The Project Gutenberg EBook of A Distributional Study of the Amphibians of
the Isthmus of Tehuantepec, Mexico, by William E. Duellman
This eBook is for the use of anyone anywhere at no cost and with
almost no restrictions whatsoever. You may copy it, give it away or
re-use it under the terms of the Project Gutenberg License included
with this eBook or online at www.gutenberg.org
Title: A Distributional Study of the Amphibians of the Isthmus of Tehuantepec, Mexico
Author: William E. Duellman
Release Date: December 30, 2011 [EBook #38440]
Language: English
Character set encoding: ISO-8859-1
*** START OF THIS PROJECT GUTENBERG EBOOK A DISTRIBUTIONAL STUDY OF ***
Produced by Chris Curnow, Joseph Cooper, Diane Monico, and
the Online Distributed Proofreading Team at
http://www.pgdp.net
University of Kansas Publications
Museum of Natural History
Volume 13, No. 2, pp. 19-72, pls. 1-8, 3 figs.
August 16, 1960
A Distributional Study of the Amphibians
of the Isthmus of Tehuantepec, México
BY
WILLIAM E. DUELLMAN
University of Kansas
Lawrence
1960
[Pg 19]
University of Kansas Publications
Museum of Natural History
Volume 13, No. 2, pp. 19-72, pls. 1-8, 3 figs.
August 16, 1960
A Distributional Study of the Amphibians
of the Isthmus of Tehuantepec, México
BY
WILLIAM E. DUELLMAN
University of Kansas
Lawrence
1960
[Pg 20]
University of Kansas Publications, Museum of Natural History
Editors: E. Raymond Hall, Chairman, Henry S. Fitch,
Robert W. Wilson
Volume 13, No. 2, pp. 19-72, pls. 1-8, 3 figs.
Published August 16, 1960
University of Kansas
Lawrence, Kansas
PRINTED IN
THE STATE PRINTING PLANT
TOPEKA, KANSAS
1960
28-3859
[Pg 21]
A Distributional Study of the Amphibians
of the Isthmus of Tehuantepec, México
BY
WILLIAM E. DUELLMAN
CONTENTS
| | PAGE |
|---|
| Introduction | 21 |
| Acknowledgments | 23 |
| Field Studies in the Isthmus of Tehuantepec | 23 |
| Sources of Material | 24 |
| Description of the Isthmus of Tehuantepec | 25 |
| Physiography | 25 |
| Climate | 28 |
| Vegetation | 29 |
| The Sierra de los Tuxtlas | 32 |
| Gazetteer | 33 |
| The Amphibian Fauna of the Lowlands | 37 |
| Composition of the Fauna | 37 |
| Ecology of the Fauna | 38 |
| Distribution of the Fauna | 42 |
| The Amphibian Fauna of the Foothills and Adjacent Highlands | 44 |
| Establishment of Present Patterns of Distribution | 45 |
| Accounts of Species | 49 |
| Summary | 68 |
| Literature Cited | 69 |
INTRODUCTION
Few regions in Middle America are so important zoogeographically as is
the Isthmus of Tehuantepec, that neck of land connecting North America
with Central America, separating the Pacific Ocean from the Gulf of
Mexico by a distance of only about 220 kilometers (airline), and
forming a low break between the highlands of México and those of
Central America. Before World War II the isthmus could be reached
readily only by railroad or by ocean vessel to Salina Cruz or
Coatzacoalcos. With the advent of roads, principally the Trans-isthmian
Highway, vast areas of the interior of the isthmus became accessible to
biologists. Nevertheless, long before roads were built in the isthmian
region collectors and biologists visited it, especially the town of
Tehuantepec, from which collections date back to the 1870's. Therefore,
it is rather[Pg 22]
surprising that no attempt has been made to present a
faunal list of the amphibians or reptiles of the isthmus. Ruthven
(1912) summarized his collections from the vicinity of Cuatotolapam,
Veracruz, and Hartweg and Oliver (1940) presented an annotated list of
the species collected by them in the vicinity of Tehuantepec. In recent
years there have been only a few papers reporting species from the
isthmus (Fugler and Webb, 1957; Langebartel and Smith, 1959). The
zoogeographic significance of the Isthmus of Tehuantepec is exemplified
by the works of Burt (1931), Duellman (1958), Gloyd (1940), Oliver
(1948), and Stuart (1941), who in their discussions of evolution and
dispersal of various genera of reptiles, pointed out that the Isthmus
of Tehuantepec was a region of zoogeographic importance.
Originally I intended to study the entire herpetofauna of the isthmus.
But I have not had opportunity to study all of the reptiles, and I have
not had the inclination to solve certain taxonomic problems concerning
them. The amphibians that I collected, together with all other known
specimens in museums, have been studied. Therefore, the present report
is concerned only with the amphibians. Only the amphibians of the
lowlands of the isthmus have been sampled adequately. Although I have
commented on the highland species in the discussion of distribution,
they are not included in the systematic section, which deals solely
with the 36 species definitely known to occur in the lowlands of the
isthmus.
Among the species of amphibians that I would expect to occur in the
isthmus, the only one not yet found there is Hyla phaeota. Sufficient
specimens of most of the species are available to show their variation
in the isthmus. Consequently, the systematics of these amphibians is on
a fairly substantial basis. Probably certain species in the isthmian
region will be found to be conspecific with others to the south, for
example Hyla ebraccata with Hyla leucophyllata and Hyla
robertmertensi with Hyla underwoodi. Nevertheless, such taxonomic
changes will not affect the distributional picture presented here. Our
greatest lack of knowledge concerning the amphibians is about their
life histories, as may be illustrated by the following questions, all
of which now are without definite answers. Where do many of the small
frogs conceal themselves during the dry season? What amount of, if any,
interspecific competition exists among several species of tree frogs,
all of which breed in the same ponds? What factors in the environment
permit certain amphibians, but not others, to live in the[Pg 23]
humid rainforests, as well as in the arid tropical scrub forest? The answers
to these questions and many others must await additional field studies.
The purpose of this paper is to make known the species of amphibians
living in the Isthmus of Tehuantepec, to describe the environments in
which they live, and to discuss their distribution in the isthmus. With
respect to the distribution of animals in the Isthmus of Tehuantepec I
will attempt to explain the present patterns of distribution with
special reference to climatic fluctuation in the Pleistocene.
Acknowledgments
My extensive field work in the Isthmus of Tehuantepec was made possible
by grants from the Penrose Fund of the American Philosophical Society
(1956) and the Bache Fund of the National Academy of Sciences (1958).
Furthermore, my field work received the hearty support of the Museum of
Zoology at the University of Michigan; for their cooperation I am
indebted to Norman Hartweg, T. H. Hubbell, and Henry van der Schalie.
In the course of my studies I received helpful suggestions from Norman
Hartweg, L. C. Stuart, and Charles F. Walker, to whom I am grateful.
For permission to examine specimens in their care I thank Doris M.
Cochran, Hobart M. Smith, and Richard G. Zweifel. I am deeply indebted
to Thomas MacDougall for many suggestions and for aid in preparing the
gazetteer. I am most grateful for the efforts of my field companions,
Richard E. Etheridge, Jerome B. Tulecke, John Wellman, and especially
my wife, Ann S. Duellman, who spent many long days and nights gathering
much of the data on which this report is based. Our work in the isthmus
was furthered by the generous help and hospitality of many residents,
especially the late Wilbur Barker of Tehuantepec, Fortunado Delgado of
Rancho Las Hojitas near Acayucan, César Fárjas of Donají, and Juan
Mayol of San Andrés Tuxtla. Profesor Jordi Juliá Z. of the Laboratorio
de Entomología, Comisión del Papaloapan, Ciudad Alemán, Veracruz,
helped make possible my field work in 1959; for this he has my sincere
thanks. In conclusion I express my gratitude to Ing. Juan Lozano
Franco, Secretaria de Agricultura y Ganadería, for providing me with
the necessary permits.
Field Studies in the Isthmus of Tehuantepec
I first visited the Isthmus of Tehuantepec and collected on the Pacific
lowlands of the isthmus in July, 1955. At that time heavy rains and
impassable roads restricted travelling. In February and March of 1956
my wife and I concentrated our efforts in the central region between
the Río Jaltepec and Matías Romero, but also made several trips across
the isthmus to gather ecological data in the dry season. In July of the
same year, accompanied by Richard E. Etheridge, we again crossed the
isthmus several times in order to gather ecological data in the wet
season, and studied especially hylid frogs, most of which had not been
seen in the dry season. Accompanied by Jerome B. Tulecke and John
Wellman, I collected again in the isthmus in July, 1958, between Salina
Cruz and Tehuantepec, and between Coatzacoalcos and[Pg 24] Cosoleacaque. In
March and April, 1959, I collected at Ciudad Alemán. Nearly 1200
specimens of 30 species of amphibians were thus collected in the
Isthmus of Tehuantepec; all specimens are now in the Museum of Zoology
at the University of Michigan. Of other species known from the isthmus,
I have had field experience with all but one (Bolitoglossa
veracrucis) in other parts of México.
Sources of Material
There are in museum collections nearly 3000 specimens of amphibians
with reliable data from the Isthmus of Tehuantepec. Among the first
herpetological specimens collected in the isthmian region are those
assembled by Francis Sumichrast in the 1870's from the vicinity of
Santa Efigenia and Tapanatepec, Oaxaca. These specimens were sent to
the United States National Museum and the Museum National d'Histoire
Naturelle in Paris; many served as the types of new species: Bufo
canaliferus Cope, Eleutherodactylus rugulosus Cope, Syrrhophus
leprus Cope, and Hylella sumichrasti Brocchi. In 1911 Alexander G.
Ruthven collected in the savanna country near Cuatotolapam, Veracruz;
the report on his collections (1912) is the first dealing with the
herpetofauna of a part of the isthmus. His specimens are in the
collection of the University of Michigan Museum of Zoology. Norman
Hartweg and James A. Oliver collected for the University of Michigan
Museum of Zoology in the vicinity of Tehuantepec, Oaxaca, during the
summer of 1936. The results of their work were published as an
annotated list of species occurring on the Pacific slopes of the
isthmus (1940). Hobart M. Smith collected in the vicinity of
Tehuantepec in January, 1940; his specimens are in the United States
National Museum. Specimens collected by Smith served as the types of
Eleutherodactylus avocalis Taylor and Smith and Diaglena reticulata
Taylor. Walter W. Dalquest collected vertebrates for the University of
Kansas in southern Veracruz in the winters of 1947 and 1948; he spent
about six months on the Gulf lowlands of the isthmus, principally in
the vicinity of Jesús Carranza. For the past two decades Thomas
MacDougall, a resident of New York City, has spent his winters
collecting biological specimens in southern México. He makes his
headquarters at Tehuantepec, but his compulsion to see the "back
country" has taken him to many remote parts of southern Oaxaca. His
earlier collections are in the American Museum of Natural History; the
later ones are in the University of Illinois Museum of Natural History.
Minor collections include those made by Matthew W. Stirling at San
Lorenzo, Veracruz, February-April, 1946 (United States National
Museum), by Fred G. Thompson on a trip across the isthmus in December,
1955 (University of Michigan Museum of Zoology), by the University of
Kansas Museum of Natural History field party under the direction of
Rollin H. Baker at Tolosita, Oaxaca, and by David A. Langebartel and
associates from southern Oaxaca in June, 1958 (University of Illinois
Museum of Natural History).
In the collections of the United States National Museum are several
species of amphibians sent to the museum from Tehuantepec by Francis
Sumichrast. These include Bolitoglossa platydactyla (USNM 30305,
30344-6, 30528), Bolitoglossa rufescens (10042), Chiropterotriton
chiropterus (30347), Lineatriton lineola (30353), Parvimolge
townsendi (30352), Pseudoeurycea cephalica (30350), Thorius
pennatulus (30348-9), Hyla miotympanum (30302-3),[Pg 25] and Hyla picta
(30304). Because of the poor condition of the specimens, determinations
of those listed as Bolitoglossa rufescens and Pseudoeurycea
cephalica are uncertain. With the exception of the Bolitoglossa
rufescens, which is stated to have come from Santa Efigenia, all of
these specimens are catalogued as having come from Tehuantepec. None of
these species has since been recorded from the Pacific slopes of the
isthmus; however, they all occur in the vicinity of Orizaba, Veracruz.
Probably Sumichrast carried the specimens with him from Orizaba, his
home before moving to Santa Efigenia, and shipped them from Tehuantepec
to the United States National Museum. These species definitely should
not be considered as inhabitants of the Pacific slopes of the Isthmus
of Tehuantepec.
DESCRIPTION OF THE ISTHMUS OF TEHUANTEPEC
The Isthmus of Tehuantepec is a strip of land forming a low pass, which
separates the mountain masses of México proper from those of Central
America, and at the same time provides a continuum of lowlands from the
Gulf of Mexico to the Pacific Ocean. This topography combines with the
climatic conditions to create extremely diverse environments, the
distribution of which can be adequately understood only after an
acquaintance with the topography and climate of the region.
Physiography
In east-central Oaxaca the mountain masses comprising the Sierra Madre
Oriental and the Sierra del Sur terminate in a series of ranges—Sierra
de Juárez, Sierra de los Míjes, and Sierra de Choapam. From lofty
peaks, such as Cerro de Zempoaltepetl (3400 meters), the highlands
diminish eastward to succeedingly lower ridges, until in the middle of
the Isthmus of Tehuantepec the continental divide is about 250 meters
above sea level. Eastward from this low divide the land rises to form
the Sierra Madre de Chiapas, which is continuous with the highland
masses of Guatemala.
For the purposes of this description, the lowlands of the isthmus may
be divided into three parts—the Gulf Coastal Plain, the central
ridges, and the Pacific Coastal Plain, which in the isthmus is called
the Plains of Tehuantepec (Figs. 1 and 2).
The Gulf Coastal Plain is broad and fairly level near the coast, but
rolling in the interior. The plain, throughout most of its length in
the isthmus, is at least 75 kilometers wide. The majority of the region
in the isthmus is drained by the Río Coatzacoalcos, which flows in a
northerly course to the Gulf of Mexico. The western part is drained by
the Río San Juan, the principal tributary of the Río Papaloapan. Behind
the coastal dunes are frequent, and sometimes[Pg 26] large, lagoons.
Immediately inland from Coatzacoalcos and along the lower stretches of
the Río Papaloapan are extensive marshes. Essentially the entire
coastal plain, with the exception of the coastal dunes, consists of
rich alluvial deposits.
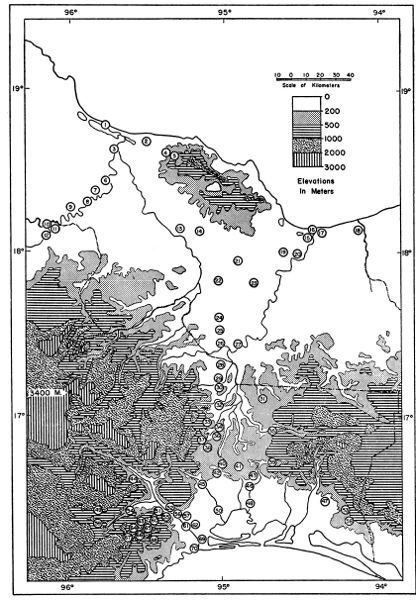 Fig. 1. Map of the Isthmus of Tehuantepec based on the
American Geographical Society's "Map of Hispanic America on the Scale
of 1:1,000,000."
Fig. 1. Map of the Isthmus of Tehuantepec based on the
American Geographical Society's "Map of Hispanic America on the Scale
of 1:1,000,000."
(Click image for larger view.)
The localities shown are numbered in the gazetteer; the numerical
sequence of localities is an arrangement whereby north takes precedence
over south and west over east. 1. Alvarado. 2. Lerdo de Tejada. 3.
Tlacotalpan. 4. Tula. 5. Tecolapan. 6. Amatitlán. 7. Cosamaloapan. 8.
Chacaltianguis. 9. Novillero. 10. Ciudad Alemán. 11. Papaloapan. 12.
Tuxtepec. 13. Cuatotolapam. 14. Hueyapan. 15. Berta. 16. Coatzacoalcos.
17. Ayentes. 18. Río de las Playas. 19. Cosoleacaque. 20. Minatitlán.
21. Acayucan. 22. Aquilera. 23. San Lorenzo. 24. Naranja. 25. Suchil.
26. Jesús Carranza. 27. La Oaxaqueña. 28. Ubero. 29. Donají. 30.
Tolosita. 31. El Modelo. 32. Sarabia, 33. Guichicovi. 34. La Princesa.
35. Santa María Chimalapa. 36. Matías Romero. 37. Santo Domingo Petapa.
38. El Barrio. 39. Palmar. 40. Chivela. 41. Santiago Chivela. 42.
Nizanda. 43. Agua Caliente. 44. Portillo Los Nanches. 45. Ixtepec. 46.
La Ventosa. 47. Zanatepec. 48. Unión Hidalgo. 49. Tres Cruces. 50.
Juchitán. 51. Escurano. 52. Salazar. 53. Santa Efigenia. 54.
Tequisistlán. 55. Cerro de Quiengola. 56. San Pablo. 57. Mixtequilla.
58. Tapanatepec. 59. Zarzamora. 60. Limón. 61. Tehuantepec. 62.
Bisilana. 63. Santa Lucía. 64. Cerro de Arenal. 65. Cerro de San Pedro.
66. La Concepción. 67. Tenango. 68. San Antonio. 69. Huilotepec. 70.
Salina Cruz.
[Pg 27]
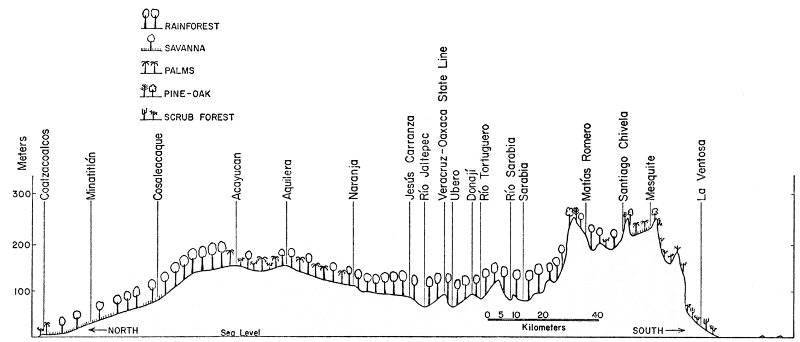 Fig. 2. Topographic profile of the Isthmus of
Tehuantepec showing major localities along the Trans-isthmian Highway
and major types of vegetation. Vertical exaggeration approximately 165
times.
Fig. 2. Topographic profile of the Isthmus of
Tehuantepec showing major localities along the Trans-isthmian Highway
and major types of vegetation. Vertical exaggeration approximately 165
times.
[Pg 28]
The central ridges extend from the Río Jaltepec southward to within 40
kilometers of the Pacific coast. It is in this area that the continuity
of the high ridges and volcanic peaks, which extend nearly the entire
length of the Americas, is interrupted at a point almost directly in
line with the shortest distance between the two oceans. The northern
part of this central region consists of hills dissected by tributaries
of the Río Coatzacoalcos; the principal ones from north to south
are—Río Jaltepec, Río Tortuguero, Río Sarabia, and Río Malatengo. The
plains of Chivela are south of these rivers and lie at an elevation of
about 200 meters; at the southern edge of these plains a range of hills
rises to 250 to 400 meters above sea level. These hills drop abruptly
to the Plains of Tehuantepec. In the northern and central parts of this
central region the rocks are granitic; the hills to the south of the
Plains of Chivela are limestone.
The Pacific Coastal Plain or Plains of Tehuantepec have a maximum width
of about 30 kilometers. From the base of the hills at an elevation of
about 75 meters the plains slope gradually to the sea. To the west of
the Río Tehuantepec and to the east of the Plains of Tehuantepec at the
base of the Sierra Madre de Chiapas, the coastal plain becomes much
narrower; in these places the continuity of the plain is frequently
interrupted by low north-south ridges extending outward from the
mountains or by isolated hills. The soil is poor, varying from volcanic
rock to gravel and sand.
Climate
The prevailing winds are from the north across the Gulf of Mexico.
These moisture-laden winds precipitate most of their moisture north of
the central ridges. This results in high rainfall on the northern
slopes and Gulf Coastal Plain and relatively little rainfall on the
southern slopes and the Pacific Coastal Plain. Precipitation is cyclic;
there is a marked wet and a dry season throughout the region, but this
is most noticeable on the Pacific lowlands (Fig. 3). At Salina Cruz on
the Pacific Ocean the average annual rainfall is 1040 mm. (Contreras,
1942); of this amount, only 15 mm. falls from November through April.
On the Gulf Coastal Plain (Minatitlán station) the average annual[Pg 29]
rainfall is 3085 mm. In this region the driest months are February
through May, during which time 236 mm. of rain falls. At Salina Cruz
the wettest month is June; at Minatitlán it is September. There is
little variation in temperature throughout the isthmus; the average
annual temperature at Salina Cruz is 26.6° C.; that at Minatitlán is
26.2° C. During the winter when masses of air from the arctic move
southward into the Great Plains of the United States, cool winds blow
across the isthmus. These are usually accompanied by overcast sky and
sometimes a slight amount of precipitation. These "nortes" may cause a
drop in temperature of about six to eight degrees in a few hours.
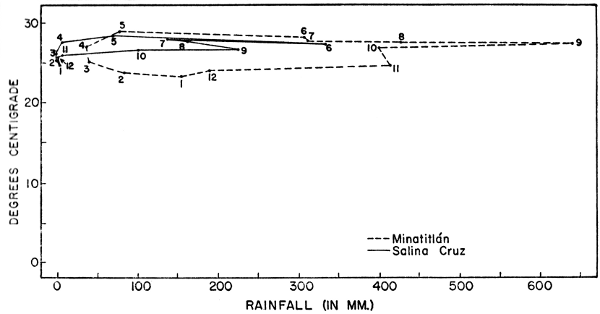 Fig. 3. Climatographs for Minatitlán, Veracruz, and
Salina Cruz, Oaxaca, based on data given by Contreras (1942). Plotted
points are for mean monthly temperatures and rainfall; months are
indicated by numbers.
Fig. 3. Climatographs for Minatitlán, Veracruz, and
Salina Cruz, Oaxaca, based on data given by Contreras (1942). Plotted
points are for mean monthly temperatures and rainfall; months are
indicated by numbers.
Vegetation
The topography and climate combine to produce drastically different
types of climax vegetation on the northern and southern lowlands of the
isthmus. The picture is somewhat complicated by the savannas on the
Gulf Coastal Plain, which, as will be shown later, are dependent upon
edaphic features more than climatic conditions. The following brief
account of the vegetation in the Isthmus of Tehuantepec is based on
data provided by Williams (1939) and Goldman (1951), supplemented by
personal observations. The purpose of this description is not to
analyze the flora of the isthmus, but to give the reader a picture of
this aspect of the biota of the major environments with which I shall
be concerned in the ensuing discourse on the amphibians of the region.[Pg 30]
The three divisions of the isthmus recognized in the account of the
physiography serve equally well in describing the vegetation. Those
divisions are as follows:
Gulf Lowlands
On the lowlands north of the continental divide and extending to the
Gulf of Mexico are three major types of vegetation—tropical
rainforest, arid tropical scrub forest, and savanna. Aside from these,
there are marshes and lagoons near the coast.
On the coastal dunes there are thickets of sea grape, patches of
Cenchrus, and clumps or scattered Opuntia. The lagoons are bordered
by mangrove thickets made up primarily of Lonchocarpus hondurensis.
In the marshes along the lower Río Coatzacoalcos and Río Papaloapan the
tall tough grass, Gynerium sagittatum, is common.
According to Beard (1953: 291) the development of savanna vegetation is
dependent upon soil, topography, and drainage. Level regions having
permeable soil horizons lying on top of an impermeable horizon provide
poor drainage. In most savanna regions in the Americas the grasslands
become waterlogged or even partly flooded during the rainy season and
desiccated in the dry season. Many ecologists and phytogeographers have
postulated that savannas are either man made or are examples of a fire
climax. Beard (op. cit.: 203) provided multitudinous evidence that
the association of savanna vegetation and certain types of edaphic and
topographic conditions was so strongly marked that grassland is the
natural vegetation in these areas.
Savannas are scattered through southern Veracruz eastward to British
Honduras. These usually are grasslands having scattered trees or clumps
of trees around depressions, which may contain water throughout the
year (Pl. 1, fig. 1). According to Williams (op. cit.), the most
common trees in the savannas in southern Veracruz are Ceiba
pentandra, Chlorophora tinctoria, and Byrsonima crassifolia.
Lying in a rain shadow cast by the Tuxtlas and on sandy and
well-drained soils is a dense xerophytic forest. The crown of this
deciduous forest usually is little more than ten to twelve meters above
the ground (Pl. 1, fig. 2). Conspicuous trees in this scrub forest are
Acacia cornigera, Bauhinia latifolia, Calliandra bijuga, Cassia
laevigata, Guazuma ulmifolia, and various species of Bursera.
The most extensive type of vegetation on the Gulf Coastal Plain is a
tall evergreen forest resembling tropical rainforest. Although[Pg 31] this
forest is made up of many species of trees that are characteristic of
true rainforest, the forest on the Gulf Coastal Plain cannot be
classified as true rainforest, neither by the climatic conditions, nor
the structure of the forest. The seasonal variation in rainfall
probably is the chief factor in hindering the development of a
rainforest climax vegetation. Usually a minimum of 65 mm. of rainfall
each month is considered essential for the development of true
rainforest. At Minatitlán the average rainfall for March (39 mm.) and
April (36 mm.) is far below this minimum. Structurally, this forest has
a crown about 30-35 meters above the ground but individual trees rising
five meters or more above the crown (Pl. 2, figs. 1-2). There is no
clear stratification within the forest; in many parts of it there are
dense growths of bushes, small trees, and palms. The forest on the Gulf
Coastal Plain, therefore, most properly might be referred to as a
quasi-rainforest, a term that has been applied to other such forests in
tropical America.
Among the abundant and dominant trees in this forest are Swietenia
macrophylla, Calophyllum brasiliense, Achras zapota, Ceiba
pentandra, Castilla elastica, Cedrela mexicana, Tabebuia
Donnell-Smithi, Calocarpum mammosum, Bombax ellipticum, and a
variety of Ficus. Epiphytes and Ilianas are abundant.
Central Ridges
The vegetation of the central ridges of the isthmus is, for the most
part, transitional between the tall rainforest of the Gulf Coastal
Plain and the low xerophytic scrub forest of the semi-arid Pacific
Coastal Plain. On the northern slopes of the ridges the rainforest is
more poorly developed than on the plains to the north. Many of the same
species of trees are present, including Ceiba pentandra, Cedrela
mexicana, Swietenia macrophylla, and Ficus sp.; nevertheless,
these seldom are as large as members of the same species in the forest
on the plains. Other species present on the forested slopes include
Tabebuia Donnell-Smithi, Zanthoxylum melanostictum, Pithecolobium
arboreum, and a species of Pterocarpus. The structure of this forest
differs from that on the Gulf Coastal Plain in that there is no
continuous upper canopy and there is a dense undergrowth (Pl. 3, fig.1).
This type of forest extends from Mogoñe southward to about Matías
Romero.
In the vicinity of Matías Romero open pine-oak forest (Pinus caribaea
and Quercus sp.) is found on some ridges as low as 250 meters above
sea level.
On the Plains of Chivela in the southern part of the central region[Pg 32]
the vegetation takes on a semi-arid appearance, especially in a savanna
on the plains. Clumps of small trees and bushes, consisting of Croton
nivea, Cordia cana, Jacquinia aurantiaca, Calycophyllum
candidissimum, and Cassia emarginata, are scattered on a grassy
plain, from which rise widely-spaced palms of an unknown species (Pl. 3, fig. 2).
Pacific Coastal Plain
The vegetation of the Pacific lowlands definitely is semi-arid in
character. Most of the trees are deciduous, thorny, and short. During
the dry season the landscape presents a barren appearance, but shortly
after the first summer rains dense green foliage appears (Pl. 4, figs. 1 and 2).
Between Juchitán and La Ventosa few trees are more than two
meters high (Pl. 5, fig. 1). In many areas the trees and bushes form an
almost impenetrable tangle, whereas on especially rocky soils or on
slopes those plants are more widely spaced. Abundant and widespread
species of trees on the Plains of Tehuantepec include Acacia
cymbispina, Prosopis chilensis, Caesalpinia coriaria, Caesalpinia
eriostachys, Celtis iguanaea, Cordia brevispicata, Jatropha
aconitifolia, and Crescentia alata.
Montane Vegetation
In order to illustrate the interruption of subtropical and temperate
types of vegetation by the lowlands of the Isthmus of Tehuantepec, it
is necessary to digress for a moment from the isthmus and consider the
types of vegetation present on the adjacent highlands. On the higher
peaks, such as Cerro de Zempoaltepetl, above about 2500 meters is fir
forest (Abies religiosa); lower on the slopes are extensive pine
forests, which on some slopes are mixed with oak or replaced entirely
by oaks. Subtropical cloud forest, characterized by relatively cool
temperatures and high humidity, is found at elevations usually between
1000 and 1800 meters on the windward slopes of the Sierra Madre
Oriental in Veracruz and northern Oaxaca and on the northern and
southern slopes of the Chiapan-Guatemalan Highlands. None of these
forest types is continuous across the Isthmus of Tehuantepec.
The Sierra de los Tuxtlas
Although actually located in the region of the Isthmus of Tehuantepec,
the Sierra de los Tuxtlas, because of its isolated position, need not
be considered in great detail in analyzing the distribution of animals
inhabiting the lowlands of the isthmus. Nevertheless[Pg 33] because some
species living in the highlands adjacent to the isthmus also live in
the Tuxtlas, this range is briefly described here. The Sierra de los
Tuxtlas is a range of volcanos lying near the Gulf Coast in southern
Veracruz between the mouths of the Río Papaloapan and the Río
Coatzacoalcos. Volcán San Martín, the highest peak, rises above 1800
meters. This range of volcanos is surrounded by lowlands, which
immediately to the south and west are covered with savanna and in
places by scrub forest. The luxuriant nature of the vegetation on these
volcanos indicates that this range receives much more rainfall than the
surrounding lowlands. Especially on the northern slopes, tropical
rainforest is well developed; this is replaced at about 1200 meters by
cloud forest. The southern and western slopes are drier, for the lower
slopes are covered with a scrubby, but evergreen, forest.
Detailed comments on the herpetofauna of the Tuxtlas have been omitted
purposefully, for the reptiles and amphibians of the region currently
are being studied by Douglas Robinson.
GAZETTEER
The following localities are those referred to in the text. The name of
the locality (listed alphabetically by states) is followed by latitude,
longitude, elevation, general description (town, ranch, etc.), and
general type of habitat. Unless otherwise noted, distances are
straight-line (airline) distances in kilometers. The localities have
been plotted from the American Geographical Society's "Map of Hispanic
America on the Scale of 1:1,000,000" (Millionth Map). Numbers in
brackets identify the position of a locality on the accompanying map
(Fig. 1).
Oaxaca
Agua Caliente.—Lat. 16° 38'; long. 94° 48'; elev. 140 m. A
hot spring, 6.9 km. north of La Ventosa on the
Trans-isthmian Highway; arid scrub forest [43].
Arenal, Cerro de.—Lat. 16° 18'; long. 95° 32'; elev. 925 m.
(crest). A ridge northeast of Tenango; scrub forest on
slopes and pine-oak forest on top [64].
Barrio, El.—Lat. 16° 38'; long. 95° 07'; elev. 314 m. A
village about 10 kilometers southwest of Matías Romero;
transition between scrub forest and broadleaf hardwood
forest [38].
Bisilana.—Lat. 16° 20'; long. 95° 13'; elev. 35 m. A place
name for a former ranch at the edge of Tehuantepec; open
arid scrub forest [62].
Chivela.—Lat. 16° 20'; long. 95° 01'; elev. 195 m. A
village on the Trans-isthmian Railroad, 26 kilometers by
rail south of Matías Romero and on the western edge of the
semi-arid Plains of Chivela [40].
Concepción.—Lat. 16° 17'; long. 95° 29'; elev. 1200 m. A
ranch on the slopes of Cerro Arenal, east-northeast of
Tenango; dry pine-oak forest [66].
[Pg 34]
Coyol.—Exact position unknown; according to Smith and
Taylor (1950: 10), Coyol is "between San Antonio and Las
Cruces."
Donají.—Lat. 17° 13'; long. 95° 02'; elev. 90 m. A village
at Km. 155 on the Trans-isthmian Highway; rainforest [29].
Escurano.—Lat. 16° 25'; long. 95° 27'; elev. 500 m. A ranch
about 25 kilometers west-northwest of Tehuantepec; arid
scrub forest [51].
Guichicovi, San Juan.—Lat. 16° 58'; long. 95° 06'; elev.
250 m. A village on the north slopes of the isthmus, 12
kilometers north-northwest of Matías Romero; cleared
hardwood forest and coffee plantations [33].
Huilotepec.—Lat. 16° 14'; long. 95° 09'; elev. 30 m. A
small village on the Río Tehuantepec, 13 kilometers
south-southeast of Tehuantepec; open arid scrub forest [69].
Ixtepec.—Lat. 16° 34'; long. 95° 06'; elev. 60 m. A town
and railroad junction on the northwestern edge of the Plains
of Tehuantepec; arid scrub forest [45].
Juchitán.—Lat. 16° 26'; long. 95° 02'; elev. 15 m. A town
on the Plains of Tehuantepec, 22 kilometers by road
east-northeast of Tehuantepec; arid scrub forest [50].
Limón.—Lat. 16° 20'; long. 95° 29'; elev. 600 m. A former
agrarian colony and now a small ranch about 27 kilometers
west of Tehuantepec; arid scrub forest [60].
Matías Romero.—Lat. 16° 53'; long. 95° 02'; elev. 200 m. A
town on the Trans-isthmian Highway and railroad in the hills
near the crest of the isthmus; broadleaf hardwood forest and
open pine-oak forest [36].
Mixtequilla.—Lat. 16° 24'; long. 95° 18'; elev. 40 m. A
village on the Río Tehuantepec, northwest of Tehuantepec;
dense scrub forest [57].
Modelo, El.—Lat. 17° 07'; long. 94° 43'; elev. 200 m. An
old rubber plantation on the Río Chalchijapa, a tributary to
the Río Coatzacoalcos; rainforest [31].
Nanches, Portillo Los.—Lat. 16° 35'; long. 95° 37'; elev.
500 m. A place name, about 4 kilometers southeast of
Totolapilla; scrub forest [44].
Nizanda.—Lat. 16° 42'; long. 95° 02'; elev. 150 m. A
village on the Trans-isthmian Railroad between Chivela and
Ixtepec; dense scrub forest [42].
Nueva Raza.—Exact location unknown; according to Thomas
MacDougall, this locality is in the lowlands of northern
Oaxaca; rainforest.
Palmar.—Lat. 16° 43'; long. 94° 40'; elev. 300 m. A small
ranch on the west base of Cerro Atravesado; scrub forest
[39].
Papaloapan.—Lat. 18° 11'; long. 96° 06'; elev. 25 m. A
small village on the Río Papaloapan in northern Oaxaca; low
evergreen forest and savanna [11].
Princesa, La.—Lat. 16° 56'; long. 95° 02'; elev. 150 m. A
ranch on the northern slopes of the isthmus, 6 kilometers by
road north of Matías Romero; poorly developed rainforest
[34].
Quiengola, Cerro de.—Lat. 16° 24'; long. 95° 22'; elev. 900
m. (crest). A hill 15 kilometers west-northwest of
Tehuantepec; dense scrub forest on slopes and scattered
pines on top [55].
Salazar.—Lat. 16° 25'; long. 95° 20'; elev. 45 m. A ranch
on the Río Tehuantepec, northwest of Tehuantepec; dense
scrub forest [52].
Salina Cruz.—Lat. 16° 10'; long. 95° 12'; sea level. A port
on the Golfo de Tehuantepec; open arid scrub forest [70].
Collections were made in the vicinity of the town and in the
open scrub forest 2.4 kilometers north at an elevation of 20
meters.
San Antonio.—Lat. 16° 15'; long. 95° 22'; elev. 40 m. A
ranch about 25 kilometers west-southwest of Tehuantepec;
arid scrub forest [68].
San Pablo.—Lat. 16° 24'; long. 95° 18'; elev. 40 m. A ranch
on the Río Tehuantepec, northwest of Tehuantepec; dense
scrub forest [56]. Cerro San Pablo probably is the hill
north of this ranch; this is shown on some maps as Cerro de
los Amates.
[Pg 35]
San Pedro, Cerro de.—Lat. 16° 18'; long. 95° 28'; elev.
about 1100 m. (crest). A ridge about 24 kilometers west of
Tehuantepec and east of Cerro Arenal; scrub forest on slopes
and pine-oak forest on top [65].
Santa Efigenia.—Lat. 16° 25'; long. 94° 13'; elev. 500 m. A
ranch on the southern slopes of the Sierra Madre de Chiapas,
8 kilometers north-northwest of Tapanatepec; scrub forest.
Former home of Francis Sumichrast [53].
Santa Lucía.—Lat. 16° 18'; long. 95° 28'; elev. 800 m. A
place name for a former ranch on the east slopes of Cerro
Arenal; scrub forest [63].
Santa María Chimalapa.—Lat. 16° 55'; long. 94° 42'; elev.
296 m. A village on the Río de los Milagros, a tributary to
the Río Coatzacoalcos; rainforest [35].
Santiago Chivela.—Lat. 16° 42'; long. 94° 53'; elev. 200 m.
A village on the Trans-isthmian Highway, 13.4 kilometers by
road south of Matías Romero; dry, grassy plains and
scattered clumps of scrubby trees and palms [41].
Collections were made in the vicinity of the village and at
a rocky stream, 11 kilometers south on the Trans-isthmian
Highway at an elevation of 230 m.
Santo Domingo (Petapa).—Lat. 16° 50'; long. 95° 08'; elev.
225 m. A village about 13 kilometers west-southwest of
Matías Romero; semi-arid scrub forest [37].
Sarabia.—Lat. 17° 04'; long. 95° 02'; elev. 100 m. A
village 25 kilometers north of Matías Romero on the
Trans-isthmian Highway; rainforest [32]. Collections were
made in the vicinity of the village and in the rainforest
along the Río Sarabia, 5 kilometers north of the village at
an elevation of 80 meters.
Tapanatepec.—Lat. 16° 32'; long. 94° 12'; elev. 90 m. A
town on the Pan-American Highway on the lower slopes of the
Sierra Madre de Chiapas; dense scrub forest [58].
Tehuantepec.—Lat. 16° 20'; long. 95° 14'; elev. 35 m. A
large town on the Plains of Tehuantepec; scrub forest [61].
Collections were made in the vicinity of the town and in the
dense scrub forest 8.6 kilometers west at an elevation of 85
meters and 14 kilometers west at an elevation of 120 meters.
Tenango.—Lat. 16° 16'; long. 95° 30'; elev. 1100 m. A town
in the mountains about 40 kilometers west-southwest of
Tehuantepec; scrub forest [67].
Tequisistlán.—Lat. 16° 24'; long. 95° 37'; elev. 190 m. A
village in the valley of the Río Tequisistlán, a tributary
to the Río Tehuantepec; dense scrub forest [54]. Most
collections were made about one kilometer north of the
village where the Pan-American Highway crosses the Río
Tequisistlán.
Tolosita.—Lat. 17° 12'; long. 95° 03'; elev. 80 m. A
village on the Río Tortuguero near the Trans-isthmian
Highway; rainforest [30].
Tres Cruces.—Lat. 16° 26'; long. 95° 51'; elev. 750 m. A
ranch near the Pan-American Highway, 70 kilometers by road
west-northwest of Tehuantepec; dense scrub forest [49].
Tuxtepec—Lat. 18° 06'; long. 96° 05'; elev. 80 m. A town on
the Río Papaloapan in northern Oaxaca; low evergreen forest
[12].
Ubero.—Lat. 17° 18'; long. 95° 00'; elev. 80 m. A lumber
camp and railroad station, 8.5 kilometers south of the Río
Jaltepec on the Trans-isthmian Highway; rainforest [28].
Unión Hidalgo.—Lat. 16° 27'; long. 94° 48'; elev. 7 m. A
village on the railroad, 20 kilometers east-northeast of
Juchitán; open scrub forest [48].
Ventosa, La.—Lat. 16° 30'; long. 94° 51'; elev. 25 m. A
village at the junction of the Pan-American and
Trans-isthmian highways; open scrub forest [46].
Zanatepec.—Lat. 16° 28'; long. 94° 22'; elev. 80 m. A
village on the Pan-American Highway at the eastern edge of
the Plains of Tehuantepec; dense scrub forest [47]. Most
collections were made in the scrub forest 5 to 8 kilometers
west-northwest of the village.
Zarzamora.—Lat. 16° 21'; long. 95° 48'; elev. 800 m. A
ranch between La Reforma (16 kilometers west of
Tequisistlán) and Santa María Ecatepec; scrub forest with
oaks on higher ridges [59].
[Pg 36]
Veracruz
Acayucan.—Lat. 17° 57'; long. 94° 55'; elev. 160 m. A large
town on the Trans-isthmian Highway; rainforest [21].
Collections were made in the vicinity of the town, but
principally at Rancho Las Hojitas, 7 kilometers northwest of
town at an elevation of 150 meters.
Alvarado.—Lat. 18° 47'; long. 95° 47'; sea level. A fishing
village at the mouth of the Río Papaloapan; coastal dunes
and marshes [1]. Most collections were made 1-3 kilometers
southeast of the village in marshes on the leeward side of
the coastal dunes.
Amatitlán.—Lat. 18° 26'; long. 95° 45'; elev. 4 m. A
village on the bank of the Río Papaloapan; savanna and sugar
plantations [6].
Aquilera.—Lat. 17° 48'; long. 95° 01'; elev. 150 m. A
village 21 kilometers southwest of Acayucan on the
Trans-isthmian Highway; rainforest [22].
Ayentes.—Lat. 18° 10'; long. 94° 26'; elev. 2 m. A railroad
station on the east bank of the Río Coatzacoalcos, across
the river from the city of Coatzacoalcos; scrub forest and
marshes [17].
Berta.—Lat. 18° 07'; long. 94° 27'; elev. 5 m. A ranch just
south of Coatzacoalcos; scrub and low evergreen forest [15].
Chacaltianguis.—Lat. 18° 18'; long. 95° 52'; elev. 5 m. A
village on the Río Papaloapan; savanna [8].
Ciudad Alemán.—Lat. 18° 13'; long. 96° 07'; elev. 30 m. A
new government town, headquarters of the Comisión del
Papaloapan; scrub and low evergreen forest [10].
Coatzacoalcos (formerly Puerto México).—Lat. 18° 10'; long.
94° 27'; elev. 2 m. A seaport at the mouth of the Río
Coatzacoalcos; scrub on coastal dunes; marshes and low
evergreen forest inland [16]. Most collections are from the
forest-savanna ecotone, 8 kilometers southwest of town.
Cosamaloapan.—Lat. 18° 22'; long. 95° 50'; elev. 4 m. An
agricultural town on the Río Papaloapan; savanna and sugar
plantations [7].
Cosoleacaque.—Lat. 17° 59'; long. 94° 38'; elev. 55 m. A
village 8 kilometers by road west of Minatitlán; savanna
[19].
Cuatotolapam.—Lat. 18° 08'; long. 95° 16'; elev. 13 m. A
village on the Trans-isthmian Railroad; savanna and low
evergreen forest along streams [13].
Hueyapan.—Lat. 18° 08'; long. 19° 09'; elev. 85 m. A town
32 kilometers by road northwest of Acayucan; savanna and low
evergreen forest [14]. Collections were made in the vicinity
of the town and from forest 10 kilometers southeast of town
at an elevation of 135 meters.
Jesús Carranza (formerly Santa Lucrecia).—Lat. 17° 27';
long. 95° 02'; elev. 80 m. A town and railroad junction in
the middle of the isthmus; rainforest [26]. Most of
Dalquest's specimens came from varying distances from Jesús
Carranza along the Río Coatzacoalcos and its tributaries.
Minatitlán.—Lat. 17° 58'; long. 94° 32'; elev. 15 m. An oil
refinery center on the Río Coatzacoalcos; savanna [20].
Naranjo.—Lat. 17° 35'; long. 95° 07'; elev. 100 m. A
village on the Trans-isthmian Highway, 45 kilometers south
of Acayucan; rainforest and palm forest [24].
Novillero.—Lat. 18° 16'; long. 95° 59'; elev. 10 m. A
village on the Río Papaloapan; scrub forest and grassland
[9].
Oaxaqueña, La.—Lat. 17° 26'; long. 94° 53'; elev. 80 m. A
hacienda on the Río Coatzacoalcos about 12 kilometers east
of Jesús Carranza; rainforest [27].
Playas, Río de las.—Lat. 18° 08'; long. 94° 07'; elev. 3 m.
The river (sometimes known as the Río Tonolá) forming the
boundary between the states of Veracruz and Tabasco;
rainforest [18].
San Lorenzo.—Lat. 17° 44'; long. 94° 42'; elev. 25 m. A
village on the Río Chiquito, about 30 kilometers southeast
of Acayucan; rainforest [23].
[Pg 37]
Suchil.—Lat. 17° 31'; long. 95° 03'; elev. 40 m. A village
on the Trans-isthmian Railroad, about 10 kilometers north of
Jesús Carranza; rainforest [25].
Tecolapan.—Lat. 18° 24'; long. 95° 18'; elev. 275 m. A
village on a small river of the same name in the western
foothills of Los Tuxtlas; rainforest [5].
Tejada, Lerdo de.—Lat. 18° 37'; long. 95° 31'; elev. 60 m.
An agricultural village, 35 kilometers by road
east-southeast of Alvarado; scrub forest, marshes, and sugar
plantations [2]. Collections were made in a marsh, 5
kilometers west-northwest of the village.
Tlacotalpan.—Lat. 18° 37'; long. 95° 42'; elev. 3 m. A town
at the confluence of the Río San Juan and Río Papaloapan;
marshes and sugar plantations [3].
Tula.—Lat. 18° 36'; long. 95° 22'; elev. 150 m. A village
near the western base of Los Tuxtlas; low evergreen forest
and marshes [4]. Collections were made in a marsh 3
kilometers northwest of the village.
THE AMPHIBIAN FAUNA OF THE LOWLANDS
In presenting an account of the amphibian fauna of the lowlands of the
Isthmus of Tehuantepec three items must be considered:
1. The composition of the fauna.
2. The ecology of the fauna.
3. The distribution of the fauna.
These items, together with similar data concerning the amphibians of
the adjacent highlands, will form the basis for the subsequent
discussion of the establishment of present patterns of distribution in
the isthmian region.
Composition of the Fauna
The amphibian fauna of the lowlands of the Isthmus of Tehuantepec
consists of 36 species definitely recorded from the area. These include
one genus and species of caecilian, one genus, including three species
of salamanders, and 14 genera and 32 species of anurans.
In comparison with the known amphibian fauna of the forested and
savanna portions of El Petén, Guatemala (Stuart, 1935 and 1958), we
find that there are more species recorded from the isthmus than from El
Petén. Stuart found only 20 species of amphibians in both forest and
savanna habitats in El Petén. Of the 36 species of amphibians known
from the isthmus, 28 occur on the Gulf lowlands and live in forest or
savanna habitats.
The geographic position of the isthmus with regard to major faunal
areas in Middle America, and the diversity of the environment are
important factors in understanding the presence of a large number of
species of amphibians in the isthmus. The large number of species
probably is a reflection of the diversity of the environment; this
diversity is the result of fluctuation of climate, and thus[Pg 38]
environments, in the not too distant past. In no individual habitat,
such as rainforest, savanna, or scrub forest, does the number of
species approach the total for the region.
Ecology of the Fauna
In the preceding section on the description of the Isthmus of
Tehuantepec I have outlined the major environments in the region. With
respect to the distribution of amphibians we may recognize three major
environments in the isthmus—rainforest, semi-arid scrub forest, and
savanna. Each of these has varying combinations of physical and biotic
factors that are important in the ecology of amphibians. Because of the
importance of moisture, not only for the maintenance of life in these
animals, but in most species their dependence on water for breeding
purposes, this environmental factor is considered the most significant
in the ecological distribution of amphibians. A second factor is the
availability of necessary shelter, especially aestivation sites. These
factors will be compared in the three major environments in the region.
Moisture is present in the environment in the form of free water or
atmospheric moisture. With respect to the latter, it is well known that
dense shaded forests have a considerably higher relative humidity than
do open plains or areas with only scattered trees. Thus, the
rainforests of the isthmus are characterized by a much higher relative
humidity than are the savannas or semi-arid scrub forests. Although
with regard to rainfall there is a pronounced dry season in the regions
supporting rainforest, there still remains considerable atmospheric
moisture in this environment throughout the year. The dense foliage
provides shade and protection from desiccating effects of wind and
sunlight; furthermore the foliage contributes moisture by
transpiration. The deep alluvial soils mixed with large quantities of
organic matter (decaying leaves and rotting logs) maintain considerable
quantities of moisture.
Conversely, the savannas and scrub forests have little atmospheric
moisture during the dry season. In the former habitat there are few
trees to provide shade or moisture through transpiration; in the latter
most of the trees lose their leaves during the dry season. Thus, these
environments are desiccated by the dry winds and direct sunlight.
Furthermore, the soils in these environments become dry and caked.
There is little or no terrestrial matter to hold moisture.
Free water in these environments is present in a variety of forms[Pg 39] at
different times of the year. During the dry season the more extensive
marshes in the savannas persist; many ponds and most of the streams in
the rainforest are permanent throughout the year. In the scrub forest
all except the largest streams become dry during the dry season, and no
ponds exist through the dry season. With the advent of the first heavy
summer rains the stream beds fill with water, marshes expand, and many
depressions become ponds (Pl. 5, fig. 2). At this time the amount of
free water in the scrub forests and savannas greatly increases, much
more so than that in the rainforests.
Environments are vertically stratified in the rainforests. There is the
deep alluvial soil, the ground litter of leaves and decaying logs, the
low bushes and small trees, and finally the tall trees of the forest.
Each of these provides certain types of shelter for amphibians. The
moist soil and litter on the forest floor is an important microhabitat
for fossorial and strictly terrestrial species. The dense foliage of
the trees, tree holes, and bromeliads growing on the trees provide
shelter for arboreal species. Arboreal and terrestrial bromeliads and
the terrestrial elephant-ear plants (Xanthosoma) contain water in the
axils of their leaves throughout the year and thus provide an important
habitat for amphibians. The low, spiny, deciduous trees of the scrub
forest and the grasses and scattered trees in the savannas provide
little shelter. In the savannas there are depressions, some of which
contain water throughout the year; these are often surrounded by trees
providing refugia for amphibians during the dry season. In the scrub
forest many species congregate along streams and in moist stream beds
during the dry season.
Now that the important ecological factors of the major environments
have been outlined, we may examine the local distribution of amphibians
in each of these. Beginning with the rainforest, we find only one
fossorial species, Gymnopis mexicanus. A large number of species are
found on the forest floor; characteristic inhabitants of the leaf
litter are: Bufo valliceps, Eleutherodactylus rhodopis,
Microbatrachylus pygmaeus, and Syrrhophus leprus. Other terrestrial
amphibians usually are not scattered throughout the rainforest, as are
those named immediately above, but instead inhabit areas of forest
adjacent to ponds or streams; these species include: Bufo marinus,
Eleutherodactylus natator, Eleutherodactylus rugulosus,
Leptodactylus labialis, Leptodactylus melanonotus, Rana palmipes
and Rana pipiens. The most striking ecological assemblage[Pg 40] of
amphibians in the rainforest is the arboreal group of species,
including:
Bolitoglossa occidentalis
Bolitoglossa platydactyla
Eleutherodactylus alfredi
Hyla baudini
Hyla ebraccata
Hyla loquax
Hyla microcephala martini
Hyla picta
Phrynohyas modesta
Phrynohyas spilomma
Phyllomedusa callidryas taylori
In the savannas Rhinophrynus dorsalis, Engystomops pustulosus, and
Gastrophryne usta are fossorial species. Bufo marinus,
Leptodactylus melanonotus, Leptodactylus labialis, Rana palmipes,
and Rana pipiens are found in the vicinity of permanent water in the
savannas. Although the savanna habitat does not provide the ecological
conditions for the existence of an arboreal fauna, many arboreal
species from the surrounding rainforest utilize the extensive marshes
and ponds in the savannas for breeding purposes. Thus, Hyla baudini,
Hyla microcephala martini, Hyla picta, and Phrynohyas spilomma
have been found breeding in savannas. In parts of savannas where clumps
of trees surround depressions containing water throughout the year,
individuals of the species named above, together with Hyla loquax and
Phyllomedusa callidryas taylori, may not only breed, but remain
throughout the year.
In the semi-arid scrub forest the same fossorial species as exist in
the savannas are found. Likewise, Bufo marinus, Leptodactylus
labialis, Leptodactylus melanonotus, and Rana pipiens are found
near permanent water. Terrestrial species in this semi-arid environment
include Bufo canaliferus, Bufo coccifer, Bufo marmoreus,
Syrrhophus pipilans, and Diaglena reticulata. Of these, Syrrhophus
pipilans sometimes inhabits low trees and bushes; the others may be
fossorial. The arboreal species in the scrub forest include Hyla
baudini, Hyla robertmertensi, Hyla staufferi, and Phyllomedusa
dacnicolor.
Eleutherodactylus rugulosus and Hylella sumichrasti live along
streams in the scrub forest. Hylella sumichrasti lays its eggs in
these streams.
In comparing the ecological differences in the amphibian assemblages in
the three major habitats, the most obvious difference is the great
percentage of arboreal species in the rainforest as compared with
savanna and scrub forest. Only four arboreal species are found in the
scrub forest, none in the savannas, but eleven in the rainforest.
Likewise, there is an absence of ground-dwelling forms in the arid
habitats; in the latter the only terrestrial[Pg 41] species are those that
are found near water. A possible exception is Syrrhophus pipilans.
From the above analysis of ecological distribution we may see that the
rainforest provides a variety of habitats for amphibians and that these
habitats are suitable for amphibian life throughout the year. On the
other hand, the savannas and scrub forests are characterized by extreme
conditions of desiccation, a factor of considerable importance in
limiting the ecological distribution of amphibians. However, there
still is a diversity of amphibians in these semi-arid environments.
Obviously, these species are adapted in various ways for survival
during the dry season, at which time environmental conditions are such
that the animals cannot carry on their normal activities.
Although there is not an abundance of data concerning the seasonal
activity of the fauna, what is available shows some interesting
correlations with the environments. During the dry season in the scrub
forest there is essentially no amphibian activity; an occasional Rana
pipiens may be seen along a river, or a Bufo marinus may be seen at
night. In the rainforest the terrestrial-breeding amphibians are active
during the dry season. Eleutherodactylus rugulosus is found at night
or by day along streams. Eleutherodactylus rhodopis,
Microbatrachylus pygmaeus, and Bufo valliceps are active during the
day; these plus Bolitoglossa occidentalis, Bolitoglossa
platydactyla, Eleutherodactylus alfredi, Eleutherodactylus
natator, and an occasional Hyla are active at night.
With the onset of the heavy summer rains and the subsequent formation
of breeding ponds, amphibian activity reaches a peak. This is
especially noticeable in the semi-arid environments, where during the
dry season there is little activity.
Among the anurans in the isthmus the four species of
Eleutherodactylus, the two species of Syrrhophus, and the one
species of Microbatrachylus are either known, or presumed, to lay
eggs on the ground; these develop directly into small frogs. All of the
other anurans deposit their eggs in water or attach them to objects
over water (Phyllomedusa); these hatch into tadpoles, which later
metamorphose into frogs. Hylella sumichrasti is known to breed only
in streams. All of the other species breed in ponds, but at times some
species deposit their eggs in streams; in this last group are Bufo
valliceps, Bufo marmoreus, Phyllomedusa callidryas taylori, and
Rana pipiens.
Although the ecological data are incomplete, they do show that[Pg 42]
ecological conditions differ greatly in the three major environments,
different species of amphibians inhabit these environments, and that
the fauna is ecologically diversified in each environment.
Distribution of the Fauna
Plotting the distributions of species of amphibians known to live in
the lowlands of the Isthmus of Tehuantepec results in an array of
geographic patterns. These may be analyzed with respect to those
species that are restricted either to the Gulf lowlands or the Pacific
lowlands, or those that occur on both the Gulf and Pacific lowlands.
Furthermore, the distributions may be analyzed with respect to those
species whose ranges extend from México across the Isthmus of
Tehuantepec into Central America, those that reach the isthmus from
Central America but do not extend into México proper, and those that
reach the isthmus from México but do not extend into Central America.
It should be kept in mind that the following analysis is of the lowland
inhabitants only. Species inhabiting the foothills and mountains will
be discussed later.
1. Species Restricted to the Gulf Lowlands. Of the 36 species of
amphibians recorded from the Isthmus of Tehuantepec, nine (25 per cent)
are in this group. Four of these (Eleutherodactylus alfredi,
Syrrhophus leprus, Hyla loquax, and Hyla picta) live in the Gulf
lowlands to the east and to the west of the isthmus. Three others
(Hyla ebraccata, Hyla microcephala martini and Phyllomedusa
callidryas taylori) are primarily Central American in their
distribution and reach the northwestern limits of their ranges in the
Gulf lowlands of the isthmus, whereas Bolitoglossa platydactyla and
Eleutherodactylus natator reach the southern limits of their
distributions in the isthmus.
2. Species Restricted to the Pacific Lowlands. This group includes six
species, or 17 per cent of the amphibian fauna of the isthmus. Two of
these (Bufo coccifer and Syrrhophus pipilans) range to the east and
to the west of the isthmus on the Pacific lowlands. Two others (Bufo
canaliferus and Hyla robertmertensi) range from the isthmus into
Central America, and Diaglena reticulata and Phyllomedusa
dacnicolor range on the Pacific lowlands of México southeastward to
the isthmus.
3. Species That Occur on the Pacific and Gulf Lowlands. This group
includes 19 species, or 53 per cent of the total amphibian fauna. Of
these, nine species (25 per cent of the entire amphibian[Pg 43] fauna) are
widespread throughout the lowlands of México and Central America; these
are:
Gymnopis mexicanus
Rhinophrynus dorsalis
Bufo marinus
Engystomops pustulosus
Leptodactylus labialis
Leptodactylus melanonotus
Hyla baudini
Hyla staufferi
Rana pipiens
Four species occur on the Gulf lowlands to the east and to the west of
the isthmus, but on the Pacific lowlands they occur only to the east;
this group includes Bufo valliceps, Eleutherodactylus rhodopis,
Phrynohyas modesta, and Phrynohyas spilomma. Three species live to
the east and to the west of the isthmus on the Pacific lowlands, but
only to the west on the Gulf lowlands; these include Eleutherodactylus
rugulosus, Microbatrachylus pygmaeus, and Gastrophryne usta.
Six species that cross the isthmus live on the humid Gulf lowlands and
on the humid lowlands of Chiapas and Guatemala, but not on the
semi-arid Plains of Tehuantepec; these include Bolitoglossa
occidentalis, Eleutherodactylus rhodopis, Microbatrachylus
pygmaeus, Phrynohyas modesta, Phrynohyas spilomma, and Rana
palmipes. Of these, Microbatrachylus pygmaeus also occurs in
scattered humid environments to the west of the isthmus on the Pacific
lowlands.
Two species are endemic to the isthmian region. Bolitoglossa
veracrucis is known only from the humid northern slopes of the
isthmus. Hylella sumichrasti occurs on the Pacific slopes of the
isthmus and extends to the east into western Chiapas.
In analyzing the distribution of the amphibians with respect to those
that are restricted to either the Pacific or Gulf lowlands or those
that cross the continental divide in the isthmus, we find that 25 per
cent of the species are restricted to the Gulf lowlands, 17 per cent
are restricted to the Pacific lowlands, and 53 per cent cross the
isthmus. In analyzing the distribution patterns with respect to those
that extend across the isthmus of Tehuantepec from east to west, we
find that 14 per cent of the species do not extend east of the isthmus
into Central America and that 19 per cent do not range west of the
isthmus into México proper; 61 per cent of the species range to the
east and to the west of the isthmus. Of the 36 species of amphibians
inhabiting the isthmus only nine species (25 per cent) range across the
isthmus, that is, occur on the Gulf and Pacific lowlands, and also
range to the east and to the west of the isthmus. To these wide-ranging
species the diversified environments of the[Pg 44] isthmus do not present a
barrier to distribution. The other 27 species (75 per cent) either do
not cross the isthmus from east to west or from north to south; thus,
probably in one way or another the isthmus presents a barrier to their
distribution.
THE AMPHIBIAN FAUNA OF THE FOOTHILLS AND ADJACENT HIGHLANDS
To amphibians inhabiting the foothills and mountains of southern México
and northern Central America, the isthmus presents a great barrier to
dispersal. For example, salamanders of the genus Thorius, the
mexicanus and augusti groups of the genus Eleutherodactylus, the
bistincta group of the genus Hyla, and the genus Tomodactylus
occur on the Mexican Plateau and southward into the mountains of
Oaxaca. Nevertheless, no members of these groups are present in the
Guatemalan-Chiapan Highlands. The genera Chiropterotriton,
Magnadigita, Pseudoeurycea, and Ptychohyla, as well as the
eximia group of Hyla are represented by different species in the
Guatemalan-Chiapan Highlands than in the mountains of México on the
other side of the isthmus. Several species of Plectrohyla occur in
the Guatemalan-Chiapan Highlands, but none is known from the Mexican
Highlands, although one species occurs in the Tuxtlas.
Living in the humid forests of the foothills are salamanders of the
genus Lineatriton, frogs of the spatulatus group of
Eleutherodactylus, Anotheca coronata, Hyla miotympanum, and
Phyllomedusa moreleti. All of these occur in the foothills of the
Sierra Madre Oriental in eastern México and in Los Tuxtlas.
Lineatriton, Hyla miotympanum, and the spatulatus group of
Eleutherodactylus do not occur in the foothills of the
Guatemalan-Chiapan Highlands; those amphibians reach the end of their
ranges at the isthmus. Phyllomedusa moreleti and Anotheca coronata
are found in the northern foothills of the Guatemalan-Chiapan
Highlands, and Phyllomedusa moreleti is found in the foothills on the
Pacific slopes of the Chiapan Highlands.
Although the above analysis is not so detailed as that of the lowland
inhabitants, it does show that all of the genera and species of
amphibians known to inhabit the foothills and highlands adjacent to the
isthmus, only two species of amphibians cross the isthmus from one
highland mass to the other. Thus, it is evident that the Isthmus of
Tehuantepec presents a great barrier to dispersal of these groups of
amphibians.
[Pg 45]
ESTABLISHMENT OF PRESENT PATTERNS OF DISTRIBUTION
From the foregoing analysis of geographical and ecological distribution
in the Isthmus of Tehuantepec we may strive for an interpretation of
the events that led to the establishment of patterns of distribution
displayed not only by the amphibians, but other terrestrial vertebrates
as well. The thesis that I am proposing below is based on the premise
that in southern México and northern Central America climatic
fluctuation during the Pleistocene was of sufficient magnitude to cause
vegetational shifts, both vertically and latitudinally, resulting in
the establishment of alternating continuous and discontinuous lowland
and highland environments, although this climatic fluctuation was not
so great as to eliminate tropical lowland environments from the region.
I feel that the present patterns of distribution of the amphibians in
the Isthmus of Tehuantepec may be explained on this premise.
Many authors dealing with the herpetofauna of Middle America have
followed Schuchert's (1935) suggestion of a seaway in the isthmus
during the Cenozoic. Thus, Burt (1931), Duellman (1956, 1958a), Gloyd
(1940), Oliver (1948), Smith and Laufe (1946), and Stuart (1941)
employed the presence of a seaway to explain distribution and
speciation in various genera. Durham, Arellano, and Peck (1952), Olson
and McGrew (1941), and Stirton (1954) have provided geological evidence
that there probably was no Cenozoic seaway in the Isthmus of
Tehuantepec. Even if there were a seaway in the Pliocene or Miocene
(the dating of this possible seaway is open to question), its presence
is not necessary to explain the present patterns of distribution in the
isthmus.
In recent years the study of natural biotic environments, palynology,
and Pleistocene chronology in Middle America has produced a wealth of
data, which although still fragmentary begins to form a picture of past
climatic events in that part of the world. Sedimentary studies by
Hutchinson, Patrick, and Deevey (1956) and Sears, Foreman, and Clisby
(1955) have provided evidence of drastic climatic shifts in México
during the Pleistocene. Further evidence of bioclimatic fluctuation is
provided by Martin and Harrell (1957) and Martin (1958); the latter has
suggested that there was a displacement of the tropical zones in
southern México and northern Central America by as much as 3000 feet
during the glacial maximum. Much of the evidence of such drastic
vertical[Pg 46] shifts in environments is based on the presence of
Pleistocene montane glaciers on Mexican volcanoes (White, 1956) and
Chirripo in Costa Rica (Weyl, 1955). Dorf (1959) supports this idea of
drastic climatic change.
In his studies of the avifauna of México and Guatemala Griscom (1932
and 1950) made an important issue of the continuity of the bird fauna
in what he called the Subtropical Life-zone, which essentially consists
of cloud forest, a widespread, but discontinuous, habitat on the Gulf
(windward) slopes of the Mexican and Central American highlands at
elevations between 1000 and 2000 meters. To account for this apparent
uniformity in the avifauna Griscom hypothesized a continuity of cloud
forest environment in the Pleistocene; this would result in the
depression of cloud forests to the coastal lowlands and the
displacement of tropical lowland environments far to the south in
Central America. Stuart (1951) objected to this displacement of lowland
tropical rainforest; he stated that a descent to sea level of a
subtropical zone would have brought about either widespread
extermination of the tropical fauna or acclimatization of that fauna to
subtropical conditions.
Although palynological studies and some faunal studies of subtropical
and temperate animals suggest a drastic climatic fluctuation that might
have eliminated tropical environments in southern México and northern
Central America, there is much biological evidence indicating the
existence of tropical environments in this region even during the
glacial maximum. Especially significant is the diversity of species
inhabiting the present tropical environments; many of these have
differentiated from related taxa to the south.
In the Pleistocene, climate fluctuated and vegetation shifted
correspondingly in southern México and northern Central America. Most
of the palynological studies and many studies of Pleistocene chronology
deal with montane regions, either the Mexican Plateau or the mountains
rising from the plateau. No such studies have been made in lowland
tropical environments. During glacial advances the tropical lowland
environments in México probably were not eliminated, for the great
diversity of animals in these environments supports the hypothesis that
they have been in existence for some time, although periodically they
may have been discontinuous.
In order to understand the nature of bioclimatological events in the
Pleistocene in lowland tropical environments of southern México,
certain factors that are of little importance in the interpretation[Pg 47] of
Pleistocene chronology in the highlands must be considered. These
factors are: 1) climatic moderation by oceans, 2) fluctuation in sea
level, and 3) fluctuation in level of the water table as affected by
sea level.
It is well-known that large bodies of water moderate the temperature on
adjacent land. Furthermore, it is known that faunas of marine
invertebrates shifted latitudinally in the Pleistocene; Trask, Phleger,
and Stetson (1947) recorded cold-water Foraminifera then as far south
as the Sigsbee Deep in the middle of the Gulf of Mexico. Large bodies
of warm water, such as the Gulf of Mexico, Caribbean Sea, and Pacific
Ocean of today, probably were not sufficiently cooled at the time of
glacial advance to affect greatly the temperature of the winds blowing
across them. Even if these bodies of water were somewhat cooler than
now, the prevailing winds blowing from them onto the lowlands of México
and northern Central America would have aided in maintaining relatively
high temperatures there. These warm winds probably counteracted the
cooling effect of glaciation in the lowlands and thereby maintained
tropical conditions near the seas.
Although no adequate studies of Pleistocene beach lines have been made
in southern México, such information is available for peninsular
Florida on the other side of the Gulf of Mexico (Cooke, 1945).
Fluctuation in sea level in the Pleistocene has been used by Hubbell
(1954), Goin (1958), and Duellman and Schwartz (1958) to explain
present patterns of distribution of animals in Florida. If Cooke's
interpretations can be applied to the western side of the Gulf of
Mexico, even generally, it would be supposed that sea level varied from
about 300 feet lower than at present during the Illinoian Glacial
Period to about 275 feet higher than at present during the Aftonian
Interglacial Period. Lowering of sea level would expand the lowlands in
the isthmus; rising sea level would restrict them, leaving only the
central ridges and many islands in the isthmus, but never forming a
seaway between the Gulf of Mexico and the Pacific Ocean.
Probably the level of the water table in the coastal lowlands and the
gradients of the streams in the lowlands and foothills was closely
correlated with fluctuation in sea level. If sea level fluctuated as
much as 575 feet in the Pleistocene, changes in the level of the water
table must have been of considerable magnitude.
During times of glacial advances the lowlands of the isthmus probably
were more extensive and had more semi-arid tropical[Pg 48] environments than
at present, with patches of rainforest existing in sheltered valleys
along the major streams. In the course of bio-climatic fluctuation the
semi-arid environments (scrub forest and/or savanna) were continuous at
times from the Pacific lowlands across the isthmus to the Gulf
lowlands. At those times such typical inhabitants of the semi-arid
environments as Rhinophrynus dorsalis, Engystomops pustulosus, and
Hyla staufferi could have made their way across the isthmus. At times
of most extensive glaciation, such as the Illinoian, temperatures in
the isthmus probably were low enough to permit the growth of pine-oak
forest and cloud forest continuously across the central ridges from the
Mexican to the Chiapan-Guatemalan highlands. At those times such
highland members of the fauna as Chiropterotriton, Pseudoeurycea,
Magnadigita, and the eximia group of Hyla could have crossed the
isthmus. During Wisconsin time, climate probably fluctuated less than
during previous glaciations; probably no montane environments, except
cloud forest, were represented in the isthmus during the Wisconsin.
Even at this relatively late date such animals as Lineatriton
lineola, Anotheca coronata, and Phyllomedusa moreleti could have
crossed the isthmus.
During the interglacial periods, which in the isthmian region were
characterized by warmer temperatures, higher sea level and consequently
more restricted areas of lowlands, and possibly more rainfall than in
the glacial periods, the continuity of pine-oak forest and cloud forest
from east to west across the isthmus was interrupted. Probably, too,
the semi-arid environments were restricted, and the rainforests were
more widespread. At those times animals now inhabiting the rainforests
of the Gulf lowlands and those inhabiting the Pacific lowlands of
Chiapas and Guatemala could have crossed the isthmus. In this group are
species such as Bolitoglossa occidentalis, Eleutherodactylus
rhodopis, Microbatrachylus pygmaeus, and Rana palmipes.
The amount of differentiation in isolated populations of amphibians in
southern México and northern Central America gives some idea of
relative lengths of time of isolation from related populations. Those
populations inhabiting high mountain environments on either side of the
isthmus are specifically distinct. Some populations inhabiting cloud
forests lower on the mountains are specifically distinct from related
populations on the other side of the isthmus; between others there is
no recognizable differentiation. Even though many populations are
isolated from other populations of the same species in the lowlands of
the isthmus, there is[Pg 49] no apparent speciation. This indicates that the
lowland environments and their inhabitants have been isolated from one
another for a shorter time than have the highland environments and
their inhabitants.
ACCOUNTS OF SPECIES
For each species of amphibian known to occur in the lowlands of the
Isthmus of Tehuantepec, localities where one or more specimens were
collected are listed, and variation, ecology, and life histories are
discussed. A total of 2833 specimens has been examined for the purposes
of this study. Individual specimens cited in the text are listed with
catalogue numbers and abbreviations of the name of the museum, as
follows:
AMNH American Museum of Natural History
KU University of Kansas Museum of Natural History
MCZ Museum of Comparative Zoology, Harvard College
UIMNH University of Illinois Museum of Natural History
UMMZ University of Michigan Museum of Zoology
USNM United States National Museum
Gymnopis mexicanus mexicanus Duméril and Bibron
Oaxaca: El Barrio (3); Matías Romero; Tehuantepec (2).
Veracruz: Cosamaloapan; Cuatotolapam (2).
The two specimens from Cuatotolapam were collected by Ruthven in an
area of mixed savanna and forest. The three specimens (USNM 30535-7)
listed above from El Barrio were collected by Sumichrast; possibly they
came from another locality. The city of Tehuantepec is divided into
seven districts called "barrios." The two specimens listed from
Tehuantepec (MCZ 1604) merely bear the data "Tehuantepec, Mexico." They
may have come from the town, the district, or from anywhere in the
isthmus. The specimen from Matías Romero has 109 primary and 67
secondary annuli, a length of 400 mm., and a diameter of 19 mm.; the
one from Cosamaloapan has 106 primary and 58 secondary annuli, a length
of 397 mm., and a diameter of 19 mm. Data on the other specimens were
recorded by Dunn (1942:475).
Bolitoglossa occidentalis Taylor
Oaxaca: Río Sarabia (2); Ubero. Veracruz: La Oaxaqueña;
14 km. E of Suchil.
The specimens from Oaxaca are only tentatively assigned to
occidentalis. All are immature and lack maxillary teeth. Taylor
(1941:147) stated that the maxillary teeth are absent in young
occidentalis. One from Río Sarabia is a male with a body-length of 29
mm. and a tail-length of 22 mm. The dorsum is reddish brown[Pg 50] streaked
with dark gray; the venter is dark gray. Two small individuals (one
from Sarabia and one from Ubero) have body-lengths of 19 and 21 mm. and
tail-lengths of 10.5 and 11 mm. In life they were pale yellowish tan
above with a brown triangular mark on the occiput, but with no
middorsal stripe. Both were found in the axils of elephant ear plants
(Xanthosoma).
This species has been noted by Goodnight and Goodnight (1956:146) on
the Atlantic lowlands at Palenque, Chiapas, and by Shannon and Werler
(1955:362) at several localities in Los Tuxtlas, Veracruz. I have
collected it at Vista Hermosa on the eastern slopes of the Sierra Madre
Oriental above Tuxtepec in northern Oaxaca. Both B. occidentalis and
B. rufescens have been reported from Palenque, Chiapas (Taylor and
Smith, 1945:547). Reëxamination of specimens from northern Chiapas and
Tabasco is needed to verify the sympatric occurrence of these two
similar species.
Bolitoglossa platydactyla Tschudi
Oaxaca: La Oaxaqueña; Tolosita (2). Veracruz: Acayucan;
Cuatotolapam; 25 km. ESE of Jesús Carranza; 14 km. E of
Suchil; 2.7 km. N of Tula.
Known only from the Gulf lowlands in the isthmian region, this species
has been taken in a variety of habitats within the humid forest area:
under outer leaves of banana plants, under a rock along a stream, under
a log in a plowed field, and on a reed in a pond at night. Three adult
males have an average snout-vent length of 44 mm. and a tail-length of
41 mm. In life the color of the dorsum varied from orange-yellow to
orange-tan, usually being more orange on the tail. The iris was a
reddish orange.
Bolitoglossa veracrucis Taylor
Veracruz: 35 km. SE of Jesús Carranza (21).
This species is known only from the type series collected at night on a
limestone cliff by Walter W. Dalquest. If this salamander is restricted
to this type of habitat, it should be found in the region of extensive
limestone outcroppings in northern Chiapas and southern Tabasco.
Rhinophrynus dorsalis Duméril and Bibron
Oaxaca: Ixtepec; Limón; Salina Cruz (18); Tehuantepec
(57); Tuxtepec (3). Veracruz: Amatitlán (3); Cosamaloapan
(5); Novillero (2); San Lorenzo.
This species inhabits the scrub forests of the Pacific coastal plain
and the savannas in southern Veracruz; apparently it does not occur in
rainforest. Consequently, its distribution in the isthmus is
discontinuous.
PLATE 1
 Fig. 1. Savanna about 75 kilometers east of
Coatzacoalcos, Veracruz. Photograph by L. C. Stuart.
Fig. 1. Savanna about 75 kilometers east of
Coatzacoalcos, Veracruz. Photograph by L. C. Stuart.
 Fig. 2. Low scrub forest near Alvarado, Veracruz.
Photograph by L. C. Stuart.
Fig. 2. Low scrub forest near Alvarado, Veracruz.
Photograph by L. C. Stuart.
PLATE 2
 Fig. 1. Rainforest near Tolosita, Oaxaca. March, 1956.
Fig. 1. Rainforest near Tolosita, Oaxaca. March, 1956.
 Fig. 2. Rainforest along the Río Sarabia, Oaxaca. March,
1956.
Fig. 2. Rainforest along the Río Sarabia, Oaxaca. March,
1956.
PLATE 3
 Fig. 1. Transition forest near La Princesa, Oaxaca.
March, 1956.
Fig. 1. Transition forest near La Princesa, Oaxaca.
March, 1956.
 Fig. 2. Palm Savanna on the Plains of Chivela, Oaxaca.
March, 1956.
Fig. 2. Palm Savanna on the Plains of Chivela, Oaxaca.
March, 1956.
PLATE 4
 Fig. 1. Scrub forest on the Plains of Tehuantepec in dry
season. March, 1956.
Fig. 1. Scrub forest on the Plains of Tehuantepec in dry
season. March, 1956.
 Fig. 2. Scrub forest on the Plains of Tehuantepec in
rainy season. View toward the north. In the distance is the Continental
Divide in the hills of the Isthmus. July, 1958.
Fig. 2. Scrub forest on the Plains of Tehuantepec in
rainy season. View toward the north. In the distance is the Continental
Divide in the hills of the Isthmus. July, 1958.
PLATE 5
 Fig. 1. Low, dense scrub forest near La Ventosa, Oaxaca.
July, 1958.
Fig. 1. Low, dense scrub forest near La Ventosa, Oaxaca.
July, 1958.
 Fig. 2. Temporary pond in scrub forest north of Salina
Cruz, Oaxaca. July 7, 1958. Rhinophrynus dorsalis, Bufo marmoreus,
and Diaglena reticulata were breeding here the previous night.
Fig. 2. Temporary pond in scrub forest north of Salina
Cruz, Oaxaca. July 7, 1958. Rhinophrynus dorsalis, Bufo marmoreus,
and Diaglena reticulata were breeding here the previous night.
PLATE 6
 Fig. 1. Calling male of Rhinophrynus dorsalis,
photographed in a pond north of Santa Cruz, Oaxaca, on July 6, 1958. ×
2/3.
Fig. 1. Calling male of Rhinophrynus dorsalis,
photographed in a pond north of Santa Cruz, Oaxaca, on July 6, 1958. ×
2/3.
 Fig. 2. Color pattern variation in two adults of Bufo
canaliferus from Juchitán, Oaxaca. × 2/3.
Fig. 2. Color pattern variation in two adults of Bufo
canaliferus from Juchitán, Oaxaca. × 2/3.
PLATE 7
 Fig. 1. Calling male of Engystomops pustulosus,
photographed in a pond west of Tehuantepec, Oaxaca, on July 5, 1956. ×
2.
Fig. 1. Calling male of Engystomops pustulosus,
photographed in a pond west of Tehuantepec, Oaxaca, on July 5, 1956. ×
2.
 Fig. 2. Foamy egg mass of Engystomops pustulosus at
the edge of a pond west of Tehuantepec, Oaxaca. July 5, 1956. × 3/8.
Fig. 2. Foamy egg mass of Engystomops pustulosus at
the edge of a pond west of Tehuantepec, Oaxaca. July 5, 1956. × 3/8.
PLATE 8
 Fig. 1. Calling male of Diaglena reticulata,
photographed at a pond north of Salina Cruz, Oaxaca, on July 6, 1958. ×
1/2.
Fig. 1. Calling male of Diaglena reticulata,
photographed at a pond north of Salina Cruz, Oaxaca, on July 6, 1958. ×
1/2.
 Fig. 2. Clasping pair of Diaglena reticulata at the
edge of a pond north of Salina Cruz, Oaxaca, on July 6, 1958. × 1.
Fig. 2. Clasping pair of Diaglena reticulata at the
edge of a pond north of Salina Cruz, Oaxaca, on July 6, 1958. × 1.
[Pg 51]
Breeding congregations were found after heavy rains at Tehuantepec on
July 5, 1956, at Cosamaloapan, Novillero, and Amatitlán on July 26,
1956, and at Salina Cruz on July 6, 1958. The call is a long "worrp"
made while the male is floating on the surface of the pond. The small
heads, small limbs, and greatly inflated bodies cause the calling males
to resemble miniature caricature balloons (Pl. 6, fig. 1). Amplexus is
inguinal. These toads are notably wary, even when calling. Often the
beam of a flashlight or the slightest disturbance of the water will
cause the males to stop calling. The body is deflated with one last
nauseous note, and the frog sinks beneath the surface of the water and
swims away with short slow kicks of the hind feet.
Bufo canaliferus Cope
Oaxaca: Chivela; Salina Cruz; Santa Efigenia; Tapanatepec
(6); Tehuantepec (10); Zanatepec (4).
This small toad apparently is restricted to the Pacific lowlands from
the Isthmus of Tehuantepec eastward to Guatemala. At Zanatepec on July
13, 1956, males were calling from a flooded field bordered by scrub
forest. The call is a rather loud nasal racket. Living individuals vary
greatly in coloration. Some have yellowish tan flanks and dorsum and an
orange middorsal stripe; others have a pale red dorsum, yellow flanks,
and a cream middorsal stripe (Pl. 6, fig. 2).
Bufo coccifer Cope
Oaxaca: Juchitán (5); Tehuantepec.
It is with some degree of hesitancy that these toads are referred to
the species coccifer. Although these and other specimens from
Guerrero and Michoacán display no striking differences from specimens
from Costa Rica, Nicaragua, and southeastern Guatemala, the ranges of
the populations are separated by a broad hiatus in Chiapas and
Guatemala. Possibly this species has utilized the sub-humid corridor
through northern Central America (Stuart, 1954) and subsequently
disappeared from the corridor in Guatemala and Chiapas. Specimens of a
coccifer-like toad collected by Stuart in the vicinity of
Jacaltenango, Departamento Huehuetenango, Guatemala, are much larger
than either the Central American or Mexican specimens of coccifer. A
final commitment on the systematic status must await a thorough study
of this group of toads.
Males of this species were calling from a grassy rain-pool in open
scrub forest at the edge of Juchitán on July 6, 1956. The call is a
low[Pg 52] "whirrr." The calling males were sitting in the shallow water at
the edge of pond, where they were hidden by the grass. None was
observed in open water, as is characteristic of calling males of Bufo
canaliferus and marmoreus.
Bufo marinus Linnaeus
Oaxaca: Agua Caliente; Guichicovi (3); Mixtequilla;
Tolosita (6); Tehuantepec (37); Tuxtepec; Unión Hidalgo.
Veracruz: Ciudad Alemán (4); Cosamaloapan; Cuatotolapam
(19); 20 km. SE of Jesús Carranza (4); 38 km. SE of Jesús
Carranza (10); 20 km. NE of Jesús Carranza (4); Novillero.
This large toad is abundant throughout the lowlands of the isthmus. The
loud rattling call of males was heard on rainy nights throughout the
summer. In March, 1956, several adults were found in a small cave back
of a spring at Agua Caliente.
Bufo marmoreus Wiegmann
Oaxaca: Cerro San Pedro (2); Chivela (5); Escurano (3);
Juchitán; Salina Cruz (101); Santa Lucía (2); 12 km. S of
Santiago Chivela (11); Santo Domingo; Tapanatepec;
Tehuantepec (100); Tequisistlán. Veracruz: Alvarado;
Coatzacoalcos.
This toad is abundant on the Pacific lowlands, where it inhabits both
open and dense scrub forest. On the Gulf lowlands its distribution
seems to be limited to xeric coastal habitats. Aside from the specimens
from Alvarado and Coatzacoalcos, it is known in Veracruz only from Boca
del Río (Langebartel and Smith, 1959:27).
The similarity in size of Bufo marmoreus and valliceps and their
almost completely allopatric ranges suggest that the two species may be
in competition at any one locality. Nevertheless, both were calling
from a small rocky stream south of Santiago Chivela on July 6, 1956.
On the night of July 6, 1958, an estimated 400 toads of this species
made up a breeding congregation near Salina Cruz. The site was a
shallow muddy pond about 20 × 40 meters located in an area cleared of
scrub forest; the banks of the pond were devoid of vegetation (Pl. 5, fig. 2).
Breeding in the same pond were Rhinophrynus dorsalis and
Diaglena reticulata. The following morning no more than a dozen
Bufo were found in the pond, but several individuals were found
beneath debris and in small burrows near the pond. On July 7, 1958,
large numbers of tadpoles and recently metamorphosed young were in a
shallow grassy pool just east of Salina Cruz.
Taylor (1943b:347) referred certain specimens from Tehuantepec to Bufo
perplexus, a species closely related to Bufo marmoreus.[Pg 53] Evidence to
be presented elsewhere shows that perplexus does not occur in the
isthmus.
Bufo valliceps valliceps Wiegmann
Oaxaca: Guichicovi (2); Matías Romero; 32 km. N of Matías
Romero (2); Nueva Raza; Río Sarabia (3); Santa María
Chimalapa (14); Santiago Chivela; 12 km. S of Santiago
Chivela (5); Santo Domingo (5); Tolosita (7). Veracruz:
Acayucan (3); Alvarado; Amatitlán; Ayentes; Cosamaloapan
(3); Cosoleacaque (6); Cuatotolapam (14); Hueyapan; 20 km.
ENE of Jesús Carranza (6); 20 km. S of Jesús Carranza; 25
km. SE of Jesús Carranza (23); 35 km. SE of Jesús Carranza;
60 km. SW of Jesús Carranza (5); La Oaxaqueña (4); Novillero
(4); San Lorenzo (5).
Individuals were found in both wet and dry seasons. In the dry season
they were most frequently found in rainforest, whereas in the rainy
season breeding congregations were found in savannas as well. This toad
occurs throughout the Gulf lowlands and on the Pacific slopes and in
the Grijalva Valley of Chiapas and Guatemala, but not on the Pacific
lowlands of the isthmus.
I have not been able to recognize individuals referrable to the race
macrocristatus. Firschein and Smith (1957:219) described
macrocristatus from the mountains of eastern Oaxaca and referred to
it specimens from the Gulf lowlands of northern Chiapas. None of the
present material shows the hypertrophied cranial crests supposedly
characteristic of macroaristatus, nor do specimens from the isthmus
resemble the population in the Grijalva Valley being described by L. C.
Stuart, who will discuss the variation in, and the validity of, the
named populations of valliceps.
Five specimens from San Lorenzo, Veracruz (USNM 123516-20), were
identified as Bufo cristatus by Smith (1947:408). Firschein (1950:83)
redefined the cristatus group of Bufo and assigned these specimens
to valliceps.
Eleutherodactylus alfredi Boulenger
Oaxaca: Tolosita (2). Veracruz: 35 km. SE of Jesús
Carranza (6).
These specimens were collected in rainforest. Shreve (1957:247) pointed
out the close resemblance between E. alfredi and E. conspicuus from
Piedras Negras, Guatemala, and treated them as subspecies. Examination
of the specimens from the isthmus, together with seven from central
Veracruz and one from Teapa, Tabasco, suggests an even closer
relationship. Eleutherodactylus conspicuus was diagnosed by Taylor
and Smith (1945:567) as differing from alfredi "in lacking a tarsal
fold, in having shorter hind legs with the tibiotarsal articulation
reaching only to the nostril instead of beyond the tip of the snout;
the vomerine teeth barely reach the posterior level of the choanae."
The specimen from[Pg 54] Teapa has the vomerine teeth reaching to the
posterior edge of the choanae; in the eight specimens from the isthmus
the teeth reach the posterior edge of the choanae in two and to the
middle of the choanae in six; in seven specimens from central Veracruz
the teeth reach the posterior edge of the choanae in two and to the
middle in five. The tibiotarsal articulation extends beyond the tip of
the snout in the specimen from Teapa and in two from central Veracruz;
in three specimens from the isthmus and in one from central Veracruz it
extends only to the nostril; in the others it extends to the snout. The
tarsal fold is absent in the specimen from Teapa, in three from the
isthmus, and in all those from central Veracruz; it is weakly present
in the others.
In the light of this evidence there seems to be little justification in
recognizing two species or even two subspecies in this group.
Consequently, Eleutherodactylus conspicuus Taylor and Smith (1945) is
here placed in the synonymy of Eleutherodactylus alfredi Boulenger
(1898), a species with a range extending from Cuautlapan and Potrero
Viejo in central Veracruz southward and eastward in forested habitats
to western El Petén, Guatemala.
Eleutherodactylus natator Taylor
Veracruz: 35 km. SE of Jesús Carranza (3); 38 km. S of
Jesús Carranza; 55 km. SE of Jesús Carranza.
The snout-vent length is 42.0 mm. in a male and averages 59.5 mm. in
three adult females. The tarsal fold is low and extends about half the
length of the tarsus; the first and second fingers are subequal in
length; the tibiotarsal articulation extends beyond the tip of the
snout. The patches of vomerine teeth lie between the posterior margins
of the choanae. The throat and belly are immaculate, and the soles of
the feet are dark. In the isthmus this species can be distinguished
from Eleutherodactylus rugulosus by less rugose skin on the dorsum
and absence of dark ventral mottling.
The specimens reported here extend the known range of natator
eastward from Camotlán, Oaxaca; northward in Veracruz the species
inhabits foothills as far north as Huatusco.
Eleutherodactylus rhodopis Cope
Oaxaca: 30 km. N of Matías Romero; Río Sarabia (5);
Tapanatepec (87); Tolosita (6); between Zanatepec and
Tapanatepec. Veracruz: 25 km. SE of Jesús Carranza; 35 km.
SE of Jesús Carranza (2); 22 km. SSW of Jesús Carranza; 20
km. ENE of Jesús Carranza (7); Minatitlán; Tapalapan (5).
For the purposes of the present study I am not recognizing
Eleutherodactylus beati, E. dorsoconcolor, and E. venustus as
specifically,[Pg 55] or even subspecifically distinct from the earlier named
E. rhodopis. Probably these are mere color varieties of a single
species.
In the dry season frogs of this species were in humid forests, where
they were most frequently found along small streams and in ravines. The
species is widespread in the Gulf lowlands, but does not occur on the
Plains of Tehuantepec. It does inhabit the Pacific slopes on the
foothills of the Sierra Madre de Chiapas, the western part of which
extends into eastern Oaxaca near Tapanatepec.
Eleutherodactylus rugulosus Cope
Oaxaca: La Princesa (30); Modelo; Santa Lucía (10);
Tapanatepec (26); Tehuantepec (6); Tres Cruces (8).
Veracruz: Tapalapan (5).
In addition to the specimens from the lowlands of the isthmus, for the
purposes of the following discussion, I have included data on two
specimens from the southern slopes of the Sierra del Sur in Oaxaca
(Mirador and Chacalapa) and on several specimens from Los Tuxtlas in
Veracruz (Los Chaneques, 67; Salto de Eyipantla, 35; and San Andrés
Tuxtla, 11).
Frogs of the Eleutherodactylus rugulosus complex occur from southern
Veracruz and Sinaloa southward through Central America. Taylor
(1940:401) described E. vocalis from Hacienda El Sabino, Michoacán;
Taylor and Smith (1945:580) described E. avocalis from Tres Cruces,
Oaxaca. These have been considered as species distinct from
rugulosus, which is known to occur in Veracruz, Guerrero, and Chiapas
southward into Central America. Although the large number of specimens
collected in the isthmus does not aid in defining the ranges of the
taxa involved, these specimens do give some idea of the variation in
certain characters in a given population.
In specimens from Los Tuxtlas the tarsal fold is well-developed and
extends two-thirds to three-fourths the length of the tarsus; the
tibiotarsal articulation reaches the nostril and sometimes slightly
beyond the tip of the snout. In males the tympanum is nearly equal to
the diameter of the eye; in females it is about one-half the diameter
of the eye. The posterior surfaces of the thighs are dark brown or
black with whitish or cream-colored spots, flecks, or irregular
mottling. The tarsal fold is dark; the throat is pale in some
individuals, but in most is mottled with dark brown or gray flecks.
Individuals from La Princesa near the continental divide in Oaxaca show
the same variation in body proportions and development of the tarsal
fold. The posterior surfaces of the thighs are dark brown[Pg 56] indistinctly
mottled with lighter brown. The throat is dark brown. Specimens from
the Pacific slopes of Oaxaca, including the Plains of Tehuantepec, have
dark brown thighs mottled with dusty cream. The tibiotarsal
articulation extends slightly beyond the tip of the snout in all
specimens. In males the tympanum is equal to about two-thirds the
diameter of the eye. Duellman (1958b:6) discussed the variation in
these characters in populations in Colima, Jalisco, and Michoacán.
Until the extent of variation of these characters is known throughout
the range of rugulosus, the recognition of populations either as
species or subspecies seems superfluous. Consequently, I have used the
oldest name; this does not necessarily imply, however, that all
populations of rugulosus (sensu lato) are conspecific.
Of the 200 specimens examined, 15 have a middorsal stripe that is red
or yellow. The iris varies from a copper to a dark golden color and
shines bright red at night. Many of the specimens are juveniles; these
were collected in the dry season, at which time they were found beneath
rocks along streams, in road culverts where there was some water, and
in holes in banks and cliffs.
Microbatrachylus pygmaeus Taylor
Oaxaca: La Princesa (5); Matías Romero (9); Río Sarabia
(41); Tolosita (2). Veracruz: Jesús Carranza; 20 km. ENE
of Jesús Carranza.
The specimens listed above vary widely in color patterns; some of the
patterns are characteristic of certain named "species": albolabris,
imitator, lineatissimus, and minimus. The large series from the
Río Sarabia contains all of the color patterns; this series was
obtained in one small ravine in the rainforest. At least in the
isthmian region, this species does not inhabit the Pacific slopes and
lowlands.
Syrrhophus leprus Cope
Oaxaca: 33 km. N of Matías Romero; Santa Efigenia.
Veracruz: San Lorenzo.
Although the type locality is stated to be Santa Efigenia on the
Pacific slopes of the Sierra Madre de Chiapas in eastern Oaxaca, the
type specimen probably came from the northern slopes of the mountains.
All other known specimens are from the Gulf slopes and lowlands, and
from several localities in Los Tuxtlas. Details concerning specimens
from the isthmus and other parts of the range were given by Duellman
(1958c:8).
Smith (1947:408) reported a specimen of Syrrhophus verruculatus
Peters from San Lorenzo, Veracruz; he stated that this specimen[Pg 57] (USNM
123530) could not be S. leprus, because it had a gray belly, nor S.
cystignathoides, because of the dark and light dorsal coloration.
Firschein (1954:57) in his review of the species of Syrrhophus in
eastern México referred the specimen to S. cystignathoides. The
specimen is in poor condition. Nevertheless, specific determination is
possible. Numerous specimens of S. leprus from Los Tuxtlas have gray
bellies; some have heavier pigmentation than the specimen from San
Lorenzo. In preservative the dorsum is dark brown with lighter
mottling. There is little doubt that the specimen from San Lorenzo is a
Syrrhophus leprus, an abundant and widespread species in the humid
Gulf lowlands of southern México, and not verruculatus, if this is a
valid species (see Firschein, op. cit.:58), and not
cystignathoides, a species known from San Luis Potosí southward to
central Veracruz.
Syrrhophus pipilans pipilans Taylor
Oaxaca: Cerro Arenal; Cerro San Pedro; 6 km. N of Chivela;
14 km. W of Tehuantepec (2).
In the isthmian region this frog is known only from the Pacific slopes
and the Plains of Tehuantepec. Males call from the ground and from
trees to heights of about four meters. The call is a single, high, long
"peep."
Engystomops pustulosus Cope
Oaxaca: Chivela; La Ventosa (3); Santo Domingo;
Tapanatepec (14); Tehuantepec (61); Unión Hidalgo (62).
Veracruz: Acayucan; Cuatotolapam (7); 10 km. SE of
Hueyapan (11).
Large congregations were breeding at Tehuantepec on July 5, at
Tapanatepec on July 13, and at Hueyapan on July 24, 1956. The frogs
were breeding in open ponds in scrub forest and savanna; none was found
in the rainforest. Males call while floating on the water (Pl. 7, fig. 1);
the call is a soft "do-ing, do-ing" with a rising tone on the last
note. Numerous individual egg masses were along the bank of a pond near
Tehuantepec; one large composite egg mass there had a surface area of
about one square meter (Pl. 7, fig. 2). The large series from Unión
Hidalgo was obtained by digging specimens out of a dry sandy river bank
in the dry season. Some of the individuals were buried to a depth of 25
centimeters.
In life individuals from the Pacific lowlands were dull brown and gray;
those from Acayucan were dark chocolate brown to black with pink or red
blotches, forearms, and dorsal stripe. Not all specimens from the
Atlantic lowlands are so colored; individuals[Pg 58] from Cordoba and
Mirador, Veracruz, are like those from Tehuantepec.
Leptodactylus labialis Cope
Oaxaca: Agua Caliente; Chivela (2); Matías Romero (12); 33
km. N of Matías Romero (4); Mixtequilla; Santa Efigenia;
Tapanatepec; Tehuantepec (38); Tolosita (2); 33 km. W of
Zanatepec (49). Veracruz: Acayucan (3); Ciudad Alemán;
Cuatotolapam (10); Hueyapan; La Oaxaqueña (4); 38 km. SE of
Jesús Carranza; 20 km. ENE of Jesús Carranza; Novillero (3);
San Lorenzo (2).
Although Leptodactylus labialis does not appear to be so abundant as
Leptodactylus melanonotus, the former was found throughout the
lowlands of the isthmus. In the dry season individuals were found along
streams, and in the rainy season breeding congregations were found in
rain pools, marshes, ponds, and even small puddles. The call is a slow
"wort, wort, wort." Males call beneath the water and from beneath rocks
and from holes in the ground. The average snout-vent length of eight
adult males is 37.2 mm. A completely metamorphosed juvenile obtained at
Hueyapan on July 24, 1956, has a snout-vent length of 11 mm.
Leptodactylus melanonotus Hallowell
Oaxaca: Agua Caliente (25); Cerro Arenal (2); Cerro
Quiengola (3); Cerro San Pedro (3); Chivela (2); Coyol;
Juchitán; Matías Romero (11); Mixtequilla (2); Papaloapan
(2); Salazar (9); Salina Cruz; 11 km. S of Santiago Chivela;
Tapanatepec (17); Tehuantepec (176); Tolosita; Unión
Hidalgo; 27 km. W of Zanatepec (6). Veracruz: Acayucan;
Cuatotolapam (9); Cosoleacaque; 20 km. ENE of Jesús Carranza
(2); 20 km. SE of Minatitlán (2); Novillero; San Lorenzo
(6).
This frog is abundant throughout the lowlands of the isthmus, where in
the dry season individuals were found along streams and beneath rocks
at a spring seepage. In the rainy season males were calling from nearly
every bit of standing water. The call is a soft clicking sound
resembling that made by striking two small stones together. The average
snout-vent length of ten adult males is 41.8 mm. There is considerable
variation in the extent of the yellowish brown glandular areas on the
belly. Some have none, whereas others have a broad area on the chest, a
band along the flanks, and a thin band across the lower abdomen.
Individuals collected in the dry season vary in the same fashion as do
those collected in the rainy season, at which time they were breeding.
The glands are equally well-developed in adults of both sexes, and were
present in some juveniles with snout-vent lengths of less than 20 mm.
Apparently the development of the glands is not associated with
maturity, sex, or size.
[Pg 59]
Diaglena reticulata Taylor
Oaxaca: Cerro Arenal; Chivela; Salina Cruz (26); San
Antonio (3); Tehuantepec (2); 8.6 km. W of Tehuantepec (11);
Zarzamora.
Breeding congregations of this rare frog were found 8.6 kilometers west
of Tehuantepec on July 5, 1956, and at Salina Cruz on July 6, 1958.
Both choruses took place immediately after torrential rains. In both
instances the frogs were in and about open muddy pools in the scrub
forest (Pl. 5, fig. 2); males called from the bank near the water, and
clasping pairs were found only on land (Pl. 8, figs. 1-2). The call is
a loud, nasal "braaa," two to three seconds in duration. Amplexus is
axillary.
The dorsal ground color is light yellowish green tending towards olive
on the head and fading to yellow on the flanks. The ventral surfaces,
including the vocal sac, are white; the iris is golden and flecked with
black. The present series agrees well with the description of
reticulata (based on two specimens) given by Taylor (1942:60). A
detailed analysis of variation, comparison with related species, and
descriptions of tadpoles are reserved for a future report.
Hyla baudini Duméril and Bibron
Oaxaca: Bisilana; Cerro Quiengola (2); Cerro San Pedro;
Coyol; Matías Romero (12); Mixtequilla; Río Sarabia (7);
Salazar; San Antonio; 11 km. S of Santiago Chivela; Santo
Domingo (3); Tapanatepec (2); Tehuantepec (23); Tolosita.
Veracruz: Acayucan; Amatitlán; Ciudad Alemán (3);
Cosamaloapan (2); Cuatotolapam (15); 10 km. SE of Hueyapan;
20 km. S of Jesús Carranza; 38 km. S of Jesús Carranza (2);
20 km. ENE of Jesús Carranza (4); La Oaxaqueña (2);
Minatitlán (2); Naranja (3); Novillero (9); Río de las
Playas (2); San Lorenzo (5); Tapalapan (2).
Commonly found on both sides of the isthmus, this large tree frog
nearly always is associated with trees; it is not found in the
savannas, although it breeds in savannas adjacent to rainforest. It
appears to be somewhat more abundant in scrub forest than in
rainforest. In the daytime individuals were found under the outer
sheaths of banana plants, in the axils of leaves of elephant ears
(Xanthosoma), in cavities in trees, and on shaded limbs in the
forest. Recently metamorphosed individuals having snout-vent lengths
slightly more than 20 mm. were found in the latter part of July.
Hyla ebraccata Cope
Oaxaca: Donají (17); 43 km. N of Matías Romero (27);
Sarabia (6); Tolosita (3); Ubero (17). Veracruz: Aquilera.
This small species was found only in forested areas, where calling
males were on bushes and trees around rain pools. The call[Pg 60] is a harsh
squawk repeated at intervals of 15 to 20 seconds, followed by a minute
or more of silence, and then repeated. Clasping pairs were found on
bushes and in the water.
The dorsum bears a dark chocolate brown hour glass-shaped mark, which
in some individuals is broken into a large mark posteriorly and a
smaller triangular one on the head and nape. The dorsal ground color
varies from pale cream or ivory to yellow or tan. The intensity of the
dorsal pigmentation is subject to rather rapid change. The flanks,
hands, and anterior part of the venter are lemon yellow; the feet,
thighs, and posterior part of the venter are golden yellow. The dorsal
surface of the shank is yellow to tan with chocolate brown bars or
spots; the heel is pale yellow. There is a dark brown bar in the loreal
region and a dark brown bar extending posteriorly from the eye to a
point above the insertion of the forelimb. The iris is a copper color.
The toes are completely webbed; the fingers, one-third webbed. There is
a small axillary web that is evident when the forelimbs are at right
angles to the body. Twenty males have an average snout-vent length of
28.1 mm.; three females, 35.3 mm. There are no nuptial tuberosities on
the pollex of breeding males.
This species has been collected at Coyame and Catemaco in Los Tuxtlas
and at various localities in Tabasco; it apparently ranges eastward
from southern Veracruz, México, in humid forests to El Petén,
Guatemala.
Hyla loquax Gaige and Stuart
Oaxaca: Donají (7); 43 km. N of Matías Romero (21).
Veracruz: 19 km. N of Acayucan (4); Aquilera (3); 8 km. SW
of Coatzacoalcos (36); Cuototolapam (11); Naranja (13); San
Lorenzo (8).
In the isthmus this species is known only from the humid forests of the
Gulf lowlands; it is also known from Boca del Río, Veracruz, and from
Teapa and Villa Hermosa, Tabasco.
Calling males were found on aquatic plants above the water in deep
ponds in the forest where it was necessary for the collector to wade
waist-deep in water to obtain them. The call is a loud "hah-onk."
Individuals, when active at night, are yellowish tan above with light
olive green spots. The flanks, belly, and vocal sac are yellow, and the
anterior and posterior surfaces of the thighs and webbing of the feet
are bright orange-red or tomato red. Individuals found during the day
are grayish brown with olive markings or reddish brown with black
markings. Sleeping individuals are ivory-gray with faint gray markings.
The iris is a bright copper color. Fifteen adult males have an average
snout-vent length of[Pg 61] 41.7 mm.; they have no horny nuptial pads on the
pollex.
The relationships of this species are with Hyla rickardsi Taylor, a
species known only from the foothills of the Sierra Madre Oriental in
the states of Puebla and Veracruz. The distinguishing characteristics
of these species are given in Table 1. Living individuals may be
distinguished immediately by the flash colors on the thighs—red in
loquax and yellow in rickardsi. The calls of the two species are
distinctly different; that of rickardsi is a high-pitched, loud
rattle continued for several seconds, notably different from the
goose-like honk of loquax.
Table 1.—Comparison of Certain Characters in Hyla loquax and Hyla rickardsi
| Character | loquax | rickardsi |
|---|
| Toe webbing | Full | Three-fourths |
| Finger webbing | Three-fourths | One-half |
| Average snout-vent length (♂) | 41.7 mm. | 37.4 mm. |
| Tympanum/eye (♂) | 63.2% | 55.8% |
| Dorsal leg pattern | Barred | Unmarked |
| Tarsal fold | Tubercular | Absent |
| Tarsal stripe | Absent or indistinct | Broad, indistinct, or absent |
| Dorsolateral stripe | Absent | Present |
| Light line over anus | Broad | Narrow |
| Flash colors | Red | Yellow |
| Iris color | Copper | Bronze |
The three specimens from San Lorenzo, Veracruz (USNM 123513-5), were
identified as Hyla rickardsi by Smith (1947:409). The flash colors
have faded in preservative, and so are of no aid in identifying these
specimens. Two are adult females with snout-vent lengths of 35 and 39
mm. In possessing a relatively large tympanum and barred thighs, and in
lacking a dorsolateral stripe they are typical of loquax, but in the
amount of webbing on the hands and feet, broad tarsal stripe, and
narrow anal stripe they are like rickardsi. The third specimen, a
juvenile, has a snout-vent length of 25 mm. In coloration it resembles
the adults; it has more distinct[Pg 62] bars on the limbs. On the basis of
geography these specimens should be loquax, for the closest known
record of rickardsi is more than 200 kilometers to the northwest,
whereas loquax is known from several localities around San Lorenzo.
Shannon and Werler (1955:383) described Hyla axillamembrana from the
lower southern slopes of Los Tuxtlas. The unique type is a small male
(27 mm. snout-vent). I have examined the type and find no great
differences between it and small specimens of loquax. It is not
possible to determine the color of the thighs, nor was this information
given in the description. Hyla axillamembrana is here considered to
be a synonym of Hyla loquax.
Hyla microcephala martini Smith
Oaxaca: Donají (15); 43 km. N of Matías Romero (19); Río
Sarabia (2); Sarabia (11); Tolosita. Veracruz: Acayucan
(17); Alvarado (41); Aquilera (21); 8 km. SW of
Coatzacoalcos (10); Cosoleacaque (26); 10 km. SE of
Hueyapan; Naranja (3); Novillero.
This frog is abundant in the Gulf lowlands of the isthmus, where large
breeding congregations were found in grassy ponds on the savannas and
in openings in the forest. Most frequently males were calling from
grasses and reeds in the ponds; many individuals were perched
precariously on thin blades as high as one meter above the water. The
call is a series of low squeaks.
Individuals found at night were pale yellow above with light brown
lines arranged in an irregular pattern on the back, but often forming a
cross or an X-shaped mark in the scapular region. There is a brown
stripe from the nostril to the eye and thence to the groin. Anteriorly
this stripe is bordered above by a thin white or cream-colored line.
Numerous small brown flecks are scattered on the back and dorsal
surface of the shank. In most specimens there are thin transverse brown
bars on the shank. The thighs and undersides of the limbs are golden
yellow; the belly and vocal sac are lemon yellow. The iris is yellowish
brown. During the day individuals assume a pale reddish tan ground
color with darker brown markings. Twenty-five adult males from Alvarado
have an average snout-vent length of 24.1 mm.
Hyla picta Günther
Oaxaca: Donají (8); Sarabia (11); Tolosita (15); Ubero
(6). Veracruz: 19 km. N of Acayucan (4); Alvarado (5);
Aquilera; 8 km. SW of Coatzacoalcos; 10 km. SE of Hueyapan
(7); Lerdo de Tejada; Tula (3).
Widespread in the forests, scrub, and savannas on the Gulf lowlands of
the isthmus, these frogs were found breeding at numerous localities.
Males call from grasses and bushes growing in and[Pg 63] about ponds. The
call is a high-pitched insect-like trill. At night these frogs are pale
yellow above; they change to light grayish tan during the day. A dark
stripe extends from the nostril to the eye and thence posteriorly to a
point between the axilla and groin. Above this dark stripe is a broader
white stripe. Scattered on the dorsum are brown flecks or spots; the
shanks are marked with poorly-defined cross-bars. The thighs are deep
yellow below and paler above with scattered dark flecks. The belly is
white, and the vocal sac is yellow. The iris is golden. Twenty males
have an average snout-vent length of 21.5 mm.; three females, 24.0 mm.
Hyla robertmertensi Taylor
Oaxaca: Tapanatepec (28); 7.5 km. NW of Tapanatepec (38);
7.2 km. WNW of Zanatepec (77).
This species was found in the isthmian region only on the Pacific
lowlands at the southern base of the western part of the Sierra Madre
de Chiapas. On July 13, 1956, many large choruses were discovered. The
calling males were on reeds and thorn scrub in and at the edge of
temporary ponds; the call is a cricket-like "creak-creack," quickly
followed by a series of notes "creak-eek-eek-eek-eek."
At night the dorsal ground color is pale yellow; this changes to
pinkish buff during the day. There is a grayish or brown dark stripe
from the nostril to the eye; the stripe continues to the groin. This
dark stripe is bordered above by a narrow white stripe. The belly is
white, and the vocal sac is yellow. The iris is dull reddish brown.
Twenty-five males have an average snout-vent length of 24.7 mm.
Hyla staufferi Cope
Oaxaca: Chivela; Huilotepec (5); Juchitán (4); Matías
Romero (4); 25 km. N of Matías Romero; Mixtequilla (4); Río
Sarabia (11); 11 km. S of Santiago Chivela; Sarabia (3);
Tapanatepec (67); Tehuantepec (66); Tolosita (2); Ubero;
Unión Hidalgo; Zanatepec (6). Veracruz: Acayucan (7);
Alvarado (3); Amatitlán; Aquilera; Ciudad Alemán (3); 8 km.
SW of Coatzacoalcos (9); Cosamaloapan (4); Cosoleacaque (8);
10 km. SE of Hueyapan; Lerdo de Tejada; Novillero (6); Tula
(2).
This is the only species of small hylid that crosses the isthmus.
Calling males were found in and about ponds on the savannas in southern
Veracruz, in ponds in open forest in northern Oaxaca (not in forest
pools), and in temporary pools in the scrub forest on the Pacific
lowlands. Individuals usually called from bushes and reeds in or at the
edge of ponds. The call is a short "braaa." Dates of breeding choruses
indicate that by the time the other small species of hylids in the Gulf
lowlands reach the peak of their[Pg 64] breeding season, that of H.
staufferi is essentially over; no large breeding congregations were
found in July. On July 8, 1956, two metamorphosing young were found
clinging to blades of grass in a pond; they had snout-vent lengths of 8
and 9 mm. and tail stumps less than 3 mm. in length. Others were found
on July 13 and 26. The juveniles are nearly unicolor olive green above
and white below.
In life the adults vary greatly in color pattern. The dorsal ground
color is yellowish tan to olive brown with olive brown or dark brown
spots, some of which in certain individuals are connected to form
longitudinal dark stripes. On the posterior surface of the thighs are
small white flecks. The belly is white, and the vocal sac is a rich
yellow. Twenty males have an average snout-vent length of 26.3 mm.;
they have no horny nuptial pads. No noticeable differences in either
color or body proportions were found between the populations on either
side of the isthmus.
Hylella sumichrasti Brocchi
Oaxaca: Cerro Arenal (5); Cerro San Pedro (2); Escurano;
La Concepción (41); Portillo Los Nanches (6); San Antonio
(16); 11 km. S of Santiago Chivela (18); Santa Lucía (7);
Tapanatepec (5); Tehuantepec (8); Tenango (49); Tres Cruces
(19).
With the exception of the series from 11 kilometers south of Santiago
Chivela, most of these specimens were found in small arboreal
bromeliads during the dry season. Males were found along a clear,
shallow, rocky stream south of Santiago Chivela on July 6, 1956. The
frogs were calling from bushes and rocks in and along the stream. When
disturbed, they jumped into the water and floated downstream until they
were able to hold onto a rock or other object. The call is a loud
"bra-a-ah." In breeding individuals the dorsum is pale yellow; the
belly is white, and the vocal sac is yellow. The iris is pale golden
yellow. Eighteen males have an average snout-vent length of 25.2 mm.
All have dark brown nuptial tuberosities on the pollex.
Certain diagnostic characters of this species as given by Taylor
(1943a:50) and Taylor and Smith (1945:598) are in need of revision.
Hylella sumichrasti has been characterized as having no vocal sac,
rarely having vomerine teeth, and as having a relatively smooth throat.
The vocal sac in breeding males is quite evident; it is single, median,
and when expanded, spherical. The openings into the vocal sac are
narrow slits along the inner posterior border of the jaw rami. Of 151
specimens studied, 74 have vomerine[Pg 65] ridges between the choanae, and 36
of these have one to three teeth on each ridge. The belly and
undersurfaces of the thighs are granular; the throat is only somewhat
less so. The granular condition may be correlated with breeding, for
specimens obtained from bromeliads in the dry season had rather smooth
throats. It seems that the vocal sac atrophys in the non-breeding
season. These seasonal changes may account for the diagnoses given by
Taylor (op. cit.) and Taylor and Smith (op. cit.); likewise, since
many of the specimens obtained by Smith in the dry season were
juveniles and subadults, the development of the vomerine ridges could
not be diagnosed properly.
The range of this species encompasses the Pacific slopes of the Isthmus
of Tehuantepec eastward to the upper Cintalapa Valley and vicinity of
Tonalá in western Chiapas. Priscilla Starrett collected tadpoles of H.
sumichrasti from a stream 19 km. N of Arriaga, Chiapas. These limited
observations on the ecology of this frog suggest that it breeds in the
fast-moving streams of the Pacific slopes, and that it seeks shelter in
arboreal bromeliads during the dry season.
Phrynohyas modesta Taylor and Smith
Oaxaca: Tuxtepec. Veracruz: 20 km. S of Jesús Carranza;
20 km. ENE of Jesús Carranza (2); Minatitlán.
I have not collected this species in the isthmus. The locality records
indicate that the range is discontinuous (Duellman, 1956:27). The
species occurs on the humid Pacific slopes from south-central Chiapas
eastward to El Salvador and on the humid Gulf lowlands from southern
Veracruz eastward into Tabasco, but is unknown from the dry Pacific
slopes and plains in the isthmus.
The acquisition of several specimens of this species in southern
Veracruz, Tabasco, and Oaxaca, together with a knowledge of the
variation displayed by Phrynohyas spilomma, suggests that modesta
may be a color variety of spilomma.
Phrynohyas spilomma Cope
Oaxaca: Tapanatepec (3). Veracruz: Amatitlán (12);
Chacaltianguis (2); Ciudad Alemán (6); Cosamaloapan;
Novillero (3).
Like the preceding species, this frog is unknown from the arid Pacific
lowlands of the isthmus; its presence at Tapanatepec, a locality
situated in more mesic conditions than prevail on the Plains of
Tehuantepec, indicates that it may have a distribution on the Pacific
slopes much like that of P. modesta. Furthermore, this frog[Pg 66] was not
detected in the rainforests of the Gulf lowlands; in that region it was
found only in scrub forest and savanna.
On July 26, 1956, numerous choruses of these frogs were heard between
Ciudad Alemán and Tlacotalpan, Veracruz. The call is a loud, nasal
"grawl" repeated continuously. The males call from the water. Several
clasping pairs were found in shallow grassy ponds amidst the scrub
forest. The ground color varies from reddish brown to tan with dark
brown dorsal markings. The iris is golden, and the vocal sacs are dark
olive brown. After a light shower during the dry season, six
individuals were found on the low branches of trees at night near
Ciudad Alemán.
Phyllomedusa callidryas taylori Funkhouser
Oaxaca: Donají (9); Sarabia (8); Tolosita (6); Ubero (27).
Veracruz: Alvarado (7); Aquilera; Berta; Coatzacoalcos
(9); 10 km. SE of Hueyapan (5); Naranja (17).
In life this frog presents a striking array of colors. The dorsum
varies from pale green to dark olive green; there may be scattered
whitish or cream-colored spots on the back. On the flanks are bright
yellow to deep cream-colored vertical bars separated by pale blue or
purple interspaces. The thighs and undersurfaces of the hind limbs are
golden orange; the belly is yellow, and the throat is cream-colored.
The iris is crimson; the transparent part of the lower eyelid has
golden reticulations. When the frog is resting, the forefeet are folded
beneath the throat, and the limbs are folded tightly against the body.
In this position and with the eyes closed and head flattened, this
gaudy frog assumes the appearance of a small elliptical green leaf.
Throughout the month of July, 1956, Phyllomedusa was breeding in
ponds in or adjacent to the rainforest in northern Oaxaca and in
southern Veracruz. Only at Alvarado was it found breeding in a grassy
pond. Males and females alike were found on bushes and trees in and
around the ponds. The call is a single "wank." Amplexing males continue
to call, but the call is softer and less nasal in quality. The eggs are
encased in pale green gelatin and attached to leaves on branches
overhanging the water. Three egg clutches contained 38, 41, and 46
eggs.
Phyllomedusa dacnicolor Cope
Oaxaca: Escurano; Tehuantepec.
Although it is abundant on the Pacific lowlands to the northwest in
Guerrero, Michoacán, and Colima, this species is known only from two
specimens from Tehuantepec. There is no apparent[Pg 67] physical barrier to
their distribution in the isthmus; in the Balsas Basin the species
lives in a hotter, more arid environment than that at Tehuantepec.
Gastrophryne usta Cope
Oaxaca: Santa Efigenia; Tehuantepec (10); 24 km. W of
Tehuantepec; Tolosita (2). Veracruz: Ayentes (6); La
Oaxaqueña; Novillero (2); San Lorenzo.
Calling males were found in open scrub forest near Tehuantepec and in
savannas near Novillero. The specimens from Tolosita were found under
cover in a clearing in the forest (Fugler and Webb, 1957:106).
Specimens from the Pacific lowlands are typical of Gastrophryne usta
gadowi Boulenger in possessing a thin line on the posterior surface of
the thighs and a thin line from the snout to the vent. Of nine
specimens from the Gulf lowlands (Ayentes, Novillero, and San Lorenzo),
seven have a middorsal line; this is narrow in four and wide in three.
Five have the stripes on the thighs. Two specimens from the middle of
the isthmus (Tolosita) have no stripes on the thighs; one has a thin
middorsal line, and the other has a broad line. The adult males have a
black throat; females have a mottled one. The brown reticulations on
the bellies of specimens from the Gulf lowlands is bolder than on
specimens from the Pacific lowlands. The presence of certain characters
supposedly diagnostic of the subspecies gadowi (line on dorsum and
thighs) in the population of usta in southern Veracruz suggests that
a redefinition of the ranges of these subspecies will be in order when
sufficient material is available to delimit them accurately. For the
present I prefer to consider all specimens from the isthmus solely as
Gastrophryne usta without referring them to subspecies.
Rana palmipes Spix
Oaxaca: Matías Romero (11); 11 km. S of Santiago Chivela;
Santo Domingo; Sarabia. Veracruz: Coatzacoalcos;
Cuatotolapam; 25 km. SE of Jesús Carranza (4); Tlacotalpan
(2); Tula.
Adults were found along streams and in marshes in savannas and
rainforest. These frogs are wary and difficult to capture, even at
night. Rana palmipes is another species that has a discontinuous
distribution in the isthmus. The species does not occur on the Pacific
lowlands of the isthmus, but does occur on the more humid Pacific
slopes of Chiapas and Guatemala.
Tadpoles were found in a small sluggish tributary to the Río Sarabia.
[Pg 68]
Rana pipiens Schreber
Oaxaca: Agua Caliente; Cerro Quiengola; Escurano (14); Río
Sarabia (2); Tapanatepec (5); Tehuantepec (24). Veracruz:
Acayucan; Cuatotolapam (15); Jesús Carranza (2); 20 km. S of
Jesús Carranza (11); 25 km. SE of Jesús Carranza; 20 km. ENE
of Jesús Carranza (10); San Lorenzo (10).
As in most other places in México and northern Central America, this
species occurs wherever there is permanent water. Males were heard
calling from woodland ponds and from savanna ponds.
SUMMARY
Investigations of the amphibians and their environments in the Isthmus
of Tehuantepec have been presented with the aim of gaining an
understanding of the present biological and of the historical events
responsible for the present patterns of distribution of amphibians in
this region.
The Isthmus of Tehuantepec embraces three major environments—savanna,
semi-arid scrub forest, and quasi-rainforest. The rainforest presents
an environment noticeably different from the other two and has a
different amphibian fauna.
Analysis of present patterns of distribution shows that certain species
are restricted to the rainforests on the Gulf lowlands; others live
only in the semi-arid scrub forests on the Pacific lowlands. A third
group of species lives on both the Gulf and Pacific lowlands; most of
these species occur only in the scrub forests or savannas on the Gulf
lowlands, but some also inhabit the rainforest. In one way or another
the isthmus presents a barrier to the distribution of 75 per cent of
the species of amphibians living in the lowlands; it is a greater
barrier still to the species inhabiting the highlands on either side.
Present patterns of distribution are attributed to bioclimatic
fluctuation in the Pleistocene. In the course of these climatic shifts,
tropical environments and their amphibian inhabitants seem to have
survived in the isthmian region.
The amphibian fauna of the lowlands of the Isthmus of Tehuantepec
consists of 16 genera and 36 species. Systematic studies of all
available specimens from the region show that Eleutherodactylus
conspicuus Taylor and Smith is a synonym of Eleutherodactylus
alfredi Boulenger and that Hyla axillamembrana Shannon and Werler is
a synonym of Hyla loquax Gaige and Stuart.
[Pg 69]
LITERATURE CITED
Beard, J. S.
1953. The savanna vegetation of northern tropical America.
Ecol. Mono., vol. 23 (2):149-215.
Boulenger, G. A.
1898. Fourth report on additions to the batrachian
collection in the Natural History Museum. Proc. Zool. Soc.
London, 1898, pp. 473-482, pls. 38-39.
Burt, C. E.
1931. A study of the teiid lizards of the genus
Cnemidophorus with special reference to their phylogenetic
relationships. Bull. U. S. Nat. Mus., No. 154, pp. viii +
286.
Contreras A., A.
1942. Mapa de las provincias climatologias de la Republica
Mexicana. México, D. F., Dirección de Geografía, Meterología
e Hidrología, pp. i-xxviii, tables 1-54, 2 maps.
Cooke, C. W.
1945. Geology of Florida. Florida Geol. Surv., Geol. Bull.,
No. 29:1-339.
Dorf, E.
1959. Climatic changes of the past and present. Contrib.
Mus. Paleo. Univ. Michigan, vol. 13 (8):181-210.
Duellman, W. E.
1956. The frogs of the hylid genus Phrynohyas Fitzinger,
1843. Miscl. Publ. Mus. Zool. Univ. Michigan, No. 96:1-47,
pls. 1-6.
1958a. A monographic study of the colubrid snake genus
Leptodeira. Bull. Amer. Mus. Nat. Hist., vol. 114:1-152,
pls. 1-31.
1958b. A preliminary analysis of the herpetofauna of Colima,
Mexico. Occas. Papers Mus. Zool. Univ. Michigan, No.
589:1-22.
1958c. A review of the frogs of the genus Syrrhophus in
western Mexico. Occas. Papers Mus. Zool. Univ. Michigan, No.
594:1-15, pls. 1-3.
Duellman, W. E., and Schwartz, A.
1958. Amphibians and reptiles of southern Florida. Bull.
Florida State Mus., vol. 3 (5):181-324.
Dunn, E. R.
1942. The American caecilians. Bull. Mus. Comp. Zool., vol.
91 (6):439-540.
Durham, J. W., Arellano, A. R. V., and Peck, Jr., J. H.
1952. No Cenozoic Tehuantepec seaways. Bull. Geol. Soc.
Amer., vol. 63:1245.
Firschein, I. L.
1950. A new toad from Mexico with a redefinition of the
cristatus group. Copeia, 1950 (2):81-87, pl. 1.
1954. Definition of some little-understood members of the
leptodactylid genus Syrrhophus, with a description of a
new species. Copeia, 1954 (1):48-58.
Firschein, I. L., and Smith, H. M.
1957. A high-crested race of toad (Bufo valliceps) and
other noteworthy reptiles and amphibians from southern
Mexico. Herpetologica, vol. 13 (3):219-222.
Fugler, C. M., and Webb, R. G.
1957. Some noteworthy reptiles and amphibians from the
states of Oaxaca and Veracruz. Herpetologica, vol. 13
(2):103-108.
Gloyd, H. K.
1940. The rattlesnakes, genera Sistrurus and Crotalus.
Chicago Acad. Sci. Special Publ., No. 4, vii + 266 pp., pls.
1-31.
[Pg 70]
Goin, C. J.
1958. Comments upon the origin of the herpetofauna of
Florida. Quart. Jour. Florida Acad. Sci., vol. 21 (1):61-70.
Goldman, E. A.
1951. Biological investigations in Mexico. Smithsonian Misc.
Publ., vol. 115, xiii + 476 pp., pls. 1-71.
Goodnight, C. J., and Goodnight, M. L.
1956. Some observations in a tropical rain forest in
Chiapas, Mexico. Ecology, vol. 37 (1):139-150.
Griscom, L.
1932. The distribution of bird-life in Guatemala. Bull.
Amer. Mus. Nat. Hist., vol. 64:1-439.
1950. Distribution and origin of the birds of Mexico. Bull.
Mus. Comp. Zool., vol. 103:341-382.
Hartweg, N., and Oliver, J. A.
1940. A contribution to the herpetology of the Isthmus of
Tehuantepec. IV. An annotated list of the amphibians and
reptiles collected on the Pacific slope during the summer of
1936. Misc. Publ. Mus. Zool. Univ. Michigan, No. 47:1-31.
Hubbell, T. H.
1954. Relationships and distribution of Mycotrupes. In The
burrowing beetles of the genus Mycotrupes, Olson, A. L.,
Hubbell, T. H., and Howden, H. F. Misc. Publ. Mus. Zool.
Univ. Michigan, No. 84:1-59, pls. 1-8.
Hutchinson, G. E., Patrick, R., and Deevey, E. S.
1956. Sediments of Lake Patzcuaro, Michoacán, Mexico. Bull.
Geol. Soc. Amer., vol. 67:1491-1504.
Langebartel, D. A., and Smith, P. W.
1959. Noteworthy records of amphibians and reptiles from
eastern Mexico. Herpetologica, vol. 15 (1):27-29.
Martin, P. S.
1958. Pleistocene ecology and biogeography of North America.
Zoogeography. Amer. Assoc. Advanc. Sci., Publ. No.
51:375-420.
Martin, P. S., and Harrell, B. E.
1957. The Pleistocene history of temperate biotas in Mexico
and eastern United States. Ecology, vol. 38:468-480.
Oliver, J. A.
1948. The relationships and zoogeography of the genus
Thalerophis Oliver. Bull. Amer. Mus. Nat. Hist., vol.
92:157-280, pls. 16-19.
Olson, E. C., and McGrew, P. O.
1941. Mammalian fauna from the Pliocene of Honduras. Bull.
Geol. Soc. Amer., vol. 52:1219-1244.
Ruthven, A. G.
1912. The amphibians and reptiles collected by the
University of Michigan—Walker Expedition in southern Vera
Cruz, Mexico. Zool. Jahrbuch, vol. 32 (4):295-332, pls.
6-11.
Schuchert, C.
1935. Historical geology of the Antillean-Caribbean region.
New York, xxvi + 811 pp.
Sears, P. B., Foreman, F., and Clisby, K. H.
1955. Palynology in southern North America. Bull. Geol. Soc.
Amer., vol. 66:471-530.
Shannon, F. A., and Werler, J.
1955. Notes on amphibians of the Los Tuxtlas Range of
Veracruz, Mexico. Trans. Kansas Acad. Sci., vol. 58
(3):360-386.
Shreve, B.
1957. Reptiles and amphibians from the Selva Lacandona. In
Biological Investigations in the Selva Lacandona, Chiapas,
Mexico. Paynter, R. A. (editor). Bull. Mus. Comp. Zool.,
vol. 116 (4):193-298.
[Pg 71]
Smith, H. M.
1947. Notes on Mexican amphibians and reptiles. Jour.
Washington Acad. Sci., vol. 37:408-412.
Smith, H. M., and Laufe, L. E.
1946. A summary of Mexican Lizards of the genus Ameiva.
Univ. Kansas Sci. Bull., vol. 31 (2):7-73.
Smith, H. M., and Taylor, E. H.
1950. An annotated checklist and key to the reptiles of
Mexico exclusive of the snakes. Bull. U. S. Natl. Mus., No.
199, v + 253 pp.
Stirton, R. A.
1954. Late Miocene mammals from Oaxaca, Mexico. Amer. Jour.
Sci., No. 252:634-638.
Stuart, L. C.
1935. A contribution to a knowledge of the herpetology of a
portion of the savanna region of central Petén, Guatemala.
Misc. Publ. Mus. Zool. Univ. Michigan, No. 29:1-56, pls.
1-4.
1941. Studies of Neotropical Colubridae VIII. A revision of
the genus Dryadophis Stuart, 1939. Misc. Publ. Mus. Zool.
Univ. Michigan, No. 49:1-106, pls. 1-4.
1951. The herpetofauna of the Guatemalan Plateau, with
special reference to its distribution on the southwestern
highlands. Contrib. Lab. Vertebrate Biol., Univ. Michigan,
No. 49:1-71, pls. 1-7.
1954. A description of a subhumid corridor across northern
Central America, with comments on its herpetofaunal
indicators. Contrib. Lab. Vertebrate Biol., Univ. Michigan,
No. 65:1-26, pls. 1-6.
1958. A study of the herpetofauna of the Uaxactun-Tikal area
of northern El Petén, Guatemala. Contrib. Lab. Vertebrate
Biol., Univ. Michigan, No. 75:1-30.
Taylor, E. H.
1940. New species of Mexican Anura. Univ. Kansas Sci. Bull.,
vol. 26 (11):385-405.
1941. New amphibians from the Hobart M. Smith Mexican
collections. Univ. Kansas Sci. Bull., vol. 27 (8):141-167.
1942. The frog genus Diaglena with a description of a new
species. Univ. Kansas Sci. Bull., vol. 28 (4):57-65.
1943a. A new Hylella from Mexico. Proc. Biol. Soc.
Washington, vol. 56:49-52.
1943b. Herpetological novelties from Mexico. Univ. Kansas
Sci. Bull., vol. 29 (8):343-361.
Taylor, E. H., and Smith, H. M.
1945. Summary of the collections of amphibians made in
Mexico under the Walter Rathbone Bacon Traveling
Scholarship. Proc. U. S. Natl. Mus., vol. 95:521-613, pls.
18-32.
Trask, P. D., Phleger, F. B., and Stetson, H. C.
1947. Recent changes in sedimentation in the Gulf of Mexico.
Science, vol. 106:460-461.
Weyl, R.
1955. Vestigios de una glaciacion del Pleistoceno en la
Cordillera de Talamanca, Costa Rica, A. C. Informe
Trimestral, Inst. Geog. de Costa Rica, pp. 9-32.
White, S. E.
1956. Probable substages of glaciation on Ixtaccihuatl,
Mexico. Jour. Geol., vol. 64:289-295.
Williams, L.
1939. Arboles y arbustos del Istmos de Tehuantepec, Mexico.
Lilloa., vol. 4:137-171.
Transmitted May 23, 1960.
[Pg 72]
UNIVERSITY OF KANSAS PUBLICATIONS MUSEUM OF NATURAL HISTORY
Institutional libraries interested in publications exchange may obtain
this series by addressing the Exchange Librarian, University of Kansas
Library, Lawrence, Kansas. Copies for individuals, persons working in a
particular field of study, may be obtained by addressing instead the
Museum of Natural History, University of Kansas, Lawrence, Kansas.
There is no provision for sale of this series by the University
Library, which meets institutional requests, or by the Museum of
Natural History, which meets the requests of individuals. However, when
individuals request copies from the Museum, 25 cents should be
included, for each separate number that is 100 pages or more in length,
for the purpose of defraying the costs of wrapping and mailing.
* An asterisk designates those numbers of which the Museum's supply
(not the Library's supply) is exhausted. Numbers published to date, in
this series, are as follows:
Vol. 1. Nos. 1-26 and index. Pp. 1-638, 1946-1950.
*Vol. 2. (Complete) Mammals of Washington. By Walter W. Dalquest.
Pp. 1-444, 140 figures in text. April 9, 1948.
Vol. 3. *1. The avifauna of Micronesia, its origin, evolution, and
distribution. By Rollin H. Baker. Pp. 1-359, 16 figures
in text. June 12, 1951.
*2. A quantitative study of the nocturnal migration of birds.
By George H. Lowery, Jr. Pp. 361-472, 47 figures in text.
June 29, 1951.
3. Phylogeny of the waxwings and allied birds. By M. Dale
Arvey. Pp. 473-530, 49 figures in text, 13 tables.
October 10, 1951.
4. Birds from the state of Veracruz, Mexico. By George H.
Lowery, Jr., and Walter W. Dalquest. Pp. 531-649, 7
figures in text, 2 tables. October 10, 1951.
Index. Pp. 651-681.
*Vol. 4. (Complete) American weasels. By E. Raymond Hall. Pp. 1-466,
41 plates, 31 figures in text. December 27, 1951.
Vol. 5. Nos. 1-37 and index. Pp. 1-676, 1951-1953.
*Vol. 6. (Complete) Mammals of Utah, taxonomy and distribution. By
Stephen D. Durrant. Pp. 1-549, 91 figures in text, 80
tables. August 10, 1952.
Vol. 7. *1. Mammals of Kansas. By E. Lendell Cockrum. Pp. 1-303, 73
figures in text, 37 tables. August 25, 1952.
2. Ecology of the opossum on a natural area in northeastern
Kansas. By Henry S. Fitch and Lewis L. Sandidge. Pp.
305-338, 5 figures in text. August 24, 1953.
3. The silky pocket mice (Perognathus flavus) of Mexico. By
Rollin H. Baker. Pp. 339-347, 1 figure in text. February
15, 1954.
4. North American jumping mice (Genus Zapus). By Philip H.
Krutzsch. Pp. 349-472, 47 figures in text, 4 tables.
April 21, 1954.
5. Mammals from Southeastern Alaska. By Rollin H. Baker and
James S. Findley. Pp. 473-477. April 21, 1954.
6. Distribution of Some Nebraskan Mammals. By J. Knox Jones,
Jr. Pp. 479-487. April 21, 1954.
7. Subspeciation in the montane meadow mouse. Microtus
montanus, in Wyoming and Colorado. By Sydney Anderson.
Pp. 489-506, 2 figures in text. July 23, 1954.
8. A new subspecies of bat (Myotis velifer) from southeastern
California and Arizona. By Terry A. Vaughan. Pp. 507-512.
July 23, 1954.
9. Mammals of the San Gabriel mountains of California. By
Terry A. Vaughan. Pp. 513-582, 1 figure in text, 12
tables. November 15, 1954.
10. A new bat (Genus Pipistrellus) from northeastern Mexico.
By Rollin H. Baker. Pp. 583-586. November 15, 1954.
11. A new subspecies of pocket mouse from Kansas. By E.
Raymond Hall. Pp. 587-590. November 15, 1954.
12. Geographic variation in the pocket gopher, Cratogeomys
castanops, in Coahuila, Mexico. By Robert J. Russell and
Rollin H. Baker. Pp. 591-608. March 15, 1955.
13. A new cottontail (Sylvilagus floridanus) from northeastern
Mexico. By Rollin H. Baker. Pp. 609-612. April 8, 1955.
14. Taxonomy and distribution of some American shrews. By
James S. Findley. Pp. 613-618. June 10, 1955.
15. The pigmy woodrat, Neotoma goldmani, its distribution and
systematic position. By Dennis G. Rainey and Rollin H.
Baker. Pp. 619-624. 2 figures in text. June 10, 1955.
Index. Pp. 625-651.
Vol. 8. 1. Life history and ecology of the five-lined skink, Eumeces
fasciatus. By Henry S. Fitch. Pp. 1-156, 26 figures in
text. September 1, 1954.
2. Myology and serology of the Avian Family Fringillidae, a
taxonomic study. By William B. Stallcup. Pp. 157-211, 23
figures in text, 4 tables. November 15, 1954.
3. An ecological study of the collared lizard (Crotaphytus
collaris). By Henry S. Fitch. Pp. 213-274, 10 figures in
text. February 10, 1956.
4. A field study of the Kansas ant-eating frog, Gastrophryne
olivacea. By Henry S. Fitch. Pp. 275-306, 9 figures in
text. February 10, 1956.
5. Check-list of the birds of Kansas. By Harrison B. Tordoff.
Pp. 307-359, 1 figure in text. March 10, 1956.
6. A population study of the prairie vole (Microtus
ochrogaster) in northeastern Kansas. By Edwin P. Martin.
Pp. 361-416, 19 figures in text. April 2, 1956.
7. Temperature responses in free-living amphibians and
reptiles of northeastern Kansas. By Henry S. Fitch. Pp.
417-476, 10 figures in text, 6 tables. June 1, 1956.
8. Food of the crow, Corvus brachyrhynchos Brehm, in
south-central Kansas. By Dwight Platt. Pp. 477-498, 4
tables. June 8, 1956.
9. Ecological observations on the woodrat, Neotoma floridana.
By Henry S. Fitch and Dennis G. Rainey. Pp. 499-533, 3
figures in text. June 12, 1956.
10. Eastern woodrat, Neotoma floridana: Life history and
ecology. By Dennis G. Rainey. Pp. 535-646, 12 plates, 13
figures in text. August 15, 1956.
Index. Pp. 647-675.
Vol. 9. 1. Speciation of the wandering shrew. By James S. Findley.
Pp. 1-68, 18 figures in text. December 10, 1955.
2. Additional records and extensions of ranges of mammals
from Utah. By Stephen D. Durrant, M. Raymond Lee, and
Richard M. Hansen. Pp. 69-80. December 10, 1955.
3. A new long-eared myotis (Myotis evotis) from northeastern
Mexico. By Rollin H. Baker and Howard J. Stains. Pp.
81-84. December 10, 1955.
4. Subspeciation in the meadow mouse, Microtus pennsylvanicus,
in Wyoming. By Sydney Anderson. Pp. 85-104, 2 figures in
text. May 10, 1956.
5. The condylarth genus Ellipsodon. By Robert W. Wilson.
Pp. 105-116, 6 figures in text. May 19, 1956.
6. Additional remains of the multituberculate genus
Eucosmodon. By Robert W. Wilson. Pp. 117-123, 10 figures
in text. May 19, 1956.
7. Mammals of Coahuila, Mexico. By Rollin H. Baker. Pp.
125-335, 75 figures in text. June 15, 1956.
8. Comments on the taxonomic status of Apodemus peninsulae,
with description of a new subspecies from North China. By
J. Knox Jones, Jr. Pp. 337-346, 1 figure in text, 1
table. August 15, 1956.
9. Extensions of known ranges of Mexican bats. By Sydney
Anderson. Pp. 347-351. August 15, 1956.
10. A new bat (Genus Leptonycteris) from Coahuila. By Howard
J. Stains. Pp. 353-356. January 21, 1957.
11. A new species of pocket gopher (Genus Pappogeomys) from
Jalisco, Mexico. By Robert J. Russell. Pp. 357-361.
January 21, 1957.
12. Geographic variation in the pocket gopher, Thomomys
bottae, in Colorado. By Phillip M. Youngman. Pp. 363-384,
7 figures in text. February 21, 1958.
13. New bog lemming (genus Synaptomys) from Nebraska. By J.
Knox Jones, Jr. Pp. 385-388. May 12, 1958.
14. Pleistocene bats from San Josecito Cave, Nuevo León,
México. By J. Knox Jones, Jr. Pp. 389-396. December 19,
1958.
15. New Subspecies of the rodent Baiomys from Central America.
By Robert L. Packard. Pp. 397-404. December 19, 1958.
16. Mammals of the Grand Mesa, Colorado. By Sydney Anderson.
Pp. 405-414, 1 figure in text. May 20, 1959.
17. Distribution, variation, and relationships of the montane
vole, Microtus montanus. By Emil K. Urban. Pp. 415-511.
12 figures in text, 2 tables. August 1, 1959.
18. Conspecificity of two pocket mice, Perognathus goldmani
and P. artus. By E. Raymond Hall and Marilyn Bailey
Ogilvie. Pp. 513-518, 1 map. January 14, 1960.
19. Records of harvest mice, Reithrodontomys, from Central
America, with description of a new subspecies from
Nicaragua. By Sydney Anderson and J. Knox Jones, Jr. Pp.
519-529. January 14, 1960.
20. Small carnivores from San Josecito Cave (Pleistocene),
Nuevo León, México. By E. Raymond Hall. Pp. 531-538, 1
figure in text. January 14, 1960.
21. Pleistocene pocket gophers from San Josecito Cave, Nuevo
León, México. By Robert J. Russell. Pp. 539-548, 1 figure
in text. January 14, 1960.
22. Review of the insectivores of Korea. By J. Knox Jones,
Jr., and David H. Johnson. Pp. 549-578. February 23, 1960.
23. Speciation and evolution of the pygmy mice, genus Baiomys.
By Robert L. Packard. Pp. 579-670, 4 plates, 12 figures in
text. June 16, 1960.
Index Pp. 671-690.
Vol. 10. 1. Studies of birds killed in nocturnal migration. By
Harrison B. Tordoff and Robert M. Mengel. Pp. 1-44, 6
figures in text, 2 tables. September 12, 1956.
2. Comparative breeding behavior of Ammospiza caudacuta and
A. maritima. By Glen E. Woolfenden. Pp. 45-75, 6 plates,
1 figure. December 20, 1956.
3. The forest habitat of the University of Kansas Natural
History Reservation. By Henry S. Fitch and Ronald R.
McGregor. Pp. 77-127, 2 plates, 7 figures in text, 4
tables. December 31, 1956.
4. Aspects of reproduction and development in the prairie
vole (Microtus ochrogaster). By Henry S. Fitch. Pp.
129-161, 8 figures in text, 4 tables. December 19, 1957.
5. Birds found on the Arctic slope of northern Alaska. By
James W. Bee. Pp. 163-211, pls. 9-10, 1 figure in text.
March 12, 1958.
6. The wood rats of Colorado: distribution and ecology. By
Robert B. Finley, Jr. Pp. 213-552, 34 plates, 8 figures
in text, 35 tables. November 7, 1958.
7. Home ranges and movements of the eastern cottontail in
Kansas. By Donald W. Janes. Pp. 553-572, 4 plates, 3
figures in text. May 4, 1959.
8. Natural history of the salamander, Aneides hardyi. By
Richard F. Johnston and Schad Gerhard. Pp. 573-585.
October 8, 1959.
9. A new subspecies of lizard, Cnemidophorus sacki, from
Michoacán, México. By William E. Duellman. Pp. 587-598,
2 figures in text. May 2, 1960.
10. A taxonomic study of the Middle American Snake, Pituophis
deppei. By William E. Duellman. Pp. 599-610, 1 plate, 1
figure in text. May 2, 1960.
Index Pp. 611-626.
Vol. 11. 1. The systematic status of the colubrid snake, Leptodeira
discolor Günther. By William E. Duellman. Pp. 1-9, 4
figs. July 14, 1958.
2. Natural history of the six-lined racerunner, Cnemidophorus
sexlineatus. By Henry S. Fitch. Pp. 11-62, 9 figs., 9
tables. September 19, 1958.
3. Home ranges, territories, and seasonal movements of
vertebrates of the Natural History Reservation. By Henry
S. Fitch. Pp. 63-326, 6 plates, 24 figures in text, 3
tables. December 12, 1958.
4. A new snake of the genus Geophis from Chihuahua, Mexico.
By John M. Legler. Pp. 327-334, 2 figures in text.
January 28, 1959.
5. A new tortoise, genus Gopherus, from north-central
Mexico. By John M. Legler. Pp. 335-343. April 24, 1959.
6. Fishes of Chautauqua, Cowley and Elk counties, Kansas. By
Artie L. Metcalf. Pp. 345-400, 2 plates, 2 figures in
text, 10 tables. May 6, 1959.
7. Fishes of the Big Blue River Basin, Kansas. By W. L.
Minckley. Pp. 401-442, 2 plates, 4 figures in text, 5
tables. May 8, 1959.
8. Birds from Coahuila, México. By Emil K. Urban. Pp.
443-516. August 1, 1959.
9. Description of a new softshell turtle from the
southeastern United States. By Robert G. Webb. Pp.
517-525, 2 pls., 1 figure in text, August 14, 1959.
10. Natural history of the ornate box turtle, Terrapene
ornata ornata Agassiz. By John M. Legler. Pp. 527-669,
16 pls., 29 figures in text. March 7, 1960.
Index will follow.
Vol. 12. 1. Functional morphology of three bats: Eumops, Myotis,
Macrotus. By Terry A. Vaughan. Pp. 1-153, 4 plates, 24
figures in text, July 8, 1959.
2. The ancestry of modern Amphibia: a review of the
evidence. By Theodore H. Eaton, Jr. Pp. 155-180, 10
figures in text. July 10, 1959.
3. The baculum in microtine rodents. By Sydney Anderson.
Pp. 181-216, 49 figures in text. February 19, 1960.
4. A new order of fishlike Amphibia from the Pennsylvanian
of Kansas. By Theodore H. Eaton, Jr., and Peggy Lou
Stewart. Pp. 217-240, 12 figures in text. May 2, 1960.
More numbers will appear in volume 12.
Vol. 13. 1. Five natural hybrid combinations in minnows (Cyprinidae).
By Frank B. Cross and W. L. Minckley. Pp. 1-18. June 1,
1960.
2. A distributional study of the amphibians of the isthmus
of Tehuantepec, México. By William E. Duellman.
Pp. 19-72, pls. 1-8, 3 figs. August 16, 1960.
More numbers will appear in volume 13.
Transcriber's Notes
Page 26: Changed "19. Cosaleacaque" to "19. Cosoleacaque".
Page 30: Changed "Brysonima crassifolia" to "Byrsonima crassifolia".
Page 34: Changed "long. 95' 29°;" to "long. 95° 29';".
Page 35: Changed "Matías Romera" to "Matías Romero".
Page 47: Changed "kown" to "known".
Pages 50 and 59: Changed "axills" to "axils".
Plate 1, Fig. 2: Changed "Veracuz" to "Veracruz".
Page 53: Changed "valadity" to "validity".
Page 67: Changed "refering" to "referring".
Page 68: Changed "survided" to "survived".
Page 71: Changed "subhimid" to "subhumid" and "Amerca" to "America".
Moved University of Kansas Publications list to end of report.
Vol. 9, No. 12: Changed pages from "363-387" to "363-384".
Vol. 10, No. 10: Changed pages from "599-612" to "599-610".
End of the Project Gutenberg EBook of A Distributional Study of the
Amphibians of the Isthmus of Tehuantepec, Mexico, by William E. Duellman
*** END OF THIS PROJECT GUTENBERG EBOOK A DISTRIBUTIONAL STUDY OF ***
***** This file should be named 38440-h.htm or 38440-h.zip *****
This and all associated files of various formats will be found in:
http://www.gutenberg.org/3/8/4/4/38440/
Produced by Chris Curnow, Joseph Cooper, Diane Monico, and
the Online Distributed Proofreading Team at
http://www.pgdp.net
Updated editions will replace the previous one--the old editions
will be renamed.
Creating the works from public domain print editions means that no
one owns a United States copyright in these works, so the Foundation
(and you!) can copy and distribute it in the United States without
permission and without paying copyright royalties. Special rules,
set forth in the General Terms of Use part of this license, apply to
copying and distributing Project Gutenberg-tm electronic works to
protect the PROJECT GUTENBERG-tm concept and trademark. Project
Gutenberg is a registered trademark, and may not be used if you
charge for the eBooks, unless you receive specific permission. If you
do not charge anything for copies of this eBook, complying with the
rules is very easy. You may use this eBook for nearly any purpose
such as creation of derivative works, reports, performances and
research. They may be modified and printed and given away--you may do
practically ANYTHING with public domain eBooks. Redistribution is
subject to the trademark license, especially commercial
redistribution.
*** START: FULL LICENSE ***
THE FULL PROJECT GUTENBERG LICENSE
PLEASE READ THIS BEFORE YOU DISTRIBUTE OR USE THIS WORK
To protect the Project Gutenberg-tm mission of promoting the free
distribution of electronic works, by using or distributing this work
(or any other work associated in any way with the phrase "Project
Gutenberg"), you agree to comply with all the terms of the Full Project
Gutenberg-tm License (available with this file or online at
http://gutenberg.org/license).
Section 1. General Terms of Use and Redistributing Project Gutenberg-tm
electronic works
1.A. By reading or using any part of this Project Gutenberg-tm
electronic work, you indicate that you have read, understand, agree to
and accept all the terms of this license and intellectual property
(trademark/copyright) agreement. If you do not agree to abide by all
the terms of this agreement, you must cease using and return or destroy
all copies of Project Gutenberg-tm electronic works in your possession.
If you paid a fee for obtaining a copy of or access to a Project
Gutenberg-tm electronic work and you do not agree to be bound by the
terms of this agreement, you may obtain a refund from the person or
entity to whom you paid the fee as set forth in paragraph 1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be
used on or associated in any way with an electronic work by people who
agree to be bound by the terms of this agreement. There are a few
things that you can do with most Project Gutenberg-tm electronic works
even without complying with the full terms of this agreement. See
paragraph 1.C below. There are a lot of things you can do with Project
Gutenberg-tm electronic works if you follow the terms of this agreement
and help preserve free future access to Project Gutenberg-tm electronic
works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the Foundation"
or PGLAF), owns a compilation copyright in the collection of Project
Gutenberg-tm electronic works. Nearly all the individual works in the
collection are in the public domain in the United States. If an
individual work is in the public domain in the United States and you are
located in the United States, we do not claim a right to prevent you from
copying, distributing, performing, displaying or creating derivative
works based on the work as long as all references to Project Gutenberg
are removed. Of course, we hope that you will support the Project
Gutenberg-tm mission of promoting free access to electronic works by
freely sharing Project Gutenberg-tm works in compliance with the terms of
this agreement for keeping the Project Gutenberg-tm name associated with
the work. You can easily comply with the terms of this agreement by
keeping this work in the same format with its attached full Project
Gutenberg-tm License when you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern
what you can do with this work. Copyright laws in most countries are in
a constant state of change. If you are outside the United States, check
the laws of your country in addition to the terms of this agreement
before downloading, copying, displaying, performing, distributing or
creating derivative works based on this work or any other Project
Gutenberg-tm work. The Foundation makes no representations concerning
the copyright status of any work in any country outside the United
States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other immediate
access to, the full Project Gutenberg-tm License must appear prominently
whenever any copy of a Project Gutenberg-tm work (any work on which the
phrase "Project Gutenberg" appears, or with which the phrase "Project
Gutenberg" is associated) is accessed, displayed, performed, viewed,
copied or distributed:
This eBook is for the use of anyone anywhere at no cost and with
almost no restrictions whatsoever. You may copy it, give it away or
re-use it under the terms of the Project Gutenberg License included
with this eBook or online at www.gutenberg.org
1.E.2. If an individual Project Gutenberg-tm electronic work is derived
from the public domain (does not contain a notice indicating that it is
posted with permission of the copyright holder), the work can be copied
and distributed to anyone in the United States without paying any fees
or charges. If you are redistributing or providing access to a work
with the phrase "Project Gutenberg" associated with or appearing on the
work, you must comply either with the requirements of paragraphs 1.E.1
through 1.E.7 or obtain permission for the use of the work and the
Project Gutenberg-tm trademark as set forth in paragraphs 1.E.8 or
1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted
with the permission of the copyright holder, your use and distribution
must comply with both paragraphs 1.E.1 through 1.E.7 and any additional
terms imposed by the copyright holder. Additional terms will be linked
to the Project Gutenberg-tm License for all works posted with the
permission of the copyright holder found at the beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm
License terms from this work, or any files containing a part of this
work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this
electronic work, or any part of this electronic work, without
prominently displaying the sentence set forth in paragraph 1.E.1 with
active links or immediate access to the full terms of the Project
Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary,
compressed, marked up, nonproprietary or proprietary form, including any
word processing or hypertext form. However, if you provide access to or
distribute copies of a Project Gutenberg-tm work in a format other than
"Plain Vanilla ASCII" or other format used in the official version
posted on the official Project Gutenberg-tm web site (www.gutenberg.org),
you must, at no additional cost, fee or expense to the user, provide a
copy, a means of exporting a copy, or a means of obtaining a copy upon
request, of the work in its original "Plain Vanilla ASCII" or other
form. Any alternate format must include the full Project Gutenberg-tm
License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying,
performing, copying or distributing any Project Gutenberg-tm works
unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing
access to or distributing Project Gutenberg-tm electronic works provided
that
- You pay a royalty fee of 20% of the gross profits you derive from
the use of Project Gutenberg-tm works calculated using the method
you already use to calculate your applicable taxes. The fee is
owed to the owner of the Project Gutenberg-tm trademark, but he
has agreed to donate royalties under this paragraph to the
Project Gutenberg Literary Archive Foundation. Royalty payments
must be paid within 60 days following each date on which you
prepare (or are legally required to prepare) your periodic tax
returns. Royalty payments should be clearly marked as such and
sent to the Project Gutenberg Literary Archive Foundation at the
address specified in Section 4, "Information about donations to
the Project Gutenberg Literary Archive Foundation."
- You provide a full refund of any money paid by a user who notifies
you in writing (or by e-mail) within 30 days of receipt that s/he
does not agree to the terms of the full Project Gutenberg-tm
License. You must require such a user to return or
destroy all copies of the works possessed in a physical medium
and discontinue all use of and all access to other copies of
Project Gutenberg-tm works.
- You provide, in accordance with paragraph 1.F.3, a full refund of any
money paid for a work or a replacement copy, if a defect in the
electronic work is discovered and reported to you within 90 days
of receipt of the work.
- You comply with all other terms of this agreement for free
distribution of Project Gutenberg-tm works.
1.E.9. If you wish to charge a fee or distribute a Project Gutenberg-tm
electronic work or group of works on different terms than are set
forth in this agreement, you must obtain permission in writing from
both the Project Gutenberg Literary Archive Foundation and Michael
Hart, the owner of the Project Gutenberg-tm trademark. Contact the
Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable
effort to identify, do copyright research on, transcribe and proofread
public domain works in creating the Project Gutenberg-tm
collection. Despite these efforts, Project Gutenberg-tm electronic
works, and the medium on which they may be stored, may contain
"Defects," such as, but not limited to, incomplete, inaccurate or
corrupt data, transcription errors, a copyright or other intellectual
property infringement, a defective or damaged disk or other medium, a
computer virus, or computer codes that damage or cannot be read by
your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right
of Replacement or Refund" described in paragraph 1.F.3, the Project
Gutenberg Literary Archive Foundation, the owner of the Project
Gutenberg-tm trademark, and any other party distributing a Project
Gutenberg-tm electronic work under this agreement, disclaim all
liability to you for damages, costs and expenses, including legal
fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT
LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE
PROVIDED IN PARAGRAPH 1.F.3. YOU AGREE THAT THE FOUNDATION, THE
TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE
LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR
INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH
DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a
defect in this electronic work within 90 days of receiving it, you can
receive a refund of the money (if any) you paid for it by sending a
written explanation to the person you received the work from. If you
received the work on a physical medium, you must return the medium with
your written explanation. The person or entity that provided you with
the defective work may elect to provide a replacement copy in lieu of a
refund. If you received the work electronically, the person or entity
providing it to you may choose to give you a second opportunity to
receive the work electronically in lieu of a refund. If the second copy
is also defective, you may demand a refund in writing without further
opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth
in paragraph 1.F.3, this work is provided to you 'AS-IS' WITH NO OTHER
WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT LIMITED TO
WARRANTIES OF MERCHANTIBILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied
warranties or the exclusion or limitation of certain types of damages.
If any disclaimer or limitation set forth in this agreement violates the
law of the state applicable to this agreement, the agreement shall be
interpreted to make the maximum disclaimer or limitation permitted by
the applicable state law. The invalidity or unenforceability of any
provision of this agreement shall not void the remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the
trademark owner, any agent or employee of the Foundation, anyone
providing copies of Project Gutenberg-tm electronic works in accordance
with this agreement, and any volunteers associated with the production,
promotion and distribution of Project Gutenberg-tm electronic works,
harmless from all liability, costs and expenses, including legal fees,
that arise directly or indirectly from any of the following which you do
or cause to occur: (a) distribution of this or any Project Gutenberg-tm
work, (b) alteration, modification, or additions or deletions to any
Project Gutenberg-tm work, and (c) any Defect you cause.
Section 2. Information about the Mission of Project Gutenberg-tm
Project Gutenberg-tm is synonymous with the free distribution of
electronic works in formats readable by the widest variety of computers
including obsolete, old, middle-aged and new computers. It exists
because of the efforts of hundreds of volunteers and donations from
people in all walks of life.
Volunteers and financial support to provide volunteers with the
assistance they need, are critical to reaching Project Gutenberg-tm's
goals and ensuring that the Project Gutenberg-tm collection will
remain freely available for generations to come. In 2001, the Project
Gutenberg Literary Archive Foundation was created to provide a secure
and permanent future for Project Gutenberg-tm and future generations.
To learn more about the Project Gutenberg Literary Archive Foundation
and how your efforts and donations can help, see Sections 3 and 4
and the Foundation web page at http://www.pglaf.org.
Section 3. Information about the Project Gutenberg Literary Archive
Foundation
The Project Gutenberg Literary Archive Foundation is a non profit
501(c)(3) educational corporation organized under the laws of the
state of Mississippi and granted tax exempt status by the Internal
Revenue Service. The Foundation's EIN or federal tax identification
number is 64-6221541. Its 501(c)(3) letter is posted at
http://pglaf.org/fundraising. Contributions to the Project Gutenberg
Literary Archive Foundation are tax deductible to the full extent
permitted by U.S. federal laws and your state's laws.
The Foundation's principal office is located at 4557 Melan Dr. S.
Fairbanks, AK, 99712., but its volunteers and employees are scattered
throughout numerous locations. Its business office is located at
809 North 1500 West, Salt Lake City, UT 84116, (801) 596-1887, email
business@pglaf.org. Email contact links and up to date contact
information can be found at the Foundation's web site and official
page at http://pglaf.org
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
gbnewby@pglaf.org
Section 4. Information about Donations to the Project Gutenberg
Literary Archive Foundation
Project Gutenberg-tm depends upon and cannot survive without wide
spread public support and donations to carry out its mission of
increasing the number of public domain and licensed works that can be
freely distributed in machine readable form accessible by the widest
array of equipment including outdated equipment. Many small donations
($1 to $5,000) are particularly important to maintaining tax exempt
status with the IRS.
The Foundation is committed to complying with the laws regulating
charities and charitable donations in all 50 states of the United
States. Compliance requirements are not uniform and it takes a
considerable effort, much paperwork and many fees to meet and keep up
with these requirements. We do not solicit donations in locations
where we have not received written confirmation of compliance. To
SEND DONATIONS or determine the status of compliance for any
particular state visit http://pglaf.org
While we cannot and do not solicit contributions from states where we
have not met the solicitation requirements, we know of no prohibition
against accepting unsolicited donations from donors in such states who
approach us with offers to donate.
International donations are gratefully accepted, but we cannot make
any statements concerning tax treatment of donations received from
outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation
methods and addresses. Donations are accepted in a number of other
ways including checks, online payments and credit card donations.
To donate, please visit: http://pglaf.org/donate
Section 5. General Information About Project Gutenberg-tm electronic
works.
Professor Michael S. Hart is the originator of the Project Gutenberg-tm
concept of a library of electronic works that could be freely shared
with anyone. For thirty years, he produced and distributed Project
Gutenberg-tm eBooks with only a loose network of volunteer support.
Project Gutenberg-tm eBooks are often created from several printed
editions, all of which are confirmed as Public Domain in the U.S.
unless a copyright notice is included. Thus, we do not necessarily
keep eBooks in compliance with any particular paper edition.
Most people start at our Web site which has the main PG search facility:
http://www.gutenberg.org
This Web site includes information about Project Gutenberg-tm,
including how to make donations to the Project Gutenberg Literary
Archive Foundation, how to help produce our new eBooks, and how to
subscribe to our email newsletter to hear about new eBooks.
 Fig. 1. Map of the Isthmus of Tehuantepec based on the
American Geographical Society's "Map of Hispanic America on the Scale
of 1:1,000,000."
Fig. 1. Map of the Isthmus of Tehuantepec based on the
American Geographical Society's "Map of Hispanic America on the Scale
of 1:1,000,000."