University of Kansas Publications
Museum of Natural History
Volume 13, No. 10, pp. 429-611, pls. 31-54, 24 figs.
![]() February 16, 1962
February 16, 1962
![]()
North American Recent Soft-shelled Turtles
(Family Trionychidae)
BY
ROBERT G. WEBB
University of Kansas
Lawrence
1962
University of Kansas Publications, Museum of Natural History
Editors: E. Raymond Hall, Chairman, Henry S. Fitch,
Robert W. Wilson
Volume 13, No. 10, pp. 429-611, pls. 31-54, 24 figs.
Published February 16, 1962
University of Kansas
Lawrence, Kansas
PRINTED IN
THE STATE PRINTING PLANT
TOPEKA, KANSAS
1962
![]()
28-7818
North American Recent Soft-shelled Turtles
(Family Trionychidae)
BY
ROBERT G. WEBB
| PAGE | |
| Contents | 431 |
| Introduction | 433 |
| Collecting Methods | 434 |
| Materials and Procedure | 437 |
| Acknowledgments | 439 |
| Taxonomy | 439 |
| Family Trionychidae Bell, 1828 | 439 |
| Genus Trionyx Geoffroy, 1809 | 443 |
| Variation | 445 |
| Secondary Sexual Variation | 446 |
| Ontogenetic Variation | 449 |
| Geographic Variation | 453 |
| Character Analysis | 460 |
| Composition of the Genus Trionyx in North America | 476 |
| Artificial Key to North American Species and Subspecies of the Genus Trionyx | 476 |
| Systematic Account of Species and Subspecies | 479 |
| Trionyx ferox | 479 |
| Trionyx spinifer | 486 |
| Trionyx spinifer spinifer | 489 |
| Trionyx spinifer hartwegi | 497 |
| Trionyx spinifer asper | 502 |
| Trionyx spinifer emoryi | 510 |
| Trionyx spinifer guadalupensis | 517 |
| Trionyx spinifer pallidus | 522 |
| Trionyx ater | 528 |
| Trionyx muticus | 531 |
| Trionyx muticus muticus | 534 |
| Trionyx muticus calvatus | 539 |
| Natural History | 541 |
| Habitat | 541 |
| Daily and Seasonal Activity | 547 |
| Diurnal Habits | 547 |
| Behavior Adaptations | 549 |
| Movement | 552 [432] |
| Nocturnal Habits | 553 |
| Seasonal Occurrence | 553 |
| Food Habits | 555 |
| Reproduction | 558 |
| Size of Males at Sexual Maturity | 558 |
| Size of Females at Sexual Maturity | 560 |
| Sexual Activity | 563 |
| Deposition of Eggs | 565 |
| Reproductive Potential | 568 |
| Eggs | 572 |
| Incubation and Hatching | 573 |
| Age and Growth | 574 |
| Mortality | 576 |
| Parasites | 576 |
| Economic Importance | 577 |
| Evolutionary History | 578 |
| Distribution | 578 |
| Relationships | 579 |
| Fossils | 582 |
| Phylogeny | 585 |
| The Importance of the Study of Turtle Populations in Relation to the History of River Systems | 588 |
| Summary | 590 |
| Literature Cited | 594 |
Is it true that the greater the degree of resemblance between two populations the shorter the time the two have been spatially isolated? Are aquatic environments more stable than terrestrial environments? These questions occurred to me while I was collecting turtles from river systems of the Gulf Coast. As a general rule, each kind of turtle seemed to occur throughout one continuous river system or large tributary, and with no barriers to dispersal therein and with the lapse of enough time for a population to reach its limits of dispersal, the question arose, "Where do subspecies and zones of intergradation occur?" It seemed logical to think that each isolated and continuous aquatic environment would not contain more than one subspecies of the same species. In terrestrial environments subspecies and transitions between them were recognizable. Terrestrial habitats were continuous for longer distances than the isolated, aquatic habitats. But, different species of turtles prefer different kinds of aquatic habitats. Also, barriers occur in large drainage systems, such as the Mississippi, where, in general, the western tributaries are sluggish, turbid and shallow, and the eastern tributaries are fast-flowing, clear and deep. But in young, relatively small, river systems that do not traverse radically different physiographic regions, and that show no gross ecological differences, habitats or microhabitats that do exist probably are only partial barriers and seem not to prevent the dispersal of most kinds of aquatic turtles. Consequently, it seemed that study of the degree of difference between closely related populations of turtles that occurred in one drainage system, or in adjacent drainage systems would indicate the length of time, respectively, that the drainage system had been continuous or the length of time that two or more systems had been isolated from one another.
Rivers or series of river systems having endemic kinds of turtles or having the most kinds of turtles that are different from those in adjacent rivers may be the oldest geologically, or may have been isolated the longest. Knowledge of the kinds of turtles and their relationships and distribution could indicate chronological changes in aquatic habitats. Of course, modifying factors such as differences between populations of turtles in rates of evolutionary change, degrees of vagility, rates of dispersal, and overland migrations need to be taken into account.
My accumulation of data on soft-shelled turtles was begun in [434] the early nineteen-fifties. Although American softshells have been discussed in a revisionary manner by Agassiz (1857), Siebenrock (1924), Stejneger (1944) and Neill (1951), the relationships of all the component populations have not hitherto been appreciated. The present account attempts to combine in one publication what is known concerning the taxonomy, geographic distribution, life history, and relationships of the Recent American species and subspecies of the genus Trionyx.
Collecting Methods
Nocturnal collecting, by hand, from a boat that was nosed among brush piles along the shore line of rivers (Chaney and Smith, 1950:323) in the early 1950's on rivers of the Gulf Coast drainage east of Texas yielded many turtles of the genus Graptemys but few softshells. Chaney and Smith (loc. cit.) reported only one softshell among 336 turtles taken in 21 collecting hours on July 5, 6 and 7 on the Sabine River; Cagle and Chaney (1950:385), however, recorded 11.6 per cent softshells of 208 turtles (collecting time not stated) taken on the Caddo Lake Spillway in Louisiana. Using hoop-nets is probably the most efficient method for collecting softshells considering the time and effort involved, and is the chief method I have used. Lagler (1943a:24) mentioned the use of watermelon rind as an effective bait. Kenneth Shain (field notes) trapped T. spinifer emoryi in hoop-nets baited with bread. I have used chopped fresh fish with most success; canned sardines have also been satisfactory. These baits seem to be more successful for trapping spinifer than they are for muticus. Hoop-nets were used to trap turtles in Lake Texoma, Oklahoma, from June 14 to July 2, 1954. The number of traps (usually four, rarely five) and trapping success varied with location. Of 156 turtles, 19 (12%) were T. spinifer and one was T. muticus.
Trotlines and set lines frequently catch softshells; sport fishermen often complain of catching these turtles on hook and line. Live worms, soft-bodied insects, small crawfish, minnows, small pieces of fish and other kinds of meat are adequate bait. Capture depends on the skill of attachment of the bait and the size of hook used. In my experience, softshells (mostly spinifer) were taken on trotlines that were set in lakes or the slower-moving parts of rivers a few inches below the surface. I have records of only two muticus taken on trotlines. Goin (1948:304) stated that commercial fishermen catch softshells on trotlines set for catfish on the bottom of river beds. Evermann and Clark (1920:595) found softshells to be caught more often than any other kind of turtle [435] in traps, on set lines, and by anglers in Lake Maxinkuckee, Indiana. Some residents of the South tell of so placing baits that turtles are lured to tread water against an object set with recurved hooks upon which the webbing of the forelimbs are impaled.
Individuals of muticus and spinifer frequently bury themselves in sand in shallow water and can be collected by hand by noting swirls or disturbances on the bottom caused by a turtle withdrawing its head (Conant, 1951:156, 159). Professional turtle collectors take them by "noodeling" (Conant, op. cit.:160); Lagler (1943a:22) elaborated on the method of "noodling." P. W. Smith (1947:39) remarked that 20 or more softshells were taken "within a few hours by probing sand bars at the water edge" near Charleston, Illinois. From a distance I observed an individual of T. s. asper bury itself in shallow water on the Escambia River, Florida. Small individuals of muticus have been taken by hand along the shore of Lake Texoma. Along the Flint River near Bainbridge, Georgia, two hatchlings that were buried in sand in shallow water emerged at my approach and scurried a few inches, then buried themselves again. Larger turtles seem to be more wary. One that was disturbed, emerged from the sand and swam toward deep water.
In clear water, water-goggling may be effective in securing softshells. Marchand (in Carr, 1952:417-18) mentioned that softshells (ferox) can be found buried in deep water with only the heads visible; the turtles are not easily frightened under water and may be captured by grasping their necks. A similar technique described by Allen and Neill (1950:3) resulted in the capture of trionychid turtles. In clear water of the White River, Arkansas, I collected a few softshells by hand as they lay on the bottom.
In shallow-water areas of large rivers, lakes and tributaries, seining often procures softshells. Methods used in fisheries investigations such as the application of rotenone and electric shockers, and even dynamiting, sometimes yield soft-shelled turtles. Carr (1952:419) wrote that numbers of ferox were incapacitated by rotenone in Florida lakes, although no other species of turtle was affected. I captured a snapping turtle (Chelydra serpentina) that was immobilized by the current from an electric shocker in a small, alga-choked tributary of Cache Creek, Comanche County, Oklahoma; presumably turtles must come in close contact with the electrodes to be affected (see discussion by Gunning and Lewis, 1957:52).
The effectiveness of gill nets in trapping turtles is indicated by information kindly supplied by Mr. Alfred Houser on gill-net operations [436] from July through December, 1952, under the direction of Mr. "Bud" Oldham, a commercial fisherman. The 4-inch mesh nets were in Lake Texoma at the mouth of Briar Creek, two miles south of Powell, Marshall County, Oklahoma, in 25 to 30 feet of water. Eighty to 90 per cent of the turtles secured were softshells; more were taken near shoreline than away from shore even though the depth was about the same. An average of only one turtle every four days was taken in July and August when the turtles presumably are most active (Table 1). One gill-net day is equivalent to one gill net, 200 yards long, operated for 24 hours.
Table 1. The Abundance of Turtles as Revealed by Gill-net Operations in Lake Texoma, 1952.
| Month | Gill-net days | Number of turtles | Gill-net days per turtle |
| July | 835 | 213 | 3.9 |
| August | 816 | 199 | 4.6 |
| September | 743 | 42 | 17.7 |
| October | 1661 | 82 | 20.3 |
| November | 1322 | 48 | 27.5 |
| December | 864 | 5 | 172.8 |
Dr. Virgil Dowell, while making fishery studies two miles east of Willis, Marshall County, Oklahoma, caught, on the average, 1.5 turtles per day. Of 75 turtles collected from July 1 through October 18, 1953, 66 were Trionyx (spinifer and muticus), five were Graptemys and four were Pseudemys scripta. No more than two gill nets were used simultaneously. The nets were moved from time to time and varied in dimensions, but those used most of the time were 200 feet long and eight feet deep with a 3-inch mesh.
The few captures by Houser probably resulted from long-continued trapping in one place; the gill nets were not moved in the entire six-month period or for some time previously. Breckenridge (1955:6) commented on the sedentary nature of spinifer (in Minnesota) and quoted a professional turtle trapper as stating that "after a section of a river has been trapped heavily for softshells, little success can be expected in that area for as much as three or four years thereafter." Both Houser's and Dowell's data indicate a higher percentage of soft-shelled turtles collected than any other [437] species. The number caught probably depends, at least partly, on the food habits of the species and is influenced by the enmeshed fish, which, serving as a food source, attract the turtles.
Materials and Procedures
In the course of this study I examined 1849 soft-shelled turtles, including some incomplete alcoholic or dried specimens, such as those represented only by skulls or by other osteological material. Material was examined from each of the collections named below (except KKA), and these are mentioned in the text by the following abbreviations:
| AMNH | American Museum of Natural History |
| ANSP | Academy of Natural Sciences, Philadelphia |
| BCB | Bryce C. Brown, private collection, Baylor University |
| CM | Carnegie Museum |
| CNHM | Chicago Natural History Museum |
| INHS | Illinois Natural History Survey, University of Illinois |
| KKA | Kraig K. Adler, private collection, data in letter dated January 8, 1960 |
| KU | Museum of Natural History, The University of Kansas |
| LSU | Louisiana State University |
| MCZ | Museum of Comparative Zoology, Harvard College |
| MSU | The Museum, Michigan State University |
| NHB | Naturhistorisches Museum Basel, Switzerland |
| OU | University of Oklahoma Museum, Division of Zoology |
| SM | Strecker Museum, Baylor University |
| TCWC | Texas Cooperative Wildlife Collection, Texas Agricultural and Mechanical College |
| TNHC | Texas Natural History Collection, The University of Texas |
| TTC | Texas Technological College |
| TU | Tulane University |
| UA | University of Alabama |
| UI | Museum of Natural History, The University of Illinois |
| UMMZ | Museum of Zoology, The University of Michigan |
| USNM | United States National Museum |
| WEB | William E. Brode, private collection, Mississippi Southern College |
| WTN | Wilfred T. Neill, private collection |
External measurements (listed under the section, "Variation") were taken by the writer by means of a Vernier caliper or a steel tape. Measurements of the skulls are in millimeters and tenths as taken by the writer with dial calipers. Partial wrinkling of the carapace at the edges of some specimens causes some error in measurements; consequently, length of plastron is used as the measurement of reference.
Scattergrams based on external measurements were constructed. Some demonstrate considerable ontogenetic variation. An inspection of the scattergrams indicated regressions essentially linear in nature, but sometimes occasioned an arbitrary separation of samples into size groups to show ontogenetic variation; no secondary sexual differences could be discerned. Several ratios were developed from the measurements. The data correspond to the regression [438] model 1A in "Statistical Methods" (Snedecor, 1956, sec. 6.13); consequently, the sample ratios indicate the slope of regression and are useful in comparisons. Sample-means and their estimated standard errors are compared graphically to show general trends in proportional characters. Comparisons of means and standard errors indicate statistical significance between populations if the sample-means plus or minus twice their standard errors do not overlap, but this method of comparison is valid only when comparing two samples (Pimentel, 1959:100).
In the section on "Variation," general features applicable to all kinds of soft-shelled turtles are discussed under the following headings: secondary sexual, ontogenetic, and geographic; individual variation is mentioned in accounts of species and subspecies. In the section "Character Analysis" external and osteological characters having taxonomic significance are discussed.
Vernacular names follow, as closely as possible, those recommended by the Committee on Herpetological Common Names (1956). The synonymy of each monotypic species or subspecies begins with the name as given in the original description. The second entry is the name-combination herein applied to the taxon. Other entries are first usages, in chronological order, of other names (synonyms) that have been applied to the taxon in question. Next, the type is briefly discussed followed by the "Range" defined in general geographic terms, and, when appropriate, in terms of river drainage systems. "Diagnosis" includes a combination of characters that facilitates quick identification. In polytypic species, the diagnosis of a subspecies is designed only to distinguish it from other subspecies of that species. The comments included under the subsection entitled "Description" pertain to individuals from an area where the taxon is most clearly differentiated. Because osteological characters are significant only at the specific level, they appear under the accounts of each species (excluding ater). Proportional characters as given in the "Diagnosis" are only in general terms; more specific data are set forth in the subsection, "Description" or in the various text figures, mostly in the section on "Variation," page 445. Proportions pertaining to the species muticus were derived only from the nominal subspecies, and appear under the account of the species. A subsection "Variation" under the accounts of some subspecies includes information concerning principally individual variation and coloration; because color is not considered to be of major taxonomic importance, color terms are used without reference to any standard color guide. The subsection "Remarks" follows the section on "Comparisons," and may include comments on nomenclature, intergradation and other information related to the distribution or taxonomy of the subspecies.
The probable geographic range of each species and subspecies is shown on one of the maps. Locality records of specimens that I have examined are shown by solid circles. Additional records of occurrence (published records or specimens otherwise not seen) are shown by hollow circles. Localities only a short distance apart share the same circle.
Under the subsection "Specimens examined," a number in parentheses following a museum number indicates the number of specimens referable to that museum number. All localities of specimens examined are indicated on one of the maps. The list of specimens is arranged alphabetically by states (Canadian provinces precede states of the United States under the account of T. spinifer spinifer, and Mexican states follow those of the United States [439] under T. s. emoryi), alphabetically by counties, and within a county alphabetically by abbreviations of museums; then, museum catalogue numbers are arranged consecutively. Records in the literature are not included if they refer to the same locality from which at least one specimen has been examined, or refer to a less restricted locality that includes the area from which at least one specimen has been examined. Localities within a county are arranged alphabetically by author; the appropriate reference may follow several localities.
All generic, specific and subspecific names (but not all the different kinds of name-combinations) that have been applied to American soft-shelled turtles are listed in a subsection entitled "Synonymy" under the heading "Genus Trionyx Geoffroy, 1809."
Acknowledgments
Completion of this study has been made possible only by the co-operation of those persons in charge of the collections listed above and I am grateful to them for the privilege of examining specimens. Also I wish to thank Dr. E. Raymond Hall for the facilities afforded by the Museum of Natural History at the University of Kansas, as well as for editorial assistance in the preparation of the manuscript, and especially Dr. Henry S. Fitch under whose guidance this research was carried out.
In addition to various staff members, graduate students, and individuals whose help is acknowledged at appropriate places in the text, Dr. Rollin H. Baker, Dr. Fred R. Cagle, Mr. J. Keever Greer, Dr. A. Byron Leonard, Dr. Carl D. Riggs, and Dr. Edward H. Taylor deserve especial mention for aid extended in the course of this study. I am indebted to Mr. J. C. Battersby, British Museum (Natural History), London, for information concerning the type of Trionyx ferox, to Dr. Jean Guibé, Museum d'Histoire Naturelle, Paris, for information concerning the types of Trionyx muticus, T. spinifer and T. carinatus, and photographs of the types of T. muticus, T. spinifer and T. ocellatus, and to Dr. Lothar Forcart of the Naturhistorisches Museum, Basel, Switzerland, for information pertaining to a published record of T. muticus.
The maps and figures are the work of Miss Lucy Jean Remple and Mrs. Lorna Cordonnier, University of Kansas. Dr. John M. Legler, University of Utah, prepared most of the photographs on plates 1-20; photographs as mentioned in the preceding paragraph were received from Dr. Guibé, one was provided through the co-operation of Roger Conant and Isabelle Hunt Conant, another was furnished by Mr. J. Keever Greer, and the others were taken by me. Field work was financed in part by funds provided by the Sigma Xi-RESA Research Fund.
Family Trionychidae Bell, 1828
Recent soft-shelled turtles comprise a well-defined assemblage of the family Trionychidae. Although the scope of this study does not involve an assay of the relationships of the soft-shelled turtles of the Old World, a brief résumé that includes some of the salient characteristics of the family is included.
Diagnosis.—Articulation between last cervical and first dorsal vertebrae by zygopophyses only; preplastra separated from hyoplastra by ʌ-shaped epiplastron, [440] entoplastron absent (Williams and McDowell, 1952:263-75); marginal bones absent or forming an incomplete series, not connected with ribs that extend beyond pleural plates; claws on only three inner digits; fourth digit having four or more phalanges; plastron united to carapace by ligamentous tissue (Smith, 1931:147).
General characters.—Size large, "… some attaining probably 5 feet in length of carapace" (Boulenger, 1890:10); body depressed; carapace and plastron lacking horny epidermal shields, covered instead with soft skin; snout ending in fleshy, tubate proboscis; jaws concealed by fleshy lips; tail short; digits well-webbed; cervical vertebrae opisthocoelous (eighth having double articulation in front); neck elongate, cervical region equaling or exceeding length of dorsal vertebral column; head and neck completely retractile, bending by means of sigmoid curve in vertical plane; ear hidden; skull elongate, having three posterior projections (median one produced by supraoccipital and two lateral projections formed chiefly by squamosals); temporal region emarginate posteriorly, forming wide shallow fossa; premaxillae fused; an intermaxillary foramen; pterygoids separated by basisphenoid that contacts palatines; vomer, if present, not separating palatines; pelvis not fused to carapace and plastron; plastron reduced, a median vacuity usually present; plastral bones developing sculpturing with increase in size, forming four to seven so-called plastral callosities; carapace with or without prenuchal bone; nuchal overlapping or overlapped by first pleural; neurals in continuous series or interrupted by pleurals; bony plates of carapace sculptured; mandible having well-developed coronoid bone; cutaneous femoral valves that conceal hind limbs present or absent; two or three pairs of scent glands; cloacal bursae absent (Smith and James, 1958:89); forelimbs having antebrachial scalation; body of hyoid apparatus formed of two or three pairs of bones; penis broad, expanded and pentifid, sulcus spermaticus quadrifid having branches in each of four lateral projections (Hoffman, 1890:298, pl. 47, fig. 2); aquatic, principally in fresh water; mainly carnivorous; flesh of many species eaten. (See Boulenger, 1889:237-41; Loveridge and Williams, 1957:412; Romer, 1956:513; Smith, op. cit.:147-54).
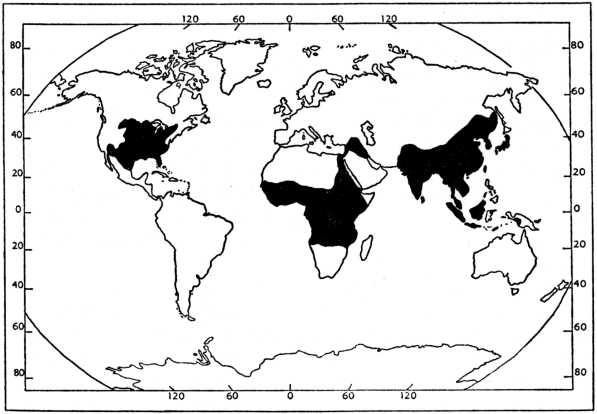
Recent distribution (Figure 1).—North America, from extreme southeastern Canada and eastern United States west to Rocky Mountains and south to northern México; introduced in southwestern United States (Conant, 1958:69-73). Africa, from Egypt and Senegal south to Angola and Zambesi River drainage (Loveridge and Williams, op. cit.:412-68); occurrence of Trionyx triunguis in Syria (Boulenger, op. cit.:255) and coastal streams of Palestine (Schmidt and Inger, 1957:36) considered accidental by Flower (1933:753-54). Southwestern Asia (Tigris and Euphrates River drainage) in eastern Turkey, Syria, Iraq and northeastern Israel (Mertens and Wermuth, 1955:388). Southeastern Asia, from Pakistan and India (Indus River drainage) and Manchuria and adjacent Siberia (Amur River drainage) to Ceylon, Japan, Formosa, Hainan, Luzon, Sumatra, Java, Borneo, Timor and southeastern New Guinea (De Rooij, 1915:325-32; Okada, 1938:108; Pope, 1935:60-64; Smith, 1931:158-79; Stejneger, 1907:514-532; Taylor, 1920:141).
Trionyx cartilagineus is questionably recorded from the Moluccas (De Rooij, op. cit.:330). T. sinensis has been introduced on Kauai Island, Hawaiian Islands (Brock, 1947:142; Oliver and Shaw, 1953:83), one of the Bonin Islands (Okada, 1930:187-94), and probably Timor (De Rooij, op. cit.:331). All insular records east of Borneo and Java are probably the result of introductions, [441] except perhaps those of Pelochelys on Luzon and New Guinea (Darlington, 1957:210).
Recent genera.—According to Mertens and Wermuth (1955:387-95), there are 21 species belonging to six genera as follows:
Chitra Gray, 1844 (1)
Cyclanorbis Gray, 1854 (2)
Cycloderma Peters, 1854 (2)
Lissemys Smith, 1931 (1)
Pelochelys Gray, 1864 (1)
Trionyx Geoffroy, 1809 (14)
Dogania is considered a synonym of Trionyx (Loveridge and Williams, op. cit.:422).
Geologic range.—Lower Cretaceous (possibly Upper Jurassic) to Recent of Asia; Upper Cretaceous to Recent of North America; Paleocene (Upper Jurassic, assuming Trionyx primoevus is a trionychid) to Pleistocene of Europe; Lower Miocene to Recent of Africa; Pleistocene to Recent in East Indies (Loveridge and Williams, op. cit.:412; Romer, 1945:594); questionable trionychid fragments from Pleistocene of Australia (Darlington, loc. cit.).
Remarks.—The genera Lissemys, Cyclanorbis and Cycloderma are distinguished from Pelochelys, Chitra and Trionyx by several characters (Loveridge and Williams, op. cit.:414). The recognition of two groups of genera caused Deraniyagala (1939:290) to erect two families, Cyclanorbidae and Trionychidae. An appraisal of fossils prompted Hummel (1929:768) to propose two corresponding subfamilies, Cyclanorbinae and Trionychinae. Williams (1950:554) considered the two groups as subfamilies (Lissemydinae and Trionychinae).
Baur (1887:97) regarded the Trionychidae as constituting a separate suborder distinct from the rest of the living turtles. Later (1891), however, he pointed out the resemblances of the Trionychidae and Carettochelyidae (having one living genus in New Guinea), and the cryptodiran affinities of Carettochelys. Bergounioux (1932:1408) mentioned the close resemblance of the Carettochelyidae to Trionyx but considered the former as having pleurodiran affinities, a view adopted by Deraniyagala (loc. cit.). Most students now consider the two families to be closely related, and conceive of both as members of the suborder Cryptodira (Hummel, 1929:768; Williams, loc. cit.; Mertens and Wermuth, 1955).
The oldest trionychid fossil, Trionyx primoevus, is from marine deposits of the Upper Jurassic (Kiméridgien) from "Cap de la Hève," and its characters do not indicate the kind of cryptodiran ancestor from which the family arose (Bergounioux, op. cit.:1409; 1937:188). Lane (1910:350) found that the entoplastron (= epiplastron) was paired in embryos of Trionyx and regarded that genus as the most primitive of the order; he also mentioned Wiedersheim's report of rudiments of teeth in embryos of Trionyx. Baur (1891:637-38) thought that the family arose directly from the Amphichelydia, that the ancestors of the Trionychidae closely resembled Carettochelys in the structure of the carapace and plastron, and that a progressive reduction in ossification of those structures occurred. Nopcsa (1926:654) also wrote that the family originated from ancestors having a well-developed plastron; he maintained that the progressive reduction in ossification of the plastron was a specialization for aquatic life, and that the more primitive trionychids had the best developed bones and callosities. Hummel (1929:772) also thought that there had been a progressive reduction in ossification. Bergounioux (1932:1408; 1936:1088, 1952:2304), on the contrary, thought that there had been a progressive increase in ossification of the marginal bones in both families as well as of the plastron (1936:1088; 1937:190). Zangerl's study of the shell elements of turtles (1939:393) indicated that Trionyx was highly specialized in having a well-developed epithecal armor (sculptured callosities, neurals and costals), and that it occurred in most aquatic turtles; the development in soft-shells suggested that members of the family had maintained an aquatic mode of life over a long period of geologic time, a view supported by Deraniyagala (1930:1066). Of interest are Stunkard's remarks (1930:214-18) concerning several Trionyx spinifer that were obtained from a commercial supply house and found to be infested with [443] pronocephalid trematodes (Opisthoporus [= Teloporia] aspidonectes). The closest relatives of that trematode (also recorded from T. ferox) live in marine turtles. Possibly, a Mesozoic ancestor of marine and essentially fresh-water soft-shelled turtles harboured ancestors of these trematodes, but possibly the parasites may have transferred relatively recently to their present hosts. Bergounioux (1937:190) judged the Trionychidae to be an ancient group of marine origin. Hummel (1929:770) wrote that the Trionychidae originated in east Asia (the region of most differentiation) in humid climates.
Baur (1891:634, 637) pointed out that the dorsal aspect of the skull of the closely related Carettochelys resembles the skull of the Dermatemydidae, Staurotypidae and Kinosternidae; the close relationship of Carettochelys and the Dermatemydidae is also mentioned by Bergounioux (1952:2304) and Hummel (1929:769). Hummel (op. cit.:771) thought that the Carettochelyidae and "die Chelydroiden" had a common ancestor, and that (op. cit.:772) the origin of the Trionychidae was older than those two groups. Dunn (1931:109) wrote that the Kinosternidae, Carettochelyidae and Dermatemydidae represented the same general ancestry. Williams (1950:552) has shown the resemblance of the cervical articulations in members of the Chelydridae (including Staurotypinae and Kinosterninae) and the Central American family Dermatemydidae. The consensus of opinion, then, is that the families Trionychidae, Carettochelyidae, Chelydridae and Dermatemydidae are relatively closely related.
Genus Trionyx Geoffroy, 1809
Testudo Linnaeus (in part), Syst. Nat., Ed. 10, 1:197, 1758; type, Testudo graeca Linnaeus by subsequent designation (Fitzinger, 1843:29).
Trionyx Geoffroy, Ann. Mus. Hist. Nat. Paris, 14:1, August, 1809; type, Trionyx aegyptiacus (= Testudo triunguis Forskål) by original designation.
Apalone Rafinesque, Atlan. Jour., Friend of Knowledge, Philadelphia, 1 (No. 2, Art. 12):64, Summer, 1832; type, Apalone hudsonica (= Trionyx spiniferus Lesueur) by monotypy.
Mesodeca Rafinesque, Atlan. Jour., Friend of Knowledge, Philadelphia, 1 (No. 2, Art. 12):64, Summer, 1832; type Mesodeca bartrami (= Testudo ferox Schneider) by monotypy.
Aspidonectes Wagler, Naturl. Syst. Amphib., p. 134, 1830; type, Aspidonectes aegyptiacus Wagler (= Testudo triunguis Forskål) by subsequent designation (Fitzinger, 1843:30).
Amyda Fitzinger, Ann. Wiener Mus. Naturg., 1:110, 120, 127, 1835; type, Amyda subplana Fitzinger by subsequent designation (Fitzinger 1843:30).
Gymnopus Duméril and Bibron, Erpét. Gén., 2:472, 1835; new (substitute) name for Aspidonectes Wagler.
Pelodiscus Fitzinger, Ann. Wiener Mus. Naturg., 1:110, 120, 127, 1835; type, Pelodiscus sinensis Fitzinger by subsequent designation (Fitzinger, 1843:30).
Platypeltis Fitzinger, Ann. Wiener Mus. Naturg., 1:109, 120, 127, 1835; type, Platypeltis ferox by subsequent designation (Fitzinger, 1843:30).
Potamochelys Fitzinger, Syst. Rept., p. 30, 1843; type, Aspidonectes javanicus Wagler (= Testudo cartilaginea Boddaert) by original designation.
Tyrse Gray, Cat. Tort. Croc. Amphis. Brit. Mus., p. 48, 1844; type, Tyrse nilotica Gray (= Testudo triunguis Forskål) by tautonomy (Tyrse, a name for the Nile River).
Callinia Gray, Proc. Zool. Soc. London, p. 222, 1869; new (substitute) name for Aspidonectes of Agassiz (1857:403); type, Callinia spicifera (mispelling for spinifera) Gray by subsequent designation (Stejneger, 1907:514).
Euamyda Stejneger, Bull. Mus. Comp. Zool., 94:7, 9, 12, 1944; new (substitute) name for Amyda mutica of Agassiz (1857:399); type, Amyda mutica Agassiz by monotypy.
Type Species.—Trionyx aegyptiacus (= Testudo triunguis Forskål).
Diagnosis.—Cutaneous femoral valves absent; width of postorbital arch of skull less than diameter of orbit; pterygoids usually not contacting opisthotics; carapace lacking prenuchal bone and marginal ossifications; nuchal bone lacking conspicuous ventral ridges; posterior margin of nuchal overlying first pair of pleurals; lateral parts of nuchal bone overlying second pair of ribs; neurals seven or eight, rarely six or nine; pleurals seven or eight pairs, posterior one or two pairs sometimes in contact medially; distinct suture usually present between hyoplastra and hypoplastra; most laterad prong of posteromedial process of hypoplastra inserted between bifid anterolateral process of xiphiplastra.
Synonomy.—Geoffroy published a synopsis of the species he recognized (1809) prior to his formal description of the genus Trionyx (1809a). Schweigger, nevertheless, probably was the first person to recognize the soft-shelled turtles as a distinct group, and he proposed for it the name Amyda in an unpublished manuscript that he sent to Geoffroy. The latter author (1809a:15) relegated the name Amyda to the synonomy of Trionyx javanicus by means of the following entry: "Amyda javanica. Schweigger, dans un manuscript communique a l'Institut." Stejneger (1944:7) maintained that this publication of Schweigger's monotypic generic name clearly established its availability for the species congeneric with Amyda javanica (= Testudo cartilaginea Boddaert, 1770). Loveridge and Williams (1957:422) contend that this mere mention of the name Amyda neither constitutes the proposal of a new name nor validates it, and that the first valid usage of the name Amyda is that of Fitzinger (1835:120), who later (1843:30) designated the type species as Amyda subplana. The name Amyda cannot date from Oken (1816:348) as Volume 3 [Zoologie] of his Lehrbuch der Naturgeschichte published in 1815-1816 has been placed on the Official Index of Rejected and Invalid Works in Zoological Nomenclature with the Title No. 33; see Opinion 417 (Hemming, 1956).
There has been considerable debate as to whether Geoffroy did or did not designate a type species of the genus Trionyx (1809a). Although not specifically designated as the type species, Trionyx aegyptiacus (= Testudo triunguis Forskål) is considered by Smith (1930:2), Schmidt (1953:108, footnote), and Loveridge and Williams (1957:422) to have been sufficiently indicated by Geoffroy as the type species. But Stejneger (1944:6), H. M. [445] Smith (1947:122), Conant and Goin (1948:11), and Mertens and Wermuth (1955) maintained that Geoffroy did not adequately designate a type species, and that Fitzinger (1843:30) designated the type species as Trionyx granosus (= Lissemys punctata), a synonym of Geoffroy's species, coromandelicus.
If Fitzinger's designation of a type species is accepted, the name Trionyx is applicable to the forms herein referred to Lissemys, and Amyda to the American forms. If Geoffroy's designation is accepted, the American forms are referable to Trionyx, and Amyda is a synonym.
The preceding includes only those generic names (listed in chronological order) that have been applied to Recent American soft-shelled turtles. Generic synonyms of the genus Trionyx applicable to Old World species are listed by Stejneger (1907:514), Smith (1931:165), and Loveridge and Williams (1957:420-21).
Trionyx is the most widespread genus of the family; most of the species occur in southeastern Asia. All North American soft-shelled turtles belong to this genus.
For quick reference, all the specific and subspecific names proposed for soft-shelled turtles in North America are listed below in alphabetical order (left hand column) with their nomenclatural status as recognized in this paper. The synonyms are listed in the account of the appropriate species or subspecies, and are discussed under the subsection entitled "Remarks."
| agassizi | Trionyx | spinifer asper |
| annulifer | Trionyx | spinifer spinifer |
| argus | Trionyx | spinifer spinifer |
| asper | Trionyx | spinifer asper |
| ater | Trionyx | ater |
| bartrami | Trionyx | ferox |
| emoryi | Trionyx | spinifer emoryi |
| calvatus | Trionyx | muticus calvatus |
| ferox | Trionyx | ferox |
| georgianus | Trionyx | ferox |
| georgicus | Trionyx | ferox |
| harlani | Trionyx | ferox |
| hartwegi | Trionyx | spinifer hartwegi |
| hudsonica | Trionyx | spinifer spinifer |
| mollis | Trionyx | ferox |
| microcephalus | Trionyx | muticus muticus |
| muticus | Trionyx | muticus muticus |
| nuchalis | Trionyx | spinifer spinifer |
| ocellatus | Trionyx | spinifer spinifer |
| olivaceus | Trionyx | spinifer spinifer |
| spiniferus | Trionyx | spinifer spinifer |
Variation
Aside from qualitative variations and comparisons of patterns of pigmentation the following external measurements (to the nearest millimeter) were used.
Length of plastron: Maximal straight-line measurement (midventrally), from the anteriormost region of the ventral surface to the posterior end of the plastron; this measurement includes an anterior cartilaginous part.
Length of carapace: Maximal, straight-line measurement (middorsally), from the nuchal region to the posteriormost region of the free edge of the carapace.
Width of carapace: Maximal, straight-line measurement between the lateral margins of the carapace.
Plane of greatest width of carapace: Maximal, straight-line measurement from the posteriormost region of the free edge of the carapace to a point on the middorsal line at the level or plane of the greatest width of the carapace; this measurement and the last two, of course, include the fringing cartilaginous parts of the dorsal bony carapace.
Width of head: Maximal measurement between the lateral margins of the head.
Length of snout: Measurement from tip of snout to interorbital region of least breadth.
Diameter of ocellus: Maximal outside diameter of largest (not conspicuously ovoid or oblong) ocellus on carapace.
The following ratios were developed from the measurements. Reference to these ratios will be made by the abbreviations within the parentheses: length of carapace/length of plastron (CL/PL); length of carapace/width of carapace (CL/CW); length of carapace/plane of width of carapace (CL/PCW); length of plastron/width of head (PL/HW); width of head/length of snout (HW/SL); diameter of ocellus/length of plastron (OD/PL).
Secondary Sexual Variation
Size
In many species of turtles, females are larger than males; the difference in size between the sexes is probably most pronounced in aquatic emydids. The ten largest individuals of each sex were selected to indicate the relative difference in size between the sexes of the three American species of Trionyx (excluding ater, Table 2). Female soft-shelled turtles attain a larger size than males. T. ferox is the largest species; muticus is the smallest. The approximate maximal size of each sex and the difference in size between the sexes are more correctly expressed for spinifer and muticus than for ferox, because fewer specimens of ferox were examined; presumably the approximate maximal size of males and females of ferox is larger than is indicated in Table 2.
Table 2. Secondary Sexual Difference in Maximal Size of North American Species of the Genus Trionyx (excluding ater) Based on the Ten Largest Specimens of Each Sex of Each Species. The Extremes Precede the Mean (in parentheses).
| Species | Plastral length (cm.) | |
| ferox | males | 17.0-26.0 (20.0) |
| females | 23.3-34.0 (27.9) | |
| spinifer | males | 13.8-16.0 (14.4) |
| females | 26.0-31.0 (28.0) | |
| muticus | males | 11.8-14.0 (12.3) |
| females | 17.7-21.5 (18.9) | |
Pattern
Secondary sexual differences in pattern are probably more pronounced in soft-shelled turtles than in other species of turtles, except perhaps for the well-known melanism and concomitant obliteration of pattern acquired by some adult males of the scripta section of the genus Pseudemys.
The difference in pattern between the sexes of American species varies with size of the individual and with the species and subspecies. The juvenal pattern of some individuals of T. spinifer asper differs according to sex. In the other species and subspecies, there are no secondary sexual differences in the juvenal pattern. That pattern in females of all species and subspecies is partly or entirely obscured by a mottled and blotched pattern as growth proceeds. This mottled and blotched pattern is present on females not yet sexually mature, and is of contrasting lichenlike figures, and in other individuals is less contrasting and a more uniform coloration. The largest males of T. spinifer retain a conspicuous juvenal pattern; in those of muticus the pattern may be well-defined or partly modified and obscured, whereas in large males of ferox the juvenal pattern is ill-defined or absent. No male normally acquires a contrasting mottled and blotched pattern on the carapace. The pattern on the carapace of many large individuals of ferox is not distinctive as to sex.
On the dorsal surface of the soft parts of the body there is a contrasting pattern in adult males and hatchlings of some forms, but in most large females the pattern is usually reduced to a near-uniform coloration; the pattern on adult males of ferox and muticus is not contrasting and resembles that on large females.
Coloration
Because most specimens examined were preserved, the detection of secondary sexual differences in coloration was difficult. There is one difference in coloration between the sexes in the subspecies T. s. emoryi. Males from the Río Grande drainage, at least those from the Big Bend region of Texas, and southwestward in the Río Conchos into Chihuahua, México, are bright orange on the side of head (postlabial and postocular pale areas); an orange tinge also occurs in pale stripes on the snout, and pale orange blotches sometimes occur on the dorsal surfaces of limbs, especially the hind limbs. The coloration of these areas on females is pale yellow, lacking orange.
Tuberculation
In all subspecies of spinifer the carapace of adult males is "sandpapery" owing to abundant, small, spiny tubercles distributed over its surface; all females lack spiny tubercles on the surface of the carapace.
Length of Tail
Elongation of the preanal region of the tail resulting in the extension of the cloacal opening beyond the posterior edge of the carapace occurs in males of several kinds of turtles, including Trionyx, at least in those from Louisiana, Texas, and Lake Texoma, Oklahoma (Webb, 1956:121). Probably this elongation is characteristic of males of all American softshells. Some females of spinifer and muticus that exceed the maximum size attained by males have the tip of the tail and cloacal opening extending a short distance beyond the posterior edge of the carapace. Some large females of ferox have more elongate tails than those of spinifer and muticus.
Width of Alveolar Surfaces of Jaws
Stejneger (1944:34-36, pl. 6) commented on a series of large skulls of ferox mostly from Kissimmee, Florida, some of which had conspicuously expanded alveolar surfaces. He suggested that the condition was confined to large males. A scattergram (Fig. 2) based on measurements obtained from 45 skulls of ferox shows widened alveolar surfaces of the upper jaws on some of the larger [448] skulls. Because the maximal size of adult males is unknown and the difference in size between the sexes of ferox is slight, such large skulls might represent either sex. The sex had been recorded for only three of the 45 skulls; none of the three exceeded 82 millimeters in basicranial length or had widened alveolar surfaces. Some of the larger skulls of approximately the same size differ markedly in width of the alveolar surfaces; this difference suggests that both sexes are included and that the sexes may be of approximately the same maximal size. On the other hand, the variation observed in skulls is possibly confined to one sex. To judge from what is known of the maximal sizes of the sexes of spinifer and muticus (see Table 2), skulls of ferox of more than 85 millimeters in basicranial length probably are of females. The largest alcoholic male (dissected) of ferox that I examined had a width of head of approximately 46.5 millimeters; that measurement corresponds to a basicranial length of 70 to 75 millimeters. The specimen of which measurements are depicted by the uppermost symbol in the scattergram (represented by KU 16528) was recorded as a female. Large females of T. s. asper from rivers emptying into the Atlantic Ocean have broadened alveolar surfaces.
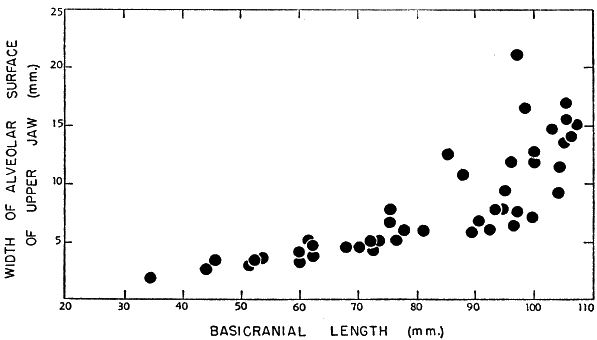
Fig. 2. Basicranial length and greatest width of alveolar surface of upper jaw on 45 skulls of T. ferox. Some skulls (sex unknown) in which the basicranial length exceeds 85 mm. develop widened alveolar surfaces of the jaws.
Length of Claw
Secondary sexual differences in length of claw on the forelimb are pronounced in some kinds of turtles. Cahn (1937:178) stated that the female of Trionyx muticus usually has long claws on the hind feet, while the male has long claws on the forefeet, but I am unable to substantiate his statement. Measurements of length of the third claw on the hind limb taken in 41 males and 45 females of spinifer from Louisiana showed no secondary sexual difference.
Ontogenetic Variation
Pattern
In all species and subspecies the juvenal pattern is replaced in females as growth proceeds by a mottled and blotched pattern that is contrasting or of nearly uniform coloration. The blotched pattern (of lichenlike figures) is evident on the carapaces of most females that have plastra so long as 8.0 centimeters. The contrasting juvenal pattern on the dorsal surfaces of the soft parts of the body is correspondingly modified in females, but at a size larger than 8.0 centimeters. Size of ocelli (OD/PL) in T. s. spinifer and hartwegi seems to vary ontogenetically (see section on Geographic Variation).
Some hatchlings have blotched patterns (T. spinifer asper, TU 16689.2, plastral length, 3.5 cm.); the largest females examined that did not show any evidence of mottling were two asper having plastrons 7.6 and 8.0 centimeters in length. Variation in color and pattern probably is modified greatly by the environment (Heude in Stejneger, 1907:518, footnote d) and the physiological condition of the individual. Smith, Nixon and Minton (1949:92) reported that a female of T. s. hartwegi developed a striking melanistic pattern in captivity and they concluded that patterns of soft-shelled turtles may be produced not only by conventional chromatophores, but also by other depositions, both intra- and extracellular. TU 16170, taken from brackish water at Delacroix Island, St. Bernard Parish, Louisiana, is the only adult male I have seen that had a blotched pattern (orange-brown in life) on the carapace in addition to the juvenal pattern. One female of muticus, KU 48229, having a plastral length 14.5 centimeters, retained a well-defined juvenal pattern, and lacked a mottled and blotched pattern (see Pl. 46).
Tuberculation
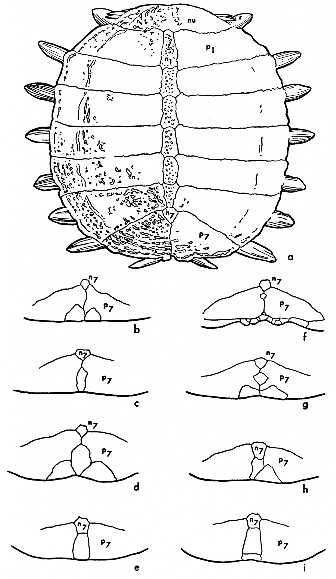
Males of the subspecies of spinifer develop small, sharp tubercles on the dorsal surface of the carapace when sexually mature. As growth proceeds, the minute prominences along the anterior edge of the carapace on hatchlings of both sexes of spinifer change in shape to conical projections or low, flattened, scarcely-elevated prominences, depending on the subspecies (Fig. 8).
Large females of spinifer and ferox acquire enlarged, flattened knobs in the nuchal region and posteriorly in the center of the carapace.
Length of Tail
The preanal region of the tail rapidly elongates in males of all soft-shells when they are sexually mature.
Width of Alveolar Surfaces of Jaws
The alveolar surfaces of the jaws are conspicuously broadened in large adults of ferox, and females of that population of T. s. asper in the Atlantic Coast drainage.
Ratios
Width of head increases at a rate slightly slower than does the length of the plastron (PL/HW, Fig. 3). The change in proportions is most pronounced [450] at a plastral length of 7.5 to 8.0 centimeters. In general, the head is narrowest in muticus and widest in ferox. T. s. asper and emoryi seemingly have the widest heads among the subspecies of spinifer. Geographically width of head increases from spinifer and hartwegi through pallidus and guadalupensis to emoryi. T. ater terminates the cline; 12 specimens, ranging in plastral length from 9.6 to 18.4 centimeters, resemble ferox and asper in having wide heads (average PL/HW of 4.93).
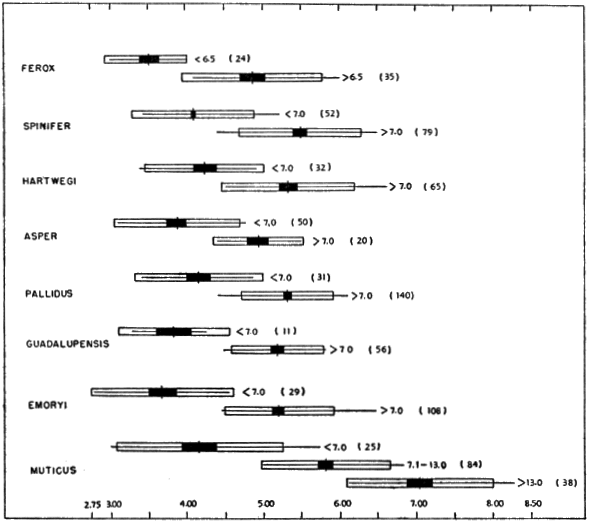
Fig. 3. Ratio of length of plastron to width of head (PL/HW) in some American species and subspecies of the genus Trionyx. The size of each sample is given in parentheses following an indication of the range (< = less than, > = greater than) in length of plastron (in cm.) of each sample. The horizontal line indicates the observed variation; the vertical line, the mean; the white rectangle, four standard deviations; and the black rectangle, four standard errors of the mean. There is some ontogenetic variation in PL/HW. The head is narrowest in muticus and widest in ferox.
The carapace increases in width more slowly than it increases in length (CL/CW, Fig. 4). The change in proportions is most pronounced when the carapace is 8.0 to 8.5 centimeters in length. Ontogenetically muticus varies least and ferox most; large specimens of ferox have narrower carapaces than muticus of corresponding size. There is also an indication of a geographical gradient that parallels the cline mentioned above for PL/HW. There is a gradual decrease in width of carapace from pallidus through guadalupensis to emoryi. Of the subspecies of spinifer, emoryi has the narrowest carapace and [451] resembles ferox. In T. ater this cline is accentuated and terminates; 12 specimens, ranging in plastral length from 9.6 to 18.4 centimeters, resemble ferox and emoryi in having narrow carapaces (average CL/CW of 1.32).
Osteological Characters
Closure of the anterior, paravertebral fontanelles on the bony carapace, and size and number of plastral callosities are subject to ontogenetic variation (see sections entitled "Carapace" and "Plastron").
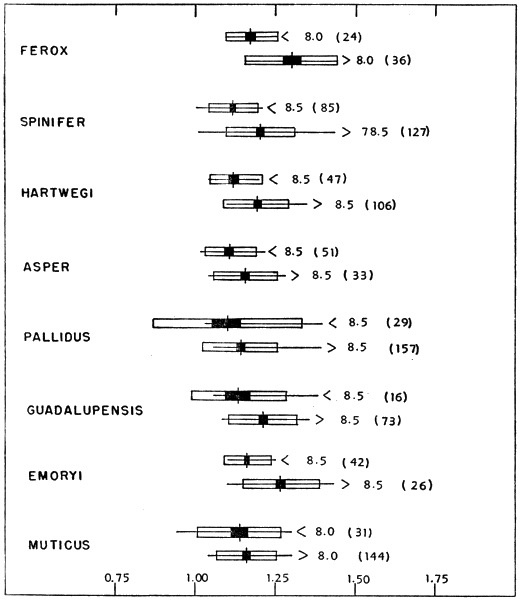
Fig. 4. Ratio of length of carapace to width of carapace (CL/CW) in some American species and subspecies of the genus Trionyx. Symbols as in Fig. 3. There is some ontogenetic variation in CL/CW (least in muticus). The carapace is narrowest in ferox and emoryi, and widest in muticus, pallidus and asper.

Fig. 5. Pattern on dorsal surface of snout of some American species and subspecies of the genus Trionyx. Note the gradual transition in pattern from that of hartwegi (b) and asper (c) to that of emoryi (h).
Geographic Variation
Geographic variation occurs in Trionyx spinifer and T. muticus. The variant populations of spinifer are segregated into six subspecies, those of muticus into two. In the subspecies of spinifer there is both group variation and clinal variation.
Group Variation
The six subspecies of spinifer can be separated into two groups on the basis of the juvenal pattern. One group (subspecies spinifer, hartwegi and asper) has a pattern of dark spots or ocelli of various sizes on the carapace, whereas the other group (subspecies pallidus, guadalupensis and emoryi) has a pattern of small white dots or tubercles on the carapace. The two groups differ also in the manner in which the mottled and blotched pattern first appears on the carapace of females. Usually, contrasting lichenlike figures initially surround the dark spots or ocelli on the carapace in females of the spinifer group (less evident in pallidus), whereas females of the emoryi group usually lack a contrasting pattern early in ontogeny. In general, the two groups differ in the degree of pigmentation. The spinifer group has larger marks and more contrasting patterns on the head and limbs, and more extensive pigmentation on the ventral surface than members of the emoryi group. T. ater is more closely related to those subspecies of the emoryi group but differs in having the ventral surface heavily speckled with black and an over-all blackish, dorsal coloration; the underlying pattern of ater resembles that of emoryi.
Clinal Variation
Several characters are arranged in a geographical gradient or cline. Some characters are relatively uniform and represent a terminus in the spinifer group. Some characters change gradually and successively through the subspecies pallidus and guadalupensis, and terminate in emoryi and T. ater. Some characters of ater, in turn, show affinity with T. muticus and T. ferox.
Pattern on Snout
The pattern (Fig. 5) on the snout usually consists of pale, dark-bordered stripes that form an acute angle in front of the eyes in spinifer, hartwegi and asper, but the corresponding marks form a dark triangle the base line of which joins the anterior margins of the orbits in emoryi and usually in guadalupensis. In pallidus, the geographic range of which is between guadalupensis and hartwegi, there are different patterns that are in various degrees intermediate between those described immediately above for hartwegi and guadalupensis.
Pattern on Side of Head
The change in pattern (Fig. 6) and its contrast with the ground color on the side of the head parallels the sequence of changes in pattern on the snout. The pattern on the side of head contrasts with the ground color and consists of dark markings below the eye and on the neck, an indication of a postlabial stripe, and a pale, dark-bordered postocular stripe that may be variously interrupted (spinifer and hartwegi; asper usually has uninterrupted postocular and postlabial stripes that unite on the side of the head). The pattern is contrasting but variable in pallidus. T. s. emoryi and usually guadalupensis have fewer dark markings, sometimes none, and an interrupted postocular pale stripe that produces a pale blotch just behind the eye.
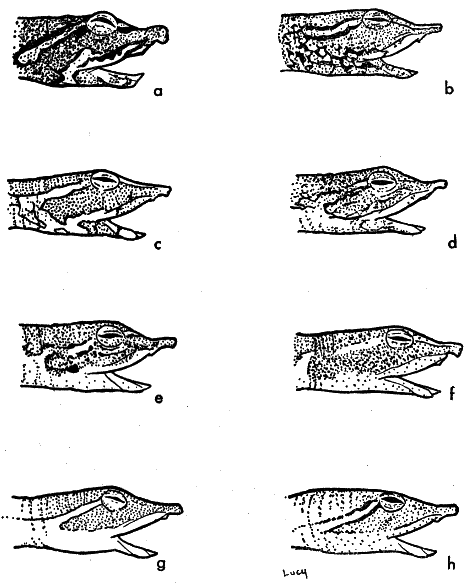
Fig. 6. Pattern on side of head of some American species and subspecies of the genus Trionyx. Note the gradual reduction in contrast of pattern and interruption of the postocular stripe from that of spinifer (b) to that of emoryi (f).
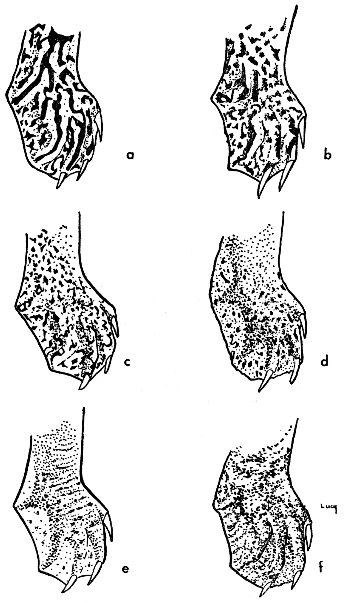
Fig. 7. Pattern on the dorsal surface of the distal part of the right hind limb of some American species and subspecies of the genus Trionyx. Note the gradual reduction in contrast of pattern from that of hartwegi (a) to that of emoryi (d).
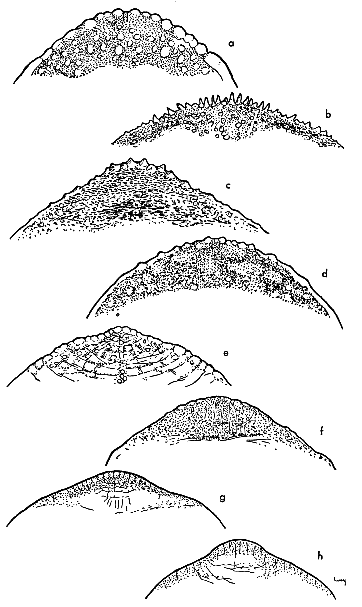
Fig. 8. Shape of tubercles on anterior edge of carapace in some American species and subspecies of the genus Trionyx (× 1/2). Note the gradual reduction in size of tubercles from that of hartwegi (b) to that of muticus (h).
Pattern on Dorsal Surface of Limbs
A corresponding sequence of change occurs in the size of dark markings on the dorsal surface of the limbs (Fig. 7). The hind limb usually has larger markings than the forelimb. The change is gradual from larger and darker markings (contrasting pattern) in hartwegi, spinifer and asper to smaller and paler markings (non-contrasting pattern) in emoryi.
Tuberculation
There is also a cline in tuberculation (Fig. 8) that parallels geographically the sequence of changes in patterns mentioned immediately above. The size of the tubercles along the anterior edge of the carapace changes in both sexes from those that are enlarged and equilateral or conical in shape in spinifer, hartwegi, asper and pallidus to those that are scarcely elevated in guadalupensis, emoryi and T. ater. Indeed, in the three kinds mentioned last, the tubercles are absent in some specimens. There seems to be a corresponding reduction in the size and number of small, sharp-tipped tubercles that cover the carapace in adult males; the carapace of T. ater is mostly smooth and has only a few small, whitish tubercles.
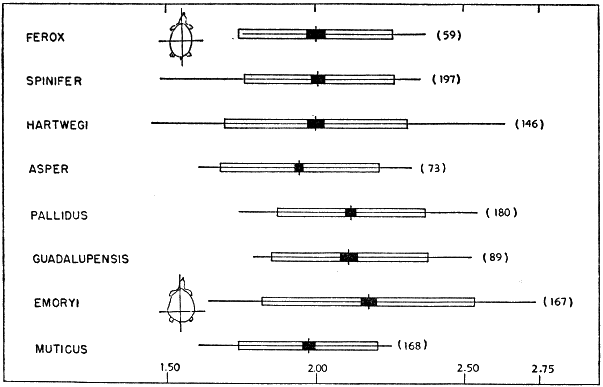
Fig. 9. Anteroposterior position of plane of greatest width of carapace (CL/PCW) in some American species and subspecies of the genus Trionyx. Symbols as in Fig. 3. The greatest width of carapace is midway between anterior and posterior ends in ferox, spinifer, hartwegi, asper and muticus, and farther posterior in the other subspecies of spinifer.
Ratios
The clinal tendencies in PL/HW (Fig. 3) and CL/CW (Fig. 4) that parallel those mentioned above for pattern and tuberculation have already been mentioned under the section "Ontogenetic Variation."
The ratio of CL/PCW (Fig. 9) was used in an effort to show further differences in the shape of the carapace, especially the plane on the carapace [459] where the greatest width occurs. Figure 9 shows the greatest width to be approximately midway between the anterior and posterior ends in the subspecies spinifer, hartwegi and asper, and in the species ferox and muticus (CL/PCW of 2.00). The greatest width of carapace is more posterior and at approximately the same plane in pallidus and guadalupensis, and farther posterior in emoryi. Calculated ratios for 12 specimens of T. ater average 2.15, a value that suggests closer affinity with pallidus, guadalupensis and emoryi than to the other species and subspecies.
Comparison of the relative lengths of snout (HW/SL, Fig. 10) in different populations of T. spinifer shows a character gradient. To facilitate a comparison utilizing large samples, the subspecies spinifer was combined with hartwegi, and pallidus with guadalupensis. The snout is longer in the subspecies spinifer and hartwegi than in emoryi; the length of the snout of emoryi resembles that of T. ferox. The snout is proportionately the longest in T. muticus. The average ratio of HW/SL for 12 individuals of T. ater is 1.37, and is nearer that of pallidus, guadalupensis, emoryi and ferox than that of muticus or the other subspecies of T. spinifer.
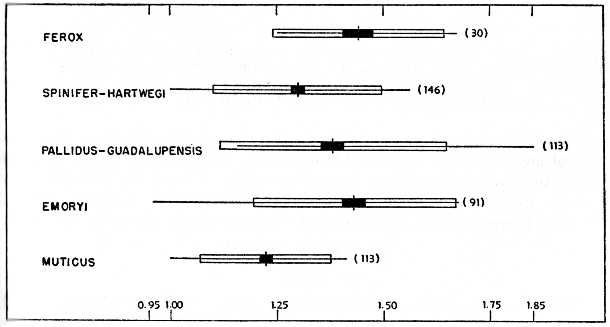
Fig. 10. Ratio of width of head to length of snout (HW/SL) in some American species and subspecies of the genus Trionyx. Symbols as in Fig. 3. Values for spinifer are combined with those of hartwegi, and those of pallidus with guadalupensis. The snout is proportionately the longest in muticus.
Size of the ocelli increases from west to east in populations of T. spinifer in the upper Mississippi River and Great Lakes drainages.
The ratio of OD/PL (Fig. 11) varies considerably but gradually increases from Kansas northeastward to Michigan. The minimal diameter of any ocellus recorded was one millimeter; solid dots on the carapace (hartwegi) were also recorded as one millimeter. Larger ratios are usually derived from measurements of larger individuals. Seemingly, there should be a clinal tendency in ontogenetic variation paralleling the size of ocelli and dependent on it; ontogenetic variation should be least in western populations in which the size of ocelli does not change appreciably with increasing size, and should be greatest in eastern populations in which the ocelli on adult males are larger than those on the carapace of young turtles. It is difficult to demonstrate [460] ontogenetic variation because specimens of corresponding size from the same general area may have ocelli of different sizes. The gradient in size of ocelli is also indicated by specimens from other states. I have the subjective impression that there is least variation in specimens from Michigan (Great Lakes-St. Lawrence River drainage), but this is not clearly shown by Figure 11.
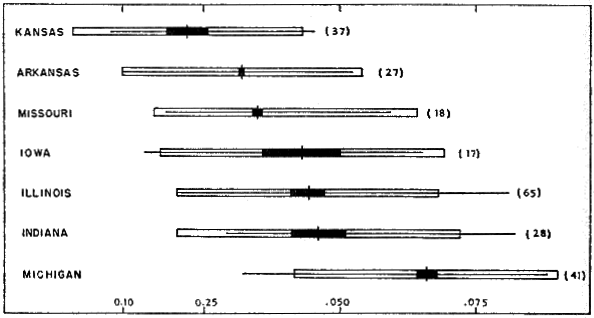
Fig. 11. Ratio of diameter of ocellus to length of plastron (OD/PL) in T. spinifer from some states in the upper Mississippi River and Great Lakes drainages. Symbols as in Fig. 3. The size of the ocelli on the carapace gradually increases from Kansas northeastward to Michigan.
Character Analysis
Snout
The snout (Fig. 12) is tubate having terminal nostrils separated by a vertical septum. One of the principal characters distinguishing T. ferox and T. spinifer from T. muticus is a lateral, whitish ridge projecting from each side of the nasal septum (hereafter referred to as septal ridges but often referred to in the literature as a papilla). The shape of the end of the snout is truncate in T. ferox and T. spinifer, and the nostrils are larger than in T. muticus. In muticus the snout usually terminates somewhat obliquely, and the nostrils tend to be slightly inferior; also, the end of the snout is usually rounded and somewhat pointed, causing the nostrils to be visible in lateral view. Some T. muticus do not differ markedly from ferox or spinifer in shape of the end of the snout. Stejneger (1944:14) mentioned indication of a septal ridge that did not reach the opening of the nostril in muticus. I have slit the outer edge of the nostril on several specimens of muticus, and have not noticed an indication of a septal ridge.
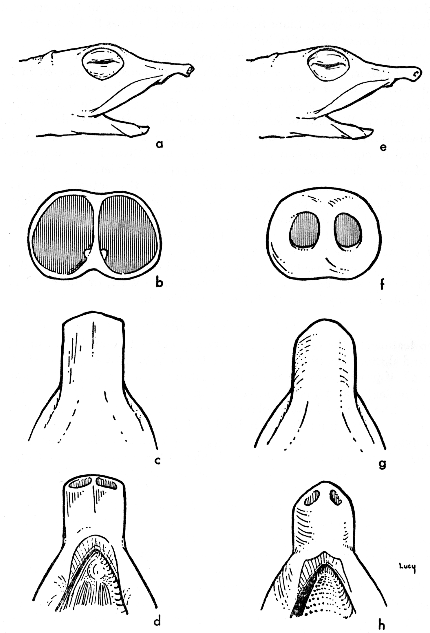
Fig. 12. Shape of snout in T. spinifer (left, a-d, from KU 46907) and T. muticus (right, e-h, from KU 48236). Lateral views—a, e (× 1); anterior views—b, f (× 5); dorsal views—c, g (× 2.5); ventral views—d, h (× 2.5).
Tuberculation
Tubercles or obtuse prominences occur on the anterior edge of the carapace (Fig. 8) or on the dorsal surface of the carapace. Trionyx muticus lacks tubercles, although some individuals show shallow, widely spaced wrinkles that suggest prominences on the anterior edge of the carapace. Both sexes of T. ferox have prominences, resembling flattened hemispheres, on the anterior edge of the carapace and in the nuchal region. Large females of ferox have obtuse prominences in the center of the carapace posteriorly, some of which are often arranged in longitudinal rows. The surface of the carapace in both sexes of T. ferox has small closely-set, blunt tubercles arranged in rows that resemble longitudinal ridges (most evident in juveniles).
Large females of T. spinifer have obtuse prominences in the center of the carapace posteriorly, some of which in many specimens are arranged in longitudinal rows; I cannot discern any correlation of number or arrangement of prominences with size in spinifer or ferox. The carapace in adult males of spinifer bears small, sharp tubercles that make the surface feel like sandpaper. The tubercles on the anterior edge of the carapace in adults of both sexes vary from round to equilateral and conical to low and flattened (see comments on tuberculation under subsection entitled "Geographic Variation"). Some large females of the same subspecies have tubercles on the anterior edge of the carapace that may be conical (higher than wide) or equilateral. The difference in shape of the tubercles seems not to be correlated with size because one T. s. pallidus, 30.5 centimeters (TU 13212) has prominent but blunted and equilateral tubercles, whereas, another female of pallidus, 20.8 centimeters (TU 13210), from the same locality has higher, conical tubercles. The blunted, equilateral tubercles may be the result of environmental wear, or the difference in shape of tubercles may be due to individual variation.
Pattern on Carapace
Two features of the pattern on the carapace are of taxonomic worth: 1) the width and distinctness of the pale rim at the periphery of the carapace (marginal rim), if present, and 2) the kind of pattern on the carapace (juvenal pattern). The marginal rim is absent in females of T. ater, and only faintly evident in males. The marginal rim is obscured or absent (adult males and females) and is not separated from the ground color of the carapace by a dark marginal line in hatchlings of T. ferox. The carapace of T. muticus has a marginal rim that is usually separated from the ground color of the carapace by an ill-defined, dark marginal line; some individuals lack the marginal dark line. The subspecies of T. spinifer have a well-defined, dark, marginal line that separates the marginal rim from the ground color of the carapace; T. s. asper has more than one dark marginal line on the carapace. The marginal rim is ill-defined and blotched, or absent, in large females of all species of Trionyx.
The marginal rim is widest at the posterior end of the carapace and lacking in the nuchal area. The width of the pale marginal rim is very narrow, almost to the degree of being absent, in juveniles of T. ferox. T. s. emoryi has a pale, marginal rim that is four or five times wider posteriorly than it is laterally, whereas posteriorly the width of the rim in the other subspecies of T. spinifer and in the species T. muticus is only two or three times wider posteriorly than it is laterally.
The juvenal pattern commonly consists of whitish tubercles or dots (T. s. emoryi, T. s. guadalupensis, T. s. pallidus, T. ater), large black ocelli (T. s. spinifer), small black dots and ocelli (T. s. hartwegi, T. s. asper), large dusky spots or ocelli (T. m. calvatus), or small dusky dots or short streaks and dashes (T. m. muticus). Some hatchlings of pallidus and emoryi have a uniform pale brown or tan carapace; hatchlings of T. ferox have a distinctive pattern (Pl. 31). [463] Further comments and illustrations pertaining to kind of pattern on the carapace are offered under the accounts of species and subspecies.
Pattern on Dorsal Surface of Snout (Fig. 5)
T. ferox has pale stripes on a dark background that unite in front of the eyes; the dark ground color becomes paler with increasing size, but the stripes retain thick black borders. T. m. muticus has ill-defined, pale stripes that are evident just in front of the eyes and do not extend anteriorly to unite in front of the eyes, whereas T. m. calvatus lacks pale stripes on the snout. The kind of pattern on the dorsal surface of the snout that is characteristic for each of the subspecies of T. spinifer has been mentioned in the discussion of clinal variation.
Pattern on Side of Head (Fig. 6)
T. ferox has a pale broad, postocular stripe in contact with the orbit or not, and other pale marks on a dark background; the ground color becomes paler with increasing size, but the stripes and other marks retain thick black borders. T. m. muticus usually has an uninterrupted, dusky-bordered, postocular stripe, whereas T. m. calvatus (in adult males only) has pale postocular stripes with thick blackish borders. The pattern on the side of head that is characteristic for each subspecies of T. spinifer has been mentioned in the discussion of clinal variation.
Pattern on Dorsal Surface of Limbs (Fig. 7)
Young specimens of T. ferox have pale marks on a blackish background. As growth proceeds the distinctive contrasting pattern is obliterated and eventually is replaced by a uniform grayish coloration in large adults. The pattern on the limbs of T. muticus is not contrasting, and is almost a uniform grayish, consisting of fine, pale markings. The clinal variation in pattern and kind of pattern on the limbs of the subspecies of T. spinifer has been mentioned in the discussion of clinal variation. Dark markings tend to form streaks that are coincident with the digits, and larger markings occur on the hind limbs than on the forelimbs.
Marginal Ridge
The anterolateral edge of the carapace in T. ferox (both sexes and all sizes) is "folded over" into a ridge having a distinct inner margin (Pls. 1 and 2), which is hereafter referred to as the marginal ridge. Siebenrock (1924:184-85) referred to this ridge as a "Hautsäume" and mentioned its occurrence in Old World species of the genus Trionyx. The marginal ridge is not present in T. muticus, T. spinifer or T. ater.
Ratios
The means of some samples (Fig. 3) differ in regard to PL/HW, but the ranges of variation overlap so much that little significance can be attributed to the difference. T. ferox, and to a lesser extent T. s. emoryi and T. s. asper, have slightly larger heads than the other forms. The width of head is proportionately the smallest in T. muticus; in most individuals of it having a plastron so long as 13.0 centimeters, the width of the head is less than 16 per cent of the length of the plastron—a percentage that is distinctive.
The visibly narrower carapace (CL/CW, Fig. 4), suggesting an ovoid or [464] oblong shape, in some large individuals of T. ferox and T. s. emoryi is indicated by the large ratio in specimens that have a plastral length of 8.0 centimeters or more. Nevertheless, the degree of overlap of the ranges of variation is such that this ratio is of relatively little use taxonomically.
The greatest width of the carapace is farther posterior in T. s. emoryi than in the other forms (CL/PCW, Fig. 9). The considerable overlap of the range of variation of this ratio for emoryi with the other forms limits its usefulness as a taxonomic character.
The snout is proportionately shortest in ferox and T. s. emoryi, and longest in muticus (HW/SL, Fig. 10). The most marked difference in this ratio is between the species muticus and ferox; the ranges of variation of those species overlap to a degree that tends to negate the taxonomic usefulness of this character.
Most adults and subadults of T. ferox show clearly in dorsal view the anterolateral portions of the plastron. This condition is much less well developed in some specimens of T. s. emoryi. T. ferox is extreme in the ratio CL/PL (relatively the longest plastron or shortest carapace, Fig. 13). T. s. asper has the shortest plastron in relation to length of carapace. Calculated ratios for 12 T. ater average 1.36, a value that suggests close affinity with some subspecies of T. spinifer (pallidus, guadalupensis, emoryi). Because of the degree of overlap of the ranges of variation in all forms, little significance can be attributed to the difference in means of ferox and asper.
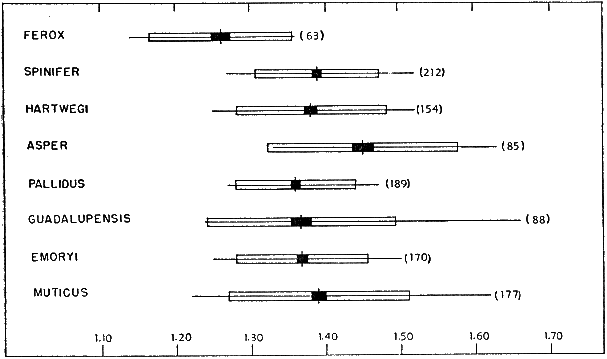
Fig. 13. Ratio of length of carapace to length of plastron (CL/PL) in some American species and subspecies of the genus Trionyx. Symbols as in Fig. 3. T. ferox has proportionately the shortest carapace.
Scalation
Cornified, smooth or cusplike areas occur on each limb, but their number and arrangement are of no taxonomic value. Normally, the anterior surface of each forelimb possesses four cornified areas for which the term antebrachial scales is proposed (Fig. 14). Two of the four scales occur in a more dorsal position; the lateral edge of the proximal one is free and cusplike along a part [465] of its length, whereas the distal scale is smooth-edged. Two scales having their lateral edges free and cusplike are ventral in position, and closer together than the two dorsad scales. Size of the scales and length of the free cusplike edges vary. Occasionally adjacent scales are fused or small additional scales are present. The number, configuration and arrangement of the two cornified areas on each hind limb are constant. One of these scales is smooth-edged and occurs posteriorly on the dorsal surface. The other scale, situated on the ventral surface posteriorly in the region of the heel and distal to the smooth-edged scale of the dorsal surface, has a pronounced, cusplike, free edge.
Fig. 14. Dorsal surface of right forelimb showing normal number and arrangement of antebrachial scales in American species of the genus Trionyx (T. spinifer hartwegi, KU 15932, × 3/4).
Choanal Papillae
This term refers to the papillate flaps of skin that project from the lateral borders of the internal nares. Webb and Legler (1960:23) noted their presence in softshells, and Parsons (1958) discussed their occurrence in sea turtles of the family Cheloniidae and in the testudinid subfamily Emydinae (1960). In preserved softshells the choanal papillae may extend laterally and partly cover the nares, or may be folded vertically against the lateral borders of the nares; in the latter position the papillae are easily overlooked. To my knowledge, choanal papillae occur in all American species and subspecies of soft-shelled turtles. The free edge of each narial flap shows various degrees of fimbriation. The fimbriated border is least developed (margin nearly entire) in T. muticus and most developed in T. ater and T. ferox. In ater at least, the anteriormost portions of the narial flaps seem wider than in the other forms and show a greater degree of fimbriation than the posteriormost parts. The choanal papillae are most easily observed in large specimens.
Skull
In general, there is less difference between the skulls of ferox and spinifer than between either of those species and muticus (Stejneger, 1944:10-11). Figure 15 shows the general differences in proportions of the skulls of spinifer and muticus; Plate 54 shows the skull of the holotype of Platypeltis agassizi (= T. s. asper), which is similar to that of ferox; Stejneger (op. cit.) provided labelled drawings of the skull of T. spinifer as well as photographs of skulls of other forms.
The total of 159 skulls examined by me include 80 of spinifer, 50 of ferox, and 29 of muticus. There are no secondary sexual differences between skulls of corresponding size, except in agassizi-form skulls mentioned under the account of T. s. asper, and possibly in ferox. Most, and possibly all, of the skulls of muticus having a basicranial length of 40.0 millimeters or more, and those of spinifer exceeding 50.0 millimeters must represent females (by correlation of known maximum size of males with greatest width of head, which is, in turn, compared with the greatest width of skull and corresponding basicranial length).
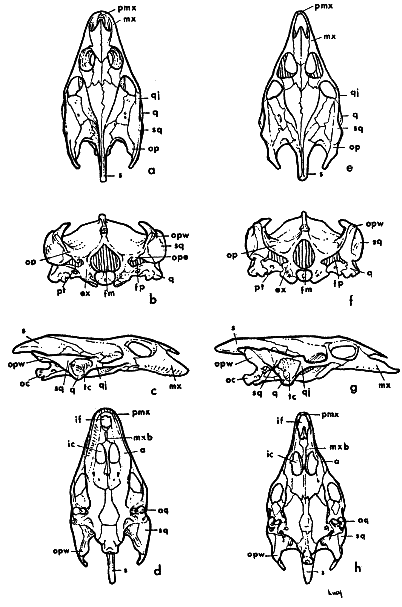
Fig. 15. Skulls of Trionyx spinifer hartwegi (left, a-d, KU 2757), and Trionyx muticus muticus (right, e-h, KU 1870). Dorsal views, a (× 1/2), e (× 3/4); occipital views, b (× 5/6), f (× 1); lateral views, c (× 1/2), g (× 3/4); ventral views, d (× 1/2), h (× 3/4).
a., alveolar surface of upper jaw
aq., articular surface of quadrate
ex., exoccipital
fp., fenestra postotica
fm., foramen magnum
if., intermaxillary foramen
ic., internal choana
mx., maxilla
mxb., maxillary bridge
oc., occipital condyle
op., opisthotic
ope., opisthotic-exoccipital spur
opw., opisthotic wing
pmx., premaxillaries (fused)
pt., pterygoid
q., quadrate
qj., quadratojugal
sq., squamosal
s., supraoccipital spine
tc., tympanic cavity
Measurements used include basicranial length (occipital condyle to tip of upper jaw), greatest width (variable in position), greatest width of alveolar surface of maxilla (taken at level immediately posterior to anterior margin of internal choanae), greatest length of internal choanae, and least breadth of maxillary bridge (separating internal choanae and intermaxillary foramen). One ratio developed from the measurements was greatest length of internal choanae/least breadth of maxillary bridge, hereafter referred to as IC/MB. This ratio is discussed under the account of T. s. asper.
Greatest Width
The position or level on the skull where the greatest width (Table 3) occurs is of some diagnostic value in distinguishing the skulls of ferox from spinifer and muticus. Skulls of ferox usually are widest at the level of the quadratojugal (immediately in front of tympanic cavity), whereas skulls of spinifer and muticus usually are widest slightly more posteriorly at a level on the squamosal immediately behind the tympanic cavity. Occasionally the width at the level of the quadratojugal and squamosal is the same, or the greatest width of skull may be ventrad between the quadrates, which are slightly flared laterally. The latter condition possibly is most prevalent in muticus.
Table 3. Variation in Position of Greatest Width of Skull of North American Species of the Genus Trionyx (excluding ater). The Number of Specimens Examined (in Parentheses) Follow the Specific Names.
| Position | Species | ||
| ferox (36) | spinifer (47) | muticus (14) | |
| Squamosal | 7 (19%) | 35 (74%) | 11 (79%) |
| Quadratojugal | 26 (72%) | 7 (15%) | 1 (7%) |
| Quadrate | 2 (6%) | ⋯ | 2 (14%) |
| Squamosal and quadratojugal of same width |
1 (3%) | 5 (11%) | ⋯ |
Supraoccipital Spine
The ventral surface of the supraoccipital spine in muticus lacks a medial ridge, and gradually increases in width anteriorly, so that it is widest proximally in the region of the roof of the foramen magnum. In ferox and spinifer, the ventral surface, usually having a medial ridge, is narrow and of the same width throughout its length or somewhat flared distally. The ventral surface of the supraoccipital spine, which is widest proximally in muticus, is always narrow proximally in ferox and spinifer. The ventral surface of the supraoccipital spine of one skull of spinifer, USNM 91311, differs little from that of muticus.
Foramen Magnum
The shape of the foramen magnum is generally rhomboidal in spinifer and ferox; the ventral angle is semicircular, the lateral angles obtuse, and the dorsal angle more acute. The shape of the foramen magnum in muticus is ovoid, higher than wide; the sides are evenly rounded.
Opisthotic-Exoccipital Spur
Skulls of spinifer normally have the fenestra postotica partly restricted by a medially-slanting, descending spur from the roof of the fenestra postotica; the spur incorporates the suture between the exoccipital and opisthotic and includes parts of those two bones. On one skull (KU 2824) the spur is displaced more medially and does not incorporate the opisthotic. The descending spur contacts the pterygoid ventrally forming a complete bony strut traversing the fenestra postotica in some skulls (KU 2228, 2666, 2762, TU 15423, MCZ 46621, TU 15415, right side only). The fenestra postotica on skulls of ferox and especially muticus is not normally restricted by an opisthotic-exoccipital spur.
Often the spur is reduced and indicated by a smooth projecting ridge. Sometimes the spur or ridge is absent on skulls of spinifer, and I have seen no well-developed spur on a skull of muticus. The development of the spur is not due to ontogenetic variation. There is some variation in development of the spur on either side of the skull; two skulls of ferox have the combination ridge/absent, and two of spinifer have the combinations ridge/spur and spur/absent. The frequency (based on counts of individual skulls) and the degree of development of the spur among the three species is indicated in Table 4.
Table 4. Frequency and Degree of Development of Opisthotic Exoccipital Spur of North American Species of the Genus Trionyx (excluding ater). The Number of Specimens Examined (in Parentheses) Follow the Specific Names.
| Development of Spur | Species | ||
| ferox (43) | spinifer (68) | muticus (29) | |
| spur (well-developed) | 1 (2%) | 45 (66%) | ⋯ |
| ridge (reduced) | 7 (16%) | 20 (30%) | 1 (3%) |
| absent | 35 (82%) | 3 (4%) | 28 (97%) |
Loveridge and Williams (1957:415, footnote) cited Siebenrock who mentioned a descending process of the opisthotic in Dogania (= Trionyx) subplana and Trionyx sinensis. I have not seen an ascending process of the pterygoids on skulls of American softshells as described by Loveridge and Williams (op. cit.:414, 429, fig. 54) for Lissemys, Cyclanorbis, Cycloderma and some Trionyx triunguis.
Opisthotic Wing
This term refers to the laterally directed, posterior part of the opisthotic that is visible in occipital, lateral and ventral views. In ventral view the opisthotic wing is most easily seen and is wider in muticus than in spinifer or ferox. In muticus the distal part is truncate, whereas in ferox and spinifer, it is more tapered and gently rounded, although somewhat unevenly flared [469] medially. Also there is more of a downward curvature (in ventral view) of the opisthotic wing in muticus than in ferox or spinifer; consequently the tip of the wing in muticus is often just visible in dorsal view (on lateral side of squamosal), certainly in lateral view. The distal part or tip of the opisthotic wing is not visible in dorsal view on skulls of ferox or spinifer.
Articular Surface of Quadrate
The ventral surface of the quadrate that articulates with the mandible is composed of a lateral condyle and a medial articular surface. The condyle and medial articular surface are separated by a furrow. On skulls of ferox and spinifer the lateral condyle, which is not conspicuously tapered posteriorly, is slightly larger than the medial articular surface, and the furrow is shallow. On skulls of muticus, the lateral condyle is conspicuously tapered posteriorly, is slightly smaller than the medial articular surface, and the furrow is deep.
Contact of Maxillaries Above Premaxillaries
The contact of the maxillaries above the premaxillaries is of diagnostic value in distinguishing skulls of ferox and spinifer from those of muticus. I have seen no skulls of muticus on which the maxillaries were in contact, and no skulls of ferox on which the maxillaries were separated. Stejneger (1944:19), however, reported a skull of muticus (USNM 102677) having the maxillaries in contact. Maxillaries are in contact (sometimes just barely) in 65 of 74 skulls of spinifer (88%); the premaxillaries are separated on nine skulls (12%).
Carapace
The dorsal surface of the bony carapace of American trionychids consists of a nuchal, seven or eight pairs of pleurals, and seven or eight, rarely nine, neurals (Fig. 16). The lateral parts of the nuchal overlie the second pair of ribs. The distal parts of the second through the ninth pair of ribs extend laterally beyond the lateral edges of the pleurals. There are no marginal ossifications. The posterior part of the bony carapace bears blunt, rounded or ovoid to linear, prominences mostly on the last pair of pleurals principally on large females of spinifer and ferox; I have seen only one adult male (stuffed, MCZ 46633) having a semblance of welts on the bony carapace. The nuchal, pleurals and neurals are sculptured.
As growth proceeds, the single, transversely-oriented, fontanelle of young turtles that separates the nuchal from the first neural and first pair of pleurals divides into two fontanelles that generally decrease in size and finally disappear. Occasionally only one (unilateral) large fontanelle is present (USNM 54734, muticus). The largest specimens noted that retain fontanelles are a ferox (USNM 029474) having a plastron 24 centimeters long, and a spinifer (USNM 54731) having a plastron 20 centimeters long. The fontanelles probably are present in some larger individuals.
Fig. 16. Carapace of Trionyx spinifer (a), and sketches of posterior parts of carapaces (b-i) of three American species, showing number and variation in arrangement of neurals and pleurals (not to scale; seventh neural, n7, and pleural, p7).
a. KU 2226, Lewisville, Lafayette County, Arkansas (× 1/3); sculpturing incompletely shown. Labels: r, ribs; nu, nuchal; n, neurals 1-7; p, pleurals 1-7.
b. ferox, USNM 60496, Auburndale, Polk County, Florida.
c. muticus, KU 1964, Doniphan Lake, Doniphan County, Kansas.
d. spinifer, USNM 100380, Plaquemine, Iberville Parish, Louisiana.
e. muticus, TCWC 7260, Red River, 8 mi. NW Ringgold, Montague County, in Clay County, Texas.
f. spinifer, USNM 59266, Homer, Winona, Minnesota.
g. muticus, KU 2840, White River, DeValls Bluff, Prairie County, Arkansas.
h. muticus, USNM 115939, Mississippi.
i. muticus, USNM 54734, Mississippi River, Fairport, Muscatine County, Iowa.
Most variation concerns the number of neurals and pairs of pleurals, and their arrangement posteriorly (H. M. Smith, 1947:121, table; Stejneger, 1944:18). Table 5 shows the frequency of occurrence of the number of neurals, pairs of pleurals, and the separation or contact of the seventh pair of pleurals; figure 16 illustrates some of the configurations of these plates posteriorly (e, g, and i not included in Table 5). The eighth pair of pleurals is reduced or absent (Loveridge and Williams, 1957:417). Eight neurals and eight pairs of pleurals occur in all three species. The seventh pleurals may contact each other in all three species, and their separation has been observed only in the species spinifer and muticus. Seven neurals and contact of the seventh pair of pleurals, or eight neurals and separation of the seventh pair of pleurals from each other occurs with approximately equal frequency in the species muticus. T. ferox and spinifer most often have seven neurals, seven pairs of pleurals, and the seventh pair of pleurals in contact. Stejneger (loc. cit.) mentioned a specimen in MCZ having nine neurals; I recorded nine neurals for USNM 54734 (Fig. 16i) for which Stejneger (loc. cit.) recorded eight. AMNH 57384 (ferox) has a small eighth pleural on the left side only, and USNM 115939 (muticus) has an eighth pleural only on the right side (Fig. 16h). Anomalous conditions observed included: an accessory bone between the first and second pleurals on the right side that contacts the first and second neurals in USNM 54733, (muticus); only six neurals in USNM 95193 (spinifer); a small accessory bony element between the first and second neurals in AMNH 57383 (ferox); and, only six pleurals (second and third fused) on the right side in USNM 54734 (muticus).
Table 5. Frequency of Occurrence of Number of Neurals, Pairs of Pleurals, and Separation or Contact of the Seventh Pair of Pleurals Among Species of American Soft-shell Turtles
| Number | Contact (+) or separation (-) of seventh pair of pleurals |
Species | |||
| Neurals | Pairs of pleurals |
ferox (16) | spinifer (60) | muticus (34) | |
| 7 | 7 | + | 9 (56%) | 50 (83%) | 13 (38%) |
| 7 | 8 | + | 5 (31%) | 2 (3%) | 2 (6%) |
| 8 | 7 | + | 2 (13%) | 3 (5%) | 3 (9%) |
| 8 | 8 | + | ⋯ | 4 (7%) | 2 (6%) |
| 8 | 7 | - | ⋯ | 1 (2%) | 14 (41%) |
Ventrally, the bony carapace shows ten thoracic vertebrae, the second through the ninth having well-developed, depressed ribs that are fused (no sutures) to the pleurals. The ribs of the first thoracic vertebra are represented by bony struts that extend posterolaterally and contact the anterior borders of the second pair of ribs. The two ribs of the ninth pair are free for most of their length and often are broken; they are slightly shorter than the eighth pair of ribs. The ribs of the tenth thoracic vertebra may be well-developed (KU 2219, 2666, 50856, spinifer, and 16528, ferox), but are usually broken off and represented only by transverse processes.
Kyphosis
Kyphosis (angular curvature of the vertebral column) or the hump-backed condition in American softshell turtles has been summarized by Nixon and Smith (1949:28). Cahn (1937:185, pl. 25e) illustrated the condition in an [473] individual of T. spinifer, and H. M. Smith (1947:119) mentioned kyphotic softshells representing the species spinifer (subspecies hartwegi and emoryi) and muticus. Neill (1951:10) mentioned two kyphotic T. s. asper and Nixon and Smith (loc. cit.) recorded the report of a kyphotic T. ferox. I have noted the condition in four muticus (subspecies muticus, KU 1959-60, 23230; INHS 2148) and seven spinifer (CNHM 22925; subspecies hartwegi, USNM 55689; subspecies spinifer, UMMZ 52948, 95615; subspecies emoryi, KU 2219, 33523, TU 16240). The smallest kyphotic specimen, a hatchling, TU 16240, has a plastral length of 3.5 centimeters. Kyphosis is to be expected in all kinds of softshells as are other abnormalities, such as albinism (reported for Lissemys by D'Abreu, 1928, and partial albinism noted in T. cartilagineus by Mohr, 1929) or congenital absence of limbs (reported by Dutta, 1931, as occurring in the genera Trionyx and Lissemys). The cause of kyphosis is not known. Smith (op. cit.:120) suggested an abnormally early fusion of the costals (= pleurals) with the ribs, and a subsequent differential rate of growth between them and the vertebral column as a hypothesis; Williams (1957:236) proposed that late retraction of the yolk mass, or retraction of an excessively large yolk mass may cause kyphosis. The cause of kyphosis may be of genetic origin or due to some environmental damage to the vertebral column prior to the cessation of growth. The variation in rate of growth of the vertebral column may produce humps of different shapes and sizes. Some of the specimens noted above (UMMZ 52948, 95615) have the carapace only slightly arched and are considered partly kyphotic. There seem to be degrees of kyphosis, a fact that should be taken into account in considering the occurrence of variation in greatest depth of shell.
Plastron
The plastron is united to the carapace by ligamentous tissue and is somewhat flexible anteriorly and posteriorly. Anteriorly the plastron is somewhat hingelike and may contact the anteriormost edge of the carapace. The bony elements are reduced. There is usually a median vacuity, which is relatively smaller in larger specimens and may be divided into two vacuities (a posteromedial and an anteromedial) by the medial juxtaposition of the hyo-hypoplastra, especially in muticus. Williams and McDowell (1952) have recommended a change in nomenclature for some of the plastral bones on the basis of reinterpretation of their homologies. The nine plastral bones include: an anterior pair of preplastra (= epiplastra, auct.); an unpaired, median bone, representing fused epiplastra (= entoplastron, auct.), hereafter referred to as the epiplastron; a pair of hyoplastra; a pair of hypoplastra; and, posteriorly, a pair of xiphiplastra (Fig. 17).
Siebenrock's (1902) synopsis of living trionychids was based entirely on plastral characters. He distinguished between muticus and spinifer principally by the shape of the epiplastron; T. ferox was not considered different from spinifer. The median angle formed by the boomerang-shaped epiplastron is obtuse and somewhat greater than 90 degrees in muticus (Fig. 17a); the angle of the epiplastron in spinifer and ferox is smaller than in muticus and forms an approximate right angle (Fig. 17b). Williams and McDowell (op. cit.:277, Pl. 1, Fig. 3) presented an illustration of the anterior plastral elements of an adult T. ferox. Siebenrock provided illustrations of the plastrons of muticus (op. cit.:823, Fig. 5) and spinifer (op. cit.:830, Fig. 10).
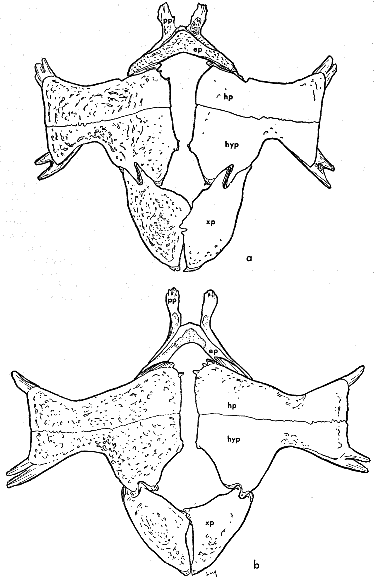
Fig. 17. Plastron of Trionyx muticus (a) and T. spinifer (b); sculpturing of callosities incompletely shown. ep, epiplastron; hp, hyoplastron; hyp, hypoplastron; pp, preplastron; xp, xiphiplastron. a—KU 1868, White River, Devall's Bluff, Prairie County, Arkansas (× 2/3); b—KU 1869, same locality (× 2/3).
Much importance has been credited to the fusion (no suture) or separation (suture present) of the hypoplastra and hyoplastra. The fusion of these bones distinguishes the genera Lissemys, Cyclanorbis and Cycloderma from Trionyx, Pelochelys, and Chitra (Siebenrock, op. cit.:815, 817; Loveridge and Williams, 1957:415). This character is also one of the criteria used by Hummel (1929: 768) in his erection of the two subfamilies Cyclanorbinae (= Lissemyinae) and Trionychinae. In my examination of specimens this character, unfortunately, was not given full attention. I have noted the fusion of the hypoplastra and hyoplastra in KU 1878 (muticus, right side only), KU 2219 (kyphotic spinifer), KU 16528 (ferox) and KU 60121 (ferox). Dr. Ernest E. Williams informs me in a letter of November 17, 1959, that of six specimens of ferox in the MCZ, the hyoplastra are fused with the hypoplastra in three (54689-90, 54686). I suspect that these bones in the three American species of the genus Trionyx, especially in ferox, fuse more often than is supposed.
In muticus the constricted part of the hyoplastron and hypoplastron is wider anteroposteriorly than in spinifer or ferox (Fig. 17).
The three American species have on the hyoplastra, hypoplastra, and xiphiplastra well-developed callosities, which enlarge with increasing size. The medial borders of the hyoplastral and hypoplastral callosities in larger specimens are rounded and closely approximated, often touching, as do the callosities of each xiphiplastron; seemingly, the callosities are relatively larger in muticus than in spinifer and ferox. I have seen one adult male muticus (KU 41380) that lacked median fontanelles or vacuities owing to the contact of the plastral elements (as viewed through overlying skin, alcoholic specimen). The bony plastron (approximately 9 cm. in maximal length) of a small muticus (KU 19460) resembles the plastron of larger individuals of muticus in having well-developed hyoplastral and hypoplastral callosities that are closely approximated medially. Large individuals of muticus usually have small, ovoid callosities on the preplastra, and a well-developed, angular callosity on the epiplastron (Fig. 17a). Siebenrock (op. cit.:823) suggests that the presence of callosities on the preplastra and epiplastron of muticus is subject to individual variation. I can not substantiate or dispute the supposition of Baur (1888:1122), Siebenrock (1924:193) and Stejneger (1944:12, 19) that the callosities are larger in males of muticus than in the females. Some individuals of spinifer have seven plastral callosities (KU 2842) as does muticus, but the callosities on the preplastra and epiplastron are less frequent and less well-developed in large specimens of spinifer than in muticus. The small epiplastral callosity in spinifer is located at the medial angle and does not extend posterolaterally to cover the entire surface of the epiplastron as it may in muticus (Fig. 17b). The epiplastron of a spinifer (KU 2826) has a medial callosity and another on the right posterolateral projection; three separate callosities occur on the epiplastron of MCZ 46615. The last specimen mentioned, a large, stuffed female, possesses a round, intercalary bone that tends to occlude the posteromedial vacuity. Seemingly, the callosity on the epiplastron appears prior to those on the preplastra; I have not seen any plastra having callosities on the preplastra and lacking a callosity on the epiplastron. I have not noted callosities on the preplastra or epiplastron of specimens of ferox.
The callosities on the plastral bones are sculptured; small, recently formed callosities on the preplastra and epiplastron lack sculpturing. The pattern [476] of sculpturing on the plastral bones as well as that of the carapace is generally of anastamosing ridges. I am unable to discern any differences in pattern of sculpturing between the three American species. Stejneger distinguished adult specimens of ferox from the other American species by the coarseness of the sculpture of the bony callosities (1944:24) and of the bony carapace (op. cit.:32). The sculpturing on the plastral callosities and carapace seems to be correlated with size; larger specimens (ferox) have coarser sculpturing than do smaller specimens (muticus). Stejneger also mentioned that the sculpturing on many specimens of ferox is specialized into prominent, longitudinal welts (loc. cit.); these welts occur also on the carapace of spinifer.
On the basis of the osteological characters examined by me, T. muticus is distinguished from spinifer and ferox by a number of characters (plastron and especially skull) whereas the species spinifer and ferox are not easily distinguished from one another.
Composition of the Genus Trionyx in North America
Analysis of the characters previously mentioned and their geographic distribution permits the recognition of ten taxa, comprising four species and eight subspecies. Two subspecies, T. spinifer pallidus and T. s. guadalupensis are described as new. The four species and the included subspecies here recognized are:
The following key is designed to permit quick identification of living individuals; therefore, ratios and osteological characters are avoided as much as possible in favor of other characters that are the least variable and most "typical." Because there is considerable variation correlated with sex and size, each taxon occurs in the key in more than one couplet. Large females having mottled and blotched patterns will be the most difficult to identify. The characters listed should be used in combination because one character alone may not be sufficient; it is advisable to read both choices of each couplet. The text, figures and illustrations should be consulted for final identification.
Artificial Key to North American Species and Subspecies of the Genus Trionyx
1. Septal ridges present; tubercles on anterior edge of carapace present or absent
2
Septal ridges absent; anterior edge of carapace lacking tubercles or raised prominences
19
2. Plastral area a uniform dark slate or blackish; soft parts of body blackish having large pale marks dorsally; carapace having large black blotches, often fused along margin, on pale background, and many well-defined longitudinal ridges
T. ferox, p. 479
Combination of characters not as above; ventral surface whitish, blackish flecks or blotches sometimes present
3
3. Carapace having pattern of white dots, or black ocelli and/or spots; carapace sometimes gritty resembling sandpaper
4
Carapace uniform pale brownish or grayish, or having mottled and blotched pattern, contrasting or not; white dots or tubercles, black ocelli and/or spots may be present; carapace not gritty
10
4. Carapace having pattern of black ocelli and/or spots; numerous, conspicuous whitish spots or tubercles absent
5
Carapace having pattern of white dots that are sometimes surrounded by small black ocelli; small black dots may be interspersed among larger white dots
7
5. Carapace having two or more marginal lines, these often diffuse and interrupted; black spots sometimes ocellate or bacilliform, or interspersed among smaller black dots; postocular and postlabial stripes usually united
spinifer asper, p. 502
Carapace having only one dark marginal line; pattern of black ocelli or spots; postocular and postlabial stripes usually not united
6
6. Carapace having prominent ocelli, which are much larger near the center than at the sides
spinifer spinifer, p. 489
Carapace having numerous small, dark spots, sometimes small ocelli, which are not much larger near the center than the sides
spinifer hartwegi, p. 497
7. White spots on anterior third of carapace; white spots on carapace often surrounded by narrow blackish ocelli; small black dots sometimes interspersed among white spots
spinifer guadalupensis, p. 517
White spots absent on anterior third of carapace, or small and inconspicuous; white spots not surrounded by narrow blackish ocelli
8
8. Pale rim of carapace narrow, partly obscured; over-all dorsal coloration (including soft parts of body) dark and lacking pattern; few, small, white tubercles confined to posterior third of carapace
ater, p. 528
Pale rim distinct, without markings; soft parts of body dorsally not uniformly dark; many white tubercles usually contrasting on pale carapace
9
9. White spots confined to posterior third of carapace; ground color of carapace usually pale brown or tan, sometimes darker; a dark, slightly curved, line connecting anterior margins of orbits; postocular stripe usually interrupted leaving pale, blotch behind eye; pale rim of carapace four or five times wider posteriorly than laterally
spinifer emoryi, p. 510
Small white spots on posterior half of carapace gradually decreasing in size anteriorly, often indistinct or absent on anterior third of carapace; pale rim of carapace no more than three times wider posteriorly than laterally
spinifer pallidus, p. 522
10. Marginal ridge present; carapace having ill-defined dark blotches on uniform grayish, lacking whitish tubercles or well-defined black spots or ocelli; pale rim of carapace absent; tubercles on anterior edge of carapace resembling flattened hemispheres; anterior parts of plastron often visible in dorsal view; postocular stripe, if present, having thick, blackish borders
ferox, p. 479
Marginal ridge absent
11
11. Carapace uniform pale brownish, lacking mottled and blotched pattern, white dots, black ocelli or spots
12
Carapace having mottled and blotched pattern, contrasting or not; white spots or tubercles, black ocelli or spots may be present
13
12. Pale rim of carapace four or five times wider posteriorly than laterally; dark, straight or slightly curved, line connecting anterior margins of orbits
spinifer emoryi, p. 510
Pale rim of carapace no more than three times wider posteriorly than laterally
spinifer pallidus, p. 522
13. Rear margin of carapace usually roughened by fine corrugations, edge often ragged; pale rim absent; carapace having dark brown-blackish, mottled and blotched pattern; anterior edge of carapace more or less smooth having scarcely elevated prominences; posterior part of plastral area and especially ventral surface of carapace having numerous black marks
ater, p. 528
Rear margin of carapace smooth, edge entire; usually some evidence of pale rim
14
14. White, rounded tubercles or spots usually evident posteriorly on carapace, sometimes indistinct; black ocelli or spots lacking in center of carapace, sometimes present at sides; shape of tubercles on anterior edge of carapace variable
15
White spots or tubercles absent; margin of carapace usually having black ocelli or spots; tubercles on anterior edge of carapace equilateral or conical, not low and flattened
17
15. White spots often present on anterior half of carapace; tubercles on anterior edge equilateral and wartlike, or less elevated, not conical
spinifer guadalupensis, p. 517
White spots usually absent on anterior half of carapace, sometimes indistinct; shape of tubercles on anterior edge of carapace variable
16
16. White spots absent on anterior half of carapace; tubercles on anterior edge of carapace low, scarcely elevated, never equilateral or conical; mottled and blotched pattern often not contrasting; ground color of carapace sometimes dark; pale rim of carapace four or five times wider posteriorly than laterally; dark, straight or slightly curved, line connecting anterior margins or orbits
spinifer emoryi, p. 510
White spots sometimes indistinct on carapace, or few, small spots present on posterior half of carapace; tubercles on anterior edge of carapace equilateral and wartlike or conical; mottled and blotched pattern usually contrasting; pale rim less than three times wider posteriorly than laterally
spinifer pallidus, p. 522
17. Carapace having evidence of more than one dark marginal line, and scattered, black spots or ocelli
spinifer asper, p. 502
Carapace having only one, dark, marginal line
18
18. Carapace having small black spots, lacking large interrupted ocelli
spinifer hartwegi, p. 497
Carapace having small black spots interspersed among larger, interrupted ocelli
spinifer spinifer, p. 489
19. Carapace having pattern of dusky spots, sometimes short lines
20
Carapace lacking pattern of dark spots or lines, having a mottled and blotched pattern
21
20. Pattern of circular spots, lacking short lines or bacilliform marks; spots sometimes slightly ocellate; no pale stripes on snout
muticus calvatus, p. 539
Pattern of dots, or dots and short lines; pale stripes on snout, at least just in front of eyes
muticus muticus, p. 534
21. Mottled and blotched pattern usually contrasting; ill-defined, blackish blotch absent behind eye
muticus muticus, p. 534
Mottled and blotched pattern usually not contrasting; ill-defined, dark blotch may be present behind eye
muticus calvatus, p. 539
Systematic Account of Species and Subspecies
Trionyx ferox (Schneider)
Florida Softshell
Plates 31 and 32
Testudo ferox Schneider, Naturg. Schildkr., p. 330, 1783 (based on Pennant, Philos. Trans. London, 61 (Pt. 1, Art. 32): 268, pl. 10 [figs. 1-3], 1772).
Trionyx ferox Schwartz, Charleston Mus. Leaflet, No. 26:17, pls. 1-3, May, 1956.
Testudo mollis Lacépède, Hist., Nat. Quadr. Ovip. Serp., 1:137, pl. 7, 1788.
Testudo (ferox?) verrucosa Schoepff, Hist. Testud., Fasc. 5 (Plag. M):90, pl. 19, 1795.
Testudo bartrami Daudin, Hist. Nat. Rept., 2:74, pl. 18, fig. 2, 1801.
Trionyx georgicus Geoffroy, Ann. Mus. Hist. Nat., Paris, 14:17, August, 1809.
Mesodeca bartrami Rafinesque, Atlan. Jour., Friend Knowledge, Philadelphia, 1 (No. 2, Art. 12):64, Summer, 1832.
Trionyx harlani Bell in Harlan, Medic. Phys. Research, p. 159, 1835.
Type.—Holotype, British Museum (Natural History) 1947.3.6.17; original number 53A, presumably that of Royal Society; stuffed adult female and skull; obtained from the Savannah River, Georgia, by Dr. Alexander Garden.
Range.—Southern South Carolina, southeastern Georgia, and all of Florida except the Keys and perhaps the western end of the panhandle (see map, Fig. 18).
Diagnosis.—Marginal ridge present; longitudinal rows of tubercles that resemble ridges on carapace of hatchlings; plastron often extending farther forward than carapace in adults; plastral area dark slate or gray in hatchlings; juvenal pattern of large slate or blackish blotches (often with pale centers) on a pale background; pale outer rim of carapace (absent on adults) narrow, not separated from ground color of carapace by distinct, dark line.
Size large; head wide; carapace relatively long and narrow; snout short; greatest width of skull at level of quadratojugal; often no suture between hypoplastra and hyoplastra; callosities on epiplastron and preplastra usually lacking.
Description.—Plastral length of smallest hatchling, 2.9 centimeters (UMMZ 95613), of largest male, 26.0 centimeters (AMNH 63642), of largest female, 34.0 centimeters (UMMZ 38123).
Septal ridges present; over-all coloration of carapace and plastron, and soft parts of body of hatchlings slate or blackish; carapace having blackish, circular blotches, usually fused at margin, often with pale centers on buff background forming coarse reticulum; pale, narrow rim of carapace not separated from ground color by dark marginal line; pale rim, coincident with marginal ridge, absent from anteriormost nuchal region; longitudinal rows of tubercles on carapace resembling ridges; undersurface blackish, usually having posterior part of carapace pale with irregular blackish marks; blackish soft parts of body dorsally having large, pale markings, most consistent of which are postocular mark that may contact orbit, postlabial mark that curves around angle of jaws, inverted Y on top of snout, and one or two streaks on side of neck.
Over-all coloration of adults grayish, paler than in hatchlings; carapace gray sometimes having slightly darker, large, irregular markings; mottled and blotched pattern on females not contrasting; sex of many large individuals not distinguishable on basis of pattern on carapace; pale rim of carapace obscure or absent; soft parts of body dorsally gray or brownish on large adults of both sexes, sometimes having slightly paler, large markings; small adult males usually having contrasting pattern on head; surface of carapace smooth (not "sandpaper") on adult males; undersurface whitish, throat often grayish; well-defined marginal ridge; anterior edge of carapace laterally to region of insertion of forelimbs studded with low, flattened tubercles resembling hemispheres, never conical; carapace usually having blunted tubercles, best developed anteriorly and posteriorly on midline, but sometimes linearly arranged, resembling ridges, especially at margins; anterolateral parts of plastron often extending farther forward than corresponding parts of carapace.
Range in length (in cm.) of plastron of ten largest specimens of each sex (mean follows extremes), males, 17.0-26.0, 20.0; females 23.3-34.0, 27.9; ontogenetic variation in PL/HW, mean PL/HW of specimens having plastral lengths 6.5 centimeters or less, 3.52, and exceeding 6.5 centimeters, 4.87; ontogenetic variation in CL/CW, mean CL/CW of specimens having plastral lengths 8.0 centimeters or less, 1.18, and exceeding 8.0 centimeters, 1.30; mean CL/PCW, 2.01; mean HW/SL, 1.44; mean CL/PL, 1.26.
Jaws of some skulls that exceed 75 millimeters in basicranial length having expanded alveolar surfaces; greatest width of skull usually at level of quadratojugal (72%); ventral surface of supraoccipital spine narrow proximally, usually having medial ridge; foramen magnum rhomboidal; opisthotic-exoccipital spur absent (82%), sometimes indicated by ridge (16%); distal part of opisthotic wing [481] tapered, not visible in dorsal view; lateral condyle of articular surface of quadrate larger than medial articular surface, not tapered posteriorly; maxillaries in contact above premaxillaries; usually a combination of seven neurals, seven pairs of pleurals, and contact of seventh pair of pleurals (56%), often eight pairs of pleurals (31%); angle of epiplastron forming approximate right angle; often no suture between hypoplastra and hyoplastra; callosities on preplastra and epiplastron usually lacking.
Variation.—Crenshaw and Hopkins (1955:19) stated that in specimens from Lake Okeechobee and southward the carapace is wider relative to the width of the head, and Neill (1951:19) quoted Allen's observations that ferox from southern Florida "average larger and darker than those collected farther north."
Carr (1952:417) reported that the pale reticulum on the carapace is yellowish olive, the markings on head are yellow on an olive ground color, some markings more orange, and the plastron slate gray. Duellman and Schwartz (1958:271) mentioned that the carapace of hatchlings is edged in orange grading to yellow posteriorly and has a pattern of bluish-black blotches on a dull brown background, whereas the carapace is dull brown or blackish on adults. Neill (op. cit.:18) wrote "that the head stripes and the marginal ring of the 'carapace' are orange rather than yellow (yellow at the time of hatching, however)."
The transition from the dark coloration of hatchlings to the paler coloration of adults is gradual and subject to individual variation. The loss of dark color ventrally occurs first on the plastral area, then the hind limbs, forelimbs, posterior part of carapace and last on the neck and throat. The soft parts of the body dorsally are gray or dark gray, and do not become so pale as the ventral surface. The smallest specimen that I have seen displaying the dark features of the hatchlings is a male, 7.7 centimeters (UMMZ 100673); a female, 9.5 centimeters (UMMZ 110987), is the smallest specimen having a whitish plastral area. The change from dark to pale coloration on the ventral surface occurs at a size of 8.0 to 9.0 centimeters. The largest specimens I have seen having indistinct, dusky blotches of the underside of the carapace are a female, 11.3 centimeters (UMMZ 100836), and a male, 16.0 centimeters (UMMZ 106322). A contrasting pattern on head and limbs, and a dark throat are still evident in a female 19.2 centimeters (UMMZ 106302).
Comparisons.—Trionyx ferox can be distinguished from all other species of the genus in North America by the presence of a marginal ridge, longitudinal ridges of tubercles on the carapace of juveniles (less evident in adults), and the unique juvenal pattern and coloration. The lack of a juvenal pattern and a smooth surface on the carapace (not gritty like sandpaper) distinguish adult males from those of T. spinifer. Most adults of both sexes can be distinguished from spinifer and muticus by the extension of the plastron farther forward than the carapace (developed to a slight degree in some specimens of T. s. emoryi). Both sexes of all ages can be distinguished from muticus by the presence of knoblike tubercles on the anterior edge of the carapace, and septal ridges.
T. ferox is the largest species in North America; the maximum size of the plastron in adult males is approximately 26.0 centimeters (16.0 in spinifer) and of adult females, 34.0 centimeters (31.0 in spinifer). The head is wider in ferox than in muticus and most subspecies of spinifer (closely approached [482] by asper, guadalupensis, emoryi and T. ater). The carapace is narrower in ferox than in muticus and most subspecies of spinifer (closely approached by emoryi and T. ater). The snout is shortest in ferox, but almost as short in T. s. emoryi and T. ater. T. ferox has proportionately the longest plastron in relation to length of carapace.
Most skulls of ferox differ from those of muticus and spinifer in having the greatest width at the level of the quadratojugal (as do some T. s. asper; see account of that subspecies). In the skull, ferox resembles spinifer but differs from muticus in having the 1) ventral surface of the supraoccipital spine narrow proximally, and usually having a medial ridge, 2) foramen magnum rhomboidal, 3) distal part of opisthotic wing tapered, 4) lateral condyle of articular surface of quadrate not tapered posteriorly, and larger than medial articular surface, and 5) maxillaries in contact above premaxillaries. T. ferox resembles muticus but differs from most individuals of spinifer in lacking a well-developed opisthotic-exoccipital spur. T. ferox resembles spinifer but differs from muticus in having the epiplastron bent at approximately a right angle; ferox differs from both muticus and spinifer in lacking a callosity on the epiplastron and probably in the more frequent fusion of the hyoplastra and hypoplastra.
Remarks.—The early taxonomic history of Trionyx ferox has been discussed in detail by Stejneger (1944:27-32), who explained that Dr. Alexander Garden of Charleston, South Carolina, sent a description and specimen of T. ferox to Thomas Pennant, and at the same time sent another specimen with drawings to a friend, John Ellis, in London. Pennant presented one of the specimens and drawings and the description to the Royal Society of London in 1771; the description was published in 1772 and included Garden's drawings. Because two specimens were involved the possibility exists that the description (text, drawings and type specimen) is a composite based on two specimens.
I have not seen the type. Garden's original description (in Pennant, 1772:268-271) leaves little doubt that the text subject is a large adult female of ferox (see especially the statements, "fore part, [of carapace] just where it covers the head and neck, is studded full of large knobs, [and] The under, or belly plate, … is … extended forward two or three inches more than the back plate, …"). I am indebted to Mr. J. C. Battersby, British Museum (Natural History), Department of Zoology (Reptiles), for information concerning the type and for comparing it with the text description and three figures published by Pennant. The carapace of the type is approximately 16 inches long, 131/2 inches wide, and has low, flattened, knoblike tubercles along the anterior edge. Some inaccuracies on the part of the artist (such as five claws on both feet on the right side of Fig. 3, and four claws on the left front foot of Fig. 2 are evident), and slight changes in the proportions of the type would have occurred after death and preservation. It is the opinion of Mr. Battersby that the type, text description and three figures represent one specimen. Figures 1 and 2, dorsal and ventral views respectively, probably represent the same specimen from life; the neck is withdrawn and the tail tip is visible in dorsal view, but concealed beneath the posterior edge of the carapace in ventral view. Presumably the same specimen (probably drawn from dried and stuffed animal) is depicted in Figure 3 (dorsal view); the neck is fully extended and a large part of the thick, pyramidal tail is visible in dorsal view. British Museum (Natural History) 1947.3.6.17 is considered a holotype. The three figures published [483] by Pennant have been duplicated by Schoepff (1795:Pl. 19) and Duméril and Bibron (1835:482). To my knowledge, the holotype was first specifically designated as the "(Type.)" of T. ferox by Boulenger (1889:259). The skull of the holotype is figured by Stejneger (1944:Pl. 5).
Garden did not list a specific locality for the two specimens that he sent to London, but did mention that the turtle was common in the Savannah and Altamaha rivers (of Georgia), and rivers in east Florida. Boulenger (loc. cit.) stated that the locality of the holotype was "Georgia." Baur (1893:220) restricted the type locality to the "Savannah river, Ga." Neill (1951:17), who believed T. ferox to be absent from the Savannah River, changed the type locality of ferox to east Florida. Schwartz (1956:8) reappraised the status of softshells in Georgia and Florida and reëstablished the Savannah River (at Savannah), Georgia, as the type locality of T. ferox.
Pennant failed to use binomial nomenclature when he published the type description of Garden. The first name-combination (Testudo ferox) was proposed by Schneider (1783:220).
Lacépède (1788:137, Pl. 7) referred to Garden's description in Pennant only as "The Molle" but on a folded paper chart entitled "Table Méthodique des Quadrupèdes ovipares," which is inserted after an introduction of 17 pages, listed T. mollis; this name is again listed on another folded chart, entitled "Synopsis methodica Quadrupedum oviparorum," which is inserted between pages 618 and 619 under the genus Testudo. The illustration (Pl. 7) was taken from Pennant (Duméril and Bibron, loc. cit.). The type locality has been designated "(following Stejneger, 1944) as eastern Florida" by Schmidt (1953:108).
Bartram failed to use a binomial name with his description of "the great soft shelled tortoise," which appeared in his Travels (1791:177-179, Pl. 4 and unnumbered plate between pages 282 and 283) and two editions of a French translation (1799 and 1801, 1:307); see Harper (1940). Recently, Bartram's Travels has been placed on the Official Index of Rejected and Invalid Works in Zoological Nomenclature, Opinion 447 (see Hemming, 1957). Bartram's description of a soft-shelled turtle has provided the basis for the proposal of at least three name-combinations. The first was Testudo (ferox?) verrucosa proposed in 1795 by Schoepff; it appeared simultaneously in The Historia Testudinum and in a German translation, Naturgeschichte der Schildkröten (see Mittleman, 1944:245). Stejneger (1944:26) listed the type locality as eastern Florida. Daudin (1801:74), also referring to Bartram's description in his Voyage (French translation), proposed the name Testudo bartrami; Harper (op. cit.:717) restricted the type locality of T. bartrami from "Halfway pond," east Florida, to southwestern Putnam County between Palatka and Gainesville, Florida. Rafinesque (1832:64-65), relying on the authenticity of the illustrations in Bartram's Travels that depict a soft-shelled turtle having five claws on each of the hind feet, tubercles on the sides of the head and neck, and ten scales in the middle of the carapace (presumably inaccuracies or a composite on the part of the artist), referred to Bartram's description as a new genus, Mesodeca bartrami, a name which Boulenger (1889:245, footnote) referred to as "mythical." Geoffroy (1809a:18-19) considered Bartram's description the basis for the recognition of a second species of Chelys (binomial nomenclature not employed), and Duméril and Bibron (loc. cit.) suggested that the description was based partly on a "Chelyde Matamata." [484] The descriptive comments of Bartram are not clearly applicable to Testudo ferox Schneider; Trionyx ferox, however, is the only species of soft-shelled turtle known to occur in the region of Bartram's observations (east Florida), and the type locality was restricted to Putnam County, Florida, by Harper. The name-combinations, Testudo (ferox?) verrucosa Schoepff, Testudo bartrami Daudin, and Mesodeca bartrami Rafinesque are junior synonyms of Testudo ferox Schneider.
Schweigger (1812:285) referred ferox to the genus Trionyx following the description of that genus by Geoffroy in 1809. Testudo ferox was listed as a synonym by Geoffroy in the description of Trionyx georgicus (1809a:17); Duméril and Bibron (1835:432) mentioned that the specific characters of georgicus were taken from Pennant. The name Trionyx georgianus presumably appears for this taxon in Geoffroy's earlier-published synopsis (1809:367). T. georgicus was listed as occurring in rivers of Georgia and the Carolinas; the type locality was restricted by Schmidt (op. cit.:109) to the Savannah River, Georgia. The two specific names georgicus and georgianus are regarded as substitute names and junior synonyms of T. ferox.
Geoffroy (1809a:14-15) also described Trionyx carinatus, a name-combination that hitherto has been considered a synonym of Trionyx ferox. There is no indication from the description that carinatus is applicable to ferox. Most comments pertain to a description of the bony carapace and plastron, which Geoffroy depicts in Plate 4. It is a young specimen judging from the small and isolated preneural; the seventh pair of pleurals is unusual in being fused (no middorsal suture), and the neurals seem large in proportion to the size of the pleurals. The anterior border of the carapace is described as having tubercles. Geoffroy listed Testudo membranacea and Testudo rostrata as synonyms of carinatus. Fitzinger (1835:127) listed T. membranacea, T. rostrata and T. carinatus as synonyms of Trionyx javanicus (= T. cartilagineus), which was also described by Geoffroy (op. cit.:15). Duméril and Bibron (op. cit.:478, 482) considered carinatus to be the young of spinifer (ferox as synonym). Gray (1844:48), however, referred T. membranacea and T. rostrata to the synonymy of T. javanicus, but considered T. carinatus to be a synonym of T. ferox (op. cit.:50), an interpretation followed by all subsequent authors. Trionyx carinatus is questionably listed as a synonym of ferox by Stejneger (1944:27). Duméril and Bibron (op. cit.:482) wrote that the young type of carinatus is in the museum at Paris. Dr. Jean Guibé informs me in letter of September 24, 1959, that the type of Geoffroy's T. carinatus cannot be found in the Natural History Museum at Paris. For the present, T. carinatus is considered a nomen dubium. According to Stejneger (1944:27), Trionyx brongniarti Schweigger is a substitute name for T. carinatus.
I am unable to add anything to Stejneger's (op. cit.:32) account of Trionyx harlani; the mention of its occurrence in east Florida indicates that it is indistinguishable from Testudo ferox Schneider.
T. ferox was considered to be indistinguishable from Lesueur's Trionyx spiniferus (described in 1827), until Agassiz (1857:401) pointed out the differences between the two species. However, Agassiz (op. cit.:402, Pl. 6, Fig. 3) regarded juveniles of T. spinifer asper as the young of ferox. Consequently, the geographic range of ferox, as envisioned by Agassiz, extended from Georgia and Florida west to Louisiana. Neill (1951:15) considered all [485] American forms conspecific. Crenshaw and Hopkins (1955) and Schwartz (1956) demonstrated that ferox is a distinct species.
Fitzinger (1843:30) designated the species ferox as the type species of his genus Platypeltis as follows: "Platypeltis. Fitz. Am[erica]. Platypelt. ferox. Fitz. Typus." If populations of soft-shelled turtles that are referable to Testudo ferox Schneider are considered to comprise a distinct genus by future workers, Platypeltis Fitzinger, 1835, is available as a generic name with Testudo ferox Schneider, 1783, as the type species (by subsequent designation).
Trionyx ferox in the northern part of its range is sympatric with T. spinifer asper. In the region of overlap, the two species are nearly always ecologically isolated; ferox inhabits lentic waters, whereas T. s. asper is partial to lotic waters (Crenshaw and Hopkins, op. cit.:16). There is no evidence of intergradation or hybridization.
Many characters of Trionyx ferox that are lacking in other North American forms are shared with some Asiatic softshells, such as the large size, longitudinal rows of tubercles that resemble ridges on the carapace, and the marginal ridge. It is thought that, of the living softshells in North America, ferox is more closely allied to Old World forms of the genus than to muticus or spinifer.
Carr (1940:107) recorded ferox from Okaloosa County, Florida, in the western end of the panhandle, whereas Crenshaw and Hopkins (1955:16) list the known westward extent of range as Leon and Wakulla counties. AMNH 6933 from west of the Apalachicola drainage in Washington County, Florida, tends to substantiate Carr's record, which is not included on the distribution map.
Specimens examined.—Total 144, as follows: Florida: Alachua: UMMZ 64178, 100969; USNM 10545, 10704, "near" Gainesville; UMMZ 56599, Levy Lake. Brevard: AMNH 12878, Canaveral. Broward: UMMZ 109441, Hugh Taylor Birch State Park; USNM 109548, 22 mi. WNW, 6 mi. SSE Fort Lauderdale. Collier: USNM 86828, Tamiami Trail, "near" Birdon. Dade: AMNH 50936, UMMZ 10183, 110981, Miami; USNM 84079, 86942, 15 mi. from (west) Miami, Tamiami Trail; UMMZ 111371, 19 mi. W, 1.3 mi. S Miami; UI 28984, 35 mi. W. (Miami) Tamiami Trail; AMNH 69932-33, UMMZ 101582, 101584, 104024, 40-45 mi. W Miami, Tamiami Trail. Glades: UMMZ 100836, mouth of Kissimmee River. Hendry: UMMZ 106302, 10.2 mi. SE Devil's Garden; UMMZ 106303-04, 106321-22, 30 mi. S Clewiston, near Devil's Garden. Hernando: TU 13624, 0.5 mi. S Citrus Co. line on US Hwy. 19. Highland: AMNH 65537, 71618, Archbold Biol. Stat., Lake Placid; AMNH 65622, Hicoria. Hillsborough: TU 13960, Hillsborough River, ca. 20 mi. NE Tampa; USNM 51184, Tampa; USNM 71156, Plant City. Indian River: USNM 55316, Vero Beach; USNM 59318, Sebastian. Lake: UMMZ 36072, USNM 20189, 029210, 029339, 38123, Eustis; UMMZ 76754-56, Lake Griffin. Lee: UMMZ 102276, 14 mi. SE Punta Gorda. Leon: CNHM 33701, USNM 95767, Lake Iamonia; USNM 103736, Silver Lake. Marion: AMNH 8294-95, UMMZ 95613 (4), USNM 52476-83, 100902-04, Eureka; AMNH 63642, near Salt Springs. Martin: TNHC 1292, 8.4 mi. N Port Mayaca. Okeechobee: AMNH 57379-84, Lake Okeechobee; AMNH 5931-32, Kissimmee Prairie. Orange: USNM 51421, 56805, Orlando; KU 16528. Osceola: USNM 029448, 029450-64, 029467-68, 029470, 029474-75, Kissimmee. Palm Beach: UMMZ 54101, Palm Beach; USNM 73199, Delray Beach. Pinellas: USNM 51417-20, St. Petersburg. Polk: AMNH 25543, Lakeland; UMMZ 112380, 6.7 mi. S Lake Wales; USNM 60496, 60532, 60534, 61083-87, Auburndale. Putnam: USNM 4373, 7651, Palatka; USNM 26035, ponds "near" Welaka. Sarasota: USNM 61352, Lake Myakka. Sumpter: UMMZ 71791, Bushnell. Volusia: UMMZ 100673, Lake Helen. Washington: AMNH 6933, Washington. County unknown: AMNH 4758; USNM 8899, St. John's River: USNM 59727-28, Lake Okeechobee, "near" mouth Taylor's Creek; USNM 84080.
Georgia: Baker: SM 2083, USNM 029619, 38980-81, 70398, Mimsville. [486] Berrien: USNM 62217, Banks Mill Pond. Charlton: AMNH 69934, Okefinokee Swamp, SW Billy's Island; UMMZ 90010, east edge Okefinokee Swamp; USNM 84603, Okefinokee Swamp, Chesser's Island. Irwin: USNM 56804. Lowndes: UMMZ 67706, 10 mi. S Valdosta. McIntosh: USNM 19621, Darien.
South Carolina: Charleston: USNM 9670, Charleston.
No Data: AMNH 22750; USNM 71608-09.
Records in the literature.—Florida: Alachua: 10 mi. ENE Gainesville (Schwartz, 1956:18). Brevard: Merritt Island (Neill, 1958:6). Broward: Fort Lauderdale (Schwartz, op. cit.:19). Charlotte: (Carr, 1940:107). Clay: Green Cove Springs (Brimley, 1910:18); St. John's River (Crenshaw and Hopkins, 1955:21); Doctor's Inlet (Schwartz, op. cit.:18). Collier: Royal Palm Hammock (Crenshaw and Hopkins, op. cit.:20); 11.2 mi. E Monroe Station (Schwartz, op. cit.: 19). Columbia: (Carr, loc. cit.). Dade: Paradise Key (Schwartz, loc. cit.); Homestead (eggs, Stejneger, 1944:43). Duval: 4-10 mi. S Jacksonville (Deckert, 1918:31). Glades: ca. 8 mi. SW Okeechobee State Park. Lake: Alexander Springs (Schwartz, op. cit.:18). Lee: 18 mi. S Fort Myers (Conant, 1930:63); 6 mi. SE Fort Myers (Hamilton, 1947:209). Levy: Gulf Hammock (Schwartz, loc. cit.); Brownson (Stejneger, op. cit.:42). Monroe and Okaloosa (Carr, loc. cit.). Okeechobee: 6 mi. E Kissimmee River; state hwy. 78 "near" Okeechobee-Glades co. line. Palm Beach: SW part of Lake Okeechobee, near Clewiston; Milton Island Cove (Schwartz, loc. cit.). Pasco: mouth Pithlachascotee River (Neill, op. cit.:26). Pinellas: Belleair (Brimley, loc. cit.); Seminole (Conant, loc. cit.); 5 mi. E Clearwater (Schwartz, op. cit.:19); Gulf Port (Stejneger, op. cit.:43). Polk: Lake Shipp, near Winter Haven (Telford, 1952:185). Sarasota: 15 mi. E Sarasota (Conant, loc. cit.); Venice (Conant, op. cit.:61). Taylor: "near" Foley. Wakulla: "near" Crawfordville (Crenshaw and Hopkins, op. cit.:15).
Georgia: Baker: 5 mi. NW Newton, 5 mi. W Newton, 4 mi. N Newton. Ben Hill: 6 mi. E Fitzgerald (Crenshaw and Hopkins, 1955:15). Bulloch: 14 mi. SE Statesboro (Schwartz, 1956:19). Decatur: "near" Bainbridge (Crenshaw and Hopkins, loc. cit.). Emanuel: "near" Midville. Evans: 8 mi. NE Manassas, Tattnall County. Ware: Laura Walker State Park (Schwartz, loc. cit.). Wilcox: 3 mi. SE Forest Glen (Crenshaw and Hopkins, op. cit.:19).
South Carolina: Beaufort: 7 mi. NE Gardens Corner (Schwartz, 1956:19). Chatham: Savannah River at Savannah (Schwartz, op. cit.:8-9). Colleton: 5 mi. from Whitehall, Combahee River (Schwartz, op. cit.:19).
Trionyx spinifer Lesueur
Spiny Softshell
Range.—In Canada, southern Ontario and Quebec; in the United States, northwestern Vermont and western New York south to northern Florida, east to central Montana, eastern Wyoming and Colorado, and New Mexico; introduced into the Colorado River system of California, Nevada, Arizona and New Mexico; in México, the northern part of the states of Tamaulipas, Nuevo León, Coahuila, and eastern Chihuahua (see map, Fig. 19).
Diagnosis.—Juvenal pattern uniform tan or brownish lacking markings, having whitish dots or spots, or having well-defined, blackish ocelli or spots; surface of carapace "sandpapery" in adult males; conical projections (in some subspecies) along anterior edge of carapace in large females; contrasting pattern of blackish marks on pale background (in some subspecies) on dorsal surface of limbs of adult males.
Opisthotic-exoccipital spur well-developed; epiplastral callosity, when present, not covering entire surface.
Description.—Septal ridges present; external and proportional characteristics variable (see accounts of subspecies); range in length of plastron (cm.) of ten [487] largest specimens of each sex (mean follows extremes), males, 13.8-16.0, 14.4; females, 26.0-31.0, 28.0.
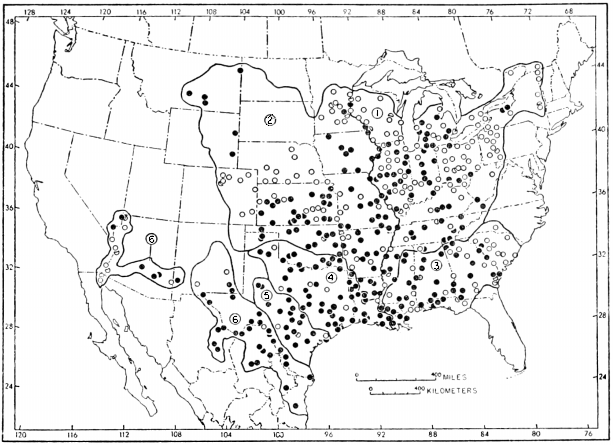
Fig. 19. Geographic distribution of Trionyx spinifer.
Guide to subspecies:
Greatest width of skull usually at level of squamosal (74%); foramen magnum rhomboidal; ventral surface of supraoccipital spine narrow proximally, usually having medial ridge; opisthotic-exoccipital spur well-developed (66%); distal part of opisthotic wing tapered, not visible in dorsal view; lateral condyle of articular surface of quadrate larger than medial articular surface, not tapered posteriorly; maxillaries in contact above premaxillaries (88%); usually a combination of seven neurals, seven pairs of pleurals and contact of seventh pair of pleurals (83%); angle of epiplastron approximately 90 degrees; callosities when present on epiplastron not covering entire surface; hyo-hypoplastral suture usually present.
Comparisons.—Trionyx spinifer can be distinguished from T. ferox and T. muticus by the presence of any one of the characters mentioned in the "Diagnosis." Both sexes and all sizes of T. spinifer resemble ferox but differ from muticus in having septal ridges. Most individuals of T. spinifer (except some large females) resemble muticus but differ from ferox and large females of ater in having a pale outer rim that is separated from the ground color of the carapace by a distinct (spinifer) or dusky (muticus) dark line. Large females of the subspecies spinifer, hartwegi, asper and pallidus may have enlarged conical projections along the anterior edge of the carapace and, unless these projections are considerably worn, are readily distinguished from large females of [488] ferox (flattened, knoblike prominences), and muticus and ater (smooth surface, no prominences). Large females of the subspecies guadalupensis and emoryi resemble muticus and ater, and to some extent ferox, in having low, scarcely elevated prominences along the anterior edge of the carapace. Some females of emoryi resemble ferox in that the plastron extends farther forward than the carapace.
T. spinifer is intermediate in size between ferox (larger) and muticus (smaller); the maximum size of the plastron in adult males is approximately 16.0 centimeters (14.0, muticus; 26.0, ferox), and of females, 31.0 centimeters (21.5, muticus; 32.5, ferox). The head for all subspecies of spinifer is proportionately narrower than in ferox but wider than in muticus.
In the skull, spinifer more closely resembles ferox than muticus, but differs from both ferox and muticus in usually having a well-developed opisthotic-exoccipital spur. Skulls of spinifer resemble those of muticus but differ from those of ferox in being widest at the level of the squamosal. Skulls of spinifer resemble those of ferox but differ from those of muticus in having the 1) ventral surface of the supraoccipital spine narrow proximally, and usually having a medial ridge, 2) foramen magnum rhomboidal, 3) distal part of opisthotic wing tapered, 4) lateral condyle of articular surface of quadrate not tapered posteriorly, and larger than medial articular surface, and 5) maxillaries in contact above premaxillaries. T. spinifer resembles ferox but differs from muticus in having the epiplastron bent at an approximate right angle. T. spinifer differs from ferox in having an epiplastral callosity, and from muticus in that the callosity does not cover the entire surface of the epiplastron. The hyo-hypoplastral suture is present more often in spinifer and muticus than in ferox.
Remarks.—Gray (1869:221) proposed the generic name Callinia as a new name for Aspidonectes as understood by Agassiz (1857:403). Gray referred Trionyx spiciferus (= spiniferus) Lesueur to Callinia. Stejneger (1907:514) designated Trionyx spiniferus Lesueur as the type species of Callinia. If Trionyx spiniferus Lesueur is considered to be generically distinct from other soft-shelled turtles, Callinia Gray, 1869, is available as a generic name with Trionyx spiniferus Lesueur, 1827, as the type species by subsequent designation.
Geographic variation.—T. spinifer is the most variable and widespread species of the genus in North America. Size of ocelli on the carapace decreases from east to west on turtles inhabiting waterways of the Upper Mississippi River drainage. The most impressive gradient, geographically oriented from western Louisiana to southwestern Texas is seen in each of several features: decrease in size of tubercles on the anterior edge of the carapace, reduction in contrast of pattern on the dorsal surface of limbs and side of head, change in pattern on the dorsal surface of the snout, and increase in the size of white spots on the carapace. But the gradient in size of white spots is reversed in T. s. emoryi, which has small white spots on the carapace. Some of the characters at the western terminus of this geographical gradient are shared with [489] T. ater and muticus. Those subspecies comprising the emoryi group also show proportional characters that correspond closely with those of T. ferox.
On the basis of tuberculation and pattern on carapace, side of head, dorsal surface of limbs and snout, Trionyx spinifer may be divided into six subspecies.
Trionyx spinifer spinifer Lesueur
Eastern Spiny Softshell
Trionyx spiniferus Lesueur, Mém. Mus. Hist. Nat. Paris, 15:258, pl. 6, December, 1827.
T[rionyx] s[pinifer] spinifer Schwartz, Charleston Mus. Leaflet, No. 26:11, May, 1956.
Trionyx ocellatus Lesueur, Mém. Mus. Hist. Nat. Paris, 15:261, December, 1827.
Apalone hudsonica Rafinesque, Atlan. Jour., Friend Knowledge, Philadelphia, 1(No. 2, Art. 12):64, Summer, 1832.
Trionyx annulifer Wied-Neuwied, Riese Nord-Amerika, 1(pt. 3):140, 1838.
Tyrse argus Gray, Cat. Tort. Croc. Amphis. Brit. Mus., p. 48, 1844.
Aspidonectes nuchalis Agassiz, Contr. Nat. Hist. United States, 1(pt. 2):406, 1857.
?G[ymnopus] olivaceus Wied-Neuwied, Nova Acta Acad. Leopold.-Carol., 32:55, pl. 5, 1865.
Type.—Lectotype, Museum d'Histoire Naturelle, Paris, No. 8808; large stuffed female obtained by C. A. Lesueur from the Wabash River, New Harmony, Posey County, Indiana (Pl. 52).
Range.—Northeastern United States and extreme southeastern Canada in tributaries flowing into the Mississippi River from the east, and the St. Lawrence River drainage; extreme southern Quebec and Ontario, Canada, east through southern Great Lakes region to Wisconsin, and south through New York, western Pennsylvania and Illinois to Tennessee and western Virginia (see map, Fig. 19).
Diagnosis.—Juvenal pattern of large, thick-bordered black ocelli, often 9-10 millimeters in diameter in center of carapace on adult males, and 2-3 millimeters in diameter on hatchlings (mean OD/PL, Michigan, .066); only one dark marginal line separating pale rim of carapace from dorsal ground color.
Description.—Plastral length of smallest hatchling, 2.7 centimeters (UMMZ 89950, INHS 3143); of largest male, 14.5 centimeters (UMMZ 72512); of largest female, 31.0 centimeters (UMMZ 40866).
Carapace olive, having large ocelli in center but smaller ocelli or spots at sides; ocelli often interrupted; pale rim of carapace not four or five times wider posteriorly than laterally, separated from darker ground color of carapace by one dark marginal line; large females often having remnants of ocelli at sides of carapace on mottled and blotched background; pattern on snout of pale, dark-bordered stripes that unite forming acute angle in front of eyes; well-defined dark markings in subocular and postlabial region; pattern contrasting [490] with ground color on side of head; postlabial stripe interrupted, diffuse; pale postocular stripe having blackish borders interrupted, not uniting with postlabial stripe; dorsal surface of soft parts of body having contrasting pattern, largest blackish marks on hind limbs; elongate tail of adult males having pale dorsolateral bands with well-defined lower blackish borders; underparts whitish, often having blackish marks, except in center of plastral area; dark marks on webbing of limbs, palms and soles; dark streaks often coincident with digits; small conical tubercles on anterior edge of carapace on adult males; conical or equilateral tubercles on anterior edge of carapace of large females; accessory knoblike tubercles in nuchal region and in middle of carapace posteriorly on large females.
Ontogenetic variation in PL/HW, mean PL/HW of specimens having plastral lengths 7.0 centimeters or less, 4.09, and exceeding 7.0 centimeters, 5.50; ontogenetic variation in CL/CW, mean CL/CW of specimens having plastral lengths 8.5 centimeters or less, 1.12, and exceeding 8.5 centimeters, 1.21; mean CL/PCW, 2.02; mean HW/SL, 1.30 (including subspecies hartwegi); mean CL/PL, 1.39.
Variation.—Variant individuals include: UMMZ 72512, an adult male, having some ocelli seven millimeters in diameter that are almost solid spots; UMMZ 89659 having postocular and postlabial stripes connected on right side of head; UMMZ 95615, 52948, 54402 having inner dark borders of pale stripes on snout represented by short dashes and dots (a ragged line connecting anterior margins of orbits on 54402); UMMZ 52948, 89659 having interrupted, black marginal lines on carapace with ends of some segments oriented inward and overlapping portion of adjacent segments; UMMZ 81699, female having plastral length of 19.0 centimeters, lacking conspicuous tubercles on anterior edge of carapace; UI 2403, CNHM 92204 having extensive dark mottling and marbling on throat and neck, undersurface of limbs and posterior portion of carapace.
Comparisons.—T. s. spinifer can be distinguished from all other subspecies of T. spinifer by the presence of large black ocelli (diameter 9-10 mm. on adult males, 2-3 mm. on hatchlings) in combination with only one dark marginal line. T. s. spinifer resembles asper in having ocelli or dots on the carapace but differs from asper in having only one dark marginal line and larger ocelli. T. s. spinifer differs from hartwegi only in the large size of the ocelli. T. s. spinifer resembles hartwegi and asper but differs from pallidus, guadalupensis and emoryi in having blackish spots and ocelli on the carapace and lacking whitish dots. T. s. spinifer resembles hartwegi, asper, and pallidus and differs from guadalupensis and emoryi in having conical or knoblike tubercles on the anterior edge of the carapace on large females.
T. s. spinifer differs from the subspecies asper, guadalupensis and emoryi in having a relatively narrower head, and from emoryi in having a relatively wider carapace. T. s. spinifer resembles hartwegi and asper but differs from the other subspecies in having the carapace widest at a plane approximately one-half way back on the carapace. The subspecies spinifer and hartwegi have longer snouts than pallidus, guadalupensis, and emoryi. T. s. spinifer differs from asper but resembles all the other subspecies in having a relatively longer plastron.
Remarks.—Lesueur's description of Trionyx spiniferus (1827:258-261, Pl. 6) seems to be based mostly, if not entirely, on a large female (length of carapace, 13 inches), which was "Le plus grand des individus observes …" [491] (op. cit.:258); an accompanying illustration depicting the dorsal surface of the bony carapace is unusual in lacking neurals (Pl. 6, E). Duméril and Bibron (1835:481) mentioned eight or nine additional specimens that Lesueur sent to the Museum of Natural History in Paris. Dr. Jean Guibé informed me under letter dated September 24, 1959, that a larger stuffed female, bearing catalog number 8808 is regarded as the holotype, and that there are seven additional specimens (1949, 4143, 8807, 8809-12) in the museum at Paris. All turtles were obtained by Lesueur from the Wabash River. To my knowledge no specimen that was available to Lesueur has been specifically designated as a type. Because the description seems to be based on one specimen, undoubtedly No. 8808, this specimen has been regarded as the holotype. However, Lesueur referred to several specimens and did not mention a type in the original description; consequently I prefer to regard No. 8808 as a lectotype.
Lesueur also described Trionyx ocellatus (op. cit.:261-263) as a variety of T. spiniferus having ocelli, or parts thereof, on the carapace and mentioned three specimens. The total number of specimens that were available to Lesueur is unknown. One young alcoholic specimen having ocelli is in the British Museum (Natural History) (Gray, 1855:69). The same letter from Dr. Guibé stated that a specimen in the Museum of Natural History, Paris, No. 6957, having a carapace 17 centimeters in length, conforms to the characters of ocellatus as mentioned by Lesueur, and was obtained from the Wabash River by Lesueur. Two of the specimens mentioned by Lesueur (loc. cit.) are stated to be females. No. 6957 is an adult male and clearly shows the juvenal pattern; it is regarded as the lectotype of T. ocellatus Lesueur, a name-combination, which is a synonym, based on a secondary sexual difference in pattern.
Rafinesque (1832:64) described a soft-shelled turtle from "the River Hudson between the falls of Hadley, Glen and Baker, and further up to the source" as Apalone hudsonica. The most outstanding characteristic was the presence of five claws on the digits of each limb. Rafinesque's recording of this characteristic was perhaps influenced by the illustration of a softshell in Bartram's Travels that showed each limb with five, clawed digits. Perhaps this was the basis for Boulenger (1889:245, footnote) regarding Apalone as "mythical." The large, yellowish, black-bordered spots, one behind and one in front of the eye presumably represent segments of the postocular stripe and the stripe on the snout; Rafinesque described the carapace as "entire … the margin is yellowish unspotted, then comes a circular black line …" and having "many round spots occulated and clouded by having a brown margin, with grey dots within." Except for five claws, the description is applicable to a softshell and referable to T. s. spinifer. To my knowledge, the only other records of the occurrence of soft-shelled turtles in the Hudson river drainage are those of Eights (in Bishop, 1923:120, Mohawk River at Cohoes), and DeKay (1842:7, Mohawk River and Hudson River near Albany); presumably these records are the basis for the comments of Holbrook (in Bishop, loc. cit.), and symbolized as an isolated locality by Conant (1958:318, map 35). The type locality of Apalone hudsonica is herein restricted to the Hudson River, near Baker's Falls, Saratoga County, New York.
Gray (1844:48) proposed the name Tyrse argus for a specimen reported to have come from Sierra Leone, West Africa; later (1855:68), he referred the species to the genus Trionyx. After comparison with a specimen of T. spiniferus Lesueur, Gray (1864:89) was "doubtful whether there must not have been some [492] confusion about the habitat of the specimen [which formed the basis of the description of Tyrse argus], and whether it is not more probably a North American species." The same author (1869:222; 1870:109) listed Tyrse argus as a synonym of Callinia spinifera (= Trionyx spiniferus Lesueur).
Agassiz (op. cit.:406-07) described Aspidonectes nuchalis on the basis of three adults from the Cumberland River and a number of young from the headwaters of the Tennessee River. Boulenger (1889:245, footnote 2) suggested that the status of A. nuchalis required further investigation. The species was not generally recognized after the turn of the century. Barbour and Loveridge (1929:226) listed MCZ 1908 (one of the juveniles) and 1623-25 as cotypes. Stejneger (1944:52) showed that nuchalis was not distinguishable from T. s. spinifer, and (op. cit.:49) listed MCZ 1623-25 as cotypes. Schmidt (1953:110) restricted the type locality to the Cumberland River, near Nashville, Tennessee.
Agassiz (loc. cit.) mentioned that nuchalis "differs strikingly from Asp. spinifer in the much more elongated form of the male, and in the great development of the marginal spines and of the tubercles upon the carapace, … But the most prominent specific character consists in the marked depressions on either side of the blunt median keel, and also in the triangular dilation of that keel behind the front margin of the carapace." These characters seem to be of no taxonomic worth. I have seen three syntypes (MCZ 1623-25) that undoubtedly correspond to the three adult specimens mentioned by Agassiz. All are females, measuring 19.5, 22.0, and 19.0 centimeters, respectively, in plastral length, and lack a contrasting mottled pattern on the carapace; the juvenal pattern is obscured, except for blackish spots at the edge of the carapace on MCZ 1625, and parts of an ocellus on MCZ 1624. The dorsal surfaces of the limbs are boldly marked. MCZ 1623, showing the diagnostic feature mentioned by Agassiz, is photographed by Stejneger (op. cit.:Pls. 14, 15), and may be regarded as the lectotype of Aspidonectes nuchalis Agassiz. MCZ 1908 is one of the young syntypes mentioned by Agassiz, and is referable to spinifer. The juvenal pattern consists of spots and ocelli; the plastron measures 3.1 centimeters in length, and the carapace 4.2 centimeters.
Wied-Neuwied (1865:55-57, Pl. 5) described the species ?G[ymnopus] olivaceus, but was uncertain whether his interpretation was based on a species, a variety or a secondary sexual difference. Wied-Neuwied mentioned that Lesueur had already named this soft-shelled turtle as Trionyx ocellatus, and agreed with Lesueur that those turtles having occulated spots on the carapace were distinguishable from T. spiniferus and T. muticus. But because Duméril and Bibron in their Erpétologie Général failed to recognize T. ocellatus, Wied-Neuwied felt obliged to bring it to the attention of his American colleagues and he renamed it. Wied-Neuwied also stated, in the context of a synonym, "Beschreibung einer Reise in Nord-America Bd. I., pag. 140." This comment presumably refers to his earlier description of T. annulifer (1838:140); seemingly Wied-Neuwied considered T. annulifer and G. olivacea as conspecific, although there is no mention of annulifer in the text proper. Stejneger (op. cit.:49) designated the type locality of T. annulifer as the Ohio River at Pittsburgh, Pennsylvania, and of Gymnopus olivacea as New Harmony, Wabash River, Illinois (lapsus for Indiana).
Trionyx spiniferus was questionably considered distinct from T. ferox by [493] Lesueur who listed "Testudo ferox Gm. Tortue de Pennant?" and "Trionyx georgicus Geoffr.?" as synonyms. Subsequently, most authors considered T. spiniferus synonymous with T. ferox until Agassiz (1857) pointed out differences between the two species.
The average size of the ocelli on the carapace of the subspecies spinifer decreases westward toward the Mississippi River; ocelli of different sizes occur on different individuals from the same state and presumably from the same population. For example, INHS 2281, plastron 9.9 centimeters in length, from Effingham County, Illinois, has some ocelli eight millimeters in diameter, whereas a larger male from the same locality, UI 1322, plastron 11.6 centimeters in length, has the largest ocelli only five millimeters in diameter. For convenience, all softshells having locality data from states east of the Mississippi River are referred to spinifer, recognizing that intergradation occurs with hartwegi over a broad area paralleling the Mississippi River. The type locality of spinifer is in an area where most turtles do not have the larger ocelli (diameter of seven to ten mm. on adult males); however, some individuals from the Wabash River (UMMZ 63523, adult male, plastron 11.5 cm. in length, ocelli diameter seven mm.) agree with more "typical" spinifer to the east. Intergradation with asper possibly occurs in that part of the Tennessee River in eastern Tennessee as exemplified by UMMZ 59198.
Published reports indicate that T. s. spinifer is not abundant in some of the northeasterly parts of its geographic range. Adams and Clark (1958:10) wrote that few softshells at Long Point on the Canadian side of Lake Erie are "ever collected and the area's game keepers report … (none) … seen in recent years. They also tell of recurrent severe stormy winters in which the muddy bottom of the marshland was repeatedly churned up and frozen. Such climatic conditions could easily destroy a large part of the Trionyx population overwintering in the mud bottom." Wright (1919:8) reported that softshells are "rarely seen" in bays on the New York side of Lake Ontario, and Babcock (1938:53) wrote that spinifer "is not common in Lake Champlain."
T. s. spinifer probably extended its geographic range into the Hudson River drainage of New York via the Erie Canal (connected Buffalo and Albany) after its completion in the early 1800's (DeKay, 1842:7). Now, the New York Barge Canal (essentially the Erie Canal, but with minor changes in course and the addition of several spurs) provides an avenue for dispersal of spinifer to the Hudson River drainage, Lake Ontario and intervening waterways in New York (Mertens, 1928:199). Netting (1944:86-87), however, suggested that spinifer occupied Lake Champlain, the Finger Lakes, Mohawk River and upper Hudson in the late stages of the formation of the Great Lakes.
A publication not seen by me is that of Mansueti and Wallace (1960). Its title suggests that Trionyx occurs in Maryland.
The unsuccessful introduction of T. s. spinifer in the Delaware drainage in New Jersey has been discussed by Fowler (1907:213), who wrote that they were found as early as the late 1860's and were introduced when young presumably to stock aquaria. Records of occurrence include Cooper's Creek, Camden County (Stone, 1906:168); Woodbury, Gloucester County (Cope, 1894:889); and Paulins Kill at Hainesburg, Warren County (Johnson, 1894:889).
Surface (1908:122) believed that soft-shelled turtles "have doubtless been [494] introduced into the eastern part of Pennsylvania through the canal from the Western and Central part of New York," and Roddy (in Neill, 1951:21) suggested that the species may be found in the Susquehanna River. Babcock (1919:420) mentioned a young specimen of spinifer in the collection of the Boston Society of Natural History that was obtained "in White River, Vermont," a tributary of the Connecticut River of the Atlantic Coast drainage; seemingly this record has not been accepted and the species is not established. To my knowledge, populations of T. s. spinifer do not occur in rivers of the Atlantic Coast drainage, except probably the Hudson-Mohawk drainage.
Stockwell (1878:401) wrote that spinifer was found "as high as Athabasca." Presumably Stockwell referred to Lake Athabaska in northern Alberta and Saskatchewan, Canada, a region where soft-shelled turtles are unknown; see also the comments by Stejneger (1944:52).
Specimens examined.—Total 250 as follows: Alabama: Morgan: UMMZ 99578, "near" Decatur.
Illinois: Adams: INHS 2150, Quincy. Bond: INHS 8345, Greenville. Carroll: CNHM 42116, Ordnance School Proving Ground. Cass: INHS 2151, Beardstown. Champaign: INHS 2273, 2311, 2413, 3142, "near" Seymour; INHS 4229, Champaign; INHS 6163, Sidney. Christian: INHS 1560, Pana. Coles: INHS 1968-69, 2 mi. W Charleston. Cumberland: INHS 2282, Greenup. De Witt: INHS 7674, Farmer City. Effingham: UI 1322, 2281, 19365, "near" Effingham. Fulton: INHS 5531, 2 mi. NE Bluff City, Schyler County; UI 23449, Liverpool; UI 24611, Spoon River, 18 mi. NW Canton. Hancock: USNM 53522, 59277, "near" Hamilton. Iroquois: INHS 6869-70, 2.5 mi. N Crescent City. Jackson: TU 1369 (12), Elkville. Kane: CNHM 42400, Aurora. Kankakee: CNHM 324, Momence. Kendall: UI 2411, Plano. Logan: INHS 7171-72, 6 mi. N Lincoln. Madison: USNM 60571. Macoupin: UI 2401-02, Beaver Dam Lake. Mason: CNHM 346, 470, INHS 1122, 1559, 5756-58, UI 42, 2404, Havana, Lake Chautauqua. Mercer: CNHM 3220, New Boston. Morgan: CNHM 2067 (2), 3290, 3303-04, 3306, INHS 2152, 2154, 5132-37, USNM 54747, Meredosia. Moultrie: INHS 8989, 2 mi. NW Lovington. Peoria: UI 2406-10, Peoria. Pope: INHS 5505, Lake Glendale. Putnam: UMMZ 81604-14, 5 mi. N Henry, Marshall County. Schuyler: UI 2405, "near" Ripley, Brown County. Scott: INHS 2149, 2153, Naples. Union: CNHM 18623, 6 mi. SW Jonesboro. Vermilion: INHS 3142, Muncie; INHS (1 untagged); UI 1970, 3209, Danville; UI 2403, 1.5 mi. E Oakwood; UI 16265, Kickapoo State Park. Wabash: USNM 12061, Mt. Carmel. Winnebago: INHS 7185, Kishwaukee Forest Preserve; INHS 7294, 1/2 mi. S Shirland. County unknown: USNM 7661.
Indiana: Bartholomew: UMMZ 61060, 10 mi. W Columbus. Carroll: USNM 42905-06, Burlington. Clark: UMMZ 110599, 14-mile Creek, 3 mi. NW Charleston. Decatur: UMMZ 55416, 3 mi. S Westport. Elkhart: UMMZ 105598, Elkhart River, south of Goshen. Gibson: UMMZ 89744, Foot's Pond. Johnson: UMMZ 108062, 2 mi. S Trafalgar. Knox: USNM 22711, Vincennes. Kosciusko: AMNH 8379, UMMZ 84287 (5), Winona Lake; UMMZ 110235, Wawasee Lake. Lake: CNHM 11019, 11021-24, Crown Point. Marion: UMMZ 103393, Ravenswood; UMMZ 110236, 1 mi. N Lawrence. Marshall: CNHM 39299; USNM 33495, Yellow River north of Burr Oak; USNM 33496-501, 35404, 42583-84, Lake Maxinkuckee. Wells: UMMZ 63523, Wabash River, Bluffton. County unknown (Lagrange or Marshall): USNM 50670, Twin Lakes.
Kentucky: Casey: UMMZ 112252, trib. of Green River, south of Yosemite. Green: UMMZ 116718, Little Barren River, 1.5 mi. E Monroe, Hart County. Rockcastle: UMMZ 98767, Rockcastle River, 5 mi. above Livingston.
Michigan: Allegan: UMMZ 42112, Kalamazoo River. Barry: UMMZ 53874, Thornapple River, 3 mi. NW Hastings. Bay: UMMZ 74670. Branch: UMMZ 95615, 1 mi. S Kinderhook; UMMZ 70748, Hog Creek. Calhoun: UMMZ 89950 (3); UMMZ 79133, near Battle Creek. Cass: UMMZ 40866-67, 53005, Diamond Lake; UMMZ 40868, 52948, Long Lake. Jackson: UMMZ [495] 72494. Kalamazoo: UMMZ 42130, 80534, Kalamazoo; UMMZ 90506, Gull Lake; UMMZ 92599, Kellogg Bird Sanctuary. Lenawee: UMMZ 72457, Devil's Lake; UMMZ 74662, Wolf Lake Park. Livingston: UMMZ 54401, 76190, Portage Lake. Monroe: UMMZ 44604-06, USNM 51213, "near" Monroe. Newaygo: UMMZ 63469. Oakland: UMMZ 64363, Hay's Creek; UMMZ 96539, Clinton River. Ottawa: UMMZ 81699. St. Joseph: UMMZ 38876, 38889, "near" White Pigeon; UMMZ 96537, Corey Lake. Van Buren: UMMZ 90003, Wolf River, west of Kalamazoo, Kalamazoo County. Washtenaw: SM 2035, 2038, 2105, UMMZ 39847, 96538, "near" Ann Arbor; UMMZ 35765, 35769, 74518 (2), Portage Lake; UMMZ 54402-03, Little Lake; UMMZ 89659, Huron River, Dexter; UMMZ 110583-85. County unknown (Washtenaw or Livingston): UMMZ 54400, Huron River near Portage Lake.
Mississippi: Adams: MCZ 46615, UMMZ 76446, "near" Natchez; MCZ 46621, 46633, USNM 01084, 01086, Washington. Coahoma: AMNH 5289, 5285-86, Moon Lake. Lafayette: MCZ 37173, Oxford; USNM 7650, Abbeville? (reported from Abbeville, South Carolina by Pickens, 1927:113; see discussion by Stejneger, 1944:50, and my comments on page 509 beyond). LeFlore: USNM 73668-69, Greenwood. Madison: USNM 95192, Big Black River. Washington: USNM 115980, Deer Creek. Yazoo: UMMZ 86669, Panther Creek west of Yazoo City; UMMZ 83304, Yazoo City.
New York: Monroe: CNHM 92001-02, Genesee River, Rochester. Wayne: AMNH 69931, CNHM 92004, Sodus Bay.
Ohio: Athens: UMMZ 111793, east branch Shade Creek. Franklin: USNM 26290. Lucas: USNM 51214, Toledo. Pike: UMMZ 99309, Morgan's Fork, Sunfish Creek. Warren: AMNH 4763, Little Miami River, 3 mi. below Morrow. County unknown: USNM 21128-29, Cuyahoga River.
Tennessee: Benton: UMMZ 113036, Eagle Creek, 1/2 mi. E Holliday. Bradley: UMMZ 59197, west branch of Chestnee Creek, 7 mi. E Cleveland. Claiborne: USNM 86677, 5 mi. SE Cumberland Gap, Powell River. Davidson: MCZ 1623-25, Cumberland River near Nashville (restricted locality); USNM 7165-67, Nashville. Decatur: KU 3000, Perryville. Hamilton: USNM 131861, Chattanooga. Monroe: TU 16058, Little Tennessee River, 10 mi. N Madisonville. Obion: UMMZ 53199, USNM 102911, Reelfoot Lake. Overton: UMMZ 69561 (2), Wirmingham. Sevier: TU 16132, UMMZ 86735, USNM 86681-82, near Sevierville; UMMZ 86734, Walden Creek "near" Gatlinburg. County unknown: MCZ 1908, headwaters of Tennessee River.
Virginia: Smythe: USNM 101386, Holston River, Seven Mile Ford.
West Virginia: McDowell: USNM 33767, Dry Fork, Perryville (county questionable, perhaps Randolph County).
Wisconsin: Chippewa: CNHM 8223, Lake Wissota, mouth of Yellow River, Anson Twp. Polk: UMMZ 72511-12, St. Croix River "near" Never's Dam. County unknown: CNHM 15971, Eau Claire River.
Records in the literature.—Ontario: Carleton: Ottawa (questionable record). Essex: Point Pelee. Haldimand: Dunville. Kent: Lake St. Clair. Norfolk: Long Point. Oxford: Beachville. Wentworth: Hamilton Bay (Logier and Toner, 1955:51).
Quebec: Iberville: Richelieu River at Iberville (Logier and Toner, 1955:51).
Alabama: Lawrence: Courtland (Stejneger, 1944:53).
Illinois: Boone: Belvidere. Bureau: Bureau. Cass: Chandlerville. Clay: Louisville (Cahn, 1937:189). Cook: Lake Michigan (Kennicott in Stejneger, 1944:44); Evanston (Necker, 1939:10); Chicago (Schmidt and Necker, 1935:76). Crawford: Robinson. Douglas: northern part of county (P. W. Smith, 1947:39). Fayette: Vandalia. Fulton: Ellisville (Cahn, loc. cit.). Grundy: Morris (Stille and Edgren, 1948:201). Jackson: Jacob (Cagle, 1942:158). Jersey: Grafton (Cahn, loc. cit.). Kane: Batavia; Dundee Game Farm (Stille and Edgren, loc. cit.). Kankakee: Kankakee River near Altort (Necker, loc. cit.). Lake: Fox Lake. LaSalle: Streator (Cahn, loc. cit.). Lawrence: (Hahn in Stejneger, 1944:44). Lee: symbol on map (Cahn, loc. cit.). McHenry: McHenry (Stille and Edgren, loc. cit.). Macon: Decatur. Macoupin: Carlinville (Cahn, loc. cit.). Ogle: Oregon (Garman in [496] Cahn, loc. cit.). Randolph: Chester, Reily Lake. Rock Island: Barstow, Hillsdale, Rock Island (Cahn, loc. cit.). Saline: Horseshoe Lake (Stein, 1954:312). Stephenson: Freeport (Cahn, loc. cit.). Union: Bluff Lake (Garman in Cahn, loc. cit.). Whiteside: Sterling, symbol on map (Cahn, loc. cit.). Williamson: Marion (Cagle, 1942:158). Winnebago: Rockton; symbol in western part of county (Cahn, loc. cit.). County unknown: Fox River (Yarrow, 1882:29).
Indiana: Brown: 1 mi. below Helmsburg (Myers, 1927:339). Clay: Eel River (Kirsch in Stejneger, 1944:45). Franklin: (Hughes in Stejneger, loc. cit.). Jasper: Jasper-Pulaski Game Preserve (Swanson, 1939:690). Jefferson: Madison (Myers, loc. cit.). Marion: Irvington (Stejneger, op. cit.:55). Marshall: 2 mi. NW Culver (KKA). Monroe: Bloomington (McLain in Stejneger, op. cit.:45). Newton: Lake Village (Stille and Edgren, loc. cit.). Posey: Wabash River at New Harmony (Lesueur, 1827:257). Starke: Grant (Stille and Edgren, loc. cit.). Steuben: Fish Creek "near" Hamilton (Stejneger, op. cit.:53). County unknown (Knox or Starke): USNM 72387, Knox (Stejneger, op. cit.:55); "White Water valley," east-central part of state (Butler, 1894:224). USNM 8359 (= Trionyx spinifer asper) has been erroneously recorded from Madison, Indiana, by Yarrow (1882:29) and Hay (1892:145); see discussion by Cahn (1937:200) and Stejneger, (op. cit.:73, 75).
Kentucky: Edmonson: Green River, Mammoth Cave National Park (Hibbard, 1936:281). Fleming: Fox Creek (Welter and Carr, 1939:130). Jefferson: (Funkhouser, 1925:71). Morgan: (Stejneger, 1944:54). County unknown: Ohio and Pond rivers (Funkhouser, loc. cit.).
Michigan: Berrien: mouth of St. Joseph River at St. Joseph (Lagler, 1943:303). Eaton: Brookfield; Olivet (Clark in Ruthven, Thompson and Thompson, 1912:133). Genesee: (Miles in Ruthven, Thompson and Thompson, loc. cit.). Iosco: (Lagler, 1943:283, symbol on map). Kent: (Lagler, loc. cit.). Montcalm: (Clark in Ruthven, Thompson and Thompson, loc. cit.). Muskegon: Muskegon River "near" Muskegon (Lagler, op. cit.:303). Van Buren: Reynolds Lake, 2.5 mi. E Lawrence (Edgren, 1942:180).
Mississippi: De Soto: Lake Cormorant (Stejneger, 1944:55). Holmes: Thornton (Cook, 1946:185). Humphreys: Belzoni (Stejneger, loc. cit.). Sunflower: Warren: Vicksburg, Eagle Lake (Cook, loc. cit.). Washington: Lake Washington (Smith and List, 1955:125); Greenville (Stejneger, loc. cit.).
New York: Albany: Hudson River at Albany (DeKay, 1842:7); Mohawk River at Cohoes (Eights in Bishop, 1923:120). Cattaraugus: Allegheny River and Red House Lake in Allegheny State Park (Eaton, 1945:115). Chautauqua: Lake Chautauqua (DeKay, loc. cit.). Monroe: Braddocks Bay and Long Pond on Lake Ontario (Wright, 1919:8). Saratoga: Hudson River near Baker's Falls (restricted locality, Rafinesque, 1832:64). County unknown: Lake Cayuga; Mohawk River (DeKay, loc. cit.).
Ohio (Conant, 1951:158-59, 264, except records from Allen, Geauga and Noble counties): Allen: Sugar Creek, 6 mi. N Lima (Adler and Dennis, 1960:27). Ashland: Long Lake, Lake Twp.; Black Fork, Sec. 27, Green Twp. Athens: Hocking River "near" Athens; "near" Fisher, Alexander Twp. Auglaize: Pusheta Creek, west of Wapakoneta. Brown: White Oak Creek, 1 mi. N Higginsport. Butler: Oxford. Champaign: Mad River, 4 mi. SW Urbana. Coshocton: Walhouding River, below dam. Defiance: Auglaize River, Shawnee Scout Camp, Defiance Twp. Erie: Huron; Sandusky. Fairfield: Buckeye Lake. Franklin: Alum Creek, Westerville; Columbus. Geauga: Chardon Twp. (Wood, 1959:8). Greene: Huffman Dam. Hamilton: Harrison; mouth of Miami River. Hardin: "near" Hepburn. Henry: Maumee River, east of Napoleon; Maumee River "near" Texas; Maumee River, 3 mi. W Texas. Highland: Little Brush Creek, 2 mi. N Sinking Spring. Huron: Huron River "near" Monroeville. Jackson: Canter's Cove, Jackson Twp.; Jackson Lake. Knox: Brinkhaven. Lake: east branch Chagrin River, Kirtland; Grand River, 4 mi. E Painesville. Lawrence: Pine Creek, Elizabeth Twp. Logan: Miami River, "near" Indian Lake. Lorain: Oberlin. Lucas: Lake Erie at Reno Beach, Jerusalem Twp.; Lake Erie, 1/2 mi. offshore from mouth of Crane Creek; Maumee River at Maumee; Swan Creek, W of Toledo; "near" Waterville; Swan Creek "near" Whitehorse. Madison: London. Medina: [497] Hinckley Lake. Meigs: Shade River, below Darwin. Miami: Miami River, above Troy. Monroe: Cranenest Fork, Green Twp. Montgomery: Mad River, Dayton; Miami River, Dayton; Stillwater River, Dayton. Morrow: Kokosing River, Franklin Twp. Noble: Jct. Sharon Twp. 1 and St. Rt. 78. (Adler and Dennis, 1960:27). Ottawa: East Harbour, Catawba Island. Pike: Chenoweth Fork, Sunfish Twp.; Scioto River, Camp Creek Twp. Ross: Paint Creek near Bainbridge. Vinton: Lake Hope; Lake Alma. Warren: Fort Ancient. Washington: Dam No. 2, Muskingum River, "near" Marietta. Williams: 1 mi. S Blakesley; St. Joseph River "near" Blakesley; West Branch, St. Joseph River, Sec. 8, Bridgewater Twp.; Edgerton. Wood: Grand Rapids; Grassy Creek, Rossford; Haskins; Maumee River opposite Toledo.
Pennsylvania: Allegheny: Monongahela River above McKeesport (Atkinson, 1901:154); Ohio River at Pittsburgh (Wied-Neuwied in Stejneger, 1944:44, 49). Armstrong: (Swanson, 1952:165). Clarion: Clarion River "near" Clarion (Allen, 1955:228); Foxburg (= Foxbury?, Boulenger, 1889:260). Crawford: Elk: Erie: Edinboro Lake. Forest: (Swanson, loc. cit.). Indiana: Plum Creek; Crooked Creek (Netting in Stejneger, 1944:48). McKean: (Swanson, loc. cit.). Somerset: Stoyestown (Surface, 1908:122). Warren: Venango: Allegheny River south of Franklin (Swanson, loc. cit.).
Tennessee: Chester: South Fork, Forked Deer River just E Henderson (Endsley, 1954:40). Clay: Mill Creek, 3 mi. from Butler's Landing; Obey River above mouth of Wolf River at Lilydale; mouth of Wolf River (Shoup, Peyton and Gentry, 1941:75); Iron Creek "near" Willow Grove (Stejneger, 1944:56). Fentress: Jackson: (Gentry, 1941:332). Lake: Reelfoot Lake (Parker, 1948:29). Obion: Walnut Log (Parker, 1937:85); east shore of Reelfoot Lake, Samburg (Rhoads, 1895:386). Overton: Medlock Branch, tributary of West Fork Obey River north of Allred (Shoup, Peyton and Gentry, loc. cit.). Roane: 2 mi. S Kingston (Stejneger, 1944:55).
Vermont: Chittenden: Lake Champlain, mouth of Winooski River; "near" Burlington; Milton (= Minton) (Babcock, 1919:420). Franklin: Swanton (Stejneger, 1944:55).
West Virginia: Randolph: Tygart River at Elkins (Green, 1937:116).
Wisconsin: Burnett: Crawford: (Pope and Dickinson, 1928:83). Dane: Lake Wingra, Madison (Noland, 1951:54). Grant: (Pope and Dickinson, loc. cit.). Green Lake: Berlin (AMNH 6840-41, listed in card file March 2, 1959). Jefferson: Lake Mills (Dickinson, 1950:75). La Crosse: West Salem (Pope, 1930:281). Oneida: Pepin: (Pope and Dickinson, loc. cit.). Racine: Eagle Lake (Edgren, 1944:498); Burlington; Rochester (Stille and Edgren, 1948:201). Sheboygan: Sheboygan (KKA). Trempealeau: Vernon: "near" Viroqua (Pope, loc. cit.). Walworth: Lake Beulah (Dickinson, loc. cit.). Washburn: (Pope and Dickinson, loc. cit.). Waukesha: Lac La Belle (Cahn, 1929:8). Winnebago: Wolfe River (Dickinson, loc. cit.).
Trionyx spinifer hartwegi (Conant and Goin)
Western Spiny Softshell
Amyda spinifera hartwegi Conant and Goin, Occas. Papers Mus. Zool. Univ. Mich., No. 510:1, pl. 1, map 1, June 15, 1948.
T[rionyx] s[pinifer] hartwegi Schwartz, Charleston Mus. Leaflet, No. 26:11, May, 1956.
Type.—Holotype, UMMZ 95365; alcoholic adult male; obtained at Wichita, Sedgwick County, Kansas, in May, 1945, by Robert Young.
Range.—Central United States in tributaries flowing into the Mississippi River from the west, except the Red River drainage; eastern Montana, North Dakota, and southern Minnesota south to eastern Colorado, northern Oklahoma and Arkansas (see map, Fig. 19).
Diagnosis.—Juvenal pattern of small ocelli, rarely as large as two millimeters [498] in diameter, or usually solid black dots that are not much larger in center of carapace than at sides (mean OD/PL, Kansas, .022); only one dark marginal line separating pale rim of carapace from dorsal ground color.
Description.—Plastral length of smallest hatchling, 2.8 centimeters (USNM 9928); of largest male, 13.1 centimeters (USNM 55687); of largest female, 25.5 centimeters (KU 2283).
Carapace olive, having small ocelli or black spots that are not much larger in the center of the carapace than at the sides; pale rim of carapace separated from darker ground color by one dark marginal line and not four or five times wider posteriorly than laterally; large females often having black dots at sides of carapace on mottled and blotched pattern; pattern on snout of pale, dark-bordered stripes that unite forming acute angle in front of eyes; well-defined dark markings in subocular and postlabial region; pattern contrasting with ground color on side of head; postlabial stripe broken, interrupted; pale postocular stripe having blackish borders interrupted, not joining with postlabial stripes; dorsal surface of soft parts of body having contrasting pattern, largest blackish marks on hind limbs; elongate tail of males having pale dorsolateral bands with well-defined, lower, blackish borders; patterns on soft parts of body usually obscured or absent on large females; underparts whitish, often having blackish marks, except in center of plastral area; dark marks on webbing of limbs, palms and soles; dark streaks often coincident with digits; tubercles along anterior edge of carapace small and conical on adult males, and conical or knoblike on large females; accessory, knoblike tubercles in nuchal region and in middle of carapace posteriorly on large females.
Ontogenetic variation in PL/HW, mean PL/HW of specimens having plastral lengths 7.0 centimeters or less, 4.24, and exceeding 7.0 centimeters, 5.33; ontogenetic variation in CL/CW, mean CL/CW of specimens having plastral lengths 8.5 centimeters or less, 1.12, and exceeding 8.5 centimeters, 1.19; mean CL/PCW, 2.00; mean SL/HW, 1.30 (including subspecies spinifer); mean CL/PL, 1.38.
Variation.—Variants include: CNHM 8949, UMMZ 72511 and TU 14591 having ocelli approximately 4 millimeters in diameter that are almost solid spots; KU 17728 having pale stripes on snout that lack black, inner borders; TTC 719 (female, plastral length 20.7 cm.), having distinct pattern on snout; USNM 14535, 17823, 55684, and 123446 (from different localities) having markings confined to margins of carapace (Stejneger, 1944:66, suggested that USNM 17823 probably came from Texas); UMMZ 92667 (female, plastral length 6.7 cm.) lacking pattern on carapace.
Comparisons.—T. s. hartwegi can be distinguished from all other subspecies of T. spinifer by the presence of small dots and ocelli on the carapace that are all of approximately the same size in combination with only one dark marginal line. T. s. hartwegi resembles asper in having small blackish ocelli or dots on the carapace but differs from asper in having only one dark marginal line. T. s. hartwegi differs from spinifer only in the small size of the ocelli. T. s. hartwegi resembles spinifer and asper, but differs from pallidus, guadalupensis and emoryi in having blackish spots and ocelli on the carapace and lacking small whitish spots. T. s. hartwegi resembles spinifer, asper and pallidus but differs from guadalupensis and emoryi in having conical or knoblike tubercles on the anterior edge of the carapace on large females.
T. s. hartwegi differs from the subspecies asper, guadalupensis and emoryi in having a narrower head, and from emoryi in having a wider carapace. T. s. hartwegi resembles spinifer and asper but differs from the other subspecies in having the carapace widest at a plane approximately one-half way back on the carapace. T. s. hartwegi and spinifer have longer snouts than do pallidus and guadalupensis or emoryi. T. s. hartwegi differs from asper but resembles the other subspecies in having a relatively longer plastron.
Remarks.—The validity of T. s. hartwegi has never been questioned. It intergrades with spinifer over a broad area paralleling the Mississippi River. For convenience, specimens occurring west of the Mississippi River are referred to the subspecies hartwegi. Figure 8 shows much variation in size of ocelli on different individuals from the same state. For example, UMMZ 92667, plastral length 6.7 centimeters has a uniform pale brown carapace lacking any dark marks, whereas UMMZ 92652, plastral length 5.9 centimeters has some ocelli three millimeters in diameter on the carapace. Both are from Iowa. One specimen from Kansas, KU 1954 (Doniphan County, plastral length 11.8 cm.), has ocelli four millimeters in diameter, and USNM 7648 captured farther west at Fort Laramie, Wyoming, an adult male having a plastral length of 11.0 centimeters, has some ocelli five millimeters in diameter on the carapace. TTC 1090, an adult male from the panhandle of Texas has some ocelli so much as 5.5 millimeters in diameter. The size of the ocelli seemingly varies in the same local population.
Specimens of T. spinifer in the lower Mississippi Valley are intergrades. Most individuals have small black dots on the carapace; some have small ocelli (TU 7216, 7501, 11912, 12123-24) interspersed with black dots (TU 5863), others have black dots confined to the edge of the carapace (TU 157, 4539, 7105), and still others have no pattern on the carapace (TU 7506, 13698.1, 10087.6). Two large males (TU 11580, 13025) have large ocelli (approximately five mm. in diameter) that have nearly black centers. In general, there is more dark pigmentation than farther north; some specimens have extensive pigmentation on the ventral surface of the carapace and soft parts of the body (TU 156, 5648). The dorsal surface of the limbs, especially the hind limbs, have a bold, black marbling and may be almost completely black (TU 5484, 5597). Many females, not exceeding plastral lengths of 7.0 centimeters, have a pale blotched pattern of lichenlike figures or have ill-defined black dots on the carapace (TU 10087, 13698.13, 13753.15).
Localities of specimens of T. spinifer occurring in the Mississippi River drainage in Mississippi are arbitrarily listed under the account of the subspecies spinifer, whereas those in Louisiana (excluding pallidus) are listed under the account of hartwegi.
Neither Over (1943) nor Wheeler (1947:169) record T. s. hartwegi, respectively, from South Dakota or North Dakota; records from the Missouri River drainage in Montana suggest the occurrence of the species in that drainage in North and South Dakota.
Specimens examined.—Total, 392 as follows: Arkansas: Clay: UMMZ 70735 (2), 7 mi. S St. Francis. Crawford: USNM 95352, Lee Creek, 7 mi. NW Natural Dam. Drew: CNHM 40785. Lafayette: KU 2225-29, 2944 (one of three specimens bearing last catalog number), 2963 (one of three specimens bearing this catalog number), 2964 (one of two specimens bearing this catalog number), Lewisville (see remarks under the account of the [500] subspecies pallidus). Lawrence: CNHM 8949; CNHM 12598-600, 12602-04, TU 5855, UI 2413, Imboden; UI 2412, Black River at Powhatan. Marion: TU 14591 (6), White River at Cotter. Prairie: KU 1867, 1869, 1879, 1949-51, 2280-83, 2285-91 (2 specimens bear catalog number 2287), 2307, 2761-62, 2666, 2826, 2842, 3346-47, White River at DeValls Bluff. Pulaski: UMMZ 96540, Little Rock. Saline: USNM 17823, Saline River at Benton. Searcy: UMMZ 92755, Little Red River, 1.5 mi. SE Leslie. Yell: TU 14565, Petit Jean Creek, 10 mi. N Casa. County unknown: CNHM 28566-67, Ouachita River.
Iowa: Allamakee: UMMZ 72556-58, 92642-49, Mississippi River "near" Lansing. Appanoose: UMMZ 92667, Chariton River, 4.3 mi. N. Centerville. Decatur: UMMZ 92651, Grand River, 3.5 mi. WSW Decatur. Dickinson: UMMZ 55249, Milford; UMMZ 92655, Spirit Lake Twp. Hamilton: USNM 9928, Webster City. Hardin: UMMZ 92650, Eldora. Louisa: UMMZ 92654, Muscatine Slough, 12 mi. SW Muscatine, Muscatine County. Muscatine: INHS 7675, 5.5 mi. SE Muscatine; USNM 54730-32, Fairport. Scott: CNHM 433, Davenport; UMMZ 92656, Steamboat Slough, 2 mi. N Princeton. Story: UMMZ 92653, Squaw Creek at Ames. Washington: UMMZ 92652, English River, 2 mi. E Riverside.
Kansas: Anderson: KU 52286-87, 31/4 mi. E, 1/2 mi. N Colony. Atchison: UMMZ 66939-41, Atchison. Barber: KU 17728, 4.5 mi. S Sun City; KU 41379, 41742, 6 mi. N, 3.5 mi. E Sharon; USNM 100580, Medicine River, 1 mi. S Lake City. Cherokee: KU 1323, Galena. Comanche: KU 18385, 3-4 mi. SE Arrington. Cowley: UMMZ 75963, USNM 90441-44, 91022, 100529-30, "near" Winfield. Doniphan: KU 1943, 1952-54, Doniphan Lake. Douglas: KU 1955-56, Wakarusa River; KU 40176-77, Kansas River at Lawrence. Franklin: KU 3290. Hamilton: KU 2990, Syracuse. Harper: KU 18159, 1 mi. N Harper. Kingman: USNM 95261, 2 mi. E Calista. Labette: KU 3339. Lane: KU 3738-41, Pendennis. Logan: KU 16531, Smoky Hill River, 3 mi. SW Elkader. Meade: KU 40210, Crooked Creek, 12.5 mi. S, 11/4 mi. W Meade. Montgomery: KU 3731-32, Independence; KU 50856, Cherryvale Lake. Neosho: UMMZ 69294, Caneville Creek, 32 mi. N. Parsons, Labette County. Osage: KU 3294-96, Appanoose Creek. Pratt: KU 15931-32, 15934, State Fish Hatchery "near" Pratt. Riley: KU 48239, McDowell Creek, WSW Manhattan; UMMZ 64434, "near" Manhattan. Russell: KU 3289. Sedgwick: UMMZ 95363-65, Wichita. Shawnee: USNM 123446, Kansas River at Topeka. Stafford: KU 3758, Little Salt Marsh; KU 41743, 13.5 mi. N, 6 mi. E Stafford. Trego: KU 2757, 3769, Smoky Hill River, 10 mi. N (NNE) Utica, Ness County; KU 51517, Saline River, 5 mi. N, 1/2 mi. E Wakeeney. Wilson: KU 56744-45, Verdigris River, 1 mi. S Altoona. Woodson: KU 55295, Neosho River, 1/2 mi. E, 11/2 mi. S Neosho Falls. County unknown: USNM 51529.
Louisiana: Catahoula: TU 12629, Ouachita River, 4 mi. N Harrisonburg. Claiborne: TU 13080, Caney Lake "near" Summerfield. Concordia: KU 50849, Tensas River at Clayton; TU 16524 (3), USNM 012349, Lake Concordia; USNM 99865, Red River "near" Shaw. East Carrol: TU 827-30, 905, 5644-45, Lake Providence. Grant: TU 12735, Big Creek at Fishville, "near" Pollock. Jefferson: TU 5592-98, 7184, 10741, 10171, Mahogany Pond. Lafourche: TU 7105, 7132, 7216, 7501, 7505-07, 10087 (14), 11828-29, 11912, 11983 (2), 12123-28, 13502, 13679 (8), 13753 (22), 13766.2, Bayou Lafourche at Raceland. Morehouse: USNM 11631 (2), Mer Rouge. Natchitoches: USNM 100420, Cane River "near" Natchitoches. Orleans: TU 16169 (3), Audubon Park, New Orleans; USNM 029310, "near" New Orleans. Ouachita: TU 12916, 12954, 12970-71, 13019, 13025, Bartholomew Bayou at Sterlington; TU 5988, Monroe. Pointe Coupee: TU 153, 156-59, 165, 5484, 5513, 5518-19, 5646, 5648, 5651, USNM 100202-12, False River at New Roads. Rapides: TU 14040, Red River at Rapides. Richland: OU 25082. St. Bernard: TU 16170, Delacroix Island. St. Charles: TU 4539, 4579, 5224, 5990, 11928 (12), 13698 (16), Bayou Gauche between Paradis and Des Allemands; TU 5863, 11580, Bonnet Carre Spillway at Norco. Tensas: TU 5762, Lake St. Joseph near Newellton. Union: USNM 138946, Meridian Creek, 1 mi. E Conway; USNM 138947, Ouachita River, Alabama Landing. Parish unknown: MCZ 1622, Lake St. John (Concordia or Tensas Parish); USNM 029266, Louisiana?
Minnesota: Hennepin: AMNH 4759-60, Fort Snelling. Lesueur: KU 46742-43, Waterville, Lake Tetonka. Winona: USNM 59263-66, Homer.
Missouri: Carter: UMMZ 70737, "near" Van Buren. Chariton: UI 17509, Triplett. Franklin: USNM 55689. Gasconade: UMMZ 95900, Bourbeuse Creek, 8 mi. S Owensville. Jefferson: USNM 95405, Glaize Creek. Lewis: USNM 59279-80, Canton. Miller: UMMZ 91929, Barren Fork Tavern Creek, 5 mi. NW Iowna. Newton: UMMZ 82822, Shoal Creek, 12 mi. W Momit. Phelps: UMMZ 91930, Bourbeuse River, 10 mi. N St. James. Reynolds: CNHM 35392, Black River at Warner Bay Spring; USNM 55688. Ripley: UMMZ 90435. Shannon: INHS 6223, Alley Spring State Park. St. Charles: USNM 93089-94, Dardenne Creek, St. Peters. St. Louis: USNM 55685-87, Mississippi River at St. Louis. Stone: USNM 55684. Washington: USNM 55690. Wayne: UI 16554, Sam A. Baker State Park; UMMZ 95879, St. Francis River at Lodi. County unknown (Wayne or Butler): UMMZ 83264, Clark National Forest, St. Francis River.
Montana: Big Horn: USNM 54421, Crow Agency. Roosevelt: USNM 58, Fort Union (locality reads "Yellowstone, Fort Union"; probably the Yellowstone River near Fort Union). Wheatland: UMMZ 92005, Musselshell River near Shawmut. Yellowstone: USNM 14535, Custer.
Oklahoma: Alfalfa: OU 9316, 2 mi. S Cherokee. Cleveland: OU 22973, Norman. Delaware: UMMZ 81476, Spavinaw. LeFlore: OU 16802, 1.5 mi. E Zoe. Osage: UMMZ 89628, Big Hominy Creek. Pottawatomie: OU 25175, 5 mi. SW Shawnee. Rogers: OU 7317, Verdigris River, 5 mi. W Claremore; UMMZ 81473-74, near Garnett, Tulsa County; UMMZ 81475, 4 mi. NE Inola. Sequoyah: OU 9008, 2 mi. NE Gore; TU 13885, Little Vian Creek, 1 mi. E Vian. Texas: OU 5005, 5 mi. SE Guymon. Tulsa: TU 17061, Bird Creek "near" Skiatook, Osage County. Woods: CHNM 11809, Waynoka; OU 9432, 2.5 mi. W Waynoka; OU 9579, 9581-82, 1 mi. S Waynoka.
Texas: Hansford: TTC 719, 10 mi. S, 2 mi. W Gruver. Hutchinson: TTC 1090, Carson Creek, Turkey Track Ranch.
Wyoming: Goshen: USNM 7648, Fort Laramie. Weston: UMMZ 78080, Beaver Creek.
No Data: CNHM 21687-88, 22925. SM 142 (locality of Waco, McLennan County, Texas, believed in error). USNM 7649, 11625, 19622-23, 36412 (Illinois River).
Records in the literature.—Arkansas: Benton: (Dowling, 1957:37). Chicot: Lake Chicot. Clark: Terre Noir Creek, 13 mi. W Arkadelphia. Garland: Ouachita River, Mountain Pine (Conant and Goin, 1948:7). Hempstead: Jefferson: (Dowling, loc. cit.). Lawrence: Black Rock (Dellinger and Black, 1938:46). Madison: Scott: St. Francis: (Dowling, loc. cit.). Washington: near Greenland (Dellinger and Black, loc. cit.).
Colorado: Boulder: Boulder Creek, E Boulder; Boulder Creek, 6 mi. S and 1 mi. E Longmont. Larimer: Cache la Poudre River. Logan: 8 mi. NE Sterling. Morgan: Platte River "near" Fort Morgan. Otero: Purgatoire River at Higbee. Prowers: Arkansas River at Lamar. Weld: Poudre River "near" Greeley; Evans. Yuma: Bonny Dam, Republican River (Maslin, 1959:24-25).
Iowa: Dickinson: Little Sioux River, Okoboji Twp. (Blanchard, 1923:24). Story: Skunk River, 5 mi. NNE Ames (Conant and Goin, 1948:9).
Kansas: Allen: Petrolia (KKA). Barber: 7 mi. S Sun City. Butler: 3 mi. SE Augusta (Burt and Hoyle, 1934:198). Chase: 10 mi. SW Olpe; 7 mi. SW Saffordville (Breukelman and Smith, 1946:112). Cherokee: tributary of Spring River, 1 mi. N Riverton (Hall and Smith, 1947:451). Coffey: (Smith, 1956:160, symbol on map). Cowley: 11 mi. SE Winfield (Stejneger, 1944:55). Crawford: Pittsburg (Hall and Smith, loc. cit.). Doniphan: "near" Geary (Linsdale, 1927:81). Elk: (Smith, loc. cit.). Ellis: Big Creek (Brennan, 1934:190); Ellis (Conant and Goin, 1948:2). Franklin: Middle Creek, SE part of county (Gloyd, 1928:135). Greenwood: (Stejneger, op. cit.:54). Leavenworth: Missouri River "near" Fort Leavenworth (Brumwell, 1951:208). Lyon: 5 mi. E Emporia (Breukelman and Smith, loc. cit.). Marion: (Smith, loc. cit.). Meade: Meade County State Park, ca. 13 mi. SW Meade (Tihen [502] and Sprague, 1939:505). Ness: 5.5 mi. NW Ness (Breukelman and Smith, loc. cit.). Osage: Marais des Cygnes River; Long and Jordan Creeks (Clarke, 1958:21). Reno: 6 mi. E Turon. Sedgwick: 2 mi. NE Cheney (Burt, 1935:321). Sheridan: State Lake 7 mi. NE Quinter, Gove County (Breukelman and Smith, loc. cit.). Wabaunsee: Dragoon Creek at Harveyville (Clarke, 1956:215). Wallace: (Burt, 1933:208). Wilson: Fall River, 1/2 mi. S Neodesha (Clarke, loc. cit.).
Minnesota: Anoka: Benton: Chisago: (Breckenridge, 1944:184, symbols on map). Crow Wing: (Breckenridge, op. cit.:185). Dakota: (Hedrick and Holmes, 1956:126). Goodhue: (Breckenridge, op. cit.:184, symbol on map). Hennepin: Minneapolis; Lake Minnetonka (Breckenridge, op. cit.:187); 5 mi. N. Minneapolis (Breckenridge, 1955:5). Houston: Root River near Hokah. Lesueur: Lake Washington (Hedrick and Holmes, loc. cit.). Meeker: Swan Lake (Breckenridge, 1957:232). Pine: (Breckenridge, 1944:185). Ramsey: Rice: Sherburne: Stearns: (Breckenridge, op. cit.:184, symbols on map). Washington: just north of Stillwater (Hedrick and Holmes, loc. cit.). Winona: Winona (Breckenridge, op. cit.:187). Yellow Medicine: (Breckenridge, op. cit.:185). County unknown (Goodhue or Wabasha): Lake Pepin (Breckenridge, op. cit.:184).
Missouri: Boone: east of Ashland (Henning, 1938:92). Jackson: Missouri River "near" Atherton (Anderson, 1942:219). Jefferson: Mississippi River "near" mouth Glaize Creek at Sulphur Springs; Glaize Creek at Barnhart (Boyer and Heinze, 1934:199). St. Clair: Osage River "near" Osceola. Vernon: Marmaton River, 7 mi. N Moundville (Conant and Goin, 1948:9).
Montana: Yellowstone River (Conant and Goin, 1948:9).
Nebraska: Adams: 1 mi. N Ayr (Hudson, 1942:101). Dawson: 2 mi. SE Gothenburg (Gehlbach and Collette, 1959:142). Franklin: 2 mi. SW Naponee. Gage: 1 mi. W Barnston. Hitchcock: 3 mi. E Stratton. Holt: Elkhorn River "near" Atkinson. Lancaster: Lincoln (Hudson, loc. cit.). Lincoln: 1 mi. S Sutherland (Gehlbach and Collette, loc. cit.). Red Willow: 14 mi. NW McCook. Richardson: 2 mi. S Rulo. Wheeler: 2 mi. W Ericson (Hudson, loc. cit.).
Oklahoma: LeFlore: Wister (Conant and Goin, 1948:9); Shady Pointe (KKA); Poteau River, 6.5 mi. W Heavener (Trowbridge, 1937:301). Tulsa: Arkansas River "near" Tulsa (Force, 1930:38).
Wyoming: Goshen: Platte River (Conant and Goin, 1948:10).
Trionyx spinifer asper (Agassiz)
Gulf Coast Spiny Softshell
Aspidonectes asper Agassiz, Contr. Nat. Hist. United States, 1(Pt. 2):405; 2(Pt. 3):pl. 6, fig. 3, 1857.
Trionyx spinifer asper Schwartz, Charleston Mus. Leaflet, No. 26:17, pls. 1-3, map 2, May, 1956.
Platypeltis agassizii Baur, Amer. Nat., 22:1121, 1888.
Type.—Lectotype, MCZ 1597; alcoholic female; locality designated as Pearl River, Columbus, Marion County, Mississippi; received from Mr. Winthrop Sargent of Natchez, Mississippi.
Range.—Southeastern United States except peninsular Florida from the Florida Parishes of Louisiana east to southern North Carolina; Gulf Coast drainage including that of Lake Pontchartrain, Louisiana, eastward to the Apalachicola River system, and Atlantic Coast drainage including that of the Altamaha River in Georgia northward to the Pee Dee River drainage in South Carolina (see map, Fig 19).
Diagnosis.—Juvenal pattern of black ocelli and spots, and two or more black, interrupted, lines paralleling rear margin of carapace; pale postocular and postlabial stripes often united on side of head; length of plastron short.
Description.—Plastral length of smallest hatchling, 2.9 centimeters (USNM 134244); of largest male, 13.2 centimeters (TU 17117); of largest female, 27.0 centimeters (TU 13474).
Blackish marginal rings on carapace number two, three or four posteriorly, but decrease in number anteriorly; segments of marginal rings may extend to nuchal region; marginal rings increasingly interrupted inwardly; pattern of hatchlings having well-defined marginal rings that are not extensively interrupted (often males), or having marginal rings broken into small segments or series of dots, and pale outer margin of carapace marked by ill-defined, hazy, inner border (often females); conspicuous marginal rings often lacking on hatchling females; pale rim of carapace not four or five times wider posteriorly than laterally; carapace having blackish dots, spots, small ocelli or a combination thereof; marks on carapace of slightly varying sizes, some occasionally barlike (usually males); some hatchling females showing pale, irregular blotching on carapace, often characterized by small lichenlike figures superimposed on blackish dots.
Striping on snout variable; pale, dark-bordered stripes usually unite in front of eyes and form right or acute angle; medial dark borders of pale stripes on snout not joined anteriorly, broken into segments or dots, reduced to single median line, united to form straight line connecting anterior margins of orbits (usually with slight medial indentation), or absent; pale postocular and postlabial stripes often joined, relationship variable and on either side of head; side of head with or without dark markings, sometimes a pale subocular blotch bordered below by a dark line; pattern on dorsal portions of soft parts of body contrasting, less so on limbs of hatchlings; pattern of irregular dark marks, dark streaks usually coincident with digits; longitudinal streaks often occur on neck; elongate tail of adult males usually having well-defined, dorsolateral, pale bands with dark lower border more diffuse than upper border.
Underparts whitish often with dusky markings on rear of carapace or in region of bridge; blackish marks often on webbing and portions of soles and palms, and chin and throat.
Small conical tubercles along anterior edge of carapace on adult males; remnants of juvenal pattern usually present on carapace of large females; conical or knoblike tubercles on anterior edge of carapace of large females; accessory knoblike tubercles in nuchal region (a paravertebral pair usually most prominent), and posteriorly in middle of carapace on large females.
Ontogenetic variation in PL/HW, mean PL/HW of specimens having plastral lengths 7.0 centimeters or less, 3.87, and exceeding 7.0 centimeters, 4.94; ontogenetic variation in CL/CW, mean CL/CW of specimens having plastral lengths 8.5 centimeters or less, 1.11, and exceeding 8.5 centimeters, 1.16; mean CL/PCW, 1.71; mean CL/PL, 1.45.
Variation.—The sex of some hatchlings can be distinguished by the pattern on the carapace (see Plate 37 for different patterns), but the sex of many hatchlings cannot be distinguished on the basis of pattern.
In the early stages of this study, I thought that the pattern on the carapace differed in eastern and western populations, and that the zone of intergradation [504] was in Alabama. Adult males from the Tombigbee-Alabama river drainage and westward were noted to have blackish spots (some slightly ocellate) intermixed with few, if any, smaller blackish dots, whereas the adult males from east of the Tombigbee-Alabama river drainage had many small, black dots intermixed with slightly larger, mostly ocellate marks (see Plate 38, left, top and bottom, for contrast); also, hatchlings from western populations were never observed to have four marginal rings. On the basis of pattern, I would have thought that the individual having many ocelli, that lacks correct locality data and that is photographed by Stejneger (1944:Pl. 26), came from Georgia or South Carolina; but, the pattern (op. cit.:Pl. 27) of a specimen, probably an adult male, from South Carolina, resembles the pattern on adult males from Louisiana. The differences noted above are probably due to individual variation rather than geographic variation.
Color notes taken from life of a freshly-killed adult male (TU 16071, Louisiana) are: carapace olive, spots blackish, outer rim buff; top of head olive, postocular and postlabial stripes yellow with blackish borders, stripes on snout buff with blackish borders; dorsal ground color of soft parts of body pale olive-green, larger marks blackish, ground color laterally toward juncture of pattern and immaculate undersurface, and toward insertions of neck and limbs becoming yellowish; webbing on hind limbs having reddish tinge; dorsolateral bands on tail yellow with blackish borders; undersurface whitish; chin and throat olive-green with blackish marks; becoming buff then whitish posteriorly.
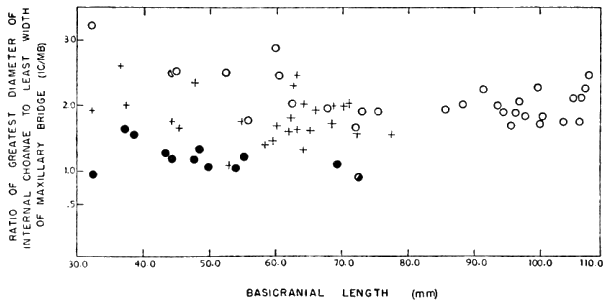
Fig. 20. Basicranial length and ratio of greatest diameter of internal choanae to least width of maxillary bridge (IC/MB) on 30 skulls of T. ferox (open circles), 26 of T. spinifer (crosses), and 12 of the agassizi-form (solid circles; half shaded circle represents holotype of agassizi). Skulls of the agassizi-form tend to have slightly smaller internal choanae than those of spinifer or ferox.
Occasional specimens have only one definite dark line paralleling the rear margin of the carapace. Schwartz (1956:16) reported that Charleston Museum No. 55.159.26 has only one solid line at the margin of the carapace, and I received an adult male (KU 47120) reported to have come from the Pearl River that is aberrant in not having more than one dark marginal line. USNM 95191, a large stuffed female from the Pearl River is mentioned by Stejneger (1944:59, Pl. 17) as having marks that "assume the form of short lines parallel [505] with the submarginal ring"; I examined this specimen and noted that it had only one dark marginal line. Stejneger (op. cit.:64) mentioned another from the Pearl River drainage, and Crenshaw and Hopkins (1955:20) wrote that some individuals from Georgia have only one dark marginal line. Presumably MCZ 1606 (now in the Albany Museum) recorded by Stejneger (op. cit.:52) as Amyda s. spinifer from Columbus, Georgia, is another specimen.
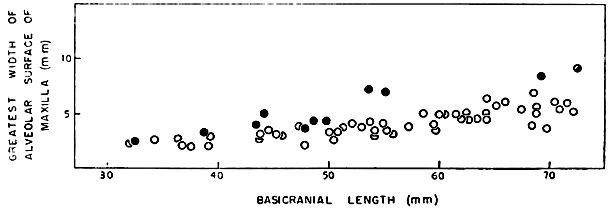
Fig. 21. Basicranial length and greatest width of alveolar surface of maxilla on 52 skulls of T. spinifer (open circles) and 11 of the agassizi-form (solid circles; half shaded circle represents holotype of agassizi). Most skulls of the agassizi-form that exceed 43 mm. in basicranial length have a more expanded, alveolar surface of the maxilla than skulls of spinifer of approximately the same size. All skulls exceeding 50 mm. are those of females.
Some skulls of soft-shelled turtles from streams of the Atlantic Coast drainage, including the skull of the holotype of Platypeltis (= Trionyx) agassizi Baur (MCZ 37172, Pl. 54), show at least two differences from other skulls of asper and from those of other subspecies of T. spinifer. Figure 20 shows that skulls of agassizi tend to have slightly smaller internal choanae (ratio IC/MB) than those of T. spinifer and T. ferox; there is seemingly little difference between skulls of ferox and spinifer, and little, if any, ontogenetic variation. Figure 21 shows that most skulls of the agassizi-form that exceed 43.0 millimeters have a more expanded, alveolar surface of the maxilla than skulls of spinifer of approximately the same size; most skulls exceeding a basicranial length of 43.0 millimeters, and certainly all skulls exceeding 50.0 millimeters are those of females. Stejneger (1944:Pl. 30) also has provided photographs of a skull of the agassizi-form. It is of interest that of the 12 agassizi-form skulls (MCZ 37172; USNM 8708, 029034, 51981, 66859, 71681, 91282, 91310-11, 92521, 92583-84) that I examined some resemble ferox (Neill, 1951:9) in having the alveolar surfaces of the jaws broadened, and the greatest width at the level of the quadratojugal (Table 3, Plate 54); also, the localities of all 12 skulls are within the geographic range of ferox. Skulls of ferox, however, have conspicuously broadened alveolar surfaces of the jaws only when they exceed in length the largest skulls of agassizi. The differences of skulls of the agassizi-form possibly reflect isolation in the Atlantic Coast drainage, and an adaptation in feeding habits. So far as I can ascertain, individuals occurring in rivers of the Atlantic Coast drainage in Georgia and South Carolina (referable to agassizi) do not differ consistently in external characters from individuals of T. s. asper that occur westward in the Apalachicola drainage.
Comparisons.—Trionyx s. asper can be distinguished from all other subspecies of T. spinifer by usually having more than one black line paralleling [506] the rear margin of the carapace. This character and the frequent fusion of the postlabial and postocular stripes on the side of the head distinguish asper from spinifer and hartwegi. T. s. asper differs from pallidus, guadalupensis and emoryi in having blackish spots and ocelli on the carapace, and lacking whitish dots or tubercles. T. s. asper resembles spinifer, hartwegi and pallidus but differs from guadalupensis and emoryi in having conical tubercles along the anterior edge of the carapace in large females. For additional differences see accounts of other subspecies.
Of the subspecies of T. spinifer, asper has a proportionately wide head that is closely approached in the subspecies guadalupensis and emoryi; T. s. asper differs from guadalupensis and emoryi in having a wider carapace, and resembles hartwegi and spinifer, but differs from the other subspecies in having the carapace widest at a plane approximately one-half way back on the carapace. T. s. asper differs from the other subspecies in having the shortest plastron.
Remarks.—Stejneger (1944:72-74) has discussed the history of Baur's Platypeltis agassizi. Briefly, Agassiz's description of Platypeltis ferox wherein he (1857:402) states that "The young ferox [Pl. 6, fig. 3] has two or three concentric black lines separating the pale margin …," was applicable to T. s. asper. Agassiz mentioned also that the young of his Aspidonectes asper (op. cit.:406) "as in Platypeltis ferox, … has … two or three black lines separating the pale rim of the posterior margin, …"; however, A. asper was distinguished chiefly by the "… prominent warts of the bony plates (loc. cit.)." Because the description of the pattern of ferox resembled that of asper, the validity of asper was not agreed upon by all workers. Boulenger (1889:245, footnote 1) referred to asper as a species that required "… further investigation."
Baur (1888:1121) realized that Agassiz's description of ferox was not that of Testudo ferox Schneider, and regarded the description of Agassiz as applying to a new species, which he named Platypeltis agassizii; Baur (op. cit.:1122) also recognized asper, referring it to the genus Aspidonectes. Baur designated a specimen from Georgia (the only individual seen by him) as the type of agassizi (Stejneger, op. cit.:73, footnote); this specimen is now MCZ 37172. Five years later (1893:218), Baur discussed generic relationships of trionychids, seemingly only on the basis of skulls (holotype of agassizi not mentioned), and referred agassizi to the resurrected genus Pelodiscus Fitzinger, 1835, which was distinguished from the other two American genera that Baur recognized (Platypeltis and Amyda) by having the "Posterior nares reduced in size by the inner and posterior extension of the maxillaries." Baur also transferred asper to the genus Platypeltis, and restricted the type locality of that species to "Lake Concordia, La." (op. cit.:220); the type locality of agassizi was restricted to "Western Georgia" (loc. cit.).
The name-combination, Pelodiscus agassizi, was not generally accepted. Hay (1892:144) and Siebenrock (1924:188) referred agassizi to the genus Trionyx. Hay regarded agassizi as a full species (see discussion by Stejneger, 1944:73), whereas Siebenrock considered it a subspecies of spiniferus; both authors regarded asper as a synonym of agassizi. Neither asper nor agassizi was mentioned in the first three editions of the Check List of North American Amphibians and Reptiles (Stejneger and Barbour, 1917, 1923, 1933); the [507] same authors in the fourth (1939:171, 172) and fifth editions (1943:212, 213) listed agassizi as a full species, and asper as a subspecies of spinifera. Stejneger (1944) used the same arrangement as set forth in the fourth and fifth editions of the Check List, and distinguished agassizi on the basis of cranial characters, namely, the small size of the internal choanae, the greater width of the alveolar surface of the lower jaw, and the position of the suture between the palatine and basisphenoid relative to the posterior edge of the temporal fossa. Neill (1951:9) regarded the peculiarities of the agassizi-type skull as inconstant, but recognized agassizi (and asper) as a subspecies of ferox. Crenshaw and Hopkins (1955) showed that asper did not intergrade with ferox. Schwartz showed that agassizi did not intergrade with ferox, and regarded agassizi as a synonym of T. s. asper (1956:17), but stated that agassizi possessed "wider crushing surfaces on the maxillae than does T. s. asper, even when skulls of the same size and sex are compared" (op. cit.:9).
The holotype of Platypeltis agassizi (MCZ 37172) is a dried adult female consisting of shell, skull and limb bones; the carapace is approximately 300 millimeters long (Schwartz, loc. cit.). I have examined only the skull of MCZ 37172 (Plate 54), and it is the largest of 12 agassizi-type skulls I have seen. The basicranial length is 72.5 millimeters, and the greatest width, which occurs at the level of the quadratojugals, is 52.9 millimeters. The agassizi-type skulls have been discussed under the subsection on variation.
The type locality of T. s. asper, Lake Concordia, Louisiana (lower Mississippi River drainage) as restricted by Baur (1893:220), is in an area of intergradation of three subspecies of Trionyx spinifer where most individuals are not typical of asper. The syntypes, the designation of MCZ 1597 as a lectotype, and Pearl River, Columbus, Marion County, Mississippi, as the type locality have been discussed elsewhere (Webb, 1960).
The range of T. s. asper overlaps that of T. ferox in Georgia and South Carolina. The two species remain distinct in the area of overlap of their geographic ranges (Crenshaw and Hopkins, 1955:16; Schwartz, op. cit.:5). Trionyx s. asper intergrades with T. s. hartwegi and T. s. spinifer in the lower Mississippi Valley (Conant and Goin, 1948:11).
However, there are few specimens available that indicate intergradation of asper with the spinifer-hartwegi complex in the lower Mississippi River drainage; this may be due to the fact that asper inhabits waterways that do not drain into the Mississippi River. Perhaps intergradation is more prevalent than the morphological basis that I have relied upon indicates; in any event, there are few specimens that have more than one dark marginal line (which is the only character that is unique for asper) from the lower Mississippi drainage. A young male (TU 11928.9) from Bayou Gauche between Paradis and Des Allemands, St. Charles Parish, Louisiana, has a pattern on the carapace resembling that of asper; several other small softshells (TU) are available from the same locality but none shows more than one dark marginal line. Another specimen (USNM 95192), a young female from a barrow pit of the Big Black River (Mississippi River drainage), Madison County, Mississippi, resembles asper in having more than one marginal ring. Of three large females from Moon Lake, an ox-bow of the Mississippi River in Coatopa County, Mississippi (AMNH 5285-86, 5289), only 5289 shows evidence of two marginal lines. USNM 73669 (Greenwood, LeFlore County, Mississippi) also indicates intergradation in that the spots tend to be linear just inside the dark marginal [508] line, but the specimen more closely resembles the hartwegi-spinifer complex rather than asper.
There seems to be little adumbration of the dark marginal lines of asper in populations from the lower Mississippi River drainage. Blackish spots and ocelli vary in size and there are many kinds of pattern on the carapace. Soft-shelled turtles inhabiting the Mississippi River and its tributaries in Louisiana and Mississippi certainly represent an intergrading population of spinifer and hartwegi, and, to a lesser extent, of asper. Soft-shelled turtles inhabiting the Pearl River drainage and rivers that drain into Lake Pontchartrain immediately adjacent to the east are predominantly asper.
Specimens having localities from the Pearl River and Lake Pontchartrain drainages are listed under the account of asper and are referred to that subspecies on the distribution map; specimens from the Mississippi drainage in Mississippi are referred to spinifer.
One specimen (UMMZ 59198, Bradley County, Tennessee), from the Tennessee River drainage where T. s. spinifer occurs, deviates markedly from spinifer and suggests intergradation. UMMZ 59198, plastral length 4.8 centimeters, has ocelli in the center of the carapace only two millimeters in diameter, a distinct but interrupted, second marginal ring consisting of spots, and the pale postlabial and postocular stripes in contact on both sides of the head.
Specimens examined.—Total 110, as follows: Alabama: Barbour: UMMZ 113038, Chattahoochee River, Eufala. Cherokee: ANSP 24592, "near" center of Terrapin Creek. Conecuh: UMMZ 70736, Murder Creek, Castleberry. Escambia: TU 15823, Escambia River, 1 mi. N Sardine; UMMZ 70734, Escambia River at Flomaton. Henry: TU 15630, 3 mi. NW jct. Echo Farm Rd. and Rt. 136 on Echo Farm Rd. Lowndes: UMMZ 67759, Pintlalla Creek. Mobile: MCZ 1608 (2), 1608A, Mobile. Sumpter: USNM 83996, 3 mi. SE Coatopa. Tuscaloosa: TU 14673 (5), Black Warrior River, 17.5 mi. SSW Tuscaloosa; UA 52-1085, Cottondale. Walker: KU 50843, 50851, TU 17137, Mulberry Fork, Black Warrior River, 9 mi. E Jasper.
Florida: Calhoun: KU 50837-38, Chipola River, 4 mi. N Scott's Ferry; TU 16689 (4), Chipola River "near" Blountstown. Escambia: TU 13474, 15869 (3), 16584, Escambia River, 1.2 mi. E Century. Okaloosa: TU 15661, Blackwater River, 4.3 mi. NW Baker on Route 4. Santa Rosa: AMNH 44621, Blackwater River, Milton. Walton: UMMZ 110421, Pond Creek, 4 mi. SW Florala, Covington County, Alabama.
Georgia: Baker: TU 15889 (3), USNM 134243-48, Flint River "near" Newton; USNM 30822. Baldwin: USNM 8708, Milledgeville. Bryan: TU 15090, Canouche River, 2.3 mi. W Groveland. Chatham: USNM 51981, 92583-84, Savannah. Chattooga: UMMZ 113037, tributary of Chattooga River, Lyerly. Decatur: KU 50839-42, Flint River, 1.5 mi. S Bainbridge. Fulton: UMMZ 53037, Roswell. Lincoln: USNM 91282-83, above Price Island, Savannah River. Murray: UMMZ 59196, 9 mi. N Spring Place. Pulaski: TU 14882, Ocmulgee River, 4.3 mi. SE Hawkinsville. Richmond: USNM 66859, Augusta. Whitfield: UMMZ 74209, Cohulla Creek, Prater's Mill "near" Dalton. County unknown: MCZ 37172; UMMZ 109864, Flint River at mouth of Dry Creek; USNM 029034.
Louisiana: East Baton Rouge: LSU 11, 1643-44, City Park Lake in Baton Rouge; TU 17237, Amite River "near" Baton Rouge. St. Tammany: TU 6356, headwater creek of Bayou Lacombe; TU 16071, USNM 66147, mouth of Tchefuncta Creek in Lake Pontchartrain. Tangipahoa: TU 13623, 3.1 mi. W Hammond; USNM 68054, Robert. Washington: KU 50840, 50846, TU 17117, Pearl River at Varnado. Parish unknown (East Baton Rouge or Tangipahoa): UMMZ 95614, Manchac.
Mississippi: Chickasaw: USNM 115981, Chookatonkchie Creek. Clarke: USNM 79350-51, 1 mi. W Melvin, Choctaw County, Alabama; USNM 100805, Enterprise. Forrest: WEB 55-586, 1 mi. S Hattiesburg. Hancock: AMNH 46780; WEB 54-651, Hickory Creek "near" Kiln. Lauderdale: UMMZ 74681, 9 mi. W Meridian; UMMZ 90130, Lake Juanita, 15 mi. W Meridian. Lawrence: KU 47120, TU 17307.1, Pearl River, 9 mi. S Monticello; USNM 7653-54, Pearl River at Monticello. Lee: CM 31904, Verona; USNM 115979, Cower's Area near Guntown. Madison: USNM 95191, 95193-94, Pearl River. Marion: MCZ 1597, Pearl River at Columbus (designated type locality). Pearl River: CM 21100, Pearl River, 20 mi. W Poplarville; TU 14362, Hobolochito Creek, 1 mi. N Picayune. Perry: WEB 55-580, Beaver Dam Creek, 1 mi. N Richton. Walthall: KU 50844, Bogue Chitto River, Dillon.
South Carolina: Abbeville: USNM 7650, Abbeville? (reported by Pickens, 1927:113; locality considered in error by Stejneger, 1944:50; USNM 7650 having only one dark marginal line paralleling rear margin of carapace is possibly an aberrant specimen—see page 495 of present account). Greenwood: USNM 71681, 73668, Greenwood. McCormick: USNM 91310-12, Savannah River, 5 mi. W Plum Branch; USNM 92521, near Parksville. Richland: AMNH 70724-25, Broad River, Columbia.
No Data: USNM 8359 (erroneously reported from Madison, Indiana by Yarrow, 1882:29 and Hay, 1892:145; see discussion by Cahn, 1937:200, and Stejneger, 1944:73-75); USNM 131859.
Records in the literature.—Alabama: Coffee: Elba (KKA). Marengo: Tombigbee River near Demopolis. Mobile: Fig Island (Löding, 1922:47).
Florida: Jackson: Chattahoochee River, 8 mi. SE Butler. Leon: Ochlocknee River, NW of Tallahassee (Goin, 1948:304).
Georgia: Bartow: Etowah River below Allatoona Dam, ca. 4 mi. ESE Cartersville (Crenshaw and Hopkins, 1955:15). Berrien: (Knepton, 1956:324). Emanuel: Ogeeche River (Schwartz, 1956:19). Fulton: Nancy Creek, Atlanta (Dunston, 1960:278). Gwinnett: Irwin: (Knepton, loc. cit.). Jenkins: Ogeeche River near Buckland Creek jct., 2.5 mi. S Millen. Liberty: Camp Stewart, 4 mi. N Hinesville. Morgan: Lake Rutledge (Schwartz, loc. cit.). Muscogee: Columbus (Stejneger, 1944:52). Wayne: Altamaha River, 5 mi. N Mt. Pleasant (Schwartz, loc. cit.). Wilcox: Ocmulgee River, 3-4 mi. SSE Abbeville (Crenshaw and Hopkins, op cit.:16, footnote; Schwartz, loc. cit.).
Mississippi: George: Whiskey Creek (Cook, 1946:185). Harrison: near Biloxi. Jackson: Pascagoula Swamp, ca. 40 mi. E. Biloxi (Corrington, 1927:101). Jones: Eastabuchie. Lee: Cain Creek Bottom. Lincoln: Old Brook Creek. Lowndes: Tombigbee River, Camp Henry Pratt and Columbus; Lake Park, Columbus. Pearl River: 21 mi. SW Poplarville; 10 mi. W Poplarville; 4 mi. W Poplarville. Wayne: Trigg Area (Cook, loc. cit.).
North Carolina: Mecklenburg: Catawba River near Charlotte (Schwartz, 1956:20).
South Carolina: Aiken: Savannah River, 10 mi. SW Jackson. Allendale: Savannah River, Fennell Hill, 2 mi. S US 301. Anderson: Pendleton. Bamberg: South Edisto River, Cannon's Bridge, 5 mi. from Bamberg. Berkeley: 2.5 mi. W Pinopolis. Charleston: Charleston. Clarendon: Upper Lake Marion at US 301; Lake Marion, 13 mi. SW Manning; 3.3 mi. S Jordan; 6.3 mi. S Jordan; Wyboo Creek, 8.5 mi. from Manning. Colleton: Edisto River (Schwartz, 1956:19-20). Darlington: Pee Dee River, Society Hill (Stejneger, 1944:72). Dorchester: Edisto River, 17 mi. from Summerville; Edisto River, 14 mi. W Summerville; Edisto River, 2.5 mi. S Hart's Bluff. Fairfield: 1 mi. N Peak, Newberry County. Georgetown: North Santee River, 1 mi. above US 17. McCormick: Little River near McCormick; Little River, 3 mi. NE Mt. Carmel. Laurens: Enoree River, 3 mi. S Cashville, Spartanburg County; Enoree River, 9.4 mi. N Clinton. Orangeburg: Edisto River, Orangeburg. Saluda: Batesburg; Lake Murray; Little Saluda River; 5 mi. from Saluda. County unknown: Upper Lake Santee (Schwartz, loc. cit.).
Trionyx spinifer emoryi (Agassiz)
Texas Spiny Softshell
Aspidonectes emoryi Agassiz (in part), Contr. Nat. Hist. United States, Vol. 1, Pt. 2, p. 407; Vol. 2, Pt. 3, pl. 6, figs. 4-5, 1857.
T[rionyx] s[pinifer] emoryi Schwartz, Charleston Mus. Leaflet, No. 26, p. 11, 1956.
Type.—Lectotype, USNM 7855; alcoholic (sex undetermined); obtained from the Río Grande near Brownsville, Texas, in the course of the Mexican Boundary Survey under the command of Colonel Wm. H. Emory.
Range.—Southwestern United States and northern México; the Río Grande drainage in Texas, New Mexico and northern México; the Río San Fernando and Río Purificación drainages in northeastern México; the Colorado River drainage in Arizona, New Mexico, and southern Nevada (see map, Fig. 19).
Diagnosis.—Juvenal pattern of white dots, not encircled with dusky or blackish ocelli, confined to posterior third of carapace; pale rim of carapace conspicuously widened, four to five times wider posteriorly than laterally; a dark triangle in front of eyes, base line connecting anterior margins of orbits; pale postocular stripe interrupted leaving conspicuous pale, usually dark-bordered, blotch just behind eye.
Description.—Plastral length of smallest hatchling, 2.5 centimeters (USNM 7632); of largest male, 13.0 centimeters (KU 2914, 3125, 3150); of largest female, 22.0 centimeters (TNHC 8023, 8104).
Carapace pale brownish or tan, lacking whitish dots on anterior half; whitish dots confined to posterior third of carapace, sometimes lacking posteriorly, especially on juveniles; small, blackish dots rarely occurring on surface of carapace, usually confined to margins when present; pale rim of carapace four to five times wider posteriorly than laterally.
Pattern on snout rarely variable, consisting of pale stripes extending forward from eyes that have only their outer borders darkened and a straight or slightly curved, dark line that connects anterior margins of orbits; few, if any, dark markings in subocular and postlabial region; pattern on side of head having few contrasting marks, often of nearly uniform coloration; postocular stripe usually interrupted; anterior segment of postocular stripe just behind eye usually dark-bordered; posterior segment usually not dark-bordered or sharply distinguished from background; pattern on dorsal parts of soft parts of body contrasting, of relatively small dark marks; dark streaks often coincident with digits.
Underparts whitish, occasionally having blackish dots or smudges on posterior part of carapace, in region of bridge, or on lateral parts of chin and throat; few dark marks often on webbing of limbs and on palms and soles.
Small, flattened or wartlike, tubercles that occasionally have sharp tips along anterior edge of carapace on adult males; tubercles flattened, scarcely elevated, never conical along anterior edge of carapace on large females; whitish, knoblike tubercles often present posteriorly in middle of carapace and in nuchal region on large females; mottled and blotched pattern sometimes contrasting on carapace of large females; whitish dots of juvenal pattern often visible through overlying blotched pattern of large females.
Ontogenetic variation in PL/HW, mean PL/HW of specimens having plastral lengths 7.0 centimeters or less, 3.68, and exceeding 7.0 centimeters, 5.19; ontogenetic variation in CL/CW, mean CL/CW of specimens having plastral lengths 8.5 centimeters or less, 1.17, and exceeding 8.5 centimeters, 1.27; mean CL/PCW, 2.18; mean HW/SL, 1.43; mean CL/PL, 1.37.
Variation.—Ten topotypes (six males, three females, one juvenile) from Brownsville, Texas (BCB 7465-73, 7564), have the following characteristics: pale rim widened posteriorly as described above; females (plastral lengths 9.8, 10.2 and 11.7 cm.) having blackish marks in pale rim, which are absent in males of corresponding size; interrupted postocular stripe with pale blotch behind eye; postocular pale blotch having blackish borders or not; dark triangular mark on snout in front of eyes; white dots present only on posterior third of carapace; carapace of females grayish, blotched pattern not contrasting; carapace of males paler, greenish-gray; undersurface immaculate except 7468 and 7472 that have blackish flecks at bridge and, on 7472, blackish marks that extend posteriorly onto ventral surface of carapace; tubercles along anterior edge of carapace flattened and rounded in adult males, more knoblike in females; largest specimen, BCB 7472, female, plastron 11.7 centimeters long.
T. s. emoryi varies more than any other subspecies of Trionyx spinifer. A large series of males and females (KU) from the Salt River (Colorado River drainage), near Phoenix, Arizona, is characterized by many adult males having indistinct white dots on posterior half of carapace; blotching on carapace of females of contrasting lichenlike figures, but usually non-contrasting and pale brownish or tan; pale rim of carapace distinct from ground color of carapace in largest female (KU 2905, plastron 21.5 cm. in length), but having dark or dusky markings: dark interorbital stripe often lacking. AMNH 58370 (Nevada) and UMMZ 92006 (Arizona) also have the dark line connecting the anterior margins of the orbits interrupted; seemingly the dark interorbital line is most often interrupted in those softshells inhabiting the Colorado River system of Nevada and Arizona.
Other variant individuals are: TU 14453.2, 14462 and 3696 having the plastron extending slightly farther forward than the carapace, thus resembling T. ferox; UMMZ 54021 and CNHM 39999, hatchlings, lacking distinct whitish dots on posterior half of carapace; UI 43509 and KU 39991 having stained (brown or blackish) claws; and, CNHM 6810, an adult male, lacking a spinose (sandpapery) carapace. I am unable to discern geographic variation in these or other characters.
The ground color of the carapace on some individuals from the Pecos River (TU, Terrell County, Texas) is grayish and in contrast with the pale rim (Pl. 44). UI 43509 from the Río Florida, La Cruz, Chihuahua, a female, has a dark brownish carapace with little evidence of a blotched pattern except on the pale rim of the carapace. A female and adult male from the Río Sabinas, Coahuila (MSU 905-06), also show considerable darkening on the dorsal surfaces; the pale rim is evident but not in sharp contrast to the coloration of the carapace. Notes taken on the freshly-killed Sabinas individuals are: male—carapace olive-gray; dorsal surface of soft parts of body olive-green to grayish, a bright yellow suffusion on limbs and neck; female—carapace and soft parts of body dark olive, laterally pale yellow; the plastron extends slightly farther forward than the carapace in both sexes.
Notes on coloration (judged to be the most common or "normal" type) [512] of living emoryi from the Río Mesquites, central Coahuila, are: Adult male (KU 53753)—pale rim butterscotch yellow; marginal line blackish; whitish dots on pale brown or tan carapace; soft parts of body olive or olive-green, slightly darker on head and paler (yellowish) on hind limbs; pale areas on side of head pale yellow, having tint of orange on neck; ventral surface white, yellow laterally on neck. Adult female (KU 53754)—carapace having contrasting blotched and mottled pattern of pale browns and tans; soft parts of body olive brown, darker brown blotching on head; dorsal surface of limbs olive-green having pale areas lemon yellow and webbing butterscotch yellow; side of neck and head, chin and throat pale lemon yellow; ventral surface white having slight red tinge to groin and soft parts posteriorly; underside of carapace near edge pale yellow.
Softshells from the Río Grande in the Big Bend region of Texas, and the Río Conchos in Chihuahua differ from other specimens of emoryi. Fifteen adult males, KU 51187-201 (no females in sample), were taken from the mouth of the Río San Pedro at Meoquí, Chihuahua (see KU 51194, Pl. 44). They are noteworthy because of a conspicuous orange or orange-yellow on the side of the head. Another relatively consistent character is the blackish tip of snout (excepting 51199), although the degree (palest on 51190) and extent of pigmentation posteriorly on the snout is variable. Eleven males, KU 51175-85, from approximately 100 miles northeastward in the Río Conchos near Ojinaga, Chihuahua, also have the bright orange on the side of the head; the tip of the snout is not blackish, although in some it is slightly darkened. Three females, KU 51174, 51186 (from Ojinaga) and 51173 (from 8 mi. S, 16 mi. W Ojinaga), lack the orange on the side of the head; KU 51186 has a plastral length of 8.0 centimeters, whereas the other two females have the same plastral length of 16.5 centimeters (larger than any male). Nineteen adult males, KU 51965-72, 51980-90, from the Río Grande near Lajitas also have the orangish coloration on the side of the head, whereas twenty females, KU 51954-64, 51973-79, 51991-92 (three smaller than largest male) lack the coloration. The tip of the snout is not blackish on any turtle in the series from Lajitas. The smallest female, from Lajitas, having a plastral length of 6.9 centimeters, has a mottled carapace.
The orange of males is most conspicuous in the pale postocular and postlabial areas; the stripes of the snout (distally) and the color of the neck at its juncture with the immaculate ventral surface are orange-yellow. The orange coloration is confined to males (all examined were sexually mature) and is probably not of seasonal occurrence (see comments under secondary sexual variation). I have not noticed this coloration in other males of the subspecies emoryi; however, long-preserved males might be expected to lack the orange color; the specimens mentioned above were initially preserved in alcohol. KU 51179 (plastral length 8.2 cm., from Ojinaga) is the smallest sexually mature male of the species spinifer that I have seen. Another character of note is the generally greater development of the plastral callosities (resembling muticus) than in other subspecies of spinifer or specimens of emoryi; three small adult males (KU 51177, 51990, 51987, plastral length 9.3, 9.9 and 9.1 cm., respectively) have large hyoplastral and hypoplastral callosities that appear to touch medially, and callosities on the epiplastron and both preplastra.
On July 8, 1953, an adult male of T. spinifer was removed from a hoop-net set in the Río Purificación at Padilla, Tamaulipas, México. I was particularly [513] impressed by the lack of whitish dots on the dark carapace; the following notes were taken from the freshly-killed specimen: carapace a uniform dark olive, lacking white dots and having a yellowish rim widest posteriorly; tubercles on anterior edge of carapace only slightly raised, inconspicuous; top of head olive with few dots and streaks; a well-defined yellowish postocular stripe not conspicuously interrupted; sharp contrast between dark olive on side of head and pale ventral coloration; yellowish-orange ventrolaterally on head; an uninterrupted slightly-curved line connecting the anterior margins of the orbits; carapace pear-shaped; underparts whitish, lacking markings. This specimen has since been destroyed. The only other specimen I have seen from this locality is a hatchling (UMMZ 69412, Pl. 43), which has a pale brownish or tan carapace that lacks whitish dots; it resembles emoryi in other characters. Although the absence of whitish dots is not distinctive, its combination with the uniform dark olive carapace in adult males and the fact that the Río Purificación is an isolated drainage system, suggests that soft-shelled turtles from that river system may warrant further taxonomic study.
Comparisons.—From all other subspecies of spinifer, T. s. emoryi can be distinguished by having a pale rim on the carapace that is four to five times wider posteriorly than it is laterally. This character, unique for emoryi, combined with patterns on the snout, side of head and carapace that are subject to little variation, permit ready identification of the subspecies emoryi. T. s. emoryi resembles pallidus, and guadalupensis and differs from spinifer, hartwegi and asper in having whitish tubercles or dots on the carapace. T. s. emoryi resembles guadalupensis but differs from pallidus, spinifer, hartwegi and asper in lacking conical tubercles along the anterior edge of the carapace on large females. For additional differences see accounts of other subspecies.
Some populations of T. s. emoryi resemble T. muticus in the size at which sexual maturity is attained and in the development of the plastral callosities. T. s. emoryi has a wide head that resembles that of T. ferox, T. ater, T. s. asper and T. s. guadalupensis; T. s. emoryi also resembles T. ferox and T. ater but differs from the other subspecies of T. spinifer and T. muticus in having a narrower carapace. T. s. emoryi resembles T. s. guadalupensis, T. s. pallidus and T. ater, and differs from the other subspecies of spinifer and T. muticus, in having the carapace widest farther posteriorly than one-half way back on the carapace. T. s. emoryi resembles T. ferox in having the shortest length of snout of the subspecies of spinifer. The plastron is shorter than in T. ferox, longer than in T. s. asper, and about the same length as in T. muticus and the other subspecies of T. spinifer.
Remarks.—Agassiz (1857, 1:407-08) did not designate a holotype in the original description of Aspidonectes emoryi; specimens are mentioned from the lower Río Grande of Texas, near Brownsville, and a stream of the Río Brazos drainage in Williamson County, Texas. The description is applicable to T. s. emoryi as herein restricted, except for the statement that the white tubercles of young specimens are "encircled by faint black lines"; that statement is presumably based on the juveniles from Williamson County. T. s. emoryi does not occur in Williamson County, Texas. Barbour and Loveridge (1929:225) listed MCZ 1909-10 and 1627 as cotypes. Stejneger (1944:65) mentioned MCZ 1909, 1913 and USNM 7855 as cotypes; the legend for Plate 20 (op. cit.) refers to a drawing that "corresponds fairly closely with the type (MCZ 1910) collected at Brownsville, Texas, by Col. Emory."
The syntypic series consists of seven specimens—MCZ 1627 (two specimens) from Williamson County, Texas; MCZ 1909 (three specimens) and 1910 from Brownsville, Texas; and USNM 7855 from Brownsville, Texas. The listing of number 1913 by Stejneger is considered a lapsus for 1910 as MCZ 1913 is catalogued as a Graptemys geographica (in letter dated November 17, 1959 from Dr. Ernest E. Williams). Stejneger's reference to MCZ 1910 as the type is considered unintentional and an inadequate designation of a lectotype.
In the "remarks" column of the USNM museum catalog, number 7855 is referred to as "Ag. Type." USNM 7855 is here designated as lectotype of Trionyx spinifer emoryi. The lectotype is a young specimen (female?) that is not easily sexed by external characters; the plastron measures (in centimeters) 6.3 in length, the carapace 8.2 in length and 7.0 in width, and the head 1.4 in width. The carapace is pale brown having inconspicuous whitish dots posteriorly and a pale rim that is approximately 6.8 times wider posteriorly (4.1 mm.) than it is laterally (0.6 mm.). The slightly curved dark line connecting the anterior margins of the orbits is dimmer than the dark lines that extend forward from the eyes. The pale postocular stripes having blackish, dotted borders are interrupted; there are no other markings on the side of the head. The ventral surface is immaculate except for a few dark dots on the right side of the carapace; the ground color is pale brown or tan, but the upper layer of skin can be scraped away revealing an underlying pale lavender-cream ground color. The tubercles along the anterior edge of the carapace resemble small rounded warts.
MCZ 1910 is an adult male T. s. emoryi having a plastron 10.7 centimeters in length. The carapace is pale brown having a relatively smooth anterior edge, inconspicuous whitish tubercles posteriorly, and a pale rim five times wider posteriorly than laterally; the pattern on the head resembles that of emoryi.
Each of three hatchlings of T. s. emoryi, 3.4, 3.5 and 3.9 centimeters in plastral length, bears an MCZ catalogue number of 1909. The carapaces are dark tan or gray having pale rims 3.7, 5.2 and 5.2 times wider posteriorly than laterally, and white dots absent or obscure posteriorly; two specimens have small blackish dots paralleling the pale rim posteriorly. The patterns on the heads are referable to emoryi.
The two juvenal syntypes (5.2 and 6.1 cm. in plastral length) from Williamson County, Texas, are both catalogued as MCZ 1627, but only one of these bears a catalogue number. The two softshells are not emoryi, and are more nearly like T. s. guadalupensis than T. s. pallidus. Actually, they are from an area of intergradation between those subspecies (see comments concerning intergradation under the accounts of the subspecies pallidus and guadalupensis). White spots occur on the carapaces anteriorly and posteriorly, the larger (more posterior) of which are encircled with dusky ocelli. The carapace of the small specimen (bearing no number) is brown having a few, small black specks intermixed with the white spots. The carapace of the large specimen is pale lavender and has a more obscure pattern than the other specimen.
After Agassiz's description, emoryi was accepted as a distinct species. Neill (1951:15) suggested that emoryi was subspecifically related to T. ferox. Crenshaw and Hopkins (1955) and Schwartz (1956), however, demonstrated that ferox was a distinct species; emoryi has since been considered a subspecies of T. spinifer.
Two specimens having blackish dots on the carapace, indicate relationship with T. s. guadalupensis. USNM 7638, a hatchling, has large whitish dots surrounded by blackish dots confined to the posterior half of the carapace, and the locality for this specimen is merely Río Bravo (= Río Grande). CNHM 47366, a hatchling from Sierra de las Palmas (Sierra de Santa Rosa, La Palma), Coahuila, has a few, small, blackish dots, irregularly spaced, on the anterior half of the carapace, but other dots more evenly distributed on the posterior half where they are intermixed with whitish dots. The drawing of the dorsal view of a hatchling emoryi (Agassiz, 1857:Pl. 6, Fig. 4) shows a sprinkling of blackish dots on the anterior half of the carapace. A hatchling from Eagle Pass (USNM 116578) does not have a noticeably widened pale rim posteriorly on the carapace, and is not distinguishable from pallidus. See account of T. s. guadalupensis for further comments on intergradation.
A soft-shelled turtle that was obtained in the Sacramento River by three fishermen, near Sacramento, California, was named Aspidonectes californiana by Rivers (1889:233). A comparison (with Aspidonectes spinifer and A. emoryi) of certain features of the skull was largely prepared by Baur and included in the description (op. cit.:234-35); seemingly, the most trenchant character of the skull of californiana was the enlarged alveolar surfaces of the jaws. This feature prompted Baur (1893:220) to refer californiana to the genus Pelodiscus, which also included agassizi (skulls also having jaws with enlarged alveolar surfaces) and several Old World species. Van Denburgh (1917) discussed the origin of the specimen that formed the basis of River's description and concluded that it was brought over from China. Siebenrock (1924:192) and Mertens and Wermuth (1955:389) listed Aspidonectes californiana as a synonym of emoryi. River's description is not that of emoryi; the enlarged alveolar surfaces of the jaws, and the dark carapace having tubercular ridges suggest a resemblance to T. ferox. The papillae on the neck are not found in any American species. Miller (1946:46, footnote 2) believed that "it obviously was introduced, apparently from China," and cited Pope (1935:61), who declared the specimen to represent Trionyx sinensis.
Schmidt (1924:64) first reported the occurrence of T. s. emoryi west of the continental divide in Arizona and suggested that it was highly probable that the species had been introduced near Phoenix in recent years. Cowles and Bogert (1936:42) mentioned a species of softshell occurring in the Boulder Dam region and presumed the species to be native to Asia and introduced by the Chinese. Linsdale and Gressitt (1937:222) determined the status of the species in the Colorado River drainage as T. s. emoryi. The discussions by Dill (1944:179-81) and Miller (1946:46) indicate that emoryi was introduced into the Gila River (Colorado River drainage) in western New Mexico near the turn of the century.
T. s. emoryi and T. ater are the only kinds of softshells occurring in México. The colloquial name for soft-shelled turtles in México is "tortuga blanca." This name is also used in reference to the Central American river turtle, Dermatemys mawei, which occurs on the east coast of México as far north as Veracruz.
Specimens examined.—Total 275, as follows: Arizona: Maricopa: CNHM 4768, KU 2214-19, 2803, 2824, 2837, 2903-07, 2909-16 (2914, 2 specimens), 2918-29, 3118-27, 3129, 3147-56, USNM 71627, Salt River, Phoenix. Pinal: UI 37713, Gila River, 6 mi. E Winkleman; UMMZ 92006-07, Gila River, 1/2 mi. [516] below Coolidge Dam; UMMZ 105824, San Pedro River about 1 mi. above confluence with Gila River.
Nevada: Clark: AMNH 58370, Boulder City boat landing, Lake Mead; TU 15802, Virgin River, Mesquite.
New Mexico: Eddy: KU 15938, Carlsbad; KU 48217-18, Black River Village. Grant: AMNH 79911, Gila River, 8 mi. NE Cliff.
Texas: Brewster: CNHM 39999, Tornillo Creek near jct. with Río Grande; KU 51954-92, Lajitas; TCWC 4291, UMMZ 66471, USNM 45545, 103678, Boquillas; INHS 7975, UMMZ 114360, Hot Springs. Cameron: BCB 7564-73, CNHM 5339-40, 6810, MCZ 1909 (3), 1910, TU 11479-80, 11561-62, UMMZ 54021, 105209-13 (Brownsville Lake), USNM 7642, 7644, 7855, Brownsville; BCB 5121, 3 mi. S Harlington. El Paso: UMMZ 85085, El Paso; USNM 7641, 7701, El Paso del Norte. Hudspeth: USNM 20846, Fort Hancock on Río Grande. Kinney: CNHM 26090, Río Pinto W of Bracketville; USNM 26426-36, Fort Clark. Loving: TTC 1143, Red Bluff Lake just below dam on Pecos River. Maverick: TU 3696-97, UMMZ 116578, Eagle Pass. Presidio: TTC 628 (2), 632 (2), 3 mi. WNW Lajitas, Brewster County. Terrell: TNHC 7997, 8022-23, Chandler Ranch, 30 mi. S Sheffield, Pecos County; TNHC 8104, Dunlap Ranch, 25 mi. SE Sheffield, Pecos County; TU 14453 (7), 14462 (2), 15415, 15423, 15586, Pecos River near jct. with Independence Creek; USNM 104240, Pecos River "near" Dryden. Val Verde: TTC 113, Pecos River. Webb: TNHC 19788, 42 mi. NW Laredo; USNM 109078-79, Laredo. Zapata: UI 19332, "near" Zapata. County unknown: MCZ 1628, USNM 7635-36, 7854; USNM 7637-38, Río Bravo (= Río Grande).
Chihuahua: KU 51173, 8 mi. S, 16 mi. W Ojinaga; KU 51174-86, 1 mi. NW Ojinaga; KU 51187-201, Río Conchos at mouth of Río San Pedro near Meoquí; UI 43508-09, Río Florida, La Cruz.
Coahuila: CNHM 26054, Sta. Helena Canyon of Río Grande; CNHM 28846, "near" Músquis; CNHM 55657, Río Alamos, Rcho. de la Gacha; CNHM 47366, Sierra de Santa Rosa, La Palma; CNHM 47367, 55661, Cuatro Ciénegas; CNHM 55658-60, Rcho. de los Borregos near Juarez; KU 33523, La Presa Don Martín; KU 39991, 39993, 8 mi. N, 2 mi. W Piedras Negras; KU 39992, 2 mi. W Jiménez; KU 46907, 16 km. S Cuatro Ciénegas; KU 46913-16, 10 km. S Cuatro Ciénegas; KU 53752-54, Río Mesquites, 8 mi. W Nadadores; KU 53757, 8.5 mi. SW Cuatro Ciénegas; MSU 905-06, Río Sabinas, 1 mi. E Sabinas.
Nuevo Leon: CNHM 1874, 2191, Rodriguez; UMMZ 69411, Río Conchos, 9 mi. N Linares.
Tamaulipas: CM 3037, Nuevo Laredo. UMMZ 7614-20, 7622-25, 7628, 7630, 7632-33, Matamoros; UMMZ 69412, Río Purificación, N of Ciudad Victoria.
No Data: MCZ 1629 (2), NHB 1032.
Records in the literature.—Arizona: Greenlee: Gila River, Duncan (Miller, 1946:46); "near" Sheldon (Dill, 1944:180). Mohave: Pierce's Ferry just below lower end of Grand Canyon (Cowles and Bogert, 1936:42); 1.5 mi. upstream (Virgin River) from Mesquite, Clarke County, Nevada (Hardy and Lamoreaux, 1945:168); Lake Havasu on Colorado River (Dill, 1944:180). Yuma: Colorado River at Headgate Rock Dam (Dill, op. cit.:179).
California: Imperial: California Lakes (Cowles and Bogert, 1936:42); Palo Verde; Colorado River at Laguna Dam (Dill, 1944:180).
Nevada: Clark: observed just north of Black Canyon (Cowles and Bogert, loc. cit.); Colorado River, 6 mi. N California line (Linsdale, 1940:255).
New Mexico: Chaves: Bitter Lakes Wildlife Refuge, 12 mi. NE Roswell (Bundy, 1951:314). Dona Ana: Río Grande near Mesilla Dam (Little and Keller, 1937:221).
Texas: Brewster: Río Grande at Castolon (Minton, 1959:38). Val Verde: mouth of Devil's River (Brown, 1950:250).
Baja California: Colorado River delta, 7 mi. E Cerro Prieto; Imperial [517] Irrigation District, Alamo Canal, 15 mi. S Internat'l Boundary and Salfatana Canal, 1 mi. N Black Butte (Linsdale and Gressitt, 1937:222).
Coahuila: San Juan (Schmidt and Owens, 1944:103).
Hitherto, soft-shelled turtles of the species Trionyx spinifer from the southern and southwestern United States having a pattern of white dots on the carapace have been relegated to the subspecies emoryi, but my examination of soft-shelled turtles from Texas has indicated that T. s. emoryi as previously conceived, is a composite of three subspecies. It is necessary, therefore, to recognize two new subspecies.
Trionyx spinifer guadalupensis new subspecies
Guadalupe Spiny Softshell
Holotype.—UMMZ 89926, alcoholic adult male; obtained 15 miles northeast Tilden, McMullen County, Texas (Pl. 41, bottom, left).
Paratypes.—Forty-two specimens: ANSP 16717 (hatchling), no data; USNM 78515-16 (hatchlings), Colleto Creek, Victoria County, Texas; TU 10143-45, 10148, 10150-59, 10161-65 (adult males), TU 10176, 10833 (immature males), TU 10147, 10149, 10155 (immature females), TU 10160 (adult female), Guadalupe River, 9 miles southeast Kerrville, Kerr County, Texas; UMMZ 89915-21, 89924-27 (adult males), UMMZ 89922-23 (immature females), same locality as holotype; UMMZ 92752 (immature female), San Antonio River, 3 miles west-northwest Goliad, Victoria County, Texas.
Description of holotype.—Carapace nearly circular, widest at level of posterior border of hypoplastra; margin entire; dorsal surface "sandpapery" to touch; pale rim separated from ground color of carapace by well-defined, blackish line that is wavy and narrowly interrupted posteriorly and anteriorly; pale rim approximately 1.8 times wider posteriorly (5.4 mm.) than laterally (3.0 mm.); pale rim increasingly narrower anteriorly, absent in nuchal region; tubercles in nuchal region low, scarcely elevated, lacking sharp tips; ground color of carapace olive having pattern of whitish spots and small tubercles; most whitish tubercles inconspicuous pinpoints; other small tubercles in center of whitish spots, mostly approximately 2 millimeters in diameter; largest white spot 3.4 millimeters in diameter; most white spots surrounded by blackish ocelli or parts thereof; whitish spots distributed over entire surface of carapace; certain features of bony carapace evident through overlying skin; carapace highest in region of second and third neurals, forming obtuse, gently sloping, vertebral, keel; undersurface of carapace butterscotch yellow, lacking markings; maximum length, 16.5 centimeters; greatest width, 13.5 centimeters.
Plastral surface butterscotch yellow, lacking markings, extending slightly farther forward than carapace; anterior and posterior lobes rounded; anterior lobe slightly truncate; certain features of bony elements of plastron visible through overlying skin; maximum length of plastron, 12.0 centimeters.
Head, extended to posterior level of eyes, terminating in flexible snout; septal ridges projecting into each rounded nostril; jaws closed, each covered by fleshy lips except anteriorly where horny portions exposed; dark triangular mark in front of eyes, base line connecting anterior margins of orbits forming series of [518] dots; pale stripes extending forward from eyes having faint inner, blackish borders; eyelids partly open having blackish dots; pale subocular blotch on right side of head having border of black dots.
Forefeet and hind feet well-webbed having five digits each; each limb having nails on first three digits; each forelimb with four antebrachial scales, three of these having free edge; each hind limb with two horny scales, one smooth on posterodorsal surface and other with free edge on posteroventral surface; pattern toward insertion of forelimbs indistinct.
Tail terminating in flexible point; penis exposed; cloacal opening extending beyond posterior edge of carapace; tail olive above bordered by blackish marks; few black dots laterally on left side.
Undersurface of soft parts of body buff, lacking markings; few dark marks posteriorly on webbing of limbs, encroaching on soles and palms.
Range.—Southcentral Texas in the drainage systems of the Nueces and Guadalupe-San Antonio rivers; the Colorado River drainage in Texas is inhabited by a population that more closely resembles guadalupensis than pallidus. See comments under subsection entitled "Remarks" and Fig. 19.
Diagnosis.—Juvenal pattern of white dots that are conspicuous on anterior half of carapace, and usually as large as those on posterior half; white dots, sometimes 3 millimeters in diameter, encircled with blackish ocelli in adult males.
Description.—Plastral length of smallest hatchling, 3.3 centimeters (ANSP 16717); of largest male, 13.5 centimeters (TU 10162); of largest female, 22.0 centimeters (TU 10160).
Hatchlings having white dots on anterior half of carapace; white dots anteriorly nearly as large as those posteriorly, encircled with blackish ocelli, and conspicuous on dark background (ANSP 16717, Pl. 41; USNM 78515-16; Stebbins, 1954:181, Pl. 26B), or smaller than those posteriorly, not encircled with dusky ocelli, and inconspicuous on pale background (TNHC 1446); pale rim of carapace less than four times as wide posteriorly as laterally.
Adult males resembling holotype; size of white tubercles on carapace variable; most, if not all, tubercles surrounded by narrow blackish ocelli, or parts thereof; largest white tubercles or dots in most specimens exceeding one millimeter and in some specimens three millimeters in diameter (TU 10163); white dots often slightly elongate (UMMZ 89917, 89920, 89926; TU 10152, 10145); juvenal pattern of white dots seemingly more contrasting in guadalupensis, owing to dark ground color of carapace, than in pallidus or emoryi that have pale brown or tan carapaces; small tubercles along anterior edge of carapace rounded, obtuse, wartlike, never conical; sharp tips often lacking (TU 10153).
Large females often having whitish spots on anterior half of carapace (TU 10160, Pl. 42, upper, right; 10142); carapace dark having ill-defined mottled and blotched pattern; tubercles along anterior edge of carapace low, rounded, rarely equilateral, never conical; small blackish dots rarely on surface of carapace (UMMZ 89923).
Pattern on side of head and snout of little diagnostic value; postocular stripe usually interrupted, but configuration variable, consisting of pale anterior, dark-bordered segment (just behind eye); posterior segment of postocular [519] stripe usually less well-defined and generally blending with adjacent ground color; pale postocular stripe sometimes uninterrupted and dark-bordered throughout its length (TU 10157, 10159, 10176); pattern on dorsal surface of snout variable; pattern usually consisting of uninterrupted dark line (slightly curved anteriorly) connecting anterior margins of orbits (TU 10161, 10164, 10159, 10143), or dark line interrupted (TU 10153, 10154, 10176), absent (TU 10163), or present in addition to dark inner borders of pale stripes that extend anteriorly from eyes (TU 10149, 10162); small, often fine, dark markings, on dorsal surface of limbs, especially forelimbs; ventral surface of plastron and soft parts of body usually whitish, lacking markings; small blackish spots occasionally in region of bridge (TU 10149); dark marks occurring on webbing of limbs and often encroaching on soles and palms.
Ontogenetic variation in PL/HW, mean PL/HW of specimens having plastral lengths 7.0 centimeters or less, 3.83, and exceeding 7.0 centimeters, 5.18; ontogenetic variation in CL/CW, mean CL/CW of specimens having plastron lengths 8.5 centimeters or less, 1.14, and exceeding 8.5 centimeters, 1.22; mean CL/PCW, 2.11; mean HW/SL, 1.38 (including subspecies pallidus); mean CL/PL, 1.37.
Variation.—Two hatchlings (ANSP 13447, Bexar County; TNHC 1446, McMullen County) more closely resemble pallidus than guadalupensis.
Some individuals from the Colorado River drainage have features suggesting those that are characteristic of pallidus. Large females have obtuse, knoblike somewhat triangular-shaped tubercles along the anterior edge of the carapace, which are never conelike (TU 14439-40, 10187, 16036.1; BCB 6010). The tubercles along the anterior edge of the carapace are more elevated than in turtles from drainage systems west of the Colorado. Whitish spots are usually absent anteriorly on the carapace, but may be evident through the mottled pattern of large females (BCB 6010, plastral length, 19.7 cm.). The pale postocular stripe is usually interrupted, whereas the dark line connecting the anterior margins of the orbits is usually not interrupted; the two characters last mentioned show alliance with guadalupensis.
The carapace of hatchlings from the Colorado River is pale having whitish dots, smaller anteriorly than posteriorly, which may be encircled with dusky ocelli (TNHC 20257) or not (ANSP 11889, BCB 5055, SM 3282). Many hatchlings are not distinguishable from pallidus (TCWC 7262, TNHC 4975, SM 4924, 6106). I have not seen hatchlings from the Colorado River that resemble ANSP 16717.
The pattern on the carapace of adult males from the Colorado River drainage resembles that of guadalupensis (Pl. 41, bottom, right) but the whitish dots are usually smaller and may not be encircled with blackish ocelli (BCB 4066, TU 14485). An adult male (TU 14476) from the South Fork of the Llano River has whitish dots three millimeters in diameter and encircled with blackish ocelli (guadalupensis), whereas another adult male (USNM 83690) from a tributary of the Colorado, the South Concho River, resembles pallidus.
Eight specimens from the San Saba River (TU 14419 [6 specimens], 14439-40), that range in plastral length from 6.8 to 17.0 centimeters are impressive because of the dark brownish coloration on the carapace. The smallest individual, which is also the only male in the series, is paler. The mottled and [520] blotched pattern on the females is therefore not contrasting; the largest females have elevated whitish prominences in the center of the carapace posteriorly. An immature male (UMMZ 70348) from the South Concho River also has a dark brown carapace, and lacks white dots. The dark coloration of the carapace of these specimens recalls the TU series of T. s. emoryi from the Pecos River, Terrell County, Texas.
Color notes taken from a freshly-killed adult female from the Llano River, two miles west Llano (TU 16036.1, Pl. 42), are: pattern on carapace of dark olive or blackish marks that form an irregular reticulum or marbling on a paler background that varies from brownish to buff and has an orange or reddish tinge in some areas; small whitish spots posteriorly; pale rim yellowish, evident only at sides of carapace; dorsal surface of soft parts of body olive-green, becoming paler with yellowish tinge toward insertions of limbs and neck; no contrasting pattern on limbs or neck and head; yellowish on sides of body; ventral surface whitish lacking dark marks, yellowish at region of bridge, axillary region and on neck; chin olive-yellow.
Comparisons.—T. s. guadalupensis can be distinguished from all other subspecies of T. spinifer in having: (1) large white dots, sometimes three millimeters in diameter, on a dark background usually surrounded with blackish ocelli and conspicuous on the anterior half of the carapace (some as large as those on posterior half) in adult males, and (2) whitish dots on the anterior half of the carapace, in hatchlings, that are often encircled with dark ocelli. T. s. guadalupensis resembles pallidus and emoryi in having white tubercles or dots on the carapace and therein differs from spinifer, hartwegi and asper. T. s. guadalupensis resembles pallidus but differs from emoryi in having a pale rim that is less than four times wider posteriorly than laterally. T. s. guadalupensis resembles emoryi but differs from pallidus, spinifer, hartwegi and asper in having along the anterior edge of the carapace tubercles that are flattened or wartlike prominences often lacking sharp tips in adult males; these tubercles are never conical in large females.
T. s. guadalupensis has a wide head, a feature shared with the subspecies asper and emoryi, but differs from emoryi in having a wider carapace. T. s. guadalupensis resembles emoryi and pallidus but differs from the other subspecies in having the carapace widest farther posterior than one-half the length of the carapace. The length of snout in pallidus and guadalupensis is shorter than in spinifer and hartwegi but is longer than in emoryi. T. s. guadalupensis differs from asper but resembles the other subspecies in having a relatively long plastron.
Remarks.—Some individuals of guadalupensis have characteristics that are applicable to emoryi. TNHC 12352 (Llano River) a hatchling, has conspicuous white dots confined to the posterior third of the carapace; the pale rim, however, is not widened posteriorly. TU 10156 (Guadalupe River) has a conspicuously widened pale rim on the carapace that is approximately 3.4 times wider posteriorly (8.5 mm.) than laterally (2.5 mm.).
T. s. guadalupensis more closely resembles pallidus than emoryi. Turtles living in rivers that drain into the Gulf of Mexico east of the Guadalupe-San Antonio river system successively show increasing resemblance to pallidus from west to east.
The expression of intergradation between guadalupensis and pallidus is of a clinal nature that involves parallel changes in the pattern on the snout, [521] side of head, limbs (to a lesser degree), tuberculation along the anterior edge of the carapace, size of whitish tubercles or dots, and the distinctness of the blackish ocelli that surround the whitish dots on the carapace. These characters form a well-marked gradation or cline that extends over a considerable area. There is, however, no continuous environmental gradient because the populations are relatively isolated by occupying adjacent drainage systems. The sharpest break in the gradation of characters mentioned above occurs between the Colorado River and Brazos River drainages. The population of softshells in the Colorado River drainage is actually an intergradient one, but more closely resembles guadalupensis, whereas the population in the Brazos River drainage more closely resembles pallidus. For convenience the turtles inhabiting the Colorado River drainage are referred to guadalupensis and those in the Brazos River drainage to pallidus. Some individuals from farther west than the Colorado River drainage will resemble pallidus, and a few individuals from father east than the Brazos River drainage will resemble guadalupensis.
The gradation of some of the characters mentioned above terminates in the subspecies emoryi. It, however, has characters not found in pallidus or guadalupensis, and is more distinct from either of those subspecies than either is from each other; the difference in characters as well as the break in the gradient of characters between guadalupensis in the Nueces River drainage and emoryi in the Río Grande drainage is greater than that between guadalupensis in the Colorado and pallidus in the Brazos River drainages.
I have refrained from designating individuals between these three subspecies (emoryi, guadalupensis and pallidus) as "intergrades" on the distribution maps, and only mention (in text) those individuals whose characters show a decided tendency toward the adjacent subspecies. For further comments on intergradation see the account of T. s. pallidus.
Specimens examined.—Total 97, as follows: Texas: Bandera: KU 50834, Hondo Creek, 4 mi. W Bandera; TNHC 797-98, 7 mi. SW Medina. Bexar: ANSP 13447, Helotes; MCZ 4587; USNM 10789, 71009, San Antonio. Borden: BCB 4066, 7 mi. N Vincent. Brown: TNHC 7262, 1 mi. E Brownwood. Comal: USNM 7700, New Braunfels. Dawson: TNHC 21594-95, 10 mi. E Lamesa. Frio: USNM 7747, Río Seco. Gillespie: TU 10185, 10187, 10205, Beaver Creek, "near" Doss. Hays: AMNH 29950-52, San Marcos. Kerr: SM 2553, headwaters Turtle Creek; TU 10142-45, 10147-65, 10176, 10833, Guadalupe River, 9 mi. SE Kerrville. Kimble: BCB 5052-55, 6010, 3 mi. SE Telegraph; TU 14476, South Fork Llano River, 1.5 mi. SE Telegraph; TU 14485, Llano River, 10 mi. W Junction. Lavaca: SM 2554-55, 2559, 3 mi. NNE Hope. Llano: TNHC 12352, TU 16036 (2), Llano River, 2 mi. W Llano. McMullen: TNHC 1446, 10 mi. W Simmons, Live Oak County; UMMZ 89915-27, 15 mi. NE Tilden. Matagorda: ANSP 11889, Matagorda. San Saba: SM 6106; TU 14419 (6), 14439-40, San Saba River, 11 mi. NNW San Saba. Tom Green: SM 3282, UMMZ 70348, USNM 83690, South Concho River at Christoval. Travis: SM 659-60, 8.5 mi. from mouth of Onion Creek in Colorado River near Austin; SM 4924, Onion Creek; TNHC 4975, Upper Bull Creek; TNHC 20257, Marshall Ford Dam. Victoria: CM 3118, Black Bayou; UMMZ 92752, San Antonio River, 3 mi. WSW Goliad; USNM 78515-17, Colleto Creek, Guadalupe River. County unknown: ANSP 16717; TNHC 1404.
Records in the literature.—Texas: Bandera: 24 mi. WNW Medina (Brown, 1950:250). Burnet: Colorado River (Strecker, 1909:8). Gillespie: 20 mi. N Harper (Brown, loc. cit.). Kendall: Cibolo Creek at Boerne (Strecker, 1926:8). Kerr: Guadalupe River, 3 mi. above Kerrville (TCWC 474, listed in card file). Mason: 12 mi. NE Mason (TCWC 3303, listed in card file). [522] Matagorda: Bay City (Brown, loc. cit.). Real: (Stejneger, 1944:66). Wilson: Cibolo River, 30 or 40 mi. N Sutherland Springs (Strecker, 1935:23).
Trionyx spinifer pallidus new subspecies
Pallid Spiny Softshell
Holotype.—TU 484, alcoholic adult male; obtained from Lake Caddo, Caddo Parish, Louisiana on June 27, 1947, by Fred R. Cagle and party (Pl. 39, lower, left).
Paratypes.—Forty-two specimens: TU 481, 490, 678 (hatchlings), TU 381, 472, 488 (immature males), TU 475, 478, 486, 1232, 1291, 10170 (adult males), TU 399, 487 (immature females), TU 469 (adult female), Caddo Lake, Caddo Parish, Louisiana; TU 15818 (immature male), TU 15819 (adult male), Cross Lake, Caddo Parish, Louisiana; TU 1253, 13211 (adult males), TU 13266 (immature female), Sabine River, 8 miles southwest Merryville, Beauregard Parish, Louisiana; TU 13281-82 (adult males), TU 13280, 13265 (immature females), TU 13303-04, 13306 (adult females), Sabine River, 8 miles southwest Negreet, Sabine Parish, Louisiana; SM 2375 (adult male), Wallace Bayou, De Soto Parish, Louisiana; TU 1122 (adult male), Lacassine Refuge, Louisiana; UMMZ 92754 (adult male), 5 miles west Iowa, Calcasieu Parish, Louisiana; KU 40174-76, OU 27297 (adult males), OU 27290 (immature female), Lake Texoma, 2 mi. E Willis, Marshall County, Oklahoma; KU 50832 (hatchling), mouth of Caney Creek, 4 miles southwest Kingston, Marshall County, Oklahoma; CNHM 15474 (immature female), Kiowa County, Oklahoma; KU 2966-67 (immature females), KU 2934, 2947 (adult males), KU 2973 (adult female) Lewisville, Lafayette County, Arkansas.
Description of holotype.—Carapace circular, widest at level of posterior edge of hyoplastra; margin entire; dorsal surface "sandpapery" to touch; pale rim separated from ground color of carapace by well-defined, slightly ragged, blackish line; pale rim approximately 2.1 times wider posteriorly (4.7 mm.) than it is laterally (2.2 mm.); pale rim increasingly narrower anteriorly, absent in nuchal region; tubercles along anterior edge of carapace triangular with sharp tips becoming flattened and inconspicuous at level of insertions of arms; ground color of carapace brownish having pattern of small whitish tubercles; most whitish tubercles inconspicuous, of pinpoint size, giving surface of carapace "sandpapery" effect; largest white tubercles posteriorly, approximately 1.2 millimeters in diameter; whitish tubercles smaller anteriorly, largest approximately 0.6 millimeters in diameter; whitish tubercles tend to form two parallel lines coincident with longitudinal sutures of neurals posteriorly in center of carapace; certain features of bony carapace evident through overlying skin; carapace highest in region of third and fourth neurals, forming obtuse, gently sloping, vertebral keel; undersurface of rear margin of carapace whitish having pinkish tinge and no markings; maximum length, 16.8 centimeters; greatest width, 14.3 centimeters.
Plastral surface extending slightly farther forward than carapace, whitish having pinkish tinge and no dark markings; anterior and posterior lobes rounded, posterior lobe more acutely; certain features of bony elements of plastron visible through overlying skin; maximum length, 12.2 centimeters.
Head extended, terminating in flexible snout; septal ridges projecting into each rounded nostril; tip of snout darkened; jaws open, each covered by fleshy lips except anteriorly where horny portions exposed; dark triangular mark in front of eyes, base line uninterrupted, slightly curved anteriorly, connecting anterior margins of orbits; eyelids having blackish dots, especially [523] upper, closing eyes; small blackish dots on dorsal surface of head; pale postocular stripe dark-bordered, interrupted; pale portion of stripe traversed by black line; pale subocular blotch margined by broken blackish border; side of head having contrasting blackish marks on pale background; postlabial stripe having lower blackish border on right side of head; chin with ill-defined marks, not contrasting on grayish background; well-defined, ragged black line on side of neck separating dorsal coloration from immaculate ventral coloration; small dark dots on dorsal surface of neck; dorsal surface of head and neck olive or brownish, becoming paler laterally and toward insertion of neck; maximum width of head, 2.1 centimeters.
Forefeet and hind feet well-webbed each having five digits; each limb having nails on first three digits; each forelimb with four antebrachial scales, three of which have free edge; each hind limb with two horny scales, one smooth on posterodorsal surface and other with free edge on posteroventral surface; contrasting pattern of blackish marks, mostly roundish, on pale background of grayish-white.
Tail terminating in flexible point; penis partly exposed; cloacal opening extending beyond posterior edge of carapace; tail having dorsal grayish band flanked by interrupted blackish lines; dark marks encroaching ventrally at tip of tail.
Undersurface of soft parts of body whitish, with pinkish tinge; dark marks lacking on soles, present on webbing and palms; dark marks arranged in linear fashion coincident with digits.
Range.—Southern Oklahoma, eastern Texas, extreme southwestern Arkansas, and the western half of Louisiana; Red River drainage and rivers that drain into the Gulf of Mexico east of the Brazos River drainage in Texas and west of the Atchafalaya River drainage in Louisiana. The Brazos River drainage is inhabited by a population that more closely resembles pallidus than guadalupensis (see comments under subsection entitled "Remarks"; see map, Fig. 19).
Diagnosis.—Juvenal pattern of white dots that are usually absent or inconspicuous, but sometimes distinct and small, on anterior third of carapace, and not surrounded with dark ocelli; white dots often absent on posterior half of carapace of hatchlings; white spots, rarely as large as two millimeters in diameter, not encircled with black ocelli on adult males; pale rim of carapace less than four times wider posteriorly than laterally.
Description.—Plastral length of smallest hatchling, 3.3 centimeters (KU 50832); of largest male, 16.0 centimeters (SM 2375); of largest female, 30.5 centimeters (TU 13213).
Surface of carapace in hatchlings uniform pale brown or tan; small white tubercles absent or inconspicuous on anterior half of carapace, but evident on posterior half of carapace, sometimes well-defined (TU 481), but usually inconspicuous (TU 678, 490); pale rim of carapace less than four times wider posteriorly than laterally.
Adult males resembling description of holotype; small whitish tubercles or dots rarely two millimeters in diameter on posterior half of carapace, smaller and usually inconspicuous on anterior half of carapace (TU 13281, 486); well-defined whitish tubercles occasionally on anterior half of carapace (KU 40174); white tubercles not surrounded with black ocelli; pattern of white dots seemingly less contrasting in pallidus than in guadalupensis, owing to pale brown or [524] tan carapace; small tubercles along anterior edge of carapace equilateral or conical having sharp tips.
Large females usually having pale brown carapaces with slightly contrasting, brownish, mottled and blotched, patterns; white prominences often evident posteriorly and anteriorly in middle of carapace and in nuchal region; tubercles along anterior edge of carapace equilateral or conical in shape.
Pattern on side of head and snout variable and of no diagnostic value; postocular stripe uninterrupted having dark borders (UMMZ 92754), or interrupted having pale segment behind eye (TU 13282); other variations in pattern shown on TU 10170 and 15818; pale stripes on snout having dark inner borders that join and form acute angle (TU 381), or lacking dark inner borders and having uninterrupted dark line connecting anterior margins of orbits (TU 13280); other variations in pattern on snout shown on TU 1232, 1291 and 15819; specimens representing illustrations of variation in pattern on snout (Fig. 5 d, e, f) all from same locality, Lewisville, Lafayette County, Arkansas; contrasting pattern on side of head of dark marks on pale background; contrasting pattern of dark marks on dorsal surface of limbs; markings on hind limbs generally larger than those on forelimbs; small or fine markings of some specimens reducing contrast in pattern (TU 478, 488); carapace sometimes having few small blackish dots confined to margin (CNHM 15474, TU 487, 1253, 13266); ventral surface of plastron and soft parts of body whitish and usually lacking dark markings; small blackish marks often occurring on flap of carapace, in region of bridge, or on chin and throat (TU 399, 469, 475, 472, 13281).
Ontogenetic variation in PL/HW, mean PL/HW of specimens having plastral lengths 7.0 centimeters or less, 4.15, and exceeding 7.0 centimeters, 5.32; ontogenetic variation in CL/CW, mean CL/CW of specimens having plastral lengths 8.5 centimeters or less, 1.10, and exceeding 8.5 centimeters, 1.14; mean CL/PCW, 2.12; mean HW/SL, 1.38 (including subspecies guadalupensis); mean CL/PL, 1.36.
Variation.—In 1953, I casually glanced at a hatchling softshell from the Calcasieu River drainage in the private collection of Mr. Wilfred T. Neill; the specimen was considered by Neill (1951:15) as "… an intergradient one (with the hartwegi-spinifer population in the lower Mississippi drainage)." The hatchling does deviate from "typical" pallidus in having darkish flecks posteriorly on the carapace.
I have seen only one adult male (USNM 94457) from the Sabine River drainage (Orange County, Texas) that shows characteristics of guadalupensis (white dots on carapace encircled with small black ocelli); another adult male (USNM 94456) from the same locality resembles pallidus. Those two USNM specimens were mentioned by Neill (1951:13) as indicating intergradation with "… the mixed spinifera-hartwegi-asper populations of Louisiana."
Two adult males (SM 2889, Pl. 40, bottom, left, and TCWC 471, Trinity River drainage) have blackish ocelli surrounding the white dots on the posterior part of the carapace; two large females (TU 14402, Pl. 40, bottom, right, plastral length, 17.5 cm., and TU 14417 plastral length, 21.3 cm., both from the Trinity River) have contrasting mottled and blotched patterns with white dots visible on the carapace. These turtles show alliance with guadalupensis.
Some individuals from the Brazos River drainage have features suggesting [525] those that are characteristic of guadalupensis. Hatchlings may have large white dots on the anterior half of the carapace (USNM 55601). Adult males may have dusky ocelli surrounding the white dots on the carapace (TU 14169, 14559.1, 14559.2). The whitish dots, rarely as large as two millimeters, are never so large as in guadalupensis (three mm. in diameter), and are usually smaller anteriorly than posteriorly; TU 14169 has white dots approximately the same size (1.2 mm.) on the anterior half as on the posterior half of the carapace. The tubercles on adult males are equilateral or subconical, usually having sharp tips (TU 14348, 14559.1, 14559.2); the tubercles on large females are subconical, resembling the end of a bullet, and, in both sexes the tubercles are less conical than those on specimens of pallidus from farther east.
Three specimens from the Brazos River drainage are particularly impressive in their alliance with guadalupensis. SM 2556, an adult male, has large white dots that are encircled with black ocelli on the posterior half of the carapace, but lacks white dots on the anterior half. TNHC 14068, a hatchling, has small black dots interspersed with the larger white dots posteriorly. CNHM 46289 has large white spots on the carapace that are surrounded with two to four black dots; scattered black dots also intermix with white spots on the surface of the carapace (less extensive anteriorly).
Color notes taken from a freshly-killed adult male (KU 47121) from the Brazos River, seven miles below Whitney Dam, Bosque-Hill county line, Texas, are: Carapace pale brown or tan bordered by black line, having pale lemon yellow rim; yellowish-cream spots on carapace faintly surrounded with black stippling; dorsal surface of soft parts of body olive having black marks and patches of grayish; webbing on limbs having golden or yellowish hue, brighter distally; interorbital region brown; black-bordered, postocular stripe orange-cream; snout and side of head olive having pale areas of orange-cream; iris cream having black stripe; yellowish at juncture of dark dorsal and pale ventral coloration with orangish tinge on forelimbs and head; tail pale brown or tan, flanked by black borders that suffuse laterally into lemon-yellow; undersurface whitish, pale yellow on neck, bluish-gray on throat.
Comparisons.—T. s. pallidus most closely resembles T. s. guadalupensis, but can be distinguished from that subspecies in having small white tubercles, rarely two millimeters in diameter, on a pale background, that are not surrounded by blackish ocelli, and are usually absent, or not conspicuous on the anterior third of the carapace in adult males; also there are usually no conspicuous white tubercles or dots on the anterior third of the carapace in hatchlings. Many adult males of pallidus from the Brazos and some from the Trinity River drainages often have dusky or black ocelli surrounding the white dots posteriorly on the carapace; males from these river systems may be distinguished from guadalupensis in having most, if not all, white dots on the anterior half of the carapace smaller than those posteriorly, and a pale brown carapace (in life, usually darker in guadalupensis). T. s. pallidus (and guadalupensis) is distinguished from emoryi in lacking a widened pale rim posteriorly, and in having small white spots on the anterior half of the carapace. T. s. pallidus resembles guadalupensis and emoryi in having white spots on the carapace in adult males. T. s. pallidus differs from spinifer, hartwegi and asper in lacking blackish dots or ocelli that occur in the center of [526] the carapace. T. s. pallidus resembles emoryi but differs from guadalupensis in lacking black ocelli surrounding the white spots. T. s. pallidus resembles spinifer, hartwegi and asper but differs from guadalupensis and emoryi in having tubercles along the anterior edge of the carapace that are conical having sharp tips in males, and conical in large females.
T. s. pallidus resembles spinifer and hartwegi but differs from the other subspecies in having a narrow head. T. s. pallidus differs from emoryi but resembles the other subspecies in having a wider carapace. T. s. pallidus resembles emoryi and guadalupensis, and differs from the other subspecies in having the carapace widest farther posterior than one-half the length of the carapace. The snout of pallidus and guadalupensis is shorter than in spinifer and hartwegi, but longer than in emoryi. T. s. pallidus differs from asper but resembles the other subspecies in having a relatively long plastron.
Remarks.—Intergradation of the subspecies pallidus and guadalupensis is of a clinal nature in which populations successively show a gradual resemblance to guadalupensis from western Louisiana and eastern Texas westward to central Texas. Because the sharpest break in this cline of characters occurs between the Colorado and Brazos River drainages, the turtles living in the Brazos River drainage and eastward are referred to pallidus, whereas those in the Colorado River drainage and westward are referred to guadalupensis. For further comments on intergradation between these two subspecies, see the account of T. s. guadalupensis.
Taylor (1935:217-18) reported on some specimens of Amyda spinifera that were obtained by Mr. R. E. McEntyre in "… the spring and summer of 1926, chiefly about Lewisville, Lafayette County (Arkansas)." Of the catalog numbers listed by Taylor from Lewisville, 58 (KU, alcoholic) represent pallidus. Three, having the same locality data, have features that are characteristic of hartwegi. KU 2944 (one of three specimens having this catalog number) is a female having a pale, mottled and blotched carapace approximately one foot in length; there are remnants of two dark ocelli, and many widely-scattered, well-defined dark spots near the periphery of the carapace. KU 2963 (one of three specimens having this catalog number) is an adult male that has solid, blackish dots on the entire surface of the carapace. KU 2964 (one of two specimens with this catalog number) is an adult male that has ocelli approximately five millimeters in diameter on the carapace (indistinct in center of carapace).
Lewisville is situated in the drainage basin of the Red River and is approximately eight miles east of the Red River and 30 miles west of the westernmost tributary of the Ouachita River drainage. T. s. pallidus occurs in the Red River drainage; hartwegi occurs in the Ouachita River drainage. Perhaps there is intergradation between pallidus and hartwegi in the intervening streams. There is no data to indicate from which river or stream each specimen obtained by McEntyre came; one would presume that all specimens came from the Red River drainage. But this is not certain. Certainly the 47 specimens designated herein as pallidus came from the Red River drainage. I suspect that KU 2944, 2963 and 2964 were obtained from tributaries of the Ouachita River drainage.
T. s. pallidus intergrades with the spinifer-hartwegi population where the Red River joins the Mississippi River in the lower Mississippi Valley in Louisiana. The majority of 13 juvenal specimens from the Red River near Shaw, Concordia [527] Parish, Louisiana (USNM 99862-69, 99871-75), resemble pallidus in having inconspicuous white tubercles on a pale brown carapace. The white tubercles are conspicuous in USNM 99871. Some specimens have a few small dark dots confined to the margin of the carapace, as do some "variant" individuals from well within the geographic range of pallidus. USNM 99865 is referred to hartwegi because the carapace is covered with dark ocelli approximately one millimeter in diameter. Some specimens from farther west in the Red River drainage are referred to hartwegi. One (USNM 100420) of three from Natchitoches Parish, Louisiana (TU 5763, USNM 100420-21), having blackish dots on the carapace, is applicable to hartwegi. Of two turtles from Grant Parish, Louisiana (TU 5647, 12735), only 12735 has dark dots and ocelli (hartwegi). One specimen from Rapides Parish, Louisiana (TU 14040), having dark dots on the entire surface of the carapace, is referred to hartwegi.
Most specimens from the lower Atchafalaya River drainage are referable to pallidus. Eastward, intergradation occurs with the spinifer-hartwegi population; USNM 100089-90 from Assumption Parish, near Napoleonville, Louisiana, are referred to pallidus. TU 11983, from Bayou Lafourche, Raceland, La Fourche Parish, and TU 13698.11, from Bayou Gauche in St. Charles Parish, Louisiana, are juvenal males that combine the characteristics of pallidus and hartwegi; the carapaces are covered with blackish spots and posteriorly have distinct whitish dots. The population in the Atchafalaya River more closely resembles pallidus than it does hartwegi or spinifer. In former times the Atchafalaya River was presumably continuous solely with the Red River (inhabited by pallidus). Now, these two rivers and the Mississippi River are interconnected in east-central Louisiana. A large volume of water of the Mississippi drainage is conveyed to the Gulf of Mexico by the Atchafalaya, and someone has said that by approximately 1975, unless man interferes, two-thirds to three-fourths of the total volume of water of the Mississippi River will be drained by the Atchafalaya. One can expect, therefore, an increase in the influence of the hartwegi-spinifer population in the Atchafalaya River drainage.
Specimens examined.—Total 270, as follows: Arkansas: Lafayette: KU 2930-37, 2939-40, 2942, 2944 (two of three specimens bear this catalog number), 2945-57, 2958 (2), 2959-61, 2963 (two of three specimens bear this catalog number), 2964 (one of two specimens bears this catalog number), 2965-73, 2987-89, 3056, Lewisville.
Louisiana: Acadia: USNM 100151-59, Mermentau River. Assumption: USNM 100089-90, Bayou Lafourche, "near" Napoleonville. Beauregard: TU 1231-32, 1253-55, 1291, 13211, 13266, Sabine River, 8 mi. SW Merryville. Bienville: TU 5649-50, Lake Bistineau. Caddo: TU 381, 397-99, 469-72, 474-90, 678, 10170, Caddo Lake: TU 15818-19, Cross Lake. Calcasieu: UMMZ 92754, 5 mi. W Iowa. Cameron: TU 1122, Lacassine Wildlife Refuge. Concordia: USNM 99862-64, 99866-69, 99871-75, Red River, "near" Shaw. De Soto: SM 2374-75, Wallace Bayou. Grant: TU 5647, Lake Iatt. Iberville: USNM 83985, 2 mi. E Mounds; USNM 100239-41, Grand Lake west of White Castle; USNM 100380, Plaquemine; USNM 100412, 100414-15, 100419, Spanish Lake, "near" St. Gabriel. Jefferson Davis: Calcasieu River drainage, WTN (no number, see page 524). Natchitoches: TU 5763, Bermuda; USNM 100421, "near" Natchitoches. Sabine: TU 13210, 13212-13, 13265, 13280-82, 13303-06, Sabine River, 8 mi. SW Negreet. St. Martin: USNM 100160, Bayou Chene; USNM 100650, Atchafalaya. St. Mary: USNM 100395-97, 100404, 100409-10, Berwick Bay near Morgan City.
Oklahoma: Atoka: OU 8966, Rock Creek, 10 mi. E Atoka; OU 8978, McGee Creek, 7 mi. SW Daisy. Caddo: ANSP 100, Washita River, Fort Cobb. [528] Choctaw: OU 27126, Mayhew Creek, 2 mi. NW Boswell. Comanche: OU 4130, 4266, 5390, 8333, 12953, 19986, Wichita Mountains Wildlife Refuge. Jackson: OU 13012, 6 mi. E El Dorado. Kiowa: CNHM 15474. Le Flore: OU 6791, Kiamichi River, 8 mi. W Arkansas State Line. McCurtain: OU 2149-50, 2152, 2155, 17126-28, 17185, 2 mi. SW Smithville; USNM 70397, Red River. Marshall: KU 40175-76, 50830-31, 50847, OU 27290, 27297, 27562-63, TU 16076 (5), 16175 (6), 16662 (5), Lake Texoma, 2 mi. E Willis; KU 50832, mouth of Caney Creek, 4 mi. SW Kingston. Pushmataha: OU 2151, 2157; OU 11365, Buffalo Creek, 5 mi. NW Tuskahoma.
Texas: Archer: TU 16174, 16668-69, Lake Diversion. Bell: SM 5667-69, Nolan Creek. Bosque: KU 47121, 7 mi. below Whitney Dam, Brazos River. Brazos: BCB 4436, 10 mi. E College Station; BCB 4437, 17 mi. S College Station; BCB 4438, 4 mi. N Bryan; KU 50833, 4 mi. W College Station; SM 2556, TCWC 472, Wickson Lake; TCWC 539, Little Brazos River; TCWC 4692, 8 mi. NE Bryan; TCWC 5121, 2 mi. S College Station; TCWC no number. Clay: TCWC 7258, 8 mi. NW Ringgold, Montague County; TU 16667.1, 3 mi. W Byers. Dallas: MCZ 3987, "near" Dallas; ANSP 13243, Dallas. Donley: ANSP 13440, S of Clarendon. Eastland: KU 3132, Cisco. Galveston: TCWC 7251, Alta Loma. Harris: UMMZ 92753, Little Cypress Creek, 1 mi. N Westfield; USNM 94335-36, "near" Houston. Harrison: USNM 95386, 16.5 mi. SE Caddo Lake. Hill: TU 14169, Richland Creek, 0.7 mi. W Mertens. Leon: CNHM 46290, 5 mi. W Marquez; TCWC 8994, 8996, 6 mi. NW Normangee. Liberty: TU 14402, 14417, Trinity River, "near" jct. with Big Creek. McLennan: BCB 4665-66, 6 mi. NNE McGregor; SM no number, 2037, 2452, 2552, 2558, 2560, 2640, 5263, 6533, Lake Waco; SM 0185, Middle Bosque River; SM 2104, 6732, Upper Bosque River; SM 5072, Bull Hide Creek; UI 2399, 1.5 mi. W China Springs; UMMZ 64063, Waco; USNM 55601. Madison: TCWC 471, 517, Twin Lakes. Montgomery: TCWC 540, 3 mi. S Conroe. Nacogdoches: TNHC 14112, Legg Creek, 5 mi. S Douglass. Orange: UMMZ 117060, 3 mi. S Orange; USNM 94456-57, Orange. Randall: TTC 576, Palo Duro Canyon, 15 mi. SE Canyon. Shackelford: TU 14547, Clear Fork Brazos River, Fort Griffin State Park. Somervell: TCWC 8995, TU 14559 (4), Brazos River, 5-6 mi. E Glen Rose. Trinity: SM 2889, Groveton. Walker: TNHC 20829, 5 mi. E New Waverly. Waller: TNHC 14068, 2.7 mi. E Brazos River on US 90. Williamson: MCZ 1627 (2); TU 14348, San Gabriel River, 6.5 mi. E Georgetown. County unknown: ANSP 13448, Wichita River; USNM 7640, Brazos River.
Records in the literature.—Louisiana: Cameron: Sabine Refuge (Cagle and Chaney, 1950:386).
Oklahoma: Le Flore: 6 mi. W Page. McCurtain: 14 mi. SE Broken Bow (Trowbridge, 1937:301).
Texas: Bosque: Bosque River, "near" Valley Mills (Strecker, 1928:6). Harris: Addicks (Brown, 1950:250). Henderson: Cedar Creek (Strecker, 1926a:7). Jefferson: 12 mi. SW Port Arthur (Guidry, 1953:56). Liberty: Daisetta (Brown, loc. cit.); San Jacinto River (Strecker, 1915:15). McLennan: "near" Crawford (Brown, loc. cit.). Orange: 1 mi. N Bridge City (Guidry, loc. cit.). Tarrant: Trinity River, Fort Worth (Stejneger, 1944:66). Taylor: Abilene (KKA). Tyler: Colmisneil (Siebenrock, 1909:603). Walker: 6 mi. E Huntsville (TCWC 329, listed in card file). Wheeler: 5 mi. N Wheeler (Brown, loc cit.).
Trionyx ater Webb and Legler
Black Softshell
Trionyx ater Webb and Legler, Univ. Kansas Sci. Bull., 40:21, pls. 1 and 2, 1960, April 20.
Type.—Holotype, KU 46903, alcoholic female; obtained 16 km. S Cuatro Ciénegas, Coahuila, México, by John M. Legler (and party), September 6, 1958.
Range.—Basin of Cuatro Ciénegas, central Coahuila, Mexico (see map, Fig. 22).
Diagnosis.—Posterior margin of carapace of some females having fine corrugations, edge often ragged, and no pale outer margin; septal ridges reduced in adult males; over-all dorsal coloration (in preservative) dark, lacking contrasting patterns.
Description.—Plastral length of adult male, 9.6 centimeters (KU 46911); of largest female, 18.4 centimeters (KU 46903).
Adult male: anterior edge of carapace smooth; septal ridges reduced; pale outer rim, and small, whitish, dots posteriorly on carapace; surface of carapace slightly gritty or sandpapery posteriorly; snout broadened; over-all dorsal coloration dark gray or slate; contrasting pattern on soft parts of body lacking; ventral surface whitish having few blackish marks posteriorly on undersurface of carapace.
Females: posterior margin of carapace usually having fine corrugations; edge of carapace posteriorly often ragged; pale rim of carapace absent; mottled and blotched pattern not contrasting on blackish carapace; dorsal surface of soft parts of body dark gray or slate, lacking contrasting pattern; ventral surface of carapace and posterior part of plastron usually having many blackish flecks and markings; tubercles lacking on anterior edge and in center of carapace posteriorly; septal ridges well developed.
Medial angle of epiplastron (as observed through overlying skin) bent at angle of approximately 90 degrees. Other osteological characters presumably as in spinifer.
Range in length of plastron (cm.) of 11 females (mean follows extremes); 10.8-18.4, 15.0; proportional measurements of 12 specimens (including adult male, mean follows extremes): PL/HW, 4.70-5.43, 4.93; CL/CW, 1.28-1.43, 1.32; CL/PCW, 1.98-2.42, 2.15; HW/SL, 1.22-1.58, 1.37; CL/PL, 1.29-1.44, 1.36; some females (especially KU 46908) have noticeably elongate carapaces.
Variation.—Corrugations best developed on two largest females (KU 46903, 46906), even present on ventral surface of carapace posteriorly and on dorsal surface of tail; development of corrugations not ontogenetic phenomenon as posterior margin relatively smooth on KU 46908 (plastral length, 16.0 cm.) but relatively rugose on KU 46909, which is smaller (plastral length, 13.9 cm.); smallest female (KU 46904) and adult male having posterior margin smooth; smallest female having indication of pale outer rim and small whitish dots posteriorly on carapace, and dark, obtusely-angular line, connecting anterior margins of orbits; blackish marks on ventral surface reduced on KU 46904, 46910, 46912, and UI 43510; UI 43510 (plastral length, 16.3 cm.) resembles T. s. emoryi in having more contrasting mottled pattern on carapace and limbs, indication of pale outer rim on carapace, and dark line connecting anterior margins of orbits; ventral surface of tail and hind limbs often tinged with red.
Color notes from life of young female, topotype (KU 53755) are: mottled carapace dark brown, pale areas buff; dorsal surface of head mottled, olive-brown, pale areas buff; iris orange-buff; upper and lower lips yellow-orange; dorsal surface of limbs olive-brown having yellow to buff suffusion and small blackish marks; pale areas on webbing yellow; ventral surface whitish having yellow at margin of carapace, on neck and limbs.
Comparisons.—T. ater most closely resembles T. spinifer (especially the subspecies emoryi) in having a gritty or "sandpapery" carapace (reduced, [530] tubercles more scattered), whitish dots on posterior third of carapace (small females and adult male) and a dark line connecting anterior margins of orbits (smallest female). Prior to acquiring the characteristic darkened, dorsal ground color, the pattern on the head and limbs seems to be that of T. s. emoryi.
T. ater resembles T. muticus in having reduced septal ridges in males, a smooth anterior edge of carapace (especially males), and no enlarged prominences on the anterior edge of the carapace or posteriorly in the center of the carapace on large females. T. ater resembles T. ferox in having an over-all dark coloration dorsally with no contrasting patterns on adults.
T. ater probably is a small species resembling T. muticus and T. spinifer emoryi. The head is wide in T. ater, resembling that of T. ferox, and closely approaching that of T. spinifer emoryi and T. s. guadalupensis. T. ater resembles T. ferox and T. s. emoryi in having a narrow carapace. T. ater resembles T. s. emoryi, T. s. guadalupensis and T. s. pallidus, but differs from T. muticus, T. ferox and the other subspecies of T. spinifer in having the carapace widest farther posterior than one-half the length of the carapace. T. ater resembles T. ferox and T. s. emoryi in shortness of snout. The plastron is short in T. ater and most closely resembles that of T. s. pallidus, T. s. guadalupensis, and T. s. emoryi.
Remarks.—T. ater is confined to permanent, clear-water ponds in the basin of Cuatro Ciénegas. The male and 11 females (KU) were taken at the type locality (a pond known locally as Tío Candido); the other female (UI 43510) was taken from a pond approximately seven miles northward (known locally as Anteojo). T. spinifer emoryi also occurs in the basin of Cuatro Ciénegas. Males and females of emoryi were collected in the Río Mesquites (Río Salado drainage) that drains the basin; two adult males of emoryi were taken from the clear-water ponds—one from the type locality of ater (KU 46907), and the other (KU 53757) from a pond (known locally as El Mojarral) from which no ater were obtained. This demonstrated sympatry indicates that the two kinds are not conspecific.
However, the nature and frequency of occurrence of characters of T. ater, suggest that it is subspecifically related to T. spinifer—in effect, a darkened race of T. s. emoryi. The diagnostic characters of fine corrugations on the posterior margin of the carapace and blackish marks on the ventral surface do not occur on every female of ater. Too, the dorsal coloration of living females (dark brown-buff) is paler than that of preserved specimens (dark gray-slate). Furthermore, a hatchling (CNHM 47367) recorded from Cuatro Ciénegas, Anteojo, is not distinguishable from emoryi.
The mention of absence of septal ridges in males of T. ater in the original description (Webb and Legler, 1960:22) should be amended. The septal ridges in the only known adult male are reduced; a small, whitish ridge is present on the medial surface of each nostril, but is not conspicuous in anterior view. The one adult male of ater is distinguished from T. s. emoryi principally on the over-all dark, dorsal coloration with concomitant loss of pattern, the noticeably broadened snout, and the reduced septal ridges. The last character mentioned possibly is variable in ater (and in emoryi in this region) in view of the variation in development of the ridge on four male emoryi from the basin: well-developed on KU 53757 (Mojarral) and KU 46907 (Tío Candido); reduced on KU 53752 (Río Mesquites), resembling development in ater; and, reduced on right side only on KU 53753 (Río Mesquites).
Presumably, the continued erosive action at the headwaters of the Río Salado has permitted the invasion of this drainage into the formerly isolated basin of Cuatro Ciénegas. In the basin, however, I know of no evidence of a direct aquatic contact between the headwater streams and the isolated, clear-water, ponds. How emoryi entered the ponds is unknown. Some of the ponds are tapped by small, man-made, irrigation canals, but, so far as I know, these are not connected to the river. The ponds have permanent water and are often separated by several miles of arid environment. Overland dispersal between waterways is possible in time of flooding. Local residents tell of the infrequent sale of softshells in Cuatro Ciénegas, which hints at their dispersal via the agency of man. The underlying gypsum substrate of the valley has been subjected to considerable erosion; the ponds observed have deep holes, and small caverns and grottos. There are conflicting reports concerning subterranean connections between ponds. Possibly there are underwater connections between some ponds and the headwater streams of the Río Mesquites. Whatever the dispersal route for emoryi into the ponds has been, it is strange that the same route has not been traversed by ater, permitting its occurrence in the Río Mesquites.
On the basis of morphological criteria, I suspect that ater and emoryi are genetically compatible. Possibly there is only sporadic entrance of emoryi into the ponds inhabited by ater, or the accessible dispersal routes for emoryi have been relatively recent and there has been insufficient time for genetic adaptation. T. ater is maintained as a full species because of the occurrence of two distinct males (KU 46907, emoryi, and KU 46911, ater) in the same pond (Tío Candido, the type locality). These two specimens are contrasted in a photograph accompanying the type description (Webb and Legler, 1960: Pl. II). The restricted distribution of ater, and its characteristics suggest a relict population derived from a ferox-like ancestor that may be in the process of becoming extinct.
There are two specimens in the CNHM recorded from Cuatro Ciénegas. One is a female (CNHM 55661) having a plastral length of 19.0 centimeters, and no specific locality other than Cuatro Ciénegas. I examined this specimen before I knew of the existence of ater, and noted no unusual features; I have not re-examined the specimen. It is considered representative of emoryi. The second is a hatchling (CNHM 47367) having a plastral length of 3.2 centimeters, recorded from Cuatro Ciénegas, Anteojo. The carapace is dark tan having small whitish dots intermixed with a few indistinct, small, blackish specks posteriorly. The specimen is indistinguishable from emoryi.
Specimens examined.—Total 12, as follows: Coahuila: KU 46903-06, 46908-12, 53755-56, 16 km. S Cuatro Ciénegas; UI 73510, 5.7 mi. W Cuatro Ciénegas.
Records in the literature.—Schmidt and Owens (1944:103) record emoryi from Cuatro Ciénegas (no museum numbers listed); presumably their reference is to CNHM 55661.
Trionyx muticus Lesueur
Smooth Softshell
Range.—United States from extreme western Pennsylvania, southern Minnesota and South Dakota south to the Gulf of Mexico in Alabama, the western end of the panhandle of Florida, and the eastern half of Texas (see map, Fig. 22.)
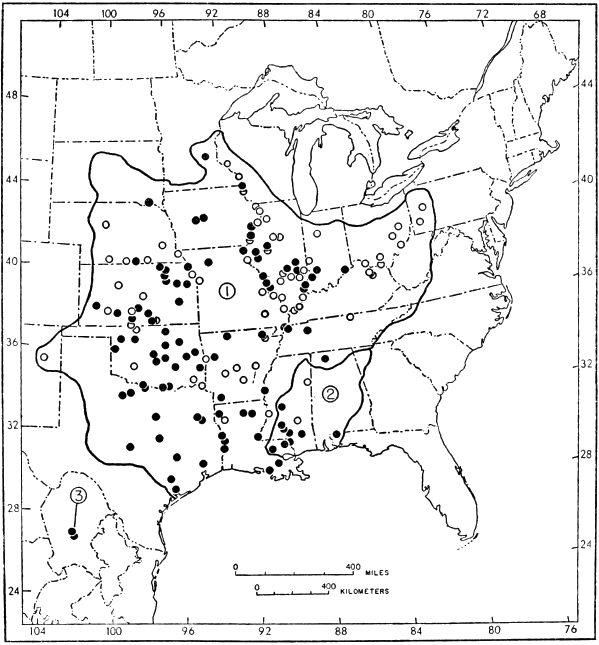
Fig. 22. Geographic distribution of Trionyx ater and Trionyx muticus. 1. T. muticus muticus. 2. T. muticus calvatus. 3. T. ater.
Diagnosis.—Septal ridges absent; anterior edge of carapace smooth, lacking prominences; juvenal pattern of large dusky spots (sometimes ocellate), or small dusky (not black), dots and short lines; side of head usually devoid of markings except for pale, usually uninterrupted, postocular stripe.
Size small; head narrow; snout long; ventral surface of supraoccipital spine broad proximally, lacking median ridge; foramen magnum evenly rounded, ovoid; opisthotic-exoccipital spur absent; distal part of opisthotic wing truncate; lateral condyle of articular surface of quadrate tapered posteriorly, smaller than medial articular surface; angle of epiplastron obtuse, approximately 100 degrees; callosity on epiplastron sometimes covering entire surface; bony bridge wide in relation to length.
Description.—Septal ridges absent; external characteristics variable (see accounts of subspecies); range in length, in centimeters, of plastron of ten largest specimens of each sex, (mean follows extremes), males, 11.8-14.0, [533] 12.3; females, 17.7-21.5, 18.9; ontogenetic variation in PL/HW, mean PL/HW of specimens having plastral lengths 7.0 centimeters or less, 4.16, ranging from 7.1 to 13.0 centimeters, 5.82, and, exceeding 13.0 centimeters, 7.04; little ontogenetic variation in CL/CW, mean CL/CW of specimens having plastral lengths 8.0 centimeters or less, 1.15, and exceeding 8.0 centimeters, 1.16; mean CL/PCW, 1.97; mean HW/SL, 1.22; mean CL/PL, 1.39.
Greatest width of skull usually at level of squamosal (79%); foramen magnum ovoid; opisthotic-exoccipital spur usually absent (97%); distal part of opisthotic wing truncate, sometimes visible in dorsal view; lateral condyle of articular surface of quadrate tapered posteriorly, smaller than medial articular surface; maxillaries not in contact above premaxillaries; combination of seven neurals, seven pairs of pleurals, and contact of seventh pair of pleurals (38%), or eight neurals, seven pairs of pleurals, and separation of seventh pair of pleurals (41%); angle of epiplastron obtuse, greater than 90 degrees; callosities well-developed, frequently on preplastra and epiplastron of adults.
Comparisons.—The absence of septal ridges distinguishes muticus from ferox, all subspecies of spinifer, and ater (ridges are reduced in males of ater). The smooth anterior edge of the carapace distinguishes muticus from all other American kinds except ater and some individuals of T. s. emoryi. T. muticus resembles only ater and ferox in usually lacking a well-defined, contrasting pattern of blackish marks on the dorsal surface of the limbs. T. muticus resembles ferox and differs from spinifer and ater in lacking a gritty or "sandpapery" carapace on adult males. Adult males of T. muticus calvatus and some individuals of T. m. muticus from the Colorado River in Texas further resemble ferox in having postocular stripes with thick black borders.
T. muticus is the smallest species in North America; the maximum size of the plastron in adult males is approximately 14.0 centimeters (16.0 cm. in spinifer) and of adult females 21.5 centimeters (31.0 cm. in spinifer). Males and females of muticus are sexually mature at approximately the same size as some T. s. emoryi; also, the great development of the plastral callosities in muticus corresponds to that in some emoryi. The head is narrower in muticus than in ferox or spinifer. The carapaces of specimens of muticus exceeding plastral lengths of 8.0 centimeters are wider than those of ferox, ater, T. s. emoryi and T. s. guadalupensis of corresponding size. T. muticus differs from ater and three subspecies of spinifer (pallidus, guadalupensis, emoryi) in having the carapace widest at a plane approximately one-half the length of the carapace. The snout is longer in muticus than in ferox and spinifer. T. muticus differs from ferox but resembles spinifer in having a relatively short plastron.
The skulls of muticus differ from those of ferox but resemble those of spinifer in usually having the skull widest at the level of the squamosals. Skulls of muticus resemble those of ferox but differ from those of spinifer in usually lacking a well-developed opisthotic-exoccipital spur. Skulls of muticus are different from those of ferox and spinifer in having the 1) ventral surface of the supraoccipital spine widest proximally, lacking a medial ridge, 2) foramen magnum ovoid, 3) distal part of opisthotic wing truncate, 4) lateral condyle of articular surface of quadrate tapered posteriorly, smaller than medial articular surface, and 5) maxillaries not in contact above premaxillaries.
Plastrons of muticus differ from those of spinifer and ferox in having an [534] obtusely-angled epiplastron, relatively large callosities in adults, and a wide hyo-hypoplastral bridge (in relation to length).
Remarks.—Agassiz (1857:399) regarded Lesueur's Trionyx muticus as the type species of the genus Amyda and the only species known to belong to the genus Amyda. Stejneger (1944:7, 9, 12) proposed the generic name Euamyda as a new name for the North American Amyda mutica as understood by Agassiz. Euamyda was proposed for use only if Agassiz's understanding was found to be correct. Actually, Stejneger thought that the Old World and New World kinds concerned were congeneric, and that the type species of the genus Amyda was the Old World species Amyda javanica Schweigger (= Testudo cartilaginea Boddaert).
If Trionyx muticus Lesueur is considered to be generically distinct from other soft-shelled turtles, Euamyda Stejneger, 1944, is available as a generic name with Trionyx muticus Lesueur, 1827, as the type species (by monotypy).
Geographical variation.—Trionyx muticus shows no obvious character gradients; the variation is mostly discontinuous and unlike that in T. spinifer. On the basis of differences in the juvenal pattern and pattern on head, T. muticus can be divided into two subspecies.
Trionyx muticus muticus Lesueur
Midland Smooth Softshell
Trionyx muticus Lesueur, Mém. Mus. Hist. Nat. Paris, 15:263, pl. 7, December, 1827.
Trionyx muticus muticus Webb, Publ. Mus. Nat. Hist. Univ. Kansas, 11:520, August 14, 1959.
Potamochelys? microcephala Gray, Proc. Zool. Soc. London, p. 87, 1864.
Type.—Lectotype, Museum d'Histoire Naturelle, Paris, No. 8813; dried carapace and plastron; obtained from the Wabash River, New Harmony, Posey County, Indiana, by C. A. Lesueur in August, 1827 (Pl. 53).
Range.—Central United States; in the Mississippi River drainage from extreme western Pennsylvania, southern Minnesota and South Dakota south to Tennessee, Louisiana and Oklahoma; streams of the Gulf Coast drainage from the Mississippi River in Louisiana westward into Texas including the Colorado River drainage (see map, Fig. 22).
Diagnosis.—Juvenal pattern of dusky dots and usually short lines or bacilliform marks; ill-defined pale stripes on snout usually evident just in front of eyes; pale postocular stripe lacking thick, black borders that are approximately one-half width of pale stripe (except some in the Colorado River drainage of Texas).
Description.—Plastral length of smallest hatchling, 2.1 centimeters (INHS 3458); of largest male, 14.0 centimeters (CNHM 92003); of largest female, 21.5 centimeters (KU 2308).
Juvenal pattern of dusky, grayish marks lacking sharp margins, and usually consisting of both small spots and short streaks or dashes, the former predominating; short streaks or dashes occasionally lacking (TU 14375, Pl. 45, bottom, left; UMMZ 92751); markings variable in number, few and widely spaced, or several and closely approximated (Pl. 45, top, topotypes); pale rim [535] separated from ground color by ill-defined, dusky margin; pattern on adult males well-defined resembling that of hatchlings (TU 16172.1, 16173), scarcely discernable (TU 13294), or absent (TU 1242); mottled and blotched pattern on carapace usually contrasting in large females.
Pale stripes extending forward from eyes usually not more than half distance to tip of snout; inner borders of pale stripes on snout usually absent or dusky and indistinct, occasionally blackish (TU 14606); outer borders of pale stripes darker than inner borders, usually blackish; pale stripes on snout occasionally absent (CNHM 7845, UMMZ 92665, TU 5989, none of these specimens being large females); pale postocular stripe having narrow, dusky or blackish borders (especially UMMZ 92751, TU 14436); pale postocular stripe usually complete, occasionally interrupted having prominent dark-bordered anterior segment just behind eye (TU 14416); lower border of postocular stripe usually in contact with dusky postlabial line; no other markings on side of head; pattern on dorsal surface of soft parts of body not contrasting, composed of closely approximated fine markings that are little darker than background, over-all coloration pale grayish; occasionally, few larger and more contrasting markings on hind limbs (UMMZ 92751, TU 14436).
Underparts white, usually lacking markings; occasional dusky markings on plastral area (UMMZ 110502), dark spots or flecks on undersurface of carapace (BCB 6043, UMMZ 92666), or markings on throat (UMMZ 95032).
Surface of carapace smooth in adult males; large females lacking prominences posteriorly in center of carapace or in nuchal region; anterior edge of carapace smooth in both sexes, but occasionally having regularly spaced furrows or wrinkles (Fig. 8g).
Variation.—Short dusky lines and streaks seem to be lacking from the juvenal pattern on the carapace more often in southern populations (Gulf Coast drainage of Texas) than in northern populations (Mississippi River drainage). I have seen one female, KU 48229 (Pl. 46, bottom, left), plastral length 14.5 centimeters that retained a well-defined juvenal pattern, and lacked a mottled and blotched pattern.
Color notes from life of 11 turtles, KU 55296-306, (eight adult males, three immature females) from the Kansas River at Lawrence, Douglas County, Kansas, are: Buff-yellow rim of carapace, sometimes having pale orange tinge; dusky, dark brown markings on pale brown or tannish carapace of males; dark and pale brown mottled and blotched pattern on carapace of females (smallest specimens having plastral length, 11.0 cm.), many having orangish or buffy hue; soft parts of body brownish to olive-green dorsally, many having small, blackish marks on hind limbs; webbing of limbs yellowish; pale orange, some yellow, laterally at juncture of dark dorsum and pale ventrum (to a lesser extent on hind limbs); pale orange in some suffusing onto dorsal surface of soft parts of body; black-bordered postocular stripes in males having orangish tinge (pattern somewhat obscured in females); whitish ventral surface in some having pale orangish tinge here and there; many having dusky, grayish flecking on plastral area and anterior ventral surface (most intense on 55306 giving appearance of grayish suffusion).
I have seen only three specimens from the Colorado River drainage in Texas. Two of these (UMMZ 92751, TU 14436) are characterized by much black pigmentation. A contrasting pattern of relatively large black marks occurs on the dorsal surface of the soft parts of the body, especially on the [536] hind limbs, and the pale postocular stripes have thick black borders. UMMZ 92751, having a plastral length of 5.5 centimeters, has a juvenal pattern of widely-spaced dark dots that lacks short lines. The other muticus from the Colorado River (CM 3055), a large female 19.0 centimeters in plastral length, has ill-defined postocular stripes lacking dark borders, although a small dusky blotch occurs on the right side of the head.
Comparisons.—T. m. muticus differs from T. m. calvatus in having pale stripes on the snout, a juvenal pattern of small dusky spots (usually lacking ocellate spots) and short lines, and a pale postocular stripe lacking thick, blackish borders (except in some turtles from the Colorado River system of Texas). One unique characteristic of muticus is the short, dusky lines in the juvenal pattern; these marks, however, are occasionally absent.
Remarks.—Trionyx muticus generally has been considered a distinct species since its description by Lesueur (1827:263-66, Pl. 7); Wied-Neuwied (1865:53), at least, questioned the identity of muticus, believing it to be based on a secondary sexual difference of T. spiniferus. Lesueur did not designate a type in the description, and mentioned that he had seen only three specimens (op. cit.:264). Stejneger (1944:17-18) discussed two mounted specimens (Nos. 787 and 788) in the Natural History Museum at Paris, and mentioned that No. 787 was designated "… as the type on the printed label (although presumably not done by Lesueur)." Dr. Jean Guibé (in litt. September 24, 1959) informed me that Nos. 787 and 788 are numbers without value and correspond, respectively, to catalog numbers 8813 and 8814. In addition, the Museum possesses an alcoholic specimen, No. 564, obtained by Lesueur from the Wabash River, that seems to have been acquired by the museum after the publication of the original description. No. 8813 is regarded as a lectotype.
Gray (1864:87) described the species microcephalus and questionably included it in the genus Potamochelys Fitzinger, 1843; the locality was stated as "Sarawak (Wallace)." Gray especially noted the small elongate head and believed that the acquisition of adult specimens would prove that it belonged to a new genus. Later, Gray (1869:221) proposed the generic name Callinia as a new name for Aspidonectes as understood by Agassiz (1857:403). Gray referred microcephala to Callinia (op. cit.:214, 222) and recognized also Amyda mutica (op. cit.:212). Baur (1888:1121) remarked that "Callinia microcephala Gray, of the British Museum, with the locality Sarawak, is Amyda mutica Les." The species microcephalus has since been considered a synonym of Trionyx muticus. Schmidt (1953:110) designated the type locality as New Harmony, Indiana.
Müller (1878:641) listed the species Trionyx muticus from México as follows: "*b. in Alcohol. Mexico. 1872. [2]." Smith and Taylor (1950:18, footnote) wrote that the record required confirmation. Webb and Legler (1960:24) questionably referred this record to the synonomy of T. ater, which resembles muticus. T. muticus is not known to occur in México. According to Dr. Lothar Forcart (in litt.) of the Naturhistorische Museum in Basel, Switzerland, only one specimen on which Müller based his record is extant. My examination of this specimen reveals that it is a hatchling T. s. emoryi, plastral length 3.5 centimeters, bearing catalog number 1032; there are no additional data of collection.
Strecker and Williams (1927:16) mentioned one specimen of muticus that was obtained at Christoval, Tom Green County, Texas, and I presume this is the basis for Pope's mention of this species from Tom Green County, Texas (1949:319). Although I do not doubt that T. muticus occurs in Tom Green County, this record possibly is based on T. spinifer because, 1) there are no specimens of muticus in the Strecker Museum from Tom Green County, but there is one specimen of spinifer (SM 3282), and in none of Strecker's publications is there any mention of spinifer from Tom Green County, and 2) Strecker had, at least once, misidentified the two species; his record of muticus from Wallace Bayou, Louisiana (Strecker and Frierson, 1926:last page, no numbers), represents T. spinifer pallidus (SM 2374-75).
Specimens examined.—Total 261, as follows: Alabama: County unknown: USNM 118167, Wheeler Reservoir, Tennessee River.
Arkansas: Franklin: KU 19459-60, Ozark. Lafayette: KU 2938, 3057, Lewisville. Lawrence: CNHM 92003, Imboden; CNHM 92005, Powhatan; USNM 59214, Black River, Black Rock. Marion: TU 14606 (2), White River at Cotter. Prairie: KU 1831, 1868, 1870, 1874-76, 1930-31, 1957-63, 2294-302, 2305-06, 2308-09, 2838-41, 3002, White River, DeVall's Bluff.
Kansas: Barber: USNM 95185-86, 1 mi. S Lake City. Doniphan: KU 1872, 1878, 1964, Doniphan Lake. Douglas: KU 2220, 16148, 23230, 40179, 50825-26, 55296-306, Kansas River, Lawrence; KU 45065-66, 1 mi. N, 1.5 mi. W Lakeview. Ford: KU 51516, Ford. Kearny: KU 48216, 4 mi. S, 1.5 mi. W Deerfield. Marshall: KU 48228, Blue Rapids. Pottawatomie: KU 48229-33, 48238, 2 mi. E Manhattan, Riley County. Reno: USNM 95260, 6 mi. E Turon. Riley: KU 46861, 48234-35, 4 mi. N Manhattan; KU 48236, 2 mi. NE Randolph. Sedgwick: UMMZ 95362, Wichita. Shawnee: UMMZ 95366-67, Topeka. Sumner: USNM 95415, 3 mi. SE Oxford. Washington: KU 48237, 8 mi. S Hanover. Woodson: KU 45064, 1 mi. E, 2 mi. S Neosho Falls. County unknown: USNM 51528.
Illinois: Cass: INHS 2146, Beardstown. Coles: INHS 1965-67, 3 mi. S Charleston. Jackson: INHS 5894, 6.5 mi. N Aldridge, Union County; UMMZ 81570, Mississippi River. Jasper: INHS 2412, Rose Hill. Jersey: INHS 2156-58, Grafton. Mason: INHS 2147, Cedar Creek. Mercer: INHS 3458, Keithsburg. Monroe: INHS 4088, 3 mi. NE Columbia. Morgan: CNHM 6028, INHS 2148, Meredosia. Pope: CNHM 2463 (30), Golconda. Schuyler: UI 40-41, Crooked Creek. Shelby: INHS 2283, Holliday. Wabash: INHS 5228, Mt. Carmel.
Indiana: Daviess: UMMZ 110234, White River, 1.5 mi. W Elnora. Jefferson: USNM 8337, Madison. Knox: UMMZ 111880-81, "near" Decker Chapel. Posey: INHS 7278-80, 7447, TTC 798, Wabash River, 2-2.5 mi. S New Harmony; UMMZ 110598, 8 mi. NW Mt. Vernon.
Iowa: Allamakee: UMMZ 92657, 1/4 mi. W Victory, Vernon County, Wisconsin; UMMZ 92658-64, Mississippi River, "near" Lansing. Boon: UMMZ 92665, Des Moines River at Ledge State Park. Greene: UMMZ 92666, 3.5 mi. N Scranton. Muscatine: USNM 53521, 54733-34, 54742, 60054-56, Fairport.
Louisiana: Beauregard: TU 1242, Sabine River, Merryville. Caddo: CNHM 7845, Gayles. Catahoula: USNM 113228, Jonesville. Concordia: USNM 99870, Red River, "near" Shaw. Ouachita: TU 5989, Monroe. Richland: USNM 100422, Rayville. Sabine: TU 13163, 13294, Sabine River, 8 miles SW Negreet. St. James: TU 7543, Vacherie. St. Mar: USNM 100406, Berwick Bay, "near" Morgan City. Vernon: KU 41380, 46777, Sabine River NW Burr Ferry.
Minnesota: Hennepin: AMNH 4761-62, Fort Snelling.
Mississippi: Washington: USNM 92605, Greenville. County unknown: USNM 115939.
Missouri: Clark: USNM 59267, 59278, Alexandria. Daviess: UMMZ 95505, Grand River, 1 mi. S Jameson. St. Louis: SM 2052, St. Louis. Wayne: UMMZ 82823, St. Francis River.
Nebraska: Webster: UMMZ 89526, Republican River, 2 mi. E Inavale.
Oklahoma: Cleveland: OU 5480-81, 6473, South Canadian River, 4 mi. SE Norman. Hughes: KU 50845, 4 mi. N Atwood. Kay: OU 9741, 8 mi. E Ponca City. Le Flore: OU 2148; OU 27390, Poteau River below Wister Dam. Love: OU 27472, Hickory Creek, 9 mi. E Marietta. Major: OU 8597, 7 mi. E Orienta. Marshall: KU 50827-29, 50848, 50853, OU 27593-94, TU 16077 (4), Lake Texoma, 2 mi. E Willis. McIntosh: OU 8993, 4 mi. W Onapa. Oklahoma: OU 10137, Lake Oberholser. Payne: UMMZ 89629, Cimarron River, 3 mi. E Ripley; UMMZ 90002, 19 mi. SE Stillwater. Pottawatomie: OU 25176-83, South Canadian River, 5 mi. SW Shawnee. Roger Mills: OU 12472. Sequoyah: OU 9006, Illinois River, 2 mi. NE Gore. Tulsa: UMMZ 95032 (4), Arkansas River at Tulsa. Woodward: CNHM 15472-73; OU 8599-600, 5 mi. E, 1 mi. N Woodward.
South Dakota: Yankton: UMMZ 110499-500, Missouri River at Fort Randall; UMMZ 110501-02, Missouri River at Yankton.
Tennessee: Benton: UMMZ 53198, Trotter's Landing. Lake: USNM 102677, Reelfoot Lake. Obion: USNM 102910, Reelfoot Lake.
Texas: Archer: TU 16173, Lake Diversion. Baylor: TU 16172 (2), Lake Kemp. Brazos: TCWC 7250, Bryan. Clay: TCWC 7248-49, 7259-61, 8 mi. NW Ringgold, Montague County; TU 16667, 3 mi. W Byers. Grayson: UI 2419, Lake Texoma. Gregg: SM 6685, near Gladewater; USNM 22629, Sabine River, 5 mi. S Longview. Liberty: TU 14416, 14375, Trinity River, "near" jct. with Big Creek. McLennan: BCB 6030, 6043, SM 2557, 2561, Lake Waco. Matagorda: CM 3055, Colorado River, Bay City. San Saba: TU 14436, San Saba River, 11 mi. NNW San Saba. Tarrant: UMMZ 92750, Worth Lake, Fort Worth. Wharton: UMMZ 92751, Colorado River, Wharton.
No Data: MCZ 1594 (erroneously recorded from Mobile, Alabama); USNM 029261, 59982.
Records in the literature.—Arkansas: Garland: Hot Springs (Combs and Hurter in Strecker, 1924:47). Jefferson: Pine Bluff. Pulaski: Little Rock. Sebastian: Fort Smith (Hurter and Strecker, 1909:21).
Illinois: Adams: Quincy (Garman in Cahn, 1937:179). Alexander: Horseshoe Lake (Cahn, loc. cit.); Cairo (Garman in Cahn, loc. cit.). Carroll: 5 mi. S Savanna (Stejneger, 1944:24). Clay: Louisville. Clinton: Carlyle. Crawford: Robinson (Cahn, loc. cit.). Cumberland: Embarrass River (Peters, 1942:183). Fayette: Vandalia. Gallatin: Shawneetown (Cahn, loc. cit.). Hancock: between Warsaw and Hamilton (Stejneger, op. cit.:23). Jackson: Murphysboro. Jasper: Newton. Marion: Centralia. Mason: Havana. Massac: symbol on map. Menard: Petersburg. Peoria: Peoria. Randolph: Chester (Cahn, loc. cit.). Richland: Olney (Stejneger, loc. cit.). Rock Island: Rock Island. St. Clair: East St. Louis (Cahn, loc. cit.). Union: (Cagle, 1942a:199). White: Carmi. Whiteside: Sterling (Cahn, loc. cit.). Woodford: Mackinaw Creek (Garman in Cahn, loc. cit.).
Indiana: Carroll: "near" Delphi (Agassiz, 1857:400). Vigo: Terre Haute (Blatchley, 1891:22).
Iowa: Des Moines: "near" Burlington (Agassiz, 1857:400). Dubuque: Mississippi River, 8 mi. S Dubuque (Goldsmith, 1945:447). Lee: Keokuk (Stejneger, 1944:23).
Kansas: Barber: 5 mi. SE Lake City; Salt River, S of Aetna (Burt, 1935:321). Cowley: symbols on map (Smith, 1956:157). Gray: Arkansas River, 1 mi. W Cimarron (Clarke, 1956:215). Leavenworth: Missouri River, Fort Leavenworth (Brumwell, 1951:207-08). McPherson: Lindsborg (Breukelman and Smith, 1946:112). Pratt: State Fish Hatchery, "near" Pratt (Taylor, 1933:269). Trego: Wakeeney (Stejneger, 1944:24).
Kentucky: Fleming: Fox. Rowan: Triplett (Welter and Carr, 1939:130). County unknown: Ohio River (Funkhouser, 1925:71).
Louisiana: De Soto: Bayou Pierre (Strecker and Frierson, 1926:last page, no numbers).
Minnesota: Houston: Brownsville (Breckenridge, 1944:183). Winona: Homer (Stejneger, 1944:23).
Mississippi: Warren: Vicksburg (Cook, 1946:185).
Missouri: Jackson: Fry's Lake (Anderson, 1942:219). Jefferson: Meramec River (Boyer and Heinze, 1934:199). County unknown: Osage River (Agassiz, 1857:400).
Nebraska: Franklin: 1/2 mi. S Franklin; 1 mi. SE Naponee. Furnas: 4 mi. E Cambridge. Lancaster: Lincoln. Nemaha: Peru. Thayer: (Hudson, 1942:102). Thomas: (Smith, 1958:36).
New Mexico: San Miguel: Conchos River above Conchos Dam (Shields and Lindeborg, 1956:120).
Ohio: Brown: mouth White Oak Creek, Higginsport. Muskingum: "near" Gaysport. Pike: Scioto River in Camp Creek, Newton and Scioto Twps.; Pike Lake. Scioto: Scioto River in Clay and Rush Twps.; Scioto River, Portsmouth; Scioto River, 3 mi. N Rushtown. Tuscarawas: Tuscarawas River, 2 mi. below Gnadenhutten; "near" Winfield. Washington: Dam No. 2, Muskingum River, northern edge of Marietta; Ohio River, 4 mi. SE Marietta (Conant, 1951:156, 264).
Oklahoma: Alfalfa: 6.5 mi. NE Ingersoll. Comanche: Camp Boulder, Wichita National Forest (Ortenburger and Freeman, 1930:188). McCurtain: Pushmataha: (Ortenburger, 1927:100).
Pennsylvania: Allegheny: Neville Island, Ohio River below Pittsburgh (Atkinson, 1901:154). Clarion: Allegheny River at Foxburg (Netting, 1944:85).
?South Dakota: County unknown: Fort Mackenzie, Missouri River, 6-8 mi. below Cedar Island (Stejneger, 1944:15).
Tennessee: Lake: Mississippi River (Parker, 1948:29). Pickett: Obey River at Eagle Creek Ford (Shoup, Peyton and Gentry, 1941:75).
Wisconsin: Crawford: Pepin: Mississippi River (Breckenridge, 1944:183; Pope and Dickinson, 1928:82).
Trionyx muticus calvatus Webb
Gulf Coast Smooth Softshell
Plate 47
Trionyx muticus calvatus Webb, Univ. Kansas Publ. Mus. Nat. Hist., 11:519, 1 fig., 2 pls., August 14, 1959.
Type.—Holotype, UI 31071, hatchling, sex undetermined, alcoholic; obtained from Pearl River, Roses Bluff, 14 miles northeast Jackson, Rankin County, Mississippi, by William F. Childers on August 25, 1952.
Range.—Southeastern United States from the Florida Parishes of Louisiana eastward to the western end of the panhandle of Florida; rivers of the Gulf Coast drainage from the Escambia River drainage, Florida, westward to Louisiana and Mississippi including the Pearl River drainage. The eastern extent of geographic range is not known (see map, Fig. 22).
Diagnosis.—Juvenal pattern of large circular spots, often ocellate; no stripes on dorsal surface of snout; pattern on dorsal surface of limbs of fine markings, not in contrast with ground color; pale postocular stripes having thick black borders approximately one half width of pale stripe on adult males.
Description.—Plastral length of smallest hatchling, 3.0 centimeters (TU 17301); of largest male, 11.8 centimeters (KU 47118); of largest female, 18.0 centimeters (TU 13473).
Juvenal pattern of dusky, circular spots, some ocellate, lacking short lines and streaks; number of spots variable; some spots on carapace of hatchlings may have maximum diameter of three millimeters (TU 17301); pale rim of carapace having dusky, ragged, inner border; juvenal pattern on adult males absent or usually evident, at least posteriorly (TU 17306.1).
Dorsal surface of snout lacking pale stripes just in front of eyes; pale postocular stripe having thick, black borders on adult males, but narrower, dusky or blackish borders on juveniles and large females; lower border of postocular stripe usually in contact with dusky postlabial line; no other markings on side of head; pattern on dorsal surface of soft parts of body of closely approximated, fine markings that are not in contrast with ground color, over-all coloration grayish; occasionally few larger and more contrasting markings, especially on hind limbs and anteriolateral surface of forelimbs.
Underparts whitish, lacking markings, occasional black flecks or dusky marks posteriorly along ventral edge of carapace (TU 17306.3).
Surface of carapace smooth in adult males; large females lacking prominences posteriorly in center of carapace or in nuchal region; anterior edge of carapace smooth in both sexes, but occasionally having regularly spaced furrows or wrinkles on hatchlings.
Comparisons.—T. m. calvatus can be distinguished from T. m. muticus by the absence of pale stripes on the snout just in front of the eyes, in having pale postocular stripes that have thick, black borders on adult males, and in having a juvenal pattern of large, circular spots that are often ocellate and three millimeters in diameter (no short lines).
Remarks.—I have not seen specimens of calvatus from the Tombigbee-Alabama river drainage; presumably Cook's record (1946:185) from Lowndes County, Mississippi, represents this subspecies.
It is still not certain that calvatus occurs in streams that drain into Lake Pontchartrain, Louisiana; TU 17236 from the Amite River that lacks a diagnostic character is questionably referred to calvatus (Webb, 1959:524). As mentioned previously T. s. asper shows little evidence of intergradation with T. spinifer in the Mississippi River drainage; asper is present in streams of the Lake Pontchartrain drainage. T. m. calvatus presumably shows a corresponding relationship with T. m. muticus in the Mississippi River drainage. There are no specimens that indicate intergradation between calvatus and muticus; calvatus is expected in streams that drain into Lake Pontchartrain, Louisiana. Probably calvatus occurs eastward in the Apalachicola drainage system.
Specimens examined.—Total, 38 as follows: Florida: Escambia: KU 47116, 50852, 50854-55, 50835-36, TU 13473, 16682, 17301, 17302 (2), Escambia River, 2 mi. E, 1 mi. N Century.
Louisiana: East Baton Rouge: TU 17236, Amite River, "near" Baton Rouge. Washington: TU 13795, Bogue Chitto River, Enon; TU 17303 (5), TU 17304 (4), Pearl River, "near" Varnado. No data: TU 17305.
Mississippi: Lawrence: KU 47117-19, TU 16956, USNM 7655, Pearl River within 4 mi. of Monticello; TU 17306 (4), Pearl River, 9 mi. S Monticello. Marion: USNM 95133-34, Pearl River, Columbia. Perry: MSC uncatalogued (3), 3 mi. SE New Augusta. Rankin: UI 31071, Pearl River, Roses Bluff, 14 mi. NE Jackson.
Records in the literature.—Mississippi: Forrest: no data. Jones: Crawford Bridge. Lowndes: Columbus, Lake Park (Cook, 1946:185).
Habitat
Most writers who describe the general habitat of soft-shelled turtles mention large rivers and streams having some current, and large permanent, quiet bodies of water having soft mud or sand bottoms, but note the general avoidance of temporary water. The impermanence of water in the ponds and "charcos" of headwaters of streams may preclude the presence of softshells from these otherwise suitable habitats. Seemingly, soft-shelled turtles are not restricted to particular local situations or microhabitats in a continuous aquatic environment as are some kinds of fish, which seem to be more or less confined to riffle areas or deep holes. Certain activities of softshells such as burying themselves in soft sand in shallow water or seeking crawfish and other food over a gravel-rock substrate or one that is débris-laden, are best carried on in different habitats. Repeated observations of turtles that are probably engaged in a specific activity in a restricted area may lead to erroneous general conclusions regarding the over-all preference for a specific habitat. Perhaps this accounts for Conant's statement (1951:156) that "In the lower portion of the Scioto River [Ohio] it appears that the present species [muticus] is abundant while spinifer is almost entirely absent."
Cagle (1954:181) wrote that softshells "inhabit the extreme headwaters and smaller tributaries." Other statements in the literature indicate the variety in kinds of habitat. In Louisiana, Beyer (1900:44) mentioned spinifer as abundant "in all inland waters, preferring, however, such bayous which have sloping and sandy banks upon which they are fond of sunning themselves." Viosca (1923:41) reported soft-shelled turtles as characteristic "of the large silt-bearing rivers … such as the Pearl, Amite, Mississippi and Atchafalaya." Cagle and Chaney (1950:386) wrote that spinifer in Louisiana was found in greatest abundance in streams having some current, but that individuals were also common in quiet areas; the habitats recorded were: False River—a lake of clear water supporting an abundance of submerged vegetation, the shallow ends having mats of water hyacinth; Lakes Iatt and Bistineau—cypress swamps having clear or muddy water; Caddo Lake—a large lake having a light oil film on the surface of the water, and vegetation toward the shore consisting of cattails, water lilies and water hyacinths, and along the bank of cypress and willow trees; Caddo Lake Spillway—muddy with swift current; Sabine River—swift current, traps set in quieter backwater areas or near cypress logs in river; Lacassine Refuge—traps set in inlets and coves of ship channel having vegetation of water hyacinth, alligator grass, and along bank, saw grass, cypress knees and snags. Stejneger (1944:59) reported spinifer taken in barrow pits in Mississippi.
In Southern Illinois, Cagle (1942:160) recorded spinifer in drainage ditches (normally having several feet of water and a lush growth of aquatic vegetation) that connect inland swamps to the Mississippi flood-plain but dry up periodically, and in Elkville Lake, an artificial lake having much aquatic vegetation in shallow areas (op. cit.:157). Myers (1927:339) recorded a spinifer from Indiana from a "tiny brook." In east-central Illinois P. W. Smith (1947:39) recorded spinifer in mud-bottomed dredge ditches, lakes, ponds, small streams and rivers, whereas muticus was found to prefer rivers having clean, sandy bottoms and was not taken from lakes or small streams. This restriction in habitat preference of muticus is again emphasized by Smith and Minton (1957:346) who wrote that in Illinois and Indiana, muticus "generally avoids lakes and minor streams." Weed (1923:48), however, recorded muticus (and spinifer) from Meredosia Bay, Illinois, presumably a broad, shallow, muddy ox-bow lake of the Illinois River.
In Minnesota, spinifer has been taken from the Mississippi River, which is described as fairly swift having a fluctuating water level, sandy islands, mud banks, a bed of pebbles and large boulders, and abundant crawfish (Breckenridge, 1955:5). In Michigan, Edgren (1942:180) recorded spinifer from a "very small muck-bottomed lake." Evans and Roecker (1951:69) recorded spinifer from Long Point, Lake Ontario, which is a "broad sand spit, straight on the lakeward side but irregular with wet flats and lagoons on the bayside."
In Kansas, Brumwell (1951:207-08) found "mostly young [muticus] … in the old ponds left during flood stages of the Missouri River" … and spinifer occasionally … "in the backwaters where stagnant ponds had been formed." In southcentral Kansas, Burt (1935:321) reported muticus from "a sandbar at junction of a small creek and Medicine River" … and … a "shallow sand-bottomed, algae-filled pasture streamlet." The same author reported spinifer from a "sand-bottomed prairie streamlet" … and … "an alga-filled pool near a stream." Burt (loc. cit.) remarked that "No ecological differences in general habitat and field behavior of mutica and spinifer are evident in Kansas." Clarke (1958:21) observed spinifer in Long Creek (Osage County, Kansas), which is a winding stream, characterized by numerous deep holes alternating with rocky riffles, and having high and wooded banks, and mostly mud bottom but occasional rock bottom.
Marr (1944:490) mentioned a spinifer that was obtained on the bank of a small, mud-bottomed stream in the Texas panhandle, and Linsdale and Gressitt (1937:222) recorded spinifer from irrigation canals in Baja California.
In southern Florida, ferox occurs in all fresh-water habitats (Duellman and Schwartz, 1958:272). Carr (1940:107) reported ferox as widely distributed in streams, lakes, big springs and canals. Judging from the numbers of turtles, "the larger canals in the Everglades must represent something like an optimum habitat" (Carr, 1952:417). Wright and Funkhouser (1915:119) wrote that in the Okefinokee Swamp, ferox was especially abundant where the water is deep and the bottom soft, and the species was found wherever there were alligators. Deckert (1918:31) wrote that young ferox were taken in springs and brooks near Jacksonville, Florida. Marchand (in Carr, 1952:417-19) observed ferox while water-goggling in Florida and noted that individuals buried themselves in deep water in white sand, mud or bubbling mud-sand springs, sometimes where there was vegetation overhead. [543] Neill (1951:16) collected ferox in marshes, "prairies," flood-plain lakes, lagoons, ox-bow lakes, mangrove swamps, rivers, creeks, calcareous spring runs, man-made lakes and lime sinks. The same author (loc. cit.) reported taking agassizi (= asper) in large muddy rivers, clear "blackwater" streams, calcareous spring runs, creeks, marshes, lagoons, ox-bow lakes, flood-plain lakes, lime sinks, man-made lakes, and smaller ponds. Crenshaw and Hopkins (1955:16), however, stated that in the area where T. ferox and T. spinifer asper overlap, "asper is nearly always an inhabitant of fluviatile situations whereas ferox is equally closely confined to non-fluviatile lakes and ponds"; in the region of sympatry, Schwartz (1956:8) reported ferox from "a moderately fast, blackwater stream [Combahee River, South Carolina]."
Carr (1952:417) wrote that ferox is not uncommon near the mouths of streams in brackish waters, where the tide must occasionally take it to sea, and cited Conant, who told of an individual found at sea in Bahaman waters; Carr (1940:25) listed ferox as occasional in the marine-littoral, mangrove swamps, as did Neill (1951:16). Neill (1958:26-27) mentioned his observance of ferox at the mouth of the Pithlachascotee River, Pasco County, Florida, where the water is sufficiently saline to favor the growth of oysters, and added that commercial fishermen had told him that these turtles are sometimes netted with loggerhead sea turtles (Caretta) in the Indian River. Neill (op. cit.:5-6) also noted the presence of ferox on Meritt Island, which supports an extensive saltwater herpetofauna, off the coast of Brevard County, Florida. Löding (1922:47) recorded spinifer from Fig Island, Mobile County, Alabama, which is probably a marine or brackish water habitat. Cagle and Chaney (1950:386) obtained one spinifer in a brackish marsh of the Sabine Wildlife Refuge, Louisiana; the poor trapping returns here (one Trionyx and one Pseudemys in 408 trap-hours) suggest that fresh-water species are not abundant in brackish habitats. Neill (1958:26-27) has summarized the occurrence of soft-shelled turtles in marine and brackish habitats.
My own observations indicate a variety of habitat preferences; the term "relatively clear" refers to waters in which visibility extends four to six inches below the surface at night using a head-light.
Individuals of spinifer have been taken in large, deep rivers having a moderate to swift current, relatively clear water, mostly sand and clay bottoms, and emergent débris intermittent along the shoreline; the banks may be steep and of mud having a sparse growth of herbs (Black Warrior River, south of Tuscaloosa, Alabama), or of low extensive, sandy bars and beaches (Escambia River, near Century, Florida, Pl. 50, Fig. 1). A juvenile spinifer was taken by hand among rocks in quiet water behind a rocky shoal in the large, deep-channeled Ocmulgee River (near Hawkinsville, Georgia). Several individuals of spinifer were seen in the Flint River (near Bainbridge, Georgia), which had a swift current in a wide, deep channel, sandy or sand-silt banks, few brush piles along shore and many oölitic, submergent snags on an otherwise sandy bottom; the water was exceedingly clear and permitted water-goggling (this habitat has been obliterated by a dam on the Apalachicola River). A large female spinifer was taken on a set line from the bottom of one of several deep holes (approximately seven feet) that were connected by shallow areas or riffles (near headwaters of Escambia River—Escambia Creek, Escambia County, Alabama). Two large females of spinifer (one escaped) were taken on a trotline set in a large, [544] deep, isolated barrow pit near the Escambia River (near Century, Florida); there was no aquatic vegetation, the water was slightly turbid, and the substrate was of a sand-silt or mud.
In Arkansas, spinifer has been taken in large deep rivers having relatively clear water, a moderate current, steep banks four to 15 feet high, and a substrate of mud with few rocks (one taken on trotline, escaped; Black River, near Black Rock, Lawrence County). Two spinifer were taken (trotline and hoop-net) from a smaller (approximately 50 feet wide) turbid river having a swift current, débris along the shoreline, and mud-gravel banks (Petit Jean Creek, Yell County). Several spinifer and muticus were taken from the White River (Marion County) having a sand-gravel or bed rock bottom and clear water; individuals were collected by hand in shallow water (approximately 31/2 feet deep) as they lay on the bottom in the main channel where the current was moderate to swift or in a quiet-water side channel having submergent vegetation.
Lake Texoma, an impoundment on the Red River, having a fluctuating water level with no permanent stand of aquatic vegetation, a mud-rock or sand-silt bottom, and turbid water (Pl. 49, Fig. 1) is a suitable habitat for spinifer and muticus. T. spinifer is found in large rivers having relatively clear water, moderate currents, emergent logs and débris, and mud or sand banks (Little River, McCurtain County, Oklahoma, Pl. 48, Fig. 1), or small, shallow, turbid creeks having sand-gravel channels of pools connected by riffle areas (Mayhew Creek, Choctaw County, Oklahoma).
Three spinifer were taken from the Llano River (near Llano, Texas) in a period of low water level in hoop-nets set in a large quiet-water pond about four feet deep and having patches of rushes encroaching into the water from the shore. The river bed of sand, gravel and large boulders consisted of narrower, swift-water channels, small pools and riffles, and large ponds.
Individuals of T. s. emoryi have been taken in large ponds having little or no current, turbid, deep water, and clay or sand-gravel banks (Río Purificación, Padilla, Tamaulipas). Two emoryi were collected from a large pond (Río Sabinas, near Sabinas, Coahuila), which was connected to an adjoining one by riffle areas and had little or no current, relatively clear, greenish water, clay or mud banks, a sand-gravel bottom, and was flanked by brush and large cypress trees. A few emoryi were trapped in hoop-nets that were set in the Río Mesquites, a stream in central Coahuila approximately 20 feet wide and six feet deep, flanked by dense stands of Phragmites, and having a moderate current, relatively clear, pea-green water and a mud-sand substrate with some gravel; the stream enlarged in some places to form quiet-water coves (Pl. 48, Fig. 2). One adult male emoryi was taken from a crystal-clear, dendritic, pond (El Mojarral, near Cuatro Ciénegas, Coahuila), having shallow areas averaging about two feet but several deep holes—in one of these at the west end of the pond the water was being emitted under pressure from an underwater cavern and "bubbling" at the surface; the vegetation consisted of scattered patches of water-lilies and stonewort; the bottom was a soft mud-marl, and in some places was carpeted with shells of small gastropods. This habitat corresponds to that of the type locality of T. ater (Pl. 49, Fig. 2); see description in Webb and Legler (1960:26). The water of the ponds is warm; at 8 p. m. on July 31, 1959, the temperature of the water at the type locality of ater was 29° C., and the air was 27° C.
An immature female spinifer was taken on a trotline in a swift, clear, cold-water [545] habitat having mud banks and an abundance of brush piles (Little Tennessee River, Monroe County, Tennessee). T. spinifer occurs also in large ox-bow lakes having relatively clear water, extensive mats of submerged vegetation, a soft mud bottom, and several emergent stumps and fallen logs (Lake Concordia, Concordia Parish, Louisiana); alligator grass and cypress trees encroached to the shoreline.
Locality data of some individuals of spinifer, hartwegi, asper, pallidus and emoryi that were examined indicated that turtles were captured in ponds, bayous, sloughs, lakes, impoundments, rivers and creeks, indicating habitation of essentially all permanent waters.
A juvenile of hartwegi was seen by Mr. Wendell L. Minckley on a gravel bar jutting into a small, shallow creek having a mud-gravel bottom (Carnahan Creek, Pottawatomie County, Kansas); the impounding of the Big Blue River by the Turtle Creek Dam will obliterate this habitat. Mr. J. Knox Jones, Jr. reported seeing a large softshell in a narrow, shallow, clear sandy creek in Holt County, Nebraska.
T. s. emoryi occurs in large rivers having generally turbid waters, a moderate to swift current and mud or sand bottoms such as the Río Grande; this habitat corresponds to that of large rivers in the western parts of the range of T. s. pallidus (Red and Washita) and T. s. hartwegi (Canadian and Cimarron). These last-named rivers, in periods of low water level, often have shallow, clear, flowing water in parts of the river bed. T. s. emoryi has also been taken from small creeks having bottoms of rocks and large boulders (Black River Village, Eddy County, New Mexico; field notes of Sydney Anderson and Kenneth Shain, June 12-14, 1958).
I received a hatchling T. s. guadalupensis that was obtained in a clear, shallow-water stream (Hondo Creek, Bandera County, Texas, on April 12, 1958). The larger streams and rivers known to be inhabited by guadalupensis are generally clear having greenish-tinted waters. The geographic distribution of guadalupensis indicates that that subspecies occurs principally in those waters that drain the limestone-mantled, Edward's Plateau off the Balcones Escarpment; the headwaters are characterized by clear, calcareous streams having occasional travertine deposits. It is probably this type of habitat to which Agassiz's statement (1857:408) of "clear, bold and rocky streams" refers.
There are a few specimens whose locality data indicate a tolerance of brackish-water habitats. An adult male spinifer was obtained at Delacroix Island, St. Bernard Parish, Louisiana, a locality said to have exceedingly brackish waters (Dr. George H. Bick, St. Mary's College, Notre Dame, Indiana); this adult male (TU 16170) is unique in having a mottled and blotched pattern. Another adult male (spinifer, TU 16071) was obtained in shallow water in Lake Pontchartrain at the mouth of Tchefuncta Creek; the salinity at the time of capture was recorded as 1.7 (datum from Dr. Royal D. Suttkus, Tulane University), indicating only slightly brackish water. Two spinifer (USNM 100409-10) and one muticus (USNM 100406) were taken at Berwick Bay, near Morgan City, St. Mary's Parish, Louisiana; the waters at this locality are probably brackish. The tolerance of brackish waters doubtless facilitates the dispersal of these turtles along coastal marshes and swamps, and into adjacent drainage systems. The greater number of records in the literature pertaining to ferox suggest that this species may be more tolerant of brackish and marine waters than are spinifer or muticus.
In summary, T. ferox occurs in all fresh-water habitats, but chiefly in lentic habitats in the northern part of its range where it and T. s. asper are sympatric. T. ferox possibly is more tolerant of brackish and marine waters than are the subspecies of spinifer and muticus.
The subspecies of T. spinifer occur in all fresh-water habitats. In the southern part of the geographic range, which overlaps that of T. ferox, T. s. asper occurs principally in running-water habitats. T. s. pallidus and T. s. asper are tolerant of brackish-water habitats. T. s. guadalupensis, known at present only from rivers and streams, occurs principally in river systems that drain the Edward's Plateau of southcentral Texas. T. ater is confined to crystal-clear ponds in central Coahuila.
The subspecies muticus occurs in large rivers and streams throughout its geographic range, but is known from lakes and impoundments principally in the southern part of its range (the northernmost record is from Reelfoot Lake, Obion County, Tennessee); there is only one record of muticus from a small, shallow, headwater creek (Reno County, Kansas), and only one from a lentic habitat (Meredosia Bay, Illinois) in the northern part of its range. T. muticus calvatus is known at present only from rivers and streams.
The seemingly greater restriction of muticus to running-water habitats suggests less vagility than in spinifer (Netting, 1944:86).
Size and coloration are adaptations to habitat. Soft-shelled turtles of large size are best adapted to mesic, essentially continuous swampy or marshy habitats, whereas small size is an adaptation to less continuous, semi-isolated habitats. A turtle of the maximum size attained by ferox in the habitat of emoryi would, in a general way, probably be more conspicuous and exposed to its enemies, both in the aquatic environment and during overland excursions; perhaps the kind and amount of food would be insufficient. In any event, small size is correlated with the more arid habitats of the southwest, and large size with mesic ones in the southeast. T. ferox, the largest species, and the smallest population of T. spinifer (resembling muticus) both occur in the southernmost part of the range of the genus. This situation does not support the corollary of Bergmann's Rule, that pertains to some groups of terrestrial reptiles, in which those subspecies occurring farther north, or in cooler climates during their season of activity, tend to be smaller.
Within the species spinifer, the emoryi group of subspecies are pallid having whitish dots on the carapace and lack extensive black pigmentation; these features seem to confer protective coloration on the inhabitants of arid, essentially sandy or muddy habitats having [547] sluggish, turbid waters, whereas the more contrasting patterns of the spinifer group of subspecies eastward seem more suited to existence in clearer, swifter waters.
The occurrence of the two clines, spinifer-hartwegi and pallidus-guadalupensis, in the species spinifer are notable in that the former occurs mostly in one large continuous drainage system, that of the Mississippi, and shows no sharp break in the one character distinguishing the two subspecies whereas populations composing the pallidus-guadalupensis cline are separated into several river drainages, and show a relatively sharp break in several characters at the Brazos-Colorado river divide. This situation seemingly supports the thesis that clines are maintained by some sort of parallel gradient in ecological or geological conditions. It is notable that streams draining the Edward's Plateau (inhabited by guadalupensis) differ in quantity (more) and quality (especially CO_{3}—, Ca++, and Mg++ ions) of their solutes, and probably pH (higher) from those farther east (Hubbs, 1957:102). The gross difference in habitats mentioned above (sandy, turbid, sluggish streams in the west vs. clear, swift streams in the east) may affect the differentiation recognizable in the spinifer-hartwegi cline.
Daily and Seasonal Activity
Diurnal Habits
Softshells bask on débris in the water or on banks close to the water; basking presumably raises the bodily temperature. In general in the southeastern and southwestern United States, I have seen softshells basking only rarely but once saw six at one time close together on logs in Bowie Creek, Hattiesburg, Mississippi (species undetermined). Surface (1908:122) saw spinifer in rows on rocks or logs in tributaries of the Ohio River. Duellman and Schwartz (1958:271-72) stated that ferox basks on banks or beds of aquatic vegetation. Deckert (1918:31) mentioned large ferox "sunning in shallow water at edge of pond." Minton (1944:447) wrote that muticus and spinifer sun on steep mud banks (Wabash River). Cahn (1937:180) stated that muticus (in Illinois) basks on banks at the water's edge but seldom on logs, and suggests that muticus is less prone to leave the water than spinifer. According to Carr (1952:438), muticus never basks on logs or rocks. In Ohio, Conant (1951:159) mentioned spinifer as occasionally basking upon a log or rock, or sometimes on steep clay banks of streams. On banks, quick escape is facilitated by directing the head toward the water, thus eliminating the time that it would [548] take to turn around on land (Conant, loc. cit.; Newman, 1906:129). Evermann and Clark (1920:593) mentioned spinifer as basking on sandy or grassy shores, and large boulders. Muller (1921:181) wrote that muticus basks four to ten feet from the water's edge on gently sloping sand and mud shores of small islands in the Mississippi River (near Fairport, Iowa). Muller stated that basking usually occurs in the morning, up until 2 p. m., and that beaches with a northern exposure were preferred; he observed 37 turtles within a 50-foot stretch of beach. In captivity, hatchlings bask on wire-mesh supports.
I have frequently observed softshells floating at the surface of the water, a habit previously mentioned by Surface (1908:122) and Pope (1949:305, 311). Individuals of Pseudemys and, to a lesser extent, Graptemys also float at the surface; those kinds of turtles and softshells at least, often appear at the surface of the water, seemingly as a result of an inquisitiveness, following repeated disturbances that cause submergence.
Newman (1906:131) described the active pursuit of food: "They crawl or swim along the bottom, thrusting their snouts under stones and into masses of aquatic vegetation, occasionally snapping up a crayfish or larva that they have succeeded in dislodging. They do not tear up their food, but swallow it whole, using the forefeet to assist in forcing it down." Surface (1908:123) suggested that softshells may feed "upon insects which may be found floating on the water," and I have had captives take insects from the surface of the water. Carr (1940:107) also wrote that ferox and numerous gars in the Tamiami Canal, often at the mouths of the tributary ditches, snap at each other furiously as floating bits of food are washed in from the Everglades. Another habit that has been mentioned as an aid in acquiring food (Breckenridge, 1944:186; Conant, 1951:156; Hudson, 1942:101) is burrowing just below the surface in a soft bottom in shallow water, to ambush passing fish, or other food. Presumably all kinds of softshells do this in both shallow and deep water of lakes or rivers having a suitable substrate; spinifer and muticus have been reported to burrow in shallow waters (no observations in deep water) by Agassiz (1857:333), Cahn (1937:180, 189), Conant (1951:159) and Weed (1923:48). Marchand (in Carr, 1952:417-19) noted that ferox burrows in deep water, and mentioned that in areas of bare white sand a group of fish invariably surrounds them, and one can locate buried softshells by observing these particular schools of fish. No mention was made of the turtles attempting to catch the fish. Other associations of soft-shelled [549] turtles and fish have been described. Kirtland (in DeKay, 1842:7) observed several large bass that closely followed large numbers of turtles floating at the surface. Newman (1906:131) reported the observations of fishermen in Lake Maxinkuckee that large-mouth black bass stay not far away from swimming softshells; the same author also mentioned the observations of Jacob Reighard, who suggested that bass may be feeding upon minnows that he noticed following softshells. Seemingly some sort of commensalistic relationship exists whereby fish acquire food that is dislodged by grubbing and scurrying of softshells. Probably food is pursued on occasion from a buried position, but this habit probably is not executed specifically for obtaining food. Newman (op. cit.:129) was of the opinion that burrowing in shallow water is a habit to facilitate "warming up."
Marchand (loc. cit.) also wrote of other notable underwater observations on ferox in Florida. He commented on this turtle's inquisitiveness in deep water and unconcern upon being touched or even upon being handled to some degree. Calf-deep in soft mud, he noted a turtle that "emerged from the mud of the bottom, headed up toward shore, circled, and when about three feet above the bottom dived suddenly and completely disappeared." Marchand wrote that some areas on the bottom (Crystal Springs), which are rooted up by the burrowing of softshells, are bare and soft, and assume a characteristic, easily recognized, appearance.
Cahn (1937:180, 189) stated that the burrowing process consists of "flipping" the loose sand or silt over the back, whereas Conant (1951:159) described the process as a rapid lateral movement of the body. My observations of captives agree essentially with Conant's observations. The initial movement, directed at a slight angle, is principally with the forelegs although complemented by lateral movements of the body. When the turtle is approximately half buried, it makes rapid lateral movements of the body, which completely bury the turtle and orient its body in a horizontal position.
Behavior and Adaptations
Some characteristics of softshells that are often mentioned in the literature are: extreme shyness or wariness, ferociousness as captives, dazzling speed and agility on land and in water, and great dependence on aquatic environment. Certainly they are wary; and this wariness may account, in part, for the scarcity of observations of basking, and statements attesting to their great speed on land. To my mind, their reported ferociousness and savage disposition [550] as captives is overrated; of the many softshells that I have collected, only a few attempted to bite. The extensibility of their long neck does warrant more careful handling than needs to be employed with other species. Holbrook (in Hay, 1892:145) even wrote that they "will sometime leap up and give a loud hiss," and Newman (1906:130) wrote that "they hiss violently and thrust out the head." Wright and Funkhouser (1915:120) reported a captive ferox that "could jump forward practically its own length." I have been bitten by individuals of Kinosternon, Sternothaerus, Pseudemys and Graptemys, and cannot support the contention that softshells are more prone to bite than those species, a view shared by LeConte (in DeKay, 1842:7); many softshells on initial capture will tend to withdraw the head completely for a short time. Newman (loc. cit.) also wrote that recently captured specimens exude a thick, yellow, semi-fluid resembling yolk of an egg from the inguinal glands; the substance, however, is odorless but "undoubtedly homologous with the emission of the inguinal glands of the musk and snapping tortoises." Perhaps there is a difference in aggressiveness associated with geographic location, the age of the turtle or individual temperament.
Smith (1956:159), referring to muticus, wrote that they are the best swimmers of all fresh-water turtles, and perhaps of any turtles. Corresponding statements of other authors attesting to their speed and agility (including spinifer and ferox) in water and on land are based principally on the published comments of Muller (1921:181), who observed that females disturbed while laying eggs "about fifty feet from water … covered the distance faster than a man can run." Cahn (1937:180) also stated that muticus on a "level, unobstructed sand beach … can outrun a man," and (op. cit.:181) can "capture fish with ease"; Cahn supported the latter statement by relating his observation of a muticus that captured a small brook trout in a large tank. Smith (op. cit.:162) wrote that spinifer is "said to overtake bass." Doubtless they are good swimmers and they do scurry rapidly on land.
Published statements relating to the strictly aquatic existence of softshells especially muticus, are based on recognition of "its drastic adaptations to aquatic existence" (Carr, 1952:428); these adaptations presumably include pharyngeal respiration and the marked depression of body form. Pharyngeal respiration was demonstrated for muticus and spinifer (Gage, 1884; Gage and Gage, 1886), and was considered the principal type of aquatic [551] respiration (some dermal and some cloacal) in Trionyx spinifer asper by Dunson (1960). Cloacal bursae (anal respiration) are lacking in trionychids (Smith and James, 1955:88). Accessory pharyngeal respiration is meaningful in light of the information furnished by Agassiz (1857:282-83), who found that Trionyx has a smaller lung capacity (weight of body in ounces/capacity of lungs in cubic inches = 16.9) than do some other genera (Pseudemys, 2.8; Testudo, 2.7; Terrapene, 1.1); corresponding values for more aquatic species were Chelydra, 9.3 and Kinosternon, 16.0. Cahn (1937:181), however, wrote that he has demonstrated pharyngeal respiration in individuals of Pseudemys, Chrysemys and Sternothaerus, and Allen and Neill (1950:13) suggested that it occurs in Macroclemys. More conclusive data are required to detect a positive correlation between small lung capacity, pharyngeal respiration, and degree of restriction to an aquatic habitat.
The depressed, soft-margined carapace of softshells has been mentioned as an adaptation to facilitate burrowing in soft sand or mud, and more suited for concealment than for speed in aquatic locomotion (Carr, 1952:429; Smith, 1956:162). Nielsen (1951:264-65), commented that in various lotic invertebrates, dorsoventral flattening of the body was no commoner than in lentic invertebrates; he wrote that a dorsoventral flattening is a disadvantage to an animal in a strong current and is an adaptation "probably … not to withstand the current directly, but to avoid it by seeking shelter in narrow crevices." Probably another aid to concealment, mentioned by Williams and McDowell (1952:272), is the plastral hinge.
Concealment of softshells is not enhanced by growths of algae on the carapace. Proctor (1958:637-38) reported that the common, epizoöphytic alga of chelonians, Basicladia, has never been reported from Trionyx; the same author recorded a large amount of filamentous algae, principally Stigeoclonium, but the algae could be easily wiped from the turtle, and Vinyard (1955:64) recorded an alga, Dermatophyton radians, attached to the skin of the legs of Trionyx. I noted a small patch of greenish scum growing near the insertion of the neck on a softshell (spinifer from Lake Texoma); cursory examination by Dr. R. H. Thompson, disclosed one of the colonial ciliate protozoans (resembling Opercularia) with enmeshed green or blue-green algae. Evermann and Clark (1920:592) mention a spinifer from Lake Maxinkuckee, Indiana, having a growth of Opercularia, covering the plastron.
Movement
The reported proclivity of softshells for a strictly aquatic existence has been over-emphasized; they are no more confined to aquatic habitats than some chelydrids (including kinosternids). In fact, there is a general parallel in habits between members of the two families, namely, a tendency toward a bottom-dwelling existence, and a burrowing habit. The alligator snapping turtle (Macroclemys) is probably the most aquatic fresh-water turtle in the United States. The common snapping turtle and some kinosternids are known to migrate overland. Kinosternids and trionychids bask frequently, and trionychids have been observed moving overland. Cox (1894:50) reported a spinifer attempting to climb a narrowly-stepped, 12-foot dam on Mud Creek at Ravenna, Nebraska; the turtle failed after repeated struggles, once reaching a height four inches shy of the brim before tumbling back into the water. Duellman and Schwartz (1958:271) commented that adults of ferox are often seen on roads bordering canals, and informants have told me verbally of similar observations. Conant (1930:61) reported an individual of ferox that was "… walking across the main street in Venice [Sarasota County, Florida]." Softshells will travel overland in search for suitable nesting sites; Newman (1906:130) wrote that spinifer will climb "steep railway embankments with considerable ease, in order to reach a sand pit some fifty yards from the water."
From an analysis of species-composition of large reservoirs and lakes and their adjacent smaller ponds in southern Illinois, Cagle (1942:162) concluded that softshells "travel overland far less often than do …" other species, but that they are "probably the first to move as the water level falls." On the basis of further observations in the same region, Cagle (1944:15) wrote that softshells rarely move overland, and once trapped in a pool of water, they bury themselves and remain there. He related instances of several individuals having been dug from dried mud where the last remnants of a water pool had evaporated and he concluded that the home range is probably confined to one body of water. That fluctuations in water level affect the movement of softshells is suggested by Mr. William E. Brode's comment that a commercial fisherman trapped numerous softshells in the Pearl River, south of Monticello, Mississippi, in unbaited hoop-nets in late May and June when the water level was receding after heavy rains.
The meager data available concerning the aquatic movements of softshells indicate that individuals wander but little. Breckenridge [553] (1955:6, table 1) found that among 30 recaptured turtles that had been marked, the greatest distance traveled was 600 yards over a two-year interval; after a three-month interval one originally captured 30 miles distant, moved only 200 yards. The statement of a professional turtle trapper mentioned by Breckenridge (loc. cit.) and data previously presented (see page 436), to the effect that over-trapping results in increasingly diminished returns, tends to support the idea that there is little aquatic movement in soft-shelled turtles.
Breckenridge (loc. cit.) mentioned methods of marking softshells and found that notching the edge of the carapace with a leather punch was satisfactory; the notches healed but were discernible as shallow sinuses. The same author mentioned a tattooing device (mentioned also by Cagle, 1939:171), but no turtles so marked were ever recognized as recovered. Tagging with a radioactive isotope and detection with suitable instruments should prove applicable to turtles (see Karlstrom, 1957).
Nocturnal Habits
Anderson (1958:212) wrote that hatchlings (muticus) leave nests within the first three hours after sunset, and are active on the surface of the sand at night. Muller (1921:183) reported hatchlings (muticus) leaving nests at night or early in the morning. Lagler (1954) stated that spinifer is nocturnal. To my knowledge there are no other published statements concerning nocturnal activity of soft-shelled turtles. I have noted them at night on only four different occasions. In two instances (Clear Fork Brazos River, Texas, and Lake Concordia, Louisiana), the turtles were resting immediately below the surface of the water on submerged branches, as one would expect Pseudemys and Graptemys to do. Another individual was seen swimming near the surface (Ocmulgee River, 11/2 mi. S Jacksonville, Georgia); this observation possibly represents nocturnal activity, or inquisitiveness owing to the disturbances caused by the motor of the boat and/or our head lights. A final observation tends to support the view of nocturnal habits. My field notes record at least four softshells collected by hand, and a few other seen in a shallow (approximately four feet deep), quiet, clear water side channel of the White River (Cotter, Arkansas); the turtles were seen resting and slowly moving on the bottom or swimming.
Seasonal Occurrence
The length of the season of activity increases with decrease in latitude. Aquatic species in general have longer periods of activity [554] than terrestrial species at the same northern temperate latitudes. The southernmost populations of all species of softshells may be active throughout the year, assuming temperature to be the limiting factor.
There are few published statements relative to the length of the annual period of activity; all records refer to spinifer. In Lake Maxinkuckee, northern Indiana, Newman (1906:128) wrote that individuals were first seen in early April on the lake shore in a weak condition with neck and legs extended, and were easily captured. Lesueur (1827:262) wrote that spinifer in Indiana appears toward the end of April. Observations of Evermann and Clark (1920:592) in Lake Maxinkuckee, and Butler (1894:224) in east central Indiana concurred in finding that of all kinds of turtles there, softshells appeared last in spring and disappeared first in fall. Evermann and Clark found small softshells, benumbed or dead, along the shore as early as March 18 and also late in fall. The earliest observation for large softshells was April 29, and the latest was September 7; Butler found that these turtles rarely appear before April 15 and sometimes not until May 1. Cahn (1937:191) stated that softshells in Illinois hibernate toward the end of October and emerge in May or the latter part of April; the same author mentioned that in southern Illinois the species might remain sluggishly active all winter. In Ohio, Conant (1951:160) wrote that individuals were collected every month from March to October, and one was even taken in December, 1929, in northwestern Ohio. Wright (1919:8) mentioned observing softshells on April 20 and September 20 (presumably these were the earliest and latest observations of them) in Monroe and Wayne counties, New York. Blatchley (1891:34) listed dates of early and late activity as March 19 and December 11 for Vigo County, Indiana. Webster (1936:22) recorded the earliest and latest dates of collection of spinifer in central Oklahoma as June 10 and November 8.
Moore and Rigney (1942:80) found an individual of muticus under six inches of ice in water about one foot deep on January 31, 1940 (Cimarron River, Payne County, Oklahoma).
The published information suggests that the length of the normal annual period of activity of spinifer in latitudes of about 40° and 43° is approximately five months, from April into September, depending upon the weather. There are numerous published statements to the effect that the period of hibernation is passed under a shallow covering of mud in deep water. Evermann and Clark (op. cit.:593) [555] found a softshell (presumably in a quiescent state) on September 6 that was "buried up to its eyes in mud at the edge of Lost Lake." Softshells possibly hibernate in shallow water or in soft mud flats. Conant (loc. cit.) found that captives would not hibernate in a pond in a zoo having a bottom of leaves.
Food Habits
Previous authors, most of whom allude to published statements preceding their own, characterize soft-shelled turtles as carnivorous and mention such food items as crawfish, insects, worms, snails, clams, frogs, tadpoles, fish, and occasional vegetable matter. Stockwell (1878:403) wrote that the relative lengths of portions of the digestive tract indicate "a purely carnivorous diet."
In an examination of the contents of 11 stomachs of spinifer from Michigan, Lagler (1943:304) found that crawfish (47%) and insects (52%), principally burrowing mayfly naiads (Hexagenia), and dragonfly naiads, comprised the bulk of the diet with cryptogams, vegetable débris, snails and fish remains present in small amounts. Breckenridge (1944:186) wrote that 18 specimens of spinifer in Minnesota contained 44 per cent crawfish, 29 per cent aquatic insects, 8 per cent fish, and 19 per cent unidentified material. Surface (1908:123) found crawfish in the only two stomachs of specimens he examined from Pennsylvania. Penn (1950) summarized the results of those authors, and estimated that crawfishes comprised 58 per cent (46% by volume) of the diet of softshells. In Indiana, three stomachs examined by Newman (1906:131) in late June contained: 1) nine crawfish, 2) four crawfish, 22 dragonfly naiads, 3) nine dragonfly naiads, few plant buds. Neill (1951a:765) found crawfishes in the stomachs of five spinifer from the Savannah River, Georgia. Evermann and Clark (1920:595) wrote that spinifer in Lake Maxinkuckee feeds principally on crawfishes. Shockley (1949:257) mentioned bottom organisms and small fishes as food. Clark and Southall (1920:16) stated that "Its principal food, to judge from a few specimens examined, consists of crayfishes."
Cahn (1937:183) wrote that the food of muticus in Illinois consists principally of crawfish, fish, frogs, tadpoles, larger insect larvae and nymphs, and aquatic mollusks. The kinds of fish eaten were Notropis heterolepis, N. spilopterus, N. hudsonius, Lepomis machrochirus, Morone chrysops, Perca flavescens, Catostomus commersonnii, and Hypentelium nigricans; Cahn (loc. cit.) also stated that the mollusks eaten by muticus are both gastropods and small, thin-shelled bivalves. In regard to the feeding habits of spinifer in Illinois, Cahn (op. cit.:193) listed the following items in decreasing order of abundance as revealed by examinations of stomachs: crawfish, minnows, fry of larger fish, frogs, tadpoles, earthworms, insects (often beetles), and mollusca (Pisidium, Viviparus, planorbids). The kinds of fish mentioned were: Notropis heterodon, N. heterolepis, N. hudsonius, Catostomus commersonnii, Lepomis humilis, L. macrochirus, Semotilus atromaculatus, Notemigonus crysoleucas, Umbra limi, and Micropterus salmoides. Cahn (loc. cit.) also found the remains of a six-inch brook trout (Salvelinus) in the stomach of a 13-inch spinifer from Wisconsin.
Agassiz (1857:399) found larvae of neuropterous insects in the stomach of one specimen of muticus, and fragments of Anodonta and Paludina (= Campeloma) in the stomach of one ferox. The expanded crushing surfaces of the jaws in some large individuals of ferox may be an adaptation to mollusc-feeding (Schmidt and Inger, 1957:36). Surface (1908:123) found spinifer to have fragments of beetles in one of two specimens examined, and large quantities of corn in another from Ohio. Webb and Legler (1960:27) reported 23 chrysomelid beetle larvae (Donacia) in one specimen of T. ater. Evermann and Clark (1920:595) reported several spinifer taken on hooks baited with grasshoppers in water 14 feet deep in Lake Maxinkuckee, Indiana. Hay (1892:144) wrote of muticus that "If there are potatoes growing near the water the turtles find their way to them and devour the stems, of which they are very fond." Wright and Funkhouser (1915:123) stated that young ferox in the Okefinokee Swamp feed on fish and frogs, and according to the natives, larger specimens take waterfowl, a statement that Smith (1956:159) was probably reiterating when he mentioned that the diet included "perhaps young birds." Parker (1939:88) wrote that of two spinifer from Reelfoot Lake, Tennessee, one contained coleopteran remains, and the other an aquatic beetle and two large tipulid larvae. Wied-Neuwied (1865:54) wrote that Lesueur found worms, snails, remains of Paludina (= Campeloma), fruits and even hard nuts in stomachs of muticus.
Holbrook (in Hay, 1892:145) mentioned that spinifer feeds on fish and such reptiles as it can secure. There are no published statements known to me that report reptiles in the diet of American softshells. Carr (1952:425) erroneously cited Strecker (1927:9) and attributed "a young lined snake" to the diet of T. s. emoryi; Strecker, however, referred to Kinosternon flavescens. In conjunction with raising softshells on turtle farms, Mitsukuri (1905:261) mentioned that first and second year-old turtles (Trionyx sinensis) must be transferred to separate ponds or they will be eaten by adults; perhaps corresponding cannibalistic tendencies exist in confined, natural habitats in American softshells.
Captives eat essentially the same things that free-living individuals do, plus scraps of meat (Strecker, 1927:9; Gloyd, 1928:135; Pope, 1949; Conant, 1951:156, 160). Lagler (1943:303) mentioned a young spinifer that fed on water fleas (Daphnia) and canned fish. Conant (op. cit.:160) wrote that no captive was observed to take vegetable matter.
Food, mostly in intestines, of two adult females of T. s. emoryi collected on June 12-14, 1959, from the Río Grande at Lajitas, Brewster County, Texas, was examined. One female, KU 51961, contained little food and mostly plant fragments; because the stomach or intestine was not full of plant fragments, this food probably was ingested incidentally to the few insects present. Another female, KU 51955, contained insects, which were identified by Dr. George W. Byers, Department of Entomology, University of Kansas, as follows: 1) Coleoptera, Dryopidae, genus Helichus, most numerous, 350 to 400 individuals; 2) Coleoptera, Scarabaeidae, genus Phyllophaga, one individual; 3) Odonata, Coenagrionidae, fragments, probably one individual; 4) Hymenoptera, Sphecidae, subfamily Bembicinae, one individual; 5) Ephemeroptera; fragments of naiad; and 6) a few plant seeds, pieces of slender roots, weed stems and a couple of fragments of tree bark. The scarab and wasp probably fell into the water and were eaten.
Table 6. Kinds of Insects Found in Stomachs and Intestines of 11 Specimens of Trionyx m. muticus (Eight Adult Males and Three Immature Females, 9.0 to 12.5 cm. in Plastral Length) From Douglas County, Kansas. Frequency of Occurrence (Approximate Number of Individual Insects/Number of Stomachs in Which Found) Is Given for Each Item Listed. Fragments of Insects Represent Adults Unless Otherwise Noted.
| Food Item | Frequency |
| Orthoptera | |
| Locustidae | 1 |
| Ephemeroptera | |
| Unknown (naiad) | |
| Odonata | |
| Anisoptera (naiad) | 3/3 |
| Zygoptera (naiad) | 4/2 |
| Plecoptera | |
| Unknown (naiad) | 2/1 |
| Homoptera | |
| Cicadellidae | 20/7 |
| Hemiptera | |
| Lygaeidae | 1 |
| Neuroptera | |
| Corydalidae (Corydalis larva) | 1 |
| Trichoptera | |
| Hydropsychidae? (incl. 18 larvae and 4 pupae) | 23/9 |
| Unknown (incl. 1 larva) | 4/4 |
| Lepidoptera | |
| Noctuidae? (larvae) | 2/1 |
| Pyralidoidea (larva) | 1 |
| Unknown | 1 |
| Coleoptera | |
| Carabidae (incl. 1 larva) | 3/3 |
| Cerambycidae? | 1 |
| Chrysomelidae | 1 |
| Cicindelidae (larva) | 1 |
| Elateridae (larva) | 1 |
| Hydrophilidae? (larvae) | 4/2 |
| Scarabaeidae (incl. Phyllophaga) | 9/6 |
| Diptera | |
| Anthomyiidae | 1 |
| Asilidae | 1 |
| Bibionidae (Bibio) | 5/2 |
| Calliphoridae (puparium) | 1 |
| Empididae | 1 |
| Mycetophilidae | 1 |
| Tipulidae (incl. Tipula bicornis and T. triplex?) | 9/4 |
| Unknown (5 muscoid, 3 acalyptrate, and 1 cyclorrhaphous types) | 9/4 |
| Hymenoptera | |
| Apoidea | 1 |
| Formicidae (incl. Camponotus) | 11/4 |
| Ichneumonidae (one questionable) | 4/3 |
| Tenthredinidae | 1 |
| Unknown (small wasps) | 3/2 |
Food from the digestive tracts of 11 specimens of T. m. muticus from the Kansas River at Lawrence, Douglas County, Kansas, were examined (Table 6). The turtles (KU 55296-306, eight adult males and three immature females, ranging in plastral length from 9.0 to 12.5 cm.) were collected in June, 1958, by Mr. Robert R. Patterson. All turtles were caught on hook and line in a period of about four or five hours at dusk. Patterson frequently fished below the bridge at Lawrence and observed that heads of softshells were often seen there about dusk and that the turtles seemed to prefer a rather shallow, quiet-water area of swirls and eddies for feeding. The stomachs, and to a lesser degree, the intestines, were nearly full. Some turtles contained plant fragments, principally elm seeds. The kinds of food in this sample were also identified by Dr. Byers and were mostly insects, the most frequent item being trichopterans; many of the insects eaten undoubtedly fell into the water. The remains of spiders were found in four stomachs and crawfish fragments in five.
Stomachs of two adults of muticus from Lake Texoma, Oklahoma, were opened. The stomach of one (OU 27593) was full of naiads of the burrowing mayfly Hexagenia; that of the other female (OU 27594) contained exoskeletal remains of crawfish. The two specimens were drowned in gill nets between the hours of 11 a. m. and 7 p. m., on July 10, 1954; the intact condition of the mayfly naiads indicated recent feeding.
The species of American softshells are mainly carnivorous. The presence of vegetable matter (mentioned in previous paragraphs) in the digestive tracts of many specimens and True's statement (1893:152) that soft-shelled turtles include a variety of vegetable matter in their food indicates omnivorous habits. Duellman and Schwartz (1958:272) stated that ferox is omnivorous and Carr (1952:430) made a similar statement for spinifer. The diet seems to be determined by the food supply available, which may vary seasonally or with adverse conditions such as flooding; under normal environmental conditions, however, vegetable matter probably is ingested incidentally to other food. There is no indication of a preference in food habits according to species and subspecies. Most of the food seems to be obtained by active foraging that is triggered primarily by movement of the prey; the sense of smell is probably secondary.
Reproduction
Size of males at Sexual Maturity
Elsewhere (1956:121) I have shown that males of spinifer from Lake Texoma, Oklahoma, and scattered localities in Texas and Louisiana are sexually mature when they reach a plastral length of 9.0-10.0 centimeters. Adult males have distinct, convoluted, non-pigmented vasa deferentia and elongate testes, the maximal measurements of which are about 10 by 30 millimeters. Testes of hatchlings are approximately 4.0 by 0.4 millimeters (TU 13698.12, [559] plastral length 3.2 cm., measured with ocular micrometer). I am not aware of seasonal changes in size of the testes.
In reading the discussion that follows, it is well to remember that males having the cloaca extending beyond the posterior edge of the carapace are regarded as sexually mature. As an indication of geographic variation in spinifer, I have listed the measurements of the 10 smallest males for only those subspecies of which there are numerous records (Table 7). Corresponding data for T. muticus muticus are also listed for comparison.
Table 7. Size at Sexual Maturity of the 10 Smallest Males of T. m. muticus and Selected Subspecies of T. spinifer. The Extremes Precede the Mean (in Parentheses).
| Species and Subspecies | Plastral length (cm.) |
| T. s. spinifer | 8.8-10.3 (9.6) |
| T. s. hartwegi | 9.6-10.5 (10.2) |
| T. s. pallidus | 9.1-11.2 (10.5) |
| T. s. guadalupensis | 9.3-10.8 (10.1) |
| T. s. emoryi | 8.2-9.0 (8.8) |
| T. m. muticus | 8.2-9.2 (8.7) |
The data indicate that the size at which sexual maturity is attained in emoryi (about 8.0-9.0 cm.) is less than in any other subspecies of T. spinifer (about 9.0-10.0 cm.), and, more importantly, corresponds to that of T. m. muticus. Although the mean for T. s. spinifer is slightly less than in the remaining subspecies, I doubt that there is any significant difference according to subspecies in the size at which sexual maturity is attained in the subspecies spinifer, hartwegi, asper, pallidus and guadalupensis. The corresponding size in T. m. muticus and T. s. emoryi heightens the morphological resemblance between these forms. The only sexually mature male of T. ater, which morphologically resembles emoryi and muticus, is 9.5 centimeters in plastral length. I do not know the size at which T. ferox attains sexual maturity. The smallest sexually mature individual examined by me was 12.0 centimeters; probably ferox attains sexual maturity at a larger size than spinifer or muticus. The relative size of attainment of sexual maturity in ferox, spinifer, and muticus corresponds to the maximum size of the three species; ferox is the largest species and muticus is the smallest (Table 2).
Size of Females at Sexual Maturity
Breckenridge (1955:6) wrote that the development of the mottled pattern "undoubtedly indicates a stage in the attainment of sexual maturity"; I have mentioned (1956:121) that the mottled pattern is apparent on females before sexual maturity is attained. To my knowledge females have no external characters which appear at the time of attainment of sexual maturity.
Sexually mature individuals of ferox have been described in various terms: 311/4 pounds (Goff and Goff, 1935:156); six pounds, lengths of carapace 101/2 and 13 inches (Hamilton, 1947:209); greatest width of head 31/2 inches (Wright and Funkhouser, 1915:120). A 101/2 inch carapace presumably represents the smallest turtle and corresponds to a plastron approximately 22.0 centimeters in length. There is no other information available concerning size at sexual maturity in T. ferox.
There is little published information concerning the size at sexual maturity in T. spinifer. Cahn (1937:193) wrote that spinifer in Illinois "must attain a carapace length of about 24 centimeters [plastral length approximately 18.0 cm.] before the females become sexually mature"; this statement is the basis for Smith's mentioning a length of 91/2 inches (1956:162). Evermann and Clark (1920:595) recorded the lengths of carapace of some females (presumably all adult) from Lake Maxinkuckee, Indiana, as 11, 113/4, 121/2, and 13 inches; the smallest measurement corresponds to a plastral length of approximately 21.0 centimeters.
The data concerning reproduction presented in succeeding paragraphs is based principally upon examinations of turtles in the TU collections; I am indebted to Dr. Fred R. Cagle for permission to dissect these turtles. Females are regarded as sexually mature when they have oviducal eggs or corpora lutea or ovarian follicles exceeding 15 millimeters in diameter. Hatchlings of spinifer have ovaries that measure approximately 6.0 × 0.3 millimeters, and straight oviducts 0.2 millimeters in width (TU 5988, plastral length 3.5 cm. measured with ocular micrometer). In the size at which sexual maturity is attained there seems to be much individual variation as well as geographic variation.
Females of T. s. emoryi from the Río Grande in the Big Bend region of Texas are sexually mature when the plastron is approximately 16.0 centimeters (16.2 cm., KU 51960), and are the smallest adult females of spinifer that I have seen; these females are representative of the population from which the smallest adult males of spinifer are known and which is unique in showing sexual differences in coloration. A female (TU 3697), having a plastral length of 16.0 centimeters, which was obtained in the Río Grande near Eagle Pass, [561] Texas, in mid-July, is immature; the ovaries are compact having the largest follicles 2.5 millimeters in diameter, and the oviduct is wrinkled and convoluted but only six millimeters wide. Of three females of emoryi from the Pecos River, Terrell County, Texas, having plastrons 17.4, 18.3 and 18.8 centimeters in length and obtained on June 11, the largest and smallest are immature, and internally resemble TU 3697. TU 14453.2 (18.3 cm.) is sexually mature having large corpora lutea and enlarged ovarian follicles. KU 53754, from the Río Salado in central Coahuila, México, having corpora lutea and a plastral length of 20.3 centimeters, is sexually mature.
Females of T. s. guadalupensis, measuring 14.5, 15.7, 16.3, 16.5, 16.8, 17.0, 19.0, and 20.0 centimeters in plastral length and obtained from June to September, are immature. The female measuring 19.0 centimeters indicates the approach of sexual maturity in having swollen and convoluted oviducts seven to ten millimeters in width, but compact ovaries having the largest follicles 4.0 millimeters. The other guadalupensis whose measurements are given above have oviducts that do not exceed four millimeters in width, and ovarian follicles that do not exceed two millimeters in diameter. TU 10187, obtained in July, plastral length 19.5 centimeters, is sexually mature having corpora lutea and enlarged follicles. Two other guadalupensis, 21.5 and 22.0 centimeters (Pl. 42, top), having ovaries with enlarged ovarian follicles (the largest in one, only 11 mm.) are considered sexually mature.
Concerning the subspecies pallidus, females (all collected in June or July) measuring 15.7, 16.3, 17.3, 17.5, 18.7, 19.5, 20.8 and 21.3 centimeters in plastral length are immature having solid, compact ovaries with the largest follicles not exceeding two millimeters in diameter; oviducts are straight not exceeding three millimeters in greatest width, except those turtles measuring 17.3 and 21.3 centimeters in which the oviducts are swollen and convoluted and, respectively, five and eight millimeters in greatest width. The smallest sexually mature pallidus is 19.8 centimeters in length; recorded lengths of other adult females are 23.5, 26.8 and 30.5 centimeters.
Of especial interest are three large female pallidus, measuring 24.8, 27.5, and 28.0 centimeters, which appear to be immature; two of these (TU 13303-04) are from the Sabine River, collected in July, and the other specimen is without data (presumably from the Sabine River). The oviducts are large, swollen and convoluted, resembling those in sexually mature individuals. The ovaries, however, are relatively solid and compact having approximate measurements of 125 × 6 millimeters (TU 13303) and 85 × 10 millimeters (TU 13304), and follicles not exceeding five millimeters in diameter.
Females of spinifer from the lower Mississippi Valley of Louisiana having plastral lengths of 15.0, 15.5, 16.7, 17.5, 18.0, 19.5, 20.0, 20.4, and 20.8 centimeters are considered immature; the ovaries are compact and solid having follicles not exceeding three millimeters in diameter, and the oviducts, swollen and convoluted in the larger individuals, do not exceed six millimeters in width. The ovaries of the specimen 19.5 centimeters in length mentioned immediately above had been removed prior to my examination; the oviducts, however, were relatively straight and only five millimeters in width. Three females 23.0, 25.5, and 25.8 centimeters in length are sexually mature. TU 5518, measuring 21.5 centimeters in length and obtained in June, indicates the onset of sexual maturity in having large convoluted oviducts, but the ovaries are solid, compact, measuring 85 × 13 millimeters, and the largest follicles are only 4.5 millimeters. A larger turtle (TU 13080), 24.5 centimeters, obtained in July, has [562] juvenal ovaries (largest follicles five mm.); the oviducts are enlarged and convoluted as in adult females.
Of two T. s. asper collected from the Escambia River in June and July, one 18.0 centimeters in plastral length is immature, whereas the other, 27.0 centimeters long, is adult. A female T. s. hartwegi, measuring 20.7 centimeters, is adult having enlarged follicles and corpora lutea (TTC 719, Pl. 36, bottom).
In summary, females of all subspecies of spinifer, except some emoryi, may be sexually mature when the plastron is 18.0 to 20.0 centimeters in length; probably all physiologically normal females are adult when 22.0 centimeters long. In general, females are sexually mature at a plastral length of approximately 20.0 centimeters, a measurement that corresponds to a length of carapace of approximately 28.0 centimeters or about 11 inches. Females representative of that population of emoryi inhabiting the Río Conchos and the Río Grande in the Big Bend region of Texas are adult when the plastron is approximately 16.0 centimeters in length, and are thus the smallest sexually mature females of the species spinifer. Oviducts are large (at least eight mm. in width, undistended), swollen and convoluted prior to the first ovulation.
Of interest are the large females (for example, TU 13303, plastral length 28.0 cm.) that seemingly have immature, relatively small, ovaries (the oviducts are large and convoluted as in adult females). Possibly such ovaries represent a regression and are in senile turtles, but I am inclined to believe that the development of these ovaries has been arrested probably owing to hormonal unbalance, and that they have never been functional.
The size of adult females of T. ater is unknown but probably approximates that of T. spinifer or is slightly less. Females of ater 15.5 and 17.2 centimeters in length are immature; the largest female, the holotype, is 18.3 centimeters in length, and was not dissected.
Females of T. muticus are sexually mature when smaller than T. spinifer. Two turtles, 13.8 and 14.0 centimeters in length, have large convoluted oviducts about 10 millimeters in width and ovarian follicles nine to twelve millimeters in diameter, and seem to be near sexual maturity. The smallest sexually mature female (subspecies muticus) is TU 14436, measuring 14.4 centimeters in plastral length and having oviducal eggs. Recorded lengths of other adult females are 16.3, 16.5, 17.2 (subspecies muticus), and 18.0 centimeters (subspecies calvatus). Two females having plastral lengths of 17.5 (subspecies muticus) and 16.0 centimeters (subspecies calvatus) seem sexually immature. These turtles collected [563] in April and May have ovarian follicles not exceeding three millimeters in diameter.
Sexual Activity
Observations by Mitsukuri (1905:263), Conant (1951:160) and Legler (1955:98), constitute the extent of our knowledge concerning courtship and copulation. Legler observed a male spinifer and a female muticus in captivity; the male was the aggressor, following at the rear or above the female, and at times nipping at the anterior part of her carapace. During these movements, the posterior edge of the female's carapace was turned up slightly whereas that of the male was turned down; the turtles frequently surfaced to breathe. Occasionally the female followed the male. On the bottom the male crawled onto the female's carapace from the rear, remaining in a somewhat posterior position as described by Conant (loc. cit.), and seemingly not clasping the female with his feet. Copulation probably occurs in this position; Mitsukuri (loc. cit.) mentioned that copulation in Trionyx sinensis occurs at the surface of the water. The male remains in the coital position for approximately 15 seconds and then slowly drifts to one side and swims away. Legler observed five coital unions in one-half hour, each preceded by courting movements.
Legler's observations indicate that the courtship patterns of muticus and spinifer are similar, and that interspecific matings are possible. I have not noted any hybrid.
Risley (1933:689) mentioned differential movements of the sexes of Sternothaerus odoratus in conjunction with the breeding cycle. Such movements are revealed by trapping procedures that yield deviations from the expected 1:1 sex ratio. That differential sexual movements probably occur in Trionyx is indicated by my trapping 17 males in a group of 19 spinifer in hoop-nets in Lake Texoma in the period June 14-July 12, 1954. On June 24-26, 1959, a field party from the University of Kansas collected 15 softshells in hoop-nets at the mouth of the Río San Pedro, near Meoquí, Chihuahua; all turtles were males. On June 17-18, 1959, the same expedition trapped 11 males in a group of 13 turtles in the Río Conchos, near Ojinaga, Chihuahua. Earlier, June 12-14, 1959, 39 softshells were trapped in the Río Grande near Lajitas, Brewster County, Texas. Of these turtles, however, 19 were adult males and 20 were females; eight females were adult (sexually mature) all having oviducal eggs (Fig. 23). One of the two females from Ojinaga, KU 51174, is sexually mature (plastral length, 16.5 cm.) having oviducal [564] eggs; the other is immature (plastral length, 8.0 cm.). The only softshell taken on June 21, 1959, 8 mi. N and 16 mi. W Ojinaga, KU 51173 (plastral length, 16.3 cm.) is a female having oviducal eggs. The two females from Lake Texoma are immature (plastral lengths, 9.8 and 12.4 cm.).
The results of trapping may indicate that females frequent shallow water for a short time before the period of deposition of eggs, but disperse to deep water after such periods or between them. The movements of immature females probably approximate those of adult males; the absence of immature females in the Meoquí series, and near absence (only one) in the Ojinaga series perhaps is due to fortuitous collecting methods or to slightly different diurnal movements between adult males and immature females. Females approaching sexual maturity and those sexually mature but not having oviducal eggs ready for deposition possibly remain relatively sedentary in deep water; such females possibly represent those absent in the 13.0-15.9 size group (Fig. 23). Certainly, factors other than those pertaining to egg deposition may cause mature egg-laden females to live in shallow water, or explain the deviations from the expected 1:1 ratio.
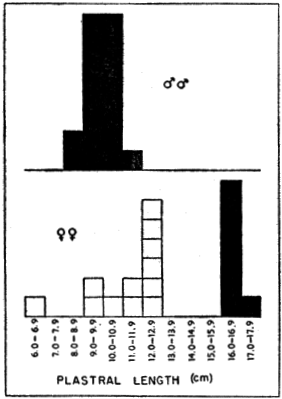
Fig. 23. Size distribution of 39 Trionyx spinifer emoryi (19 males and 20 females) collected in the period June 12 through June 14, 1959, from the Río Grande, near Lajitas, Brewster County, Texas. Solid squares represent sexually mature specimens. Females approaching sexual maturity or those not ready for egg deposition (13.0-15.9 cm. size group) are possibly sedentary in deep water.
One of the immature softshells (KU 51979, plastral length, 9.7 cm.) of the series from Lajitas is considered to be a female. It combines characteristics of both sexes. It resembles a male in having a carapace gritty to the touch, in having prominent white dots posteriorly and in not having a faint mottled and blotched pattern as do females of the same size. The postocular and postlabial [565] markings are mostly yellow (female), but a small patch of the postocular stripe near the junction with the pale ventral coloration laterally is tinted with orange (male); the morphological characters and secondary sexual difference in coloration of this series of softshells has been mentioned on page 512. The tail is short and pyramidal resembling that of a female. Internally, there are a pair of ovaries and oviducts; KU 51979 is functionally a female. An over-production of androgens probably is responsible for the external masculine characteristics (orange color, gritty carapace and absence of mottling on carapace).
Deposition of Eggs
Concerning T. ferox, Wright and Funkhouser (1915:122-23) wrote that deposition of eggs occurred in June and July in the Okefinokee Swamp on the sandy parts of the islands or in sandy fields in places exposed to the direct rays of the sun. The same authors recorded a gravid female taken on June 22 (op. cit.:120), and a nest with eggs on June 26. Harper (1926:415) reported egg-laying in June in the Okefinokee Swamp. Goff and Goff (1935:156) found a female in search of a nesting site crawling toward a cleared area within a hammock at 11 a. m. on May 19, about 25 yards from the western shore of Lake Griffin, Florida. Carr (1940:107) stated that eggs in Florida "are laid from March to July 10. One individual laid her eggs on a block of ice which we had buried in the sand." Hamilton (1947:209) observed deposition of eggs near Fort Myers, Florida, in "a sandy roadbed slightly above the cypress swamp and ditch levels on either side of the road." … either in … "the ruts formed by cars or the slope of the roadbed"; dates of deposition of eggs recorded are March 30 at 11 a. m. in bright sun, and March 31 (from context, the date given as March 21 is considered an error) at 5 p. m. following a heavy rain. The daily temperatures at the time of Hamilton's observations "averaged 85° F., the first really warm spell of the season."
Eigenmann (1896:262) reported egg-laying of spinifer in sand and gravel in June and July at Turkey Lake (= Lake Wawasee), Indiana. A turtle was seen digging a nest on June 26, and fresh nests of eggs were found on June 27 and July 9. Hedrick and Holmes (1956:126) wrote that a clutch of eggs of spinifer in Minnesota was found about ten inches deep in sand about one foot from the river; a steep gravel bank was also cited as a favorite nesting site. Surface (1908:123) stated that eggs of spinifer in Pennsylvania are laid in May, and the young hatch in August. Gehlbach and Collette (1959:142) found eggs of spinifer on June 19 on a sand bank 15 feet from the edge of the Platte River in Nebraska. Breckenridge (1944:187) wrote that spinifer in Minnesota nests on sandy beaches from June 14 to July 6. Cahn (1937:193) stated that deposition of eggs in Illinois occurs in "June or early July: earlier in the southern part of the state, later in the northern portion." Force (1930:38) mentioned a gravid female from Oklahoma obtained on May 20. Evermann and Clark (1920:593) were of the opinion that spinifer began laying about mid-June and continued until perhaps late July at Lake Maxinkuckee, Indiana; a female opened on June 14 had oviducal eggs, and the first nest was found [566] on June 18. Nests were usually at the edge of an abrupt ascent in sand; one nest was found in black, mucky soil (op. cit.:595). Newman (1906:128) wrote that spinifer in the same lake nests later than the other species of turtles, as a rule not earlier than the middle of June (but as early as June 10, op. cit.:132), and rarely later than the middle of July; he observed deposition of eggs on June 22. Sites of deposition of eggs were mostly in soft sand not more than six feet from water; other sites found by Newman (op. cit.:132-33) were a sandy, abandoned road bed separated from the shore by a strip of tall grass, a rock pile (the eggs being dropped into crevices and sand packed around them), among roots of a tree (the eggs being deposited between the roots and under them in a very irregular fashion), and in clay "so hard packed that one could scarcely break it with the fingers." Natural nest sites in hard clay and a rock pile seem incongruous with nesting habits of softshells. I note that Newman's study was not begun until 1902 (op. cit.:127), and it was that year that the water level of the lake was high, flooding the surrounding lowlands (Evermann and Clark, 1920:49-53). Perhaps some of the nests found by Newman were old and not natural because of conditions resulting from the receding water level.
Newman (op. cit.:134-35) mentioned that in small sandy areas nests were frequently in contact and overlapped; he found one nest containing nine small eggs contiguous with 23 large eggs. Breckenridge (1944:187) reported a nest of 56 eggs of two slightly differing sizes, and probably from two females. Evermann and Clark (1920:594) discovered "probably 10 or 12 nests in a distance of a few yards" and mentioned one nest containing 25 eggs "that evidently belonged to two different sets … In the bottom were 10 eggs that looked old … and … separated from them by sand, were 15 other eggs."
Nesting sites of muticus were mentioned by Muller (1921:181) on one of several small islands having "gently sloping sand and mud shores, and interior areas of open sand and densely growing willows" in the Mississippi River, near Fairport, Iowa. The same author wrote that the egg-laying season is from late June to early July, and that the female selected a place 10 to 60 feet inland "with an unobstructed view of the open water." Farther north on the Mississippi River near Dubuque, Iowa, Goldsmith (1945:447) found that muticus preferred "clean, somewhat level sandbars and sandy shores free from trash, weeds … and exposed to open view." The same species, however, may "make unsatisfactory nests … in any place they can find sand, even in the weeds and bushes … when the river is high, covering the sandy plots …" Sometimes nests, which were "seldom nearer than six feet or more than twenty-five feet from water …," were submerged by a rise in water level. In Missouri, Hurter (1911:251) found that individuals of muticus came "… out on the sandbars in the Mississippi River to deposit their eggs … At the end of May up to the middle of June …" Cahn (1937:182) wrote that the nesting season of muticus is early July near Meredosia, Illinois. Anderson (1958:212, Fig. 1) found nests of muticus along the Pearl River in Louisiana on an open sandbar (not in gravel, fine sand or silt), whereas nests of Graptemys were confined to the landward margin of the sandbar.
The onset and length of the breeding season seems to be geared to the climatic conditions under which the species occurs, and, as would be expected, [567] begins earlier and lasts longer in southern latitudes than in northern latitudes. The period of deposition of eggs in T. ferox (Florida) is from late March to mid-July, whereas that of northern populations of spinifer and muticus (southern Great Lakes region) is usually from mid-June to mid-July.
Seemingly there is little difference between species in preference of nesting sites; a sandy substrate is probably preferred. Gravid females of ferox and spinifer may wander overland some distance and select places where the view of the water is obstructed by vegetation; both species may wander little and nest in full view of the water. Concerning muticus, it is of interest that of the many nests discovered by Anderson (loc. cit.) on an open sandbar, all were those of muticus and none was a nest of spinifer. The nests of muticus mentioned by Muller (loc. cit.) and Goldsmith (loc. cit.) were on open sandbars. On June 4, 1953, six clutches of eggs were found on an open sandbar of the Escambia River, Florida; all hatchlings from those eggs that were successfully incubated were muticus. On June 1, 1954, three nests were found on an open sandbar of the same river (Pl. 50); the temperature within the nests at 6:30 a. m. was approximately 25° C. Two nests were dug in a sand substrate on the level portion of the bar (Pl. 51, Fig. 1). The third clutch of eggs was deposited in a sand-gravel substrate at the brim of the incline from the shore (approximately 30 degrees and about five feet above the water); the eggs of this clutch were arranged rather symmetrically (Pl. 51, Fig. 2). Unfortunately, most of the eggs from these three clutches failed to hatch. Although the data are far from conclusive, I have the impression that muticus limits its sites of egg deposition to the open regions of sand bars and does not lay inland where it must traverse vegetated areas unless preferred nesting sites are submerged or otherwise unsatisfactory. Females of spinifer may utilize open sandbars for deposition of eggs but not areas where muticus occurs. In areas where both muticus and spinifer occur, the latter probably lays farther inland or on the landward margins of sandbars.
Excavation of nests has been observed in ferox (Hamilton, 1947:209), muticus (Muller, 1921:181-82; Goldsmith, 1945:448), and spinifer (Newman, 1906:132-33; Cahn, 1937:191-92; Breckenridge, 1960:284). Turtles leaving the water are cautious, usually stopping and extending the neck to its greatest length, holding the head high, and sometimes returning to the water for a short time. Depending on the condition of the substrate and wariness of the female, nest construction may begin immediately, or several holes may be dug and then abandoned. The excavation on level ground or a slight incline is made by means of the hind feet (Muller mentions digging with the forefeet; I agree with Pope, 1949:321, and Conant, 1951:264, who consider Muller in error); the forefeet are firmly planted and not moved during the excavation, deposition of eggs or the filling of the nest cavity. The hind feet are used alternately; cloacal water may be used to facilitate digging or to provide a suitable substrate for the eggs. Cahn mentioned that some sand may be flung four or five feet, and that during the digging the head is held high. Breckenridge (loc. cit.) reported that sand was thrown a distance of ten feet. The nest may be completed in 16 minutes (Cahn, loc. cit.) or less than 40 minutes (Newman, loc. cit.). Breckenridge recorded 17 eggs laid in six minutes, Cahn recorded 12 eggs laid in eight minutes, and Hamilton recorded four eggs laid in three minutes. The hind feet are used to arrange the eggs and are used alternately to fill the nest cavity; sometimes a little sand is scraped in [568] before all the eggs are deposited. Muller recorded the nest cavity as five inches in diameter and ten inches deep, the finished nest appearing "as a small crater … about a foot in diameter, or where the surface is covered with pebbles, as a circular area of clear sand." Goldsmith reported that the nest cavity was six to nine inches in depth, and that after deposition and filling with sand "By certain twisting movements with all four legs, she drags the plastron around over the sand, making a perfect camouflage." Newman found the nest flask-shaped having a depth of about six inches, and diameters of about three inches at the bottom and one and one-half inches in the neck. Hamilton described a flask-shaped nest, the entrance of which would "barely permit the passage of an egg … the bottom, at a depth of five inches, being about the width of a quart milk bottle." Cahn related that the "hole descended at an angle of about 60°," and the eggs thus rolled down an inclined plane.
Possibly the nests of ferox and spinifer differ from those of muticus in being flask-shaped. A nest of spinifer was reported by Gehlbach and Collette (loc. cit.) as having a neck three inches across, a depth of six inches and a width of five inches at the bottom. The nests of muticus that I discovered on the Escambia River were not flask-shaped; the eggs were five to seven inches below the surface. Evermann and Clark (1920:594) reported eggs of spinifer "generally at a depth of four to ten inches," and Breckenridge (loc. cit.) found the topmost eggs about five inches below the surface. There may be behavioral differences between ferox and spinifer and muticus. Hamilton (loc. cit.) mentioned that ferox proceeded with its reproductive duties even when he stood only a few yards away. Muller (op. cit.:181) found that muticus would run to the water if disturbed, without completing deposition of eggs; the same behavior was described by Cahn (op. cit.:191) for spinifer. Newman (1906:133) wrote that spinifer will abandon nesting activities if surprised before egg deposition begins, but will wait to cover the eggs if interrupted while laying eggs. Goldsmith (1945:448) found that an observer did not disturb females of muticus when they were laying eggs (females "could be approached and even touched"), but that, in the presence of an observer, they would scurry toward the water without covering the eggs and would not return to cover them. Turtles frightened in the process of the construction of the nests would not return to complete the original nest. Harper (1926:415) wrote that ferox, after completing nesting activities, will crawl a few feet from the nest and scuffle up the surface, presumably to decoy predators that might otherwise destroy the eggs; this observation has not been corroborated by other authors. Harper (op. cit.:416) recorded the observation of Allen Chesser, who says that females, after egg deposition, often "… bury themselves, before they go ter the water, an' stay there ten er twelve hours."
Reproductive Potential
Estimates of reproductive potentials are subject to variation of one kind or another. Counts of oviducal eggs or those in nests may be misleading, as in some individuals one or more eggs may have been deposited previously. Mitsukuri (1905:263), Newman (1906:135), Muller (1921:182), and Cahn (1937:183) have mentioned that the number of eggs per clutch corresponds to the size of the female. Females of northern populations may have larger clutches than females of the same size from southern populations.
Table 8. Records in the Literature Pertaining to Number and Size of Eggs of Three American Species of Trionyx.
| Species | Number of eggs per clutch; oviducal (o), nest (n); ave. = average | Size of eggs; ave. = average | Authority and remarks |
| ferox | 24 mm. | Agassiz (1857, pl. 7, fig. 20); nat. size. | |
| 22 (n) | ave. 31 mm. | Wright and Funkhouser (1915:120) | |
| some (o) | 32 mm. | " | |
| 20 (o) | ave. 25 mm., and 12 gms. | Goff and Goff (1935:156) | |
| 17 (o) | ave. 27 mm. | Hamilton (1947:209) | |
| 21 (o) | " | ||
| 7 (o) | " (egg deposition probably interrupted) | ||
| spinifer | 29 mm., 26.5 mm. | Agassiz (1857, pl. 7, figs. 20 and 23, respectively); nat. size. | |
| 9, 12, 17, 18, 27 and 32 | Eigenmann (1896:263); northern Indiana | ||
| 9 to 24, ave. 18 | Newman (1906:135); northern Indiana | ||
| about 30 (n), 4 (n), 3 (n) | 1.09 × 1 inch | Evermann and Clark (1920:593-94); northern Indiana | |
| 21 (n and o) | (o) and some (n) .93 × .93 inches; rest of (n) 1.07 × 1.07 inches | " | |
| 32 (o) | ave. 11/4 inches | Force (1930:38); Oklahoma | |
| 9, 12, 13, 15, 17, 19, 19, 21, 22, 23, and 25; ave. 18 | ave. 28.3 mm. (217 eggs) | Cahn (1937:193); Illinois | |
| 12 (o), 26 (o), 24 (n), and 30 (n) | 22.0 to 28.5 mm. | Breckenridge (1944:187); Minnesota | |
| 21 (o)[570] | 24 to 27.8 (ave. 25.6 mm.) × 25.8 to 29 (ave. 27 mm.) | Conant (1951:160); Michigan | |
| 22 (n), 22 or 23 (n) | Hedrick and Holmes (1956:126); Minnesota | ||
| 25 (n) | ave. 24 × 25.2 mm. | Gehlbach and Collette (1959:142); Nebraska | |
| 17 (n) | Breckenridge (1960:284); Minnesota | ||
| muticus | about 22 mm. | Agassiz (1857, pl. 7, fig. 21); nat. size. | |
| 21 | about 20 mm. | Hurter (1911:249); Missouri | |
| 4, 12, 13, 16, 21, 22, 26, and 33, all (n); ave. 22 | ave. 2.3 cm. and 7 gms. | Muller (1921:182); Iowa | |
| 18 to 22, maximum 31 | ave. 22.6 mm. (116 eggs) | Cahn (1937:183); Illinois | |
| 93 from 5 nests, ave. 18.6; 10, 10, 16, 17, 17, 19, 21, 21, 22, 22, 31, all (n), ave. 18.7 | variable—largest ca. 13/8 inches, smallest less than one inch. | Goldsmith (1945:449); Iowa |
Additional records of size of clutch are provided by data from dissected females (Table 9). All females were collected from May through September from localities south of latitude 36.5°. The number of eggs includes those in both oviducts, and the number of ovarian follicles those in both ovaries. The number and range in size of only the largest group of follicles is listed; in some instances the size of follicles formed a graded series, and the designation of a group was arbitrary.
Table 9. Length, Number of Oviducal Eggs, and Condition of Ovaries in Adult Females of T. spinifer and T. muticus.
| Species | Size of female (plastral length, cm.) |
Eggs (total) |
Ovarian follicles (total) | |
| Number | Size (mm.) | |||
| muticus | 14.4 | 6 | 14 | 15-18 |
| 16.3 | 9 | 4 | 15-17 | |
| 16.5 | …… | 3 | 16 | |
| 16.5 | 3 | 4 | 14-18 | |
| 17.2 | …… | 13 | 14-21 | |
| 27.0 | …… | 25 | 18-21 | |
| spinifer | 16.2 | 7 | 4 | 16-20 |
| 16.2 | 7 | 5 | 18-20 | |
| 16.2 | 7 | 1 | 18 | |
| 16.3 | 6 | 5 | 16-18 | |
| 16.3 | 4 | 5 | 15-19 | |
| 16.8 | 6 | 1 | 18 | |
| 17.3 | 3 | 2 | 17 | |
| 18.3 | …… | 13 | 19-20 | |
| 19.5 | …… | 2 | 17 | |
| 19.8 | …… | 4 | 20 | |
| 20.7 | …… | 11 | 15-18 | |
| 21.5 | …… | 6 | 8-11 | |
| 22.0 | …… | 13 | 11-14 | |
| 23.5 | 8 | 12 | 20-24 | |
| 25.5 | 11 | several | 18-22 | |
| 25.8 | 13 | ? | 18-21 | |
| 26.8 | 10 | 5 | 18-20 | |
| 30.5 | 13 | 5 | 20-21 | |
| 16 | 16 | 16-21 | ||
| 11 | 19 | 15-20 | ||
| 17 | 23 | 18-22 | ||
| 17 | 22 | 14-20 | ||
| 8 | 15 | 18-22 | ||
Published data (Table 8) indicate that the average number of eggs per clutch for the three American species is about 20, although the number of eggs may exceed 30 in spinifer and muticus. Except for those of ferox, most of these records are based on observations in northern latitudes (approximately 40°). My examination of females from southern latitudes (below 36.5°) reveals no oviducal egg count greater than 17 and an average number of eggs per clutch of 9.6 per spinifer (Table 9); that of muticus is 7.3, as based on data given in Table 9 as well as on egg-nest counts of 15, 6, 6, 6, 6, [572] 5, 9, 8, and 8. Ovarian follicles larger than 15 millimeters in diameter are arbitrarily considered to comprise the next clutch that will be deposited in the current season. Follicles of this size possibly are retained until the following year or some may undergo regression; some of the included follicles may not be representative of the succeeding egg complement. The average number of follicles of the most enlarged groups is 9.0 for spinifer and 10.5 for muticus. Females in northern latitudes probably have a greater reproductive potential than those in southern latitudes if it is assumed that there is only one laying per season for an individual; the maximum number of eggs laid at any one time probably does not exceed 35. There is an indication that larger females deposit more eggs than smaller females (Table 9). Muller (1921:184) mentioned two double eggs (each having two yolks) in the complement of 33, indicating an abnormally large number and excessive crowding of eggs in the oviducts. Simkins (1925:188) also mentioned some eggs of a clutch (form and locality unknown) that were five or six millimeters larger (about 31-32 mm.) than the rest, and which "invariably bore twins." The largest number of eggs in a single nest mentioned by Simkins is 22. If the presence of double-yolked eggs is indicative of crowding of eggs in the oviducts, the egg complements of 22 and 33 indicate the approximate maximal number of eggs per clutch. In the species spinifer, the average size of sexually mature females is slightly smaller at some places in the south than in the north. Therefore, smaller clutches are to be expected in the south.
Many of the females collected in June or July contained corpora lutea four to eight millimeters in diameter in addition to enlarged ovarian follicles. Presumably the corpora lutea indicate clutches deposited earlier in the current season, and the enlarged follicles represent clutches to be deposited in the current season. One female of muticus (OU 27593) obtained on July 10, contains oviducal eggs, ovarian follicles 15-17 millimeters in diameter, and corpora lutea of different sizes that exceed the number of oviducal eggs; possibly this female was capable of laying three clutches each season. Corpora lutea, representing ovulation points of eggs in the oviducts, are approximately eight millimeters in diameter. In order to establish definitely the reproductive potentials of any species of turtle, it is desirable to know the approximate size of ovarian follicles that are retained by sexually inactive females, and the rate of regression of the corpora lutea. The data suggest that, in southern populations at least, two and possibly three clutches of eggs are deposited in the annual breeding season. Mitsukuri (in Cagle, 1950:38) found that T. sinensis deposited four groups of eggs each season.
It is suggested that the seasonal reproductive potential of northern populations (averaging about 20 eggs per clutch, and probably one clutch per season) is less than that for southern populations (averaging about 10 eggs per clutch, but three clutches per season). But owing to variation, there may be no great discrepancy between the actual potentials of northern and southern populations.
Eggs
The eggs of Trionyx are white and spherical having a brittle shell. Some eggs are occasionally abnormal in shape and size; overcrowding of eggs in the oviducts may result in small, irregular-shaped eggs, or large double-yolked eggs. Presumably enlargement of the eggs occurs in the oviducts and within [573] a short period after deposition prior to complete hardening of the brittle shell; therefore some eggs in the oviducts are smaller than those in nests.
The data concerning ferox (Table 8) suggest that the maximum size of eggs is 31 to 32 millimeters, whereas oviducal eggs are slightly smaller, about 25 to 27 millimeters. Eggs of spinifer from northern latitudes (most from approximately 40°, Table 8) also vary in size, oviducal eggs being as small as 22 millimeters in diameter and the maximal size about 29 millimeters. Average extreme measurements (in mm.) of oviducal eggs (number of eggs in parentheses) from females taken in latitudes of 33 degrees or less are: 25 × 29 (11), 29 × 30 (11), 28 × 30 (13), 28 × 30 (10), 29 × 30 (5), 29 × 29 (8), 25 × 26 (17), 29 × 30 (5), and 28 × 29 (8). The average size of these eggs is slightly larger than the oviducal eggs of which measurements are given in Table 8, and suggest larger eggs from more southern latitudes. Eggs of muticus are smaller than those of spinifer (Cahn, 1937:183) or ferox; the average size of eggs from nests found in Iowa and Illinois is 22 to 23 millimeters (Table 8). Nine oviducal eggs from a female obtained in Lake Texoma, Oklahoma, averaged 22 × 23 millimeters. The largest eggs of muticus are from the southernmost locality; eight eggs from a nest found along the Escambia River, Florida, averaged 26 × 27 millimeters.
In general, the data suggest that at each laying slightly smaller eggs but larger numbers are laid by females in northern latitudes, whereas larger but fewer eggs are laid by females from farther south.
Incubation and Hatching
Length of the incubation period seems to depend upon conditions of heat and moisture, and, in general, to be geared to the prevailing climatic conditions. Goff and Goff (1935:156) artificially incubated some eggs of ferox at temperatures varying from 82.3 to 89.2° F., and found that the incubation period was 64 days. Muller (1921:184) wrote that the period of incubation of eggs of muticus (natural nests at temperatures about 90°., op. cit.:182, and artificial nests) in Iowa is from 70 to 75 days. Breckenridge (1944:187) stated that spinifer makes nests in Minnesota from June 14 to July 6, and cited reports that indicate hatching in September. Hedrick and Holmes (1956:126) discovered a nest of eggs in Minnesota on September 5; the eggs were artificially incubated and some hatched on October 29. Eigenmann (1896:263) found eggs as late as September in northern Indiana that "contained young which would have been ready to hatch about a month later." Cahn (1937:193) wrote that spinifer in Illinois lays in June or early July and that "young-of-the-year are taken in late August and September." Some recently deposited eggs of muticus (as indicated by fresh turtle tracks, Pl. 50, Fig. 2) that I obtained on June 1 were artificially incubated and hatched on August 4, indicating an approximate incubation period of 65 days. Dr. Paul K. Anderson in the course of field work on the Pearl River, Louisiana (1958:211), found that eggs collected on June 13 from a nest excavated three to five days before, hatched on August 15, indicating an incubation period of approximately 67 days. Eggs collected on May 17 to 25 (three clutches) hatched on August 4 to 6, indicating an incubation period of approximately 77 days. In any latitude the incubation period probably is at least 60 days. Eigenmann (loc. cit.), however, mentioned empty nests that were found in July; this indicates early hatching or more probably the action of predators.
In northern latitudes eggs or young turtles may over-winter in the nest if deposition is late in the season. In northern Indiana Evermann and Clark (1920:595) found a nest on November 16 that contained "well-formed young" and believed that the turtles would have wintered in the nest. Conant (1951:160) was of the opinion that most eggs probably hatch in early fall, but that some do not hatch until spring.
The hatching of eggs of muticus has been described by Muller (1921:183). According to him, the forelimbs first emerge through the shell and enlarge the opening. There is an "egg tooth below the flexible proboscis" but "it does not seem to be used in escape from the eggs, and is dropped a week after hatching." Hatchlings burrow almost straight upward through the sand leaving the egg shell below the surface and a hole in the sand about an inch in diameter. Muller found that young turtles emerge from the nests in the night or early morning and always go downhill probably influenced in their movements by the open sky and sloping beach. Anderson (1958:212-15) found that hatchlings of muticus leave nests within the first three hours after sunset and travel a direct route to the water. He discovered that hatchlings are active on the surface of the sand at night and generally show a positive reaction to light (moonlight, flashlight), whereas, in daytime, there is a negative reaction to bright sunlight (causing the turtles to bury themselves in sand). Anderson believed that the positive response to light at night is not correlated with the water-approach behavior of hatchlings, but that movements to water are possibly influenced by a negative reaction to dark masses of environment (such as shadows formed by landward forests).
Age and Growth
Goff and Goff (1935:156) found that hatchlings of ferox average 8.82 grams (extremes, 8.50 to 9.25); one of these, UMMZ 76755, is illustrated in Plate 31. Muller (1921:184) recorded measurements of five hatchlings of muticus; the average measurements (in cm., extremes in parentheses) were: length of carapace, 3.54 (3.43 to 3.67); width of carapace, 3.20 (3.10 to 3.25); length of plastron, 2.54 (2.47 to 2.60). I recorded measurements of 32 hatchlings (three clutches combined) of muticus on August 16; the turtles hatched on August 4 to 6 from eggs collected along the Pearl River, Louisiana. The average measurements (in mm., extremes in parentheses) of the 32 turtles were: length of carapace, 41.3 (34.0 to 45.0); width of carapace, 38.6 (31.0 to 40.0); length of plastron, 30.1 (25.0 to 32.0). These turtles have circular umbilical scars averaging approximately two millimeters in diameter. The smallest hatchling that I have seen measures 21.0 millimeters in plastral length (T. m. muticus, INHS 3458). There are no data to indicate a difference in size of hatchlings among the American species of soft-shelled turtles. The average plastral length of most hatchlings probably is 28.0 to 30.0 millimeters.
Owing to the lack of a horny epidermal covering of the carapace and plastron, soft-shelled turtles are not so well suited to studies of age and growth as are the "hard-shelled" species, which have visible impressions of growth annuli on the epidermal scutes. Mattox (1936:255) found annular rings in the long bones of specimens of Chrysemys and suggested that it is tenable to correlate the number of rings with the age of the turtle.
Mitsukuri (1905:265) reported that in hatchlings of Trionyx sinensis the [575] length of the carapace averages 2.7 centimeters (hatchlings of sinensis seem to average smaller than any American species), and that the average length of carapace (cm.) at the end of the first year is 4.5, second year 10.5, third year 12.5, fourth year 16.0, and end of fifth year 17.5; he stated also that females of sinensis are sexually mature in their sixth year. Breckenridge (1955:7-9) computed a growth curve based on 11 recaptures of females of spinifer in Minnesota; his data on rate of growth for the first five years do not differ appreciably from those of Mitsukuri. As most females of spinifer are sexually mature when the carapace is about 11 inches long, the age at sexual maturity is approximately 12 years according to Breckenridge (op. cit.:8, Fig. 4). The discrepancy in age of females at the size of attainment of sexual maturity (Mitsukuri—six years; Breckenridge—12 years) is, in part, rectified by the fact that Trionyx sinensis probably is a smaller species. Also, Breckenridge's computation of the growth curve is based on continuously decreasing increments of growth and seemingly eliminates consideration of the probable marked decrease in rate of growth that occurs when sexual maturity is attained—a phenomenon noted in other species of turtles. I think that increments of growth of immature turtles are, on the average, larger than those of sexually mature turtles. Judging from these criteria, the age of a female of spinifer at sexual maturity is less than 12 years, and turtles having carapaces 17 to 18 inches in length (maximal size for spinifer) would be older than 53 years (op. cit.:9). Occasional individuals, however, may greatly exceed the usual growth rate in which event large adults may be younger than 50 years.
Females of muticus are sexually mature when the plastron is 14.0 to 16.0 centimeters long, which corresponds to a carapace 19.6 to 22.4 centimeters (about 73/4 to 83/4 inches) long (average CL/PL approximately 1.4, see Fig. 13). The smaller adult females probably mature sexually in their sixth year, but most probably do so when seven years old. Accordingly, some T. spinifer emoryi, which are sexually mature at a plastral length of 16.0 centimeters, are also sexually mature in their seventh year, whereas most spinifer (sexually mature at a plastral length of 18.0 to 20.0 cm., corresponding to a length of carapace of 25.2 to 28.0 cm. or about 10 to 11 inches) probably become sexually mature in their ninth year, and some when eight years old. Most males of spinifer are sexually mature when the plastron is 9.0 to 10.0 centimeters long (length of carapace 12.6 to 14.0 cm. or 5 to 51/2 inches), whereas males of muticus and some T. spinifer emoryi are sexually mature at a plastral length of 8.0 to 9.0 centimeters (length of carapace 11.2 to 12.6 cm. or 41/2 to 5 inches). The smaller adult males are probably sexually mature in their fourth growing season. Breckenridge (op. cit.:7, Tab. II) commented on the abundance of females between five and 12 inches in length, and males that ranged in length from five to seven inches. The abundance of turtles in these size ranges is probably due, in part, to a slowing of the rate of growth indicating the approach of sexual maturity; the abundance of the smallest males is especially in accord with the size at sexual maturity of males (about five inches).
The largest acceptable record of size of spinifer is 18 inches in length of carapace (Breckenridge, 1957:232). Stockwell (1878:402), however, wrote that females of spinifer attain "an extreme length of from twenty-four to twenty-eight, and, in rare instances, thirty inches, with an average length of carapace of fifteen to eighteen," and True (1893:152) mentioned lengths of two feet or even more. Turtles 17 to 18 inches long are doubtless rare and probably [576] about 60 years old. A specimen of ferox lived the longest time in captivity—25 years (Pope, 1949:304). Individuals of ferox probably exceed the maximum recorded length of carapace of 181/2 inches (Agassiz, 1857:401). The head of a ferox having a width of 31/2 inches (Wright and Funkhouser, 1915:120) corresponds to a length of carapace of approximately 221/2 inches (PL/HW == 4.9; CL/PL == 1.3). De Sola and Abrams (1933:12) wrote that ferox in the Okefinokee Swamp, Georgia, attains a length of two feet. The largest female of muticus of which I have record is 21.5 centimeters in plastral length (KU 2308), a measurement corresponding to a carapace about 13 inches long.
Mortality
Man, in one sense or another, is a great enemy of soft-shelled turtles. Those caught by fishermen are destroyed because of the erroneous belief that they are harmful to fish populations. Some are drowned in hoop-nets or gill nets used by commercial fishermen. Many softshells are used by man for food. Herald (1949:118-19) reported the results of spraying an area with DDT and mentioned a 10-inch individual of ferox that was eating a dead bluegill, and which "probably died as a result of ingesting contaminated food."
Predation on eggs probably accounts for most mortality. Hamilton (1947:209) reported tracks of spotted skunks, raccoons and foxes seen about destroyed nests, and Cahn (1937:183) incriminated skunks and raccoons. Goldsmith (1945:449) reported a raccoon that unearthed seven nests in one night. Little and Keller (1937:221) wrote of egg shells found in the sand (probably not as a result of hatching), and Muller (1921:182) reported egg shells around dug-up nests, suggesting such predators as "ground moles," raccoons and crows. Chesser (in Harper, 1926:416) said that in the Okefinokee Swamp the jackdaw (fish crow), raccoon, bear and domestic dogs will eat the eggs. Wright and Funkhouser (1915:122) recorded a young ferox in the stomach of a water moccasin (Agkistrodon piscivorus), and suggested that young soft-shells probably are food of larger snakes. Kellogg (1929:26) wrote that stomachs of two alligators each contained one soft-shelled turtle. Newman (1906:136) found that young captives were eaten by individuals of Chrysemys and Sternothaerus, and I found that they were eaten by Kinosternon. Mitsukuri (1905:261-62) stated that first- and second-year individuals of T. sinensis are eaten by the adults.
Breckenridge (1960) wrote that a clutch of eggs probably failed to develop because of an "… unusually cool season." Evermann and Clark (1920:595) stated that "many young appear to perish during the first winter." They (op. cit.:594) found two eggs submerged in two feet of water and it is supposed that they never hatched. Dundee (1950:139) reported remains of soft-shelled turtles left on the mud of a dried swamp.
Parasites
Muller (1921:182) found maggots in a few eggs of a clutch, but thought that only the infertile and decomposing eggs were infested. I removed a hard, spherical cyst from the hind leg of a preserved softshell (TU). A captive hatchling (TU 17304) died as the result of a continuously enlarging and deepening hole on the top of its head; I could not discern a visible parasite with the naked eye. I found 25 leeches (Placobdella parasitica, largest about 13 mm.; identified by Dr. Kenneth B. Armitage, Department of Zoology, University [577] of Kansas) in association with 11 T. m. muticus (number per turtle not known) that were collected from the Kansas River at Lawrence, Douglas County, Kansas. Evermann and Clark (1920:596) reported a few nematodes in the stomachs of some spinifer, and three nematodes are listed by Harwood (1932:46, 60, 62, 66) in the same species. Hughes, Higginbotham and Clary (1941) have listed the known reptilian hosts of parasitic trematodes, and Hughes, Baker and Dawson (1941) have done the same for tapeworms. The species of parasites and their trionychid hosts are listed below.
| Trematoda | ||
| Trionyx ferox: | Neopolystoma orbiculare | Vasotrema amydae |
| Neopolystoma rugosa | Vasotrema attenuatum | |
| Polystomoides coronatus | Vasotrema robustum | |
| Teloporia aspidonectes | ||
Trionyx muticus: |
Crepidostomum cooperi |
Opisthorchis ovalis |
Trionyx spinifer: |
Hapalorhynchus evaginatus |
Vasotrema amydae |
| Opisthorchis ovalis | Vasotrema attenuatum | |
| Polystomoides coronatus | Vasotrema longitestis | |
| Teloporia aspidonectes | Vasotrema robustum | |
Cestoda |
||
| Trionyx ferox: | Proteocephalus trionychinus | |
| Trionyx spinifer: | Proteocephalus testudo | |
Nematoda |
||
| Trionyx spinifer: | Camallanus trispinosus | Spiroxys amydae |
| Falcaustra chelydrae | ||
Economic Importance
Several authors have mentioned softshells as a food item much sought after by man. The commercial value of these turtles has been summarized by Clark and Southall (1920:15-16). Softshells are consumed in quantity only in small towns near the place of capture. They are found only occasionally in the markets of large cities because the turtles are little known and the palatability of their flesh is unappreciated. Also, they do not stand shipment so well as other turtles, and they are "not so meaty as the snapper; so there is more waste" (Clark and Southall, loc. cit.). Little and Keller (1937:221) reported living individuals for sale at the market in Ciudad Juarez, Chihuahua; however my inquiry at markets in Juarez in the summer of 1959 disclosed no evidence of recent sale of soft-shelled turtles. In the southeastern United States the demand is perhaps greater than in other regions. I have noted softshells in the market at New Orleans, and Oliver (1955:19) has [578] mentioned the sale of "some 146,600 pounds" in one recent year in Florida. Over most of their range, however, there probably is no general demand for softshells and no special efforts are made to capture them. Softshells have been raised successfully on "turtle farms" in Japan (Mitsukuri, 1905). True (1893:152) wrote that "The eggs also are considered very excellent."
Softshells generally are condemned by fishermen because of the mistaken belief that they are detrimental to fish populations. Food of softshells is principally crawfish and insects. Fish comprise a small proportion of the diet (frequency 1.9% in Michigan, Lagler, 1943: Tab. 9). Most of the fishes eaten seem to be small minnows. Probably fish would comprise a larger percentage of the diet if they could be caught. I doubt that a softshell can pursue and capture a healthy fish in natural waters. Recently dead fish are eaten and perhaps fish eggs, and senile and decrepit fishes. There is no evidence that soft-shelled turtles are active predators on any kind of fish. Of course in congested areas such as ponds of fish hatcheries, it is desirable to eliminate the turtles. The known food habits of soft-shelled turtles suggest that they compete with game fishes for food, but there is no information on the intensity of competition (Lagler, op. cit.:305).
The combined statements of many authors in their general accounts of food habits (for instance, Babcock, 1919:425) have tended to create the erroneous belief that soft-shelled turtles harm waterfowl. To my knowledge the only basis for this belief is the statement of Wright and Funkhouser (1915:123) that according to the natives of the Okefinokee Swamp, the larger turtles "devour also such waterfowl as are unfortunate enough to be taken unaware by these reptiles." Perhaps an occasional waterfowl is eaten, but the present information on kinds of food eaten certainly does not warrant the destruction of soft-shelled turtles. There may be some mortality in congested areas such as game refuges where young birds crowd the surface of the water.
The kind of bait successfully used in trapping softshell turtles suggests that they are of some value as scavengers.
Before attempting to reconstruct the history of soft-shelled turtles in North America, it will be helpful to summarize the salient facts concerning the distribution and relationships of the living forms, and to comment on fossils.
Distribution
The geographic range of the family Trionychidae in North America is principally in the eastern two-thirds of the continent and contributes to the well-known floral and faunal resemblance of eastern North America to that of eastern Asia (Schmidt, 1946:149) because Trionyx ferox (see Fig. 18) resembles the species of the genus in Asia more closely than it does any North American species. The Recent distribution in America does not include the Neotropical region, whereas the geographical range in the Old World extends south of the equator (Fig. 1; Dunn, 1931:109, fig. 2; Gadow, 1909:333, fig. 72; Hay, 1908:35, fig. 16).
American softshells occur in all river systems in the United States and the two adjacent river systems on the east coast of México that drain into the Gulf of México. Softshells inhabit streams of the Great Plains and occur westward to the foothills of the Rocky Mountains in the western tributaries of the [579] Mississippi River. Only T. s. spinifer occurs in the southern part of the Great Lakes-St. Lawrence drainage. Softshells are absent from the Atlantic Coast drainage except the Hudson River and those rivers at least south of (and including) the Pee Dee River in South Carolina.
T. s. emoryi is not known to be indigenous west of the Río Grande drainage, and has probably been introduced across the Continental Divide via the Gila River in western New Mexico into the Colorado River drainage of Arizona (Miller, 1946:46); the species undoubtedly occurs in México on the Sonoran side of the Colorado River opposite Baja California (Bogert and Oliver, 1945:417).
In the summer of 1959, I trapped turtles and with a specimen in hand inquired about softshells occurring in the inland drainages of northern México. From two collecting stations on the Río Nazas in Durango, only specimens of Pseudemys and Kinosternon were obtained; local inhabitants had neither seen nor heard of softshells. Flooded conditions in August of 1959 permitted trapping in only one of the inland drainages of northwestern Chihuahua, the Río Santa María; only specimens of Kinosternon were obtained. Local residents near that river as well as those living near the Río Casa Grandes and Río del Carmen had not seen or heard of softshells. A person that I judge to be a competent observer reported seeing a softshell in June of 1958 in the Río Alamos (Arroyo Cuchujáqui) near Alamos, Sonora, in the Río del Fuerte drainage on the west coast of México. I was a member of a field party from the University of Kansas that visited that locality in late January of 1959; only specimens of Pseudemys and Kinosternon were collected. Possibly isolated populations occur in streams of the Pacific Coast drainage of northern México. If so, they may have entered Pacific Coast drainages by stream capture across the Continental Divide. Several species of fish that are characteristic of the Río Grande traversed the Sierra Madre Occidental at some former time (presumably via the Río Conchos and Río Papigochic) and occur in the Yáqui River drainage (Meek, 1904:xxxviii, xlvii; Miller, 1959:214-15, 217). Because of the probability that the Río Nazas at some former time flowed north into the Río Grande (Meek, op. cit.:xxxiv), it is notable that softshells are absent in the Río Nazas drainage; the Big Bend turtle, Pseudemys scripta gaigeae, occurs in both drainages.
Relationships
Characters of Trionyx ferox suggesting a closer resemblance to some Old World members of the family than to the other three American species are: large size; marked difference between juvenal and adult patterns on the carapace; the marginal ridge; and the longitudinal ridgelike prominences on the carapace, especially in juveniles. Other characters of ferox suggesting a corresponding, but less marked resemblance to Old World species of Trionyx are: the large size of the eighth pair of pleurals; the absence of callosities on the epiplastron and preplastra; frequent fusion of the hyoplastra and hypoplastra (more than in spinifer or muticus); and tolerance of marine waters (more than muticus or spinifer). Some fossils also suggest alliance with ferox and some Old World members of [580] the genus in their large size, large eighth pair of pleurals, and occurrence in marine deposits; several Old World species have been reported at sea (Pelochelys, T. triunguis, T. sinensis). T. ferox is monotypic and has the most southeasterly displaced, geographic range.
Because ferox resembles softshells from the Old World more closely than it does any American species, ferox is assumed to be more closely related to Old World softshells than to any American species, and, because of resemblance to some fossils, ferox is assumed to resemble most closely the primitive, ancestral stock of softshells that occupied North America. T. spinifer, T. muticus and T. ater, which resemble each other more closely than any of them resembles T. ferox or any Old World species, are considered autochthonous in North America. T. spinifer and T. muticus are distinct, sympatric species. Burt (1935:321) suggested that the two species "may be variants of the same species." T. ater is weakly differentiated from T. spinifer emoryi. The species, ferox, spinifer and muticus are well-differentiated and were considered by Agassiz (1857), Gray (1869) and Baur (1893) as belonging to three different genera.
In the widely distributed T. spinifer, the subspecies spinifer, hartwegi and asper closely resemble one another; asper seems most distinct, whereas spinifer and hartwegi are terminal populations of an east-west cline in one character. The subspecies pallidus, guadalupensis and emoryi resemble one another more closely than any resembles any of the subspecies mentioned immediately above; T. s. pallidus, however, is annectent. T. s. pallidus and guadalupensis represent terminal populations of clines in several characters, some of which occur in emoryi, but that subspecies is more distinct from pallidus and guadalupensis than those subspecies are from each other. T. s. emoryi is the most variable subspecies. T. ater, known only from a restricted area in central Coahuila, is most closely related to T. s. emoryi, and possesses some characters judged to represent the attenuation of the geographic cline in pallidus, guadalupensis and emoryi mentioned above. Some characters of ater show alliance to the species muticus. Of T. muticus, whose geographic range is removed from that of ater, there are two subspecies. Four subspecies of spinifer (spinifer, hartwegi, pallidus and asper) intergrade in the Mississippi River drainage of Louisiana; few specimens, however, are typical of asper. The subspecies of muticus do not show definite evidence of intergradation. To facilitate quick reference, the occurrence of some characters that are shared by, or are approximated in, two or more forms are listed in Table 10. In addition to external characters, some ratios emphasize the clinal relationship between T. s. pallidus, guadalupensis, and emoryi mentioned above. Of especial interest is the frequent resemblance of those subspecies and T. ater to T. ferox (dorsal pattern on limbs of adults, reduction in anterior tuberculation, wide head, narrow carapace, and short snout), and the less marked resemblance of T. muticus to T. ferox; not shown in Table 10 is the resemblance of ferox to T. muticus calvatus in having thick, black-bordered postocular stripes. Some populations of T. s. emoryi resemble T. muticus in the corresponding size at sexual maturity and in having well-developed plastral callosities. It is notable that the occurrence of ater, and to a lesser extent that of T. s. emoryi, which resembles ferox (and muticus), is in the southwestern United States and northern México.
Table 10. Frequency of Selected Characters Among Species and Subspecies of Trionyx in North America. Characters of muticus Refer to the Typical Subspecies; Horizontal Dashes Connecting X's Indicate that Computations for Those Subspecies Were Combined; Vertical Dashes Indicate that the Subspecies Is Intermediate Between the Adjacent Subspecies.
| Characters | Species and subspecies | ||||||||
| ferox | spinifer | hartwegi | asper | pallidus | guadalupensis | emoryi | ater | muticus | |
| Juvenal pattern: | |||||||||
| black spots, ocelli | X | X | X | ||||||
| white dots | X | X | X | X | |||||
| Pattern on snout: | |||||||||
| acute angle (reduced in muticus) | X | X | X | X | X | X | |||
| | | |||||||||
| triangular | X | X | X | X | |||||
| Pattern on side of head: | |||||||||
| contrasting marks | X | X | X | X | X | X | |||
| | | |||||||||
| non-contrasting marks (distinct stripe in muticus) |
X | X | X | X | |||||
| Pattern on limbs of adults: | |||||||||
| contrasting | X | X | X | X | X | ||||
| | | |||||||||
| reduced or absent | X | X | X | X | X | ||||
| Tuberculation (anterior edge of carapace): |
|||||||||
| conical, equilateral | X | X | X | X | |||||
| reduced or absent | X | X | X | X | X | ||||
| Head (PL/HW, fig. 3): | |||||||||
| wide | X | X | X | X | X | ||||
| narrow | X | X | X | X | |||||
| Carapace (CL/CW, fig. 4): | |||||||||
| wide | X | X | X | X | X | X | |||
| | | |||||||||
| narrow | X | X | X | X | |||||
| Level of Carapace Width (CL/PCW, fig. 5): |
|||||||||
| middle of carapace | X | X | X | X | X | ||||
| farther posteriorly | X | X | X | X | |||||
| Snout (HW/SL, fig. 6): | |||||||||
| long | X —— | —— X | X | X —— | —— X | X | |||
| | | | | ||||||||
| short | X | X —— | —— X | X | X | ||||
Fossils
The known occurrence of fossil trionychids throughout the world indicates a former distribution more widespread than the family has today; the principal difference in the former and present distributions is the lack of living softshells in Europe.
I have not studied in detail the many fossil remains but such examination as I have made of them suggests that many of the characters used as a basis for distinguishing fossil forms in North America are subject to individual variation or are of no diagnostic value in the living species (Hummel, 1929:769). Knowledge of the variation in the living species of the Old World would aid in adequately appraising the North American fossils. Some osteological characters of the three living American species (excluding ater) together with data on variation within a given species are mentioned below. Some differences in skulls of the three species already were mentioned in the section "Osteological Characters." Because most fossil remains are those of the carapace and plastron, attention is here given to those structures.
Widened alveolar surfaces of jaws.—An ontogenetic variation affecting large skulls of T. ferox and some individuals of T. spinifer asper; presumably confined to females. Of especial interest is its presence in some populations of asper that are not otherwise distinguishable (external characters) from the rest of the individuals comprising that subspecies.
Sculpturing.—No differences in pattern (generally of anastomosing ridges) on carapace or plastron; fineness or coarseness seemingly correlated with size; frequency and kind (knoblike or ridgelike) of bony prominences on carapace variable; bony prominences confined to species spinifer and ferox, occurring principally on large females.
Fontanelles of carapace.—Closure more or less correlated with increasing size, although much variation noted between individuals of same size; small individuals have fontanelles confluent (medially), thus separating nuchal from contact with first neural and first pair of pleurals.
Number and arrangement of neurals and pleurals.—Neurals number six to nine, usually seven or eight; pleurals number seven or eight pairs, and may or may not be in contact with each other posteriorly; eighth pair of pleurals when present reduced, never contacting seventh neural; arrangement posteriorly variable (see Fig. 16 and Tab. 5).
Plastral callosities.—Increase in size with advancing age causing corresponding reduction in size of plastral vacuity; relatively best developed in muticus (all elements touching medially on KU 41380 leaving no plastral vacuity); probably no callosities on preplastra or epiplastron of ferox; callosity on epiplastron of spinifer not covering entire surface (as it may in muticus).
Epiplastron.—Obtusely-angled (greater than 90 degrees) in muticus; acutely-angled (90 degrees or less) in ferox and spinifer.
Hyo-hypoplastral suture.—Usually present, but occasionally absent, in all species.
The fossil turtles of North America have been treated monographically by Hay (1908), who apportioned fossil trionychid remains into eight genera (three living) of two families. Recently, Romer (1956:514) relegated all trionychid fossils to the genus Trionyx. Characters, as gleaned from Hay's synopsis (op. cit.:465-548, Pls. 85-113), that seem especially worthy of taxonomic consideration are: (1) The presence of a preneural, which is not known to occur in the living American species (seemingly the preneural is fused with the first neural and represents the elongate first neural in living species); (2) The large eighth pair of pleurals, especially when they contact the seventh neural; (3) The thickness of the costal plates, a condition probably correlated with the size of some fossils, which are larger than any living species (for example, Hay, op. cit.:518, mentioned the greatest dimension of a nuchal bone as approximately 300 mm.).
The approximate extent of the known horizontal distribution of fossils is indicated in Figure 24. A comparison of known localities of fossils and the distribution of living softshells (introduced population of T. s. emoryi in Colorado River drainage omitted) shows that the distribution was more widespread in former times. Localities of fossils are centered on the Atlantic Coast from New Jersey to North Carolina and in the Rocky Mountain-Great Plains region from Alberta and Saskatchewan to northwestern New Mexico; the oldest fossils, which occur in each region, are found in Upper Cretaceous deposits. Many fossils occur in marine and [584] brackish water deposits. Most localities depicted on the map are mentioned by Hay (1908:36-37, 465-548). Other localities included on the map are in southern Alberta (Russell, 1929:164; 1930:27; Sternberg, 1926:104), southern Saskatchewan (Russell, 1934:109), northern South Dakota (Hay, 1910:324), central Utah (Gilmore, 1946), western Colorado (Schmidt, 1945), southwestern Kansas (Galbreath, 1948:284), southeastern Texas (Hay in Stejneger, 1944:65), southern California (Brattstrom, 1958:5), and northeastern Coahuila, México (Mullerried, 1943:623). Hay's record of the living Platypeltis (= Trionyx) ferox and other remains from the Peace Creek formation in Hillsborough County, Florida (op. cit.:548), presumably is the same record mentioned by Pope (1949:305).
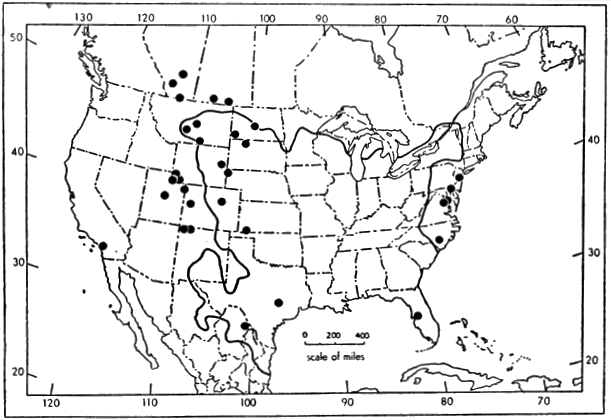
Fig. 24. Geographic distribution of Recent soft-shelled turtles (bordered by heavy black line) and fossil trionychids (black circles) in North America. The introduced population of T. s. emoryi in the southwestern United States is not shown.
Ameghino (in Hay, op. cit.:35) recorded specimens of a trionychid from the Cretaceous of Patagonia, a record that, at present, cannot be accepted (Simpson, 1943:423). Mullerried (loc. cit.) also mentioned some trionychid remains that were housed in Tuxtla Gutierrez, Chiapas, México, (material now lost), but their geographical provenance was unknown. The former extent of range [585] southward is not known; it is improbable that trionychids occurred in South America (Simpson, 1943:423).
Phylogeny
The occurrence of T. ferox in Florida and the suggestion of ferox-like characters in turtles from southwestern Texas and northern Mexico presents a distributional pattern that resembles the disjunct ranges of many other pairs of closely related taxa. The clear-water ponds in central Coahuila, which are inhabited by ater, correspond to aquatic habitats supporting ferox in Florida. The splitting of the geographic range into eastern and western parts possibly resulted from a southward shift of colder climates in glacial stages of the Pleistocene, or from the development of an intervening arid region in the late Miocene and Pliocene (see discussions in Martin and Harrell, 1957, and Blair, 1959). An initial separation of range by an arid environment in the Pliocene may have been terminated by the colder climates in the Pleistocene.
The degree of morphological difference between ferox and the forms in southwestern Texas and northern México, suggests that the time of separation antedated the Pleistocene.
Trionychid turtles may have traversed the Bering land bridge between Asia and North America in late Mesozoic times for they occur as fossils on the Atlantic Coast and in the Rocky Mountain-Great Plains region in Upper Cretaceous deposits. Shallow, inland seas may have afforded no barrier to the dispersal of softshells which presumably were tolerant of saline waters. The orogeny and volcanic action with subsequent erosion and sedimentation of the Rocky Mountain system, which was later accompanied by drier climates, tended to obliterate suitable habitats in the western United States; softshells persisted at least until the Upper Eocene on the west coast (Brattstrom, 1958:5). The factors responsible for the disappearance of softshells on the Atlantic Coast probably were related to the glacial advances in the Pleistocene; the most recent fossils known occur in Miocene deposits.
The relationships of the living species and subspecies were probably correlated with geologic change in aquatic environments and drainage patterns. These changes probably included stream capture, flooding, drought, uplifting and planation. A hypothetical, evolutionary history is presented in the phylogenetic diagram where letter symbols represent species and subspecies, and grouped symbols (referred to in subsequent paragraphs) represent ancestral stocks.
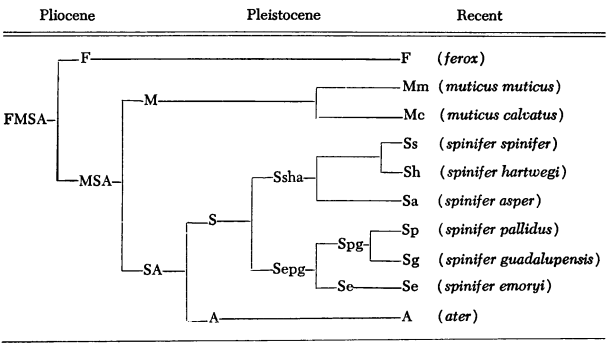
Click here to see text version of above chart.
An arid environment in the central and southern United States and northern Mexico may have increased in area especially southward from Miocene times into the Pliocene (Dorf, 1959:189, 191). The combination of physiographic changes and aridity, which modified the mesic, essentially continuous, aquatic habitats, may have isolated and aided in the differentiation of the ferox, muticus and spinifer stocks. Encroachment of the Eocene seas, the maximal extent of which corresponded to the Gulf Coastal Plain and included a northerly extension as far as Cairo in southern Illinois (Mississippi embayment), possibly was an initial barrier isolating the ferox stock of the east.
In the late Miocene or early Pliocene, the MSA (muticus-spinifer-ater) stock presumably occupied a large region of the central United States, which extended southward into northern México and along the Gulf Coast at least as far as Alabama. Farther eastward, the ferox stock was isolated in more mesic, probably swampy, marshy habitats.
Later, in the southwestern part of the range of the MSA stock (southern Texas and northern México), the SA and muticus stocks were separated. The muticus stock occurred to the northeastward, and presumably no farther south than the area included within the present drainage basin of the Colorado River. Southward, the SA stock was isolated into several populations that are today represented by ater and T. s. emoryi, the most variable subspecies; the distribution of the most distinctive population of emoryi indicates [587] a former isolated inland drainage. The multiple fragmentation of the SA stock presumably terminated by the end of the Pliocene. The progenitors of T. ater probably closely resembled ferox. Trionyx ater and T. ferox resemble each other morphologically and in habitat. Therefore, the progenitors of ater are considered to have undergone comparatively little differentiation.
The spinifer stock, occurring principally in the area included within the present drainage basin of the Río Grande, extended its geographic range eastward and became sympatric with muticus and ferox. An expansion of range necessarily demands more mesic conditions; these were perhaps afforded by the pluvials (wet, rainy ages) that were coincident with the glacial periods in the Pleistocene (Antevs, 1948:168). The pluvials permitted the isolated populations of the spinifer stock to unite, and permitted that stock to extend its range eastward. The concurrent continental glaciation permitted the spinifer stock to extend its range eastward only in a belt approximately 300 miles wide along the Gulf Coast, and also displaced the ranges of ferox and muticus to southern latitudes. Perhaps ferox was less tolerant of decreased temperatures or changes in habitat than was the spinifer stock but, for some unknown reason, ferox did not extend its range westward. Because T. ater closely resembles T. s. emoryi, continued isolation of ater since the beginning of the Pleistocene seems unlikely and ater may have been reunited in subsequent pluvial periods with the spinifer (emoryi) stock. A climatic fluctuation between relatively wet and dry periods is corroborated by studies of soil profiles in Trans-Pecos Texas (Bryan and Albritton, 1943).
The separation of the range of spinifer in the general region of western Louisiana, resulting in the differentiation of the spinifer group of subspecies to the east and the emoryi group of subspecies to the west, and the differentiation of T. s. asper and T. m. calvatus, both having corresponding western limits of distribution (Mississippi River drainage), are associated with the activities of the Mississippi River and its flood-plain. The combined effects of the pluvials and interpluvials seem responsible for changes in the lower Mississippi Valley. Great volumes of summer melt-water in the glacial stages greatly increased the breadth of the channel of the lower Mississippi River (corresponding to the northern extent of the Mississippi embayment; Hobbs, 1950), and this, coupled with the encroachment of Pleistocene seas (especially in the Mississippi embayment) in the interglacial periods, perhaps separated populations eastward represented today by T. m. calvatus and T. s. asper. The [588] spinifer-hartwegi stock probably developed in southern Louisiana in association with the meandering of the Mississippi River and its tributaries, and its broad alluvial plain. The biota of that plain differed from that adjacent to the east or west (see discussion in Viosca, 1944) and constituted a barrier, of a sort, to free communication between the east and west. Westward the emoryi group of subspecies differentiated, its eastern limit probably being the Red River, which followed its own course to the Gulf along the lowlands on the west side of the Mississippi Valley and did not empty directly into the Mississippi until Recent times (Holland, 1944:20). There was not an equally-marked, corresponding separation of the range of muticus. However, the juvenal pattern of the subspecies muticus that inhabits the Gulf Coast streams is slightly different (having less short lines) from that of muticus elsewhere.
The Río Grande (inhabited by emoryi) presumably had its own exit to the Gulf whereas rivers westward to (and including) the Red River (inhabited by pallidus-guadalupensis cline) probably were joined near their mouths forming a large drainage system. Hubbs (1957:93) pointed out that the Río Grande-Nueces divide also limits a large number of species of fish. The differentiation of pallidus and guadalupensis is possibly due to a difference in the salt content of waters that drain the Edward's Plateau (see page 547), or to isolation of those subspecies in separate drainage systems that had their own exits to the Gulf.
In the lower Mississippi drainage, the spinifer-hartwegi stock extended its range northward following the retreat of the last glacial stage, and differentiated into those two subspecies in the upper Mississippi drainage and Great Lakes-St. Lawrence drainage system.
I have seen one specimen (UMMZ 59198) from the eastern part of the Tennessee drainage (inhabited by T. s. spinifer) that resembles T. s. asper (occupying the Gulf Coast drainages of the southeast). This resemblance tends to support the thesis of a former confluence of the Coosa (Alabama River system) and Tennessee drainages as believed by some malacologists to explain resemblances in molluscan fauna and as corroborated by physiographical evidence (see discussion in van der Schalie, 1945).
The Importance of the Study of Turtle Populations in Relation to the History of River Systems
In the Río Grande drainage the geographic distribution of the population of emoryi having orange color in males is approximately the same as that of Pseudemys scripta gaigeae; the corresponding [589] distributions suggest that a part of the Río Grande drainage consisting of the Río Conchos in Chihuahua and the Big Bend region of Texas was isolated in former times. Accordingly, the known aquatic chelonian fauna in the basin of Cuatro Ciénegas in central Coahuila, México, is endemic (except T. s. emoryi). And the coincidence of the geographic ranges of T. muticus calvatus and Graptemys pulchra in the southeast suggest a former association of the included (Pearl to Escambia) river systems. The occurrence of T. s. pallidus in the Red River drainage indicates that the Red River was formerly associated with the Gulf Coast streams of eastern Texas and western Louisiana (inhabited by pallidus) and not with the Mississippi River drainage. The lower Mississippi River valley forms a prominent barrier to the eastern and western dispersal of many kinds of species and subspecies of turtles. T. m. calvatus and T. s. asper, which occur in rivers of the Gulf Coast drainage east of the Mississippi, are well-differentiated subspecies showing little or no evidence of intergradation with their relatives in the Mississippi River. The large faunal break provided by the Mississippi River would seem to indicate greater age for that river than for other rivers of the Gulf Coast drainage.
A comparison of the distributions of Trionyx and Graptemys in Texas suggests a faunal break between the drainage systems of the Brazos and Colorado rivers. Graptemys versa occurs in the Colorado and Guadalupe-San Antonio drainages. To my knowledge versa hitherto has not been recorded from the latter drainage system. I have seen one specimen of Graptemys (custody of Gerald Raun, University of Texas) from the Guadalupe River drainage, which I judge to be representative of versa, and Olson (1959:48) has reported Graptemys (probably versa) in the San Antonio River. The distribution of G. versa parallels in a general way, the distribution of T. s. guadalupensis. G. kohni and T. s. pallidus occur in the Brazos River and eastward. Also, it is notable that the population of T. m. muticus occurring in the Colorado River drainage differs slightly (more black pigmentation) from the same subspecies in the adjacent Brazos River system.
There is much difference in the patterns of distribution and degree of differentiation of different genera of aquatic turtles in the eastern United States. Tinkle (1958:41-43, Figs. 49-55) concluded that a general resemblance in the patterns of distribution of the different genera of turtles was evidence that the rates of evolution were essentially the same, assuming that each genus had had a [590] similar time interval for differentiation (op. cit.:42). If this is true, corresponding patterns of distribution might indicate the same relative age of the population of turtles concerned. Generally, the genera of turtles that on morphological grounds are considered the oldest and most primitive (Macroclemys, Chelydra) show less differentiation into species and subspecies than those considered younger and more recently evolved (Graptemys, Pseudemys). In the genus Graptemys, much differentiation occurs in the geologically, recently formed, Gulf Coast drainage systems of the southeastern United States. It would seem then, that faster rates of differentiation denote more recent genera, whereas older genera are endowed with a "genetic senility" and are less subject to change.
Evidence of the relative age of two genera of turtles, as suggested by their degree of differentiation into minor taxa, and the degree of difference between populations of two genera that inhabit adjacent drainage systems, may indicate the relative ages of particular river systems. For example, the slight resemblance of G. versa to kohni and the close resemblance of T. s. guadalupensis to pallidus in Texas may reflect the age of the genus Trionyx and the youth of the genus Graptemys. Remembering that the genus Graptemys is relatively recently evolved and assuming G. versa to be the most primitive and ancestral species of the genus (at least it is monotypic, the most aberrant species, and unlike any other species of the genus), it seems logical to suppose that the physiographic changes responsible for the Colorado-Brazos divide and the isolation of versa occurred early in the evolutionary history of the genus Graptemys. The degree of differentiation of Trionyx suggests that that genus is, comparatively, much older, and that the same physiographic changes responsible for the Colorado-Brazos divide and differentiation of the subspecies pallidus and guadalupensis occurred late in the evolutionary history of the genus Trionyx.
In general, patterns of distribution of turtle populations support physiographic evidence concerning changes in stream confluence and relative age of river systems.
In North America, soft-shelled turtles (genus Trionyx) occur in northern México, the eastern two-thirds of the United States, and extreme southeastern Canada. The genus fits the well-known Sino-American distributional pattern. In North America there are four species. Three (ferox, spinifer and muticus) are well-differentiated [591] and one (ater) is not well-differentiated from spinifer. Characters of taxonomic worth are provided by the following: size; proportions of snout, head and shell; pattern on carapace, snout, side of head, and limbs; tuberculation; sizes of parts of skull; number of parts of carapaces; and, shape and number of some parts of plastra. Many features show geographical gradients or clines. T. ferox is the largest species and muticus is the smallest. Females of all species are larger than males. With increasing size of individual, the juvenal pattern is replaced by a mottled and blotched pattern in females of all species; adult males of spinifer retain a conspicuous juvenal pattern, whereas the juvenal pattern is sometimes obscured or lost on those of ferox and muticus. The elongation of the preanal region in all males, and the acquisition of a "sandpapery" carapace in males of spinifer occur at sexual maturity. There is a marked secondary sexual difference in coloration in a population of T. s. emoryi (side of head bright orange in males and yellow in females). The sex of many hatchlings of T. s. asper can be distinguished by the pattern on the carapace. Slight ontogenetic variation occurs in some proportional measurements. Large skulls of ferox and some asper (those in Atlantic Coast drainages) have expanded crushing surfaces on the jaws. Considering osteological characters, muticus is most distinct; there is less difference between ferox and spinifer than between those species and muticus.
T. ferox is monotypic, confined to the southeastern United States, and resembles Old World softshells more than it does any American species. The northern part of the geographic range of ferox overlaps that of T. s. asper; there, the two species are ecologically isolated. T. spinifer is polytypic, has the largest geographic range, and is composed of six subspecies, of which two are described as new (pallidus and guadalupensis). The subspecies are divisible into two groups. One, the spinifer group (spinifer, hartwegi and asper) is recognized by a juvenal pattern having black spots or ocelli; asper is the most distinctive and shows little evidence of intergradation in the lower Mississippi River drainage with the spinifer-hartwegi complex, which, northward, is differentiated into two subspecies in which there is an east-west cline in size of the ocelli on the carapace. The emoryi group (pallidus, guadalupensis, emoryi) is recognized by a pattern of white spots; emoryi is most distinctive. Each of several characters behaves as a cline if traced from east to west through the three subspecies. T. s. pallidus intergrades with [592] the spinifer-hartwegi complex in the lower Mississippi River drainage. T. s. emoryi is the most variable subspecies; in its most notable population the males have orange coloration. T. s. emoryi has been introduced into the Colorado River drainage of Arizona. T. ater most closely resembles T. s. emoryi, but shows alliance with T. muticus and T. ferox. T. ater is confined to ponds of crystal-clear water in central Coahuila, México. T. muticus is completely sympatric with spinifer, and is composed of two subspecies (muticus and calvatus). T. m. calvatus shows no evidence of intergradation in the lower Mississippi River drainage with T. m. muticus, corresponding somewhat to the relationship of T. s. asper with the intergradient population of T. spinifer in the Mississippi River.
Softshells have pharyngeal respiration and probably are incapacitated by rotenone. T. ferox and the subspecies of spinifer occur in a wide variety of fresh-water habitats; muticus is more nearly restricted to running water (especially in the northern parts of its range) than spinifer, and may be less vagile than spinifer. T. ferox is more tolerant of marine and brackish waters than are muticus or spinifer. Small size and pallid coloration seem correlated with arid environments. The largest species (ferox) and the smallest population of spinifer (resembling muticus) both occur in the southernmost part of the range of the genus. Diurnal habits include basking on shores or débris in water, floating at the surface, procuring food, and burrowing in shallow and deep water (no observations for spinifer and muticus in deep water). Softshells are principally carnivorous; the food consists mostly of crawfish and insects; there is evidence of cannibalism involving predation on first- and second-year-old turtles. The capture of food is triggered primarily by movement of prey; sight seems to be more important than smell to Trionyx in capturing food. There is no indication of a food preference between species; enlarged crushing surfaces of jaws in some ferox and asper may be an adaptation for feeding on mollusks. Schools of fish are reported to follow softshells, and presumably acquire food that is dislodged by the grubbing and scurrying of the turtles on the bottom. Softshells are wary. They are good swimmers, and travel rapidly on land. The depressed body is an adaptation for burrowing and concealment. Permanent growths of algae do not occur on the dorsal surface of softshells. There is evidence of some nocturnal activity, and a general parallel in habits between trionychids and chelydrids. Softshells sometimes move overland; they move little in aquatic habitats. The normal annual period of [593] activity of spinifer in latitudes 40° to 43° is approximately five months from April into September, depending on the weather; they hibernate under a shallow covering of mud in deep water. The southernmost populations may be active throughout the year.
Males of spinifer are sexually mature when the plastron is 9.0 to 10.0 centimeters in length (some when 8.0 long), whereas those of muticus are sexually mature at 8.0 to 9.0 centimeters. In the mentioned size range, the smaller adult males are probably in their fourth growing season, and the larger males in their fifth. Most females of spinifer are sexually mature at a plastral length of 18.0 to 20.0 centimeters and are probably in their ninth year; the smaller individuals probably are in their eighth. Females of muticus are sexually mature when the plastron is 14.0 to 16.0 centimeters long. Most of these are seven years old but some are only six years old. Some large females contain immature ovaries. The near-maximum length of carapace of spinifer is 18 inches, and such turtles are perhaps 60 years old; ferox perhaps attains a length of two feet.
T. ferox deposits eggs from late March to mid-July, whereas northern populations of spinifer and muticus usually deposit theirs from mid-June to mid-July. Sandy sites are preferred for nests, although movement to other sites occurs if the preferred sandy sites are submerged or otherwise rendered unusable. T. muticus limits its nest sites to the open areas of sand bars and does not lay inland where it must traverse vegetated areas, as does spinifer. Nests of ferox and spinifer seem to differ from those of muticus in being flask-shaped.
The seasonal reproductive potential is perhaps less in northern populations (averaging 20 eggs per clutch and only one clutch per season) than in southern populations (averaging about 10 eggs per clutch, but three clutches per season). Larger females deposit more eggs than smaller females. Eggs laid in northern latitudes are slightly smaller than those laid farther south. In any latitude the incubation period probably is at least 60 days. Hatchlings presumably leave nests at dusk, nighttime or dawn, and may winter over in eggs or nests.
Man is a great enemy of softshells. Predation on eggs probably accounts for most mortality. Physical conditions of the environment (overcrowding of nest sites, inadequate hibernation sites) and probably some kinds of parasitism contribute to mortality. Softshells are eaten locally and sometimes appear in the market of large cities, but over most of their range, there probably is no general [594] demand and no special efforts are made to capture them. Fish, mostly minnows, comprise a small proportion of the diet. There is no evidence that softshells are active predators on any kind of fish, but their known food habits suggest that they compete with game fishes for food. Softshells are scavengers.
Fossil material was not studied in detail. The fossil softshells indicate a more widespread, former distribution. Some osteological characters and their variation in the living species are mentioned as an aid to future workers concerned with an assay of fossil remains. Fossils occur in marine, brackish and fresh-water deposits, and many are much larger than the living species; the oldest American fossils are of Upper Cretaceous age.
The interrelationships of the living species and subspecies suggest that the species spinifer, ater, and muticus are derivatives of a ferox-like ancestor, and that they differentiated in North America; most differentiation occurs in southwestern Texas and northern México where characters of some populations indicate alliance with ferox. It is hypothesized that aridity in the late Tertiary effected specific differentiation by the modification and isolation of aquatic habitats. Pluvial periods in the Pleistocene provided for confluence of aquatic habitats and expansion of geographic ranges, and coupled with physiographic changes, conceivably caused or enhanced some of the subspecific variation.
References marked with an asterisk were not seen by the author.
1958. A herpetofaunal survey of Long Point, Ontario, Canada. Herpetologica, 14(1):8-10, April 25.
1960. New herpetological records from Ohio. Jour. Ohio Herp. Soc., 2(4):25-27.
1857. Contributions to the natural history of the United States of America. Vol. I. Part II. North American Testudinata. Vol. II. Part III. Embryology of the turtle. Little, Brown and Co., Boston, pp. 233-452d; pp. 449-643, 27 pls.
1950. The alligator snapping turtle Macrochelys temminckii in Florida. Spec. Publ. Ross Allen's Rept. Inst., 4:1-15, 6 figs., May.
1955. Some notes on reptiles. Herpetologica, 11(Pt. 3):228, November 30.
1942. Amphibians and reptiles of Jackson County, Missouri. Bull. Chicago Acad. Sci., 6(11):203-220.
1958. The photic responses and water-approach behavior of hatchling turtles. Copeia, 1958(3):211-215, 5 figs., August 28.
1948. Climatic changes and pre-white man. In The Great Basin with emphasis on glacial and postglacial times. Bull. Univ. Utah, 38(20), Biol. Ser., 10(7):168-191, 1 fig., 1 table, June 30.
1901. The reptiles of Allegheny County, Pennsylvania. Ann. Carnegie Mus., 1901-1902, 1(4):145-157.
1919. The turtles of New England. Mem. Boston Soc. Nat. Hist., 8(3): 325-431, 16 pls., April.
1938. Field guide to New England turtles. New England Mus. Nat. Hist., Nat. Hist. Guides, 2:1-56, 9 pls., November.
1929. Typical reptiles and amphibians. Bull. Mus. Comp. Zool., 69(10): 205-360, June.
1791. Travels through North and South Carolina, Georgia, east and west Florida, the Cherokee country, the extensive territories of the Muscogulges, or Creek Confederacy, and the country of the Chactaws; containing an account of the soil and natural productions of those regions, together with observations on the manners of the Indians. First Edition, Philadelphia, xxxiv+522 pp., 8 pls. (including frontispiece), 1 map. Plates unnumbered except "Plate IV" (dorsal view of head of softshell), inserted between pages 282 and 283.
1887. Über die stellung der Trionychidae zu den übrigen Testudinata. Zool. Anz., 10(242):96-102, January 17.
1888. Notes on the American Trionychidae. Amer. Nat., 22(264): 1121-22, December.
1891. On the relations of Carettochelys Ramsay. Amer. Nat., 25:631-639, 3 pls., July.
1893. Notes on the classification and taxonomy of the Testudinata. III. The genera of the Trionychidae. Proc. Amer. Phil. Soc., 31:213-221, May 5.
1932. Sur la place des Trionyx dans la classification des chéloniens. Compt. Rend. Acad. Sci., 195(26):1407-09, December 27.
1936. Sur l'origine du groupe des Trionychoidés. Compt. Rend. Acad. Sci., 203(21):1087-88, November 23.
1937. Chéloniens fossiles du Kiméridgien du Cap de la Hève. Bull. Soc. Nat. Hist. de Toulouse, 71(1-2):180-91, 2 figs., 2 pls., June 30.
1952. Remarques due les chéloniens fossiles de la famille des Carettochelyidae. Compt. Rend. Acad. Sci., 234(23):2302-04, June 4.
1900. Louisiana herpetology with a check-list of the batrachians and reptiles of the state. Proc. Louisiana Soc. Nat., 1897-1899, pp. 25-46.
1923. Notes on the herpetology of Albany County, New York, III. Copeia, 1923(125):115-20, December 31.
1959. Distributional patterns of vertebrates in the southern United States in relation to past and present environments. In Zoogeography, edit. C. L. Hubbs. Amer. Assoc. Adv. Sci., Publ. No. 51, Washington, pp. 433-468, 11 figs., January 16.
1923. The amphibians and reptiles of Dickinson County, Iowa. Univ. Iowa Stud. Nat. Hist., 10:19-26.
1891. Notes on the batrachians and reptiles of Vigo County, Indiana. Jour. Cincinnati Soc. Nat. Hist., 14:22-35 (from Carr, 1952:467).
1945. A preliminary analysis of the herpetofauna of Sonora. Bull. Amer. Mus. Nat. Hist., 83(6):297-426, 13 figs., 8 pls., 1 table, 2 maps, March 30.
1889. Catalogue of the chelonians, rhynchocephalians, and crocodiles in the British Museum (Natural History). Taylor and Francis, London, x + 311 pp., 73 figs., 6 pls.
1890. The fauna of British India, including Ceylon and Burma. Reptilia and Batrachia. Taylor and Francis, London, xviii + 541 pp., 142 figs.
1934. An annotated list of the amphibians and reptiles of Jefferson County, Missouri. Trans. Acad. Sci. St. Louis, 28(3):185-202, 1 fig., April 1.
1958. New records of Cenozoic amphibians and reptiles from California. Bull. Southern California Acad. Sci., 57(1):5-12, 2 pls.
1944. Reptiles and amphibians of Minnesota. Univ. Minnesota Press, Minneapolis, xiii + 202 pp., 52 figs., 45 maps.
1955. Observations on the life history of the soft-shelled turtle Trionyx ferox, with especial reference to growth. Copeia, 1955(1):5-9, 4 figs., 3 tables, February 18.
1957. A large spiny soft-shelled turtle. Copeia, 1957(3):232, August 26.
1960. A spiny soft-shelled turtle nest study. Herpetologica, 16(4): 284-285, December 30.
1934. A check list of the amphibians and reptiles of Ellis County, Kansas. Trans. Kansas Acad. Sci., 37:189-191.
1946. Selected records of reptiles and amphibians from Kansas. Univ. Kansas Publ. Mus. Nat. Hist., 1(5):101-112, August 15.
1910. Records of some reptiles and batrachians from the southeastern United States. Proc. Biol. Soc. Washington, 23:9-18, March 23.
1947. The establishment of Trionyx sinensis in Hawaii. Copeia, 1947 (2):142, June 30.
1950. An annotated check list of the reptiles and amphibians of Texas. Baylor Univ. Press, Waco, xii + 257 pp.
1951. An ecological survey of the Fort Leavenworth Military Reservation. Amer. Midl. Nat., 45(1):187-231, 6 figs., January.
1943. Soil phenomena as evidence of climatic changes. Amer. Jour. Sci., 241(8):469-490, August.
1951. New locality records of reptiles in New Mexico. Copeia, 1951(4): 314, December 28.
1933. Some distributional and ecological records of Kansas reptiles. Trans. Kansas Acad. Sci., 36:186-208.
1935. Further records of the ecology and distribution of amphibians and reptiles in the middle west. Amer. Midl. Nat., 16(3):311-66, May.
1934. Additional records of the reptiles of the central prairie region of the United States. Trans. Kansas Acad. Sci., 37:193-216.
1894. On the habits of turtles. Proc. Indiana Acad. Sci., 1893, p. 224, August.
1939. A system of marking turtles for future identification. Copeia, 1939(3):170-73, 1 fig., September 9.
1942. Turtle populations in southern Illinois. Copeia, 1942(3):155-162, 1 pl., October 8.
1942a. Herpetological fauna of Jackson and Union counties, Illinois. Amer. Midl. Nat., 28(1):164-200, July.
1944. Home range, homing behavior, and migration in turtles. Misc. Publ. Mus. Zool., Univ. Michigan, 61:1-34, 4 figs., 2 pls., 1 map, November 21.
1950. 100 The life history of the slider turtle, Pseudemys scripta troostii (Holbrook). Ecol. Monog., 20(1):31-54, 18 figs., January.
1954. Two new species of the genus Graptemys. Tulane Stud. Zool., 1(11):167-86, 15 figs., 1 table, August 26.
1950. Turtle populations in Louisiana. Amer. Midl. Nat., 43(2):383-88, 2 figs., March.
1929. The herpetology of Waukesha County, Wisconsin. Copeia, 1929 (170):4-8, April 30.
1937. The turtles of Illinois. Illinois Biol. Monog., 16(1-2):1-218, 31 pls., 20 tables, 20 maps, 15 figs., August 31.
1940. A contribution to the herpetology of Florida. Univ. Florida Publ., Biol. Sci. Ser., 3(1):1-118, January.
1952. Handbook of turtles. The turtles of the United States, Canada and Baja California. Comstock Publ. Assoc., Cornell Univ. Press, xv + 542 pp., 37 figs., 82 pls., 15 tables, 23 maps.
1950. Methods for collecting map turtles. Copeia, 1950(4):323-24, December 22.
1920. Fresh-water turtles: a source of meat supply. Rep't. U. S. Comm. Fish., 1919, Appendix 7, U. S. Bur. Fish. Doc., 889:1-20, 8 pls.
1956. Distributional notes on some amphibians and reptiles in Kansas. Trans. Kansas Acad. Sci., 59:213-19.
1958. An ecological study of reptiles and amphibians in Osage County, Kansas. Emporia St. Res. Stud., 7(1):1-52, 15 figs., 4 tables, September.
1956. Common names for North American amphibians and reptiles. Copeia, 1956(3):172-185, August 29.
1930. Field notes of a collecting trip. Bull. Antivenin Inst. Amer., 4(3): 60-64, December.
1951. The reptiles of Ohio. Second edition (with revisionary addenda). Amer. Midl. Nat., Univ. Notre Dame Press, 284 pp., 27 pls., 77 maps.
1958. A field guide to reptiles and amphibians of eastern North America. Houghton Mifflin Co., Boston, xviii + 366 pp., 62 figs., 40 pls., 248 maps.
1948. A new subspecies of soft-shelled turtle from the central United States, with comments on the application of the name Amyda. Occas. Pap. Mus. Zool., Univ. Michigan, No. 510:1-19, 2 pls.
1946. Distribution of species of Amyda in Mississippi. Jour. Mississippi Acad. Sci., 1946, pp. 185-190.
1894. Note on the above (Trionyches in the Delaware drainage by C. W. Johnson). Amer. Nat., 28:889, October.
1927. Field notes on some amphibians and reptiles at Biloxi, Mississippi. Copeia, 1927(165):98-102, December 23.
1936. The herpetology of the Boulder Dam region (Nev., Ariz., Utah). Herpetologica, 1 (Pt. 2):33-42, December 30.
1894. The climbing habits of the softshell turtle (Aspidonectes spinifer). Science, 23(573):50, January 26.
1955. The relationships of the soft-shelled turtles Trionyx ferox ferox and Trionyx ferox aspera. Copeia, 1955 (1):13-23, 4 figs., February 18.
1928. An albino turtle. Jour. Bombay Nat. Hist. Soc., 32(3):608.
1957. Zoogeography: The geographical distribution of animals. John Wiley and Sons, Inc., New York, xiv + 675 pp., 80 figs., 21 tables, frontispiece.
1801. Histoire naturelle, générale et particuliére des Reptiles. Paris, 2:1-432, 13 pls. (from Loveridge and Williams, 1957:511).
1918. A list of reptiles from Jacksonville, Florida. Copeia, 1918 (54):30-33, February 17.
1842. Zoology of New-York. Part III. Reptiles and Amphibia. Albany, vi + 98 pp., 23 pls.
1938. Herpetology of Arkansas. Part One. The Reptiles Occas. Pap. Univ. Arkansas Mus., No. 1:1-47 pp., June 11.
1930. Testudinate evolution. Proc. Zool. Soc. London, 1930, pp. 1057-70, 3 pls., January 21, 1931.
1939. The tetrapod reptiles of Ceylon. Volume 1. Testudinates and crocodilians. Ceylon Jour. Sci., Columbo Mus. Nat. Hist. Ser., xxxii + 412 pp., 137 figs., 24 pls., 62 tables.
1915. The reptiles of the Indo-Australian Archipelago. I. Lacertilia, Chelonia, Emydosauria. E. J. Brill Ltd., Leiden, xiv + 384 pp., 132 figs.
1933. Testudinata from south-eastern Georgia, including the Okefinokee swamp. Copeia, 1933(1):10-12, April 3.
1950. Recent additions to the records of the distribution of the reptiles in Wisconsin. Trans. Wisconsin Acad. Sci. Arts Letters, 40(1):71-77, August 15.
1944. The fishery of the lower Colorado River. California Fish and Game, 30(3):109-211, 38 figs., 7 tables, July.
1959. Climatic changes of the past and present. Contr. Mus. Paleont., Univ. Michigan, 13(8):181-210, 3 figs., 1 pl., 7 maps, April 17.
1957. A review of the amphibians and reptiles of Arkansas. Occas. Pap. Univ. Arkansas Mus., No. 3:1-51, 1 map, July 15.
1958. Amphibians and reptiles of southern Florida. Bull. Florida St. Mus., 3(5):81-324, 28 figs.
1835. Erpétologie générale ou histoire naturelle complète des reptiles. Paris, 2:iv + 682 pp., 12 pls., 2 folding tables.
1950. Kinosternon subrubrum hippocrepis (Gray) in Oklahoma. Herpetologica, 6(Pt. 5):138-39, November 20.
1931. The herpetological fauna of the Americas. Copeia, 1931(3):106-119, 6 figs., October 30.
1960. Aquatic respiration in Trionyx spinifer asper. Herpetologica, 16 (4):277-83, 2 figs., December 30.
1931. Congenital absence of limbs in tortoises of the genera Trionyx and Emyda. Allahabad Univ. Stud., Sci. Sect., 8(2):1-8 (from Biol. Absts.).
1945. Herpetological notes from Allegany State Park, New York. Copeia, 1945(2):115, June 30.
1942. Amphibians and reptiles from Van Buren County, Michigan. Copeia, 1942(3):180, October 8.
1944. Notes on amphibians and reptiles from Wisconsin. Amer. Midl. Nat., 32(2):495-498, September.
1896. Testudinata (of Turkey Lake, Indiana). Proc. Indiana Acad. Sci., 1895, 5:262-64.
1954. An annotated listing of a herpetological collection mainly from Tennessee. Jour. Tennessee Acad. Sci., 29(1):36-41, January.
1951. Notes on the herpetology of Ontario, Canada. Herpetologica, 7(Pt. 2):69-71, June 5.
1920. Lake Maxinkuckee: A physical and biological survey. Vol. 1. Dep't. Conserv. St. Indiana, Publ. No. 7, 660 pp., 23 figs., 8 pls., 36 color pls., frontispiece, 1 folding map. Contains "The turtles and batrachians of the Lake Maxinkuckee region," Proc. Indiana Acad. Sci., 1917 (1916), pp. 472-518, with minor changes.
1835. Entwurf einer systematischen Anordnung der Schildkröten nach den Grundsätzen der natürlichen Methode. Ann. Wiener Mus. Naturg., 1(1):103-128.
1843. Systema reptilium. Fasc. I. Ambyglossae. Vindobonae, vi + 106 pp.
1933. Notes on recent reptiles and amphibians. Proc. Zool. Soc. London, 1933, pp. 735-851, 1 fig., 1 map, September 20.
1930. The amphibians and reptiles of Tulsa County, Oklahoma, and vicinity. Copeia, 1930(2):25-39, June 30.
1907. The amphibians and reptiles of New Jersey. Ann. Rep't. New Jersey St. Museum, 1906, 2:23-250.
1925. Wild life in Kentucky. Kentucky Geol. Surv., 16(ser. 6):xiv + 385, 89 figs., frontispiece.
1909. Amphibia and reptiles. Macmillan and Co., Ltd., London, xiii + 668 pp., 181 figs.
1884. Pharyngeal respiration in the soft-shelled turtle (Aspidonectes spinifer). Proc. Amer. Assoc. Adv. Sci., 1883, 32:316-18.
1886. Aquatic respiration in soft-shelled turtles: a contribution to the physiology of respiration in vertebrates. Amer. Nat., 20:233-236, March.
1948. Pliocene and Pleistocene records of fossil turtles from western Kansas and Oklahoma. Univ. Kansas Publ. Mus. Nat. Hist., 1(17):283-284, August 16.
1959. Distributional and biological notes on the Nebraska herpetofauna. Herpetologica, 15(Pt. 3):141-143, September 10.
1941. Herpetological collections from counties in the vicinity of the Obey River drainage of Tennessee. Jour. Tennessee Acad. Sci., 16(3):329-332, July.
* 1809. Mémoire sur les tortues molles. Nouv. Bull. Sci. Philom., Paris, 1(22):363-67, July (from Stejneger, 1944:27, and Loveridge and Williams, 1957:515).
1809a. Mémoire sur les tortues molles, nouveau genre sous le nom de Trionyx, et sur la formation des carapaces. Ann. Mus. Hist. Nat., Paris, 14:1-20 pp., 5 pls., August.
1946. Reptilian fauna of the North Horn Formation of central Utah. Prof. Pap. U. S. Geol. Surv., 210-C:28-53 (from Biol. Absts.).
1928. The amphibians and reptiles of Franklin County, Kansas. Trans. Kansas Acad. Sci., 31:115-141.
1935. On the incubation of a clutch of eggs of Amyda ferox (Schneider). Copeia, 1935(3):156, October 15.
1948. The occurrence of Amyda spinifer aspera in Florida. Copeia, 1948 (4):304, December 31.
1945. Notes on the egg laying habits of the softshell turtles. Proc. Iowa Acad. Sci., 1944, 51:447-49, December.
1844. Catalogue of the tortoises, crocodiles, and amphisbaenians in the collection of the British Museum. London, viii + 80 pp.
1855. Catalogue of shield reptiles in the collection of the British Museum. Part I. Testudinata (Tortoises). London, 79 pp., 42 pls.
1864. Revision of the species of Trionychidae found in Asia and Africa, with descriptions of some new species. Proc. Zool. Soc. London, 1864, pp. 76-98, 21 figs., February 23.
1869. Notes on the families and genera of tortoises (Testudinata) and on the characters afforded by the study of their skulls. Proc. Zool. Soc. London, 1869, pp. 165-225, 20 figs., 1 pl., March 11.
1870. Supplement to the catalogue of shield reptiles in the collection of the British Museum. Part I. Testudinata (Tortoises). With figures of skulls of 36 genera. London, 120 pp., 40 figs.
1937. The amphibians and reptiles of Randolph County, West Virginia. Herpetologica, 1(Pt. 4):113-16, November 16.
1953. Herpetological notes from southeastern Texas. Herpetologica, 9 (Pt. 1):49-56, June 1.
1957. An electrical shocker for the collection of amphibians and reptiles in the aquatic environment. Copeia, 1957 (1):52, April 5.
1947. Selected records of reptiles and amphibians from southeastern Kansas. Trans. Kansas Acad. Sci., 49(4):447-54.
1947. Egg laying of Trionyx ferox. Copeia, 1947(3):209, September 12.
1945. Emory's turtle in the Virgin River drainage of northwestern Arizona. Copeia, 1945(3):168, October 15.
1926. Tales of the Okefinokee. Amer. Speech, 1(8):407-20, May.
1940. Some works of Bartram, Daudin, Latreille, and Sonnini, and their bearing upon North American herpetological nomenclature. Amer. Midl. Nat., 23(3):692-723, 1 fig., May.
1932. The helminths parasitic in the Amphibia and Reptilia of Houston, Texas and vicinity. Proc. U. S. Nat. Mus., 81(2940):1-71, 5 pls.
1892. The batrachians and reptiles of the state of Indiana. Ann. Rep't Indiana Dep't Geol. Nat. Res., 17:1-204 pp., 3 pls.
1908. The fossil turtles of North America. Publ. Carnegie Inst. Washington, 75:iv + 568, 704 figs., 113 pls.
1910. Descriptions of eight new species of fossil turtles from west of the one-hundredth meridian. Proc. U. S. Nat. Mus., 38(1747):307-26, 23 figs., 3 pls., June 29.
1956. Additional Minnesota herpetological notes. Flicker, 28(3):123-26, September.
1956. Opinion 417. Ops. Decs. Internat. Comm. Zool. Nomencl., 14(1):1-42.
1957. Opinion 447. Ops. Decs. Internat. Comm. Zool. Nomencl., 15(12):211-224.
1938. Amphibians and reptiles of a 2,200-acre tract in central Missouri. Copeia, 1938(2):91-92, June 30.
1949. Effects of DDT-oil solutions upon amphibians and reptiles. Herpetologica, 5(Pt. 6):117-20, December 15.
1936. The amphibians and reptiles of Mammoth Cave National Park proposed. Trans. Kansas Acad., Sci., 39:277-81.
1950. The Pleistocene history of the Mississippi River. Science, N. S., 111:260-62, 2 figs., March 10.
1890. Reptilian. I. Schildkröten. In Bronn's Klassen und Ordnungen des Thier-Reichs. Leipzig, 6(3):1-442, 48 pls.
1944. Physiographic divisions of the Quarternary lowlands of Louisiana. Proc. Louisiana Acad. Sci., 8:11-24, 1 fig., December.
1957. Distributional patterns of Texas fresh-water fishes. So'western Nat., 2(2-3):89-104, 2 figs., December 27.
1942. The amphibians and reptiles of Nebraska. Nebraska Conserv. Bull., No. 24:1-145, 20 pls., 32 maps, June.
1941. The tapeworms of reptiles. Part II. Host catalogue. Wasmann Collector, 4(3):97-104, April.
1941. The trematodes of reptiles. Part II. Host catalogue. Proc. Oklahoma Acad. Sci., 1940, 21:35-43.
1929. Die fossilen weichschildkröten (Trionychia). Eine morphologisch-systematische und stammesgeschichtliche studie. Geol. und Paläont. Abh. N. F., 16(5):357-487, 34 figs. Abstract by K. Staesche in Neu. Jahrb. Min., Geol. und Paläont., Referate III, 1929, pp. 768-72.
1911. Herpetology of Missouri. Trans. Acad. Sci. St. Louis, 20(5):59-274, 7 pls., July 28.
1909. Amphibians and reptiles of Arkansas. Trans. Acad. Sci. St. Louis, 18(2):11-27, May 14.
1894. Trionyches in the Delaware drainage. Amer. Nat., 28:889, October.
1957. The use of CO^{60} as a tag for recovering amphibians in the field. Ecology, 38(2):187-195, 6 figs., 1 table, April.
1929. The habits and economic importance of alligators. U. S. Dep't. Agric., Tech. Bull. 147:1-36, December.
1956. County records of Testudinata collected in Georgia. Jour. Tennessee Acad. Sci., 31(4):322-24, October.
1788. Histoire naturelle des quadrupèdes ovipares et des serpens. Volume I. Paris, 17 pp., 1 folding table, 651 pp., 1 folding table, 41 pls.
1943. Food habits and economic relations of the turtles of Michigan with special reference to fish management. Amer. Midl. Nat., 29(2): 257-312, 9 figs., 9 tables, March.
1943a. Methods of collecting fresh-water turtles. Copeia, 1943(1):21-25 1 fig., March 31.
1954. Michigan turtles. Reprint from Michigan Conserv., 23(3):no page numbers, May-June.
1910. A paired entoplastron in Trionyx and its significance. Proc. Indiana Acad. Sci., 1909, pp. 345-50.
1955. Observations on the sexual behavior of captive turtles. Lloydia, 18(2):95-99, June.
1827. Note sur deux especes de tortues du genre Trionyx Gffr. St. H. Mém. Mus. Hist. Nat. Paris, 15:257-68, 2 pls.
1927. Amphibians and reptiles of Doniphan County, Kansas. Copeia, 1927(164):75-81, July-September.
1940. Amphibians and reptiles in Nevada. Proc. Amer. Acad. Arts Sci., 73(8): 197-257, 29 figs., May.
1937. Soft-shelled turtles in the Colorado River basin. Copeia, 1937 (4):222-225, 3 figs., December 31.
1937. Amphibians and reptiles of the Jornada Experimental Range, New Mexico. Copeia, 1937(4):216-22, December 31.
1922. A preliminary catalogue of Alabama amphibians and reptiles. Geol. Surv. Alabama, Alabama Mus. Nat. Hist., No. 5:1-59 pp., September.
1955. Check-list of the amphibians and reptiles of Canada and Alaska. Contr. Roy. Ontario Mus. Zool. Palaeont., No. 41:v + 88 pp., 77 maps, August 31.
1957. Revision of the African tortoises and turtles of the suborder Cryptodira. Bull. Mus. Comp. Zool., 115(6):163-557, 62 figs., 18 pls., February.
1960. Notes on the soft-shell turtle (Trionyx) in Maryland waters. Chesapeake Sci., 1(1):71-72.
1944. Notes on amphibians and reptiles from the central United States. Amer. Midl. Nat., 32(2):478-90, September.
1957. The Pleistocene history of temperate biotas in Mexico and eastern United States. Ecology, 38(3):468-80, 15 figs., 2 tables, July.
1959. An annotated check list of the amphibians and reptiles of Colorado. Univ. Colorado Stud., Ser. Biol., 6:vi + 98 pp., November.
1936. Annular rings in the long bones of turtles and their correlation with size. Trans. Illinois St. Acad. Sci., 1935, 28(2):255-56, March.
1904. The fresh-water fishes of Mexico north of the Isthmus of Tehuantepec. Field Columbian Mus. Publ. 93, Zool. Ser., 5:lxiii + 252, 72 figs., 17 pls., 1 map, August.
1928. Über die einwirkung der kulturlandschaft auf die verbreitung der amphibien und reptilien. Zool. Garten Leipzig, N. S., 1(Pt. 5-6): 195-203 (from Biol. Absts., and Stejneger, 1944:47).
1955. Die rezenten schildkröten, krokodile und brückenechsen. Zool. Jahrb., 83(5):323-440, October 31.
1946. The probable origin of the soft-shelled turtle in the Colorado River basin. Copeia, 1946(1):46, April 30.
1959. Origin and affinities of the fresh-water fish fauna of western North America. In Zoogeography, edit. C. L. Hubbs. Amer. Assoc. Adv. Sci., Publ. No. 51, Washington, pp. 187-222, 19 figs., 2 tables, January 16.
1944. Introduction to the study of the reptiles of Indiana. Amer. Midl. Nat., 32(2):438-477, September.
1959. Observations on amphibians and reptiles of the Big Bend region of Texas. So'western Nat., 1958, 3(1-4):28-54, 6 tables, 1 map, May 29.
1905. The cultivation of marine and fresh-water animals in Japan. The snapping turtle, or soft-shell tortoise, "suppon." Bull. U. S. Bur. Fish., 24:60-266, 2 figs., 1 pl.
1944. The status of Testudo terrapin Schoepf. Copeia, 1944(4):245-250, December 26.
1929. A case of partial albinism in Trionyx cartilagineus (Bodd.). Misc. Zool. Sumatrana, 40:1-2 (from Biol. Absts.).
1942. Notes on the herpetology of Payne County, Oklahoma. Proc. Oklahoma Acad. Sci., 22:77-80.
1878. Katalog der im museum und universitätskabinet zu Basel aufgestellten amphibien und reptilien nebst anmerkungen. Verh. der Natur. Ges. Basel, 6:557-709, 3 pls.
1921. Notes on the habits of the soft-shell turtle, Amyda mutica. Amer. Midl. Nat., 7(6):180-83, November.
1943. Fosiles raros de Mexico III.—Una tortuga fosil del estado de Chiapas. An. Inst. Biol. Mexico, 14(2):623-24.
1927. Notes on Indiana amphibians and reptiles. Proc. Indiana Acad. Sci., 1926, 36:337-40.
1939. Records of amphibians and reptiles of the Chicago region, 1935-1938. Bull. Chicago Acad. Sci., 6(1):1-10.
1951. Taxonomy of North American soft-shelled turtles, genus Amyda. Publ. Res. Div. Ross Allen's Rept. Inst., 1(2):7-24, 1 fig., January 31.
1951a. Notes on the role of crawfishes in the ecology of reptiles, amphibians, and fishes. Ecology, 32(4):764-66, October.
1958. The occurrence of amphibians and reptiles in saltwater areas, and a bibliography. Bull. Marine Sci. Gulf and Caribbean, 8(1):97 pp., March.
1944. The spineless soft-shelled turtle, Amyda mutica (Le Sueur), in Pennsylvania. Ann. Carnegie Mus., 30:85-88.
1906. The habits of certain tortoises. Jour. Comp. Neurol. Psychol., 16(2):126-52.
1951. Is dorsoventral flattening of the body an adaptation to torrential life? Proc. Internat'l. Assoc. Limnology, 11:264-67.
1949. The occurrence of kyphosis in turtles. Turtox News, 27(1):28-29, 3 figs., January.
1951. The hydrography, fish, and turtle population of Lake Wingra. Trans. Wisconsin Acad. Sci. Arts Letters, 40(2):5-58, 1 map, April 27.
1926. Heredity and evolution. Proc. Zool. Soc. London, 1926, pp. 633-65, 9 figs.
* 1930. Notes on the herpetology of the Chichijima, one of the Bonin Islands. Bull. Biogeog. Soc. Japan, 1(3):187-91 (Japanese), pp. 193-94 (English summary) (from Biol. Absts.).
1938. A catalogue of vertebrates of Japan. Maruzen Co., Ltd., Tokyo, iv + 412 pp.
1816. Lehrbuch Zoologie (Naturgeschichte III, 2). Leipzig, xvi + 1270 pp. (from Conant and Goin, 1948:18).
1955. The natural history of North American amphibians and reptiles. Van Nostrand Co., Inc., Princeton, New Jersey, ix + 359 pp., 74 figs., 15 tables, 12 pls.
1953. The amphibians and reptiles of the Hawaiian Islands. Zoologica, 38(5):65-95, 19 figs.
1959. Notes on some Texas herptiles. Herpetologica, 15(Pt. 1):48, February 25.
1927. A report on the amphibians and reptiles of Oklahoma. Proc. Oklahoma Acad. Sci., 1926, 6(Pt. 1):89-100.
1930. Notes on some reptiles and amphibians from western Oklahoma. Publ. Univ. Oklahoma Biol. Surv., 2(4):175-88, 2 maps.
1943. Amphibians and reptiles of South Dakota. Nat. Hist. Stud., Univ. So. Dakota, 6:1-31, 20 figs., July.
1937. Some amphibians and reptiles from Reelfoot Lake. Jour. Tennessee Acad. Sci., 12(1):60-86, 18 figs., January.
1939. The amphibians and reptiles of Reelfoot Lake and vicinity, with a key for the separation of species and subspecies. Jour. Tennessee Acad. Sci., 14(1):72-101, 14 figs., January.
1948. A contribution to the herpetology of western Tennessee. Jour. Tennessee Acad. Sci., 23(1):20-30, January.
1958. The choanal papillae of the Cheloniidae. Breviora, 85:1-5, 2 pls., January 31.
1960. The structure of the choanae of the Emydinae (Testudines, Testudinidae). Bull. Mus. Comp. Zool., 123(4):113-127, 4 figs., September.
1950. Utilization of crawfishes by cold-blooded vertebrates in the eastern United States. Amer. Midl. Nat., 44(3):643-58, 3 figs., 4 tables, November.
1772. An account of two new tortoises: in a letter to Matthew Maty, M. D. Sci. R. S. Philos. Trans. London, 1771, 61:266-73, 1 pl.
1942. Reptiles and amphibians of Cumberland County, Illinois. Copeia, 1942(3):182-83, October 8.
1927. Reptiles of upper South Carolina. Copeia, 1927(165):110-13, December 23.
1959. On the analysis of biological data. Turtox News, 37(4):98-101, 1 fig., April.
1935. The reptiles of China. Natural history of central Asia. Vol. X. Amer. Mus. Nat. Hist., lii + 604 pp., 78 figs., 64 tables, 27 pls.
1949. Turtles of the United States and Canada. A. A. Knopf, New York, 3rd printing, xviii + 343 + v pp., 99 figs., September.
1930. Wisconsin herpetological notes. Trans. Wisconsin Acad. Sci. Arts Letters, 25:273-84.
1928. The amphibians and reptiles of Wisconsin. Bull. Public Mus. City Milwaukee, 8(1):1-138, 28 figs., 21 pls., April 3.
1958. The growth of Basicladia on turtles. Ecology, 39(4):634-45, 3 figs., October.
1832. Description of two new genera of softshell turtles of North America. Atlantic Jour. and Friend of Knowledge, Philadelphia, 1(2):64-65, Summer.
1933. Observations on the natural history of the common musk turtle, Sternotherus odoratus (Latreille). Pap. Michigan Acad. Sci. Arts Letters, 1932, 17:685-711, 2 figs., 1 table.
1889. Description of a new turtle from the Sacramento River, belonging to the family of Trionychidae. Proc. California Acad. Sci., 2nd Ser., 2:233-36, December 20.
1895. Contributions to the zoology of Tennessee. No. 1. Reptiles and batrachians. Proc. Acad. Nat. Sci. Philadelphia, 47:376-407.
1945. Vertebrate paleontology. Univ. Chicago Press, 2nd Ed. (5th printing, 1953), x + 687 pp., 377 figs., 4 tables.
1956. Osteology of the reptiles. Univ. Chicago Press, xxi + 772 pp., 248 figs.
1929. Paleocene vertebrates from Alberta. Amer. Jour. Sci., 17(98):162-178, 5 figs., February.
1930. A new species of Aspideretes from the Paskapoo Formation of Alberta. Amer. Jour. Sci., 20(115):27-32, 3 figs., July.
1934. Fossil turtles from Saskatchewan and Alberta. Trans. Roy. Soc. Canada, 28:101-11, 6 pls., 1 table, May.
1912. The herpetology of Michigan. Michigan Geol. Biol. Serv., Publ. 10, Biol. Ser. 3, 161 pp., 55 figs., 16 pls.
1924. Emory's soft-shelled turtle in Arizona. Copeia, 1924(131):64, June 30.
1945. A new turtle from the Paleocene of Colorado. Fieldiana Geol., 10(1):1-4, 1 fig., September 19.
1946. On the zoogeography of the Holarctic Region. Copeia, 1946 (3):144-152, 1 map, October 20.
1953. A check list of North American amphibians and reptiles. Sixth edition. Amer. Soc. Ichth. Herp., Univ. Chicago Press, viii + 280 pp.
1957. Living reptiles of the world. Hanover House, Garden City, New York, 287 pp., 266 illus.
1935. Amphibians and reptiles of the Chicago region. Bull. Chicago Acad. Sci., 5(4):57-77.
1944. Amphibians and reptiles of northern Coahuila, Mexico. Zool. Ser. Field Mus. Nat. Hist., 29(6):97-115, February 23.
1783. Allgemeine Naturgeschichte der Schildkröten, nebst einem system. Verzeichnisse der einzelnen arten. Leipzig, "2 Kpftaf. (wovon 1 illum.) gr. 8" (from Engelmann, W., 1846, Bibliotheca Historico-naturalis. 1700-1846, p. 422).
1795. The Historia Testudinum iconibus illustrata. Fasc. 5. Erlangen, Plag. L-O (32 pp.), 6 pls.
1956. The relationships and nomenclature of the soft-shelled turtles (genus Trionyx) of the southeastern United States. Charleston Mus. Leaflet, 26:1-21, 1 fig., 2 maps, 3 pls., May.
1812. Monographiae cheloniorum. Königsberger Arch. Naturwiss. Math., 1:271-368, 406-458 (from Loveridge and Williams, 1957:533).
1956. Records of the spineless soft-shelled turtle and the snapping turtle from New Mexico. Copeia, 1956(2):120-21, May 29.
1949. Fish and invertebrate populations of an Indiana bass stream. Invest. Indiana Lakes Streams, 3(5):247-70, 4 figs., 4 tables.
1941. A limited biological survey of the Obey River and adjacent streams in Tennessee. Jour. Tennessee Acad. Sci., 16(1):48-76, 9 tables, January.
1902. Zur systematik der schildkrötenfamilie Trionychidae Bell, nebst der beschreibung einer neuen Cyclanorbis-art. Sitzungsb. Akad. Wiss. Wien, 91(1):807-46, 18 figs., October.
1909. Synopsis der rezenten schildkröten, mit berücksichtigung der in historischer zeit ausgestorbenen arten. Zool. Jahrb., Suppl. 10, Pt. 3, pp. 427-618.
1924. Die nearktischen Trionychidae. Verh. Zool.-Bot. Ges. Wien, 1923, 73:180-94.
1925. Origin of the germ cells in Trionyx. Amer. Jour. Anat., 36(2): 185-213, 4 pls., November 15.
1943. Turtles and the origin of the fauna of Latin America. Amer. Jour. Sci., 241(7):413-29, July.
1958. Natural History of Thomas County, Nebraska. Privately published, 99 pp.
1947. Kyphosis and other variations in soft-shelled turtles. Univ. Kansas Publ. Mus. Nat. Hist., 1(6):117-24, 1 table, July 7.
1956. Handbook of amphibians and reptiles of Kansas. Misc. Publ. Univ. Kansas Mus. Nat. Hist., 2nd ed., No. 9:1-356, 253 figs., April 20.
1958. Taxonomic significance of cloacal bursae in turtles. Trans. Kansas Acad. Sci., 61(1):86-98.
1949. Observations on constancy of color and pattern in soft-shelled turtles. Trans. Kansas Acad. Sci., 56(1):92-98, 3 figs.
1950. An annotated checklist and key to the reptiles of Mexico exclusive of the snakes. Bull. U. S. Nat. Mus., 199:v + 253.
1930. The Reptilia and Amphibia of the Malay peninsula. A supplement to G. A. Boulenger's Reptilia and Batrachia, 1912. Bull. Raffles Mus., 3:xviii + 149 pp., 13 figs., April.
1931. The fauna of British India including Ceylon and Burma. Reptilia and Amphibia. Vol. I.—Loricata, Testudines. Taylor and Francis, London, xxviii + 185 pp., 2 pls., 42 figs., 1 map (frontispiece), March.
1947. The reptiles and amphibians of eastern central Illinois. Bull. Chicago Acad. Sci., 8(2):21-40.
1955. Notes on Mississippi amphibians and reptiles. Amer. Midl. Nat., 53(1):115-25, January.
1957. A distributional summary of the herpetofauna of Indiana and Illinois. Amer. Midl. Nat., 58(2):341-51, 13 figs.
1956. Statistical methods. Iowa St. College Press, Ames, Iowa, 5th ed., xiii + 534 pp.
1954. Amphibians and reptiles of western North America. McGraw-Hill Book Co., New York, vii + 528 pp., 104 pls., 52 figs.
1954. Additional records of amphibians and reptiles in southern Illinois. Amer. Midl. Nat., 51(1):311-12.
1907. Herpetology of Japan and adjacent territory. Bull. U. S. Nat. Mus., 58:xx + 577 pp., 409 figs., 35 pls.
1944. Notes on the American soft-shell turtles, with special reference to Amyda agassizii. Bull. Mus. Comp. Zool., 94(1):1-75 pp., 30 pls., 10 tables, May.
1917. A check list of North American amphibians and reptiles. Harvard Univ. Press, Cambridge, Mass., 1st ed., iv + 125 pp.
1923. A check list of North American amphibians and reptiles. Harvard Univ. Press, Cambridge, Mass., 2d ed., x + 171 pp.
1933. A check list of North American amphibians and reptiles. Harvard Univ. Press, Cambridge, Mass., 3rd ed., xiv + 185 pp.
1939. A check list of North American amphibians and reptiles. Harvard Univ. Press, Cambridge, Mass., 4th ed., xvi + 207 pp.
1943. A check list of North American amphibians and reptiles. Fifth edition. Bull. Mus. Comp. Zool., 93(1):xix + 260 pp.
1926. Notes on the Edmonton Formation of Alberta. Canadian Field Nat., 40(5):102-04, May.
1948. New records for amphibians and reptiles in the Chicago area, 1939-1947. Bull. Chicago Acad. Sci., 8(7):195-202.
1878. On some peculiarities in the anatomy of soft-shelled turtles. The Zoologist, 3rd Ser., 2(23):401-07, November.
1906. Notes on reptiles and batrachians of Pennsylvania, New Jersey and Delaware. Amer. Nat., 40(471):159-70, March.
1909. Notes on the herpetology of Burnet County, Texas. Bull. Baylor Univ., 12(1):1-9, January.
1915. Reptiles and amphibians of Texas. Bull. Baylor Univ., 18(4):1-82, August.
1924. Notes on the herpetology of Hot Springs, Arkansas. Bull. Baylor Univ., 27(3):29-47, September.
1926. A list of reptiles and amphibians collected by Louis Garni in the vicinity of Boerne, Texas. Contr. Baylor Univ. Mus., No. 6:1-9.
1926a. Notes on the herpetology of the east Texas timber belt. 2. Henderson County amphibians and reptiles. Contr. Baylor Univ. Mus., 7:3-7, July 15.
1927. Observations on the food habits of Texas amphibians and reptiles. Copeia, 1927(162):6-9, January-March.
1928. Occurrence of the spotted night snake (Hypsiglena ochrorhynchus Cope) in Central Texas, with other Bosque County herpetological notes. Contr. Baylor Univ. Mus., 15:1-6, July 10.
1935. Notes on the zoology of Texas from the unpublished manuscripts of John Kern Strecker (assembled and edited by Walter J. Williams). Bull. Baylor Univ., 38(3):vii + 69 pp.
1926. The herpetology of Caddo and De Soto Parishes, Louisiana. Contr. Baylor Univ. Mus., 5:3-10, May 15.
1927. Herpetological records from the vicinity of San Marcos, Texas, with distributional data on the amphibians and reptiles of the Edwards Plateau region and central Texas. Contr. Baylor Univ. Mus., 12:1-16, December.
1930. Morphology and relationships of the trematode Opisthoporus aspidonectes (MacCallum, 1917) Fukui, 1929. Trans. Amer. Micros. Soc., 49(3):210-19, 1 pl., July.
1908. First report on the economic features of turtles of Pennsylvania. Zool. Bull. Div. Zool. Pennsylvania St. Dep't. Agric., 6(4-5):105-96.
1939. Herpetological notes from Indiana. Amer. Midl. Nat., 22(3):684-95, November.
1952. The reptiles of Venango County, Pennsylvania. Amer. Midl. Nat., 47(1):161-82, 1 fig., January.
1920. Philippine turtles. Philippine Jour. Sci., 16(2):111-44, 7 pls.
1933. Observations on the courtship of turtles. Univ. Kansas Sci. Bull., 21(9):269-71, March 15.
1935. Arkansas amphibians and reptiles in the Kansas University Museum. Univ. Kansas Sci. Bull., 22(10):207-18, April 15.
1952. A herpetological survey in the vicinity of Lake Shipp, Polk County, Florida. Quart. Jour. Florida Acad. Sci., 15(3):175-85.
1939. Amphibians, reptiles, and mammals of the Meade County State Park. Trans. Kansas Acad. Sci., 42:499-512, 5 figs., 3 pls.
1958. The systematics and ecology of the Sternothaerus carinatus complex (Testudinata, Chelydridae). Tulane Stud. Zool., 6(1):1-56, 57 figs., March 31.
1937. Ecological observations on amphibians and reptiles collected in southeastern Oklahoma during the summer of 1934. Amer. Midl. Nat., 18(2):285-303, 1 fig., March.
1893. Useful aquatic reptiles and batrachians of the United States. In The fisheries and fishery industries of the United States by G. B. Goode, Sect. I, Part II, pp. 137-162.
1917. Concerning the origin of the soft-shelled turtle, Aspidonectes californiana Rivers. Proc. Calif. Acad. Sci. 4th Ser., 7(2):33-35.
1945. The value of mussel distribution in tracing stream confluence. Pap. Michigan Acad. Sci., Arts Letters, 1944, 30:355-73, 4 figs., September.
1955. Epizoophytic algae from mollusks, turtles, and fish in Oklahoma. Proc. Oklahoma Acad. Sci., 1953, 34:63-65, January.
1923. An ecological study of the cold-blooded vertebrates of southeastern Louisiana. Copeia, 1923(115):35-44, February 1.
1944. Distribution of certain cold-blooded animals in Louisiana in relationship to the geology and physiography of the state. Proc. Louisiana Acad. Sci., 8:47-62, 3 figs., December.
1956. Size at sexual maturity in the male softshell turtle, Trionyx ferox emoryi. Copeia, 1956(2):121-22, 1 fig., May 29.
1959. Description of a new softshell turtle from the southeastern United States. Univ. Kansas Publ. Mus. Nat. Hist., 11(9):517-25, 1 fig., 2 pls., August 14.
1960. Type and type locality of the Gulf Coast spiny softshell turtle, Trionyx spinifer asper (Agassiz). Breviora, No. 129:1-8, 2 pls., December 21.
1960. A new softshell turtle (genus Trionyx) from Coahuila, Mexico. Univ. Kansas Sci. Bull., 40(2):21-30, 2 pls., April 20.
1936. A preliminary list of the reptiles and amphibians of Pottawatomie County, Oklahoma, near Shawnee. Proc. Oklahoma Acad. Sci., 1935, 16:20-22.
1923. Notes on reptiles and batrachians of central Illinois. Copeia, 1923(116):47-50, March 15.
1939. Amphibians and reptiles of northeastern Kentucky. Copeia, 1939 (3):128-30, September 9.
1947. The amphibians and reptiles of North Dakota. Amer. Midl. Nat., 38(1):162-90, July.
* 1838. Reise in das innere Nord-America in den Jahren 1832 bis 1834. Reise von Bethlehem nach Pittsburgh über die Alleghanys, vom 17. September bis zum 7. October. Coblenz, pp. 121-42 (from Carr, 1952:527).
1865. Verziechniss der reptilien welche auf einer reise im nordlichen America beobachtet wurden. Nova Acta Acad. Leopold.—Carol., 32:viii + 146, 7 pls.
1950. Variation and selection in the cervical central articulations of living turtles. Bull. Amer. Mus. Nat. Hist., 94(9):509-61, 20 figs., 10 tables, March 24.
1952. The plastron of soft-shelled turtles (Testudinata, Trionychidae): a new interpretation. Jour. Morph., 90(2):263-75, 2 pls., March.
1957. Yolk retraction as a possible cause of kyphosis in turtles. Herpetologica, 13(Pt. 3):236, October 31.
1959. New Ohio county records in the herpetology collection of the Cleveland Museum of Natural History. Jour. Ohio Herp. Soc., 2(2):8, September.
1919. The turtles and the lizard of Monroe and Wayne counties, New York. Copeia, 1919(66):6-8, February 25.
1915. A biological reconnaissance of the Okefinokee Swamp in Georgia. The reptiles. Proc. Acad. Nat. Sci., Phila., 67:107-92, 14 figs., 3 pls., April 23.
1882. Check list of North American Reptilia and Batrachia, with catalogue of specimens in the U. S. National Museum. Bull. U. S. Nat. Mus., 24:vi + 249 pp.
1939. The homology of the shell elements in turtles. Jour. Morph., 65(3):383-407, 9 figs., 2 pls., November.
Transmitted June 8, 1961.
PLATE 31
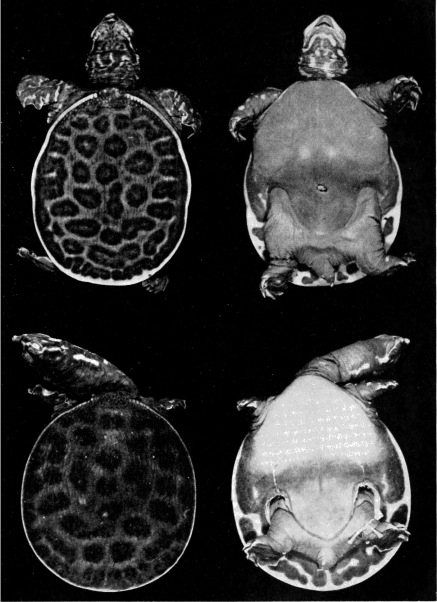
Trionyx ferox, juveniles. Top—UMMZ 76755 (× 1) dorsal and ventral views; Lake Griffin, Lake County, Florida. Bottom—TU 13960 (× 3/4), dorsal and ventral views; Hillsborough River, ca. 20 mi. NE Tampa, Hillsborough County, Florida.
PLATE 32

Top—Trionyx ferox, female, UMMZ 90010 (× 2/9); east edge Okefinokee Swamp, Charlton County, Georgia. Bottom—Left, Trionyx ferox, adult male, UMMZ 102276 (× 1/5), 14 mi. SE Punta Gorda, Lee County, Florida; right, Trionyx sinensis, female, KU 39417 (× 3/10), 5 mi. ESE Seoul, Korea. All dorsal views; note resemblance of two species in having longitudinal ridging and marginal ridge of carapace.
PLATE 33

Trionyx spinifer spinifer, juveniles, dorsal views. Top—UMMZ 74518 (× 12/5); Portage Lake, Washtenaw County, Michigan. Bottom—TU 16132 (× 11/5); Sevierville, Sevier County, Tennessee.
PLATE 34

Trionyx spinifer spinifer, dorsal views. Top—Adult male, UMMZ 54401 (× 3/7), Portage Lake, Livingston County, Michigan. Bottom—Female, UMMZ 81699 (× 2/7), Ottawa County, Michigan.
PLATE 35
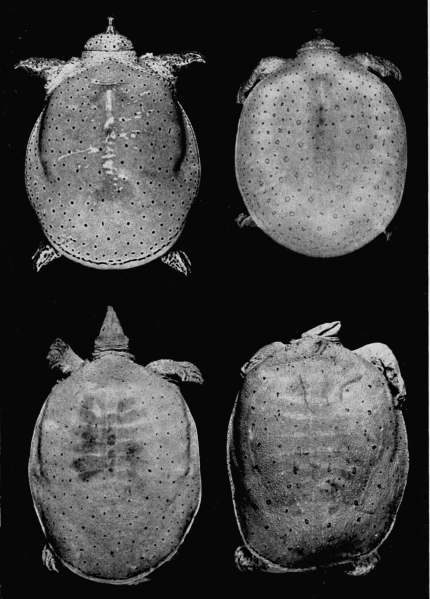
Trionyx spinifer hartwegi, dorsal views. Top—Juveniles; left, KU 40210 (× 9/10), 121/2 mi. S, 11/4 mi. W Meade, Meade County, Kansas; right, KU 16531 (× 1), Smoky Hill River, 3 mi. SW Elkader, Logan County, Kansas. Bottom—Adult Males; left, KU 18385 (× 2/5), Arrington, Comanche County, Kansas; right, KU 3758 (× 3/10), Little Salt Marsh, Stafford County, Kansas.
PLATE 36

Trionyx spinifer hartwegi. Top—Juveniles; left, TU 13885, dorsal view (× 3/4), Little Vian Creek, 1 mi. E Vian, Sequoyah County, Oklahoma; right, KU 3732, ventral view (× 5/7), Independence, Montgomery County, Kansas. Bottom—Adult female, TTC 719, dorsal view (× 2/7), 10 mi. S, 2 mi. W Gruver, Hansford County, Texas.
PLATE 37

Trionyx spinifer asper, juveniles, dorsal views. Top—Left, male, KU 50839 (× 9/10), Flint River, 11/2 mi. S Bainbridge, Decatur County, Georgia; right, female, TU 15661 (× 9/10), Blackwater River, 4.3 mi. NW Baker, Okaloosa County, Florida. Bottom—Left, male, TU 13623 (× 7/9), Yellow River, 3.1 mi. W Hammond, Tangipahoa Parish, Louisiana; right, female, TU 14362 (× 4/5), Hobolochito Creek, 1 mi. N Picayune, Pearl River County, Mississippi.
PLATE 38

Trionyx spinifer asper, dorsal views. Top—Left, adult male, TU 15869 (× 1/2), Escambia River, 1.2 mi. E Century, Escambia County, Florida; right, female, TU 14673.3 (× 1/2), Black Warrior River, 171/2 mi. SSW Tuscaloosa, Tuscaloosa County, Alabama. Bottom—Left, adult male, TU 17117 (× 1/4), Pearl River, Varnado, Washington Parish, Louisiana; right, female, TU 16584 (× 1/5), locality same as TU 15869.
PLATE 39

Trionyx spinifer pallidus, new subspecies, dorsal views. Top—Juveniles; left, TU 481 (× 2/3), Caddo Lake, Caddo Parish, Louisiana; right, KU 50832 (× 9/10), mouth of Caney Creek, 4 mi. SW Kingston, Marshall County, Oklahoma. Bottom—Adult males; left, holotype, TU 484 (× 1/3), locality same as TU 481; right, TU 1122 (× 2/9), Lacassine Refuge, Cameron Parish, Louisiana.
PLATE 40

Trionyx spinifer pallidus, new subspecies, dorsal views. Top—Females; left, TU 13213 (× 1/4), Sabine River, 8 mi. SW Negreet, Sabine Parish, Louisiana; right, TU 13266 (× 2/9), Sabine River, 8 mi. SW Merryville, Beauregard Parish, Louisiana. Bottom—Left, adult male, SM 2889 (× 1/4), Groveton, Trinity County, Texas; right, female, TU 14402 (× 1/5), Trinity River, near junction with Big Creek, Liberty County, Texas.
PLATE 41

Trionyx spinifer guadalupensis, new subspecies, dorsal views. Top—Juveniles; left, ANSP 16717 (× 1), no data; right, KU 50834 (× 11/10), Hondo Creek, 4 mi. W Bandera, Bandera County, Texas. Bottom—Adult males; left, holotype UMMZ 89926 (× 1/3), 15 mi. NE Tilden, McMullen County, Texas; right, SM 659 (× 3/10), Colorado River, near Austin, Travis County, Texas.
PLATE 42

Trionyx spinifer guadalupensis, new subspecies, dorsal views. Top—Adult females; left, TU 16036.1 (× 1/5), Llano River, 2 mi. W Llano, Llano County, Texas; right, TU 10160 (× 1/5), Guadalupe River, 9 mi. SE Kerrville, Kerr County, Texas. Bottom—Left, female, CM 3118 (× 3/4), Black Bayou, Victoria County, Texas; right, male, TU 14419.6 (× 5/9), San Saba River, 11 mi. NNW San Saba, San Saba County, Texas.
PLATE 43
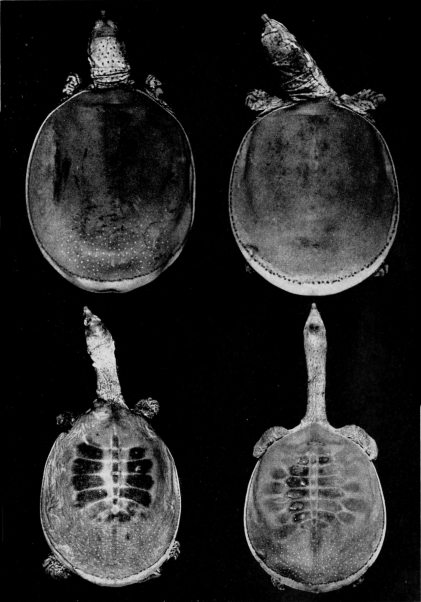
Trionyx spinifer emoryi, dorsal views. Top—Juveniles; left, UMMZ 69411 (× 3/4), Río Conchos, 9 mi. N Linares, Nuevo León, México; right, UMMZ 69412 (× 5/6), Río Purificación, north Ciudad Victoria, Tamaulipas, México. Bottom—Adult males; left, topotype, TU 11561 (× 1/3), Brownsville, Cameron County, Texas; right, KU 48217 (× 1/3), Black River Village, Eddy County, New Mexico.
PLATE 44
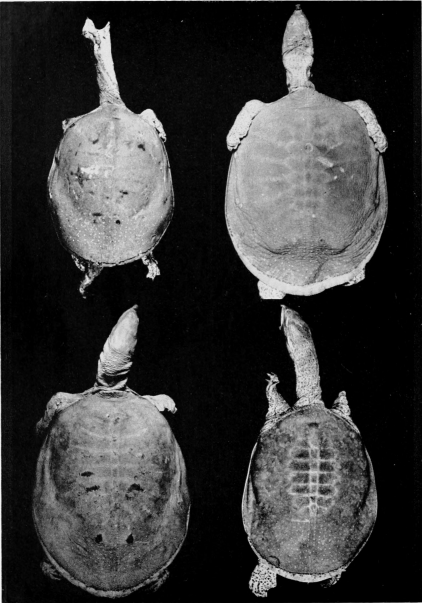
Trionyx spinifer emoryi, dorsal views. Top—Left, adult male, KU 51194 (× 2/7), Río Conchos, near Meoquí, Chihuahua, México; right, female, KU 3119 (× 4/9), Salt River, Phoenix, Maricopa County, Arizona. Bottom—Females; left, KU 3118 (× 1/5), locality same as KU 3119; right, TU 14453 (× 3/10), Pecos River, near junction with Independence Creek, Terrell County, Texas.
PLATE 45

Trionyx muticus muticus, juveniles, dorsal views. Top—Topotypes (× 1), Wabash River, 2 mi. S New Harmony, Posey County, Indiana; left, INHS 7278; right, INHS 7279. Bottom—Left, TU 14375 (× 3/4), Trinity River near junction with Big Creek, Liberty County, Texas; right, KU 50845 (× 12/5), 4 mi. N Atwood, Hughes County, Oklahoma.
PLATE 46
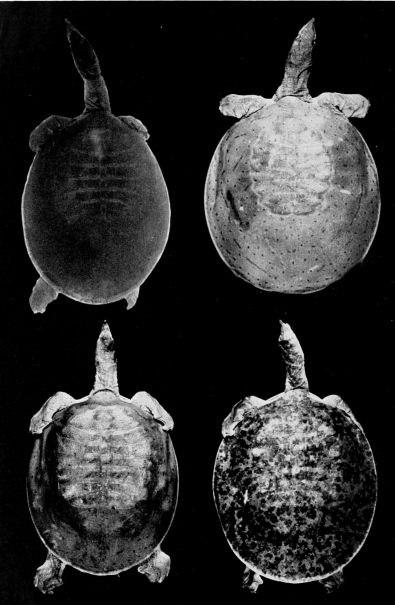
Trionyx muticus muticus, dorsal views. Top—Adult males; left, TU 14606 (× 3/10), White River, Cotter, Marion County, Arkansas; right, KU 48237 (× 1/3), 8 mi. S Hanover, Washington County, Kansas. Bottom—Females (× 1/4), 2 mi. E Manhattan, in Pottawatomie County, Kansas; left, KU 48229; right, KU 48238.
PLATE 47

Trionyx muticus calvatus, dorsal views. Top—Juvenile, TU 17303 (× 12/5), Pearl River, Varnado, Washington Parish, Louisiana. Bottom—Left, adult male, KU 47118 (× 3/10), Pearl River within 4 mi. of Monticello, Lawrence County, Mississippi; right, adult female, TU 17306 (× 2/9), Pearl River, 9 mi. S Monticello, Lawrence County, Mississippi.
PLATE 48

Fig. 1. Habitat of T. s. pallidus, Little River, 6.5 mi. S Broken Bow, McCurtain County, Oklahoma, September 7, 1953.

Fig. 2. Habitat of T. s. emoryi, Río Mesquites, 2 mi. W Nadadores, Coahuila, México, July 27, 1959. Two emoryi were trapped in hoop nets set in quiet water to left of what is believed to be a muskrat house.
PLATE 49

Fig. 1. General habitat of T. s. pallidus and T. m. muticus, Lake Texoma, in a period of low water, 2 mi. E Willis, Marshall County, Oklahoma, February 24, 1951.

Fig. 2. Type locality of T. ater, Tío Candido, 16 km. S Cuatro Ciénegas, Coahuila, México, July 30, 1959. An adult male of T. s. emoryi was also netted here.
PLATE 50

Fig. 1. General habitat of T. s. asper and T. m. calvatus, Escambia River, 2 mi. E, 1/2 mi. N Century, Escambia County, Florida, June 1, 1954. Three nests of calvatus found on sand bar in foreground.
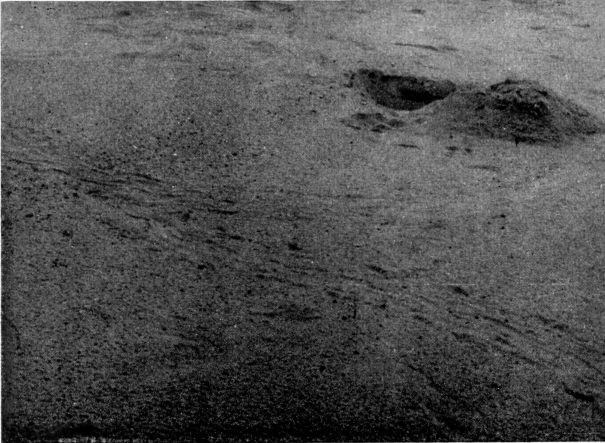
Fig. 2. Nest site of T. m. calvatus (excavated by investigator) on open sand bar shown above in Fig. 1, June 1, 1954. Note tracks of turtle in foreground leading toward and away from disturbed area at left.
PLATE 51
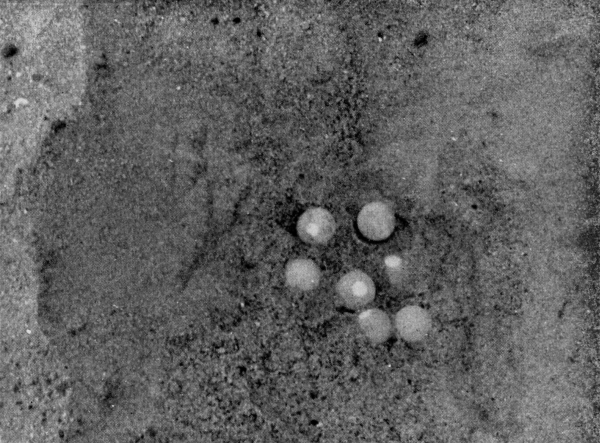
Fig. 1. Eggs of T. m. calvatus in situ, June 1, 1954, approximately six inches below surface, from nest shown in Fig. 2, Pl. 50. Note sandy substrate and seemingly irregular arrangement of eggs.
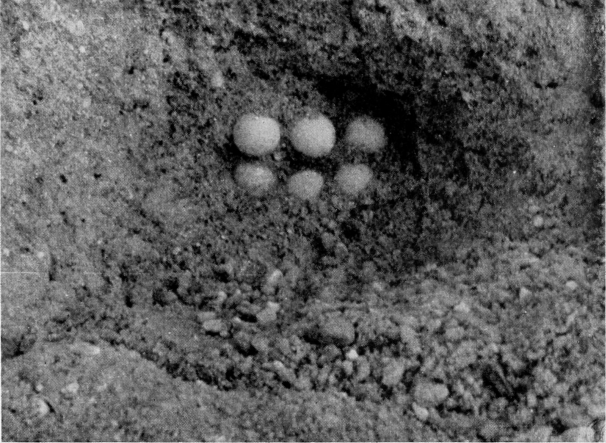
Fig. 2. Eggs of T. m. calvatus in situ, June 1, 1954; nest located at brim of incline shown in foreground of Fig. 1, Pl. 50. Note gravelly substrate (in foreground) and symmetrical arrangement of eggs.
PLATE 52
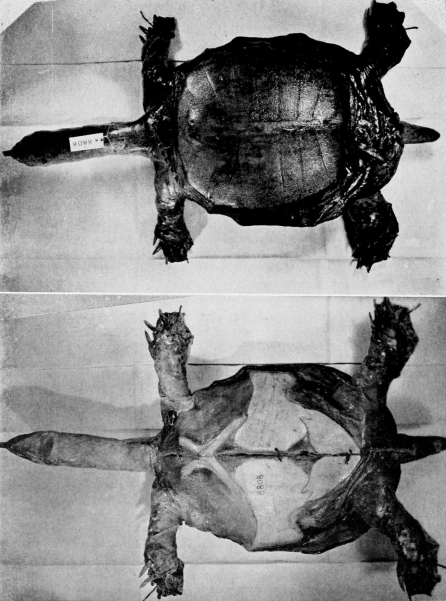
Lectotype of Trionyx spinifer Lesueur, Museum d'Histoire Naturelle, Paris, No. 8808 (× 1/5); obtained by C. A. Lesueur from the Wabash River, New Harmony, Posey County, Indiana. Top—Dorsal view. Bottom—Ventral view.
PLATE 53
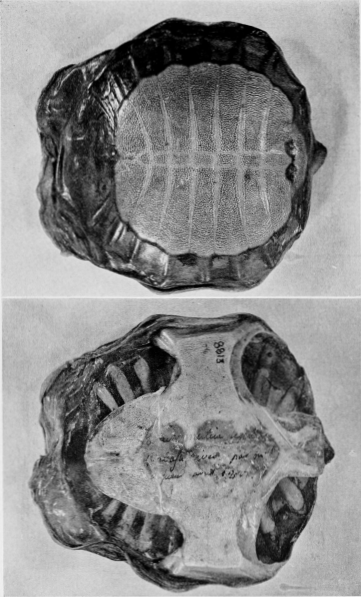
Lectotype of Trionyx muticus Lesueur, Museum d'Histoire Naturelle, Paris, No. 8813 (× 1/2); obtained by C. A. Lesueur from the Wabash River, New Harmony, Posey County, Indiana. Top—Dorsal view. Bottom—Ventral view.
PLATE 54
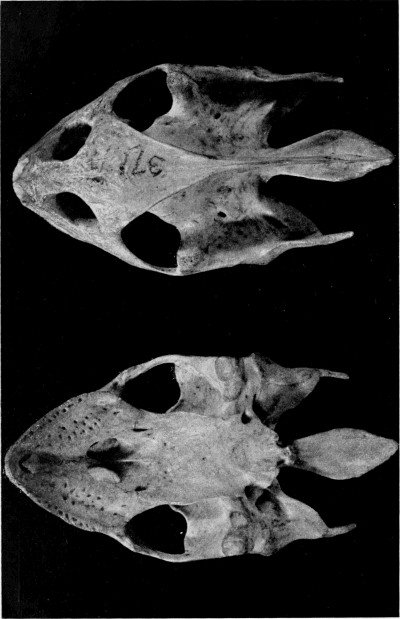
Skull of holotype of Platypeltis agassizi Baur (= T. s. asper), MCZ 37172 (× 1), Savannah River, Georgia. Top—Dorsal view. Bottom—Ventral view.
28-7818
UNIVERSITY OF KANSAS PUBLICATIONS
MUSEUM OF NATURAL HISTORY
Institutional libraries interested in publications exchange may obtain this series by addressing the Exchange Librarian, University of Kansas Library, Lawrence, Kansas. Copies for individuals, persons working in a particular field of study, may be obtained by addressing instead the Museum of Natural History, University of Kansas, Lawrence, Kansas. There is no provision for sale of this series by the University Library, which meets institutional requests, or by the Museum of Natural History, which meets the requests of individuals. Nevertheless, when individuals request copies from the Museum, 25 cents should be included, for each separate number that is 100 pages or more in length, for the purpose of defraying the costs of wrapping and mailing.
* An asterisk designates those numbers of which the Museum's supply (not the Library's supply) is exhausted. Numbers published to date, in this series, are as follows:
| Vol. 1. | Nos. 1-26 and index. Pp. 1-638, 1946-1950. | |
| *Vol. 2. | (Complete) Mammals of Washington. By Walter W. Dalquest. Pp. 1-444, 140 figures in text. April 9, 1948. | |
| Vol. 3. | *1. | The avifauna of Micronesia, its origin, evolution, and distribution. By Rollin H. Baker. Pp. 1-359, 16 figures in text. June 12, 1951. |
| *2. | A quantitative study of the nocturnal migration of birds. By George H. Lowery, Jr. Pp. 361-472, 47 figures in text. June 29, 1951. | |
| 3. | Phylogeny of the waxwings and allied birds. By M. Dale Arvey. Pp. 473-530, 49 figures in text, 13 tables. October 10, 1951. | |
| 4. | Birds from the state of Veracruz, Mexico. By George H. Lowery, Jr., and Walter W. Dalquest. Pp. 531-649, 7 figures in text, 2 tables. October 10, 1951. | |
| Index. Pp. 651-681. | ||
| *Vol. 4. | (Complete) American weasels. By E. Raymond Hall. Pp. 1-466, 41 plates, 31 figures in text. December 27, 1951. | |
| Vol. 5. | Nos. 1-37 and index. Pp. 1-676, 1951-1953. | |
| *Vol. 6. | (Complete) Mammals of Utah, taxonomy and distribution. By Stephen D. Durrant. Pp. 1-549, 91 figures in text, 30 tables. August 10, 1952. | |
| Vol. 7. | *1. | Mammals of Kansas. By E. Lendell Cockrum. Pp. 1-303, 73 figures in text, 37 tables. August 25, 1952. |
| 2. | Ecology of the opossum on a natural area in northeastern Kansas. By Henry S. Fitch and Lewis L. Sandidge. Pp. 305-338, 5 figures in text. August 24, 1953. | |
| 3. | The silky pocket mice (Perognathus flavus) of Mexico. By Rollin H. Baker. Pp. 339-347, 1 figure in text. February 15, 1954. | |
| 4. | North American jumping mice (Genus Zapus). By Philip H. Krutzsch. Pp. 349-472, 47 figures in text, 4 tables. April 21, 1954. | |
| 5. | Mammals from Southeastern Alaska. By Rollin H. Baker and James S. Findley. Pp. 473-477. April 21, 1954. | |
| 6. | Distribution of Some Nebraskan Mammals. By J. Knox Jones, Jr. Pp. 479-487. April 21, 1954. | |
| 7. | Subspeciation in the montane meadow mouse, Microtus montanus, in Wyoming and Colorado. By Sydney Anderson. Pp. 489-506, 2 figures in text. July 23, 1954. | |
| 8. | A new subspecies of bat (Myotis velifer) from southeastern California and Arizona. By Terry A. Vaughn. Pp. 507-512. July 23, 1954. | |
| 9. | Mammals of the San Gabriel mountains of California. By Terry A. Vaughn. Pp. 513-582, 1 figure in text, 12 tables. November 15, 1954. | |
| 10. | A new bat (Genus Pipistrellus) from northeastern Mexico. By Rollin H. Baker. Pp. 583-586. November 15, 1954. | |
| 11. | A new subspecies of pocket mouse from Kansas. By E. Raymond Hall. Pp. 587-590. November 15, 1954. | |
| 12. | Geographic variation in the pocket gopher, Cratogeomys castanops, in Coahuila, Mexico. By Robert J. Russell and Rollin H. Baker. Pp. 591-608. March 15, 1955. | |
| 13. | A new cottontail (Sylvilagus floridanus) from northeastern Mexico. By Rollin H. Baker. Pp. 609-612. April 8, 1955. | |
| 14. | Taxonomy and distribution of some American shrews. By James S. Findley. Pp. 613-618. June 10, 1955. | |
| 15. | The pigmy woodrat, Neotoma goldmani, its distribution and systematic position. By Dennis G. Rainey and Rollin H. Baker. Pp. 619-624, 2 figures in text. June 10, 1955. | |
| Index. Pp. 625-651. | ||
| Vol. 8. | Nos. 1-10 and index. Pp. 1-675, 1954-1956. | |
| Vol. 9. [ii] | 1. | Speciation of the wandering shrew. By James S. Findley. Pp. 1-68, 18 figures in text. December 10, 1955. |
| 2. | Additional records and extension of ranges of mammals from Utah. By Stephen D. Durrant, M. Raymond Lee, and Richard M. Hansen. Pp. 69-80. December 10, 1955. | |
| 3. | A new long-eared myotis (Myotis evotis) from northeastern Mexico. By Rollin H. Baker and Howard J. Stains. Pp. 81-84. December 10, 1955. | |
| 4. | Subspeciation in the meadow mouse, Microtus pennsylvanicus, in Wyoming. By Sydney Anderson. Pp. 85-104, 2 figures in text. May 10, 1956. | |
| 5. | The condylarth genus Ellipsodon. By Robert W. Wilson. Pp. 105-116, 6 figures in text. May 19, 1956. | |
| 6. | Additional remains of the multituberculate genus Eucosmodon. By Robert W. Wilson. Pp. 117-123, 10 figures in text. May 19, 1956. | |
| 7. | Mammals of Coahulia, Mexico. By Rollin H. Baker. Pp. 125-335, 75 figuresin text. June 15, 1956. | |
| 8. | Comments on the taxonomic status of Apodemus peninsulae, with description of a new subspecies from North China. By J. Knox Jones, Jr. Pp. 337-346, 1 figure in text, 1 table. August 15, 1956. | |
| 9. | Extensions of known ranges of Mexican bats. By Sydney Anderson. Pp. 347-351. August 15, 1956. | |
| 10. | A new bat (Genus Leptonycteris) from Coahulia. By Howard J. Stains. Pp. 353-356. January 21, 1957. | |
| 11. | A new species of pocket gopher (Genus Pappogeomys) from Jalisco, Mexico. By Robert J. Russell. Pp. 357-361. January 21, 1957. | |
| 12. | Geographic variation in the pocket gopher, Thomomys bottae, in Colorado. By Phillip M. Youngman. Pp. 363-387, 7 figures in text. February 21, 1958. | |
| 13. | New bog lemming (genus Synaptomys) from Nebraska. By J. Knox Jones, Jr. Pp. 385-388. May 12, 1958. | |
| 14. | Pleistocene bats from San Josecito Cave, Nuevo León, México. By J. Knox Jones, Jr. Pp. 389-396. December 19, 1958. | |
| 15. | New subspecies of the rodent Baiomys from Central America. By Robert L. Packard. Pp. 397-404. December 19, 1958. | |
| 16. | Mammals of the Grand Mesa, Colorado. By Sydney Anderson. Pp. 405-414, 1 figure in text, May 20, 1959. | |
| 17. | Distribution, variation, and relationships of the montane vole, Microtus montanus. By Sydney Anderson. Pp. 415-511, 12 figures in text, 2 tables. August 1, 1959. | |
| 18. | Conspecificity of two pocket mice, Perognathus goldmani and P. artus. By E. Raymond Hall and Marilyn Bailey Ogilvie. Pp. 513-518, 1 map. January 14 1960. | |
| 19. | Records of harvest mice, Reithrodontomys, from Central America, with description of a new subspecies from Nicaragua. By Sydney Anderson and J. Knox Jones, Jr. Pp. 519-529. January 14, 1960. | |
| 20. | Small carnivores from San Josecito Cave (Pleistocene), Nuevo León, México. By E. Raymond Hall. Pp. 531-538, 1 figure in text. January 14, 1960. | |
| 21. | Pleistocene pocket gophers from San Josecito Cave, Nuevo León, México. By Robert J. Russell. Pp. 539-548, 1 figure in text. January 14, 1960. | |
| 22. | Review of the insectivores of Korea. By J. Knox Jones, Jr., and David H. Johnson. Pp. 549-578. February 23, 1960. | |
| 23. | Speciation and evolution of the pygmy mice, genus Baimoys. By Robert L. Packard. Pp. 579-670, 4 plates, 12 figures in text. June 16, 1960. | |
| Index. Pp. 671-690 | ||
| Vol. 10. | 1. | Studies of birds killed in nocturnal migration. By Harrison B. Tordoff and Robert M. Mengel. Pp. 1-44, 6 figures in text, 2 tables. September 12, 1956. |
| 2. | Comparative breeding behavior of Ammospiza caudacuta and A. maritima. By Glen E. Woolfenden. Pp. 45-75, 6 plates, 1 figure. December 20, 1956. | |
| 3. | The forest habitat of the University of Kansas Natural History Reservation. By Henry S. Fitch and Ronald R. McGregor. Pp. 77-127, 2 plates, 7 figures in text, 4 tables. December 31, 1956. | |
| 4. | Aspects of reproduction and development in the prairie vole (Microtus ochrogaster). By Henry S. Fitch. Pp. 129-161, 8 figures in text, 4 tables. December 19, 1957. | |
| 5. | Birds found on the Arctic slope of northern Alaska. By James W. Bee. Pp. 163-211, plates 9-10, 1 figure in text. March 12, 1958. | |
| 6. | The wood rats of Colorado: distribution and ecology. By Robert B. Finley, Jr. Pp. 213-552, 34 plates, 8 figures in text, 35 tables. November 7, 1958. | |
| 7. | Home ranges and movements of the eastern cottontail in Kansas. By Donald W. Janes. Pp. 553-572, 4 plates, 3 figures in text. May 4, 1959. | |
| 8. | Natural history of the salamander, Aneides hardyi. By Richard F. Johnston and Gerhard A. Schad. Pp. 573-585. October 8, 1959. | |
| 9. | A new subspecies of lizard, Cnemidophorus sacki, from Michoacán, México. By William E. Duellman. Pp. 587-598, 2 figures in text. May 2, 1960. | |
| 10. | A taxonomic study of the middle American snake, Pituophis deppei. By William E. Duellman. Pp. 599-610, 1 plate, 1 figure in text. May 2, 1960. | |
| Index. Pp. 611-626. | ||
| Vol. 11. [iii] | 1. | The systematic status of the colubrid snake, Leptodeira discolor Günther. By William E. Duellman. Pp. 1-9, 4 figures. July 14, 1958. |
| 2. | Natural history of the six-lined racerunner, Cnemidophorus sexlineatus. By Henry S. Fitch. Pp. 11-62, 9 figures, 9 tables. September 19, 1958. | |
| 3. | Home ranges, territories, and seasonal movements of vertebrates of the Natural History Reservation. By Henry S. Fitch. Pp. 63-326, 6 plates, 24 figures in text, 3 tables. December 12, 1958. | |
| 4. | A new snake of the genus Geophis from Chihuahua, Mexico. By John M. Legler. Pp. 327-334, 2 figures in text. January 28, 1959. | |
| 5. | A new tortoise, genus Gopherus, from north-central Mexico. By John M. Legler. Pp. 335-343. April 24, 1959. | |
| 6. | Fishes of Chautauqua, Cowley and Elk counties, Kansas. By Artie L. Metcalf. Pp. 345-400, 2 plates, 2 figures in text, 10 tables. May 6, 1959. | |
| 7. | Fishes of the Big Blue river basin, Kansas. By W. L. Minckley. Pp. 401-442, 2 plates, 4 figures in text, 5 tables. May 8, 1959. | |
| 8. | Birds from Coahuila, México. By Emil K. Urban. Pp. 443-516. August 1, 1959. | |
| 9. | Description of a new softshell turtle from the southeastern United States. By Robert G. Webb. Pp. 517-525, 2 plates, 1 figure in text. August 14, 1959. | |
| 10. | Natural history of the ornate box turtle, Terrapene ornata ornata Agassiz. By John M. Legler. Pp. 527-669, 16 pls., 29 figures in text. March 7, 1960. | |
| Index Pp. 671-703. | ||
| Vol. 12. | 1. | Functional morphology of three bats: Sumops, Myotis, Macrotus. By Terry A. Vaughan. Pp. 1-153, 4 plates, 24 figures in text. July 8, 1959. |
| 2. | The ancestry of modern Amphibia: a review of the evidence. By Theodore H. Eaton, Jr. Pp. 155-180, 10 figures in text. July 10, 1959. | |
| 3. | The baculum in microtine rodents. By Sydney Anderson. Pp. 181-216, 49 figures in text. February 19, 1960. | |
| 4. | A new order of fishlike Amphibia from the Pennsylvanian of Kansas. By Theodore H. Eaton, Jr., and Peggy Lou Stewart. Pp. 217-240, 12 figures in text. May 2, 1960. | |
| More numbers will appear in volume 12. | ||
| Vol. 13. | 1. | Five natural hybrid combinations in minnows (Cyprinidae). By Frank B. Cross and W. L. Minckley. Pp. 1-18. June 1, 1960. |
| 2. | A distributional study of the amphibians of the Isthmus of Tehuantepec, México. By William E. Duellman. Pp. 19-72, pls. 1-8, 3 figures in text. August 16, 1960. | |
| 3. | A new subspecies of the slider turtle (Pseudemys scripta) from Coahulia, México. By John M. Legler. Pp. 73-84, pls. 9-12, 3 figures in text. August 16, 1960. | |
| 4. | Autecology of the copperhead. By Henry S. Fitch. Pp. 85-288, pls. 13-20, 26 figures in text. November 30, 1960. | |
| 5. | Occurrence of the garter snake, Thamnophis sirtalis, in the Great Plains and Rocky Mountains. By Henry S. Fitch and T. Paul Maslin. Pp. 289-308, 4 figures in text. February 10, 1961. | |
| 6. | Fishes of the Wakarusa river in Kansas. By James E. Deacon and Artie L. Metcalf. Pp. 309-322, 1 figure in text. February 10, 1961. | |
| 7. | Geographic variation in the North American cyprinid fish, Hybopsis gracilis. By Leonard J. Olund and Frank B. Cross. Pp. 323-348, pls. 21-24, 2 figures in text. February 10, 1961. | |
| 8. | Descriptions of two species of frogs, genus Ptychohyla; studies of American hylid frogs, V. By William E. Duellman. Pp. 349-357, pl. 25, 2 figures in text. April 27, 1961. | |
| 9. | Fish populations, following a drought, in the Neosho and Marais des Cygnes rivers of Kansas. By James Everett Deacon. Pp. 359-427, pls. 26-30, 3 figs. August 11, 1961. | |
| 10. | Recent soft-shelled turtles of North America (family Trionychidae). By Robert G. Webb. Pp. 429-611, pls. 31-54, 24 figures in text, February 16, 1962. | |
| Vol. 14. | 1. | Neotropical bats from western México. By Sydney Anderson. Pp. 1-8. October 24, 1960. |
| 2. | Geographic variation in the harvest mouse. Reithrodontomys megalotis, on the central Great Plains and in adjacent regions. By J. Knox Jones, Jr., and B. Mursaloglu. Pp. 9-27, 1 figure in text. July 24, 1961. | |
| 3. | Mammals of Mesa Verde National Park, Colorado. By Sydney Anderson. Pp. 29-67, pls. 1 and 2, 3 figures in text. July 24, 1961. | |
| 4. | A new subspecies of the black myotis (bat) from eastern Mexico. By E. Raymond Hall and Ticul Alvarez. Pp. 69-72, 1 figure in text. December 29, 1961. | |
| 5. | North American yellow bats, "Dasypterus," and a list of the named kinds of the genus Lasiurus Gray. By E. Raymond Hall and J. Knox Jones, Jr. Pp. 73-98, 4 figures in text. December 29, 1961. | |
| 6. | Natural history of the brush mouse (Peromyscus boylii) in Kansas with description of a new subspecies. By Charles A. Long. Pp. 99-111, 1 figure in text. December 29, 1961. | |
| 7. | Taxonomic status of some mice of the Peromyscus boylii group in eastern Mexico, with description of a new subspecies. By Ticul Alvarez. Pp. 113-120, 1 figure in text. December 29, 1961. | |
| More numbers will appear in volume 14. | ||
| Vol. 15. | 1. | The amphibians and reptiles of Michoacán, México. By William E. Duellman. Pp. 1-148, pls. 1-6, 11 figures in text. December 20, 1961. |
| 2. | Some reptiles and amphibians from Korea. By Robert G. Webb, J. Knox Jones, Jr., and George W. Byers. Pp. 149-173. January 31, 1962. | |
| More numbers will appear in volume 15. | ||
All obvious typographical error were corrected. Due to variant usage of hyphens, "soft-shelled" and "crystal-clear" were used as the standard for variants of those terms. All other variant spellings of "softshell(s)" and where variant spellings occur in quoted text or literature titles they were retained. Variants in accented and non-accented words may exist. Paragraphs split by tables or illustrations were rejoined.
For those using screen readers, here is a text version of graphic on page 586.
Pliocene Pleistocene Recent
==========================================================================
+--F-----------------------------------F (ferox)
|
| +---------Mm (muticus muticus)
| +--M------------------+
| | +---------Mc (muticus calvatus)
| |
FMSA-+ | +--Ss (spinifer spinifer)
| | +------+
| | | +--Sh (spinifer hartwegi)
| | +--Ssha-+
+--MSA-+ | |
| | +---------Sa (spinifer asper)
| |
| +--S---+ +--Sp (spinifer pallidus)
| | | +--Spg-+
| | | | +--Sg (spinifer guadalupensis)
+--SA--+ +--Sepg-+
| |
| +--Se-----Se (spinifer emoryi)
|
+--A---------------------A (ater)
--------------------------------------------------------------------------
Typographical Corrections
| Page | Correction |
| 494 | Ordinance School Proving Ground → Ordnance |
| 494 | Pl. 12, top → Pl. 42, top |
| 567 | Pl. 52, Fig. 2 → Pl. 51, Fig. 2 |
| 576 | Agkistrodon piscivorous → piscivorus |
| 595 | Carettochlydae → Carettochelyidae |