Project Gutenberg's The Baculum in Microtine Rodents, by Sydney Anderson This eBook is for the use of anyone anywhere at no cost and with almost no restrictions whatsoever. You may copy it, give it away or re-use it under the terms of the Project Gutenberg License included with this eBook or online at www.gutenberg.org Title: The Baculum in Microtine Rodents Author: Sydney Anderson Release Date: November 15, 2011 [EBook #38021] Language: English Character set encoding: ISO-8859-1 *** START OF THIS PROJECT GUTENBERG EBOOK THE BACULUM IN MICROTINE RODENTS *** Produced by Chris Curnow, Alex Gam, Joseph Cooper and the Online Distributed Proofreading Team at http://www.pgdp.net
University of Kansas Publications
Museum of Natural History
Volume 12, No. 3, pp. 181-216, 49 figs.
February 19, 1960
BY
SYDNEY ANDERSON
University of Kansas
Lawrence
1960
University of Kansas Publications, Museum of Natural History
Editors: E. Raymond Hall, Chairman, Henry S. Fitch, Robert W. Wilson
Volume 12, No. 3, pp. 181-216, 49 figs.
Published February 19, 1960
University of Kansas
Lawrence, Kansas
PRINTED IN
THE STATE PRINTING PLANT
TOPEKA, KANSAS
1960
28-774
The Baculum in Microtine Rodents
BY
SYDNEY ANDERSON
Didier (1943, 1954) has described the bacula of several Old World microtines, and other rodents. Argyropulo studied (1933a, 1933b) five species of Cricetinae and Microtus socialis. Ognev (1950) illustrated numerous species of Eurasian microtines. Hamilton (1946) figured and described the baculum of 11 species of North American microtines. Hibbard and Rinker (1942, 1943) figured the baculum of Synaptomys cooperi paludis and of Microtus ochrogaster taylori. Dearden (1958) studied the baculum in two Asiatic species of Lagurus, in six subspecies of Lagurus curtatus of North America, and in six other species of microtines of other genera.
The baculum can be preserved easily with standard study skins, and is potentially useful in interpreting relationships on any taxonomic level, and especially in determining the relationships of species within a genus, if used together with other structures.
The anatomical orientation of the baculum needs comment because some confusion exists in the literature, especially concerning the use of the terms ventral and dorsal. The urethra lies on the anatomically ventral side of the penis, and of the baculum. In the center of the penis lies a single corpus cavernosum penis, shown in cross section proximal to the baculum in Figure 1c. Dorsally an artery, thinner walled than the ventral urethra, ends in a somewhat reticulate sinus surrounding primarily the middle part of the baculum within the bulbous glans penis. The corpus cavernosum penis (the structure has no median septum, at least distally) terminates with the baculum and is closely knit to it. The site of this bond is evident in the tuberosities and sculpturing of the base of the baculum.
The part of the penis enclosing the baculum, when not erect, is folded back as shown in Figures 1a and 1b. As a result the anatomically ventral surface faces upwards, or at least posterodorsally. The use of the term ventral in this account refers to the anatomically ventral side, that is to say to the side of the baculum facing the urethra.
The baculum in microtines consists of an elongate stalk, having a laterally, and to a lesser extent dorsoventrally, expanded base and [Pg 184] an attenuate distal shaft. Usually, three digitate processes of cartilaginous material in which additional ossifications may occur arise from the terminus of the shaft. The proportions and curvature of the stalk vary as do the proportions of the terminal ossifications to each other and to the stalk. In some species one or more of the digital processes are frequently completely unossified.
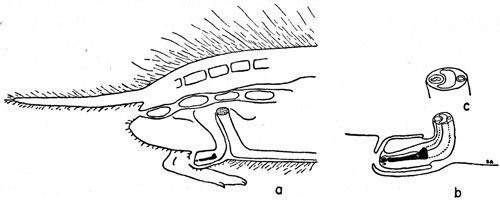
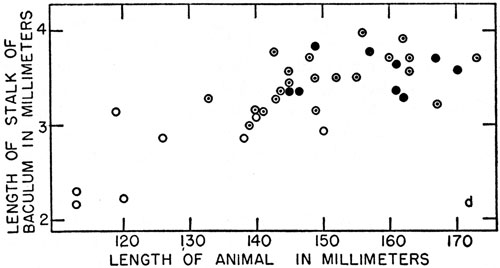
Figure 1. The baculum in Microtus ochrogaster—orientation and variation with age. a. Diagram of a sagittal section of the posterior half of a vole, natural size. The penis, containing the baculum (in black), extends ventrally from a point posterior to the pubic symphysis (stippled), along the body wall, and bends posteriorly at the distal end. b. Distal end of penis (× 2) showing baculum (in black), the urethra (solid lines) adjacent to the baculum, and the corpus cavernosum (broken lines) proximal to the baculum. c. Oblique view of the cross section of penis (× 4) shown in Figure 1 b. The thick-walled urethra lies ventral to the curved corpus cavernosum. A thinner-walled blood-vessel lies dorsal to the corpus cavernosum. The anatomically ventral side of the baculum, in the normal non-erect penis shown, is seen to face dorsally. d. Graph showing the relationship between size of baculum, size of animal, and development of digital ossifications. Circles show presence of ossification in stalk only; circles enclosing dots indicate presence of secondary ossification in median process also; large dots indicate the addition of tertiary ossification in one or both of the lateral digitate processes.
Preserved specimens of Microtus arvalis, Microtus agrestis, Microtus orcadensis, Microtus nivalis, Microtus guentheri, Microtus subterraneus, Clethrionomys glareolus, and Ellobius lutescens were provided by Prof. Robert Matthey of Lausanne, Switzerland. J. Knox Jones, Jr. carefully saved the bacula with specimens of Microtus fortis and Clethrionomys rufocanus from Korea. Dr. W. B. Quay, Department of Zoology, University of California, supplied specimens of Synaptomys cooperi, Phenacomys intermedius, and Microtus oregoni. Dr. Franklin Sturges and Mr. John W. Goertz, Museum of Natural History, Oregon State College, Corvallis, have provided specimens including bacula of Clethrionomys occidentalis, Microtus oregoni, and Microtus townsendii. Dr. Randolph L. Peterson and Mr. Bristol Foster, Royal Ontario Museum of Zoology, Toronto, Canada, provided specimens of Phenacomys intermedius. Dr. J. N. Layne, University of Florida, Gainsville, Florida, presented me with a baculum of Microtus parvulus.
I am indebted to all of these persons for their aid, and to various collectors for the Museum of Natural History, who preserved bacula with specimens. Many of these specimens were obtained through the assistance of the University of Kansas Endowment Association and the National Science Foundation.
Bacula were obtained from fresh specimens, specimens preserved in alcohol or formalin, and dried study skins. The processing of bacula has been discussed by Hamilton (1946), Friley (1947), White (1951), and Dearden (1958). The methods used to preserve bacula for my study differed some from any of those reported. The terminal part of each penis including the baculum imbedded in the glans penis was removed in its entirety and placed in a vial. The catalogue number was kept with each specimen at all times. A two per cent solution of potassium hydroxide was added. All specimens were examined at least once a day. If tissues other than the glans penis were present they were removed with forceps when softened usually at the end of one day. Several drops of Alizarin red-S stain in a saturated alcoholic solution were added to the 3 to 5 ccs. of KOH solution in each vial. Solutions were replaced if they became turbid enough to obstruct observation of the clearing penis. After one day the solution containing stain was removed and replaced with two per cent KOH solution without stain. When the glans became sufficiently cleared that the stained baculum could be seen easily, the solution was replaced by glycerin in which clearing was completed. The time required for the entire process varied from one day to more than two weeks depending on the size of the specimen and on its condition. Fresh specimens clear more rapidly than dried specimens, and those that are dried more rapidly than those that are preserved. A three or four per cent solution of hydroxide will hasten the process, but more frequent observation is required to prevent excessive maceration.
Specimens were then examined in a shallow dish containing glycerin under a binocular microscope. The baculum can be viewed from any desired direction. The method described above leaves the baculum intact within the glans penis; therefore its orientation can be determined relative to the thick walled urethra and the thin walled dorsal artery that extends onto the dorsal side of the baculum. The ventral curvature of the penis proximal to the baculum, and the distal extension, characteristic of most species, of the dorsal border of [Pg 186] the glans (both shown in Figure 1) are other features aiding in correctly orienting cleared specimens. The digitate processes are not so often injured, lost, or displaced when the method described above is used as they are when the penis is dissected. Specimens were stored in glycerin in glass shell vials having polyethylene stoppers. A small card bearing the name, number, locality, and other data was placed in each vial. A specimen thus enclosed can be kept indefinitely, or removed and mounted in balsam as described by White (1951:631) or in plastic as described by Dearden (1958:541) and thus stored in the vial containing the skull of the specimen.
Drawings were made on millimeter ruled paper while the baculum was viewed under a binocular microscope with a square ruled eyepiece.
Unless otherwise noted all specimens listed are in the University of Kansas Museum of Natural History. Catalogue numbers are cited. Measurements are accurate to within less than one-tenth of a millimeter. Proportions as stated in the text are approximations, accurate to within one-twelfth (8.33 per cent). The range of variation is unknown for some species. Mention is made if maturity is known or suspected to differ in specimens being compared.
The development of the baculum has been studied by Callery (1951) in Mesocricetus auratus and by Ruth (1934) in the laboratory rat. In the rat (Rattus norvegicus) the bone is of endoblastemal origin being laid down by a condensation of undifferentiated mesenchymal cells. At the distal end of the bone dense fibrous tissue is then differentiated and at the proximal end hyaline cartilage. Growth is by substitution at the proximal end and by subperiosteal lamellation circumferentially. A marrow cavity is formed by resorption. In the baculum of the hamster the primary center of ossification is in the stalk, and is present at the age of three days; the secondary centers are in lateral processes and are present at 80 days and enlarge subsequently. A tertiary center, in each median process, may or may not develop later. Maximum development of the baculum is reached late in the reproductive life of the hamster.
The early ossification of the baculum noted in the rat and the hamster occurs in Microtus also. A specimen of Microtus montanus fusus (76831, from 5 mi. N, 26 mi. W Saguache, 9600 ft., Saguache County, Colorado) only 74 mm. in total length and weighing only 6.6 grams, had a slender ossified baculum having enlarged ends. This vole was one-half of the average length and less than one-fifth of the average weight of an adult, and of approximately the size at which weaning takes place.
The development of the baculum in Microtus ochrogaster was studied in 32 specimens of various ages. The specimens (between Nos. 74994 and 75074) were collected between August 15 and September 4, 1957, at localities on the Great Plains. These specimens were from breeding populations, as evidenced by pregnancy of females and by large size of testes of males. The length and width of the stalk of the baculum, the presence of digital ossifications, the [Pg 187] total length of the animal, and the size of the testes were noted. Variability in length of testes is greatest when voles are from 140 to 150 mm. in total length. Sexual maturity is reached rather abruptly when the total length of most individuals is 140 to 150 millimeters. If the baculum likewise underwent more rapid growth at the onset of sexual maturity, greater variability should be evident in the length of the baculum of voles 140 to 150 mm. in total length than in bacula of voles of other sizes. This was the case (see Figure 1d). The baculum does not, however, suddenly reach its maximum maturity.
The primary ossification is in the stalk. The secondary ossification is in the median process except in Lagurus (Dearden, 1958:551) and some individuals of Neofiber (see account on page 258). Tertiary centers of ossification are in the lateral processes. The primary ossification is present at an early age and subsequently increases in size and solidity. The secondary and tertiary ossifications are progressively more common in older voles. The increase in degree of ossification of all parts continues after sexual maturity is reached. Individual variation and variation with age in the baculum of Microtus pennsylvanicus have been illustrated by Hamilton (1946:380). Figures 14, 15, and 17 illustrate variation with size, which is correlated with age, and also illustrate individual variation. The three bacula are from adult voles having testes that measured 15, 16 and 16 mm. in length, respectively. Each vole was trapped in late June. The total lengths in millimeters of the three voles are 172, 167, and 181; weights are 55, 52.4, and 65.5 grams. I judge that the greater size of the stalk and the better developed base shown in Figure 17 than in Figure 15 are illustrative of age variation; the difference in the size of the lateral digitate processes is, in this case, attributable to individual variation. Differences in the distal end of the baculum in Figures 42 and 43, show individual variation also. Figures 35 and 36 represent two different subspecies; different individuals of M. mexicanus mogollonensis, however, exhibit individual variation of the same degree.
Hall and Cockrum (1953) list 44 species of microtines in North America. At least twelve of these are insular or local forms perhaps derived from some other species; for example Microtus coronarius, an insular form derived from Microtus longicaudus; Microtus provectus, considered by Chamberlain (1954:587) and by Wheeler (1956:176) as a subspecies of Microtus pennsylvanicus; and Microtus ludovicianus, a close relative of Microtus ochrogaster.
All North American genera have been studied. Of the genus Microtus in North America, all subgenera but Orthriomys and all species but the following nine, have been studied: M. (Orthriomys) umbrosus, the insular M. (Stenocranius) abbreviatus, M. (Microtus) breweri, M. (Microtus) nesophilus, M.
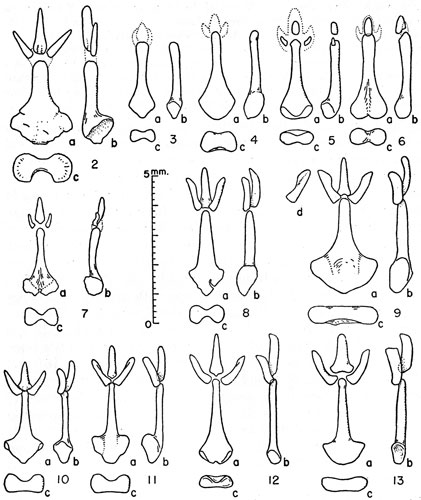
Figures 2-13. Bacula of microtines. Unless indicated otherwise views are (a) of the dorsum, (b) the right side, and (c) the proximal end with the dorsal surface upward. Exact localities are given in accounts of species concerned.
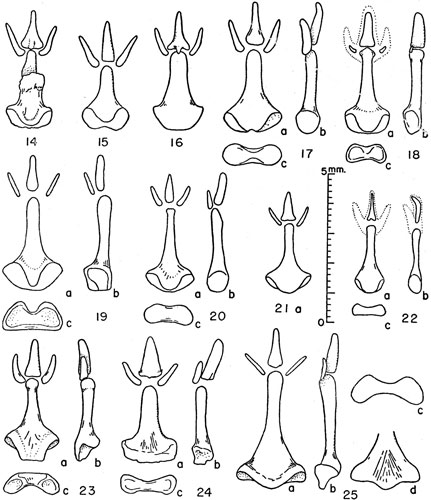
Figures 14-25. Bacula of Microtus. Unless indicated otherwise views are (a) of the dorsum, (b) the right side, and (c) the proximal end with dorsal surface upward.
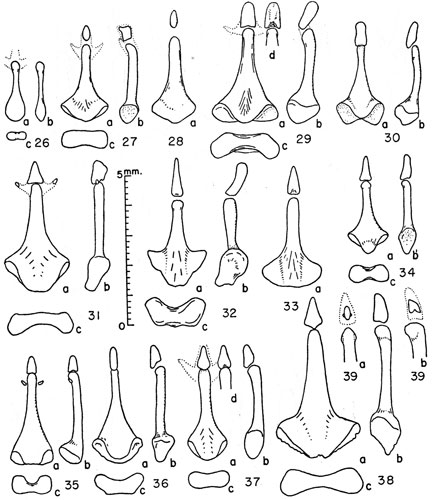
Figures 26-39. Bacula of microtines. Unless indicated otherwise views are (a) of the dorsum, (b) the right side, and (c) the proximal end with the dorsal surface upward.
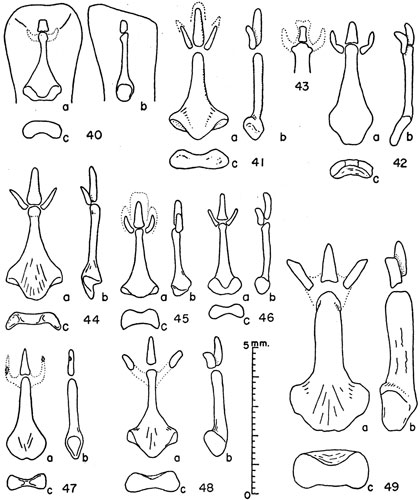
Figures 40-49. Bacula of microtines. Unless indicated otherwise views are (a) of the dorsum, (b) the right side, and (c) the proximal end with the dorsal surface upward.
(Microtus) provectus (the last three are probably insular derivatives of M. pennsylvanicus), M. (Microtus) fulviventer (perhaps derived from the same stock as Microtus mexicanus), M. (Microtus) xanthognathus (perhaps related to Microtus chrotorrhinus), M. (Microtus) coronarius, and M. (Pedomys) ludovicianus.
| Species of Which Bacula Were Examined | |
|---|---|
| Subfamily: Microtinae | Number of Specimens |
| Tribe: Lemmi | |
| Dicrostonyx groenlandicus (Traill) | 4 |
| Lemmus trimucronatus (Richardson) | 6 |
| Synaptomys cooperi Baird | 5 |
| Tribe: Microti | |
| Genus: Clethrionomys Tilesius, 1850 | |
| Clethrionomys rutilus Pallas | 4 |
| Clethrionomys gapperi (Vigors) | 9 |
| Clethrionomys occidentalis (Merriam) | 1 |
| Clethrionomys glareolus Schreber | 1 |
| Clethrionomys rufocanus Sundevall | 1 |
| Genus: Phenacomys Merriam, 1897 | |
| Phenacomys intermedius Merriam | 5 |
| Genus: Ondatra Link, 1795 | |
| Ondatra zibethicus (Linnaeus) | 1 |
| Genus: Microtus Schrank, 1798 | |
| (Herpetomys) guatemalensis Merriam | 3 |
| (Arvicola) richardsoni (DeKay) | 2 |
| (Chilotus) oregoni (Bachman) | 3 |
| (Stenocranius) gregalis (Pallas) | 1 |
| (Stenocranius) miurus Osgood | 9 |
| (Chionomys) longicaudus (Merriam) | 6 |
| (Chionomys) nivalis Martins | 2 |
| (Microtus) arvalis (Pallas) | 1 |
| (Microtus) orcadensis Millais | 1 |
| (Microtus) guentheri Danford and Alston | 1 |
| (Microtus) fortis Büchner | 2 |
| (Microtus) montanus (Peale) | 15 |
| (Microtus) townsendii (Bachman) | 3 |
| (Microtus) oeconomus (Pallas) | 10 |
| (Microtus) mexicanus (Saussure) | 13 |
| (Microtus) californicus (Peale) | 2 |
| (Microtus) pennsylvanicus (Ord) | 13 |
| (Microtus) agrestis (Linnaeus) | 1 |
| (Pedomys) ochrogaster (Wagner) | 41 |
| (Pitymys) pinetorum (LeConte) | 2 |
| (Pitymys) parvulus (Howell) | 1 |
| (Pitymys) quasiater (Coues) | 5 |
| (Pitymys) fatioi Mottaz | 1 |
| Genus: Neofiber True, 1884 | |
| Neofiber alleni True | 2 |
| Genus: Lagurus Gloger, 1841 | |
| Lagurus curtatus (Cope) | 7 |
| Total number examined | 184 |
Figs. 3 and 4
Baculum: stalk elongate, greatest length (3.1 mm.) 2 1/5 to 2½ times greatest breadth, and 4½ times greatest depth; digitate processes usually cartilaginous, occasionally lateral processes partly ossified; basal tuberosities weakly to moderately developed, medially confluent; posterior profile in dorsal view rounded with rounded posterior apex or shallow notch; dorsal concavity in end-view shallower and not so wide as ventral concavity; median constriction approximately 2/3 greatest depth; ventral part of base in end-view wider than dorsal part; shaft straight or slightly curved; base of stalk placed dorsally relative to axis of shaft; stalk spatulate, sometimes with distal enlargement; at mid-point stalk wider than high; lateral profile in dorsal view sloping gradually without abrupt curvature anterior to point of greatest width.
The baculum of Dicrostonyx torquatus figured by Ognev (1948:476) agrees with that of D. groenlandicus in shape of stalk, and in lateral digitate processes that are small relative to size of median process; but differs in more elongate, terminally enlarged, bulbar shape of median process. None of my specimens showed ossification in the lateral processes, observed by Hamilton (1946:381) in Dicrostonyx rubricatus richardsoni [ = D. groenlandicus richardsoni]. In all of my specimens the cartilaginous median process was larger than that figured by Hamilton, or by Dearden (1958:542).
Specimens examined: Four from; Point Barrow, Alaska, 52524 (Barrow Village), 67264 (died in captivity); Brooks Range, Alaska, 50536 (Wahoo Lake, 69°08', 146°58'), 50539 (Porcupine Lake, 68°51'57", 146°29'50", 3140 ft.).
Fig. 2
Baculum: Stalk heavy, broad, greatest length (2.8 mm.) in mature individuals (Fig. 2) as little as 11/3 times greatest breadth, greatest length no less than 22/3 times greatest depth of base; three ossified processes, median one from as long as to ½ longer than the lateral processes, and approximately 2/3 wider and twice as deep as lateral processes; length of median process almost 3½ times its breadth, approximately ½ length of stalk; basal fossae broadly confluent; posterior profile in dorsal view evenly rounded; in end-view ventral concavity deeper than dorsal concavity, constriction as little as ½ greatest depth in mature specimens; shaft straight, bluntly rounded, or slightly decurved and laterally inflated terminally; lateral profile in dorsal view a gradual slope from widest point of stalk anteriorly onto shaft; in younger individuals stalk slenderer, otherwise as described above.
Five specimens examined by me differ from one figured and described by Hamilton (1946:379) in that stalk is better developed, larger relative to size of processes, length of stalk in my specimen (Fig. 2) 2.8 as opposed to 2.1 mm. in Hamilton's specimen; median process shorter, 1.5 as opposed to 1.8 mm., proximal end rounded rather than concave, not partially enclosing tip of shaft; proportion of and relative sizes of median and lateral processes approximately same as in Hamilton's Lemmus helvolus [= Lemmus trimucronatus helvolus]. [Pg 194] A specimen figured by Dearden (1958:542) has a basally trilobed median process.
The baculum of the Asiatic Lemmus lemmus figured by Ognev (1948:413) agrees with my specimens in the ossification of three processes, the relative sizes of these processes to each other and to the stalk, the well-developed base of the stalk and heavy bluntly rounded shaft; the baculum of Lemmus lemmus differs in greater anterolateral extent of basal tuberosities, in proximal notch seemingly separating these tuberosities, and in median process being slenderer.
Specimens examined: Five, of two subspecies; Lemmus trimucronatus alascensis, Point Barrow, Alaska, numbers 50591, 50678, 50731, 50758; Lemmus trimucronatus subarcticus, Wahoo Lake, 69°08', 146°58', 2350 ft., Brooks Range, Alaska, 50948.
Figs. 5 and 6
Baculum: Stalk elongate, greatest length (2.7 to 2.8 mm.) 2 1/3 to 2½ times greatest breadth, 4 to 5 times greatest depth; three processes ossified or lateral processes unossified, ossifications relatively small (in 78380, median ossification less than ¼ as large as lateral ossifications although median cartilaginous process is larger), length of median process 1/5 to 1/6 of length of stalk, cartilaginous part of median process larger; posterior profile in dorsal view convex throughout or bilobate; tuberosities moderately developed, deflected dorsal to axis of shaft; in end-view medial construction 3/5 greatest depth of tuberosities; shaft tapered from point of greatest width, slightly inflated terminally.
The specimen (KU 13716) figured by Hibbard and Rinker (1942:29) has been restudied. It was first cleared and stained to soften the dry cartilage binding the digital processes together and to differentiate bone and cartilage. The lateral processes are small and cartilaginous (Fig. 6) and seem intact. The differences between this specimen and others examined by Hamilton (1946:381), Dearden (1958:542), and myself, namely the relatively larger median ossification, the absence of ossification in lateral processes, and the distinctly bilobate base and larger size, may represent geographic differences, or individual variation. The proportions of length, width, and depth of the stalk, and the appearance in lateral view do not differ greatly from others examined by Hamilton, by Dearden (1958:546), and by me.
Specimens examined: Five, representing four subspecies; S. cooperi gossii, 6 mi. N Midway, Holt Co., Nebraska 78379, 78380; S. cooperi relictus, 5 mi. N, 2 mi. W Parks, Dundy Co., Nebraska, 72601 (immature); S. cooperi saturatus, 3 mi. S Demotte, Jasper Co., Indiana, 3-C-454, collection of W. B. Quay; S. cooperi paludis, Meade County State Park, Kansas, 13716.
Fig. 11
Baculum: Stalk elongate, and proximally enlarged, greatest length (2.7 mm.) 2 times greatest breadth; less than 4 times greatest depth; three well-developed ossified processes; length of stalk 21/3 times length of median process; median process with basal (and ventral) protuberence and lateral lobes, arched in dorsoventral plane; lateral processes as large as median process, flattened distally, having ventromedial vane on distal half; basal tuberosities of stalk well developed, medially confluent; posterior profile in dorsal view trilobate or convex [Pg 195] throughout with rounded posterior apex; dorsal concavity well developed, ventral surface but slightly concave, medial constriction of base as little as ½ greatest depth; shaft straight, slender, at mid-point of stalk but slightly wider than high; basal tuberosities largely dorsal to axis of shaft in lateral view; lateral profile in dorsal view with an abrupt curvature separating the gently sloping sides of the shaft from the basal part at its greatest breadth.
The specimen of Clethrionomys rutilus figured by Ognev (1950:120) is essentially like the North American specimens examined by me in the relative sizes of the ossifications and the general shape of the stalk.
Specimens examined: Four, of one subspecies; C. r. dawsoni, west bank Gakona River, 1700 ft., 5 mi. NNE Gulkana, Alaska, 42865, 42866; SW end Dezadeash Lake, 2400 ft., Yukon Territory, 42910, 42921.
Fig. 10
Baculum: Stalk elongate, greatest length (2.8 mm.) 1¾ times greatest breadth, and 3¾ times greatest depth; proximally enlarged, greatest depth ½ greatest breadth; three well-developed ossified processes; length of stalk 2 1/3 times length of median process; median process arched in dorsoventral plane, with basiventral protuberence or spine and lateral lobes; lateral processes as large as median process, flattened distally, arched; basal tuberosities of stalk well developed, medially confluent; posterior profile in dorsal view trilobate or convex throughout with a rounded posterior apex; dorsal concavity well developed, ventral surface but slightly concave, or in some cases slightly convex; medial constriction of base 3/5 greatest depth; shaft straight, slender, at mid-point of stalk twice as wide as high; basal tuberosities dorsally placed relative to axis of shaft; lateral profile in dorsal view abruptly curved anterior to point of greatest width; slender stalk distinct from angular enlarged base.
The most noticeable difference between the baculum of C. rutilus and C. gapperi is size. The proportions of the four ossifications are approximately the same. Ventral vanes on the lateral processes are not developed in C. gapperi. C. gapperi and C. rutilus are more nearly alike in their bacula than any other two species of Clethrionomys examined. Clethrionomys occidentalis, the other New World species, is also much like C. gapperi and C. rutilus. The differences are of a magnitude comparable to those between the bacula in subspecies of Microtus montanus (Figs. 19-21) for example, or in subspecies of Lagurus curtatus (Dearden, 1958:542).
Specimens examined: Nine, of two subspecies; Clethrionomys gapperi athabascae, British Columbia, 42922 (Indian Creek, Mile Post 234 of Alaskan Highway), 64281 (West bank Racing River, 89 mi. W Muskwa), 64287 (North bank Tetsa River, 56 mi. W, 11 mi. S Muskwa), 64290 (44 mi. W, 9 mi. S Muskwa), 64310 (32 mi. W, 2 mi. S Muskwa); Clethrionomys gapperi galei, 31 mi. N Pinedale, Sublette Co., Wyoming, 42108; Grand Mesa, Delta Co., Colorado, 60014 and 60015 (5½ mi. E, 12 mi. S Collbran), 60022 (8 mi. E, 1/2 mi. S Skyway).
Fig. 12
Baculum: Stalk elongate, greatest length (2.8 mm.) 2½ times greatest breadth, 6 times greatest depth; three well-developed ossified processes; median process larger than lateral processes, ½ the length of stalk, curved, basally [Pg 196] broad, ventrally keeled, trilobate posteriorly; lateral ossifications large, flattened distally, curved; posterior profile of stalk posteriorly slightly emarginate, thus bilobate in outline; in end-view dorsal concavity deeper than ventral, constriction less than ½ greatest depth, tuberosities confluent, visible in dorsal view at each side; shaft slender, especially in depth, straight; at mid-point of stalk almost twice as wide as deep, slight terminal inflation.
The general proportions of the stalk and the relatively large, uniquely shaped processes, are characteristic of most specimens of the genus Clethrionomys examined.
Specimen examined: C. occidentalis californicus, one from Mary's Peak, Benton Co., Oregon, 30, F. W. Sturges' collection.
Fig. 13
Baculum: Stalk elongate, greatest length (2.9 mm.) twice the greatest breadth in the specimen examined, flattened proximally, greatest length almost 6 times greatest depth of base; three well-developed ossified processes; median process arched in a dorsoventral plane, with basal notch and lateral lobes; lateral processes as long as median process, bowed in dorsal view, flattened distally, with ventromedial vane; basal tuberosities of stalk weakly developed, medially confluent; posterior profile in dorsal view evenly rounded; in end-view dorsal concavity shallow in comparison to most species but deeper than ventral concavity, constriction ¾ greatest depth; shaft straight, at mid-point slightly wider than high, elongate, widest point of stalk less than ¼ of total length from proximal end, slight lateral inflation at tip; lateral profile in dorsal view sloping at first abruptly and then gradually from widest point of stalk anteriorly onto shaft.
The specimen of Clethrionomys glareolus figured by Ognev (1950:31) in dorsal view as I interpret it, resembles my specimen in the rounded base; in the elongate, distally inflated shaft; in the initially abrupt slope of the lateral profile in dorsal view from the greatest width of stalk anteriorly; and in the presence of three well ossified processes. Ognev's specimen differs from mine in the median process being more elongate relative to its width, and rounded proximally, lacking lateral lobes and basal notch; in lateral processes being less curved; in the greater terminal inflation of the shaft; and in the closer approximation of the terminal processes to the shaft. The baculum of Clethrionomys glareolus as described and figured by Didier (1954:243-244) resembles my specimen in general proportions, but is more pointed proximally and more curved in dorsoventral plane. Didier states that the baculum is rather variable in form in this species, in different regions, but that a large number of specimens must be examined to assess the geographic nature of this variation.
Specimen examined: One from Zermatt, Valais, Switzerland, 67100.
Fig. 9
Baculum: Base of stalk broad but relatively flattened dorsoventrally, greatest length (3.2 mm.) less than 1½ greatest width, 4 times greatest depth; three well-developed ossified processes; median process arched in dorsoventral plane, having basal notch and lateral lobes; lateral processes as long as median process, [Pg 197] flattened distally, with ventromedial vane; basal tuberosities of stalk weakly developed, medially confluent; posterior profile in dorsal view convex with rounded posterior apex; dorsal surface of base almost flat, ventral concavity broad and shallow; constriction ¾ greatest depth (not including an unusual irregularity on the ventral surface of the base); shaft straight, at mid-point of stalk distinctly wider than high, slender at distal end, widest point of stalk almost 1/3 of total length from proximal end, tip of shaft rounded; lateral profile in dorsal view gradually sloping from widest point anteriorly onto shaft.
The specimen of Clethrionomys rufocanus figured by Ognev (1950:97) resembles my specimen in the presence of three well ossified processes. Ognev's specimen differs however in the lack of a proximal notch on the median process, the lesser proportion of the stalk included in the basal enlargement, the more posterior position of the point of greatest width, and the presence of a concavity in the posterior profile of the stalk in dorsal view. These differences in the stalk may be owing to a difference in age (my specimen perhaps being older).
Specimen examined: One from 1 mi. NW Oho-ri, 6 M., Korea, 60438.
Figs. 7 and 8
Baculum: Stalk slender, greatest length (2.9 mm.) 2¼ to 2½ times greatest breadth, 4 times greatest depth; three well-developed ossified processes, median one almost ½ length of stalk, curved, broad basally and slightly larger in all dimensions than either lateral process; lateral processes flattened distally, curved; base of stalk well developed, basal tuberosities medially confluent or separated by medial emargination, posterolateral faces flattened or rough; emarginations in the four adults examined; posterior profile in dorsal view bluntly pointed or flattened except for emargination posterially, abruptly curved at point of greatest width; shaft arising broadly from distal side of base of stalk; in end-view hour-glass shaped, medial constriction pronounced, both dorsal and ventral concavities deep; shaft having relatively straight but distally convergent sides; at mid-point of stalk, 1 to 1½ times as wide as deep; tip bluntly rounded, or slightly inflated.
The specimens from Quebec differ from the one from Wyoming in smaller size, relatively smaller lateral digital processes, larger more medial basal emargination, and slender shafts. The baculum of Phenacomys intermedius differs much from that of Phenacomys longicaudus, described by Hamilton (1946:381) and by Dearden (1958:547). Dearden states that the three bacula examined by him of Phenacomys longicaudus differ markedly from the specimen described by Hamilton. It seems to me that in major features the resemblance is greater between the specimens of Phenacomys longicaudus examined by these two authors than between their specimens and specimens of other microtines, including Phenacomys intermedius. Neither Hamilton nor Dearden record the exact localities of capture, the collections in which the specimens are deposited, or the catalogue numbers of specimens. Consequently verification of identifications and observations is difficult.
Specimens examined: Five, of two subspecies; P. intermedius intermedius, 5.4 mi. S Moran, Teton Co., Wyoming, 3-C-309, collection of W. B. Quay; P. intermedius celatus, four (including one immature specimen) from Authiernord, [Pg 198] Abitibi-ouest Co., Quebec, specimens in collection of Bristol Foster designated by numbers 2041-2044 of S. Anderson's field catalogue. Smith and Foster (1957:107) were of the view that Phenacomys ungava (including the above specimens from Quebec) may be specifically distinct from Phenacomys intermedius.
Not figured
Baculum: In the single specimen examined, less mature than that figured by Hamilton (1946:384), the digitate processes are cartilaginous, the basal tuberosities are less well developed, and the shaft is slenderer throughout. The cartilaginous processes are of the same proportions as ossified processes in the figure mentioned. The shaft is also convex ventrally in lateral profile. The view of the side here considered to be anatomically the ventral side (adjacent to the urethra) is labelled dorsal view in Hamilton's specimen.
Specimen examined: One, from Reserve, Brown Co., Kansas, 72405.
Figs. 42 and 43
Baculum: Stalk moderately elongate, greatest length (3.5 mm.) 21/3 times greatest breadth, spatulate, flattened throughout, greatest thickness 1/3 millimeter; three ossified processes; median process having three cornered base, curved dorsally, wider than high, ¼ to 1/5 greatest length of stalk; each lateral process bent at middle, as long as median process, compressed laterally; base of stalk curved dorsally, tuberosities marginal, hence narrow, lateral excavations of tuberous margin not confluent medially; in end-view ventral concavity broad, no dorsal concavity, medial constriction but slightly less than greatest thickness (not depth); shaft wider than high throughout, at mid-point more than 3 times as wide as high; tip of shaft slightly inflated both laterally and dorsoventrally; lateral profile gradually sloping anteriorly from widest point of stalk.
Specimen number 65921 (Fig. 43) differs from number 65895 (Fig. 42) described above. Terminus of shaft of number 65921 has lateral lobes from which arise lateral cartilaginous processes; median terminal ossification irregular in shape, smaller, imbedded in terminally bilobate cartilage. In the spatulate flattened stalk these two specimens are much alike. An immature specimen, number 65908, is smaller (length of stalk 2.6 mm.) also flattened and spatulate, has the terminal processes cartilaginous, the lateral processes bent medially, and proportions as in the adult.
The baculum shows no noteworthy resemblance to that of any other species of North American Microtus; on the other hand the differences between M. guatemalensis and some other species are no greater than the differences between certain species included in the subgenus Microtus. The baculum neither strengthens nor weakens the case for subgeneric rank for M. (Herpetomys) guatemalensis.
Specimens examined: Three from Guatemala; 65895 (2 mi. S San Juan Ixcoy), 65908, (3-1/2 mi. SW San Juan Ixcoy), 65921 (10 mi. E, 4 mi. S Totonicapán).
Figs. 38 and 39
Baculum: Stalk broad, greatest length (3.7 to 4.3 mm.) 1½ times greatest breadth, relatively flattened, greatest depth 1/3 greatest breadth; single median ossified process, in smaller of two specimens this ossification incomplete and of unusual shape (Fig. 39); length of stalk 4 times length of median process; concavities of basal tuberosities medially confluent, constriction less than ½ greatest depth; widest point of shaft less than ¼ length of shaft from posteriormost point; shaft wider than high except at distal end that is inflated dorsally and sometimes laterally; both ventral and dorsal concavities of base of stalk broad and moderately deep; posterior profile in dorsal view evenly rounded or having marginal notch.
In the absence of ossified lateral processes my two specimens differ from bacula of Microtus (Arvicola) terrestris figured by Didier (1943:79, 1954:245, 247, 248) and by Ognev (1950:591). The median process relative to the size of the shaft is smaller, and the shaft relative to its length is wider in M. richardsoni than in M. terrestris. The stalk of M. (Arvicola) amphibius figured by Didier is like that of M. richardsoni in its greater breadth and median notch on posterior border.
The relationship of the New World water rat, M. richardsoni, to the Old World water rats (genus Arvicola of some European authors) is uncertain. Miller (1896:66) placed all of them in the subgenus Arvicola. Subsequent authors, stressing differences in the teeth, have placed M. richardsoni in the subgenus Aulacomys of Rhoads. Zimmerman (1955) has shown that teeth in some Arvicola approach the more complex pattern of M. richardsoni. He argues also that Arvicola is generically distinct from Microtus on the grounds that the two groups have separate origins, Arvicola having descended from the genus Mimomys and Microtus from some other group of microtines. This argument also was advanced by Hinton (1926:47-48). Pending further studies of the possible polyphyletic origin of other subgenera of the genus Microtus, I refer both M. richardsoni and M. terrestris to the subgenus Arvicola.
The evidence afforded by the bacula available is not conclusive as to relations of Old World and New World water rats. No general agreement on the number of species in this Palaearctic group has been reached, and bacula of only three or four of the numerous Old World subspecies have been figured. I have examined none.
Specimens examined: Two, from Wyoming; 42454 (31 mi. N Pinedale, 8025 ft., Sublette Co.), 37903 (23-1/2 mi. S, 5 mi. W Lander, 8600 ft., Fremont Co.).
Fig. 45
Baculum: Stalk broad, greatest length (2.2 mm.) 1¾ times greatest breadth, 3½ times greatest depth; three well-developed ossified processes; median process 2/5 length of stalk, rounded or tapered terminally, proximal end opposed to tip of stalk and flattened obliquely; lateral processes 2/3 length of median process, deeper than wide, curved; tuberosities of stalk well developed, confluent medially, visible in dorsal view; in end-view dorsal concavity narrow, moderately [Pg 200] deep, rounded, ventral concavity wide, deep, flattened; base wider ventrally than dorsally; shaft tapering more or less uniformly, terminally inflated.
In the relative sizes, to each other and to the stalk, of the three digitate ossifications M. oregoni resembles closely the Old World representative of the same subgenus, M. (Chilotus) socialis, as figured by Argyropulo (1933b:181). In M. oregoni the greatest width of the baculum is more proximal on the stalk than in the M. socialis figured by Argyropulo but closely resembles the baculum of the M. socialis figured by Didier (1954:242). In possessing a shallow emargination in the base of the stalk and in possessing a median process that is smaller than the lateral processes, M. socialis, as figured by Didier, differs from M. oregoni. The baculum figured by Argyropulo (loc. cit.) of Sumeriomys colchicus schidlovskii [ = Microtus (Chilotus) socialis schidlovskii according to Ognev, 1950:392] differs from other Chilotus that have been studied in having an unusually elongate median process and a more distal placement of the widest part of the stalk.
Specimens examined: Three, of the subspecies M. oregoni oregoni, from 5 mi. N Orick, Humboldt Co., California, 3-C-248, collection of W. B. Quay; from Mary's Peak, Benton Co., Oregon, 66, collection of F. W. Sturges; and from Sec. 3, T. 11S, R. 5W, Benton Co., Oregon, 79183.
Fig. 34
Baculum: Length of stalk (2.4 mm.) 1¾, times greatest breadth, 41/3 times greatest depth; median ossified process well developed, more than 1/3 length of stalk, higher than wide, slightly bowed, closely appressed to terminus of shaft; basal tuberosities of stalk moderately developed, confluent medially, posterior profile of medial apex rounded in dorsal view, lateral indentations present, hence trilobate outline; in proximal end-view base wider ventrally, ventral concavity broader than dorsal concavity but of equal depth, medial constriction 2/3 greatest depth; shaft slender in distal part, inflated terminally, and wider than high at mid-point of stalk; lateral profile a smooth slope of gradually decreasing curvature from point of greatest width to near distal end.
The baculum of this species figured by Ognev (1950:461) differs in having lateral ossified processes, and a more rounded base of the stalk. Resemblance to the New World Stenocranius is discussed below.
Specimen examined: One from "Eastern Europe," 8059.
Figs. 32 and 33
Baculum: Length of stalk (2.8 mm.) 1½ times greatest breadth, 3½ times greatest depth; median process ossified, 2/5 to 3/5 length of stalk, laterally compressed, sometimes arched in dorsoventral plane; lateral processes cartilaginous, slender; basal tuberosities well developed, averaging less enlarged than shown in Figure 32, but more angular in lateral outline than shown in Figure 33; tuberosities confluent posteriorly; posterior profile smoothly rounded to trilobate, curvature at point of greatest breadth usually acute; in proximal end-view base wider dorsally, deep dorsal concavity, shallow ventral concavity, medial constriction 3/5 of greatest depth; shaft slender anteriorly, at mid-point of stalk [Pg 201] twice as wide as high, at tip higher than wide, laterally inflated; lateral profile in most specimens abruptly curved anterior to point of greatest breadth.
The single specimen of the Old World M. (Stenocranius) gregalis examined resembles the New World M. (Stenocranius) miurus in the angular lateral profile at the point of greatest breadth of the stalk, slender shaft in comparison to broad base of stalk, and presence of a single well-developed laterally compressed median process. The base of the stalk in the baculum of M. gregalis is less well developed and smaller than in the baculum of M. miurus.
Specimens examined: Nine, all of the subspecies Microtus miurus muriei, from the Brooks Range, Alaska; 51077 (Lake Schrader, 145°09'50", 69°24'28", 2900 ft., Romanzof Mts.); 51151, 51152, 51154, 51164, 51166, 51169 (last 6 from Wahoo Lake, 69°08', 146°58', 2350 ft.); 51210, 51213 (last 2 from Porcupine Lake, 68°51'57", 146°29'50", 3140 ft.).
Fig. 47
Baculum: Greatest length of stalk (2.7 mm.) 2¼ times greatest breadth, 4½ times greatest depth; three digitate processes, lateral processes mostly cartilaginous in single adult examined; median process well ossified, approximately 1/3 length of stalk, basally notched, not arched, laterally compressed distally; base of stalk broad and flat, basal tuberosities well developed, separate; posterior profile in dorsal view rounded, convex except for medial notch separating tuberosities; dorsal and ventral concavities deep, broad, equal; medial constriction less than ½ greatest depth; in dorsal view shaft tapering gradually from widest point, terminally rounded; at mid-point of stalk almost twice as wide as high.
In the elongate, largely cartilaginous lateral processes of the baculum, the specimen described above resembles M. longicaudus. The size of the median process in comparison to the size of the stalk is also the same. The lateral processes have larger ossifications and the base of the stalk is more robust in M. longicaudus than in M. nivalis.
The well ossified lateral processes and enlarged base of Didier's (1954:240) specimen suggest that it is of a more mature individual than the one described above. These specimens of M. nivalis, as well as the specimens of M. longicaudus, exhibit dorso-ventral flattening of the mid-part of the base of the stalk.
The baculum of a specimen from Switzerland is weakly developed, of small size (shaft 2.0 mm. in length), slender, thin, spatulate, and terminally inflated. Digital processes were not observed, perhaps owing to excessive maceration in preparation. The general appearance of the baculum is that of an immature individual, although the animal was not small (165 mm. total length in preservative).
Specimens examined: Two Microtus nivalis nivalis; Zermatt, Valais, Switzerland, 67105; Wetterstein, Germany, 65127.
Fig. 48
Baculum: Base of stalk well developed, greatest length (3 mm.) 1¾ times greatest breadth, 32/3 times greatest depth; three ossified processes; base of median process rounded; median process slightly curved in dorsoventral plane, in length almost 1/3 greatest length of stalk; ossifications in lateral processes [Pg 202] variable in size, frequently widely separated from shaft by cartilage, rarely as large as median ossification; basal tuberosities usually well-developed, medially confluent; profile of base in dorsal view trilobate or irregularly convex throughout; constriction ½ greatest depth; shaft relatively straight or slightly bowed ventrally or dorsally, shaft at mid-point of stalk wider than high; tip of shaft laterally inflated; widest point of stalk approximately ¼ length of stalk from proximal end; lateral profile in dorsal view tapers gradually onto shaft anteriorly from point of greatest width of stalk; shaft variable, from slender terminally and nearly parallel sided (Fig. 48), to broad distally and tapered.
In many of the features that distinguish M. longicaudus (and the closely related insular species M. coronarius) from other North American Microtus, longicaudus resembles the Old World species of the subgenus Chionomys (that is to say, M. nivalis, M. gud, and M. roberti). These features are medium size, long tail, grayish color, montane habitat, relatively short molar tooth-row, moderate sized and unconstricted incisive foramen, relatively decurved upper incisors, elongate nasals, relatively broad interorbital region without well-developed median ridge, and similar chromosomes (Matthey, 1955:178). For these reasons I am here referring Microtus longicaudus to the subgenus Chionomys; previously it has not been referred to that subgenus.
Specimens examined: Six, of three subspecies; Microtus longicaudus littoralis, Sullivan Island, Alaska, 42972, 42969; M. l. mordax, 3/4 mi. N, 2 mi. W Allenspark, 8400 ft., Boulder Co., Colorado, 50335, 76829; M. l. sierrae, Crane Flat, Mariposa Co., California, 50252, 50253.
Fig. 22
Baculum: In the single specimen examined, stalk small, greatest length (2.3 mm.) 21/3 times greatest width, almost 6 times greatest depth, flattened proximally; three well-developed digitate processes, the median one ossified, the lateral processes cartilaginous; median ossification laterally compressed and decurved at tip, bilobate at base; basal tuberosities of stalk weakly developed, medially confluent; posterior profile in dorsal view evenly rounded; ventral concavity deeper and narrower than dorsal concavity, but both comparatively shallow; medial constriction 2/3 greatest depth; shaft straight, at mid-point twice as wide as deep; lateral profile tapering from greatest width gradually to parallel sides of distal third of stalk.
From the baculum of Microtus arvalis figured by Ognev (1950:173), and from the baculum figured by Didier (1954:238) my specimen differs in the absence of lateral ossifications in the digitate processes, smaller and slenderer median ossification, and weaker base. These differences in part may be owing to a difference in age, my specimen being the less mature.
Specimen examined: One from Vidy, Switzerland, 67101.
Fig. 24
Baculum: In the one specimen examined, stalk broad, greatest length (2.6 mm.) 1½ times greatest breadth, 3½ times greatest depth; three digitate processes ossified; median process relatively broad, in length more than ½ length of stalk, triangular in dorsal view, with small spurs posterolaterally, [Pg 203] middorsal ridge posteriorly; lateral ossifications slightly curved, slenderer, less than ½ depth and less than ½ transverse thickness of median process; basal tuberosities well-developed, confluent medially; in end-view base wider dorsally than ventrally, dorsal concavity broader and more abruptly curved at mid-point than ventral concavity; constriction ½ greatest depth; posterior profile in dorsal view notched, setting off a posterior shelf; stalk including shaft wider than deep throughout, at mid-point width twice depth; lateral profile abruptly curved anterior to point of greatest width, sides of shaft tapering gradually anteriorly to rounded uninflated tip.
The baculum of this insular species, placed in the "arvalis" group by Ellerman (1941:595), resembles the baculum of both Microtus agrestis and Microtus guentheri more than it resembles the baculum of Microtus arvalis. Similarities in the chromosomes of M. arvalis and M. orcadensis were noted by Matthey (1953:254, 279), who was of the opinion that M. orcadensis is an insular derivative of the arvalis-group.
Specimen examined: One from the Orkney Islands, 67106.
Fig. 23
Baculum: In the one specimen examined, stalk broad, greatest length (2.9 mm.) 1½ times greatest breadth, 3½ times greatest depth; three digitate processes ossified; median process slightly less than ½ length of stalk, broad, dorsally curved; curved lateral ossifications shorter and more slender than median ossification; basal tuberosities well developed, angular, confluent across posterior border of projecting shelf; in end-view tuberosities projecting ventrolaterally from central shelf; dorsal surface at medial constriction flat, ventral surface broadly and deeply concave; posterior profile in dorsal view trilobate, central lobe formed by posteriorly flattened shelf, surface of attachment visible only on lateral lobes; at mid-point stalk almost twice as wide as deep, depth of shaft greater than width proximal to inflated terminus.
Specimen examined: One from Palestine, 67104.
Fig. 25
Baculum: Stalk large, greatest length (3.8 mm.) 14/5 times greatest breadth, 4½ times greatest depth; three digitate processes ossified; median ossification almost 1/3 length of stalk; lateral ossifications slender, smaller than median ossification; posterior profile of stalk in dorsal view trilobate, basal tuberosities well developed, confluent medially; in end-view dorsal concavity broader and deeper than ventral concavity; medial constriction pronounced (less than ½ greatest depth); lateral profile at widest point of stalk convex, becoming abruptly concave as the flange of the basal tuberosities grades into the shaft, then gradually converging to narrowest point 1/3 of length of stalk from the terminus; stalk wider than deep in proximal 2/3, circular in cross section in terminal 1/3, slight terminal inflation.
A specimen figured by Ognev (1950:297) has the same general proportions, slender lateral processes, and proximal placement of the point of greatest breadth.
Specimens examined: Two from Chipo-ri, Korea, 60443, 63841.
Figs. 19, 20 and 21
Baculum: Stalk broad, greatest length (varying with subspecies from 2.3 to 3.1 mm.) 1½ to 1¾ times greatest breadth, 31/3 to 41/3 times greatest depth; three ossified processes, median one largest, more than twice as wide and as deep as shorter, slenderer, lateral processes; median process laterally compressed distally except in one specimen in which moderately inflated distally, proximally enlarged in some specimens (Fig. 21) and 1/3 to 2/5 length of stalk; base broad, posterior profile in dorsal view evenly convex throughout, at widest point of stalk abruptly incurved; basal tuberosities moderately to strongly developed, medially confluent; in end-view base wider ventrally than dorsally, dorsal concavity slightly to much deeper than the nearly flattened ventral concavity; medial constriction 2/3 to 4/5 of greatest depth; shaft relatively slender, at mid-point of stalk slightly wider than high and ¼ as wide as base of stalk, terminally rounded or slightly inflated; lateral profile in dorsal view a gradual curve from point of greatest width anteriorly onto shaft.
The different subspecies figured show the essential characteristics of the species, differing primarily in size.
Specimens examined: Fourteen, of three subspecies; Microtus montanus amosus, ½ mi. E Soldier Summit, Wasatch Co., Utah, 62241; M. montanus fusus, La Manga Pass, Conejos Co., Colorado, 42164; 5 mi. N, 26 mi. W Saguache, 9500 ft., Saguache Co., Colorado, 42307, 42315; 5 mi. N, 27 mi. W Saguache, 9350 ft., Saguache Co., Colorado, 42308; 5 mi. N, 28 mi. W Saguache, 9325 ft., Saguache Co., Colorado, 42309; 5 mi. S, 24 mi. W Antonito, 9600 ft., Conejos Co., Colorado, 42327, 42330; Prater Canyon, Mesa Verde National Park, Montezuma Co., Colorado, 69456, 69457, 69463; Microtus montanus nanus, 2 mi. N, 2 mi. W Pocatello, Bannock Co., Idaho, 57470, 57472; ¼ mi. N, 2 mi. W Allenspark, 8400 ft., Boulder Co., Colorado, 50330.
Fig. 41
Baculum: Stalk broad, greatest length (3.0 mm.) 1½ times greatest breadth, 4½ times greatest depth; three ossified processes, median one largest, deeper and more than twice as wide as curved, shorter, compressed lateral processes and more than 2/5 as long as stalk; base broad, in dorsal view posterior profile trilobate, basal tuberosities visible; basal tuberosities well developed, medially confluent; in end-view base wider ventrally than dorsally, dorsal concavity deeper than ventral concavity; medial constriction 3/5 of greatest depth; shaft broad, at mid-point more than twice as wide as high and 1/3 as wide as base of stalk, terminally rounded.
Specimens examined: Three, all M. t. townsendii; Fort Lewis, Pierce Co., Washington, 57998, subadult; Sec. 33, T. 11S, R. 5W, Benton Co., Oregon, 79186; Sec. 5, T. 12S, R. 4W, Benton Co., Oregon, 79188.
Fig. 44
Baculum: Stalk broad and flattened, greatest length (3.5 mm.) 12/3 to 2 times greatest width, 4 to 5½ times greatest depth; three ossified processes, median one largest, lateral processes slender, relatively small; length of median process 3/8 length of stalk; median process decurved, dorsoventrally flattened in [Pg 205] some specimens, widened at base; attachment of processes to shaft displaced ventrally; base of stalk widened, posterior profile in dorsal view usually trilobate, in a few cases rounded, median lobe forming posterior shelf, lateral lobes dorsally raised and forming margins of lateral tuberosities; in end-view thickness frequently more or less uniform throughout central part, broad depression dorsally, ventral concavity narrower and shallower (as figured); base, and occasionally shaft, flattened, width at mid-point of stalk 2 to 3 times depth, narrowest point posterior to terminal inflation of shaft in terminal 1/3 of shaft.
The baculum of M. oeconomus (Old World) figured by Ognev (1950:257) resembles but exceeds that of M. oeconomus (New World) in the relatively large median process and slender lateral processes, but differs noticeably in the presence of a deep median notch in the base of the stalk. A specimen from Hungary is intermediate between Ognev's specimen and those from the New World in both size of median process and size of lateral processes, and has an unnotched base resembling that in Figure 44.
Specimens examined: Ten, of three subspecies; M. oeconomus gilmorei, Umiat, Alaska, 51354, 51361, 51399, 51408; Lake Schrader, Brooks Range, Alaska, 51422; M. o. macfarlani, 5 mi. NNE Gulkana, Alaska, 43039, 43041; 20 mi. NE Anchorage, Alaska, 43044; Kelsall Lake, British Columbia, 43048; M. o. mehelyi, Kisbalatan, Hungary, 75159.
Figs. 35 and 36
Baculum: Stalk attenuate, greatest breadth relatively near proximal end; greatest length (3.1 to 3.4 mm.) more or less twice greatest breadth, 4 to 5 times greatest depth; usually a single process ossified; lateral processes relatively small, cartilaginous or (in three specimens, 63094, 69453, 68019) with small ossifications; median process relatively small, sometimes appressed to tip of shaft, in length less than ¼ length of stalk; posterior profile in dorsal view rounded, flattened posteriorly, or in some specimens trilobate with angular edges; in end-view relative depths of dorsal and ventral concavities variable, dorsal usually deeper than ventral; distal end of stalk frequently bowed dorsally; shaft slender distally, sometimes slightly inflated terminally, or (in one specimen, 63085) near tip small lateral projections that are perhaps fused lateral ossifications; lateral profile in dorsal view a gradual slope anteriorly from point of greatest width to slender tip.
Specimens examined: Thirteen, of four subspecies; Microtus mexicanus mexicanus, Las Vigas, Veracruz, 30692; Nevada de Toluca, México, 63101; Valle de Bravo, México, 63094; Microtus mexicanus mogollonensis, Mt. Taylor, Valencia Co., New Mexico, 63298, 76830; Park Well, Mesa Verde National Park, Montezuma Co., Colorado, 69448, 69453; Upper Nutria, McKinley Co., New Mexico, 69997, 70000; Microtus mexicanus phaeus, Sierra Patamba, 9000 ft., Michoacán, 63085; Microtus mexicanus subsimus, 2 mi. E Mesa de Tablas, Coahuila, 58916; 13 mi. E San Antonio de las Alazanas, Coahuila, 68019, 68021.
Fig. 37
Baculum: Stalk elongate, greatest length (3.0 mm.) 21/3 times greatest breadth, 4½ times greatest depth; median process ossified, ¼ length of stalk, basally broadened, flattened and shallowly grooved ventrally to fit tip of shaft, to which the process is closely appressed; lateral processes cartilaginous; ends [Pg 206] of stalk bowed upwardly; posterior profile of base of stalk rounded or slightly trilobate if posterolateral concavities form in tuberosities; moderate development of tuberosities, in end-view dorsal concavity slightly deeper and narrower than ventral concavity, both comparatively shallow, median constriction 4/5 greatest depth; shaft curved, more or less terete at mid-point of stalk, terminally inflated dorsally; lateral profile in dorsal view gradually curved from point of greatest width anteriorly onto shaft.
Specimens examined: Two, of two subspecies; Microtus californicus californicus, 1 mi. NE Berkeley, in Contra Costa Co., California, 76828; Microtus californicus mohavensis, ½ mi. SE Victorville, San Bernardino Co., California, 63745.
Figs. 14, 15, 16 and 17
Baculum: Stalk heavy, broad, greatest length (2.2 to 3.0 mm.) 11/3 to 12/3 times greatest breadth, up to 3¾ times greatest depth; three ossified processes, median one largest, usually not twice so deep as lateral ossifications; median process usually distinctly widened basally, in length approximately ½ length of stalk; base broad, frequently angular laterally and basally, sometimes bilobate; basal tuberosities well developed, medially confluent; in end-view more or less uniformly biconvex or ventral surface more flattened than dorsal surface, medial constriction ½ to 2/3 greatest depth; shaft relatively heavy, at mid-point stalk almost twice as wide as deep and 1/3 as wide as base of stalk; shaft terminally rounded and sometimes slightly inflated; lateral profile in dorsal view abruptly or gradually curved anterior to point of greatest width and then gradually curved anteriorly.
Specimens examined averaged slightly smaller and were more variable than those described by Hamilton (1946:382). The greater variation may be in part geographic, as five subspecies are represented. Lateral processes are the last to ossify. One specimen (75082) with well-ossified median process lacks any lateral ossification. Four bacula of M. pennsylvanicus (locality not specified) studied by Dearden (1958:547) agree in general with the description above.
One specimen shows a break, perhaps resulting from injury, in the shaft (Fig. 14). One specimen has a posteromedian spine on the median digital ossification (Fig. 16). Comparison with M. agrestis is included with the description of M. agrestis.
Specimens examined: Thirteen, of six subspecies; Microtus pennsylvanicus alcorni, 20 mi. NE Anchorage, Alaska, 43043; Microtus pennsylvanicus finitus, Laird, Yuma Co., Colorado, 68544; Microtus pennsylvanicus modestus, 5 mi. N, 26 mi. W Saguache, 9500 ft., Saguache Co., Colorado, 42306; 3 mi. N, 16 mi. W Saguache, 8500 ft., Saguache Co., Colorado, 42416, 42417, 42418; 1 mi. S, 2 mi. E Eagle Nest, 8100 ft., Colfax Co., New Mexico, 42430, 42439; Microtus pennsylvanicus pennsylvanicus, 2 mi. S, 3 mi. E Ft. Thompson, 1370 ft., Buffalo Co., South Dakota, 42379; Vermillion, Clay Co., South Dakota, 37070; Microtus pennsylvanicus pullatus, 12 mi. S, 5 mi. E Butte, Silver Bow Co., Montana, 57501, 57503; Microtus pennsylvanicus uligocola, Muir Springs, 2 mi. N, 2½mi. W Ft. Morgan, Morgan Co., Colorado, 75082.
Fig. 18
Baculum: Greatest length of stalk (2.9 mm.) twice greatest breadth, 4½ times greatest depth; stalk well developed, shaft not flattened dorsoventrally; large median ossified process, minute lateral ossifications in single specimen [Pg 207] examined; length of stalk 2½ times length of median ossification which is higher than wide, slightly decurved, sagittate in dorsal view, with three-cornered base; basal tuberosities of stalk moderately well developed, medially joined; posterior profile in dorsal view evenly rounded; ventral concavity broader than, but of comparable depth to, dorsal concavity in end-view, base of stalk wider ventrally, constriction ¾ greatest depth; at mid-point of stalk shaft is but slightly wider than high; pronounced terminal inflation of shaft; lateral profile in dorsal view sloping abruptly from widest point of stalk anteriorly onto stalk which then tapers more gradually to terminal inflation.
From the baculum of its New World counterpart, namely Microtus pennsylvanicus, my specimen of Microtus agrestis and the specimen figured by Didier (1954:239) differ in their minute lateral processes, relatively larger median processes, and more elongate, less dorsoventrally flattened shafts.
The specimen of M. agrestis figured by Ognev (1950:320), in dorsal view has lateral concavities producing a somewhat trilobate outline in the base of the stalk, and the lateral processes are well developed; the median process is larger and bulbous, wider distally than proximally. Without larger numbers of bacula of M. agrestis I am unable to reconcile these differences. The differences between M. agrestis and M. pennsylvanicus seem comparable to the differences between some other species of Microtus.
Specimen examined: One, from Gryon, Switzerland, 67102.
Fig. 31
Baculum: Stalk broad, greatest length (3.2-4.0 mm.) 12/3 to 2 times greatest breadth, 2½ to 4 times greatest depth; median process ossified, relatively small, less than 3/10 length of stalk; lateral processes arising from subterminal part of stalk, cartilaginous or with small ossifications; posterior profile in dorsal view broadly rounded or slightly angular, widest point of stalk 1/6 to ¼ the length of stalk from base; basal tuberosities well developed and medially confluent, in end-view dorsally convex, or at least less deeply concave than ventrally; shaft straight, base bent ventrally or more commonly dorsally; at mid-point of stalk wider than high, often twice as wide as high; viewed from above, lateral profile from point of greatest breadth to middle of shaft a gradual sigmoid curve; slight terminal inflation of shaft.
Specimens examined: Forty-one, of three subspecies; Microtus ochrogaster haydeni, Muir Springs, 2 mi. N, 2½ mi. W Ft. Morgan, Morgan Co., Colorado, 74995, 74998, 74999, 75002; 1 mi. W Laird, Yuma Co., Colorado, 57304, 76833; 2 mi. N, 2 mi. W Haigler, Dundy Co., Nebraska, 75016; 2 mi. S Franklin, Franklin Co., Nebraska, 75043, 75044; Atwood, Rawlins Co., Kansas, 75020, 75023, 75025, 75027, 75028; 1 mi. N, 2 mi. E Oberlin, Decatur Co., Kansas, 75030, 75032, 75034, 75035, 75036; 1½ mi. N, ¼ mi. E Norton, Norton Co., Kansas, 68327; 1 mi. SW Norton, Norton Co., Kansas, 75037; 2 mi. S, 1 mi. W Norton, Norton Co., Kansas, 75038; M. ochrogaster ochrogaster, Rydal, Republic Co., Kansas, 75047-75053, 75060, 75062, 75063, 75066, 75070, 75071, 75073; 1 mi. N, 1 mi. W Holton, Jackson Co., Kansas, 75077; 2 mi. W Court House, Lawrence, Douglas Co., Kansas, 76832; Univ. Kansas Natural History Reservation, Douglas Co., Kansas, 68536; M. ochrogaster taylori, Meade County State Park, Kansas, 68539, 68542.
Figs. 27 and 28
Baculum: Stalk broad, greatest length (2.5 to 2.7 mm.) 12/3 times greatest breadth, 4 times greatest depth; median process ossified, size small, 1/5 length of stalk, higher than wide, having small anterodorsal prominence in both specimens examined; lateral processes cartilaginous, relatively small, displaced posteriorly, attenuate; posterior margin in dorsal view broadly rounded, or having blunt median apex, convex throughout; basal tuberosities moderately well developed, medially confluent, barely visible in dorsal view when mature; in end-view median constriction 4/5 greatest depth, ventral concavity deeper than dorsal concavity, both comparatively shallow; stalk at mid-point 1½ times as wide as deep; shaft relatively slender, bowed dorsally at tip, relatively straight otherwise; lateral profile in dorsal view a gradual concave slope from point of greatest width anteriorly to distal part of shaft.
Specimens examined: Two, from Douglas Co., Kansas, 76834 (2 mi. N Baldwin), 68545 (1 mi. NE Pleasant Grove).
Fig. 40
Baculum: Stalk broad, greatest length (2.4 mm. in specimen examined) 1¾ times greatest breadth, 4 times greatest depth; median process ossified, size small, less than ¼ length of stalk, wider than high, terminally flattened; lateral processes cartilaginous, relatively small, attenuate; posterior margin in dorsal view flattened, irregularly curved with concavities medially and laterally; basal tuberosities well developed, medially confluent; visible in dorsal view; in end-view median constriction 2/3 greatest depth, ventral concavity well-formed, no dorsal concavity; stalk at mid-point twice as wide as deep; shaft relatively slender, bowed dorsally toward tip; in dorsal view lateral profile a gradual concave slope from point of greatest width anteriorly to distal part of shaft; tip of shaft enlarged.
The baculum of M. parvulus resembles that of M. pinetorum more than it resembles the baculum of any other microtine studied, differing primarily in smaller size.
Specimen examined: One, from 1 mi. W Micanopy, Alachua Co., Florida, Univ. Florida No. 1508.
Figs. 29 and 30
Baculum: Stalk broad, greatest length (2.6-3.2 mm.) 11/3 to 12/3 times greatest breadth, 31/3 to 32/3 times greatest depth; median process ossified, with ventral depression, process ¼ to 1/3 length of stalk, appressed to tip of shaft, wider than high proximally, relatively broad terminally; lateral processes cartilaginous, small, attenuate; posterior profile of stalk in dorsal view broadly rounded, bilobate, or trilobate, median lobe formed by posterior projection of dorsal shelf between enlarged lateral tuberosities that form outer lobes, posterolateral faces of these tuberosities visible in dorsal view of stalk; in end-view dorsal surface slightly concave, ventral concavity broad and deep, median constriction [Pg 209] ½ greatest depth; shaft flattened except tip that is more terete, and bowed dorsally; at mid-point, stalk twice as wide as high; shaft relatively slender terminally, narrower than median ossification.
The baculum of M. quasiater is the largest and has the best developed base and median process of the three American species of the subgenus Pitymys. The three species closely resemble each other in basic form.
Specimens examined: Five, all from Veracruz; Teocelo, 4500 ft., 30709, 30711; 4 km. N Tlapacoyán, 1700 ft., 24466; 5 km. N Jalapa, 4500 ft., 19869, 19878.
Fig. 26
The baculum of a single specimen (KU 67103) of M. fatioi from Zermatt, Valais, Switzerland, was examined. The baculum is immature, as evidenced by its small size, slender stalk and absence of ossified processes, therefore no characterization is included.
The baculum of another Old World species of the subgenus Pitymys, M. pyrenaicus from France, figured and described by Didier (1954:242-243), differs from all New World Pitymys examined in processing ossified lateral processes.
The status of Pitymys, as a genus or as a subgenus, is uncertain. Hall and Cockrum (1953:448) considered the North American Pitymys and Pedomys as subgenera of Microtus. They did not state specifically the basis for this point of view, but mention the fact that these two subgenera (Pitymys and Pedomys) closely resemble each other cranially. These authors did not study nor comment upon the status of the Old World Pitymys. It may be asked whether the Old World and New World Pitymys have developed as fossorial Microtus independently, or from an ancestor common to both groups and not common to any other Microtus. Matthey (1955:202) found 62 chromosomes (2N) in both the New World Pitymys pinetorum and the Old World Pitymys duodecimcostatus. This suggests, but does not prove, common ancestry.
Fig. 49
Baculum: Stalk massive, greatest length (4.7 mm.) 1¾ times greatest breadth, 4 times greatest depth; ossification in digitate processes variable; in one (KU 27123) of two specimens examined lateral processes ossified and median process unossified, as in two specimens examined by Hamilton (1946:379) from "southern Florida"; in my other specimen (KU 27268) that is possibly more mature, median process ossified although less deeply stained than lateral ossifications or stalk; posterior profile in probable dorsal view roughly rounded; in end-view probable dorsal concavity deep, ventral concavity broad but shallow, and with center convex; median constriction 3/5 greatest depth; shaft heavy, least depth 2/3 greatest depth of base; stalk, at mid-point, slightly wider than deep and more than 1/3 width of base; lateral profile in dorsal view sharply incurved distal to point of greatest breadth, shaft therefore relatively distinct from basal part of stalk; slight subterminal constriction; tip less reduced in the two specimens examined than in two figured by Hamilton. In preparation, the tissues that make it possible to distinguish [Pg 210] with certainty the dorsal and ventral surfaces of the baculum were removed in both specimens.
Specimens examined: Two, of the subspecies Neofiber alleni alleni, 2 mi. S Gainesville, Alachua Co., Florida, 27268; 1 mi. E Courtenay, Merritt Island, Brevard Co., Florida, 27123.
Fig. 46
Baculum: Stalk slender, greatest length (2.5 mm.) 2 to 22/3 times greatest breadth, 4 to 5 times greatest depth; three ossified processes; median one more than 1/3 length of stalk, curved dorsally toward tip, proximally flattened and having acute lateral angles in dorsal view, wider than deep except in distal half; lateral processes smaller than median one, slenderer, shorter, of approximately same depth, also curved dorsally; base of stalk well developed, basal tuberosities medially confluent, in part visible in dorsal view, in end-view wider ventrally than dorsally, dorsal and ventral concavities of equal depth and both wide, medial constriction ½ greatest depth; posterior profile in dorsal view broadly bilobate; lateral profile with abrupt transition from basal tuberosities to gradually converging, slightly curved sides of shaft; shaft terminally inflated.
Dearden (1958:543) described and figured the bacula of six subspecies of Lagurus curtatus and two Asiatic species, Lagurus lagurus and Lagurus luteus. He examined at least 34 specimens of L. curtatus and found geographic variation in size, breadth of shaft distally, and proportions of digital ossifications to each other and to the stalk. The description that I have given above pertains to L. c. levidensis.
The baculum of the Asiatic Lagurus (Lagurus) lagurus figured by Ognev (1950:554) agrees with that of Lagurus (Lemmiscus) curtatus, described here, in the relatively elongate shaft and slender stalk, the proportions of the processes, and the well-formed and moderately enlarged base of the stalk. The bacula of three Lagurus lagurus examined by Dearden (1958:545) were of older individuals than the specimen that Ognev figures and differ from it and from bacula of Lagurus curtatus (all subspecies) in the unusual, almost heart shaped, median process, and in larger size. Lagurus luteus examined by Dearden (1958:545) differs from both Lagurus lagurus and Lagurus curtatus in lacking lateral digital ossifications and in having shorter median digital ossifications and wider base of stalk.
Specimens examined: Seven Lagurus curtatus levidensis from Wyoming; 9 mi. S Robertson, Uinta Co., 26045, 26053; 8 mi. S, 2½ mi. E Robertson, Uinta Co., 26049; Farson, Sweetwater Co., 37906; 16 mi. S, 11 mi. W Waltman, Natrona Co., 42457; 32 mi. S, 22 mi. E Rock Springs, 42465, 42466.
The following key to the bacula in some adult North American Microtinae is intended to help point out some of the most important differences. It should be noted that not all species can be keyed out on the basis of the baculum. The most difficult group in this respect includes the species of Microtus that have small or no ossified lateral processes, especially species of the subgenera Pedomys and Pitymys, and the two species Microtus californicus and Microtus mexicanus of the subgenus Microtus. Another complicating factor is the variability of bacula evident in some species even in the small samples available. [Pg 211] It is to be expected that additional specimens will show variations not yet observed.
| 1. Length of lateral digital ossifications more than 1/3 breadth of stalk | 2 |
| 1´. Length of lateral digital ossifications less than 1/3 breadth of stalk or absent | 15 |
| 2. Size small (total length of baculum less than 5.5 mm.) | 3 |
| 2´. Size large (total length of baculum more than 5.5 mm.) | 14 |
| 3. Width at mid-point of stalk more than 1/3 greatest breadth of stalk | 4 |
| 2´. Size large (total length of baculum more than 5.5 mm.) | 14 |
| 3´. Width at mid-point of stalk less than 1/3 greatest breadth of stalk, | 8 |
| 4. Stalk, viewed from proximal end hour-glass shaped, and width of stalk less than ½ length of stalk.... Phenacomys intermedius, | p. 197 |
| 4´. Stalk not both hour-glass shaped when viewed from proximal end, and with width less than ½ length of stalk | 5 |
| 5. Shaft thin basally, thickness less than 1/3 of greatest breadth | 6 |
| 5´. Shaft thick basally, thickness 1/3 or more of greatest breadth | 7 |
| 6. Stalk more or less straight, base not deflected. Microtus oeconomus, | p. 204 |
| 6´. Stalk spatulate, and base deflected from axis of shaft.... Microtus guatemalensis, | p. 198 |
| 7. Base enlarged, depth nearly ½ of breadth.... Lemmus trimucronatus, | p. 193 |
| 7´. Base moderately enlarged, depth near 1/3 of breadth.... Microtus pennsylvanicus, p. 206, or Microtus townsendii, | p. 204 |
| 8. Base hour-glass shaped as viewed from proximal end.... Phenacomys intermedius, | p. 197 |
| 8´. Not so | 9 |
| 9. Lateral processes separated from tip of shaft by more than the thickness of the lateral process | 10 |
| 9´. Lateral processes separated from tip of shaft by less than the thickness of the lateral process | 11 |
| 10. Lateral processes more than ½ the width of median process.... Microtus longicaudus, | p. 201 |
| 10´. Lateral processes slender, less than ½ the width of median process.... Microtus montanus, | p. 204 |
| 11. Lateral ossifications equal in length to median ossification.... Clethrionomys, | p. 194 |
| 11´. Lateral ossifications shorter than median ossification | 12 |
| 12. Size small, less than 3.4 mm. in total length.... Microtus oregoni, | p. 199 |
| 12´. Size medium, more than 3.4 mm. in total length | 13 |
| 13. Greatest width of stalk at a point about 1/3 of length of stalk from base.... Microtus chrotorrhinus (Hamilton, 1946:382). | |
| 13´. Greatest width of stalk at a point less than 1/3 of length of stalk from base.... Lagurus curtatus, | p. 210 |
| 14. Size of baculum larger, base more than 3 mm. wide, processes all well developed.... Ondatra zibethicus, | p. 198 |
| 14´. Size of baculum smaller, base less than 3 mm. wide, processes poorly developed in some animals.... Neofiber alleni, | p. 209 |
| 15. At least one digital ossification present | 16 |
| [Pg 212] 15´. Digital ossifications not present.... Dicrostonyx groenlandicus, | p. 193 |
| 16. Breadth of stalk at least ½ length of stalk | 17 |
| 16´. Breadth of stalk less than ½ length of stalk | 19 |
| 17. Length of stalk greater than 3.6 mm. and less than 1½ times its greatest breadth.... Microtus richardsoni, | p. 199 |
| 17´. Length of stalk usually less than 3.6 mm., or if more than 3.6 mm. (up to 4.0 mm.) then length 1½ times or more its greatest breadth | 18 |
| 18. Median process attenuate distally in dorsal view, and relatively long (more than twice its own breadth), 1/5 to 3/5 the length of stalk; breadth of stalk usually 2/3 or more length of stalk.... Microtus miurus, | p. 200 |
| 18´. Median process relatively blunt distally in dorsal view, relatively short (usually less than ¼ length of stalk), breadth of stalk usually less than 2/3 length of stalk.... Pitymys, p. 208, Pedomys, p. 207, or Microtus mexicanus, | p. 205 |
| 19. Distal processes small and firmly ankylosed to distal end of shaft.... Phenacomys longicaudus, | p. 197 |
| 19´. Distal processes if present not firmly ankylosed to distal end of shaft | 20 |
| 20. Dorsal concavity of base as viewed from proximal end usually deeper than ventral concavity.... Microtus mexicanus, | p. 205 |
| 20´. Dorsal and ventral concavities of base equal in depth or ventral one the deeper | 21 |
| 21. Total length of baculum more than 3.6 mm.... Microtus californicus, | p. 205 |
| 21´. Total length of baculum less than 3.6 mm.... Synaptomys cooperi, | p. 194 |
Owing to shortness of lower incisors and present geographic distribution of the species, Hinton (1926:35) considered the Tribe Lemmi (lemmings) to be more primitive than the Tribe Microti (voles). The surviving lemmings are specialized in many features and therefore are considered as advanced end-products of an evolutionary radiation of a primitive microtine stock, of which all earlier stages are extinct.
Hinton regarded Dicrostonyx as the most primitive of the genera of lemmings on account of its more complex molar teeth (complexity was considered to be primitive), and on account of the presence of three primitive longitudinal rows of tubercles in unworn molars. The other three genera were arranged in order of increasing specialization as follows: Synaptomys, Myopus, Lemmus.
If the baculum tended to retain its primitive character while specializations in the external anatomy developed, and if the above arrangement is correct the most primitive bacula would be found in Dicrostonyx and in Synaptomys. The baculum in these two genera in comparison to that in Myopus (as figured by Ognev, 1948:512) [Pg 213] and Lemmus has a slenderer stalk and smaller digital ossifications or none at all. The baculum in the genera of lemmings increases in robustness and the development of processes from Dicrostonyx, to Synaptomys, to Myopus, to Lemmus—the same order outlined above for total of specialization. The two extremes in this series are near the extremes of variation in bacula to be found in all microtines. The baculum in lemmings as a group cannot then be considered more primitive than in voles as a group, although the voles are usually considered to be more advanced. The situation in the voles, as we shall see, casts a different light on the matter.
The voles, Tribe Microti, were considered by Hinton (1926:40) to be more advanced than the lemmings because the incisors of the voles are longer and the root of their last lower molar is lingual to the root of the incisor. Hinton thought also that the murine ancestors of microtines had shorter incisors and that the backward extension of the incisors in the voles is a more ancient feature than the hypsodonty of the molars. A trend in the molar teeth has been toward greater hypsodonty. The voles in which the molars are least hypsodont are thus considered primitive. These include the living genera Clethrionomys, Phenacomys, Ondatra, Dolomys, Ellobius, and Prometheomys. Therefore, the baculum, in these assumedly primitive genera, would be expected to resemble the baculum in the lemmings or at least the most primitive lemmings. This is not the case.
The bacula that I have examined of Clethrionomys and Phenacomys have well-developed digital ossifications. In this they resemble the baculum of the genus Lemmus, the most advanced genus of lemmings according to Hinton. The baculum of Dolomys has not been studied. The baculum in Ondatra, and in Prometheomys as illustrated by Ognev (1948:552), also possesses well-developed processes. The baculum of Ellobius is small and lacks processes (as figured by Ognev, 1950:662). No ossification was found in a single specimen of Ellobius examined by me although the entire glans penis was removed and cleared without dissection. So far as known then, with the exception of Ellobius and Phenacomys longicaudus (Dearden, 1958:547), the primitive microtines having rooted molars possess bacula having three well-developed ossified processes.
Voles of the genus Microtus vary in the structure of the baculum almost as much as do the lemmings. Within the single subgenus Microtus some individuals of Microtus mexicanus, for example, have minute ossified lateral processes and other individuals lack these [Pg 214] processes; Microtus pennsylvanicus and some other species have proportionately large lateral ossifications. If the well-developed condition of the baculum in the microtines having rooted molars is primitive, then within the genus Microtus those species having well-developed bacula may be considered primitive.
The genera Lagurus and Neofiber have moderately developed or well-developed lateral processes. Neofiber exhibits a tendency, not prominent elsewhere, to have a proportionately smaller median process rather than reduced lateral processes.
American species of Microtus (genus and subgenus) that have moderately- to well-developed ossified lateral processes are M. townsendii, M. oeconomus, M. pennsylvanicus, M. montanus, and M. chrotorrhinus. Microtus of other subgenera having this type of baculum include M. (Herpetomys) guatemalensis, M. (Chilotus) oregoni, and M. (Chionomys) longicaudus.
American species of Microtus (genus and subgenus) in which the lateral ossifications are weakly developed or absent (although cartilaginous lateral processes are present) include M. mexicanus and M. californicus. In other subgenera, species of Microtus having reduced lateral ossifications are M. (Pedomys) ochrogaster, M. (Pitymys) pinetorum, M. (Pitymys) parvulus, M. (Pitymys) quasiater, M. (Arvicola) richardsoni, and M. (Stenocranius) miurus.
The microtines are essentially holarctic in distribution. Both of the tribes, the lemmings and the voles, as well as primitive representatives of each tribe (not considering Ellobius) occur in both the Old World and New World. It is not certain on which continent (or continents) the Microtinae first differentiated. It is certain, however, that at various times, both early and late in the evolution of the subfamily, representatives have crossed from Eurasia to North America or vice versa. Each of 10 or more microtines in the New World is more closely related to some microtine in the Old World than to any other microtine in the New World.
The similarities or differences in the baculum in Old World and New World representatives placed in the same genus or subgenus, or thought to be "companion species" have been commented upon in accounts of Lemmus, Dicrostonyx, Clethrionomys, Lagurus, Arvicola, Stenocranius, Chilotus, Chionomys, Pitymys, and in accounts of Microtus agrestis as compared with M. pennsylvanicus, and Microtus oeconomus (both Old World and New World).
The baculum in the Microtinae more closely resembles the baculum in the Cricetinae of the Old World than in the Murinae, or than in any other rodents known to me. This resemblance suggests relationship between Microtinae and Cricetinae.
| Argyropulo, A. I. | |
| 1933a. | Die Gattungen und Arten der Hamster (Cricetinae Murray, 1866) der Paläarktik. Zeitschr. f. Säugetierkunde, 8:129-149, 27 figs. in text. |
| 1933b. | über zwei neue paläarktische Wühlmäuse. Zeitschr. f. Säugetierkunde, 8:180-183, 3 figs. in text. |
| Callery, R. | |
| 1951. | Development of the os genitale in the golden hamster, Mesocricetus (Cricetus) auratus. Jour. Mamm., 32:204-207, 1 fig. in text. |
| Chamberlain, J. L. | |
| 1954. | The Block Island meadow mouse, Microtus provectus. Jour. Mamm., 35:587-589, 2 tables in text. |
| Dearden, L. C. | |
| 1958. | The baculum in Lagurus and related microtines. Jour. Mamm., 39:541-553, 1 fig. in text. |
| Didier, R. | |
| 1943. | L'os pénien des Campagnols de France du Genre Arvicola. Mammalia, 7:74-79, 10 figs. in text. |
| 1954. | Etude systématique de l'os pénien des Mammifères (suite), Rongeurs: Muridés. Mammalia, 18:237-256, 14 figs. in text. |
| Ellerman, J. R. | |
| 1941. | The families and genera of living rodents. Vol. II. Family Muridae. The British Museum (Natural History), London, pp. xii + 690, 50 figs. |
| Friley, Charles E. | |
| 1947. | Preparation and preservation of the baculum of mammals. Jour. Mamm., 28:395-397, 1 fig. in text. |
| Hall, E. R., and E. L. Cockrum. | |
| 1953. | A synopsis of the North American microtine rodents. Univ. Kansas Publ., Mus. Nat. Hist., 5:373-498, 149 figs. in text. |
| Hamilton, W. J., Jr. | |
| 1946. | A study of the baculum in some North American Microtinae. Jour. Mamm., 27:378-387, 3 figs. in text. |
| Hibbard, C. W., and G. C. Rinker. | |
| 1942. | A new bog-lemming (Synaptomys) from Meade County, Kansas. Univ. Kansas Sci. Bull., 28:25-35, 3 figs. in text. |
| 1943. | A new meadow mouse (Microtus ochrogaster taylori) from Meade County, Kansas. Univ. Kansas Sci. Bull., 29:255-268, 5 figs. in text. |
| Hinton, M. A. C. | |
| 1926. | Monograph of the voles and lemmings (Microtinae), living and extinct, Vol. I. British Museum (Natural History), London, pp. xvi + 488, plus 15 plates, 110 figs. in text. |
| Matthey, R. | |
| 1953. | Les Chromosomes des Muridae. Revue Suisse de Zoologie, 60:225-283, avec les planches 1 à 4 groupant 84 photomicrographies, 98 figures et 5 schemas dans le texte. |
| 1955. [Pg 216] | Nouveaux documents sur les chromosomes des Muridae. Problèmes de cytologie comparée et de taxonomie chez les Microtinae. Revue Suisse de Zoologie, 62:163-206, avec 114 figures. |
| Miller, G. S. | |
| 1896. | Genera and subgenera of voles and lemmings. North American Fauna No. 12, pp. 1-85, 40 figs. and 3 plates in text. |
| Ognev, S. I. | |
| 1948. | The mammals of Russia (USSR) and adjacent countries (The mammals of Eastern Europe and Northern Asia), Vol. 6. Publ. Acad. Sci. USSR, pp. 1-587, 260 figs., 12 maps, and 11 color plates in text (in Russian). |
| 1950. | The mammals of Russia (USSR) and adjacent countries (The mammals of Eastern Europe and Northern Asia), Vol. 7. Publ. Acad. Sci. USSR, pp. 1-736, 347 figs., 15 maps, and 10 color plates in text (in Russian). |
| Ruth, E. B. | |
| 1934. | The os priapi: A study in bone development. Anat. Rec., 60:231-249, 16 figs. in 3 plates. |
| Smith, D. A., and J. B. Foster. | |
| 1957. | Notes on the small mammals of Churchill, Manitoba. Jour. Mamm., 38:98-115, 3 figs. and 3 tables in text. |
| Wheeler, B. | |
| 1956. | Comparison of the Block Island "species" of Microtus with M. pennsylvanicus. Evolution, 10:176-186, 4 figs. and 2 tables in text. |
| White, J. A. | |
| 1951. | A practical method for mounting the bacula of small mammals. Jour. Mamm., 32:125. |
| Zimmerman, K. | |
| 1955. | Die Gattung Arvicola Lac. im System der Microtinae. Säugetierkundliche Mitteilungen, 3:110-112, 2 figs. in text. |
| Transmitted August 14, 1959. | |
28-774
Institutional libraries interested in publications exchange may obtain this series by addressing the Exchange Librarian, University of Kansas Library, Lawrence, Kansas. Copies for individuals, persons working in a particular field of study, may be obtained by addressing instead the Museum of Natural History, University of Kansas, Lawrence, Kansas. There is no provision for sale of this series by the University Library which meets institutional requests, or by the Museum of Natural History which meets the requests of individuals. However, when individuals request copies from the Museum, 25 cents should be included, for each separate number that is 100 pages or more in length, for the purpose of defraying the costs of wrapping and mailing.
* An asterisk designates those numbers of which the Museum's supply (not the Library's supply) is exhausted. Numbers published to date, in this series, are as follows:
End of the Project Gutenberg EBook of The Baculum in Microtine Rodents, by
Sydney Anderson
*** END OF THIS PROJECT GUTENBERG EBOOK THE BACULUM IN MICROTINE RODENTS ***
***** This file should be named 38021-h.htm or 38021-h.zip *****
This and all associated files of various formats will be found in:
http://www.gutenberg.org/3/8/0/2/38021/
Produced by Chris Curnow, Alex Gam, Joseph Cooper and the
Online Distributed Proofreading Team at http://www.pgdp.net
Updated editions will replace the previous one--the old editions
will be renamed.
Creating the works from public domain print editions means that no
one owns a United States copyright in these works, so the Foundation
(and you!) can copy and distribute it in the United States without
permission and without paying copyright royalties. Special rules,
set forth in the General Terms of Use part of this license, apply to
copying and distributing Project Gutenberg-tm electronic works to
protect the PROJECT GUTENBERG-tm concept and trademark. Project
Gutenberg is a registered trademark, and may not be used if you
charge for the eBooks, unless you receive specific permission. If you
do not charge anything for copies of this eBook, complying with the
rules is very easy. You may use this eBook for nearly any purpose
such as creation of derivative works, reports, performances and
research. They may be modified and printed and given away--you may do
practically ANYTHING with public domain eBooks. Redistribution is
subject to the trademark license, especially commercial
redistribution.
*** START: FULL LICENSE ***
THE FULL PROJECT GUTENBERG LICENSE
PLEASE READ THIS BEFORE YOU DISTRIBUTE OR USE THIS WORK
To protect the Project Gutenberg-tm mission of promoting the free
distribution of electronic works, by using or distributing this work
(or any other work associated in any way with the phrase "Project
Gutenberg"), you agree to comply with all the terms of the Full Project
Gutenberg-tm License (available with this file or online at
http://gutenberg.org/license).
Section 1. General Terms of Use and Redistributing Project Gutenberg-tm
electronic works
1.A. By reading or using any part of this Project Gutenberg-tm
electronic work, you indicate that you have read, understand, agree to
and accept all the terms of this license and intellectual property
(trademark/copyright) agreement. If you do not agree to abide by all
the terms of this agreement, you must cease using and return or destroy
all copies of Project Gutenberg-tm electronic works in your possession.
If you paid a fee for obtaining a copy of or access to a Project
Gutenberg-tm electronic work and you do not agree to be bound by the
terms of this agreement, you may obtain a refund from the person or
entity to whom you paid the fee as set forth in paragraph 1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be
used on or associated in any way with an electronic work by people who
agree to be bound by the terms of this agreement. There are a few
things that you can do with most Project Gutenberg-tm electronic works
even without complying with the full terms of this agreement. See
paragraph 1.C below. There are a lot of things you can do with Project
Gutenberg-tm electronic works if you follow the terms of this agreement
and help preserve free future access to Project Gutenberg-tm electronic
works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the Foundation"
or PGLAF), owns a compilation copyright in the collection of Project
Gutenberg-tm electronic works. Nearly all the individual works in the
collection are in the public domain in the United States. If an
individual work is in the public domain in the United States and you are
located in the United States, we do not claim a right to prevent you from
copying, distributing, performing, displaying or creating derivative
works based on the work as long as all references to Project Gutenberg
are removed. Of course, we hope that you will support the Project
Gutenberg-tm mission of promoting free access to electronic works by
freely sharing Project Gutenberg-tm works in compliance with the terms of
this agreement for keeping the Project Gutenberg-tm name associated with
the work. You can easily comply with the terms of this agreement by
keeping this work in the same format with its attached full Project
Gutenberg-tm License when you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern
what you can do with this work. Copyright laws in most countries are in
a constant state of change. If you are outside the United States, check
the laws of your country in addition to the terms of this agreement
before downloading, copying, displaying, performing, distributing or
creating derivative works based on this work or any other Project
Gutenberg-tm work. The Foundation makes no representations concerning
the copyright status of any work in any country outside the United
States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other immediate
access to, the full Project Gutenberg-tm License must appear prominently
whenever any copy of a Project Gutenberg-tm work (any work on which the
phrase "Project Gutenberg" appears, or with which the phrase "Project
Gutenberg" is associated) is accessed, displayed, performed, viewed,
copied or distributed:
This eBook is for the use of anyone anywhere at no cost and with
almost no restrictions whatsoever. You may copy it, give it away or
re-use it under the terms of the Project Gutenberg License included
with this eBook or online at www.gutenberg.org
1.E.2. If an individual Project Gutenberg-tm electronic work is derived
from the public domain (does not contain a notice indicating that it is
posted with permission of the copyright holder), the work can be copied
and distributed to anyone in the United States without paying any fees
or charges. If you are redistributing or providing access to a work
with the phrase "Project Gutenberg" associated with or appearing on the
work, you must comply either with the requirements of paragraphs 1.E.1
through 1.E.7 or obtain permission for the use of the work and the
Project Gutenberg-tm trademark as set forth in paragraphs 1.E.8 or
1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted
with the permission of the copyright holder, your use and distribution
must comply with both paragraphs 1.E.1 through 1.E.7 and any additional
terms imposed by the copyright holder. Additional terms will be linked
to the Project Gutenberg-tm License for all works posted with the
permission of the copyright holder found at the beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm
License terms from this work, or any files containing a part of this
work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this
electronic work, or any part of this electronic work, without
prominently displaying the sentence set forth in paragraph 1.E.1 with
active links or immediate access to the full terms of the Project
Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary,
compressed, marked up, nonproprietary or proprietary form, including any
word processing or hypertext form. However, if you provide access to or
distribute copies of a Project Gutenberg-tm work in a format other than
"Plain Vanilla ASCII" or other format used in the official version
posted on the official Project Gutenberg-tm web site (www.gutenberg.org),
you must, at no additional cost, fee or expense to the user, provide a
copy, a means of exporting a copy, or a means of obtaining a copy upon
request, of the work in its original "Plain Vanilla ASCII" or other
form. Any alternate format must include the full Project Gutenberg-tm
License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying,
performing, copying or distributing any Project Gutenberg-tm works
unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing
access to or distributing Project Gutenberg-tm electronic works provided
that
- You pay a royalty fee of 20% of the gross profits you derive from
the use of Project Gutenberg-tm works calculated using the method
you already use to calculate your applicable taxes. The fee is
owed to the owner of the Project Gutenberg-tm trademark, but he
has agreed to donate royalties under this paragraph to the
Project Gutenberg Literary Archive Foundation. Royalty payments
must be paid within 60 days following each date on which you
prepare (or are legally required to prepare) your periodic tax
returns. Royalty payments should be clearly marked as such and
sent to the Project Gutenberg Literary Archive Foundation at the
address specified in Section 4, "Information about donations to
the Project Gutenberg Literary Archive Foundation."
- You provide a full refund of any money paid by a user who notifies
you in writing (or by e-mail) within 30 days of receipt that s/he
does not agree to the terms of the full Project Gutenberg-tm
License. You must require such a user to return or
destroy all copies of the works possessed in a physical medium
and discontinue all use of and all access to other copies of
Project Gutenberg-tm works.
- You provide, in accordance with paragraph 1.F.3, a full refund of any
money paid for a work or a replacement copy, if a defect in the
electronic work is discovered and reported to you within 90 days
of receipt of the work.
- You comply with all other terms of this agreement for free
distribution of Project Gutenberg-tm works.
1.E.9. If you wish to charge a fee or distribute a Project Gutenberg-tm
electronic work or group of works on different terms than are set
forth in this agreement, you must obtain permission in writing from
both the Project Gutenberg Literary Archive Foundation and Michael
Hart, the owner of the Project Gutenberg-tm trademark. Contact the
Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable
effort to identify, do copyright research on, transcribe and proofread
public domain works in creating the Project Gutenberg-tm
collection. Despite these efforts, Project Gutenberg-tm electronic
works, and the medium on which they may be stored, may contain
"Defects," such as, but not limited to, incomplete, inaccurate or
corrupt data, transcription errors, a copyright or other intellectual
property infringement, a defective or damaged disk or other medium, a
computer virus, or computer codes that damage or cannot be read by
your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right
of Replacement or Refund" described in paragraph 1.F.3, the Project
Gutenberg Literary Archive Foundation, the owner of the Project
Gutenberg-tm trademark, and any other party distributing a Project
Gutenberg-tm electronic work under this agreement, disclaim all
liability to you for damages, costs and expenses, including legal
fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT
LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE
PROVIDED IN PARAGRAPH 1.F.3. YOU AGREE THAT THE FOUNDATION, THE
TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE
LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR
INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH
DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a
defect in this electronic work within 90 days of receiving it, you can
receive a refund of the money (if any) you paid for it by sending a
written explanation to the person you received the work from. If you
received the work on a physical medium, you must return the medium with
your written explanation. The person or entity that provided you with
the defective work may elect to provide a replacement copy in lieu of a
refund. If you received the work electronically, the person or entity
providing it to you may choose to give you a second opportunity to
receive the work electronically in lieu of a refund. If the second copy
is also defective, you may demand a refund in writing without further
opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth
in paragraph 1.F.3, this work is provided to you 'AS-IS' WITH NO OTHER
WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT LIMITED TO
WARRANTIES OF MERCHANTIBILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied
warranties or the exclusion or limitation of certain types of damages.
If any disclaimer or limitation set forth in this agreement violates the
law of the state applicable to this agreement, the agreement shall be
interpreted to make the maximum disclaimer or limitation permitted by
the applicable state law. The invalidity or unenforceability of any
provision of this agreement shall not void the remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the
trademark owner, any agent or employee of the Foundation, anyone
providing copies of Project Gutenberg-tm electronic works in accordance
with this agreement, and any volunteers associated with the production,
promotion and distribution of Project Gutenberg-tm electronic works,
harmless from all liability, costs and expenses, including legal fees,
that arise directly or indirectly from any of the following which you do
or cause to occur: (a) distribution of this or any Project Gutenberg-tm
work, (b) alteration, modification, or additions or deletions to any
Project Gutenberg-tm work, and (c) any Defect you cause.
Section 2. Information about the Mission of Project Gutenberg-tm
Project Gutenberg-tm is synonymous with the free distribution of
electronic works in formats readable by the widest variety of computers
including obsolete, old, middle-aged and new computers. It exists
because of the efforts of hundreds of volunteers and donations from
people in all walks of life.
Volunteers and financial support to provide volunteers with the
assistance they need, are critical to reaching Project Gutenberg-tm's
goals and ensuring that the Project Gutenberg-tm collection will
remain freely available for generations to come. In 2001, the Project
Gutenberg Literary Archive Foundation was created to provide a secure
and permanent future for Project Gutenberg-tm and future generations.
To learn more about the Project Gutenberg Literary Archive Foundation
and how your efforts and donations can help, see Sections 3 and 4
and the Foundation web page at http://www.pglaf.org.
Section 3. Information about the Project Gutenberg Literary Archive
Foundation
The Project Gutenberg Literary Archive Foundation is a non profit
501(c)(3) educational corporation organized under the laws of the
state of Mississippi and granted tax exempt status by the Internal
Revenue Service. The Foundation's EIN or federal tax identification
number is 64-6221541. Its 501(c)(3) letter is posted at
http://pglaf.org/fundraising. Contributions to the Project Gutenberg
Literary Archive Foundation are tax deductible to the full extent
permitted by U.S. federal laws and your state's laws.
The Foundation's principal office is located at 4557 Melan Dr. S.
Fairbanks, AK, 99712., but its volunteers and employees are scattered
throughout numerous locations. Its business office is located at
809 North 1500 West, Salt Lake City, UT 84116, (801) 596-1887, email
business@pglaf.org. Email contact links and up to date contact
information can be found at the Foundation's web site and official
page at http://pglaf.org
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
gbnewby@pglaf.org
Section 4. Information about Donations to the Project Gutenberg
Literary Archive Foundation
Project Gutenberg-tm depends upon and cannot survive without wide
spread public support and donations to carry out its mission of
increasing the number of public domain and licensed works that can be
freely distributed in machine readable form accessible by the widest
array of equipment including outdated equipment. Many small donations
($1 to $5,000) are particularly important to maintaining tax exempt
status with the IRS.
The Foundation is committed to complying with the laws regulating
charities and charitable donations in all 50 states of the United
States. Compliance requirements are not uniform and it takes a
considerable effort, much paperwork and many fees to meet and keep up
with these requirements. We do not solicit donations in locations
where we have not received written confirmation of compliance. To
SEND DONATIONS or determine the status of compliance for any
particular state visit http://pglaf.org
While we cannot and do not solicit contributions from states where we
have not met the solicitation requirements, we know of no prohibition
against accepting unsolicited donations from donors in such states who
approach us with offers to donate.
International donations are gratefully accepted, but we cannot make
any statements concerning tax treatment of donations received from
outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation
methods and addresses. Donations are accepted in a number of other
ways including checks, online payments and credit card donations.
To donate, please visit: http://pglaf.org/donate
Section 5. General Information About Project Gutenberg-tm electronic
works.
Professor Michael S. Hart is the originator of the Project Gutenberg-tm
concept of a library of electronic works that could be freely shared
with anyone. For thirty years, he produced and distributed Project
Gutenberg-tm eBooks with only a loose network of volunteer support.
Project Gutenberg-tm eBooks are often created from several printed
editions, all of which are confirmed as Public Domain in the U.S.
unless a copyright notice is included. Thus, we do not necessarily
keep eBooks in compliance with any particular paper edition.
Most people start at our Web site which has the main PG search facility:
http://www.gutenberg.org
This Web site includes information about Project Gutenberg-tm,
including how to make donations to the Project Gutenberg Literary
Archive Foundation, how to help produce our new eBooks, and how to
subscribe to our email newsletter to hear about new eBooks.