University of Kansas Publications
Museum of Natural History
Vol. 12, No. 9, pp. 363-474, 20 figs.
October 25, 1963
Variation in the Muscles and Nerves
of the Leg in Two Genera of Grouse
(Tympanuchus and Pedioecetes)
BY
E. BRUCE HOLMES
University of Kansas
Lawrence
1963
UNIVERSITY OF KANSAS PUBLICATIONS
MUSEUM OF NATURAL HISTORY
Institutional libraries interested in publications exchange may obtain this series by addressing the Exchange Librarian, University of Kansas Library, Lawrence, Kansas. Copies for individuals, persons working in a particular field of study, may be obtained by addressing instead the Museum of Natural History, University of Kansas, Lawrence, Kansas. There is no provision for sale of this series by the University Library, which meets institutional requests, or by the Museum of Natural History, which meets the requests of individuals. Nevertheless, when individuals request copies from the Museum, 25 cents should be included, for each separate number that is 100 pages or more in length, for the purpose of defraying the costs of wrapping and mailing.
* An asterisk designates those numbers of which the Museum's supply (not the Library's supply) is exhausted. Numbers published to date, in this series, are as follows:
| Vol. | 1. | Nos. 1-26 and index. Pp. 1-638, 1946-1950. | ||
|---|---|---|---|---|
| * | Vol. | 2. | (Complete) Mammals of Washington. By Walter W. Dalquest. Pp. 1-444, 140 figures in text. April 9, 1948. | |
| Vol. | 3. | *1. | The avifauna of Micronesia, its origin, evolution, and distribution. By Rollin H. Baker. Pp. 1-359, 16 figures in text. June 12, 1951. | |
| *2. | A quantitative study of the nocturnal migration of birds. By George H. Lowery, Jr. Pp. 361-472, 47 figures in text. June 29, 1951. | |||
| 3. | Phylogeny of the waxwings and allied birds. By M. Dale Arvey. Pp. 473-530, 49 figures in text, 13 tables. October 10, 1951. | |||
| *4. | Birds from the state of Veracruz, Mexico. By George H. Lowery, Jr., and Walter W. Dalquest. Pp. 531-649, 7 figures in text, 2 tables. October 10, 1951. | |||
| Index. Pp. 651-681. | ||||
| * | Vol. | 4. | (Complete) American weasels. By E. Raymond Hall. Pp. 1-466, 41 plates, 31 figures in text. December 27, 1951. | |
| Vol. | 5. | Nos. 1-37 and index. Pp. 1-676, 1951-1953. | ||
| * | Vol. | 6. | (Complete) Mammals of Utah, taxonomy and distribution. By Stephen D. Durrant. Pp. 1-549, 91 figures in text, 30 tables. August 10, 1952. | |
| Vol. | 7. | Nos. 1-15 and index. Pp. 1-651, 1952-1955. | ||
| Vol. | 8. | Nos. 1-10 and index. Pp. 1-675, 1954-1956. | ||
| Vol. | 9. | *1. | Speciation of the wandering shrew. By James S. Findley. Pp. 1-68, 18 figures in text. December 10, 1955. | |
| 2. | Additional records and extension of ranges of mammals from Utah. By tephen D. Durrant, M. Raymond Lee, and Richard M. Hansen, Pp. 69-80. December 10, 1955. | |||
| 3. | A new long-eared myotis (Myotis evotis) from northeastern Mexico. By Rollin H. Baker and Howard J. Stains. Pp. 81-84. December 10, 1955. | |||
| 4. | Subspeciation in the meadow mouse, Microtus pennsylvanicus, in Wyoming. By Sydney Anderson. Pp. 85-104, 2 figures in text. May 10, 1956. | |||
| 5. | The condylarth genus Ellipsodon. By Robert W. Wilson. Pp. 105-116, 6 figures in text. May 19, 1956. | |||
| 6. | Additional remains of the multituberculate genus Eucosmodon. By Robert W. Wilson. Pp. 117-123, 10 figures in text. May 19, 1956. | |||
| 7. | Mammals of Coahulia, Mexico. By Rollin H. Baker. Pp. 125-335, 75 figures in text. June 15, 1956. | |||
| 8. | Comments on the taxonomic status of Apodemus peninsulae, with description of a new subspecies from North China. By J. Knox Jones, Jr. Pp. 337-346, 1 figure in text, 1 table. August 15, 1956.. | |||
| 9. | Extensions of known ranges of Mexican bats. By Sydney Anderson. Pp. 347-351. August 15, 1956. | |||
| 10. | A new bat (Genus Leptonycteris) from Coahulia. By Howard J. Stains. Pp. 353-356. January 21, 1957. | |||
| 11. | A new species of pocket gopher (Genus Pappogeomys) from Jalisco, Mexico. By Robert J. Russell. Pp. 357-361. January 21, 1957. | |||
| 12. | Geographic variation in the pocket gopher, Thomomys bottae, in Colorado. By Phillip M. Youngman. Pp. 363-384, 7 figures in text. February 21, 1958. | |||
| 13. | New bog lemming (genus Synaptomys) from Nebraska. By J. Knox Jones, Jr. Pp. 385-388. May 12, 1958. | |||
| 14. | Pleistocene bats from San Josecito Cave, Nuevo León, México. By J. Knox Jones, Jr. Pp. 389-396. December 19, 1958. | |||
| 15. | New subspecies of the rodent Baiomys from Central America. By Robert L. Packard. Pp. 397-404. December 19, 1958. | |||
| 16. | Mammals of the Grand Mesa, Colorado. By Sydney Anderson. Pp. 405-414, 1 figure in text. May 20, 1959. | |||
| 17. | Distribution, variation, and relationships of the montane vole, Microtus montanus. By Sydney Anderson. Pp. 415-511, 12 figures in text, 2 tables. August 1, 1959. | |||
| (Continued on inside of back cover) | ||||
University of Kansas Publications
Museum of Natural History
Vol. 12, No. 9, pp. 363-474, 20 figs.
October 25, 1963
Variation in the Muscles and Nerves
of the Leg in Two Genera of Grouse
(Tympanuchus and Pedioecetes)
BY
E. BRUCE HOLMES
University of Kansas
Lawrence
1963
University of Kansas Publications, Museum of Natural History
Editors: E. Raymond Hall, Chairman, Henry S. Fitch,
Theodore H. Eaton, Jr.
Volume 12, No. 9, pp. 363-474, 20 figs.
Published October 25, 1963
University of Kansas
Lawrence, Kansas
PRINTED BY
JEAN M. NEIBARGER, STATE PRINTER
TOPEKA, KANSAS
1963
29-5835
BY
E. BRUCE HOLMES
| PAGE | |
| Introduction | 367 |
| Materials and Methods | 368 |
| Terminology | 369 |
| Acknowledgments | 375 |
| Skeleton | 375 |
| Nerves | 376 |
| Lumbosacral Plexus | 376 |
| Femoral Nerve | 377 |
| Obturator Nerve | 379 |
| Sciatic Nerve | 379 |
| Peroneal Nerve | 382 |
| Tibial Nerve | 384 |
| Muscles | 396 |
| M. Extensor Iliotibialis Lateralis | 398 |
| M. Extensor Iliotibialis Anticus | 405 |
| M. Ambiens | 408 |
| M. Vastus Lateralis | 408 |
| M. Vastus Medialis | 410 |
| M. Piriformis | 412 |
| M. Gluteus Profundus | 413 |
| M. Iliacus | 414 |
| M. Iliotrochantericus Medius | 415 |
| M. Psoas | 416 |
| M. Flexor Cruris Lateralis | 416 |
| M. Flexor Cruris Medialis | 417 |
| M. Caudofemoralis | 418 |
| M. Flexor Ischiofemorali | 420 |
| M. Adductor Superficialis | 420 |
| M. Adductor Profundus | 421 |
| [Pg 366]M. Obturator | 422 |
| M. Femorocruralis | 425 |
| M. Gastrocnemius | 426 |
| M. Flexor Perforans et Perforatus Digiti II | 427 |
| M. Flexor Perforans et Perforatus Digiti III | 429 |
| M. Flexor Perforatus Digiti IV | 430 |
| M. Flexor Perforatus Digiti III | 432 |
| M. Flexor Perforatus Digiti II | 433 |
| M. Flexor Hallucis Longus | 435 |
| M. Plantaris | 435 |
| M. Flexor Digitorum Longus | 436 |
| M. Popliteus | 438 |
| M. Peroneus Longus | 438 |
| M. Tibialis Anticus | 439 |
| M. Extensor Digitorum Longus | 440 |
| M. Peroneus Brevis | 441 |
| M. Extensor Hallucis Longus | 442 |
| M. Abductor Digiti II | 443 |
| M. Extensor Brevis Digiti III | 444 |
| M. Extensor Proprius Digiti III | 444 |
| M. Extensor Brevis Digiti IV | 445 |
| M. Lumbricalis | 445 |
| M. Abductor Digiti IV | 446 |
| M. Flexor Hallucis Brevis | 446 |
| Discussion and Conclusions | 446 |
| Analysis of Individual Variation | 446 |
| Muscles | 447 |
| Nerves | 449 |
| Analysis of Variation Between Species | 451 |
| Comparison with Other Studies of Innervation | 452 |
| Summary | 457 |
| Literature Cited | 473 |
| PAGE | ||
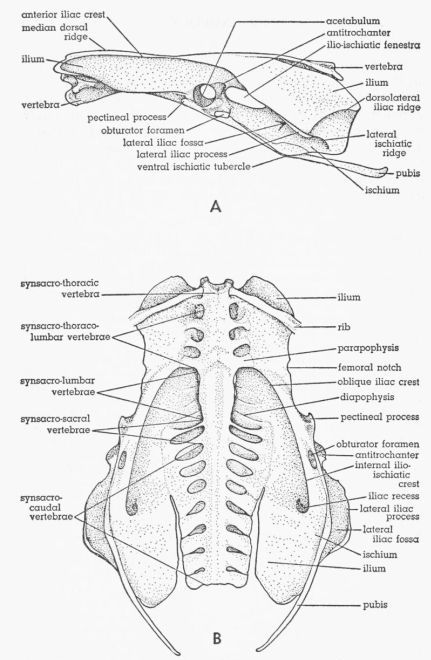
| Fig. 1. | Pelvis of Tympanuchus pallidicinctus. A. Lateral view. × 1. B. Ventral view. × 1⅛. | 370 |
| Fig. 2. | Ventral views of the lumbosacral plexus of Tympanuchus pallidicinctus. Sympathetic ganglionated chain removed. Numbers indicate synsacral spinal nerves. × 2. A. T.p. 1L. B. T.p. 2L. | 386 |
| Fig. 3. | Ventral views of the lumbosacral plexus. Sympathetic ganglionated chain removed. Numbers indicate synsacral spinal nerves. × 2. A. Tympanuchus cupido pinnatus 3L. B. Pedioecetes phasianellus jamesi 4L. | 387 |
| Fig. 4. | Semidiagrammatic ventral views of the femoral nerve, showing the distribution of the branches. × 3. 1,2, M. extensor iliotibialis anticus; 3, cutaneous; 4-6, M. extensor iliotibialis lateralis; 7,8, M. iliacus; 9, M. gluteus profundus; 10-12, fused Mm. vastus lateralis and vastus medialis; 13,14, M. vastus medialis; 15, M. ambiens; 16, M. femoritibialis internus; 17, nonmuscular; 18, M. psoas; 19, M. iliotrochantericus medius. A. Tympanuchus cupido pinnatus 3L. B. Pedioecetes phasianellus jamesi 3L. | 388 |
| Fig. 5. | Semidiagrammatic ventral views of the femoral nerve, showing the distribution of the branches. × 3. 1,2, M. extensor iliotibialis anticus; 3, cutaneous; 5,6, M. extensor iliotibialis lateralis; 7,8, M. iliacus; 9, M. gluteus profundus; 10,11, fused Mm. vastus lateralis and vastus medialis; 13, M. vastus medialis; 15, M. ambiens; 16, M. femoritibialis internus; 17, nonmuscular; 18, M. psoas; 19, M. iliotrochantericus medius. A. Tympanuchus pallidicinctus 2L. B. Tympanuchus cupido attwateri 1R | 389 |
| Fig. 6. | Semidiagrammatic dorsolateral view of the sciatic nerve of Pedioecetes phasianellus jamesi 3R, showing the distribution of the branches. × 2½. 1, M. gluteus profundus; 2, M. piriformis; 3, M. extensor iliotibialis lateralis; 4-7, M. extensor iliofibularis; 8, M. flexor cruris medialis; 9, cutaneous; 10, to pudendal plexus; 11, M. flexor cruris lateralis; 12, M. caudofemoralis pars caudifemoralis; 13-15, M. caudofemoralis pars iliofemoralis; 16,17, M. flexor ischiofemoralis; 18,19, M. femorocruralis (branch of tibial nerve); 20, cutaneous; 21, M. gastrocnemius pars media (branch of tibial nerve); 22, cutaneous. | 390 |
| Fig. 7. | Semidiagrammatic dorsolateral view of the sciatic nerve of Tympanuchus pallidicinctus 2L, showing the distribution of the branches. × 2½. 1, M. gluteus profundus; 2, M. piriformis; 3, M. extensor iliotibialis lateralis; 4, 7, M. extensor iliofibularis; 8, M. flexor cruris medialis; 9, cutaneous; 10, to pudendal plexus; 11, M. flexor cruris lateralis; 12, M. caudofemoralis pars caudifemoralis; 13-15, M. caudofemoralis pars iliofemoralis; 17, M. flexor ischiofemoralis; 18, M. femorocruralis (branch of tibial nerve); 22, cutaneous; 23, nonmuscular (branch of peroneal nerve). | 391 |
| Fig. 8. | Semidiagrammatic dorsolateral view of the sciatic nerve of Tympanuchus cupido pinnatus 3L, showing the distribution of the branches. × 2½. 1, M. gluteus profundus; 2, M. piriformis; 3, M. extensor iliotibialis lateralis; 4,7, M. extensor iliofibularis; 8, M. flexor cruris medialis; 9, cutaneous; 11, M. flexor cruris lateralis; 12, M. caudofemoralis pars caudifemoralis; 13, M. caudofemoralis pars iliofemoralis; 17, M. flexor ischiofemoralis; 18, M. femorocruralis (branch of tibial nerve); 20, cutaneous; 22, cutaneous. | 392 |
| Fig. 9. | Semidiagrammatic dorsolateral view of the sciatic nerve of Pedioecetes phasianellus jamesi 3L, showing the distribution of the branches. × 2½. 1, M. gluteus profundus; 2, M. piriformis; 3, M. extensor iliotibialis lateralis; 4,5,7, M. extensor iliofibularis; 8, M. flexor cruris medialis; 9, cutaneous; 11, M. flexor cruris lateralis; 13,14, M. caudofemoralis pars iliofemoralis; 16,17, M. flexor ischiofemoralis; 18,19, M. femorocruralis (branch of tibial nerve); 20, cutaneous; 22, cutaneous. | 393 |
| Fig. 10. | A,B. Semidiagrammatic drawings of the peroneal nerve of Tympanuchus pallidicinctus 1L, showing the distribution of the branches. × 2. C. Semidiagrammatic drawing of the distal part of the peroneal nerve of Tympanuchus cupido attwateri 1R, showing the distribution of the branches. × 2. 1,2, M. tibialis anticus (tibial head); 3,4, M. tibialis anticus (femoral head); 5, M. extensor digitorum longus; 6, nonmuscular; 7,8, M. peroneus longus; 9, M. peroneus brevis; 10,11, M. extensor hallucis longus (proximal head); 12, M. extensor hallucis longus (distal head); 13-15, nonmuscular (to toes); 16, M. abductor digiti II; 17, M. extensor brevis digiti III; 18, M. extensor brevis digiti IV. | 394 |
| Fig. 11. | A,B. Semidiagrammatic drawings of the tibial nerve (excluding the paraperoneal branch) of Tympanuchus pallidicinctus, showing the distribution of the branches. × 2. A. T.p. 1L. B. T.p. 3R. C. Semidiagrammatic drawing of the distal part of the paraperoneal branch of the tibial nerve of Pedioecetes phasianellus jamesi 2L, showing the distribution of the branches. × 2. 1, M. femorocruralis; 2, M. gastrocnemius pars media; 3, M. popliteus; 4, M. plantaris; 5, M. flexor digitorum longus; 6-8, nonmuscular; 9-11, M. gastrocnemius pars interna; 12,13, M. flexor hallucis longus; 14-16, M. flexor perforatus digiti IV (medial head); 17, M. flexor perforatus digiti III (medial head); 18-20, M. flexor perforatus digiti II; 21, M. flexor perforatus digiti IV (lateral head); 22-24, M. flexor perforatus digiti IV (anterolateral head); 25, M. flexor perforatus digiti III (anterolateral head); 26, M. flexor perforans et perforatus digiti III; 27,28, M. flexor perforans et perforatus digiti II; 29, M. gastrocnemius pars externa; 30,31, M. abductor digiti IV; 32,33, M. flexor hallucis brevis; 34,35, nonmuscular (to toes). | 395 |
| Fig. 12. | Tympanuchus pallidicinctus 2L. Lateral view of the superficial muscles of the left leg. × 1. | 397 |
| Fig. 13. | Tympanuchus pallidicinctus 2L. Medial view of the superficial muscles of the left leg. × 1. Articular capsule shown by concentrically arranged dashes. | 398 |
| Fig. 14. | Tympanuchus pallidicinctus 2L. Lateral view of the muscles of the left leg. The following muscles have been removed: extensor iliotibialis lateralis, extensor iliotibialis anticus, gastrocnemius pars externa and pars interna, and peroneus longus. × 1. | 399 |
| Fig. 15. | Tympanuchus pallidicinctus 2L. Medial view of the muscles of the left leg. The following muscles have been removed: extensor iliotibialis lateralis, extensor iliotibialis anticus, ambiens, flexor cruris lateralis (in part), flexor cruris medialis (in part), gastrocnemius pars externa and pars interna, and peroneus longus. × 1. | 400 |
| Fig. 16. | Tympanuchus pallidicinctus 2L. Lateral view of the muscles of the left leg. The following muscles, in addition to those listed for Fig. 14, have been removed: ambiens, vastus lateralis pars lateralis, vastus medialis (except for part of patellar tendon), extensor iliofibularis, flexor cruris lateralis (in part), flexor perforans et perforatus digiti II, and flexor perforans et perforatus digiti III. × 1. | 401 |
| Fig. 17. | Tympanuchus pallidicinctus 2L. Lateral view of the muscles of the left leg. The following muscles, in addition to those listed for Fig. 16, have been removed: vastus lateralis pars postica, gluteus profundus, flexor cruris medialis (in part), caudofemoralis, flexor perforatus digiti IV, and tibialis anticus. × 1. | 402 |
| Fig. 18. | Tympanuchus pallidicinctus 2L. Lateral view of the muscles of the left leg. The following muscles, in addition to those listed for Fig. 17, have been removed: patellar tendon, iliacus, iliotrochantericus medius, flexor cruris lateralis, flexor cruris medialis, flexor ischiofemoralis, adductor superficialis, femorocruralis, gastrocnemius pars media, flexor perforatus digiti III, flexor perforatus digiti II, flexor hallucis longus, plantaris, flexor digitorum longus, popliteus, and extensor digitorum longus. × 1. | 403 |
| Fig. 19. | Tympanuchus pallidicinctus 2L. A. Posterior view of the muscles of the left shank. The following shank muscles, in addition to those listed for Fig. 17, have been removed: gastrocnemius pars media, flexor perforatus digiti III, and flexor perforatus digiti II. × 1. B. Posterior view of the proximal end of the shank, showing the most deeply situated muscle. × 1. C. Lateral view of the head of the left femur and the middle part of the pelvis, showing the deepest part of M. obturator. × 1. D. Medial view of the posteroventral part of the left side of the pelvis, showing the intrapelvic part of M. obturator. × 1. E. Anterior view of the left tarsometatarsus, showing the dorsal intrinsic muscles of the foot. × 1½. F. Posterior view of the left tarsometatarsus, showing the ventral intrinsic muscles of the foot. × 1½. | 404 |
| Fig. 20. |
A-D. Dorsal views of M. iliotrochantericus
medius, showing its relationship to femoral notch. × 1. In D,
note absence of femoral notch and location of branch of femoral
nerve. A. Tympanuchus pallidicinctus 2L. B. T. cupido
pinnatus 4L. C. Pedioecetes phasianellus jamesi 1L.
D. T. pallidicinctus 3L. E. Medial view of distal end of M. flexor cruris medialis of P. p. jamesi 4L. × 1. Part of insertion is covered by medial collateral ligament. F,G. Lateral views of posteroproximal corner of M. extensor iliotibialis lateralis (removed from specimen). × 1. F. T. pallidicinctus 2L. G. P. p. jamesi 3L. H,I. Dorsolateral views of M. piriformis. × 1. H. P. p. jamesi 1L. I. T. cupido attwateri 1L. J. Lateral view of M. caudofemoralis pars caudifemoralis (removed from specimen) of T. c. pinnatus 4L. × 1. K. Lateral view of extrapelvic part of M. obturator of T. pallidicinctus 3L (bones not shown). × 2. L,M. Region surrounding obturator foramen of T. pallidicinctus 3L, showing points of attachment of three parts of M. obturator (muscles removed). × 3. L. Lateral view. M. Medial view. N. Anterior view of left tarsometatarsus of P. p. jamesi 4L, showing dorsal intrinsic muscles of foot. × 1½. Tendon of M. extensor digitorum longus has been removed. |
406 |
The purposes of this study were: (1) to obtain information on individual variation in the anatomy of the muscles and nerves of the leg of Tympanuchus cupido pinnatus (Greater Prairie Chicken), T. c. attwateri (Attwater's Prairie Chicken), T. pallidicinctus (Lesser Prairie Chicken), and Pedioecetes phasianellus jamesi (Sharp-tailed Grouse); (2) to determine whether or not the two species of the genus Tympanuchus differ constantly in the myology of the leg; and (3) to determine what constant differences in the myology of the leg exist between the two closely related genera Tympanuchus and Pedioecetes.
These particular birds were chosen because they are closely related, and closely resemble one another in habitats occupied and in patterns of behavior. It was desired to study examples that showed as few adaptive differences as possible among the grouse. Series of each of the three species of grouse were readily obtainable, making it possible to draw comparisons at the level of individuals, subspecies, species, and genera.
The study here reported on was begun in the spring of 1957 and was completed in the autumn of 1961.
Prior work on the muscles of the leg of birds has been reviewed by Hudson (1937) and Hudson, et al. (1959). Only papers dealing with the innervation of the leg in birds are reviewed below.
DeMan (1873) treated the nerves of Paradisea papuana, Corvus monedula, and the chicken; he also commented briefly on a few other species. Jhering (Ihering, 1873) briefly described the lumbosacral plexus in approximately a dozen birds, but illustrated only two. Gadow (1880) described the nerves in Struthio, Rhea, and Casuarius; his paper contains some excellent illustrations of nerves. Unfortunately, the text is marred by numerous confusing typographical errors. Carlsson (1884) described the nerves of Eudyptes chrysolopha, Alca torda, Mergulus alle, and Mormon arcticus. Gadow (1891) described the nerves in a study that included a large variety of birds, but published few illustrations. DuToit (1913) described the lumbosacral plexus of the chicken. Romer (1927) gave the innervation of the hip and thigh muscles in the chicken, but did not cover the lumbosacral plexus. Appleton (1928) gave the innervation, in various birds, only of those muscles of the hip and thigh that are supplied by the tibial and peroneal nerves; he did not include the lumbosacral plexus. Sudilovskaya (1931) described the nerves of Struthio, Rhea, and Dromaeus (Dromiceius). Unfortunately, his illustrations are almost useless as far as the nerves are concerned. Boas (1933) described the lumbosacral plexus in a large number of birds. His extensive account includes numerous good illustrations. Howell (1938) listed the innervation of the hip and thigh muscles in the chicken; he did not include the lumbosacral plexus. Fisher (1946) listed the innervation of the muscles of vultures, but did not include the lumbosacral plexus. Wilcox (1948) gave the innervation of the muscles [Pg 368] of Gavia immer, but did not include the lumbosacral plexus. Fisher and Goodman (1955) described the nerves in the Whooping Crane. Papers by Chomiak (1950) and Yasuda, et al. (1959), both dealing with the chicken, were not examined.
Complete dissections of the muscles and nerves were made in eight legs (of five specimens) of the Lesser Prairie Chicken (Tympanuchus pallidicinctus), six legs (of four specimens) of the Greater Prairie Chicken (T. cupido pinnatus), three legs (of two specimens) of Attwater's Prairie Chicken (T. cupido attwateri), and six legs (of four specimens) of the Sharp-tailed Grouse (Pedioecetes phasianellus jamesi).
For convenience and simplicity of reference, each specimen has been designated by a symbol consisting of the first letter of the genus and of the species (and also of the subspecies in T. cupido) plus a number. The letter "L" or "R" is added to indicate the left or right leg. Thus the symbol T.p. 1L refers to the left leg of specimen number one of T. pallidicinctus.
All specimens are in the University of Kansas Museum of Natural History. The catalogue number of each specimen, and the legs of it that were dissected, are listed below.
| T.p. 1L,R | KU38520 | T.c.p. 4L | KU38518 | |
| T.p. 2L,R | KU38521 | T.c.a. 1L,R | KU36617 | |
| T.p. 3L,R | KU38522 | T.c.a. 2L | KU36618 | |
| T.p. 4L | KU38523 | P.p. 1L,R | KU38526 | |
| T.p. 5R | KU38527 | P.p. 2L | KU38518 | |
| T.c.p. 1L,R | KU38515 | P.p. 3L,R | KU38528 | |
| T.c.p. 2L,R | KU38516 | P.p. 4L | KU38529 | |
| T.c.p. 3L | KU38517 |
The specimens were injected in the field either with formalin (10%) or embalming fluid, except for those of T. c. attwateri, which were frozen; the latter were later injected with embalming fluid. Injection in all the birds was by hypodermic syringe into all major muscle masses, into the body cavities, and subcutaneously in the neck, wings, and feet. In those specimens injected with embalming fluid, the body cavities were injected with formalin. The embalming fluid consisted of 70 per cent alcohol, glycerin (or propylene glycol), and formalin (full strength) in the approximate ratio of 78:20:2, respectively. This fluid gave good preservation; these specimens had the advantages of lacking almost entirely the irritating odor of formalin and of having pliable tissues. The skin of those specimens originally injected with formalin was slit in several places and they were transferred to crocks containing embalming fluid (without the formalin). After a period of many weeks, with two changes of fluid, most of the formalin odor was eliminated and the muscles were sufficiently pliable to be easily dissected. All specimens were kept in containers filled with embalming fluid. No mold ever appeared, even though no phenol or other chemical was added.
To facilitate comparison, two or three specimens were frequently dissected simultaneously. The nerves and smaller muscles were dissected with the aid of a stereoscopic microscope mounted on a long movable arm. In order satisfactorily to expose the lumbosacral plexus the posterior half of the sternum and pectoral muscles, as well as the abdominal viscera, were removed.
To insure more nearly accurate proportions, drawings of the pelvis and of some of the muscles were made with the aid of photographs of the several specimens listed above.
Skeleton
The majority of the osteological terms used in the present paper are those used by Howard (1929); however, many skeletal features are not named by Howard. Since names for most of these parts were not found in the other literature examined, it was necessary for me to propose terms for them. Most of this new terminology pertains to the pelvis. All of the osteological terms used in the present paper, whether used by Howard or not, are briefly defined below. Those of the pelvis are illustrated in Fig. 1. Most of the remaining terms are illustrated by Howard (1929).
Pelvis
The median dorsal ridge is the blunt ridge in the midline of the anterior part of the synsacrum formed by the neural spines of the vertebrae. The antitrochanter, on the posterodorsal rim of the acetabulum, is a pyramid-shaped projection that articulates with the proximal end of the femur. The anterior iliac crest is a ridge along the dorsomedial border of the ilium, beginning almost at the anterior end of that bone; the crest curves laterally as it extends posteriorly and (for purposes of the present definition) ends at the level of the posterior edge of the antitrochanter, where the crest is continuous with the lateral iliac process. The lateral iliac process is a pronounced, laterally or ventrolaterally, projecting ridge on the ventrolateral surface of the ilium posterior to the level of the antitrochanter; the process does not extend as far as the posterior end of the ilium. The lateral ischiatic ridge is a relatively slight ridge continuous with the posterior end of the lateral iliac process and curves posteroventrally across the lateral surface of the posterior part of the ischium; the ridge extends to the ventral edge of the ischium in some individuals and not in others. The dorsolateral iliac ridge begins at the lateral edge of the ilium near the posterior end of the lateral iliac process and curves posteromedially and somewhat dorsally, extending to the posterior edge of the ilium. The lateral iliac fossa is the concavity below the overhanging lateral iliac process. The ilio-ischiatic fenestra is a large oblong opening behind the acetabulum between the ilium and the ischium. The obturator foramen is a small oval opening posteroventral to the acetabulum between the ischium and the pubis. The ventral ischiatic tubercle is the angle formed by the ventrally projecting ischium at the point (near its midlength) where the ischium overlaps and lies lateral to (and fused to) the pubis. The pectineal process is an anterolaterally directed projection of the ventrolateral edge of the ilium anteroventral to the acetabulum. The femoral notch of the ilium is a shallow notch in the ventrolateral edge of the ilium approximately halfway between the last rib and the pectineal process. The oblique iliac crest is a pronounced blunt ridge on the ventral surface of the ilium and extends from the posterolateral corner of the last synsacro-thoraco-lumbar vertebra to near the anteroventral border of the ilio-ischiatic fenestra. The internal ilio-ischiatic crest is more or less continuous with the oblique iliac crest and extends posteriorly along the dorsal border of the ischium (forming the ventral border of the ilio-ischiatic fenestra), and then curves sharply dorsomedially onto the ventral surface of the ilium. The iliac recess is a concavity dorsolateral to the sharply curving posterior end of the internal ilio-ischiatic crest.
The terminology applied to the synsacral vertebrae by different authors varies. The terminology proposed by DuToit (1913) is employed in the present account. See my Fig. 1B. This terminology differs considerably from that used by Howard (1929). DuToit divides the fused synsacral vertebrae into the following five groups, listed in anteroposterior sequence: (1) synsacro-thoracic, which bear movable ribs; (2) synsacro-thoraco-lumbar, which lack movable ribs but possess well developed laterally directed parapophyses, in addition to the more dorsally directed diapophyses; (3) synsacro-lumbar, which lack parapophyses, although possessing inconspicuous diapophyses; these vertebrae are shortened anteroposteriorly and are so firmly fused together that often the number present can be determined only by counting the intervertebral foramina; (4) synsacro-sacral, which have much more pronounced transverse processes than do the synsacro-lumbar vertebrae; these transverse processes are expanded distally where they fuse with the ilium and represent both parapophyses and diapophyses partly or completely fused together plus sacral ribs (detectable only in the embryo); there are considered to be two of these vertebrae; they are situated at approximately the level of the acetabulum; (5) synsacro-caudal, which include the remainder of the fused vertebrae; no marked gross morphological features differentiate the synsacro-sacral and the synsacro-caudal groups of vertebrae. The boundaries between all but the last two groups of vertebrae are usually, but not always, easily determined. It may be difficult to determine whether a vertebra with rudimentary parapophyses belongs to the synsacro-thoraco-lumbar or the synsacro-lumbar group. Sometimes a parapophysis will be better developed on one side of a vertebra than on the other.
Femur
The trochanter is a large squarish tuberosity on the lateral surface of the proximal end of the femur. The trochanteric ridge is a sharp, longitudinal (relative to the femur) ridge forming the anterior edge of the trochanter. The obturator ridge is a short, blunt, longitudinal ridge forming the posterior edge of the trochanter. The anterior intermuscular line is a slight ridge extending distally from the trochanteric ridge. The posterolateral intermuscular line is a slight ridge extending distally from the obturator ridge. The posterior intermuscular line is a slight, longitudinal ridge on the mid-posterior surface of the femur. The internal condyle is a large rounded articular prominence on the medial side of the distal end of the femur. On the lateral side of the distal end of the femur are two articular prominences—the lateralmost, smaller one is the fibular condyle, separated by the fibular groove (visible from posterior aspect only) from the larger and more medial external condyle. The popliteal area is a depression on the posterior surface of the distal part of the femur immediately proximal to the condyles.
Tibiotarsus and Fibula
The inner cnemial crest is pronounced and directed anteriorly on the anterior surface of the proximal end of the tibiotarsus. The outer cnemial crest is pronounced and directed anterolaterally on the anterolateral surface of the proximal end of the tibiotarsus. The rotular crest is transverse and forms the anterior border of the proximal end of the tibiotarsus; the crest extends between the dorsal ends of the two cnemial crests and also extends medial[Pg 372] to the inner cnemial crest. The fibular crest is longitudinal on the lateral surface of the tibiotarsus and fuses with the middle part of the fibula. The fibular tubercle is small and on the lateral surface of the fibula near the level of the middle of the fibular crest. The anteromedial intermuscular line is a slight ridge extending from the inner cnemial crest down the anteromedial surface of the tibiotarsus. The anterolateral intermuscular line is a slight ridge extending from the fibular crest down the anterolateral surface of the tibiotarsus. The supratendinal bridge is a transverse bony arch over a longitudinal groove near the distal end of the anterior surface of the tibiotarsus.
Tarsometatarsus
The hypotarsus is a large, pronounced, squarish protuberance on the posterior surface of the proximal end of the tarsometatarsus and contains grooves and canals for the passage of the flexor tendons. The longitudinal ridges forming the lateral and medial edges of the posterior surface of the hypotarsus are termed calcaneal ridges. The posterior metatarsal crest is long and sharp; it is continuous with the medial calcaneal ridge that extends most of the way down the posterior surface of the tarsometatarsus medial to the midline; there is an opening between this crest and the tarsometatarsus immediately distal to the hypotarsus. The medial metatarsal depression is large; it is on the medial surface of the proximal end of the tarsometatarsus. The anterior metatarsal groove is a longitudinal groove in the midline of the proximal part of the anterior surface of the tarsometatarsus. The three trochleae are large rounded articular prominences at the distal end of the tarsometatarsus; there is one at the base of each of the digits II, III, and IV. The term distal foramen (as used by Howard) refers to a short, anteroposteriorly directed canal that perforates the tarsometatarsus a short distance proximal to the intertrochlear notch between the trochleae for digits III and IV. Beginning at the middle of this canal and extending distally at a right angle to it is the intertrochlear canal, which opens via the terminal foramen into the intertrochlear notch between the trochleae for digits III and IV.
Nerves
For ease of description I have coined terms for the major divisions of the femoral and sciatic nerves.
Muscles
My terminology follows that of Fisher (1946) and Fisher and Goodman (1955) except for Mm. femoritibialis externus, flexor cruris lateralis (accessory head), and obturator internus et externus. Fisher (1946:547) states that most of his names for the hip and thigh muscles are those of Howell (1938) and the names for the shank and foot muscles are those of Hudson (1937). Fisher deviates, without explanation, from Howell's terminology in respect to Mm. vastus medialis and femoritibialis internus, M. caudofemoralis, M. flexor cruris lateralis, and Mm. obturator internus and obturator externus. Fisher's synonymy of these muscles (1946: table 42) [Pg 373] is in error. Fisher understandably deviates from Hudson in respect to Mm. extensor brevis digiti III and extensor proprius digiti III (see Holmes, 1962), although Fisher's synonymy is in error here. See my table 1.
I am not using Fisher and Goodman's term femoritibialis externus; this muscle is here considered as a part of M. vastus lateralis. A great deal of confusion surrounds the terminology of the muscle complex here termed Mm. vastus lateralis and vastus medialis. Hudson (1937), Hudson, et al. (1959), Fisher (1946), and Fisher and Goodman (1955) have used different terminology for this complex. Most of the confusion stems from Gadow's (1891) unclear description of this complex, which he subdivided into two units termed Mm. femori-tibialis externus and femori-tibialis medius. Many birds have three parts to this complex. It is difficult to determine how to apply Gadow's two terms to these three parts. As nearly as I can determine, the correct method is that of Hudson, et al. (1959); but because Gadow's terms have been used in different ways (even by the same worker), it seems best to abandon these terms. Berger (1956:272) believes that the muscle unit that Fisher and Goodman term M. femoritibialis externus represents a head of M. vastus lateralis; I am accepting his opinion. For the three parts of the complex under discussion, I am using the terms M. vastus medialis and M. vastus lateralis pars lateralis and pars postica.
Fisher (Fisher, 1946; Fisher and Goodman, 1955) considers the muscle here termed M. femorocruralis as an accessory head of M. flexor cruris lateralis. The two muscle units in question are closely associated; they insert broadly on opposite sides of a common tendinous raphe. Howell (1938:73) considers this to be a secondary fusion of unrelated muscles. Romer (1927:366) states that in the chick embryo M. femorocruralis is in reality a shank muscle that migrates into the thigh during development. Therefore, Fisher's usage of a single name for these two unrelated muscles is unsatisfactory. I am using Howell's terminology in which the name flexor cruris lateralis represents the main head only of Fisher's M. flexor cruris lateralis and the name femorocruralis represents Fisher's accessory head.
Gadow (1891) divides the obturator complex into two muscles (or muscle groups), which he terms M. obturator and Mm. accessorii M. obturatoris. He states that the former is homologous with the mammalian obturator internus and the latter with the obturator externus. Hudson (1937), accepting Gadow's homologies, renamed these muscles M. obturator internus and M. obturator externus. Nearly all subsequent workers have followed Hudson's terminology, with its implication that these muscles are homologous with the mammalian muscles of the same name. Howell (1938) is an exception. He points out (pp. 78, 79) that the obturator internus of Hudson is homologous with the obturator externus of mammals. His evidence is convincing: "In origin the obturator is somewhat suggestive of the mammalian obturator internus, for which it has uniformly been mistaken. That the latter interpretation is incorrect, however, is attested by the facts that it receives twigs of n. obturatorius within the pelvis, passes through the obturator foramen rather than dorsal to the border of the ischium, and it is segregated from any muscle with tibial innervation. Insertion has shifted only to a slight and unimportant degree as compared with that of the [Pg 374] mammalian obturator externus, and beyond question it is the equivalent of that muscle. The stimulus for a longer muscle, has been the same, resulting in the extension of origin to within the pelvis of the externus in birds and the internus in mammals, but the obturator internus is an extension of a part of the gemellus mass and this does not occur in any vertebrate class but Mammalia." Howell applies the term M. obturator to the entire obturator complex.
Romer (1927), studying the development of the thigh musculature in chick embryos, concluded that the entire obturator complex is homologous with the mammalian obturator externus plus quadratus femoris. He considered the avian M. flexor ischiofemoralis to be the homologue of the mammalian obturator internus.
Gadow, in his work on the ratites (1880:34), states that M. obturator (obturator internus of Hudson) cannot be homologous to the mammalian obturator internus, but must represent the obturator externus. His reasoning is as follows: "Als M. pectineus kann man diesen Muskel nicht auffassen, da er auf der Aussenfläche des Trochanter major inserirt, ferner auch nicht als M. obturator internus der menschlichen Anatomie, da er nicht vom Plexus ischiadicus, sondern vom Plexus cruralis aus innervirt wird. Seiner Innervation und Insertion nach wäre er nur mit dem M. obturator externus zu vergleichen, wobei er seinen Ursprung im Verhältniss zum Menschen nur bedeutend weiter auf das Os ischii und Os pubis distalwärts ausgedehnt hätte und so allerdings der Lage nach mit Ausnahme seines Insertionsdrittels ein 'internus' geworden wäre."
Since Gadow gives different homologues for M. obturator in two of his works (1880 and 1891), one would suspect that he had changed his opinion in the interim; however, there is no evidence that he did so. In 1880 he gives supporting evidence (quoted above) for his view; in 1891 he does not. After describing (1891:173) how the origin of M. obturator in bird ancestors presumably migrated from a location outside the pelvis to a position inside the pelvis prior to the meeting of the pubis and ischium external to the muscle, he states: "Eine ähnliche Entwicklung ist für den Obturator internus der Säugethiere anzunehmen, welchem der M. obturator der Vögel entspricht." A similar development in mammals is impossible, owing to the different relationship of the muscle to the pelvic bones in this class. Gadow says nothing more about the mammalian homologue of M. obturator. In view of this discrepancy, Gadow can hardly be considered as a supporter of the idea that the avian M. obturator is homologous with the mammalian obturator internus.
The evidence is conclusive, it seems to me, that the obturator internus of Hudson is not homologous with the mammalian obturator internus. Therefore, the term obturator internus is inappropriate for the avian muscle and must be abandoned. I shall follow Howell (1938) in naming the entire obturator complex M. obturator. This term, of course, is not used in the sense in which it is used by Gadow. The use of the term obturator externus for the entire complex is avoided because it may not correspond exactly to the mammalian obturator externus. As mentioned previously, Romer considers the avian muscle to be homologous not only with the mammalian obturator externus but also with the quadratus femoris.
I am following the policy of Wilcox (1948) and Berger (1952) in latinizing the term anterior, changing it to anticus. When preceded by the feminine word pars, the feminine ending is used (antica).
In table 1 my terminology is compared with that of Fisher and Goodman (1955), Howell (1938), Hudson (1937), and Gadow (1891). The terminology of Fisher (1946) is identical with that of Fisher and Goodman (1955) except that in his earlier work Fisher did not describe or name M. femoritibialis externus, and M. lumbricales of his earlier work is not mentioned in his later work. The terminology of Hudson, et al. (1959) is identical with that of Hudson (1937) except that the manner in which the femoritibialis complex is subdivided is identical with that of Gadow (1891) and different from that in Hudson's earlier work; also the abbreviations p. ext. and p. int. are substituted in his later paper for pars anterior and pars posterior, respectively, of M. adductor longus et brevis.
I gratefully acknowledge the generous help of Professor A. Byron Leonard, under whose guidance this study was conducted and thank Professor E. Raymond Hall, Professor Howard A. Matzke, and Dr. Irwin Baird for numerous helpful suggestions and criticisms.
For help in collecting specimens I thank J. R. Alcorn, W. C. Glazener (through the courtesy of the Texas Game and Fish Commission), Dr. Harrison B. Tordoff, Jerry Tash, William Brecheisen, and Louis Brecheisen. I thank also Edwin Gebhard of the Kansas Forestry, Fish and Game Commission for help in locating the Lesser Prairie Chickens.
I am grateful for the assistance of Mrs. Chester Alexander and Dr. L. C. Dahl in translating a Russian and a Dutch reference, and thank George Young and James Bee for making equipment used in my study.
All of the original drawings except Fig. 1 were made by me, although the final inking of Figs. 12 through 19 was done by Bret Waller. Fig. 1 was drawn by Kay Swearingen.
I was aided in this study during the summer of 1960 by a research grant from the University of Kansas.
Although no special study was made of the skeleton, certain conspicuous variations are discussed here.
There are a few pronounced differences between the pelvis of Tympanuchus and that of Pedioecetes. Whereas in the former the thick lateral iliac process has a pronounced overhang with the ventral edge lateral to the ischium (Fig. 1), in Pedioecetes there is no overhang at all and the edge of this process is much thinner. The ischium in Pedioecetes is wider (in dorsoventral extent), especially posteriorly, than in Tympanuchus. In Tympanuchus the posteroventral margin of the ischium is rounded and is free from the pubis, whereas in Pedioecetes it is pointed and fused with the pubis.
In Tympanuchus cupido (both subspecies) the lateral iliac process extends farther ventrally than in T. pallidicinctus, approaching or extending ventral to the level of the pubis in the former species; also the edge of this process is thicker in T. cupido.
All specimens studied have a single synsacro-thoracic vertebra. The number of combined synsacro-thoraco-lumbar and synsacro-lumbar vertebrae is eight in each specimen of Tympanuchus and in one specimen of Pedioecetes phasianellus jamesi and is seven in three specimens of the latter. In most specimens of Tympanuchus there are three synsacro-thoraco-lumbar and five synsacro-lumbar vertebrae, although in two specimens (T. pallidicinctus) there are four of each group; in one of these latter two specimens the parapophysis on one side of the fourth synsacro-thoraco-lumbar vertebra is small. The first (of five) synsacro-lumbar vertebra has a rudimentary parapophysis on one side in one specimen of Tympanuchus and on both sides in another specimen. One specimen of Pedioecetes phasianellus jamesi has five synsacro-lumbar vertebrae and the others have four; all have three synsacro-thoraco-lumbar vertebrae.
For each nerve (or plexus) the condition found in most specimens of the Lesser Prairie Chicken (T. pallidicinctus) is described first. Following this, variations from the typical T. pallidicinctus condition are given for T. pallidicinctus, then for T. cupido (both subspecies considered together), and finally for P. p. jamesi.
Lumbosacral Plexus, Figs. 2, 3
T. pallidicinctus
Description.—Eight spinal nerves contribute to the lumbosacral plexus. These are the second through the ninth synsacral spinal nerves (S2 to S9). The entire ventral ramus of each of these nerves, excepting S2 and S9, contributes to this plexus. The ventral ramus of S2 divides into two branches, only the posterior of which contributes to the plexus; the anterior branch directly innervates muscles of the abdominal wall (as does the entire ventral ramus of S1). The ventral ramus of S9 divides into two branches, only the anterior of which contributes to this plexus; the posterior branch contributes to the more posteriorly situated pudendal plexus.
Each root of the plexus corresponds to a single spinal nerve except one spinal nerve (S5—the furcal) that contributes a root to both the femoral nerve and the sciatic nerve; thus typically the plexus has nine roots (but see below). The four anteriormost roots (S2 to S5) contribute to the femoral nerve, although the contribution from S2 is small. S3 and S4 contribute to the obturator nerve. The five posteriormost roots (S5 to S9) contribute to the sciatic nerve, although the contribution from S9 is relatively small.
Individual Variation.—In all specimens (of all species) examined, the right and left sides of the plexus in any one individual were practically identical. In T.p. 2 (Fig. 2B), there appear to be two furcal nerves; S5 is typical, but a small branch of S4 apparently also contributes to the sciatic nerve. In T.p. 5, S9 is unique in dividing into three branches; the anterior two join the sciatic nerve separately; the posterior one joins the pudendal plexus as usual.
T. cupido
Individual Variation.—S2 or S5, or both, may contribute to a limited extent to the obturator nerve. In T.c.p. 3 (Fig. 3A) and T.c.a. 1 and 2, much of the plexus has shifted one segment anteriorly, relative to the synsacral vertebrae (the so-called prefixed condition); the roots of the femoral nerve are S2, S3, and S4 (all large); the furcal nerve is S4 (in T.c.a. 1, S5 gives an extremely small root to the femoral nerve, thus making two furcal nerves); six roots (S4 to S9) contribute to the sciatic nerve; S3 and S4 remain as the main contributors to the obturator nerve except in T.c.a. 2 in which only S2 and S3 contribute to it.
P. p. jamesi
Individual Variation.—In P.p. 1, the plexus resembles the typical condition in T. pallidicinctus. In P.p. 2, 3, and 4, the plexus is prefixed. P.p. 2 resembles T.c.p. 3. In P.p. 3 and 4 (Fig. 3B) there are two furcal nerves (S4 and S5); S2 to S4 are the main contributors to the femoral nerve; only S2 and S3 contribute to the obturator nerve; S4 to S9 contribute to the sciatic nerve (the anteriormost and posteriormost roots are small).
T. pallidicinctus
Description.—The femoral nerve is short, dividing inside the pelvis into six major divisions—anterior, middle, posterior, anterodorsal, dorsal, and posterodorsal. The anterodorsal and posterodorsal divisions are short, failing to extend so far laterally as the inguinal ligament; the posterodorsal division is also small and is usually covered by other divisions and is not visible when viewed from the ventral side.
The anterior division passes ventral to Mm. iliotrochantericus medius and iliacus and dorsal to the anterior end of the inguinal ligament. The division branches into two parts. The anterior part extends around the posterior border of M. extensor iliotibialis anticus and sends several twigs to the lateral surface of this muscle. The posterior part passes between the proximal parts of Mm. extensor iliotibialis anticus and extensor iliotibialis lateralis and supplies the skin.
The middle division passes ventral to Mm. iliotrochantericus medius and iliacus and dorsal to the inguinal ligament. The division branches into a large but variable number of parts. A variable number of branches (usually two) pass posterior to M. extensor iliotibialis anticus and penetrate the medial surface of M. extensor iliotibialis lateralis. Several branches supply the fused Mm. vastus lateralis and vastus medialis. The posteriormost branch of this division passes between Mm. ambiens and vastus medialis, [Pg 378] giving twigs to the lateral surface of M. ambiens, and sometimes also to the medial surface of M. vastus medialis, and terminates in M. femoritibialis internus.
The posterior division, which does not subdivide, spirals completely around M. psoas (passing in turn anterior, dorsal, posterior, and ventral to it) and gives twigs into this muscle. This nerve then extends distally into the proximal part of the shank and there has a nonmuscular termination.
The short, thick anterodorsal division, partly covered by the anterior division, turns dorsally and passes through the femoral notch of the ilium and penetrates the deep surface of M. gluteus profundus.
The slender dorsal division passes ventral to M. iliotrochantericus medius and dorsal to the inguinal ligament and penetrates the ventral surface of M. iliacus.
The small, short posterodorsal division penetrates the ventral surface of M. iliotrochantericus medius.
Individual Variation.—In two legs the anterior division gives a twig or two twigs to M. extensor iliotibialis lateralis. The dorsal division may fuse proximally with either the anterior or middle division, thus appearing to be a branch of one of these divisions. In one leg (Fig. 5A), there are two separate branches (both fused with the middle division) to M. iliacus. On both sides of one specimen (Fig. 5A), the anteriormost branch of the middle division, which supplies M. extensor iliotibialis lateralis, gives off a twig that anastomoses with the branch of the anterior division that supplies M. extensor iliotibialis anticus. On both sides of another specimen, the anterodorsal division passes lateral to the anterior end of M. iliotrochantericus medius instead of through the femoral notch, which is lacking.
T. cupido
Individual Variation.—In three legs, the anterior division gives twigs into M. extensor iliotibialis lateralis. The dorsal division is fused proximally with the middle division in one instance. In three cases, a twig from the middle division anastomoses with the branch of the anterior division supplying M. extensor iliotibialis anticus. In the example shown in Fig. 5B, a twig comes off the cutaneous branch of the anterior division, perforates the ventral part of M. iliacus, and rejoins the cutaneous branch. In both legs of one specimen, the cutaneous branch of the anterior division perforates the anterior edge of M. extensor iliotibialis lateralis instead of passing between the latter and M. extensor iliotibialis anticus. The posteriormost branch of the middle division, which terminates in M. femoritibialis internus, perforates the medial part of M. vastus medialis in one leg. In another leg, one of the branches to the fused Mm. vastus lateralis and vastus medialis sends a twig into M. extensor iliotibialis lateralis.
P. p. jamesi
Individual Variation.—In three legs, the anterior branch of the anterior division is cutaneous and the posterior branch supplies M. extensor iliotibialis anticus. The dorsal division may fuse proximally with either the anterior or middle division. In one leg (Fig. 4B), there are two branches to M. iliacus, one associated with the anterior division and one with the middle division.
Obturator Nerve
T. pallidicinctus
Description.—The long slender obturator nerve passes along the oblique iliac crest and divides into several branches immediately before reaching the obturator foramen. One or two branches, which do not pass through the foramen, penetrate the superficial surface of M. obturator pars postica. Several small branches (variable in number and arrangement) pass through the obturator foramen and supply pars ventralis, pars dorsalis, and pars antica of M. obturator. When pars ventralis and pars dorsalis are fused, one branch perforates the proximal end of this mass and reaches pars antica. One large branch passes through the obturator foramen dorsal to the tendon of M. obturator pars postica, then turns ventrally, passing lateral to the latter; the branch passes between Mm. adductor superficialis and adductor profundus and gives twigs to each of these two muscles.
Individual Variation.—None of significance in any of the three species.
Sciatic Nerve, Figs. 6, 7, 8, 9
T. pallidicinctus
Description.—The sciatic nerve passes through the anterior part of the ilio-ischiatic fenestra. Several branches diverge from the nerve immediately after it emerges from the fenestra. The main trunk of the nerve then extends distally through the thigh deep to M. extensor iliofibularis and superficial (lateral) to Mm. flexor ischiofemoralis, caudofemoralis, adductor superficialis, and femorocruralis. The main trunk subdivides into two large nerves—peroneal and tibial—that are adjacent and bound to each other throughout the thigh; the peroneal nerve lies anterior to the tibial. At the distal end of the thigh the main trunk splits grossly into two large branches that diverge and enter the shank. This division does not represent the separation between peroneal and tibial nerves, as is sometimes assumed; the anterior branch includes a part of the tibial nerve as well as the entire peroneal nerve.
A longitudinal groove is visible grossly on the lateral surface of the main trunk, except at the proximal end; distally a second groove is visible posterior to the first one (Fig. 6). The long anterior groove indicates the boundary between the peroneal and tibial nerves; this groove may disappear distally, although the posterior groove is always visible distally. The posterior groove, which is continuous with the division of the sciatic nerve into anterior and posterior branches, represents the boundary between two divisions of the tibial nerve. (This is discussed in detail below.) In the middle of the thigh the peroneal and tibial nerves are enclosed in separate connective tissue sheaths, although the two sheaths are fused together; the point of fusion is marked by the anterior groove. If the two sheaths are slit open, the two nerves can be removed and can be seen to be entirely separate. In the proximal part of the main trunk the peroneal and tibial components are enclosed in a single sheath and appear as an undivided trunk; but if the sheath is removed, the two components can be pulled apart rather easily, although there may be some intermingling of a few fibers. This separation can be extended to a point proximal to the origin of all the branches of the sciatic nerve; thus it can be determined which branches arise from the [Pg 380] peroneal component and which from the tibial. (These branches arise from the sciatic nerve as, or immediately before, the nerve passes through the ilio-ischiatic fenestra; since this level of the intact nerve could not be adequately observed, it was necessary to cut the nerve inside the pelvis and pull the intrapelvic part of the nerve out through the ilio-ischiatic fenestra. In doing this, care had to be taken to avoid damaging the most proximal branches.)
Three main branches arise from the peroneal component (apart from the main trunk) and two from the tibial. Including the peroneal and tibial components of the main trunk, the sciatic nerve can be divided into seven major divisions—anterior peroneal, middle peroneal, dorsal peroneal, posterior or main peroneal (contributes to main trunk), anterior or main tibial (contributes to main trunk), middle tibial, and posterior tibial. Farther distally, the posterior peroneal division becomes the peroneal nerve and the anterior tibial division becomes the tibial nerve. For descriptive purposes, the term peroneal (or tibial) nerve will be applied only where the nerve is enclosed in its own sheath, but regardless of whether or not the sheath is fused with another; proximal to this, where the separation may not be precise, the terms peroneal (or tibial) division or component will be used.
The small anterior peroneal division arises from the anterior edge of the sciatic nerve. Immediately after emerging from the ilio-ischiatic fenestra, the division turns anteriorly and passes deep to M. piriformis, to which the division gives a twig (in some cases more than one twig), then continues forward to supply the posterior part of M. gluteus profundus.
The middle peroneal division branches into two parts. One part penetrates the deep surface of the anteroproximal part of M. extensor iliofibularis. The other part emerges between the proximal ends of Mm. extensor iliofibularis and vastus lateralis and penetrates the deep surface of M. extensor iliotibialis lateralis.
The dorsal peroneal division arises from the posterodorsal part of the peroneal component, then angles posteriorly, crossing the dorsal surface of the anterior tibial division and superficially appears to arise from the tibial component. The dorsal peroneal division usually subdivides into two unequal branches, both of which penetrate the deep surface of the proximal end of M. extensor iliofibularis.
The large middle tibial division soon subdivides into two branches that pass posterodistally lateral to M. flexor ischiofemoralis. One branch (usually the anterior one) passes lateral to M. caudofemoralis (both heads) and emerges between Mm. extensor iliofibularis and flexor cruris lateralis and enters the skin. The other branch passes deep to M. caudofemoralis pars iliofemoralis, and divides into several branches. Several tiny branches penetrate the deep surface of M. caudofemoralis pars iliofemoralis. Another branch also enters the substance of the latter and emerges from the ventral edge of it, giving a twig to pars caudifemoralis, then passes lateral to pars caudifemoralis and enters M. flexor cruris lateralis. Still another branch passes deep to both heads of M. caudofemoralis and enters the anterior part of M. flexor cruris medialis.
The small posterior tibial division arises from the posterior edge of the sciatic nerve. The division diverges from the remainder of the nerve, as the [Pg 381] latter passes through the ilio-ischiatic fenestra, and penetrates the dorsal surface of M. flexor ischiofemoralis.
Below the middle of the main trunk a bundle of fibers of moderate size separates from the anterior edge of the tibial nerve, leaves the tibial sheath, and enters its own sheath, lying superficially between the tibial and peroneal sheaths (Fig. 6). At the distal end of the thigh the sheath enclosing this bundle of fibers remains fused with the posterior edge of the peroneal nerve and passes with the latter (diverging from the remainder of the tibial nerve) through the tendinous guide loop for M. extensor iliofibularis, and then diverges from the peroneal nerve. Since this bundle of fibers is distributed with the peroneal nerve, and since the origin of the bundle may be easily overlooked, it has sometimes been misinterpreted as a branch of the peroneal nerve, whereas it almost certainly is a branch of the tibial nerve; this bundle will here be termed the paraperoneal branch of the tibial nerve.
A small but long branch separates from the posterior edge of the proximal end of the tibial nerve or from the tibial component proximal to this and extends distally for some distance adjacent to the tibial nerve, then passes posterodistally between Mm. extensor iliofibularis and flexor cruris lateralis and supplies the skin.
A small branch separates from the anterior edge of the peroneal nerve a short distance above the distal end of the main trunk and passes distolaterally between Mm. extensor iliotibialis lateralis and extensor iliofibularis and supplies the skin.
A twig comes off the medial surface of the tibial nerve near the distal end of the main trunk, passes anteriorly deep to the peroneal nerve, and penetrates the lateral surface of M. femorocruralis; in some cases two twigs enter this muscle.
Individual Variation.—In one leg (Fig. 7), the twig to M. caudofemoralis pars caudifemoralis arises more proximally than usual and perforates pars iliofemoralis independently of the branch to M. flexor cruris lateralis. The nerve supplying M. flexor cruris lateralis does not perforate M. caudofemoralis pars iliofemoralis, but passes deep to it in three legs. In half the legs, the paraperoneal branch of the tibial nerve, after extending a short distance in its own sheath, enters the sheath of the peroneal nerve and appears grossly to unite with it; if, however, the sheath is slit open, the paraperoneal branch can be easily pulled apart from the posterior edge of the peroneal nerve; the paraperoneal branch is again enclosed in its own sheath at the distal end of the thigh. In one leg, the cutaneous branch of the peroneal nerve perforates the posteroproximal part of M. gastrocnemius pars externa; in three others, this branch is absent. In one of these last three legs (Fig. 7), the distal cutaneous branch of the tibial nerve is also absent. In three legs (of different specimens), a minute twig from the middle tibial division passes posteriorly deep to M. caudofemoralis pars caudifemoralis toward the tail (Fig. 7); this twig joins the pudendal plexus in one leg; in the other two the twig could not be traced to its termination. Minute twigs come off the peroneal nerve near the middle of the thigh and enter M. extensor iliofibularis in some legs. In a few cases, a minute nonmuscular twig arises from the peroneal nerve near the distal end of the main trunk and passes anteriorly deep to M. vastus lateralis pars postica (Fig. 7).
T. cupido
Individual Variation.—In several legs, the nerve supplying M. flexor cruris lateralis does not perforate M. caudofemoralis pars iliofemoralis, but passes deep to it. The branch to M. flexor cruris medialis arises from the posterior (rather than the middle) tibial division in one instance (Fig. 8). In one leg, a minute twig from the middle tibial division passes posteriorly and joins the pudendal plexus; in another, a similar twig is present but could not be traced to its termination. In some specimens, minute twigs come off the peroneal nerve near the middle of the thigh and enter M. extensor iliofibularis. In one leg, a nonmuscular twig arises from the base of the cutaneous branch of the peroneal nerve and passes anteriorly deep to M. vastus lateralis pars postica. In another leg (Fig. 8), a tiny additional twig arises from the posterior edge of the tibial nerve and subdivides, one branch joining the cutaneous branch of the middle tibial division and the other joining the distal cutaneous branch of the tibial nerve.
P. p. jamesi
Individual Variation.—In both legs of one specimen, the branch to M. flexor cruris medialis arises from the posterior (rather than the middle) tibial division; in three legs, this branch arises as an independent division of the tibial nerve (Fig. 6). (Only in one leg does this branch arise as in T. pallidicinctus.) The branch to M. flexor cruris medialis perforates the lateral part of M. flexor ischiofemoralis in one instance. In all legs except one (nerve possibly destroyed), a second twig to M. flexor ischiofemoralis arises from the branch to M. flexor cruris medialis (Fig. 6). In one leg (Fig. 9), an additional branch, arising as an independent division of the sciatic nerve, enters M. extensor iliofibularis distal to the point of entrance of the dorsal peroneal division; this extra branch arises posterior (adjacent) to the dorsal peroneal division, but it could not be determined with certainty whether it arises from the peroneal or tibial component. A minute twig from the branch to M. flexor cruris medialis passes posteriorly and joins the pudendal plexus in one leg (Fig. 6); in another, a similar twig is present but could not be traced to its termination. In nearly all the legs, minute twigs come off the peroneal nerve near the middle of the thigh and enter M. extensor iliofibularis (Fig. 6). In both legs of one specimen, the paraperoneal branch enters the peroneal sheath (although separable from the peroneal nerve). The distal branch to M. femorocruralis gives off a long twig to M. gastrocnemius pars media in one instance (Fig. 6).
Peroneal Nerve, Fig. 10
T. pallidicinctus
Description.—The branch that is given off in the thigh has been discussed above. The peroneal nerve passes, with the paraperoneal branch of the tibial nerve, through the guide loop for M. extensor iliofibularis. The peroneal nerve diverges from the paraperoneal branch and passes along the anterior (proximal) edge of the tendon of M. extensor iliofibularis medial to the common tendon of the lateral heads of Mm. flexor perforatus digiti IV [Pg 383] and flexor perforatus digiti II and lateral to the common tendon of the anterolateral heads of Mm. flexor perforatus digiti IV, flexor perforatus digiti II, and flexor perforatus digiti III.
The peroneal nerve soon gives off a spray of branches that supplies the following: femoral head of M. tibialis anticus, tibial head of M. tibialis anticus (branch passes deep to femoral head), M. extensor digitorum longus (branch passes deep to tibial head of M. tibialis anticus), and M. peroneus longus. A part of the nerve may or may not pass through a notch in the proximal end of the lateral head of M. flexor digitorum longus. The nerve then extends distally along the anterolateral edge of the latter muscle and subdivides into two long branches. Gadow (1891) termed these branches the superficial peroneal and the deep peroneal; his terminology will be used here.
The superficial peroneal branch, after giving off, near its proximal end, one or two twigs into M. peroneus brevis, passes lateral to the retinaculum for the tendon of M. tibialis anticus, then across the intratarsal joint lateral to the latter, then lateral to the insertion of M. tibialis anticus, where the branch subdivides. One of the two resulting branches gives one or two twigs into M. extensor brevis digiti IV, then terminates nonmuscularly in the digits. The other branch passes between the main and accessory insertions of M. tibialis anticus and joins the branch of the deep peroneal which supplies M. abductor digiti II. (See next paragraph.)
The deep peroneal branch passes through the retinaculum for the tendon of M. tibialis anticus, lying lateral, then deep, then medial to the latter; it crosses the intratarsal joint medial to the latter. Immediately above the insertion of M. tibialis anticus, the deep peroneal branch divides, one branch passing on each side of the main insertion. The branch passing lateral to the main insertion passes between the latter and the accessory insertion (medial to the latter) and is joined by a branch of the superficial peroneal nerve. This fused branch extends distally between Mm. extensor hallucis longus and extensor brevis digiti IV and medial to M. extensor brevis digiti III, giving twigs into the latter and into M. abductor digiti II before terminating nonmuscularly in the digits. The branch of the deep peroneal nerve that passes medial to the main insertion of M. tibialis anticus gives one or two twigs into the proximal head of M. extensor hallucis longus, then terminates nonmuscularly in the digits.
Individual Variation.—In four legs, the branch of the superficial peroneal nerve that usually joins the lateral branch of the deep peroneal nerve is lacking (Fig. 10B). In these legs it can be seen that Mm. extensor brevis digiti III and abductor digiti II are supplied by the deep peroneal nerve.
T. cupido
Individual Variation.—In two legs, the same branch that gives twigs into the proximal head of M. extensor hallucis longus also sends a twig into the distal head of this muscle (Fig. 10C).
P. p. jamesi
Individual Variation.—None of significance.
Tibial Nerve, Fig. 11
T. pallidicinctus
Description.—The branches given off in the thigh have been discussed in the account of the sciatic nerve. At the distal end of the thigh the peroneal nerve and the paraperoneal branch of the tibial nerve diverge from the remainder of the tibial nerve and pass through the tendinous guide loop for M. extensor iliofibularis whereas the remainder of the tibial nerve does not. This main part of the tibial nerve immediately divides into three main divisions—lateral, posterior, and medial.
The lateral division passes between Mm. flexor perforatus digiti IV and gastrocnemius pars externa and subdivides into two branches, one of which penetrates the medial surface of M. gastrocnemius pars externa. The other branch passes deep to the latter and sends twigs into the posterior head of M. flexor perforans et perforatus digiti II, then passes deep to the latter and enters M. flexor perforans et perforatus digiti III.
The posterior division sends a branch into the medial head of M. flexor perforatus digiti IV, then passes between the latter and the medial head of M. flexor perforatus digiti III, and extends distally giving off twigs to each of the three heads of M. flexor perforatus digiti IV, to each of the two heads of M. flexor perforatus digiti III, and to each of the three heads of M. flexor perforatus digiti II. The number and arrangement of these twigs is variable.
The medial division passes medial to the medial head of M. flexor perforatus digiti III, sends a twig to the lateral surface of M. gastrocnemius pars media, then passes into the shank musculature between Mm. plantaris and flexor hallucis longus, and sends a branch along the medial edge of M. flexor hallucis longus that gives several twigs into this muscle before terminating nonmuscularly. A small branch extends to M. popliteus, another to M. plantaris, and another to the posterior head of M. flexor digitorum longus. A nonmuscular branch passes between the medial and posterior heads of M. flexor digitorum longus and extends distally deep to this muscle. A long branch gives off near its proximal end a variable number of twigs that pass deep to M. plantaris and enter M. gastrocnemius pars interna; the branch then extends distally along the lateral edge of M. plantaris and terminates nonmuscularly.
The paraperoneal branch diverges from the peroneal nerve, passing medial and then distal to the insertion of M. extensor iliofibularis, whereas the peroneal nerve passes proximal and then lateral to this insertion. The paraperoneal branch passes deep to the lateral heads of Mm. flexor perforatus digiti IV and flexor perforatus digiti II and superficial to the tendon of the anterolateral head of M. flexor perforatus digiti IV and then passes distally along the anterolateral borders of the latter and the lateral head of M. flexor perforatus digiti III and the posterolateral border of M. flexor digitorum longus. This branch is thus separated from the peroneal nerve by M. flexor digitorum longus and by the fibula; the branch passes along the lateral surface of the tibial cartilage, continues lateral to the hypotarsus, then turns medially before extending distally between Mm. abductor digiti IV and flexor hallucis brevis, sending twigs into each of these muscles and a long twig into M. lumbricalis before terminating nonmuscularly.
Individual Variation.—In T.p. 3L,R (Fig. 11B), an extra branch arises from the tibial nerve as a separate (fourth) division; it enters the medial head of M. flexor perforatus digiti IV and also gives off a twig that anastomoses with the posterior division (left leg) or with the first branch of the posterior division (right leg). In T.p. 3R (Fig. 11B), a large extra branch arises from the proximal part of the medial division and passes medial and then deep to the medial head of M. flexor perforatus digiti III, perforates the tendinous part of the medial head of M. flexor perforatus digiti II, and joins the posterior division (lateral to the medial head of M. flexor perforatus digiti III). A similar branch is found in T.p. 3L except that it arises from the proximal part of the posterior (rather than the medial) division. In T.p. 3R (Fig. 11B), the branch to M. gastrocnemius pars externa arises so far proximally that it appears as a separate (fifth) division of the tibial nerve. In two legs, the branch of the medial division that supplies M. gastrocnemius pars media sends a twig into the distal end of M. femorocruralis (Fig. 11A).
T. cupido
Individual Variation.—In one leg, an extra branch of the medial division arises immediately distal to the branch to M. gastrocnemius pars media and enters the proximal end of the medial head of M. flexor perforatus digiti III. In one instance, the branch to M. gastrocnemius pars interna passes through a gap in the origin of M. plantaris rather than distal to the origin of the latter.
P. p. jamesi
Individual Variation.—The branch to M. gastrocnemius pars interna gives a minute twig to the deep surface of the free belly of M. plantaris in one leg.
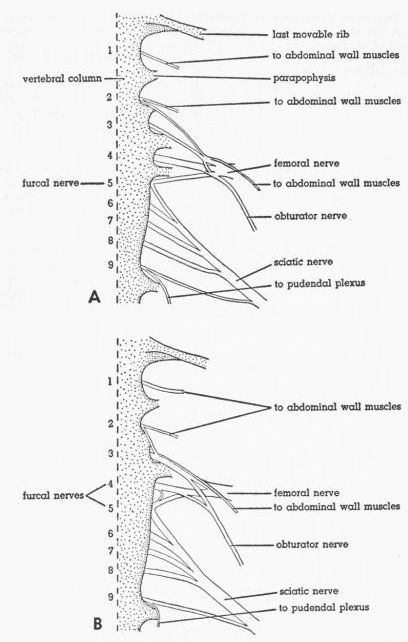 Fig. 2.
Ventral views of the lumbosacral plexus of
Tympanuchus pallidicinctus.
Sympathetic ganglionated chain removed. Numbers indicate
synsacral spinal nerves. × 2. A. T.p. 1L. B. T.p. 2L.
Fig. 2.
Ventral views of the lumbosacral plexus of
Tympanuchus pallidicinctus.
Sympathetic ganglionated chain removed. Numbers indicate
synsacral spinal nerves. × 2. A. T.p. 1L. B. T.p. 2L.
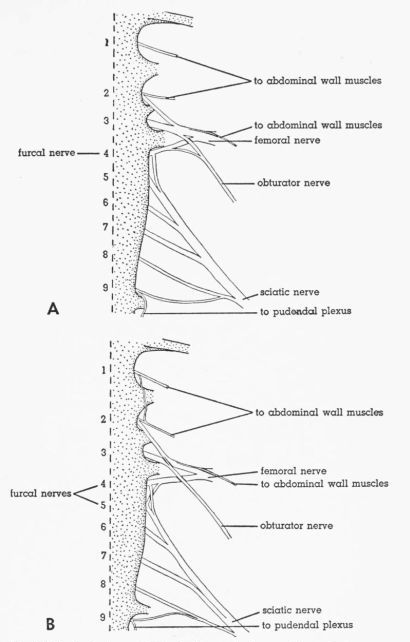 Fig. 3.
Ventral views of the lumbosacral plexus.
Sympathetic ganglionated chain removed. Numbers indicate
synsacral spinal nerves. × 2. A. Tympanuchus cupido pinnatus
3L. B. Pedioecetes phasianellus jamesi 4L.
Fig. 3.
Ventral views of the lumbosacral plexus.
Sympathetic ganglionated chain removed. Numbers indicate
synsacral spinal nerves. × 2. A. Tympanuchus cupido pinnatus
3L. B. Pedioecetes phasianellus jamesi 4L.
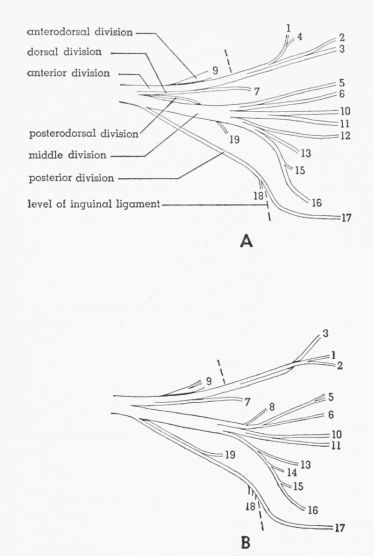 Fig. 4.
Semidiagrammatic ventral views of the femoral
nerve, showing the distribution of the branches. × 3. 1,2,
M. extensor iliotibialis anticus; 3, cutaneous; 4-6, M. extensor
iliotibialis lateralis; 7,8, M. iliacus; 9, M. gluteus
profundus; 10-12, fused Mm. vastus lateralis and vastus
medialis; 13,14, M. vastus medialis; 15, M. ambiens;
16, M. femoritibialis internus; 17, nonmuscular; 18, M.
psoas; 19, M. iliotrochantericus medius. A. Tympanuchus
cupido pinnatus 3L. B. Pedioecetes phasianellus
jamesi 3L.
Fig. 4.
Semidiagrammatic ventral views of the femoral
nerve, showing the distribution of the branches. × 3. 1,2,
M. extensor iliotibialis anticus; 3, cutaneous; 4-6, M. extensor
iliotibialis lateralis; 7,8, M. iliacus; 9, M. gluteus
profundus; 10-12, fused Mm. vastus lateralis and vastus
medialis; 13,14, M. vastus medialis; 15, M. ambiens;
16, M. femoritibialis internus; 17, nonmuscular; 18, M.
psoas; 19, M. iliotrochantericus medius. A. Tympanuchus
cupido pinnatus 3L. B. Pedioecetes phasianellus
jamesi 3L.
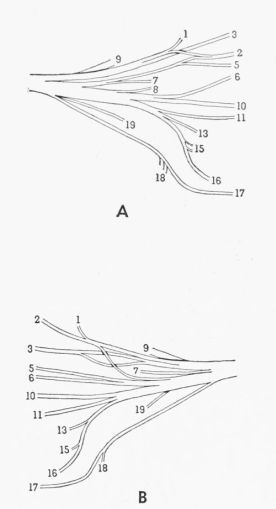 Fig. 5.
Semidiagrammatic ventral views of the femoral
nerve, showing the distribution of the branches. × 3. 1,2, M.
extensor iliotibialis anticus; 3, cutaneous; 5,6, M. extensor
iliotibialis lateralis; 7,8, M. iliacus; 9, M. gluteus
profundus; 10,11, fused Mm. vastus lateralis and vastus
medialis; 13, M. vastus medialis; 15, M. ambiens; 16,
M. femoritibialis internus; 17, nonmuscular; 18, M. psoas;
19, M. iliotrochantericus medius. A. Tympanuchus
pallidicinctus 2L. B. Tympanuchus cupido attwateri 1R.
Fig. 5.
Semidiagrammatic ventral views of the femoral
nerve, showing the distribution of the branches. × 3. 1,2, M.
extensor iliotibialis anticus; 3, cutaneous; 5,6, M. extensor
iliotibialis lateralis; 7,8, M. iliacus; 9, M. gluteus
profundus; 10,11, fused Mm. vastus lateralis and vastus
medialis; 13, M. vastus medialis; 15, M. ambiens; 16,
M. femoritibialis internus; 17, nonmuscular; 18, M. psoas;
19, M. iliotrochantericus medius. A. Tympanuchus
pallidicinctus 2L. B. Tympanuchus cupido attwateri 1R.
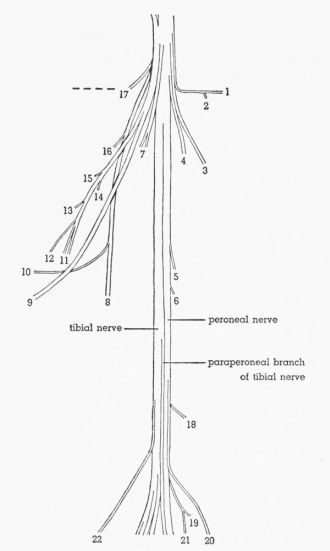 Fig. 6.
Semidiagrammatic dorsolateral view of the sciatic
nerve of Pedioecetes phasianellus jamesi 3R, showing
the distribution of the branches. × 2½. 1, M. gluteus
profundus; 2, M. piriformis; 3, M. extensor iliotibialis
lateralis; 4-7, M. extensor iliofibularis; 8, M. flexor cruris
medialis; 9, cutaneous; 10, to pudendal plexus; 11, M. flexor
cruris lateralis; 12, M. caudofemoralis pars caudifemoralis;
13-15, M. caudofemoralis pars iliofemoralis; 16,17, M. flexor
ischiofemoralis; 18,19, M. femorocruralis (branch of tibial
nerve); 20, cutaneous; 21, M. gastrocnemius pars media (branch
of tibial nerve); 22, cutaneous.
Fig. 6.
Semidiagrammatic dorsolateral view of the sciatic
nerve of Pedioecetes phasianellus jamesi 3R, showing
the distribution of the branches. × 2½. 1, M. gluteus
profundus; 2, M. piriformis; 3, M. extensor iliotibialis
lateralis; 4-7, M. extensor iliofibularis; 8, M. flexor cruris
medialis; 9, cutaneous; 10, to pudendal plexus; 11, M. flexor
cruris lateralis; 12, M. caudofemoralis pars caudifemoralis;
13-15, M. caudofemoralis pars iliofemoralis; 16,17, M. flexor
ischiofemoralis; 18,19, M. femorocruralis (branch of tibial
nerve); 20, cutaneous; 21, M. gastrocnemius pars media (branch
of tibial nerve); 22, cutaneous.
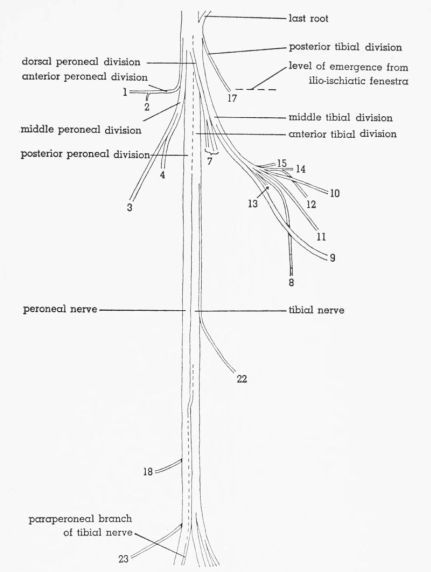 Fig. 7.
Semidiagrammatic dorsolateral view of the sciatic
nerve of Tympanuchus pallidicinctus 2L, showing the
distribution of the branches. × 2½. 1, M. gluteus profundus;
2, M. piriformis; 3, M. extensor iliotibialis lateralis; 4, 7,
M. extensor iliofibularis; 8, M. flexor cruris medialis; 9,
cutaneous; 10, to pudendal plexus; 11, M. flexor cruris
lateralis; 12, M. caudofemoralis pars caudifemoralis; 13-15,
M. caudofemoralis pars iliofemoralis; 17, M. flexor
ischiofemoralis; 18, M. femorocruralis (branch of tibial nerve);
22, cutaneous; 23, nonmuscular (branch of peroneal nerve).
Fig. 7.
Semidiagrammatic dorsolateral view of the sciatic
nerve of Tympanuchus pallidicinctus 2L, showing the
distribution of the branches. × 2½. 1, M. gluteus profundus;
2, M. piriformis; 3, M. extensor iliotibialis lateralis; 4, 7,
M. extensor iliofibularis; 8, M. flexor cruris medialis; 9,
cutaneous; 10, to pudendal plexus; 11, M. flexor cruris
lateralis; 12, M. caudofemoralis pars caudifemoralis; 13-15,
M. caudofemoralis pars iliofemoralis; 17, M. flexor
ischiofemoralis; 18, M. femorocruralis (branch of tibial nerve);
22, cutaneous; 23, nonmuscular (branch of peroneal nerve).
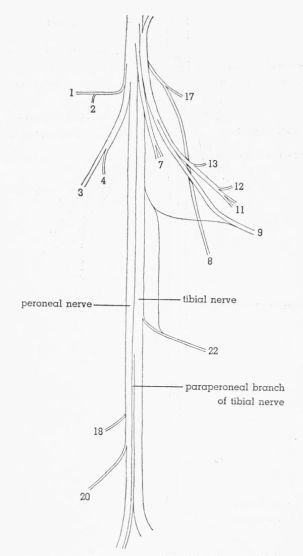 Fig. 8.
Semidiagrammatic dorsolateral view of the
sciatic nerve of Tympanuchus cupido pinnatus 3L,
showing the distribution of the branches. × 2½. 1,
M. gluteus profundus; 2, M. piriformis; 3, M. extensor
iliotibialis lateralis; 4,7, M. extensor iliofibularis;
8, M. flexor cruris medialis; 9, cutaneous; 11, M. flexor
cruris lateralis; 12, M. caudofemoralis pars caudifemoralis;
13, M. caudofemoralis pars iliofemoralis; 17, M. flexor
ischiofemoralis; 18, M. femorocruralis (branch of tibial
nerve); 20, cutaneous; 22, cutaneous.
Fig. 8.
Semidiagrammatic dorsolateral view of the
sciatic nerve of Tympanuchus cupido pinnatus 3L,
showing the distribution of the branches. × 2½. 1,
M. gluteus profundus; 2, M. piriformis; 3, M. extensor
iliotibialis lateralis; 4,7, M. extensor iliofibularis;
8, M. flexor cruris medialis; 9, cutaneous; 11, M. flexor
cruris lateralis; 12, M. caudofemoralis pars caudifemoralis;
13, M. caudofemoralis pars iliofemoralis; 17, M. flexor
ischiofemoralis; 18, M. femorocruralis (branch of tibial
nerve); 20, cutaneous; 22, cutaneous.
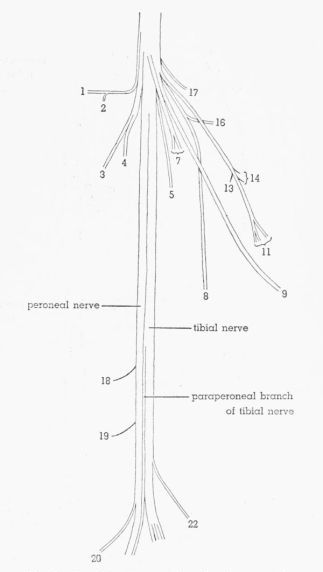 Fig. 9.
Semidiagrammatic dorsolateral view of the
sciatic nerve of Pedioecetes phasianellus jamesi 3L,
showing the distribution of the branches. × 2½. 1,
M. gluteus profundus; 2, M. piriformis; 3, M. extensor
iliotibialis lateralis; 4,5,7, M. extensor iliofibularis;
8, M. flexor cruris medialis; 9, cutaneous; 11, M. flexor
cruris lateralis; 13,14, M. caudofemoralis pars iliofemoralis;
16,17, M. flexor ischiofemoralis; 18,19, M. femorocruralis
(branch of tibial nerve); 20, cutaneous; 22, cutaneous.
Fig. 9.
Semidiagrammatic dorsolateral view of the
sciatic nerve of Pedioecetes phasianellus jamesi 3L,
showing the distribution of the branches. × 2½. 1,
M. gluteus profundus; 2, M. piriformis; 3, M. extensor
iliotibialis lateralis; 4,5,7, M. extensor iliofibularis;
8, M. flexor cruris medialis; 9, cutaneous; 11, M. flexor
cruris lateralis; 13,14, M. caudofemoralis pars iliofemoralis;
16,17, M. flexor ischiofemoralis; 18,19, M. femorocruralis
(branch of tibial nerve); 20, cutaneous; 22, cutaneous.
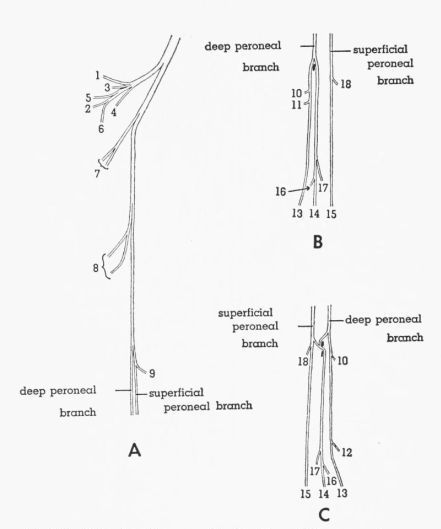 Fig. 10.
A,B. Semidiagrammatic drawings of the peroneal
nerve of Tympanuchus pallidicinctus 1L, showing the
distribution of the branches. × 2. C. Semidiagrammatic drawing
of the distal part of the peroneal nerve of Tympanuchus
cupido attwateri 1R, showing the distribution of the
branches. × 2. 1,2, M. tibialis anticus (tibial head); 3,4,
M. tibialis anticus (femoral head); 5, M. extensor digitorum
longus; 6, nonmuscular; 7,8, M. peroneus longus; 9, M. peroneus
brevis; 10,11, M. extensor hallucis longus (proximal head);
12, M. extensor hallucis longus (distal head); 13-15, nonmuscular
(to toes); 16, M. abductor digiti II; 17, M. extensor brevis
digiti III; 18, M. extensor brevis digiti IV.
Fig. 10.
A,B. Semidiagrammatic drawings of the peroneal
nerve of Tympanuchus pallidicinctus 1L, showing the
distribution of the branches. × 2. C. Semidiagrammatic drawing
of the distal part of the peroneal nerve of Tympanuchus
cupido attwateri 1R, showing the distribution of the
branches. × 2. 1,2, M. tibialis anticus (tibial head); 3,4,
M. tibialis anticus (femoral head); 5, M. extensor digitorum
longus; 6, nonmuscular; 7,8, M. peroneus longus; 9, M. peroneus
brevis; 10,11, M. extensor hallucis longus (proximal head);
12, M. extensor hallucis longus (distal head); 13-15, nonmuscular
(to toes); 16, M. abductor digiti II; 17, M. extensor brevis
digiti III; 18, M. extensor brevis digiti IV.
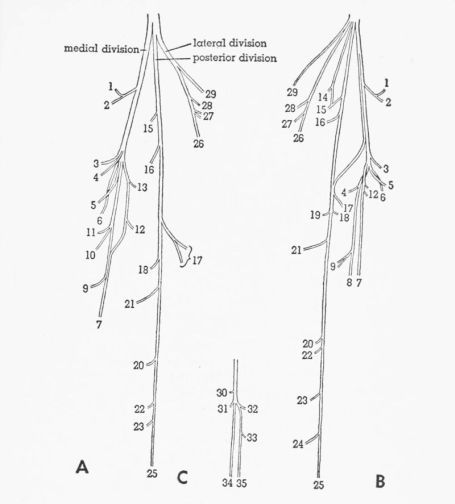 Fig. 11.
A,B. Semidiagrammatic drawings of the tibial
nerve (excluding the paraperoneal branch) of Tympanuchus
pallidicinctus, showing the distribution of the branches.
× 2. A. T.p. 1L. B. T.p. 3R. C. Semidiagrammatic drawing
of the distal part of the paraperoneal branch of the tibial
nerve of Pedioecetes phasianellus jamesi 2L, showing
the distribution of the branches. × 2. 1, M. femorocruralis;
2, M. gastrocnemius pars media; 3, M. popliteus; 4, M. plantaris;
5, M. flexor digitorum longus; 6-8, nonmuscular; 9-11, M.
gastrocnemius pars interna; 12,13, M. flexor hallucis longus;
14-16, M. flexor perforatus digiti IV (medial head); 17, M.
flexor perforatus digiti III (medial head); 18-20, M. flexor
perforatus digiti II; 21, M. flexor perforatus digiti IV
(lateral head); 22-24, M. flexor perforatus digiti IV
(anterolateral head); 25, M. flexor perforatus digiti III
(anterolateral head); 26, M. flexor perforans et perforatus
digiti III; 27,28, M. flexor perforans et perforatus digiti II;
29, M. gastrocnemius pars externa; 30,31, M. abductor digiti IV;
32,33, M. flexor hallucis brevis; 34,35, nonmuscular (to toes).
Fig. 11.
A,B. Semidiagrammatic drawings of the tibial
nerve (excluding the paraperoneal branch) of Tympanuchus
pallidicinctus, showing the distribution of the branches.
× 2. A. T.p. 1L. B. T.p. 3R. C. Semidiagrammatic drawing
of the distal part of the paraperoneal branch of the tibial
nerve of Pedioecetes phasianellus jamesi 2L, showing
the distribution of the branches. × 2. 1, M. femorocruralis;
2, M. gastrocnemius pars media; 3, M. popliteus; 4, M. plantaris;
5, M. flexor digitorum longus; 6-8, nonmuscular; 9-11, M.
gastrocnemius pars interna; 12,13, M. flexor hallucis longus;
14-16, M. flexor perforatus digiti IV (medial head); 17, M.
flexor perforatus digiti III (medial head); 18-20, M. flexor
perforatus digiti II; 21, M. flexor perforatus digiti IV
(lateral head); 22-24, M. flexor perforatus digiti IV
(anterolateral head); 25, M. flexor perforatus digiti III
(anterolateral head); 26, M. flexor perforans et perforatus
digiti III; 27,28, M. flexor perforans et perforatus digiti II;
29, M. gastrocnemius pars externa; 30,31, M. abductor digiti IV;
32,33, M. flexor hallucis brevis; 34,35, nonmuscular (to toes).
In the accounts of the muscles the name used by Hudson, et al. (1959) for each muscle is given in parentheses after the name used by me if the two differ.
In the account of each muscle, the description of the condition found in most specimens of the Lesser Prairie Chicken (T. pallidicinctus) is given first. This is hereafter referred to as the typical condition for T. pallidicinctus. Then any individual variations found within this species are given. Under the heading T. cupido any constant differences between this species and typical T. pallidicinctus are given first, and any individual variations found within the species T. cupido (both subspecies considered together) are given second. Under the heading P. p. jamesi any constant differences between this subspecies and the typical condition for T. pallidicinctus (thus these differences are not necessarily constant between the two genera) are given first, and any individual variations found within the subspecies P. p. jamesi are given second.
In the bird embryo, according to the studies of Romer (1927) and Wortham (1948), the muscles within each segment of the leg differentiate from distinct dorsal or ventral mesenchymal masses. Presumably these represent the primitive dorsal extensor and ventral flexor muscle masses. The list below indicates the ontogenetic origin of the avian leg muscles, according to the studies of Romer and Wortham. The individual muscles are discussed in the order in which they are listed below.
| M. extensor iliotibialis lateralis | M. extensor iliofibularis | |
| M. extensor iliotibialis anticus | M. piriformis | |
| M. ambiens | M. gluteus profundus | |
| M. vastus lateralis | M. iliacus | |
| M. vastus medialis | M. iliotrochantericus medius | |
| M. femoritibialis internus | M. psoas |
| M. flexor cruris lateralis | M. adductor superficialis | |
| M. flexor cruris medialis | M. adductor profundus | |
| M. caudofemoralis | M. obturator | |
| M. flexor ischiofemoralis | M. femorocruralis |
| M. gastrocnemius | M. flexor perforatus digiti III | |
| M. flexor perforans et perforatus digiti II | M. flexor perforatus digiti II | |
| M. flexor perforans et perforatus digiti III | M. flexor hallucis longus | |
| M. flexor perforatus digiti IV | M. plantaris | |
| M. flexor digitorum longus | ||
| M. popliteus |
| M. peroneus longus | M. extensor digitorum longus | |
| M. tibialis anticus | M. peroneus brevis |
| M. extensor hallucis longus | M. extensor proprius digiti III | |
| M. abductor digiti II | M. extensor brevis digiti IV | |
| M. extensor brevis digiti III |
| M. lumbricalis | (M. adductor digiti II—not present) | |
| M. abductor digiti IV | (M. adductor digiti IV—not present) | |
| M. flexor hallucis brevis |
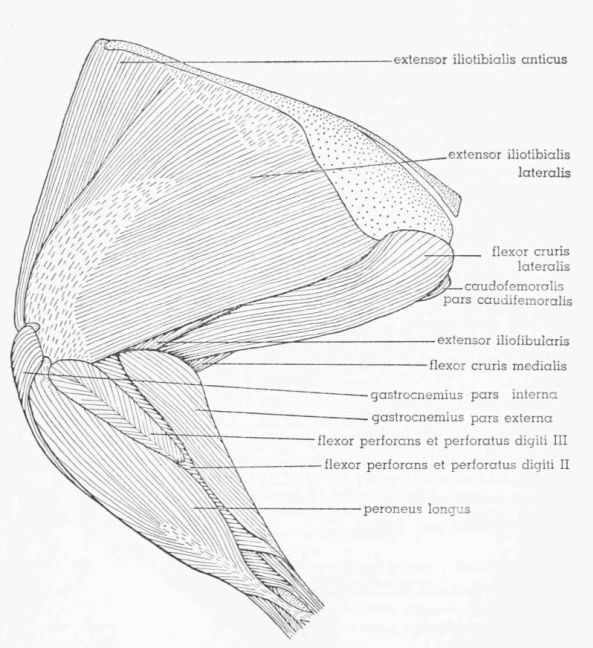 Fig. 12.
Tympanuchus pallidicinctus 2L. Lateral
view of the superficial muscles of the left leg. × 1.
Fig. 12.
Tympanuchus pallidicinctus 2L. Lateral
view of the superficial muscles of the left leg. × 1.
M. Extensor Iliotibialis Lateralis (M. iliotibialis), Figs. 12, 13, 20F, G
T. pallidicinctus
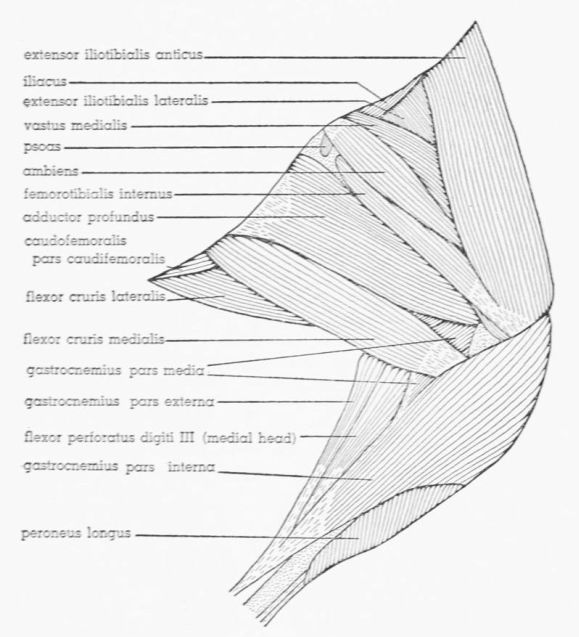 Fig. 13.
Tympanuchus pallidicinctus 2L. Medial
view of the superficial muscles of the left leg. × 1. Articular
capsule shown by concentrically arranged dashes.
Fig. 13.
Tympanuchus pallidicinctus 2L. Medial
view of the superficial muscles of the left leg. × 1. Articular
capsule shown by concentrically arranged dashes.
General Description and Relations.—Most superficial muscle on lateral surface of thigh; broad, flat, and triangular; bounded anteriorly by M. extensor iliotibialis anticus and posteriorly by M. flexor cruris lateralis; posterior part considerably thicker than anterior part; anteroproximal and centrodistal parts aponeurotic; extreme posteroproximal corner also aponeurotic (could be considered tough sheet of connective tissue intimately fused with M. extensor iliotibialis lateralis, rather than part of muscle itself; see Fig. 20F); latter aponeurosis, as well as adjacent fleshy fibers, overlapped by M. flexor cruris lateralis; this aponeurosis fused with posterior end of underlying M. caudofemoralis pars iliofemoralis; centrodistal aponeurosis tightly fused to underlying Mm. vastus lateralis and vastus medialis; fleshy fibers posterior to this aponeurosis also fused with M. vastus lateralis, although posterior third of [Pg 399] muscle free; fleshy part anterior to this aponeurosis bound by tough connective tissue to underlying M. vastus medialis, although no fusion of fibers; anterior edge tightly bound by strong connective tissue to M. extensor iliotibialis anticus, with some fusion of fibers (proximally); posteroproximal corner bound by tough connective tissue to adjacent muscles; anteroproximal aponeurosis fused with aponeurotic anteroproximal part of underlying M. extensor iliofibularis. Continuous proximal aponeurosis of M. extensor iliotibialis anticus and of M. extensor iliotibialis lateralis underlain by tough fascial sheet overlying M. gluteus profundus; anterior part of this fascia tightly fused to latter muscle but free from overlying aponeurosis; posterior part of this fascia tightly fused to overlying aponeurosis but free from M. gluteus profundus; middle part of fascia fused to both aponeurosis and M. gluteus profundus.
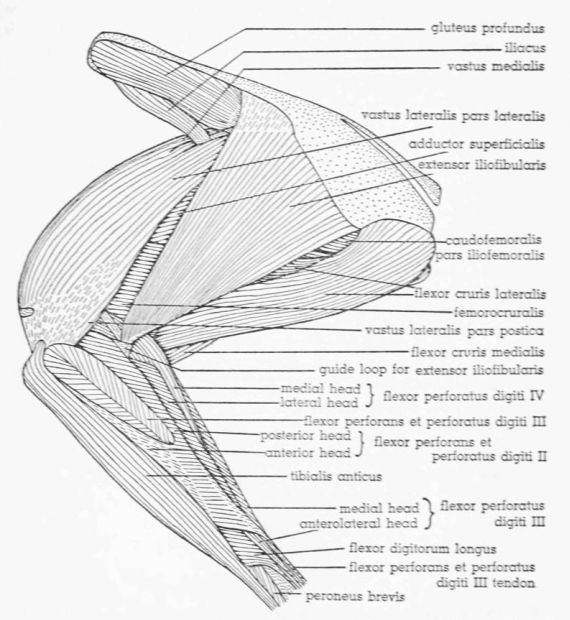 Fig. 14.
Tympanuchus pallidicinctus 2L. Lateral
view of the muscles of the left leg. The following muscles have
been removed: extensor iliotibialis lateralis, extensor
iliotibialis anticus, gastrocnemius pars externa and pars
interna, and peroneus longus. × 1.
Fig. 14.
Tympanuchus pallidicinctus 2L. Lateral
view of the muscles of the left leg. The following muscles have
been removed: extensor iliotibialis lateralis, extensor
iliotibialis anticus, gastrocnemius pars externa and pars
interna, and peroneus longus. × 1.
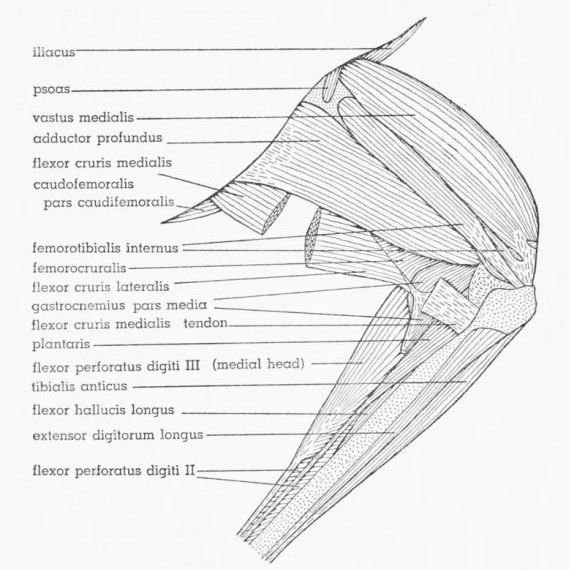 Fig. 15.
Tympanuchus pallidicinctus 2L. Medial
view of the muscles of the left leg. The following muscles have
been removed: extensor iliotibialis lateralis, extensor
iliotibialis anticus, ambiens, flexor cruris lateralis (in part),
flexor cruris medialis (in part), gastrocnemius pars externa
and pars interna, and peroneus longus. × 1.
Fig. 15.
Tympanuchus pallidicinctus 2L. Medial
view of the muscles of the left leg. The following muscles have
been removed: extensor iliotibialis lateralis, extensor
iliotibialis anticus, ambiens, flexor cruris lateralis (in part),
flexor cruris medialis (in part), gastrocnemius pars externa
and pars interna, and peroneus longus. × 1.
Origin.—Approximately the anterior half attaches by an extensive aponeurosis, which is continuous anteriorly with that of M. extensor iliotibialis anticus, to the anterior iliac crest, ending posteriorly at the anterior end of the lateral iliac process; the posterior part attaches fleshily to the edge of the entire lateral iliac process and (posterior few mm.) aponeurotically to the entire lateral ischiatic ridge. The proximal part of the belly is much thicker than the fleshy origin. Two accessory aponeuroses associate with the anterior part of the muscle; the proximal one of these comes off the deep surface several mm. distal to the proximal end of the fleshy belly and passes medially between Mm. gluteus profundus and iliacus, fusing to both these muscles, and attaches to the lateral edge of M. iliotrochantericus medius and to the lateral edge of the ilium anterior to the latter; the aponeurosis actually splits into two sheets at the edge of M. iliotrochantericus medius; these sheets fuse to the dorsal and ventral surfaces of the latter muscle, enclosing it; the part of this aponeurosis between Mm. iliacus and iliotrochantericus medius is strongly fused with the underlying body wall. The distal accessory aponeurosis [Pg 401] (sometimes weak) comes off the deep surface several mm. distal to the proximal one and passes medially along the ventral surface of M. iliacus, fusing with the latter, then joining the proximal accessory aponeurosis medial to M. iliacus.
Insertion.—The muscle inserts by a broad aponeurosis strongly fused to the underlying Mm. vastus lateralis and vastus medialis; the aponeurosis contributes superficially to the patellar tendon, attaching to the lateral half of the rotular crest.
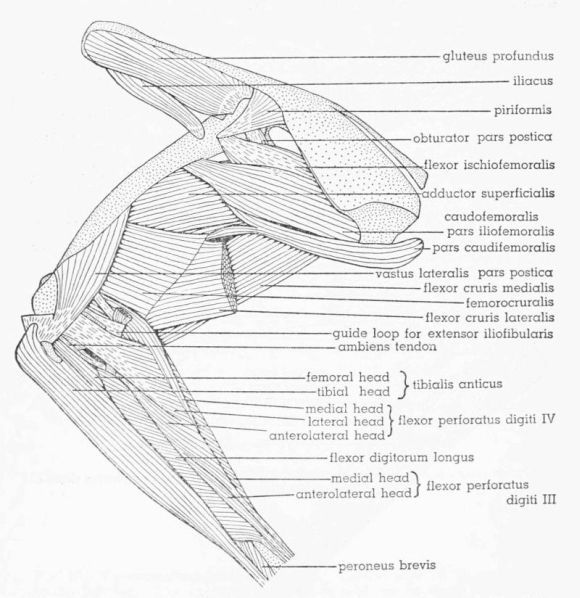 Fig. 16.
Tympanuchus pallidicinctus 2L. Lateral
view of the muscles of the left leg. The following muscles,
in addition to those listed for Fig. 14, have been removed:
ambiens, vastus lateralis pars lateralis, vastus medialis (except
for part of patellar tendon), extensor iliofibularis, flexor
cruris lateralis (in part), flexor perforans et perforatus
digiti II, and flexor perforans et perforatus digiti III. × 1.
Fig. 16.
Tympanuchus pallidicinctus 2L. Lateral
view of the muscles of the left leg. The following muscles,
in addition to those listed for Fig. 14, have been removed:
ambiens, vastus lateralis pars lateralis, vastus medialis (except
for part of patellar tendon), extensor iliofibularis, flexor
cruris lateralis (in part), flexor perforans et perforatus
digiti II, and flexor perforans et perforatus digiti III. × 1.
Innervation.—A variable number of branches (usually two) of the middle division of the femoral nerve pass ventral to M. iliacus and between Mm. extensor iliotibialis anticus and vastus medialis and enter the deep surface of the anteroproximal part of the muscle. The branch of the middle peroneal [Pg 402] division of the sciatic nerve emerges between the proximal ends of Mm. extensor iliofibularis and vastus lateralis and sends twigs into the deep surface of M. extensor iliotibialis lateralis.
Individual Variation.—In two legs, the nerve supplying M. extensor iliotibialis anticus gives twigs into M. extensor iliotibialis lateralis.
T. cupido
Differences From T. pallidicinctus.—The fleshy origin from the lateral iliac process is considerably thicker (reflected in a thicker lateral iliac process).
Individual Variation.—In three legs the nerve supplying M. extensor iliotibialis anticus gives twigs into M. extensor iliotibialis lateralis. In another leg one of the branches to the fused Mm. vastus lateralis and vastus medialis sends a twig into M. extensor iliotibialis lateralis.
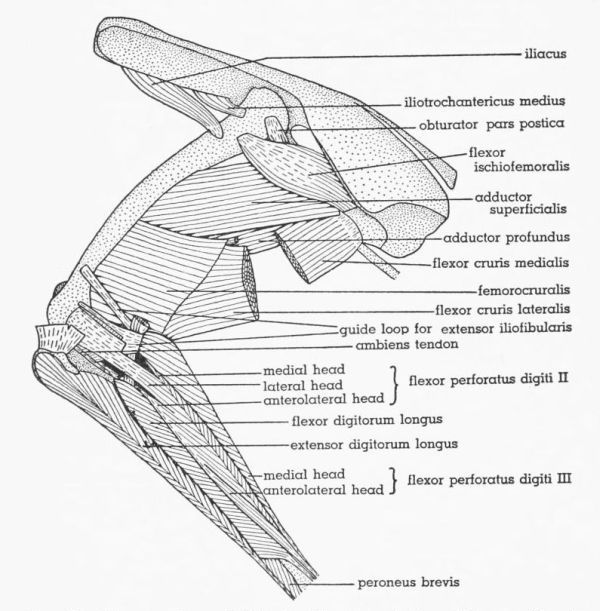 Fig. 17.
Tympanuchus pallidicinctus 2L. Lateral
view of the muscles of the left leg. The following muscles, in
addition to those listed for Fig. 16, have been removed: vastus
lateralis pars postica, gluteus profundus, flexor cruris medialis
(in part), caudofemoralis, flexor perforatus digiti IV, and
tibialis anticus. × 1.
Fig. 17.
Tympanuchus pallidicinctus 2L. Lateral
view of the muscles of the left leg. The following muscles, in
addition to those listed for Fig. 16, have been removed: vastus
lateralis pars postica, gluteus profundus, flexor cruris medialis
(in part), caudofemoralis, flexor perforatus digiti IV, and
tibialis anticus. × 1.
P. p. jamesi
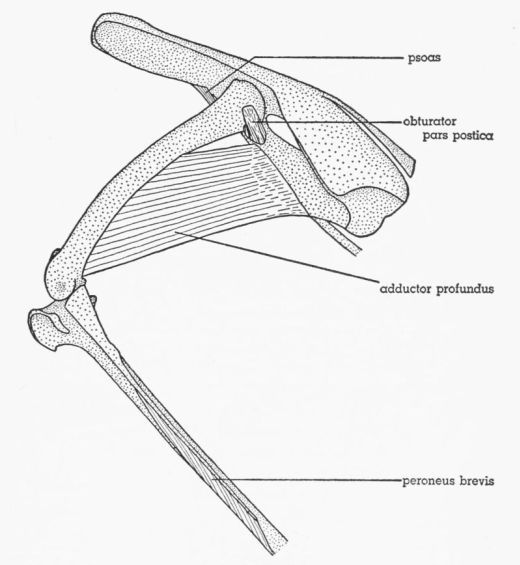 Fig. 18.
Tympanuchus pallidicinctus 2L. Lateral
view of the muscles of the left leg. The following muscles, in
addition to those listed for Fig. 17, have been removed: patellar
tendon, iliacus, iliotrochantericus medius, flexor cruris
lateralis, flexor cruris medialis, flexor ischiofemoralis,
adductor superficialis, femorocruralis, gastrocnemius pars media,
flexor perforatus digiti III, flexor perforatus digiti II, flexor
hallucis longus, plantaris, flexor digitorum longus, popliteus,
and extensor digitorum longus. × 1.
Fig. 18.
Tympanuchus pallidicinctus 2L. Lateral
view of the muscles of the left leg. The following muscles, in
addition to those listed for Fig. 17, have been removed: patellar
tendon, iliacus, iliotrochantericus medius, flexor cruris
lateralis, flexor cruris medialis, flexor ischiofemoralis,
adductor superficialis, femorocruralis, gastrocnemius pars media,
flexor perforatus digiti III, flexor perforatus digiti II, flexor
hallucis longus, plantaris, flexor digitorum longus, popliteus,
and extensor digitorum longus. × 1.
Differences From Typical T. pallidicinctus.—The posteroproximal aponeurosis is more extensive, resulting in a narrower proximal fleshy end (Fig. 20G); the fleshy fibers adjacent to this aponeurosis are not overlapped by M. flexor cruris lateralis. There is a fusion of fibers between the anterodistal fleshy part of M. extensor iliotibialis lateralis and the underlying M. vastus medialis, but there is no fusion of fibers between the anterior edge of M. extensor iliotibialis lateralis and M. extensor iliotibialis anticus. The connective tissue binding the posteroproximal corner to adjacent muscles is stronger. The fleshy part of the origin is narrower, partly tendinous, and much thinner (reflected in a thin lateral iliac process). The proximal border is much more nearly straight, owing to a less pronounced lateral iliac process. The distal accessory aponeurosis is absent.
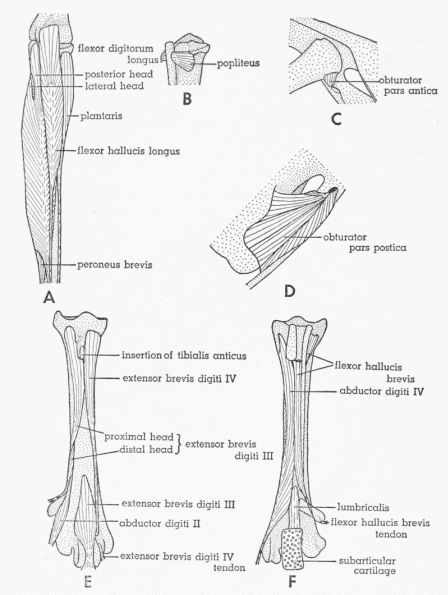 Fig. 19.
Tympanuchus pallidicinctus 2L. A.
Posterior view of the muscles of the left shank. The following
shank muscles, in addition to those listed for Fig. 17, have
been removed: gastrocnemius pars media, flexor perforatus
digiti III, and flexor perforatus digiti II. × 1. B. Posterior
view of the proximal end of the shank, showing the most deeply
situated muscle. × 1. C. Lateral view of the head of the left
femur and the middle part of the pelvis, showing the deepest
part of M. obturator. × 1. D. Medial view of the posteroventral
part of the left side of the pelvis, showing the intrapelvic part
of M. obturator. × 1. E. Anterior view of the left
tarsometatarsus, showing the dorsal intrinsic muscles of the
foot. × 1½. F. Posterior view of the left tarsometatarsus,
showing the ventral intrinsic muscles of the foot. × 1½.
Fig. 19.
Tympanuchus pallidicinctus 2L. A.
Posterior view of the muscles of the left shank. The following
shank muscles, in addition to those listed for Fig. 17, have
been removed: gastrocnemius pars media, flexor perforatus
digiti III, and flexor perforatus digiti II. × 1. B. Posterior
view of the proximal end of the shank, showing the most deeply
situated muscle. × 1. C. Lateral view of the head of the left
femur and the middle part of the pelvis, showing the deepest
part of M. obturator. × 1. D. Medial view of the posteroventral
part of the left side of the pelvis, showing the intrapelvic part
of M. obturator. × 1. E. Anterior view of the left
tarsometatarsus, showing the dorsal intrinsic muscles of the
foot. × 1½. F. Posterior view of the left tarsometatarsus,
showing the ventral intrinsic muscles of the foot. × 1½.
Individual Variation.—The muscle is usually somewhat fused to the posteroproximal and anteroproximal fleshy corners of the underlying M. extensor iliofibularis.
M. Extensor Iliotibialis Anticus (M. sartorius), Figs. 12, 13
T. pallidicinctus
General Description and Relations.—Anteriormost muscle of thigh; long and strap-shaped; proximal part entirely anterior (adjacent) to M. extensor iliotibialis lateralis; posterior edge of middle part medial to latter muscle; distal part mostly medial to Mm. extensor iliotibialis lateralis and vastus medialis; proximal part aponeurotic, continuous posteriorly with anteroproximal aponeurosis of M. extensor iliotibialis lateralis; anterior edge of M. extensor iliotibialis lateralis bound by strong connective tissue to adjacent part of M. extensor iliotibialis anticus; some fusion of fibers (proximally) between these two muscles; anteroproximal corner of fleshy part of muscle sometimes fused to underlying anterior edge of ilium and fascia covering body wall musculature adjacent (anterior) to ilium.
Origin.—The muscle arises aponeurotically from the anterior part of the anterior iliac crest and (anteroproximal corner) from the anterior end of the median dorsal ridge.
Insertion.—The flat tendon, continuous posteriorly with the superficial tendon of M. femoritibialis internus, fuses to the tendon of M. vastus medialis, contributing superficially to the medial part of the patellar tendon, which attaches to the medial half of the rotular crest; most of the tendon is overlapped by the edge of M. gastrocnemius pars interna.
Innervation.—A branch of the anterior division of the femoral nerve gives twigs into the lateral surface of the posterior part.
Individual Variation.—In two legs, a twig from the anteriormost branch of the middle division of the femoral nerve anastomoses with the typical branch to M. extensor iliotibialis anticus.
T. cupido
Individual Variation.—In several legs, the anterior edge of origin extends forward onto the neural spine of the last free thoracic vertebra. A twig from the middle division of the femoral nerve anastomoses with the typical branch to M. extensor iliotibialis anticus in three legs.
P. p. jamesi
Differences From Typical T. pallidicinctus.—There is no fusion of fibers between M. extensor iliotibialis anticus and M. extensor iliotibialis lateralis.
Individual Variation.—The anterior edge of origin extends forward onto the neural spine of the last free thoracic vertebra in some legs.
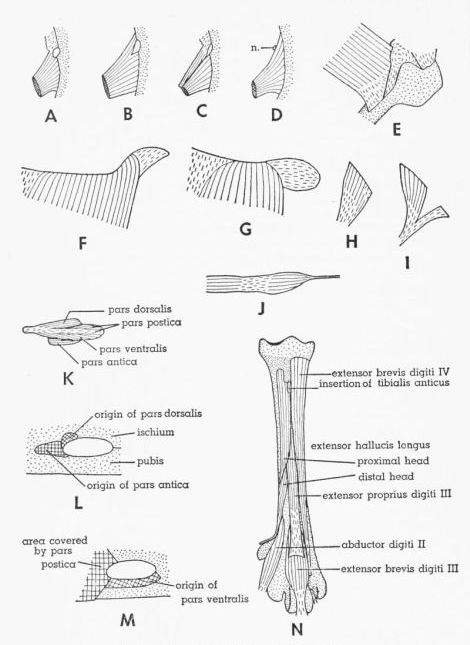 Fig. 20.
A-D. Dorsal views of M. iliotrochantericus
medius, showing its relationship to femoral notch. × 1. In D,
note absence of femoral notch and location of branch of femoral
nerve. A. Tympanuchus pallidicinctus 2L. B. T. cupido
pinnatus 4L. C. Pedioecetes phasianellus jamesi 1L.
D. T. pallidicinctus 3L.
Fig. 20.
A-D. Dorsal views of M. iliotrochantericus
medius, showing its relationship to femoral notch. × 1. In D,
note absence of femoral notch and location of branch of femoral
nerve. A. Tympanuchus pallidicinctus 2L. B. T. cupido
pinnatus 4L. C. Pedioecetes phasianellus jamesi 1L.
D. T. pallidicinctus 3L.T. pallidicinctus
General Description and Relations.—Thin and elongate; on medial surface of thigh; broadest above middle of belly; belly narrowed distally, forming long slender tendon passing lateral to distal part of M. extensor iliotibialis anticus; bounded anterolaterally by M. vastus medialis and posterolaterally by Mm. femoritibialis internus and psoas (proximally).
Origin.—The muscle arises by a short flat tendon from the pectineal process.
Insertion.—The long slender tendon enters an elongate channel within the patellar tendon; the point of entrance is at the proximal end of the latter tendon just medial to the patella; the tendon passes distolaterally (within the channel) below the patella and emerges from the distolateral edge of the patellar tendon and then extends distally along the anterolateral surface of the head of the fibula, superficial to the fibular arm of the guide loop for M. extensor iliofibularis, and joins the anterolateral surface of the common tendon of origin of the anterolateral heads of Mm. flexor perforatus digiti III, flexor perforatus digiti IV, and flexor perforatus digiti II; the point of junction is usually immediately proximal to the proximal end of the lateral head of M. flexor digitorum longus.
Innervation.—The branch of the middle division of the femoral nerve that supplies M. femoritibialis internus gives off a tiny twig or twigs that penetrate the lateral surface of the proximal part of M. ambiens.
Individual Variation.—None of significance in T. pallidicinctus or in P. p. jamesi; in T. cupido the origin is partly fleshy in one leg.
M. Vastus Lateralis (M. femoritibialis externus + part of M. femoritibialis medius), Figs. 14, 16
Fisher and Goodman (1955) apply the name femoritibialis externus to the muscle unit that I here term the pars postica of M. vastus lateralis. The reasons for this change are discussed in the section on terminology.
T. pallidicinctus
General Description and Relations.—Thick; on lateral surface of femur deep to M. extensor iliotibialis lateralis; anterior to M. extensor iliofibularis and lateral to M. vastus medialis; much of lateral surface, except proximal part, fused with overlying M. extensor iliotibialis lateralis; deep surface of anterior half fused with M. vastus medialis; proximal part overlapping, but usually not fusing with, insertions of Mm. iliacus and caudofemoralis; partially separable into two parts—pars lateralis and pars postica, former constituting main part of muscle; latter considerably smaller and situated deep to posterodistal part of pars lateralis, except for posterodistal part extending posterior to edge of pars lateralis; proximal part of pars postica strongly fused with pars lateralis; posterodistal tendinous edge of pars lateralis fused or not fused with lateral surface of pars postica; proximal end (narrow) of pars postica tendinous and variable in length.
Origin.—Pars lateralis: This arises fleshily from most of the lateral surface and (distally) from the anterior surface of the femur, extending anteriorly to [Pg 409] the anterior intermuscular line, fusing with M. vastus medialis, and extending posteriorly to the posterolateral intermuscular line (proximally) and the origin of pars postica (distally); the proximal end begins at the level of the distal edge of the insertion of M. iliotrochantericus medius, contacting the insertions of Mm. iliotrochantericus medius, piriformis, and flexor ischiofemoralis, and terminates distally at the level of the proximal ends of the femoral condyles.
Pars postica: This arises fleshily and tendinously (proximal end and deep surface) from the posterolateral surface of approximately the distal half of the femur, extends posteromedially to the posterolateral intermuscular line where it contacts the origin of M. femorocruralis, and extends anteriorly to the level of a line drawn diagonally across the femur from the proximal end of the origin (at the posterolateral intermuscular line) to the proximal end of the external condyle; the distal end is anterior (adjacent) to the attachment of the proximal arm of the tendinous guide loop for M. extensor iliofibularis; the origin is adjacent to, but distinct from, the origin of pars lateralis.
Insertion.—Pars lateralis is fused indistinguishably with M. vastus medialis; these two muscles form the main (middle) part of the patellar tendon, which also receives contributions from pars postica and Mm. femoritibialis internus, extensor iliotibialis lateralis, and extensor iliotibialis anticus; the patellar tendon attaches to the entire rotular crest of the tibia; the patella is situated in the proximal part of this tendon; some deep fleshy fibers of M. vastus lateralis pars lateralis and M. vastus medialis attach to the proximal edge of the patella. Pars postica forms a short narrow tendon that fuses to the lateral part of the tendon of pars lateralis, forming the lateralmost part of the patellar tendon. A broad flat vinculum extends from the lateral surface of the femorofibular fascia (defined under M. flexor perforans et perforatus digiti II) to the deep surface of the lateral part of the patellar tendon; a similar vinculum extends from the medial surface of the internal condyle to the deep surface of the medial part of the patellar tendon.
Innervation.—Two or more branches of the middle division of the femoral nerve penetrate the anterior surface of the fused Mm. vastus lateralis (pars lateralis) and vastus medialis; short twigs emerge from the deep surface of pars lateralis and penetrate the superficial surface of the anteroproximal part of pars postica.
Individual Variation.—The proximal ends of M. vastus medialis and M. vastus lateralis are usually separated by a deep notch. In some legs, a small bundle of fibers forming the anteroproximal part of M. vastus lateralis attaches to the lateral surface of M. vastus medialis anterior to this notch.
T. cupido
Individual Variation.—One leg shows the same variation found in T. pallidicinctus (see above). In several legs, pars lateralis does not extend so far proximally as usual, but begins at the level of insertion of M. piriformis (does not contact the insertion of M. iliotrochantericus medius) and may not overlap M. iliacus. In a few legs, no vincula are associated with the patellar tendon.
P. p. jamesi
Individual Variation.—Pars lateralis often begins proximally at the level of the insertion of M. piriformis.
M. Vastus Medialis (Part of M. femoritibialis medius), Figs. 13, 14, 15
T. pallidicinctus
General Description and Relations.—Thick; on anteromedial surface of femur medial to anterior part of M. vastus lateralis pars lateralis; bounded medially by Mm. ambiens and extensor iliotibialis anticus (distally); bounded posteromedially by M. femoritibialis internus; proximal part medial to posterior ends of Mm. iliacus, iliotrochantericus medius, and gluteus profundus; lateral surface, except proximal part, fused with anterior part of M. vastus lateralis pars lateralis; part of lateral surface of M. vastus medialis covered by sheet of fascia attaching to anterior intermuscular line; M. vastus lateralis separable from this fascia, but fascia absent anteriorly and distally and these two muscles indistinguishably fused.
Origin.—The proximal third is attached narrowly by its lateral edge; the distal two thirds is attached broadly by its entire deep surface. The proximal third arises tendinously from the trochanteric ridge and the proximal end of the anterior intermuscular line and fleshily from a narrow area of the femur adjacent (medial) to the latter; the distal part arises tendinously from the anterior intermuscular line and fleshily from a broad adjacent area on the anteromedial surface of the femur, terminating distally at the level of the proximal end of the internal condyle; the posterior edge contacts the origin of M. femoritibialis internus.
Insertion.—Attachment is in common with M. vastus lateralis pars lateralis, which see.
Innervation.—Two or more branches of the middle division of the femoral nerve penetrate the anterior surface of the fused Mm. vastus medialis and vastus lateralis pars lateralis; a variable number of branches of the same division penetrate the medial surface of the proximal part of M. vastus medialis.
Individual Variation.—None of significance in any of the three species studied.
M. Femoritibialis Internus, Figs. 13, 15
T. pallidicinctus
General Description and Relations.—Elongate; on posteromedial surface of femur; bounded anteriorly by M. vastus medialis and posteriorly by M. adductor profundus (overlapping anterior edge of latter); anteroproximal part lateral to M. ambiens; anterodistal corner deep to distal end of M. extensor iliotibialis anticus; distal part of muscle split into superficial and deep layers; superficial layer thin, narrow, and tendinous except for proximal end; deep layer wider, much thicker, and fleshy except for distal end taking form of flat tendon; anterior edge of latter somewhat fused to medial edge of tendon of M. vastus medialis; deep layer widest near distal end of fleshy part; posterior edge of superficial layer fused to underlying deep layer, and anterior edge fused to (continuous with) posterior edge of tendon of M. extensor iliotibialis anticus.
Origin.—The origin is mostly fleshy from the posteromedial surface of the femur between the origin of M. vastus medialis and the posterior intermuscular line, terminating immediately proximal to the internal condyle.
Insertion.—The tendons of both superficial and deep layers attach to the medial part of the rotular crest, forming the medialmost part of the patellar tendon.
Innervation.—The posteriormost branch of the middle division of the femoral nerve penetrates the medial surface of the muscle near the proximal end.
Individual Variation.—None of significance in any of the three species studied.
M. Extensor Iliofibularis (M. biceps femoris), Figs. 12, 14, 16, 17
The term extensor in the name of this muscle does not refer to its function. Howell (1938) used the term extensor to indicate derivation of the muscle from the primitive dorsal extensor muscle mass. (Likewise he used the term flexor to indicate derivation from the primitive ventral flexor muscle mass.)
T. pallidicinctus
General Description and Relations.—Deep to M. extensor iliotibialis lateralis and posterior to femur; broad proximally and narrow distally; posterior to M. vastus lateralis and anterior to proximal part of M. flexor cruris lateralis (superficial to distal part of latter); anteroproximal part aponeurotic, fused to deep surface of aponeurosis of M. extensor iliotibialis lateralis; proximal part of aponeurosis of M. extensor iliofibularis also fused to dorsal edges of underlying Mm. gluteus profundus and piriformis.
Origin.—The posterior part is fleshy from the ventromedial surface of the entire lateral iliac process; the anterior part is aponeurotic from the posterior part of the anterior iliac crest.
Insertion.—The tendon forms along the posterodistal edge of the belly and continues beyond the end of the belly as a cylindrical tendon that passes through the tendinous guide loop (the belly terminates approximately at the level of the guide loop), then extends anterodistally into the shank musculature; the tendon passes between the medial and lateral heads of M. flexor perforatus digiti IV, between the medial and lateral heads of M. flexor perforatus digiti II, lateral to the common tendon of the anterolateral heads of Mm. flexor perforatus digiti IV, flexor perforatus digiti II, and flexor perforatus digiti III, and between the posterior and lateral heads of M. flexor digitorum longus, attaching to the fibular tubercle.
The tendinous guide loop has three arms—proximal femoral, distal femoral, and fibular; the proximal and distal femoral arms unite posterior to the tendon of M. extensor iliofibularis; the proximal arm is medial to, and the distal arm is lateral to, the latter; the fibular arm joins the distal edge of the distal arm lateral to the tendon of M. extensor iliofibularis. The proximal arm extends anteroproximally lateral to the medial head of M. flexor perforatus digiti IV and medial to M. vastus lateralis pars postica, attaching to a narrow line on the anterolateral surface of the femur a short distance proximal to the external condyle and adjacent (posterior) to the origin of M. vastus lateralis pars postica. The distal arm extends anteriorly medial to the posterior head of M. flexor perforans et perforatus digiti II and medial to M. vastus lateralis pars postica, attaching in common with the tendon of origin of M. gastrocnemius pars externa to a small oval area on the posterolateral surface[Pg 412] of the femur a short distance proximal to the fibular groove; the arm is also fused to the underlying articular capsule. The fibular arm (broadest of the three) passes deep to, and fused with, the common tendon of origin of the lateral heads of Mm. flexor perforatus digiti IV and flexor perforatus digiti II, superficial to the common tendon of origin of the anterolateral heads of Mm. flexor perforatus digiti IV, flexor perforatus digiti II, and flexor perforatus digiti III, and deep to the tendon of M. ambiens, attaching broadly to a narrow line on the anterolateral surface of the proximal part of the fibula; the arm is also fused to the underlying articular capsule.
Innervation.—A branch of the middle peroneal division of the sciatic nerve sends twigs to the deep surface of the anteroproximal part; the dorsal peroneal division of the sciatic nerve penetrates the deep surface of the proximal end.
Individual Variation.—In some instances a variable number of twigs arises from the peroneal nerve near the middle of the thigh and enters the deep surface of the muscle. They are difficult to expose without breaking and may have been overlooked in some specimens.
T. cupido
Individual Variation.—The same variation is found as in T. pallidicinctus (see above). In one leg, the tendon of insertion bifurcates into proximal and distal arms before attaching.
P. p. jamesi
Differences from Typical T. pallidicinctus.—It arises from the ventral rather than the ventromedial surface of the lateral iliac process (there is no ventromedial surface to this process).
Individual Variation.—In nearly all of the legs, minute twigs to M. extensor iliofibularis come off the peroneal nerve near the middle of the thigh. The insertional tendon tends toward doubleness in two legs.
M. Piriformis (M. gluteus medius et minimus), Figs. 16, 20H, I
T. pallidicinctus
General Description and Relations.—Small, thin, and triangular; lateral to antitrochanter and posterior part of trochanter; deep to M. extensor iliofibularis and posterior (adjacent) to M. gluteus profundus; distal half (or more) tendinous.
Origin.—The muscle arises fleshily from the posterior end of the anterior iliac crest (ventral to the origins of Mm. extensor iliotibialis lateralis and extensor iliofibularis) beginning adjacent to the posterior end of M. gluteus profundus.
Insertion.—The flat tendon narrows, overlaps the anteroproximal corner of insertion of M. flexor ischiofemoralis, and attaches to the lateral surface of the proximal part of the femur immediately anterior to the insertion of M. flexor ischiofemoralis and posterior to the proximal end of M. vastus lateralis; the attachment is posterodistal to the insertion of M. iliotrochantericus medius and posteroproximal to the insertion of M. iliacus.
Innervation.—The small anterior peroneal division of the sciatic nerve turns anteriorly immediately after emerging from the ilio-ischiatic fenestra and passes deep to M. piriformis, giving twigs to the deep surface.
Individual Variation.—In both legs of one specimen, the insertion does not overlap the insertion of M. flexor ischiofemoralis. The posteroproximal corner of the muscle is tendinous in one leg.
T. cupido
Individual Variation.—The anterior border is somewhat fused with the posterior edge of M. gluteus profundus in one leg, while in another there is a slight gap between the origins of M. gluteus profundus and M. piriformis. In one leg, the posterior edge of the origin is aponeurotic. On both sides of one specimen, an accessory tendinous band arises several mm. posterior to the main part of M. piriformis and joins the proximal part of the insertional tendon, thus forming a Y-shaped unit (Fig. 20I); the accessory tendon arises from the anterior end of the lateral iliac process (left side) or from the anterior part of the lateral iliac fossa (right side). The insertion may be proximal (rather than posterior) to the proximal end of M. vastus lateralis. In one leg, the insertional tendon is partly fused to the insertional tendon of M. flexor ischiofemoralis.
P. p. jamesi
Individual Variation.—There is often a gap between the origins of M. gluteus profundus and M. piriformis. In one leg (Fig. 20H), the posteroproximal corner of the muscle is aponeurotic. The insertion is often proximal (rather than posterior) to the proximal end of M. vastus lateralis. In one instance, the insertion does not overlap the insertion of M. flexor ischiofemoralis.
M. Gluteus Profundus (M. iliotrochantericus posterior), Figs. 14, 16
T. pallidicinctus
General Description and Relations.—Large and thick; covering dorsolateral surface of entire preacetabular part of ilium; deep to Mm. extensor iliotibialis lateralis and extensor iliotibialis anticus; bounded posteriorly by M. piriformis and ventrally by M. iliacus; ventral edge fused with anterior part of latter and with proximal accessory aponeurosis of M. extensor iliotibialis lateralis; tough sheet of fascia strongly fused to anterior two thirds of lateral surface; posterior to this, fascia overlying muscle but not attaching to it; posterior half of fascia fused to overlying aponeurosis of M. extensor iliotibialis lateralis; deep surface of muscle somewhat fused to proximal part of M. iliotrochantericus medius.
Origin.—The superficial surface is tendinous from the entire anterior iliac crest except the posterior end and from the crest forming the anterior and anterolateral edges of the ilium; the muscle arises fleshily from the entire dorsolateral surface of the preacetabular ilium as far posteriorly as the level of the pectineal process; the dorsal edge is adjacent (anterior) to the origin of M. piriformis.
Insertion.—The attachment is by a short, wide, thick tendon to a curved line (convex anteriorly) on the lateral surface of the femoral trochanter.
Innervation.—The anterodorsal division of the femoral nerve turns dorsally[Pg 414] through the femoral notch of the ilium and penetrates the deep surface of the ventral part of the muscle midway of its length; the anterior peroneal division of the sciatic nerve passes deep to M. piriformis and terminates near the posterodorsal edge of M. gluteus profundus.
Individual Variation.—On both sides of one specimen, the branch from the femoral nerve passes lateral to the extreme anteroproximal corner of M. iliotrochantericus medius instead of through the femoral notch.
T. cupido
Individual Variation.—In one leg, the insertional tendon is strongly fused to the insertional tendon of M. iliotrochantericus medius.
P. p. jamesi
Individual Variation.—None of significance.
M. Iliacus (M. iliotrochantericus anterior), Figs. 13, 14, 15, 16, 17
T. pallidicinctus
General Description and Relations.—Adjacent ventrally to ventrolateral edge of M. gluteus profundus; lateral edge much thicker than medial edge; deep to M. extensor iliotibialis lateralis and anterolateral to M. iliotrochantericus medius; distal (posterior) end passing between proximal ends of Mm. vastus medialis and vastus lateralis pars lateralis; insertion overlapped by latter; dorsal surface of anterior part fused with ventrolateral edge of M. gluteus profundus and with ventral surface of proximal accessory aponeurosis of M. extensor iliotibialis lateralis; ventral surface partly fused with distal accessory aponeurosis of latter muscle.
Origin.—The origin is fleshy and tendinous from the lateral edge of the anterior part of the ilium.
Insertion.—The attachment is by a short flat tendon to the lateral surface of the femur distal to the trochanter and anterodistal to the insertion of M. piriformis and deep to the proximal part of M. vastus lateralis pars lateralis.
Innervation.—The dorsal division of the femoral nerve penetrates the ventral surface.
Individual Variation.—The dorsal division of the femoral nerve may fuse proximally with either the anterior or middle division. In one leg, there are two separate branches to the muscle.
T. cupido
Individual Variation.—The insertion may not be overlapped by M. vastus lateralis. The dorsal division of the femoral nerve is fused proximally with the middle division in one leg.
P. p. jamesi
Differences from Typical T. pallidicinctus.—The fleshy origin is wider.
Individual Variation.—The dorsal division of the femoral nerve may fuse proximally with either the anterior or middle division. In one leg, there are two branches to M. iliacus, one fused with the anterior division and the other with the middle division.
M. Iliotrochantericus Medius, Figs. 17, 20A, B, C, D
T. pallidicinctus
General Description and Relations.—Small and triangular; ventral to posterior half of M. gluteus profundus; all but posteroventral corner deep to latter; posteromedial to M. iliacus, anterior to neck of femur, and dorsolateral (adjacent proximally) to M. psoas; proximal end notched at level of femoral notch for passage of anterodorsal division of femoral nerve; part anterior to femoral notch mainly tendinous; dorsal surface of proximal part somewhat fused to M. gluteus profundus, proximal accessory aponeurosis of M. extensor iliotibialis lateralis split into two sheets enclosing and fusing with M. iliotrochantericus medius, ultimately attaching to lateral edge of ilium in common with origin of latter muscle.
Origin.—The muscle arises from the ventrolateral surface of the ilium anterior to the acetabulum and posterior to the origin of M. iliacus; the anterior part attaches to the ventrolateral edge of the ilium and the posterior part attaches just above the ventral edge. The muscle is not attached to the concavity of the femoral notch (the origin is notched here). The part attaching anterior to the femoral notch is narrow, tendinous, and continuous anteriorly with the accessory aponeurosis of M. extensor iliotibialis lateralis (thus the anterior border of the muscle cannot be exactly delimited). The part attaching posterior to the femoral notch is wider and fleshy (Fig. 20A).
Insertion.—The short flat tendon attaches to the lateral surface of the distal end of the trochanter slightly anterior and immediately distal to the insertion of M. gluteus profundus; the attachment is proximal to the origin of M. vastus lateralis, anteroproximal to the insertion of M. piriformis, and several mm. proximal to the insertion of M. iliacus.
Innervation.—The small posterodorsal division of the femoral nerve penetrates the ventral surface.
Individual Variation.—On both sides of one specimen, the femoral notch is absent and the proximal end of the muscle is not notched; the proximal part is entirely fleshy and the anterior border is well defined (Fig. 20D).
T. cupido
Individual Variation.—The part attaching anterior to the femoral notch has a fleshy origin in one leg (Fig. 20B), but in another, no part attaches anterior to the femoral notch (thus the muscle is not notched). In one leg, the insertional tendon is strongly fused to, and continuous with, the ventral edge of the insertional tendon of M. gluteus profundus.
P. p. jamesi
Individual Variation.—The part attaching anterior to the femoral notch may be mainly or entirely fleshy. In one leg, the part attaching anterior to the femoral notch is entirely separate from, although overlapped by, the main part of the muscle for the entire length of the fleshy belly (Fig. 20C); both parts have a common insertional tendon.
M. Psoas (M. iliacus), Figs. 13, 15, 18
T. pallidicinctus
General Description and Relations.—Small and slender; on medial aspect of proximal end of thigh lateral to proximal end of M. ambiens; ventromedial to M. iliotrochantericus medius; proximal end visible from inside pelvis (medial to inguinal ligament); passes dorsolateral to inguinal ligament.
Origin.—The muscle arises fleshily from the ventrolateral edge of the ilium posterior to the femoral notch and ventral (adjacent) to the origin of M. iliotrochantericus medius.
Insertion.—The attachment is tendinous to the medial surface of the femur a short distance proximal to the origin of M. femoritibialis internus.
Innervation.—The posterior division of the femoral nerve, which spirals completely around M. psoas, gives several twigs into the proximal part.
Individual Variation.—None of significance.
T. cupido
Individual Variation.—In two legs the insertion is partly fleshy.
P. p. jamesi
Individual Variation.—In one leg the insertion is partly fleshy. The posterior division of the femoral nerve perforates the muscle in one instance.
M. Flexor Cruris Lateralis (M. semitendinosus), Figs. 12, 13, 14, 15, 16, 17
This muscle represents only the main head of the muscle for which Fisher and Goodman (1955) used the same name. Their accessory head of M. flexor cruris lateralis is here termed M. femorocruralis.
T. pallidicinctus
General Description and Relations.—Large, thick, and strap-shaped; on posterior surface of thigh; proximal part bounded anteriorly by Mm. extensor iliotibialis lateralis and extensor iliofibularis; anterodistal part deep to latter; bounded medially by Mm. caudofemoralis (proximally) and flexor cruris medialis (distally); proximal end much narrower than remainder and posterior to ilium; fused to underlying tough membrane, which forms body wall posterior to ilium; proximal half of narrow part aponeurotic; distal part of muscle posterior to M. femorocruralis; separated from latter by common raphe to which both attach; caudal muscle (M. transversoanalis) attached aponeurotically to superficial surface of posteroproximal fleshy part of M. flexor cruris lateralis.
Origin.—The origin is tendinous (superficial surface) and fleshy from the entire dorsolateral iliac ridge and fleshy from an area of the ilium below this ridge, also tendinous from the posterior edge of the ilium medial to the dorsolateral iliac ridge, and also tendinous from the transverse processes of the first free caudal vertebra and the vertebra either anterior or posterior to the latter.
Insertion.—M. flexor cruris lateralis and M. femorocruralis insert broadly on opposite sides of a long tendinous raphe that extends parallel to, but some distance posterior to, the distal half of the femur; the distal end of this tendon[Pg 417] broadens somewhat and fuses to the medial surface of M. gastrocnemius pars media (continuous with the tendon of the latter); the superficial part of this tendon continues toward the tibiotarsus, soon fusing to the deep surface of the overlying tendon of M. flexor cruris medialis; thus the common tendon of M. flexor cruris lateralis and M. femorocruralis insert in common with both M. flexor cruris medialis and M. gastrocnemius pars media.
Innervation.—A branch of the middle tibial division of the sciatic nerve enters the substance of M. caudofemoralis pars iliofemoralis, and emerges near its ventral edge, then passes lateral to M. caudofemoralis pars caudifemoralis and enters the anterior part of M. flexor cruris lateralis.
Individual Variation.—In three legs, the nerve does not perforate M. caudofemoralis pars iliofemoralis, but passes deep to it.
T. cupido
Individual Variation.—In one leg, a small accessory slip arises from the ventrolateral surface of the caudal musculature and joins the posterior edge of the main part of M. flexor cruris lateralis a short distance dorsal to the pubis. In several legs, the nerve does not perforate M. caudofemoralis pars iliofemoralis, but passes deep to it.
P. p. jamesi
Differences from Typical T. pallidicinctus.—The muscle is wider. The extreme proximal end is fleshy up to its origin, which is fleshy and tendinous from the vertebrae. The common insertional tendon of M. flexor cruris lateralis and M. femorocruralis fuses with the distal end of the fleshy part (instead of tendon) of M. flexor cruris medialis.
Individual Variation.—None of significance.
M. Flexor Cruris Medialis (M. semimembranosus), Figs. 12, 13, 14, 15, 16, 17, 20E
T. pallidicinctus
General Description and Relations.—Most posterior muscle on medial surface of thigh; long and strap-shaped; bounded anteriorly by M. adductor profundus; posteroproximal corner of latter medial to anteroproximal part of M. flexor cruris medialis; bounded laterally by Mm. caudofemoralis (proximally) and flexor cruris lateralis (distally); anteroproximal corner adjacent to posteroventral corner of M. flexor ischiofemoralis and lateral to extreme posteroproximal corner of M. adductor superficialis; distal end tendinous, extending into proximal part of shank; bounded medially by M. gastrocnemius pars interna and laterally by Mm. gastrocnemius pars media and plantaris.
Origin.—The muscle arises by a wide flat tendon from a narrow line on the lateral surface of the ischium dorsal to the ventral ischiatic tubercle.
Insertion.—The wide flat tendon attaches to a narrow line on the medial surface of the proximal part of the tibiotarsus a short distance anterior to the proximal part of M. plantaris and deep to M. gastrocnemius pars interna; the proximal end attaches immediately anterior to the distal end of the medial collateral ligament. Part of the common tendon of Mm. flexor cruris lateralis and femorocruralis fuses with the lateral surface of the tendon of M. flexor cruris medialis, inserting in common with it.
Innervation.—A branch of the middle tibial division of the sciatic nerve passes deep to both heads of M. caudofemoralis and enters the anterior part of M. flexor cruris medialis.
Individual Variation.—In several legs, the anterior edge of the proximal part fits into a deep longitudinal groove in the posterior edge of the proximal part of M. adductor superficialis; the two muscles fuse slightly at this point.
T. cupido
Individual Variation.—In two legs, the extreme posterior end of the origin is from the pubis. In two others, the proximal end is separated by a slight gap from M. adductor superficialis. The nerve arises from the posterior (rather than middle) tibial division in one leg.
P. p. jamesi
Differences from Typical T. pallidicinctus.—The origin is wider; the posterior third to half of the origin is fleshy. The entire origin is from a strongly curved line, the middle part of which attaches to the ventral edge of the ischium posterior to the ventral ischiatic tubercle. The insertion is wider. The insertional tendon attaches posterior (rather than anterior) to the distal end of the medial collateral ligament; the proximal end of the insertion attaches to the articular capsule (Fig. 20E). The insertional tendon is shorter; as a result, the common tendon of Mm. flexor cruris lateralis and femorocruralis fuses with the distal end of the fleshy belly (instead of the tendon) of M. flexor cruris medialis.
Individual Variation.—In two thirds of the legs, the proximal part of the insertion is fleshy rather than tendinous. In one leg, the middle part of the insertional tendon splits into two sheets, one attaching anterior to and one attaching posterior to the distal end of the medial collateral ligament. The nerve may arise from the posterior tibial division (two legs), from the middle tibial division (one leg), or as an independent division of the tibial nerve (three legs). In one leg, the nerve perforates the lateral part of M. flexor ischiofemoralis.
M. Caudofemoralis (M. piriformis), Figs. 12, 13, 14, 15, 16, 20J
T. pallidicinctus
General Description and Relations.—Posterior to proximal part of shaft of femur and deep to M. extensor iliofibularis; posterior part deep to M. flexor cruris lateralis; bounded medially by Mm. flexor ischiofemoralis (dorsally), flexor cruris medialis (posteriorly), and adductor superficialis (anteroventrally); anterior end distal to anterior end of M. flexor ischiofemoralis; two distinct heads—pars iliofemoralis and pars caudifemoralis; pars iliofemoralis dorsal to pars caudifemoralis; posteroventral corner of former overlapped by latter; pars iliofemoralis wider and much shorter than pars caudifemoralis; extreme posterior end of pars iliofemoralis fused to overlying posteroproximal aponeurosis of M. extensor iliotibialis lateralis; small part of ventral edge sometimes fused with underlying tendinous posteroproximal corner of M. flexor cruris medialis; entirely fleshy except for small triangular tendinous area along dorsal margin at point where branch of middle tibial division of sciatic nerve passes deep to muscle; pars caudifemoralis long, thin,[Pg 419] narrow, and strap-shaped; overlapping posteroventral corner of ischium; posterior end of fleshy belly narrowed and forming long slender tendon passing into caudal musculature; anterior end forming short narrow tendon fused to deep surface of ventral edge of pars iliofemoralis relatively near insertion; tendon continuous to insertion; fleshy anterodorsal corner of pars caudifemoralis slightly overlapped by ventral edge of pars iliofemoralis; some form of connection usually present between anterior part of M. caudofemoralis pars caudifemoralis and dorsal end of raphe between Mm. flexor cruris lateralis and femorocruralis, most often consisting of narrow weak tendon.
Origin.—Pars iliofemoralis: This arises fleshily from the ventromedial surface of the posterior part of the lateral iliac process, from the entire lateral ischiatic ridge, and from the lateral surface of the ischium anterior to this ridge nearly as far forward as the posterior edge of origin of M. flexor ischiofemoralis; the posteroventral corner reaches the ventral edge of the ischium and usually attaches to the ischiopubic membrane posterior to M. flexor cruris medialis. Pars caudifemoralis: This arises by a narrow tendon from the ventral surface of a broad, thick, tendinous sheet ventral to the pygostyle, which, in turn, attaches to the ventral surface of the pygostyle.
Insertion.—The common belly formed by the union of the two heads narrows (width variable) and attaches to the posterolateral surface of the femur distal to the level of insertion of M. iliacus and in contact with the posterior edge of origin of M. vastus lateralis pars lateralis; the dorsal part is fleshy and the ventral part is tendinous.
Innervation.—A branch of the middle tibial division of the sciatic nerve gives several twigs to the deep surface of pars iliofemoralis; another twig enters the substance of pars iliofemoralis and emerges from the ventral edge of the latter, then enters the dorsal edge of pars caudifemoralis. The latter twig was not found in all legs, but was probably destroyed during dissection.
Individual Variation.—The tendinous area in the dorsal margin of pars iliofemoralis is lacking in one leg and extremely small in some others. In both legs of one specimen, the connection between M. caudofemoralis pars caudifemoralis and the raphe between Mm. flexor cruris lateralis and femorocruralis consists of a small (11 × 2 mm.) but well developed and entirely fleshy muscle slip (Fig. 16). In one leg, the ventral third of this connection is fleshy, the remainder tendinous; in another, this connection is completely lacking.
T. cupido
Individual Variation.—The tendinous area in the dorsal margin of pars iliofemoralis is lacking in one leg. The connection between pars caudifemoralis and the raphe between Mm. flexor cruris lateralis and femorocruralis is lacking in several legs. A conspicuous variation occurring in three legs is the presence of a tendinous area in the belly of pars caudifemoralis, dividing the latter into proximal and distal parts (Fig. 20J). In one leg, the posteroventral corner of pars iliofemoralis arises from the pubis. The origin of pars caudifemoralis in three legs is directly from the anteroventral surface of the pygostyle. In one instance, the insertional tendon of pars caudifemoralis is long and extremely slender and extends for some distance in a groove on the medial surface of pars iliofemoralis before fusing with the latter.
P. p. jamesi
Differences from Typical T. pallidicinctus.—There is no connection at all between pars caudifemoralis and the raphe between Mm. flexor cruris lateralis and femorocruralis. The posteroventral corner of pars iliofemoralis is some distance dorsal to the ventral edge of the ischium and, therefore, does not attach to the ischiopubic membrane.
Individual Variation.—The insertion (narrow) is entirely tendinous in one leg.
M. Flexor Ischiofemoralis (M. ischiofemoralis), Figs. 16, 17
T. pallidicinctus
General Description and Relations.—Thick; on lateral surface of anterior part of ischium; posterior end in lateral iliac fossa; deep to Mm. extensor iliofibularis and caudofemoralis pars iliofemoralis; overlapping ventral extrapelvic part of M. obturator and anteroproximal part of M. adductor superficialis (slightly fused to proximal edge of latter); posteroventral corner contacting anteroproximal corner of M. flexor cruris medialis; extreme anterodorsal corner usually overlapped by tendon of M. piriformis.
Origin.—The muscle arises fleshily from a large area on the lateral surface of the ischium extending ventrally to the origin of M. adductor superficialis, anteriorly to the level of the posterior end of the obturator foramen, dorsally to the ventral border of the ilio-ischiatic fenestra and to the depth of the lateral iliac fossa, and posteriorly approximately to the level of the ventral ischiatic tubercle.
Insertion.—The short flat tendon attaches to the lateral surface of the femur immediately posterior to the insertion of M. piriformis.
Innervation.—The posterior tibial division of the sciatic nerve penetrates the dorsal surface.
Individual Variation.—The ventral part of the insertion may be fleshy.
T. cupido
Individual Variation.—None of significance.
P. p. jamesi
Individual Variation.—In all the legs except one, an additional twig arises from the branch to M. flexor cruris medialis and penetrates the lateral surface of M. flexor ischiofemoralis. The ventral part of the insertion is fleshy in one leg.
M. Adductor Superficialis (M. adductor longus et brevis, pars externa), Figs. 14, 16, 17
T. pallidicinctus
General Description and Relations.—Posterior to femur, lateral to M. adductor profundus, and medial to Mm. flexor ischiofemoralis, caudofemoralis, and femorocruralis; proximal end (fleshy) fused to proximal tendinous end of M. adductor profundus.
Origin.—The origin is fleshy and tendinous from the proximal end of the lateral surface of M. adductor profundus and from a narrow line on the ischium[Pg 421] adjacent (dorsal) to the origin of the latter; the posterior part of the origin sometimes extends farther dorsally on the lateral surface of the ischium; the origin does not extend so far anteriorly nor so far posteriorly as the origin of M. adductor profundus; the anterior edge is at the posterior border of the obturator foramen.
Insertion.—The attachment is fleshy and thick (distal end thin) to the posterior surface of the middle part of the femur between the posterior and posterolateral intermuscular lines; the attachment is adjacent (lateral) to the insertion of M. adductor profundus and adjacent (medial) to the origins of Mm. vastus lateralis (proximally) and femorocruralis (distally); the proximal edge is approximately at the level of the distal edge of the insertion of M. caudofemoralis.
Innervation.—A branch of the obturator nerve emerges from the obturator foramen dorsal to the tendon of insertion of M. obturator pars postica, turns ventrally (crossing latter), and passes deep to the anteroproximal corner of M. adductor superficialis, extending posterodistally between the adductor muscles and giving twigs to the medial surface of M. adductor superficialis and to the lateral surface of M. adductor profundus.
Individual Variation.—The anterior edges of the two adductor muscles are so firmly fused together in some cases that the boundaries cannot be identified at this point. In several legs, there is a deep longitudinal groove in the posterior edge of the proximal part of the muscle into which the anterior edge of M. flexor cruris medialis fits.
T. cupido
Individual Variation.—In some cases, the anterior edges of the two adductor muscles are firmly fused together.
P. p. jamesi
Differences from Typical T. pallidicinctus.—The origin is narrower.
Individual Variation.—The anterior edges of the two adductor muscles may be fused together. In one leg, the entire muscle is indistinguishably fused with M. adductor profundus and they appear as a single muscle.
M. Adductor Profundus (M. adductor longus et brevis, pars interna), Figs. 13, 15, 17, 18
T. pallidicinctus
General Description and Relations.—Broad; on medial surface of thigh immediately posterior to femur; bounded posteriorly by M. flexor cruris medialis (medial to anteroproximal corner of latter), anteriorly by M. femoritibialis internus (anterior edge overlapped by latter), and laterally by Mm. adductor superficialis and femorocruralis; proximal end tendinous (except anterior edge), fused to proximal fleshy end of M. adductor superficialis.
Origin.—The muscle arises tendinously from the ventral edge of the ischium extending from the posterior border of the obturator foramen to the ventral ischiatic tubercle and (anterior edge) fleshily from the lateral surface of the pubis ventral to the obturator foramen; the origin is adjacent (ventral) to the origin of M. adductor superficialis.
Insertion.—The attachment is fleshy and tendinous from the posterior intermuscular line and (proximally and distally) from a narrow adjacent area. Proximally there are often two approximately parallel lines a short distance apart, representing points of attachment of the lateral and medial edges of the muscle; if there is only one line proximally, it may represent the attachment of either the lateral or medial edge of the muscle; distally there is usually only one line, representing the lateral edge of the muscle. The distal end extends onto the posterior surface of the proximal part of the internal condyle, and is adjacent (lateral) to the origin of M. femoritibialis internus, adjacent (medial) to Mm. adductor superficialis and femorocruralis, and adjacent (proximal) to M. gastrocnemius pars media.
Innervation.—See M. adductor superficialis.
Individual Variation.—The anterior edges of the two adductor muscles are strongly fused together in some cases.
T. cupido
Individual Variation.—The anterior edge may be fused with that of M. adductor superficialis. The distal end is sometimes slightly fused with M. gastrocnemius pars media. In one leg, the proximal two thirds of the insertion is entirely tendinous, whereas in another the distal end of the insertion is tendinous.
P. p. jamesi
Individual Variation.—The anterior edge (in one leg the entire muscle) in some legs fuses with that of M. adductor superficialis.
M. Obturator (M. obturator externus + M. obturator internus), Figs. 16, 17, 18, 19C, D, 20K, L, M
I am adopting the single name M. obturator for the complex that Fisher (Fisher, 1946; Fisher and Goodman, 1955) subdivides into Mm. obturator externus and obturator internus. The reasons for this change are given in the section on terminology.
For ease of description, it is desirable to apply names to the subdivisions of M. obturator. It has been customary to divide the obturator complex into two parts—an obturator internus and an obturator externus; the latter has often been further subdivided. The evidence given below demonstrates that a primary division of the complex into only two parts is unsatisfactory.
I strongly suspect that comparable parts of the obturator complex have been considered a part of the "internus" in some birds and a part of the "externus" in others. In their work on the Galliformes, Hudson, et al. (1959) subdivide the obturator complex into only two divisions—obturator externus and obturator internus. The extrapelvic part of this complex that arises from the rim of the obturator foramen and inserts in common with the stout tendon of the main intrapelvic part of the obturator internus is considered by them to be a part of the obturator internus. Their obturator externus lies anterior and deep to the extrapelvic part of the obturator internus and inserts separately from the latter. (I also have found this same arrangement in Tympanuchus and Pedioecetes.)
Berger (1952), in his description of the Black-billed Cuckoo (Coccyzus erythrophthalmus), also divides the obturator complex into an obturator internus[Pg 423] and an obturator externus; the latter he subdivides into a dorsal and a ventral part. He states (p. 530) that he did not find any measurable differences in myology between C. erythrophthalmus and C. americanus. In order better to compare this arrangement with that in Tympanuchus, I have examined two specimens of C. americanus. My findings in the latter differ from Berger's description (p. 541) in one respect. Whereas Berger states that the dorsal and ventral parts of M. obturator externus are distinct except at their origin, I find them fused for their entire length; the muscle fibers that connect these two parts lie deep to the tendon of M. obturator internus. The origin of all parts of the complex in Coccyzus is similar to that in Tympanuchus. The only notable difference in configuration is that the part in Coccyzus that appears to correspond to the obturator externus of Hudson, et al. (1959) is not separate from the remainder of the extrapelvic part of the muscle. Berger (1952) considers all parts of the muscle having an extrapelvic origin to make up the obturator externus. It appears to me that the dorsal part and a part of the ventral part of the obturator externus of Berger correspond to the extrapelvic fleshy part of the obturator internus of Hudson, et al.
From my limited study, it seems to me to be desirable to recognize four subdivisions of the obturator complex, for which I propose the terms pars antica, pars dorsalis, pars ventralis, and pars postica. These parts exhibit various degrees of fusion in different groups of birds and some parts appear to be absent in certain birds. A study of a wide variety of birds will be required to determine whether or not a subdivision into the four parts proposed here is suitable for birds as a whole.
Applying these terms to Coccyzus, pars postica is equivalent to the entire obturator internus of Berger (1952). Pars dorsalis is apparently equivalent to the dorsal part of Berger's obturator externus. The ventral part of the obturator externus of Berger represents the fused pars antica and pars ventralis.
The main parts of the obturator muscle appear to be pars postica and pars antica. Pars dorsalis and pars ventralis are more variable; in Coccyzus these two parts are closely associated with pars antica whereas in Tympanuchus they are most closely associated with pars postica. Apparently pars dorsalis and pars ventralis may be absent in some birds.
T. pallidicinctus
General Description and Relations.—Deeply situated immediately posterior to head of femur; part extending through obturator foramen and lying inside pelvis; extrapelvic part deep to Mm. flexor ischiofemoralis and piriformis; muscle partially divisible into four parts—pars antica, pars dorsalis, pars ventralis, and pars postica (Fig. 20K); pars postica: mostly inside pelvis; much larger than other parts; broad (narrow anteriorly); on medial surface of ischium; composed of several fascicles; anterior end forming narrow, heavy tendon (with some fleshy fibers on posterior part of deep surface) passing through obturator foramen; anteriormost fleshy fibers of ventralmost fascicle fused with pars ventralis; pars ventralis: essentially extrapelvic (see origin); mostly ventral to tendon of pars postica; superficial to pars antica; fused to anterior fleshy part of pars postica; anterodorsal edge usually adjacent to, and often slightly fused with, ventral edge of pars dorsalis (deep to tendon of pars postica); pars dorsalis: entirely extrapelvic; mostly dorsal to tendon of pars[Pg 424] postica; superficial to dorsal part of pars antica; pars antica: extremely short but relatively thick; entirely fleshy; entirely extrapelvic; between obturator foramen and head of femur; anterior surface adjacent to articular capsule; almost completely covered by other parts of muscle; proximal end of posterior surface often slightly fused with adjacent parts of pars ventralis and pars dorsalis.
Origin.—Pars postica: This arises fleshily from the medial surface of the entire ischium except the posterior end, from the dorsomedial and medial surfaces of the anterior half of the pubis as far forward as the obturator foramen, from the internal ilio-ischiatic crest, from the medial surface of the ilium for a short distance posterior to this crest, and from the iliac recess; the posteroventral corner usually arises from the medial surface of the ischiopubic membrane. Pars ventralis: This arises fleshily from the dorsomedial edge of the ventral border of the obturator foramen (Fig. 20M) and (narrowly) from the anterior border of the foramen; this part may or may not arise from the lateral surface of the anteroventral border of the foramen and is usually adjacent along the anterior border of the foramen to pars dorsalis; pars ventralis is continuous along the ventral border of the foramen with the intrapelvic origin of pars postica. Pars dorsalis: This arises fleshily from the lateral surface of the anterodorsal border of the foramen (Fig. 20L) and may extend posteriorly along the dorsal border of the foramen. Pars antica: This arises fleshily from the depresssed area anterior to the obturator foramen (adjacent to pars dorsalis and pars ventralis); the posteroventral corner may arise from the lateral surface of the anteroventral border of the obturator foramen (ventral to the anterior end of pars ventralis; Fig. 20L).
Insertion.—Pars postica: Several tendinous bands (intrapelvic) converge and coalesce, forming a single strong tendon that passes through the obturator foramen and attaches to the lateral surface of the femoral trochanter a short distance posterior to the insertion of M. gluteus profundus and proximal to the insertion of M. flexor ischiofemoralis. Pars ventralis: The attachment is fleshy and tendinous to the ventral edge and the deep surface of the tendon of pars postica. Pars dorsalis: The attachment is fleshy and tendinous to the dorsal edge of the tendon of pars postica. Pars antica: The attachment is fleshy to the posterior surface of the proximal end of the femur several mm. posterior to the insertion of pars postica; the lateral edge attaches to the obturator ridge.
Innervation.—The muscle is supplied by the obturator nerve; several twigs, which do not pass through the obturator foramen, penetrate the anterior part of the medial surface of pars postica; several twigs pass through the obturator foramen and supply pars dorsalis, pars ventralis, and pars antica.
Individual Variation.—In some cases the origin of pars postica does not include the dorsal end of the internal ilio-ischiatic crest nor the ilium posterior to it. Tiny but distinct accessory slips are sometimes present. In one leg a tendinous slip of pars antica extends beyond the remainder of the muscle and inserts independently on the trochanter close to the insertion of pars postica. In another leg, a fleshy and tendinous slip of pars antica attaches to the deep surface of the insertional tendon of pars postica. In still another leg, a fleshy and tendinous slip of pars dorsalis inserts adjacent (anterior) to the dorsal edge of the insertion of pars antica.
T. cupido
Individual Variation.—The variations are similar to those given above for T. pallidicinctus except that there is no slip of pars antica attaching to the tendon of pars postica.
P. p. jamesi.
Individual Variation.—There are variations similar to those given above for T. pallidicinctus except that there is no independent slip of pars antica attaching on the trochanter close to the insertion of pars postica. Pars dorsalis may be quite small. In several legs, pars dorsalis is more closely associated with pars antica than with pars postica; in one of these, pars dorsalis is indistinguishably fused with pars antica (inserting with the latter) except for a few fibers which insert with pars postica.
M. Femorocruralis (M. accessorius semitendinosi), Figs. 14, 15, 16, 17
Fisher (Fisher, 1946; Fisher and Goodman, 1955) considers this muscle as an accessory head of M. flexor cruris lateralis. The reasons for this change in terminology are given in the section on terminology.
T. pallidicinctus
General Description and Relations.—Short and broad; posterior to distal part of femur; deep to Mm. extensor iliofibularis and vastus lateralis pars postica; bounded posteriorly by M. flexor cruris lateralis, medially by Mm. adductor superficialis and adductor profundus, and distally by M. gastrocnemius pars media; fused to a variable degree with the latter (in some cases these two muscles fused firmly together, appearing as single muscle); distal and medial to proximal end of M. flexor perforatus digiti IV.
Origin.—The muscle arises fleshily (thin proximally, thick distally) from the posterior surface of approximately the distal half of the femur between the posterior and posterolateral intermuscular lines. The ventral end is continuous with the origin of M. gastrocnemius pars media, adjacent (medial) to the origin of M. vastus lateralis pars postica, and adjacent (lateral) to the insertions of Mm. adductor superficialis and adductor profundus.
Insertion.—The attachment is to the tendinous raphe in common with M. flexor cruris lateralis (which see).
Innervation.—One or two tiny branches come off the tibial nerve near the distal end of the main trunk of the sciatic nerve, pass anteriorly deep to the peroneal nerve, and penetrate the lateral surface.
Individual Variation.—In two legs, the branch of the medial division of the tibial nerve which supplies M. gastrocnemius pars media sends a twig to the lateral surface of the distal end of M. femorocruralis (in addition to the usual innervation).
T. cupido
Individual Variation.—None of significance.
P. p. jamesi
Differences from Typical T. pallidicinctus.—The muscle is much wider, extending farther proximally on the femur.
Individual Variation.—None of significance.
M. Gastrocnemius, Figs. 12, 13, 15
T. pallidicinctus
General Description and Relations.—Divided into three distinct, widely separated parts—pars externa, pars interna, and pars media; pars externa: large; on posterolateral surface of shank; narrow proximally and distally; bounded anterolaterally by M. flexor perforans et perforatus digiti II and anteromedially by medial head of M. flexor perforatus digiti III; completely separate from pars interna and media except for common tendon of insertion; pars interna: large; on anteromedial surface of shank; narrow distally; bounded anterolaterally by M. peroneus longus and posteromedially by pars media (proximally) and medial head of M. flexor perforatus digiti III; broad sheet of tough connective tissue extending between distal parts of pars externa and pars interna; covering underlying M. flexor perforatus digiti III (medial head), somewhat fused with anteroproximal edge of M. peroneus longus; pars media: small and short; on medial surface of proximal part of shank; deep to tendon of insertion of M. flexor cruris medialis; bounded anteromedially by pars interna, posterolaterally by medial head of M. flexor perforatus digiti III, and proximally by M. femorocruralis; fused to latter, and boundary between the two difficult to locate.
Origin.—Pars externa: The short cylindrical tendon fuses with the anterior half of the distal arm of the tendinous guide loop for M. extensor iliofibularis and attaches in common with the latter to the posterolateral surface of the femur immediately proximal to the fibular condyle; the attachment is proximal (adjacent) to the origin of M. flexor perforans et perforatus digiti II and distal (adjacent) to the origin of M. flexor perforatus digiti IV and is fused to the articular capsule.
Pars interna: The proximal end is partly separable into two layers—a superficial longer one and a deep shorter one. The superficial layer attaches fleshily to the ventral part of the anterior surface of the patella and to the medial half of the superficial surface of the patellar tendon; this layer slightly overlaps the distal fleshy end of M. extensor iliotibialis anticus. The deep layer (overlapped by the superficial layer) attaches to the medial surface of the inner cnemial crest, to the rotular crest medial to the latter, to the medial surface of the proximal part of the tibiotarsus, and (posteroproximal corner) to the distomedial edge of the patellar tendon and to the articular capsule posteromedial to the rotular crest; the entire ventral edge is tendinous, the remainder fleshy.
Pars media: This arises fleshily from an oblique line beginning at the distal end of the origin of M. femorocruralis (continuous with the latter) and extending distomedially across the proximal part of the popliteal area to the proximal edge of the internal condyle, then attaching to the adjacent part of the articular capsule; this part is adjacent (distal) to the insertion of M. adductor profundus and adjacent (proximomedial) to the medial head of M. flexor perforatus digiti IV.
Insertion.—Pars media narrows distally with a narrow tendon along the posterior edge of the fleshy belly; approximately one third of the way down the tibiotarsus the fleshy part terminates and the tendon joins the posterior edge of pars interna, continuing distally in this position. The ossified tendon on the superficial surface of the distal part of pars interna, continuous posteriorly with the tendon of pars media, is joined approximately two thirds of the way down the tibiotarsus by the tendon of pars externa; the fleshy belly of pars interna ends just below the junction. The ossified tendon on the superficial surface of the distal part of pars externa extends beyond the fleshy belly and becomes flexible before joining the tendon of pars interna and media. The common tendon (partly ossified) extends along the posterior surface of the tibiotarsus and widens as it passes posterior to the tibial cartilage, bound to the latter by a thin tough sheet of connective tissue which attaches to the edges of the tibial cartilage, thus forming a sheath for the tendon; the tendon attaches by its edges to the posterior edges of the calcaneal ridges of the hypotarsus, then continues distally (much reduced in thickness) along the posterior surface of the tarsometatarsus, enclosing the flexor tendons; the lateral edge of the tendon attaches to the posterolateral edge of the tarsometatarsus, terminating immediately above the level of the hallux; the medial edge attaches to the edge of the posterior metatarsal crest; the tendon terminates as a thin sheet that attaches to the fascia on the sole of the foot. (Hudson, et al., 1959 consider the posterior metatarsal crest to be an ossified part of the tendon of M. gastrocnemius.)
Innervation.—A branch of the lateral division of the tibial nerve penetrates the proximal part of the medial surface of pars externa. One or two branches of the medial division of the tibial nerve pass deep to M. plantaris and penetrate the deep surface of the posterior part of pars interna. The most proximal branch of the medial division of the tibial nerve penetrates the lateral surface of pars media.
Individual Variation.—None of significance.
T. cupido
Individual Variation.—In one leg, the lateral edge of pars interna overlaps the proximomedial edge of M. peroneus longus; some fibers attach to the lateral surface of the inner cnemial crest.
P. p. jamesi
Differences from Typical T. pallidicinctus.—The proximal end of pars interna does not reach the patella.
Individual Variation.—In one leg, an additional twig to pars media arises from the distal branch to M. femorocruralis.
M. Flexor Perforans et Perforatus Digiti II, Figs. 12, 14
T. pallidicinctus
General Description and Relations.—Long, slender, and Y-shaped; on lateral surface of shank; the two heads enclosing M. flexor perforans et perforatus digiti III; posterior head bounded posteriorly by M. gastrocnemius pars externa; extreme proximal end deep to M. vastus lateralis pars postica; anterior surface fused to posterior surface of M. flexor perforans et perforatus digiti[Pg 428] III; deep surface fused to tendinous part of lateral head of M. flexor perforatus digiti IV; anterior head tendinous except for extreme distal end; covered by, and fused to, posterior edge of M. peroneus longus; fused to anterior surface of M. flexor perforans et perforatus digiti III; two heads join above middle of shank; anteroproximal and posterodistal parts of common belly usually tendinous.
Origin.—Anterior head: This arises by a narrow tendon (partly ossified) from the distal tip of the outer cnemial crest. The tendon is so intimately fused with a connective tissue sheet fused to the deep and posterior surfaces of M. peroneus longus and to the anterior surface of M. flexor perforans et perforatus digiti III that M. flexor perforans et perforatus digiti II could be considered to arise from these two muscles. Posterior head: This arises mostly fleshily from the lateral surface of a compound sheet of tough connective tissue formed by the fusion of the tendinous posteroproximal corner of M. flexor perforans et perforatus digiti III, the proximal parts of the tendons of origin of the lateral heads of Mm. flexor perforatus digiti IV and flexor perforatus digiti II, the fibular and distal arms of the guide loop for M. extensor iliofibularis, and the lateral part of the articular capsule; a part of the common tendon of origin of the anterolateral heads of Mm. flexor perforatus digiti III, flexor perforatus digiti IV, and flexor perforatus digiti II also contributes to this sheet, which attaches to the lateral surface of the external condyle of the femur and to the anterolateral surface of the head of the fibula; for convenience in description, this complex connective tissue sheet will hereafter be termed the femorofibular fascia. The anteroproximal corner of the posterior head of M. flexor perforans et perforatus digiti II often attaches to the lateral surface of the vinculum that passes from the femorofibular fascia to the deep surface of the patellar tendon; the extreme proximal end usually attaches fleshily to a small area on the femur immediately proximal to the fibular condyle and adjacent (distal) to the attachment of the distal arm of the guide loop for M. extensor iliofibularis.
Insertion.—The common belly terminates approximately two thirds of the way down the shank; the slender ossified tendon begins along the posteromedial edge of the common belly, continues distally along the posterior surface of the shank, and becomes flexible before passing through the canal in the tibial cartilage that lies posteromedial to the canal for M. flexor digitorum longus. The tendon passes with the tendon of M. flexor perforatus digiti II (medial to the latter) through a canal in the hypotarsus (see M. flexor perforatus digiti II); just below the hypotarsus, the tendon becomes superficial to the tendon of M. flexor perforatus digiti II and farther distally becomes lateral and finally deep to the latter; the tendon is ossified for most of the length of the tarsometatarsus. At the distal end of this bone, the tendon expands before passing onto the ventral surface of digit II between the tendons of Mm. flexor perforatus digiti II and flexor digitorum longus; at the level of the first phalanx, the edges of the tendon extend dorsally around the tendon of M. flexor digitorum longus and fuse, forming a sheath around the latter; the latter emerges from the sheath near the distal end of the first phalanx; the tendon attaches to the proximal end of the subarticular cartilage ventral to the first interphalangeal joint (the strongest attachment is on the medial side).
Innervation.—The lateral division of the tibial nerve sends twigs into the posteromedial edge of the posterior head.
Individual Variation.—In one leg, the fleshy part of the anterior head is unusually long. In another leg, the anterior head is entirely tendinous. In one leg, a bundle of fibers of the posterior head attaches to the deep surface of the distal part of the patellar tendon. In one leg, near the middle of the tarsometatarsus a rather long and narrow but thick and strong vinculum arises from the tendon of M. flexor perforatus digiti II and, farther distally, joins the tendon of M. flexor perforans et perforatus digiti II.
T. cupido
Individual Variation.—In one leg, the posterior head arises in part from the distolateral edge of the patellar tendon and in another, in part from the superficial surface of the distolateral corner of the patellar tendon.
P. p. jamesi
Individual Variation.—None of significance.
M. Flexor Perforans et Perforatus Digiti III, Figs. 12, 14
T. pallidicinctus
General Description and Relations.—Thick, bipinnate; on lateral surface of proximal part of shank between two heads of M. flexor perforans et perforatus digiti II; bounded anteriorly by M. peroneus longus; anterior surface fused with tendinous anterior head of M. flexor perforans et perforatus digiti II; anterolateral edge somewhat fused to posterior edge of M. peroneus longus superficial to latter tendon; posterior surface fused to posterior head of M. flexor perforans et perforatus digiti II; distal part of belly covered by common belly of latter muscle; posteromedial edge fused to underlying lateral head of M. flexor perforatus digiti IV; anteromedial edge usually somewhat fused to underlying M. flexor digitorum longus.
Origin.—The origin is fleshy and tendinous from the edge of the outer cnemial crest and fleshy from the superficial surface of the distolateral part of the patellar tendon; the posteroproximal corner arises tendinously from the femorofibular fascia.
Insertion.—The belly narrows abruptly, terminating approximately at the middle of the shank; the slender ossified tendon extends posterodistally along the shank, becoming flexible before passing posterior to the tibial cartilage deep to the tendon of M. gastrocnemius, medial to the tendon of M. flexor perforatus digiti IV, and superficial to the medial half of the tendon of M. flexor perforatus digiti III; a thin sheet of connective tissue covers the tendon and attaches by its edges to the underlying tendon of M. flexor perforatus digiti III (thus the latter tendon forms a sheath for the tendon of M. flexor perforans et perforatus digiti II); the tendon is ossified for most of the length of the tarsometatarsus; at midlength of the latter, the tendon lies between the tendons of Mm. flexor perforatus digiti IV and flexor perforatus digiti III; near the distal end of the tarsometatarsus, the tendon becomes lateral and then deep to the tendon of M. flexor perforatus digiti III and is connected by a vinculum to the latter (which see). The tendon enters the ventral surface of digiti III between the tendons of Mm. flexor perforatus digiti III and flexor digitorum longus; after sending a dorsal slip (lateral to the tendon of M. flexor digitorum longus) to the subarticular cartilage ventral to the first[Pg 430] interphalangeal joint, the tendon divides into two branches, between which emerges the tendon of M. flexor digitorum longus; the lateral branch attaches to the subarticular cartilage of the second interphalangeal joint and to the lateral surface of the distal end of the second phalanx; the medial branch has similar attachments on the medial side of the digit.
Innervation.—A branch of the lateral division of the tibial nerve passes deep to the posterior head of M. flexor perforans et perforatus digiti II and enters the posteromedial edge of M. flexor perforans et perforatus digiti III.
Individual Variation.—In both legs of one specimen, the part arising from the femorofibular fascia appears as a distinct but short accessory head. There is no significant individual variation in T. cupido or P. p. jamesi.
M. Flexor Perforatus Digiti IV, Figs. 14, 16
T. pallidicinctus
General Description and Relations.—On posterolateral aspect of shank deep to M. gastrocnemius pars externa; bounded medially by medial head of M. flexor perforatus digiti III, anterolaterally by posterior head of M. flexor perforans et perforatus digiti II, and anteriorly by M. flexor digitorum longus; divided into three heads—medial (largest), lateral, and anterolateral (smallest); tendon of insertion of M. extensor iliofibularis passing between medial and lateral heads; proximal and anteroproximal parts of lateral head an extremely thin, flat tendon; anterodistal part of tendon fused to lateral surface of fleshy part of underlying lateral head of M. flexor perforatus digiti II; proximal part of tendon fused indistinguishably to tendinous part of underlying lateral head of M. flexor perforatus digiti II; fleshy part of anterolateral head anterodistal to lateral head; proximal part of former a long slender tendon anterior to lateral head; anterior surface of anterolateral head (both fleshy and tendinous parts) fused to tendon of anterolateral head of M. flexor perforatus digiti III; deep surface fused to underlying anterolateral head (fleshy) of M. flexor perforatus digiti II; common tendon of anterolateral heads of M. flexor perforatus digiti IV and M. flexor perforatus digiti III passing medial to tendon of insertion of M. extensor iliofibularis, to peroneal nerve, and to fibular arm of guide loop for M. extensor iliofibularis; tendon of M. ambiens inserting on anterolateral surface of this common tendon; medial head entirely fleshy; medial surface fused to medial head of M. flexor perforatus digiti III; deep surface fused to medial edge of underlying medial head of M. flexor perforatus digiti II; medial and lateral heads joined, forming bipinnate belly (pinnate structure most evident on deep surface); anterolateral head joined to distolateral part of belly.
Origin.—The medial head attaches fleshily to the proximal part of the popliteal area proximal (adjacent) to the origin of M. flexor hallucis longus and distolateral to the distal end of the origin of M. femorocruralis; the attachment extends laterally onto the posterolateral surface of the femur proximal (adjacent) to the common attachment of M. gastrocnemius pars externa and the distal arm of the guide loop for M. extensor iliofibularis; the medial edge of the origin is fused with part of the tendinous origin of the medial head of M. flexor perforatus digiti III.
The broad flat common tendon of the lateral head and the lateral head of[Pg 431] M. flexor perforatus digiti II fuses to the superficial surface of the fibular arm of the guide loop for M. extensor iliofibularis and contributes to the femorofibular fascia; consequently the ultimate origin would be the external femoral condyle and the head of the fibula.
The slender common tendon of the anterolateral head and the anterolateral heads of Mm. flexor perforatus digiti II and flexor perforatus digiti III passes deep to the insertional tendon of M. extensor iliofibularis and to the fibular arm of the guide loop for the latter muscle (to which it partly fuses); the tendon attaches to a narrow line on the head of the fibula adjacent to the attachment of the fibular arm of the guide loop and to the deep part of the femorofibular fascia.
Insertion.—The slender ossified tendon becomes flexible before it passes posterior to the tibial cartilage deep to the tendon of M. gastrocnemius, lateral to the tendon of M. flexor perforans et perforatus digiti III, and superficial to the lateral half of the tendon of M. flexor perforatus digiti III; a thin sheet of connective tissue covers the tendon and attaches by its edges to the underlying tendon of M. flexor perforatus digiti III (thus the latter tendon forms a sheath for the tendon of M. flexor perforatus digiti IV; this sheath is separate from a similar sheath surrounding the tendon of M. flexor perforans et perforatus digiti III); the tendon is again ossified where it passes along the posterolateral surface of the tarsometatarsus posterolateral to the tendon of M. flexor perforans et perforatus digiti III; near the distal end of the tarsometatarsus the tendon becomes flexible and expands greatly in width and thickness, and sends a small slip dorsally, medial to the underlying tendons, that attaches to the subarticular cartilage ventral to the trochlea for digit IV; sometimes this slip is continuous with the retinaculum ventral to the tendon at the level of the proximal end of the digit. Several more or less distinct sheets of tough connective tissue lie ventral to all of the flexor tendons at the level of the trochleae and the proximal end of the digits, holding them in place. The tendon narrows as it passes onto the ventral surface of digit IV and soon divides into three branches; the tendon of M. flexor digitorum longus emerges between the medial and middle branches. The lateral branch attaches to the subarticular cartilage ventral to the first interphalangeal joint and is also bound by connective tissue to the ventrolateral surface of the first phalanx. A dorsal slip arises at the point of divergence of the lateral and middle branches and attaches to the subarticular cartilage of the first interphalangeal joint. The middle branch attaches to the subarticular cartilage of the second joint. The medial branch, after sending dorsal slips to each of the first two subarticular cartilages, attaches to the subarticular cartilage of the third interphalangeal joint.
Innervation.—The posterior division of the tibial nerve sends a branch into the posterior edge of the medial head, then passes between the latter and the medial head of M. flexor perforatus digiti III; as it extends distally it gives off twigs to the medial surface of the medial head, to the deep surface of the lateral head, and to the deep surface of the anterolateral head.
Individual Variation.—In one leg, an additional branch arises from the tibial nerve at the level of origin of the posterior division and enters the posterior surface of the medial head; a twig from this branch anastomoses with the first twig of the posterior division to the same head; a branch of the[Pg 432] medial division joins the posterior division distal to the origin of the twigs to the medial head but proximal to the origin of the twigs to the other heads.
T. cupido
Individual Variation.—None of significance.
P. p. jamesi
Individual Variation.—In four legs, a tiny vinculum connects with the tendon of M. flexor digitorum longus (which see).
M. Flexor Perforatus Digiti III, Figs. 13, 14, 15, 16, 17
T. pallidicinctus
General Description and Relations.—Divided into two widely separated heads—medial and anterolateral—with completely separate bellies but with common insertional tendon; small anterolateral head on lateral aspect of thigh deep to M. flexor perforans et perforatus digiti II and posterior to M. flexor digitorum longus; fleshy part of head distolateral to belly of M. flexor perforatus digiti IV; fleshy part fused to lateral edge of belly of M. flexor perforatus digiti II; proximal part of head a slender ossified tendon fused to anterior edge of both fleshy and tendinous parts of anterolateral head of M. flexor perforatus digiti IV and to lateral edge of anterolateral head of M. flexor perforatus digiti II; this tendon passing deep to tendon of insertion of M. extensor iliofibularis and to peroneal nerve; large medial head on posteromedial surface of thigh anterior to medial edge of M. gastrocnemius pars externa, lateral to M. gastrocnemius pars media, and medial to M. flexor perforatus digiti IV; fused to medial surface of medial head of latter and to medial edges of Mm. flexor perforatus digiti II and flexor hallucis longus; proximal end of head tendinous.
Origin.—The medial head attaches tendinously to the medial part of the popliteal area in common with the medial head of M. flexor perforatus digiti II and with the medial edges of Mm. flexor perforatus digiti IV (medial head) and flexor hallucis longus; and is also fused to the articular capsule. The anterolateral head arises in common with the anterolateral heads of Mm. flexor perforatus digiti II and flexor perforatus digiti IV (see account of latter).
Insertion.—The short unossified tendon of the anterolateral head and the longer ossified tendon of the medial head join (after the latter becomes flexible) a short distance above the tibial cartilage, forming a broad flat common tendon that passes posterior to the tibial cartilage (in a shallow groove of the latter); the main part of the tendon is deep to the tendons of Mm. flexor perforatus digiti IV and flexor perforans et perforatus digiti III, but forms separate thin sheaths around these two tendons at the level of the tibial cartilage. A thin sheet of connective tissue covers these three tendons and attaches by its edges to the tibial cartilage, forming a sheath for them. These three tendons pass through the superficial groove in the hypotarsus deep to the tendon of M. gastrocnemius; the tendon of M. flexor perforatus digiti III is ossified for most of the length of the tarsometatarsus; a short distance below the hypotarsus, the anterior branch of the tendon of M. peroneus longus attaches broadly to the lateral edge of the tendon of M. flexor perforatus digiti III. In the proximal[Pg 433] part of the tarsometatarsus the tendon is deep to the tendon of M. flexor perforans et perforatus digiti III, but farther distally becomes medial and then superficial to the latter and lateral to the tendon of M. flexor perforans et perforatus digiti II; near the distal end of the tarsometatarsus a narrow but strong vinculum extends from the lateral edge of the tendon somewhat distally to the lateral edge of the tendon of M. flexor perforans et perforatus digiti III. At the distal end of the tarsometatarsus the tendon expands before entering the ventral surface of digit III where it soon divides into two branches, between which emerge the tendons of Mm. flexor perforans et perforatus digiti III and flexor digitorum longus; the lateral branch attaches to the subarticular cartilage ventral to the first interphalangeal joint and to the lateral surface of the distal end of the first phalanx; the medial branch has similar attachments on the medial side of the digit.
Innervation.—The posterior division of the tibial nerve passes between the medial heads of M. flexor perforatus digiti III and M. flexor perforatus digiti IV and sends a twig to the lateral surface of the former, then passes deep to the common belly of M. flexor perforatus digiti IV and sends a twig to the posterior surface of the anterolateral head of M. flexor perforatus digiti III.
Individual Variation.—None of significance.
T. cupido
Individual Variation.—In one leg, an extra branch (immediately distal to the branch to M. gastrocnemius pars media) of the medial division of the tibial nerve penetrates the medial surface of the proximal end of the medial head.
P. p. jamesi
Individual Variation.—None of significance.
M. Flexor Perforatus Digiti II,Figs. 15, 17
T. pallidicinctus
General Description and Relations.—Bipinnate; on posterior aspect of shank deep to M. flexor perforatus digiti IV and between two heads of M. flexor perforatus digiti III; bounded anteriorly by Mm. flexor digitorum longus and flexor hallucis longus; proximal part divided into three small heads—medial, lateral, and anterolateral; medial and proximal parts of medial head tendinous and extremely thin except for ossified medial edge; proximal part of lateral head tendinous and lateral to insertional tendon of M. extensor iliofibularis; both tendinous and fleshy parts fused to overlying tendon of M. flexor perforatus digiti IV; narrow anterolateral head fused to overlying anterolateral head of latter muscle and (anterolateral edge) to ossified tendon of anterolateral head of M. flexor perforatus digiti III; lateral edge of common belly fused to latter head; medial edge of muscle fused to medial heads of Mm. flexor perforatus digiti IV and flexor perforatus digiti III and to M. flexor hallucis longus.
Origin.—The medial head attaches by a slender ossified tendon to the medial part of the popliteal area in common with the medial head of M. flexor perforatus digiti III and with the medial edges of Mm. flexor perforatus[Pg 434] digiti IV (medial head) and flexor hallucis longus; this head is also fused to the articular capsule. The above-mentioned ossified part of the tendon is situated at the junction of M. flexor perforatus digiti II and M. flexor perforatus digiti III (medial head) and could be considered to be a part of the latter rather than the former. The flat tendon of the lateral head arises in common with the lateral head of M. flexor perforatus digiti IV (which see). The anterolateral head arises in common with the anterolateral heads of Mm. flexor perforatus digiti IV and flexor perforatus digiti III (see former).
Insertion.—The short, slender, ossified tendon becomes flexible and passes through the canal in the tibial cartilage that lies medial to the canal for M. flexor hallucis longus and lateral to the canals for Mm. flexor digitorum longus and flexor perforans et perforatus digiti II. The tendon passes with the tendon of M. flexor perforans et perforatus digiti II (lateral to latter) through the canal in the hypotarsus that is deep to the groove for M. flexor perforatus digiti III and superficial to the canal for M. flexor digitorum longus; the former canal has a bony floor and sides but a fibrous roof; a fibrous partition subdivides the proximal half of this canal, forming a separate channel for each tendon. The tendon is ossified for most of the length of the tarsometatarsus and is situated lateral (adjacent) to the posterior metatarsal crest; immediately below the hypotarsus, the tendon becomes situated deep to the tendon of M. flexor perforans et perforatus digiti II and farther distally becomes situated medial and finally superficial to the latter; at the distal end of the tarsometatarsus the tendon expands greatly and its edges (thick) pass dorsally around the underlying flexor tendons and become continuous with the subarticular cartilage ventral to the trochlea for digit II. The tendon extends onto the ventral surface of digit II and attaches by its edges to the ventromedial and ventrolateral surfaces of the proximal part of the first phalanx (the lateral edge extending farthest distally); the tendons of Mm. flexor perforans et perforatus digiti II and flexor digitorum longus emerge from the distal end of the tendon of M. flexor perforatus digiti II.
Innervation.—The posterior division of the tibial nerve passes between the medial heads of Mm. flexor perforatus digiti III and flexor perforatus digiti IV and gives a twig to the superficial surface of each of the three heads of M. flexor perforatus digiti II and sometimes gives another twig to the superficial surface of the distal part of the common belly.
Individual Variation.—In one leg, a vinculum connects the tendon with that of M. flexor perforans et perforatus digiti II (which see).
T. cupido
Individual Variation.—The canal in the hypotarsus through which the tendon passes has a bony (instead of fibrous) roof in one leg.
P. p. jamesi
Individual Variation.—The variation given above for T. cupido is found in both legs of one specimen.
M. Flexor Hallucis Longus, Figs. 15, 19A
T. pallidicinctus
General Description and Relations.—Elongate and tapering; on posterior aspect of shank deep to M. flexor perforatus digiti II and to proximal end of medial head of M. flexor perforatus digiti IV; bounded anterolaterally by M. flexor digitorum longus and anteromedially by M. plantaris; tendinous anteromedial surface of proximal end fused to common tendon of origin of medial heads of Mm. flexor perforatus digiti III and flexor perforatus digiti II; belly ending approximately halfway down shank.
Origin.—The origin is fleshy and tendinous (anteromedial surface) from the popliteal area immediately distal to the origin of the medial head of M. flexor perforatus digiti IV, extending laterally to the area immediately proximal to the external femoral condyle (medial to the origin of M. gastrocnemius pars externa); the muscle also arises from the proximal end of the posterior part of the articular capsule.
Insertion.—The slender ossified tendon becomes flexible and passes through the canal in the tibial cartilage that lies lateral to the canal for M. flexor perforatus digiti II, then passes through a slight groove in the lateral surface of the hypotarsus and becomes ossified again; midway of the tarsometatarsus, the tendon becomes superficial to the tendon of M. flexor digitorum longus and is connected with the latter by an extensive vinculum, which extends from the deep surface and lateral edge of the tendon of M. flexor hallucis longus distally to the superficial surface of the tendon of M. flexor digitorum longus; the tendon continues, unossified and considerably reduced in size, distally medial to the tendon of M. flexor digitorum longus, and passes through the flexor groove of the first metatarsal anterolateral (adjacent) to the tendon of M. flexor hallucis brevis, then passes deep to the terminal expansion of the latter onto the ventral surface of the hallux; the tendon emerges from under the end of the tendon of M. flexor hallucis brevis and attaches to the ventral surface of the ungual phalanx; a weak dorsal slip attaching to the ventral surface of the distal end of the first phalanx is usually present.
Innervation.—A branch of the medial division of the tibial nerve passes along the medial edge of the muscle, giving several twigs into it.
Individual Variation.—None of significance in any of the three species studied.
T. pallidicinctus
General Description and Relations.—Elongate and tapering; on posteromedial surface of tibiotarsus; bounded medially by M. gastrocnemius pars interna and tendon of M. flexor cruris medialis, posteriorly by M. gastrocnemius pars media and medial head of M. flexor perforatus digiti III, posterolaterally by M. flexor hallucis longus; medial to M. flexor digitorum longus; anterolateral surface of proximal end often slightly overlapping and fused to posterior surface of medial end of M. popliteus; belly terminating above middle of shank.
Origin.—The origin is fleshy and tendinous (distal edge only) from an elongate area on the posteromedial surface of the proximal end of the tibiotarsus adjacent to the insertion of M. popliteus.
Insertion.—The long, slender, ossified tendon extends along the posteromedial aspect of the tibiotarsus and becomes flexible just before attaching to the proximomedial part of the tibial cartilage. The tibial cartilage is a large, mostly cartilaginous pad fitting closely over the posterior surface of the intratarsal joint; the distomedial corner is ossified. This cartilage is perforated by the tendons of several flexor muscles; the distal end of the cartilage attaches to the posteroproximal corner of the tarsometatarsus.
Innervation.—A branch of the medial division of the tibial nerve penetrates the lateral surface.
Individual Variation.—In one leg, a small bundle of fibers separates from the proximal end of the muscle, forming a short accessory head which attaches, separately from the remainder, to the articular capsule posteroproximal to the main origin; a blood vessel passes between the main and accessory heads.
T. cupido
Individual Variation.—In one leg, a small bundle of fibers arises from the medial collateral ligament. In another leg, the nerve to M. gastrocnemius pars interna passes through a gap in the origin of M. plantaris rather than distal to its origin.
P. p. jamesi
Individual Variation.—The nerve branch supplying M. gastrocnemius pars interna gives a minute twig to the deep surface of the free belly of M. plantaris in one instance.
M. Flexor Digitorum Longus, Figs. 14, 16, 17, 19A
T. pallidicinctus
General Description and Relations.—Relatively broad; bipinnate; on posterolateral surface of tibiotarsus; bounded posteromedially by M. flexor hallucis longus, posteriorly by M. flexor perforatus digiti II and anterolateral head of M. flexor perforatus digiti III, laterally by Mm. flexor perforans et perforatus digiti III and flexor perforans et perforatus digiti II, and anterolaterally by Mm. peroneus brevis and tibialis anticus; anterior surface of lateral part of distal half of common belly fused to M. peroneus brevis; divided into three heads—posterior (largest), lateral, and medial; posterior head on posterior surface of head of fibula; overlapping and fused to lateral end of M. popliteus; proximomedial corner deep to latter; lateral head on lateral surface of fibula; lateral and posterior heads separated by insertion of M. extensor iliofibularis; these two heads joined immediately distal to insertion of latter; medial head on posterior surface of tibiotarsus; group of blood vessels and nerves passing between medial and posterior heads; these two heads joined several mm. distal to junction of lateral and posterior heads; deep surface of insertional tendon near distal end of tarsometatarsus serving as origin for M. lumbricalis.
Origin.—Posterior head: This arises fleshily from the posterior surface of the fibula beginning almost at the proximal end and from the medial surface of the fibula beginning deep to the distal part of M. popliteus. Lateral head: This arises fleshily (sometimes partly tendinously) from the lateral surface of the fibula proximal to the fibular tubercle. Some fibers arise from the distal[Pg 437] edge of the tendon of insertion of M. extensor iliofibularis. Medial head: This arises fleshily from the posterior surface of the tibiotarsus just medial to the distal part of the posterior head, distal to M. popliteus, and either lateral or distolateral to the origin of M. plantaris. Distal to the junction of the three heads, the muscle arises fleshily from the posterior surface of the tibiotarsus (except the distal part) and from the medial and posterior surfaces of the fibula.
Insertion.—The slender ossified tendon becomes flexible and passes through the canal in the tibial cartilage that lies anterolateral to the canal for M. flexor perforans et perforatus digiti II and anteromedial to the canal for M. flexor perforatus digiti II, then passes through the bony canal of the hypotarsus that is deep to all the other flexor tendons; the tendon ossifies again and lies adjacent (lateral) to the posterior metatarsal crest; the vinculum from the tendon of M. flexor hallucis longus fuses extensively to the superficial surface of the present tendon a short distance below the midpoint of the tarsometatarsus; the tendon is considerably broader below this point than above it. At the level of the first metatarsal, the tendon divides into three branches (unossified) that diverge, each passing through a groove on the ventral surface of the subarticular cartilages ventral to the trochleae, then pass onto the ventral surfaces of digits II, III, and IV. On digit IV the tendon gives off two dorsal fibro-elastic slips before attaching to the ventral surface of the ungual phalanx; one slip attaches to the subarticular cartilage ventral to the third interphalangeal joint, the other to the subarticular cartilage of the fourth joint and may also attach in part to the distal end of the fourth phalanx. On digit III the tendon gives off two dorsal slips before attaching to the ventral surface of the ungual phalanx; one slip attaches to the subarticular cartilage of the second interphalangeal joint, the other to the subarticular cartilage of the third joint and may also attach in part to the distal end of the third phalanx. On digit II the tendon gives off one dorsal slip before attaching to the ventral surface of the ungual phalanx; the slip attaches to the subarticular cartilage of the second interphalangeal joint and may also attach in part to the distal end of the second phalanx.
Innervation.—A branch of the medial division of the tibial nerve penetrates the medial surface of the posterior head.
Individual Variation.—In half the legs, the proximal end of the lateral head is notched for the passage of the peroneal nerve; the main part of the head lies medial to this nerve; the short fleshy slip lateral to this nerve arises by a long, slender, and extremely weak tendon from connective tissue surrounding the femorotibiotarsal joint. In one leg, a bundle of fibers separates from the lateral head and attaches to the terminal four mm. of the anterior (proximal) edge of the tendon of M. extensor iliofibularis. Each of the following variations occurs in several legs: a third dorsal slip on digit IV attaches to the distal end of the fourth phalanx in some legs and to the subarticular cartilage of the fourth joint in other legs; a third dorsal slip on digit III attaches to the distal end of the third phalanx in some legs and to the subarticular cartilage of the third joint in other legs; a second dorsal slip on digit II attaches to the distal end of the second phalanx in some legs and to the subarticular cartilage of the second joint in other legs.
T. cupido
Individual Variation.—The dorsal slips of insertion show variations similar to those noted above for T. pallidicinctus.
P. p. jamesi
Individual Variation.—In one leg, the proximal end of the lateral head is notched for the passage of the peroneal nerve. The dorsal slips of insertion show variations similiar to those given above for T. pallidicinctus. In four legs, a tiny vinculum extends from the lateral edge of the branch of the tendon on digit IV to the lateral edge of the underlying medial branch of the tendon of M. flexor perforatus digiti IV at the level of the second phalanx.
M. Popliteus, Fig. 19B
T. pallidicinctus
General Description and Relations.—Extremely short but relatively broad and thick; on posterior surface of proximal end of tibiotarsus; extending distomedially from proximal part of fibula; deep to M. flexor hallucis longus; lateral end overlapped by, and fused to, posterior head of M. flexor digitorum longus; medial end often slightly overlapped by, and fused to, M. plantaris; medial end (insertion) much wider than lateral end (origin).
Origin.—The origin is fleshy and tendinous (superficial surface) from the medial surface of the fibula near the proximal end.
Insertion.—The attachment is fleshy to the posterior surface of the proximal end of the tibiotarsus adjacent (lateral) to the origin of M. plantaris.
Innervation.—A branch of the medial division of the tibial nerve penetrates the posterior surface.
Individual Variation.—None of significance in any of the three species studied.
M. Peroneus Longus, Figs. 12, 13
T. pallidicinctus
General Description and Relations.—Large; on anterolateral surface of shank; bounded medially by M. gastrocnemius pars interna and posterolaterally by Mm. flexor perforans et perforatus digiti III and flexor perforans et perforatus digiti II; proximal three fourths of posteromedial part (covered by M. gastrocnemius pars interna) aponeurotic and tightly fused to medial surfaces of underlying Mm. tibialis anticus and extensor digitorum longus; proximal part of fleshy belly somewhat fused to anterior surface of underlying M. tibialis anticus; posterolateral surface strongly fused to aponeurotic medial head of M. flexor perforans et perforatus digiti II and slightly fused to anterolateral edge of M. flexor perforans et perforatus digiti III.
Origin.—The muscle arises by fleshy and tendinous fibers from the edges of the inner and outer cnemial crests; the extreme proximal end arises either fleshily or aponeurotically from the rotular crest between the cnemial crests; the posteromedial edge (aponeurotic except distal one fourth fleshy) arises from the anteromedial intermuscular line.
Insertion.—The narrow ossified tendon on the superficial surface of the distal part of the fleshy belly extends several mm. beyond the belly where it[Pg 439] becomes flexible and divides into two branches. The short, broad posterior branch attaches broadly to the proximolateral corner of the tibial cartilage. The narrow anterior branch passes along the lateral surface of the tibiotarsus, through a strong retinaculum immediately proximal to the external condyle, and crosses the lateral surface of the joint, where it is covered by connective tissue nearly as tough as, and continuous with, the retinaculum; the tendon attaches broadly to the lateral edge of the ossified tendon of M. flexor perforatus digiti III a short distance below the hypotarsus.
Innervation.—The peroneal nerve sends twigs to the deep surface.
Individual Variation.—In both legs of two specimens, the extreme proximal end extends proximal to the rotular crest and attaches fleshily to the superficial surface of the distal end of the patellar tendon.
T. cupido
Individual Variation.—None of significance.
P. p. jamesi
Individual Variation.—One leg shows the variation described above for T. pallidicinctus.
M. Tibialis Anticus, Figs. 14, 15, 16, 19E, 20N
T. pallidicinctus
General Description and Relations.—Thick; on anterior aspect of thigh deep to M. peroneus longus; bounded posteriorly by M. extensor digitorum longus and posterolaterally by Mm. flexor digitorum longus and peroneus brevis; divided into two heads—tibial and femoral; small femoral head adjacent to posterolateral surface of much larger tibial head; two heads joined near midpoint of fleshy part of muscle, forming bipinnate belly (pinnate structure most evident on deep surface); proximal part of femoral head situated between outer cnemial crest and head of fibula; proximal part of anterior surface of tibial head somewhat fused to overlying M. peroneus longus; medial surface fused to aponeurosis of latter.
Origin.—Tibial head: This arises by fleshy and tendinous fibers from the edge of the inner cnemial crest, from the rotular crest between the inner and outer cnemial crests, and from the anterior surface, distal edge, and posterior surface of the outer cnemial crest; the attachment may or may not extend onto the superficial surface of the distal part of the patellar tendon; the attachment is adjacent to the origin of the underlying M. extensor digitorum longus. Femoral head: This arises by a slender tendon from the notch in the distal end of the external condyle of the femur.
Insertion.—The slender ossified tendon extends along the anterior surface of the distal end of the tibiotarsus and passes through a large, strong, oblique retinaculum (superficial to the supratendinal bridge); the lateral end of the retinaculum attaches to the lateral end of the supratendinal bridge; the medial end attaches immediately proximal to the medial end of the bridge. The tendon widens and becomes flexible as it passes across the anterior surface of the intratarsal joint, then narrows and attaches to the tubercle on the anterior surface of the proximal part of the tarsometatarsus between Mm. extensor hallucis longus and extensor brevis digiti IV. The[Pg 440] distalmost bundle of tendinous fibers does not attach to the tubercle, but extends distally along the anterior surface of the tarsometatarsus and attaches to the latter a few mm. distal to the tubercle, forming an accessory insertion. A part of the peroneal nerve passes between the main and accessory insertions.
Innervation.—A variable number of branches of the peroneal nerve penetrate the lateral surface of the femoral head; a variable number of branches of the same division pass deep to the femoral head and enter the posterior edge of the tibial head.
Individual Variation.—- In one leg, the accessory insertion is absent.
T. cupido
Individual Variation.—None of significance.
P. p. jamesi
Differences from Typical T. pallidicinctus.—The origin of the tibial head does not extend onto the patellar tendon.
Individual Variation.—The accessory insertion is absent in one leg.
M. Extensor Digitorum Longus, Figs. 15, 17
T. pallidicinctus
General Description and Relations.—Bipinnate; on anterior surface of tibiotarsus deep to M. tibialis anticus; bounded laterally by M. peroneus brevis; lateral edge usually slightly fused to proximal half of latter; medial surface fused to aponeurosis of M. peroneus longus.
Origin.—The muscle arises fleshily from the lateral surface of the inner cnemial crest, from the rotular crest between the cnemial crests (deep to the attachment of M. tibialis anticus), from the basal (medial) half of the anterior surface of the outer cnemial crest, and from the anterior surface of the tibiotarsus (except the distal part) between the anteromedial and anterolateral intermuscular lines; proximal to the anterolateral intermuscular line, the origin usually extends almost to the lateral edge of the tibiotarsus.
Insertion.—The ossified tendon extends along the mid-anterior surface of the distal part of the tibiotarsus deep to the tendon of M. tibialis anticus and passes under the supratendinal bridge, becoming flexible and widening slightly as it crosses the anterior surface of the intratarsal joint; the tendon narrows again and passes through a small but strong retinaculum on the anterior surface (medial to midline) of the proximal part of the tarsometatarsus; the retinaculum is immediately proximal and medial to the insertion of M. tibialis anticus. The tendon ossifies again as it passes down the anterior surface of the tarsometatarsus and bifurcates near the midpoint of the latter; the lateral branch soon bifurcates again; of these three branches, which are ossified for some distance, the lateral one passes onto the dorsal surface of digit IV, the middle one passes onto the dorsolateral surface of digit III, and the medial one subdivides (at the level of the trochleae) into three branches—one passing onto the dorsal surface of digit III and two passing onto the dorsal surface of digit II. At the level of the metatarsophalangeal joints, all of these tendons are interconnected by strong sheets of connective tissue and it is often difficult exactly to delimit the tendons at this level. On[Pg 441] the digits, tough connective tissue binds the tendons to the phalanges; this is most pronounced at the interphalangeal joints. The tendons are distinct on the first phalanx of each digit, but are often poorly defined farther distally. On digit IV the tendon subdivides into branches that attach to the proximal ends of the ungual, fourth, third, and (usually) second phalanges. On digit III the lateralmost tendon bifurcates, with one branch attaching to the ungual phalanx and the other to the proximal end of the third phalanx; the medial tendon attaches to the proximal end of the second phalanx. On digit II the originally medial tendon passes underneath and then lateral to the other tendon and attaches to the ungual phalanx; the other tendon attaches to the proximal end of the second phalanx.
Innervation.—One or more branches of the peroneal nerve enter the lateral edge.
Individual Variation.—In four legs, the lateral branch of the trifurcated tendon is not ossified at all.
T. cupido
Individual Variation.—In a few cases, the muscle does not come in contact with M. peroneus brevis.
P. p. jamesi
Differences from Typical T. pallidicinctus.—The belly is shorter. The lateral branch of the tendon on the tarsometatarsus is not ossified (true also of some legs of Tympanuchus).
Individual Variation.—In several legs, the muscle also arises from the distal part of the posterior surface of the outer cnemial crest.
M. Peroneus Brevis, Figs. 14, 16, 17, 188, 19A
T. pallidicinctus
General Description and Relations.—Small; on lateral surface of distal part of tibiotarsus; mainly anterior to fibula; bounded posteriorly and laterally by M. flexor digitorum longus (fused with latter), anteriorly by M. tibialis anticus, and anteromedially by M. extensor digitorum longus (usually slightly fused to latter).
Origin.—The muscle arises by fleshy and tendinous fibers from the medial and anterior surfaces of the fibula beginning a short distance below the distal end of the fibular crest and from the anterolateral surface of the tibiotarsus anterior to the fibula; the anteromedial edge attaches to the anterolateral intermuscular line.
Insertion.—The short, slender, ossified tendon passes along the anterolateral surface of the tibiotarsus and through a retinaculum immediately proximal and anteromedial to the retinaculum for the anterior branch of the tendon of M. peroneus longus; the tendon becomes flexible and widens as it passes across the lateral surface of the intratarsal joint deep to the tendon of M. peroneus longus, turning posteriorly and attaching to the proximolateral corner of the hypotarsus.
Innervation.—The superficial peroneal branch of the peroneal nerve gives one or two twigs to the anterior surface of the proximal part.
Individual Variation.—None of significance.
T. cupido
Individual Variation.—In a few legs, the muscle does not come in contact with M. extensor digitorum longus.
P. p. jamesi
Individual Variation.—None of significance.
M. Extensor Hallucis Longus, Figs. 19E, 20N
T. pallidicinctus
General Description and Relations.—Slender and elongate; proximal part on anterior surface of tarsometatarsus medial to anterior metatarsal groove; near midlength of tarsometatarsus, muscle twisted onto medial surface of latter; divisible into two heads—proximal and distal; belly of proximal head (largest) ending at level of twisting onto medial surface of bone; short distal head beginning at this point deep to tendon of proximal head and soon joining latter tendon.
Origin.—Proximal head: This arises fleshily from the anterior surface of approximately the proximal half of the tarsometatarsus medial to the anterior metatarsal groove; the proximal end is partly medial to and partly deep to the retinaculum for M. extensor digitorum longus; some fibers arise from the extreme distal edge of the main insertion of M. tibialis anticus; the distal end of the belly is unattached. Distal head: This arises fleshily from the medial surface of the tarsometatarsus proximal to the first metatarsal and deep to the tendon of the proximal head.
Insertion.—The slender tendon of the proximal head, which begins along the medial edge of the distal part of the belly, soon fuses with the superficial surface of the distal head (ossified here); the common tendon (unossified) passes onto the dorsal (proximal) surface of the first metatarsal, where it passes through a retinaculum, then passes along the dorsal surface of the hallux (bound by strong connective tissue to the metatarsophalangeal joint), attaching to the dorsal surface of the ungual phalanx.
Innervation.—The branch of the deep peroneal nerve that passes medial to the main insertion of M. tibialis anticus gives one or two twigs into the proximal part of the proximal head. No supply to the distal head was found, but see below.
Individual Variation.—In one leg, the proximal end of the distal head is fused to the distal end of the belly of the proximal head, whereas in three legs, a distinct gap separates the fleshy parts of the two heads. The following variations, each found in one leg, pertain to the relationship of the origin of the proximal head to the retinaculum for M. extensor digitorum longus: the origin does not extend proximally medial to the retinaculum; the origin does not extend proximally deep to this retinaculum; a part of the proximal end extends proximally lateral to this retinaculum (in this instance there is an unusually wide gap between the retinaculum and the insertion of M. tibialis anticus). In one leg, the distalmost fibers of the distal head do not join the common tendon but insert independently on the articular capsule of the metatarsophalangeal joint (deep to the common tendon).
T. cupido
Individual Variation.—The relationship between the two heads varies as follows: the proximal end of the distal head may be fused to the distal end of the belly of the proximal head; the proximal end of the distal head may begin anterior (adjacent) to the distal end of the belly of the proximal head; there may be a distinct gap between the fleshy parts of the two heads. In two legs, there is no origin from the insertion of M. tibialis anticus. In one leg, a small accessory bundle of fleshy fibers arises from the proximal end of the first metatarsal (widely separated from the origin of the distal head), passes through the retinaculum deep to the common tendon and attaches to the dorsal surface of the articular capsule of the metatarsophalangeal joint; thus this bundle is completely separate from the remainder of the muscle. In two legs, the same nerve branch that gives twigs into the proximal head also gives off (much farther distally) a twig that enters the distal head.
P. p. jamesi
Individual Variation.—The proximal end of the distal head may begin anterior (adjacent) to the distal end of the belly of the proximal head. In four legs, the origin of the proximal head does not extend proximally medial to the retinaculum for M. extensor digitorum longus; in one of these legs, a part of the proximal end extends proximally lateral to this retinaculum. The distalmost fibers of the distal head do not join the common tendon but insert independently on the dorsal surface of the articular capsule of the metatarsophalangeal joint in four legs; in another leg, the entire distal head has the latter insertion (consequently the two heads are completely separate).
M. Abductor Digiti II Figs. 19E, 20N
T. pallidicinctus
General Description and Relations.—Short; on medial surface of distal part of tarsometatarsus; proximal end adjacent (anterior) to distal head of M. extensor hallucis longus.
Origin.—The origin is fleshy from the medial surface of the distal part of the tarsometatarsus anterior (adjacent) to the first metatarsal and from the anteromedial surface of the basal half of the first metatarsal.
Insertion.—The flat tendon passes over the medial surface of the trochlea for digit II and attaches to the medial surface of the proximal end of the first phalanx of digit II; the tendon is fused with the articular capsule.
Innervation.—The compound nerve formed by the fusion of a branch of the superficial peroneal nerve with a branch of the deep peroneal nerve gives a twig to the anterolateral edge of the muscle.
Individual Variation.—In some cases, the twig arises from the deep peroneal branch alone (which is not joined by the superficial peroneal nerve).
T. cupido
Individual Variation.—In one leg, some of the fleshy fibers arising from the first metatarsal insert independently on the medial surface of the trochlea for digit II (deep to the main part of the muscle).
P. p. jamesi
Individual Variation.—None of significance.
M. Extensor Brevis Digiti III (M. extensor proprius digiti III), Figs. 19E, 20N
T. pallidicinctus
General Description and Relations.—Short and relatively broad (narrow proximally); on mid-anterior surface of distal part of tarsometatarsus; tendon of insertion fused with articular capsule.
Origin.—The origin is fleshy from the mid-anterior surface of the distal part of the tarsometatarsus ending a short distance proximal to the trochlea for digit III.
Insertion.—The flat tendon passes over the trochlea for digit III and attaches to the dorsal surface of the proximal end of the first phalanx of digit III.
Innervation.—The compound nerve formed by the fusion of a branch of the superficial peroneal nerve with a branch of the deep peroneal nerve gives a twig to the proximal end of the muscle.
Individual Variation.—In some cases, the twig arises from the deep peroneal branch alone (which is not joined by the superficial peroneal nerve). The individual variation is insignificant in T. cupido and P. p. jamesi.
M. Extensor Proprius Digiti III (Not found by Hudson, et al.), Fig. 20N
T. pallidicinctus and T. cupido
Absent in both species.
P. p. jamesi
This atypical muscle was found in only two legs (P.p. 1L and 4L). The following description applies to P.p. 4L (Fig. 20N).
General Description and Relations.—Small but well developed; fleshy part 1½ × 13 mm.; proximal end narrower; on mid-anterior surface of tarsometatarsus between Mm. extensor brevis digiti IV and extensor hallucis longus and mostly proximal to M. extensor brevis digiti III; tendinous distal part superficial to latter; fleshy belly ending immediately distal to proximal end of latter.
Origin.—The origin is fleshy from a narrow elongate area on the mid-anterior surface of the tarsometatarsus between Mm. extensor brevis digiti IV and extensor hallucis longus, beginning at the distal end (bony) of the elongate accessory insertion of M. tibialis anticus. The distal part of the belly is free.
Insertion.—The attachment is by a thin, wide (relative to belly) tendon to the superficial surface of M. extensor brevis digiti III.
Innervation.—Not found.
Individual Variation.—In P.p. 1L, the muscle is less well developed. The fleshy belly is 1 × 5 mm. It arises from the lateral edge of M. extensor hallucis longus. The extremely slender insertional tendon attaches as above.
M. Extensor Brevis Digiti IV, Figs. 19E, 20N
T. pallidicinctus
General Description and Relations.—Slender and tapering; on lateral part of anterior surface of tarsometatarsus; length of belly variable; middle of medial edge in contact with M. extensor hallucis longus.
Origin.—The origin is fleshy from the lateral part of the anterior surface of the tarsometatarsus, including the anterior metatarsal groove.
Insertion.—The long slender tendon enters the anterior aperture of the distal foramen, passes through the intertrochlear canal, emerges from the terminal foramen, and attaches to the medial surface of the proximal end of the first phalanx of digit IV.
Innervation.—The superficial peroneal branch of the peroneal nerve sends a twig into the proximal part of the muscle.
Individual Variation.—None of significance in any of the three species studied.
M. Lumbricalis, Fig. 19F
T. pallidicinctus
General Description and Relations.—Small, thin, and strap-shaped; on mid-posterior surface of distal end of tarsometatarsus deep to tendon of M. flexor digitorum longus; belly partly fleshy and partly elastic connective tissue.
Origin.—The muscle arises from the deep (anterior) surface of the tendon of M. flexor digitorum longus a short distance proximal to the trifurcation of the latter.
Insertion.—The muscle attaches to the proximal end of the subarticular cartilage ventral to the trochlea for digit III.
Innervation.—A long but extremely small twig arises from the paraperoneal branch of the tibial nerve a short distance distal to the hypotarsus and extends distally along the mid-posterior surface of the tarsometatarsus (parallel to a larger nonmuscular branch) and enters the deep surface distal to the middle. It was possible to follow this twig in only two legs; it was presumably destroyed in the course of dissection in the others.
Individual Variation.—In some cases, the "muscle" appears grossly to be entirely connective tissue, although a distinct entity.
T. cupido
Individual Variation.—In some cases, the "muscle" appears grossly to be entirely connective tissue. The innervation was found in only one leg, in which the twig arises more distally than in T. pallidicinctus.
P. p. jamesi
The innervation was not found.
M. Abductor Digiti IV, Fig. 19F
T. pallidicinctus
General Description and Relations.—Slender and elongate; on posterior surface of tarsometatarsus lateral to midline; in contact with M. flexor hallucis brevis in midline.
Origin.—The origin is fleshy from the posterior surface of the tarsometatarsus lateral to the midline beginning near the proximal end (lateral to the hypotarsus) and ending at the level of the first metatarsal.
Insertion.—The slender tendon, which begins along the lateral edge of the distal part of the belly, passes through a retinaculum on the posterolateral surface of the tarsometatarsus immediately above the outer trochlea and attaches to the lateral surface of the proximal end of the first phalanx of digit IV.
Innervation.—The paraperoneal branch of the tibial nerve gives one or two twigs to the proximal part of the muscle.
Individual Variation.—None of significance in any of the three species studied.
M. Flexor Hallucis Brevis, Fig. 19F
T. pallidicinctus
General Description and Relations.—Slender and elongate; on posterior surface of tarsometatarsus medial to midline; belly (except proximal end) adjacent (lateral) to posterior metatarsal crest; proximal end passing under latter (immediately distal to hypotarsus) and lying anteromedial to hypotarsus.
Origin.—The origin is fleshy from the medial metatarsal depression and from the posterior surface of the tarsometatarsus between the midline and the posterior metatarsal crest beginning immediately below the hypotarsus and ending a short distance above the first metatarsal (sometimes more proximally).
Insertion.—The slender tendon, which begins along the medial edge of the distal part of the belly, passes through the groove on the posterodistal surface of the first metatarsal and onto the proximal end of the ventral surface of the hallux; the tendon widens considerably and attaches by its edges to the ventral surface of the proximal end of the first phalanx, forming a short "tunnel" through which the tendon of M. flexor hallucis longus passes.
Innervation.—The paraperoneal branch of the tibial nerve sends one or two twigs into the proximal part of the muscle (but distal to the hypotarsus).
Individual Variation.—In two legs, the muscle arises in part from the distal end of the lateral calcaneal ridge. The individual variation is insignificant in T. cupido and P. p. jamesi.
FOOTNOTES:
Analysis of Individual Variation
Considerable individual variation occurs in both the muscles and the nerves of the leg of the three species studied. The amount of variation reported by a worker depends in large part on the degree of variation that he considers significant.
Individual variation in the muscles and in the nerves will be discussed separately; that of the muscles (excluding innervation) will be considered first.
Muscles
Considering the number, rather than degree, of variations, the most variable muscles are: Mm. flexor digitorum longus, obturator, caudofemoralis, and extensor hallucis longus. The first-mentioned muscle exhibits 14 different variations in the specimens studied. Mm. vastus lateralis, flexor perforans et perforatus digiti II, and piriformis also showed a considerable number of variations. The following muscles did not exhibit any variations considered significant in this study: Mm. vastus medialis, femoritibialis internus, flexor perforatus digiti III, extensor brevis digiti III, and abductor digiti IV.
Muscles showing a great degree of individual variation included the following: M. extensor proprius digiti III was present in two legs of Pedioecetes but absent in the other legs studied. A fleshy muscle slip connected M. caudofemoralis pars caudifemoralis with the tendinous raphe between Mm. flexor cruris lateralis and femorocruralis in two legs, whereas in others this connection was tendinous or even absent altogether. M. caudofemoralis pars caudifemoralis had a tendinous area within the belly in only three legs. A vinculum connected the insertional tendons of Mm. flexor perforans et perforatus digiti II and flexor perforatus digiti II in only one leg. The fleshy belly of M. iliotrochantericus medius was completely split into two parts in one leg. M. flexor cruris lateralis had an accessory slip arising from the caudal musculature in one leg.
Certain individual variations reported in the accounts of the muscles formed a graduated series, as far as degree is concerned, from the typical to the extreme condition. Therefore it was difficult or impossible in some cases to state whether or not certain specimens exhibited such a variation. Elimination of the doubtful instances of variation leaves a total of 50 different variations (excluding variations between species) which can be attributed to a definite number of specimens. The remainder of the discussion of individual variation in the muscles concerns these 50 variations. See table 3.
The typical condition of any structure is considered to be the condition of that structure in the majority of the legs studied. Some conditions considered as typical in the present study might not be so considered if a larger number of specimens had been studied. If exactly half of the legs of one species shows a particular condition[Pg 448] of a structure, the condition typical for this species is considered (for purposes of the following discussion) to be that found in the majority of the legs of the other species.
In all instances except two (of 50) the typical condition of the muscles in T. pallidicinctus was also the typical condition in T. cupido. The majority of the legs in T. cupido had an additional dorsal slip on the tendon of M. flexor digitorum longus in digits II and III. In all instances except seven the typical condition in T. pallidicinctus was also the typical condition in Pedioecetes. In these seven instances a variation in the former was the typical condition in the latter. These were: an additional dorsal slip on the tendon of M. flexor digitorum longus in each of three digits, a vinculum between the latter and M. flexor perforatus digiti IV, a partly fleshy insertion of M. flexor cruris medialis, an unossified lateral branch of the insertional tendon of M. extensor digitorum longus, and an independent insertion of the distalmost fibers of the distal head of M. extensor hallucis longus. For all characters except the number of the dorsal slips on the tendon of M. flexor digitorum longus in digits II and III, the typical condition in T. pallidicinctus was also the typical condition for all species considered together. To facilitate comparison, in the following discussion all of the above-mentioned characters are considered in all species as variants from the typical condition.
Certain legs showed a greater number of variations from the typical condition than did others. The majority of legs showed from four to seven variations in the muscles of the leg. The extremes were P.p. 1L, which showed 11, and T.c.p. 2L, which exhibited only one variation.
Twenty-three of the 50 variations were found in only one leg (out of 23). It would be expected that if additional specimens were studied, more kinds of variations would be found. Nine variations were found in only two legs, five in three legs, five in four legs, and four in five legs. One variation was found in nine legs, one in ten legs, and two in 12 legs; the last four variations were in the number of dorsal slips of the insertional tendon of M. flexor digitorum longus in digits II, III, and IV and in the ossification of the insertional tendon of M. extensor digitorum longus.
Five of the variations were found only in specimens in which only one leg was dissected. Considering only those eight specimens in which both legs were dissected, five of the 45 variations were found in both legs of each specimen exhibiting the variation;[Pg 449] 28 variations were found in only one leg of each specimen exhibiting the variation; 12 variations were found in both legs of some specimens but in only one leg of other specimens. Of the six muscle features showing the greatest degree of individual variation (described previously), only two (both pertaining to M. caudofemoralis) were found in both legs of the specimens exhibiting the variation.
For one leg (the one showing the most variations) of each specimen of which both legs were studied, the number of variations that this leg had in common with every other leg (of all species) was determined. Then the number of variations in common between the two legs of one individual was compared with the number of variations in common between one leg of this individual and each leg of every other individual. See table 4. One leg of six of the eight specimens showed at least as many variations in common with a leg of another individual as with the other leg of the same individual. The two exceptions were T.p. 2R and T.c.a. 1R. Thus for most specimens there was as much variation in the muscles between the right and left legs of one individual as there was between individuals.
Of the 50 muscle variations seven were found only in T. pallidicinctus (eight legs), 16 were found only in T. cupido (nine legs), and ten were found only in Pedioecetes (six legs). Two were found in both species of Tympanuchus (but not in Pedioecetes). Fifteen were found in both Tympanuchus and Pedioecetes; of these, five were found in all three species studied, eight were shared by T. pallidicinctus and Pedioecetes, and two occurred in T. cupido and Pedioecetes.
Nerves
The lumbosacral plexus, femoral nerve, sciatic nerve, and tibial nerve all showed numerous individual variations. The peroneal nerve, however, was relatively constant. Variations in the obturator nerve were considered to be insignificant. See table 5.
In all instances except one (of 40) the typical condition in T. pallidicinctus was also the typical condition in T. cupido. In most of the legs of the latter the nerve to M. flexor cruris lateralis did not perforate M. caudofemoralis. In all instances except four the typical condition in T. pallidicinctus was also the typical condition in Pedioecetes. These exceptions were: prefixation of the lumbosacral plexus, six roots of the sciatic nerve, femoral nerve formed mainly from S2 to S4 and two twigs to M. flexor ischiofemoralis.[Pg 450] In all instances the typical condition in T. pallidicinctus was also the typical condition for all species considered together.
Certain legs showed a greater number of variations from the typical condition of the nerves than did others. The greatest number of variations was shown by P.p. 3L, which had 12. T.p. 1R and T.c.p. 1L both showed only one.
All six variations in the lumbosacral plexus were found on both sides of each specimen exhibiting the variation. In marked contrast to the other nerves, there was no significant variation in the lumbosacral plexus between the right and left sides of one individual. (This might not always be true, however, if a larger number of specimens were studied.) Of the variations in the lumbosacral plexus, one was found in only one specimen (of 15), one was found in three specimens, one in four specimens, two in six specimens, and one in seven specimens. Of the 34 variations found in the other nerves, 14 were found in only one leg (of 23), six occurred in two legs, four in three legs, three in four legs, three in five legs, two in six legs, one in seven legs, and one in nine legs.
Four of the variations were found only in specimens in which only one leg was dissected. Considering only those eight specimens in which both legs were dissected, and excluding the lumbosacral plexus, ten of the 30 variations were found in both legs of each specimen exhibiting the variation; 16 variations were found in only one leg of each specimen exhibiting the variation; four variations were found in both legs of some specimens but in only one leg of other specimens.
The number of variations in common between the two legs of one individual was compared with the number between individuals in the same manner as for the muscles; the lumbosacral plexus was excluded from consideration. See table 6. One leg of six of the eight specimens showed at least as many variations in common with a leg of another individual as with the other leg of the same individual. The two exceptions were T.p. 2L and T.p. 3R. Thus for most specimens there was as much variation in the nerves other than the lumbosacral plexus between the right and left legs of one individual as there was between individuals.
Of the 40 nerve variations (including the lumbosacral plexus) 11 were found only in T. pallidicinctus, seven were found only in T. cupido, and seven were found only in Pedioecetes. Four were found in both species of Tympanuchus (but not in Pedioecetes). Eleven were found in both Tympanuchus and Pedioecetes; of[Pg 451] these, four were found in all three species, three were shared by T. pallidicinctus and Pedioecetes and four occurred in T. cupido and Pedioecetes.
The average number of variations per leg in both muscles and nerves was 11 in T. pallidicinctus, nine in T. cupido, and 16 in Pedioecetes. The high number in the last is in part the result of these being variations from the typical condition of T. pallidicinctus (rather than from Pedioecetes).
Analysis of Variation Between Species
No constant differences in the muscles or nerves was found between T. cupido pinnatus and T. cupido attwateri. Only one constant difference was found between T. cupido and T. pallidicinctus: a thicker fleshy origin of M. extensor iliotibialis lateralis in T. cupido (associated with a thicker edge of the lateral iliac process).
Although no constant differences in the nerves were found between Pedioecetes and Tympanuchus (both species), 17 constant differences in the muscles were found between these two genera. Seven of these differences pertain to features of a single muscle—M. flexor cruris medialis. Compared with the condition in Tympanuchus, M. flexor cruris medialis in Pedioecetes has a wider origin, a partly fleshy (instead of entirely tendinous) origin, a more pronounced curvature of the line of origin, a wider insertion, an insertion posterior (rather than anterior) to the medial collateral ligament, an insertion that attaches in part to the articular capsule, and a shorter tendon of insertion (resulting in the fusion of the common insertional tendon of Mm. flexor cruris lateralis and femorocruralis with the fleshy belly rather than with the insertional tendon). Other differences include the following. A more extensive posteroproximal aponeurosis of M. extensor iliotibialis lateralis in Pedioecetes (resulting in a narrower fleshy origin); a more nearly straight line of origin of this muscle (associated with a less pronounced lateral iliac process); a thinner fleshy origin of this muscle (associated with a thinner edge of the lateral iliac process); a wider M. flexor cruris lateralis that is fleshy up to the origin from the vertebrae; a wider fleshy origin of M. iliacus; the origin of M. caudofemoralis pars iliofemoralis not reaching the ventral edge of the ischium; a narrower origin of M. adductor superficialis; a wider M. femorocruralis; and a shorter belly of M. extensor digitorum longus. Some additional differences between these two genera, which are slight in degree, are given in the accounts[Pg 452] of the muscles. If additional specimens were studied, some of the differences listed above possibly would prove to be subject to individual variation and so could not properly be listed as constant differences between the two genera.
The picture of the differences between Tympanuchus and Pedioecetes that the present study presents is radically different from that presented by the study of Hudson, et al. (1959). These authors reported the following differences between these two genera. (I am using my terminology.) The origin of M. piriformis is narrower in Pedioecetes and is more posteriorly situated; the belly of M. extensor iliotibialis anticus is broader in Pedioecetes; the belly of M. tibialis anticus is longer; the belly of M. peroneus brevis is shorter; the insertional tendon of the anterolateral head of M. flexor perforatus digiti III is shorter; the belly of M. flexor digitorum longus is shorter; only two (rather than three) of the branches of M. extensor digitorum longus on the tarsometatarsus are ossified; the posterior metatarsal crest is shorter; M. flexor perforans et perforatus digiti II has two heads in Pedioecetes but only one in Tympanuchus; the roof over the hypotarsal canal enclosing the tendon of M. flexor digitorum longus is bony in Pedioecetes but fibrous in Tympanuchus; M. flexor cruris lateralis is wider in Pedioecetes; and the origin of M. femorocruralis is wider. I paid particular attention in my study to these 13 features given by Hudson, et al.; of these the only differences that I found to be constant were the last two. The apparent reason for this great discrepancy is the small number of legs of Tympanuchus studied by Hudson, et al. They studied eight legs of Pedioecetes but only two legs of Tympanuchus. This emphasizes the danger of making comparisons based on a very small number of specimens (a criticism which may prove to apply to the present study as well). The reason why Hudson, et al. did not report most of the differences found by me is not so apparent. Either the specimens studied by the former workers showed a greater variation in these characters than did my specimens or else those workers overlooked the differences. Probably both factors are involved. It remains to be determined how many specimens need to be studied in order to obtain a fairly accurate picture of variation.
Comparison with Other Studies of Innervation
I accept the following concept of muscle-nerve relationship. All muscles of the pelvic limb of birds have developed phylogenetically[Pg 453] from either the dorsal extensor muscle mass or the ventral flexor muscle mass. The former was (at least originally) supplied by only the femoral and peroneal nerves ("dorsal" nerves), the latter by only the obturator and tibial nerves ("ventral" nerves). The best guide for determining which muscles are phylogenetically dorsal and which are ventral seems to be their embryogeny (as shown in the studies of Romer, 1927, and Wortham, 1948). In the phylogenetic changes undergone by the muscles under consideration, the innervation may have changed in some instances, although this is less likely to occur than changes in the attachment or function of the muscles. If a change in innervation has occurred, it would be more likely to be a change from one dorsal nerve to the other or from one ventral nerve to the other rather than from a dorsal nerve to a ventral one or vice versa.
Thus, in my opinion, a report of a dorsal muscle supplied by a ventral nerve, or vice versa, should be viewed with suspicion until it is verified. I suspect that many previous workers have ignored this concept of muscle-nerve relationship, or else do not accept it, since they report, without comment, dorsal muscles (as determined embryologically) innervated by ventral nerves, or vice versa. Owing to the intimate association between the proximal parts of the tibial and peroneal nerves, the true relationship may be difficult to determine. I suspect that this relationship has been misinterpreted by a number of workers. I found in Tympanuchus and Pedioecetes a branch of the tibial nerve that is closely associated with, and distributed with, the peroneal nerve and has been mistakenly considered a part of the peroneal nerve by some workers. In the study here reported on, I have found no definite exceptions to the expected innervation. The only possible exception is an extra branch, which could not be traced to its origin, supplying M. extensor iliofibularis in one leg. Thus my study of innervation agrees with the embryological determination of the (phylogenetic) dorsal and ventral muscles and lends strong support to the above-stated concept of muscle-nerve relationship.
I have compared my findings on the nerves with those of other workers, who have studied the nerves with a varying degree of thoroughness. The important differences in innervation between these studies and the present one are discussed below.
In neither of Gadow's works did he distinguish tibial and peroneal components in the thigh. In his later work (1891), covering a wide variety of birds, he found that M. piriformis sometimes has[Pg 454] a femoral innervation in addition to the constant sciatic one and that M. gluteus profundus may or may not have a sciatic supply in addition to the femoral one. A comparison of Gadow's terminology of the sciatic nerve branches in the shank and foot (in both works) with mine shows that his branch I represents my peroneal nerve plus my paraperoneal branch of the tibial nerve (Ic); his branch II represents my medial division of the tibial nerve; and his branch III represents my posterior (IIIa) and lateral (IIIb) divisions of the tibial nerve.
Gadow's study (1880) on the ratites included Struthio, Rhea, and Casuarius. Only in Casuarius did Gadow find a branch (IIe) of the sciatic nerve supplying Mm. lumbricalis, adductor digiti II, and abductor digiti II. The two former muscles are typically supplied (as in Rhea) by the paraperoneal branch of the tibial nerve; Gadow's branch IIe presumably represents a segregated branch of this nerve. More surprising is his finding that M. abductor digiti II is innervated in Casuarius by both the deep peroneal nerve and branch IIe and in Rhea by branch Ic (paraperoneal branch of tibial nerve). The deep peroneal innervation is typical. Also unexpected is his finding that the posterior division of the femoral nerve gives minute twigs into M. gastrocnemius pars interna in Struthio and Casuarius. Since the other terminal branches of this nerve in these birds are nonmuscular, since this muscle is chiefly supplied by other nerves, and since the innervation from the femoral nerve is apparently atypical for most birds, the possibility should be considered that the femoral twigs are sensory rather than motor.
Sudilovskaya (1931), studying Struthio, Rhea, and Dromaeus (Dromiceius), used the same terminology as Gadow except that he designates as branch III Gadow's branch Ic. Sudilovskaya's discussion of the main branches of the sciatic nerve is confusing. He states that in Struthio, branches I, II, and III all pass through the tendinous guide loop for M. extensor iliofibularis; this is hard to believe. As near as I can determine, he has mistakenly given the same designation (branch III) to two separate branches (Gadow's Ic and III). There is no problem, however, in determining to which of these two branches he is referring when he is describing the innervation of a particular muscle, since one supplies only muscles of the shank and the other only intrinsic foot muscles. Sudilovskaya found M. abductor digiti II to be innervated by branch III (Ic of Gadow); thus the innervation of this muscle corresponds to that found in Rhea by Gadow. Although M. adductor digiti II had the expected innervation from branch III (paraperoneal branch of[Pg 455] tibial nerve) in Dromaeus, that muscle was found to be supplied by branch II in Rhea. (Gadow, on the other hand, reports a typical innervation for this muscle in Rhea.) Sudilovskaya found M. peroneus brevis to be supplied by the deep peroneal branch (in contrast to the superficial peroneal supply that I found in Tympanuchus and Pedioecetes). He found M. gastrocnemius pars interna to be supplied in Struthio by twigs of the femoral nerve in addition to its typical innervation from branch II of the sciatic nerve; this agrees with Gadow's findings in the same genus. Sudilovskaya reports that M. gastrocnemius pars externa was innervated by branches II and III in Struthio and Rhea and by branches I and III in Dromaeus. (Gadow found only the typical innervation—branch III.)
In the Whooping Crane, Fisher and Goodman (1955) found a peroneal, rather than a femoral, nerve supply for pars postica of M. vastus lateralis. They also report a peroneal nerve supply for M. flexor ischiofemoralis (in contrast to the usual tibial nerve supply) and for M. adductor superficialis (in addition to the usual supply from the obturator nerve). The innervation was not given for the intrinsic foot musculature.
Fisher (1946), studying vultures, reports the following: tibial branches, in addition to the main sciatic branch, supplying M. extensor iliofibularis (typically supplied by the peroneal nerve); an obturator supply, in addition to the usual tibial supply, to M. flexor cruris medialis; a tibial supply, in addition to the typical obturator supply, to M. obturator pars postica; a possible obturator supply, in addition to the typical femoral supply, to M. ambiens; a possible peroneal supply, in addition to the typical tibial supply, to M. flexor digitorum longus; and a peroneal supply to Mm. abductor digiti IV, flexor hallucis brevis, and adductor digiti II (which are typically supplied by the paraperoneal branch of the tibial nerve). Fisher's postfibular branch of the peroneal nerve, which supplies the latter three muscles, apparently represents the paraperoneal branch of the tibial nerve.
Carlsson (1884) did not find a femoral nerve supply for M. gluteus profundus. He found an obturator supply, in addition to the usual sciatic supply, to M. flexor ischiofemoralis in Eudyptes chrysolopha and Mergulus alle but not in the other two forms studied. He reported a peroneal supply, rather than the expected tibial (paraperoneal) supply, to Mm. abductor digiti IV and adductor digiti IV.
DeMan (1873) found a twig of the obturator nerve supplying[Pg 456] M. flexor ischiofemoralis, in addition to the typical innervation, in Corvus monedula, but not in the few other forms studied. He did not distinguish tibial and peroneal components in the thigh.
Wilcox (1948), studying a loon, did not find any peroneal supply to M. extensor iliotibialis lateralis or to M. gluteus profundus. He found a femoral, rather than a peroneal, supply to M. piriformis. He found an obturator, instead of a tibial, supply to M. flexor ischiofemoralis. (In some of my specimens I found a tiny blood vessel, appearing much like a nerve, emerging from the obturator foramen and entering M. flexor ischiofemoralis.) Wilcox reports an innervation of M. caudofemoralis pars caudifemoralis from the pudendal plexus, in addition to the usual sciatic one. Wilcox did not distinguish tibial and peroneal components in the thigh. In the shank and foot he misidentified the peroneal nerve as the tibial nerve and therefore gave erroneous innervations for all the muscles supplied by this nerve, except for M. adductor digiti IV, which actually should be supplied by the tibial nerve.
Howell (1938) studied only the hip and thigh musculature of the chicken. He overlooked the femoral nerve supply for M. gluteus profundus.
Romer (1927) studied only the hip and thigh muscles of the chick. He did not distinguish tibial and peroneal components in the thigh. He did not mention any sciatic supply for M. gluteus profundus.
Appleton (1928), studied (in various birds) only those muscles of the hip and thigh that are innervated by the tibial and peroneal nerves. He terms the former "ischiadicus ventralis" and the latter "ischiadicus dorsalis." His findings did not differ from mine.
Many differences in the innervation of specific muscles are reported in the literature, even in the same species (by different workers). Some of these differences may be real; others are probably misinterpretations. Consequently more work needs to be done before a complete understanding can be obtained of the innervation of the leg muscles of birds. Especially needed are studies of the tibial-peroneal nerve relationship, perhaps approached by a method other than gross dissection.
FOOTNOTES:
The muscles and nerves were dissected in eight legs of the Lesser Prairie Chicken (Tympanuchus pallidicinctus), six legs of the Greater Prairie Chicken (T. cupido pinnatus), three legs of Attwater's Prairie Chicken (T. c. attwateri), and six legs of the Sharp-tailed Grouse (Pedioecetes phasianellus jamesi) for the purpose of obtaining information on individual variation as well as variation between these closely related species. Relatively little information is available regarding the nerves of the leg of birds and little is known about individual variation and variation between closely related forms in the muscles of the leg of birds.
All osteological terms used in the present paper are defined and those of the pelvis are illustrated. New terms were coined for some structures for which no names could be found in the literature. Terms were also coined for the major divisions of the femoral and sciatic nerves. With three exceptions, my muscle terminology follows that of Fisher (1946) and Fisher and Goodman (1955). Their term femoritibialis externus is not used here; the muscle so named is considered to be a part of M. vastus lateralis. Fisher's accessory head of M. flexor cruris lateralis is considered to be a distinct muscle—M. femorocruralis. Usage of the term obturator internus is avoided because the muscle so named is considered not to be homologous with the mammalian muscle of the same name; the entire obturator complex is called M. obturator, and is subdivided into four parts.
The typical (most common) condition of the nerves and muscles in Tympanuchus pallidicinctus is described in detail. Variations from this condition among the other birds studied are then described. All muscles of one leg of T. pallidicinctus are illustrated. Several variations in the muscles are also illustrated. The lumbosacral plexus and nerves of the leg in several specimens that show variations are illustrated.
Considerable individual variation was found in both the muscles and the nerves of the leg of the species studied. Certain muscles were more variable than others. Mm. flexor digitorum longus, obturator, caudofemoralis, and extensor hallucis longus showed the greatest number of variations. Mm. vastus medialis, femoritibialis internus, flexor perforatus digiti III, extensor brevis digiti III, and abductor digiti IV did not exhibit any variations considered significant. Certain legs showed a greater number of variations from the typical condition than did others.
Although most of the variations were minor, some were major. M. extensor proprius digiti III was present in two legs of Pedioecetes but absent in the other legs studied. A fleshy muscle slip connected M. caudofemoralis pars caudifemoralis with the tendinous raphe between Mm. flexor cruris lateralis and femorocruralis in two legs, whereas in others this connection was tendinous or even absent altogether. M. flexor cruris lateralis had an accessory slip arising from the caudal musculature in one leg. A vinculum connected the insertional tendons of Mm. flexor perforans et perforatus digiti II and flexor perforatus digiti II in one leg.
In most specimens there was as much variation between the muscles of the right and left legs of one individual as there was between individuals. The same was true for the nerves, except for the lumbosacral plexus, in which there was no significant variation between the right and left sides of any individual. The peroneal and obturator nerves varied less than the other nerves.
No constant differences in the muscles or nerves was found between T. cupido pinnatus and T. c. attwateri. One constant difference was found between T. cupido and T. pallidicinctus: the fleshy origin of M. extensor iliotibialis lateralis in T. cupido was thicker (associated with a thicker edge of the lateral iliac process).
Although no constant differences in the nerves were found between Pedioecetes and Tympanuchus (both species), 17 constant differences in the muscles were found between these two genera. Study of additional specimens possibly would show enough individual variation in some of these differences to reduce the number of constant differences to fewer than 17. Seven of these differences pertain to features of a single muscle—M. flexor cruris medialis. Some of the other differences are associated with the thinner and much less pronounced lateral iliac process in Pedioecetes. The picture of the differences between Tympanuchus and Pedioecetes that this study presents is radically different from that presented by the study of Hudson, et al. (1959).
The important differences in innervation between previous studies and the present one are discussed.
All of the muscles under consideration have been grouped as either dorsal or ventral muscles, according to their embryonic origin, as described by Romer (1927) and Wortham (1948). This grouping probably represents accurately the phylogenetic origin of these muscles. The dorsal muscles probably were originally supplied by dorsal nerves—the femoral and peroneal—and the ventral[Pg 459] muscles probably were originally supplied by ventral nerves—the obturator and tibial. This primitive muscle-nerve relationship has been relatively constant.
Several previous workers have reported some dorsal muscles supplied by ventral nerves and vice versa. Those findings should be viewed with suspicion until verified, because the proximal parts of the tibial and peroneal nerves are intimately associated and their relationship is easily misinterpreted. I found a branch of the tibial nerve that is closely associated with, and distributed with, the peroneal nerve. That branch of the tibial nerve has been mistakenly considered a part of the peroneal nerve by some workers. My study revealed no definite exceptions to the expected innervation.
| Gadow (1891) | Hudson (1937) | Howell (1938) | Fisher & Goodman (1955) | Holmes |
| ilio-tibialis | ilio-tibialis | extensor iliotibialis lateralis |
extensor ilio-tibialis lateralis |
extensor iliotibialis lateralis |
| ilio-tibialis internus s. sartorius |
sartorius | extensor iliotibialis anterior |
extensor ilio-tibialis anterior |
extensor iliotibialis anticus |
| ambiens | ambiens | ambiens | ambiens | ambiens |
femori-tibialis externus |
............ femori-tibialis externus |
............ vastus lateralis |
femoritibialis externus | vastus lateralis (a) pars postica (b) pars lateralis |
|---|---|---|---|---|
| { femori-tibialis { medius { { |
vastus lateralis | |||
| femori-tibialis medius |
} { } vastus medialis { } { |
vastus medialis | vastus medialis | |
| femori-tibialis internus | femori-tibialis internus | femoritibialis internus | femoritibialis internus | |
| ilio-fibularis | biceps fernoris | extensor iliofibularis | extensor ilio-fibularis | extensor iliofibularis |
| ilio-femoralis externus | glutaeus medius et minimus | piriformis | piriformis | piriformis |
| ilio-trochantericus posterior | ilio-trochantericus posterior | gluteus profundus | gluteus profundus | gluteus profundus |
| [Pg 461] ilio-trochantericus anterior | ilio-trochantericus anterior | iliacus | iliacus | iliacus |
| ilio-trochantericus medius | ilio-trochantericus medius | ............ | ilio-trochantericus medius | iliotrochantericus medius |
| ilio-femoralis internus | iliacus | psoas | psoas | psoas |
| caud ilio flexorius | semitendinosus | flexor cruris lateralis | flexor cruris lateralis (a) main head (b) accessory heads |
flexor cruris lateralis |
| accessorius semitendinosi | accessorius semitendinosi | femorocruralis | femorocruralis | |
| ischio-flexorius | semimembranosus | flexor cruris medialis | flexor cruris medialis | flexor cruris medialis |
| caud-ilio-femoralis (a) pars caudi-femoralis (b) pars ilio-femoralis |
piriformis (a) pars caudi-femoralis (b) pars ilio-femoralis |
caudofemoralis |
caudofemoralis (a) pars caudi-femoralis (b) pars ilio-femoralis |
caudofemoralis (a) pars caudifemoralis (b) pars iliofemoralis |
| flexor iliofemoralis | ||||
| ischio-femoralis | ischio-femoralis | flexor ischiofemoralis | flexor ischiofemoralis | flexor ischiofemoralis |
| { pub-ischio-femoralis { { |
adductor longus et brevis (a) pars anterior (b) pars posterior |
adductor superficialis |
adductor superficialis |
adductor superficialis |
| adductor profundus | adductor profundus | adductor profundus | ||
| [Pg 462] obturator | obturator internus |
} { } obturator { } { |
obturator internus |
} } obturator } |
| accessorii M. obturatoris | obturator externus | obturator externus | ||
| gastrocnemius | gastrocnemius | gastrocnemius | gastrocnemius | |
| flexor perforans et perforatus digiti II |
flexor perforans et perforatus digiti II |
flexor perforans et perforatus digiti II |
flexor perforans et perforatus digiti II |
|
| flexor perforans et perforatus digiti III |
flexor perforans et perforatus digiti III |
flexor perforans et perforatus digiti III |
flexor perforans et perforatus digiti III |
|
| flexor perforatus digiti IV |
flexor perforatus digiti IV |
flexor perforatus digiti IV |
flexor perforatus digiti IV |
|
| flexor perforatus digiti III |
flexor perforatus digiti III |
flexor perforatus digiti III |
flexor perforatus digiti III |
|
| flexor perforatus digiti II |
flexor perforatus digiti II |
flexor perforatus digiti II |
flexor perforatus digiti II |
|
| flexor hallucis longus | flexor hallucis longus | flexor hallucis longus | flexor hallucis longus | |
| plantaris | plantaris | plantaris | plantaris | |
| flexor profundus s. perforans | flexor digitorum longus | flexor digitorum longus | flexor digitorum longus | |
| [Pg 463] popliteus | popliteus | popliteus | popliteus | |
| peroneus superficialis | peronaeus longus | peroneus longus | peroneus longus | |
| tibialis anticus | tibialis anterior | tibialis anterior | tibialis anticus | |
| extensor digitorum communis | extensor digitorum longus | extensor digitorum longus | extensor digitorum longus | |
| peroneus profundus | peronaeus brevis | peroneus brevis | peroneus brevis | |
| extensor hallucis brevis | extensor hallucis longus | extensor hallucis longus | extensor hallucis longus | |
| abductor digiti II | abductor digiti II | abductor digiti II | abductor digiti II— | |
| extensor brevis digiti III |
} { } extensor proprius { } digiti III { } { |
extensor brevis digiti III | extensor brevis digiti III | |
| extensor brevis digiti IV | extensor brevis digiti IV | extensor brevis digiti IV | ||
| flexor brevis digiti III | lumbricalis | ............ | lumbricalis | |
| abductor digiti IV | abductor digiti IV | abductor digiti IV | abductor digiti IV | |
| flexor hallucis brevis | flexor hallucis brevis | flexor hallucis brevis | flexor hallucis brevis | |
| adductor digiti II | adductor digiti II | adductor digiti II | ............ | |
| adductor digiti IV | adductor digiti IV | ............ | ............ |
| Muscle | Tympanuchus | Pedioecetes | ||||
|---|---|---|---|---|---|---|
| Ave. |
Range | No.[1] | Ave. | Range | No.[1] | |
|
Iliacus: width of fleshy origin (divided by length of ilium)
|
.10 | .08-.11 | 13 | .19 | .17-.19 | 6 |
|
Flexor cruris lateralis: maximum width of exposed part (divided by length of ilium)
|
.22 | .19-.27 | 13 | .31 | .27-.36 | 6 |
|
Flexor cruris medialis: width of origin (divided by length of ilium)
|
.11 | .08-.16 | 13 | .22 | .19-.23 | 6 |
|
Flexor cruris medialis: width of insertion (divided by length of tibiotarsus)
|
.09 | .08-.13 | 13 | .17 | .15-.17 | 4 |
|
Adductor superficialis: width of origin (divided by length of ilium)
|
.20 | .17-.23 | 13 | .13 | .10-.16 | 5 |
|
Femorocruralis: distance of proximal end of origin from proximal end of femur (divided by length of femur)
|
.59 | .55-.63 | 13 | .40 | .38-.43 | 6 |
|
Extensor digitorum longus: length of fleshy belly (divided by length of tibiotarsus)
|
.73 | .64-.83 | 13 | .59 | .50-.62 | 4 |
FOOTNOTES:
[1] No. = number of legs.
| T.p. | T.c.p. | T.c.a. | P.p. | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 L |
1 R |
2 L |
2 R |
3 L |
3 R |
4 L |
5 R |
1 L |
1 R |
2 L |
2 R |
3 L |
4 L |
1 L |
1 R |
2 L |
1 L |
1 R |
2 L |
3 L |
3 R |
4 L |
|
| Ambiens | |||||||||||||||||||||||
|
origin partly fleshy
|
x | ||||||||||||||||||||||
| Vastus lateralis | |||||||||||||||||||||||
|
no vincula
|
x | x | |||||||||||||||||||||
| Extensor iliofibularis | |||||||||||||||||||||||
|
insertional tendon double
|
x | ||||||||||||||||||||||
| Piriformis | |||||||||||||||||||||||
|
posteroproximal corner tendinous
|
x | x | x | x | x | ||||||||||||||||||
|
insertion fused to flexor ischiofemoralis
|
x | ||||||||||||||||||||||
| Iliotrochantericus medius | |||||||||||||||||||||||
|
not notched
|
x | x | x | ||||||||||||||||||||
|
anterior part with fleshy origin
|
x | x | x | x | |||||||||||||||||||
|
insertion fused to gluteus profundus
|
x | ||||||||||||||||||||||
|
muscle split
|
x | ||||||||||||||||||||||
| Flexor cruris lateralis | |||||||||||||||||||||||
|
accessory slip present
|
x | ||||||||||||||||||||||
| Flexor cruris medialis | |||||||||||||||||||||||
|
origin from pubis
|
x | x | |||||||||||||||||||||
|
insertion partly fleshy
|
x | x | x | x | |||||||||||||||||||
|
insertional tendon split
|
x | ||||||||||||||||||||||
| Caudofemoralis | |||||||||||||||||||||||
|
accessory slip fleshy
|
x | x | |||||||||||||||||||||
|
tendinous area in belly of pars caudifemoralis
|
x | x | x | ||||||||||||||||||||
|
origin from pubis
|
x | ||||||||||||||||||||||
|
insertion entirely tendinous
|
x | ||||||||||||||||||||||
| [Pg 466] Flexor ischiofemoralis | |||||||||||||||||||||||
|
insertion partly fleshy
|
x | x | x | ||||||||||||||||||||
| Adductor superficialis | |||||||||||||||||||||||
|
groove for flexor cruris medialis present
|
x | x | x | x | |||||||||||||||||||
|
completely fused with adductor profundus
|
x | ||||||||||||||||||||||
| Adductor profundus | |||||||||||||||||||||||
|
proximal part of insertion tendinous
|
x | ||||||||||||||||||||||
|
distal end of insertion tendinous
|
x | ||||||||||||||||||||||
| Obturator | |||||||||||||||||||||||
|
independent slip of pars antica present
|
x | x | |||||||||||||||||||||
|
slip of pars antica fused to pars postica
|
x | x | |||||||||||||||||||||
|
independent slip of pars dorsalis present
|
x | x | x | x | |||||||||||||||||||
|
pars dorsalis fused with pars antica
|
x | ||||||||||||||||||||||
| Gastrocnemius | |||||||||||||||||||||||
|
pars interna overlaps peroneus longus
|
x | ||||||||||||||||||||||
| Flexor perforans et perforatus digiti II | |||||||||||||||||||||||
|
anterior head entirely tendinous
|
x | ||||||||||||||||||||||
|
vinculum joins flexor perforatus digiti II
|
x | ||||||||||||||||||||||
|
origin from superficial surface of patellar tendon
|
x | ||||||||||||||||||||||
| Flexor perforans et perforatus digiti III | |||||||||||||||||||||||
|
accessory head present
|
x | x | |||||||||||||||||||||
| Flexor perforatus digiti II | |||||||||||||||||||||||
|
roof of hypotarsal canal bony
|
x | x | x | ||||||||||||||||||||
| [Pg 467] Plantaris | |||||||||||||||||||||||
|
accessory head present
|
x | ||||||||||||||||||||||
|
origin from medial collateral ligament
|
x | ||||||||||||||||||||||
| Flexor digitorum longus | |||||||||||||||||||||||
|
notched for peroneal nerve
|
x | x | x | x | x | ||||||||||||||||||
|
origin from tendon of extensor iliofibularis
|
x | ||||||||||||||||||||||
|
third dorsal slip present in digiti IV
|
x | x | x | x | x | x | x | x | x | ||||||||||||||
|
third dorsal slip present in digiti III
|
x | x | x | x | x | x | x | x | x | x | x | x | |||||||||||
|
second dorsal slip present in digiti II
|
x | x | x | x | x | x | x | x | x | x | x | x | |||||||||||
|
vinculum joins flexor perforatus digiti IV
|
x | x | x | x | |||||||||||||||||||
| Peroneus longus | |||||||||||||||||||||||
|
origin from patellar tendon
|
x | x | x | x | x | ||||||||||||||||||
| Tibialis anticus | |||||||||||||||||||||||
|
accessory insertion absent
|
x | x | |||||||||||||||||||||
| Extensor digitorum longus | |||||||||||||||||||||||
|
lateral branch of tendon not ossified
|
x | x | x | x | x | x | x | x | x | x | |||||||||||||
|
origin from posterior surface of outer cnemial crest
|
x | x | x | ||||||||||||||||||||
| Extensor hallucis longus | |||||||||||||||||||||||
|
origin lateral to retinaculum
|
x | x | |||||||||||||||||||||
|
distal fibers of distal head insert independently
|
x | x | x | x | x | ||||||||||||||||||
|
accessory bundle present
|
x | ||||||||||||||||||||||
|
entire distal head inserts independently
|
x | ||||||||||||||||||||||
| Abductor digiti II | |||||||||||||||||||||||
|
accessory insertion present
|
x | ||||||||||||||||||||||
| Extensor proprius digiti III | |||||||||||||||||||||||
|
present
|
x | x | |||||||||||||||||||||
| Leg | Other leg of same specimen |
T.p. | T.c.p. | T.c.a. | P.p. | No other legs |
|||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 L |
1 R |
2 L |
2 R |
3 L |
3 R |
4 L |
4 R |
1 L |
1 R |
2 L |
2 R |
3 L |
4 L |
1 L |
1 R |
2 L |
1 L |
1 R |
2 L |
3 L |
3 R |
4 L |
|||
| T.p. 1 L | 1 | — | — | 2 | 2 | 1 | 1 | 1 | 1 | 2 | 1 | 1 | 2 | 1 | |||||||||||
| T.p. 2 R | 6 | — | — | 2 | 1 | 1 | 2 | 1 | 1 | 1 | 2 | 3 | 3 | 3 | 2 | 3 | 1 | 1 | 4 | 2 | 0 | ||||
| T.p. 3 L | 4 | 2 | 1 | 2 | 2 | — | — | 1 | 1 | 1 | 2 | 3 | 2 | 2 | 4 | 4 | 1 | 2 | 3 | 5 | 1 | ||||
| T.c.p. 1 R | 1 | 1 | — | — | 1 | 2 | |||||||||||||||||||
| T.c.p. 2 R | 0 | 1 | 1 | 1 | — | — | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 3 | ||||||||||
| T.c.a. 1 R | 5 | 1 | 3 | 3 | 2 | 1 | 1 | 1 | 2 | 2 | — | — | 3 | 3 | 4 | 1 | 1 | 3 | 2 | 3 | |||||
| P.p. 1 L | 4 | 2 | 2 | 2 | 3 | 1 | 1 | 1 | 3 | 3 | 3 | 2 | — | — | 3 | 2 | 3 | 7 | 2 | ||||||
| P.p. 3 R | 4 | 4 | 4 | 2 | 2 | 1 | 1 | 1 | 1 | 2 | 3 | 3 | 3 | 3 | 5 | 4 | — | — | 5 | 1 | |||||
| T.p. | T.c.p. | T.c.a. | P.p. | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 L |
1 R |
2 L |
2 R |
3 L |
3 R |
4 L |
5 R |
1 L |
1 R |
2 L |
2 R |
3 L |
4 L |
1 L |
1 R |
2 L |
1 L |
1 R |
2 L |
3 L |
3 R |
4 L |
|
| Lumbosacral plexus | |||||||||||||||||||||||
|
two fureal nerves
|
x | x | x | x | x | x | x | ||||||||||||||||
|
S9 with three branches
|
x | ||||||||||||||||||||||
|
prefixed
|
x | x | x | x | x | x | x | x | |||||||||||||||
|
sciatic nerve with six roots
|
x | x | x | x | x | x | x | x | x | x | |||||||||||||
|
obturator nerve from S2 and S3 only
|
x | x | x | x | |||||||||||||||||||
|
femoral nerve mainly from S2-S4
|
x | x | x | x | x | x | x | x | |||||||||||||||
| Femoral nerve | |||||||||||||||||||||||
|
anterior division innervates extensor iliotibialis lateralis
|
x | x | x | x | x | ||||||||||||||||||
|
dorsal division fused with anterior division
|
x | x | x | x | |||||||||||||||||||
|
dorsal division fused with middle division
|
x | x | x | x | x | x | |||||||||||||||||
|
two branches to iliacus
|
x | x | |||||||||||||||||||||
|
middle division anastomoses with anterior division
|
x | x | x | x | x | ||||||||||||||||||
|
anterodorsal division does not go through femoral notch
|
x | x | |||||||||||||||||||||
|
branch of anterior division perforates iliacus
|
x | ||||||||||||||||||||||
|
cutaneous branch perforates extensor iliotibialis lateralis
|
x | x | |||||||||||||||||||||
|
branch of middle division perforates vastus medialis
|
x | ||||||||||||||||||||||
|
[Pg 470]
branch to vasti innervates extensor iliotibialis lateralis
|
x | ||||||||||||||||||||||
|
anterior branch of anterior division cutaneous
|
x | x | x | ||||||||||||||||||||
| Sciatic nerve | |||||||||||||||||||||||
|
twig to pars caudifemoralis independent
|
x | ||||||||||||||||||||||
|
branch to flexor cruris lateralis does not perforate caudofemoralis
|
x | x | x | x | x | x | x | x | x | ||||||||||||||
|
paraperoneal nerve enters peroneal sheath
|
x | x | x | x | x | x | |||||||||||||||||
|
cutaneous peroneal branch perforates gastrocnemius pars externa
|
x | x | x | ||||||||||||||||||||
|
cutaneous peroneal branch absent
|
x | x | x | ||||||||||||||||||||
|
distal cutaneous tibial branch absent
|
x | ||||||||||||||||||||||
|
twig to tail present
|
x | x | x | x | x | x | x | ||||||||||||||||
|
nonmuscular peroneal twig deep to vastus lateralis pars postica
|
x | x | x | x | |||||||||||||||||||
|
branch to flexor cruris medialis from posterior tibial division
|
x | x | x | ||||||||||||||||||||
|
extra twigs join cutaneous tibial branches
|
x | ||||||||||||||||||||||
|
branch to flexor cruris medialis an independent division
|
x | x | x | ||||||||||||||||||||
|
[Pg 471]
branch to flexor cruris medialis perforates flexor ischiofemoralis
|
x | ||||||||||||||||||||||
|
two twigs to flexor ischiofemoralis
|
x | x | x | x | x | ||||||||||||||||||
|
independent extra branch innervates extensor iliofibularis
|
x | ||||||||||||||||||||||
|
branch to femorocruralis innervates gastrocnemius pars media
|
x | ||||||||||||||||||||||
| Peroneal nerve | |||||||||||||||||||||||
|
superficial and deep peroneal nerves do not join
|
x | x | x | x | |||||||||||||||||||
| Tibial nerve | |||||||||||||||||||||||
|
independent extra branch innervates flexor perforatus digiti IV
|
x | x | |||||||||||||||||||||
|
anastomosis involving posterior division
|
x | x | |||||||||||||||||||||
|
branch to gastrocnemius pars externa an independent division
|
x | ||||||||||||||||||||||
|
branch to gastrocnemius pars media innervates femorocruralis
|
x | x | |||||||||||||||||||||
|
extra branch innervates flexor perforatus digiti III
|
x | x | |||||||||||||||||||||
|
branch to gastrocnemius pars interna perforates plantaris
|
x | ||||||||||||||||||||||
|
branch to gastrocnemius pars interna innervates plantaris
|
x | ||||||||||||||||||||||
| Leg | Other leg of same specimen |
T.p. | T.c.p. | T.c.a. | P.p. | No other legs |
|||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 L |
1 R |
2 L |
2 R |
3 L |
3 R |
4 L |
5 R |
1 L |
1 R |
2 L |
2 R |
3 L |
4 L |
1 L |
1 R |
2 L |
1 L |
1 R |
2 L |
3 L |
3 R |
4 L |
|||
| T.p. 1 L | 1 | — | — | 1 | 2 | 1 | 1 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | 1 | 2 | 0 | |||||
| T.p. 2 L | 5 | 1 | — | — | 2 | 1 | 3 | 1 | 2 | 2 | 2 | 1 | 2 | 1 | 2 | 1 | 1 | ||||||||
| T.p. 3 R | 3 | 1 | 2 | 1 | — | — | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | ||||||||||
| T.c.p. 1 R | 1 | 1 | 1 | 1 | 1 | 1 | — | — | 2 | 1 | 2 | 1 | 0 | ||||||||||||
| T.c.p. 2 L | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 2 | — | — | 2 | 1 | 1 | ||||||||||||
| T.c.a. 1 R | 1 | 2 | 1 | 1 | 1 | 1 | — | — | 1 | 1 | 1 | 2 | |||||||||||||
| P.p. 1 R | 3 | 2 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | — | — | 1 | 2 | 2 | 3 | 1 | ||||||||
| P.p. 3 L | 2 | 1 | 2 | 1 | 1 | 1 | 2 | 3 | — | — | 2 | 2 | |||||||||||||
| Appleton, A. B. | ||
|---|---|---|
| 1928. | The muscles and nerves of the post-axial region of the tetrapod thigh. Parts I and II. Jour. Anat., 62(3,4):364-438. | |
| Berger, A. J. | ||
| 1952. | The comparative functional morphology of the pelvic appendage in three genera of Cuculidae. Amer. Midl. Nat., 47(3):513-605. | |
| Berger, A. J. | ||
| 1956. | The appendicular myology of the Sandhill Crane, with comparative remarks on the Whooping Crane. Wilson Bull., 68(4):282-304. | |
| Boas, J. E. V. | ||
| 1933. | Kreuzbein, Becken und Plexus Lumbosacralis der Vögel. Det Kongelige Danske Videnskabernes Selskabs Skrifter. Naturvidenskabelig og Mathematisk Afdeling. 9. række, 5(l):l-74, 15 pls. | |
| Carlsson, A. | ||
| 1884. | Beiträge zur Kenntniss der Anatomie der Schwimmvögel. Bihang till K. Svenska Vetenskapsakad. Handlingar, 9(3):l-44, 5 pls. | |
| Chomiak, M. | ||
| 1950. | [Studies on the plexus lumbalis et sacralis in the domestic hen.] Ann. Univ. Mariae Curie-Sklodowska, Lublin, Sect. DD-Vet. Med., 5(3):29-45. | |
| Fisher, H. I. | ||
| 1946. | Adaptations and comparative anatomy of the locomotor apparatus of new world vultures. Amer. Midi. Nat., 35(3):545-727, 13 pls. | |
| Fisher, H. I., and Goodman, D. C. | ||
| 1955. | The myology of the Whooping Crane, _Grus americana_. Ill. Biol. Mono., 24(2):viii + 1-127. | |
| Gadow, H. | ||
| 1880. | Zur vergleichenden Anatomie der Muskulatur des Beckens und der hinteren Gliedmasse der Ratiten. Fischer, Jena, 56 pp., 5 pls. | |
| Gadow, H. (with E. Selenka). | ||
| 1891. | Vögel. I. Anatomischer Theil. _In_ Bronn's Klassen und Ordnungen des Their-Reichs, 6(4):1-1008. Winter, Leipzig. | |
| Holmes, E. B. | ||
| 1962. | The terminology of the short extensor muscles of the third toe in birds. Auk, 79(3):485-488. | |
| Howard, H. | ||
| 1929. | The avifauna of Emeryville shellmound. Univ. Calif. Publ. Zool., 32(2):301-394, 4 pls. | |
| Howell, A. B. | ||
| 1938. | Muscles of the avian hip and thigh. Auk, 55(1):71-81. | |
| Hudson, G. E. | ||
| 1937. | Studies on the muscles of the pelvic appendage in birds. Amer. Midl. Nat., 18(1):1-108, incl. 26 pls. | |
| Hudson, G. E., et al. | ||
| 1959. | Muscles of the pelvic limb in galliform birds. Amer. Midl. Nat., 61(1):l-67. | |
| Jhering (Ihering), H. V. | ||
| 1878. | Das peripherische Nervensystem der Wirbelthiere. Vogel, Leipzig, xiv-238 pp., 5 pls. | |
| Man, J. G. de | ||
| 1873. | Vergelijkende myologische en neurologische Studien over Amphibien en Vögels. van Doesburgh, Leiden, 148 pp., 4 pls. | |
| Romer, A. S. | ||
| 1927. | The development of the thigh musculature of the chick. Jour. Morph., 43(2):347-385. | |
| Sudilovskaya, A. M. | ||
|---|---|---|
| 1931. | [Study on the comparative anatomy of the musculature and innervation of the pelvic region and the hind appendages of the Ratitae (Struthio, Rhea, Dromaeus).] Acad. Sci. U.S.S.R., Leningrad, 84 pp. (In Russian.) | |
| Toit, P. J. du | ||
| 1913. | Untersuchungen über das Synsacrum und den Schwanz von Gallus domesticus nebst Beobachtungen über Schwanzlosigkeit bei Kaulhühnern. Jenaische Zeitschr. Naturw., 49:149-312, 3 pls. | |
| Wilcox, H. H., Jr. | ||
| 1948. | The pelvic musculature of the loon (Gavia immer). Univ. Microfilms, Ann Arbor, 95 pp., 26 pls. | |
| Wortham, R. A. | ||
| 1948. | The development of the muscles and tendons in the lower leg and foot of chick embryos. Jour. Morph., 83(1):105-148. | |
| Yasuda, M. | ||
| 1959. | [Comparative and topographical anatomy of the fowl. XI. On the nervous supply of the hind limb.] In Proc. of 47th Meeting of Jap. Soc. of Vet. Sci. Jap. Jour. Vet. Sci., 21(6):36. (Japanese abstract.) | |
|
Transmitted October 30, 1962. |
||
|
(Continued from inside of front cover) |
||||
| |
||||
| 18. | Conspecificity of two pocket mice, Perognathus goldmani and P. artus. By E. Raymond Hall and Marilyn Bailey Ogilvie. Pp. 513-518, 1 map. January 14, 1960. | |||
| 19. | Records of harvest mice, Reithrodontomys, from Central America, with description of a new subspecies from Nicaragua. By Sydney Anderson and J. Knox Jones, Jr. Pp. 519-529. January 14, 1960. | |||
| 20. | Small carnivores from San Josecito Cave (Pleistocene), Nuevo León, México. By E. Raymond Hall. Pp. 531-538, 1 figure in text. January 14, 1960. | |||
| 21. | Pleistocene pocket gophers from San Josecito Cave, Nuevo León, México. By Robert J. Russell. Pp. 539-548, 1 figure in text. January 14, 1960. | |||
| 22. | Review of the insectivores of Korea. By J. Knox Jones, Jr., and David H. Johnson. Pp. 549-578, February 23, 1960. | |||
| 23. | Speciation and evolution of the pygmy mice, genus Baimoys. By Robert L. Packard. Pp. 579-670, 4 plates, 12 figures in text. June 16, 1960. | |||
| Index. Pp. 671-690. | ||||
| Vol. | 10. | 1. | Speciation and evolution of the pygmy mice, genus Baimoys. By Robert L. Packard. Pp. 579-670, 4 plates, 12 figures in text. June 16, 1960. | |
| 2. | Comparative breeding behavior of Ammospiza caudacuta and A. maritima. By Glen E. Woolfenden. Pp. 45-75, 6 plates, 1 figure. December 20, 1956. | |||
| 3. | The forest habitat of the University of Kansas Natural History Reservation. By Henry S. Fitch and Ronald R. McGregor. Pp. 77-127, 2 plates, 7 figures in text, 4 tables. December 31, 1956. | |||
| 4. | Aspects of reproduction and development in the prairie vole (Microtus ochrogaster). By Henry S. Fitch. Pp. 129-161, 8 figures in text, 4 tables. December 19, 1957. | |||
| 5. | Birds found on the Arctic slope of northern Alaska. By James W. Bee. Pp. 163-211, plates 9-10, 1 figure in text. March 12, 1958.. | |||
| *6. | The wood rats of Colorado: distribution and ecology. By Robert B. Finley, Jr. Pp. 213-552, 34 plates, 8 figures in text, 35 tables. November 7, 1958. | |||
| 7. | Home ranges and movements of the eastern cottontail in Kansas. By Donald W. Janes. Pp. 553-572, 4 plates, 3 figures in text. May 4, 1959. | |||
| 8. | Natural history of the salamander, Aneides hardyi. By Richard F. Johnston and Gerhard A. Schad. Pp. 573-585. October 8, 1959. | |||
| 9. | A new subspecies of lizard, Cnemidophorus sacki, from Michoacán, México. By William E. Duellman. Pp. 587-598, 2 figures in text. May 2, 1960. | |||
| 10. | A taxonomic study of the middle American snake, Pituophis deppei. By William E. Duellman. Pp. 599-610, 1 plate, 1 figure in text. May 2, 1960. | |||
| Index. Pp. 611-626. | ||||
| Vol. | 11. | Nos. 1-10 and index. Pp. 1-703, 1958-1960. | ||
| Vol. | 12. | 1. | Functional morphology of three bats: Sumops, Myotis, Macrotus. By Terry A. Vaughan. Pp. 1-153, 4 plates, 24 figures in text. July 8, 1959. | |
| *2. | The ancestry of modern Amphibia: a review of the evidence. By Theodore H. Eaton, Jr. Pp. 155-180, 10 figures in text. July 10, 1959. | |||
| 3. | The baculum in microtine rodents. By Sydney Anderson. Pp. 181-216, 49 figures in text. February 19, 1960. | |||
| *4. | A new order of fishlike Amphibia from the Pennsylvanian of Kansas. By Theodore H. Eaton, Jr., and Peggy Lou Stewart. Pp. 217-240, 12 figures in text. May 2, 1960. | |||
| 5. | Natural history of the bell vireo. By Jon C. Barlow. Pp. 241-296, 6 figures in text. March 7, 1962. | |||
| 6. | Two new pelycosaurs from the lower Permian of Oklahoma. By Richard C. Fox. Pp. 297-307, 6 figures in text. May 21, 1962. | |||
| 7. | Vertebrates from the barrier island of Tamaulipas, México. By Robert K. Selander, Richard F. Johnston, B. J. Wilks, and Gerald G. Raun. Pp. 309-345, pls. 5-8. June 18, 1962. | |||
| 8. | Teeth of Edestid sharks. By Theodore H. Eaton, Jr. Pp. 347-362, 10 figures in text. October 1, 1962. | |||
| 9. | Variation in the muscles and nerves of the leg in two genera of grouse (Tympanuchus and Pedioecetes). By E. Bruce Holmes. Pp. 363-474, 20 figs. October 25, 1963. | |||
| More numbers will appear in volume 12. | ||||
| Vol. | 13. | 1. | Five natural hybrid combinations in minnows (Cyprinidae). By Frank B. Cross and W. L. Minckley. Pp. 1-18. June 1, 1960.. | |
| 2. | A distributional study of the amphibians of the Isthmus of Tehuantepec, México. By William E. Duellman. Pp. 19-72, pls. 1-8, 3 figures in text. August 16, 1960. | |||
| 3. | A new subspecies of the slider turtle (Pseudemys scripta) from Coahulia, México. By John M. Legler. Pp. 73-84, pls. 9-12, 3 figures in text. August 16, 1960. | |||
| 4. | Autecology of the copperhead. By Henry S. Fitch. Pp. 85-288, pls. 13-20, 26 figures in text. November 30, 1960. | |||
| 5. | Occurrence of the garter snake, Thamnophis sirtalis, in the Great Plains and Rocky Mountains. By Henry S. Fitch and T. Paul Maslin. Pp. 289-308, 4 figures in text. February 10, 1961. | |||
| 6. | Fishes of the Wakarusa river in Kansas. By James E. Deacon and Artie L. Metcalf. Pp. 309-322, 1 figure in text. February 10, 1961. | |||
| 7. | Geographic variation in the North American cyprinid fish, Hybopsis gracilis. By Leonard J. Olund and Frank B. Cross. Pp. 323-348, pls. 21-24, 2 figures in text. February 10, 1961. | |||
| (Continued on outside of back cover) | ||||
|
(Continued from inside of back cover) |
||||
| |
||||
| 8. | Decriptions of two species of frogs, genus Ptychohyla; studies of American hylid frogs, V. By William E. Duellman. Pp. 349-357, pl. 25, 2 figures in text. April 27, 1961. | |||
| 9. | Fish populations, following a drought, in the Neosho and Marais des Cygnes rivers of Kansas. By James Everett Deacon. Pp. 359-427, pls. 26-30, 8 figs. August 11, 1961. | |||
| 10. | Recent soft-shelled turtles of North America (family Trionychidae). By obert G. Webb. Pp. 429-611, pls. 31-54, 24 figures in text. February 16, 1962. | |||
| Index. Pp. 613-624. | ||||
|---|---|---|---|---|
| Vol. | 14. | 1. | Neotropical bats, from western Mexico. By Sydney Anderson. Pp. 1-8. October 24, 1960. | |
| 2. | Geographic variation in the harvest mouse, Reithrodontomys megalotis, on the central Great Plains and in adjacent regions. By J. Knox Jones, Jr., and B. Mursaloglu. Pp. 9-27, 1 figure in text. July 24, 1961 | |||
| 3. | Mammals of Mesa Verde National Park, Colorado. By Sydney Anderson. Pp. 29-67, pls. 1 and 2, 3 figures in text. July 24, 1961. | |||
| 4. | A new subspecies of the black myotis (bat) from eastern Mexico. By E. Raymond Hall and Ticul Alvarez. Pp. 69-72, 1 figure in text. December 29, 1961. | |||
| 5. | North American yellow bats, "Dasypterus," and a list of the named kinds of the genus Lasiurus Gray. By E. Raymond Hall and J. Knox Jones, Jr. Pp. 73-98, 4 figures in text. December 29, 1961. | |||
| 6. | Natural history of the brush mouse (Peromyscus boylii) in Kansas with description of a new subspecies. By Charles A. Long. Pp. 99-111, 1 figure in text. December 29, 1961. | |||
| 7. | Taxonomic status of some mice of the Peromyscus boylii group in eastern Mexico, with description of a new subspecies. By Ticul Alvarez. Pp. 113-120, 1 figure in text. December 29, 1961. | |||
| 8. | A new subspecies of ground squirrel (Spermophilus spilosoma) from Tamaulipas, Mexico. By Ticul Alvarez. Pp. 121-124. March 7, 1962 | |||
| 9. | Taxonomic status of the free-tailed bat, Tadarida yucatanica Miller. By J. Knox Jones, Jr., and Ticul Alvarez. Pp. 125-133,1 figure in text. March 7, 1962. | |||
| 10. | A new doglike carnivore, genus Cynaretus, from the Clarendonian Pliocene, of Texas. By E. Raymond Hall and Walter W. Dalquest. Pp. 135-138, 2 figures in text. April 30, 1962. | |||
| 11. | A new subspecies of wood rat (Neotoma) from northeastern Mexico. By Ticul Alvarez. Pp. 139-143. April 30, 1962. | |||
| 12. | Noteworthy mammals from Sinaloa, Mexico. By J. Knox Jones, Jr., Ticul Alvarez, and M. Raymond Lee. Pp. 145-159, 1 figure in text. May 18, 1962. | |||
| 13. | A new bat (Myotis) from Mexico. By E. Raymond Hall. Pp. 161-164, 1 figure in text. May 21, 1962. | |||
| 14. | The mammals of Veracruz. By E. Raymond Hall and Walter W. Dalquest. Pp. 165-362, 2 figures. May 20, 1963. | |||
| 15. | The recent mammals of Tamaulipas, Mexico. By Ticul Alvarez. Pp. 363-473, 5 figures in text. May 20, 1963. | |||
| More numbers will appear in volume 14. | ||||
| Vol. | 15. | 1. | The amphibians and reptiles of Michoacán, Mexico. By William E. Duellman. Pp. 1-148, pls. 1-6, 11 figures in text. December 20, 1961. | |
| 2. | Some reptiles and amphibians from Korea. By Robert G. Webb, J. Knox Jones, Jr., and George W. Byers. Pp. 149-173. January 31, 1962. | |||
| 3. | A new species of frog (Genus Tomodactylus) from western Mexico. By Robert G. Webb. Pp. 175-181, 1 figure in text. March 7, 1962. | |||
| 4. | Type specimens of amphibians and reptiles in the Museum of Natural History, the University of Kansas. By William E. Duellman and Barbara Berg. Pp. 183-204. October 26, 1962 | |||
| 5. | Amphibians and Reptiles of the Rainforests of Southern El Petén, Guatemala. By William E. Duellman. Pp. 205-249, pls. 7-10, 6 figures in text. October 4, 1963. | |||
| 6. | A revision of snakes of the genus Conophis (Family Colubridae, from Middle America). By John Wellman. Pp. 251-295, 9 figures in text. October 4, 1963. | |||
| 7. | A review of the Middle American tree frogs of the genus Ptychohyla. By William E. Duellman. Pp. 297-349, pls. 11-18, 7 figures in text. October 18, 1963. | |||
| More numbers will appear in volume 15. | ||||