with lead acetate, glucose or fructose, and washed zymin can be prepared which scarcely undergoes any fermentation unless phosphate be added. If now arsenates or arsenites can replace phosphate, they should be capable of setting up fermentation in such a mixture. Experiment shows that they do not possess this power. For fermentation to proceed phosphate must be present and it cannot be replaced either by arsenate or arsenite [Harden and Young, 1911, 1].
The effect of these salts on the action of the hexosephosphatase can also be ascertained by a modification of the foregoing experiment. If a hexosephosphate be made the sole source of phosphate in such a mixture as that described above, in which it must be remembered abundance of sugar is present, the rate at which fermentation can proceed will be controlled by the rate at which the hexosephosphate is decomposed with formation of phosphate. Experiment shows that in the presence of added arsenate or arsenite the rate of fermentation is largely increased, so that the effect of these salts must be to increase the rate of liberation of phosphate, or in other words, to accelerate the hydrolytic action of the hexosephosphatase.
This conclusion is even more strikingly confirmed by a comparison of the direct action of yeast-juice on hexosephosphate in presence and in absence of arsenate, as measured by the actual production of free phosphate. In a particular experiment this gave rise to 0·0707 gram of Mg2P2O7 in the absence of arsenate and 0·6136 gram of Mg2P2O7 in the presence of arsenate.
The results obtained with arsenite are throughout very similar to those given by arsenate, but are not quite so striking. It may therefore be affirmed with some confidence that the chief action of arsenates [p080] and arsenites in accelerating the rate of fermentation of sugars by yeast-juice or zymin, consists in an acceleration of the rate at which phosphate is produced from the hexosephosphate by the action of the hexosephosphatase.
It has further been found that arsenates, and to a less degree arsenites, also produce an acceleration of the rate of autofermentation of yeast-juice and of the rate at which glycogen is fermented. This turns out to be due in all probability to an increase in the activity of the glycogenase by the action of which the sugar is supplied which is the direct subject of fermentation. Thus in one case an initial rate of fermentation of glycogen of 1·9 c.c. per five minutes was increased by 0·05 molar arsenate to 9·7 and the amount of carbon dioxide evolved in two hours from 38 to 158 c.c. Even this enhanced production of glucose from glycogen, however, is not nearly sufficient for the complete utilisation of the phosphate also being liberated by the action on the hexosephosphatase, for the addition of an excess of sugar produces a much higher rate, in this case 36 c.c. per five minutes. The effect of arsenate on the rate of action of the glycogenase seems therefore to be much smaller than on that of the hexosephosphatase.
No other substances have yet been found which share these interesting properties with arsenates and arsenites, and no advance has been made towards an understanding of the mechanism of the accelerating action of these salts on the specific enzymes which are affected by them.
An observation of remarkable interest, which promises to throw light on several important features of the biochemistry of yeast, was made in 1911, and has since then formed the subject of detailed investigation by Neuberg and a number of co-workers.
It was found that yeast had the power of rapidly decomposing a large number of hydroxy-and keto-acids [Neuberg and Hildesheimer, 1911; Neuberg and Tir, 1911; see also Karczag, 1912, 1, 2]. The most important among these are pyruvic acid, CH3·CO·COOH, and a considerable number of other aliphatic a-keto-acids which are decomposed with evolution of carbon dioxide and formation of the corresponding aldehyde:—
The reaction is produced by all races of brewer's yeast which have been tried, as well as by active yeast preparations and extracts and by wine yeasts [Neuberg and Karczag, 1911, 4; Neuberg and Kerb, 1912, 2]. The phenomenon can readily be exhibited as a lecture experiment by shaking up 2 g. of pressed yeast with 12 c.c. of 1 per cent. pyruvic acid, placing the mixture in a Schrötter's fermentation tube, closing the open limb by means of a rubber stopper carrying a long glass tube and plunging the whole in water of 38–40°. Comparison tubes of yeast and water and yeast and 1 per cent. glucose may be started at the same time, and it is then seen that glucose and pyruvic acid are fermented at approximately the same rate [Neuberg and Karczag, 1911, 1]. If English top yeast be used it is well to take 0·5 per cent. pyruvic acid solution and to saturate the liquids with carbon dioxide before commencing the experiment. The production of acetaldehyde can be readily demonstrated by distilling the mixture at the close of fermentation and testing for the aldehyde either by Rimini's reaction (a blue coloration with diethylamine and sodium nitroprusside) or by means of p-nitrophenylhydrazine which precipitates the hydrazone, melting at 128·5° [Neuberg and Karczag, 1911, 2, 3]. [p082]
As the result of quantitative experiments it has been shown that 80 per cent. of the theoretical amount of acetaldehyde can be recovered. The salts of the acids are also attacked, the carbonate of the metal, which may be strongly alkaline, being formed. Thus taking the case of pyruvic acid, the salts are decomposed according to the following equation:—
Under these conditions a considerable portion of the aldehyde undergoes condensation to aldol [Neuberg, 1912]:—
This change appears to be due entirely to the alkali and not to an enzyme since the aldol obtained yields inactive β-hydroxybutyric acid on oxidation [Neuberg and Karczag, 1911, 3; Neuberg, 1912]. The various preparations derived from yeast which are capable of producing alcoholic fermentation also effect the decomposition of pyruvic acid in the same manner as living yeast. They are, however, more sensitive to the acidity of the pyruvic acid, and it is therefore advisable to employ a salt of the acid in presence of excess of a weak acid, such as boric or arsenious acid, which decomposes the carbonate formed but has no inhibiting action on the enzyme [Harden, 1913; Neuberg and Rosenthal, 1913].
As already mentioned the action is exerted on α-ketonic acids as a class and proceeds with great readiness with oxalacetic acid, COOH·CH2·CO·COOH, all the three forms of which are decomposed, with α-ketoglutaric acid, and with α-ketobutyric acid. Hydroxypyruvic acid CH2(OH)·CO·COOH is slowly decomposed yielding glycolaldehyde, CH2(OH)·CHO, and this condenses to a sugar [Neuberg and Kerb, 1912, 3; 1913, 1]. Positive results have also been obtained with diketobutyric, phenylpyruvic, p-hydroxyphenylpyruvic, phenylglyoxylic and acetonedicarboxylic acids [Neuberg and Karczag, 1911, 5].
With regard to the relation of carboxylase to the process of alcoholic fermentation, nothing definite is yet known. As Neuberg points out [see Neuberg and Kerb, 1913, 1] the universal presence of the enzyme in yeasts capable of producing alcoholic fermentation, and the extreme readiness with which the fermentation of pyruvic acid takes place create a [p083] strong presumption that the decomposition of pyruvic acid actually forms a stage in the process of the alcoholic fermentation of the sugars. On the other hand Ehrlich's alcoholic fermentation of the amino-acids (p. 87) provides another function for carboxylase—that of decomposing the α-ketonic acids produced by the deaminisation of the amino-acids. It must be remembered in this connection that carboxylase is not specific in its action, but catalyses the decomposition not only of pyruvic acid but also of a large number of other α-ketonic acids, including many of those which correspond to the amino-acids of proteins and are doubtless formed in the characteristic decomposition of these amino-acids by yeast. Carboxylase undoubtedly effects one stage in the production of alcohols from amino-acids, whether it is also the agent by which one stage in the alcoholic fermentation of sugar is brought about still remains to be proved.
A comparison of the conditions of action of carboxylase and zymase has revealed several interesting points of difference. Neuberg and Rosenthal [1913] have observed that the fermentation of pyruvic acid by maceration extract commences much more rapidly than that of glucose and interpret this to mean that in the fermentation of glucose a long preliminary process occurs before sufficient pyruvic acid has been produced to yield a perceptible amount of carbon dioxide. The long delay (3 hours) which they sometimes observed in the action of maceration juice on glucose is however by no means invariable (see p. 46), but in any case indicates that the sugar fermentation can be affected by conditions which are without influence on the pyruvic fermentation. A similar conclusion is to be drawn from the fact that the pyruvic acid fermentation is less affected by antiseptics than the glucose fermentation [Neuberg and Karczag, 1911, 4; Neuberg and Rosenthal, 1913], chloroform sufficient to stop the glucose fermentation brought about by yeast or dried yeast being usually without effect on the fermentation of the pyruvates either alone or in presence of boric or arsenious acid. A more important difference is that carboxylase decomposes pyruvic acid in the absence of the co-enzyme which is necessary for the fermentation of glucose [Harden, 1913; Neuberg and Rosenthal, 1913]. This can be demonstrated experimentally by washing dried yeast or zymin with water (see p. 63) until it is no longer capable of decomposing glucose (Harden), or by allowing maceration extract to autolyse or dialyse until it is free from co-enzyme (Neuberg and Rosenthal). The zymase of maceration extract is moreover inactivated in 10 minutes at 50–51°, whereas after this treatment the carboxylase is still active. [p084]
The only conclusion that can be legitimately drawn from these highly interesting facts is that if the decomposition of pyruvic acid actually be a stage in the alcoholic fermentation of glucose the soluble co-enzyme is required for some change precedent to this, so that in its absence the production of pyruvic acid cannot be effected.
When pure yeast is allowed to develop in a solution of sugar containing a suitable nitrogenous diet and the proper mineral salts, the liquid at the close of the fermentation contains not only alcohol and some carbon dioxide but also a considerable number of other substances, some arising from the carbonaceous and others from the nitrogenous metabolism of the cell. Prominent among the non-nitrogenous substances which are thus found in fermented sugar solutions are fusel oil, succinic acid, glycerol, acetic acid, aldehyde, formic acid, esters, and traces of many other aldehydes and acids. In addition to these substances which are found in the liquid, there are also the carbonaceous constituents of the newly formed cells of the organism, comprising the material of the cell walls, yeast gum, glycogen, complex organic phosphates, as well as other substances.
The attention of chemists has been directed to these compounds since Pasteur first emphasised their importance as essential products of the alcoholic fermentation of sugar, and his example was generally followed in attributing their origin to the sugar.
The study of cell-free fermentation by means of yeast-juice or zymin has, however, revealed the facts that certain of these substances are not formed in the absence of living cells, and that their origin is to be sought in the metabolic processes which accompany the life of the cell. Their source, moreover, has been traced not to the sugar but to the amino-acids, formed by the hydrolysis of the proteins, which occur in all such liquids as beer wort, grape juice, etc., which are usually submitted to alcoholic fermentation. This has so far been proved with certainty for the fusel oil and succinic acid, and rendered highly probable for all the various aldehydes and acids of which traces have been detected.
All forms of alcohol prepared by fermentation contain a fraction of high boiling-point, which is termed fusel oil, and amounts to about [p086] 0·1 to 0·7 per cent. of the crude spirit obtained by distillation. This material is not an individual substance, but consists of a mixture of very varied compounds, all occurring in small amount relatively to the ethyl alcohol from which they have been separated. The chief constituents of the mixture are the two amyl alcohols, isoamyl alcohol,
and d-amyl alcohol,
which contains an asymmetric carbon atom and is optically active. In addition to these, much smaller amounts of propyl alcohol and isobutyl alcohol are present, together with traces of fatty acids, aldehydes, and other substances.
The origin of these purely non-nitrogenous compounds was usually sought in the sugar of the liquid fermented, from which they were thought to be formed by the yeast itself or by the agency of bacteria [Emmerling, 1904, 1905; Pringsheim, 1905, 1907, 1908, 1909], whilst others traced their formation to the direct reduction of fatty acids. Felix Ehrlich has, however, conclusively shown in a series of masterly researches that the alcohols, and probably also the aldehydes, contained in fusel oil are in reality derived from the amino-acids which are formed by the hydrolysis of the proteins.
The close relationship between the composition of leucine,
and isoamyl alcohol,
had previously led to the surmise that a genetic relation might exist between these substances, but the idea had not been experimentally confirmed. In 1903 Ehrlich discovered [1903; 1904, 1, 2; 1907, 2; 1908; Ehrlich and Wendel, 1908, 2] that proteins also yield on hydrolysis an isomeride of leucine known as isoleucine, which has the constitution
and therefore stands to d-amyl alcohol,
in precisely the same relation as leucine to isoamyl alcohol. This suggestive fact at once directed his attention to the problem of the origin of the amyl alcohols in alcoholic fermentation. Using a pure culture of yeast, and thus excluding the participation of bacteria in the change, he found that leucine readily yielded isoamyl alcohol, and isoleucine d-amyl alcohol when these amino-acids were added in the pure state [p087] to a solution of sugar and treated with a considerable proportion of yeast [1905; 1906, 2, 3; 1907, 1, 3]. The chemical reactions involved are simple ones and are represented by the following equations:—
The experiments by which these important changes were demonstrated were of a very simple and convincing character [Ehrlich, 1907, 1]. Two hundred grams of sugar and 3 to 10 grams of the nitrogenous substance to be examined were dissolved in 2 to 2·5 litres of tap water in a 3 to 4 litre flask, the liquid was sterilised by being boiled for several hours, and after cooling 40 to 60 grams of fresh yeast were added and the flask allowed to stand at room temperature until the whole of the sugar had been decomposed by fermentation. In the earlier experiments the amyl alcohols were isolated and identified by conversion into the corresponding valerianic acids, but as a rule the fusel oil as a whole was quantitatively estimated in the filtrate by the Röse-Herzfeld method [Lunge, 1905, p. 571].
The following are typical results. (1) An experiment carried out as above without any addition of leucine gave 97·32 grams of alcohol containing 0·40 per cent. of fusel oil. (2) When 6 grams of synthetic, optically inactive leucine were added, 97·26 grams of alcohol were obtained, containing 2·11 per cent. of fusel oil, which was also optically inactive; 2·5 grams of leucine were recovered, so that 87 per cent. of the theoretical yield of isoamyl alcohol was obtained from the 3·5 grams of leucine decomposed. (3) In the presence of 2·5 grams of d-isoleucine (prepared from molasses residues), 200 grams of sugar gave 93·99 grams of alcohol, containing 1·44 per cent. of fusel oil, which was lævo-rotatory. This corresponds with 80 per cent. of the theoretical yield of d-amyl alcohol from the isoleucine added.
This change, which Ehrlich has termed the alcoholic fermentation of the amino-acids, although brought about by living yeast, does not appear to occur at all when zymin [Ehrlich, 1906, 4; Pringsheim, 1906] or yeast-juice [Buchner and Meisenheimer, 1906] is substituted for the intact organism, nor is it effected even by living yeast in the absence of a fermentable sugar [Ehrlich, 1907, 1]. The reaction appears indeed to be intimately connected with the nitrogenous metabolism of the cell, and the whole of the ammonia produced is at once assimilated and does not appear in the fermented liquid. Other amino-acids [p088] undergo a corresponding change, and the reaction appears to be a general one. Thus tyrosine, OH·C6H4·CH2·CH(NH2)·COOH, yields p-hydroxyphenylethyl alcohol, or tyrosol [Ehrlich, 1911, 1; Ehrlich and Pistschimucka, 1912, 2], OH·C6H4·CH2·CH2OH, a substance of intensely bitter taste, which was first prepared in this way and is probably one of the most important factors in determining the flavour of beers, etc. Phenylalanine, C6H5·CH2·CH(NH2)·COOH, in a similar way yields phenylethyl alcohol, C6H5·CH2·CH2OH, one of the constituents of oil of roses, whilst tryptophane,
| C6H4 | ||||
| ╱ | │ | |||
| HN | │ | |||
| ╲ | │ | |||
| H | C═══ | C·CH2·CH(NH2)·COOH |
yields tryptophol,
| C6H4 | ||||
| ╱ | │ | |||
| HN | │ | |||
| ╲ | │ | |||
| H | C═══ | C·CH2·CH2OH |
which was also first prepared in this way [Ehrlich, 1912] and has a very faintly bitter, somewhat biting taste.
The extent to which the amino-acids of a medium in which yeast is producing fermentation are decomposed in this sense depends on the amount of the available nitrogen and on the form in which it is present. Thus the addition of ammonium carbonate to a mixture of yeast and sugar was found to lower the production of fusel oil from 0·7 to 0·33 per cent. of the alcohol produced. The addition of leucine alone raised the percentage from 0·7 to 2·78, but the addition of both leucine and ammonium carbonate resulted in the formation of only 0·78 per cent. of fusel oil, The production of fusel oil therefore and the character of the constituents of the fusel oil alike depend on the composition of the medium in which fermentation occurs. This affords a ready explanation of the fact that molasses, which contains almost equal amounts of leucine and isoleucine, yields a fusel oil also containing approximately equal amounts of isoamyl alcohol and d-amyl alcohol [Marckwald, 1902], whilst corn and potatoes, in which leucine preponderates over isoleucine, yield fusel oils containing a relatively large amount of the inactive alcohol. The subject is, in fact, one of great interest to the technologist, for as Ehrlich points out "the great variety of the bouquets of wine and aromas of brandy, cognac, arrak, rum, etc., may be very simply referred to the manifold variety of the proteins of the raw materials (grapes, corn, rice, sugar cane, etc.) from which they are derived".
Yeast can also form fusel oil at the expense of its own protein, but this only occurs to any considerable extent when the external [p089] supply of nitrogen is insufficient. Under these circumstances the amino-acids formed by autolysis may be decomposed and their nitrogen employed over again for the construction of the protein of the cell.
The yield is also influenced by the condition of the yeast employed with regard to nitrogen, a yeast poor in nitrogen being more efficacious in decomposing amino-acids than one which is already well supplied with nitrogenous materials. The nature of the carbonaceous nutriment and finally the species of yeast are also of great importance [see Ehrlich, 1911, 2; Ehrlich and Jacobsen, 1911].
A very important characteristic of the action of yeast on the amino-acids is that the two stereo-isomerides of these optically active compounds are fermented at different rates. When inactive, racemic leucine is treated with yeast and sugar, the naturally occurring component, the l-leucine, is more rapidly attacked, so that if the experiment be interrupted at the proper moment the other component, the d-leucine, alone is present and may be isolated in the pure state. In an actual experiment 3·8 grams of this component were obtained in the pure state from 10 grams of dl-leucine [Ehrlich, 1906, 1], so that the whole of the l-leucine (5 grams) had been decomposed but only 1·2 grams of the d-leucine. This mode of action has been found to be characteristic of the alcoholic fermentation of the amino-acids by yeast. In all the instances so far observed, both components of the inactive amino-acid are attacked, but usually the naturally occurring isomeride is the more rapidly decomposed, although in the case of β-aminobutyric acid both components disappear at the same rate [Ehrlich and Wendel, 1908, 1]. This reaction therefore must be classed along with the action of moulds on hydroxy-acids [McKenzie and Harden, 1903], and the action of lipase on inactive esters [Dakin, 1903, 1905], in which both isomerides are attacked but at unequal rates, and differs sharply from the action of yeast itself on sugars [Fischer and Thierfelder, 1894], and of emulsin, maltase, etc., which only act on one isomeride and leave the other entirely untouched [see Bayliss, 1914, pp. 55, 77, 117].
The origin of the succinic acid formed in fermentation has also been traced by Ehrlich [1909] to the alcoholic fermentation of the amino-acids. It was shown by Buchner and by Kunz [1906] that succinic acid like fusel oil is not formed during fermentation by yeast-juice or zymin, and, in the light of Ehrlich's work on fusel oil, several [p090] modes of formation appeared possible for this substance [Ehrlich, 1906, 3]. The dibasic amino-acids might, for example, undergo simple reduction, the NH2 group being removed as ammonia and replaced by hydrogen. Aspartic acid would thus pass into succinic acid:—
This change can be effected in the laboratory only by heating with hydriodic acid. Biologically it has been observed [E. and H. Salkowski, 1879] when aspartic acid is submitted to the action of putrefactive bacteria, and almost quantitatively when Bacillus coli communis is cultivated in a mixture of aspartic acid and glucose [Harden, 1901]. In this case a well-defined source of hydrogen exists in the glucose, which when acted on by this bacillus yields a large volume of gaseous hydrogen, which is not evolved in the presence of aspartic acid. Some such source is also available in the case of yeast, although it cannot be chemically defined, for this organism is known to produce many reducing actions, which are usually ascribed to the presence of reducing ferments or reductases in the cell.
A similar action would convert glutamic acid,
into glutaric acid,
which also is found among the products of fermentation, whilst the monamino-acids would pass into the simple fatty acids.
On submitting these ideas to the test of experiment, however, Erhlich found that the addition of aspartic acid did not in any way increase the yield of succinic acid, and that of all the amino-acids which were tried only glutamic acid, COOH·CH2·CH2·CH(NH2)·COOH, produced a definite increase in the amount of this substance. Further experiments showed that glutamic acid was actually the source of the succinic acid, the relations being quite similar to those which exist for the production of fusel oil.
Succinic acid is formed whenever sugar is fermented by yeast, even in the absence of added nitrogenous matter, and amounts to 0·2 to 0·6 per cent. of the weight of the sugar decomposed, its origin in this case being the glutamic acid formed by the autolysis of the yeast protein. When some other source of nitrogen is present, such as asparagine or an ammonium salt, the amount falls to 0·05 to 0·1. If glutamic acid be added it rises to about 1 to 1·5 per cent. but falls again to about 0·05 to 0·1 when other sources of nitrogen, such as asparagine or ammonium salts, are simultaneously available, either in the presence or [p091] absence of added glutamic acid. As in the case of fusel oil, the production does not occur in the absence of sugar, and is not effected by yeast-juice or zymin.
The chemical reaction involved in the production of succinic acid differs to some extent from that by which fusel oil is formed, inasmuch as an oxidation is involved:—
From analogy with the production of amyl alcohol from leucine, glutamic acid would be expected to yield γ-hydroxybutyric acid:—
As a matter of fact this substance cannot be detected among the products of fermentation, but succinic acid as already explained is formed. This acid might, however, possibly be formed by the oxidation of the γ-hydroxybutyric acid:—
although this change is on biological grounds improbable.
The conversion of the group —CH(NH2)— into the terminal CH2·OH in fusel oil, or COOH in succinic acid, may possibly be effected in several different ways, the most probable of which are the following:—
I. Direct elimination of carbon dioxide, followed by hydrolysis of the resulting amine:—
The reaction (1) is actually effected by many bacteria and has been employed for the preparation of bases from amino-acids [cf. Barger, 1914, p. 7], although there is no direct evidence that it can be brought about by yeast. On the other hand reaction (2) has actually been observed with some yeasts. Thus it has been found [Ehrlich and Pistschimuka, 1912, 1] that many "wild" yeasts produce this change with great readiness in presence of sugar, glycerol or ethyl alcohol as sources of carbon and grow well in media in which amines, such as p-hydroxyphenylethylamine or iso-amylamine, form the only source of nitrogen. Willia anomala (Hansen), a yeast which forms surface growths, succeeds admirably under these conditions, whereas culture yeasts are much less active in this way, although they produce a certain amount of change. It is therefore possible that this mode of decomposition plays some part in the production of fusel oil, but in the case of culture yeasts it is entirely subordinated to the mode next to be discussed. [p092]
II. Oxidative removal of the –NH2 group with formation of an α-ketonic acid:—
followed by the decomposition of the ketonic acid into carbon dioxide and an aldehyde and the subsequent reduction or oxidation of the aldehyde:—
The evidence for the occurrence of reaction (1) is supplied by the experiments of Neubauer and Fromherz [1911]. Having previously found that amino-acids undergo a change of this kind in the animal body, Neubauer investigated their behaviour towards yeast. Taking dl-phenylaminoacetic acid, C6H5·CH(NH2)·COOH, it was found that the changes produced were essentially the same as in the animal body. The l-component of the acid was partly acetylated and partly unchanged, whereas the d-component of the acid yielded benzyl alcohol, C6H5·CH2·OH, phenylglyoxylic acid, C6H5·CO·COOH, and the hydroxy-acid C6H5·CH(OH)·COOH. Since however this hydroxy-acid was produced in the l-form it probably arose by the asymmetric reduction of phenylglyoxylic acid, a reaction which can be effected by yeast as was also found to be the case in the animal body [see Dakin, 1912, pp. 52, 78]. Moreover it was shown that when the effects of yeast on a ketonic acid and the corresponding hydroxy-acid were compared, the alcohol was formed in much better yield from the ketonic acid (70 per cent.) than from the hydroxy-acid (3–4 per cent.), the actual example being the production of tyrosol (p-hydroxyphenylethyl alcohol), OH·C6H4·CH2·CH2OH, from p-hydroxyphenylpyruvic acid, OH·C6H4·CH2·CO·COOH, and p-hydroxyphenyl-lactic acid, OH·C6H4·CH2·CH(OH)·COOH respectively.
Neubauer by these experiments established two extremely important points. 1. That the amino-acids actually yield the corresponding α-ketonic acids when treated with yeast and sugar solution. 2. That the a-ketonic acids under similar conditions give the alcohol containing one carbon atom less in good yield, whereas the corresponding hydroxy-acids only give an extremely small amount of these alcohols.
It is therefore probable that at an early stage in the decomposition of the amino-acids by yeast a ketonic acid is produced, which then undergoes further change.
| O | H | |
| │ | ||
| R· | C | —COOH |
| │ | ||
| N | H2 |
The spontaneous production of ketonic aldehydes from amino-acids and from hydroxy-acids in aqueous solution, which has been demonstrated by Dakin and Dudley [1913], points however to the possibility that the ketonic acid may be a secondary product derived from the corresponding ketonic aldehyde [see also Dakin, 1908; Neuberg, 1908, 1909]. This itself may either arise directly from the amino-acid or from a previously formed hydroxy-acid, the latter alternative being, however, improbable in view of the small yield of alcohol obtained from hydroxy-acids by the action of yeast in the experiments of Neubauer and Fromherz.
| R·CH(NH2)·COOH | → | R·CH(OH)·COOH |
| ⇅ | ⇅ | |
| R·CO·CHO | ||
| ↓ + Oxygen | ||
| R·CO·COOH | ||
(2) Whatever be the exact mode by which the ketonic acid is formed, it appears most probable that a compound of this nature forms the starting-point for the next stage in the production of the alcohols. The researches of Neuberg, which have already been discussed on p. 81, have revealed a mechanism in yeast—the enzyme carboxylase—by which these α-ketonic acids are rapidly broken up into an aldehyde and carbon dioxide:
and it can scarcely be doubted that this is the actual course of the reaction.
(3) The final conversion of the aldehyde into the corresponding alcohol is also a change which it has been proved can be effected by yeast [Neuberg and Rosenthal, 1913] probably by the aid of the reductase which is one of the weapons in its armoury of enzymes.
Yeast is capable of producing many vigorous reducing actions and rapidly reduces methylene blue and sodium selenite. It is in all probability due to a reaction of this kind that the iso-amylaldehyde and isovaleraldehyde were reduced to the alcohols in Neuberg and Steenbock's experiments [1913, 1914], and that considerable quantities of ethyl alcohol are formed in the sugar free fermentation of pyruvic acid [Neuberg and Kerb, 1913, 1] (see later p. 110 for a discussion of this question).
A further possibility exists that in some cases the aldehyde may [p094] be simultaneously oxidised and reduced or the molecule of one aldehyde reduced and that of another oxidised with production of the corresponding acid and alcohol by an "aldehydo-mutase," similar to that which has been observed by Parnas [1910] in many animal tissues. Finally the aldehyde may simply be converted into the corresponding acid by oxidation as appears to take place in the formation of succinic acid.
The intermediate production of an aldehyde would thus be consistent both with the production of alcohols and acids from amino-acids.
Fusel oil would be formed by the reduction of the aldehydes arising from the simple monobasic amino-acids, succinic acid would be produced by oxidation of the aldehyde derived from the dibasic glutamic acid.
In favour of this view is to be adduced the fact that aldehydes such as isobutyraldehyde and valeraldehyde have been found in crude spirit, whilst acetaldehyde is a regular product of alcoholic fermentation [see Ashdown and Hewitt, 1910]. Benzaldehyde, moreover, has been actually detected as a product of the alcoholic fermentation of phenylaminoacetic acid, C6H5·CH(NH2)·COOH [Ehrlich, 1907, 1]. Further, the aldehydes so produced would readily pass by oxidation into the corresponding fatty acids, small quantities of which are invariably produced in fermentation.
This view of the nature of the alcoholic fermentation of the amino-acids is undoubtedly to be preferred to that previously suggested by Ehrlich [1906, 3] according to which a hydroxy-acid is first formed and then either directly decomposed into an alcohol and carbon dioxide or into an aldehyde and formic acid, the aldehyde being reduced and the formic acid destroyed (see p. 115).
| R·CH(NH2)·COOH → | R·CH( | OH)·COOH | |
| ↓ | or | ↓ | |
| R·CH2OH + CO2 | R·CHO + H·CO2H | ||
| ↓ | |||
| R·CH2OH | |||
The most probable course of the decomposition by which isoamyl alcohol and succinic acid are produced from leucine and glutamic acid respectively is therefore the following:—
Of the three chief by-products of alcoholic fermentation, only glycerol remains at present referable directly to the sugar. This substance, as shown by the careful experiments of Buchner and Meisenheimer [1906], is formed by the action both of yeast-juice and zymin to the extent of 3·8 per cent. of the sugar decomposed, and no other source for its production has so far been experimentally demonstrated. If it be true that during the decomposition of sugar into alcohol and carbon dioxide, substances containing three carbon atoms are formed as intermediate compounds (see p. 100), it is obvious that these might by reduction be converted into glycerol which would thus be a true by-product of the alcoholic fermentation of sugar. [See Oppenheimer, 1914, 2.] It has, however, been suggested that it may in reality be a product of decomposition of lipoid substances or of the nuclein of the cell (Ehrlich).
The effect of Ehrlich's work has been clearly to distinguish the chemical changes involved in the production of fusel oil and succinic acid from those concerned in the decomposition of sugar into alcohol and carbon dioxide, and to bring to light a most important series of reactions by means of which the yeast-cell is able to supply itself with nitrogen, one of the indispensable conditions of life.
It has long been the opinion of chemists that the remarkable and almost quantitative conversion of sugar into alcohol and carbon dioxide during the process of fermentation is most probably the result of a series of reactions, during which various intermediate products are momentarily formed and then used up in the succeeding stage of the process. No very good ground can be adduced for this belief except the contrast between the chemical complexity of the sugar molecule and the comparative simplicity of the constitution of the products. Many attempts have, however, been made to obtain evidence of such a series of reactions, and numerous suggestions have been made of probable directions in which such changes might proceed. In making these suggestions, investigators have been guided mainly by the changes which are produced in the hexoses by reagents of known composition. The fermentable hexoses, glucose, fructose, mannose, and galactose, appear to be relatively stable in the presence of dilute acids at the ordinary temperature, and are only slowly decomposed at 100°, more rapidly by concentrated acids, with formation of ketonic acids, such as levulinic acid, and of coloured substances of complex and unknown constitution.
In the presence of alkalis, on the other hand, the sugar molecule is extremely susceptible of change. In the first place, as was discovered by Lobry de Bruyn [1895; Bruyn and Ekenstein, 1895; 1896; 1897, 1, 2, 3, 4], each of the three hexoses, glucose, fructose, and mannose is converted by dilute alkalis into an optically almost inactive mixture containing all three, and probably ultimately of the same composition whichever hexose is employed as the starting-point.
This interesting phenomenon is most simply explained on the assumption that in the aqueous solution of any one of these hexoses, along with the molecules of the hexose itself, there exists a small proportion of those of an enolic form which is common to all the three hexoses, as illustrated by the following formulæ, the aldehyde formulæ [p097] being employed instead of the γ-oxide formulæ for the sake of simplicity:—
| C | HO | C | HO | C | H2(OH) | C | H(OH) | ||||||
| │ | │ | │ | ║ | ||||||||||
| H | C | OH | HO | C | H | C | O | C | OH | ||||
| │ | │ | │ | │ | ||||||||||
| HO | C | H | HO | C | H | HO | C | H | HO | C | H | ||
| │ | │ | │ | │ | ||||||||||
| H | C | OH | H | C | OH | H | C | OH | H | C | OH | ||
| │ | │ | │ | │ | ||||||||||
| H | C | OH | H | C | OH | H | C | OH | H | C | OH | ||
| │ | │ | │ | │ | ||||||||||
| C | H2(OH) | C | H2(OH) | C | H2(OH) | C | H2(OH) | ||||||
| Glucose | Mannose | Fructose | Enolic form |
||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
This enolic form is capable of giving rise to all three hexoses, and the change by which the enolic form is produced and converted into an equilibrium mixture of the three corresponding hexoses is catalytically accelerated by alkalis, or rather by hydroxyl ions. In neutral solution the change is so slow that it has never been experimentally observed; in the presence of decinormal caustic soda solution at 70° the conversion is complete in three hours. Precisely similar effects are produced with galactose, which yields an equilibrium mixture containing talose and tagatose, sugars which appear not to be fermentable.
The continued action even of dilute alkaline solutions carries the change much further and brings about a complex decomposition which is much more rapidly effected by more concentrated alkalis and at higher temperatures. This change has been the subject of very numerous investigations [for an account of these see E. v. Lippmann, 1904, pp. 328, 713, 835], but for the present purpose the results recently obtained by Meisenheimer [1908] may be quoted as typical. Using normal solutions of caustic soda and concentrations of from 2 to 5 grams of hexose per 100 c.c., it was found that at air temperature in 27 to 139 days from 30 to 54 per cent. of the hexose was converted into inactive lactic acid, C3H6O3, from 0·5 to 2 per cent. into formic acid, CH2O2, and about 40 per cent. into a complex mixture of hydroxy-acids, containing six and four carbon atoms in the molecule. Usually only about 74 to 90 per cent. of the sugar which had disappeared was accounted for, but in one case the products amounted to 97 per cent. of the sugar. About 1 per cent. of the sugar was probably converted into alcohol and carbon dioxide. No glycollic acid, oxalic acid, glycol, or glycerol was produced.
The fact that alcohol is actually formed by the action of alkalis on sugar was established by Buchner and Meisenheimer [1905], who obtained small quantities of alcohol (1·8 to 2·8 grams from 3 kilos. of cane sugar) by acting on cane sugar with boiling concentrated caustic soda [p098] solution. It is evident that under these conditions an extremely complex series of reactions occurs, but the formation of alcohol and carbon dioxide and of a large proportion of lactic acid deserves more particular attention.
The direct formation of alcohol from sugar by the action of alkalis appears first to have been observed by Duclaux [1886], who exposed a solution of glucose and caustic potash to sunlight and obtained both alcohol and carbon dioxide. As much as 2·6 per cent. of the sugar was converted into alcohol in a similar experiment made by Buchner and Meisenheimer [1904]. When the weaker alkalis, lime water or baryta water, were employed instead of caustic potash, however, no alcohol was formed, but 50 per cent. of the sugar was converted into inactive lactic acid [Duclaux, 1893, 1896]. Duclaux therefore regarded the alcohol and carbon dioxide as secondary products of the action of a comparatively strong alkali on preformed lactic acid. Ethyl alcohol can, in fact, be produced from lactic acid both by the action of bacteria [Fitz, 1880] and of moulds [Mazé, 1902], and also by chemical means. Thus Duclaux [1886] found that calcium lactate solution exposed to sunlight underwent decomposition, yielding alcohol and calcium carbonate and acetate, whilst Hanriot [1885, 1886], by heating calcium lactate with slaked lime obtained a considerable quantity of a liquid which he regarded as ethyl alcohol, but which was shown by Buchner and Meisenheimer [1905] to be a mixture of ethyl alcohol with isopropyl alcohol.
It appears, therefore, that inactive lactic acid can be quite readily obtained in large proportion from the sugars by the action of alkalis, whilst alcohol can only be prepared in comparatively small amount and probably only as a secondary product of the decomposition of lactic acid.
The study of the action of alkalis on sugar has, however, yielded still further information as regards the mechanism of the reaction by which lactic acid is formed. A considerable body of evidence has accumulated, tending to show that some intermediate product of the nature of an aldehyde or ketone containing three carbon atoms is first formed.
Thus Pinkus [1898] and subsequently Nef [1904, 1907], by acting on glucose with alkali in presence of phenylhydrazine obtained the osazone of methylglyoxal, CH3·CO·CHO. This osazone may be formed either from methylglyoxal itself, from acetol, CH3·CO·CH2·OH, or from lactic aldehyde, CH3·CH(OH)·CHO [Wohl, 1908]. Methylglyoxal itself may also be regarded as a secondary [p099] product derived from glyceraldehyde, CH2(OH)·CH(OH)·CHO, or dihydroxyacetone, CH2(OH)·CO·CH2(OH), by a process of intramolecular dehydration, so that the osazone might also be derived indirectly from either of these compounds [see also Neuberg and Oertel, 1913]. Methylglyoxal itself readily passes into lactic acid when it is treated with alkalis, a molecule of water being taken up:—
Further evidence in the same direction is afforded by the interesting discovery of Windaus and Knoop [1905], that glucose is converted by ammonia in presence of zinc hydroxide into methyliminoazole,
| CH3· | C | ·NH· | C | H | |
| ║ | ║ | , | |||
| H | C | ──── | N |
a substance which is a derivative of methylglyoxal.
The idea suggested by Pinkus that acetol is the first product of the action of alkalis on sugar has been rendered very improbable by the experiments of Nef, and the prevailing view (Nef, Windaus and Knoop, Buchner and Meisenheimer) is that the first product is glyceraldehyde, which then passes into methylglyoxal, and finally into lactic acid:—
All these changes may occur at ordinary temperatures in the presence of a catalyst, and in so far resemble the processes of fermentation by yeasts and bacteria.
The first attempt to suggest a scheme of chemical reactions by which the changes brought about by living organisms might be effected was made in 1870 by Baeyer [1870], who pointed out that these decompositions might be produced by the successive removal and re-addition of the elements of water. The result of this would be to cause an accumulation of oxygen atoms towards the centre of the chain of six carbon atoms, which, in accordance with general experience, would render the chain more easily broken. Baeyer formulated the changes characteristic of the alcoholic and lactic fermentations as follows, the intermediate stages being derived from the hydrated aldehyde formula of glucose by the successive removal and addition of the elements of water: [p100]
| I. | |
|---|---|
| C | H2·OH |
| │ | |
| C | H·OH |
| │ | |
| C | H·OH |
| │ | |
| C | H·OH |
| │ | |
| C | H·OH |
| │ | |
| C | H(OH)2 |
| II. | |
|---|---|
| C | H2 . . . OH |
| │ | |
| C | OH . . H |
| │ | |
| C | . . OH . . H |
| │ | |
| C | OH . . . H |
| │ | |
| C | OH . . . H |
| │ | |
| C | H . . . (OH)2 |
| III. | |
|---|---|
| C | H3 |
| │ | |
| C | H . OH |
| │ | |
| C | (OH)2 |
| │ | |
| C | (OH)2 |
| │ | |
| C | (OH)2 |
| │ | |
| C | H3 |
| IV. | |||
|---|---|---|---|
| C | H3 | ||
| │ | |||
| C | H(OH) | ||
| │ | |||
| C | O | ||
| ╱ | |||
| O | |||
| ╲ | |||
| C | O | ||
| │ | |||
| C | H(OH) | ||
| │ | |||
| C | H3 | ||
| V. | |||
|---|---|---|---|
| C | H3 | ||
| │ | |||
| C | H2 | ||
| ╱ | |||
| O | |||
| ╲ | |||
| C | O | ||
| ╱ | |||
| O | |||
| ╲ | |||
| C | O | ||
| ╱ | |||
| O | |||
| ╲ | |||
| C | H2 | ||
| │ | |||
| C | H3 | ||
The immediate precursor of alcohol and carbon dioxide is here seen to be the anhydride of ethoxycarboxylic acid (V), whilst that of lactic acid is lactic anhydride (IV). (Baeyer does not appear, as recently stated by Meisenheimer [1907, p. 8], Wohl [1907, 2], and Buchner and Meisenheimer [1909] to have suggested that lactic acid was an intermediate product in alcoholic fermentation, but rather to have represented independently the course of the two different kinds of fermentation, the alcoholic and the lactic.)
It was subsequently pointed out by Buchner and Meisenheimer [1904] that Baeyer's principle of oxygen accumulation might be applied in a different way, so that a ketonic acid would be produced, the decomposition of which, in a manner analogous to that of acetoacetic acid, would lead to the formation of two molecules of lactic acid, from which the final products alcohol and carbon dioxide might be directly derived, as shown in the following formulæ:—
| C | HO |
| · | |
| C | H(OH) |
| · | |
| C | H(OH) |
| · | |
| C | H(OH) |
| · | |
| C | H(OH) |
| · | |
| C | H2(OH) |
| C | OOH |
| · | |
| C | H(OH) |
| · | |
| C | H2 |
| · | |
| C | O |
| · | |
| C | H(OH) |
| · | |
| C | H3 |
| C | OOH |
| · | |
| C | H(OH) |
| · | |
| C | H3 |
| ─ | ─── |
| C | OOH |
| · | |
| C | H(OH) |
| · | |
| C | H3 |
| C | O2 |
| ─ | ─── |
| C | H2·OH |
| · | |
| C | H3 |
| ─ | ─── |
| C | O2 |
| ─ | ─── |
| C | H2·OH |
| · | |
| C | H3 |
A scheme based on somewhat different principles has been propounded by Wohl [Lippmann, 1904, p. 1891], and has been accepted by Buchner and Meisenheimer [1905] as more probable than that quoted above. Wohl and Oesterlin [1901] were able to trace experimentally the various stages of the conversion of tartaric acid (I) into oxalacetic acid (III), which can be carried out by reactions taking place at the ordinary temperature, and they found that the first stage consisted in the removal of the elements of water leaving an unsaturated hydroxy derivative (II) which in the second stage underwent intramolecular change into the corresponding keto-compound (III): [p101]
| C | OOH | C | OOH | C | OOH | |||||
| · | · | · | ||||||||
| C | H(OH) | H | C | (OH) | C | O | ||||
| · | − | · | = | ║ | ⇌ | · | ||||
| C | H(OH) | OH | C | H | C | H3 | ||||
| · | · | · | ||||||||
| C | OOH | C | OOH | C | OOH | |||||
| I. Tartaric acid |
II. | III. Oxalacetic acid. |
||||||||
|---|---|---|---|---|---|---|---|---|---|---|
This change differs in principle from that assumed by Baeyer, inasmuch as the second stage is not effected by the re-addition of water, but by the keto-enol transformation, which is now usually ascribed to the migration of the hydrogen atom, although the same result can theoretically be arrived at by the addition and removal of the elements of water. The analogy of this process to what might be supposed to occur in the conversion of sugar into carbon dioxide and alcohol was pointed out by Wohl and Oesterlin, and subsequently Wohl developed a theoretical scheme of reactions by which the process of alcoholic fermentation could be represented. In the first place the elements of water are removed from the α and β carbon atoms of glucose (I) and the resulting enol (II) undergoes conversion into the corresponding ketone (III), which has the constitution of a condensation product of methylglyoxal and glyceraldehyde, and hence is readily resolved by hydrolysis into these compounds (IV). The glyceraldehyde passes by a similar series of changes (V, VI) into methylglyoxal, and this is then converted by addition of water into lactic acid (VII), a reaction which is common to all ketoaldehydes of this kind. Finally, the lactic acid is split up into alcohol and carbon dioxide (VIII):—
| C | HO | C | HO | C | HO | |||
| │ | │ | │ | ||||||
| C | H(OH) | C | (OH) | C | O | |||
| │ | H | ║ | │ | |||||
| C | H(OH) | − | OH | C | H | ⇌ | C | H2 |
| │ | │ | │ | ||||||
| C | H(OH) | C | H(OH) | C | H(OH) | |||
| │ | │ | │ | ||||||
| C | H(OH) | C | H(OH) | C | H(OH) | |||
| │ | │ | │ | ||||||
| C | H2(OH) | C | H2(OH) | C | H2(OH) | |||
| I. Glucose. |
II. | III. | ||||||
|---|---|---|---|---|---|---|---|---|
| Methyl- glyoxal |
||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| C | HO | C | OOH | C | O2 | |||||||
| │ | │ | ─ | ─── | |||||||||
| C | O | +H2O | C | H(OH) | C | H2OH | ||||||
| │ | │ | │ | ||||||||||
| C | H3 | C | H3 | C | H3 | |||||||
| C | HO | C | HO | C | HO | C | OOH | C | O2 | |||
| │ | │ | │ | │ | ─ | ──── | |||||||
| C | H(OH) | H | C | (OH) | ⇌ | C | O + H2O | C | H(OH) | C | H2OH | |
| │ | · | ║ | │ | │ | │ | |||||||
| C | H2(OH) | − HO | C | H2 | C | H3 | C | H3 | C | H3 | ||
| IV. Glyceral- dehyde. |
V. | VI. Methyl- glyoxal. |
VII. Lactic acid. |
VIII. Alcohol and carbon dioxide. |
||||||||
This scheme agrees well with the current ideas as to the formation of lactic acid from glucose under the influence of alkalis (p. 99). It postulates the formation as intermediate products of no less than three compounds containing a chain of three carbon atoms—glyceraldehyde, methylglyoxal, and lactic acid.
A practical interest was given to these various schemes by the fact that Buchner and Meisenheimer adduced experimental evidence in favour of the view that lactic acid is an intermediate product in the formation of alcohol and carbon dioxide from sugar by fermentation [1904, 1905, 1906, 1909].
These observers proved by a series of very careful analyses that yeast-juice frequently, but not invariably, contains small quantities of lactic acid, not exceeding 0·2 per cent. When yeast-juice is incubated alone or with sugar the amount of lactic acid may either increase or decrease. Moreover, lactic acid added to the juice is sometimes diminished and sometimes increased in quantity. On the whole it appears that the addition of a considerable quantity of sugar or of some lactic acid favours the disappearance of lactic acid. Juices of low fermenting power produce a diminution in the lactic acid present, those of high fermenting power an increase.
In all cases the amounts of lactic acid either produced or destroyed are very small in relation to the volume of the yeast-juice employed.
Throughout the whole series of experiments the greatest increase amounted to 0·47 per cent. on the juice employed, and the greatest decrease to 0·3 per cent. [See also Oppenheimer, 1914, 1.] Buchner and Meisenheimer at one time regarded these facts as strong evidence that lactic acid is an intermediate product of alcoholic fermentation. It was thought probable that the production of alcohol and carbon dioxide from sugar occurred in at least two stages and under the influence of two distinct enzymes. The first stage consisted in the conversion of sugar into lactic acid, and for the enzyme which brought about this decomposition was reserved the name zymase or yeast-zymase. The lactic acid was then broken down into alcohol and carbon dioxide by the second enzyme, lactacidase.
This theory, which is quite in harmony with the current ideas as to the mode of decomposition of sugars by alkalis, and is also consistent with Wohl's scheme of reactions, is open to adverse criticism from several points of view. In the first place, it is noticeable that the total amount of lactic acid used up by the juice is extremely small, even [p103] in the most favourable cases, relatively to the amount of the juice [Harden, 1905], and it may be added to the sugar-fermenting power of the juice. Moreover, as pointed out by Buchner and Meisenheimer themselves [1909], no proof is afforded that the lactic acid which disappears is converted into alcohol and carbon dioxide. It is not even certain, although doubtless probable, that the lactic acid which occurs or is produced in the juice is really derived from sugar.
The most weighty criticism of the theory is that of Slator [1906, 1907; 1908, 1, 2], which is based on the consideration that if lactic acid be an intermediate product of alcoholic fermentation the reaction by which it is fermented must proceed at least as rapidly as that by which it is formed, in order to prevent accumulation of lactic acid. The fermentation of lactic acid by yeast should therefore proceed at least as rapidly as that of glucose. So far is that from being the case that it has been experimentally demonstrated that lactic acid is not fermented at all by living yeast. This conclusion was rendered extremely probable by Slator, who showed that lactic acid, even in concentrations insufficient to prevent the fermentation of glucose, is not fermented to any considerable extent. The final proof that lactic acid is neither formed nor fermented by pure yeast has been brought by Buchner and Meisenheimer in a series of very careful quantitative experiments carried out with a pure yeast and with strict precautions against bacterial contamination [1909, 1910].
At first sight this fact appears decisive against the validity of the lactic acid theory, and it is recognised as such by Buchner and Meisenheimer. Wohl has, however, suggested that the non-fermentability of lactic acid by yeast is not really conclusive [1907, 1; see also Franzen and Steppuhn, 1912, 1]. The production of lactic acid from glucose is attended by the evolution of a considerable amount of heat (22 cal.), and it is possible that at the moment of production the molecule of the acid is in a condition of activity corresponding with a much higher temperature than the average temperature of the fermenting liquid. Under these circumstances the molecule would be much more susceptible of chemical change than at a later period when temperature equilibrium had been attained. It has, however, been pointed out by Tafel [1907], that such a decomposition of the lactic acid would occur at the very instant of formation of the molecule, so that no ground remains even on this view for assuming the actual existence of lactic acid as a definite intermediate product. It has also been suggested by Luther [1907] that an unknown isomeride of lactic acid is formed as an intermediate product and fermented, and that traces of lactic [p104] acid are formed by a secondary reaction from this, but no satisfactory evidence for this view is forthcoming. There still remains a doubt as to whether the living yeast-cell is permeable to lactic acid, a fact which would of course afford a very simple explanation of the non-fermentability of the acid. Apart from this, however, it is difficult, in face of the evidence just quoted, to believe that lactic acid is in reality an intermediate product in alcoholic fermentation.
As regards the fermentability by yeast of compounds containing three carbon atoms, which may possibly appear as intermediate products in the transformation of sugar into carbon dioxide and alcohol, many experiments have been carried out, with somewhat uncertain results. Care has to be taken that the substance to be tested is not added in such quantity as to inhibit the fermenting power of the yeast or yeast-juice, and further that the conditions are such that the substance in question, often of a very unstable nature, is not converted by some chemical change into a different fermentable compound. It is also possible that the substance to be tested may accelerate the rate of autofermentation in a similar manner to arsenates (pp. 80, 126) and many other substances. These are all points which have not up to the present received sufficient attention. In the case of living yeast the further question arises of the permeability of the cell.
Methylglyoxal, CH3·CO·CHO, has been tested by Mayer [1907] and Wohl [1907, 2] with yeast, and by Buchner and Meisenheimer both with acetone-yeast [1906] and yeast-juice [1910], in every case with negative results, but it may be noted that the concentration employed in the last mentioned of these experiments was such as considerably to diminish the autofermentation of the juice.
Glyceraldehyde, CH2(OH)·CH(OH)·CHO, was also tested with yeast with negative results by Wohl [1898] and by Emmerling [1899], who employed a number of different yeasts. The same negative result attended the experiments of Piloty [1897] and Emmerling [1899] with pure dihydroxyacetone. Fischer and Tafel [1888, 1889], however, had previously found that glycerose, a mixture of glyceraldehyde and dihydroxyacetone prepared by the oxidation of glycerol, was readily fermented by yeast, agreeing in this respect with the still older observations of Van Deen and of Grimaux. The reason for this diversity of result has not been definitely ascertained, but it has been supposed by Emmerling to lie in the formation of some fermentable sugar from [p105] glycerose when the latter is subjected to too high a temperature during its preparation.
On the other hand, Bertrand [1904] succeeded in fermenting pure dihydroxyacetone by treating a solution of 1 gram in 30 c.c. of liquid with a small quantity of yeast for ten days at 30°, the best result being a fermentation of 25 per cent. of the substance taken. Moreover, Boysen-Jensen [1908, 1910, 1914] states that he has also observed both the formation from glucose and the fermentation of this substance by living yeast, but the amounts of alcohol and carbon dioxide produced were so minute and the evidence for the production of dihydroxyacetone so inconclusive that the experiments cannot be regarded as in any way decisive [see Chick, 1912; Euler and Fodor, 1911; Karauschanoff, 1911; Buchner and Meisenheimer, 1912]. A careful investigation by Buchner [1910] and Buchner and Meisenheimer [1910] has led them to the conclusion that both glyceraldehyde and dihydroxyacetone are fermentable. Glyceraldehyde exerts a powerful inhibiting action both on yeast and yeast-juice, and was only found to give rise to a very limited amount of carbon dioxide, quantities of 0·15 to 0·025 gram being treated with 1 gram of yeast or 5 c.c. of yeast-juice and a production of 4 to 12 c.c. of carbon dioxide being attained.
When 0·1 gram of dihydroxyacetone in 5 c.c. of water was brought in contact with 1 gram of living yeast, about half was fermented, 17 c.c. of carbon dioxide (at 20° and 600 mm.) being evolved in excess of the autofermentation of the yeast (13 c.c.). A much greater effect was obtained by the aid of yeast-juice, and the remarkable observation was made that whilst yeast-juice alone produced comparatively little action a mixture of yeast-juice and boiled yeast-juice was much more effective, quantities of 20 to 50 c.c. of yeast-juice mixed with an equal volume of boiled juice, which in some experiments was concentrated, yielding with 0·4, 1, and 2 grams of dihydroxyacetone almost the theoretical amount of carbon dioxide and alcohol in excess of that evolved in the absence of this substance. It was further observed that the fermentation of this substance commenced much more slowly than that of glucose. No explanation of either of these facts has at present been offered. The conclusion drawn from their experiments by Buchner and Meisenheimer that dihydroxyacetone is readily fermentable, was confirmed by Lebedeff [1911, 1], who further made the important observation that during the fermentation of dihydroxyacetone the same hexosephosphoric acid is produced as is formed during the fermentation of the hexoses. Lebedeff accordingly propounded a scheme of alcoholic fermentation according to which the hexose [p106] was first converted into two molecules of triose, the latter being first esterified to triosephosphoric acid and then condensed to hexosediphosphoric acid, which then underwent fermentation, after being hydrolysed to phosphoric acid, and some unidentified substance, probably an unstable modification of a hexose, much more readily attacked by an appropriate enzyme than the original glucose or fructose [1911, 1, pp. 2941–2].
The idea that the sugar is first converted into triose and this into triosemonophosphoric acid had been previously suggested by Iwanoff who postulated the agency of a special enzyme termed synthease [1909, 1], and supposed that this triosemonophosphoric acid was then directly fermented to alcohol, carbon dioxide and phosphoric acid. According both to Iwanoff and Lebedeff the phosphoric ester is an intermediate product and its decomposition provides this sole source of carbon dioxide and alcohol. This is quite inconsistent with the facts recounted above (Chap. III), which prove that the formation of the hexosephosphate is accompanied by an amount of alcoholic fermentation exactly equivalent to the quantity of hexosephosphate produced, and that the rate of fermentation rapidly falls as soon as the free phosphate has disappeared, in spite of the fact that at that moment the concentration of the hexosephosphate is at its highest, whereas according to Iwanoff's theory it is precisely under these conditions that the maximum rate of fermentation should be maintained.
It has also been shown that the arguments adduced by Iwanoff in favour of the existence of his synthease are not valid [Harden and Young, 1910, 1].
The fermentation of dihydroxyacetone was moreover proved by Harden and Young [1912] to be effected by yeast-juice and maceration extract at a much slower rate than that of the sugars, in spite of the fact that the addition of dihydroxyacetone did not inhibit the sugar fermentation. The same thing has been shown for living yeast by Slator [1912] in agreement with the earlier results of Buchner [1910] and Buchner and Meisenheimer [1910].
The logical conclusion from Lebedeff's experiments would appear rather to be that dihydroxyacetone is slowly condensed to a hexose and that this is then fermented in the normal manner [Harden and Young, 1912; Buchner and Meisenheimer, 1912; Kostytscheff, 1912, 2]. Buchner and Meisenheimer, however, regard this as improbable on the ground that dihydroxyacetone, being symmetric in constitution, would yield an inactive hexose of which only at most 50 per cent. would be fermentable. Against this it may be urged, however, [p107] that enzymic condensation of dihydroxyacetone might very probably occur asymmetrically yielding an active and completely fermentable hexose. Buchner and Meisenheimer, however, still support the view that dihydroxyacetone forms an intermediate stage in the fermentation of glucose and adduce as confirmatory evidence of the probability of such a change the observation of Fernbach [1910] that this compound is produced from glucose by a bacillus, Tyrothrix tenuis, which effects the change both when living and after treatment with acetone.
The balance of evidence, however, appears to be in favour of the opinion that dihydroxyacetone does not fulfil the conditions laid down by Slator (see p. 103) as essential for an intermediate product in the process of fermentation [see also Löb, 1910].
Lebedeff subsequently [1912, 4; Lebedeff and Griaznoff, 1912] extended his experiments to glyceraldehyde and modified his theory very considerably. Using maceration extract it was found in general agreement with the results of Buchner and Meisenheimer (p. 105) that 20 c.c. of juice were capable of producing about half the theoretical amount of carbon dioxide from 0·2 gram of glyceraldehyde, whereas 0·4 gram caused coagulation of the extract and a diminished evolution of carbon dioxide. The addition of phosphate diminished rather than increased the fermentation. Even in the most favourable concentration however (0·2 gram per 20 c.c.) the glyceraldehyde is fermented much more slowly than dihydroxyacetone or saccharose, as is shown by the following figures:—
| 20 c.c. Extract + 0·2 gram. |
CO2 in grams in successive periods of | Duration of fer- ment- ation |
Total CO2 | ||
|---|---|---|---|---|---|
| 6 hours. | 18 hours. | 24 hours. | |||
| Cane sugar | 0·050 | 0·000 | 0·000 | 6 | 0·05 |
| Dihydroxy- acetone |
0·042 | 0·000 | 0·000 | 6 | 0·042 |
| Glycer- aldehyde |
0·008 | 0·022 | 0·005 | 48 | 0·035 |
Further, during an experiment in which 0·129 gram of CO2 was evolved in 22·5 hours from 0·9 gram of glyceraldehyde in presence of phosphate, no change in free phosphate was observed, whereas in a similar experiment with glucose a loss of about 0·2 gram of P2O5 would have occurred. Hence the fermentation takes place without formation of hexosediphosphate. This was confirmed by the fact that the osazone of hexosephosphoric acid was readily isolated from the products of fermentation of dihydroxyacetone (0·259 gram of CO2 having been evolved in twenty hours) but could not be obtained from those of glyceraldehyde (0·138 gram CO2 in twenty hours). [p108]
This result is extremely interesting, although it is not impossible that the rate of fermentation of the glyceraldehyde is so slow that any phosphoric ester produced is hydrolysed as rapidly as it is formed.
Lebedeff regards the experiments as proof that phosphate takes no part in the fermentation of glyceraldehyde and bases on this conclusion and his other work the following theory of alcoholic fermentation.
1. The sugar is split up into equimolecular proportions of glyceraldehyde and dihydroxyacetone:—
2. The dihydroxyacetone then passes through the stages previously postulated (p. 106).
After which the hexose, C6H12O6 re-enters the cycle at (a).
3. The fermentation of the glyceraldehyde occurs according to the scheme developed by Kostytscheff (p. 109), pyruvic acid being formed along with hydrogen and then decomposed into carbon dioxide and acetaldehyde, which is reduced by the hydrogen. Lebedeff, however, suggests [1914, 1, 2] that glyceric acid is first formed (1) and then converted by an enzyme, which he terms dehydratase into pyruvic acid (2):—
The experimental basis for this idea is the fact that glyceric acid is fermented by dried yeast and maceration juice [compare Neuberg and Tir, 1911].
This scheme has the merit of recognising the fact that the carbon dioxide does not wholly arise from the products of decomposition of hexosephosphate, nor from its direct fermentation. The function assigned to the phosphate is that of removing dihydroxyacetone and thus preventing it from inhibiting further conversion of hexose into triose, according to the reversible reaction
This however appears to be quite inadequate, since, on the one hand, the fermentation of glucose proceeds quite freely in presence of as much as 5 grams per 100 c.c. of dihydroxyacetone [Harden and Young, 1912], and on the other hand alcoholic fermentation appears not to proceed at all in the absence of phosphate (see p. 55). This forms the chief objection to the theory in its present form. The slow rate at which [p109] glyceraldehyde is fermented also affords an argument against the validity of Lebedeff's view, but this may possibly be accounted for to some extent by the fact that glyceraldehyde is a strong inhibiting agent so that it might be more rapidly fermented if added in a more dilute condition.
The unfermented glyceraldehyde cannot be recovered from the solution and nothing is known as to its fate except that it readily gives rise both to lactic acid and glycerol [Oppenheimer, 1914, 1, 2]. Evidently the reaction between glyceraldehyde and yeast-juice is by no means a simple one.
The third stage of Lebedeff's theory postulates the intermediate formation of pyruvic acid. This idea immediately suggested itself when it became known that yeast was capable of rapidly decomposing a-ketonic acids with evolution of carbon dioxide [see Neubauer and Fromherz, 1911, p. 350; Neuberg and Kerb, 1912, 4; Kostytscheff, 1912, 2].
This scheme has been differently elaborated by different workers. According to Kostytscheff it involves (1) the production of pyruvic acid from the hexoses, a process accompanied by loss of hydrogen; (2) the decomposition of pyruvic acid into acetaldehyde and carbon dioxide; and (3) the reduction of the acetaldehyde to ethyl alcohol.
1. As regards the production of pyruvic acid from the hexoses by yeast, the only direct evidence is afforded by the experiments of Fernbach and Schoen [1913] who have obtained a calcium salt having the qualitative properties of a pyruvate by carrying out alcoholic fermentation by yeast in presence of calcium carbonate, but have not yet definitely settled either the identity of the acid or its origin from sugar. Pyruvic acid is, however, very closely related to several substances which are intimately connected both chemically and biochemically with the hexoses. Thus lactic acid is its reduction product,
glyceraldehyde can readily be converted into it by oxidation to glyceric acid followed by abstraction of water (Erlenmeyer), [p110]
and finally methylglyoxal CH3·CO·CHO is its aldehyde.
2. The decomposition of pyruvic acid into acetaldehyde and carbon dioxide has already been fully discussed (Chapter VI). The universality of the enzyme carboxylase in yeasts and the rapidity of its action on pyruvic acid form the strongest evidence at present available in favour of the pyruvic acid theory. Given the pyruvic acid, there is no doubt that yeast is provided with a mechanism capable of decomposing it at the same rate as an equivalent amount of sugar.
3. The final step postulated by the pyruvic acid theory is the quantitative reduction to ethyl alcohol of the acetaldehyde formed from the pyruvic acid.
The idea that acetaldehyde is an intermediate product in the various fermentations of sugar has frequently been entertained [Magnus Levy, 1902; Leathes, 1906; Buchner and Meisenheimer, 1908; Harden and Norris, D., 1912] although no very definite experimental foundation exists for the belief. It is, however, a well-known fact that traces of acetaldehyde are invariably formed during alcoholic fermentation [see Ashdown and Hewitt, 1910], and this is of course consistent with the occurrence of acetaldehyde as an intermediate product. Important evidence as to the specific capability of yeast to reduce acetaldehyde to alcohol has been obtained by several workers. Thus Kostytscheff [1912, 3; Kostytscheff and Hübbenet, 1913] found that pressed yeast, dried yeast and zymin all reduced acetaldehyde to alcohol, 50 grams of yeast in 10 hours producing from 660 mg. of aldehyde 265 mg. of alcohol in excess of the amount produced by autofermentation in absence of added aldehyde. Maceration extract was found to reduce both in absence and in presence of sugar, whereas Lebedeff and Griaznoff [1912] obtained no reduction in presence of sugar, and observed that the power of reduction was lost by the extract on digestion, a circumstance which suggests the co-operation of a co-enzyme in the process. Neuberg and Kerb [1912, 4; 1913, 1] have also been able to show by large scale experiments that alcohol is produced in considerable quantity by the fermentation of pyruvic acid by living yeast in absence of sugar and that the yield is increased by the presence of glycerol. When treated with 22 kilos, of yeast, 1 kilo, of pyruvic acid yielded 241 grams of alcohol in excess of that given by the yeast alone, whilst in presence of glycerol the amount was 360 grams, the amount theoretically obtainable being 523 grams. The function of the glycerol is not understood but is probably that of lessening the rate of destruction of the yeast enzymes. [p111]
That yeast possesses powerful reducing properties has long been known and many investigations have been made as to the relation of these properties to the process of alcoholic fermentation. Thus Hahn (Buchner, E. and H., and Hahn, 1903, p. 343) found that the power of reducing methylene blue was possessed both by yeast and zymin and on the whole ran parallel to the fermenting power in the process of alcoholic fermentation. The intervention of a reducing enzyme was suggested by Grüss [1904, 1908, 1, 2] and was supported by Palladin [1908]. The latter observed that zymin which reduces sodium selenite and methylene blue in absence of sugar almost ceases to do so in presence of a fermentable sugar, and concluded that the great diminution of reduction during fermentation was due to the fact that the reducing enzyme was largely combined with a different substrate arising from the sugar, the reduction of which was necessary for alcoholic fermentation. Grüss, however, found that with living yeast the reduction is greatly increased in presence of a fermentable sugar, while Harden and Norris, R. V. [1914] confirmed the observation of Grüss, but found that the reducing power of zymin is not seriously affected by the presence of a fermentable sugar in concentration less then 20 grams per 100 c.c., whilst its fermenting power for glucose is inhibited by 1 per cent. sodium selenite. Hence Palladin's conclusion cannot be regarded as proved.
Interesting attempts have been made by Kostytscheff and later by Lvoff to obtain evidence of the participation of a reductase in alcoholic fermentation by adding some substance which would be capable either of taking up hydrogen and thus preventing the reduction of the acetaldehyde or of converting the aldehyde into some compound less liable to reduction.
Kostytscheff [1912, 1; 1913, 1, 2; 1914; Kostytscheff and Hübbenet, 1913; Kostytscheff and Scheloumoff, 1913; Kostytscheff and Brilliant, 1913] has examined the effect of the addition of zinc chloride, chosen with the idea that it might polymerise the aldehyde and thus remove it from the sphere of action. As pointed out by Neuberg and Kerb [1912, 1] this action is not very probable, and it was subsequently found [Kostytscheff and Scheloumoff, 1913] that the effect of added zinc salts was more probably specifically due to the zinc ion. Fermentation of sugar by dried yeast still proceeds when 0·6 gram of ZnCl2 is added to 10 grams of the yeast and 50 c.c. of water, whereas it ceases in the presence of 1·2 gram of ZnCl2. Even the addition of 0·075 gram however greatly diminishes the rate of fermentation and the total amount of sugar decomposed. The most noteworthy effect is that the production of acetaldehyde is increased both in autofermentation and [p112] in sugar fermentation. The course of the reaction is further modified in the sense that the percentage of sugar used up which can be accounted for in the products decreases, in other words the "disappearing sugar" (p. 31) increases. In long continued fermentations moreover and particularly with high concentrations of zinc chloride less alcohol is produced than is equivalent to the carbon dioxide evolved. The interpretation of these results is difficult. Kostytscheff takes them to mean (1) that the zinc salt modifies one stage of the reaction so that a higher concentration of intermediate products is obtained, and (2) that the carbon dioxide and alcohol must be produced at different stages or their ratio, in the absence of secondary changes, would be unalterable.
Alternative interpretations are, however, by no means excluded. Thus Neuberg and Kerb [1912, 1; 1913, 2] do not regard it as conclusively proved that the aldehyde really arises from the sugar since they have observed its production in maceration extract free from autofermentation. The method used by Kostytscheff for the separation of alcohol and aldehyde (treatment with bisulphite) has also proved unsatisfactory in their hands and the results obtained as to the reduction of acetaldehyde by yeast, etc., are not accepted. They also consider that in any case the small amounts produced (less than 0·2 per cent. of the sugar used) would not afford convincing evidence that the aldehyde is an intermediate product, although it must be admitted that no large accumulation of an intermediate product could be reasonably expected. It may also be pointed out that the increase in "disappearing sugar" may be simply due to the fact that in the controls the whole of the sugar was fermented, so that any polysaccharide formed at an earlier stage would have been hydrolysed and fermented, whereas in the presence of zinc chloride excess of sugar was present throughout the whole experiment.
Lvoff [1913, 1, 2, 3] has made quantitative experiments on the effect of methylene blue both on the sugar fermentation and autofermentation of dried yeast and maceration extract. In presence of sugar the methylene blue causes a decrease in the extent of fermentation, the difference during the time required for reduction of the methylene blue being represented by an amount of glucose equimolecular to the latter. In the absence of sugar on the other hand an excess of carbon dioxide equimolecular to the methylene blue is evolved but no corresponding increase in the alcohol production occurs. The effect of methylene blue is evidently complex and it is impossible at present to say whether Lvoff's contention is correct that the methylene blue actually [p113] interferes with the fermentation by taking up hydrogen (2 atoms per molecule of glucose) destined for the subsequent reduction of some intermediate product or whether the effect is one of general depression of the fermenting power which would be presumably proportional to the concentration of methylene blue and inversely proportional to that of the fermenting complex [see Harden and Norris, R. V., 1914]. In any case it will be noticed that Lvoff s interpretation of the results is at variance with the requirements of Kostytscheff's theory (p. 109) according to which 4 atoms of hydrogen should be given off by a molecule of glucose.
Kostytscheff [1913, 2; Kostytscheff and Scheloumoff, 1913] has also observed a depression of the extent of fermentation by methylene blue without any serious alteration in the ratio of CO2 to alcohol, although an increase occurs in the production of acetaldehyde.
On the whole it cannot be said that the evidence gathered from experiments on the reduction of acetaldehyde and methylene blue is very convincing. All that is established beyond doubt seems to be that yeast possesses a reducing mechanism for many aldehydes [see also in this connection Lintner and Luers, 1913; Lintner and von Liebig, 1911; as well as Neuberg and Steenbock, 1913, 1914] and colouring matters. This mechanism appears to be capable of activity in the absence of sugar and it is to be supposed that in accordance with the views of Bach [1913] the necessary hydrogen is derived from water and that some acceptor for the oxygen simultaneously liberated is also present. There seems however at the moment to be no sufficient reason to suppose that this mode of reduction is in any way altered by the presence of sugar and until the production of intermediate products equivalent to the amount of substance reduced is actually demonstrated, the conclusions of these workers may be regarded as not fully justified.
Neuberg and Kerb [1913, 2] themselves tentatively propose a complicated scheme possessing some novel features according to which methylglyoxal is the starting-point for the later stages of the change.
(a) A small portion of this is converted by a reaction which may be variously interpreted as a Cannizzaro transformation or a reductase reaction into glycerol and pyruvic acid.
| CH2:C(OH)·CHO + H2O | H2 + │ = O |
CH2(OH)·CHOH·CH2(OH) (glycerol) + |
| CH2:C(OH)·CHO | CH2:C(OH)·COOH (Pyruvic acid) |
(b) The pyruvic acid is then decomposed by carboxylase yielding aldehyde and carbon dioxide (equation 2, p. 109). [p114]
(c) The aldehyde and a molecule of glyoxal then undergo a Cannizzaro reaction and yield alcohol and pyruvic acid,
| CH3·CO·CHO | O | CH3·CO·COOH |
| + │ = | + | |
| CH3·CHO | H2 | CH3CH2(OH) |
and the latter then undergoes reaction (b).
A small amount of glycerol is thus necessarily formed, as is actually found to be the case.
The experimental foundation for stages (a) and (c) will be awaited with great interest, as well as the proof that methylglyoxal is readily fermentable (see p. 104).
An interesting interpretation of the phenomena of fermentation was attempted by Schade [1906] based upon the conception that glucose under the influence of catalytic agents readily decomposes into acetaldehyde and formic acid. It was subsequently found that the experimental evidence upon which this conclusion was founded had been wrongly interpreted [Buchner, Meisenheimer, and Schade, 1906; Schade, 1907], but Schade has succeeded in devising an interesting series of reactions by means of which alcohol and carbon dioxide can be obtained from sugar by the successive action of various catalysts. The following are the stages of this series: (1) Glucose, fructose, and mannose are converted by alkalis into lactic acid along with other products. (2) Lactic acid when heated with dilute sulphuric acid yields a mixture of acetaldehyde and formic acid:—
(3) It has long been known that formic acid is catalysed by metallic rhodium at the ordinary temperature into hydrogen and carbon dioxide, and Schade has found that when a mixture of acetaldehyde and formic acid is submitted to the action of rhodium the acetaldehyde is reduced to alcohol at the expense of the hydrogen and the carbon dioxide is evolved:—
Schade suggests [1908] that the fermentation of sugar may proceed by a similar series of reactions catalysed by enzymes, the acetaldehyde and formic acid being derived not from the relatively stable lactic acid but more probably from a labile substance capable of undergoing change either into lactic acid or into aldehyde and formic acid.
It will be noticed that this theory resembles the pyruvic acid [p115] theory in postulating the immediate formation of acetaldehyde but differs from it by supposing that the reduction is effected at the expense of formic acid produced at the same time.
The acetaldehyde question has already been discussed. In view of the fact that formic acid is a regular product of the action of many bacteria on glucose [see Harden, 1901], Schade's theory of alcoholic fermentation may be said to be a possible interpretation of the facts. Formic acid is known to be present in small amounts in fermented sugar solutions and the actual behaviour of yeast towards this substance has been investigated in some detail by Franzen and Steppuhn [1911; 1912, 1, 2], who have obtained results strongly reminiscent of those obtained with lactic acid by Buchner and Meisenheimer (p. 102). Many yeasts when grown in presence of sodium formate decompose a certain proportion of it, whereas in absence of formate they actually produce a small amount of formic acid—the absolute quantities being usually of the order of 0·0005 gram molecule (0·023 gram) per 100 c.c. of medium in 4 to 5 days. Only in the case of S. validus did the consumption of formic acid in 5 days reach 0·0017 gram molecule (0·08 gram). Somewhat similar but rather smaller results were given by yeast-juice, a small consumption of formic acid being usually observed. The possibility thus exists that formic acid may be an intermediate product of alcoholic fermentation and Franzen argues strongly in favour of this view.
Direct experiment, on the other hand, shows that yeast-juice cannot ferment a mixture of acetaldehyde and formic acid, even when these are gradually produced in molecular proportions in the liquid by the slow hydrolysis of a compound of the two, ethylideneoxyformate, OHC·O·CH(CH3)·O·CH(CH3)·O·CHO, this method being adopted to avoid the inhibiting effect of free acetaldehyde and formic acid [Buchner and Meisenheimer, 1910]. Nor is the reduction of acetaldehyde assisted by the presence of formate [Neuberg and Kerb, 1912, 4; Kostytscheff and Hübbenet, 1912].
A modified form of Schade's theory has been suggested by Ashdown and Hewitt [1910], who have found that when brewer's yeast is cultivated in presence of sodium formate the yield of aldehyde, as a rule, becomes less. They regard the aldehyde as derived from alanine, CH3·CH(NH2)·COOH, one of the amino-acids formed from the proteins by hydrolysis, which is known to be attacked by yeast in the characteristic manner (p. 87), forming alcohol, carbon dioxide, and ammonia. Fermentation is supposed to proceed in such a way that the sugar is first decomposed into two smaller molecules, C3H6O3 [p116] (equation i), and that these react with formamide to produce alanine and formic acid (ii). The alanine then enters into reaction with formic acid, producing alcohol, carbon dioxide, and formamide (iii):—
According to this scheme all the sugar fermented passes through the form of alanine, and the formic acid acts along with the enzyme as catalyst, passing into formamide in reaction (iii) and being regenerated in (ii). The alanine is in the first place derived from the hydrolysis of proteins, or possibly by the reaction of the C3H6O3 group with one of the higher amino-acids:—
There is as little positive evidence for this course of events as for that postulated by Schade, and the theory suffers from the additional disability that the chemical reactions involved have not been realised in the laboratory. Direct experiments with yeast-juice, moreover, show that a mixture of alanine with formic acid or a formate is not fermented, whilst neither the added mixture nor formamide seriously effects the action of the juice on glucose.
Among other suggestions may be mentioned that of Kohl [1909] who asserts that sodium lactate is readily fermented, whilst Kusseroff [1910] holds the view that the glucose is first reduced to sorbitol and the latter fermented, in spite of the fact that sorbitol itself in the free state is not fermented by yeast.
The rapid appearance and disappearance of glycogen in the yeast cell at various stages of fermentation [see Pavy and Bywaters, 1907; Wager and Peniston, 1910] has led to the suggestion [Grüss, 1904; Kohl, 1907] that this substance is of great importance in fermentation, and represents a stage through which all the sugar must pass before being fermented. The fact that the formation of glycogen has been observed in yeast-juice by Cremer [1899], and that complex carbohydrates are also undoubtedly formed (p. 31), are consistent with this theory. The low rate of autofermentation of living yeast, which is only a few per cent. of the rate of sugar fermentation, renders this supposition very improbable (Slator), as does the fact that the fermentation of glycogen by yeast-juice is usually slower than that of glucose [see also Euler, 1914].
An entirely different explanation of the chemical changes attendant on alcoholic fermentation has been suggested by Löb [1906; [p117] 1908, 1, 2; 1909, 1, 2, 3, 4; 1910; Löb and Pulvermacher, 1909], founded on the idea that the various decompositions of the sugar molecule both by chemical and biological agents are to be explained by a reversal of the synthesis of sugar from formaldehyde. As the sugar molecule can be built up by the condensation of formaldehyde, so it tends to break down again into this substance, and the products observed in any particular case are formed either by partial depolymerisation in this sense or by partial re-synthesis following on depolymerisation.
Löb has adduced many striking facts in favour of this view, and has shown that very dilute alkalis produce no lactic acid but formaldehyde and a pentose as primary products. These substances represent the first stage of depolymerisation and are also formed by the electrolysis of glucose.
Löb has himself been unable to detect definite intermediate products of fermentation by adding reagents, such as aniline, ammonia, and phloroglucinol, which would combine with such substances and prevent their further decomposition [1906].
The occurrence of traces of formaldehyde as a product of alcoholic fermentation by yeast-juice [Lebedeff, 1908] is at least consistent with this theory, but no decisive evidence has so far been obtained either for or against it.
In all the foregoing attempts to indicate the probable stages in the production of alcohol and carbon dioxide from sugar, a single molecule of the sugar forms the starting-point. The facts recounted in Chapter III as to the function of phosphates in alcoholic fermentation, which are summed up in the equation:—
render it in the highest degree probable that two molecules of the sugar are concerned. The most reasonable interpretation of this equation appears to be that in the presence of phosphate and of the complicated machinery of enzyme and co-enzyme two molecules of the hexose, or possibly of the enolic form, are each decomposed primarily into two groups.
Of the four groups thus produced, two go to form alcohol and carbon dioxide and the other two are synthesised to a new chain of six carbon atoms, which forms the carbohydrate residue of the hexosephosphate. The introduction of the phosphoric acid groups may possibly occur before the rupture of the original molecules, and may even be the determining factor of this rupture, or again this introduction may take place during or after the formation of the new carbon [p118] chain. Sufficient information is not yet available for the exact formulation of a scheme for this reaction. Such a scheme, it may be noted, would not necessarily be inconsistent with the views of Wohl and of Buchner as to the way in which the carbon chain of a hexose is broken in the process of fermentation, but would interpret differently the subsequent changes which are undergone by the simpler groups which are the result of this rupture. The reaction might thus proceed without the formation of definite intermediate products, whilst opportunity would be afforded for the production of a small quantity of by-products such as formaldehyde, glycerol, lactic acid, acetic acid, etc., by secondary reactions.
A symmetrical scheme can readily be constructed for such a change, but much further information is required before any decisive conclusion can be drawn as to the precise course of the reaction which actually occurs in alcoholic fermentation.
The analysis of the process of alcoholic fermentation by yeast-juice and other preparations from yeast which has been carried out in the preceding chapters has shown that the phenomenon is one of a very complex character. The principal substances directly concerned in the change appear to be the enzyme and co-enzyme of the juice, a second enzyme, hexosephosphatase, and, in addition, sugar, phosphate, and the hexosephosphate formed from these. During autofermentation two other factors are involved, the complex carbohydrates of the juice, including glycogen and dextrins, and the diastatic ferment by which these are converted into fermentable sugars. It is also possible that the supply of free phosphate is partially provided by the action of proteoclastic ferments on phosphoproteins. Under special circumstances the rate at which fermentation proceeds may be controlled by the available amount of any one of these numerous substances.
When the juice from well-washed yeast is incubated, the phenomenon of autofermentation is observed. The juice contains an abundant supply of enzyme, co-enzyme, and phosphate or hexosephosphate, and in this case the controlling factor is usually the supply of sugar, which is conditioned by the concentration of the diastatic enzyme or of the complex carbohydrates as the case may be. When this is the case the measured rate of fermentation is the rate at which sugar is being produced in the juice, this being the slowest of the various reactions which are proceeding under these circumstances. If sugar be now added, an entirely different state of affairs is set up. As soon as any accumulated phosphate has been converted into hexosephosphate, the normal rate of fermentation which is usually higher than that of autofermentation is attained, and, provided that excess of sugar be present, fermentation continues for a considerable period at a slowly diminishing rate and finally ceases. During the first part of this fermentation the rate is controlled entirely by the supply of free phosphate, and this depends mainly on the concentration of the hexosephosphatase and of the hexosephosphate, and only in a secondary degree on the decomposition [p120] of other phosphorus compounds by other enzymes and on the concentration of the sugar. The amount of hexosephosphate in yeast-juice is usually such that an increase in its concentration does not greatly affect the rate of fermentation, and hence the measured rate during this period represents the rate at which hexosephosphate is being decomposed, and this in its turn depends on the concentration of hexosephosphatase, which is therefore the controlling factor. As fermentation proceeds, the concentration of both enzyme and co-enzyme steadily diminishes, as already explained, probably owing to the action of other enzymes, so that at an advanced stage of the fermentation, the controlling factor may be the concentration of either of these, or the product of the two concentrations (see p. 122). The hexosephosphatase appears invariably to outlast the enzyme and co-enzyme. The condition at any moment could be determined experimentally if it were possible to add enzyme, co-enzyme and hexosephosphatase at will and so ascertain which of these produced an acceleration of the rate.
Unfortunately this can at present be only very imperfectly accomplished, owing to the impossibility of separating these substances from each other and from accompanying matter which interferes with the interpretation of the result.
A third condition can also be established by adding to the fermenting mixture of the juice and sugar a solution of phosphate. The supply of phosphate is now almost independent of the action of the hexosephosphatase, and the measured rate represents the rate at which reaction (1), p. 51, can occur between sugar and phosphate in the presence of the fermenting complex consisting of enzyme and co-enzyme. This change is controlled, so long as sugar and phosphate are present in the proper amounts, by the concentration of the fermenting complex or possibly of either the enzyme or the co-enzyme. If only a single addition of a small quantity of phosphate be made, the rate falls as soon as the whole of this has been converted into hexosephosphate and the reaction then passes into the stage just considered, in which the rate is controlled by the production of free phosphate.
Although these varying reactions have not yet been exhaustively studied from the kinetic point of view, owing to the experimental difficulties to which allusion has already been made, investigations have nevertheless been carried out on the effect of the variation of concentration of yeast-juice and zymin as a whole, as well as of the carbohydrate. Herzog [1902, 1904] has made experiments of this kind with zymin, and Euler [1905] with yeast-juice, whilst many of the results [p121] obtained by Buchner and by Harden and Young are also available.
The actual observations made by these authors show that the initial velocity of fermentation is almost independent of the concentration of sugar within certain limits, but decreases slowly as the concentration increases. When the velocity constant is calculated on the assumption that the reaction is monomolecular [see Bayliss, 1914, Chap. VI], approximate constancy is found for the first period of the fermentation. This method of dealing with the results is, however, as pointed out by Slator, misleading, the apparent agreement with the law of monomolecular reactions being probably due to the gradual destruction of the fermenting complex.
Experiments with low concentrations of sugar are difficult to interpret, the influence of the hydrolysis of glycogen and of dextrins on the one hand, and the synthesis of sugar to more complex carbohydrates on the other (p. 31), having a relatively great effect on the concentration of the sugar. Unpublished experiments (Harden and Young) indicate, however, that the velocity of fermentation remains approximately constant, until a certain very low limit of sugar concentration is reached, and then falls rapidly. The fall in rate, however, only continues over a small interval of concentration, after which the velocity again becomes approximately constant and equal to the rate of autofermentation. During this last phase, as already indicated, the velocity is generally controlled by the rate of production of sugar and no longer by that of phosphate, this substance being now present in excess. In other words, the rate of fermentation of sugar by yeast-juice and zymin is not proportional to the concentration of the sugar present as required by the law of mass, but, after a certain low limit of sugar concentration, is independent of this and is actually slightly decreased by increase in the concentration of the sugar.
The relations here are very similar to those shown to exist by Duclaux [1899] and Adrian Brown [1902] for the action of invertase on cane sugar and are probably to be explained in the manner suggested by the latter. According to this investigator, the enzyme unites with the fermentable material, or as it is now termed, the substrate or zymolyte, forming a compound which only slowly decomposes so that it remains in existence for a perceptible interval of time. The rate of fermentation depends on the rate of decomposition of this compound and hence varies with its concentration. This conception leads to the result that the rate of fermentation will increase with the concentration of the substrate up to a certain limit and will then remain [p122] constant, unless interfered with by secondary actions. This limit of concentration is that at which there is just sufficient of the material in question present to combine with practically the whole of the enzyme, so that no further increase in its amount can cause a corresponding increase in the quantity of its compound with the enzyme or in the rate of fermentation which depends on the concentration of that compound.
The curve relating the rate of action of such an enzyme with the concentration of the zymolyte therefore consists of two portions, one in which the rate at any moment is proportional to the concentration of the zymolyte, according to the well-known law of the action of mass, and a second in which the rate at any moment is almost independent of that concentration, approximately equal amounts being decomposed in equal times whatever the concentration of the substrate.
The results of the experiments with yeast-juice therefore indicate that what is being measured is a typical enzyme action, but afford no information as to which of the many possible actions is the controlling one, a fact which must be ascertained for each particular case in the manner indicated above.
Clowes [1909], using washed zymin free from fermenting power and adding various volumes of boiled yeast extract, found that the velocity of reaction was proportional to the product of the concentrations of zymin and yeast extract up to a certain optimum concentration. He interprets these concentrations as representing the concentrations of zymase and co-enzyme, but they also represent the concentrations of hexosephosphatase (present in the zymin) and phosphate (present in the yeast extract), so that at least four factors were being altered instead of only two.
It has already been mentioned that Euler and Kullberg [1911, 3] found the conversion of phosphate into hexosephosphate in presence of excess of glucose to proceed according to a monomolecular reaction (p. 58).
The rate of fermentation is diminished by dilution of the yeast-juice, but less rapidly than the concentration of the juice. Herzog found that when the relation between concentration of enzyme and the velocity constant of the reaction is expressed by the formula K1/K2 = (C1/C2)n where K1 and K2 are the velocity constants corresponding with the enzyme concentrations C1 and C2, the value for n is 2 for zymin, whilst Euler working with yeast-juice obtained values varying from 1·29 to 1·67 and decreasing as K increased.
The temperature coefficient of fermentation by zymin was found [p123] by Herzog to be K24·5°/K14·5° = 2·88, which agrees well with the value found by Slator for yeast-cells (p. 129).
When we endeavour to apply the results of the investigations of the fermentation of sugar by yeast-juice, zymin, etc., to the process which goes on in the living cell, considerable difficulties present themselves. A scheme of fermentation in the living cell can, however, easily be imagined, which is in harmony with these results. According to the most simple form of this ideal scheme, the sugar which has diffused into the cell unites with the fermenting complex and undergoes the characteristic reaction with phosphate, already present in the cell, yielding carbon dioxide, alcohol, and hexosephosphate. The latter is then decomposed, just as it is in yeast-juice, but more rapidly, and the liberated phosphate again enters into reaction, partly with the sugar formed from the hexosephosphate and partly with fresh sugar supplied from outside the cell. The main difference between fermentation by yeast-juice and by the living cell would then consist in the rate of decomposition of the hexosephosphate, for it has been shown that yeast-juice in presence of sufficient phosphate can ferment sugar at a rate of the same order of magnitude (from 30 to 50 per cent.) as that attained by living yeast.
The difference between the two therefore would appear to lie not so much in their content of fermenting complex as in their very different capacity for liberating phosphate from hexosephosphate and thus supplying the necessary conditions for fermentation.
A simple calculation based on the phosphorus content of living yeast [Buchner and Haehn, 1910, 2] shows that the whole of this phosphate must pass through the stage of hexosephosphate every five or six minutes in order to maintain the normal rate of fermentation, whereas in an average sample of yeast-juice the cycle, calculated in the same way, would last nearly two hours.
Wherein this difference resides is a difficult question, which cannot at present be answered with certainty.
In the first place it must be remembered that a very great acceleration of the action of the hexosephosphatase is produced by arsenates (p. 79), and this suggests the possibility that some substance possessing a similar accelerating power is present in the yeast-cell and is lost or destroyed in the various processes involved in rendering the yeast susceptible to phosphate. The great variety of these processes—extraction of yeast-juice by grinding and pressing, drying and macerating, heating, treating with acetone and with toluene—renders this somewhat improbable, and so far no such substance has been detected. [p124]
A comparison of living yeast, zymin, and yeast-juice shows that these are situated on an ascending scale with respect to their response to phosphate. Taking fructose as the substrate in each case, yeast does not respond to phosphate at all (Slator), the rate of fermentation by zymin is approximately doubled (p. 46), and that by yeast-juice increased ten to forty times, whilst the maximum rates are in each case of the same order of magnitude. Euler and Kullberg, however, have observed an acceleration of about 25 per cent. in the rate of fermentation of yeast in presence of a 2 per cent. solution of monosodium phosphate, NaH2PO4 [1911, 1, 2].
The high rate of fermentation by living yeast and its lack of response to phosphate may possibly be explained by supposing that the balance of enzymes in the living cell is such that the supply of phosphate is maintained at the optimum, and the rate of fermentation cannot therefore be increased by a further supply.
A further difference lies in the fact that yeast-juice and zymin respond to phosphate more strongly in presence of fructose than of glucose, whereas yeast ferments both sugars at the same rate (p. 131), and this property has been shown to be connected with the specific relations of fructose to the fermenting complex. It seems possible that these differences are associated with the gradual passage from the complete living cell of yeast, through the dead and partially disorganised cell of zymin to yeast-juice in which the last trace of cellular organisation has disappeared and the contents of the cell are uniformly diffused throughout the liquid. Living yeast is, moreover, not only unaffected by phosphate but only decomposes hexosephosphate extremely slowly (Iwanoff).
Some light is thrown on these interesting problems by the effect of antiseptics on fermentation by yeast-cells and by yeast-juice. The action of toluene has hitherto been most completely studied, and this substance is an extremely suitable one for the purpose since it has practically no action whatever on fermentation by yeast-juice. The experiments of Buchner have, in fact, shown that the normal rate of fermentation and the total fermentation produced, are almost unaffected by the presence of toluene even in the proportion of 1 c.c. to 20 c.c. of yeast-juice. What then is the effect of toluene on the living yeast-cell? When toluene in large excess is agitated with a fermenting mixture of yeast and sugar, the rate of fermentation falls rapidly at first and then more slowly until a relatively constant rate is attained which gradually decreases in a similar manner to the rate of fermentation by yeast-juice. Thus at air temperature (16°) 10 grams of [p125] yeast suspended in 50 c.c. of 6 per cent. glucose solution gave the following results when agitated with toluene:—
| Time after Addition of Toluene, Minutes |
C.c. of CO2 per Minute. |
Time. | C.c. per Minute. |
|---|---|---|---|
| 0 | 4·6 | 6 | 1·6 |
| 1 | 4 | 8 | 1·2 |
| 2 | 3·3 | 12 | 0·85 |
| 3 | 2·6 | 24 | 0·8 |
| 4 | 2 | 32 | 0·5 |
| 5 | 1·8 | constant |
Simultaneously with this, the yeast acquires the property of decomposing and fermenting hexosephosphate and of responding to the addition of phosphate. This last property is only acquired to a small degree in this way but it becomes much more strongly developed if the pressed yeast be washed with toluene on the filter pump. Thus 10 grams of yeast after this treatment fermented fructose at 1·2 c.c. per three minutes; after the addition of phosphate (5 c.c. of 0·6 molar phosphate) the rate rose to 6·9 and then gradually fell in the typical manner [Harden, 1910; see also Euler and Johansson, 1912, 3].
The current explanation of the great decrease in rate of fermentation which attends the action of toluene and other antiseptics on living yeast, and also follows upon the disintegration of the cell, appears to be that in living yeast the high rate of fermentation is maintained by the continued production of relatively large fresh supplies of fermenting complex, and that when the power of producing this catalytic agent is destroyed by the poison, the rate of fermentation falls to a low value, corresponding to the store of zymase still present in the cell (cf. Buchner, E. and H., and Hahn, 1903, pp. 176, 180).
This explanation implies that the rate of fermentation after the action of the toluene represents the amount of fermenting complex present, a supposition which has been shown (p. 53) to be highly improbable. It further necessitates, as also pointed out independently by Euler and Ugglas [1911], a rapid destruction of the fermenting complex both in the process of fermentation and by the action of the antiseptic, as otherwise the store of zymase remaining in the dead cell would be practically the same as that contained in the living cell at the moment when it was subjected to the antiseptic, and this store would therefore suffice to carry out fermentation at the same rate in the dead as in the living cell. No such rapid destruction, however, occurs in yeast-juice, as judged by the rate of fermentation, which falls off [p126] slowly and to about the same extent in the presence or absence of toluene. Moreover, as shown above, it is highly probable that the actual amount of fermenting complex in yeast-juice is a large fraction of that present at any moment in the cell, and is capable under suitable conditions of producing fermentation at a rate comparable with that of the living cell.
This last criticism also applies to the view expressed by Euler [Euler and Ugglas, 1911; Euler and Kullberg, 1911, 1, 2] that in the living cell the zymase is partly free and partly combined with the protoplasm; when the vital activity of the cell is interfered with, the combined portion of the zymase is thrown out of action and only that which was free remains active.
The suggestion made by Rubner [1913] that the action of yeast on sugar is in reality chiefly a vital act, but that a small proportion of the change is due to enzyme action, is similar in its consequences to that of Euler and may be met by the same arguments. Buchner and Skraup [1914] have moreover shown that the effects of sodium chloride and toluene on the fermenting power of yeast which were observed by Rubner, can be explained in other ways.
Some other explanation must therefore be sought for this phenomenon. Great significance must be attached in this connection to the relation noted above between the degree of disintegration and disorganisation of the cell and the fall in the normal rate of fermentation. It seems not impossible that fermentation may be associated in the living cell with some special structure, or carried on in some special portion of the cell, perhaps the nuclear vacuole described by Janssens and Leblanc [1898], Wager [1898, 1911; Wager and Peniston, 1910] and others which undergoes remarkable changes both during fermentation and autofermentation [Harden and Rowland, 1901]. The disorganisation of the cell might lead to many modifications of the conditions, among others to the dilution of the various catalytic agents by diffusion throughout the whole volume of the cell. As a matter of observation the dilution of yeast-juice leads to a considerable diminution of the rate of fermentation of sugar, and it is possible that this is one of the chief factors concerned. That phenomena of this kind may be involved is shown by the remarkable effect of toluene on the autofermentation of yeast. Whereas the fermentation of sugar is greatly diminished by the action of toluene, the rate of autofermentation, which is carried on at the expense of the glycogen of the cell, is greatly increased. In a typical case, for example, the autofermentation of 10 grams of yeast suspended in 20 c.c. of water amounted to 28 c.c. in 4·8 hours [p127] at 25°, whereas the same amount of yeast in presence of 2 c.c. of toluene gave 97·6 c.c. in the same time.
Many salts produce a similar effect on English top yeasts (in which the autofermentation is large) [Harden and Paine, 1912], whereas Neuberg and Karczag in Berlin [1911, 2] were unable to observe this phenomenon.
A necessary preliminary of the fermentation of glycogen is its conversion by a diastatic enzyme into a fermentable sugar, and it is probable that the effect of the disorganisation of the cell by toluene is that this enzyme finds more ready access to the glycogen, which is stored in the plasma of the cell. No such acceleration of autofermentation is effected by the addition of toluene to yeast-juice, and hence the result is not due to an acceleration of the action of the diastatic enzyme on the glycogen.
This effect of toluene is similar in character to the action of anæsthetics on the leaves of many plants containing glucosides and enzymes, whereby an immediate decomposition of the glucoside is initiated [see H. E. and E. F. Armstrong, 1910].
Although as indicated above Euler's theory cannot apply to zymase itself, if applied to the hexosephosphatase it would afford a consistent explanation of the facts. According to this modified view it would be the hexosephosphatase of yeast which existed largely in the combined form, so that in extracts, in dried yeast and in presence of toluene only the small fraction which was free would remain active. The zymase on the other hand would have to be regarded as existing to a large extent in the free state so that it would pass into extracts comparatively unimpaired in amount and capable under proper conditions (i.e. when supplied with sufficient phosphate) of bringing about a very vigorous fermentation. The theory of combined and free enzymes is undoubtedly of considerable value, although it cannot be considered as fully established.
Much important information as to the nature of the processes involved in fermentation has been acquired by the direct experimental study of the action of living yeast on different sugars.
This phenomenon has formed the subject of several investigations from the kinetic point of view, and its general features may now be regarded as well established.
The difficulty, which must as far as possible be avoided in quantitative experiments of this sort with living yeast, is the alteration [p128] in the amount or properties of the yeast, due to growth or to some change in the cells. This has been obviated in the work of Slator [1906] by determining in every case the initial rate of fermentation, so that the process only continues for a very short period, during which any change in the amount or constitution of the yeast is negligible. The method has the further advantage that interference of the products of the reaction is to a large extent avoided. The pressure apparatus already described (p. 29) was employed by Slator, the rate of production of carbon dioxide being measured by the increase of pressure in the experimental vessel.
With regard to this important factor it is found that the action of living yeast follows the same law as that of most enzymes (p. 121); within certain wide limits the rate of fermentation is almost independent of the concentration of the sugar. This conclusion has been drawn by many previous investigators from their experiments [Dumas, 1874; Tammann, 1889; Adrian Brown, 1892; O'Sullivan, 1898, 1899] and is implicitly contained in the results of Aberson [1903], although he himself regarded the reaction as monomolecular.
Slator, working with a suspension of ten to twelve yeast-cells per 1/4000 cubic millimetre at 30°, obtained the results which are embodied in the curve (Fig. 8).
This shows that, for the amount of yeast in question, the rate of fermentation is almost constant for concentrations of glucose between [p129] 1 and 10 grams per 100 c.c., but gradually decreases as the concentration increases. Below 1 gram per 100 c.c. the rate decreases very rapidly with the concentration.
It follows from this, in the light of what has already been said (p. 121), that the action of living yeast on sugar follows the same course as a typical enzyme reaction, although in this case, as in that of yeast-juice, no information is given as to the exact nature of this reaction.
It appears to be well established that, when changes in the quantity and constitution of the yeast employed are eliminated, the rate of fermentation is exactly proportional to the number of the yeast-cells present (Aberson, Slator). This result might be anticipated, as pointed out by Slator, from the fact that the fermentation takes place within the cell, each cell acting as an independent individual.
The diffusion of sugar into the yeast-cell which necessarily precedes the act of fermentation has been shown by Slator and Sand [1910] to occur at such a rate that the supply of sugar is always in excess of the amount which can be fermented by the cell.
The temperature coefficient of fermentation by living yeast has been carefully determined by Slator by measurements of the initial rates at a series of temperatures from 5° to 40° C. The coefficient is found to be of the same order as that for many chemical reactions, but to vary considerably with the temperature, a rise in temperature corresponding with a diminution in the coefficient. The following values were obtained for glucose; they are independent of the concentration of yeast and glucose, the class of yeast, and presence or absence of nutrient salts, and remain the same when inhibiting agents are present. Almost precisely the same ratios are obtained for fructose and mannose:—
| t. | V(t+5)/Vt. | V(t+10)/Vt. |
|---|---|---|
| 5 | 2·65 | 5·6 |
| 10 | 2·11 | 3·8 |
| 15 | 1·80 | 2·8 |
| 20 | 1·57 | 2·25 |
| 25 | 1·43 | 1·95 |
| 30 | 1·35 | 1·6 |
| 35 | 1·20 |
Aberson's result, K(t+10)/Kt = 2·72, which represents the mean coefficient for 10° between 12° and 33°, agrees well with this. [p130]
Slator [1908, 1] was unable to find any agent which greatly accelerated the rate of fermentation of living yeast. Small concentrations of various inhibiting agents which are often supposed to act in this way were quite ineffective, and phosphates, which produce such a striking change in yeast-juice, were almost without action (cp. p. 124).
Euler and Bäckström [1912], however, have made the important observation that sodium hexosephosphate causes a considerable acceleration although it is itself neither fermented nor hydrolysed under these conditions. The extent of this is evident from the following numbers:—
| 20 c.c. of 20 per cent. glucose solution. 0·25 g. yeast [Yeast H of St. Erik's brewery]. | |||
|---|---|---|---|
| Without addition. | + 0·5 g. Na hexosephosphate. | ||
| Time. Min. |
CO2. | Time. Min. |
CO2. |
| 46 | 10·5 | 37 | 8 |
| 76 | 17·5 | 73 | 19 |
| 197 | 45 | 188 | 52·5 |
| 347 | 74·5 | 321 | 123 |
| 488 | 95 | 450 | 193·5 |
The observation has been confirmed with English top yeast (Harden and Young, unpublished experiments), but no explanation of the phenomenon is at present forthcoming.
Euler has also found [Euler and Cassel, 1913; Euler and Berggren, 1912] that yeast extract, sodium nucleinate and ammonium formate also increase the rate of fermentation of glucose by yeast, but these results have been criticised by Harden and Young [1913] on the ground that the possibility of growth of the yeast during the experiment has not been excluded.
Many valuable ideas as to the nature of fermentation have been obtained by a consideration of the phenomena presented by the action of yeast on the different hexoses. Of these only glucose, fructose, mannose, and galactose are susceptible of alcoholic fermentation by yeast, the stereoisomeric hexoses prepared in the laboratory being unfermentable, as are also the pentoses, tetroses, and the alcohols corresponding to all the sugars. The yeast-cell is therefore much more limited in its power of producing fermentation than such an organism as, for example, Bacillus coli communis, which attacks substances as [p131] diverse as arabinose, glucose, glycerol and mannitol, and yields with all of them products of the same chemical character, although in varying proportions.
A careful examination of a number of different genera and species of the Saccharomycetaceæ and allied organisms by E. F. Armstrong [1905] has shown that all yeasts which ferment glucose also ferment fructose and mannose. Armstrong grew his yeasts in a nutrient solution containing the sugar to be investigated, and his experiments are open to the criticism that the organisms were hereby afforded an opportunity for becoming acclimatised to the sugar. His results, therefore, only demonstrate the fact that the organisms in question when cultivated in presence of the sugars examined brought about their fermentation, and do not exclude the possibility that the same organism when grown in presence of a different sugar might not be capable of fermenting the one to which it had in the other type of experiment become acclimatised.
This has actually been shown to be the case for galactose by Slator [1908, 1], and it is possible that this circumstance explains the negative results obtained by Lindner [1905] with S. exiguus and Schizosaccharomyces Pombe upon mannose, a sugar which, according to Armstrong, is fermented by both these organisms.
The same problem has been attacked quantitatively by Slator, who has shown that living yeast of various species and genera ferments glucose and fructose at approximately the same rate. Moreover, when the yeast is acted upon by various inhibiting agents, such as heat, iodine, alcohol, or alkalis, the crippled yeast also ferments glucose and fructose at the same rate.
With mannose the relations are somewhat different. The relative rate of fermentation of mannose and glucose by yeast is dependent on the variety of the yeast and the treatment which it has received. Fresh samples of yeast ferment mannose more quickly than glucose, but by older samples the glucose is the more rapidly decomposed. This is especially the case with yeast, the activity of which has been partly destroyed by heat, the relative fermenting power to mannose being sometimes reduced by this treatment from 120 per cent. of that of glucose to only 12 per cent. (Slator).
A further difference consists in the fact that with certain yeasts the rate of fermentation of glucose is somewhat increased by monosodium phosphate whilst that of mannose is unaffected [Euler and Lundeqvist, 1911].
Mixtures of glucose and fructose are fermented by yeast at the [p132] same rate as either the glucose or the fructose contained in the mixture would be alone. When, however, mannose and glucose are fermented simultaneously interference between the reactions takes place, and this is especially evident when the yeast has comparatively little action on mannose. The following are the results obtained by Slator:—
| Yeast. | Relative Rates. | ||
|---|---|---|---|
| 2·5 per cent. Glucose. | 2·5 per cent. Mannose. | 2·5 per cent. Glucose + 2·5 per cent. Mannose. | |
| S. Thermantitonum | 100 | 105 | 92 |
| Brewery yeast, 53 per cent. activity destroyed by heat | 100 | 21 | 33 |
| Brewery yeast, 60 per cent. activity destroyed by heat | 100 | 12 | 42 |
The case of galactose merits special attention. Previous investigations [see Lippmann, 1904, p. 734] have shown that the fermentation of galactose by yeast differs greatly from that of the other hexoses. The subject has been re-investigated by E. F. Armstrong [1905], and by Slator [1908, 1]. Armstrong carried out his experiments in the manner already described (p. 131), and found that some yeasts had, and others had not, the power of fermenting galactose, although all were capable of fermenting glucose, fructose, and mannose.
Slator made quantitative experiments on the same subject. He was able to confirm the statement which had previously been made, that certain yeasts which have the property of fermenting galactose possess it only after the yeast has become acclimatised by culture in presence of the sugar. This was shown for brewery yeast and for the species mentioned below. This phenomenon is one of great interest and is strictly analogous to the adaptation of bacteria which has now been quite conclusively established [Neisser, 1906].
| Yeast. | Mode of Culture. Grown in: |
Relative Rates. | |
|---|---|---|---|
| Glucose. | Galactose. | ||
| S. Carlsbergensis | wort | 100 | <1 |
| " | hydrolysed lactose | 100 | 86, 83, 85, 25, 46, 51, 69, 54, 155 |
| S. Cerevisiæ | wort | 100 | <1 |
| " | hydrolysed lactose | 100 | 21, 26, 29 |
| S. Thermantitonum. | wort | 100 | <1 |
| " | hydrolysed lactose | 100 | 77, 53, 35 |
| S. Ludwigii | wort | 100 | <1 |
| " | hydrolysed lactose | 100 | <1 |
It will be seen that in one case the rate of fermentation of galactose was considerably greater than that of glucose. S. Ludwigii did not respond to the cultivation in hydrolysed lactose, but, as Slator points out, it is quite possible that repeated cultivation in this medium might effect the change, and this would be strictly analogous to the results obtained with bacteria. Slator's results have been confirmed by Harden and Norris, R. V. [1910], and by Euler and Johansson [1912, 2] who have made an exceedingly interesting study of the progress of the adaptation. As in the case of mannose the rates of fermentation of glucose and galactose are differently affected by agents such as heat and alcohol; moreover, the rate of fermentation of mixtures of dextrose and galactose is in no case either the sum or the mean of the rates obtained with the separate sugars. The temperature coefficient of the fermentation of galactose also differs slightly from that of the other hexoses.
| Yeast. | Relative Rates. | ||
|---|---|---|---|
| Glucose. | Galactose. | Glucose + Galactose. |
|
| S. Cerevisiæ | 100 | 0 | 103 |
| " | 100 | 34 | 103 |
| S. Carlsbergensis | 100 | 155 | 119 |
| S. Thermantitonum | 100 | 76 | 124 |
Assuming that his conclusion that all yeasts which ferment glucose also ferment fructose and mannose is correct, Armstrong has drawn attention to the fact that these three hexoses are also related by the possession of a common enolic form (p. 97) and has suggested that this enolic form is the substance actually fermented to carbon dioxide and alcohol [1904].
The idea that such an intermediate form is the direct subject of fermentation has much to recommend it. In the first place it is almost certain, as already pointed out, that the sugars in aqueous solution do exist, although to a very small extent, in this enolic form. The slow rate at which equilibrium is established in aqueous solution, however, must be taken as definite evidence that under these circumstances the enolic form is only produced very slowly [compare Lowry, 1903]. This has been used by Slator [1908, 1] as an argument against the probability of the preliminary conversion of the sugars into the enolic form before fermentation. It appears, however, quite possible that under the influence of the fermenting complex of the yeast-cell, or of special enzymes, this change might occur much more rapidly, [p134] and at different rates with the different sugars. This reaction might in fact control the observed rate of fermentation. This conception affords a simple explanation of the different rates of fermentation of mannose and glucose, and also of galactose, the enolic form of which is quite different, by yeast under different circumstances, but does not explain the uniformity of rate observed by Slator for glucose and fructose nor the results with mixtures of sugars. The direct fermentation of a common enolic form is also consistent with the fact that the same hexosephosphate is produced from all three hexoses.
Slator himself prefers the view that the first stage of fermentation consists in the rapid combination of the sugar with the enzyme, producing a compound, which then breaks up at a rate which determines the observed rate of fermentation. This rate will of course vary with the nature of the compound, so that if two sugars form the same compound they will be fermented at the same rate; if they form different compounds, different rates may result. Slator supposes that glucose and fructose form the same compound with the enzyme. This, however, appears to involve an intramolecular change of the same order as the production of the enolic form, and moreover is not absolutely essential, as it is probably sufficient to suppose that the two compounds derived from glucose and fructose are very similar, although possibly not absolutely identical. Mannose and galactose, on the other hand, form stereoisomeric compounds, and the capacity of the fermenting complex to form these compounds may be affected by various agents to a different extent from its capacity for combining with glucose or fructose.
A third theory has also been suggested to explain these phenomena, according to which the various sugars are fermented by different enzymes [see Slator, 1908, 1]. The uniformity of the result obtained with glucose and fructose suggests that these two sugars are fermented by the same enzyme (glucozymase), mannose and galactose by different ones (mannozymase and galactozymase). This would afford a simple explanation of the different rates of fermentation for different sugars and of different degrees of sensitiveness towards reagents.
If, however, a separate and independent mechanism were present for each sugar, the rate of fermentation of mixtures should be the sum of the rates for the constituents. This, as shown above, is not found to be the case, and it is therefore necessary to suppose, either that one sugar influences the fermentation of another in some unknown way, or that only a part of the mechanism of fermentation is specific for the particular sugar. Thus the enzyme may be specific and, the co-enzyme [p135] non-specific, so that only a certain maximum rate is attainable, or again, the supply of free phosphate may be the controlling factor.
In the prevailing state of ignorance as to the exact function of the co-enzyme and of the conditions upon which the velocity of fermentation in the cell depends, it is at present impossible to decide between these various theories, but they all offer points of attack which justify the hope that much further information can be obtained by experimental inquiry.
It will be seen from the foregoing that Buchner's discovery of zymase has opened a chapter in the history of alcoholic fermentation which is yet far from being completed. In every direction fresh problems present themselves, and it cannot be doubted that as in the past, the investigation of the action of the yeast-cell will still prove to be of fundamental importance for our knowledge of the mode in which chemical change is brought about by living organisms. [p136]
| Page of Text on which reference is made. | |
|---|---|
Abeles, Hans (1898), Zur Frage der alkoholischen Gährung ohne Hefezellen Ber., 31, 2261–2267. |
19 |
Aberson, J. H. (1903), La fermentation alcoolique Rec. trav. chim., 22, 78–132. |
128 |
Ahrens, Felix B. (1900), Ein Beitrag zur zellenfreien Gärung Zeitsch. angew. Chemie, 483. |
19 |
Ahrens, Felix B. (1902), Das Gärungsproblem (F. Enke, Stuttgart.) |
9 |
Albert, R. (1899, 1), Ueber künstliche Anreicherung der Hefe an Zymase Ber., 32, 2372–2374. |
24 |
Albert, R. (1899, 2), Erfahrungen bei der Herstellung von Hefepresssaft aus untergähriger Bierhefe der Versuchs- und Lehrbrauerei zu Berlin Wochensch. Brauerei, 16, 485–488. |
36 |
Albert, R. (1900), Einfacher Versuch zur Veranschaulichung der Zymase-Wirkung Ber., 33, 3775–3778. |
21, 39 |
Albert, R. (1901), Verfahren zur Herstellung gärwirksamer steriler Dauerhefe mittels Aceton German Patent Kl. 6a, Nr. 135535. |
21 |
Albert, R. and W. (1901), Chemische Vorgänge in der abgetöteten Hefezelle Centr. Bakt. Par., Abt. II., 7, 737–742. |
39 |
Albert, R., und E. Buchner (1900, 1), Hefepresssaft und Fällungsmittel Ber., 33, 266–271. |
38 |
Albert, R., und E. Buchner (1900, 2), Hefepresssaft und Fällungsmittel Ber., 33, 971–975. |
38 |
Albert, R., E. Buchner, und R. Rapp (1902), Herstellung von Dauerhefe mittels Aceton Ber., 35, 2376–2382. |
21, 39 |
Armstrong, Edward Frankland (1904), Studies on enzyme action, III. The influence of the products of change on the rate of change conditioned by sucro-clastic enzymes Proc. Roy. Soc., 73, 516–526. |
133 |
Armstrong, Edward Frankland (1905), Studies on enzyme action, VIII. The mechanism of fermentation Proc. Roy. Soc., B., 76, 600–605. |
131, 132 |
Armstrong, H. E., and E. F. Armstrong (1910), The origin of osmotic effects, III. The function of hormones in stimulating enzymic change in relation to narcosis and the phenomena of degenerative and regenerative change in living structures Proc. Roy. Soc., B., 82, 588–602. |
127 |
Ashdown, O. E., and J. T. Hewitt (1910), The by-products of alcoholic fermentation J. Chem. Soc., 1636–1648. |
94, 110, 115 |
Bach, A. (1913), Oxydationsprozesse in der lebenden Substanz. Handbuch der Biochemie, Oppenheimer. Ergänzungsband (G. Fischer, Jena), 133–182. |
113 |
Baeyer, Adolf (1870), Ueber die Wasserentziehung und ihre Bedeutung für das Pflanzenleben und die Gärung Ber., 3, 63–75. |
99 |
| 90 | |
Bayliss, W. M. (1914), The nature of enzyme action (Longmans, Green & Co., London.) |
89, 121 |
Beijerinck, M. W. (1897), Weitere Beobachtungen über die Octosporushefe Centr. Bakt. Par., Abt. II., 3, 449–454. |
19 |
Beijerinck, M. W. (1900), Ueber Chinonbildung durch Streptothrix chromogena und Lebensweise dieses Mikroben Centr. Bakt. Par., Abt. II., 6, 2–12. |
19 |
Berthelot, M. (1857), Sur la fermentation alcoolique Compt. rend., 44, 702–706. |
14 |
Berthelot, M. (1860), Sur la fermentation glucosique du sucre de canne Compt. rend., 50, 980–984. |
14 |
Bertrand, G. (1897), Sur l'intervention du manganèse dans les oxydations provoquées par la laccase Compt. rend., 124, 1032–1035. |
63 |
Bertrand, G. (1904), Étude Biochimique de la Bacterie du Sorbose Ann. Chim. Phys., (8), 3, 181–288. |
105 |
Berzelius, Jacob (1836), Einige Ideen über eine bei der Bildung organischer Verbindungen in der lebenden Natur wirksame, aber bisher nicht bemerkte Kraft Berz. Jahresbericht, 15, 237–245. |
8 |
|
Berz. Jahresbericht, 18, 400–403. |
7 |
|
Berz. Jahresbericht, 27, 500. |
9 |
Bokorny, Th. (1906), Ueber die Trennung von Leben und Gärkraft in der Hefe Pflüger's Arch., 114, 535–544. |
19 |
Boysen-Jensen, P. (1908), Die Zersetzung des Zuckers während des Respirationsprocesses. (Vorläufige Mitteilung) Ber. deut. bot. Ges., 26a, 666–667. |
105 |
Boysen-Jensen, P. (1910), Sukkersönderdelingen under Respirationsprocessen hos höjere Planter Dissertation, Köbenhavn. (H. H. Hagerup.) |
105 |
Boysen-Jensen, P. (1914), Die Zersetzung des Zuckers bei der alkoholischen Gärung Biochem. Zeitsch., 58, 451–466. |
105 |
Bredig, Georg (1901), Anorganische Fermente (Engelmann, Leipzig), 68. |
37 |
Brown, Adrian, J. (1892), Influence of oxygen and concentration on alcoholic fermentation J. Chem. Soc., 61, 369–385. |
128 |
Brown, Adrian, J. (1902), Enzyme action J. Chem. Soc., 81, 373–388. |
121 |
Bruyn, C. A. Lobry de (1895), Action des alcalis sur les hydrates de carbone, I. (Expériences provisoires) Rec. trav. chim., 14, 156–165. |
96 |
Bruyn, C. A. Lobry de, and W. A. van Ekenstein (1895), Action des alcalis sur les sucres, II. Transformation réciproque des uns dans les autres des sucres glucose, fructose et mannose Rec. trav. chim., 14, 203–216. |
96 |
Bruyn, C. A. Lobry de, and W. A. van Ekenstein (1896), Action des alcalis sur les sucres, III. Transformations des sucres sous l'influence de l'hydroxyde de plomb Rec. trav. chim., 15, 92–96. |
96 |
Bruyn, C. A. Lobry de, and W. A. van Ekenstein (1897, 1), Action des alcalis sur les sucres, IV. Remarques générales Rec. trav. chim., 16, 257–261. |
96 |
Bruyn, C. A. Lobry de, and W. A. van Ekenstein (1897, 2), Action des alcalis sur les sucres, V. Transformation de la galactose, les tagatoses et la galtose Rec. trav. chim., 16, 262–273. |
96 |
Bruyn, C. A. Lobry de, and W. A. van Ekenstein (1897, 3), Action des alcalis sur les sucres, VI. La glutose et la pseudo-fructose Rec. trav. chim., 16, 274–281. | 96 |
Bruyn, C. A. Lobry de, and W. A. van Ekenstein (1897, 4), Action de l'eau bouillante sur la fructose Rec. trav. chim., 16, 282–283. | 96 |
Buchner, Eduard (1897, 1), Alkoholische Gärung ohne Hefezellen. [Vorläuf. Mitt.] Ber., 30, 117–124. |
16, 18 |
Buchner, Eduard (1897, 2), Alkoholische Gärung ohne Hefezellen. Zweite Mitt. Ber., 30, 1110–1113. |
18, 20, 21 |
Buchner, Eduard (1898), Ueber zellenfreie Gärung Ber., 31, 568–574. |
18 |
Buchner, Eduard (1900, 1), Zymase aus getöteter Hefe Ber., 33, 3307–3310. |
21 |
Buchner, Eduard (1900, 2), Bemerkungen zur Arbeit von A. Macfadyen, G. H. Morris and S. Rowland "Ueber ausgepresstes Hefezellplasma (Buchner's Zymase)" Ber., 33, 3311–3315. |
21, 33 |
Buchner, Eduard (1904), Zur Geschichte der Gärungstheorien Wochensch. Brauerei, 21, 507–510. |
9 |
Buchner, Eduard (1910), Sur la fermentation alcoolique du Sucre Bull. Soc. Chim. |
105, 106 |
Buchner, Eduard, und Wilhelm Antoni (1905, 1), Weitere Versuche über die zellfreie Gärung Zeitsch. physiol. Chem., 44, 206–228. | 36, 40 |
Buchner, Eduard, und Wilhelm Antoni (1905, 2), Existiert ein Coenzym für die Zymase? Zeitsch. physiol. Chem., 46, 136–154. |
62 |
Buchner, Eduard, Hans Buchner, und Martin Hahn (1903), Die Zymasegärung. Untersuchungen über den Inhalt der Hefezellen und die biologische Seite des Gärungsproblems [R. Oldenbourg, München, pp. viii, 416.] |
18, 20, 21, 22, 30, 34, 35, 36, 37, 38, 39, 42, 77, 78, 111, 125 |
Buchner, Eduard, und Franz Duchaček (1909), Ueber fraktionierte Fällung des Hefepresssaftes Biochem. Zeitsch., 15, 221–253. |
38, 66 |
Buchner, Eduard, und Hugo Haehn (1909), Ueber das Spiel der Enzyme im Hefepresssaft Biochem. Zeitsch., 19, 191–218. | 62, 68 |
Buchner, Eduard, und Hugo Haehn (1910, 1), Ueber eine Antiprotease im Hefepresssaft Biochem. Zeitsch., 26, 171–198. |
42 |
Buchner, Eduard, und Hugo Haehn (1910, 2), Studien über den Phosphorgehalt der Hefe und einiger Hefepräparate Biochem. Zeitsch., 27, 418–426. |
65, 123 |
Buchner, Eduard, und Robert Hoffmann (1907), Einige Versuche mit Hefepresssaft Biochem. Zeitsch., 4, 215–234. |
36 |
Buchner, Eduard, und F. Klatte (1908), Ueber das Koenzym des Hefepresssaftes Biochem. Zeitsch., 8, 520–557. |
64, 67 |
Buchner, Eduard, und Jakob Meisenheimer (1904), Die chemischen Vorgänge bei der alkoholischen Gärung Ber., 37, 417–428. |
98, 100, 102 |
Buchner, Eduard, und Jakob Meisenheimer (1905), Die chemischen Vorgänge bei der alkoholischen Gärung. (Zweite Mitteilung) Ber., 38, 620–630. |
97, 98, 100, 102 |
Buchner, Eduard, und Jakob Meisenheimer (1906), Die chemischen Vorgänge bei der alkoholischen Gärung. (Dritte Mitteilung) Ber., 39, 3201–3218. |
31, 87, 95, 102, 104 |
Buchner, Eduard, und Jakob Meisenheimer (1908), Ueber Buttersäuregärung Ber., 41, 1410–1419. |
110 |
Buchner, Eduard, und Jakob Meisenheimer (1909), Ueber die Rolle der Milchsäure bei der alkoholischen Gärung des Zuckers Zeitsch. wiss. Landwirtschaft, 38, Ergänzungsband V., 265–288. |
100, 102, 103 |
Buchner, Eduard, und Jakob Meisenheimer (1910), Die chemischen Vorgänge bei der alkoholischen Gärung. (IV. Mitteilung) Ber., 43, 1773–1795. |
103, 104, 105, 106, 115 |
Buchner, Eduard, und Jakob Meisenheimer (1912), Die chemischen Vorgänge bei der alkoholischen Gärung Ber., 45, 1633–1643. |
105, 106 |
Buchner, Eduard, J. Meisenheimer, und H. Schade (1906), Zur Vergärung des Zuckers ohne Enzyme Ber., 39, 4217–4231. |
114 |
Buchner, Eduard, und Sigurd Mitscherlich (1904), Herstellung glykogenarmer Hefe und deren Anwendung zum Zuckernachweis im Harn Zeitsch. physiol. Chem., 42, 554–562. |
39 |
Buchner, Eduard, und Rudolf Rapp (1897), Alkoholische Gärung ohne Hefezellen Ber., 30, 2668–2678. |
19, 34, 36, 77 |
Buchner, Eduard, und Rudolph Rapp (1898, 1), Alkoholische Gärung ohne Hefezellen Ber., 31, 209–217. |
15, 59, 77 |
Buchner, Eduard, und Rudolf Rapp (1898, 2), Alkoholische Gärung ohne Hefezellen. (5 Mitteilung) Ber., 31, 1084–1090. |
36, 77 |
Buchner, Eduard, und Rudolf Rapp (1898, 3), Alkoholische Gärung ohne Hefezellen. (6 Mitteilung) Ber., 31, 1090–1094. |
32, 36, 77 |
Buchner, Eduard, und Rudolf Rapp (1898, 4), Alkoholische Gärung ohne Hefezellen. (7 Mitteilung) Ber., 31, 1531–1533. |
38 |
Buchner, Eduard, und Rudolf Rapp (1899, 1), Alkoholische Gärung ohne Hefezellen. (8 Mitteilung) Ber., 32, 127–137. |
36 |
Buchner, Eduard, und Rudolf Rapp (1899, 2), Alkoholische Gärung ohne Hefezellen. (9 Mitteilung) Ber., 32, 2086–2094. |
32, 42, 77 |
Buchner, Eduard, und Rudolf Rapp (1901), Alkoholische Gärung ohne Hefezellen. (10 Mitteilung) Ber., 34, 1523–1530. |
38 |
Buchner, Eduard, und Siegfried Skraup (1914), Ist die Enzym-Theorie der Gärung einzuschränken? Ber., 1914, 47, 853–870. |
127 |
Buchner, Eduard, and Albert Spitta (1902), Zymasebildung in der Hefe Ber., 35, 1703–1706. |
38 |
Buchner, Hans (1897), Die Bedeutung der aktiven löslichen Zellprodukte für den Chemismus der Zelle München. med. Wochensch. (No. 12), 44, 300–302, and 322. |
18 |
Cagniard-Latour (1838), Mémoire sur la Fermentation vineuse. (Présenté à l'Académie des Sciences, le 12 juin, 1837) Ann. Chim. Phys., 68, 206–222. |
5 |
Cavendish, Henry (1766), Three papers containing experiments on factitious air. Part III. Containing experiments on the air, produced by fermentation and putrefaction. (Read 13 Nov., 1766) Phil. Trans., 175–184. |
2 |
Chick, Frances (1912), Die vermeintliche Dioxyacetonbildung während der alkoholischen Gärung und die Wirkung von Tierkohle und von Methylphenylhydrazin auf Dioxyaceton Biochem. Zeitsch., 40, 479–485. |
105 |
Clowes, George H. A. (1909), A critical study of the conditions under which zymase and its associated coenzyme bring about alcoholic fermentation Proc. Exper. Biol. and Medicine, 6, 44–46. |
122 |
Colin (1825), Mémoire sur la Fermentation du Sucre. (Lu à l'Académie des Sciences, le 31st janvier, 1825) Ann. Chim. Phys., 28, 128–142. |
5 |
Cremer, M. (1899), Ueber Glykogenbildung im Hefepresssaft Ber., 32, 2062–2064. |
116 |
Dakin, H. D. (1903), The hydrolysis of ethyl mandelate by lipase Proc. Chem. Soc., 19, 161. |
89 |
Dakin, H. D. (1905), The fractional hydrolysis of optically inactive esters by lipase Part II. J. Physiol., 32, 199–206. |
89 |
Dakin, H. D. (1908), The oxidation of leucin, α-amido-isovaleric acid and of α-amido-n-valeric acid with hydrogen peroxide J. Biol. Chem., 4, 63–76. |
93 |
Dakin, H. D. (1912), Oxidations and reductions in the animal body (this series) |
92 |
Dakin, H. D., and H. W. Dudley (1913), The interconversion of α-amino-acids, α-hydroxy-acids, and α-ketonic aldehydes. Part II. J. Biol. Chem., 15, 127–143. |
93 |
Delbrück, Max (1897), Alkoholische Gärung ohne Hefezellen Wochensch. Brauerei, 14, 363–364. |
19 |
|
Annales des Sciences naturelles, 10, 42–67. |
5 |
Dixon, Henry H., and W. R. G. Atkins (1913), The extraction of zymase by means of liquid air. (Preliminary Note) Sci. Proc. Roy. Dublin Soc., 14, 1–8. |
25, 26 |
Duchaček, Franz (1909), Einwirkung verschiedener Antiseptika auf die Enzyme des Hefepresssaftes Biochem. Zeitsch., 18, 211–227. |
36 |
Duclaux, E. (1886), Sur les transformations chimiques provoquées par la lumière solaire Compt. rend., 103, 881–882. |
98 |
Duclaux, E. (1893), Sur les analogies entre les procès de fermentation et de combustion solaire Ann. Inst. Pasteur, 7, 751–754. |
98 |
Duclaux, E. (1896), Études sur l'action solaire. (Premier Mémoire) Ann. Inst. Pasteur, 10, 129–168. |
98 |
|
Mikrobiologie, 2, 142. |
121 |
Dumas, Jean Baptiste (1874), Recherches sur la fermentation alcoolique Ann. Chim. Phys., 3, 57–108. |
128 |
Ehrlich, Felix (1903), Ueber neue stickstoffhaltige Bestandteile der Zuckerabläufe Zeitsch. Verein. Rübenzucker-Ind., 809–829. |
86 |
Ehrlich, Felix (1904,1), Ueber das natürliche Isomere des Leucins Ber., 37, 1809–1840. |
86 |
Ehrlich, Felix (1904, 2), Ueber den neuen optisch-aktiven Nichtzucker, das Isoleucin Zeitsch. Verein. Rübenzucker-Ind., 775–803. |
86 |
Ehrlich, Felix (1905), Ueber die Entstehung des Fuselöles Zeitsch. Verein. Rübenzucker-Ind., 539–567. |
87 |
Ehrlich, Felix (1906, 1), Ueber eine Methode zur Spaltung racemischer Aminosäuren mittels Hefe Biochem. Zeitsch., 1, 8–31; Zeitsch. Verein. Rübenzucker-Ind., 840–860. |
89 |
Ehrlich, Felix (1906, 2), Verfahren zur Gewinnung von Fuselölen und dessen Bestandteilen German Patent Kl. 120, Nr. 177174, vom. 1, 4, 1905 (17 Nov., 1906). |
87 |
Ehrlich, Felix (1906, 3), Die chemischen Vorgänge bei der Hefegärung Biochem. Zeitsch., 2, 52–80; Zeitsch. Verein. Rübenzucker-Ind., 1145–1168. |
87, 90, 94 |
Ehrlich, Felix (1906, 4), Zur Frage der Fuselölbildung der Hefe Ber., 39, 4072–4075. |
87 |
Ehrlich, Felix (1907, 1), Ueber die Bedingungen der Fuselölbildung und über ihren Zusammenhang mit dem Eiweissaufbau der Hefe Ber., 40, 1027–1047; Zeitsch. Verein. Rübenzucker-Ind., 1907, 461. |
87, 94 |
Ehrlich, Felix (1907, 2), Ueber das natürliche Isomere des Leucins Ber., 40, 2538–2562. |
86 |
Ehrlich, Felix (1907, 3), Die Rolle des Eiweisses und der Eiweissabbauprodukte bei der Gärung Jahrb. d. Versuchs u. Lehranstalt f. Brauerei in Berlin, 10, 515–529. |
87 |
Ehrlich, Felix (1908), Ueber eine Synthese des Isoleucins Ber., 41, 1453–1458; Zeitsch. Verein. deutsch. Zuckerind., 1908, 528–533. |
86 |
Ehrlich, Felix (1909), Ueber die Entstehung der Bernsteinsäure bei der alkoholischen Gärung Biochem. Zeitsch., 18, 391–423. |
89 |
Ehrlich, Felix (1911, 1), Ueber die Vergärung des Tyrosins zu p-Oxyphenylethyl alkohol. (Tyrosol) Ber., 44, 139–147. |
88 |
Ehrlich, Felix (1911, 2), Ueber die Bildung des Plasmaeiweisses bei Hefen und Schimmelpilzen Biochem. Zeitsch., 36, 477–497. |
89 |
Ehrlich, Felix (1912), Ueber Tryptophol (β-Indolylæthylalkohol), ein neues Gärprodukt der Hefe aus Aminosäuren Ber., 45, 883–889. |
88 |
Ehrlich, Felix, and K. A. Jacobsen (1911), Ueber die Umwandlung von Aminosäuren in Oxysäuren durch Schimmelpilze Ber., 44, 888–897. |
89 |
Ehrlich, Felix, and P. Pistschimuka (1912, 1), Ueberfuhrung von Aminen in Alkohole durch Hefe und Schimmelpilze Ber., 45, 1006–1012. |
91 |
Ehrlich, Felix, and P. Pistschimuka (1912, 2), Synthesen des Tyrosols und seine Umwandlung in Hordenin Ber., 45, 2428–2437. |
88 |
Ehrlich, Felix (mit A. Wendel) (1908, 1), Ueber die Spaltung racemischer Amidosäuren mittels Hefe, II. Biochem. Zeitsch., 8, 438–466. |
89 |
Ehrlich, Felix, und Adolph Wendel (1908, 2), Zur Kenntnis der Leucinfraktion des Eiweisses Biochem. Zeitsch., 8, 399–437. |
86 |
Emmerling, O. (1899), Das Verhalten von Glycerin-aldehyd und Dioxyaceton gegen Hefe Ber., 32, 542–544. |
104 |
Emmerling, O. (1904), Ueber den Ursprung der Fuselöle Ber., 37, 3535–3538. |
86 |
Emmerling, O. (1905), Ueber den Ursprung der Fuselöle Ber., 38, 953–956. |
86 |
Euler, Hans (1905), Chemische Dynamik der zellfreien Gärung Zeitsch. physiol. Chem., 44, 53–73. |
120 |
Euler, Hans (1912, 1), Ueber die Wirkungsweise der Phosphatese. (3 Mitteilung) Biochem. Zeitsch., 41, 215–223. |
47, 58 |
Euler, Hans (1912, 2), Verhalten der Kohlenhydratphosphorsäure-ester im Tierkörper. Nach Versuchen von E. Thorin und D. Johansson Zeitsch. physiol. Chem., 79, 375–397. |
51 |
Euler, Hans (1914), Ueber die Rolle des Glykogens bei der Gärung durch lebende Hefe Zeitsch. physiol. Chem., 89, 337–344. |
116 |
Euler, Hans, and Bäckström, Helma (1912), Zum Kenntnis der Hefegärung. (2 Mitteilung) Zeitsch. physiol. Chem., 77, 394–401. |
47, 68, 130 |
Euler, Hans, and Th. Berggren (1912), Ueber die primäre Umwandlung der Hexosen bei der alkoholischen Gärung Zeitsch. Gärungsphysiol., 1, 203–218. |
32, 130 |
Euler, Hans, and Henry Cassel (1913), Ueber Katalysatoren der alkoholischen Gärung. Vorläufige Mitteilung Zeitsch. physiol. Chem., 86, 122–129. |
130 |
Euler, Hans, and A. Fodor (1911), Ueber ein Zwischenprodukt der alkoholischen Gärung Biochem. Zeitsch., 36, 401–410. |
47, 48, 49, 50, 105 |
Euler, Hans, and Yngve Funke (1912), Ueber die Spaltung der Kohlenhydratphosphorsäure-ester Zeitsch. physiol. Chem., 77, 488–496. |
51 |
Euler, Hans, and David Johansson (1912, 1), Umwandlung des Zuckers und Bildung der Kohlensäure bei der alkoholischen Gärung Zeitsch. physiol. Chem., 76, 347–354. |
32 |
Euler, Hans, and David Johansson (1912, 2), Untersuchungen über die chemische Zusammensetzung und Bildung der Enzyme. IV. Ueber die Anpassung einer Hefe an Galaktose Zeitsch. physiol. Chem., 78, 246–265. |
133 |
Euler, Hans, and David Johannson (1912, 3), Ueber den Einfluss des Toluols auf die Zymase und auf die Phosphatese Zeitsch. physiol. Chem., 80, 175–181. |
125 |
Euler, Hans, and David Johansson (1912, 4), Versuche über die enzymatische Phosphatbindung Zeitsch. physiol. Chem., 80, 205–211. |
47, 57 |
Euler, Hans, and David Johansson (1913), Ueber die Reaktionsphasen der alkoholischen Gärung Zeitsch. physiol. Chem., 85, 192–208. |
47, 52, 54, 73 |
Euler, Hans, and Sixten Kullberg (1911, 1), Untersuchungen über die chemische Zusammensetzung und Bildung der Enzyme Zeitsch. physiol. Chem., 71, 14–30. |
124, 126 |
Euler, Hans, and Sixten Kullberg (1911, 2), Ueber das Verhalten freier und an Protoplasma gebundener Hefenenzyme Zeitsch. physiol. Chem., 73, 85–100 and partly in Arkiv. Kem. Min. Geol., 4, No. 13, 1–11. |
124, 126 |
Euler, Hans, and Sixten Kullberg (1911, 3), Ueber die Wirkungsweise der Phosphatese Zeitsch. physiol. Chem., 74, 15–28. |
47, 58, 122 |
Euler, Hans, and Gunnar Lundeqvist (1911), Zur Kenntnis der Hefegärung Zeitsch. physiol. Chem., 72, 97–112. |
131 |
Euler, Hans, and Hjalmar Ohlsén (1911), Ueber den Einfluss der Temperatur auf die Wirkung der Phosphatese Biochem. Zeitsch., 37, 313–320. |
47, 57, 58 |
Euler, Hans, and Hjalmar Ohlsén (1912), Ueber die Wirkungsweise der Phosphatese, II Zeitsch. physiol. Chem., 76, 468–477. |
47, 57 |
Euler, Hans, and Beth af. Ugglas (1911), Untersuchungen über die chemische Zusammensetzung und Bildung der Enzyme. (2 Mitteilung) Zeitsch. physiol. Chem., 70, 279–290. |
125, 126 |
Fernbach, A. (1910), Sur la dégradation biologique des hydrates de carbone Compt. rend., 151, 1004–1006. |
107 |
Fernbach, A., and Schoen M. (1913), L'acide pyruvique, produit de la vie de la levure Compt. rend., 157, 1478–1480. |
109 |
Fischer, Emil, und Julius Tafel (1888), Oxydation des Glycerins Ber., 21, 2634–2637. |
104 |
Fischer, Emil, und Julius Tafel (1889), Oxydation des Glycerins, II. Ber., 22, 106–110. |
104 |
Fischer, Emil, und Hans Thierfelder (1894), Verhalten der verschiedenen Zucker gegen reine Hefen Ber., 27, 2031–2037. |
89 |
Fischer, Hugo (1903), Ueber Enzymwirkung und Gärung Centr. Bakt. Par., Abt. II., 10, 547–8. |
19 |
Fitz, Alb. (1880), Ueber Spaltpilzgärungen. (6 Mitteilung) Ber., 13, 1309–1312. |
98 |
Franzen, Hartwig, and O. Steppuhn (1911), Ein Beitrag zur Kenntnis der alkoholischen Gärung Ber., 44, 2915–2919. |
115 |
Franzen, Hartwig, and O. Steppuhn (1912, 1), Vergärung und Bildung der Ameisensäure durch Hefen Zeitsch. physiol. Chem., 77, 129–182. |
103, 115 |
Franzen, Hartwig, and O. Steppuhn (1912, 2), Berichtigung zu der Abhandlung: Ueber die Vergärung und Bildung der Ameisensäure durch Hefen Zeitsch. physiol. Chem., 78, 164. |
115 |
Gay-lussac, Louis Joseph (1810), Extrait d'un Mémoire sur la Fermentation. (Lu à l'Inst., 3 Dec, 1810) Ann. Chim. Phys., 76, 245–259. |
4 |
Geret, L., und M. Hahn (1898, 1), Zum Nachweis des im Hefepresssaft enthaltenen proteolytischen Enzyms Ber., 31, 202–205. |
20 |
Geret, L., und M. Hahn (1898, 2), Weitere Mitteilungen über das im Hefepresssaft enthaltene proteolytische Enzym Ber., 31, 2335–2344. |
20 |
Geret, L., und M. Hahn (1900), Ueber das Hefe-endotrypsin Zeitsch. Biologie, 40, 117–172. |
20 |
Gerhardt, Charles (1856), Traité de Chimie Organique, 4, 537–546 |
10, 15 |
Giglioli, J. (1911), Della probabile funzione degli olii essenziali e di altri prodotti volatili delle piante, quale causa di movemento dei succhi nei tessuli viventi Atti. R. Accad. Lincei, 20, II., 349–361. |
26 |
Green, J. Reynolds (1897), The supposed alcoholic enzyme in yeast Annals of Botany, 11, 555–562. |
19 |
Green, J. Reynolds (1898), The alcohol-producing enzyme of yeast Annals of Botany, 12, 491–497. |
19 |
Gromoff, T., und O. Grigorieff (1904), Die Arbeit der Zymase und der Endotryptase in den abgetöteten Hefezellen unter verschiedenen Verhältnissen Zeitsch. physiol. Chem., 42, 299–329. |
36 |
Gruber, M. (1908), Eduard Buchner München, med. Wochensch., 342. |
18 |
Grüss, J. (1904), Untersuchungen über die Atmung und Atmungsenzyme der Hefe Zeitsch. Ges. Brauwesen, 27, 689. |
111 |
Grüss, J. (1908, 1), Ueber den Nachweis mittelst Chromogramm-Methode dass die Hydrogenase aktiv bei der Alkoholgärung beteiligt ist Ber. deutsch, botan. Ges., 26a, 191–196. |
111 |
Grüss, J. (1908, 2), Hydrogenase oder Reduktase? Ber. deutsch, botan. Ges., 26a, 627–630, Abstract J. Inst. Brewing, 1909, 344. |
111 |
Hahn, Martin (1898), Das proteolytische Enzym des Hefepresssaftes Ber., 31, 200–201. |
20 |
Hahn, Martin (1908), Zur Geschichte der Zymaseentdeckung München. med. Wochensch., 515. |
18 |
Hanriot, M. (1885, 1886), Sur la décomposition pyrogénée des acides de la série grasse Bull. Soc. Chim., 43, 417; 45, 79–80. | 98 |
Harden, Arthur (1901), The chemical action of Bacillus coli communis and similar organisms on carbohydrates and allied compounds J. Chem. Soc., 79, 612–628. |
90, 115 |
Harden, Arthur (1903), Ueber alkoholische Gärung mit Hefepresssaft (Buchner's "Zymase") bei Gegenwart von Blutserum Ber., 36, 715–716. |
20, 41 |
Harden, Arthur (1905), Zymase and alcoholic fermentation J. Inst. Brewing, 11, No. 1. |
42, 103 |
Harden, Arthur (1910), Recent researches on alcoholic fermentation J. Inst. Brewing, 16, 623–639. |
125 |
Harden, Arthur (1913), The enzymes of washed zymin and dried yeast (Lebedeff). I. Carboxylase Biochem. J., 7, 214–217. |
82, 83 |
Harden, Arthur, and Dorothy Norris (1912), The bacterial production of acetylmethylcarbinol and 2–3 butylene glycol from various substances Proc. Roy. Soc., B., 84, 492–499. |
110 |
Harden, Arthur, and Roland V. Norris (1910), The fermentation of galactose by yeast and yeast-juice. (Preliminary Communication) Proc. Roy. Soc., B., 82, 645–649. |
32, 133 |
Harden, Arthur, and Roland V. Norris (1914), The enzymes of washed zymin and dried yeast (Lebedeff). II. Reductase Biochem. J., 8, 100–106. |
68, 111, 113 |
Harden, Arthur, and S. G. Paine (1912), The action of dissolved substances on the autofermentation of yeast Proc. Roy. Soc., B., 84, 448–459. |
127 |
Harden, Arthur, and Robert Robison (1914), A new phosphoric ester obtained by the aid of yeast-juice. (Preliminary Note) Proc. Chem. Soc., 30, 16–17. |
48 |
Harden, Arthur, and Sydney Rowland (1901), Autofermentation and liquefaction of pressed yeast J. Chem. Soc., 79, 1227–1235. |
126 |
Harden, Arthur, J. Thompson, and W. J. Young (1910), Apparatus for the collection of gases evolved in fermentation Biochem. J., 5, 230–235. |
28 |
Harden, Arthur, and W. J. Young (1902), Glycogen from yeast J. Chem. Soc., 81, 1224–1233. |
33 |
Harden, Arthur, and W. J. Young (1904), Gärversuche mit Presssaft aus obergäriger Hefe Ber., 37, 1052–1070. |
30, 33, 35 |
Harden, Arthur, and W. J. Young (1905, 1), The alcoholic ferment of yeast-juice J. Physiol., 32. Proceedings of 12 November, 1904. |
41, 42, 59 |
Harden, Arthur, and W. J. Young (1905, 2), The influence of phosphates on the fermentation of glucose by yeast-juice. (Preliminary communication, 1 June, 1905) Proc. Chem. Soc., 21, 189–191. |
42, 47 |
Harden, Arthur, and W. J. Young (1906, 1), The alcoholic ferment of yeast-juice Proc. Roy. Soc., B., 77, 405–420. |
43 |
Harden, Arthur, and W. J. Young (1906, 2), The alcoholic ferment of yeast-juice. Part II. The co-ferment of yeast-juice Proc. Roy. Soc, B., 78, 369–375. |
59, 62 |
Harden, Arthur, and W. J. Young (1906, 3), Influence of sodium arsenate on the fermentation of glucose by yeast-juice. (Preliminary notice) Proc. Chem. Soc, 22, 283–284. |
37, 75 |
Harden, Arthur, and W. J. Young (1907) Biochem. Centr., 6, 888. |
65 |
Harden, Arthur, and W. J. Young (1908, 1), The alcoholic ferment of yeast-juice. Part III. The function of phosphates in the fermentation of glucose by yeast-juice Proc. Roy. Soc., B., 80, 299–311. |
47, 53, 71 |
Harden, Arthur, and W. J. Young (1908, 2), The fermentation of mannose and lævulose by yeast-juice. (Preliminary note) Proc. Chem. Soc., 24, 115–117. |
73 |
Harden, Arthur, and W. J. Young (1909), The alcoholic ferment of yeast-juice. Part IV. The fermentation of glucose, mannose, and fructose by yeast-juice Proc. Roy. Soc., B., 81, 336–347. |
32, 44, 47, 73 |
Harden, Arthur, and W. J. Young (1910, 1), The function of phosphates in alcoholic fermentation Centr. Bakt. Par., Abt. II., 26, 178–184. |
46, 106 |
Harden, Arthur, and W. J. Young (1910, 2), The alcoholic ferment of yeast-juice. Part V. The function of phosphates in alcoholic fermentation Proc. Roy. Soc., B., 82, 321–330. |
44, 52, 56 |
Harden, Arthur, and W. J. Young (1911, 1), The alcoholic ferment of yeast-juice. Part VI. The effect of arsenates and arsenites on the fermentation of the sugars by yeast-juice Proc. Roy. Soc., B., 83, 451–475. |
56, 75, 77, 79 |
Harden, Arthur, and W. J. Young (1911, 2), Ueber die Zusammensetzung der durch Hefepresssaft gebildeten Hexosephosphorsäure I. Biochem. Zeitsch., 32, 173–176. |
47 |
Harden, Arthur, and W. J. Young (1912), Der Mechanismus der alkoholischen Gärung Biochem. Zeitsch., 40, 458–478. |
46, 106, 108 |
Harden, Arthur, and W. J. Young (1913), The enzymatic formation of polysaccharides by yeast preparations Biochem. J., 7, 630–636. |
31, 130 |
Harding, Victor J. (1912), The action of enzymes on hexosephosphate Proc. Roy. Soc., B., 85, 418–422. |
51 |
Helmholtz, Hermann Ludwig (1843), Ueber das Wesen der Fäulnis und Gärung Arch. Anat. Physiol. Joh. Müller, 5, 453–462. |
10 |
Herzog, R. O. (1902), Ueber alkoholische Gärung, I. Zeitsch. physiol. Chem., 37, 149–160. |
120 |
Herzog, R. O. (1904), Ueber die Geschwindigkeit enzymatischer Reaktionen Zeitsch, physiol. Chem., 41, 416. |
120 |
Hoppe-Seyler, F. (1876), Ueber die Processe der Gärungen und ihre Beziehungen zum Leben der Organismen Pflüger's Archiv, 12, 1–17. |
14 |
Hoppe-Seyler, F. (1877), Ueber Gärungen. Antwort auf einen Angriff des Herrn Moritz Traube Ber., 10, 693–695. |
14 |
Iwanoff, Leonid (1905), Ueber Umwandlungen des Phosphors in der Pflanze S. Travaux de la Soc. des Naturalistes de St. Petersburg, 34. |
47 |
Iwanoff, Leonid (1907), Ueber die Synthese der phospho-organischen Verbindungen in abgetöteten Hefezellen Zeitsch. physiol. Chem., 50, 281–288. |
47 |
Iwanoff, Leonid (1909, 1), Ueber die Bildung der phospho-organischen Verbindung und ihre Rolle bei der Zymasegärung Centr. Bakt. Par., Abt. II., 24, 1–12. |
47, 49, 56, 106 |
Iwanoff, Leonid (1909, 2), Ueber einen neuen Apparat für Gärungsversuche Centr. Bakt. Par., Abt. II., 24, 429–432. |
29 |
Janssens, A., and A. Leblanc (1898), Recherches cytologiques sous la cellule de levure La Cellule, 14, 203–241. |
126 |
Karauschanoff, S. (1911), Zur Frage nach der Bedeutung des Dioxyacetons als eines intermediären Produktes der alkoholischen Gärung Ber. deutsch, bot. Ges., 29, 322. |
105 |
Karczag, L. (1912, 1), Ueber die Gärung der verschiedenen Weinsäuren Biochem. Zeitsch., 38, 516–518. |
81 |
Karczag, L. (1912, 2), In welcher Weise wird die Weinsäure durch Hefe angegriffen? Biochem. Zeitsch., 43, 44–46. |
81 |
Kayser, E. (1911), Sur le suc de levure de bière Compt. rend., 152, 1279–1280. |
26 |
Knoop, F. (1910), Ueber den physiologischen Abbau der Säuren und die Synthese einer Aminosäure im Tierkorper Zeitsch. physiol. Chem., 67, 487–502. |
93 |
Kohl, F. G. (1907), Ueber das Glykogen und einige Erscheinungen bei der Sporulation der Hefe Ber. deut. bot. Ges., 25, 74–85. |
116 |
Kohl, F. G. (1909), Alkoholische Gärung Inaug. Diss. Leipzig Abstr. in Zeitsch. Brauereiwesen, 32, 406–7, and J. Inst. Brewing, 15, 710–711. |
116 |
Kostytscheff, S. (1912, 1), Bildung von Acetaldehyd bei der alkoholischen Zuckergärung. (Vorläufige Mitteilung) Ber., 45, 1289–1293. |
111 |
Kostytscheff, S. (1912, 2), Ueber Alkoholgärung. (1 Mitteilung.) Ueber die Bildung von Acetaldehyd bei der alkoholischen Zuckergärung Zeitsch. physiol. Chem., 79, 130–145. |
106, 109 |
Kostytscheff, S. (1912, 3), Ueber Alkoholgärung. (2 Mitteilung.) Ueber die Bildung von Äthylalkohol aus Acetaldehyd durch lebende und getötete Hefe. Von S. Kostytscheff und E. Hübbenet Zeitsch. physiol. Chem., 79, 359–374. |
110 |
Kostytscheff, S. (1913, 1), Ueber den Mechanismus der alkoholischen Gärung Ber., 46, 339. |
111 |
Kostytscheff, S. (1913, 2), Ueber Alkoholgärung. III. Die Bedingungen der Bildung von Acetaldehyd bei der Gärung von Dauerhefe Zeitsch. physiol. Chem., 83, 93–111. |
111, 113 |
Kostytscheff, S. (1914), Ueber Alkoholgärung (6 Mitteilung) Das Wesen der reduktion von Acetaldehyd durch lebenden Hefe Zeitsch. physiol. Chem., 89, 367–372. |
111 |
Kostytscheff, S., and E. Hübbenet (1913), Zur Frage der Reduktion von Acetaldehyd durch Hefesaft Zeitsch. physiol. Chem., 85, 408–411. |
110, 115 |
Kostytscheff, S., and W. Brilliant (1913), Ueber Alkoholgärung. V. Ueber Eiweissspaltung durch Dauerhefe in Gegenwart von Zinkchlorid Zeitsch. physiol. Chem., 85, 507–516. |
111 |
Kostytscheff, S., and A. Scheloumoff (1913), Ueber Alkoholgärung. IV. Ueber Zuckerspaltung durch Dauerhefe in Gegenwart von Zinkchlorid Zeitsch. physiol. Chem., 85, 493–506. |
111, 113 |
Kunz, R. (1906), Ist die bei der alkoholischen Hefegärung entstehende Bernsteinsäure als Spaltungsprodukt des Zuckers anzusehen? Zeitsch. Unters. Nahr. Genussmittel, 12, 641–645. |
89 |
|
München, med. Wochensch., 44, 321–322. |
19 |
Kusseroff, R. (1910), Eine neue Theorie der alkoholischen Gärung Centr. Bakt. Par., Abt. II., 26, 184–187. |
116 |
Kützing, Friedrich (1837), Mikroskopische Unter-suchungen über die Hefe und Essigmutter, nebst mehreren anderen dazugehörigen vegetabilischen Gebilden. (Im Auszuge vorgetragen in der Vers. d. Naturhistor. Vereins des Harzes, in Alexisbad am 26 Juli, 1837) J. pr. Chem., 11, 385–409. |
7 |
Lange, H. (1898), Beitrag zur alkoholischen Gärung ohne Hefezellen Wochensch. Brauerei, 15, 377–378. |
19 |
Lavoisier, A. (1789), Traité Élémentaire de Chymie, Chap. XIII |
2 |
Leathes, J. B. (1906), Problems in Animal Metabolism London, 81–86. |
110 |
Lebedeff, Alex. v. (1908), Auftreten von Formaldehyd bei der zellfreien Gärung Biochem. Zeitsch., 10, 454–457. |
117 |
Lebedeff, Alex. v. (1909), Versuche zur Aufklärung des zellenfreien Gärungsprocesses mit Hilfe des Ultrafilters Biochem. Zeitsch., 20, 114–125. |
29, 31, 47, 49, 50 |
Lebedeff, Alex. v. (1910), Ueber Hexosephosphorsäureester. (1 Mitteilung) Biochem. Zeitsch., 28, 213–229. |
31, 47, 48, 50 |
Lebedeff, Alex. v. (1911, 1), Ueber den Mechanismus der alkoholischen Gärung Ber., 44, 2932–2942. |
105, 106 |
Lebedeff, Alex. v. (1911, 2), Extraction de la zymase par simple macération Compt. rend., 152, 49–51. |
24 |
Lebedeff, Alex. v. (1911, 3), Sur l'extraction de la zymase Compt. rend., 152, 1129. |
24 |
Lebedeff, Alex. v. (1911, 4), La zymase est-elle une diastase? Ann. Inst. Past., 25, 682–694. (Bull. Soc. Chim., IV., 9, 672–682.) |
34, 35 |
Lebedeff, Alex. v. (1911, 5), Bemerkungen zu der Arbeit von Hans Euler und Sixten Kullberg. Ueber die Wirkungsweise der Phosphatese Zeitsch. physiol. Chem., 75, 499–500. |
47 |
Lebedeff, Alex. v. (1911, 6), Ueber Hexosephosphorsäureester. II. Biochem. Zeitsch., 36, 248–260. |
47, 50 |
Lebedeff, Alex. v. (1911, 7), Sur l'extraction de la zymase Bull. Soc. Chim., IV., 9, 744–750. |
24 |
Lebedeff, Alex. v. (1912, 1), Ueber die Extraktion der Zymase Chem. Zeit., 36, 365. |
24 |
Lebedeff, Alex. v. (1912, 2), Extraction de la Zymase par simple macération Ann. Inst. Past., 26, 8–37. |
24 |
Lebedeff, Alex. v. (1912, 3), Notiz über "Phosphatese" Biochem. Zeitsch., 39, 155–157. |
47 |
Lebedeff, Alex. v. (1912, 4), Ueber den Mechanismus der alkoholischen Gärung. Biochem. Zeitsch., 46, 483–489 (and Bull. Soc. Chim., IV., 11, 1039–1041). |
107 |
Lebedeff, Alex. v. (1913, 1), Ueber die Veresterung von Dioxyaceton mit Phosphaten Zeitsch. physiol. Chem., 84, 305. |
47 |
Lebedeff, Alex. v. (1913, 2), Ueber den kinetischen Verlauf der alkoholischen Gärung Zeitsch. Gärungsphysiol., 2, 104–106. |
31 |
Lebedeff, Alex. v. (1914, 1), Ueber den Mechanismus der alkoholischen Gärung III. Zellenfreie Gärung der Polyoxy-carbonsäuren Ber., 1914, 47, 660–672. |
108 |
Lebedeff, Alex. v. (1914, 2), Notij über zellenfreie Gärung der Poly-oxycarbonsäuren Ber., 1914, 47, 965–967. |
108 |
Lebedeff, Alex. v., and N. Griaznoff (1912), Ueber den Mechanismus der alkoholischen Gärung Ber., 45, 3256–3272. |
107, 110 |
Liebig, Justus v. (1839), Ueber die Erscheinungen der Gärung, Fäulnis und Verwesung und ihre Ursachen Annalen, 30, 250–287. |
8 |
Liebig, Justus v. (1870), Ueber die Gärung und die Quelle der Muskelkraft Annalen, 153, 1–47; 137–228. |
13 |
Lindner, Paul (1905), Mikroskopische Betriebskontrolle in den Gärungsgewerben [Berlin, 4th edition], 234. |
131 |
|
Chem. Zeit, 23, 851. |
19 |
Lintner, C. J., and H. J. v. Liebig (1911), Ueber die Reduktion des Furfurols durch Hefe bei der alkoholischen Gärung Zeitsch. physiol. Chem., 72, 449–454. |
113 |
Lintner, C. J., and H. Lüers (1913), Ueber die Reduktion des Chloralhydrates durch Hefe bei der alkoholischen Gärung Zeitsch. physiol. Chem., 88, 122–123. |
113 |
Lippmann, E. v. (1904), Die Chemie der Zuckerarten Braunschweig. |
97, 100, 132 |
Löb, Walter (1906), Zur chemischen Theorie der alkoholischen Gärung Landwirtsch. Jahrb., 35, 541; Chem. Zeit., 42, 540; Zeitsch. Elektrochem., 1906, 12, 282; 1907, 13, 311–516. |
116, 117 |
Löb, Walter (1908, 1), Zur Kenntnis der Zuckerspaltungen. I. Mitteilung. Die Einwirkung von Zinkcarbonat auf Formaldehydlösungen Biochem. Zeitsch., 12, 78–96. |
117 |
Löb, Walter (1908, 2), Zur Kenntnis der Zuckerspaltungen. II. Mitteilung. Die Einwirkung von Zinkstaub und Eisen auf Formaldehydlösungen; die Einwirkung von Zinkstaub auf Traubenzucker Biochem. Zeitsch., 12, 466–472. |
117 |
Löb, Walter (1909, 1), Zur Kenntnis der Zuckerspaltungen. III. Mitteilung. Die Elektrolyse des Traubenzuckers Biochem. Zeitsch., 17, 132–144. |
117 |
Löb, Walter (1909, 2), Zur Kenntnis der Zuckerspaltungen. V. (Vorläufige) Mitteilung. Die Umkehrung der Zuckersynthese Biochem. Zeitsch., 20, 516–522. |
117 |
Löb, Walter (1909, 3), Zur Kenntnis der Zuckerspaltungen. VI. Mitteilung. Die elektrolytische Reduktion des Traubenzuckers Biochem. Zeitsch., 22, 103–105. |
117 |
Löb, Walter (1909, 4), Zur Kenntnis der Zuckerspaltungen. VII. Mitteilung. Die Umkehrung der Zuckersynthese (von Walter Löb und Georg Pulvermacher) Biochem. Zeitsch., 23, 10–26. |
117 |
Löb, Walter (1910), Zur geschichte der chemischen Gärungshypothesen Biochem. Zeitsch., 29, 311–315. |
107 |
Löb, Walter, und Georg Pulvermacher (1909), Zur Kenntnis der Zuckerspaltung. IV. Mitteilung. Die Elektrolyse des glycerins und des Glykols Biochem. Zeitsch., 17, 343–355. |
117 |
Lowry, T. Martin (1903), Studies of dynamic isomerism. I. The mutarotation of glucose J. Chem. Soc, 83, 1314–1323. |
133 |
Lüdersdorff, F. v. (1846), Ueber die Natur der Hefe Ann. Physik., 76, 408–411. |
15 |
Lunge, G. (1905), Chemisch-technische Untersuchungsmethoden (Berlin, 5te Aufgabe), 3, 571. |
87 |
|
Zeitsch. Elektrochem., 13, 517. |
103 |
Lvoff, Sergius (1913, 1), Zymase und Reduktase in ihren gegenseitigen Beziehungen Ber. deut. bot. Ges., 31, 141–147. |
112 |
Lvoff, Sergius (1913, 2), Sur le rôle de la reductase dans la fermentation alcoolique Bull. Acad. Sci. St. Pétersbourg, 501–532. |
112 |
Lvoff, Sergius (1913, 3), Hefegärung und Wasserstoff Zeitsch. Gärungsphysiol., 3, 289–320. |
112 |
Macfadyen, Allan, G. Harris Morris, and Sydney Rowland (1900), Ueber ausgepresstes Hefezellplasma (Buchner's "Zymase") Ber., 33, 2764–2790. |
19, 24, 27, 30 |
Mckenzie, Alex., and Arthur Harden (1903), The biological method for resolving inactive acids into their optically active components J. Chem. Soc, 83, 424–438. |
89 |
Magnus-levy, A. (1902), Ueber den Aufbau der hohen Fettsäuren aus Zucker Arch. Anat. Physiol., 365–368. |
110 |
Manasseïn, Marie v. (1872), Mikroskopische Untersuchungen. Herausgegeben von Wiesner [Stuttgart], 128. |
15 |
Manasseïn, Marie v. (1897), Zur Frage von der alkoholischen Gärung ohne lebende Hefezellen Ber., 30, 3061–3062. |
15 |
Marckwald, W. (1902), Ueber die Trennung der Amylalkohole des Fuselöles. III. Ber., 35, 1595–1601. |
88 |
Martin, C. J. (1896), A rapid method of separating colloids from crystalloids in solutions containing both J. Physiol., 20, 364–371. |
59 |
Martin, C. J., and H. G. Chapman (1898), An endeavour to procure an alcoholic ferment from yeast-cells Proc. physiol. Soc., 11 June, ii. |
19 |
Mayer, Adolf (1879), Lehrbuch der Gärungschemie 3. Ausgabe Heidelberg, Carl Winters' Univ.-buchh. |
15 |
Mayer, P. (1907), Zur Frage der Vergärbarkeit von Methylglyoxal Biochem. Zeitsch., 2, 435–437. |
104 |
Mazé, P. (1902), Recherches sur les modes d'utilisaion du carbone térnaire par les végétaux et les microbes Ann. Inst. Pasteur, 16, 446; Compt. rend., 134, 241. |
98 |
Meisenheimer, Jakob (1903), Neue Versuche mit Hefepresssaft Zeitsch. physiol. Chem., 37, 518–526. |
35 |
Meisenheimer, Jakob (1907), Ueber die chemischen Vorgänge bei den als Enzymreaktionen erkannten Gärungen Biochem. Centr., 6, 1–13. |
100 |
Meisenheimer, Jakob (1908), Ueber das Verhalten der Glukose, Fructose und Galaktose gegen verdünnte Natronlauge Ber., 41, 1009–1019. |
97 |
Mitscherlich, Eilhard (1841), Ueber die chemische Zersetzung und Verbindung mittels Contactsubstanzen Monatsber. K. Akad. d. Wissensch. Berlin, 1841, 392; Ann. Physik., 1842, 55, 209–229. |
10 |
Nägeli, C. v. (1879), Theorie der Gärung [München. R. Oldenbourg,] 156. |
15 |
Nāgeli, C., und O. Loew (1878), Ueber die chemische Zusammensetzung der Hefe Annalen, 193, 322–348. |
15 |
Nef, J. U. (1904), Dissoziationsvorgänge in der Glycolglycerinreihe Annalen, 335, 247–333. |
98 |
Nef, J. U. (1907), Dissoziationsvorgänge in der Zuckergruppe. (I. Abhandlung.) Ueber das Verhalten der Zuckerarten gegen die Fehling'sche Lösung sowie gegen andere Oxydationsmittel Annalen, 357, 214–312. |
98 |
Neisser, M. (1906), Ein Fall von Mutation nach de Vries bei Bakterien und andere Demonstrationen Cent. Bakt. Par., Abt. I., Ref. 38, Append., 98–102. |
132 |
Neubauer, Otto and Konrad Fromherz (1911), Ueber den Abbau der Aminosäuren bei der Hefegärung Zeitsch. physiol. Chem., 70, 326–350. |
92, 109 |
Neuberg, Carl (1908), Chemische Umwandlungen durch Strahlenarten. I. Mitteilung. Katalytische Reaktionen des Sonnenlichtes Biochem. Zeitsch., 13, 305–320. |
93 |
Neuberg, Carl (1909), Chemische Umwandlungen durch Strahlenarten. II. Mitteilung. Wirkungen des elektrischen Gleichstroms Biochem. Zeitsch., 17, 270–292. |
93 |
Neuberg, Carl (1912), Ueber zuckerfreie Hefegärungen, VII. Bildung von β-Oxybuttersäurealdehyd (Aldol) bei der Vergärung von Brenztraubensäure Biochem. Zeitsch., 43, 491–493. |
82 |
Neuberg, Carl, and A. Hildesheimer (1911), Ueber zuckerfreie Hefegärungen, I. Biochem. Zeitsch., 31, 170–176. |
81 |
Neuberg, Carl, and L. Karczag (1911, 1), Die Gärung der Brenztraubensäure und Oxalessigsäure als Vorlesungsversuch Ber., 44, 2477–2479. |
81 |
Neuberg, Carl, and L. Karczag (1911, 2), Ueber zuckerfreie Hefegärungen, III. Biochem. Zeitsch., 36, 60–67. |
82, 127 |
Neuberg, Carl, and L. Karczag (1911, 3), Ueber zuckerfreie Hefegärungen, IV. Carboxylase ein neues Enzym der Hefe Biochem. Zeitsch., 36, 68–75. |
82 |
Neuberg, Carl, and L. Karczag (1911, 4), Ueber zuckerfreie Hefegärungen, V. Zur Kenntnis der Carboxylase Biochem. Zeitsch., 36, 76–81. |
81, 83 |
Neuberg, Carl, and L. Karczag (1911, 5), Ueber zuckerfreie Hefegärungen, VI. Biochem. Zeitsch., 37, 170–176. |
82 |
Neuberg, Carl, and Johannes Kerb (1912, 1), Ueber zuckerfreie Hefegärungen, VIII. Enstehung von Acetaldehyd bei der sog. Selbstgärung Biochem. Zeitsch., 43, 494–499. |
111, 112 |
Neuberg, Carl, and Johannes Kerb (1912, 2), Ueber zuckerfreie Hefegärungen, IX. Vergärung von Ketosäuren durch Weinhefen Biochem. Zeitsch., 47, 405–412. |
81 |
Neuberg, Carl, and Johannes Kerb (1912, 3), Ueber zuckerfreie Hefegärungen, X. Die Gärung der α-Ketobuttersäure Biochem. Zeitsch., 47, 413–420. |
82 |
Neuberg, Carl, and Johannes Kerb (1912, 4), Entsteht bei zuckerfreien Hefegärungen Æthylalkohol? Zeitsch. Gärungphysiol., 1, 114–120. |
109, 110, 115 |
Neuberg, Carl, and Johannes Kerb (1913, 1), Ueber zuckerfreie Hefegärungen, XII. Ueber die Vorgänge bei der Hefegärung Biochem. Zeitsch., 53, 406–419; Ber., 46, 2225–2228. |
82, 93, 110 |
Neuberg, Carl, and Johannes Kerb (1913, 2), Ueber zuckerfreie Hefegärungen, XIII. Zur Frage der Aldehydbildung bei der Gärung von Hexosen sowie bei der sog. Selbstgärung Biochem. Zeitsch., 58, 158–170. |
112, 113 |
Neuberg, Carl, and W. Oertel (1913), Studien über Methylglyoxalbildung Biochem. Zeitsch., 55, 495–503. |
99 |
Neuberg, Carl, and P. Rosenthal (1913), Ueber zuckerfreie Hefegärungen, XI. Weiteres zur Kenntnis der Carboxylase Biochem. Zeitsch., 51, 128–142. |
46, 82, 83, 93 |
Neuberg, Carl, and H. Steenbock (1913), Ueber die Bildung höherer Alkohole aus Aldehyden durch Hefe, I. Uebergang von Valeraldehyd in Amyl Alkohol Biochem. Zeitsch., 52, 494–503. |
93, 113 |
Neuberg, Carl, and H. Steenbock (1914), Ueber die Bildung höherer Alkohole aus Aldehyden durch Hefe, II. Biochem. Zeitsch., 59, 188–192. |
93, 113 |
Neuberg, Carl, and L. Tir (1911), Ueber zuckerfreie Hefegärungen, II. Biochem. Zeitsch., 32, 323–331. |
81 |
Neumeister, R. (1897), Bemerkungen zu Eduard Buchner's Mitteilungen über Zymase Ber., 30, 2963–2966. |
19 |
Oppenheimer, Max (1914, 1), Ueber die Bildung von Milchsäure bei der alkoholischen Gärung Zeitsch. physiol. Chem., 89, 45–62. |
102, 109 |
Oppenheimer, Max (1914, 2), Ueber die Bildung von Glycerin bei der alkoholischen Gärung Zeitsch. physiol. Chem., 89, 63–77. |
33, 95, 102, 109 |
O'Sullivan, James (1898), On the rate of alcoholic fermentation J. Soc. Chem. Ind., 17, 559–560. |
128 |
O'Sullivan, James (1899), The hydrolytic and fermentative functions of yeast J. Inst. Brewing (3), 5, 161–175. |
128 |
Parnas, Jakob (1910), Ueber fermentative Beschleunigung der Cannizzaroschen Aldehydumlagerung durch Gewebesäfte Biochem. Zeitsch., 28, 274–294. |
94 |
Paine, Sydney, G. (1911), The permeability of the yeast cell Proc. Roy. Soc,, B., 84, 289–307. |
51 |
Palladin, W. (1908), Beteiligung der Reduktase im Prozesse der Alkoholgärung Zeitsch. physiol. Chem., 56, 81–88. |
111 |
Pasteur, Louis (1857), Mémoire sur la fermentation appelée lactique Compt. rend., 45, 913–916. |
11 |
Pasteur, Louis (1860), Mémoire sur la fermentation alcoolique Ann. Chim. Phys., (3), 58, 323–426. |
11, 12 |
Pasteur, Louis (1872), Note sur la mémoire de M. Liebig relatif aux fermentations Ann. Chim. Phys., (4), 25, 145–151. |
13 |
Pasteur, Louis (1875), Nouvelles observations sur la nature de la fermentation alcoolique Compt. rend., 80, 452–457. |
13, 14 |
Pavy, F. W., and H. W. Bywaters (1907), On glycogen formation by yeast J. Physiol., 36, 149–163. |
116 |
Payen, Anselme et Persoz (1833), Mémoire sur la diastase, les principaux produits de ses reactions et leur applications aux arts industriels Ann. Chim. Phys., 53, 73–92. |
14 |
Piloty, Oscar (1897), Ueber eine neue Totalsynthese des glycerins and des Dioxyacetons Ber., 30, 3161–3169. |
104 |
Pinkus, Georg (1898), Ueber die Einwirkung von Benzhydrazid auf Glukose Ber., 31, 31–37. |
98 |
Plimmer, Robert Henry Aders (1913), The metabolism of organic phosphorus compounds. Their hydrolysis by the action of enzymes Biochem. J., 7, 43–71. |
51 |
Pringsheim, Hans H. (1905), Zur Fuselölfrage Ber., 38, 486–487. |
86 |
Pringsheim, Hans H. (1906), Ueber die Bildung von Fuselöl bei Acetondauerhefe-Gärung Ber., 39, 3713–3715. |
87 |
Pringsheim, Hans H. (1907), Ueber die Stickstoffernährung der Hefe. (Ein Beitrag zur Physiologie der Hefe) Biochem. Zeitsch., 3, 121–286. |
86 |
Pringsheim, Hans H. (1908), Ueber die Unterdrückung der Fuselölbildung und die Mitwirkung von Bakterien an der Bildung höherer Alkohole bei der Gärung Biochem. Zeitsch., 10, 490–497. |
86 |
Pringsheim, Hans H. (1909), Bemerkungen zur Mitwirkung von Bakterien an der Fuselölbildung Biochem. Zeitsch., 16, 243–245. |
86 |
Resenscheck, Friederich (1908, 1), Einwirkung des elektrischen Stromes auf den Hefepresssaft Biochem. Zeitsch., 9, 255–263. |
67 |
Resenscheck, Friederich (1908, 2), Einwirkung von kolloidalem Eisenhydroxyd auf den Hefepresssaft Biochem. Zeitsch., 15, 1–11. |
67 |
Rinckleben, P. (1911), Gewinnung von Zymase aus frischer Brauereihefe durch Plasmolyse Chem. Zeit., 35, 1149–1150. |
26 |
Rowland, Sydney (1901), A method of obtaining intracellular juices J. Physiol., 27, 53–56. |
24 |
Rubner, Max (1913), Die Ernährungsphysiologie der Hefezelle bei alkoholischer Gärung Rubner's Archiv. Physiologie Suppl. Bd., 1–369. |
126 |
Salkowski, E. und H. (1879), Weitere Beiträge zur Kenntnis der Fäulnisprodukte des Eiweiss Ber., 12, 648–655. |
90 |
Schade, H. (1906), Ueber die Vergärung des Zuckers ohne Enzyme Zeitsch. physikal. Chem., 57, 1–46. |
114 |
Schade., H. (1907), Berichtigung und Nachtrag zu der Arbeit "Ueber die Vergärung des Zuckers ohne Fermente" Zeitsch. physikal. Chem., 60, 510–512. |
114 |
Schade, H. (1908), Ueber die Vorgänge der Gärung vom Standpunkt der Katalyse Biochem. Zeitsch., 7, 299–326. |
114 |
Schlossberger, J. (1844), Ueber die Natur der Hefe, mit Rücksicht auf die Gärungserscheinungen Annalen, 51, 193–212. |
10 |
Schmidt, C. (1847), Gärungsversuche Annalen, 61, 168–174. |
15 |
Schroeder, H. (1859, 1861), Ueber Filtration der Luft in Beziehung auf Fäulnis und Gärung Annalen, 109, 35–52; 117, 273–295. |
10 |
Schroeder, H., und Th. v. Dusch (1854), Ueber Filtration der Luft in Beziehung auf Fäulnis und Gärung Annalen, 89, 232–343. |
10 |
Schrohe, A. (1904), Eilhard Mitscherlich und die vitalistische Gärungstheorie in der deutschen Literatur vor Pasteur Hefe, Gärung und Fäulnis, 208–229 [Parey, Berlin]. |
9, 11 |
Schulze, Franz (1836), Vorläufige Mitteilung der Resultate einer experimentellen Beobachtung über Generatio aequivoca Ann. Physik., 39, 487–489. |
6 |
Schwann, Theodor (1837), Vorläufige Mitteilung, betreffend Versuche über die Weingärung und Fäulnis Ann. Physik., 41, 184–193. |
6 |
Slator, Arthur (1906), Studies in fermentation. Part I. The chemical dynamics of alcoholic fermentation by yeast J. Chem. Soc., 89, 128–142. |
29, 103, 128 |
Slator, Arthur (1907), Ueber Zwischenprodukte der alkoholischen Gärung Ber., 40, 123–126. |
103 |
Slator, Arthur (1908, 1), Studies in Fermentation. Part II. The mechanism of alcoholic fermentation J. Chem. Soc., 93, 217–242. |
103, 130, 131, 132, 133, 134 |
Slator, Arthur (1908, 2), Factors which influence fermentation Chem. News, 98, 175, and Brit. Ass. Reports, 1908. | 103 |
Slator, Arthur (1912), Ueber Dioxyaceton als zwischenstufe der alkoholischen Gärung Ber., 45, 43–46. |
106 |
Slator, Arthur, and H. J. S. Sand (1910), Studies in fermentation. Part III. The rôle of diffusion in fermentation by yeast-cells J. Chem. Soc, 922–927. |
129 |
Stahl, Georg Ernst (1697), Zymotechnia fundamentalis (Franckfurth, 1734), 304. |
2 |
Stavenhagen, A. (1897), Zur Kenntnis der Gärungserscheinungen Ber., 30, 2422–2423 and 2963. |
19 |
Tafel, Julius (1907), Ueber Zwischenprodukte bei chemischen Reaktionen Ber., 40, 3318–3321. |
103 |
Tammann, G. (1889), Ueber die Wirkung der Fermente Zeitsch. physikal. Chem., 3, 25–37. |
128 |
Thenard, Louis Jacques (1803), Mémoire sur la fermentation vineuse Ann. Chim. Phys., 46, 294–320. |
4 |
Traube, Moritz (1858), Theorie der Ferment-wirkungen [Berlin, Ferd. Dummler's Verlagsbuchh.], 119. |
14 |
Traube, Moritz (1877), Die chemische Theorie der Fermentwirkungen und der Chemismus der Respiration. Antwort auf die Ausserungen des Herrn Hoppe-Seyler Ber., 10, 1984–1992. |
14 |
Trommsdorff, Richard (1902), Ueber die Beziehungen der Gram'schen Färbung zu chemischen Vorgängen in der abgetöteten Hefezelle Centr. Bakt. Par., Abt. II., 8, 82. |
39 |
Turpin (1838), Mémoire sur la cause et les effets de la fermentation alcoolique et acéteuse Compt. rend., 7, 369–402. |
8 |
|
München, med. Wochensch., 44, 321. |
19 |
Volhard, Jakob (1909), Justus von Liebig [Johann Ambrosius Barth. Leipzig.] |
13 |
Wager, Harold (1898), The nucleus of the yeast plant Annals of Botany, 12, 449. |
126 |
Wager, Harold (1911), The yeast cell J. Inst. Brewing, 2–22. |
126 |
Wager, Harold, and Annie Peniston (1910), Cytological observations on the yeast plant Annals of Botany, 24, 45–83. |
116, 126 |
Walton, James Henri, Jr. (1904), Die Jodionenkatalyse des Wasserstoffsuperoxyds Zeitsch. physikal. Chem., 47, 185–222. |
29 |
|
Botan. Zeit., 53. |
19 |
Will, H. (1897), Alkoholische Gärung ohne Hefezellen Zeitsch. ges. Brauwesen., 20, 363–364. |
19 |
Will, H. (1898), Zur Frage der alkoholischen Gärung ohne Hefezellen Zeitsch. ges. Brauwesen., 21, 291. |
19 |
Windaus, A., und F. Knoop (1905), Ueberführung von Traubenzucker in Methylimidazol Ber., 38, 1166–1170. |
99 |
Wohl, A. (1898), Ueber die Acetale des Akroleïns und des Glycerinaldehyds Ber., 31, 1796–1801. |
104 |
Wohl, A. (1907, 1), Ueber Oxyfumar- und Oxymaleïnsäure Ber., 40, 2282–2300. |
103 |
Wohl, A. (1907, 2), Die neueren Ansichten über den chemischen Verlauf der Gärung Biochem. Zeitsch., 5, 45–65, and Zeitsch. angew. Chem., 20, 1169–1177. |
100, 104 |
Wohl, A. (1908), Zur Kenntnis der Dreikohlenstoffreihe Ber., 41, 3599–3612. |
98 |
Wohl, A., und C. Oesterlin (1901), Ueberführung der Weinsäure in Oxalessigsäure durch Wasserabspaltung bei niederer Temperatur Ber., 34, 1139–1148. |
100 |
[Wöhler] (1839), Das enträtselte Geheimniss der geistigen Gärung. (Vorläufige briefliche Mitteilung.) Annalen, 29, 100–104. |
8 |
Wroblewski, A. (1898), Gärung ohne Hefezellen Ber., 31, 3218–3225; Centr. Physiol., 12, 697–701. |
19 |
Wroblewski, A. (1899), Ueber den Buchner'schen Hefepresssaft Centr. Physiol., 13, 284–297. |
19 |
Wroblewski, A. (1901), Ueber den Buchner'schen Hefepresssaft J. pr. Chem., (2), 64, 1–70. |
19, 42 |
Young, W. J. (1909), The hexosephosphate formed by yeast-juice from hexose and phosphate (Prel. Note, Proc. Chem. Soc., 1907, 65.) Proc. Roy. Soc., B., 81, 528–545. |
47, 56 |
Young, W. J. (1911), Ueber die Zusammensetzung der durch Hefepresssaft gebildeten Hexosephosphorsäure II. Biochem. Zeitsch., 32, 178–188. |
47, 50 |
With a few exceptions, original spelling and grammar were retained. The cover image is a modified version of the original scanned image from archive.org. This book contains several uncommon Unicode characters, including "→", "↓", "⇌", "⇅", "│", "╱", "╲", "═", "║". A font and ebook reader software capable of rendering these is necessary for reading this book.
Full stops, middle dots "·", or even "˙"—Dot Above"—were variably (randomly?) used in the original as decimal points and for denoting chemical bonds. These have been rendered as middle dots herein.
The reference to Colin's paper on page 5 has been changed from 1826 to 1825, to agree with the corresponding entry in the Bibliography. The reference to Turpin's paper on page 8 was changed to 1838 from 1839, for the same reason.
Tables and formulas have been edited for clarity and readability, while honoring the original form. For example, the two sequential equations at the top of page 110 originally had "+O" and "−H2O" written under the two arrows, and the two equations appeared on one line. These have been converted into two equations on two lines, with the arrow subtext moved into the equations.
The incorrect formula for the enol II. in the equation for Glucose dehydration near bottom of page 101 was corrected.