The Project Gutenberg EBook of Systematics of Megachiropteran Bats in the Solomon Islands, by Carleton J. Phillips This eBook is for the use of anyone anywhere at no cost and with almost no restrictions whatsoever. You may copy it, give it away or re-use it under the terms of the Project Gutenberg License included with this eBook or online at www.gutenberg.org Title: Systematics of Megachiropteran Bats in the Solomon Islands Author: Carleton J. Phillips Release Date: July 1, 2012 [EBook #40112] Language: English Character set encoding: ISO-8859-1 *** START OF THIS PROJECT GUTENBERG EBOOK SYSTEMATICS OF MEGACHIROPTERAN BATS *** Produced by Chris Curnow, Joseph Cooper, Tom Cosmas and the Online Distributed Proofreading Team at http://www.pgdp.net
University of Kansas Publications, Museum of Natural History
Editorial Committee: E. Raymond Hall, Chairman; Frank B. Cross, Editor;
Henry S. Fitch; J. Knox Jones, Jr.
Volume 16, No. 8, pp. 777-837, 17 figs.
Published December 16, 1968
University of Kansas
Lawrence, Kansas
PRINTED BY
ROBERT R. (BOB) SANDERS, STATE PRINTER
TOPEKA, KANSAS
1968
![]()
31-9490
Systematics of Megachiropteran Bats
in the Solomon Islands
BY
CARLETON J. PHILLIPS
The Solomon Islands constitute an archipelago east of the large island of New Guinea and more than a thousand miles off the northeastern coast of Australia. This archipelago, which is principally of volcanic origin although sedimentary layers of calcareous rocks occur on many islands (Lever, 1934; Belkin, 1962), consists of a double chain of islands having a northwest-southeast axis of more than 600 miles. The archipelago is more or less an extension of New Guinea and in fact is connected to it in stepping-stone fashion by New Britain, New Ireland, and numerous smaller islands (see Fig. 1).
Australia and New Guinea have many kinds of mammals but the only terrestrial mammals in the Solomon Islands are a species of the genus Phalanger (order Marsupialia), and several species of four genera of rodents, one genus of which probably was introduced by man. Additionally, several kinds of bats have reached and colonized the Solomon Islands.
In the past 100 years at least 43 species and subspecies of Chiroptera of 16 genera have been recorded from the Solomon Islands; of these 27 species and subspecies of seven genera are in the suborder [782] Megachiroptera. At least one genus of Megachiroptera is endemic as are numerous species of other genera, and subspecies of still other species.
In 1963 and 1964, the Bernice P. Bishop Museum sent several collecting parties to the British Solomon Islands Protectorate and the Australian Trust Territory of New Guinea. In the Solomons, J. Linsley Gressitt, Philip Temple, Peter Shanahan, and Ray Straatmann visited many of the larger and more accessible islands and collected a wealth of zoological materials. I have had the opportunity to study and report on specimens of mammals, especially bats, collected by the persons named and deposited in the Bishop Museum. This report is the third in a series on bats from the Solomons (Phillips, 1966; 1967). Other specimens, mostly obtained in 1944 by personnel of United States military units, are stored in the United States National Museum and have been available for study. Aims of the following report are to (1) identify the megachiropteran bats to species and subspecies and (2) discuss distribution of these bats in the Solomon Islands.
In all, 27 kinds (subspecies and monotypic species) of the order Megachiroptera are known from the Solomon Islands. These pertain to three subfamilies of the one family Pteropodidae.
The 43 Solomon Islands, having a total land area of more than 15,300 square miles (see Belkin, 1962:42-43), are listed in the gazetteer (see also Figure 2). Politically, all of the Solomon Islands except Buka and Bougainville, which are included in New Guinea Trust Territory under mandate to Australia, are in the British Solomon Islands Protectorate.
The Solomons are within 300 to 700 miles of the equator and have a fairly constant tropical climate, except at high elevations. The temperature varies little; monthly mean temperature is between 81° and 83° F. and at sea level ranges from about 70° to 93° F. yearly (Belkin, 1962:42).
Southeast tradewinds are relatively constant from May to October and this period, in general, is a dry season except at higher elevations on windward coasts. From December to March prevailing winds are from the north and precipitation throughout the island group is especially heavy. Rainfall on the island of Tulagi averages about 120 inches per year (Bryan, Edwin H., 1941; MS, p. 2, at Pacific Sci. Information Center, Bishop Museum) and up to 300 inches have been recorded on the north coast of Guadalcanal (Belkin, 1962:42-43). Occasional dry periods occur even in the period of December to March.
Most islands of the Solomon Group support dense tropical rain forest. Much of it has been modified by man. Some clearings and scattered coconut plantations are found along coasts. On some of the larger islands (for example, Guadalcanal) coastal scrub (especially on leeward coasts) and extensive grassy areas are to be found. Additional notes on vegetation are in the gazetteer.
The 165,000 persons living on the Solomon Islands are mostly Melanesians [783] but some are mixed Papuan, Malay, and Polynesian. These native peoples are notorious for their cannibalistic tendencies; the eating of human flesh usually was related to warfare, although malefactors and human sacrifices accounted for some of the cannibalism (Cranstone, 1961:29). Prior to the Second World War few Europeans visited the Solomons and several islands still remain beyond reach of modern-day technology. For example, Rennell and Bellona islands, south of the main part of the archipelago, are visited only rarely, and then only by a medical officer or the Resident Commissioner. According to Troughton (1936:341), the islanders in the interior of Bougainville as late as 1935, were prone to kill and feast upon strangers. In 1932, Lewis (1951:37) felt that the natives of Malaita Island were especially resistant to outside interference by Caucasians and reported that no "white man or foreigner" was safe on Malaita.
Troughton (1936), who listed Melanesian names for mammals, indicated that the native peoples distinguished between kinds of bats that closely resembled one another. Of these, the only bats that seem to be used as food belong to the genus Pteropus.
In the following list, currently-used names of islands are given; when available, older names and variant spellings are indicated in parentheses. For certain islands, especially those visited by field parties from the Bishop Museum or those frequently mentioned in previous literature on bats, some descriptive and ecological information also is provided.
Latitude and longitude of islands are from publication no. 881 of the Hydrographic Office of the United States Navy Department (Anonymous, 1944); names of islands were checked against a list by Brigham (1900); descriptive information mostly is from reports by Temple and Straatmann (1964, field notes, at the Department of Entomology, Bishop Museum).
ALU.—7° 07' S, 155° 54' E.
BANIKA.—9° 05' S, 155° 13' E.
BARA (Gera).—9° 31' S, 160° 31' E.
BELLONA (Bello).—11° 18' S, 159° 48' E.
BOUGAINVILLE (Mamamolimo).—6° 12' S, 155° 15' E. This is the largest island in the Solomon Group, being 127 miles long (northwest to southeast) and about 59 miles across at the widest place. The highest elevations are 9850 and 10171 feet, at the tops of active volcanoes. Ecologically, Bougainville is mostly dense rain forest, which is less dense on the summits of higher mountains.
BUKA.—5° 15' S, 154° 38' E.
CHOISEUL.—7° 04' S, 157° 01' E. This island, formed along a northwest-southeast line of low mountains (maximum elevation of 3500 feet), is about 90 miles long and 20 miles wide. Most collecting was at Malangona (Sasamunga on some maps) on the southwestern coast.
FAURO.—6° 55' S, 156° 07' E. This small island, about 14 miles long (north-south) and six miles wide (east-west), lies about 10 miles south and east of Bougainville. Fauro is formed around a volcanic cone having a maximum elevation of 1925 feet; it has considerable dense mangrove swamp along the west coast, and mature rain forest with little understory growth. Most collecting was at Toumoa, on one of two southern peninsulas.
FLORIDA (Nggela).—9° 05' S, 160° 16' E. Florida, the main island in the Nggela Island Subgroup, is mountainous and except for some small grassy areas, supports dense rain forest. It is nearly 25 miles long (east-west) and [784] nine miles wide (north-south), with a maximum elevation, at Mount Barnett, of about 1366 feet. Most collecting was at Haleta, on the southwestern coast. At this locality there were scattered mangrove swamps, rain forest, and gardens inland.
GANONGGA (Ronogo, Ronongo).—8° 03' S, 156° 35' E.
GATUKAI.—8° 47' S, 158° 12' E.
GHIZO (Gizo, Keso).—8° 05' S, 156° 59' E.
GOWER (N'dai).—7° 54' S, 160° 34' E.
GUADALCANAL (Guadalcanar).—9° 15' S, 159° 35' E. Guadalcanal is mostly of volcanic origin and has an irregular chain of mountains along the southern coast. The highest elevation is 8005 feet at Mount Popomanasiu. This large island is nearly 80 miles long (east-west) and 25 miles wide (north-south). Most of the northwestern part of Guadalcanal supports alang-alang grass. The remainder of the island is heavily wooded.
KILINAILAU (Cartaret).—4° 44' S, 155° 28' E.
KOLOMBANGARA (Duki, Kulambangara).—8° 00' S, 157° 05' E. Kolombangara, formed from an extinct volcano, is about 18 miles in diameter and nearly circular. The highest peaks, rising as precipitous cliffs in some places, reach a maximum elevation of about 5000 feet. The vegetation is mostly virgin rain forest. Mangrove swamp and small coconut groves occur along the coast. Field parties from the Bishop Museum were able to reach the highest elevations, and concentrated their work along the southwestern side of the island.
MALAITA (Mala, Malanta, Malayta).—9° 00' S, 161° 00' E. This long (104 miles northwest to southeast), narrow (about 23 miles at its widest spot) island, between Santa Ysabel and San Cristobal islands, is basically of volcanic origin with some limestone (coral) deposits along the coast. Mount Kolovrat, [785] having an elevation of 4275 feet, is the highest point. The Bishop Museum field party lived at Dala, in dense rain forest about 12 miles north of Auki on the northwestern coast of Malaita.
MALAPA.—9° 49' S, 160° 53' E.
MONO (Treasury).—7° 22' S, 155° 35' E. This is a small island (maximum elevation 1150 feet) in the Treasury Island Subgroup just south of Bougainville. Mono is about nine miles long (east-west) and five and one half miles wide (north-south). The basic volcanic core is described in field notes as topped with coral limestone.
NEW GEORGIA (Kausagi).—8° 20' S, 157° 30' E. The New Georgia Subgroup is composed of 11 moderate-sized islands and islets. New Georgia Island, the main member of the subgroup, is 50 miles long (northwest to southeast) and from five to 30 miles wide. On the northern side several volcanic peaks attain an elevation of about 3000 feet. The entire island is forested.
NGGELA (Florida Islands).—4° 31' S, 154° 11' E. This subgroup consists of several small to medium-sized islands between Guadalcanal and Malaita. Florida is the main island.
NISSAN (Green, Sir Charles Hardy's).—4° 31' S, 154° 11' E.
NUKUMANU (Le Maira, Tasman).—4° 32' S, 159° 25' E.
ONTONG JAVA (Lord Howe Atoll, Liuniuwu).—5° 25' S, 159° 30' E.
PAVUVO.—9° 04' S, 159° 08' E.
RAMOS.—8° 16' S, 160° 11' E.
RENNELL.—11° 38' S, 160° 14' E. This island, of limestone (coral) origin, along with Bellona, is nearly 100 miles southwest of any other member of the Solomons and has been regarded, because of this distance, as an oceanic island instead of a continental island. It is about 50 miles long (east-west) and nine miles wide (north-south); its highest elevation is 500 feet.
ROVIANA (Rendova, Rovianna, Rubiana).—8° 21' S, 157° 20' E.
RUSSELL.—9° 04' S, 159° 12' E.
SAN CRISTOBAL (San Christoval, Bauro, Makira, Arussi).—11° 33' S, 161° 43' E. This island is composed mostly of ancient volcanic rock, has a maximum elevation of 4100 feet, is nearly 70 miles long (northwest to southeast) and 24 miles wide, and supports a dense rain forest.
SANTA YSABEL (George, Ysabel, San Isabel, Isbel, Mahaga).—8° 00' S, 159° 07' E. Santa Ysabel is a long (90 miles from northwest to southeast), narrow (19 miles at the widest spot), forested island, consisting of a single chain of volcanic mountains. The numerous bays and mouths of rivers provide excellent anchorages. Collecting was at Tatamba approximately two miles south of Tanambuli where the considerable area of forest was dense and bamboo thickets were abundant.
SAVO (Savu).—9° 08' S, 159° 49' E.
SHORTLAND.—7° 03' S, 155° 47' E.
SIKAIANA (Stewart).—8° 22' S, 162° 44' E.
SIMBO (Narovo, Naorovo, Naravo, Navoro, Sembo).—8° 16' S, 156° 31' E.
STIRLING.—7° 25' S, 155° 35' E.
TANABULI (Tanambuli, Tunnibili, Tunnibilis, Tunnibul, Tunnivula).—8° 24' S, 159° 35' E.
TAUU (Marqueen, Mortlock).—4° 48' S, 157° 32' E.
TELIPARI.—8° 15' S, 157° 32' E.
UGI.—10° 14' S, 161° 44' E.
VANGUNO (Vangunu).—8° 39' S, 158° 00' E.
VELLA LAVELLA.—7° 43' S, 156° 40' E. The coastline is rugged and indented by numerous small bays. Some peaks are 3000 feet high. The southeastern half of Vella Lavella is said to consist of uplifted coral, and to be thickly planted to coconut palms. The native population is concentrated here. [786] The northwestern half of the island is rain forest and is nearly uninhabited. Most of the collecting was at Pusisama, on the southern beach and on Ulo Crater, an extinct volcano at the middle of the island.
YANNTA.—10° 20' S, 161° 20' E.
The phylogenetic arrangement and nomenclature in the text beyond are mainly that of Laurie and Hill (1954). The synonymies for accounts of genera are as follows: (1) first use of the generic name employed along with the original description, and (2) original proposals, in chronological order, of other generic names subsequently applied to the bat in the Solomons. The synonymies in accounts of species and subspecies are as follows: (1) first use of the accepted name, followed by its type locality, followed, in chronological order, by other references to the first name-combination, (2) first use of the name-combination employed herein (if different from the original combination), followed, in chronological order, by other references to the present name-combination, and (3) other name-combinations, in chronological order, employed for the bat in the Solomons. The word "part" is used in parentheses after a name if some specimens listed under that name are from the Solomon Islands and are referable to the species or subspecies being written about.
Unless noted otherwise, specimens listed as examined were prepared originally as museum skins with skulls. Approximately 70 per cent of bats collected in the Solomons were preserved in formalin and now are stored in alcohol. Because it was necessary to obtain dimensions and examine various morphological characteristics of skulls, many crania were extracted from bats preserved in alcohol.
Although all specimens in the Bishop Museum from the Solomon Islands have been catalogued with the prefix BBM-BSIP, catalogue numbers without prefixes in the lists of specimens examined refer to this museum. Catalogue numbers with the prefix USNM refer to specimens in the U. S. National Museum and those with the prefix AM-M refer to specimens in the Australian Museum.
Unless indicated otherwise, all measurements in this paper are in millimeters and are of adults. Cranial measurements, and external measurements of specimens stored in alcohol, were taken by me. The cranial measurements were taken with dial calipers using techniques described by Hall (1946:672-685). External measurements (except length of forearm) of specimens originally prepared as dried study skins, were transcribed from specimen labels.
Capitalized color nomenclature is from Ridgway (1912). Noncapitalized color terms are from published reports that did not use Ridgway's terminology.
Financial support for this investigation was from (1) a United States Army Medical Research and Development Command grant (DA-MD-49-193-62-G65) to the Entomology Department of the Bernice P. Bishop Museum, and (2) a National Science Foundation grant (2185-4703) to the author, through the Committee on Systematics and Evolutionary Biology of The University of Kansas. I am grateful to many individuals who have helped me in various ways throughout the course of this study. Dr. J. Linsley Gressitt, Chairman of the Entomology Department, Bernice Bishop Museum, allowed me to study specimens collected by his expeditions; Professors E. Raymond Hall and J. Knox Jones, Jr., of the Museum of Natural History and the Department of Zoology, The University of Kansas, offered advice and guidance and constructively [787] reviewed the manuscript. Other persons who have given me assistance and, in some cases, arranged for loans of comparative materials, are: Dr. David H. Johnson, Division of Mammals, United States National Museum; Mr. Hobart M. Van Deusen and Dr. Richard G. Van Gelder, Archbold Expeditions and Department of Mammalogy, American Museum of Natural History; Messrs. Ellis LeG. Troughton and Basil Marlow, Mammal Department, The Australian Museum; Dr. Joseph Curtis Moore, Department of Mammalogy, Field Museum of Natural History; Mr. John Edwards Hill, Mammal Room, British Museum (Natural History); Prof. William B. Davis, Department of Zoology, Texas A & M University; Miss Barbara Lawrence, Museum of Comparative Zoology, Harvard University. Messrs. Jerry R. Choate and H. H. Genoways, two colleagues in zoology at The University of Kansas, have assisted me in many ways, for which I am grateful. Linda Anne Phillips, my wife, prepared many of the figures and tables used herein. I thank also Setsuko Nakata, Edwin H. Bryan, Robert Bowan, and Ilse Koehler, who, as staff members of the Bishop Museum, were especially helpful to me. Most of the specimens reported herein were collected by Philip Temple and Peter Shanahan.
| 1. | Uropatagium lacking, or, if present, deeply indented in center; tail vertebrae absent, or if present, free |
| 2 | |
| 1'. | Uropatagium present, not indented; tail vertebrae present, free or in uropatagium |
| MICROCHIROPTERA 1 | |
| 2(1). | External tail-vertebrae lacking, or, if present, less than 3 mm long |
| 3 | |
| 2'. | External tail-vertebrae more than 3 mm long |
| 6 | |
| 3(2). | Small or medium-sized (forearm less than 50); tongue long, extensile |
| 4 | |
| 3'. | Large (forearm more than 80); tongue not long and extensile |
| 5 | |
| 4(3). | Uropatagium present; small claw present on second phalanx of second digit; tail short (about 3 mm) |
| Macroglossus, p. 812 | |
| 4'. | Uropatagium absent; no claw on second phalanx of second digit; no tail |
| Melonycteris, p. 814 | |
| 5(3'). | Entire back set with hair; wing membranes not meeting at middle of back |
| Pteropus, p. 793 | |
| 5'. | Back naked; wing membranes meeting at middle of back, |
| Pteralopex, p. 790 | |
| 6(2'). | Nostrils having definite tubelike extensions |
| Nyctimene, p. 817 | |
| 6'. | Nostrils lacking tubelike extensions |
| 7 | |
| 7(6'). | Forearm less than 80; large, sharp claw on second phalanx of second digit; four upper incisors |
| Rousettus, p. 787 | |
| 7'. | Forearm more than 90; small, blunt claw on second phalanx of second digit; two upper incisors |
| Dobsonia, p. 807 | |
Family PTEROPODIDAE
Subfamily Pteropodinae
Rousettus Gray
1821. Rousettus Gray, London Medical Repository, 15:299, April 1.
1843. Xantharpyia Gray, List of species ... British Museum, p. 37.
1852. Cynonycteris Peters, Reise nach Mossambique, p. 25.
The genus Rousettus occurs throughout the tropical regions of the Old World, and in the Solomons is readily distinguished from all other megachiropteran genera by having both a small claw on the second digit and free caudal vertebrae. The oriental species have been divided into two groups on the basis of size (Tate, 1942:344). The subspecies Rousettus amplexicaudatus hedigeri appears to be the sole representative of this genus in the Solomon [788] Islands. Prior to 1953, several workers (Thomas, 1887b:323, 1888b:475; Matschie, 1899:68; Sanborn, 1931:11) used the name Rousettus amplexicaudatus brachyotis for it, but Pohle (1953) suggested that the specimens from the Solomons recorded by earlier workers were R. a. hedigeri named by him on the basis of the specimen that he saw from Bougainville.
Rousettus amplexicaudatus
Rousettus amplexicaudatus has at least three subspecies, one of which is endemic to the Solomon Islands. The species is wide-ranging, being known from as far west as Thailand (Ellerman and Morrison-Scott, 1966:93) and as far east as the Solomons.
Rousettus amplexicaudatus hedigeri Pohle
1953. Rousettus amplexicaudatus hedigeri Pohle, Z. Säugetierk., 17:127, October 27, type from Bougainville.
1887. Cynonycteris brachyotis, Thomas, Proc. Zool. Soc. London, p. 323, March 15; 1888, Thomas, Proc. Zool. Soc. London, p. 475, December 4, from Fauro.
1889. Xantharpyia brachyotis, Matschie, Die Megachiroptera ... naturkunde, p. 68, from Guadalcanal.
1912. Rousettus brachyotis, Andersen, Catalogue of the Chiroptera ... British Museum, 1:809; 1931, Sanborn, Publ. Field Mus. Nat. Hist., Zool. Ser., 18:11, February 12, from Santa Ysabel.
Specimens examined (20 males and 21 females; all in alcohol; ten crania extracted and cleaned).—Guadalcanal in May, 23863, 23915; Fauro in April, 23804-5; Malaita in June, 24079; Choiseul in March, 23563-4, 23616, 23627, 23630, 23632-3, 23642, 23658, 23663-4, 23680, 23692-3, 23713, 23722; Kolombangara in January and February, 23343, 23366, 23382-4, 23389-90, 23408-9, 23424, 23455, 23471-4, 23501.
Measurements.—Average and extreme external measurements of 13 males and 18 females are, respectively, as follows: Length of head and body, 104.4 (99-118), 108.6 (104-117); tail vertebrae, 16.8 (13-19), 17.6 (15-24); hind foot, 18.0 (16-19), 16.2 (12-18); ear, 15.9 (15-17), 15.0 (14-16); length of forearm, 70.1 (66.0-74.1), 68.1 (65.0-69.1). Average and extreme measurements of skulls of five males and five females are, respectively, as follows: Greatest length of skull, 33.2 (33.0-33.7), 31.5 (30.9-32.1); condylobasal length, 31.3 (30.9-31.9), 30.1 (29.3-30.8); palatal length, 14.0 (13.3-14.8), 13.3 (13.0-13.7); zygomatic breadth, 20.8 (19.8-21.8), 19.4 (18.7-20.8); length of maxillary tooth-row, 11.0 (10.9-11.3), 10.3 (10.1-10.6); length of mandibular tooth-row, 12.6 (12.4-12.9), 11.8 (11.7-12.2).
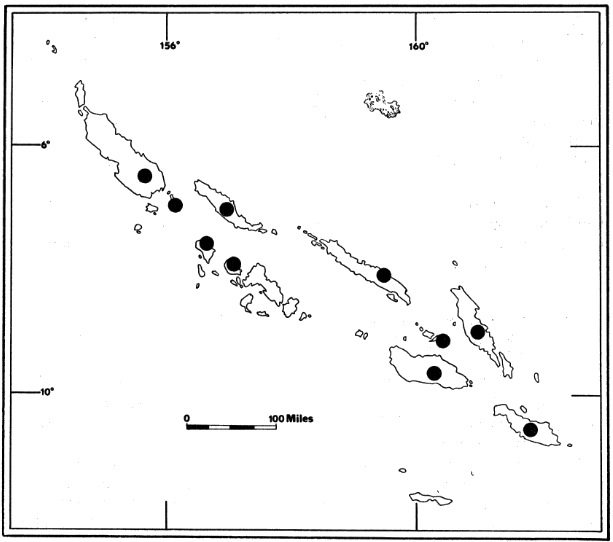
Remarks.—The specimens from Choiseul, Kolombangara, and Malaita islands provide new records of distribution for Rousettus amplexicaudatus hedigeri (Fig. 3). It was described as smaller than R. a. brachyotis Dobson, which is known from New Guinea, Amboina, and the Bismarck Archipelago (Pohle, 1953:127-128). Andersen (1912:809) gave the range of length of forearm in R. a. brachyotis as 73-81, whereas Pohle (1953:127) gave the length of forearm of the type specimen of R. a. hedigeri (adult male) as 67. Measurements of specimens examined by me indicate that hedigeri occurs throughout the Solomon Islands. Cranial measurements of my specimens and Pohle's type are less than those of R. a. brachyotis (see Andersen, 1912:48).
Sanborn (1931:11) noted that the forearms of three males examined by him were longer than that of a female. Mean and range for length of forearm of males and females listed herein, respectively, are 70.1 (66.0-74.1) and 68.1 (65.0-69.1). Also, each of seven cranial measurements taken by me averaged more in males than in females. Sagittal and lambdoidal crests are more prominent in males than in females.
Table 1. A Summary of Breeding Data for Females of Rousettus amplexicaudatus hedigeri Collected December to June.
| Month | Total number collected | Number adult ♀ ♀ collected |
Number lactating | Number of immature individuals |
| December | 3 | 3 | 3 | 0 |
| January | 11 | 11 | 8 | 0 |
| February | 6 | 0 | — | 1 |
| March | 16 | 1 | 0 | 9 |
| April | 2 | 2 | 0 | 0 |
| June | 1 | 1 | 0 | 0 |
As shown in Table 1, adult females obtained in December and January were lactating when captured whereas those obtained in March, April, and [790] June were not. More than half of the individuals collected in March were immature (judging from small size, unfused epiphyses, and lack of wear on teeth). The immature individuals probably had been nursing in December and January.
Pteralopex Thomas
1888. Pteralopex Thomas, Ann. Mag. Nat. Hist., ser. 6, 1:155, February 1.
1762. Pteropus Brisson, Regnum animale ..., ed. 2, p. 153.
Pteralopex, with one species and two subspecies, is the only megachiropteran genus endemic to the Solomons. Thomas (1888b:475) considered this unusual bat a relic, isolated from the time when pteropodids had cuspidate cheek-teeth. Although two workers (Matschie, 1899:11; Simpson, 1945:54) have synonymized Pteralopex with Pteropus, I regard Pteralopex as a morphologically distinct genus.
Individuals of Pteralopex can be distinguished from all species of Pteropus in the Solomon Islands by the following features: wing membranes originate along dorsal midline; braincase diminutive relative to rest of skull; sagittal crest pronounced; cheek-teeth cuspidate, broad and massive; i2 about 10 times larger than i1; upper canines with well-developed secondary cusp; postorbital process fused with zygomatic arch, forming complete bony ring around orbit.
Andersen (1909a:216; 1912:436) considered the relationships of Pteralopex and Pteropus and concluded that Pteropus pselaphon Lay, 1829, from the Sulphur Islands east of Taiwan, and Pteropus samoensis Peale, 1848, from the Samoan Islands, were the "closest" living relatives of Pteralopex. He stated further that Pteralopex "presents in fact scarcely a single character which is not either developed to a certain extent or at least distinctly foreshadowed in Pteropus pselaphon, pilosus, tuberculatus, or leucopterus." In summary, Andersen thought several species of Pteropus had undergone evolutionary development resembling that in Pteralopex, and that the latter, with its massive, cuspidate cheek-teeth, could be considered a highly modified Pteropus. For this hypothesis to be plausible, one must assume that the originally complex cheek-teeth of pteropodids became simple and, at least in the case of Pteralopex, secondarily became complex once again. According to present-day theory of evolutionary development, his hypothesis is improbable. Thomas (1888b:475) probably was correct when he considered Pteralopex an isolated relic.
Although Pteralopex usually is listed after Pteropus in phylogenetic arrangements (see, for example, Sanborn, 1931:21; Pohle, 1953:129; Laurie and Hill, 1954:40), I have placed Pteralopex before Pteropus.
Pteralopex atrata
Two subspecies of Pteralopex atrata (P. a. atrata and P. a. anceps) have been named; specimens of both are rare in museum collections. Thomas (1888a:155) described adults of atrata. Sanborn (1931:21) examined the one additional specimen known to me and reported that it agreed with Thomas' description.
Andersen (1909b:266) used a subadult female ("nearly fully grown") as the holotype of anceps. At least five additional specimens, all adults, of anceps now are housed in various collections. Judging from these individuals, [791] the holotype of anceps was only four-fifths grown and because he used an immature individual, Andersen's (1912:437) criteria for distinguishing the two subspecies mostly are invalid.

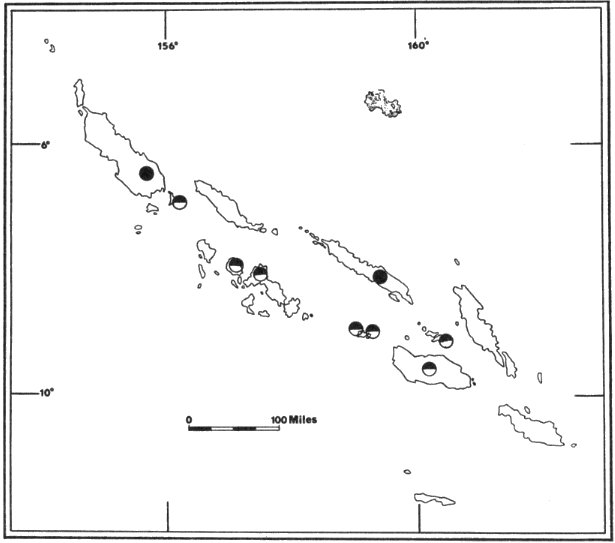
Fig. 4. Distribution of Pteralopex atrata;
P. atrata atrata (![]() ) and
P. atrata anceps (
) and
P. atrata anceps (![]() ).
For names of islands see Fig. 2.
).
For names of islands see Fig. 2.
Key to Subspecies of Pteralopex atrata
| 1. | Length of forearm 139-144 mm.; dorsal surface of distal one-fourth of tibia and entire metatarsus naked; known only from Guadalcanal and Santa Ysabel islands |
| Pteralopex atrata atrata | |
| 1'. | Length of forearm 162-166 mm.; dorsal surface of distal one-fourth of tibia and entire metatarsus furred; known only from Bougainville and Choiseul islands |
| Pteralopex atrata anceps | |
Pteralopex atrata atrata Thomas
1888. Pteralopex atrata Thomas, Ann. Mag. Nat. Hist., ser. 6, 1:155, February, type from Guadalcanal; 1888, Thomas, Proc. Zool. Soc. London, p. 475, December 4; 1896, Heude, Mém. Hist. Nat. Emp. China, 3:179; 1897, Trouessart, Catalogus Mammalium ..., 1:83; 1907, Miller, Bull. U. S. Nat. Mus., 57:60, June 29; 1912, Andersen, Catalogue of the Chiroptera ... British Museum, 1:439; 1931, Sanborn, Publ. Field Mus. Nat. Hist., Zool. Ser., 18:21, February 12, from Santa Ysabel.
1954. Pteralopex atrata atrata, Laurie and Hill, List of land mammals of New Guinea, Celebes and adjacent islands, p. 40, June 30.
1899. Pteropus (Pteralopex) atrata, Matschie, Die Megachiroptera ... naturkunde, p. 11; 1904, Trouessart, Catalogus Mammalium ..., Suppl., p. 49.
Specimens examined.—None.
Remarks.—Pteralopex atrata atrata is known from four specimens from Guadalcanal and one from Santa Ysabel (Sanborn, 1931:21).
Sanborn (loc. cit.) reported that a specimen wounded at night, while feeding on young green coconuts, was the only fruit bat that attempted to attack the collectors. Troughton (1936:348) has suggested, on the basis of his experiences with Pteropus, that this behavior probably was a reaction from fear rather than an indication of general aggressiveness on the part of Pteralopex.
Pteralopex atrata anceps Andersen
1909. Pteralopex anceps Andersen, Ann. Mag. Nat. Hist., ser. 8, 3:266, March, type from Bougainville; 1912, Andersen, Catalogue of the Chiroptera ... British Museum, 1:437; 1936, Troughton, Rec. Australian Mus., 14:348, April 7; 1953, Pohle, Z. Säugetierk., 17:129, October 27.
1954. Pteralopex atrata anceps, Laurie and Hill, List of land mammals of New Guinea, Celebes and adjacent islands, p. 40, June 30.
Specimens examined (three males, two females; one skull-only and one in alcohol).—Choiseul in March, 23682; Bougainville in July, USNM 276973-74, USNM 276928, USNM 277112.
Measurements.—Measurements of three males and one female are, respectively, as follows: Length of head and body, 280, 271, 261, 255; hind foot, 50, 54, 52, 59; ear, 23, 23, 26, 22; length of forearm, 160, 162, 166, 171; greatest length of skull, 77.6, 77.9, 78.9, 77.0; condylobasal length, 74.3, 74.3, 75.5, 73.8; zygomatic breadth, 42.2, 45.4, 43.1, 42.6; breadth across upper canines, 18.7, 21.1, 19.0, 19.0; breadth across first upper molars, 22.2, 25.3, 22.9, 22.0; length of maxillary tooth-row, 29.3, 29.8, 28.9, 28.2; length of mandibular tooth-row, 32.8, 32.8, 32.1, 31.4.
Remarks.—Heretofore, Pteralopex atrata anceps was not known from Choiseul. The specimen from that island agrees well with specimens in the U. S. National Museum from Cape Torokina, Bougainville.
The type specimen of this subspecies is a subadult and is smaller than the specimens examined by me; Andersen (1912:440) gave length of forearm of the type as 137 (as opposed to 164 in adults). He (1912:438) figured the dentition of anceps and described the ways in which it differed from the dentition of atrata. Although he (1912:437) concluded that anceps and atrata represented "two stages of specialization of ... dentition," there apparently are few, if any, dental differences between the two subspecies. Teeth of adults of anceps differ from teeth of the immature type of anceps as follows: in adults the anterior basal ledge of P4 extends onto the labial surface, whereas in the type it does not; and maxillary and mandibular teeth in adults are spaced as in the subspecies atrata (see Andersen, 1912:438, fig. 22) and not crowded as in the type of anceps. Distance between individual cheek-teeth apparently increases with growth of the cranium and mandible.
Adults of P. a. anceps that I examined are darker than the subadult type. The mantle in these adults is black, whereas it is seal-brown in the type (Andersen, 1912:439).
An adult female was lactating when obtained on Bougainville in July (USNM 276928).
Key to Pteropus in the Solomon Islands
| 1. | Premolars having distinct basal ledges; molars 2.5-4.0 wide | |
| 2 | ||
| 1'. | Premolars lacking definite basal ledges; molars 1.0-2.4 wide | |
| 14 | ||
| 2(1). | Rostrum unshortened (orbit to anterior tip of nasals about one-third greatest length of skull); dorsal surface of tibiae nearly naked | |
| 3 | ||
| 2'. | Rostrum shortened (orbit to anterior tip of nasals less than one-third greatest length of skull); dorsal surface of tibiae usually at least partially furred | |
| 8 | ||
| 3(2). | Forearm more than 128 | |
| 4 | ||
| 3'. | Forearm less than 128 | |
| 5 | ||
| 4(3). | Forearm about 155; venter and dorsum nearly black, mantle pale yellow | |
| P. tonganus geddiei, p. 798 | ||
| 4'. | Forearm 128-136; venter and dorsum near Mars Brown, mantle Ochraceous or Cream-Buff | |
| P. hypomelanus luteus, p. 796 | ||
| 5(3'). | Mantle dark, russet or cinnamon, not strongly contrasting with color of back | |
| P. admiralitatum solomonis, p. 796 | ||
| 5'. | Mantle pale, Ochraceous-Buff or Cream-Buff, strongly contrasting with color of back | |
| 6 | ||
| 6(5'). | Mantle Ochraceous-Orange to Ochraceous-Buff, hairs pale basally; forearm 108-111 | |
| P. admiralitatum goweri, p. 797 | ||
| 6'. | Mantle Ochraceous to Cream-Buff, but hairs dark brown basally; forearm 110-122 | |
| 7 | ||
| 7(6'). | Length of forearm 110-112 | |
| P. admiralitatum colonus, p. 796 | ||
| 7'. | Length of forearm about 122 | |
| P. howensis, p. 797 | ||
| 8(2'). | Forearm more than 145 | |
| 9 | ||
| 8'. | Forearm less than 144 | |
| 12 | ||
| 9(8). | Forearm more than 162 | |
| 10 | ||
| 9'. | Forearm less than 162 | |
| 11 | ||
| 10(9). | Forearm 167-173 | |
| P. rayneri grandis, p. 801 | ||
| 10'. | Forearm about 164 | |
| P. rayneri rubianus, p. 802 | ||
| 11(9'). | Flanks and lower belly brightly colored, Burnt Sienna to Sanford's Brown; forearm less than 150 | |
| P. rayneri monoensis, p. 803 | ||
| 11'. | Flanks and lower belly darker, near tawny; forearm more than 150, | |
| P. rayneri lavellanus, p. 802 | ||
| 12(8'). | Pelage of dorsum tricolored; rump brightly colored; forearm 139-141, | |
| P. rayneri rayneri, p. 800 | ||
| 12'. | Pelage of dorsum bicolored; rump dark; forearm less than 135 | |
| 13 | ||
| 13(12'). | Mantle tawny with some Ochraceous-Buff; forearm about 130, | |
| P. rayneri rennelli, p. 804 | ||
| 13'. | Mantle russet, lacking Ochraceous-Buff; forearm about 121, | |
| P. rayneri cognatus, p. 803 | ||
| 14(1'). | Forearm more than 131; dorsum Tawny Olive | |
| P. mahaganus, p. 806 | ||
| 14'. | Forearm less than 100; dorsum dark brown | |
| P. woodfordi, p. 804 | ||
Pteropus Brisson
1762. Pteropus Brisson, Regnum animale ..., ed. 2, p. 153.
Remarks.—More species (seven) and subspecies (12) of Pteropus occur in the Solomon Islands than of any other chiropteran genus. Other kinds of Pteropus, as yet unknown, may live there.
The relationships among the species of these large fruit-eating bats, commonly termed "flying foxes," are obscure and the genus is in need of revision. The basic, definitive work is still that of Andersen (1912). Tate (1942) and Felten (1964a, 1964b) have offered some additional remarks but groupings and suggested relationships of species of Pteropus almost entirely are the products of Kund Andersen. According to present-day concepts of variation and speciation, Andersen's criteria are artificial.
Basically, there are three "species-groups" of Pteropus in the Solomon Islands. The first is composed of species in which the rostrum is "unshortened" (its length about one third of greatest length of skull), and the cheek-teeth are of moderate size (M1 is 2.8-3.2 wide). The species are P. hypomelanus, P. admiralitatum, P. tonganus, and P. howensis. The first and second species were placed in the Pteropus hypomelanus group by Andersen (1912:98).
In the second group the rostrum is "shortened" (its length less than one third of greatest length of skull) and the cheek-teeth are of moderate to large size (M1 3.3-4.1 wide). Pteropus rayneri, endemic to the Solomons and represented there by at least seven subspecies, fits into this category.
The third group is represented by P. mahaganus and P. woodfordi. Both species are endemic to the Solomon Islands. In these species the rostrum is unshortened but the cheek-teeth are greatly reduced, especially in width (M1 is 1.0-2.2 wide). Both P. mahaganus and P. woodfordi can be included in the Pteropus scapulatus group of Andersen (1912:402).
Pteropus hypomelanus
Pteropus hypomelanus is a wide-ranging species of flying fox having at least seven subspecies; three occur in southeastern Asia, two on and near Celebes, and two in New Guinea and islands adjacent to the southeastern coast of New Guinea, including one island in the Solomons (Ellerman and Morrison-Scott, 1966:95; Laurie and Hill, 1954:32-33).

Fig. 5. Distribution of
Pteropus hypomelanus luteus (![]() ),
Pteropus admiralitatum solomonis (
),
Pteropus admiralitatum solomonis (![]() ),
Pteropus a. colonus (
),
Pteropus a. colonus (![]() ),
Pteropus a. goweri (
),
Pteropus a. goweri (![]() ),
Pteropus tonganus geddiei (
),
Pteropus tonganus geddiei (![]() ),
and Pteropus howensis (
),
and Pteropus howensis (![]() ).
For names of islands see Fig. 2.
).
For names of islands see Fig. 2.
Pteropus hypomelanus luteus Andersen
1908. Pteropus hypomelanus luteus Andersen, Ann. Mag. Nat. Hist., ser. 8, 2:362, October, type from Kiriwini Island, Trobriand Islands; 1912, Andersen, Catalogue of the Chiroptera ... British Museum, 1:128; 1947, Sanborn and Beecher, Jour. Mamm., 28:388, November 19, from Banika Island, Russell Islands.
Specimens examined.—None.
Remarks.—Andersen (1908:362) identified specimens of Pteropus hypomelanus from eastern New Guinea and three nearby islands (Conflict Islands, Trobriand Islands, and Woodlark Island) as P. hypomelanus luteus. Sanborn and Beecher (1947:388) identified a female from Banika Island in the Solomons as of this subspecies although this specimen was darker and had a slightly smaller skull than typical P. hypomelanus luteus. They noted that the pelage of the venter of the female was uniformly dark rather than the typical Ochraceous-Buff to Cream-Buff; the specimen was regarded as a dark phase of the subspecies. Although not recorded previously for luteus, other subspecies of P. hypomelanus were known in dark phase as well as pale and intermediate phases of coloration (Andersen, 1912:122). The reported occurrence of P. h. luteus on Banika Island extended the known geographic range about 450 miles eastward from Woodlark Island.
Pteropus admiralitatum
Three subspecies, all about the same size but differing in coloration, have been described from the Solomon Islands. P. a. goweri is known only from Gower (Ndai) Island, notably removed from the western chain of islands inhabited by P. a. colonus and P. a. solomonis. Only one other subspecies, from the Admiralty Islands, is known.
Pteropus admiralitatum solomonis Thomas
1904. Pteropus solomonis Thomas, Novit. Zool., 11:597, type from Ghizo Island; 1912, Andersen, Catalogue of the Chiroptera ... British Museum, 1:149; 1931, Sanborn, Publ. Field Mus. Nat. Hist., Zool. Ser., 18:12, February 12, from Ronongo (Ganongga), Vella Lavella, and Narovo (Simbo) islands; 1947, Sanborn and Beecher, Jour. Mamm., 28:389, November 19, from Banika and Guadalcanal islands.
1954. Pteropus admiralitatum solomonis, Laurie and Hill, List of land mammals of New Guinea, Celebes and adjacent islands, p. 33, June 30.
Specimens examined.—None.
Remarks.—Andersen (1912:149) considered Pteropus admiralitatum, and especially the subspecies P. a. solomonis, to be the easternmost "representative" of Pteropus hypomelanus. In comparison with P. hypomelanus luteus, P. a. solomonis differs mostly in size, being much smaller (length of forearm about 110 rather than 134). It is now known that both species occur on Banika Island in the Solomons.
The subspecies P. a. solomonis has been recorded from a "chain" of islands that included Vella Lavella, Simbo, Ghizo, Ganongga, Banika, and Guadalcanal (see Fig. 5).
Pteropus admiralitatum colonus Andersen
1908. Pteropus colonus Andersen, Ann. Mag. Nat. Hist., ser. 8, 2:363, October, type from Shortland Island; 1912, Andersen, Catalogue of the Chiroptera ... British Museum, 1:150; 1931, Sanborn, Publ. Field Mus. Nat. Hist., Zool. Ser., 18:12, February 12, from Mono Island.
1954. Pteropus admiralitatum colonus, Laurie and Hill, List of land mammals of New Guinea, Celebes and adjacent islands, p. 33, June 30.
1887. Pteropus hypomelanus (part), Thomas, Proc. Zool. Soc. London, p. 471, December 4; 1898, Trouessart, Catalogus Mammalium ..., 1:82, from "I. Salomonis."
1899. Pteropus (Spectrum) hypomelanus (part), Matschie, Die Megachiroptera ... naturkunde, p. 24.
Specimens examined.—None.
Remarks.—Pteropus admiralitatum colonus is the largest of the three subspecies that occur in the Solomon Islands. It closely resembles P. hypomelanus luteus, except in being smaller throughout (see Andersen, 1912:151-152, for measurements) and darker on the underparts.
This bat has been found in a group of small islands (Alu, Mono, and Shortland) about 30 miles south of Bougainville. Because of this proximity and because yet another subspecies of this species occurs northward of Bougainville, it is interesting that neither Troughton (1936) nor Pohle (1953) included the species in their faunal lists for Bougainville.
Andersen (1912:152) indicated that the M1 in P. admiralitatum colonus is smaller than in P. a. solomonis, the subspecies found in islands to the southeast (4.4-4.5 and 5.2, respectively), but Sanborn (1931:13) studied specimens of these two subspecies that overlapped in size of M1.
Pteropus admiralitatum goweri Tate
1934. Pteropus goweri Tate, Amer. Mus. Novit., 718:1, May 4, type from Gower (Ndai) Island.
1954. Pteropus admiralitatum goweri, Laurie and Hill, List of land mammals of New Guinea, Celebes and adjacent islands, p. 33, June 30.
Specimens examined.—None.
Remarks.—Pteropus admiralitatum goweri was described from six specimens collected in 1930 by the Whitney South Sea Expedition (Tate, 1934:1). This subspecies closely resembles the other two subspecies of P. admiralitatum (colonus and solomonis) found in the Solomon Islands. Color and length of forearm (see key on p. 793) seem to be the only reliable criteria for distinguishing between these subspecies. The longitude of Gower Island, 160° 34' E, was incorrectly listed in Laurie and Hill (1954:152) as 159° 34' E.
1931. Pteropus howensis Troughton, Proc. Linn. Soc. New South Wales, 56:204, June 24, type from Lord Howe Islands (Ontong Java); 1950, Sanborn and Nicholson, Fieldiana:Zool., 31:329, August 31.
Specimens examined (one male, three females, and two sex unknown; two embryos in alcohol).—Liuniuwu, Lord Howe Islands (Ontong Java) in August, USNM 278703-6, USNM 279715-6.
Measurements.—Average and extreme measurements of one male and three females are as follows: Length of head and body, 185.2 (176-196); hind foot, 34.5 (33-36); ear, 21.5 (21-23); forearm not measured [broken in all specimens examined]. Cranial measurements of a male and a female are, respectively, as follows: Greatest length of skull, 55.3, 53.8; condylobasal length, [798] 54.2, 52.8; palatal length, 26.7, 26.0; zygomatic breadth, 30.6, 29.9; breadth of braincase, 19.9, 19.2; breadth across first upper molars, 14.3, 14.3; length of maxillary tooth-row, 20.7, 19.6; length of mandibular tooth-row, 23.1, —.
Remarks.—Apparently Pteropus howensis is confined to Ontong Java (Lord Howe Islands) located northeastward of the main body of islands that constitute the Solomon Archipelago (see Fig. 5). According to A. J. Nicholson, who collected the specimens listed above, P. howensis is not abundant in Ontong Java. He related this circumstance to the fact that these small islands are nothing more than parts of a coral atoll used almost entirely for the production of coconuts (see Sanborn and Nicholson, 1950:329).
Specimens of Pteropus howensis deposited in the U. S. National Museum agree well in most ways with the original description of the species by Troughton (1931:204-205). Slight variation in color is evident; in two specimens, the mantle, just posterior to the ears, is Ochraceous-Buff.
The relationship of this species to other kinds of Pteropus known from Melanesia is not clear. Troughton (1931:204, 206) compared P. howensis with P. hypomelanus and P. admiralitatum and found that it resembled each of them. Tate (1934:2) noted that the skull of P. admiralitatum goweri was similar to that of P. howensis in structure. The latter species is, however, larger (length of forearm 122 according to Troughton, 1931:205) than any subspecies of P. admiralitatum (length of forearm 108-112). Also, the cheek-teeth of P. howensis that I have studied are relatively larger than those of either P. hypomelanus or P. admiralitatum. Furthermore, in P. howensis there is a small but distinct cusp located medio-posteriorly on P4 (most noticeable in young individuals) that is more reduced or undeveloped in specimens of the other two species. Cheek-teeth of P. howensis resemble those in a dull-colored specimen of P. tonganus from Fiji Island with which I compared the specimens listed above.
Weights and crown-rump lengths of the two embryos (in an advanced stage of development) examined were 20 and 29 grams and 43 and 51 mm. (apparently these are the specimens listed by Sanborn and Nicholson, 1950:329).
Pteropus tonganus
Pteropus tonganus has at least three subspecies, one of which has been recorded from the Solomons. The species ranges from a small island off the eastern coast of New Guinea, where there is an endemic subspecies, eastward to Tonga and the New Hebrides (Laurie and Hill, 1954:33-34). Felten (1964a) recently has reported on the species in the New Hebrides.
Pteropus tonganus geddiei MacGillivary
1860. Pteropus geddiei MacGillivary, Zoologist, 18:7134, September, type from Aneitum Island, New Hebrides; 1912, Andersen, Catalogue of the Chiroptera ... British Museum, 1:189; 1931, Sanborn, Publ. Field Mus. Nat. Hist., Zool. Ser., 18:13, February 12, from Rennell Island in the Solomons.
1914. Pteropus tonganus geddiei, Revilliod, in Sarasin and Roux, Nova Caledonia (A), 1:341; 1954, Laurie and Hill, List of land mammals of New Guinea, Celebes and adjacent islands, p. 34, June 30.
Specimens examined.—None.
Remarks.—Pteropus tonganus geddiei, as far as is known, is the widest ranging subspecies of this genus. It is the only megachiropteran in the Solomon Islands having affinities with bats to the southeast (the New Hebrides, Santa [799] Cruz Islands, Samoan Islands and Fiji Islands) rather than with those to the west (New Guinea). The subspecies P. tonganus geddiei, which ranges from the Solomons to the New Hebrides (about 500 miles straight-line distance), is said to be remarkably uniform throughout its range. Sanborn (1931:14) compared color and size in specimens from the Solomon Islands and the New Hebrides and found little variation. Another subspecies, P. t. bascilicus Thomas 1915, apparently closely related to geddiei, is known from Dampier [= Kar-kar] Island off the northeastern coast of New Guinea and therefore farther westward from the New Hebrides than are the Solomon Islands. Additional remarks on the distribution of this species are in the section on Zoogeography and Speciation.
Pteropus rayneri
Pteropus rayneri is endemic to the Solomon Islands. It is divisible into seven subspecies (see Fig. 6), which, excepting P. r. rennelli and P. r. cognatus, are strikingly colored—the mantle, back, and rump being of different colors. Differences in color and size provide characters differentiating the subspecies (see key, p. 793). Recorded lengths of forearms do not overlap between any two subspecies. P. r. grandis, northernmost in distribution, has the longest (about 170) forearm and P. r. cognatus, known from two of the southernmost islands, has the shortest (about 121).

Fig. 6. Distribution of
Pteropus rayneri: P. r. rayneri (![]() );
P. r. grandis (
);
P. r. grandis (![]() );
P. r. lavellanus (
);
P. r. lavellanus (![]() );
P. r. monoensis (
);
P. r. monoensis (![]() );
P. r. rubianus (
);
P. r. rubianus (![]() );
P. r. cognatus (
);
P. r. cognatus (![]() );
P. r. rennelli (
);
P. r. rennelli (![]() ).
For names of islands see Fig. 2.
).
For names of islands see Fig. 2.
Adult males of Pteropus rayneri have well-developed tufts of hair on each [800] side of the neck where a gland is located (see Andersen, 1912:259). Apparently these glands are not present in females as none were found in specimens studied by me or those reported by Sanborn (1931:16). Evidently, these glands are associated with sexual maturity in males because neither Sanborn nor I found them in subadult males.
1870. Pteropus rayneri (part), Gray, Catalogue of monkeys, lemurs and fruit-eating bats ... British Museum, p. 108, cotypes from Guadalcanal; 1878, Dobson, Catalogue of the Chiroptera ... British Museum, p. 33; 1879, Trouessart, Rev. Mag. Zool., 6:204; 1879, Trouessart, Ann. Sci. Nat. Zool, 8:16; 1887, Thomas, Proc. Zool. Soc. London, p. 322, March 15; 1888, Thomas, Proc. Zool. Soc. London, p. 472, December 4; 1898, Trouessart, Catalogus Mammalium ..., 1:78; 1912, Andersen, Catalogue of the Chiroptera ... British Museum, p. 254; 1931, Sanborn, Publ. Field Mus. Nat. Hist., Zool. Ser., 18:15, February 12, from Guadalcanal and Malaita.
1954. Pteropus rayneri rayneri, Laurie and Hill, List of land mammals of New Guinea, Celebes and adjacent islands, p. 35, June 30.
1899. Pteropus (Spectrum) rayneri (part), Matschie, Die Megachiroptera ... naturkunde, p. 22; 1904, Trouessart, Catalogus Mammalium ..., Suppl., p. 51.
Specimens examined (four males and one female; one embryo in alcohol).—Guadalcanal in July and November, USNM 278700-02, USNM 278142, USNM 278714.
Measurements.—Measurements of three males and one female are, respectively, as follows: Length of head and body, —, 210, 214, 215; hind foot, —, 33, 39, 42; ear, —, 23, 23, 23; length of forearm, —, 138, 136, 134; greatest length of skull, 61.5, 59.2, 61.6, 61.2; condylobasal length, 61.4, 58.2, 60.3, 60.0; zygomatic breadth, 36.6, 35.3, 35.4, 36.5; breadth of braincase, 23.7, 22.5, 22.6, 24.1; breadth across first upper molars, —, 16.9, 16.7, 16.8; width of M1, 3.4, 3.5, 3.5, 3.5; length of maxillary tooth-row, 22.4, 22.1, 23.6, 23.2; length of mandibular tooth-row, 26.4, 25.5, 25.9, 25.6.
Remarks.—Pteropus rayneri was named on the basis of two specimens (cotypes) obtained on Guadalcanal and listed as "male" and "female"; according to Andersen (1912:254), however, both are females.
P. r. rayneri is known from Guadalcanal and Malaita (see Fig. 6), and is of almost the same size as P. r. cognatus, which is known from San Cristobal and Ugi, only about 40 miles to the southeast. In the latter subspecies the back and rump are the same color (Prouts Brown), whereas in P. r. rayneri the rump is brightly colored and therefore contrasts strongly with the dark brown back. A specimen of rayneri from Malaita was reported by Sanborn (1931:15) as unusually small and having a dark-colored rump patch. In the specimens examined from Guadalcanal, there is noticeable variation in color of the mantle that does not seem related to age or sex. In two specimens (adult male and female) the mantle is Cinnamon-Rufous tinged with Russet, strongly contrasting with the crown, which is Ochraceous-Tawny and has scattered silvery hairs. Another specimen has a darker mantle (near Chestnut-Brown) and a crown of about the same color, but with a few scattered Ochraceous-Tawny hairs.
The skull of one adult male bears an extra peglike tooth posterior to M3 on the right side.
An embryo, in an advanced stage of development, in the collection of the U. S. National Museum, measures: Length of head and body, 98; hind foot, 30; ear, 8.5; length of forearm, 48 (this may be the same specimen listed by Sanborn and Nicholson, 1950:329).
Pteropus rayneri grandis Thomas
1887. Pteropus grandis Thomas, Ann. Mag. Nat. Hist., ser. 5, 19:147, March, type from Shortland; 1887, Thomas, Proc. Zool. Soc. London, p. 320, March 15, from Alu and Shortland; 1897, Trouessart, Catalogus Mammalium ..., 1:80, from "I. Salomonis"; 1899, Matschie, Die Megachiroptera ... naturkunde, p. 15; 1904, Trouessart, Catalogus Mammalium ..., Suppl., p. 49; 1907, Miller, Bull. U. S. Nat. Mus., 57:58, June 29; 1912, Andersen, Catalogue of the Chiroptera ... British Museum, 1:259, from Bougainville; 1931, Sanborn, Publ. Field Mus. Nat. Hist., Zool. Ser., 18:16, February 12, from Choiseul, and Santa Ysabel; 1936, Troughton, Rec. Australian Mus., 19:348, April 7; 1953, Pohle, Z. Säugetierk., 17:128, October 27.
1954. Pteropus rayneri grandis, Laurie and Hill, List of land mammals of New Guinea, Celebes and adjacent islands, p. 35, June 30.
Specimens examined (six males and 10 females; five in alcohol).—Choiseul in March, 23580, 23644, 23593; Bougainville in July, August, September, and October, USNM 276926-7, USNM 276968, USNM 277091-9.
Measurements.—Average and extreme measurements of four males and seven females are as follows: Length of head and body, 281 (260-302); hind foot, 52.3 (50-58); ear, 33.1 (31-37); length of forearm, 173 (168-180). Average and extreme measurements of skulls of three males and six females are as follows: Greatest length of skull, 73.7 (71.3-77.7); condylobasal length, 73.1 (70.5-77.4); zygomatic breadth, 40 (36.4-41.5); breadth across first-upper molars, 20.9 (18.3-22.1); length of maxillary tooth-row, 28.1 (26.9-29.9); length of mandibular tooth-row, 31.8 (29.7-32.7).
Remarks.—Pteropus rayneri grandis is the largest subspecies of the species. It is also the widest ranging subspecies, being found on six islands (see Fig. 6).
Although the specimens listed above agree well with descriptions of color given by Thomas (1887a:147) and Andersen (1912:259, 263-264), some individual variation is noticeable. In bats not yet fully grown (judging from small size, unfused epiphyses, and lack of wear on teeth), numerous scattered hairs on the sides of the face and crown are buffy. In adults the face and crown are blackish. With regard to individual variation in color of mantle and rump patch, specimens with the following combinations were noted (1) mantle Brick Red, rump patch bright, basal three-quarters of hairs white, tips Warm Buff (2) mantle darker, near Hessian Brown, rump patch dark, Chestnut along edges, center Ochraceous-Tawny (3) mantle Brick Red, rump patch intermediate between the two other types. Size of rump patch also is variable. In some specimens it extends onto the upper parts of the thighs whereas in other specimens it does not.
Sanborn (1931:16) reported an extra tooth, behind the last lower molar, in a specimen from Choiseul. In one of three specimens in the Bishop Museum, m3 is lacking. Judging from Troughton's (1936:346) remarks, size of individuals varies considerably. Specimens that he examined from Bougainville had longer forearms (up to 177) and larger hind feet (54-57) than those examined by me from Choiseul. On the other hand, specimens listed above from Bougainville agree well with those from Choiseul. In many specimens in [802] the U. S. National Museum, length of the right- and left-forearm differ. For example, in No. 276926 the right forearm measures 180 whereas the left is 174; in No. 277098 the right is 172 and the left is 167. Troughton (1936:346) gave standard ear measurement in P. r. grandis as ranging from 29.5 to 31.5. Ears of specimens that I examined varied from 31.0 to 37.0.
Pteropus rayneri rubianus Andersen
1908. Pteropus rubianus Andersen, Ann. Mag. Nat. Hist., ser. 8, 2:366, October, type from Rubiana; 1912, Andersen, Catalogue of the Chiroptera ... British Museum, 1:255; 1931, Sanborn, Publ. Field Mus. Nat. Hist., Zool. Ser., 18:15, February 12, from Narovo (Simbo).
1954. Pteropus rayneri rubianus, Laurie and Hill, List of land mammals of New Guinea, Celebes and adjacent islands, p. 35, June 30.
1888. Pteropus grandis (part), Thomas, Proc. Zool. Soc. London, p. 470, December 4, from Rubiana; 1899, Matschie, Die Megachiroptera ... naturkunde, p. 15; 1904, Trouessart, Catalogus Mammalium ..., Suppl., p. 49.
Specimens examined (two males and one female).—Kolombangara, in February, 23458-60.
Measurements.—Measurements of two males and one female are, respectively, as follows: Length of head and body, 253, 265, 251; hind foot, 53, 50, 50; ear, 30, 31, 32; length of forearm, 158, 161, 160; greatest length of skull, 70.2, 67.4, —; condylobasal length, 67.0, —, 68.4; zygomatic breadth, 40.0, 39.4, 40.7; breadth across first upper molars, 19.4, 20.4, 19.9; length of mandible, 53.9, 49.4, 51.3.
Remarks.—Kolombangara Island is a new locality for Pteropus rayneri rubianus; heretofore this subspecies was known only from Rubiana and Narovo islands (Andersen, 1908:366; Sanborn, 1931:15). The coloration of a specimen from Narovo Island was described as between that of P. r. rubianus and P. r. lavellanus. Sanborn (1931:16) allocated it to the subspecies rubianus on the basis of length of forearm.
Andersen's descriptions (1908:366; 1912:256) of rubianus were of a specimen stored in alcohol. Coloration of the museum skins examined by me is as follows: Dorsum from shoulders to rump near Vandyke Brown; crown and mantle Brick Red; face close to Mummy Brown; rump patch and thighs close to Warm Buff, strongly contrasting with back and mantle; base of hairs dark, Seal Brown; venter dark; chest about same as back but paler laterally (to Ochraceous Tawny); throat Brick Red.
Pteropus rayneri lavellanus Andersen
1908. Pteropus lavellanus Andersen, Ann. Mag. Nat. Hist., ser. 8, 2:366, October, type from Vella Lavella; 1912, Andersen, Catalogue of the Chiroptera ... British Museum, 1:259; 1931, Sanborn, Publ. Field Mus. Nat. Hist., Zool. Ser., 18:16, February 12, from Ghizo and Ronongo.
1954. Pteropus rayneri lavellanus, Laurie and Hill, List of land mammals of New Guinea, Celebes and adjacent islands, p. 36, June 30.
Specimens examined (one male and one female).—Vella Lavella in November, 23192, 23142.
Measurements.—Measurements of a male and a female are, respectively, as follows: Length of head and body, 286, 282; hind foot, 55, 56; ear, 30, 30; [803] length of forearm, 156, 155; greatest length of skull, 72.9, 67.6; condylobasal length, 71.8, 64.2; zygomatic breadth, 38.4, 37.9; breadth across first upper molars, 19.9, 19.8; length of mandible, 54.6, 50.8.
Remarks.—Pteropus rayneri lavellanus inhabits islands geographically near those from which P. r. rubianus is known (see Fig. 6) and in most respects the two subspecies closely resemble each other. P. r. lavellanus is slightly the smaller (average length of forearm about 156 instead of 160) and darker. A bat from Narovo [Simbo] Island, only a few miles from Vella Lavella, identified by Sanborn (1931:16) on basis of its size as P. r. rubianus, resembled the subspecies lavellanus in color and probably represents an intergrade between the two populations.
The color of P. r. lavellanus is close to that of P. r. rubianus except that the crown, mantle, and foreneck are near Chestnut-Brown, the basal portions of hair black, and the fur of the venter, from sternum to pectoral region, is dark, almost black (compare with description of P. r. rubianus under account of that subspecies).
Measurements of the male examined are greater than those of the female studied. Andersen (1912:259) noted that the canine teeth are heavier in males than in females.
Pteropus rayneri monoensis Lawrence
1945. Pteropus rayneri monoensis Lawrence, Proc. New England Zool. Club, 23:63, March 26, type from Mono (Treasury); 1954, Laurie and Hill, List of land mammals of New Guinea, Celebes and adjacent islands, p. 36, June 30.
Specimens examined.—None.
Remarks.—Pteropus rayneri monoensis is the most recently described subspecies of P. rayneri. Lawrence (1945:63) judged that in most ways this bat is intermediate between P. r. grandis and P. r. lavellanus. Coloration of monoensis indicates affinity with the former, whereas length of forearm (145-148) approaches that in the latter. The small skull, narrow palate, and whitish rump patch of monoensis are differences that distinguish it from grandis and lavellanus. The relatively isolated position of Mono Island may have been important in establishment of the distinctive features of this bat.
Lawrence (1945:65) quoted a collector as stating: "They [individuals of P. r. monoensis] rest quietly during the day in the tops of heavy-leaved, tall jungle trees, and start flying about dusk, looking for feeding spots. There is usually quite a flight for fifteen to twenty minutes at twilight...."
No additional specimens of this subspecies have been collected on small adjacent islands and monoensis may therefore be confined to Mono Island.
Pteropus rayneri cognatus Andersen
1908. Pteropus cognatus Andersen, Ann. Mag. Nat. Hist., ser. 8, 2:365, October 1, type from San Cristobal; 1912, Andersen, Catalogue of the Chiroptera ... British Museum, 1:251; 1931, Sanborn, Publ. Field Mus. Nat. Hist., Zool. Ser., 18:15, February 12, from San Cristobal and Ugi; 1954, Laurie and Hill, List of land mammals of New Guinea, Celebes and adjacent islands, p. 35, June 30.
1962. Pteropus rayneri cognatus, Hill, The natural history of Rennell Island, British Solomon Islands, 4:9, February.
1870. Pteropus rayneri (part), Gray, Catalogue of monkeys, lemurs and fruit-eating bats ... British Museum, p. 108, from San Cristobal; 1878, Dobson, Catalogue of the Chiroptera ... British Museum, p. 33.
1904. Pteropus (Spectrum) rayneri (part), Trouessart, Catalogus Mammalium ..., Suppl., p. 51.
Specimens examined.—None.
Remarks.—Specimens of Pteropus rayneri cognatus first were reported under the name Pteropus rayneri based on three specimens (one from San Cristobal and two from Guadalcanal). Because the description was based mostly on the two specimens from Guadalcanal, the name rayneri is applicable to the bats from that island. Andersen (1908:365) thought that specimens that he studied, from San Cristobal, were specifically distinct from P. rayneri and he proposed the name Pteropus cognatus for them. Later, Hill (1962:9) reduced cognatus to subspecific status under P. rayneri.
Presently P. r. cognatus is known only from San Cristobal and the small adjacent island of Ugi (see Fig. 6).
Pteropus rayneri rennelli Troughton
1929. Pteropus rennelli Troughton, Rec. Australian Mus., 17:193, September 4, type from Rennell Island; 1954, Laurie and Hill, List of land mammals of New Guinea, Celebes and adjacent islands, p. 35, June 30.
1962. Pteropus rayneri rennelli, Hill, The natural history of Rennell Island, British Solomon Islands, 4:7, February.
Specimens examined.—None.
Remarks.—Until recently, Pteropus rayneri rennelli was known from but a single specimen. Hill (1962:7) reported two additional specimens and pointed out that P. r. cognatus and P. r. rennelli probably represent the extremes of an east-west cline in size. P. r. rennelli and P. r. cognatus differ from other subspecies of the species in lacking tricolored pelage on the dorsum, but their short rostrum clearly indicates affinity with other members of this complex group in the Solomon Islands (Hill, 1962:8).
The relationship of the subspecies rennelli and cognatus is close, both geographically and genetically. Longer forearm, longer metacarpals, and longer mandibular tooth-row serve to differentiate rennelli from cognatus.
1888. Pteropus woodfordi Thomas, Ann. Mag. Nat. Hist., ser. 6, 1:156, February, type from Guadalcanal; 1888, Thomas, Proc. Zool. Soc. London, p. 472, December 4; 1898, Trouessart, Catalogus Mammalium ..., 1:78; 1907, Elliot, Field Columbian Mus., Zool. Ser., 8:491; 1912, Andersen, Catalogue of the Chiroptera ... British Museum, 1:410, from New Georgia and Guadalcanal; 1931, Sanborn, Publ. Field Mus. Nat. Hist., Zool. Ser., 18:19, February 12, from Kolombangara; 1947, Sanborn and Beecher, Jour. Mamm., 28:389, November 19, from Banika and Guadalcanal; 1954, Laurie and Hill, List of land mammals of New Guinea, Celebes and adjacent islands, p. 39, June 30.
1899. Pteropus (Sericonycteris) woodfordi, Matschie, Die Megachiroptera ... naturkunde, p. 83; 1904, Trouessart, Catalogus Mammalium ..., Suppl., p. 54.
1945. Pteropus austini Lawrence, Proc. New England Zool. Club, 23:59, March 26, from Florida.
Specimens examined (four males and three females; five in alcohol and two skin-onlys).—Fauro, in April, 23727, 23790; Guadalcanal in May and June, 23823, 23931; Pavuvo (Russell Islands) in August and October, USNM 277887, USNM 283872-3.
Measurements.—External measurements of two males and two females are, respectively, as follows: Length of head and body, 152, 128, 132, 155; hind foot, 29, 26, 31, 28; ear, 16, 14, 14, 17; length of forearm, 79, 76, 86, 90.
Remarks.—Heretofore, Pteropus woodfordi was known from New Georgia, Guadalcanal, Kolombangara, and Banika (see Fig. 7); specimens from Fauro and Pavuvo islands, listed above, provide new northern localities of record for this species.
Judging by small size and unfused epiphyses, a bat obtained in April and another obtained in June are subadults. Specimens of adults, examined by me, agree well with the descriptions of P. woodfordi by Thomas (1888a:156) and Andersen (1912:407-409), but are slightly smaller than specimens listed by Sanborn and Beecher (1947:389). Color of pelage in this species seems to vary. Adults seen have a pale head and mantle, contrasting strongly with the dark back. Andersen (1912:409) and Lawrence (1945:61) discussed individuals that had scattered silvery hairs mixed with dark fur dorsally and darker mantles that did not contrast noticeably with the rest of the dorsum.
Lawrence (1945:389) named Pteropus austini as a new species closely related to P. woodfordi and other species of the P. scapulatus group of Andersen (1912:402) and Tate (1942:336). Sanborn and Beecher (1947:389), studied a series of P. woodfordi from Banika and Guadalcanal and found that skulls of two subadults agreed well with cranial characteristics ascribed to P. austini, which was based on two subadults. Lawrence (1945:61) stated also that "the interfemoral membrane is entirely absent medially in austini, while in woodfordi it is present as a barely discernible ridge 8 mm. wide." Andersen (1912:408) had earlier reported that in the type of woodfordi the interfemoral membrane was "undeveloped in [the] centre." In 13 adults (in alcohol) studied by Sanborn and Beecher (1947:389), as well as in adults examined by me, the uropatagium is not present. In size, however, these specimens agree with dimensions given for woodfordi by Thomas (1888a:156) and Andersen (1912:410); for example, length of forearm is 93-99. According to Lawrence (1945:59) austini, in which the interfemoral membrane is lacking, is smaller than woodfordi and has a forearm of about 84. In two juveniles of P. woodfordi in the U. S. National Museum, the medially-developed interfemoral membrane is about 7 wide. One specimen has small but distinct calcars whereas the other (slightly larger) apparently lacks calcars. This suggests individual variation in the presence or absence, as well as in the size, of the uropatagium in Pteropus woodfordi.
Sanborn and Beecher (1947:389) decided that "until fully adult specimens showing the characters of austini are available, it best be considered a synonym of woodfordi." For the following reasons I agree with these authors: (1) austini is known from only two specimens, both of which are apparently subadults; (2) austini is reported to have a forearm 84 long and no interfemoral membrane, whereas woodfordi has a forearm about 96 long and an interfemoral membrane that is only slightly developed; (3) specimens that agree in size and cranial characters with the type of woodfordi but that lack an interfemoral membrane have been obtained; and (4) skulls of subadults of woodfordi agree with the description of skulls of austini.
Sanborn (1931:19) reported that specimens of Pteropus woodfordi were obtained at night, while feeding on young green coconuts. Lawrence (1945:62) reported that in the late afternoon a collector found individuals of austini [= woodfordi] in the fronds of a coconut tree, apparently feeding on pollen shoots. Sanborn and Beecher (1947:388) have reported malaria (Plasmodium) in P. woodfordi obtained on Guadalcanal. They suggested that malaria might have rendered one individual helpless because when it was found, on the ground, no wounds were evident and parasites were present in the blood.
1931. Pteropus mahaganus Sanborn, Publ. Field Mus. Nat. Hist., Zool. Ser., 18:19, February 12, type from Santa Ysabel, also reported from Bougainville; 1954, Laurie and Hill, List of land mammals of New Guinea, Celebes and adjacent islands, p. 39, June 30.
Specimens examined (one male and two females; one in alcohol).—Bougainville, in August and October, USNM 276972, USNM 277104-5.
Measurements.—Measurements of one male and two females are, respectively, as follows: Length of head and body, 180, 204, 198; hind foot, 42, 38, 44; ear, 25, 23, 22; length of forearm, 134, 138, 140. Measurements of the skull of the male and one female are, respectively, as follows: Greatest length of skull, 52.5, 55.8; condylobasal length, 50.9, 54.3; palatal length, 24.1, 26.0; [807] zygomatic breadth, 28.9, 32.5; breadth across first upper molars, 14.4, 15.0; width of M1, 2.2, 2.2; length of maxillary tooth-row, 17.4, 18.4; length of mandibular tooth-row, 20.1, 21.4.
Remarks.—Sanborn (1931:19-21) described Pteropus mahaganus on basis of six specimens, five from Santa Ysabel and one from Bougainville. The latter was in poor condition and only provisionally allocated to this species. The specimens examined by me (listed above) confirm the occurrence of P. mahaganus on Bougainville.
Sanborn (1931:20) described mahaganus as "similar to and about the size of [Pteropus scapulatus] from Australia, but lighter in color," and considered it, along with P. woodfordi, a member of the Pteropus scapulatus group of Andersen (1912:402) and Tate (1942:336). I would judge, however, that P. mahaganus and P. woodfordi are much more closely related to one another than to P. scapulatus of Australia. The only significant characteristic that the latter has in common with the two species from the Solomons is small cheek-teeth. In fact, teeth of scapulatus are relatively smaller than teeth of either mahaganus or woodfordi. Also, in scapulatus the upper canines are widely separated due to lateral expansion of the palate at that point, whereas in mahaganus and woodfordi the width across the upper canines is relatively much less.
Dobsonia Palmer
1898. Dobsonia Palmer, Proc. Biol. Soc. Washington, 12:114, April 30.
1810. Cephalotes (part) É. Geoffroy, Ann. du Mus. d'Hist. Nat., 15:104.
Dobsonia, a genus of large to medium-sized fruit bats, occurring from Celebes to the Solomon Islands, contains at least nine species. One species and its two subspecies are endemic to the Solomons.
Dobsonia differs from all other genera of megachiropteran bats in the Solomons by combining absence of a small claw on the second digit and presence of external tail vertebrae.
The cranium of Dobsonia resembles, in some ways, the cranium of Rousettus as well as that of Pteropus. Even so, in Dobsonia the rostrum is shorter and the cheek-teeth, especially in the upper jaw, are more crowded. The anterior part of the mandible is narrow and the lower incisors are diminutive and often concealed by the flesh of the gum.
Dobsonia inermis
In a review of the genus Dobsonia, Andersen (1909c:532) named and described D. inermis and D. nesea from the Solomons. Specimens of Dobsonia inermis from San Cristobal and Ugi were said to differ from specimens of D. nesea from Alu, Shortland, and Rubiana in having perpendicular as opposed to anteriorly slanted upper canines. Andersen (1909c:532) reported that the two species were of "... the same general size." Troughton (1936:348-349) studied specimens of Dobsonia from Bougainville and Santa Ysabel and, because of individual variation in proclivity of the upper canines, concluded that D. nesea was conspecific with D. inermis. He (p. 349) noted that the ears were shorter in inermis than in nesea, but the size of teeth showed insular variation and a "... confusing amount of intergradation ... [that obscures] ... diagnostic importance."
Specimens of Dobsonia from Choiseul are smaller (externally and cranially) than those from Alu, Shortland, Rubiana, Bougainville, Fauro, Vella Lavella, Guadalcanal, Florida, Ugi, San Cristobal, and Rennell. Specimens from Santa Ysabel (see Fig. 8) are intermediate in size between those from Choiseul and the other islands listed. Judging from available specimens, two subspecies of Dobsonia inermis occur in the Solomons. Specimens from Choiseul (see A, Fig. 8), which are smaller than those from other islands, represent one subspecies (heretofore unrecognized), whereas specimens from other islands (except Santa Ysabel) represent a second subspecies. Specimens from Santa Ysabel are slightly larger than those on Choiseul and are regarded as intergrades between the two subspecies. Specimens from Rennell, Ugi, San Cristobal, Florida, Fauro, and Guadalcanal are slightly smaller than those from Bougainville, Vella Lavella, Shortland, and Rubiana, but the differences are not great enough to warrant recognition of two subspecies. Therefore, the subspecific name nesea is arranged as a synonym of inermis, which has priority, and the latter name is used for specimens of Dobsonia inermis from the Solomon islands other than Choiseul and Santa Ysabel. Additional remarks on the distribution of this species are in the section on Zoogeography and Speciation.
Pohle (1953:130) suggested that Dobsonia inermis (as well as D. crenulata and D. praedatrix) is conspecific with D. viridis, but Laurie and Hill (1954:41) did not adopt his suggestion. I have not seen adequate series of crenulata, praedatrix, and viridis (none of which occurs in the Solomons) to judge systematic relationships of these kinds; therefore I follow Laurie and Hill.
Dobsonia inermis inermis Andersen
1909. Dobsonia inermis Andersen, Ann. Mag. Nat. Hist., ser. 8, 4:532, December, type from San Cristobal; 1912, Andersen, Catalogue of the Chiroptera ... British Museum, 1:475.
1936. Dobsonia inermis inermis, Troughton, Rec. Australian Mus., 14:349, April 7, from Santa Ysabel; 1954, Laurie and Hill, List of land mammals of New Guinea, Celebes and adjacent islands, p. 41, June 30; 1956, Hill, The natural history of Rennell Island, British Solomon Islands, 1:74, November 28, from Rennell Island.
1878. Cephalotes peroni (part), Dobson, Catalogue of the Chiroptera ... British Museum, p. 91; 1879, Trouessart, Rev. Mag. Zool., 3:208; 1887, Thomas, Proc. Zool. Soc. London, p. 323, March 15, from Ugi and San Cristobal; 1888, Thomas, Proc. Zool. Soc. London, p. 476, December 4; 1897, Trouessart, Catalogus Mammalium ..., 1:87.
1899. Dobsonia peroni (part), Trouessart, Catalogus Mammalium ..., 2:1278.
1909. Dobsonia nesea Andersen, Ann. Mag. Nat. Hist., ser. 8, 4:532, December 1, type from Shortland Island; 1912, Andersen, Catalogue of the Chiroptera ... British Museum, 1:476, from Shortland and Rubiana; 1931, Sanborn, Publ. Field Mus. Nat. Hist., Zool. Ser., 18:22, February 12, from San Cristobal.
1936. Dobsonia inermis nesea, Troughton, Rec. Australian Mus., 14:348, April 7, from Bougainville; 1953, Pohle, Z. Säugetierk., 17:130, October 27; 1954, Laurie and Hill, List of land mammals of New Guinea, Celebes and adjacent islands, p. 41, June 30, from New Georgia.
Specimens examined (13 males and three females; three in alcohol, crania extracted and cleaned).—Fauro in April, 23728, 23740, 23751; Vella Lavella in November, 23134-36, 23141, 23145, 23147, 23149, 23151, 23153; Guadalcanal in May and June, 23865, 23914, 24008; Florida in October, 24416.
Measurements.—See tables 2 and 3.
Remarks.—Heretofore, Dobsonia inermis inermis was unreported from Fauro, Vella Lavella, Guadalcanal, and Florida. Apparently the subspecies occurs on most islands of the archipelago (see Fig. 9).
In coloration and most cranial dimensions the specimens listed above agree with specimens of D. i. inermis from Alu, Shortland, and Rubiana (Andersen, 1909c:532; 1912:475, 476), Bougainville (Troughton, 1936:348, 349), and Rennell (Hill, 1963:74). The forearm in the adult male holotype of "nesea," from Shortland, is 109.5 as opposed to 109.0 in an adult female topotype of inermis from Ugi (Andersen, 1912:478) in the southeastern part of the archipelago (see Fig. 9). Forearms of specimens of D. i. inermis from Vella Lavella are 107 to 112 (measurements from labels because forearms of these specimens were broken and therefore could not be remeasured). Forearms of specimens from Fauro, Florida, Guadalcanal, and Rennell are 103.6 to 110.0 (see Hill, 1956:74). Variation in length of forearm probably is not significant because no cline is evident (see Fig. 9 and Table 2).
In 1964, 11 specimens of Dobsonia inermis were collected on Choiseul. They are smaller, externally and cranially, than specimens of D. inermis from Sun Cristobal, Ugi, Rennell, Guadalcanal, Florida, Rubiana, Vella Lavella, Shortland, Alu, Bougainville, and Fauro, and may be named and described as follows:
Dobsonia inermis minimus, new subspecies
Type.—Adult male skin and skull, in good condition (originally stored in 70 per cent alcohol for about one year), no. BBM-BSIP 23716, Bernice P. Bishop Museum; from Choiseul Island, British Solomon Islands Protectorate; obtained on 20 March 1964 by Philip Temple, original number 1524.
Distribution.—Choiseul Island (type locality); intergrades from Santa Ysabel also assigned to this subspecies.
Diagnosis.—Size small for species; wing membranes, feet, and ears black; dorsal surface of interfemoral membrane sparsely set with silvery hairs, other membranes naked; hair soft, medium length (10 on mantle, 5 on crown), black hairs and scattered white hairs on face and crown; fur of dorsal surface of mantle composed of whitish hairs having faint olive cast imparting general color of Buffy-Citrine; hair of venter short (about 5), soft, and fine; general coloration Buffy-Citrine; cranium delicate; rostrum narrow in dorsal aspect (nasals not expanded laterally); forehead (junction of nasals and frontals) pronounced in lateral aspect; teeth resembling those of other subspecies of D. inermis but slightly smaller.
Comparisons.—From adults of Dobsonia inermis inermis, which occurs on Rennell, San Cristobal, Ugi, Malaita, Florida, Guadalcanal, Rubiana, Vella Lavella, Shortland, Alu, Bougainville, and Fauro, minimus differs in being smaller. Average length of mandible 31.2 and 33.4. For other measurements see Table 2.
From Dobsonia praedatrix, which occurs on New Britain, New Ireland, and Duke of York (northward of the Solomons), minimus differs in being smaller in all dimensions; length of forearm averaging 100.5 as opposed to 116.0, and greatest length of skull 42.4 as opposed to 50.0.

Fig. 8. Greatest length of
skull plotted against zygomatic breadth for two subspecies of Dobsonia inermis.
Symbols represent D. i. inermis (![]() ),
D. i. minimus (
),
D. i. minimus (![]() ),
and intergrades assigned to minimus
(
),
and intergrades assigned to minimus
(![]() ). Capital letters are
used to relate groups of specimens to the island or islands from which they
were collected; spatial distribution of specimens indicated in the scatter diagram
thus is shown in the inset map. Specimens from Santa Ysabel and Bougainville
are deposited in the Australian Museum. The type specimen of D. i. inermis is
labeled "E." For names of islands see Fig. 2.
). Capital letters are
used to relate groups of specimens to the island or islands from which they
were collected; spatial distribution of specimens indicated in the scatter diagram
thus is shown in the inset map. Specimens from Santa Ysabel and Bougainville
are deposited in the Australian Museum. The type specimen of D. i. inermis is
labeled "E." For names of islands see Fig. 2.
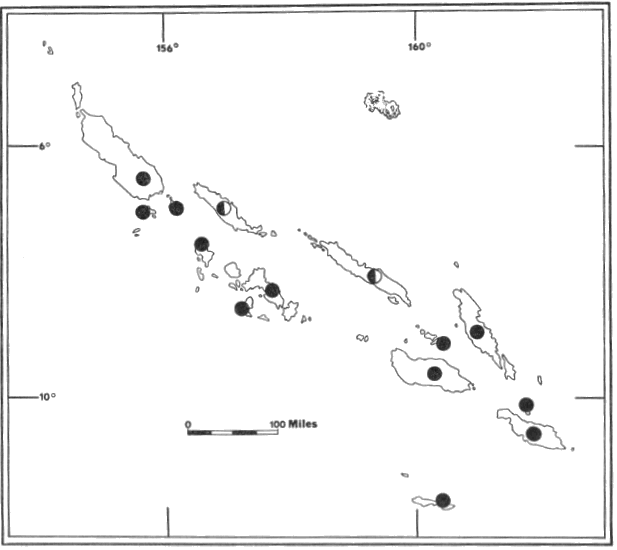
Fig. 9. Distribution of
Dobsonia inermis inermis (![]() ) and
D. inermis minimus (
) and
D. inermis minimus (![]() ).
For names of islands see Fig. 2.
).
For names of islands see Fig. 2.
Table 2. Average and Extreme Measurements of Two Subspecies of Dobsonia inermis.
| Measurement | D. i. minimus | D. i. inermis | ||||||||||
| Choiseul 4 ♂, 2 ♀ |
Santa Ysabel 1 ♂, 2 ♀ |
Guadalcanal, Fauro, Vella Lavella, Florida 9 ♂, 2 ♀ |
||||||||||
| Length of head and body | 174.5 | (170 | - | 180) | .......... | 174.5 | (160 | - | 190) | |||
| Tail vertebrae | 28.5 | ( 24 | - | 33) | 23.0 | (21.5 | - | 24.0) | 30.4 | (25 | - | 35) |
| Hind foot | 25.3 | (25.1 | - | 25.9) | 23.5 | (23.5 | - | 24.0) | 29.3 | (26.0 | - | 31.6) |
| Ear | 21.5 | (21.0 | - | 22.9) | 21.1 | (21.0 | - | 21.5) | 23.1 | (19 | - | 25) |
| Length of forearm | 100.5 | (98.1 | - | 104.0) | 105.3 | (104 | - | 107) | 108.4 | (105 | - | 112) |
| 2nd metacarpal | 43.6 | (42.6 | - | 45.2) | .......... | 48.1 | (45.9 | - | 50.9) | |||
| 3rd metacarpal | 61.5 | (59.8 | - | 62.9) | .......... | 67.6 | (65.2 | - | 68.5) | |||
| 4th metacarpal | 57.5 | (56.5 | - | 58.5) | .......... | 62.5 | (58.7 | - | 65.5) | |||
| 5th metacarpal | 59.0 | (57.0 | - | 60.5) | .......... | 64.4 | (61.8 | - | 66.0) | |||
| Greatest length of skull | 42.4 | (42.1 | - | 43.5) | 44.0 | (43.0 | - | 45.6) | 45.9 | (45.2 | - | 47.4) |
| Condylobasal length | 40.4 | (39.5 | - | 41.3) | 41.7 | (41.1 | - | 42.6) | 43.6 | (43.1 | - | 45.0) |
| Zygomatic breadth | 25.6 | (24.9 | - | 26.8) | 26.5 | (25.7 | - | 27.6) | 27.9 | (27.2 | - | 28.5) |
| Breadth of braincase | 16.8 | (16.5 | - | 17.4) | 18.0 | (17.1 | - | 19.9) | 17.9 | (16.7 | - | 19.0) |
| Breadth across upper canines | 8.4 | (8.1 | - | 8.7) | .......... | 9.2 | (9.2 | - | 9.5) | |||
| Breadth across first upper molars | 12.1 | (11.8 | - | 12.6) | .......... | 13.1 | (12.6 | - | 13.3) | |||
| Length of maxillary tooth-row | 15.6 | (15.5 | - | 15.8) | 16.4 | (16.0 | - | 17.2) | 16.4 | (15.9 | - | 17.0) |
| Length of mandibular tooth-row | 17.1 | (16.8 | - | 17.6) | 17.8 | (17.3 | - | 18.4) | 18.2 | (17.8 | - | 19.4) |
Measurements.—Comparative measurements of the subspecies inermis and minimus are given in Table 2. Some measurements of the type are as follows: Length of head and body, 147; tail vertebrae, 31; hind foot, 25; ear, 21; length of forearm, 99.5; 2nd metacarpal, 42.8; 3rd metacarpal, 62.7; 4th metacarpal, 58.5; 5th metacarpal, 59.1; greatest length of skull, 42.2; condylobasal length, 40.6; zygomatic breadth, 25.8; breadth of braincase, 16.8; length of maxillary tooth-row, 15.8; length of mandible, 31.2.
Remarks.—Dobsonia inermis minimus is the smallest subspecies of Dobsonia inermis. Specimens from Santa Ysabel, southeastward of Choiseul, are slightly larger than the type and paratypes of minimus. As can be seen in the scatter diagram (Fig. 8), a male from Santa Ysabel is as large as one male and most females of D. i. inermis. The other three specimens from Santa Ysabel also are slightly larger than specimens of minimus from Choiseul, but are much smaller than specimens of D. i. inermis, and, therefore, are referred to D. i. minimus.
Although there is a cline in size of Dobsonia inermis from Choiseul to Florida (generally southward; Fig. 9), no cline in size is apparent between Choiseul and Fauro (generally westward). Specimens of D. inermis from Fauro are average for the subspecies inermis; there is no evidence, in the small series available, of intergradation between minimus on Choiseul and inermis on Fauro.
Specimens examined (eight males and three females, all originally in alcohol; seven crania, all adults, extracted and cleaned).—Choiseul in March, 23565, 23628, 23637, 23665-67, 23640, 23714, 23716 (holotype), 23717, 23720. Ellis LeG. Troughton kindly examined and measured nos. AM-M. 3693 ♂, AM-M. 3694 ♂, AM-M. 3937 ♀, and AM-M. 3940 ♀, from Santa Ysabel in the Australian Museum.
Subfamily Macroglossinae
Macroglossus F. Cuvier
1824. Macroglossus F. Cuvier, Des dents des mammiferes ... zoologiques, p. 248.
1840. Kiodotus Blyth, in Cuvier's animal kingdom ..., p. 69.
1891. Carponycteris Lydekker, in Flower and Lydekker, mammals living and extinct, p. 654.
1902. Odontonycteris Jentink, Notes Leyden Mus., 23:140, July 15.
Macroglossus, the widest-ranging genus of macroglossine bats, occurs from southeastern Asia to the southern islands of the Solomon Archipelago (see Ellerman and Morrison-Scott, 1966:101; Laurie and Hill, 1954:44). One species, known also from Celebes and New Guinea, occurs in the Solomons and is represented there by an endemic subspecies.
Numerous generic names have been applied, at one time or another, to bats now considered as Macroglossus. Trouessart (1904:65) and Miller (1907:70) listed the one bat of this genus occurring in the Solomons under Carponycteris and Kiodotus, respectively. Andersen (1911:642; 1912:767) and, later, Sanborn (1931:22) identified this bat as Macroglossus lagochilus microtus. Troughton (1936:350), reporting an extension of range of this species in the Solomons, used the generic name Odontonycteris without explanation. Andersen (1912:754) pointed out that Jentink originally established the name Odontonycteris on the basis of an extra premolar in each upper jaw as opposed to the usual two in Macroglossus, and arranged Odontonycteris as a synonym of Macroglossus because "in no genus of Megachiroptera are dental anomalies of so frequent occurrence as in Macroglossus, and on no point of the jaws are these [813] anomalies ... so often met with as on that occupied by the molar series." Sanborn (1931:22) and Phillips (1966:27) noted variation in number of incisors in Macroglossus as well as in Melonycteris, another macroglossine genus. All of the more recent workers (Ellerman and Morrison-Scott, 1966; Pohle, 1953; Laurie and Hill, 1954) use the name Macroglossus.
Macroglossus lagochilus
Macroglossus lagochilus has at least three subspecies, one of which is endemic to the Solomons. The species ranges from Celebes on the west to the Solomon Islands on the east, occurring not only in New Guinea but also on many of the small adjacent islands (see Laurie and Hill, 1954:44).
Macroglossus lagochilus microtus Andersen
1911. Macroglossus lagochilus microtus Andersen, Ann. Mag. Nat. Hist., Ser. 8, 7:642, June, type from Guadalcanal, additional specimens from Florida; 1912, Andersen, Catalogue of the Chiroptera ... British Museum, 1:767; 1931, Sanborn, Publ. Field Mus. Nat. Hist., Zool. Ser., 18:22, February 12, from San Cristobal; 1953, Pohle, Z. Säugetierk., 17:130, October 27, from Bougainville; 1954, Laurie and Hill, List of land mammals of New Guinea, Celebes and adjacent islands, p. 44, June 30.
1888. Macroglossus australis (part). Thomas, Proc. Zool. Soc. London, p. 476, December 4, from Guadalcanal.
1904. Carponycteris nana (part), Trouessart, Catalogus Mammalium ..., Suppl., p. 65.
1907. Kiodotus sp., Miller, Bull. U. S. Nat. Mus., 57:70, June 29.
1936. Odontonycteris lagochilus microtus, Troughton, Rec. Australian Mus., 14:350, April 7, from Bougainville.
Specimens examined (14 males and 16 females; in alcohol).—Choiseul in March, 23654-57, 23614, 23629, 23643, 23645, 23647, 23677-79, 23684; Vella Lavella in December, 23277-79, 23283-84; Fauro in April, 23765; Guadalcanal in May and June, 23830, 23864, 23935; Kolombangara in January, 23385, 23399, 23397, 23407, 23420-21; Santa Ysabel in June, 24067; Malaita in June, 24067.
Measurements.—Average and extreme external measurements of 14 males and 15 females are as follows: Length of head and body, 68.3 (63-72); tail vertebrae present but scarcely perceptible and therefore not measured; hind foot, 11.4 (9.0-12.9); ear, 12.0 (10.0-12.9); length of forearm, 37.6 (36.2-39.9).
Remarks.—The distribution of Macroglossus lagochilus microtus has not been well known. Specimens herein reported from Choiseul, Fauro, and Vella Lavella provide new records of distribution. As shown on Figure 10, the subspecies occurs throughout the Solomon Islands.
Macroglossus lagochilus microtus differs slightly from M. l. nanus Matschie, the subspecies of the Bismarck Archipelago and Admiralty Islands to the north of the Solomons. M. l. nanus averages slightly larger than microtus (see Andersen, 1912:768-769, for comparative measurements) but otherwise closely resembles it.
Individual variation is evident in several measurements of the specimens at hand (in length of forearm, for example) but no clines are apparent. Four females obtained in March were lactating, as was one taken in December and one taken in January.
Melonycteris Dobson
1877. Melonycteris Dobson, Proc. Zool. Soc. London, p. 119, June 1.
1877. Cheiropteruges Ramsay, Proc. Linn. Soc. New South Wales, 2:19, July.
1887. Nesonycteris Thomas, Ann. Mag. Nat. Hist., ser. 5, 14:147, February.
The genus Melonycteris is known from three species, two apparently endemic to the Solomon Islands and the third occurring in eastern New Guinea and the Bismarck Archipelago (Laurie and Hill, 1954:45).
Heretofore, the generic name Nesonycteris has been applied to the species in the Solomons, whereas Melonycteris has been restricted to the one species in the Bismarck Archipelago and New Guinea. Andersen (1912:792) judged that Nesonycteris was clearly distinct from Melonycteris on the basis of two characters (loss of a claw on the second digit and loss of the inner, lower incisors). On the other hand, he noted striking similarities in general cranial features, dentition, palatal ridges, tongue, and external appearance of the two genera. Pohle (1953:131) synonymized the two but Laurie and Hill (1954:45) considered them distinct. I have suggested previously (Phillips, 1966:26, 27) that characteristics used to distinguish between Melonycteris and Nesonycteris are of less than generic value. Variability of number of incisors in the upper jaw of specimens of Melonycteris (and in other macroglossine genera, as well) indicates a lack of selective pressure for either increase or decrease in number of incisors. Furthermore, the loss of the small claw on the second digit might not be important because, as Bader and Hall (1960:15) have pointed out, limbs of bats vary more in phenotypic expression than do other parts of the skeletal structure.
The discovery of a new species (Melonycteris aurantius) in the Solomon Islands sheds additional light on the problem. Although M. aurantius possesses the distinguishing characteristics of the genus "Nesonycteris," the species closely resembles Melonycteris in other features. Similarity in structure of hair of Melonycteris and Nesonycteris, as first reported by Benedict (1957:293), also supports the argument for synonymy (see Phillips, 1966:26).
Melonycteris aurantius lacks a small claw on the second digit and has only two lower incisors. In these ways this species is like woodfordi, which also is restricted to the Solomons. On the other hand, the structure of the skull of M. aurantius is like that of M. melanops, which is the species found in the Bismarck Archipelago.
Although melanops is not yet known from the Solomon Islands, I have included it in the following key.
Key to Known Species of Melonycteris
| 1. | Ventral surface darker than dorsum, but not strongly contrasting with it; lacking a small claw on the second digit, |
| 2 | |
| 1´. | Ventral surface nearly black, strongly contrasting with dorsum; small claw on second digit, |
| Melonycteris melanops | |
| 2(1´). | Pelage bright, Cinnamon-Rufous; postorbital region of skull expanded (about 8.3 wide), |
| Melonycteris aurantius, p. 816 | |
| 2´. | Pelage dark, near Wood-Brown or Cinnamon; postorbital region of skull constricted (about 7.5), |
| Melonycteris woodfordi, p. 816 | |
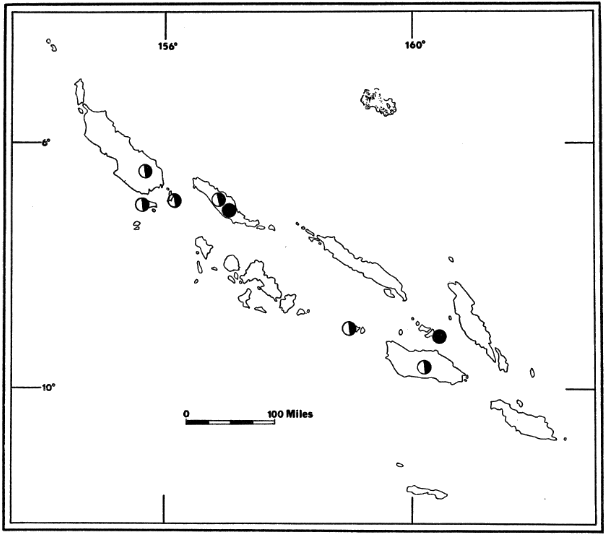
Fig. 11. Distribution of
Melonycteris aurantius (![]() ) and
M. woodfordi (
) and
M. woodfordi (![]() ).
For names of islands see Fig. 2.
).
For names of islands see Fig. 2.
Melonycteris aurantius Phillips
1966. Melonycteris aurantius Phillips, Jour. Mamm., 47:23-27, March 12, type from Florida Island, additional specimens from Choiseul Island.
Specimens examined (six females; three in alcohol).—Florida in October, 24440; Choiseul in March, 23615, 23617, 23558, 23694, 23681.
Measurements.—Average and extreme measurements of six females are as follows: Length of head and body, 80.8 (77-106); hind foot, 17.2 (16.0-18.7); ear, 12.7 (11.5-14.0); length of forearm, 49.3 (42.9-53.8). Average and extreme measurements of skulls of five females are as follows: Greatest length of skull, 31.8 (30.8-33.3); condylobasal length, 29.7 (28.6-32.4); zygomatic breadth, 18 (17.2-20.0); breadth of braincase, 12.6 (12.4-13.2); postorbital breadth, 8.3 (8.0-8.9); length of maxillary tooth-row, 10.1 (9.4-10.4); length of mandibular tooth-row, 11.7 (10.8-12.2).
Remarks.—On Choiseul Island Melonycteris aurantius was taken at the same locality as its congener, Melonycteris woodfordi.
Externally, M. aurantius resembles M. woodfordi. These species are the same size, but the former is brighter in color (nearly orange in adults) than the latter, which is Wood-Brown dorsally. Internally, differences between M. aurantius and M. woodfordi are more obvious. In the skull of M. aurantius, the postorbital region is expanded (measuring about 8.3), whereas in M. woodfordi the postorbital region is constricted. Furthermore, in lateral aspect the posterior portion of the skull of M. aurantius is down-turned and the angle of the facial axis with the basicranial axis is much more acute than in M. woodfordi.
The number of upper incisors is highly variable in the six specimens of M. aurantius that I have examined. In two specimens an extra tooth has erupted just anterior to I2 and there is a total of six upper incisors. In two other specimens an extra tooth has erupted in front of I2 on one side but not the other. I could find no trace of an extra tooth in the remaining two specimens.
Practically nothing is known about the natural history of M. aurantius, or, indeed, that of either of the other two species of this genus. One field collector (Temple, in litt.) for the Bishop Museum reported that he obtained both M. aurantius and M. woodfordi in the same mist net in one night. The holotype, an adult female, was lactating when obtained in October.
Melonycteris woodfordi (Thomas)
1887. Nesonycteris woodfordi Thomas, Ann. Mag. Nat. Hist., ser. 5, 14:147, February, type from Shortland Island; 1887, Thomas, Proc. Zool. Soc. London, p. 324, March 15; 1888, Thomas, Proc. Zool. Soc. London, p. 476, December 4; 1898, Trouessart, Catalogus Mammalium ..., 1:90; 1899, Matschie, Die Megachiroptera ... naturkunde, p. 91; 1904, Trouessart, Catalogus Mammalium .., Suppl., p. 66; 1907, Miller, Bull. U. S. Nat. Mus., 57:74, June 29; 1912, Andersen, Catalogue of the Chiroptera ... British Museum, 1:792, from Alu, Shortland, Fauro, and Guadalcanal; 1931, Sanborn, Publ. Field Mus. Nat. Hist., Zool. Ser., 18:23, February 12, from Russell Island (Pavuvo); 1954, Laurie and Hill, List of land mammals of New Guinea, Celebes and adjacent islands, p. 45, June 30.
1953. Melonycteris woodfordi, Pohle, Z. Säugetierk., 17:130, October 27, from Bougainville Island; 1966, Phillips, Jour. Mamm., 47:23, March 12, from Choiseul.
Specimens examined (three males and one female; in alcohol).—Choiseul, in April, 23413-14, 23434, 23275.
Measurements.—Average and extreme measurements of three males and one female are as follows: Length of head and body, 86.1 (83.1-91.0); hind foot, 19.6 (17.2-22.2); ear, 11.3 (10.8-11.7); length of forearm, 54.4 (52.1-57.7).
Remarks.—Specimens of Melonycteris woodfordi from Choiseul constitute a new locality of occurrence for the species. Apparently M. woodfordi occurs throughout the Solomons (see Fig. 11).
Thomas (1887a:147) named Nesonycteris woodfordi in a preliminary report that appeared before the publication of the more detailed description of the genus and species (1887b:323-324). In the second paper he stated that the anterior projections of the premaxillary bones are separated distinctly in both Nesonycteris and Melonycteris. According to Thomas (1887b:323), it was by some "accident" that Dobson (1878:4) reported the anterior projections of the premaxillary bones in Melonycteris melanops to be united. Writing at a later date, Andersen (1912:785) reported that in Melonycteris melanops the premaxillary bones have "simple contact with each other." Furthermore, in Andersen's (1912:791) illustration of M. woodfordi the premaxillary bones are in contact anteriorly. In specimens of woodfordi and melanops examined by me, the premaxillary bones are in contact. In M. aurantius the premaxillary bones are not in contact, and it differs from woodfordi in several other respects.
In M. woodfordi, as in other macroglossine bats, there is variability in dentition. One specimen examined has a total of three upper incisors, and another had an extra peglike tooth just anterior to I1.
Subfamily Nyctimeninae
Nyctimene Borkhausen
1797. Nyctimene Borkhausen, Deutsche fauna ..., 1:86.
1810. Cephalotes É. Geoffroy, Ann. du Mus. d'Hist. Nat., 15:104.
1811. Harpyia Illiger, Prodr. Syst. Mamm. et Avium, p. 118.
1837. Gelasinus Temminck, Monographe de Mammalia ..., 2:100.
Tube-nosed bats of the genus Nyctimene occur from Celebes on the west to the Santa Cruz Islands on the east. Heretofore, two species (N. albiventer and N. major), each with an endemic subspecies, were known from the Solomon Islands. Both species occur also in New Guinea and on many adjacent islands. A new species of Nyctimene, apparently endemic to the Solomons, and a new subspecies of N. albiventer are named beyond.
Nyctimene is related closely to Cynopterus and the "Cynopterus group" of Andersen (1912:691). Because Nyctimene is a highly specialized bat, Miller (1907:75) placed it in a subfamily separate from that of Cynopterus and its allies.
Andersen (1912:696, 697) placed the species of Nyctimene previously known from the Solomons in two groups, the "papuanus" group and the "cephalotes" group, on the basis of difference in length of forearm and length of maxillary tooth-row. Because of its short forearm (about 58), N. albiventer is in the papuanus group; and N. major, because of its long forearm (about 74), is in the cephalotes group.
Key to Species of Nyctimene in the Solomons
| 1. | Forearm longer than 70; males grayish-brown, females pale gray, |
| N. major scitulus, p. 825 | |
| 1'. | Forearm shorter than 70; males dark brown, females pale brown, |
| 2 | |
| 2(1'). | Forearm about 65, |
| N. malaitensis, p. 822 | |
| 2'. | Forearm less than 61, |
| N. albiventer, p. 818 | |
Nyctimene albiventer
This species occurs throughout New Guinea and on many adjacent islands, including the Bismarck Archipelago and the Admiralty and Solomon islands. The species varies geographically and five subspecies are recognized. The two subspecies in the Solomons resemble N. albiventer papuanus, the subspecies that ranges from eastern New Guinea to New Britain. N. albiventer bougainville occurs in the western chain of islands of the Solomons, whereas another subspecies, named as new beyond, occurs in the eastern chain of islands (see Fig. 12).
Sexual dichromatism is striking. As Andersen (1912:690) previously reported, females generally are paler, more brownish than males, which are dark and have a better defined black dorsal stripe.
Nyctimene albiventer bougainville Troughton
1936. Nyctimene bougainville Troughton, Rec. Australian Mus., 19:349, April 7, type from Bougainville.
1954. Nyctimene albiventer bougainville, Laurie and Hill, List of land mammals of New Guinea, Celebes and adjacent islands, p. 46, June 30.
1953. Nyctimene papuanus bougainville, Pohle, Z. Säugetierk., 17:130, October 27.
Specimens examined (nine males, one female; nine in alcohol, nine crania extracted and cleaned).—Bougainville in December, AM-M. 5786 (paratype); Guadalcanal in May, 23812, 23815, 23827; Kolombangara in January and February, 23369, 23381, 23388, 23406, 23444, 23456.
Measurements.—See Table 3.
Remarks.—Heretofore, Nyctimene albiventer bougainville was not known from Kolombangara and Guadalcanal. The subspecies apparently ranges throughout the western chain of the Solomons.
Troughton (1936:350) considered Nyctimene bougainville specifically distinct from its nearest ally, N. papuanus. Pohle (1953:130) did not examine specimens of either kind, but on the basis of Troughton's description decided that N. bougainville differed only subspecifically from N. papuanus. Laurie and Hill (1954:46) synonymized bougainville and papuanus with N. albiventer. However, Troughton (1936:350) pointed out that in addition to size bougainville differed from papuanus by having narrower and longer pm3 and pm4. Judging from specimens examined by me, such is the case, and the difference is even more pronounced in m1.
Specimens of N. a. bougainville from Kolombangara and Guadalcanal agree with a paratype of this subspecies from Bougainville. Geographic variation, if present in the population in the western chain of islands (see Fig. 12), is slight and not notable in the series available. Some individual variation was found, especially in the shape of the interorbital region of the skull. An adult [819] male from Kolombangara is unusually dark, almost black; color of the other specimens (all in alcohol) is consistent according to sex.
Nyctimene albiventer from Choiseul and Santa Ysabel is smaller, in all respects, than N. albiventer from Bougainville, Kolombangara, and Guadalcanal (see Table 3), and therefore may be named and described as follows:
Nyctimene albiventer minor, new subspecies
Type.—Adult male, skin and skull, in good condition (originally stored in alcohol for about one year), no. BSIP 23636, Bernice P. Bishop Museum; from Choiseul Island, British Solomon Islands Protectorate; obtained on 11 March 1964, by Philip Temple, original number 1441.
Distribution.—Known only from Choiseul and Santa Ysabel islands (see Fig. 12).
Diagnosis.—Small for Nyctimene; wing membranes brown with scattered yellow spots (dried specimens); uropatagium, feet, and ears brown; dorsum of tibia set with hair, ventral surface naked; dorsum of uropatagium sparsely set with pale brown hairs, ventral surface almost bare; fringe of hairs along two centimeters of dorsal and ventral surfaces of trailing edge of wing membrane; proximal third of dorsal surface of forearm sparsely set with hairs; pelage of back soft and thick, of medium length (about 7); hair on crown and nape short (about 4); well-defined black dorsal stripe, extending from uropatagium to shoulders; skull resembling that of other subspecies of N. albiventer but relatively smaller; zygomatic arch delicate, slender anteriorly; P2 small (see Fig. 14). Sexually dichromatic as follows: male—dorsum Hair-Brown, bases of hairs darker; hair on throat sparse, medium length (about 6), Hair-Brown; fur along sides of abdomen Drab; female—dorsum having Buffy-Brown cast, some individual hairs Hair-Brown; shoulders Sayal-Brown; hair on throat sparse, Hair-Brown on throat and midline of abdomen; sides of abdomen Sayal-Brown.
Comparisons.—From Nyctimene major scitulus, the largest member of this genus in the Solomons, N. a. minor differs in being smaller in all measurements taken; forearm averaging 54.8 as opposed to 73.5; greatest length of skull 28.2 as opposed to 37.0, and females pale brown instead of pale gray.
From nine adults of Nyctimene albiventer bougainville from Bougainville, Kolombangara, and Guadalcanal, minor differs as follows: averaging slightly smaller in all dimensions; forearm averaging 54.8 as opposed to 57.9; second metacarpal averaging 27.4 as opposed to 28.3; 5th metacarpal averaging 38.5 as opposed to 40.0; condylobasal length 26.7 as opposed to 28.0; length of mandibular tooth-row 10.3 as opposed to 10.9; mandible smaller (see Fig. 14); dorsal stripe fainter.
From Nyctimene albiventer papuanus, known from eastern New Guinea, New Britain, and the Admiralty Islands, minor differs as follows: slightly smaller in most dimensions; forearm averaging 54.8 as opposed to 57.0; length of maxillary tooth-row 8.9 as opposed to 9.8; length of mandibular tooth-row 10.3 as opposed to 11.0; breadth across upper third premolars notably less (7.5 as opposed to 8.4).
N. a. minor differs from N. albiventer albiventer Gray, which occurs about 800 miles to the west of minor, in ways made apparent by the description by Andersen (1912:700-701). N. a. minor occurs about 1500 miles eastward of the place from which N. a. draconilla Thomas, a subspecies essentially unknown to me, was named (see Laurie and Hill, 1954:46).
From Nyctimene sanctacrucis, known from the Santa Cruz Islands, minor differs as follows: much smaller in all dimensions; forearm averaging 54.8 as opposed to 75; greatest length of skull 28.2 as opposed to 34.5; length of maxillary tooth-row 8.9 as opposed to 12.9.

Fig. 12. Distribution of
Nyctimene albiventer bougainville (![]() )
and N. albiventer minor (
)
and N. albiventer minor (![]() ).
For names of islands see Fig. 2.
).
For names of islands see Fig. 2.
Fig. 13. Scatter diagram comparing
two subspecies of Nyctimene albiventer. One individual of specimens thought
to be intergrades is as large as specimens of Nyctimene a. bougainville,
whereas the other three intergrades are about the same size as specimens of
N. a. minor. Symbols represent N. a. bougainville
(![]() ),
N. a. minor (
),
N. a. minor (![]() ),
and intergrades assigned to minor (
),
and intergrades assigned to minor (![]() ).
For names of islands see Fig. 2.
).
For names of islands see Fig. 2.
Table 3. Average and Extreme Measurements of Nyctimene albiventer bougainville and N. a. minor.
| Measurement | N. a. minor Choiseul, Santa Ysabel 4 ♂, 1 ♀ |
Intergrades Fauro 1 ♂, 3 ♀ |
N. a. bougainville Kolombangara, Guadalcanal 8 ♂, 1 ♀ |
|||||||||
| Length of head and body | 107.0 | (105 | - | 109) | 109.2 | (105 | - | 112) | 110.0 | (106 | - | 117) |
| Tail vertebrae | 20.0 | (19.3 | - | 20.5) | 21.0 | ( 20 | - | 22) | 19.2 | ( 15.5 | - | 23.0) |
| Hind foot | 14.2 | (13.5 | - | 15.0) | .......... | 14.3 | ( 13.0 | - | 15.9) | |||
| Ear | 11.9 | (11.0 | - | 13.0) | .......... | 12.8 | ( 11.8 | - | 14.5) | |||
| Length of forearm | 54.8 | (54.0 | - | 55.8) | 57.1 | (55.9 | - | 59.0) | 57.9 | ( 55.8 | - | 59.8) |
| Greatest length of skull | 28.2 | (27.2 | - | 28.9) | 28.6 | (28.3 | - | 29.7) | 29.7 | ( 28.6 | - | 30.1) |
| Condylobasal length | 26.7 | (26.2 | - | 27.5) | 27.4 | (26.6 | - | 28.0) | 28.0 | ( 27.8 | - | 28.9) |
| Palatal length | 11.2 | (10.9 | - | 11.9) | 11.6 | (11.3 | - | 11.8) | 11.7 | ( 11.0 | - | 12.5) |
| Breadth of braincase | 12.0 | (11.5 | - | 12.4) | 12.0 | (11.7 | - | 12.2) | 12.3 | ( 12.1 | - | 12.8) |
| Zygomatic breadth | 18.9 | (18.4 | - | 19.7) | 18.6 | (18.4 | - | 19.2) | 19.2 | ( 18.7 | - | 20.0) |
| Interorbital breadth | 5.0 | ( 4.7 | - | 5.6) | 5.3 | ( 5.0 | - | 5.6) | 5.1 | ( 4.7 | - | 5.5) |
| Breadth across first upper molars | 8.6 | ( 8.4 | - | 8.9) | 8.9 | ( 8.7 | - | 9.1) | 9.1 | ( 8.8 | - | 9.6) |
| Maxillary tooth-row | 8.9 | ( 8.7 | - | 9.3) | 9.3 | ( 9.1 | - | 9.5) | 9.5 | ( 9.2 | - | 9.8) |
| Mandibular tooth-row | 10.3 | (10.0 | - | 10.6) | 10.5 | (10.2 | - | 11.1) | 10.9 | ( 10.7 | - | 11.4) |
Measurements.—Measurements of the two subspecies from the Solomons are given in Table 3. Some measurements of the type are as follows: Length of head and body, 108; tail vertebrae, 20.5; hind foot, 14.7; ear, 11.3; length of forearm, 55.1; 2nd metacarpal, 27.4; 3rd metacarpal, 39.0; 4th metacarpal, 37.5; 5th metacarpal, 39.1; greatest length of skull, 28.6; condylobasal length, 27.5; zygomatic breadth, 18.4; length of maxillary tooth-row, 9.0; length of mandibular tooth-row, 10.4.
Remarks.—Nyctimene albiventer minor closely resembles N. albiventer bougainville, differing from the latter mostly in size. Although adults of minor average only slightly smaller than adults of bougainville (see Table 3), there is only slight overlap (about 0.2 at most) in most minimum dimensions of external and cranial features of bougainville and corresponding maximum dimensions of externals and crania of minor. The difference in size is clearly shown in Figs. 13 and 14.
Four specimens of Nyctimene albiventer from Fauro herein are considered to be intergrades between N. a. bougainville and N. a. minor. As shown in Table 3, the specimens from Fauro average slightly larger than those of minor from Choiseul and Santa Ysabel and slightly smaller than specimens of bougainville from Kolombangara and Guadalcanal. I have assigned the specimens from Fauro to N. a. minor because they generally are closer to minor in size (see Fig. 13).
Specimens examined (five males and four females; seven in alcohol; seven crania extracted and cleaned).—Choiseul in February and March, 23636 (holotype), 23631, 23540, 23646; Santa Ysabel in February, 23539; Fauro in April, 23742, 23743, 23763, 23764.
One specimen of Nyctimene from Malaita Island is smaller than Nyctimene major, which is known from Shortland, Alu, Florida, New Georgia, Guadalcanal, Choiseul, and Malapa (see Fig. 15) and is larger than either of the two subspecies of Nyctimene albiventer known from Bougainville, Fauro, Kolombangara, Guadalcanal, Choiseul, and Santa Ysabel. This specimen represents a previously unknown species and may be named and described as follows:
Nyctimene malaitensis, new species
Type.—Adult female, skin and skull, in good condition (originally stored in alcohol for about one year), no. BSIP 24103, Bernice P. Bishop Museum; from Malaita Island, British Solomon Islands Protectorate; obtained on 1 July 1964, by Peter Shanahan, original no. unknown.
Distribution.—Known only from Malaita (see Fig. 16).
Diagnosis.—Size average for genus but larger than closest relative, Nyctimene albiventer; wing membranes brown with scattered yellow spots (dried specimen); uropatagium, ears, and feet brown; dorsal surface of tibia set with hair, ventral surface bare; dorsal surface of uropatagium sparsely set with hair, ventral surface having few, scattered hairs; dorsal surface of trailing edge of wing membrane sparsely set with hairs, ventral surface bare; proximal third of upper- and under-surface of forearm set with hair; pelage of back luxuriant and soft (about 10 long); hair on crown and nape shorter than on back (4 to 8); well-defined black dorsal stripe from shoulders to rump (about 2 wide); basal half of most hairs on dorsum Deep Mouse Gray, distal half Light Buff, tips Ochraceous-Tawny; some hairs on back entirely Light Buff; hairs of crown Light Ochraceous Buff tipped with Ochraceous-Tawny; hair on throat and along sides of abdomen Light Ochraceous Buff; hairs of ventral midline Smoke Gray; braincase narrow; zygomatic breadth relatively narrow; well-developed lambdoidal crest in female; rostrum short, wide; upper canines slanted posteriorly; upper incisors large; foramen ovale large (see Fig. 14).
Fig. 14. Dorsal and ventral views of skulls of (A) Nyctimene albiventer minor [specimen 23631 ♂, (B) N. a. bougainville [specimen 23381 ♂], and (C) N. malaitensis [specimen 24103 ♀].
Comparisons.—From Nyctimene major scitulus, the largest kind of Nyctimene in the Solomons, malaitensis differs as follows: smaller in all dimensions (forearm 65 as opposed to 73.5); greatest length of skull 32.4 as opposed to 37.0; length of maxillary tooth-row 10.5 as opposed to 13.0; length of mandibular tooth-row 11.8 as opposed to 14.2.
From nine adults of Nyctimene albiventer bougainville from Bougainville, Kolombangara, and Guadalcanal, malaitensis differs as follows: larger in all dimensions: forearm 65 as opposed to 57.9; greatest length of skull 32.4 as opposed to 29.7; zygomatic breadth 20.4 as opposed to 19.2; and length of maxillary tooth-row 10.5 as opposed to 9.5; length of mandibular tooth-row 11.8 as opposed to 11.1.
From five adults of Nyctimene albiventer minor, from Choiseul and Santa Ysabel, malaitensis differs in the same ways it differs from N. a. bougainville, but the contrast is even greater when malaitensis and minor are compared.
From Nyctimene sanctacrucis, known only from the Santa Cruz Islands, malaitensis differs in being smaller in all dimensions: forearm 65 as opposed to 75; greatest length of skull 32.4 as opposed to 34.5; and length of maxillary tooth-row 10.5 as opposed to 12.9.
Measurements of the holotype.—Length of head and body, 118; tail vertebrae, 23.0; hind foot, 16.0; ear, 14.0; length of forearm, 65.0; 2nd metacarpal, 33.2; 3rd metacarpal, 46.4; 4th metacarpal, 44.3; 5th metacarpal, 46.0; greatest length of skull, 32.4; condylobasal length, 30.6; palatal length, 13.0; breadth of braincase, 12.5; zygomatic breadth, 20.4; interorbital breadth, 5.5; breadth across first upper molars, 9.5; length of maxillary tooth-row, 10.5; length of mandibular tooth-row, 11.8.
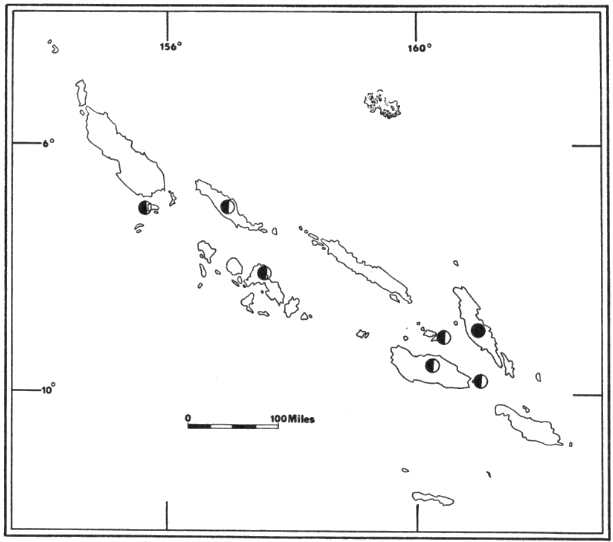
Fig. 15. Distribution of
Nyctimene malaitensis (![]() )
and N. major scitulus (
)
and N. major scitulus (![]() ).
For names of islands see Fig. 2.
).
For names of islands see Fig. 2.
Remarks.—In size, Nyctimene malaitensis is intermediate between N. albiventer and N. major. Because the type of malaitensis is brown and not pale gray, as are females of major, N. malaitensis most likely is more closely related to N. albiventer, in which the females are brown. The teeth of the holotype and only known specimen of malaitensis are too worn to be useful in determining the relationships between these species.
When more specimens are available, N. malaitensis may prove to be a subspecies of N. albiventer. At present, malaitensis is accorded specific rank in order not to obscure the apparent relationships of N. albiventer bougainville and N. a. minor. Additionally, N. malaitensis is given specific rank because (1) it is larger (especially in external dimensions) than the largest subspecies of N. albiventer (compare above measurements with those in Table 3), and (2) malaitensis does not form a cline with either of the two subspecies of N. albiventer.
Specimen examined (one female).—Malaita in July, 24103 (holotype).
Nyctimene major
This large species of tube-nosed bat has at least four subspecies, one of which (N. major scitulus) is endemic to the Solomons. The species occurs throughout eastern New Guinea and on many of the islands adjacent to the [825] eastern coast of New Guinea, including the Trobriand Islands, the Bismarck Archipelago, and the Solomons (see Laurie and Hill, 1954:47). The geographic distribution of the species generally is the same as that of N. albiventer.
In Nyctimene major, as in N. albiventer, most males are grayish-brown, whereas most females are pale gray.
Nyctimene major scitulus Andersen
1910. Nyctimene scitulus Andersen, Ann. Mag. Nat. Hist., ser. 8, 6:623, December 1, type from Shortland; 1912, Andersen, Catalogue of the Chiroptera ... British Museum, 1:711, from Shortland, New Georgia, Florida, Guadalcanal; 1931, Troughton, Proc. Linnean Soc. New South Wales, 56:206, July 15; 1931, Sanborn, Publ. Field Mus. Nat. Hist., 18:22, February 12, from Choiseul and Malapa; 1942, Tate, Bull. Amer. Mus. Nat. Hist., 80:342, December 31.
1954. Nyctimene major scitulus, Laurie and Hill, List of land mammals of New Guinea, Celebes and adjacent islands, p. 47, June 30.
1862. Harpyia pallasi, Gerrard, Catalogue of the bones ... British Museum, p. 58.
1870. Harpyia cephalotes, Gray, Catalogue of monkeys, lemurs and fruit-eating bats in the British Museum, p. 121.
1878. Harpyia major, Dobson, Catalogue of the Chiroptera ... British Museum, p. 90; 1879, Trouessart, Rev. Mag. Zool., 3:207; 1887, Thomas, Proc. Zool. Soc. London, p. 323; 1888, Thomas, Proc. Zool. Soc. London, p. 476; 1897, Trouessart, Catalogus Mammalium ..., 1:87.
1899. Cephalotes major, Trouessart, Catalogus Mammalium ..., 2:1277.
1899. Gelasinus major, Matschie, Die Megachiroptera ... naturkunde, p. 84; 1904, Trouessart, Catalogus Mammalium ..., Suppl., p. 64.
Specimens examined (four males and one female; dried skins with skulls inside).—Florida in October, 24397, 24413, 24418, 24419.
Measurements.—External measurements of four males and one female are, respectively, as follows: Length of head and body, 134, 128, 134, 134, 136; tail vertebrae, 28, 23, 27, 26, 21; hind foot, 20, 16, 19, 16, 21; ear, 17, 17, 17, 17, 18; length of forearm, 73.8, 68.0, 74.0, 73.6, 78.0.
Remarks.—Nyctimene major scitulus has been recorded only from the western chain of islands in the Solomons (see Fig. 15). Specimens examined by me agree well in external dimensions and color with specimens described by Andersen (1912:712) and Troughton (1931:206-207).
De Beaufort (1951:113) considered bats of "less zoogeographical importance" than other mammals because the ocean is not an "absolute barrier to their dispersal." Volant animals are ecologically terrestrial and therefore are more nearly earthbound than De Beaufort's remarks would suggest (see Miller, 1966:10). Indeed, many kinds of volant animals are endemic to the Solomons. Birds, for example, are well adapted for flight but pose some of the most complex zoogeographic problems in the area of New Guinea and the Solomon Islands (Mayr, 1940:198; 1942:81-83; Koopman, [826] 1957). Rapid speciation can take place in any situation where there is a high degree of isolation (Wright, 1931; Lack, 1947). In fact, isolation is a most important factor in speciation of insular populations (Baker, 1951:55). The one genus, nine species, and 19 subspecies of megachiropterans that are endemic to the Solomons (Table 4) obviously indicate that bats, although volant, can be restricted to one or more islands long enough for new taxa to evolve.
Table 4. A Summary of the Kinds of Megachiropteran Bats in the Solomon Islands and Their Affinities with Faunas of Adjacent Islands.
| Totals | Endemic to Solomons | Common only to Solomons and Bismarcks | Common to Solomons, Bismarcks, and New Guinea | Common to Solomons, New Hebrides, and New Caledonia | |
| Genera | 7 | 1 | 0 | 6 | 0 |
| Species | 16 | 9 | 1 | 6 | 1 |
| Subspecies | 20 | 19 | 0 | 0 | 1 |
The megachiropteran bats of the Solomons have their affinities with the fauna of New Guinea (Table 4); the Solomons and New Guinea have six genera and six species in common. Because the two areas never have been connected (via the Bismarck Archipelago) by dry land, bats probably have reached the Solomons by flying from island to island (see Durham, 1963:357, 359, 361, 363). Deignan (1963:266) has dismissed voluntary or involuntary flight as possible explanations for distributions of bats and birds on islands of the Pacific.
The taxonomic level of endemism can be used as an indicator of antiquity (Dobzhansky, 1941; Koopman, 1958:429-430). The one megachiropteran genus (Pteralopex) endemic to the Solomons apparently is an ancient relic. Bats of this monotypic genus occur on Bougainville, Choiseul, Santa Ysabel, and Guadalcanal (see Fig. 4). These four islands probably were contiguous during the maximum lowering of sea level in the Pleistocene (see Durham, 1963:362-363). Bats of the genus Pteralopex are the only kind in the Solomons having a distribution that can be correlated with former land connections between islands.
The distributions of 16 species of megachiropterans known from the Solomons are summarized in Table 5 and in Figure 16. The larger islands (in terms of surface area and elevation) in general [827] have the highest number of species (Guadalcanal 10, Choiseul 9, and Bougainville 8). But Fauro, one of the smallest islands for which data are available, has six species of megachiropterans whereas San Cristobal and Malaita, two of the larger islands, have only three and four species, respectively. Possibly this difference signals the need for additional collecting.
Bougainville and Choiseul, about 60 miles apart, have seven species of megachiropterans in common (Table 5). Fauro, 25 miles southeast of Bougainville and 35 miles west of Choiseul, shares five species with each of these islands (Fig. 16). Pteralopex atrata and Pteropus rayneri occur on Choiseul and on Bougainville, but not on Fauro. Individuals of these species are the largest fruit bats in the Solomons, and their absence on Fauro suggests, therefore, that this small island is ecologically unsuitable, at least in some months, for the support of populations of bats that require relatively large amounts of food. The small size of the island is consistent with this hypothesis, but several other islands as small as Fauro do support populations of the large kinds of Pteropus, at least in some months.
Table 5. A Summary of Distribution of All Species of Megachiropteran Bats Known from the Solomons. Only Islands Well Known Faunistically Are Listed.
| Subspecies | ||||||||||||||||
| R. amplexicaudatus | X | X | X | X | X | X | X | |||||||||
| P. atrata | X | X | X | X | ||||||||||||
| Pt. hypomelanus | X | |||||||||||||||
| Pt. admiralitatum | X | X | X | X | X | |||||||||||
| Pt. tonganus | X | |||||||||||||||
| Pt. howensis | X | |||||||||||||||
| Pt. rayneri | X | X | X | X | X | X | X | X | X | X | ||||||
| Pt. woodfordi | X | X | X | X | ||||||||||||
| Pt. mahaganus | X | X | ||||||||||||||
| D. inermis | X | X | X | X | X | X | X | X | X | X | X | X | ||||
| M. lagochilus | X | X | X | X | X | X | X | |||||||||
| M. woodfordi | X | X | X | X | X | X | ||||||||||
| M. aurantius | X | X | ||||||||||||||
| N. albiventer | X | X | X | X | X | X | ||||||||||
| N. major | X | X | X | X | ||||||||||||
| N. malaitensis | X | |||||||||||||||
| Totals | 8 | 9 | 6 | 1 | 4 | 4 | 6 | 4 | 3 | 5 | 4 | 10 | 3 | 2 | 3 | 1 |
Santa Ysabel has six species of megachiropterans and 10 occur on Guadalcanal (Table 5). These two islands, separated by about 100 [828] miles of water, share five species (Rousettus amplexicaudatus, Pteralopex atrata, Pteropus rayneri, Dobsonia inermis, and Nyctimene albiventer). The Nggela Group, in which Florida is the largest island and the only one from which bats have been collected, is 50 miles southeast of Santa Ysabel and 30 miles north of Guadalcanal (Fig. 16). Four species of megachiropterans are known from Florida (Dobsonia inermis, Macroglossus lagochilus, Melonycteris aurantius, and Nyctimene major). Three of these are known from Guadalcanal and one occurs on Santa Ysabel. This situation resembles the one involving Fauro, Bougainville, and Choiseul because none of the large bats (Pteropus and Pteralopex) is known from Florida, even though two species of large bats that occur on Santa Ysabel to the northwest occur also on Guadalcanal to the south. Possibly Florida and the smaller islands that comprise the Nggela Group are ecologically unsuitable for large bats, or perhaps these small islands can support only limited numbers of individuals during part of a year.
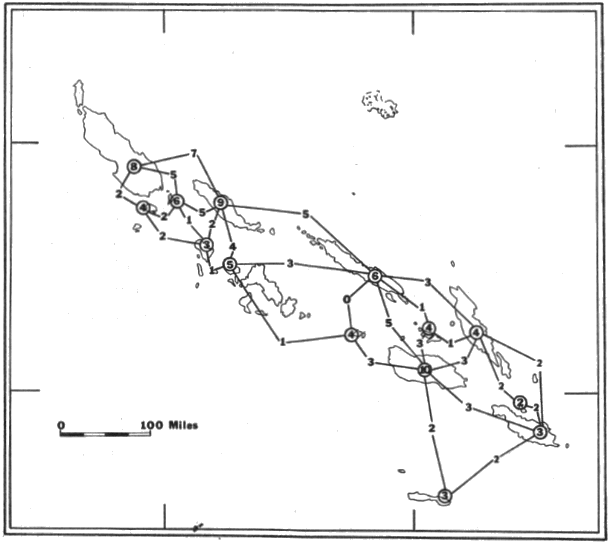
Fig. 16. The number of megachiropteran species known from individual islands (number within a circle) is compared with the number of species common to two different islands (number without a circle). For names of islands see Fig. 2.
Some of the small islands in the Solomons have populations of large fruit bats. For example, Pteropus admiralitatum and P. hypomelanus have been reported from the small islands in the Russell Group (Table 5). Possibly these species do not live concurrently in the Russells; specimens of the two were obtained in different years. Two small megachiropterans, P. woodfordi and Melonycteris woodfordi, also inhabit the Russells. Shortland, a small island about 15 miles south of Bougainville, supports one large bat, P. admiralitatum, as well as smaller megachiropterans.
Kolombangara and Vella Lavella are about the same size and are separated by about 15 miles of water. Rousettus amplexicaudatus, Pteropus rayneri, P. woodfordi, Macroglossus lagochilus, and Nyctimene albiventer have been collected on Kolombangara but only P. admiralitatum, P. rayneri, and Dobsonia inermis have been found on Vella Lavella. The difference in the known megachiropteran faunas is more striking when one compares each island with adjacent islands. Two species on Vella Lavella occur also on Choiseul, which is about 35 miles northeastward, and two species occur also on Shortland, which is 120 miles northwestward (Fig. 16). Four of the five megachiropterans on Kolombangara also have been found on Choiseul, about 50 miles northward (Table 5). Pteropus rayneri is the only megachiropteran known from both Kolombangara and Vella Lavella, even though the islands are separated by only a few miles of water. Inadequate data possibly account for the differences in the megachiropteran fauna, but I suspect that some other factors are involved. Although Vella Lavella and Kolombangara do have one species (P. rayneri) in common, a different subspecies occurs on each island—rubianus on Kolombangara and lavellanus on Vella Lavella (Fig. 17 and Table 6). This indicates that some factor or factors are operating to keep megachiropterans from moving frequently or easily from one island to the other.
Each of several subspecies of species in the genus Pteropus are known from one or two small islands separated by only a few miles from other islands on which different subspecies occur (see Fig. 6). Judging from this kind of distribution, these bats do not move frequently from island to island. Possibly this is because they cannot easily cross water barriers, or are not inclined to do so because food is abundantly available throughout the year on their home [830] island. Because "flying foxes" frequently are seen in flight over water several hundred yards from shore, the first factor probably is unimportant—at least where short distances are involved. It seems most likely that when abundant food is available these bats have no reason to move even moderate distances.
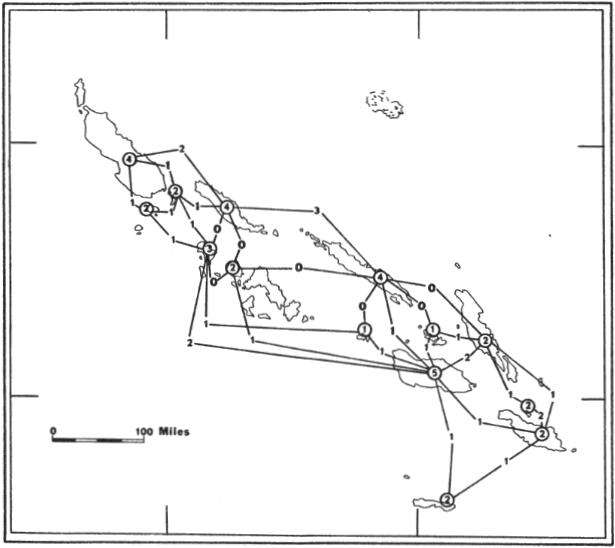
Fig. 17. The number of subspecies of megachiropterans known from individual islands (number within a circle) is compared with the number of subspecies common to different islands (number without a circle). For names of islands see Fig. 2.
Distributions of subspecies of polytypic species are summarized in Table 6 and Figure 17. Generally, more subspecies are known from the larger islands than from the smaller islands (Guadalcanal with 5, Bougainville, Choiseul, and Santa Ysabel with 4, Fauro with 2.) The distributions of some subspecies can be used to judge the differential effectiveness of water gaps between islands. The distribution of Pteropus rayneri lavellanus and P. rayneri rubianus is an example.
Choiseul and Santa Ysabel are separated by about 50 miles of water (see Fig. 17) but have three subspecies in common (Pteropus rayneri grandis, Dobsonia inermis minimus, and Nyctimene albiventer [831] minor.) Choiseul is about 50 miles from Kolombangara and about 35 miles from Vella Lavella, but shares no subspecies with these smaller islands although some species are shared (Tables 5 and 6). From these data one can conclude that exchange of genes between populations on Choiseul and populations on Santa Ysabel is frequent but for some reason exchange of genes between populations on Vella Lavella and Choiseul and Kolombangara and Choiseul is infrequent. A series of small islands (Rob Roy, Wagina, and the Arnavon Islands, not named on the maps) connect Choiseul and Santa Ysabel in stepping-stone fashion (see Fig. 17). Possibly these small islands enhance movement of megachiropterans between Choiseul and Santa Ysabel.
Table 6. A Summary of Distribution of Polytypic Species of Megachiropteran Bats in the Solomon Islands. Only Islands Well Known Faunistically Are Listed.
| Subspecies | ||||||||||||||||
| P. atrata atrata | X | X | ||||||||||||||
| P. atrata anceps | X | X | ||||||||||||||
| Pt. a. solomonis | X | X | X | |||||||||||||
| Pt. a. colonus | X | |||||||||||||||
| Pt. a. grandis | X | |||||||||||||||
| Pt. r. rayneri | X | X | ||||||||||||||
| Pt. r. grandis | X | X | X | |||||||||||||
| Pt. r. rubianus | X | |||||||||||||||
| Pt. r. lavellanus | X | |||||||||||||||
| Pt. r. monoensis | X | |||||||||||||||
| Pt. r. cognatus | X | X | ||||||||||||||
| Pt. r. rennelli | X | |||||||||||||||
| D. i. inermis | X | X | X | X | X | X | X | X | X | X | ||||||
| D. i. minimus | X | X | ||||||||||||||
| N. a. bougainville | X | X | X | |||||||||||||
| N. a. minor | X | X | X | |||||||||||||
| Totals | 4 | 4 | 4 | 1 | 2 | 1 | 2 | 2 | 3 | 2 | 1 | 5 | 2 | 2 | 2 | 1 |
Florida, of the Nggela Group, is approximately halfway between Santa Ysabel and Guadalcanal. Pteralopex atrata anceps occurs on Santa Ysabel and on Guadalcanal but is unknown from Florida. Fauro lies between Bougainville and Choiseul. Pteralopex atrata atrata and Pteropus rayneri grandis occur on Choiseul and on Bougainville but are unknown from Fauro. As suggested earlier, small islands like Fauro and Florida possibly cannot support large fruit bats, although they probably would utilize these small islands when in transit between larger islands.
Fauro apparently is important to the distribution of the two subspecies of Dobsonia inermis and Nyctimene albiventer in the Solomons (see Figs. 9 and 13). In both species, one subspecies is found in the eastern chain of islands and one subspecies is found in the western chain. Specimens of Dobsonia inermis from Fauro and Bougainville can be identified as the subspecies inermis whereas those from Choiseul are assignable to the subspecies minimus. Nyctimene albiventer bougainville occurs on Bougainville but specimens of N. albiventer from Fauro and Choiseul can be identified as the subspecies minor. Although interchange of genes occurs between populations on Bougainville and Fauro in the case of D. inermis, the population of N. albiventer on Fauro is at least partially isolated from the population on Bougainville.
Rennell and Ontong Java are relatively isolated from other islands in the Solomons (see Fig. 17). Only one kind of bat (Pteropus howensis) is known from Ontong Java and apparently is endemic to that atoll. Pteropus tonganus geddiei, one of the megachiropterans that occurs on Rennell (Table 5), also is found in the New Hebrides and on New Caledonia (Table 4). This makes P. t. geddiei the only megachiropteran bat in the Solomons that is more closely related to bats on islands to the southeast of the Solomons than to bats on other islands of the Solomons, the Bismarcks, or New Guinea, to the north and west. The other species of megachiropterans (Dobsonia inermis and Pteropus rayneri) on Rennell are found also on other islands in the Solomons. It is to be noted that Mayr (1931) regarded the avifauna of Rennell as most nearly like that of the New Hebrides and New Caledonia. He suggested that the prevailing winds from the southeast have been important for birds that have reached Rennell. The New Hebrides and New Caledonia are four and a half times farther from Rennell than are San Cristobal and Guadalcanal. On first consideration a person might doubt that the winds would be favorable enough to compensate for the great distance between Rennell and the New Hebrides and New Caledonia. Darlington (1938) has used the formula X n/m to obtain a comparison of barriers of different widths. [X = the probability of an individual crossing a barrier of width m; the probability of an individual crossing a similar barrier of width n is the ratio n/m.] If this formula is applied here, one finds that winds from the southeast (that is, from the New Hebrides and New Caledonia) would have to be more than 100 times more [833] favorable than winds from the northeast (from Guadalcanal and San Cristobal) in order to compensate for the distance of Rennell from the New Hebrides and New Caledonia. Even so, tropical storms with unusually strong winds, frequent during some parts of the year, possibly account for the present distributional pattern of bats and birds that live on Rennell.
Whatever the means by which bats of the species P. tonganus reached Rennell, the fact remains that specimens from Rennell cannot be distinguished from specimens of P. tonganus geddiei from the New Hebrides and New Caledonia, more than 500 miles to the southeast.
Note: An important and interesting paper on zoogeography of bats, which was published too late to be included here, is: Krzanowski, A., 1967, The magnitude of islands and the size of bats (Chiroptera), Acta Zool. Cracoviensia, 12:281-348.
LITERATURE CITED
1944. Gazetteer of Solomon Islands, Bismarck Archipelago, and Islands of the south-eastern end of New Guinea. Hydrographic office of the United States Navy Department, No. 881.
1908. Twenty new forms of Pteropus. Ann. Mag. Nat. Hist., ser. 8, 2:361-370, October.
1909a. On the characters and affinities of "Desmalopex" and Pteralopex. Ann. Mag. Nat. Hist., ser. 8, 3:213-222, February.
1909b. Two new bats from the Solomon Islands. Ann. Mag. Nat. Hist., ser. 8, 3:266-270, March.
1909c. On the fruit-bats of the genus Dobsonia. Ann. Mag. Nat. Hist., ser. 8, 4:528-533, December.
1911. Six new fruit-bats of the genera Macroglossus and Syconycteris. Ann. Mag. Nat. Hist., ser. 8, 7:641-643, June.
1912. Catalogue of the Chiroptera in the collection of the British Museum, British Mus. (Nat. Hist.), London, 1:ci + 1-854, 79 figs.
1960. Osteometric variation and function in bats. Evol., 14:8-17, 3 figs., March 21.
1951. The avifauna of Micronesia, its origin, evolution, and distribution. Univ. Kansas Publ., Mus. Nat. Hist., 3:1-359, 16 figs., June 12.
1951. Zoogeography of the land and inland waters. Sidgwick and Jackson, London, viii + 208 pp., maps.
1962. The mosquitoes of the South Pacific (Diptera, Culicidae). Univ. California Press, Berkeley, 1:xii + 1-608, July 18.
1957. Hair structure as a generic character in bats. Univ. California Publ. Zool., 59:285-548, 9 pls., 4 figs., October 10.
1900. An index to the islands of the Pacific Ocean. Mem. Bernice P. Bishop Mus., Honolulu, 2:1-170, maps.
1961. Melanesia: a short ethnography. British Mus., London, 115 pp., 26 pls., 43 figs., map.
1938. The origin of the fauna of the Greater Antilles, with discussion of dispersal of animals over water and through the air. Quart. Rev. Biol., 13:274-300, 5 figs.
1963. Birds in the tropical Pacific, pp. 263-269, in Pacific Basin biogeography (J. L. Gressitt, ed.). Bernice P. Bishop Mus. Press, Honolulu, xi + 563 pp.
1878. Catalogue of the Chiroptera in the ... British Museum. British Mus., London, xlii + 567 pp., 30 pls.
1941. Genetics and the origin of species. Columbia Univ. Press, New York, 2nd. ed., rev., ix + 466 pp., 24 figs.
1963. Paleogeographic conclusions in light of biological data, pp. 355-365, 4 figs., in Pacific Basin biogeography (J. L. Gressitt, ed.). Bernice P. Bishop Mus. Press, Honolulu, xi + 563 pp.
1966. Checklist of Palaearctic and Indian mammals, 1758-1946. British Mus. (Nat. Hist.), London, 2nd. ed., 810 pp., July.
1964a. Flughunde der Gattung Pteropus von den Neuen Hebriden (Mammalia, Chiroptera). Senck. biol., 45:87-92, 6 figs., May 15.
1964b. Flughunde der Gattung Pteropus von Neukaledonien und den Loyalty-Inseln (Mammalia, Chiroptera). Senck. biol., 45:671-683, December 21.
1946. Mammals of Nevada. Univ. California Press, Berkeley, xi + 710 pp., frontispiece, 11 pls., 485 figs., July 1.
1956. The mammals of Rennell Island. The natural history of Rennell Island, British Solomon Islands, Copenhagen, 1:73-84, November 28.
1962. A little-known fruit-bat from Rennell Island. The natural history of Rennell Island, British Solomon Islands, Copenhagen, 4:7-9, February 15.
1963. A revision of the genus Hipposideros. Bull. British Mus. (Nat. Hist.), 2:1-129, October.
1957. Evolution in the genus Myzomela (Aves: Meliphagidae). The Auk, 74:49-72, 5 figs., January.
1958. Land bridges and ecology in bat distribution on islands off the northern coast of South America. Evol., 12:429-439, 2 figs., December.
1947. Darwin's finches. Cambridge Univ. Press, Cambridge, x + 209 pp., 8 pls., 27 figs.
1954. List of land mammals of New Guinea, Celebes, and adjacent islands. British Mus. (Nat. Hist.), London, 175 pp., June 30.
1945. Three new Pteropus from New Caledonia and the Solomons. Proc. New England Zool. Club, 23:59-69, March 26.
1934. Notes on mosquitoes of the British Solomon Islands. British Solomon Islands Agr. Gaz., 2:16.
1951. The Melanesians: People of the South Pacific. Chicago Nat. Hist. Mus. Press, Chicago, 259 pp., 56 figs., maps, September.
1899. Die Megachiroptera des Berliner Museums für Naturkunde. Druck und verlag von George Reimer, Berlin, viii + 102 pp., 14 figs.
1931. A systematic list of the birds of Rennell Island with descriptions of new species and subspecies. Amer. Mus. Novit., 486:1-29, August 29.
1940. The origin and the history of the bird fauna of Polynesia. Proc. 6th Pacific Sci. Cong., 1939, 4:197-216.
1942. Systematics and the origin of species from the viewpoint of a zoologist. Columbia Univ. Press, New York, xiv + 334 pp., 29 figs.
1966. Animal evolution on islands, pp. 10-16, in The Galapagos (R. I. Bowman, ed.), Univ. California Press, Berkeley.
Miller, G. S.
1907. The families and genera of bats. Bull. U. S. Nat. Mus., 57:xvii + 282, 14 pls., 49 figs., June 29.
1966. A new species of bat of the genus Melonycteris from the Solomon Islands. Jour. Mamm., 47:23-27, 1 fig., March 12.
1967. A new subspecies of horseshoe bat (Hipposideros diadema) from the Solomon Islands. Proc. Biol. Soc. Washington, 80:35-40, 2 figs., March 24.
1953. Über die Fledertiere von Bougainville. Z. Säugetierk., 17:127-137, October 27.
1912. Color standards and color nomenclature. Washington, D. C., iv + 44 pp., 53 pls.
1931. Bats from Polynesia, Melanesia, and Malaysia. Publ. Field Mus. Nat. Hist., Zool. Ser., 18:7-29, February 12.
1947. Bats from the Solomon Islands. Jour. Mamm., 28:387-391, November 19.
1950. Bats from New Caledonia, the Solomon Islands, and New Hebrides. Fieldiana (Zool.), 31:313-338, 6 figs., August 31.
1945. The principles of classification and a classification of mammals. Bull. Amer. Mus. Nat. Hist., 85:xvi + 350, October 5.
1934. An apparently new fruit bat of the Pteropus hypomelanus group from Gower Island, Solomon Islands. Amer. Mus. Novit., 718:1-2, May 4.
1942. Pteropodidae (Chiroptera) of the Archbold Collections. Bull. Amer. Mus. Nat. Hist., 80:331-347, December 31.
1887a. Diagnoses of two new fruit-eating bats from the Solomon Islands. Ann. Mag. Nat. Hist., ser. 5, 60:147, February.
1887b. On the bats collected by Mr. C. M. Woodford in the Solomon Islands. Proc. Zool. Soc. London, 23:320-328, 3 figs., 2 pls., March 15.
1888a. Diagnoses of six new mammals from the Solomon Islands. Ann. Mag. Nat. Hist., ser. 6, 2:155-158, February.
1888b. The mammals of the Solomon Islands, based on the collections made by Mr. C. M. Woodford during his second expedition to the Archipelago. Proc. Zool. Soc. London, 33:470-484, 3 pls., December 4.
1904. Catalogus Mammalium tam viventium quam fossilium. Quinquennale Supplementum, Berlin, vii + 929 pp.
1931. Three new bats of the genera Pteropus Nyctimene, and Chaerephon from Melanesia. Proc. Linn. Soc. New South Wales, 56:204-209, June 15.
1936. The mammalian fauna of Bougainville Island, Solomons Group. Rec. Australian Mus., 14:341-353, April 7.
1931. Evolution in Mendelian populations. Genetics, 16:97-159, 21 figs.
Transmitted August 10, 1967.
Obvious typographical and punctuation errors repaired. The "Key to Pteropus in the Solomon Islands" was moved above the beginning of the listing for Pteropus Brisson. Where figures or tables split paragraphs, they were moved above or below the split.
| Page | Correction | |
| 797 | Liuinuwu → Liuniuwu | |
| 809 | intermis → inermis | |
| 824 | adbiventer → albiventer | |
| 832 | Gaudalcanal → Guadalcanal |
End of the Project Gutenberg EBook of Systematics of Megachiropteran Bats in
the Solomon Islands, by Carleton J. Phillips
*** END OF THIS PROJECT GUTENBERG EBOOK SYSTEMATICS OF MEGACHIROPTERAN BATS ***
***** This file should be named 40112-h.htm or 40112-h.zip *****
This and all associated files of various formats will be found in:
http://www.gutenberg.org/4/0/1/1/40112/
Produced by Chris Curnow, Joseph Cooper, Tom Cosmas and
the Online Distributed Proofreading Team at
http://www.pgdp.net
Updated editions will replace the previous one--the old editions
will be renamed.
Creating the works from public domain print editions means that no
one owns a United States copyright in these works, so the Foundation
(and you!) can copy and distribute it in the United States without
permission and without paying copyright royalties. Special rules,
set forth in the General Terms of Use part of this license, apply to
copying and distributing Project Gutenberg-tm electronic works to
protect the PROJECT GUTENBERG-tm concept and trademark. Project
Gutenberg is a registered trademark, and may not be used if you
charge for the eBooks, unless you receive specific permission. If you
do not charge anything for copies of this eBook, complying with the
rules is very easy. You may use this eBook for nearly any purpose
such as creation of derivative works, reports, performances and
research. They may be modified and printed and given away--you may do
practically ANYTHING with public domain eBooks. Redistribution is
subject to the trademark license, especially commercial
redistribution.
*** START: FULL LICENSE ***
THE FULL PROJECT GUTENBERG LICENSE
PLEASE READ THIS BEFORE YOU DISTRIBUTE OR USE THIS WORK
To protect the Project Gutenberg-tm mission of promoting the free
distribution of electronic works, by using or distributing this work
(or any other work associated in any way with the phrase "Project
Gutenberg"), you agree to comply with all the terms of the Full Project
Gutenberg-tm License available with this file or online at
www.gutenberg.org/license.
Section 1. General Terms of Use and Redistributing Project Gutenberg-tm
electronic works
1.A. By reading or using any part of this Project Gutenberg-tm
electronic work, you indicate that you have read, understand, agree to
and accept all the terms of this license and intellectual property
(trademark/copyright) agreement. If you do not agree to abide by all
the terms of this agreement, you must cease using and return or destroy
all copies of Project Gutenberg-tm electronic works in your possession.
If you paid a fee for obtaining a copy of or access to a Project
Gutenberg-tm electronic work and you do not agree to be bound by the
terms of this agreement, you may obtain a refund from the person or
entity to whom you paid the fee as set forth in paragraph 1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be
used on or associated in any way with an electronic work by people who
agree to be bound by the terms of this agreement. There are a few
things that you can do with most Project Gutenberg-tm electronic works
even without complying with the full terms of this agreement. See
paragraph 1.C below. There are a lot of things you can do with Project
Gutenberg-tm electronic works if you follow the terms of this agreement
and help preserve free future access to Project Gutenberg-tm electronic
works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the Foundation"
or PGLAF), owns a compilation copyright in the collection of Project
Gutenberg-tm electronic works. Nearly all the individual works in the
collection are in the public domain in the United States. If an
individual work is in the public domain in the United States and you are
located in the United States, we do not claim a right to prevent you from
copying, distributing, performing, displaying or creating derivative
works based on the work as long as all references to Project Gutenberg
are removed. Of course, we hope that you will support the Project
Gutenberg-tm mission of promoting free access to electronic works by
freely sharing Project Gutenberg-tm works in compliance with the terms of
this agreement for keeping the Project Gutenberg-tm name associated with
the work. You can easily comply with the terms of this agreement by
keeping this work in the same format with its attached full Project
Gutenberg-tm License when you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern
what you can do with this work. Copyright laws in most countries are in
a constant state of change. If you are outside the United States, check
the laws of your country in addition to the terms of this agreement
before downloading, copying, displaying, performing, distributing or
creating derivative works based on this work or any other Project
Gutenberg-tm work. The Foundation makes no representations concerning
the copyright status of any work in any country outside the United
States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other immediate
access to, the full Project Gutenberg-tm License must appear prominently
whenever any copy of a Project Gutenberg-tm work (any work on which the
phrase "Project Gutenberg" appears, or with which the phrase "Project
Gutenberg" is associated) is accessed, displayed, performed, viewed,
copied or distributed:
This eBook is for the use of anyone anywhere at no cost and with
almost no restrictions whatsoever. You may copy it, give it away or
re-use it under the terms of the Project Gutenberg License included
with this eBook or online at www.gutenberg.org
1.E.2. If an individual Project Gutenberg-tm electronic work is derived
from the public domain (does not contain a notice indicating that it is
posted with permission of the copyright holder), the work can be copied
and distributed to anyone in the United States without paying any fees
or charges. If you are redistributing or providing access to a work
with the phrase "Project Gutenberg" associated with or appearing on the
work, you must comply either with the requirements of paragraphs 1.E.1
through 1.E.7 or obtain permission for the use of the work and the
Project Gutenberg-tm trademark as set forth in paragraphs 1.E.8 or
1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted
with the permission of the copyright holder, your use and distribution
must comply with both paragraphs 1.E.1 through 1.E.7 and any additional
terms imposed by the copyright holder. Additional terms will be linked
to the Project Gutenberg-tm License for all works posted with the
permission of the copyright holder found at the beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm
License terms from this work, or any files containing a part of this
work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this
electronic work, or any part of this electronic work, without
prominently displaying the sentence set forth in paragraph 1.E.1 with
active links or immediate access to the full terms of the Project
Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary,
compressed, marked up, nonproprietary or proprietary form, including any
word processing or hypertext form. However, if you provide access to or
distribute copies of a Project Gutenberg-tm work in a format other than
"Plain Vanilla ASCII" or other format used in the official version
posted on the official Project Gutenberg-tm web site (www.gutenberg.org),
you must, at no additional cost, fee or expense to the user, provide a
copy, a means of exporting a copy, or a means of obtaining a copy upon
request, of the work in its original "Plain Vanilla ASCII" or other
form. Any alternate format must include the full Project Gutenberg-tm
License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying,
performing, copying or distributing any Project Gutenberg-tm works
unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing
access to or distributing Project Gutenberg-tm electronic works provided
that
- You pay a royalty fee of 20% of the gross profits you derive from
the use of Project Gutenberg-tm works calculated using the method
you already use to calculate your applicable taxes. The fee is
owed to the owner of the Project Gutenberg-tm trademark, but he
has agreed to donate royalties under this paragraph to the
Project Gutenberg Literary Archive Foundation. Royalty payments
must be paid within 60 days following each date on which you
prepare (or are legally required to prepare) your periodic tax
returns. Royalty payments should be clearly marked as such and
sent to the Project Gutenberg Literary Archive Foundation at the
address specified in Section 4, "Information about donations to
the Project Gutenberg Literary Archive Foundation."
- You provide a full refund of any money paid by a user who notifies
you in writing (or by e-mail) within 30 days of receipt that s/he
does not agree to the terms of the full Project Gutenberg-tm
License. You must require such a user to return or
destroy all copies of the works possessed in a physical medium
and discontinue all use of and all access to other copies of
Project Gutenberg-tm works.
- You provide, in accordance with paragraph 1.F.3, a full refund of any
money paid for a work or a replacement copy, if a defect in the
electronic work is discovered and reported to you within 90 days
of receipt of the work.
- You comply with all other terms of this agreement for free
distribution of Project Gutenberg-tm works.
1.E.9. If you wish to charge a fee or distribute a Project Gutenberg-tm
electronic work or group of works on different terms than are set
forth in this agreement, you must obtain permission in writing from
both the Project Gutenberg Literary Archive Foundation and Michael
Hart, the owner of the Project Gutenberg-tm trademark. Contact the
Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable
effort to identify, do copyright research on, transcribe and proofread
public domain works in creating the Project Gutenberg-tm
collection. Despite these efforts, Project Gutenberg-tm electronic
works, and the medium on which they may be stored, may contain
"Defects," such as, but not limited to, incomplete, inaccurate or
corrupt data, transcription errors, a copyright or other intellectual
property infringement, a defective or damaged disk or other medium, a
computer virus, or computer codes that damage or cannot be read by
your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right
of Replacement or Refund" described in paragraph 1.F.3, the Project
Gutenberg Literary Archive Foundation, the owner of the Project
Gutenberg-tm trademark, and any other party distributing a Project
Gutenberg-tm electronic work under this agreement, disclaim all
liability to you for damages, costs and expenses, including legal
fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT
LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE
PROVIDED IN PARAGRAPH 1.F.3. YOU AGREE THAT THE FOUNDATION, THE
TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE
LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR
INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH
DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a
defect in this electronic work within 90 days of receiving it, you can
receive a refund of the money (if any) you paid for it by sending a
written explanation to the person you received the work from. If you
received the work on a physical medium, you must return the medium with
your written explanation. The person or entity that provided you with
the defective work may elect to provide a replacement copy in lieu of a
refund. If you received the work electronically, the person or entity
providing it to you may choose to give you a second opportunity to
receive the work electronically in lieu of a refund. If the second copy
is also defective, you may demand a refund in writing without further
opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth
in paragraph 1.F.3, this work is provided to you 'AS-IS', WITH NO OTHER
WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT LIMITED TO
WARRANTIES OF MERCHANTABILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied
warranties or the exclusion or limitation of certain types of damages.
If any disclaimer or limitation set forth in this agreement violates the
law of the state applicable to this agreement, the agreement shall be
interpreted to make the maximum disclaimer or limitation permitted by
the applicable state law. The invalidity or unenforceability of any
provision of this agreement shall not void the remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the
trademark owner, any agent or employee of the Foundation, anyone
providing copies of Project Gutenberg-tm electronic works in accordance
with this agreement, and any volunteers associated with the production,
promotion and distribution of Project Gutenberg-tm electronic works,
harmless from all liability, costs and expenses, including legal fees,
that arise directly or indirectly from any of the following which you do
or cause to occur: (a) distribution of this or any Project Gutenberg-tm
work, (b) alteration, modification, or additions or deletions to any
Project Gutenberg-tm work, and (c) any Defect you cause.
Section 2. Information about the Mission of Project Gutenberg-tm
Project Gutenberg-tm is synonymous with the free distribution of
electronic works in formats readable by the widest variety of computers
including obsolete, old, middle-aged and new computers. It exists
because of the efforts of hundreds of volunteers and donations from
people in all walks of life.
Volunteers and financial support to provide volunteers with the
assistance they need are critical to reaching Project Gutenberg-tm's
goals and ensuring that the Project Gutenberg-tm collection will
remain freely available for generations to come. In 2001, the Project
Gutenberg Literary Archive Foundation was created to provide a secure
and permanent future for Project Gutenberg-tm and future generations.
To learn more about the Project Gutenberg Literary Archive Foundation
and how your efforts and donations can help, see Sections 3 and 4
and the Foundation information page at www.gutenberg.org
Section 3. Information about the Project Gutenberg Literary Archive
Foundation
The Project Gutenberg Literary Archive Foundation is a non profit
501(c)(3) educational corporation organized under the laws of the
state of Mississippi and granted tax exempt status by the Internal
Revenue Service. The Foundation's EIN or federal tax identification
number is 64-6221541. Contributions to the Project Gutenberg
Literary Archive Foundation are tax deductible to the full extent
permitted by U.S. federal laws and your state's laws.
The Foundation's principal office is located at 4557 Melan Dr. S.
Fairbanks, AK, 99712., but its volunteers and employees are scattered
throughout numerous locations. Its business office is located at 809
North 1500 West, Salt Lake City, UT 84116, (801) 596-1887. Email
contact links and up to date contact information can be found at the
Foundation's web site and official page at www.gutenberg.org/contact
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
gbnewby@pglaf.org
Section 4. Information about Donations to the Project Gutenberg
Literary Archive Foundation
Project Gutenberg-tm depends upon and cannot survive without wide
spread public support and donations to carry out its mission of
increasing the number of public domain and licensed works that can be
freely distributed in machine readable form accessible by the widest
array of equipment including outdated equipment. Many small donations
($1 to $5,000) are particularly important to maintaining tax exempt
status with the IRS.
The Foundation is committed to complying with the laws regulating
charities and charitable donations in all 50 states of the United
States. Compliance requirements are not uniform and it takes a
considerable effort, much paperwork and many fees to meet and keep up
with these requirements. We do not solicit donations in locations
where we have not received written confirmation of compliance. To
SEND DONATIONS or determine the status of compliance for any
particular state visit www.gutenberg.org/donate
While we cannot and do not solicit contributions from states where we
have not met the solicitation requirements, we know of no prohibition
against accepting unsolicited donations from donors in such states who
approach us with offers to donate.
International donations are gratefully accepted, but we cannot make
any statements concerning tax treatment of donations received from
outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation
methods and addresses. Donations are accepted in a number of other
ways including checks, online payments and credit card donations.
To donate, please visit: www.gutenberg.org/donate
Section 5. General Information About Project Gutenberg-tm electronic
works.
Professor Michael S. Hart was the originator of the Project Gutenberg-tm
concept of a library of electronic works that could be freely shared
with anyone. For forty years, he produced and distributed Project
Gutenberg-tm eBooks with only a loose network of volunteer support.
Project Gutenberg-tm eBooks are often created from several printed
editions, all of which are confirmed as Public Domain in the U.S.
unless a copyright notice is included. Thus, we do not necessarily
keep eBooks in compliance with any particular paper edition.
Most people start at our Web site which has the main PG search facility:
www.gutenberg.org
This Web site includes information about Project Gutenberg-tm,
including how to make donations to the Project Gutenberg Literary
Archive Foundation, how to help produce our new eBooks, and how to
subscribe to our email newsletter to hear about new eBooks.