The Project Gutenberg EBook of Natural History of Cottonmouth Moccasin, Agkistrodon piscovorus (Reptilia), by Ray D. Burkett This eBook is for the use of anyone anywhere at no cost and with almost no restrictions whatsoever. You may copy it, give it away or re-use it under the terms of the Project Gutenberg License included with this eBook or online at www.gutenberg.org Title: Natural History of Cottonmouth Moccasin, Agkistrodon piscovorus (Reptilia) Author: Ray D. Burkett Release Date: December 15, 2011 [EBook #38308] Language: English Character set encoding: UTF-8 *** START OF THIS PROJECT GUTENBERG EBOOK NATURAL HISTORY OF *** Produced by Chris Curnow, Joseph Cooper and the Online Distributed Proofreading Team at http://www.pgdp.net
| PAGE | ||
| Introduction | 439 | |
| Acknowledgments | 440 | |
| Systematic Relationships and Distribution | 441 | |
| Description | 444 | |
| Color and Pattern | 444 | |
| Scutellation | 444 | |
| Dentition | 449 | |
| Habitat and Limiting Factors | 450 | |
| Reproduction | 452 | |
| Courtship and Mating | 452 | |
| Reproductive Cycles | 452 | |
| Embryonic Development | 454 | |
| Birth of Young | 454 | |
| Number of Young per Litter | 454 | |
| Population Composition | 455 | |
| Reproductive Potential | 455 | |
| Growth and Development | 456 | |
| Size at Birth and Early Growth | 456 | |
| The Umbilical Scar | 457 | |
| Later Growth and Bodily Proportions | 457 | |
| Shedding | 459 | |
| The Shedding Operation | 459 | |
| Frequency of Shedding | 460 | |
| Food Habits | 461 | |
| Methods of Obtaining Prey | 461 | |
| Food and Food Preferences | 462 | |
| Mortality Factors | 465 | |
| Natural Enemies and Predators | 465 | |
| Parasites and Diseases | 465 | |
| Miscellaneous Causes of Death | 466 | |
| Behavior | 466 | |
| Annual and Diel Cycles of Activity | 466 | |
| Basking | 469 | |
| Coiling | 469 | |
| Locomotion | 470 | |
| Disposition | 470 | |
| Defense and Escape | 471 | |
| "Head Bobbing" | 471 | |
| Combat Dance | 472 | |
| The Venom | 473 | |
| Properties of the Venom | 473 | |
| Venom Yield and Toxicity | 473 | |
| Susceptibility of Snakes | 475 | |
| The Bite | 476 | |
| Effects of the Bite | 476 | |
| Treatment | 477 | |
| Case History of a Bite | 479 | |
| Snakebite in the United States | 480 | |
| Summary | 480 | |
| Literature Cited | 485 | |
Objectives of the study here reported on were to: (1) learn as much as possible concerning the natural history and economic importance of the cottonmouth; (2) determine what factors limit its geographic distribution; (3) determine the role of the cottonmouth in its ecological community; and (4) compare the cottonmouth's life history with that of other crotalid snakes, especially the kinds that are most closely related to it.
Twenty-five live cottonmouths were kept in the laboratory for the purpose of studying behavior and fang shedding and for comparison of measurements with those of preserved specimens. Live snakes were obtained in Brazoria and Nacogdoches counties, Texas, from Hermann Park Zoo, Houston, Texas, and from the late Paul Anderson of Independence, Missouri. Preserved western cottonmouths were examined for the purpose of determining variation, distribution, food habits, body proportions, embryonic development, and reproductive cycles. The cottonmouths examined include: 221 from Texas; 33 from Arkansas; 22 from Louisiana; 2 from Illinois; and 1 each from Kansas, Mississippi, and Oklahoma.
In the preparation of this report I have examined all available literature pertaining to the cottonmouth and have drawn from these sources for comparative or additional material. Some of the more noteworthy contributions to knowledge of the cottonmouth are the general accounts of the life history by Allen and Swindell (1948), Barbour (1956), and Wright and Wright (1957); the publications by Gloyd and Conant (1943) concerning taxonomy; Klimstra (1959) concerning food habits; and Allen (1937), Parrish and Pollard (1959), Swanson (1946), and Wolff and Githens (1939b) concerning the venom. Numerous other publications, although brief, contain worthwhile contributions. Also of special interest as a source of material for comparison of cottonmouths with other crotalids are the works of Fitch (1960) on the copperhead and of Klauber (1956) on the rattlesnakes.
The cottonmouth has been well known for nearly 200 years. Wright and Wright (1957) listed the following vernacular names that are applied to the cottonmouth: black moccasin, black snake, blunt-tail moccasin, congo, copperhead, cottonmouth water moccasin, cotton-mouthed snake, gapper, highland moccasin, lowland moccasin, mangrove rattler, moccasin, North American cottonmouth snake, North American water moccasin, North American water viper, pilot, rusty moccasin, salt-water rattler, stubtail, stump [440] (-tail) moccasin, stump-tail viper, swamp lion, Texas Moccasin, trapjaw, Troost's moccasin, true horn snake, true water moccasin, viper, water mokeson, water pilot, water rattlesnake, and water viper.
Some of the names listed above are based upon superstition and folklore prevailing in pioneer times, and others are based upon the behavior or appearance of the snake at various ages. Names like "stump-tail moccasin" are derived from the appearance of females which have short tails or snakes that have lost part of the tail. Names like "gapper" and "trapjaw" came to be applied because of the habit of the snake's lying with its mouth open when approached. The name "cottonmouth" also was derived from this behavior, although the lining of the mouth is whitish in most other snakes. The term "rattlesnake" may have come from the fact that the cottonmouth vibrates its tail vigorously when nervous as do many other snakes, or it may have been confused with rattlesnakes. Because of the general public's fear of snakes and their reluctance to learn to discriminate between the poisonous and harmless species, numerous kinds of snakes seen in or near water have been called moccasins. The general appearance, pugnacious behavior, and whitish mouth of water-snakes (Natrix) have earned them a bad reputation. In fact, a great majority of the "cottonmouths" reported in many areas are found to be water-snakes.
The cottonmouth is economically important mainly because of the injurious or fatal effects of its bite and the psychological effect that its actual or suspected presence has upon many persons. The species eats a wide variety of prey items and helps to prevent overabundance of certain kinds of organisms. The venom has been used in the therapeutic treatment of blood clots owing to its anticoagulant properties (Didisheim and Lewis, 1956). It also is employed in the treatment of haemorrhagic conditions and rheumatoid arthritis, as well as in the production of antivenin (Allen and Swindell, op cit.:13). None of these uses of venom has become widely accepted, and its value is questionable.
For guidance in the course of my study, I am especially indebted to Professor Henry S. Fitch. For suggestions concerning the preparation of the manuscript, I thank Professor E. Raymond Hall. I am grateful to my wife, Janis, for her invaluable assistance and for typing the manuscript.
For use of specimens in their care, I thank Professors William E. Duellman, University of Kansas; Robert L. Packard, formerly of Stephen F. Austin State College; W. Frank Blair, University of Texas; and William B. Davis and
Richard J. Baldauf, Texas Agricultural and Mechanical College. Mr. John E. Werler of the Hermann Park Zoo, Houston, Texas, contributed live individuals; Mr. Richard S. Funk contributed information on the birth of a brood of cottonmouths; and Dr. Henry M. Parrish contributed information on the incidence of snakebite. To numerous other persons at leading museums throughout the United States for information on the cottonmouths in their collections, to all who helped with the field work in various ways, and to others at the University of Kansas for their help and suggestions I am grateful.
Snakes of the genus Agkistrodon are relatively primitive members of the Crotalidae, which is one of the most specialized families of snakes. A majority of the pit-vipers are found in the Americas, but close relatives are found from extreme southeastern Europe through temperate Asia to Japan (A. halys) and southeastern Asia including Indonesia (Agkistrodon and Trimeresurus). Familial characters include: vertical pupil of the eye; facial pit present between the preoculars and loreal; scales usually keeled; short, rotatable maxilla bearing a large hollow fang; toothless premaxilla; chiefly hematoxic venom; and undivided anal plate.
The genus Agkistrodon includes about nine species in the Old World and three in North and Central America. Some of the primitive characters of the genus are: head covered with nine enlarged shields or having the internasals and prefrontals broken up into small scales; subcaudals on proximal part of tail undivided; fangs relatively short; tail lacking rattles. In one species, A. rhodostoma, the scales are smooth; and the female is oviparous and guards her eggs until they hatch. Other species have keeled scales and are ovo-viviparous.
There is little paleontological evidence illustrating evolution of the cottonmouth or for that matter of crotalids in general. Brattstrom (1954) summarized the current knowledge of fossil pit-vipers in North America. The few fossils found of the cottonmouth are from Alacha, Brevard, Citrus, Levy, Pasco, and Pinellas counties, Florida (Brattstrom, op. cit.:35; Auffenberg, 1963:202). All are of late Pleistocene Age and well within the present geographic range of the cottonmouth.
Of crotalid genera only Agkistrodon occurs in both the Old World and the New World, suggesting that this genus is relatively old. Schmidt (1946: 149-150) mentioned several other closely related groups of animals found in both eastern Asia and eastern North America, including the reptilian genera: Natrix, Opheodrys, Elaphe, Ophisaurus, Leiolopisma (= Lygosoma), Eumeces, Clemmys, Emmys, and Alligator. Of the groups of animals now confined to these two regions the most important are the cryptobranchid salamanders, the genus Alligator, and the spoon-bills (Psephurus in China and Polyodon in the Mississippi drainage). Fossil evidence for these groups indicates that existing forms common to eastern Asia and eastern North America are remnants of a late Cretaceous or early Tertiary Holarctic fauna which was forced southward as the climate became gradually cooler to the north. "Other clues suggest that both Agkistrodon and Trimeresurus (Bothrops) moved from Asia to America, one of these presumably giving rise to the rattlesnakes." (Darlington, 1957:228).
The named, American kinds of Agkistrodon currently are arranged as three species: the copperhead, the cantil and the cottonmouth. The copperhead (A. contortrix) is divided into four subspecies, all of which are terrestrial. This species occurs from southern New England to eastern Kansas and along the Atlantic and Gulf Coastal plains, exclusive of peninsular Florida and the delta of the Mississippi River in Louisiana. It extends southwest from Kansas through the Edwards Plateau of west-central Texas. Isolated populations occur in the Chisos and Davis mountains of Trans-Pecos Texas. The cantil or Mexican moccasin (A. bilineatus), probably the nearest relative of the cottonmouth (A. piscivorus), is divisible into two subspecies and occupies a nearly complementary range from Mexico south to Nicaragua. The cottonmouth occurs throughout the coastal plains of the southeastern United States, usually at altitudes of 500 feet or less. Two subspecies are recognized, the eastern A. p. piscivorus and the western A. p. leucostoma. A revision of the genus is underway by Professor Howard K. Gloyd.
The basic pattern and various behavioral traits are common to all three species. The young are more nearly alike in appearance than adults, the copperhead and the cottonmouth being easily confused. Adults differ in color, size, body proportions, habitat, and habits. In range and habitat preference the cottonmouth more closely resembles the southern subspecies of the copperhead, A. c. contortrix, which is usually found in lowlands, near swamps and streams, but seldom in water.
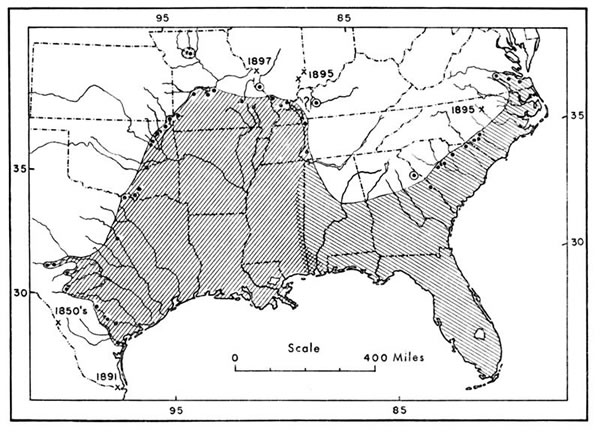 Fig. 1.
Fig. 1. Fig. 1. Geographic range of the cottonmouth, showing marginal and near-marginal records, based largely upon maps by Gloyd and Conant (1943:165) and Conant (1958:336) but including additional records. The more important of these records (from east to west) are discussed in the following paragraphs. Crosshatching indicates the area of intergradation between the eastern and western subspecies. Old records, indicated by dates, and their sources are as follows: 1850's and 1891—U. S. National Museum numbers 4263 and 32753 respectively; 1897—Hurter (1897); and 1895—Stejneger (1895:408).
The northernmost record for the eastern subspecies is in the Petersburg area, Prince George County, Virginia (Anon., 1953:24). A sight record (Hickman, 1922:39) near Bristol, West Virginia, probably was based on a water-snake (Natrix sp.), since the stream in which the snake was seen flows north into the Ohio River rather than southeast through Virginia. In North Carolina the most inland record is from the Neuse River, six to eight miles east of Raleigh (Stejneger, 1895:408). Neill (1947:205) reported a population in the vicinity of Dry Fork Creek on the boundary line of Wilkes and Oglethorpe counties, Georgia. Distribution of cottonmouths in Florida is statewide, including the Keys and other offshore islands.
The ranges of the two subspecies, piscivorus and leucostoma, meet near the eastern border of Mississippi. A. p. piscivorus has been reported from Tishomingo County to the Gulf and east of the Loess Bluff area in central Mississippi, and A. p. leucostoma has been reported from this area westward. A few specimens from along the Coast indicate intergradation (Cook, 1962:33) between the two subspecies.
Barbour (1956:33) reported one specimen from Cypress Creek, in the Green River drainage, Muhlenberg County, Kentucky, and stated that suitable habitat can be found in several areas east of Kentucky Lake. Hence, cottonmouths may have entered this area via the Ohio River. Stejneger (loc. cit.) reported the species in the Wabash River at Mount Carmel, Wabash County, Illinois, and mentioned a former occurrence at Vincennes, Knox County, Indiana; but there are no recent records at these localities. Hurter (1897) reported having seen cottonmouths in Illinois, opposite St. Louis; Smith (1961:265) believes that this and a population in Monroe County, Illinois, are isolated relicts, since no specimens have been found within 50 miles to the south of Monroe County. The specimens reported by Anderson (1941:178; 1945:274) near Chillicothe (three miles southwest and seven miles northwest, respectively), Livingston County, Missouri, also are thought to represent a relict population. Hall and Smith (1947:453) reported one specimen from Jasper County, Missouri, in the Spring River which flows through extreme southeastern Kansas and into Oklahoma and another in the Neosho River at Chetopa, Kansas. Both of these specimens were taken after a flood, and no additional specimens have been taken in this region. Nevertheless, sufficient habitat is probably available along the Neosho and Verdigris rivers in the southeastern part of Kansas.
In Texas the cottonmouth has penetrated marginal habitat perhaps farther than anywhere else in its range. Formerly it was thought to be limited to the country east of the Balcones Escarpment (Smith and Buechner, 1947:8), but semiarid areas of the state have been invaded primarily via the Colorado and Brazos River systems up to altitudes of 2300 feet. Two additional specimens are said to have been collected along the Rio Grande. Dr. Howard K. Gloyd (in litt.) stated that the specimen reported from Eagle Pass, Maverick County, is believed to have been taken in the 1850's; and the one said to have come from the mouth of the Devil's River is actually marked "near Santa Rosa, Cameron County, September 30, 1891." No additional specimens have been taken in that area; and the range now probably extends no farther south than Corpus Christi, Texas. Brown's (1903:554) knowledge of the extension of the range of the cottonmouth west of longitude 98° is probably based upon the records along the Rio Grande reported in the nineteenth century.
Three extensions of the known range in Texas are reported herein. One specimen was captured by Mr. Harry Green (HWG 346) along the San Saba River, 8.1 miles west of Menard, Menard County. The other two specimens (KU 84375 and 84376) were taken by the late Paul Anderson one and one-half miles north of Pecan Crossing, South Concho River, Tom Green County, and one mile west of Mertzon, Irion County.
In the hypsithermal period following Pleistocene glaciation, cottonmouths gradually moved northward occupying areas beyond their present range. The distributional records since the 1850's and the apparent relict populations now in existence indicate that the range of this species has since receded.
Color predominantly brown, ranging through pale reddish-brown or dark reddish-brown, brownish-green, to almost black; 10 to 17 irregular dark brown bands on paler brown ground color; young paler (some nearly salmon pink), retaining a vivid pattern throughout first year; pattern of most individuals nearly obliterated by third year; brilliance and dullness of predominant color correlated with molting cycle (skin especially bright and shiny immediately following shedding; tip of tail yellowish in juveniles; posterior part of venter and tail uniformly black in some adult individuals, especially females; secondary sexual differences in dorsal coloration, such as found in copperhead by Fitch (1960:102), not noted.
The eastern subspecies, A. p. piscivorus, has the more brilliant pattern in which the centers of the dark cross-bands are invaded by the ground color. The cross-bands are slightly constricted in the mid-line and may or may not be bilaterally symmetrical. One-half of the cross-band may be displaced anteriorly or posteriorly to a slight degree or may even be completely absent. From one to several dark spots may be present within the cross-bands.
The western subspecies, A. p. leucostoma, has a comparatively dull pattern in which the ground color does not invade the center of the cross-bands. In many instances the bands are outlined by white scales, as in the Mexican moccasin (this character is not so prominent in A. p. piscivorus because of the paler ground color). A large, dark blotch usually occurs at the base of the cross-band and may completely cross the ventral scales. The characteristic variations found in piscivorus are also present in leucostoma.
The number of bands is often difficult to count because of the dark color of some specimens. Gloyd and Conant (1943:168) reported averages of 12.5 (11 to 16) and 12.2 (10 to 16) in males and females, respectively, of leucostoma and ranges of 10 to 17 for males and 10 to 16 for females with averages of 13 in both sexes of piscivorus. On 20 specimens of leucostoma from Texas the average number of bands was 12.7 (11 to 15). If the number of bands differed on the two sides of an animal, the total number of the two sides was divided by two.
Scutellation
The scutellation of the cottonmouth closely resembles that of the other species of Agkistrodon. For example, the nine cephalic shields are characteristic of most species of Agkistrodon, as well as most other primitive crotalids and viperids, and most colubrids. Most individuals have an additional pair of large scales behind the parietals.
The numbers of postoculars, supralabials, and infralabials are variable. On either side the postoculars (three in most specimens) are reduced to two in some specimens. The supralabials (eight in most specimens) frequently vary (usually on one side only) from seven to nine. The number of infralabials is somewhat more variable than the number of supralabials, the usual number being 11, but 10 is also common; 8, 9, and 12 are more rare (Table 1). In 102 snakes in which these characters were examined, four different combinations of supralabials and seven combinations of infralabials were found. Both characters together yielded 16 combinations, considering only the actual number of scales and not taking into account the side of the head on which they occurred (Table 2). The combinations found in a brood of seven young from Houston, Texas, are shown in Table 3 to illustrate the variability of this character. Gloyd and Conant (1943:168) found a variation of 6 to 11 (8) and 7 to 9 (8) supralabials and 8 to 13 (11) and 8 to 12 (10.4) infralabials in samples of 301 leucostoma and 119 piscivorus, respectively (numbers in parentheses represent average). Also of interest is the variability of the scales themselves. In one instance a scale was found that had not completely divided. In another specimen the last supralabial and last infralabial were one scale that completely lined the angle of the jaw. Instances of one scale almost [445] crowding out another were common. In still other instances one or two supralabials were divided horizontally into two scales. Individual variation rather than geographical variation occurs in these characters.
TABLE 1.—Frequency of Occurrence of Various Numbers of Supralabial and
Infralabial Scales in 102 Cottonmouths.
| Number of scales | Specimens having number on both sides | Specimens having number on one side | Total | Percentage |
| Supralabials | ||||
| 7 | 11 | 24 | 35 | 25.2 |
| 8 | 64 | 27 | 91 | 65.5 |
| 9 | 0 | 3 | 3 | 2.2 |
| Infralabials | ||||
| 8 | 0 | 2 | 2 | 1.5 |
| 9 | 3 | 10 | 13 | 9.6 |
| 10 | 12 | 32 | 44 | 32.4 |
| 11 | 53 | 22 | 75 | 55.1 |
| 12 | 0 | 2 | 2 | 1.5 |
TABLE 2.—Numbers of Supralabials and Infralabials of 102 Cottonmouths.
| Number of individuals |
Number of supralabials |
Number of infralabials |
| 37 | 8 | 11 |
| 15 | 8 | 10-11 |
| 12 | 7-8 | 11 |
| 6 | 7-8 | 10-11 |
| 5 | 8 | 10 |
| 5 | 8 | 9-10 |
| 4 | 7 | 11 |
| 3 | 7 | 9-10 |
| 3 | 7-8 | 10 |
| 2 | 7 | 9 |
| 2 | 7 | 10 |
| 2 | 8 | 10-12 |
| 2 | 8-9 | 10 |
| 2 | 7-8 | 8-9 |
| 1 | 7-8 | 9 |
| 1 | 8-9 | 10-11 |
The dorsal scales of cottonmouths are strongly keeled except that those of the two lower scale-rows on each side are weakly keeled. Also they are slightly larger than the others. Two apical pits are present on each dorsal scale. The shape of the scales and number of scale rows vary depending upon the position on the body. Scales on the neck are considerably smaller than those elsewhere on the body and are arranged in two or three more rows than those at mid-body. The skin in the region of the throat, neck, and fore-body is [446] especially elastic and allows the swallowing of large prey. Posteriorly from the mid-body the scales decrease in size and become more angular, those on the tail tending to be rhomboidal and wider than long. In the region of the anus the number of scale rows diminishes rapidly, leaving only 12 to 14 rows at the base of the tail and only three rows immediately ahead of the tail tip. The tail ends in a spine composed of two scales: one scale covers the bottom, lower parts of the sides, and tip of the spine; and a shorter dorsal scale covers the top and upper parts of the sides of the basal two-thirds of the spine. The spine of embryos and young cottonmouths is blunt, but is pointed in most adults.
TABLE 3.—Variation in Numbers of Supralabials and Infralabials in a Brood
of Seven Cottonmouths.
| Number of individuals |
Number of supralabials |
Number of infralabials |
| 1 | 7 | 9 |
| 1 | 7 | 9-10 |
| 2 | 7-8 | 8-9 |
| 1 | 7-8 | 9 |
| 1 | 8 | 9-10 |
| 1 | 8-9 | 10 |
TABLE 4.—Analysis of Number of Scale Rows at Three Parts of the Body
in 81 Cottonmouths.
| Number of scales per row |
Neck | Mid-body | Anterior to anus | |||
| Number of individuals |
Percentage | Number of individuals |
Percentage | Number of individuals |
Percentage | |
| 29 | 1 | 1.2 | ||||
| 28 | 3 | 3.7 | ||||
| 27 | 52 | 64.2 | ||||
| 26 | 16 | 18.0 | 2 | 2.5 | ||
| 25 | 8 | 9.9 | 67 | 82.7 | ||
| 24 | 1 | 1.2 | 4 | 4.9 | ||
| 23 | 8 | 9.9 | 4 | 4.9 | ||
| 22 | 4 | 4.9 | ||||
| 21 | 68 | 84.0 | ||||
| 20 | 5 | 6.2 | ||||
The number of scale rows on the neck, at mid-body, and just anterior to the anus is relatively constant at 27-25-21, respectively; but some individual variation is evident (Table 4). Since the rows are diagonally arranged, it is necessary in counting scales to proceed either anteriorly or posteriorly across the back; or the row may be counted in either direction up to the center of the back and then reversed on the other side of the snake. In order to count the scale rows in a position where no scale reduction or addition was occurring and to avoid as much error as possible, I counted from anterior to center and back on the neck, in any direction at mid-body, and from posterior to center and back near the anus. Because females generally are the larger in circumference posteriorly, they could have more scale rows than males just anterior [447] to the anus. The few snakes having more than 21 scale rows in the posterior region offer no conclusive evidence as to tendencies, but in both instances in which this occurred the females outnumbered the males three to one. An odd, rather than an even, number of scale rows occurs on most of the length of the snakes examined, because there is a mid-dorsal row and scale rows tend to be lost on both sides at about the same level. An example of scale reduction of one snake was as follows:
| 6 + 7 (13) | 6 + 7 (96) | |||
| 27 ————— | 25 ————— | 24 ————— | 23 ————— | 22 ————— |
| 5 + 6 (13) | 5 + 6 (90) | 7 + 8 (111) | 7 + 8 (114) | |
| 6 + 7 (122) | + 7, -5 (125) | |||
| 23 ————— | 22 ————— | 23 ————— | 21 ————— | 22 ————— |
| -6 (118) | + 6 (119) | 6 + 7 (121) | + 6 (123) | |
| -6 (126) | ||||
| 22 ————— | 21 (130). | |||
This scale reduction follows the method proposed by Dowling (1951b: 133) in which the numbers on the mid-line represent the number of scale rows, upper figures refer to the right side of the snake, and figures in parentheses indicate the number of the ventral scale (counted from the anterior end of the series), thus marking the position of the addition or reduction. Addition of a row is shown by a plus sign and the number of the row, whereas reductions are shown by a minus sign and the number of the row that is lost or by a plus sign between the number of two rows that join. According to Dowling, variation in number of dorsal scales characterizes the few genera and species of snakes in which it has been studied. The time and difficulty involved in ascertaining the number of scales explain why it has not been widely used in classification.
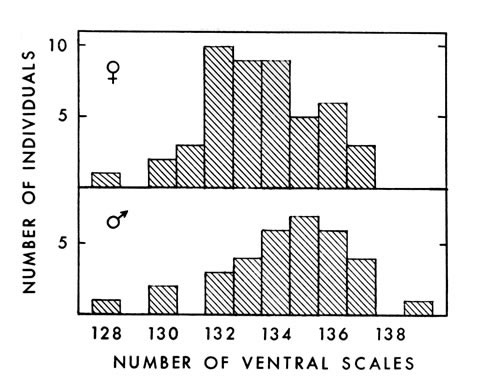 Fig. 2. Number of ventral scales in 48 female and 34
male A. p. leucostoma.
Fig. 2. Number of ventral scales in 48 female and 34
male A. p. leucostoma. Ventral scales on 34 males averaged 134.4 (128 to 139), and on 48 females 133.5 (128 to 137) (Fig. 2.). Barbour (1956:34) found an average of 135.3 ventral scales on 64 males and 44 females, and Gloyd and Conant (loc. cit.) found an average of 134 for both males and females. The average for the eastern cottonmouth obtained by Gloyd and Conant, however, was 137 ventrals in both sexes. Some of my counts were made before I knew of the standard system of counting ventrals proposed by Dowling (1951a:97-99), in which [448] the first ventral plate is defined as the most anterior one bordered on both sides by the first row of dorsals. Therefore, some inconsistencies may exist in my counts. Where differences occur, Dowling's method probably will indicate the presence of an additional scale, since it appears to begin farther anteriorly on the average, than I began counting.
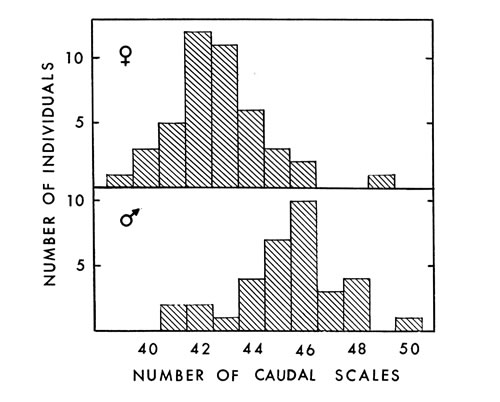 Fig. 3. Number of caudal scales in 44 female and 34
male A. p. leucostoma.
Fig. 3. Number of caudal scales in 44 female and 34
male A. p. leucostoma.
TABLE 5.—Caudal Scale Combinations in 95 Cottonmouths. U = Undivided;
D = Divided.
Number |
Number of scales | ||||||||||||||||
| D | U | D | U | D | U | D | U | D | U | D | U | D | U | D | U | D | |
| 25 | 13-35 | 10-32 | |||||||||||||||
| 11 | 1-2 | 12-33 | 14-28 | ||||||||||||||
| 20 | 16-39 | 1-9 | 1-3 | 3-24 | |||||||||||||
| 20 | 1-4 | 3-37 | 1-21 | 1-5 | 1-29 | ||||||||||||
| 4 | 14-30 | 1-8 | 1-7 | 1-8 | 1-4 | 2-10 | |||||||||||
| 3 | 1 | 18-23 | 1-2 | 1-2 | 6-11 | 1-3 | 6-9 | ||||||||||
| 4 | 1-17 | 1 | 1-3 | 1-8 | 1-4 | 1-3 | 1-4 | 13-22 | |||||||||
| 2 | 1-2 | 4-16 | 1 | 1-4 | 2 | 1 | 1-4 | 1 | 18-21 | ||||||||
| 1 | 20 | 1 | 1 | 1 | 1 | 6 | 1 | 3 | 1 | 11 | |||||||
| 1 | 10 | 2 | 3 | 2 | 10 | 1 | 2 | 2 | 1 | 4 | 4 | ||||||
| 1 | 20 | 1 | 1 | 2 | 1 | 1 | 4 | 4 | 2 | 4 | 1 | 3 | |||||
| 1 | 1 | 13 | 1 | 1 | 1 | 3 | 1 | 1 | 1 | 4 | 2 | 4 | 13 | ||||
| 1 | 17 | 1 | 1 | 2 | 1 | 1 | 6 | 2 | 1 | 2 | 3 | 2 | 7 | ||||
| 1 | 9 | 1 | 1 | 8 | 1 | 3 | 1 | 1 | 3 | 1 | 1 | 2 | 1 | 1 | 1 | 6 | |
Analysis of caudal scales revealed sexual dimorphism. In the six specimens from Tennessee, Blanchard (1922:16) found the same thing. Caudals averaged 45.4 (41 to 50) on 34 males and 42.6 (39 to 49) on 44 females (Fig. 3). Barbour (loc. cit.) found an average of 45.7 (30 to 54) caudals in males and 43 (17 to 56) in females. Caudal scale counts by Gloyd and Conant (loc. cit.) averaged 44 (38 to 49) in males and 42 (37 to 48) in females of leucostoma; in piscivorus they averaged 48 (42 to 53) in males and 44 (41 to 49) in females. Another seldom-mentioned, unusual characteristic of the caudal [449] scales of copperheads and cottonmouths is that some are single (usually those at the base of the tail) and others divided (Table 5). To my knowledge, all other species have either single or divided scales the entire length of the tail. See Klauber (1941:73) and Fox (1948:252) concerning correlation of few scales with warm environment.
Cottonmouths, like other pit-vipers, have their teeth reduced in number and have enlarged, highly specialized fangs. Small teeth occur on the palatine and the pterygoid in the upper jaw and on the dentary in the lower jaw. The dentary bone bears 17 curved teeth that decrease in size posteriorly. The palatine bears five small, strongly curved teeth, and the pterygoid bears 16 to 18 strongly curved teeth decreasing in size posteriorly. The numbers of teeth mentioned above in each instance refer to the number of sockets rather than the actual number of teeth, because teeth are frequently shed, leaving some of the sockets empty at any one time.
The maxillary bone has two sockets side by side which bear the poison fangs, usually one at a time. During the period shortly before a fang is to be shed, however, its replacement becomes attached in the alternate socket; and both fangs may be functional for a short time. The old fang then becomes weakened at its base, eventually breaks off, and is swallowed. At any one time four or five replacement fangs in various stages of development are found in the gum behind the functional fang. These replacement fangs, which are arranged in alternate rows, gradually enlarge as they move forward in their development and, in juveniles, are generally slightly longer than the fangs that they replace.
In 1963 I examined the fangs of 14 cottonmouths at four- to seven-day intervals for a period of six weeks. The fang-shedding cycle was found to be highly irregular, with a double condition (on one or both sides) occurring one-third of the time. Approximately the same proportion of double fangs was found in preserved individuals. A replacement period of at least five days was observed in one snake. One-half the cycle (from replacement on one side to replacement on the other) varied from five to twenty days, indicating that the cycles for each fang are independent of one another. Bogert (1943:324) found that young rattlesnakes are born with functional fangs in the two inner sockets. Nonsynchronous use of the sockets on opposite sides of the head in rattlesnakes is a later development which results from accidents or other conditions leading to a longer retention of the fang on one side than on the other (Klauber, 1956:723). I found a double set of fangs in cottonmouths only twice in the six-week period. A complete cycle was recorded in ten instances in a period of 19 to 23 days and in two instances in 32 days. One cottonmouth was examined periodically over a 34-day period by Allen and Swindell (1948:12), but a complete fang-shedding cycle was not observed. Fitch (1960:110) reported a 33-day cycle in copperheads; Klauber (1956:726) estimated the normal active life of each fang of an adult rattlesnake to be from six to ten weeks, but he made no observations to confirm his estimation.
Fangs measured from the tip of the notch of the basal lumen to the end of the fang vary from about 1.3 per cent of the snout-vent length in juveniles to about 1.0 per cent in large adults (Table 6). The fangs are longer than those of copperheads (Fitch, 1960:111). Klauber's (1956:736) figures on fang-lengths in all species of rattlesnakes are percentages of total length rather than of the snout-vent length. The fangs of various species of rattlesnakes range from nearly the same proportionate length as those of cottonmouths to some much longer.
From patterns of bites of venomous snakes, Pope and Perkins (1944:333-335) attempted to correlate number, size, and patterns of tooth marks with size and generic identity of the snake responsible for the bite. Distance between fangs is relatively constant for snakes of a particular size (Table 6) regardless of genus, but the fangs of a cottonmouth are directed outward to variable degrees, and puncture wounds could easily resemble those of a much larger snake (Table 7). Also there is no direct relationship between size of [450] snake and toxicity or amount of venom injected. Consequently information of this kind is of little or no value from a medical standpoint.
TABLE 6.—Correlation of Relative Fang-length and Distance Between Fangs
at Base with Snout-vent Length of Cottonmouths.
| Snout-vent length (millimeters) |
Number
in |
Average |
Number
|
Average ratio of distance between fangs to snout-vent length (percent) |
| 200-299 | 3 | 1.33 | 3 | 2.57 |
| 300-399 | 7 | 1.30 | 5 | 2.48 |
| 400-499 | 13 | 1.21 | 9 | 2.21 |
| 500-599 | 12 | 1.22 | 8 | 2.19 |
| 600-699 | 7 | 1.17 | 1 | 2.10 |
| 700-799 | 5 | 1.07 | 4 | 1.65 |
| 800-899 | 1 | 1.00 | 1 | 2.00 |
TABLE 7.—Contrast in Measurements Between the Base of the Fangs and
Between Fang Punctures of Nine Cottonmouths (in millimeters).
| Distance between base of fangs |
Distance between fang punctures |
Snout-vent length |
| 7.7 | 13.0 | 400 |
| 8.7 | 14.0 | 575 |
| 10.0 | 22.5 | 526 |
| 11.0 | 18.0-19.0 | 590 |
| 12.0 | 18.0 | 793 |
| 13.0 | 17.0, 20.0 | 558, 612 |
| 15.5 | 23.5 | 800 |
| 16.0 | 24.0 | 800 |
Although usually associated with swamps and lowlands along river bottoms, the cottonmouth lives in a variety of habitats ranging from salt marshes to cool, clear streams and from sea level to an altitude of 2300 feet. Shaded, moist areas either in or beside shallow waters are preferred, but cottonmouths occasionally wander as far as a mile from water.
In the pine-oak forests of Nacogdoches County in eastern Texas cottonmouths and copperheads are probably the most abundant species of snakes. Specimens have been collected near Nacogdoches in ponds, swamps, clear and fast-running streams with rock bottoms, and sluggish muddy streams. On the Stephen F. Austin Experimental Forest numerous cottonmouths live in a swamp until around mid-July, when it becomes dry. A small stream west of [451] the swamp seems to be used as a migration route to and from the swamp. Slightly more than a mile downstream cottonmouths are common in a bottomland area. The ground is always moist and no undergrowth occurs; a few small clear springs produce shallow trickles that run into a swamp. Cottonmouths can often be found here, lying in or beside the small trickles.
I have seen cottonmouths in various types of aquatic habitats in Brazoria County. In most places in this area, cottonmouths are found in association with one or more species of water-snakes (including Natrix cyclopion, N. erythrogaster, N. rhombifera, and N. confluens), which greatly outnumber the cottonmouth. Interspecific competition may be reduced somewhat by cottonmouths sometimes feeding on water-snakes.
The numerous statements in the literature concerning the habitat of the cottonmouth can be summarized most easily by the following short quotations:
Agkistrodon piscivorus piscivorus—"Marshes and lakes; ponds and streams with wooded shores; low country near water; roadside ponds; drainage ditches; coastal 'banks'; keys; some Gulf coast islands; mangrove swamps." (Wright and Wright, 1957:919.)
Agkistrodon piscivorus leucostoma—"Cypress, gum, river swamps; alluvial swamps wooded or not wooded; water courses of the south such as rivers, bayous, backwaters of small branches; hill streams in the north; ... marshy places in prairies ... rice fields, bottomland pools; margins of above habitats, pools, shallow lakes, swampy places, temporary flood lands. ... In, under, or on fallen timber, in holes in banks, rocky bluffs, crayfish burrows. In short it is very aquatic." (Wright and Wright, op. cit.:923.)
Geographically cottonmouths differ somewhat in their ecological requirements, but are basically much alike in most respects. The areas of greatest abundance are those having 40 inches or more of annual rainfall. The northern edge of the range has a mean temperature of approximately 38° F. in January in Virginia and 30° F. in Missouri, although the lowest temperature reached in these areas is more important as a limiting factor. The annual rainfall in both Virginia and Missouri amounts to approximately 40 inches. Moisture, as well as temperature, may play an important role in the northward distribution of the species. The eastern cottonmouth seems to be less tolerant of low temperatures than the western subspecies. Mean January temperatures equal to those along the northern limits of the western cottonmouth's distribution are reached in the vicinity of Connecticut, which is north of the geographic range of the eastern subspecies.
The depths to which cottonmouths penetrate into their dens may have a limiting influence upon the geographic range, especially in the northern extremes. Bailey (1948:215) discussed the possibility that populations of snakes may be significantly depressed because of winter kill of individuals that "hibernate" at shallow depths. He speculated also that the short growing season does not allow enough time for the essentials of existence to be carried out, and the prolonged period of inactivity overtaxes the energy reserve of the species.
Available food does not seem to be of much importance as a limiting factor, for the cottonmouth is remarkably indiscriminate in its choice of prey, feeding upon almost any vertebrate animal that happens to come within reach. Competition for food, however, may play an important role.
A review of available literature indicates no records of courtship of the cottonmouth other than statements that breeding occurs in early spring. In a close relative, the copperhead (see Fitch, 1960:159-160), mating occurs almost any time in the season of activity but is mainly concentrated in the few weeks after spring emergence, at about the time when females are ovulating. Klauber (1956:692) concluded that along the southern border of the United States rattlesnakes normally mate in spring soon after coming out of their winter retreats; but farther north where broods are produced biennially, the mating times may be more widely dispersed, and summer and fall matings may even predominate.
The only record of copulation in the cottonmouth was reported by Allen and Swindell (1948:11), who observed a pair copulating for three hours on October 19, 1946, at the Ross Allen Reptile Institute. Davis (1936:267-268) stated that courtship in cottonmouths is violent and prolonged but did not note any nervous, jerky motions or nudging of the female along her back and sides as had been observed in other genera of snakes. Carr (1936:90) saw a male cottonmouth seize a female in his mouth and hold her, but no courtship followed.
Many persons have assumed that gestation periods in snakes are the intervals between mating and parturition, and that mating and ovulation occur at approximately the same time. However, retention of spermatozoa and delayed fertilization indicate that copulation is not a stimulus for ovulation.
A biennial reproductive cycle was found for the copperhead in Kansas (Fitch, 1960:162), the prairie rattler in Wyoming (Rahn, 1942:239) and in South Dakota (Klauber, 1956:688), the great basin rattler in Utah (Glissmeyer, 1951:24), and the western diamondback rattler in northwestern Texas (Tinkle, 1962:309). Klauber's (1956:687) belief that the reproductive cycle of rattlesnakes varies with climate, being biennial in the north and annual in the south, is supported by similar climatic variation in the reproductive cycle of the European viper which was discussed by Volsøe (1944:18, 149).
If data for a large number of females were arranged as are those in Table 8, they might reveal whether the breeding cycle is annual or biennial. The figures presented in Table 8 are misleading if viewed separately because of the small number of individuals included in some of the size classes.
The smallest reproductive female found measured 455 millimeters in snout-vent length. Conant (1933:43) reported that a female raised in captivity gave birth to two young at an age of two years and ten months. The size classes represented by gravid females found by Barbour (1956:38) in Kentucky indicate that breeding occurs at least by the third year.
The ovaries of female cottonmouths examined revealed ova in various stages of development. In individuals less than 300 millimeters in snout-vent length the ovaries are almost completely undeveloped; in immature individuals from 300 to 450 millimeters in length the follicles are from one to two millimeters in length; in post-post females follicles vary in size, the largest being about seven millimeters. Reproductive females also contain follicles of various sizes. [453] One or two sets are less than three millimeters in length, and large ova that soon are to be ovulated are present. Ovarian ova found in April ranged in length from 23 to 35 millimeters. No embryonic development was observed in most individuals until June or later.
TABLE 8.—Percentage of Gravid Females of A. p. leucostoma in 50 Millimeter Size Classes.
| Snout-vent length |
Number of gravid females |
Total number in size class |
Percentage gravid |
| 450-499 | 3 | 14 | 21.4 |
| 500-549 | 7 | 17 | 41.2 |
| 550-599 | 8 | 17 | 47.1 |
| 600-649 | 5 | 7 | 71.4 |
| 650-699 | 2 | 9 | 22.2 |
| 700-749 | 2 | 3 | 66.7 |
| 750-799 | 1 | 1 | 100.0 |
| 850-899 | 1 | 1 | 100.0 |
| Totals | 29 | 69 | 42.0 |
Increase in length of testes appears to be correlated with length of the individual rather than cyclic reproductive periods (Fig. 4).
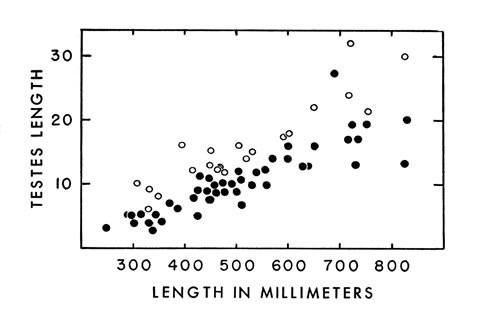 Fig. 4. Length of testes in cottonmouths of various sizes
Fig. 4. Length of testes in cottonmouths of various sizes The reproductive cycle in cottonmouths resembles that illustrated by Rahn (op. cit.:237), in which the ovarian follicles of post-partum females begin to enlarge in late summer and autumn, with ovulation occurring the following spring. By means of retaining sperm successive broods possibly are produced after only one mating. In captivity, at least, some females may not follow this biennial cycle; Stanley Roth (M.S.), biology teacher in high school at Lawrence, Kansas, had a female of A. p. piscivorus, from Florida, that produced broods of 14 and 12 young in two consecutive years.
After ova are fertilized a three and one-half to four-month period of development begins which varies somewhat depending on the temperature. In almost every instance the ova in the right uterus outnumber those in the left. Embryos usually assume the serpentine form in the latter part of June and are coiled in a counterclockwise spiral with the head on the outside of the coil. At this time the head is relatively large and birdlike in appearance with conspicuous protruding eyes. Sex is easily noted because the hemipenes of males are everted. By late July scales are well developed and the embryo is more snakelike in appearance, but pigmentation is still absent. By mid-August the color and pattern are well developed, the egg tooth is present, the snake shows a considerable increase in size over that of the previous month, and much of the yolk has been consumed. Some females that contain well developed embryos also contain eggs that fail to develop. Sizes of ova vary irrespective of size of female and stage of embryonic development. Lengths of ova ranged from 22 to 51 millimeters in May to 35 to 49 millimeters in July and August. A two-yolked egg was found in one female.
Accounts in the literature of 15 litters of cottonmouths fix the time of birth as August and September. Conant (1933:43) reported the birth of a litter in mid-July by a female that had been raised in captivity, and one female that I had kept in captivity for two months gave birth to a litter between October 19 and October 25. The conditions of captivity undoubtedly affected the time of birth in both instances.
Wharton (1960:125-126) reported the birth and behavior of a brood of seven cottonmouths in Florida. I was given notes of a similar nature by Richard S. Funk of Junction City, Kansas, on a brood of five cottonmouths. The mother of the brood was caught in June, 1962, in Tarrant County, Texas, by Richard E. Smith, and was 705 millimeters in snout-vent length. The first young was found dead in an extended position a few inches from the fetal membranes at 11:05 p.m. on August 22. The second young was born at 11:07 p.m. The intervals between the successive births were three, seven, and four minutes; and time until the sac was ruptured in each instance was six, five, eight, and 11 minutes. The time interval between the rupture of the sac and emergence of each individual was 41, 92, 154, and 34 minutes. The mother's actions in giving birth to the last four young were essentially as described by Wharton (loc. cit.), except that the intervals between successive births did not increase. Within one minute after rupturing the sac and while its head was protruding, each of the four living young opened its mouth widely from three to seven times, then took its first breath. Breaths for the first three hours were steady at three or four per minute but then decreased to two or three per minute. Pulse rate for the four averaged 38 per minute while at rest but increased to 44 per minute after voluntarily crawling.
Records of from one to 16 young per litter have been reported (Ditmars, 1945:330; Clark, 1949:259), but the average is probably between six or seven. Most accounts in the literature present information on number of ova or embryos [455] per female rather than the number of young. Size and age of the mother (Table 9) influence the number of ova produced. Allen and Swindell (1948:11) recorded three to 12 embryos in 31 cottonmouths varying in total length from 26 to 44 inches. An average of 6.5 embryos per female was found.
TABLE 9.—Number of Ova Produced by Fecund Cottonmouths.
| Snout-vent length in millimeters |
Number in sample |
Number of ova, average and extremes |
| 450-549 | 10 | 4.1 (2 to 7) |
| 550-649 | 11 | 4.9 (1 to 8) |
| 650-749 | 4 | 6.3 (4 to 8) |
| 750-849 | 1 | 5 |
| 850-949 | 1 | 14 |
Mortality at birth has been recorded for almost every litter born in captivity (see Allen and Swindell, loc. cit.; Conant, 1933:43; Wharton, 1960:125). A female that I kept in captivity gave birth to seven young. Three never ruptured their sacs, and another died soon after leaving the sac. The effects of captivity on females may result in higher rates of deformity and mortality in young than is common in nature. Klauber (1956:699-700) estimated that the defects brought about by conditions of captivity on rattlesnakes eliminate about three young per litter.
No investigator has yet analyzed the composition of a population of cottonmouths according to age, sex and snout-vent length. Barbour (1956:35) did sort 167 snakes into size classes, but did not determine sex ratio, size at sexual maturity, reproductive cycles, or snout-vent length. He recorded total lengths from which snout-vent lengths cannot be computed because of differential growth rates and different bodily proportions of the two sexes. I judge from my findings that he included immature individuals in his three smallest size classes (45.5 per cent of the population). I found at least 32.5 per cent immature individuals (Fig. 5) in my material, but it was not a natural population.
The sex ratios of several small collections from natural populations varied, and no conclusions could be drawn. Females comprised 53 per cent of the specimens included in Fig. 5 and in a group of 48 embryos which represented eight broods. That percentage may not be the percentage in a natural population but is used in making assumptions because I lack better information.
If data in Fig. 5 are representative of a natural population and if 61 per cent of the females are sexually mature, the reproductive potential can be estimated as follows: assuming a cohort of 1000 cottonmouths contains 530 females, 61 per cent of the females (323 individuals) probably are adults. If 42 per cent of these females produce 6.5 young per female in any season (Tables 8 and 9), 136 females will produce 884 young. But if 50 per cent [456] of the adult females are reproductive (as would be assumed if reproduction is biennial), 1050 young will be produced. Actually the number of young required per year to sustain a population is unknown, because mortality rates at any age are unknown.
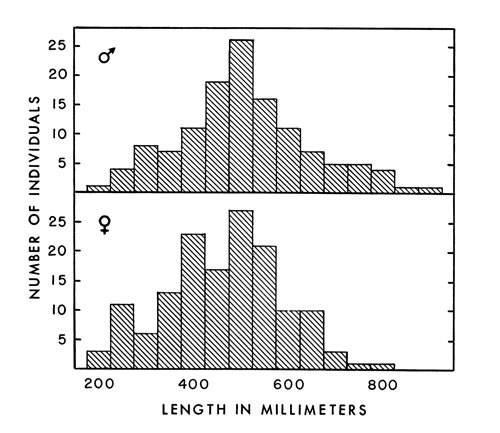 Fig. 5. Composition of a group of cottonmouths examined in this
study. Individuals less than 450 millimeters in snout-vent length
are considered as immature. Specimens from 200 to 249 millimeters
in length are included in the 200-millimeter class, etc.
Fig. 5. Composition of a group of cottonmouths examined in this
study. Individuals less than 450 millimeters in snout-vent length
are considered as immature. Specimens from 200 to 249 millimeters
in length are included in the 200-millimeter class, etc. Size at birth depends on the health of the mother. According to Fitch (1960:182), many litters of copperheads born in captivity are stunted. Seven young cottonmouths (two males and five females) born in captivity were each 185 millimeters in snout-vent length and 40 millimeters in tail length. Weights of the three living young were 10.0, 10.1, and 11.1 grams. Another litter of five young measured by Richard S. Funk were larger, and differences in the proportions of the tail length and snout-vent length suggest the sexual dimorphism found in larger individuals. However, sex of these young snakes was not recorded. Snout-vent length and tail length in millimeters were 232, 41; 243, 47; 229, 40; 240, 48; and 225, 40 in the order of their birth. These snakes are considerably smaller than the nine young of A. p. piscivorus reported by Wharton (1960:127) that averaged 338 millimeters total length and 28.7 grams. The yolk of one young piscivorus was 11.7 per cent of the total weight. [457] Yolk is used up in about two weeks if its rate of utilization resembles that of the copperhead as reported by Gloyd (1934:600).
Early rates of growth of three living young are shown in Table 10. On the 56th day after birth, each was fed one minnow less than two inches long. Between the 80th and 120th days three additional small minnows were fed to each snake. Young cottonmouths increase nearly 50 millimeters in length by the first spring if they inhabit warm areas and feed in autumn or winter.
Variation in size of newborn cottonmouths may be less in nature than in captivity. Average size at birth can be determined accurately by the size of young captured in early spring, at least in northern parts of the range where winter feeding and growth do not occur at all or are negligible. Total lengths of 19 juveniles thought by Barbour (1956:38) to be seven to eight months old do not differ markedly from lengths of the five newly-born young measured by Funk.
TABLE 10.—Rate of Growth of Three Young Cottonmouths.
| Age in days |
Snout-vent length / tail length—weight in grams | ||
| Female No. 1 | Female No. 2 | Male | |
| 2 | 185/40—11.1 | 185/40—10.1 | 185/40—10.0 |
| 7 | 192/40— | 190/40— | 189/40— |
| 22 | 195/40—10.3 | 200/41.5—10.6 | 197/40— |
| 80 | 204/40—11.7 | 203/42—10.4 | 218/48—14.3 |
| 88 | .... | 204/44— | .... |
| 143 | 215/40.5—13.3 | .... | 225/48—15.1 |
The umbilical cord is broken at birth and the navel closes within a few days; but the scar, involving from two to four ventral scales, remains throughout life. Position of the scar was found by Edgren (1951:1) to be sexually dimorphic in the eastern hog-nose snake (Heterodon platyrhinos), but nothing has been published on this matter concerning the cottonmouth. Consequently, I counted the scales of several individuals from the anal plate, and there was no marked difference in the position of the scar in males and females; it varied in position from the 10th to the 18th scale. When counted from the anterior end, the scar ranged from ventral number 115 to 122 (average, 119) in 28 females and from number 117 to 126 (average, 121) in 14 males. The difference between male and female cottonmouths is not nearly so great as in Heterodon.
The only records of growth increments in a natural population of cottonmouths are those in Table 11. The period of growth is mostly the period of activity, and differences are expected between northern and southern populations. As size increases, determination of growth rate becomes more difficult because age classes overlap in size. Growth of any individual depends not only on climate and food but also on disease and parasitism and the innate [458] size potential. Stabler (1951:91) showed weight and length relationships in two cottonmouths for a period of six and one-half years.
TABLE 11.—Growth Increments in Cottonmouths (Barbour, 1956:38-39).
| Number of individuals |
Total length in millimeters |
Estimated age in months |
Estimated growth from preceding year in millimeters |
| 19 | 260-298 | 7-8 | 25 |
| 11 | 312-337 | 19-20 | 45 |
| 40 | 355-485 | 31-32 | 95± |
| 83 | 500-1000 | 43-44+ | ? |
My study failed to reveal any secondary sexual difference in growth rate and maximum size. Of the 306 cottonmouths measured by me, 16 males and five females exceeded 700 millimeters in snout-vent length. Two males were more than 850 millimeters long. One cottonmouth lived in captivity for 18 years and 11 months (Perkins, 1955:262). The maximum total lengths were reported by Conant (1958:186-187) to be 74 inches (1876 mm.) in A. p. piscivorus and 54 inches (1370 mm.) in A. p. leucostoma.
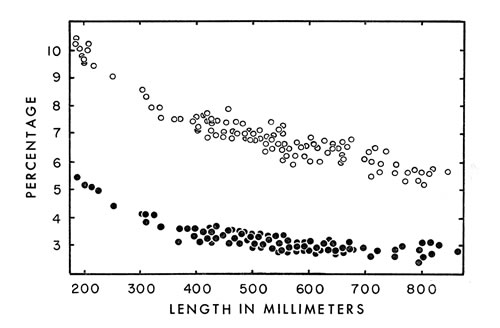 Fig. 6. Head length ( ° ) and head width ( · ) expressed as a percentage
of snout-vent length of living and preserved cottonmouths. Head length
was measured from the tip of the snout to the posterior end of the mandible.
Head width was measured across the supraocular scales, since accuracy was
greater than if measured at the posterior edge of the jaw. No sexual dimorphism
or geographical variation occurs in these characters.
Fig. 6. Head length ( ° ) and head width ( · ) expressed as a percentage
of snout-vent length of living and preserved cottonmouths. Head length
was measured from the tip of the snout to the posterior end of the mandible.
Head width was measured across the supraocular scales, since accuracy was
greater than if measured at the posterior edge of the jaw. No sexual dimorphism
or geographical variation occurs in these characters. Proportions of various parts of the body vary considerably depending on age, size and, in some instances, sex. Heads are proportionately larger in young than in adults (Fig. 6), as is true of vertebrates in general. This larger head has survival value for the cottonmouth in permitting more venom [459] to be produced and in permitting it to be injected deeper than would be the case if the proportions were the same as in adults. Relative to the remainder of the snake the head is considerably larger than in the copperhead (Fitch, 1960:108) and slightly larger than in the rattlesnake, Crotalus ruber (Klauber, 1956:152).
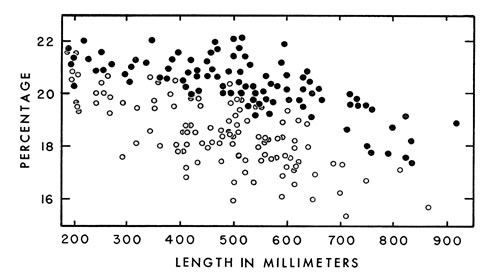 Fig. 7. Tail length expressed as a percentage of snout-vent length of living and preserved cottonmouths ( ·—males; °—females ).
Fig. 7. Tail length expressed as a percentage of snout-vent length of living and preserved cottonmouths ( ·—males; °—females ). In general, tails are relatively longer in males than in females of the same size (Fig. 7), except that there is little or no difference at birth. Growth of the tail in males proceeds at a more rapid rate. In certain individuals sex cannot be recognized from length of the tail relative to snout-vent length because overlapping occurs, especially in medium-sized individuals. Similar changes of proportions with increase in age occur in copperheads (Fitch, 1960:106) and rattlesnakes (Klauber, 1956:158-159), but the tail of the cottonmouth is relatively much longer.
Shedding of the skin is necessary to provide for growth and wear in snakes. The milkiness or bluing of the eyes, which causes partial blindness, marks the initial stage of shedding and is caused by a discharge of the exuvial glands that loosens the old stratum corneum from the layer below. In four to seven days the opaqueness disappears, and the snake sheds after an additional three to six days (Table 12). Young snakes first shed within a few days after birth and generally shed more frequently than adults, but the interval is variable. The eyes of three young cottonmouths observed by Wharton (1960:126) became milky on the fourth day but cleared on the seventh day, and the skin was shed on the eighth day. The eyes of three young kept by me became milky two to three days after birth, cleared on the seventh to tenth days, and the skin was shed on the thirteenth day. Possibly the relatively long interval in [460] this instance resulted from low relative humidity in the room where the snakes were kept. According to Fitch (1960:134), litters of young copperheads usually shed within three to ten days after birth; but under unusually dry conditions shedding did not occur for several weeks.
TABLE 12.—Duration of Preparatory Period (in days) to Shedding in 11 Cottonmouths.
| Duration of cloudiness of eyes |
Time between clearing and shedding |
Time from beginning of cloudiness until shedding |
| 5 | 6 | 11 |
| 7 | 3 | 10 |
| - | - | 6 |
| - | - | 6 |
| 5 | 3 | 8 |
| 4 | 6 | 10 |
| 7 | 3 | 10 |
| 5 | 6 | 11 |
| 5 | 3 | 8 |
| 7 | - | - |
| 7 | 3 | 10 |
| X 5.4 | X 3.8 | X 9.0 |
Cottonmouths as well as other snakes usually do not feed until after the skin is shed and are generally quiescent during the period preceding shedding, except that immediately before shedding they become active and rub their snouts on some rough object and may yawn several times seemingly in an attempt to loosen the skin along the edges of the lips. After the skin is loosened from the head, more rubbing against rough surfaces and writhing serves to pull the old skin off, turning it inside out. Once the old skin has passed over the thick mid-body, the snake often crawls forward using rectilinear locomotion until the skin is completely shed. It normally comes off in one piece; but, if the snake is unhealthy or has not had sufficient food or water, the skin may come off in patches. Frequently one or both of the lens coverings are not shed immediately and impair the sight. Bathing or swimming ordinarily causes dried skin to peel off; and, because of the cottonmouth's aquatic habits, its chances of shedding successfully are much greater than those of less aquatic snakes. Cottonmouths that have recently shed have bright and glossy patterns, in contrast to the dull and dark appearance of those that are preparing to shed.
Most of our knowledge concerning the frequency of shedding is based upon observations of captives. It is known that the intervals between exuviations are largely dependent upon the amount of food taken and the rate of growth. Unless laboratory conditions closely resemble those in the field, shedding frequencies in captives probably differ much from those of free-living snakes.
Only two of my captives shed twice. The intervals between exuviations [461] in the two snakes were eight and five months, lasting from August to April and from December to May, respectively. Ten other snakes shed once in the period from January through July. Stabler (1951:91) presented data concerning shedding of two cottonmouths kept 12 and 14 years in captivity. One shed 25 times in 12 years and the other shed 37 times in 14 years, giving an average of 2.1 and 2.6 per year, respectively. Neither of the snakes shed from December through March, but the period of shedding corresponded to the period of greatest activity and growth. In Florida, cottonmouths shed four to six times a year, according to rate of growth (Allen and Swindell, 1948:7).
Food is obtained by a variety of methods depending on the type of food, age of the cottonmouth, and possibly other factors. Some captives lie in ambush and others crawl slowly in active search. At the first cue of possible prey, either by sight, scent, or differential temperature detection by the pit, the snake appears to become alert and flicks its tongue out at fairly rapid intervals.
By means of the facial or loreal pit found in all crotalids, the snake is able to detect objects having temperatures different from that of the surroundings of the objects. In detecting prey the tongue acts to sharpen the sense of "smell" by conveying particles to Jacobson's organs in the roof of the mouth. On many occasions cottonmouths appeared to rely solely on sight; they passed within a few inches of prey, apparently unaware of its presence until it moved. When pools of water begin to dry up toward the end of summer, cottonmouths often congregate and feed on dying fish. In these instances the fish are usually taken as they come to the surface. In laboratory observations moccasins seize live fish and some moccasins carry the fish until they have received lethal doses of venom; afterward the fish are swallowed. But grasping and manipulation of the prey occurs without the fangs' being employed, especially in the case of dead fish. On one occasion a cottonmouth was observed to grasp the edge of a glass dish that had contained fish and apparently retained the odor. On another occasion I placed several fish in a bowl, rubbed a stick on the fish, and then touched each snake lightly on the nose with the stick. The snakes crawled directly to the bowl and began feeding. At other times these same snakes crawled around the cage in an apparent attempt to locate the food but paid little attention to fish held in front of them. If the catching of prey under natural conditions were as uncoordinated as it sometimes is in captivity, the snakes probably would not be able to survive.
Wharton (1960:127-129) described tail-luring in one individual of a 76-day-old brood of cottonmouths. The snake lay loosely coiled with the tail held about six centimeters from the ground; a constant waving motion passed posteriorly through the terminal inch of the tail. These movements ceased at 7:20 p.m. but were resumed at 7:40 a.m. the following day. All observations were under artificial light. The "caudal lure" as a means of obtaining prey has been described in other species and related genera by Neill (1960:194) and Ditmars (1915:424).
Various authors have suggested that the method of capture differs according to the kind of prey. Allen and Swindell (1948:5) stated that cottonmouths retain their hold after striking fish or frogs but will release a mouse after delivering a bite and are timid in striking at larger rodents. Neill (1947:203) noted that a cottonmouth always waited several minutes after biting a large rat before approaching its prey. This same type of behavior has been reported for copperheads (Fitch, 1960:194) and rattlesnakes (Klauber, 1956:618). [462] Cottonmouths observed by me retained a strong hold on fish, frogs, and sometimes mice, but almost always released large mice and baby chicks, which were not eaten until after death.
Different behavior according to type of prey is correlated with ability of prey to retaliate, although some animals may not be released because they could easily escape. For instance, a frog could hop far enough to escape in a matter of seconds if released. A 73-millimeter Rana pipiens that I observed was bitten twice within one and a quarter hours and died 45 minutes after the last bite. Its movement was uncoordinated by the time of the second bite, but it could have escaped had the frog not been confined. Although it is doubtful that normal, healthy fish are frequently captured by cottonmouths, Allen (1932:17) reported that a cottonmouth was seen pushing a small, dead pike about on the surface of a stream. A wound on the belly of the fish indicated that it had been bitten. A 17-gram creek chub (Semotilus) and a 13.7-gram bass (Micropterus) were injected by me with one-fourth cubic centimeter of fresh venom near the base of the tail in order to determine whether the fish could escape after being bitten and released. The creek chub flipped onto its back after a minute and 45 seconds and gill movements stopped in eight minutes and 35 seconds; the bass flipped over after 50 seconds and died in two minutes and 10 seconds. The venom immediately affected both fish, and it is unlikely that either could have swum more than a few feet.
After its prey has been killed, a cottonmouth examines the body from end to end by touching it with the tongue. Then the animal is grasped in the mouth without the use of the fangs and is slowly manipulated until one end (usually the head) is held in the mouth. The lengthy process of swallowing then takes place, the fangs and lower jaws alternately pushing the prey down the throat.
The cottonmouth seems to be an opportunistic omni-carnivore, because it eats almost any type of flesh that is available, including carrion. It feeds primarily upon vertebrates found in or near water; but invertebrates and eggs have also been found in the diet. The only potential prey items that seem not to be normally eaten are bufonid toads and tadpoles. I have occasionally offered tadpoles and frogs to cottonmouths, but only the frogs were accepted. But, Stanley Roth kept a cottonmouth in captivity that ate both toads and tadpoles. If tadpoles are commonly eaten, their probable rapid digestion would make identification almost impossible.
Following is a list of known foods of the cottonmouth:
Captivity: "... rattlesnake.... The same moccasin also killed and ate a smaller snake of its own species...." (Conant, 1934:382.)
Florida: "3 heron feathers, bird bone, Eumeces inexpectatus, 3 fish all under one inch in length, 1 heron egg shell" (Carr, 1936:89). According to Allen and Swindell (1948:5), "the food included other moccasins, prairie rattlesnakes, king-snakes, black snakes, water snakes, garter snakes, ribbon snakes, and horn snakes ... most of the species of frogs, baby alligators, mice, rats, guinea pigs, young rabbits, birds, bats, squirrels, and lizards ... a mud turtle ... a case of a four footer eating ten to twelve chicken eggs. The most common food appears to be fish and frogs. Catfish are included on this list...." Yerger (1953:115) mentions "an adult yellow bullhead, Ameiurus natalis ... 306 mm. in standard length [from a 63-inch cottonmouth]."
Georgia: "... full grown Rana catesbeiana, several foot-long pickerel ... dead fish if placed in a pan of water.... Natrix sipedon fasciata and Masticophis flagellum ... rats.... Toads and large Eumeces laticeps were always ignored." (Neill, 1947:203.) "Natrix, Heterodon, Kinosternon, Rana, Hyla cinerea, Microhyla, Microtine [Pitymys pinetorum]." (Hamilton and Pollack, 1955:3.)
Mississippi: "... Hyla gratiosa.... In captivity specimens have eaten frogs, mice, birds, dead fish, pigmy rattlers and copperheads. Toads ... were refused" (Allen, 1932:17). One moccasin "disgorged a smaller decapitated moccasin ... killed the day before by boys" (Smith and List, 1955:123).
Tennessee: "Beetles in one stomach; lizard (Eumeces) in another stomach; small snake (Natrix) in one intestine, and hair in another intestine. One stomach contained numerous bits of wood, up to four inches in length...." (Goodman, 1958:149.)
Kentucky: "Siren intermedia was the most abundant food item in both volume and occurrence. Frogs of the genus Rana ranked second. Together, these two items comprised almost 2/3 of the food of the snakes. The other food items were distributed among the fishes, reptiles, and other amphibians [one Rana tadpole included]." (Based on 42 samples—Barbour, 1956:37.)
Illinois: (Based on 84 samples—Klimstra, 1959:5.)
Food Item Per cent Frequency
of OccurrencePer cent
VolumePisces 39.3 31.9 Amphibia 36.9 26.0 Reptilia 25.0 18.2 Mammalia 30.9 17.9 Gastropoda 17.8 1.0 Miscellaneous 25.0 5.0 (Algae, Arachnida, Aves, Insecta) Louisiana: Penn (1943:59) mentions that a "female had just eaten two young cottonmouths...." Clark (1949:259) mentions "100 specimens—34 fish; 25 Rana pipiens; 16 Rana clamitans; 7 Acris; 4 Natrix sipedon confluens; 8 birds; 5 squirrels ... catfish thirteen and one-half inches in length ... small-mouth black bass [eleven inches]."
Oklahoma: Force (1930:37) remarks that the moccasin "eats bullfrogs ... but refuses leopard frogs." Trowbridge (1937:299) writes: "several sun perch.... Another had eaten six catfish six to ten inches long ... a water snake (Natrix s. transversa) about 18 inches long ... frogs, mostly Rana sphenocephala." Carpenter (1958:115) mentions "a juvenile woodthrush.... Seven last instar cicadas ... a young cottontail." According to Laughlin (1959:84), one moccasin "contained the following items: 18 contour feathers of a duck, probably a teal; one juvenile cooter turtle, Pseudemys floridana; and a large mass of odd-looking unidentifiable material. The other cottonmouth contained one juvenile pond turtle, Pseudemys scripta...."
Texas: "... several ... feeding on frogs.... One ... found DOR was found to contain a large catfish." (Guidry, 1953:54.)
Of 246 cottonmouths that I examined for food items, only 46 contained prey in their digestive tracts. Almost all of the snakes examined were museum specimens that had been collected at many places over a period of about 40 years. It was not known how long each had been kept alive before being preserved. Therefore it was impossible to determine what proportion of any population of cottonmouths could be expected to contain food. The food items were not analyzed numerically because the scales and hair, by means of which many food items in the intestine were identified, yielded no clue as to the number of individuals actually present unless several [464] distinct kinds were found. Each occurrence of scales or hair was thus recorded as a single individual, although some such occurrences may have represented more than one animal. The contents of some stomachs were so well digested that it was difficult to determine the number of items present. As a rule only one food item was present in a digestive tract, but a few tracts contained several items of the same or different species. Three frogs (Acris crepitans) were in one snake and three hylas (Hyla versicolor) in another. Still another individual captured beside a drying pond contained six individuals of Lepomis each about three inches long and two pikes (Esox) about six inches long.
TABLE 13.—Analysis of Food Items of 46 Cottonmouths Collected in Arkansas, Louisiana, and Texas (1922-1962).
| Food Items | Number of samples in which item occurred |
Percent frequency of occurrence |
Estimated weight in grams |
Estimated percentage by bulk |
|
| Fish | (7) | 13.2 | 18.4 | ||
| Esox sp. | 1 | 20 | |||
| Lepomis sp. | 2 | 15 | |||
| Unidentified | 4 | ||||
| Amphibians | (12) | 23.0 | 20.4 | ||
| Scaphiopus hurteri | 1 | 13 | |||
| Acris crepitans | 2 | 4 | |||
| Hyla cinerea | 2 | 12 | |||
| Hyla versicolor | 1 | 12 | |||
| Rana catesbeiana | 1 | 20 | |||
| Rana pipiens | 3 | 15 | |||
| Unidentified | 2 | ||||
| Reptiles | (15) | 28.4 | 29.9 | ||
| Pseudemys scripta | 2 | 15 | |||
| Anolis carolinensis | 1 | 6 | |||
| Eumeces fasciatus | 1 | 7 | |||
| Lygosoma laterale | 2 | 5 | |||
| Natrix sp. | 1 | 10 | |||
| Natrix erythrogaster | 2 | 10 | |||
| Agkistrodon piscivorus | 2 | 20 | |||
| Crotalus sp. | 1 | 30 | |||
| Unidentified snakes | 3 | ||||
| Birds | (4) | 7.6 | 18.6 | ||
| Anhinga anhinga (juv.) | 1 | 60 | |||
| Egret (head and neck) | 1 | 20 | |||
| Passeriformes | 2 | 20 | |||
| Mammals | (6) | 11.3 | 12.7 | ||
| Blarina brevicauda | 1 | 12 | |||
| Cricetinae | 5 | 18 | |||
| Unidentified | (9) | 17.0 | |||
The "unidentified" category (Table 13) refers to jellylike masses in the stomach or material in the intestine in which no scales, feathers, hair, or bones [465] could be found. Most of the unidentifiable matter could be assumed to consist of remains of amphibians, since they leave no hard parts. If this assumption is correct, amphibians comprise about 40 per cent of the diet. Since intestinal contents were included, a volumetric analysis was not feasible. Therefore, the weight of each type of food item was estimated and the percentage by bulk calculated from it (Table 13).
Pieces of dead leaves and small sticks constituted most of the plant material found and presumably were ingested secondarily because they adhered to the moist skin of the prey, especially to fish and amphibians. However, some plant materials probably are eaten because they have acquired the odor of the prey. One cottonmouth contained a Hyla cinerea, several leaves, and five sticks from 37 to 95 millimeters long and from 12 to 14 millimeters in diameter.
Most reports in the literature state that gravid females do not feed, but four gravid females examined by me containing large, well-developed embryos also contained evidences of having recently fed. Two of them had scales of snakes in the stomach or intestine, one contained a six-inch Lepomis, and the other had hair in the intestine and the head and neck of an adult egret in the stomach.
Published records of other animals preying on cottonmouths or killing them are few. Reptiles more often than other classes of vertebrates prey on the cottonmouth. McIlhenny (1935:44) reported on the scarcity of snakes in areas where alligators were present. Predation on cottonmouths by indigo snakes (Drymarchon corais) was reported by Conant (1958:153) and Lee (1964:32). Allen and Swindell (1948:6) obtained a photograph of a king-snake (Lampropeltis getulus) killing a cottonmouth but thought that moccasins are not eaten by L. getulus. However, one occasion reported herein shows that cottonmouths are eaten by king-snakes; and Clark (1949:252) reported finding 13 cottonmouths, along with other prey, in the stomach contents of 301 king-snakes (L. g. holbrooki) from northwestern Louisiana. Cannibalism is also common among cottonmouths. Klauber (1956:1058;1079) cited predation on cottonmouths by a blue heron (Ardea herodias) and a largemouth bass (Micropterus salmoides). Man is probably the greatest enemy of the cottonmouth. Intentional killing, capturing, road kills, and alteration of the environment destroy large numbers.
Allen and Swindell (1948:12) listed several diseases and parasites of snakes and stated that "some moccasins captured in the woods are so poor and weak from parasitic infection that they can barely crawl." The only kind of ectoparasite found on captive cottonmouths in the course of my study was a snake mite, Ophionyssus natricus. An infestation of that mite was thought to be partly responsible for the death of one captive moccasin. Other moccasins spent increasing amounts of time in their water dish after they became infected with mites. Under natural conditions frequent swimming probably keeps cottonmouths nearly free of mites.
Endoparasites found included lung flukes, stomach nematodes, and tapeworms. Lung flukes (Ochetosoma sp.) were found in 16 of 20 captive cottonmouths. [466] Snails and frogs serve as intermediate hosts for various stages in the life cycle of these flukes. The high percentage of cottonmouths infested with flukes is indicative of the use of frogs as a major source of food. Less than ten flukes were usually observed in the snakes' mouths but occasionally more were seen. One snake was observed thrashing about in its cage for nearly an hour, after which time it died. Upon examination of the mouth, 32 flukes were found, most of which were located in the Jacobson's organs. Whether or not flukes caused the death is not known. Nematodes (Kalicephalus sp.) were found in the stomach of each of several preserved specimens; most of these snakes had no food in their digestive tracts. In a high percentage of the moccasins, tapeworms (Ophiotaenia sp.) were in the duodenum, in many instances so tightly packed as seemingly to prevent passage of food. The importance of fish in the diet is reflected by the high percentage of snakes containing tapeworms. An unidentified cyst (?) about an inch long and containing two hooks on one end was found attached to the outer wall of the stomach of a cottonmouth. Yamaguti (1958) listed all the kinds of helminths known from cottonmouths.
Munro (1949:71-72) reported on the lethal effect of 10 per cent DDT powder on two young cottonmouths which were dusted with it to kill mites. Herald (1949:117) reported an equal effect caused by spraying a five per cent DDT solution in a room with several snakes. All but three large cottonmouths, which were under shelter at the time of spraying, were killed.
One individual that refused to eat was dissected soon after death, and a short piece of a branch on which two large thorns were located at 90° angles was found blocking the intestine at the posterior end of the stomach.
An unexpected and probably unusual circumstance caused the death of two captives. After cleaning a cage containing five cottonmouths and placing several mice in the cage for food, I noticed two of the snakes lying stretched out, partially on one side, and almost unable to move. At first I thought they had been bitten by other snakes which were in pursuit of the mice. The two died after two days. When a similar incident occurred in another cage, I removed the "bitten" snake and it fully recovered after 11 days. When the same symptoms were observed in a garter snake in another cage, I realized that in each instance the cage had been cleaned and fresh cedar chips placed in it immediately prior to observation of these symptoms. Fine cedar dust on the chips had evidently poisoned the snakes.
In the days following emergence in spring, cottonmouths often endure uncomfortable and even dangerous temperatures in order to obtain food and mates. They are more sluggish at this time and more vulnerable to predation than later in the season when temperatures are optimal. Fitch (1956:463) found that copperheads in northeastern Kansas begin their annual cycle of activity in the latter part of April, when the daily maximum temperature is about 22° C. and the minimum is about 4° C., and become dormant in late [467] October or early November, at which time the daily maximum temperature is about 15° and the minimum is about 0°. Indications are that in the northern part of its range the annual activity cycle of the cottonmouth resembles that of the copperhead in northeastern Kansas. Klimstra (1959:2) captured cottonmouths from April to October in southern Illinois. Barbour (1956:36) collected large numbers of them in early April in Kentucky and stated that they migrate from swamps to wooded hillsides in late August and early September. Spring migrations begin after a few consecutive warm days in March. In northern Oklahoma cottonmouths have been found along the Verdigris River as early as March, suggesting that a few winter in crayfish holes and mammal burrows. The majority of individuals found in this area were at denning sites along cliffs above the river and emerged later than those near the river (Dundee and Burger, 1948:1-2). In Virginia cottonmouths have been seen as early as March 5 (Martin and Wood, 1955:237) and as late as December 4. They have been observed in migration from the swamps of the barrier beach to the mainland in late October and early November in southeastern Virginia (Wood, 1954a:159). According to Neill (1947:204), the cottonmouth tolerates lower temperatures than do most snakes in Georgia and is one of the last to go into hibernation. Allen and Swindell (1948:4) stated that cottonmouths usually bask during the mornings of the cooler months in Florida, but they mentioned nothing of denning such as occurs farther north. Although winter aggregations occur in the northern parts of the range, I have never seen such aggregations in the South. However, in one instance related to me by a reliable observer, seven cottonmouths were found together on a creek bank near the Gulf Coast in early spring.
During late summer and early autumn, fat is deposited in lobes in the lower abdomen in preparation for the period of winter quiescence. Gravid females usually do not feed so frequently or so much as other snakes, because they tend to become inactive as the ova develop. Whether or not females feed heavily after parturition and previous to denning is not known. Peaks of activity in autumn may be caused by final attempts to feed before denning and by the appearance of large numbers of newborn young. The young usually have from one to two months in which to feed before the advent of cold weather. According to Barbour's (op. cit.:38) findings, the young probably feed before hibernation because they grow substantially in winter. For those that do not feed, the rate of survival is perhaps much lower.
In preparation for winter, cottonmouths migrate inland, usually to dry forested hillsides where they den, commonly among rocks at the tops of bluffs, along with several other species of snakes. In such aggregations there is no hostility and each individual may derive benefit from contact with others by which favorable conditions of temperature and humidity are maintained.
Neill (1947:204) has found many specimens in winter by tearing bark from rotting pine stumps on hillsides overlooking lakes or streams. On cold days they evidently retreat below the surface, while on warm days they lie just below the bark or emerge and bask. Neill believes that the use of stumps by cottonmouths is an innate pattern of behavior, because of the large number of young-of-the-year found in such surroundings. Cottonmouths were observed in winter also under logs and stumps by Allen (1932:17). I have twice observed cottonmouths crawling into crayfish burrows along the Gulf Coast of Texas, and suppose they are used as denning sites to some extent.
The diel cycle of activity of cottonmouths is of necessity closely related to the seasonal cycle. Since optimal temperatures determine activity, the diel cycle varies greatly from time to time. It has been well established that cottonmouths, like most other crotalids and many snakes of other families, prefer nocturnal to diurnal activity, even though the temperature may be less favorable at night. This preference is correlated with increased nocturnal activity of frogs and reptiles that constitute the principal food supply.
During spring and autumn, activity is more restricted to the day and long periods of basking occur. However, as hot weather approaches, basking occurs mainly in the morning and evening and activity becomes primarily nocturnal. But, in well shaded, moist forests, cottonmouths feed actively in the daytime.
Availability of food also has an important influence upon activity. Allen and Swindell (op. cit.:5) stated that moccasins congregate around drying ponds and feed on dying fish until the moccasins can hold no more. They then usually stay nearby as long as food remains. In an area of the Stephen F. Austin Experimental Forest near Nacogdoches, Texas, many cottonmouths journey daily to and from a swamp and a dry field, evidently to feed on rodents inhabiting the area. Ten individuals captured along a snake-proof fence that was built 30 yards from the swamp were found lying coiled along the fence after 4:30 p.m., at which time the area was shaded. On another occasion, I captured a large cottonmouth that was feeding upon dying fish in a drying pool about 10:30 a.m. on August 19, 1962.
Because of the aquatic habits of the cottonmouth, relative humidity probably has little influence on the snake's activity. However, cottonmouths are more restricted to the vicinity of water in dry weather than during rains or muggy weather when many of their natural prey species also move about more freely. Increased activity on cloudy days may result from protection from long exposure to sunshine. Torrential rains and floods, such as those following hurricanes along the Gulf and Atlantic coasts of the southeastern United States, bring out quantities of snakes of all species. Rattlesnakes and cottonmouths in particular are killed by the thousands at these times because they seek shelter in human habitations. However, these are unusual circumstances and do not reflect voluntary activity as a result of preferences.
Thermal reactions of reptiles were classified by Cowles and Bogert (1944) into several categories. For each species there is a basking and normal activity range limited by the voluntary minimum and voluntary maximum at which the animal seeks shelter. Beyond this normal range are the critical thermal minimum and critical thermal maximum (C. T. M.) at which effective locomotion is prevented. The lethal minimum and maximum are those temperatures at which short exposure produces irreparable damage, and death inevitably results. These classifications are modified somewhat by seasonal or laboratory acclimation or by the physiological state of the animal. The C. T. M. of five cottonmouths was determined by placing each individual in an enclosed area and heating it with an infrared lamp. Cloacal temperatures were taken with a Schultheis quick-recording thermometer as soon as the snake could no longer right itself when placed on its back. All temperatures were in degrees Celcius. The C. T. M. averaged 39.2° (38.0° to 40.0°). A temperature of 38.0° was lethal to one individual. These cottonmouths had [469] been in captivity for nine months. The behavior of the snakes during heating resembled those instances described by Klauber (1956:382-387) for rattlesnakes. As the body temperature of the snakes rose past the optimum, each individual became disturbed and tried to escape from the enclosure. The snakes soon became frantic in their efforts to escape. After about five minutes the mouth was opened and heavy, slow breathing was begun, accompanied by a loss of coordination and a slowing down of movements. The snakes writhed spasmodically for a few seconds and then lay still, usually with the mouth open. Recovery was begun by rolling on the belly and flicking the tongue, followed by movements of the head and then the body. Cottonmouths are rarely exposed to dangerously high temperatures owing to their semi-aquatic habits, but there are probably occasions when individuals reach the C. T. M. for the species.
Since activity, digestion, and gestation depend upon adequate internal temperatures, there must be a process by which these temperatures are attained and for an appropriate time maintained. Basking is important in this respect. The cottonmouths prefer to lie in a coiled position and, during basking, can usually be found beside bodies of water or on branches of dead trees overhanging the water. They are good climbers and have a prehensile tail that is frequently employed in descending from small branches. Since cottonmouths are semi-aquatic and are often exposed to temperatures that are lower than those of the air, they either must bask more often than terrestrial snakes or tolerate lower temperatures. Length of the period of basking is determined not only by amounts of insolation and temperature but also by the size of the snake. A smaller snake can reach its optimum temperature more rapidly because of a higher surface-to-volume ratio. Another factor that may play a minor role in the rate of temperature change is the color of the snake. The wide variation in color of cottonmouths probably affects rates of heat increase and loss due to direct radiation. Slight hormonal control of melanophores described in snakes by Neill and Allen (1955) also may exert some influence on the length of time spent basking. No rates of temperature increase or decrease are available for cottonmouths.
While inactive the cottonmouth spends most of its time lying in a coiled position with the tail outermost, with the body usually wound into about one and one-half cycles, and the head and neck in a reversed direction forming a U- or S-shaped loop. From this position the snake is able to make a short strike or a hasty getaway if necessary. In my opinion this position is used primarily for basking or resting and only secondarily for feeding. Most individuals appear to pursue their prey actively, not lying in ambush for the approaching prey to the extent that most other crotalids do.
Many of the cottonmouths that I kept in captivity were observed in a coiled position for periods up to three or four days. Under natural conditions, however, they are more active. Young cottonmouths are inclined to remain in a coiled position for longer periods than older individuals.
Four distinct types of locomotion have been described in snakes: horizontal undulatory, rectilinear, sidewinding, and concertina (Klauber, 1956: 331-350). Most snakes are capable of employing two or more of these types of progression, at least to a certain degree; but horizontal undulatory locomotion is the most common method used by the majority of snakes, including the cottonmouth. In this method the snake's body is thrown into lateral undulations that conform with irregularities in the substrate. Pressure is exerted on the outside and posterior surface of each curve, thus forcing the body forward.
Rectilinear locomotion is more useful to large, thick-bodied snakes which use this method of progression, chiefly when they are prowling and unhurried. This method depends upon the movement of alternate sections of the venter forward and drawing the body over the ventral scales resting on the substratum by means of muscular action. This mode of locomotion was most frequently observed in captive cottonmouths when they were crawling along the edge of their cages, especially when they were first introduced to the cages and toward the end of the shedding process. The other two types of locomotion, sidewinding and concertina, have not, to my knowledge, been observed in the cottonmouth.
Both the cottonmouth and the cantil have definite affinities for water and are as likely to be found in water as out of it. Copperheads and rattlesnakes, although not aquatic, are good swimmers. When swimming, a motion resembling horizontal undulatory progression is used.
The number of different opinions expressed in the literature concerning the cottonmouth's disposition is not at all surprising. As with any species there is a wide range of individual temperament, which is affected by many factors. The cottonmouth is considered by some writers to be docile while others consider it to be highly dangerous. Allen and Swindell (1948:7) described the variability in temperament, even among individuals. They wrote: "On rare occasions, moccasins are found which will attack. A perfectly docile snake will turn and bite viciously without any apparent reason." They also recounted a case in which a cottonmouth was kept as a pet for six years, being allowed the freedom of the house. Smith and List (1955:123) found them "... surprisingly docile in the gulf region [Mississippi], displaying none of the pugnacity of more northern cottonmouths." Smith (1956:310) stated: "Unlike the copperhead, cottonmouths are pugnacious; their powerful jaws, long fangs, vicious disposition and potent venom make them a very dangerous animal."
My own observations are in general agreement with the statements of Allen and Swindell (loc. cit.). In my encounters with cottonmouths, I have never found any aggressive individuals except for three juveniles that were born in captivity. In their first three days in the laboratory these juveniles were observed to strike repeatedly whenever anyone entered the room. After this short period of aggressiveness, however, they slowly became more docile. The disposition shown by the newborn young is clearly an innate behavioral pattern that undoubtedly has a direct relationship to survival. The majority of cottonmouths [471] that I have approached in the field have moved swiftly to seek refuge in nearby water; a few have remained motionless as I approached, and one showed the typical threat display. Upon capture and handling, they react similarly to other pit-vipers by opening and closing the mouth and erecting the fangs in an attempt to bite. They often bite through the lower jaw and eject venom at this time as well as when the mouth is open. Of more than a dozen individuals kept in captivity, four were particularly difficult to handle whereas another was extremely docile. It was almost never found in aggregations with the other snakes and did not struggle or attempt to bite when handled. The majority remained unpredictable in disposition, usually appearing docile and lazy but capable of extremely rapid movements when disturbed.
The typical threatening posture of rattlesnakes is all but lacking in the cottonmouth, which relies primarily on concealing coloration or nearness to water for escape. When approached, it usually plunges into nearby water or remains motionless with the head held up at a 45° angle and the mouth opened widely exposing the white interior. The tail is sometimes vibrated rapidly and musk is expelled. This threat display is unique to cottonmouths; although it does not attract as much attention as the display of rattlesnakes, it is probably an effective warning to most intruders at close range.
Neill (1947:205) reported one case in which a cottonmouth used the "body blow" defense, described for Crotalus by Cowles (1938:13), when approached by a king-snake, Lampropeltis getulus. In this unusual posture the anterior and posterior portions of the body are held against the ground and the middle one-fourth to one-third of the body is lifted up and used in striking the intruder. This same defense posture also was observed in rattlesnakes when presented with the odor of the spotted skunk, Spilogale phenax. However, the "king-snake defense posture" is probably not a well-established behavioral pattern in the cottonmouth, for it sometimes feeds upon king-snakes. I observed the killing and devouring of a cottonmouth by a speckled king-snake, L. g. holbrooki; the only attempts to escape were by rapid crawling and biting.
Cottonmouths often squirt musk as a defensive action. The tail is switched back and forth, and musk is emitted from glands on each side of the base of the tail. The fine jets of musk are sprayed upward at about 45° angles for a distance of nearly five feet. How often this defense mechanism is used against other animals is not known, but the musky odor can frequently be detected in areas where cottonmouths are common. The odor is repulsive and, if concentrated, can cause nausea in some individuals. To me, the scent is indistinguishable from that of the copperhead.
"Head bobbing" in snakes has been described frequently in the literature, and many interpretations have been advanced to explain its occurrence. One of the earlier accounts was that of Corrington (1929:72) describing behavior of the corn snake, Elaphe guttata. Characteristic bobbing occurred when the snake was cornered, and seemingly the purpose was to warn or frighten foes. Neill (1949:114-115) mentioned the jerking or bobbing of the head in several species of snakes including the cottonmouth, and remarked that "it is apparently [472] connected with courtship and with the recognition of individuals." According to Munro (1950:88), "head bobbing" appears to be a sign of annoyance in some instances but is usually concerned with reproduction and individual recognition. Richmond (1952:38) thought that many types of head movements among not only reptiles but also birds and some mammals are a result of poor vision and serve "to delimit and orient an object that for lack of motion is otherwise invisible." Head movements undoubtedly occur in animals to facilitate accommodation, but it is obvious from Richmond's conclusions that he has never observed "head bobbing" in snakes. The term itself is grossly misleading and should be discarded. Mansueti (1946:98) correctly described the movements as spastic contractions of the body. I have observed numerous instances of these movements in cottonmouths, copperheads, and rat snakes (Elaphe obsoleta); and in no case has the movement resembled a head bob as is described in lizards and other animals. The movement appears to be a result of a nervous or sexually excited state and consists of highly spastic contractions confined to the anterior part of the snake most of the time but affecting the entire body on some occasions.
I found the response to be most common among cottonmouths in confinement when food was introduced to a cage containing several individuals (increasing the tendency to strike at a moving object) and when an individual was placed back in the cage after being handled. At these times the snakes that were inactive began to jerk for a few seconds. When the snake is in this seemingly nervous state, the same response is elicited by another snake crawling over it. At other times the movement of one individual causes no such response. The jerking movements appear to be released by the recognition of a nervous state in another individual and may serve to protect the jerking individual from aggressive advances of the former.
Where courtship is involved, the jerking motions are made in conjunction with writhing of the male and do not result from the same type of releaser described above.
The so-called combat dance between male snakes has long been known, but its significance is still poorly understood. It was for many years believed to be courtship behavior until the participants were examined and found to be males. Carr and Carr (1942:1-6) described one such instance in two cottonmouths as courtship. In their observations, as well as those of others, copulation was never observed following the "dance" but was assumed to be the ultimate goal. After the discovery that only males participated, it was suggested that combat involved competition for mates, but the "dance" has been observed at times other than the breeding season (Ramsey, 1948:228).
Shaw (1948:137-145) discussed the combat of crotalids in some detail but drew no conclusions as to the cause of the behavior. Lowe (1948:134) concluded with little actual evidence that combat among male snakes is solely for territorial purposes. Shaw (1951:167) stated that combat may occur as a possible defense against homosexuality. One case of homosexual mating among cottonmouths was reported (Lederer, 1931:651-653), but the incomplete description seems to be of normal courtship procedure except that the "female" tried to avoid the male. Two instances of combat observed between timber rattlesnakes (C. h. horridus) by Sutherland (1958:23-24) were definitely [473] initiated because of competition for food. More observations are needed before the significance of the combat can be fully understood.
The venom and poison apparatus have developed primarily as a means of causing rapid death in small animals that are the usual prey. As a protective device against larger enemies, including man, the venom may have some value; but this was probably unimportant in the evolution of the poison mechanism. A secondary function of the venom is to begin digestion of tissues of the prey. Since food is swallowed whole, injection of digestive enzymes into the body cavity enhances digestion of the prey.
Kellogg (1925:5) described venom as a somewhat viscid fluid of a yellowish color and composed of 50 to 70 per cent proteins, the chief remaining components being water and carbohydrates, with occasional admixtures of abraded epithelial cells or saprophytic microorganisms. Salts, such as chlorides, phosphates of calcium, magnesium, and ammonium, occur in small quantities. Each of the components of snake venom has a different effect on the body of the victim. It was at first believed that there were two types of venoms: neurotoxic, which acts upon nervous tissue; and haemotoxic, which acts on blood and other tissues. It has since been found that venoms are composed of varying mixtures of both types. Fairley (1929:301) described the constituents of venom as: (1) neurotoxic elements that act on the bulbar and spinal ganglion cells of the central nervous system; (2) hemorrhagins that destroy the lining of the walls of blood vessels; (3) thrombose, producing clots within blood vessels; (4) hemolysins, destroying red blood corpuscles; (5) cytolysins that act on leucocytes and on cells of other tissues; (6) elements that retard coagulation of the blood; (7) antibactericidal substances; and (8) ferments that prepare food for pancreatic digestion. Elapid snakes tend to have more of elements 1, 4, and 6 in their venoms, while viperids and crotalids, of which the cottonmouth is one, have higher quantities of elements 2, 3, and 5. Kellogg (loc. cit.) stated that venom of cottonmouths contains more neurotoxin than that of rattlesnakes and not only breaks down the nuclei of ganglion cells but also produces granular disintegration of the myelin sheath and fragmentation of the conducting portions of nerve fibers.
Thus, venoms contain both toxic elements and non-toxic substances that promote rapid spreading of the venom through the body of the victim. Jacques (1956:291) attributed this rapid spreading to the hyaluronidase content of venoms.
One of the most important yet undeterminable factors of the gravity of snakebite is the amount of venom injected into the victim. Since this volume varies considerably in every bite, attempts have been made to determine the amount and toxicity of venom produced by each species of poisonous snake. Individual yield is so variable that a large number of snakes must be milked in order to determine the average yield. Even then there remains an uncertainty as to how this amount may compare with that injected by a biting snake.
Wolff and Githens (1939b:234) made 16 venom extractions from a group [474] of cottonmouths in a two-year period. The average yield per snake fluctuated between 80 and 237 milligrams (actual weight), and toxicity measured as the minimum lethal dose for pigeons varied from 0.05 to 0.16 milligrams (dry weight). No decrease in yield or toxicity was evident during this period. Another group of cottonmouths from which venom was extracted over a period of five years also showed no decrease in yield or toxicity. Of 315 individual extractions the average amount obtained from each individual was 0.55 cubic centimeters of liquid or 0.158 grams of dried venom (28.0 per cent solids). The minimum lethal dosage (M. L. D.) which was determined by injecting intravenously into 350-gram pigeons was found to be 0.09 milligrams (dry weight). Each snake carried approximately 1755 M. L. D.'s of venom.
The record venom extraction for the cottonmouth was 4.0 cubic centimeters (1.094 grams dried venom) taken from a five-foot snake which had been in captivity for 11 weeks and milked five weeks earlier (Wolff and Githens, 1939a:52). The average yield of venom of cottonmouths is about three times the average yield reported for copperheads by Fitch (1960:256), a difference correlated with the greater bulk and relatively large head of the cottonmouth.
Allen and Swindell (1948:13) stated that cottonmouth venom rates third in potency, compared drop for drop to that of Micrurus fulvius and Crotalus adamanteus. Freshly dried cottonmouth venom tested on young white rats showed the lethal dose to be from 23 to 29 milligrams per kilogram of body weight. The venom of 11 one-week-old cottonmouths was found to be more potent than that of adult males. Githens (1935:171) rated C. adamanteus venom as being weaker than that of the copperhead (A. contortrix), which he rated only slightly lower than cottonmouth venom. The crotalids which he ranked more toxic than cottonmouths are: the Pacific rattlesnake (C. viridis oreganus) and the massasauga (S. catenatus). He found A. bilineatus, C. durissus, and C. v. lutosus to have the same toxicity as cottonmouths. Minton (1953:214) found that the intraperitoneal "lethal dose 50" (the dose capable of killing half the experimental mice receiving injections of it) was 6.36 milligrams per kilogram for copperheads. However, in later publications Minton (1954:1079; 1956:146) reported that the "lethal dose 50" for copperheads was 25.65 milligrams. Approximately the same potency was determined for cottonmouths. Several rattlesnakes that he tested showed a higher toxicity than copperheads or cottonmouths.
Criley (1956:378) found the venom of copperheads to be 6.95, nearer Minton's earlier estimate, and rated cottonmouth venom as being twice as toxic as that of copperheads. The relative toxicities of other crotalids tested, considering the cottonmouth to be one unit, were: C. basiliscus, 0.3; A. contortrix, 0.5; C. viridis oreganus, 1.4; A. bilineatus, 2.2; C. adamanteus, 2.3; C. v. viridis, 3.2; C. durissus terrificus, 27.5.
It can be seen from the above examples that toxicity of venoms and the resistance of the animal receiving an injection of venom is highly variable. Possibly the venom of each species of snake has greatest effect on animals of the particular group relied on for food by the snake. If that is so, the venom of cottonmouths would be expected to be more toxic when tested on fish, reptiles, and amphibians than on birds and mammals. Likewise, the venom of most species of rattlesnakes would be expected to be more virulent when injected into mammals than when injected into lower vertebrates. But, according to Netting (1929:108), species of rattlesnakes that prey on cold-blooded [475] animals, which are less susceptible to venoms than warm-blooded animals, are thought to have highly toxic venoms. This explanation accounts for the powerful venom of Sistrurus catenatus; and, in this respect, venom of cottonmouths should be highly toxic also. However, no clear-cut trends have been shown in most cases. Allen (1937) injected 250-gram guinea pigs with 4 milligrams of venom of various poisonous snakes. Survival time was recorded in order to indicate the relative potency of the venoms. Of 16 such tests C. adamanteus held places 1, 2, 3, 12, and 16; Bothrops atrox held places 4, 9, 10, and 13; and A. piscivorus held places 5, 7, 8, and 15. Places 6, 11, and 14 were held by three individuals of different species. No relationship to size or sex was indicated by the results of this experiment.
Numerous experiments have been conducted to determine the susceptibility of various snakes to venom. The majority of these experiments were performed to learn whether or not venomous snakes were immune to their own poison. Conant (1934:382) reported on a 30-inch cottonmouth that killed two Pacific rattlesnakes and another cottonmouth. One rattlesnake was bitten on the tail and the other on or near the head and partially swallowed. Gloyd (1933:13-14) recorded fatal effects to a rattlesnake from the bite of a cottonmouth. He also reported on the observations of three other crotalids bitten by themselves or other snakes, from which no harmful effects were observed. Allen (1937) injected several snakes with dried cottonmouth venom which was diluted with distilled water just before each injection. Four cottonmouths receiving 9, 18, 19, and 20 milligrams of venom per ounce of body weight survived, while another receiving 18.7 milligrams per ounce died after three hours. A specimen of S. miliarius receiving 8.3 milligrams per ounce died in about ten hours, while a C. durissus receiving 12.5 milligrams per ounce succumbed in 45 minutes. An alligator receiving 6 milligrams per ounce died in 14 hours. Even the snakes that survived showed some degree of swelling.
The studies of Keegan and Andrews (1942:252) show that king-snakes are sometimes killed by poisonous snakes. A Lampropeltis calligaster injected with A. contortrix venom (0.767 milligrams per gram) died five days following the injection. This amount was more than twice the amount of A. piscivorus venom injected into a L. getulus by Allen (1937) in which the snake showed no ill effects. Keegan and Andrews (loc. cit.) stated that success in overpowering and eating poisonous snakes by Lampropeltis and Drymarchon may be due to the ability to avoid bites rather than to immunity to the venom. However, Rosenfeld and Glass (1940) demonstrated that the plasma of L. g. getulus had an inhibiting effect on the hemorrhagic action on mice of the venoms of several vipers.
One of the more extensive studies on effects of venoms on snakes is that by Swanson (1946:242-249). In his studies freshly extracted liquid venom was used. His studies indicated that snakes are not immune to venom of their own kind or to closely related species. Copperhead venom killed copperheads faster than did other venoms but took more time to kill massasaugas, cottonmouths, and timber rattlers. However, most of the snakes were able to survive normal or average doses of venom although they are not necessarily immune to it.
One of the major problems in comparing the data on toxicity of venom in studies of this type is that no standard method of estimating toxicity has been used. Swanson's (loc. cit.) amount of venom equalling one minim (M.L.D.?) ranged from 0.058 to 0.065 cubic centimeters. There were no different values given for each species, but the time that elapsed from injection of the venom to death represented the toxicity. There also was no attempt in his study to convert the amount of venom used into a ratio of the volume of venom per weight of snake, making the results somewhat difficult to interpret. Additional work in this field should provide for many injections into many individuals of several size classes. The studies to date have been on far too few individuals to allow statistical analyses to be accurate.
Factors determining the outcome of snakebite are: size, health, and species of snake; individual variation of venom toxicity of the species; age and size of the victim; allergic or immune responses; location of the bite; and the amount of venom injected and the depth to which it is injected. The last factor is one of the most variable, owing to (1) character and thickness of clothing between the snake and the victim's skin, (2) accuracy of the snake's strike, and (3) size of the snake, since a large snake can deliver more venom and at a greater depth than can a small snake.
Pope and Perkins (1944) demonstrated that pit-vipers of the United States bite as effectively as most innocuous snakes and that a careful study of the bite may reveal the location of the pocket of venom, size of the snake, and possibly its generic identity (see Dentition). The bite pattern of the cottonmouth as well as the other crotalids showed the typical fang punctures plus punctures of teeth on both the pterygoid and mandible. Even so, a varying picture may be presented because from one to four fang marks may be present. At times in the fang-shedding cycle three and even four fangs can be in operation simultaneously.
Various authors have attributed death of the prey to the following causes: paralysis of the central nervous system, paralysis of the respiratory center, asphyxiation from clotting of the blood, stoppage of the heart, urine suppression due to crystallized hemoglobin in the kidney tubules, dehydration of the body following edema in the area of the bite, or tissue damage. Mouths of snakes are reservoirs for infectious bacteria, which are especially prolific in damaged tissue. Bacterial growth is aided by the venom which blocks the bactericidal power of the blood.
Three grades in the severity of snakebite (I, minimal; II, moderate; and III, severe) were described by Wood, Hoback, and Green (1955). Parrish (1959:396) added a zero classification to describe the bite of a poisonous snake in which no envenomation occurred. Grade IV (very severe) was added by McCollough and Gennaro (1963:961) to account for many bites of the eastern and western diamondback rattlesnakes.
The first symptom of poisonous snakebite is an immediate burning sensation at the site of the bite. Within a few minutes the loss of blood into the tissues causes discoloration. Swelling proceeds rapidly and can become so great as to rupture the skin. Pain is soon felt in the lymph ducts and glands. Weakness, [477] nausea, and vomiting may ensue at a relatively early stage. Loss of blood into tissues may spread to the internal organs. In conjunction with a rapid pulse, the blood pressure and body temperature can drop. Some difficulty in breathing can occur, especially if large amounts of neurotoxin are present in the venom. In severe cases the tension due to edema obstructs venous and even arterial flow, in which case bacteria may multiply rapidly in the necrotic tissue and gangrene can occur. Blindness due to retinal hemorrhages may occur. Symptoms of shock may be present after any bite.
Perhaps one of the most important factors in the outcome of snakebite is the treatment. Because of the variable reactions to snakebite, treatment should vary accordingly. Many methods have been proposed for treating snakebite, and there is disagreement as to which is the best. The list of remedies that have been used in cases of snakebite includes many that add additional injury or that possibly increase the action of the venom. The use of poultices made by splitting open living chickens and the use of alcohol, potassium permanganate, strychnine, caffeine, or injection of ammonia have no known therapeutic value, and may cause serious complications. The most important steps in the treatment of snakebite are to prevent the spread of lethal doses of venom, to remove as much venom as possible, and to neutralize the venom as quickly as possible.
It is generally agreed that the first step in snakebite treatment should be to place a ligature above the bite to restrict the flow of venom, and also to immobilize the patient as much as possible. The ligature should be loosened at least every fifteen minutes. The next steps are sterilization of the skin and the making of an incision through the fang punctures. As pointed out by Stahnke (1954:8), the incision should be made in line with the snake's body at the time of the bite, so as to account for the rearward curvature of the fangs and possibly to reach the deposition of venom. Many instruction booklets and first-aid guides have specified the length and depth of incision to be made, but the actual size and depth of the cut should depend upon the location of the bite. An "X" cut or connection of the fang punctures is likely to facilitate the spread of the venom. No cut should be made that would sever a large blood vessel or ligament.
Extensive damage is often caused by well-meaning individuals whose attempts at first aid result in brutally deep incisions and tourniquets applied too tightly and for too long a period of time; the resultant damage in many instances exceeds that of the bite itself (Stimson and Engelhardt, 1960:165). Stimson and Engelhardt also think that time should be sacrificed to surgical cleanliness, and incisions should not be made if a hospital can be reached within an hour.
The ligature-cryotherapy (L-C) method proposed by Stahnke (1953) has been severely criticized by other workers. He stated that the ligature should be tight enough to restrict completely the flow of venom until the temperature of the area can be lowered sufficiently to prevent any action of the venom. After 10 minutes the ligature may be removed and the bitten area kept immersed in a vessel of crushed ice and water. If the envenomized member is to be treated for more than four hours (which is the case with almost all [478] pit-viper bites), it should be protected by placing it in a plastic bag. The venom action should be tested after 12 or more hours. This consists of a brief warming period to determine whether or not the action of the venom can be felt. The patient should be kept warm at all times; and the warming at the termination of treatment should be done gradually, preferably by allowing the water to warm slowly to room temperature.
Advocates of the L-C method warn against making incisions unless they are absolutely necessary, the theory being that each cut permits additional bacterial infection and does little good in removing venom. However, McCollough and Gennaro (1963:963) demonstrated that, in bites where the fangs had only slightly penetrated the skin, more than 50 per cent of the venom was removed in some instances if suction was started within three minutes after the injection. With deeper injection the amount of venom recovered sometimes reached 20 per cent of the dose. Stahnke suggested that an incision be made at the site of the bite only after the site has been refrigerated for at least 30 minutes.
Stimson and Engelhardt (loc. cit.) stated that two constricting bands should be used between the bite and the body and that cracked ice in a cloth should be applied to the bite before reaching a hospital. In addition, they suggested the following procedure. Rings of incisions should follow the swelling, and suction should continue for several hours. After the edema has receded, the limb should be wrapped in a towel containing crushed ice. Antivenin should be given only in severe cases. Calcium gluconate and gas gangrene antitoxin as well as antibiotics are helpful.
The most recent and up-to-date summary of snakebite treatment is that by McCollough and Gennaro (1963). Following is a brief summary of their suggestions:
1. Immobilization—Systemic immobilization is effected by body rest and locally by splinting the bitten area.
2. Tourniquet—A lightly occlusive tourniquet during a 30- to 60-minute period of incision and suction would seem to possess some advantages. In severe cases where medical attention is hours away, a completely occlusive tourniquet may be necessary to prevent death. Sacrifice of the extremity may be necessary for the preservation of life.
3. Incision and suction—Suction should begin three to five minutes after injection of venom if symptoms of poisoning are present. Incisions one-fourth inch to an inch long across each fang mark should be made in order to open the wound for more efficient suction. Multiple incisions are not useful for the removal of venom but may be employed under hospital conditions to reduce subcutaneous tensions and ischemia.
4. Cryotherapy—An ice cap over the site of the bite for relief of pain would seem to be permissible, especially prior to the administration of antivenin. It must be remembered that cooling during the administration of the antivenin radically reduces the access of the antiserum to the bite area.
5. Antivenin—Antiserum is the keystone to the therapy of snakebite. Careful evaluation of the severity of the bite and the patient's sensitivity should be made before the use of antivenin. In Grade II (moderate) bites, the intramuscular injection on the side of the bite may suffice. In Grades III (severe) and IV (very severe), shock and systemic effects require intravenous injection. In bites producing symptoms of this severity, antivenin must be given in [479] amounts large enough to produce clinical improvement. Ten to 20 units may be necessary to prevent the relapse that sometimes occurs after small doses of antivenin. Permanent remission of swelling and interruption of necrosis are the therapeutic end point in the clinical use of the antiserum.
In all cases of snakebite where there is any doubt as to the snake's identity, it should be killed if possible and taken to the hospital for positive identification. In many instances of actual bites by poisonous snakes the only treatment needed was an injection of tetanus antitoxin or toxoid and sedation, because physical examination revealed no indication of poisoning (Stimson and Engelhardt, loc. cit.).
On July 29, 1963, at 8:20 a.m., I was treating a nine-month-old cottonmouth for mites. As I dropped the snake into a sink, it twisted its head and bit the tip of my right middle finger with one fang. The fang entered just under the fingernail and was directed downward, the venom being injected about five millimeters below the site of fang penetration. After placing the snake back in its cage, I squeezed the finger once to promote bleeding, wrapped a string around the base of the finger, and drove to Watkins Memorial Hospital on the University of Kansas campus. I began to feel a burning sensation in the tip of the finger almost immediately. Upon my arrival at the hospital, an additional ligature was placed around my wrist. At 8:30 a.m. a small incision was made in the end of the finger, which by this time was beginning to darken at the point of venom deposition. I sucked on the finger until 8:35 a.m., when a pan of ice water that I had requested was brought to me. No pain was felt except that caused by the ice. Fresh ice was added as needed to keep the temperature low. By 9:30 a.m. the finger had swollen and stiffened. At 10:00 a.m. the swelling had progressed to the index finger and back of the hand. I experienced difficulty in opening and closing the hand. Blood oozed slowly from the incision. A dull ache persisted and about every two to four minutes a sharp throb could be felt until nearly 11:00 a.m., when the pain diminished. The rate and intensity of throbbing increased whenever the hand was removed from the ice bath for more than a few seconds. Although only the hand was immersed, the entire forearm was cold. Pain was felt along the lymphatics on top of the arm when it was touched, and by 1:00 p.m. a slight pain could be felt in the armpit. Since swelling and pain were almost nonexistent by 2:00 p.m., I was permitted to leave. After walking to a nearby building, I again felt a burning sensation as the hand warmed. I made another ice bath and again immersed the hand in it until 4:10 p.m., at which time it was removed from the water. The pain and swelling began anew, and the hand was placed back in an ice bath from 5:30 p.m. until about 7:30 p.m. At this time cryotherapy was discontinued. From 10:00 p.m. to 12:00 midnight my legs twitched periodically, and pain could be felt in both armpits. A slight difficulty in breathing also was experienced for a short time. The acute pain and burning sensation remained in the finger until the following morning, but swelling progressed only as far as the wrist. The only discomfort that day was in the finger. The tip was darkened, the entire first digit red and feverish, and the lymphatics still painful when touched. By the third day the swelling had regressed. The incision itself was the main cause of discomfort, and the soreness at the site of the bite persisted for at least four days.
Although the L-C method of snakebite treatment has been vigorously attacked by many, there is still need of much more data before it can be unequivocally condemned or praised. It was preferred in the treatment of this bite because: I knew that envenomation was minimal and that there would be no need for antivenin; only one fang of a snake less than one foot long had entered the tip of the finger; the snake had bitten three frogs in the previous [480] two days and had possibly used up a considerable amount of its venom; the venom was deposited at such a shallow depth that at least a portion of it could be removed by suction; and the wound bled freely even before suction was applied. The ice water was uncomfortably cold but was not cold enough to cause frostbite, a major objection to the L-C method. Ideally, fresh ice should be added little by little to replace that which is melting, and the immersed area should be protected from the water by a plastic bag. Pain and swelling can be minimized by cryotherapy, but I would recommend its use only in cases of mild poisoning such as the one described herein.
Many estimates have been made of the number of bites of poisonous snakes that occur annually in the United States. The occurrence of poisonous snakebite has been nearly as badly underestimated as fatal results of their envenomations have been overrated. For important data on number of persons bitten by poisonous snakes in the United States, see the following: Allen and Swindell (1948:15); Githens (1935:172); Klauber (1956:811); Parrish (1963); Sowder and Gehres (1963:973); Stimson and Engelhardt (1960:153); Swaroop and Grab (1956:441); Swartzwelder (1950:579); Willson (1908:530); and Wood (1954b:937).
Judging from estimates made in several states, the number of poisonous snakebites in the United States would be about 5000 per year. In the region where the cottonmouth occurs there are approximately 2000 persons bitten annually by poisonous snakes. Of these approximately 39 per cent are copperhead bites, 30 per cent each are cottonmouth and rattlesnake bites, and I per cent are coral snake bites. These percentages vary considerably from place to place, because of the distribution and abundance of the eight species of poisonous snakes whose ranges overlap that of the cottonmouth.
According to Parrish (1963), about 14 people die of snakebite each year in the United States. Of these deaths, about 6.6 per cent are attributable to cottonmouths, 77.0 per cent to rattlesnakes, and 1.6 per cent to coral snakes; 14.8 per cent are unidentified. Almost half of the fatalities are in persons less than 20 years of age, the high mortality rate being partially due to the greater ratio of venom to body weight.
In my study, 306 living and preserved cottonmouths were examined. This species occurs throughout the coastal plains of the southeastern United States, usually at altitudes of less than 500 feet but occasionally up to altitudes of more than 2000 feet.
Two subspecies are recognized: the eastern cottonmouth, A. p. piscivorus, occurring from extreme eastern Mississippi to southeastern Virginia and Florida; and the western cottonmouth, A. p. leucostoma, occurring from eastern Mississippi northward to southern Illinois and Missouri and westward to central Texas. Intergradation occurs in eastern Mississippi.
The northern edge of the range is probably limited by low temperatures in winter, and the western edge by lack of available habitat resulting from insufficient precipitation. Old records of occurrence indicate that the range has decreased in the last 100 years. The species inhabits mostly areas where water is found, but at times wanders a mile or more from the nearest water.
The ground color is predominantly a brown, but varies from a brownish-green to almost black with a pattern of 10 to 17 irregular bands of a darker shade of brown. The pattern is better defined in the eastern subspecies than in the western.
The scutellation resembles that of other species of Agkistrodon. In the specimens examined supralabials ranged from 7 to 9, and infralabials from 8 to 12. The number of dorsal scale rows on the neck, at mid-body, and immediately anterior to the anus is relatively constant at 27-25-21, respectively. Ventral scales of 34 males averaged 134.4 (128 to 139), and those of 48 females 133.5 (128 to 137). The number of caudal scales showed some degree of sexual dimorphism; the average was 45.4 (41 to 50) in 34 males and 42.6 (39 to 49) in 44 females. In general, caudal scales on the basal half of the tail are undivided, whereas those on the distal half are divided. No marked geographical variation was found in any scale character.
The poison fangs vary in length from 1.3 per cent of snout-vent length in juveniles to 1.0 per cent in large adults. Fangs of captive cottonmouths were shed and replaced at intervals of about 21 days, but the interval was variable. Relationships in distance between the base of fangs and between fang punctures in an actual bite indicate that examination of the wound does not provide a good basis for judging accurately the size of the snake that inflicted the bite.
In general, females less than 450 millimeters in snout-vent length were juveniles; those more than 450 millimeters were classified as post partum or reproductive on the basis of sizes of ovarian follicles. Since about half the adult females were fecund, it was concluded that a biennial reproductive cycle occurs in this species. An annual cycle may occur in areas where temperature permits year-round activity. It was estimated that females become sexually mature at an age of approximately two and one-half years. Mating is probably most concentrated in early spring at about the time when females ovulate, but copulation is not a stimulus for ovulation. Sperm retention and delayed fertilization allow young to be produced without [482] copulation occurring in each breeding season. The testes increase in size gradually rather than rapidly at maturity or in each breeding season, but seasonal cycles in sperm production occur.
The gestation period is three and one-half to four months. Determination of sex in the embryos is possible by late June, because the hemipenes of males are evaginated until the time of birth. Parturition generally occurs in August or September, but captivity may delay birth for a month or more. From one to 16 young per litter are born, depending on size of the mother and other factors; but the average is between six and seven. Mortality rate at birth is high in captive individuals but has not been determined in natural populations. The sex ratio in embryos and adults examined revealed about 53 per cent females. Because sufficient information on population composition is not available, an estimate of the percentage of adults in a natural population was based upon the number found in my study. The reproductive potential was estimated from these figures.
Normal young at birth are 230 to 240 millimeters in snout-vent length, but their size is influenced by the condition of the mother. Comparison of newborn young with those captured in spring indicates that little growth occurs during winter. Early growth is largely dependent upon feeding before winter quiescence.
The umbilical cord is broken at birth and the navel closes within a few days, but the scar remains throughout life. Sexual dimorphism in the position of the scar is characteristic of some snakes but is minimal in cottonmouths.
In those snakes more than 700 millimeters in length, males outnumber females three to one. The maximum age of cottonmouths in nature is unknown, but one has been kept in captivity for more than 18 years.
Allometric growth is striking in cottonmouths. The head and tail are proportionately longer in young individuals than in adults; and in males the tail is, on the average, slightly longer than in females of the same size.
Shedding of the skin provides for growth and wear in snakes. The young shed within a few days after birth and generally shed more frequently than adults. Frequency of shedding depends mostly on amount of food consumed, and there is some evidence that injuries on the head and neck increase the frequency of shedding. Before shedding, the eyes become cloudy for about five [483] and one-half days, then clear up again for about four days before the skin is shed.
The food of cottonmouths consists mainly of small vertebrates and occasionally invertebrates that are found near water. Fish, amphibians, and reptiles make up nearly 70 per cent of the diet. Carrion is also eaten and cannibalism occurs occasionally. Food is obtained by lying in ambush or by active searching. The young are known to lure their prey within striking range by waving their yellow tails in a manner suggestive of writhing grubs. The method of obtaining prey differs according to the kind of prey. Generally, cottonmouths retain their hold on fish or frogs but release mice and larger prey after delivering a bite.
The major causes of mortality of cottonmouths are obscure. Predators are known to include alligators, indigo snakes, king-snakes, largemouth bass, and blue herons; there are probably numerous others. Heavy parasitic infestations were found among the snakes examined. Snake mites, Ophionyssus natricus, became increasingly abundant on almost all captive snakes in April and May of 1963. Lung flukes (Ochetosoma sp.) were in 16 of 20 captive snakes, and many preserved specimens contained nematodes (Kalicephalus sp.) in the stomach and/or tapeworms (Ophiotaenia sp.) in the intestine. Although parasitic infestation causes discomfort and may lower resistance to other detrimental factors, it is difficult to attribute death to the effect of any particular kind of parasite. Miscellaneous causes of death of some captive snakes also were discussed.
The maximal body temperatures tolerated by four cottonmouths were between 38° and 40° C., but a temperature of 38° was lethal to a fifth individual. Cottonmouths have been found on occasion when other snakes were inactive because of low temperatures, but minimal temperatures tolerated by this species are not known. The annual cycle of activity is dependent upon temperature and thus varies from north to south. Cottonmouths generally migrate inland in autumn, usually to dry forested hillsides, where they den along with other species of snakes. After a few warm days in spring they migrate back to the water's edge. The diel activity cycle likewise depends upon temperatures but is influenced by other factors as well. In spring and autumn, the snakes are active mostly on warm, sunny days, whereas in summer they are active mostly at night. In order to maintain adequate internal temperatures, much [484] time is spent basking mostly in a characteristic flat, resting coil either beside a body of water or above water on limbs of dead trees. In this position the snake is ready either for a short strike or a hasty getaway.
Juveniles appear particularly aggressive and strike repeatedly when approached, a behavioral pattern definitely favoring survival. Adults vary in disposition, usually appearing sluggish and lazy, but they are capable of striking rapidly when disturbed. The typical threat display consists of lying in a coiled position with the mouth opened widely, exposing the white interior, and with the tail vibrating rapidly. The striking posture resembles the resting coil except that the anterior part of the body is raised off the ground and the mouth is sometimes opened. Musk is often ejected in a fine spray from glands in the tail as a further defensive action.
"Head bobbing," more properly described as spastic contractions of the body, was observed in captives when food was introduced into a cage containing several individuals or when one of the snakes was returned to the cage after being handled. Reports in the literature also have connected these jerking movements with courtship. The response appears to be elicited whenever a nervous state is recognized in another individual and may serve to protect the jerking individual from aggressive advances of the former.
The relatively heavy appearance of the body, sluggish habits, and cryptic coloration are correlated with the development of venom and fangs. The poison apparatus has developed primarily as a means of causing rapid death in prey and secondarily, perhaps, to begin the digestion of small animals that are the usual prey, but it is also important as a defensive device. The venom contains at least eight constituents that aid in its action on prey. Toxicity of the venom is difficult to determine because of numerous variables, but cottonmouth venom is generally believed to be less potent than that of most rattlesnakes and more potent than that of the copperhead. Snakes in general are more resistant to snake venoms than other vertebrates of similar size, but there is no immunity even to their own venom.
About ten per cent of the approximately 5000 bites of poisonous snakes per year in the United States are attributable to cottonmouths, and about seven per cent of the approximately 14 deaths per year are caused by cottonmouths.
Allen, E. R.
1937. Florida snake venom experiments. Florida Acad. Sci., 2:7 pp.
Allen, E. R., and Swindell, D.
1948. Cottonmouth moccasin of Florida. Herpetologica, 4:1-16 (first supplement).
Allen, M. J.
1932. A survey of the amphibians and reptiles of Harrison County, Mississippi. Amer. Mus. Novit., 542:1-20.
Anderson, P.
1941. The cottonmouth in northern Missouri. Copeia, 1941(3):178.
1945. New herpetological records for Missouri. Bull. Chicago Acad. Sci., 7(5):271-275.
Anon.
1953. Snake collecting hobby of Colonial Heights lad. Virginia Wildlife, 14-24.
Auffenberg, W.
1963. The fossil snakes of Florida. Tulane Studies Zool., 10(3):131-216, 51 figs.
Bailey, R. M.
1948. Winter mortality in the snake Storeria dekayi. Copeia, 1948(3): 215.
Barbour, R. W.
1956. A study of the cottonmouth, Agkistrodon piscivorus leucostoma Troost, in Kentucky. Trans. Kentucky Acad. Sci., 17(1):33-41, 1 fig.
Blanchard, F. N.
1922. The amphibians and reptiles of western Tennessee. Occ. Pap. Mus. Zool.; Univ. Michigan, 117:1-18.
Bogert, C. M.
1943. Dentitional phenomena in cobras and other elapids with notes on adaptive modifications of fangs. Bull. Amer. Mus. Nat. Hist., 81(3):285-360.
Brattstrom, B. H.
1954. The fossil pitvipers (Reptilia: Crotalidae) of North America. Trans. San Diego Soc. Nat. Hist., 12(3):31-46, 2 figs.
Brown, A. E.
1903. Texas reptiles and their faunal relations. Proc. Acad. Nat. Sci. Philadelphia, pp. 543-558.
Carpenter, C. C.
1958. Reproduction, young, eggs, and food of Oklahoma snakes. Herpetologica, 14:113-115.
Carr, A. F., Jr.
1936. The gulf-island cottonmouths. Proc. Florida Acad. Sci., 1:86-90.
Carr, A. F., Jr., and Carr, M. H.
1942. Notes on the courtship of the cottonmouth moccasin. Proc. New England Zool. Club, July 18, 20:1-6.
Clark, R. F.
1949. Snakes of the hill parishes of Louisiana. Jour. Tennessee Acad. Sci., 24(4):244-261.
Conant, R.
1933. Three generations of cottonmouths, Agkistrodon piscivorus (Lacépède). Copeia, 1933(1):43.
1934. Two rattlesnakes killed by a cottonmouth. Science, 80(2078):382.
1958. A field guide to reptiles and amphibians. Houghton Mifflin Co., Boston, 366 pp.
Cook, F. A.
1962. Snakes of Mississippi. Mississippi Game and Fish Comm., Surv. Bull., ii + 45 pp.
Corrington, J. D.
1929. Herpetology of the Columbia, South Carolina region. Copeia, 1929:58-83.
Cowles, R. B.
1938. Unusual defense postures assumed by rattlesnakes. Copeia, 1938(1):13.
Cowles, R. B., and Bogert, C. M.
1944. A preliminary study of the thermal requirements of desert reptiles. Bull. Amer. Mus. Nat. Hist., 83(5):263-296.
Criley, B. R.
1956. Development of a multivalent antivenin for the family Crotalidae. Pp. 373-380 in Venoms (ed. Buckley, E. E., and Porges, N., Amer. Assoc. Adv. Sci., Publ. No. 44).
Darlington, P. J.
1957. Zoogeography: the geographical distribution of animals. John Wiley and Sons, Inc., New York, xi + 675 pp.
Davis, D. D.
1936. Courtship and mating behavior in snakes. Zool. Series Field Mus. Nat. Hist., 20:257-290, figs. 28-34.
Didisheim, P., and Lewis, J. H.
1956. Fibrinolytic and coagulent activities of certain snake venoms and proteases. Proc. Soc. Exp. Biol. Med., 93(1):10-13.
Ditmars, R. L.
1915. The reptile book. Doubleday, Page and Co., Garden City, N. Y., xxxii + 472 pp.
1945. The reptiles of North America. Doubleday, Doran and Co., Inc., Garden City, N. Y., xvi + 476 pp., 135 pls.
Dowling, H. G.
1951a. A proposed standard system of counting ventrals in snakes. British Jour. Herp., 1(5):97-98, 1 fig.
1951b. A proposed method of expressing scale reductions in snakes. Copeia, 1951(2):131-134.
Dundee, H. A., and Burger, W. L., Jr.
1948. A denning aggregation of the western cottonmouth. Nat. Hist. Misc., Chicago Acad. Sci., 21:1-2.
Edgren, R. A.
1951. The umbilical scar, a sexually dimorphic character in Heterodon platyrhinos. Nat. Hist. Misc., 83:1-2.
Fairley, N. H.
1929. The present position of snake bite and the snake bitten in Australia. Med. Jour. Australia, 1(10):296-313.
Fitch, H. S.
1956. Temperature responses in free-living amphibians and reptiles in northeastern Kansas. Univ. Kansas Publ. Mus. Nat. Hist., 8(7):417-476, 10 figs., 6 tables.
1960. Autecology of the copperhead. Univ. Kansas Publ. Mus. Nat. Hist., 13(4):85-288, pls. 13-20, 26 figs.
Force, E. R.
1930. The amphibians and reptiles of Tulsa County, Oklahoma, and vicinity. Copeia, 1930(2):25-39.
Fox, W.
1948. Effect of temperature on development of scutulation in the garter snake, Thamnophis elegans atratus. Copeia, 1948(4):252-262.
Githens, T. S.
1935. Studies on the venoms of North American pit vipers. Jour. Immun., 29(2):165-173.
Glissmeyer, H. R.
1951. Symposium. A snake den in Tooele County, Utah. Egg production of the great basin rattlesnake. Herpetologica, 7(1):24-27.
Gloyd, H. K.
1933. On the effects of moccasin venom upon a rattlesnake. Science, 78(2010):13-14.
1934. Studies on the breeding habits and young of the copperhead, Agkistrodon mokasen Beauvois. Papers Michigan Acad. Sci., Arts and Letters, 19:587-604.
Gloyd, H. K., and Conant, R.
1943. A synopsis of the American forms of Agkistrodon (copperheads and moccasins). Bull. Chicago Acad. Sci., 7(2):147-170, 16 figs.
Goodman, J. D.
1958. Material ingested by the cottonmouth, Agkistrodon piscivorus, at Reelfoot Lake, Tennessee. Copeia, 1958(2):149.
Guidry, E. V.
1953. Herpetological notes from southeastern Texas. Herpetologica, 9:49-56.
Hall, H. H., and Smith, H. M.
1947. Selected records of reptiles and amphibians from southeastern Kansas. Trans. Kansas Acad. Sci., 49(4):447-454.
Hamilton, W. J., Jr., and Pollack, J. A.
1955. The food of some crotalid snakes from Fort Benning, Georgia. Nat. Hist. Misc., 140:1-4.
Herald, E. S.
1949. Effects of DDT oil solution upon amphibians and reptiles. Herpetologica, 5(6):117-120.
Hickman, C. P.
1922. A northern record for the water moccasin. Copeia, 1922(106):39.
Hurter, J. H.
1897. A contribution to the herpetology of Missouri. Trans. Acad. Sci. St. Louis, 7:499-503.
Jacques, R.
1956. The hyaluronidase content of animal venoms. Pp. 291-293 in Venoms (ed. Buckley, E. E., and Porges, N., Amer. Assoc. Adv. Sci., Publ. No. 44).
Keegan, H. L., and Andrews, T. F.
1942. Effects of crotalid venom on North American snakes. Copeia, 1942:251-254.
Kellogg, R.
1925. Poisonous snakes of the United States. U. S. D. A. Papers (371), 13 pp.
Klauber, L. M.
1941. Four papers on the application of statistical methods to herpetological problems. Bull. Zool. Soc. San Diego, 17:1-95.
1956. Rattlesnakes. Univ. California Press, 2 vols., xxix + 1476 pp.
Klimstra, W. D.
1959. Food habits of the cottonmouth in southern Illinois. Chicago Acad. Sci., Nat. Hist. Misc., 168:1-8.
Laughlin, H. E.
1959. Stomach contents of some aquatic snakes from Lake McAlester, Pittsburgh County, Oklahoma. Texas Jour. Sci., 11(1):83-85.
Lederer, G.
1931. Aus meinem Tagebuch. Wachenschrift fur Aquar. und Terr'kde., 28:651-653.
Lee, H. T.
1964. Letters to the Editor, Texas Game and Fish Mag., 22(3):32.
Lowe, C. H., Jr.
1948. Territorial behavior in snakes and the so-called courtship dance. Herpetologica, 4(4):129-145.
Mansueti, R.
1946. Mating of the pilot blacksnake. Herpetologica, 3:98-100, 1 fig.
Martin, J. R., and Wood, J. T.
1955. Notes on the poisonous snakes of the Dismal Swamp area. Herpetologica, 11(3):237-238.
McCollough, N. C., and Gennaro, J. F., Jr.
1963. Evaluation of venomous snake bite in the southern United States from parallel clinical and laboratory investigations. Jour. Florida Med. Assn., 49(12):959-967.
McIlhenny, E. A.
1935. The alligator's life history. The Christopher Publ. House, Boston, Massachusetts, 117 pp.
Minton, S. A., Jr.
1953. Variation in venom samples from copperheads (Agkistrodon contortrix mokeson) and timber rattlesnakes (Crotalus horridus horridus). Copeia, 1953:212-215.
1954. Polyvalent antivenin in the treatment of experimental snake venom poisoning. Amer. Jour. Trop. Med. and Hyg., 3:1077-1082.
1956. Some properties of North American pit viper venoms and their correlation with phylogeny. Pp. 145-151 in Venoms (ed. Buckley, E. E., and Porges, N., Amer. Assoc. Adv. Sci., Publ. No. 44).
Munro, D. F.
1949. Effect of DDT powder on small cottonmouths. Herpetologica, 5:71-72.
1950. Additional observations on head bobbing by snakes. Herpetologica, 6:88.
Neill, W. T.
1947. Size and habits of the cottonmouth moccasin. Herpetologica, 3:203-205.
1949. Head bobbing, a widespread habit of snakes. Herpetologica, 5:114-115.
1960. The caudal lure of various juvenile snakes. Quart. Jour. Florida Acad. Sci., 23(3):173-200, 2 figs.
Neill, W. T., and Allen, E. R.
1955. Metachrosis in snakes. Quart. Jour. Florida Acad. Sci., 18(3):207-215.
Netting, M. G.
1929. The venom of Sistrurus catenatus. Bull. Antivenin Inst. Amer., 2(4):108-109.
Parrish, H. M.
1959. Poisonous snakebites resulting in lack of venom poisoning. Virginia Med. Month, 86:396-401.
1963. Analysis of 460 fatalities from venomous animals in the United States. Amer. Jour. Med. Sci., 245(2):35-47.
Parrish, H. M., and Pollard, C. B.
1959. Effects of repeated poisonous snakebite in man. Amer. Jour. Med. Sci., 237(3):277-286.
Penn, G. H.
1943. Herpetological notes from Cameron Parish, Louisiana. Copeia, 1943(1):58-59.
Perkins, C. B.
1955. Longevity of snakes in captivity in the United States. Copeia, 1955(3):262.
Pope, C. H., and Perkins, R. M.
1944. Differences in the patterns of bites of venomous and of harmless snakes. Archives of Surgery, 49:331-336.
Rahn, H.
1942. The reproductive cycle of the prairie rattler. Copeia, 1942(4):233-240.
Ramsey, L. W.
1948. Combat dance and range extension of Agkistrodon piscivorus leucostoma. Herpetologica, 4:228.
Richmond, M. D.
1952. Head bobbing in reptiles. Herpetologica, 8:38.
Rosenfeld, S., and Glass, S.
1940. The inhibiting effect of snake bloods upon the hemorrhagic action of viper venoms on mice. Amer. Jour. Med. Sci., 199:482-486.
Schmidt, K. P.
1946. On the zoogeography of the Holarctic region. Copeia, 1946:144-152.
Shaw, C. E.
1948. The male combat "dance" of some crotalid snakes. Herpetologica, 4:137-145.
1951. Male combat in American colubrid snakes with remarks on combat in other colubrid and elapid snakes. Herpetologica, 7(4):149-168.
Smith, H. M.
1956. Handbook of amphibians and reptiles of Kansas. 2nd Edition. Univ. Kansas Mus. Nat. Hist., Misc. Publ., 9:1-356, 253 figs.
Smith, H. M., and Buechner, H. K.
1947. The influence of the Balcones Escarpment on the distribution of amphibians and reptiles in Texas. Bull. Chicago Acad. Sci., pp. 1-16.
Smith, P. W.
1961. The amphibians and reptiles of Illinois. Illinois Nat. Hist. Survey, 28(1):1-298.
Smith, P. W., and List, J. C.
1955. Notes on Mississippi amphibians and reptiles. Amer. Midl. Nat., 53(1):115-125.
Sowder, W. T., and Gehres, G. W.
1963. Snakebites in Florida. Jour. Florida Med. Assn., 49(12):973-976.
Stabler, R. M.
1951. Some observations on two cottonmouth moccasins made during 12 and 14 years of captivity. Herpetologica, 7:89-92.
Stahnke, H. M.
1953. The L-C treatment of venomous bites and stings. Amer. Jour. Trop. Med. and Hyg., 2(1):142-143.
1954. The L-C method of treating venomous bites and stings. Pois. Anim. Res. Lab., Arizona State Coll., 28 pp.
Stejneger, L.
1895. The poisonous snakes of North America. Smithsonian Inst., U. S. Nat. Mus., 1893:337-487, pls. 1-19, figs. 1-70.
Stimson, A. C., and Engelhardt, T. H.
1960. The treatment of snakebite. Jour. Occ. Med., 2(4):163-168.
Sutherland, I. D.
1958. The "combat dance" of the timber rattlesnake. Herpetologica, 14(1):23-24.
Swanson, P. L.
1946. Effects of snake venoms on snakes. Copeia, 1946(4):242-249.
Swaroop, S., and Grab, B.
1956. The snakebite mortality problem in the world. Pp. 439-466 in Venoms (ed. Buckley, E. E., and Porges, N., Amer. Assoc. Adv. Sci., Publ. No. 44).
Swartzwelder, J. C.
1950. Snake-bite accidents in Louisiana with data on 306 cases. Amer. Jour. Trop. Med., 30(4):575-589.
Tinkle, D. W.
1962. Reproductive potential and cycles in female Crotalus atrox from northwestern Texas. Copeia, 1962(2):306-313.
Trowbridge, A. H.
1937. Ecological observations on amphibians and reptiles collected in southeastern Oklahoma during the summer of 1934. Amer. Midl. Nat., 18(2):285-303.
Volsøe, H.
1944. Structure and seasonal variation of the male reproductive organs in Vipera berus (L.). Spolia Zool. Mus. Hauniensis V. Reprint, Copenhagen, pp. 1-172.
Wharton, C. H.
1960. Birth and behavior of a brood of cottonmouths, Agkistrodon piscivorus piscivorus, with notes on tail-luring. Herpetologica, 16:125-129.
Willson, P.
1908. Snake poisoning in the United States: a study based on an analysis of 740 cases. Arch. Int. Med., 1(5):516-570.
Wolff, N. O., and Githens, T. S.
1939a. Record venom extraction from water moccasin. Copeia, 1939(1):52.
1939b. Yield and toxicity of venom from snakes extracted over a period of two years. Copeia, 1939(4):234.
Wood, J. T.
1954a. The distribution of poisonous snakes in Virginia. Virginia Jour. Sci., 5(3):152-167, 4 maps.
1954b. A survey of 200 cases of snake-bite in Virginia. Amer. Jour. Trop. Med. and Hyg., 3(5):936-943.
Wood, J. T., Hoback, W. W., and Green, T. W.
1955. Treatment of snake venom poisoning with ACTH and cortisone. Virginia Med. Month, 82:130-135.
Wright, A. H., and Wright, A. A.
1957. Handbook of snakes of the United States and Canada. Comstock Publ. Assoc., Cornell Univ. Press, 2:ix + 565-1106 pp.
Yamaguti, S.
1958. Systema helminthum. Interscience Publ., Inc., New York, 3 vols., 5 parts, 1:xi + 1575 pp., 2:vii + 860 pp., 3:1261 pp.
Yerger, R. W.
1953. Yellow bullhead preyed upon by cottonmouth moccasin. Copeia, 1953(2):115.
Transmitted June 20, 1966.
Institutional libraries interested in publications exchange may obtain this series by addressing the Exchange Librarian, University of Kansas Library, Lawrence, Kansas. Copies for individuals, persons working in a particular field of study, may be obtained by addressing instead the Museum of Natural History, University of Kansas, Lawrence, Kansas. When copies are requested from the Museum, 25 cents should be included (for each 100 pages or part thereof) for the purpose of defraying the costs of wrapping and mailing. For certain longer papers an additional amount indicated below, toward the cost of production, is to be included. Materials published to date in this series are as follows.
* An asterisk designates those numbers of which the Museum's supply (not necessarily
the Library's supply) is exhausted. Materials published to date, in this series, are as
follows:
Vol. 1. Nos. 1-26 and index. Pp. 1-638, 1946-1950.
*Vol. 2. (Complete) Mammals of Washington. By Walter W. Dalquest. Pp. 1-444, 140 figures in text. April 9, 1948.
*Vol. 3. Nos. 1-4 and index. Pp. 1-681. 1951.
*Vol. 4. (Complete) American weasels. By E. Raymond Hall. Pp. 1-466, 41 plates, 31 figures in text. December 27, 1951.
Vol. 5. Nos. 1-37 and index. Pp. 1-676, 1951-1953.
*Vol. 6. (Complete) Mammals of Utah, taxonomy and distribution. By Stephen D. Durrant. Pp. 1-549, 91 figures in text, 30 tables. August 10, 1952.
Vol. 7. Nos. 1-15 and index. Pp. 1-651, 1952-1955.
Vol. 8. Nos. 1-10 and index. Pp. 1-675, 1954-1956.
Vol. 9. Nos. 1-23 and index. Pp. 1-690, 1955-1960.
Vol. 10. Nos. 1-10 and index. Pp. 1-626, 1956-1960.
Vol. 11. Nos. 1-10 and index. Pp. 1-703, 1958-1960.
Vol. 12. *1. Functional morphology of three bats: Eumops, Myotis, Macrotus. By Terry
A. Vaughan. Pp. 1-153, 4 plates, 24 figures in text. July 8, 1959.
*2. The ancestry of modern Amphibia: a review of the evidence. By Theodore
H. Eaton, Jr. Pp. 155-180, 10 figures in text. July 10, 1959.
3. The baculum in microtine rodents. By Sydney Anderson. Pp. 181-216, 49
figures in text. February 19, 1960.
*4. A new order of fishlike Amphibia from the Pennsylvanian of Kansas. By
Theodore H. Eaton, Jr., and Peggy Lou Stewart. Pp. 217-240, 12 figures in
text. May 2, 1960.
5. Natural history of the Bell Vireo. By Jon C. Barlow. Pp. 241-296, 6 figures
in text. March 7, 1962.
6. Two new pelycosaurs from the lower Permian of Oklahoma. By Richard C.
Fox. Pp. 297-307, 6 figures in text. May 21, 1962.
7. Vertebrates from the barrier island of Tamaulipas, México. By Robert K.
Selander, Richard F. Johnston, B. J. Wilks, and Gerald G. Raun. Pp. 309-345,
plates 5-8. June 18, 1962.
8. Teeth of edestid sharks. By Theodore H. Eaton, Jr. Pp. 347-362, 10 figures
in text. October 1, 1962.
9. Variation in the muscles and nerves of the leg in two genera of grouse
(Tympanuchus and Pedioecetes). By E. Bruce Holmes. Pp. 363-474, 20
figures. October 25, 1962. $1.00.
10. A new genus of Pennsylvanian fish (Crossopterygii, Coelacanthiformes) from
Kansas. By Joan Echols. Pp. 475-501, 7 figures. October 25, 1963.
11. Observations on the Mississippi Kite in southwestern Kansas. By Henry S.
Fitch. Pp. 503-519. October 25, 1963.
12. Jaw musculature of the Mourning and White-winged doves. By Robert L.
Merz. Pp. 521-551, 22 figures. October 25, 1963.
13. Thoracic and coracoid arteries in two families of birds, Columbidae and
Hirundinidae. By Marion Anne Jenkinson. Pp. 553-573, 7 figures. March
2, 1964.
14. The breeding birds of Kansas. By Richard F. Johnston. Pp. 575-655, 10
figures. May 18, 1964. 75 cents.
15. The adductor muscles of the jaw in some primitive reptiles. By Richard C.
Fox. Pp. 657-680, 11 figures in text. May 18, 1964.
Index. Pp. 681-694.
Vol. 13. 1. Five natural hybrid combinations in minnows (Cyprinidae). By Frank B.
Cross and W. L. Minckley. Pp. 1-18. June 1, 1960.
2. A distributional study of the amphibians of the Isthmus of Tehuantepec,
México. By William E. Duellman. Pp. 19-72, plates 1-8, 3 figures in text.
August 16, 1960. 50 cents.
3. A new subspecies of the slider turtle (Pseudemys scripta) from Coahuila,
México. By John M. Legler. Pp. 73-84, plates 9-12, 3 figures in text.
August 16, 1960.
*4. Autecology of the copperhead. By Henry S. Fitch. Pp. 85-288, plates 13-20,
26 figures in text. November 30, 1960.
5. Occurrence of the garter snake, Thamnophis sirtalis, in the Great Plains and
Rocky Mountains. By Henry S. Fitch and T. Paul Maslin. Pp. 289-308,
4 figures in text. February 10, 1961.
6. Fishes of the Wakarusa River in Kansas. By James E. Deacon and Artie L.
Metcalf. Pp. 309-322, 1 figure in text. February 10, 1961.
7. Geographic variation in the North American cyprinid fish, Hybopsis gracilis.
By Leonard J. Olund and Frank B. Cross. Pp. 323-348, plates 21-24, 2
figures in text. February 10, 1961.
8. Descriptions of two species of frogs, genus Ptychohyla; studies of American
hylid frogs, V. By William E. Duellman. Pp. 349-357, plate 25, 2
figures in text. April 27, 1961.
9. Fish populations, following a drought, in the Neosho and Marais des Cygnes
rivers of Kansas. By James Everett Deacon. Pp. 359-427, plates 26-30,
3 figures. August 11, 1961. 75 cents.
10. Recent soft-shelled turtles of North America (family Trionychidae). By
Robert G. Webb. Pp. 429-611, plates 31-54, 24 figures in text. February
16, 1962. $2.00.
Index. Pp. 613-624.
Vol. 14. 1. Neotropical bats from western México. By Sydney Anderson. Pp. 1-8.
October 24, 1960.
2. Geographic variation in the harvest mouse, Reithrodontomys megalotis, on
the central Great Plains and in adjacent regions. By J. Knox Jones, Jr.,
and B. Mursaloglu. Pp. 9-27, 1 figure in text. July 24, 1961.
3. Mammals of Mesa Verde National Pork, Colorado. By Sydney Anderson.
Pp. 29-67, plates 1 and 2, 3 figures in text. July 24, 1961.
4. A new subspecies of the black myotis (bat) from eastern Mexico. By E.
Raymond Hall and Ticul Alvarez. Pp. 69-72, 1 figure in text. December
29, 1961.
5. North American yellow bats, "Dasypterus," and a list of the named kinds
of the genus Lasiurus Gray. By E. Raymond Hall and J. Knox Jones, Jr.
Pp. 73-98, 4 figures in text. December 29, 1961.
6. Natural history of the brush mouse (Peromyscus boylii) in Kansas with
description of a new subspecies. By Charles A. Long. Pp. 99-111, 1 figure
in text. December 29, 1961.
7. Taxonomic status of some mice of the Peromyscus boylii group in eastern
Mexico, with description of a new subspecies. By Ticul Alvarez. Pp. 113-120,
1 figure in text. December 29, 1961.
8. A new subspecies of ground squirrel (Spermophilus spilosoma) from Tamaulipas,
Mexico. By Ticul Alvarez. Pp. 121-124. March 7, 1962.
9. Taxonomic status of the free-tailed bat, Tadarida yucatanica Miller. By J.
Knox Jones, Jr., and Ticul Alvarez. Pp. 125-133, 1 figure in text. March 7,
1962.
10. A new doglike carnivore, genus Cynaretus, from the Clarendonian Pliocene,
of Texas. By E. Raymond Hall and Walter W. Dalquest. Pp. 135-138,
2 figures in text. April 30, 1962.
11. A new subspecies of wood rat (Neotoma) from northeastern Mexico. By
Ticul Alvarez. Pp. 139-143, April 30, 1962.
12. Noteworthy mammals from Sinaloa, Mexico. By J. Knox Jones, Jr., Ticul
Alvarez, and M. Raymond Lee. Pp. 145-159. 1 figure in text. May 18,
1962.
13. A new bat (Myotis) from Mexico. By E. Raymond Hall. Pp. 161-164,
1 figure in text. May 21, 1962.
*14. The mammals of Veracruz. By E. Raymond Hall and Walter W. Dalquest.
Pp. 165-362, 2 figures. May 20, 1963. $2.00.
15. The recent mammals of Tamaulipas, México. By Ticul Alvarez. Pp. 363-473,
5 figures in text. May 20, 1963. $1.00.
16. A new subspecies of the fruit-eating bat, Sturnira ludovici, from western
Mexico. By J. Knox Jones, Jr., and Gary L. Phillips. Pp. 475-481, 1 figure
in text. March 2, 1964.
17. Records of the fossil mammal Sinclairella, Family Apatemyidae, from the
Chadronian and Orellan. By William A. Clemens. Pp. 483-491. 2 figures
in text. March 2, 1964.
18. The mammals of Wyoming. By Charles A. Long. Pp. 493-758, 82 figs.
July 6, 1965. $3.00.
Index. Pp. 759-784.
Vol. 15. 1. The amphibians and reptiles of Michoacán, México. By William E. Duellman.
Pp. 1-148, plates 1-6, 11 figures in text. December 20, 1961. $1.50.
2. Some reptiles and amphibians from Korea. By Robert G. Webb, J. Knox
Jones, Jr., and George W. Byers. Pp. 149-173. January 31, 1962.
3. A new species of frog (Genus Tomodactylus) from western México. By
Robert G. Webb. Pp. 175-181, 1 figure in text. March 7, 1962.
4. Type specimens of amphibians and reptiles in the Museum of Natural History,
the University of Kansas. By William E. Duellman and Barbara Berg.
Pp. 183-204. October 26, 1962.
5. Amphibians and Reptiles of the Rainforests of Southern El Petén, Guatemala.
By William E. Duellman. Pp. 205-249, plates 7-10, 6 figures in text. October
4, 1963.
6. A revision of snakes of the genus Conophis (Family Colubridae, from Middle
America). By John Wellman. Pp. 251-295, 9 figures in text. October 4,
1963.
7. A review of the Middle American tree frogs of the genus Ptychohyla. By
William E. Duellman. Pp. 297-349, plates 11-18, 7 figures in text. October
18, 1963. 50 cents.
*8. Natural history of the racer Coluber constrictor. By Henry S. Fitch. Pp.
351-468, plates 19-22, 20 figures in text. December 30, 1963. $1.00.
9. A review of the frogs of the Hyla bistincta group. By William E. Duellman.
Pp. 469-491, 4 figures in text. March 2, 1964.
10. An ecological study of the garter snake, Thamnophis sirtalis. By Henry S.
Fitch. Pp. 493-564, plates 23-25, 14 figures in text. May 17, 1965.
11. Breeding cycle in the ground skink, Lygosoma laterale. By Henry S. Fitch
and Harry W. Greene. Pp. 565-575, 3 figures in text. May 17, 1965.
12. Amphibians and reptiles from the Yucatan Peninsula, México. By William
E. Duellman. Pp. 577-614, 1 figure in text. June 22, 1965.
13. A new species of turtle, Genus Kinosternon, from Central America, by John
M. Legler. Pp. 615-625, pls. 26-28, 2 figures in text. July 20, 1965.
14. A biogeographic account of the herpetofauna of Michoacán, México. By
William E. Duellman. Pp. 627-709, pls. 29-36, 5 figures in text. December
30, 1965.
15. Amphibians and reptiles of Mesa Verde National Park, Colorado. By Charles
L. Douglas. Pp. 711-744, pls. 37, 38, 6 figures in text. March 7, 1966.
Index. Pp. 745-770.
Vol. 16. 1. Distribution and taxonomy of Mammals of Nebraska. By J. Knox Jones, Jr.
Pp. 1-356, pls. 1-4, 82 figures in text. October 1, 1964. $3.50.
2. Synopsis of the lagomorphs and rodents of Korea. By J. Knox Jones, Jr.,
and David H. Johnson. Pp. 357-407. February 12, 1965.
3. Mammals from Isla Cozumel, Mexico, with description of a new species of
harvest mouse. By J. Knox Jones, Jr., and Timothy E. Lawlor. Pp. 409-419,
1 figure in text. April 13, 1965.
4. The Yucatan deer mouse, Peromyscus yucatanicus. By Timothy E. Lawlor.
Pp. 421-438, 2 figures in text. July 20, 1965.
5. Bats from Guatemala. By J. Knox Jones, Jr. Pp. 439-472. April 18, 1966.
More numbers will appear in volume 16.
Vol. 17. 1. Localities of fossil vertebrates obtained from the Niobrara Formation (Cretaceous)
of Kansas. By David Bardack. Pp. 1-14. January 22, 1965.
2. Chorda tympani branch of the facial nerve in the middle ear of tetrapods.
By Richard C. Fox. Pp. 15-21, May 22, 1965.
3. Fishes of the Kansas River System in relation to zoogeography of the Great
Plains. By Artie L. Metcalf. Pp. 23-189, 4 figures in text, 51 maps.
March 24, 1966.
4. Factors affecting growth and reproduction of channel catfish, Ictalurus punctatus.
By Bill A. Simco and Frank B. Cross. Pp. 191-256, 13 figures in
text. June 6, 1966.
5. A new species of fringe-limbed tree frog, genus Hyla, from Darién, Panamá.
By William E. Duellman. Pp. 257-262, 1 figure in text. June 17, 1966.
6. Taxonomic notes on some Mexican and Central American hylid frogs. By
William E. Duellman. Pp. 263-279. June 17, 1966.
7. Neotropical hylid frogs, genus Smilisca. By William E. Duellman and
Linda Trueb. Pp. 281-375, pls. 1-12, 17 figures in text. July 14, 1966.
8. Birds from North Borneo. By Max C. Thompson. Pp. 377-433, 1 figure in
text. October 27, 1966.
9. Natural history of cottonmouth moccasin, Agkistrodon piscivorus (Reptilia).
By Ray D. Burkett. Pp. 435-491, 7 figures in text. October 27, 1966.
More numbers will appear in volume 17.
End of the Project Gutenberg EBook of Natural History of Cottonmouth
Moccasin, Agkistrodon piscovor, by Ray D. Burkett
*** END OF THIS PROJECT GUTENBERG EBOOK NATURAL HISTORY OF ***
***** This file should be named 38308-h.htm or 38308-h.zip *****
This and all associated files of various formats will be found in:
http://www.gutenberg.org/3/8/3/0/38308/
Produced by Chris Curnow, Joseph Cooper and the Online
Distributed Proofreading Team at http://www.pgdp.net
Updated editions will replace the previous one--the old editions
will be renamed.
Creating the works from public domain print editions means that no
one owns a United States copyright in these works, so the Foundation
(and you!) can copy and distribute it in the United States without
permission and without paying copyright royalties. Special rules,
set forth in the General Terms of Use part of this license, apply to
copying and distributing Project Gutenberg-tm electronic works to
protect the PROJECT GUTENBERG-tm concept and trademark. Project
Gutenberg is a registered trademark, and may not be used if you
charge for the eBooks, unless you receive specific permission. If you
do not charge anything for copies of this eBook, complying with the
rules is very easy. You may use this eBook for nearly any purpose
such as creation of derivative works, reports, performances and
research. They may be modified and printed and given away--you may do
practically ANYTHING with public domain eBooks. Redistribution is
subject to the trademark license, especially commercial
redistribution.
*** START: FULL LICENSE ***
THE FULL PROJECT GUTENBERG LICENSE
PLEASE READ THIS BEFORE YOU DISTRIBUTE OR USE THIS WORK
To protect the Project Gutenberg-tm mission of promoting the free
distribution of electronic works, by using or distributing this work
(or any other work associated in any way with the phrase "Project
Gutenberg"), you agree to comply with all the terms of the Full Project
Gutenberg-tm License (available with this file or online at
http://gutenberg.org/license).
Section 1. General Terms of Use and Redistributing Project Gutenberg-tm
electronic works
1.A. By reading or using any part of this Project Gutenberg-tm
electronic work, you indicate that you have read, understand, agree to
and accept all the terms of this license and intellectual property
(trademark/copyright) agreement. If you do not agree to abide by all
the terms of this agreement, you must cease using and return or destroy
all copies of Project Gutenberg-tm electronic works in your possession.
If you paid a fee for obtaining a copy of or access to a Project
Gutenberg-tm electronic work and you do not agree to be bound by the
terms of this agreement, you may obtain a refund from the person or
entity to whom you paid the fee as set forth in paragraph 1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be
used on or associated in any way with an electronic work by people who
agree to be bound by the terms of this agreement. There are a few
things that you can do with most Project Gutenberg-tm electronic works
even without complying with the full terms of this agreement. See
paragraph 1.C below. There are a lot of things you can do with Project
Gutenberg-tm electronic works if you follow the terms of this agreement
and help preserve free future access to Project Gutenberg-tm electronic
works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the Foundation"
or PGLAF), owns a compilation copyright in the collection of Project
Gutenberg-tm electronic works. Nearly all the individual works in the
collection are in the public domain in the United States. If an
individual work is in the public domain in the United States and you are
located in the United States, we do not claim a right to prevent you from
copying, distributing, performing, displaying or creating derivative
works based on the work as long as all references to Project Gutenberg
are removed. Of course, we hope that you will support the Project
Gutenberg-tm mission of promoting free access to electronic works by
freely sharing Project Gutenberg-tm works in compliance with the terms of
this agreement for keeping the Project Gutenberg-tm name associated with
the work. You can easily comply with the terms of this agreement by
keeping this work in the same format with its attached full Project
Gutenberg-tm License when you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern
what you can do with this work. Copyright laws in most countries are in
a constant state of change. If you are outside the United States, check
the laws of your country in addition to the terms of this agreement
before downloading, copying, displaying, performing, distributing or
creating derivative works based on this work or any other Project
Gutenberg-tm work. The Foundation makes no representations concerning
the copyright status of any work in any country outside the United
States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other immediate
access to, the full Project Gutenberg-tm License must appear prominently
whenever any copy of a Project Gutenberg-tm work (any work on which the
phrase "Project Gutenberg" appears, or with which the phrase "Project
Gutenberg" is associated) is accessed, displayed, performed, viewed,
copied or distributed:
This eBook is for the use of anyone anywhere at no cost and with
almost no restrictions whatsoever. You may copy it, give it away or
re-use it under the terms of the Project Gutenberg License included
with this eBook or online at www.gutenberg.org
1.E.2. If an individual Project Gutenberg-tm electronic work is derived
from the public domain (does not contain a notice indicating that it is
posted with permission of the copyright holder), the work can be copied
and distributed to anyone in the United States without paying any fees
or charges. If you are redistributing or providing access to a work
with the phrase "Project Gutenberg" associated with or appearing on the
work, you must comply either with the requirements of paragraphs 1.E.1
through 1.E.7 or obtain permission for the use of the work and the
Project Gutenberg-tm trademark as set forth in paragraphs 1.E.8 or
1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted
with the permission of the copyright holder, your use and distribution
must comply with both paragraphs 1.E.1 through 1.E.7 and any additional
terms imposed by the copyright holder. Additional terms will be linked
to the Project Gutenberg-tm License for all works posted with the
permission of the copyright holder found at the beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm
License terms from this work, or any files containing a part of this
work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this
electronic work, or any part of this electronic work, without
prominently displaying the sentence set forth in paragraph 1.E.1 with
active links or immediate access to the full terms of the Project
Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary,
compressed, marked up, nonproprietary or proprietary form, including any
word processing or hypertext form. However, if you provide access to or
distribute copies of a Project Gutenberg-tm work in a format other than
"Plain Vanilla ASCII" or other format used in the official version
posted on the official Project Gutenberg-tm web site (www.gutenberg.org),
you must, at no additional cost, fee or expense to the user, provide a
copy, a means of exporting a copy, or a means of obtaining a copy upon
request, of the work in its original "Plain Vanilla ASCII" or other
form. Any alternate format must include the full Project Gutenberg-tm
License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying,
performing, copying or distributing any Project Gutenberg-tm works
unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing
access to or distributing Project Gutenberg-tm electronic works provided
that
- You pay a royalty fee of 20% of the gross profits you derive from
the use of Project Gutenberg-tm works calculated using the method
you already use to calculate your applicable taxes. The fee is
owed to the owner of the Project Gutenberg-tm trademark, but he
has agreed to donate royalties under this paragraph to the
Project Gutenberg Literary Archive Foundation. Royalty payments
must be paid within 60 days following each date on which you
prepare (or are legally required to prepare) your periodic tax
returns. Royalty payments should be clearly marked as such and
sent to the Project Gutenberg Literary Archive Foundation at the
address specified in Section 4, "Information about donations to
the Project Gutenberg Literary Archive Foundation."
- You provide a full refund of any money paid by a user who notifies
you in writing (or by e-mail) within 30 days of receipt that s/he
does not agree to the terms of the full Project Gutenberg-tm
License. You must require such a user to return or
destroy all copies of the works possessed in a physical medium
and discontinue all use of and all access to other copies of
Project Gutenberg-tm works.
- You provide, in accordance with paragraph 1.F.3, a full refund of any
money paid for a work or a replacement copy, if a defect in the
electronic work is discovered and reported to you within 90 days
of receipt of the work.
- You comply with all other terms of this agreement for free
distribution of Project Gutenberg-tm works.
1.E.9. If you wish to charge a fee or distribute a Project Gutenberg-tm
electronic work or group of works on different terms than are set
forth in this agreement, you must obtain permission in writing from
both the Project Gutenberg Literary Archive Foundation and Michael
Hart, the owner of the Project Gutenberg-tm trademark. Contact the
Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable
effort to identify, do copyright research on, transcribe and proofread
public domain works in creating the Project Gutenberg-tm
collection. Despite these efforts, Project Gutenberg-tm electronic
works, and the medium on which they may be stored, may contain
"Defects," such as, but not limited to, incomplete, inaccurate or
corrupt data, transcription errors, a copyright or other intellectual
property infringement, a defective or damaged disk or other medium, a
computer virus, or computer codes that damage or cannot be read by
your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right
of Replacement or Refund" described in paragraph 1.F.3, the Project
Gutenberg Literary Archive Foundation, the owner of the Project
Gutenberg-tm trademark, and any other party distributing a Project
Gutenberg-tm electronic work under this agreement, disclaim all
liability to you for damages, costs and expenses, including legal
fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT
LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE
PROVIDED IN PARAGRAPH 1.F.3. YOU AGREE THAT THE FOUNDATION, THE
TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE
LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR
INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH
DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a
defect in this electronic work within 90 days of receiving it, you can
receive a refund of the money (if any) you paid for it by sending a
written explanation to the person you received the work from. If you
received the work on a physical medium, you must return the medium with
your written explanation. The person or entity that provided you with
the defective work may elect to provide a replacement copy in lieu of a
refund. If you received the work electronically, the person or entity
providing it to you may choose to give you a second opportunity to
receive the work electronically in lieu of a refund. If the second copy
is also defective, you may demand a refund in writing without further
opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth
in paragraph 1.F.3, this work is provided to you 'AS-IS' WITH NO OTHER
WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT LIMITED TO
WARRANTIES OF MERCHANTIBILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied
warranties or the exclusion or limitation of certain types of damages.
If any disclaimer or limitation set forth in this agreement violates the
law of the state applicable to this agreement, the agreement shall be
interpreted to make the maximum disclaimer or limitation permitted by
the applicable state law. The invalidity or unenforceability of any
provision of this agreement shall not void the remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the
trademark owner, any agent or employee of the Foundation, anyone
providing copies of Project Gutenberg-tm electronic works in accordance
with this agreement, and any volunteers associated with the production,
promotion and distribution of Project Gutenberg-tm electronic works,
harmless from all liability, costs and expenses, including legal fees,
that arise directly or indirectly from any of the following which you do
or cause to occur: (a) distribution of this or any Project Gutenberg-tm
work, (b) alteration, modification, or additions or deletions to any
Project Gutenberg-tm work, and (c) any Defect you cause.
Section 2. Information about the Mission of Project Gutenberg-tm
Project Gutenberg-tm is synonymous with the free distribution of
electronic works in formats readable by the widest variety of computers
including obsolete, old, middle-aged and new computers. It exists
because of the efforts of hundreds of volunteers and donations from
people in all walks of life.
Volunteers and financial support to provide volunteers with the
assistance they need, are critical to reaching Project Gutenberg-tm's
goals and ensuring that the Project Gutenberg-tm collection will
remain freely available for generations to come. In 2001, the Project
Gutenberg Literary Archive Foundation was created to provide a secure
and permanent future for Project Gutenberg-tm and future generations.
To learn more about the Project Gutenberg Literary Archive Foundation
and how your efforts and donations can help, see Sections 3 and 4
and the Foundation web page at http://www.pglaf.org.
Section 3. Information about the Project Gutenberg Literary Archive
Foundation
The Project Gutenberg Literary Archive Foundation is a non profit
501(c)(3) educational corporation organized under the laws of the
state of Mississippi and granted tax exempt status by the Internal
Revenue Service. The Foundation's EIN or federal tax identification
number is 64-6221541. Its 501(c)(3) letter is posted at
http://pglaf.org/fundraising. Contributions to the Project Gutenberg
Literary Archive Foundation are tax deductible to the full extent
permitted by U.S. federal laws and your state's laws.
The Foundation's principal office is located at 4557 Melan Dr. S.
Fairbanks, AK, 99712., but its volunteers and employees are scattered
throughout numerous locations. Its business office is located at
809 North 1500 West, Salt Lake City, UT 84116, (801) 596-1887, email
business@pglaf.org. Email contact links and up to date contact
information can be found at the Foundation's web site and official
page at http://pglaf.org
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
gbnewby@pglaf.org
Section 4. Information about Donations to the Project Gutenberg
Literary Archive Foundation
Project Gutenberg-tm depends upon and cannot survive without wide
spread public support and donations to carry out its mission of
increasing the number of public domain and licensed works that can be
freely distributed in machine readable form accessible by the widest
array of equipment including outdated equipment. Many small donations
($1 to $5,000) are particularly important to maintaining tax exempt
status with the IRS.
The Foundation is committed to complying with the laws regulating
charities and charitable donations in all 50 states of the United
States. Compliance requirements are not uniform and it takes a
considerable effort, much paperwork and many fees to meet and keep up
with these requirements. We do not solicit donations in locations
where we have not received written confirmation of compliance. To
SEND DONATIONS or determine the status of compliance for any
particular state visit http://pglaf.org
While we cannot and do not solicit contributions from states where we
have not met the solicitation requirements, we know of no prohibition
against accepting unsolicited donations from donors in such states who
approach us with offers to donate.
International donations are gratefully accepted, but we cannot make
any statements concerning tax treatment of donations received from
outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation
methods and addresses. Donations are accepted in a number of other
ways including checks, online payments and credit card donations.
To donate, please visit: http://pglaf.org/donate
Section 5. General Information About Project Gutenberg-tm electronic
works.
Professor Michael S. Hart is the originator of the Project Gutenberg-tm
concept of a library of electronic works that could be freely shared
with anyone. For thirty years, he produced and distributed Project
Gutenberg-tm eBooks with only a loose network of volunteer support.
Project Gutenberg-tm eBooks are often created from several printed
editions, all of which are confirmed as Public Domain in the U.S.
unless a copyright notice is included. Thus, we do not necessarily
keep eBooks in compliance with any particular paper edition.
Most people start at our Web site which has the main PG search facility:
http://www.gutenberg.org
This Web site includes information about Project Gutenberg-tm,
including how to make donations to the Project Gutenberg Literary
Archive Foundation, how to help produce our new eBooks, and how to
subscribe to our email newsletter to hear about new eBooks.