BY
JOÃO MOOJEN
University of Kansas Publications
Museum of Natural History
Volume 1, No. 19, pp. 301-406, 140 figures in text
December 10, 1948
University of Kansas
LAWRENCE
1948
University of Kansas Publications, Museum of Natural History
Editors: E. Raymond Hall, Chairman, A. Byron Leonard,
Edward H. Taylor
Volume 1, No. 19, pp. 301-406, 1 plate, 140 figures in text
Published December 10, 1948
University of Kansas
Lawrence, Kansas
PRINTED BY
FERD VOILAND, JR., STATE PRINTER
TOPEKA, KANSAS
1948
22-3343
Speciation in the Brazilian Spiny Rats
(Genus Proechimys, Family Echimyidae)
By
JOÃO MOOJEN
| PAGE | |||
| Introduction | 305 | ||
| Methods and Terminology | 305 | ||
| Acknowledgments | 308 | ||
| Paleontology | 308 | ||
| Speciation | 311 | ||
| Subgeneric variation | 312 | ||
| Specific variation in the subgenus Proechimys | 314 | ||
| Subspecific variation in the subgenus Proechimys | 317 | ||
| Specific variation in the subgenus Trinomys | 320 | ||
| Subspecific variation in the subgenus Trinomys | 322 | ||
| Taxonomic Characters | 323 | ||
| Size and proportions of external parts | 323 | ||
| Pelage | 324 | ||
| Skull | 326 | ||
| Incisive foramen | 326 | ||
| Teeth | 327 | ||
| Habits | 330 | ||
| Changes with Age | 331 | ||
| Genus Proechimys | 333 | ||
| Artificial Key to Subgenera and Species | 334 | ||
| Subgenus Proechimys | 338 | ||
| Proechimys goeldii | 338 | ||
| Proechimys goeldii steerei | 338 | ||
| Proechimys goeldii goeldii | 340 | ||
| Proechimys semispinosus | 342 | ||
| Proechimys semispinosus liminalis | 343 | ||
| Proechimys semispinosus amphichoricus | 344 | ||
| Proechimys semispinosus kermiti | 345 | ||
| [Pg 304] | Proechimys longicaudatus | 346 | |
| Proechimys semispinosus brevicauda | 349 | ||
| Proechimys semispinosus boimensis | 350 | ||
| Proechimys semispinosus longicaudatus | 351 | ||
| Proechimys semispinosus leucomystax | 352 | ||
| Proechimys semispinosus roberti | 353 | ||
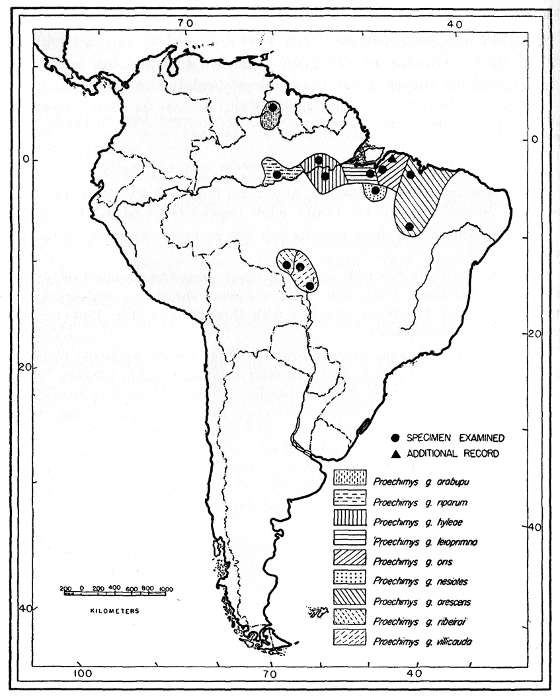
| Proechimys guyannensis | 355 | ||
| Proechimys guyannensis villicauda | 355 | ||
| Proechimys guyannensis ribeiroi | 361 | ||
| Proechimys guyannensis hyleae | 361 | ||
| Proechimys guyannensis nesiotes | 363 | ||
| Proechimys guyannensis leioprimna | 364 | ||
| Proechimys guyannensis oris | 365 | ||
| Proechimys guyannensis arescens | 366 | ||
| Proechimys guyannensis riparum | 367 | ||
| Proechimys guyannensis arabupu | 369 | ||
| Subgenus Trinomys | 369 | ||
| Proechimys dimidiatus | 371 | ||
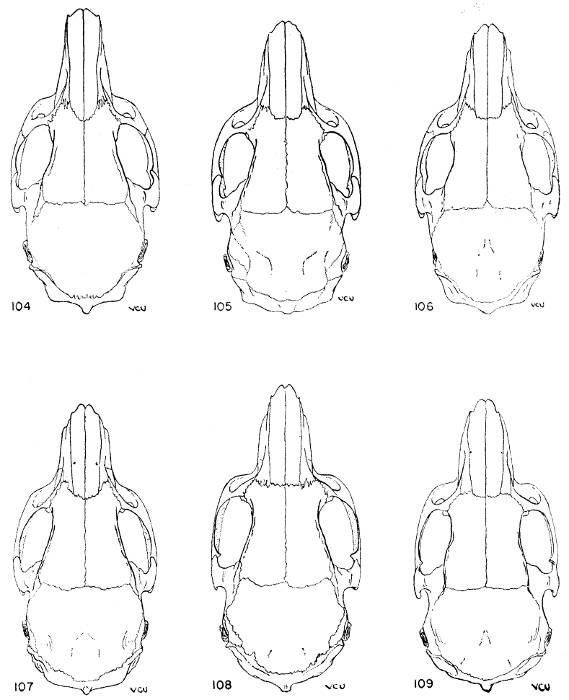
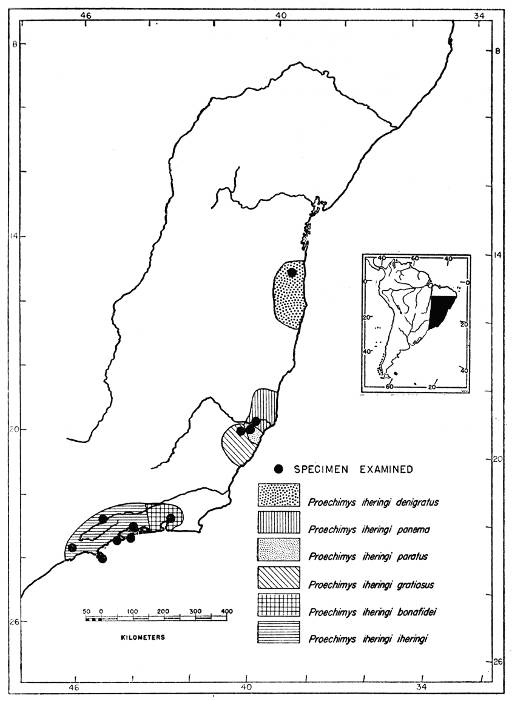
| Proechimys iheringi | 373 | ||
| Proechimys iheringi iheringi | 378 | ||
| Proechimys iheringi bonafidei | 378 | ||
| Proechimys iheringi gratiosus | 379 | ||
| Proechimys iheringi panema | 380 | ||
| Proechimys iheringi denigratus | 381 | ||
| Proechimys iheringi paratus | 382 | ||
| Proechimys setosus | 384 | ||
| Proechimys setosus setosus | 385 | ||
| Proechimys setosus elegans | 387 | ||
| Proechimys albispinus | 388 | ||
| Proechimys albispinus albispinus | 390 | ||
| Proechimys albispinus sertonius | 391 | ||
| Incerta Sedis | 392 | ||
| Proechimys myosuros | 392 | ||
| Conclusions | 393 | ||
| Table of Measurements | 395 | ||
| Literature Cited | 400 | ||
 Fig. 1. Proechimys dimidiatus (Günther). Live female on left and male on
right. × ½. From Tingua, Nova Iguassú, Rio de Janeiro, Brazil. Photographed
in spring (August or September) of 1942 by author.
Fig. 1. Proechimys dimidiatus (Günther). Live female on left and male on
right. × ½. From Tingua, Nova Iguassú, Rio de Janeiro, Brazil. Photographed
in spring (August or September) of 1942 by author.The spiny-rats included in the genus Proechimys are common in almost every forest of South America above the Tropic of Capricorn, and in Central America northward to approximately 12° N, in Nicaragua. In size and proportions they are similar to the brown rat Rattus norvegicus but actually they belong to a very different suborder of rodents—the Hystricomorpha. The hystricomorphs are represented in South America by a large variety of animals, of which capybaras, agoutis and cavies are common representatives.
The pelage of the spiny-rats has a large number of flattened, spinelike hairs, especially on the back. The color ranges through different tints and shades of reddish-brown more or less evenly distributed on the upper parts; the underparts are usually pure white, sharply contrasting with the brown color above. The tail is bicolored, brown above and white below.
The spiny-rats live in forests of different types, generally in the proximity of water. Shelter is usually procured under boulders, stumps or masses of roots. The reproductive rate is low; on the average, there are only two young per litter and only two litters per year.
Sixty-odd names have been given to species and subspecies of Proechimys in the last hundred and fifty years and no serious revision of the taxonomy of the genus was undertaken in the last century. The purpose of the present work is to provide means of understanding species and subspecies within the genus and to describe the different kinds known to occur within the confines of Brazil.
Pelage.—It was found advisable to use a standardized nomenclature for hairs. The names here proposed are a choice of those used in the literature, with the suffix "form" as an element of uniformity. I feel that it would be advantageous if everyone adopted a similar universal system in mammalogy.
The names listed below are used as nouns and are considered as English versions which could easily be adapted to different languages. These names may be complemented with adjectives as needed. Examples are lanceolate aristiforms, spinous aristiforms, and woolly setiforms.
Aristiforms: The most conspicuously developed hairs in a three-layered pelage or the corresponding hairs in a simpler pelage. Names previously used for these hairs are: guard hair, leithaar and jarre.
Setiforms: Common to all species and most numerous throughout the pelage; second in conspicuousness, being the dominant hairs in the middle layer. Synonyms are: over hairs, grannenhaare and soies.
Villiforms: The smallest hairs in the three-layered pelage. Synonyms are: underfur, wollhaar and duvet.
Vibrissiforms: The vibrissae proper, or any typically sensory hair.
Teeth.—The tritubercular nomenclature was abandoned because of overwhelming difficulties; more research on the Hystricomorpha is certainly needed before the tritubercular nomenclature can be applied with confidence. The following names are used for features of the molariform teeth:
Main fold: The inner or lingual fold in the upper molariform teeth and outer or labial fold in the lower molariform teeth.
Counterfold: Any outer or labial fold in the upper or any inner or lingual fold in the lower molariform teeth.
For incisors Thomas (1921:141) is followed: opisthodont, orthodont and proodont depending on the angle between the exposed part of incisors and the ventral surface of the rostrum.
The capital letters P and M designate premolars and molars, respectively, of the upper jaws; lower case letters p and m designate corresponding teeth in the lower jaws.
Measurements.—Measurements of skins were used only when provided by the collector. The length of the hind-foot is intended to be always cum unguis, but in a few instances it is impossible to be sure whether the collector included the nail. Length of tail was used only when the tail seemed not to be mutilated. Ear measurements taken by collectors are scarce. In spite of the apparent usefulness of length of ear, it was found to be inadvisable to take the measurement on the dry skins.
The following measurements of the skull are used in the tables:
Greatest length: From the anteriormost part of the nasals to the posteriormost part of the supraoccipital.
Condylo-incisive length: From the anterior face of one incisor, at the alveolus, to the posteriormost part of the exoccipital condyle of the same side.
Zygomatic breadth: Maximum distance across zygomata in a plane perpendicular to longitudinal axis of the skull.
Length of nasals: Maximum length of one or both, whichever is the greater.
Interorbital constriction: Least width between the orbits on top of the skull.
Palatilar length: From the posterior face of an incisor, at the alveolus, to the nearest part of the posterior edge of the palatine bone.
Crown length of cheekteeth: From the anterior border of P4 to the posterior border of M3.
In the accounts of species, measurements of aristiforms and setiforms are used. The hairs measured were taken from the middorsal region and outer thighs, and the measurements are means.
All specimens of which measurements are here recorded, as for example in [Pg 307] the tables, are fully adult; each specimen shows some wear on each of the four upper molariform teeth unless otherwise indicated.
Capitalized color terms are after Ridgway "Color Standards and Color Nomenclature," Washington, D. C., U. S. A., 1912. One setiform was taken from the animal and placed over the rectangles in Ridgway's charts and the examination made under a microscope with low (×7) magnification and natural light. This method was found to give the most satisfactory results.
The following abbreviations are used for names of institutions:
AMNH—American Museum of Natural History.
CNHM—Chicago Natural History Museum.
DZ—Departamento de Zoologia da Secretaria de Agricultura, São Paulo, Brazil.
MCZ—Museum of Comparative Zoology at Harvard College.
MN—Museu Nacional, Brazil.
MZ—Museum of Zoology, University of Michigan.
SEPFA—**Serviço de Estudos e Pesquisas sobre a Febre Amarela, Brazil.
USNM—United States National Museum.
UZM—Universitets Zoologiske Museum, Copenhagen.
Approximately two thousand skins and skulls were assembled at the Museum of Natural History, University of Kansas, through the coöperation of the authorities in the various institutions of North America, Brazil and Denmark, as listed immediately above. This comprehensive material was used to obtain a more complete understanding of the group, and for the loan of these specimens I am extremely grateful to the authorities of each of the institutions.
First of all I acknowledge the encouragement given me in the Proechimys project by Heloisa Alberto Torres, Director of the Museu Nacional, Rio de Janeiro. I extend my thanks also to Stephen D. Durrant, of the University of Utah, for helpful corrections in the preparation of the manuscript; to Mrs. Virginia Cassell Unruh, for the preparation of the drawings of the skulls; to Miss Alice M. Bruce for assistance in drawing the maps; and to my daughter, Julieta, for help in assembling data and for typing.
Dr. Remington Kellogg, Curator of Mammals in the United States National Museum, and the late Dr. Wilfred H. Osgood, formerly Curator Emeritus of the Department of Zoölogy in the Chicago Natural History Museum, generously permitted me to use their private lists of South American mammals. These lists contain much unpublished data, as for example, proof, in Kellogg's list, that Proechimys guyannensis (E. Geoffroy Saint-Hilaire, 1803) antedates P. cayennensis (Desmarest, 1817). I register here my gratitude to both these zoölogists and acknowledge other critical assistance from Dr. Kellogg.
The John Simon Guggenheim Memorial Foundation awarded me a fellowship for which I am deeply grateful. This expression of the Foundation's interest in education and good neighborliness made possible the completion of the present paper.
Finally I desire to express my deepest gratitude to Professor E. Raymond Hall, Director of the Museum of Natural History and Chairman of the Department of Zoölogy at the University of Kansas whose untiring aid and guidance has enabled me to terminate this study.
The only known, significant, fossil Proechimys comes from deposits in the limestone caves of Lagoa Santa, Minas Gerais, Brazil. These deposits, of Late Pleistocene or Recent age, were extensively studied by P. W. Lund and the results published in a series of French and Danish papers. F. Ameghino (1934:110) studied another fauna from a deposit of similar age in the cave of Iporanga, São Paulo, Brazil. Proechimys is recorded in his account under the inclusive specific name fuliginosus.
The molariform teeth of the fossil described by Lund (1841:pl. 21, fig. 14) shows its close relationship to the living form P. s. [Pg 309] elegans (Lund) which still inhabits the same region. It belongs in the more specialized subgenus Trinomys which seems to have been derived from Proechimys. Trinomys has the main fold in the molars always greatly developed and the fold tends to set apart one lamina in the occlusal surface. The Lagoa Santa fossil, like some specimens of the living subspecies, has a small main fold in P4. However, the main fold is large in all upper molars and in the lower molariform teeth which are notably specialized in the extreme reduction of the number of counterfolds to only one.
One hypothesis concerning the evolution of the genus is that a more primitive group of Proechimys lived in all of the Central Plateau of Brazil in the Pleistocene Time. The climatic conditions at that time might have been such as to support large forests but, since the Pleistocene, these climatic conditions may have changed from humid to the present drier conditions, which support the dominant, savanna, floral climax. Actually the extinct fauna from the caves includes animals which have disappeared from the area and now live only in more humid areas, as for example Myocastor, which has shifted to the lowlands to the west and south.
Possibly climatic changes were responsible for the faunal shift from the region that is now a plateau in Central Brazil. This climatic change may have resulted from the gradual uplift of the eastern part of the continent. This uplift prevents part of the trade winds which come from the east from carrying the same amount of moisture inland as they did previously. In fact, the Andean revolution, even if it occurred as late as Late Tertiary, would have had no perceptible influence on the amount of water precipitated on the more eastern parts of the continent. Oliveira and Leonardos (1943:617) point out that after a Cretaceous submersion of the central part of Brazil, there was a general uplift. The authors (op. cit.:689) mention the presence of continental Cretaceous deposits in the Central Plateau of Brazil, in support of these changes, and state that "pelo menos em certas zonas do litoral a elevação do continente prolongou-se até o Pleistoceno."
Berry (1942:373) concluded, among other things, that there was a southward extension "in South America of equatorial floras in the lower Miocene," and (op. cit.:372) that ... "east of the Andean Axis in the south temperate zone there was a normal mesophytic flora ... instead ... of present day large steppes."
My idea is that a tropical forest still covered the Central Plateau of Brazil in (early?) Pleistocene times and that populations of [Pg 310] Proechimys of a primitive type, similar to P. g. steerei, for example, lived in that extensive forest-climax. The gradual uplift of the plateau, however, gradually brought about drier conditions in this region. As a result a large cliseral change was initiated, which shifted the forest-climax to the more humid eastern escarpments and lowlands that were gradually being developed, while the savanna climax was being established on the plateau. Eventually the effect of the decreasing moisture was locally accentuated by the erosion of the sandstones (Oliveira and Leonardos, 1943:690) in northeastern Brazil, thus depriving it of a natural reservoir of rain water. An arid belt was developed which now constitutes an efficient geographic barrier to the distribution of many kinds of animals.
One marginal species may have shifted eastward with the forest-climax to effect the Recent distribution. The eastern species became completely isolated from the main group, accumulated mutations, and evolved into the subgeneric type Trinomys. The generic trend that gave rise to Trinomys probably remained more stable as far as supraspecific changes are concerned. The lack of barriers in the distributional area of the original group favored the dispersal and submergence of mutations and, therefore, there was but little further supraspecific evolution. The speciation in both subgenera finally resulted from gradual differentiation of varying populations since they show combinations of the generic biotypes and possess few truly qualitative characters.
The cliseral changes in the Central Plateau, which developed the dry belt, a barrier, might explain the evolution of a few more supraspecific groups of mammals, as indicated by the presence of similar forms in the Amazonian region and in Southeastern Brazil. Among these Echimys and Phyllomys, in the same family with Proechimys, show differences that are parallel to those observed in Proechimys. One of these parallel changes is the increased lamination of the cheekteeth. Although Echimys, from the Amazonian region, has upper molariform teeth with the four laminae fused, Phyllomys has the four laminae completely separated.
None of the genera known from the Upper Oligocene and Miocene of Argentine deposits seems to be directly ancestral to Proechimys.
The detection of differences of systematic worth between populations of animals, represented by skins and skulls, is a step preliminary to deducing the factors responsible for the differences. Ordinarily the factors which cause heritable differences have to do with geographic isolation and adaptation to ecological conditions. When differences in the structure of the animal are known, a person is led to speculate on the factors which could cause them. For one thing, does the observed degree of difference tend to isolate animals possessing the "new" character from the other animals? It would seem to me that the isolation once started by one of these differences tends to be accentuated with time and the difference itself thus then becomes a factor responsible for further differentiation.
Whether or not transition from one character to another occurs gradually, in its geographic expression, and thus whether or not intergradation occurs between two subspecies, can be ascertained by the analysis of a series of population-samples appropriately distributed geographically. If two characters of systematic worth are known to blend in one part of the geographic range of a subgenus, and if the same two characters are seen in two other populations, far removed geographically from each other and without any samples of annectent populations to provide actual evidence of intergradation, then such intergradation is to be inferred.
The available collections of Proechimys mostly were made haphazardly with the result that there are extensive areas from which no specimens as yet are available. Thus, actual proof of intergradation is often lacking in areas where it almost certainly occurs. In some extensive areas, however, many samples, from relatively regular intervals, have been available and they provide genuine proof of intergradation. These instances have served as a guide for estimating whether other samples should be considered to be full species or instead merely subspecies of the same species.
Lack of intergradation in any of the characters may be accepted as the criterion of full species. Where two populations occupying the same range (sympatric populations) show different qualitative characters, they almost certainly do not crossbreed. Furthermore the characters that distinguish such kinds of nonintergrading animals are likely to be considered as of full specific value when detected in far distant parts of the range of the subgenus.
In a genus that is widespread and continuously distributed, it is useful to know which characters always distinguish full species and which ones, sometimes or always, distinguish only subspecies, since in a population from a small island, there is, ordinarily, less individual variation than in a corresponding population from the mainland or a larger island; under certain circumstances a person might be tempted to give specific rank to the population when its characters actually are analogous to those separating subspecies elsewhere.
Sometimes it is convenient to recognize species-groups, a systematic category without nomenclatural status, intermediate between the species and the subgenus. When there are two groups of species not sharply separated, including one species whose characters overlap those of each of the two groups, it would seem most appropriate to recognize only species-groups instead of subgenera. When, on the other hand, the two groups of species have mutually exclusive characters and a species with intermediate characters is unknown, the two groups of species can conveniently be accorded separate subgeneric rank.
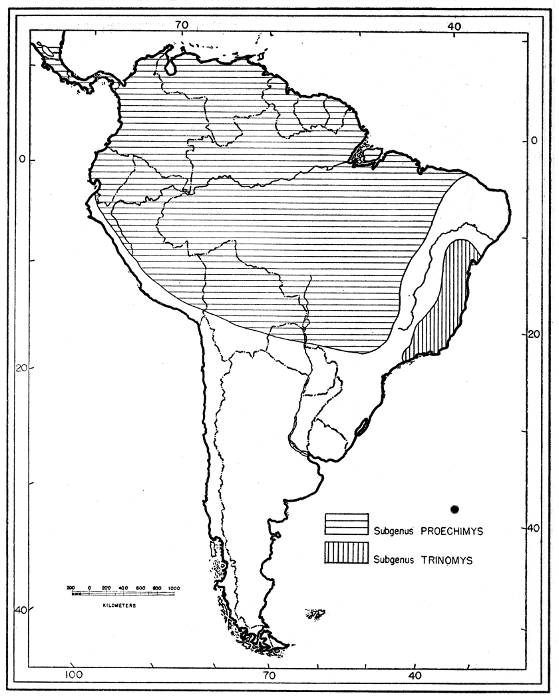
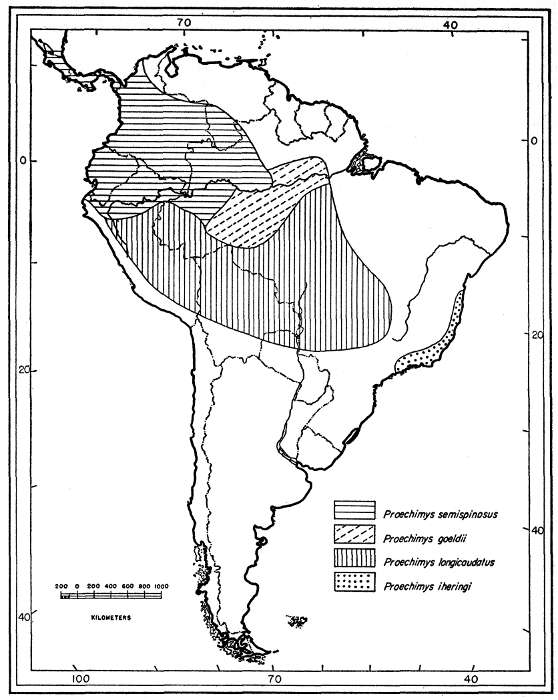
A few characters are common to one group of species and other features are common to a second group. The most striking of these features is the character of the main fold in the molariform teeth. In one group the fold transversely crosses the crown of the tooth and in the other it extends scarcely halfway across. No specimen is intermediate in this respect. These two groups, furthermore, are separated geographically by an important barrier, the arid belt that starts in the northeastern littoral of Brazil (Ceará), and that extends south and southwesterly, more or less accompanying the São Francisco River in the Plateau, to about 20° S. Proechimys is thought not to inhabit this arid belt. At the latitude of 20° S the conditions become more suitable for Proechimys, especially along the rivers which flow eastward, but there the Plateau is replaced by mountains: the Serra Geral at the west, and Serra da Mantiqueira at the south; these ranges are bare of forests at higher elevations. Two groups of species of Proechimys are, therefore, kept geographically isolated: one group lives in southeastern Brazil, and the other lives in a large area to the west which starts at 21° S in Paraguay and Brazil and widens northward and includes, farther west, central and northern Brazil and all the South American countries above [Pg 313] 21° S, as well as Central America northward to southern Nicaragua.
The two groups which are here treated as subgenera may be designated as follows:
Trinomys—main fold deep: aristiforms well-developed on the rump and outer thighs; tail no less than 75 per cent of length of head and body; skull without ridges across the parietals; no conspicuous groove for transmission of nerve inside infraorbital foramen; molariform teeth decreasing in size from premolar to third molar; 1 to 3 counterfolds in the molariform teeth.
Proechimys—main fold shallow: aristiforms not developed on rump and outer thighs; tail less than 75 per cent of length of head and body; groove for transmission of nerve present in infraorbital foramen of several subspecies; molariform teeth increasing in size from premolar to second molar; 2 to 5 counterfolds in molariform teeth.
Most of these characters vary but do not overlap. Subgeneric rank is here accorded to the two groups of Proechimys characterized immediately above.
The primary cause of the subgeneric differentiation is thought to have been geologic changes in the continental area. As already pointed out (see Paleontology), decreasing humidity in the Central Plateau of Brazil may have caused a migration southwestward of one or more of the species along with the forests. Once isolated geographically, the species probably differentiated at an accelerated rate.
The fact that a much larger number of subspecies occupies the larger geographic range of the subgenus Proechimys would not be sufficient to prove that this subgenus, Proechimys, is nearer to the primitive group than Trinomys, the subgenus occupying the smaller range with fewer subspecies. The paleontological evolution of the rodents, however, consistently points to teeth with a larger number of counterfolds (as seen in Proechimys) as the primitive condition. The extension of the main fold, tending to set apart one lamina in each upper molariform tooth, seems to be a specialization; reduction in the size of the head and body, increase in length of tail and decreasing size of molars posteriorly also may be specializations. The main point, however, is to establish if Trinomys is a relic group rather than a "differentiated" one. If an intermediate form were known which connected Trinomys with one species of Proechimys more than with another or even if Trinomys itself more closely resembled one of the groups of species of the subgenus Proechimys than [Pg 314] it did another, we would assume that divergence and selection accounted for the subgeneric variation. The lack of any such connecting link favors the first idea, namely that Trinomys differentiated rapidly with the aid of geographic variation.
If Trinomys is, as I am inclined to consider it, the result of "differentiation," its subgeneric features are to be admitted as "new" and therefore the most primitive species in the genus should be found in the subgenus Proechimys.
It is a matter of common sense to admit the two groups considered above as subgenera rather than genera. Since the two structural plans were established they would, and do, act as different sources of variation. On the other hand, the morphological differences do not give the two groups an amount of morphological differences that would justify full generic rank for each.
Most of the described forms in the subgenus were initially named as distinct full species. More recently, however, in accordance with the ideas now prevalent in systematic work, many of the named kinds were reduced to the rank of subspecies. Tate first made a geographic arrangement (1935:399-400) and later (1939:177-178) provisionally synonymised several named kinds of Proechimys with Proechimys "cayennensis cayennensis." A similar tendency was clearly displayed by Ellerman (1940:115-122) who allocated 29 names, out of 33 (in the subgenus, as here understood), to the species Proechimys guyannensis and gave full specific rank to four other named kinds. Osgood (1944) also had the same viewpoint; that is to say, he appeared to have the idea that there were only two full species in the subgenus in Brazil—admitting this orally—and consequently he synonymised some full species where two or more occurred in the same place, thinking that he was dealing with individual, rather than specific, differences. Evidently the number of species in the subgenus cannot be great because the known kinds show few patterns worthy of specific designation and therefore the majority of the existing names should be suspected of having no more than subspecific value. Nevertheless none of the above writers presented real evidence in support of his arrangement.
Criteria for the recognition of full species are most easily recognized where two or more different species live together. In the [Pg 315] literature, P. goeldii and P. "oris" were mentioned by Thomas (1912:89) as having been collected in the same place; P. mincae and P. canicollis, by H. H. Smith (in Allen, 1904:440); P. "leucomystax," from Utiarití, by Miranda Ribeiro (1914:42) and P. "longicaudatus," from the same place, by Allen (1916:569) were other examples. In these, and other alleged instances of two or more kinds occurring together, detailed study of the specimens concerned was necessary to learn the true facts. Also with the opportunity to compare collections from several different places, new facts emerged. P. longicaudatus, as it was conceived of by Allen, was a composite species, but in one locality, Utiarití, Ribeiro and Allen actually were dealing with two distinct species.
The species, or subspecies belonging to different species, living together are: goeldii and hyleae, at Fazenda Paraiso; goeldii and riparum in Manaus; boimensis and hyleae in Tauarí; leucomystax and villicauda in Utiarití; mincae and canicollis in Bonda; gularis and hendeei on the banks of Rio Napo ("same trap lines," according to P. Hershkovitz, In Litt.). Study of samples of the above named pairs of kinds of Proechimys showed the following specific differences: goeldii is large with narrow aristiforms, has a large and strongly built skull, with four counterfolds in one or more upper molars: hyleae is smaller, has wide aristiforms, smaller skull with less pronounced ridges, and never has more than three counterfolds in the upper molariform teeth; riparum closely resembles hyleae; boimensis has thin aristiforms, small skull and no more than three counterfolds in the upper molariform teeth in contrast to hyleae, already discussed; leucomystax closely resembles boimensis; villicauda closely resembles both hyleae and riparum; mincae is similar to hyleae-riparum-villicauda; canicollis has the number of counterfolds in all molars reduced to two; gularis is large, has a strongly built and ridged skull, some upper molariform teeth with four counterfolds and wide aristiforms; hendeei closely resembles leucomystax and boimensis.
The evidence obtained from study of specimens where two or more species occurred together was applied to the remaining samples and the geographic distribution was worked out. As a result the arrangement below was made, including all valid kinds already named and those here newly named from Brazil. The names of kinds I do not consider as belonging to the subgenus (and genus) are excluded. These are Echimys macrourus Jentink, not seen, and Proechimys cayennensis hoplomyoides Tate (= genus Hoplomys). The application of names is tentative, however, because the types [Pg 316] deposited in Europe have not been seen. An asterisk denotes the forms not seen by me.
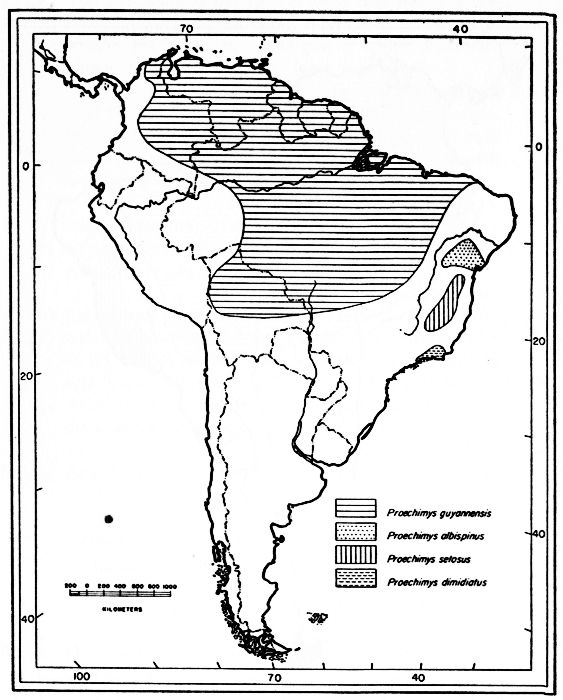
Proechimys guyannensis: arabupu, arescens, bolivianus, cherriei, chrysaeolus, guairae, o'connelli, guyannensis*, hyleae, leioprimna, mincae, nesiotes, ochraceus, oris, poliopus, ribeiroi, riparum, trinitatis, urichi, vacillator*, villicauda, warreni.
Proechimys longicaudatus: boimensis, brevicauda, elassopus, hendeei, leucomystax, longicaudatus, nigrofulvus, pachita, rattinus*, roberti, securus, simonsi.
Proechimys semispinosus: amphichoricus, burrus, calidior, centralis, chiriquinus, colombianus, decumanus, goldmani*, gorgonae, gularis, hilda*, ignotus, kermiti, liminalis, panamensis, rosa*, rubellus, semispinosus.
Proechimys goeldii: goeldii, steerei.
Proechimys canicollis.
Proechimys guyannensis appears to be more plastic than any other species. In size of animal, width of aristiforms, color and number of counterfolds in the cheekteeth, it shows marked response to variations in geographic conditions. Proechimys longicaudatus is apparently less plastic; only the number of counterfolds shows marked variation. Proechimys semispinosus varies much within its range. Proechimys goeldii seems to be relatively uniform. Proechimys canicollis shows relatively little variation throughout its range but probably is divisible into two or more subspecies.
The primitive Proechimys probably was large with a short tail, narrow aristiforms, strongly built skull, and five counterfolds in each molariform tooth. Primitiveness here is inferred from characters which now are of general occurrence in the whole group as opposed to those restricted in geographic occurrence.
It is a curious fact that in this genus, populations from small islands are more primitive than populations on the mainland. Apparently a small population restricted to a small island tends to revert to the primitive type. The homozygous condition will tend toward a generalized genotype and the disappearance of secondary biotypes. P. i. iheringi on the Island of São Sebastião averages larger, has thinner aristiforms, and a stronger skull than the same subspecies on the mainland, and the cheekteeth usually have two and three counterfolds. The same subspecies on the mainland has no more than two counterfolds. Proechimys semispinosus gorgonae and Proechimys semispinosus ignotus, living on Gorgona and San José islands, respectively, are both characterized by large size, short tails, strong and conspicuously ridged skulls, and cheekteeth frequently with four and five counterfolds. On the mainland, closely related subspecies, like P. s. panamensis, chiriquinus and gularis, far [Pg 317] less frequently have four counterfolds in more than one or two teeth. More striking still is the population-sample of gularis from the island of Llunchi, in the Rio Napo, eastern Ecuador. In it there is a higher ratio of cheekteeth with four counterfolds than there is in the samples from the banks of the river.
The two insular forms, P. s. gorgonae and P. s. ignotus, referred to as primitive in the discussion above, have wide aristiforms, which is contrary to what would be expected in a primitive Proechimys. Supposing, however, as actually seems to be the fact, that narrowness of the aristiforms depends on an increased number of genes, we deduce that the population from the mainland, that gave rise to the populations of the islands, did not have all of the genes necessary to make the aristiforms narrow. In fact the subspecies known on the mainland, near the aforementioned islands, have wide aristiforms.
Another point which favors the idea that narrow aristiforms result from an increased number of genes is that, generally, the aristiforms are narrow in any species whose geographic range is extensive and relatively uniform.
Proechimys goeldii is the species which has the largest number of characters that are judged to be primitive, and it may be the oldest stock. P. semispinosus, P. longicaudatus and P. guyannensis may have been derived from an early splitting of the genus or they may have branched off the main stem at different times. P. canicollis, however, seems clearly to be an offshoot of P. guyannensis; canicollis shows greater resemblance to guyannensis than to any other species. P. g. vacillator is another close relative of P. guyannensis with the number of counterfolds almost as much reduced as in P. canicollis. Conceivably, vacillator is a full species, but the reduction in number of counterfolds in the teeth more probably expresses only one extreme of a gradient, as will be discussed below.
In spite of the lack of specimens from areas in which Proechimys certainly occurs, it is evident that the genus has great plasticity and that the number of subspecies will be greatly increased as additional material is studied. Only perfunctory examination of samples from outside the area of Brazil shows me that there are several unnamed subspecies there. My impression is that Allen's trinitatis, of Trinidad, the genotype of Proechimys, will eventually be split.
There are two main lines of subspeciation in Proechimys guyannensis. The one south of the Amazon River includes P. g. bolivianus, [Pg 318] in Bolivia, P. g. villicauda, and P. g. ribeiroi occurring on the divide of the headwaters of the Amazon and Parana rivers, in Brazil, and P. g. hyleae in the lower Tapajoz and P. g. nesiotes in the lower Tocantins. All six subspecies have a large number of counterfolds in the molariform teeth. In these six subspecies, p4 has four counterfolds and the lower molars have three each. Toward the northeastern coast the number of counterfolds decreases to three in p4 and to two in the lower molars, as in P. g. arescens, P. g. leioprimna and P. g. oris.
In northern South America, north of the Amazon River, the subspecies with the greatest number of counterfolds is P. guyannensis warreni (known from only the Demerara River area); p4 has four counterfolds and the lower molars have three each. The number decreases in all the adjacent populations: P. g. guyannensis, in the Guianas, P. g. trinitatis, and P. g. urichi (going westward from the Guianas to Venezuela) have the counterfolds reduced to three in p4, but the lower molars still have the same number of counterfolds, namely, three, although there is a tendency for them to coalesce; farther west, on the coast, the number decreases to three counterfolds in p4 and to only two in the lower molars as in P. g. guairae and P. g. mincae. Subspecies south of the coast show the same reduction of counterfolds, P. g. cherriei and P. g. o'connelli being examples; P. g. ochraceus and P. g. poliopus have the reduction carried to the upper molars, M3 having usually only two counterfolds; P. g. chrysaeolus in the valley between the Madalena and the Cauca rivers seems to be somewhat isolated and shows reversion to three counterfolds in the lower molariform teeth; directly southward of the range of P. g. warreni the number of counterfolds decreases to three in all lower cheekteeth (population at Ayan-Tepuy, southern Venezuela), and then to three in p4 and to two in the lower molars, as in P. g. arabupu on the Brazilian side of Mount Roraima, and the reduction is extended to the upper molars in P. g. vacillator.
On the north bank of the Amazon, the only population of P. g. hyleae known to me (from Obidos) has four counterfolds in p4 and three in the lower molars; P. g. riparum, from Manaus, also on the north bank of the Amazon, has three counterfolds in p4 and two counterfolds in the lower molars. P. g. hyleae occurs also on the south bank of the Amazon. P. g. riparum, therefore, may be the northern part of the southern cline, instead of the southern end of the northern cline.
The whole picture, as outlined above, may be explained by assuming that the species P. guyannensis differentiated somewhere on the Central Plateau of South America, with three counterfolds in each upper molariform tooth, four counterfolds in the lower premolar and three counterfolds in the lower molars. The species might have extended its range to the Guianas and then all the biotypes with reduced number of counterfolds might have slowly developed by natural selection. The gradient is, broadly, from subspecies with greater number of counterfolds in more humid areas, to a gradually lessening number of counterfolds in less humid areas.
Proechimys longicaudatus is limited in the south to the headwaters of the Parana River drainage, where the subspecies P. l. roberti and P. l. longicaudatus are found. The species ranges northward through the Tapajoz drainage, with P. l. leucomystax in the headwaters and P. l. boimensis in the lower course. To the northwest and west the species is represented in Bolivia by P. l. securus; P. l. elassopus, P. l. simonsi, P. l. pachita, and P. l. hendeei occur in Peru and P. l. brevicauda in Peru and Brazil; and P. l. nigrofulvus occurs in Ecuador. Again in P. longicaudatus it seems that the number of counterfolds follows a gradient from more humid areas with four counterfolds in p4, as seen in nigrofulvus, pachita, simonsi, elassopus and brevicauda, decreasing to three or four in securus, to three in longicaudatus, but with m3 having only two counterfolds in leucomystax and roberti. P. l. boimensis, widely separated in the lower Tapajoz (no samples being known from the intervening range) may be the end of a cline started by leucomystax with only 2 counterfolds in m3 and ending to the northward with four counterfolds in m3. Over the same area the counterfolds in p4 increase from 3 to 4.
Of Proechimys goeldii I have had inadequate material but there seems to be a similar gradient in it which may be traced from P. g. steerei to P. g. goeldii. P. g. steerei has four counterfolds in more upper molars than occurs in the other subspecies.
Proechimys semispinosus has its wide range in the mountainous, western area of South America, the headwaters of the Amazon drainage and northward in Central America and the nearby Pacific Islands. In these populations a gradient may exist in the number of counterfolds which is varied in every population. The highest number seems to occur in the populations from northern Peru and Ecuador, decreasing from there in all directions, except in the Panamanian and Columbian islands. In gross examination, it seems that the size of the animals increases to the northwards.
Some specific characters are duplicated in each of the two subgenera; that is to say, there are some parallel developments and they give the common generic stock its biotypical variability. Among these parallel developments are the width of the aristiforms, the amount of pigment in the agouti-colored setiforms, and the shape of the nasal bones. Other characters, however, appear in one subgeneric group and not in the other. The specific variation will be discussed separately for each subgenus.
The aristiforms are narrow and soft in P. dimidiatus and in the other species are wide and stiff, and on the outer thighs and rump some are light-colored. P. albispinus has the maximum number of light-colored aristiforms; they are present over the sides and back. This species has, however, a type of aristiforms unique in the genus—the clavate type. The tail is longer in P. iheringi and P. setosus than in P. dimidiatus and P. albispinus; the longer type is associated with a penicillate tip suggesting an adaptation to arboreal habit. The skull and nasals are longer in P. dimidiatus and P. iheringi than in P. setosus and P. albispinus. In the latter two species the longitudinal dorsal outline of the skull is conspicuously convex as opposed to slightly convex in the other two species. The palate is longest in P. dimidiatus and P. iheringi extending posteriorly to the level of the second molars; it is slightly shorter in P. setosus and shortest in P. albispinus where it does not extend behind the level of the first molars. The incisors are opisthodont in P. dimidiatus and P. iheringi and orthodont in P. setosus and P. albispinus and even proodont in one part of the last species.
The molariform teeth have a large number of counterfolds in both P. dimidiatus and P. iheringi, although the number varies but little in the first species and much in the second. The variation in P. iheringi decreases in populations of increasingly more northern geographic distribution; in both P. setosus and P. albispinus the number of counterfolds is greatly reduced; there is only one in most specimens of P. albispinus. The incisive foramen is small and nearly round in P. dimidiatus, larger and elongate in P. iheringi, very narrow and fissurelike in both P. setosus and P. albispinus.
The characters of Trinomys, as briefly outlined above, seem to be the result of one original species having split first into four species which provide a gradient for certain characters. Subsequently one of these four species, P. iheringi, split into six subspecies and another [Pg 321] gradient, parallel to the first, and involving the same characters, is to be seen.
The interrelationship among the species is evident, not only because they have the same subgeneric characters, but because the full species themselves provide successive steps in a stairway of increasing specialization from P. dimidiatus to P. albispinus.
Morphologically P. dimidiatus and P. iheringi are sometimes difficult to distinguish, especially on the basis of cranial features. Nevertheless close attention to the small, nearly round, incisive foramen of P. dimidiatus versus the larger, more elongate foramen in P. iheringi will permit separation of the two. However, the two species live in the same place and one is led to infer that there may be greater differences in their physiology than in their morphology. In fact Dr. H. W. Laemmert, from the Serviço de Estudos e Pesquisas Sobre a Febre Amarela in Brazil, informs me that while P. dimidiatus was highly susceptible to the virus of yellow fever (18 out of 24 with virus in circulation), P. iheringi showed a lower rate of susceptibility (3 out of 25 with virus in circulation). P. longicaudatus roberti, belonging in the other subgenus, showed no susceptibility at all.
At Teresópolis, Estado do Rio de Janeiro, the two species were found in two different forests, only a few kilometers apart, but dimidiatus lived at a higher elevation, where the humidity was remarkably higher. Naturally the plant associations were different in the two forests. This seeming ecological adaptation of the two kinds of Proechimys may explain why P. iheringi ranges farther north; the forests to the northward are less humid.
One of the four species, P. setosus, subspecies elegans, was used by Winge (1941:80, 82) as representative of the genus Proechimys when he was estimating the relationships of that genus. Because Cercomys, with four crests in each of its cheekteeth, was, on other grounds, regarded by him (op. cit.:80) as "... the most primitive genus within the group.", and because he noted in P. s. elegans 4 crests in P4 and in some first molars, he concluded that Proechimys was "very closely related to Cercomys." His conclusion seems to be correct, but actually other species of Proechimys (subgenus Trinomys), for example, P. dimidiatus, have four or more crests in each cheektooth, and, therefore, may be considered as more closely related to Cercomys than is P. setosus. If a large number of crests indicates primitiveness, P. dimidiatus, always with four, is more primitive than any other species in the subgenus Trinomys. Also, the large skull, long hind foot, short tail and thin aristiforms of P. dimidiatus, in my opinion, are primitive characters.
One of the species of Trinomys, Proechimys iheringi, is here subdivided into six subspecies which show a clinal variation. P. i. iheringi, in the southernmost part of the range of the species (Ilha de São Sebastião), has three counterfolds in the upper cheekteeth of almost every young specimen but one of these counterfolds, since it is small, very shallow, and disappears after little wear, is probably in the process of disappearance; all lower cheekteeth have two counterfolds or, rarely, m3 has only one. P. i. bonafidei is the next subspecies northward, where it was collected at 850 m altitude (Fazenda Boa Fé). This subspecies still has two counterfolds in all the upper cheekteeth; only 3 out of 16 specimens fail to have these counterfolds coalesced in one or more of the teeth. In the lower cheekteeth the coalescence is evident in 18 per cent of the specimens. P. i. gratiosus, from Floresta da Caixa Dagua (alt. 750 m), geographically is well removed from bonafidei (more than two degrees north), and no samples were obtained from the intervening area. It shows such great reduction in the counterfolds that the existence of intermediate populations is clearly suggested. Every upper cheektooth of this subspecies has the two counterfolds coalesced and in 40 per cent of the specimens M3 has only one counterfold; in the lower cheekteeth 60 per cent of the specimens have only one counterfold in m3. P. i. panema, occurring approximately 100 kilometers to the northward of P. i. gratiosus (lowland form), has one counterfold in M3 in only 20 per cent of the specimens but the lower third molar has only one counterfold in 80 per cent of the specimens. In P. i. denigratus, from about 3 degrees north of the range of P. i. panema, the reduction is proportionately greater: P4 now is the only upper cheektooth with two counterfolds in every specimen; all molars tend to have only one; p4 has also two counterfolds but all lower molars have only one.
The relative size of the tail also varies in a cline from south to north. Its length is approximately 87 per cent of the length of the head and body in P. iheringi; 88 per cent in bonafidei; 99 per cent in gratiosus; 100 per cent in panema; and 103 in denigratus.
One of the subspecies, P. i. paratus, however, seems to be completely out of the dental cline. It was collected in the near proximity of the type locality of P. i. gratiosus, at an elevation of 120 m lower. This subspecies has two counterfolds in all molariform teeth [Pg 323] and only one of the two specimens known shows these counterfolds coalesced in P4 and M1. The sample, 2 specimens, is too small to be trustworthy; hence it is impossible satisfactorily to account for the break in the clinal variation. Conceivably two full species are involved, but I prefer at present to defer decision on this problem until such time as more evidence is accumulated.
P. setosus is poorly represented, both of the available skins being faded. Furthermore, no type locality is known for the subspecies P. s. setosus.
P. albispinus has only two known subspecies: P. a. albispinus, living in a region of higher humidity, is slightly the darker and has subapical zones of the setiforms on the sides Ochraceous-Tawny; P. a. sertonius, living in a much drier region, has the same subapical zone Ochraceous-Buff. The number of specimens of P. a. sertonius is so few that no gradient can be detected, even if one exists.
Absolute size of head and body, tail, hind-foot and ear are useful in distinguishing subgenera and subspecies and to some extent in differentiating species.
The length of head and body is large to medium in Proechimys and medium to small in Trinomys. The tail is long to medium in Trinomys and short in Proechimys. The longest tail, 242 mm, is found in P. i. denigratus, and the shortest tail, 123 mm, in P. g. steerei. The relative length of tail also provides gradients or clines.
In every species, males surpass females in average size. Nevertheless, the largest animals are usually females. How this paradoxal fact is to be accounted for, I am not sure, but it may be that the animals grow as long as they live and that females have more chances to survive longer since the care of the young keeps them closer to shelter.
Color.—Upper parts vary from Buckthorn Brown to Ochraceous-Buff. Dark color ordinarily is correlated with an environment of higher degree of humidity and light color with lower humidity. However, species may be found in similar conditions of humidity but differing in color. Proechimys albispinus albispinus, for example, a light-colored form, is found in areas where the rainfall averages 1,000 to 1,500 mm of annual precipitation, in the isohygra of 80 per cent relative humidity. These conditions actually are similar to those where P. dimidiatus, of darker color, is found. The [Pg 324] subspecies albispinus, however, ranges mostly over a dry area and the fact that it occurs also in a moist area without appreciable change in color is difficult to explain.
Insular populations are usually darker or richer in color than corresponding continental populations. On a small island, uniformity of environment and inbreeding may be responsible for an accumulation of characters for richness of color.
The pelage provides most useful taxonomic characters. Excepting the vibrissiform hairs, all of the elements of the pelage have a common feature, the flattened shape. The hair constellation (cf. Toldt, 1935) on the upper and lateral surfaces is composed of hairs of two main types: aristiforms (guard hairs) and setiforms (over hairs).
The aristiforms are wide, strong, and have the dorsal (= anterior) margins raised, forming a wide shallow longitudinal groove on the dorsal face of the hair. The tip is a filament that usually is lacking in aristiforms which are especially strong. Wear probably removes these tips. The aristiforms have the bases whitish or grayish and the amount of pigment gradually increases distally to a dark brown or blackish shade. On the dorsal and lateral surfaces of the head the aristiforms are small and narrow but gradually increase in length and width caudad on the animal. The maximum development is reached in the middorsal region, from where they decrease in size and number toward the lateral surfaces or caudad. This decrease in the development of the aristiforms, however, is not uniformly gradual. Generally, the aristiforms become increasingly conspicuous in a middorsal band, but they extend to the sides and onto the outer sides of the thighs; the band narrows rapidly on the rump. In the subgenus Trinomys, where the aristiforms attain their maximum development, they are still strong and conspicuous on the rump and sometimes around the base of the tail. In Proechimys the aristiforms do not extend caudad from the hips. Also, in Trinomys, besides the ordinary lanceolate type, there are some aristiforms on the dorsal surface with a clavate shape; the base is wide and the distal part narrow. This parallels the conditions in the pelage of the most spiny species in the genus Echimys, Echimys chrysurus (Lichtenstein).
The recently named subspecies Proechimys cayennensis hoplomyoides Tate, 1939, shows an extraordinary development of the [Pg 325] aristiforms on the back and sides such as occurs in the genus Hoplomys. Actually the small bulla, wide basisphenoid and tooth structure add to the possibility of hoplomyoides being a true Hoplomys, and worn teeth might have been responsible for the difficulty which Tate had in allocating the form to the proper genus. However, the narrow braincase is more nearly like that of Proechimys than that of Hoplomys. The intermediate nature of hoplomyoides argues for including the genus Hoplomys as a subgenus of Proechimys.
Species with narrow aristiforms have a rather soft and flexible pelage, while those with wide aristiforms have harsh, spiny pelage. The aristiforms vary in width from 0.45 to 1.3 mm, depending upon the species or subspecies.
Animals with narrow aristiforms tend to have a more or less uniform coloration throughout the dorsal parts. The blackish distal parts of the aristiforms regularly interline the ground color made by the subapical zone of the setiforms. If, on the contrary, wide aristiforms occur, the dorsal surface is conspicuously marked by the wide blackish lines among spots of color formed by the subapical zones of the setiforms. No clinal variation was detected in width of aristiforms but geographic variation in width was noted; for example, the subspecies of P. iheringi differ in this respect.
The setiforms are narrow and flattened but are without pronouncedly raised margins. The setiforms are usually bicolored on the dorsal and lateral surfaces of the animals, with a subapical zone of some reddish-brown color, like Ochraceous-Orange or Ochraceous-Buff. They are whitish or gray on the basal parts and gradually blacken toward the tip, but have a reddish subapical zone. Common exceptions to this pattern are setiforms without subapical zones; these appear on the dorsal surface among setiforms which are normal in possessing distinctive subapical zones. Also there are setiforms without blackened tips on the lateral surfaces. Due to their relative abundance and subapical color, these setiforms are responsible for the dominant color on the upper parts. Like the aristiforms, they are longer and wider in the middorsal region of the animal and are gradually less developed on the remainder of the upper parts. Actually there is more than one type of setiform in the hair constellation; they vary in length, width and color. Attention was not given, however, to every type of setiform.
The ventral surface of the body and the inner sides of the legs are uniformly covered by short setiforms, thinner and more sparsely [Pg 326] distributed on the inner side of the legs. These setiforms are usually uniformly white in color or, sometimes, the distal parts are buff or more richly colored.
Vibrissiforms are scattered on the dorsal and lateral surfaces of the body, and in penicillate arrangements on the head. They are longer than the pelage proper, have a nearly circular cross-section and are blackish in color.
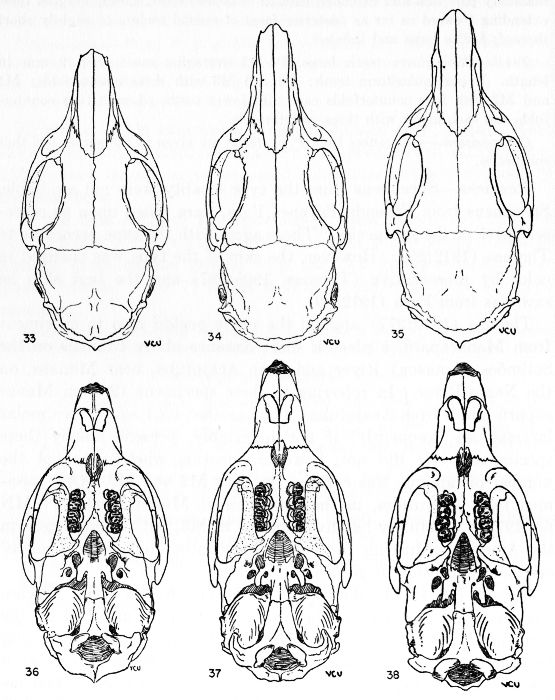
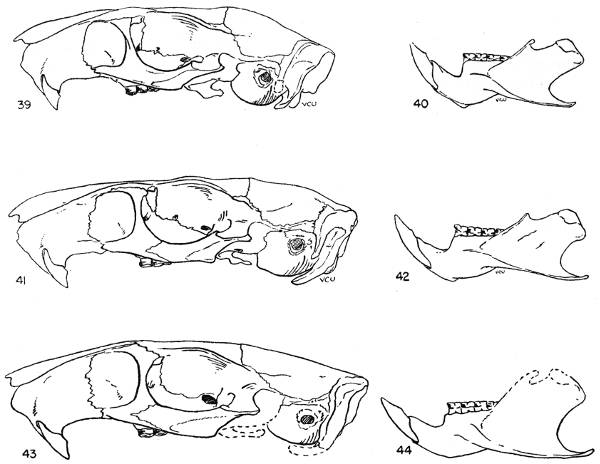
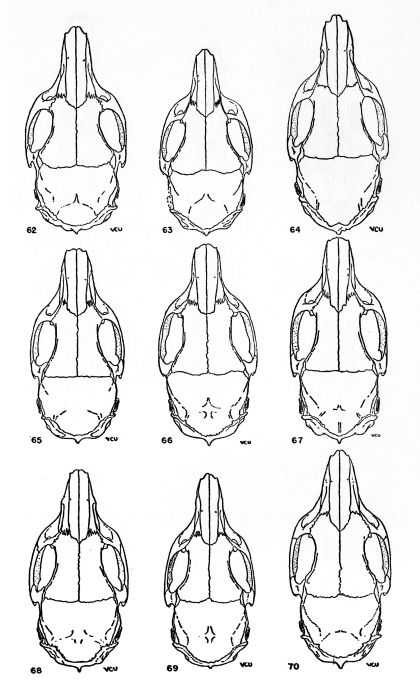
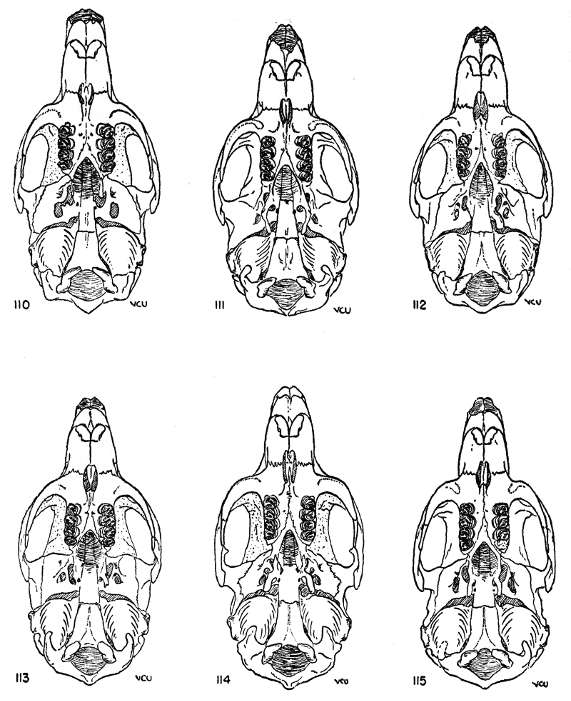
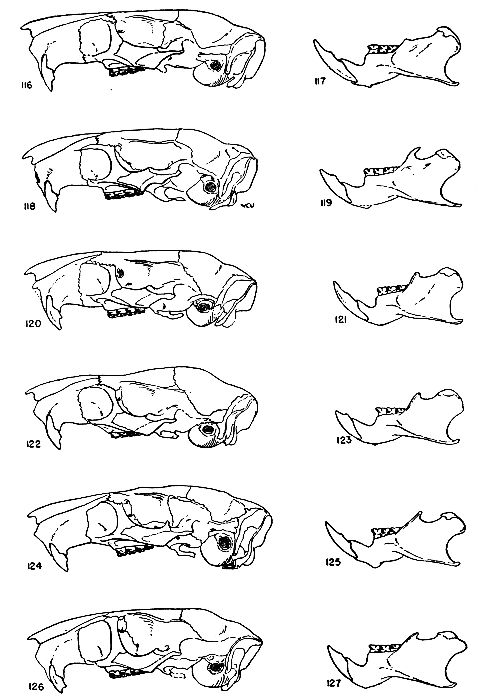
The absolute size of the skull is proportionate to bulk of the body. The supraorbital and parietal ridges are especially developed in the P. semispinosus group, where they extend across the parietals to the interparietals. In all members of the subgenus Proechimys, these ridges extend onto the parietal region. In Trinomys, however, they do not extend so far posteriorly as the parietal, but only onto the squamosal.
The rostrum varies from slender to stout. Elongate rostra are common in Proechimys; Trinomys has a short blunt rostrum.
The infraorbital foramen commonly has a ventral groove for nerve transmission in many forms of Proechimys but Trinomys almost always lacks this groove. Presence or absence of the groove is a subspecific character in the subgenus Proechimys.
The jugals are dorso-ventrally wide in Trinomys except in the species P. setosus. In Proechimys a dorso-ventrally narrow jugal is the rule, but P. canicollis has an especially wide jugal. A postorbital process appears on the jugo-squamosal suture and is here called postorbital process of the zygoma. In Proechimys it is more or less weakly developed and shows no variation of systematic worth. In Trinomys, on the other hand, this process varies in a clinal way (P. iheringi) and stages of the gradient characterize populations of subspecific rank.
Linear and spatulate shape of the humular process of the pterygoid constituted specific characters for Thomas, but there is so much individual variation in the shape of this process in almost every population that it has not been used in the present account.
The mesopterygoid (interpterygoid) fossa in almost every specimen extends anteriorly to the level of M1 or M2 in Trinomys, and to M3 in Proechimys. Exceptions may occur, as in P. hendeei, where the fossa extends to the level of M2.
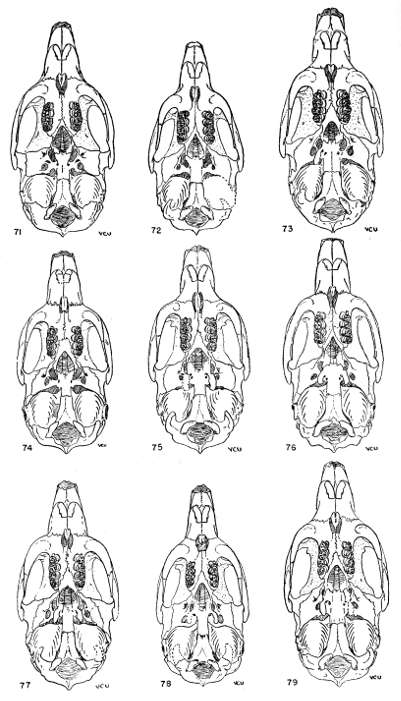
The shape and dimensions of the incisive foramen long have been recognized as providing specific characters. Large size of the foramen is probably correlated with the requirement for a large amount [Pg 327] of moisture reaching Jacobson's organ in the nasopalatine space; the moistening of the sensory epithelium is certainly involved. There seems to be a certain correlation between small size of the incisive foramen and high degree of humidity in the environment. Shapes and dimensions of the foramen appear as simple or multiple biotypes and provide characters which can be employed to differentiate subspecies, species and even subgenera. Usually a character, say a general shape, occurs in nearly all populations of a given subspecies but the particular shape seems to be more closely correlated with ecological conditions, especially humidity. Animals which live far away from large rivers usually have larger foramina than animals which live close to rivers.
Both the premaxilla and the maxilla develop processes which form a sheath for the vomer. This vomerine sheath forms a bridge which longitudinally crosses the incisive foramen; the structure of this bridge varies widely. Sometimes the maxillary part is not developed and the sheath is incomplete posteriorly; sometimes this maxillary part is very slender and merely touches the premaxillary part. The premaxillary part, however, is always well developed.
Considered by itself the variation in the tooth pattern can lead to erroneous conclusions as to differentiation of species, because the number of folds on the occlusal face of a tooth and the depth of certain folds may be subject to great individual variation as shown by examination of more than one large series of specimens of the same kind, age and sex from a single locality. Also there are geographic gradients or clines, in number of folds. Nevertheless the variation in number of folds, when measured at sufficient intervals along a cline, may provide quantitative characters useful in differentiating subspecies.
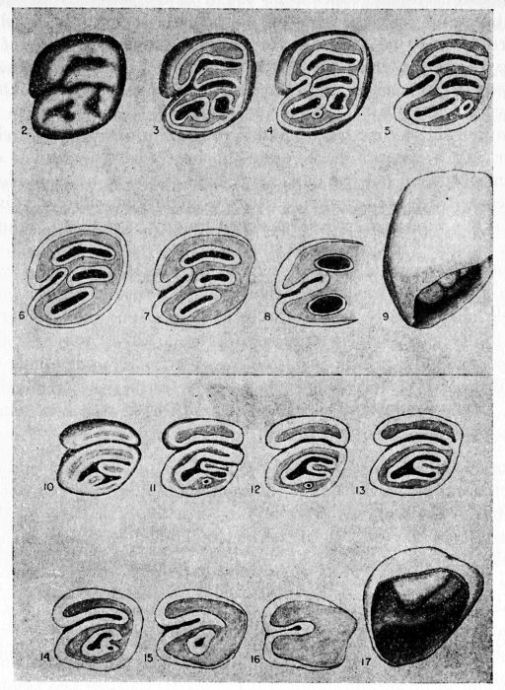
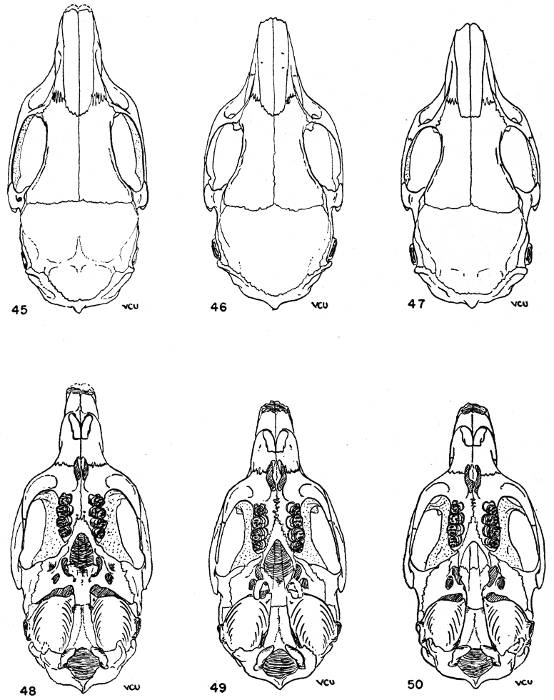
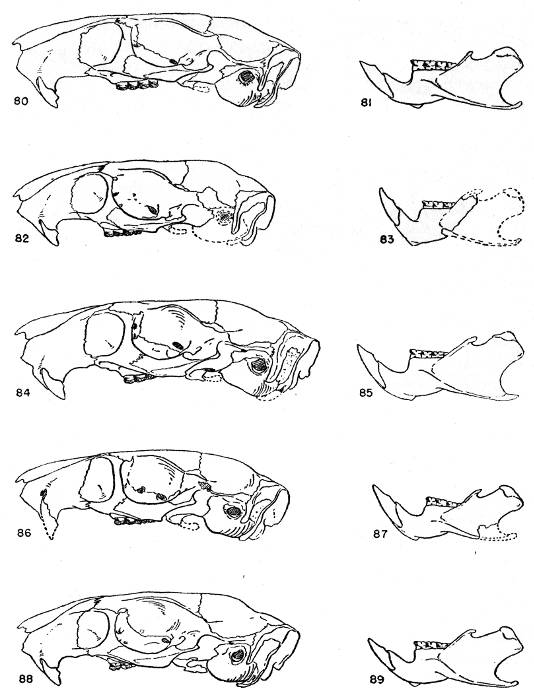
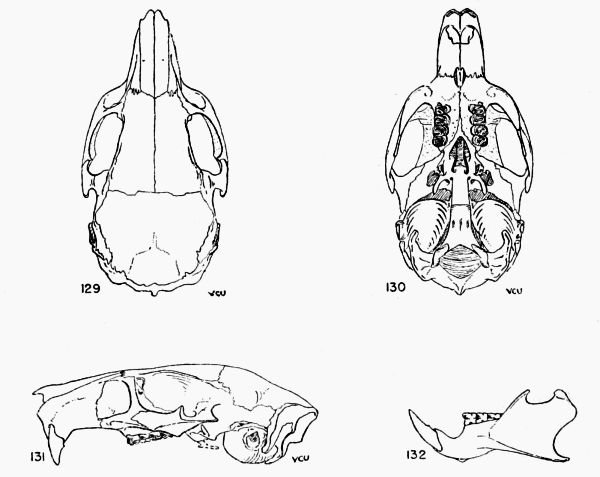
Figs. 2-17. Second left upper molar of the two subgenera Proechimys and Trinomys. All × 8. Anterior border of tooth is at the top of each figure (Nos. 9 and 17 excepted). Note especially that main fold is short in Proechimys and long in Trinomys.
Figs. 2-9. Proechimys (Proechimys) semispinosus liminalis, female, MN no. 6243, Rio Quichito. Fig. 2, unworn crown. Figs. 3-8, cross sections at 0.5 mm. intervals, showing changes in the main fold and counterfolds at increasing depths as the tooth was ground down. Fig. 8 is 3 mm. below surface shown in fig. 2. Fig. 9, posterior view with proximal end of the tooth open showing basal ends of folds. Later in life the proximal end closes and three roots are formed.
Figs. 10-17. Proechimys (Trinomys) iheringi denigratus, female, SEPFA no. 17060, Mata do Ribeirão da Fortuna. Figs. 10-16 corresponding to figs. 2-8. Fig. 17, posterioventral view with proximal end of the tooth open and part of walls cut away, showing basal ends of folds. Later in life, as in Proechimys, the proximal end closes and three roots are formed.
The main fold involves both the occlusal face of the tooth and the side wall. The counterfolds, which are smaller counterparts of the main fold, in most instances also implicate the wall of the tooth opposite to that marked by the main fold, but are to be seen mostly on only the occlusal face of the tooth. Unerupted teeth with the crowns unworn and other teeth which had barely broken through the gums were ground down to permit the making of drawings of the surfaces at different levels. This study revealed that the main fold is deepest in the wall of the tooth. The development of the main fold varies in two different ways: in all samples from southeastern and eastern Brazil it is strongly developed, deeply grooves the tooth through its crown and, in younger individuals, completely divides the occlusal surface of the tooth. As use wears down the crown, the main fold soon becomes separated from the opposite wall and then gradually shortens toward its basal portion. In the other type, common to animals of all the remaining part of the range of the genus, the main fold is rather short, never reaching the opposite wall. In this case, however, one of the counterfolds usually appears almost opposite the main fold in such a way that in non-erupted or just-erupted teeth the main fold and one counterfold may be connected by a shallow groove that may give the impression of extension of the main fold and, therefore, lead to false interpretations. Closer examination shows that the counterfold which apparently meets the main fold is really situated anteriorly or posteriorly to it, in upper or lower teeth respectively. One subspecies in the subgenus Trinomys differs from the general characteristics of the subgenus in sometimes showing a small main fold in P4 whereas it is large in all other cheekteeth. The structural differences here mentioned in the main fold were never before recognized. Therefore, the meaning of "quadrilaminate" pattern, "three outer folds" or "three inner folds" (of authors) is not consistent insofar as the two groups are concerned.
Writers have more or less tacitly admitted three as the usual number of counterfolds present in the upper molariform teeth. Ellerman, for example (1940:117), states: "Upper cheekteeth normally with three outer and one inner folds each, these soon becoming isolated as islands. A few species, which will be discussed below, vary slightly in pattern." Thomas (1921:140) erected the subgenus Trinomys on the basis of the upper molariform teeth having only three laminae. Actually a meticulous study of widely varying samples shows that the number of counterfolds may vary from one to five, the usual number being three or four. One of the most important facts to record on this subject is that young specimens with slightly worn molariform teeth are more apt than either adult or younger specimens to show the maximum number of counterfolds. Usually nonworn teeth show rounded crests and valleys of different depth. For example, it is common to see one continuous groove giving the impression of a main fold crossing the occlusal surface transversely. The slightest wear of the occlusal surface, however, shows that really there are two valleys instead of one. The two are the main fold and one counterfold. In this case, it would be easy to confuse the two types of teeth, one with the main [Pg 330] fold short and the other with the main fold extending almost all the way across the occlusal surface.
Some of the counterfolds are especially shallow and tend to disappear in an early stage of wear, and adult individuals may have these folds completely worn away. Advanced wear usually develops a cuplike occlusal surface with only the remains of the main fold and also remains of one or more counterfolds represented by small enamel islands (Figs. 2 to 17). In the form Proechimys iheringi iheringi, for example, every tooth shows three counterfolds in the upper molariform teeth of individuals in which the wear is not advanced. This number, however, is less in all or part of the molariform teeth of older individuals.
Adjacent counterfolds may appear to be coalesced in many instances. Coalescence is more likely to be seen in species where a wider variation in the number of the counterfolds is involved and it appears as a gradient in the reduction of the number of counterfolds.
Of great importance, as a general feature of molariform teeth, is the relative size as related to the geographical distribution, showing, again, a natural division in the genus. In all forms of southeastern Brazil the premolars are larger than the first molars, the first molars are larger than the second molars, and the second molars are larger than the third molars. The forms from central and northern Bahia, Brazil, have the molariform teeth more or less the same size. The forms from the remaining part of the area occupied by the genus have premolars smaller than the first molars, the first molars smaller than the second molars, but the second molars larger than third ones.
P. dimidiatus was studied in the field and laboratory. P. dimidiatus in captivity showed regular diurnal activity, coming out of the nest for food at intervals. Individuals were fed a cereal mixture and nuts of small size. The animals usually buried the nuts in the sand of the outer cage. While holding the nut with the mouth and front feet, the animal patted the sand rapidly, thus burying the nut, and it then pushed more sand over the place with the front feet.
Sometimes the emergence from the nest is followed by a long yawning and stretching ceremony. The animal spreads the fore and hind legs widely apart, while the back is curved down and the head and tail turn upward. Then one of the hind legs is stretched backward and, at the same time, the mouth is opened widely and the tail is moved in an undulatory fashion. The operation may be repeated using the other legs, or not.
P. dimidiatus was regularly found in climax forest. The best shelter and nesting ground was usually under boulders, commonly not farther than 10 meters from water. The entrance to the nest was kept clean. No more than two adult animals (male and female) were captured in the same shelter, and only a few times were young captured in the same place with adults. Nesting places were located also at the bases of trees and near fallen logs where litter accumulates.
Records of animals kept in captivity show that the species dimidiatus survives more than two years. Specimen MN no. 5448 [M] was adult when captured by the Serviços de Estudos e Pesquisas sobre a Febre Amarela on December 5, 1938, and died on January 17, 1942. Therefore, it lived for more than 1,139 days.
Proechimys dimidiatus, in Rio de Janeiro, as well as P. i. bonafidei and P. i. iheringi which live in the same region, were found breeding from September to November and from March to May. Proechimys longicaudatus roberti, in Anapolis, Goiaz, was found breeding from July to November and from January to March. P. g. hyleae and P. g. leioprimna in the lower Tapajoz and lower Tocantins rivers, Pará, were found breeding in January.
It seems that in the Central Plateau and southeastern Brazil, Proechimys has two litters per year, one in the early spring and a second in the late summer. The number of young per litter varies from 1 to 5, although 2 is the usual number.
Juveniles.—The animals are born with a thick pelage of thin aristiforms and thin setiforms. The color is uniformly blackish brown. The nose, hands, feet, ears and tail are pinkish; P4 and M1 are already erupted and the second molars are included in the bony alveoli. The incisors are orthodont; the rostrum is short and the braincase is wide. The posterior part of the skull is greatly curved dorsally. No change is noticed in the pelage before the second molars erupt and become functional.
Adolescents.—As soon as the second molars become functional, the pelage starts molting on the back. The thin aristiforms are still in place but the aristiforms of the adult pelage may be noticed growing under them in an oval patch which extends from behind the shoulders caudad to the hips. At that age the first, agouti-colored aristiforms appear on the mystacial region, immediately behind the vibrissiforms. The rostrum gradually lengthens and the braincase appears to become less inflated.
By the time the third molars erupt, the aristiforms start showing among the setiforms which are now changing to agouti color in the same area on the back, while the thin aristiforms of the juvenal stage disappear. The agouti setiforms are appearing also posteriorly from the mystacial region to the sides of the head and neck and, at the same time, on the frontal region. The patch of glossy aristiforms and setiforms on the back is sharply differentiated from the dull juvenal pelage of the sides and rump. In a later stage the area of agouti setiforms on the sides of the neck extends to the outer sides of the arms and finally reaches the area on the back where the agouti setiforms were already developed.
Adults.—When the third molars become functional, the agouti setiforms are in place except for those on the upper sides of the neck. The aristiforms have now extended over their normal area of distribution. As soon as the third molars show wear, the premolars and first molars have the counterfolds isolated in the occlusal surfaces as enamel islands. Wearing gradually isolates all counterfolds.
Senile individuals.—Progression of wear soon eliminates the signs of the shallowest counterfolds from the occlusal surface and finally the tooth is reduced to a short crown with a cuplike occlusal surface completely filled with dentine. Only the main fold usually remains; it is more or less reduced in size.
From the records of the Serviços de Estudos e Pesquisas sobre a Febre Amarela, the following data for males of Proechimys longicaudatus roberti were obtained:
| Age | Number of cheek-teeth | Weight in grams | Length of head and body | Length of tail | Length of hind foot |
|---|---|---|---|---|---|
| One day old | 2 | 20.5 | 70 | 53 | 24 |
| 17 days old | 2 | 26.0 | 110 | 60 | 28 |
| Juvenile | 2 | 85.0 | 150 | 105 | 39 |
| Adolescent | 3 | 115.0 | 175 | 120 | 45 |
| Adolescent (older) | 3 | 180.0 | 195 | 135 | 45 |
| Adult | 4 | 200.0 | 223 | 158 | 48 |
| Senile individual | 4 | 360.0 | 230 | 170 | 48 |
The weights and measurements (except for one- and 17-day-old animals) represent averages of specimens of the different ages.
Genotype.—Echimys trinitatis Allen and Chapman, by original designation.
Proechimys Allen and Chapman, 26 December 1899, Bull. Amer. Mus. Nat. Hist., 12(20):264, orig. description; Tate, 1935, Bull. Amer. Mus. Nat. Hist., 68(5):398; Ellerman, 1940, The families and genera of living rodents, Brit. Mus. (Nat. Hist.), 1:115.
General characters.—Muriform echimyids of medium size; pelage with flattened and lanceolate and sometimes clavate aristiforms, varying greatly in width and distributed over most of the dorsal surface from shoulders to hips or base of tail; setiforms also flattened, evenly distributed throughout; entire ventral surface and inner sides of legs white or, rarely, invaded by some buffy color; tail shorter than, equal to, or slightly longer than, head and body, bicolored and with a few bristles between scales, sometimes heavily penicillated; feet long and narrow; pollex rudimentary; hallux shorter than fifth toe; ears wide and long; mammae 2-1=6.
Skull.—Generally elongate and strongly built, with supraorbital ridges well developed, frequently extending across parietals toward occipital region; zygomatic arches variable in depth, always with postorbital process; infraorbital foramen with or without lower groove for transmission of nerve; incisive foramen usually large; vomerine sheath complete or incomplete; mesopterygoid fossa extending forward at least to plane of third molars; bullae large; angular process of mandible turned upward.
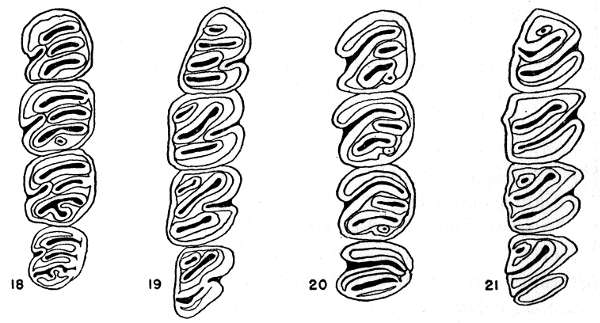
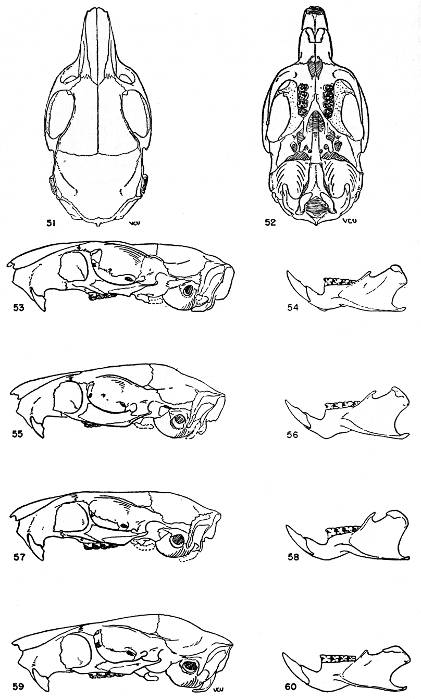
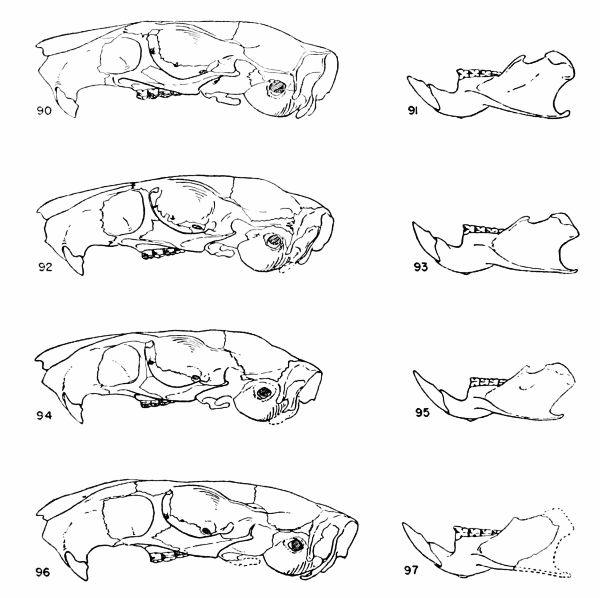
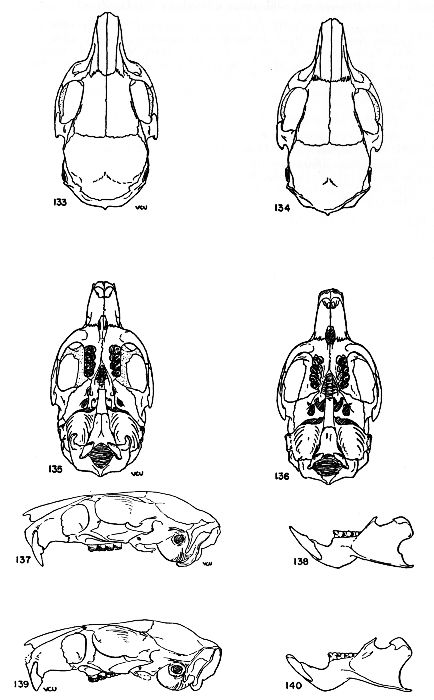
Figs. 18-21. Occlusal views of the upper left and lower right molariform teeth of the two subgenera of the genus Proechimys. Anterior end of the tooth row at the top of drawing. All × 6.
Figs. 18-19. Proechimys (Proechimys) goeldii steerei, sex ?, USNM no. 105537, "Hyutanaham." Upper teeth at left (fig. 18).
Figs. 20-21. Proechimys (Trinomys) dimidiatus, male, MN no. 6256, Pedra Branca.
Teeth.—Incisors opisthodont, orthodont or proodont, not grooved; upper molariform teeth with a main internal fold and one to five external counterfolds which usually appear as enamel islands in worn teeth, these counterfolds barely implicating the lateral wall; lower molariform teeth with folds as in the upper molariform teeth except that they are reversed and the number of internal counterfolds is usually fewer in the molars.
| 1. | (a) | Tail less than 90 per cent of head and body; aristiforms not evident on outer thighs and rump; skull with ridges across parietals; size of upper cheekteeth increasing from P4 to M2; main fold small. | ||
| subgenus Proechimys, | 2 | |||
| (b) | Tail 90 per cent or more of head and body; aristiforms evident on outer thighs and rump; skull with no ridges across parietals; size of upper cheekteeth decreasing from P4 to M3; main fold large. | |||
| subgenus Trinomys, | 5 | |||
| 2. | (a) | One or more upper molars with four counterfolds, | 3 | |
| (b) | Upper molars with no more than three counterfolds, | 4 | ||
| 3. | (a) | Aristiforms wide (more than 0.7 mm). | ||
| P. semispinosus, | p. 342 | |||
| (b) | Aristiforms narrow (less than 0.7 mm). | |||
| P. goeldii, | p. 338 | |||
| 4. | (a) | Aristiforms wide (0.9 mm or more), or narrow (0.6 to 0.7 mm) but then with only two counterfolds in each lower molar. | ||
| P. guyannensis, | p. 355 | |||
| (b) | Aristiforms narrow (0.5 to 0.65 mm) but with one or more lower molars having three counterfold. | |||
| P. longicaudatus, | p. 346 | |||
| 5. | (a) | Aristiforms narrow (0.5 mm) and limber; no differentiated light-colored aristiforms on outer thighs and rump; incisive foramen short and widest posteriorly. | ||
| P. dimidiatus, | p. 371 | |||
| (b) | Aristiforms 0.6 mm or more and stiff; differentiated light-colored aristiforms on outer thighs and rump; incisive foramen elongated and constricted posteriorly. | 6 | ||
| 6. | (a) | Skull large, more than 50 mm in total length, incisors opisthodont. | ||
| P. iheringi, | p. 373 | |||
| (b) | Skull small, less than 49 mm in total length, incisors orthodont or proodont. | 7 | ||
| 7. | (a) | No clavate aristiforms, tail with white tip, | ||
| P. setosus, | p. 384 | |||
| (b) | Clavate aristiforms among the ordinary ones, tail without white tip, | |||
| P. albispinus, | p. 388 | |||
General characters.—Pelage with lanceolate aristiforms limited to an area on the dorsal surface between the shoulders and the hips; length of tail less than 90 per cent of length of head and body; skull with conspicuous ridges; extension of supraorbital ridges always evident on parietals; infraorbital foramen usually with separate groove for transmission of nerve; palate usually extended posteriorly as far as third molars; incisors opisthodont; molariform teeth with a small main fold, never extended transversely to opposite wall in occlusal surface of tooth; usually one counterfold anterior to main fold in upper molariform teeth and posterior to main fold in lower molariform teeth; premolars usually smaller than first molars, first molars smaller than second molars but second molars larger than third molars.
General characters.—Size large; tail short; aristiforms narrow and soft, usually concealed in pelage by setiforms; general color of upper parts some tint of orange, gradually becoming lighter on sides with no conspicuous, dark longitudinal band on back; feet dark; ventral surface of body and inner side of legs white but sometimes with some buff locally; skull broad and strongly built but not conspicuously ridged; zygomatic expanse great and rostrum not elongate; incisive foramen narrow; bullae large and inflated; upper molariform teeth with three to four counterfolds, M3 ordinarily with four; lower premolars with four, and molars with three, counterfolds.
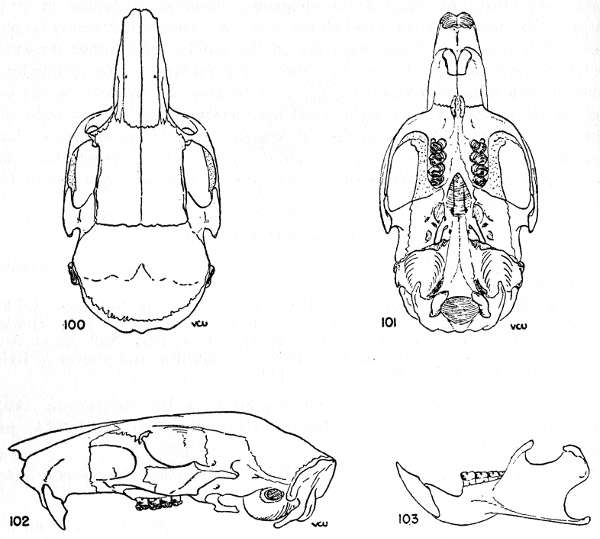
Proechimys steerei Goldman, Proc. Biol. Soc. Washington, 24:238, 28 November 1911 (original description); Goldman, 1912, Proc. Biol. Soc. Washington, 25:186; Tate, 1935, Bull. Amer. Mus. Nat. Hist., 68(5):400; Ellerman, 1940, The families and genera of living rodents, Brit. Mus. (Nat. Hist.), 1:119; Osgood, 1944, Zool. Ser. Field Mus. Nat. Hist., 29(13):204.
Type locality.—Hyutanaham, Upper Purus, Lábrea, Amazonas, Brazil. Type: United States National Museum, no. 105535, adult male; collected in 1901 by Prof. J. B. Steere.
Range.—Known only from the type locality and Porto Velho.
Diagnosis.—Upper parts Mars Orange on back, grading to Ochraceous-Tawny on sides; zygomatic breadth narrow; nasals short; incisive foramen narrow and short; vomerine sheath complete and thick; upper molars usually with four counterfolds.
Pelage.—Aristiforms on middorsal region: Grayish basally, gradually blackening toward tip; total length, 16 to 19 mm; maximum width, 0.5 mm. Setiforms on middorsal region: Grayish on basal third, gradually blackening toward tip but interrupted by a Mars Orange, subapical zone 1.5 mm long; total length, 16 to 19 mm; maximum width, 0.06 mm. Setiforms on outer thighs: Whitish basally, gradually blackening toward tip but interrupted by Orange Rufous or Ochraceous-Tawny, subapical zone; total length, 14 to 16 mm; maximum width, 0.05 mm.
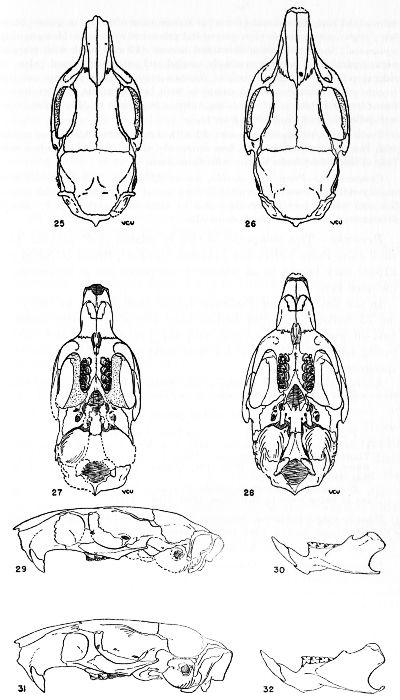
Figs. 25, 27. Proechimys goeldii steerei, sex ?, "Hyutanaham," USNM no. 105537. × 1.
Figs. 26, 28. Proechimys goeldii goeldii, female, AMNH no. 37488. × 1.
Figs. 29, 30. Proechimys goeldii steerei, sex ?, USNM no. 105537, "Hyutanaham." × 1.
Figs. 31, 32. Proechimys goeldii goeldii, female, AMNH no. 37488, Fazenda Paraiso. × 1.
Skull.—Large and strong; rostrum rather pointed posteriorly; supraorbital ridges not much expanded and extending across anterior half of parietals; infraorbital foramen without groove for transmission of nerve, or groove obsolete; zygomatic arches slender; postorbital process of zygoma involving mostly squamosal; incisive foramen short and narrow (4.5 × 2.5 mm) with margins almost parallel or tapering gradually caudad and extending toward palate as ridges; posterior margin of incisive foramen approximately 2.5 mm anterior to premolars; vomerine sheath complete, with both elements well-developed; mesopterygoid fossa never extending anterior to middle of M3; bullae large, well inflated and with shallow grooves.
Teeth.—Upper molariform teeth: P4 with three counterfolds; upper molars with four counterfolds each or, less commonly, three. Lower premolars with four counterfolds; lower molars with three each.
Comparisons.—From P. g. goeldii, steerei differs in: Back and sides with more reddish; narrower interorbitally and across zygomata; palatilar length less and nasals shorter; maxillary part of vomerine sheath thicker; usually four instead of three counterfolds in M3.
Remarks.—This subspecies is clearly related to P. goeldii. One skull from Porto Velho, Rio Madeira, Guaporé, Brazil (CNHM no. 21558) may belong to an unnamed subspecies but is provisionally included here.
In the field notes of Professor Joseph Beal Steere, an entry for no. 72 reads: "Big white bellied wood rats x two young found in nest of grass on the ground with the two young—much darker young female." No. 77 in his field notes corresponds to the type specimen.
Specimens examined.—Total number, 4, from Brazil, as follows: Amazonas. Lábrea, Hyutanaham, 3 (USNM); Territ. Guaporé, Porto Velho, 1 (CNHM).
Proechimys goeldii Thomas, June 1905, Ann. Mag. Nat. Hist., 15 (ser. 7):587, (orig. descr.); Thomas, 1912, Ann. Mag. Nat. Hist., 9 (ser. 8):89; Thomas, 1920, Ann. Mag. Nat. Hist., 6 (ser. 9):277, Tate, 1935, Bull. Amer. Mus. Nat. Hist., 68(5):400; Osgood, 1944, Zool. Ser. Field Mus. Nat. Hist., 29(13):199.
Proechimys cayennensis goeldii, Ellerman, 1940, The families and genera of living rodents, Brit. Mus. (Nat. Hist.), 1:121.
Type locality.—Santarem, Santarem, Pará, Brazil. Type: British Museum (Nat. Hist.), no. 5.1.25.6, adult female; presented by Dr. E. A. Goeldi.
Range.—Margins of the Amazon, between Jamundá and Tapajoz rivers.
Diagnosis.—Upper parts Ochraceous-Tawny; wide across zygomata; nasals of moderate length; incisive foramen long and narrow; vomerine sheath complete but maxillary part slender; first and second upper molars with four counterfolds.
Pelage.—Aristiforms on middorsal region: Whitish basally and gradually blackening toward tip which is extended as long, thin filament; total length, 22 to 24 mm; maximum width, 0.5 mm. Setiforms on middorsal region: Gray basally, gradually blackening toward tip but interrupted by Ochraceous-Tawny, subapical zone 3.3 mm long; total length, 19 to 21 mm; maximum width, 0.06 mm. Setiforms on outer thighs: Whitish basally, gradually blackening toward tip but interrupted by Ochraceous-Tawny, subapical zone 3 mm long; total length 14 to 16 mm; maximum width, 0.04 mm.
Skull.—Large and strong; nasals pointed posteriorly; supraorbital ridges moderately developed and extended caudad across anterior third of parietals; zygomatic arches strong; postorbital process of zygoma involving mostly squamosal; incisive foramen elongate and narrow (5 to 6.5 × 2.3 mm) with margins more or less parallel and raised to form ridges which extend posteriorly to within 3 mm of plane of premolars; vomerine sheath complete, with maxillary part thin and extended caudad as medial crest; mesopterygoid fossa extending forward as far as posterior faces of second molars or slightly short thereof; bullae large and inflated.
Teeth.—Molariform teeth large, P4-M3 averaging more than 9 mm in length. Upper molariform teeth: P4 and M3 with three counterfolds; M1 and M2 with four counterfolds each. In lower teeth, p4 with four counterfolds and each molar with three counterfolds.
Comparisons.—Differences from P. g. steerei are given in the account of that subspecies.
Remarks.—Specimens from the type locality were not available. Specimens from Fazenda Paraiso, Faro, were relied upon as representative of the subspecies. These agree with the type according to Thomas (1912:89). However, the skin of the type was changed in color by preservative (Thomas, 1905:587) and the best skin he saw was from Faro (1912:89).
Thomas (1920:277) applied the name goeldii also to specimens from Manacaparú, a place a short distance above Manaus on the Solimões (Amazon) River and from Acajutuba, near Manaus, on the Negro River. In referring to these specimens (2 from Manacaparú and 2 from Acajutuba) Thomas (loc. cit.) said "Five molar laminae are frequently, if not invariably, present among these specimens." He did not, however, mention whether or not the number of laminae was constant in both M2 and M3. One specimen from Acajutuba, in the collection of Museu Nacional (MN no. 1973 [M]), actually has five laminae in M3, but the specimens in the American Museum from Faro agree absolutely with Thomas' original description of goeldii.
Osgood (1944:199) doubted that goeldii was a valid species. Evidence that Osgood's doubt was unjustified is furnished by the fact that Thomas (1912:89) pointed out that his specimen from Faro agrees with the type. Likewise, my two specimens from Faro agree with the type insofar as it has been described. Thomas (1912:89) mentioned two additional skulls from the type locality which, he stated, agree with the type which was received from the Museu Goeldi, Pará.
Specimens examined.—Total number, 4, from Brazil as follows: Pará, Faro, Faro, Fazenda Paraiso, 2 (AMNH); Amazonas, Manaus, Manaus, 1 skull (AMNH); Amazonas, Manaus, Acajutuba, 1 (MN).
Additional records.—Total number, 7 (British Museum), from Brazil, as follows: Pará, Santarem, Santarem (Thomas, 1912:89; 1920:277), 3; Amazonas, Manaus, Acajutuba (Thomas, 1920:277), 2; Manacaparú, Manacaparú (Thomas, 1920:277), 2.
Figs. 33, 36. Proechimys semispinosus liminalis, female, MN no. 6253, Rio Quichito. Type. × 1.
Figs. 34, 37. Proechimys semispinosus amphichoricus, male, AMNH no. 77020, Mount Duida. Type. × 1.
Figs. 35, 38. Proechimys semispinosus kermiti, female, AMNH no. 37124, Lower Rio Solimões. Type. × 1.2 (from photograph).
General characters.—Size large; tail short and hairy; aristiforms wide and stiff, especially well-developed on back; general color on upper parts some shade of ochraceous, usually much darker on back and forming a conspicuous dorsal band; feet dark; ventral surfaces and inner sides of legs white; skull elongate and strong with ridges well developed; incisive foramen long and narrow; bullae large; usually four counterfolds in M3 and M2; usually three but sometimes four counterfolds in M1 and even P4; lower premolar with four and lower molars with three counterfolds.
Figs. 39, 40. Proechimys semispinosus liminalis, female, MN no. 6253, Rio Quichito. Type. × 1.
Figs. 41, 42. Proechimys semispinosus amphichoricus, male, AMNH no. 77020, Mount Duida. Type. × 1.
Figs. 43, 44. Proechimys semispinosus kermiti, female, AMNH no. 37124, Lower Rio Solimões. Type. × 1.2 (from photograph).
Type locality.—Rio Quichito, affluent from the south of the Javarí River, near Benjamin Constant, Benjamin Constant, Amazonas, Brazil. Type: Museu Nacional, no. 6253, adult female, collected in August, 1942, by E. Parko.
Range.—Known only from the type locality.
Diagnosis.—Color uniformly dark, setiforms marked with Ochraceous-Tawny; skull wide across zygomata; nasals short; prepalatilar part of skull long; incisive foramen long and narrow; vomerine sheath incomplete or complete; M2 and M3 almost always with four counterfolds; M1 more rarely with four counterfolds.
Pelage.—Aristiforms on middorsal region: Gray basally, gradually blackening toward tip which is generally extended as a filament; total length, 21 to 23 mm; maximum width, 0.9 to 1 mm. Setiforms on middorsal region: Gray basally, gradually blackening toward tip but interrupted by Ochraceous-subapical [Pg 344]Tawny, zone 3 mm long; total length, 22 to 24 mm; maximum width, 0.06 mm. Setiforms on outer thighs: Whitish basally, gradually blackening toward tip but interrupted by Ochraceous-Buff, subapical zone 2.5 mm long; total length, 13 to 15 mm; maximum width, 0.08 mm; some with gray base, blackening gradually toward tip, without any subapical zone; some with Light Ochraceous-Tawny, subapical zone.
Skull.—Large and strongly built throughout; supraorbital ridges expanded and thick, extending, in old specimens, across parietals to anterior angles of interparietals; interparietal ridges always conspicuous; rostrum elongated; nasals blunt posteriorly; zygomatic arches strong; infraorbital foramen with weakly-developed groove for transmission of nerve; postorbital process of zygoma involving mostly squamosal; incisive foramen averaging 6 x 2.7 mm, widest in middle part and posteriorly constricted, with raised margins which do not extend across maxillae as ridges; posterior margin of incisive foramen approximately 1.5 mm anterior to plane of premolars; vomerine sheath incomplete or, sometimes, complete but always with maxillary part slender; mesopterygoid fossa not extending forward past centers of third molars; bullae moderately developed.
Teeth.—Upper molariform teeth: P4 always with three counterfolds; M1 with three counterfolds in 9 of 10 specimens and four counterfolds in remainder; M2 with four counterfolds in 7 specimens, three counterfolds in remainder; M3 with four counterfolds in 6 specimens, three counterfolds in remainder. Lower premolar always with four, and molars with three, counterfolds.
Comparisons.—From P. s. semispinosus, liminalis differs in: darker color; wider aristiforms; greater percentage of upper molars with four counterfolds. From P. s. amphichoricus, liminalis differs in: lighter upper parts of almost uniform color instead of with conspicuous, blackish, middorsal, longitudinal band; more strongly built skull; longer incisive foramen; vomerine sheath usually incomplete instead of always complete.
Specimens examined.—Total number, 10 (MN) from the type locality.
Type locality.—Mount Duida, Esmeralda, Amazonas, Venezuela; altitude 325 m. Type: American Museum of Natural History, no. 77020, adult male; collected 16 October 1920 by Olalla Bros.
Range.—Headwaters of Negro and Orinoco rivers, along boundary of Brazil and Venezuela.
Diagnosis.—Color dark, blackish on middorsal area; subapical zone of setiforms on back Buckthorn Brown, but many with distal parts black; skull broad across zygomata; nasals long; prepalatilar area of skull long; incisive foramen long and narrow; vomerine sheath complete; upper molars usually with four counterfolds but P4 usually with only three.
Pelage.—Aristiforms on middorsal region: Grayish basally, gradually blackening toward tip; total length, 18 to 20 mm; maximum width, 0.8 to 1.0 mm. Setiforms on middorsal region: Gray basally, gradually blackening toward tip but interrupted by a light (16 i), Buckthorn Brown, subapical zone 2 mm long; total length, 18 to 22 mm; maximum width, 0.03 mm. Most of them, [Pg 345] however, whitish basally, gradually blackening toward tip without any distinctively-colored, subapical zone; total length, 24 to 26 mm; maximum width, 0.5 mm. Setiforms on outer thighs: Whitish basally, gradually blackening toward tip but interrupted by an Ochraceous-Buff, subapical zone 3.5 mm long; black tip short; total length, 17 to 19 mm; maximum width, 0.05 mm.
Skull.—Large and slender; rostrum elongate; nasals bluntly pointed posteriorly; supraorbital ridges thick (but not expanded) and extending across parietals but almost obsolete in middle part of parietals; infraorbital foramen with weakly-developed groove for transmission of nerve; postorbital process of zygoma involving mostly squamosal; incisive foramen 5.5 x 2.8 mm wide in anterior third, with margins constricted posteriorly and extending as ridges approximately 2 mm beyond posterior margin of incisive foramen; posterior margin of incisive foramen approximately 2.5 mm anterior to premolars; vomerine sheath complete with maxillary part weak and premaxillary part extending posteriorly beyond middle of incisive foramen; mesopterygoid fossa extending forward as far as middle of M3; bullae well inflated and elongated.
Teeth.—P4 with four counterfolds in one of five specimens and with three in remainder; M1 with four counterfolds in three of five specimens and with three in remainder; M2 with three counterfolds in one specimen and with four in all four remaining specimens; M3 always with four counterfolds. Lower premolars with four counterfolds and lower molars with only three.
Comparisons.—The subspecies is easily distinguishable from P. s. angularis by: larger number of black setiforms on back, forming an almost black longitudinal band; more elongate skull; larger and longer bulla; longer incisive foramen which is more constricted posteriorly.
Specimens examined.—Total number, 6 (AMNH), as follows: Venezuela, territ. Amazonas, Esmeralda, Mt. Duida, altitude 325 m., 4; Venezuela, territ. Amazonas, Rio Cassiquiare, Quemapuré, 1; Brazil, Amazonas, São Gabriel, Rio Uaupés or Caiari, Tatú, 1.
Proechimys kermiti Allen, 30 December 1915, Bull. Amer. Mus. Nat. Hist., 34(22):629 (orig. descr.); Allen, 1916, Bull. Amer. Mus. Nat. Hist., 35(30):569; Tate, 1935, Bull. Amer. Mus. Nat. Hist., 68(5):400; Ellerman, 1940, The families and genera of living rodents, Brit. Mus. (Nat. Hist.), 1:119.
Type locality.—Lower Rio Solimões (up the Solimões 50 to 60 miles on the north bank of the river), Manacaparú, Amazonas, Brazil. Type: American Museum of Natural History, no. 37124, adult female; collected 20 April, 1914, by Leo E. Miller (Roosevelt Brazilian Expedition).
Range.—Known only from type locality.
Diagnosis.—Upper parts Tawny, with darker longitudinal band on back, gradually becoming Ochraceous-Buff on sides; zygomata widely spread; nasals long; incisive foramen long; vomerine sheath incomplete; only M3 with four counterfolds.
Pelage.—Aristiforms on middorsal region: Grayish basally, gradually blackening toward tip; total length, 18 to 20 mm; maximum width, 0.8 mm. Setiforms on middorsal region: Grayish basally, gradually blackening toward tip but interrupted by Tawny, subapical zone 2 mm long; total length, 18 to 20 [Pg 346] mm; maximum width, 0.06 mm; some blackened toward tip without subapical zone. Setiforms on outer thighs: Whitish basally, gradually blackening toward tip but interrupted by Ochraceous-Buff, subapical zone 2.5 mm long; total length, 18 to 20 mm; maximum width, 0.05 mm.
Skull.—Large, elongate, and strongly built; rostrum not conspicuously elongated; nasals bluntly pointed posteriorly; supraorbital ridges wide and extending posteriorly across parietals almost to level of interparietal; infraorbital foramen with moderate development of groove for transmission of nerve; zygomatic arches slender; postorbital process of zygoma involving mostly squamosal; incisive foramen 6.5 mm long and 2.7 mm wide, wider in anterior third and gradually constricted posteriorly, with margins extended toward palate as ridges; vomerine sheath incomplete, maxillary part threadlike; mesopterygoid fossa extending forward as far as anterior third of m3; bullae large and well inflated.
Teeth.—P4 with three counterfolds; M3 with four counterfolds; M1 and M2 with three counterfolds. Lower premolars with four counterfolds; lower molars with three counterfolds.
Comparisons.—From P. s. amphichoricus, kermiti differs in: upper parts Tawny instead of Buckthorn Brown; incisive foramen longer and wider; vomerine sheath incomplete; only M3 instead of usually all molars, with four counterfolds. From P. s. liminalis, kermiti differs in: upper parts Tawny instead of Ochraceous-Tawny; aristiforms narrower; M3 only, instead of usually M2 and M3, with four counterfolds.
Specimens examined.—Only the type.
General characters.—Size medium to large; tail short; aristiforms long and narrow; general color on upper parts Ochraceous-Buff to Ochraceous-Orange, finely and uniformly lined with blackish and not forming evident dark band on back; feet dorsally white or gray; underparts of body and inner sides of legs white; skull elongate and slender with moderate ridges; incisive foramen of medium size; vomerine sheath complete or incomplete; bullae large and elongate; upper molariform teeth with three counterfolds; lower molariform teeth with three counterfolds but commonly one or two molars have only two although premolar may have four.
Remarks.—The identity of "Echimys longicaudatus Rengger" can be ascertained only after samples have been collected in the area indicated by Rengger: "unter dem ein und zwansigsten Breitengrade" in Paraguay. Of the samples available to me, those from Urucum, in western Brazil, are geographically nearest the type locality. North of Urucum, both in Brazil and Bolivia, two species of Proechimys live together and one of them is the same species as that at Urucum. Of the two species found to the northward in Brazil and Bolivia, the one that ranges farther south probably will occur at the locality indicated by Rengger. Provisionally, therefore, the name longicaudatus is allocated to the Urucum sample (see Osgood, 1944:198). In fact, the lack of a type specimen and the [Pg 347] general nature of Rengger's description make "Echimys longicaudatus" a nomen vanum. If two species are found living together in the region of northern Paraguay indicated by Rengger it probably will be impossible to be sure to which one his vague description applies.
The form from Urucum, to which the name Proechimys longicaudatus is here applied, is undoubtedly closely related to Proechimys leucomystax Ribeiro, from Utiarití, on the Rio Papagaio and also to P. roberti and P. boimensis, all from Brazil. P. longicaudatus is used as the name of the species because it is the oldest of the four names.
Figs. 45, 48. Proechimys longicaudatus boimensis, male, MCZ no. 30881, Boim. × 1.
Figs. 41, 42. Proechimys longicaudatus longicaudatus, male, AMNH no. 37085, Urucum. × 1.
Figs. 43, 44. Proechimys longicaudatus leucomystax, male, AMNH no. 37509, Tapirapoa. × 1.
Figs. 51, 52. Proechimys longicaudatus roberti, male, MN no. 6233, Pouso Alto, Goiaz. × 1.
Figs. 53, 54. Proechimys longicaudatus boimensis, male, MCZ no. 30881, Boim. × 1.
Figs. 55, 56. Proechimys longicaudatus longicaudatus, male, AMNH no. 37085, Urucum. × 1.
Figs. 57, 58. Proechimys longicaudatus leucomystax, male, AMNH no. 37509, Tapirapoa. × 1.
Figs. 59, 60. Proechimys longicaudatus roberti, male, MN no. 6233, Pouso Alto, Goiaz. × 1.
Echimys brevicauda Günther, 1 April 1877, Proc. Zool. Soc. London for 1876, (49):748, fig. 9.
Proechimys brevicauda Ihering, 1904, Rev. Mus. Paulista, S. Paulo, 6:422; Osgood, 1914, Zool. Ser. Field Mus. Nat. Hist., 10(12):168; Thomas, 1924, Ann. Mag. Nat. Hist., 13 (ser. 9):534; Thomas, 1927, Ann. Mag. Nat. Hist., 19 (ser. 9):553; Thomas, 1927, Ann. Mag. Nat. Hist., 20 (ser. 9):604; Thomas, 1928, Ann. Mag. Nat. Hist., 2 (ser. 10):262; Thomas, 1928, Ann. Mag. Nat. Hist., 2 (ser. 10):292; Tate, 1935, Bull. Amer. Mus. Nat. Hist., 68 (5):399; Osgood, 1944, Zool. Ser. Field Mus. Nat. Hist., 29 (13):201.
Proechimys cayennensis brevicauda Ellerman, 1940, The families and genera of living rodents, Brit. Mus. (Nat. Hist.), 1:120.
Type locality.—Chamicuros, Rio Huallaga, Peru. Type: British Museum (Nat. Hist.), no. 69.3.31.7,[F]; Lectoparatype: British Museum (Nat. Hist.), no. 66.1.29.8, from Upper Amazons (E. Barttet), selected by Thomas (1900:301).
Range.—Region of the headwaters and upper courses of the Juruá and Ucaiali rivers, eastern Peru and northwestern Brazil.
Diagnosis.—Upper parts Tawny and blackish without marked longitudinal band on back; underparts buffy or white; aristiforms narrow; skull slender; incisive foramen wide; vomerine sheath complete; molariform teeth with three counterfolds, except p4 with four and m3 with only two.
Pelage.—Aristiforms on middorsal region: Blackish basally, gradually blackening toward tip which is extended as a long filament; total length, 18 to 20 mm; maximum width, 0.65 mm. Setiforms on middorsal region: Gray basally, gradually blackening toward tip but interrupted by Tawny, subapical zone 1.2 mm long; total length, 19 to 21 mm; maximum width, 0.1 mm. Setiforms on outer thighs: Whitish basally, gradually blackening toward tip but interrupted by Ochraceous-Tawny, subapical zone 3 mm long.
Skull.—Slender but not elongated; nasals tapering posteriorly; interparietals wide; supraorbital ridges not much extended and faintly shown across parietals; jugals dorso-ventrally "wide" (3.5 mm); postorbital process of zygoma weakly developed; incisive foramen 5.5 x 3 mm, oval, with posterior borders raised to form ridges which extend toward premolars; vomerine sheath complete, with maxillary part laterally compressed and extended toward palate as ridge; mesopterygoid fossa extending forward as far as third molars; bullae large and well inflated.
Teeth.—Molariform teeth with three counterfolds, except p4 which has four and m3 which has only two counterfolds.
Comparisons.—From P. l. longicaudatus, brevicauda differs in: upper parts Tawny instead of Ochraceous-Buff; lower premolar with four instead of three counterfolds; m3 only, instead of both m1 and m3, with two counterfolds. From P. l. boimensis, brevicauda differs in: upper parts Tawny instead of Ochraceous-Orange; aristiforms wider; m3 with two instead of three counterfolds.
Specimens examined.—Total number, 3 (DZ), from Brazil, Amazonas, João Pessoa, Rio Juruá.
Proechimys boimensis Allen, 24 July, 1916, Bull. Amer. Mus. Nat. Hist., 35(27):523; Tate, 1935, Bull. Amer. Mus. Nat. Hist., 68(5):400; Ellerman, 1940, The families and genera of living rodents, Brit. Mus. (Nat. Hist.), 1:119.
Proechimys cayennensis Osgood, 1944, Zool. Ser. Field Mus. Nat. Hist., 29(13):199.
Type locality.—Boim, Rio Tapajoz, Santarem, Pará, Brazil. Type: American Museum of Natural History, no. 37486, adult male; "October 10, 1911 (ex Museu Goeldi)."
Range.—Along lower course of Tapajoz River.
Diagnosis.—Upper parts Ochraceous-Orange; incisive foramen posteriorly constricted; mesopterygoid fossa sharply pointed anteriorly; p4 with four counterfolds, remaining molariform teeth with three counterfolds.
Pelage.—Aristiforms on middorsal region: Gray basally, gradually blackening toward tip which is extended as a long filament; total length, 16 to 18 mm; maximum width, 0.5 mm. Setiforms on middorsal region: Gray basally, gradually blackening toward tip, but interrupted by short (1.5 mm), Ochraceous-Orange, subapical zone; total length 18 to 20 mm; maximum width 0.09 mm. Setiforms on outer thighs: White basally, gradually blackening toward tip but interrupted by Ochraceous-Orange, or Ochraceous-Buff, subapical zone 3.5 mm long; total length 15 to 17 mm; maximum width, 0.06 mm.
Skull.—Elongate and slender; rostrum slender and long; nasals evenly pointed posteriorly; supraorbital ridges weak and barely reaching anteriormost part of parietals; zygomatic arches slender; infraorbital foramen with groove for nerve-transmission obsolete; postorbital process of zygoma weak and involving mostly squamosal; incisive foramen 5 mm long and 2.5 to 3 mm wide, oval, with margins not much extended caudad as ridges; posterior margin of incisive foramen approximately 2 mm anterior to premolars; vomerine sheath complete with maxillary part slender; mesopterygoid fossa sharply pointed anteriorly and extending forward as far as anterior part of third molar; bullae of medium size, smooth and more or less triangular in contour.
Teeth.—Crown length of upper molariform teeth 7 to 7.5 mm; all upper molariform teeth with three counterfolds; lower premolar with four counterfolds; lower molars with three counterfolds each.
Comparisons.—From three of the other four subspecies found in Brazil, boimensis differs in having four, instead of three, counterfolds in the lower premolars. Differences from P. l. brevicauda are indicated in the account of that subspecies.
Remarks.—The material available from Boim, Tapajoz, is rather poor and of no great significance. The Museu Nacional has one specimen from Boim which agrees with the type in the American Museum of Natural History. The Museum of Comparative Zoölogy at Harvard College, however, has one specimen in its collection (MCZ no. 30888 [M]), adult, also from Boim, in which the color pattern is different although agreement with the type specimen is shown in cranial characters and width and length of hairs. This [Pg 351] specimen is much darker than the other specimens (of orange-tint) assigned to this subspecies. However, in other samples of this species, similarly dark animals have been noted. It seems that the orange tint is gained only in old age. Another specimen (MCZ no. 30878 [F]) agrees in all respects with the type of the subspecies but the locality recorded on the label is Cametá, on the Tocantins River. Possibly the subspecies has a range so wide as to include Cametá but I suspect that the data on the label is incorrect as to locality. The collector marked [M] on the label although the specimen is a [F]. The mistake might have occurred through labeling of specimens at a time later than that at which they were collected. The collector was in both localities on more than one occasion.
Specimens examined.—Total number, 5, from Brazil, Pará, as follows: Santarem, Rio Tapajoz, Boim, 3 (AMNH type, MCZ 1, MN 1); Porto de Moz, Tauarí, 1 (MCZ); Cametá (?) 1 (MCZ).
Echimys longicaudatus Rengger, 1830, Naturgeschichte der Saeugethiere von Paraguay, p. 236.
Loncheres myosuros Lichtenstein, 1832, Darstellung neuer oder wenig bekannter Säugethiere, pl. 36 and text.
Echimys myosuros Is. Geoffroy Saint-Hilaire, 1840, Mag. Zool., Ann. 2 (ser. 2):15, 17; Allen, 1899, Bull. Amer. Mus. Nat. Hist., 12(20):261.
Echimys cayennensis Pictet, 1841, Mém. Soc. Phys. Hist. Nat., Genève, 9:145; Waterhouse, 1848, Nat. Hist. Mammalia, 2:334.
Proechimys longicaudatus Thomas, 1901, Ann. Mag. Nat. Hist., 8 (ser. 7):532; Thomas, 1904, Proc. Zool. Soc. London, p. 240; Allen, 1916, Bull. Amer. Mus. Nat. Hist., 35(30):569; Tate, 1935, Bull. Amer. Mus. Nat. Hist., 68(5):400.
Proechimys cayennensis longicaudatus Ellerman, 1940, The families and genera of living rodents, Brit. Mus. (Nat. Hist.), 1:121; Osgood, 1944, Zool. Ser. Field Mus. Nat. Hist, 29(13):198.
Type locality.—Northern Paraguay ("unter dem ein und zwansigsten Breitengrade"). Type: Apparently no type specimen was preserved.
Range.—Western Mato Grosso, Brazil, and northern Paraguay.
Diagnosis.—Upper parts almost uniformly Ochraceous-Buff; incisive foramen widest posteriorly; vomerine sheath complete; p4 and m2 with three counterfolds; m1 and m3 usually with two counterfolds.
Pelage.—Aristiforms on middorsal region: Dark gray, gradually blackening toward tip that has long filament; total length 20 to 22 mm; maximum width 0.6 mm. Setiforms on middorsal region: Whitish basally, gradually blackening toward tip but interrupted by Ochraceous-Buff, subapical zone; blackish tip extended (3.5 mm) and thin; total length 19 to 20 mm; maximum width 0.06 mm. Setiforms on outer thighs: Whitish basally, progressively grayish or blackish toward tip but interrupted by Light Ochraceous-Buff or Ochraceous-Buff, subapical zone; total length 18 to 20 mm; maximum width 0.06 mm.
Skull.—Slender; supraorbital ridge notably raised; bullae large, elongate, smooth and inflated; jugals narrow; postorbital process of zygoma of medium size and constructed entirely of jugal; incisive foramen wide and large (5.5 x [Pg 352] 3 mm), being wider posteriorly than anteriorly and with posterior margins raised; vomerine sheath complete, maxillary part slender and laterally flattened; mesopterygoid fossa extending forward as far as centers, or anterior margins, of third molars; posterior palatine foramina on plane with posterior faces, or centers, of second molars.
Teeth.—Upper molariform teeth always with three counterfolds. Lower molariform teeth: p4 and m2 always with three counterfolds, sometimes the two anterior ones coalesced in m2; m1 with three counterfolds in one specimen (33 per cent); m3 with only two counterfolds in all specimens.
Comparisons.—Differences from P. l. leucomystax and P. l. roberti are given in the accounts of those subspecies.
Specimens examined.—Total number, 3 (2 AMNH, 1 CNHM), from Brazil, Mato Grosso, Corumbá, Urucum.
Proechimys leucomystax Ribeiro, May, 1914, Commissão de linhas telegraphicas estrategicas de Matto Grosso ao Amazonas, Annexo no. 5. Hit. Nat., Zool., Mammiferos, p. 42, pl. 24 (orig. descr.); Tate, 1935, Bull. Amer. Mus. Nat. Hist., 68(5):400; Ellerman, 1940, The families and genera of living rodents, Brit. Mus. (Nat. Hist.), 1:119.
Type locality.—Utiarití, Rio Papagaio, Diamantion, Mato Grosso, Brazil. Type: Museu Nacional, no. 2212, adult, skull only, collected on 5 May 1909, by Prof. A. Miaranda Ribeiro, is here designated lectotype. See remarks.
Range.—Serra dos Parecís, Mato Grosso, Brazil.
Diagnosis.—Ochraceous-Buff, richly lined with blackish, on upper parts; some setiforms completely blackened distally; incisive foramen regularly ovoid; vomerine sheath incomplete; upper molariform teeth and lower premolar with three counterfolds; m2 with three counterfolds but m1 and m3 usually with two counterfolds.
Pelage.—Aristiforms on middorsal region: Grayish basally, gradually blackening toward tip which is extended as a long filament; total length 21 to 22 mm; maximum width 0.65 mm. Setiforms on middorsal region: Whitish basally, gradually blackening toward tip but interrupted by Ochraceous-Buff, subapical zone; some setiforms grayish basally and gradually blackening toward distal portion without any colored, subapical zone; total length 20 to 22 mm, maximum width 0.06 mm. Setiforms on outer thigh: White basally, gradually becoming gray toward tip but interrupted by Light Ochraceous-Buff, subapical zone; some setiforms gray basally and blackening toward tip, but interrupted by Ochraceous-Buff, subapical zone; total length 15 to 18 mm; maximum width 0.08 mm.
Skull.—Slender; supraorbital ridges conspicuous; bullae large, smooth and inflated, with slight, transverse groove; jugals narrow; postorbital processes of zygomata small and involving only squamosal; incisive foramen of medium size but narrow (5 x 2.5 mm), regularly oval and with margins uplifted posteriorly; vomerine sheath incomplete but maxillary part projecting forward and sometimes almost reaching premaxillary part; mesopterygoid fossa reaching forward as far as centers of third molars; posterior palatine foramina at plane of posterior faces of second molars or slightly anterior thereto.
Teeth.—Upper molariform teeth with three counterfolds. Lower molariform teeth: p4 and m2 always with 3 counterfolds; m1 and m3 with 2 counterfolds.
Comparisons.—From P. l. longicaudatus, leucomystax differs in: upper parts richly lined, instead of scarcely lined, with blackish; incisive foramen narrower, and regularly oval instead of widest anteriorly.
Remarks.—Proechimys leucomystax was described mainly on the basis of the Utiarití specimen, here designated lectotype of the species. The specimen from the Juina River is younger, as stated by Ribeiro in his description. Ribeiro mentions the skin of the specimen from Utiarití as "em muito mao estado" and I presume it was discarded as it has not been found in the collection of the Museu Nacional, Brazil.
Specimens examined.—Total number, 6, from Brazil, Mato Grosso: Cáceres, Salto Sepotube, 2 (MN); Cáceres, Tapirapoã, Rio Sepotuba, 2 (AMNH); Diamantino, Utiarití, Rio Papagaio, 1 skull (MN); Diamantino, Rio Juina, 1 (MN).
Proechimys roberti Thomas, December, 1901, Ann. Mag. Nat. Hist., 8 (ser. 7):531 (orig. descr.); Thomas, 1904, Ann. Mag. Nat. Hist., 14 (ser. 7):195; Thomas, 1921, Ann. Mag. Nat. Hist., 8 (ser. 9):140; Tate, 1935, Bull. Amer. Mus. Nat. Hist., 68(5):400.
Proechimys cayennensis roberti Ellerman, 1940, The families and genera of living rodents, Brit. Mus. (Nat. Hist.), 1:121.
Type locality.—Rio Jordão, Araguarí, Minas Gerais, Brazil; 960 meters alt. Type: British Museum (Nat. Hist.), no. 1.11.3.62, old male, collected 8 August, 1902, by A. Robert; original number, 705.
Range.—Western Minas Gerais and southern Goiaz.
Diagnosis.—Color almost uniformly Ochraceous-Orange on upper parts; setiforms long and narrow; incisive foramen long and wide; vomerine sheath usually complete; upper molariform teeth and lower premolar with three counterfolds; m3 with two, and m1 and m2 with two or three, counterfolds.
Pelage.—Aristiforms on middorsal region: Whitish basally, gradually blackening toward tip; total length, 21 to 23 mm; maximum width, 0.6 mm. Setiforms on middorsal region: Whitish on basal half, gradually blackening toward tip but interrupted by an Ochraceous-Orange, subapical zone 6 to 7 mm long; total length 25 to 30 mm; maximum width, 0.05 mm. Setiforms on outer thighs: Whitish on basal half, gradually becoming gray and then blackish toward tip but interrupted by wide, Ochraceous-Buff, subapical zone, the tip being brownish or blackish; total length, 22 to 23 mm; maximum width, 0.04 mm.
Skull.—Slender; supraorbital ridges bent outward and making sharp angle at point of frontosquamosal suture, then continuing backward parallel to each other and extending across parietals; squamosal taking small part in supraorbital ridges; bullae large (11 x 8 mm), inflated, with shallow depressions; incisive foramen not especially long but wide (5 x 3 to 3.5 mm), widest in anterior third and constricted posteriorly; vomerine sheath usually complete, with maxillary part reduced to slender threadlike process or, less commonly, missing; mesopterygoid fossa extending forward as far as centers of third molars; zygomatic arches strong with jugals of medium dorsoventral width (approximately 3.5 mm); postorbital process of zygoma weakly developed and involving only squamosal; posterior palatine foramina on plane of posterior margins of first molars or slightly anterior thereto.
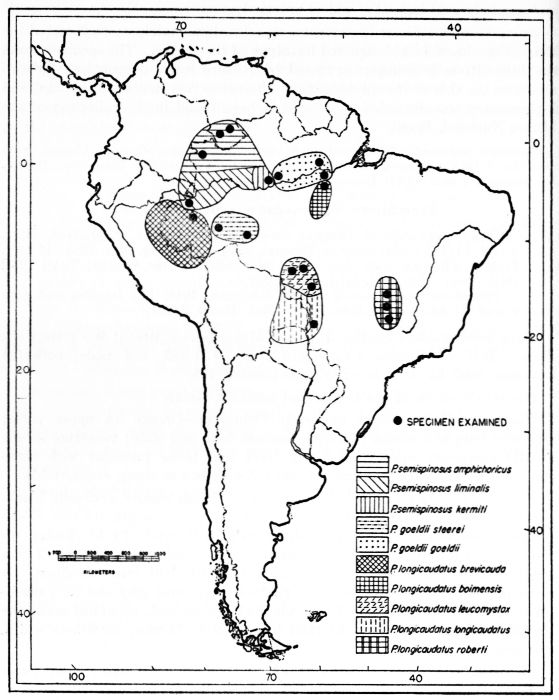 Fig. 61. Map showing the geographic ranges of the subspecies of three species of the
subgenus Proechimys in Brazil.
Fig. 61. Map showing the geographic ranges of the subspecies of three species of the
subgenus Proechimys in Brazil.Teeth.—Upper molariform teeth usually with three counterfolds (92.5 per cent of 42 specimens); M2 with four counterfolds in 5 per cent of specimens; M3 with two counterfolds in 2.5 per cent of specimens. Lower molariform teeth: p4 usually with three counterfolds (97.5 per cent of 39 specimens); rarely with four (2.5 per cent); m1 with three counterfolds in 58 per cent and two counterfolds in 42 per cent of specimens; m2 with three counterfolds in 61 per cent and two counterfolds in 39 per cent of specimens; m3 always with only two counterfolds.
Comparisons.—From P. l. boimensis, roberti differs in: wider incisive foramen; lower premolar with three, and one or two lower molars with only two, counterfolds, instead of lower premolar with four and all lower molars with three counterfolds. From P. longicaudatus, roberti differs in: upper parts Ochraceous-Orange instead of Ochraceous-Buff; incisive foramen wider in posterior third than in anterior third.
Remarks.—This subspecies seems to be adapted to forests of post-climactic conditions which is probably typical of most valleys and margins of the rivers in southern Goiaz and western Minas Gerais. It was found in Goiaz usually in riparian forests with climactic associations or in some advanced stage of the sere. The animals also make incursions into nearby open areas or crops of corn.
Specimens examined.—Total number, 52, from Brazil, as follows: Minas Gerais, Araguarí, Rio Jordão (effluent of Parnaiba), 960 meters alt., 2 (1 CNHM, 1 DZ); Goiaz, Anapolis, 1010 meters alt., 38 (MN); Goiaz, Pouso Alto, 768 meters alt., 11 (MN); Goiaz, Tio São Miguel, 2 (MN).
Mus guyannensis E. Geoffroy Saint-Hilaire, 1803, Catalogue des mammifères du Museum d'Histoire Naturelle, Paris, p. 194.
Echimys cayennensis Desmarest, 1817, Nouv. Dict. Hist. Nat., Paris, nouv. ed., 10:59.
Proechimys cayennensis Allen, 1899, Bull. Amer. Mus. Nat. Hist., 12(20):261, 264; Tate, 1935, Bull. Amer. Mus. Nat. Hist., 68(5):399; Ellerman, 1940, The families and genera of living rodents, Brit. Mus. (Nat. Hist.), 1:120.
General characters.—Size medium to large; tail usually short; aristiforms narrow to wide; general color of setiforms on back ranging from Tawny to Ochraceous-Buff and becoming gradually lighter on sides; no conspicuous dark longitudinal band on back; upper parts of hands and feet white to light brown; underparts white, including inner sides of legs; skull elongate and not conspicuously ridged; vomerine sheath complete or incomplete; upper premolar with three counterfolds and molars with two or three; lower premolar with three or four counterfolds, and lower molars with two or three.
Type locality.—Tapirapoã, Rio Sepotuba, Cáceres, Mato Grosso, Brazil. Type: Museu Nacional, no. 1932, adult male (color faded); collected on 2 February, 1909, by Prof. A. Miranda Ribeiro; original number, 788 A.
Range.—Serra dos Parecís, headwaters of Paraguai and Tapajoz rivers.
Diagnosis.—Aristiforms wide and stiff; general color on upper parts Ochraceous-Orange; incisive foramen long; vomerine sheath incomplete or complete; lower premolar with four counterfolds, remaining molariform teeth with three counterfolds.
Pelage.—Aristiforms on middorsal region: Whitish basally, gradually becoming gray toward tip, with distal fourth blackish and ending as a long filament; total length, 22 to 23 mm; maximum width, 1 mm. Setiforms on middorsal region: Whitish on basal half, gradually blackening toward tip but interrupted by Ochraceous-Orange, subapical zone 5 mm long; total length, 26 to 30 [Pg 356] mm; width, 0.04 to 0.12 mm. Setiforms on outer thighs: Whitish on basal half, gradually blackening toward tip but interrupted by Ochraceous-Buff to Ochraceous-Orange subapical zone; total length 20 to 23 mm; width, 0.03 to 0.18 mm.
Skull.—Strong; supraorbital ridges raised and extending across anterior fourth of parietals; nasals elongate; bullae rounded, inflated, with shallow grooves; postorbital process of zygoma weakly developed and constructed entirely of jugal; incisive foramen elongate and narrow (5.5 x 2.5 mm), posteriorly constricted with posterior margins elevated above surface of bones; vomerine sheath incomplete or complete but, when complete, with maxillary part filiform and delicate; mesopterygoid fossa extending forward as far as middle, or even anterior, parts of third molars; posterior palatine foramina on plane of centers, or even anterior faces, of second molars.
Teeth.—Upper molariform teeth with three counterfolds. Lower molariform teeth: premolar with four counterfolds, molars with three counterfolds, sometimes with two folds coalesced in center of tooth.
Comparison.—From P. g. ribeiroi, and P. g. bolivianus, villicauda differs in wider aristiforms. From P. g. ribeiroi, villicauda further differs in: larger and wider incisive foramen; vomerine sheath incomplete or complete instead of always complete and thick. From P. g. bolivianus, villicauda differs in: lower premolars always, instead of rarely, with four counterfolds, and m3 always with three counterfolds instead of usually with only two counterfolds.
Specimens examined.—Total number, 4, from Brazil, Mato Grosso, as follows: Cáceres, Tapirapoã, 3 (MN); Diamantino, Rio Papagaio, Utiarití, 1 (AMNH).
Fig. 62. Proechimys guyannensis villicauda, male, MN no. 1932, Tapirapoã. Type. × 1. × 1.
Fig. 63. Proechimys guyannensis ribeiroi, male, MN no. 1935, Rio Doze de Outubro. Type. × 1.
Fig. 64. Proechimys guyannensis hyleae, male, MCZ no. 30887, Tauarí. Type. × 1.
Fig. 65. Proechimys guyannensis nesiotes, male, CNHM no. 19496, Ilha de Manapirí. Type. × 1.
Fig. 66. Proechimys guyannensis leioprimna, female, CNHM no. 19503, Cametá. Type. × 1.
Fig. 67. Proechimys guyannensis oris, male, CNHM no. 19495, Providencia. × 1.
Fig. 68. Proechimys guyannensis arescens, male, CNHM no. 26440, Fazenda Inhuma. Paratype. × 1.
Fig. 69. Proechimys guyannensis riparum, female, AMNH no. 143018, Manaus. Type. × 1.
Fig. 70. Proechimys guyannensis arabupu, male, AMNH no. 75816, Arabupu. Type. × 1.
Fig. 71. Proechimys guyannensis villicauda, male, MN no. 1932, Tapirapoã. Type. × 1.
Fig. 72. Proechimys guyannensis ribeiroi, male, MN no. 1935, Rio Doze de Outubro. Type. × 1.
Fig. 73. Proechimys guyannensis hyleae, male, MCZ no. 30887, Tauarí. Type. × 1.
Fig. 74. Proechimys guyannensis nesiotes, male, CNHM no. 19496, Ilha de Manapirí. Type. × 1.
Fig. 75. Proechimys guyannensis leioprimna, female, CNHM no. 19503, Cametá. Type. × 1.
Fig. 76. Proechimys guyannensis oris, male, CNHM no. 19495, Providencia. × 1.
Fig. 77. Proechimys guyannensis arescens, male, CNHM no. 26440, Fazenda Inhuma. Paratype. × 1.
Fig. 78. Proechimys guyannensis riparum, female, AMNH no. 143018, Manaus. Type. × 1.
Fig. 79. Proechimys guyannensis arabupu, male, AMNH no. 75816, Arabupu. Type. × 1.
Figs. 80, 81. Proechimys guyannensis villicauda, male, MN no. 1932, Tapirapoã. Type. × 1.
Figs. 82, 83. Proechimys guyannensis ribeiroi, male, MN no. 1935, Rio Doze de Outubro. Type. × 1.
Figs. 84, 85. Proechimys guyannensis hyleae, male, MCZ no. 30887, Tauarí. Type. × 1.
Figs. 86, 87. Proechimys guyannensis nesiotes, male, CNHM no. 19496, Ilha de Manapirí. Type. × 1.
Figs. 88, 89. Proechimys guyannensis leioprimna, female, CNHM no. 19503. Type. × 1.
Figs. 90, 91. Proechimys guyannensis oris, male, CNHM no. 19495, Providencia. × 1.
Figs. 92, 93. Proechimys guyannensis arescens, male, CNHM no. 26440, Fazenda Inhuma. Paratype. × 1.
Figs. 94, 95. Proechimys guyannensis riparum, female, AMNH no. 143018, Manaus. Type. × 1.
Figs. 96, 97. Proechimys guyannensis arabupu, male, AMNH no. 75816, Arabupu. Type. × 1.
Type locality.—Rio 12 de Outubro, affluent of the Camararé, Mato Grosso, Mato Grosso, Brazil; about 190 kilometers west of Utiarití; altitude 414 meters. Type: Museu Nacional, no. 1935, adult male (colors faded); collected on 20 June, 1909, by Prof. A. Miranda Ribeiro; original number G.
Range.—Known only from the type locality.
Diagnosis.—Aristiforms wide and stiff; incisive foramen small and narrow; vomerine sheath complete and thick; p4 with four counterfolds; remaining molariform teeth with three counterfolds.
Pelage.—Aristiforms on middorsal region: Whitish basally, gradually blackening toward tip; total length, 19 to 22 mm; maximum width, 0.8 mm. Setiforms on middorsal region: Whitish on basal half, gradually blackening toward tip but interrupted by subapical zone probably of some tint of ochraceous; total length, 22 to 24 mm; maximum width, 0.06 mm. Setiforms on outer thighs: Whitish basally, gradually blackening toward tip but interrupted by probably light ochraceous, subapical zone; total length 14 to 16 mm; maximum width, 0.03 mm.
Skull.—Slender; supraorbital ridges low; bullae ovate with shallow grooves; postorbital process of zygoma almost obsolete and involving mostly jugal; incisive foramen short and narrow (4 × 2 mm), constricted posteriorly and with posterior margins raised; vomerine sheath complete and thick; mesopterygoid fossa extending forward as far as posterior faces of second molars; posterior palatine foramina on plane with centers of second molars.
Teeth.—Upper molariform teeth with three counterfolds. Lower molariform teeth: p4 with four counterfolds; molars with three counterfolds which sometimes are fused.
Comparisons.—From P. g. bolivianus, ribeiroi differs in: aristiforms wider; incisive foramen shorter and narrower; vomerine sheath complete and thick, instead of complete or incomplete and not thick; p4 always with four, instead of usually only three, counterfolds and lower molars always with three, instead of sometimes with only two, counterfolds in m3. Differences from P. g. villicauda are given in the account of that subspecies.
Remarks.—The name ribeiroi is given in honor of the late Professor Alipio Miranda Ribeiro, in recognition of his important work in Brazilian vertebrate zoology.
Specimens examined.—Total number, 2 (MN), from Brazil, Mato Grosso, Mato Grosso, Rio 12 de Outubro; altitude, 414 meters.
Type locality.—Tauarí, Rio Tapajoz, Porto de Moz, Pará, Brazil; approximately 87 kilometers south of Santarem. Type: Museum of Comparative Zoology at Harvard College, no. 30887, adult male; collected on 19 January, 1934, by A. M. Olalla; original number 7288.
Range.—Region of lower Tapajoz River and banks of Amazon up to the Jamundá River.
Diagnosis.—Aristiforms conspicuously wide and stiff; general color on upper parts Tawny; incisive foramen long and oval; vomerine sheath complete [Pg 362] but with maxillary part slender or, sometimes, incomplete; p4 with four counterfolds, rarely three; remaining molariform teeth with three counterfolds.
Pelage.—Aristiforms on middorsal region: Whitish basally, gradually becoming blackish toward tip; total length, 19 to 21 mm; maximum width, 1.1 mm. Setiforms on middorsal region: a. Gray basally, gradually blackening toward tip but interrupted by wide (5 to 6 mm) Tawny, subapical zone; some are whitish basally and gradually become sooty brown toward tip except for same type of subapical zone (tip only slightly darker than subapical zone); b. With the same type described above and some completely blackish, with the base gray; total length 22 to 25 mm; maximum width, 0.1 mm. Setiforms on outer thighs: Whitish basally, gradually becoming gray and then blackish toward tip but interrupted by long, Ochraceous-Tawny, subapical zone; tip, itself, blackish brown; sometimes this type appears with some lighter ones and sometimes with completely blackish setiforms; total length 16 to 18 mm; maximum width 0.3 mm.
Skull.—Medium in size and slender; cranium narrow and not increasing much in breadth posteriorly; rostrum stout, laterally thick, with masseteric crest well-developed; nasals pointed posteriorly; supraorbital ridges broad but barely extended across parietals; zygomatic arches strong; postorbital process of zygoma involving mostly squamosal; incisive foramen long and narrow (5.5 to 6 × 2.5 mm), oval and extending posteriorly to point only 2 mm anterior to premolars; vomerine sheath complete with maxillary part usually slender; mesopterygoid fossa extending forward as far as centers of third molars; bulla of medium size, well inflated and with shallow grooves on surface.
Teeth.—Upper molariform teeth with three counterfolds. Lower premolar with four counterfolds or, sometimes (20 per cent of 15 specimens), with only three; lower molars with three counterfolds.
Comparisons.—From P. g. oris and P. g. nesiotes, hyleae differs in: wider aristiforms; general color on upper parts Tawny, instead of Ochraceous-Orange; vomerine sheath not always complete, instead of always complete. From P. g. oris, hyleae differs in: p4 usually with four, instead of only three counterfolds and all molars with three, instead of only lower molars with three counterfolds. From P. g. nesiotes, hyleae differs in: p4 usually, instead of always, with four counterfolds; color on back Tawny instead of Ochraceous-Orange; aristiforms wider.
Remarks.—This subspecies shows greater variability than any other in this species. There are two types of coloration. The most common type of coloration is dark, with Tawny, subapical zones in the setiforms of the middorsal region and many completely black setiforms; in the other type the subapical zone is still Tawny but there are no black setiforms. One specimen from Obidos, on the north bank of the Amazon, completely agrees in the characteristics of color and skull with the reddish type and suggests either that there is an extension of the range of the subspecies along the lower [Pg 363] course of the Tapajoz or that there are two subspecies, in which event the animals from Tauarí are intergrades between hyleae and an unnamed, tawny-colored subspecies occurring to the southward.
Between 13 and 23 January, 1934, A. M. Olalla collected 10 adult females, 6 of which contained embryos. Three of the females had 2 embryos each, two had 3 embryos each and one had only 1 embryo. At this same time and place only ten per cent of specimens obtained were not fully adult.
Specimens examined.—Total number, 21, from Brazil, Pará, as follows: Porto de Moz, Tauarí, right bank of Tapajoz, approximately 85 kilometers south of Santarem, 20 (19 MCZ, 1 CNHM); Obidos, Obidos, 1 (MCZ).
Type locality.—Ilha de Manapirí, Rio Tocantins, Pará, Brazil. Type: Chicago Natural History Museum, no. 19496, adult male; collected on 9 December, 1910, by Dr. Emilia Snethlage; original number, 12.
Range.—Known only from the type locality.
Diagnosis.—Aristiforms wide and stiff; general color on upper parts Ochraceous-Orange; incisive foramen long, with parallel borders; vomerine sheath complete and thick; p4 with four counterfolds, remaining molariform teeth with three counterfolds.
Pelage.—Aristiforms on middorsal region: Gray basally, gradually blackening toward tip; total length, 18 to 19 mm; maximum width, 0.9 mm. Setiforms on middorsal region: Gray basally, gradually blackening toward tip but interrupted by Ochraceous-Orange, subapical zone 4 mm long; total length 16 to 19 mm; maximum width, 0.06 mm. Setiforms on outer thighs: Whitish basally, gradually becoming gray and blackish toward tip but interrupted by Ochraceous-Buff, subapical zone 3 mm long; total length, 13 to 15 mm; maximum width, 0.03 mm.
Skull.—Of medium size; rostrum short; nasals pointed posteriorly; postorbital ridges extending caudad across anterior fifth of parietals; zygomatic arches strong; jugal with process in posterior part of masseteric fossa; postorbital process of zygoma involving mostly squamosal; incisive foramen elongate, narrow (5 x 2.3 mm), and parallel sided; posterior margin of incisive foramen approximately 3 mm anterior to premolars; margins of foramen raised to form ridges; vomerine sheath complete, of almost uniform width and set deeply in foramen; mesopterygoid fossa extending forward as far as centers of third molars; bullae of medium size and inflated.
Teeth.—Upper molariform teeth with three counterfolds; p4 with four counterfolds; m1-3 with three counterfolds.
Comparison.—From P. g. oris, nesiotes differs in: Aristiforms conspicuously wider; incisive foramen shorter and narrower, with borders parallel instead of posteriorly constricted; posterior margin of incisive foramen farther from premolars; p4 with four, instead of three, counterfolds; lower molars with three instead of two counterfolds. From P. g. leioprimna, nesiotes differs in: Incisive foramen with parallel borders instead of oval; p4 with four instead of three counterfolds; m3 always with three instead of two counterfolds.
Remarks.—Dr. E. Snethlage mentions the type as having been collected at night in the forest.
Specimens examined.—Total number, 8 (MCZ, CNHM, MN), from Brazil, Pará, Tocantins River, Ilha de Manapirí.
Type locality.—Cametá, left bank of Tocantins River, near its mouth, Cametá, Pará, Brazil. Type: Chicago Natural History Museum, no. 19503, adult female; collected on 21 January, 1911, by Dr. Emilia Snethlage; original number, 35.
Range.—Known only from type locality but probably extending westward toward Xingú River.
Diagnosis.—Aristiforms wide and stiff; general color on upper parts Ochraceous-Orange, incisive foramen moderately long; oval; vomerine sheath complete; all molariform teeth with three counterfolds, except lower, third molar which has only two.
Pelage.—Aristiforms on middorsal region: Gray basally, gradually blackening toward tip which is extended as a short filament; total length, 19 to 21 mm; maximum width, 0.8 to 1 mm. Setiforms on middorsal region: Whitish basally, gradually blackening toward tip but interrupted by Ochraceous-Orange, subapical zone 2 to 3 mm long; total length, 17 to 20 mm; maximum width 0.1 mm. Setiforms on outer thighs: Whitish basally, becoming gradually gray and then blackish toward tip but interrupted by Ochraceous-Buff, subapical zone; blackish tip short; total length 13 to 15 mm; maximum width 0.06 mm.
Skull.—Of medium size; rostrum relatively short; nasals with posterior borders rounded; postorbital ridges extending across anterior fourth of parietals; zygomatic arches moderately strong; postorbital process of zygoma involving both jugal and squamosal; incisive foramen of medium length (4 to 5 mm) and narrow (about 2.5 mm), oval and extending caudad to a plane approximately 2 mm anterior to premolars; vomerine sheath complete, with premaxillae forming approximately anterior 3/4 of sheath; maxillary part of sheath short but well-developed; mesopterygoid fossa extending forward as far as centers of third molars; bullae of medium size but well-inflated.
Teeth.—Upper molariform teeth with three counterfolds. Lower molariform teeth with three counterfolds, except third molar which has only two.
Comparison.—From P. g. oris, leioprimna differs in: conspicuously wider aristiforms; shorter and narrower incisive foramen; lower molariform teeth with three counterfolds (except m3 with only two), instead of lower molars with only two counterfolds. Differences from P. g. nesiotes are given in the account of that subspecies.
Remarks.—The paratype was collected in an "igarapé," depression usually invaded by the river waters; the paratype, collected on 18 January, 1911, had two large embryos.
Specimens examined.—Total number, 4 (2 CNHM, 2 AMNH), from Brazil, Pará, Cametá.
Proechimys oris Thomas, September, 1904, Ann. Mag. Nat. Hist., 14 (ser. 7): 195; Thomas, 1905, Ann. Mag. Nat. Hist., 15 (ser. 7):587; Thomas, 1912, Ann. Mag. Nat. Hist., 9 (ser. 8):89; Tate, 1935, Bull. Amer. Mus. Nat. Hist., 68:400; Osgood, 1944, Zool. Ser. Field Mus. Nat. Hist., 29:199.
Proechimys cayennensis oris Ellerman, 1940, The families and genera of living rodents, Brit. Mus. (Nat. Hist.), 1:121.
Type locality.—Igarapé-assú, E. F. B., near Belem, Igarapé-assú, Pará, Brazil. Type: British Museum (Nat. Hist.), no. 4.7.4.78, old male; collected on 6 March, 1904, by Alphonse Robert; original number, 1818.
Range.—Probably most of the region on south bank of Amazon River, between Tocantins (west) and Gurupí River (south).
Diagnosis.—Aristiforms narrow but somewhat stiff; color on upper parts Ochraceous-Orange; incisive foramen long and wide, conspicuously constricted posteriorly; posterior margin of incisive foramen close to plane of premolars; vomerine sheath complete but maxillary part threadlike; upper molariform teeth and lower premolar with three counterfolds; lower molars with only two counterfolds.
Pelage.—Aristiforms on middorsal region: Gray basally, gradually blackening toward tip, which is extended as a filament; total length, 16 to 17 mm; maximum width, 0.6 to 0.7 mm. Setiforms on middorsal region: Gray basally, gradually becoming blackish toward tip but interrupted by Ochraceous-Orange, subapical zone 2 to 4 mm long; total length, 18 to 20 mm; maximum width 0.06 mm. Setiforms on outer thighs: Whitish basally, gradually becoming blackish toward tip but interrupted by Ochraceous-Buff, subapical zone; total length, 15 to 16 mm; maximum width, 0.04 mm.
Skull.—Of medium size; supraorbital ridges well developed and extending across anterior fifth of parietals; zygomatic arches strong; jugal with masseteric fossa deep and with well-developed posterior process; postorbital zygomatic process involving mostly squamosal; incisive foramen long (6 to 7 mm), widest anteriorly (2.5 to 3.2 mm); but narrowing posteriorly to less than 1 mm and extending caudad almost to plane of premolars; vomerine sheath complete but maxillary part delicate and threadlike; mesopterygoid fossa extending forward as far as third molars or posterior parts of second molars; bullae large and inflated.
Teeth.—Each upper molariform tooth with three counterfolds. In lower jaw, premolar with three, and molars with only two, counterfolds.
Comparisons.—Differences from the subspecies with adjoining ranges are given in the accounts of those subspecies.
Remarks.—Thomas (1912:89) extended the known range of the subspecies to Faro, on the Jamundá River, on the left bank of the Amazon, and to Boim, on the Tapajoz River, as well as to Benevides, E. F. Braganca, near Belem. It seems to me that the specimens from Faro should be referred provisionally to Proechimys guyannensis hyleae; the specimens from Boim are "more brightly [Pg 366] rufous" (Thomas, loc. cit.) and could be referred to Allen's P. boimensis, described in 1914, but P. guyannensis hyleae probably lives in the same place and only an examination of the specimens, which I have not seen, would permit of certainly allocating the specimens to their correct species. The specimens from Benevides are more certainly P. g. oris.
E. Snethlage collected one specimen in a garden (Providencia, E. F. B.). However, according to the personnel of the Brazilian Health Service, the animals are strictly forest dwellers although they do make excursions into more open places.
Specimens examined.—Total number, 3, from Brazil, Pará, as follows: Providencia, E. F. B., approximately 15 kilometers east from Belem, 1 (CNHM); Tanaquará, near Belem, 1 (MN); Rio Guamá, near Belem, 1 (AMNH).
Additional record.—Brazil, Pará, Benevides, E. F. B., approximately 100 kilometers north-east of Belem (Thomas, 1912:89).
Proechimys cayennensis arescens Osgood, 12 July 1944, Zool. Ser. Field Mus. Nat. Hist., 29(13): 198.
Type locality.—Fazenda Inhuma, below Santa Filomena, upper Rio Parnaiba, Vitoria do Alto Parnaiba, Maranhão, Brazil. Type: Chicago Natural History Museum, no. 26441, adult male; collected on 5 August, 1925, by Heinrich E. Snethlage.
Range.—Region including the valleys of the Turí-assú and Parnaiba rivers, Maranhão, Brazil.
Diagnosis.—Aristiforms moderately wide and not conspicuously stiff; general color of upper parts near (15'a) Ochraceous-Orange; incisive foramen long and wide; vomerine sheath complete or incomplete; upper molariform teeth and lower premolar with three counterfolds; lower molars with only two.
Pelage.—Aristiforms on middorsal region: Whitish basally, gradually blackening toward tip; total length, 19 to 21 mm; maximum width, 0.7 mm. Setiforms on middorsal region: Whitish basally or on basal half, gradually becoming gray and then blackish toward tip, but interrupted by long (5 to 6 mm) subapical zone near (15'a) Ochraceous-Orange; total length, 15 to 16 mm; maximum width 0.05 mm. Setiforms on outer thighs: Whitish on basal half, gradually becoming gray and then blackish toward tip but interrupted by Ochraceous-Buff, subapical zone; tip sometimes not conspicuously darker than subapical zone; total length 18 to 25 mm; maximum width, 0.03 mm.
Skull.—Medium in size, not elongated; nasals pointed posteriorly; supraorbital ridges strong and thick, extending caudad across anterior third of parietals; zygomatic arches strong; postorbital process of zygoma involving only squamosal; incisive foramen 5 by 2.7 mm, oval and extending caudad to plane approximately 2 mm anterior to premolars; posterior margins of incisive foramen not forming a ridge; vomerine sheath complete and with maxillary part slender and threadlike, or incomplete, in which event, maxillary part not extended enough to join premaxillary process; mesopterygoid fossa [Pg 367] extending forward as far as centers of third molars; bulla large and more or less triangular in its peripheral outline.
Teeth.—Upper molariform teeth with three counterfolds each. Lower premolar with three counterfolds; lower molars with two counterfolds.
Comparisons.—From P. g. oris, arescens differs in: Color of upper parts lighter and more uniform; incisive foramen oval instead of conspicuously constricted posteriorly; posterior margin of incisive foramen farther from premolars.
Remarks.—One specimen from Turí-assú (MN) has been identified by O. Thomas as "P. oris" (his own handwriting is on the label) and the subspecies is really closely related to oris.
Specimens examined.—Total number, 3, from Brazil, Maranhão, as follows: Vitoria do Alto Parnaiba, Fazenda Inhuma (below Santa Filomena), 2 (CNHM); Alto da Alegria, Turí-assú, 1 (MN).
Type locality.—Manaus, Manaus, Amazonas, Brazil. Type: American Museum of Natural History, no. 143018, adult female; collected 6 March, 1943.
Range.—Known only from type locality but probably extending northward and eastward.
Diagnosis.—Aristiforms wide and stiff; upper parts Ochraceous-Tawny; incisive foramen short, wide, and oval; vomerine sheath incomplete; upper molariform teeth and lower premolar with three counterfolds; lower molars with only two counterfolds.
Pelage.—Aristiforms on middorsal region: Gray basally gradually blackening toward tip; total length, 18 to 20 mm; maximum width, 0.9 mm. Setiforms on middorsal region: Whitish basally, gradually blackening toward tip but interrupted by 2 mm long, Ochraceous-Tawny, subapical zone; total length 20 to 22 mm; maximum width, 0.04 mm. Some are whitish basally and gradually become black toward tip with no subapical zone. Setiforms on outer thighs: Whitish basally, gradually becoming gray and then blackish toward tip but interrupted by Ochraceous-Buff, subapical zone 3 to 4 mm long; tip not conspicuously dark; total length, 17 to 19 mm; maximum width, 0.03 mm.
Skull.—Of medium size and slender; rostrum slender; nasals rounded posteriorly; supraorbital ridges well developed and barely extended onto anteriormost part of parietals; zygomatic arches slender; infraorbital foramen with well-developed groove for nerve transmission; postorbital process of zygoma almost obsolete and involving mostly squamosal; incisive foramen short and wide (4.5 × 3.3 mm), oval and with posterior margins raised to form ridges which extend toward premolars; posterior margin of incisive foramen approximately 2.5 mm anterior to premolars; vomerine sheath incomplete, with only short, premaxillary part; mesopterygoid fossa extending forward as far as anterior parts of third molars; bullae large and inflated, with more or less triangular outline.
Teeth.—Crown length of well worn P4-M3, 6.8 mm; upper molariform [Pg 368] teeth with three counterfolds each. Lower premolar with three counterfolds; lower molars with two counterfolds.
Comparisons.—From P. g. oris and P. g. hyleae, riparum differs in: Shorter and wider incisive foramen; vomerine sheath incomplete, instead of sometimes incomplete. From P. g. oris, riparum differs in: Upper parts Ochraceous-Tawny instead of Ochraceous-Orange; aristiforms conspicuously wider. From P. g. hyleae, riparum differs in: Aristiforms narrower; upper parts Ochraceous-Tawny instead of Tawny; lower premolars with three, instead of four, counterfolds; lower molars with two, instead of three, counterfolds.
Specimens examined.—Type only.
Type locality.—Arabupu, Mount Roraima, Boa Vista, Territ. Rio Branco; about 1540 meters altitude. Type: American Museum of Natural History, no. 75816, adult male; collected by Dr. G. H. H. Tate on 30 December, 1927; original number, 4716.
Range.—Known only from the type locality.
Diagnosis.—Aristiforms conspicuously wide and stiff; color on upper parts dark, near (15'j) Ochraceous-Tawny; incisive foramen widest in anterior third; vomerine sheath complete, sometimes incomplete; upper molariform teeth and lower premolar with three counterfolds; lower molars with two counterfolds.
Pelage.—Aristiforms on middorsal region: Whitish basally, gradually blackening toward tip; total length, 19 to 22 mm; maximum width, 1.1 mm. Setiforms on middorsal region: Gray basally, gradually blackening toward tip but interrupted by dark (15'j) Ochraceous-Tawny, subapical zone 3 mm long; some completely blackish on distal parts; total length, 20 to 23 mm; maximum width, 0.07. Setiforms on outer thighs: Whitish basally, gradually blackening toward tip but interrupted by Ochraceous-Buff or Light Ochraceous-Buff, subapical zone; total length, 20 to 23 mm; maximum width, 0.07 mm.
Skull.—Size medium; nasals pointed posteriorly; supraorbital ridges prominent and slightly extended caudad onto anterior half of parietals; groove for transmission of nerve in infraorbital foramen weakly developed; zygomatic arches strong; postorbital process of zygoma involving mostly squamosal; incisive foramen approximately 5.5 × 2.7 mm, widest in anterior third and constricted posteriorly, with posterior margin about 1 mm anterior to plane of premolars; vomerine sheath complete, with premaxillary part expanded and maxillary part notably slender and sometimes lacking; mesopterygoid fossa in some specimens extending forward as far as middle parts of second molars; bullae large and inflated.
Teeth.—Upper molariform teeth with three counterfolds each. Lower premolar with three counterfolds; molars with only two.
Comparisons.—From P. g. warreni, arabupu differs in: Narrower aristiforms; narrower incisive foramen; lower premolar with three instead of four counterfolds; lower molars with two, instead of three, counterfolds. From P. g. oris, arabupu differs in: Aristiforms wider; posterior margin of incisive foramen farther from plane of premolars; upper parts dark (15'j) Ochraceous-Tawny, instead of Ochraceous-Orange.
Remarks.—The sample is fairly uniform.
Specimens examined.—Total number, 6 (AMNH), from Brazil, Territorio do Rio Branco, Boa Vista, Mount Roraima, Arabupu; approximately 1540 m. altitude.
Genotype.—Echimys albispinus Is. Geoffroy Saint-Hilaire, 1838; by original designation.
Trinomys Thomas, July 1921, Ann. Mag. Nat. Hist., 8 (ser. 9):140 (orig. descr.); Tate, 1935, Bull. Amer. Mus. Nat. Hist., 68(5):401; Ellerman, 1940, The families and genera of living rodents, Brit. Mus. (Nat. Hist.), 1:115.
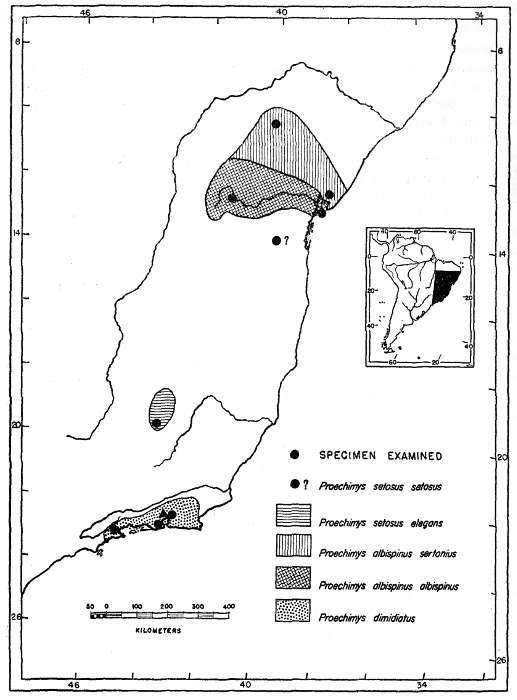 Fig. 99. Map showing the geographic ranges of the subspecies of three species of the subgenus
Trinomys.
Fig. 99. Map showing the geographic ranges of the subspecies of three species of the subgenus
Trinomys.General characters.—Pelage of upper parts with lanceolate and, sometimes, clavate aristiforms extending over most of rump and onto thighs; tail 86 to 103 per cent of length of head and body; tail sometimes white-tipped and sometimes penicillate; skull small, with ridges moderately developed; supraorbital ridges involving no part of parietals; infraorbital foramen with no separate groove for transmission of nerve; mesopterygoid fossa extending forward to level of second or first molars; incisors opisthodont, orthodont or proodont; molariform teeth, in occlusal view, with main fold large and usually [Pg 371] reaching opposite wall; no counterfold anterior to main fold in upper molariform teeth and usually no counterfold posterior to main fold in lower molariform teeth; premolars larger than first molars, first molars larger than second molars and second molars larger than third molars; four molariform teeth of nearly equal size in some animals.
Thomas (1921:140) erected the subgenus Trinomys, including in it the species albispinus and setosus and stated that "the primary distinction between these [Trinomys and Proechimys] lies in the number of laminae present in the cheekteeth—four in Proechimys, three in Trinomys." The distinction is valueless as a subgeneric character, not only because the character is not constant in the species in the subgenus but also because there is subspecific variation in number of laminae in the cheekteeth. Proechimys albispinus, however, shares with three other species common characters, as listed above, and the name Trinomys will, therefore, apply to this group of species, since Proechimys albispinus is the genotype.
Echimys dimidiatus Günther, 1 April 1877, Proc. Zool. Soc. London, 1876(4):747.
Proechimys dimidiatus Allen, 1899, Bull. Amer. Mus. Nat. Hist., 12(20):264; Ribeiro, 1905, Arch. Mus. Nac. Rio de Janeiro, 13:187; Thomas, 1921, Ann. Mag. Nat. Hist., 8 (ser. 9):141; Tate, 1935, Bull. Amer. Mus. Nat. Hist., 68(5):400; Ellerman, 1940, The families and genera of living rodents, Brit. Mus. (Nat. Hist.), 1:122.
Type locality.—Unknown; probably southwestern Rio de Janeiro, Brazil (see Remarks). Type: British Museum (Nat. Hist.), no. 51.7.21.24; presented by Lord Derby.
Range.—Rio de Janeiro, from the southern limit of the state northward to and including the Distrito Federal.
General characters.—Size large; tail averaging 80 per cent of head and body; aristiforms narrow and soft (0.4 to 0.5 mm wide), imparting a non-spiny character to the pelage; general color of upper parts Ochraceous-Buff, finely lined with blackish brown, gradually becoming lighter on sides; ventral surface of body and inner sides of legs white; feet dorsally white but with a sepia-colored stripe along outer margin; tail brownish-black above and white below, but white sometimes extended to upper side in distal part; skull broad with no conspicuous ridges; jugals deep with transverse ridge usually conspicuous; postorbital process of zygoma involving only squamosal; incisive foramen short and wide posteriorly; vomerine sheath complete in 95 per cent of specimens and with maxillary part thick; posterior palatine foramina at plane of first molars or slightly anterior to them; bullae moderately developed; in juvenal specimens, each upper molariform tooth with three counterfolds, but posteriormost counterfold small; in adult specimens, posteriormost counterfold disappearing in 50 per cent of fourth premolars and first molars, in 20 per cent of second molars, and in 15 per cent of third molars; lower molariform teeth with two counterfolds in almost every juvenal specimen, this number, in adult animals, decreasing in m3 to one in 20 per cent of specimens but rarely being reduced in other teeth.
General characters.—Aristiforms soft and narrow, ranging from 15 to 19 mm in total length and 0.4 to 0.5 mm in maximum width; pelage generally non-spiny and not harsh; length of tail ranging from 20 per cent shorter than head and body to as long as, or slightly longer than, head and body; ears rather small (23 to 25 mm).
Color.—General color of back and sides results from uniform mixture of black distal parts of aristiforms with Ochraceous-Buff of subapical zone of setiforms. Dorsally, from nose caudad to rump, mixture appears brownish-black, lined with Ochraceous-Buff; toward sides, amount of Ochraceous-Buff gradually increases and resultant color is much lighter brown than on back. On outer parts of arms and legs, color turns gradually to sepia toward distal parts and finally to uniform sepia on wrists and ankles, this color extending to outer dorsal parts of hands and feet; on ankles, sepia forms complete ring, as usual in the genus. Tail blackish-brown on upper parts, this stripe gradually tapering toward tip where dark brown hairs form small pencil; white of under side of tail sometimes seen also entirely around distal part, short of tip which remains dark brown. Ventral surfaces wholly white, from upper lips caudad including inner surfaces of legs.
Hairs.—Aristiforms on middorsal region: Gray basally, gradually blackening toward tip that has long, fine filament; total length 16 to 19 mm; maximum width 0.5 mm. On outer thigh whitish basally, gradually blackening toward tip; some with Ochraceous-Buff, subapical zone; total length 13 to [Pg 373] 15 mm; maximum width 0.25 mm. Setiforms on middorsal region: Whitish on basal half, gradually blackening toward tip, but interrupted by Ochraceous-Buff, subapical zone; some with Light Ochraceous-Buff, subapical zone and short, blackish zone on tip; total length 12 to 14 mm; maximum width 0.02 mm. Setiforms on outer thighs: Whitish on basal half, then gradually becoming gray on middle part and finally Light Ochraceous-Buff on distal third, or with tip blackish and Ochraceous-Buff, subapical zone.
Skull.—Elongate and broad with no conspicuous crests; rostrum rather stout; jugals deep with transverse crest usually well-developed; zygomatic postorbital process conspicuous and formed entirely of squamosal; incisive foramen short and wide posteriorly; vomerine sheath complete in great majority of specimens, its maxillary part wide and strong; posterior palatine foramina on plane with front of M1 or slightly farther forward; bullae rather small and elongate.
Teeth.—P4 with three secondary folds in all juvenal specimens, but posteriormost fold small and disappearing in 50 per cent of adult specimens; M1 with 3 outer folds in juveniles and also disappearing in 50 per cent of adults; M2 with three outer folds in juveniles, but only 20 per cent remaining in adults; M3 with 3 outer folds in 50 per cent of juveniles, decreasing to 15 per cent in adults. Lower molariform teeth: p4 with 2 secondary folds; m1 with 2 secondary folds in 90 per cent of adults and in all juveniles; m2 with 2 secondary folds in 98 per cent of adults and in all juveniles; m3 with 2 secondary folds in 81 per cent of adults, remaining percentage with only one counterfold, and with 2 secondary folds in all juveniles.
Remarks.—Samples studied of P. dimidiatus are notably uniform throughout the geographic range of the species. The few biotypes detected seemed unworthy of subspecific rank.
In discussing the type locality of the species, Thomas (1921:141) states: "We know that its donor did obtain a number of specimens from Rio Janeiro, and the skull agrees so closely with those of two examples from Itatiaia, near to the Rio-Minas frontier, collected and presented by Prof. J. P. Hill, that I have no hesitation in referring the latter to Günther's species."
Specimens examined.—Total number, 211 (MN), from Brazil as follows: Rio de Janeiro; Parati, Pedra Branca (400 m.), 113; Mangaratiba, Fazenda do Rubião (750 m.), 3; Fazenda do Tenente (700 m.), 4; Fazenda da Lapa (450 m.), 13; Teresópolis, Fazenda Guinle (960 m.), 61; Nova Iguassú, Barro Branco (20 m.), 16; Distrito Federal, Tijuca, 1.
Additional records.—Rio de Janeiro, Itatiaia (Thomas, 1921:141); Rio de Janeiro, Zona da mata, Mont-Serrat, Serra do Itatiaia (Ribeiro, 1905:187).
General characters.—Size large; tail long; aristiforms generally wide and stiff; general color on upper parts and sides a combination of blackish from tips of aristiforms with cinnamon ground color from subapical zones of setiforms; darker band on middorsal line; differentiated light-colored aristiforms conspicuous on outer sides of thighs and rump; usually rufous tint on neck and postauricular region; underparts white; tail with white tip, usually accentuated by white brush; feet white on dorsal surface; hind feet slightly [Pg 374] darker on outer sides; skull elongate and smooth; jugals wide dorso-ventrally; incisive foramen elongate; upper molariform teeth usually with one to five counterfolds, number varying with subspecies; lower premolar always with two counterfolds and lower molars always with one or two counterfolds.
Remarks.—As a whole, the samples of the populations of the species do not afford a satisfactory record of the distribution; my concept of the group may be changed when further collections are made in localities geographically intermediate between those from which specimens now are known. If some of the forms prove to be physiologically isolated, they may deserve treatment as full species according to the conventional standards of systematic zoology. P. panema, for example, does not seem to be geographically isolated from P. gratiosus. P. denigratus, at the northernmost known occurrence of the species, actually represents a striking jump in the cline, although collections from intermediate regions may provide intermediate structural stages. Further collecting may also prove that the southern form, P. iheringi iheringi, is completely isolated from the rest of the group. However, these samples are certainly more related to each other than any one of them is to that of the other species found in the same range, namely P. dimidiatus, and all the forms in question, therefore, seem best arranged as subspecies of one full species. A clinal variation certainly exists among these forms and the most striking differences correspond to larger geographical distances.
Fig. 104. Proechimys iheringi iheringi, female, MN no. 6453, Ilha de São Sebastião. × 1.
Fig. 105. Proechimys iheringi bonafidei, male, MN no. 6183, Fazenda Bõa Fé. Type. × 1.
Fig. 106. Proechimys iheringi gratiosus, male, MN no. 4024, Floresta da Caixa Dagua. Type. × 1.
Fig. 107. Proechimys iheringi panema, female, MN no. 8288, Campinho. Type. × 1.
Fig. 108. Proechimys iheringi denigratus, male, MN no. 8500, Mata do Ribeirão da Fortuna. Type. × 1.
Fig. 109. Proechimys iheringi paratus, female, MN no. 4012, Floresta da Capela de São Braz. Type. × 1.
Fig. 110. Proechimys iheringi iheringi, female, MN no. 6453, Ilha de São Sebastião. × 1.
Fig. 111. Proechimys iheringi bonafidei, male, MN no. 6183, Fazenda Bõa Fé. Type. × 1.
Fig. 112. Proechimys iheringi gratiosus, male, MN no. 4024, Floresta da Caixa Dagua. Type. × 1.
Fig. 113. Proechimys iheringi panema, female, MN no. 8288, Campinho. Type. × 1.
Fig. 114. Proechimys iheringi denigratus, male, MN no. 8500, Mata do Ribeirão da Fortuna. Type. × 1.
Fig. 115. Proechimys iheringi paratus, female, MN no. 4012, Floresta da Capela de São Braz. Type. × 1.
Figs. 116, 117. Proechimys iheringi iheringi, female, MN no. 6453, Ilha de São Sebastião. × 1.
Figs. 118, 119. Proechimys iheringi bonafidei, male, MN no. 6183, Fazenda Bõa Fé. Type. × 1.
Figs. 120, 121. Proechimys iheringi gratiosus, male, MN no. 4024, Floresta da Caixa Dagua. Type. × 1.
Figs. 122, 123. Proechimys iheringi panema, female, MN no. 8288, Campinho. Type. × 1.
Figs. 124, 125. Proechimys iheringi denigratus, male, MN no. 8500, Mata do Ribeirão da Fortuna. Type. × 1.
Figs. 126, 127. Proechimys iheringi paratus, female, MN no. 4012, Floresta da Capela de São Braz. Type. × 1.
Proechimys iheringi Thomas, August, 1911, Ann. Mag. Nat. Hist., 8 (ser. 8):252 (orig. descr.); Thomas, 1921, Ann. Mag. Nat. Hist., 8 (ser. 9):141; Tate, 1935, Bull. Amer. Mus. Nat. Hist., 68(5):400; Ellerman, 1940, The families and genera of living rodents, Brit. Mus. (Nat. Hist.), 1:122.
Type locality.—Island of São Sebastião (off São Paulo), Formosa, São Paulo, Brazil. Type: British Museum (Nat. Hist.), no. 2.8.25.5, adult male, presented by the São Paulo Museum.
Range.—Littoral and islands of São Paulo and Rio de Janeiro.
Diagnosis.—Aristiforms narrow; tail shorter than head and body; setiforms Cinnamon-Buff; incisive foramen short; vomerine sheath complete; upper molariform teeth with two or three counterfolds; lower molariform teeth with two counterfolds, rarely one in m3.
Pelage.—Aristiforms on middorsal region: Gray basally, gradually blackening toward tip; total length 18 to 23 mm; maximum width, 0.6 mm. Aristiforms on outer thighs: Gray basally, blackening distally toward tip; some differentiated with Cinnamon-Buff tip. Setiforms on middorsal region: Gray basally, gradually blackening toward tip but interrupted by a Cinnamon-Buff, subapical zone 3 mm long; total length, 16 to 20 mm; maximum width, 0.06 mm. Setiforms on outer thighs: Gray basally, gradually blackening toward tip but interrupted by Cinnamon-Buff, subapical zone or with Cinnamon-Buff continuous to tip.
Skull.—Slender; bullae small and well inflated; jugal dorso-ventrally wide with transverse ridge inconspicuous; incisive foramen short, 3.5 × 2.5 mm; vomerine sheath complete; mesopterygoid fossa extending forward as far as middle parts of second molars; postorbital process of zygoma small, formed by both jugal and squamosal; posterior palatine foramina at plane of premolars; interorbital breadth narrow.
Teeth.—Upper molariform teeth with two or three counterfolds (when unworn usually three and rarely four); sometimes only one counterfold in M3 and sometimes counterfolds fused in molars. Lower molariform teeth with two counterfolds, rarely one in m3.
Comparisons.—From P. i. bonafidei and P. i. gratiosus, iheringi differs in: Incisive foramen shorter; vomerine sheath complete, instead of usually incomplete; setiforms Cinnamon-Buff, instead of Ochraceous-Buff; upper molariform teeth with two or three separate counterfolds, instead of having counterfolds fused or reduced to one or two; aristiforms narrower in iheringi than in bonafidei.
Specimens examined.—Total number, 25, from Brazil, as follows: São Paulo, Formosa, Ilha de São Sebastião, 9 (DZ 6, MN 2, MCZ 1); São Paulo, Mogi das Cruzes, Alto da Serra, alt. 900 m., 2 (DZ); São Paulo, Ubatuba, alt. 10 m., 4 (2 DZ, 2 MN); Rio de Janeiro, Angra dos Reis, 2 (MN); Rio de Janeiro, Angra dos Reis, Ilha Grande, 7 (5 DZ, 1 MCZ, 1 MN).
Type locality.—Fazenda Bõa Fé, Teresópolis, Rio de Janeiro, Brazil; alt. 850 meters. Type: Museu Nacional, no. 6183, adult male; collected on 18 August, 1942, by G. Pereira; SEPFA no. M 14663.
Range.—Known only from the type locality.
Diagnosis.—Aristiforms wide and stiff; tail shorter than head and body; setiforms Ochraceous-Buff; incisive foramen long; vomerine sheath incomplete, or rarely complete; molariform teeth with two counterfolds usually fused.
Pelage.—Aristiforms on middorsal region: Gray basally, gradually blackening toward tip; total length, 22 to 26 mm; maximum width, 0.8 mm. Aristiforms on outer thighs: Gray basally, gradually blackening toward tip but interrupted by Ochraceous-Buff subapical zone; some Ochraceous-Buff to tip; total length, 18 to 20 mm; maximum width, 0.7 mm. Setiforms on middorsal region: Gray basally, gradually blackening toward tip but interrupted by Ochraceous-Buff, subapical zone; total length, 17 to 20 mm; maximum width, 0.06 mm. Setiforms on outer thighs: Gray basally, gradually blackening toward tip but interrupted by Ochraceous-Buff, subapical zone; only a short blackened tip.
Skull.—Large, with elongate rostrum; bullae large and well inflated; jugals with transverse ridge inconspicuous; postorbital process of zygoma small, formed mostly by squamosal; incisive foramen elongated (5.5 × 2.5 mm); vomerine sheath incomplete or, if complete, with maxillary part thin and delicate; posterior palatine foramen at plane of first molars; mesopterygoid fossa extending forward as far as middle parts of second molars.
Teeth.—Upper molariform teeth with two counterfolds; these completely separated in 3 of 16 specimens; two counterfolds coalesced in all three molars in 6 specimens; counterfolds coalesced in only two molars in 3 specimens; counterfolds coalesced in only one molar in 4 specimens. Lower molariform teeth with two counterfolds which are completely separated in 13 of 16 specimens; counterfolds coalesced in only one molar in 2 specimens; counterfolds coalesced in all three molars in one specimen.
Comparisons.—From P. i. gratiosus, bonafidei differs in: Aristiforms wider; tail shorter; molariform teeth with two counterfolds instead of one or two. Differences from P. i. iheringi are given in the account of that subspecies.
Remarks.—Of females with embryos two were captured in April and one in September. The embryos number 2, 1, 2. Young were captured mostly in April, but two were taken in July. Male gonads seemed to be most active in March, April and September. The animals lived in a second growth forest, approaching the climax. The rainfall was more than 1600 mm annually, and the mean annual temperature was 18.5° centigrade.
Specimens examined.—Total number, 18 (MN), from Brazil, Rio de Janeiro, Teresópolis, Fazenda Bõa Fé.
Type locality.—Floresta da Caixa Dagua, Santa Teresa, Espirito Santo, Brazil; altitude 750 meters. Type: Museu Nacional, no. 4024, adult male; collected on 25 May, 1940, by C. Lako; SEPFA no. M 6911.
Range.—Known only from the type locality.
Diagnosis.—Aristiforms narrow; tail of same length as head and body; [Pg 380] setiforms Ochraceous-Buff; incisive foramen long; vomerine sheath usually incomplete; upper molariform teeth with one or two counterfolds; lower molariform teeth with two counterfolds, except that m3 usually has only one.
Pelage.—Aristiforms on middorsal region: Gray basally, gradually blackening toward tip; total length, 21 to 27 mm; maximum width, 0.6 mm. Aristiforms on outer thighs: Gray basally, gradually blackening toward middle, and Ochraceous-Buff on distal half; total length, 18 to 21 mm; maximum width, 0.5 mm. Setiforms on middorsal region: Gray basally, gradually blackening toward tip but interrupted by short, Ochraceous-Buff, subapical zone; total length, 18 to 20 mm; maximum width, 0.06 mm. Setiforms on outer thighs: Gray basally, gradually blackening toward middle, and distal part Ochraceous-Buff or with only tip blackened; total length, 14 to 16 mm; maximum width, 0.05 mm.
Skull.—Slender; bullae small but well-inflated; upper edge of jugals deeply concave; transverse ridge of jugals conspicuous; postorbital process of zygoma small, involving only squamosal; incisive foramen elongate (5 x 2.5 mm); vomerine sheath almost always incomplete, and maxillary part lacking or, when present, slender; mesopterygoid fossa extending forward as far as middle of second molars; posterior palatine foramina at plane of front border of first molars or slightly anterior thereto.
Teeth.—Upper molariform teeth with two counterfolds in 10 of 16 specimens and only one in remainder; these folds commonly coalesced; M3 with only one counterfold in 6 specimens, and 2 counterfolds in remainder. Lower molariform teeth with two counterfolds in 6 specimens and in 10 of them m3 has only one counterfold.
Comparisons.—From P. i. panema, gratiosus differs in: Lower molariform teeth with only one counterfold in smaller percentage of specimens; incisive foramen shorter; aristiforms narrower; setiforms Ochraceous-Buff instead of Cinnamon. Differences from iheringi and paratus are given in the accounts of those subspecies.
Remarks.—All the animals were captured in climax forest.
Specimens examined.—Total number, 16 (MN), from Brazil, Espirito Santo, Santa Teresa, Floresta da Caixa Dagua, altitude 750 meters.
Type locality.—Campinho, Colatina, Espirito Santo, Brazil; altitude 500 meters. Type: Museu Nacional, no. 8288, adult female; collected on 15 July, 1942, by C. Lako.
Range.—Known only from the type locality.
Diagnosis.—Aristiforms moderately wide; tail of approximately same length as head and body; setiforms Cinnamon; incisive foramen moderately long and narrow; vomerine sheath incomplete; upper molariform teeth with two counterfolds, but m3 most frequently with one.
Pelage.—Aristiforms on middorsal region: Gray basally, gradually blackening toward tip; total length, 21 to 23 mm; maximum width, 0.8 mm. Aristiforms on outer thighs: Gray, some gradually blackening toward tip and others with distal part Cinnamon; total length, 17 to 19 mm; maximum width, 0.7 mm. Setiforms on middorsal region: Gray, gradually blackening [Pg 381] toward tip, but interrupted by Cinnamon, subapical zone; total length, 18 to 20 mm; maximum width, 0.06 mm. Setiforms on outer thighs: Gray, gradually blackening toward middle, and Cinnamon on all of distal parts or with tip blackish; total length, 13 to 15 mm; maximum width, 0.09 mm.
Skull.—Strong, with jugals dorso-ventrally wide; interorbital region and cranium wide; bullae well inflated; transverse ridge of jugals not well-developed; postorbital process of zygoma small and formed only of squamosal; incisive foramen 4.7 × 2.2 mm; vomerine sheath always incomplete, with maxillary part reduced to small process; mesopterygoid fossa extending forward as far as middle of second molars or only slightly short thereof; posterior palatine foramina at plane of front of first molars.
Teeth.—All upper molariform teeth with two counterfolds in 4 specimens; one having only one counterfold in M3; 3 with counterfolds coalesced in one or two molars. Lower molariform teeth with two counterfolds in one specimen, these counterfolds not coalesced; m3 with one counterfold in 4 specimens and with two in one specimen.
Comparisons.—Differences from P. denigratus and P. i. paratus are given in the accounts of those animals.
Specimens examined.—Total number, 5 (MN), from Brazil, Espirito Santo, Colatina, Campinho; altitude 500 meters.
Type locality.—Mata do Ribeirão da Fortuna, 40 kilometers west of Ilheus, Itabuna, Bahia, Brazil. Type: Museu Nacional, no. 8500, adult male; collected 16 March, 1945.
Range.—Known only from the type locality.
Diagnosis.—Aristiforms wide and stiff; tail longer than head and body; setiforms near (15"a) Cinnamon; incisive foramen long and narrow; vomerine sheath complete; premolars with two counterfolds, upper molars with one or two, and lower molars with only one.
Pelage.—Aristiforms on middorsal region: Gray basally, gradually blackening toward tip; total length, 20 to 22 mm; maximum width, 1.1 mm. Aristiforms on outer thighs: Gray basally, gradually blackening toward tip or with distal part near (15"a) Cinnamon; total length, 14 to 16 mm; maximum width, 0.5 mm. Setiforms on middorsal region: Gray basally, gradually blackening toward tip but interrupted by near (15"a) Cinnamon, subapical zone 4 mm wide; total length, 18 to 20 mm; maximum width, 0.05 mm. Setiforms on outer thighs: Gray basally, gradually blackening toward tip but interrupted by wide, near (15"a) Cinnamon, subapical zone.
Skull.—Slender; nasals short; bullae large and well-inflated; jugals with conspicuous transverse ridge; postorbital process of zygoma conspicuous, spiniform and formed almost exclusively by jugal; incisive foramen elongated and narrow (5 × 1.8 mm); vomerine sheath complete and formed almost exclusively by premaxillae; maxillary part of this sheath short and in most specimens the two parts of sheath completed by vomer itself; mesopterygoid fossa extending forward as far as middle of second molars and in some skulls as far as anterior border of second molars; posterior palatine foramina at anterior plane of first molars.
Teeth.—Upper molariform teeth: P4 always with two counterfolds; M1 with two counterfolds in 65 per cent of specimens but anterior counterfold poorly developed; rest of specimens with only one counterfold in M1; M2 with two counterfolds in 50 per cent of specimens and only one in remainder; M3 with two counterfolds in only 17 per cent of specimens, and remainder with only one. Lower molariform teeth: p4 always with two counterfolds; molars always with only one counterfold.
Comparisons.—From P. i. panema, denigratus differs in: Each lower molar with only one, instead of with more than one, counterfold; incisive foramen longer and narrower; vomerine sheath complete instead of incomplete; aristiforms conspicuously wider; tail longer.
Remarks.—One female (SEPFA no. M 17060) captured on 9 January, 1944, gave birth to two females on 26 January, 1944. Each of these young measured 177 mm in total length and weighed 27.8 g. On 4 March, 1944, their measurements were: head and body, 120,120; tail, 120,130; hind foot, 32,33; ear, 21,22; skull:—total length, 36.0,35.0; condyloincisive length, 29.0,29.1; zygomatic breadth, 19.1,18.5; length of nasals, 12.5,11.6; interorbital constriction, 9.3,8.8; cranial breadth, 16.4,16.9; palatilar length, 11.5,10.5; crown length of P4 and M1, 4.3,4.3 mm.
The forest where the animals were captured has a high percentage of deciduous trees in spite of the heavy rainfall in this region. All of the animals were trapped near water. Young were captured from January to May. Most animals have a conspicuous Cinnamon patch on the nuchal region.
Specimens examined.—Total number, 34 (SEPFA 33, MN 1), from Brazil, Bahia, Itabuna, Mata do Ribeirão da Fortuna.
Type locality.—Floresta da Capela de São Braz, Santa Teresa, Espirito Santo, Brazil; altitude 630 meters. Type: Museu Nacional, no. 4012, adult female; collected on 24 September, 1940, by Dr. H. W. Laemmert; SEPFA no. M 8447.
Range.—Known only from the type locality.
Diagnosis.—Aristiforms wide and stiff; tail 96 per cent of head and body; color on setiform Cinnamon-Buff; incisive foramen short and moderately wide; vomerine sheath complete; all molariform teeth with two counterfolds.
Pelage.—Aristiforms on middorsal region: Gray basally, gradually blackening toward tip; total length, 24 to 26 mm; maximum width, 1.3 mm. Aristiforms on outer thighs: Gray basally, gradually blackening toward middle, and distal parts near (15"c) Pinkish Cinnamon; total length, 18 to 20 mm; maximum width 0.8 mm. Setiforms on middorsal region: Gray basally, gradually blackening toward tip but interrupted by Cinnamon-Buff, subapical zone; total length 14 to 16 mm; maximum width, 0.06 mm.
Skull.—Slender; bullae large and well-inflated; jugals with conspicuous, [Pg 383] transverse ridge; postorbital process of zygoma moderately developed and involving only squamosal; incisive foramen short and narrow (4.1 × 2.1 mm); vomerine sheath complete, with maxillary part short and thick; mesopterygoid fossa extending forward as far as posterior parts of second molars; posterior palatine foramina at plane of premolars.
Teeth.—Upper and lower molariform teeth with two counterfolds. Counterfolds coalesced in P4 and M1 of one specimen.
Comparisons.—From P. i. gratiosus and P. i. panema, paratus differs in: all molariform teeth with two, instead of some with fewer, counterfolds; vomerine sheath complete and thick instead of usually incomplete; incisive foramen shorter and narrower; aristiforms conspicuously wider; setiforms Cinnamon-Buff instead of Ochraceous-Buff and Cinnamon, respectively. Tail 96 per cent of head and body in paratus instead of 100 per cent as in panema.
Remarks.—The animals were captured in climax forest.
Specimens examined.—Total number, 3 (MN), from Brazil, Espirito Santo, Santa Teresa, Floresta da Capela de São Braz; altitude 630 meters.
General characters.—Size medium; tail approximately same length as head and body; aristiforms moderately wide; feet rather large; ears of medium size; color on upper parts and sides sepia gradually changing to Ochraceous-Tawny; few differentiated, light-colored aristiforms present on outer thighs and rump; under surface of body and inner sides of legs white; tail with white tip and conspicuous, white pencil; feet white dorsally; skull short and smooth, somewhat flattened in interorbital region; jugals narrow dorso-ventrally; incisive foramen moderately long and notably narrow; vomerine sheath complete and slender; postorbital process of zygoma spinelike and involving mostly jugal; premolars usually with two counterfolds; molars with only one counterfold, rarely two in M1 or in M3.
Remarks.—The specimens available are undoubtedly faded and, therefore, the colors mentioned above for the upper parts and sides may not correspond to the colors of unfaded pelages. Desmarest (1817:59) describes the color of setosus as similar to that of the "Echimys de Cayenne" (Proechimys guyannensis) but being more "rousse." Is. Geoffroy Saint-Hilaire (1840:52) describes the same animal as being "d'un brun roussâtre" on the upper parts.
The Proechimys from Lagoa Santa, Minas Gerais, "Echimys" elegans Lund, is certainly related to P. iheringi as well as to P. albispinus. From P. iheringi, elegans differs in having a smaller skull with shorter rostrum, narrower incisive foramen, and orthodont incisors. On the other hand the restricted distribution of the aristiforms in the pelage and the white, penicillate tail are points of resemblance to iheringi. From P. albispinus, elegans differs in having a less spinous pelage and longer tail with white pencil instead of a brown pencil. The skulls, however, are similar, except for the fact that elegans does not have proodont incisors as albispinus sometimes does. Thomas (1921:141) states, after describing the skull of the type of setosus, that "Specimens corresponding to this animal have been obtained at Lagoa Santa, Minas, by Lund and others, and at Bahia." Thomas, however, would not have referred to specimens from "Bahia" as being comparable to elegans had they not [Pg 385] been different from albispinus which he discussed in the same paper. Also, he would not have confused "specimens comparable to elegans" with a subspecies of P. iheringi (P. i. denigratus, from southern Bahia) which has opisthodont instead of orthodont incisors. Since French collectors sent material to Europe at the beginning of the 19th century from (southern?) Bahia, possibly setosus came from there.
In the collection of the American Museum of Natural History there is one specimen (AMNH no. 16140) of Proechimys, included in the so-called Maximilian Collection. The characters of this specimen agree closely with those of the specimens from Lagoa Santa. The locality of capture of specimen no. 16140 is unknown, but it is reasonable to assume that Prince Maximilian zu Wied obtained it somewhere along his route of travel through southeastern Bahia. Wied (1826:445) mentions "L[oncheres]. myosuros Licht." as "am Parahyba, am Peruhype und Belmonte," which greatly increases the possibility of its having come from southern Bahia. The close similarity to elegans of Wied's specimen indicates that the locality of capture possibly was in the region of the less humid, low escarpments of southern Bahia.
My conclusion is that Wied's specimen corresponds closely to setosus and, tentatively, I identify it as such. "Echimys elegans," due to the relationships mentioned above is here considered to be a subspecies of setosus.
Among the species described in earlier times, and whose identity was never ascertained, "Echinomys" fuliginosus Wagner seems to be synonymous with setosus. Wagner describes the animal as having a tail "apicis versus pilis albidis vestita" and the figure of the cheekteeth (1844, pl. 239 D) shows a typical trilaminate condition which occurs commonly in elegans. Moreover, the tail of fuliginosus is only 9 per cent shorter than the head and body and the aristiforms of this subspecies are moderately wide.
Echimys setosus Desmarest (Geoffroy's MS), 1817, Nouv. Dict. Hist. Nat. nouv. ed., 10:59 (orig. descr.); Is. Geoffroy Saint-Hilaire, 1838, Comptes Rendus Acad. Sci., Paris, 6(26):886; Is. Geoffroy Saint-Hilaire, 1840, Mag. Zool., Paris, (ser. 2, année 2):12, 33, 52; Allen, 1899, Bull. Amer. Mus. Nat. Hist., 12(20):257, 261.
Echimys cayennensis Pictet, 1841, Mém. Soc. phys. Hist. Nat., Genève, 9:145; Waterhouse, 1848, Nat. Hist. Mammalia, 2:334.
Echinomys fuliginosus Wagner, 1843, Schreber's Säugethiere, suppl. 3:343; Wagner, 1844, Schreber's Säugethiere, suppl. 4, pl. 39 D.
Proechimys setosus Allen, 1899, Bull. Amer. Mus. Nat. Hist., 12(20):264; Thomas, 1921, Ann. Mag. Nat. Hist., 8 (ser. 9):141; Tate, 1935, Bull.
Amer. Mus. Nat. Hist., 68(5):401; Ellerman, 1940, The families and genera of living rodents, Brit. Mus. (Nat. Hist.), 1:122.
Proechimys fuliginosus Tate, 1935, Bull. Amer. Mus. Nat. Hist., 68(5): 400; Ellerman, 1940, The families and genera of living rodents, Brit. Mus. (Nat. Hist.), 1:119.
Type locality.—Unknown; see remarks under P. setosus. Type: Museum d'Histoire Naturelle, Paris, no. A. 7787 (Thomas, 1921:141), "very imperfect."
Diagnosis.—Aristiforms wide; P4 and M1 with two counterfolds; p4 with two counterfolds, one anterior to main fold.
Pelage.—Aristiforms on middorsal region: Gray basally, gradually blackening toward tip; total length, 18 to 20 mm; maximum width, 0.8 mm. Aristiforms on outer thighs: Color much faded; total length, 15 to 17 mm; maximum width, 0.3 mm. Setiforms on middorsal region: Color faded; total length, 16 to 18 mm; maximum width, 0.04 mm. Setiforms on outer thighs: Color faded; total length, 10 to 13 mm; maximum width, 0.03 mm.
Figs. 129-132. Proechimys setosus elegans, sex ?, UZM no. L 104, Lagoa Santa. × 1.
Skull.—Short; rostrum short and stout; length of nasals approximately 15 mm (broken); bullae roundish, smooth and well-inflated; jugals dorso-ventrally narrow (3.1 mm) with strong transverse ridge; postorbital process of zygoma spiniform, slender and involving mostly jugal; incisive foramen narrow (3.8 x 1.7 mm) and narrowest in posterior part; vomerine sheath complete; posterior palatine foramina obsolete; mesopterygoid fossa extending forward as far as middle of second molars.
Teeth.—Incisors orthodont. P4 with two counterfolds; M1 with two counterfolds but anterior one notably small; M2 and M3 with only one counterfold each. In lower jaw: p4 with two counterfolds, one anterior to main fold; molars with only one counterfold.
Comparisons.—From P. s. elegans, setosus differs in: M1 with two counterfolds as opposed to only one; M3 with one counterfold instead of sometimes with two counterfolds; p4 with one counterfold anterior to main fold and another posterior, instead of both counterfolds posterior.
Remarks.—The measurements above were taken from the Maximilian specimen mentioned above. Measurements of the type were given by Desmarest as: head and body, 5-1/2 inches, tail about 6-1/2 inches. Is. Geoffroy Saint-Hilaire (1838:886) corrects these measurements to: head and body 195 mm; tail (part missing), 170 mm.
E[chimys]. elegans Lund, 1841, Kong. Danske Videnskab. Selsk. natur-vidensk. math. Afhandl., Kjöbenhavn, 8:99 (orig. descr.).
Loncheres elegans Lund, 1841, Kong. Danske Videnskab. Selsk, natur-vidensk. math. Afhandl., Kjöbenhavn, 8:245, 266, 294; Wagner, 1843, Wiegman's Archiv f. Naturg., Berlin, 2 (Jahrg. 9):47.
Echimys cayennensis Waterhouse, 1848, Nat. Hist., Mammalia, 2:337.
Echinomys cajennensis Winge, 1888, Jordfundne og nulevende Gnavere (Rodentia), E Museo Lundii, Kjöbenhavn, 1(3):71, pl. 6, figs. 5-6, pl. 7, fig. 1.
Proechimys setosus Thomas, 1921, Ann. Mag. Nat. Hist., 8 (ser. 9):141.
Proechimys elegans Tate, 1935, Bull. Amer. Mus. Nat. Hist., 68(5):400; Ellerman, 1940, The families and genera of living rodents, Brit. Mus. (Nat. Hist.), 1:119.
Type locality.—Lagoa Santa, Nova Lima, Minas Gerais, Brazil. Type: Syntypes in Universitets Zoologiske Museum, Kjöbenhavn; collected by P. W. Lund.
Range.—Known only from the type locality.
Diagnosis.—Aristiforms wide; P4 usually with two counterfolds; M3 sometimes with two counterfolds; p4 with two counterfolds anterior to main fold.
Pelage.—Aristiforms on middorsal region: Gray basally, gradually blackening toward tip; total length, 18 to 20 mm; maximum width, 0.7 mm. Aristiforms on outer thighs: Gray basally, gradually blackening toward tip which is Cinnamon; total length, 15 to 17 mm; maximum width, 0.3 mm. Setiforms on middorsal region: Whitish basally, gradually blackening toward tip, but interrupted by Cinnamon, subapical zone; total length, 17 to 19 mm; maximum width, 0.04 mm. Setiforms on outer thighs: Whitish basally, gradually blackening toward tip but interrupted by near (15"a) Cinnamon, subapical zone; total length, 10 to 12 mm; maximum width, 0.03 mm.
Skull.—Short; rostrum short but not stout; length of nasals 17 mm; bullae large, smooth, and well-inflated; jugals with conspicuous, transverse ridge; postorbital process of zygoma long, spiniform and constructed entirely of jugal; incisive foramen narrow (4 × 1.7 mm); vomerine sheath complete and slender; posterior palatine foramina obsolete; mesopterygoid fossa extending anteriorly as far as middle parts of second molars.
Teeth.—Incisors orthodont. P4 usually with two counterfolds, rarely with three; upper molars with only one counterfold, but M3 sometimes with two, posterior one being vestigial. Lower molariform teeth: p4 with two counterfolds, both being anterior to main fold; molars with only one counterfold.
Comparisons.—Differences from P. s. setosus are given in the account of that subspecies.
Remarks.—According to Lund, these animals are found in the vicinity of small pools, swim well in spite of not having webbed toes, at night go after food and climb the corn stalks, and have their nests in the grass at the margins of the pools.
Specimens examined.—Total number, 2 (UZM), from Brazil, Minas Gerais, Nova Lima, Lagoa Santa.
General characters.—Size small; tail of same length as head and body or slightly less; feet small; ears of medium size; color of upper parts Ochraceous-Tawny gradually changing to Ochraceous-Buff on sides; differentiated, light-colored aristiforms on back, sides, rump and at base of tail; clavate aristiforms on back with Ochraceous-Tawny or Ochraceous-Buff, subapical zone; underparts of body and inner sides of legs white; tail blackish above, white below, with no white tip; hands and feet white on dorsal parts and some specimens darker on outer margins of feet; skull short and smooth, somewhat flattened in frontal region; jugal dorso-ventrally wide and with moderately conspicuous transverse ridge; postorbital process of zygoma well developed and involving both jugal and squamosal; bullae large and smooth; incisive foramen short and narrow; vomerine sheath incomplete or complete; molariform teeth with only one counterfold; incisors orthodont or proodont.
Remarks.—A good series from Macaco Seco, Andaraí, Bahia, agrees closely with the form first described (albispinus) from the Island Madre de Deus, in Todos os Santos Bay, Bahia. Compared with topotypes of P. albispinus sertonius, the animal from Macaco Seco in general color is more Ochraceous-Tawny and has a narrower skull with orthodont incisors. Specimens from Bonfim, northeastern Bahia, on the other hand, agree with Thomas' albispinus sertonius, from Lamarão, being browner and having broader skulls than P. a. albispinus and having proodont, instead of orthodont, incisors. The range of each of the two subspecies is, therefore, fairly extensive. The insular form extends to the less rainy, continental area and the form from Lamarão ranges northward (NNW) in the same type of highly deciduous forest, the "caatinga."
The species albispinus is certainly the most specialized form of the entire genus for drier habitats. In addition to the general adaptations described above, it is noteworthy for having both lanceolate and clavate aristiforms. The latter type has a wide basal part and an abruptly narrowed, distal part. The same development is seen in the genus Echimys, where highly spinous forms, like Echimys paleacea (Lichtenstein), show the same two types of aristiforms.
Figs. 133, 135. Proechimys albispinus albispinus, male, CNHM no. 20409, Macaco Seco. × 1.
Figs. 134, 136. Proechimys albispinus sertonius, male, MN no. 6454, Bonfim. × 1
Figs. 137, 138. Proechimys albispinus albispinus, male, CNHM no. 20409, Macaco Seco. × 1.
Figs. 139, 140. Proechimys albispinus sertonius, male, MN no. 6454, Bonfim. × 1.
Echimys albispinus Is. Geoffroy Saint-Hilaire, 25 June 1838, Comptes Rendus Acad. Sci., Paris, 6(26):886; Is. Geoffroy, August, 1838, Ann. Sci. Nat., Paris, 10 (ser. 2):125; Is. Geoffroy, 1840, Mag. Zool., Paris (ser. 2, année 2, livr. 13):33, 53, pl. 26, pl. 29 (figs. 1, 2, 3); Allen, 1899, Bull. Amer. Mus. Nat. Hist., 12(20):261.
Echinomys fuliginosus Wagner, 1843, Schreber's Säugethiere, suppl., 3:343.
Echimys albispinosus Waterhouse, Nat. Hist., Mammalia, 2:341.
Proechimys albispinus Allen, 1899, Bull. Amer. Mus. Nat. Hist., 12(20):264; Thomas, 1911, Ann. Mag. Nat. Hist., 8 (ser. 8):252.
Proechimys albispinus albispinus Thomas, 1921, Ann. Mag. Nat. Hist., 8 (ser. 9):141; Tate, 1935, Bull. Amer. Mus. Nat. Hist., 68(5):401; Ellerman, 1940, The families and genera of living rodents, Brit. Mus. (Nat. Hist.), 1:122.
Type locality.—Ilha Madre de Deus, Itaparica (near Salvador), Bahia, Brazil. Type: Museum d'Histoire Naturelle, Paris, no. A 7669, "skull ... practically perfect" (Thomas, 1921:142).
Range.—Island Madre de Deus, Macaco Seco, Andaraí and probably islands of the bay of Todos os Santos and valley of the Paraguassú River.
Diagnosis.—Aristiforms wide; color on setiforms Ochraceous-Tawny on upper parts and sides; incisors orthodont; molariform teeth with one counterfold, p4 rarely with two.
Pelage.—Aristiforms on middorsal region: Lanceolate aristiforms, with basal part whitish, gradually blackening toward tip; total length, 25 to 28 mm; maximum width, 1.2 mm; clavate aristiforms with base whitish, gradually blackening toward tip but interrupted by Ochraceous-Tawny, subapical zone. Aristiforms on outer thighs: Whitish on basal half, gradually blackening toward tip; total length, 24 to 26 mm; maximum width, 0.9 mm. Some are whitish basally, gradually blackening toward distal part but distal fourth or fifth near (15'j) Ochraceous-Tawny. Setiforms on middorsal region: Whitish basally, gradually blackening toward tip but interrupted by Ochraceous-Tawny, subapical zone; total length, 20 to 23 mm; maximum width, 0.1 mm. Some setiforms almost completely whitish. Setiforms on outer thighs: Whitish basally, gradually blackening toward tip but interrupted by near (15'j) Ochraceous-Tawny, subapical zone; total length, 18 to 20 mm; maximum width, 0.06 mm.
Skull.—Narrow; bullae small and smooth; jugals dorso-ventrally wide with conspicuous transverse ridge; postorbital process of zygoma well-developed and formed by jugal and squamosal; posterior palatine foramina obsolete; incisive foramen narrow and short; vomerine sheath complete or incomplete but premaxillary part at a level lower than that of maxillary part (when skull is viewed from ventral face); mesopterygoid fossa extending forward as far as anterior borders of second molars.
Teeth.—Incisors orthodont; molariform teeth with only one counterfold, except that p4 rarely has two counterfolds.
Comparisons.—From P. a. sertonius, albispinus differs in: sides of body darker; incisors orthodont as opposed to proodont; p4 rarely with two counterfolds instead of always with one counterfold.
Remarks.—The localities where P. a. albispinus has been collected have a forest climax with a moderate percentage of deciduous trees.
Specimens examined.—Total number, 19 (18 CMNH, 1 MCZ), from Brazil, Bahia, Andaraí, Macaco Seco.
Proechimys albispinus sertonius Thomas, July, 1921, Ann. Mag. Nat. Hist., 8 (ser. 9):142 (orig. descr.); Tate, 1935, Bull. Amer. Mus. Nat. Hist., 68(5):401; Ellerman, 1940, The families and genera of living rodents, Brit. Mus. (Nat. Hist.), 1:122.
Type locality.—Lamarão ("about 70 miles north of Bahia City"), Ituiutaba, Bahia, Brazil; altitude 300 meters. Type: British Museum (Nat. Hist.), no. 3.9.5.86, adult male; collected on 16 June, 1903, by Alphonse Robert; original number, 1508.
Range.—Known from the type locality and Bonfim; probably occupies valleys of Jacuipe and the Itapicurú rivers and littoral between them.
Diagnosis.—Aristiforms wide; color of setiforms Ochraceous-Tawny on back, grading to Ochraceous-Buff on sides; incisors proodont; no molariform tooth with more than one counterfold.
Pelage.—Aristiforms on middorsal region: Lanceolate aristiforms whitish basally, gradually blackening toward tip; total length, 23 to 27 mm; maximum width, 1.3 mm. Clavate aristiforms, and some lanceolate ones, whitish basally, gradually blackening toward tip but interrupted by Ochraceous-Tawny, subapical zone. Some clavate aristiforms without subapical zone but blackened in distal part; total length, 23 to 24 mm; maximum width, 0.7 mm. Aristiforms on outer thighs: Whitish basally, gradually blackening toward tip but interrupted by Light Ochraceous-Buff, subapical zone; tip slightly darker; some whitish basally, grayish in middle and light yellowish toward tip; total length, 20 to 22 mm; maximum width, 0.9 mm. Setiforms on middorsal region: Whitish basal part succeeded by grayish, then by long, light, yellowish band, which becomes Light Ochraceous-Buff, and blackish tip; total length, 26 to 29 mm; maximum width, 0.15 mm. Setiforms on outer thighs: Whitish basal part succeeded by grayish, then Light Ochraceous-Buff, subapical zone and blackish tip; total length, 18 to 20 mm; maximum width, 0.13 mm.
Skull.—Broad; bullae small and smooth; jugals dorso-ventrally "wide," with conspicuous transverse ridge; postorbital process of zygoma well-developed and formed by both jugal and squamosal; incisive foramen narrow and short; vomerine sheath incomplete or complete but premaxillary part on a lower level than maxillary part (when skull is viewed from ventral face); mesopterygoid fossa extending forward as far as anterior faces of second molars.
Teeth.—Incisors proodont; molariform teeth with only one counterfold.
Comparisons.—Differences from P. a. albispinus are given in the account of that subspecies.
Remarks.—Localities where samples were collected are typical "caatinga" forest, a climax of mainly deciduous trees; cacti are also common in the region.
Specimens examined.—Total number, 10, from Brazil, Bahia, as follows: Ituiutaba, Lamarão, 4 (1 DZ, 1 CMNH, 1 MCZ, 1 USNM); Bonfim, Bonfim, 6 (5 DZ, 1 MN).
Loncheres myosuros Lichtenstein, 1818, Das zoologische Museum der Universität zu Berlin, (2):18 (nomen nudum); Lichtenstein, 1820, Abhandl. K. Akad. Wissensch., Berlin (1818-1819):192, pl. 1, fig. 2 (orig. descr.); Wied, 1826, Beiträge zur Naturgeschichte von Brasilien, 2:445.
Mus leptosoma Brants, 1827, Het geslacht der Muizen door Linnaeus opgesteld ..., Berlyn, p. 150; Lichtenstein, 1830, Darstellung neuer oder wenig bekannter Säugethiere, Berlin, Heft 7, pl. 36, fig. and text pages.
Mus cinnamoneus Lichtenstein, 1830, Darstellung neuer oder wenig bekannter Säugethiere, Berlin, Heft 7, pl. 36.
Echimys myosuros Is. Geoffroy Saint-Hilaire, 1838, Comptes Rendus Acad. Sci., 6(26):886, and 1840, Mag. Zool., Paris (ser. 2, année 2):15, 33, 53; Allen, 1899, Bull. Amer. Mus. Nat. Hist., 12(20):261.
Echinomys leptosoma Wagner, 1843, Schreber's Säugethiere, suppl., 3:341.
Echinomys myosuros Burmeister, 1854, Syst. ubersicht Thiere Brasiliens, p. 199.
Proechimys setosus Thomas, 1921, Ann. Mag. Nat. Hist., 8 (ser. 9):141.
Proechimys myosuros Tate, 1935, Bull. Amer. Mus. Nat. Hist., 68(5):400; Ellerman, 1940, The families and genera of living rodents, Brit. Mus. (Nat. Hist.), 1:119.
Proechimys leptosoma Tate, 1935, Bull. Amer. Mus. Nat. Hist., 68(5): 400; Ellerman, 1940, The families and genera of living rodents, Brit. Mus. (Nat. Hist.), 1:119.
General characters.—Aristiforms wide (1''') and numerous on dorsal parts of body; tail longer (9") than head and body (8"); hind feet short (1"6''').
Color (According to Lichtenstein and Brants' descriptions).—Black between ears; dark brown on middorsal line with reddish tinge on front and upper side of neck; posteriorly from shoulders there is a greasy shine added to color; this dark, dorsal band widens posteriorly, there encroaching on sides of body; sides lighter brown, sparsely marked with dark brown lines, from sides of head caudad to, and including, outer surfaces of hind legs; outer sides of forelegs colored like outer sides of hind legs; ankles ringed with brown. Tail blackish above, whitish below. Upper surfaces of hands and feet white.
Skull.—No description of skull or teeth found.
Remarks.—Thomas (1921:140-143) summarized the available information on the forms from southeastern Brazil and synonymised myosuros with setosus. The description of leptosoma by Brants applies to this same species except in a few features. However, neither Brants nor Lichtenstein described the tail of myosuros as having a white tip or even as having a heavily pencilled tip, although Wagner (1843:342) in redescribing Lichtenstein's species indicated that the tail had a white pencil. He gave also measurements of the head and body, and tail, which do not agree with the original measurements given by either Lichtenstein or Brants. Lichtenstein, in the [Pg 393] original description, gave the type locality of myosuros as Bahia, and stated that it was collected by Freireiss. Brants also gave Bahia as the type locality for leptosoma. The names leptosoma Brants and cinnamoneus Lichtenstein are evidently no more than duplicate names for myosuros, as pointed out by Thomas (1921:141) and subsequent writers. Specimens referred to any of these forms, therefore, could take the earlier name myosuros until identified otherwise. Lichtenstein later (1830, text for plate 36, fig. 2) added São Paulo State to the known range of the species, mentioning specimens collected there by Sello. Probably, therefore, Wagner redescribed leptosoma using a composite sample; the white tip on the tail could occur in any race of P. iheringi from São Paulo.
Considering the short hind feet and the wide aristiforms, Proechimys myosuros probably will eventually prove to be related to albispinus; perhaps it will prove to be a synonym of albispinus.
1. The genus Proechimys is divisible into two subgenera. In all Brazil there are four full species of each subgenus, or 8 species in all. All but one of these are divisible into subspecies of which there are 29, making a total of 30 kinds in Brazil; 14 of these are here newly named.
2. It is new information, I think, that: (1) One main fold extending entirely across the worn crown of the molariform tooth is peculiar to Trinomys; in the subgenus Proechimys, apparent complete division of the crown surface is accomplished by a short main fold meeting a counterfold originating on the opposite side of the tooth; (2) progressive decrease in size of molariform teeth from P4 to M3 is peculiar to the subgenus Trinomys; in the subgenus Proechimys, M2 is largest and the teeth are progressively smaller anteriorly.
3. In the one species, Proechimys albispinus, which has the widest distribution of aristiforms on the body of any species in the genus, some of the aristiforms are clavate. Clavate aristiforms occur in the most spiny species of the related genus Echimys.
4. In subspecies of any one full species the incisive foramen is larger in animals which inhabit arid areas than in those which inhabit humid areas. Possibly increased area of moist mucosa associated with Jacobson's organ is required in arid areas for maintenance of the necessary keenness of smell.
5. The number of counterfolds in the molariform teeth vary in clinal fashion. Their variation is in response to humidity. Increasing humidity is correlated with increasing number of folds, and decreasing humidity is correlated with decreasing number of folds.
6. Clinal variation correlated with increasing humidity is to be seen also in longer tail and darker color of pelage.
7. The primitive Proechimys probably was large, with a short tail, narrow aristiforms, strongly built skull, and five counterfolds in each molariform tooth.
8. Geographic isolation appears to have been a factor in the establishment of the two subgenera; the arid belt along the São Francisco River and northward to Ceará appears to be uninhabited by Proechimys and constitutes a barrier separating the two subgenera, Proechimys and Trinomys.
9. This arid belt probably developed relatively early, since in deposits of late Pleistocene age, remains of the subgenus Trinomys have been found in the area where the subgenus still occurs.
10. The most primitive types occur at the periphery of the range of the genus.
11. Populations from small islands tend to be more primitive than populations on the mainland. Insular populations develop a homozygous condition with resultant disappearance of secondary biotypes.
12. Insular animals ordinarily are larger than their mainland counterpart.
Measurements (in Millimeters) of adults of Proechimys
| Length of head and body | Length of tail | Length of hind foot | Length of ear from notch | Greatest length of skull | Condylo-incisive length | Zygomatic breadth | Length of nasals | Interorbital constriction | Palatilar length | Crown length of cheekteeth | |
|---|---|---|---|---|---|---|---|---|---|---|---|
P. g. steerei, ♂ ♂ Hyutanaham |
|||||||||||
USNM 105535 |
218 |
123 |
48 |
17 |
53.5 |
44.0 |
25.2 |
19.3 |
11.7 |
18.2 |
8.2 |
USNM 105536 |
217 |
135 |
50 |
|
55.2 |
45.3 |
25.7 |
20.7 |
11.4 |
19.2 |
8.0 |
? |
|||||||||||
USNM 105537 |
|
|
|
|
56.32 |
44.9 |
24.0 |
20.4 |
11.4 |
19.3 |
8.7 |
P. g. goeldii, ♂ Fazenda Paraiso |
|||||||||||
AMNH 37489 |
218 |
|
52 |
20 |
55.1 |
44.9 |
27.0 |
22.1 |
12.0 |
18.6 |
9.4 |
♀ |
|||||||||||
AMNH 37488 |
228 |
157 |
49 |
22 |
57.3 |
47.6 |
27.9 |
22.1 |
12.4 |
20.3 |
9.6 |
P. s. liminalis, ♂ ♂ Rio Quichito |
|||||||||||
Mean |
229 |
145 |
43 |
21 |
58.4 |
47.6 |
27.3 |
21.8 |
12.9 |
20.1 |
8.3 |
Maximum |
250 |
|
45 |
24 |
61.7 |
50.0 |
28.9 |
23.0 |
13.8 |
21.2 |
8.8 |
Minimum |
210 |
|
40 |
18 |
53.3 |
44.3 |
25.5 |
19.4 |
12.0 |
18.5 |
7.9 |
No. of specimens |
5 |
1 |
5 |
5 |
5 |
5 |
5 |
5 |
5 |
5 |
5 |
♀ ♀ |
|||||||||||
MN 6253 |
215 |
150 |
43 |
20 |
57.5 |
46.5 |
28.3 |
21.5 |
12.0 |
19.4 |
8.7 |
MN 6250 |
213 |
|
42 |
20 |
60.8 |
49.7 |
|
22.0 |
13.7 |
21.0 |
9.1 |
P. s. amphichoricus, ♂ ♂ Esmeralda |
|||||||||||
AMNH 77000 |
252 |
163 |
53 |
|
60.8 |
50.3 |
27.5 |
24.4 |
13.7 |
20.7 |
9.4 |
AMNH 76994 |
260 |
160 |
54 |
|
|
|
25.5 |
22.8 |
12.6 |
|
9.5 |
AMNH 77020 |
250 |
181 |
57 |
|
62.0 |
52.0 |
27.3 |
25.9 |
13.8 |
21.5 |
9.6 |
♀ |
|||||||||||
AMNH 76999 |
235 |
149 |
50 |
|
|
|
|
24.0 |
|
19.0 |
9.3 |
P. s. kermiti, ♀ Lower Solimões |
|||||||||||
AMNH 37124 |
210 |
|
55 |
|
65.2 |
53.7 |
29.2 |
27.6 |
13.5 |
21.4 |
9.0 |
P. l. brevicauda, ♂ João Pessoa |
|||||||||||
DZ 900 |
245 |
147 |
48 |
|
58.3 |
46.6 |
26.5 |
22.5 |
11.6 |
18.2 |
8.2 |
P. l. boimensis, ♂ Boim |
|||||||||||
MCZ 30881 |
220 |
160 |
50 |
|
54.6 |
44.2 |
24.8 |
21.0 |
11.7 |
17.3 |
7.5 |
♀ |
|||||||||||
MN 1976 |
182 |
140 |
45 |
|
52.6 |
42.2 |
24.3 |
20.8 |
11.4 |
16.8 |
7.6 |
♀ Cametá |
|||||||||||
MCZ 30878 |
240 |
|
48 |
|
55.1 |
45.0 |
25.2 |
22.4 |
11.5 |
18.4 |
7.6 |
P. l. longicaudatus, ♂ Urucum |
|||||||||||
CNHM 26732 |
229 |
121 |
48 |
|
|
|
|
21.5 |
11.5 |
18.5 |
8.3 |
♀ ♀ |
|||||||||||
AMNH 37085 |
210 |
150 |
44 |
|
51.4 |
42.5 |
24.3 |
19.5 |
10.8 |
17.1 |
8.9 |
AMNH 37086 |
210 |
|
50 |
|
48.5 |
40.9 |
23.7 |
17.1 |
10.2 |
17.1 |
8.3 |
| [Pg 396]
P. l. leucomystax, ♂ Tapirapoã |
|||||||||||
AMNH 37509 |
230 |
150 |
43 |
|
50.7 |
42.2 |
24.5 |
18.4 |
10.3 |
17.5 |
8.1 |
♀ |
|||||||||||
AMNH 37510 |
210 |
|
42 |
|
48.1 |
41.3 |
23.0 |
16.9 |
9.9 |
17.2 |
8.1 |
♀ Utiarití |
|||||||||||
MN 2212 |
|
|
|
|
48.0 |
39.9 |
|
18.3 |
11.2 |
15.9 |
8.0 |
♀ Salto Sepotuba |
|||||||||||
MN 1936 |
202 |
147 |
44 |
|
52.6 |
43.0 |
23.6 |
19.2 |
10.7 |
17.8 |
7.8 |
P. l. roberti, ♂ ♂ Anapolis |
|||||||||||
Mean |
208 |
159 |
45 |
21 |
52.7 |
43.5 |
24.9 |
20.7 |
13.1 |
17.2 |
7.9 |
Maximum |
235 |
190 |
55 |
25 |
56.1 |
47.8 |
27.1 |
23.7 |
12.0 |
19.1 |
8.2 |
Minimum |
170 |
135 |
36 |
18 |
48.1 |
40.0 |
22.8 |
18.2 |
10.6 |
15.6 |
7.6 |
No. of specimens |
16 |
14 |
16 |
16 |
11 |
11 |
11 |
11 |
11 |
11 |
11 |
♀ ♀ |
|||||||||||
Mean |
219 |
149 |
44 |
20 |
51.1 |
42.3 |
24.1 |
20.0 |
10.7 |
17.2 |
8.0 |
Maximum |
290 |
155 |
48 |
24 |
55.5 |
45.4 |
25.8 |
21.5 |
11.1 |
17.7 |
8.7 |
Minimum |
195 |
125 |
40 |
18 |
48.9 |
40.3 |
23.1 |
19.1 |
10.5 |
16.6 |
7.7 |
No. of specimens |
10 |
8 |
10 |
10 |
7 |
7 |
7 |
7 |
7 |
7 |
7 |
P. g. villicauda, ♂ ♂ Tapirapoã |
|||||||||||
MN 1932 |
225 |
145 |
47 |
|
55.6 |
45.5 |
26.8 |
24.0 |
12.0 |
18.1 |
8.9 |
MN 1934 |
215 |
162 |
50 |
|
56.2 |
46.0 |
26.1 |
21.3 |
12.0 |
18.6 |
8.4 |
♂ Utiarití |
|||||||||||
AMNH 57544 |
250 |
200 |
55 |
|
|
|
|
24.3 |
13.1 |
19.9 |
9.1 |
P. g. ribeiroi, ♂ Rio 12 de Outubro |
|||||||||||
MN 1935 |
190 |
134 |
47 |
|
50.1 |
41.0 |
24.3 |
20.0 |
11.5 |
15.9 |
8.1 |
P. g. hyleae, ♂ ♂ Tauarí |
|||||||||||
Mean |
248 |
146 |
52 |
|
58.1 |
47.6 |
27.1 |
22.9 |
12.1 |
19.4 |
8.5 |
Maximum |
260 |
174 |
53 |
|
59.0 |
47.8 |
... |
23.4 |
13.2 |
20.0 |
8.8 |
Minimum |
217 |
143 |
51 |
|
57.2 |
46.5 |
... |
22.4 |
11.1 |
18.8 |
8.3 |
No. of specimens |
4 |
3 |
4 |
|
2 |
2 |
1 |
2 |
2 |
2 |
2 |
♀ ♀ |
|||||||||||
Mean |
229 |
149 |
50 |
|
54.3 |
44.9 |
25.8 |
21.0 |
11.8 |
18.1 |
8.5 |
Maximum |
270 |
168 |
54 |
|
56.1 |
46.3 |
27.4 |
23.0 |
13.4 |
19.3 |
9.0 |
Minimum |
190 |
132 |
49 |
|
51.5 |
42.9 |
24.5 |
19.1 |
11.1 |
17.1 |
7.9 |
No. of specimens |
10 |
9 |
10 |
|
9 |
11 |
11 |
10 |
10 |
10 |
10 |
P. g. nesiotes, ♂ Ilha de Manapirí |
|||||||||||
CNHM 19496 |
201 |
133 |
47 |
20 |
52.7 |
42.7 |
25.1 |
19.5 |
11.1 |
17.8 |
8.0 |
♀ |
|||||||||||
MN 1975 |
200 |
152 |
47 |
21 |
52.1 |
42.6 |
25.8 |
19.5 |
12.3 |
18.3 |
8.0 |
| [Pg 397]
P. g. leioprimna, ♀ ♀ Cametá |
|||||||||||
AMNH 37484 |
192 |
151 |
41 |
22 |
54.8 |
44.9 |
25.4 |
19.2 |
12.6 |
18.4 |
8.4 |
CNHM 19503 |
189 |
164 |
47 |
22 |
54.4 |
43.7 |
26.2 |
20.5 |
12.6 |
18.2 |
8.2 |
CNHM 19536 |
189 |
|
43 |
22 |
|
|
|
|
|
|
|
P. g. oris, ♂ Providencia |
|||||||||||
CNHM 19495 |
230 |
170 |
45 |
24 |
56.1 |
47.1 |
25.8 |
22.0 |
11.4 |
17.4 |
8.3 |
♂ Tanaquará |
|||||||||||
MN 1974 |
230 |
175 |
49 |
23 |
53.2 |
43.0 |
23.8 |
20.1 |
10.5 |
17.9 |
7.7 |
♂ Rio Guamá |
|||||||||||
AMNH 37487 |
205 |
142 |
42 |
21 |
|
|
|
|
|
|
|
P. g. arescens, ♂ ♂ Fazenda Inhuma |
|||||||||||
CNHM 26440 |
206 |
149 |
51 |
24 |
54.7 |
44.1 |
26.3 |
21.4 |
12.1 |
18.7 |
8.3 |
CNHM 26441 |
191 |
164 |
51 |
|
55.6 |
45.0 |
25.7 |
22.4 |
11.7 |
18.7 |
8.7 |
P. g. riparum, ♀ Manaus |
|||||||||||
AMNH 143018 |
225 |
|
44 |
20 |
52.6 |
43.2 |
24.0 |
20.5 |
11.0 |
17.2 |
6.7 |
P. g. riparum, ♀ Manaus |
|||||||||||
AMNH 143018 |
225 |
|
44 |
20 |
52.6 |
43.2 |
24.0 |
20.5 |
11.0 |
17.2 |
6.7 |
P. g. arabupu, ♂ ♂ Arabupu |
|||||||||||
AMNH 75816 |
243 |
220 |
56 |
|
59.2 |
48.7 |
27.0 |
23.8 |
12.9 |
17.8 |
8.7 |
AMNH 75819 |
230 |
181 |
52 |
|
55.0 |
46.0 |
26.0 |
22.5 |
12.3 |
16.9 |
8.3 |
AMNH 75815 |
228 |
198 |
52 |
|
|
|
|
|
|
|
|
♀ ♀ |
|||||||||||
AMNH 75810 |
226 |
170 |
48 |
|
53.9 |
45.6 |
25.6 |
21.0 |
12.0 |
16.1 |
8.3 |
AMNH 75823 |
209 |
188 |
48 |
|
53.4 |
43.9 |
24.5 |
21.5 |
11.7 |
16.4 |
8.3 |
AMNH 75817 |
204 |
167 |
47 |
|
51.1 |
43.1 |
... |
21.6 |
11.3 |
16.6 |
8.2 |
P. dimidiatus, ♂ ♂ Pedra Branca |
|||||||||||
Mean |
199 |
170 |
46 |
|
52.4 |
43.5 |
26.2 |
19.5 |
12.1 |
16.4 |
8.3 |
Maximum |
220 |
195 |
50 |
|
56.4 |
47.1 |
27.5 |
21.5 |
13.6 |
18.0 |
8.7 |
Minimum |
180 |
150 |
44 |
|
48.1 |
40.4 |
24.6 |
17.5 |
11.0 |
14.4 |
7.4 |
No. of specimens |
19 |
18 |
19 |
|
45 |
46 |
45 |
45 |
46 |
46 |
46 |
♀ ♀ |
|||||||||||
Mean |
197 |
162 |
44 |
|
51.8 |
42.9 |
25.8 |
19.4 |
11.8 |
16.3 |
8.3 |
Maximum |
230 |
180 |
46 |
|
55.1 |
45.9 |
27.4 |
22.0 |
13.0 |
18.4 |
8.9 |
Minimum |
165 |
145 |
42 |
|
48.6 |
39.5 |
23.8 |
17.6 |
10.7 |
14.8 |
7.7 |
No. of specimens |
14 |
12 |
14 |
|
42 |
44 |
44 |
42 |
44 |
44 |
44 |
P. i. iheringi, ♂ ♂ Ilha de São Sebastião |
|||||||||||
Mean |
207 |
197 |
48 |
|
54.5 |
44.6 |
26.2 |
19.9 |
12.0 |
18.3 |
8.3 |
Maximum |
2207 |
205 |
50 |
|
55.0 |
45.2 |
27.1 |
20.4 |
12.8 |
18.7 |
8.5 |
Minimum |
196 |
190 |
46 |
|
53.5 |
43.7 |
25.9 |
19.3 |
10.9 |
17.5 |
8.0 |
No. of specimens |
5 |
2 |
5 |
|
5 |
5 |
4 |
4 |
5 |
5 |
5 |
♀ ♀ |
|||||||||||
MN 6453 |
228 |
185 |
46 |
|
54.3 |
44.5 |
25.9 |
20.5 |
11.0 |
18.9 |
8.2 |
DZ 2095 |
205 |
180 |
46 |
|
53.2 |
42.9 |
26.4 |
18.9 |
11.1 |
17.0 |
8.2 |
DZ 2525 |
205 |
|
46 |
|
56.9 |
45.8 |
27.8 |
20.9 |
12.5 |
18.9 |
8.2 |
| [Pg 398]
P. i. bonafidei, ♂ ♂ Fazenda Bõa Fé |
|||||||||||
Mean |
211 |
186 |
50 |
25 |
53.3 |
43.4 |
25.6 |
20.2 |
12.1 |
16.3 |
8.5 |
Maximum |
220 |
194 |
54 |
26 |
55.8 |
45.2 |
26.3 |
21.7 |
13.2 |
17.4 |
9.1 |
Minimum |
200 |
176 |
47 |
24 |
50.7 |
41.0 |
24.2 |
19.1 |
11.0 |
14.8 |
8.1 |
No. of specimens |
7 |
5 |
8 |
8 |
4 |
5 |
5 |
7 |
7 |
7 |
7 |
♀ ♀ |
|||||||||||
Mean |
209 |
185 |
52 |
25 |
52.6 |
44.4 |
26.6 |
20.0 |
12.4 |
16.4 |
8.6 |
Maximum |
226 |
203 |
55 |
27 |
56.9 |
46.4 |
28.6 |
21.4 |
13.2 |
17.0 |
9.2 |
Minimum |
185 |
153 |
50 |
22 |
44.4 |
42.3 |
24.9 |
18.0 |
11.4 |
15.5 |
8.2 |
No. of specimens |
7 |
7 |
7 |
7 |
6 |
6 |
7 |
7 |
7 |
7 |
7 |
P. i. gratiosus, ♂ ♂ Floresta da Caixa Dagua |
|||||||||||
Mean |
193 |
191 |
48 |
|
51.2 |
41.4 |
25.5 |
18.5 |
11.7 |
16.3 |
8.0 |
Maximum |
200 |
200 |
49 |
|
51.7 |
42.2 |
27.0 |
18.9 |
12.0 |
16.7 |
8.2 |
Minimum |
185 |
175 |
47 |
|
50.5 |
40.4 |
24.6 |
18.0 |
11.1 |
15.6 |
7.9 |
No. of specimens |
5 |
5 |
5 |
5 |
5 |
5 |
5 |
5 |
5 |
5 |
5 |
♀ ♀ |
|||||||||||
Mean |
204 |
175 |
49 |
24 |
50.5 |
42.1 |
26.0 |
18.4 |
11.4 |
16.3 |
7.9 |
Maximum |
220 |
190 |
50 |
26 |
52.6 |
43.1 |
26.6 |
19.5 |
12.1 |
17.5 |
8.2 |
Minimum |
195 |
160 |
47 |
22 |
48.4 |
41.0 |
25.3 |
17.5 |
10.7 |
15.3 |
7.6 |
No. of specimens |
5 |
2 |
5 |
4 |
4 |
4 |
4 |
4 |
4 |
4 |
4 |
P. i. paratus, ♂ ♂ Floresta da Capela de São Braz |
|||||||||||
MN 4023 |
203 |
195 |
54 |
28 |
51.2 |
41.7 |
25.4 |
18.3 |
10.4 |
16.2 |
8.4 |
MN 5458 |
190 |
170 |
51 |
27 |
|
|
|
|
|
|
|
♀ |
|||||||||||
MN 4012 |
220 |
210 |
54 |
29 |
52.2 |
42.3 |
25.4 |
19.1 |
12.3 |
17.5 |
8.7 |
P. i. panema, ♂ ♂ Campinho |
|||||||||||
MN 8286 |
215 |
|
45 |
23 |
54.0 |
45.1 |
27.7 |
19.5 |
13.4 |
16.4 |
8.1 |
MN 8284 |
195 |
180 |
43 |
23 |
51.5 |
41.8 |
24.5 |
18.1 |
11.4 |
16.0 |
7.6 |
♀ ♀ |
|||||||||||
MN 8288 |
190 |
190 |
46 |
21 |
51.6 |
42.8 |
25.3 |
18.1 |
11.7 |
15.7 |
7.9 |
MN 8287 |
200 |
190 |
46 |
23 |
52.6 |
43.6 |
27.2 |
18.4 |
12.5 |
16.7 |
8.3 |
MN 8285 |
190 |
|
45 |
24 |
50.0 |
41.1 |
26.6 |
19.4 |
12.3 |
16.1 |
8.1 |
P. i. denigratus, ♂ ♂ Mata do Ribeirão da Fortuna |
|||||||||||
Mean |
197 |
218 |
52 |
24 |
51.5 |
42.2 |
25.7 |
18.3 |
11.3 |
16.0 |
8.2 |
Maximum |
217 |
242 |
54 |
28 |
55.4 |
45.3 |
27.0 |
20.3 |
12.4 |
17.6 |
8.5 |
Minimum |
190 |
204 |
50 |
21 |
48.7 |
39.5 |
23.7 |
16.8 |
10.4 |
15.0 |
8.0 |
No. of specimens |
10 |
9 |
9 |
10 |
8 |
8 |
8 |
8 |
8 |
8 |
8 |
♀ ♀ |
|||||||||||
Mean |
201 |
207 |
52 |
24 |
49.1 |
41.2 |
25.0 |
18.3 |
11.2 |
16.0 |
7.8 |
Maximum |
225 |
225 |
54 |
28 |
54.1 |
44.6 |
25.7 |
21.5 |
11.8 |
17.1 |
8.3 |
Minimum |
180 |
175 |
49 |
20 |
48.2 |
39.5 |
23.5 |
17.4 |
10.5 |
15.5 |
7.5 |
No. of specimens |
12 |
12 |
12 |
12 |
8 |
8 |
8 |
8 |
8 |
8 |
7 |
| [Pg 399]
P. s. setosus, ? No locality |
|||||||||||
AMNH 16140 |
|
|
|
|
|
39.8 |
25.2 |
|
11.0 |
15.9 |
7.6 |
P. s. elegans, ♂ Lagoa Santa |
|||||||||||
UZM H82 |
190 |
190 |
47 |
25 |
|
|
24.3 |
17.5 |
11.2 |
16.6 |
7.7 |
? |
|||||||||||
UZM L104 |
|
|
|
|
48.6 |
40.8 |
24.8 |
16.6 |
10.3 |
15.9 |
7.7 |
P. a. albispinus, ♂ ♂ Macaco Seco |
|||||||||||
Mean |
175 |
162 |
40 |
|
45.9 |
39.7 |
23.6 |
15.6 |
10.6 |
15.7 |
7.6 |
Maximum |
193 |
173 |
47 |
|
48.2 |
42.0 |
24.9 |
17.1 |
11.6 |
17.5 |
8.2 |
Minimum |
165 |
147 |
38 |
|
44.2 |
38.1 |
22.7 |
14.8 |
9.7 |
14.3 |
7.2 |
No. of specimens |
9 |
8 |
9 |
|
11 |
12 |
12 |
14 |
13 |
14 |
13 |
♀ ♀ |
|||||||||||
CNHM 20402 |
187 |
171 |
40 |
|
46.0 |
40.8 |
23.8 |
14.9 |
11.3 |
16.0 |
7.4 |
CNHM 20408 |
|
|
|
|
47.6 |
40.7 |
24.6 |
17.1 |
11.2 |
15.9 |
8.2 |
P. a. sertonius, ♂ Lamarão |
|||||||||||
CNHM 18196 |
190 |
160 |
36 |
25 |
|
|
|
|
11.7 |
|
|
♂ Bonfim |
|||||||||||
MN 6454 |
185 |
175 |
37 |
|
45.7 |
39.8 |
24.2 |
15.5 |
10.7 |
16.1 |
7.5 |
♀ ♀ |
|||||||||||
DZ 2636 |
190 |
150 |
35 |
|
43.8 |
37.6 |
22.8 |
14.7 |
9.6 |
14.5 |
|
DZ 2635 |
175 |
155 |
38 |
|
46.7 |
40.6 |
23.3 |
16.9 |
10.2 |
15.1 |
7.4 |
Allen, J. A. 1904. Mammals from the District of Santa Marta, Colombia, collected by Mr. Herbert H. Smith, with field notes by Mr. Smith. Bull. Amer. Mus. Nat. Hist., 20:407-468, November 28, 1904. 1916. Mammals collected on the Roosevelt Brazilian Expedition, with field notes by Leo E. Miller. Bull. Amer. Mus. Nat. Hist., 35:559-610, August 9, 1916. Ameghino, F. 1934. Notas sobre una pequéna coleccion de huesos de mamiferos, procedentes de las grutas calcáreas de Iporanga en el Estado São Paulo (Brasil), Obras completas y Correspondencia Cientifica de Florentino Ameghino, La Plata, 17:103-153. Berry, E. W. 1942. Mesozoic and Cenozoic plants of South America, Central America and the Antilles. Proc. Eighth Amer. Sci. Congress, 4:365-373. Desmarest, A. G. 1817. Nouveau Dictionaire d'Histoire Naturelle, ed. 2, 10:59. Ellerman, J. R. 1940. The families and genera of living Rodents. Vol. 1, Rodents other than Muridae, pp. xxvi + 689, 189 text figs. British Museum (Natural History). June 8, 1940. Geoffroy Saint-Hilaire, Isidore 1840. Notice sur les rongeurs épineux, désignés par lés auteurs sous les noms d'Echimys, Loncheres Heteromys et Nelomys. Mag. Zool. (Guerin-Meneville), s. 2, Ann. 2, pp. 1-57, pls. 20-29. Lichtenstein, M. H. C. 1830. Darstellung neuer oder wenig bekannter Säugethiere, 65 Arten, etc., pp. 118, 50 pls. colored. Lund, P. W. 1841. Blik paa Brasiliens Dyreverden för sidste Jordomvaeltning, af Dr. Lund. Kongelige Danske Videnskabernes Selskabs naturvidenskabelige og mathematiske. Fortsaettelse af Pattedyrene. Kjöbenhavn, 2:217-272, pls. 14-24. Oliveira, A. I. and Leonardos, O. H. 1943. Geologia do Brasil. Min. da Agricultura. Rio de Janeiro, 26 + 813, 202 figs., photographs, 1 map. Osgood, Wilfred H. 1944. Nine new South American rodents. Zool. Ser. Field Mus. Nat. Hist., 29:191-204, July 12, 1944. Ribeiro, Alipio de Miranda 1914. Historia Natural. Zoologia. Mammiferos. Commissão de Linhas Telegraficas, Estrategicas de Matto Grosso ao Amazonas. Annexo no. 5, Rio de Janeiro, pp. 1-49 + 1-3, pls. 25, May, 1914. [Pg 401] Ridgway, Robert 1912. Color standards and color nomenclature, iv + 44 pp., 53 pls. Published by the author, Washington, D. C. Tate, G. H. H. 1935. The taxonomy of the genera of Neotropical hystricoid rodents. Bull. Amer. Mus. Nat. Hist., 68:295-447, June 12, 1935. 1939. The mammals of the Guiana Region. Bull. Amer. Mus. Nat. Hist., 76:151-229, October 20, 1939. Thomas, Oldfield 1900. Descriptions of new rodents from western South America. Ann. and Mag. Nat. Hist., 6(ser. 7):294-302, September, 1900. 1905. New Neotropical Molossus, Conepatus, Nectomys, Proechimys, and Agouti, with a note on the genus Mesomys. Ann. and Mag. Nat. Hist., 15(ser. 7):584-591, June, 1905. 1912. On small mammals from the Lower Amazon. Ann. and Mag. Nat. Hist., 9(ser. 8):84-90, January, 1912. 1920. On mammals from the Lower Amazons in the Goeldi Museum, Pará. Ann. and Mag. Nat. Hist., 6(ser. 9):266-283, September, 1920. 1921. On the spiny rats of the Proechimys group from Southeastern Brazil. Ann. and Mag. Nat. Hist., 8(ser. 9):140-143, July, 1921. Wagner, Andreas 1843. Beschreibung einiger neuer Nager, welche auf der Reise des Herrn Hofrats v. Schubert gesammelt wurden, mit Bezugnahme auf einige andere verwandte Formen. Abhandl. Akad. Wiss. math.-phys. Cl., 3(4):173-216, pl. 2. Wagner, J. A. 1844. (1774-1846). In Schreber's Die Säugethiere ..., 7 pts. including text and pls., colored (347). Wied, Maximilian zu 1826. Beiträge zur Naturgeschichte von Brazilien, vol. 2, Mammalia, 600 pp., 6 pls., Weimar. Winge, Herluf 1888. 1888. Jordfundne og nulevende Gnavere (Rodentia) fra Lagoa Santa, Minas Geraes, Brasilien. E Museo Lundii Afhandlinger, 1(3):1-178, pls. 1-8. 1941. The interrelationships of the mammalian genera. Translated from Danish by E. Deichmarm and G. M. Allen. København, 3 vols. (vol. 1, xii + 418, 3 pls., 1941; vol. 2, 376 pp., 1941; vol. 3, 308 pp., 1942.
Transmitted December 1, 1947.
22-3343