The Project Gutenberg EBook of The Cambridge Natural History, Vol X.,
Mammalia, by Frank Evers Beddard
This eBook is for the use of anyone anywhere at no cost and with
almost no restrictions whatsoever. You may copy it, give it away or
re-use it under the terms of the Project Gutenberg License included
with this eBook or online at www.gutenberg.org
Title: The Cambridge Natural History, Vol X., Mammalia
Author: Frank Evers Beddard
Release Date: June 1, 2012 [EBook #39887]
Language: English
Character set encoding: ISO-8859-1
*** START OF THIS PROJECT GUTENBERG EBOOK CAMBRIDGE NATURAL HISTORY ***
Produced by Chris Curnow, Joseph Cooper, Keith Edkins and
the Online Distributed Proofreading Team at
http://www.pgdp.net
|
Transcriber's note:
|
A few typographical errors have been corrected. They
appear in the text like this, and the
explanation will appear when the mouse pointer is moved over the marked
passage.
|
THE
CAMBRIDGE NATURAL HISTORY
EDITED BY
S. F. Harmer and A. E. Shipley
VOLUME X
MAMMALIA
by F. BEDDARD
Reprint Edition
1958
CODICOTE, ENGLAND
WHELDON & WESLEY, LTD.
| |
WEINHEIM, GERMANY
H. R. ENGELMANN (J. CRAMER)
|
|
NEW YORK
HAFNER PUBLISHING CO.
|
© 1902, by Macmillan & Co., Limited
Authorized reprint by
Wheldon & Wesley, Ltd. and H. R. Engelmann (J. Cramer)
Printed in Germany
{iii}
PREFACE
Inasmuch as Sir W. H. Flower and Mr. Lydekker could not profess to
treat the Mammalia exhaustively within the limits of nearly 800 pages, in
their Introduction to the Study of Mammals, it is obvious that the
present volume, which appears ten years later and is of rather less size,
can contain but a selection of the enormous mass of facts at the disposal
of the student of this group. Thus the chief question for myself was what
to select and what to leave aside. It will be observed that I have
reduced the pages of this book to conformity with those of other volumes
of the series by treating some groups more briefly than others. It has
appeared to me to be desirable to treat fully such groups as the Edentata
and the Marsupialia, and permissible to be more brief in dealing with
such huge Orders as those of the Rodentia and Chiroptera. Lengthy
disquisitions upon such familiar and comparatively uninteresting animals
as the Lion and Leopard have been curtailed, and the space thus saved has
been devoted to shorter and more numerous accounts of other creatures. As
there are nearly six hundred genera of living Mammals known to science,
omission as well as compression became an absolute necessity. I have
given, I hope, adequate treatment from the standpoint of a necessarily
limited treatise to the majority of the more important genera of Mammals
both living and extinct; but the length of this part of the book had to
be increased by the discoveries, which give me at once an advantage and a
disadvantage as compared with the two authors whose names I have quoted,
of a considerable number of important new types in the last ten years.
{iv}Such forms as Notoryctes,
Romerolagus, Caenolestes, "Neomylodon," and
Ocapia could not possibly have been omitted.
In preparing my accounts of both living and extinct forms I have
nearly invariably consulted the original authorities, and have often
supplemented or verified these accounts by my own dissections at the
Zoological Society's Gardens. My rule has not, however, been invariable
in this matter, inasmuch as there exist two recent and trustworthy
text-books of Mammalian Palaeontology—Professor Zittel's
Handbuch der Palaeontologie, and Dr. A. Smith Woodward's manual,
Outlines of Vertebrate Palaeontology, in the Cambridge Biological
Series. Where the name of a genus only or its range, or merely one or two
facts about it, are mentioned, I have not thought it necessary to go
further than these two works. But a good deal has been done even since
the appearance of these two volumes which it will be found that I have
not ignored.
I have to thank my editors for the trouble which they have taken in
the revision of the proofs and for many suggestions. To Professor Osborn,
of Columbia University, New York, I am indebted for some kind
suggestions. My daughter Iris has assisted me in various ways. Finally, I
desire to express my indebtedness to Mr. Dixon and to Mr. M. P. Parker
for the care which they have taken in the preparation of the figures
which were drawn by them especially for this work.
Frank E. Beddard.
London, February 28, 1902.
{v}
CONTENTS
| |
Page |
| Preface |
iii |
| Scheme of the Classification adopted in this Book |
ix |
| CHAPTER I |
| Introductory |
1 |
| CHAPTER II |
| Structure and Present Distribution of the Mammalia |
5 |
| CHAPTER III |
| The Possible Forerunners of the Mammalia |
90 |
| CHAPTER IV |
| The Dawn of Mammalian Life |
96 |
| CHAPTER V |
| The Existing Orders of Mammals: Prototheria—Monotremata |
105 |
| CHAPTER VI |
| Introduction to the Sub-Class Eutheria |
116 |
| CHAPTER VII |
| Eutheria—Marsupialia |
122 |
|
{vi}
CHAPTER VIII |
| Edentata—Ganodonta |
161 |
| CHAPTER IX |
Ungulata—Condylarthra—Amblypoda—Ancylopoda—Typotheria—
Toxodontia—Proboscidea—Hyracoidea |
195 |
| CHAPTER X |
| Ungulata (continued)—Perissodactyla (Odd-toed Ungulates)—Litopterna |
235 |
| CHAPTER XI |
| Ungulata (continued)—Artiodactyla (Even-toed Ungulates)—Sirenia |
269 |
| CHAPTER XII |
| Cetacea—Whales and Dolphins |
339 |
| CHAPTER XIII |
| Carnivora—Fissipedia |
386 |
| CHAPTER XIV |
| Carnivora (continued)—Pinnipedia (Seals and Walruses)—Creodonta |
446 |
| CHAPTER XV |
| Rodentia—Tillodontia |
458 |
| CHAPTER XVI |
| Insectivora—Chiroptera |
508 |
| CHAPTER XVII |
| Primates |
533 |
| Index |
591 |
{vii}
Scheme of the Classification Adopted In This Book
| Sub-Class Prototheria (p. 105). |
|---|
| Order. |
|
Sub-order. |
|
Family. |
|
Sub-family. |
|---|
| MONOTREMATA (p. 106) |
 |
|
|
Echidnidae (p. 110).
Ornithorhynchidae (p. 112). |
| ?ALLOTHERIA (p. 96). |
| Sub-class Eutheria (p. 116) |
|---|
| MARSUPIALIA (p. 122) |
|
Diprotodontia (p. 128) |
|
Macropodidae (p. 129) |
|
Macropodinae (p. 132).
Potoroinae (p. 137).
Hypsiprymnodontinae (p. 138).
|
| Phalangeridae (p. 138) |
|
Phalangerinae (p. 140).
Phascolarctinae (p. 142).
Phascolomyinae (p. 144).
Tarsipedinae (p. 145).
|
| Epanorthidae (p. 145). |
| Polyprotodontia (p. 149) |
|
Dasyuridae (p. 149).
Didelphyidae (p. 155).
Peramelidae (p. 156).
Notoryctidae (p. 158). |
| EDENTATA (p. 161) |
|
Xenarthra (p. 166) |
|
Myrmecophagidae (p. 166).
Bradypodidae (p. 170).
Dasypodidae (p. 173).
Mylodontidae (p. 179).
Megalonychidae (p. 183).
Megatheriidae (p. 183).
Glyptodontidae (p. 184).
|
| Nomarthra (p. 186) |
|
Orycteropodidae (p. 187).
Manidae (p. 188). |
| GANODONTA (p. 190) |
|
|
|
Stylinodontidae (p. 191).
Conoryctidae (p. 193). |
|
{viii}
UNGULATA (p. 195) |
|
Condylarthra (p. 202).
Amblypoda (p. 205).
Ancylopoda (p. 211).
Typotheria (p. 212).
Toxodontia (p. 214). |
| Proboscidea (p. 216) |
|
Elephantidae (p. 217).
Dinotheriidae (p. 231). |
| Hyracoidea (p. 232). |
| Perissodactyla (p. 235) |
|
Equidae (p. 237).
Lophiodontidae (p. 247).
Palaeotheriidae (p. 247).
Tapiridae (p. 260).
Rhinocerotidae (p. 253).
Titanotheriidae (p. 264). |
| Litopterna (p. 267) |
|
Macraucheniidae (p. 267). |
| Artiodactyla (p. 269) |
|
Hippopotamidae (p. 273).
Suidae (p. 275).
Dicotylidae (p. 278).
Tragulidae (p. 282).
Proceratidae (p. 284).
Camelidae (p. 285). |
| Cervidae (p. 291) |
|
Cervinae (p. 293).
Moschinae (p. 299). |
Giraffidae (p. 301).
Antilocapridae (p. 306).
Bovidae (p. 307).
Anthracotheriidae (p. 328).
Caenotheriidae (p. 329).
Xiphodontidae (p. 329).
Oreodontidae (p. 330).
Anoplotheriidae (p. 332). |
| SIRENIA (p. 333). |
| CETACEA (p. 339) |
|
Mystacoceti (p. 353) |
|
Balaenopteridae (p. 355).
Balaenidae (p. 358). |
| Odontoceti (p. 362) |
|
Physeteridae (p. 362) |
|
Physeterinae (p. 363).
Ziphiinae (p. 367). |
|
Delphinidae (p. 372).
Platanistidae (p. 380).
Squalodontidae (p. 384). |
| Archaeoceti (p. 384) |
|
Zeuglodontidae (p. 384). |
| CARNIVORA (p. 386) |
|
Fissipedia (p. 387) |
|
Felidae (p. 390).
Machaerodontidae (p. 401). |
| Viverridae (p. 403) |
|
Euplerinae (p. 403).
Galidictiinae (p. 404).
Cryptoproctinae (p. 404).
Viverrinae (p. 405).
Herpestinae (p. 409). |
{ix}
Hyaenidae (p. 411).
Canidae (p. 413).
Procyonidae (p. 426). |
| Mustelidae (p. 431) |
|
Melinae (p. 432).
Mustelinae (p. 433).
Lutrinae (p. 439). |
| Ursidae (p. 442). |
| Pinnipedia (p. 446) |
|
Otariidae (p. 450).
Trichechidae (p. 451).
Phocidae (p. 452). |
| CREODONTA (p. 455). |
| RODENTIA (p. 458) |
|
Simplicidentata (p. 462) |
|
Anomaluridae (p. 462).
Soiuridae (p. 463).
Castoridae (p. 467).
Haplodontidae (p. 469).
Gliridae (p. 470). |
| Muridae (p. 471) |
|
Murinae (p. 471).
Phlaeomyinae (p. 473).
Hydromyinae (p. 474).
Rhynchomyinae (p. 474).
Gerbillinae (p. 475).
Otomyinae (p. 475).
Dendromyinae (p. 476).
Lophiomyinae (p. 476).
Microtinae (p. 477).
Sigmodontinae (p. 479).
Neotominae (p. 480). |
Bathyergidae (p. 480).
Spalacidae (p. 482).
Geomyidae (p. 483).
Heteromyidae (p. 484).
Dipodidae (p. 484).
Pedetidae (p. 486). |
| Octodontidae (p. 487) |
|
Octodontinae (p. 487).
Loncherinae (p. 488).
Capromyinae (p. 489). |
Ctenodactylidae (p. 490).
Caviidae (p. 491).
Dasyproctidae (p. 493).
Dinomyidae (p. 495).
Chinchillidae (p. 496).
Cercolabidae (p. 497).
Hystricidae (p. 499). |
| Duplicidentata (p. 502) |
|
Leporidae (p. 502).
Lagomyidae (p. 505). |
| TILLODONTIA (p. 506). |
| INSECTIVORA (p. 508) |
|
Insectivora Vera (p. 509) |
|
Erinaceidae (p. 509).
Tupaiidae (p. 511).
Centetidae (p. 511).
Potamogalidae (p. 513).
Solenodontidae (p. 513).
Chrysochloridae (p. 514).
Macroscelidae (p. 515).
Talpidae (p. 516).
Soricidae (p. 518). |
| Dermoptera (p. 520) |
|
Galeopithecidae (p. 520). |
|
{x}
CHIROPTERA (p. 521) |
|
Megachiroptera (p. 524) |
|
Pteropodidae (p. 524). |
| Microchiroptera (p. 526) |
|
Rhinolophidae (p. 527).
Nycteridae (p. 527).
Vespertilionidae (p. 528).
Emballonuridae (p. 530).
Phyllostomatidae (p. 531). |
| PRIMATES (p. 533) |
|
Lemuroidea (p. 534) |
|
Lemuridae (p. 538) |
|
Indrisinae (p. 538).
Lemurinae (p. 540).
Galagininae (p. 542).
Lorisinae (p. 545). |
Chiromyidae (p. 548).
Tarsiidae (p. 550).
Anaptomorphidae (p. 552).
Chriacidae (p. 552).
Megaladapididae (p. 554). |
| Anthropoidea (p. 554) |
|
Hapalidae (p. 556).
Cebidae (p. 557).
Cercopithecidae (p. 562).
Simiidae (p. 570).
Hominidae (p. 585). |
{1}
CHAPTER I
INTRODUCTORY
The Mammalia form a group of vertebrated animals which roughly
correspond with what are termed in popular language "quadrupeds," or with
the still more vernacular terms of "beasts" or "animals." The name
"Mammal" is derived from the most salient characteristic of the group,
i.e. the possession of teats; but if the term were used in an
absolutely strict etymological sense, it could not include the
Monotremes, which, though they have mammary glands, have not
fully-differentiated teats (see p. 16). There are,
however, as will be seen shortly, other characters which necessitate the
inclusion of these egg-laying quadrupeds within the class Mammalia.
The Mammalia are unquestionably the highest of the Vertebrata. This
statement, however, though generally acceptable, needs some explanation
and justification. "Highest" implies perfection, or, at any rate,
relative perfection. It might be said with perfect truth that a serpent
is in its way an example of perfection of structure: not incommoded with
limbs it can slip rapidly through the grass, swim like a fish, climb like
a monkey, and dart upon its prey with rapidity and accuracy. It is an
example of an extremely specialised reptile, the loss of the limbs being
the most obvious way in which it is specialised from more generalised
reptilian types. Specialisation in fact is often synonymous with
degradation, and, this being the case, implies a restricted life. On the
other hand, simplification is not always to be read as degeneration. The
lower jaw, for instance, of mammals has fewer bones in it than that of
reptiles, and is more concisely articulated to the skull; this implies
greater efficiency {2}as a biting organ. The term highest, however,
includes increased complexity as well as simplification, the two series
of modifications being interwoven to form a more efficient organism. It
cannot be doubted that the increased complexity of the brain of mammals
raises them in the scale, as does also the complex and delicately
adjusted series of bonelets which form the organ for the transmission of
sound to the internal ear. The separation of the cavity containing the
lungs, and the investment of the partition so formed with muscular
fibres, renders the action of the lungs more effective; and there are
other instances among the Mammalia of greater complexity of the various
parts and organs of the body when compared with lower forms, which help
to justify the term "highest" generally applied to these creatures.
Complexity and finish of structure are often accompanied by large
size; and the Mammalia are, on the whole, larger than any other
Vertebrates, and also contain the most colossal species. The huge
Dinosaurs of the Mesozoic epoch, though among the largest of animals, are
exceeded by the Whales; and the latter group includes the mightiest
creature that exists or has ever existed, the eighty-five-feet-long
Sibbald's Rorqual. Confining ourselves rigidly to facts, and avoiding all
theorising on the possible relation between complexity and nicety of
build and the capacity for increase in bulk, it is plain from the history
of more than one group of mammals that increase in bulk accompanies
specialisation of structure. The huge Dinocerata when compared with the
ancestral Pantolambda teach us this, as do many similar examples.
Within the mammalian group, as in the case of other Vertebrates,
difference of size has a certain rough correspondence with difference of
habitat. The Whales not only contain the largest of animals, but their
average size is great; so too with the equally aquatic Sirenia and very
aquatic Pinnipedia. Here the support offered by the water and the
consequent decreased need for muscular power to neutralise the effects of
gravity permit of an increase in bulk. Purely terrestrial animals come
next; and finally arboreal, and, still more, "flying" mammals are of
small size, since the maintenance of the position when moving and feeding
needs enormous muscular effort.
The Mammals are more easily to be separated from the Vertebrates lying
lower in the series than any of the latter are from each other in
ascending order. A large number of {3}characters might be used in
addition to those which will be made use of in the following brief
catalogue of essential mammalian features, were it not for the low-placed
Monotremata on the one hand and the highly specialised Whales on the
other. Including those forms, the Mammalia are to be distinguished from
all other Vertebrates by the following series of structural features,
which will be expanded later into a short disquisition upon the general
structure of the Mammalia. The class Mammalia may, in fact, be thus
defined:—
Hair-clad Vertebrates, with cutaneous glands in the female, secreting
milk for the nourishment of the young. Skull without prefrontal,
postfrontal, quadrato-jugal, and some other bones, and with two occipital
condyles formed entirely by the exoccipitals. Lower jaw composed of
dentary bone only, articulating only with the squamosal. Ear bones a
chain of three or four separate bonelets. Cervical vertebrae sharply
distinguished from the dorsals, and if with free ribs, showing no
transition between these and the thoracic ribs. Brain with four optic
lobes. Lungs and heart separated from abdominal cavity by a muscular
diaphragm. Heart with a single left aortic arch. Red blood-corpuscles
non-nucleate.
The following characters are also very nearly universal, and in any
case absolutely distinctive:—Cervical
vertebrae, seven; vertebrae with epiphyses. Ankle-joint "cruro-tarsal,"
i.e. between the leg and the ankle, and not in the middle of the
ankle.[1] Attachment of the
pelvis to the vertebral column pre-acetabular in position.
The Mammalia since they are hot-blooded creatures are more independent
of temperature than reptiles; they are thus found spread over a wider
area of the earth's surface. As however, though hot-blooded, they have
not the powers of locomotion possessed by birds, they are not quite so
widely distributed as are those animals. The Mammalia range up into the
extreme north, but, excepting only forms mainly aquatic, such as the Sea
Lions, are not known to occur on the Antarctic continent. With the
exception of the flying Bats, indigenous mammals are totally absent from
New Zealand; and it seems to be doubtful whether those supposed oceanic
islands which have a mammalian fauna are really {4}oceanic in origin. The
continents and oceans are peopled by rather over three thousand species
of Mammalia, a number which is considerably less than that of either
birds or reptiles. It seems clear that, so far at any rate as concerns
the numbers of families and genera, the mammalian fauna of to-day is less
varied than it was during the Mid-tertiary period, the heyday of
mammalian life. It is rather remarkable to contrast in this way the
mammals and the birds. The two classes of the animal kingdom seem to have
come into being at about the same period; but the birds either have
reached their culminating point to-day, or have not yet reached it. The
Mammalia, on the other hand, multiplied to an extraordinary extent during
the Eocene and the Miocene periods, and have since dwindled. The break is
most marked at the close of the Pleistocene, and may be in part due to
the direct influence of man. At present man exercises so enormous an
effect, both directly and indirectly, that the future history of the
Mammalia is probably foreshadowed by the instances of the White
Rhinoceros and the Quagga. On the other hand, the economic usefulness of
the Mammalia is greater than that of any other animals; and the next most
important era in their history will be probably that of domesticity and
"preservation."
{5}
CHAPTER II
STRUCTURE AND PRESENT DISTRIBUTION OF THE MAMMALIA
External Form.—It would be quite impossible for any one
to confuse any other quadrupedal animal with a mammal. The body of a
reptile is, as it were, slung between its limbs, like the body of an
eighteenth century chariot between its four wheels; in the mammal the
body is raised entirely above, and is supported by, the four limbs. The
axes of these limbs too, as a general rule, are parallel with the
vertical axis of the body of their possessor. There is thus a greater
perfection of the relations of the limbs to the trunk from the point of
view of a terrestrial creature, which has to use those limbs for rapid
movement. The same perfection in these relations is to be seen, it should
be observed, in such running forms among the lower Vertebrata as the
Birds and the Dinosaurs, where the actual angulation of the limbs is as
in the purely running Mammalia. These relations are of course absolutely
lost in the aquatic Cetacea, and not marked in various burrowing
creatures. The way in which the fore- and hind-limbs are angulated is
considerably different in the two cases. In the latter, which are most
used and, as it were, push on the anterior part of the body, the femur
has its lower end directed forwards, the tibia and the fibula project
backwards at the lower end, while the ankle and foot are again inclined
in the same direction as the femur. With the fore-limbs there is not this
regular alternation. The humerus is directed backwards, the fore-arm
forwards, and the hand still more forwards. This angulation seems to
facilitate movement, inasmuch as it is seen in even the Amphibia and the
lower Reptiles, in which, however, the differences between the fore- and
hind-limbs are less marked, {6}indicating therefore a less specialised
condition of the limbs. It is an interesting fact that the angulation of
the limbs is to some extent obliterated in very bulky creatures, and
almost entirely so in the elephants (see p. 217),
which seem to need strong and straight pillars for the due support of
their huge bodies.
The alertness and general intellectual superiority of mammals to all
animals lying below them in the series (with the exception of the birds,
which are in their way almost on a level with the Mammalia) are seen by
their active and continuous movements. The lengthy periods of absolute
motionlessness, so familiar to everybody in such a creature as the
Crocodile, are unknown among the more typical Mammalia except indeed
during sleep. This mental condition is clearly shown by the proportionate
development of the external parts of all the organs of the higher senses.
The Mammalia as a rule have well-developed, often extremely large, flaps
of skin surrounding the entrance to the organ of hearing, often called
"ears," but better termed "pinnae." These are provided with special
muscles, and can be often moved and in many directions. The nose is
always, or nearly always, very conspicuous by its naked character; by the
large surface, often moist, which surrounds the nostrils; and again by
the muscles, which enable this tract of the integument to be moved at
will. The eyes, perhaps, are less marked in their predominance over the
eyes of lower Vertebrates than are the ears and nose; but they are
provided as a rule with upper and lower eyelids, as well as by a
nictitating membrane as in lower Vertebrates. The apparent predominance
of the senses of smell and hearing over that of sight appears to be
marked in the Mammalia, and may account for their diversity of voice as
well as of odour, and for the general sameness of coloration which
distinguishes this group from the brilliantly-coloured birds and
reptiles. The head, too, which bears these organs of special sense, is
more obviously marked out from the neck and body than is the case with
the duller creatures occupying the lower branches of the Vertebrate
stem.
|
Fig. 1.—A, Section of human skin. Co,
Dermis; D, sebaceous glands; F, fat
in dermis; G, vessels in dermis; GP,
vascular papillae; H, hair; N, nerves
in dermis; NP, nervous papillae; Sc,
horny layer of epidermis; SD, sweat
gland; SD1, duct of sweat gland; SM,
Malpighian layer. B, Longitudinal section
through a hair (diagrammatic). Ap,
Band of muscular fibres inserted into the
hair-follicle; Co, corium (dermis); F,
external longitudinal; F1, internal circular,
fibrous layer of follicle; Ft, fatty
tissue in the dermis; GH, hyaline
membrane between the root-sheath and
the follicle; HBD, sebaceous gland;
HP, hair-papilla with vessels in its interior;
M, medullary substance (pith)
of the hair; O, cuticle of root-sheath;
R, cortical layer; Sc, horny layer of
epidermis; Sch, Hair shaft; SM, Malpighian
layer of epidermis; WS, WS1,
outer and inner layers of root-sheath. (From Wiedersheim's Comparative
Anatomy.)
|
|
The Hair.—The Mammalia are absolutely distinguished from
all other Vertebrates (or, for the matter of that, Invertebrates) by the
possession of hair. To define a mammal as a Vertebrate with hair would be
an entirely exclusive definition; even in the smooth Whales a few hairs
at least are present, which may be {7}reduced to as few as two
bristles on the lips. The term "hair," however, is apt to be somewhat
loosely applied; it has been made use of to describe, for example, the
slender processes of the chitinous skin of the Crustacea. It will be
necessary, therefore, to enter into the microscopical structure and
development of the mammalian hair. Hair is found in every mammal. The
first appearance of a hair is a slight thickening of the stratum
Malpighii of the epidermis, the cells taking part in this being {8}elongated and
converging slightly above and below. Dr. Maurer has called attention to
the remarkable likeness between the embryonic hair when at this stage and
the simple sense-organs of lower Vertebrates. Later there is formed below
this a denser aggregation of the corium, which ultimately becomes the
papilla of the hair. This is the apparent homologue of the first formed
part of a feather, which projects as a papilla before the epidermis has
undergone any modification. Hence there is from the very first a
difference between feathers and hairs—a difference which must be
carefully borne in mind, especially when we consider the strong
superficial resemblance between hairs and the simple barbless feathers.
Still later the knob of epidermic cells becomes depressed into a tubular
structure, which is lined with cells also derived from the stratum
Malpighii, but is filled with a continuation of the more superficial
cells of the epidermis. This is the hair-follicle, and from the epidermic
cells arises the hair by direct metamorphosis of those cells; there is no
excretion of the hair by the cells, but the cells become the hair. From
the hair-follicle also grows out a pair of sebaceous glands, which serve
to keep the fully-formed hair moist.

Fig. 2.—Four diagrams of stages in the development
of a hair. A, Earliest stage in one of those mammals
in which the dermal papilla appears first; B, C, D,
three stages in the development of the hair in the
human embryo. blb, Hair-bulb; crn, horny layer
of the epidermis; foll, hair-follicle; grm, hair-germ;
h, hair, in D, projecting on the surface;
muc, Malpighian layer of epidermis; pp,
dermal papilla; seb, developing sebaceous glands;
sh.1, sh.2, inner and outer root-sheaths. (After
Hertwig.)
Dr. Meijerle[2] has lately
described in some detail the {9}particular arrangement of the individual hairs
among mammals; they are not by any manner of means scattered without
order, but show a definite and regular arrangement, which varies with the
animal. For instance, in an American Monkey (Midas), the hairs
arise in threes—three hairs of equal size springing from the
epidermis close together; in the Paca (Coelogenys) there are in
each group three stout hairs alternating with three slender hairs. In
some forms a number of hairs spring from a common point: in the Jerboa
(Dipus) twelve or thirteen arise from a single hole; in Ursus
arctos there is the same general plan, but there is one stout hair
and four or five slender ones. There are numerous other complications and
modifications, but the facts, although interesting, do not appear to
throw any light upon the mutual affinities of the animals. Allied forms
may have a very different arrangement, while in forms which have no near
relationship the plan may be very similar, as is shown by the examples
cited from Dr. Meijerle's paper. The groups of hairs, moreover, have
themselves a definite placing, which the same anatomist has compared with
the disposition of the bundles of hairs behind and between the scales of
the Armadillo, and which has led him to the view that the ancestors of
mammals were scaly creatures—a view also supported by Professor Max
Weber,[3]and not in itself
unreasonable when we consider the numerous points of affinity between the
primitive Mammalia and certain extinct forms of reptiles.[4]
The hairs are greatly modified in form in different mammals and in
different parts of their bodies. It is very commonly the case that a soft
under-fur can be distinguished from the longer and coarser hairs, which
to some extent hide the latter. Thus the "sealskin" of commerce is the
under-fur of the Otaria ursina of the North. The coarser hairs may
be further differentiated into bristles; these again into spines, such as
those of the Hedgehog and of the Porcupine. Again, the flattening and
agglutination of hairs seems to be responsible for the scales of the
Manis {10}and for the horns of the Rhinoceros.
It is a matter of common knowledge that upon the head of various animals,
e.g. the Domestic Cat, long and sensitive hairs are developed,
which are connected with the terminations of nerves, and perform a
sensory, probably tactile function. These occur on the snout, above the
eyes, and in the neighbourhood of the ears. It is an interesting fact
that a tuft of quite similar hairs occurs on the hand of many mammals
close to the wrist, which, at least in the case of Bassaricyon,
are connected with a strong branch from the arm-nerve. These tufts also
occur in Lemurs, in the Cat, various Rodents and Marsupials, and are
probably quite general in mammals who "feel" with their
fore-limbs;—in which, in fact, the fore-limbs are not exclusively
running organs. That the last remaining hairs of the Cetacea are found
upon the muzzle, is perhaps significant of the importance of these
sensory bristles. The entire absence of hairs is quite common in this
order, although traces of them are sometimes found in the embryo. The
Sirenia, too, are comparatively hairless, as are also many Ungulates.
Whether the presence of blubber in the former case and the existence of a
very thick skin in the latter animals are facts which have had anything
to do with the disappearance of hair or not, is a matter for further
inquiry.
The intimate structure of the hair varies considerably. The variations
concern the form of the hair, which may be round in transverse section,
or so oval as to appear quite flat when the hair is examined in its
entirety. The substance of the hair is made up of a central medulla or
pith with a peripheral cortex; the latter is scaled, and the scales are
often imbricated and with prominent edges. The amount of the two
constituents also differs, and the cortex may be reduced to a series of
bands surrounding only tracts of the enclosed pith. In the hair is
contained the pigment to which the colour of mammals is chiefly due.
Tracts of brightly-coloured skin may exist, as in the Apes of certain
genera; but such structures are not general. The pigment of the hair
seems to consist of those pigmentary substances known as melanins. It is
remarkable to find such a uniform cause of coloration, when we consider the great variety
of feather-pigments found in birds. The variations of colour of the hair
of mammals are due to the unequal distribution of these brown pigments.
There are very few mammals which can {11}be called brightly
coloured. The Bats of the genus Kerivoula have been compared to
large butterflies, and some of the Flying Squirrels have strongly-marked
contrasts of reddish brown, white, and yellow. The same may be said of
the spines of certain Porcupines. But we find in the hair no bright
blues, greens, and reds such as are common among birds.
There are certain general facts about the coloration of mammals which
require some notice here. Next to the usually sombre hues of these
animals the general absence of secondary sexual coloration is noteworthy.
In but a few cases among the Lemurs and Bats do we find any marked
divergences in hues between males and females. Secondary sexual
characters in mammals are, it is true, often exhibited by the great
length of certain hair-tracts in the male, such as the mane of the Lion,
the throat- and leg-tufts of the Barbary Sheep, and so forth; but apart
from these, the secondary sexual characters of mammals are chiefly shown
in size, e.g. the Gorilla, or in the presence of tusks,
e.g. various Boars, or of horns, as in the Deer, etc. The
coloration of mammals frequently exhibits conspicuous patterns of
marking. These are in the form of longitudinal stripes, of cross-stripes,
or of spots; the latter may be "solid" spots, or broken up, as in the
Leopard and Jaguar, into groups of smaller spots arranged in a
rosette-fashion. We never find in mammals the complicated "eyes" and
other markings which occur in so many birds and in other lower
Vertebrates. It is important to note that in the Mammalia whose sense of
sight is quite keen there should be a practical absence of secondary
sexual colours. As to the relationship of the various forms of marking
that do occur, it seems clear that there has been a progression from a
striped or spotted condition to uniform coloration. For we find that many
Deer have spotted young; that the young Tapir of the New World is
spotted, while its parents are uniform blackish brown; the
strongly-marked spotting of the young Puma contrasts with the uniform
brown of the adult; and the Lion cub, as every one knows, is also
spotted, the adult lioness showing considerable traces of the spots.
The seasonal change in the colours of certain mammals is a subject
upon which much has been written. The extreme of this is seen in those
creatures, such as the Polar Hare and the Arctic Fox, which become
entirely blanched in the winter, recovering {12}their darker coat in the
spring. This is, however, only an extreme case of a change which is
general. Most animals get a thicker fur in winter and exchange it for a
lighter one in summer. And the hues of the coat change in
correspondence.
Glands of the Skin.—The great variety of integumental
glands possessed by the Mammalia distinguishes them from any group of
lower Vertebrates. This variability, however, only concerns the
anatomical structure of the glands in question. Histologically they are
all of them apparently to be referred to one of two types, the
sudoriparous or sweat gland and the sebaceous gland. Simple sweat and
sebaceous glands are abundant in mammals, with but a few exceptions. The
structures that we are now concerned with are agglomerations of these
glands. The mammary glands will be treated of in connexion with the
marsupium; they are either masses of sweat glands, or of sebaceous glands
whose secretion has been converted into milk.
Many Carnivora possess glands opening to the exterior, near the anus,
by a large orifice. These secrete various odoriferous substances, of
which the well-known "civet" is an example. Other odoriferous glands are
the musk glands of the Musk-deer and of the Beaver; the suborbital gland
of many Antelopes; the dorsal gland of the Peccary, which has given the
name of Dicotyles to the genus on account of its resemblance in
form to a navel. This gland may be seen to secrete a clear watery fluid.
The Elephant has a gland situated on the temple, which is said to secrete
during certain periods only, and to be a warning to leave the animal
alone. Very remarkable are the foot glands of certain species of
Rhinoceros; they are not universally present in those animals, and
are therefore useful as specific distinctions. On the back of the root of
the tail in many Dogs are similar glands. The Gentle Lemur
(Hapalemur) has a peculiar gland upon the arm, about the size of
an almond, which in the male underlies a patch of spiny outgrowths. In
Lemur varius is a hard patch of black skin which may be the
remnants of such a gland. It is thought that the callosities on the legs
of Horses and Asses are remnants of glands.
One of the most complex of these structures which has been examined
microscopically exists in the Marsupial Myrmecobius.[5] On the skin of the anterior
part of the chest, just in front of the {13}sternum, is a naked patch
of skin which is seen to be perforated by numerous pores. Besides the
ordinary sebaceous and sweat glands there are a series of masses of
glands, opening by larger orifices, which present the appearance of
groups of sebaceous glands, and are of a racemose character; but the
existence of muscular fibres in their coats seems to show that they
should be referred rather to the sudoriparous series. Beneath the
integument is a large compound tubular gland quite half an inch in
diameter.
In Didelphys dimidiata there is a precisely similar glandular
area and large underlying gland, the correspondence being remarkable in
two Marsupials so distant in geographical position and affinities. Even
among the Diprotodont genera there is something of the kind; for in
Dorcopsis luctuosa and D. muelleri is a collection of four
unusually large sebaceous follicles upon the throat, and in the Tree
Kangaroo (Dendrolagus bennettii) there is the same collection of
enlarged hair-follicles, though they are apparently somewhat reduced as
compared with those of Dorcopsis. These are of course a few
examples out of many.
It seems to be possible that the functions of these various glands is
at least twofold. In the first place, they may serve, where predominant
in one sex, to attract the sexes together. In the second place, the
glands may be useful to enable a strayed animal of a gregarious species
to regain the herd. It is perfectly conceivable too that in other cases
the glands may be a protection, as they most undoubtedly are in the
Skunk, from attacks. In connexion with the first, and more especially the
second, of the possible uses of these glands, it is interesting to note
that in purely terrestrial creatures, such as the Rhinoceros, the glands
are situated on the feet, and would therefore taint the grass and herbage
as the animal passed, and thus leave a track for the benefit of its mate.
The same may be said of the rudimentary glands of Horses if they are
really glands. The secretion of the "crumen" of Antelopes is sometimes
deposited deliberately by Oreotragus upon surrounding objects, a
proceeding which would attain the same end. One may even perhaps detect
"mimicry" in the similar odours of certain animals. Prey may be lured to
their destruction, or enemies frightened away. The defenceless Musk-deer
may escape its foes by the suggestion of the musky odour of a crocodile.
It is at any rate perfectly conceivable that the variety of odours among
mammals may play a very {14}important part in their life, and it is
perhaps worthy of note that birds with highly-variegated plumage are
provided only with the uropygial gland, while mammals with usually dull
and similar coloration have a great variety of skin glands. Scent is no
doubt a sense of higher importance in mammals than in birds. The subject
is one which will bear further study.
Nails and Claws.—Except for the Cetacea (where rudiments
have been found in the foetus), the extremities of the fingers and of the
toes of mammals are covered by, or encased in, horny epidermic plates,
known as nails, claws, and hoofs.
The variety in the shape and development of these corneous sheaths to
the digits is highly characteristic of mammals as opposed to lower
Vertebrates. If we take extreme cases, such as the nail of the thumb in
Man, the hoof of a Horse, and the claw of a Cat, it is easy to
distinguish the three kinds of phalangeal horny coverings. But the
differences become extinguished as we pass from these to related types.
The nail of the little finger in Man approaches the claw-like form; and
the hoofs of the Lama are almost claws in the sharpness of their
extremities. On the whole it may be said that claws and hoofs embrace the
bone which they cover, while nails lie only upon its dorsal surface. The
form of the distal phalanx which bears the nail shows, however, two kinds
of modification which do not support such a classification. When those
phalanges are clad with hoofs or covered by a nail they end in a rounded
and flattened termination. On the other hand, when they bear a claw they
are themselves sharpened at the extremity and often grooved above.
The Marsupium.—It may appear to be unnecessary at this
juncture to speak of the marsupial pouch, which is so usually believed to
be a characteristic of the group Marsupialia. Rudiments of this structure
have, however, been recently discovered in the higher mammals, and, as
Dr. Klaatsch[6] has remarked,
all researches into the "history of the mammals culminate in the question
whether the placental mammals pass through a marsupial stage or not." We
cannot, therefore, look upon the marsupial pouch as a matter affecting
only the Marsupials, though it is true that this organ is at present
functional only in them and in the Monotremata. {15}
Fig. 3.—Echidna hystrix. A, Lower surface of brooding female; B; dissection showing
a dorsal view of the pouch and mammary glands; ††, the two tufts of hair in the
lateral folds of the mammary pouch from which the secretion flows, b.m, Pouch;
cl, cloaca; g.m, groups of mammary glands. (From Wiedersheim's Comparative
Anatomy, after W. Haacke.)
In the Marsupials the pouch shelters the young, which are born in an
exceedingly imperfect state, minute, nude, and blind, with a "larval"
mouth fitted only to grasp in a permanent fashion the teat, upon which
they are carefully fixed by the parent. But even later the pouch is made
use of as a temporary harbour of refuge: from the pouch of female
Kangaroos at the Zoological Gardens may frequently be observed to
protrude the tail and hind-legs of a young Kangaroo as big as a Cat, and
perfectly well able to take care of itself.
In the Monotremata (in Echidna) there is a deep fold of the
skin which lodges the unhatched egg, and into which the mammary glands
open, one on either side. This structure is only periodically developed,
and arises from two rudiments, one corresponding to each mammary area;
but in the female with eggs or young there is but a single deep
depression, which occupies the same region of the body as the marsupial
pouch of the {16}Marsupials.[7] It is usually held that this structure is
not of precisely the same morphological value as the pouch of the
Marsupial; and the difference is expressed by terming the one (that of
Echidna) the mammary pouch, and the other the marsupium. At first
sight it may appear to be an unnecessary refinement to separate two
structures which have so many and such obvious likenesses. It is not
quite certain, however, that the difference is not even more profound
than later opinions seem to indicate. The Monotremata not only have no
teats, as has already been pointed out, but the mammary glands themselves
are of a perfectly different nature to those of the higher mammals,
including the Marsupials. There is therefore no a priori objection
to the view that the accessory parts developed in connexion with the
mammary glands should also be different. The teat of the higher Mammalia
grows up round the area upon which the ducts of the mammary glands open;
it is a fold of skin which eventually assumes the cylindrical form of the
adult teat, and which includes the ducts of the milk glands. It has been
suggested that the two folds of skin which form the mammary pouch of
Echidna are to be looked upon as the equivalent of the commencing
teat of the higher mammal.[8]
In this case it is clear that the marsupial folds of the Marsupial cannot
correspond accurately with the apparently similar folds of
Echidna, because there are teats as well. It is the teats which
correspond to the marsupial folds of Echidna. This view is in
apparent contradiction to an interesting discovery in a specimen of a
Phalanger by Dr. Klaatsch.[9]
This Marsupial, like most others, has a well-developed marsupial pouch,
in which the young are lodged at birth; but round two of the teats is
another distinct fold on either side, the outer wall of which forms the
general wall of the pouch. Dr. Klaatsch thinks that these smaller and
included pouches are the equivalents of the mammary pouches of
Echidna. They contain teats, but this comparison does not do away
with the validity of Gegenbaur's suggestion already referred to, because
the teats are (see above) {17}secondary. If this fact be fairly to be
interpreted in the sense which Dr. Klaatsch attaches to it, we have an
interesting case of the growth of a new organ out of and partly replacing
an old organ. In the Monotremes there is a pouch which facilitates or
performs both nutritive and protective functions; in the Phalanger these
two functions are carried on in separate pouches; finally, in other
Marsupials, there is a return to the undifferentiated state of affairs
found in the Monotremata, but with the help of a new organ not found in
them.

Fig. 4.—Diagram of the development of the nipple
(in vertical section). A, Indifferent stage, glandular
area flat; B, elevation of the glandular area
with the nipple; C, elevation of the periphery
of the glandular area into the false teat, a,
Periphery of the glandular area; b, glandular
area; gl, glands. (From Gegenbaur.)
Though so characteristic of Marsupials, the marsupial pouch is not
always developed in them. It is present in all the Kangaroos, Wallabies,
and Wombats, in fact in the Diprotodonts. It is also present in a number
of the carnivorous Polyprotodont Marsupials; but in Phascologale
it is only present in rudiment, and in Myrmecobius it is entirely
obsolete. In the American Opossums the state of the pouch is variable.
"Generally absent, sometimes merely composed of two lateral folds of skin
separate at each end, rarely complete," is Mr. Thomas' summary in his
definition of the family Didelphyidae.[10] Another curious feature of the pouch in
the Marsupials is the variability in the position of the mouth of the
pouch: in all the Diprotodonts it looks forward; but in many
Polyprotodonts it looks backward. This, however, has some connexion with
the habitual attitude of the possessor: in the Kangaroo, leaping along on
its hind-legs, it is requisite that the pouch should open forwards; but
in the dog-like Thylacine, going on all fours, the fact that the pouch
{18}opens backwards is less disadvantageous to
the contained young.
The male Thylacine has a pouch which is quite or very nearly as well
formed as in the female. There are also rudiments of a pouch in the male
foetuses of many Marsupials, especially of those belonging to the
Polyprotodont section of the order, though these rudiments are by no
means confined to that subdivision. Up to so late a period as the age of
four months (length 19.8 cm.) the male Dasyurus ursinus has a
pouch.
We have now to consider the interesting series of facts relative to
the permanence—in a rudimentary condition it is true—of the
mammary pouch in the higher Mammalia, facts which seem to be an
additional proof that they have been derived from an ancestor in which
the pouch was an organ of functional importance. The first definite proof
of the occurrence of a pouch in any mammal not a Marsupial or a Monotreme
was made by Malkmus, who found this structure in a Sheep. It seems,
however, that the structures found in the higher mammals are not always
comparable to the marsupium of the Marsupials, but sometimes to the
mammary pouch of the Monotreme. That the Marsupials are a side line, and
not involved in the ancestry of the Eutheria, is an opinion which is at
present widely held. At the same time it is reasonable to suppose that
the original stock lying between the Prototheria and the Metatheria,
whence the latter and the Eutheria have arisen, preserved both the
mammary pouch of the lower mammal and the marsupium of the
further-developed stage, as does Phalangista occasionally at the
present day. Hence to find remnants of both structures in existing
mammals would not so incredible. This is what Dr. Klaatsch believes to be
the case. In certain Ungulates, including two species of Antelope, Dr.
Klaatsch found very considerable rudiments of folds provided with
unstriated muscular fibre; there were in the adult Cervicapra
isabellina a pair of pouches, one on each side, and a rudiment of a
second on either side; possibly this multiplication of the pouches has
relation to the number of young. That there is more than one pouch makes
a comparison with the mammary pouch rather than with the marsupium
probable. The Ungulate teat, it must be remembered (see p. 16), is a secondary teat; hence there is no difficulty
in the comparison from this point of view. A pouch containing a primary
{19}teat would of course be absolutely
incomparable with a mammary pouch, because in that case the wall of the
teat itself would be the pouch.
Mammals belonging to quite different Orders show traces more or less
marked of a marsupium. In young Dogs the teats are borne upon an area
where the skin is thinner, the covering of hair less dense than
elsewhere—all points of resemblance to the inside of the pouch of a
Marsupial; in addition to this there are traces of the sphincter marsupii
muscle. In other Carnivora there are similar vestiges. In Lemur
catta a more complete rudiment of a marsupial pouch is to be met
with. In this Lemur the teats are both inguinal and pectoral; the skin in
these regions is thin and but slightly hairy, and extends forwards as two
bands of the same thinness and smoothness on each side of the densely
hairy skin covering the sternum. This area is sharply separated from the
rest of the integument by a fold which runs parallel to the longitudinal
axis of the body, and can be comparable with nothing save the rudiment of
the marsupial fold.
One is tempted to wonder how far the habit which certain Lemurs have
of carrying their young across the abdomen with the tail wrapped round
the body of the mother is a reminiscence of a marsupial pouch.
Skeleton.
The skeleton of the Mammalia consists almost solely of the
endoskeleton. It is only among the Edentata that an exoskeleton of bony
plates in the skin is met with. As in other Vertebrates, the skeleton is
divisible into an axial portion, the skull and vertebral column, and an
appendicular skeleton, that of the limbs. The bones of mammals are well
ossified, and in the adult there are but few and small tracts of
cartilage left.
Vertebral Column.—The vertebral column of the mammals,
like that of the higher Vertebrata, consists of a number of separate and
fully-ossified vertebrae.
The constitution of a vertebra upon which all the usual processes are
marked is as follows:—There is first of all
the body or centrum of the vertebra, a massive piece of bone shaped like
a disc or a cylinder. The centra of contiguous vertebrae {20}are separated by
a certain amount of fibrous tissue forming the intervertebral disc, and
the apposed surfaces of the centra are as a rule nearly flat. In this
last feature, and in the important fact that the centra are ossified from
three distinct centres, the anterior and posterior pieces ("epiphyses")
remaining distinct for a time, even for a long time (as in the Whales),
the centra in the mammals differ from those of reptiles and birds. The
epiphyses are not found throughout the vertebral column of the
lowly-organised Monotremata, and they do not appear to exist in the
Sirenia.
|
|
|
|
Fig. 5.—Anterior surface of
Human thoracic vertebra
(fourth), × ⅔. az, Anterior
zygapophysis; c, body or
centrum; l, lamina, and
p, pedicle, of the neural
arch; nc, neural canal; t,
transverse process. (From
Flower's Osteology of the
Mammalia.)
|
Fig. 6.—Side view of first
lumbar vertebra of Dog
(Canis familiaris). × ¾.
a, Anapophysis; az, anterior
zygapophysis; m,
metapophysis; pz, posterior
zygapophysis; s,
spinous process; t, transverse
process. (From
Flower's Osteology.)
|
From each side of the centrum on the dorsal side arises a process of
bone which meets its fellow in the middle line above, and is from there
often prolonged into a spine. A canal is thus formed which lodges the
spinal cord. This arch of bone is known as the neural arch, and the
dorsal process of the same as the spinous process. The sides of the
neural arch bear oval facets, by which successive vertebrae articulate
with one another: those situated anteriorly are the anterior
zygapophyses, while those on the posterior aspect of the arch are the
posterior zygapophyses; these articular facets do not exist in the
tail-region of many mammals, e.g. Whales.
In addition to the dorsal median spinous process of the {21}vertebra there
may be a ventral median process, arising of course from the centrum,
termed the hypapophysis.
From the sides of the neural arch, or from the centrum itself, there
is commonly a longer or shorter process on each side, known as the
transverse process. This is sometimes formed of two distinct processes,
one above the other; in such cases the upper part is called a
diapophysis, the lower a parapophysis.
The neural arch may also bear other lateral processes, of which one
directed forwards is the metapophysis, the other directed backwards the
anapophysis.
The series of bones which constitute the vertebral column can be
divided into regions. It is possible to recognise cervical, dorsal,
lumbar, sacral, and caudal vertebrae. In the case of animals with only
rudimentary hind-limbs, such as the Whales, there is no recognisable
sacral region. The neck or cervical vertebrae are nearly always seven in
number. The well-known exceptions are the Manatee, where there are six,
and certain Sloths, where there are six, eight, or nine. These rare
exceptions only accentuate the very remarkable constancy in number, which
is very distinctive of the mammals as compared with lower Vertebrata.
There are of course abnormalities, the last cervical, and sometimes the
last two, assuming the characters of the ensuing dorsals, by developing a
more or less complete rib. There are also recorded examples of
Bradypus, in which the number of cervicals is increased to ten.
The characteristics, then, of the cervical vertebrae are, in the first
place, that they do not normally bear free ribs, and that there is a
break as a rule between the last cervical and the first dorsal on this
account. In birds, for example, the cervicals, differing in number in
different families and genera, gradually approach the dorsals by the
gradually lengthening ribs. The transverse processes of the vertebrae are
commonly perforated by a canal for the vertebral artery, and are bifid at
their extremities. In some Ungulates these vertebrae, moreover,
approximate to the vertebrae of lower Vertebrata in the fact that there
are ball and socket joints between the centra, instead of only the
fibrous discs of the remaining vertebrae.
The first two vertebrae of the series are always very different from
those which follow. The first is termed the {22}atlas, and articulates
with the skull. The most remarkable fact about this bone (shared,
however, by lower Vertebrates) is that its centrum is detached from it
and attached to the next vertebra, in connexion with which it will be
referred to immediately. The whole bone thus gets a ring-like form, and
the salient processes of other vertebrae are but little developed, with
the exception of the transverse processes, which are wide and wing-like.
In many Marsupials, such as the Wombat and Kangaroo, the arch of the
atlas is open below, there being no centre of ossification. In others,
such as Thylacinus, there is a distinct nodule of bone in this
situation not concrescent with the rest of the arch.
|
|
|
|
Fig. 7.—Human atlas (young), showing development.
× ¾. as, Articular surface for
occiput; g, groove for first spinal nerve
and vertebral artery; i a, inferior arch;
t, transverse process. (From Flower's
Osteology.)
|
Fig. 8.—Inferior surface of atlas of
Dog. × ½. sn, Foramen for first
spinal nerve; v, vertebrarterial
canal. (From Flower's Osteology.)
|
Fig. 9.—Atlas of Kangaroo.... (From
Parker and Haswell's Zoology.)
The second vertebra, which is known as the axis or epistropheus, is a
compound structure, the anterior "odontoid process," which fits into the
ring of the atlas, being in reality the detached centrum of that
vertebra.[11] It is a
curious fact about that process that it has independently become
spoon-shaped in two divisions of Ungulates; that it has become so seems
to be shown by the fact that in the earlier types of both it has the
simple peg-like form, which is the prevailing form. The cervical {23}vertebrae are occasionally wholly (Right
Whales) or partially (many Whales, Jerboa, certain Edentates) welded into
a combined mass. Indications of this have even been recorded in the human
subject.
|
|
|
|
Fig. 10.—Side view of axis of Dog.
× ⅔. o, Odontoid process; pz,
posterior zygapophysis; s, spinous
process; t, transverse process;
v, vertebrarterial canal.
(From Flower's Osteology.)
|
Fig. 11.—Anterior surface of axis
of Red Deer. × ⅔. o, Odontoid
process; pz, posterior
zygapophysis; sn, foramen for
second spinal nerve. (From
Flower's Osteology.)
|
The dorsal vertebrae vary greatly in number: nine (Hyperoodon)
seems to be the lowest number existing normally; while there may be as
many as nineteen, as in Centetes, or twenty-two, as in
Hyrax. These vertebrae are to be defined by the fact that they
carry ribs, and the first one or two lumbars are often "converted into"
dorsals by the appearance of a small supernumerary rib. The spinous
processes of these vertebrae are commonly long, and sometimes very long.
It is only among the Glyptodons that any of these vertebrae are fused
together into a mass.
The lumbar vertebrae, which follow the dorsal, vary greatly in number.
There are as few as two in the whale Neobalaena, as many as
seventeen in Tursiops; this group, the Cetacea, contains the
extremes. Nine lumbars are found in the Lemurs Indris and
Loris. As a rule the number of lumbars is to some extent dependent
upon that of the dorsals. It often happens that the number of
thoraco-lumbar vertebrae is constant for a given group. Thus the
Artiodactyles have nineteen of these vertebrae, and the Perissodactyles
as a rule twenty-three. A greater number of dorsals implies a smaller
number of lumbars, and of course vice versa. The existence of a
sacral region formed of a {24}number of vertebrae fused together and
supported by the pelvic girdle is characteristic of the mammals, but is
not found in the Cetacea and the Sirenia, where functional hind-limbs are
wanting. Strictly speaking, the sacrum is limited to the two or three
vertebrae whose expanded transverse processes meet the ilia. But to these
are or may be added a variable number of vertebrae withdrawn from both
the lumbar and the caudal series, which unite with each other to form the
massive piece of bone which constitutes the sacrum of the adult.
|
|
|
|
Fig. 12.—Lepus cuniculus. Innominate
bones and sacrum, ventral aspect.
acet, Acetabulum; il, ilium; isch,
ischium; obt, obturator foramen;
pub, pubis; sacr, sacrum; sy, symphysis.
(From Parker and Haswell's
Zoology.)
|
Fig. 13.—Anterior surface of fourth
caudal vertebra of Porpoise (Phocoena
communis), × ½. h, Chevron
bone; m, metapophysis; s, spinous
process; t, transverse process.
(From Flower's Osteology.)
|
The caudal vertebrae complete the series. They begin in as fully
developed a condition as the lumbars, with well-marked transverse
processes, etc.; but they end as no more than centra, from which
sometimes tiny outgrowths represent in a rudimentary way the neural
arches, etc. Very often the caudal vertebrae are furnished with ventral,
generally 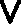 -shaped, appendages, the chevron bones or intercentra.[12] These are {25}particularly conspicuous
in the Whales and in the Edentates. In the former group the occurrence of
the first intercentrum serves to mark the separation of the caudal from
the lumbar series. The number of caudals varies from three in
Man—and those quite rudimentary—to nearly fifty in Manis
macrura and Microgale longicaudata.
-shaped, appendages, the chevron bones or intercentra.[12] These are {25}particularly conspicuous
in the Whales and in the Edentates. In the former group the occurrence of
the first intercentrum serves to mark the separation of the caudal from
the lumbar series. The number of caudals varies from three in
Man—and those quite rudimentary—to nearly fifty in Manis
macrura and Microgale longicaudata.

Fig. 14.—Lateral view of skull of a Dog. C.occ, Occipital condyle; F, frontal;
F.inf, infra-orbital foramen; Jg, jugal; Jm, premaxilla; L, lachrymal; M,
maxilla; Maud, external auditory meatus; Md, mandible; N, nasal; P,
parietal; Pal, palatine; Pjt, process of squamosal; Pt, pterygoid; Sph, alisphenoid;
Sq, squamosal; Sq.occ, supraoccipital; T, tympanic. (From Wiedersheim's
Comparative Anatomy.)
The Skull.—The skull in the Mammalia differs from that of
the lower Vertebrata in a number of important features, which will be
enumerated in the following brief sketch of its structure. In the first
place, the skull is a more consolidated whole than in reptiles; the
number of elements entering into its formation is less, and they are on
the whole more firmly welded together than in Vertebrates standing below
the Mammalia in the series. Thus in the cranial region the post- and
pre-frontals, the post-orbitals and the supra-orbitals have disappeared,
though now and again we are reminded of their occurrence in the ancestors
of the Mammalia by a separate ossification corresponding to some of the
bones. Nowhere is this consolidation seen with greater clearness than in
the lower jaw. That bone, or rather each half of it, is in mammals formed
of one bone, the dentary (to which occasionally, as it appears, a
separate mento-Meckelian {26}ossification may be added). The angular,
splenial, and all the other elements of the reptilian jaw have vanished,
though the numerous points from which the mammalian dentary ossifies is a
reminiscence of a former state of affairs; and here again an occasional
continuance of the separation is preserved, as the case observed by
Professor Albrecht of a separate supra-angular bone in a Rorqual attests.
Among other reptilian bones that are not to be found in the mammalian
skull are the basipterygoids, quadrato-jugal, and supratemporal. A few of
these bones, however, though no longer traceable in the adult skull save
in cases of what we term abnormalities, do find their representatives in
the foetal skull. Professor Parker, for example, has described a
supra-orbital in the embryo Hedgehog; a supratemporal also appears to be
occasionally independent.

Fig. 15.—Head of a Human embryo of the fourth month. Dissected to show the
auditory ossicles, tympanic ring, and Meckel's cartilage, with the hyoid and thyroid
apparatus. All these parts are delineated on a larger scale than the rest of the
skull. an, Tympanic ring; b.hy, basihyal element; hy, so-called hyoid bone; in,
incus; md, bony mandible; ml, malleus; st, stapes; tp, tympanum; tr, trachea;
I. (mk), first skeletal (mandibular) arch (Meckel's cartilage); II. second skeletal
(hyoid) arch; III. third (first branchial) arch; IV. V. fourth and fifth arches
(thyroid cartilage). (From Wiedersheim's Structure of Man.)
In the mode of the articulation of the lower jaw to the skull the
Mammalia apparently, perhaps really, differ from other Vertebrates. In
the Amphibia and Reptilia, with which groups alone any comparisons are
profitable, the lower jaw articulates by means of a quadrate bone, which
may be movably or firmly attached to the skull. In the mammals the
articulation of the lower jaw is with the squamosal. The nature of this
articulation is one of the most debated points in comparative anatomy.
Seeing that Professor Kingsley[13] in the most recent contribution to the
subject quotes no less than fifty-two different views, many of which are
more or less convergent, it will be obvious that in a work like the
present the matter cannot be treated exhaustively. As, however, Professor
Kingsley justly says that "no single bone occupies a more important
position in the discussion of the origin of the Mammalia than does the
quadrate," and with equal justice adds that "upon the answer given as to
its fate in this group depends, in large measure, the broader problem of
the phylogeny of the Mammalia," it becomes, or indeed has long been, a
matter which cannot be ignored in any work dealing with the mammals. A
simple view, due to the late Dr. Baur and to Professor Dollo, commends
itself at first sight as meeting the case. The last-named author holds,
or held, that in all the higher Vertebrates it is at least on
a priori grounds likely that two such characteristically
vertebrate features as the lower jaw and the chain of bones bringing the
outer world {27}into communication with the internal organ
of hearing would be homologous throughout the series. He believed,
therefore, that the entire chain of ossicula auditus in the mammal is
equal to the columella of the reptile, since their relations are the same
to the tympanum on the one hand and to the foramen ovale on the other;
and that the lower jaw articulates in the same way in both. It follows,
therefore, that the glenoid part of the squamosal must be the quadrate
which has become ankylosed with it after the fashion of concentration in
the mammalian skull that has already been referred to. The fact that
occasionally the glenoid part of the squamosal is a separate bone[14] appeared to confirm this
way of looking at the {28}matter. But the hall-mark of truth is not
always simplicity; indeed the converse appears to be frequently the case.
And on the whole this view does not commend itself to zoologists at
present. For it must be borne in mind that the lower jaw of the mammal is
not the precise equivalent of that of the reptiles. Apart from the
membrane bones, which may be collectively the equivalents of the dentary
of the mammal, there is the cartilaginous articular bone to be
considered, which forms the connexion between the rest of the jaw and the
quadrate in reptiles. Even in the Anomodontia, whose relations to the
Mammalia are considered elsewhere, there is this bone. But in these
reptiles the articular bone articulates not only with the quadrate, but
also to a large extent with the squamosal, the quadrate shrinking in size
and developing processes which give to it very much the look of either
the incus or the malleus of the mammalian ear. In fact it seems on the
whole to fit in with the views of the majority, as well as with a fair
interpretation of the facts of embryology, to consider that the chain of
ear bones in the mammal is not the equivalent of the columella of the
reptile, but that the stapes of the mammal is the columella, and that the
articulare is represented by the malleus and the quadrate by the incus.
It is very interesting to note this entire change of function in the
bones in question. Bones which in the reptile serve as a means of
attachment of the lower jaw to the skull are used in the mammal to convey
the waves of sound from the tympanum of the ear to the internal organ of
hearing.
Another important and diagnostic feature in the mammalian skull is
that the first vertebra of the vertebral column always articulates with
two separate occipital condyles, which are borne by the exoccipital bones
and formed mainly though not entirely by them. Certain Anomodontia form
the nearest approach to the mammals in this particular. The two condyles
of Amphibia are purely exoccipital in origin.
In the Mammalia, unlike what is found in lower Vertebrates (but here
again the Anomodontia form at least a partial exception), the jugal arch
does not connect the face with the quadrate, for, as already said, that
bone does not exist, in the Sauropsidan form, in mammals. This arch
passes from the squamosal to the maxillary, and has but one separate bone
in addition to those two, viz. the jugal or malar. {29}

Fig. 16.—Under surface of the cranium of a Dog.
× ½. apf, Anterior palatine foramen; as, posterior
opening of alisphenoid canal; AS,
alisphenoid; BO, basioccipital; BS, basisphenoid;
cf, condylar foramen; eam, external
auditory meatus; Ex.O, exoccipital;
flm, foramen lacerum medium; flp, foramen
lacerum posterius; fm, foramen magnum;
fo, foramen ovale; fr, foramen rotundum;
Fr, frontal; gf, glenoid fossa; gp, post-glenoid
process; Ma, malar; Mx, maxilla;
oc, occipital condyle; op, optic foramen;
Per, mastoid portion of periotic; pgf, post-glenoid
fossa; Pl, palatine; PMx, premaxilla;
pp, paroccipital process; ppf, posterior
palatine foramen; PS, presphenoid;
Pt, pterygoid; sf, sphenoidal fissure or foramen
lacerum anterius; sm, stylomastoid foramen;
SO, supraoccipital; Sq, zygomatic
process of squamosal; Ty, tympanic bulla;
Vo, vomer. (From Flower's Osteology.)
In connexion with the elaboration of the chain of auditory ossicles it
is very usual for mammals to possess a thin inflated bone, sometimes
partly or entirely formed out of the tympanic bone, and known as the
tympanic bulla. Whether this structure is thin and inflated or thick and
depressed in form it is characteristic of the mammals, and does not occur
below them in the series. But it is not present in all mammals. It is
absent, for example, in the Monotremes. When it is present it is
sometimes formed from other bones, as, for instance, from the
alisphenoids. The tympanic ring has been held to be the equivalent of the
quadrate. It is more probably the quadrato-jugal.[15]

Fig. 17.—A, First thoracic skeletal segment for comparison with B, fifth cervical vertebra
(Man), b.v. Body of vertebra; c, first thoracic rib; c′, cervical rib (which has become
united with the transverse process, tr), the two enclosing the costo-transverse
foramen (f.c.t); st, sternum; zy, articular process of the arch (zygapophysis).
(From Wiedersheim's Structure of Man.)
Ribs.—All mammals are furnished with ribs, of which the
number of pairs differs considerably from group to group, or it may be
even from species to species. The ribs are attached as a rule by two
heads, of which one, the capitulum, arises as a rule between two centra
of successive vertebrae. The other, the tuberculum, springs from the
transverse process. Only in the Monotremes {30}are there ribs with but
one, the capitular, head. In the posterior part of the series the two
heads often gradually coalesce, so that there comes to be but one, the
capitular, head. The Whales also, at least the Whalebone Whales, are
exceptional in possessing but one head to the ribs, which is the
capitular. The first rib joins the sternum below, and a variable number
after this have the same attachment. There are always a number of ribs,
sometimes called floating ribs, which have no sternal attachment. In the
Whalebone Whales it is the first rib alone which is so attached. As a
rule, to which the Whales mentioned are again an exception, the rib is
divided into at least two regions—the vertebral portion which is
always ossified, and the sternal moiety which is usually cartilaginous.
This is, however, often very short in the first rib. They are, however,
ossified in the Armadillos and in some other animals. Between the
vertebral and sternal portions an intermediate tract is separated off and
ossified in the Monotremata. The ribs of existing mammals belong only to
the dorsal region of the vertebral column, but there are traces of lumbar
ribs and also of cervical ribs. In the Monotremata, indeed, these latter
{31}are
persistently free for a very long period, and in some cases never become
ankylosed with their vertebrae. But it should be noted that in this group
there is no approximation to the state of affairs which exists in many
lower Vertebrates, where there is a gradual transition between the ribs
of the cervical and those of the dorsal region of the vertebral column;
for that of the seventh ribs in Monotremes is smaller than those which
precede it.
|
|
|
|
Fig. 18.—Sternum and sternal ribs of the
Common Mole (Talpa europaea), with
the clavicles (cl) and humeri (H); M,
manubrium sterni. Nat. size. (From
Flower's Osteology.)
|
Fig. 19.—Sternum
of the Pig (Sus
scrofa). × ¼. ms,
Mesosternum;
ps, presternum;
xs, xiphisternum.
(From Flower's
Osteology.)
|
The Sternum.—All the Mammalia so far as is known possess
a sternum. This is the bone, or series of bones (sternebrae), which lies
upon the ventral surface of the chest, and to which the ribs are attached
below. The development of the sternum has been shown to take place from
the fusion of the ribs below into two lateral bands, one on each side;
the approximation of these bands forms the single and unpaired sternum of
most mammals. Very considerable traces, however, of the paired state of
the sternal bones often exist; thus in the Sperm Whale the first piece of
the sternum is divided into two by a longitudinal division, and the
second piece is longitudinally grooved. The development of the sternum
out of the fused ends of ribs is shown in a more complete condition in
some species of Manis than in many other mammals. Thus in M.
tricuspis the last ribs of those which are attached to the sternum
are completely fused together into a single piece on each side.[16] As a general rule the
last ribs which come into relation with the sternum do so only in an
imperfect way, being simply firmly attached at their sides to, but not
fused with, the last ribs which are definitely articulated with the
sternum. Contrary to what is found in lower {32}Vertebrates, the sternum
of the Mammalia consists of a series of pieces, as many as eight or nine
or even sixteen in Choloepus, of which the first is called the
manubrium sterni, and the last the ensiform cartilage, xiphisternum, or
xiphoid process. The latter often remains largely cartilaginous
throughout life; in fact this is generally but not universally the case
with that part of the breastbone. The most extraordinary modification of
the xiphoid process is seen in the African species of the genus
Manis, where it diverges into two long cartilages, which run back
to the pelvis and then, curving round, run forwards and fuse together in
the middle line anteriorly. These processes serve for the attachment of
certain tongue-muscles. They were looked upon by Professor Parker as the
equivalents of the "abdominal ribs" of reptiles elsewhere non-existent
among mammals. This view is not, however, usually held. The manubrium
sterni is often keeled in the middle line below; this is so with the
Bats, which thus approach the birds, and probably for the same reason,
i.e. the need of an enlarged origin for the pectoral muscle, which is
concerned in the movements of flight. In many forms this part of the
sternum is much broader than the pieces which follow; this is so with the
Viscacha. In the Pig the precise reverse is seen, the manubrium being
narrower than the rest of the sternal bonelets. It will be noticed,
however, that in this and similar cases there are no clavicles. Ribs are
attached between the successive pieces of the sternum. When the sternum
is reduced, as it is in the Cetacea and in the Sirenia, it is the
intermediate part of the series of bones which becomes abbreviated or
vanishes. The Sperm Whale has only a manubrium sterni and a following
piece belonging to the mesosternum. It is fair to say that the xiphoid
process and the rest of the sternum have disappeared, since among the
Toothed Whales a progressive shortening of the sternum can be seen. In
the Whalebone Whales the sternum is still further reduced; the manubrium
is alone left, and to it are attached but a single pair of ribs. In
Balaena, however, a rudimentary {33}piece, apparently
comparable to a xiphoid process, has been detected.
|
|
|
|
Fig. 20.—Sternum of Rudolphi's Whale (Balaenoptera
borealis), showing its relation to
the inferior extremities of the first pair of
ribs. × 1⁄10. (From Flower's Osteology.)
|
Fig. 21.—Sternum of a young
Dugong (Halicore indicus).
× ¼. From a specimen in
the Leyden Museum, ps,
Presternum; xs, xiphisternum.
(From Flower's Osteology).
|
From the instances which have been described, as well as from the mode
of development of the sternum and from the number of free ribs,
i.e. ribs which are not attached to it, it would seem that the
sternum has undergone a considerable reduction in its size. This
reduction may be possibly accounted for by the need for respiratory
activity, which is clearly increased by a less-marked fixity of the walls
of the thoracic cavity. In the case of the Whales one can hardly help
coming to that conclusion. The arrangement in the Monotremata does not,
however, point in the same direction; for these animals are precisely
like the higher Mammalia in the reduction of the sternum and of the
number of ribs which reach it.
Fig. 22.—Shoulder girdle of Ornithorhynchus. c1, c2, c3, First, second, third ribs; cl,
clavicle; e.c, epicoracoid; es′ and es″, interclavicle (episternum); m.c, metacoracoid;
m.s, manubrium sterni; sc, scapula; st, sternebra. (From Wiedersheim's Structure
of Man.)
Fig. 23.—Episternum of an embryo
Mole. (After A. Götte.)
cl, Clavicle; es′, central portion
of the episternum; es″,
lateral portion of the same;
r.c, costal ribs; st, sternum.
(The figure was constructed
from two consecutive horizontal
sections.) (From
Wiedersheim's Structure of
Man.)
The Episternum.—The Mammalia are as a rule to be
distinguished from lower Vertebrates by the absence of an episternum, or
interclavicle as it is also called. In the Monotremata, however, there is
a large  -shaped bone which does not overlie the sternum as in reptiles, but is
anterior to it. The relations of this bone to the clavicles seem to leave
no doubt that it is the equivalent of the Lacertilian interclavicle or
episternum. The Monotremata are not, however, the only mammals in which
this structure is to be seen. The Mole in the embryonic condition is {34}provided
with pieces of bone which overlie the manubrium sterni and are attached
to the clavicles, and are no doubt to be regarded as the same structure.
Probably in many mammals the manubrium will be found to be partly made up
of corresponding rudiments. In any case, vestiges of an episternum in the
shape of two minute ossicles have been discovered in Man, lying in front
of the manubrium. They have been termed ossa suprasternalia. In Man and
in the Mole the paired nature of the episternum is clearly apparent. It
has been suggested that this structure in its entirety belongs to the
clavicles, just as the sternum belongs to the ribs; i.e. that it
formed out of the approximated and fused ends of the clavicles. Dr.
Mivart[17] figured a good
many years since a pair of ossicles in Mycetes, lying in one case
between the ends of the clavicles and the manubrium sterni, and in
another example anterior to the ventral ends of the clavicles. Gegenbaur
has figured a {35}pair of similar bones in the Hamster.[18] It is possible that these
are to be referred to the same category. It has also been suggested that
these supposed episternal rudiments are the vestiges of a pair of
cervical ribs.
-shaped bone which does not overlie the sternum as in reptiles, but is
anterior to it. The relations of this bone to the clavicles seem to leave
no doubt that it is the equivalent of the Lacertilian interclavicle or
episternum. The Monotremata are not, however, the only mammals in which
this structure is to be seen. The Mole in the embryonic condition is {34}provided
with pieces of bone which overlie the manubrium sterni and are attached
to the clavicles, and are no doubt to be regarded as the same structure.
Probably in many mammals the manubrium will be found to be partly made up
of corresponding rudiments. In any case, vestiges of an episternum in the
shape of two minute ossicles have been discovered in Man, lying in front
of the manubrium. They have been termed ossa suprasternalia. In Man and
in the Mole the paired nature of the episternum is clearly apparent. It
has been suggested that this structure in its entirety belongs to the
clavicles, just as the sternum belongs to the ribs; i.e. that it
formed out of the approximated and fused ends of the clavicles. Dr.
Mivart[17] figured a good
many years since a pair of ossicles in Mycetes, lying in one case
between the ends of the clavicles and the manubrium sterni, and in
another example anterior to the ventral ends of the clavicles. Gegenbaur
has figured a {35}pair of similar bones in the Hamster.[18] It is possible that these
are to be referred to the same category. It has also been suggested that
these supposed episternal rudiments are the vestiges of a pair of
cervical ribs.

Fig. 24.—Episternal
vestiges in Man.
cl, Clavicle, sawn
through; es, "episternum"
(sternoclavicular
cartilage);
l′, interclavicular
ligament; l″, costoclavicular
ligament;
m.s, manubrium
sterni; o.s, ossa
suprasternalia; r.c,
first rib; st, sternum.
(From Wiedersheim's
Structure
of Man.)
The Pectoral Girdle.—The skeleton by which the fore-limb
is connected with the trunk is known as the Pectoral Girdle. The main
part of this girdle is formed by the large scapula, or blade-bone as it
is often termed. The coracoidal elements will be dealt with later. The
scapula is not firmly connected with the backbone; it is attached merely
by muscles, thus presenting a great difference from the corresponding
pelvic girdle. The reason for this difference is not easy to understand.
On the one hand it may be pointed out that in all running animals at any
rate there is a greater need for the fixation in a particularly firm way
of the hind-limbs; but, again, in the climbing creatures both limbs
would, one might suppose, be bettered by a firm fixation. It must be
remembered, however, that in the latter case the same result is at least
partly brought about by a well-developed clavicle, which fixes the girdle
to the sternum and so to the vertebral column by means of the ribs.
Broadly speaking, too, the fore-limbs require a greater freedom and
variety of movement than the hind-limbs, which are supports {36}for or serve to
push along the rapidly-moving body. Stronger fixation is therefore a
greater necessity posteriorly than anteriorly. In any case, whatever the
explanation, this important difference exists.
|
Fig. 25.—Right scapula of
Dog (Canis familiaris).
× ¼. a, Acromion; af,
prescapular fossa; c,
coracoid; cb, coracoid or
anterior border; css, indicates
the position of
the coraco-scapular suture,
obliterated in adult
animals by the complete
ankylosis of the two
bones; gb, glenoid or
posterior border; gc,
glenoid cavity; pf, postscapular
fossa; s, spine;
ss, suprascapular border.
(From Flower's Osteology.)
|
Fig. 26.—Right scapula of
Red Deer (Cervus elaphus).
× ¼. a, Acromion; af, anterior
or prescapular fossa;
c, coracoid; gc, glenoid
cavity; pf, postscapular
fossa; ss, partially ossified
suprascapular border.
(From Flower's Osteology.)
|
The shoulder-blade of mammals is as a rule a much-flattened bone with
a ridge on the outer surface known as the spine; this ridge ends in a
freely-projecting process, the acromion, from which a branch often arises
known as the metacromion. This gives a bifurcate appearance to the end of
the ridge. The spine is less developed and the scapula is narrower in
such animals as the Dog and the Deer which simply run, and whose
fore-limbs therefore are not endowed with the complexity of movement
seen, for instance, in the Apes.
{37}
Fig. 27.—Right scapula of Dolphin (Tursiops tursio).
× ¼. a, Acromion; af, prescapular fossa; c, coracoid;
gc, glenoid cavity; pf, postscapular fossa.
(From Flower's Osteology.)
Fig. 28.—Side view of right half of
shoulder girdle of a young Echidna
(Echidna hystrix). × ⅔. a, Acromion;
c, coracoid; cb, coracoid
border; cl, clavicle; css, coraco-scapular
suture; ec, epicoracoid;
gb, glenoid border; gc, glenoid
cavity; ic, interclavicle; pf, postscapular
fossa; ps, presternum; s,
spine; ss, suprascapular epiphysis;
ssf, subscapular fossa. (From
Flower's Osteology.)
It has been pointed out that the area which lies in front of the
spine, the prescapular lamina, is most extensively developed in such
animals as perform complex movements with the fore-limbs. The Sea Lion
and the Great Anteater are cited by Professor G. B. Howes as examples of
this preponderance of the anterior portion of the scapula over that which
lies behind the spine. The general shape of the scapula varies
considerably among the different orders of mammals; but it always
presents the characters mentioned, which are nowhere seen among the
Sauropsida except among certain Anomodonts, which will be duly referred
to (see p. 90). The most conspicuous divergences
from the normal are to be found in the Cetacea and the Monotremata. In
the former the acromion is approximated so nearly to the anterior border
of the blade-bone that the prescapular fossa is reduced to a very small
area; and in Platanista the acromion actually coincides with the
anterior border, so that that fossa actually disappears. In the Whales,
too, the scapula is as a rule very broad, especially above; it has
frequently a fan-like contour. In the Monotremata the acromion also
coincides with the anterior border of the scapula; but the sameness of
appearance which it thus presents (in this feature) to the Cetacean
scapula is {38}apparently not due to real resemblance. What
has happened in the Monotremata is, that the prescapular fossa is so
enormously expanded that it occupies the whole of the inner side of the
blade-bone, while the subscapular fossa which, so to speak, should occupy
that situation, has been thus pushed round to the front, where it is
divided from the postscapular fossa by a slight ridge only.
The clavicle is a bone which varies much in mammals. It is sometimes
indeed, as in the Ungulata, entirely absent; in other forms it shows
varying degrees of retrocession in importance; it is only in climbing,
burrowing, digging, and flying mammals that it is really well
developed.
Fig. 29.—Shoulder girdle, with upper end of sternum (inner
surface) of Shrew (Sorex), after Parker, × 7. a, Acromion;
c, coracoid; cl, clavicle; ec, partially ossified
"epicoracoid" of Parker, or rudiment of the sternal
extremity of the coracoid; ''ma'', metacromial process;
mss, ossified "mesoscapular segment"; ost, omosternum;
pc, rudiment of precoracoid (Parker); ps, presternum;
sr1, first sternal rib; sr2, second sternal rib. (From
Flower's Osteology.)
In the higher Mammalia the coracoid[19] is present, but does not reach the
sternum as in the Monotremata. It is known to human anatomists as the
coracoid process of the scapula. It has been found, however, by Professor
Howes[20] and others, that
this process really consists of two separate centres of ossification,
forming two separate bonelets, which in the adult become firmly ankylosed
to each other and to the scapula. These two separate bones have been met
with in the embryo of Lepus, Sciurus, and the young of various
other mammals belonging to very diverse orders, such as Edentates and
Primates. The separation even occasionally persists in the adult. The
question is, What is the relation of these bonelets to the coracoid of
the Monotremata and to the corresponding regions of reptiles? Professor
Howes terms the lower patch of bone the metacoracoid and the upper the
epicoracoid; {39}the former is alone concerned with the
glenoid cavity. It must therefore, one would suppose, correspond to the
"coracoid" of the Monotremata, while the upper piece of bone is the
epicoracoid process of that mammal. The Mammalia, therefore, higher as
well as lower, differ from the reptiles in that the coracoid is formed of
two bones, the exceptions being, among some other extinct forms, certain
of the Anomodontia, a group which it will be recollected is the nearest
of all reptiles to the mammals.

Fig. 30.—Distal extremity
of the humerus to
show Epicondylar Foramina.
A, In Hatteria;
B, in a Lizard (Lacerta
ocellata); C, in the
Domestic Cat; D, in
Man. c.e, External condyle;
c.i, internal condyle.
In A the two
foramina are developed
(at i, the entepicondylar;
at ii, the ectepicondylar).
The only
canal (†) present in the
Lizard (B) is on the
external ulnar side, in
the cartilaginous distal
extremity. In Man (D)
an entepicondylar process
(pr) is sometimes
developed and continued
as a fibrous band.
(From Wiedersheim's
Anatomy of Man.)
The Fore-limb.—The humerus is of varying length among
mammals. A feature which it sometimes shares with the humerus of lower
forms is the presence of an entepicondylar foramen, a defect of
ossification situated above the inner condyle of that bone which
transmits a nerve. The same foramen and an additional ectepicondylar
foramen are found in the ancient reptilian type Hatteria
(Sphenodon); it occurs also in the Anomodont reptiles. It is as a
rule only the lower forms among mammals which show this foramen; thus it
is present in the Mole and absent in the {40}Horse. The fact that it
is occasionally met with in Man is an additional proof of the, in many
respects, ancient structure of the highest type of Primate.
The radius and the ulna, which together constitute the fore-arm, are
both present in a large number of mammals, but the ulna tends to vanish
in the purely walking and digitigrade Ungulates, being present, however,
in the more ancient forms of these Ungulates. In Man and in many other
mammals the radius can be moved from its normal position and crossed over
the ulna; this movement of pronation has been permanently fixed in the
Elephant, where the bones are crossed but cannot be altered in position
by the contractions of any muscles. Other types agree with the Elephant
in this fixation of the two bones.
Fig. 31.—Bones of fore-arm
and manus of Mole (Talpa
europaea). × 2. c, Cuneiform;
ce, centrale; l, lunar;
m, magnum; p, pisiform;
R, radius; rs, radial sesamoid
(falciform); s, scaphoid;
td, trapezoid; tm,
trapezium; U, ulna; u,
unciform; I-V, the digits.
(From Flower's Osteology.)
The bones of the wrist show great variation among mammals. The
greatest number present are to be seen in such a type as the Mole. Here
we have a proximal row, consisting of the scaphoid, lunar, cuneiform, and
pisiform, which are arranged in their proper order, beginning with that
on the radial side of the limb, that side which bears the first digit. A
second row articulates proximally with these bonelets and distally with
the metacarpals; the bones composing it are, mentioning them in the same
order, trapezium, trapezoid, centrale, magnum, unciform.
The centrale does not, however, really belong to the distal carpal
row, and is as a rule situated in the middle of the carpus away from
articulation with the metacarpals. It is a bone which is not commonly
present in the mammalian hand, but is present in various lower forms,
such as the Beaver and Hyrax. It also occurs in such high types as the
majority of Monkeys; it is to be found in the Human foetal carpus. Many
extinct forms possessed a separate centrale. Its importance in the
formation of the interlocking condition of the Ungulate foot is referred
to later, {41}on p. 196. The only
mammal which appears to have the proper five bones in the distal row of
the carpus corresponding to the five metacarpals is Hyperoodon,
where this state of affairs at least occasionally occurs. The final bone
of that series, the unciform, seems to represent two bones fused. Very
often the carpus is reduced by the fusion of certain of the carpal bones;
thus among the Carnivora it is usual for the scaphoid and the lunar to be
fused. It is interestingly significant that these bones retain their
distinctness in the ancestral Creodonts. In many Ungulates the trapezium
vanishes. The reduction of the toes in fact implies a reduction of the
separate elements of the carpus.
As to the digits of the mammalian hand, the greatest number is five,
the various supplementary bonelets known as prepollex and postminimus
being, it is now generally held, merely supplementary ossifications not
representing the rudiments of pre-existing fingers. They may, however,
bear claws.[21] The number
of phalanges which follow upon the metacarpals is almost constantly three
in the mammals, excepting for the thumb, which has only two. This is
highly characteristic of the group as opposed to reptiles and birds, and
the increase in the number of these bones in the Whales and to a very
faint degree in the Sirenia is a special reduplication, which will be
mentioned when those animals are treated of.
The Pelvic Girdle.—The pelvic girdle or hip girdle is the
combined set of bones which are attached on the one hand to the sacrum
and on the other articulate with the hind-limb. Four distinct elements
are to be recognised in each "os innominatum," the name given to the
conjoined bones of each half of the entire pelvis. These are:—the ilium, which articulates with the sacrum;
the ischium, which is posterior; the pubis, which is anterior; and
finally, a small element, the cotyloid, which lies within the acetabular
cavity where the femur articulates. The epipubes of the Monotreme and the
Marsupial are dealt with elsewhere (see p. 116) as
they are peculiar to those groups.
Professor Huxley pointed out many years since that while the Eutherian
Mammalia differ from the reptiles in the fact that the axis of the ilium
lies at a less angle with that of the sacrum, {42}Ornithorhynchus
comes nearest to the reptile in the fact that this axis is nearly at
right angles to that of the sacrum. It is particularly interesting to
find that this peculiarity of Ornithorhynchus is only acquired
later in life, and that the pelvis of the foetus conforms in these angles
to the adults of other mammalian groups. In any case, the backward
rotation of the pelvis is a mammalian characteristic, and it is most
nearly approached among reptiles by the extinct Anomodontia, whose
affinities to mammals will be dealt with on a later page (p. 90). Another peculiarity of the mammalian pelvis
appears to be the cotyloid bone already referred to. In the Rabbit this
bone completely shuts out the pubis from any share in the acetabular
cavity; later it ankyloses with that bone. In Ornithorhynchus the
cotyloid or os acetabuli is a larger element of the girdle than is the
pubis. In other mammals, therefore, it seems to be a rudimentary
structure. But it seems to be a bone peculiar to and thus distinctive of
the mammals as compared with other vertebrates. The acetabular cavity is
perforated in Echidna as in birds; but in certain Rodents the same
region is very thin and only closed by membrane, as in Circolabes
villosus.
The number and the arrangement of the bones in the hind-limb
correspond exactly to those of the fore-limb. The femur, which
corresponds to the humerus, shows some diversities of form. The neck,
which follows upon the almost globular head, the surface of articulation
to the acetabular cavity of the pelvis, has two roughened areas or
tuberosities for the insertions of muscles. A third such area, known as
the third trochanter, is present or absent as the case may be, and its
presence or absence is of systematic import. As a general rule the
thigh-bones of the ancient types of mammals are smoother and less
roughened by the presence of these three trochanters than in their modern
representatives. The radius and the ulna are represented in the hind-leg
by the tibia and the fibula. These bones are not crossed, and do not
allow of rotation as is the case with the radius and the ulna. In
Ungulate animals there is the same tendency to the shortening and
rudimentary character of the fibula that occurs in the case of the ulna,
but it is more marked. It has been shown in tracing the history of fossil
Ungulates that the hind-limbs in their degree of degeneration are as a
rule ahead of the fore-limbs. This is natural when we reflect that {43}the
hind-limbs must have preceded the fore-limbs in their thorough adaptation
to the cursorial mode of progression. In the Mammalia the ankle-joint is
always what is termed cruro-tarsal, i.e. between the ends of the
limb-bones and the proximal row of tarsals; not in the middle of the
tarsus as in some Sauropsida (reptiles and birds). The bones of the ankle
are much like those of the hand; but there are never more than two bones
in the proximal row, which are the astragalus and the calcaneum. The
former is perhaps to be looked upon as the equivalent of the cuneiform
and lunar together. But the views as to the homologies of the tarsal
bones differ widely. Below these is the navicular, regarded as a
centrale. The distal row of the tarsus has four bones, three cuneiforms
and a cuboid. Reduction is effected by the soldering together of two
cuneiforms as in the Horse, by the fusion of the navicular and cuboid as
in the Deer. No mammal has more than five toes, and the number tends to
become reduced in cursorial animals (Rodents, Ungulates, Kangaroos).

Fig. 32.—Anterior aspect of
right femur of Rhinoceros
(Rhinoceros indicus). × ½.
h, Head; t, great trochanter;
t′, third trochanter.
(From Flower's Osteology.)
Teeth.—The teeth of the Mammalia[22] differ from those of other vertebrated
animals in a number of important points. These, however, entirely concern
the form of the adult teeth, their position in the mouth, and the
succession of the series of teeth. Developmentally and histologically
there are no fundamental divergences from the teeth of vertebrates lower
in the scale.
In mammals, as for example in the Dog, the teeth consist of three
kinds of tissue—the enamel, the dentine, and the cement. The enamel
is derived from the epidermis of the mouth cavity, and the two remaining
constituents from the underlying dermis. The teeth originate quite
independently of the jaws, with which they are later so intimately
connected; the independence of origin being one of the facts upon which
the current theory {44}of the nature of teeth is founded. It has
been pointed out that the scales of the Elasmobranch fishes consist of a
cap of enamel upon a base of dentine, the former being derived from the
epidermis and modelled upon a papilla of the dermis whose cells secrete
the dentine. The fact that similar structures arise within the mouth
(i.e. the teeth) is explicable when it is remembered that the
mouth itself is a late invagination from the outside of the body, and
that therefore the retention by its tissues of the capacity to produce
such structures is not remarkable.

Fig. 33.—Diagrammatic sections of various forms
of teeth. I, Incisor or tusk of Elephant,
with pulp cavity persistently open at base;
II, Human incisor during development, with
root imperfectly formed, and pulp cavity
widely open at base; III, completely formed
Human incisor, with pulp cavity opening by
a contracted aperture at base of root; IV,
Human molar with broad crown and two
roots; V, molar of the Ox, with the enamel
covering the crown deeply folded, and the
depressions filled up with cement; the surface
is worn by use, otherwise the enamel
coating would be continuous at the top of the
ridges. In all the figures the enamel is black,
the pulp white; the dentine represented by
horizontal lines, and the cement by dots.
(After Flower and Lydekker.)
The relations of the three constituents of the tooth in its simplest
form is shown in the accompanying diagram, where the intimate structure
of the enamel, dentine, and cement (or crusta petrosa as it is sometimes
called) is not indicated. The latter has the closest resemblance to bone.
The dentine is traversed by fine canals which run parallel to each other
and anastomose here and there. The enamel is formed of long prismatic
fibres, and is excessively hard in structure, containing less animal
matter than the other tooth tissues. To this fact is frequently {45}due the
complicated patterns upon the grinding teeth of Ungulates, which are
produced by the wearing away of the dentine and the cement, and the
resistance of the enamel.
The centre of the tooth papilla remains soft and forms the pulp of the
tooth, which is continuous with the underlying tissues of the gum by a
fine canal or a wide cavity as the case may be. In teeth which
persistently grow throughout the lifetime of the animal, as for example
the incisors of the Rodents, there is a wide intercommunication between
the cavity of the tooth and the tissues of the gum; only a narrow canal
exists in, for instance, the teeth of Man, and in fact in the vast
majority of cases. The three constituents of the typical teeth are not,
however, found in all mammals; the layer which is sometimes wanting is
the enamel. This is the case with most Edentates; but the interesting
discovery has been made (by Tomes) that in the Armadillo there is a
downgrowth of the epidermis similar to that which forms the enamel in
other mammals, a rudimentary "enamel organ."
Teeth are present in nearly all the Mammalia; and where they are
absent there is frequently some evidence to show that the loss is a
recent one. The Whalebone Whales, the Monotremata, Manis, and the
American Anteaters among the Edentata are devoid of teeth in the adult
state. In several of these instances, however, more or less rudimentary
teeth have been found, which either never cut the gums or else become
lost early in life. The latter is the case with Ornithorhynchus,
where there are teeth up to maturity (see p. 113).
Kükenthal has found germs of teeth in Whales, and Röse in the Oriental
Manis. The loss of the teeth in these cases seems to have some
relation to the nature of the food. In ant-eating mammals, as in the
Anteaters and Echidna, the ants are licked up by the long and
viscid tongue, and require no mastication. Yet it must be remembered that
Orycteropus is also an anteater, like the Marsupial
Myrmecobius, both of which genera have teeth.
The first of the essential peculiarities of the mammalian teeth as
compared with those of other vertebrates concerns the position of the
teeth in the mouth. There is no undoubted mammal extinct or living in
which the teeth are attached to any bones other than the dentary, the
maxilla, and the {46}premaxilla. There are no vomerine, palatine,
or pterygoid teeth, such as are met with in Amphibia and Reptilia.
The other peculiarities of the mammalian teeth, though true of the
great majority of cases, are none of them absolutely universal.
But it is necessary to go into the subject at some length on account
of the great importance which has been laid upon the teeth in deciding
questions of relationship; moreover, largely no doubt on account of their
hardness and imperishability, our knowledge of certain extinct forms of
Mammalia is entirely based upon a few scattered teeth; while of some
others, notably of the Triassic and Jurassic genera, there is not a great
deal of evidence except that which is furnished by the teeth. Indeed the
important place which odontography holds in comparative anatomy is from
many points of view to be regretted, though inevitable. "In hardly any
other system of organs of vertebrated animals," remarks Dr. Leche, "is
there so much danger of confounding the results of convergence of
development with true homologies, for scarcely any other set of organs is
less conservative and more completely subservient to the lightest impulse
from without." Affinities as indicated by the teeth are sometimes in
direct contradiction to those afforded by other organs; or, as in the
case of the simple Toothed Whales, no evidence of any kind is
forthcoming. Dr. Leche has pointed out that, judged merely from its
teeth, Arctictis would be referred to the Raccoons, though it is
really a Viverrid; while Bassariscus, which Sir W. Flower showed
to be a Raccoon, is in its teeth a Viverrid. Mr. Bateson has been obliged
to hamper the subject with another difficulty.
In dealing with the variations of teeth,[23] Mr. Bateson has brought together an
immense number of facts, which tend to prove that the variability of
these structures is much greater than had been previously recognised;
that this variability is often symmetrical; and that in some animals, as
in "Canis cancrivorus, a South American fox, the majority showed
some abnormality." When we learn from Mr. Bateson that "of Felis
fontanieri, an aberrant leopard, two skulls only are known, both
showing dental abnormalities," it seems dangerous to rear too lofty a
superstructure upon a single fossil jaw. It must be noted too that, {47}contrary
to the prevailing superstition, it is not domestic animals which show the
greatest amount of tooth variation. As to special homologies between
tooth and tooth, with which we shall deal on a later page, Mr. Bateson
has urged almost insuperable difficulties.

Fig. 34.—Skull of Dasyurus (lateral view). al.sph, Alisphenoid; ang, angular process
of mandible; fr, frontal; ju, jugal; lcr, lachrymal; max, maxilla; nas, nasal;
oc.cond, occipital condyle; par, parietal; par.oc, paroccipital process; p.max,
premaxilla; s.oc, supraoccipital; sq, squamosal; sq′, zygomatic process of squamosal.
(From Parker and Haswell's Zoology.)
Fig. 35.—Upper and lower teeth of one side of the mouth of a Dolphin (Lagenorhynchus),
illustrating the homodont type of dentition in a mammal. (After Flower
and Lydekker.)
The teeth of the Mammalia are almost without exception "heterodont,"
i.e. they show differences of structure in different parts of the
mouth. As a general rule, teeth can be grouped into cutting incisors,
sharp conical canines, and molars, with a surface which is in the
majority of cases suited for grinding. In this they contrast with the
majority of the lower vertebrates, where the teeth are "homodont" (or,
better, homoeodont), i.e. all more or less similar and not
fitted by change of form to perform different duties. But there are
exceptions on both sides. In {48}the Toothed Whales the teeth are homodont,
as they are in the frog and in most reptiles; on the other hand, some of
the remarkable reptiles belonging to Professor Huxley's order of the
Anomodontia have distinct canines, and show other differentiations in
their teeth.
A second characteristic of the mammalian dentition is the limited
number of the teeth, which rarely exceeds fifty-four. Here again the
Toothed Whales are an exception, the number of their teeth being as great
as in many reptiles. In the Mammalia the number of the teeth is fixed
(excepting of course for abnormalities), while in reptiles there is
frequently no precise normal. Two regions may be distinguished in every
tooth—the crown and the root; the latter, as its name denotes, is
imbedded in the gum, while the crown is the freely-projecting summit of
the tooth. The varying proportions of these two regions of the tooth
enables us to divide teeth into two series—the brachyodont and the
hypselodont; in the latter the crown is developed at the expense of the
root, which is small; the hypselodont tooth is one that grows from a
persistent pulp or, at any rate, one that is long open. Brachyodont teeth
on the contrary have narrow canals running into the dentine. The
primitive form of the tooth seems undoubtedly to be a conical
single-rooted tooth, such as is now preserved in the Toothed Whales and
in the canine teeth of nearly all animals. The development of the teeth,
that is, the simple bell-shaped form of the enamel organ, seems to go
some way towards proving this; but it is quite another question whether
we can fairly regard the Whales as having retained this early form of
tooth. In their case the simplification, as is so often the case where
organs are simplified, seems to be rather degeneration than retention of
primitive characters. But this is a matter which must be deferred for the
present.
The incisor teeth are generally of simple structure and nearly always
single rooted. In the Rodents, in the extinct Tillodontia and in
Diprotodont Marsupials, they have grown large, and, as has been already
stated, they increase in size continuously from the growing pulp. These
teeth have a layer of enamel only on the anterior face, which keeps a
sharp chisel-like edge upon them by reason of the fact that the harder
enamel is worn away more slowly than the comparatively soft dentine. The
{49}"horn" of the Narwhal is another
modification of an incisor, as are the tusks of Elephants. Among the
Lemurs the incisors are denticulate, and serve to clean the fur in a
comb-like fashion. This is markedly the case in Galeopithecus. The
incisors are sometimes totally absent, as in the Sloths, sometimes
partially absent, as in many Artiodactyles, where the lower incisors bite
against a callous pad in the upper jaw, in which no trace of incisors has
been found.
Canine teeth are present in the majority of mammals, but are absent
without a single exception from the jaws of the Rodentia. The canine
tooth of the upper jaw is that tooth which comes immediately after the
suture dividing the premaxillary from the maxillary bone. The canines are
as a rule simple conical teeth, with but a single root; indeed they
resemble what we may presume to have been the first kind of tooth
developed in mammals. In this they resemble also as a general rule the
foregoing incisors. But instances are known where the canines are
implanted by two roots. This is to be seen in Triconodon, in the
pig Hyotherium, in the Mole and some other Insectivores, and in
Galeopithecus, where the incisors also may be thus implanted in
the jaw. Furthermore, the simple condition of the crown of the tooth may
be departed from. This is the case with a Fruit Bat belonging to the
genus Pteralopex. In the more primitive Mammalia it is common to
find no great difference between the canines and incisors; such is the
case with the early Ungulate types of Eocene times, such as
Xiphodon. In modern mammals, however, especially among the
Carnivora, the canines tend to become larger and stronger than the
incisors, and in some of the Cats and in the Walrus these teeth are
represented by enormous offensive tusks. It is not rare for the canines
of male animals to be larger than those of their mates. There are also
cases such as the Musk-deer and the Kanchil where the male alone
possesses these teeth, but only in the upper jaw. The teeth which follow
the canines are known as the grinders or cheek teeth, or more technically
as premolars and molars. These two latter terms separate teeth which
arise at different periods, and their use will be explained later. In the
meantime it may be pointed out that the cheek teeth are the teeth which
show the greatest amount of variation in their structure; this is shown
by the number and variety of the cusps in which {50}the biting surface ends.
The grinding teeth vary from simple one-cusped teeth, precisely like
canines, to teeth with an enormous number of separate tubercles. In the
former case it is hard to distinguish between incisors, canines, and
cheek teeth in the lower jaw, where no suture separates the bone.
Moreover it is quite common for the first cheek tooth in the lower jaw to
have the characters of a canine, while the true canine approximates in
its form to the antecedent incisors. This is so, for instance, with the
Lemurs, where the first premolar is caniniform, and the canine shares in
the curious procumbent attitude which distinguishes the lower incisors of
many of those animals.
A variable number of the anterior cheek teeth may be little more than
simple conical teeth; but the rest of the set are commonly more
complicated. No definite laws can be laid down as to the complication of
the posterior as compared with the anterior set. Broadly speaking, it is
purely herbivorous creatures in which the least difference can be
detected at the two extremities, and which are at the same time the most
elaborately decorated with tubercles and ridges. The converse is true
that in purely carnivorous animals, including insect- and fish-eating
forms, there is the greatest difference between the anterior set of
grinding teeth and those which follow. In these two respects such animals
as a Lemur and a Rhinoceros occupy the extremes. Furthermore, it may be
said that omnivorous creatures lie, as their diet would suggest, in an
intermediate position. Generally speaking, when there is a marked
difference between the first premolar and molars at the end of the
series, there is a gradual approximation in structure of a progressive
kind. The tubercles become more numerous in successive teeth; but the
corollary which is apparently deducible from this, i.e. that the
last molar is the most elaborate of the series, is by no means always
true. The last cheek tooth indeed is often degenerate. On the other hand,
it is very markedly the largest of the series in such diverse types as
the Elephant, the hog Phacochoerus, and the Rodent
Hydrochoerus. It is a rule that the cheek teeth of the upper jaw
are more complicated than the corresponding teeth of the lower jaw.
The structure of the cheek teeth is very diverse among the Mammalia.
Broadly, two types are to be recognised. There are {51}teeth in which the
grinding surface is raised into a series of two, to many, tubercles
sharper or blunter as the case may be;—sharper and fewer at the
same time in carnivorous and especially in insectivorous types, more
abundant in omnivorous animals. To this form of tooth the term "bunodont"
is applied. There is no doubt that this is the earliest type of tooth;
but whether the fewer or the more cusped condition is the primitive one
is a question that is reserved for consideration at the end of the
present chapter. The other type of grinding tooth is known as
"lophodont." This is exemplified by such types as the Perissodactyla and
Ungulates generally, and by the Rodents. The tooth is traversed by ridges
which have generally a transverse direction to the long axis of the jaw
in which the tooth lies. The ridges may be regarded as having been
developed between tubercles which they connect and whose distinctness as
tubercles is thereby destroyed. Lophodont teeth are only found in
vegetable-feeding animals.

Fig. 36.—Molar teeth of Aceratherium platycephalum. × ½. m.1-m.3., Molars; mh, metaloph;
p.1-p.4, premolars; ph, protoloph; ps.f, parastyle fossa; te, tetartocone.
(After Osborn.)
The special characteristics of the teeth of various groups of animals
will be considered further under the accounts of the several orders of
recent and fossil Mammalia.
Fig. 37.—Two stages in the development of the teeth of a Mammal (diagrammatic sections).
alv, Bone of alveolus; dent, dentine; dent.s, dental sac; en, enamel;
en.m, enamel membrane; en.m2, enamel membrane of permanent tooth; en.plp,
enamel pulp; gr, dental groove; lam, dental lamina; lam′, part of dental lamina
which grows downwards below the tooth germ; n, neck connecting germs of milk
and permanent tooth; pap, dental papilla; pap2, dental papilla of permanent tooth.
(After O. Hertwig.)
A very general feature of the teeth of the Mammalia is what is usually
termed the diphyodont dentition. In the majority of cases there are two
sets of teeth developed, of which the first lasts for a comparatively
short time, and is termed on account of its usual time of appearance the
"milk dentition"; this is replaced later by the permanent dentition. In
lower vertebrates the teeth are replaced as worn away. There is not,
however, so great an antithesis in this matter between the Mammalia {52}and
other vertebrates as was at one time assumed. But in order to explain
this very important part of the subject it will be necessary to give some
account of the development of the teeth. The type selected is the
Hedgehog, which has been recently and carefully described by Dr. Leche of
Stockholm, which type has furthermore the advantage of being a "central"
type of mammal. The first step in the formation of the teeth is a
continuous invagination of the epithelium covering the jaw to form a
deepish wall of tissue running in the thickness of the jaw; this is
perfectly continuous from end to end of the lower jaw. From this "common
enamel germ" (Schmelzleiste of the Germans[24]) "special enamel germs"
(Schmelzorgane, enamel organs) are developed here and there as
thickenings in the form of buds {53}which arise on the outer side of the fold of
epithelium and some way above its lower termination. These ultimately
acquire a bell-like form, and are as it were moulded on to a thickened
concentration of the dermis beneath; they then become separate from the
downgrowth of the epithelium whence they have arisen. Finally, each of
the eight germs becomes one of the milk teeth of the animal. The lower
end of the sheet of invaginated epithelium, the common enamel germ, is
the seat of the formation of the second set of teeth, of which, however,
in the animal under consideration, there are only two in each jaw. But
corresponding to each of the enamel germs of the milk dentition, with the
exception of the first two molars, there is a slight thickening of the
end of the common enamel germ, which at a certain stage is
indistinguishable from the thickening which will become one of the
permanent teeth. We have thus the diphyodont arrangement. But this does
not exhaust the series of rudimentary teeth, though no more come to
maturity than those whose development has already been touched upon. In
the upper jaw a small outgrowth of the common enamel germ arises above
and to the outer side of the enamel germ of the third milk incisor; this
does not develop any further, but its resemblance to the commencing germ
of a tooth seems to indicate that it is the remnant of a tooth series
antecedent to the milk series. Furthermore, there are indications in the
fourth premolar of a fourth series of teeth posterior in appearance to
the permanent dentition. We arrive therefore at the important conclusion
that although here as elsewhere there are only two sets of calcified
teeth ever developed, there are feeble though unmistakable remains of two
other series, one antecedent to and the other posterior to the diphyodont
dentition. The gap therefore which separates the mammalian dentition from
that of reptiles is less than has hitherto appeared. Dr. Leche also
carefully studied the tooth development of Iguana; he found that
in this lizard there are four series of teeth which come to maturity, and
a rudimentary series antecedent to these which never produces fully
formed teeth.
In a few mammals there is a kind of dentition known as the
monophyodont, in which only one series of teeth reaches maturity; where
in fact there is no replacement of a milk series by a permanent
dentition. Of the monophyodont dentition Whales form an example. The
Marsupials are very nearly an instance of the {54}same phenomenon; for Sir
W. Flower showed, and Mr. Thomas confirmed his discovery, that only one
tooth, according to Mr. Thomas the fourth premolar, is replaced in that
group. But even the purely monophyodont dentition of the Toothed Whales
is a more apparent than real contrast to the diphyodont dentition
elsewhere prevalent. An investigation of the embryos of various Toothed
Whales by Dr. Kükenthal and by Dr. Leche has brought to light the highly
important fact that two dentitions are present, but that one only comes
to maturity; from this fact obviously follows the interesting
question:—To which of the two dentitions of
more normal Mammalia does the monophyodont dentition of the Whales and
Marsupials belong? To this question a clear answer is fortunately
possible. As has been pointed out in the foregoing sketch of tooth
development, and has been illustrated in the figures, the milk teeth
develop as lateral outgrowths of the common enamel germ, while the
permanent teeth arise from the end of the same band of tissue. This fact
enables it to be stated apparently beyond a doubt that in the Whales and
in the Marsupials it is the milk dentition which is the only one to
arrive at maturity. Thus the earlier theoretical conclusion that the
Marsupial dentition "is a secondary dentition with only one tooth of the
primary set left," is proved on embryological grounds to be untrue. But
there are other monophyodont animals than those already mentioned.[25] Orycteropus, the
Cape Anteater, is an example. Mr. Thomas has lately discovered that in
this Edentate there is a set of minute though calcified milk teeth which
probably never cut the gum; here we have a different sort of
monophyodontism, in which the teeth belong to the second and not to the
first set. Between the latter condition and the diphyodont state are
intermediate stages. Thus in the Sea Lions the milk teeth are developed
but disappear early, probably before the animal is born.
In the typical diphyodont dentition, such as is exhibited for example
in Man and the vast majority of mammals, the milk teeth eventually
completely disappear and are entirely replaced by the permanent set of
teeth, with the exception, of course, of the molars, which though they
are developed late belong to the milk series. {55}
Their correspondence with the milk series is shown in an interesting
way by the close resemblance which the last milk premolar often bears to
the first molar. These two extremes of dentition, i.e. purely
monophyodont and, excepting for the molars, purely diphyodont, are
however connected by an intermediate state of affairs, which is
represented by more than one stage. In Borhyaena (probably a
Sparassodont) the incisors and the canines and two out of the four
premolars belong to the permanent dentition, while the two remaining
premolars and of course the three molars are of the milk series.
Prothylacinus, a genus belonging to the same group, has a
dentition which is a step or two further advanced in the direction of the
recent Marsupials. We find, according to Ameghino,[26] whose conclusions are accepted by Mr.
Lydekker, that the incisors, canines, and two premolars belong to the
milk series, while the permanent series is represented only by the two
remaining premolars. We can tabulate this series as follows:—
(1) Purely monophyodont, with teeth only of the first
set—Toothed Whales.
(2) Incompletely monophyodont, as in the Marsupials, where there is a
milk dentition with only one tooth replaced.[27]
(3) Incompletely diphyodont, with the dentition made up partly of
milk, partly of permanent teeth, as in Borhyaena.
(4) Diphyodont, where all the teeth except the molars are of the
second set; this characterises nearly all the mammals.
As we pass from older forms to their more recent representatives there
is as a rule a progressive development of the form of the teeth. This is
especially marked among the Ungulata. The extremely complicated type of
tooth found in such a form as the existing Horse can be traced back
through a series of stages to a tooth in which the crown is marked by a
few separated tubercles or cusps. Arrived at this point, the differences
between the teeth of ancestral Horses and ancestral Rhinoceroses and
Tapirs are hard to distinguish with accuracy; and the same difficulty is
experienced in attempting to give a definition of other large orders by
the characters of the teeth, such as will apply to the Eocene or {56}even
earlier representatives of these families. Fig. 36 (p. 51) illustrating a series of mammalian teeth will
illustrate the above remarks. That there is such a convergence in tooth
structure shows that it is, theoretically at least, possible to determine
the ancestral form of the mammalian tooth. Practically, however, the
difficulties which beset such theorising are great; that there are such
divergent and such strongly-held antithetical views is sufficient proof
of this. Two main views hold the field: one, which has found most favour
in America, and is due chiefly to the labours and persuasiveness of
Professors Cope, Scott, Osborn, and others, is known as "trituberculy."[28] The alternative view, as
urged by Forsyth Major, Woodward, and Goodrich, attempts to show that the
dentition of the original mammal included grinding teeth which were
multicuspidate or "multitubercular." There is much to be said for both
views, and something to be said against both.

Fig. 38.—Molar teeth of A, Phenacodus, and
B, the Creodont Palaeonictis. End, endoconid;
hld, hypoconulid; hyd, hypoconid;
med, metaconid; prd, protoconid.
(After Osborn and Wortman.)
This question is, however, wrapped up in a wider one. Its solution
depends upon the ancestry of mammals. If the Mammalia are to be derived
from reptiles with simple conical teeth, then the first stage in the
development of trituberculy is proved. On the other hand, however, the
evidence is gradually growing that the Theromorpha represent more nearly
than any non-mammalian group with which we are acquainted the probable
ancestral form of the mammals. These animals offer some support to both
the leading views. Cynognathus had triconodont teeth which, as
will be pointed out later, are a theoretically intermediate stage in the
evolution of tritubercular teeth; on the other hand, the teeth of
Diademodon and some others are multituberculate, and have been
very properly compared to the multitubercular teeth of such primitive
mammalia as the Ornithorhynchus. Professor Osborn is no doubt correct in
italicising a remark of an anonymous writer in Science to the
effect that in Diademodon the teeth, though multitubercular, show
the prevalence of three cusps arranged in the tritubercular fashion. {57}But this
may be only a proof that the multitubercular antedates the tritubercular.
It may be, indeed, that the mammalian tooth was already differentiated
among the mammal-like Saurians and that from such a form as
Cynognathus the Eutheria and other forms in which a tritubercular
arrangement can be detected were evolved, and from such form as
Tritylodon the Monotrematous branch of the mammals. This way of
looking at the matter harmonises a much-disputed question, but involves a
diphyletic origin of the mammals—an origin which for other reasons
is not without its supporters.
We shall now attempt to give a general idea of the facts and arguments
which support or tend to support "trituberculy." As a matter of fact the
name is inaccurate; for the holders of this view do not derive the
mammalian molar from a trituberculate condition, but in the first place
from a simple cone such as that of a crocodile!
To this main and at first only cusp came as a reinforcement an
additional cusp at each side, or rather at each end, having regard to
their position with reference to the long axis of the jaw. This stage is
the "triconodont" stage, and teeth exist among living as well as extinct
mammals which show this early form of tooth. We have, indeed, the genus
Triconodon, so named on that very account. Among living mammals
the Seals and the Thylacine all show some triconodont teeth. A Toothed
Whale, it may be remarked, is a living example of a mammal with
monoconodont teeth. The three primary cusps, as the supporters of Cope's
theory of trituberculism denominate them, are termed respectively the
protocone, paracone, and metacone, or, if they are in the teeth of the
lower jaw, protoconid, paraconid, and metaconid. At a slightly later
stage, or coincidently, a rim partly surrounded the crown of the tooth;
the rim is known as the cingulum, and from a prominent elevation of this
rim a fourth cusp, the hypocone, was developed. The three main cones then
moved, or rather two of them moved, so as to form a triangle; this is the
tritubercular stage. Teeth of this pattern are common, and occur in such
ancient forms as Insectivora and Lemurs, besides numerous extinct groups.
An amendment has been suggested, and that is to term the teeth with the
simple primitive triangle "trigonodont," and to reserve the term
tritubercular for those teeth in which the hypocone has appeared. The
platform bearing the hypocone widened into the {58}"talon"; and this ledge
became produced into two additional cusps, the hypoconule or hypoconulid,
and the ectocone or ectoconid. Thus the typical sextuberculate tooth of
the primitive Ungulate, and indeed of many primitive Eutherians, is
arrived at. From this the still further complicated teeth of modern
Ungulates can be derived by further additions or fusions, etc.[29] On the other hand, the
development of the Primate molar stops short at the stage of four
cusps.

Fig. 39.—Epitome of the evolution of a cusped tooth. 1, Reptile; 2, Dromatherium;
3, Microconodon; 4, Spalacotherium; me, metaconid; pa, paraconid; pr, protoconid;
5, Amphitherium. (After Osborn.)
That such a series can be traced is an undoubted fact. Every stage
exists, or has existed. But whether the stages can be connected or not is
quite another question. It is by three main lines of argument that the
view here sketched out in brief is supported. In the first place, the
tracing of the pedigrees of many groups of mammals has met with very
considerable success; and it is clear that as we pass from the living
Horse and Rhinoceros, with their complicated molars, to their
forerunners, we find that both can be referred to a primitive Ungulate
molar with but six cusps. Going still further back to the lowest Eocene
and ancestral type as it appears, Euprotogonia, we still find in
the molar tooth the sextubercular plan of structure. We can hardly get
further back in the evolution of the Perissodactyles with any probability
of security. On the other hand, many facts point to a fundamental
relationship between the primitive Ungulates and the early Creodonts. The
latter frequently show plainly tritubercular molars. Such Ungulates as
Euprotogonia and Protogonodon, though sex- or
quinque-tubercular as to their molars, have a distinctly prevailing
trituberculism, when the size and importance of three of the cusps
is taken into account. But this {59}lacks finality as a convincing proof of the
tritubercular tooth as a primitive Ungulate tooth.
Professor Osborn has ingeniously utilised certain deviations from the
normal type of tooth structure (for the group) in favour of his
strongly-urged opinions. If the stages of development have been as he
suggests, a retrogression would naturally be in the inverse order; thus
the "apparently 'triconodont' lower molar of Thylacinus" may be
interpreted as a retrogression from a tritubercular tooth. In the same
way may be explained the triconodont teeth of Seals and of the Cetacean
Zeuglodon. Finally, the modern Toothed Whales have retrograded
into "haplodonty."
Embryological evidence has also been called in, and with some success,
to contribute towards the proof of the tritubercular theory of teeth.
Taeker has shown that in the Horse and the Pig, and some other Ungulates,
there is first of all a single hillock or cusp, and that later the
additional cones arise separately. An apparent stumbling-block raised by
these investigations is that it is not always the protocone or its
equivalent in the upper jaw which arises first, as it obviously ought to
do phylogenetically. This, however, is not a final argument in either
direction. We know from plenty of examples that ontogenetic processes
sometimes do not correspond in their order with phylogenetic changes.
Thus in the mammalian heart the ventricle divides before the auricle; and
of coarse, phylogenetically, the reverse ought to occur, since a divided
auricle precedes a divided ventricle. This method of development has,
moreover, been interpreted otherwise. It has been held to signify that
the complex teeth of mammals are indeed derived from simple cones but by
the fusion of a number of those cones.
On the other hand there are the claims of the multitubercular theory
of the origin of mammalian teeth to be considered. The palaeontological
evidence has been already, to some extent, utilised. The occurrence of
such teeth among the possible forerunners of mammals, and in some of the
most primitive types of Mammalia, has been referred to. Señor Ameghino
dwells upon the sextubercular condition of many primitive mammals even
belonging to the Eutheria. In a recent communication[30] he attempts to identify six tubercles
in the molars of types belonging to a {60}variety of Orders. The
same condition, as has been noted, characterises that ancient Ungulate
form Euprotogonia. Even where the teeth seem at first sight to be
tritubercular a detailed study shows traces of otherwise vanished
cusps.
It must be remembered in basing arguments upon the early Jurassic and
Cretaceous mammals, that our knowledge of them mainly depends upon lower
jaws, the teeth of which are usually simpler in pattern than those of the
upper jaws. Moreover, another fact, not always insisted upon, must not be
lost sight of. In many of those creatures the jaws were of small size,
and yet accommodated a large series of molar teeth. Amphitherium,
for example, had six molar teeth, and five is a number frequently met
with. As the teeth are so numerous and the jaws so small it seems
reasonable to connect the simplicity of the structure of the teeth with
the need for crowding a number together. The same argument may partly
account for the superabundant teeth of many Toothed Whales. It is true
that the Manatee has very numerous grinders which are yet complex; but
then in this animal there is a succession, and the jaw does not hold at a
given time the entire series, with which it is provided in relays. On the
other hand, where there are few molars they are often of the
multitubercular type, or at least approach it; of this the
Multituberculate Polymastodon is a good example; so, too, the
molars of Hydrochoerus, and of many other Rodents.
It is well known that the fourth deciduous molar of the upper jaw,
which is replaced by a permanent premolar in the fully adult animal, is
of a more complex structure than its successor. This may indeed be
extended to premolars earlier in the series. In the Dog "the second and
first milk molars closely resemble the third and second premolars"; now
the milk premolars belong evidently to the same dentition as the
permanent molars, and they are earlier teeth than the later-developed
replacing teeth. It is therefore significant that these earlier teeth
should be more cuspidate than the later teeth. It tells distinctly in
favour of the simplification as opposed to the complication of teeth in
time, in the groups concerned.
These facts may possibly be applied in explanation of the simple teeth
of some of the Jurassic and Cretaceous mammals. It has been mentioned
that absolute trituberculy is exceedingly rare among those ancient
creatures; more generally there are to {61}be found at least traces
of more cusps. Now in some of them we may be dealing with instances of a
complete tooth change; the suppression, save for one tooth, which is
found in Marsupials, was probably not developed in at least some of these
early mammals. The simplicity may therefore have been preceded by
complexity, and may have been merely an adaptation to an insectivorous
diet.
Alimentary Canal.—The mouth of the Mammalia is
remarkable for the fact that with a few exceptions, such as the Whales,
there are thick and fleshy lips. The office of these is to seize the
food. The roof of the mouth is formed by the "hard palate" in front,
which covers over the maxillary and palatine regions. This region is
often covered with raised ridges, which have a symmetrical disposition,
and are particularly strong in Ruminant animals. They are much reduced in
the Rodents, where the anterior part of the palate is ill-defined owing
to the way in which its sides fade into the lateral surface of the face.
It has been shown that these ridges, in the Cat at least, develop as
separate papilliform outgrowths, and it has been suggested that these
papillae, which later become united to form the ridges, are the last
remnant of palatine teeth such as occur in lower vertebrates.

Fig. 40.—Palatal folds of the Raccoon
(Procyon lotor). p.p, Papilla palatina;
r.p, palatal folds. (From
Wiedersheim's Structure of Man.)
The tongue is a well-developed organ, usually playing a double
part. It acts as an organ of prehension, especially in such animals as
the Giraffe and the Anteater, where it is long and protrusible beyond the
mouth for a considerable distance. It also carries gustatory organs,
which serve for the discrimination of the nature of the food. Beneath the
tongue there may be a hardish plate, known as the sublingua. This is
especially prominent in the Lemurs, where it projects as a horny
structure below the tongue, and has an independent and free tip. It is
supported in some of these animals by a cartilaginous {62}structure. It is
held by Gegenbaur that this organ is the equivalent of the reptilian
tongue, and that in the skeletal vestiges which it contains are to be
found the equivalents of the hyoid skeletal cartilages which support the
tongue in lizards. In this case the tongue of mammals is a subsequently
added structure.
The oesophagus leads from the mouth cavity to the
stomach. The latter organ has commonly a distinctive shape in
mammals. This is well shown in Man. The orifices of the oesophagus and
intestine are somewhat approximated; and this causes a bulging of the
lower border of the organ, usually spoken of as the greater curvature. A
stomach of this typical form is found in many orders of mammals, and is
unlike the stomach in any of the groups of lower vertebrates in shape.
Sometimes the shape of the organ is greatly altered: it may be drawn out,
sacculated, or divided, as in the Ruminants and Whales, into a series of
differentiated chambers, each of which plays some special part in the
phenomena of digestion.
The intestine of mammals is always long and much coiled, though
the length and consequent degree of coiling naturally varies. On the
whole it is perhaps safe to say that it is shorter in carnivorous than in
vegetable-feeding beasts. Thus the Paca has an intestine of 39 inches
total length, while the Cat, an animal of about the same size, has an
intestine which is only 36 inches long. A fish diet, however, to judge
from the Seals, is associated with a long intestinal tract. The intestine
is divisible in the vast majority of mammals into a small and a large
intestine. The two are separated by a valvular constriction save in
certain Carnivores; and in the majority of cases the distinction is also
emphasised by the presence at the junction of a blindly-ending
diverticulum, the caecum. This latter organ varies greatly in
length, being very short in the Cat-tribe and exceedingly long in
Rodents. Its size is, to some extent, dependent upon the flesh-eating or
grass-eating propensities of the animal in which it occurs. One of the
longest caeca is possessed by the Vulpine Phalanger, in which the organ
is one-fifth of the length of the small intestine; while the opposite
extremity is reached by Felis macroscelis, which has a small
intestine one hundred times the length of the caecum.
{63}
Fig. 41.—Different forms of the stomach in Mammals. A, Dog; B, Mus decumanus;
C, Mus musculus; D, Weasel; E, scheme of the Ruminant stomach, the arrow
with the dotted line showing the course taken by the food; F, Human stomach.
a, Minor curvature; b, major curvature; c, cardiac end. G, Camel; H, Echidna
aculeata. Cma, Major curvature; Cmi, minor curvature. I, Bradypus tridactylus.
Du, Duodenum; MB, coecal diverticulum; **, outgrowths of duodenum; †, reticulum;
††, rumen. A (in E and G), Abomasum; Ca, cardiac division; O,
psalterium; Oe, oesophagus; P, pylorus; R (to the right in E and to the left in G),
rumen; R (to the left in E and to the right in G), reticulum; Sc, cardiac
division; Sp, pyloric division; WZ, water-cells. (From Wiedersheim's Comparative
Anatomy.)
An interesting point in connexion with the gut of mammals is the
varying proportion of the small to the large intestine. As a general rule
the former is very considerably longer than the latter; in
Paradoxurus, for instance, the small intestine may be fifteen
times the length of the large. The excess of length of one section over
the other is not generally so marked {64}as this. In Phalanger
maculatus the two sections of the gut are as nearly as possible equal
in length, while in Phaseolarctos the large intestine is
considerably longer than the small, the lengths being respectively 160
inches and 111 inches. It is common among the Marsupials and also among
the Rodents for these proportions to exist, i.e. for the large
intestine to be as long as, or longer than, the small. But there are so
many exceptions that no general statements can be extracted from the
facts.
Some few details will be found in the systematic part of this book.
Mr. Chalmers Mitchell has brought forward some reasons for associating a
great length of large intestine with an archaic systematic position, in
the birds at any rate. The facts here briefly touched upon are not at
variance with the extension of such a view to the mammals.
Fig. 42.—Diagrammatic plan of the liver of a Mammal
(posterior surface). c, Caudate lobe; cf, cystic
fissure; dv, ductus venosus; g, gall-bladder; lc,
left central lobe; ll, left lateral lobe; llf, left
lateral fissure; p, portal vein entering transverse
fissure; rc, right central lobe; rl, right lateral lobe;
rlf, right lateral fissure; s, Spigelian lobe; u, umbilical
vein; vc, post-caval vein. (After Flower
and Lydekker.)
Appended to the alimentary tract are three glands or sets of glands.
Opening into the mouth cavity are the salivary glands, which are
of enormous size in Anteaters, and small or absent in Whales. In their
number and position these glands are characteristic of mammals. Into the
intestine open the ducts of the pancreas and liver, two glands which the
mammals share with lower vertebrates. The form of the liver is,
however, generally characteristic of mammals. It is divided as a rule
into a right and a left half, the line of division being marked by the
insertion of the umbilical ligament, a vestige of the primitive ventral
mesentery. Each half is again commonly subdivided into central and
lateral lobes. In addition to these, two other divisions are often to be
seen—the Spigelian and the caudate lobe. The liver is less divided
in Cetacea and {65}some others, very much subdivided in Rodents
and other groups. The degree of subdivision and the proportions of the
several lobes frequently offer valuable systematic characters. The
gall-bladder may be present or absent; it is always a diverticulum of the
hepatic duct. The two are never separate, as in birds, for instance.
Organs of Circulation.—The heart of all mammals is a
completely four-chambered organ. In the adult heart there is no
communication between the right and left halves. The auricles are
comparatively thin-walled, the ventricles thick-walled, in relation to
the amount of work that they have severally to perform. The right
ventricle, moreover, which has only to drive the blood into the lungs, is
much thinner-walled than the left ventricle, which is concerned with the
entire systemic circulation. The exits of the arteries and the
auriculo-ventricular orifices are guarded by valves, which are so
arranged as only to permit the blood to flow in the proper direction. But
these valves have a morphological as well as a physiological interest. At
the origin of each artery, the aorta and the pulmonary, there is a row of
three watch-pocket valves, as they have been generally termed on account
of their form. These three valves meet accurately in the middle of the
lumen of the arterial tube when liquid is poured into them from above,
and thus completely occlude the orifice. The auriculo-ventricular valves
differ in structure in the two ventricles. That of the left ventricle has
only two flaps, and is therefore often spoken of as the bicuspid or
mitral valve. Both these flaps are membranous, and together they
completely surround the exit from the auricle into the ventricle. The
edges of the valve are bound down to the parietes of the heart by
numerous branching tendinous threads, the chordae tendineae, which often
take their origin from pillar-like muscles arising from the walls of the
heart, the so-called musculi papillares. The valve of the right ventricle
is composed of three flaps, and is therefore often spoken of as the
tricuspid valve; it is in the same way membranous, and has chordae
tendineae and musculi papillares connected with it. The disposition of
the musculi papillares and their number differ in different mammals, but
no exhaustive study has as yet been made of the arrangements in different
groups; the amount of individual variation even is not known, though it
is certainly considerable in some cases, for {66}instance in the heart of
the Rabbit. The heart of the Monotremata presents differences of some
importance from those of other Mammalia; the modern knowledge of the
Monotrematous heart is mainly due to Gegenbaur[31] and Lankester,[32] in whose memoirs references to the
older literature will be found. The principal features of interest in
which the heart of the Monotremata differs from that of the higher
Mammalia are these. When the two ventricles are cut across transversely,
the cavity of the right is seen to be wrapped round that of the left in a
fashion precisely like that of the bird's heart; on the other hand in the
higher mammal the two cavities lie side by side. The main difference
between Monotremes and other Mammals concerns the right
auriculo-ventricular valve. The differences which it presents from the
corresponding structure of the rest of the Mammalia are two: in the first
place, the valve itself does not completely surround the ostium; it is
only developed on one side; the septal half (i.e. that turned
towards the interventricular septum) is either entirely absent or more
generally represented by a small bit of membrane; nevertheless I found[33] recently in an
Ornithorhynchus heart a complete septal half to the right
auriculo-ventricular valve. The second point of interest in connexion
with this valve is, that the musculi papillares instead of ending in
chordae tendineae attached to the free edge of the valve are directly
attached to the valve, and in some cases pass through its membranous
flap, to be attached to its origin at the boundary of the auricle and of
the ventricle. The invading of the valve-flap by muscle in this way is
highly interesting, as it recalls the heart of the bird and of the
crocodile. The imperfect condition of the valve (from which, as has
already been stated, the septal half is as a rule nearly absent) is a
point of resemblance to the heart of the bird; the corresponding valve of
the crocodile's heart being complete.
{67}

Fig. 43.—Lepus cuniculus. Ventral
view of the vascular system.
The heart is somewhat
displaced towards the left of
the subject; the arteries of
the right and the veins of the
left side are in great measure
removed. a.epg, internal mammary
artery; a.f, anterior facial
vein; a.m, anterior mesenteric
artery; a.ph, anterior phrenic
vein; az.v, azygos vein; br,
brachial artery; c.il.a, common
iliac artery; c.il.v, common
iliac vein; cœ, coeliac artery;
d.ao, dorsal aorta: e.c, external
carotid artery; e.il.a, external
iliac artery; e.il.v, external
iliac vein; e.ju, external jugular
vein; fm.a, femoral artery;
fm.v, femoral vein; h.v, hepatic
veins; i.c, internal carotid
artery; i.cs, intercostal vessels;
i.il.a, internal iliac artery; i.il.v,
internal iliac vein; i.ju, internal
jugular vein; i.l, iliolumbar
artery and vein; in, innominate
artery; l.au, left auricle;
l.c.c; left common carotid artery;
l.pr.c, left pre-caval vein;
l.v, left ventricle; m.sc, median
sacral artery; p.a, pulmonary
artery; p.epg, epigastric artery
and vein; p.f, posterior facial
vein; p.m, posterior mesenteric
artery; p.ph, posterior
phrenic veins; pt.c, post-caval
vein; p.v, pulmonary vein; r,
renal artery and vein; r.au,
right auricle; r.c.c, right common
carotid artery; r.prc, right
pre-caval vein; r.v, right ventricle;
s.cl.a, right subclavian
artery; s.cl.v, subclavian vein;
spm, spermatic artery; s.vs,
vesical artery; ut, uterine artery
and vein; vr, vertebral artery.
(From Parker's Zootomy.)
There are also features in the system of arteries and veins which are
eminently distinctive of mammals. In the first place, the aorta leaving
the heart and conveying blood to the body is only a half arch, and bends
to the left side as seen in Fig. 43. The right and left halves are
present in reptiles, and meet behind the heart. In the bird the right
half alone has remained. This fact, therefore, shows that the mammal
cannot have been derived from a bird-like ancestor, but that both must
have independently come from an ancestor with both halves of the aortic
arch present, of which one half has disappeared in one group, and the
other half in the other. It is an interesting fact, too, to notice that
the four {68}cavities of the mammal's heart, which
fourfold division it shares with birds alone, do not exactly correspond
compartment for compartment with those of the bird's heart, at least in
so far as concerns the ventricles. For the reptilian heart is provided
with only one ventricle, and therefore the division of that cavity must
have been independently accomplished in mammals and in birds.
There are two features in the venous system which distinguish all the
Mammalia (with the exception of Echidna in one of these points)
from vertebrates standing lower in the series. The hepatic portal system
is limited to a vein which conveys to the liver blood derived from the
alimentary tract; in no mammal except in Echidna is there any
representative of the anterior abdominal vein of lower vertebrates. In
that animal there is such a vein, which apparently arises from a
capillary network upon the bladder and passes up, supported by a
membrane, along the ventral wall of the abdomen to the liver, thus
emptying blood into that organ exactly as does the anterior abdominal
vein of the frog. In no mammal is there any trace of a renal portal
system. The kidneys derive their blood from the renal arteries only.
Many mammals have two superior venae cavae; this is the case, for
instance, in the Elephant and the Rodents and other types lying
comparatively far down in the series. In most if not in all mammals there
are considerable remains of one of the posterior cardinals, in the form
of the azygos vein, which opens into the vena cava superior or pre-caval
vein, i.e. the superior cardinal just before the latter debouches
into the heart. This one posterior cardinal is usually on the right side;
but it may be on the left side, for instance in Trichosurus
vulpecula. In Halmaturus bennettii there are two azygos veins,
one left and one right, of which the left is rather the larger.[34]
Urinary Organs.—The kidneys in the Mammalia have a
compact form, which contrasts with the somewhat diffuse and
vaguely-outlined kidneys of the Sauropsida. In mammals the organ is as a
rule of that peculiar shape which is called "kidney-shaped"; a depression
termed the hilum, which receives the ducts of the glands, indenting the
border of an otherwise oval-shaped gland. In some few mammals the kidney
is broken up {69}into lobules; this is the case with the
Whales, the Bears, the Oxen, and a few other forms. A curious fact about
the kidneys of the Mammalia is their very general asymmetry of position.
One of them usually lies in a more advanced position than the other. The
ureters lead from the kidneys to the urinary bladder, which in its form
and relations is quite distinctive of the Mammalia. The bladder is formed
out of the remains of the allantois, and is therefore not the exact
homologue of the bladder of the frog, which is the equivalent of the
entire sac which grows out of the cloaca in the mammal, and is the foetal
allantois. The ureters open into the bladder in the higher Mammalia, but
lower down in the urino-genital passage in the more primitive
mammals.
The Body Cavity.—The Mammalia differ from all other
living vertebrates by the arrangement of the body cavity in which lie the
viscera. That cavity is divided into two by a partly muscular and partly
tendinous partition, the diaphragm. No other vertebrate has this precise
disposition of the coelom. The diaphragm lies usually transversely to the
longitudinal axis of the body, but gets a much more oblique arrangement
in the Cetacea and the Sirenia, whose needs demand a more expanded
chamber for the lungs. For in front of the diaphragm lie the lungs and
heart; behind it the stomach, liver, intestines, and the organs of
reproduction and excretion. The diaphragm is used in respiration; when
its muscles contract, the surface directed toward the pleural cavity
becomes less convex, and the cavity of the lungs is thus increased,
allowing them to expand under the pressure of the entering air.
The Lungs.—The lungs of the Mammalia differ from those of
animals lying lower in the series by the fact, just referred to, that
they occupy a pleural cavity completely shut off from the abdomen by the
diaphragm. As a rule the lungs of the Mammalia are to be distinguished by
their more or less extensive lobation. In the Whales, however, and in the
Sirenia, they are not much divided, but present the appearance of the
simple sac-like lungs of the reptiles. In some mammals there is a median
and posterior unpaired lobe of the lung, which lies in the
post-pericardial cavity behind the pericardium. This is not universally
present. The lungs are very frequently not symmetrical in their lobation,
the number of separate lobes on the right side {70}and on the left being
different. The lungs of mammals agree with those of the lower reptiles in
being freely suspended within their coelomic cavity, and in not being, as
in birds, crocodiles, and the Varanidae among lizards, tied down to the
dorsal surface of that cavity by a sheet of peritoneum covering them.

Fig. 44.—Part of a sagittal section of an ovary of a child just born. bl.v, Blood-vessels;
foll, strings and groups of cells derived from the germinal epithelium becoming
developed into follicles; g.ep, germinal epithelium; in, ingrowing cord of cells
from the germinal epithelium; pr.ov, primitive ova. (From Hertwig, after
Waldeyer.)
The Gonads (Ovaries and Testes).—The ovary in the
Mammalia is always paired; there is never a partial or complete abortion
of one gonad as in birds—except of course in pathological cases.
The ovaries are small, and lie in the abdominal cavity behind the
kidneys. In the immense majority of the Mammalia the ova which are
produced within the ovaries are of minute size; those of even the
colossal Rorqual are, so far as we know, not markedly larger than the ova
of a Mouse. The smallness of size of these reproductive elements implies
necessarily an absence of much nutritive yolk; and as a consequence the
developing embryo, since it is not hatched in an early stage as a free
living larva, has to be nourished by the mother, to whose tissues it is
attached through the intermediary of the placenta, a structure partly
composed of foetal structures derived from the embryo, and partly of
portions of the lining membranes of the uterus of the mother. The ova of
the {71}Eutherian mammals, including the Marsupials,
are very small as compared with those of any other vertebrates, excepting
only Amphioxus, where the young are hatched early as free swimming
larvae. They also differ in a highly characteristic way in the mode of
their development within the ovary. These processes are to some extent
illustrated in Fig. 44. The main framework of the ovary is formed of the
so-called "stroma," which is a mass of tissue formed of more or less
connective-tissue-like cells. Within this are numerous cavities, the
Graafian follicles. The very young follicles consist of but a single
layer of follicular cells surrounding the ovum, which lies centrally. The
follicular cells gradually increase in number until the ovum lies in the
midst of several layers of cells. At this period a vacuity is formed
between some of these cells, and grows into a large cell-free cavity; the
ovum does not lie loosely in this space, but is connected at one side
with the follicular cells, which still line the interior of the Graafian
follicle by the so-called discus or cumulus proligerus. The egg or ovum
has, moreover, a layer of cells immediately surrounding itself. All these
facts can be gathered by an inspection of Fig. 45. It has been shown
that, as in lower vertebrates, the cells immediately surrounding the ovum
are connected with it directly by delicate processes which penetrate the
actual membrane of the egg.

Fig. 45.—Two stages in the development of the Graafian follicle. A, With the follicular
fluid beginning to appear; B, after the space has largely increased. caps,
Capsule; disc, cumulus proligerus; memb, membrana granulosa; ov, ovum; sp,
space containing fluid. (After Hertwig.)
Fig. 46.—Ovarian egg of Echidna. b, Basilar membrane; fe, follicular epithelium;
o, oil globules; vm, vitelline membrane; y1, y2, yolk. (Partly after Caldwell.)
The only ova which depart at all in structure from that above
described are those of the Monotremata. The credit of this {72}discovery rests
with Owen and with Professor Poulton, who pointed out in 1884,[35] that the ovum of
Ornithorhynchus is very large as compared with those of other
Mammalia (6 mm. as against .2 mm.), that it is filled with yolk, and that
it completely fills the follicle, being surrounded by two layers of
follicular cells only. This latter fact was proved by Caldwell.
Subsequently Gyldberg[36]
and I[37] described the
ovarian ovum of Echidna, showing it to be identical with that of
Ornithorhynchus. Later still a more elaborate and beautifully
illustrated paper was published by Caldwell[38] upon the early stages of development in
the Monotremata and Marsupials, in which the ovum of the former was
accurately described (see Fig. 46). In the particulars mentioned above,
the ovum of the Monotremata is practically identical with that of the
large-yolked ova of the Sauropsida. {73}

Fig. 47.—Lepus cuniculus. The anterior end of the vagina,
with the right uterus, Fallopian tube, and ovary. (Nat.
size.) Part of the ventral wall of the vagina is removed,
and the proximal end of the left uterus is shown in
longitudinal section, fl.t, Fallopian tube; fl.t′, its peritoneal
aperture; l.ut, left uterus; l.ut′, left os uteri;
ov, ovary; r.ut, right uterus; r.ut′, right os uteri; s,
vaginal septum; va, vagina. (From Parker's Zootomy.)
It is the general rule among vertebrate animals that the ovaries are
completely independent of the ducts which convey their products to the
exterior. In certain fishes, however, there is an absolute continuity
between the two structures, which is believed to be due to a simple
concrescence between the originally distinct ovary and oviduct. The
latter has grown round the former, an obvious advantage in preventing the
eggs from wandering into the abdominal cavity and becoming lost. In the
Mammalia we find discontinuity as a general rule. But in quite a number
of forms folds of the lining membrane of the abdominal cavity are
developed, which practically ensure the passage of the ova into the
oviduct when they are extruded from the ovaries. The oviduct, moreover,
has a large and fimbriated mouth, called in human anatomy—which is
provided with a number of fanciful names—the morsus diaboli. This
almost wraps round the ovary, and thus prevents the ova from straying in
the wrong direction. Moreover, the ovary itself is often so arranged that
it can easily be withdrawn into a pocket of the peritoneum, from which
the obvious exit is by the gaping mouth of the oviduct. This disposition
of the generative parts is still further modified in a few animals, such
as the Rat[39] and the
Kinkajou.[40] In these
animals the mouth of the oviduct actually opens into the interior of a
closed chamber which contains the ovary. In this case there is but one
route for the {74}extruded ova to follow. This series of steps
in the perfecting of the mode of safe extrusion of the ova is highly
interesting, and is a piece of evidence in favour of the high position of
the mammals.

Fig. 48.—Female urino-genital
apparatus of various Marsupials.
A, Didelphys dorsigera
(young); B, Trichosurus;
C, Phascolomys wombat. B,
Urinary bladder; Cl,
"cloaca"; Fim, fimbriae;
g, clitoris; N, kidney; Od,
Fallopian tube; Ot, aperture
of Fallopian tube; Ov,
ovary; r, rectum; Sp, septum
dividing vagina; Sug,
urino-genital sinus; Ur,
ureter; Ut, uterus; Ut′,
opening of the uterus into
the median vagina (VgB);
Vg, lateral vagina; Vg′, its
opening into the urino-genital
sinus; † (in B), point of
approximation of uteri; † (in
C) and *, rectal glands. (From
Wiedersheim's Comparative
Anatomy.)
{75}
The oviducal apparatus of the mammal is more specialised than that of
lower vertebrates. It is most simple, as might be imagined, in the
egg-laying Monotremes, where, indeed, it is on the same level as that of
reptiles. But in the Eutheria the fimbriated mouth of the oviduct passes
into a narrow and winding tube, the Fallopian tube; this widens into a
uterus, and the two uteri combine into a single tube in the higher forms.
They are called the Monodelphia on this account. In the Marsupials the
uteri are distinct though they often join above, and from this junction
depends a median "uterus." After the uterus or the uteri follows in every
case a single vagina.
The testes of the Mammalia, like those of other vertebrates, occupy
primitively a position within the body cavity precisely corresponding to
that of the ovaries. And in the lowly-organised Monotremata, and some
other forms, such as the Whales, they retain that primitive position
within the body. It is, however, distinctive of the Mammalia as opposed
to lower vertebrates that the testes descend later into a scrotum, which
is simply a protrusion of the skin of the body surrounded by muscles,
and, of course, containing a section of the body cavity in which lie the
testes. The penis of the Mammalia, represented by the clitoris and
associated structures in the female, is of a structure entirely peculiar
to this group.

Fig. 49.—Brain of Dog. A, ventral; B, dorsal; C, lateral aspect. B.ol, Olfactory lobe;
Cr.ce, crura cerebri; Fi.p, great longitudinal fissure; HH, HH1, lateral lobes of
cerebellum; Hyp, hypophysis; Med, spinal cord; NH, medulla oblongata; Po,
pons Varolii; VH, cerebral hemispheres; Wu, middle lobe (vermis) of cerebellum;
I-XII, cerebral nerves. (From Wiedersheim's Comparative Anatomy.)
The Brain.—Inasmuch as Professor Wiedersheim has said
with perfect truth that "the brain of the extinct Ungulate
Dinoceras shows so striking a likeness to that of a lizard that
one would be compelled to explain it as that of a lizard without a
knowledge of the skeleton," it is clear that to define the mammalian
brain is a difficult matter. The existing Mammalia, however, all possess
brains which can be readily distinguished from those of vertebrates lying
lower in the scale. They are of relatively large size, brought about
mainly by the dimensions of the cerebral hemispheres, which have an
importance in this class of vertebrates that they have not elsewhere.
Coupled with this large size of the hemispheres is a more elaborate
system of transverse commissures uniting the two; and this culminates in
the higher Mammalia, where the corpus callosum attains a large size and
great physiological importance. A {76}very marked feature,
moreover, of the mammal's brain is the development of regular fissures
upon its surface, which fissures are only absent from
Ornithorhynchus, various small Rodents, Bats, and Insectivores,
among living mammals. It is sometimes, but erroneously, said that the
more complicated the fissures of the brain are, the higher in
intelligence and "zoological position" is the possessor of that brain.
Instances can undoubtedly be quoted to support such a view; but they are
{77}merely selected cases, which do not indicate
a wide applicability of such a generalisation. Thus it is true that the
brain of a Man is more elaborate in its furrows and convolutions than is
that of a Cat. The real fact of the matter is, that the complexity of the
brain from this point of view increases with the size of the animal
within the group.

Fig. 50.—Lepus cuniculus. Longitudinal vertical section of the brain. (Nat. size.)
a.co, Anterior commissure; b.fo, body of the fornix; cb, cerebellum, showing arbor
vitae; c.c, crus cerebri; c.h1, parencephalon or cerebral hemisphere; c.h2, temporal
lobe; c.ma, corpus mammillare; cp.cl, corpus callosum; f.m, foramen of Monro;
inf, infundibulum; l.t, lamina terminalis; ly, lyra; m.co, middle commissure;
m.o, medulla oblongata; o.ch, optic chiasma; o.l1, o.l2, corpora quadrigemina or
optic lobes; olf, olfactory lobe; p.co, posterior commissure; pd.pn, peduncle of the
pineal "gland," pn; p.fo, anterior pillar of the fornix; pty, pituitary body; pv.a,
pons Varolii; sp.lu, septum lucidum; v4, fourth ventricle; vl.ip, velum interpositum;
v.vn, valve of Vicussens; II, optic nerve. (From Parker's Zootomy.)
The Gorilla and the Chimpanzee have a more furrowed brain than has the
little Marmoset; the Bear a more complicated brain than the Weasel, etc.
The most highly-convoluted brains of all mammals are those of the
Elephants, and there does not seem in the Ungulates to be so marked a
relation between size and abundance of fissures as there is among other
mammals. A regular plan of the fissures can be detected with certainty
for each group considered by itself; but it is not so easy to homologise
the details of arrangement from group to group. This is so far in accord
with the view that the existing groups of mammals have diverged from each
other ab initio.
Another marked characteristic of the mammalian as opposed to other
brains is the relatively small importance in size and yet the fourfold
nature of the optic lobes. What was the case with the optic lobes of the
early Ungulates is difficult to understand, on account of the fact that
the casts are necessarily imperfect. {78}Altogether the enormous
progress in the complexity of the brain from the early Tertiary mammals
down to the present, is one of the most remarkable revelations of
palaeontology. It goes perhaps some way in explaining the remarkable
diversity in mode of life exhibited by the mammals as compared, for
example, with the birds, whose brains have not diverged so much or in so
many directions from the primitive form.
The present Distribution of the Mammalia.—In the
following pages some of the principal facts in the geographical range of
the orders, families, and many of the genera of Mammalia will be given.
It has been justly observed by Mr. Sclater that the habitat of an animal
is as much a part of its definition as is its structure or external form.
No systematic account of the Mammalia would therefore be complete without
such geographical facts. But that branch of zoology which is concerned
with the past and present distribution of animals is wider in scope than
this. Zoogeography deals not only with the actual facts in the range of
animals, but with the inferences as to past changes in the relations of
land and sea which the facts seem to indicate, and with speculations as
to the place of origin of the different groups, of which more than hints
are sometimes given by their past and present distribution. In addition
to this, the earth can be mapped out into provinces and regions which are
definable by their animal inhabitants. In the present volume, dealing
only with the Mammalia, it will be obviously impossible to enter fully
into the entire subject of zoogeography. All that will be attempted is a
brief general survey of the science so far as it can be illustrated by
the Mammalia. For fuller knowledge the reader is referred to the
treatises mentioned below.[41]
There are certain facts in the distribution of animals which are
commonplaces of knowledge, but which may be set forth with definiteness.
Everybody knows that an animal has a given range: Elephants, for example,
are found in India and certain adjacent parts of Asia, and again in
Africa; the Rhinoceroses have roughly the same range; the Tiger is
limited to Asia; the {79}Jaguar to America, and so forth. The entire
expanse of country which is inhabited by an animal is called its area of
distribution. Such areas are larger or smaller. The Lion ranges over the
whole of Africa, a small part of India, and some neighbouring countries;
on the other hand, the Insectivore Solenodon is limited to Cuba
and Hayti, a separate species to each. Among other groups of animals are
instances of an even more restricted range. There are humming-birds
confined to the slopes of a single mountain, and fishes limited in their
range to a single small lake.
A species may be found everywhere within the area of its distribution,
or it may be confined to a number of limited tracts within that area. In
this case it is usual to speak of "stations." In such cases the species
in question is generally suited to some particular kind of environment.
Thus the Otter and other aquatic mammals will only be found where there
is water; and intervening tracts of waterless country will contain no
Otters. Goats and Chamois live only upon mountains; the intervening
plains are destitute of them. This discontinuity of distribution within
the area is very general. But a discontinuity of area is also
seen—not so commonly however; and, indeed, when it does occur, it
is a matter of a genus and not of a species. Thus the Tapir is found in
the East Indies on the one hand and in South and Central America on the
other, being absent in the intermediate tracts.
It is clear that tracts of country eminently suitable for the housing
of a particular mammal do not always possess that kind, or even an allied
form. Africa, for example, possesses no arboreal Anteaters; there are no
Anteaters at all (of the order Edentata) in Australia, though there are
plenty of ants for them to feed upon, and tropical conditions of climate
prevail. But as in these cases the inference may be denied on the grounds
that no experiments exist to prove or to disprove the assertion, the
matter may be better emphasised by such cases as the introduction of the
Rabbit into Australia, and various mammals, such as Goats, into oceanic
islands. The plague caused by the former is a matter of notoriety. But
although climate and conditions and animal inhabitants do not march
accurately together, there is certainly some connexion between
temperature and the range of animals. Mr. Lydekker writes on this point
as follows: "The llama-like animals, respectively known as vicunas and
guanacos, are met with in {80}company on the highlands of the Cordillera
in Peru and Ecuador, but as we go farther south the latter are found on
the plains of southern Argentina and Patagonia, as well as on the island
of Tierra del Fuego at the sea level. Here then is a clear proof of the
intimate connexion existing between temperature and station; the guanaco
being an animal which can only live in cold or temperate climates, finds
suitable conditions for its existence in tropical latitudes solely at a
height of so many thousands of feet, although farther south it is able to
thrive at the sea level." This, however, cannot be pushed too
far—the world cannot be mapped out into areas bounded by parallels
of temperature as was once attempted—since there are plenty of
cases like that of the Tiger, which is as much at home in a tropical
jungle as on the icy plains of Northern Asia.
Seeing that there are in many cases no climatic barriers to the
spreading of a given race of animals over a larger area of distribution
than it actually occupies, it becomes important to inquire why there are
so many cases of restriction in range.
It is possible to see, at any rate, three causes which are responsible
for a large number of such cases. In the first place, a given species of
animal must have originated at a certain spot; its multiplication in
individuals must always be a slow matter, since enemies, and untoward
events generally, would conspire to check the natural multiplication by
geometrical progression. A long time might therefore elapse before the
species greatly extended its range. A restricted distribution may
therefore, in some cases, mean a modern race. In the second place, there
are definite physical barriers which check the migration of species. The
terrestrial Mammalia cannot cross wide arms of the sea; that they can and
do swim for considerable distances has been proved in several instances;
but, as has been pointed out, it is unlikely that a purely terrestrial
mammal would voluntarily swim out into an unknown sea. And then if it
did, and successfully reached the opposite side, nothing would happen
unless it were a pregnant female; or, if not pregnant, till a male swam
very soon afterwards in exactly the same direction. Many travellers have
told of floating islands, formed of torn-up trees and brushwood, which
have been seen at the mouths of large rivers, with animal passengers upon
them. These are, however, so much at the mercy of currents and storms,
{81}that but little reliance can be placed on
them as a means of transit; besides, here again, two individuals, or a
pregnant female, would be required to effect a settlement on a foreign
shore. The existence of oceanic islands is often urged as a proof of this
inability to cross tracts of sea; even those which are comparatively near
an extensive continent, such as, for example, Fernando Noronha in the
Atlantic, are destitute of mammals (except, indeed, the ubiquitous Mouse,
which is believed to have been carried there, often in company with the
equally widely-spread Rat, in ships). This argument, however, is not so
conclusive as might appear; it doubtless is in the case of far-distant
islands. But the size of the islands has to be taken into account. For
there are islands, such as the Galapagos, or, to take a less contested
instance, some of the islands of the Malagasy Archipelago, undoubtedly
continental, which have an exceedingly reduced number of mammals. An area
of a certain size seems to be a necessity.
The converse of this is in many cases easy to show, that is, the wide
range of animals when there are no marine barriers to stop their
spreading. John Hunter, the celebrated anatomist and surgeon (not often
quoted, however, as an authority upon geographical distribution),
observes: "It is a curious circumstance in the natural history of animals
to find most of the northern animals the same both on the continent of
America and what is called the Old World, while those of the warmer parts
of both continents are not so. Thus we find the bear, fox, wolf, elk,
reindeer, ptarmigan, etc., in the northern parts of both.... The reason
why the same animals are to be found in the northern parts is the
nearness of the two continents. They are so near as to be within the
power of accident to bring the animals, especially the large ones, from
one continent to the other either on the ice or even by water. But the
continents diverging from each other southward, so as to be at a very
considerable distance from each other even beyond the flight of birds, is
the reason why the quadrupeds are not the same."
There is no doubt, in fact, that the ocean is the most insuperable of
all barriers to the dispersal of mammals. In a less degree mountain
ranges and deserts are also barriers. The Desert of Sahara is a striking
instance to the point; it separates two exceedingly different faunas.
{82}
A third cause of more or less limited range is the barrier due to
competition. If the ground is already taken up, there is no room for new
immigrants. There is obviously a limit to the number of Antelopes or Deer
that can graze upon a given tract of grassy plain. These two groups of
Ungulates illustrate the matter well: the Antelopes are African and
Indian, especially the former, while Africa has no Deer at all; America,
on the other hand, has plenty of Deer but no Antelopes, save the
Prong-horn. The more nearly akin the two species or groups of species
are, the fiercer will be the competition; for a near kinship will at
least often imply similar habits, the need for similar food, and other
likenesses which will prevent both from successfully occupying the same
tract of country. The remarkable fauna of Australia is believed to afford
an example of this. In that country the prevalent inhabitants are the
Marsupials. The Monotremes are found there also, and nowhere else save in
New Guinea and Tasmania. The remaining mammals are inconspicuous; they
embrace a few Rodents and Bats, and the doubtfully indigenous Dingo-dog.
Now the Marsupials are fitted to every variety of life. We have the
grazing Kangaroos and Wallabies, the burrowing Wombats, the arboreal
Phalangers, and the carnivorous Dasyures. In the second place, it is an
unquestioned fact that the Marsupials are an older race than are the
existing Eutherian mammals; they were the dominant mammals during the
Secondary epoch. At that time they were more widely distributed than at
present. In most parts of the world they are now absent, since they have
been successfully ousted by the more highly organised groups of Eutheria.
But at that period, when the higher Eutheria were in the ascendant,
Australia and the islands to the north became cut off from Asia, and thus
became freed from inroads of Eutheria, which were partly prevented by the
physical barrier of the sea from effecting a settlement, and partly
perhaps prevented owing to the ground being already taken up by the
Marsupials. Likeness of habit gave the older inhabitants victory in the
struggle for existence.
The general statements that have been here made are in accord with
current opinion upon the factors of geographical distribution. But the
past range of animals appears to be less consonant with the received
views. In the Tertiary {83}period, groups of animals had often a far
wider range than at present. To-day the Rhinoceroses are limited to Asia
and Africa, and to quite limited parts of the former continent. In the
past, these animals were abundant in Europe and North America. Wild
Horses now have a range which is not widely different from that of the
Rhinoceroses, save that they extend into the more northern regions of
Asia. Their remains are abundant both in North and South America. The
Hippopotamus, now confined to Africa, once ranged over Europe,
Madagascar, and India. There were plenty of American and European Lemurs.
Elephants were nearly world-wide in their range; and, in short,
restricted distribution seems to be on the whole a characteristic of
animals of the present day.
These statements, however, though perfectly true, must not lead to
erroneous inferences. It is rather impressed upon the reader, in books
which contain sections dealing with geographical distribution, that
animals on the whole occupy more restricted areas at present than in the
past. There are, however, plenty of examples of groups of extinct
creatures which had, so far as we know, quite a restricted range. Thus
the Toxodonts were purely South American, as were the Glyptodonts and
some other forms. And, on the other hand, the Cervidae of to-day are as
widely, if not more widely, distributed than at any other time. The Hares
and Rabbits are now nearly universal in range; the Cats almost so. We
meet with Bovidae, even excluding the Sheep and Goats, in all the four
quarters of the globe, excluding only South America and, of course,
Australia. The Camelidae are still common to both the Old and the New
Worlds.
During certain periods of the Tertiary epoch it is true that there was
more similarity between Europe and North America than there is at
present. It would have been quite necessary to unite both into a
Holarctic area, such as is now insisted upon by many; but the reasons for
this union would then have been stronger. The fact is, however, that the
closer resemblances were due to the larger number of families of animals
which existed then than now; these have decayed away from both
continents, and allowed the unlikenesses between the mammalian fauna of
both to become evident. But the likenesses which still survive have led
many to associate the two regions closely together.
So far as the history of a genus or family or larger division {84}can be
traced, it results as a conclusion that from a given area of origin the
group in question migrated in all directions where possible to a varying
degree; it then died out in intervening tracts, or was left only in a
certain part of its former and more extensive area of range.
Zoological Regions.—Seeing that each species of animal
has its own definite range, it is clear that the earth's surface can be
apportioned into divisions which are characterised by their animal
inhabitants. We shall divide the earth into realms, which are the largest
divisions; then into regions; and finally into subregions. It must be
borne in mind that the various groups of the animal kingdom are of
different ages, geologically speaking, and have therefore had less or
more time, as the case may be, to settle down into their present
distribution, and that different animals differ greatly in their rate of
multiplication, their power of migration, and their susceptibility to the
effectiveness of various natural and other barriers to distribution. It
is not, therefore, possible to divide the world into realms and regions
which shall express the facts of distribution of the entire animal
kingdom. Such divisions, which are common in text-books of zoology having
but a small section devoted to zoogeography, are at best mere
approximations and averages; no good is gained by taking such a
comprehensive view of the matter, as the essential object of subdividing
the earth's surface is thereby lost sight of. The zoogeographical
division of the earth which will be adopted here is that originally
recommended by Dr. Blanford, and now accepted by a number of authorities.
There are three "realms," to which a fourth may perhaps be
added—though on negative grounds, and merely for the purpose of
emphasising the parts of the world to which mammals have not gained
access. The realms are again divisible into regions, at least in the case
of one of them, and the regions may be again separated into more or less
distinct subregions or provinces. The three primary divisions or realms
which contain mammals are the Notogaean, including Australia and certain
islands to the north of it; the Neogaean, or the South American continent
and Central America; the Arctogaean, including the continents of North
America, Europe, Asia, and Africa, together with the adjacent islands,
such as the West Indies, East Indies (exclusive of those which fall
within {85}the realm of Notogaea), and Madagascar; and
finally, the realm of Antarctogaea or Atheriogaea, which embraces New
Zealand, the Antarctic continent, and a series of islands such as South
Georgia and Kerguelen, and possibly even the extreme south of Patagonia.
This latter quarter of the globe will need no further reference, as it
has no truly indigenous terrestrial mammalian inhabitants. We cannot
include the Bats in this statement, as their distribution is due to
different powers of extending their range, and to different barriers from
those which govern the range of other groups of mammals.
(1) Notogaea.[42] This
realm is characterised by the exclusive possession of the Monotremes:—that is to say, one of the two primary
divisions of the Mammalia is absolutely restricted to this area. It
contains, moreover, the vast majority of the Marsupials. Further, the
realm of Notogaea is to be distinguished by the entire absence of the
higher mammals, with the exception of a few small Rodents. (The Bats are
ignored for the reasons stated, and the Dingo is believed to have been an
importation.) It cannot be disputed that this is a very distinctly-marked
area of the earth's surface.
(2) Neogaea. The continent of South America has no Monotremes and only
a few Marsupials, all of which, with the exception of Caenolestes,
belong to the Polyprotodont division of that order, and to a peculiar
family, Didelphyidae. The recent discovery of other fossil Marsupials,
however, to some extent favours Huxley's view that Neogaea and Notogaea
form one realm as opposed to the rest of the world. Besides this, Neogaea
possesses the Edentata, which are found nowhere else;—that is, the
division of the Edentata to which the name is now restricted by some
authorities. It is also characterised by the nearly entire absence of the
important order of Insectivora; and, as minor marks of distinction, by
the absence of Antelopes, Oxen and Sheep, of the Ichneumon tribe, of
Horses, and of Lemurs. It has the exclusive possession of the Hapalidae
and Cebidae, and of several families of Rodents.
(3) Arctogaea. This vast realm is clearly capable of subdivision into
four regions, which will be considered in detail later. In the meantime
the points of likeness between these subdivisions is more marked than are
either the resemblances or the {86}differences of any one of them to either of
the two realms which have just been defined. The two realms that have
been discussed retain their distinctness from each other and from
Arctogaea for a considerable way back into the Tertiary period. It is not
until we reach very early Tertiary times that Edentates are met with in
North America; and then it cannot be regarded as absolutely settled that
the Ganodonta are really the forerunners of the Armadillos, Sloths, etc.
Nor do we find Marsupials in Europe until far back in time, and at a
corresponding period in North America. Indeed the fauna of South America
in late Tertiary times was even more distinct than it is now; for then we
had confined to that region the Toxodonts, Glyptodonts,
Macrauchenia, and other forms, while in Australia there were still
Marsupials. In late Tertiary times Europe and India were by no means so
distinct from Africa as they are to-day. North America does not resemble
the Old World quite so much as the subdivisions of the Old World resemble
each other; but, as will be pointed out later, there are and were very
substantial agreements. The Elephants, Rhinoceroses, Giraffe,
Hippopotamus, Orycteropus, are now distinctively African or Indian
animals; but all these genera, or at least families (in the case of the
Giraffe), have occurred in Europe during quite recent times.
Lycaon indeed, now confined to Africa, is thought to have had a
European origin from its occurrence in caves there. The Hyaena and the
Lion, certain members of the Horse tribe, Apes, and other animals, were
also but are not now European.
India again, and the Oriental region generally, once possessed the
Hippopotamus, the Chimpanzee, Giraffidae, the Antelopes, Cobus,
Hippotragus, Strepsiceros, and Orias, which are now
purely African animals. It shares at present with the Ethiopian region
the Catarhines, including the Anthropoid Apes, the Lemurs, Tragulina (the
genus Dorcatherium is also known from fossils in India),
Manis, Hyaena, the Cheetah, Elephant, Rhinoceros, and the
Ratel. There is, in fact, no order of mammals which is now absent from
one of these three regions though present in the others, save the Lemurs,
and they occurred in past times in Europe. The Tapir of India is known
fossil in Europe, and the latter continent had its Monkeys and even
Anthropoids. On the other hand, North America is more distinct. It has no
Lemurs, Apes, Elephants, Rhinoceroses, Tapirs, Old World Edentates {87}(Effodientia), Viverridae, Horses, or
Antelopes, excepting Antilocapra, a type of a separate division of
Bovidae. But since several of these groups have been represented in
recent times, no primary line of division can be profitably drawn.
Arctogaea as a whole may be characterised by both negative and
positive characters. As negative features may be mentioned;—the
entire absence of Edentates (Necrodasypus of Filhol is rather
doubtful, see p. 164, n.), though a few crept up
into the Nearctic region from Neogaea during past times; and of
Hapalidae, Cebidae, and Marsupials, except an Opossum in North America.
This realm has, on the other hand, all the Lemurs, all the Insectivores
with the exception of the West Indian Solenodon, all the
Proboscidea, Rhinoceroses, Horses, Deer, Antelopes, the last group
including the Oxen and a variety of other important families. It is in
fact the headquarters of all the Eutheria with the exception of the
Edentata and Marsupials.
The subdivisions of this realm have been variously effected. The
classical subdivisions are of course those of Mr. Sclater, who would
recognise (1) the Nearctic, North America; (2) the Palaearctic, including
Europe, Northern Asia, and Japan; (3) the Oriental, including Asia south
of the Himalayas and the islands of the Malay Archipelago as far east as
the Australian region; and (4) the Ethiopian, i.e. tropical Africa
and Madagascar. Some would alter this by uniting America and the north of
the Old World into a Holarctic region, separating off the southern parts
of the North American continent into a Sonoran region. To some, the
claims of Madagascar to form a separate region are convincing. To
distinguish the boundaries of the several regions is a difficult task;
they dovetail into each other on the frontiers with the complex curves of
a puzzle-map. The difficulty has been grappled with by the suggestion of
intermediate transitional areas; but this proceeding really doubles the
difficulty, for there are then two frontiers to delimit in each case
instead of only one. The animal inhabitants must be expected to mingle
somewhat at the lines of junction of one region with another.
The Sonoran region does not appear to us to have great claims to
recognition. It shows a mingling of southern with northern forms exactly
as might be expected. An Armadillo and Didelphys have, as it is
believed, invaded it from the Neogaeic realm; it possesses also the South
American genera, Dicotyles, Nasua, {88}Conepatus,
Sigmodon. On the other hand, the Sonoran genera
Antilocapra, Cynomys, Procyon, and the Insectivora
Blarina and Scapanus, extend further north. Peculiar to
this region are only six genera of Rodents, which seems an insufficient
reason for raising the Sonoran province to the dignity of a region.
Considered from the point of view of numbers of peculiar forms, the
Thibetan subregion has more claims to distinction as a region; for
confined to that area we have the genera Nectogale,
Aeluropus, Eupetaurus, Pantholops, Budorcas;
while by slightly extending its limits, a number of other peculiar forms
might be added. Madagascar has distinctly more claims to regional
division. Absolutely confined to it are eleven of the seventeen existing
genera of Lemurs, the family Centetidae among the Insectivora, which
contains seven genera, and another recently discovered and peculiar
genus, Geogale; it has six peculiar genera of Viverridae; it has
five peculiar genera of Rodents. In addition to this it is negatively
characterised by the absence of the following typical African animals,
Felidae, Proboscidea, Rhinocerotidae, Equidae, Monkeys, etc. It seems to
be impossible to avoid allowing the rank of a region to this part of the
world.
In separating the Nearctic from the Palaearctic region, stress must be
laid rather upon the absence of Asiatic and European forms from North
America than upon the existence in the northern half of the New World of
many peculiar forms. Peculiar to the Nearctic are the Goat genus
Haploceros, the Rodents Erethizon, Zapus, and the
family Haplodontidae. The Mole genus Condylura is also restricted
to this part of the New World. Even so it has more peculiar forms than
the Sonoran. If we add to this the absence of Horses, Antelopes except
Antilocapra, Pigs, Hyaenas, etc., there are strong grounds for
retaining this division. It must be agreed, however, that it comes rather
nearer to the Eurasian district than the latter does to the Oriental.
The Oriental region has many characteristic animals. It has among the
Anthropoid Apes the Orangs and Gibbons; of Old World Apes it has confined
to its own area the genera Semnopithecus and Nasalis. Of
Lemurs there are Loris and Nycticebus, and Tarsius,
representing a family of that order, or even a sub-order. The
Galeopithecidae are entirely Malayan. There are many Rodent, Carnivorous,
and Insectivorous genera; the Rhinoceroses and the Elephant of this
region differ from those of Africa. {89}Tragulus concludes
a sample from a very rich list of peculiar forms.
The Ethiopian region has also its Anthropoids, the Gorilla and the
Chimpanzee, but they belong to genera or a genus different from those
which include the Oriental forms. There are five peculiar genera of
Cercopithecidae. The Lemurs restricted to this region are Galago,
Perodicticus and Arctocebus. The peculiar Insectivorous
families Macroscelidae and Chrysochloridae are only found here, besides
many other peculiar genera. Africa is especially the home of Antelopes,
and the Giraffe is not found now outside its borders. The Elephant and
the Rhinoceroses are of different species from those of India. There are
many peculiar Rodents and Ungulates.
{90}
CHAPTER III
THE POSSIBLE FORERUNNERS OF THE MAMMALIA
The relationship of Mammals to Vertebrates lying below them in the
scale, their origin in fact, is a much-debated question, with many
attempted solutions. To enter into this large question in detail would
involve a great deal of useless statement of arguments founded upon
misleading or upon quite inaccurate "facts." It will perhaps be
sufficient if we reflect here the current view most in vogue at the
present, i.e. that which would refer the Mammalia to reptiles
belonging to the extinct Permian and Triassic group of the Theromorpha
(also called Anomodontia). These have been explored lately to a very
large extent, and chiefly by Professor Seeley.[43] The very fact that a genus
Tritylodon, only known by the forepart of the skull, has been
called Mammalian and Anomodont by various authors, shows at least the
difficulty of differentiating the two groups when the material for study
is imperfect. As a matter of fact these Theromorpha are without doubt
reptiles; they show, for example, a lower jaw formed out of several
distinct pieces, of which the articular articulates with a fixed quadrate
on the skull. They possess the characteristic reptilian bones, the
"transverse," the pre- and post-frontals, and there are various other
points of structure which leave no room for doubt as to their truly
reptilian nature. There are, however, numerous indications of an
evolution in the mammalian direction in all parts of the skeleton, to the
more important of which some reference will be made here. It may be as
well to clear the {91}ground by mentioning the fact that among the
Theromorpha four distinct types of reptiles are included, which are
considered to form four orders, i.e. the Pareiasauri, the
Theriodontia, the Anomodontia (Dicynodontia), and the Placodontia.
The first of these divisions includes what seem to be basal forms.
These reptiles show numerous points of likeness to the Amphibian
Labyrinthodonts.[44] On the
other hand the third division, that of the Dicynodontia, are
highly-specialised Theromorpha, from which no further evolution would
appear to have been possible. Thus the dentition was either completely
lost, or reduced to tusks as in Dicynodon. We need not therefore
concern ourselves in the present volume with these Anomodonts. It is with
the Theriodonts that our business lies. The very name, be it observed, is
aptly chosen on the hypothesis to be explained here; but it is not only
in the teeth that these reptiles show likenesses to the Theria or
Mammals, but in almost every feature of their organisation. Unlike other
reptiles, the Theromorpha in general were lifted comparatively high above
the ground on legs of fair length and of mammalian relationship in the
position of the segments of the limbs. The typical reptile grovels upon
the earth with legs sprawling out, as indeed the very name suggests. One
bar to the Theriodonts being on the direct line of mammalian ancestry has
been urged as a preliminary difficulty, and that is their large size. The
earliest undoubted mammals were small creatures, comparable to a Rat or a
Mouse in size; whereas a good-sized Bear or a Wolf is a better standard
of size for some of the best-known genera of Theriodonts. It has,
however, been quite permissibly suggested that living in company with
these large Theriodonts were less obtrusive genera, from which the
mammals might have sprung. It is so familiar a fact that a given group of
animals generally contains giants, dwarfs, and members of intermediate
size, that this suggestion may almost be accepted as a fact. It need at
least present no difficulties to us in our comparisons.
The most salient "mammalian" feature of the Theriodonts is the
heterodonty of the teeth, the pattern of the "molars," and the limited
number which constitute the series. The fact, too, that they are limited
to the dentary bones below and to the {92}maxillae and the
premaxillae above, is a sine qua non for mammalian comparison. In
the more basal Theromorpha the teeth are not so limited in position.
Finally, to complete the remarkable mammalian resemblance of the teeth of
these reptiles, it must be mentioned that in Tritylodon and
Diademodon the roots of the molars, as we may fairly term them,
though not actually divided after the mammalian fashion, were deeply
marked by a groove, which suggests an incipient division or a fusion of
two distinct roots. Some of these facts of structure may now be
considered in further detail. As to the incisors and canines, it is
sufficient to say that the numbers of the former, and the shape of the
latter, are in perfect consonance with a derivation of the Mammalia from
this group. The molar series can be divided into premolars and molars, at
least in so far as regards their shape; for the anterior teeth are often
smaller and less complicated than those which follow, as is often the
case with the two series in mammals. The molar series also consist of
teeth in close apposition to each other and separated from the canines by
a diastema, which is a character of mammalian teeth. The fact that in the
reptile Cynognathus and the mammal Myrmecobius there are
nine of these molar teeth in each half of each jaw is perhaps not a point
upon which it is desirable to dwell with too much weight; but the general
fact that the molars are further reduced in some genera of Theriodontia
than in that which has been mentioned, is clearly a matter of
significance when the ancestry of the mammals is under consideration.
The most interesting fact about the molar series in the Theriodontia
is that we meet with the two types of molars that occur in the mammals.
Cynognathus and other genera have molars which consist of a main
cusp, and of one cusp before and one after the main cusp; in fact these
teeth are triconodont as in certain early mammals, a state of affairs
which is believed by the "trituberculists" (see p. 56) to have preceded the tritubercular tooth. There
are also "multitubercular" teeth, especially well developed in
Tritylodon, where they exactly resemble those of certain
Multituberculata, and whose structure originally led to the placing of
Tritylodon among the mammals of that group. If there is any
question about the mammalian nature of this fossil, there remain several
other Theriodontia in which the multituberculism is well marked. It is so
in Trirhackodon {93}and in Diademodon for instance. This
incidentally lends some support to the idea that the Mammalia have been
evolved from two sources, a way of looking at the origin of the group
that will coincide with the views of some authors like the late Dr.
Mivart, and will at the same time reconcile the trituberculists and the
multituberculists. For we should then assume that the Eutheria and
Triconodontia had originated from some such form as Cynognathus;
and the Multituberculata and the existing Monotremes from some form like
Diademodon. It is not of great use to point out that
Diademodon is really of the trituberculate pattern, because in its
molars, though multituberculate, the trituberculate main cones can be
recognised; for that state of affairs could just as well have been
brought about by a reduction from the multituberculate type. The skull of
these Theriodonts shows some well-marked approximations to the mammalian
type. There is in the first place a commencing consolidation and
reduction of the individual bones, which is so distinguishing a feature
of the mammalian skull as opposed to the skull of lower vertebrates. In
Cynognathus the postorbital is fused with the jugal, and the
supratemporal with the squamosal, forming apparently one bone. In the
lower jaw the splenial is often reduced to the thinness of paper, thus
indicating a commencing disappearance. In many Theromorpha the squamosal
shares largely in the formation of the articular facet for the lower jaw,
obviously an important mammalian characteristic; this is brought about by
the reduction of the quadrate, which latter bone, moreover, acquires in
certain particulars the appearance of the mammalian malleus, with which
it is, according to many, homologous. But this subject has been already
dealt with on page 26. A very pronounced likeness
to the mammalian skull is that there are two occipital condyles. That
this has been brought about by the further development of a tripartite
condyle such as occurs in tortoises, and that by the suppression of the
basi-occipital part, does not affect the resemblance to the mammalian
skull; in fact it explains the origin of two condyles from the typical
reptilian single condyle, and disposes of the necessity for believing,
with Huxley and others, the Amphibia to be on the main line of mammalian
evolution on account of their two condyles. The general aspect of the
skull in Cynognathus has been {94}compared to that "of
Thylacinus or Dissacus." No one can examine the actual
sketches of the skull of that Theriodont without endorsing that opinion.
As a curious detailed point of likeness to certain Mammalia may be
mentioned "a small descending process of the malar bone, which may be a
diminutive representative of the descending element of the malar seen in
Elotherium, Nototherium, Diprotodon,
Macropus, certain Edentata, such as Glyptodon,
Megatherium, Mylodon, Bradypus, but unparalleled so
far as I am aware in fossil reptiles." (Osborn.) The zoologist cannot
help being impressed with the significance of small details of
similarity, which do not seem to be due in any way to surrounding
conditions of life, and thus referable to mere convergence, like the
fish-like form of Whales and Seals.
The rest of the skeleton of the Theriodontia is by no means so well
known as the skull and teeth. But from what is known, other mammalian
characters can be pointed out. Perhaps the most striking mammalian
feature is to be found in the scapula of Cynognathus. It is in
this creature somewhat narrow and elongated; but it has a well-marked
spine, ending in a hooked acromion. Now it is to be noted in support, so
far, of the diphyletic origin of mammals, that in the Monotreme, as in
Whales indeed, the spine forms the anterior border of the scapula, and is
coincident with it, there being thus no prescapula at all in the
Monotreme, and only a trace of it in certain Whales.[45] Whether the multituberculate
Tritylodon or Diademodon had a scapula after the Monotreme
pattern is not known; but it is clear that the scapula of the triconodont
Cynognathus is quite after the pattern of the Eutherian scapula.
Furthermore, Professor Seeley is of opinion that the coracoid was
relatively small, and indeed smaller than the same bone in Edentates, and
a fortiori than in Monotremes. Another fact of structure which
points also, possibly, in the direction of a diphyletic origin for the
Mammalia, is the double-headed ribs of Cynognathus. As is well
known, the ribs of the Monotremata have only the central head, the
capitulum.
As a general mark of affinity with mammals the reduction of the
intercentra in Cynognathus may be noted, and also the existence of
a small though perfectly obvious obturator-foramen, separating the pubis
from the ischium. There are further details {95}which tend in the same
direction. And we shall probably not go far wrong in the present state of
our knowledge if we assign the origin of the mammals to some type which
would be included in the order Theriodontia or at least in the sub-class
Theromorpha.
{96}
CHAPTER IV
THE DAWN OF MAMMALIAN LIFE
The animals that we considered in the last chapter, though showing
certain unmistakable likenesses to the mammals, are nevertheless
unquestionably not mammals but reptiles. In the Triassic strata, however,
we first meet with the remains of undoubted mammals. The Mammalia first
appeared upon the earth in a tentative and hesitating way: they had not
cast off many of the characters of their supposed reptilian forefathers;
they shrank from observation and destruction by their small size, and
apparently, so far at any rate as their teeth afford a clue, by an
omnivorous diet. The world abounded at that period in large and
carnivorous reptiles, which may indeed have been the principal enemies
with which the first mammals had to cope. These early mammals lingered on
to so late a period as the Eocene; but the majority of the genera were
Triassic, Jurassic, and Cretaceous. Certain of the primitive mammalian
forms have been referred to the Marsupials, and their resemblances to the
Monotremata have also been pointed out. The current view of the present
time, however, is that they form a special order, which may possibly have
embraced the ancestors of both Marsupials and Monotremes; for it is
reasonable to explain in this way the combination of characters of these
two orders which they present. For this group the name Allotheria has
been proposed by Marsh, and Multituberculata by Cope; the latter term is
the less suitable, in that the Monotremata (Ornithorhynchus) are
also "multituberculate." The group is known in a very imperfect fashion.
The remains are but few and fragmentary; and for the most part we have
only a few teeth to speculate upon. This is natural enough, for the
harder teeth might easily be supposed to {97}have resisted the decay
which would more readily affect the softer bones. Where there are bones
it is frequently the lower jaw alone which has been preserved for
us—a bone which has also been preserved in the case of some of the
contemporary Marsupials.
It has been pointed out (from the observation of dead dogs floating in
canals) that the lower jaw is occasionally detached from the carcase. It
is the most readily separable part which contains a skeleton. It may be,
therefore, that the remains of these early mammals, floating down some
river to the sea, may have lost their jaws while in the river, or at
furthest in the shallow waters of the sea, the rest of the carcase
floating out to a greater distance, and being finally entombed in the
stomach of some carnivorous fish, or in the mud at the bottom of a deep
ocean, which has never since seen the light.
The characters of this group are really more those of the Monotremata
than of the Marsupials. The undoubted likeness which their molar teeth
show to the temporary teeth of the Platypus have already been commented
upon. Like the Monotremes the Allotheria appear to have possessed a large
and independent coracoid; the evidence for this rests upon the discovery
of the lower end of a scapula of Camptomus, a Cretaceous genus
from North America upon which there is a distinct facet for the
articulation of what can have been nothing else than a coracoid. On the
other hand they differ from the Monotremata by the presence of incisor
teeth which were Rodent-like in form, and not very different from those
of certain Marsupials. This point of difference cannot be regarded as of
very first-rate importance; no one would relegate the Sloth and the
Armadillo to different orders on account of their tooth differences,
which are about on a par with those to which we have just referred. It
seems indeed likely that it will be ultimately necessary to rub out the
boundary line which now divides the Allotheria and the Monotremata.
The Plagiaulacidae are unquestionably mammals, and they are placed by
most naturalists in this at present uncertain group of Multituberculata,
which will be retained here in deference to the distinguished authorities
who have instituted the group, though there are but few characters by
which it can be defined. This family though appearing in the Trias,
extends down in time to the Eocene. The type-genus, that which has given
its name to {98}the family, is Plagiaulax. As it is
not Triassic, the consideration of its characters will be deferred until
later. Microlestes is a Rhaetic genus, known from rocks in Germany
and England; but it is entirely based upon molar teeth. M.
antiquus has a two-rooted molar of an elongated form with a row of
tubercles on either side of a median groove, which traverses the long
axis of the tooth. To some extent the teeth of the ancient form resemble
those of Ornithorhynchus. Microlestes has been sometimes
spoken of as a Marsupial, but Mr. Tomes[46] has found that it does not show one
very universal character of the Marsupial teeth: it has not those
continuations of the dentinal tubes which traverse the enamel in all
Marsupials that have been examined with the sole exception of the
Wombat.
The rarity of the remains of mammals in these earliest rocks of the
Secondary epoch has been accounted for in another way from that which has
been suggested above. It may be that the group Mammalia was not evolved
in Europe at all, and that the stray remains which have been found in
that continent represent the fragmentary remnants of a few scattered
immigrants which heralded the later invasion of more numerous genera
during the Jurassic period.
The Mammals of the Jurassic Period.—Some of the
Allotheria or Multituberculata described in the last section occur in the
rocks of this early part of the Secondary epoch. They are doubtful in
position, as already stated; some of them indeed, as for instance
Tritylodon and Dromatherium, are possibly not mammals at
all, while the remainder probably belong to a non-existent order of
mammals. Along with these dubious creatures are the fragmentary remains
of small animals which are not merely mammals, but in all probability
definitely Marsupials. It is true that here again we have little beyond
lower jaws and teeth to deal with; so that there may be less certainty in
referring them to the Marsupials than appears to be the opinion of the
majority of Palaeontologists.
Professor Osborn in fact considers that the Mesozoic mammals consist
of three groups: (1) The Multituberculata, including the Bolodontidae,
Stereognathidae, Plagiaulacidae, Polymastodontidae, and possibly the
Tritylodontidae (which, however, are regarded by him and by others as
more probably reptiles of the {99}Theromorphous group). (2) The Triconodonta,
which were Marsupials, though in all probability with a complete
succession of teeth and with an allantoic placentation. This group will
include the genera Phascolotherium and Amphilestes, as well
as Triconodon and Spalacotherium. Finally we have (3) the
Trituberculata (or Insectivora Primitiva) with the genera
Amphitherium, Peramus, Amblotherium,
Stylacodon, and Dryolestes.
We shall take these three groups in order. The Multituberculata have
already been to some extent defined, if such a word can be used to
express the summation of the very scanty information at our disposal. Of
this group, Plagiaulax is a genus which occurs in the Purbeck
beds; it is only known by lower jaws implying an animal of the size of a
Rat or rather smaller. The jaws have in front a large incisor which looks
Rodent-like, and also like those of the Diprotodont Marsupials; but it is
held that these teeth did not grow from persistent pulps, and there is in
any case no anterior thickened coating of enamel. Canines are absent; the
diastema is followed by four premolars increasing progressively in size
and possessing somewhat complicated grinding surfaces. These surfaces are
formed by several obliquely-set ridges. The succeeding teeth are termed
molars on account of their difference in structure, and there are but two
of them on each side. The molars are of a pattern common in the
Multituberculata; the centre is hollowed, and the raised rim is beset by
tubercles. Other Jurassic genera of Multituberculates are Bolodon,
Allodon, and Stereognathus. All of these possess the same
multituberculate molars.
Of the Triconodonta the type-genus is Triconodon. This genus is
better known than most Jurassic mammals, since both the upper and the
lower dentition have been described. It appears to have possessed the
typical Eutherian dentition of forty-four teeth, to which a fourth molar
is added in some species. The great difference between the molars and
premolars argues a complete tooth-change. The genus is American as well
as European.
Spalacotherium has more molars, five or six.
Phascolotherium bucklandi, on the other hand, is a much older
type in the form of its teeth. There are, however, not so many of them as
in Amphitherium; Phascolotherium has but two premolars and
five molars, making a total of forty-eight teeth. The teeth are of the
triconodont form, the three cusps being in line, and the middle one the
largest. {100}
Amphilestes has teeth of the same pattern but has more of them,
the premolars and molars being respectively four and five. All these
animals had the lower jaw inflected. Whether they are all Marsupials or
not, it is clear that Phascolotherium and Amphilestes
should be united and placed away from Amphitherium on account of
the more primitive form of their teeth.
We next come to the Trituberculata.
Among the most celebrated of these remains are a few jaws discovered
in the Stonesfield slates near Oxford, and examined by Buckland, Cuvier,
and some of the most eminent naturalists of the beginning of the last
century. These jaws have been lately submitted to a careful
re-examination by Mr. Goodrich,[47] who has increased our knowledge of the
subject by exposing from the rocky matrix in which the jaws lie fresh
details of their structure; it is probable therefore that now all that
there is to be learnt from these specimens has been recorded.
Amphitherium prevostii was a creature about the size of a Rat.
Its jaw was first brought to Dean Buckland about the year 1814, and
described six years later. Buckland thought the jaw to be that of an
Opossum, an opinion in which Cuvier concurred. The jaw, however, is
marked by a groove running along its length, and this groove was regarded
by de Blainville as evidence of the composition of the jaw out of more
than one element, which would naturally lead to its being regarded as the
jaw of a reptile.[48] This
species and another named after Sir Richard Owen have a dental formula
which, like that of the Marsupials, is large as compared with that of the
Placental mammals; it runs: I 4, C 1, Pm 5, M 6—i.e. 64
teeth altogether. This is a larger number than we find in any existing
Marsupial. But as in Marsupials, and in certain Insectivora also, the
angle of the jaw is inflected. These teeth are of the tritubercular
pattern with a "heel." They are in fact closely like those of the living
Myrmecobius; but not, it should be remarked, unlike those of
certain Insectivora.
The Mammals of the Cretaceous Period.—At one time there
was a totally inexplicable gap between the Jurassic and the basal Eocene,
a series of strata which occupy an enormous expanse of time in the
history of the earth having appeared to {101}be devoid of mammalian
remains. This gap, however, has been filled up by the discovery of
mammalian remains in the North American Laramic formation, which seems to
be clearly of Cretaceous age. Furthermore, it is held by some that the
Purbeck beds are more properly to be placed with the Cretaceous, which
would then necessitate the consideration under the present heading of
some of the types already dealt with; and if, as is suggested in the
following section, the lowest so-called Eocene beds are really referable
to the Cretaceous, there is no lack of mammalian remains in that period.
And, moreover, it was in that case the Cretaceous period which witnessed
the evolution of the existing orders of Placental mammals. Otherwise the
mammalian remains of the Cretaceous agree with those of the Jurassic. We
find remains of the Multituberculata in fragments of Plagiaulacidae and
Polymastodontidae. Ptilodus is a genus which has two premolars;
and Meniscoessus is another multituberculate from the same Laramic
formation. The other detached fragments of mammals are thought by Osborn
to represent both Placentals and Marsupials.
The Mammals of the Tertiary Period.—Unless the lowest
beds of the earliest Tertiary period, the Eocene, such as the Torrejon of
North America, should be in reality referred to the Cretaceous, there is
no evidence that the modern groups of Mammalia existed before the present
epoch of the earth's history. It is probable, however, that the Eutheria
as a group were Mesozoic. The fossil jaws that have been considered in
the last chapter may quite probably be primitive Eutherians, or even
divisible, as believed by Professor Osborn, into Marsupials and
Insectivores. In the Tertiary, however, apart from the question as to the
nature of the Puerco and Torrejon formations, and as to certain South
American strata whose fossil contents have been investigated by Professor
Ameghino, we find the first traces of mammals definitely referable to
existing orders, or to be distinctly compared with existing orders.
Since, however, representatives of types which have obvious relationships
to modern types appear in considerable profusion in the very earliest
strata of the Eocene, it seems clear that much remains to be discovered
in beds earlier than these. Confining ourselves, however, to facts and to
comparisons which can be made on more than a few lower jaws and scattered
teeth, which is practically all that we {102}possess of earlier
mammals, we must arrive at the general conclusion that two of the
existing larger groups of the Eutherian, non-Marsupial, mammals were
differentiated at quite the beginning of the Eocene, and were represented
by forms from which it is possible to derive at least the existing
Carnivora, Insectivora, Artiodactyla, and Perissodactyla. These were the
Creodonta and the Ungulate Condylarthra. In addition to these we may
enumerate as very early types the Lemuroidea, represented by such forms
as Indrodon in the New World, and (though later) by
Necrolemur, etc., in the Old World, and the Edentata, if we are to
allow as their ancestors the Ganodonta.
The early Eocene strata also contain representatives of at least one
order, the Amblypoda, which increased subsequently, but has died out
without descendants, unless we are to believe with some that the
Elephants are to be derived from these Eocene "pachyderms." In later
Eocene times the great majority of the existing orders, and even
subdivisions of orders, are to be met with; and there are in addition
such totally extinct orders as the Typotheria, Ancylopoda, and
Tillodontia. Coupled with this gradual specialisation in the orders of
Eutherian mammals, there is naturally a vast increase in the number of
generic and family types. This culminates perhaps in the Miocene, from
which time there has been a gradual decline in mammalian variety, so that
it is justly said that we live now in an epoch which is impoverished of
mammals. This gradual decay has persisted until to-day, as is witnessed
by the extinction of the Rhytina and the Quagga, and the growing rarity
of the White Rhinoceros and the American Bison.
The early Eutherian stock consisted of small mammals with small heads
and slender, long tails. The limbs were pentadactyle, ensheathed in claws
or broader hoofs. The fore-limbs may have been partly prehensile. The
teeth were forty-four, completely differentiated into incisors, canines,
molars, and premolars; and there appears to have been a complete
diphyodontism. The canines were not greatly enlarged, and no diastema
separated any of the teeth. The molars were bunodont or of a more cutting
pattern, with some five or six tubercles. These animals were, moreover,
very small-brained. This early stock is represented by Creodont and
Condylarthrous animals, the exact boundaries between which are hardly
marked in the {103}very early types. Professor Osborn has
argued that from this early Eutherian stock there were two waves of
progress, or, as he expresses it, "two great centres of functional
radiation."[49]
The first was largely ineffective, the second has produced all the
Eutherian orders of to-day. These two divisions are termed by him
"Mesoplacentalia" and "Cenoplacentalia." The first division embraces the
Amblypoda and their descendants the Coryphodonts and Dinocerata, many of
the Condylarthra, the bulk of the Creodonts and the Tillodonts. These
creatures persisted for a time, but died out in the Miocene. They were
mainly distinguished by the smallness of their brain; the great
specialisation of structure which they exhibit having left that organ
unaffected, and therefore tending in the long run to render them unable
to cope with changes in the inorganic and organic world. The successful
division of the primitive Eutheria comprises the groups which exist at
the present day, and is not connected directly with those small-brained
Mesoplacentals; it has apparently originated, however, from the least
specialised of their ancestors. Professor Osborn thinks, moreover, that
the Lemurs and the Insectivores are persistent descendants of the earlier
wave of Eutherian life. It appears in fact as if Nature had created the
existing Ungulate, Unguiculate, and other types on a defective plan, and,
instead of mending them to suit more modern requirements, had evolved an
entirely new set of similarly-organised types from some of the more
ancient and plastic forms remaining over. The Marsupials may be the only
group of the early wave remaining, and they have been able to hold their
own for the geological reason that Australia was early cut off from
communication with the rest of the world. That they are disappearing
seems to be shown by their gradual diminution as we pass from Australia
towards the continent of Asia, through the islands of the Malay
Archipelago. Competition has here decimated them, as it may do in the
remote future in Australia.
It is often said, but with some looseness of statement, that ancient
quadrupeds are huger than their modern representatives. This statement is
partly true in fact, but largely wrong in implication. For it suggests
that—and the suggestion is often expressed in books that are not
authoritative—huge animals {104}have left a dwarfish
offspring; that there were giants of old, and that there is a puny race
to-day. As a matter of fact, the study of the gradual evolution of the
early Tertiary Mammalia into their descendants of later times shows very
plainly the truth of this interesting generalisation: That the primitive
types were all small creatures, and that in those instances where we can
trace a pedigree, there was a gradual increase in size up to a point
where greater increase led to extinction. We point out later on a number
of facts illustrating this matter in detail. It has been ascertained, for
instance, that the pedigree of the Horses, the Camels, the Rhinoceroses,
and many other groups, commences with small forms and culminates in large
ones. It may be urged that such animals as the Tapir are to-day smallish
forms, and that related to them in the past were the gigantic
Titanotheres; but in this and similar cases it will be found that the
extinct giants were not in the direct line of pedigree, but represented
side-branches which waxed huge on their own account and then
disappeared.
{105}
CHAPTER V
THE EXISTING ORDERS OF MAMMALS
Prototheria—Monotremata
Apart from those creatures whose fragmentary remains have been
considered in the last chapter, and which belong to the earliest of
mammaliferous strata, the remains of Mammalia are all referable to
existing orders. In the pages which follow we shall therefore deal with
the actual representatives of living families side by side with their
extinct relatives. The existing orders of Mammalia, together with those
of their fossil allies, can be plainly divided into two great
subdivisions, or, as we shall term them, sub-classes; the Mammalia as a
whole being termed a class of the Vertebrata comparable with the class
Reptilia, etc. It has been usual, owing to the initiative of Professor
Huxley, to divide the Mammalia into three divisions of primary
importance. We shall adduce reasons later for not accepting this mode of
division, but that which allows of only two primary divisions. These two
divisions are (1) Prototheria and (2) Eutheria. Whether the
Multituberculata, Trituberculata, and Triconodonta, considered in the
last chapter, are really to be distributed among these two sub-classes is
a matter upon which it is possible to form an opinion, but not to
dogmatise. The Prototheria stand at the base of the mammalian series, and
present many likenesses to the Sauropsida; the Eutheria are the animals
which are most fully differentiated as mammals. We shall commence
with
Sub-Class I.—PROTOTHERIA.
To this group belongs the order Monotremata, and possibly also the
so-called Allotheria or Multituberculata. As, however, {106}the latter are
only known from very fragmentary remains, which are not sufficient to
determine the systematic position of the animals of which they are
fragments, I have not thought it worth while to attempt a serious
definition of the order Multituberculata. I have introduced a short
account of the principal facts which are known concerning the creatures
grouped together under this name into the historical sketch of the
progress of mammalian life in Chapter IV. As to the Monotremata, there is
no question that they are entitled to rank in a group equivalent to that
including all other mammals of which we have sufficient knowledge to
construct a classificatory scheme. There have been, indeed, naturalists,
such as Meckel, who would altogether deny the mammalian rank of these
creatures.
The Monotremata or Ornithodelphia may be thus defined:—
Mammalia with no teats, but with a temporary pouch in which the young
are hatched, or to which they are transferred after hatching, and into
which open the ducts of the mammary glands. An anterior abdominal vein,
or at least the membrane supporting it, persists throughout the abdominal
cavity. Heart with an incomplete and largely fleshy right
auriculo-ventricular valve. Brain without a corpus callosum. Shoulder
girdle with a large coracoid reaching the sternum; clavicles and an
interclavicle present. There are "marsupial" or epipubic bones attached
to the pelvis. Vertebrae with no epiphyses for the most part. Ribs with
only capitulum and no tuberculum. Mammary glands of the sudoriparous and
not the sebaceous type of epidermic gland.[50] Oviparous, with a large-yolked and
meroblastic ovum, enclosed within a follicle of two rows of cells.
To call these animals Mammalia is of course an abuse of the meaning of
that word in one sense, but it is not in another; since the pouch of
these Monotremes is, as has been explained elsewhere (p. 16), the real equivalent of a teat, and not of the
pouch of the Marsupials.
The most salient characteristic of this group of mammals in the
estimation of their position in the vertebrate series is not so much the
fact that they are oviparous as that the eggs are large-yolked, and
develop therefore, so far as regards their early stages, after the
fashion of the egg of a reptile. The laying {107}of eggs, or at least
ovoviviparity, would follow from the structure of the egg, since the
abundance of yolk would do away with the necessity for a placenta. That
the eggs had this Saurian characteristic was first definitely made known
by Professor Poulton[51]
for Ornithorhynchus, and his results were confirmed later for
Echidna.[52] The
structure of the eggs has, however, already been dealt with on p. 72. The fact that these animals lay eggs appears to
have been known for a very long time, though rediscovered so lately as
1884 by Mr. Caldwell.[53]
In connexion with the structure of the ova, the ovaries themselves and
the oviducts are built upon the Sauropsidan plan. In the male the testes
retain the primitive abdominal position. The fact that the urinary and
genital products escape by means of their ducts into a chamber which also
receives the end of the alimentary tract is not a distinctive feature of
this group, inasmuch as it is seen in the Marsupials, and also in certain
low Eutheria, such as the Beaver and other Rodents, and a few
Insectivores. As to external features, the Monotremata show certain
archaic characters. The unspecialised arrangement of the mammary glands
has already been described. These animals are plantigrade, if the term
may be used also to describe the aquatic Ornithorhynchus. The ears
are absolutely destitute of a conch. The remarkable spur upon the
hind-legs furnished with a gland, which is more marked in the male, and
indeed disappears in the female of Ornithorhynchus, is a structure
which argues the specialised condition of these two modern
representatives of what must have been a large order in the past.
{108}
Fig. 51.—Ventral view of skull of Echidna aculeata, and right half of mandible, ang,
Angle of mandible; aud.oss, auditory ossicles; cond, condyle of mandible; cor,
coronoid process; max, maxilla; oc.cond, occipital condyle; pal, palatine; p.max,
premaxilla; pt, pterygoid; sq, squamosal; ty, tympanic ring. (After Parker and
Haswell.)
The skeleton shows numerous ancient characteristics. In the skull
there is no demarcation of the orbit from the temporal fossa, a feature
widely found in archaic mammals. The tympanic remains as a slender ring,
there being no auditory bulla formed either from this or from any other
bone. The malleus and incus are large, and thus reminiscent of the
quadrate and articular bone of reptiles. In the lower jaw the absence of
a marked coronoid process, and the absence of a firm ossification at the
meeting of the two rami, may be a primitive state of affairs. It must be
remembered, however, that the Cetacea show the same characters, though it
is possible that they too are developed from a low mammalian stock. In
the vertebral column we find the typical mammalian seven cervicals; but
those characteristically mammalian structures the epiphyses are totally
absent in Echidna, and only to be seen in the tail-region in
Ornithorhynchus. In having only the capitular head to the ribs,
these mammals are evidently far removed from all other mammals, and are
even more reptilian than the Theromorphous reptiles. The large clavicles
and the interclavicle (Fig. 52, {109}p. 109) are characteristic of the group,
and the latter bone is peculiar to the Monotremata among mammals. So,
too, is the large coracoid. In the scapula there is a spine which
coincides with the anterior border of that bone. The arrangement of the
muscles in this region proves conclusively that this projection is the
homologue of the spine and the acromion of other mammals. Here, again, we
have a point of likeness to the Cetacea.[54] In the pelvis the acetabulum is
perforate (in Echidna), as in Sauropsida.

Fig. 52.—Side view of right half of the
shoulder girdle of a young Echidna
(Echidna aculeata). × 1. a, Acromion;
c, coracoid; cb, coracoid border;
cl, clavicle; css, coraco-scapular
suture; ec, epicoracoid; gb, glenoid
border; gc, glenoid cavity; ic, interclavicle;
pf, postscapular fossa; ps,
presternum; s, spine; ss, suprascapular
epiphysis; ssf, subscapular fossa.
(From Flower's Osteology.)
Considering the numerous very archaic features which the general
structure of this group displays, it is surprising to find how typically
mammalian they are in certain other peculiarities. The mammalian
diaphragm, one of the distinguishing features of the class, is perfectly
normal in the Monotremata. The alimentary canal shows no great
divergences from the normal structure. The stomach is almost globular,
with a projecting pyloric region in Ornithorhynchus; the intestine
is divided into a "small" and "large" intestine by a slender caecum. The
liver has the subdivisions that this organ usually shows in the Mammalia.
However, the presence of the ventral mesentery and of the abdominal vein
in Echidna and Ornithorhynchus has already been mentioned
as a distinctive character. The peculiar and apparently partly primitive
valve of the right ventricle has been described above (see p. 66). The brain is in most respects mammalian in its
characters, but naturally shows some important differences. Dr. Elliot
Smith, who has most recently studied this question,[55] is of opinion that the size of the
cerebral hemispheres is not at all reptilian; indeed, it "greatly exceeds
that of {110}many other mammals." In Echidna,
too, but not in Ornithorhynchus, the hemispheres are well
convoluted, though the arrangement of these convolutions cannot be
brought into line with what is known concerning the convolutions upon the
hemispheres of other mammals. It had been stated that in these animals,
at least in Echidna, there were only two optic lobes, as in lower
vertebrates, instead of the mammalian four. The late Sir W. H. Flower set
this matter at rest,[56]
and showed that Echidna was in this respect typically mammalian.
The absence of the corpus callosum is one of the principal features
separating the Monotremes from other mammals.
The Monotremata are represented to-day by two types,
Ornithorhynchus and Echidna, which are no doubt worthy of
being placed in separate families. Fossil remains of the group (apart
from the problematical Multituberculata) are only known from Pleistocene
times in Australia, and consist of the bones of a large species of
Echidna, and some fragments of Ornithorhynchus, indicating
a smaller animal than the living Platypus.
Fig. 53.—Brain of Echidna aculeata,
dorsal view. (Nat. size.) (From
Parker and Haswell's Zoology.)
Fam. 1. Echidnidae.—This family contains two genera, of
which Echidna is the older and much the better known. The skin is
abundantly covered with spines, with which are mingled hairs. The snout
is tapering, the tail rudimentary, and the fingers and toes five in
number. The spur and gland upon the calcaneum are smaller than in
Ornithorhynchus. The claws are very strong, serving to tear open
the ants' nests, upon the inhabitants of which the Echidna feeds, licking
them up with a long extensile tongue like that of Myrmecophaga. In
relation to this habit the salivary glands are enormously developed, and
indeed the animal has been confounded with Myrmecophaga,[57] as the vernacular name
"Australian Anteater" exemplifies. {111}
In the skull the Echidna differs from Ornithorhynchus in the
greater extension backwards of the palatines, and the larger size of the
pterygoids. The extent and relations of these bones to each other is not
at all unlike that which obtains in many Whales. The premaxillae show
traces of the same divergence followed by convergence of their ends that
is seen in the Platypus. There are only sixteen pairs of ribs, and either
three or four lumbar vertebrae. Echidna has no trace of teeth, and
there are no horny pads which take their place; the mouth is as
edentulous as in the true American Anteaters. The brain (Fig. 53) is
marked by sulci, contrary to what we find in Ornithorhynchus. The
genus has been divided into three species, but it is doubtful whether
more than one can be allowed, which ranges from Australia through the
Papuan region. While there is but one species of true Echidna, a
New Guinea species must clearly be referred to a distinct genus
Proechidna.[58] This
animal is to be distinguished by the fact that there are usually but
three toes on each foot. But there are copious rudiments of the other
phalanges, upon which claws are sometimes developed. The beak is curved
downwards, and the back is rather arched; the whole animal has the most
singular likeness to an Elephant! The ribs are increased by one pair, and
there are four lumbar vertebrae. The one species is named P.
bruijnii. The Hon. W. Rothschild[59] distinguishes a form P.
nigroaculeata, which is allowed by Mr. Lydekker.

Fig. 54.—Australian Anteater. Echidna aculeata. × 1⁄6.
{112}
The Echidna feeds like anteaters, by thrusting its tongue into an
ant-hill, and waiting until it is covered with indignant and marauding
ants, which are then swallowed. But this animal also devours worms and
insects, which are extracted from their hiding-places by the tongue. It
is mainly nocturnal, and prefers the seclusion of the densest scrubs of
the bush, or rocky spots where it is free from intrusion. Dr. Semon did
not find that the spur of this animal was used at all in self-defence;
but he thinks that possibly the weapon may be used, in the breeding
season only, in the combats of the males for the females, when perhaps,
as has been shown to be the case in Ornithorhynchus, the gland
attached to it produces a poisonous secretion.
The egg, as it appears, is transferred to the pouch by the mouth of
the mother; the shell is broken by the emerging young one, which has an
egg-breaking tubercle on its snout for this purpose; the mother removes
the shell. When the young has attained a certain size, the mother removes
it from the pouch, but takes it in from time to time to suckle it. When
on her nightly rambles the young one is left in a burrow dug for the
purpose. Dr. Semon was able, from his own observations, to substantiate
this act of intelligence on the part of the Echidna. It is well known
that the temperature of the Monotremes is less than that of higher
mammals; in addition to this fact Dr. Semon found that the range of
variation of temperature in the Echidna was as much as 13 degrees or
more. It is thus intermediate between the "poikilothermal" reptiles and
the "homoeothermal" mammals.
Fam. 2. Ornithorhynchidae.—There is no need to attempt to
define this family, since it contains but one genus
Ornithorhynchus, with but one species, O. anatinus. The
general aspect of the animal is well known. It is covered with dense fur
of a blackish brown colour; the limbs are short and five-toed, the toes
being webbed. The tail is longish and broad, being flattened from above
downwards. The webbing on the anterior toes considerably outdistances the
tips of the claws, as in the Seals. But this is not the case with the
hind-feet. The "beak," which is broad and flat, and does actually suggest
that of a duck, is not covered with horn, as is often stated, but with a
fine, soft, sensitive, naked skin, which abounds in sense-organs of a
tactile nature. As to characters derived from the skeleton,
Ornithorhynchus has {113}seventeen pairs of ribs and only two
lumbar vertebrae. The skull is expanded in front, and the bill is
supported by two, at first diverging, and then converging, premaxillae.
Between them is the famous "dumb-bell shaped bone," which is believed to
be the representative of the reptilian prevomer. The pterygoids are
smaller than in Echidna, and the hard palate does not extend so
far back as in that genus. The brain of this genus is smooth.

Fig. 55.—Duck-billed Platypus. Ornithorhynchus anatinus. × 1⁄6.
The discovery of the real teeth of Ornithorhynchus only dates
from the year 1888, when they were found by Professor Poulton[60] in an embryo. Later Mr.
Thomas found[61] that the
teeth persist for a considerable portion of the animal's life, and are
only shed, like milk teeth, "after being worn down by friction with food
and sand." We have already (p. 98) called attention
to the general similarity of these teeth to those of certain of the
earliest Mammalia and of mammal-like reptiles. The teeth are all molars,
and they are either eight or ten in number. They are replaced by the
horny plates of the adult animal; but the mode of replacement is curious.
The plates are developed from the epithelium of the mouth, but round and
under the true teeth; the epithelium of the mouth grows gradually under
the calcified teeth, a method of growth which has possibly something to
do with the shedding of the latter. The hollows and {114}grooves in the
plates are the remains of the original alveoli of the teeth.

Fig. 56.—Skeleton of
male Ornithorhynchus.
Ventral view.
The right fore-limb
has been separated
and turned round so
as to bring into view
the dorsal surface of
the manus. The
lower jaw is removed.
acc.tars, Accessory
tarsal bone supporting
the spur; ant.pal.for,
anterior palatine foramen;
ast, astragalus;
atl, atlas; ax, axis;
bs.oc, basi-occipital;
bs.sph, basi-sphenoid;
calc, calcaneum; cbd,
cuboid; cerv.rb, cervical
rib; clac, clavicle;
cond.for, foramen
above inner condyle
of humerus; cor,
coracoid; cun, cuneiform
of carpus; dent,
horny dental plate;
ect.cun, ecto-cuneiform;
ent.cun, ento-cuneiform;
ep.co,
epicoracoid;
epist, episternum;
ep.pb, epipubis; fb,
fibula; fem, femur;
for.mag, foramen magnum;
glen, glenoid
cavity of shoulder-joint;
glen, glenoid
cavity for mandible;
hum, humerus;
in.cond, inner condyle
of humerus;
inf.orb.for, points to
position of infra-orbital
foramen; infr.proc,
inferior processes of
caudal vertebrae;
int.rbs, intermediate
ribs; isch, ischium;
mag, magnum of carpus;
max, maxilla;
max.for, maxillary
foramen; metat.I,
first metatarsal;
metat.V, fifth metatarsal;
nas.cart, nasal
cartilage; obt, obturator
foramen; ol,
olecranon; out.cond, outer condyle of humerus; pal, palatine; pat, patella;
post.pal.for, posterior palatine foramen; pr.max, premaxilla; pr.st, presternum;
pter, pterygoid; pub, pubis; rad, radius; scap, scapula; scaph, scaphoid of tarsus;
scaph.lun, scapho-lunar; ses, sesamoid bones of wrist and ankle; sp, tarsal horny spur;
sq, squamosal; tib, tibia; trd, trapezoid; trm, trapezium; tym.c, tympanic cavity;
uln, ulna; unc, unciform; vom, vomer; x, dumb-bell shaped bone; zyg, zygomatic
arch; I-V, digits of manus; V, foramen for fifth nerve. (From Parker's Zoology.)
{115}
The Duck-billed Platypus is, as every one knows, an aquatic animal. It
is not found all over Australia, but is limited to the southern and
eastern parts of that continent, and to Tasmania. The animal excavates a
burrow for itself in the bank of the slow streams which it frequents. The
burrow has one opening below the water and one above; and it is of some
length, twenty to fifty feet. The Platypus feeds upon animal food,
chiefly "grubs, worms, snails, and, most of all, mussels." These it stows
away when captured into its capacious cheek-pouches. The food is then
chewed and swallowed above the surface as the animal drifts slowly along.
Dr. Semon, from whose work, In the Australian Bush, this account
of the animal's habits is quoted, thinks that in the nature of the food
of the creature the explanation of the loss of the teeth is to be found.
He is of opinion that for cracking the hard shells of the mollusc
Corbicula nepeanensis, upon which Ornithorhynchus mainly
feeds, the horny plates are preferable to brittle teeth.
Ornithorhynchus is apparently not eaten by the natives by reason
of its ancient and fish-like smell. Besides, it is hard to catch on
account of its diving capacities, which are aided by an acute sense of
sight and of hearing. When the Duck-bill was first brought to this
country it was believed to be a deliberate fraud, analogous to the
mermaids produced by neatly stitching together the forepart of a monkey
and the tail of a salmon.
{116}
CHAPTER VI
INTRODUCTION TO THE SUB-CLASS EUTHERIA
Sub-Class II.—EUTHERIA
Definition.—Mammalia with teats. Mammary glands of
sebaceous type. Heart with entirely membranous and complete right
auriculo-ventricular valve. Brain generally with a corpus callosum.
Coracoid much reduced and not reaching sternum. No interclavicle.
Vertebrae with epiphyses. Ribs double-headed. Viviparous, with a small
ovum.
In this group are included not only the Eutheria in the sense of
Huxley, but also his Metatheria. Though the Metatheria, or Marsupials as
we shall term them, undoubtedly form a most distinct order of mammals,
perhaps even a trifle more distinct than most others, their differences
from the remaining tribes are not by any means so great as those which
separate Ornithorhynchus and Echidna from all other
mammals. In his well-known memoir upon the arrangement of the Mammalia,[62] Professor Huxley
enumerated eleven characters as distinguishing the Metatheria either from
the Prototheria or from the Eutheria. Of these only three were characters
in which they approach the lower mammals. According to his showing,
therefore, the preponderance of marsupial features are Eutherian. The
three characters of Prototherian type are (1) the presence of epipubes;
(2) the small corpus callosum; (3) the absence of an allantoic
placenta.
The last of these can be dismissed, in consequence of the recent
discovery of an allantoic placenta in Perameles. The first
character is apparently a valid distinction between the Marsupials {117}and
their mammalian relatives higher in the series; but it is not a character
that should have been made use of by Huxley, since he believed in the
existence of a corresponding element in the Dog. As to the corpus
callosum (Fig. 50, p. 77) being small, that seems
to be not more than a slight difference of degree.[63] A number of other characters of
secondary importance were added by Huxley to the weight of evidence which
led him to form a group Metatheria for the Marsupials. Some of these,
however, are now known to be not evidence in that direction. For instance
he observed that no Marsupial had more than a single successional tooth.
It seems at the present moment to be fairly clear that Marsupials have a
milk dentition like other Eutherians, but that only one of these teeth,
the fourth premolar, comes to functional maturity. That it is really one
of a complete milk series is evidenced by the fact that this tooth is
differentiated contemporaneously with another series formerly held to
belong to the so-called prelacteal dentition.[64] There still remains, of course, the
actual fact that the milk dentition is not for the most part functional,
but its significance breaks down with these fresh discoveries. Of this
Professor Osborn has remarked: "The discovery of the complete double
series seems to have removed the last straw from the theory of the
marsupial ancestry of the Placentals." But Huxley did not lay much stress
upon this matter of the teeth, since he observed that similar
suppressions of the milk dentition were to be found in many other mammals
admittedly Eutherian.

Fig. 57.—Brain of Echidna aculeata; sagittal
section. ant.com, Anterior commissure;
cbl, cerebellum; c.mam, corpus mammillare;
col.forn, column of the fornix; c.qu,
corpora quadrigemina; gang.hab, ganglion
habenulare; hip.com, hippocampal commissure;
med, medulla oblongata; mid.com,
middle commissure; olf, olfactory lobe;
opt, optic chiasma; tub.olf, tuberculum
olfactorium; vent. 3, third ventricle. (From
Parker and Haswell's Zoology.)
Huxley regarded the peculiarities in the reproductive organs {118}of the
Marsupials as "singularly specialised characters," in no way intermediate
in character. This view applies also to the pouch, which, as already
stated, distinguishes the adults of that group. But the impossibility of
using this last character as one of any importance has been shown by the
discovery of rudiments of it in embryos of undoubtedly Eutherian mammals
(see p. 18).

Fig. 58.—Sagittal section of brain of Rock Wallaby
(Petrogale penicillata). ant.com, Anterior commissure;
cbl, cerebellum; c.mam, corpus mammillare;
c.qu, corpora quadrigemina; crur, crura cerebri;
epi, epiphysis, with the posterior commissure immediately
behind it; f.mon, position of foramen of
Monro; hip.com, hippocampal commissure, consisting
here of two layers continuous behind at the
spleneium, somewhat divergent in front where the
septum lucidum extends between them; hypo, hypophysis;
med, medulla oblongata; mid.com, middle
commissure; olf, olfactory lobe; opt, optic chiasma;
vent. 3, third ventricle. (From Parker and Haswell's
Zoology.)
Less stress is laid now upon the existence of four molars in the
Marsupials as dividing them from the higher mammals than was formerly the
case. The total dentition of the group is on the whole composed of more
numerous individual teeth than in the typical Eutheria; but we have
exceptions like the Whales, the Armadillo Priodontes, and the
Manatee; or better, because free from the suspicion of secondary
multiplication, Otocyon and occasionally (according to Mr. Thomas)
Centetes. In the last two there are at least sometimes four
molars.
On the other hand, a few archaic characters of some importance crop up
here and there among the Marsupials, which are sometimes held to point to
a primitive ancestry. It has been remarked that in Marsupials it is the
fourth toe which is dominant in size, whereas in Ungulates it is the
third. An attempt has been made to explain this on the view (reasonable
enough in itself) of a tree-living ancestry for the group. A greater
development of the fourth toe is, however, by no means a necessary
character of arboreal creatures; the Primates themselves are an
exception. Nor is this prevalence universal among the Marsupials; {119}in
Myrmecobius (alone) is the third toe the longest; and no great
difference can be detected between the third and fourth toes in the case
of the genera Phascologale, Didelphys, and some others.
Professor Leche compares the predominance of the fourth toe with the
hyperphalangeal condition in the fourth toe of the embryo Crocodile, and
considers it an archaic feature, not surpassed by the ancient
characteristics of the Monotremata. Again it has been pointed out that in
Phascologale and Perameles, the epistropheus (axis
vertebra) has a separate rib as in Ornithorhynchus. In the third
place, the likeness of the teeth of Myrmecobius to those of
Ornithorhynchus is an argument in the same direction, which is
furthermore supported by the great age (Mesozoic) of the Metatherian
group, if we are right in regarding those extinct creatures as
Marsupials.
We may now mention certain facts which are not so generally used. The
partly primitive structure of the right auriculo-ventricular valve in the
Monotremata has no counterpart in any Marsupial which has been dissected;
but there are traces in the latter of the characteristic "ventral
mesentery" of Ornithorhynchus and Echidna.[65] Mr. Caldwell's interesting observation
upon the segmenting egg of the Marsupial, the incompleteness of the first
segmentation furrow (reminding us of the meroblastic ovum of the
Monotreme), may possibly not turn out to be so exclusively Marsupial a
feature as has been thought.
The balance of evidence thus points to the nearer relationship of the
Marsupials to the Eutherian mammals; and their great specialisation
combined with certain evidences of degeneration (disappearance in part of
the milk dentition), and their age, point to the fact that they are, at
any rate, the descendants of an early form of Eutherian. But they must
have separated from the Eutherian stock after it had acquired a definite
diphyodonty and the allantoic placenta, the two principal features of the
Eutherian as opposed to the Prototherian mammals.
Nevertheless it seems probable that the Marsupial tribe is derived
from some of the earliest Eutherians. And on this view may be explained
the retention of Prototherian characters.
The remaining Eutheria are obviously all to be referred to one great
division with the possible exception of the Whales, whose affinities form
one of the principal difficulties to the student {120}of this group. A short
résumé of what is at present thought of the systematic position of this
anomalous order is appropriate here. Albrecht went so far as to regard
the Cetacea as the nearest group of animals to the hypothetical
Promammalia.[66] But
discounting his arguments by the removal of such of them as relate to
structure plainly altered by the singular mode of life of these
creatures, there is really a great deal to be said in favour of his
view.
The chief facts which argue a primitive position among mammals for the
Cetacea are perhaps: (1) the slight union of the rami of the lower jaw;
(2) the occasionally rather marked traces of the double constitution of
the sternum; (3) the long and simple lungs; (4) the retention of the
testes within the body-cavity; (5) the occasional presence (in
Balaenoptera) of a separate supra-angular bone. These points,
however, are but few, and are not of such great weight as those which
ought to be present to establish a claim to separate treatment for the
Cetacea as opposed to the Eutheria. If this group of mammals can be
tacked on anywhere, it appears to us that the nearest relatives are not,
as is sometimes put forward, the Ungulata or the Carnivora, but the
Edentata. There are quite a number of rather striking features in which a
likeness is shown between these apparently diverse orders of mammals. The
chief ones are these: (1) the existence of traces of a hard exoskeleton,
of which vestiges remain in the Porpoise; (2) the double articulation of
the rib of the Balaenopterids to the sternum, with which compare the
conditions obtaining in the Great Anteater; (3) the concrescence of some
of the cervical vertebrae; (4) the share which the pterygoids may take in
the formation of the hard palate; (5) the fact that in the Porpoise, at
any rate, as in many Edentates, the vena cava, instead of increasing in
size as it approaches the liver, diminishes.
Another group which is perfectly isolated is that of the Sirenia. The
alliance advocated by some with the Cetacea, and quite recently renewed
by Professor Haeckel, is contradicted by so many important features that
it seems necessary to abandon it. The recent discovery of a fossil
Sirenian jaw by Dr. Lydekker with teeth highly suggestive of those of
Artiodactyla, may prove a clue. A third group which is so isolated as to
have been placed in a {121}primary division, proposed to be called
Paratheria, is that of the Edentates. Probably the group so called should
really be divided into the Edentata and the Effodientia, the latter
containing the Old World forms. Whether or not it be ultimately shown
that the Ganodonta are ancestral Edentates (sensu strictiori), the
connexion of the group with others is not at present plain. The same is
the case with the extensive order of Rodents. It is true that the extinct
order of the Tillodontia shows certain Rodent-like characters on the one
hand, and likenesses to Ungulates on the other. Certain likenesses shown
by such apparently diverse animals as the Rabbit and the Elephant used to
be insisted upon by Professor Huxley. For the present, however, the
Rodents must remain as an isolated group with only very dubious
affinities to others. The remaining groups of existing mammals are easier
to connect. At first the differences between a Cat and a Horse seem to be
quite as wide as those which separate any two of the higher Eutherian
orders. But it seems to become clearer and clearer, as palaeontological
investigation proceeds, that the bulk of the Ungulate and the
Carnivorous, Insectivorous, and perhaps Lemuroid stocks converge into the
early Eocene Creodonta. From the Lemuroid branch the higher Primates can
be derived. The only "Ungulates" which cannot be fitted in with some
reasonable probability is the group of the Proboscidea. But of the early
forms of this division we have at present no knowledge.
{122}
CHAPTER VII
EUTHERIA—MARSUPIALIA
Order I. MARSUPIALIA[67]
The Marsupials may be thus defined:—Terrestrial, arboreal, or burrowing (rarely
aquatic) mammals, with furry integuments; palate generally somewhat
imperfectly ossified; jugal bone reaching as far as the glenoid cavity;
angle of lower jaw nearly always inflected. The clavicle is developed.
Arising from the pubes are well developed and ossified epipubic bones.
Fourth toe usually the most pronounced. Teeth often exceed the typical
Eutherian number of forty-four; molars generally four on each side of
each jaw. As a rule but one tooth of the milk set is functional, which is
(according to many) the fourth premolar. Teats lying within a pouch, in
which the young are placed. Young born in an imperfect condition, and
showing certain larval characters. There is a shallow cloaca. The testes
are extra-abdominal, but hang in front of the penis. In the brain the
cerebellum is completely exposed; the hemispheres are furrowed, but the
corpus callosum is rudimentary. An allantoic placenta is rarely
present.
Structurally the Marsupials are somewhat intermediate between the
Prototheria and the more typical Eutheria, with a greater resemblance to
the latter.
Fig. 59.—Rock Wallaby (Petrogale xanthopus), with young in pouch. × 1⁄7.
(After Vogt and Specht.)
The name Marsupial indicates what is perhaps the most salient
character of this order. The pouch in which the young are carried is
almost universally present. It is less developed {123}on the whole in the
Polyprotodont forms, such as the Thylacine, Dasyures, etc., but is found
in so many of them that the two divisions of the Marsupials, the
Diprotodonts and the Polyprotodonts, cannot be raised to distinct orders
on this and other grounds. The marsupial pouch of the Marsupials must
not, as has been already pointed out, be confounded with the pouch of the
Monotreme mammals. Distinct teats are found in the marsupium of the
Marsupials, while there are none in the mammary pouch of the Monotreme,
the pouch itself indeed representing an undifferentiated teat, of which
the walls have not closed up. The pouch opens forward in the Kangaroos,
and backwards in the Phalangers and in the Polyprotodonts. Its walls are
supported by a pair of bones diverging from each other in a -shaped
manner; these are cartilaginous and vestigial in the Thylacine. They {124}are
the precise equivalents of similar bones in the Monotremata. It has been
held, but apparently erroneously, that these bones are mere ossifications
in the tendons of the external oblique muscle of the abdomen, or of the
pyramidalis of the same region; and vestiges have been asserted to exist
in the Dog. Such bonelets are undoubtedly present in the Dog; but it
seems clear from their development in Marsupials, as structures actually
continuous with the median unossified portion of the symphysis pubis,
that the "marsupial bones" belong to that part of the skeleton, and that
they correspond with the epipubis of certain amphibians and reptiles. The
pouch, it may be remarked, exists in a rudimentary form in the males of
many Marsupials.

Fig. 60.—Ventral surface of
innominate bone of Kangaroo
(Macropus major).
× ⅓. a, Acetabulum; ab,
acetabular border of ilium;
is, iliac surface; m, "marsupial"
bone; pb, pubic
border; pt, pectineal tubercle;
s, symphysis; si,
supra-iliac border; ss,
sacral surface; thf, thyroid
foramen; ti, tuberosity
of ischium. (From
Flower's Osteology.)
Fig. 61.—Mammary foetus
of Kangaroo attached to
the teat. (Nat. size.)
(From Parker and Haswell's
Zoology.)
The most salient feature in the life-history of the Marsupials is the
imperfect condition in which the young are born. The egg is no longer
laid, as in the Monotremes; but curiously enough the ovum, which has the
small size of that of the Eutheria, divides incompletely at the first
division (as Mr. Caldwell has shown), and this developmental feature may
perhaps be looked upon as a reminiscence of a former large-yolked
condition. The young when born are small and nude; the newly born young
of a large Kangaroo is perhaps as large as the little finger. The young
are transferred by the lips of the mother to the pouch, where they are
placed upon a teat. It is an interesting fact that they are not merely
imperfect foetuses, but that they are actual larvae. They possess in fact
at any rate one larval organ in the shape of {125}a special sucking
mouth. This sucking mouth is an extra-uterine production, and is of
course an adaptation to the particular needs of the young, just as are
other larval organs, such as the chin-suckers of the tadpole, or the
regular ciliated bands of the larvae of various marine invertebrate
organisms.
There are a number of other features which distinguish the Marsupials
from other mammals.
The cloaca of the Marsupials is somewhat reduced, but is still
recognisable. Its margins in Tarsipes are even raised into a wall,
which projects from the body.
The tooth series of the Marsupials was once held to consist of one
dentition only, with the exception of the last premolar, which has a
forerunner. The interpretation of the teeth of Marsupials are various.
Perhaps most authorities regard the teeth as being of the milk dentition,
with the exception of course of the single tooth that has an obvious
forerunner. But there are some who hold that the teeth are of the
permanent dentition. In any case it is proved that a set of rudimentary
teeth are developed before those which persist. Those who believe in the
persisting milk dentition describe these as prelacteal. Another matter of
importance about the teeth of this order of mammals is that their numbers
are sometimes in excess of the typical Eutherian 44. This, however, holds
good of the Polyprotodonts only.
It was for a long time held that the Marsupials differed from all
other mammals in having no allantoic placenta. But quite recently this
supposed difference has been proved to be not universal by the discovery
in Perameles of a true allantoic placenta. The Marsupials have
been sometimes called the Didelphia. This is on account of the fact that
the uterus and the vagina are double. Very frequently the two uteri fuse
above, and from the point of junction an unpaired descending passage is
formed (see Fig. 48 on p. 74).
A character of the brain of Marsupials has been the subject of some
controversy. Sir Richard Owen stated many years ago that they were to be
distinguished from the higher mammals by the absence of the corpus
callosum. Later still it was urged that a true corpus callosum, though a
small one, was present; while, finally, Professor Symington[68] seems to have shown that
{126}the original statement of Owen was
correct, at least in part. It is at most feebly developed (see Fig. 58,
p. 118).
As to skeletal characters, the Marsupial skull has on the whole a
tendency towards a permanent separation of bones usually firmly
ankylosed. Thus the orbitosphenoids remain distinct from the presphenoid.
The palate is largely fenestrated, a return as it were—says
Professor Parker—to the Schizognathous palate of the bird. The
mandible is inflected; this familiar character of the Marsupials goes
back to the earliest representatives of the order in Mesozoic times (see
p. 96); but it is not absolutely universal, being
absent from the much weakened skull of Tarsipes. On the other
hand, the inflection is nearly as great in certain Insectivores, in
Otocyon, etc. The malar always extends back to form part of the
glenoid cavity. The shoulder girdle has lost the large coracoid of
Monotremes; this bone has the vestigial character that it possesses in
other Eutheria. The clavicle is present except in the Peramelidae. A
third trochanter upon the femur seems to be never present.

Fig. 62.—Skull of Rock Wallaby (Petrogale
penicillata). (Ventral view.) ali, Alisphenoid;
bas.oc, basi-occipital; bas.sph, basi-sphenoid;
ex.oc, ex-occipital; ju, jugal; max, maxilla;
pal, palatine; par.oc, paroccipital; p.max,
premaxilla; pr.sph, presphenoid; pt, pterygoid;
sq, squamosal; ty, tympanic. (From
Parker and Haswell's Zoology.)
The Marsupials cannot be regarded as an intermediate stage in the
origin of the Eutheria for a number of reasons. In the first place, the
nature of their teeth shows them to be degenerate animals; one set,
whether we regard it as the milk or permanent dentition, has become
vestigial. The recent discovery of a true allantoic placenta in
Perameles removes one reason for regarding {127}the Marsupials as
primitive creatures. It implies on the whole that the Marsupials have
sprung from a stock with an allantoic placenta. The alternative is to
assume the independent development of an allantoic placenta in both
groups of the Mammalia; unless indeed the genus Perameles is to be
held to be the most primitive race of Marsupials living, a hypothesis
which does not appear on the face of it likely. So long as it was
believed that the mammary pouch of the Monotremes was the equivalent of
the marsupium of the Marsupials, the persistence of this structure seemed
to be a bond of union between the groups. But it is now known that the
marsupium is a special organ confined to the Marsupials, an argument
which is rather in favour of their being a lateral development of the
mammalian stem. It is to be remarked also that the marsupium is feeblest
in the Polyprotodonts, which may perhaps be looked upon as the most
primitive of the Marsupials, owing to their more numerous teeth and other
points to be referred to immediately.
Not only are the Marsupials interesting from the point of view of
their structure; their present and past distribution is of equal
interest. During the Mesozoic epoch they occurred in Europe and North
America; but not, so far as negative evidence means anything, in
Australia, which is now their headquarters. In Europe Marsupials lingered
on into the Tertiary period, when they finally became extinct. In
America, of course, the group has persisted to the present day. Now it is
important to notice that the two main subdivisions of the Marsupials, the
Polyprotodontia and the Diprotodontia, exist to-day in both Australia and
South America. These two divisions, it should be explained, differ
principally in that one has numerous, the other rarely more than two,[69] incisors in the lower
jaw. It is perhaps the more widely distributed opinion that the
Polyprotodontia are the more archaic group; this opinion rests upon one
or two facts in addition to the absence of specialisation in the incisor
teeth. Among the Polyprotodontia the total number of teeth is
greater—a clearly primitive character; secondly, the general form
of the body of these animals, with four subequal limbs and carnivorous or
omnivorous diet, contrasts with the purely vegetarian and much
specialised Kangaroos at any rate. Finally—and sufficient stress
{128}has perhaps not been laid upon this
matter—the brain among the Polyprotodonts is less convoluted than
among the genera of the other division. This statement is of course made
with due regard to parallelism in size (see p. 77).
It is well known that the complexity of a brain bears a distinct relation
to the size of its possessor within the group. Now the most ancient
Marsupials are decidedly more Polyprotodont-like. No European form from
the earlier periods is distinctly to be referred to the Diprotodonts. But
both divisions now exist in America and Australia.
We must assume, therefore, one of three hypotheses. Either the
differentiation into the two great divisions occurred in Jurassic or
Cretaceous times before the migration of the order southwards; or the
Diprotodont type is only a type, and not a natural group, i.e. it
has been separately evolved in America and Australia; or, finally, there
was formerly a land-connexion in the Antarctic hemisphere, along which
the Diprotodonts of Australia wandered into South America. The middle
hypothesis has this to commend it, that syndactylism occurs in both
divisions, and that in some Diprotodonts the pouch opens backwards as it
does in the Polyprotodonts. So great are the resemblances that but little
difference is really left—of great importance that is to say. Hence
it is not difficult to imagine the reduction of the incisors having taken
place twice. In favour of the first hypothesis there are no positive
facts. Finally, in favour of the last, which is so strongly supported by
the facts of distribution derived from the study of other groups of
animals,[70] there is at
least this striking fact or rather series of facts: that some of the
South American fossil Polyprotodonts have a "strictly Dasyurine
relationship."[71] If there
has not been a direct migration, then the Dasyurine type has been twice
evolved, an improbability that few will attempt to explain away. In any
case we shall adopt here the usual division of the Marsupials into
Diprotodontia and Polyprotodontia.
Sub-Order 1. DIPROTODONTIA.
This group includes the herbivorous Marsupials. The incisors are as a
rule three above, but one only in the Wombats. Below {129}is one strong
pair, with occasionally one or two rudimentary incisors. The upper
canines, if present, are not large. The molars are tuberculate or ridged.
All Marsupials (except the Wombats) to some extent, and the Macropods
especially, are characterised by the prolongation of the tubes of the
dentine into the clear enamel. The significance of this fact is, however,
lessened by the fact that the same penetration of the enamel by dentinal
tubes occurs in the Jerboa, the Hyrax, and some Shrews. The feet have two
syndactylous toes,[72] less
marked in the Wombats than in the Kangaroos and Phalangers.

Fig. 63.—Skull of Wombat (Phascolomys wombat). (Lateral view.) ang, Angular process;
cond, condyle of mandible; ext.aud, opening of bony auditory meatus;
ex.oc, exoccipital; ju, jugal; lcr, lachrymal; max, maxilla; nas, nasal; p.max,
premaxilla; sq, squamosal; ty, tympanic. (From Parker and Haswell's Zoology.)
This order is mainly Australian at the present day, using the term of
course in the "regional" sense (see p. 84); the
only exception indeed to this statement is the occurrence of the genus
Caenolestes in South America. But it is now known that Diprotodont
Marsupials formerly existed in the same part of the world.
Fig. 64.—Bones of right
foot of Kangaroo (Macropus
bennetti). a, Astragalus;
c, calcaneum;
cb, cuboid; e3, ento-cuneiform;
n, navicular;
II-V, second to fifth toes.
(From Flower's Osteology.)
{130}
Fig. 65.—Skeleton of Wallaby (Macropus ualabatus). The scapula is raised somewhat
higher than in nature. The end of the tail is omitted. The head of
the femur has been separated from the acetabulum. acet, Acetabulum; acr,
acromion process; ast, astragalus; cal, calcaneum; cbd, cuboid; chev, chevron-bones;
cl, clavicle; cun, cuneiform of carpus; epi, epipubis; fb, fibula; fem,
femur; hd, head of femur; hu, humerus; il, ilium; isch, ischium; obt, obturator-foramen;
orb, orbit; pis, pisiform; pub, pubis; rad, radius; rb1, first rib; rb13,
last rib; sc, scapula; st, sternum; tb, tibia; troch, great trochanter of femur; uln,
ulna; unc, unciform; IV, fourth toe. (From Parker and Haswell's Zoology.)
Fam. 1. Macropodidae.—This family contains the Kangaroos,
Wallabies, Rat-Kangaroos, and Tree-Kangaroos. With the exception of
Dendrolagus the family is terrestrial, and its numerous species
progress by leaps effected by the long hind-limbs, which are decidedly,
often greatly, longer than the fore-limbs. In the hind-limb the fourth
toe is very long and strong; the fifth moderately so; the second and
third are slender and united by skin. The tail is always long, but
differs in its characters from {131}genus to genus. The stomach is much
sacculated. The dental formula is I 3/1 C (1 or 0)/0 P 2/2 M 4/4. The
atlas is often open below, forming thus an incomplete ring.
Though the number of the incisor teeth in the adult Diprotodonts is
never more than three on each side in each jaw, more numerous rudiments
are present. Mr. M. Woodward[73] has lately investigated the subject
with interesting results. He finds that many species present decided
traces of two additional incisors, raising the total to that which
characterises the Polyprotodontia; but in two cases, viz. Macropus
giganteus and Petrogale penicillata, a sixth is present, the
total number being thus in excess of that found in any other Marsupial.
This, as the author himself admits, proves too much. No mammal is known
which in the adult condition has so many incisors; nor do the fossil
Mammalia help us to get over the difficulty; even among reptiles it is
not usual for so many teeth to occur upon the premaxillaries.
It is a curious fact that the two long lower incisors can be used
after the fashion of a pair of scissors, or rather a pair of shears.
Their inner edges are sharpened, and they are capable of some motion
towards and away from each other; by their means grass is cropped.
The stomach of Macropus (and of other allied genera) is
peculiar by reason of its long and sacculated character; the oesophagus
enters it very near the cardiac end, which is bifid. Messrs. Schäfer and
Williams[74] have shown
that the squamous, non-glandular epithelium of the oesophagus extends
over the greater part of the stomach, only the pyloric extremity and one
of the two cardiac caeca being lined with columnar epithelium.
The Macropodidae are clearly divisible into three sub-families, which
are distinguished by marked anatomical characters.
In the sub-family Macropodinae (including the
genera Macropus, Petrogale, Lagorchestes,
Dorcopsis, Dendrolagus, Onychogale, and
Lagostrophus) there is no hallux, and the tail is hairy. The
oesophagus enters the stomach near the cardiac end. The caecum when short
has no longitudinal bands; the liver has a Spigelian lobe.
The second sub-family, Potoroinae or Hypsiprymninae (including the genera Potorous,
Aepyprymnus, Bettongia, and {132}Caloprymnus),
consists of smaller animals than the Macropodinae, which, however,
resemble them in having no hallux, but a hairy tail. The oesophagus
enters the stomach near the pyloric end of that organ. The caecum, though
short, has lateral longitudinal bands. The liver has no special Spigelian
lobe. The canines are always present, being rarely so in Macropodinae,
and are usually well developed.
The third sub-family, that of the Hypsiprymnodontidae, is doubtfully referable to the
family; it consists of but one genus Hypsiprymnodon, which is in
many points more like a Phalanger than a Kangaroo. It has an opposable
hallux and a non-hairy, but scaly, tail. It has canines in the upper
jaw.
Fig. 66.—Red Kangaroo. Macropus rufus. × 1⁄18.
Sub-Fam. 1. Macropodinae.—The genus Macropus
includes not only the Kangaroos but also the Wallabies, which are really
indistinguishable, though they have sometimes been placed in a separate
genus Halmaturus. The genus thus enlarged contains twenty-three
species. It may be thus characterised: the ears are long, the rhinarium
is usually naked, but in M. giganteus and others a band of hairs
descends to the upper lip; a naked band extends from the ankle to the
pads on the digits, which is interrupted in M. rufus by a band of
hairs just in front of the digits. The mammae are four. The tail is not
bushy, {133}but is crested in M. irma. They are
for the most part found on the Australian continent, but some species are
found in the islands to the north which belong to the Australian region.
Thus M. brunii, which is of interest as the first Kangaroo seen by
a European, is a native of the Aru islands. A specimen of this animal,
which was then living in the garden of the Dutch governor of Batavia, was
described by Bruyn in the year 1711. M. rufus, the largest member
of the group, is remarkable for the red secretion which adorns the neck
of the male. It is caused by particles which have the appearance and
colour of carmine. M. giganteus is not, as its specific name might
imply, the "giant" of the race; its dimensions are given as 5 feet, while
M. rufus is said to attain a length of 5 feet 5 inches, exclusive
(in both cases) of the tail.
The account which Sir Joseph Banks gives[75] in his diary of the Kangaroo is
interesting, since he was one of the first naturalists to see that
creature. In July 1770 it was reported to him that an "animal as large as
a greyhound, of a mouse colour, and very swift" had been seen by his
people. A little later he was surprised to observe that the animal "went
only upon two legs, making vast bounds just as the jerboa does." The
second lieutenant killed one of these Kangaroos, of which Sir Joseph
Banks wrote that "to compare it to any European animal would be
impossible, as it has not the least resemblance to any one I have seen.
Its fore-limbs are extremely short and of no use to it in walking; its
hind, again, as disproportionately long; with these it hops seven or
eight feet at a time, in the same manner as the jerboa, to which animal
indeed it bears much resemblance, except in size, this being in weight 38
lbs., and the jerboa no larger than a common rat." The beast was killed
and eaten, and proved excellent meat. Sir Joseph Banks' observations upon
the leaping of the Kangaroo are of interest, because it is often asserted
that the tail is largely made use of as a third foot or as a support. Mr.
Aflalo declares in the most positive way that after repeatedly examining
the tracks upon soft sand immediately after the animal had passed, not
the very faintest trace of the impression of the tail could be
discovered. The leaps of a big Kangaroo seem to be somewhat greater than
is recorded {134}by Banks. It is said that 15 or even 20
feet are covered at a bound, and in bound after bound. But in walking
slowly it can be readily seen from an inspection of Kangaroos at the
Zoological Society's Gardens that the animal does rest upon its tail,
which with the hind-legs forms a tripod.
Petrogale with six species comes next to Macropus, and
is indeed only to be differentiated from it by the thickly-haired and
more slender tail, which is not used, as it is sometimes in the
Kangaroos, as an extra hind-limb. The Rock-Kangaroos live among rocks,
which they climb, and from which they leap; and the tail acts rather as a
balancing pole. The most elaborate account of the anatomy of
Petrogale known to me is by Mr. Parsons.[76] The dentition as given by Mr. Thomas is
I 3/1 C 0/0 Pm 2/2 M 4/4—that of Macropus without the
occasionally occurring canine of the upper jaw. The osteological
characters which separate it from Macropus are quite
insignificant. Mr. Parsons mentions a wormian bone, "os epilepticum," at
the junction of the coronal and sagittal sutures. It was found to occur
in two out of five skulls examined, and appears not to occur in other
Kangaroos. The palatine foramina of Petrogale are so large that
the posterior part of the bone is only a narrow thickened ridge. The
small intestine of P. xanthopus is 102 inches long, the large
intestine 44 inches. The caecum has a length of 6 inches, and is not
sacculated, differing in this from the caecum of Macropus major.
The best known species are P. xanthopus and P. penicillata.
The genus is confined to Australia itself, and does not enter
Tasmania.
Onychogale includes the so-called "Nail-tailed Wallabies,"
which have a thorn at the end of the tail, reminding one of the Lion and
the Leopard, whose tails have a similar armature. The muffle is hairy.
Three species are allowed by Mr. Thomas.
Lagorchestes has, like the last genus, the rhinarium,
i.e. that part of the nose immediately surrounding the nostrils,
hairy instead of smooth as in the Kangaroos proper. It is distinguished
from Onychogale by the absence of the terminal callosity to the
tail, which is rather short. The name Hare-Kangaroo is given to the
members of this genus (three species) on account of their exceeding
fleetness. This genus is limited to Australia itself. L.
conspicillatus is said to present "a remarkable {135}resemblance to
the English hare," and L. leporoides was so called by Gould on
account of general appearance as well as face.
Dorcopsis has shorter hind-legs than Macropus, and a
naked muffle. The ears are small. The structure of D. luctuosa has
been studied by Garrod,[77]
who pointed out the existence of four enlarged hair follicles on the neck
near the mandibular symphysis. These are, however, represented in the
next genus Dendrolagus, and occur also in Petrogale. The
limbs are not so disproportionate as in Macropus, and the tail is
naked at the tip.
Dorcopsis and the next genus to be described,
Dendrolagus, differ from Macropus and its immediate allies,
Petrogale and Lagorchestes, in a number of anatomical
points. In the first place, the premolars are twice the size of those of
Macropus, and they have a characteristic pattern not observable in
the Kangaroos. This consists of a median ridge (the whole tooth being
rather prismatic in shape), with lateral ridges at right angles to it.
The upper canines are developed, but are minute.
The stomach is not quite like that of Macropus, though built
upon a similar plan. The blind cardiac extremity is a single, not a
double cul-de-sac; in this it is like that of Petrogale. The
distribution of the squamous, white, oesophageal epithelium is very much
like that of Dendrolagus. In both genera the orifice of the
oesophagus into the stomach is guarded by two strong longitudinal folds,
which run for some distance towards the pylorus. In Dendrolagus,
at any rate, this tract is bordered on each side by glandular patches. In
Dendrolagus, moreover, the squamous epithelium does not extend
into the cardiac cul-de-sac. This latter is separated from the rest of
the stomach by two slightly diverging folds, which are faintly
represented in Petrogale and in Halmaturus. In the last two
genera the folds surrounding the oesophageal orifice are but slightly
represented; better in Halmaturus than in Petrogale. But
there are not the patches of glands already referred to. The small
intestine of Dorcopsis is 97 inches in length, the large being 32,
i.e. proportionately long, as in Marsupials generally. The small
caecum (2½ inches) is not sacculated.
The spleen is Macropodine, being -shaped or  -shaped.
The differences between Dorcopsis and the evidently closely allied
Dendrolagus will be further considered under the description of
{136}the latter. Dorcopsis is confined
to New Guinea, and contains three species, viz. D. muelleri, D.
luctuosa, and D. macleani. D. muelleri has a striking
resemblance to Macropus brunii, with which it has been confounded.
Though intermediate between Macropus and Dendrolagus, these
Kangaroos are not arboreal.
-shaped.
The differences between Dorcopsis and the evidently closely allied
Dendrolagus will be further considered under the description of
{136}the latter. Dorcopsis is confined
to New Guinea, and contains three species, viz. D. muelleri, D.
luctuosa, and D. macleani. D. muelleri has a striking
resemblance to Macropus brunii, with which it has been confounded.
Though intermediate between Macropus and Dendrolagus, these
Kangaroos are not arboreal.
The genus Dendrolagus is remarkable for its un-kangaroo-like
habit of living in trees. In accordance with this change of habit is a
relative shortening of the hind-limbs, a feature which begins to be
observable in Dorcopsis. "The general build," writes Mr. Thomas,
"is of the ordinary mammalian proportions, not macropodiform at all." The
muffle is not naked for the greater part, though the shortness of the
hairs gives that effect. As in Dorcopsis, but not as in
Macropus, the bulla tympani is not swollen. There are altogether
five species, the fifth, D. bennetti, having been lately described
from specimens living in the Zoological Society's Gardens.
Fig. 67.—Tree-Kangaroo. Dendrolagus bennetti. × 1⁄12.
The anatomy of this genus has been described by Owen for D.
inustus,[78] and by
myself for D. bennetti. The stomach, which {137}has a single, not
bifid, cul-de-sac, is sacculated by two principal bands and other
subsidiary ones. Its internal structure has already been to some extent
described. The spleen of D. bennetti is remarkable for the fact
that it is not -shaped, whereas D. inustus agrees with other Macropodines in
the form of this organ. The small intestine of D. bennetti is 95
inches long, the large 38. The caecum appears to differ in the two
species; it is smaller in D. bennetti, where it is only 2 inches
in length. The most remarkable feature of the liver is the large size of
the left lateral lobe and the bilobed condition of the Spigelian lobe;
this at least was the case with D. bennetti. A recently-described
species[79] has been
attentively studied in its native haunts by Dr. Lumholtz.[80] It lives in the highest parts of the
mountainous scrubs of Queensland, where it moves quickly on the ground as
well as among the trees. It is hunted with Dingos by the "blacks," and is
eaten by them.[81]
Lagostrophus is a generic name that has been proposed by Mr.
Thomas for a small Wallaby 18 inches in length, which is distinguished by
the fact that the long claws of the hind-limbs are entirely hidden by
long and bristly hairs; the muffle is naked; there is no canine. The
bullae are swollen. There is but one species of the genus, L.
fasciatus, a native of West Australia.
Sub-Fam. 2. Potoroinae.—Aepyprymnus and the other
genera placed in this sub-family are known by the vernacular name of
Rat-Kangaroos, or sometimes Kangaroo-Rats. The latter term has been
called "incorrect," though it is just as good as the former, both of them
in fact being inaccurate as implying some likeness to or relation with a
Rat. The present genus has a partially hairy rhinarium; the auditory
bullae are not swollen. It contains but one species, Ae.
rufescens, a native of Eastern Australia, which is distinguished by
its very long hind-feet.
Bettongia has long hind-feet as in Aepyprymnus, but the
rhinarium is entirely naked instead of being partially hairy, while the
ears are much shorter. The genus, which contains four species, is
remarkable as being the only ground-living mammal with a prehensile tail,
which it uses to carry grass, etc. {138}B. lesueuri
burrows in the ground, often to so great a depth as 10 feet. The genus
occurs in Tasmania as well as in Australia.
Caloprymnus, with one species, is a genus instituted by Mr.
Thomas in his Catalogue of Marsupials for a form (C. campestris)
which combines in a remarkable way the characters of Aepyprymnus,
Bettongia, and Potorous. The external characters and the
general shape of the skull are as in Bettongia, while the molars
have the structure of those of Aepyprymnus. The last premolar is
as in Potorous.
Of the genus Potorous there are three species, which are
Tasmanian as well as Australian. Unlike the other Rat-Kangaroos, the
hind-feet are comparatively short, and the animal is therefore less
addicted to jumping than its relatives. The rhinarium is naked, and the
ears are of fair length.
Sub-Fam. 3. Hypsiprymnodontinae.—The Musk-Kangaroo,
Hypsiprymnodon, is the last genus of the present family, and the
only genus of this sub-family. It is intermediate between the
Macropodidae and the Phalangeridae, the annectant character being mainly
the hind-feet, which though they have the same long fourth digit as the
Kangaroos, have it more feebly developed, and possess also an opposable
hallux, which is one of the salient features in the structure of the
Phalangeridae. The tail is naked and scaly; the rhinarium is entirely
naked. The ears are large and not furry. The single species, H.
moschatus, appears to feed upon insects as well as vegetables.
"Its habits are chiefly diurnal, and its actions when not disturbed by
no means ungraceful. It progresses in much the same manner as the
Kangaroo-Rats (Potorous), to which it is closely allied, but
procures its food by turning over the débris in the scrubs in search of
insects, worms, and tuberous roots, frequently eating the palm berries,
which it holds in its fore-paws after the manner of the Phalangers,
sitting up on its haunches, or sometimes digging like the bandicoots."
This is Mr. Ramsay's description of the animal, which he was the first to
discover.[82]
Fam. 2. Phalangeridae.—The genus Hypsiprymnodon
bridges over the not very wide gap which separates the Kangaroos from the
Phalangers. The Phalangers are Marsupials with five fingers and toes; the
second and third toes are bound together by a {139}common integument as in
the Macropodidae. The hallux is opposable and nailless. The tail is
nearly always long and prehensile. The pouch is well developed; the
stomach not sacculated; a caecum is present (except in Tarsipes).
These are really the principal distinctions between the two families. In
addition, it may be mentioned that the lower incisors have not a
scissor-like action as in the Kangaroos.
The Phalangers may be divided into four sub-families.
The first of these, that of the Phalangerinae,
contains the genera Phalanger (including Cuscus),
Acrobates, Distaechurus, Dromicia,
Gymnobelideus, Petaurus, Petauroides,
Dactylopsila, Pseudochirus, and Trichosurus.
These genera agree in the following generalities:—Tail well developed, often very long; three
incisors above, and at least two premolars both above and below; caecum
long and simple; stomach without a cardiac gland; liver not very
complicated by secondary furrows, with a distinct caudate lobe; the
vaginal median culs-de-sac often coalesced; lungs with an azygos
lobe.
Fig. 68.—Bones of leg and foot of Phalanger. ast,
Astragalus; calc, calcaneum; cub, cuboid;
ect.cun, ecto-cuneiform; ent.cun, ento-cuneiform;
fb, fibula; mes.cun, meso-cuneiform;
nav, navicular; tib, tibia; I-V, first to fifth
toes. (After Owen.)
The second sub-family, Phascolarctinae (with
the Koala only), is thus characterised:—Tail rudimentary; cheek-pouches present;
superior incisors three, but only one premolar above and below; {140}caecum
extraordinarily long; stomach with a cardiac gland; liver complicated by
additional furrows, without a free caudate lobe; no azygos lobe to lungs;
vaginal culs-de-sac free.
The third sub-family, Phascolomyinae,
contrasts with the others as follows:—Tail
rudimentary; cheek-pouches present, but rudimentary; one incisor on each
side above, but no additional premolars; all the teeth rootless; caecum
not peculiar in shape; stomach with a cardiac gland; liver complicated by
secondary furrows, without a free caudate lobe; lung with an azygos lobe;
vaginal culs-de-sac free.
The last sub-family, Tarsipedinae, is thus
defined:—Tail long; tongue extensile; only
one premolar; molars reduced; caecum absent.
Fig. 69.—Vulpine Phalanger. Trichosurus vulpecula. × 1⁄6.
Sub-Fam. 1. Phalangerinae.—The genus Phalanger
embraces five species, sometimes called by the generic name of
Cuscus. They are largish animals with short ears; only the end of
the tail is naked. Of these animals only one species is found in
Australia itself, the rest inhabiting the islands lying to the north. The
Spotted Cuscus, Ph. maculatus, is in spite of its vegetarian diet,
and perhaps on account of its spots, spoken of as the "Tiger Cat." Mr.
Aflalo remarks of it that though provided with a prehensile tail, it is
little better as a climber than the tailless Koala.
Trichosurus, including the "True Phalangers," includes largish
species, which can be distinguished from the last genus by a chest-gland
similar to that which occurs in Myrmecobius and some other
Marsupials of the present group. There are but two species, which are
purely Australian. The "Brush-tailed Opossum," T. vulpecula
(perhaps better known as Phalangista {141}vulpina), like
its American pseudo-namesake (a true Opossum, genus Didelphys),
"plays 'possum" on occasions. The dental formula is I 3/2 C 1/0 Pm 2/3
M 4/4. The ears are shortish.
The Ring-tailed Phalangers, Pseudochirus, are more widely
distributed than the last two genera; they range from Tasmania in the
south to New Guinea in the north. They are not, however, ring-tailed,
though the tip of the tail is generally white. As in the last genera,
which have prehensile tails, the end of this appendage is naked. The
mammae are four. The tooth formula is I 3/2 C 1/0 Pm 3/3 M 4/4. There are
some ten species of the genus.
The Striped Phalanger, Dactylopsila trivirgata, is an animal
about a foot long, whose identity can be ascertained by its striped,
black and white skin. It is an arboreal creature that lives apparently
both on leaves and grubs like so many arboreal creatures of quite
different groups—Squirrels, for instance, and New-World Monkeys.
The tooth formula is I 3/3 C 1/6 Pm 3/2 M 4/4.
Gymnobelideus leadbeateri is a small creature with a body 6
inches in length. It is restricted to the colony of Victoria. The general
look is that of Petaurus; the ears are naked.
Dromicia is a genus of Phalangers which although devoid of a
parachute, such as is possessed by certain genera that will be considered
immediately, is able to leap with great agility from branch to branch.
The ears are large and thin and almost naked; the tooth formula is I 3/2
C 1/0 Pm 3/3 M 4/4. They are minute creatures, the longest measuring,
with the tail, but 10 inches. Dormouse-Phalanger is a name sometimes
given to them. There are four species, ranging from Tasmania to New
Guinea. The name Dormouse as applied to the genus seems to be owing to
the way in which they hold a nut in the paws when feeding. D. nana
is 4 inches long, with a tail of nearly the same length. It is thick at
the base.
Distaechurus is the last genus of non-flying Phalangers. Its
name refers to the arrangement of the hairs on the tail, which are
disposed on either side in a row like the vane of a feather. The tooth
formula is I 3/2 C 1/0 Pm 3/2 M 3/3, very nearly as in Acrobates.
The ears are as in that genus.
Petaurus is the first genus of the Flying Phalangers, all of
which are provided with a parachute-like expansion of the skin between
the fore- and hind-limbs; the ears are large and naked; and the tooth
formula is I 3/2 C 1/0 Pm 3/3 M 4/4. There are three {142}species of the
genus, which extend through pretty well the entire Australian region. The
term "flying" as applied to these and the other "flying" genera is of
course an exaggeration. The animals cannot fly upwards; they can only
descend in a skimming fashion, the folds of skin breaking their fall.
P. breviceps is perhaps the best-known species. The body is 8, the
tail 9 inches long.
Petauroides seems to be chiefly distinguished from
Petaurus by the fact that, as in its ally Dactylopsila, the
tail is partly naked terminally. In Petaurus and
Gymnobelideus the tail is bushy to the very end, including its
extreme tip below.
A third genus of Flying Phalangers is the minute Acrobates,
which has a distichous tail like that of Distaechurus. It is not
more than 6 inches in length including the tail. As to these Flying
Phalangers it is exceedingly instructive to observe that the same method
of "flight" has been apparently evolved three times; for the three genera
are each of them specially related to a separate type of non-flying
Phalanger. The same observation can be made about the Flying Squirrels,
Anomalurus and Sciuropterus. The dental formula is I 3/2
C 1/0 Pm 3/3 M 3/3. The ears are thinly clad with hair. There are four
teats.
Sub-Fam. 2. Phascolarctinae.—The Koala, or Native Bear,
Phascolarctos cinereus, is the only representative of its
sub-family. It is, like the Wombat, aberrant in the lack of an obvious
tail. The absence of this appendage is curious in an arboreal creature
whose near allies have a long and prehensile one. The structure of the
Koala was investigated by the late Mr. W. A. Forbes.[83] There are some unexpected points of
likeness to the Wombat: thus they agree in the absence of the tail, in
the structure of the stomach, and in the great subdivision of the lobes
of the liver. The brain, however, is smooth, and the caecum is
exceedingly large and complicated in structure, that of the Wombat being
short. That both animals have cheek-pouches is perhaps due to similar
habits of temporarily storing masses of food. This animal has only eleven
pairs of ribs. The tail has only seven or eight vertebra, and these have
no chevron-bones.
A peculiarity of the skull is seen in the great size of the
alisphenoid bulla, which is comparable in size and appearance with that
of the Pig. As in the Kangaroos, the atlas is incomplete below. {143}
The tooth formula of the genus is I 3/1 C 1/0 Pm 1/1 M 4/(4 or 5). The
additional lower molar seems to be exceptional, and has been found in one
specimen only.
In the alimentary tract the most remarkable structure is the large
intestine, which is very capacious for the first 28 inches or so of its
course. This section of the colon is lined with rugae precisely like
those which are found in the caecum. These folds, which at first are some
twelve in number, fuse lower down, and by the time that the colon
approaches the external orifice are reduced to five. Similar folds, as
already stated, occur in the caecum, but do not extend as far as its
blind end. The caecum is proportionately and actually larger than in any
other Marsupial. The gall-bladder is unusually elongated.
Fig. 70.—Koala. Phascolarctos cinereus. × 1⁄9.
The Koala is mainly crepuscular or nocturnal in its habits. It feeds
so exclusively upon the leaves of the gum-tree (Eucalyptus) that
it is impossible to keep the creature long in captivity in lands where
that particular kind of food is not available.
The female, though she seems to bear but a single young one, which is
carried on the back after the fashion of some Opossums, has two nipples.
The animal's slow habits seem to require a nocturnal and retired life. It
is about as lethargic as the Sloth, and it is said to further resemble
that animal in clinging firmly to a branch even after it is shot. {144}
Fig. 71.—Wombat. Phascolomys wombat. × 1⁄12.
Sub-Fam. 3. Phascolomyinae.—Phascolomys, the
Wombat, is the only genus of this sub-family. This animal has the
appearance of a heavily-built Marmot, like which it has a mere stump for
a tail, and a pair of strong chisel-shaped and Rodent-like incisors,
which, however, differ from those of Rodents in having a complete coating
of cement. All the teeth of the animal are rootless, and there are no
canines. The incisors have enamel on the front and lateral faces only.
The dental formula is I 1/1 C 0/0 Pm 1/1 M 4/4. The affinities with other
Diprotodont Marsupials are shown by the commencing syndactyly of the
second and third toes. The rhinarium is naked or hairy. There is a
rudimentary cheek-pouch, as in Phascolarctos. The Wombat has, like
the Koala, and also the Beaver—which does away with some of the
value of the comparison—a peculiar gland-patch in the stomach, a
raised area of collected glands. In no other Marsupial is such a
structure found, "whilst in the two forms under consideration its
identity is almost precise. That such a unique structure should have been
independently developed in two forms unrelated to each other, appears to
me to be in the highest degree improbable." This is Mr. Forbes' opinion.
It might be strengthened by adding the observation that, as there are
other points of likeness between the Wombat and the Koala, it seems more
unlikely that a structure so nearly identical should have been twice {145}developed in two not very distant forms.
As in the Kangaroos, the atlas is open below. Ph. ursinus has 15
ribs; the other species the normal (for Marsupials) 13. Other points of
likeness will be mentioned under the description of the Koala. These
animals mainly feed upon roots; they live in companies in burrows. There
are three species—Ph. ursinus, Ph. latifrons, and
Ph. mitchelli. Ph. ursinus is Tasmanian in range, the other
two species South Australian.

Fig. 72.—Skull of Wombat. Phascolomys wombat. (Lateral view.) ang, Angular process;
cond, condyle of mandible; ex.oc, exoccipital; ext.aud, opening of bony
auditory meatus; ju, jugal; lcr, lachrymal; max, maxilla; nas, nasal; p.max,
premaxilla; sq, squamosal; ty, tympanic. (From Parker and Haswell's Zoology.)
Sub-Fam. 4. Tarsipedinae.—The genus Tarsipes ought
perhaps to be removed from the present family. There is but a single
species, which is a small creature of 7 inches in total length, of which
the tail measures 4 inches. The teeth are much dwindled, the formula
being I 2/1 C 1/0 Pm 1/0 M 3/3 = 22. The lower incisors are procumbent.
The lower jaw, moreover, has not the characteristic Marsupial inflection.
The intestinal canal is without the caecum present in the remaining
Phalangeridae. It is a curious fact that this aberrant little Phalanger
should come from Western Australia, like the even more aberrant
Myrmecobius. Like the latter also, Tarsipes has a long
exsertile tongue, with which, however, it extracts honey from flowers.
Probably it also catches minute insects in the corollas of the flowers.
It has been proved, in fact, that in captivity at any rate the animal is
insectivorous; for it has been known to eat moths.
Fam. 3. Epanorthidae.—The extinct Epanorthidae of {146}Patagonia are represented to-day by a
small Marsupial which has been rediscovered within the last two or three
years. This little animal, formerly called Hyracodon (a
pre-occupied name), is now termed Caenolestes, and is a native of
Colombia and Ecuador. There are two species, and of these C.
obscurus is called by the inhabitants "Raton runcho," which means
opossum-rat. It lives apparently upon bird's eggs and small birds, though
it belongs to the Diprotodont division of the Marsupials.
Caenolestes, however, although diprotodont, has not the
syndactylous character of the digits of the feet already referred to in
the Kangaroos and their allies. The pouch is small and rudimentary. The
dentition is I 4/3 C 1/1 Pm 3/3 M 4/4 = 46, and the teeth are said by Mr.
Thomas to be much like those of the Australian Dromicia.[84]
In the skull a peculiarity which does not bear upon its affinities to
other Marsupials, but is still interesting, is mentioned by Mr. Thomas.
The nasals are not sufficiently prolonged to meet the upper edge of the
maxillae, and so a vacuity is left, as in the skulls of many Ruminants
(e.g. the Sable Antelope). The palate is very imperfect; the
foramina, which render it so, reach as far forward as the last premolar.
The lower jaw has quite the appearance of that of a Macropus or
Phalanger, with long and forwardly projecting incisors.
Extinct Diprotodonts.—The great Diprotodon is a
creature with a skull a yard long, which must have been of the size of a
large Rhinoceros. Though closely allied to Macropus, it seems that
this great beast did not hop after the fashion of a Kangaroo, its limbs
being of a more equal size than in the Kangaroo. Recently some further
remains of Diprotodon have been discovered in a lake known as Lake
Mulligan, where they had apparently been bogged. Professor Stirling has
contributed an account of these remains, which fills up a considerable
gap in our knowledge. He has been able to state the structure of the
fore- and hind-limbs. Both limbs are pentadactyle, the fingers of the
fore-limb being approximately equal in length and general development. In
the hind-limb the hallux is small, and consists of the metatarsal only.
This bone is fixed in the position of "extreme abduction," and is
suggestive of an arboreal limb. Digits two and three may have {147}been
syndactylous, and the authors of the account[85] of these bones think that the fourth
toe may have shared in this syndactyly. The metatarsal of the fifth digit
is enormously expanded at its edge, and seems to have furnished a strong
support to the creature; this is also seen in the metacarpal of the
fore-limb. Probably, therefore, Diprotodon was quadrupedal in its
mode of progression, with the emphasis laid upon the little finger and
the little toe instead of, as in ourselves, the first toe. The hind-foot
of the Diprotodon could not be more unlike that of a Kangaroo than
it actually is.
|
|
|
|
Fig. 73.—Diprotodon australis. (After Owen.)
|
Fig. 74.—Thylacoleo carnifex. Side view of skull.
(After Flower.)
|
Another giant among these Marsupials was the genus Thylacoleo,
whose name was given to it by Sir Richard Owen on the view that it was a
Marsupial Tiger. Sir W. Flower has, however, controverted this opinion,
and the genus is in fact, in spite of its large size, closely allied to
the Phalangers and {148}Cuscuses.[86] The dental formula is I 3/1 C 1/0
Pm 3/1 M 1/2; the last premolar is a great blade-shaped tooth like that
of Potorous.
Nototherium was a creature smaller than Diprotodon, but
still of large size; it is believed to have been a burrowing creature,
and to connect the Wombats with Diprotodon. More certainly allied
to the existing Wombat was Phascolonus, a Wombat as big as a
Tapir.
Fig. 75.—Nototherium mitchelli. Side view of skull. × 1⁄6. (After Owen.)
Of extinct American Diprotodonts the Epanorthidae, already referred to
in connexion with the living Caenolestes, were the most prominent
forms. The genus Epanorthus occurs in the Santa Cruz formation of
Patagonia, which is believed to be Miocene. The incisors are three in the
upper jaw; and the single incisor of each ramus of the lower jaw is a
great chisel-shaped, cutting instrument.
Abderites is also typically Diprotodont by reason of the large
projecting incisors of the lower jaw. It has a large cutting tooth in the
lower jaw, which appears to be the last premolar, and is thus comparable
to the great cutting tooth of the lower jaw and of the upper jaw of the
extinct Phalanger, Thylacoleo. {149}It may also be
comparable to the great premolar of such Multituberculata as
Ptilodus and Plagiaulax. It is, moreover, marked with
vertical grooves.
An interesting form, which is unfortunately but little known, is the
Australian and Pleistocene genus Triclis, with one species, T.
oscillans. In having a minute canine tooth in the lower jaw it agrees
with some Phalangeridae, and being otherwise closely allied to
Hypsiprymnodon, it unites the Macropodidae with the
Phalangeridae.
Sub-Order 2. POLYPROTODONTIA.
In this mainly carnivorous or insectivorous division of the Marsupials
the incisors are four or five on each side of the upper jaw, and one or
two fewer in the lower jaw. Figs. 76 and 77 illustrate the Polyprotodont
and Diprotodont dentitions. The canines are those of flesh-eaters and so
are the molars, being as a rule sharply cuspidate. As a rule, which has
an exception in the Peramelidae, there is no syndactylism of toes in the
hind-foot. This sub-order is at the present day Australian and American
in its range.
Fig. 76.—Front view of the skull of Tasmanian Devil (Sarcophilus ursinus), showing
Polyprotodont and carnivorous dentition. (After Flower.)
Fam. 1. Dasyuridae.—This family consists of Marsupials
which are generally pentadactylous, but with occasionally the hallux
missing. The tail is long but not prehensile. The pouch is present or
absent. The teeth vary in the different genera, but {150}the upper
incisors are never less than three, and may be as many as five in the
upper jaw and six in the lower. The canines are trenchant. There is no
caecum.
Fig. 77.—Front view of skull of Koala (Phascolarctos
cinereus), illustrating Diprotodont
and herbivorous dentition. (From Flower.)
Fig. 78.—Longitudinal section of the skull of the Thylacine (Thylacinus cynocephalus).
× ½. a, Angular process of mandible; AS, alisphenoid; BO, basioccipital; BS,
basisphenoid; cd, condyle of mandible; ET, ethmoturbinal; Ex.O, exoccipital;
Fr, frontal; ME, ossified portion of mesethmoid; MT, maxilloturbinal; Mx,
maxilla; Na, nasal; OS, orbitosphenoid; Pa, parietal; Per, periotic; Pl, palatine;
PMx, premaxilla; PS, presphenoid; Pt, pterygoid; SO, supraoccipital;
Sq, squamosal; Vo, vomer. (From Flower's Osteology.)
The genus Thylacinus contains but a single species, which is
now limited to Tasmania, and is generally known as the Tasmanian Wolf. It
has the build of an ordinary Wolf, and is of about the same size. The
hinder part of the body is marked with a series of black transverse
bands. The hallux is entirely wanting; the pouch opens backwards. The
marsupial bones are minute and unossified. The dental formula is I 4/3
C 1/1 Pm 3/3 M 4/4 = 46. There are four mammae. This animal, now confined
to Tasmania, {151}is getting rarer on account of its
sheep-killing propensities, and the consequent war of extermination
declared upon it by the colonists. It will, however, feed upon other
animals; and it is related that the first specimen ever captured had in
its stomach the remains of an Echidna! Mr. Thomas thinks that the
persistence of this and of some of the other larger carnivorous
Marsupials in Tasmania after their extinction in Australia is not
unconnected with the advent of the Dingo. But it is stated that the
Thylacine is quite capable of keeping even a pack of dogs at bay.

Fig. 79.—Tasmanian Devil. Sarcophilus ursinus. × 1⁄10.
The genus Sarcophilus has been frequently confounded with the
next, but it is kept apart by Mr. Thomas, who follows Cuvier in this. An
alternative generic name is Diabolus, which, like the first name,
refers to the habits and character of the single species which this genus
contains. The genus is more like Thylacinus than is
Dasyurus. The hallux is wanting, and the teeth, though fewer in
number (42), resemble those of the Thylacine more closely than do those
of the Dasyure. The species is called S. ursinus, the popular name
being Tasmanian Devil. It is black with a variable number of white
patches on the body. It is of about the size of a Badger, and is, like
the Thylacine, a nocturnal animal. The Tasmanian Devil is said to be one
of the most ferocious of animals, and to express its ferocity by a
"yelling growl."
{152}
Fig. 80.—Skull of Dasyurus. (Lateral view.) al.sph, Alisphenoid; ang, angular process
of mandible; fr, frontal; ju, jugal; lcr, lachrymal; max, maxilla; nas, nasal;
oc.cond, occipital condyle; par, parietal; par.oc, paroccipital process; p.max,
premaxilla; s.oc, supraoccipital; sq, squamosal; sq′, zygomatic process of squamosal.
(From Parker and Haswell's Zoology.)
Fig. 81.—Dasyure. Dasyurus viverrinus. × 1⁄5. (After Vogt and Specht.)
The next genus of this family, Dasyurus, comprises five
species, which range over the whole of the Papuan and Australian
sub-regions. The general form is Viverrine, and the hallux is sometimes
present though small. The dental formula is as in the last genus, but the
teeth "are more insectivorous in their character." There are six or eight
mammae. The members of this genus are grey or brown, and spotted with
white; they are all arboreal, and feed largely upon birds and their eggs.
Mr. Thomas has pointed out that in two species, D. viverrinus and
D. geoffroyi, the striae upon the foot-pads are absent, and that
therefore these at least are probably not so purely arboreal as the rest.
The animals are not diurnal, and during the day hide themselves in the
hollow trunks of trees. They are spoken of as "Native Cats," but have the
general habits of Martens. D. maculatus is common in Tasmania, but
is rare in Australia, thus "approaching the condition now exhibited by
the Thylacine and {153}Tasmanian Devil, namely, complete
extermination in Australia, where both once lived." D. hallucatus
shows an approach to Phascologale in its five-toed hind-feet and
slender build.
Phascologale is a genus which, like the last, is usually
arboreal (although not P. virginiae of North Queensland), but is
of much smaller size, the species not exceeding the dimensions of a rat.
They have no spots, but there is sometimes a stripe down the back. There
are thirteen species, which have the same range as the last genus. The
hallux is present though small, but the pouch is "practically obsolete,"
though there is a small fold of skin behind the teats. The rhinarium is
naked; the tail is long, "bushy, crested, or nearly naked." The mammae
are four to ten in number. The dental formula is as in Dasyurus,
and the teeth are not very different in form; sometimes the last premolar
is wanting. "The members of this genus," remarks Mr. Thomas, "evidently
take the place in the Australian region filled in the Oriental by the
Tupaiae, and in the Neotropical by the smaller Opossums."
The genus Sminthopsis comprises not more than four species,
even smaller than the last. The largest species, S. virginiae, is
only 125 mm. in length. The hallux is present, and there is a
well-developed pouch. There are forty-six teeth, as in the Dasyures. The
feet are narrow with granulated or hairy soles, whereas in
Phascologale they are broad with smooth soles. The mammae are
eight or ten. The genus ranges through Australia and Tasmania.
The genus Antechinomys has but a single species, which is a
native of Queensland and New South Wales. The build is Jerboa-like, and
the animal is, as might be inferred, terrestrial. The ears are very long,
and the limbs elongated; the hallux is absent; the teeth are exactly as
in Sminthopsis.
Antechinomys has thirteen dorsal and seven lumbar vertebrae;
three sacrals and twenty-five caudals, the latter number being in excess
of that of its allies. The stomach is nearly globular, with approximated
orifices; the intestine measured 6.8 inches, a little more than twice the
length of the animal itself. A. lanigera is a native of East
Central Australia, and appears to be entirely terrestrial in habit, and
to progress by a series of leaps—at any rate when going at full
speed.
Professor Spencer, who found examples of this rare species, gives {154}an
interesting description of its habits. Antechinomys has much the
look of the Australian Rat, Hapalotis mitchelli; and as the two
animals lead a similar kind of life, the resemblance is not unexpected.
Professor Spencer wonders why these creatures are saltatory in habit. The
country which they inhabit is arid, but with patches of grass and shrubs.
For a big kangaroo the advantage of the power of leaping over such
obstacles may be obvious, but not for the small and slender
Antechinomys. The chief foes of this rare Marsupial appear to be
predatory birds; and Professor Spencer thinks that the saltatory mode of
progression may be more baffling to such pursuers than even a rapid
run.
The genus Dasyuroides has been lately instituted by Professor
Spencer for a Marsupial from Central Australia somewhat intermediate
between Sminthopsis and Phascologale. As there is but one
species, the generic will be considered with the specific characters.
D. byrnei is an animal of about the size of the Common Rat. The
hallux is absent. The tail is fairly thick, but not "incrassated." There
are six mammae, and the pouch is but slightly developed, with two low
lateral folds. The dentition is I 4/3 C 1/1 Pm 3/2 M 4/4. This Marsupial
is nocturnal, and burrowing in habit. Its food consists of insects.[87]
Myrmecobius is so different from the last-described genera
(Dasyurinae) that it is usually separated from
them as a sub-family Myrmecobiinae. The animal is
of a bright rufous colour, banded posteriorly with white. There is no
hallux, though the metatarsal belonging to that digit is present. There
are four mammae.[88] On the
chest is a naked patch of some extent, upon which open the ducts of a
complex gland, which has been described and figured by myself.[89] There is no pouch, but a
tract of skin shows indications of a pouch-like structure. The teeth are
extraordinarily numerous, fifty to fifty-four; the formula being I 4/3(4)
C 1/1 Pm 3/3 M 5/6. Their resemblance to those of certain Jurassic
Marsupials is dealt with on p. 100.[90] In this matter lies of
{155}course the chief interest of the genus,
which may be "an unmodified survivor from Mesozoic times, and therefore
from a time long before the Didelphyidae, Peramelidae, and Dasyuridae
were differentiated one from the other." Another ancient feature (found
in Jurassic mammals) is a mylo-hyoid groove upon the lower jaw, which,
however, is not always present, and its existence has therefore been
denied. The single species, M. fasciatus, is partly arboreal and
partly terrestrial in habit, and feeds upon ants. It is a Western and
Southern Australian form.

Fig. 82.—Banded Australian Anteater. Myrmecobius fasciatus. × 1⁄5.
Fam. 2. Didelphyidae.—All the members of this family are
pentadactylous. The teeth are fifty in number, arranged thus: I 5/4 C 1/1
Pm 3/3 M 4/4. The caecum is small; the pouch is generally absent; the
tail generally long and prehensile.
Fig. 83.—Virginian Opossum. Didelphys virginiana.
× 1⁄5. (After Vogt and Specht.)
The genus Didelphys contains most of the forms belonging to
this family, including as it does some twenty-three species. The Opossums
are mainly arboreal animals, insectivorous in their food; but the larger
species eat reptiles, birds, and their eggs. Several of the small species
carry their young, when able to leave the teats, on {156}their back,
the tails of the young being wrapped round that of the mother. It is not
only the pouched species which carry their young in something of this
fashion. Azara's Opossum, an animal as big as a cat, is said to carry its
eleven young ones (themselves as large as rats) on the back, though their
foothold does not appear to be strengthened by intertwining the tails.
Even with this huge family on her back, the mother can climb trees with
considerable alacrity. The mammae are seven to twenty-five in number. The
genus has been lately split up into a number of genera, Marmosa,
Dromiciops, Peramys, etc.

Fig. 84.—Thick tailed Opossum. Didelphys crassicaudata. × 1⁄5.
Chironectes is hardly different from Didelphys. It has
webbed hind-feet, and is aquatic in habit. The one species of the genus
is known as the Yapock, and is a Central and South American form. It is
of about the size of a large rat, and appears to be an expert diver after
the fish upon which it lives.
Fam. 3. Peramelidae.—The Bandicoots, although clearly
belonging to the Polyprotodont Marsupials, yet agree with the
Diprotodonts in the fact that the second and third toes of the feet are
bound up in a common integument, which is not the case with the
Diprotodont Caenolestes. The hind-feet are longer than the front;
of the former limb, two or three of the fingers alone are long and
functional; the others are rudimentary or absent. Tail long, hairy, and
non-prehensile. Dentition I 5/3 C 1/1 Pm 3/3 M 4/4 = 48, or sometimes,
owing to the absence of a pair of upper incisors, 46. There is a
caecum.
{157}
Fig. 85.—Bones of manus.
A, of Choeropus castanotis.
× 2. B, of
Bandicoot (Perameles).
× 1½. c, Cuneiform;
l, lunar; m, magnum;
R, radius; s, scaphoid;
td, trapezoid; tm,
trapezium; u, unciform;
U, ulna; I-V,
digits. (From Flower's
Osteology.)
Fig. 86.—Rabbit Bandicoot. Peragale lagotis. × 1⁄5.
The genus Peragale, the Rabbit-Bandicoots, consists of two
species entirely Australian in range. The enormous ears (whence "Rabbit"
Bandicoot) distinguish this genus from Perameles. The pouch opens
backwards, and there are eight mammae. P. lagotis, the only
species about whose ways of life anything is {158}known, burrows in the
soil, whence it extracts grubs; it is also a grass-feeder, and it is said
that its likeness to a Rabbit in appearance is strengthened by its
similarity in flavour!
Perameles is a genus consisting of twelve species, which are
found in Tasmania, Australia, and New Guinea. Like the last genus, from
which it does not widely differ in other points, Perameles
consists of species which combine insectivorous and vegetarian habits.
One species is said to become in captivity an expert in catching mice.
The pouch opens backwards, and there are six or eight mammae.
Fig. 87.—Pig-footed Bandicoot. Choeropus castanotis. × ⅓.
The last genus of this family is Choeropus, containing but one
species, Ch. castanotis. It is confined to the Australian
continent. It is to be distinguished from the last two by the fact that
there are only two functional digits, the second and third, in the
fore-limb; the fourth is rudimentary; the other two are absent. It
burrows, and is omnivorous like its allies. The two metacarpals that are
developed are very long and closely apposed; they have hence a remarkably
pig-like aspect, and justify its name. The pouch opens backwards, and
there are eight mammae.
Fam. 4. Notoryctidae.—This family contains but a single
genus and species, the recently-discovered Notoryctes typhlops.[91]
{159}
We may regard as family-characters the pentadactyle limbs, the
existence of three pairs of incisors in the lower and four in the upper
jaw; and the tritubercular nature of the upper molars. Notoryctes
typhlops, the "Marsupial Mole" as it has been termed, was originally
discovered by Professor Stirling in Central South Australia. It is a
burrowing creature, clothed in a silky fur of a pale golden red, without
external ears. It has been compared in appearance with
Chrysochloris, the Cape Golden Mole, and the eminent
palaeontologist, Professor Cope, has even insisted upon a real genetic
affinity. Edentate affinities have also been suggested. But
Notoryctes has a small pouch opening backwards as in other
Polyprotodonts,[92] and as
it also possesses marsupial bones it must undoubtedly be referred to the
Marsupialia. The animal shows many curious adaptations to its underground
mode of life. Certain of the vertebrae in the neck and in the lumbar
region are firmly welded together, giving of course a strength of push,
and suggesting the Armadillos; the claws of the third and fourth
front-toes are greatly enlarged, and must be efficient digging organs.
The track of the animal is like that of a railway in mountainous country;
it burrows for a short distance, emerges, and then descending beneath the
surface re-emerges. The red colour of the fur is said to be in harmony
with the arid soil in which it lives. The native name of the creature is
"Urquamata." It feeds upon ants and other insects.

Fig. 88.—Australian Marsupial Mole. Notoryctes typhlops. × ¼.
Extinct Polyprotodonts.—Of extinct Polyprotodonts (apart
from those Mesozoic forms which are considered on p. 100) extinct species of Thylacinus and
Dasyurus are known from {160}Australia. The most interesting fact in
connexion with the Tertiary Polyprotodonts is the existence in South
America of such genera as Prothylacinus and
Amphiproviverra, which are not merely Polyprotodonts but
definitely Dasyures, and not referable to the Didelphyidae.
These forms have been included in an order, Sparassodonta. But it is not by any means certain
whether these forms are rightly placed in the neighbourhood of the
carnivorous Marsupials; it is possible that they ought to be relegated to
the Creodonta or to their allies. Their structure is in fact somewhat
intermediate between those two groups. The teeth seem to be carnivorous
and Marsupial-like in form; but as already mentioned, in connexion with
the general structure of teeth, more than a single premolar is replaced.
These animals in fact, in so far as regards their teeth, are midway
between the Marsupials and the typical Eutheria. The angle of the lower
jaw is inflected, but the palate is not marked by deficient ossification.
At least this is not the case with all the members of the group. Whether
the small Microbiotherium, which is made the type of a family, is
rightly referred here is not certain. This animal had palatine vacuities
as well as an inflected angle to the lower jaw.
{161}
CHAPTER VIII
EDENTATA—GANODONTA
Order II. EDENTATA
Terrestrial, partly subterranean, or arboreal creatures of quite small
to gigantic size (some extinct genera), with frequently a covering of
scales or bony scutes. Limbs clawed. Teeth either totally absent or, if
present, imperfect in structure, being without enamel, and not forming a
complete series; incisors and canines being as a rule absent. Teats
axillary, pectoral, or inguinal.[93] Retia mirabilia very common in the
extremities.
To this group the name of Bruta was given by Linnaeus, but then it
included not only the families which we now place in the modern order
Edentata, but also the Elephant and the genus Trichechus. Mr.
Thomas has proposed to change the name into Paratheria, which name is
suggestive of what he and some others think concerning the systematic
position of the group, i.e. that it is not to be placed in the
Eutherian group of mammals at all, but represents a separate twig which
has arisen with the Eutheria from a low mammalian stock. This view can
hardly be accepted if the Ganodonta—which will be treated of
presently—be really ancestral Edentates, for they are not in any
way a Prototherian mammalian group, so far as their remains enable us to
judge.
The Edentata contain the Sloths, Ant-bears, Armadillos, Manis
and Orycteropus, among living forms. The great Ground-Sloths,
Megatherium, etc., and Armadillos, Glyptodon, etc.,
represent the extinct forms.
The name that has been applied to this group is inappropriate {162}inasmuch as many Edentates have teeth. It
is, however, by a number of small tooth-characters that the order can be
defined. Thus if teeth are present they are simple in structure, without
enamel in the adult condition, though a rudimentary enamel-organ has been
discovered in an Armadillo. The teeth, moreover, are not found in the
anterior part of the mouth, and they grow from persistent pulps; neither
is there much differentiation among them. It is not possible, however, to
speak of the Edentates as quite homodont, since in Orycteropus
there are large cheek-teeth; but there is at any rate not a marked
heterodonty in that or in any other Edentate. It used to be said that the
Edentates were monophyodont. But the Armadillo Tatusia was
subsequently found to possess a second suppressed dentition, and after
this discovery Mr. Thomas proved that Orycteropus is also
diphyodont. Since then other Armadillos have been shown to be diphyodont;
and the whole group therefore, so far as concerns those members that have
teeth, may in all probability be regarded as typically mammalian in this
respect.
These characters are slender enough, but there seem to be no others by
means of which the members of this order can be satisfactorily linked
together. The fact is, that we have here a polymorphic order which
contains in all probability representatives of at least two separate
orders. We have at present a very few, and these perhaps highly modified,
descendants of a large and diverse group of mammals. For convenience'
sake they will be all treated of under the head of Edentata.
Although for the probable reasons already stated it is a hard matter
to frame such a definition as will include all existing Edentates, it is
easy enough to define two groups in this heterogeneous order; to define
one group we should say, rather, and then to regard the leavings as
forming another not so easily definable a group.
The perfectly-definable group is that which includes the American
Anteaters, the Armadillos, and the Sloths. In all these creatures, which
may certainly be regarded as representing on their own account as many
family types, there are a number of important and highly-characteristic
anatomical features which they share in common. So exceedingly different
are these three types in general appearance and (correlated with that) in
way of life that these common characters acquire increased importance.
{163}

Fig. 89.—Great Anteater (Myrmecophaga jubata). A, Side view of twelfth and thirteenth
thoracic vertebrae. B, Posterior surface of second lumbar vertebra. C, Anterior
surface of third lumbar vertebra, × ⅔. az, Anterior zygapophysis; az1, az2, az3,
additional anterior, articular facets; cc, facet for capitulum of rib; m, metapophysis;
pz, posterior zygapophysis; pz1, pz2, pz3, additional posterior articular
facets; t, transverse process; tc, facet for articulation of tubercle of rib. (From
Flower's Osteology.)
The first of these characters is the series of additional zygapophyses
on the posterior dorsal and lumbar vertebrae; these are very clear in the
Anteaters and Armadillos; less clear, but still obviously represented, in
the Sloths. In the second place, they all possess a clavicle,
rudimentary, it is true, in the Great Ant-bear, but still present.
Thirdly, the testes are abdominal throughout life, a character which they
share with such lowly-organised animals as the Monotremata and the
Whales. Finally, and this is by no means a matter to be overlooked, not
only are all the existing members of this group American in range, but
there is no evidence to prove that they have ever existed elsewhere. No
European or Old-World {164}representatives have as yet been
discovered which can be referred to the Anteater, Armadillo, or Sloth
type with certainty.[94]
Of these American forms, which will be treated of first, the
Armadillos are further apart from either Sloths or Anteaters than the
last two are from each other. The name Xenarthra
has been suggested for the American Edentates with "abnormal" vertebral
articulations; the corresponding Nomarthra
includes the Old-World forms.
Fig. 90.—Right scapula and clavicle
of Two-toed Sloth (Choloepus
hoffmanni). × 1⅔. a, Acromion;
af, prescapular fossa;
c, coracoid; cl, clavicle; csf,
coraco-scapular foramen; gc,
glenoid cavity; pf, postscapular
fossa. (From Flower's Osteology.)
Between the Sloths and Anteaters the extinct Megatherium and
some of its allies are to a certain extent intermediate. But it may be
pointed out in the first place that there are certain important
resemblances between the living forms. In both, retia mirabilia are
developed in the tail (in spite of its reduction in the Sloths) and in
the limbs. But, as is well known, retia are also found in other mammals
far removed in the series from these under consideration. The
reproductive organs generally are very similar, and they have both a
dome-shaped and deciduate placenta. The latter character they share with
the Armadillos and with the Aard Vark; Manis having a
non-deciduate placenta which is, like that of the Carnivora, zonary in
form. The Edentates, at any rate the American forms, have a double vena
cava posterior and no azygos vein. This condition is also met with among
Whales.
Osteologically the Sloths and Anteaters are united by the fact that
the coracoid becomes fused with the coracoid border of the scapula, thus
forming a foramen; the importance of this character is, however,
discounted by its occurrence in three genera of Cebidae.
The above facts embody the views of Sir William Flower.[95]
{165}
A subsequent study of the brain and of the muscles of these animals
has led to results not entirely in harmony with these views.
Dr. Elliot Smith is of opinion,[96] after an exhaustive study of the
Edentate brain, that in this region of the body the present group shows
very decided points of likeness to the Carnivora; that is, so far as
concerns the Anteaters. On the other hand, Orycteropus is as
distinctly comparable with a primitive Ungulate type, such as is
exemplified by Moschus. "If the brain of Orycteropus," he
remarks, "were given to an anatomist acquainted with all the other
variations of the mammalian type of brain, there is probably only one
feature which would lead him to hesitate in describing it as an
exceedingly simple Ungulate brain. That one feature is the high degree of
macrosmatism.[97]
Manis, on the other hand, does not come especially near to
Orycteropus. The brain of Manis conforms to a simple type
of architecture, which agrees in many points with both those of
Orycteropus and the American Edentates; there is not sufficient
evidence to show which type it really favours." Elliot Smith would, in
fact, agree with Max Weber that it is better, if a division is to be
made, to divide the group into three orders:—the Xenarthra (Sloths, Anteaters, and
Armadillos), Tubulidentata (Orycteropus), and Squamata
(Manis), instead of into Xenarthra and Nomarthra.
Messrs. Windle and Parsons[98] are disposed to see in muscular
similarities reasons for uniting Manis with the American
Edentates, though they confess to being unable to place
Orycteropus; in this animal, they say, "we are more struck by the
generalised mammalian arrangement of its muscles than by any special
Edentate characters. There are, however, two muscles in
Orycteropus which show peculiarities not found elsewhere than in
the Edentates";—the triceps, which has more than one scapular head,
and the tibialis posticus, which is double. They conclude that
Orycteropus "presents some feeble claims to be taken into the
order."
We shall here adopt the following divisions. {166}
Sub-Order 1. XENARTHRA.
Fam. 1. Myrmecophagidae.—The family Myrmecophagidae
contains three genera, all South American in range. These genera,
Myrmecophaga, Tamandua, and Cycloturus, agree
greatly in their outward form. They are all without teeth, and have long
snouts and long protrusible tongues. The fur is thick, and they have
powerful claws wherewith to break down the strong ant-hills upon whose
inhabitants they feed. Tamandua and Cycloturus are
arboreal, Myrmecophaga is terrestrial in habit. The claws of the
arboreal forms are useful to destroy the bark, and thus bring to light
insects which lurk in such situations.
Fig. 91.—Great Anteater. Myrmecophaga jubata. × 1⁄10.
The genus Myrmecophaga contains but one species, the Great
Anteater, Myrmecophaga jubata. It is a large and handsome animal,
with long, shaggy, greyish-black hair and a broad white stripe across the
shoulder. The coloration is similar in the two sexes. Including the long
and bushy tail it reaches a length of over 7 feet. It is on account of
its long tongue and greatly developed salivary glands that this and the
allied genera were originally placed with Manis. It is the
submaxillary glands which are so enormous; they extend back over the
chest, and open by three distinct ducts, of which two unite just before
the external orifice. {167}Along their course these ducts are
provided with a sphincter muscle, which squeezes the secretion towards
the external orifice into the mouth-cavity. The stomach is somewhat
gizzard-like. The intestine has no caecum.[99]
The Anteater's great claws are not only serviceable in tearing up the
ground to get at its food; armed with them he does not fear, as Mr.
Waterton remarked, "the fatal pressure of the serpent's fold or the teeth
of the famished jaguar." An Anteater, too, is more than a match for a big
dog, and will rip open its belly with the claws while the dog is vainly
trying to make an impression with its teeth upon the shaggy hair.
Tamandua is a smaller animal than Myrmecophaga, and, as
has been stated, is arboreal; associated with this habit is a prehensile
tail. Like the last genus, Tamandua has a rudimentary clavicle,
this bone being well developed in the little Cycloturus.
Fig. 92.—Skull of Anteater (Myrmecophaga). Lateral view, al.sph, Alisphenoid; cond,
condyle of mandible; cor, coronoid process of mandible; ex.oc, exoccipital;
ext.aud, external auditory meatus; fr, frontal; ju, jugal; lcr, lachrymal; max,
maxilla; nas, nasal; occ.cond, occipital condyle; pal, palatine; par, parietal;
p.max, premaxilla; s.oc, supraoccipital; sq, squamosal; ty, tympanic. (From
Parker and Haswell's Zoology).
The skull of the Anteater[100] is very long and low; the fore-part
is tubular, and there appear to be no traces of teeth. The premaxilla is
very small; the zygomatic arch is imperfect, and does not reach the
squamosal behind. A curious feature of this genus, which it shares with
some Dolphins and other Whales, is that the pterygoid bones develop
palatine plates which meet each other in the middle line, and thus shift
the opening of the {168}posterior nares backwards. This is also,
of course, a character of various lower vertebrates. Another Whale-like
character in the skull is the weak character of the mandible, which does
not give off a marked coronoid process. But then in neither group is
there much mastication. The tympanic, periotic and squamosal are
ankylosed together. A peculiarity of the cervical vertebrae is that (as
in the Camels) the vertebrarterial canal of several of the vertebrae
perforates the pedicle obliquely. There are fifteen or sixteen dorsal and
three or two lumbar vertebrae. The additional zygapophyses upon the
former have been already referred to. The mode of articulation of the
ribs is highly singular.
|
Fig. 93.—Skull of Anteater (Myrmecophaga).
Ventral view. Letters
as in Fig. 92. In addition, b.oc,
basioccipital; glen, glenoid surface
for mandible; pter, pterygoid.
(From Parker and Haswell's
Zoology.)
|
Fig. 94.—Side view of three
mesosternal segments of a
young Anteater (Tamandua),
showing the mode of
articulation of the sternal
rib (sr). mst, The upper
or inner surface of the
mesosternal segment; sy,
the synovial articulation
between the segments.
(From Flower's Osteology,
after Parker.)
|
Each segment of the sternum (of which there are eight) is separated
from the next by a synovial membrane: and it has on either side two
facets for articulation with the ribs. The way in {169}which these latter
bones are connected with the sternum is curiously like their mode of
connexion with the spinal column at their other end. With this may be
possibly compared the double articulation of the single rib (which
articulates with the sternum) in the Rorquals. In Cycloturus this
mode of articulation does not occur.
The manus of Myrmecophaga is five-fingered. Of these the third
digit (as in Perissodactyles) is the most prominent; it is at least
double the width of the second or third finger; the pollex is very
slender. In the little Cycloturus this is carried to a greater
extent: the third digit is relatively enormous; the first and the fourth
have become quite rudimentary; while the fifth is only just recognisable
as a minute ossification.
Fig. 95.—A, Manus of Great Anteater (Myrmecophaga jubata). × ⅓. B, Manus of
Little Anteater (Cycloturus didactylus). × 2. c, Cuneiform; l, lunar; m, magnum;
p, pisiform; s, scaphoid; td, trapezoid; tm, trapezium; u, unciform; I-V, digits.
(From Flower's Osteology.)
The chevron-bones in the tail surround a well-developed rete mirabile,
a rete being found in precisely the same position in the Eastern
Manis. Tamandua has also retia, which are also found in the
Spider-monkeys.
Cycloturus is by far the smallest of the Anteaters. It has {170}only
two toes on the fore-feet. It is to be distinguished, anatomically, from
its larger relatives by the complete clavicle, and by the fact that the
pterygoids do not meet in the middle line of the skull. The ribs, too,
are unusually wide, as in the Whale Neobalaena, and form a bony
encasement for the body. It has two small caeca. Of fossil Anteaters but
little is known. The most interesting form is Scotaeops,
interesting because it has two small back teeth, which are totally lost
in its living allies. The huge Patagonian extinct bird
Phororhacos, first known by a lower jaw, was at one time regarded
as a member of this group on account of the form and edentulous character
of the jaw.
Fig. 96.—Unau, or Two-toed Sloth. Choloepus didactylus. × 1⁄5.
(After Vogt and Specht.)
{171}
Fig. 97.—Skull of Three-toed Sloth. Bradypus tridactylus. Lateral view. fr, Frontal;
ju, jugal; lcr, lachrymal; max, maxilla; nas, nasal; par, parietal; s.oc, supra-occipital;
ty, tympanic. (From Parker and Haswell's Zoology.)
Fam. 2. Bradypodidae.—The Sloths, genera Bradypus
and Choloepus, come, as already stated, very near to the
Anteaters, in spite of their striking difference in appearance. The
Sloths are purely arboreal creatures, with strong recurved claws, which
serve as hooks to keep them suspended from the lower side of a branch.
The three-toed sloth, Bradypus (or "Ai"), has the exceptional
number of nine cervical vertebrae; the two-toed sloth, Choloepus
hoffmanni (or "Unau"), has the equally exceptional number of six. The
hair is long and shaggy, and gets an adventitious green colour from the
presence of minute algae.[101] This gives to the animal the
appearance of a lichen-covered bough, a resemblance which is increased in
one species by an oval mark upon the back, which suggests forcibly a
broken end of such a branch. The likeness of a Sloth to its surroundings
is pointed out by Dr. Siemann,[102] who observed that a species occurring
in Nicaragua "has almost exactly the same greyish-green colour as
Tillandsia usneoides, the so-called 'Vegetable Horsehair' common
in the district.... If it could be shown that it frequented trees covered
with that plant ... there would be a curious case of mimicry between the
sloth's hair and the Tillandsia, and a good reason why so few of
these Sloths are seen." The stomach in the Sloths is complicated in
structure, with several chambers; one of these gives off a long
crescent-shaped caecum. The skull of the Sloths agrees in a number of
particulars with that of the Anteaters. {172}

Fig. 98.—Skeleton of Three-toed Sloth. Bradypus tridactylus. (After de Blainville.)
The zygoma is incomplete, though the part connected with the frontal
has a strong downward process like that found in Diprotodon and
some other mammals. There is, moreover, a process from the squamosal,
though it does not reach the anterior part and thus {173}complete the
arcade. The premaxillaries are very small, and are usually lost in dried
skulls. Coupled with these points of likeness are some differences. The
lower jaw, for instance, has a well-marked coronoid process. The
pterygoids do not meet in the middle line. The teeth are five or four in
each half of each jaw. There is no trace of a second set.
A peculiarity of the Sloths is the enormous number of dorsal
vertebrae. There are twenty-three of these in Choloepus hoffmanni,
but only fifteen to seventeen in the Three-toed Sloth, Bradypus.
As in other American Edentates, the acromion joins the coracoid. This
connexion occurs in both the Two-toed and the Three-toed species. The
limbs of these creatures are very long, a concomitant of an arboreal
life. The femur has no third trochanter. The genus Bradypus, which
by reason of the fact that it has not lost the third toe on the manus
seems to be more primitive than Choloepus, shows another
structural feature which does not bear out this conclusion. The trapezoid
and the os magnum of the carpus are united, while in Choloepus
they are perfectly distinct bones.
The intestine has no caecum.
There are several species of Sloths. Eminently perfect though the
organisation of the Sloth in relation to its particular surroundings
appears to us, Buffon selected the animal as the very type of
imperfection in nature. "One more defect," he wrote, "they could not have
existed."
Fam. 3. Dasypodidae.—The family Dasypodidae or Armadillos
contains a considerable number of genera. Tatusia,
Tolypeutes, Dasypus, Xenurus, Priodon,[103] and
Chlamydophorus. They have all a more or less rigid covering of
bony plates imbedded in the skin, which are not in the least comparable
with the scales of the Manis. Save the Whales, in one or two genera of
which traces of a dermal armature exist, the Armadillos are unique among
existing mammals in this particular. The term "Edentate" is especially
inapplicable to the Armadillos; the genus Priodon may have more
than forty teeth in each jaw; a total of ninety was found in one specimen
examined by Professor Kükenthal. In the tendency of the teeth to
multiply, we have another example of a state of affairs which
characterises so many Whales. Generally, however, seven to nine is the
number of teeth in each {174}half jaw, of which one is often implanted
in the premaxilla. The Armadillos show their alliance with the other
American Edentates in the points enumerated above. Their teeth specially
ally them to the Sloths, while the salivary and digestive organs
generally are on the Anteater plan, but present a less extreme
development. There are, however, caeca, paired as in birds, in the genera
Dasypus and Chlamydophorus. The others have none. But there
is a dilatation at the commencement of the large intestine, which is not
very different from the slightly-developed caeca of Dasypus.
There are certain peculiarities in the skeleton, which distinguish
this family.
Fig. 99.—Skull of Armadillo. Dasypus sexcinctus. × ⅔. ex.oc, Exoccipital; fr,
frontal; max, maxilla; nas, nasal; par, parietal; peri, periotic; p.max, premaxilla;
s.oc, supraoccipital; sq, squamosal; ty, tympanic. (From Parker and
Haswell's Zoology.)
{175}
|
|
|
|
Fig. 100.—Bones of the right
manus of the Hairy Armadillo.
Dasypus villosus. × ⅔. c,
Cuneiform; l, lunar; m, magnum;
p, pisiform; R, radius;
s, scaphoid; td, trapezoid; tm,
trapezium; u, unciform; U,
ulna; I-V, digits. (From
Flower's Osteology.)
|
Fig. 101.—Bones of the manus of
the Great Armadillo. Priodon
giganteus. × ⅓. a, An accessory
carpal ossicle in front of
the pisiform, which is not seen
in the figure. Other letters as
in Fig. 100. (From Flower's
Osteology.)
|
The skull in the Armadillos presents a number of likenesses to the
other American Edentates.[104] The premaxillaries are small, but are
larger in Dasypus than in Tatusia. On the other hand the
lachrymals are larger in the latter. The zygomatic arch is complete, but
there is no downward process as in the Sloths. In Tatusia (but not
in Dasypus) the "short thick pterygoids add somewhat to the hard
palate." This is clearly a beginning or a remnant of the quite
crocodilian character of the palate of Myrmecophaga. In the
cervical vertebrae we see the Whale-like character of fusion between
individual vertebrae; and also, as in the Whales, the degree to which
this fusion is carried out varies; two to four may be thus united. The
additional articular facets upon the dorsal vertebrae have been already
commented upon as a point of important likeness to other American
Edentates. The dorsal vertebrae are commonly eleven in number, the lumbar
being three. But in Priodon the numbers are twelve and two
respectively. There are traces to be observed of the double-headed
attachment of the ribs to the sternum. The shoulder girdle of the
Armadillos is somewhat diverse in form in different genera; the acromion
is always large, and is remarkable in Priodon for the fact that
the humerus also articulates with it, its extremity being recurved, and
forming a socket for this purpose. As in some other Edentates there is a
second spine on the scapula behind the first. The clavicle is strong.
There is some variation in the form of the manus. It is five-fingered in
Dasypus; in Tolypeutes the first digit has vanished; on the
other hand, in Priodon, the fifth has become rudimentary {176}and the
third enormously enlarged. This latter fact recalls the arrangement
characteristic of Myrmecophaga. The pelvis is greatly attached by
the ischium to the vertebral column. The femur has a third
trochanter.
The various forms of Armadillos are largely distinguished by the
number of movable thin bands of scutes lying between the large anterior
and posterior shields. Thus we have Dasypus sexcinctus,
Tolypeutes tricinctus, etc.
Fig. 102.—Pelvis and sacrum of Armadillo. Dasypus
sexcinctus. ac, Acetabulum; il, ilium; isch, ischium;
obt.for, obturator-foramen; pect.tub, pectineal tubercle;
pub, pubis. (From Parker and Haswell's
Zoology.)
The little Pichi-chago (or, more correctly, Pichy-ciego),
Chlamydophorus, which only grows to about 5 inches in length, has
no movable bands at all. It is covered with a uniform series of plates,
which, moreover, are not discontinuous at the neck. It differs, too, from
the prevailing Armadillo-type by the absence of conspicuous external
ears. In the anterior part of the body the armature consists of little
more than the horny plates, which in other Armadillos overlie the bony
dermal plates. In the hinder region the bony plates are strong. In this
animal, therefore, we have the dermal armature reduced to a minimum; but
it must be noticed that, like the extinct Glyptodons, the armature is
continuous and nowhere ringed.
The genus Tolypeutes, of which the best-known species is T.
tricinctus, the Apar (there are two other species in the genus), can
roll itself up into a ball like the Pill-Millipede (Glomeris),
and, protected by its armour, roll away from its enemies like the
Arthropod under similar circumstances. This mode of protection, be it
observed, is also adopted by the Pangolin and by the {177}Hedgehog. The
genus has only three movable bands. The tail is short, and is covered
with large tubercles. This genus is very markedly digitigrade when
running.
Fig. 103.—Three-banded Armadillo or Apar. Tolypeutes tricinctus. × ¼.
Fig. 104.—Peludo Armadillo. Dasypus sexcinctus. × ¼. (After Vogt and Specht.)
The Peludo, Dasypus sexcinctus, is, like other Armadillos, an
omnivorous creature, and appears to be particularly fond of carrion. It
will burrow up to a decaying carcase like the ground-beetles. {178}Mr. W. H.
Hudson has described the way in which this Armadillo will kill a snake by
holding it down and literally sawing the reptile in half by help of the
sharp and serrated edges of the carapace. Dasypus has a very short
tail, which is shielded by distinct rings near the base.
Tatusia novemcincta is a species with nine movable bands. The
genus has four teats; the ears are near together. There are no caeca and
no azygos lobe to the lung. A species apparently belonging to this genus,
but described under the generic names of Cryptophractus and
Praopus, is remarkable for the thick covering of hair, not
entirely wanting but usually thin in other Armadillos. In this particular
species the coat of hair is so thick as to conceal the underlying plates
of the carapace. The individual hairs are stiff, and one inch and a half
in length.[105]
The genus Xenurus contains several species, the best known of
which is inaptly named X. unicinctus. As a matter of fact the
characteristic feature of the genus is the existence of twelve or
thirteen movable plates between the two ends of the body. X.
unicinctus has twelve dorsal and three lumbar vertebrae. This
Armadillo, known by the vernacular name of the Cabassou, has one of the
most modified hands that are found in the family. The first two digits
are slender and elongated; but are quite normal in the number of their
phalanges. In the remaining three digits the metacarpal is short and
broad, while the proximal phalanx is either suppressed altogether or
fused with the metacarpal, the middle phalanx is present but short, while
the third phalanx is very large indeed. As in Dasypus, but not as
in Tatusia, which is in so many other respects divergent from
these genera, the lungs have an azygos lobe. As a small point of
difference, tending to show an alliance between the genera Xenurus
and Dasypus and their difference from Tatusia, is the
deeply-imbedded gall-bladder; this sac is not nearly so deeply plunged
into the hepatic tissue in Tatusia. Xenurus has no caecal
dilatations. The brain "is intermediate in its form and surface markings
between Dasypus and Tolypeutes." The small intestine is
nearly eighteen times the length of the large. But these intestinal
measurements are not of much avail in this group as marks of affinity,
since in three species of Dasypus Garrod gives the following
widely-divergent lengths:—D.
villosus, 11.5 feet and 1.25; D. minutus {179}5.1, with a
large intestine of no less than 7 feet; D. vellerosus 4.3 and
.66.
Priodon is the giant of its race. This Armadillo may reach a
length of 3 feet to the base of the tail. The tail is some 20 inches
long. The large number of teeth has been already noticed. There are
twelve or thirteen bands. Other points in the structure of this genus
have already been mentioned, and need not be recapitulated. This
Armadillo feeds upon termites and carrion.
Scleropleura is unfortunately but imperfectly known. The single
species, named by Milne-Edwards[106] S. bruneti, is apparently a
very rare inhabitant of Brazil. It is known by a single skin, which was
tanned by the hunter who obtained it. Thus the hair, if any, has dropped
out. The plates in the skin are deficient along the back and even upon
the top of the head, and are barely represented upon the tail
posteriorly. The ears are small and distant from each other. The tail is
longish, about one-third of the length of the body. The total length of
the creature including the tail is rather more than a foot and a half.
The hunter who obtained it regarded it as a hybrid between an Armadillo
and an Anteater.
Extinct Xenarthra.—There are a good many extinct forms of
Armadillo, apart of course from the Glyptodons. Peltephilus is
referred to later (p. 186). Dasypus was
represented by a large form, 6 feet long, with a skull of one foot in
length. The genus Eutatus was also large. The carapace was formed
of thirty-three distinct bands, of which the last twelve are soldered
together, but not fused into a shield as in Dasypus, etc.
An extinct group of American Edentates, termed the Gravigrada,[107] are somewhat intermediate between the
Sloths and the Anteaters. A number of the genera are well known from
complete skeletons.
One of the typical forms of this group is Mylodon, which,
together with its immediate allies, is often placed in a separate family,
Mylodontidae.
Mylodon itself was a large creature, as big as a Rhinoceros. It
was covered externally by armour in the skin, which did not form a
massive armature as in the Glyptodonts, but was in the {180}form of
scattered plates, small and not fused together. The general aspect of the
skull is decidedly Sloth-like. As in that animal, the malar bone is bifid
posteriorly, and between the bifurcation is embraced the process of the
squamosal. This latter is thus more developed than in the Sloth, but
there is no actual union between it and the malar. The premaxilla is
small. The lower jaw has both coronoid and ascending processes, and is
massive. There are five teeth on each side above, and four on each side
below, as in the Sloths. There are the normal seven cervical vertebrae
and sixteen dorsals. The limbs are not long and slender,but short and
strong, the animal having been terrestrial. The fore-feet were five-toed,
of which the three inner toes had claws. The hind-feet were only
four-toed, and the two inner only were clawed.

Fig. 105.—Mylodon robustus. (Restoration, after Owen.)
Scelidotherium is a genus which is a trifle smaller than the
last. It has only four properly-developed toes in the fore-foot, the
thumb being rudimentary; of these, the first two bear claws. The
hind-feet are also four-toed. Like Mylodon, Scelidotherium
is a Pleistocene genus.
Glossotherium has a skull very much like the last two genera;
but it is remarkable for the fact that the nostrils instead of being
unprotected with bone anteriorly are there closed by a plate of bone
formed by the well-developed premaxillae, the nostrils appearing at the
sides, and giving the skull a curious likeness to that of a Chelonian.
From a series of recent and most important observations it appears to be
clear that this genus has survived into quite modern times.[108]
{181}
The well-known naturalist of La Plata, Señor Moreno, engaged in
studies connected with the political boundary line between Chili and the
Argentine, had occasion to visit Consuelo Cove on Last Hope Inlet in
Patagonia. Hanging from a tree he noticed a piece of dried skin, which at
once struck him as looking more like the remains of a Mylodon than of any
living animal. The inhabitants regarded this piece of skin as a great
curiosity, but were of opinion that it was the hide of a cow encrusted
with pebbles! This fragment from a bygone age was originally described
by Professor Ameghino, who had apparently seen some of the bonelets
imbedded in it, as Neomylodon listai, "a living representative of
the ancient Gravigrade Edentates of Argentina." That this piece of skin
is of quite recent date seems to be proved by a number of considerations.
In the first place it is covered by long hair of a light yellowish-brown
colour; it does not seem likely that hair would preserve its character
for geological epochs. The nearest corresponding case is that of the
remains of Moas in New Zealand, whose feathers, dried skin, and tendons
are known. Now the Moa was unquestionably contemporaneous with man, as
abundant surviving legends prove, and indeed it cannot have been long
extinct. Still, hair is a resisting structure, and in a dry cave, with no
possibility of irruptions of floods, might retain its characters for long
periods. The evidence, however, of more recent date is stronger than
this. The skin shows patches of reddish colour, suggestive of course of
blood-stains. A small piece of the outside of the skin at the cut edge,
which presented the appearance of freshly or comparatively freshly dried
fluid, was submitted to a chemical examination and shown to be serum! Dr.
Lönnberg examined chemically a bit of the skin itself and found in it,
after boiling, glue, "which proves that the collagen and gelatinous
substances are perfectly preserved." After this it seems impossible to
suppose that the skin can be of any very great age; for bacteria would
have finished their work upon the serum and gelatine long ago. Combined
with the fresh appearance of the skin is the very fresh appearance of the
skull. In fact it is impossible to believe that the animal was not alive
quite a few years since, relatively speaking. It is admitted that this
animal was contemporaneous with man. There are actually legends of a
creature which may have been this Glossotherium. "Ancient
chroniclers inform us that the indigenous inhabitants recorded the
existence of a {182}strange, huge, ugly monster, which had its
abode in the Cordillera to the south of latitude 37. The Tehuelches and
the Gennakens have mentioned similar animals to me, of whose existence
their ancestors had transmitted the remembrance; and in the neighbourhood
of Rio Negro, the aged Cacique Sinchel, in 1875, pointed out to me a
cave, the supposed lair of one of these monsters, called 'Ellengassen';
but I must add that none of the many Indians with whom I have conversed
in Patagonia have ever referred to the actual existence of animals to
which we can attribute the skin in question."
A rude painting in a cavern, in red ochre, seems to Dr. Moreno (whose
words we have just quoted) to be somewhat suggestive of a
Glyptodon. There are some reasons for believing that this
quadruped was kept by man as a domestic creature. In the cave are two
walls of rough pieces of stone which seem to have dropped down owing to
the wearing away of the roof; they also seem to have been loosely piled
together to form two walls, within which enclosure an imperfect skull of
the animal was found. This skull shows clearly that the so-called
"Neomylodon" must be referred to Glossotherium or
Grypotherium, as it is sometimes termed. This skull is perforated
on the roof in such a way as could only have been effected (in the
opinion of experts) by a weapon in the hand of a man. A hole in the skin
has been even compared to a bullet-wound. But this it is perhaps
unnecessary to discuss. The skin of Glossotherium is, like that of
other extinct "Ground-sloths" (e.g. Mylodon), filled with small
and irregular ossicles. But in Mylodon, the sculptured appearance
of the dermal ossicles appears to indicate that they reached the surface
of the body and were covered by epidermis alone, which is not the case
with the animal now under consideration. The microscopic characters of
the ossicles, too, show differences in the two. Glossotherium
being "precisely intermediate between Mylodon and the existing
Armadillo (Dasypus)." Now Glossotherium and Mylodon
are regarded as forms which lie between the existing Anteaters and the
Sloths of the same part of the world. We have already pointed out the
facts of structure which lead to this conclusion. It might therefore be
reasonably surmised that the hair of Glossotherium would be also
intermediate, or at least like that of one of the two genera
Myrmecophaga and Bradypus. But microscopical investigation
has {183}negatived this supposition. It has shown
that the Armadillos are in this matter the nearest relatives of
Glossotherium. This result is important as tending further to
confirm the close interrelationship of all the American Edentates as
contrasted with the Old-World forms—a matter which has already been
emphasised. It is suggested, however, that the absence of under fur,
which is so well developed in the Sloth, and the difference shown in
transverse sections from the hair of Myrmecophaga, may be
explained by difference in habitat. Glossotherium lived under
conditions similar to those under which the Armadillos live to-day. Thus
the outer covering of the body became alike in the two cases, the same
needs supervening in both genera.
Lestodon is another allied genus, which seems to possess
canines. At any rate, in front of the four molars, and separated from
them by a diastema, is a smallish, somewhat canine-like tooth, in both
jaws.
Megalonyx and its allies are sometimes placed in a distinct
family, Megalonychidae. Megalonyx itself had a skull very
like that of Bradypus, being shorter and not so elongated as in
the Mylodontidae. There is a strong tusk anteriorly, which is separated
by a considerable space from the three molars lying behind it. Both pairs
of limbs seem to have possessed five toes. This is a North American
genus. It differs from the bulk of the American Edentates in having a
complete jugal arch.
Megatherium is the type of yet a third family,
Megatheriidae, of the Gravigrade Edentates. This creature is
familiar from the many restorations which have been built up, and from
its huge bulk, little short of that of an elephant. The skull, which is
small for the size of the creature, has a complete jugal arch, from the
middle of which depends a downward process as in other allied forms. The
teeth grow to an extraordinary depth, and there are five of them in the
upper and four in the lower jaw—on each side of course. The
fore-limbs of the Megatherium are very much more slender than the
enormously bulky hind-limbs, upon which and the equally massive tail the
animal seems to have supported itself while tearing down branches of
trees, upon whose leaves it fed. In the scapula the acromion joins the
coracoid as in Bradypus; the clavicle is large. The fore-limb is
four-toed, and the hind-limb three-toed. The latter has but one clawed
digit (the third, i.e. the inner). {184}On the manus, the three
inner digits have powerful claws. This animal, too, was Pleistocene in
time. The Megatheriidae had, however, small as well as gigantic
forms.
The genus Zamicrus had a skull no bigger than that of a Sloth,
while Nothrotherium was also a comparatively small creature; the
teeth of the latter genus are reduced to 4/3.
The extinct group of the Glyptodontidae comprises large
creatures with a dense covering of bony scutes which are arranged in a
tesselated fashion, and thus form an immobile armature of immense
strength. In correspondence with this massive carapace the dorsal
vertebrae have fused together, and the lumbar vertebrae form a series
ankylosed to each other and to the following sacrals. These creatures are
all South American.
Fig. 106.—Glyptodon clavipes. × 1⁄12. (After Owen.)
Glyptodon, the genus which gives its name to the family, is
known from numerous remains in South America, and also from so far north
as Texas and Mexico. It grew to be as long as 16 or 17 feet. In the skull
there is an exceedingly long downward process of the zygomatic arch, as
in Sloths, the arch itself being complete. The process extends so far
down as to reach a point about on a level with the middle of the lower
jaw. The nasals are short or rudimentary. As in Myrmecophaga, the
pterygoids enter into the formation of the bony palate. The lower jaw has
a spout-shaped extremity, and, behind, it rises into an enormous vertical
branch as high as the front part of the jaw is long. There are eight
teeth in each half of each jaw. As in {185}some Armadillos, the
cervical vertebrae are at least partly fused. The atlas is free, but the
rest, or at any rate five of them, are united. The last cervical is
sometimes fused with the succeeding dorsals; the latter are twelve in
number, and are fused together so far as concerns their centra and neural
processes. The succeeding region of the vertebral column includes seven
to nine lumbars, which are fused with the eight sacrals; in this region
the neural processes are high, and there is thus produced a strong and
lofty ridge along the back, which forms a powerful support for the
carapace. The fore-limbs are shorter than the hind-limbs, which latter
are attached to an unusually massive pelvis. The claws of the limbs are blunt and
almost hoof-like.
The heavy carapace consists of sculptured, five or six-sided plates,
which have no particular arrangement in the middle, but towards the
margins show indications of an arrangement in transverse rows. The
moderately long tail is also encircled by bony skin-plates which are
thorny above, or at least provided each with a blunt upstanding process.
It appears that outside this bony system of scutes were horny epidermic
scales, corresponding exactly with the tesserae which they cover. There
are apparently a good many species of Glyptodon.
In the allied genus Panochthus the tail is rather longer, and
the bony rings which surround it, instead of being all movable as in
Glyptodon, are at first so, but later, i.e. towards the end
of the tail, become welded into a single and massive piece. Both feet are
here four-toed, while in Glyptodon the hind-feet are five-toed and
the fore-feet four-toed.
Daedicurus shows a further specialisation, in that the feet
have three and four digits respectively. The orbit too shows a
specialisation in being separated from the temporal fossa. The descending
process of the zygomatic arch is not so extraordinarily exaggerated as it
is in Glyptodon. It has the same terminal tube of osseous scutes
upon the tail. This creature seems to have reached a length of about
twelve feet.
Propalaeohoplophorus is, unlike the great Armadillos that we
have hitherto dealt with, a small animal, not exceeding 2 feet or so in
length of carapace. A small alveolus on each side of the premaxillae
seems to suggest the former presence of an incisor tooth; and it seems
that the animal possesses both true molars and premolars; for the first
four of the eight teeth are much {186}simpler in structure
than those which follow. The dorsal vertebrae again are not fused
together; the hind-limbs are five-toed. All the plates of the carapace
are arranged in definite transverse rows; it has been observed, too, that
some of the anterior scutes overlap like those of the Armadillos, to
which this animal possesses further likenesses in the exclusion of the
maxillae from the border of the nostril (a Glyptodont character), and the
comparative feebleness of the scutes.
A primitive genus also appears to be Peltephilus, which is
perhaps rather an Armadillo than a Glyptodon. However, it comes
somewhat between the two, like Propalaeohoplophorus, with which it
may therefore be treated. A most singular feature of this genus has been
mentioned on p. 27 in connexion with the skull in
the Mammalia generally. That is the fact that a portion of the squamosal
surrounding the articular facet for the lower jaw is separated by a
suture from the rest of that bone, and is therefore obviously suggestive
of the quadrate in the lower Vertebrates. As in certain Armadillos and
Glyptodons, etc., the pterygoids appear in this genus to have taken a
share in the formation of the hard palate. The plates of the carapace
were movable, as is shown by the fact that they sometimes slightly
overlap. In view of the possible origin of the Edentates from
lowly-organised Mammalia, it is noteworthy that the humerus has been
especially compared to that of the Monotreme. Peltephilus differs
from other Armadillos in having teeth in the front of the jaws. The total
number of teeth is twenty-eight, i.e. seven in each half of each
jaw.
Sub-Order 2. NOMARTHRA.
As already explained, the Old-World Edentates differ from the
New-World forms in having normal dorsal vertebrae, that is to say,
without additional zygapophyses. That negative feature, however, though
combined with the positive fact that both the Old-World forms feed upon
ants, is hardly sufficient to outweigh the many structural differences
which distinguish the Orycteropodidae from the Manidae; which will be
placed therefore in different groups. To that containing the Aard Vark,
the name Tubulidentata may be applied. {187}
This group contains but one family, the Orycteropodidae, of
which there is but a single genus.
The Aard Vark (earth-pig), genus Orycteropus, is characterised
by its heavy build, the body being covered by rather coarse and not very
abundant hair; the snout is long and pig-like, with round nostrils at its
end; the ears are long, erect, and pointed; the tail is very thick at
first, so that it has been aptly described as "a tapering of the body to
a point." The fore-limbs are four-toed, the hind five-toed.
Fig. 107.—Aard Vark, or Cape Anteater. Orycteropus capensis. × 1⁄16.
In the skull there is a complete though slender zygoma; the
premaxillaries, though small, are not so rudimentary as in the American
Edentates. The annular tympanic is not ankylosed to the surrounding
bones, a character found in other low mammals. Contrary to what is found
in Manis, Orycteropus has a huge lachrymal. There are
thirteen dorsal and seven lumbar vertebrae. The clavicle is well
developed. Orycteropus is peculiar among Edentates in that the
ischia do not unite with the vertebral column. The femur has a third
trochanter.
As mentioned on p. 162, the Aard Vark is
diphyodont like normal mammals. The permanent teeth consist of five
molars and premolars on each side of each jaw; the first two of these are
premolars, and are simpler in their form than the succeeding two teeth,
which are partly divided by a median furrow into two halves. These teeth
are also peculiar in that they consist entirely of vaso-dentine. They
have been compared in minute structure to those of the Ray
Myliobates. According to Mr. Oldfield {188}Thomas[109] there are seven milk teeth on each
side of the upper jaw (limited to the maxillae, and thus not incisors).
An eighth tooth was discovered on one side of one of the specimens
examined by Thomas. In the lower jaw there are only four milk teeth on
each side. It is interesting to note that the histological structure of
these milk teeth agrees with that of the permanent teeth. There are two
species of this genus found in Africa: the southern, O. capensis,
is more hairy than the northern, O. aethiopicus. O. gaudryi
is a Pliocene species from the Island of Samos and from Persia, described
by Dr. Forsyth Major and Dr. Andrews.[110] It closely resembles the existing
O. aethiopicus.

Fig. 108.—Section of lower jaw with the teeth of Orycteropus. × 2. (After Owen.)
Of the Scaly Anteaters, Group Squamata or
Manidae, there is really but one genus, though Phatagin,
Pholidotus, Smutsia, and Pangolin have been used to
distinguish various forms. The genus Manis is African and Oriental
in range. Dr. Jentink, who has lately revised the species, allows
seven.[111] The external
form of these animals is fairly well known, the remarkable scales
distinguishing the Pangolins from other animals. Between the scales lie
hairs, which seem to be absent in the adults of the African species,
though present in the young, thus affording a convenient method of
distinguishing the Ethiopian from the Oriental forms. The scales have
been compared to agglutinated hairs. That they are not "merely mimetic of
the Lizards' scales" is held by Weber,[112] who compares them directly with those
{189}structures, as he does the scales of other
mammals, such as those upon the tail of Anomalurus, etc. This,
however, is not a universal opinion. It is true that these scales occur
chiefly in the lower forms of mammals such as those under consideration,
Marsupials, Rodents, and Insectivores; but the fact that the hairs are
developed before the scales shows, or seems to show, that the former are
the older structures, and to lead to the inference that the scales of
mammals are new structures. The scattered hairs of the Pangolin have no
sebaceous glands excepting on the snout. This, again, looks as if they
were degenerate structures, and emphasises the non-archaic character of
the scales. These animals have no trace of teeth except possibly some
slight epithelial thickenings which have been interpreted as a last
remnant; the tongue is suited for the capture of ants, and is therefore
much like that of the not nearly-related American Anteaters. The stomach
is of simple form; it is characterised by a large gland, which suggests
that of the Koala (see p. 144); the intestine has
no caecum. Retia mirabilia occur on the limb arteries. The placenta is
non-deciduate and diffuse; it is specially compared by Weber with that of
the Horse. Considering the many adaptive resemblances between this genus
and the American Anteaters, especially in the mouth cavity, it is
remarkable that in Manis the pterygoids are not joined as they are
in Myrmecophaga. In spite of statements to the contrary, it
appears that there is sometimes a distinct lachrymal.
A remarkable feature in the skeleton of Manis is the singular
sternum. The xiphoid cartilage is extraordinarily elongated into thin
strips, which reach the pelvis and return. This state of affairs is to be
found in the African species only. This structure is not comparable, as
it has been said to be, with abdominal ribs such as those of the reptile
Hatteria.
These animals are mainly anteaters. The Japanese have a curious legend
as to the method adopted for the capture of ants, which is related by Dr.
Jentink in his monograph of the genus. The Manis "erects his scales and
feigns to be dead; the ants creep between the erected scales, after which
the anteater again closes its scales and enters the water; he now again
erects the scales, the ants are set floating, and are then swallowed by
the anteaters"! The same story is related by Mr. Stanley Flower on the
authority of the Malays.
Though it seems clear that the likenesses which Manis shows
{190}to the Anteaters of the New World are
chiefly adaptive and have nothing to do with real affinity, being merely
an expression of a similar mode of life, it is curious to note that here
and there we do find certain resemblances which do not seem to be
susceptible of the latter explanation. The jugal bone, absent in
Manis, is small in Myrmecophaga; the clavicle is absent and
again small or rudimentary in the Anteaters; it is large in other
Edentates. The third trochanter is absent, as in Myrmecophaga (and
the Sloths). There are many scales on the body; in Myrmecophaga
there are traces of these structures on the tail, as also in
Tamandua. In the features mentioned, the Myrmecophagidae differ
from either or from both of the two other American families (i.e.
Dasypodidae, Bradypodidae) and agree with Manis. The facts are not
a little remarkable.
Fig. 109.—Manis. Manis gigantea. × 1⁄12.
Order III. GANODONTA.[113]
Allied to the Edentata, and apparently representing the ancestral
forms from which they, at any rate the Xenarthra were derived, is the
order of the Ganodonta. Of this order a number of genera are now known,
which can be ranged in a series which more and more approaches the
Edentata as we pass from the older to the newer forms. This interesting
and transitional series will be made manifest by a description of the
characters of the various genera taken in their proper {191}chronological
order. The following genera are included by Wortman in his family
Stylinodontidae.
The earliest type of the Ganodonta is the genus Hemiganus, with
but one species, H. otariidens. This animal lived during the
deposition of the lowest Eocene strata, the Puerco beds of North America.
It was about as big as a fair-sized Dog, and had powerful jaws. There
were at least two pairs of incisors in the upper jaw, together with
powerful canines and the full premolar and molar formula. In the lower
jaw the canines were also strong, but the incisors are not certainly
known to be more than two pairs. The enamel upon the posterior surface of
the canine is thin, and in the case of the incisors the enamel seems to
be limited to the anterior face. The lower molars are quadritubercular.
It is believed from the presence of a suture on the upper surface of the
premaxillary that the snout of the creature was tubular. The cervical
vertebrae, only known by their centra, are like those of the Armadillos
(and for the matter of that of the Whales) in the great transverse as
opposed to the antero-posterior diameter. The feet are especially
compared with those of the Ground Sloths. The single ungual phalanx is
marked by a large subungual process, which is pierced by a considerable
foramen. The tibia again is to be compared with that of the
Armadillos.
In the Upper Puerco (Torrejon) beds the remains of
Psittacotherium are found. This genus, when first discovered, was
referred to the Tillodontia by some and to the Ungulates, the latter
being a refuge for indeterminate Eocene mammals, just as the
"Multituberculata" is for similarly-placed Secondary mammals. It is now
known to be clearly a member of the order Ganodonta. Wortman thinks that
there is but one species, P. multifragum. It seems to have had a
general aspect much like that of Hemiganus—that is judging
from the skull—and was not very greatly different in size. The
facial portion of the skull is short, and the zygoma is deep. The
infra-orbital canal is double, a feature which crops up in the Sloth, and
has been mentioned in the later form of Ground Sloth, Megalonyx
(but it must be remembered that the same characteristic is not unknown in
Rodents). The dentition is reduced as compared with that of
Hemiganus, that is to say, as far as concerns the molars and the
incisors. There is but a single pair of incisors in each jaw; the canines
are strong; the premolar and molar series seem to have been complete in
the lower jaw, {192}but reduced by one premolar at least in
the upper jaw. It is very important to notice that the incisors have
enamel only on their anterior faces, and that the same is the case with
the canines, the slender layer present behind the tooth in
Hemiganus having vanished in this later form. The tooth pattern of
the molars is like that of Hemiganus. The fore-limb is decidedly
Edentate-like; but it is the foot which presents the strongest likenesses
to that order. "If an anatomist," remarks Dr. Wortman, "had no other part
of the skeleton than that of the foot to guide his judgment, and he
should fail to detect a most striking similarity between it and that of
the Edentata, especially the Ground Sloths, he would not only lay himself
open to the criticism of being lacking in the ordinary powers of
observation and comparison, but would be suspected of placing the matter
upon a basis other than that established by such a method." It is not
certain how many toes upon the fore-limbs were possessed by
Psittacotherium, but the close resemblance to Mylodon is
indeed striking, the third digit being in both forms the most pronounced.
Some vertebrae of this Ganodont have been discovered which do not show
the complex articular arrangements of later American Edentates. The
sacrum, on the other hand, is very like that of the Sloth, and there is a
foreshadowing of the attachment of the ilia to the sacrum by
co-ossification which is met with in later Edentates. A still later type
is the genus Calamodon, which has been shown to occur in Europe as
well as in America. C. simplex was a larger beast than either of
the genera that have already been treated of, thus affording another
example of the increase in size of later as compared with earlier members
of the same group, so pronounced among the Ungulata. The lower jaw has
the same massive structure that characterises that bone in
Hemiganus and Psittacotherium. There is but one incisor,
but the premolar and molar series are complete. The canine is Rodent-like
in appearance, being imbedded throughout the greater part of the lower
jaw; it evidently grew from a persistent pulp. It is enamelled upon the
anterior face only. The premolar and molar teeth are in this genus
commencing to lose their enamel, which is distributed in the form of
vertical bands, leaving interspaces which are not covered by enamel.
These teeth, moreover, are decidedly hypselodont, more decidedly so than
in Psittacotherium; they are, when unworn, quadricuspidate, with
accessory cusps; when more worn, the teeth {193}are double-ridged, and
that transversely to the long axis of the jaw; finally, the much-worn
teeth have flattish crowns more or less surrounded by a ring of
enamel.
A still later form, coming from the Lower and Middle Eocene strata, is
the genus Stylinodon. S. cylindrifer, which is the more
archaic of the two described species, is only known from a single molar,
fragments of a canine, and "some inconsiderable pieces of the skull." The
molar is interesting on account of the fact that the enamel is still
further reduced; it is represented only by narrow vertical strips, which
are much narrower than those of older forms of Ganodonts. It is also
hypselodont, and has a persistent pulp. So, too, the canine which had a
thick anterior facing of enamel. The later species, S. mirus, is
more fully known. The teeth seem to have been much the same as in the
last-described species; the premolars and molars were seven in all in the
lower jaw, and the canine was imbedded in the bone for a long distance,
as in Calamodon. The cervical vertebrae have short centra as in
Hemiganus. The clavicles were well developed. The humerus
possessed an entepicondylar foramen, and its head displays the pyriform
pattern so characteristic of later Edentates. The foot is clearly like
that of Psittacotherium.
In reviewing the series, therefore, we see a gradual diminution of the
incisors, a gradual loss of enamel on the teeth generally, and the
production of hypselodont teeth growing from persistent pulps; all of
which are features of the later Edentates. The progression is so gradual
that the forms enumerated and described seem to have been part of a
continuous series culminating in the Ground Sloths of later times. The
other points of similarity will be gathered from the facts given in the
foregoing pages.
There is another family belonging to the Ganodonta whose position with
regard to the Edentata is not so clear. This is the family
Conoryctidae, of which two genera are known. The earliest of
these, from the Lower Puerco, is Onychodectes. In O.
tissonensis the skull is long and narrow, thus contrasting with that
of the last family. The facial part is also long. The lower jaw is much
more slender. The molar formula was complete, but there is some doubt as
to the incisors. The molars are tritubercular.
The other known genus is Conoryctes. Its skull has a shorter
{194}facial portion, and is thus more like that
of Stylinodontidae than that of Onychodectes. The dental formula
is known, and is complete save for the loss of one incisor above and
below, and one premolar above. The relationship of these Ganodonts to any
later forms is uncertain; but their skeletal structure is as yet by no
means fully known.
{195}
CHAPTER IX
UNGULATA—CONDYLARTHRA—AMBLYPODA—ANCYLOPODA—TYPOTHERIA—TOXODONTIA—PROBOSCIDEA—HYRACOIDEA
Order IV. UNGULATA
The existing members of this order can be readily grouped into the
Hyracoidea, Proboscidea, Perissodactyla, and Artiodactyla, each of which
divisions has quite the value of an order, and all of which are sharply
marked off from each other. But as the discovery of so many fossil forms
has to a great extent rendered these demarcations less sharp, it is
better to regard all these groups as not more than sub-orders of a larger
"Order" Ungulata. Even when this conclusion has been necessarily arrived
at from a consideration of the more ancient groups of Ungulate animals,
the definition of such an order remains a difficult matter for the
systematist. For the earliest of these forms, more particularly the
Ancylopoda, the Amblypoda, and the Condylarthra, whose peculiarities will
be dealt with at length subsequently, are not by any means easily
differentiated from the primitive Carnivorous mammals of that date, the
Creodonta; these latter, moreover, fade into the Marsupials through the
so-called Sparassodonta of Professor Ameghino. To confine ourselves to
the Ungulates, we may perhaps define them as terrestrial animals with
hoofs rather than claws or nails, and chiefly, if not entirely,
vegetarian in habit. The teeth are bunodont or lophodont, the tendency to
the production of the latter type being always marked. The walk, although
plantigrade in the older types, becomes more and more digitigrade, except
in such survivals from antiquity as Hyrax. There is, too, as we
pass from the ancient types to the modern, a gradual perfection of the
limbs as running {196}and not climbing or grasping organs; the
number of toes becomes reduced, and culminates twice (in the horse and in
the Litopterna) in one toe on each foot; at the same time the ulna
becomes rudimentary and fuses with the radius, and the fibula in the
hind-limb undergoes a like reduction. The clavicle is absent even in some
of the oldest types; its presence in Typotherium[114] is highly remarkable. The tail too,
an organ which is long in some of the early forms, gets short in their
modern derivatives.

Fig. 110.—An early Ungulate. Phenacodus primaevus. × 1⁄12. (After Osborn.)
Fig. 111.—Series of metacarpals and metatarsals of Camelidae, to show secular and
progressive increase in size. From left to right the species are Protylopus
petersoni, Poebrotherium labiatum, Gomphotherium sternbergi, Procamelus occidentalis.
F, Fore-foot; H, hind-foot; III, IV, third and fourth metapodials.
(After Wortman.)
Coupled with the increasing perfection of the foot as an organ used
merely for the support of the body, certain interesting changes have
taken place in the arrangement with regard to each other of the several
bonelets of the wrist and ankle. It has been held by Cope and others that
the truly primitive disposition of these bones was that presented to us
by certain early types, such as Meniscotherium or the existing
elephant or Hyrax. In these animals there is (see Fig. 112) a
serial {197}arrangement of these bones, the distal
bones only, or very nearly only, articulating with the corresponding
bones in the upper series. In the modern types (cf. Fig. 113) there is,
on the other hand, an interlocking, so that the bones of the distal
series articulate with two of those of the proximal series. By this is
produced, as it would appear, a much firmer foot, less liable to "give"
under pressure, and thus more fitted for an animal that runs. It is the
same principle as that adopted in the laying of bricks. The actual stress
and strain of impact has been held responsible for those changes. An
equally ingenious and possibly truer explanation of the undoubted facts
has lately been advanced by Mr. W. D. {198}Matthew.[115] He has pointed out
that in some ancient Ungulates the carpus is not serial but interlocking,
even in forms which belong to the earliest Eocene groups, such as the
genus Protolambda among the Amblypoda. Now in the fore-foot of
Meniscotherium and the living Hyrax there is a separate
centrale which is wanting in the greater number of Ungulates. The
absorption, that is the practical dropping out of this bone, would
restore to an interlocking carpus the serial arrangement; while on the
other hand, by the fusion of this bone with the scaphoid, the
interlocking disposition would be maintained.

Fig. 112.—Bones of the manus A, of the Indian Elephant, Elephas indicus. × ⅛.
B, of the Cape Hyrax, Hyrax capensis. × 1. c, Cuneiform; cc, centrale; l,
lunar; m, magnum; p, pisiform; R, radius; td, trapezoid; tm, trapezium; s,
scaphoid; u, unciform; U, ulna. (From Flower's Osteology.)
Fig. 113.—Bones of the manus A, of Rhinoceros, Rhinoceros sumatrensis. × 1⁄5. B, of
Pig, Sus scrofa. × ⅓. Letters as in Fig. 112. (From Flower's Osteology.)
The gradual perfecting of the fore- and hind-limbs as running organs
has been put down to the advent of the grasses, and the formation of
large plains covered with this herbage. The same reason would also be in
harmony with the equally gradual change in the shape of the molar teeth,
from a tubercular form calculated for a mixed or even a carnivorous diet,
to the flatter crushing surfaces exhibited by the lophodont teeth of
later Ungulates. Strong {199}canines would in the same way cease to be
useful, and even become encumbrances to such grazing creatures; and their
disappearance is one of the salient features in the history of the
Ungulata, that is of the modern representatives of the order. The
extraordinary hypertrophy of these teeth in such a line as that of the
Amblypoda, which has left no descendants, was one of the reasons perhaps
for the decay of those great pachyderms of mid-Tertiary times; their
excessive armature became an encumbrance, since it was not accompanied by
improvements in other necessary directions. Some of the features of the
Tertiary Ungulates have, however, been dealt with in our general sketch
of the mammalian life during that epoch, and need not be again referred
to here. Of existing Ungulates there are no clear indications of the
descent of the Elephants or of the Hyracoidea. Their structure proclaims
these two divisions to be of ancient descent, and not to be modern twigs
of the Ungulate stem. As to the Perissodactyla and the Artiodactyla we
cannot bring them together nearer than in quite early Tertiary times. The
order Condylarthra seems to be the starting-point of both these
sub-divisions. Euprotogonia has been considered to be an ancestor
of the Perissodactyle branch, and Protogonodon or
Protoselene of the Artiodactyla. If this be true, {200}the likenesses
which Titanotherium shows to the Artiodactyla must be either
purely superficial and secondary, or a cropping out of ancient characters
which had been dormant for many generations.
Horns.—The Ungulata are the only order of mammals which
possess horns; as they are on the whole a more defenceless group than the
Carnivora, it may be that the horns are a counterpoise to the teeth and
claws of the latter; need for defence and for armature in the combats
with their own kind for the favours of the does has led to a different
kind of protective and aggressive mechanism. Horns as weapons are,
however, particularly effective in this group wherever they exist. A
Ruminant is most frequently a large and heavy animal without the agility
and litheness of the Carnivore. It is precisely to this sort of animal,
where weight is an important consideration, that horns are the most
suitable weapons. This is further shown by the fact that although the
general term horn is used to describe the weapons of the Ungulate
mammals, there is more than one kind of structure included under this
general term; it is indeed probable that the extreme terms in the series
of horns have been independently acquired by their possessors. There is
but little in common between the horns of a Giraffe and of a Rhinoceros.
In the Rhinoceros we have one or two horns, in the latter case one placed
behind the other, which are purely epidermic growths; they may indeed be
regarded as matted masses of hair, borne, it is true, upon a boss of
bone, which however is not a separate structure. The Giraffe supplies us
with the simplest term in that series of horns which are partly epidermal
and partly bony. The paired horns of this animal have often been
contrasted with those of the Deer, for example; but there is no
fundamental difference between them. In the Giraffe a pair of bony
outgrowths, originally separate from the skull which bears them, but
ultimately ankylosed to it, are covered by a layer of entirely unmodified
skin. A distinction of undoubtedly practical importance is usually drawn
between the Hollow-horned Ruminants, i.e. Oxen, Goats and
Antelopes, and the Deer tribe. There is nevertheless no fundamental
distinction. In the Antelopes there is a core of bone, the "os cornu" as
it has been termed, which is covered by a horny layer, the horn proper,
variously modified in shape and size according to the genus or species.
In the Deer there is the {201}same os cornu, which may however be
branched, but which is in the same way covered by a layer of modified
integument; this is known as the "velvet"; it only lasts for a certain
period, and is then torn off by the exertions of the animal itself,
leaving behind the bony core, which is popularly termed the horn. It will
be clear that here is only a difference of comparative unimportance; the
same essential features are present in both groups of animals, but the
modification of the epidermis has progressed along different lines. Both
can be referred back to the primitive conditions seen in the paired horns
of the Giraffe. Even the difference, such as it is, is bridged over by
the Antelope Antilocapra, where the os cornu is bifid and the horn
is periodically shed, as is the velvet of the stag; but in the stag the
bony part of the horn is also shed, a state of affairs which has no
parallel in the Hollow-horned Ruminants. The great Sivatherium may
conceivably be an annectant form between the two types of compound horns,
i.e. those of the Antelope and those of the Deer. This creature
had two pairs of horns, of which, naturally, only the bony cores remain;
the hinder pair of these were branched. But although so far they resemble
the Deer's horns rather than the Antelope's, Dr. Murie has thought that
they were covered by a horny sheath and not by soft skin as in the Deer.
In any case these horns were apparently never shed, which is a point of
likeness with the Antelope and of difference from the Deer. Apart
therefore from the nature of the covering of the bony cores, there are
good grounds for looking upon them as intermediate between those of the
Deer and those of the Antelopes.
The horns of the Ruminants are frequently a secondary sexual
character; this is especially the case with the Deer. The Reindeer is,
however, an exception, both the stags and the does having horns. That
they are associated with the reproductive function is shown by their
being shed after the period of rut, the destruction of the velvet at that
period, and also by the effect upon the horns which any injury to the
reproductive glands produces. Some useful facts upon this latter head
have been amassed by Dr. G. H. Fowler,[116] who noticed in a series of stags,
horns showing various degrees of degeneration in the antlers produced by
varying degrees and periods of gelding. From the facts {202}here collected
it is clear that a direct effect is produced. If we are to regard horns
as secondary sexual appendages which have been subsequently handed on to
the female by heredity, we should expect to meet with examples of animals
now horned in both sexes, of which the earlier representatives had the
horns confined to one sex. This is most interestingly shown by the
extinct and Miocene Giraffe, Samotherium, of which the male alone
had a pair of short horns, while the skull of the female was entirely
hornless; the modern Giraffa, as is well known, has horns in both
sexes.
It is interesting to note that the existing Perissodactyles and
Artiodactyles are to be distinguished by their unpaired or paired horns.
But while there are no Artiodactyles with unpaired horns (save occasional
sports) the Perissodactyles have more than once tried, so to speak,
paired horns, which ultimately proved fatal to them. The Rhinoceros
Diceratherium apparently inherited and improved upon the small
paired horns of Aceratherium, but it has left no descendant. The
paired horned Titanotheria offer another instance of the same apparent
incompatibility between the Perissodactyle structure and the persistence
of paired horns.
Sub-Order 1. CONDYLARTHRA.
This group is characterised by the following assemblage of characters.
Extinct, often plantigrade Ungulates, with five-toed limbs. Bones of
carpus and tarsus not always interlocking, but sometimes lying above each
other in corresponding positions. The humerus has an entepicondylar
foramen. Dental formula quite complete; the molars brachyodont and
bunodont. The premolars are simpler than the molars. The canines are
small. As with other early types, the zygapophyses are flat and do not
interlock. The astragalus is like that of the Creodonta. This group was
American and European in range, the remains of its rather numerous genera
being of Eocene time. The best-known genus is Phenacodus, of which
some account will be given before discussing the, in many cases, more
fragmentary remains of other allied forms.
The genus Phenacodus was first described so long ago as 1872,
from a few scattered teeth. Since then several nearly complete skeletons
have been obtained, and we are in full possession of {203}the details of
its osteology. It was not a large creature (see Fig. 110, p. 196), about 6 feet in length, with a small head. The
feet were more or less plantigrade, and five-toed. The last phalanges of
the toes show that they carried hoofs and not claws; yet the fore-feet
look a little as if they could be used as grasping organs. The third
digit of both hind- and fore-feet exceeds the others, and thus a
Perissodactyle-like foot characterised this Eocene creature. The tail is
exceedingly long, and must have reached the ground as the animal walked.
This is of course by no means an Ungulate character. Still, in the
totality of its organisation the animal was decidedly Ungulate, though
Professor Cope spoke of Phenacodus as not merely an ancestral
Ungulate but as the parent form of Insectivores, Carnivores, Lemurs,
Monkeys, and Man himself! The scapula indeed is from its breadth and oval
contour rather like that of a Carnivore. The clavicles as in other
Ungulates are absent. The femur is Perissodactyle rather than
Artiodactyle in the presence of a third trochanter. The creature had
fifteen pairs of ribs and five or six lumbar vertebrae. The two bones of
the leg which lie below the femur are perfectly distinct and separate. A
cast of the brain-case shows that the cerebral hemispheres were smooth
and small, the cerebellum of course completely uncovered and nearly as
large as the cerebrum. The olfactory lobes were also large. The complete
skeleton of Phenacodus has lately been excavated more fully from
the enveloping matrix by Professor Osborn,[117] and mounted in what is regarded as
the natural position of the beast. It appears that though five-toed it
went upon the three middle toes only, and furthermore that of these the
middle one was the more prevailing, so that Phenacodus was
distinctly "Perissodactyle," at least in habit. Moreover its "long
hind-quarters, the long powerful tail ... are reminiscent of Creodont
ancestry." The genus was European and American in range.
Meniscotherium ( = Hyracops[118]) comprises several forms of about the
size of a fox; they are both European and American in range. The teeth
are more distinctly Ungulate in form than those of Phenacodus,
with a  -shaped outer wall. The skull is described as possessing "indifferent,
primitive characters," permitting a comparison with those of Opossums,
Insectivores, and {204}Creodonta. It has, as in
Phenacodus, no orbital ring. The humerus resembles that of a
Carnivore rather than that of an Ungulate. The carpus and tarsus are
serial. The fibula articulates with both the calcaneum and the
astragalus, which is not the case with Phenacodus. It is suggested
that these animals are ancestral forms of the Chalicotheres. In the brain
the hemispheres do not cover the cerebellum.
-shaped outer wall. The skull is described as possessing "indifferent,
primitive characters," permitting a comparison with those of Opossums,
Insectivores, and {204}Creodonta. It has, as in
Phenacodus, no orbital ring. The humerus resembles that of a
Carnivore rather than that of an Ungulate. The carpus and tarsus are
serial. The fibula articulates with both the calcaneum and the
astragalus, which is not the case with Phenacodus. It is suggested
that these animals are ancestral forms of the Chalicotheres. In the brain
the hemispheres do not cover the cerebellum.
More primitive apparently than Phenacodus was the less-known
genus Euprotogonia, or Protogonia[119] as it has been called. The best-known
species is E. puercensis, so called from its occurrence in the
Puerco beds of the American Eocene. It was a slender, long-limbed
creature, smaller than Phenacodus, with a long and heavy tail as
in that animal. Like Phenacodus it was semiplantigrade, and shows
more likenesses to the Creodonta. The skull is only known by a part of
the lower jaw with teeth, and by the teeth of the upper jaw. The
vertebrae are not entirely preserved, but enough remain to show that the
animal had a tail of 16 or 17 inches, which is a considerable length when
compared to its height, about a foot at the rump. In the fore-limb the
most noteworthy point is that the ulna has a convex posterior border as
in the Creodonts, the same border in Phenacodus being concave. The
humerus is slender, with less-marked tuberosities. The fifth digit seems
to have been less reduced. The phalanges seem to have borne horny sheaths
somewhat intermediate between hoofs and claws. The pelvis is described as
being, as is also that of Phenacodus, rather like that of the
Creodonta. The right hind-limb is known in all its details. It appears
that the bones are not serial but interlocking; this, however, on the
views with regard to the relations of these two forms of tarsus mentioned
on p. 198, does not militate against regarding
Euprotogonia as the ancestor of the genus Phenacodus. The
third toe is the pre-eminent one, the animal thus being Perissodactyle.
The lateral digits are larger than in Phenacodus, and the
metatarsals and the phalanges are slightly curved, which is again a
Creodont character as compared to the perfectly straight corresponding
bones of Phenacodus. It seems evident that this animal is to be
looked upon as a more ancient type than Phenacodus, even if not as
its actual ancestor.
Another group of the Condylarthra contains the genus
Pertipychus and some others. Periptychus has the full
dentition {205}of forty-four teeth, the molars being of
course bunodont, with the three chief tubercles most developed. The bones
of the tarsus interlock and are not serial, as they are in many other
members of the Condylarthra. The astragalus has a shorter neck than in
Meniscotherium, for example. It has in this a likeness to the same
bone in the Amblypoda, to the primitive members of which, such as
Pantolambda, this animal bears much resemblance. "Astragali and
many skeletal bones of Periptychus rhabdodon and Pantolambda
bathmodon are almost indistinguishable," observes Mr. Matthew. The
fore-feet of this genus are unknown, but it would seem that it was
plantigrade from the evidence of the hind-feet. There are several species
of the genus.
Possibly, but not at all certainly, the Mioclaenidae, with the genera
Mioclaenus and Protoselene, are to be referred to this same
order of primitive Ungulates. It is only necessary to mention them here,
because they show very clearly the primitive form of dentition of these
early Eocene mammals. The teeth are quite complete and unbroken by a
diastema. The canines are but little pronounced. The molars are not
strictly tritubercular, but have a prevailing trituberculy. The nature of
the feet is not known. Since the genus Protoselene, as its name
denotes, shows an indication of a commencing selenodonty, it has been
suggested that this group is the stock whence the Artiodactyles have been
derived.
In any case, whether the particular comparisons that have been made as
to the relationship of various forms of Condylarthra are valid or not, it
seems to be plain that this group represents the earliest Ungulate stock,
but little differentiated from the contemporaneous Creodonts.
Sub-Order 2. AMBLYPODA.
This group of extinct mammals has the following principal
characteristics:—
They are large, semiplantigrade Ungulates, of heavy build and
apparently elephantine gait. The dentition is for the most part complete
as in other ancient groups, and the canines are in the later forms big
tusks. The back teeth are brachyodont and ridged (lophodont). Both radius
and ulna in the fore-limb, and tibia and fibula in the hind-limb, are
well developed. The bones {206}in the carpus are alternating in position.
The toes are five in both feet, and are very short. There is a hint of
commencing "perissodactylism" in the fore-feet at any rate. The brain is
small and the hemispheres smooth.
The Amblypoda, or Amblydactyla, are so called on account of their
short and stumpy feet and toes. They were held by Professor Cope to be on
the direct line of ancestry of both Perissodactyles and Artiodactyles, a
view which is on the whole not accepted at present.
Fig. 114.—Skull of Protolambda bathmodon. × ¾. e.a.m, External auditory meatus;
m, mastoid; m.f, mastoid foramen. (After Osborn.)
As is the case with other groups, the Amblypoda commenced existence as
a sub-order with relatively small forms such as Pantolambda, the
most ancient type known, which is in many respects a transition between
the later forms and other groups of mammals such as the Creodonta.[120] The race culminated
and ended in the giant Dinoceras and Coryphodon, and spread
into the Old World. In spite of their smooth and diminutive brain, these
mammals were able to hold their own and to multiply into many species and
genera; in this they were perhaps aided by their formidable tusks and by
the horns which many of them possessed. The teeth seem to imply an
omnivorous diet, which was quite possibly an additional advantage in the
struggle for existence. It does not seem to be necessary to divide off
the Dinoceratidae into a sub-order equivalent to the Coryphodontidae as
was done {207}by Professor Marsh; the numerous points in
common possessed by the members of both families forbid their separation
more widely than as families.
The earliest types of Amblypoda belong to the genus
Pantolambda, of which the species P. bathmodon was about
four feet in length. As restored it seems to have had proportionately
short fore- and hind-limbs, and it had a long tail. It was apparently
plantigrade, and would have had not a little likeness to a carnivorous
type. The skull has no air cavities, such as are developed in the later
types from the Lower Eocene, e.g. Coryphodon; Pantolambda
is from the basal Eocene. The frontal bones show no trace of the horns
that are developed in subsequent forms; the nasals are comparatively
long; the zygomatic arch is slender. The molar teeth are in the primitive
form of trituberculy, and the premolars, as is so often the case with
primitive animals, are unlike the molars in form, being less markedly
selenodont. As to the vertebral column, the dorsal vertebrae appear to
have had short spines, which argues, as it does also in the case of the
larger and heavier Coryphodon, a feebleness in the development of
ligaments and muscles supporting and moving the head. The scapula seems
to have the same peculiar leaf-like form that it has in the later
Coryphodon.[121]
This primitive type shows an entepicondylar foramen in the humerus. It is
interesting to observe that the posterior border of the ulna is convex,
as in the Creodonts, and in the early Condylarthrous form
Euprotogonia. In the subsequently-developed Amblypoda, as in the
later Condylarthra, that bone acquires a concave outer border. In the
carpus the os centrale is distinct. In the femur the third trochanter is
well formed; it gradually dies out in later Amblypoda. The fibula
articulates with the calcaneum. This species, according to Osborn,
"typifies the hypothetical Protungulate, being more primitive than either
Euprotogonia or Phenacodus."[122]

Fig. 115.—Skeleton of Coryphodon radians. × 1⁄10. (After Osborn.)
The genus Coryphodon is known by a large number of species, of
which the first was discovered in this country, and was represented
merely by a jaw with some teeth. This was named by Sir R. Owen C.
eocaenus, and was dredged up from the bottom of the sea off the Essex
coast. A second specimen consisted of a single {209}canine tooth only, and
was brought up from a depth of 160 feet during the making of a well at
Camberwell. More abundant remains have since been found in North
America.
This genus had a large head, and in some specimens traces of the "horn
cores," so marked in the related Dinoceras, are to be noticed. The
skull is broad behind and narrowed in front; the roofing bones show the
cellular spaces so characteristic of the Elephant. The jugal bone,
however, is not, as it is in the Elephant, placed in the middle of the
somewhat massive zygomatic arch. As in some other primitive Ungulates
(e.g. Phenacodus) there are twenty dorso-lumbar vertebrae, of
which fifteen bore ribs.
The scapula seems to have possessed a peculiar leaf-like form,
swelling in the middle and ending almost in a point above. It has a
well-marked spine, and the acromion projects much. The fore- as well as
the hind-feet are in a state of transition between plantigradism and
digitigradism. It was at one time held that the animal was digitigrade as
to the fore-feet and plantigrade as to the hind-feet. Though, as has been
pointed out, it is a fact that the hind-feet are often on a different
plane of evolution from the fore-feet, it seems that this amount of
difference does not characterise any Ungulate, not excepting the genus
now under consideration.
The toes are very spreading. The pelvic girdle is of great strength
and broadness. The femur, as in the Perissodactyles, has a well-developed
third trochanter; but whereas in this particular the hind-limb is
Perissodactyle, it is Artiodactyle in the fact that the tibia and the
fibula articulate with the astragalus and calcaneum. The ridged teeth
have given the name to the genus.
A curious feature in the structure of the genus are the slender spines
of the dorsal vertebrae, which contrast with the enormous ones of some
other Ungulates—more curious in this genus, which is of heavy
build, than in the lighter Pantolambda. The back of the animal is
short, and the limbs are very spreading, so that the gait was doubtless
shuffling. The large head, and short and heavy limbs and limb girdles
added probably to its cumbrous walk or trot. The canines are great tusks,
and spread out on both sides of the mouth.[123]
The late Professor Cope, in 1874, described the probable appearance of
the Coryphodon in the following words:—"The general appearance of the Coryphodons, as
determined by the skeleton {210}probably resembled the bears more than any
living animals, with the important exception that in their feet they were
much like the elephant. To the general proportions of the bears must be
added the tail of medium length. Whether they were covered with hair or
not is of course uncertain. Of their nearest living allies, the
elephants, some were hairy and others naked.... The movements of the
Coryphodons doubtless resembled those of the elephant in its shuffling
and ambling gait, and may have been even more awkward from the
inflexibility of the ankle."
The most recent members of this sub-order come from the Middle Eocene
beds, and are chiefly referable to the genus Dinoceras, with which
Tinoceras and Uintatherium are at least very nearly
related, if not identical. These creatures were of great size, larger
than the earlier types which have been considered. They show a certain
superficial resemblance to the Titanotheriidae, on account of the massive
horn cores upon the skull. These horn cores are large upon the maxillae
and the parietals, and are paired; on the nasals are smaller horns. The
bones of the skull have air cavities. The incisors of the upper jaw are
absent; the canines are enormous tusks, and the lower jaws are flanged
downwards near the symphysis where these tusks border them. Contrary to
what is found in the older types, where the position of the condyle of
the lower jaw is normal, this prominence faces backwards in the
Dinocerata. The same shortness of the spines of the dorsal vertebrae
prevails in this group as in the other Amblypoda, though it is perhaps
hardly so marked. The scapula has not the peculiar acuminate form that
exists in Coryphodon, but is triangular and broad above. The limbs
are elephantine, in that the angle between the humerus and the femur
respectively, and the bones which follow, is not marked. The hind-limbs
are especially straight. The tail is short as compared with that of the
primitive Amblypoda. The Dinocerata are purely digitigrade. The
entepicondylar foramen has, as in the Coryphodonts, disappeared. The os
centrale of the carpus has become fused, and no longer exists as a
separate bone. The fibula no longer articulates with the calcaneum, but
both that bone and the ulna are well developed. The genus
Astrapotherium is placed among the Amblypoda by some
authorities.[124]
{211}
Sub-Order 3. ANCYLOPODA.
The history of the discovery of the members of this order is very
instructive as illustrating the dangers of laying too much classificatory
importance upon detached fragments of animals. So long ago as 1825
terminal phalanges of a new creature were found in the Miocene of
Eppelsheim, and sent to Cuvier. Cuvier named them "Pangolin gigantesque,"
deeming them, on account of their general form and cleft terminations, to
pertain to the Edentata. In the same bed some seven years later were
found certain teeth clearly of an Ungulate character, to which the
generic name of Chalicotherium was applied. It was subsequently
discovered that the teeth and the claws belonged to the same animal, and,
later, further remains turned up which disclosed a creature having the
anomalous composition of an Ungulate with decisively Ungulate teeth, but
with the feet to a large extent like those of an unguiculate animal. The
same confusion of characters occurs also, it will be remembered, in the
distinctly Artiodactyle Agriochoerus (see p. 331). Indeed the feet of the latter when first
discovered were erroneously, as it now appears, referred to the present
order of Ungulates under the name of Artionyx. It is probable that
the genus Moropus of North America is a member of this group, and
that it is probably congeneric with a somewhat different type of
Ancylopod known as Macrotherium. It is also clear that
Anisodon, Schizotherium, and Ancylotherium, if not
congeneric with either of the two recognised genera, are at least very
close to them.
Chalicotherium has a skull which recalls that of some of the
earlier Ungulates; it has, however, no incisors at all, and no canines in
the upper jaw; this feature has led to the belief that the animal is
related to the Edentata, and that it is in fact a link between them and
the Ungulata. The molars, like those of the Perissodactyla, are of the
buno-selenodont type. It also agrees with that group (to which it has
been approximated by several writers) in the. tridactyl manus and pes,
and in the characters of the tarsus. But although tridactyl, the axis of
the limb passes through the fourth digit. Chalicotherium is not
mesaxonic, as are the Perissodactyles. Moreover, it has no third {212}trochanter, and the unguiculate claws have
already been referred to. As to the latter, which are short, it is not
the end phalanx but the first which is retracted; thus
Chalicotherium differs markedly from both Carnivorous and Edentate
types; for in the former it is the last phalanx which is retracted, while
in the Edentates the same phalanx is flexed downwards. The limbs of
Chalicotherium are nearly of the same size, and the animal seems
to have been stout and quadrupedal.[125]
Macrotherium, like the last genus, seems to have been common to
both New and Old Worlds. It is to be distinguished by a number of
characters. It is supposed to have been "semi-arboreal and fossorial";
the fore-limbs are much longer than the hind, the relative proportions of
the radius and tibia being 70 to 29. The ulna was distinct from the
radius, whereas in Chalicotherium the two are coalesced, or nearly
so. Young specimens appear to possess a full set of incisors; whether
this is the case or not with Chalicotherium is not known.[126]
Homalodontotherium is sometimes placed in the group.
Sub-Order 4. TYPOTHERIA.
It is a little difficult to be confident that the Typotheria are
rightly referred to the Ungulata, since they contradict two important
Ungulate rules. They have clavicles, which are elsewhere missing, and the
thumb looks as if it were opposable.[127] An Ungulate is essentially a running
animal, and has no need of a grasping finger. Still Typotheria are placed
by most within the Ungulate series, though their undoubted likenesses to
other groups, especially to the Rodentia, are admitted, and indeed
emphasised. Cope places them definitely with the Toxodonts.
The Typotheria are an extinct group of smallish beasts, confined, like
the Toxodontia, to South America, a region which during the Tertiary
period, and into the Pleistocene, abounded with strange and varied types
of Ungulate animals.
The earlier forms of Typotheria may be exemplified by some {213}account of
the genus Protypotherium. This animal was of about the size of a
Hyrax, which indeed it resembles in several points of structure.
The teeth have the primitive number of forty-four, and they are close
set, leaving no diastema; the molars are rootless and grow persistently;
they are simple and Rodent-like in surface pattern. The shape of the
lower jaw is like that of Hyrax, being rounded in outline
posteriorly; there is no projecting angle as in the Rodents, and this
remark applies to the Typotheria in general. The aspect of the Rodent
lower jaw is characteristically different from that of Hyrax and
the forms under consideration.
Some other characters of these early forms of Typotheria can be
gathered from an inspection of other genera. In Icochilus both
hand and foot were five-toed, and, as in ancient Ungulates generally, the
bones of the wrist and of the ankle are serially and not alternately
arranged. Moreover, an os centrale is present in the carpus. Both thumb
and big toe were opposable. The skull has a remarkably Rodent-like
appearance, but the palate is not so narrowed as in these animals.
In the more recent forms of Typotheria the dentition has become
reduced. The canines are lost, and as the incisors are reduced also, to
one on each side of the upper, and two on each side of the lower jaw, the
likeness to a Rodent skull is increased. There is also evidence of a
modification from the more primitive forms in the loss of one premolar or
even more, in the alternating bones of the carpus, in the disappearance
of the centrale, and in the loss of a toe upon the hind-foot. In these
more recent forms the fibula articulates with the astragalus instead of
with the calcaneum. Typotheria of these more recent forms may be
illustrated by the typical genus Typotherium. It has the reduced
dental formula I 1/2 C 0/0 Pm 2/1 M 3/3; the molars are simple in
pattern, and much like those of Toxodon. The upper incisors are
powerful and curved, but are surrounded by a layer of enamel, which is
not limited to the anterior face, as it practically is in Rodents. The
sacrum is composed of a large number of vertebrae—some
seven—a state of affairs which recalls the Edentata. The shoulder
blade is not Ungulate in form. It has a strong spine, with an acromion
and a well-developed metacromion. The terminal phalanges are enlarged and
hoof-like.
In the genus Pachyrucos there are three premolars, otherwise
{214}the formula is the same as in
Typotherium. The animal seems to have had nails rather than hoofs.
The thumb was opposable. The fibula is fused below with the tibia,
whereas in the last genus these two bones are quite separate from each
other.
Sub-Order 5. TOXODONTIA
The group Toxodontia,[128] like so many others, is exceedingly
hard to define. Nor are its limits any easier to mark out than many
others of the groups of Ungulates. It will be best perhaps to give an
account of Toxodon, and of a few types which seem to lie near it
in the system, and then to indicate how far they resemble or depart in
structure from other Ungulates. Toxodon itself is known from
complete skeletons. It lived in Argentina during the "Pampean" period,
which seems to be of the Pleistocene age. A large number of species,
however, have been described, some of which seem to go farther back in
time, and to have existed during the Miocene period further south in
Patagonia.
The size of this creature was about that of a large Rhinoceros; it has
a bulky body and a large head, which was borne low down, on account of
the bending downwards of the anterior vertebrae; in this aspect the
figures of the skeletons recall Glyptodon and similar Edentates.
The beast was discovered by Darwin, and originally described by Owen.
"During his (Mr. Darwin's) sojourn in Banda Oriental," writes the Rev. H.
Hutchinson, "having heard of some 'giants' bones' at a farmhouse on the
Sarandis, a small stream entering the Rio Negro, he rode there, and
purchased for the sum of eighteenpence the skull which has been described
by Sir R. Owen. The people at the farm-house told Mr. Darwin that the
remains were exposed by a flood having washed down part of a bank of
earth. When found, the head was quite perfect, but the boys knocked the
teeth out with stones, and then set up the head as a mark to throw at."
The whole of the Pampean area is a valley of dry bones, and the remains
of Toxodon are abundant there. The skull of Toxodon is not
unlike that of a horse in general aspect; but the orbit is not separated
from the temporal fossa. The premaxillae are furnished above with a
slight protuberance directed towards {215}the free end of the
nasals, which may be related to the presence of a short proboscis. The
zygomatic arch is strong and broad: the mandibles are provided with a
long symphysis. The dental formula is I 2/3 C (0-1)/1 Pm 4/(3-4) M 3/3.
The teeth are prismatic and hypselodont, growing from persistent pulps.
The molar teeth are slightly arched in form, whence the name of
Toxodon, "bow teeth." The strong chisel-shaped incisors suggest
the Rodents and Hyrax. The cheek teeth, moreover, are by no means
unlike those of Rodents in their pattern. They are at any rate not at all
like those of existing Ungulates. The small size of the canine and of the
first premolar produces a diastema in the tooth series. The sacrum
consists of five vertebrae, and the ischium does not articulate with
it.
The shoulder blade has a strong spine, but only a rudimentary
acromion; nor is the coracoid well developed. The radius crosses the
ulna, as in the Elephant; the whole fore-limb is shorter than the
hind-limb, which must have exaggerated the hang-dog expression of the
creature when alive. The elements of the carpus interlock in the modern
fashion. Those of the tarsus, however, are primitive in lying below each
other without alternation. The carpus has a centrale. The fibula
articulates with the calcaneum. The femur has no third trochanter. There
are three toes to all the limbs. It is clear that this assemblage of
characters will not allow the placing of Toxodon in any living
Ungulate order. If the middle toes appear by their slight pre-eminence to
approach the Perissodactyle form, the peculiar surface contour of the
molar teeth, letting alone the absence of a third trochanter on the
femur, will not permit this classification.
Allied to Toxodon is the genus Nesodon. It was so named
from an "island lobe" on the inner side of the upper molars. This
creature, smaller than Toxodon, also differs from it in the fact
that the dentition is complete, and in the pattern of the molars, which
is rather more complex. There is still the slight projection upon the
premaxillary bones, but the nostril is directed more forwards than in
Toxodon. The zygoma, too, is massive. The second pair of incisors
in the upper jaw and the outer (third) pair in the lower jaw form biggish
tusks in the adult. These and the molar teeth are, however, finally
rooted, and do not grow, as in Toxodon, from persistent pulps. The
genus is from the older Tertiary of Patagonia. Five or six species have
been described. Some are as large as a Rhinoceros, others as small as a
Sheep. {216}
There is no doubt about the close alliance of the two genera just
referred to. It is more doubtful whether Homalodontotherium and
its allies should be placed, as they often are, in the neighbourhood of
the Toxodonts. Homalodontotherium owes its name to its even row of
teeth without a diastema. It was a creature of equally large size with
Toxodon, and also came from the Tertiary strata of Patagonia. The
teeth are the typical forty-four, and the molars like those of a
Rhinoceros; they are, however, brachyodont and not hypselodont as in
Toxodon. This genus, however, shows an important difference from
the Rhinocerotidae and from the other Toxodontia in the fact that it was
five-toed, and that the bones of the carpus and tarsus are set in
relation to each other in the linear serial fashion.
Undoubtedly a near relative of Homalodontotherium is
Astrapotherium. This creature was of equal bulk, and was also
Patagonian in range. The teeth are reduced in number, but the animal was
provided, like a Wild Boar, with great tusks, which were, however, formed
by the incisors. This animal is very imperfectly known; it is the form of
the molars and the large size of the incisors which have led to its
association with the Toxodontia. As to the resemblance of the teeth of
this genus and of Homalodontotherium to those of
Rhinoceros, it is difficult to regard it as evidence of near
affinity. The likeness is probably to be looked upon as a case of
parallelism in development. Exactly the same explanation is possibly to
be given to the likeness which the teeth of Toxodon and
Nesodon show to Rodents, or even to Edentates. As to their
affinities Zittel observes:—
"The entirety of their osteological characters argues for the Toxodon
a separate position in the neighbourhood of the Perissodactyla,
Proboscidea, Typotheria, and Hyracoidea. The relations to the Rodentia
rest mainly upon the converging development of the teeth, not upon true
relationship."
Sub-Order 6. PROBOSCIDEA.
Large vegetable-feeding animals, usually scantily covered with hair,
and with the nostrils and upper lip drawn out into a long proboscis.
Digits five on both limbs. Femur and humerus not bent upon lower leg and
fore-arm in a position of rest. Skull {217}with abundant air
cavities in the roofing and other bones. The incisors are developed into
long tusks, which exist in the upper jaw alone, in the lower jaw alone,
or in both jaws. There are no canines. The molars are lophodont. The
clavicle is absent. The femur has no third trochanter. The bones of the
carpus are serially arranged and do not interlock. The stomach is simple.
The brain has much convoluted cerebral hemispheres, but the cerebellum is
completely uncovered by them. The intestine is provided with a wide
caecum. The testes are abdominal. The teats are pectoral in position. The
placenta is non-deciduate and zonary. There are two venae cavae
superiores.
The position of the limbs in the Elephant tribe is unique among living
animals: their straightness that is to say, and the absence or very
slight development of angulation at the joints of the limb bones. This
same feature has been observed in the extinct Dinocerata and in the
Titanotheria. It must not, however, be assumed from the resemblance to
these ancient forms that there is much affinity between them and the
Proboscidea, or that the latter have retained an ancient feature of
organisation. The oldest Ungulates for the most part, and the Creodonts
to which they are undoubtedly related, have much bent limbs. It must be
considered, therefore, that the arrangement obtaining in the Elephants is
purely secondary. Professor Osborn has put forward the reasonable view[129] that the vertical
limbs of all these colossal creatures are due to "an adaptation designed
to transmit the increasing weight" of these animals. The huge bulk of the
body is better borne by vertical pillars than by an angulated limb. Other
points, however, such as the exposure of the cerebellum, the two venae
cavae, the five digits, and the absence of a third trochanter, argue a
low position for the Proboscidea in the Eutherian group.
The group can be readily divided into two families, the Elephantidae
and the Dinotheriidae. We will commence with the former.
The Elephants proper, Elephantidae, differ from the
Dinotheriidae in, and are characterised by, a number of anatomical
features. They possess long tusks (incisors) either in both jaws, or, if
only in one jaw, in the upper. The molar teeth are very large—so
large that only a few of them are simultaneously in use. There are but
three definable genera of Elephantidae, of which {218}Elephas alone
survives. This genus also includes many extinct forms, both American and
European, as well as Asiatic and African. The entirely extinct genera are
Stegodon and Mastodon. The group is clearly one dwindling
towards extermination. From the Middle Miocene downwards these great
"pachyderms" have existed; and from the Miocene up to Pleistocene times
they were almost world-wide in range and numerous in species.
The genus Elephas comprises usually large, but occasionally
(the pygmy Elephant of Malta) quite small forms. The external features of
the genus differ slightly in different species, and will therefore be
described in relation to those species which we shall notice here. The
vertebral formula is C 7, D 19-20, L 3-5, Sa 4-5, Ca 24-30, or even
more.
The bodies of the vertebrae are remarkable for their shortness and for
the very flattened articular surfaces.
Fig. 116.—A section of the cranium of a full-grown African Elephant, taken to the left
of the middle line, and including the vomer (Vo) and the mesethmoid (ME); an,
anterior, and pn, posterior narial aperture. × 1⁄12. (From Flower's Osteology.)
The skull is large and massive. Its large and heavy character is, as
has been stated in the definition of the sub-order, due to the immense
development of air cavities in the diploe; the diameter of the wall of
the skull is actually greater than that of the cranial cavity. These
cavities are not obvious in the young animal. They are most conspicuous
in the roofing bones of the skull, but are seen elsewhere, and thicken
the basis cranii, {219}the maxillae, and so forth. This state of
affairs, together with the presence of the huge tusks, has, as it were,
pushed back the nasal orifices to near the top of the skull in a very
Whale-like fashion. As in the Cetacea, the nasal bones are limited in
size, and the premaxillae send up processes to join the frontals and the
nasals. There is a straight and somewhat slender zygomatic arch, but the
orbit is not separated from the temporal fossa. The malar bone is small,
and, as in Rodents, forms the middle part of the zygoma. This is not the
case with most Ungulata. The symphysis of the mandibles forms a
spout-like rim. The scapula has a narrow prescapular, but a very wide
postscapular region. The spine has a strong process projecting backwards
from near its middle; this is a point of likeness to certain Rodents. No
Elephant has a clavicle. The most remarkable feature about the fore-limb
is the separation and crossing of the radius and ulna. The arms of these
animals are permanently fixed in the position of pronation. The foot is
short, and the bones of the carpus are serially arranged. There are,
however, traces of a commencing interlocking of these bones in many
forms. The hind-feet are somewhat smaller than the fore-feet, and the
tibia and fibula are both developed.
As to the teeth, this genus is to be distinguished from allied forms
by the presence of tusks in the upper jaw only. These tusks have no bands
of enamel such as characterise those of Mastodon. They are
incisors. There is, however, a trace of the former enamelling in the
shape of a patch at the tip, which soon wears away. The molar teeth of
Elephas are so large that the jaws cannot accommodate more than at
the most two and a part of a third at a time. These are gradually
replaced by others to the number of three, the replacement of teeth
suggesting that of the Manatee. Each molar is deeply ridged, the
interstices between the ridges being filled up with cement. As the tooth
wears away, therefore, the surface continues to be flat. Each ridge
consists of a core of dentine surrounded by a coat of enamel. The number
of these ridges varies greatly from species to species. The Indian
Elephant is one of those which have the greatest number of plates in a
single tooth, as many as twenty-seven.[130] Of the six molars which {220}eventually
appear, the first three are considered to correspond to premolars. But
successional teeth are rare in the genus; that is to say as far as
concerns the molars, for the tusks have their milk forerunners. As to the
molars it is apparently only E. planifrons which certainly shows a
milk dentition. In Mastodon and older types a milk dentition is
commoner.
The viscera of the Elephant have been examined by many zoologists. The
latest paper, dealing chiefly with the African species, but containing
facts about its Indian congener also, is quoted below.[131] The Elephant is remarkable in
possessing, in addition to the three usual pairs of salivary glands
present in mammals, a fourth, situated in the molar region, and opening
on to the cheek by many pores. This gland is especially well developed in
Rodents. There is a gland which may be mentioned in this connexion,
though it opens externally between the eye and ear, known as the temporal
gland; its use does not seem clear. The thoracic cavity of the Elephant,
as may be inferred from the large number of ribs, is very large as
compared with the abdominal.
The stomach is simple in form, and the epithelium of the oesophagus
does not extend into it as is the case with the Horse and Rhinoceros. A
gland or a collection of smaller glands occurs in the stomach, and
recalls the "cardiac gland" of the Wombat and the Beaver, also that of
the Giraffe. The large intestine is long, rather more than half the
length of the small intestine. The caecum is well developed in these
animals. The liver has a very simple form, being but slightly lobulated.
It is actually only bilobed, but it is important to notice that this
division does not correspond to the two halves of the liver. As shown by
the attachment of the suspensory ligament, one half consists of the left
lateral lobe alone, the other half embracing the remaining primary lobes.
The simplicity of the liver looks like an archaic character. No Elephant
has a gall-bladder. The lungs again are simple in form through their
slight lobulation. Each half in fact is without subdivisions, and is of a
triangular form. In this the Elephants resemble the Whales, as in the
simple liver. In both cases probably the likeness is due to the
permanence of primitive features of organisation. The brain[132] of the Elephant {221}has
hemispheres which are extremely well convoluted; but they leave the
cerebellum entirely uncovered. This suggests a brain which is a great
specialisation of a low type. The brain has been particularly compared
with that of the Carnivora, with which group the Elephants agree in the
characters of the placenta. It is, however, always a matter of the very
greatest difficulty to compare the brains of mammals belonging to
different orders.
There are but two living species of Elephant, of both of which we
shall now proceed to give some account. Only a few of the rather numerous
fossil forms can be touched upon here.
The African Elephant, E. africanus, has been sometimes referred
to a distinct genus or sub-genus, Loxodon, by reason of the
lozenge-shaped areas on the worn grinding-teeth. It lives, as its name
denotes, in Africa. This species has a number of external features which
enable it to be distinguished from the Oriental Elephant. The head slopes
back more, and has not the two rounded bosses which give so wise a
countenance to the Indian species. The ears are very much larger. The tip
of the trunk has a slight triangular projection on both the lower and the
upper part of the circumference of the aperture. There are four nails on
the fore-feet and three on the hind. As in the Indian form, the toes are
all bound together, and do not appear for any part as free digits. A
thick pad of fat, etc., makes the animal when alive look as if
plantigrade, whereas it is, as a matter of fact, digitigrade. In internal
features the most prominent difference from E. indicus is in the
molar teeth, which are ridged by much fewer ridges. The outside number
for a single tooth in the present species is 10 or 11. In Elephas
indicus on the other hand there are as many as 27.
The African Elephant, thinks Sir Samuel Baker, reaches a height of
about 12 feet, and it will be remembered that the notorious "Jumbo" was
found to be 11 feet high at the shoulder. The tusks are found in both
sexes, as in the Indian beast, but are relatively larger in the female in
the species now under consideration. It is also a rather more active
creature, and is more savage;[133] however it can be tamed, as is shown
by several {222}specimens which have been and are in the
possession of the Zoological Society, and other proprietors. It was
apparently used in the past. Certain Carthaginian coins are stamped with
a figure of the African Elephant; but in Africa no attempts are now made
to utilise this creature except for food and ivory.
Fig. 117.—African Elephant. Elephas africanus. × 1⁄56. (After Sir Samuel Baker.)
The meaning of an Elephant as an emblem. upon a coin appears to be
eternity, and there is no question but that the {223}Elephant is a
long-lived animal. It is said that it hardly reaches proper maturity
before forty, and that 150 years is not beyond probability in the way of
longevity. Even longer periods have been assigned to it.
The tusks of the Elephant are by no means necessarily sexual
adornments, used for fighting purposes only. The African Elephant is a
most "industrious digger," and grubs up innumerable roots as food. It
appears to be a fact that during these operations the right tusk is
mainly used, and in consequence that tusk is shorter as well as thinner
than the other. Two average tusks would weigh respectively 75 and 65
lbs., the latter of course being the weight of the more worn right tusk.
These weights, it should be observed, by no means indicate the limits to
which finely-developed tusks can attain. The very heaviest tusk known to
Sir Samuel Baker[134]
weighed 188 lbs. This was sold at an ivory sale in London in the year
1874. The pace of the African Elephant, says the same authority, is at
most at the rate of fifteen miles an hour at first, and of course in a
furious rush. This pace cannot be kept up for more than two or three
hundred yards, after which ten miles an hour is a better approximation to
the rate which can be kept up for long distances.
The Indian Elephant, Elephas indicus (or Euelephas
indicus, if the genus Loxodon is to be accepted), is better
known and has been longer known than the African. It occurs in India and
Ceylon, and in some of the Malayan islands, the Elephants of which latter
parts of the world have been regarded as a distinct form, an apparently
unnecessary procedure.
Fig. 118.—Indian Elephant. Elephas indicus. × 1⁄54. (After Sir Samuel Baker.)
This species does not stand so high at the shoulder as the African;
its back is more rounded in the middle. The trunk has but one pointed
tip; there are five nails on the fore- and four on the hind-feet. As this
species comes from India and the East, it has been longer as well as
better known than the African form. Thus many of the stories and legends
that have congregated round Elephants apply really to this form. As is
well known, the Indian Elephant is much used as a beast of burden, and
for other purposes where its huge strength renders it invaluable. But its
great drawback as a servant of man is its great independability. On the
one hand we have furious, vicious, and generally unreliable {224}Elephants, and
on the other perfectly docile creatures, who obey the slightest hint from
their driver. Huge though the Elephant is, it is frequently a timid
beast. Sir Samuel Baker relates how one which he was riding fairly bolted
at the sight of a Hare. To {225}be bolted with by an Elephant is far from
pleasing, though a rather exciting event. It makes for the nearest jungle
at once, being, much more than the African species, an inhabitant of
forest. And in rushing through the dense undergrowth, the occupiers of
the Elephant's back are apt to be swept off or cut to pieces by
innumerable thorns.
Elephants, no doubt of the Indian species, were used by the Persians
in battle, and from fifteen which were captured at the battle of Arbela
some notes were drawn up by Aristotle. In stating that the animal reaches
an age of 200 years, the naturalist and philosopher was probably not very
far out. The mode of Elephant-catching as related by Aristotle is that
pursued at the present day. Then, as now, tame Elephants were made use of
as decoys. Pliny,[135]
who was apt to confound fact and fiction in a somewhat inseparable
tangle, had something to say about Elephants, both Indian and African.
Serpents, he thought, were their chief enemies, which slew them by
coiling round them and thrusting their heads into the trunk, and so
stopping respiration. In Europe Elephants were first seen in the year
B.C. 280. Pyrrhus used them in his invasion, and
copying his example the Romans themselves learnt to use Elephants. The
first Elephant seen in England arrived in the year 1257, presented by the
King of France to Henry III. It was kept in the Tower (for long
afterwards a menagerie), and died at twelve years of age. Much use of the
Elephant has been made in symbols. We have spoken of the African Elephant
on Carthaginian coins as an emblem of eternity. The Oriental Elephant
resting on the back of a tortoise and supporting the world is the same
idea; and it is instructive to note that remains have been found in the
Siwalik Hills of a tortoise which would have been actually big enough to
support the creature, even "Jumbo," who weighed 6½ tons. Another symbol
is that of an Elephant upon whose back is a child with arrows; this
occurs on a medal of the Emperor Philip. It can perhaps hardly signify
the eternity of a strong human feeling!
The intelligence of the Elephant has been both exaggerated and
minimised. Perhaps the most elaborate attempt to endow the beast with
unusual mental perceptions is that of Aelian, who related that an
Elephant carefully watching his keeper, wrote after him with his trunk
letters upon a board. That the animal does {226}possess a good deal of
brains, seems to be shown by the way in which a well-trained animal will
obey the slightest sign of the mahout in India. According to Sir Samuel
Baker, localities which produce in abundance particular kinds of fruit
are remembered, as well as the time at which the fruit will be at its
best. Stories of revenge, which are numerous enough, attest, so far as
their data are to be accepted as accurate, the power of memory possessed
by the Elephant.
In spite of their longevity, however, Elephants, unlike Rome, have not
been built for eternity. We can only find two living species; but in past
times Elephants were very numerous. They commenced, so far as we know, in
the Miocene.
The existing forms are known in a fossil, or at least sub-fossil
state, from diluvial deposits; and it is interesting to note that the
African Elephant had formerly a wider range than now. Its bones
(described as E. priscus) have been met with in Spain and
Sicily.
One of the best known of completely-extinct Elephants is the Mammoth,
E. primigenius. This great Elephant in most respects more nearly
approached the existing Indian Elephant. The teeth have quite as numerous
plates. The tusks were enormous, reaching a maximum length of 15 feet;
they were much curved upwards as well as outwards. A large tusk weighs as
much as 250 lbs. The Mammoth was of exceedingly wide range. Not only was
it found in various parts of Europe, but it was especially abundant in
Siberia, as is exemplified by the fact that for the last two hundred
years as many or more than 100 pairs of tusks annually have been sold
from that region. It also occurred in America together with forms at
least not far removed from it, such as E. columbianus. Mammoths
have been more than once found as entire carcases in the frozen soil of
Siberia. The first was discovered in the year 1799, and rescued some
years later for the St. Petersburg Museum. This example showed that the
Mammoth, unlike existing Elephants, was covered with thick wool mingled
with long and more bristly hairs of some 10 inches in length. The softer
wool formed a kind of mane beneath the neck, which hung down as far as
the knees. Another carcase was discovered later by Lieut. Benkendorf, who
did not save it, but was nearly swept along with it into the sea by a
flood. These creatures died in the position in which they were found by
being bogged when in search of vegetation or water. {227}
How primeval man, with his inferior weapons, slew the Mammoth is not
easy to understand; but that they were contemporaneous is clearly shown
by associated remains, and by the notorious sketch of the Mammoth on a
piece of its own ivory, in which curved tusks and a forehead like that of
an Indian Elephant are plainly to be seen. Although it was only so
recently as the year 1799 that an example of this great creature was
actually studied on the spot, and removed to St. Petersburg, the
existence of Mammoths and of ivory is a matter of much more ancient
knowledge. M. Trouessart relates[136] that fossil ivory was known to the
Greeks. Theophrastus spoke of ivory imbedded in the soil, and the tusks
were recovered by the Chinese. It is a curious fact that the Chinese
described and figured the Mammoth as a kind of gigantic Rat. The likeness
between the elephantine molar and that of Rodents has been commented
upon; but the existence of its tusks below the level of the ground led
the Chinese Natural Historians to consider that the ways of life of the
Mammoth were those of the Mole. As to the carcases themselves, the
Chinese said that the flesh was cold, but very healthy to eat. This
expression can hardly be explained, except upon the view that fresh
carcases were known to that people long before they were known to us of
the Western world. The value of the Mammoth ivory was known to antiquity;
the famous Haroun-al-Raschid gave to King Charlemagne not only a pair of
living Elephants, but a "horn of Licorne," which seems undoubtedly to
have been a name for the tusks of the Mammoth. For in an account of the
sacred treasures of Saint Denis, published in the year 1646, the author
states this to be the fact.
The causes of the disappearance of the Mammoth are not easy to
understand. Some held that it was a naked animal like the existing
Elephants, and that the lowering of the temperature in Siberia proved
fatal; it is, of course, now certain that it was clothed with dense
woolly hair. Along with the bogged corpses of the great pachyderm,
numerous trunks of pine-trees have been found, together with associated
remains of other animals now extinct in that neighbourhood. Thus it is
plain that Siberia was once covered by mighty forests, through which the
Mammoth roamed. The decay of these forests, upon whose branches the
Elephant fed, as is attested by the remains of pine leaves found {228}in
the interstices of its teeth, was the signal for the disappearance of
their most colossal inhabitant.
The large number of remains of this and of other extinct species of
Elephas in this country gave rise to the supposition that they
were Elephants brought over by Caesar to aid in the subjugation of these
islands. The Rev. J. Coleridge (father of the poet) pointed out that
though Caesar in his Commentaries made no mention of any such
importation of Elephants, a passage in the Stratagems of Polyaenus
expressly mentions that Cassivelaunus was confronted by the Romans with
an Elephant clad in a coat of mail, by whose aid the crossing of the
Thames was effected. At the time that attention was called to this (1757)
it was not popular to hint at the possibility of fossils. So that fact,
conveniently historical, served to explain away a difficulty. It is
remarkable that the Elephant, common enough of course in Asiatic
monuments, actually occurs in English architecture. Mr. Watkins, from
whose interesting work (Natural History of the Ancients) a good
many of the facts detailed here are drawn, tells us that the church of
Ottery St. Mary has an Elephant's head sculptured on one of its pillars.
The same ornament appears in Gosberton Church, Lincolnshire. Whether this
has anything to do with a reminiscence of formerly existing Elephants is
a hard question to answer. In this figure of an Elephant the trunk has a
spiral representation, and the trunk of an Elephant is believed by some
to be intended by the common "so-called Pictish ornamentation" in
Scotland; this spiral alone is to be seen constantly. If it is a
reduction of an Elephant to its simplest terms, it is highly interesting
as an almost undoubted survival of remembrance of Elephants. For at such
a period we cannot use the memories of Crusaders or others who may have
visited the East to explain the facts. The sculptured Elephants' heads
might conceivably be so explained.
The name Mammoth, thinks Mr. Watkins, may be derivable from the Arabic
word Behemoth. He quotes a writer, who first described the beast in 1694,
as using the two words indifferently. The Arabs, moreover, were then as
they are now great ivory traders; and in the ninth and the two succeeding
centuries explored the confines of Siberia, as they now do the forests of
Africa, for ivory. The "Behemoth" of Job "eateth grass as an ox.... He
moveth his tail like a cedar" (the Hippopotamus has a much more {229}stumpy
appendage). "Behold, he drinketh up a river, and hasteth not" is surely
much more suggestive of the copious draughts of an Elephant than the
possibly equally copious but not so visible libations of a
Hippopotamus.
The most ancient of the true Elephants (genus Elephas) is E.
meridionalis. It is of the African type, i.e. the plates of
the molar teeth are not abundant, and are not so many as in the existing
E. africanus. It seems to have been one of the largest of
Elephants, standing 4 metres high. Its remains are abundant in Europe,
and are known also from England. Like this species E. antiquus is
also of the African type. It was contemporary with man. Certain dwarf or
"pony" races found in caves in Malta, and called Elephas
melitensis or E. falconeri, are believed to belong to this
species. Mr. Leith Adams, who described these[137] remains, placed them in two dwarf
species called by the names used above, and found associated with them a
larger form, which he referred to E. antiquus. The existence of
these animals in Malta seems to argue at least its former larger
dimensions, and the presence of more abundant fresh water. The remarkable
swimming capabilities of the Elephant do not necessarily imply either a
former absence of land connexion or, on the other hand, its existence.
Nor as a third possibility can it be suggested that the dwarf size argues
an island of limited dimensions, when we bear in mind the huge tortoises
of the Galapagos and some other islands. It is important to notice that
Elephants of the African type (Loxodon) were not formerly absent
from India. E. planifrons was one of these.
The genus Stegodon is so called from the fact that the molar
teeth, seen in longitudinal section, present a series of roof-shaped
folds, the interstices between which are not, or are, imperfectly filled
up with the cement which in Elephas reduces the surface of the
teeth to a level plane. This genus is exclusively Asiatic, and is Miocene
to Pleistocene in time range. The number of ridges on the molars is
small, not more than two. The incisors (tusks) have no enamel; the
skeleton generally is like that of Elephas, between which and
Mastodon the present genus is intermediate. Among the four or five
species is S. ganesa (called after the Indian Elephant-headed
divinity), with tusks 10 feet long, to be seen at the British Museum of
Natural History. {230}
The last genus of the family Elephantidae is Mastodon, so
called from the structure of the molar teeth. These are provided with but
few transverse ridges, not more than five, so that their structure is
intermediate between those of Dinotherium and those of
Stegodon. Between the ridges are sometimes isolated, boss-like
protuberances (whence the name of Mastodon), produced by a
subdivision of the ridges. There is either but little or no cement
between the ridges. This genus differs from nearly all other Elephantidae
by the possession of milk molars, which occasionally persist throughout
life, the permanent dentition in those cases being a mixture of milk and
permanent teeth, as has been (erroneously) stated of the Hedgehog.[138]
The tusks (incisors) are sometimes present in both jaws, and as they
have, during youth at any rate, a coating of enamel, the likeness to the
chisel-shaped incisors of Rodents is patent. In connexion with the
implantation of incisors in the lower jaw, many species have a
prolongation of the bones of that part of the skeleton. In the bones,
generally, there is not very much difference from Elephas, but the
forehead is a little less pronounced. The genus existed from the Miocene
and became extinct in the Pleistocene. It was nearly world-wide in range,
being known from all four continents. Naturally with this very wide range
was associated a large number of species. Zittel enumerates no less than
thirty-two.
This genus is the only one of the Elephantidae which extended its
range into South America, where the remains of two species occur. The
bones of these great Elephants have attracted attention for some
centuries. They were often held to be the bones of giants (as they
actually were!), and in one case were ascribed to a deceased monarch,
Teutobochus. The American Indians considered that equally gigantic men
lived who were able to combat these great Proboscideans. There are
legends of the Mastodons as living animals, which is quite probable,
considering their geological age. There is a curious parallelism between
the legends of two such widely-separated localities as North America and
Greece. Buffon relates how among the Indians of Canada there was a belief
that the Great Being destroyed both Mastodons and men of equal
proportions, with thunderbolts. With this we may perhaps compare the
story of the destruction of Typhoeus by Zeus, who {231}also used thunderbolts.
One of the giants was not slain, but was compelled to stand and bear up
the heavens. Atlas holds thus the position of the Elephant supporting the
globe of Indian mythology.

Fig. 119.—Dinotherium giganteum. Side view of skull, 1⁄15th natural size.
Miocene, Germany. (After Kaup.)
The genus Dinotherium, sole representative of the family
Dinotheriidae, differs in a number of important particulars from
the true Elephants. In the Elephants, if there is but a single pair of
incisors, these are found in the upper jaw; in Dinotherium there
is apparently but a single pair, but these are implanted in the lower
jaw, the symphysis of which is much prolonged and greatly bent downwards,
so that the tusks emerge at right angles to the long axis of the head,
and are even bent backwards. The molar teeth are five in number on each
side of each jaw and are bi- or tri-lophodont, not unlike those of the
Tapir. There is no cement in the valleys between the ridges of these
teeth, and there is a regular succession, the premolars being two and the
molars three.[139] All
the teeth are in use at the same time, {232}their small size
enabling them to be accommodated in the jaw together. The skull of
Dinotherium is lower than that of Elephas or
Mastodon. The bones of the skeleton generally are like those of
Elephas.
Though a suggestion of marsupial bones attached to the pelvis has been
discredited, there is no doubt that Dinotherium occupies the most
primitive position among the Proboscidea; but at the same time it cannot
be regarded as the ancestor of Elephants, as it is so much specialised in
various ways. The incisors for one thing forbid this way of looking at
the creature. It is an ancient genus found in beds of Miocene age in
Europe and Asia. It is not known from America. The creature was larger
than any Elephant. Eighteen feet in length has been assigned to it. The
enormous weight of the lower jaw and tusks seems to argue that it was at
least partially aquatic in habit, and that it may have used these tusks
for grubbing up aquatic roots or for mooring itself to the bank. At first
there were naturalists who considered it as an ally of the Manatee, and
the skull is not unsuggestive of that of the Sirenia.
Pyrotherium has been referred to the Proboscidea; but our
knowledge of that form is limited to a few teeth from Patagonian rocks of
an uncertain age.[140]
They are simple bilophodont molars, very like those of
Dinotherium. A tusk has been found in the neighbourhood of these
teeth which may possibly belong to the same animal; but it is
uncertain.
Sub-Order 7. HYRACOIDEA.
This group of small mammals contains only one well-marked genus which
is usually named Hyrax, although Procavia seems to be the
accurate term. Popularly these creatures are known as Coneys. They have a
singular resemblance to Rodents, the short ears and much reduced tail,
besides the squatting attitude adopted, contributing to this merely
skin-deep likeness. They agree with other Ungulates in the structure of
the molar teeth, which are much like those of Rhinoceros; in the
absence of a clavicle; in the absence of an acromion; in the reduction of
the digits of the limbs to four digits in the manus and three in the pes.
On the {233}other hand they differ from most Ungulates
in the incisors growing from persistent pulps, a point in which they
resemble the Rodentia. The muffle also is split as in those animals. The
Hyracoidea are peculiar in the fact that in addition to the caecum at the
junction of the small and large intestines, there are a pair of caeca
(bird-like in being paired) some way down the large intestine. The dorsal
vertebrae are unusually numerous, 22. The adult dentition according to
Woodward,[141] who has
recently examined the matter, is I 1/2 C (1/0) Pm 4/4 M 3/3, while the
milk dentition is I 3/2 C 1/1 Pm 4/4.

Fig. 120.—Cape Hyrax. Hyrax capensis. × ⅛.
The inclusion of the canine of the permanent set of teeth in brackets
signifies that it is the milk canine which occasionally persists. It
should further be remarked about the teeth that they are both hypselodont
and brachyodont, the extremes being connected by intermediate forms.
Another peculiarity of the genus is the dorsal gland, which is covered
with hair of a different colour to that covering the body generally. This
is present in all species.
The genus Hyrax (the most recent authority on the subject, Mr.
Oldfield Thomas,[142]
only allows one genus) is limited in its range to Ethiopian Africa and to
Arabia, including Palestine, It does not reach Madagascar. Mr. Thomas
allows fourteen species with two or three sub-species. {234}
Some of the Coneys live in rocky ground, while others, formerly placed
in the genus Dendrohyrax, frequent trees, in holes in which they
sleep. The Coney of the Scriptures is familiar, who is "exceeding wise,"
though a "feeble folk." But the further observation that he "cheweth the
cud but divided not the hoof," is obviously entirely wrong. As to the
wisdom, it is said that this beast is too wary to be taken in traps;
while the suggestion of chewing the cud is, according to Canon Tristram,
to be interpreted in the light of a habit of working and moving its jaws
which the animal has. The traveller Bruce kept one in captivity to see if
it did really chew the cud, and found that it did!
{235}
CHAPTER X
UNGULATA (continued)—PERISSODACTYLA (ODD-TOED
UNGULATES)—LITOPTERNA
Sub-Order 8. PERISSODACTYLA
Fig. 121.—Bones of the manus A, of Tapir (Tapirus indicus). × 1⁄5. B, of Rhinoceros
(Rhinoceros sumatrensis). × 1⁄5. C, of Horse (Equus caballus). × ⅛. c, Cuneiform;
l, lunar; m, magnum; p, pisiform; R, radius; s, scaphoid; td, trapezoid;
tm, trapezium; u, unciform; U, ulna; II-V, second to fifth digits; V in B, and II
and IV in C, represented by rudimentary metacarpals. (From Flower's Osteology.)
These Ungulates derive their name, which is that given by the late Sir
Richard Owen, from the fact that the middle digit of the hand and foot is
pre-eminent. As will be seen from Fig. 121, the axis of {236}the limb
passes through the third finger, which is larger than any of the others,
and is symmetrical in itself. In this the present group contrasts with
the Artiodactyla, where the axis is not "mesaxonic," but where there are
two digits, on either side of the axis, which are symmetrical with each
other. This arrangement of the limbs is highly characteristic, but
appears to be not quite universal. In the Titanotheres, which form a
group of the Perissodactyles, the fore-limbs are not quite accurately
mesaxonic. Nor on the other hand can all Ungulates which show the
Perissodactyle condition be safely included in the present group. The
ancient Condylarthra and the Litopterna show precisely the same state of
affairs. But other features in their organisation lead to their
separation from the Perissodactyles, of which, however, the Condylarthra
are probably ancestors. The Litopterna on the other hand, which possess
even one-toed members like Equus, are believed to represent a case
of parallelism in development. The number of functional toes varies from
four to one. In the ankle joint the astragalus either does not, or does
only to a comparatively slight extent, articulate with the cuboid as well
as with the navicular bone. Moreover the fibula when present does not as
a rule articulate with the calcaneum. In the opposed group of
Artiodactyles the precise reverse of these conditions obtains. It is
usually stated as part of the definition of this group that they do not
possess horns of the type of those met with in the Cervicornia and
Cavicornia. But the strong bony bosses on the skull of many Titanotheres,
so curiously reminiscent of those of the not nearly related
Dinoceras and Protoceras, may well have supported horns of
the Ox and Antelope pattern.

Fig. 122.—Bones of the
manus of Camel (Camelus
bactrianus). × ⅛. c,
Cuneiform; l, lunar; m,
magnum; R, radius; s,
scaphoid; td, trapezoid;
u, unciform. (From
Flower's Osteology.)
The teeth of the Perissodactyles are lophodont, more rarely bunodont.
The selenodont Artiodactyle form of molar is not met with. The dental
formula, moreover, is at least near the {237}complete one, the more
modern forms as usual being the more deficient in numbers of teeth.
The dorso-lumbar vertebrae are as a rule twenty-three; but the extinct
Titanotheres are again an exception; for, at least in
Titanotherium, there are but twenty of these vertebrae—an
Artiodactyle character. The femur has a third trochanter. There are so
few recent Perissodactyles that an enumeration of the distinguishing
characters of the viscera may very probably be useless for purposes of
classification. But the living genera at any rate are to be separated
from the living Artiodactyles by the invariable simplicity of the stomach
coupled with a very large and sacculated caecum. The liver is simple and
not much broken up into lobes, and the gall-bladder is always absent. The
brain is well convoluted. The teats are in the inguinal region. The
placenta in this group is of the diffused kind.
Fig. 123.—Anterior aspect of
right femur of Rhinoceros
(Rhinoceros indicus). × ½.
h, Head; t, great trochanter;
t′, third trochanter. (From
Flower's Osteology.)
The living Perissodactyles belong to three types only, indeed to three
genera only (in the estimation of most), which are the Horses, Tapirs,
and Rhinoceroses. But taking into account the extinct forms, they may be
divided primarily (according to Professor Osborn) into the four following
groups:—(1) Titanotherioidea, including but
one family, Titanotheriidae; (2) Hippoidea, including the families
Equidae and Palaeotheriidae; (3) Tapiroidea, with two families, Tapiridae
and Lophiodontidae; and (4) Rhinocerotoidea with families Hyracodontidae,
Amynodontidae, and Rhinocerotidae. It is conceivable, according to the
same writer, that the Chalicotheres (here treated of as a separate
sub-order, Ancylopoda) should be added to the Perissodactyle series.

Fig. 124.—Side view of skull of Horse with the bone removed so as to expose the whole
of the teeth. c, Canine; Fr, frontal; i1, i2, i3, incisors; L, lachrymal;
m1, m2, m3,
molars; Ma, malar or jugal; Mx, maxilla; Na, nasal; oc, occipital condyle; Pa,
parietal; pm1, situation of the vestigial first premolar, which has been lost in the
lower, but is present in the upper jaw; pm2, pm3, pm4, remaining premolars; PMx,
premaxilla; pp, paroccipital process; Sq, squamosal. (After Flower and Lydekker.)
Fam. 1. Equidae.—This family, which includes the living
Horse, Zebras, and Asses, as well as a number of extinct genera agreeing
with those types in structure, may be defined by the possession of but
one functional toe, the two lateral ones being mere splints, or but
little more. The molar teeth are hypselodont, and {238}the premolars, with the
exception of the first, resemble the molars in their pattern. The orbit
is completely surrounded by bone. The incisors are chisel-shaped, with a
pit on the free surface. The canines are rudimentary if present. The
radius and ulna are fused, as are the tibia and fibula. Although for the
sake of uniformity a family, Equidae, is here separated from its allies,
it is quite impossible owing to the full state of our knowledge of this
group to draw a really hard-and-fast line between this family and the
Palaeotheriidae. We shall deal presently with the conjectured pedigree of
the Horse, which naturally involves that family, and which presents an
unbroken series from four-toed Perissodactyles to the present one-toed
Horse, the various bones and teeth becoming modified in the course of the
descent "with the regularity of clockwork." We are compelled to draw the
line at functional second and third toes; directly these are no longer
used the animal is a Horse in the strict sense! This is irrational and
regrettable, but necessary for practical purposes, if {239}we are to
continue the plan of defining the various families of Mammalia.
The genus Equus[143] contains not only the Horse, but the
Asses and Zebras. The genus is to be distinguished as regards external
characters by the following features:—The
body is thickly clothed with hair; there is a more or less bushy tail and
mane; the colours are apt to be disposed in stripes of black or blackish
upon a yellowish brown ground; this is of course best seen in the Zebras,
but the wild Asses also have some traces of it, if only in the single
cross-bar of the African Wild Ass, and it is even "reversionary" in the
domestic Horse at times. There are no horns upon the forehead or
elsewhere; the fore-limbs or both pairs have a callous pad upon the
inside, which is possibly to be looked upon as an aborted gland, possibly
originally of use as secreting some odorous substance calculated to
enable strayed members of the herd to regain their companions. The
terminal phalanx of each of the (functionally) single digits is enclosed
in a large horny hoof.
The main internal features of structure which divide this genus of
Perissodactyles from the Rhinoceros or the Tapir, or from both, are: the
existence of strong incisors, three on each side of each jaw; there are
canines, but these are small and do not always persist in the full-grown
mare. They are popularly known as "tusks" or "tushes." The first of the
four premolars (the "wolf tooth") is small and quite rudimentary; it is
often absent. As there are three molars, the present genus has the
"typical" number of the Eutherian dentition, i.e. forty-four. In
the skull the orbit is—as it is not in Tapirs and
Rhinoceroses—completely encircled by bone. There is but one
functional finger and toe on each hand (Fig. 121 C) and foot; the second
and third digits are represented by mere splints, one of which may as an
abnormality be enlarged, and reach nearly as far as the well-developed
digit. There are even occasionally traces of digit number two.
The Horse, E. caballus, is to be distinguished from its
congeners by the small callosities on the hind-limbs which it possesses
in addition to the larger ones on the fore-limbs. The hairy covering of
the tail is more abundant, as is also the mane. The head too is
proportionately smaller, and the general contour {240}more graceful. Though
Zebra markings are not usual upon E. caballus, there are plenty of
examples of—what we may perhaps in this case term—a
"reversion" to a striped state. The celebrated "Lord Morton's mare,"[144] whose portrait hangs
in the Royal College of Surgeons, is an interesting case of this. It was
as a matter of fact thought to be an example of that rather
doubtfully-occurring phenomenon, "telegony." Its history is briefly this.
The animal was the offspring of a mare that had previously produced to a
male Quagga a hybrid foal. Afterwards a second foal was produced by the
same mare to an Arab sire. This foal, the one in question, was striped,
and hence was thought to be an example of male prepotency. But instances
are known of unquestioned Horses which show the same stripes, such as a
Norway pony which had not even seen a Zebra!
A last remnant of the naked palm of the hand and sole of the foot is
left in the shape of a small bare area, smaller in the Horse than in the
Asses, known technically as the "ergot," the term being that of the
French veterinarians. As already mentioned, the Horse differs from the
Asses and Zebras in the fact that the hind-limbs have callosities on the
inner side. They are known as "chestnuts," and their nature has been much
disputed. It has been suggested that they are the last rudiment of a
vanished toe; but in all probability they are, as already suggested,
traces of glandular structures, which are common, upon the limbs in many
animals (see above, p. 12).
It is a singular fact that there are apparently no wild Horses of this
species. The case is curiously analogous to that of the Camel, which also
is only known as feral or domesticated. Why the Horse should have become
extinct as a wild animal, considering that when it does run wild it can
thrive abundantly, is impossible to understand. Sir W. Flower thinks[145] that "the nearest
approach to truly wild horses existing at present are the so-called
Tarpans, which occur in the Steppe country north of the sea of Azov
between the river Dnieper and the Caspian. They are described as being of
small size, dun colour, with short mane and rounded obtuse nose." But he
adds that there is no evidence to prove whether they are really wild. In
favour, however, of their possibly being wild and indigenous European
Horses, may be {241}mentioned the fact that their general
build and appearance is highly suggestive of the wild Horses sketched by
primitive man upon ivory.
A really wild Horse, and possibly the ancestor of the European
domestic Horse, is E. przewalskii of the sandy deserts of Central
Asia. This animal has been believed to be a mule between the Wild Ass and
a feral Horse; but if a distinct form, and probability seems to urge that
view, it is interesting as breaking down the distinctions between Horses
and Asses. The species possesses the four callosities of the Horse, but
has a poorer mane and an asinine tail.
There is no question that the Horse has been a domestic animal for
very many centuries. Hieroglyphics appear to show that the Egyptians had
not originally domesticated the Horse; it seems to have been first
introduced among them by the Hyksos or Shepherd Kings.[146] Whatever the date may be, it is
certain that considerably anterior to the Egyptians the Assyrians and
Phoenicians possessed Horses. In Western Europe the date of the
introduction of the Horse seems to have been during the bronze epoch.
Lord Avebury[147] has
pointed out that out of eighteen cases of graves in which the remains of
Horse were found, twelve contained metal implements, i.e. 66 per
cent. This does not of course prove that the Horse was domesticated at
that period, but it throws doubt upon the earlier occurrence of the Horse
in abundance. The Horse, however, does occur on the Continent associated
with the remains of man during the Quaternary period.[148]
Messrs. Cuyer and Alix enumerate between fifty and sixty domesticated
races of Horse, not counting the supposed wild varieties which have been
already referred to. These may be further subdivided; for instance, under
the race "pony" we may distinguish the Irish, Scotch, and Shetland
varieties, all of which, however, according to Sanson, have originated in
Ireland. They are used, remark the authors above quoted, "par les jeunes
filles des lords pour leurs promenades." The Arab, the Barb, the Suffolk
Punch, etc., are among the numerous races of domestic Horses, into which
to enter properly would require another volume, and that of large size.
{242}
The Asses and Zebras differ from the Horse in the characters mentioned
under the description of Equus caballus. In addition to these may
be pointed out a feature to which attention has been directed by Mr.
Tegetmeier.[149]
According to him the period of gestation in the Horse is only eleven
months; in the others more than twelve.
Fig. 125.—Asiatic Wild Ass. Equus onager. × 1⁄20.
Opinions as to the number of species of Asses differ. On the most
liberal estimate there are three Asiatic and two African species. The
best known of the Asiatic Wild Asses is the Onager, E. onager. It
is of a uniform yellowish, "desert" colour, with a dark stripe along the
middle of the back, and is found in Persia, the Punjab, and the country
of Cutch. The creature is of great swiftness; it has been stated to be
untameable, but Mr. Tegetmeier makes the absolutely opposite statement
that the Ass occasionally "becomes so tame as to be troublesome"! The
Syrian Wild Ass, E. hemippus, hardly, if at all, differs from
this.
Fig. 126.—Nubian Wild Ass. Equus africanus. × 1⁄20.
The Kiang, E. hemionus, seems to have more claims to
distinctness. In the first place it has a more limited and a {243}different
distribution; it is confined to the high tablelands of Thibet at an
elevation of 15,000 feet and upwards. In correlation with this habitat it
has a thicker and more "furry" coat, which is, moreover, of a darker
shade than that of the Onager. This coat is shed in the summer, and
replaced by one which is not so dark in hue. It is an interesting fact
that the African Wild Asses approach to the zebra type in having at least
traces of stripings. There are apparently two species. The best known,
the Nubian Ass, E. africanus, is probably the parent of the
domestic donkey. It has a dorsal longitudinal stripe, and another across
the shoulder—in legend the marks of the Saviour. The matter of the
name of this Ass seems difficult to decide. It has been called also E.
asinus and E. taeniopus. It has been observed that this animal
has a great aversion to water, and a delight in rolling in the
dust—both of which characteristics argue a desert existence. But on
the other hand the Kiang will plunge boldly into streams, yet it would
seem to be the descendant of a purely desert form. The Ass is a
longer-lived {244}animal than the Horse. Mr. Tegetmeier
calls attention to a donkey living in 1893 which had been ridden
fifty-five years previously. The Horse, on the other hand, lives not much
more than twenty-five years.
A second species of African Wild Ass, E. somalicus,[150] is distinguished by
its greyer colour, by the absence of the shoulder stripe, by the very
faint development of the dorsal stripe, and by the presence of numerous
cross stripes upon the legs. It has, too, smaller ears, and a longer and
more flowing mane. Mr. Lort Phillips, an experienced naturalist and
traveller, saw a herd of these Wild Asses in Somaliland, which he
regarded as being of quite a new species. A living example in the
Zoological Society's Gardens led Mr. Sclater to an identical conclusion,
which was supported, as he pointed out, by the fact that this Ass has a
different range to the African or Nubian Wild Ass.
Of the Zebras three species are usually allowed; these are E.
zebra, the "Mountain" or "Common" Zebra, E. burchelli, E.
grevyi, as well as E. quagga. Professor Ewart thinks that the
Common Zebra, Burchell's, and the Quagga are not very distinctly marked
off from each other. No one, however, has any doubt of the distinctness
of E. grevyi. This latter differs from the rest in its larger
size, in the large head and ears, and in the marked hairiness of the
ears. It would seem to be a primitive type of Zebra, if the fact that the
occasional reversion of hybrids to a parent form be allowed; for
Professor Ewart found a cross-bred Zebra to present several
characteristics in the face-marking of this, the finest of the Zebra
tribe. Only four specimens of E. grevyi have been exhibited alive
in Europe—two in Paris, and two in the Zoological Society's Gardens
in London. The latter were presented to Queen Victoria by King Menelek of
Abyssinia. The species was named by Professor A. Milne-Edwards in honour
of a late President of the French Republic, from an example also sent by
King Menelek.
The Common Zebra has closer and darker stripes than Burchell's, but
not quite so close as in E. grevyi. It has also a very
characteristic arrangement of stripes on the withers in the form of a
gridiron. This latter is wanting in both the other species. In E.
grevyi, in fact, this part of the back is white. E. zebra has
also a dewlap in front. E. burchelli has fewer and broader {245}stripes, and between them lie in many
cases shadow-stripes of a faint brown.
Fig. 127.—Burchell's Zebra. Equus burchelli. × 1⁄20.
All these animals, and the Quagga too, are absolutely confined to
Africa. Mr. R Crawshay,[151] in describing what he considered to
be a new variety, remarked upon the curiosity of E. burchelli.
"They remain out in the sun on the plains all day long, not retiring into
covert at all. They are then an intolerable nuisance to any one in
pursuit of other game; indeed this may be said of them at all times. If
once they notice you, they draw in and mob you in their
curiosity—only, however, when one takes no interest in them, for
when they fancy they are the object of the intruder's attention, no
animals are more watchful and cunning in safeguarding themselves. If only
their curiosity were manifested in silence it would not so much matter,
but it vents itself in snorts and thundering stampedes, which puts every
beast within earshot on the qui vive."
Whether Burchell's Zebra[152] can be further subdivided into
species or sub-species appears to be doubtful. Dr. Matschie considers
that Equus boehmi may be regarded as a valid form, and in addition
to this two sub-species, E. burchelli granti and {246}E.
burchelli selousi, have been proposed for what are at most local
races. But it is at present far from certain whether their distribution
favours this subdivision.
The Quagga was more striped than is sometimes represented in
illustrations. According to Dr. Noack, from whose paper[153] upon the animal I quote here, the
transverse stripes reached back as far as the buttocks; they were,
however, completely absent from the legs. The animal is, as every one
knows, probably completely extinct. In the year 1836 it was still
abundant; in 1864 the last specimen ever exhibited was received by the
Zoological Society. Mr. W. L. Sclater thinks that it may have survived in
the Orange River Colony as late as 1878, but admits that any certainty is
difficult, as it was frequently confounded by the Boers with Burchell's
Zebra. Its rarity is emphasised by the fact that it is not mentioned in
the recent work of that most skilful of hunters, Mr. F. Selous. Gaudry
places the Quagga nearest of all living Equidae to the Hipparion
gracile of Pikermi.
Fossil Equidae.—The existing Equidae all belong to the
genus Equus, though there are some who would (quite unnecessarily)
divide off the Zebras as a genus Hippotigris. The genus
Equus itself goes back in time to the Pliocene, during which epoch
there lived in India E. sivalensis, the same species according to
some with the E. stenonis of Europe. None of these species, Old
World or New, are easily to be separated from E. caballus. But
many names have been given to them. It is of course perfectly conceivable
that they may have differed among themselves as much as do the existing
Zebras and Asses, the separation of which would be hardly possible did we
know their bones only. There are, however, extinct genera, undoubtedly
related so closely to Equus as to be placed in the same family,
though clearly separable as genera. Hipparion is one of these
genera; its remains are known from Europe, Asia, and North Africa, from
beds of Miocene and Pliocene times. A large number of different species
have been described. It was a beast of about the size of a Zebra. The
principal characters are that each foot has three toes, of which,
however, the two side ones are smaller than the central toe. There is a
marked round fossa on the maxillary bone, a feature shared by the South
American Onohippidium.[154] The pattern of {247}the molar teeth is,
too, a little different from that of Equus. Protohippus of
the North American Pliocene is also three-toed, but the two
additionally-developed toes are smaller than in Hipparion. Other
forms are dealt with below in connexion with the ancestry of
Perissodactyles. It is a curious fact about Hipparion, which is
not now regarded as on the direct line of equine descent, that the edges
of the enamel plates of the molars may show a complicated folding very
like that presented by that clearly terminal form of Perissodactyle life,
the gigantic Elasmotherium. This is indicative of high
specialisation, which ended in extinction.
Ancestry of the Horses.—The Lophiodontidae and the
Palaeotheriidae are two of the most interesting extinct families
of Perissodactyles; for among them we find what would appear to be the
ancestral forms of both the existing Tapirs and Horses. The Rhinoceroses
also would seem to be derivable from the Palaeotheriidae. The very
vagueness of the characters of these creatures, considered from a
classificatory point of view, has led to much diversity in their placing.
This though gratifying to the evolutionist is tiresome to the writer who
wishes to give a methodical account of their various characters. It will
be best perhaps not to attempt an accurate placing or to reconcile
conflicting opinion, but to give some salient features of osteology which
lead to the belief in their relationship to existing groups of
Perissodactyles. A book upon the history of mammals would be incomplete
without some account of that well-ascertained series of forms which seem
to connect these primitive Perissodactyles with the modern Horse.
Equus, in fact, is not only the "show horse" of the doctrine of
evolution, but also the "stalking horse."
In the Eocene of both Europe and America are met with a number of
forms from which we may start. Hyracotherium, regarded on the one
hand as the type of a sub-family of the Equidae themselves, and on the
other as a member of the family Lophiodontidae, was a small-sized animal,
three feet or so in length; it possesses the complete Eutherian dentition
with a slight diastema. The orbits are not separated from the temporal
fossa; the fore-limbs were four-toed, the hind three-toed, with
moderately long metapodia, especially on the hind-feet. The shoulder
blade {248}has a well-marked coracoid process. The
radius and ulna are separate; so too are the tibia and fibula.
Eohippus, belonging to the same sub-family, is slightly more
primitive; for the hind-feet have a rudiment of digit I. Orohippus
is a little nearer to the Horses in that the molar teeth have acquired a
little further advance towards the equine type. Instead of the tubercles
of the teeth remaining for the most part separate, they have fused into a
set of ridges, of which, however, the pattern is less complex than in the
modern Horses. In other respects Orohippus is much like
Hyracotherium. Pachynolophus seems to be but a synonym.
The next stage is shown by Mesohippus, a Lower Miocene form,
usually referred to the neighbourhood of Palaeotherium. It has
nearly lost one of the toes of the fore-foot, a rudiment only remaining;
the metapodials, at any rate of the fore-feet, seem to be slightly
increased in length. The orbit is not encircled by bone, but there is a
strong process from the frontal, which nearly meets the zygomatic
arch.
Anchitherium, from the Upper Miocene, is not far removed in
structure from the last-mentioned form; it is a trifle nearer the
existing Horse in several points. The ulna is further reduced and fused
with the radius below: the rudiment of digit V is still more rudimentary;
the two lateral digits are smaller in proportion to the central one than
they are in Mesohippus; the fibula is fused below with the tibia.
From this form to Equus is a small series of steps, characterised
by the still further reduction of all the digits except III, by the still
further reduction of the already rudimentary ulna and fibula, and by the
increasing depth of the molar teeth, which are of course, in
Equus, hypselodont.
Another interesting conclusion may seem to follow when we consider the
geographical range of the ancestral Horses. Hyracotherium and
Pachynolophus occurred both in the Old and New World. From them
may have arisen the Horses of both hemispheres. After that point there is
a division. Mesohippus is American, and we get at Equus in
that continent through Desmatippus and Protohippus. On the
other hand there are no remains known of Mesohippus in Europe; and
unless subsequent researches prove the existence of Mesohippus, we
have to rely upon forms which are placed with Anchitherium and
Hipparion.
It seems that in America the next genus in the direct line of equine
descent to Mesohippus is Miohippus. It is smaller in {249}size
than Anchitherium, to be considered immediately. The odontoid
process of the axis is just beginning to assume the characteristic
spout-like shape of that of the existing Horse and many modern Ungulates.
The median digit of both fore- and hind-limbs has become greatly enlarged
as compared with the corresponding digit of earlier forms.
It is held, however, that Anchitherium is not on the direct
line of descent either in America or in Europe, in both of which it
occurs. Its teeth are in some respects less Horse-like than in some of
the more ancient genera, to which the converse would be expected on the
descent theory. Its hoofs are much elongated and flattened, a mark of
specialisation and not appropriate to a creature holding an intermediate
position in the equine series. Both the American (A. equinum) and
the European species (A. aureliense) are of very large size,
larger than its successors, and such "alternations in bulk are
unlikely."
The genus Desmatippus of Professor Scott[155] fills in the gap between
Miohippus and Protohippus. The molars and premolars are
brachyodont, but there is a thin deposit of cement in the tooth valleys,
leading towards the more complete filling of these valleys with cement,
which is found in Protohippus. This genus of Horses, of which
there is at present but one species, D. crenidens, was three-toed,
and "the lateral digits, so far as can be judged by fragmentary remains,
were still fairly developed, and though much more reduced than in
Miohippus, appear to be somewhat less so than in
Protohippus."
To recapitulate, the following is the probable series of equines in
America—Mesohippus, Miohippus, Desmatippus,
Protohippus.
The development of the limbs of the Horse shows a most interesting
series, of stages, which correspond in part to the ancestral forms which
palaeontology seems to prove to be the line of the descent of our
existing Equidae. This matter has recently been elucidated by Professor
Ewart, who details the following facts and comparisons:—
In the youngest embryo (about 20 mm. in length) the humerus is
somewhat curved, and considerably longer than the radius and carpus taken
together. The first-named bone is shorter in the adult, and the
proportions of that bone in the young as well as its curvature are
suggestive of that ancient {250}Ungulate Phenacodus (see p. 202). In the next stage (an embryo of 25 mm.) the
humerus has slightly decreased in proportionate length, and has come to
be more like that of Hipparion. In both of these embryos it should
be noted that the ulna is complete and separate from the radius. In the
second of the two it has more distinctly acquired the form which it will
possess in the adult. The second metacarpal—one of the splint bones
of the adult—is tipped with a small nodule of cartilage, which is
clearly the representative of one or more of the phalanges belonging to
that digit.
Fam. 2. Tapiridae.—The Tapirs may be distinguished from
the Horse and from the Rhinoceros tribe by a few characters, which are as
follows:—
The dentition is generally the full one of forty-four teeth. The
premolars in the more ancient forms are unlike the molars, but like them
in more recent forms. The molars of the upper jaw have two crests
parallel and united by an outer crest. The fore-feet have four, the
hind-feet three toes.
The family is fully as ancient as that of the Equidae, but the
specialisation of the toes never advances so far. The modern
representatives of the order are, so far as the feet are concerned, in
the condition of very early representatives of the equine stock. Nor do
the teeth of the Tapirs ever reach the complicated pattern of that
presented by at least the modern Horses, or indeed of the Palaeotheres.
Apart from this it is not an easy matter to distinguish accurately
between these several families, including the Lophiodontidae, which, as
already mentioned, is placed nearer to the Tapiridae than to the
Palaeotheriidae. Indeed the differentiation of these two families, the
Tapiridae and the Lophiodontidae, seems to be a matter of the greatest
difficulty. The difficulty is well emphasised by the fact that
naturalists disagree most profoundly as to the relations of various
genera of extinct Tapir-like animals. For Mr. Lydekker the genus
Lophiodon includes also the American genera Isectolophus
and Systemodon, which are placed by Zittel in the sub-family
Tapirinae as opposed to Lophiodontinae, which contains Lophiodon
and Helaletes. The existing Tapirs can be differentiated from the
existing Horses with great ease, as the following account of the existing
genera will show.
{251}
Fig. 128.—American Tapir. Tapirus terrestris. × 1⁄10.
The genus Tapirus is now met with only in South and Central
America, and in the Malay Peninsula and the islands of Java and Sumatra.
This animal is in many respects the most ancient of existing forms
referable to the Perissodactyle order. It has four toes on the
front-feet, though only three on the hind-feet. The number of teeth is
42—nearly the typical Eutherian number. The Tapirs are always
moderately-sized animals, entirely covered with hair, and usually of a
brownish-black colour. The Malayan Tapir is, however, banded broadly with
white—a single band; the young of the Tapir is spotted, and striped
with white. The nose and upper lip conjoined are produced into a short
trunk, precisely comparable with that of the Elephant. As in the
Rhinoceros—and in this both contrast with the other existing
Perissodactyle genus Equus—the temporal fossa is not
separated from the orbit by bone. Of existing Tapirs there are at any
rate T. terrestris,[156] T. roulini (the "Tapir
Pinchaque" of Cuvier), T. dowi and T. bairdi in America
(the last two being sometimes separated into a distinct genus,
Elasmognathus, on account of the prolongation of the ossified
mesethmoid), and T. indicus in the East. The tapir, probably T.
terrestris, is described by Buffon as "a dull and gloomy animal." It
is certainly mainly nocturnal in habit. The name terrestris was
given by Linnaeus, who placed it in the same genus as {252}Hippopotamus amphibius; hence the
epithet applied to the Tapir. But as a matter of fact it loves marshy
neighbourhoods, and is in a way amphibious. This does not of course apply
to the Andesian T. roulini, which inhabits the cordillera of
Ecuador and Colombia. The distribution of existing Tapirs is, as is so
often the case, restricted when compared with that of their extinct
congeners and allies. In Europe the remains of the genus Tapirus
are abundant from Pliocene strata, and its remains are there known from
as far back as the Miocene. The genus is thus one of the very oldest
forms of Mammalia at present inhabiting the earth.

Fig. 129.—Malayan Tapir. Tapirus indicus, young. × 1⁄10. (From Nature.)
The Malayan Tapir is to be distinguished from the American (T.
terrestris—the other species have not been dissected) by the
greater development of the valvulae conniventes in the intestine, the
absence of a moderator band in the heart, and the less elongated caecum,
which is sacculated by only three bands, there being four in T.
terrestris.[157] The
animal frequents the most retired spots among the hill woods, by which
habit it seems {253}largely to escape the Tiger, its most
formidable foe in those regions of the world. Its quickness of senses
enables it also to slip away with rapidity. It can proceed at a great
pace when disturbed, and can readily push its way through obstacles. The
young animal, like that of the American species, is dark brown with
yellowish spots. It is stated by Mr. H. N. Ridley that the young animal
lies during the hot part of the day under bushes, in which situation "its
coat is so exactly like a patch of ground flecked with sunlight that it
is quite invisible." It is interesting to note that here, as with some
other animals, it is the young that are especially protected by such
mechanisms. Moreover, some of the spots are round and some are more
elongated, so that the resemblance to spots of sunlight which come in a
direct and in a slanting direction is greatly increased. Even the colours
of the adult are not so conspicuous when it is in its native haunts as
might be supposed. The breaking up of the ground colour into tracts of
two different colours prevent it from striking the eye so plainly as if
it were of one colour throughout. "When lying down during the day it
exactly resembles a grey boulder, and as it often lives near the rocky
streams of the hill jungles, it is really nearly as invisible then as it
was when it was speckled."[158]
Fam. 3. Rhinocerotidae.—This family is to be
distinguished from the preceding by a number of characters, which though
not universal are general. In the first place, there are commonly horns,
or a horn, consisting of what appears to be an agglomeration of hair-like
structures fixed upon a roughened patch of bone on the surface of the
nasals. The incisors are diminished or defective, and the upper canines
are often wanting. The molars and premolars are alike. The fore-feet are
four- or three-toed, but are functionally tridactyle; the hind-feet are
three-toed. The skeleton in this family is massive, and the limbs
relatively short. The skull, as in the Tapirs, has a confluent orbit and
temporal fossa. The upper lip is generally more or less prehensile; the
body is as a rule—to which the Pleistocene Hairy Rhinoceros is of
course an exception—rather sparsely covered with hair. In this
feature the Rhinocerotidae contrast both with the Tapiridae and the
Equidae. The family in reality contains but one existing genus, though
three have been instituted, viz. {254}Rhinoceros,
Ceratorhinus, and Atelodus. As there are so few existing
species the subdivision of animals which agree in so many and such
highly-characteristic features seems to be an unnecessary procedure. The
existing Rhinoceroses are but a fragment of the total number of known
forms from past epochs. The family is very markedly on the wane.
The genus Rhinoceros is characterised by its heavy build and
thick, almost smooth, skin—smooth, that is to say, so far as
concerns the slight development of hair—which is often thrown into
folds. There is one or there are two horns on the fore-part of the head,
which are, as has already been pointed out, structures sui
generis, and not exactly comparable with the horns of other living
Ungulates. There are three nearly equal toes on both fore- and
hind-limbs. The canine teeth of existing species have disappeared; the
incisors are, or are not, present; the molars and premolars are three and
four in each half of each jaw.
The visceral anatomy of the Rhinoceros has been much investigated so
far as concerns the Asiatic forms. A curious feature, which serves to
discriminate some of the Asiatic species from others, is to be seen in
the small intestine. In Rh. indicus[159] this gut is furnished with numerous
long cylindrical narrow outgrowths "like tags of worsted"; in the allied
Rh. sondaicus these tags are present, but are flatter and broader;
while in the two-horned Rh. sumatrensis there are no tags at all,
but only smooth valve-like folds. Another mark by which these species can
be distinguished depends upon the variation in the presence or absence of
certain glands imbedded in the integument of the foot—the so-called
"hoof glands." These occur in Rh. indicus and Rh.
sondaicus, but are absent in Rh. sumatrensis.
Sir W. Flower[160]
studied some years since the skull features which serve to differentiate
the existing forms.
In Rh. sumatrensis the two long downward processes of the
squamosal bone, termed respectively post-glenoid and post-tympanic, do
not unite below the auditory meatus. In this the species in question
agrees with the African forms but not with the one-horned Asiatic
species, where the two processes completely fuse. Again, another
character, though perhaps less important, {255}is the sloping
backwards instead of forward of the occipital crest in all two-horned
species, whether African or Asiatic.
The Asiatic Rhinoceroses have, what the African animals have not,
functional incisor teeth throughout life. It has been proposed on these
and other grounds to separate generically the African and Asiatic
forms.
Fig. 130.—Indian Rhinoceros. Rhinoceros indicus. × 1⁄40.
The Asiatic Rhinoceroses include three well-differentiated species, in
all of which the skin is much thrown into folds. Rh. indicus is
the largest form. It is one horned, and has enormous folds of skin at the
neck and hanging over the limbs. So like artificial armour is this thick
plating, that Albrecht Dürer may be excused for having given the beast
the appearance of being actually mail-plated in a sketch which he made of
a specimen sent over to the King of Portugal in 1513. This particular
beast, one of if not the first sent over to Europe, proved so intractable
in disposition that the king sent it as a present to the Pope. But "in an
access of fury it sunk the vessel on its passage"! The horn of this and
of other species was held until almost our times to have medicinal and
other more curious values. So recently as 1763 it was gravely asserted
that a cup made of its horn would fall to pieces if poison were poured
into it. "When wine is poured therein," wrote Dr. Brookes in the year
referred to, "it will rise, ferment, and seem to boil; but when {256}mixed with
poison it cleaves in two, which experiment has been seen by thousands of
people." John Evelyn also wrote of a well in Italy which was kept sweet
by a Rhinoceros' horn. This species seems to be long-lived, even in
captivity; a specimen now to be seen in the Zoological Society's Gardens
has been there since the year 1864.
Rhinoceros sondaicus, the Rhinoceros of the Sunderbunds, has a
much wider range than the last species or Indian Rhinoceros. This is
unknown out of India itself, and is there limited to a small region; the
Sondaic form is found in Bengal and in the Malayan Islands. It is a
smaller species, and the armour has a tesselated appearance. The female
generally, if not always, is hornless.
Fig. 131.—Sumatran Rhinoceros. Rhinoceros sumatrensis. × 1⁄15. (From Nature.)
The Sumatran species, Rhinoceros sumatrensis, is to be
distinguished from the last two by its two horns. It is also covered {257}by a
much thicker coat of hairs, which are sometimes blacker and sometimes
redder. On account of its two horns it has been proposed to separate it
from the other Oriental species into a distinct genus,
Ceratorhinus. The animal has much the same range as the last
species, but extends to Borneo. A variety of this species with hairy
ears, from Assam, has been separated as a distinct form, under the name
of Rh. lasiotis, by Mr. Sclater. The animal upon which that
species was founded was until quite recently living in the Zoological
Society's Gardens.
Fig. 132.—Hairy-eared Rhinoceros. Rhinoceros lasiotis. × 1⁄30.
There are only two certainly-known species of Rhinoceros in Africa.
These are the White Rhinoceros (Rh. simus) and the Black
Rhinoceros (Rh. bicornis). The origin of the names is not easy to
understand, since the "white" animal is, if anything, darker in colour
than the Black Rhinoceros. It is stated, however, that in past years the
specimens of Rh. simus found in the south-west of Cape Colony were
"paler and whiter in colour than those in the north-east." At present
there are no grounds for distinguishing the species by their colour
characters. But they are plainly distinguishable on other grounds.
Rhinoceros simus has a square upper lip, and in relation to this
crops the herbage upon the ground. Rh. bicornis has a prehensile
upper lip projecting beyond the lower, and in a corresponding fashion
feeds principally upon the branches of shrubs, It has been pointed out by
Mr. {258}Coryndon[161] that the calf of Rh. simus
"always runs in front of the cow, while the calf of Rh. bicornis
invariably follows its mother." Both animals of course have two horns,
and upon the varying proportions of the horns a large number of "species"
have been made in the past. It is stated that the longest horn of the
"White Rhinoceros" known measures 56½ inches; while that of R.
bicornis is shorter, 40 inches being apparently the maximum. But the
animal is smaller.

Fig. 133.—Head of Rhinoceros bicornis.
The possible third African species of Rhinoceros[162] has been provisionally
named after Mr. Holmwood, and is based upon two horns 41 and 42 inches
long, which may be abnormal horns of Rh. bicornis; but they are
thinner and have a smaller pedicel.
Extinct Rhinocerotidae.—The existing Rhinoceroses are
thus confined to Africa, to certain parts of the continent of Asia, and
to some of the large islands lying to the south of that continent. But
formerly the genus, and allied genera, had a wider range. As far back as
the Miocene we meet with remains of Rhinoceroses closely allied to
existing forms. The more ancient forms have, as is natural, more ancient
characters. Thus in Rh. schleiermacheri of the Miocene, canines
appear to have been present. The Miocene Aceratherium, primitive
in the absence of horns as its {259}name denotes,[163] had also canines and, in one species,
six incisors in the lower jaw. This Aceratherium had, moreover,
four toes in the fore-feet. In the Miocene and later the Rhinoceros
existed in Europe and America. There was even a purely northern form, the
Rh. tichorhinus, which possessed a woolly covering and had the
same range as the Mammoth. This Rhinoceros was two-horned.
The post-Pliocene and European Elasmotherium was a colossal
rhinocerotine creature. This great beast had two horns and a body 15 feet
long. Its limbs are not known, and as the teeth are different from those
of Rhinoceroses in general, it may not have belonged to this group at
all, though Osborn is inclined to derive it from Aceratherium,
admitting at the same time that the evidence is "decidedly slender." The
teeth in fact are like those of a Horse in being hypselodont and
prismatic in form. As to the two horns, they were apparently not exactly
like those of typical Rhinoceroses; there was an enormous horn
posteriorly, supported on a huge boss of bone, and in front of this a
roughened spot suggests a smaller or at least a much more slender
horn.
It is important to notice that fossil Rhinoceroses belonging to the
restricted genus Rhinoceros were in Europe invariably two-horned;
it is only in India, where they still exist, that one-horned forms are
met with in a fossil state.
Fig. 134.—Skeleton of Hyracodon nebrascensis. × 1⁄12 (After Scott.)
The Rhinoceroses of America were mostly hornless. Diceratherium
is an exception; but in many cases it had two parallel not successive
horns, and these were, to judge from the slight prominences, but feeble
in development, and perhaps hardly exactly comparable with the formidable
weapons of the Old-World forms. Aceratherium tridactylum, with
indications of paired horns, may be ancestral to Diceratherium.
The American forms have weak and slender nasals in correspondence with
the absence of horns; the sagittal crest is retained in contradistinction
to the great flattened surface of the skull in the horned Rhinoceroses.
Aceratherium of both divisions of the globe probably represents
the ancestral group of the horned and the hornless forms. This being the
case it is highly interesting to note a distinct convergence in the quite
{260}separate American genera towards the
European horned genera. A genus sometimes united with
Aceratherium, but still differing from it in some points, is
Aphelops (Teleoceras).[164] This animal is more nearly
approximated to "the modern standard" of Rhinoceroses than is its
possible ancestor Aceratherium. The skeleton in general is more
robust, even surpassing that of modern forms, and approaching the
Hippopotamus. There is a reduction in the upper incisors, which
are limited to two pairs, and the lower molars {262}are
reduced to five. The lower incisors are only two. The sagittal crest is
less marked; the fifth digit is reduced to a tiny nodule representing the
metacarpus. It had a small nasal horn. There are numerous other details
of likeness to modern Rhinoceroses in this creature, which has only
community of descent with them from the older hornless forms, such as
Aceratherium and Caenopus. In the genus Peraceras
the upper incisors are as completely gone as in the living African
Rhinoceroses.

Fig. 135.—Skeleton of Aphelops (Teleoceras) fossiger. × 1⁄15. (After Osborn.)
The most ancient rhinocerotine types[165] are the Hyracodonts and the
Amynodonts. They both date from the Eocene, and became extinct in the
succeeding Oligocene. Hyracodon[166] (Fig. 134) was "an agile,
light-chested, and rather long-necked" type, resembling a Horse in build.
There were no horns present, but the hoofs were more like those of the
Horses than of the existing Rhinoceroses. These animals were apparently
plain dwellers and defenceless, which is held to account for their
compact hoofs and outward similarity to a Horse. The genus is Oligocene.
The dental formula is I 3/3 C 1/1 Pm 4/3 M 3/3.
It is surmised by Professor Scott that the number of dorso-lumbar
vertebrae was twenty-three or twenty-four. The radius and ulna are
complete and separate bones, but the latter is somewhat reduced. There
are four metacarpal bones, of which, however, the fifth is much reduced.
The animal is only three-fingered. The tibia and the fibula are distinct,
and show no tendencies towards fusion; but the fibula is much reduced.
There are only three metatarsals and three toes. Had this line, which is
to be regarded as a side branch of the Rhinoceros stem, not died out, it
would probably have resulted, thinks Professor Scott, in
monodactyle—very Horse-like types. It is later than the next genus
to be described, Hyrachyus, of which it is possibly a descendant.
An intermediate type, Triplopus, appears to bind together
Hyracodon and Hyrachyus.
In Hyrachyus agrarius the skull is long and narrow, the facial
region being markedly longer than in existing Rhinoceroses. The mastoid
portion of the periotic bone is widely exposed upon the outer face of the
skull, which is, as has been said, not the case with the existing genus
Rhinoceros. The dentition is the complete Eutherian dentition of
forty-four teeth. The upper {263}molar teeth are strikingly like those of
the genus Rhinoceros. The fore-feet are pentadactyle, but
functionally tetradactyle; the hind-feet tridactyle. The ulna is less
reduced than in Hyracodon, and the dorso-lumbar vertebrae are
twenty-five.
Fig. 136.—Skeleton of Metamynodon planifrons. × 1⁄22. (After Osborn and Wortman.)
The Amynodonts were short, heavy types, probably marsh-haunting in
habit, and possibly with a proboscis like that of the Tapir. The orbit is
higher than it is in the purely terrestrial {264}Hyracodonts, and it is
suggested that when swimming it was raised above the surface as with the
Hippopotamus. "This feature," observes Professor Osborn, "with the long
curved tusks, undoubtedly used in uprooting, suggests the resemblance
between the habits of these animals and those of the hippopotami." There
were no horns in the Amynodonts. The face is shorter than in the
Hyracodonts, and the mastoid is covered as in recent Rhinoceroses. The
canines are very strongly developed into tusks, but the incisors show
signs of disappearance. We know of the genera Amynodon,
Metamynodon, and Cadurcotherium. All except the last, which
is European, are American in range.
Fam. 4. Titanotheriidae.—These Oligocene Ungulates, often
attaining to large dimensions, are nearly peculiar, so far as is at
present known, to the North American Continent, and are at least most
abundant in it.[167] Many
generic names, such as Titanotherium, Brontotherium,
Brontops, Titanops, and Menodus, have been given to
them; but a recent study of the entire material accessible for
description or already described has led Professor Osborn to the opinion
that there was but a single genus, to which the name Titanotherium
must be applied. Of this genus there are some thirty well-characterised
species, of which the gradual evolution can be traced from the lowest
strata of the White River beds where their remains occur. An entire
skeleton of T. robustum enables us to understand the osteology of
these forms and to compare them with other Perissodactyles. This animal
was more than 13 feet long, standing some 7 feet 7 inches in height. It
seems to have presented during life the aspect of a Rhinoceros with
perhaps a touch of Elephant. The skull is not unlike that of a Rhinoceros
in general dimensions and shape; but there are a pair of apparent horn
cores anteriorly, which are smaller in the more ancient forms and acquire
a large size, a forward direction with a divergence of the two in the
later forms. A glance at the accompanying figures of skulls (Fig. 137) of
early and later Titanotheres will exhibit the changes in this particular
which the skulls underwent in the lapse of time occupied by the
deposition of these Oligocene beds. The nasals are short in the later,
longer in the more early species, such as T. heloceras and T.
coloradense. The zygomatic arch projects much, and is "shelf-like" in
the later forms, the skull thus getting an immense breadth, which, {266}together with the long and divergent horn
cores, must have given to the living animal a most bizarre appearance. It
is an interesting fact that this animal, though a Perissodactyle, agrees
with the Artiodactyla in the nineteen dorso-lumbar vertebrae, of which
seventeen bear ribs.

Fig. 137.—Three figures showing the cranial evolution of Titanotherium. Upper figure,
T. trigonoceras; middle figure, T. elatum; lower figure, T. platyceras. (After Osborn.)
The genus further agrees with the Artiodactyles in the structure of
the carpus. The toes of the fore-limb are four, those of the hind-limb
three; but while the hind-limb is undoubtedly Perissodactyle in the
arrangement of its component parts, the fore-limb shows a hint of an
Artiodactyle mode of structure. This limb is paraxonic, the axis of the
limb passing between the two middle digits. It may be that this genus
represents more nearly than any other Perissodactyle or Artiodactyle the
primitive stem from which both have diverged, though, of course, it is
not old enough to be very near to the actual ancestor. The molar
dentition is the typical one; the incisors seem to vary as to their
presence or absence, and, if present, in their numbers. In comparing the
older with the more recent forms it is noteworthy that there has been an
increase of size exactly as there has been during the evolution of the
Camels and some other groups of Ungulates. As already mentioned, the size
of the horn cores also increases until it culminates in the extraordinary
species, T. platyceras and T. ramosum, in which these are
half as long as the skull, flattened in form, and connected at their
bases by a "web" of bone. Arrived at this amount of specialisation the
genus Titanotherium apparently exhausted its capacities for
modification and ceased to be. The many generic names may be explained by
sexual differences on the one hand and an incomplete knowledge of
connecting links on the other.[168]
Palaeosyops is somewhat like a Tapir in build, the skull
especially resembling that of the Tapir. As in Titanotherium the
molar teeth, instead of having an outer wall formed by fused cusps, have
a -shaped outer wall on one side and two or one cusps on the opposite
side. It is, moreover, an Eocene form, and in correspondence with its
greater age is more primitive in some points of structure, for example,
in the absence of horns and in the full dental formula. The fore-limbs
are four-toed, the hind {267}three-toed. It was intermediate between a
Tapir and a Rhinoceros in size. It has been shown, too, from casts of the
interior of the skull, that the cerebral hemispheres are much less
convoluted than were those of Titanotherium.
Related to Palaeosyops is another primitive Titanothere, the
genus Telmatotherium. This is also Eocene, from the Uinta Basin,
the uppermost of Eocene strata. The skull of these creatures was rather
elongated, and not unlike that of a Titanothere in general aspect. The
dentition was complete and the canines not very large. The horns, which
acquire so prodigious a development in the later Titanotheres, are just
recognisable in at any rate many species of this genus
Telmatotherium, the name being thus by no means an apt one. Better
was that proposed by Dr. Wortman, of Manteoceras or "prophet
horned." The horns are small elevations upon the frontals just at the
junction of these with the nasals, and, indeed, lying partly upon the
latter bones. In T. cornutum the horns are chiefly borne upon the
very long nasals, whose size contrasts with the same bones in the more
highly-developed Titanotherium. It appears to be quite possible
that Titanotherium was evolved from the genus
Telmatotherium.[169]
Sub-Order 9. LITOPTERNA.
Whether the Macraucheniidae should be considered as a separate
group of Ungulata is a matter of dispute. Cope placed them in a special
order of Ungulates which he called Litopterna. Zittel, on the other hand,
regards them as definitely Perissodactyles. One curious point of
resemblance to existing Horses is shown—that is the presence of a
pit in the incisor teeth. This matter seems to be so important as to need
a placing of these forms in the neighbourhood of the Perissodactyles,
even of the Equidae; it is so peculiar a character, and apparently so
little related to any obvious similarity in way of life, that it seems to
mark a special affinity. Not so the fact that in Macrauchenia at
any rate the orbit was entirely surrounded by bone as in the Horse. We
find that condition so frequently acquired in many groups,—a
development from an earlier condition where the cavity for the lodgment
of the eye is in continuity with the temporal {268}fossa, that it cannot
be regarded as anything more than a mark of specialisation. It is, in
fact, the case that the Macraucheniidae are in many points specialised,
while retaining many primitive features of structure.
The chief primitive features are: the non-alternating positions of the
wrist- and ankle-bones; these, of course, interlock in the
Perissodactyles of to-day and in many extinct families. Then the absence
of a diastema in the tooth series, coupled with the presence in
Macrauchenia of a complete dentition. The small brain may be
referred to the same category. Macrauchenia must have been a
strange-looking animal. It walked upon three toes on each limb; the skull
was Horse-like in general form, but the nostrils are removed to a point
about as far back as in the Whales or nearly so, the nasal bones being
correspondingly reduced. This it is thought argues a proboscis. The
humerus is particularly compared by Burmeister[170] to that of a Horse. The radius and
ulna though both well developed are fused. The neck is long, and, as in
the Camel, the vertebral arteries run inside the neural arches. Since the
fore-legs seem to have been rather longer than the hind-legs, though only
very slightly, and the neck was long, the animal may have presented some
likeness to the Giraffe. It is interesting to note that in the
proportions of humerus to ulna this animal is more Lama-like than
Horse-like. On the other hand, the proportions of femur to tibia are more
Horse-like. The remains of the creature are limited to South America, and
to quite superficial deposits. It is evidently a specialised type, and
has pursued a course parallel to that of the Horse. Much nearer to the
Horse however, but apparently by convergence only, is the genus
Thoatherium, usually placed in a separate family, the
Protorotheriidae. In this creature, which has many archaic
characters, the toes are reduced to one in each foot. In an allied form,
Protorotherium, we have the two lateral toes diminishing just as
in Anchitherium.
{269}
CHAPTER XI
UNGULATA (continued)—ARTIODACTYLA (EVEN-TOED
UNGULATES)—SIRENIA
Sub-Order 10. ARTIODACTYLA.
Fig. 138.—Bones of the Manus—A, of Pig (Sus scrofa). × ⅓. B, of Red Deer (Cervus
elaphus). × ½. C, of Camel (Camelus bactrianus). × ⅛. c, Cuneiform; l, lunar;
m, magnum; m2, m5, second and fifth metacarpals; R, radius; s, scaphoid; td,
trapezoid; u, unciform; U, ulna; II-V, second to fifth fingers. (From Flower's
Osteology.)
The Artiodactyle or "Even-toed" Ungulates are to be distinguished from
the Perissodactyla, and from other Ungulate groups, by a number of
trenchant characters. The most salient {270}of these, and that
which has given its name to the group, concerns the arrangement of the
digits. Instead of there being but one prevailing digit—the third,
in the hand and foot, through which the axis of the foot passes, there
are two, numbers three and four, between which the same axis passes, and
which are perfectly symmetrical with each other. This type of foot has
been termed "paraxonic," as opposed to the "mesaxonic" Perissodactyle
foot (see Fig. 121 B, p. 235). It has been
attempted to prove that the single prevailing digit of the Horse's foot
is a fused pair of digits, and the state of affairs which characterises
the Camel, where the two metacarpals or metatarsals are to an almost
complete extent united, has been urged in proof; so, too, certain
abnormalities, such as those called "solid-hoofed pigs."[171] These latter are simply Pigs in which
the two central metacarpals and the terminal hoofs are completely fused
with one another. In some of such cases there is not the slightest trace
of the union of the separate metacarpals and phalanges. Even the sesamoid
bones, attached behind to the toes, are two in number instead of four.
And, furthermore, the tendon supplying the bones is single, though
showing traces of its double origin. Such Pigs often show the abnormality
from generation to generation, and they proved convenient for those whose
scruples would not allow them to eat the flesh of a beast "dividing the
hoof" and not chewing the cud. More singular still, as showing a
pathological approach from another side to the Perissodactyle condition
in an Artiodactyle, is a calf, where the foot ended in three equi-sized
digits, of which the middle one lay in the longitudinal axis of the limb.
From the opposite side cases are known of a Horse with a split hoof and
phalanges, thus presenting the most striking likeness to a Camel.
There is, furthermore, in certain groups of Artiodactyles (e.g.
the Tragulidae) a tendency for the two middle metacarpals to unite, quite
apart from such "sports" as those illustrated by the cases just set
forth. And, as already mentioned, the union of the two middle metacarpals
culminates in the Camel, Ox, etc. There is, however, absolutely no trace
of such a fusion in the series of Perissodactyle animals known to us; and
it would be by fusion rather than dismemberment that, as it would appear
on this theory, the modern Ungulate foot has been arrived at. Of course
{271}the facts of Ungulate descent are
absolutely destructive of any such comparisons.
As is the case with the Perissodactyles, the Artiodactyles show a
historical series, the primitive five-toed condition being almost
preserved in Oreodon, up to the most modern modification
exemplified by the Ox, Sheep, etc., in which animals there are not even
vestiges of the fourth and fifth toes. It has been stated, however, that
the foetal Sheep has traces of those rudiments. The so-called cannon bone
(the fused third and fourth metapodia) is accompanied in its fusion by an
increase in length. At the same time the functional middle metacarpals
push aside the rudiments and, forming a broad surface for that purpose,
articulate with the magnum and unciform bones to the exclusion of the
rudiments. This has been termed an "adaptive reduction." In the
"inadaptive reduction" there is the same reduction of the metacarpals,
but the rudiments still articulate as in the primitive Artiodactyle foot,
i.e. Mc II with trapezium, trapezoid, and magnum; Mc III with
magnum and unciform; Mc IV and V with unciform. This would appear to give
greater solidity and consequently greater strength to the foot.

Fig. 139.—Dorsal surface of
right tarsus of Red Deer
(Cervus elaphus). × ⅓.
a, Astragalus; c, calcaneum;
c3, cuneiform;
cb, cuboid; mIII, mIV,
metatarsals; n, navicular.
(From Flower's
Osteology.)
The carpal bones of the Artiodactyla alternate in their articulation;
the primitive state of affairs[172] is not retained even in the earliest
types. The femur has no third trochanter, so prevalent in the
Perissodactyles. In the hind-foot the calcaneum has an articular facet
for the fibula, which is not characteristic of the Perissodactyla. In the
more modern forms, e.g. the Cervidae, the navicular and cuboid
become fused into one bone; and there are even further fusions which will
be referred to later as characteristic features of different groups. It
is interesting to notice that the reduction begins earlier and is clearer
in the hind-foot than in the fore. One {272}can see how this may be
purely adaptive, the push of the hind-legs in running needing a firmer
support. In Hyomoschus this is the case. The hind-limbs are
provided with a cannon bone, while the metacarpals of the fore-feet are
still free.
The number of dorso-lumbar vertebrae is less in the Artiodactyle than
in the Perissodactyle Ungulates. Whereas the former have but nineteen,
the latter have, as a rule, twenty-three such vertebrae.[173] The number of ribs varies from twelve
(Camelus, Hydropotes) through thirteen (Cervus,
Gazella) to fourteen in Dicotyles, Giraffa, etc.
The curious form of teeth known as "selenodont" is characteristic of
the Artiodactyla, though only found well developed in the modern forms,
and of those only in the Pecora. The more primitive forms had "bunodont"
teeth with typically four tubercles (if we except the tritubercular and
but little-known Pantolestes); and the intermediate
"buno-selenodont" type characterises such groups as the
Anthracotheriidae.
While the stomach of the Perissodactyles is always a simple sac, it is
complicated, or shows signs of complication, in the Artiodactyles. That
of the Hippopotamus is divided into two chambers; there are three in
Tragulus, and four in the typical Ruminants such as Cervus,
Ovis, etc.
Had we to deal only with the still living genera of Artiodactyles, it
would be easy to sort them into two groups on the characters of the
teeth; for the Pigs and Hippopotamus are provided with tubercular molars;
they are bunodont. The Deer, Camels, Oxen, Giraffes, etc., have
selenodont molars. Besides, the latter are "Ruminants," and have a more
complicated stomach. The existing Chevrotains forbid a more trenchant
division, since they are, as will be pointed out in due course, somewhat
intermediate in structure; the feet are more Pig-like, and the stomach is
not so typically Ruminant. In any case such a division is prevented by
certain extinct families which are perhaps ancestral to both. They have
teeth which are not quite bunodont and not quite selenodont. These teeth
have been termed buno-selenodont or buno-lophodont.
The distribution of the living Artiodactyles presents us with some
interesting facts. The vast preponderance of species occurs in the Old
World—34 in America as against over 250 species {273}in Europe,
Asia, and Africa. The Neotropical region has no Oxen, or Sheep, or
Antelopes. The latter are confined to Africa, Asia, and certain parts of
the Palaearctic region; they are vastly more prevalent in Africa, where
they take the place of the totally absent Deer. The Pig tribe is almost
entirely Oriental and Ethiopian in distribution, only one form, the
European Wild Boar, ranging into the Palaearctic region; and the two
species of Peccary are found in both North and South America. Broadly
speaking, the Ethiopian region is the headquarters of the Artiodactyla.
But the great island of Madagascar has but one form of Artiodactyle, a
Pig of the genus Potamochoerus.[174]
Group I.—SUINA.
Fam. 1. Hippopotamidae.—The family Hippopotamidae
contains of existing genera only Hippopotamus, for the Liberian
dwarf Hippopotamus is not now regarded, as it was formerly, as the type
of another genus, Choeropsis. The reasons for its former
separation were the loss of the outer pair of incisors and the different
proportions of various parts of the skull. This little Liberian animal
has, however, been shown by Sir W. Flower[175] to possess the missing incisors
occasionally; and as to the proportions of the skull, it is exceedingly
common for small animals to vary from larger relatives in this way.
Hence, considering the characteristic features of the Hippopotamus and
the fewness of species, it seems unnecessary to divide it up further. We
shall therefore only recognise one genus.
The Hippopotamus at present is African in range, and confined to that
continent. But quite recently it inhabited Madagascar; and further back
still in time the existing African species, H. amphibius, ranged
into Europe; there were also Indian forms, which were contemporary with
the Stone-age man. The Common Hippopotamus is a great thick-skinned beast
with but few hairs. It has four toes on each foot, a complex stomach, but
no caecum. The strong incisors continue growing through life, as do the
great canines. The number of incisors is two on each side of each jaw.
Some of the extinct species had six in each {274}jaw, and they were
distinguished as a genus Hexaprotodon, contrasting with
Tetraprotodon, until intermediate conditions were observed.
Choeropsis, as already observed, was a still further reduction of
the tetraprotodont type. The molars (the formula is Pm 4/4 M 3/3) when
worn show a double trefoil pattern. The orbital cavity is encircled by
bone. As with many other aquatic mammals the kidneys are lobulated.

Fig. 140.—Hippopotamus. Hippopotamus amphibius. × 1⁄40.
A very singular fact about the Hippopotamus is the production of a
"bloody sweat," a carmine-coloured secretion, containing small crystals
and corpuscles, from the skin. This coloured fluid has of course nothing
to do with blood.[176]
The animal grows to a length of at any rate 14 feet. The limbs and the
tail are short. Like other aquatic animals the nostrils are on the
surface of the head, and can be closed when the animal is under water.
When it reaches the surface of the water after a prolonged immersion, it
spouts like a Whale. Sir Samuel Baker says that ten minutes is the
longest time that the Hippopotamus can remain below the water. It is
frequently a dangerous animal to encounter, as it will capsize boats, and
even bite large pieces out of their bottoms; with its huge teeth it {275}can
and does attack and destroy human beings. The Hippopotamus not only
swims, but can walk along the bottom of a river with great rapidity. It
occasionally puts out to sea from the mouths of rivers frequented by it;
and it is supposed that in this way Madagascar was populated with
Hippopotamuses, whose remains are now found in swamps in that island.
Fig. 141.—Wild Boar. Sus scrofa. × 1⁄12.
Fam. 2. Suidae.—The Pig family, Suidae, differ from the
last in their smaller size, in the terminal nostrils and mobile snout,
which is not grooved, except faintly as in Babirusa. They are
generally hairy, but the Babyroussa is an exception, while
Phacochoerus is but slightly haired. Though there are four digits,
as in the Hippopotamus, only two reach the ground in walking. The
stomach, furthermore, is simple, and (except in Dicotyles) there
is a caecum. The kidneys are smooth, and the liver is more lobate than in
Hippopotamus. The orbital cavity is confluent with the temporal
fossa. The typical genus, Sus, is distributed over Europe, Asia,
and the islands of the Malay Archipelago, reaching as far as Borneo and
Celebes. The dentition[177] is complete. A single species, the
so-called S. sennaariensis, is from Ethiopian Africa, but it is
not certain how far this animal may be an escaped species introduced by
man. A very large number of "species" of Sus have been described,
but Dr. Forsyth {276}Major is disposed to reduce them to four
if not to fewer species. He allows the widely-ranging S. scrofa,
S. vittatus, and the eastern Malayan S. verrucosus and
S. barbatus.

Fig. 142.—Pygmy Hog (from Nature). Sus salvania. × 1⁄6.
The Pygmy Hog of the Bhotans seems to be not entitled to specific
rank, certainly not to generic (in the opinion of some), though it has
been termed Porcula salvania.[178] The Wild Boar of Europe is Sus
scrofa. It was formerly quite abundant in this country; not merely
are its remains exhumed from fens, caves and peat bogs, but there is
ample evidence of its continuance down to a comparatively late historic
period. Enactments are on record as to the hunting of these animals;
there are places, such as Boarstall, whose names are clearly derived from
the name of the animal, presumably once a native of the locality; and
various documents all show the presence of the Wild Boar {277}in this
country down to so late a period as the end of the sixteenth century.
Fig. 143.—Wart Hog. Phacochoerus aethiopicus. × 1⁄6.
The African Wart Hog, genus Phacochoerus, is usually regarded
as the type of a distinct genus of Pigs. This animal, "superlatively
ugly" with its huge tusks and great protuberances upon the face, is
chiefly to be distinguished from the genus Sus by these
characters, and by the complexity of the last molar, which, with the
tusks, are sometimes in aged animals the only teeth left. The complete
formula is Pm 2/2 M 3/3. There are two species of this genus, P.
aethiopicus and P. africanus. When enraged the Wart Hog is
said to carry its tail directly up, and to present a ludicrous as well as
ferocious appearance.
Fig. 144.—Head of Wart Hog.
The Celebesian Babyroussa, genus Babirusa, is an almost
hairless hog with enormously upturned tusks in both jaws of the {278}male. In
the Wild Boar there is a hint of this, which is carried still further in
Phacochoerus; but in Babirusa the upper tusks turn upwards
before they leave the substance of the jaw, for which reason they appear
to arise on its dorsal surface; the lower tusks are nearly as long. It
has been found that the young of this Pig are not striped as are those of
other Pigs. By means of the curved upper tusks this animal has been said
by old writers to suspend itself to branches of trees, just as does by
his downwardly-projecting tusks the male Chevrotain! There is but one
species, B. alfurus.
From Sus proper the African and Malagasy Potamochoerus,
including the Red River Hog, is barely separable generically. Their
principal claim to generic distinction lies in the existence of a horny
outgrowth arising from a bony apophysis above the canine in the male.
These have been compared to the osseous "horn cores" in the extinct
Dinocerata. But the Javan Sus verrucosus shows at least the
beginning of a similar modification. The popular name of the animal is
derived from the fine rufous colour of its pelage, not seen, however, in
all the species. Dr. Forsyth Major[179] recognises five species, of which
only one is from Madagascar.
Fig. 145.—Peccary. Dicotyles tajaçu. × 1⁄6.
Fam. 3. Dicotylidae.—The Peccaries are generally placed
in a different family from that of the other Pigs. This family, {279}Dicotylidae, contains but one genus,
Dicotyles, with at most two species. The name of the animal is
connected with the dorsal gland; the animal thus appeared to possess two
navels. The Peccaries, exclusively confined to the New World, differ from
the Old-World Pigs in one or more important characters. They have only
three toes on the hind-feet, and the stomach is complicated. Though the
Peccaries have but small tusks they hunt in packs and are very dangerous
animals to meet with. They owe, too, their safety from many foes to their
sociable habits. Being nocturnal animals they are liable to the attacks
of the Jaguar, which will speedily overpower and devour a Peccary that
has strayed from its herd.
Fossil Swine.—The existing genera of the Pig tribe are
also known in a fossil condition. Sus itself goes back as far as
the Upper Miocene. Sus erymanthius, the Erymanthine Boar, is known
from beds of that age in Greece, England, and Germany. This genus is not
known to have had a wider distribution in the past than it has in the
present. Dicotyles occurs in the Pleistocene of both North and
South America, the regions which it inhabits at the present day. The
genus Listriodon, also Miocene, is remarkable for having lophodont
instead of bunodont teeth, that is so far as concerns the molars, which
resemble those of the Tapir. It was European and Indian in range. A
number of genera, more remote from the existing Pigs than those which
have just been dealt with, are placed together in a special sub-family,
Achaenodontinae. The type genus, Achaenodon, had a somewhat short
skull for a Pig; and it is in general aspect and in the characters of the
canine teeth highly suggestive of that of a Carnivore. The bunodont
molars, however, are Suine, as is the form of the lower jaw with a
rounded angle. This is an Eocene animal found in Wyoming.
Elotherium[180]
occurs chiefly in the Miocene of both North America and Europe; but E.
uintense is Eocene. The orbits are completely encircled by bone in
the more modern forms; this is not the case in the last-described genus,
with which E. uintense agrees. The skull is also longer and more
Pig-like. The zygomatic arch is powerful, with sometimes a large
descending process, such as is found in Diprotodon, more faintly
in Kangaroos, and in Sloths and certain extinct Edentates. The lower jaw
has a pair {280}of dependent processes near the symphysis,
which suggest processes occupying a corresponding position in
Dinoceras. The skull and body are heavy, but the two-toed limbs
are slender. There is a smaller pair of toes behind these. The dentition
is complete, and the canines are not inordinately developed. The brain is
very diminutive. Perhaps E. uintense should be separated as a
distinct genus, Protelotherium.[181]
Hyotherium (which is regarded as identical with
Palaeochoerus) has a sharp sagittal crest; the orbit is nearly but
not quite closed. The canines are not strongly developed. The upper
canines have double fangs as in Triconodon among extinct mammals,
and as in the Hedgehog and other forms among living Mammalia. The
premolars have the cutting and serrated edge of those of some other Pigs,
a feature which gives them a curious resemblance to the "grinding" teeth
of Seals. The molars are tuberculate, and like those of living Pigs. It
is European and Indian in range, and Miocene.
The genus Choeropotamus has a complete dental formula save for
the loss of a premolar in the lower jaw. Though it has lost this tooth,
it is from an older stratum than some of those forms which have retained
that premolar; it has been found in the Upper Eocene of the Isle of Wight
and of the neighbourhood of Paris.
The American and Miocene Chaenohyus has lost the corresponding
teeth of the upper jaw.
Homacodon[182]
is a genus consisting of several species, which has a bunodont and
complete dentition. The molars are sextubercular in the upper jaw. H.
vagans was of about the size of a Rabbit, and it appears to have had
a curved neck. The limbs had five digits, as is so generally the case
with Eocene Ungulates. It is known from the Middle Eocene of Wyoming.
Group II.—RUMINANTIA.
The Selenodontia or Ruminantia form the second division of existing
Artiodactyles. The characters of the teeth, which give them their name,
have already been referred to. They also differ in that there are never
more than a single pair of incisors {281}in the upper jaw, and
very usually there are none. As a general rule the third and fourth
metacarpals and metatarsals become united to form a cannon bone. To this
there is but one exception, the African Hyomoschus. Moreover, the
second and fifth digits are nearly always rudimentary, and may
practically disappear altogether. Here again the Tragulidae are an
exception. The Ruminantia are so-called on account of the fact that they
"ruminate," that is, after the food has been rapidly swallowed, it is
forced back up the gullet and more thoroughly masticated. Associated with
this is a complex stomach, which is divided into several compartments.
This stomach has at least three compartments, as in the Tragulidae; but
it has usually four. Its characters are illustrated in Fig. 146. The
majority of the Selenodontia possess horns, which are partly formed of
solid protuberances of the frontal bones. In the Giraffe they are
somewhat different.

Fig. 146.—Stomach of Ruminant opened to show the internal structure. a, Oesophagus;
b, rumen; c, reticulum; d, psalterium; e, abomasum; f, duodenum. (After
Flower and Lydekker.)
This group may be divided into—A. Tragulina, Chevrotains; B. Tylopoda, Camels, Lamas; and C. Pecora, Deer, Antelopes, Oxen, Giraffes, Goats,
Sheep.
A. TRAGULINA.
As the Tragulina are undoubtedly the most ancient of the Selenodontia
it will be logical to commence with an account of them. {282}
Fam. 4. Tragulidae.—This family comprises a number of
small Deer-like animals, which are really in many points more related to
the Pigs than to the true Deer. They are known as Chevrotains; and the
term "Deerlet," introduced by Professor Garrod, is certainly appropriate,
since they have the aspect of very small and hornless Deer. If it were
not for their Artiodactyle feet one might at a glance confuse these
creatures with some Marsupial type. The family is Oriental and West
African in range. The two genera (whose individual peculiarities will be
considered later) differ from other Artiodactyles in a number of rather
important characters.
Fig. 147.—Indian Chevrotain. Tragulus meminna. × ¼.
They are absolutely hornless in both sexes. The canines are present in
both jaws, and are especially well developed in the upper jaw. The dental
formula is I 0/3 C 1/1 Pm 3/3 M 3/3. In the skull the tympanic bulla is
usually, as in the non-ruminating Artiodactyles, filled with loose bony
tissue. The feet (usually) have the four toes of the Suina, and are
therefore in a more primitive condition than in Deer and Antelopes. But
as the middle metacarpals are fused in Tragulus (though separate
in Hyomoschus) they are a stage further than are the Pigs, in the
direction of the typical Ruminants.
The stomach is comparatively simple, thus offering {283}intermediate
characters between the Pigs and the Ruminants; there are but three
separate compartments. A highly-interesting character is afforded by the
placenta. This is in the present family of the diffuse kind, not
presenting the separated and tufted cotyledons of the Ruminant placenta.
This we may fairly assume is a further proof of the less-specialised
characters of this group[183] as compared with the Ruminantia, a
view, however, which is not universally accepted. While the molars have
the selenodont character of other Pecora, the premolars are more adapted
for cutting, with sharp edges.
The genus Tragulus consists of several species (e.g. T.
stanleyanus, T. napu, etc.), which have been aptly compared in
external appearance to certain Rodents such as the Agoutis. The legs are
delicate and slender, hardly "thicker than an ordinary cedar pencil."
These creatures have got among the Malays a considerable reputation for
astuteness, embodied in the saying, "Cunning as a kanchil." The
male has tusks, which greatly contributed to the confusion of this
creature with the totally different Musk Deer, Moschus
moschiferus. It is even said to suspend itself by their aid to the
branches of trees, and so avoid danger.
Hyomoschus (or Dorcatherium as it should properly be
called) is West African. Its rich brown colour, with spots and stripes,
is much like that of the Chevrotains, but it has shorter limbs. The only
species is D. aquaticum, which is sometimes called, on account of
its frequenting the banks of streams, the Water Chevrotain. Remains of
this genus occur in Miocene and Pliocene strata of Europe.
The separate metacarpals, comparatively simple stomach, absence of
horns, diffuse placenta, and spotted pelage are features which argue the
primitive position of these animals among existing Artiodactyles.
Besides the two existing genera which have just been treated of, there
are a number of extinct genera undoubtedly belonging to the same
group.
Gelocus (Eocene and Oligocene in range) is a European genus
known from France. It differs from the living members of the group by the
fact that the second and fifth toes on both hind- and fore-feet are
represented, as in certain Deer, {284}by rudiments at the
upper and at the lower end only; they are deficient in the middle. The
middle large metacarpals, though closely applied, are not fused. The
metatarsals, on the other hand, are, or are not fused, according to the
species. A later form is the genus Leptomeryx from the Miocene of
North America. This genus departs from the typical Traguline structure in
more than one point. The tympanic bulla is hollow instead of being filled
with cancellated bone; the cuneiform is not fused with the cuboid and
navicular, though the latter are with each other; the lateral digits of
the hind-feet are rudimentary. The magnum and trapezoid, however, are
fused. In the fore-feet the middle metacarpals are separate, and the
lateral less perfect metacarpals have toes. The metatarsals are
fused.
Not definitely referable to the Tragulidae, but coming near to them,
are the Protoceratidae. Of this family there is but one well-known
genus, Protoceras,[184] from the Miocene of North
America.
The skull is singularly reminiscent of Dinoceras, with which
this quite Artiodactyle genus has, of course, nothing to do. It merely
exemplifies the phenomenon of "parallelism." In general form it is
peculiarly long and low. There are three pairs of bony protuberances:
one, the largest, pair are situated on the maxillae rising up just behind
the implantation of the canine teeth; the parietals have a second pair;
and a third much more diminutive pair of bosses are upon the frontals,
near their junction with the nasals. This description refers to the male;
the female has only traces of the parietal bosses. These were all
possibly tipped or sheathed with horn or roughened skin. The dentition of
this genus is precisely that of the Tragulidae, i.e. I 0/3 C 1/1
Pm 4/4 M 3/3. The orbit is completely encircled by bone; the auditory
bulla is not swollen; the premaxillae are small.
The nasal cavity is very large and open, the end of the nasal bones
anteriorly being situated at about the middle of the skull; this would
seem to indicate at least a flexible and long nose like that of the Saiga
Antelope, if not a trunk.
The brain was of good size, and quite well convoluted.
The limbs are constituted on the Traguline plan; in the fore-limbs the
middle metacarpals are quite free from each other, and the more
diminutive lateral digits are complete. The {285}metatarsals are free,
but with a tendency to fusion; the lateral toes are only represented at
the upper extremity. The carpal bones are separated.
This animal, which was about the size of a Sheep, though of more
delicate proportions, was allied not only to the Tragulidae but to the
Giraffidae; it is impossible to refer it definitely to either family.
B. TYLOPODA.
Fam. 5. Camelidae.—This small group of Selenodonts
includes only the Camels and Lamas. The limbs are long and have no traces
of the second and fifth toes. The fused metacarpals and metatarsals
diverge somewhat at their distal ends. In the upper jaw is a single pair
of incisors. The stomach differs from that of the typical Ruminants. The
rumen has smooth and not papillose walls, and from it are developed the
"water cells," diverticula with narrow mouths provided with a closing
sphincter muscle. The psalterium is reduced to a mere vestige, and so the
stomach has, as in the Tragulina, but three chambers. This, so far
ancient, character in the structure of the Camel tribe is associated with
another, also seen in the more primitive Ungulates, viz. the diffuse
character of the placenta. A very singular peculiarity of this group is
the fact that the blood corpuscles instead of showing the ordinary
mammalian round contours are elliptical.
Fig. 148.—Bactrian Camel. Camelus bactrianus. × 1⁄30.
The genus Camelus, confined to the Old World, is made up of two
quite distinct species, the Bactrian Camel, C. bactrianus, with
two humps, and the Dromedary, C. dromedarius, with only one. The
former species is Asiatic. It is a singular fact that neither of the
species is known to occur in a genuinely wild condition. The so-called
"wild" Camels appear to be invariably feral. The two species will
interbreed; and there is at the Zoological Society's Gardens such a
hybrid, which has the general appearance and shaggy brown hair[185] of the Bactrian
animal, but the one hump of the Dromedary. It may be that the Bactrian
Camels of Lob-nor are really wild; but the desert contains so many
remains of cities destroyed by sand-storms that these reputed wild {286}Camels may be the descendants of animals
belonging to the inhabitants of those cities. A strayed herd of Camels
has established itself in a feral state in Spain. Otherwise the genus
does not occur in Europe. The Camels are also represented in the New
World. The genus Lama (Auchenia of many authors) belongs to
this family. These Camels differ from their allies in the Old World by
their smaller size, by the absence of the characteristic hump, and by the
dropping of one premolar, the dental formula being otherwise similar. A
variety of names, Lama, Alpaca, Huanaco, Vicuña, have been applied to
these animals; but it appears that the names are in excess of the number
of the species. Mr. Thomas, who has lately inquired into the matter, will
only allow two, the Huanaco, Lama huanacos, of which there are two
domestic races, the Llama and the Alpaca, and the Vicuña, Lama
vicugna. They are both South American in range. Not only is there a
herd of escaped Camels in Spain, but the Spaniards attempted to introduce
and acclimatise the useful Lama. The first Lama ever seen in Europe was
brought in the year 1558 to {287}the town of Middelburg in Holland; it was
purchased and presented to the Emperor of Germany. Gesner gives a curious
figure of it, representing the animal as a comparatively colossal beast
submitting itself to the guidance of a dwarfish man. The habit of
"spitting" of the Lama is well known. Augustin de Zarate and Buffon speak
of the Lama as having no protection save this habit, which is more than a
mere ejection of saliva: the contents of the stomach are forcibly shot at
the object of its annoyance. It can also kick and bite. In the intestines
(as in those of some other mammals) are found Bezoar stones, or Bezards
as they are variously spelt. These were once valued in medicine, and even
so lately as 1847 were, according to Gay, the historian of Chili, in
vogue; these concretions, comparable to the ambergris of the Whales, were
supposed to be an antidote to poison.

Fig. 149.—Lama. Lama huanacos. × 1⁄12.
Extinct Camels.—The earliest cameloid type is the genus
Protylopus,[186]
of which we are acquainted with an imperfect skull {288}and the greater part of
a radius and ulna belonging to one individual, and most portions of the
hind-limbs in other specimens. The one species, P. petersoni, was
about the size of a "jack rabbit," and is late Eocene (Uinta formation)
and American in range. The teeth of this mammal are the typical
forty-four, and the canines are not pronounced, being incisiform in
shape. In the skull the nasals overhang, as in the genus
Poebrotherium. The orbit is not closed by bone. There is in this
ancient Camel a trace of the supra-orbital notch so characteristic of the
Camel tribe. "The vertebrae resemble those of the modern Lamas closely in
their general proportions." The lumbars have the usually Cameloid formula
of 7. This genus has but two functional toes on the hind-feet, the second
and fifth being reduced to vestiges. It is interesting to note that the
radius and ulna appear to remain distinct, except in very old animals, in
which they come to be co-ossified in the middle only, thus foreshadowing
their complete union in the next genus, Poebrotherium. The present
genus, moreover, as well as Poebrotherium, was distinctly
unguligrade; it has not acquired the characteristic phalangigrade mode of
progression of the modern types of Camels.

Fig. 150.—Skull of Poebrotherium wilsoni. i1, i2, i3, Incisors 1-3. × ½.
(After Wortman.)
The American and Oligocene Poebrotherium has been recently and
exhaustively studied by Professor Scott.[187] It was considerably smaller than a
Lama. Its neck was long as compared with other Artiodactyles, but still
shorter than that of the Lama. It was a lightly-built, graceful creature,
with apparently some external likeness to a Lama. It is an important fact
to notice that at this {289}period, and for a long time after, there
were no types referable to the Camelidae in the Old World. Though a Camel
in many features of its organisation, Poebrotherium was
"generalised" in many ways. Thus the metacarpals and metatarsals were not
fused to form a cannon bone, and the two lateral digits were represented
by splint rudiments of metacarpals and metatarsals. The dentition was
complete. The skull though distinctly Tylopodan, also shows more
generalised characters. Thus the orbit is not quite, though nearly,
completed by bone. In the Camel it is quite closed. The nasal bones are
much longer, reaching nearly to the end of the snout. The odontoid
process of the axis vertebra is not spout-like as in existing forms, but
cylindrical, though slightly flattened upon the upper surface. The
scapula is described as being more like that of the Lama than of the
Camel, though variations occur which approximate to the Camel. The brain,
judging of course from casts, has those sulci "which are common to the
whole series of Ungulates, and closely resemble those of a foetal
Sheep."

Fig. 151.—Anterior surface of
axis of Red Deer, × ⅔. o,
Odontoid process; pz, posterior
zygapophysis; sn, foramen
for second spinal nerve.
(From Flower's Osteology.)
Later in historical sequence than Poebrotherium, and
structurally intermediate between it and Protolabis, is the
Miocene genus Gomphotherium. It shows an advance in structure upon
Poebrotherium, in that the orbit is completely encircled by bone,
though the posterior wall is thin; the lower canines instead of being
incisiform are curved back as in later Camels, and separated by a wide
diastema from the preceding and the succeeding teeth.
Later in age than Poebrotherium is Protolabis, a
Tylopod in which the full number of teeth is still retained; its
skull presents no particular changes from the Poebrotherine type; the
nasals, however, are somewhat shortened.
Later still in point of time is Procamelus. In this form we
have apparently an ancestral stock, whence both Camels and Lamas were
derived. The upper incisors are as in existing forms, but the first and
second persist for a somewhat longer time. The skull shows two
well-marked types of structure; in P. occidentalis {290}there are more
points of likeness to the Lama, in P. angustidens to the Camel. In
both, the orbits are completely encircled by bone. The nasals are much
shortened. The odontoid process of the axis is still more concave than in
Poebrotherium, but not spout-like as in existing forms. This fact
shows that the spout-like character of the Camels' odontoid process is
not a point of affinity to other Artiodactyles—in fact the
occurrence of the same form of odontoid process in Perissodactyles is
enough proof of this. We must come to the conclusion that the form is
adaptive in all cases. If we were not obliged on palaeontological
evidence to come to this conclusion, the structure in question is just
one which would be fastened upon as evidence of genetic affinity; for it
is a resemblance in a small though distinctive point of structure having
no obvious relation to utility. The metacarpals and metatarsals have
coalesced to form the cannon bones, though a rudiment of one metacarpal
seems to remain. The genera referred to appear to be on the direct line
of descent of the modern representatives of the family. But there are
other forms which are offshoots of the main stem. Such are
Homocamelus, Eschatia, and Holomeniscus. The last
two are Pliocene and American; the teeth are much reduced.
C. PECORA.
The Pecora are a group which possess so many characters in common that
it is not an easy task further to subdivide them.
In all there are but two functional digits on the feet, and the
metacarpals and metatarsals of these are fused. There are no upper
incisors, and canines in the upper jaw are not universal, and generally
small. Horns are confined to this group of the Selenodontia.[188] The premolar teeth are
of a simpler form than the molars. The stomach has four chambers, of
which two may be regarded as belonging to its cardiac half and two to the
pyloric. The former are, in the first place, a large paunch or rumen,
followed by a smaller reticulum, so called on account of the network
arrangement of the folds of its lining membrane. Connected with the
latter, and constituting the first part of the pyloric half of the
stomach, is the psalterium or "manyplies," so called on account of the
longitudinal folds, like the leaves of a {291}book, into which its
lining membrane is raised. Finally there is the abomasum, out of which
proceeds the small intestine. Garrod has observed that the chamber of the
stomach which varies most among the Pecora is the psalterium. This
chamber is specially large in Bos, and particularly small in the
Antelopes Nannotragus and Cephalophus. But its variation
relates more especially to the folds of its mucous membrane. These folds
are of varying lengths and have a definite arrangement There may be as
many as five sets of laminae of regular depths. The most simple
psalterium is that of Cephalophus, where there are only two sets
of laminae of different sizes, a deeper set and a very much shallower
set; this form is termed by Garrod "duplicate." Most common is the
"quadruplicate" arrangement, with four sets of laminae of differing
depths. In all Pecora the liver is but little divided by fissures.
Fam. 6. Cervidae.—The Deer tribe is a very extensive one,
and, with the exception of Africa and Australia, world-wide in
distribution.[189]
The Deer are absolutely distinguished from all other Ruminant animals
by the existence of antlers, which are invariably present in the male
sex, save in the aberrant genera Moschus and Hydropotes; in
the Reindeer alone are antlers present in both sexes. The general
characters of these appendages have been dealt with on a former page (p.
200), where they are compared to, or rather
contrasted with, the horns of the Bovidae. These antlers, so
characteristic of the Cervidae, are very variously developed among the
members of the family. Thus in Elaphodus the antlers are very
small and entirely unbranched. In the Muntjacs, Cervulus, the
antlers are hardly larger, but they have a small anterior branch arising
from near the pedicel, the "brow tine." In Cariacus antisiensis
only one branch, the brow tine, is present, but it is nearly as long as
the main stem of the antler, the "beam." In Capreolus capraea the
beam bears two tines; in Cervus sika three; in C. duvauceli
two of the three tines present bear secondary branches. There are other
complications (some of which are illustrated in Figs. 152-157) of the
simple antler which culminate in the complex antlers with their expanded
"palms" of the Elk and the Fallow Deer. {292}
Another highly-interesting fact concerning these same antlers is their
gradual increase in complexity of tines and palm from the Miocene
Cervus matheroni to the great Irish Elk of post-Tertiary
times.
Beyond the antlers there seems to be no character of universal
applicability which distinguishes the Cervidae from the nearly-related
Antelopes. There are, however, a number of structural features which are
nearly universally characteristic. Excepting Moschus (which
Professor Garrod would not allow to be a "Deer"), no Cervine has a
gall-bladder[190] to its
liver. All Bovidae (including Antelopes) have, with the exception of
Cephalophus.
A small but constant character of the Deer is the existence of two
orifices to the lachrymal duct. The genus Tragelaphus alone among
Antelopes shows this character.
So far as is known the placenta of the Deer has but few cotyledons,
that of the Bovidae many. But not many types are known.
The navicular, cuboid and ectocuneiform are often united. This is
never the case in the Bovidae.
The first and second phalanges of the lateral (imperfectly developed)
digits are always present except in the Muntjacs; they are never found in
Bovidae. The Deer always present a light brown to a darker brown
coloration. Elaphodus michianus is almost black. There is commonly
white on the under parts and beneath the short tail. Some Deer, such as
the Fallow Deer, are spotted; and the young of others that are uniformly
coloured when adult are spotted. In some cases a winter coat, darker than
the summer coat, is developed.
Altogether some sixty species of Deer are known, of which the
preponderance are Old-World forms. The Deer of the Old World are
distributed among the genera[191] Cervus (all Europe and Asia);
Cervulus, the Muntjacs (India, Burmah, China, etc.);
Hydropotes (Eastern China); Capreolus (Europe and Central
Asia); Elaphodus (Eastern China); there is one American
Cervus, the Wapiti. The American genera are Cariacus and
Pudua. The Elk (Alces) and the Reindeer (Rangifer)
are circumpolar. The principal structural modification which occurs
within {293}the family Cervidae concerns the
rudimentary fifth and second toes. In Capreolus,
Hydropotes, Moschus, Alces, Rangifer, and
Pudua there are considerable remains of the lower parts of
metacarpals II. and V.; in the other genera smaller traces of the upper
ends of the same bones.
The two most abnormal genera are Moschus and Hydropotes,
more particularly the former, which neither Sir V. Brooke nor Professor
Garrod allow to be members of the family at all. Moschus is
usually placed in a special sub-family by itself, Moschinae, the
remaining Deer being referred to another sub-family, Cervinae.
Sub-Fam. 1. Cervinae.—The genus Cervus comprises
rather over twenty existing species, which, except the Wapiti (C.
canadensis), are exclusively Old World in distribution. The principal
features of variation in the genus, in accordance with which it has been
divided up into sub-genera, are (1) palmated (Fallow Deer, Dama)
or non-palmated antlers; (2) adults spotted with white at all ages and
seasons (Axis), or in summer only (Pseudaxis), or not at
all; (3) spotted or unspotted young; (4) existence or absence of
rudimentary canines in the upper jaw.
Among the members of this genus, Cervus (Elaphurus) davidianus
is interesting as having been first observed by the missionary Père David
in a park belonging to the Emperor of China near Pekin. Its horns are
remarkable for dividing early into two branches of equal length, of which
the anterior again branches into two. Specimens of this Deer were
ultimately obtained for the Zoological Society's Gardens.
The species of Cervus are fairly distributed between the
Palaearctic and the Indian regions. The Palaearctic species, such as
Lühdorff's Deer (Fig. 152), are mainly Asiatic. Cervus elaphus and
Cervus dama alone are European and British. The former of course
is the Red Deer, the latter the Fallow Deer. The Red Deer is
reddish-brown in summer and greyish-brown in winter, with the white patch
on the rump so common in the Deer tribe. The Red Deer is genuinely wild
in Scotland, in certain parts of Devonshire and Westmoreland, and in the
New Forest. At the beginning of the last century, according to Gilbert
White, there were 500 head of deer in Wolmer Forest, which were inspected
by Queen Anne. The antlers may have as many as forty-eight points; and a
stag with more than the three anterior tines is termed a "Royal Hart."
The Fallow Deer has {294}palmated antlers, and is usually spotted.
It seems to be an introduced species, common report pointing to the
Romans as the introducers. It would be more correct to say
"re-introduced," for fossil remains of this Deer have been met with.

Fig. 152.—Lühdorff's Deer. Cervus luehdorffi. × 1⁄15. (From Nature.)
Elaphodus[192]
contains probably two species, E. cephalophus of Milne-Edwards and
E michianus of Swinhoe, both from China. The antlers are small and
unbranched; the canines in the male are massive; it differs from
Cervulus, to which it is closely allied, principally in the
absence of frontal glands. The second {295}species has a dark
iron-grey pelage, and the late Mr. Consul Swinhoe described it as very
Goat-like in aspect.
Capreolus.—The Roe Deer has fairly complex antlers. It is
a small Deer and has spotted young. The common Roe Deer, C.
capraea, is a native of this country. It is the smallest of our Deer,
and its antlers only have three tines in stags of the third year. It is a
singular fact about this Deer that though the pairing season is in July
and August, the young are not born until the following May or June, a
period which does not represent that of gestation. The germ remains
dormant for some time before developing.
Fig. 153.—Mule Deer. Cariacus macrotis. × 1⁄15. (From Nature.)
The Muntjacs, Cervulus, form a distinct generic type confined
to the Indian and the South-Eastern Palaearctic region. They are small
Deer with spotted young, and short one-branched antlers placed upon
pedicels as long as themselves. The canines are strongly developed in the
males. There are about half-a-dozen species.
Cariacus is exclusively American in range, and contains about
twenty species. There are or are not upper canines. The young {296}are
spotted. The antlers are occasionally very simple; in C. rufus and
a few allies (placed in a special sub-genus Coassus) they are
simple spikes without branches. In this genus, and in the nearly allied
and also New-World Pudua, the vomer is prolonged backwards and
divides the posterior nares into two. The bulk of the species are South
American.
Fig. 154.—Chilian Deer. Cariacus chilensis. × 1⁄12. (From Nature.)
Pudua, just mentioned, comes from the Chilian Andes. It is a
small Deer without canines and with minute antlers. Other generic names
have been proposed for various species of American deer.
Hydropotes inermis is a small perfectly hornless Deer, living
on the islands of the Yang-tse-kiang. The male has tusks; the young are
spotted. Though, like other deer, Hydropotes has no gall-bladder,
both Mr. Garrod[193] and
Mr. Forbes[194] found the
{297}rudiments of one in the shape of a white
ligamentous cord. Mr. Forbes has especially dwelt upon the likeness of
the brain to that of Capreolus. The female has four teats, and
produces three to six young at a time.
Fig. 155.—Water Deer. Hydropotes inermis. × 1⁄10. (From Nature.)
Alces machlis, the Elk or Moose, is a circumpolar species with
palmated antlers and is of large size. The young are unspotted. This
animal is the largest of the Deer tribe. The aspect of this creature is
by no means that of a Deer, the long, thick, and rather prehensile upper
lip not by any means suggesting the family to which it belongs; the legs,
too, are ungainly through their unusual length. The Moose has a curious
method of protecting himself from Wolves. Instead of moving about during
heavy snowstorms, and being thus on the heavy ground an easy prey for
these agile enemies, the animal forms what is known as a "Moose yard." An
area of ground is kept well {298}trampled down, and the animal contents
itself with browsing upon the adjacent stems. The well-trampled ground
gives an easy footing, and by his powerful horns the great stag is able
to keep his enemies at bay.
Fig. 156.—Moose. Alces machlis. × 1⁄20.
Rangifer tarandus, the Reindeer, is unique among Deer by reason
of the fact that both sexes wear antlers. These antlers are palmated. The
brow tine and the next or bez tine are also palmated and are directed
forwards and a little downwards. The young are unspotted. The pelage
alters in winter. Like the Moose, the Reindeer is circumpolar. As is well
known, during the Pleistocene period the Reindeer extended its range as
far as the South of France. Even in the historic period it is said to
have been hunted in Caithness.
Reindeer, like so many other particularly Arctic animals, have regular
migrations. In Spitzbergen, for instance, the animal migrates in the
summer to the inland region of the island, and in {299}the autumn back again
to the sea coast to browse upon the seaweed. These migrating herds have
been stated to be led by a large female.
Fig. 157.—Reindeer. Rangifer tarandus. × 1⁄15.
Sub-Fam. 2. Moschinae.—Moschus moschiferus[195] is a native of the
Asiatic Highlands. It is 3 feet or so high, perfectly hornless, and with
very large canines in the male. It is noteworthy that in
Hydropotes, where the canines are also very large, horns are
absent. These are examples, perhaps, of correlation. The musk sac (whence
the name) is present on the abdomen of the male only. There is no crumen
or suborbital gland, which is so generally (though by no means
universally) present in Cervidae. But the male has, in addition to the
musk glands, glands near the tail and on the outside of the thigh. Unlike
other Deer, the lachrymal bone of Moschus bears but one orifice.
The feet, so far as concerns the preservation of the outer rudimentary
{300}metacarpals, are of the more ancient type
represented in Alces, Hydropotes, etc. A gall-bladder is
present. The young, as in so many Cervidae are spotted; but the adult is
of a greyish-brown colour.

Fig. 158.—Musk Deer. Moschus moschiferus. × 1⁄6. (From Nature.)
There is no doubt that Moschus is more nearly related to the
Cervidae than to any other Ruminants. It is regarded by Sir W. Flower as
"an undeveloped deer—an animal which in most points (absence of
horns, smooth brain, retention of gall-bladder, etc.) has ceased to
progress with the rest of the group, while in some few (musk gland,
mobile feet) it has taken a special line of advance of its own."
The musk itself, which gives its name to the creature, is found in a
gland on the belly, about the size of a hen's egg. The whole gland is cut
out and sold in this condition. Such quantities of musk deer have been
and are killed for this purpose that the rarity of the animal is
increasing. In the seventeenth {301}century it was so common that the
traveller Tavernier purchased 7673 musk "pods" in one journey, or,
according to Buffon, 1663. The tusks, which recall those of
Hydropotes, to which Moschus is not nearly allied, and of
Tragulus, with which it has of course still less connexion, are
said to be used for the digging up of roots. Its feet, in relation to its
mountain-ranging habits, are very mobile.
Extinct Species of Deer.—It has been already mentioned
that the most primitive kinds of Deer had no horns at all, resembling in
this the modern Moschus and Hydropotes, and that with lapse
of time went hand in hand an increasing complexity of antler; the facts
of palaeontology harmonising in the most striking manner with the facts
of individual development from year to year. The oldest forms seem to be
more nearly akin to the living Muntjacs, and their remains occur in the
lowest Miocene beds of both Europe and America. At present the group is
confined to the warmer parts of Asia and some of the islands belonging to
that continent.
One of the oldest types is Amphitragulus. This genus, which
consists of several species, inhabited Europe, and differed from living
Muntjacs in being totally hornless in both sexes; the skull had no
lachrymal fossa or deficient lateral ossification.
Nearly allied is Dremotherium of similar age and range.
The Middle Miocene has furnished the remains of the genus
Dicroceras. This is the earliest Deer in which horns have been
found. The horns are, as the name of the genus implies, bifid, and have,
like those of the living Muntjac, a very long pedicel. This is also a
European genus like the last. From this period we come across true Deer,
which commence in the Upper Miocene and have branched horns. Moreover
they belong, at least for the most part, to the existing genera. One of
the most remarkable forms is Cervus sedgwicki (sometimes placed in
a separate genus, Polycladus) from the Forest Bed of Norfolk and
from the Upper Pliocene of the Val d'Arno. This creature was remarkable
for its multitudinously-branched antlers. These end in no less than
twelve points. No Deer exists or has existed in which the horns are so
completely branched. They are like those of a Red Deer exaggerated.
Fig. 159.—Giraffe. Giraffa camelopardalis. × 1⁄40.
Fam. 7. Giraffidae.—Undoubtedly the type of a distinct
family, Giraffidae, is the genus Giraffa. It is characterised by
{302}the long neck, which, nevertheless,
consists of only the normal seven vertebrae, and by the "horns" which
differ from those of all other Ruminants; they are small bony prominences
of the frontal bones, which become fused with the skull, and which are
covered with unmodified skin. They are not shed. Between them is a median
prominence. This cranial armature is present in the female as well as in
the male, and is well developed even in the {303}new-born young. The
orbits are completely encircled by bone, and there is no lachrymal fossa,
so common in Deer and Antelopes. There are no canines above; but these
are present in the lower jaw. The rudimentary digits of other Ruminants
have disappeared in this genus. There are fourteen pairs of ribs as in
many other Artiodactyla. The liver of the Giraffe[196] is, as in many, but not all,
Ruminants, devoid of a gall-bladder; neither has it a caudate or a
Spigelian lobe. The caecum is actually largish (2½ feet in length), but
is relatively very small, as the small and the large intestines measure
196 and 75 feet in length respectively. The Giraffe has a well-marked
"ileo-caecal" gland, found in many Ruminants; its appearance in
Giraffa is especially compared by Garrod with its appearance in
Alces.
Considered by itself, Giraffa forms a very isolated type of
Ruminant. But after we have dealt with certain facts concerning extinct
forms clearly allied to Giraffa, the isolation of the family will
be found to be less marked.
The Giraffe ("one who walks swiftly," the word means in Arabic) is, as
every one knows, limited in its range to the African continent. It is
not, however, so familiar a fact that there are two quite distinct
species of Giraffe, one a northern form from Somaliland, and the other
South African. The distinctness of these two, G. camelopardalis
and G. australis, has been lately worked out in some detail by Mr.
de Winton.[197] The
principal point of difference between them consists in the large size of
the median horn in the Cape species, which is represented by the merest
excrescence in the other species. The Giraffe of West Africa is held to
differ from the northern and southern species, coming nearer to the
former. It appears in the first place to be a larger animal, and slight
differences in the skull have been pointed out. This series of
peculiarities may be expressed, for those who do not object to trinomial
nomenclature, by calling this novel western form Giraffa
camelopardalis peralta. The existence of the three horns covered with
unaltered skin is the main characteristic of this Ungulate. But the
Giraffe also differs from other Artiodactyles by its enormously long
neck, which enables it to browse upon trees inaccessible to the common
herd {304}of Ruminants. The neck is often supposed
to have some relation to this method of feeding. But a more ingenious
explanation of its inordinate length is that it serves as a watch-tower.
The long grass of the districts inhabited by the animal swarms with Lions
and Leopards, which must be foes. The long neck allows of a wide look out
being kept, and it is noteworthy that the Ostrich, living under similar
conditions, is also renowned for its length of neck. It is the spots upon
the Giraffe which have given it its name of Cameleopard; these spots
present in the southern form a series of chocolate-coloured areas,
sharply marked off by white spaces. Of these spots it is asserted that
they serve as a means of concealing their possessor. Sir Samuel Baker[198] wrote of it in the
following words: "The red-barked mimosa, which is its favourite food,
seldom grows higher than 14 or 15 feet. Many woods are almost entirely
composed of these trees, upon the flat heads of which the giraffe can
feed when looking downwards. I have frequently been mistaken when
remarking some particular dead tree-stem at a distance that appeared like
a decayed relic of the forest, until upon nearer approach I have been
struck by the peculiar inclination of the trunk; suddenly it has started
into movement and disappeared."
The Giraffe, remarked Pliny, "is as quiet as a sheep." The Roman
public, to whom the first Giraffe ever brought into Europe was exhibited,
expected from its name "to find in it a combination of the size of the
camel and the ferocity of a panther." As a matter of fact, Giraffes in
captivity are not always sheep-like in temper. They will kick with
viciousness and vigour, and will even initiate an attack upon their
keeper. At the same time they are singularly nervous creatures, and have
been known to die from a shock. In moving, the Giraffe uses the fore- and
hind-limb of each side simultaneously; this gives to its gait a peculiar
rocking motion, the singularity of which is heightened by the curving
movements of the long neck, which even describes now and then a figure of
eight in the air. Giraffa camelopardalis and the species (?)
already referred to are the only existing Giraffes (of the genus
Giraffa), and they are not found out of Africa. Sir Harry Johnston
has lately given a brief account of a larger and more brilliantly
coloured species from Uganda {305}which will probably prove to belong to a
distinct genus. It has five horns, the additional pair being placed above
the ears.
Sir Harry Johnston has quite recently made known another genus of
Giraffidae living in the Semliki forest, Belgian Congo district. The skin
and two skulls, as well as the bones of the feet, are known from
specimens sent by Sir Harry Johnston to the Natural History Museum, and
briefly described to the Zoological Society by Professor Ray Lankester.[199] This creature, of
which the native name is "Okapi," is proposed to be called Ocapia
johnstoni. The first actual specimens which reached this country were
two bandoliers made from the skin of the flanks, which were striped black
and white, and were not unnaturally held to be portions of the skin of a
new species of Zebra. The animal is of about the size of a Sable
Antelope, and the back and sides are of a rich brown colour; it is only
the fore- and hind-limbs which are striped, the striping being
longitudinal, i.e. parallel with the long axis of the body. The
head is Giraffe-like, but there are no external horns; wisps of curled
hairs seem to represent the vestiges of the horns of other Giraffes. The
tail is rather short, and the neck is rather thick and short. The skull
is clearly Giraffine. The basicranial axis is straight, and the
fontanelle in the lachrymal region is very large. Upon the frontal bones
near their parietal border is a large boss on either side, which
presumably represents the horn core or "os cornu." On the mandible the
great length of the diastema between the incisors and premolars is a
Giraffine characteristic. The Okapi lives in pairs in the deepest
recesses of the forest.
We are acquainted with a few extinct forms, belonging to
Giraffa, which are extra-African in range. G. sivalensis is
from the Pliocene of the Siwalik Hills in India, G. attica from
Greece. These remains, however, do not include the top of the skull, so
that it is doubtful whether their horns were as in G.
camelopardalis.
A closely-allied genus is the extinct Samotherium. This
flourished in Miocene times, and its remains have been found in the Greek
island of Samos. The neck and limbs are shorter than in the Giraffe, and
the horns, longer than in Giraffa, are placed just above the orbit
upon the frontal bones alone, instead of upon the boundary line of
frontals and parietals as in Giraffa. In several ways, therefore,
the existing Giraffe is a more modified or {306}specialised animal than
its forerunner of the Miocene. In the latter, the male alone carried
horns, and in neither sex does the unpaired median bony excrescence
appear. The remains of this genus (probably even the same species, S.
boissieri) also occur in Persia.
Helladotherium (there is but one species, H. duvernoyi)
has its four limbs of nearly the same length; the skull of the only known
example is hornless; the neck is shorter than in Giraffa. It is
known from the Miocene deposits of Pikermi in Greece.
Palaeotragus is a genus which is not referred to the Giraffidae
by all systematists. Its very name, given to it by the eminent French
palaeontologist M. Gaudry, indicates his opinion as to its Antelopine
affinities. The chief and indeed (according to Forsyth Major[200]) the only reason for
placing this Ruminant with the Antelopes is the large size of the horns.
They undoubtedly suggest the horn cores of Antelopes. But they are placed
wider apart than in those animals. It is thought that the hornless
Camelopardalis parva is the female of this species, which is from
Pikermi.
Rather more different from Giraffa is the extinct genus
Sivatherium, from the Siwalik deposits of India. Here again there
has been some discussion as to its affinities. Some place it in the
neighbourhood of Antilocapra, but most palaeontologists now regard
it as a Giraffe. The main peculiarity of this large beast was the
existence of two pairs of horn cores; the larger are upon the parietal
bones, and are of a palmated form, with a few short tines, which are
highly suggestive of those of the Elk (Alces). The shorter
anterior pair are upon the frontal bones. The neck is short, the limbs of
equal length, and there are no additional toes upon the limbs.
Sivatherium was almost as large as an Elephant, and in
restorations it is depicted as having a fleshy dilated nose like the
Saiga Antelope; this view is based upon the position and size of the
nasal bones. Hornless skulls have been identified as the female of
Sivatherium.
Vishnutherium, Hydraspotherium, and Bramatherium
are allied genera.
Fam. 8. Antilocapridae.—This family contains but one
genus and species, the N. American "Pronghorn," Antilocapra
americana. This animal deserves a family to itself on account of the
singular structure of the horns, which are intermediate in character {307}between those of the Deer and those of the
Antelopes. They are unquestionably "hollow-horned" Ruminants, in that
there is an osseous horn core, upon which lies the actual horn. This,
however, is softer than in Bovidae, and is semicorneous. It is, indeed,
more like the velvet of the stag's horn. Moreover the horn is branched,
and there are sometimes even three prongs. Furthermore, it is now
certainly known that the Pronghorn sheds its horns not merely
occasionally, but with definite annual periodicity. It so far resembles
the Deer. But it must be borne in mind that in the Deer the horn shedding
is a twofold process. There is first of all the stripping off of the
velvet, and secondly the shedding of a portion of the horn core down to
the burr. What happens in the Prongbuck is the shedding of the true horn
only ( = the shedding of the velvet), not of the horn core. It
appears, however, that occasionally (once in their lifetime?) certain
undoubted Antelopes may cast their horns.[201] Another external character of this
animal is the total absence of "false hoofs," the last vestiges of the
second and fifth digits. The Pronghorn is a gregarious creature running
in bands of six up to hundreds.
Fam. 9. Bovidae.—This family, more extensive than that of
the Cervidae, contains not only the Oxen, Sheep, and Goats, but also the
Antelopes, save only Antilocapra, which must be placed in a family
by itself. The only two points which distinguish all Bovidae from all
Cervidae[202] are the
nature of the horns already described, and the polycotyledonary condition
of the placenta. Moreover the horns are usually present in both sexes,
though there are exceptions, such as the Sheep and Goats, and various
genera of Antelopes (Tragelaphus, Tetraceros, etc.). There
are never the first two phalanges belonging to the rudimentary digits
II., V., as there are in all Deer excepting Cervulus. There is as
a rule but one orifice to the lachrymal duct. There are never persistent
upper canine teeth in either sex.
It is exceedingly difficult to separate the Antelopes from the Sheep,
Oxen, and Goats. Their inclusion along with these creatures in one
family, Bovidae, shows that no differences of an important character
exist. The term Antelope is rather of popular than {308}of zoological
significance. As a rule there are horns in both sexes; but this rule is
not without exceptions, of which one is the genus Strepsiceros,
the Koodoo. Many other Bovidae are horned in the males only, e.g.
Saiga, Tragelaphus. The Antelopes further differ from the
true Oxen in their more graceful build, and in the fact that the horns,
if they curve at all, generally curve backwards towards the neck. In the
Oxen, on the other hand, the build is stouter, and the horns usually
curve outwards. The same remarks apply to the Sheep. Such an Antelope,
however, as the Eland (Orias) is very Ox-like in habit. Another
feature which may be remarked upon, though not of absolute differential
value, is that while the Antelopes are as a rule smooth and sleek in
their skins, the Oxen tend to be rough and shaggy. The Zebu, however, in
this, in its hump, and in general aspect, is far from being unlike an
Eland. But then the Zebu is a domestic race, and we do not know what the
wild stock was like. It is perhaps with the Goats that the Antelopes have
the nearest affinities, and it is difficult to place such a form as
Nemorrhaedus, and indeed some others. In the Antelopes as a rule
the middle lower incisors are larger than the lateral ones; in the Sheep
and Goats they are alike in size. The parietal bones, too, in the
Antelopes are moderately large and are much shortened in the remaining
Cavicornia, especially in the Oxen. As the Antelopes are the oldest, so
far as we know, of all bovine animals, one would expect to find them
combining the characters of the rest. But they do this so effectually
that a disentanglement is really impossible. They date from the Miocene.
Antelopes are now limited to Europe, Asia, and Africa; they have always
had the same range, though more abundant in former times in Europe. They
preponderate now in tropical Africa, and abound in genera and species.
Messrs. Sclater and Thomas[203] allow altogether thirty-five genera,
of which twenty-four are exclusively Ethiopian in range.
In the following summary of the group Messrs. Sclater and Thomas's
work is followed. They commence with a section or sub-family of which the
type is the Hartebeest.
Bubalis, or Alcelaphus as it is sometimes called, is an
African genus, ranging however into Arabia. These Antelopes are
characterised by the long skull and the doubly-curved horns. There are
eight species of the genus, of which B. caama is the {309}best known;
this is the animal known as the Hartebeest. The Bontebok and Blessbok
belong to a closely-allied genus, Damaliscus, distinguished mainly
by the fact that the bony base of the horn cores is not extended upwards,
and therefore the parietal bones are visible when the skull is viewed
from in front, which is not the case in Bubalis.
Fig. 160.—Brindled Gnu. Connochaetes taurinus. × 1⁄20.
The Gnus, Connochaetes, are familiar owing to their curious
aspect. The hairy face, and rump and tail like those of a pony are highly
characteristic. The horns are bovine in appearance, standing outwards and
then curving upwards.[204] There are three species of Gnu, all
from South Africa. They are C. gnu, C. taurinus, and C.
albogulatus.
Of the Cephalophine section there are two genera:—
Cephalophus is an African genus. These animals are known as
Duikerboks; they are small, and have short non-curved horns in the male
sex only. Their general aspect is not unlike that of certain Deer with
simple horns, such as Cervulus. Messrs. Sclater and Thomas allow
thirty-eight species. The {310}smallest species do not exceed the
dimensions of a Hare. None are really large.
Tetraceros is an Indian genus characterised, as its name
denotes, by the fact that it possesses four horns. It is the posterior
pair which correspond to the single pair of Cephalophus. The
anterior pair, which are much smaller and are sometimes absent, are a new
pair. The female of this Antelope is hornless. Sheep are occasionally
four-horned, and there is indeed a breed of such in Kashmir. A
four-horned Chamois was described by the late Mr. Alston.
The Klipspringer, Oreotragus saltator, is the first type of a
third section; as its name denotes, it is an Antelope with Goat-like
habits, being found particularly among rocks. The horns are short and
straight. This, the only species of the genus, is African in range, of
which its Dutch name gives evidence. A specimen in the Zoological
Society's Gardens (as has been pointed out to me by Mr. Mercer) had the
habit of depositing the secretion of the tear gland upon a mass of
concrete in its enclosure, the secretion thus exuded forming a pointed
heap of hardish matter. It may be that the object of this is to guide its
fellows to its whereabouts.
Ourebia is a less-known genus, larger in size, but with horns
of the same character, though longer.
The Grysbok and the Steinbok, genus Raphiceros, have similar
horns. This as well as the last two genera have horns in the male
only.
One of the smallest of Antelopes belongs to an allied genus; this is
Neotragus pygmaeus. It is known as the Royal Antelope, a name
apparently derived from Bosman's statement that the negroes called it
"the king of the harts." Its horns are very small. The height of the
animal is only 10 inches. Horns are present in the male alone. The last
three genera are African.
The Cervicaprine series, which is also African, includes the
Waterbucks and Reedbucks, so called on account of their water-loving
propensities. As in the last series, from which they are separated by
Sclater and Thomas, but with which they are united by Flower, there are
horns in the male only. These horns, though not twisted, are long. The
typical genus is Cobus, of which there are eleven species. The
Waterbuck, C. ellipsiprymnus, and the Sing-sing, C.
unctuosus, are perhaps the best-known species; the former is {311}blackish grey, the latter browner in
colour. In C. maria and one or two other species the horns are
more curved backwards and again forwards than in some of the others,
where their form is sublyrate.
The Reedbucks, Cervicapra, are closely allied to Cobus;
they are, however, of smaller size. Here, as in that genus, the females
are hornless, and the horns of the males are of medium size. Five species
are referred to the genus. They are all of a brownish fawn colour. A
genus Pelea, with but one species, P. capreolus, has been
separated on account of the fact that the horns are nearly straight and
that there is no naked patch of skin beneath the ears. This animal has
received its name on account of its resemblance to the Roebuck.
The Antilopine section includes a number of genera.
The genus Antilope is Indian in range. It includes but one
species, A. cervicapra. This Antelope is of medium size, with a
brown pelage getting blacker with years; it is thus known as the
Black-buck. The female, which is hornless, is lighter brown. The horns
are long, spirally twisted, and closely ringed.
Aepyceros, with two species, is African. The Palla (Ae.
melampus) is a large Antelope, with longish lyrate horns in the male,
which are half-ringed.
The Saiga Antelope, genus Saiga, is one of the most remarkable
types of Antelope in its outward appearance. Its nose is very large and
inflated, the two nostrils being quite widely separated, a depression
indeed lying between them dorsally. The horns are lyrate in the male,
absent in the female. The "ovine expression" of this bovine animal is
more pronounced in the female. Corresponding with the clumsy nose are
very short nostrils, the commencement of the narial aperture being
therefore very far back. It is almost suggestive of Macrauchenia
in this respect. The fleece is also Sheep-like. The genus occurred in
this country during the Pleistocene. It is now an inhabitant of Eastern
Europe and Western Asia. The only species is S. tartarica.
The Chiru, Pantholops, is allied to the Saiga. The horns of the
male are long and nearly straight; they are ringed in front. The muzzle
is swollen in the male; the nostrils are large, and provided with
extensive sacs internally. The colour of this animal, which is
exclusively Thibetan in range, is a pale fawn. The hair, in accord with
its habitat, is very woolly. No living specimens have ever been brought
to Europe. This creature has accumulated much {312}legend. Its blood is
believed by the Mongols to possess virtues, and by means of the rings on
the horns fortunes are told. Naturally the animal is on these grounds
hard to stalk and shoot.
Fig. 161.—Loder's Gazelle. Gazella loderi. × 1⁄10.
The Gazelles, genus Gazella, are fairly numerous in species,
which are both Palaearctic and Ethiopian. There are altogether
twenty-five of them. The genus as a whole is characterised by the small
or moderate size, the sandy coloration with white belly, the presence of
dark and light stripes on the face and on the flanks. These streaks,
however, are not always present, and their presence or absence serves to
differentiate some of the species. The horns are usually present in both
sexes. The horns are of fair length, ringed, and of lyrate form.
The Springbok is separated from the rest of the Gazelles, to which
genus it is clearly most nearly related, as a genus Antidorcas.
This genus differs from Gazella by having only two lower premolars
as in Saiga. Otherwise it resembles the Gazelles; there is but a
single species, A. euchore, which is African. {313}
Ammodorcas is closely allied to the Gazelles, but differs from
them in having an elongated neck and also a long tail. A. clarkei,
the only species, is limited to Somaliland.
Lithocranius, not unlike the last, has a still longer neck,
which makes it almost Giraffe-like; its tail, however, is short. The
scientific name is derived from the "solid stony character of the
cranium." In running, this Gazelle carries the head forward in a straight
line with the body. It is African.
Dorcotragus with one species, D. megalotis, is a pigmy
Gazelle restricted to Somaliland. Its likeness, on account of size and in
some other superficial features, to the Klipspringer, led to its original
confusion with that genus (Oreotragus).
Fig. 162.—Sable Antelope. Hippotragus niger. × 1⁄20. The horns of the specimen
figured have not nearly reached their full dimensions.
A sub-family Hippotraginae, or Hippotragine section, includes a number
of Antelopes which agree in the possession of four mammae, and of molars
more like those of the true Oxen, of {314}horns of some length,
present in both sexes, and of a longish tail. They are all African in
range.
The type genus Hippotragus has its horns placed above the
orbits; they are not twisted, but curved backwards. There are three
species in the genus. Of these the best known is H. niger, the
beautiful Sable Antelope. Its general colour is a rich, dark, glossy
brown with white stripes on the face, and with a white belly. The other
species are the Roan Antelope, H. equinus, and the Blaaubok, H.
leucophaeus, of which the last specimen was probably killed in
1799.[205]
Fig. 163.—Beatrix Antelope. Oryx beatrix. × 1⁄16. (From Nature.)
The genus Oryx (chiefly African, but also Arabian and Syrian)
also contains a number of species, which are fairly familiar through the
fact that several of them are always on view in the Zoological Society's
Gardens. The genus differs from Hippotragus in that the horns,
present in both sexes, are placed behind the orbits, and slant backwards
in a line with the face. They are annulated. The Leucoryx (O.
leucoryx) is of a pale colour, but {315}this is not so marked
as in O. beatrix, which is largely white with, however, brown
legs. The Gemsbok is a handsome creature with greyish tawny colour, much
darker on the legs, and with a Gazelle-like, dark, side stripe. It has
received its vernacular name on account of its supposed likeness to the
Chamois ("Gemse"), just as the Rehbok was so-called from its supposed
likeness to the Roe Deer, and the Eland to the Elk. The Beisa (O.
beisa) is of a similar tawny colour to the last, and also with darker
stripes.
The Addax (Addax) of North Africa, Arabia and Syria, has but
one species (A. nasomaculatus). The horns are spirally
twisted.
Fig. 164.—Speke's Antelope. Tragelaphus spekii (♀). × 1⁄16.
The Tragelaphine section includes the Kudus, Elands, Nilgais, and
Harnessed Antelopes. They are all long-horned (when the horns are present
in both sexes), the horns being twisted; the nose is naked with a slight
median groove, and all are Ethiopian or Oriental in range.
The genus Tragelaphus includes the Harnessed Antelopes, so
called on account of the direction of the stripes suggesting harness. The
females are hornless, and the colours of the two sexes are different. The
hoofs are long and the toes rather unusually separable, which state of
affairs is in accord with the {316}swampy country affected by many. T.
gratus and T. spekei are larger forms; the Boschbok, T.
sylvaticus, is smaller.
The Kudus, genus Strepsiceros, have more markedly twisted
horns, which are absent in the female. The body is vertically striped
with white. The largest species is S. kudu; a smaller form, S.
imberbis, is from Somaliland.
Fig. 165.—Eland. Orias canna. × 1⁄25.
The last genus of this section or sub-family is the African Eland,
genus Oreas[206]
(which it appears should be spelt Orias). The Elands are perhaps
more Ox-like in appearance than the other members of this group, and in
both sexes have horns, in which the spiral twisting is more close.
Orias canna is the name of the common Eland. O.
livingstonii has been applied to an East African variety, which has
thin and faint lateral stripes like the other members of the group to
which it belongs.
The genus Boselaphus includes only B. tragocamelus, the
Nilgai, which is purely Indian in range. The female is hornless, and the
horns of the male are smooth and not long. {317}
The members of the Bovine section or Oxen are to be distinguished from
other hollow-horned Ruminants by their stouter build and by the fact that
the horns stand out from the sides of the skull and are simply curved,
not twisted; and smooth, not annulate like those of other Ruminants. The
muffle is naked, broad, and moist. The Oxen are widely distributed; but
are entirely absent from the Australian region and from South America and
Madagascar.
The true Oxen are perhaps best considered to form but a single genus,
Bos. They have, however, been divided into a number of genera.
Even the supposed aberrant Anoa depressicornis of Celebes hardly
differs sufficiently to warrant its separation. In favour of this view,
too, is the extraordinary ease with which different "genera" will cross
with each other and produce fertile offspring. The following is the
pedigree of an animal lately living in the Zoological Society's Gardens.
The female offspring of a male Zebu and a female Gayal was mated with a
male Bison. The female calf was again mated with a Bison and produced a
calf, also a female, which contained therefore the three species, Bos
indicus, Bibos frontalis, and Bison americanus. It is
clearly unwise in view of this fact to insist too much upon generic
distinctions in any of those types.[207]
Of this genus the Oriental Gaur (Bos gaurus), the Gayal (B.
frontalis), and the Banteng (B. sondaicus) form a well-marked
section, characterised by their dark coloration and by the somewhat
flattened horns.
The Gaur, Bos gaurus, has a more concave forehead than its
allies; the horns are less curved than those of the Banteng, and less so
than the horns of the Gayal (Bos frontalis). It inhabits the
Indian Peninsula; and extends through Burmah to the extremity of the
Malay Peninsula. The Malay name of this animal is Sakiutan, which simply
means wild cattle. It chiefly frequents wooded hills and is an excellent
mountain climber.
Bos frontalis, the Indian Gayal, has a white caudal disc like
the last species, but the forehead is flat and the horns curve but
little. It is chiefly known as a tame animal, and its occurrence in the
wild state has been doubted. It has furthermore been suggested that it is
merely a tame race of the Gaur altered {318}slightly through
domestication. It is, however, said not to cross in a state of nature
with the Gaur.[208]
Fig. 166.—Gayal. Bos frontalis. × 1⁄20.
The Banteng, B. sondaicus, is distributed through Chittagong,
Tenasserim, and the Malay Peninsula to Java and Borneo. There are
apparently two races of this animal. The species differs from the others
by the fact that the horns are smaller and more curved; there is a white
caudal disc; the forehead is narrower and the skull longer than in the
others.
The American Bison and the European Aurochs form another section; they
are indeed extremely alike, specific differences being hardly
recognisable. The Bison of America, formerly present in such numbers that
the prairies were black with countless herds, has now diminished to about
a thousand head.
One of the largest of existing Bovidae is the Aurochs, Wisent, or
European Bison, Bos bonasus (or Bison europaeus). It is
exceedingly like its American relative. Formerly the animal was much more
widely spread than it is now, extending its range from Europe into North
America. It is now limited to certain districts on the Urals, in the
Caucasus, and a herd of them are kept up through the fostering care of
the Emperor of Russia in the forest of Bielovege in Lithuania. The term
"Aurochs" should not really be applied to this species but to the Wild
Cattle, Bos taurus. It is, however, so generally used for the
Wisent (which is the German name) that it {319}is not necessary to
change it. The Sclavonic name is Zubr or Suber. It is a great beast,
standing 6 feet or so in height at the shoulder. It ranged further over
Europe well within the historic period. In the days of Charlemagne it was
spread over Germany and was a beast of the chase. In the year 1848 the
Emperor of Russia presented a pair of these Oxen to the Zoological
Society of London. At the time of their presentation an interesting
communication was made to the Society by M. Dolmatoff, on the method of
the capture of these two examples. The creature is not easy to capture
and is alarming to confront. "The eyes," says an old writer, "are red and
fiery; the looks are furious and commanding." It has of course the shaggy
mane and hump of the American animal. The herd in Lithuania was said to
be 1900 in the year 1856. Mr. E. N. Buxton,[209] who has lately visited the forest,
quotes M. Neverli to the effect that at present the numbers are not more
than 700.

Fig. 167.—Bison. Bison americanus. × 1⁄25.
Allied to this animal, and apparently still nearer to the American
Bison, is the extinct B. priscus of Europe. The Pleistocene Bisons
of North America, B. antiquus and B. latifrons, are not
remote from the living forms. Finally, the Miocene B. sivalensis
from India, and the Pliocene B. ferox and B. alleni of
North America, take back this group to as remote a period as any other
genus of Oxen. {320}
Fig. 168.—Yak. Bos grunniens. × 1⁄15.
Fig. 169.—British Wild Ox. Bos taurus. From Vaynol Park, Bangor. × 1⁄20.
The Yak, Bos grunniens, is a long-haired peculiar type,
confined to the Thibetan plateau. B. (Anoa)
depressicornis of Celebes is characterised by its straight horns;
allied to it is B. mindorensis (Philippine Islands), supposed,
however, to be a hybrid between {321}it and some other species. Africa has at
least two Buffaloes. We may finally mention the Wild Ox of Europe, B.
primigenius, the supposed progenitor of our domestic cattle, believed
to be still surviving in the herds at Chillingham, Chartley, and
elsewhere. This animal is sometimes called the Aurochs. The Romans spoke
of it as the Urus, and it appears to have formerly attained to more
gigantic proportions than at present. It is the small size of the present
race that is the chief objection to tracing them back to the large Oxen
existing near London in 1174, and found sub-fossil in the Cambridgeshire
fens.
Fig. 170.—Punjab Wild Sheep. Ovis vignei. × 1⁄10.
Of the true sheep, genus Ovis, there are a considerable number
of species. The Sheep are to be distinguished from the Goats by their
rather stouter build and by the absence of the beard in the male. The
horns are developed in both sexes, and are usually twisted and often of
large size.
The Sheep are almost entirely Palaearctic and Nearctic. They only just
get into the Oriental region. One of the finest species is the great
Pamir Sheep, O. poli, whose length reaches 6 feet {322}7 inches, and
height 3 feet 10 inches. The horns of this fine Sheep may measure more
than five feet round the curves. The Rocky Mountain Bighorn (O.
montana) is a Sheep ranging along the Rockies as far south as New
Mexico, and also to the far north; they are not confined to the chain of
mountains mentioned, but occur also on the mountains of British Columbia
down to those of California. The horns are not quite as large as those of
the last species, but measurements give a length (along the curve) of 32
to 40 inches.
Fig. 171.—Himalayan Burrhel Sheep. Ovis burrhel. × 1⁄12. (From Nature.)
Fig. 172.—Blanford's Sheep. Ovis blanfordi. × 1⁄10. (From Nature.)
Just as the Goats are often limited to islands and small stretches of
country, so are the Sheep. Thus Cyprus has a species, O. ophion,
peculiar to itself. This, which is known as the Cyprus Mouflon, is
limited to a range of mountains, the Troodos, in that island. In 1878 it
was believed that the animal was nearly exterminated, a flock of
twenty-five members alone surviving. They have, however, since increased.
Confined {324}to the Thibetan plateau are O.
hodgsoni and O. nahura. Corsica has the Mouflon, O.
musimon; and the Barbary Sheep or Arui, O. tragelaphus, is
found only in Northern Africa. Ovis burrhel and O.
blanfordi are Indian forms.
Fig. 173.—Barbary Sheep. Ovis tragelaphus. × 1⁄10.
Fig. 174.—Thar. Capra jemlaica. × 1⁄10. (From Nature.)
Ovis nahura is chiefly responsible for the impossibility of
strictly separating the Sheep and Goats. It has no suborbital glands or
lachrymal fossae, which are as a rule present in the Sheep and absent
from the Goats. On the other hand interdigital glands are present, which
is the case with Sheep. Its habits, too, are a blending of those of the
Sheep and the Goat. It lives largely on undulating ground like Sheep, and
frequently lies down during the day on its feeding ground. On the other
hand it is, like the Goats, a splendid climber.
The Goats, genus Capra, differ from the Sheep in their slighter
build and in the fact that the horns are not spirally curved, but arched
over the back. There is also the characteristic beard, {325}and the male
is odorous. The true Goats are almost exclusively Palaearctic in range.
They show the limited distribution of the Sheep, a distribution which
follows from their mountain-loving habits.
Fig. 175.—Sinaitic Ibex. Capra sinaitica. × 1⁄10.
Thus we have the Spanish Ibex (C. pyrenaica), limited to the
Pyrenees and other mountain ranges of the peninsula; C. ibex, the
Steinbok of the Alps and the Tyrol; the Markhoor, C. falconeri, of
certain mountain ranges of Afghanistan; the Caucasian, Sinaitic, and
Cretan Ibexes, and the Thar.
Capra aegagrus, the Persian Wild Goat, ranges from the Caucasus
to Sind. It is this animal which produces the true "bezoar stone." The
substance in question is a secretion apparently found in the stomach. It
is still, according to Mr. Blanford, regarded as an antidote to poison in
Persia. Buffon called this Goat the "Pasan," which is evidently a
corruption of the word bezoar. When the substance was in repute as a
medicine of the "alexipharmic" kind, the supply naturally came up to the
demand. Thus the bezoar stones of the Lama in South America gained
repute, and there were "Oriental bezoar, cow {326}bezoar, hog bezoar, and
monkey bezoar"! As concretions of one kind or another are not uncommon
objects in the alimentary tract of mammals it was easy enough to obtain a
fair amount of some substance which was sure to sell well. It is said
that a stone weighing four ounces was once sold in this country (or at
any rate in Europe) for £200.
"There can be no doubt," observes Mr. Blanford, "that C.
aegagrus is one of the species, and probably the principal, from
which tame goats are derived."
Fig. 176.—Japanese Goat Antelope. Nemorrhaedus crispus. × 1⁄12. (From Nature.)
The Chamois (Rupricapra) and the Goral (Nemorrhaedus)
are best described as Goat-like Antelopes; but, as already said, it is
difficult to split up the Bovidae satisfactorily. The Rocky Mountain
Goat, Haploceros montanus, is a large Goat-like creature, {327}which
has the peculiarity of having the shortest cannon bones of any Ruminant.
Its name denotes its range.
Fig. 177.—Goral. Nemorrhaedus goral. × 1⁄12. (From Nature.)
The Musk Ox, Ovibos moschatus, has been thought to be on the
borderland between the Sheep and Oxen, as indeed expressed in its
scientific name. It is a purely Arctic creature, now confined to the
Nearctic region; but it formerly existed in the Arctic regions of
Europe.
The anatomy of the "soft parts" of this genus has lately been
investigated by Dr. Lönnberg.[210] The animal has no foot glands such as
occur in Ovis. Its kidneys, however, are non-lobate, and it has
orbital glands. The cotyledons of the placenta are unusually large, and
the cow has the "primary four" teats. It cannot, in fact, be definitely
referred to either the Caprine or the Bovine section of the Cavicornia,
and while possibly most allied to Budorcas, it may be regarded, at
least for the present, as entitled to form a separate sub-family of its
own. The muzzle {328}has a slight naked strip above the
nostrils, as in the Sheep, but there is no fissure of the upper lip.
Extinct Families of Artiodactyla.
The origin of the Artiodactyla is placed by Cope in the family
Pantolestidae,[211] allied to the genus
Protogonodon of the Condylarthra. As, however, this family is
represented by but a few back teeth and a fragment of the hind-foot, it
seems premature to regard it as the necessary starting-point of the
Bunodont and Ruminant groups.
Fam. Anthracotheriidae.—This well-known and ancient
family consists of creatures of for the most part a Pig-like form, with
teeth approaching the selenodont shape, and a complete dentition. The
carpals, tarsals, metacarpals, and metatarsals are all free. The toes are
four (or five) to each foot, with the outermost beginning to be reduced.
These of course are all generalised and primitive characters, pointing
nowhere in particular, except, of course, to an Artiodactyle stock, on
account of the teeth and the two predominating toes.
The type genus of the family, Anthracotherium, is not, as its
name might seem to denote, a relic of the Carboniferous period; its
remains were found in lignite, which may also show that it was at least
semi-aquatic in habit. Its form, however, must have been Pig-like, so at
least one would presume from the elongated skull and shortish legs. There
were species as great as a Rhinoceros, and smaller forms. The genus began
in the Oligocene and continued down to the Pliocene. It is known from
Europe, Asia, and America.
The skull is long with a prominent sagittal crest. The facial part is
also very long, and the orbits are not closed by a bony ring. The
premolars are simple teeth; the molars distinctly bunodont with a
tendency in one or two to the selenodont condition. The canines are
powerful, as are also the incisors. The scapula has been specially
compared with that of the Camel. It has no acromion, which is usually
though not always absent in Ungulates. An ally of the present animal, for
instance, the Hippopotamus, has the acromion developed. The radius and
ulna, the tibia and fibula, are all fully developed. {329}
Ancodus (or Hyopotamus, as it has been called) is also
Oligocene in range, and its remains have been found in the same countries
as have those of Anthracotherium. Both genera are indeed closely
allied. Ancodus seems to be a more slightly-built creature. The
skull looks weaker, but presents much the same features of organisation.
In A. velaunus, a species found in French rocks, a metacarpal of
digit I. was present in the manus, while A. brachyrhynchus had a
completely five-fingered manus.
The Miocene genus Merycopotamus (from the lower layers of the
Siwalik formation in India) is more distinctly selenodont than the forms
already discussed. On this ground it has been placed in a separate
sub-family. As, however, in other respects it does not depart from the
Anthracotherian type of structure, this proceeding seems to be hardly
necessary. There are two species known, of which one, M. nanus,
is, as its name denotes, a dwarf form.
Fam. Caenotheriidae.—While the last family consisted of
animals rather more akin to the Pigs, the present is more Pecorine in its
characters. The molars are selenodont; but as in the Tragulidae the
premolars are more of the nature of cutting teeth. The dentition, like
that of so many of these early Ungulates, is complete, and the canines
are not prominent. The feet are four-toed, the lateral toes not reaching
the ground.
The principal genus is the Eocene and Miocene Caenotherium. Of
this genus there were a considerable number of species all European in
range, and of small size—not more than a foot or so in length.
Their small size is suggestive of the Chevrotains. In the skull the
orbital cavity is nearly or quite surrounded by bone, and the tympanic
bulla is large and inflated. A common feature of Artiodactyles, a failure
of the nasals and maxillae to meet at the side of the face, is to be seen
in this ancient forerunner of the Pecora.
Plesiomeryx, also European, and from the same geological
horizon, is a very closely allied form.
Fam. Xiphodontidae.—This family consists of slender,
small Artiodactyles which are, like the Caenotheriidae, related to the
Pecora. They are confined in their range to Europe.
The type genus Xiphodon has selenodont molars and elongated,
slender, cutting premolars. The dentition was complete and the canines
not highly developed. Like Caenotherium, {330}Xiphodon was a
hornless creature, but with only two toes, the two lateral digits being
represented by the merest rudiments of metacarpals. The other metacarpals
were unusually long.
Amphimeryx (also called Xiphodontotherium) is much more
imperfectly known, but belongs to this family or to that of the
Caenotheriidae. Dichodon is another member of the same family.
Fam. Oreodontidae.—This family, consisting of numerous
genera, is limited to the North American continent. Its range in time is
from the Eocene to the Lower Pliocene. The family as a whole is to be
distinguished by a number of primitive characters. The dentition is
complete; the feet are four- or even five-toed; the orbit is sometimes
open behind. The canines of the lower jaw are not more pronounced than
the incisors. The characteristics of the group will be further developed
by a consideration of some of the principal genera which are included in
this family.
Oreodon is a Miocene form about as large as a Peccary. The
skull has a short face with a completely-closed orbital cavity. In front
of the orbit is a deep pit, not a mere deficiency of ossification, such
as occurs in many Artiodactyles. This is placed on the lachrymal bone,
and is in fact a lachrymal fossa, such as occurs in other forms. The
odontoid process of the axis vertebra is somewhat cheese-taster shaped,
as in recent Artiodactyles. There are fourteen dorsal vertebrae and a
very large number of caudals. The radius and the ulna are completely
separated, as are the carpals. There are five digits to the fore-limbs.
The fibula is complete and independent. The hind-foot is four-toed.
Several species of the genus are known.
Merycochoerus is an allied Miocene genus. It is more massive in
form than the last, but otherwise does not present differences of
importance.
Mesoreodon is another genus of this family which presents some
curious features of organisation. In the skull and teeth there is nothing
very noteworthy, but the hyoid is remarkable. This appendage of the skull
is by no means always preserved, and when it is, it might be denied that
it belonged to any particular skull. In the present case there appears to
be no doubt as to the identity of the bones, which resemble the
corresponding bones of the Perissodactyla much more than they do those of
other Artiodactyles. Associated with the bones an ossified {331}thyroid
cartilage of the larynx was found. As the skull was that of a male, this
character may be a sexual one. It is quite comparable to the ossification
of the same cartilage in the American monkey Callithrix. "The
function of the bone," observes Professor Scott,[212] "was probably similar to that
performed by the enormously-inflated basihyal of the howling monkeys, and
must have given to these animals most unusual powers of voice." Another
important anatomical fact about Mesoreodon is the apparent
existence of a clavicle. It is of course conceivable that the remains of
some other animal have got mixed up with that of the individuals upon
which the present genus is founded; but failing that, here is a clavicle
in an Ungulate. The spine of the scapula possesses a metacromion. This
greater development of the spine of the scapula in Artiodactyles than in
Perissodactyles is, it is suggested, to be correlated with the earlier
loss of the clavicle in the latter group of Ungulates.
Cyclopidius (synonymous with Brachymeryx) is a kind of
pug form of Oreodon. The skull is short and broad, and the end of
the snout a little turned up. The upper incisors are small and drop out
early. On each side of the nasals is a large oval vacuity which is
perhaps to be compared to the lateral deficiency to be found in other
Artiodactyles. One species of this singular-looking form is appropriately
called C. simus.
Other allied genera are Merychyus and Leptauchenia. The
former extends as far down as the Lower Pliocene, and is thus one of the
newest forms of Oreodontidae.
Agriochoerus[213] (Fig. 178) is placed in a separate
sub-family from the types which have just been considered. It is Miocene
in range. It differs from Oreodon and its closer relatives by the
fact that the orbit is open behind and not closed. The most remarkable
fact about this creature is that the terminal phalanges of the digits
(five in the fore- and four in the hind-feet) being pointed, seem to
suggest their encasement with claws rather than hoofs. The pollex, though
small, seems to have been opposable. As with other Oreodonts, the molars
are selenodont. The premaxillae are toothless—at least in adults,
for two teeth are present in the {332}young. There are
several species. Agriochoerus, like Oreodon and primitive
Ungulates in general, had a long tail. The genus thus shows a mixture of
ancient and specialised characters.
Fig. 178.—Skeleton of Agriochoerus latifrons. × ⅛. (After Wortman.)
The most ancient form of Oreodont is Protoreodon. This is
Eocene, and became extinct during that period. It had a complete
dentition, open orbit, and no lachrymal fossa. The fore-feet were
five-toed, the hind four-toed.
Fam. Anoplotheriidae.—This family is entirely Eocene in
point of time, and is unknown outside Europe. The dentition of the group
is complete; the molars are seleno-bunodont, like those of the
Anthracotheriidae. The bones of the carpus, tarsus, metacarpus and
metatarsus are all free; the toes are four to two in number on each foot.
The orbit is widely open behind. The tail is long, as in Xiphodon,
etc. {333}
These general characters only just serve to differentiate the family;
but they illustrate its archaic character, in which it resembles the
Xiphodontidae, and even more the Anthracotheriidae. A survey of some of
the genera which have been assigned to the family will bring out other
features in the organisation of these very ancient Artiodactyles.
Anoplotherium is so called on account of the fact that it is,
like all ancient Artiodactyles, without horns or claws. Tusks it might
have, but as a matter of fact has not. There are, as in Artiodactyles
generally, nineteen dorso-lumbar vertebrae; the long tail has numerous
chevrons. The shoulder blade has a well-marked acromion and a distinct
coracoid process; it is wide proximally. The bones of the fore-arm and
fore-leg are, as is usual in primitive Artiodactyles, separate.
In the skull the chief features, in addition to that mentioned in the
definition of the family, are the large size of the paroccipital
processes; there is no fossa lachrymalis or deficiency in the side of the
face. The animal is three-toed, both in the fore- and hind-limbs. The
second toe is nearly as large as the Artiodactyle third and fourth. There
are tiny rudiments of the two remaining fingers. The hind-foot is also
three-toed, and there is a trace of the hallux. The fingers are so widely
separated and divergent from each other that it has been suggested that
the animal had webbed feet and inhabited marshes, in which it swam by the
aid of its long tail. The creature was the size of a Tapir.
Closely resembling Anoplotherium are a number of other
genera.
Diplobune ( = Hyracodontotherium) was much like the
last, but was a more delicately-formed animal. The fingers and toes
(three of each) end in such sharply-pointed phalanges that claws seem to
be almost suggested. There are several species of this genus.
Dacrytherium differs by the presence of a lachrymal fossa.
Dichobune has four-toed extremities, of which the lateral ones
are more slender and shorter than the two middle ones. As in other
Anoplotheriidae, the anterior premolars are furnished with a sharp
cutting edge.
Order V. SIRENIA.
Aquatic Mammalia, with but few scattered hairs; hind-limbs absent;
fore-limbs paddle-shaped; tail flattened, and either {334}Whale-like or
rhomboidal to circular in form. Nostrils on upper surface of not
specially-elongated snout. Clavicles are absent. The scapula has the
normal mammalian form, with a well-developed and roughly median spine.
The bones of the arm and hand articulate together, as in land animals;
the phalanges show at most traces of increase in number above the normal.
Pelvis represented by a vestige, more highly developed in some fossil
than in recent forms. Stomach complex, consisting of several chambers.
Lungs simple and not lobulated. Diaphragm oblique and very muscular.
Brain peculiar in form and but slightly convoluted. Testes abdominal.
Teats two, and pectoral in position. Placenta non-deciduous and zonary.[214]
This limited group consists of purely aquatic forms, which are both
marine and fresh-water in their proclivities. They have been placed in
the immediate vicinity of the Whales; but it is now believed by most
zoologists that the likenesses which they undoubtedly show to the Cetacea
are of an adaptive kind and related to their similar mode of life. The
group is a readily-definable one. Externally they are marked by their
dark coloration, somewhat Whale-like though of clumsier build, and by the
total absence of external ears and hind-limbs; the latter are, however,
as will be pointed out shortly, marked by certain rudimentary bones.
There is a flattened tail, which in the Dugong and Rhytina is
precisely like that of a Whale. It is interesting to note that the former
genus, whose tail is, judging it at least by the standard of the Whales,
more completely modified for the aquatic life, should also show other
features which indicate their longer life as marine creatures. For the
flippers are more Whale-like in that the fore-arm is completely enclosed
within the body, or nearly so, and the nostrils have a more decidedly
superior position than in the Manatee. The fore-limbs of this group, as
may be inferred from what has just been said, are flipper-like; but,
contrary to what we find in Whales, the phalanges do not as a rule show
any traces of multiplication, so characteristic a feature of the Cetacean
hand, and the individual bones are connected by well-formed joints.
Beneath the thick skin, which is sparsely provided with stout hairs in
the Dugong, is a layer of blubber. Dr. Murie has called attention to the
fact that this layer in the {335}Manatee[215] differs from the blubber of the Whale
in that there is no free oil anywhere.[216]
The skeleton of the Sirenia is strong and massive, thus contrasting
with the loosely-textured bones of the Cetacea. The cervical vertebrae
are, as a rule, free, but the second and third are fused in
Manatus and the extinct Halitherium. It is noteworthy that
in Rhytina the cervical vertebrae have the exceedingly thin centra
that characterises the neck vertebrae in Whales. The ribs are most of
them firmly articulated by two heads. The breastbone is generally
reduced, as in Whales; and but few ribs are attached thereto. The
vertebrae, moreover, are well locked together by zygapophyses, and not
loosely attached as in Whales.
The shoulder blade is long and narrow, and not unlike that of the
Seals. It is totally unlike the peculiarly-modified scapula of the Whale
tribe. But, as in the latter, there are no clavicles.
The hind-limbs are only represented by the pelvis; and this is a
rudimentary structure, varying, however, in the degree of its
degeneration. That of the extinct Halitherium recalls the pelvis
of the Rorqual. There is a single triradiate bone with an acetabular
cavity for the rudiment of the femur in the centre; it suggests that here
the three normal elements of the pelvis have become fused into a single
bone. In the Dugong there are two small bones on each side.
The Manatees (Manatus)[217] are found in the fresh-waters and
along the Atlantic coasts of South America and Africa. It appears that
there are four species, of which one only is African, the others
American. Report asserts the former occurrence of this genus on the
shores of St. Helena.
The Manatee is provided with only six cervical vertebrae, a fact which
distinguishes it from the other existing genera of its group. A
remarkable feature which it exhibits is the large number of molar teeth.
These apparently go on increasing indefinitely during its life, the
suggestion being that they are worn away by the nature of the
food—algae with much sand intermixed. As many as twenty molar teeth
have been counted in one half of the jaw, and there is no reason to
forbid the assumption that they {336}may get still more numerous. This large
number of grinding teeth is obviously suggestive of the Whales, with
which the Sirenia are believed by some to be allied. It is at least a
remarkable coincidence that these two aquatic groups of mammals should
both have assumed the same indefinite tooth formula. It is correct to say
assumed, since extinct forms of Manatees, such as Halitherium and
Prorastoma, have not a continuous succession of molars. The brain
of the Manatee is, contrary to the usual arrangement among aquatic
mammals, smooth, and only marked by one or two fissures.
The Manatee[218] is
black in colour, its thick skin being wrinkled. The animal is assisted in
feeding by a curious mechanism of the upper lip; this is split in two,
and the two halves, which are furnished with strong bristles, can play
upon each other like the points of a pair of forceps. The flippers are
furnished with nails, save in M. inunguis, but in the nailed forms
it is not every finger which is thus armed.
Fig. 179.—Skeleton of Dugong. Halicore australis. (After de Blainville.)
Halicore,[219]
the Dugong, is an entirely Oriental and Australian {337}form; there appears to
be but a single species, though more than one name has been given to
supposed distinct species. As already mentioned, it differs from the
Manatee in the possession of a Whale-like tail; the nostrils, too, are
more upon the upper surface of the head, and there are no nails upon the
flipper. The peculiar cleft lip of the Manatee is not so well developed
in the Dugong, but there are traces of it; and in the foetus the likeness
to the Manatee in this respect is very striking. It would thus appear
that Halicore is a stage in advance upon Manatus; that the
remarkable mechanism of the lip of the latter has been possessed, but has
been lost, by the Dugong. The skull of the Dugong is distinguished by the
stout premaxillary bones, which bear a tusk in the male. In the female
the tooth is there, but is lodged within the bone. This incisor has a
milk forerunner. The back teeth of the Dugong (there are no canines) are
few in number (four or five, even six), thus showing a gradual reduction
when compared with Manatus; and this culminates in the toothless
Rhytina. It is also interesting to notice that in the massive
lower jaw there are traces of an incisor. Were this to be developed into
a tusk, the jaw would present a curious resemblance to that of
Dinotherium.
The Dugong, H. dugong, has the reputation of being the original
of the mermaid legends, since the young is held to the
pectorally-situated breast with one flipper. "But it should be
remembered," justly observes Dr. Blanford, "that stories of beings half
man or woman, half fish, are as common in temperate as in tropical seas,
and that some of them are more ancient than any European knowledge of the
Dugong."
Extinct Sirenians.—The earliest genus that can be with
certainty referred to this order is the Oligocene Prorastoma. This
genus, though offering no particular skull-characters that assist in the
determination of the much-debated affinities of the Sirenia, shows a
remarkable condition of the teeth that may afford a clue. The species
P. veronense, recently described by Mr. Lydekker,[220] is founded upon a fragment of the
skull which contains two teeth apparently representing the third and
fourth upper milk molars. The interest attaching to these teeth lies in
the fact that they clearly exhibit the buno-selenodont condition
characteristic of certain early Artiodactyles, e.g.
Merycopotamus.
Halitherium is a later genus, which is known by the nearly {338}complete skeleton. The skull is like that
of other Sirenia, with the down-turned premaxillary region. But the nasal
bones, lost, or at least rudimentary, in recent forms, are well
developed; the likeness of ancient to living forms in this respect being
exactly paralleled by the Zeuglodonts, when compared with recent Whales.
The vertebral centra exhibit distinct epiphyses, which have disappeared
in living Sirenians. The cervical vertebrae are seven, of which the
second and third are occasionally fused. There are nineteen pairs of
ribs, and there are three lumbar vertebrae. The sternum consists of three
separate pieces. There is a rudimentary femur.
The recently-extinct Steller's Sea-cow, belonging to the genus
Rhytina, was a huge beast, seen in the flesh up to nearly the end
of the last century. It frequented the shores of Bering's Straits. Its
remains occur in the peat on the shores of those seas. It reaches a
length of some 20 to 30 feet. The external characters were much like
those of other recent Sirenians. The nostrils were above the fore part of
the snout, the latter being truncated and obtuse. The tail was of the
Cetacean pattern, and thus like that of Halicore. The head of this
Sirenian was small, and the teeth had entirely vanished save for the
apparent existence as transitory structures of two small incisors in the
upper jaw. The absence of teeth was compensated by the presence of a
horny palate for the trituration of the sea-weeds which constituted the
food of Steller's Sea-cow. The fore-limbs seem to have possessed no
nails, but were covered at the extremity with short, bristly hairs, no
doubt serving the purpose of keeping the animal moored in safety to the
slippery beds of Fucus upon which it browsed.
There are nineteen pairs of ribs. The vertebrae of the cervical region
are the customary seven, and the centra are thin and plate-like as in the
Cetacea, the animal being thus short-necked like those marine
creatures.
{339}
CHAPTER XII
CETACEA—WHALES AND DOLPHINS
Order VI. CETACEA.[221]
Aquatic Mammalia of fish-like form; tail expanded into horizontal
flukes; a fatty dorsal "fin" present in most species; anterior limbs
converted into fin-like paddles; posterior limbs only represented by
skeletal rudiments. Hairy covering reduced to a few isolated hairs in the
neighbourhood of the muzzle. Nostrils represented by the single or double
blow-hole, nearly always situated far back upon the skull. Bones of loose
texture and much impregnated with oil. The skull has a greatly-developed
facial portion; supra-occipital bones meeting the frontal by overgrowing,
or growing in between the parietals; bones surrounding the organ of
hearing loosely attached to the skull, the tympanics of peculiar
cowrie-shell form. Coronoid process of mandible absent, or very feebly
developed. Teeth, when present, few or numerous, always of simple conical
form, with at most traces of additional cusps (Inia); if absent
their place taken by whalebone. Cervical vertebrae of short
antero-posterior diameter, often more or less completely welded together
into a single mass. Articulations between dorsal and other vertebrae
feeble. Scapula peculiarly flattened; acromion strongly developed as a
rule, but arising from a slightly-marked spine; coracoid process
generally strongly developed. Phalanges of digits always more numerous
than in other mammals. Clavicles absent. Stomach complex, consisting of
at least four and often more chambers. Lungs simple and non-lobulated.
Diaphragm obliquely set and very {340}muscular. Brain much
expanded transversely and well convoluted. Testes abdominal. Teats two,
inguinal in position. Placenta diffuse and non-deciduate.
The Whales and Dolphins, which constitute this order, form an
assemblage which is easily characterised by reason of the fact that their
affinities to other groups of Mammalia are so doubtful that they furnish
matter rather for speculation than for authoritative statement. Some hold
that they resemble in certain points the Ungulata; while others again see
in them the culminating term of a series which commences with such a form
as the Otter, and of which the Seals and Sea-lions are intermediate
stages. A third opinion is that the Whales have arisen from some low
mammalian stock, too primitive to be assigned to any existing order of
mammals. Palaeontology, as will be seen later, throws no light whatever
upon their origin. This matter has already been referred to (see p. 120) in considering the position of the Cetacea.
The Whales include the most gigantic of all the orders of vertebrated
animals. No creature living or extinct is so large as the Sibbald's
Rorqual, which attains to a length of some 85 feet, or perhaps even
rather more. On the other hand we have what are by comparison minute
forms. Apart from the possibly problematical Delphinus minutus,
stated to be only 2 feet in length, we have as a minimum 3 or 4 feet. The
size of the Cetacea has been subjected to much exaggeration. The first
duty of a Whale, observed the late Sir William Flower, is to be large;
and Natural Historians, in the recent as well as in the remote past, have
not hesitated to put very round numbers upon the dimensions of the larger
members of the order. We may perhaps pass over Pliny's "fish called
balaena or whirlpool, which is so long and broad as to take up more in
length and breadth than two acres of ground," and a number of analogous
exaggerations, which gradually dwindled down to the dimensions just
stated of the great Rorqual. M. Pouchet has made the ingenious suggestion
that the statements of the ancients may have been nearer the truth than
observations of to-day would have us believe; he pointed out justly that
in former times Whales were not so relentlessly pursued as during the
last century; the inference being that they may have lived to a greater
age, and attained a more colossal bulk. The more modern exaggerations in
the {341}dimensions of the bigger Whales are
probably due to the fact that measurements have been taken, not in a
straight line from snout to tail, but along the bulging sides of the
Cetacean, rendered even more convex than in nature by decomposition, and
by pressure due to the immense tonnage of the creature.
The Cetacea are the most perfectly aquatic of all mammals; they never
leave the waters which they inhabit. It is true that legends have
represented them as pasturing upon the shore—Aelian spoke of
Dolphins basking in the sun's rays upon the sand; and the "Devil Fish" of
California, Rhachianectes (see p. 357) has
given rise to improbable stories—but they are apparently only
legends. Indeed a stranded Whale cannot live long, for it is unable to
breathe, the comparatively feeble breast being crushed by its own weight.
In accordance with the purely aquatic habit, we find a modification of
the outward form of the body (and as we shall see later of many of the
internal organs), which renders the Cetacea externally unlike all other
mammals. The form is fish-like, the fore-limbs are paddles, the tail is
expanded into two horizontal flukes, which serve to propel the creature
through the water.
Fig. 180.—Killer. Orca gladiator. × 1⁄40 (After True.)
The skin is smooth and shiny, so smooth and so shiny that it has often
been compared to coach leather. But nevertheless they are not entirely
without that most essential character of the class Mammalia, a coating of
hair. The hairy covering is, however, reduced to the very smallest
proportions; it is represented {342}by a few hairs only—so few that they
can be counted with ease—in the neighbourhood of the muzzle. These
hairs are not present in all Whales; they are absent, for example, in the
White Whale or Beluga. When present they are not furnished either with
sebaceous glands or with muscular fibres, which are such universal
concomitants of the hair follicles in the Mammalia generally. This
appears to be conclusive evidence that the hairs, few as they are, are
still undergoing degeneration. The need for a furry coat is removed by
the presence of a thick coating of fat immediately underlying the skin.
This is known as the blubber, and is the main incentive to the pursuit of
Whales. It must not, however, be assumed without further argument that
the hair is absent because its place is taken, as a mechanism for
retaining the heat, by the blubber; for the Seal tribe possess both fur
and blubber. Another conceivable explanation is quite at variance with
such a view of economy. It may be noticed that among Ungulates there is a
tendency to lose hair, particularly among more or less aquatic forms.
Thus the Hippopotamus is almost naked (as is indeed the Walrus); the
Rhinoceros, too, often a frequenter of marshy soil, is almost as denuded
as is the Hippopotamus. It is not, however, settled that the Whales have
anything to do with the Ungulata; otherwise an additional argument might
be used, that is, the secular loss of hair in some members of this group.
The Hairy Rhinoceros, Rh. tichorhinus, was, as its name denotes, a
hairy beast; the Mammoth was equally so. The descendants, or at least the
modern representatives of both these creatures, are but scantily clad
with hairs.
A final reason for the naked character of the skin in existing Cetacea
is closely connected with a feature in the organisation of three or four
living species which must first be described.
Some years ago the late Dr. J. E. Gray of the British Museum described
from the sea, off Margate, what he considered to be a new species of
Porpoise, characterised by the presence on the dorsal fin of a row of
stony tubercles. As a matter of fact it was subsequently shown that the
Common Porpoise has the same structures, so that there was no need for a
Margate species, Phocaena tuberculifera. Moreover, in the Indian
Neomeris, a close ally of the Porpoise, a more abundant calcified
covering of scales exists along the whole back of the animal. These
plates, {343}it has been discovered, are larger in the
foetus, a fact which naturally points to their being an inheritance from
the past, now undergoing retrogressive changes. Such a way of looking
upon the facts is confirmed by the finding, many years ago, by the
naturalist and physiologist Johannes Müller, of bony plates in connexion
with the remains of a Zeuglodont Cetacean. It looks, therefore, very much
as if the Eocene ancestors of the modern Cetacea had a skin studded with
bony plates, as have the armadillos. This being the case, the
disappearance of hair is not surprising. The room would be taken up by
the calcified plates, and when the latter disappeared, as they have in
the vast majority of existing Whales, the naked skin alone would be
left.
Whales possess no externally-visible hind-limbs; rudiments of these
appendages are present, which will be dealt with under the description of
the principal features of the skeleton. But it has been discovered that
in the Porpoise, external vestiges of hind-limbs do appear in the foetus,
a fact which, be it observed, does away with the old view that the flukes
of the Whale are the last term in the series of vanishing hind-limbs, of
which the Seals, with their hind-limbs and tail bound up together, offer
an intermediate step.
The tail is fish-like in form, but the flukes are horizontal instead
of vertical as in fishes and Ichthyosaurus. This arrangement is no
doubt associated with the need for rapid return to the surface waters
after a prolonged immersion in search of food. A downward stroke, such as
is given by the powerful and large tail flukes, would naturally bring
about this result rapidly. The tail, moreover, is under all circumstances
the swimming organ. Its motion has been stated to be slightly rotatory,
like that of a screw, and it is the case that the two flukes are often
alternate in shape like the flanges of a screw; one being convex upwards,
the other convex downwards.
The fore-limbs are in the form of paddles, but they do not apparently
serve as organs of locomotion so much as balancers. When a Whale is
killed, it falls over on to one side, the office of the flippers being to
maintain the proper position. It is believed, however, from the fact that
the embryo often shows a relatively larger pectoral fin than that of the
adult—the difference being due to a reduction in the adult of the
number of phalanges—that the fin was once an organ of progression.
{344}
The pectoral fin of Whales exists in two forms. In the Toothed Whales
it is shorter and rounder; in the Whalebone Whales longer and narrower.
Structural differences accompany these outward dissimilarities. In the
first-named group the humerus and the beginning of the radius and ulna
are within the body, and do not form a part of the fin. In the Whalebone
Whales, on the other hand, the fin contains all the bones of the
fore-limb. Another remarkable contrast between the hand in the two groups
of Whales is that while the Toothed Whales have five fingers, thus
justifying the prevailing opinion that they are the more primitive of the
two groups, the Whalebone Whales have only four fingers. Actually the
Right Whale, Balaena, seems to have five fingers; and, indeed, the
fact that it has, is often used to distinguish it from the Humpback,
which has undoubtedly only four. But a careful consideration of the state
of affairs which prevails in the foetus of Balaenoptera dispels
this idea. Between what are apparently the second and third fingers, a
rudimentary finger, consisting of four phalanges, appears. This is not
produced, as is an additional finger found in the White Whale or Beluga,
by a splitting of a finger. Accordingly the four-fingered condition of
the Whalebone Whales is produced by the dropping out of a finger in the
middle of the series,—a very remarkable fact. When fingers
disappear, as, for instance, in the Horse, etc., it is at the two ends of the series
that the digits vanish. If this view of Professor Kükenthal's[222] be accepted, it
follows that the presumed thumb of the Right Whale is what has been
termed the prepollex.
The hand of the Whales, like those of some other aquatic creatures,
e.g. the reptile Ichthyosaurus, has a larger number of
phalanges than have terrestrial animals. The result of this is, of
course, to increase the length of the fin and its utility as a paddle. It
is commonly not all the fingers that have developed this great number of
accessory phalanges. Rudimentary nails have been found upon the Cetacean
hand; but in no case are they functionally developed. In the Manatees we
have the disappearance of the nails still imperfectly accomplished. In
M. latirostris there are nails; these have vanished, apart from
possible traces to be seen with a microscope, in M. inunguis.
A very characteristic feature of certain Whales are the furrows {345}to be
seen on the throat. This is especially the case with the Rorquals, in
which group the Humpback Whale, Megaptera, is to be included. The
whales of these two genera (Balaenoptera and Megaptera)
have a large number of the throat furrows—as many as sixty have
been counted. Some other Whales have a smaller number; thus
Rhachianectes has but two on each side, and the Physeteridae have
not many more. These furrows are absent in very young embryos. It is
thought by Professor Kükenthal that they allow of a wide opening of the
mouth.
Fig. 181.—Dorsal surface of bones of
right anterior limb of Round-headed
Dolphin (Globicephalus melas). × 1⁄10.
The shaded portions of the digits
are cartilaginous. c, Cuneiform; H,
humerus; l, lunar; R, radius; s,
scaphoid; td, trapezoid or magnum;
U, ulna; u, unciform; II-V, digits.
(From Flower's Osteology.)
The blow-hole of Whales is, of course, the aperture of the nostrils,
which are not so far back in the foetus as in the adult. By the
characters of the nostrils the Toothed Whales can be distinguished from
the Baleen Whales; in the latter the orifice is double, in the former
single. In embryos of Dolphins, however, the two apertures are quite
independent. The phenomena of spouting have often been misinterpreted.[223] When the Whale
breathes, the expired air rushes out through the nostrils. The water
vapour in the breath condenses into drops of water in the cold Arctic
regions where the phenomenon has been mainly observed. Hence the idea
that water taken in at the mouth is expelled through the blow-hole. As
the Whale approaches the surface to breathe, it may be that some of the
water of the sea is {346}driven upwards by the forcible expulsion
of air from the lungs. But for the most part the water which is spouted
is simply condensed breath.
Like some, but not all, other aquatic Mammalia the Whales have
apparently no external ear. Indeed the opening of the ear is excessively
small. In a huge Rorqual it will "admit a quill"; and although "a quill"
is rather vague, we may fairly allow any sized quill without proving that
the orifice of the auditory passage is anything but exceedingly minute.
As a proof, added to so many, that the Whales are the progeny of
terrestrial creatures, we have the occasional traces of external ears.[224]
Fig. 182.—Left lower jaw of foetus of Balaenoptera rostrata. Inner aspect,
natural size, showing teeth. (After Julin.)
Whalebone Whales never possess permanent teeth as well as the baleen;
but in the foetus are more than traces of true teeth, which, however,
never arrive at maturity. The whalebone itself is described later (p. 354). That the Whalebone Whales possess teeth while
in the foetal condition was discovered so long ago as 1807. It has since
been confirmed by many observers. Not only is there one set of teeth
developed in the foetal Balaenoptera but two, of which one comes
to a greater maturity; the other, in fact, remaining at a very early
stage of development. The more complete dentition belongs to the milk
series, as is the case with the Toothed Whales. A very interesting
conclusion with regard to the derivation of the simple conical teeth of
Whales seems to follow from the development of these structures in
Balaenoptera. There are in the young foetus fewer teeth than in the more
advanced embryo. Now in the younger embryo some of the teeth are
furnished with more than one cusp; they are bi- or even tri-conodont. As
Sir R. Owen observed, the teeth—some of {347}them—are
literally double teeth. This is a suggestion of the more complicated
teeth of the Zeuglodonts, and shows so far that the simple conical teeth
of existing Whales (cf. however the Platanistidae) are not by any manner
of means so primitive as their actual structure would undoubtedly lead
one to believe. Further than this, the greater number of teeth in the
older embryo coincided with the disappearance of these double teeth,
which seem to split up into the simple conical teeth.
The Toothed Whales are not furnished with baleen, but with teeth only.
These teeth are more or less numerous, their arrangement being of value
in the classification of the group; a matter which is dealt with
later.
In the Narwhal, whose dentition in the adult is reduced to the
well-known tusk or tusks (properly developed only in the male), there is
a complete foetal dentition. A very curious fact has been elucidated by
Professor Kükenthal about the dentition of the Common Porpoise. It
appears that in this Cetacean the two teeth corresponding to each other
of the two dentitions may fuse into a single tooth, which has in
consequence a double crown. It may be that this is the case with the
Platanistid Inia, and that its diconodont teeth are not,
therefore, a reminiscence of the comparatively complicated teeth of the
ancient Zeuglodonts.
The internal organs of Whales which show the greatest peculiarities as
compared with other mammals are the stomach, the lungs, and the
diaphragm. Whales always possess a complicated stomach divided into many,
but into a variable number of, chambers: there are as few as four in
some, as many as fourteen in Ziphioids.
On account of its complication the stomach[225] has been compared to that of
Ruminants—it has even been alleged that Whales "ruminate"—but
the comparison will not hold good. Nor, on the other hand, is there a
very close resemblance to the equally-complicated stomach of the
Sirenians.
The Rorqual has a stomach with as few compartments as any. The only
Whale which appears to have fewer is Balaena mysticetus, where
there are but three. In the Rorqual the oesophagus opens into a more or
less globular sac; from the upper end of this, i.e. close to the
entry of the oesophagus, arises the second chamber, long and narrowish;
then follows an extremely short third sac, {348}then a larger fourth,
after which comes the dilated commencement of the small intestine. The
latter might be regarded as a chamber of the stomach were it not for the
fact that the ducts of the liver and the pancreas open into it. This
represents one type of the Cetacean stomach, which seems to be found in
all Whales except the Ziphioids. In the latter, the oesophagus opens into
the first compartment as usual; but the second division of the stomach
arises not close to the entrance of the oesophagus, but at the opposite
end. It would seem, therefore, as if the first division of the stomach,
found in most Whales, were missing in Ziphioids. This way of looking at
the matter is confirmed by the fact that in Hyperoodon a remnant
of the missing first stomach is found in the shape of a small
diverticulum of the oesophagus just before it enters the stomach.
The essential difference between the Whale's and the Ruminant's
stomach is this: in the latter the stomach is primarily divided into two
portions, of which the first is non-digestive and is clothed with
oesophageal epithelium. The second, the abomasum, is the digestive
region. The first part is again divided into three compartments. In the
Whales, on the other hand, it is the digestive part which is again
subdivided, while if the first part is divided it is not markedly so as
in the Ruminants.
The lungs are remarkable for their unlobulated character; in this they
agree with the lungs of the Sirenia. The thoracic cavity in which they
lie is barrel-shaped, and not, as is usual in terrestrial mammals,
boat-shaped, i.e. narrower sternally than above. The alteration of
the shape of the thoracic cavity is associated with the aquatic life; so
at any rate the fact that it is also marked in Seals and even in the
Otter seems to show. The Whales are also characterised by the great
obliquity of the diaphragm, which is extremely muscular. In this
character again we find an agreement with the Sirenia, and also with
other aquatic mammals; it is not therefore a character of Whales so much
as evidence of an adaptation to the aquatic life. The advantage is, it
appears, in the increased capacity of the thoracic cavity, and the
consequent greater possibilities of expansion of the lungs, which it must
be remembered serve as hydrostatic as well as breathing organs.
Some of the internal arteries of Whales break up into retia mirabilia.
Their kidneys are lobulated; whether this has {349}anything to do with the
aquatic life is not so clear. It also characterises the Sirenia, more or
less, and the Otters; but, on the other hand, the terrestrial Bears show
the same structure as do also some Ungulates. It must be borne in mind,
too, that the kidneys of foetal Man are lobulated.
The liver is a compact organ not showing such lobulation as is common,
but not universal, among mammals.
The bones of Whales have a somewhat loose structure, and are much
impregnated with oil. In many features the skeleton of Whales is highly
distinctive of the order.
Fig. 183.—Skeleton of Porpoise (Phocoena communis), acr, Acromion process of scapula; cerv, united cervical vertebrae; chev, chevron
bones; cor, coracoid process; hu, humerus; hy, hyoid; jug, jugal; lumb.trans, lumbar transverse processes; max, maxilla; nas, nasal;
orb, orbit; par, parietal; pelv, vestige of pelvis; per.ot, periotic; pr.max, premaxilla; rad, radius; rb1, first rib; rb12, twelfth rib; sc,
scapula; s.occ, supra-occipital; st, sternum; uln, ulna; zyg, prezygapophysis. (From Parker and Haswell's Zoology.)

Fig. 184.—Under surface of the cranium of a young Caa'ing Whale (Globicephalus
melas). × 1⁄5. AS, Alisphenoid; BO, basioccipital; cf, condylar foramen; ExO,
exoccipital; Fr, supra-orbital process of frontal; gf, glenoid fossa of squamosal;
Ma, body of malar; Mx, maxilla; OS, orbitosphenoid; Per, posterior (mastoid)
process of periotic; Pl, palatine; PMx, premaxilla; Pt, pterygoid; Sq, squamosal;
tg, deep groove on squamosal for meatus auditorius externus, leading to tympanic
cavity; Ty, tympanic; Vo, vomer; ZM, zygomatic process of malar. (From
Flower's Osteology.)
The brain case is small proportionately and rounded. The "face" is
therefore long, and in some cases, especially among the fossil forms of
Platanistidae, the rostrum is extraordinarily elongated. The asymmetry of
the Whale's skull is one of its most remarkable features; this, however,
is entirely limited to the Toothed Whales, and among them is more
pronounced in some forms than in others. Thus the Platanistidae and many
Ziphioids are not nearly so asymmetrical as the {350}Dolphins and,
especially, Physeter. This asymmetry affects particularly the
premaxillae, the maxillae, and the nasals. The base of the skull is
symmetrical. The Whale's skull has very long premaxillae which, however,
do not, except in the extinct Zeuglodonts, bear any teeth. The nasal
bones, whether symmetrical or the reverse, are very small in existing
Whales, which arrangement, together with the long and broad maxillary
bones, removes the anterior nostrils, the blow-hole, far backwards. The
roof of the skull is not at all formed by the parietals externally. These
bones form a portion of the side of the cranium, but are replaced or
covered by the {351}enormously-developed supra-occipital in
the adult. Here again the Zeuglodonts are more typically Mammalian, for
in them the parietals have a normal development and situation, rising
even into a median crest as in so many quadrupeds. The bones related to
the organ of hearing, the tympanis and petrous bones, are very solid and
dense in structure. Moreover they are but loosely attached to surrounding
bones, and are thus easily and frequently lost. Nearly the only mammals
which resemble the Whales in the fact that the pterygoids sometimes meet
in the middle line below are the Edentata (Anteater and Armadillo, see p.
167). But in both groups this peculiarity is not
universal.

Fig. 185.—A section of a skull of a young Caa'ing Whale (Globicephalus melas). × 1⁄5. a,
Angle; an, anterior nares; AS, alisphenoid; bh, basihyal; BO, basioccipital; BS,
basispnenoid; cd, condyle; cp, coronoid process; ExO, exoccipital; Fr, frontal;
id, inferior dental canal; IP, interparietal; ME, ossified portion of the mesethmoid;
Mx, maxilla; Na, nasal; Pa, parietal; Per, periotic; Pl, palatine; PMx,
premaxilla; pn, posterior nares; PS, presphenoid; Pt, pterygoid; s, symphysis
of mandible; sh, stylohyal; SO, supra-occipital; Sq, squamosal; th, thyrohyal;
Vo, vomer. (From Flower's Osteology.)
The vertebral column is remarkable for the fact that more or fewer of
the cervical vertebrae may be fused together into a short and compact
mass. This is seen at its maximum in the genera Balaena and
Neobalaena. The odontoid process of the second {352}vertebra,
though hardly at all marked, is nevertheless really present and developed
from a bony centre of its own, as in other mammals. The dorsal and lumbar
vertebrae are, of course, to be distinguished by the presence of ribs
attached to the former; but as there is only a rudimentary pelvis, not
attached to the vertebral column, no sacral region can be detected. The
caudal vertebrae are to be recognised by the -shaped chevron bones
below.

Fig. 186.—Section through middle
line of united cervical vertebrae
of Greenland Right Whale (Balaena
mysticetus). × 1⁄9. a, Articular
surface for occipital condyle;
e, epiphysis on posterior end of
body of seventh cervical vertebra;
sn, foramen in arch of atlas for
first spinal nerve; 1, arch of atlas;
2, 3, 4, 5, 6, conjoined arches of
the axis and four following vertebrae;
7, arch of seventh vertebra.
(From Flower's Osteology.)
Fig. 187.—A, Sternum of Greenland Right Whale (Balaena mysticetus). × 1⁄15. B,
Sternum of Common Rorqual or Fin Whale (Balaenoptera musculus). × 1⁄10.
(From Flower's Osteology.)
The sternum in the Whale tribe is much more modified in the Whalebone
Whales than in the Odontocetes. In the latter it is made up of several
pieces, as in other mammals, which often, however, become coalesced. In
the Mystacoceti this bone is a single piece, to which only one pair of
ribs is attached, and its {353}form is characteristic of the genus. It is
heart-shaped, more or less, in Balaena, and somewhat cross- or
-shaped in the genus Balaenoptera. In the Odontocetes the ribs
have, some of them, the normal attachment by capitulum and tuberculum. In
the Mystacocetes the attachment, where it exists, is very loose, and the
tuberculum alone is attached to its vertebra. This allows of the freer
play of the ribs during respiration. The scapula has a very
characteristic form in these animals. The acromion, where it exists, is
placed near the anterior margin of the shoulder blade, and overlaps the
generally long coracoid process. Clavicles are totally absent. The pelvis
is very rudimentary, consisting merely of a single bonelet, to which are
attached the rudiments (in some cases) of a femur, and, in Balaena
(Fig. 188), of a tibia also.

Fig. 188.—Side view of bones of posterior extremity of
Greenland Right Whale (Balaena mysticetus). × ⅛.
i, Ischium; f, femur; t, accessory ossicle representing
the tibia. (After Eschricht and Reinhardt)
(from Flower's Osteology.)
Whales are to be divided into three great groups:—(1) the Whalebone Whales or Mystacoceti; (2)
the Toothed Whales or Odontoceti; and (3) the entirely-extinct
Archaeoceti or Zeuglodonts.
Sub-Order 1. MYSTACOCETI.
This division is thus characterised:—Teeth are never functionally developed; they
are present in the young, but replaced in the adult by the baleen or
whalebone; the external respiratory aperture is double; the skull is
perfectly symmetrical; the rami of the mandible are arched outwards and
do not form a true symphysis; the sternum is always composed of a single
piece of bone; the ribs articulate only with the transverse processes of
the vertebrae.
The Mystacoceti are nearly invariably huge creatures, the sole
exceptions being the Pygmy Right Whale, Neobalaena, and {354}a small
Rorqual. But even these are larger than the majority of Toothed
Whales.
The most characteristic feature by which the Whalebone Whales are to
be distinguished from other Whales is that which gives to them their
name, the presence of whalebone. Whalebone is a horny product of the
epithelium lining the mouth, and is comparable to an exaggeration of the
transverse ridges which are found in the mouths of all mammals upon the
palate. In non-Cetacean mammals these ridges vary in depth, and are
arranged as a rule transversely, but with an oblique inclination. This is
precisely how the plates of baleen are disposed in the mouth of a Whale.
Each piece of "bone" is triangular in shape, the broader end being that
of attachment while it narrows gradually; the inner side of the blades is
frayed out into a number of threads which form the straining apparatus.
The plates vary in length up to as great an extreme length as 13 feet,
which occurs in the Right Whale at times. The colour is black or paler,
even white. The number of these plates in the mouth is very great. As
many as 370 blades have been counted. They diminish in length towards
both ends of the series. Though whalebone has been in use for a long
period, whence the whalebone came was formerly one of those things not
generally known.

Fig. 189.—Section of upper jaw, with
baleen plates, of Balaenoptera. a,
Bone of jaw; b, gum; c, straight
edge of baleen plate; d, e, frayed
out surface of baleen plates. (After
Owen.)
A very prevalent notion was that the whalebone formed the eyelids or
perhaps the eyelashes of the creature. Scaliger, commenting upon
Aristotle, held that the whale had "lamellae upon the eyebrows, which,
when the head is plunged below the surface, were raised by the water; but
when the animal raised its head {355}above the waves the lamellae fell and
covered the eyes." Whalebone, too, has been often spoken of as "the fin
of a whale," "the finnes that stand forth of their mouths." The value of
whalebone is still great, in spite of various substitutes which are now
used in its place. In the year 1897, for example, the value of this
article was £2000 per ton. As a single Whale may produce several tons of
this material, it is not surprising to find that the results of a whaling
voyage may be very profitable.
Fam. 1. Balaenopteridae.—This genus Balaenoptera
includes the Rorquals, which are Whalebone Whales of large size,
differing from the Right Whales in three important external characters:
the head is comparatively small; there is a dorsal fin; the throat is
marked by numerous longitudinal furrows. The bones of the cranium are not
so arched as in the Right Whales, and as a consequence the plates of
baleen are shorter. The hand is only four-fingered. The cervical
vertebrae are for the most part all free. One of the earliest records of
a Whale stranded in the Thames was probably of a species of this genus in
the year 1658, and is thus described by John Evelyn:—"A large whale was taken betwixt my land
butting on the Thames and Greenewich, which drew an infinite concourse to
see it, by water, horse, coach, and on foot, from London and all
parts.... It was killed with a harping yron, struck in the head, out of
which spouted blood and water by two tunnells, and after an horrid grone
it ran quite on shore and died. Its length was 58 foot, heighth 16; black
skinn'd like coach leather, very small eyes, greate taile, onely two
small finns, a picked snout, and a mouth so wide that divers men might
have stood upright in it; no teeth, but suck'd slime onely as thro' a
grate of that bone which we call whalebone, the throate yet so narrow as
would not have admitted the least of fishes ... all of it prodigious, but
in nothing more wonderful that an animal of so greate a bulk should be
nourished onely by slime thro' those grates."
Professor Collett has recently given[226] an elaborate account of the
characters and habits of this great Whale (Balaenoptera musculus).
Though a large beast (44 to 67 feet in length) it is exceeded by other
Rorquals; it is of a dark grey blue colour above, white, for the most
part, below. The dorsal fin is large and high; the flippers relatively
slender and small. The whole throat from the {356}symphysis of the jaws
to the middle of the belly is, as in other species, marked by furrows,
forty to fifty-eight in number. The hairy covering is reduced (in an
adult female) to thirteen hairs on each side of the lower jaw; in a
foetus there were also seven hairs on each side of the upper jaw, as well
as rather more on the lower jaw—altogether, forty-eight. This Whale
appears to feed chiefly upon small Crustacea, especially the Copepod,
Calanus finmarchicus. The number of baleen plates is about 330 on
each side of the jaw. This Whale sometimes swims singly, but usually in
schools of even as many as fifty.
Rudolphi's Rorqual (B. borealis) seems to be a perfectly
inoffensive beast; it is said to be able to stay under water for as long
a time as twelve hours.
A smaller species than the last is B. rostrata—at the
outside 33 feet in length. Here the hairy covering is reduced[227] to "two small hairs on
the integument covering the apex of the lower maxilla." The colour is
greyish black above, the underside white. On the other hand, B.
sibbaldii, the Blue Whale, is the giant of its race, reaching a
length of 85 feet. Its colour is a dark bluish grey, with small whitish
spots on the breast. The dorsal fin is small and low with straight
margins.
B. musculus, the Finner, is intermediate in size—not more
than 70 feet. It seems doubtful whether the "sulphur bottom," B.
australis, of Antarctica and B. patachonica differ
specifically from this.[228]
The genus Megaptera is very near Balaenoptera, but
differs from it mainly in the following external and internal characters.
The dorsal fin is not very prominent, and its place is taken by a lowish
hump, whence, indeed, the common name of this Whale, "Humpback." The
pectoral fin is unusually long, and the creature uses it to beat itself,
the surrounding water, and, more playfully, its mates. The general
outline of this Cetacean is more clumsy than that of Balaenoptera.
The most important internal difference is in the form of the scapula,
which has at most a slight acromion and coracoid process. These are
rather more pronounced, according to Messrs. van Beneden and Gervais,[229] {357}in the southern form of
the genus, which is known as M. lalandii. The head, it should also
be remarked, is studded with large tubercles about the size of an orange,
which seem to be hyper-trophied rudiments of the hairs, which should be
present in this region of the body. As is the case with other Whales,
numerous species have been made out of individuals of Megaptera.
Captain Scammon, who observed many "gams" or herds of these Whales,
remarked[230] that he had
extreme difficulty in finding any two individuals precisely alike! The
best-known species in any case is the northern M. longimana, which
occurs on our own coasts. The genus is, like so many Cetaceans,
world-wide in range; and it is possible that the difference in the
scapula already referred to may justify the separation of a southern
M. lalandii (with which in that case, perhaps, M. capensis
and M. novae zelandiae will be synonymous). Quite recently M.
Gervais has insisted upon a Megaptera indica from the Persian
Gulf. Megaptera grows to a length of 50 to 60 feet. Seventy-five
feet have been stated, but measurements of Whales have usually to be
received with caution.
Rhachianectes, with but one species, R. glaucus,[231] the "Californian Grey
Whale," is the last genus of the family Balaenopteridae. This Whale is
but imperfectly known anatomically; but quite sufficient has been
ascertained to show its great divergence from Balaenoptera or
Megaptera. The dorsal fin is completely absent, and the throat
pleats, so characteristic of the typical Balaenopteridae, are reduced to
two. It has, however, the general outline of a Rorqual, with a relatively
small head. In osteological characters it tends to unite the two families
Balaenopteridae and Balaenidae (if they are really necessary
subdivisions). The skull is on the whole Rorqual-like; but its fore-part
is narrow as in the Greenland Whale, and the premaxillaries are pinched
up in the middle line so as to be visible from the side; this again is a
Balaenid character. The cervical vertebrae are free as in Rorquals, and
the sternum is quite as in that group. The scapula has more the shape of
that of Balaena.
Rhachianectes glaucus is confined to the Pacific, and has been
extensively hunted from the shore. It is not, however, a very valuable
Whale, since the baleen is short as in Rorquals, and the {358}beast,
moreover, appears to be fierce, a somewhat rare attribute of Whales. It
has been spoken of, indeed, as "a cunning, courageous, and vicious"
animal. Rhachianectes is essentially a coast Whale, and loves to
lie in the surf in quite shallow water waiting for the tide to float it
off. This Whale varies much in colour from black to mottled grey and
black, and reaches a length of about 40 feet.
Fam. 2. Balaenidae.—The Right Whales of the genus
Balaena are to be distinguished from Neobalaena and from
the Rorquals by the following characters:—
The size is large, 50 to 60 feet. There is no dorsal fin. The head is
more than or nearly one-fourth of the entire length of the animal. The
baleen is very long. The throat is not grooved. The orbital process of
the frontal is not wider than the downward process of the maxilla. The
cervical vertebrae are all fused. The scapula is rather high. The
hind-limb has the rudiment of a tibia. The intestine has no caecum.
A vast number of different genera have been founded on detached bones,
bits of whalebone, and more or less complete skeletons of Right Whales
coming from different parts of the world. In Dr. Gray's catalogues we
find the following allowed, viz. Balaena, Eubalaena,
Hunterius, Caperea, Macleayius. The number of
"species" distributed among the genera is some thirteen or more, with
whose names we shall not trouble the reader. As a matter of fact there
are not more than two species which can with certainty be identified and
distinguished, both of which are so close that they cannot possibly be
placed but in the same genus, Balaena. In no group of
Whales—in no group of animals probably—has imagination run
riot to so terrible an extent in the formation of genera and species as
in these Right Whales. This multiplication or rather division of genera
has arisen from an old idea that Whales coming from different seas must
be of different kinds, a notion now thoroughly exploded.
The term "Right Whale" simply means that the Whales of this genus are
the right kind of Whale for the whaler to pursue. Their whalebone is
longer and more valuable, while the oil is not only more abundant but of
a superior quality. The two species demand a separate account.
The Greenland Whale, Balaena mysticetus, is one of the rare
instances of a Whale which has an exceedingly limited range in {359}space. It
is absolutely confined to the Arctic Ocean, and reported occurrences on
our coasts are due to a confusion with B. australis, to be
presently described. At the "Devil's Dyke," near Brighton, there is, or
was, the skull of a most flagrant Rorqual, which is carefully labelled
"Greenland Whale." This Whale grows to a length of 50, 60, rarely 70
feet. It is black in colour, save for a white patch on the under side of
the jaw. The head is quite one-third of the body in length. There are a
few scattered hairs at the extremity of the jaws. The length of time
which this Whale can endure immersion has been variously stated. The
utmost limit of endurance is stated by Scammon to be one hour and twenty
minutes. The pursuit of this Whale is attended by dangers, not in the
least because the animal is itself fierce and ready to attack, but simply
on account of the velocity with which, and the great depth to which, it
will dive, and also to the huge muscular force which is exerted in its
struggles to free itself from the harpoons. It is indeed an extremely
timid beast. It has been remarked that "a bird alighting upon its back
sometimes sets it off in great agitation and terror." Combined with this
timidity of disposition is an intense affection for its young, "which
would do honour," observed Scoresby, "to the superior intelligence of
human beings." Yet that trader and observer goes on to remark that "the
value of the prize ... cannot be sacrificed to feelings of compassion"!
The fact that this Whale and its congener, B. australis, feed
among swarms of minute pelagic creatures, which they engulf in their huge
mouths, led the ancients to believe and assert that they fed upon water
only. When the Whale feeds it moves along with some velocity, taking in
huge mouthfuls of sea water with the contained organisms, which are then
strained off by the whalebone and left stranded upon the tongue.
Unlike its congener, the southern Right Whale, B. australis,[232] is world-wide in
distribution, avoiding only the Arctic regions. Where the Greenland Whale
is found B. australis does not exist. The principal differences
which it shows from B. mysticetus are firstly in the relatively
shorter head and shorter and coarser whalebone. In the second place it
has more ribs, fifteen pairs as against thirteen; but there is apparently
some little confusion in the matter of ribs. An additional rib at the end
of the series {360}is apt to get lost, and in the skeleton of
so huge and unmanageable a beast there is nothing more unwise than to
insist upon, as specific characters, what may be due merely to defective
preparation. This Whale has often, and the Greenland Whale also, a rough
horny protuberance upon the snout known as the "bonnet." The causation of
this is not clear. It has been spoken of as "a rudimentary frontal horn."
But this suggestion of an Ungulate affinity can hardly be accepted. It
seems to be more like a kind of corn.
This Whale was once more abundant on the coasts of Europe than it is
to-day; it was much hunted by the Basques in past time. The Whale which
frequented the Bay of Biscay was usually called the Biscayan Whale or
B. biscayensis; but there is probably no specific difference.
Among the small towns which fringe the Bay, it is very common to find the
Whale incorporated into the armorial bearings. "Over the portal of the
first old house in the steep street of Guetaria," writes Sir Clements
Markham,[233] "there is a
shield of arms consisting of Whales amid waves of the sea. At Motrico the
town arms consist of a Whale in the sea harpooned, and with a boat with
men holding the line." Plenty of other such examples testify to the
prevalence of the whaling industry on these adjoining coasts of Spain and
France. It appears that though the fishery began much earlier—even
in the ninth century—the first actual document relating to it dates
from the year 1150. It is in the shape of privileges granted by Sancho
the Wise to the city of San Sebastian. The trade was still very
flourishing in the sixteenth century. Rondeletius the naturalist
described Bayonne as the centre of the trade, and tells us that the
flesh, especially of the tongue, was exposed for sale as food in the
markets.
M. Fischer,[234] who,
as well as Sir Clements Markham, has given an important account of the
whaling industry on the Basque shores, quotes an account of the methods
pursued in the sixteenth century. It was at Biarritz—or as Ambroise
Pare, from whom Fischer quotes, spelt it, Biaris—that the main
fisheries were undertaken. The inhabitants set upon a hill a tower from
which they could see "the Balaines which pass, and perceiving them coming
partly by the loud noise they make, and {361}partly by the water
which they throw out by a conduit which they possess in the middle of the
forehead." Several boats then set out in pursuit, some of which were
reserved for men whose sole duty it was to pick out of the water their
comrades who had overbalanced themselves in their excitement. The
harpoons bore a mark by which their respective owners could recognise
them, and the carcase of the animal was shared in accordance with the
numbers and owners of the harpoons found sticking in the dead body of the
Whale. At this period the fishery was at its height. But it continued to
be an occupation along those shores until the beginning of the eighteenth
century, after which it gradually declined. The fishery of Whales began
to be carried farther afield than the shore, and for a long time the
Basques furnished expert harpooners to whaling vessels proceeding to the
Arctic seas. A curious example of the continuance of the fishery until at
least 1712 is given by Sir C. Markham. In the parish records of Lequeito
for that year, it is noted that a couple were married who possessed
between them all the necessary outfit for a whaling cruise.
The genus Neobalaena is interesting from more than one point of
view. Its size compared with its gigantic relatives is small, some 16 or
17 feet. The genus bears the same kind of proportion to Balaena
that Kogia does to Physeter among the Physeteridae. It is
one of those Whales which are very restricted in habitat; up to the
present it is only known from the Antarctic region in the neighbourhood
of New Zealand and South Australia. Structurally it is in a few points
intermediate between the Right Whales and the Rorquals. The head is
proportionately (as well as, of course, actually) not so large as in
Balaena. There is a falcate dorsal fin; but the head in outline is
not Rorqual-like in spite of its similar proportions. The whalebone is
long. The throat is not grooved. Neobalaena has forty-three
vertebrae, of which the cervicals are all fused. There are as many as
seventeen or eighteen dorsal vertebrae, the largest number in any
Cetacean as far as is known. With these are articulated not eighteen but
only seventeen ribs. The first dorsal vertebra appears to be without a
rib. The ribs are very broad and flat. The body thus gets an appearance
of a Sirenian. The lumbar vertebrae are fewer than in any other Cetacean,
being only two. The scapula is more like that of the Rorquals than that
of the Right Whales; {362}that is to say, it is long and not very
high. The skull is most like that of Balaena, but the process of
the frontal arching over the eye is broader relatively than in
Balaena, and thus approaches Balaenoptera. Nothing is known
of the viscera of this Whale. The whalebone is white, and the animal was
first described by Dr. Gray from pieces of "bone." It is not always that
so fortunate a diagnosis of specific or generic difference has been made
from a structure which apparently offers so little aid for
discrimination.
There is but a single species of the genus which is named
Neobalaena marginata.[235]
Sub-Order 2. ODONTOCETI.
The Odontoceti have teeth but no whalebone; the blow-hole is
single; the skull is not symmetrical; some of the ribs are
two-headed.
Fam. 1. Physeteridae.—This family of the Odontocetes may
be thus defined:—All or most of the
cervical vertebrae are fused together. The costal cartilages are not
ossified. In the skull the pterygoids are thick and meet in the middle
line; the symphysis of the mandible is long. Teeth, more or fewer, are
found in both jaws, but those of the mandible are alone functional (?
exc. Kogia). The pectoral limb is smallish. The throat is grooved
by two or four furrows.
This family of Whales is again susceptible of division into the two
sub-families—Physeterinae or Sperm Whales and the Ziphiinae or
Beaked Whales. Professor P. J. van Beneden was strongly against any
subdivision of what is here regarded as a perfectly natural family,
embracing the Physeters and the Beaked Whales. There are, however, some
reasons for the subdivision. The Ziphiinae have a reduced series of
teeth, never exceeding two on each mandible, which contrasts with the
fully-toothed mandibles of both Physeter and Kogia. The
stomach of the Ziphioids is extraordinarily complicated even for a
Cetacean. The small head of the latter group, which recalls in a curious
way that of Mosasauroid reptiles and some Dinosaurs, is in contrast to
the {363}enormous head of the Cachalot and the very
fairly-developed skull of the "Pygmy Sperm Whale." Both, however, furnish
spermaceti, and in various osteological details come near together. On
the whole we incline towards separating the Cachalots from the Ziphioids,
and shall therefore commence with the former as being in some respects
the more primitive members of the family Physeteridae.
Sub-Fam. 1. Physeterinae.—This sub-family may be thus
defined:—Teeth in lower jaw numerous. No
distinct lachrymal bone. Stomach with only four compartments (? as to
Kogia).
Of this sub-family the best-known genus is Physeter, including
the Sperm Whale or Cachalot. Of other reputed species we shall speak
later. The genus is characterised in the first place by its large
size—as much as 82 feet of length have been assigned to Physeter
macrocephalus; but Sir William Flower thought that 55 or possibly 60
feet might be a better approximation to the greatest length of the
Cachalot. The head is enormous, a third of the length of the body, and
terminates in a massive and bluntish snout. This is, however, not so
abruptly truncated as is often represented in figures. According to
Messrs. Pouchet and Chaves,[236] it slopes forward two metres beyond
the end of the lower jaw; the mouth is thus ventral and almost shark-like
in position, as is the case also with the Pygmy Sperm Whale, to be
considered later. In connexion with this peculiar position of the mouth,
it has been asserted—Mr. F. T. Bullen figures it[237]—that the Sperm Whale turns over
upon its back to bite. The blow-hole is single, and shaped like the
sound-hole of a violin; it lies upon one side, and is not median in
position. The throat is grooved as in the Ziphioids by two grooves. The
dorsal fin is represented by a whole series of lowish humps, decreasing
in elevation from before backwards. The pectoral fins are not large
relatively speaking. The great square head is not occupied entirely by
the skull; the cavity lying above, which is of course traversed by the
tube ending in the blow-hole, is filled with the spermaceti, which is
fluid fat during the life of the animal. Spermaceti also occurs in other
Whales; and that of Hyperoodon, whence it has been extracted for
commercial purposes, is said to offer no differences of importance from
the spermaceti of the {364}Sperm Whale. Spermaceti as a drug appears
to have been first mentioned in the pharmacopoeias of the famous medical
school of Salerno towards the year 1100. But it was confounded with a
totally distinct substance, viz. ambergris. The confusion was also made
by the famous alchemist Albertus Magnus, and by the observant Archbishop
of Upsala, Olaus Magnus, in his work De gentibus
septentrionalibus. It was supposed in fact by these writers to be the
liberated sperm of the Whale, hence obviously the name. Later on, the
substance in question was regarded as the brain of the Cachalot, in fact
as late as the middle of the eighteenth century. It was Hunter and Camper
who really discovered the true nature of the substance, oil of course, in
the cavities of the skull.[238] The huge skull of Physeter "is
perhaps the most modified from the ordinary type" of skull in the whole
mammalian class.
The top of the skull rises into a huge crest lying transversely, and
from it slope forward two lateral crests formed from the maxillary bones;
in this great basin lies the spermaceti already referred to. The skull,
as in Toothed Whales generally, is exceedingly asymmetrical. The right
premaxillary and the left nasal bones are much larger than their fellows;
indeed the right nasal is hardly present as a separate bone. The parietal
if present is fused with the supra-occipital. The jugal is large, and is
not divided into two pieces as it is in the Ziphioids. The pterygoids
meet below for a considerable distance, as in many Dolphins, and in the
Edentata among other mammals. The symphysis of the lower jaw is very
long, but the bones do not appear to be ankylosed. The length of the
symphysis recalls that of the Gangetic Dolphin, Platanista.
In the vertebral column the atlas alone is free, the remaining
cervicals being fused. There are only eleven dorsal vertebrae, eight
lumbars, and twenty-four caudals. The breastbone of this Whale is a
roughly-triangular bone made up of three pieces. Four cartilaginous
sternal ribs are attached to this bone. The scapula is remarkable for the
fact that it is concave on the outer and convex on the inner surface;
otherwise it is quite typically Cetacean in form. The shortness of the
pectoral limb is shown by the phalangeal formula, which is as
follows:—I 1, II 5, III 5, IV 4, V 3. {365}
One of the reasons for the pursuit of the Sperm Whale is the desire to
obtain that extremely valuable product ambergris. This substance has long
been known; but its true nature was for centuries in dispute. In Dr.
Johnson's Dictionary (so recently as the edition of 1818!)
ambergris is provided with alternative definitions; it is either the
excrement of birds washed off rocks, or honeycombs that have fallen into
the sea!
An old writer asserted of ambergris that it was "not the scum or
excrement of the whale, but issues out of the root of a tree, which tree,
howsoever it stands on the land, alwaies shoots forth its roots towards
the sea, seaking the warmth of it, thereby to deliver the fattest gum
that coms out of it, which tree otherwise by its copious fatness might be
burnt and destroyed." These "explanations" were caused by the fact that
ambergris is sometimes found floating in the sea. Ambergris is, of
course, a product of the intestinal canal of the Sperm Whale; it seems to
be of the nature of cholesterin, and its place of origin was conclusively
proved by finding the beaks of cuttle-fish imbedded in it. When first
extracted from the alimentary canal it is of greasy feel and consistency;
later it hardens, and acquires its characteristic sweet earthy odour.
Ambergris is used mainly as a vehicle for scents, and is a costly
substance. A piece weighing 130 lbs. was valued at £500. Though now
entirely used in connexion with perfumery, it was held by the ancients to
be of great value as a specific in certain diseases.
The Sperm Whale is chiefly a tropical animal. Examples that have been
cast up on our shores are strayed individuals. It often goes about in
herds, which seem to be composed of females. Its food is chiefly
cuttle-fishes, and it is said to have a predilection for those colossal
cuttle-fishes whose existence has until recently been doubted. Mr. Bullen
has sketched a conflict between these two giants of the deep. On the
other hand it is said that its large throat, more than big enough to
swallow a man (the Whale is credited with being that which swallowed
Jonah), does not usually admit fishes larger than Bonitos and
Albacores.
The ferocity of the Cachalot has been denied and affirmed. It
certainly has great strength, for it can throw itself completely out of
the water. Captain Scammon thinks that ships which are mysteriously lost
at sea, with no obviously assignable cause, are sometimes the victims of
the furious rushes of a bull {366}Sperm Whale. Marco Polo took much the same
view, but suggested that the Whale did not deliberately attack the ship,
but was deceived by the foam following in its wake into thinking "there
is something to eat afloat, and makes a rush forward, whereby it shall
often stave in some part of the ship."[239]
Sir W. Flower and many others are of opinion that there is but one
species of Cachalot. But many names have been given to supposed other
forms. The genus itself has even been divided, and to a set of vertebrae
from the south Dr. Gray gave the perfectly superfluous name of
Meganeuron kreffti. The "High-finned Cachalot" rests mainly upon
the suggestions of Sir Robert Sibbald. It is supposed to have a high
dorsal fin, and teeth in the upper as well as in the lower jaw. Common
though it was asserted by its describer to be, there is not a bone, not a
fragment even of a bone, alleged to belong to Physeter tursio in
any museum in the world! It seems premature, therefore, to include this
mysterious creature in any list of Cetacea, though that was done by no
less a naturalist than the late Mr. Thomas Bell. It is this creature
round which most of the stories of ferocity congregate. It is held to be
the monster from which Perseus delivered Andromeda, and which was about
to devour Angelica upon the shore of Brittany. The fact of the matter is,
that the Sperm Whale, like so very many other Whales, is world-wide in
range; and those naturalists who did not believe in so wide a
distribution found themselves obliged, in order to satisfy their own
views, to create new species for those of distant localities. Hence the
dozen or so of synonyms which refer to what is to be called Physeter
macrocephalus.
The genus Kogia (sometimes written Cogia), the so-called
"Pygmy Sperm Whale," is a southern form of much smaller dimensions than
its gigantic ally just described. Kogia does not exceed 15 feet or
so in length. It differs from Physeter also in the well-marked and
falcate dorsal fin, in its generally delphinoid form, in the short snout,
and the more normal (for a Whale) shape of the blow-hole, which is
crescentic.
There are also a number of osteological characters in which the two
Physeterines differ from each other. In Kogia all the cervical
vertebrae are ankylosed together; the skull is short, though equally
asymmetrical; the ribs are as many as twelve or {367}fourteen; the scapula
has not the concave face that it has in Physeter. The functional
teeth of the lower jaw seem to be reinforced by two on each side of the
upper jaw. Moreover, the articulation of the ribs with the vertebrae does
not show the very anomalous state of affairs that characterises
Physeter, where the two heads of a rib may be upon one
vertebra.
While there is no doubt as to the generic distinctness of
Kogia, there is again the same difficulty that is met with
throughout the whole of the order in settling into how many species the
genus requires dividing.
We can dismiss, as unnecessary, additional generic names
(Euphysetes, Callignathus), but there do appear to be
reasons for allowing two species, if the accounts of their osteology are
to be depended upon. One of these is K. breviceps, with thirteen
pairs of ribs, no teeth in the upper jaw, fourteen or fifteen on each
side of the lower jaw, vertebral formula C 7, D 13, L 9, Ca 25, and
phalangeal formula I 2, II 8, III 8, IV 8, V 7.
The other will then be K. simus, with fourteen pairs of ribs,
two teeth in the upper jaw, nine in each ramus of the lower jaw,
vertebral formula C 7, D 14, L 5, Ca 24, and phalangeal formula I 2,
II 5, III 4, IV 4, V 2.
A Californian species has been called K. floweri, whose teeth
seem to be particularly long and recurved. And the New Zealand K.
pottsi has been held to be also a distinct form. There seems to be
nothing of special interest to record about the way of life of these
Cetaceans, which are but imperfectly known.
Sub-Fam. 2. Ziphiinae.—Teeth in the lower jaw not more
than two on each side. A distinct lachrymal bone. Stomach with very
numerous compartments.
These Whales are all of moderate size, not exceeding 30 feet or so in
length. They have a falcate dorsal fin rather near the end of the body;
the muzzle is prolonged, hence the name often given to them of "Beaked
Whales." The throat is grooved; the blow-hole is single and median,
crescentic in form, with the concavity pointing forwards. A character
possibly differentiating the Ziphioids from other Whales is the fact that
the body ends in a rounded projection between the flukes of the tail.
This has at any rate been noted in Mesoplodon, Ziphius, and
Hyperoodon. The Ziphioid Whales are by no means common; indeed of
Berardius but four or five specimens have ever been {368}met with. Most
of them are southern in range, and the vast stretches of desolate coast
which occur in these regions of the world account possibly for the rarity
of their remains. These Whales have done duty more than once for the "Sea
Serpent." Quite recently an alleged sea serpent turned out to be a couple
of Mesoplodon lying head to tail! The head in these Whales is
small compared to the body. The skull is characterised by the strong
maxillary crests, enormously developed in the male Hyperoodon. The
vertex of the skull too is raised, forming a pronounced prominence behind
the aperture of the nares (blow-hole); in many forms the rostrum is made
of very dense bone, and is thus relatively abundant in rock strata. The
pterygoids meet in the middle line as in the Cachalot. In addition to the
few functional teeth in the lower jaw there are more numerous but small
teeth in the upper jaw. These are not always to be recognised, as they
are not attached to the bone, but merely imbedded in the gums, so that
they come away when the skull is prepared.
The genus Berardius[240] differs from Mesoplodon by its
rather more symmetrical skull, of which the vertex is formed by the
nasals. The mesethmoid is only partly ossified. The teeth are two on each
side of the mandible, with their apices directed forwards. The vertebral
formula is C 7, D 10, L 12, Ca 19.
B. arnouxi, from the seas of New Zealand, is the only species
of this genus which is well known. It is 30 or 32 feet in length, and is
of a velvety black colour, with a greyish belly. Instead of lowing like a
cow, this Whale has been described as "bellowing like a bull"! A singular
and somewhat inexplicable fact has been stated of this species. The teeth
were said to be protrusible, and Sir James Hector stated that the teeth
were imbedded "in a tough cartilaginous sac which adheres loosely in the
socket of the jaw, and is moved by a series of muscular bundles that
elevate or depress it." Sir William Flower justly observed that these
statements "accord so little with anything hitherto known in mammalian
anatomy that further observations on the subject are extremely
desirable." Like other Ziphioids, Berardius feeds mainly, if not
entirely, upon cuttle-fish, a prey eminently suited to their almost
toothless mouths. It is not known whether Berardius has the {369}Ziphioid grooves upon the throat. Nothing
is known of the structure of the internal viscera of this Whale. It
appears not to be really limited to the region of New Zealand, as is
often stated, for Malm has lately described a skull (Berardius
vegae) from Bering's Straits.[241]
Mesoplodon[242]
is a world-wide genus embracing a number of species; on the lowest
estimate seven species can be distinguished, and Sir W. Flower would add
two more. These are moderate-sized Whales, 15 to 17 feet in length. In
the skull the mesethmoid is ossified; the nasals are sunk between the
upper ends of the premaxillae. There are but a single pair of teeth in
the mandible attached to nearly the middle of its length (whence the
generic name). The vertebral formula is C 7, D 9 or 10, L 10 or 11, Ca 19
or 20. The sternum consists of four or five pieces. The amount to which
the cervical vertebrae are fused varies; but some are always fused.
The only species which has ever been stranded on the shores of this
country is M. bidens, an example of which was described many years
ago as the "Toothless Whale of Havre"; it was an old animal which had
probably lost its teeth. Nevertheless it received the separate generic
and specific name of Aodon dalei. The animal lived for two days
out of the water, and made a sound like the "lowing of a cow." An
instance of the rarity of the Whales of this genus is afforded by M.
europaeus, of which only a single skull is known; this was extracted
from a dead body, found floating, about the year 1840. It has never
appeared since. M. layardi is remarkable on account of the very
large size of its strap-shaped teeth; these curve over the upper jaw in
such a way as to prevent the animal from fully opening its jaws. The case
is curiously paralleled by the Sabre-toothed Tiger. This species is
antarctic in range. From the opposite extremity of the globe comes M.
stejnegeri, again known by but a single skull. It is singular on
account of the large size of the brain case, and is a native of Bering's
Straits. M. hectori has its two teeth situated quite at the
extremity of the mandible, and in this feature approximates to the genus
Berardius. It was, indeed, confounded with that genus by one
naturalist. {370}
Ziphius is a genus which is also of world-wide range. Here
again the number of species is at present merely a matter of opinion. The
prevalent impression, however, is that but a single species exists, which
will therefore have the name of Z. cavirostris. The genus (and for
the matter of that the species too) may be thus characterised in
comparison with its allies. The mesethmoid is ossified as in
Mesoplodon, but the nasals joined together form the vertex of the
skull. There are two teeth near the symphysis of the mandible, besides
the usual small and "functionless" teeth in the upper jaw. The vertebral
formula is C 7, D 9 or 10, L 11, Ca 21.
The throat of a Ziphius from New Zealand was described by
Messrs. Scott and Parker[243] as having three grooves on each side.
Whether this form is the same as von Haast's Z. novae zelandiae is
a matter of doubt; but the individual to which his name has been applied
was 26 feet long, and had but a single groove on each side. Even in the
external characters of many Whales many points require clearing up. Our
knowledge of Ziphius dates from the year 1804, when a skull
"completely petrified in appearance" was picked up upon the Mediterranean
coast of France, and described by the great Cuvier. It was forty years
before another specimen was found. In the New Zealand specimen of von
Haast already referred to, the body was scored by numerous lacerations.
These wounds may have been due to fights among the Whales themselves; the
forwardly-situated teeth would be capable of inflicting such wounds. But
it has also been stated that the armed suckers of gigantic cuttle-fish
are responsible for these scratches.
Hyperoodon is the most easily-distinguishable genus of Ziphioid
Whales. Its characters are the following:—The skull has enormously-developed maxillary
crests in the adult male; the mesethmoid is not fully ossified. There is
but a single tooth to each ramus of the lower jaw, besides, of course,
the usual small teeth in the upper jaw. The vertebral formula is C 7,
D 9, L 9, Ca 18. The cervicals are fused into one mass, more or fewer
being free in other Ziphioids. The sternum consists of three pieces only,
the last of which is bifid posteriorly.
The name Hyperoodon was given to this Whale by Colonel Lacepède
on account of the rough papillae upon the palate, which {371}were mistaken
by that observer for teeth. It is curious that the name is really
appropriate in spite of this mistake, though of course it would be so to
all the Ziphioids. In more than one feature this genus comes nearest of
all the Ziphiinae to Physeter. Its enormous maxillary crests are
paralleled in that Whale; but in Hyperoodon their great thickness
contrasts with the thinness of those of the Cachalot. The correspondence
in the attachment of a rib to its vertebra by both heads is noteworthy.
It is remarkable that in this particular Hyperoodon is more like
Physeter than the supposed nearest ally of the
latter—Kogia.
Of this genus two species are known. The best known is the common
northern H. rostratum (with many aliases); the second species from
the southern hemisphere, H. planifrons, is only known from a
single water- and pebble-worn skull. Its identification, however, depends
upon the known accuracy of the late Sir William Flower.
The northern species (Hyperoodon rostratum) has often been
recorded upon our own coasts; the first record of the stranding of this
Whale was in the year 1717. In that year an example was found at Maldon,
in Essex. Like the Beluga, Hyperoodon rostratum gets lighter in
colour with advancing years. The young are black; the old animals pale
brown with some white about them. The under surface, however, is always
greyish white. The length of this Whale reaches to at any rate 30 feet.
But John Hunter had a specimen which he believed to be 40 feet in length.
The specimen, however, consisted only of a skull, so that error might
have crept in. It has already been mentioned that the old males have
enormous maxillary crests. According to M. Bouvier, who has lately made
an exhaustive examination of the anatomy of this Whale,[244] the females occasionally exhibit the
same crests, which are thus presumably of the nature of spurs sometimes
seen in old females among the Gallinaceous birds. The number of grooves
upon the throat is in dispute in this Whale as in Ziphius. One
pair is the usual allowance; but Kükenthal found four in some embryos
studied by him. Attention has already been called to the voice of
Ziphioid Whales. Hyperoodon neither "lows" nor "bellows," but
"sobs"! Hyperoodon rostratum is a gregarious Whale, going about in
herds, or "gams" as they should technically be termed, of four to ten or
even fifteen. This Whale {372}can leap right out of the water, and while
in the air can turn its head from side to side, a capability which has
not been mentioned in any other Whale. It can also stay under water for
an unusually long period. Captain Gray,[245] who has made an accurate study of
this species, states that so long a period as two hours is the limit of
endurance; this event occurred in the case of a harpooned Whale.
Fam. 2. Delphinidae.—This family, which includes the
greater number of Cetacea, may thus be characterised:—Whales of small to moderate size. Teeth as a
rule numerous, and present in the upper as well as in the lower jaw.
Maxillae without large crests; the pterygoids, often meeting in the
middle line, enclose an air space open behind. The anterior (five to
eight) ribs are two headed, the posterior with tubercular head only. The
sternal ribs are ossified.
The Dolphins and Porpoises, as already stated, embrace the greater
number of existing species of Whales. Sir W. Flower and others who have
followed him, allow nineteen genera. But as to the exact number of known
species there is much uncertainty. That very careful observer, Mr. True,
considers[246] that there
are fifty which demand recognition. As many as one hundred have received
names. The matter is one which is perhaps barely ripe for decision. All
the Dolphin tribe are, for Whales, smallish animals. The Killer Whale,
Orca, is the only genus (or species?) which usually attains to
more than moderate bulk. The rather mysterious Delphinus
coronatus, 36 feet in length, of M. de Fréminville, would seem to be
a Ziphioid; it was described as having a very pointed beak, and as having
the dorsal fin situated near the tail; such characters suggest a
Mesoplodon.
The genus Delphinapterus, the Beluga or White Whale, consists
of but a single species, though as usual more than one name has been
given to supposed different species. It is characterised as a genus by
the following assemblage of structural features:—It has only eight to ten teeth occupying the
anterior part of the jaws only. All the cervical vertebrae are free and
unjoined. The vertebral formula is C 7, D 11 (or 12), L 9, Ca 23. The
pterygoids are wide apart, though they converge as if about to meet at
their posterior ends. There is no dorsal fin. The colour is white. {373}
The Beluga is a northern species purely. The reputed form, D.
kingii, was said to come from Australian seas; but there seems to
have been an error in this statement. It is interesting to note that the
white colour, so characteristic of the genus and species, is not found in
the young, which are blackish. They gradually pale as they advance
towards maturity. Delphinapterus leucas reaches a length of 10
feet, and like other Porpoises will ascend rivers in search of food. It
is said to be specially addicted to salmon. Among the contents of the
stomach have been found quantities of sand. But this habit of swallowing
sand or pebbles has been noted in other Whales. Whether it is or is not
accidental (taken in with ground-living food), it seems hardly likely
that it is used for purposes of ballast! The Beluga has a voice; but the
name "Sea Canary" is hardly suitable to it. A specimen of this species,
recently described from the shores of Scotland (it is often thrown up
upon our coasts), which had got entangled in the stakes of a new net, was
regarded by the natives, on account of its white colour, as a ghost.
Externally, besides its colour, the Beluga is remarkable for possessing a
distinct neck, which is correlated of course with the freedom of the
cervical vertebrae, and is also seen in Platanistidae.
The Narwhal (Monodon) is closely allied in structure to the
last genus. It has the following anatomical characters:—The teeth are reduced to a single "horn" in the
upper jaw, which is rudimentary in the female. The neck vertebrae are
free. The vertebral formula is C 7, D 11, L 6, Ca 26. The pterygoids are
as in Delphinapterus, and, as in that genus, there are no hairs
upon the face or dorsal fin.
This genus is of course most obviously characterised by the twisted
tusk of the male, which is occasionally double. This tusk has given to
the only species of the genus, M. monoceros, both its generic and
specific name. The animal has a spotted colour; but, as in the case of
the Beluga, old animals tend to become white. The use of its horn to
Monodon has been debated. In the first place it is clearly a
secondary sexual character. The males have been observed to cross their
horns like rapiers in a fencing match. It may be that they are used in
more serious combats. An ingenious suggestion is that the long and strong
tusk enables its possessor to break the thick ice and {374}make a
breathing hole. A third suggestion is due to Scoresby, who was led to
make it from having taken out of the stomach of a Narwhal a large skate.
He held that with its tusk the Whale empaled the fish and then swallowed
it. The Narwhal is not large, 15 feet or so in length. But Lacepède, who
was apt to compile with lack of discrimination, speaks of 60 feet long
Narwhals. Monodon is purely Arctic, and but three or four
specimens have ever been cast up on our shores.
Of true Porpoises, genus Phocaena, there are apparently several
species. The genus itself has the following characters:—The teeth are sixteen to twenty-six on each
half of each jaw; their crowns are compressed and lobed. The pterygoids
do not meet. The dorsal fin has a row of tubercles along its margin.
The Porpoise of our coasts, P. communis, is a smallish species
6 to 8 feet in length. There are two to four hairs present in the young;
its colour is black, generally lighter on the belly. The first six
cervical vertebrae are fused. The ribs vary in number from twelve to
fourteen pairs. It is a gregarious Whale, and will ascend rivers; it has
been seen for example in the Seine at Paris. The name Porpoise is often
written Porkpisce, which of course shows its origin. Very conveniently it
was regarded as a fish, and therefore allowed to be eaten in Lent. The
celebrated Dr. Caius, a gourmet as well as a physician and the refounder
of a college, invented a particular sauce wherewith to dress this royal
dish. Some time since Dr. Gray described a Porpoise from Margate as a
distinct species (see p. 342) on account of the
tubercles, which are now known to be a generic character.
Dr. Burmeister's P. spinipennis seems, however, to be really
distinct. It was captured near the mouth of the Rio de la Plata. It is
more tuberculated on the fin and back, and has fewer teeth (sixteen as
against twenty-six).
Mr. True's P. dallii of the Pacific (where the Common Porpoise
also occurs) is characterised chiefly by its very long vertebral column,
consisting of ninety-eight vertebrae; there are only sixty-eight in the
other species. The Eastern genus Neomeris is placed with
Phocaena by Dr. Blanford. It practically only differs by the
absence of a dorsal fin. It is only about 4 feet long, and inhabits the
seas of India, Cape of Good Hope, and Japan. The one species is called
N. phocaenoides.
The genus Globicephalus is to be defined thus:—Teeth {375}seven to twelve on each side, confined to
anterior end of jaws. Skull raised into a prominence behind the
blow-hole; pterygoids large and in contact. Pectoral fin long and
falcate; dorsal fin present. No beak. Vertebral formula C 7, D 11, L 11
to 14, Ca 27 to 29. Six pairs of the ribs are two-headed.
The best known species of the genus is the Ca'ing Whale, G.
melas.[247] This
animal reaches a length of 20 feet, and is thus one of the largest of the
Delphinidae. It is gregarious and was, even is now, much hunted in the
Faeroe Islands. Its sheep-like habits (embodied in one scientific name
deductor) enable it to be easily driven on shore in herds, which
are then harpooned. The foetus of this Whale has a few hairs; the number
of phalanges in the two middle digits is very great, as many as eleven to
fourteen. G. scammoni, G. brachypterus, and G.
indicus are other reputed species of the genus allowed by True and
Blanford.
Grampus is a genus allied to the last. It has no teeth in the
upper jaw, and but three to seven in the lower jaw, near the symphysis of
the mandible. The pterygoids are in contact. There is no beak, and the
pectoral fin is long. There are twelve pairs of ribs, of which six are
two-headed. Apparently there is but one species, G. griseus, known
as "Risso's Dolphin." It is a Mediterranean and Atlantic form, and is not
common.
The genus Orca has as characters:—Teeth ten to thirteen, long and strong.
Pterygoids not quite meeting. Vertebrae C 7, D 11 to 12, L 10, Ca 23. The
first two or three fused. The dorsal fin is long and pointed.
Of this genus there may be more than one species; but the best known
is the Killer Whale, O. gladiator (Fig. 180, p. 341), often spoken of as the "Grampus."[248] It is marked with
contrasting bands of white or yellow upon a black body-colour. The animal
grows to a considerable length, as much as 30 feet. Orca is a
powerful and rapacious Whale; and Eschricht has stated that from the
stomach of one, thirteen Porpoises and fourteen Seals were extracted.
They will also combine to attack larger Whales, and Scammon has related
how he witnessed such an onslaught upon a Californian {376}Grey Whale.
"Belua truculenta dentibus," observed Olaus Magnus of this Cetacean. The
high dorsal fin has been much exaggerated in old drawings; it has been
even represented as strong and sharpened at the end, so as to be capable
of ripping open the belly of a Whale. The fact that it sometimes lies
over a little to one side is responsible for another anecdote: that an
example of this Whale was seen to retire with a couple of Seals tucked
away under the flippers, another grasped by the dorsal fin, and a fourth
in the mouth! "When an Orca pursues a whale," wrote Dr. Frangius, "the
latter makes a terrible bellowing like a bull when bitten by a dog." It
is probable, according to F. Cuvier, that this Whale is the "Aries
marinus" of the ancients, certain bands of white upon the head giving an
impression of curved horns. It may also be the "horrible Sea-satyre" of
Edmund Spencer.
Allied to Orca, but distinguishable from it by some rather
minute peculiarities, is Pseudorca. It may be thus defined:—Teeth eight to ten, much like those of
Orca. Dorsal fin rather small, falcate. Vertebral formula C 7,
D 10, L 9, Ca 24. Six or all the cervicals united. The curious fact about
this Whale, which embraces only a single species, P. crassidens,
is that it was first known in the fossil condition from remains
discovered in the fens of Lincolnshire. An important day for cetologists
was that on which a whole herd entered the Baltic and furnished material
for a better study of this Whale. It is not, any more than its near ally
Orca, confined to northern seas; for several examples, at first
relegated to a distinct species (P. meridionalis), have been
obtained from the seas round Tasmania.
Orcella (which has been written Orcaella) has fourteen
to nineteen small sharp teeth in each half of each jaw. The pterygoids
are widely separate. The dorsal fin is small and falcate. The vertebral
formula is C 7, D 14, L 14, Ca 26. Seven ribs are two-headed, and five of
them reach the sternum.
This genus contains but a single species, O. brevirostris,
which is both marine and fresh-water in habit; it occurs in the Indian
seas, and in the Irrawaddy even as far up as 900 miles from the sea. Some
regard the fresh-water individuals as a distinct form, O.
fluminalis.
Sagmatias is a genus known only from a skull, which is
remarkable for the elevation of the premaxillae into a crest; the {377}pterygoids are short, and there are
thirty-two teeth in each half of each jaw.
Feresia is known from two skulls which are provided with ten to
twelve teeth in each half of each jaw. It is intermediate between
Globicephalus, Grampus, and Lagenorhynchus,
according to Sir W. Flower.
The genus Delphinus contains the Dolphin, D. delphis.[249] The genus may be
characterised as follows:—Teeth small and
numerous, forty-seven to sixty-five. Vertebral formula C 7, D 14 or 15,
L 21 or 22, Ca 30 to 32. The atlas and axis are fused, the rest free. The
palatal border of the maxillae is deeply grooved. The fins are falcate;
the beak long and distinct.
The Common Dolphin of the Mediterranean shows so many variations of
colour, slight differences in the proportions of the bones of the skull,
and in the number of the teeth, that it has been divided up into at least
seventeen "species." But M. Fischer, who has studied many of these forms,
does not admit them, and most students of this group of mammals follow
him in the matter. The Dolphin is and has been the most familiar of
Cetaceans; in consequence it has accumulated much anecdote of a mythical
character. The extreme intelligence and goodwill towards man assigned to
this creature by the ancients are possibly due to the anomaly of a
creature ostensibly a fish showing many of the characters of higher
animals. Its unfishlike intelligence baffled the early observers, who at
once endowed it with especially advanced attributes. Hence the stories of
Arion and others. The leaping of the Dolphin out of the water is
exemplified in many Mediterranean coins and coats of arms; the heraldic
dolphin is represented with an arched back as in leaping. The Dolphin
reaches a length of some 7 feet, and appears to be world-wide in range.
Possibly distinct is D. longirostris, characterised, as the name
denotes, by the very long beak; it has also more teeth and is a native of
Malabar. D. roseiventris again may be a third species of
Delphinus. It comes from Torres Straits, and has the under parts
rosy in colour.
The genus Prodelphinus has, like Delphinus, a distinct
beak; but it has not the grooved maxillaries. No other character of
importance appears to separate it from Delphinus. {378}
The genus consists of some eight widely distributed species, which are
none of them large Dolphins.
Lagenorhynchus has the following assemblage of characters:—Head with short, not very distinct beak. Dorsal
and pectoral fins falcate. Teeth small, twenty-two to forty-five in each
half jaw. Vertebrae ranging in number from seventy-three to ninety-two.
Pterygoid bones either in contact or separate. There are fifteen or
sixteen pairs of ribs, of which six are two-headed. Of this genus Mr.
True allows eight species, which have been increased by a ninth since the
publication of his "Revision."[250]
Two species of Lagenorhynchus are known from our coasts; the
rest are mainly southern in range. The British species are, firstly,
L. albirostris, a Dolphin of some 9 feet in length. It has a large
number of vertebrae, ninety-two in number. L. albirostris is a
rare species, the first record of its occurrence on these shores being in
1834. Since that date some eighteen individuals have been shot or
stranded on the shores of the British Isles. The second British species,
L. acutus, differs in colour from the first. As in the last, the
upper parts are black and the under parts white; but in L. acutus
there is also a stripe on the flanks, brownish in colour. It has fewer
vertebrae, not more than eighty-two.
The next genus of Dolphins, Sotalia, is characterised
by—Teeth tolerably large, twenty-six to thirty-five. The vertebral
formula is C 7, D 11 or 12, L 10 to 14, Ca 22. The pterygoids are not in
contact in the middle line. It has a distinct beak.
Of this genus there are some six species (the exact number, as in so
many other genera, cannot be positively asserted), most of which are
fluviatile or estuarine in habit. They are also on the whole
characterised by their pale, if not actually white, coloration. S.
sinensis of the Amoy is white with pinkish fins. Sotalia
guianensis is American as its name denotes. It is figured by van
Beneden as of a pale brown colour. It is very abundant in the Bay of Rio
de Janeiro, and has the reputation of being a friend of man like some
other Dolphins. The natives hold that it will bring to shore the bodies
of drowned persons. The most singular species of the genus is that
recently described by Professor Kükenthal as S. teüszii.[251] This animal is purely
fresh-water, being found in {379}the Camaroon river, where it is extremely
rare. The nostrils (blow-hole) are prolonged into a snout-like process, a
fact which is of interest in connexion with the assertion that in
Balaenoptera the blow-hole is puffed out during spouting. What is
temporary in the Rorqual appears to be permanent in the Sotalia.
More remarkable still, perhaps, is the assertion that it is a
vegetable-feeding Dolphin. This is not a mere assertion except that it
may not apply universally; for in the stomach of a specimen nothing but
vegetable débris was found. But in the stomachs of other Whales (e.g.
Rhachianectes) vegetable matter has also been found, which may
perhaps have been taken in accidentally with the food.
Steno comes near Sotalia, and Dr. Blanford has
transferred to it (under the one name of Steno perniger) the two
species, Sotalia gadamu and Sotalia lentiginosa. It is,
however, to be distinguished from Sotalia by the following
characters:—Teeth large and few, twenty to
twenty-seven on each side of each jaw, with furrowed surfaces to crowns.
Vertebrae C 7, D 12 or 13, L 15, Ca 30 to 32. Pterygoids in contact.
There are but two species apparently (not counting Dr. Blanford's).
Tursiops is not a very easily definable genus. These are its
chief features:—Teeth large, twenty-two to
twenty-six in number in each half of each jaw. Vertebral formula C 7,
D 12 or 13, L 16 or 17, Ca 27. Pterygoids in contact. Beak distinct. Some
five species are allowed; but it seems to be difficult to differentiate
the others from Tursiops tursio. This, the best-known form, is
quite or nearly world-wide in range, and occurs, though not abundantly,
on our own coasts. Mr. True has observed that the eyelids of this Whale,
which is largely hunted on the American coast, are as mobile as those of
a terrestrial mammal. The name "tursio" is derived from Pliny. Belon
would also derive from this word the French vernacular "marsouin." The
latter term is sometimes regarded as a corruption of "Meerschwein," but
it would seem to be more probably derivable from "marinum suem," from the
Latin direct. T. tursio has the back black to lead-colour; the
under parts white. In the reputed species, T. abusalam, from the
Red Sea, the back is a dark sea-green. T. tursio reaches a length
of 12 feet, but is more usually smaller.
The genus Tursio must be carefully distinguished from {380}Tursiops. It has no dorsal fin, the
teeth are small and numerous (forty-four), and the pterygoids are
separate. There are two species, T. borealis and T.
peronii, the former being northern and the latter more widely
spread.
The genus Cephalorhynchus has for its chief characters the
following:—Teeth twenty-five to thirty-one,
small and sharp. Pterygoids widely separated. Dorsal fin not falcate, but
triangular or ovate in form. Beak not well marked off from the head. The
species of this genus are all southern in range; four are perhaps to be
allowed.
Fam. 3. Platanistidae.—This family of Odontocetes may be
distinguished from the Dolphins by the following assemblage of structural
features:—Cervical vertebrae all free, and
each one of some length (for a Cetacean). Jaws long and narrow, with a
considerable length of symphysis. Teeth very numerous.
This very meagre series of differential characters is largely due to
Pontoporia on the Platanistid side, and to Monodon and
Delphinapterus upon the Delphinid side. Otherwise the family
Platanistidae would be extremely distinct. The two last-named genera have
separate cervical vertebrae, and in the Beluga at any rate this is
expressed externally by a quite distinct neck. Moreover, as Mr. True has
pointed out, the pterygoid bones have not the involuted cavity below
which characterises other Dolphins; and they have, what other Dolphins
have not, an articulation outwards with the roofing bones of the skull.
Sir W. Flower described the fact that in Inia (and the same occurs in
Pontoporia) the palatines are separated from each other by the
intervention of the vomer. In this feature they resemble certain
Ziphioids, Berardius, Oulodon (= Mesoplodon)
grayi, and Hyperoodon. The true Dolphins also appear to
show the same intervention of the vomer in a few cases. There is nothing,
therefore, distinctive from the Delphinidae in this feature.
The existence of cartilaginous sternal ribs in Inia and
Platanista shows affinity between these two genera and the
Physeteridae. Pontoporia is Dolphin-like in this particular, as it
is also in the mode of articulation of the ribs with the vertebral
column. But this last matter has already been dealt with. The principal
reason for placing Pontoporia with the other two genera is the
close resemblance which its skull bears to that of Inia.
The first genus of this family which will be noticed is
Platanista. {381}The following are its main characters:—Dorsal fin absent. Eyes rudimentary. Pectoral
fins large and truncated at the extremity. Teeth, about twenty-nine in
each half of each jaw. Scapula with the acromion coinciding with its
anterior edge. Skull with enormous maxillary crests, and with the
palatines entirely concealed by the pterygoids. The length of the above
definition will serve to indicate how anomalous in many particulars is
the structure of this "Dolphin."
There is apparently but one species, P. gangetica, the "Susu."
The Indian vernacular name is derived from the sound that the animal
makes when spouting. It is an inhabitant of the Ganges and the Indus,
together with their tributaries, and ascends very high up its streams. It
is also thought to be purely fluviatile and never to desert the rivers
for the sea. Platanista lives chiefly by grubbing in the mud for
prawns and fish. Grains of rice have also been found in the stomach, but
this would seem to be accidental. The long snout of the Susu has been
compared to the long snout of the Gharial, a native of the same region.
This Whale grows to a length of over 9 feet, but this length is
exceptional. Its anatomy has been elaborately described by Dr.
Anderson.[252]
The next genus, Inia, is thus to be characterised:—Dorsal fin rudimentary; pectorals large and
ovate. Teeth, as many as thirty-two on each side, often with an
additional tubercle. Skull without large maxillary crests; palatines not
hidden by pterygoids, but divided by vomer. The vertebrae of this genus
are few in number, only forty-one in all, which are thus distributed:
C 7, D 13, L 3, Ca 18. The peculiarities of the vertebral column are
several. In the first place, as has been mentioned in the definition of
the family, all the cervicals are separate and individually of some
length. Secondly, the axis has a better trace of an odontoid process than
in any other Whale except Platanista, where it is even more
obvious. The lumbar region is remarkable on account of its restriction to
three vertebrae. The sternum, by what we must regard as convergence, is
somewhat like that of the Whalebone whales. It consists of one piece
only, of a roughly-oval form, to which apparently only two pairs of
(cartilaginous) sternal ribs are attached. In the fore-limb the
proportions between the humerus and the radius are more like {382}those of
terrestrial mammals; i.e. the humerus is distinctly the longer,
the converse usually obtaining among Whales. But Platanista again
agrees with Inia. The teeth are remarkable for the fact that the
hindermost ones of the series have an additional lobe; they are not
purely conical as are those of Whales generally.
There is but one species, Inia geoffrensis, which inhabits the
Amazons, and grows to a length of 8 feet. Its colour variations are
rather extraordinary, unless they can be set down to sex, which has been
denied. Some individuals are wholly pink; others are black above and pink
beneath. This Whale is believed by the Indians to attack a man in the
water, and it is added that the Sotalia of the same streams will
defend him from these attacks! Naturally, therefore, superstitious
reverence attaches to this Dolphin, which is tiresome to the naturalist
who wants specimens, as Professor Louis Agassiz found.
In the genus Pontoporia[253] the dorsal fin is well developed and
falcate. The teeth are very numerous, 200 in all. The ribs articulate as
in Dolphins. The skull closely resembles that of Inia, and the scapula
is, as in that genus, "normal."
The proper name for Pontoporia is really Stenodelphis,
which name was first used by Gervais a month or two before Gray, who
separated it from the vague Delphinus of its original discoverer,
Gervais himself. It has a longer snout than Inia, which, being
bent towards the extremity in a downward direction, curiously suggests
the skull of a Curlew. In details, however, the skull is exceedingly like
that of Inia. It is nearly symmetrical. The vertebral formula
appears to be the following:—C 7, D 10,
L 5, Ca 20 = 42, just one over the number of the vertebrae in
Inia. The sternum is in two pieces. Of the ten pairs of ribs the
first three are double-headed. These and the next have sternal moieties
joining the sternum, of which the first three are ossified, the last
being apparently merely a ligament.
There is a single species of the genus, P. blainvillii. This
Whale is described by Mr. Lydekker as being of a clear brown colour,
harmonising with the waters of the estuary of the Amazons and the La
Plata which it inhabits. The same colour characterises Sotalia
pallida of those parts of the world, and {383}may be a colour
adaptation. But the extant accounts of the colour of this Dolphin
vary—quite possibly in accordance with real variations, such as are
exhibited by Inia already spoken of. Pontoporia blainvillii
is a smallish Dolphin some 4 feet in length.
Fossil Odontocetes.—Several of the existing genera of
Dolphins are also known in a fossil condition, as well as Ziphioid Whales
closely related to existing forms. We shall deal here only with a few
genera of fossil Odontocetes which depart in their structure from
existing forms.
The genus Physodon is Miocene, and has been found in Patagonia.
It appears to be most nearly allied to the Physeteridae, but should
probably form a distinct family. Physodon was not so large as
Physeter, the skull measuring only some 10 feet. It thus comes
nearer in point of size to Kogia, and it is interesting to note
that its relatively-shorter snout is also suggestive of the dwarf
Cachalot. The general outline of the skull is, however, more like that of
Physeter, and there is the same deep cavity for the lodgment of
spermaceti. The main feature of interest in the skull is the presence of
teeth in both jaws, and the fact that two or three are lodged in the
premaxillae. This is precisely what is found in the most ancient Whales,
the Zeuglodonts.
Extinct Dolphins, apparently referable to the Platanistidae, are the
most numerous among the earlier forms of Cetaceans, and it is significant
that the earliest known forms of these go back to the Eocene.
The genus Iniopsis of Mr. Lydekker,[254] with one species, I.
caucasica, comes from rocks which seem to be of that age. The back
part of the skull of this animal, the only part of the skull known, has
the same squarish excavation of the maxillaries that characterises
Inia and Pontoporia. Its lower jaw was slender and
possessed numerous teeth.
The long snout and jaws of Platanistids, especially exaggerated in
Pontoporia among living forms, are constantly found in these
Tertiary Platanistids.
Eurhinodelphis had a beak three and a half times the length of
the cranium, whereas in Pontoporia the proportions are as 2:1. The
teeth too were very numerous.
The genus Argyrocetus, from Patagonian Tertiary strata, was an
animal about as large as the existing Dolphin. It had the {384}slender
rostrum and numerous teeth of the Platanistids and the squared
excavations of the maxillaries. Argyrocetus patagonicus possessed
also archaic characters, suggesting earlier affinities still. The two
condyles of the skull instead of being closely adpressed to the skull
stood out in a way more like that met with in terrestrial mammals. The
nasal bones instead of being abbreviated rudiments are well developed as
in the archaic Zeuglodonts. The cervical vertebrae of this Whale are all
perfectly free from each other and individually long. The skull is on the
whole bilaterally symmetrical; this again is a feature more pronounced
among the Platanistidae than among other Odontocetes. Accompanying these
generalised Cetacean characters are some which show that the animal was
too specialised to be the direct ancestor of any existing forms. The end
of the mandible was upturned and without teeth, its form being quite
unique among Cetacea. Other allied forms, such as Zarrhachis and
Priscodelphinus, showed the same length of the cervical
vertebrae.
A very distinct family of extinct Whales is that of the
Squalodontidae. They to some extent bridge over the gap between
the existing Odontoceti and the Eocene Archaeoceti (Zeuglodonts).
The skull of these Whales was on the whole Dolphin-like. But they
possessed teeth which were distinctly specialised into incisors, canines,
and molars. The molars have a coarsely-serrated cutting edge as in the
Zeuglodonts, and are also to some extent two-rooted. But they are more
numerous, and so far approximate to the conditions which characterise the
more typical modern Odontocetes. Squalodon was a long-beaked form,
and Prosqualodon had a skull whose proportions are nearer those of
Kogia.
Sub-Order 3. ARCHAEOCETI.
This division of the Whale tribe embraces but a single family,
Zeuglodontidae, of which but a single genus, Zeuglodon, can
with certainty be discriminated.
Zeuglodon is an Eocene form of large size, with teeth which are
limited in number and disposed in three series as incisors, canines, and
molars. The molars are double-rooted, a fact which has given to the genus
its name. The nasal bones being long {385}instead of rudimentary
like those of other Whales, the blow-hole lies more in the middle of the
face. The skull, too, is not Whale-like in a number of other points. Thus
the premaxillaries take their fair share in the outline of the upper jaw;
and, furthermore, bear the incisor teeth. The parietals meet above in a
crest and are not excluded from the roof of the skull. The vertebrae of
the neck are in no way shortened; neither are they fused together. The
ribs are double-headed, and the sternum is made up of several pieces.
Some naturalists, particularly Professor D'Arcy Thompson,[255] have assigned a
relationship to the Seals to these ancient Cetacea; but others[256] have disputed this
view chiefly on the grounds that the characters which appear to be
Seal-like are simply characters which are generalised and so far at most
not Whale-like. Thus the long neck and the serrated character of the
teeth may be accepted as Seal-like on the one hand; but on the other, a
simple serrated tooth and a long neck are not by any means features of
organisation which we should consider out of the way in an ancient form
of Cetacean which probably preyed upon fish. The humerus of
Zeuglodon, according to Mr. Lydekker, puts out of court any
possible near relationship to the Seals. But the matter under dispute can
be further studied by reference to the three memoirs quoted below.
{386}
CHAPTER XIII
CARNIVORA[257]—FISSIPEDIA
Order VII. CARNIVORA
This order may be thus defined:—Small to
large quadrupeds, terrestrial, arboreal, or aquatic, of usually
carnivorous habits. The teeth have generally sharp and cutting edges, and
the canines are well developed; the incisors are small, and four to six
in number. The number of toes is never less than four. There are usually
strong and sharp claws. The clavicles are incomplete or absent. In the
hand the scaphoid and lunar bones are always united. The brain is well
developed, and the hemispheres are well convoluted. The stomach is always
simple, while the caecum, if present, is always small. The members of
this group have a deciduate and zonary placenta.
The fewness of the characters used in the above definition is chiefly
owing to the fact that the Seals and Sea-lions, although they are
referable without a doubt to this order, have undergone in their
metamorphosis into aquatic animals so many changes that some of the main
features in the structure of their terrestrial relatives have been lost.
This group will, however, be again characterised. We shall deal at
present with the land division of the Carnivora, the Carnivora Fissipedia as they are generally termed. The
name is of course given to them to distinguish them from the
corresponding division of the Pinnipedia. In the
latter group the feet and hands are modified into "fins"; in the other
the fingers and toes are cleft, as with terrestrial beasts generally.
{387}
Sub-Order 1. FISSIPEDIA.
Fig. 190.—Under surface of the
cranium of a Dog. × ½. apf,
Anterior palatine foramen; AS,
alisphenoid; as, posterior opening
of alisphenoid canal; BO,
basioccipital; BS, basisphenoid;
cf, condylar foramen; eam, external
auditory meatus; ExO,
exoccipital; flm, flp, foramen
lacerum medium and posterius;
fm, foramen magnum; fo, foramen
ovale; Fr, frontal; fr,
foramen rotundum; gf, glenoid
fossa; gp, post-glenoid process;
Ma, malar; Mx, maxilla; oc,
occipital condyle; op, optic foramen;
Per, mastoid portion of
periotic; pgf, post-glenoid foramen;
Pl, palatine; PMx, premaxilla;
pp, paroccipital process;
ppf, posterior palatine
foramen; PS, presphenoid; Pt,
pterygoid; sf, sphenoidal fissure
or foramen lacerum anterius;
sm, stylomastoid foramen; SO,
supraoccipital; Sq, zygomatic
process of squamosal; Ty, tympanic
bulla; Vo, vomer. (From
Flower's Osteology.)
A very marked feature of the terrestrial Carnivora is to be found in
the structure of the teeth. The incisors are nearly always six, and are
somewhat feebly developed in many cases. The canines are almost
invariably very large strong teeth, and are always present. In some of
the extinct Cats they reached enormous dimensions. The number of cheek
teeth is not always identical; but the last premolar in the upper jaw and
the first true molar in the lower jaw, known as the "carnassial" or
"sectorial" teeth, mark a difference in structure between the anterior
and the posterior crushing teeth; those in front of the carnassial tooth
have cutting edges, and are often merely small, conical teeth; those
behind have broader crowns and are tuberculate; those of simpler forms
often trituberculate; those of others {388}with numerous
tubercles. The carnassial tooth is often, but by no means always, very
much larger and especially longer than the rest of the molar and premolar
series. It is less pronounced in some of the omnivorous Arctoidea. The
skull of the Carnivora is longer in the more primitive types, such as the
Canidae, and shorter in the more specialised Felidae. The orbit is hardly
ever completely shut off by bone, though the postorbital process of the
frontal sometimes approaches the corresponding upward process of the
zygomatic arch. The palate, which is completely ossified, sometimes
reaches back for some distance behind the teeth; it always extends as far
as the last molar. The tympanic bulla is often very inflated, and if
flatter, as in the Bears, is at any rate large and conspicuous. The lower
jaw has a high coronoid process, and the condyle is transversely
elongated, this part of the bone being rolled into an almost cylindrical
form; it fits very closely into the glenoid cavity, and the articulation
is thereby very strict—an obvious advantage in a creature with so
great a need for power of jaw.

Fig. 191.—A, Atlas of Dog. Ventral view, × ½. B, Axis of Dog. Side view, × ⅔.
o, Odontoid process; pz, posterior zygapophysis; s, spinous process; sn, foramen
for first spinal nerve; t, transverse process; v, vertebrarterial canal. (From
Flower's Osteology.)
In the vertebral column the atlas always has large wing-like
processes; the spine of the axis vertebra has a long antero-posteriorly
elongated form. The transverse processes of the fourth to the sixth
cervicals are, as a rule, double. These features, however, though
characteristic of the Carnivora are not by any means distinctive. The
true sacrum consists of but a single vertebra to which the ilia are
attached; but at most two other vertebrae are fused with this. The
clavicle is always small and sometimes quite rudimentary, or even absent.
The spine of the scapula is well developed, and almost equally divides
the {389}surface of that bone. The digits of the
Carnivora are mostly five, and are never less than four. The mode of
progression may be digitigrade or plantigrade, and the intermediate
semidigitigrade mode of walking also occurs. The brain in all Carnivora
is large and well convoluted. The arrangement of the convolutions is
characteristic. There are three or four gyri disposed round each other,
of which the lowest surrounds the Sylvian fissure. The stomach in these
creatures is always simple in form, without {390}subdivisions. The
caecum is never large, and may be, as in the Bear tribe, completely
absent.

Fig. 192.—Brain of Dog. A, Ventral; B, dorsal; C, lateral aspect. B.ol, Olfactory
lobe; Cr.ce, crura cerebri; Fi.p, great longitudinal fissure; HH, HH1, lateral
lobes of cerebellum; Hyp, hypophysis; Med, spinal cord; NH, medulla oblongata;
Po, pons Varolii; VH, cerebral hemispheres; Wu, middle lobe (vermis) of cerebellum;
I-XII, cerebral nerves. (From Wiedersheim's Comparative Anatomy.)
The distribution of the Carnivora is world-wide, excluding only the
Australian region, if, as seems probable, the Dingo of that region is an
introduced species. The most striking features in their distribution are
perhaps the following:—There are no Bears
in the Ethiopian region or in Madagascar, and but a single species in the
Neotropical. The only Carnivora in Madagascar are the Viverridae, and of
the seven genera there found six are peculiar. The Procyonidae are nearly
entirely New World in range; out of sixteen genera of Mustelidae only
five are New World, and only two of those are peculiar to the American
continent. The Hyaenidae are limited to the Old World.
The classification of the Carnivora is a matter which is difficult,
and which has therefore been very variously effected. It is unfortunate
that the classification of Flower (based upon the researches of H. N.
Turner as well as his own, and accepted by Mivart) should fail when
applied to fossil forms. For it separates with great clearness the
existing genera into three great divisions, the Cynoidea, Aeluroidea, and
Arctoidea, definable by visceral as well as by osteological characters.
The apparent anomaly, too, of a single supposed Viverrine genus, to wit
Bassariscus, existing in America, while all the rest of its kin
are Old-World forms, was shown by his characters to be neither an anomaly
nor a fact. It will be better, therefore, to divide the Carnivora into
the families, Felidae, Machaerodontidae, Viverridae, Hyaenidae, Canidae,
Ursidae, Procyonidae, and Mustelidae, indicating at the same time the
reasons for and against retaining the three divisions of Sir W.
Flower.
Fam. 1. Felidae.[258]—This family includes only the
Cats (i.e. Lions, Tigers, "Cats," Hunting Leopard, etc.), and is
to be distinguished by the following characters:—In the skull the auditory bulla is much
inflated, and there is an internal septum; the paroccipital processes are
flattened against the bullae. There is no alisphenoidal canal. The dental
formula is I 3, C 1, Pm 3 to 2, M 1. The carnassial tooth of the upper
jaw has three lobes to the blade; that of the lower jaw is without an
inner cusp. {391}The digits are five on the fore-feet, four
on the hind. The caecum is present and small. This family contains but
two genera, Felis and Cynaelurus.
Fig. 193.—Section of auditory bulla of Tiger. am, Auditory
meatus; BO, basioccipital; e, Eustachian canal;
ic, oc, two chambers of bulla divided by s, septum;
*, their aperture of communication; Pt, periotic;
Sq, squamosal; t, tympanic ring. (From Flower's
Osteology.)
The genus Felis is very wide in its distribution, being common
to both the Old and the New Worlds. Its distinctive characters, as
opposed to Cynaelurus, are mainly the following:—The claws are retractile, and the retractility
is more markedly developed than in the Cheetah. The molar is not so
nearly in a line with the other teeth; the upper carnassial, moreover,
has an inner tubercle. The legs are relatively shorter.
The complete retractility of the claws is a very distinctive feature
of the true Cats. It is brought about in this way: the terminal joint of
the toe, which is clad with the claw, folds back into a sheath by the
outer side of or above the middle phalanx. It is held in this position by
a strong ligament. The flexor muscles straighten the phalanx which bears
the claw, so that the natural position for the animal is to be in a state
of retracted claws, which of course preserves them from friction; when
wanted for aggressive purposes, they are pulled into sight by the action
of the muscles already mentioned.
Fig. 194.—The phalanges of the middle
digit of the manus of the Lion (Felis
leo). × ½. a, The central portion
forming the internal support to the
horny claw; b, the bony lamina reflected
around the base of the claw;
ph1, proximal phalanx; ph2, middle
phalanx; ph3, ungual phalanx. (From
Flower's Osteology.)
Much has been written as to the shape of the pupil of the Cat's eye.
Some careful observations upon the matter have been {392}made by Dr.
Lindsay Johnson,[259] who
found that out of 180 Domestic Cats 111 had round pupils, and that in 19
the shape was a pointed oval, intermediate conditions being offered by
the rest. These 180 comprised males and females of many varieties. When
the pupil of the Cat's eye contracts, it forms a vertical slit with two
pin holes, one at each end, through which alone light appears to enter.
In the Genet and the Civet the contraction of the pupil is as in the Cat.
In the Lion, Tiger—in fact apparently in all the large
Cats—the pupil retains its circular shape even when contraction is
fully effected. Dr. Johnson has, furthermore,[260] made some interesting experiments
upon the Seal's eye—a creature which has, of course, to exert its
powers of vision in two media, and from one to the other. This is
effected by dilatation of the pupil when in the water, and its
contraction to a vertical slit with parallel margins and rounded ends
when in the air, the contraction being to some extent at least under the
influence of the animal's will.
The coloration of these creatures is very varied: spots of black, or
bordered with black upon a more or less tawny ground-colour, is the
prevailing pattern. Stripes are also met with, as in the Tiger, but these
are usually cross stripes,[261] while in the related Viverridae there
are many examples of longitudinal stripes. Finally, many Cats, as for
instance the Puma and the Eyra, are "self-coloured"—have, that is
to say, a uniform tint. Just as the unstriped Horse sometimes shows
traces of the former existence of stripes, so the self-coloured Cats are
occasionally spotted when young; this is markedly so in the case of the
Puma; while the Lion is spotted as a cub, and in the
adult—particularly in the lioness—there are distinct
indications of these spots. It is evident, therefore, that there are
grounds for regarding a spotted condition to be antecedent, at least in
some cases, to a uniform colour. There are divers explanations of these
hues and of these changes. It is held by many that the coloration has a
relation to the habits of the creature: the spotted Cats, it is pointed
out, are largely arboreal; this is eminently so with the Jaguar at any
rate; and in an arboreal {393}creature the spots, it is said, give the
impression of flecks of sunlight broken up by foliage. On the other hand,
the self-coloured Cats of a sandy to earthen hue assimilate in tint with
a sandy or stony soil. The stripes of the Tiger, it is thought,
approximate to the tall parallel stems of grasses and other plants in the
dense cover in which it lives. In favour of these views is undoubtedly
the fact that in other mammals and other animals belonging to quite
different groups the same four plans of coloration are met with. Spots
and cross stripes are found in the Marsupials; the young Tapir is spotted
while the adult is self-coloured, and so forth. This last fact, however,
serves to illustrate another view which has been put forward in
explanation of these characteristic markings of the Felidae. Eimer has
come to the conclusion that there is and has been a regular series of
steps in the evolution of these markings. The primitive condition was, he
thinks, a longitudinally striped one; the stripes then broke up into
spots, and the spots rearranged themselves as transverse stripes; the
self-coloured Puma and Lion are a final stage in this gradual evolution.
In support of this is the fact that spots precede self-coloration in the
individual growth of these animals. The exact sequence of these markings
is, however, contradicted by Dr. Haacke's observations upon a certain
Australian fish which is cross striped when young and longitudinally
striped when adult, a precise reversal of what ought to occur on Eimer's
view.
The Felidae are almost universally distributed with the exception, of
course, of Australia and a good deal of the Australian region; the
headquarters of the group are undoubtedly in the tropics of the Old
World.
The characteristics of a few species of the Cat tribe will now be
given. As there are at any rate forty-five species, this survey will have
to be somewhat incomplete.
The Lion, F. leo, differs from all other species by the mane of
the male. It is an inhabitant of Africa, India, and certain parts of
Western Asia. Within the historic period it ranged into Europe. According
to Sir Samuel Baker those of us who have not seen the Lion in his native
haunts have never seen a really magnificent specimen of the brute; but
other travellers disagree, and state that a captive Lion is often a finer
animal—by reason, of course, of good feeding. Unlike the majority
of Cats, the Lion {394}cannot climb. His roar (which is so
suggestive, towards its end, of that animal who once dressed himself up
in his skin) is literally after his prey. The Lion, it is stated,
does not roar except upon a full stomach. The Lion is mainly nocturnal in
its habits, and is said to be not in the least dangerous if unprovoked in
the daytime; but here again opinions differ. The tail of the animal is
provided at the extremity with a slight claw, but it can hardly be
sufficient for the animal to lash itself into a fury with it. A Lion will
live for thirty or forty years, and will breed freely in captivity. The
Gardens of the Zoological Society of Dublin have been famed for their
success in breeding Lions; but more surprising still, this has been
successfully accomplished in travelling menageries. The "desert" colour
of the Lion is familiar to all. It is stated that the likeness to the
parched soil of certain parts of Africa is greatly heightened by black
patches in the mane, for in certain regions of that continent the arid
yellow of the general environment is diversified by pieces of black lava.
It is apparently a popular delusion to speak of the Maneless Lion of
Guzerat. No doubt maneless Lions do come from there, but so do young and
maneless Lions from other places; in short, it is simply a question of
age, and old Lions from the Asiatic continent are as fully maned as those
from Africa.
The Tiger, F. tigris, is an animal of about the same size as
the Lion, distinguished, of course, by the stripes. The skeletons are
much like those of other Cats; but the skull of the Tiger may be
distinguished from that of the Lion by the fact that the nasal bones
reach back beyond the frontal processes of the maxillae. The Tiger is an
exclusively Asiatic beast, ranging northward into icy Siberia. The
northern individuals have a closer fur, and have been quite unnecessarily
separated as a distinct variety. Nine feet six inches is the size of the
average full-grown Tiger; but the skins will stretch, a fact of which the
sportsman will sometimes take advantage. A "man-eater" is a Tiger which
has discovered "that it is far easier to kill a native than to hunt for
the scarce jungle game." As with the Lion, the accounts of travellers
differ enormously, particularly with regard to the strength of the
creature. Some have said that a Tiger can easily lift a full-grown
bullock and leap with it in the mouth over a considerable obstacle, a
statement which is ridiculed by Sir Samuel Baker. Unlike the Lion, the
Tiger can climb trees; {395}it will also voluntarily enter the water,
and can swim considerable rivers.
Mr. H. N. Ridley[262]
observes that Tigers "habitually swim over to Singapore across the Johore
Strait, usually by way of the intermediate islands of Pulau Ubin and
Pulau Tekong. They make the passage at night, landing in the early
morning. As so much of the coast is mangrove swamp, and the animals do
not risk going through the mud, they are only able to cross where the
shores are sandy, and thus they have regular starting- and
landing-places."
The Tiger is mainly nocturnal, but begins its depredations towards
five o'clock in the afternoon, before which it remains sleeping in shady
thickets. If the weather is rainy and windy it becomes restless and
wanders about earlier. Under the provocation of extreme hunger it will
hunt during the daytime. Hunger, too, naturally produces extreme
boldness. Mr. Ridley relates a story of four Tigers who walked up the
steps of a house in search of the master of the house or his dog, and
broke into it, the inhabitants retiring in their favour. The Malays have
superstitions about Tigers, which are precisely paralleled by the
man-and-wolf stories of Europe. "Certain people are supposed to have the
power of turning into tigers for a short time, and resuming their human
form at pleasure. The transformation commences tail first, and the human
tiger is so completely changed that not only has it all the actions and
appearance of the tiger, but on resuming its human form it is quite
unconscious of what it has been doing in the tiger state." Mr. Ridley
disputes the common stories as to man-eaters. If a Tiger has once tasted
human flesh it does not always confine itself afterwards to that article
of diet, nor is it only aged and comparatively toothless animals which
hunt man. That they do take a large toll of coolies is an undoubted fact,
and many are the artifices to prevent the rest from knowing the fate of
one of their fellow-workmen, or of becoming acquainted with the presence
in the neighbourhood of one of the dreaded beasts.
The Leopard or Panther, F. pardus, is, like the Lion, African
and Asiatic in range. The animal is spotted with rosettes of black spots
surrounding a central field of the tawny colour of the body generally.
Some of the spots are solid and black. "The {396}pantere like unto the
smaragdyne" seems to be an inapt description of this Cat, unless indeed
the eyes be referred to. The ancients ascribed to it a most fragrant
odour. As with the Tiger, a northern variety of this Carnivore has a
closer and longer fur. There is a tendency towards melanism in this
animal, the black Leopard being comparatively common, particularly, it
appears, in high lands. Several other variations in colour are known.
These have received different specific names; but it seems that there is
in reality but one species of Leopard. The Leopard can climb with the
agility of any Cat. Sir S. Baker reserves the name Panther for large
Leopards, which reach a length of 7 feet 6 inches. But there is no valid
distinction between any two such varieties. The Leopard is as ferocious
as the Tiger; and Sir Samuel Baker advises that the power of the human
eye be not experimented with when meeting unarmed one of these
brutes.

Fig. 195.—Snow Leopard. Felis uncia. × 1⁄20.
The Snow Leopard or Ounce, F. uncia, is a beautiful creature,
confined to the highlands of Central Asia. The ground-colour is white,
and the spots are larger than those of the ordinary Leopard. Two examples
of this rather rare Carnivore have been recently on view in the
Zoological Society's Gardens, London. The Clouded Leopard, F.
nebulosa, is an animal of considerable size (6 feet total length).
{397}
The Fishing Cat, F. viverrina, of India and China, is about 3
feet 6 inches including the tail. Its black spots upon a grey-brown
ground have a tendency to form longitudinal lines. It is in fact, on
Eimer's theory, a case of longitudinal stripes breaking up into spots. It
differs from the bulk of Cats by preying upon fish, though it is not
known how it catches them. It also feeds upon the large snail
Ampullaria. In addition to these there are twenty-four species of
Cats found in the Old World, mainly in the Oriental region, of small to
moderate size.
Fig. 196.—European Lynx. Felis lynx. × 1⁄12.
The European Lynx, F. lynx, has rather long legs, a short tail,
and tufted, pointed ears. It has only two premolars in the upper jaw
instead of the usual three. It seems to be doubtful whether the Asiatic
Lynx can be distinguished from the European, but the Spanish form, F.
pardina, does appear to be distinct. The Common Lynx, sometimes
called F. canadensis, also ranges into America, where some other
forms exist, known by the specific names of F. rufa and F.
baileyi.
In America there are altogether sixteen species of Cats, if we allow
three species of Lynx, none of which, however, does Dr. Mivart allow to
be different from the European and Asiatic Lynx (F. lynx). {398}
The largest of American Cats is the Jaguar, F. onca. This is an
arboreal creature with a long, heavy body and short limbs. Its pelage is
much like that of the Leopard, but the spots are larger and more
definitely arranged in groups. There are a number of distinct rows of
spots. The length of the body alone is not greater than 4 feet. They prey
very largely on the Capybara, and upon turtles, which they surprise upon
the sand when about to lay their eggs; the reptiles are turned upon their
backs, so as to be incapable of escaping, and the Jaguar then easily
devours them. The Jaguar will even pursue the turtle into the water, and
will devour its eggs and the newly-hatched young.
Fig. 197.—Jaguar. Felis onca. × 1⁄15.
The Ocelot is another spotted American Cat. F. pardalis[263] ranges from Arkansas
in North America southwards, its range corresponding with that of the
Jaguar. Although small for one of the "larger cats," the Ocelot inspired
with considerable respect Captain Dampier, who remarked of it: "The
Tigre-cat is about the bigness of a bull-dog, with short truss, body
shaped much like a mastiff, but in all things else, its head, the colour
of its hair, the manner of its preying, much resembling the {399}tigre, only
somewhat less.... But I have wisht them farther off when I have met them
in the woods; because their aspect appears so very stately and
fierce."
Fig. 198.—Ocelot. Felis pardalis. × 1⁄10.
The Puma, F. concolor, the American Lion as it is called in the
north, is a rather smaller animal than the last, and of a uniform tawny
colour, tending to white on the abdomen and to a dark stripe along the
back. The young, as already mentioned, are very distinctly spotted. Like
the Tiger, the Puma can endure extremes of heat and cold; it is equally
at home in the snow of North America and among the tropical forests and
swamps of the south. It is a ferocious creature so far as concerns Deer,
Lamas, Raccoons, even Skunks and Rheas, but, according to Mr. W. H.
Hudson, will not attack man, and will even defend him against the
Jaguar.[264] In captivity
the Puma will purr like a Cat.
The Eyra, F. eyra, is another self-coloured American cat, which
has a curious likeness to the totally distinct Cryptoprocta of
Madagascar.
The Wild Cat of Europe, F. catus, is found over the greater
part of Europe, and also in Northern Asia. It was undoubtedly common at
one time in this country, though it appears never to have extended its
range into Ireland. But the real Wild Cat is now rare in this island, and
is confined to certain districts in {400}Scotland. Plenty of
alleged wild Cats have been seen and even shot; but these are too
frequently merely feral Cats, i.e. domestic tabbies which have
reverted to a hunting life. The real Wild Cat differs from the domestic
races by the proportionately longer body and limbs, the shorter and
thicker tail; the pads of the toes are not quite black. The period of the
gestation of the Wild Cat, according to Mr. Cocks, is a week or so longer
than that of any domestic Cat.
The Domestic Cat is in fact regarded as the descendant of the Eastern
F. caffra, or (perhaps and) the closely-allied F.
maniculata. It is highly probable, however, that after introduction
into this country as a domestic animal it has interbred with the Wild
Cat. Many allied species of Cats will interbreed, even two so far apart
as the Lion and the Tiger. There are interesting archaeological and
linguistic reasons for regarding the Domestic Cat as an importation. The
legend of Dick Whittington's Cat points to it being a rare and valuable
animal, which a tamed F. catus would not at that time have been.
There was an enactment in Wales of a penalty against him who should kill
the king's Cat, again suggestive of its rarity and consequent value. The
very name "Puss" is a hint of a foreign origin. Some would derive it from
Perse, and upon this is based the notion that the Cat is from Persia. But
it seems that Puss is the same as Pasht and Bubastis, showing so far an
Egyptian origin for the animal. The ancestral Cats mentioned above are
natives of Egypt.[265]
The genus Cynaelurus, which includes but a single species,
C. jubatus, the Cheetah or Hunting Leopard, is separated from
Felis by a number of characters. In the first place the claws are
non-retractile, or at least less retractile than those of the true Cats.
It is, moreover, longer legged. The molar is more in a line with the
other teeth of the jaw, and the upper carnassial tooth has no inner
tubercle. Messrs. Windle and Parsons have lately pointed out many
Dog-like features in the muscles. This animal is about as large as a
Leopard, but has plain black spots. As its vernacular name implies it is
used for sport, and is quite easily tameable. It will purr like the Puma.
The Cheetah occurs in India, Persia, Turkestan, and also in Africa; the
latter {401}form is sometimes, though quite
unnecessarily, separated as C. lanigera. The genus occurs fossil
in the Siwalik deposits of India, the species being known as C.
brachygnatha.
Fam. 2. Machaerodontidae.—This is a family of
totally-extinct Cats which range from the Eocene down to the Pleistocene.
Their general structure is like that of the Felidae; but they differ in a
number of skeletal features. Thus there is an alisphenoid canal, and, as
in Bears, there is a postglenoid foramen. There is also a distinct
carotid foramen, which does not occur in the true Cats. The teeth are
often distinguished by the huge size of the superior canines, which are
"weapons for penetrating wounds, without rival among carnivorous
animals." These must have been displayed at the sides of the chin when
the mouth was closed, and it has even been suggested that the animal
possessing these exaggerated canines could hardly have properly closed
its mouth. The lower canines were often on the contrary much reduced, and
in fact incisor-like. In tracing the series of these Cats we find a
gradual reduction of the teeth from a more nearly complete number down to
the specialised dentition of the existing Cats. The genus
Proaelurus, Miocene in range, had four premolars in each jaw, and
two molars in the lower and one in the upper. This is the greatest number
of teeth found in any member of the group.
The resemblance of this genus to Cryptoprocta has been insisted
upon. Archaelurus has suffered a reduction, since one premolar in
the lower jaw has disappeared, its formula being thus I 3/3 C 1/1 Pm 4/3
M 1/2. The next stage is shown by Dinictis with three premolars in
both jaws. There are a good many species of this genus which are all
American and Miocene. This genus has five toes upon the hind-feet, and
was probably plantigrade. It had retractile claws.
In the genus Nimravus the dental formula is still further
reduced. Another premolar of the lower jaw has gone, the formula being
thus I 3/3 C 1/1 Pm 3/2 M 1/2. Nimravus gomphodus was a Carnivore
about the size of a Panther. It has no third trochanter upon the femur,
which process is present in the corresponding bone of Dinictis.
Pogonodon was an equally large animal in which the premolars were
three in each jaw, but the molars have become reduced to one in the
lower, as they have in this and other genera in the upper {402}jaw. Finally,
Hoplophoneus has acquired the dentition of existing Cats.
The Machaerodons, however, show examples with a yet more reduced
dentition than that of the most reduced existing Cat, viz. the Lynx,
which has only two premolars in each jaw and one molar. In
Eusmilus the molar in both jaws is single, and there is but one
premolar in the lower jaw.
The genus Machaerodus itself, which appears to include
Smilodon, is referred by Cope to the true Cats, and not to the
Nimravidae, as he terms the family which we have called here the
Machaerodontidae. These creatures are known as "Sabre-toothed Tigers,"
and were of very wide distribution, occurring in South America as well as
in Europe and North America. "As nothing," remarks Professor Cope, "but
the characters of the canine teeth distinguished these from typical
felines, it is to these that we must look for the cause of their failure
to continue. Professor Flower's suggestion appears to be a good one, viz.
that the length of these teeth became an inconvenience and a hindrance to
their possessors. I think there can be no doubt that the huge canines in
the Smilodons must have prevented the biting off of flesh from large
pieces, so as to greatly interfere with feeding, and to keep the animals
in poor condition. The size of the canines is such as to prevent their
use as cutting instruments excepting with the mouth closed; for the
latter could not have been opened sufficiently to allow any object to
enter it from the front. Even when it opens so far as to allow the
mandible to pass behind the apices of the canines, there would appear to
be some risk of the latter being caught on the point of one or the other
canine, and forced to remain open, causing early starvation. Such may
have been the fate of the fine individual of the S. neogaeus,
Lund, whose skull was found in Brazil by Lund, and which is familiar to
us through the figures of de Blainville."
Machaerodus is placed among the Felidae on account of the fact
that the condyloid and carotid foramina unite with the foramen lacerum
posterius. But as in at least one species, M. palmidens, there is
an alisphenoid canal, which, however, has disappeared in the more recent
American forms, it seems permissible to retain the genus in the family
Machaerodontidae though its existence reduces the differential character
of that family to a minimum. The genus goes back to the Eocene. {403}
Fam. 3. Viverridae.—The Civets, Genets, and their kind
differ from the Cats in a number of points. They form, however, by no
means so uniform an assemblage as do the Cats; so that the difficulty is,
as Dr. Mivart has remarked, not to divide them into sub-families, but to
avoid making too many. But before proceeding to subdivide the family we
shall describe the characters of the family and contrast them with those
of the Felidae.
All the Viverridae are comparatively small creatures. The head and
body are more elongate than in the Cats. The fingers and toes are
generally five; but there are some (e.g. Cynictis) where the
formula of the toes is as in the Cats, i.e. four on the hind-foot.
In the Suricate the fingers are also reduced to four. The claws are
perhaps never completely retractile,[266] and often are not at all so. The
dental formulae of the genera differ considerably; but in the majority
there are more teeth than in the Felidae. The well-known sharp-pointed,
conical papillae of the Cat's tongue are not present. The majority have a
scent gland beneath the tail, from which the perfume civet is derived.
There are a number of osteological characters which differentiate the two
families; thus the alisphenoid canal is sometimes present. The bulla is
divided, as in the Cats, but is externally constricted.
It seems clear from some at any rate of the characters, i.e.
the more complete dentition, the five-fingered hands and feet, the
non-retractile claws, etc., that the Civets are on a lower level of
specialisation than are the Cats.
Sub-Fam. 1. Euplerinae.—The genus Eupleres is in
many ways the most aberrant type of Viverrid, and is placed in a
sub-family, Euplerinae. Its salient feature is the very peculiar
dentition: peculiar in the small size of the canines, the canine-like
character of the anterior premolars, and the resemblance of the premolars
to molars. In some of the characters of the teeth, Eupleres is
Insectivore-like, and was formerly grouped with that family. There are
four premolars and two molars in each jaw on each side. It has five toes
upon both fore- and hind-limbs; the skull is very slender. It has no
alisphenoid canal. The only species, E. goudotii, is of an
olive-grey colour, with dark bands across the shoulders in the young. The
nose and upper lip are grooved. There are no scent glands. It appears to
burrow in the ground, and possibly contents itself with a diet of worms.
Eupleres is a {404}native of Madagascar, where all the most
peculiar Viverridae live.
Sub-Fam. 2. Galidictiinae.—Mivart has placed in this
sub-family the three Mascarene genera, Galidia,
Hemigalidia, and Galidictis. In them the orbit is not
enclosed by bone; there is no alisphenoid canal, and there are five toes
and fingers.
Galidia consists of but one species, G. elegans, of a
chestnut brown colour, with a tail ringed with black. The claws are not
retractile. The scent gland is absent. There are five digits upon both
hand and foot. There are three premolars and two molars on each side of
each jaw. The caecum is (for an Aeluroid) long, and pointed at the apex;
it is quite twice the length of that of Genetta.
Closely allied to Galidia is the genus Hemigalidia, of
which there are two species. It is distinguished from the last genus by
the non-annulated tail. It also differs in the dental formula, which is
for the molars Pm 4/3 M 2/1. This animal is termed by Buffon the Vansire.
He correctly enumerates its grinders, and distinguishes it from the
Ferret!
Galidictis is a third genus from Madagascar containing two
species, one of which has been unfortunately named G. vittata,
leading perhaps to some confusion with the totally distinct Galictis
vittata. As in the last two genera the digits are five. The dental
formula is that of Galidia. It is distinguished from the other two
genera of its sub-family by the longitudinal brown striping of the upper
part of the greyish body.
Sub-Fam. 3. Cryptoproctinae.—Cryptoprocta[267] represents a special
sub-family, Cryptoproctinae, and includes only a single species, the
Fossa (C. ferox) of Madagascar. It is the largest Carnivore of
Madagascar, being about twice the size of a Cat, but with an elongated
body; the colour is a tawny brown with no striping. The animal is active
and lithe in its motions, and is said to be of almost unexampled ferocity
in disposition. Its exact systematic position has been much discussed. By
Zittel it is placed in a sub-family (including the extinct
Proaelurus and Pseudaelurus) of the Felidae. Mivart and
Lydekker, on the other hand, regard it as a genus of the Viverridae. The
dental formula of the molars, Pm 3/3 M 1/1, is {405}more like that of the
Felidae than of the Viverridae, and the teeth are more
Feline in structure. The claws of the feet are retractile. As to internal
structure the Fossa agrees largely with the Viverridae, but then this
family has no very marked points of difference from the Felidae; but
where the anatomy does diverge from that of the Felidae it approaches the
Viverridae, especially in the muscular system.

Fig. 199.—Fossa. Cryptoprocta ferox. × 1⁄6.
The remaining and by far the larger number of genera of Civets are
grouped by Professor Mivart in two sub-families: the Viverrinae, including the genera Viverra,
Viverricula, Fossa, Genetta, Prionodon,
Poiana, Paradoxurus, Arctogale, Hemigale,
Arctictis, Nandinia, and Cynogale; and the Herpestinae, including the genera Herpestes,
Helogale, Cynictis, and probably Bdeogale and
Rhynchogale. In the Viverrinae the digits are always five, the
claws are more or less retractile, the prescrotal scent glands are
usually present, and the anus does not open into a sac. On the other
hand, the Herpestinae are characterised by the non-retractility of the
claws, the absence of the glands in question, and the fact that the anus
does open into a terminal sac.
Sub-Fam. 4. Viverrinae.—Viverra includes the true
Civets. The genus, save for one African species, is Oriental in range.
The molar formula is the complete one for the Viverridae, viz. {406}Pm 4/4
M 2/2. The secretion of the prescrotal gland of V. civetta yields
the civet of commerce.
The "Rasse," genus Viverricula, has been separated generically
from the true Civets. It is, remarkably enough, common to both
Madagascar[268] and many
parts of the Oriental region. It is, moreover, capable of climbing trees,
which its relatives are not. It has no mane like Viverra and is of
slighter build.
Fig. 200.—Civet Cat. Viverra civetta. × 1⁄6.
Prionodon or Linsang differs from the last two genera in
the loss of an upper molar. It thus approaches the Cats, with which it
also agrees in the furry feet. It is a purely Oriental genus. It also
resembles the Cats in that the claws are apparently quite retractile, a
feature not common among the group. There are three species of the genus.
P. pardicolor has large black spots and a ringed tail. Its body is
some 15 inches in length. Dr. Mivart has commented upon the particularly
small caecum, which, like that of Arctictis, seems to be on the
verge of disappearance.
Genetta, including the Genets, is almost purely African. It has
the full tooth formula of Viverra; but is to be distinguished by
the absence of a scent pouch, and by a naked strip of skin running up the
metatarsus. These animals are all brownish yellowish to greyish with
darker spots. The Common Genet, G. vulgaris, is South European,
and just gets into Asia; it is also North African. The Genet, an animal
"with an appetite for petty carnage," is one of those smaller Carnivora
which are possibly to be regarded as meant by the word γαλῆ, and appear to have "functioned"
as Cats among the Greeks. So recently as {407}the times of Belon we
are told (by him) that Genets were common and tame at Constantinople.
Poiana, containing a single African species, a spotted and
entirely Genet-like animal, has been separated as a distinct genus. Dr.
Mivart, however, holds it to be a Prionodon which has acquired a
Genet-like tarsus.
Arctictis, containing but one species, A. binturong, the
Binturong, is in some ways an exceptional form. It is a black arboreal
creature of not very wide range in the Oriental region, with a fully
prehensile tail. This feature and its plantigrade foot with naked sole
have led to its being regarded as more allied to the Arctoidea. It is,
however, undoubtedly an ally of Paradoxurus. The caecum is small,
or may be quite absent. The dentition is I 3/3 C 1/1 Pm 4/3 M 2/2. The
structure of the animal has been investigated by Garrod.[269]
The genus Fossa is a Viverrine confined to Madagascar. There is
but one species, F. daubentoni, the "Fossane." It is distinguished
from Viverra by the presence of two bare spots on the under
surface of the metatarsus in the hind-limb, and by the absence of a scent
pouch. The animal is not much spotted and striped, but the striping in
the young is much more marked.
Of the genus Paradoxurus there are some ten or a dozen species,
belonging entirely to the Oriental region. The teeth are as in
Viverra, but occasionally the molars are reduced to one. The
pupils are vertical. The tail though long is not prehensile, "but the
animal appears to have the power of coiling it to some extent, and in
caged specimens the coiled condition not unfrequently becomes confirmed
and permanent" (Blanford). This fact accounts for the name
Paradoxurus; for a prehensile tail is hardly to be expected in an
animal of the zoological position of the Palm Civets, and yet its
occasional twisting led originally to the view that it was so. The genus
has scent glands. The dentition is I 3/3 C 1/1 Pm 4/4 M 2/2. P.
niger, the Indian Palm Civet, is, like other species, not often to be
seen in a wild condition. It is arboreal, and, like other members of the
genus, feeds upon a mixed diet, consisting of all kinds of small
Vertebrata and insects, varied by fruit. Another species, P.
grayi, is so distinctly vegetarian in its habits that it makes
considerable havoc in pine-apple beds in the Andaman Islands.
{408}
Arctogale is another Oriental genus with very small teeth,
those of the molar series being hardly in contact. The soles of the feet
are more naked than in the last genus, and the scent glands, if present,
appear to be small and ill developed. It has also a long tail, and is
arboreal in way of life. There is "nothing particular recorded" as to its
habits. The species are A. leucotis and A. stigmatica.
Fig. 201.—Hardwicke's Civet Cat. Hemigale hardwicki. × 1⁄5. (From Nature.)
Closely allied to both the last genera is Hemigale, also an
Oriental genus. It is to be distinguished from Paradoxurus by
having the soles of the feet much less naked, though they are more so
than in Viverra or Prionodon. The coloration of the
species, H. hardwicki (a Malayan animal), is very peculiar. The
body is banded with five or six broad transverse stripes, and the basal
portion of the tail is also ringed, an uncommon feature in the group. A
second species of this genus is H. hosei, from Borneo. It is
blackish in colour, but is not a melanic variety of the last.
Nandinia appears never to possess a caecum.[270] It is also peculiar among Carnivora
in the non-ossification of the hinder {409}part of the bulla. It
is an African genus, containing two species which are spotted. The tail
is ringed.
Cynogale is at any rate a partially aquatic, short-tailed,
web-footed, reddish brown-coloured Civet, which lives upon fish and
Crustacea, and inhabits the Malay Peninsula, Sumatra, and Borneo. It has
long "moustaches," and is said to have a head bearing a singular
resemblance to the head of the Insectivorous "Otter" Potamogale.
The metatarsus is bald, and the pollex and hallux are very well
developed.
Sub-Fam. 5. Herpestinae.—There are over twenty species of
Herpestes (Mongooses) divided between the Ethiopian and Oriental
regions, one species, H. ichneumon, being also found in Europe.
The fur has a "pepper and salt" appearance; the feet are plantigrade.
There are five fingers and toes. The pollex and hallux are small; the
tail is long. The tarsus and metatarsus are usually naked. The Egyptian
species "has been injudiciously denominated the Cat of Pharaoh." It is
perhaps better known as Pharaoh's Mouse. The beast is so far Cat-like
that it will destroy Rats and Mice; and it has been exported to sugar
plantations for that very purpose. More famous are its combats with
venomous serpents. According to Aristotle and Pliny the Ichneumon first
coats its body with a coating of mud, in which it wallows, and then with
this armour can defy the serpent. Topsell tells the tale better. The
Ichneumon burrows in the sand, and "when the aspe espyeth her threatening
rage, presently turning about her taile, provoketh the ichneumon to
combate, and with an open mouth and lofty head doth enter the list, to
her owne perdition. For the ichneumon being nothing afraid of this great
bravado, receiveth the encounter, and taking the head of the aspe in his
mouth biteth that off to prevent the casting out of her poison." In the
West Indies the animal has been described as fearlessly attacking the
deadly Fer de Lance and receiving its bites with impunity; it is also
added that it will eat the leaves of a particular plant as an antidote!
The real explanation of the result of these encounters is of course the
agility of the Ichneumon[271]—fort cauteleuse beste,
as Belon says.
Another species, H. albicauda, is distinguished, as the name
denotes, by its white tail. A species of this genus, H. urva, {410}sometimes raised to generic rank as
Urva, is partly aquatic in habit; it feeds upon crabs and frogs,
but is quite willing to take to poultry and their eggs.
Helogale is a genus whose validity appears doubtful (to Dr.
Mivart). It is African, and contains two species.
Fig. 202.—White-tailed Ichneumon. Herpestes albicauda. × 1⁄5.
Cynictis is an African genus, with five digits on the
fore-limbs and four on the hind. As in Herpestes, the orbit is
completely encircled by bone. There is but a single species, C.
penicillata, which is of a reddish colour and has a bushy tail.
Bdeogale, also African, has the toes still further reduced;
there are only four on both limbs. The tarsus is hairy and the tail
bushy. They are "very rare animals, and nothing is known of their
habits." It is known, however, that they will kill poisonous snakes, for
Dr. Peters took a Rhinoceros Viper out of the stomach of one.
Rhynchogale[272] differs from all other genera of
Viverridae, except Crossarchus and Suricata, in having no
groove upon the muzzle. There are five digits. There is the full
Viverrine dentition, with five premolars in the upper jaw; but this may
be an abnormality.[273]
Crossarchus differs from the last in only having three
premolars on each side of each jaw. It is also African, and there are
several species.
Suricata is the last genus of Viverridae; it is also African,
and contains a single species, Suricata tetradactyla, the
"Meerkat" of the Cape. The Suricate has but four toes on each foot; the
tarsus and the metatarsus are naked below. The body is banded
posteriorly. There are fifteen dorsal vertebrae, and the orbit is {411}closed by bone. The Suricate lives in
caves and rock crevices, and will dig burrows. It is distinctly a diurnal
animal, and sits upon its hind-legs after the fashion of a Marmot. As
Buffon noticed in a tame specimen (thought by him to be a native of
Surinam), the animal barks like a dog. The Suricate is largely
vegetarian, living upon roots.
Fig. 203.—Suricate. Suricata tetradactyla. × ¼.
Fam. 4. Hyaenidae.—Unlike though the Hyaenas appear to be
to the last family—mainly perhaps on account of size—they
are, nevertheless, very nearly akin to them, more so than to the Cat
tribe. It will be remembered that the striping and spotting of the
Hyaenas is very Genet- and Suricate-like.
There are admittedly two genera among the Hyaenidae, Hyaena
itself with three species,[274] and the Aard Wolf, Proteles,
with but one. But Dr. Mivart considers that the Spotted Hyaena should
form a genus apart, Crocuta—a proceeding which was initiated
by the late Dr. Gray of the British Museum. The Hyaenidae are to be
distinguished by the following characters:—There are generally four toes, always so in the
hind-foot. The claws are non-retractile. The nose and upper lip are
grooved. The molar formula is Pm 4/3 M 1/1. The soles of the feet are
covered with hairs upon the tarsus and metatarsus. No scent glands. Tail
short. Dorsal vertebrae more numerous than in other Aeluroids,
i.e. fifteen. The bulla is divided by a rudimentary septum
only.
{412}
Fig. 204.—Spotted Hyaena. Crocuta maculata. × 1⁄12.
The genera Hyaena and Crocuta, the Striped and Spotted
Hyaena respectively, are African and Asiatic in range, Crocuta
being limited to South Africa. There is neither hallux nor pollex.
Fig. 205.—Striped Hyaena. Hyaena striata. × 1⁄12.
The Hyaenas, stigmatised by Sir Samuel Baker as "low-caste creatures,"
are mainly carrion feeders. Much Arab superstition is associated with
them. Certain peculiarities in the structure {413}of the organs of
reproduction have led to the belief that a Hyaena changes its sex every
year. Its almost human-sounding howls are supposed to be a deliberate
trap for the unwary traveller. There is also a legend that in the eye of
the Hyaena is a stone which if placed under the tongue of a man endows
him with the gift of prophecy.
Proteles presents many resemblances to the Hyaenas, but also
certain differences; by many it is placed in a separate family. There is
but one species, P. cristata, the Aard Wolf of South Africa. In
outward aspect it is very Hyaena-like, the coat being striped, and the
ears, though longer, resembling those of a Hyaena. There is also a mane.
There are, however, five toes on the fore-feet. The teeth are feebler,
particularly the molars, which are also reduced in number. The skull, as
in Hyaena, has no alisphenoid canal, but the bulla tympani is
divided by a septum. The animal seems to feed largely upon insects,
particularly Termites, and also upon carrion.[275]
Of extinct Hyaenoids Ictitherium seems to be transitional
between them and the Viverridae. Its dentition, 3/3, 1/1, 4/3, 2/1, is
that of a Viverrid, and the feet are five-toed. The upper carnassial
tooth, however, is like that of Hyaena in having a strong inner
cusp. Other extinct genera of Hyaenas are Lycyaena and
Hyaenictis. The genus Hyaena itself goes back as far as to
the Miocene, and occurred in Europe until the Pleistocene. The Cave
Hyaena of this country seems to be indistinguishable from Crocuta
maculata, though it has received the name of H. spelaea.
Fam. 5. Canidae.[276]—This family cannot be divided
into more than five genera, and is universally distributed with the
exception of New Zealand. The auditory bulla is smooth and rounded, and
has internally a very incomplete septum, extending through about
one-fourth or one-third of the cavity. The meatus has a fairly prominent
under lip. The paroccipital process is long and prominent. The mastoid is
distinct, though but slightly developed. The glenoid foramen is large;
the condyloid foramen is conspicuous, and the carotid canal is deep
within the foramen lacerum posterius. The last three characters are
Bear-like; the {414}form of the bulla is Aeluroid. The teeth
vary somewhat in number, and the following table will serve to indicate
the gradual reduction observable in the number of molars:—
| Otocyon |
I 3/3 C 1/1 Pm 4/4 M (3 or 4)/4 |
| Canis generally |
I 3/3 C 1/1 Pm 4/4 M (3 or 2)/(4 or 3) |
| Cyon |
I 3/3 C 1/1 Pm 4/4 M 2/2 |
| Icticyon |
I 3/3 C 1/1 Pm 4/4 M (2 or 1)/2 |
All the Dogs have a caecum[277] of simple cylindrical form. In C.
cancrivorus, C. jubatus, and Nyctereutes procyonides
this organ is straight or only very faintly curved; in other Dogs it is
coiled into an  -like form, sometimes with an additional twist. The Dogs have, as a
rule, five toes, one being dropped in Lycaon. The tail is fairly
long and distinctly bushy. There is in a number of species a gland at the
root of the tail, the presence of which can frequently be detected by the
wet appearance due to the oozing secretion. The great majority of
existing Canidae belong to the genus Canis. But certainly three,
and more doubtfully four, other genera can be distinguished.
-like form, sometimes with an additional twist. The Dogs have, as a
rule, five toes, one being dropped in Lycaon. The tail is fairly
long and distinctly bushy. There is in a number of species a gland at the
root of the tail, the presence of which can frequently be detected by the
wet appearance due to the oozing secretion. The great majority of
existing Canidae belong to the genus Canis. But certainly three,
and more doubtfully four, other genera can be distinguished.
The genus Icticyon contains but one recent species, the Bush
Dog (I. venaticus, Lund) of British Guiana. The animal has a
somewhat Paradoxure-like, at any rate a distinctly un-dog-like, aspect,
being longish in the body (some 2 feet long), shortish in the legs, and
big-headed. It is blackish in colour, verging towards golden brown on the
head and back. Sir W. Flower, to whom we owe our chief knowledge of its
structure, characterises it as like a young Fox, and with the playful
manners of a puppy. The animal appears to hunt in packs and by scent, and
has a reputation for ferocity. Icticyon differs from Canis
and agrees with the Indian Cuon in having but forty teeth, the
last molar having disappeared from the upper and lower jaws. The caecum,
unlike that of the majority of Canidae, is only slightly curved. The
brain, oddly enough, shows a Cat-like peculiarity. It has been pointed
out that in their long bodies and short legs the genera Cuon and
Icticyon resemble the primitive dogs.[278]
A genus Nyctereutes is usually separated from Canis for
the inclusion of N. procyonides only. The separation is based upon
{415}the strikingly unusual coloration of this
Dog. It is a small animal, with numerous long white hairs dorsally. The
face, chest, and much of the belly are black. Its aspect distinctly
recalls that of a Raccoon,[279] especially in the black patches below
the eyes, whence of course the scientific name and the pseudo-vernacular
"Raccoon-like Dog." It inhabits China and Japan. As to structure, there
is hardly anything that justifies its exclusion from the genus
Canis. Garrod, however, mentions the unusually large size of the
Spigelian lobe of the liver.
Fig. 206.—Raccoon-like Dog. Nyctereutes procyonides. × 1⁄6.
Wortman and Malkens[280] have instituted a genus
Nothocyon for Dr. Mivart's species C. urostictus[281] and C.
parvidens, which are both South American forms.
The genus Otocyon contains but one species, O.
megalotis, an African species, ranging pretty widely in that
continent (from the Cape to Somaliland, in sandy districts), and
sometimes confused with the Fennec on account of its long ears. Its
principal structural difference from other Dogs is that there is an
additional molar in each jaw, the molar formula being thus M 3/4 or even
4/4. Moreover the carnassial teeth are not so pronounced, and Professor
Huxley laid especial stress upon the {416}likeness of some of the
cheek teeth to those of the more primitive Arctoids. The angle of the
lower jaw is inflected, a character, however, which seems to be more
general than is usually allowed among animals not referable to the
Marsupials. It is possible that Otocyon is a persistent
Creodont-like form which has developed in a direction curiously, and in a
most detailed fashion, parallel to the Dogs. If, however, we may assume
the addition of the molar, then this anomalous but not necessarily
untenable conclusion is obviated.
The genus Cuon, or Cyon, has been instituted for the two
or three species of Eastern Dogs (C. primaevus, C.
dukkunensis, etc.) which agree with each other in the constant loss
of a molar in the lower jaw, or, it should be said, almost constant loss,
for the missing tooth is occasionally represented. The latter of the two
species mentioned, the Dhole, is, like its congeners, an animal which
hunts in packs; it is said to hunt even the ferocious Tiger, and to be
thus one of the few animals which can face the largest and fiercest of
the Carnivora.
The genus Lycaon is a very distinct type, being differentiated
from other Dogs by the possession of only four toes on both fore- and
hind-limbs, and by the dental formula, which is Pm 4/4 M 2/3. The one
species is L. pictus, the Cape Hunting Dog. It is singularly like
a Hyaena[282] in general
appearance; the ochraceous grey ground-colour with black markings and the
long ears produce this likeness. The animal has got its vernacular name
from the habit of hunting in packs. Its range is over a good part of
Africa. The occurrence of this species (or at least genus, for the name
L. anglicus has been used) in caves in Glamorganshire seems to
show that it is a comparatively recent immigrant into Africa. As to its
visceral structures, Lycaon[283] does not differ widely from other
Dogs. It has, however, no lytta beneath the tongue. The intestines are
thus divided: large, 9 feet 1 inch; small, 1 foot 3 inches. This
contrasts with the proportions observable in some other Dogs. While other
Dogs have but a cartilaginous rudiment of the clavicle, Lycaon has
a considerably larger representative of this bone.

Fig. 207.—Fennec Fox. Canis zerda. × 1⁄5.
Fig. 208.—Prairie Wolf or Coyote. Canis latrans. × ⅛.
The bulk of the Dogs, Wolves, Foxes, and Jackals are thus left over
for inclusion in the genus Canis. But the numerous {417}members of
this genus can, according to Professor Huxley, be sorted into two series
by certain cranial characters. The two series he termed the "Alopecoid"
or Fox-like, and the "Thooid" or Wolf-like. It was suggested that the
generic name Vulpes be used for the former, and Canis for
the second. The characters which will be dealt with immediately are also
to be noted among {418}the Dogs belonging to genera that have
already been separated off. Thus Lycaon is distinctly Thooid. The
characters in question are these:—In the
Fox series, the frontal air-sinus of the Thooids is absent; the cranial
cavity is pear-shaped, without an abrupt angle coinciding with the
supra-orbital sulcus, such as exists in the other group; the coronoid
process of the mandible is rather higher and more turned back in the
Foxes, while the depth of the mandible at the level of the first molar is
greater.

Fig. 209.—Japanese Wolf. Canis hodophylax. × ⅛ (From Nature.)
To the Fox series belong among others the species C. lagopus
(Arctic Fox), C. zerda (the Fennec), C. chama (the
Silver-backed Fox of Africa), C. virginianus (the Virginian Fox),
C. velox (the Kit Fox), and of course the Common Fox of this
country. On the other hand, the Dogs proper (such as C. dingo),
the Wolves (C. lupus, C. pallipes, C. niger), the
Japanese Wolf (C. hodophylax), the Red Wolf of America (C.
jubatus), the Jackals (C. aureus, C. anthus, etc.), the
Prairie Wolf (C. latrans), and a number of American forms, such as
C. azarae, its close ally C. cancrivorus ( = C.
rudis), C. antarcticus, C. magellanicus, etc., are
decidedly Wolves rather than Foxes. {419}
The Arctic Fox, Canis lagopus, is known by its bluish summer
and pure white winter dress as "Blue Fox" and "White Fox" respectively.
It is an inhabitant of the Arctic north; but in former days, as its
remains show, it descended to such southern latitudes as Germany and this
country. The most southern point which it now inhabits is Iceland. This
small Fox is well known as being one of the few animals which change
their dress to a complete white in winter. This change is, however, not
absolutely universal; and M. Trouessart has even stated that the supposed
change does not exist, but that the colours are a question of age and
sex. This Fox feeds on birds and cast-up carcases of Whales and Seals; it
is also said to devour shell-fish, and actually to store up food when
abundant for seasons of scarcity. A Fox has been observed to "carry off
eggs in his mouth from an eider duck's nest, one at a time, until the
whole were removed"; and in winter to "scratch a hole down through very
deep snow to a cache of eggs beneath." These anecdotes are told by
Sir Leopold M‘Clintock; but others have also asserted the storing
habits of this Fox, which really has only a short time of the year in
which it can catch suitable living food.
Canis vulpes, the Fox, is not only a native of England, but
extends as far to the east as Egypt, the so-called C. aegyptiacus
being at most a mere variety. Varieties indeed occur in these islands;
the English Fox being redder, the Scotch greyer. Not only is the Fox a
truly indigenous English beast, but its remains go back a very long way
into past time. Its bones occur in the Red Crag, a deposit of Pliocene
times. Its prevalence now is no doubt due to its preservation as a beast
of chase. It lives in burrows, either excavating them itself or taking
possession of those of some other animal; the Badger suffers in this way,
and is said to be vanquished not by the teeth of the burglarious Fox, but
by its far fouler habits! It is curious that the expression "foxing" is
not so suitable to this animal as to many others. The habit of "shamming
death" is a widely-spread one in the animal world, but at least not
common with our Fox. The sagacity of the Fox appears to be a little more
proverbial than actual; literature teems with its accomplishments. The
worthy Archbishop of Upsala, Olaus Magnus, figured Foxes dipping their
tails in the streams, and then pulling out inquisitive crayfishes {420}which
had seized upon them. "It is a crafty, lively, and libidinous creature,"
observed a writer of the last century.
Of Jackals there are many species, both African and Oriental. Mr. de
Winton allows the following list of African species[284]:—C.
anthus, C. variegatus, C. mesomelas, C.
lateralis. C. mesomelas is distinguished by the broad black
patch in the middle of the back. These animals do not appear to go in
packs as so many Canidae do; they live upon carrion, but also rob
hen-roosts, and commit other depredations upon the live stock of farmers.
The "Quaha," C. lateralis, is distinguished from the last by its
sharp bark, and by the obvious side stripe which has given to it its
name. It is curious that it should live in apparent amity with C.
mesomelas, since the habits of the two are identical and would lead,
one might suppose, to a severe struggle for existence, in which one of
the two would disappear. Of Indian Jackals C. aureus is the most
familiar type.
Fig. 210.—Wolf. Canis lupus. × ⅛.
The European Wolf, Canis lupus, was once, but is no longer, an
inhabitant of the British Islands. Their former prevalence is indicated
by many names of towns and villages, such as Ulceby and Usselby in
Lincolnshire, the town of Wolverton, and Woolmer Forest. In Saxon times
Wolves were very abundant; and even so recently as the reign of Elizabeth
they were to be seen on {421}Dartmoor and in the Forest of Dean. In the
New Forest they were hunted in the twelfth century. It would seem that
the last English Wolf was slain some time during the reign of Henry VII.
In Scotland, however, they persisted very much longer. So recently as
1743 was the last killed. But before this period they had begun to get
exceedingly scarce, for the price of a skin in 1620 is quoted at £6:13:4.
In Ireland Wolves lingered yet longer; about 1770 is believed to be the
date of their final extinction in that island. The Wolf nowadays is
distributed over the greater part of Europe, Northern Asia, and North
America, the American form not being considered to be distinct from its
European ally. Much legend has collected round this fierce Carnivore.
Aristotle, usually accurate in the main, still "states more of wolves
than experience warranted." Pliny, unable to sift truth from falsehood,
was in this matter "an eager listener to all old women's tales." Aelian
added to his marvels and asserted that the Wolf cannot bend its head
back; if it should happen to tread on the flower of the squill it at once
becomes torpid. So the wily fox, fearing his more powerful enemy, takes
care to strew his path with squills! The conversion of men into Wolves
was a well-known superstition, dating from Grecian and Roman times; it
formed the basis of much of the witchcraft persecutions of the Middle
Ages and onwards, and has left its mark in folklore, e.g. the Wolf
in "Red Riding Hood."
The Indian Wolves, C. pallipes, C. chanco, and C.
laniger, are hardly, if at all, different from C. lupus.
Professor Huxley has remarked upon the likeness of C. pallipes to
a Jackal, thus bridging over the very inconsiderable gap that may be held
to divide Jackals and Wolves.
The Dingo, Canis dingo, is an interesting and somewhat
mysterious species of Dog or Wolf. As is well known, it is an Australian
species; but it does not seem to be certain whether it was tamed and
brought over to Australia by the native races, or is a true and
indigenous Australian species.
The colour of this species varies, but is usually of a reddish brown;
it is, however, often grey and indeed almost black. Whether indigenous or
introduced, the Dingo is a plague to Australian settlers, devouring
Sheep, which it generally destroys by tearing out the paunch. It does not
as a rule hunt in packs. The Dingo is stated to feign death with so much
persistence that {422}an individual has been known to be partly
flayed before moving. Dingo remains have been found in river-gravels in
Australia where no human remains have been detected. This argues for its
indigeneity; but, on the other hand, it has been pointed out that man
himself in the Australian continent goes back a very long way into time,
and may thus still have imported this companion with him. Anyhow it is
quite a wild creature now. Dr. Nehring, an expert investigator into the
subject of domestic animals, has stated that the skeleton of the Dingo
does not suggest a feral animal at all but a purely wild race.

Fig. 211.—Dingo. Canis dingo. × ⅛.
The Domestic Dog is usually spoken of as Canis familiaris; but
to remains in bone caverns the name of C. ferus or C. mikii
has been given. There seems to be no doubt that the Dog was the "friend
of man" in very early times. Its remains have been met with in Danish
kitchen-middens, in the lake-dwellings of the Swiss lakes, and during the
Bronze Age in Europe generally. But "there are few more vexed questions
in the archaeology of natural history than the origin of the dog." Its
remains already referred to may in many cases have argued its use as
food. But in a Neolithic barrow a Dog was found buried with a woman, the
{423}skeletons of both being in situ;
this animal was about the size of a Shepherd Dog. The actual Dog of
to-day is divisible into more than 180 different breeds; but in a work
upon "Natural History" it would seem out of place to enumerate and
characterise these artificial products. Authors vary in their opinion as
to what stock gave rise to the domestic races of the past and of to-day.
The Jackal, the Bunasu (C. primaevus), the Indian Wolf (C.
pallipes), have been proposed as likely ancestors. It is more
probable that there is much admixture, and that various wild types have
been selected by man in various countries.
Extinct Canidae.—Many of the existing species of Canidae
are also to be found in Pleistocene deposits of the countries which they
now inhabit. A few show a wider range in the immediate past than in the
present. Thus Lycaon (L. anglicus) has been met with in
caves in Glamorganshire, while Icticyon of South America appears
to be congeneric with Speothos of the Brazilian caves. The African
Otocyon seems to occur in deposits in India. There are also
numerous extinct species belonging to the genus Canis, which
extend as far back as the Pliocene.
The earlier types of Dogs have been placed in different genera.
Cynodictis is an Eocene form from European strata. The skull is
decidedly Civet-like, with a short snout. The fore- and hind-feet were
five-toed, with well-developed pollex and hallux. The dentition was that
of modern Dogs, the molars being two in the upper and three in the lower
jaw. The general aspect of the creature and the form of the skeleton was
much like that of the Viverrine genus Paradoxurus, of which, as
well as of the Dogs, Cynodictis might have been an ancestor.
Simocyon of the Upper Miocene serves as the type of a separate
sub-family of Dogs, Simocyoninae. The skull is short, broad, and high;
the shortening of the skull affecting the jaws has reduced the teeth
greatly; the first three premolars are very small, fall out soon, and are
thus often deficient. There are only two molars in each jaw. This type is
of course nowhere near the ancestral Dog. It is a much-specialised branch
of an early type. Cephalogale is less specialised; there are the
usual four premolars. Enhydrocyon is an intermediate form; it has
lost one premolar in each jaw.
Amphicyon, forming the type of another sub-family,
Amphicyoninae, though usually placed among the Dogs, presents us with
{424}many Bear-like features in its
organisation. The feet, for instance, were plantigrade and five-toed. The
ulna and the radius are specially compared with the same bones in the
Bear tribe. The skull on the other hand is as distinctly Dog-like in
form. The molars are large, broad, and crushing, and Bear-like. The
largest known species, A. giganteus, is of about the size of the
Brown Bear. Amphicyon is a Miocene genus. Eocene and allied to it
is Pseudamphicyon. This genus has, like Amphicyon, the
complete dentition of forty-four teeth. In the Amphicyoninae generally
the feet are five-toed, the humerus has an entepicondylar foramen and the
femur a third trochanter. The upper molars are large.
The closely allied and American genus Daphaenus has also
plantigrade feet, and has in its structure many reminiscences of the
Creodonts. So, too, has the Eocene Uintacyon.
Cynodesmus is closely allied to Cynodictis. It has
ancient features combined with quite modern ones. The skull is described
as being Creodont-like, but the dentition is that of the microdont modern
Dogs. In accordance with its age the cerebral convolutions of this Dog
are much simpler than in existing Dogs, and the hemispheres do not cover
the cerebellum so much.
The Bear-like Carnivora or Arctoidea.—That division of
the Carnivora which is typically represented by the Bears embraces three
recent families, which are united by a number of characters. These
Carnivora are always plantigrade or nearly so. They have nearly always
five toes. The claws are not retractile, or at most semi-retractile as in
the Panda. In the skull the tympanic bulla is often depressed, and is not
so globular and obvious as in the Cats. Its cavity is not divided by a
septum. The paroccipital processes are not applied to it. The carnassial
tooth is less emphasised in this group than in the Cats.
These characters, however, have to be used with caution, as they are
hardly universally applicable. A fairly typical Arctoid bulla is seen in
such a form as Cercoleptes. The bulla itself is a little more
swollen than in Ursus, but it is flattened off in the same way
towards the bony meatus. The paroccipital processes, slightly developed,
are at a distance of ¼-inch from the posterior margin of the bulla. In
the Raccoon the bullae are much more swollen, and the paroccipital
processes are closer to them. In the Marbled Polecat, Putorius
sarmaticus, the bullae are fairly {425}swollen, and there is
but little flattening towards the meatus: the paroccipital processes,
though slight, are in contact with the bullae basally, though their free
tips are turned away from them. Finally, in Ictonyx the bullae are
much swollen; there is but little flattening towards the meatus, and the
paroccipital processes, themselves much swollen, are pressed closely
against the bullae. The Mustelidae, therefore, in this as in other
characters, approach the Aeluroids.

Fig. 212.—Section of the left auditory bulla and surrounding bones of a Bear (Ursus
ferox). am, External auditory meatus; BO, basioccipital; Car, carotid canal;
e, Eustachian canal; g, glenoid canal; Sq, squamosal; T, tympanic; t, tympanic
ring. (From Flower, Proc. Zool. Soc. 1869.)
There is no caecum, a feature which marks off the Arctoidea from all
Carnivora except the Viverrids Nandinia and Arctictis
(occasionally). The brain is characterised by the possession of what Dr.
Mivart has described as the "ursine lozenge," a tract about the middle of
the hemispheres, defined posteriorly by the crucial sulcus, and formed by
the emergence on to the surface of the brain of the hippocampal
gyrus.
The Arctoidea are very widely distributed. But there are some curious
exceptions. Thus there are no representatives of the group (as might be
expected) in the Australian region; they are completely absent from
Madagascar; while the true Bears (family Ursidae) are totally absent from
Ethiopian Africa, and are only represented by a single species, Ursus
ornatus, in the Neotropical region.
It is noteworthy that the Arctoidea never show spots or {426}cross stripes
(save rings on the tail), which are so common a feature of the coloration
of the Cat-like forms.
In bracketing together the three families which are described in the
following pages, emphasis is laid upon a number of undoubtedly common
features. Palaeontology seems, however, to suggest that the Mustelidae
come nearer to the Viverridae. That the Bears and Dogs are connected by
extinct annectent genera does not interfere with their present
distinctness.
The systematic arrangement of these Carnivora is not easy. It may be
useful, however, to give a method of arrangement for the convenient
placing of the genera.
The most primitive group is perhaps that of the true Bears, family
Ursidae; for in them the molars are two above and three below, and have
thus not become diminished in number as in some of the other members of
the order. Moreover, the Bears have lobate kidneys, which character,
often occurring in the young of animals which when adult have smooth
kidneys, may be looked upon as a primitive character. The feet
furthermore are completely plantigrade. This family will contain only
three genera, Ursus, Melursus, and Aeluropus.
Next comes the family Procyonidae, in several members of which one
molar is lost below, though in others the more archaic formula is
retained. The kidneys are simple. This family contains the American
genera Procyon, Nasua, Bassariscus,
Bassaricyon, Cercoleptes, and the Old-World form
Aelurus.
The third family, Mustelidae, has the molar formula reduced to 1/2 or
1/1. The kidneys are simple except in the Otters. To this family are
assigned the following genera:—Arctonyx, Conepatus,
Meles, Mephitis, Taxidea, Mydaus,
Mellivora, Helictis, Ictonyx, Mustela,
Galictis, Grisonia, Putorius, Gulo, and the
aquatic Lutra, Enhydris, and Aonyx.
Fam. 6. Procyonidae.—This family is mainly American in
range, the genus Aelurus alone being a native of the Old World.
But Zittel would include with the genera of this family the Viverrine and
Oriental genus Arctictis, a proceeding which is perhaps hardly
admissible, though the occasional absence of a caecum in that animal is
so far in favour of such an alliance. The largely vegetable nature of its
food and its arboreal habits cause a certain amount of likeness to some
of the members of the present group of Carnivores. The Procyonidae have
two {427}molars in either half of each jaw. The
carnassial teeth are not typically developed, and the molars are broad
and tuberculate. The tail is long, often prehensile, and often ringed in
the disposition of its colour pattern. The alisphenoid canal is absent
save in the aberrant Aelurus. Both condyloid and postglenoid
foramina are present. The members of this family are plantigrade.
Fig. 213.—Raccoon. Procyon lotor. × 1⁄5.
The genus Procyon includes at least two species of Raccoon, the
northern form, P. lotor, and the South American, P.
cancrivorus. To these may possibly be added a third, P.
nigripes. This genus is characterised by the length and the mobility
of the fingers, and indeed it uses its hands greatly. It has no median
groove upon the muzzle, which is found in many other Arctoids; the ears
are moderately large; the tail is not long, being about one-third of the
entire length of the animal, including the tail. The soles of the feet
are naked. Its limbs are very long (for an Arctoid), and this gives to
the animal a bunched-up appearance when walking. There are four premolars
and two molars on each side of each jaw. There are fourteen pairs of
ribs, of which ten pairs reach the sternum. The latter is composed of
nine pieces.
The first-named species has received its name from the fact—of
which there is abundant proof—that it dips its food into water. As
a matter of fact, the animal frequents the margins of streams, and hunts
in the shallow water beneath stones for {428}crayfish, and it also
captures fish. Not only is this animal partially aquatic, but it can
climb well—"they make their homes in trees, but carry on their
business elsewhere." The animal can be readily tamed, but is a tiresome
pet on account of its insatiable curiosity and its skill in the use of
its hands, which enables it to unlatch doors and generally to pry about
everywhere. The Raccoons are mostly nocturnal creatures.
The genus Bassaricyon[285] includes two species, both American,
B. alleni being from Ecuador, and B. gabbii from Costa
Rica. They have so much the aspect of a Kinkajou that a specimen, which
arrived at the Zoological Gardens, was presented and entered as one of
those animals. Nevertheless there are many differences between the two
genera. The tail of Bassaricyon is not prehensile, and the animal,
as will be seen from Fig. 214, has a sharper snout; the brain is more
like that of Bassariscus. The likeness to Cercoleptes can
hardly perhaps be regarded as an example of "mimicry" since the forms are
so nearly related, and the advantage of such an imitation remains to be
proved. The muzzle of Bassaricyon is grooved; the ears are fairly
large; the soles of the feet are naked; there is but a single pair of
teats. There are two molars and four premolars to each half jaw.
Fig. 214.—Bassaricyon. Bassaricyon alleni. × 1⁄5.
The dorsal vertebrae are thirteen in number; nine of the ribs reach
the sternum. The slenderness and convexity of the lower margin of the
lower jaw, as well as the feeble angular process, distinguish this genus
from its undoubtedly near ally Cercoleptes. The dental formula
also is different.
Bassariscus has a ringed tail like a Raccoon, and is also
American in range; it furthermore agrees with the Raccoon in {429}being
nocturnal and mainly arboreal in habit. There are apparently three
species, of which B. astutus is the best known, having been on
several occasions exhibited at the Zoological Society's Gardens, the last
examples so lately as 1900. The animal was for a long time believed to be
allied to the Oriental Paradoxures, and its occurrence in America was
therefore puzzling. The real affinities of the creature were, however,
definitely set at rest by Sir W. Flower, and later accounts of its
anatomy have confirmed this opinion.[286] The vertebrae are more numerous than
in Procyon, and the teeth are slightly different; otherwise it
presents many likenesses to its nearest ally. The ears are long; the nose
is grooved; and the palms and soles are naked.
Fig. 215.—Cunning Bassarisc. Bassariscus astutus. × 1⁄5. (From Nature.)
The Kinkajou, Cercoleptes, is likewise an American Arctoid. It
ranges from Central Mexico down to the Rio Negro in Brazil. It was at one
time confounded, and, considering its external appearance, not
unnaturally, with the Lemurs. Sir R. Owen dispelled this view by a
careful dissection of the creature. Nevertheless, there are certain
anatomical features in which it differs {430}from Carnivora and
resembles Lemuroids.[287]
It has been pointed out that the form of the lower jaw "much resembles
that of the Lemuroid Microrhynchus." There is, however, no doubt
that it is rightly placed in the present group. The tail is very
prehensile, and the animal is therefore, as might be supposed from this
circumstance, purely arboreal. It has some twenty-eight vertebrae. This
genus has a median groove upon the nose. The claws are long and sharply
pointed, and the palms and soles of the feet are naked. The premolars are
three, the molars two. There are fourteen dorsal vertebrae, of which nine
are united to the nine-jointed sternum by ribs. There is but one species,
C. caudivolvulus, of a uniform yellowish-brown colour.

Fig. 216.—Kinkajou. Cercoleptes caudivolvulus. × 1⁄6.
Fig. 217.—Coati. Nasua rufa. × 1⁄6.
Nasua, the Coati, ranges from Texas to Paraguay, and has two
species. In Guatemala it reaches a height of 9000 feet on the mountains.
The nose is produced into a short and very {431}mobile proboscis, hence
its name. The native Mexican name for the creature is "Quanhpecotl."
The Coati is largely arboreal, and hunts iguanas in large bands, some
of them being on the trees and some on the ground beneath. It also grubs
up worms and larvae, for which purpose its long snout is suited. The
molars of the genus resemble those of Procyon.
There is not a median groove upon the nose. The palms and soles are
naked. Six teats occur. There are thirteen dorsal vertebrae. Nasua
nasica[288] and N.
rufa are the best known and perhaps the only species. The colour of
the fur varies a good deal, and has led to the use of other names for
supposed species.
Aelurus, the Panda, is a largish animal found in the
south-eastern Himalayas up to a height of 12,000 feet. It has a glossy
fur of a reddish colour, and a "white somewhat cat-like face." The molar
formula which distinguishes it from the New-World Arctoids belonging to
the Procyonidae, as well as from its possible ally Aeluropus, is
Pm 3/4 M 2/2. The anatomy of the animal has been described by Sir W.
Flower.[289] Dr. Mivart
has pointed out that the muzzle though short is upturned in a way
distinctly recalling that of Nasua. The animal inhabits forests,
and feeds almost entirely upon vegetable food. It eats eggs, however, and
insects. Though living to a great extent upon the ground, it is also
arboreal, and has sharp semi-retractile claws. It is said to be dull of
sight, hearing, and smell, and yet with these disadvantages is also
unprovided with cunning or ferocity. Its habits have been compared with
those of a Kinkajou.
Fossil Procyonidae.—In addition to several of the
existing genera, the remains are known of various extinct forms of
Procyonidae. Leptarctus, with one species, L. primaevus, is
of Pliocene age, but is known only by one ramus of the lower jaw. It
appears to "offer a number of transitional characters between the more
typical Procyonidae and the aberrant Cercoleptes."[290]
Fam. 7. Mustelidae.—Contrary to what has been stated with
regard to the habits of the Procyonidae, the Mustelidae are for the most
part "bloodthirsty robbers," and are spread over the {432}whole surface
of the world, with the exception of Australia and Madagascar. The molar
teeth are generally reduced to one in the upper jaw, and sometimes to one
in the lower jaw, which thus gives "a sort of prima facie
resemblance to the feline dentition." There is no alisphenoid canal;
postglenoid and condyloid foramina are found.
Sub-Fam. 1. Melinae.—Of this sub-family there are
representatives both in the Old and New Worlds.
Fig. 218.—Badger. Meles taxus. × 1⁄6.
Meles, the Badger, is exclusively Palaearctic in range.[291] Dr. Mivart says that
Meles has a relatively longer dorsal region than any other
Carnivore, and that it is most nearly approached by its allies
Ictonyx and Conepatus. The molar formula is, as in
Arctonyx, Mydaus, and Helictis, Pm 4/4 M 1/2. The
molars differ from those of any other Carnivore in the much greater size
of the first molars than of the last premolars. The nose is not grooved;
the soles of the feet are naked. The claws of the fore-feet are much
longer than those of the hind-feet.
The genus Arctonyx is a "pig-like badger" from Hindostan,
Assam, and North China. The epithet "pig-like" is derived from the long
and mobile snout, which is truncated and has terminal nostrils. It is
remarkable for having a part of the palate formed by the pterygoids, as
in Whales and certain Edentata (e.g. Myrmecophaga). There are
sixteen dorsal vertebrae. A. collaris lives in {433}fissures of
rocks, or in holes dug by itself. It is a purely nocturnal beast.
The singular genus Mydaus, containing the species M.
meliceps, the Teledu or Javanese Skunk, is an inhabitant of Java and
Sumatra. It frequents the mountains of these islands, into the soil of
which it burrows in search of worms and larvae. There is but one species,
which is "like a miniature badger, of rather eccentric colours." It is
blackish brown, with a yellowish-white top to its head, and a stripe of
the same colour down the back. It may be distinguished by its elongated
snout, obliquely truncated, and with inferiorly-placed nostrils. As to
osteological characters, it has a more oblique symphysis of the mandible
than in any other Carnivore. The secretion of the anal glands is said to
rival that of the Skunk in offensiveness and in the distance to which it
can be propelled.
Sub-Fam. 2. Mustelinae.—Representatives occur in both the
Old and New Worlds; but the genera and even the species are in one or two
cases common to both.
Fig. 219.—Tayra. Galictis barbara. × 1⁄7.
Galictis barbara,[292] the Tayra, is a brown, elongated, and
Weasel-like animal from Mexico and South America. As is the case with the
Weasel, it is sometimes gregarious, a herd of twenty having been
observed. The soles of the feet are naked, and the molar formula is
Pm 3/3 M 1/2. In these characters the Grison (G. vittata) agrees
with G. barbara; but it has been referred to a different genus,
Grisonia.
The Grison, "this savage and diabolical-looking weasel," as {434}Mr. Aplin
terms it,[293] is known
also as the "Hurón." It almost rivals the Skunk in the power of the odour
which it can emit when enraged. A trapped specimen was placed in a cage
50 yards or so from the house, and even at this distance it was
disagreeably easy to tell when any one visited the animal—at least
when the wind set in the right direction. It is greyish yellow above and
blackish beneath, presenting, as has been remarked, a curious similarity
to the Ratel. The nose of this animal is destitute of a median groove,
which is present in the Tayra; the soles of the feet, however, are naked
as in that animal, and it is nearly plantigrade in walk. It differs also
from Galictis in having sixteen[294] instead of fourteen dorsal vertebrae.
Eleven of the ribs reach the sternum. Considering the differences that
exist between some other genera of Arctoids, it may be fairly allowed
that a genus Grisonia is tenable.

Fig. 220.—Grison. Grisonia vittata. × 1⁄7.
G. allamandi is darker coloured than the Grison, with a white
band from the forehead to the neck. Mr. T. Bell described a tame
individual as eating eggs, frogs, and even a young alligator.
A third genus of this group has recently been founded by Mr. Oldfield
Thomas[295] for a small
African animal, which is Grison-like in its coloration. The name given to
the genus, Galeriscus, is intended to suggest its likeness to the
Grison (Galera or Grisonia). The chief distinctive feature
of this genus, whose skeleton is not yet known, is the presence of only
four digits on each limb; the pollex and the hallux being entirely
absent. The ears of this Grison are short. {435}
The genus Mustela includes the Martens and Sables, which are
distinguished from the following genus by the molar formula, which is
Pm 4/4 M 1/2. The same character separates them from Galictis, and
also the generally hairy under surface of the feet. In more southern
latitudes, however, the palms are sometimes naked. The nose is grooved,
and the ears are short and broad. The genus is widely distributed, being
common to the Old and New Worlds. In the Old World it extends from Europe
to Java, Sumatra, and Borneo. The largest species of the genus is the
American Pekan, an animal which may be 46 inches in length, including the
tail. There are two species of Sable, one European (M. zibellina),
the other American.
The only British species of the genus is the Pine Marten, M.
martes. It is dark brown, with a brownish-yellow throat, and reaches
a length of some 17 inches, with an eight-inch tail. It is getting rare,
but is still fairly common in the Lake country. The animal is largely
arboreal in habit, whence the vernacular name. It is also called Marten
Cat. The allied M. foina, the Beech Marten, has been stated to be,
but apparently is not, an inhabitant of these islands. The colour of the
animal is a rich brown. It has small eyes and ears and a short tail. The
palms of the hands and the soles of the feet are hairy; the muzzle is
naked, and has a groove as in Cercoleptes, etc.
The Glutton, Gulo, is a well-marked genus, containing but one
species, which is circumpolar in range. The dentition is Pm 4/4 M 1/2.
The ferocity but not the voracity of this animal appears to have been
exaggerated. It mainly feeds on carcases, and is not really a successful
hunter. As to the carcases, Olaus Magnus tells in straightforward
language the way in which the animal dilates in size during a meal, and
presently, after following the practice of the ancient Romans, returns to
the banquet: "Creditur a natura creatum ad ruborem hominum qui vorando
bibendoque vomunt redeuntque ad mensam"!
This is one of the few land animals which ranges completely round the
pole. There is no difference to be noted between the Old-World and the
New-World specimens. It is now an entirely northern form, but in
Pleistocene times it reached as far south as this country. The fossil
species seems to be Gulo luscus, and to be quite indistinguishable
from the living forms.
Putorius, the genus which embraces the Weasel tribe, contains
{436}many species known popularly as Weasels,
Ermines, Stoats, Ferrets, Polecats, Minks, and Vison. Not only is the
genus common to both Old and New Worlds, but in a few cases the species
(e.g. P. erminea) range from Asia to America. The molar formula is
Pm 3/3 M 1/2. The form of the body is an exaggerated one, the length of
the trunk to the limbs being very great. The feet are more or less hairy
beneath, and the animals are digitigrade. The nose is grooved. The dorsal
vertebrae vary from thirteen to sixteen.
Fig. 221.—Polecat. Mustela putorius. × 1⁄6.
There are four British representatives of this genus:—
The Polecat, P. foetidus, is a dark brown-coloured animal. Its
total length is about 2 feet, of which the tail occupies some 7 inches.
It is a species banned by the gamekeeper, and hence is approaching
extinction in this country. It is excessively bloodthirsty, as are
apparently all the members of this genus, and kills out of mere
wantonness. The Ferret is simply a domesticated variety of the
Polecat.
The Stoat or Ermine, P. erminea, is reddish brown above, white
beneath. In winter, in certain localities, it becomes white with the
exception of the black tip of the tail. This colour-change bears some
relation to the degree of latitude. It is universal in the north of
Scotland, rare in the south of England. As is the case with some other
animals that generally change {437}their colour in the winter, there are
individuals which seem to have lost the power of change, and others which
change in an apparently capricious manner, not influenced by season or
cold. Like so many other animals, the Stoat appears at times to migrate,
which it does in large parties. Such parties are said to be dangerous,
and will attack a man who crosses their path.
The Weasel, P. vulgaris, has much the same colour as the Stoat,
but is a smaller animal; it differs also by undergoing no seasonal
change. It is equally agile and ferocious, and ought to be encouraged, as
it vents its ferocity largely upon Voles and Moles, which it can pursue
underground. Like other species of Putorius, it seems to kill its
prey by biting through the brain-case.
The fourth British species is the recently-described Irish Stoat,
P. hibernicus. It is somewhat intermediate between the last
two.
Poecilogale is a genus recently instituted by Mr. Thomas for a
small South African Weasel, P. albinucha, coloured like the
Zorilla, i.e. with whitish stripes upon black, but differing in
its reduced molar formula, which is Pm 2/2 M 1/1 or 1/2.
Lyncodon[296]
is thought to be more doubtful; it is South American (Patagonian), with
the same molar formula as the most reduced forms of the last genus,
i.e. Pm 2/2 M 1/1. The ears are short and almost invisible; the
claws of the anterior limbs are long, those of the hind limbs short. It
is not quite certain that it is not "an aberrant southern form of
Putorius brasiliensis." That its distinction is justifiable
appears to be shown by the discovery in the same region of a fossil
species, L. luganensis. Matschie places it near
Galictis.
The Ratel, Mellivora, is common to India and West and South
Africa. It is a black animal with a grey back and grey on the top of the
head, the contrast of colour suggesting a dorsal carapace. It runs with a
swift trot. The animal lives much on the ground, but can climb trees. It
is exclusively nocturnal in its habits. It has the reputation in India of
feeding upon dead bodies, a view which has probably no foundation in fact
save that it can burrow. The molar formula is Pm 3/3 M 1/1. There are
fourteen dorsal vertebrae. The African and Indian species are {438}hardly to
be distinguished from each other. The ears are very minute. The tail is
short. The muzzle is rather pointed, and the soles and palms are
naked.
Fig. 222.—Ratel. Mellivora capensis. × ⅛.
The structure of Helictis has been described by the late
Professor Garrod,[297] as
well as by Sir W. Flower in his general account of the Carnivorous
skeleton. The animal, which is a native of East Asia, is sometimes gaily
coloured. H. subaurantiaca, the species dissected and figured by
Garrod, is a varied black and orange. The genus is arboreal, and the tail
may be moderately long and bushy. The ears are small; the nose is
grooved; the palms are naked, but the soles of the feet are hairy. There
are fourteen dorsal vertebrae. The molar formula is Pm 4/4 M 1/2.
The Zorilla, Ictonyx, is the last of the Old-World genera of
Melinae. It is African, ranging from the tropical parts of the continent
to the Cape. "In colour and markings," remarks Dr. Mivart, "as well as in
the odour of the secretion of its anal glands, the one or two species
which form this genus resemble the skunks; so much so that did they
inhabit the same region, and were they devoid of an offensive secretion,
they would certainly be said to mimic the skunks." The molar formula of
the genus is Pm 3/3 M 1/2. There are fifteen dorsal vertebrae. The nose
is grooved and the soles partly hairy.
The American Badger, Taxidea, is a burrower of omnivorous
tastes, and correlated with the former habit are the immense {439}claws of
the fore-paws. It is North American, but gets into Mexico. The molar
formula is as in the American genera Mephitis and
Conepatus, and as in the Old-World Ictonyx, and it thus
differs from that of Meles. Besides the great size of the claws
upon the hand, which are larger relatively than those of any other
Carnivore, the genus Taxidea is to be distinguished from all
Arctoids (indeed, from all Carnivora) except Mydaus, by the fact
that the pelvic limb is of the same length as the pectoral. The muzzle is
furry except at the very extremity; this is grooved. The animal is
carnivorous, subsisting upon the following very varied kinds of
food—"Spermophiles, Arvicolas, birds' eggs, and snails, also
honey-comb, wax, and bees."
The Skunk, Mephitis, is an American animal with several
species, which range from North to Central America. The black-and-white
colour distinguishes the genus, which is furthermore marked by the fact
that the third digit of the hand is relatively longer than in any other
Carnivore except Taxidea. The soles are partly hairy. It is a
terrestrial fossorial animal with well-known powers of protecting itself
from aggression. But nevertheless the Skunk has its enemies, and is not
quite so unmolested as is sometimes popularly supposed. The Puma, Harpy
Eagle, and the Great Horned Owl will at least occasionally attack and
devour it. The molar formula is Pm 3/3 M 1/2. There are sixteen dorsal
vertebrae.
Conepatus is a more southern form of Skunk, extending down into
South America. Its dentition is like that of Mephitis save for the
loss of an upper premolar. This genus, which has been further subdivided,
differs from Mephitis in the fact that the soles of the feet are
wholly naked, whereas in Mephitis those of the hind-limbs are partially
hairy. It has no groove on the nose. Its tail is shorter than that of
Mephitis. This Skunk has the same habits as the last. In certain
parts of South America the animals are so abundant and their odour so
powerful that in the evening there is generally a recognisable smell
about. This is said to be good for the headache!
Sub-Fam. 3. Lutrinae.—Of this sub-family there are at
least two genera. Enhydris (Latax),[298] the Sea-Otter, is confined to the
shores of the North Pacific. It is more purely aquatic than are {440}other
Otters. Specimens have been seen swimming fifteen miles from land. The
gait of the creature when on land is suggestive of a marine animal; the
webbed hind-feet are doubled back upon the knuckles during progression
upon land, and locomotion is effected by a series of short springs from
these feet; the Otter does not walk "in ordinary acceptance of the term."
The tail is flattened, being twice as broad as it is thick, and ends in a
bluntish point. Enhydris feeds mainly upon crabs and sea-urchins,
but also upon fish. Its dental formula is peculiar by reason chiefly of
the reduction of the lower incisors. The formula runs as follows: I 3/2
C 1/1 Pm 3/3 M 1/2.
The molar teeth of this creature, in accordance with its diet, have
lost the sharp points of the Mustelidae in general; the crowns are
flattened, and the tubercles very blunt. In this it contrasts with
Lutra, and presents some resemblance to the Crab-eating Raccoon,
Procyon cancrivorus; but the teeth are still further blunted.
Enhydris feeds largely upon sea-urchins and shell-fish, and needs
blunt teeth for the crushing of the hard shells of its prey. It is
interesting to notice that the habits of this animal have been altered by
the interference of man. The creature has been hotly pursued for a long
time on account of its valuable fur. Instead of feeding and breeding upon
the shore in places readily accessible to its pursuers, the Sea-Otter has
now taken to the open sea in a greater degree. It utilises masses of
floating seaweed for those purposes, and hunts for its food in the deeper
water at a greater distance from the shore. In conjunction with the
increasing rarity of the Sea-Otter the price of its skin has enormously
increased: whereas in 1888 the average price per skin was £21:10s., the
value of a fine skin now is at least £100, and as much as £200 and even
£250 has been given. The animal is captured by netting and by clubbing
and spearing.[299] From
the Miocene Siwalik beds remains of an allied form, Enhydridon,
have been obtained, whose teeth are somewhat intermediate in their crowns
between Lutra and Enhydris.
Lutra, including the Otters, is widely distributed. Both manus
and pes are webbed. The ears are small and hairy. The nose is not
grooved, and the naked part is very circumscribed; {441}the claws upon the
hind-feet are flattened and somewhat nail-like. There are about ten
species, but of course, as is so universally the case, a great many more
names have been given. The molar formula is like that of Enhydris
save that there is an extra premolar in the upper jaw. There are fourteen
pairs of ribs, of which eleven pairs reach the ten-jointed sternum. The
caudals are twenty-three. The Cape Otter, the "clawless" Otter, has been
separated as a genus Aonyx. So too has the South American
Pteronura brasiliensis. But in neither case is the separation
allowed by Mr. Thomas in a recent revision of the genus.[300] The latter species has the reputation
of being very fierce, and is known in Uruguay by the name of "Lobo de
pecho blanco." The British species, L. vulgaris, reaches a length
of 2 feet or so, with a tail of 16 inches; it ranges over the whole of
Europe and a large portion of Asia. This Otter often burrows in the banks
of the streams which it frequents; and in the burrow in March or April
the female brings forth her young, three to five in number. It will also
frequent the sea-coast.

Fig. 223.—Otter. Lutra vulgaris. × 1⁄6.
Fossil Mustelidae.—Besides a number of the existing
genera there are fossil members of this family which cannot be referred
to existing genera. These latter extend back into time as far as the
Eocene. Stenoplesictis, one of these Eocene forms referable to the
sub-family Mustelinae, is to be distinguished {442}from living Mustelines
by its comparatively long legs. In this genus as in several others there
are two upper molars.
Fam. 8. Ursidae.—This family is nearly universal in
distribution, and consists of but three genera, Ursus,
Melursus, and Aeluropus.
Ursus has the palms and soles naked except in the Polar Bear,
which needs a furry sole to walk with ease upon ice surfaces. The ears
are fairly large, and the nose may or may not be traversed by a median
groove.[301] The molar
formula[302] is Pm 4/4
M 2/3. The brain is naturally (because of the size of the animals of this
genus) richly convoluted. The lobate kidneys have already been mentioned
in defining this family (see p. 426).
Fig. 224.—Himalayan Bear. Ursus tibetanus. × 1⁄15.
A very large number of species of Bears have been described. But it is
the opinion of Mr. Lydekker[303] and of others that many of these are
really to be referred to the European Brown Bear; in this event the
Grizzly of North America, the Isabelline Bear, the Syrian Bear, a Bear
from Algeria, the Kamschatkan and Japanese Bears, besides the extinct
Ursus fossilis of Pleistocene caves, are to be regarded as slight
modifications of Ursus arctos. On the other hand, the great Cave
Bear, U. spelaeus, {443}and the Thibetan Blue Bear (U.
pruinosus) are distinct species, not to be confounded with U.
arctos. Neither, of course, are the Peruvian U. ornatus and
the Sun Bear, U. malayanus.
Fig. 225.—Malayan Bear. Ursus malayanus. × 1⁄12.
The Polar Bear has even been placed in a separate genus,
Thalassarctos, a proceeding which is quite unnecessary. The white
colour of this Bear tends to become browner with age. It is one of the
few mammals which extend right round the pole; the Polar Bear is of
course a purely Arctic animal. The chief food of the Polar Bear is Seal.
Out of thirty Bears examined, Mr. Koettlitz found that only fifteen had
animal remains in their stomachs, and these remains were invariably Seal.
The animal apparently hunts by scent rather than by sight or hearing,
both of which senses seem to be somewhat dull. The males and females
wander separately, except of course during the breeding season. The Bears
dig holes in which they may remain for some time, but there is no
hibernation. In Pleistocene times, the Polar Bear extended as far south
as Hamburg. The female has four mammae, pectoral in position.
Melursus includes only M. labiatus, the Sloth Bear of
India. This animal has an upturned snout, which is described as closely
resembling that of Mydaus, the Teledu. The snout has no groove.
{444}All Bears are largely vegetarian and
insect feeders; but this Bear is especially so. It delights in the nests
of Termites, and its energy in destroying these hills for the sake of
their inhabitants is so great that the name of "sloth" appeared to Sir
Samuel Baker to be an entire misnomer.
Aeluropus, a rare Carnivore with but one species, A.
melanoleucus, is not inferior in size to the Brown Bear, and is
distinguished by its largely white coloration. It was discovered in the
mountains of East Thibet by Père David, and described by Milne-Edwards[304] as a distinct and new
genus, the discoverer himself having named it as a species of
Ursus. It is a vegetable-feeding creature and bulky in form, with
a rudimentary tail and a short broad head; in fact, more like a Bear than
a Procyonid (with which group it is placed by some). The width of the
head, however, is greater than in any other Carnivore; it is most closely
approached in this by Aelurus and by Hyaena. The molar
formula is Pm 4/3 M 2/3. The soles are hairy. There is no alisphenoid
canal. The molars are especially large and multicuspid.
Fig. 226.—Aeluropus melanoleucus. × 1⁄12.
Fossil Ursidae.—The genus Ursus itself goes back
to Pliocene times. The well-known Cave Bear, Ursus spelaeus of
{445}Pleistocene times, was one of the
commonest of Carnivorous creatures during the very early times of the
present era. It was as huge as a Polar Bear or a Grizzly. The skull is
remarkable for the fact that the first three premolars, which are small
in all Bears, dropped out early in life. An immense number of names have
been given to what are in all probability the same species as this Cave
Bear of remote antiquity.
Hyaenarctos is the oldest genus of true Ursidae. It goes back
into Middle Miocene times, and ranged over Europe and North Africa.
Arctotherium is an American genus of Pleistocene times. The
likeness of some of the extinct Canidae to Bears has been already
commented upon.
{446}
CHAPTER XIV
CARNIVORA (CONTINUED)—PINNIPEDIA (SEALS AND WALRUSES)—CREODONTA
Sub-Order 2. PINNIPEDIA
This group includes the Seals, Sea-Lions, and Walruses,[305] all aquatic and, for
the larger part, marine creatures. Being aquatic they have to some extent
acquired a fish-like form, though not so completely as have the Whales
and even the Sirenia. This is most complete so far as the group is
concerned in the Seals, where the hind-limbs have become soldered to the
tail and are inefficient as walking legs, where the external ears have
vanished, and where the general shape of the body is tapering and thus
fish-like. The Walruses and Sea-Lions are less modified in this
direction; in the latter (not in the former) the external ear, though
small, is persistent, and the hind-limbs are capable of being used as
organs of progression upon dry land. The general characters applicable to
the Carnivora, given upon a previous page, apply to the Pinnipedia.
Fig. 227.—Skeleton of Seal. Phoca vitulina. (After de Blainville.)
The characters confined to the Pinnipedia as a whole are mainly
these:—The greater part of the limbs are
enclosed within the skin, the hands and feet are fully webbed, and there
is a tendency for the nails to disappear, and for the phalanges to
increase in number—characters which are clearly not diagnostic of
the order but correlated with an aquatic life, since they reappear, and
are indeed exaggerated, in the Cetacea. The teeth are peculiar in that
the milk dentition is feeble and is early shed. This, as it were, undue
emphasis upon one of the two sets of teeth is another likeness to the
Whales, {447}where, however, it is the milk dentition
that is most pronounced, the "permanent" being feeble and very early
shed. But the dentition of the Pinnipedes presents other likenesses to
the Cetacea, which are, it must be remembered, regarded by some as a
modification of the Carnivorous stock, in which case, of course, the
likenesses may be genetic rather than due to adaptation in the two cases.
There is a distinct tendency towards a homodont series, the grinding
teeth being often very simple, and the very distinct carnassial tooth of
many terrestrial Carnivores being absent. Finally, the number of the back
teeth shows some signs of being on the increase; and Professor Kükenthal
has found that this increase is due to the division of existing teeth.
Here is a point of likeness to the many teeth of the typical Toothed
Whales. Dr. Nehring found in several examples of Halichoerus
grypus the normal five back teeth increased to {448}six, and the additional
molar was at the end of the series, thus suggesting a lengthening of jaw
coupled with an increase in number of teeth.
The incisor teeth of the Pinnipedia differ from those of the land
Carnivora in that there are nearly always fewer than 3/3, at least in the
adult animal. In possessing lobulated kidneys the Pinnipedia differ from
all terrestrial Carnivores except the Otters and Bears—a
significant fact.
In the characters of the skeleton the Pinnipedia show many
peculiarities. The cranial part of the skull is proportionately to the
facial part greater than in terrestrial Carnivora; there is no lachrymal
bone, and the orbit is to some extent defective in ossification. The
alisphenoid canal, so important a feature in the Carnivora, may be
present or absent. It is present, for example, in Otaria jubata.[306] This genus also has
the more primitive small and rugged tympanic bullae, which are inflated
and more Cat-like in others. The vertebrae show an interesting Creodont
peculiarity in the complex interlocking arrangements of the zygapophyses
of the dorsal vertebrae. The ossicula auditus differ from those of their
terrestrial allies in their large size and massive growth. In this they
have come to be like those of the Whales and Sirenians.
Fig. 228.—Patagonian Sea-Lion. Otaria jubata. × 1⁄20.
There is no doubt about their close resemblance to the {449}terrestrial
Carnivora, but the question is, to which group of Carnivora have they the
most likeness. The semiaquatic Otter, and the still more thoroughly
aquatic (marine) Enhydris, suggest an affinity in that direction.
The long body and short legs of the Otter, which is more thoroughly at
home pursuing fish in the streams than in waddling clumsily upon the
banks of the streams, seem to require but little external change to
convert it into a small Seal, while the long and completely webbed hind
digits of Enhydris are even more like those of a Pinniped. The
Sea-Lions, in which the external ear has been preserved, and in which the
limbs have not become so entirely useless for progression on the land as
they have in the Seals, seem to be the intermediate step in the evolution
of the latter. This, however, is not the opinion of Dr. Mivart, who,
without definitely committing himself on the point, presents some
evidence for the assumption that the marine Carnivora are diphyletic.
This double origin, however, is not from two groups of the terrestrial
Carnivora. Dr. Mivart, in common with many others, holds that the
Pinnipedia as a whole are undoubtedly nearer to the Arctoidea than to
either of the two remaining sections of the sub-order. One of the most
striking structural characters in which they show this resemblance is the
brain; the peculiar Ursine lozenge, already treated of as so distinctive
a character of the Arctoidea, is repeated in the Pinnipedia.
There are, however, other points of likeness which seem rather to
point to a Creodont origin. Patriofelis is a genus that from more
than one side may be looked upon as a possible ancestor of these animals.
The Creodont peculiarity of the vertebrae has already been referred to.
It may be added that the facial part of the skull is small in
Patriofelis, which appears, moreover, to have had an alisphenoid
canal. A very remarkable resemblance lies in the structure of the
astragalus. This is not deeply grooved on the tibial facet as it is in
Fissiped Carnivora. This might be held to be an instance of degeneration
in the aquatic Seals, which do not use their limbs as walking organs. But
Professor Wortman[307]
has pointed out that in the Sea-Otter, which is entirely aquatic, the
groove exists and is plain. The likeness offered to the Seals by the
spreading feet of Patriofelis is noticed under the description of
that genus.[308]
{450}
Fig. 229.—Cape Sea-Lion. Otaria pusilla. × 1⁄16.
Fam. 1. Otariidae.—The family Otariidae[309] is no doubt the least modified of the
aquatic Carnivora. It is rational, therefore, to commence the survey of
the group with this family. They have preserved, as already noted, the
independence of the hind-limbs; the external ear is present, though
small; there is an obvious neck, and the nostrils are at the end of the
snout, as in terrestrial creatures generally. The nails are small and
rudimentary, save those upon the three middle digits of the foot. It is a
singular fact that among the Otaries the angle of the lower jaw is
"inflected as much as in any Marsupial." The literature relating to this
family is great, and it seems difficult to reconcile the very varying
opinions as to how many genera ought to be admitted. Mr. Allen arranged
the nine species which he allowed {451}in six genera; but more
generic names have been proposed. At the other extreme stands Dr. Mivart,
who speaks of only one genus, Otaria; of this genus the number of
species is by no means agreed upon. There can, however, be no doubt of
the distinctness of the Northern Fur Seal, O. ursina (the "Seal"
of commerce and the cause of international complications), of the
Patagonian Maned Sea-Lion, O. jubata,[310] of O. pusilla of the Cape, of
the Californian O. gillespiei, of O. hookeri from the
Auckland Islands, and of four or five others. The range of the genus is
wide, but is mainly Antarctic. It is usual to speak of "Hair Seals" and
"Fur Seals," the latter being the species which produce the "sealskin" of
commerce. The difference is that in the Fur Seals there is a dense, soft
under-fur, which is wanting in the other group. It is, however,
impossible to make this character the basis of a generic subdivision.
There is a Fur Seal, O. nigrescens, in South America as well as
the more widely-known northern form.
Fam. 2. Trichechidae.—This family contains but one genus,
Trichechus, the Walrus or Morse, or Odobaenus, as the more
correct term seems to be. It is a tiresome result of accurate conformity
with the rules of priority in nomenclature that the name
Trichechus should be applied to the Manatee. There is but one
species of Walrus, though it has been attempted to show that the Pacific
and Eastern forms are different. The animal is Arctic and circumpolar.
The Walrus is characterised by the enormous canines of its upper jaw,
which form the well-known tusks and reach a length of 30 inches. The
animal can progress on land like the Sea-Lions; but, as in the Seals,
there are no external ears, though there is a slight protuberance above
the meatus auditorius. The strong bristles upon the upper lip are as
thick as crow quills. The pectoral limb has nails, but these are small,
as in the Sea-Lions. The under surface of the manus has a warty pad,
which cannot but assist[311] in maintaining a foothold upon
slippery ice. The hind-limbs have longer nails, which are still
diminutive and subequal in size. There is no free tail. The liver of this
animal is much furrowed, but not so much so as in Otaria, though
more so than in Phoca. The kidneys are of course lobulate, as in
the other aquatic Carnivores. The milk dental formula appears to be I 3/3
C 1/1 Pm + M 5/4. In the adult the formula[312] is I 1/0 C 1/1 M 3/3.
{452}
Fig. 230.—Common Seal. Phoca vitulina. × ⅛. (From Parker and Haswell's Zoology.)
Fam. 3. Phocidae.—The true Seals have no external ears,
and the nostrils are quite dorsal in position as in other aquatic
animals, such as the Crocodile. There is obviously an approach to the
conditions characteristic of the Whales. The hind-limbs are useless for
locomotion on land. They are bound up with the tail, and form
functionally merely a part of the tail. In this family there are, at any
rate, eight genera.
Phoca and Halichoerus are not very wide apart from each
other. In both there are five well-developed claws on feet and hands.
They are British, and generally Arctic and temperate in range. For some
reason or other the late Dr. Gray placed Halichoerus in the same
sub-family with the Walrus! Phoca is not only marine, but is found
in the Caspian and in Lake Baikal. Their existence in those inland seas
is believed to be a vestige of a former connexion with the sea.
Halichoerus grypus is a large seal 8 feet in length when full
grown. Its colour is yellowish grey, with darker grey spots and blotches.
It is not uncommon on the shores of our islands, particularly of the
Hebrides and Argyllshire. The commonest Seal is Phoca vitulina,
not more than 4 to 5 feet long, and of the same spotted coloration as the
last. This Seal has, however, a much wider distribution, being Arctic as
well as British, American, and North Pacific. A curious fact about this
Seal is that it is not impatient of fresh water; not only will it ascend
rivers, but it will live in inland lakes. It is said to be especially
sensitive to musical sounds. P. hispida is British, but a rare
visitor to our islands. It is essentially an Arctic species. The Harp
Seal, P. groenlandica, is so called on account of a harp-shaped
black bar in the males, which starts at {453}the shoulders and
extends to the thighs. Like the other Seals mentioned, the young are
white when first born. As may be inferred from its scientific name this
species is also Arctic in range. It is also a rare visitor to these
shores.
The genus Cystophora is the only other genus of which there is
a British representative. It is called the Hooded Seal on account of an
inflatable sac upon the face, with which it is said to attempt to terrify
its enemies. The genus has an incisor less in each half of each jaw than
Phoca and Halichoerus. Its formula is I 2/1 while these
genera are both 3/2. C. cristata is a large species reaching a
length of 10 feet. The colour of the back is dark grey with deeper
coloured spots. A few individuals only have been recorded from our
coasts.
Stenorhynchus ( = Ogmorhinus) is an Antarctic genus. The
hind-feet are clawless. The incisors are 2/2. The molars have an
additional cusp, i.e. three in all.
The genus Leptonyx with but one species, L. weddelli, is
purely Antarctic in range. Like the last genus it has two incisors, and
has but rudimentary claws upon the hind-feet; the first and fifth toes
moreover are the longest. The genus chiefly differs from the last in the
simple conical crowns of the molars, which have not the additional cusps
of Stenorhynchus.
Ommatophoca is another Antarctic genus with but a single
species, O. rossi. In this genus the hind-feet have no claws, and
the first and fifth toes are longer than the others. The claws of the
fore-feet are rudimentary. The immense size of the orbits gives the name
to the genus. There are two incisors, and the molars are all very
small.
Monachus is a northern genus inhabiting the Mediterranean and
the Atlantic in the vicinity of Madeira and the Canary Islands. It has
rudimentary nails upon both pairs of feet. The first and fifth toe of the
hind-feet are longer than the others. As with the preceding genera, the
incisors are two in each jaw. The species are M. albiventer, the
Monk-Seal, and M. tropicalis, the Jamaica Seal.
Allied to Cystophora is the genus Macrorhinus, with
(possibly) two species, of which one is Antarctic, the other frequents or
frequented the coast of California. The incisors are two in the upper
jaw, and but one in the lower. The premolars are four and the molar one;
all the teeth are small and simple, but {454}have long roots. The
nose of the male has a dilatable proboscis. The southern Elephant Seal is
M. leoninus, and reaches a length of some 20 feet. It occurs on
the shores of Kerguelen and some other more or less remote islands. Its
habits have been studied and described by several observers, beginning
with Anson in the last century. The late Professor Moseley gave a good
account of this marine monster in his book on the voyage of the
"Challenger." When the animal is enraged, the end of the snout is
dilated; but when this happens there is no long and hanging proboscis
such as has sometimes been described. The inflation affects the skin on
the top of the snout, which thus rises rather upwards during inflation.
The inflated region, according to Mr. Vallentin, quoted by Mr. J. T.
Cunningham, is about 1 foot long in an individual of 17 feet. It has been
stated that this proboscis is a temporary structure, only appearing in
the breeding season; but recent observations have shown that this
statement is inaccurate; it persists all the year round. The males fight
greatly during the breeding season, and produce a roar which has been
compared to the "noise made by a man when gargling." The females and the
young males bellow like a bull. The males fight of course with their
teeth, literally falling upon one another with their whole weight. Mr.
Cunningham thinks that the use of the proboscis is to protect the nose
from injury; or that it may be merely the result of "emotional
excitement." In any case the Bladder-nosed Seal, Cystophora, is
undoubtedly protected from injury by the possession of a corresponding
hood. The nose is the most vulnerable place, and the existence of this
hood would stave off the effects of a blow in that region. Moseley,
however, has said of Macrorhinus that it cannot be stunned by
blows on the nose as other Seals can; but he attributes this, not to the
dilated snout, but to the bony crest on the skull, and to the strength of
the bones about the nose. This Seal crawls with difficulty on the land,
and as the animals move "the vast body trembles like a great bag of
jelly, owing to the mass of blubber by which the whole animal is
invested, and which is as thick as it is in a whale."[313] When lying on the shore, these
animals scrape sand and throw it over themselves, apparently to prevent
themselves from being {455}incommoded by the direct rays of the sun,
to the effects of which they are very susceptible. The Elephant Seal is
mild and inoffensive, unless enraged, and, of course, during the breeding
season.
Order VIII. CREODONTA.
This entirely extinct group of Mammalia may be thus characterised:—Small to large carnivorous mammals, with skull
on the whole like that of the Carnivora and with trenchant teeth; digits
with unguiculate phalanges; tail long; extremities usually with five,
sometimes with four digits. In the carpus a centrale is present, and the
scaphoid and lunar are separate. Interlocking of posterior dorsal and
lumbar zygapophyses very perfect. Brain small but convoluted.
This group, which corresponds with the Carnivora
Primigenia of Mr. Lydekker, is not easy to separate absolutely
from the existing and more especially from some of the extinct members of
the Carnivora Vera. They also come exceedingly
near the Condylarthra, the presumed ancestors of the Ungulata, and like
them begin in the earliest Tertiary deposits. Their likeness to the
carnivorous Marsupials has also been insisted upon; but it would seem
that the succession of teeth in the Creodonta is typically Eutherian.
The characteristics of the group may be exemplified by an account of
the genus Hyaenodon, after which some of the more important
deviations in structure shown by other genera will be referred to.
Hyaenodon is both American and European, and ranges through the
Eocene and the Upper Miocene. It is a much-specialised Creodont, and
therefore exhibits well the distinctive characters of the group. About a
dozen species have been described. One of the best-known is the American
H. cruentus, and the following description refers to it. The back
part of the skull is low and broad, and is compared by Professor Scott
(who has described this and other species) as being "somewhat like that
of an opossum."[314] The
whole skull is {456}long, and the top has a great sagittal
crest. The paroccipital processes are short and are closely applied to
the mastoid processes. The mesethmoid is larger than in the carnivorous
Marsupials, and the frontals are very large. The palate has a peculiar
structure; in most species the hinder ends of the palatines are separated
by a narrow fissure which broadens gradually, thus forming the posterior
nares. In H. leptocephalus the posterior nares are brought very
far back by the meeting of the alisphenoids. The presphenoid, contrary to
what we find in the Dog, for example, is chiefly concealed by the vomer,
which covers it. The mandible has a long and strong symphysis, and its
angle is not inflected. The fore-limb is described as being "weak when
compared with the modern Carnivora." The scaphoid and lunar are separate,
and there is a centrale. The teeth present us with nearly the typical
formula. There is only one molar missing in the upper jaw. The canines
are enlarged. It has been suggested from a consideration of its palate
that Hyaenodon was a semiaquatic animal; the deep cleaving at the
extremities of the phalanges seems to point in the same direction, since
they resemble in this the genus Patriofelis, which there are other
reasons to regard as aquatic. This latter genus has a fore-limb which is
very like that of the Pinnipedia, the digits are much spread out, and
would seem to have supported a kind of paddle. In any case it certainly
fed upon aquatic tortoises, for their remains have been found in its
coprolites. The name Limnofelis, also applied to what appear to
have been members of this genus, is suggestive of their habits.
Patriofelis, at least one species, seems to have been of about the
size of a Lion.
Mesonyx has a brain case which is actually smaller than that of
the Marsupial Thylacinus. The lachrymal bone is very large, and
extends a little way over the face, as is also the case with
Hyaenodon; this condition is also found in Insectivora and in
Thylacinus. The axis vertebra has a curiously-shaped spine, which
is very different from the hatchet-shaped process of that vertebra usual
in the Carnivora, but is not unlike what exists in the Arctoid genera
Meles and Mydaus. The limbs show much disparity in length,
and seem to argue a much-arched back when the creature progressed. The
carpus is stated to be strikingly like that of the Insectivora. There is
as in other Creodonts a separation between the scaphoid and lunar; {457}the
centrale appears to be present. The pelvis "is most like that of the
bear," the metacarpals and the tibia, and some other bones, resemble
those of the Hyaena. In fact this animal shows those combined characters
which are common in archaic forms.
{458}
CHAPTER XV
RODENTIA—TILLODONTIA
Order IX. RODENTIA[315]
Small to moderately large animals, furry, sometimes with spines. Toes
with nails of a claw-like character, or sometimes approaching hoofs.
Usually plantigrade, and only occasionally and partly carnivorous. Canine
teeth absent; incisors long and strong, growing from persistent pulps,
and with enamel only or chiefly on the anterior face, producing a
chisel-shaped edge; molars few (two to six), separated from the incisors
by a wide diastema. Caecum (nearly always present) very large, and often
complicated in structure. Brain, if not smooth, with few furrows, the
hemispheres not overlapping the cerebellum. Surface of skull rather flat;
orbits not separated from temporal fossae; malar bone in middle of
zygomatic arch; palate very narrow, with elongated incisive foramina;
articular surface for lower jaw antero-posteriorly elongated. Clavicles
generally present. Testes generally abdominal. Placenta deciduate, and
discoidal in form.
The Rodents are a very large assemblage of usually small, sometimes
quite minute, creatures, embracing an enormous number of living generic
types. They are distributed all over the world, including the Australian
region, and, being small and often nocturnal, and by no means particular
in their diet, have managed to thrive and multiply to a greater extent
than any other group of living mammals. They are chiefly terrestrial
creatures, and often burrow or live in ready-made burrows. {459}Some, however,
such as the Voles, are aquatic; others, e.g. the Squirrels, are
arboreal, and there are "flying" Rodents exemplified by the genus
Anomalurus. Their range of habitat is in fact as wide as that of
any other Order of mammals, and wider than that of most.

Fig. 231.—Side view of skull of Cape Jumping Hare
(Pedetes caffer). × 3⁄5. AS, Alisphenoid; Ex.O,
exoccipital; Fr, frontal; L, lachrymal; Ma,
malar; Mx, maxilla; Na, nasal; OS, orbito-sphenoid;
Pa, parietal; Per points to the large
supratympanic or mastoid bulla; PMx, premaxilla;
Sq, squamosal; Ty, tympanic. (From
Flower's Osteology.)
The most distinct anatomical characteristic of the Rodents concerns
the teeth. They are without exception entirely deprived of canines. Thus
there is a long diastema between the incisors and the molars. Another
peculiarity is, that in many cases the dentition is absolutely
monophyodont. In such forms as the Muridae there seems to be no milk
dentition at all. In that family there are only three molars; but in
other types where there are four, five, or six molars, the first one,
two, or three, as the case may be, have milk predecessors, and may thus
be termed premolars. This has been definitely proved to be the case in
the common Rabbit, which has the unusually large number of six grinding
teeth in each half of the upper jaw when adult. The first three of these
have milk forerunners. On the other hand the existence of four molars
does not apparently always argue that the first is a premolar; for Sir W.
Flower found that in Hydrochoerus,[316] none of the teeth had any
forerunners, at any rate so far as could be detected from the examination
of a very young animal. The Rabbit appears to be also exceptional, in
that the second incisor of the upper jaw and the incisor of the lower jaw
have milk forerunners. In any case the tendency towards monophyodontism
is peculiarly well-marked in this group of mammals. The incisors of
Rodents are as a rule in each jaw a single pair of long and strong teeth,
which grow from persistent pulps, and {460}grow to a very great
length, extending back within the jaw to near the hinder part of the
skull. These teeth are reinforced in the upper jaw by a small second pair
in the Lagomorpha only. The incisors are chisel-shaped, and often brown
or yellow upon the outer face, as is the case also with some
Insectivores. This peculiar shape, and their strength, renders them
especially capable of the gnawing action which characterises the Rodents.
It has been pointed out that where the incisors are wider than thick, the
gnawing powers are feebly developed; and that on the contrary, where
these teeth are thicker than wide, the animals are good gnawers. The
incisors have often an anterior groove, or it may be grooves.

Fig. 232.—Molar teeth of Rodents. A, of Capybara (Hydrochoerus); B, of Squirrel
(Sciurus); C, of Ctenodactylus. (After Tullberg.)
The cheek teeth vary in number from two (Hydromys) to six
(Rabbit) on each side of the two jaws. Four is the prevailing number
outside the large division of the Rat-like Rodents. They are often set at
an angle to the horizontal plane of the jaw, looking outwards and
obliquely to its longitudinal axis; the individual teeth too are not
unfrequently bowed in form, reminding us of those of Toxodon. This
of course only occurs in those genera which have hypselodont teeth. The
pattern of the teeth varies much, and the different forms recall the
teeth of more than one other group of mammals. They are either bunodont
or lophodont. In many cases the tooth is encircled with a ridge of
enamel, which is either almost simple or has a more complicated contour;
such teeth are by no means unsuggestive of the Toxodonts. Some of the
lophodont molars are by no means unlike those of the Proboscidea. In
Sciurus vulgaris the {461}encircling ridge is broken up into
tubercles, which gives to the tooth a striking likeness to those of
Ornithorhynchus. Other genera have teeth like those of many
Ungulates. It has been shown by Sir J. Tomes[317] that the minute structure of the
enamel differs in different groups of Rodents.
The skull shows certain primitive characters. In the first place there
is no distinction between the orbital and the temporal fossa.[318] The sutures between
the bones retain their distinctness for very long. Other characteristic
features are the following:—The nasals are
large, and so are the paroccipital processes. The palate in front of the
molars is not distinct from the sides of the skull, its edge gradually
becoming rounded off above. It is also very narrow. The premaxillae are
large in relation to the great incisors. There is often a very much
enlarged infra-orbital foramen through which passes a part of the
masseter muscle. The jugal bone lies in the middle of the zygomatic arch,
which is complete and enormously enlarged in the Spotted Cavy
(Coelogenys paca). As in many Marsupials, the jugal bone sometimes
extends backwards to the glenoid cavity, where the lower jaw articulates.
It is usually said with an absolute want of accuracy that the cerebral
hemispheres of the Rodents are smooth and without convolutions. This
error has been repeated again and again in text-books. As a matter of
fact the cerebral hemispheres of many forms are quite well convoluted,[319] the degree of
furrowing corresponding, as in so many groups of mammals, with the size
of the animal. This at any rate is generally true, though the large
Beaver with its scant convolutions is an exception. The smaller forms,
such as Mus, Sciurus, Dipus, and Cricetus are
quite smooth-brained. The best furrowed brain of any Rodent which has
been examined is that of the huge Hydrochoerus. The Sylvian
fissure is very generally not pronounced; but is particularly well-marked
in Lagostomus. In all, or in most, Rodents the hemispheres are
separated by an interval from the cerebellum, the optic lobes being
visible between the two.
The mouth cavity of this group of mammals is divided into two chambers
by a hairy ingrowth behind the incisors; this arrangement is useful for
animals which use their strong incisors {462}as gnawing and
excavating tools as well as for the purposes of alimentation; for it
allows of substances being gnawed away without the products of the
chisel-like action being taken into the hinder cavity of the mouth. The
Rodents have for the most part a simple stomach of normal form; but in a
few this is complicated by a marked constriction, which divides the
cardiac from the pyloric portions. The Hamster, for example, is thus
characterised. In all the members of the order, with the exception of the
Dormice and some allied forms, the caecum is large and often sacculated.
In some forms (e.g. Arvicola, Myodes, Cuniculus) the
large intestine is coiled upon itself in a spiral way—a state of
affairs strongly suggestive of Ruminants.
The Rodents are primarily divisible into two great groups, the
Simplicidentata and the Duplicidentata, characterised mainly by the upper
incisor teeth. In the former there is but one pair of these teeth; in the
latter a second smaller pair lie behind the former.
Sub-Order 1. SIMPLICIDENTATA.
Section 1. Sciuromorpha.
The Anomaluri are separated by Thomas and others from this section as
an equal section, while by Tullberg they are grouped with
Pedetes.
Fam. 1. Anomaluridae.—The genus Anomalurus
suggests at first sight the Flying Squirrels of Asia, Pteromys. It
is, however, an entirely African genus, and is to be distinguished from
the Asiatic Rodents by a series of scales at the root of the tail,
imbricated, keeled, and forming possibly a "climbing organ." This
character serves also to distinguish the present genus from
Sciuropterus. The cartilage, moreover, which supports the patagium
springs from the elbow. There are four molars in each half of each jaw.
The eyes and ears are large. There are five fingers and toes, but the
thumb is small, though provided with a nail. The sternum has seven
joints, and nine ribs reach it. The clavicle is strong. Huet, who has
recently monographed the genus,[320] allows six species. The species vary
in size. {463}
Anomalurus peli appears, according to Mr. W. H. Adams,[321] to be a common species
on the Gold Coast; it is coloured black and white, but in spite of the
warning which this colour should convey, is considered by the perhaps
rather omnivorous native as "the greatest delicacy." The animal is
nocturnal, but affects only bright moonlight nights. Their "flying"
consists of a jump from a high branch to a lower one, after which they
reascend the tree to a point of vantage for another jump. They are said
to feed upon nuts; but Tullberg only found the remains of leaves in the
stomach.
Idiurus is a lately-described genus allied to
Anomalurus. There are at any rate two species, I. zenkeri
and I. macrotis. The thumb is more reduced than in
Anomalurus, and the fibula, contrary to what is found in that
genus and in most Sciuromorphs, is fused with the tibia below.
A third genus, very recently described and allied to both the
foregoing, is Aëthurus. It is a native of the French Congo,[322] and differs by the
absence of flying membranes. It has, however, the pad of large scales.
There is but one species, A. glirinus. It has a black bushy tail.
The postorbital processes of the frontals are totally wanting—there
are not even the traces to be seen in Anomalurus. The thumb has
vanished. If we are to compare Anomalurus with the Squirrels then,
thinks Mr. de Winton, the present genus is probably diurnal by reason of
the want of flying membranes.
Fam. 2. Sciuridae.—The Squirrels, genus Sciurus,
are world-wide in range, the Australian region and the island of
Madagascar being alone excepted.
The eyes and the ears are large; the tail is of course long and bushy.
The fore-feet have an inconspicuous thumb; the hind-feet have four toes.
The soles of the fore-feet are naked or furry, those of the hind-feet are
hairy. There are twelve or thirteen dorsal vertebrae, and in
correspondence seven or six lumbars. The caudal vertebrae may be as many
as twenty-five. In the skull the frontals are broad, and there are long
{464}postorbital processes. The infra-orbital
foramen is, as a rule, not large, but is increased in size in a few
forms. The number of separate pieces of bone in the sternum is five. The
molars of the upper jaw are five, but the first is very small and soon
drops out.
The Squirrels are often rather brilliantly coloured. The Chinese
Sc. castaneiventris has grey fur with a rich chestnut-coloured
under surface. The Malabar Squirrel, Sc. maximus, as its name
implies, a large animal, has a deep reddish or chestnut-coloured fur
above, which becomes yellow below. The "Common Squirrel," "the lytill
squerell full of besynesse," which is the Squirrel of this country, is
brownish red on the upper parts and white below. It ranges from this
country as far east as Japan. Like many other Rodents the Squirrel likes
animal food and will eat both eggs and young birds. "Camel's hair"
brushes are made from this animal. The genus Tamias, almost
exclusively North American in range, is included by Dr. Forsyth Major[323] in this genus, which
then consists of considerably over one hundred species.
The Ethiopian Ground Squirrels, genus Xerus, have a more
elongated skull than Sciurus, and the postorbital processes are
shorter. The feet are not hairy.
Nannosciurus forms a perfectly distinct genus of Squirrels.
These "Pygmy Squirrels" differ in possessing a very elongated "face" and
in the very broad frontal region. The teeth are unlike those of
Sciurus in certain features, and have been especially compared by
Forsyth Major to those of the Dormice. Four species of this genus are
Malayan; one is West African.
The Bornean Rheithrosciurus macrotis is the only species of its
genus. The genus may be distinguished by the exceedingly brachyodont
molars, this feature being more marked in this genus than in all other
Squirrels. It is called the "Groove-toothed Squirrel," from the "seven to
ten minute parallel vertical grooves running down the front face of its
incisors."[324]
The genus Spermophilus includes a large number (forty or so) of
Palaearctic and Nearctic animals known as Sousliks. The ears are small;
there are cheek pouches as in Tamias. The general aspect of the
animal is like that of a Marmot, and they bridge over the exceedingly
narrow gap which separates the Marmots from the true Squirrels.
Anatomically the skull is like that of {465}Arctomys; the
molars are five in the upper and four in the lower jaw. The caecum is
relatively speaking very small; the measurements in a specimen of S.
tredecimlineatus, dissected by Dr. Tullberg, were: small intestine,
580 mm.; large intestine, 170 mm.; and caecum, 27 mm. In Tamias
also the caecum is not greatly developed. These animals are burrowing in
habit.
The Prairie-dogs, genus Cynomys, of which the best-known
species is perhaps C. ludovicianus, are very like the Squirrels,
but they are not arboreal creatures; they live in burrows on the ground,
as their vernacular name denotes. The genus is entirely North American,
and four species have been differentiated.
The Prairie-dog or Prairie-marmot is some 10 inches to one foot in
length. The tail is no more than 2 inches. The ears are very small; the
thumb is fully developed and bears a claw. The measurements of the
various sections of the intestine are the following:—Small intestine, 860 mm.; large intestine, 690
mm.; caecum, 75 mm. Thus the caecum is not large comparatively speaking.
These animals dig burrows on grassy plains which they share with the
Ground Owl (Speotyto cunicularis) and with Rattlesnakes, all three
species appearing to live in perfect amity. Probably the Owls use the
conveniently-constructed burrows, and the Rattlesnakes come there to look
after the young of both.
Fig. 233.—Long-tailed Marmot. Arctomys caudatus. × 1⁄7.
Closely allied to the last are the Marmots, genus Arctomys.
They differ in the rudimentary character of the thumb and in the longer
tail. The eyes and ears are small. The distribution of the genus is
Nearctic and Palaearctic. There are ten species of {466}the genus. The Alpine
Marmot, A. marmotta, is familiar to most persons. The animal lives
high up in the Alps, and when danger threatens it gives vent to a shrill
whistle. It hibernates in the winter, and as many as ten to fifteen
animals may be found closely packed together in a single, carefully-lined
burrow.
The only other European species is A. bobac, the Siberian
Marmot, which occurs in the extreme east of Europe, and is also Asiatic.
There are four North American species, including the Quebec Marmot, A.
monax.
Fig. 234.—Flying Squirrel. Pteromys alborufus. × 1⁄5.
The genus Pteromys (of which the proper name, antedating
Pteromys by five years, appears to be Petaurista) is
confined to the Oriental region, where there are a dozen species or so.
The limbs are united by a parachute extending to the toes, and supported
anteriorly by a cartilage attached to the wrists. There are also
membranes anteriorly uniting the fore-limbs to the neck, and posteriorly
uniting the hind-limbs to the root of the tail and a trifle beyond. The
skull and the dental formula are as in Sciurus, but the pattern of the
molars, which is much complicated, seems to argue a different mode of
nutrition. There are twelve pairs of ribs. The large intestine (in P.
petaurista) is very nearly as long as the small, and the caecum is
also "colossal"; the measurements in an individual of the species named
were: small intestine, 670 mm.; caecum, 320 mm.; large intestine, 650 mm.
{467}The caecum is disposed in a spiral. The
teats are three pairs, non-inguinal in position.
The size of these squirrels is 16 to 18 inches exclusive of the tail,
which is longer. These animals can make an exceedingly long jump with the
help of their flying membrane. Nearly eighty yards is the longest
distance given for these aerial excursions. It is stated that they are
able to steer themselves to some extent while in the air. As appears to
be the case with so many Rodents, these animals feed largely upon beetles
and other insects, besides bark, nuts, etc.
The allied genus Sciuropterus has a much wider range. It
extends into the Palaearctic region and into North America, besides being
found in India. There is here no membrane reaching to the tail. The palms
and soles are furry. The caecum is very much shorter, and so is the large
intestine. The latter, in S. volucella, is not more than one-third
of the length of the small intestine. In other features there are no
remarkable differences in structure, save that the mammae, always three
pairs, may be inguinal.
Of the genus Eupetaurus[325] but a single species is known, which
is limited to high altitudes at Gilgit and perhaps in Thibet. Its
principal difference from the other genera of Flying Squirrels is that
the molars are hypselodont instead of brachyodont. The interfemoral
membrane is rudimentary or wanting. The one species is E.
cinereus. It is thought to live "on rocks, perhaps among precipices."
Dr. Tullberg attributes the hypselodont teeth to the fact that the mosses
upon which it is believed to feed may have much sand and earth
intermingled, which would naturally lead to a more rapid wearing away of
the teeth, and hence a need for a good supply of dental tissue to meet
this destruction.
Fam. 3. Castoridae.—This, the third family of the
Sciuromorpha, contains but one genus, Castor, the Beaver, with at
most two species, one North American, the other European. This large
Rodent has small eyes and ears, as befits an aquatic animal, and the tail
is exceedingly broad and covered with scales; the transverse processes of
the caudal vertebrae, in order better to support the thick tissues lying
outside them, are divided in the middle of the series into two. The
hind-feet are much larger than the fore-feet, and are more webbed than in
any other aquatic Rodent. {468}
In the skull the infraorbital foramen is small as in Squirrels. The
postorbital process has practically vanished. The four molars stand out
laterally from the jaws. The incisors, as might be surmised from the
habits, are particularly strong. The stomach has near the entrance of the
oesophagus a glandular patch, which seems to be like that of the Wombat
(see p. 144). In both sexes the cloaca is very
distinct and comparatively deep.
The two species of the genus are C. canadensis and C.
fiber. The latter is of course the European species, which is now
found in several of the large rivers of Europe, such as the Danube and
the Rhone. But it is everywhere getting scarce, and limited to quite
small and isolated colonies.
In this country it is absolutely extinct and has been since before the
historic period. There is apparently no documentary evidence of its
survival down to this period. But the numerous names of places which are
called from this animal illustrate its former prevalence. Examples of
such names are Beverley in Yorkshire, and Barbourne or Beaverbourne in
Worcestershire. In Wales, however, Beavers seem to have persisted longer.
But they were rare in the Principality for a hundred years or so before
the Norman Conquest. The king Howel Dda, who died in 948 A.D., fixed the price of a Beaver skin at 120 pence,
the skins of Stag, Wolf, and Fox being worth only 8 pence apiece. The
Beaver was called by the Welsh "Llost-llyddan," which means "broad-tail."
Its existence in the country is handed down in the name of
Llyn-ar-afange, which means Beaver lake. The last positive record of the
Beaver in Wales seems to be the statement of Giraldus Cambrensis that in
1188 the animal was still to be found in the river Teivy in
Cardiganshire. In Scotland the Beaver is said to have continued down to a
later date. Ireland it never reached. The remains of this animal by their
abundance show the former prevalence of C. fiber in this country.
It is known from the fens of Cambridgeshire, and from superficial
deposits elsewhere. The Thames formerly had its Beavers, and apparently
it was widely spread through the country generally.
The Beaver not only furnishes collars and cuffs for coats; it was
used, as every one knows, to provide hats. But the usefulness of the
animal by no means ended here in the eyes of our {469}forefathers. The Rev.
Edward Topsell observed that "for giving great ease unto the gowt the
skinnes of beavers burned with drie oynions" are excellent. Castorein as
a drug, if not in actual use, has quite recently been a part of the
pharmacopoeia. It is derived from the anal glands common to this and
other Rodents, and indeed many other mammals.
A large extinct form of Beaver is Trogontherium,[326] found in the
"Forest-bed" of Cromer. The skull is about one-fourth longer than that of
Castor. It has a less inflated bulla, and slightly more pronounced
postorbital processes than Castor. The third molar (fourth
grinding tooth) is relatively larger than in Castor, and has a
rather more folded crown. The foramen magnum is more triangular.
Fam. 4. Haplodontidae.—A separate family seems to be
required for the genus Haplodon, whose characters will therefore
be merged with those of its family. It is to be distinguished from most
other Squirrel-like creatures by the fact that there is no postorbital
process to the frontal. The molar teeth are five in the upper and four in
the lower jaw. The Sewellel, H. rufus, like the other species of
the genus (H. major), is found in North America west of the Rocky
Mountains. It has the habit of the Prairie-marmot, and has a short tail,
only moderately long ears, and five-toed feet. Tullberg is of opinion
that this animal nearly represents the ancestral form of the Squirrel
tribe.
Section 2. Myomorpha.
This subdivision of the Rodents contains, according to Mr. Thomas's
recent estimate,[327] no
less than 102 genera. It is therefore obviously impossible to do more
than refer to some of the more interesting of these. This group is again
divided into the following families:—
(1) Gliridae, including the Dormice.
(2) Muridae, the Rats, Mice, Gerbilles, Australian Water-rats,
Hamster.
(3) Bathyergidae, Cape Mole, etc.
(4) Spalacidae, Bamboo Rats. {470}
(5) Geomyidae, Pouched Rats.
(6) Heteromyidae, Kangaroo Rats.
(7) Dipodidae, Jerboas.
(8) Pedetidae.
The Gliridae have no caecum, so usual in the Rodentia. It is true that
all the genera have not been dissected, but it is known that in the true
Dormice, as well as in the genus Platacanthomys, a caecum is
absent.
Apart from these few exceptions the Mouse-like Rodents all possess a
caecum, though it is often not very large. They are all smallish animals,
and are modified to a great variety of habit and habitat. There are
burrowing, swimming, and climbing forms. The group is universal in range,
even including the Australian region, in which they are the only
Rodents.
Fam. 1. Gliridae.—This family, also called Myoxidae,[328] includes the Dormice,
and is entirely an Old-World family, absent only from the Malagasy
region. Its most important differential character is the total absence of
the caecum and of any sharp boundary between the small and large
intestine. The molars are usually four. The eyes and ears are well
developed.
The genus Muscardinus includes only the Common Dormouse, M.
arellanarius. This small creature, 3 inches long with a tail of 2½
inches, is, of course, a well-known inhabitant of this country. It is
also found all over Europe. It is not particularly abundant in this
country, and a good specimen is said to be worth half a guinea. As the
specific name denotes, it lives largely on hazel nuts; but it will also
suck eggs and devour insects. The animal makes a "nest" in the form of a
hollow ball. Its hibernation is well known, and has also given rise to
the German name ("Schläfer") of the group. It was well known to
Aristotle, who gave or adopted the name Ἐλειὸς for the animal.
Its winter sleep—suggestive of death—and its revivification
in the spring gave the Bishop of Salamis, Epiphanius, an argument for the
resurrection of man. The fur was reckoned in Pliny's time a remedy for
paralysis and also for disease of the ears.
The genus Myoxus includes also but a single species, M.
glis, the so-called "Fat Dormouse" of the Continent. It has no {471}glandular swelling at the base of the
oesophagus, such as occurs in the last genus and in Graphiurus. Of
Graphiurus there are thirteen species, all African in range. The
genus does not differ widely from the last. There is, however, a
glandular region of the oesophagus. Eliomys is the last genus of
typical Dormice. It is Palaearctic in range.
Platacanthomys, of a Dormouse-like form, has like other Dormice
a long tail, on which the long coarse hairs are arranged in two rows on
opposite sides towards the tip; it is represented by a single species,
P. lasiurus, from the Malabar coast. It is arboreal in habit. The
fur is mingled with flattened spines. The molars are reduced to three on
each side of each jaw. This form has been bandied about between the
"Mice" and the "Dormice"; but Mr. Thomas's discovery of the absence of
the caecum argues strongly in favour of its correct location among the
Gliridae. Typhlomys is an allied genus, also from the Oriental
region. This and the last are placed in a special sub-family of the
Gliridae, Platacanthomyinae, by Mr. Thomas.
Fam. 2. Muridae.—This family, that of the Rats and Mice
in a wide sense, is the most extensive family of Rodents. In it Mr.
Thomas includes no less than seventy-six genera. The molars are generally
three. The tail is fairly long, or very long, and the soles of the feet
are naked.
Sub-Fam. 1. Murinae.—The true Rats and Mice may be
considered to form a sub-family, Murinae. The genus Mus, including
the Rats and Mice in the limited sense of the word, contains about 130
species. They are exclusively Old World in range, being only absent from
the Island of Madagascar. In the New World there are no species of the
restricted genus Mus. The eyes and the ears are large; the pollex
is rudimentary, and bears a nail instead of a claw. The tail is largely
scaly. All the members of the genus are small animals, some quite minute.
In this country there are five species[329] of the genus, viz. the Harvest Mouse,
M. minutus, which has a body only 2½ inches long with an equally
long tail. It is the smallest of British quadrupeds with the exception of
the Lesser Shrew. The Wood Mouse, M. sylvaticus, is about twice
the size; it differs also from the last species in that it {472}frequents
barns, and is thus sometimes mistaken for the Common Mouse, from which,
however, it is to be distinguished by its coloration and longer ears. The
latter, M. musculus, is too familiar to need much description. A
curious variety of it has occurred. This has a thickened and a folded
skin like that of a Rhinoceros, and the hair has disappeared. The Black
Rat, M. rattus, is like a large Mouse, and is smaller and blacker
in colour than the "Hanoverian Rat." It is sometimes called the "Old
English Rat," but seems nevertheless to be not a truly indigenous Rodent.
It has been so defeated by competition with the Hanoverian Rat that it is
now not a common species in this country.
The Hanoverian or Brown Rat, M. decumanus, is a larger and a
browner animal than the last. It is very widely distributed through the
globe, no doubt largely on account of the fact that it is readily
transported by man. The same is the case with the Common Mouse, whose
real origin must be a matter of doubt. The original home of the Brown Eat
is thought by Dr. Blanford to be Mongolia. There is so far a
justification for the name "Hanoverian Rat" that the animal seems to have
reached this country about the year 1728. But there is no reason for
calling it, as is sometimes done, the Norway Rat.
Some members of this genus, whose fur is interspersed with spines, or
which are quite spiny, have been separated as a genus, Leggada,
which, however, is not generally allowed.
Closely allied again is Chiruromys, which has a strongly
prehensile tail, a feature which is not common among the Myomorpha,
though Dendromys, a tree-frequenting form, and Mus minutus,
already spoken of, show the same character. Many Mice seem to have
prehensile tails, which they can curl round branches; but it is not so
fully developed as in the species just named.
A number of other genera are referable to the true Mice, the
sub-family Murinae of Thomas's classification. The Syrian and African
Acomys has very spiny fur, so much so that "when it has its spines
erected it is almost indistinguishable at the first glance from a
diminutive hedgehog." The genera Cricetomys, Malacomys,
Lophuromys, Saccostomus, Dasymys are restricted to
the Ethiopian region. Nesokia is Oriental, reaching {473}also the
Palaearctic region. Vandeleuria, Chiropodomys,
Batomys, Carpomys are Oriental, the last two being confined
to the Philippines.
Another peculiar Philippine genus is Phlaeomys, of large size,
and allied to it is Crateromys, originally confounded with it.
Batomys granti is also confined to Luzon. Its molars are three,
like those of the also restricted and Philippine Carpomys
melanurus, which is an arboreal form. There is a second species,
C. phaeurus.
Phlaeomys is placed, however, by Mr. Thomas in a distinct
sub-family of its own, Phlaeomyinae, and is removed from the
Murinae.
Hapalomys, with but one species, is Burmese.
Pithecochirus is Javanese and Sumatran. Conilurus (also
known as Hapalotis) is a genus containing species which are termed
Jerboa Rats, on account of their mode of progression. They are desert and
Australian forms. There are sixteen species.
Fig. 235.—Spiny Mouse. Acomys cahirinus. × ½.
Mastacomys, with one species, is limited to Tasmania.
Uromys, with some eight species, is from Queensland, and inhabits
also the Aru Islands and the Solomon Islands. The Celebesian
Echiothrix, or Craurothrix as it should apparently properly
be called, is another genus containing but a single species.
Golunda is both Oriental and Ethiopian, one species occurring in
each region. The beautiful little striped Barbary Mice,
Arviacanthis (or Isomys), are African, north as well as
tropical.
The genus Saccostomus resembles the Hamsters in the presence of
cheek pouches. Its teeth, however, are Murine. It agrees with
Steatomys in the comparatively short tail. The caecum is rather
long. {474}
Sub-Fam. 2. Hydromyinae.—The genus Hydromys,[330] of which there are
several species, the best known being H. chrysogaster, is an
exclusively Australian form, and is aquatic in habit. It is a foot or so
in length, and has a fairly long tail. The fore- and hind-limbs are
webbed, in correspondence with its habits. The Australian Water-Rat is
black, with an admixture of golden-coloured hairs dorsally and golden
colour below, with a lighter median stripe. The thumb is small, and the
webbing of the hands is not so marked as is that of the feet. The molars
are only two in each half of each jaw. The caecum is rather small, the
measurements of the alimentary canal being: small intestine, 895 mm.;
large intestine, 278 mm.; caecum, 70 mm. Allied to the last is
Xeromys, a genus which is also Australian, but limited to
Queensland. It has been established by Mr. Thomas,[331] who discovered that it has the same
reduced formula as Hydromys. Xeromys, however, is not an
aquatic animal, and has unwebbed feet.
In the Luzon highlands Mr. Whitehead has discovered, and Mr. Thomas
quite recently described,[332] a number of peculiar Rodents. Of
these the genera Chrotomys, Celaenomys, and Crunomys
are allied to the Australian and New Guinea Hydromys.
Chrotomys whiteheadi is unusual among Muridae, in its
coloration being marked by a pale stripe down the back. The creature is
the size of the Black Rat (Mus rattus). It is terrestrial not
aquatic in habit, in spite of its likeness to Hydromys. The
molars, however, are 3/3.
Crunomys fallax is more like Hydromys. It has, however,
three molars, as in the last genus. But the skull has the flattened form
characteristic of Hydromys as opposed to Mus.
Like Batomys, Celaenomys silaceus is also somewhat
intermediate between Hydromys and Mus. It is described as
very Shrew-like in appearance, and has a very pointed muzzle. Its habits
Mr. Whitehead is "quite unable even to guess at." Like Hydromys
and Xeromys this Rodent has but two molars.
Sub-Fam. 3. Rhynchomyinae.—The genus Rhynchomys,
containing but one species, Rh. soricioides (of Thomas), is also,
as both its generic and specific names imply, a somewhat Shrew-like form
in external aspect. The skull, too, is Insectivore-like in its {475}elongation, and the lower incisors are
worn to needle-like points. The two molars are excessively minute, and
thus the always large gap in the jaws is greatly exaggerated. It is
suggested that this Rat is an insect-eater, but nothing positive is
known.
Sub-Fam. 4. Gerbillinae.—The Gerbilles form another
sub-family, Gerbillinae, of the Muridae, or a family, according to some.
The best-known genus is Gerbillus, including the Gerbilles proper.
These animals are Old World in range, belonging to the three regions of
that part of the world. There are a large number of species in the genus,
over thirty. They have a Jerboa-like form, with rather long hind-limbs
and a long and hairy tail. But the hind- as well as the fore-feet are
five-toed. The molar teeth have no trace of tubercles, but only
transverse lamellae of enamel. The incisors are orange; they are white in
Dipus. Gerbillus pyramidum is 90 mm. long, with a tail of
125 mm. The ears are long, 13 mm. The tail has longer hairs at the
tip.
Fig. 236.—Gerbille. Gerbillus aegyptius. × ½.
Psammomys is in some respects different. The tail is shorter
than in Gerbillus; its length in an individual of 165 mm. was 130
mm. As in Gerbillus there are four pairs of teats, two pectoral
and two inguinal. This genus is exclusively Palaearctic in range.
Meriones has a range co-extensive with that of
Gerbillus.
Pachyuromys is an Ethiopian genus with a short tail. As the
generic name denotes, the tail is not only short but thick and
fleshy.
Sub-Fam. 5. Otomyinae.—The allied genera, Otomys
and Oreinomys, are Ethiopian. Otomys unisulcatus has a tail
shorter than the body, the measurements of a female of this species being
137 mm. with a tail of 87 mm. The ear is long, whence the name; it
measured in this specimen 20 mm. {476}
Sub-Fam. 6. Dendromyinae.—The genus Deomys is an
African form, consisting of only one species from the Congo region. D.
ferrugineus has a reddish colour as its name implies; the soles are
quite naked and the tail is long and slender. It is considerably longer
than the body, measuring (minus a fragment of the tip) 172 mm., while the
body is 125 mm. long. The characters of the molar teeth, which are three,
are intermediate in their form between those of the true Rats and those
of the Hamsters.
Dendromys is also Ethiopian in range. There are several
species. D. mesomelas is a smallish creature, 60 mm. long, with a
tail of 90 mm.
Steatomys is another African genus, allied to the last. Its
tail, however, is only half the length of the body. The two remaining
genera are Malacothrix and Limacomys. Their range is
African.
Sub-Fam. 7. Lophiomyinae.—Allied to the Hamsters is the
singular East African genus Lophiomys, with only one species,
L. imhausi, of Milne-Edwards.[333] The size is between that of a Rabbit
and of a Guinea-pig. The stomach is curved and somewhat intestiniform. It
has been termed the Crested Rat on account of the "prominent crest of
stiff hair running down the back." The fingers and toes are five, and the
very long tail is clad with hair longer than that upon the body
generally. The pollex is rudimentary, and the hallux is opposable.
The most remarkable structural feature in this genus concerns the
skull, and on account of this it has been regarded as the type of a
separate family. The temporal fossa behind the eye is covered over by a
complete bony plate, formed by a downgrowth of the parietal, meeting an
upgrowth from the malar; this singular arrangement of the bones recalls
the conditions which obtain in turtles. The whole skull, moreover, is
covered with symmetrically disposed granulations, such as are found in no
other mammal; it suggests rather the skull of certain fish. It is
believed that the bony plate already referred to is not really a portion
of the bones of which it appears to be a prolongation, but merely an
ossification of fasciae in this region. The atlas is granulated like the
skull; there are sixteen pairs of ribs and a feeble clavicle. The molars
are three, and of a peculiar form. {477}They have, in the case
of the first three, transverse ridges, from which stand up two sharp and
long tubercles. The other teeth have two ridges. The incisors are pale
yellow. The shape of the teeth and the smallness of the caecum suggest
that this Rodent is not so purely a vegetarian as others, and that it
nourishes itself largely upon insects.
Sub-Fam. 8. Microtinae.—The Voles or Water-Rats form a
distinct group of Murine animals, to which the sub-family name of
Microtinae has been applied from the genus Microtus (more
generally known as Arvicola), a genus which includes the Water-Rat
and Field-Voles of this country. This genus has short ears, and a short
and hairy tail. Its build is stouter and clumsier than that of the Rats.
The genus is confined to the Palaearctic and the Nearctic regions. In
this country there are three species. The best known is the Water-Vole or
Water-Rat, M. amphibius, which has been seen by most people, and
which frequents streams, ponds, and canals. The feet, curiously enough,
are not webbed, which seems to argue the recent adoption of an aquatic
life. Mr. Trevor-Battye has remarked that this animal, when swimming at
leisure, uses its hind-limbs only, carrying the fore pair at the sides
like a Seal. The Bank-Vole, M. glareolus, is rather a local
species in this country. It is a terrestrial Vole, and burrows. The
Field-Vole, M. agrestis, has become notorious on account of the
"plagues," to which its immense numbers have on occasions given rise. It
is the smallest species, and has a greyish-brown fur like the Water-Vole,
the Bank-Vole being redder. To give an idea of the cost of the
depredations of this animal, Mr. Scherren quotes[334] a farmer who gave evidence before the
Agricultural Commission to the effect that, putting the damage of one
Vole at two pence, the amount of loss suffered on a farm of 6500 acres in
two years would be £50,000!
The genus Fiber comes very near the last. It is a North
American genus. The hind-feet are slightly webbed; the tail is a trifle
shorter than the body, and is compressed and scaly, with scattered hairs.
The thumb is short, but with a fully-developed claw. As in the last
genus, the small and large intestines are roughly of the same length, and
the caecum is about one-fourth of the length of either. It is known as
the "Musquash." {478}
Of Fiber zibethicus, or rather a closely-allied form, F.
osoyoosensis, from Lake Osoyoos near the Rockies, Mr. Lord writes[335] that it constructs for
itself a house of bulrushes built up from the bottom in 3 or 4 feet of
water. It is dome-shaped, and rises about a foot out of the water. "If a
dead or badly-wounded duck be left on the pool, it is at once seized on,
towed into the house, and doomed." Thus it appears that this Rodent, like
so many others, is largely carnivorous. It has also been asserted that it
eats fish.
Neofiber is an allied genus, North American in range. The
species, N. alleni, is compared, as regards outward form, with the
Water-Vole, M. amphibius. It has, however, a shorter tail.
Another very well-known member of this sub-family is the Lemming. The
name, however, applies to two quite distinct genera. The genus
Cuniculus, including the Banded Lemming, C. torquatus, is
an inhabitant of North America, Siberia, and Greenland. The tail is
short, its length being 12 mm. as against a body length of 101 mm. The
feet are furred beneath, a not unusual state of affairs in Arctic
mammals. The ears are very slight. The thumb is well developed, and bears
a claw.
In Myodes, on the other hand, which is not so markedly an
Arctic animal, though occurring in both Palaearctic and Nearctic regions,
the ears are rather bigger, though still smaller than those of
Microtus. The under surfaces of the feet are similarly furred. The
tail is also short. It is commonly said that the two genera are to be
distinguished by the furred feet of Cuniculus, and by the absence
of fur in the present genus. That, according to Tullberg, does not appear
to be the case. The differences are thus so much reduced that it seems
almost unnecessary to retain the two genera. The best known species of
Myodes is of course the Scandinavian Lemming, M. lemmus.
This animal used to occur in this country in Pleistocene times (as did
also C. torquatus), and recently Dr. Gadow has found remains with
skins attached in caves in Portugal. It may still survive in the
mountains of the Peninsula.
The actual habitat of the Lemming in Scandinavia is the great
tablelands, 3000 feet high in the centre. The migrations do not take
place with regularity; even twenty years may elapse before the appearance
in cultivated lands of those countless {479}hordes so familiar (as
far as their description is concerned) to everybody. The Lemmings do not
return from their exodus. They die from various causes, including combats
with one another. Their chief foes, however, are Wolves and Gluttons,
Buzzards and Ravens, Owls and Skuas, which batten on the migrant hordes.
Their sudden increase in numbers recalls the similar increase at times of
the Field-Vole, to which reference has already been made.
Ellobius is an Old-World genus, which leads a "Talpine" life,
and has in consequence rudimentary external ears and very small eyes. The
tail is short. Contrary to what might be expected from its mode of life,
the claws upon the digits are not strong.
The remaining genera of Vole-like Murines are Phenacomys and
Synaptomys from North America, and Siphneus from Palaearctic Asia.
Evotomys is one of those genera which are common to both the
Palaearctic and the Nearctic regions, but the bulk of the species are
North American.
Sub-Fam. 9. Sigmodontinae.—This is the name given to
another sub-family of Murine Rodents, a group which includes the Hamsters
in the Old World as well as a large number of South American genera of
Rat-like animals. Of these latter there are a very large number, the bulk
of the group being American.
The Hamsters, genus Cricetus, as it is usually called, although
apparently the correct name is Hamster, are Old-World forms of Pouched
Rats. The Common Hamster, C. frumentarius, is about 210 mm. long,
with a tail of 58 mm. It has cheek pouches. The small and the large
intestines are not very unequal in length, and the caecum is fairly
large, being about one-sixth to one-seventh of the length of either. It
is a purely vegetable-feeding creature, and in Germany where it occurs
(and from which language its vernacular name is derived), hibernates
during the winter in its burrow, having previously surrounded itself with
a great accumulation of food carried thither.
To North America are peculiar the genera Onychomys,
Sigmodon, and Peromyscus. The genus Sigmodon, the
Cotton Rats, reaches Central America, and even gets a little farther
south. The other two genera, though mainly North American, also extend
their range to the south. Onychomys has hairy {480}foot-pads, a
state of affairs which characterises a number of these Rodents.
The genera Megalomys, Chilomys, Reithrodontomys,
Eligmodontia, Nectomys, Rhipidomys, Tylomys,
Holochilus, Reithrodon, Phyllotis,
Scapteromys, Acodon, Oxymycterus, Ichthyomys,
Blarinomys, Notiomys are South American forms.
Oryzomys and Rheithrodontomys are common to both parts of
the New World.
The genus Ichthyomys is remarkable on account of its
un-Rodent-like habits and of certain associated structural changes. I.
stolzmanni was obtained from Mount Chanchamays in Peru at an altitude
of 3000 feet; it is a habitual fish-eater, and lives in streams. Another
species, I. hydrobates, was formerly referred to
Habrothrix. The skull shows likenesses to that of the Australian
Hydromys; but the most marked characters of adaptation are those
of the teeth and caecum. The cutting edges of the upper incisors form a
reversed of obvious use in holding a slippery fish. The caecum is much reduced,
short, and narrow. The general Otter-like shape of the creature is
largely due to its flattened head, though its "size and general
proportions are much as in the common Black Rat."[336]
This sub-family contains a number of genera from Madagascar, viz.
Brachytarsomys, Nesomys, Hallomys,
Brachyuromys, Hypogeomys, Gymnuromys, and
Eliurus.
Sub-Fam. 10. Neotominae.—The last sub-family of the
Muridae is that of the Neotominae, containing the North American genera
Neotoma, Xenomys, Hodomys, and Nelsonia.
Fam. 3. Bathyergidae.—This family contains several genera
which consist of subterranean forms. All these Rodents agree in a number
of characters, of which the principal are as follows:—
The eyes are very small, and the external ears are reduced to the
merest fringe of skin round the aural aperture. The legs are short, as is
the tail; the hair-covering is reduced—a reduction which finds its
culmination in the nearly nude Heterocephalus. Being burrowing
creatures, a number of other modifications in accordance with this mode
of life are to be seen in their structure. The upper incisors stand out
in front of the closed lips, and prevent the entrance of earth. For the
same reason {481}the nostrils are small, and the forehead
but little expanded between them.
The genus Bathyergus contains but a single species, the Cape
Mole-Rat, which is found in Southern Africa; it is of moderate size, not
exceeding a small Rabbit in dimensions. On the fore-limbs are exceedingly
long claws, of which that borne by the second finger is the longest, and
the claw of the thumb the shortest. The hind-feet have by no means such
long claws. The scratching and burrowing is naturally chiefly effected by
the fore-limbs. The small and large intestines are of equal length, and
each is rather more than six times the length of the caecum; in these
measurements the present genus differs from the next.
Georhychus.—Of this African genus there are about ten
species. The claws are not so long as in the last genus, but there are,
as in Bathyergus, four molar teeth on each half of each jaw. The
intestinal measurements in an example of G. capensis were: small
intestine, 25 inches; caecum, 4 inches; large intestine, 15 inches.
The genus Myoscalops or Heliphohius (also with an
African range) has six back teeth on each side. A number of species
sometimes referred to the last genus are placed here by Mr. Thomas. The
claws are small.
One of the most remarkable genera of this family is the little
Heterocephalus from Abyssinia and Somaliland. As Mr. Thomas justly
remarks,[337] it "is a
peculiar-looking little creature, about the size of the Common Mouse, but
looking almost more like a tiny hairless puppy on account of its nearly
naked skin, small eyes, and peculiar physiognomy." Though apparently
naked, there are numerous scattered hairs over the entire body, and the
toes are fringed with stiffish hairs, which must be advantageous to a
burrowing animal. There are two species, H. glaber (originally
described by Rüppell), and H. phillipsii, of which our knowledge
is due to Mr. Thomas. The length of the entire creature including the
tail is not more than 134 mm., both species being approximately of the
same dimensions. Mr. Lort Phillips, the discoverer of the species which
bears his name, writes "that this little creature, called 'Farumfer' by
the Somali, throws up in places groups of miniature craters, which
exactly resemble volcanoes in active eruption. When the little beasts
were at {482}work, I used frequently to watch them, and
found that the loose earth from their excavations was brought to the
bottom of the crater, and sent with great force into the air in a
succession of rapid jerks, and that they themselves never ventured forth
from the shelter of the burrows."[338]
Fam. 4. Spalacidae.—"The Spalacidae," observes Dr.
Blanford, "are sometimes called rodent moles, and resemble a mole in
general aspect, having cylindrical bodies, short limbs, small eyes and
ears, large claws, and a short or rudimentary tail." The existence of a
spiral valve in the caecum may perhaps characterise this family; but it
has at present only been found in the two genera Spalax and
Rhizomys.
Spalax has inconspicuous eyes and external ears. The tail is
totally absent. The lower incisors are more developed than in other
Rodents; they project in a bony sheath beyond the posterior end of the
ramus of the lower jaw. The scapula is long and narrow. The large
intestine is half the length of the small intestine. The animal seems to
have only two pairs of teats, one pectoral the other inguinal.
Spalax typhlus of Egypt, which is probably not different from
the European form, makes extensive burrows, some of the branches being
even 30 to 40 yards in length. In a "domical chamber," situated along the
course of one of these burrows, Dr. Anderson found no less than 68 bulbs
stored up. Its eyes are mere black specks among the muscles, but they
appear, however, to have a proper organisation. There are altogether
eight species of the genus, which is entirely Palaearctic in its
range.
The genus Rhizomys, including a number of species known as
Bamboo Rats, is purely Oriental in range. Rh. sumatrensis reaches
a length of 19 inches; the better-known species, Rh. badius, is at
most only 9 inches in length—in both cases the measurements are
exclusive of the tail, which is a quarter to one-third of the length of
the body, and is not scaly but nearly naked, with a few scattered hairs.
The molars are three, and the incisors usually orange in colour; but
sometimes the upper incisors are white as in Rh. badius. There are
thirteen dorsal vertebrae. In Rh. pruinosus the large intestine is
considerably longer than the small intestine; the lengths of the two
sections of the gut are 42 and 30 inches respectively. In another {483}species the large intestine is slightly
shorter than the small intestine. In Rh. badius the two parts of
the gut are almost exactly equal in length. There are three pairs of
inguinal and two pairs of pectoral teats. The name Rhizomys
appears to have been given to the animals of this genus for the reason
that they feed largely on roots. They burrow, and, like many other
burrowing animals, feed in the evening. As is the case with other forms,
Rhizomys is said to burrow with the assistance of its teeth as
well as of its claws.

Fig. 237.—Bamboo Rat. Rhizomys badius. × ¼.
Tachyoryctes is an African genus closely allied to the last.
There are three Ethiopian species. It is mainly to be distinguished by
the different pattern upon the grinding surface of the molars.
Fam. 5. Geomyidae.—This family of burrowing Rodents is
limited to North and Central America. The animals have cheek pouches, and
small eyes and ears, in accordance with their mode of life. The claws of
the fore-limbs are very strongly developed.
The genus Geomys contains some eight species, which are Central
and North American, not extending, however, far north. The incisors of
the upper jaw are grooved with two grooves. There are three pairs of
teats—one axillary, and the two remaining inguinal.
Thomomys, without grooves on the incisors, reaches to Canada in
the north, and does not extend as far south as the last genus. {484}
Allied to this family, and indeed united with it by Tullberg, but kept
separate by Thomas, is the
Fam. 6. Heteromyidae.—The members of this family are also
American, but are not confined to the northern-central regions of that
continent, for the genus Heteromys extends into South America.
The genus Dipodomys, with twelve species, is of a Jerboa-like
form, as the following measurements of an example of D. merriami
will show. The length of the head and body was 85 mm.; of the tail 127
mm.; the hind-foot is 32 mm. It has but four toes. The hind-limb is
longer than the front-limbs.
In Perodipus the same form is exhibited. There are, however,
five toes, and the sole of the foot is hairy. The axis vertebra and the
two following vertebrae are fused together.
Perognathus is a third genus. It has the same general slender
form, but the tail is not so long, being but little longer than the body.
The hind-limbs, too, are shorter. The teats of this and of
Perodipus are as in Geomys. The two remaining genera of the
family are Heteromys and Microdipodops.
Fig. 238.—Jerboa. Dipus hirtipes. × ⅓. Eastern Europe.
Fam. 7. Dipodidae.—This family consists of small,
plain-living, and leaping or arboreal creatures, commonly known as
Jerboas. The main anatomical characters of the family are the
following:—There is a large infra-orbital
foramen. The molars are always reduced, the premolar being either absent
in the lower jaw alone or in both jaws. This family presents an obvious
likeness to Dipodomys (hence the name of the latter) and to some
other members of the American family Heteromyidae {485}There is even the same
ankylosis of the neck vertebrae. We find, moreover, the same association
of long-legged and shorter-legged forms that characterises the
Heteromyidae.
Fig. 239.—Bones of right pes
of Jerboa, Dipus aegyptius.
× ¾. a, Astragalus; c,
calcaneum; c2, middle
cuneiform; c3, outer
cuneiform; cb, cuboid;
n, navicular; I-IV, first
to fourth toes. (From
Flower's Osteology.)
The typical genus Dipus is a smallish quadruped with long naked
ears and a long tail. The ten species are all Palaearctic in range. The
fore-limbs are short and five fingered, and the short pollex has no claw;
the hind-limbs are excessively long and only three-toed. The bony
structure of these limbs is remarkable. The three metatarsals are
elongated almost like those of a bird, and are ankylosed together. The
digits have long phalanges which alone reach the ground as the animal
hops. It is a curious fact, and one not so easily identifiable with the
way of life, that the neck vertebrae of this genus are ankylosed together
with the exception of the atlas, which is free; the arrangement is
precisely like that of the Sperm Whale. The last vertebra is, however,
sometimes free. The Jerboas not only leap but they burrow, and their
strong incisors are said to be used in burrowing through stony ground.
They are eaten by the Arabs, and are, or have been, called Daman Israel,
i.e. Lamb of Israel. In D. hirtipes the body and tail
measure respectively 4½ and 7 inches. The hind-feet have a tuft of long
hairs below. Mr. W. L. Sclater's newly-founded genus Euchoreutes[339] is somewhat more
primitive in its characters than is Dipus. The general form is the
same, with long ears and a long tail. But there are five toes to the
hind-limb, the two lateral ones though nailed being much shorter than the
middle three. It has a "long pig-like snout," and the tail is cylindrical
as in most other Jerboas, with a tuft of longer hairs at the end. The
incisor teeth, grooved in Dipus, are here smooth, as in
Alactaga. The species was probably obtained "in the sandy plains
round the city of Yarkand." {486}
Alactaga is much like Euchoreutes; it has five toes, a
cylindrical tufted tail, the hairs at the end distichous, smooth
incisors, and a premolar present in the upper jaw. It also differs from
Euchoreutes by the much smaller auditory bulla as well as in the
fact that the infra-orbital foramen has no separate passage for the
nerve, which passage is to be distinguished in both Dipus and
Euchoreutes. The best-known species is the Siberian Jumping
Rabbit, A. jaculus. Beneath the ends of the three main toes of the
feet are remarkable fan-shaped pads. In A. decumana the body and
tail measure 7 and 10 inches respectively, the ears 2 inches.
Platycercomys, a fourth genus of the family, is much less known
and is to be differentiated from the last three genera by the fact that
it has no premolars at all, the grinding tooth formula being thus 3/3.
The tail too is flattened and "lancet shaped." It extends from Siberia to
Nubia, and thus just enters the Ethiopian region.
The above are the more typical Jerboas. There remain several forms
which are not at all Jerboa-like in their way of life, but are
nevertheless, on anatomical grounds, placed with them. Zapus, an
American genus, with the exception of one Palaearctic species, is
transitional in that its hind-legs are rather long, but there is not so
much difference between them as in the typical Dipodidae. Sminthus
is at the opposite extreme to Dipus. Its feet are short and of
equal length; it climbs in trees, and may perhaps be looked upon as
nearest of all Dipodidae to the ancestral form of the group.
Fam. 8. Pedetidae.—The genus Pedetes contains but
one species, P. caffer, the Cape Jumping Hare. The animal suggests
a large Jerboa in appearance on account of its jumping habits, the long
hind-limbs, and the long tail. The length of a fair-sized example is some
17 inches, with a tail of the same length. The eyes and ears are large.
The hands are five-fingered and the feet only four-toed, the hallux being
of course the absent digit. In the skeleton it is interesting to note
that the second and third cervical vertebrae are so close together that
there can be no free movement; interesting because in Dipus the
cervicals are actually ankylosed. The dorsal vertebrae are twelve. The
small intestine is long, measuring 7 feet 4 inches, while the caecum is
short, being only 8 inches long. The large intestine is 3 feet 10 inches
long. The gall-bladder appears to be {487}absent,[340] an exceptional state
of affairs in Rodents. A singular fact in the anatomy of this animal is
the existence of a septum dividing the lower part of the trachea. This is
sometimes met with in birds. As might be supposed from its large eyes,
the Spring Haas, as the animal is sometimes called, is nocturnal. Its
long hind-limbs permit it to leap enormous distances. It is a burrowing
Rodent.
Section 3. Hystricomorpha.
Fam. 1. Octodontidae.—The Rodents of this family are of
small to moderate size, the only, relatively speaking, giant in the
family being the "Water-Rat," Myocastor. The toes are with but one
exception not reduced; the tail is long in the majority of the genera.
The teats are placed high up on the sides of the body. The clavicle is
fully ossified. All the genera are South or Central American in range
with the exception of Petromys.[341]
Sub-Fam. 1. Octodontinae.—Octodon has four
species, which are all Chilian, Peruvian, and Bolivian in distribution.
The Degu, O. degus, has a length of 160 mm., with a tail 105 mm.
long. The ears are 18 mm. long. At the roots of the claws are longish and
stiff hairs which appear to serve as "combs." The tail has long but
sparsely scattered hairs. There are twelve pairs of ribs. The lengths of
the various sections of the intestine are as follows: small intestine,
680 mm.; caecum, 90 mm.; large intestine, 390 mm. These animals live in
large companies. Closely allied is the genus Habrocoma (more
correctly, as it appears, to be written Abrocoma), with two
species. H. bennetti is 204 mm. long, with a tail of 103 mm. The
ears are long, 22 mm. The fore-feet have no outward trace of the thumb.
Stiff hairs like those that characterise Octodon are found also in
this genus. The fur is very soft. The furring of the tail is much thicker
than in Octodon.
Spalacopus with but a single species, S. poeppigi, is a
burrowing animal, from which indeed, and on account of its resemblance to
Spalax, it has received its name. The ears in accordance with the
underground life are short, only 5 mm. in length in an {488}example of 120
mm. The tail too is reduced, being in the same example only 42 mm. in
length. As in the last two genera the large intestine is about one half
of the length of the small intestine.
The "Tuco-tuco," genus Ctenomys, has also short ears and tail.
The claws of the fore-feet are longer than those of the hind-feet.
A related form is Aconaemys (better known as Schizodon),
with similar external characters; it inhabits high localities on the
Andes.
Petromys is the only genus of the sub-family which is not
American in habitat. It is an African form and there is but one species.
Its anatomy conforms to that of the genera already considered. The main
difference in structure is shown by the teeth. Their surface is uneven,
and differs from that of other Hystricomorphs "in that the enamel to the
inside of each upper jaw-tooth and outside on each lower jaw-tooth forms
two tubercles, to which correspond grooves in the reverse position of the
applied teeth."
Sub-Fam. 2. Loncherinae.—The genus Echinomys with
thirteen species belongs to the Neotropical region. The members of the
genus are entitled "Spiny Rats" since they have spines mixed with the
fur. The tail is long and the ears are very well developed. Both feet are
five-toed. The tail is scaly as well as haired. Trichomys (also
called Nelomys) is very close to the above, and is also from the
same part of the world.
The genus Cannabateomys contains but one species, C.
amblyonyx, which was formerly included in the genus
Dactylomys, but has lately been separated by Dr. Jentink.[342] The animal is
Brazilian and has a total length of 520 mm., of which 320 mm. belong to
the tail. It is a climbing rat, and in accordance with that way of life
has undergone some modifications. The fore-feet are four-toed, the two
middle toes being markedly longer than the outer ones. The hind-feet are
five-toed with the same greater development of the two middle toes. The
claws are small and somewhat nail-like.
Dactylomys, also Brazilian, and with but one species, D.
dactylinus, differs from the last in the fact that the molars are
simpler in form; they are divided into two lobes, each of which {489}has but a
single enamel fold, whereas in Cannabateomys these teeth have
several enamel folds. The tail, moreover, is but slightly hairy.
Loncheres with eighteen species is another Neotropical genus
allied to the foregoing. Small spines are, as in many of these genera,
intermingled with the fur. This genus has as many as seventeen dorsal
vertebrae, which is an unusually large number. L. guianae is known
as the "Porcupine Rat." Allied genera, also South American, and without
spines in their fur, are Mesomys, Cercomys, and
Carterodon.
The South American Thrinacodus is also known by one species,[343] T. albicauda,
which has rather more than the distal half of the long tail of a white
colour. The fore-feet have four toes. The ears are broad and short.
Sub-Fam. 3. Capromyinae.—A third sub-family of the
Octodontidae is formed by the genera Myocastor, Capromys,
Plagiodontia, and Thrynomys, which are all Neotropical
forms with the exception of the last, which is African.
Thrynomys (better known perhaps as Aulacodus) is a genus
of African Rodent, containing some four species. The best-known of these
is T. swindernianus, the Ground-Rat of West and South Africa. Its
structure has been investigated by Garrod,[344] by Tullberg,[345] and by myself.[346] The fur is mingled with flattish
bristles; the tail is moderately long, about half as long as the body.
The fore-feet are five-toed, but the two toes at each end of the series
are quite small. The hind-feet are only four-toed, the hallux being
absent. The claws of the hind-feet are stronger than those of the
fore-feet. The ears are not long. The limbs are decidedly short, hence
the name of "Ground-Pig" sometimes applied to this animal. The molars are
four in number in both jaws. The incisors of the upper jaw are twice
grooved. There are thirteen dorsal vertebrae. The length of the small
intestine is 60½ inches, that of the large 49; the caecum is short, being
only 8 inches long. It is a remarkable fact that the acromion is joined
to the rest of the spine of the scapula by a joint.
Myocastor, a name which seems to have the rights of priority
over the more familiar Myopotamus, applies to a large South
American aquatic Rodent. The general aspect of the animal {490}suggests a
Water-Rat of large size (it has been exhibited in shows as a phenomenal
product of London sewers!); the tail is nearly as long as the body. The
ears are small. The limbs are short. The tail is naked. The hind-feet are
webbed, but not so much so as in Hydromys. A small thumb is
present. The animal has thirteen pairs of ribs; the molars are four in
each jaw. The large intestine is more than three times the length of the
small, and the caecum is, as in the last genus, relatively short.
Capromys is a genus[347] which is remarkable on account of its
restricted distribution. It is found only in the islands of Cuba and
Jamaica. There are four species, of which C. melanurus is a dark
brown-coloured animal with a blacker tail, nearly as large as a Rabbit.
The native name of this Rodent is "hutia." It is also remarkable
for having a stomach more complicated than is the rule among the mammals
of this group. The organ is divided by two constrictions into three
compartments. In C. pilorides the liver is occasionally divided up
in an extraordinary fashion into small lobules. Capromys has the
large number of sixteen dorsal vertebrae.
Fam. 2. Ctenodactylidae.—For these African genera it
seems admissible to form a distinct family, though Thomas, and Flower and
Lydekker, only allow to the genera Ctenodactylus,
Pectinator, and Massoutiera sub-family rank. On the other
hand, Tullberg removed these genera entirely from the Hystricomorph
section and placed them as a section of the sub-tribe Myomorphi of the
tribe Sciurognathi. It was chiefly the form of the mandible which led to
this placing, for in these Rodents, as in all Squirrel- and Rat-like
Rodents, and unlike what is found in the Hystriciform genera, the angular
process of the mandible is not bent sideways.
The genus Ctenodactylus derives its name from the peculiar
strong bristles which form a comb-like structure upon the hind-feet and
hide the claws; these are stated to be for the purpose of dressing the
fur. The Gundi of North Africa, C. gundi, has a length of 190 mm.,
with a short tail of 17 mm. The ears are only moderate in size. The
dental formula of the molars is 4/3. The incisors are white. The feet
have four digits, and the hind-limbs are the longer. The large intestine
is distinctly longer than the small intestine. {491}
Pectinator spekii is the only representative of a genus not far
removed from Ctenodactylus; it is a smallish Rodent, 6 inches in
length, exclusive of a rather bushy tail nearly 3 inches long. It comes
from Abyssinia. It has somewhat the appearance of a Squirrel, which is
heightened by the fact that when sitting the tail is arched over the
back; when running the tail is carried out straight. There are only four
toes visible externally on both fore- and hind-limbs, but pollex and
hallux exist in the skeleton, with a single phalanx each. There is only a
single pair of mammae, and in correspondence with this but two or three
young are produced at a time. The hind-feet have bristles very much like
those of Ctenodactylus. The molars, however, are 4/4. There are
twelve ribs, of which six reach the sternum. The latter is made up of six
pieces, and the manubrium in its breadth anteriorly suggests that of the
Vizcachas. The clavicles are present.[348]

Fig. 240.—Carpincho. Hydrochoerus capybara. × 1⁄12.
Fam. 3. Caviidae.—This family, which includes the Cavies
and the Capybara, is entirely South American and West Indian in
distribution. It embraces animals of fair to large size, the Capybara
{492}(or Carpincho) being the greatest of
existing Rodents. The ears are well developed. The toes are commonly
reduced, and the members of this family possess only a rudimentary tail.
The hair though rough is not spiny. Other characters had best be deferred
until the several genera are treated of. We shall begin with the giant of
the family, the genus Hydrochoerus. This genus contains but a
single species, H. capybara of South America. It reaches a length
of some 4 or 5 feet. The ears are not large; the tail is completely
absent. The fore-feet are four-toed, the hind-feet three-toed; the digits
are webbed, though not to a very great degree, and the nails have the
appearance of hoofs. There are fourteen dorsal vertebrae; the clavicle is
absent. In the skull the paroccipital processes are of great length. The
infra-orbital foramen is large. The most remarkable fact about the teeth
is the great size of the posterior molar of the upper jaw; it has
fourteen folds of enamel, more than all the anterior teeth possess
collectively. The incisors are white and grooved in front. The
measurements of the alimentary tract as given by Tullberg are: small
intestine, 4350 mm.; caecum, 450 mm.; large intestine, 1500 mm.

Fig. 241.—Patagonian Cavy. Dolichotis patachonica. × 1⁄10.
The Capybara or Carpincho is largely aquatic in its habits. Their
"favourite locality," writes Mr. Aplin,[349] "is a broad laguna {493}in the river,
furnished with open water, and also beds of 'camelotes,'—a sloping
open grassy bank on one side, where the Carpinchos can lie in the daytime
in the cooler weather, sleeping and basking in the sunshine; on the other
a low shelving bank, clothed with 'Sarandi' scrub growing out into the
black reeking mud and shallow water beyond." They always take to the
water when alarmed, at a rate and with a gait which reminded Mr. Aplin of
a Pig. When in the water they swim slowly with the upper part of the
head, including nose, eyes, and ears, above the surface. But they can
dive for a considerable time and distance, and baffle their enemies by
seeking the shelter of a mass of water-plants, and lying there with their
noses only just above the surface.
The genus Dolichotis[350] has long ears, and generally
resembles a rather long-legged Hare in appearance. The front-feet are
four-toed, the hind three-toed. The Patagonian Cavy, as this animal is
called, has twelve dorsal vertebrae, and rudimentary clavicles.[351] The paroccipital
processes are long; the incisors are white, and are not grooved in front.
The sternum has six pieces, and seven ribs reach it.
Cavia, including the species C. porcellus, the
Guinea-pig (which name is a corruption apparently of Guiana pig), has the
same number of toes on its hind- and fore-feet as has the Capybara. The
name applied to the wild stock whence our Guinea-pig is derived is the
Restless Cavy. The fur is greyish; of the domestic animals the colour is
too well known to need description.
Fam. 4. Dasyproctidae.—The genus Coelogenys
includes but two species. C. paca, known as the "Spotted Cavy" or
"Paca," has a brown body, with white spots like those of a Dasyure; it is
one of the largest of Rodents, and has a quite short tail. The hand and
foot are both provided with five digits; but the thumb is small, and in
the foot the three middle toes considerably exceed the others in length.
The hind-foot is practically three-toed. The fibula is not nearly so
reduced as in Dolichotis. The skull of the animal is remarkable
for the extraordinary development in breadth of the jugal arch, which is
sculptured externally. There is a large cavity formed below, at the
maxillary end of this huge {494}arch, by the curving inwards of the bone,
which lodges a cavity continuous with the mouth. The palate has
anteriorly a ridge on either side, and is thus divided from the sides of
the face in a way which is not found[352] in the allies of Coelogenys.
Clavicles are present. There are thirteen dorsal vertebrae. The incisors
are coloured red in front. The animal is South American, and in that
continent is limited to the Brazilian sub-region. This, the best-known
species of Paca, is called the Gualilla by the natives of Ecuador; in the
same district another form is met with which the natives term Sachacui
(signifying Forest Cavy). It is very often the case that a different
native name expresses a real specific difference; and to the latter form
M. T. Stolzmann has given the name of C. taczanowskii.[353] This form, unlike the
common Paca, which is fond of forests and low-lying ground in the
neighbourhood of water, is alpine in habitat, living upon mountains of
6000 to 10,000 feet. It burrows in much the same way as its congener, and
is greatly sought after as food, its meat possessing an "exquisite
taste." It is pursued by dogs, by whose aid one of the two entrances to
the burrow is guarded, and the creature is smoked out and killed with a
stick.
The genus Dasyprocta, containing those Rodents known as
Agoutis, is divisible into several species, apparently about twelve, all
of which are, like the Pacas, confined to the Neotropical region. They
have, however, a much wider range within that region, and occur as far
north as in Central America and in some of the West Indian Islands. They
are of rather smaller size than the Paca, and are without spots. The
colour is of a golden brown in some forms, but usually has a freckled,
grizzled, greenish kind of appearance. The tail is stumpy, the hind-limbs
are distinctly longer than those of the Paca, and the two lateral toes
have disappeared from the feet—a concomitant as it seems of the
Agouti's greater powers of running. The three metatarsals are closely
pressed together, and the foot is as it were on the way towards the
highly-modified foot of the Jerboa. The fore-feet are, however,
five-toed. The clavicle is rudimentary,[354] whereas it is well developed in the
Paca. The skull has not the peculiar modifications of that of the
last-mentioned type. The sternum has seven {495}pieces, and eight ribs
reach it. A curious difference between this genus and the last is in the
relative proportions of the regions of the intestine. The figures given
by Tullberg for the two animals are—for Coelogenys, small
intestine, 4800 mm.; caecum, 230 mm.; large intestine, 21,000
mm.;—for Dasyprocta aguti the same author gives: small
intestine, 4200 mm.; caecum, 200 mm.; large intestine, 1000 mm. The
Agouti, says Mr. Rodway,[355] is as wily as the Fox. "If chased he
will run along the shallows of a creek to hide his scent from the dogs,
or swim over and back again several times for the same purpose. He never
runs straight when pursued, but doubles, often hiding until a dog has
passed, and then making off in a different direction. Like the fox he has
been hunted for a very long period, and, like Reynard, has grown wiser
with every generation."

Fig. 242.—Agouti. Dasyprocta aguti. × 1⁄10.
Fam. 5. Dinomyidae.—The genus Dinomys of Dr.
Peters[356] is a very
little known and remarkable form from South America allied to the
Capybara, the Chinchilla, and other South American Rodents. It is only
known by a single example found wandering about a courtyard in a town of
Peru. It is externally like, and of about the same size as the Paca, but
has a hairy tail. The animal is four-toed and plantigrade; the ears are
short, and the nostrils are -shaped. It is usually regarded as belonging {496}to a separate family
which will include but the one species, D. branickii.
Fam. 6. Chinchillidae.—This family, likewise South
American, contains three genera,[357] all of which agree in having long
limbs, especially the hind-limbs, and a bushy and well-developed tail.
The hair is exceedingly soft, hence the commercial value of
"chinchilla."
The genus Chinchilla, containing but a single species, C.
laniger, is a small and squirrel-like creature, living at
considerable heights in the Andes. The eyes, as it is a nocturnal
creature, are naturally large; and so also are the ears. The fore-feet
have five toes, the hind-feet only four; they are furnished with feeble
nails. The innermost toe of the hind-foot has a flat and nail-like claw.
There are thirteen dorsal vertebrae, and the long tail has more than
twenty. The clavicle is well developed, as in the other genera of this
family. The large intestine of this animal is extraordinarily long; the
proportions of the different regions of the gut are shown by the
following measurements: small intestine, 820 mm.; caecum, 125 mm.; large
intestine, 1340 mm. Such a disproportion between the large intestine and
the small, to the advantage of the former, is a very strange fact in the
anatomy of this Rodent.
The genus Lagidium (also called Lagotis), which includes
"Cuvier's Chinchilla," is also a mountain dweller. There are several
species of this genus, which differs from Chinchilla by the
complete abortion of the thumb and of the great toe. The intestinal
proportions are those of Chinchilla. The ears and tail are long.
L. cuvieri measures 1½ feet in length.
Lagostomus, again, has but one species, L.
trichodactylus. The animal has a tail about half the length of the
body. The digits are reduced as compared with Chinchilla, there
being but four on the fore- and three on the hind-feet. There are only
twelve dorsal vertebrae, and seven ribs reach the sternum. In the skull a
distinguishing mark from the last two genera is the separation of the
infra-orbital foramen into two by a thin lamella of bone. The large
intestine is between one-half and one-third the length of the small
intestine, and thus differs much from that of Chinchilla. {497}
Fig. 243.—Vizcacha. Lagostomus trichodactylus. × 1⁄10.
The Vizcacha lives in societies of twenty to thirty members,[358] in a "village"
("Vizcachera"), a dozen or so of burrows, which intercommunicate. They
lie at home during the day and come out in the evening. Their burrows,
like those of the Prairie Marmot, harbour other creatures, which
apparently live on amicable terms with the Vizcachas; such are the
burrowing owl, a small swallow, and a Geositta. The Fox also
affects these burrows, but then he ejects the rightful owner of the
particular burrow which he selects. When the young Foxes are born the
vixen hunts the Vizcachas for food. The Vizcacha has a most varied voice,
producing "guttural, sighing, shrill, and deep tones," and Mr. Hudson
doubts if there is "any other four-footed beast so loquacious or with a
dialect so extensive." These animals are very friendly, and pay visits
from village to village; they will attempt to rescue their friends if
attacked by a Weasel or a Peccary, and to disinter those covered up in
their burrows by man.
Fam. 7. Cercolabidae.—A number of the characters which
differentiate this family from the Hystricidae or Ground Porcupines of
the Old World are given under the description of the latter. The
principal external characters are the prehensile tail, the admixture of
spines with hairs, and the nature of the sole of the foot. In these
points the New-World Cercolabidae differ from the Old-World Hystricidae.
It is interesting to notice that {498}in both families we
have long-tailed and short-tailed forms. Cercolabes corresponds to
Atherura or Trichys, and Erethizon to
Hystrix.
The genus Erethizon, the "Urson" of Canada, has a short, stumpy
tail. Its spines are almost hidden by enveloping hair. The fore-feet have
four, the hind-feet five toes. The short tail of this creature is
remarkable when we reflect upon its climbing habits. It appears, however,
to be a weapon with which it strikes sideways at the enemy.
Fig. 244.—Brazilian Tree Porcupine. Sphingurus prehensilis. × 1⁄6.
Of the Neotropical genus Cercolabes (sometimes called
Sphingurus, Synetheres, or Coendou) there are some
eight or nine species, all found in Central and South America. The animal
is arboreal, and has in correspondence with that habit a prehensile tail.
The spines are not so stout as in the Ground Porcupines, and are often
coloured yellowish or reddish. In correlation with its tree-frequenting
habits the bones of Cercolabes show certain differences from those
of the Ground Porcupines. The scapula is broader and rounder in front
than is that of Hystrix; the phalanges of the thumb (which is
rudimentary) are fused together as in the Canadian Erethizon; but
those of the very small hallux are also fused, whereas in
Erethizon, as in Hystrix, they are separate. In one
species, C. insidiosus, Sir W. Flower states that there are as
many as seventeen dorsal vertebrae and thirty-six caudals. The tail is
thus very long. In C. villosus there are fifteen dorsals and
twenty-seven caudals; eight ribs reach the sternum, which is composed of
seven pieces, the {499}sixth being very small. The clavicles are
well developed. A curious fact about C. villosus is that the
acetabular cavity is perforate (on both sides), or at least only closed
by membrane. In many forms of Rodents the bone is very thin in this
region. This fact perhaps lessens the significance of the perforation of
the acetabulum of Echidna (see p. 109).
Of the allied genus Chaetomys, also Neotropical, there is but a
single species, which inhabits Brazil. It has a nearly completely closed
orbit, a feature which differentiates it from the last animal, and one
which also shows it to be a more modified form. The spiny covering is
less pronounced than in its allies.
Fam. 8. Hystricidae.—This family is characterised by the
fact that all its members possess spines; but the tail, if at all long,
is not prehensile, and the soles of the feet are smooth and not covered
with rough tubercles, as in the Tree Porcupines of the next family,
Erethizontidae. The clavicle is less developed than in the arboreal
forms. In the organs of digestion there are points of a family difference
between the two groups of spiny Rodents. The tongue has serrated scales
arranged in transverse rows, which are directed backwards. A
gall-bladder, though not always present, is sometimes found; it
apparently never exists in the arboreal Porcupines and in
Erethizon. The lungs show a great tendency to subdivision, which
appears to be especially marked in the genus Atherura. The caecum
seems also to be shorter in the Ground Porcupines. In Hystrix
cristata the small intestine measures 15 feet 7 inches; the caecum, 8
inches; the large intestine, 4 feet 4 inches:—in Atherura africana the caecum measures
7½ inches; the large intestine, 1 foot 10 inches. The corresponding
measurements of Synetheres villosus were: small intestine, 7 feet
3 inches; caecum, 1 foot 4 inches; large intestine, 2 feet 7 inches. In
Erethizon the caecum is 2 feet 4 inches in length. These
differences are too large and too constant in a number of presumedly
allied forms to be overlooked.
Mr. Parsons has directed attention[359] also to a number of muscular
differences, such indeed as might be expected to occur between animals of
such different habits.
The genus Hystrix embraces the better-known Porcupines. It is a
genus of wide range, extending from the East Indies to Africa, {500}and even
occurring in Europe. There are several species, of which the common
Hystrix cristata is the best known, and is the one which is to be
found in Europe.
Fig. 245.—Common Porcupine. Hystrix cristata. × 1⁄10.
The spines of the common form and of the others are solid in the
middle of the body, but on the tail they are expanded into hollow quills,
which make much rattling. They are as a rule black and white, the middle
of the spine being banded with black. A great crest of coarse long hairs
on the head is responsible for the scientific name of the well-known
form. Sometimes in this genus, as in the Tree Porcupines of Brazil, the
spines are orange or yellow; but it is said that the colour is soon lost
in this country. As a matter of fact it is the easiest thing in the world
to wash out with ordinary tap-water much of the yellow colour of the
spines of the South American Sphingurus. The same may be the case
with the pigment of the Old-World Porcupines. There are fourteen to
fifteen dorsal vertebrae and four or five lumbars. The tail varies in
length, but is shorter than the long tail of the arboreal New-World
forms. It seems impossible when mentioning the Porcupine to escape from
some observations about its alleged habit of shooting its quills. For
some reason or other Buffon has got the credit of inventing, or at {501}least
promulgating, this legend, which has even grown so in the telling that
the quills are said to be capable of penetrating planks of wood. What
Buffon said apropos of this matter is, "The marvellous commonly is
pleasingly believed, and increases in proportion to the number of hands
it passes through." It is of course the rattling of the spines and the
occasional falling out of loose ones which has started the legend. They
are, however, excellent weapons of offence, and the animal charges
somewhat backwards to make the best use of them against the foe. The
spines, however, are by no means an absolute protection, since, as Mr.
Ridley informs us,[360]
Tigers will kill and eat these animals just as the Thylacine is
apparently indifferent to the spiny armature of Echidna.
Of the Brush-tail Porcupine, Atherura[361] there are at any rate two species,
the West African A. africana and the Malayan A.
fasciculata. It is interesting that the gap in the present
distribution is partially filled by the discovery of fossil teeth near
Madras. The genus does not differ widely in external appearance from
Hystrix; it has, however, a rather longer tail; there are fewer
large spines, and there is a tuft of them at the end of the tail, whence
is derived the name of the genus. The frontal bones project a little
distance between the nasals, a feature which does not seem to appear in
the true Porcupines. There are fourteen dorsal vertebrae and five
lumbars. The twenty-four caudal vertebrae of this Porcupine shows how
much longer is its tail than that of Hystrix; for in the latter
twelve is about the number.
A third genus of Old-World Porcupine is the singular Trichys.[362] Of this there is but
one species, T. lipura. It is a curious fact that out of three
examples, all from Borneo, two were quite without a tail. But this
appears to be merely a mutilation, though it is singular that the natives
state it to be without a tail. One cannot help thinking of the way in
which lizards sometimes shed their tails when pecked at. The tail of this
genus is more than half the length of the body and head. Trichys
has sixteen dorsal and six lumbar vertebrae. There is a tuft of quills at
the end of the tail, which are thin and compressed, {502}though
truncate at the free extremity and hollow; they represent in a more
rudimentary way the much stronger tuft at the end of the tail of other
Porcupines. It is a curious fact that this and other Porcupines possess a
mechanism for warning their foes precisely comparable to that of the
rattlesnake. There are sixteen dorsal vertebrae.
Sub-Order 2. DUPLICIDENTATA.
The chief feature of this group is the existence of two pairs of
incisor teeth in the upper jaw, of which the inner are very small and lie
behind the outer. In the skull the infra-orbital foramen is small; the
incisive foramina are very large. The tail is short or absent.
Fam. 1. Leporidae.—This family is distinguished from the
Lagomyidae by the long ears, by the tail, which is present, though short,
and by the longer limbs. There are six teeth belonging to the molar
series in the upper jaw, and five of the same in the lower. The clavicle
is imperfect.
The longest known genus of this family, Lepus, was, until the
quite recent discovery of Romerolagus, the only genus. It is of
universal range, excepting Australasia and Madagascar, and consists of
about sixty species. These are the Hares and Rabbits, to the former being
assigned the longer-limbed forms.
As every text-book of zoology contains a more or less elaborate
account of the structure of the Common Rabbit, and as there is but little
structural difference between the members of the genus, a short account
of the generic peculiarities of Lepus will suffice here. The
fore-feet are five-toed, the hind-limbs four-toed. The hairy integument
enters the mouth cavity, and the inside of the cheeks have a hairy
covering. The soles of the feet are, moreover, hairy. The maxillary bones
are curiously sculptured.

Fig. 246.—Lepus cuniculus. Skull. A, Lateral view; B, ventral view. ang.pro,
Angular process of mandible; as, alisphenoid (external pterygoid process); aud.me,
external auditory meatus; b.oc, basioccipital; b.sph, basisphenoid; cond, condyle;
cor, coronoid process; fr, frontal; int.pa, inter-parietal; ju, jugal; lcr, lachrymal;
max, maxilla; nas, nasal; opt.fo, optic foramen; o.sph, orbitosphenoid; pa, parietal;
pal, palatine; pal.max, palatine plate of maxilla; pal.p.max, palatine process of
premaxilla; par.oc, paroccipital process; peri, periotic; p.max, premaxilla; pt,
pterygoid; p.t.sq, post-tympanic process of squamosal; s.oc, supraoccipital; sq,
squamosal; ty.bul, tympanic bulla; vo, vomer; zyg.max, zygomatic process of
maxilla. (From Parker and Haswell's Zoology.)
The Common Rabbit, L. cuniculus, differs from the Common Hare
in the comparatively shorter ears and legs. The ears have not, to so
marked a degree, the black tips of those of the Hare. The animal,
moreover, produces naked young, and lives in burrows of its own
excavation. A difference in the structure of the caecum, which
distinguishes the Rabbit from the Hare, has been {504}pointed out by
Professor W. N. Parker.[363] These differences have led some to
approve of its separation from the Hares into a genus Oryctolagus.
This animal is believed to be an introduced species, and to have been
brought by man into these islands. Its original home is the Spanish
Peninsula, the south of France, Algiers, and some of the Mediterranean
islands. Mr. Lydekker thinks that the only other species of Lepus
which can be considered to be a "Rabbit" is the Asiatic L.
hispidus.
Of Hares there are two species in this country. The Common Hare, L.
europaeus (the name L. timidus seems to be really applicable
to another species to be referred to presently), extends all over Europe
excepting the extreme north of Russia and Scandinavia. It is not known in
Ireland, and, curiously enough, attempts to acclimatise this animal in
that island have failed—a state of affairs which contrasts with the
fatal ease with which the Rabbit has been introduced into Australia.
Ireland has, however, the Variable Hare, L. timidus (also called
L. variabilis), a species which is common in other parts of
Europe, and which extends as far east as Japan. This species differs from
its ally by the fact that it often turns white in winter with the
exception of the black tips to the ears. In Ireland this change does not
always occur; but Mr. Barrett-Hamilton has commented upon the fact that
Hares of this species do change on Irish mountains. It appears that in
this animal the change from the winter to the summer dress is
accomplished by the actual casting off of the white hairs and their
replacement by a fresh growth of "blue" hairs. A similar change occurs in
the American L. americanus.
Dr. Forsyth Major has noted the fact that the various species of Hares
can be distinguished by the condition of the furrows upon the upper
incisors. Thus two African species, L. crawshayi and L.
whytei, are to be separated by the fact that in the former the
incisors are quite flat, whereas in L. whytei the groove is more
prominent and there is a second shallow furrow.
The genus Romerolagus[364] is quite a recent discovery. It
occurs on the slopes of Popocatepetl in Mexico; it has the general aspect
of the last genus, and is spoken of as a "Rabbit." It inhabits runs in
the long grass which clothes the sides of the {505}mountain. Externally it
is something like the Pikas, since it has no tail visible. The ears, too,
are short, and the hind-legs comparatively short. The skull is very like
that of the Rabbit; but in other osteological details it is aberrant.
Thus the clavicle is quite complete, and only six ribs articulate with
the sternum, instead of the seven that we find in the Rabbit.
Fam. 2. Lagomyidae.—The animals of this family are
smaller than the Hares and Rabbits; they have short Vole-like ears and no
external tail. The limbs also appear to be shorter. As there is but a
single genus, the characters of the family may be described in connexion
with those of the genus, which is known as Lagomys (apparently
more correctly Ochotona). Of this genus there are about sixteen
species, which are mainly Asiatic; one species extends its range into
Eastern Europe, and three are North American.
The skull has not the supra-orbital grooves of the Rabbits, and has a
well-marked backward process of the zygomatic arch. There are eighteen
dorsal vertebrae. The molars and premolars are five.
The vernacular names of "Pika" and "Piping Hares" have been applied to
the members of this genus, the latter on account of their peculiar call.
They live among rocks in companies and they burrow. They are usually
found at considerable altitudes: thus L. roylei, the "Himalayan
Mouse Hare," is found at elevations as high as 16,000 feet; while L.
ladacensis gets even higher, 19,000 feet having been recorded. With
the habits of a Marmot, so far as concerns living in burrows and at great
altitudes, the animals of this genus, with their squat form and short
ears, are not unlike those animals. In the past this genus occurred more
generally over Europe. Species from Miocene beds have been met with in
England, France, Germany, and Italy.
Fossil Rodents.—Quite a large number of existing genera
of Rodents are known from even the earlier strata of the Tertiary period.
The Squirrels (and even the genus Sciurus itself) occur in the
Upper Eocene. So, too, do the genera Myoxus, and (in South
America) Lagostomus. Spermophilus, Acomys,
Hystrix, Lagomys, Lepus, Hesperomys are known
from Miocene rocks. Rhizomys, Castor, Cricetus,
Mus, Microtus, and some others appear to have originated so
far as we know in the Pliocene, while a still larger series of existing
genera are Pleistocene. It is interesting {506}to note that some of
the extinct genera were much larger than recent forms. At present,
Hydrochoerus is the biggest Rodent; but the genus Megamys
from the Pampas formation of Argentina was "nearly as large as an ox."
The wider range of genera in the past is illustrated by Hystrix,
which, now an Old-World form, is represented by remains in the Miocene
and Pliocene of America.
It is a significant fact that of living genera Sciurus is the
oldest; for it has been pointed out that in a number of features the
Squirrels are among the most primitive of Rodents. The zygomatic arch is
slender, and has thus not acquired the specialisation that is to be found
in that part of the skull in other Rodents; moreover, the "jugal bone is
not supported by any process from the maxilla exactly as in the primitive
Ungulata." The feet, too, are unspecialised, though that is the case with
many other genera. It may also be pointed out that the teeth bear not a
little likeness to those of Ornithorhynchus in their
multituberculate character.
Some few fossil forms have already been dealt with in the preceding
pages.
The two genera Castoroides and Amblyrhiza, from the
Pleistocene of North America and the West Indies, are usually regarded as
forming a family. The skull of the former genus indicates an animal of
the size of a Bear. It is compared to that of Castor, but it has a
wide infra-orbital foramen. The teeth are four in each jaw, and are
formed of three to five lamellae; the incisors of this animal are
powerful but short. Amblyrhiza, on the other hand, has long
incisors which are longitudinally grooved anteriorly. It has a free
fibula. This latter as well as other characters have led Tullberg to
remove it from association with Castoroides.
Order X. TILLODONTIA.
This group of Eocene mammals is to be defined by a number of
characters, of which the more important are the following:—The incisors are enlarged, grow from persistent
pulps, and are coated with enamel upon the outer surface only; they are
those of the second pair only, the first and third having disappeared or
become small. The canines are reduced in the later forms. {507}
Fig. 247.—Tillotherium fodiens. Left lateral view of skull. (From Flower,
after Marsh.)
These animals have been regarded as ancestral Rodents, to which the
tooth characters just mentioned clearly show likenesses. The earliest
known form is Esthonyx. This genus shows such primitive
characters, compared with its later representatives, as the existence of
all three pairs of incisors in the upper jaw, but only two in the lower
jaw. The enlarged incisors of both jaws do not seem to have grown from
persistent pulps.
Anchippodus, a later form, still preserves the upper pair of
first incisors in a vestigial form; the strong second incisors grew from
persistent pulps. The most recent genus, Tillotherium, shows the
characteristics of the group at their height. The strong Rodent-like,
chisel-shaped incisors, which are reinforced by a small additional pair
in the upper jaws only, are persistent. The grinding teeth are of the
tritubercular pattern; there are three of each kind in the upper jaw, but
in the lower jaw only two premolars on each side. This is at any rate the
case with some, while others have three. The canine, though present in
both jaws, is insignificant. As in many ancient types, there is an
entepicondylar foramen in the humerus. The feet were five-toed, and bore
sharp, laterally-compressed claws. The skull has been compared in general
aspect to that of a Bear.
{508}
CHAPTER XVI
INSECTIVORA—CHIROPTERA
Order XI. INSECTIVORA.
The Insectivora[365]
are an order of mammals to which it is (to quote Professor Huxley)
"exceedingly difficult to give a definition." They are, however, none of
them large animals, and most of them are nocturnal in habit—two
circumstances which may have had something to do with their survival from
past ages, as may have also their modification to so many and diverse
modes of life; for everything points to the antiquity of the group. They
are, for instance, more or less plantigrade. The snout is generally long,
and is often prolonged into a short proboscis.[366] There is a tendency for the teeth to
be of a generalised type, and their number is often the typical mammalian
forty-four. Moreover, trituberculate teeth, which are certainly an
ancient form of tooth, are common; and indeed the Insectivora of the
southern regions of the globe, e.g. Centetidae, Solenodontidae,
and Chrysochloridae, have the most prevalent trituberculism, a fact which
is of importance in considering the age of the animal life of these
regions of the world. The limbs are, as a rule, provided with five
digits. The hemispheres of the brain are usually smooth, and do not
extend over the cerebellum. The palate is often fenestrate as in the
Marsupials, and as in that group the lower jaw is sometimes inflected.
But the latter character also occurs in the Sea-lions and elsewhere.
Clavicles are present, as a rule, but not in Potamogale. {509}
There is, furthermore, a distinct tendency towards a disappearance of
functional milk teeth, which is best seen in Sorex, where there
are only seven milk teeth, none of which ever cut the gum. This
suppression of the milk dentition is like that of the Marsupials,
Edentates, and Whales, all of which appear to be—the first
certainly are—ancient forms of mammalian life.
There is also a fairly well-defined, though shallow, cloaca in many
genera. Finally, the testes are purely abdominal in some, and in none is
there a full descent into a scrotum, as in the more highly-developed
Eutheria.
Sub-Order 1. INSECTIVORA VERA.
Fam. 1. Erinaceidae.—This family contains the genera
Erinaceus, Hylomys, and Gymnura.
Hylomys, considered by Dobson to fall within Gymnura, is
kept separate by Leche.[367] H. suillus is a Malayan
animal, small in size, about 5 inches long, with a short tail. Like
Gymnura it is spineless. The ears are decidedly large and nude.
There is one pair of inguinal and one pair of thoracic teats. The colour
above is a rusty brown with yellowish-white under parts. The palms and
soles are quite naked. In its general form it recalls Tupaia very
much more than its own immediate relatives. There is no doubt, however,
of its systematic position when the skeleton and teeth are examined. A
variety has been described from altitudes of 3000 to 8000 feet on Mount
Kina Balu in Borneo. It has the complete dentition of forty-four teeth.
There are fourteen pairs of ribs. As in Gymnura the tibia and
fibula are united below. The genus is considered by Leche to be the
oldest existing type of Erinaceidae.
Gymnura[368] is
also a Malayan form with the complete dentition of the last, but with
fifteen pairs of ribs and a longer tail, consisting of twenty-three
vertebrae as against fourteen. There is, as with Hylomys, but one
species, G. rafflesii. This animal has a peculiar odour,
resembling decomposed cooked vegetables. {510}The under surface of
the tail is rough, and it is thought by Dr. Blanford that it may be of
use to the animal in climbing. Its compressed terminal third and the
fringe of stiff bristles on the under surface of this indicate, according
to Dr. Dobson, powers of swimming, or at any rate a not very remote
ancestry of swimming creatures. It is purely insectivorous in diet.
Erinaceus, including the Hedgehogs, is a widely distributed
genus—Palaearctic, Oriental, and Ethiopian in range. There are
about twenty species. The familiar spines distinguish the Hedgehogs from
their allies, as also the fact that they possess but thirty-six teeth,
the formula being I 3/2 C 1/1 Pm 3/2 M 3/3. There are fifteen or fourteen
ribs, and the tail is very short, consisting of only twelve vertebrae. As
in Gymnura there is no caecum. The upper canine has usually, as in
other Erinaceidae, two roots, but not in E. europaeus, which is
one of the most modified of Hedgehogs.
The Hedgehog is a more omnivorous creature than Gymnura. It
eats not only insects and slugs, but also chickens and young game birds,
and lastly vipers. Four, or in some cases as many as five or six, young
are produced at a birth; they are blind, with soft and flexible white
spines. In hot and dry weather Hedgehogs disappear; they come forth in
rainy weather. The English Hedgehog, as is well known, hibernates. The
Indian species do not. The Hedgehog is occasionally spineless, which
condition may be regarded as an atavistic reversion.[369]
The Hedgehog has acquired the reputation of carrying off apples
transfixed upon its spines. Blumenbach thus quaintly describes this and
other habits of the animal, whose English name he gives as "hedgidog":
"Il se nourrit des productions des deux règnes organisés, miaule comme un
chat, et peut avaler une quantité énorme de mouches cantharides. Il est
certain qu'il pique les fruits avec les épines de son dos, et les porte
ainsi dans son terrier."[370]
The Miocene Palaeoerinaceus is so little different from
Erinaceus that it is really hardly generically separable.
Erinaceus is therefore clearly one of the oldest living genera of
mammals.
Necrogymnura of the same epoch and the same beds (Quercy
Phosphorites) is doubtless an ancestral form. The palate is {511}perforated as
in Erinaceus (it is not so in Gymnura and Hylomys),
but on the whole it comes nearest to Hylomys.
Fam. 2. Tupaiidae.—This family contains the genera
Tupaia and Ptilocercus. Tupaia is Oriental in range,
extending as far east as Borneo. There are a dozen or so of species,
which are generally arboreal and have the outward aspect of Squirrels. It
has been suggested that this is a case of mimicry, the animal gaining
some advantage by its likeness to the Rodent. The name Tupaia, it should
be added, means Squirrel, and the long-nosed Squirrel, Sciurus
laticaudatus, is so extraordinarily like it that "one has to look at
the teeth" to distinguish them. Moreover this Squirrel, like some
Tupaias, lives largely on the ground among fallen logs. Tupaia
resembles a Lemur in the complete orbit. The dental formula is I 2/3
C 1/1 Pm 3/3 M 3/3 = 38. The sublingua, too, is stated by Garrod to be
like that of Chiromys. There is a minute caecum in T.
belangeri, none in T. tana.
Ptilocercus[371] has a pen-like posterior portion to
the tail, a modification which is found in other groups of animals. The
tail of certain Phalangers, for instance, shows this same modification.
The rest of the tail is scaly. The animal, as was pointed out by Dr.
Gray,[372] looks very
much like a Phalanger. The orbit is entire as in Tupaia. The
fingers and toes are five. The one species, called after Sir Hugh Low,
G.C.M.G., P. lowi, is a Bornean animal.
Fam. 3. Centetidae.—This family is entirely confined to
the Island of Madagascar. It includes some seven genera. The best-known
genus is Centetes. C. ecaudatus, the Tanrec, Tenrec, or
Tendrac, is an animal a foot or so in length, without a tail, and with
forty-four teeth.[373]
The immature animal is so different from the parent as to appear quite a
different form. It has three narrow rows of spines along the back, which
do not wholly disappear until the permanent dentition has been acquired.
Even then the hairs are of a rather spiny character, particularly those
upon the back of the head, which are erected when the animal is {512}annoyed.
The Tanrec feeds mainly upon earthworms. It is "probably the most
prolific of all animals," since as many as twenty-one young are said to
have been brought forth at a birth. Some Opossums, however, have
twenty-five teats.

Fig. 248.—Skull of Tenrec. Centetes ecaudatus. fr, Frontal; max, maxilla; pa,
parietal; p.max, premaxilla; sq, squamosal. (After Dobson.)
Hemicentetes[374] is a genus with two species. These
have spines mixed with the fur of the back. There is no caecum in this or
in other Centetidae. The teeth are forty in number, there being only
three molars.
Ericulus setosus is a small Insectivore, resembling externally
a small Hedgehog. It is covered with close-set spines which, unlike what
is found in Erinaceus, extend over the short tail. The total
number of teeth is thirty-six, the formula being I 2/2 C 1/1 Pm 3/3
M 3/3.
Echinops[375]
is another spiny genus which is a stage in advance of Ericulus,
for still another molar has been lost, reducing the total number of teeth
to thirty-two. The dental formula is thus I 2/2 C 1/1 Pm 3/3 M 2/2. The
zygomata are reduced to mere threads.
Microgale, a genus recently instituted by Mr. Thomas, is a
small furry Insectivore with a long tail, which is more than double the
length of the head and body. There are no less than forty-seven vertebrae
in the tail, which is relatively longer than that of any other
mammal.
Limnogale, discovered by Forsyth Major, is an aquatic genus,
also furry and not spiny, which has departed from the Centetid type and
taken to an aquatic life. The single species, {513}L. mergulus, is
about the size of Mus rattus; it has webbed toes and a powerful
laterally-compressed tail. Clavicles are present, which is not the case
with Potamogale.
Oryzoryctes is a Mole-like Centetid. It has fossorial
fore-limbs, but a fairly long tail. This genus is furry like the last
two. It is said to burrow in the rice-fields and to do much harm. The
teeth are forty in number, three incisors and three molars in each half
of each jaw.
Fam. 4. Potamogalidae.—This family contains two genera,
Potamogale and Geogale.
Potamogale velox is a West African animal, which though an
Insectivore has the habits of an Otter. It is "somewhat larger than a
stoat." The upper surface of the body is dark brown, the belly brownish
yellow. It has a flat head and a long tail like the Stoat, but the tail
is laterally compressed and very thick. The eyes are very small; the
nostril has valves. The toes are not webbed; but the second and third
toes are united for the whole length of their first phalanges. Along the
outer side of the foot is a thin extension of the integument. In swimming
the feet are drawn up along the body, hence webbing would be of no use;
but the thin flattening prevents the edge of the foot from acting as a
hindrance to the motion of the animal. M. du Chaillu describes it as
catching fish, which it pursues with extreme rapidity in the clear
mountain streams it frequents; but Dr. Dobson, remarking that no stomachs
have been examined, thinks that water insects are more probably its prey.
It is not known whether the animal possesses a caecum. The tooth
formula[376] is I 3/3
C 1/1 Pm 3/3 M 3/3. The animal is exceptional among the Insectivora in
having no clavicles.[377]
There are sixteen ribs; there is no zygomatic arch, and the pterygoids
converge posteriorly.
Geogale, with one species, G. aurita, is a small
representative of this family from Madagascar. It has only thirty-four
teeth. When better known it may be necessary, thinks Mr. Lydekker, to
make this animal the type of a separate family. The tibia and fibula are
distinct, not confluent with one another as in Potamogale.
Fam. 5. Solenodontidae.—This family contains but a single
genus. {514}
Solenodon. This genus, including two species, one from Cuba,
the other from Hayti, was at one time referred to the Centetidae. It
offers, however, numerous points of difference from the members of that
family with some general points of agreement. Possibly its isolation in
the two West Indian islands mentioned is comparable to the isolation of
the Centetidae in Madagascar; they are both survivors of an ancient group
of Insectivores extinct elsewhere. Solenodon has nearly the
complete dentition. It has lost only one premolar, and has therefore
forty teeth in all. The formula is thus I 3/3 C 1/1 Pm 3/3 M 3/3. It also
differs from the Centetidae in having only two inguinal mammae instead of
both inguinal and thoracic; the penis of the male does not project from a
cloaca, but lies forward. On the other hand, the molars have their cusps
arranged in the -fashion of the Centetidae, a fact, however, which, in the opinion of
some, merely points to an ancient trituberculism not indicative of
special affinity. It has, moreover, no zygoma in the skull, and there is
no caecum. Dr. Dobson has furthermore tabulated a number of differences
in muscular anatomy between the two families. Solenodon has a long
naked tail. The snout, always developed in Insectivores, is
extraordinarily long in this genus. It is a furry, not a spiny animal.
S. cubanus is liable to fits of rage when irritated, a feature
which it has in common with Shrews and Moles; it is also stated to have
the ostrich-like way of concealing its head in a crevice, "apparently
thinking itself then secure." But nothing is known of the genus in a wild
state.
Fam. 6. Chrysochloridae.—This family contains only the
genus Chrysochloris, comprising some five species, all natives of
Africa south of the equator. The scientific name of the genus, and also
the vernacular name Cape Golden Mole, are derived from the beautiful
iridescent hairs which are intermingled with softer and non-iridescent
fur. Chrysochloris has -shaped cusped teeth like those that are possessed by the Centetidae
and Solenodontidae. In the skull as in the Macroscelidae, etc., but not
in the Centetidae, there are complete zygomata. They are Moles in habit,
and the eyes are covered with skin; the ears, moreover, have no conches.
The teeth are forty or thirty-six in number, the reduction being caused
by the losing of a molar in those forms which possess the smaller
number.[378] It is
interesting to notice that the {515}adaptation to a digging life is brought
about in quite a different way from that of the true Moles
(Talpa). In the latter the fore-limbs are changed in position by
the elongation of the manubrium sterni, carrying with it the clavicles,
which are extraordinarily shortened (Fig. 251). In Chrysochloris,
on the other hand, the same need (i.e. that the limbs project as
little as possible from the sides of the body, while the length of the
limbs is retained, and the leverage of the muscles unaffected) is
provided for by a hollowing out of the walls of the thorax, the ribs and
the sternum being here convex inwards. The sternum and the clavicles are
not modified. The tibia and fibula are ankylosed below. In the manus,
moreover, there are but four digits, of which the two middle ones are
greatly enlarged. In the Moles there are five fingers, and all are
enlarged; there is, too, a great radial sesamoid bone, which is as good
as a sixth finger (which, indeed, it is considered to be, in common with
similar structures in other animals, by some anatomists). The foot has
only four toes.

Fig. 249.—Golden Mole. Chrysochloris trevelyani. A, Lower surface of fore-foot.
× ½. (After Günther.)
Fam. 7. Macroscelidae.[379]—This family contains three
genera, all of them African in range, and mainly Ethiopian.
Macroscelides, the Elephant Shrews, are jumping creatures of
Shrew-like appearance, combined with a Marsupial look. Both radius and
ulna, and tibia and fibula, are ankylosed. There {516}are five fingers and
toes. There is a caecum as in but few Insectivores. The tooth formula, as
revised by Thomas,[380]
is I 3/3 C 1/1 Pm 4/4 M 2/(2 or 3), the total number being thus forty or
forty-two. There are several species of this genus.
Fig. 250.—Rhynchocyon chrysopygus. × ¼. (After Günther.)
Rhynchocyon and Petrodromus differ from
Macroscelides in not having such long hind-legs. The dental
formula of the first is I (1 or 0)/3 C 1/1 Pm 4/4 M 2/2 = 34 or 36, of
the latter I 3/3 C 1/1 Pm 4/4 M 2/2 = 40. In Petrodromus the toes
are reduced to four; in Rhynchocyon there are only four digits in
the manus as well as in the pes. This animal, as its name implies, has a
longish proboscis, which can be bent, and is really very like a miniature
Elephant's trunk, and also like that of the Desman (Myogale). It
has thirteen pairs of ribs, and a well-developed caecum. Dr. Günther has
pointed out that in Petrodromus tetradactylus the hairs of the
lower part of the tail are stiff elastic bristles 5 mm. long, with a
swelling at the free tip. The use of this singular modification is not at
all apparent. Pseudorhynchocyon, of European Oligocene, is
believed to be related to this family.
|
|
|
|
Fig. 251.—Sternum and sternal ribs of the
Common Mole (Talpa europaea), with the
clavicles (cl) and humeri (H); M, Manubrium
sterni. Nat. size. (From Flower's
Osteology.)
|
Fig. 252.—Bones of fore-arm and
manus of Mole (Talpa europaea).
× 2. C, Cuneiform; ce, centrale;
l, lunar; m, magnum; p, pisiform;
R, radius; rs, radial sesamoid
(falciform); s, scaphoid; td,
trapezoid; tm, trapezium; U,
ulna; u, unciform; I-V, the
digits. (From Flower's Osteology.)
|
Fam. 8. Talpidae.—This family is confined to the
Palaearctic and Nearctic regions, or practically so, being fairly equally
{517}distributed as regards genera; a Mole just
gets over the boundary into the Oriental region. The genus Talpa
is entirely Old World in range, and includes several species, of which
the Common Mole, T. europaea, is the best known. There are forty
teeth, one of the molars of the full mammalian dentition not being
represented. In the milk dentition there is an additional premolar, not
represented by a successor in the permanent dentition. The formula is
thus I 3/3 C 1/1 Pm 4/4 M 3/3. There are no external ears, and the eyes
are rudimentary; the soft silky fur is familiar to everybody. The sternum
has a strong crest, associated with a powerful development of the
pectoral muscles, so necessary to a burrowing animal. The animal, it is
hardly necessary to state, lives underground in burrows excavated by
itself, which have not, it has been stated, the elaborate and, it
appears, fanciful shape assigned to them by many writers. At times Moles
appear above ground. {518}Their principal food consists of
earthworms, and it may not be out of place to quote Topsell's quaint
account of their pursuit of the annelids: "When the wormes are followed
by molds (for by digging and heaving they foreknow their owne perdition)
they fly to the superficies and very toppe of the earth, the silly beast
knowing that the molde, their adversary, dare not followe them into the
light, so that their wit in flying their enemy is greater than in turning
againe when they are troade upon." It has lately been said[381] that Moles store up
earthworms for consumption during the winter, biting off their heads to
prevent their crawling away.
Scalops, an American genus, is a Mole-like creature of largely
aquatic habits, as its webbed hind-feet show; it has a short, naked tail.
Apparently, like the Shrews, it has no lower canines.
Condylura, another American genus, is called the Star-nosed
Mole on account of a curious radiating structure at the end of the
snout.
Myogale, the Desman, is still more aquatic in habit, and
connects the Moles with the Shrews, though, as in many of the former, it
has lower canines. It has webbed hind-feet and a long tail. One species
occurs in the Pyrenees, the other in Russia. A few other genera
(Urotrichus, Uropsilus, Scaptonyx, Dymecodon,
Scapasius, Perascalops) belong to the same family.
Fam. 9. Soricidae.—The true Shrews have a much wider
range than other families of the present order. In the Palaearctic region
are found Sorex, Crossopus, Crocidura,
Nectogale, Chimarrogale. The first is also Nearctic, and
reaches Central America. In the Ethiopian region is the single peculiar
genus Myosorex, but Crocidura occurs there also.
Blarina and Notiosorex are "Sonoran" in range;
Soriculus Oriental. Crocidura, Anurosorex, and
Chimarrogale also enter this region. Sorex has teeth tipped
with reddish colour, its dental formula being, according to Mr.
Woodward's recent researches, I 3/(2 or 3) C 1/0 Pm 3/1 M 3/3 = 32 or
34.
As compared with other Insectivores, therefore, the most remarkable
fact found throughout the family is the absence of the lower canines. In
addition to this the genus may be known—the family indeed—by
the large size of the first pair of incisors. In the above formula it is
possible, thinks Mr. Woodward, that there may be errors; he is not
certain whether the supposed {519}upper canine may not be a fourth incisor,
and whether the first premolar may not be really the canine. Another
peculiar feature about the dentition of Sorex is the suppression
of the teeth of the milk dentition, which are functionless, and probably
uncalcified. The genus Sorex is terrestrial. The tail is long and
covered with hairs. There are two species in this country, S.
vulgaris and S. minutus. The former is the Shrew of legend and
superstition; and it is no doubt the species that has lent its name to
the more untameable members of the softer sex, though it is the males
which are especially pugnacious. As to legend, everybody has heard of the
shrew ash whose leaves, after a Shrew has been inserted living into a
hole cleft in the tree, are a specific for diseases of cattle, caused by
the Shrew itself creeping over them.
The Rev. Edward Topsell, author of The Historie of Four-footed
Beastes, who defends his veracity by asserting that he does not write
"for the rude and vulgar sort, who being utterly ignorant of the
operation of learning, do presently condemne al strange things," says of
the Shrew that "it is a ravening beast, feigning itself gentle and tame,
but, being touched, it biteth deep and poysoneth deadly. It beareth a
cruel minde, desiring to hurt anything, neither is there any creature
that it loveth, or it loveth him, because it is feared of all." It is
probable that all this rustic feeling is due to the powerful effluvium
which the Shrew undoubtedly emits.
S. minutus has the distinction of being the smallest British
mammal; it is scarcer than the last. This form is found upon the Alps, as
is also the peculiarly Alpine species S. alpinus, which inhabits
the Alps, Pyrenees, Carpathians, and the Hartz.
Crossopus fodiens, the Water Shrew, has also brown-stained
teeth. It is not uncommon in this country, and lives in burrows excavated
by the sides of the streams which it affects.
Besides these two genera, Soriculus, Blarina, and
Notiosorex have red-tipped teeth. In Crocidura,
Myosorex, Diplomesodon, Anurosorex,
Chimarrogale, and Nectogale the teeth are white-tipped.
These are all the genera of the family allowed by the late Dr. Dobson in
a review of that family.[382]
Chimarrogale and Nectogale are aquatic genera. The
former {520}consists of a Himalayan and Bornean, and
of a Japanese species, which have not webbed feet, but have a tail with a
fringe of elongated hairs.
Nectogale elegans is one of the characteristic animals of the
Thibetan plateau. It has webbed feet. The teeth are as in
Chimarrogale I 3/2 C 1/0 Pm 1/1 M 3/3.
The other genera are terrestrial in habit.
Sub-Order 2. DERMOPTERA.
Fig. 253.—Galeopithecus volans. × ⅓. (After Vogt and Specht.)
The family Galeopithecidae contains but one genus, which has
been at times referred to the Lemurs, to the Bats, or has been made the
type of a special order of mammals. It is better to regard it as an
aberrant Insectivore—so different indeed from other forms that it
requires a special sub-order for its reception. {521}
Galeopithecus[383] inhabits the Oriental region. It is a
larger animal than any other Insectivore, about the size of a Cat, and
has a patagium extending between the neck and the fore-limb, between the
fore-limb and the hind-limb, and between the hind-limb and the tail. This
patagium is abundantly supplied with musculature, but the fingers are not
elongated as in the Bats for its support. In the degree of its
development, however, the patagium of this creature is midway between
that of Sciuropterus on the one hand, and the Bats on the other.
It presents many remarkable features in its organisation. The brain is
like that of the Insectivora in the exposure of the corpora quadrigemina
by the slight extension backward of the cerebral hemispheres; but its
upper surface is marked by two longitudinal furrows on each side, a state
of affairs (in combination) which is unparalleled among the Mammalia. The
teeth are peculiar by reason of the singular "comb-like" structure of the
lower incisors. This, however, is an exaggeration of what is to be found
in Rhynchocyon and Petrodromus, while the same style of
tooth, though not so highly developed, characterises certain Bats. The
Tupaiidae and certain Lemurs show what Dr. Leche regards as the beginning
of the same thing. As in Tupaia also there is an indication of the
characteristically Lemurine sublingua. The stomach is more specialised
than in other Insectivores, the pyloric region being extended as a
narrowish tube. There is a caecum. A peculiarity of the intestinal tract
is that the large intestine is longer than the small.
Order XII. CHIROPTERA.
We may thus define the Bats:—Flying
mammals, with the phalanges of the four digits of the hand following the
pollex greatly elongated, and supporting between themselves and the
hind-limbs and tail a thin integumental membrane, which forms the wing.
The radius is long and curved; the ulna rudimentary. The knee is directed
backwards, owing to the rotation of the limb outward by the wing
membrane. From the inner side of the ankle-joint arises a cartilaginous
process, the calcar, which supports the interfemoral part of the wing
{522}membrane. The mammae are thoracic; the
placenta discoidal and deciduate. The cerebral hemispheres, which are
smooth, do not extend over the cerebellum.
Fig. 254.—Barbastelle. Synotus barbastellus. × ½. (After Vogt and Specht.)
This large order of mammals was once placed with the Primates. There
is no doubt, however, that they form a perfectly distinct order; no
knowledge of fossil forms in any way bridges over the gap which
distinguishes them from the highest mammals. The most salient feature in
their organisation is clearly the wings. These consist of membrane, an
expansion of the integument, provided with nerves, blood-vessels, etc.,
which mainly lie stretched between the digits 2 to 5. These digits
themselves, which are enormously elongated, act like the ribs of an
umbrella, and when the wing is folded they come into contact. Besides
this part of the flying apparatus there is a tract of membrane lying in
front of the arm, which corresponds to the wing membrane of the bird, but
which in the Bats takes quite a subordinate place. In the bird, on the
other hand, there is a metapatagium, which is the main part of the wing
of the Bat. It seems just possible that in Archaeopteryx the
metapatagium was more Bat-like. Furthermore, a steering membrane, like
that which fringes the tail in some Pterosaurians, lies interfemorally in
Bats, and includes the whole or a part of the tail. The pollex takes no
share in the wing, but projects, strongly armed with a claw, from the
upper margin.
The bones of this order of mammals are slender and marrowy; they are
thus light, and subserve the function of flight. A most remarkable
feature among the external characters of the Bat tribe is the
extraordinary and often highly complicated membranes which surround the
nostrils. These are at least often {523}more strongly developed
in males than in females, and may perhaps be partly relegated to the
category of secondary sexual characters. But it seems that they have also
an important tactile function, and enable the creatures to fly without
touching bodies which intrude themselves upon their way. The ears, too,
are frequently very large, and it may be supposed that the sense of
hearing is correspondingly acute. In the common Long-eared Bat of this
country, the ears are not greatly inferior in length to the head and body
of the animal combined. The ears are of every variety of shape, and offer
characters which are valuable in the systematic arrangement of the
members of the order.

Fig. 255.—Skeleton of Flying Fox. Pteropus jubatus. × ⅛. (After de Blainville.)
In the skull of Bats there is very rarely a complete separation
between the orbital and temporal fossae; the lachrymal duct is outside
the orbit. The tympanics are annular, and in a rudimentary condition. The
centra {524}of the vertebrae tend to become ankylosed
in old individuals; the caudals have no processes, but are like those
quite at the end of the series in long-tailed animals. The sternum is
keeled for the better attachment of the pectoral muscles, the chief
muscles of flight. The ribs, which are much flattened, are occasionally
ankylosed together by their margins. There is a well-developed clavicle.
In the carpus the scaphoid, lunar, and cuneiform are all fused together.
In the hind-limb the fibula is rarely fully developed.
The Bats are divisible into two primary groups, which are those of the
Megachiroptera and the Microchiroptera.
Sub-Order 1. MEGACHIROPTERA.
Fig. 256.—Skull of Pteropus fuscus. × 3⁄2. (After de Blainville.)
The Pteropodidae are frugivorous Bats, usually of large size.
The chief distinguishing feature is the fact that the molars are not
tubercular, but marked with a longitudinal furrow, which is, however,
concealed in the genus Pteralopex by cusps. The palate is
continued back behind the molars. The index finger has three phalanges,
and is usually clawed. The ears are oval, and the two edges are in
contact at the base of the ear. The tail, if present, has nothing to do
with the interfemoral membrane. This group is entirely Old World in
range. The genus Pteropus embraces the creatures known as Flying
Foxes. They are the largest forms in the sub-order, sometimes having an
expanse of wing of 5 feet (this is the case with P. edulis). The
muzzle is long, and the face therefore "foxy" in appearance. {525}The inner
margin of the nostrils projects, a preparation for the tubular nostrils
of Harpyia. The tail is absent. The premolars are three and the
molars two. The pyloric region of the stomach is extended and twisted
upon itself. Of this genus there are nearly sixty species, extending from
Madagascar to Queensland. Thirty species inhabit the Australian, twenty
the Oriental region. Madagascar has seven, and one species just enters
the Palaearctic. The occurrence of this genus in India and in Madagascar
is one of those facts which favour the view supported, on these and other
grounds, by Dr. Dobson and Dr. Blanford that a connexion between India
and Madagascar must once have existed; for these slow-flying creatures
could hardly be believed capable of traversing vast stretches of ocean by
their unaided efforts.[384]

Fig. 257.—Flying Fox. Pteropus poliocephalus. × ⅓.
Pteropus is represented in the Ethiopian region by the allied
genus Epomophorus. Of this there are perhaps a dozen species. The
teeth are reduced to two premolars in the upper jaw, {526}three
remaining below; while there is but one molar in each upper jaw, and two
in each lower. Dr. Dobson has studied the structure of the remarkable
pharyngeal sacs which exist in the neck of the male, and are capable of
inflation.
Pteralopex of the Solomon Islands has shorter ears than have
many Pteropus, otherwise its external characters are the same. As
in Pteropus nicobaricus, this genus has the orbits shut off by a
bony ring, an extremely rare phenomenon in Bats. The canines have two
cusps. The characters of the grinding teeth have already been mentioned.
It is uncertain whether the only species of this genus, P. atrata,
is, or is not, a vegetable feeder. Harpyia has shortish ears and
extraordinarily prolonged and tubular nostrils. There is a hint of the
accessory cusp to the canines mentioned above in Pteralopex. The
incisors are reduced to one on each upper jaw, and none below.
Cynopterus has also often bituberculate canines. It is an Oriental
genus with several species.
Nesonycteris, with one species from the Solomon Islands, N.
woodfordi, has the dental formula I 2/1 C 1/1 Pm 3/3 M 2/3. The index
finger has no claw; the tail is absent. The premaxillae are separated
anteriorly.
Eonycteris, with a single cave-dwelling species from Burmah,
E. spelaea, has also no claw upon the index; the tooth formula is
fuller by reason of the presence of an additional incisor below. The
tongue is very long and is armed with papillae. There is a short but
distinct tail.
Notopteris, from New Guinea and the Fiji Islands, is
distinguished from the related genera by its long tail.
The remaining genera of Fruit Bats are Boneia,
Harpyionycteris, Cephalotes, Callinycteris, and
Macroglossus, from the Oriental region, and Scotonycteris,
Liponyx, and Megaloglossus from the Ethiopian region;
finally, there is the Australian Melonycteris.
Sub-Order 2. MICROCHIROPTERA.
The members of this sub-order are mostly insectivorous though
occasionally "frugivorous or sanguivorous" Bats. The molars are
multicuspid with sharp cusps. The palate is not continued back behind the
last molar. The second finger has but one phalanx, or {527}none;
occasionally there are two. It has no claw. The ear has its two sides
separate from their point of origin upon the head. The group is of
Old-World distribution.
Fam. 1. Rhinolophidae.—The Bats of this family possess
the leafy outgrowths around the nostrils. The ears are large, but have no
tragus. The index finger has no phalanx at all. The premaxillary bones
are quite rudimentary, and are suspended from the nasal cartilages. In
addition to the pectoral mammae they have two teat-like processes
situated abdominally. The tail is long, and extends to the end of the
interfemoral membrane.
The genus Rhinolophus has a large nose leaf, and an antitragus
to the ear. The first toe has two joints, the remaining toes have three
joints each. The dentition is I 1/2 C 1/1 Pm 2/3 M 3/3. There are nearly
thirty species of the genus, which are restricted to the Old World. Two
species occur in this country, viz. R. ferrum equinum, the Great
Horse-shoe Bat, and the Lesser Horse-shoe Bat, R. hipposiderus.
The name is of course derived from the shape of the nose leaf.
The genus Hipposiderus and some allied forms are placed away
from Rhinolophus and its immediate allies in a sub-family
Hipposiderinae. The type genus Hipposiderus, or, as it
ought apparently to be called, Phyllorhina, is Old World in range,
like all the other members of the family.
The nose leaf is complicated, and there are only two phalanges in all
the toes; there is no antitragus to the ear. A curious feature in the
osteology of the genus, and indeed of the sub-family, is the fact that
the ileo-pectineal process is connected with the ilium by a bony bridge;
this arrangement is unique among mammals.
The genus Anthops, only known from the Solomon Islands, and
represented there by but a single species (A. ornatus), has an
extraordinarily complicated nose leaf. The tail, like that of the
Oriental Coelops, likewise represented by a single species (C.
frithii), is rudimentary.
Triaenops, Ethiopian and Malagasy, has, like the Australian
Rhinonycteris, a well-developed tail. Triaenops has also a
highly-complicated nose leaf.
Fam. 2. Nycteridae.—This family is to be distinguished
from the Rhinolophidae by the fact that the ear has a small tragus, and
by the small and cartilaginous premaxillae. In addition to {528}these two
characters it may be added that the nose leaf is well developed, but is
not so complicated as in the last family. The type genus Nycteris
is Ethiopian and Oriental, nine species being African, and only one,
N. javanica, being, as the specific name denotes, from the East.
Megaderma is to be distinguished by the loss of the upper
incisors. There is no tail, and the ears are particularly large. They are
carnivorous Bats, and M. lyra, called the "Indian Vampire Bat,"
chiefly affects frogs as an article of diet.
Fam. 3. Vespertilionidae.—This family has not the nose
leaf of other families. The apertures of the nostrils are simple, round,
or crescentic apertures. The ear has a tragus, and the tail is not
produced to any great degree behind the interfemoral membrane. There are
two phalanges to the index digit.
This family in numbers of species is vastly in excess of any other
family of Bats. The most recent estimate, that of P. L. and W. L.
Sclater, allows 190. But the generic types are by no means so numerous as
in the Phyllostomatidae. This is a significant fact when we reflect upon
the geographical range of the two families. The Vespertilionidae range
over the whole earth, while the Phyllostomatidae are practically limited
to the South American continent, only just getting into the Nearctic
region. They inhabit, therefore, a more restricted area, and, in
consequence of competition, have specialised more freely than the
widely-spread and therefore not crowded Vespertilionidae.
The genus Vesperugo is by far the largest genus of this family,
embracing no less than seventy species. The tail is shorter than the head
and body together; the ears are separate, and moderate or short in size;
the tragus is generally short and obtuse. The dentition is I 2, C 1, Pm 2
or 1, M 3. It is a remarkable fact that this genus, unlike most Bats,
produces two young at a time. The genus is universal in range, and one
species, the Serotine Bat, known in this country, even ranges from the
New World to the Old; but with so small a creature the possibility of
accidental transportation by man must not be left out of sight. The
British species are—V. serotinus, the Serotine already
mentioned; V. discolor, a single example only of which has
occurred, and may have been introduced; V. noctula, the habits of
which were described by Gilbert White; V. leisleri; and the
Pipistrelle, V. pipistrellus, which is the best-known member of
the genus in this country.
{529}
The genus Vespertilio contains some forty-five species, and is
world-wide in range. It has one more premolar in the upper jaw than has
Vesperugo. There are no less than six British species, of which
V. murinus is the largest species of Bat recorded from this
country, but is not quite certainly indigenous.
Plecotus has very long ears. The dentition is I 2/3 C 1/1
Pm 2/3 M 3/3. The tragus is very large. There are but two or possibly
three species, of which one is North American, and the other is the
Long-eared Bat, P. auritus, of this country, but ranging as far as
India. The shrill voice, inaudible to some ears, of this Bat has been
heard of by everybody.
Synotus includes the British Barbastelle, S.
barbastellus, as well as an Eastern form. It differs from the last
genus principally by the loss of a lower premolar. The ears, too, are not
so large. Otonycteris, Nyctophilus, and Antrozous
are allied genera; the last is Californian, the others Old-World
forms.
Kerivoula (or Cerivoula) has a long, pointed, narrow
tragus. The tail is as long as or longer than the head and body. The
dentition is as in Vespertilio; but the upper incisors are
parallel instead of divergent as in that genus. The brilliantly-coloured
K. picta is, on account of this very fact, the best-known species.
The name Kerivoula, a corruption of the Cinghalese "Kehel vulha,"
signifies plantain bat. This Bat has been described as looking, when
disturbed in the daytime, more like a huge butterfly than a Bat, which is
naturally associated with sombre hues. Other species occur in the
Oriental, Australian, and Ethiopian regions.
Miniopterus has a premolar less in the upper jaw; it has a long
tail as in the last genus. One species, M. scheibersi, has almost
the widest range of any Bat, it being found from South Europe to Africa,
Asia, Madagascar, and Australia.
Natalus is an allied form from Tropical America and the West
Indies. It is chiefly to be separated from Kerivoula by the short
tragus to the ear.
Thyroptera is also South American. It is distinguished by the
curious sucker-like discs upon the thumb and foot. These "resemble in
miniature the sucking cups of cuttle-fishes." The Madagascar genus,
Myxopoda, with but one species, has also an adhesive but
horse-shoe-shaped pad upon the thumb and foot.
Scotophilus has shortish ears with a tapering tragus. The tail
is shorter than the head and body, and is nearly contained within {530}the
interfemoral membrane. The dentition is I 1/3 C 1/1 Pm 1/2 M 3/3, with
another upper incisor in the young. It is African, Asiatic, and
Australian.
This genus appears to be connected with Vesperugo by Mr.
Dobson's proposed genus, or sub-genus as it is generally held to be,
Scotozous.[385]
The genus Nycticejus, founded for the inclusion of Scotozous
dormeri, an Indian species, should, according to Dr. Blanford,
replace on grounds of priority the name Scotophilus. But as this
name (Nycticejus) is one introduced by Rafinesque, whose work was
so uncertain and untrustworthy, it seems preferable to retain the
better-known name of Scotophilus, introduced by William Elford
Leach.
The genus Chalinolobus[386] has short, broad ears with an
expanded tragus. A distinct fleshy lobule projects from the lower lip on
either side of the mouth. The tail is as long as the head and the body.
The dental formula is I 2/3 C 1/1 Pm 2/2 or 1/2 M 3/3. The genus occurs
in Africa, Australia, and New Zealand; but the African species, with
diminished premolars and pale coloration, have been distinguished as
Glauconycteris.
Fam. 4. Emballonuridae.—The Bats belonging to this family
have no nose leaf. The tragus is present, but often very small. The ears
in this family are often united. There are two phalanges in the middle
finger. The tail is partly free, either perforating the interfemoral
membrane and appearing upon its upper surface, or prolonged beyond its
end. The face is obliquely truncated in front, the nostrils appearing
beyond the lower lip.
Emballonura is Australian, Oriental, and Mascarene in range.
The ears arise separately, and there is a fairly developed and narrow
tragus. The tail perforates the interfemoral membrane. The dental formula
is I 2/3 C 1/1 Pm 2/2 M 3/3.
Rhinopoma has the ears united; the incisors are reduced by one
on each side of each jaw, and the premolars are similarly reduced, but
only in the upper jaw.
Noctilio is an American genus of two or three species, which
has one pair of markedly large upper incisors, which completely conceal
the outer pair. On these grounds this Bat was removed from its allies and
placed by Linnaeus among the Rodents, an instance of the disadvantage of
the artificial scheme of classification. The species named N.
leporinus has been shown to feed upon fish. {531}
Furia, Amorphochilus, Rhynchonycteris,
Saccopteryx, Cormura, and Diclidurus are other
Neotropical genera of the same family.
The genus Taphozous[387] has a tail which perforates the
interfemoral membrane, appearing on its upper surface; it is capable of
being withdrawn. The premaxillaries are cartilaginous. The dentition is
I 1/2 C 1/1 Pm 2/2 M 3/3. The upper incisors often disappear. Many
species of the genus have a gular sac, opening anteriorly between the
jaws. This is better developed in the males. The genus ranges from Africa
through Asia to New Guinea and Australia. There are some twelve
species.
The genus Molossus[388] has short legs and well-developed
fibulae. The tail is thick and fleshy, and is prolonged far beyond the
margin of the interfemoral membrane. The ears are united together above
the nose; the tragus is minute. The dentition is I 1/1 or 1/2 C 1/1
Pm 1/2 or 2/2 M 3/3. This genus, which is confined to the tropical and
subtropical portions of America, has long and narrow wings. The Bats can
thus fly rapidly, twist about with ease, and capture strongly-flying
insects. There are a large number of species.
Nyctinomus is an allied genus, and also has many species. These
range through both hemispheres. The chief differences from
Molossus are that the premaxillary bones are separate in front or
united by cartilage, and that the incisors may be three in the lower
jaw.
Fam. 5. Phyllostomatidae.—The Bats of this family are
extremely numerous and almost entirely confined to South America. None of
them occur outside the New World. There are some thirty-five genera. The
members of the family are to be distinguished by the presence of the nose
leaf, by the well-developed premaxillae, and by the possession of three
phalanges by the middle finger. They are large, and the tragus of the ear
is well developed.
Vampyrus of South America contains the large species V.
spectrum, which, mainly on account as it seems of its "forbidding
aspect," was supposed to be a bloodsucker. This genus has two incisors on
each side of the upper jaw.
The genus Glossophaga represents another type of structure in
this family. The tongue is long and extensile, and is much attenuated
towards the tip, where it is covered with strong and recurved papillae.
This structure was at one time thought to indicate a {532}bloodsucking
habit; but its use appears to be merely that of scooping out the soft
insides of fruits, upon which the Bat mainly lives. The incisors are only
one on each side of the upper jaw. The really bloodsucking Bats of this
family belong to the genera Desmodus and Diphylla. The
former is the Vampire, the species being known as Desmodus rufus.
These Bats have no tail; there is no true molar tooth; the canines are
large, and the single pair of upper incisors quite caniniform, and very
sharp and strong. These are the main teeth for aggression. In accordance
with its diet of blood, the Vampire has a peculiarly modified intestine.
The gullet is provided with a bore so small that nothing but fluid food
could pass down it; the stomach is intestiniform in shape.
{533}
CHAPTER XVII
PRIMATES
Order XIII. PRIMATES.
The highest of mammals, the Primates,[389] may be thus differentiated from other
groups:—Completely hairy, generally
arboreal mammals, with five digits on fore- and hind-limbs, provided with
flat nails (except in the case of certain Lemurs and the Marmosets), the
phalanges that bear these being flattened at the extremity and expanded
rather than diminished in size. The fore-feet are grasping hands as a
rule, and the hind-feet walking as well as (generally) grasping organs,
and the mode of progression is plantigrade. The teats, except in
Chiromys, are thoracic, and even axillary in position. The skull
is characterised by the fact that the orbital and the temporal vacuities
are, at least partly, separated by bone. The clavicles are always
present. The carpus has separate lunar and scaphoid bones, and the
centrale is often present. There is rarely an entepicondylar foramen in
the humerus, except in some archaic Lemurs. The femur has no third
trochanter. The stomach is usually simple, being sacculated only in
Semnopithecinae. The caecum is always present, and often large.
This great group could be easily divided into two separate orders, the
Apes and the Lemurs, if it were not for certain fossil types. As will be
seen from the description of Nesopithecus and of Tarsius,
the actual hard and fast lines between all Apes and all
Lemurs are very few. On the other hand, it is a little difficult to draw
a hard and fast line between the Primates as a whole—or at least
between the Lemurine section—and the Creodonta, a {534}group to which
so many others appear to converge. It is disputed, for example, whether
the Chriacidae among extinct Lemurs are rightly placed, or whether they
should be referred to the Creodonta. The number of primitive characters
seen among the Primates, even in Man himself, is remarkable. Of these the
more important are the five digits of both limbs and the plantigrade
walk, the presence of clavicles and of a centrale, and the absence of a
third trochanter. All these features distinguish the early Eutheria.
Sub-Order 1. LEMUROIDEA.[390]
The animals known as Lemurs, from their nocturnal and ghostly habits,
are on a lower level of organisation than the other division of the
Primates. Even the external form enables the members of the present
sub-order to be readily distinguished from the higher Anthropoidea. The
head is more like that of a Fox, with a sharp muzzle; it lacks the human
expression of the face of even the lower among the Apes. The long tail is
never prehensile, and there is never any trace of cheek pouches or of
integumental callosities, which are frequently so characteristic of the
Apes. The Lemurs agree with the remainder of the Primates in having
pectoral mammae (sometimes abdominal ones are present in addition, and in
Hapalemur—in the male at least—there is a mamma upon
each shoulder), in having opposable thumbs and toes, and in the flattened
digits. The tail varies from complete absence (in the Loris) to a
great length and bushiness in the Aye-aye. The pectoral limbs are always
shorter than the hind-limbs; the reverse is occasionally the case in the
Anthropoidea. A curious contrast between the two divisions of the
Primates concerns the digits of the hands and feet. In the Anthropoidea
it is the hallux or pollex which is subject to great variation. In the
Lemurs, on the contrary, the thumb and great toe are always well
developed, but the second or the third digit constantly shows some
abnormality; thus the singular elongation of the third digit of the hand
in Chiromys and the absence of the index in the Potto.[391] In all Lemurs the {535}second toe is furnished with a sharp nail,
unlike the flattened nails of the other fingers and toes, and in
Tarsius the third also is thus provided. As to osteology, the
shape of the head, already referred to, indicates some of the differences
in the skull which mark off the Lemurs from the Anthropoidea. The brain
case is small relatively to the face; the orbital and temporal fossae are
in communication, though the frontal and jugal bones are united behind
the orbit. The two halves of the lower jaw are not invariably ossified to
form one piece, as is the case with most Apes. The lachrymal foramen lies
upon the face in front of the orbit. The teeth are characteristic, not so
much in their number (the dental formula is usually I 2, C 1, Pm 3, M 3 =
36) as in the disposition of the incisors. The incisors of the lower jaw
and the canines project forwards in a way only found in a few American
Monkeys; as in the Apes there are four incisors in each jaw, but, with
the exception of the highly aberrant Chiromys, there is a space in
the upper jaw between the incisors of the two sides. The canines of the
lower jaw, moreover, are often incisiform. There is a well-developed
sublingua beneath the tongue (see p. 61). The
stomach is perfectly simple; and the caecum, always present and varying
in length, never has a vermiform appendix. The gall-bladder is always
present. The brain differs from that of the Anthropoidea in that the
cerebellum is, as in the lower Mammalia, exposed. The convolutions upon
the cerebral hemispheres are not greatly developed, a circumstance,
however, which (see p. 77) may have more relation
to the size of the animals than to their mental development. Though the
brain in its general outlines is not like that of the other Primates,
there are certain resemblances; the most striking of these is perhaps the
presence, though in rather a rudimentary condition, of the "Simian
fissure."
The Lemurine brain has been chiefly studied by Flower,[392] by Milne-Edwards,[393] and by myself.[394] There are also a
number of scattered papers dealing with particular types, such as the
memoirs of Owen[395] and
Oudemans,[396] upon the
brain (and the general anatomy) of Chiromys. Without going into
great {536}detail it may be stated generally that the
anatomy of the brain of this group confirms the classification which is
adopted in this work.
A curious feature in the anatomy of the Lemurs, which they share with
animals so remote from them in the system as the Edentata, is the
breaking up of some of the arteries of the limbs to form retia mirabilia;
nothing of the kind is known among the other Primates.
Perhaps the most remarkable difference between the Lemurs and the
Anthropoidea, which are really in many respects more closely allied than
might be inferred from the above summary of differences, is in the
structure of the placenta. The Lemurs agree with the Ungulates in having
a non-deciduate placenta.
A curious feature confined to the sub-family Lemurinae was first
discovered by myself in Hapalemur griseus.[397] On the forearm (see Fig. 258) is an
area of hardened skin, which is raised into spine-like processes. Fully
developed, this organ is characteristic of the male, the area being
marked off in the female, but without the spiny outgrowths. On removing
the skin a gland about the size and shape of an almond is brought into
view. In other Lemurs there is no modified skin, but a small tuft of
particularly long hairs, which are also present in Hapalemur, and
a small gland beneath the skin. The gland of Hapalemur may be
comparable with a tract of hardened skin in Lemur catta, which
projects to a large extent and has been spoken of as a "climbing
organ."
An almost exactly similar tuft of spine-like outgrowths exists upon
the lower end of the ankle of Galago garnetti. The spines are
black and bent, just as they are in Hapalemur. There appears also
to be a gland. This structure is not universal in the genus Galago
any more than is the patch of spines in the genus Hapalemur.
In addition to this gland and to the patch of spines which cover it,
the same Lemur as well as Chirogaleus and certain species of
Lemur possess to the inner side of it a bundle of long and stiff
bristles associated with unusually large sebaceous glands; these
structures are, of course, not homologous with the gland of the arm of
Hapalemur, as they coexist in the same {537}species. They are,
moreover, not peculiar to the Lemurs, but exist in the Squirrel, in the
Domestic Cat, in the Leopard, in Bassaricyon,[398] the Otter, various Marsupials, and
doubtless in many mammals which require a tactile organ, for these hairs
are associated with a large branch of the radial nerve.
Fig. 258.—A, left arm of Hapalemur griseus ♂. a, Teat; b, spines on arm gland; c,
tactile bristle. B, left foot of Nycticebus tardigradus. 1 to 5, Pads upon sole of
foot. (After Sutton, and Mivart and Murie.[399])
The Lemurs have at the present time a most remarkable distribution.
There are altogether about fifty species, referable to seventeen genera.
Thirty-six species are confined to Madagascar {538}and to some small
neighbouring islands. The rest occur in the Ethiopian and in the Oriental
region. The rest of the world is at present totally without Lemurs,
though, as will be seen in the sequel, the order was more widely spread
over the globe in past times.
Fam. 1. Lemuridae.—This family can be usefully subdivided
into four sub-families.
Sub-Fam. 1. Indrisinae.—This sub-family is limited to
Madagascar, and has been exhaustively treated of by M. Grandidier and
Professor Milne-Edwards in the Histoire de Madagascar. These
Lemurs contrast with others by the large size of the hind- as compared
with the fore-limbs. The ears are short. The tail varies in length. The
thumb is but slightly opposable, and the toes are webbed. Correlated with
the first two of these characters, these Lemurs when upon the ground
progress by means of the hind-limbs, holding their arms above their
heads. The number of teeth is reduced, the total being thirty. The
formula[400] is I 2/2
C 1/0 Pm 2/2 M 3/3. The colon or large intestine, as figured by
Milne-Edwards, has a remarkable watch-spring-like coil, highly suggestive
of the Ruminants and of certain Rodents. This, however, is only in
Propithecus and Avahis. The caecum in this sub-family is
specially large. The brain is characterised by the comparatively slight
development of the angular fissure in Propithecus and
Avahis; it is in them anterior in position. In Indris it is
more -shaped and larger as in Lemur. The parieto-occipital fissure is
fairly well developed, so too is the antero-temporal.
The genus Indris has more pronounced external ears than have
the two other genera of the sub-family. The tail is rudimentary. The
incisors of the upper jaw are sub-equal and set close together, those of
the lower jaw have marked longitudinal ridges upon the outer surface,
which suggests Galeopithecus (see p. 520).
The molars are quadricuspidate. There is but a single species, I.
brevicaudata, which is of a black colour, diversified with white upon
the rump and the limbs. The term "Indri"[401] means, as does "Aye-aye," "look." One
of the native names for the {539}animal, "Amboanala," signifies "dog of the
forest," and is derived not only from the woeful howls of the creature,
but from the fact that in certain parts of the island it is used as a dog
to chase birds.
These howls are largely effected by means of a laryngeal pouch, which
is described as different from that of Apes; the mechanism must also
differ from that of Megaladapis, inasmuch as the lower jaw is not
deep as in that extinct Lemur. The Indri is the largest of Lemurs,
measuring about two feet in length. It is arboreal and social, travelling
in large companies. As is the case with the Propithecus, the
natives of Madagascar hold the Indri in awe and veneration. It is curious
that the name Lemur or ghost is peculiarly applicable to the Indri or
Babakote in another sense from that which led to its adoption by
Linnaeus. The natives, in fact, believe that men after death become
Indris. Naturally, therefore, these Lemurs have reaped the advantage of
this superstition in almost perfect immunity from destruction. Their
"long-drawn-out, melancholy cries" are probably at the root of much of
the ghostly terrors which they inspire.
The genus Avahis[402] has but a single species, A.
laniger, which is the smallest of this sub-family. It is a foot long
without the tail. The Avahi has a long tail (15 inches in length) like
Propithecus. The outer incisors are larger than the inner, thus
differentiating the genus from Propithecus. The molars of the
upper jaw are quadricuspidate, of the lower jaw five cusped. This genus
has only eleven pairs of ribs instead of the twelve of Indris and
Propithecus. The Avahis, unlike the Sifakas and Indrinas, lead a
solitary life, or go about in pairs. They are, moreover, completely
nocturnal.
The genus Propithecus is characterised by the fur being rather
silky than woolly, which latter is the kind of fur found in the two other
genera of the sub-family. They are also rather larger animals, the body
reaching a length of nearly 2 feet. The tail is long as in Avahis;
the inner incisors are larger than the outer. The "Sifakas," as these
Lemurs are termed, have a reputation for gentleness of character, but, as
is the case with other animals, the males fight for the possession of the
females at the breeding season. They are mainly vegetarian in habit, and
travel in large companies. There are at least three species, and {540}several varieties are allowed. The colours
of these Lemurs are bright, and distributed so as to form contrasting
bands; thus P. coquereli, a variety of P. verreauxi, has a
black face and a body mainly white, with splashes of a rich maroon upon
the limbs and upon the chest.
These Lemurs are diurnal, and are especially active in the early
morning and evening, sleeping, or at any rate remaining quiet, during the
heat of the day. Their fitness for an arboreal life is shown by the
existence of a parachute-like fold of skin between the arms and the body,
which suggests a commencement of the more complete parachute of Flying
Foxes, etc. These Lemurs are said to be reverenced and therefore shielded
from injury by the natives of Madagascar.
Sub-Fam. 2. Lemurinae.—The "True Lemurs" are all
inhabitants of Madagascar and of the Comoro Islands. They have not such
long hind-limbs as have the members of the last sub-family, nor are the
toes webbed. The tooth formula differs from that of the Indrisinae in
that there is one more premolar on each side of the upper jaw, and often
one more incisor in the lower jaw, making thus a total of thirty-six
teeth. Sometimes, however, the incisors of the upper jaw are totally
wanting.
The Hattock, genus Mixocebus, is a scarce creature, only known
from a single species, M. caniceps. As it is rare, nothing is
known of its habits. It has one pair of upper incisors. The creature is
one foot and half an inch long, exclusive of the tail, which is an inch
longer than the body.
Genus Lepilemur.—The Lemurs belonging to this genus,
entirely confined to Madagascar, as are all the Lemurinae, have received
the perfectly unnecessary and pseudo-vernacular name of "Sportive
Lemurs"; an equally inappropriate and not at all ingenious name of
"Gentle Lemurs" being bestowed upon the allied genus Hapalemur. In
Lepilemur there are seven species, which are to be distinguished
from Mixocebus in having the tail shorter than the body. There are
no incisors in the upper jaw. The last molar is tricuspidate in the upper
jaw; that of the lower jaw has five cusps. They are nocturnal creatures,
and but little is known of their habits. Previously to Dr. Forsyth
Major's visit to Madagascar only two species of the genus were known; he
has added five others. The length of the body is 14 inches, and that of
the tail 10 inches, in L. mustelinus, which is the largest
species. {541}
The genus Hapalemur[403] has a shorter muzzle than
Lemur, and shorter ears. There are two pairs of mammae instead of
only one; these are upon the breast and abdomen. In the male there is a
pair upon the shoulder. The incisors are small, sub-equal, and placed one
behind the other; the last one is at the inside of the canines. The
molars of the upper jaw and the last premolar have only three well-marked
cusps; in the lower jaw they have four. The caecum is blunter and is not
so long as in Lemur; it differs from that of other Lemurinae in
having only two supporting mesenteries, which are both furnished with
blood-vessels. As in Lepilemur and the Indrisinae the carpus has
no os centrale.
The genus, which is confined to the island of Madagascar, has two
species, of which one, H. simus, is the larger and has a broader
muzzle, and does not possess the peculiar arm gland (Fig. 258) already
described in H. griseus. The former species is stated by Mr. Shaw
to be chiefly a grass-eater, and to dislike berries and fruits, which are
usually so popular with Lemurs. It is, however, believed by some that
there is but one species of Hapalemur. H. griseus is 15
inches long, and has a tail of the same length. Its native name is
"Bokombouli." It is nocturnal, and is especially addicted to bamboos,
upon the shoots of which it feeds and among which it lives. It is often
exhibited in the Zoological Society's Gardens; but the specimens seem to
be always males. This Lemur is of a dark iron-grey colour with a tinge of
yellow, more marked in individuals which have received the separate
specific name of H. olivaceus.
The genus Lemur is distinguished by the long tail, half as long
as the body at the least, by the elongated face, and by the Fox-like
muzzle; the teeth are present to the full number of the family, viz.
thirty-six; the incisors are small and equal in size, and are separated
from each other and from the canines by spaces. The molars of the upper
jaw have five cusps, but there are only four in the lower jaw.
This genus is entirely confined to Madagascar and the Comoro Islands,
and consists of several species, the exact number of which is doubtful.
Wallace in his Geographical Distribution allows fifteen; Dr.
Forbes only eight, with a plentiful allowance of varieties. One of the
best-known species is Lemur catta, the {542}Ring-tailed Lemur, or
the "Madagascar Cat" of sailors. Lemur macaco shows a remarkable
sexual dimorphism, the male being black, and the female—formerly
described as a distinct species, L. leucomystax—being
reddish brown with white whiskers and ear tufts. This led to a confusion
with a totally distinct species, L. rufipes, of which the male
(regarded as distinct and called L. nigerrimus) is entirely black.
This latter identification is, however, considered by Dr. Forsyth Major[404] to be not quite
certain at present.
The young Lemur is at least sometimes carried by the mother across her
belly; its tail passes round her back and then round its own neck.
Fig. 259.—Ruffed Lemur. Lemur varius. × 1⁄9.
The Lemurs of this genus agree with those of some other genera in the
loudness of their voice, which is constantly exercised. Some move about
by day and others by night. They are insectivorous and carnivorous as
well as vegetarian; and Mr. Lydekker suggests[405] that their abundance and hardiness is
to be traced to this fondness for a mixed diet. Lemur catta seems
to be the only member of the genus that is not arboreal. It lives among
rocks where but few trees, and those much stunted, occur. Many species of
Lemur are always to be seen in the Zoological Society's Gardens.
Fourteen "species" have at one time or another been exhibited.
Sub-Fam. 3. Galagininae.—This sub-family is found on the
continent of Africa as well as in Madagascar; but the genera are {543}different in the two districts. In
Madagascar we have Opolemur, Microcebus, and
Chirogale; on the continent, Galago. The members of this
sub-family have markedly large ears, which are but little furry; the tail
is long. A very marked skeletal character distinguishes this sub-family
from other Lemuridae, and allies them to Tarsius, that is the
lengthening of the calcaneum and naviculare in the ankle. The dental
formula is as in Lemur. The supporting bands of the caecum are in
this sub-family as in the genus Lemur. There are but two folds, of
which one is median and non-vascular; the lateral fold bears a
blood-vessel, and is joined by the median frenum. The brain is but little
known. The only figure of the brain of Galago is one by myself.
There are four mammae, two on the breast and two upon the abdomen.
The genus Galago comprises at any rate six distinct species.
They are all African, and range right across the continent from Abyssinia
as far south as Natal, and to Senegambia in the west. The incisors of the
upper jaw are small and equal; there is a gap between the canine and the
first premolar. The molars and the last premolar have four cusps; the
last molar of the lower jaw has an additional fifth cusp as in
Macacus, etc. The Galagos are chiefly nocturnal, and are more or
less omnivorous. Owing to their long hind-legs these animals when they
leave the trees advance upon the ground by hops like a Kangaroo.
Galago senegalensis makes a nest in the fork of two branches,
where it sleeps during the day. The Great Galago (G.
crassicaudatus) is named by the Portuguese "Rat of the Cocoa-nut
Palm." Sir John Kirk, after whom a variety of this species is called,
relates that it is incapable of resisting the fascinations of palm wine,
upon which it will readily intoxicate itself, and as a consequence brave
probable captivity. I have referred above (p. 536)
to the patch of spines upon the tarsus of G. garnetti.
The genus Chirogale is entirely confined to Madagascar. It is
to be distinguished from Galago by the fact that the inner
incisors are larger than the outer. There are five species of the genus
known: four previously to Dr. Forsyth Major's recent visit to Madagascar,
and a fifth brought back by him.[406] In connexion with this genus the
naturalist just mentioned has observed that all the Lemurs of Madagascar,
including the aberrant {544}Chiromys, differ from the African
forms by the fact that the tympanic ring "is completely enclosed by the
bulla ossea, but without osseous connexion with the same." This character
he thinks so important as to justify the inclusion of all the Mascarene
forms in one group as opposed to another group consisting of the
continental Lemurs. In this event Chirogale will have to be
separated from its close association with Galago. For the present,
however, it is left in the more generally accepted position.
Fig. 260.—Smith's Dwarf Lemur. Microcebus smithii. × ¾.
Fig. 261.—Mouse Lemur. Chirogale coquereli. × ½.
Microcebus contains the most minute among the Lemurs. M.
smithii has a body only 5 inches long, the tail being another 6
inches. It occurs in Madagascar, and includes five species.
Opolemur, the Fat-tailed Lemur, was so called on account of a
{545}deposit of fat formed chiefly at the root
of the tail, and intended to tide over the time of the creature's
hibernation. But, as a matter of fact, this peculiarity also exists in
Chirogale. Of Opolemur but two species are known, and of
one of these, named after Mr. Thomas of the British Museum, only three
examples are in existence in museums, that is to say in one
museum—our own at South Kensington. Many of these dwarf Lemurs are
exceedingly rare. In this genus and in the last two the palate has a pair
of posterior fenestrae, of which there are also traces in other Lemurs,
but which are particularly large in Microcebus. This is, of
course, a well-known character of the Marsupials, and also, which is more
important in the present connexion, of certain Insectivores.
Sub-Fam. 4. Lorisinae.—This sub-family is the only one
with a wide distribution, and it contains, with the exception of
Tarsius, the only Asiatic members of the group. Correlated with
its wide distribution there is more divergence in anatomical characters
than is the case with the other sub-families of the Lemuridae. In
external features all the three genera of this sub-family agree in their
small size, their short or entirely deficient tail, large staring eyes,
and the rudimentary character, or absence, of the index finger, which is
never provided with a nail; in all of them the thumb diverges widely from
the other fingers, and the great toe is so divergent as to be directed
backwards. In the brain there is one character common to all three
genera, and that is the small length of the angular fissure. The caecum,
which is long, is supported by three folds, of which the median is
anangious, and is sometimes attached to the longer of the two lateral
folds, which are vascular. The members of this sub-family have more
dorsal vertebrae than are found in other Lemurs; the range is from
fourteen in Loris, to sixteen in Nycticebus.
The genus Nycticebus contains only a single species, N.
tardigradus, though four other names have been given to supposed
varieties. Moreover, the genus itself has been named Stenops, as
also the next genus Loris. The body of this animal is stouter than
that of the next to be described. Professor Mivart has pointed out that,
though Asiatic like the Loris, it presents more resemblances to the
African Potto. The index finger is small; the inner of the two incisors
is smaller than the outer, but both of one side are close together. They
may be reduced to one on each side of the upper jaw. {546}
Fig. 262.—Slow Loris. Nycticebus tardigradus. × ⅓.
The animal has a wide distribution in the East, occurring in Assam and
Burmah, the Malay Peninsula, Siam, and Cochin-China, Sumatra, Java,
Borneo, and the Philippines. Its vernacular names signify "Bashful Cat"
and "Bashful Monkey" in allusion to its nocturnal and shy habits. It
lives among trees, which it does not voluntarily leave. Its movements are
deliberate, as its popular name, Slow Loris, implies; but it makes up for
this by a vigorous tenacity of grasp. The animals "make a curious
chattering when angry, and when pleased at night they utter a short
though tuneful whistle of one unvaried note, which is thought by Chinese
sailors to presage wind." Much superstition has collected round this
harmless though rather weird-looking creature. Its influence over human
beings is as active when it is dead as when it is alive. "Thus," writes
Mr. Stanley Flower,[407]
"a Malay may commit a crime he did not premeditate, and then find that an
enemy had buried a particular part of a loris under his threshold, which
had, unknown to him, compelled him to act to his own disadvantage." The
life of the Loris, adds Mr. Flower, {547}"is not a happy one,
for it is continually seeing ghosts; and that is why it hides its face in
its hands!"
The genus Perodicticus contains two quite recognisable species,
known respectively as the Angwantibo and Bosman's Potto. The former has
been regarded as referable to a distinct genus, Arctocebus. A
curious internal character of the Potto which is visible, or at least can
be felt, externally, is the long neural processes of the cervical
vertebrae, which project beyond the level of the skin. The index finger
is rudimentary and so is the tail, being only just visible (about an inch
in length) in the Potto. The colour of both genera is a reddish grey,
redder in the Potto. The incisors are equal and minute. Both species are
confined in their range to West Africa, and are arboreal like the other
members of the sub-family. The Potto seems to share the leisurely mode of
progression of its Asiatic relatives, if Bosman, its original describer,
is to be trusted. He says: "By the negroes called Potto, but known to us
by the name of Sluggard, doubtless from its lazy, sluggish nature; a
whole day being little enough for it to advance ten steps forward." The
same writer did not at all appreciate his addition to zoological
knowledge, for he remarked that the Potto "hath nothing very particular
but his odious ugliness." The Angwantibo is rare and but little known.
Our knowledge of its anatomy is derived from a paper by Huxley.[408] It is an animal
measuring about 10½ inches in total length to the end of the tail, which
is only a quarter of an inch long. The hands and feet are smaller than
those of Perodicticus. The index finger is rudimentary and has but
two phalanges, and it has no trace of a nail. In this it agrees with the
Potto, but "the spinous processes of the cervical vertebrae do not
project in the manner described by van der Hoeven in the Potto, though
they can be readily felt through the skin." The dental formula of this
genus as of the last is I 2/2 C 1/1 Pm 3/3 M 3/3. The last lower molar
has a fifth cusp, which is wanting in the Potto. The last upper molar is
tricuspid. It is bicuspid in the Potto. It seems impossible to avoid
agreeing with Professor Huxley that the Angwantibo is entitled to generic
separation.
The genus Loris also contains but a single species, L.
gracilis, and is, as its name denotes, an animal of more slender
build than the Slow Loris. Its eyes are very large, and the limbs
excessively {548}slender. The index finger is much as in
Nycticebus. The colour, too, is not widely different, being of a
yellowish grey, but it lacks the dorsal stripe which distinguishes its
relative. The incisor teeth are equal and very small. The last upper
molar has four cusps instead of the three of Nycticebus. This
Lemur is confined to Southern India and Ceylon, and has much the same
habits as the last. But it is rather more active, and can capture small
birds when sleeping upon the trees; its diet, however, is mixed, and is
vegetarian as well as animal.
A mysterious Lemur, which we conveniently place as a kind of appendix
to the present family on account of its locality, has been shortly
described by Nachtrieb from the Philippines. The tail is rudimentary;
there are two upper incisors, but as many as six lower. It is doubtful
what the beast really is.
Fig. 263.—Aye-aye. Chiromys madagascariensis. × 1⁄10.
Fam. 2. Chiromyidae.—This family contains but a single
genus and species, the Aye-aye, Chiromys madagascariensis, whose
characters therefore are for the present those of the family as well as
of the genus and species. The external features of this extraordinary
animal will be gathered from an inspection of Fig. 263, from which it
will be seen that the earlier name of Sciurus given to the
creature was not by any means a misnomer. The Squirrel-like appearance is
due, of course, chiefly to the strong and long incisor teeth. As to the
external characters, which are of systematic importance, {549}attention may
be called to the long and bushy tail, to the greater length of the
hind-limbs, to the abdominal teats (one pair) in the female, and above
all to the singular third digit of the hand, which is thin and elongated.
The thumb is, as in other Lemurs, opposable, and has a flat nail; the
remaining digits have claws, as have also the toes with the exception of
the great toe, which has a flat nail like the thumb.
The anatomy of this animal has occupied the attention of a
considerable number of observers, dating from Sir R. Owen, who was the
first to give a connected account of its entire organisation. The most
recent paper of importance is by Dr. Oudemans.[409] The teeth are very unlike those of
other Lemurs. The most remarkable divergence is in the incisors, which
are present to the number of but a single pair in each jaw, and are
shaped like those of the Rodentia, and in the same way as in that group
grow from persistent pulps. There are likewise, as in the Rodents, no
canines. There are two premolars in the upper jaw (none in the lower) and
altogether twelve molars, so that there is a total of eighteen teeth. The
intestine has a moderately long caecum. The brain has been most fully
described by Oudemans, who had fresh material to work with, the brain
described by Owen having been extracted from a spirit-preserved carcase.
The angular fissure is well developed, as in Lemur and the Indri; but it
does not join the infero-frontal. The antero-temporal fissure is also
well developed.
"The name of Aye-aye," wrote Sonnerat, the discoverer of the animal,
"which I have retained for it, is a cry of surprise of the inhabitants of
Madagascar." It is, however, usually said that the animal itself makes a
sound which may be written in the same way (or with an initial H). It is
an arboreal and nocturnal animal, which accounts for its excessive rarity
at one time. In one of his many eloquent essays upon natural history the
late Mr. P. H. Gosse adduced the Aye-aye as an example of a creature on
the verge of extinction. It is, however, now more frequently met with,
though the superstition of the natives renders its capture a matter of
some difficulty. There is a specimen at the moment of writing in the
Zoological Society's Gardens. There has been some discussion as to the
use of the slender middle finger: it is stated that it can thrust it into
the {550}borings of the larva of a certain beetle
of which the Lemur is particularly fond, and can extract the insect, or
at any rate discover its position, when it may be extracted by the
powerful chisel-shaped teeth. The partiality of the Aye-aye for animal
food of any kind including insects has been both reaffirmed and denied;
and Mr. Bartlett has seen the creature use its slender finger for combing
out its hair, and for other purposes of the "toilet." Dr. Oudemans has
figured in his paper an apple which has been largely eaten by the
Chiromys; the fleshy pulp has been entirely excavated, leaving
only the core and the skin, which are untouched. The Rev. Mr. Baron is
one of the latest writers upon the ways of life of Chiromys.[410] He states that it
inhabits the most dense parts of the forests. It has the habit of
prowling about in pairs, and the female produces but a single young one
at a birth. A nest, which is about 2 feet across, is made of twigs in
lofty branches. This is occupied during the day, and entered by a hole in
the side. With regard to the superstitious veneration in which the animal
is held, it is said that if a person sleeps in the forest the Aye-aye
will bring him a pillow. "If a pillow for the head, the person will
become rich; if for the feet, he will shortly succumb to the creature's
fatal power, or at least will become bewitched." But a counter-charm may
be obtained. It is said that the reverence for this beast leads the
natives to bury carefully a specimen found dead.
Fam. 3. Tarsiidae.—This family also consists of but a
single genus, Tarsius, to which it is the general opinion that but
a single species belongs; there are, however, at least four different
specific names on record. The general aspect of the animal is not unlike
that of a Galago, with which it also agrees in the elongation of the
ankle; but the elongation is more pronounced in the present genus. The
ears are large, and the eyes are extraordinarily developed. The fingers
and toes terminate in large expanded discs, and are furnished with
flattened nails except on the second and third toes, which have claws.
The tail is longer than the body and is tufted at the end. The skull is
more like that of the Anthropoidea than is the skull of any other Lemur.
The resemblance is by reason of the almost complete separation of the
orbit and the temporal fossa by bone; {551}there is, however, a
gap left to mark the Lemurine characters of the animal. The placenta,
too, has been compared to that of the Apes. The dental formula is that of
the genus Lemur, save for the absence of an incisor on each side
of the lower jaw; the number of teeth is therefore thirty-four. The
incisors of the lower jaw are upright, and not procumbent as in other
Lemurs. The caecum is of moderate length. The brain is almost smooth, but
there is a Sylvian fissure and an antero-temporal, which latter does not
reach the lower margin of the brain, but divides the middle part of the
temporal lobe. The name Tarsier, as may be inferred, was originally given
to this creature by Buffon on account of the abnormal ankle, and it was
compared by him with the Jerboa, like which animal the Tarsier leaps when
it descends to the ground. The genus is Malayan, but its range extends to
the Philippines and to Celebes and Borneo. The Tarsiers are nocturnal and
particularly arboreal; they live in pairs, in holes in tree stems, and
are mainly insectivorous in their food. One, rarely two young are
produced at a birth. Contrary to what is found in many Lemurs, the
Tarsier is a silent creature, and at most emits a "sharp, shrill call."
Dr. Charles Hose, who has studied this creature, has noticed that the
mother often carries her young one about in her mouth like a Cat. Like so
many Lemurs this animal is held in superstitious dread, which no doubt is
the result of its most weird appearance.[411]

Fig. 264.—Right pes of Tarsius
spectrum. (Nat. size.) a, Astragalus;
c, calcaneum; c1, internal
cuneiform; c2, middle cuneiform;
c3, external cuneiform;
cb, cuboid; n, navicular; I-V,
the digits. (From Flower's
Osteology.)
Fossil Lemurs.—The Lemuroids are a very ancient race;
they extend back to the very earliest strata of the Eocene, the Torrejon
and Puerco beds, which, as already said, are thought to be more referable
to the Cretaceous than to the Tertiary epoch. {552}One of these early
forms is referred to the genus Mixodectes, a genus which has been
placed, though with a query, in the order Rodentia. It appears, however,
to be a Lemuroid, and is of American range. The incisor teeth have been
held to argue that it lies on the direct track of Chiromys; but
other features, more especially the form of the astragalus, have been
used to argue the justice of the inclusion of this type within the order
Rodentia. Allied, as it is supposed, to this form is Indrodon,
also of the lowest Eocene deposits of the United States. Indrodon
malaris is known from fragments of nearly all parts of the skeleton.
They indicate the existence of a creature of about one-half the size of
Lemur varius. It had slender limbs and a long and powerful tail.
The humerus, as in so many archaic beasts, has an entepicondylar foramen.
The femur has three trochanters, and the fibula articulates with the
astragalus. It is not always easy to distinguish these primitive mammals
from each other, so that the minutest of characters have to be called in
to our assistance. One of the contemporaneous groups with which these
early Lemurs might be confused is that of the Condylarthra; it is
important, therefore, to note that in Indrodon the
calcaneo-cuboidal articulation is nearly flat, and not bent as it is in
the former group. The teeth are of the tritubercular pattern. The
incisors are not known, but the molars and premolars are each three. To
the same family, which has been termed Anaptomorphidae, is
referred the genus Anaptomorphus, which has been specially
compared to Tarsius. This small animal has a Lemurine face with
huge orbits. It has a premolar less than Indrodon. It has been
ascertained that A. homunculus had an external lachrymal
foramen.[412]
Another family, that of the Chriacidae, appear to hover on the
border line of Lemurs and Creodonts, having been referred to both by
various palaeontologists. Professor Scott suggests their Lemurine or at
least Primate relationships, while Cope urged their Creodont affinities.
A difficulty raised by Scott was, that in Chriacus the premolars
of the lower jaw were spaced. But it appears that this is not fatal to
their inclusion in the Primates, since Tomitherium, an "undoubted
Primate," shows the same feature. If Chriacus is a Lemur it is an
earlier type than those {553}which have been considered; for it has the
typical Eutherian dentition of four premolars and three molars. These
teeth, especially the superior molars, are particularly compared to the
corresponding teeth of Lemur and Galago. Of this and the
allied genus, Protochriacus, several species are known.
Adapis, a representative of another family, is one of the best
known of ancient Lemuroids. It has the typical mammalian dentition of
forty-four teeth in a close series without diastemata. The orbits are
completely separated from the temporal cavity, the eyes looking forwards.
The canines are large and caniniform. The skull is deeply ridged behind
with the usual sagittal crest. This genus is European, and corresponds to
the already mentioned American Eocene Tomitherium, perhaps
belonging to the same family.
Nesopithecus is an extinct genus from Madagascar, lately
described by Dr. Forsyth Major.[413] There are two species, N.
roberti and N. australis. The dental formula is I 2, C 1,
Pm 3, M 3, for the upper jaw, the lower jaw having but a single pair of
incisors. The lachrymal foramen is just inside, or on the edge, of the
orbit, so that one distinctive Lemurine character is lost. The genus is
also Ape-like in the form of the canines and incisors, these having been
especially compared by Dr. Forsyth Major with those of the
Cercopithecidae. The molars, too, agree with those of the same family.
There is, however, one important feature in which Nesopithecus
resembles not only the Lemurs as opposed to the Apes, but the Malagasy
Lemurs. As already mentioned (p. 544), Dr. Major
has shown that in the Malagasy Lemurs, even including the aberrant
Chiromys, and in the Tertiary and European Adapis, the
bulla tympani is not produced by an ossified extension of the annulus
tympanicus, but from the adjacent periotic bone, the annulus remaining
separate and lying within the fully-formed bulla. This feature shows
conclusively that Adapis is a Lemur, and that Nesopithecus,
originally supposed to be a Monkey, cannot be removed from the
Lemuroidea, many though its likenesses to the higher Primates undoubtedly
are. However, this feature, combined with the fact that the orbital and
temporal cavities are in communication, shows the Lemuroid position of
Nesopithecus, though it is quite conceivable that it is on the way
to become an Ape. {554}
A family, Megaladapididae, has been quite lately founded by Dr.
Forsyth Major[414] to
include the remains of a gigantic extinct Lemur from Madagascar, which
when alive, so far as we can judge from the skull, must have been three
or four times the size of the Common Cat. The name Megaladapis
madagascariensis was given to the fossil on account of certain
resemblances to the also extinct Adapis. It differs from other
Lemurs in a number of characters which jointly warrant its inclusion in a
distinct family. The small size of the orbits suggest a diurnal life; the
deep mandibles, which, unlike what is found in other Lemurs, are
completely blended at the suture, point to the existence of a howling
apparatus, as in Mycetes. The low brain-case is a character which
is found in so many extinct Mammalia belonging to many different orders
that it weighs neither one way nor the other in considering the
systematic position of the animal. The shape of the molars, which are
three in each half of each jaw as in other Lemurs, is, according to the
discoverer, like that of the genus Lepilemur. The incisors and the
canines are not known. Of a still larger form, M. insignis, the
molar teeth are known.[415]
Sub-Order 2. ANTHROPOIDEA.
The Apes differ from the Lemurs in that the teats are always
restricted to the thoracic region; the orbit, though surrounded by bone
as in the Lemurs (and in Tupaia, a very Lemur-like Insectivore),
does not open freely behind into the temporal fossa as in Lemurs (except
Tarsius). The lachrymal opening is inside the orbit instead of
outside; the cerebral hemispheres are more highly developed, and conceal,
or nearly conceal, the cerebellum; the upper incisors are in close
contact; a few other points are mentioned under the description of the
characters of the Lemurs. There are altogether about 212 species of
Monkeys and Apes. They are tropical and subtropical in range, and, with
but few exceptions, are impatient of cold.
The Monkeys are primarily divisible into two great divisions, which
have been termed, on account of the characters of the nose, {555}the
Catarrhines and Platyrrhines. In the former the nostrils look downward
and are close together; in the latter they are separated by a broad
cartilaginous septum, and the apertures are directed outwards. But
numerous other points of difference separate these two groups of the
Monkey tribe. The Catarrhines often have those remarkable ischial
callosities, patches of hard skin brightly coloured; the tail may be
totally wanting as a distinct organ, as is the case, for instance, with
the Anthropoid Apes; there are often cheek pouches, so that, as Mr.
Lydekker has remarked, if a Monkey be observed to stow nuts away in its
cheeks for future reference, we may be certain that its home is in the
Old World, for the Catarrhines are exclusively denizens of the Old World,
while the Platyrrhines are as exclusively New World in range. Again,
those of the Catarrhines which do possess a long tail, such as the
members of the genus Cercocebus, never show the least sign of
prehensility in that tail. The teeth of the Catarrhines are invariably
thirty-two in number, the formula being I 2/2 C 1/1 Pm 2/2 M 3/3 =
32.
In the Old-World Apes there is a bony external auditory meatus, which
is wanting (as a bony structure) in the Platyrrhines. The late Mr. W. A.
Forbes pointed out that in most of the New-World forms the parietals and
the malars come into contact; in the Monkeys of the Old World they are
hindered from coming into contact by the frontals and the alisphenoids.
The Platyrrhines may have the same number of teeth; this is the case with
the Marmosets, but in them there are three premolars and two molars; in
the remaining New-World Monkeys there are thirty-six teeth, but of these
three are premolars and three molars.
Not only are these two groups of the Primates absolutely distinct at
the present day, but they have been, so far as we know, for a very long
time, since no fossil remains of Monkeys at all intermediate have been so
far discovered. This has led to the suggestion that the Monkeys are what
is termed diphyletic, i.e. that they have originated from two
separate stocks of ancestors. It is hard, however, to understand on this
view the very great similarities which underlie the divergences that have
just been mentioned. But, on the other hand, it is equally hard to
understand how it is that, having been separated from each other for so
long a period, they have not diverged further in {556}structure than they
have. The Platyrrhines seem to stand at the base of the series. This is
another example of the existence of archaic creatures in South
America.
Group I. PLATYRRHINA.
Fam. 1. Hapalidae.—We may begin the account of the
Platyrrhine Monkeys with the Hapalidae or Marmosets; for this family is
structurally lower than the rest. They have thirty-two teeth, arranged as
in the following formula: I 2/2 C 1/1 Pm 3/3 M 2/2 = 32. The molars have
three main tubercles, and not four as in the higher forms. The digits are
for the most part clawed, not nailed, as in the higher types; the great
toe alone bears a flat nail. The tail, too, is ringed, a condition which
is characteristic of many of the lower groups of mammals, but not of the
higher Apes. The cerebral hemispheres are smooth, but this is a matter
rather connected with their small size than with low zoological position.
The tails of the Marmosets, unlike those of so many other American
Monkeys, are not prehensile though they are long.
The genus Hapale is broadly distinguished from the other genus,
Midas, by the fact that the lower incisors slant forwards as in
the Lemurs. They are small, soft-furred, long-tailed Monkeys, familiar to
every one. There are some seven species, which are entirely restricted in
range to Brazil, Bolivia, and Colombia, one species only, H.
pygmaea, extending northward into Mexico.
Of Tamarins, genus Midas, there are rather more
species—about fourteen. They are South and Central American in
distribution. Since both these genera are arboreal in habit, it is
extraordinary that they have not the prehensile tails of their American
allies. As, however, the late Mr. Bates observed an individual of the
species M. nigricollis fall head-foremost from a height of at
least 50 feet, alight on its feet, and run off as if nothing in
particular had occurred, it is evident that no extra prehensile powers
are absolutely necessary. Some of the Tamarins have a long mane; this is
well seen in M. rosalia, or rather in M. leoninus, which,
if not identical with it, is at least very closely allied to it. The name
is obviously derived from the character {557}referred to, and the
Monkey, originally described by the traveller von Humboldt, is said to
have "the appearance of a diminutive lion." M. bicolor is an
example of the species with no mane, but with a patch of white round the
mouth, looking like "a ball of snow-white cotton" held in the teeth.
Fam. 2. Cebidae.—The remaining American Monkeys are
comprised in the family Cebidae. This is to be distinguished from the
last by the fact that there is an additional molar, thus making
thirty-six teeth in all. The tail, sometimes very short, is more
generally long and highly prehensile, being nude at the extremity, which
part is therefore especially prehensile; this state of affairs is often
to be seen in animals with prehensile tails. The Cebidae, though for the
most part larger than the Marmosets, never approach in size the Old-World
Apes.
Typical of the family is the genus Cebus, including the
"Capuchin" Monkeys, and consisting of nearly twenty species; the tail,
though prehensile, is covered with hair to the tip, a fact which is
indicative of a less perfect prehensility than is exhibited in some
Monkeys with a naked under surface to the tip of the tail. The thumb is
well developed. The genus ranges from Costa Rica to Paraguay. The
commonest Monkey which accompanies the street organs of this country is a
Cebus. It is a popular delusion that these and other monkeys are
purely vegetable-feeding animals. Cebus is in fact particularly
fond of caterpillars, as are also the Marmosets.
Allied to Cebus is Lagothrix, the Woolly Monkey, of
which L. humboldti is the best-known species, there being indeed
but one other. It is a larger and heavier animal than any species of
Cebus; and the Hare-like woolliness of the fur suggested its
scientific name to its original describer, von Humboldt. It has a
perfectly prehensile tail, naked at the tip. The thumb and great toe are
well developed. These are purely fruit-eating Monkeys, and are known as
"Barrigudos" by the Portuguese of the Amazon country on account of their
prominent belly, due apparently to the immense amount of fruit consumed.
They are, or were, much eaten by natives.
Brachyteles is a little-known genus, connecting the last with
the next genus. The under fur is woolly; the thumb is small or absent.
The tail is naked below.
The Spider monkeys, Ateles or Coaitas, have been described as
{558}the most typically arboreal of American
monkeys. The use of the prehensile tail can frequently be studied in
living examples in the Zoological Society's Gardens. With this "fifth
hand" the Monkey feels for a place to grasp, and securely twists its tail
round, moving it with the greatest ease from point to point. When the
tail is being thus used it is carried erect over the head. The fact that
this genus possesses no functional thumb is thought to be associated with
the extreme perfection of its adaptability to an exclusively arboreal
life. The hand without a thumb can act as an equally efficient hook for
suspending the body; and what is useless in nature tends to disappear.
These Monkeys have a wide range, extending from Mexico in the north to
Uruguay in the south. There are ten species. The flesh of many Monkeys is
eaten not only by natives but by Europeans; but the Spider Monkeys are
said to furnish the most sapid food of all.

Fig. 265.—Spider Monkey. Ateles ater. × 1⁄12.
The Howling Monkeys, genus Mycetes, have also received the
appropriate generic names of Alouatta and Stentor. The
former of these two names, indeed, is that which should properly be
applied to the genus. But Mycetes is perhaps better known. The
"howling" is produced by saccular diverticula of the larynx, larger than
those of other American Monkeys, such as Ateles, where, however,
they are also developed. The hyoid bones, too, {559}are enormously enlarged
and cavernous, while the jaw—in order to accommodate and protect
these various structures—is unusually large and deep. The Howlers
are furnished with a fully prehensile tail. The thumb is present. They
are described as being the most hideous in aspect of the American
Monkeys, and of the lowest intelligence, with which latter characteristic
is associated a less convoluted brain than in Ateles, for example.
The noise produced by these Monkeys is audible for miles, and is said not
to be due to emulation, i.e. not to be comparable to singing or
talking, but to serve to intimidate their enemies. The story told of
these and other Monkeys with prehensile tails, that they cross rivers by
means of a bridge of intertwined Monkeys, is apparently devoid of truth.
There are six species, which are Central and South American in range.
The Squirrel Monkeys, genus Chrysothrix, are small creatures
with a long head, the occiput projecting. Their tail, though long, has no
naked area at the extremity and is non-prehensile. It is a remarkable
fact that the proportions of the cranium as compared with the face are
greater, not only than in other Monkeys, but than in Man himself. The
thumb is short, but not so short as in the Spider Monkeys. The cerebral
hemispheres are very smooth; but, as already remarked, this is a matter
of size, and not of low position in the series. It may appear at first
sight that this statement contradicts the one made concerning the
Howlers. But the latter are large Monkeys, and therefore ought, so to
speak, to have a more complex brain; but they have not. Like so many of
the American Monkeys, the Squirrel Monkeys are gregarious, and, in spite
of their tails, arboreal. They are largely insect-feeders, and also catch
small birds and devour eggs. There are four species, of which C.
sciurea is the commonest, and is constantly an inmate of the
Zoological Society's Gardens. Humboldt asserted of it that when vexed its
eyes filled with tears; but Darwin did not succeed in seeing this very
human expression of an emotion.
Callithrix is a genus not far removed from the last, and, like
it, occurs both in Central and in South America. It is chiefly to be
distinguished from Chrysothrix by the non-extension backwards of
the head, and by the more furry character of the tail. The lower jaw is
rather deep, as in the Howlers; but there is not, or there has not been
discovered, a howling apparatus like {560}that of Mycetes.
Nevertheless Professor Weldon[416] has found in a female of C.
gigot a patch of ossification on the thyroid cartilage of the larynx
which may be an indication of something more in the male. There are
eleven species.
Nyctipithecus, the Doroucouli Monkeys, is a genus of somewhat
Lemurine appearance, caused by their large eyes. But they reminded Bates
of an Owl or a Tiger-cat! They have a long, but not prehensile tail. As
in the Marmosets, the lower incisors project forwards in a Lemurine
fashion. The thumb is very short. A peculiarity of this genus is the
twenty-two dorso-lumbar vertebrae. As in Chrysothrix, but not as
in Callithrix, the hemispheres of the brain are smooth. There are
five species, of which one occurs so far north as Nicaragua; the rest are
Brazilian, extending down to the Argentine.
Fig. 266.—Red-faced Ouakari. Brachyurus rubicundus. × 1⁄5.
The Ouakari Monkeys, Brachyurus,[417] are, as the name denotes,
short-tailed forms. Two species, B. rubicundus and B.
calvus, have bright red faces; B. melanocephalus has a black
one. There is a small thumb. The brain is fairly convoluted, and is to be
specially compared with that of Cebus and Pithecia. The
{561}species B. rubicundus at any rate
has an absolutely as well as a relatively greater length of intestines
and caecum than any other American Monkey known.
Fig. 267.—White-nosed Saki. Pithecia albinasa. × 1⁄5. (From Nature.)
Not the least remarkable fact about these Ouakari Monkeys is their
distribution in South America. We cannot do better than quote the summary
given by Messrs. P. L. and W. L. Sclater in their Geography of
Mammals, which is as follows: "Each of them, as first shown by Bates
and afterwards further explained by Forbes, is limited to a comparatively
small tract of forest on the banks of the Amazon and its affluents. The
Black-headed Ouakari (B. melanocephalus) ... is met with only in a
tract {562}traversed by the Rio Negro; the
Bald-headed Ouakari appears to be confined to the triangle formed by the
union of the Amazon with another affluent, the Japura; and the Red
Ouakari to the forests on the north bank of the Amazon opposite Olivença,
and lying between the main stream and the River Iça. Each of them
evidently takes the place of the others in its particular district. Of
this peculiar kind of distribution few instances are known amongst
mammals, but many somewhat similar cases have been observed in birds,
reptiles, and insects."
The genus Pithecia, the Sakis, consists of five species with
long bushy tails, which are non-prehensile. They are bearded and have a
thumb. Like the last genus, Pithecia does not extend into Central
America. The incisors project forwards, and the lower jaw is deep, though
the howling apparatus of Mycetes is wanting. The thin,
closely-set, and projecting incisors are very suggestive of those of the
Lemurs. Brachyurus is much like Pithecia in this respect,
and both differ markedly from such a genus as Cebus, where the
lower incisors are vertical. An anatomical peculiarity of Pithecia
is the breadth of the ribs. P. satanas is perhaps the best-known
species, but all five have been exhibited at the Zoological Society's
Gardens. As its name suggests, P. satanas is entirely black; it
shows a curious point of difference from P. cheiropotes in its way
of drinking. The latter species, as its name denotes, uses its hand to
drink, while P. satanas puts its mouth to the water. P.
albinasa is black with a red patch on the nose, within which again is
a small white patch.
Group II. CATARRHINA.
The Catarrhine Apes are divisible into three or perhaps only two
families, the Cercopithecidae and the Simiidae, to which must be added
the Hominidae. The Simiidae are sometimes spoken of as the Anthropoid
Apes.
Fam. 1. Cercopithecidae.—Of the Cercopithecidae there are
eight genera (perhaps nine) to be recognised, which may be distributed
into two sub-families. The first of these two sub-families, that of the
Cercopithecinae, has the following characters:—There are cheek pouches in which the animals
store food temporarily. {563}The stomach is simple and globular; this
corresponds with a mixed diet. The tail is long or short, or practically
absent.
Fig. 268.—Tcheli Monkey. Macacus tcheliensis. × 1⁄6. (From Nature.)
The most familiar genus is undoubtedly Macacus. This includes
all the common so-called Macaques, the Bonnet Monkey, the Pig-tailed
Monkey, etc. In this genus we find that the males are larger than the
females, and have stronger canine teeth. Ischial callosities are well
developed. The genus is purely Asiatic, reaching as far east as Japan,
with the exception of the Barbary Ape, M. inuus, also known as the
Gibraltar Ape. There are altogether some seventeen species.
Macacus inuus is doubtfully indigenous to Gibraltar. It is,
however, definitely established there at present, and is carefully
fostered. It is a large Ape with no external tail, in which {564}particular it
is unique among the members of its genus. At one time its extinction on
the "Rock" was nearly accomplished, but three individuals being known. In
1893 the Governor of Gibraltar informed Mr. Sclater that he had himself
counted as many as thirty in one herd. Its depredations seem to have led
to the expression of a wish in some quarters that the numbers should be
thinned; but feeling on the opposite side appears to be stronger, so that
whatever was the actual mode of its introduction on to the "Rock" it will
at any rate remain there unmolested for the present.
M. tcheliensis is a species found in the Yung-ling Mountains in
North China. It is, with the possible exception of M. speciosus,
the most northerly form of Monkey. It is interesting on account of the
fact that like the Tiger of those regions it has put on an extra coating
of fur to enable it to combat with the bitter winters. It is doubtful
whether it is more than a variety of the Rhesus Monkey (M.
rhesus).
M. nemestrinus, "the Pig-tailed Macaque," is trained by the
natives of the east to climb cocoa-nut palms and to carefully select and
throw down only the ripe fruit. Sir Stamford Raffles apparently was the
first to report upon this useful intelligence of the animal, and Dr.
Charles Hose of Borneo has confirmed him.
The Japanese Macaque (M. speciosus) is well known from the work
of Japanese artists. It is the only species of Monkey found in Japan, and
goes very far north.
A rather rare form is M. leoninus. It has a short tail, and
occurs in Burmah. M. silenus is distinguished by a ruff of long
light-coloured hair surrounding the face. It is sometimes called the
Wanderoo; but this is apparently quite inaccurate, since that term is
used by the Ceylonese for a Semnopithecus. For those who wish a
"pseudo-vernacular" name Dr. Blanford suggests Pennant's name of
"Lion-tailed Monkey."
The commonest species of the genus are M. cynomolgus, M.
sinicus, and M. rhesus.
The genus Cercocebus, including those Monkeys known as
Mangabeys, is confined to West Africa. They have always a long tail,
quite as long as the body. The upper eyelids are pure white in colour.
The ischial callosities are more pronounced than in the Macaques. In the
Mangabeys also the hairs are not ringed with differently coloured bars,
as is the case with both {565}Macaques and Cercopithecus, giving
to them the greenish hue which characterises so many of the last two
genera. There are no laryngeal air sacs as in the Macaques. There are not
more than seven species.
The genus Cercopithecus (the Guenons) represents in Africa the
Oriental and Palaearctic Macaques; the genus has a long tail. The cheek
pouches are larger than in the genus Macacus. The ischial
callosities are less extensive than in that genus. A tooth character also
distinguishes this genus from Macacus; the last molar of the lower
jaw has, as a very general rule, only four cusps instead of the five
which are found in Macacus. The supraciliary ridges in the skull
are by no means so marked as in the allied genera.
One species, the Talapoin, C. talapoin, has been separated into
a distinct genus, Miopithecus, on account of the fact that the
lower molars have only three tubercles instead of the usual four. But if
this be done, then Cercopithecus moloneyi, which has a lower molar
with five tubercles, should also be separated.
Fig. 269.—Diana Monkey. Cercopithecus diana. × 1⁄6.
The genus Cercopithecus is limited to Africa, and its numerous
species have often a very limited range. They are frequently rather
brightly coloured, with blue and white patches on the face. The Diana
Monkey has a pointed white beard. Of the Vervet Monkey (C.
lalandii) a curious fact was noticed at the Zoological {566}Society's
Gardens a year or two back: the young was observed to take both teats of
the mother into its mouth at once. Mr. Sclater[418] in a recent list of the group allows
forty-seven species, of which thirty-three were examined by himself.
Subsequently, however, the list has been reduced to forty by the same
authority. One of the rarest species is C. stairsi, first
described from a skin stripped from a specimen which lived for a short
time at the Zoological Gardens.
The genus Cynocephalus (or Papio) includes the Baboons;
and the scientific name indicates the Dog-like aspect of these animals,
due to the projecting snout. Cynocephalus is confined to Africa
and Arabia. Several of the species of the genus are well known. The
Mandrill, C. mormon (or maimon), has blue ridges on the
muzzle, the bridge of the nose being red. The animal lives in herds, and
is ferocious and omnivorous. The Chacma Baboon, C. porcarius, is
the largest of Baboons. It lives in South Africa in large herds. The
Arabian Baboon, C. hamadryas, is the Sacred Baboon of the
Egyptians. The names of two other species, C. thoth and C.
anubis, serve also to remind us of the ancient Egyptians. There are
altogether eleven species of Cynocephalus.
Gelada (or Theropithecus) is separated as a distinct
genus. Though regarded as a Baboon, Garrod has pointed out many points of
likeness to Cercopithecus.[419] The two species are, like the other
Baboons, African.
Cynopithecus niger is a small black Baboon from Celebes. It has
swellings on the muzzle as in other Baboons, but differs from them in
being a more amiable creature as well as in its smaller size. It has a
rudimentary tail, smaller even than the small tail of the typical
Baboons. It has, like them, ischial callosities.
In the second sub-family, Semnopithecinae, the following
characters are distinctive:—All the Apes of
this group are slender in form, with a long tail. There are no cheek
pouches. The stomach is sacculated; it is divided into three portions.
This is accompanied by an apparently more exclusively vegetarian diet
than characterises other Apes, which mingle with their diet of fruit a
large proportion of insects, eggs, etc. {567}
Fig. 270.—Black Celebesian Ape. Cynopithecus niger. × 1⁄5.
The first with which we shall deal is Colobus, containing the
Monkeys known as Guerezas. These creatures are entirely confined to the
African continent, and they are arboreal in habit. It has been attempted
to show that their affinities are more with the Platyrrhines than with
the group in which they are really to be placed. In favour of regarding
them as nearer akin to the American monkeys are only two facts of
importance: the first is the practical absence of the thumb, which of
course recalls the condition characteristic of Ateles; in the
second place, the nostrils in their wideness somewhat resemble those of
the Platyrrhines. They are slender Monkeys with well-marked callosities.
They have a complex sacculated stomach, resembling the large intestine of
some other animals; it is not divided into distinct chambers like the
stomach of a Ruminant or of a Whale. Correlated apparently with this
large stomach is the small development of the cheek pouches. This genus,
of which there are about ten species, is characterised by beautiful
skins, which are largely collected. The Arabs have a legend to the effect
that one species, when wounded, and seeing its capture and the removal of
its skin {568}inevitable, carefully tears the latter,
that its captors may not profit by it. The species of this genus are most
abundant on the west coast of Africa. It is interesting that one species,
C. kirki, is limited to the Island of Zanzibar, where, however, it
is nearly extinct.
The "Holy Apes," or Langurs, genus Semnopithecus, are allied to
the last, but they are Asiatic in range. The thumb is better developed,
but still shorter than in other Cercopithecidae; the callosities are
small, and the cheek pouches are absent. There is a single large
laryngeal sac, and the stomach is complex.
This genus is, like the Tiger, often quoted as an example of a race
supposed to be characteristically tropical, existing habitually in the
coldest climate. A species of Semnopithecus has been observed
climbing snow-laden branches at a height of 11,000 feet in the Himalayas.
There are some thirty species, which extend as far east as Borneo.
Fig. 271.—Entellus Monkey or Hanuman. Semnopithecus entellus. × 1⁄6.
The name Semnopithecus is derived from the fact that the
Hanuman is regarded as sacred by the Hindus. The best-known species of
Semnopithecus is this Langur or Hanuman, S. entellus. Being
regarded as a sacred animal, and with the advantage thus gained, it has
become a fell nuisance in gardens and to crops. Though the veneration
with which the Hindoos regard these animals will not allow them to slay
them, they are exceedingly thankful to a European who will enable them to
{569}commit a sin vicariously. This Ape has
immense powers of leaping—a space of 20 to 30 feet can be cleared
by them if one side, that from which the leap is taken, be considerably
higher than the other. They are useful to the Tiger hunter, as they
follow and hoot at this, their deadly enemy. S. schistaceus is a
species which lives at great heights, not less than 5000 feet, in the
Himalayas.
The genus Nasalis is hardly separable from the genus
Semnopithecus. It is a Bornean animal, and is distinguished by a
comical long nose, which not only suggests, but goes beyond, the aquiline
nose of the human species. It is no doubt on this account that the
Borneans, unconsciously imitating our habit of comparing "natives" in
general to Monkeys, call it by a name which signifies "white man."
Rhinopithecus has also a long, but a more definitely upturned
nose.
Fossil Monkeys.—Several of the existing genera of
Old-World Apes are also known to have existed in past times; in some
cases their past distribution indicates a greater range. Thus
Macacus is now represented—and that doubtfully—in
Europe by the Barbary Ape alone. But from Montpellier have been unearthed
the remains of M. priscus, from Pliocene beds. The Asiatic
Semnopithecus is known to have lived during the Pliocene period;
its remains are discovered in France and Italy, as well as in Asia. In
addition to these existing forms, a number of totally extinct Old-World
genera are known. The rich formation at Pikermi near Athens has produced
Mesopithecus pentelici; this Monkey has a skull which recalls that
of Semnopithecus, while the stout limbs are rather Macaque-like.
As is the case with many living Catarrhines, the males have stronger
canines. The animal had a long tail.
An analogous annectent character is shown by the Italian fossil,
Oreopithecus bambolii. This animal was referred by one
palaeontologist to the Man-like Apes, by another to the Cercopithecidae.
It suggests a common ancestral form, and is Middle Miocene in
horizon.
Just as there are no Platyrrhine Apes in the Old World so there are no
Catarrhines met with in a fossil condition in the New World; the two
great divisions of the Apes were as distinct in the past, so far as we
know, as they are now—a strong argument in favour of those who
would derive them from two sources. The {570}existing genera,
Cebus, Mycetes, and Callithrix, now living in South
America, are also known in a fossil state. The extinct genus
Homunculus is known from the Tertiary strata of Patagonia, and an
apparently allied form is Anthropops. These creatures, however,
are at present far from exhaustively known.
Fam. 2. Simiidae.—The Anthropoid, or Man-like Apes,[420] may be separated from
the lower Apes as a group, Simiae, or perhaps better, on account of the
after all slender points of difference, a family Simiidae, which has the
following distinctive characters.
Though arboreal creatures for the most part, these Apes, when they
come to the ground, progress in at least a semi-erect fashion. Moreover,
when they, as is usually the case, put their hands upon the ground to aid
in walking, they do not rest as do the lower Apes upon the flat of the
hand, but upon the back of the fingers. None of the Anthropoids has a
tail, or cheek pouches. Ischial callosities are only seen in the Gibbons.
There is commonly a laryngeal pouch, which is of large size, and aids in
the production of the generally loud voice of these creatures. The hair
is rather more scanty than in the Cercopithecidae, which is an approach
to Man. The placenta differs in detail from that of the lower Apes, and
is exactly like that of Man. These Apes show as further differences from
the underlying Cercopithecidae, the greater length of the arms as
compared with the legs, and the presence of a vermiform appendix to the
caecum. In the latter but not the former character they agree with Man,
whom we shall place in a separate family, Hominidae. The Anthropoid Apes
are entirely Old World and intratropical in range at the present
time.
The Gibbons, genus Hylobates, stand quite at the base of the
series of existing Anthropoid Apes. They are the smallest and the most
purely tree-frequenting of all the members of that group. Connected with
this habit is the structural peculiarity that their arms are
proportionately longer than in the other Anthropoids. The affinity of the
Gibbons to the Catarrhines is proved by the presence of distinct but
small ischial callosities. The arms are so long that when walking upright
the hands reach the ground. The hallux is well developed. The ribs are
thirteen pairs. In the skull the chief noteworthy character as {571}compared
with other Anthropoids is the fact of the large size of the canines,
which are of equal or nearly equal size in the two sexes. The molars on
the other hand have been particularly compared to those of Man. The brain
is simpler than in the higher forms. But it is not clear that this may
not be a case of diminished complexity of convolution going hand in hand
with smallness of size.

Fig. 272.—Hoolock. Hylobates hoolock. × 1⁄6.
The Gibbons range through south-eastern Asia from Assam and Burmah to
Hainan. The number of species is a little doubtful. It is clear that in
the first place we may distinguish the Siamang, H. syndactylus,
which indeed some regard as a separate genus. It is mainly to be defined
by the syndactylous character of the second and third toes; they are
united by skin as far as the last joint. The Hainan species, H.
hainanus, is probably distinct, and the following names have been
given to various other species or races, viz. H. agilis, H.
leuciscus, H. leucogenys, H. lar, H. hoolock.
These animals can walk erect; and when they do so, the big toe is
separated as in unsophisticated or at least unbooted man. The voice is
well known to be loud, {572}and it is a curious fact that the Siamang,
which has a large laryngeal pouch, is not excelled in this respect by
species in which this sac is not developed.

Fig. 273.—Cerebrum of the Gibbon (Hylobates). (Lateral aspect.) c.c′, c.c″, Anterior
and posterior central convolution; fi, interparietal fissure; fr, frontal lobe;
f.s, Sylvian fissure; oc, occipital lobe; pa, parietal lobe; s.c, fissure of Rolando;
tp, temporal lobe; *, fronto-orbital fissure. (From Wiedersheim's Structure of
Man.)
Of Gorillas, genus Gorilla, there is but one species, which
must apparently and rather unfortunately be called Gorilla
gorilla.
The misfortune is double: in the first place the repetition of the
same word as both generic and specific appellation is tiresome to the
ears and barbarous in its suggestion; in the second place it is now well
known that the "Gorilla" of Hanno, observed by that Carthaginian voyager
on an island off the African coast, was not a Gorilla at all as the word
is now understood, but probably a Baboon. The external aspect of this
great Anthropoid is familiar from many reproductions. The male, as is
usual, is larger than the female, and his characters are more
pronounced.
The face is naked and black, and the skin generally is deep black,
even at birth. The ear is comparatively small, and is adpressed to the
side of the head; it is altogether more human in form than that of the
Chimpanzee, and this statement applies also to the rudimentary condition
of the muscles of the ear, which are more rudimentary than in the
Chimpanzee. The nose has an obvious median ridge, and is thus pronounced
as an external feature; the nostrils are very wide. The hands and feet
are short, thick, and broad; the digits are webbed. In the foot the heel
is more apparent than in other Anthropoids. It is not, {573}however, so
marked as in Man, and the phrase "Ex pede Herculem" has been aptly
supplemented by "Ex calce hominem." The hair upon the head forms a kind
of crest, which can be elevated when the animal is enraged. The neck is
thick and short, and the beast has massive shoulders and a broad
chest.
Fig. 274.—Gorilla. Gorilla gorilla, ♀. × ⅛.
If it were not for the fewness of the Anthropoid Apes, and their
nearness to Man, it is doubtful whether the Gorilla would be ranked as a
distinct genus,[421] for
in internal structure it is very near the Chimpanzee. The microscopic
character of the investigations into the anatomy of Man have somewhat
dimmed the proper sense of perspective, and have tended to throw into
greater prominence than seems necessary the divergences of structure seen
in the Gorilla. Dr. Keith[422] has recently summed up and commented
upon these divergences, and the following account of this Anthropoid is
mainly deduced from his memoir.
The cranial capacity of the Gorilla is greater than that of the
Chimpanzee. It is not possible, however, to decide from this point of
view whether a given skull is that of one or of the other of these Apes.
Some Chimpanzees are higher in capacity {574}than some Gorillas. But
the average is undoubtedly as stated. It is to be noted that there is a
correspondence between cranial capacity and size of palate, the
correspondence being converse, i.e. the greater the brain the
smaller the palate. This applies to Man as compared with his Ape-like
relatives, but does not apply so accurately to the Gorilla, which has a
more extensive palate than the Chimpanzee; its "brute development" is
much greater than that of the Chimpanzee. Not only is the palate larger,
but the molar teeth, slightly different in form, are also larger and
stronger. This is so plainly marked that "one may say almost with
certainty, that any upper molar tooth over 12 mm. in length is that of a
Gorilla, and under 12 mm. is that of a Chimpanzee." In the skeleton
generally it may be said that the crests for muscular attachments upon
the bones are greater in the Gorilla. The nasal bones are more like those
of lower Apes in their length, and they have a sharp ridge more marked
than in the Chimpanzee, which, however, disappears in aged animals. It is
a curious fact that Gorillas often have a "cleft palate," owing to the
failure of the palatal part of the palatine bones to meet completely. The
general conformation of the skull is less brachycephalic in the
Gorilla.
The limbs show a number of small differences, which are associated
with a more completely arboreal life in the Chimpanzee as compared with
the Gorilla. The latter is approaching the human way of life. In spite,
however, of these differences, no hard and fast lines of divergence can
be laid down between the two African Anthropoids, for it appears from the
many memoirs that have been written upon both that "there is scarcely a
feature in any muscle or bone found in one animal which is not also found
in the other." The heel of the Gorilla has already been referred to. This
is, of course, associated with a plantigrade and therefore non-arboreal
mode of progression. Certain of the muscles of the calf of the leg
attached to the heel show a more human arrangement in the Gorilla than in
the Chimpanzee. It is interesting to find that the muscles of the little
toe are diminishing in the Gorilla as in Man. This is most clearly due to
terrestrial progression and we may apply the same explanation to Man and
ignore tight boots! The arm of the Gorilla is less adapted to arboreal
progression. Its proportions differ from those of the arm of the
Chimpanzee in that the fore-arm is shorter. In {575}both animals the thumb
is not of much use, and this digit is more retrograde in the Gorilla, not
only in proportionate length but in its muscular supply. The hip girdle
tells the same tale. It is broader in the Gorilla, and the glutaei
muscles are more prominent, all these features being connected with the
more erect gait.
The brain of both animals have been studied, but not in the case of
the Gorilla from a sufficiently large number of examples to make any
generalisations of great value. On the whole, the Gorilla has the larger
brain, but this must be discounted by the fact that it also has the
larger body. It is a remarkable fact that the Gorilla's liver is much
more like that of lower Apes than the liver of other Anthropoids. It has,
as has the Chimpanzee, laryngeal sacs. The general conclusion concerning
the relative position of the two African Anthropoids seems to be that the
Gorilla is the more primitive; and as thus it must approach more nearly
to the original parent than does the Chimpanzee, it may be said that it
also comes rather nearer to Man, since the Chimpanzee has travelled away
from the common stock on another line. The detailed likenesses to Man,
however, are not to be unduly dwelt upon; for they mainly come from a
tendency to assume the plantigrade mode of progression.
In mental characteristics there is the widest difference between the
two Apes that we are considering. The Chimpanzee is lively, and—at
least when young—teachable and tameable. The Gorilla, on the other
hand, is gloomy and ferocious, and quite untameable. When angry the
Gorilla beats its breast, a statement that was originally made, we
believe, by M. du Chaillu, but which has been disputed, though it appears
to be perfectly true. A young Gorilla, exhibited some time since in the
Gardens of the Zoological Society, could be observed to do so. The cry of
the Chimpanzee is different from the "howl" of the Gorilla. An immense
amount has been written upon the ways of this animal in its own home,
including much that is legendary. The Gorilla has been said to lurk in
the depths of the forest, and to stretch down a prehensile foot to grasp
and strangle an unfortunate black man passing below. It is said, too, to
vanquish the Elephant by hitting it hard upon the trunk with a stout
stick, and to crumple up the barrel of a rifle with its powerful teeth.
{576}
Apart from the doubtful "Pongo" and "Engeco" of Andrew Battel, our
first intelligence concerning the Gorilla is due to Dr. Savage, after
whom, indeed, the late Sir Richard Owen called the animal Troglodytes
savagei, a name which has to be abandoned in favour of an earlier
name.
The Gorilla is limited in its distribution to the forest tract of the
Gaboon. It goes about in families, with but one adult male, who later has
to dispute his position as leader of the band with another male, whom he
kills or drives away, or by whom he is killed or driven away. The animal
is said to make a nest in a tree like the Orang; but this statement has
been questioned.
It feeds upon the berries of various plants, and upon other vegetable
substances; there is apparently not so marked an inclination for animal
food as is exhibited by the Chimpanzee. In search of their food they
wander through the forest, walking partly upon the bent hand, and
progressing with a shuffling gait. It is noteworthy that the Gorilla has
been said to walk upon the palm of the hand and not upon the back, as is
the case with the Chimpanzee. It can readily assume the upright posture,
and, in this case, balances itself largely with its arms. Professor
Hartmann, however, states that the back of the hand is also used. Unlike
most or many wild beasts, the Gorilla exhibits no desire to run away when
he views a human enemy. Dr. Savage remarks that "when the male is first
seen, he gives a terrific yell, that resounds far and wide through the
forest, something like kh-ah! kh-ah! prolonged and shrill." This is
accompanied by offensive tactics, which the natives do not willingly
encounter. When making an attack the Gorilla rises to his feet, and as a
full-grown animal reaches a height of some five feet, he is a most
formidable antagonist. The attack of one of these animals is said to be
made with the hand, with which he strikes his adversary to the ground,
and then uses the powerful canines. The beating of the breast which
heralds an attack is a statement made by M. du Chaillu. It has been
denied with a vigour and asperity quite incommensurate with the
importance of the matter.[423]

Fig. 275.—A, Cerebrum of a female Chimpanzee two years old. × ½. (Dorsal aspect,
showing asymmetrical development.) c.c′, c.c″, Anterior and posterior central convolutions;
f.i, interparietal fissure; f.l, the longitudinal fissure; f.po, parieto-occipital
fissure; fr, frontal lobes; oc, occipital lobes; s.c, sulcus centralis. B,
Brain of a female Chimpanzee two years old. × ½. (Lateral aspect.) cb, Cerebellum;
c.c′, c.c″, anterior and posterior central convolutions; fr, frontal lobe; f.s,
fissura Sylvii; is, island of Reil; md, medulla oblongata; oc, occipital lobe; pa,
parietal lobe; s.c, sulcus centralis; tp, temporal lobe. (From Wiedersheim's
Structure of Man.)
The Chimpanzees, genus Anthropopithecus (or
Troglodytes), are {577}to be distinguished from the Gorilla by
the characters mentioned in the account of the latter animal. Briefly
summed up they are mainly as follows:—The
ears are large, and generally stand out from the head; but there are
exceptions to be noted {578}presently. The pigmentation of the body is
not always so pronounced as in the Gorilla. The nasal bones are shorter.
The skull as a whole is more brachycephalic, and the molar teeth are
smaller. The hands and feet are much longer, the animal being more purely
arboreal than the Gorilla. The female Chimpanzee is slightly smaller than
the male, but the great disparity observable in the Gorilla does not
characterise its ally. The animal, like the Gorilla, has large air
sacs.
Fig. 276.—Skull of Chimpanzee. Anthropopithecus troglodytes. × ⅓.
(After de Blainville.)
Chimpanzees are entirely restricted to Africa, and though they appear
to extend rather farther east than the Gorilla, the forest-clad region of
the equatorial belt is their home.
It has been mentioned in treating of the Gorilla that the main feature
of this animal, which affords a constant difference from the Chimpanzee,
is its gloomy and ferocious manner. The Chimpanzee, on the other hand, is
lively and playful, though often maliciously so, and quite tameable, as
many instances—particularly the notorious "Sally" of the Zoological
Gardens—show. The earliest mention of animals that are probably
Chimpanzees is to be found in a work upon the Kingdom of Congo, published
in 1598. In a cut illustrating that work, and of {579}which a part is
reproduced in Professor Huxley's essay referred to below,[424] the Apes, which
correspond roughly in their appearance to Chimpanzees, are represented as
being captured by the device of limed boots, which the Apes are putting
on. This idea has been subsequently imitated and acted upon. A little
later, Andrew Battel wrote of the Pongo and of another creature the
Engeco. This latter, whatever may be the case with the former, is in all
probability the Chimpanzee, since the word 'Nchego, now applied to those
creatures, seems to be the same word. From this seems also to be derived
the sailor's term "Jacko." Whether there are or are not more than one
species of Chimpanzee, is a matter which has exercised and perplexed
naturalists. That there are plain differences of external features, at
any rate between individuals, is perfectly clear. We are justified in
recognising three forms, but the question of their specific distinctness
may for the present be held in reserve. The commonest of these is the
variety known as A. troglodytes. This is frequent in menageries,
though the specimens on view are nearly always young and small. The face
and the hands are flesh-coloured, and the ears are very large. The black
hair gets a reddish tinge on the flanks. The second variety is that which
was termed by du Chaillu Troglodytes kooloo-kamba. This animal
appears to be also the T. aubryi of MM. Gratiolet and Alix,[425] and to be identical
with two Apes known by the names of "Mafuca" and "Johanna."[426] The former of these
was exhibited in Dresden, the latter at Messrs. Barnum and Bailey's show.
The two animals have been carefully studied. They differ from the common
Chimpanzee by the dark colour of the face, and in the case of Mafuca the
ear was Gorilline in form. So too was the ear of A. aubryi, while
Johanna has a larger one. These features have led to the suggestion that
the Kooloo-kamba was the result of a mésalliance between a Gorilla and a
common Chimpanzee.
It has at any rate been stated that the two Anthropoids do go about in
company; but there seems to be little doubt that there is no question
here of a hybrid. Dr. Keith's careful studies[427] upon Johanna have demonstrated the
impossibility of {580}regarding this Ape as anything but a
Chimpanzee. The animal has the ways and manners of the Chimpanzee; has a
cry exactly like that of A. troglodytes; does not beat her breast
like a Gorilla when annoyed. Anatomical knowledge, however, of this
specimen is at present wanting.
Fig. 277.—Young Orang-Utan. Simia satyrus. Zeitschrift für Ethnologie (Anthropolog.
Gesellschaft), Bd. viii. (From Wiedersheim's Structure of Man.)
Anthropopithecus calvus[428] seems to be at least as much entitled
to distinction as the last. It was originally described by du Chaillu;
but Dr. Gray who examined the skins thought that the baldness was
accidental, and then after this wise caution proceeded to describe, under
the name of A. vellerosus, perhaps the "worst" species of
Chimpanzee that has been added to the unnecessarily long list of
"species" of Chimpanzees. To this variety belonged "Sally"[429] of the Zoo, whose
intelligence has been celebrated by the late Dr. Romanes. The form is
characterised {581}by its intense blackness, the red
reflection of other Chimpanzees not being visible; also by the bald head,
whence of course the name. The nostrils of this Ape, as of Johanna, were
somewhat expanded, and thus present a certain likeness to the Gorilla.
But there can be no suggestion that A. calvus is the product of a
union between the two African Anthropoids. As is the case with Johanna,
Sally was given and enjoyed animal food on occasions. It is a curious
fact that both Sally and Johanna appear to have been colour-blind.

Fig. 278.—Young Orang-Utan. Simia satyrus. Zeitschrift für Ethnologie (Anthropolog.
Gesellschaft), Bd. viii. (From Wiedersheim's Structure of Man.)
The Orang Utan, genus Simia, has but one definable species,
viz. S. satyrus. The supposed species of Owen, S. morio,
cannot be satisfactorily defined. Plenty of other specific names have
also been given to what is in all probability but a single species of
large Anthropoid Ape inhabiting the islands of Borneo and Sumatra. {582}
Fig. 279.—Skeleton of Orang. Simia satyrus. (After de Blainville.)
The name Orang-Utan, now applied exclusively to the subject of the
present description, was formerly applied also to the Chimpanzee, and to
that animal, moreover, under the latinised version of Homo
sylvestris. The Orang is a large and heavy Ape with a particularly
protuberant belly and a melancholy expression. The face of the old male
is broadened by a kind of callous expansion of naked skin at the sides.
The colour of the animal is a yellow brown, varying in the exact shade.
The ears are particularly small and graceful in appearance, pressed
closely to the sides of the head. The head is very brachycephalic. The
arms are very long, and when the animal is in the erect posture they
reach as far as the ankle. The hallux is very short and usually destitute
of a nail. It is a curious fact that the head of the thigh bone is
unattached by a ligament to the socket of the pelvis in which it
articulates, a state of affairs which may give the limb greater freedom
in movement, but does not add to its strength; {583}indeed, the Orang has
been described as moving with laborious caution.
This Ape inhabits flat and forest-clad ground, and lives mainly in the
trees. The male leads a solitary life except at the pairing season, but
the female goes about with her family. On the ground the Orang walks with
no great ease, and uses his arms as crutches to swing the body along.
Even on trees the rate of progress is not rapid, and is accomplished with
careful investigations as to the capabilities of the branches to bear his
weight. The "Man of the Woods" has been stated to build a hut in trees.
This is an exaggeration of the fact that it constructs a temporary
nest.
Fig. 280.—A, Skull of a young Orang-Utan. Simia satyrus. (One-third natural size.)
B, Skull of an adult Orang-Utan. (One-third natural size.) (From Wiedersheim's
Structure of Man.)
One of these nests has lately been described elaborately by Dr.
Moebius. It was found (by Dr. Selenka) on the fork of a tree at a height
of 11 metres from the ground. Every night, as it appears, or every second
night, the animal constructs a new nest for himself, abandoning the old
one. So numerous, therefore, are these nests in localities frequented by
Orangs, that a dozen can be readily found in a day. The particular nest
which Dr. Moebius examined was 1.42 metres long, and at most .80 metre
broad. It was built of about twenty-five branches, broken off and laid
for {584}the most part parallel to each other.
Above this framework a number of loose leaves lay. There is no doubt,
therefore, that these nests are not by any means elaborate structures,
and that they only serve as sleeping-places, and not as nurseries for the
upbringing of the young, as has been asserted.
The Orang seems to be usually of a fairly mild disposition; it will
rarely attack a man unprovoked. But Dr. Wallace, who has accumulated a
large number of observations upon these animals, describes a female Orang
who "on a durian tree kept up for at least ten minutes a continuous
shower of branches, and of the heavy-spined fruits as large as
32-pounders, which most effectually kept us clear of the tree she was on.
She could be seen breaking them off and throwing them down with every
appearance of rage, uttering at intervals a loud pumping grunt, and
evidently meaning mischief." The name given by the Dyaks to the Orang is
Mias Pappan.[430]
Fossil Anthropoid Apes.—Undoubtedly the most interesting
of fossil Anthropoids is the now famous Pithecanthropus erectus.
Our knowledge of it is due in the first place to Dubois.[431] But there is hardly an anatomist or
an anthropologist who has not had his say upon this regrettably very
incomplete remnant. The creature is only known by a calvarium, two
separate teeth, and a femur. And the femur, moreover, is diseased. M.
Dubois discovered these remains in the island of Java in andesite tufa of
Pliocene or at least early Pleistocene age. The remains were found in
company with Stegodon, which is now extinct, and
Hippopotamus, which is no longer found in that part of the world.
The name Pithecanthropus was given to it by the discoverer in
order to furnish with a definite habitation and a name the theoretical
Pithecanthropus of Haeckel. Even the most particular of students
of mammalian nomenclature will hardly object to the utilisation of a name
for a second time which is with some clearness a nomen nudum! The
animal when erect must have stood 5 feet 6 inches high. The contents of
the cranium must have been 1000 cm., that is to say 400 cm. more than the
cranial capacity of any Anthropoid {585}Ape, and quite as great
as or even a trifle greater than the cranial capacity of some female
Australians and Veddahs. But as these latter are not 5 feet in height,
the Ape-like man had really a less capacious cerebral cavity. The skull
in its profile outline stands roughly midway between that of a young
Chimpanzee (young in order to do away with the secondary modifications
caused by the crest) and the lowest human skull, that of Neanderthal Man.
This creature is truly, as Professor Haeckel put it, "the long searched
for 'missing link,'" in other words represents "the commencement of
humanity."
The remains of Apes, more distinctly Apes than Pithecanthropus,
are known from Miocene strata of France. Two genera, Pliopithecus
and Dryopithecus, are known. The former appears to be close to
Hylobates. Dryopithecus is more Man-like than any other,
and seems to have been as large as a Chimpanzee. The incisors are human
in their relatively small size. But it has been pointed out that the long
and narrow symphysis of the lower jaw is a point of likeness to the
Cercopithecidae.
Fam. 3. Hominidae.—Apart from Pithecanthropus,
which perhaps is a member of this family, but whose remains permit us to
leave it among the Simiidae, at least for the present, the family
Hominidae contains but one genus, Homo, and probably but one
species, H. sapiens. The characters of the family may therefore be
merged in those of the genus.[432]
Though it is easy enough to distinguish a Man from an Ape, it is by no
means easy to find absolutely distinctive characters which are other than
"relative." As Professor Haeckel has pointed out, there are really only
four characters which differentiate Man: these are the erect walk, and
the consequent modification of the fore- and hind-limbs to that position;
the existence of articulate speech; the faculty of reason. Whether one
body of psychologists are right who argue that reason is a distinctive
human attribute, not to be confused with the apparent reasoning powers of
lower animals, or whether others are justified in separating Man only in
degree from the lower animals, it is clear that this very diversity of
opinion prevents us for the present from utilising such characters as
absolute differences. In any case the discussion of these matters is
beyond the scope of the present book. {586}

Fig. 281.—Skull of Immanuel Kant. (After C. von Kupffer.) The great size of the
cranium is a noteworthy feature. (From Wiedersheim's Structure of Man.)
Anatomically there are a number of small points which distinguish Man;
but they are mainly due to the erect gait. It is sometimes attempted to
divide Man as a naked animal. But this is an apparent difference only;
the hair is not so much developed upon the body as in the Apes, save in
occasional abnormalities, such as the various hairy men and women who can
be seen in travelling shows, and to a less extent the Japanese Ainos, but
it is present everywhere, as is shown by microscopical investigation of
the skin. The skull in Man "is a smooth and imposing, rounded or oval
bony case," which contrasts with the smaller and deeply ridged skull of
the Anthropoid Apes. The shape of the skull is largely in accord with the
large brain. The face does not project so much as in the Anthropoid Apes,
though this character must not {587}be insisted upon too strongly, as in some
American Monkeys the face is as little projecting. Still we are now
comparing Man with his undoubtedly nearest relatives the Simiidae. In the
lower jaw the anterior line at the symphysis is an approximately straight
one, that is at right angles to the long axis of the jaw, while Apes have
a more retreating chin. The "beautiful sigmoid curve formed by the lumbar
and dorsal vertebrae" is more pronounced in Man, but exists not only in
the Anthropoids, but in other Apes.[433]

Fig. 282.—Foot of Man, Gorilla, and Orang of the same absolute length, to show the
difference in proportions. The line a′a′ indicates the boundary between the tarsus
and metatarsus; b′b′, that between the latter and the proximal phalanges; and c′c′
bounds the ends of the distal phalanges. as, Astragalus; ca, calcaneum; sc,
scaphoid. (After Huxley.)
Fig. 283.—Skeleton of the left pes of a Chimpanzee. (Dorsal aspect.) as, Astragalus;
cb, cuboid; cl, calcaneum; ec, ectocuneiform; en, endocuneiform; ms, mesocuneiform;
nv, navicular; I-V, digits. (From Wiedersheim's Structure of Man.)
The fore-limbs are relatively short, the extreme length of the arm
being such that the outstretched hand does not reach the knee. The thumb
is a large and useful digit in Man, much more so than in the Anthropoids.
On the other hand the hallux is not opposable. This is, of course,
correlated with the upright attitude, as is also the greater relative
thickness of that digit, upon which {588}the greatest stress is
laid in walking. As to muscles, the glutaeus maximus is more developed in
Man—the Ape which most nearly approaches him being the Gorilla, in
which animal the life is less thoroughly arboreal than in some others.
The so-called "scansorius" is only present in Man as an occasional
occurrence. The rudimentary character of the ear muscles for the movement
of the external ear in Man has often been insisted upon, as also their
occasional functional activity. But here and elsewhere, so numerous are
the abnormalities, that "the gap which usually separates the muscular
system of Man from that of the Anthropoids appears to be completely
bridged over." These are words of Professor Wiedersheim quoted from
Testut, and express a final summary of the matter of muscles in Man and
the Apes. {589}

Fig. 284.—The hard palate, A, of a Caucasian; B, of a Negro; C, of an adult Orang-Utan,
showing the differences in shape of the bones. The palate of the Negro
represents a type transitional between that of the Caucasian and that of the Orang.
mx, Maxilla; pl, palatine; p.mx, premaxilla. (From Wiedersheim's Structure of
Man.)
In his teeth Man differs by the small exaggeration of the canines,
which hardly, if at all, differ in the two sexes. There is also a
complete absence of a diastema. The teeth are also on the whole weaker
than in the Anthropoids, though Hylobates is very human in this
particular.
Fig. 285.—Human Larynx in frontal
section. cr, Cricoid cartilage; sn,
sinus of Morgagni; t.c, first
tracheal cartilage; th, thyroid
cartilage. (From Wiedersheim's
Structure of Man.)
There is a tendency in Man towards the disappearance of the upper
outer incisors, and more markedly still of the wisdom teeth, which appear
very late, and are often imperfect. In a large number of cases the tooth
does not appear at all. In the larynx there is no great development of
the great throat pouches of the Anthropoids. The minute diverticula of
that organ, known to human anatomists as the ventricles of Morgagni,
alone remain to testify to a former howling apparatus in the ancestors of
Man.
{590}
{591}
INDEX
Every reference is to the page: words in italics are names of genera
or species; figures in thick type refer to an illustration; f. = and in
following page or pages; n. = note.
{592}
Aard Vark, 187
Aard Wolf, 413
Abderites, 148
Aceratherium, 258;
A. incisivum, 259 n.;
A. platycephalum, molar teeth of, 51;
A. tridactylum, 259
Achaenodon, 279
Acodon, 480
Acomys, 472;
A. cahirinus, 473
Aconaemys, 488
Acrobates, 142
Adapis, 553
Addax, 315;
A. nasomaculata, 315
Aeluroidea, 390
Aeluropus, 444;
Ae. melanoleucus, 444
Aelurus, 431
Aepyceros, 311;
Ae. melampus, 311
Aepyprymnus, 137;
Ae. rufescens, 137
Aëthurus, 463;
A. glirinus, 463
Agouti, 495
Agriochoerus, 331;
skeleton of, 332;
A. latifrons, 332
Ai, 171
Alactaga, 486;
A. decumana, 486;
A. jaculus, 486
Alcelaphus, 308
Alces machlis, 297, 298
Alimentary canal, 61
Allodon, 99
Allotheria, 105
Alouatta, 558
Alpaca, 286
Ambergris, 364
Amblotherium, 99
Amblydactyla, 206
Amblypoda, 205 f.
Amblyrhiza, 506
Ammodorcas, 313;
A. clarkei, 313
Amorphochilus, 531
Amphicyon, 423;
A. giganteus, 424
Amphilestes, 99, 100
{593}
Amphimeryx, 330
Amphiproviverra, 160
Amphitherium, 99;
A. prevostii, 100
Amphitragulus, 301
Anaptomorphidae, 552
Anaptomorphus, 552;
A. homunculus, 552
Anchippodus, 507
Anchitherium, 248;
A. aureliense, 249;
A. equinum, 249
Ancodus, 329;
A. brachyrhynchus, 329;
A. velaunus, 329
Ancylopoda, 211 f.
Ancylotherium, 211
Angwantibo, 547
Anisodon, 211
Ankle, 43
Anoa, 317;
A. depressicornis, 317
Anomaluridae, 462
Anomalurus, 462;
A. peli, 463
Anoplotheriidae, 332
Anoplotherium, 333
Ant-eater, 166
Antechinomys, 153;
A. lanigera, 153
Antelopes, 307 f.
Anthops, 527;
A. ornatus, 527
Anthracotherium, 328
Anthropoid Apes, 570 f.
Anthropoidea, 554 f.
Anthropopithecus, 580;
A. calvus, 580;
A. troglodytes, 579
Anthropops, 570
Antidorcas, 312;
A. euchore, 312
Antilocapra americana, 306
Antilocapridae, 306
Antilope, 311;
A. cervicapra, 311
Antlers; see Horns
Antrozous, 529
Anurosorex, 518, 519
Aodon dalei, 369
Aonyx, 441
Apar, 177
{594}
Apes, 554 f.
Aphelops, 260
Archaelurus, 401
Archaeoceti, 384
Arctic region, mammals of, 81
Arctictis, 407;
A. binturong, 407
Arctocebus, 547
Arctogaea, 85
Arctogale, 408;
A. leucotis, 408;
A. stigmatica, 408
Arctoidea, 424
Arctomys, 465;
A. bobac, 466;
A. caudatus, 465;
A. marmotta, 466;
A. monax, 466
Arctonyx, 432;
A. collaris, 432
Arctotherium, 445
Argyrocetus, 383;
A. patagonicus, 384
Armadillo, 177
Arteries, 66
Artiodactyla, 209
Artionyx, 211;
five-toed, 271;
extinct;
forms of, 328
Arviacanthis, 473
Arvicola, 477
Ass, Wild, 242
Astrapotherium, 210, 216
Ateles, 557;
A. ater, 558
Atelodus, 254
Atheriogaea, 85
Atherura, 499;
A. africana, 501;
A. fasciculata, 501
Auchenia, 286
Aulacodus, 489
Aurochs, 318, 321
Avahis, 539;
A. laniger, 539
Aye-aye, 548;
legends relating to, 550
Babirusa, 277;
B. alfurus, 278
Baboon, 566
Badger, 432
Balaena, 358;
B. australis, 359;
B. biscayensis, 360;
B. glacialis, 359;
B. mysticetus, 359;
bones of hind-limbs of, 353;
cervical vertebrae of, 352;
sternum of, 352
Balaenidae, 358
Balaenoptera, 355;
B. australis, 356;
B. borealis, 356;
sternum of, 33;
B. musculus, 356;
sternum of, 352;
B. patachonica, 356;
B. rostrata, 356;
B. sibbaldii, 356
Balaenopteridae, 355
Baleen, 354
Bandicoot, 156
Banteng, 317, 318
Bassaricyon, 428;
B. alleni, 428;
B. gabbii, 428
Bassariscus, 428;
B. astutus, 429
Bat, 521 f.; wings of, 522
Bathyergus, 481
Batomys, 473, 474;
B. granti, 473
Bdeogale, 410
{595}
Bear, 442;
Grizzly, 442;
Himalayan, 442;
Malayan, 443;
Polar, 443;
Sloth-, 443
Beatrix Antelope, 314
Beaver, 467
Behemoth, equivalent to Mammoth, 228
Beisa, 315
Beluga, 372
Berardius, 368;
B. arnouxi, 368;
B. vegae, 369
Bettongia, 137;
B. lesueuri, 138
Bezoar stones, 287, 325
Bibos frontalis, 317, 318
Binturong, 407
Bison, 318;
B. americanus, 319;
B. europaeus, 318
Blaaubok, 314
Blarina, 518
Blarinomys, 480
Blessbok, 309
Boar, Wild, of Europe, 275, 276
Body cavity, 69
Bolodon, 99
Boneia, 526
Bontebok, 309
Bos, 317;
B. alleni, 319;
B. antiquus, 319;
B. bonasus, 318;
B. depressicornis, 320;
B. ferox, 319;
B. frontalis, 318;
B. gaurus, 317;
B. grunniens, 320;
B. latifrons, 319;
B. mindorensis, 320;
B. primigenius, 321;
B. sondaicus, 318;
B. taurus, 320
Boselaphus, 316;
B. tragocamelus, 316
Bovidae, 307
Brachymeryx, 331
Brachyodont, 48
Brachytarsomys, 480
Brachyuromys, 480
Brachyurus, 560;
B. calvus, 560;
B. melanocephalus, 561;
B. rubicundus, 560
Bradypodidae, 170
Bradypus, 170;
B. tridactylus, 171;
skull of, 171;
skeleton of, 172
Brain, 75
of Dog, 76;
of Rabbit, 77;
of Echidna, 110, 117;
of Wallaby, 118
Bramatherium, 306
Brontops, 264
Brontotherium, 264
Bruta, 161
Bubalis, 308;
B. caama, 308
Buffalo, 321
Bunodont, 272
Bush Dog, 414
Cachalot, 363;
ferocity of, 365;
food of, 365
Cadurcotherium, 264
Caenolestes, 146;
C. obscurus, 146
Caenopus, 262
Caenotheriidae, 329
Caenotherium, 329
{596}
Calamodon, 192;
C. simplex, 192
Callignathus, 367
Callinycteris, 526
Callithrix, 559;
C. gigot, 560
Caloprymnus, 138;
C. campestris, 138
Camel, 285
Camelidae, 285;
ancestry of, 289
Camelus, 285;
C. bactrianus, 285, 286;
C. dromedarius, 285
Canidae, 413
Canine teeth, 49
Canis, 416;
C. aegyptiacus, 419;
C. antarcticus, 418;
C. anthus, 420;
C. aureus, 420;
C. azarae, 418;
C. cancrivorus, 414;
C. chama, 418;
C. chanco, 421;
C. dingo, 421, 422;
C. familiaris, 422;
C. ferus, 422;
C. hodophylax, 418;
C. jubatus, 414;
C. lagopus, 419;
C. laniger, 421;
C. lateralis, 420;
C. latrans, 417;
C. lupus, 420;
C. magellanicus, 418;
C. mesomelas, 420;
C. mikii, 422;
C. niger, 418;
C. pallipes, 418;
C. parvidens, 415;
C. primaevus, 423;
C. rudis, 418;
C. urostictus, 415;
C. variegatus, 420;
C. velox, 418;
C. virginianus, 418;
C. vulpes, 419;
C. zerda, 417;
Cannabateomys, 488;
C. amblyonyx, 488
Caperea, 358
Capra, 324
C. aegagrus, 325;
C. falconeri, 325;
C. ibex, 325;
C. jemlaica, 324;
C. pyrenaica, 325;
C. sinaitica, 325
Capreolus, 295;
C. capraea, 295
Capromys, 489, 490;
C. melanurus, 490;
C. pilorides, 490
Capuchin Monkey, 557
Capybara, 491
Cardinal vein, 68
Cariacus, 295;
C. chilensis, 296;
C. macrotis, 295;
C. rufus, 296
Carnivora, 386 f.;
C. Fissipedia, 387 f.;
C. Pinnipedia, 446 f.
Carpomys, 473;
C. melanurus, 473;
C. phaeurus, 473
Carterodon, 489
Castor, 467;
C. canadensis, 468;
C. fiber, 468
Castoridae, 467
Castoroides, 506
Cat, Domestic, 10, 14, 392, 400;
-tribe, 390;
coloration of, 392;
Wild, 399
Catarrhines, 555, 562
Cavia, 493;
C. porcellus, 493
Caviidae, 491
Cavy, 493;
Patagonian, 492
Cebidae, 557
Cebus, 557
Celaenomys, 474;
C. silaceus, 474
Cenoplacentalia, 103
{597}
Centetes, 511;
C. ecaudatus, 511;
skull of, 512
Centetidae, 511
Cephalogale, 423
Cephalophus, 309
Cephalorhynchus, 380
Cephalotes, 526
Ceratorhinus, 254, 257
Cercocebus, 564
Cercolabes, 498;
C. villosus, 42, 499;
C. insidiosus, 498
Cercolabidae, 497
Cercoleptes, 429;
C. caudivolvula, 430
Cercomys, 489
Cercopithecidae, 562
Cercopithecus, 565;
C. diana, 565;
C. lalandii, 565;
C. moloneyi, 565;
C. stairsi, 566;
C. talapoin, 565
Cervicapra, 311;
C. isabellina, 18
Cervidae, 291
Cervulus, 295
Cervus, 293;
C. dama, 293;
C. davidianus, 293;
C. duvauceli, 291;
C. elaphus, 293;
C. luehdorffi, 294;
C. sedgwicki, 301;
C. sika, 291
Cetacea, 339 f.;
primitive position of, 120
Chaenohyus, 280
Chaetomys, 499
Chalicotherium, 211
Chalinolobus, 530
Chamois, 326
Cheetah, 400
Chestnuts of Horse, 240
Chevrotain, 282
Chillingham Cattle, 321
Chilomys, 480
Chimarrogale, 518, 519
Chimpanzee, 576;
brain of, 577;
species of, 579
Chinchilla, 496;
C. lanigera, 496
Chinchillidae, 496
Chirogale, 543;
C. coquereli, 544
Chiromyidae, 548
Chiromys, 548;
C. madagascariensis, 548
Chironectes, 156
Chiropodomys, 473
Chiroptera, 521 f.
Chiru, 311
Chiruromys, 472
Chlamydophorus, 173, 176
Choeropotamus, 280
Choeropsis, 273
Choeropus, 158;
C. castanotis, 158;
manus of, 157
Choloepus, 170;
C. didactylus, 170;
C. hoffmanni, 171;
scapula of, 164
Chriacidae, 552
Chriacus, 552
Chrotomys, 474;
C. whiteheadi, 474
Chrysochloridae, 514
Chrysochloris, 514;
C. trevelyani, 515
Chrysothrix, 559;
C. sciurea, 559
{598}
Civet Cat, 406
Clavicle, 38
Claws, on tip of tail of Lion and Leopard, 41;
see also Nails
Coaitas, 557
Coassus, 296
Coati, 430
Cobus, 310;
C. ellipsiprymnus, 310;
C. maria, 311;
C. unctuosus, 310
Coelogenys, 493;
C. paca, 493;
C. taczanowskii, 494
Coelops, 527;
C. frithii, 527
Coendou, 498
Cogia;
see Kogia
Colobus, 567
Condylarthra, 202 f.
Condylura, 518
Conepatus, 439
Conilurus, 473
Connochaetes, 309;
C. albogulatus, 309;
C. gnu, 309;
C. taurinus, 309
Conoryctes, 193
Conoryctidae, 193
Coracoid, 38
Cormura, 531
Corpus callosum, 77, 125
Coryphodon, 207;
C. eocaenus, 207;
C. radians, skeleton of, 208
Cotton Rats, 479
Coypu;
see Myopotamus
Crateromys, 474
Craurothrix, 473
Creodonta, 449, 455 f.
Cricetomys, 472
Cricetus, 479;
C. frumentarius, 479
Crocidura, 518
Crocuta, 411;
C. maculata, 412
Crossarchus, 410
Crossopus, 518;
C. fodiens, 519
Crumen, of Antelopes, 13;
of Deer, 299
Crunomys, 474;
C. fallax, 474
Cryptophractus, 178
Cryptoprocta, 404;
C. ferox, 405
Ctenodactylidae, 490
Ctenodactylus, 490;
C. gundi, 490
Ctenomys, 488
Cuniculus, 478;
C. torquatus, 478
Cuscus, 140
Cyclopidius, 331;
C. simus, 331
Cycloturus, 167;
C. didactylus, bones of manus of, 169
Cynaelurus, 400;
C. brachygnatha, 401;
C. jubatus, 400;
C. lanigera, 401
Cynictis, 410;
C. penicillata, 410
Cynocephalus, 566;
C. anubis, 566;
C. hamadryas, 566;
C. maimon, 566;
C. mormon, 566;
C. porcarius, 566;
C. thoth, 566
Cynodesmus, 424
Cynodictis, 423
Cynogale, 409
Cynomys, 465;
C. ludovicianus, 465
{599}
Cynopithecus niger, 566, 567
Cynopterus, 526
Cyon, 416;
C. dukkunensis, 416;
C. primaevus, 416
Cystophora, 453;
C. cristata, 453
Dacrytherium, 333
Dactylomys, 488;
D. dactylinus, 488
Dactylopsila trivirgata, 141
Daedicurus, 185
Damaliscus, 309
Daphaenus, 424
Dasymys, 472
Dasypodidae, 173
Dasyprocta, 494;
D. aguti, 495
Dasyproctidae, 493
Dasypus, 173;
D. minutus, 178;
D. sexcinctus, skull of, 174;
pelvis of, 176;
D. villosus, 178;
manus of, 175
Dasyure, 151
Dasyuridae, 149
Dasyuroides, 154;
D. byrnei, 154
Dasyurus, 151;
skull of, 152;
D. geoffroyi, 152;
D. hallucatus, 153;
D. maculatus, 152;
D. ursinus, 18;
D. viverrinus, 152
Deer, 11, 291 f.
Delphinapterus, 372;
D. kingii, 373;
D. leucas, 373
Delphinidae, 372
Delphinus, 377;
D. delphis, 377;
D. longirostris, 377;
D. roseiventris, 377
Dendrohyrax, 234
Dendrolagus, 135;
D. bennetti, 136;
D. inustus, 136
Dendromys, 472, 476;
D. mesomelas, 476
Deomys, 476;
D. ferrugineus, 476
Dermal plates, in Armadillos, 173;
in Whales, 342
Dermoptera, 520
Desman, 518
Desmatippus, 248
Desmodus, 532;
D. rufus, 532
Diabolus, 151
Diaphragm, 69
Diceratherium, 259
Dichobune, 333
Dichodon, 330
Diclidurus, 531
Dicotyles, 279;
D. tajaçu, 278
Dicotylidae, 278
Dicroceras, 301
Didelphyidae, 155
Didelphys, 155;
D. crassicaudata, 156;
D. dimidiata, 13;
D. virginiana, 155
Dinictis, 401
Dinoceras, 210
Dinocerata, 210
Dinomyidae, 495
Dinomys, 495;
D. branickii, 496
Dinotheriidae, 231
{600}
Dinotherium, 231;
D. giganteum, skull of, 231
Diphylla, 532
Diphyodonty, definition of, 51
Diplobune, 333
Diplomesodon, 519
Dipodidae, 484
Dipodomys, 484;
D. merriami, 484
Diprotodon, 146;
D. australis, 147
Diprotodontia, 128 f.
Dipus, 485;
D. aegyptius, 485;
pes of, 485;
D. hirtipes, 484
Distaechurus, 141
Distribution, geographical, 78;
in the past 83
Dogs, 413;
Raccoon-like, 415
Dolichotis, 493;
D. patachonica, 492
Dolphin, 372
Dorcatherium; see Hyomoschus
Dorcopsis, 135;
D. luctuosa, 135, 136;
D. macleani, 136;
D. muelleri, 136
Dorcotragus, 313;
D. megalotis, 313
Dormouse, 470
Doroucouli, 560
Dremotherium, 301
Dromatherium, 98
Dromicia, 141;
D. nana, 141
Dromiciops, 156
Dryolestes, 99
Dryopithecus, 585
Duck-billed Platypus; see Platypus and Ornithorhynchus
Dugong, 336;
skeleton of, 336
Duplicidentata, 502 f.
Dymecodon, 518
Echidna, 110;
egg of, 72;
E. aculeata, 111;
skull of, 108;
shoulder girdle of, 109;
brain of, 110;
E. hystrix (= E. aculeata), 15;
shoulder girdle of, 37
Echidnidae, 110
Echinomys, 488
Echinops, 512
Echiothrix, 473
Edentata, 161 f.
Eland, 316
Elaphodus, 294;
E. cephalophus, 294;
E. michianus, 294
Elasmognathus, 251
Elasmotherium, 259
Elephant, 217;
observations of Pliny and Aristotle upon, 225
Elephantidae, 217
Elephas, 218;
E. africanus, 221, 222;
skull of, 218;
E. antiquus, 229;
E. falconeri, 229;
E. indicus, 221, 223, 224;
foot of, 198;
E. melitensis, 229;
E. meridionalis, 229;
E. planifrons, 229;
E. primigenius, 226;
E. priscus, 226
Eligmodontia, 480
Eliomys, 471
Eliurus, 480
{601}
Elk, 297
Ellobius, 479
Elotherium uintense, 279
Emballonura, 530
Emballonuridae, 530
Enhydridon, 440
Enhydris, 439
Enhydrocyon, 423
Eohippus, 248
Eonycteris, 526;
E. spelaea, 526
Epanorthidae, 145
Epanorthus, 148
Epicondylar foramina, 39
Epipubes of Monotremes and Marsupials, 116
Episternum, 33, 34, 35
Epomophorus, 525
Equidae, 237
Equus, 239;
E. africanus, 243;
E. asinus, 243;
E. boehmi, 245;
E. burchelli, 244, 245;
E. caballus, 239;
E. grevyi, 244;
E. hemionus, 242;
E. hemippus, 242;
E. onager, 242;
E. przewalskii, 241;
E. quagga, 244;
E. sivalensis, 246;
E. somalicus, 244;
E. stenonis, 246;
E. taeniopus, 243;
E. zebra, 244
Erethizon, 498
Ergot, of Horse, 240
Ericulus, 512;
E. setosus, 512
Erinaceidae, 509
Erinaceus, 510;
E. europaeus, 510
Ermine, 436
Eschatia, 290
Esthonyx, 507
Eubalaena, 358
Euchoreutes, 485
Eupetaurus, 467;
E. cinereus, 467
Euphyseter, 367
Eupleres, 403;
E. goudotii, 403
Euprotogonia, 204;
E. puercensis, 204
Eurhinodelphis, 383
Eusmilus, 402
Eutatus, 179
Eutheria, 116;
earliest forms of, 102
Evotomys, 479
Fallow Deer, 293
Felidae, 390
Felis, 391;
F. baileyi, 397;
F. caffra, 400;
F. canadensis, 397;
F. catus, 399;
F. concolor, 399;
F. eyra, 399;
F. leo, 39;3
F. lynx, 397;
F. maniculata, 400;
F. nebulosa, 396;
F. onca, 398;
F. pardalis, 398, 399;
F. pardina, 397;
F. pardus, 395;
F. rufa, 397;
F. tigris, 394;
F. uncia 396;
F. viverrina, 397
Feresia, 377
Fiber, 477;
F. osoyoosensis, 478;
F. zibethicus, 478
Flying Fox, 524;
skeleton of, 523
Flying Squirrel, 466
{602}
Fore-limb, 39
Fossa, 405
Fossa, 407;
F. daubentoni, 407
Fossil mammals, 96
Fox, 419;
Arctic, 419
Fur Seals, 451
Furia, 531
Galago, 543;
G. crassicaudatus, 543;
G. garnetti, 543;
G. senegalensis, 543
Galeopithecidae, 520
Galeopithecus, 521;
G. volans, 520
Galera, 434
Galeriscus, 434
Galictis, 433;
G. barbara, 433
Galidia, 404;
G. elegans, 404
Galidictis, 404;
G. vittata, 404
Ganodonta, 190 f.
Gaur, 317
Gayal, 318
Gazella, 312;
G. loderi, 312
Gelada, 566
Gelocus, 283
Gemsbok, 315
Genet, 406
Genetta, 406;
G. vulgaris, 406
Geogale, 513;
G. aurita, 513
Geomyidae, 483
Geomys, 483
Georhychus, 481
Gerbillus, 475;
G. aegyptius, 475;
G. pyramidum, 475
Gibbon, 570
Giraffa, 301;
G. attica, 305;
G. australis, 303;
G. camelopardalis, 302;
G. sivalensis, 305
Giraffidae, 301
Glands, of skin, 12;
of Lemurs, 12, 537;
of Marsupials, 13;
of Skunk, 13, 439;
on feet of Rhinoceros, 13, 254;
temporal,of Elephant, 12;
musk, 13, 300;
mammary, 16
Glauconycteris, 530
Gliridae, 470
Globicephalus, 374;
G. brachypterus, 375;
G. indicus, 375;
G. melas, 375;
hand of, 345;
G. scammoni, 375
Glossophaga, 531
Glossotherium, 180
Glutton, 435
Glyptodon, 184;
G. clavipes, skeleton of, 184
Glyptodontidae, 184
Gnu, 309
Goats, species of, 324
Golunda, 473
Gomphotherium sternbergi, foot of, 197
Goral, 327
Gorilla, 572, 573;
brain of, 575;
habits of, 576
Gorilla, 572;
G. gorilla, 572
Grampus, 375
{603}
Grampus, 375;
G. griseus, 375
Graphiurus, 471
Gravigrada, 179
Greenland Whale, 358
Grisonia, 433;
G. allamandi, 434;
G. vittata, 434
Ground Sloths, 179
Grypotherium, 182
Grysbok, 310
Guinea-pig, 493
Gulo, 435;
G. luscus, 435
Gymnobelideus, 141
Gymnura, 509;
G. rafflesii, 509
Gymnuromys, 480
Habrocoma, 487;
H. bennetti, 487
Hair, structure of, 6;
sensory, 10;
tuft of elongated, on wrist, 10;
of Whales, 341
Halichoerus, 452;
H. grypus, 452
Halicore, 336;
H. australis, skeleton of, 336;
H. dugong, 337;
H. indicus, 33
Halitherium, 335
Hallomys, 480
Hamster, 479
Hapale, 556;
H. pygmaea, 556
Hapalemur, 541;
H. griseus, 541;
arm of, 537;
H. olivaceus, 541;
H. simus, 541
Hapalidae, 556
Hapalomys, 473
Hapalotis, 473
Haploceros montanus, 326
Haplodon, 469;
H. major, 469;
H. rufus, 469
Haplodontidae, 469
Hare, 504;
Cape Jumping, 486;
-Kangaroo, 134;
Variable, 504
Harnessed Antelopes, 315
Harpyia, 526
Harpyionycteris, 526
Hartebeest, 309
Heart, 65;
of Monotremata, 66
Hedgehog, 510
Helaletes, 250
Helictis, 438;
H. subaurantiaca, 438
Heliphobius, 481
Helladotherium, 306;
H. duvernoyi, 306
Helogale, 410
Hemicentetes, 512
Hemigale, 408;
H. hardwicki, 408;
H. hosei, 408
Hemigalidia, 404
Hemiganus, 191;
H. otariidens, 191
Herpestes, 409;
H. albicauda, 409, 410;
H. ichneumon, 409;
H. urva, 409
Heterocephalus, 481;
H. glaber, 481;
H. phillipsii, 481
Heterodont, 47
Heteromyidae, 484
Heteromys, 484
Hexaprotodon, 274
{604}
Hind-limb, 42
Hipparion, 250
Hippopotamidae, 273
Hippopotamus, 273;
H. amphibius, 273, 274
Hipposiderus, 527
Hippotigris, 246
Hippotragus, 314;
H. equinus, 314;
H. leucophaeus, 314;
H. niger, 313 f.
Hodomys, 480
Holochilus, 480
Holomeniscus, 290
Homacodon, 280;
H. vagans, 280
Homalodontotherium, 212, 216
Hominidae, 585
Homo, 585;
skull of, 586;
foot of, 587;
palate of, 589;
H. sapiens, 585
Homocamelus, 290
Homodont, 47
Homunculus, 570
Hoolock, 571
Hoplophoneus, 402
Horns, 200, 307
Horse, 239;
Wild, 240;
striping of, 240;
Domestic, 241;
ancestry of, 247
Howling Monkeys, 558
Humpback Whale, 356
Hunterius, 358
Hunting Dog, 416
Hutia, 490
Hyaena, 411;
H. crocuta, 412;
H. spelaea, 413;
H. striata, 412
Hyaenarctos, 445
Hyaenictis, 413
Hyaenidae, 411
Hyaenodon, 455
Hybrid Oxen, 317
Hydraspotherium, 306
Hydrochoerus, 492;
H. capybara, 491
Hydromys, 474;
H. chrysogaster, 474
Hydropotes, 296;
H. inermis, 296, 297
Hylobates, 570;
H. agilis, 571;
H. hainanus, 571;
H. hoolock, 571
Hylomys, 509;
H. suillus, 509
Hyomoschus, 283;
H. aquaticus, 283
Hyopotamus, 329
Hyotherium, 280
Hyperoodon, 23, 41, 370;
H. planifrons, 371;
H. rostratum, 371
Hypogeomys, 480
Hypselodont, 48
Hypsiprymninae, 131
Hypsiprymnodon, 138;
H. moschatus, 138
Hypsiprymnodonitinae, 138
Hyrachyus, 262;
H. agrarius, 262
Hyracodon, 146, 262;
H. nebrascensis, 260
Hyracodontotherium, 333
Hyracoidea, 232 f.
Hyracops, 203
Hyracotherium, 247
{605}
Hyrax, 232;
H. capensis, 233;
manus of, 198
Hystricidae, 499
Hystricomorpha, 487
Hystrix, 499;
H. cristata, 499, 500
Ibex, 325
Ichneumon, 409
Ichthyomys, 480;
I. hydrobates, 480;
I. stolzmanni, 480
Icticyon, 414;
I. venaticus, 414
Ictonyx, 438
Idiurus, 463;
I. macrotis, 463;
I. zenkeri, 463
Incisor teeth, 48
Indris, 538;
I. brevicaudata, 538
Indrodon, 552;
I. malaris, 552
Inia, 381;
I. geoffrensis, 382
Iniopsis caucasica, 383
Insectivora, 508 f.
Intercentra, 24
Intestine, 62;
large and small, 64
Isectolophus, 250
Isomys, 473
Ivory, of Elephant, 227
Kanchil, 283
Kangaroo, 132;
Tree-, 136
Kerivoula, 529;
K. picta, 529
Kiang, 242
Kidney, 68
Killer Whale, 341, 375
Kinkajou, 430
Klipspringer, 310
Koala, 142, 143
Kogia, 366;
K. breviceps, 367;
K. floweri, 367;
K. pottsi, 367;
K. simus, 367
Kudu, 316
Lagenorhynchus, 378;
L. acutus, 378;
L. albirostris, 378
Lagidium, 496;
L. cuvieri, 496
Lagomyidae, 505
Lagomys, 505;
L. ladacensis, 505;
L. roylei, 505
Lagorchestes, 134;
L. conspicillatus, 134;
L. leporoides, 135
Lagostomus, 496;
L. trichodactylus, 496, 497
Lagostrophus, 137;
L. fasciatus, 137
Lagothrix, 557;
L. humboldti, 557
Lagotis, 496
Lama, 286;
L. huanacos, 286, 287;
L. vicugna, 286
Langur, 568
Latax, 439
Leggada, 472
{606}
Lemming, 478
Lemur, 541;
L. catta, 542;
N. leucomystax, 542;
L. macaco, 542;
L. nigerrimus, 542;
L. rufipes, 542;
L. varius, 12, 542
Lemur, Gentle, 540
Lemuridae, 538
Lemuroidea, 534
Leopard, 395;
Snow, 396;
Hunting, 400
Lepilemur, 540;
L. mustelinus, 540
Leporidae, 502
Leptarctus, 431;
L. primaevus, 431
Leptauchenia, 331
Leptomeryx, 284
Leptonyx, 453;
L. weddelli, 453
Lepus, 502;
L. americanus, 504;
L. crawshayi, 504;
L. cuniculus, 502;
brain of, 77;
L. europaeus, 504;
L. timidus, 504;
L. whytei, 504
Lestodon, 183
Limacomys, 476
Limnofelis, 456
Limnogale, 512;
L. mergulus, 513
Linsang, 406
Lion, 393
Liponyx, 526
Listriodon, 279
Lithocranius, 313
Litopterna, 267
Liver, 64
Loncheres, 489;
L. guianae, 489
Lophiodon, 250
Lophiodontidae, 247
Lophiomys, 476;
L. imhausi, 476
Lophodont teeth, definition of, 51
Lophotragus michianus, 294 n.
Lophuromys, 472
Loris, 23, 547;
L. gracilis, 547
Lungs, 69
Lutra, 440;
L. vulgaris, 441
Lycaon, 416;
L. anglicus, 416;
L. pictus, 416
Lycyaena, 413
Lyncodon, 437;
L. luganensis, 437
Lynx, 397
Lytta, a rod formed of connective tissue within the tongue, 416
Macacus, 563;
M. cynomolgus, 564;
M. inuus, 563;
M. leoninus, 564;
M. nemestrinus, 564;
M. priscus, 569;
M. rhesus, 564;
M. silenus, 564;
M. sinicus, 564;
M. speciosus, 564;
M. tcheliensis, 563, 564
Machaerodontidae, 401
Machaerodus, 402;
M. palmidens, 402
Macleayius, 358
Macrauchenia, 267
Macraucheniidae, 267
Macroglossus, 526
Macropodidae, 129
Macropodinae, 131, 132
{607}
Macropus, 132;
M. brunii, 133;
M. giganteus, 132;
M. irma, 133;
M. rufus, 132
Macrorhinus, 453;
M. leoninus, 454
Macroscelidae, 515
Macroscelides, 515
Macrotherium, 211
Malacomys, 472
Malacothrix, 476
Mammals, position in Vertebrate series, 1;
characters of, 3;
coloration of, 10, 11;
Reptilian ancestors of, 90;
earliest known forms, 96
Mammoth, 226
Man, 585
Manatee, 335
Manatus, 335;
M. inunguis, 336
Mandrill, 566
Mangabeys, 564
Manidae, 188
Manis, 188;
M. gigantea, 190;
M. macrura, 25;
M. tricuspis, 31
Manteoceras, 267
Marine Mammals; see Sirenia, Cetacea
Marmosa, 156
Marmoset, 556
Marmot, 465;
Prairie-, 465
Marsupial Mole, 159
Marsupials, 122;
imperfectly born young of, 124
Marsupium or Pouch, 14;
of Monotremata, 15;
of Marsupials, 17;
rudiments of, in higher Mammals, 23, 18
Marten, 435
Massoutiera, 490
Mastacomys, 473
Mastodon, 230
Meerkat, 410, 411
Megachiroptera, 524 f.
Megaderma, 528;
M. lyra, 528
Megaladapididae, 554
Megaladapis insignis, 554;
M. madagascariensis, 554
Megaloglossus, 526
Megalomys, 480
Megalonychidae, 183
Megalonyx, 183
Megamys, 506
Meganeuron kreffti, 366
Megaptera, 356;
M. capensis, 357;
M. indica, 357;
M. lalandii, 357;
M. longimana, 357
Megatherium, 183
Meles, 432;
M. anakuma, 432 n.;
M. taxus, 432
Mellivora, 437;
M. capensis, 438
Melonycteris, 526
Melursus, 443;
M. labiatus, 443
Meniscoessus, 101
Meniscotherium, 203
Menodus, 264
Mephitis, 439
{608}
Meriones, 475
Mermaids, 337
Merychyus, 331
Merycochoerus, 330
Merycopotamus, 329;
M. nanus, 329
Mesohippus, 248
Mesomys, 489
Mesonyx, 456
Mesopithecus pentelici, 569
Mesoplacentalia, 103
Mesoplodon, 369;
M. bidens, 369;
M. europaeus, 369;
M. hectori, 369;
M. layardi, 369;
M. stejnegeri, 369
Mesoreodon, 330
Metamynodon, 264;
M. planifrons, skeleton of, 263
Metatheria, 116
Microbiotherium, 160
Microcebus, 544;
M. smithii, 564
Microchiroptera, 526 f.
Microdipodops, 484
Microgale, 512;
M. longicaudata, 25
Microlestes, 98;
M. antiquus, 98
Microtus, 477;
M. agrestis, 477;
M. amphibius, 477;
M. glareolus, 477
Midas, 556;
M. bicolor, 557;
M. leoninus, 556;
M. nigricollis, 556;
M. rosalia, 556
Mimicry, in Bassaricyon, 428;
in Tupaia, 511
Minopterus, 529;
M. scheibersi, 529
Mioclaenidae, 205
Mioclaenus, 205
Miohippus, 248
Miopithecus, 565
Mixocebus, 540;
M. caniceps, 540
Molar teeth, 51
Mole, 517;
sternum of, 31;
fore-foot of, 40;
Golden, 515
Mole Rat, 481
Molossus, 531
Monachus, 453;
M. albiventer, 453;
M. tropicalis, 453
Mongoose, 409
Monkeys, 554 f.
Monodon, 373;
M. monoceros, 373
Monophyodonty, definition of, 53
Monotremata, 105 f.
Moose, 297, 298
Moropus, 211
Morse, 451
Moschinae, 299
Moschus, 299;
M. moschiferus, 299, 300
Mouflon, 324;
Cyprus, 322
Mouse, 472;
Pharaoh's, 409
Multituberculata, 98
Multituberculy, definition of, 59
Muntjac, 295
Muridae, 471
Mus, 471;
M. decumanus, 472;
M. musculus, 472;
M. minutus, 472;
M. rattus, 472
{609}
Muscardinus, 470;
M. avellanarius, 470
Musk, 300;
Deer, 12, 13, 299;
Kangaroo, 138;
Ox, 327
Musquash, 477
Mustela, 435;
M. foina, 435;
M. martes, 435;
M. putorius (= Putorius foetidus), 436;
M. zibellina, 435
Mustelidae, 431
Mycetes, 34, 558
Mydaus, 433;
M. meliceps, 433
Mylodon, 179;
M. robustus, 179;
skeleton of, 180
Mylodontidae, 179
Myocastor, 489
Myodes, 478;
M. lemmus, 478
Myogale, 518
Myomorpha, 469
Myopotamus, 489
Myoscalops, 481
Myosorex, 518
Myoxus, 470;
M. glis, 470
Myrmecobius, 154;
M. fasciatus, 155
Myrmecophaga, 166;
M. jubata, 166;
skull of, 167, 168;
hand of, 169;
vertebrae of, 163
Myrmecophagidae, 166
Mystacoceti, 353 f.
Myxopoda, 529
Nail-tailed Wallaby, 134
Nails, 14
Nandinia, 408
Nannosciurus, 464
Narwhal, 373;
"horn" of, 49
Nasalis, 569
Nasua, 430;
N. nasica, 431;
N. rufa, 430, 431
Natalus, 529
Native, Bear, 142;
Cat, 152
Necrodasypus, 164 n.
Necrogymnura, 510
Nectogale, 518;
N. elegans, 520
Nectomys, 480
Nelomys, 488
Nelsonia, 480
Nemorrhaedus, 326;
N. crispus, 326;
N. goral, 327
Neobalaena, 361;
N. marginata, 362
Neofiber, 478;
N. alleni, 478
Neogaea, 85
Neomeris, 374;
N. phocaenoides, 374
Neomylodon listai, 181
Neotoma, 480
Neotragus pygmaeus, 310
Nesodon, 215
Nesokia, 472
Nesonycteris, 526
Nesopithecus, 553;
N. australis, 553
New Zealand, absence of terrestrial mammals from, 85
Nilgai, 316
Nimravus, 401;
N. gomphodus, 401
{610}
Noctilio, 530;
N. leporinus, 530
Nomarthra, 186 f.
Nothocyon, 415
Notiomys, 480
Notiosorex, 518
Notogaea, 85
Notopteris, 526
Notoryctes typhlops, 159
Notoryctidae, 158
Nototherium, 148;
N. mitchelli, 148;
skull of, 148
Nyctereutes, 414;
N. procyonides, 415
Nycteridae, 527
Nycteris, 528;
N. javanica, 528
Nycticebus, 545;
N. tardigradus, 546;
foot of, 537
Nycticejus, 530
Nyctinomus, 531
Nyctipithecus, 560
Nyctophilus, 529
Ocapia johnstoni, 305
Oceanic islands, mammalian fauna of, 81
Ocelot, 398, 399
Ochotona, 505
Octodon, 487;
O. degus, 487
Octodontidae, 487
Odobaenus, 451
Odontoceti, 362 f.;
fossil forms of, 383
Ogmorhinus, 453
Okapi, 305
Ommatophoca, 453;
O. rossi, 453
Onohippidium, 247 n.
Onychodectes, 193;
O. tissonensis, 193
Onychogale, 134
Onychomys, 479
Opolemur, 544
Opossum, 155;
Thick-tailed, 156;
Virginian, 155
Orang Utan, 580, 581, 582, 583, 587, 589
Orca, 375;
O. gladiator, 341, 375
Orcella, 376;
O. brevirostris, 376;
O. fluminalis, 376
Oreinomys, 475
Oreodon, 330
Oreodontidae, 330
Oreopithecus bambolii, 569
Oreotragus saltator, 310
Orias, 316;
O. canna, 316;
O. livingstonii, 316
Ornithodelphia, 106
Ornithorhynchidae, 112
Ornithorhynchus, 112;
O. anatinus, 112;
shoulder girdle of, 34;
skeleton of, 114
Orohippus, 248
Orycteropodidae, 187
Orycteropus, 187;
teeth of, 188;
O. aethiopicus, 188;
O. capensis, 187;
O. gaudryi, 188
Oryx, 314;
O. beatrix, 314;
O. beisa, 315;
O. leucoryx, 314
Oryzomys, 480
{611}
Oryzoryctes, 513
Otaria, 451;
O. gillespiei, 451;
O. hookeri, 451;
O. jubata, 448;
O. nigrescens, 451;
O. pusilla, 450;
O. ursina, 451
Otariidae, 450
Otocyon, 415;
O. megalotis, 415
Otomys, 475;
O. unisulcatus, 475
Otonycteris, 529
Otter, 441;
Sea, 439
Ouakari Monkey, 560;
Red-faced, 560
Oulodon grayi, 380
Ounce, 396
Ourebia, 310
Ova, 71;
of Monotremes, 72
Ovaries, 70
Ovibos moschatus, 327
Oviduct, 73;
of Marsupials, 74
Ovis, 321;
O. blanfordi, 323;
O. burrhel, 322;
O. montana, 322;
O. musimon, 324;
O. nahura, 324;
O. ophion, 322;
O. poli, 321;
O. tragelaphus, 323;
O. vignei, 321
Oxen, 317
Oxymycterus, 480
Paca, 493
Pachynolophus, 248
Pachyrucos, 213
Pachyuromys, 475
Palaeochoerus, 280
Palaeoerinaceus, 510
Palaeonictis, molar teeth of, 56
Palaeosyops, 266
Palaeotragus, 306
Palate, 61
Palm Civet, 407
Pancreas, 64
Panda, 431
Pangolin, 188
Pangolin gigantesque, 211
Panochthus, 185
Panther, 395
Pantholops, 311
Pantolambda, 206;
P. bathmodon, 207;
skull of, 206
Pantolestidae, 328
Paradoxurus, 407;
P. grayi, 407;
P. niger, 407
Patriofelis, 449, 456
Peccary, 278
Pecora, 290 f.
Pectinator, 490;
P. spekii, 491
Pectoral girdle, 35
Pedetes, 486;
P. caffer, 486
Pedetidae, 486
Pelea, 311;
P. capreolus, 311
Peltephilus, 186
Peludo, 177
Pelvis, 41
Peraceras, 262
Peragale, 156;
P. lagotis, 157
Perameles, 158
{612}
Peramelidae, 156
Peramus, 99
Peramys, 156
Perascalops, 518
Periptychus, 204;
P. rhabdodon, 205
Perissodactyla, 235 f.
Perodicticus, 547
Perodipus, 484
Perognathus, 484
Peromyscus, 479
Petauroides, 142
Petaurus, 141;
P. breviceps, 142
Petrodromus, 516;
P. tetradactylus, 516
Petrogale, 134;
P. penicillata, 134;
P. xanthopus, 134
Petromys, 488
Phacochoerus, 277;
P. aethiopicus, 277;
P. africanus, 277
Phalanger, 140;
P. maculatus, 140
Phalanger, 138;
Flying, 141;
Striped, 141;
Ring-tailed, 141
Phalangeridae, 138
Phalangista vulpina, 140
Phascolarctos cinereus, 142, 143
Phascologale, 153;
P. virginia, 153
Phascolomys, 144;
skull of, 145;
P. latifrons, 145;
P. mitchelli, 145;
P. ursinus, 145;
P. wombat, 144
Phascolonus, 148
Phascolotherium, 99;
P. bucklandi, 99
Phatagin, 188
Phenacodus, 202;
P. primaevus, skeleton of, 196;
molar teeth of, 56
Phenacomys, 479
Phlaeomys, 473
Phoca, 452;
P. groenlandica, 452;
P. hispidia, 452;
P. vitulina, 452;
skeleton of, 447
Phocaena, 374;
P. communis, 374;
P. dallii, 374;
P. spinipennis, 374;
P. tuberculifera, 342
Phocidae, 452
Pholidotus, 188
Phyllorhina, 527
Phyllostomatidae, 531
Phyllotis, 480
Physeter, 363;
P. macrocephalus, 363;
P. tursio, 366
Physeteridae, 362
Physodon, 383
Pichy-ciego, 176
Pig, 275;
sternum of, 32;
foot of, 199;
solid-hoofed, 270
Pika, 505
Pinnipedia, 446 f.
Pithecanthropus erectus, 584
Pithechirus, 473
Pithecia, 562;
P. albinasa, 561;
P. cheiropotes, 562;
P. satanas, 562
Placenta, 125
Plagiaulax, 99
Plagiodontia, 489
{613}
Platacanthomys, 471;
P. lasiurus, 471
Platanista, 380;
P. gangetica 381
Platanistidae, 380
Platycercomys, 486
Platypus, 113
Platyrrhines, 555, 556
Plecotus, 529;
P. auritus, 529
Plesiomeryx, 329
Pliopithecus, 585
Poebrotherium, 288;
P. labiatum, foot of, 197;
P. wilsoni, skull of, 288
Poecilogale, 437;
P. albinucha, 437
Poephagus grunniens; see Bos grunniens
Pogonodon, 401
Poiana, 407
Polecat, 436
Polycladus, 301
Polyprotodontia, 149 f.
Pontoporia, 382;
P. blainvillii, 382
Porcula salvania, 276
Porcupine, 499, 500;
Tree, 498;
Brush-tailed, 501
Porpoise, 374
Potamochoerus, 278
Potamogale, 513;
P. velox, 513
Potamogalidae, 513
Potoroinae, 131, 137
Potorous, 138
Potto, 547
Pouch; see Marsupium
Prairie-dog, 465
Praopus, 178
Prepollex, 41
Primates, 533 f.
Priodon, 173, 179;
P. giganteus, hand of 175
Prionodon, 406;
P. pardicolor, 406
Priscodelphinus, 384
Proaelurus, 401
Proboscidea, 216 f.
Procamelus, 289;
P. angustidens, 290;
P. occidentalis, foot of, 197
Procavia, 232
Procyon, 427;
P. cancrivorus, 427;
P. lotor, 427;
P. nigripes, 427
Procyonidae, 426
Prodelphinus, 377
Proechidna, 111;
P. bruijnii, 111;
P. nigroaculeata, 111
Pronghorn, 306
Propalaehoplophorus, 185
Propithecus, 539;
P. coquereli, 540;
P. verreauxi, 540
Prorastoma, 336;
P. veronense, 337
Prosqualodon, 384
Proteles, 413;
P. cristata, 413
Protelotherium, 280
Prothylacinus, 160
Protoceras, 284
Protoceratidae, 284
Protochriacus, 553
Protogonia, 204
{614}
Protohippus, 249
Protolabis, 289
Protoreodon, 332
Protorotheriidae, 268
Protorotherium, 268
Protoselene, 205
Prototheria, 105 f.
Protylopus, 287;
P. petersoni, feet of, 197
Protypotherium, 213
Psammomys, 475
Pseudamphicyon, 424
Pseudochirus, 141
Pseudorca, 376;
P. crassidens, 376;
P. meridionalis, 376
Pseudorhynchocyon, 516
Psittacotherium, 191;
P. multifragum, 191
Pteralopex, 526;
P. atrata, 526
Pteromys, 466;
P. alborufus, 466;
P. petaurista, 466
Pteroneura brasiliensis, 441
Pteropodidae, 524
Pteropus, 524;
P. edulis, 524;
P. fuscus, skull of, 524;
P. nicobaricus, 526;
P. poliocephalus, 525
Ptilocercus, 511;
P. lowi, 511
Ptilodus, 101
Pudua, 296
Puma, 399
Putorius, 435;
P. brasiliensis, 437;
P. erminea, 436;
P. foetidus (= Mustela putorius), 436;
P. hibernicus, 437;
P. vulgaris, 437
Pygmy, Hog, 276;
Right Whale, 361
Pyrotherium, 232
Rabbit, 502;
skull of, 503;
oviduct of, 73;
brain of, 77
Raccoon, 427
Rangifer tarandus, 298, 299
Raphiceros, 310
Rasse, 406
Rat, 472;
-Kangaroo, 137
Ratel, 437, 438
Red Deer, 293
Reed Buck, 311
Reindeer, 292, 298, 299
Reithrodon, 480
Reithrodontomys, 480
Reithrosciurus macrotis, 464
Reproductive organs, 70
Respiratory organs; see Lungs
Rhachianectes, 357;
Rh. glaucus, 357
Rhinoceros, 254;
Rh. bicornis, 257, 258;
Rh. indicus (= Rh. unicornis), 255;
femur of, 43;
Rh. lasiotis, 257;
Rh. schleiermacheri, 258;
Rh. simus, 257;
Rh. sondaicus, 256;
Rh. sumatrensis, 256;
hand of, 199;
Rh. tichorhinus, 259
{615}
Rhinoceros, White, 257;
Black, 257
Rhinocerotidae, 253
Rhinogale, 410 n.
Rhinolophidae, 527
Rhinolophus, 527;
Rh. ferrum equinum, 527;
Rh. hipposiderus, 527
Rhinonycteris, 527
Rhinopithecus, 569
Rhinopoma, 530
Rhipidomys, 480
Rhizomys, 482;
Rh. badius, 483;
Rh. pruinosus, 482;
Rh. sumatrensis, 482
Rhynchocyon, 516;
Rh. chrysopygus, 516
Rhynchogale, 410
Rhynchomys, 474;
Rh. soricioides, 474
Rhynchonycteris, 531
Rhytina, 338
Ribs, 29;
single-headed, of Whales and Monotremes, 30
Risso's Dolphin, 375
River Hog, Red, 278
Roan Antelope, 314
Rodentia, 458 f.;
brain of, 461;
teeth of, 459
Roe Deer, 295
Romerolagus, 504
Rorqual, 355
Ruminantia, 280 f.;
stomach of, 281 f.
Rupicapra, 326
Sable, 435;
Antelope, 313
Saccopteryx, 531
Saccostomus, 472
Sacrum, 24
Sagmatias, 376
Saiga, 311;
S. tartarica, 311
Saki monkey, 562
Salivary glands, 64
Samotherium, 305;
S. boissieri, 306
Sarcophilus ursinus, 151;
skull of, 149
Scales, 9;
of Anomaluridae, 463;
of Manidae, 188
Scalops, 518
Scapasius, 518
Scapteromys, 480
Scaptonyx, 518
Scapula, 36;
of Whales, 37
Scelidotherium, 180
Schizodon, 488
Schizotherium, 211
Sciuridae, 463
Sciuropterus, 467;
S. volucella, 467
Sciurus, 463;
S. castaneiventris, 464;
S. laticaudatus, 511;
S. maximus, 464
Scleropleura, 179;
S. bruneti, 179
Scotophilus, 529
Scotozous, 530;
S. dormeri, 530
Seal, 452;
Bladder-nosed, 454;
compared with Zeuglodonts, 385;
Elephant, 454;
Eared, 450;
hooded, 453
Sea-Lion, 450;
see Otaria
{616}
Sealskin, 9
Sea Serpent, 368
Selenodont, 272
Selenodontia, 280
Semnopithecus, 568;
S. entellus, 568;
S. schistaceus, 569
Sewellel, 469
Sheep, Barbary, Blanford's, and other species of, 321 f.
Shoulder girdle, 35;
of Ornithorhynchus, 34;
of Echidna, 37
Shrew, 518;
Elephant-, 515
Siamang, 571
Sibbald's Rorqual, 2, 356
Sigmodon, 479
Simia satyrus, 580, 581;
skeleton of, 582;
skull of, 583;
nest of, 583
Simiidae, 570
Simocyon, 423
Simplicidentata, 462 f.
Sing-sing Antelope, 310
Siphneus, 479
Sirenia, 333 f.
Sivatherium, 306
Size, of Mammals, 2;
progressive secular increase in, 103;
of Whales, 342
Skull, 25;
of Human embryo, 27;
of adult, 586;
of Dog, 25, 29;
comparison of, with that of Anomodontia, 28
Skunk, 439
Sloth, 170;
Ground-, 180
Smilodon, 402;
S. neogaeus, 402
Sminthopsis, 153;
S. virginiae, 153
Sminthus, 486
Smutsia, 188
Solenodon, 514;
S. cubanus, 514
Solenodontidae, 513
Sorex, 518;
S. alpinus, 519;
S. minutus, 519;
S. vulgaris, 519
Soricidae, 518
Soriculus, 518
Sotalia, 378;
S. gadamu, 379;
S. guianensis, 378;
S. lentiginosa, 379;
S. pallida, 382;
S. sinensis, 378;
S. teüszii, 378
Souslik, 464
Spalacidae, 482
Spalacopus, 487;
S. poeppigi, 487
Spalacotherium, 99
Spalax, 482;
S. typhlus, 482
Sparassodonta, 160
Speke's Antelope, 315 f.
Speothos, 423
Sperm Whale, 363
Spermaceti, 363
Spermophilus, 464;
S. tredecimlineatus, 465
Sphingurus, 498;
S. prehensilis, 498 f.
Spider Monkey, 558
Spiny Anteater; see Echidna
Springbok, 312
Squalodon, 384
{617}
Squalodontidae, 384
Squamata, 188
Squirrel, 463;
Pygmy, 464;
Ground, 464;
Flying, 466
Steatomys, 473, 476
Stegodon, 229;
S. ganesa, 229
Steinbok, 310
Steller's Sea-cow, 338
Steno, 379;
S. perniger, 379
Stenodelphis, 382
Stenoplesictis, 441
Stenops, 545
Stenorhynchus, 453
Stentor, 558
Stereognathus, 99
Sternum, 31;
of Whales, 32, 33, 352;
of Dugong, 33
Stoat, 436
Stomach, 62;
of various mammals, 63;
of Whales, 347
Strepsiceros, 316;
S. imberbis, 316;
S. kudu, 316
Stylacodon, 99
Stylinodon, 193;
S. cylindrifer, 193;
S. mirus, 193
Stylinodontidae, 191
Sublingua, 61
Suidae, 275
Suricata, 410;
S. tetradactyla, 411
Sus, 275;
S. barbatus, 276;
S. erymanthius, 279;
S. salvania, 276;
S. scrofa, 275;
S. sennaariensis, 275;
S. verrucosus, 276;
S. vittatus, 276
Synaptomys, 479
Synetheres, 498
Synotus, 529;
S. barbastellus, 522, 529
Systemodon, 250
Tachyoryctes, 483
Talpa, 517;
T. europaea, 517
Talpidae, 516
Tamandua, 167;
sternum of, 168
Tamarin, 556
Tamias, 464
Taphozous, 531
Tapir, 11
Tapiridae, 250
Tapirus, 250;
T. bairdi, 251;
T. dowi, 251;
T. ecuadorensis, 251 n.;
T. indicus, 252;
T. leucogenys, 251 n.;
T. roulini, 251;
T. terrestris, 251
Tarsiidae, 550
Tarsipedinae, 140, 145
Tarsipes, 145
Tarsius, 550;
T. spectrum,foot of, 551
Tasmanian, Devil, 151;
Wolf, 150
Tatusia, 173;
T. novemcincta, 178
Taurotragus oryx, 316 n.
Taxidea, 438
Tayra, 433
Teeth, 43;
minute structure of, 44;
development of, 52;
brachyodont (with short roots), 48;
{618}
bunodont, 272;
diphyodont, 51;
heterodont, 47;
homodont, 47;
hypselodont (with long roots), 48;
lophodont, 51;
monophyodont, 53;
selenodont, 272
Teledu, 433
Teleoceras, 259 n.;
T. fossiger, 261
Telmatotherium, 267;
T. cornutum, 267
Temperature of Monotremata, 112
Tenrec, 511
Testes, 75
Tetraceros, 310
Tetraprotodon, 274
Thalassarctos, 443
Theropithecus, 566
Thoatherium, 268
Thomomys, 483
Thrinacodus, 489;
T. albicauda, 489
Thrynomys, 489;
T. swindernianus, 489
Thylacinus, 150;
T. cynocephalus, skull of, 150
Thylacoleo, 147;
T. carnifex, skull of, 147
Thyroptera, 529
Tiger, 394
Tillodontia, 506 f.
Tillotherium, 507;
T. fodiens, 507;
skull of, 507
Tinoceras, 210
Titanops, 264
Titanotheriidae, 264
Titanotherium, 264;
T. coloradense, 264;
T. elatum, 265;
T. heloceras, 264;
T. platyceras, 265, 266;
T. ramosum, 266;
T. trigonoceras, 265
Tolypeutes, 173;
T. tricinctus, 176, 177
Tomitherium, 552
Tongue, 61
Toxodon, 214
Toxodontia, 214 f.
Tragelaphus, 315;
T. gratus, 316;
T. spekii, 315;
T. sulvaticus, 316
Tragulidae, 282
Tragulina, 281
Tragulus, 282;
T. meminna, 282;
T. napu, 283;
T. stanleyanus, 283
Tree- Porcupines, 498;
Kangaroos, 136
Triaenops, 527
Trichechidae, 451
Trichechus, 451
Trichomys, 488
Trichosurus, 140;
T. vulpecula, 140
Trichys, 501;
T. lipura, 501
Triclis, 149;
T. oscillans, 149
Triconodon, 99
Trituberculata, 99
Trituberculy, definition of, 56
Tritylodon, 98
Troglodytes, 576;
T. aubryi, 579;
T. kooloo-kamba, 579;
T. savagei, 576
{619}
Trogontherium, 469
Tubulidentata, 186
Tuco-tuco, 488
Tupaia, 511;
T. belangeri, 511;
T. tana, 511
Tupaiidae, 511
Tursio, 379;
T. borealis, 380:
T. peronii, 380
Tursiops, 379;
T. abusalam, 379;
T. tursio, 379
Tusks, 49
Tylomys, 480
Tylopoda, 285 f.
Typhlomys, 471
Typotheria, 212 f.
Typotherium, 213
Uintacyon, 424
Uintatherium, 210
Unau, 170, 171
Ungulata, 195 f.
Urinary organs, 68
Uromys, 473
Uropsilus, 518
Urotrichus, 518
Ursidae, 442
Urson, 498
Ursus, 442;
U. arctos, 442;
U. fossilis, 442;
U. malayanus, 443;
U. ornatus, 443;
U. pruinosus, 443;
U. spelaeus, 442, 444;
U. tibetanus, 442
Urus, 321
Vampire, 532
Vampyrus, 531;
V. spectrum, 531
Vandeleuria, 473
Vansire, 404
Veins, 67;
anterior abdominal, in Echidna, 68
Vena cava, double in Elephants, 68
Vertebrae, 19;
cervical, 21, 22;
dorsal, 20, 23;
lumbar, 20, 23;
caudal, 24
Vespertilio, 529;
V. murinus, 529
Vespertilionidae, 528
Vesperugo, 528;
V. discolor, 528;
V. leisleri, 528;
V. noctula, 528;
V. pipistrellus, 528;
V. serotinus, 528
Vicuna, 286
Vishnutherium, 306
Viverra, 405;
V. civetta, 406
Viverricula, 406
Viverridae, 403
Vizcacha, 497
Vole, 477
Vulpine Phalanger, 140
Wallaby, 129;
skeleton of, 130;
Nail-tailed, 134
Walrus, 451
Wapiti, 292
Wart Hog, 277
Waterbuck, 310
{620}
Water Chevrotain, 283
Weasel, 437
Whalebone, 354
Whalebone Whales, 354
Whales, 339;
hunting of, 360
White Whale, 372
Wild Boar of Europe, 276
Wild Cat of Europe, 399
Wisent, 318
Wolf, 420
Wombat, 144;
skull of, 145
Wrist, 40;
tactile hairs of, 10
Xenarthra, 166 f.
Xenomys, 480
Xenurus, 173, 178;
X. unicinctus, 178
Xeromys, 474
Xerus, 464
Xiphodon, 329
{621}
Xiphodontidae, 329
Xiphodontotherium, 330
Zaglossus, 111 n.
Zamicrus, 184
Zapus, 486
Zarrhachis, 384
Zebra, 244;
Burchell's, 245;
Common, 244;
Grevy's, 244
Zenkerella, 463 n.
Zeuglodon, 384
Zeuglodontidae, 384
Ziphius, 367, 370;
Z. cavirostris, 370;
Z. novae zelandiae, 370
Zoological regions, 84
Zorilla, 438
{622}
{623}
THE CAMBRIDGE NATURAL HISTORY
Edited by Sir S. F. Harmer, K.B.E., Sc.D., F.R.S., and A. E. Shipley, G.B.E., M.A., F.R.S.
In Ten Volumes. Fully Illustrated. Medium 8vo. 21s. net each.
FIELD.—"The Cambridge Natural History series of volumes
is one of very great value to all students of biological science. The
books are not intended for popular reading, but for utilisation by those
who are desirous of making themselves thoroughly acquainted with the
branches of zoology of which they treat."
VOLUME I.
Protozoa, by Marcus Hartog, M.A., D.Sc.; Porifera (Sponges), by
Igerna B. J. Sollas, B.Sc; Coelenterata and Ctenophora, by S. J. Hickson,
M.A., F.R.S.; Echinodermata, by E. W. MacBride, M.A, F.R.S.
FIELD.—"The book can be in the strongest manner
recommended to those for whose benefit it has been written. We know of no
work from which a more truly scientific account of the Protozoa,
Echinodermata, and other lower forms of animal life could be gained."
VOLUME II.
Flatworms and Mesozoa, by F. W. Gamble, D.Sc.; Nemertines, by Miss
L. Sheldon; Threadworms and Sagitta, by Sir A. E. Shipley, G.B.E.,
M.A., F.R.S.; Rotifers, by Marcus Hartog, M.A., D.Sc.; Polychaet
Worms, by W. Blaxland Benham, D.Sc., M.A.; Earthworms and
Leeches, by F. E. Beddard, M.A., F.R.S.; Gephyrea and Phoronis,
by Sir A. E. Shipley, G.B.E., M.A, F.R.S.; Polyzoa, by Sir S. F.
Harmer, K.B.E., Sc.D., F.R.S.
CAMBRIDGE REVIEW.—"Most of the articles are of a very
high order of merit—taken as a whole, it may be said that they are
by far the best which have as yet been published.... We may say with
confidence that the same amount of information, within the same compass,
is to be had in no other zoological work."
VOLUME III.
Molluscs, by the Rev. A. H. Cooke, M.A.; Brachiopods (Recent), by Sir
A. E. Shipley, M.A., F.R.S.; Brachiopods (Fossil), by F. R. C. Reed,
M.A.
TIMES.—"There are very many, not only among educated
people who take an interest in science, but even among specialists, who
will welcome a work of reasonable compass and handy form containing a
trustworthy treatment of the various departments of Natural History by
men who are familiar with, and competent to deal with, the latest results
of scientific research. Altogether, to judge from this first volume, the
Cambridge Natural History promises to fulfil all the expectations that
its prospectus holds out."
VOLUME IV.
Crustacea, by Geoffrey W. Smith, M.A., and the late W. F. R. Weldon,
M.A.; Trilobites, by Henry Woods, M.A.; Introduction to
Arachnida, and King-Crabs, by Sir A. E. Shipley, G.B.E., M.A.,
F.R.S.; Eurypterida, by Henry Woods, M.A.; Scorpions, Spiders,
Mites, Ticks, etc., by Cecil Warburton, M.A.; Tardigrada
(Water-Bears), by Sir A. E. Shipley, G.B.E., M.A., F.R.S.;
Pentastomida, by Sir A. E. Shipley, G.B.E., M.A., F.R.S.;
Pycnogonida, by D'Arcy W. Thompson, C.B., M.A.
{624}
VOLUME V.
Peripatus, by Adam Sedgwick, M.A., F.R.S.; Myriapods, by
F. G. Sinclair, M.A.; Insects, Part I., Introduction, Aptera, Orthoptera,
Neuroptera, and a portion of Hymenoptera (Sessiliventres and
Parasitica), by David Sharp, M.A., M.B., F.R.S.
Prof. RAPHAEL MELDOLA, F.R.S., F.C.S., in his Presidential Address
to the Entomological Society of London, said:—"The authors of this volume are certainly to be
congratulated upon having furnished such a valuable contribution to our
literature. When its successor appears, and I will venture to express the
hope that this will be at no very distant period, we shall be in
possession of a treatise on the natural history of insects which, from
the point of view of the general reader, will compare most favourably
with any similar work that has been published in the English
language."
VOLUME VI.
Hymenoptera (continued) (Tubulifera and Aculeata), Coleoptera,
Strepsiptera, Lepidoptera, Diptera, Aphaniptera, Thysanoptera,
Hemiptera, Anoplura, by David Sharp, M.A., M.B., F.R.S.
SATURDAY REVIEW.—"Dr. Sharp's treatment is altogether
worthy of the series and of his own high scientific reputation....
Certainly this is a book that should be in every entomologist's
library."
VOLUME VII.
Hemichordata, by Sir S. F. Harmer, K.B.E., Sc.D., F.R.S.; Ascidians and
Amphioxus, by W. A. Herdman, D.Sc., F.R.S.; Fishes (exclusive
of the Systematic Account of Teleostei), by T. W. Bridge, Sc.D.,
F.R.S.; Fishes (Systematic Account of Teleostei), by G. A. Boulenger, F.R.S.
ATHENÆUM.—"All who take a serious interest in the advance
of ichthyology will find this a fascinating book."
VOLUME VIII.
Amphibia and Reptiles, by Hans Gadow,
M.A., F.R.S.
NATURE.—"In concluding the review we would express the
opinion that by this handsome volume a very important addition to science
has been made; that the beautiful illustrations, together with the clear
and charming accounts of the life-histories which it contains, will do
much to popularise the study of a rather neglected section of zoology;
and that lovers of Reptiles, of which there are more than one generally
thinks, will feel that the new knowledge imparted to them emanates from
one who is thoroughly in sympathy with their enthusiasm."
VOLUME IX.
Birds, by A. H. Evans, M.A.
IBIS.—"Mr. Evans has produced a book full of concentrated
essence of information on birds, especially as regards their outer
structure and habits, and one that we can cordially recommend as a work
of reference to all students of ornithology."
VOLUME X.
Mammalia, by Frank Evers Beddard, M.A.,
F.R.S.
NATURE.—"Cannot fail to be of very high value to all
students of the Mammalia, especially from the standpoints of morphology
and palæontology."
LONDON: MACMILLAN AND CO., Ltd.
NOTES
[1] The degeneration of the hind-limb
in Whales and Sirenia forbids the use of this character as a distinctive
one on the principles advocated by the selection of the above list. But
it would be absurd to leave out hair.
[2] "Über die Haare der Säugethiere,"
Morph. Jahrb. xxi. 1894, p. 312.
[3] "Bemerkungen über den Ursprung
der Haare," Anat. Anz. 1893, p. 413.
[4] See for this matter, p. 90. Dr. Bonavia has recently advanced (Studies in
Evolution, London, 1895) the somewhat fantastic view that the
pigment-patches of Carnivorous and other mammals are a reminiscence of an
earlier scaly condition. There is no direct evidence that the primitive
mammals were scaly, nor are the Monotremata or Marsupials furnished with
any more traces of such a condition than are other mammals; and they are
the most lowly organised of existing Mammalia.
[5] Proc. Zool. Soc. 1887, p.
527.
[6] "Über Marsupialrudimente bei
Placentaliern," Morph. Jahrb. xx. 1893, p. 276.
[7] See Haacke, "On the Marsupial
Ovum, the Mammary Pouch, etc., of the Echidna," Proc. Roy. Soc.
1885, p. 72; and "Über die Entstehung der Säugetiere," Biol.
Centralbl. viii. 1889, p. 8.
[8] See Gegenbaur's Elements of
Comp. Anat. Transl. by Bell, 1878, p. 421.
[9] "Über die Beziehungen zwischen
Mammartasche u. Marsupium," Morph. Jahrb. xvii. 1891, p. 483.
[10] Catalogue of Marsupials in
British Museum, 1886.
[11] Its independence from the
epistropheus is emphasised in Monotremes and some Marsupials by its late
fusion with that vertebra.
[12] Intercentra are but rarely met
with anterior to the caudal series. Mr. Parsons has, however, recorded
their occurrence in the lumbar vertebrae of Atherura.
[13] Tufts College Studies,
No. 6, 1900.
[14] Cf. the Armadillo
Peltephilus, p. 186.
[15] Gegenbaur, Vergl. Anat.
Wirbelth. Leipzig, 1898, p. 404.
[16] Ehler's Zool.
Miscellen, i. 1894.
[17] Proc. Zool. Soc. 1865,
p. 567.
[18] Vergl. Anat. der
Wirbelth. Leipzig, 1898, p. 497.
[19] To this category are perhaps
to be referred cartilaginous pieces occurring in the Rabbit, Mus
and Sorex (see Fig. 29 above).
[20] "On the Coracoid of the
Terrestrial Vertebrates," P.Z.S. 1893, p. 585.
[21] Horny matter is apt to be
formed upon extremities; instances which are well known are the "claws"
upon the tail of the Lion and Leopard and the Kangaroo Onychogale.
For an account of the first see Proc. Zool. Soc. 1832, p. 146.
[22] Cf. Tomes, A Manual of
Dental Anatomy, 5th ed. London, 1898.
[23] Materials for the Study of
Variation, London, 1894.
[24] Morph. Jahrb. xix.
1892, p. 502.
[25] It would be of the greatest
interest in relation to this and many other problems to ascertain the
precise meaning of the monophyodont dentition of
Ornithorhynchus.
[26] Proc. Zool. Soc. 1899,
p. 922.
[27] Mr. M. Woodward, however
(P.Z.S. 1893, p. 467), is disposed to think that in some
Macropodidae at any rate the supposed tooth of the second set really
belongs to the milk dentition, arising late between Pm_{3} and
Pm_{4}.
[28] See for a summary, Osborn,
American Nat. Dec. 1897, p. 993.
[29] e.g. the "protoloph,"
"metaloph," etc. (see Fig. 36, p. 51), of the
modern Ungulate form of tooth.
[30] "On the Primitive Type of the
Plexodont Molars of Mammals," Proc. Zool. Soc. 1899, p. 555.
[31] Jen. Zeitschr. ii.
1866, p. 365.
[32] Proc. Zool. Soc. 1883,
p. 8.
[33] Proc. Zool. Soc. 1894,
p. 715.
[34] Beddard, Proc. Zool.
Soc. 1895, p. 136.
[35] Quart. Journ. Micr.
Sci. xxiv. 1884, p. 9.
[36] S.B. Jen. Gesells.
1885, p. 1.
[37] Proc. Roy. Phys. Soc.
Edin. viii. 1885, p. 354.
[38] Phil. Trans. clxxviii.
1887, p. 463.
[39] Robinson, Studies Biol.
Lab. Owens Coll. ii. 1890, p. 35.
[40] Beddard, Proc. Zool.
Soc. 1900, p. 667.
[41] Wallace, The Geographical
Distribution of Animals, 1876. Heilprin, The Distribution of
Animals, Internat. Scientific Series, 1887. Beddard, A Text-book
of Zoogeography, Cambridge Natural Science Manuals, 1895. Lydekker,
Geographical History of Mammals, Cambridge Geographical Series,
1896. W. L. and P. L. Sclater, The Geography of Mammals, Kegan
Paul and Co. 1899.
[42] This term is sometimes used in
a wider sense; cf. vol. viii. p. 74.
[43] A series of papers in the
Phil. Trans. for 1888-96, of which a useful abstract by Professor
Osborn was published in the American Naturalist, 1898, p. 309; see
also Cambr. Nat. Hist. viii. 1901, p. 303.
[44] Cf. vol. viii. p. 82.
[45] It may be necessary to exclude
the Whales from the comparison.
[46] Dental Anatomy, 5th ed.
1898, p. 304.
[47] "On the Fossil Mammalia from
the Stonesfield Slate," Quart. Journ. Micr. Sci. xxxv. 1894, p.
407.
[48] This groove has been found in
the existing Myrmecobius, see p. 154.
[49] Trans. New York Acad.
Sci. xiii. 1894, p. 234.
[50] Gegenbaur, Zur Kenntniss
der Mammarorgane der Monotremen, Leipzig, 1886.
[51] Quart. Journ. Micr.
Sci. xxiv. 1884, p. 124.
[52] Beddard, Proc. Roy. Phys.
Soc. Edinb. viii. 1885, p. 354.
[53] See Phil. Trans.
clxxviii. 1887, where the literature of the subject is fully cited.
[54] Muscular insertions and
attachments do not, however, altogether support the comparison.
[55] Journ. Anat. Phys.
1899, p. 309.
[56] Proc. Zool. Soc. 1864,
p. 18.
[57] Myrmecophaga aculeata
was the name given by Shaw.
[58] Zaglossus has
apparently priority as a name; but Proechidna is better known.
[59] Proc. Zool. Soc. 1892,
p. 545.
[60] Quart. J. Micr. Sci.
xxix. 1888, p. 353.
[61] Proc. Roy. Soc. xlvi.
1889, p. 127. See also Stewart, Quart. J. Micr. Sci. xxxiii. 1892,
p. 229.
[62] Proc. Zool. Soc. 1880,
p. 649.
[63] Moreover, the "corpus callosum
and the anterior commissure ... in ... Erinaceus and
Dasypus are almost Monotreme-like."
[64] See Wilson and Hill, Quart.
J. Micr. Sci. xxxix. 1899, p. 427.
[65] In Dendrolagus at any
rate. See Proc. Zool. Soc. 1895, p. 132.
[66] Anat. Anz. i. 1886, p.
338; and see Weber, ibid. ii. 1887, p. 42.
[67] Works dealing exclusively with
the Marsupials are: Lydekker, in Allen's Naturalists' Library,
1894; Aflalo, Natural History of Australia, Macmillan and Co.
1896; Waterhouse, Natural History of Mammalia, i. London, 1848;
Oldfield Thomas, British Museum Catalogue of Marsupialia and
Monotremata, 1888.
[68] "The Cerebral Commissures in
the Marsupialia and Monotremata," Journ. Anat. Phys. xxvii. 1893,
p. 69.
[69] When there are more than two,
two are especially developed. See Figs. 76, 77 (pp. 149, 150).
[70] See for a further discussion
of this subject the zoogeographical handbooks of Mr. Lydekker and myself,
quoted on p. 78 (footnote).
[71] To this may be added Mr.
Thomas' observation that the family of American Opossums is "very closely
allied to the Dasyuridae, from which, were it not for its isolated
geographical position, it would be very doubtfully separable."
[72] Except in the South American
Diprotodonts.
[73] Proc. Zool. Soc. 1893,
p. 450.
[74] Ibid. 1876, p. 165.
[75] Journal of the Rt. Hon. Sir
Joseph Banks, Bart., K.B., P.R.S., edited by Sir Joseph Hooker,
London, 1896.
[76] Proc. Zool. Soc. 1896,
p. 683.
[77] Proc. Zool. Soc. 1875,
p. 48.
[78] Proc. Zool. Soc. 1852,
p. 103.
[79] Proc. Zool. Soc. 1895,
p. 131.
[80] Ibid. 1884, p. 387.
[81] Ibid. 1884, p. 407.
[82] Proc. Linn. Soc. N.S.
Wales, i. 1877, p. 34.
[83] "On some Points in the Anatomy
of the Koala," Proc. Zool. Soc. 1881, p. 180.
[84] Thomas, "On
Caenolestes, a still existing survivor of the Epanorthidae of
Ameghino, and the representative of a new family of recent Marsupials,"
P.Z.S. 1895, p. 870.
[85] Stirling and Zietz, Mem.
Roy. Soc. South Australia, i.; see also a notice in Nature,
January 18, 1900.
[86] Quite recently (Proc. Linn.
Soc. N.S.W. 1898, p. 1) the carnivorous character of
Thylacoleo has been reasserted by Mr. Broom.
[87] Horn Scientific
Expedition, pt. ii. Zoology, 1896, p. 36.
[88] Leche found five, and
Waterhouse stated eight to be the number.
[89] Proc. Zool. Soc. 1887,
p. 527. See also Leche, Biol. Fören. Förhandl. 1891, p. 136, and
literature quoted.
[90] Traces of horny pads, like
those of the Duck-bill, have been asserted to exist in this animal. This
is exceedingly interesting when regarded in conjunction with its
multituberculate molars.
[91] See for an account of this
animal, Professor Stirling's Memoir in Trans. Roy. Soc. S.
Australia, 1891, p. 154, and Gadow, Proc. Zool. Soc. 1892, p.
361.
[92] The male, according to
Professor Spencer, has a rudimentary pouch.
[93] Pectoral and abdominal in the
Armadillo Tatusia.
[94] A rather problematical
Armadillo, Necrodasypus, has been recorded from French strata. It
consists of a few scutes only.
[95] Proc. Zool. Soc. 1882,
p. 358.
[96] Trans. Linn. Soc. (2)
vii. 1898, p. 277.
[97] i.e. large olfactory
lobes.
[98] Proc. Zool. Soc. 1899,
p. 1014.
[99] See for anatomy Owen,
Trans. Zool. Soc. iv. 1862, p. 117, and Forbes, Proc. Zool.
Soc. 1882, p. 287.
[100] For the skull of Edentates
generally see Parker, Phil. Trans. clxxvi. 1885, pt. i. p.
121.
[101] The colour fades in
captivity owing to the disappearance of the algae.
[102] In a letter addressed to
Dr. Gray, quoted by the latter in a revision of the Sloths, Proc.
Zool. Soc. 1871, p. 428.
[103] This name is written
"Prionodos" by Gray, which might lead to a confusion with the
Carnivore Prionodon.
[104] For the anatomy of several
forms, see Garrod, Proc. Zool. Soc. 1878, p. 222, who quotes other
memoirs.
[105] Flower, Proc. Zool.
Soc. 1886, p. 419.
[106] Milne-Edwards, Nouv.
Arch. Mus. vii. 1871, p. 177.
[107] See especially Lydekker,
An. Mus. La Plata, Pal. Arg. iii. 1894.
[108] Dr. Moreno and Mr. A. Smith
Woodward in Proc. Zool. Soc. 1899, p. 144; Wiss. Ergeb. Schwed.
Exped. Magellansländ. ii. 1899, p. 149.
[109] Proc. Roy. Soc.
xlvii. 1890, p. 246.
[110] Proc. Zool. Soc.
1893, p. 239, and 1896, p. 296.
[111] "Revision of the Manidae in
the Leyden Museum," Notes Leyd. Mus. iv. 1882, p. 193.
[112] Weber, Zool. Ergebnisse
einer Reise in Niederl. Ost Indien, 1892. See also Römer, in Jen.
Zeitschr. xxxi. 1896, p. 604, and Reh, ibid. xxx. 1895, p.
137.
[113] See Wortman, "The Ganodonta
and their Relationship to the Edentata," Bull. Am. Mus. Nat. Hist.
ix. 1897, p. 59.
[114] This creature is, however,
sometimes referred to the neighbourhood of the Rodents.
[115] Bull. Amer. Mus. Nat.
Hist. ix. 1897, p. 321.
[116] "Notes on some Specimens of
Antlers of the Fallow Deer, etc.," Proc. Zool. Soc. 1894, p.
485.
[117] Bull. Amer. Mus. Nat.
Hist. x. 1898, p. 159.
[118] Marsh, Amer. Journ.
Sci. xliii. 1892, p. 447.
[119] See W. D. Matthew, Bull.
Amer. Mus. Nat. Hist. ix. 1897, p. 303.
[120] Or perhaps rather to the
primitive Ungulates Condylarthra. It is especially compared with
Periptychus of that group.
[121] The scapula of P.
bathmodon is unknown.
[122] For the structure of this
genus and of Coryphodon, see Osborn, Bull. Amer. Mus. Nat.
Hist. x. 1898, p. 169.
[123] Osborn, Bull. Amer. Mus.
Nat. Hist. x. 1898, p. 81.
[124] Gadow, A Classification
of Vertebrata, Recent and Extinct, London, 1898.
[125] See Osborn, American
Naturalist, February 1893, p. 118.
[126] It is not absolutely clear
whether both or only one genus ranged into America. Different opinions
have been expressed.
[127] It must be remembered,
however, that there is a suggestion of a prehensile character in the hand
of Phenacodus (see p. 203).
[128] Cope, American
Naturalist, xxxi. 1897, p. 485.
[129] American Nat.
February 1900, p. 89.
[130] It must be borne in mind
that the teeth increase in complexity, those first pushed up having the
fewest plates. The first has only four transverse plates.
[131] Forbes, Proc. Zool.
Soc. 1879, p. 420.
[132] See Krueg, Zeitschr.
wiss. Zool. xxxiii. 1881, p. 652, and Beddard, Proc. Zool.
Soc. 1893, p. 311.
[133] So convinced are some
persons of the untameable character of the African Elephant, that it has
even been suggested that the animals with which Hannibal crossed the Alps
were not E. africanus, but a now extinct species!
[134] Wild Beasts and their
Ways, London, 1890.
[135] See Natural History of
the Ancients, by Rev. M. G. Watkins, London, 1896.
[136] Bull. Soc. Nat.
d'Acclimat. xlv. 1898, p. 41.
[137] Trans. Zool. Soc.
ix. 1874, p. 1.
[138] See Busk in Trans. Zool.
Soc. vi. 1868, p. 227.
[139] There are, however, three
milk forerunners of the premolars, of which one has no successor.
[140] Lydekker, An. Mus. La
Plata, Pal. Arg. iii. 1894.
[141] M. F. Woodward "On the Milk
Dentition of Procavia (Hyrax) capensis, etc," Proc. Zool.
Soc. 1892, p. 38.
[142] "On the Species of the
Hyracoidea," Proc. Zool. Soc. 1892, p. 50.
[143] Sir W. H. Flower, The
Horse, London, 1890.
[144] See Ewart, The Penicuik
Experiments, Constable and Co., 1899.
[145] The Horse, London,
1890.
[146] Cuyer and Alix, Le
Cheval, Paris, 1886.
[147] Lubbock, Prehistoric
Times, London, 1865.
[148] J. Geikie, Prehistoric
Europe, London, 1881.
[149] Horses, Asses, and
Zebras, London, 1895.
[150] Proc. Zool. Soc.
1884, p. 540.
[151] Proc. Zool. Soc.
1895, p. 688.
[152] See Pocock, Ann. Nat.
Hist. (6) xx. 1897, p. 33.
[153] "Das Quagga," Zool.
Garten, 1893, p. 289.
[154] Of this Horse, remains have
been lately discovered (see Lönnberg, Proc. Zool. Soc. 1900, p.
379) in the cave which produced the remains of Glossotherium. A
piece of skin covered with Fox-red hair, possibly spotted with paler
areas, is believed to be a relic of Onohippidium.
[155] Trans. American Phil.
Soc. xviii. 1896, p. 55.
[156] T. leucogenys and
T. ecuadorensis are probably not distinct, the latter being in
reality T. terrestris, the former T. roulini.
[157] See Beddard, Proc. Zool.
Soc. 1889, p. 252, and other papers there cited, for the anatomy of
the Tapir.
[158] Natural Science, vi.
1895, p. 161.
[159] Garrod, Proc. Zool.
Soc. 1873, p. 92; ibid. 1877, p. 707. Beddard and Treves,
Trans. Zool. Soc. xii. 1887, p. 183.
[160] Proc. Zool. Soc.
1876, p. 443.
[161] Proc. Zool. Soc.
1894, p. 329. See also Mr. Selous' paper in Proc. Zool. Soc. 1881,
p. 275.
[162] P. L. Sclater, Proc.
Zool. Soc. 1893, p. 514.
[163] Quite recently, however, a
species, A. incisivum, preserved at Darmstadt, has been found by
Professor Osborn to possess a slight rugosity upon the frontal bones,
which probably indicates the presence of a rudimentary horn, and the same
author is apparently inclined to place in Aceratherium the horned
Teleoceras (see p. 261).
[164] Osborn, Bull. Amer. Mus.
Nat. Hist. x. 1898, p. 51.
[165] See Osborn, Mem.
American Mus. Nat. Hist. vol. i. pt. iii. 1898.
[166] Scott, in Gegenbaur's
Festschrift, ii. 1896, p. 351.
[167] Remains of the genus have
been met with in the Balkans.
[168] See especially Osborn and
Wortman, Bull. Amer. Mus. Nat. Hist. vii. 1895, p. 333, and
Osborn, ibid. viii. 1896, p. 157.
[169] See Osborn, Bull. Amer.
Mus. Nat. Hist. vii. 1895, p. 82.
[170] N. Acta Acad. Caes.
Leop. Car. xxvii. 1885, p. 238.
[171] See Bateson, Materials
for the Study of Variation, London, 1894, p. 387.
[172] See, however, p. 196, for a discussion as to which is the more
primitive arrangement.
[173] Titanotherium (see
p. 266) is exceptional.
[174] Bones of
Hippopotamus, however, indicate the very recent occurrence of that
animal in Madagascar.
[175] "On the Pygmy Hippopotamus
of Liberia," Proc. Zool. Soc. 1887, p. 612.
[176] Tomes, Proc. Zool.
Soc. 1850, p. 160.
[177] There is, however, some
doubt about the first premolars.
[178] Dr. Garson has investigated
its anatomy, Proc. Zool. Soc. 1883, p. 413, and states that its
differences from Sus are "unimportant and few."
[179] "On the Species of
Potamochoerus," Proc. Zool. Soc. 1897, p. 359.
[180] Marsh, Amer. Journ.
Sci. xlvii. 1894, p. 407.
[181] Osborn, Bull. Amer. Mus.
Nat. Hist. vii. 1895, p. 102.
[182] Marsh, Amer. Journ.
Sci. xlviii. 1894, p. 262.
[183] For the structure of
Tragulus, see Milne-Edwards, Ann. Sci. Nat. (5) ii. 1864,
p. 49.
[184] Marsh, Amer. Journ.
Sci. 1897, p. 165.
[185] This is the winter dress.
In the summer both camels lose their long rough hair.
[186] See Wortman, Bull. Amer.
Mus. Nat. Hist. x. 1898, p. 93.
[187] "Osteology of
Poebrotherium," Journ. Morph. v. 1891, p. 1.
[188] Unless Protoceras
(see p. 284) was furnished with horns.
[189] Sir Victor Brooke, "On the
Classification of the Cervidae," Proc. Zool. Soc. 1878, p.
883.
[190] It has been occasionally
recorded in an Axis Deer, and in another species, Cariacus
superciliaris.
[191] It is not every one that
admits so many genera. I follow Sir Victor Brooke.
[192] Garrod, "On the Chinese
Deer named Lophotragus michianus by Mr. Swinhoe," Proc. Zool.
Soc. 1876, p. 757.
[193] Proc. Zool. Soc.
1877, p. 789.
[194] Proc. Zool. Soc.
1882, p. 636.
[195] Sir W. Flower "On the
Structure and Affinities of the Musk Deer (Moschus moschiferus),"
Proc. Zool. Soc. 1875, p. 159; Garrod, loc. cit. 1877, p.
287; and F. Jeffrey Bell, Proc. Zool. Soc. 1876, p. 182.
[196] For the viscera, see
Garrod, Proc. Zool. Soc. 1877, p. 5, etc.; and ibid. p.
289, etc.
[197] Proc. Zool. Soc.
1897, p. 273.
[198] Wild Beasts and their
Ways, 1890, p. 151.
[199] See also Sclater, Proc.
Zool. Soc., 1901, ii. p. 3.
[200] Forsyth Major. Proc.
Zool. Soc. 1891, p. 315.
[201] "On the Shedding of the
Horns in the Prongbuck," see Bartlett, Proc. Zool. Soc. 1865, p.
718; Canfield, ibid. 1866, p. 105; Murie, ibid. 1870, p.
334; and Forbes, ibid. 1880, p. 540.
[202] The distinction between the
two families has been called "fanciful." It may be admitted that it is
not great.
[203] The Book of
Antelopes, London, Porter, 1894-1900.
[204] They are straight in the
young.
[205] W. L. Sclater, The Fauna
of South Africa, Mammals, i. 1900.
[206] Taurotragus oryx has
unfortunately been discovered to be the correct name for the Eland.
[207] A. D. Bartlett, "On some
Hybrid Bovine Animals bred in the Society's Gardens," Proc. Zool.
Soc. 1884, p. 399.
[208] See Proc. Zool. Soc.
1890, p. 592.
[209] Proc. Zool. Soc.
1899, p 64.
[210] Proc. Zool. Soc.
1900, p. 142.
[211] The name
Trigonolestes has to be substituted for Pantolestes.
[212] Trans. American Phil.
Soc. xviii. 1896, p. 125.
[213] For complete osteology see
Wortman, Bull. Amer. Mus. Nat. Hist. vii. 1895, p. 145.
[214] In Halicore;
probably also in Manatus. See Turner, Trans. Roy. Soc.
Edinb. xxxv. 1889, p. 641.
[215] Kükenthal has discovered a
thick coating of rudimentary hairs in the foetus of the Manatee, thus
showing that it is the descendant of an animal furry like a Seal.
[216] "On the Manatee," in
Trans. Zool. Soc. vol. viii. 1872, p. 127.
[217] Hartlaub, "Beiträge zur
Kenntnis der Manatus-Arten," Zool. Jahrb. 1886, p. 1.
[218] Beddard, "Notes upon the
Anatomy of a Manatee (Manatus inunguis)," Proc. Zool. Soc.
1897, p. 47.
[219] See Kükenthal in Semon's
"Zoolog. Forschungen," Denkschr. Jen. 1897; Langkavel, "Der
Dugong," Zool. Garten, 1896, p. 337.
[220] Proc. Zool. Soc.
1892, p. 77.
[221] See van Beneden and
Gervais, Ostéographie des Cétacés; and for a more general account
Beddard, A Book of Whales, London, Murray, 1900.
[222] Vergleichend-anatomische
Untersuchungen an Walthiere, Jena, 1889-93.
[223] "And at his gills draws in,
and at his trunk spouts out, a sea," wrote Milton, and think many
others.
[224] These have been recorded by
Professor Howes in the Porpoise.
[225] For details and literature
see Jungklaus; Jen. Zeitschr. xxxii. 1898, p. 1.
[226] In Proc. Zool. Soc.
1886, p. 243.
[227] Perrin, "Notes on the
Anatomy of B. rostrata," Proc. Zool. Soc. 1870, p. 805.
[228] von Haast, "Notes on a
Skeleton of Balaenoptera australis," Proc. Zool. Soc. 1883,
p. 592.
[229] Ostéographie des
Cétacés, Paris, 1880, p. 130.
[230] Marine Mammals of the
North-West Coast of North America, 1874.
[231] Cf. Scammon, loc.
cit.
[232] The name that has priority
seems to be glacialis.
[233] Proc. Zool. Soc.
1881, p. 969.
[234] Actes Linn. Soc.
Bordeaux, 1881.
[235] For osteology see Hector,
Trans. New Zeal. Inst. vii. 1876, p. 251; and Beddard, Trans.
Zool. Soc. xv. 1901, p. 87.
[236] Journ. de l'Anat.
xxvi. 1890, p. 270.
[237] The Cruise of the
Cachalot, London, 1900.
[238] See Pouchet, "Contribution
a l'histoire du spermaceti," Bergens Museums Aarbog for 1893, No.
I.
[239] Yule, Travels of Marco
Polo, ii. London, 1874, p. 231.
[240] See Flower, Trans. Zool.
Soc. viii. 1872, p. 203.
[241] Bihang Svensk. Akad.
Handl. viii. 1883.
[242] Flower, Trans. Zool.
Soc. x. 1878, p. 415; and H. O. Forbes, Proc. Zool. Soc. 1893,
p. 216.
[243] Trans. Zool. Soc.
xii. 1889, p. 241.
[244] Ann. Sci. Nat. (7),
xiii. 1892, p. 259.
[245] Proc. Zool. Soc.
1882, pp. 722, 726.
[246] Bull. U.S. Nat. Mus.
No. 36, 1889, p. 7.
[247] See an essay on the hunting
of this Whale, by S. H. C. Müller, in Fish and Fisheries,
Edinburgh (Blackwood), 1883.
[248] Grampus being a contraction
of grand poisson is an obvious name to apply to any Whale.
[249] See Actes Soc. Linn.
Bordeaux, 1881; and for another figure, also coloured, Flower, in
Trans. Zool. Soc. xi. 1880, pl. i.
[250] Bull. U.S. Nat. Mus.
No. 36, 1889.
[251] Zool. Jahrb. Syst.
Theil, vi. 1892, p. 442.
[252] Anatomical Researches
Yunnan Exp. 1878, p. 417.
[253] Flower, Trans. Zool.
Soc. vi. 1867, p. 106; and Burmeister, Proc. Zool. Soc. 1867,
p. 484.
[254] Proc. Zool. Soc.
1892, p. 558.
[255] Thompson, Studies Mus.
Dundee, i. 1890; and C. R. Congrès de Zoologie, 1889, p.
225.
[256] Lydekker, Proc. Zool.
Soc. 1892, p. 560.
[257] For a general account of
the osteology, see Flower, Proc. Zool. Soc. 1869, p. 4; and for
muscular anatomy, Windle and Parsons, Proc. Zool. Soc. 1897, p.
370, and 1898, p. 152.
[258] See St. G. Mivart "On the
Aeluroidea," Proc. Zool. Soc. 1882, p. 135: and The Cat,
London, J. Murray, 1881.
[259] "On the Pupils of the
Felidae," Proc. Zool. Soc. 1894, p. 481.
[260] "Observations ... on the
Seal's Eye," Proc. Zool. Soc. 1893, p. 719.
[261] It is noteworthy that in
the Tiger some of the stripes have pale centres and are thus like spots
pulled out, while there are also small black spots.
[262] Natural Science, vi.
1895, p. 89.
[263] For an account of this and
of other mammals which occur in Central America, see Alston in Messrs.
Godman and Salvin's Biologia Centrali-Americana, 1879-1882.
[264] But Mr. Belt says that the
"Tigre" never attacks man unless it be provoked.
[265] See E. Hamilton, The
Wild Cat of Europe, London, Porter, 1896; and M. G. Watkins,
Gleanings from the Natural History of the Ancients, London, Elliot
Stock, 1896.
[266] The retractility is most
marked in the Linsangs.
[267] Beddard in Proc. Zool.
Soc. 1895, p. 430.
[268] Where it has probably been
introduced.
[269] Proc. Zool. Soc.
1873, p. 196.
[270] Flower, Proc. Zool.
Soc. 1872, p. 683.
[271] See also vol. viii. p.
591.
[272] The original name was
Rhinogale.
[273] That it is an abnormality
has been recently stated.
[274] For the anatomy of Hyaenas
see Morrison Watson in Proc. Zool. Soc. 1877, p. 369; 1878, p.
416; and 1879, p. 79.
[275] Flower, Proc. Zool.
Soc. 1869, p. 457.
[276] For a general account of
the Canidae see Mivart, A Monograph of the Canidae, London,
1890.
[277] Flower, Proc. Zool.
Soc. 1879, p. 766.
[278] Proc. Zool. Soc.
1880, p. 70.
[279] The relationship between
the Canidae and the Procyonidae must not be lost sight of in considering
this point of external likeness.
[280] Bull. Amer. Mus. Nat.
Hist. xii. 1900, p. 109.
[281] Proc. Zool. Soc.
1890, p. 98.
[282] Temminck, its original
describer, placed it in the genus Hyaena.
[283] See Garrod, Proc. Zool.
Soc. 1878, p. 373.
[284] Proc. Zool. Soc.
1899, p. 533.
[285] See Beddard, Proc. Zool.
Soc. 1900, p. 661, for anatomy.
[286] Beddard, Proc. Zool.
Soc. 1898, p. 129.
[287] It is a curious fact that a
native name for the creature is "Pottos" (cf. of course Potto);
and indeed the generic name Potos seems to have the priority over
Cercoleptes.
[288] "Narica" is
generally written, after Linnaeus. But this was, according to Mr. Alston,
probably an error for nasica.
[289] Proc. Zool. Soc.
1870, p. 752.
[290] See Wortman, Bull. Amer.
Mus. Nat. Hist. vi. 1894, p. 229.
[291] As a small point of
likeness between this Mustelid and the Procyonidae may be mentioned the
colours of the face. M. anakuma is particularly Raccoon-like.
[292] See Trans. Zool.
Soc. ii. 1841, p. 201.
[293] Proc. Zool. Soc.
1894, p. 306.
[294] I found fifteen.
[295] Ann. Nat. Hist. (6)
xiii. 1893, p. 522.
[296] See Matschie, SB. Ges.
Naturf. Berlin, 1895, p. 171.
[297] Proc. Zool. Soc.
1879, p. 305.
[298] Lydekker, "Note on the
Structure and Habits of the Sea-Otter (Latax lutris)," Proc.
Zool. Soc. 1895, p. 421; and ibid. 1896, p. 235.
[299] See an article by Mr.
Lydekker in Knowledge, April 1898, from which many of the above
facts have been taken.
[300] "Preliminary Notes on the
Characters and Synonymy of the different Species of Otter," Proc.
Zool. Soc. 1889, p. 190.
[301] Even apparently in the same
species.
[302] The number of premolars is
reduced in the Polar Bear.
[303] "The Blue Bear of Thibet,"
etc., Proc. Zool. Soc. 1897, p. 412.
[304] Nouv. Arch. Mus.
vii. 1872, Bull. p. 92; and Recherches pour servir à l'histoire
naturelle des Mammifères, 1868-1874, p. 321. This genus has quite
recently (Lankester, Trans. Linn. Soc. viii. 1901, p. 163) been
definitely referred to the Procyonidae.
[305] For the genera of
Pinnipedia see Mivart, Proc. Zool. Soc. 1885, p. 484.
[306] Murie, Trans. Zool.
Soc. viii. 1874, p. 501.
[307] Bull. Amer. Mus. Nat.
Hist. vi. 1894, p. 129.
[308] P. 456 below.
[309] See especially Allen,
North American Pinnipedes, 1880.
[310] Murie, Trans. Zool.
Soc. vii. 1894, p. 411.
[311] Cf. the Dugong, p. 336.
[312] Kükenthal, Jen.
Zeitschr. xxviii. 1894, p. 76.
[313] Cunningham, "Sexual
Dimorphism in the Animal Kingdom," London, 1900; see also Flower,
Proc. Zool. Soc. 1881, p. 145.
[314] Journ. Ac. Sci.
Philadelphia, ix. 1886, p. 175.
[315] See especially Tullberg,
"Ueber das System der Nagethiere," Act. Ak. Upsala, 1899; and
Alston, Proc. Zool. Soc. 1875, p. 61; and for nomenclature,
Thomas, Proc. Zool. Soc. 1896, p. 1012; and Palmer, Proc. Biol.
Soc. Washington; xi. 1897, p. 241.
[316] Proc. Zool. Soc.
1884, p. 252.
[317] Phil. Trans. 1850,
pt. ii. p. 529.
[318] Seen, however, in
Chaetomys.
[319] See Beddard, Proc. Zool.
Soc. 1892, p. 596, and Gervais, Journ. Zool. i. 1872, p.
450.
[320] "Observations sur le genre
Anomalurus," Nouv. Arch. Mus. (2), vi. 1883, p. 277.
[321] "On the Habits of the
Flying Squirrels of the genus Anomalurus," Proc. Zool. Soc.
1894, p. 243.
[322] W. E. de Winton, "On a New
Genus and Species of Rodents," etc., Proc. Zool. Soc. 1898, p.
450. Apparently just at the time of the publication of this paper
Matschie described the same animal as Zenkerella.
[323] Proc. Zool. Soc.
1893, p. 179.
[324] Flower and Lydekker.
[325] Thomas, J. Asiat. Soc.
Bengal, lvii. 1888, p. 256.
[326] E. T. Newton, Trans.
Zool. Soc. xiii. 1892, p. 165.
[327] Proc. Zool. Soc.
1896, p. 1016.
[328] Reuvens, "Die Myoxidae oder
Schläfer," Leyden, 1890, allows but one genus, Myoxus, the other
genera adopted here being termed subgenera.
[329] To which a sixth, the
"Yellow-necked Mouse," Mus flavicollis, may perhaps be added.
[330] For anatomy see Windle,
Proc. Zool. Soc. 1887, p. 53.
[331] Proc. Zool. Soc.
1889, p. 247.
[332] Trans. Zool. Soc.
xiv. 1898, p. 377.
[333] Nouv. Arch. Mus.
iii. 1867, p. 81.
[334] Popular Natural History
of Animals, London, 1898.
[335] Proc. Zool. Soc.
1863, p. 95.
[336] See O. Thomas, "On some
Mammals from Central Peru," Proc. Zool. Soc. 1893, p. 333.
[337] "Notes on the Rodent genus
Heterocephalus," Proc. Zool. Soc. 1885, p. 845.
[338] Proc. Zool. Soc.
1885, p. 611.
[339] Proc. Zool. Soc.
1890, p. 610.
[340] Parsons, Proc. Zool.
Soc. 1898, p. 858.
[341] Very probably this form
should be rather, as it is by Thomas, referred to the neighbourhood of
Pectinator, which would clear up the geographical anomaly.
[342] Notes Leyd. Mus.
1891, p. 105.
[343] Günther, Proc. Zool.
Soc. 1879, p. 144.
[344] Proc. Zool. Soc.
1873, p. 786.
[345] Loc. cit. (on p. 458), p. 123.
[346] Proc. Zool. Soc.
1892, p. 520.
[347] See Dobson, Proc. Zool.
Soc. 1884, p. 233.
[348] Peters, Trans. Zool.
Soc. vii. 1871, p. 397.
[349] "Field Notes on the Mammals
of Uruguay," Proc. Zool. Soc. 1894, p. 297.
[350] Beddard, Proc. Zool.
Soc. 1891, p. 236.
[351] These are stated by
Tullberg to be absent. I have found them, but they are very small bones,
not more than half an inch long.
[352] There is a faint
development of these ridges, but behind the palatine foramina in
Dasyprocta aguti.
[353] Proc. Zool. Soc.
1885, p. 161.
[354] Or absent?
[355] In the Guiana
Forest, London, 1894.
[356] MB. Ak. Berlin,
1873, p. 551.
[357] An account of the three
genera is to be found in Trans. Zool. Soc. i, 1833, p. 35, by Mr.
E. T. Bennett.
[358] Hudson, "On the Habits of
the Vizcacha," Proc. Zool. Soc. 1872, p. 822.
[359] Proc. Zool. Soc.
1894, pp. 251, 680.
[360] Nat. Science, vi.
1895, p. 94.
[361] See Parsons. Proc. Zool.
Soc. 1894, p. 675.
[362] Günther, Proc. Zool.
Soc. 1876, p. 739, and 1889, p. 75; and Cederblom, Zool. Jahrb.
Syst. Abth. xi. 1897-98, p. 497.
[363] Proc. Zool. Soc.
1881, p. 624.
[364] Proc. Biol. Soc.
Washington, x. 1896, p. 169.
[365] See especially Dobson, A
Monograph of the Insectivora, London, 1886-90.
[366] Even in the Otter-like
Potamogale the upper jaw, though broad and flat, projects
considerably beyond the lower.
[367] "Bemerkungen über die
Genealogie der Erinaceen." In Festschrift f. Liljeborg, 1896. See
also Anderson, Trans. Zool. Soc. viii. 1874, p. 453.
[368] Dobson, "Notes on the
Anatomy of the Erinaceidae," Proc. Zool. Soc. 1881, p. 389.
[369] See Natural Science,
xiii. 1898, p. 156.
[370] Manuel d'Hist. Nat.
French trans. by Artaud, 1803.
[371] "Notes on the Visceral
Anatomy of the Tupaia of Burmah," Proc. Zool. Soc. 1879, p.
301.
[372] Proc. Zool. Soc.
1848, p. 23.
[373] I quote Woodward, Proc.
Zool. Soc. 1896, for this dentition. The fourth molar of the lower
jaw is not always present. It comes late, and only old animals
possess it.
[374] Mivart in Proc. Zool.
Soc. 1871, p. 58.
[375] Thomas, Proc. Zool.
Soc. 1892, p. 500.
[376] Allman states the canines
to be absent. I follow Flower and Lydekker.
[377] See Allman in Trans.
Zool. Soc. vi. 1869, p. 1.
[378] The generic name of
Chalcochloris was proposed by Dr. Mivart for these.
[379] See Peters, Reise nach
Mosambique, 1852, for external characters and anatomy.
[380] "Mammals collected by Dr.
Emin Pasha," in Proc. Zool. Soc. 1890, p. 446.
[381] Ritsema Bos, Biol.
Centralbl. xviii. 1898, p. 63.
[382] "A Synopsis of the Genera
of the Family Soricidae," Proc. Zool. Soc. 1890, p. 49.
[383] Leche, "Über
Galeopithecus," K. Svensk. Ak. Handl. 1886.
[384] See Dobson, Ann. Nat.
Hist. (5) xiv. 1884, p. 153.
[385] Dobson, Proc. Zool.
Soc. 1875, p. 370.
[386] Ibid. p. 381.
[387] Dobson, Proc. Zool.
Soc. 1875, p. 546.
[388] Ibid. 1876, p.
701.
[389] For a general account of
the Primates, see Forbes in Allen's Naturalists' Library, London,
1894.
[390] See Dr. Mivart's papers in
Proc. Zool. Soc. 1864, -65, -66, -67, and -73 for osteology and teeth.
[391] Murie and Mivart, Trans.
Zool. Soc. vii. 1869, p. 1.
[392] Trans. Zool. Soc. v.
1863, p. 103.
[393] Hist. Nat. de
Madagascar, Mamm. 1875.
[394] Proc. Zool. Soc.
1895, p. 142.
[395] Trans. Zool. Soc. v.
1863, p. 33.
[396] Verh. Ak. Amsterdam,
xxvii. 1890, Art. 2.
[397] "On some Points in the
Structure of Hapalemur griseus" Proc. Zool. Soc. 1884, p.
301.
[398] Beddard, Proc. Zool.
Soc. 1900, p. 661.
[399] On the Arm Glands of the
Lemurs, Proc. Zool. Soc. 1887, p. 369.
[400] So at least the formula has
been given; but it is very possible that the supposed second incisor is
really, judging from the other Lemurs, a canine.
[401] The Malagasy, however, must
be vague in definition, or their interpreters not well grounded in the
rudiments of the language; for Sonnerat states that Indri signifies
"homme des bois."
[402] Syn.
Microrhynchus.
[403] Beddard, Proc. Zool.
Soc. 1884, p. 391, and 1891, p. 449; and Jentink, Notes Leyd.
Mus. 1885, p. 33.
[404] Proc. Zool. Soc.
1899, p. 554.
[405] Royal Natural
History, London, 1894, p. 211.
[406] See Novitates
Zoologicae, vol. i. 1894, p. 2.
[407] Proc. Zool. Soc.
1900, p. 321.
[408] "On the Angwantibo,"
Proc. Zool. Soc. 1864, p. 314.
[409] Verh. Ak. Amsterdam,
xxvii. 1890.
[410] Proc. Zool. Soc.
1882, p. 639; see also Rev. G. A. Shaw, Proc. Zool. Soc. 1883, p.
44, 2nd Art.
[411] For a survey of the
position of Tarsius, see Earle, Amer. Naturalist, xxxi.
1897, p. 569; and Nat. Science, x. 1897, p. 309.
[412] See Schlosser, Beiträge
Pal. Osterr. Hung. 1888; also Osborn and Earle, Bull. Amer. Mus.
Nat. Hist. vii. 1895, p. 16.
[413] Proc. Zool. Soc.
1899, p. 987.
[414] Phil. Trans. clxxxv.
B, 1894, p. 15.
[415] It seems to be possible
that this great Lemur was extant so lately as 1658, when a creature
possibly answering to it was described by de Flacourt.
[416] "Notes on Callithrix
gigot," Proc. Zool. Soc. 1884, p. 6.
[417] Forbes, Proc. Zool.
Soc. 1880, p. 639.
[418] "On a new African Monkey of
the genus Cercopithecus, with a List of the known Species,"
Proc. Zool. Soc. 1893, p. 243; see also p. 441.
[419] Proc. Zool. Soc.
1879, p. 451.
[420] See the books quoted on p.
576 (footnote).
[421] It is not so ranked by
everybody.
[422] Proc. Zool. Soc.
1899, p. 296.
[423] For accounts of the habits
of the Gorilla, compiled from various sources, see Hartmann's "Anthropoid
Apes," International Scient. Ser. London, 1885; H. O. Forbes,
"Monkeys," in Allen's Naturalists' Series, London, 1894; and
Huxley, "Man's Place in Nature," vol. vii. of Collected Essays,
London, 1894.
[424] "Man's Place in Nature,"
vol. vii. of Collected Essays, London, 1894.
[425] Hartmann's "Anthropoid
Apes," in International Sci. Ser. London, 1885.
[426] Nouv. Arch. Mus. Hist.
Nat. Paris, ii. 1866.
[427] Proc. Zool. Soc.
1899, p. 296.
[428] See also Duckworth,
Proc. Zool. Soc. 1898, p. 989.
[429] For the structure of this
Ape see Beddard, Trans. Zool. Soc. xiii. 1893, p. 177; and for
experiments on her intelligence, Romanes, Proc. Zool. Soc. 1889,
p. 316.
[430] For the external appearance
of the Orang see Hermes, Zeitschr. f. Ethn. 1876, a paper which
has coloured plates.
[431] Pithecanthropus erectus.
Eine menschenähnliche Uebergangsform aus Java, Batavia, 1894. See
also Ernst Haeckel, The Last Link (with notes by H. Gadow),
London, 1898; Manouvrier, Amer. Journ. Sci. 1897, p. 213
(extracts); and Klaatsch, Zoolog. Centralbl. vi. 1899, p. 217.
[432] See especially Wiedersheim,
The Structure of Man, transl. by Howes, London, 1895.
[433] Cunningham, "Cunningham
Memoirs," No. II. Royal Irish Acad. 1886.
End of the Project Gutenberg EBook of The Cambridge Natural History, Vol X.,
Mammalia, by Frank Evers Beddard
*** END OF THIS PROJECT GUTENBERG EBOOK CAMBRIDGE NATURAL HISTORY ***
***** This file should be named 39887-h.htm or 39887-h.zip *****
This and all associated files of various formats will be found in:
http://www.gutenberg.org/3/9/8/8/39887/
Produced by Chris Curnow, Joseph Cooper, Keith Edkins and
the Online Distributed Proofreading Team at
http://www.pgdp.net
Updated editions will replace the previous one--the old editions
will be renamed.
Creating the works from public domain print editions means that no
one owns a United States copyright in these works, so the Foundation
(and you!) can copy and distribute it in the United States without
permission and without paying copyright royalties. Special rules,
set forth in the General Terms of Use part of this license, apply to
copying and distributing Project Gutenberg-tm electronic works to
protect the PROJECT GUTENBERG-tm concept and trademark. Project
Gutenberg is a registered trademark, and may not be used if you
charge for the eBooks, unless you receive specific permission. If you
do not charge anything for copies of this eBook, complying with the
rules is very easy. You may use this eBook for nearly any purpose
such as creation of derivative works, reports, performances and
research. They may be modified and printed and given away--you may do
practically ANYTHING with public domain eBooks. Redistribution is
subject to the trademark license, especially commercial
redistribution.
*** START: FULL LICENSE ***
THE FULL PROJECT GUTENBERG LICENSE
PLEASE READ THIS BEFORE YOU DISTRIBUTE OR USE THIS WORK
To protect the Project Gutenberg-tm mission of promoting the free
distribution of electronic works, by using or distributing this work
(or any other work associated in any way with the phrase "Project
Gutenberg"), you agree to comply with all the terms of the Full Project
Gutenberg-tm License available with this file or online at
www.gutenberg.org/license.
Section 1. General Terms of Use and Redistributing Project Gutenberg-tm
electronic works
1.A. By reading or using any part of this Project Gutenberg-tm
electronic work, you indicate that you have read, understand, agree to
and accept all the terms of this license and intellectual property
(trademark/copyright) agreement. If you do not agree to abide by all
the terms of this agreement, you must cease using and return or destroy
all copies of Project Gutenberg-tm electronic works in your possession.
If you paid a fee for obtaining a copy of or access to a Project
Gutenberg-tm electronic work and you do not agree to be bound by the
terms of this agreement, you may obtain a refund from the person or
entity to whom you paid the fee as set forth in paragraph 1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be
used on or associated in any way with an electronic work by people who
agree to be bound by the terms of this agreement. There are a few
things that you can do with most Project Gutenberg-tm electronic works
even without complying with the full terms of this agreement. See
paragraph 1.C below. There are a lot of things you can do with Project
Gutenberg-tm electronic works if you follow the terms of this agreement
and help preserve free future access to Project Gutenberg-tm electronic
works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the Foundation"
or PGLAF), owns a compilation copyright in the collection of Project
Gutenberg-tm electronic works. Nearly all the individual works in the
collection are in the public domain in the United States. If an
individual work is in the public domain in the United States and you are
located in the United States, we do not claim a right to prevent you from
copying, distributing, performing, displaying or creating derivative
works based on the work as long as all references to Project Gutenberg
are removed. Of course, we hope that you will support the Project
Gutenberg-tm mission of promoting free access to electronic works by
freely sharing Project Gutenberg-tm works in compliance with the terms of
this agreement for keeping the Project Gutenberg-tm name associated with
the work. You can easily comply with the terms of this agreement by
keeping this work in the same format with its attached full Project
Gutenberg-tm License when you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern
what you can do with this work. Copyright laws in most countries are in
a constant state of change. If you are outside the United States, check
the laws of your country in addition to the terms of this agreement
before downloading, copying, displaying, performing, distributing or
creating derivative works based on this work or any other Project
Gutenberg-tm work. The Foundation makes no representations concerning
the copyright status of any work in any country outside the United
States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other immediate
access to, the full Project Gutenberg-tm License must appear prominently
whenever any copy of a Project Gutenberg-tm work (any work on which the
phrase "Project Gutenberg" appears, or with which the phrase "Project
Gutenberg" is associated) is accessed, displayed, performed, viewed,
copied or distributed:
This eBook is for the use of anyone anywhere at no cost and with
almost no restrictions whatsoever. You may copy it, give it away or
re-use it under the terms of the Project Gutenberg License included
with this eBook or online at www.gutenberg.org
1.E.2. If an individual Project Gutenberg-tm electronic work is derived
from the public domain (does not contain a notice indicating that it is
posted with permission of the copyright holder), the work can be copied
and distributed to anyone in the United States without paying any fees
or charges. If you are redistributing or providing access to a work
with the phrase "Project Gutenberg" associated with or appearing on the
work, you must comply either with the requirements of paragraphs 1.E.1
through 1.E.7 or obtain permission for the use of the work and the
Project Gutenberg-tm trademark as set forth in paragraphs 1.E.8 or
1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted
with the permission of the copyright holder, your use and distribution
must comply with both paragraphs 1.E.1 through 1.E.7 and any additional
terms imposed by the copyright holder. Additional terms will be linked
to the Project Gutenberg-tm License for all works posted with the
permission of the copyright holder found at the beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm
License terms from this work, or any files containing a part of this
work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this
electronic work, or any part of this electronic work, without
prominently displaying the sentence set forth in paragraph 1.E.1 with
active links or immediate access to the full terms of the Project
Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary,
compressed, marked up, nonproprietary or proprietary form, including any
word processing or hypertext form. However, if you provide access to or
distribute copies of a Project Gutenberg-tm work in a format other than
"Plain Vanilla ASCII" or other format used in the official version
posted on the official Project Gutenberg-tm web site (www.gutenberg.org),
you must, at no additional cost, fee or expense to the user, provide a
copy, a means of exporting a copy, or a means of obtaining a copy upon
request, of the work in its original "Plain Vanilla ASCII" or other
form. Any alternate format must include the full Project Gutenberg-tm
License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying,
performing, copying or distributing any Project Gutenberg-tm works
unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing
access to or distributing Project Gutenberg-tm electronic works provided
that
- You pay a royalty fee of 20% of the gross profits you derive from
the use of Project Gutenberg-tm works calculated using the method
you already use to calculate your applicable taxes. The fee is
owed to the owner of the Project Gutenberg-tm trademark, but he
has agreed to donate royalties under this paragraph to the
Project Gutenberg Literary Archive Foundation. Royalty payments
must be paid within 60 days following each date on which you
prepare (or are legally required to prepare) your periodic tax
returns. Royalty payments should be clearly marked as such and
sent to the Project Gutenberg Literary Archive Foundation at the
address specified in Section 4, "Information about donations to
the Project Gutenberg Literary Archive Foundation."
- You provide a full refund of any money paid by a user who notifies
you in writing (or by e-mail) within 30 days of receipt that s/he
does not agree to the terms of the full Project Gutenberg-tm
License. You must require such a user to return or
destroy all copies of the works possessed in a physical medium
and discontinue all use of and all access to other copies of
Project Gutenberg-tm works.
- You provide, in accordance with paragraph 1.F.3, a full refund of any
money paid for a work or a replacement copy, if a defect in the
electronic work is discovered and reported to you within 90 days
of receipt of the work.
- You comply with all other terms of this agreement for free
distribution of Project Gutenberg-tm works.
1.E.9. If you wish to charge a fee or distribute a Project Gutenberg-tm
electronic work or group of works on different terms than are set
forth in this agreement, you must obtain permission in writing from
both the Project Gutenberg Literary Archive Foundation and Michael
Hart, the owner of the Project Gutenberg-tm trademark. Contact the
Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable
effort to identify, do copyright research on, transcribe and proofread
public domain works in creating the Project Gutenberg-tm
collection. Despite these efforts, Project Gutenberg-tm electronic
works, and the medium on which they may be stored, may contain
"Defects," such as, but not limited to, incomplete, inaccurate or
corrupt data, transcription errors, a copyright or other intellectual
property infringement, a defective or damaged disk or other medium, a
computer virus, or computer codes that damage or cannot be read by
your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right
of Replacement or Refund" described in paragraph 1.F.3, the Project
Gutenberg Literary Archive Foundation, the owner of the Project
Gutenberg-tm trademark, and any other party distributing a Project
Gutenberg-tm electronic work under this agreement, disclaim all
liability to you for damages, costs and expenses, including legal
fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT
LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE
PROVIDED IN PARAGRAPH 1.F.3. YOU AGREE THAT THE FOUNDATION, THE
TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE
LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR
INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH
DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a
defect in this electronic work within 90 days of receiving it, you can
receive a refund of the money (if any) you paid for it by sending a
written explanation to the person you received the work from. If you
received the work on a physical medium, you must return the medium with
your written explanation. The person or entity that provided you with
the defective work may elect to provide a replacement copy in lieu of a
refund. If you received the work electronically, the person or entity
providing it to you may choose to give you a second opportunity to
receive the work electronically in lieu of a refund. If the second copy
is also defective, you may demand a refund in writing without further
opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth
in paragraph 1.F.3, this work is provided to you 'AS-IS', WITH NO OTHER
WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT LIMITED TO
WARRANTIES OF MERCHANTABILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied
warranties or the exclusion or limitation of certain types of damages.
If any disclaimer or limitation set forth in this agreement violates the
law of the state applicable to this agreement, the agreement shall be
interpreted to make the maximum disclaimer or limitation permitted by
the applicable state law. The invalidity or unenforceability of any
provision of this agreement shall not void the remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the
trademark owner, any agent or employee of the Foundation, anyone
providing copies of Project Gutenberg-tm electronic works in accordance
with this agreement, and any volunteers associated with the production,
promotion and distribution of Project Gutenberg-tm electronic works,
harmless from all liability, costs and expenses, including legal fees,
that arise directly or indirectly from any of the following which you do
or cause to occur: (a) distribution of this or any Project Gutenberg-tm
work, (b) alteration, modification, or additions or deletions to any
Project Gutenberg-tm work, and (c) any Defect you cause.
Section 2. Information about the Mission of Project Gutenberg-tm
Project Gutenberg-tm is synonymous with the free distribution of
electronic works in formats readable by the widest variety of computers
including obsolete, old, middle-aged and new computers. It exists
because of the efforts of hundreds of volunteers and donations from
people in all walks of life.
Volunteers and financial support to provide volunteers with the
assistance they need are critical to reaching Project Gutenberg-tm's
goals and ensuring that the Project Gutenberg-tm collection will
remain freely available for generations to come. In 2001, the Project
Gutenberg Literary Archive Foundation was created to provide a secure
and permanent future for Project Gutenberg-tm and future generations.
To learn more about the Project Gutenberg Literary Archive Foundation
and how your efforts and donations can help, see Sections 3 and 4
and the Foundation information page at www.gutenberg.org
Section 3. Information about the Project Gutenberg Literary Archive
Foundation
The Project Gutenberg Literary Archive Foundation is a non profit
501(c)(3) educational corporation organized under the laws of the
state of Mississippi and granted tax exempt status by the Internal
Revenue Service. The Foundation's EIN or federal tax identification
number is 64-6221541. Contributions to the Project Gutenberg
Literary Archive Foundation are tax deductible to the full extent
permitted by U.S. federal laws and your state's laws.
The Foundation's principal office is located at 4557 Melan Dr. S.
Fairbanks, AK, 99712., but its volunteers and employees are scattered
throughout numerous locations. Its business office is located at 809
North 1500 West, Salt Lake City, UT 84116, (801) 596-1887. Email
contact links and up to date contact information can be found at the
Foundation's web site and official page at www.gutenberg.org/contact
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
gbnewby@pglaf.org
Section 4. Information about Donations to the Project Gutenberg
Literary Archive Foundation
Project Gutenberg-tm depends upon and cannot survive without wide
spread public support and donations to carry out its mission of
increasing the number of public domain and licensed works that can be
freely distributed in machine readable form accessible by the widest
array of equipment including outdated equipment. Many small donations
($1 to $5,000) are particularly important to maintaining tax exempt
status with the IRS.
The Foundation is committed to complying with the laws regulating
charities and charitable donations in all 50 states of the United
States. Compliance requirements are not uniform and it takes a
considerable effort, much paperwork and many fees to meet and keep up
with these requirements. We do not solicit donations in locations
where we have not received written confirmation of compliance. To
SEND DONATIONS or determine the status of compliance for any
particular state visit www.gutenberg.org/donate
While we cannot and do not solicit contributions from states where we
have not met the solicitation requirements, we know of no prohibition
against accepting unsolicited donations from donors in such states who
approach us with offers to donate.
International donations are gratefully accepted, but we cannot make
any statements concerning tax treatment of donations received from
outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation
methods and addresses. Donations are accepted in a number of other
ways including checks, online payments and credit card donations.
To donate, please visit: www.gutenberg.org/donate
Section 5. General Information About Project Gutenberg-tm electronic
works.
Professor Michael S. Hart was the originator of the Project Gutenberg-tm
concept of a library of electronic works that could be freely shared
with anyone. For forty years, he produced and distributed Project
Gutenberg-tm eBooks with only a loose network of volunteer support.
Project Gutenberg-tm eBooks are often created from several printed
editions, all of which are confirmed as Public Domain in the U.S.
unless a copyright notice is included. Thus, we do not necessarily
keep eBooks in compliance with any particular paper edition.
Most people start at our Web site which has the main PG search facility:
www.gutenberg.org
This Web site includes information about Project Gutenberg-tm,
including how to make donations to the Project Gutenberg Literary
Archive Foundation, how to help produce our new eBooks, and how to
subscribe to our email newsletter to hear about new eBooks.




























































































































































































