
Museum of Natural History
Volume 17, No. 7, pp. 281-375, pls. 1-12, 17 figs.
July 14, 1966
Editors: E. Raymond Hall, Chairman, Henry S. Fitch, Frank B. Cross
Volume 17, No. 7, pp. 281-375, pls. 1-12, 17 figs.
Published July 14, 1966
ROBERT R. (BOB) SANDERS, STATE PRINTER
TOPEKA, KANSAS
1966
31-3430
The family Hylidae, as currently recognized, is composed of about 34 genera and more than 400 species. Most genera (30) and about 350 species live in the American tropics. Hyla and 10 other genera inhabit Central America; four of those 10 genera (Gastrotheca, Hemiphractus, Phrynohyas, and Phyllomedusa) are widely distributed in South America. The other six genera are either restricted to Central America or have their greatest differentiation there. Plectrohyla and Ptychohyla inhabit streams in the highlands of southern Mexico and northern Central America; Diaglena and Triprion are casque-headed inhabitants of arid regions in México and northern Central America. Anotheca is a tree-hole breeder in cloud forests in Middle America. The genus Smilisca is the most widespread geographically and diverse ecologically of the Central American genera.
The definition of genera in the family Hylidae is difficult owing to the vast array of species, most of which are poorly known as regards their osteology, colors in life, and modes of life history. The genera Diaglena, Triprion, Tetraprion, Osteocephalus, Trachycephalus, Aparasphenodon, Corythomantis, Hemiphractus, Pternohyla, and Anotheca have been recognized as distinct from one another and from the genus Hyla on the basis of various modifications of dermal bones of the cranium. Phyllomedusa is recognized on the basis of a vertical pupil and opposable thumb; Plectrohyla is characterized by the presence of a bony prepollex and the absence of a quadratojugal. Gastrotheca is distinguished from other hylids by the presence of a pouch in the back of females. A pair of lateral vocal sacs behind the angles of the jaws and the well-developed dermal glands were used by Duellman (1956) to distinguish Phrynohyas from Hyla. He (1963a) cited the ventrolateral glands in breeding males as diagnostic of Ptychohyla. Some species groups within the vaguely defined genus Hyla have equally distinctive characters. The Hyla septentrionalis group is characterized by a casque-head, not much different from that in the genus Osteocephalus (Trueb, MS). Males in the Hyla maxima group have a protruding bony prepollex like that characteristically found in Plectrohyla.
Ontogenetic development, osteology, breeding call, behavior, and ecology are important in the recognition of species. By utilizing [Pg 286] the combination of many morphological and biological factors, the genus Smilisca can be defined reasonably well as a natural, phyletic assemblage of species. Because the wealth of data pertaining to the morphology and biology of Smilisca is lacking for most other tree frogs in Middle America it is not possible at present to compare Smilisca with related groups in more than a general way.
Smilisca is an excellent example of an Autochthonous Middle American genus. As defined by Stuart (1950) the Autochthonous Middle American fauna originated from "hanging relicts" left in Central America by the ancestral fauna that moved into South America and differentiated there at a time when South America was isolated from North and Middle America. The genus Smilisca, as we define it, consists of six species, all of which occur in Central America. One species ranges northward to southern Texas, and one extends southward on the Pacific lowlands of South America to Ecuador. We consider the genus Smilisca to be composed of rather generalized hylids. Consequently, an understanding of the systematics and zoogeography of the genus can be expected to be of aid in studying more specialized members of the family.
Examination of many of the specimens used in our study was possible only because of the cooperation of the curators of many systematic collections. For lending specimens or providing working space in their respective institutions we are grateful to Doris M. Cochran, Alice G. C. Grandison, Jean Guibe, Robert F. Inger, Günther Peters, Gerald Raun, William J. Riemer, Jay M. Savage, Hobart M. Smith, Wilmer W. Tanner, Charles F. Walker, Ernest E. Williams, and Richard G. Zweifel.
We are indebted to Charles J. Cole and Charles W. Myers for able assistance in the field. The cooperation of Martin H. Moynihan at Barro Colorado Island, Charles M. Keenan of Corozal, Canal Zone, and Robert Hunter of San José, Costa Rica, is gratefully acknowledged. Jay M. Savage turned over to us many Costa Rican specimens and aided greatly in our work in Costa Rica. James A. Peters helped us locate sites of collections in Ecuador and Coleman J. Goin provided a list of localities for the genus in Colombia.
We especially thank Charles J. Cole for contributing the information on the chromosomes, and Robert R. Patterson for preparing osteological specimens. We thank M. J. Fouquette, Jr., who read the section on breeding calls and offered constructive criticism.
Permits for collecting were generously provided by Ing. Rodolfo Hernandez Corzo in México, Sr. Jorge A. Ibarra in Guatemala, and Ing. Milton Lopez in Costa Rica. This report was made possible by support from the National Science Foundation (Grants G-9827 and GB-1441) and the cooperation of the Museum of Natural History at the University of Kansas. Some of the field studies were carried out in Panamá under the auspices of a grant from the National Institutes of Health (NIH GM-12020) in cooperation with the Gorgas Memorial Laboratory in Panamá.
In our study we examined 4151 preserved frogs, 93 skeletal preparations, 88 lots of tadpoles and young, and six lots of eggs. We have collected specimens in the field of all of the species. Observations on behavior and life history were begun by the senior author in México in 1956 and completed by us in Central America in 1964 and 1965.
Osteological data were obtained from dried skeletons and cleaned and stained specimens of all species, plus serial sections of the skull of Smilisca baudini. Developmental stages to which tadpoles are assigned are in accordance with the table of development published by Gosner (1960). Breeding calls were recorded in the field on tape using Magnemite and Uher portable tape recorders. Audiospectrographs were made by means of a Vibralyzer (Kay Electric Company). External morphological features were measured in the manner described by Duellman (1956). In the accounts of the species we have attempted to give a complete synonymy. At the end of each species account the localities from which specimens were examined are listed alphabetically within each state, province, or department, which in turn are listed alphabetically within each country. The countries are arranged from north to south. Abbreviations for museum specimens are listed below:
| AMNH—American Museum of Natural History BMNH—British Museum (Natural History) BYU—Brigham Young University CNHM—Chicago Natural History Museum KU—University of Kansas Museum of Natural History MCZ—Museum of Comparative Zoology MNHN—Museu National d'Histoire Naturelle, Paris UF—University of Florida Collections UIMNH—University of Illinois Museum of Natural History UMMZ—University of Michigan Museum of Zoology USC—University of Southern California USNM—United States National Museum TNHC—Texas Natural History Collection, University of Texas ZMB—Zoologisches Museum Berlin |
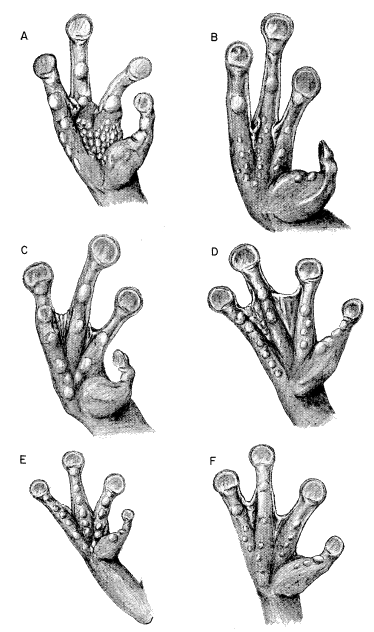
Definition.—Medium to large tree frogs having: (1) broad, well ossified skull (consisting of a minimum amount of cartilage and/or secondarily ossified cartilage), (2) no dermal co-ossification, (3) quadratojugal and internasal septum present, (4) large ethmoid, (5) M. depressor mandibulae consisting of two parts, one arising from dorsal fascia and other from posterior arm of squamosal, (6) divided M. adductor mandibulae, (7) paired subgular vocal sacs in males, (8) no dermal appendages, (9) pupil horizontally elliptical (10) small amounts of amines and other active substances in skin, (11) chromosome number of N = 12 and 2N = 24, (12) breeding call consisting of poorly modulated, explosive notes, and (13) 2/3 tooth-rows in tadpoles.
Composition of genus.—As defined here the genus Smilisca contains six recognizable species. An alphabetical list of the specific and subspecific names [Pg 288] that we consider to be applicable to species of Smilisca recognized herein is given below.
| Names proposed | Valid names | |
| Hyla baudini Duméril and Bibron, 1841 | = S. baudini | |
| Hyla baudini dolomedes Barbour, 1923 | = S. phaeota | |
| Hyla beltrani Taylor, 1942 | = S. baudini | |
| Hyla gabbi Cope, 1876 | = S. sordida | |
| Hyla labialis Peters, 1863 | = S. phaeota | |
| Hyla manisorum Taylor, 1954 | = S. baudini | |
| Hyla muricolor Cope, 1862 | = S. baudini | |
| Hyla nigripes Cope, 1876 | = S. sordida | |
| Hyla pansosana Brocchi, 1877 | = S. baudini | |
| Hyla phaeota Cope, 1862 | = S. phaeota | |
| Hyla phaeota cyanosticta Smith, 1953 | = S. cyanosticta | |
| Hyla puma Cope, 1885 | = S. puma | |
| Hyla salvini Boulenger, 1882 | = S. sordida | |
| Hyla sordida Peters, 1863 | = S. sordida | |
| Hyla vanvlietii Baird, 1854 | = S. baudini | |
| Hyla vociferans Baird, 1859 | = S. baudini | |
| Hyla wellmanorum Taylor, 1952 | = S. puma |
Distribution of genus.—Most of lowlands of México and Central America, in some places to elevations of nearly 2000 meters, southward from southern Sonora and Río Grande Embayment of Texas, including such continental islands as Isla Cozumel, México, and Isla Popa and Isla Cebaco, Panamá, to northern South America, where known from Caribbean coastal regions and valleys of Río Cauca and Río Magdalena in Colombia, and Pacific slopes of Colombia and northern Ecuador.
| 1. | Larger frogs ([M] to 76 mm., [F] to 90 mm.) having broad flat heads and a dark brown or black postorbital mark encompassing tympanum |
| 2 | |
| Smaller frogs ([M] to 45 mm., [F] to 64 mm.) having narrower heads and lacking a dark brown or black postorbital mark encompassing tympanum | |
| 4 | |
| 2. | Lips barred; flanks cream-colored with bold brown or black mottling in groin; posterior surfaces of thighs brown with cream-colored flecks |
| S. baudini, p. 289 | |
| Lips not barred; narrow white labial stripe present; flanks not cream-colored with bold brown or black mottling in groin; posterior surfaces of thighs variable | |
| 3 | |
| 3. | Flanks and anterior and posterior surfaces of thighs dark brown with large pale blue spots on flanks and small blue spots on thighs |
| S. cyanosticta, p. 303 | |
| Flanks cream-colored with fine black venation; posterior surfaces of thighs pale brown with or without darker flecks or small cream-colored spots | |
| S. phaeota, p. 308 | |
| 4. | Fingers having only vestige of web; diameter of tympanum two-thirds that of eye; dorsum pale yellowish tan with pair of broad dark brown stripes |
| S. puma, p. 314 | |
| Fingers about one-half webbed; diameter of tympanum about one-half that of eye; dorsum variously marked with spots or blotches | |
| 5 | |
| 5. | Snout short, truncate; vocal sacs in breeding males dark gray or brown; blue spots on flanks and posterior surfaces of thighs |
| S. sila, p. 318 | |
| Snout long, sloping, rounded; vocal sacs in breeding males white; cream-colored or pale blue flecks on flanks and posterior surfaces of thighs | |
| S. sordida, p. 323 | |
| 1. | Pond tadpoles; tail about half again as long as body; mouth anteroventral |
| 2 | |
| Stream tadpoles; tail about twice as long as body; mouth ventral | |
| 5 | |
| 2. | Labial papillae in two rows |
| 3 | |
| Labial papillae in one row | |
| 4 | |
| 3. | First upper tooth row strongly arched medially; third lower tooth row much shorter than other rows; dorsal fin deepest at about two-thirds length of tail; tail cream-colored with dense gray reticulations |
| S. puma, p. 314 | |
| First upper tooth row not arched medially; third lower tooth row nearly as long as others; dorsal fin deepest at about one-third length of tail; tail tan with brown flecks and blotches | |
| S. baudini, p. 289 | |
| 4. | Dorsal fin extending onto body |
| S. phaeota, p. 308 | |
| Dorsal fin not extending onto body | |
| S. cyanosticta, p. 303 | |
| 5. | Mouth completely bordered by two rows of papillae; inner margin of upper beak not forming continuous arch with lateral processes; red or reddish brown markings on tail |
| S. sordida, p. 323 | |
| Median part of upper lip bare; rest of mouth bordered by one row of papillae; inner margin of upper beak forming continuous arch with lateral processes; dark brown markings on tail | |
| S. sila, p. 318 | |
Diagnosis.—Size large ([M] 76 mm., [F] 90 mm.); skull noticeably wider than long, having small frontoparietal fontanelle (roofed with bone in large individuals); postorbital processes long, pointed, curving along posterior border of orbit; squamosal large, contacting maxillary; tarsal fold strong, full length of tarsus; inner metatarsal tubercle large, high, elliptical; hind limbs relatively short, tibia length less than 55 per cent snout-vent length; lips strongly barred with brown and creamy tan; flanks pale cream with bold brown or black reticulations in groin; posterior surfaces of thighs brown with cream-colored flecks; dorsal surfaces of limbs marked with dark brown transverse bands. (Foregoing combination of characters distinguishing S. baudini from any other species in genus.)
Description and Variation.—Considerable variation in size, and in certain proportions and structural characters was observed; variation in some characters seems to show geographic trends, whereas variation in other characters apparently is random. Noticeable variation is evident in coloration, but this will be discussed later.
In order to analyze geographic variation in size and proportions, ten adult males from each of 14 samples from various localities throughout the range of the species were measured. Snout-vent length, length of the tibia in relation to snout-vent length, and relative size of the tympanum to the eye are the only measurements and proportions that vary noticeably (Table 1). The largest specimens are from southern Sinaloa; individuals from the Atlantic lowlands of Alta Verapaz in Guatemala, Honduras, and Costa Rica are somewhat smaller, and most specimens from the Pacific lowlands of Central America are slightly smaller than those from the Atlantic lowlands. The smallest males are from the Atlantic lowlands of México, including Tamaulipas, Veracruz, the Yucatán Peninsula, and British Honduras.
| Table 1.—Geographic Variation in Size and Proportions in Males of Smilisca baudini. (Means in Parentheses Below Observed Ranges; Data Based on 10 Specimens From Each Locality.) | |||
| Locality | Snout-vent length |
Tibia length/ snout-vent |
Tympanum/ eye |
| Southern Sinaloa | 62.3-75.9 | 43.2-46.7 | 84.2-94.4 |
| (68.6) | (44.9) | (87.8 | |
| Ocotito, Guerrero | 55.6-64.0 | 46.1-51.2 | 66.7-82.8 |
| (58.7) | (47.8) | (74.6) | |
| Pochutla, Oaxaca | 56.1-65.1 | 44.7-49.4 | 73.0-84.2 |
| (60.2) | (47.5) | (77.4) | |
| San Salvador, El Salvador | 57.0-68.0 | 42.1-46.1 | 74.6-83.3 |
| (62.1) | (44.9) | (77.6) | |
| Managua, Nicaragua | 52.9-63.6 | 45.6-49.4 | 73.7-89.7 |
| (57.3) | (47.5) | (79.4) | |
| Esparta, Costa Rica | 57.6-66.0 | 44.6-49.3 | 65.5-83.6 |
| (61.3) | (47.3) | (75.2) | |
| Ciudad Victoria, Tamaulipas | 50.6-56.9 | 44.5-48.7 | 67.2-84.3 |
| (53.7) | (46.6) | (73.9) | |
| Córdoba, Veracruz | 53.8-63.4 | 43.9-48.4 | 66.1-75.9 |
| (57.5) | (45.6) | (70.0) | |
| Isla del Carmen, Campeche | 47.3-56.6 | 44.7-48.9 | 61.5-72.6 |
| (50.9) | (47.6) | (65.7) | |
| Chichén-Itzá, Yucatán | 49.6-57.1 | 45.2-53.4 | 62.7-80.7 |
| (53.8) | (49.5) | (72.6) | |
| British Honduras | 49.0-59.6 | 47.5-50.7 | 67.9-76.8 |
| (54.9) | (49.1) | (72.2) | |
| Chinajá, Guatemala | 56.8-67.6 | 47.0-51.0 | 70.0-82. |
| (63.2) | (49.5) | (73.6 | |
| Atlantidad, Honduras | 52.5-65.1 | 49.8-53.6 | 56.1-76.5 |
| (57.6) | (51.5) | (67.0) | |
| Limón, Costa Rica | 57.7-71.3 | 50.4-52.3 | 63.9-73.0 |
| (62.4) | (51.2) | (68.5) | |
The ratio of the tibia to the snout-vent length varies from 42.1 to 53.6 in the 14 samples analyzed. The average ratio in samples from the Pacific lowlands varies from 44.9 in Sinaloa and El Salvador to 47.8 in Guerrero; on the Gulf lowlands of México the average ratio varies from 45.6 in Veracruz to 47.6 on Isla del Carmen, Campeche. Specimens from the Yucatán Peninsula and the Caribbean lowlands have relatively longer legs; the variation in average ratios ranges from 49.1 in British Honduras to 51.2 in Costa Rica and 51.5 in Honduras.
Specimens from southern Sinaloa are outstanding in the large size of the [Pg 293] tympanum; the tympanum/eye ratio varies from 84.2 to 94.4 (average 87.8). In most other samples the variation in average ratios ranges from 72.2 to 79.3, but specimens from Veracruz have an average ratio of 70.0; Campeche, 65.7; Honduras, 67.0; and Limón, Costa Rica, 68.5.
No noticeable geographic trends in size and proportions are evident. Specimens from southern Sinaloa are extreme in their large size, relatively short tibia, and large tympani, but in size and relative length of the tibia the Sinaloan frogs are approached by specimens from such far-removed localities as San Salvador, El Salvador, and Chinajá, Guatemala. Frogs from the Caribbean lowlands of Honduras and Costa Rica are relatively large and have relatively long tibiae and small tympani.
The inner metatarsal tubercle is large and high and its shape varies. The tubercle is most pronounced in specimens from northwestern México, Tamaulipas, and the Pacific lowlands of Central America. Possibly the large tubercle is associated with drier habitats, where perhaps the frogs use the tubercles for digging.
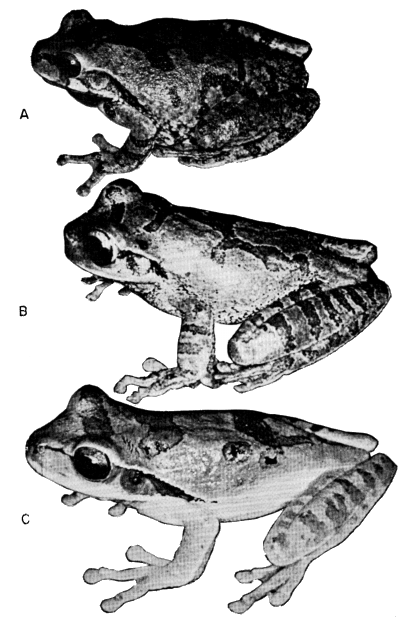
The ground color of Smilisca baudini is pale green to brown dorsally and white to creamy yellow ventrally. The dorsum is variously marked with dark brown or dark olive-green spots or blotches (Pl. 6A). In most specimens a dark interorbital bar extends across the head to the lateral edges of the eyelid; usually this bar is connected medially to a large dorsal blotch. There is no tendency for the markings on the dorsum to form transverse bands or longitudinal bars. In specimens from the southern part of the range the dorsal dark markings are often fragmented into small spots, especially posteriorly. The limbs are marked by dark transverse bands, usually three on the forearm, three on the thigh, and three or four on the shank. Transverse bands also are present on the tarsi and proximal segments of the fingers and toes. The webbing on the hands and feet is pale grayish brown. The loreal region and upper lip are pale green or tan; the lip usually is boldly marked with broad vertical dark brown bars, especially evident is the bar beneath the eye. A dark brown or black mark extends from the tympanum to a point above the insertion of the forearm; in some specimens this black mark is narrow or indistinct, but in most individuals it is quite evident. The flanks are pale gray to creamy white with brown or black mottling, which sometimes forms reticulations enclosing white spots. The anterior surfaces of the thighs usually are creamy white with brown mottling, whereas the posterior surfaces of the thighs usually are brown with small cream-colored flecks. A distinct creamy white anal stripe usually is present. Usually, there are no white stripes on the outer edges of the tarsi and forearms. In breeding males the throat is gray.
Most variation in coloration does not seem to be correlated with geography. The lips are strongly barred in specimens from throughout the range of the species, except that in some specimens from southern Nicaragua and Costa Rica the lips are pale and in some specimens the vertical bars are indistinct. Six specimens from 7.3 kilometers southwest of Matatán, Sinaloa, are distinctively marked. The dorsum is uniformly grayish green with the only dorsal marks being on the tarsi; canthal and post-tympanic dark marks absent. A broad white labial stripe is present and interrupted by a single vertical dark mark below the eye. A white stripe is present on the outer edge of the foot. The flanks and posterior surfaces of the thighs are creamy white, boldly marked with black. Two specimens from Alta Verapaz, Guatemala (CNHM 21006 [Pg 294] from Cobán and UMMZ 90908 from Finca Canihor), are distinctive in having many narrow transverse bands on the limbs and fine reticulations on the flanks. Two specimens from Limón Province, Costa Rica (KU 34927 from Batán and 36789 from Suretka), lack a dorsal pattern; instead these specimens are nearly uniform brown above and have only a few small dark brown spots on the back and lack transverse bands on the limbs. The post-tympanic dark marks and dark mottling on the flanks are absent. Specimens lacking the usual dorsal markings are known from scattered localities on the Caribbean lowlands from Guatemala to Costa Rica.
The coloration in life is highly variable; much of the apparent variation is due to metachrosis, for individuals of Smilisca baudini are capable of undergoing drastic and rapid change in coloration. When active at night the frogs usually are pale bright green with olive-green markings, olive-green with brown markings, or pale brown with dark brown markings. The dark markings on the back and dorsal surfaces of the limbs are narrowly outlined by black. The pale area below the eye and just posterior to the broad suborbital dark bar is creamy white, pale green, or ashy gray in life. The presence of this mark is an excellent character by which to identify juveniles of the species. The flanks are creamy yellow, or yellow with brown or black mottling. In most individuals the belly is white, but in specimens from southern El Petén and northern Alta Verapaz, Guatemala, the belly is yellow, especially posteriorly. The iris varies from golden bronze to dull bronze with black reticulations, somewhat darker ventrally.
Natural History.—Throughout most of its range Smilisca baudini occurs in sub-humid habitats; consequently the activity is controlled by the seasonal nature of the rainfall and usually extends from May or June through September. Throughout México and Central America the species is known to call and breed in June, July, and August. Several records indicate that the breeding season in Central America is more lengthy. Gaige, Hartweg, and Stuart (1937:4) noted gravid females collected at El Recreo, Nicaragua, in August and September. Schmidt (1941:486) reported calling males in February in British Honduras. Stuart (1958:17) stated that tadpoles were found in mid-February, juveniles in February and March and half-grown individuals from mid-March to mid-May at Tikal, El Petén, Guatemala. Stuart (1961:74) reported juveniles from Tikal in July, and that individuals were active at night when there had been light rain in the dry season in February and March in El Petén, Guatemala. Smilisca baudini seeks daytime retreats in bromeliads, elephant-ear plants (Xanthosoma), and beneath bark or in holes in trees. By far the most utilized retreat in the dry season in parts of the range is beneath the outer sheaths of banana plants. Large numbers of these frogs were found in banana plants at Cuautlapan, Veracruz, in March, 1956, in March and December, 1959.
Large breeding congregations of this frog are often found at the time of the first heavy rains in the wet season. Gadow (1908:76) estimated 45,000 frogs at one breeding site in Veracruz. In the vicinity of Tehuantepec, Oaxaca, large numbers of individuals were found around rain pools and roadside ditches in July, 1956, and July, 1958; large concentrations were found near Chinajá, Guatemala, in June, 1960, and near Esparta, Costa Rica in July, 1961. Usually males call from the ground at the edge of the water or not infrequently sit in shallow water, but sometimes males call from bushes and low trees [Pg 295] around the water. Stuart (1935:38) recorded individuals calling and breeding throughout the day at La Libertad, Guatemala. Smilisca baudini usually is absent from breeding congregations of hylids; frequently S. baudini breeds alone in small temporary pools separated from large ponds where numerous other species are breeding. In Guerrero and Oaxaca, México, S. baudini breeds in the same ponds with Rhinophrynus dorsalis, Bufo marmoreus, Engystomops pustulosus, and Diaglena reticulata, and in the vicinity of Esparta, Costa Rica, S. baudini breeds in ponds with Bufo coccifer, Hyla staufferi, and Phrynohyas venulosa. In nearly all instances the breeding sites of S. baudini are shallow, temporary pools.
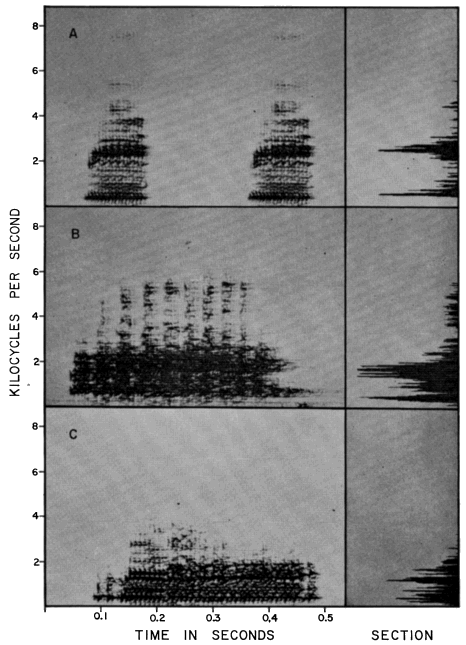
The breeding call of Smilisca baudini consists of a series of short explosive notes. Each note has a duration of 0.09 to 0.13 seconds; two to 15 notes make up a call group. Individual call groups are spaced from about 15 seconds to several minutes apart. The notes are moderately high-pitched and resemble "wonk-wonk-wonk." Little vibration is discernible in the notes, which have 140 to 195 pulses per second and a dominant frequency of 2400 to 2725 cycles per second (Pl. 10A).
The eggs are laid as a surface film on the water in temporary pools. The only membrane enclosing the individual eggs is the vitelline membrane. In ten eggs (KU 62154 from San Salvador, El Salvador) the average diameter of the embryos in first cleavage is 1.3 mm. and of the vitelline membranes, 1.5 mm. Hatchling tadpoles have body lengths of 2.6 to 2.7 mm. and total lengths of 5.1 to 5.4 mm. The body and caudal musculature is brown; the fins are densely flecked with brown. The gills are long and filamentous. Growth and development of tadpoles are summarized in Table 9.
A typical tadpole in stage 30 of development (KU 60018 from Chinajá, Alta Verapaz, Guatemala) has a body length of 8.7 mm., a tail length of 13.6 mm., and a total length of 22.3 mm.; body slightly wider than deep; snout rounded dorsally and laterally; eyes widely separated, directed dorsolaterally; nostril about midway between eye and tip of snout; mouth anteroventral; spiracle sinistral, located about midway on length of body and slightly below midline; anal tube dextral; caudal musculature slender, slightly curved upward distally; dorsal fin extending onto body, deepest at about one-third length of tail; depth of dorsal fin slightly more than that of ventral fin at mid-length of tail; dorsal part of body dark brown; pale crescent-shaped mark on posterior part of body; ventral surfaces transparent with scattered brown pigment ventrolaterally, especially below eye; caudal musculature pale tan with a dark brown longitudinal streak on middle of anterior one-third of tail; dorsum of anterior one-third of tail dark brown; brown flecks and blotches on rest of caudal musculature, on all of dorsal fin, and on posterior two-thirds of ventral fin; iris bronze in life (Fig. 11). Mouth small; median third of upper lip bare; rest of mouth bordered by two rows of conical papillae; lateral fold present; tooth rows 2/3; two upper rows about equal in length; second row broadly interrupted medially, three lower rows complete, first and second equal in length, slightly shorter than upper rows; third lower row shortest; first upper row sharply curved anteriorly in midline; upper beak moderately deep, forming a board arch with slender lateral processes; lower beak more slender, broadly V-shaped; both beaks bearing blunt serrations (Fig. 15A).
In tadpoles having fully developed mouthparts the tooth-row formula of 2/3 is invariable, but the coloration is highly variable. The color and pattern [Pg 296] described above is about average. Some tadpoles are much darker, such as those from 11 kilometers north of Vista Hermosa, Oaxaca, (KU 87639-44), 3.5 kilometers east of Yokdzonot, Yucatán (KU 71720), and 4 kilometers west-southwest Puerto Juárez, Quintana Roo, México (KU 71721), whereas others, notably from 17 kilometers northeast of Juchatengo, Oaxaca, México (KU 87645), are much paler and lack the dark markings on the caudal musculature. The variation in intensity of pigmentation possibly can be correlated with environmental conditions, especially the amount of light. In general, tadpoles that were found in open, sunlit pools are pallid by comparison with those from shaded forest pools. These subjective comparisons were made with preserved specimens; detailed comparative data on living tadpoles are not available.
The relative length and depth of the tail are variable; in some individuals the greatest depth of the tail is about at mid-length of the tail, whereas in most specimens the tail is deepest at about one-third its length. The length of the tail relative to the total length is usually 58 to 64 per cent in tadpoles in stages 29 and 30 of development. In some individuals the tail is about 70 per cent of the total length. On the basis of the material examined, these variations in proportions do not show geographical trends. Probably the proportions are a reflection of crowding of the tadpoles in the pools where they are developing or possibly due to water currents or other environmental factors.
Stuart (1948:26) described and illustrated the tadpole of Smilisca baudini from Finca Chejel, Alta Verapaz, Guatemala. The description and figures agree with ours, except that the first lower tooth row does not have a sharp angle medially in Stuart's figure. He (1948:27) stated that color in tadpoles from different localities probably varies with soil color and turbidity of water. Maslin (1963:125) described and illustrated tadpoles of S. baudini from Pisté, Yucatán, México. These specimens are heavily pigmented like specimens that we have examined from the Yucatán Peninsula and from other places in the range of the species. Maslin stated that the anal tube is median in the specimens that he examined; we have not studied Maslin's specimens, but all tadpoles of Smilisca that we have examined have a dextral anal tube.
Newly metamorphosed young have snout-vent lengths of 12.0 to 15.5 mm. (average 13.4 in 23 specimens). The largest young are from La Libertad, El Petén, Guatemala; these have snout-vent lengths of 14.0 to 15.5 mm. (average 14.5 in five specimens). Young from 11 kilometers north of Vista Hermosa, Oaxaca, México, are the smallest and have snout-vent lengths of 12.0 to 12.5 mm. (average 12.3 in three specimens). Recently metamorphosed young usually are dull olive green above and white below; brown transverse bands are visible on the hind limbs. The labial markings characteristic of the adults are represented only by a creamy white suborbital spot, which is a good diagnostic mark for young of this species. In life the iris is pale gold.
Remarks: The considerable variation in color and the extensive geographic distribution of Smilisca baudini have resulted in the proposal of eight specific names for the frogs that we consider to represent one species. Duméril and Bibron (1841:564) proposed the name Hyla baudini for a specimen (MNHN 4798) from México. Smith and Taylor (1950:347) restricted the type locality to Córdoba, Veracruz, México, an area where the species occurs in abundance. Baird (1854:61) named Hyla vanvlieti from Brownsville, Texas, and (1859:35) labelled the figures of Hyla vanvlieti [= Hyla baudini] on plate 38 as Hyla vociferans, a nomen nudum. Cope (1862:359) named Hyla muricolor from Mirador, Veracruz, México, and (1865:194) used the name Smilisca daulinia for a skeleton that he employed as the basis for the cranial characters diagnostic of the genus Smilisca, as defined by him. Although we cannot be certain, Cope apparently inadvertently used daulinia for baudini, just as he used daudinii for baudini (1871:205). Brocchi (1877:125) named Hyla pansosana from Panzos, Alta Verapaz, Guatemala.











Aside from the skeleton referred to as Smilisca daulinia by Cope (1865:194), we have examined each of the types of the species synonymized with S. baudini. All unquestionably are representatives of S. baudini.
Taylor (1942:306) named Hyla beltrani from Tapachula, Chiapas. This specimen (UIMNH 25046) is a small female (snout-vent length, 44 mm.) of S. baudini. Taylor (1954:630) named Hyla manisorum from Batán, Limón, Costa Rica. The type (KU 34927) is a large female (snout-vent length, 75.3 mm.) S. baudini. In this specimen and a male from Suretka, Costa Rica, the usual dorsal color pattern is absent, but the distinctive curved supraorbital processes, together with other structural features, show that the two specimens are S. baudini.
Hyla baudini dolomedes Barbour (1923:11), as shown by Dunn (1931a:413), was based on a specimen of Smilisca phaeota from Río Esnápe, Darién, Panamá.
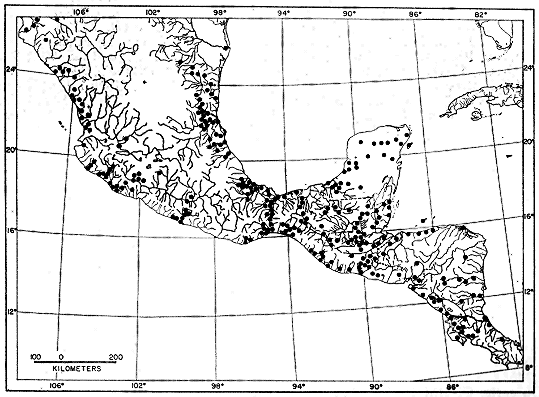
Distribution.—Smilisca baudini inhabits lowlands and foothills usually covered by xerophytic vegetation or savannas, but in the southern part of its range baudini inhabits tropical evergreen forest. The species ranges throughout the Pacific and Atlantic lowlands of México from southern Sonora and the Río Grande embayment of Texas southward to Costa Rica, where on the Pacific lowlands the range terminates at the southern limits of the arid tropical forest in the vicinity of Esparta; on the Caribbean lowlands the distribution [Pg 298] seems to be discontinuous southward to Suretka (Fig. 1). Most localities where the species has been collected are at elevations of less than 1000 meters. Three localities are notably higher; calling males were found at small temporary ponds in pine-oak forest at Linda Vista, 2 kilometers northwest of Pueblo Nuevo Solistahuacán, Chiapas, elevation 1675 meters, and 10 kilometers northwest of Comitán, Chiapas, at an elevation of 1925 meters. Tadpoles and metamorphosing young were obtained from a pond in arid scrub forest, 17 kilometers northeast of Juchatengo, Oaxaca, elevation 1600 meters. Stuart (1954:46) recorded the species at elevations up to 1400 meters in the south-eastern highlands of Guatemala.
Specimens examined.—3006, as follows: United States: Texas: Cameron County, Brownsville, CNHM 5412-3, 6869, UMMZ 54036, USNM 3256.
Mexico: Campeche: Balchacaj, CNHM 102285, 102288, 102291, 102311, UIMNH 30709-22, 30726; Champotón, UMMZ 73172 (2), 73176, 73180; 16 km. E Champotón, UMMZ 73181; 5 km. S Champotón, KU 71369-75; 9 km. S Champotón, KU 71367-8; 10.5 km. S Champotón, KU 71365-6, 71722 (tadpoles), 71723 (yg.); 24 km. S Champotón, UMMZ 73177 (2); Chuina, KU 75101-3; Ciudad del Carmen, UIMNH 30703-8; Dzibalchén, KU 75413-31; Encarnación, CNHM 102282, 102289, 102294-5, 102300, 102306-8, 102312, 102314, 102316-7, 102319, 102322, UIMNH 30727-40, 30836-7; 1 km. W Escárcega, KU 71391-6; 6 km. W Escárcega, KU 71397-403; 7.5 km. W Escárcega, KU 71376-89; 14 km. W Escárcega, KU 71390; 13 km. W, 1 km. N Escárcega, KU 71404; 3 km. N Hopelchén, KU 75410-11; 2 km. NE Hopelchén, KU 75412; Matamoras, CNHM 36573; Pitál, UIMNH 30741; 1 km. SW Puerto Real, Isla del Carmen, KU 71345-64; San José Carpizo, UMMZ 99879; Tres Brazos, CNHM 102284, UIMNH 30723-5; Tuxpeña Camp, UMMZ 73239.
Chiapas: Acacoyagua, USNM 114487-92; 2 km. W Acacoyagua, USNM 114493-4; 5 km. E Arroyo Minas, UIMNH 9533-7; Berriozabal, UMMZ 119186 (7); Chiapa de Corzo, UMMZ 119185 (2); Cintalapa, UIMNH 50077; Colonia Soconusco, USNM 114495-9; 5 km. W Colonia Soconusco, UMMZ 87885 (7); Comitán, UMMZ 94438; 10 km. NW Comitán, KU 57185; El Suspiro, UMMZ 118819 (11); Escuintla, UMMZ 88271 (7), 88278, 88327, 109233; 6 km. NE Escuintla, UMMZ 87856 (26); 3 km. E Finca Juárez, UIMNH 9538; Finca Prussia, UMMZ 95167; Honduras, UMMZ 94434-7; La Grada, UMMZ 87862; 21 km. S La Trinitaria, UIMNH 9540-1; 14.4 km. SW Las Cruces, KU 64239-44; Palenque, UIMNH 49286, USNM 114473-84; 2 km. NW Pueblo Nuevo Solistahuacán, KU 57182-4, UMMZ 119948 (8), 121514; 1.3 km. N Puerto Madero, KU 57186-9; 4 km. N Puerto Madero, KU 57190-1; 8 km. N Puerto Madero, UMMZ 118379 (2); 12 km. N Puerto Madero, KU 57192; 17.6 km. N Puerto Madero, UMMZ 118378; Rancho Monserrata, UIMNH 9531-2, UMMZ 102266-7; Region Soconusco, UIMNH 33542-56; San Bartola, UIMNH 9519-30; San Gerónimo, UIMNH 30804; San Juanito, USNM 114485-6; San Ricardo, CNHM 102406; Solosuchiapa, KU 75432-3; Tapachula, CNHM 102208, 102219, 102239, 102405, UIMNH 25046, 30802-3; Tonolá, AMNH 531, CNHM 102232, 102416, UIMNH 30805-9, USNM 46760; Tuina, KU 41593 (skeleton); Tuxtla Gutierrez, CNHM 102231, 102248; 6 km. E Tuxtla Gutierrez, UIMNH 9539; 10 km. E Tuxtla Gutierrez, UMMZ 119949.
Chihuahua: 2.4 km. SW Toquina, KU 47226-7; Riito, KU 47228.
Coahuila: mountain near Saltillo, UIMNH 30833-4.
Colima: No specific locality, CNHM 1632; Colima, AMNH 510-11; Hacienda Albarradito, UMMZ 80029 (2); Hacienda del Colomo, AMNH 6208; Los Mezcales, UMMZ 80028; Manzanillo, AMNH 6207, 6209; Paso del Río, CNHM 102207, 102229-30, UIMNH 30819-21, UMMZ 110875 (3); Periquillo, UMMZ 80025 (3), 80026 (14); 1.6 km. SW Pueblo Juárez, UMMZ 115564; Queseria, CNHM 102204, 102216-7, 102224, UIMNH 30816-8, UMMZ 80023 (7), 80024 (7); Santiago, UMMZ 80027; 7.2 km. SW Tecolapa, UMMZ 115184.
[Pg 299] Guerrero: Acahuizotla, UF 1338 (2), 1339-40, UMMZ 119182 (2), 119184; 3 km. S Acahuizotla, KU 87183-7; Acapulco, AMNH 55276, UMMZ 121879 (4), USNM 47909; 3 km. N Acapulco, UMMZ 110127; 8 km. NW Acapulco, UF 11203 (7); 27 km. NE Acapulco, UIMNH 26597-610; Agua del Obispo, CNHM 102214, 102290, 102293, 102310, 102413, KU 60413, 87180-2, UIMNH 30764-6; Atoyca, KU 87175-8; Buena Vista, CNHM 102279, 102304, 102313, 102315, UIMNH 30774; Caculutla, KU 87179; 20 km. S Chilpancingo, CNHM 102242, 102401, 102410-1, 102415; Colonia Buenas Aires, UMMZ 119189; El Limoncito, CNHM 102292, 102303, 102321, 102414; El Treinte, CNHM 102212, 102221, 102237, 102240-1, UIMNH 30783-5, USNM 114508-10; Laguna Coyuca, UMMZ 80960 (2); 3 km. N Mazatlán, UIMNH 30777-9; 9 km. S Mazatlán, CNHM 102209, 102215, 102234, 102246, UIMNH 30781-2; Mexcala, CNHM 102399, 102403, 102409, 106539-40, UIMNH 30775-6; Ocotito, KU 60414-23; 5.4 km. N Ocotito, UMMZ 119181 (4); 1.6 km. N Organos, UIMNH 30752-63; Palo Blanco, CNHM 102283, 102286, 102305, 102320, 102404, UIMNH 30767-70; Pie de la Cuesta, AMNH 55275, 59202-5; Puerto Marquéz, AMNH 59200-1 (13); 5.6 km. S San Andreas de la Cruz, KU 87173-4; San Vincente, KU 87172; Zaculapán, UMMZ 119183.
Hidalgo: Below Tianguistengo, CNHM 102318.
Jalisco: Atenqueque, KU 91435-6; 5 km. NE Autlán, UIMNH 30810; 5 km. E Barro de Navidad, UMMZ 110900; Charco Hondo, UMMZ 95247; Puerto Vallarta, UIMNH 41346; between La Huerta and Tecomates, KU 91437; 3 km. SE La Resolana, KU 27619, 27620 (skeleton); 11 km. S, 1.6 km. E Yahualica, KU 29039; Zapotilitic, CNHM 102238.
Michoacán: Aguililla, UMMZ 119179 (5); Apatzingán, CNHM 38766-90, KU 69101 (skeleton); 7 km. E Apatzingán, UMMZ 112843; 11 km. E Apatzingán, UMMZ 112841 (3); 27 km. S Apatzingán, KU 37621-3; 1.6 km. N Arteaga, UMMZ 119180; Charapendo, UMMZ 112840; Coahuayana, UMMZ 104458; El Sabino, CNHM 102205-6, 102210-1, 102220, 102228, 102233, UIMNH 30822-3; La Placita, UMMZ 104456; La Playa, UMMZ 105163; 30 km. E Nueva Italia, UMMZ 120255 (2); 4 km. S Nueva Italia, UMMZ 112842; Ostula, UMMZ 104457 (4); Salitre de Estopilas, UMMZ 104459; San José de la Montaña, UMMZ 104461 (2); 11 km. S Tumbiscatio, KU 37626; 12 km. S Tzitzio, UMMZ 119178.
Morelos: 3.5 km. W Cuautlixco, KU 87188-90; 1 km. NE Puente de Ixtla, KU 60393-4; 20 km. S Puente de Ixtla, CNHM 102400, UIMNH 30832; Tequesquitengo, AMNH 52036-9.
Nayarit: 3 km. S Acaponeta, UMMZ 123030 (4); 56 km. S Esquinapa (Sinaloa), KU 73909; Jesús María, AMNH 58239; San Blas, KU 28087, 37624, 62360-2, USNM 51408; 8.6 km. E San Blas, UMMZ 115185; Tepic, UIMNH 30812-5; 4 km. E Tuxpan, KU 67786; 11 km. SE Tuxpan, UIMNH 7329-31, 7335-59.
Nuevo León: Galeana, CNHM 34389; Salto de Cola de Caballo, CNHM 30628-31, 30632 (40), 30633-7, 34454-67.
Oaxaca: 11 km. S Candelaria, UIMNH 9515-8; Cerro San Pedro, 24 km. SW Tehuantepec, UMMZ 82156; Chachalapa, KU 38199; 8 km. S Chiltepec, KU 87191; 12 km. S Chivela, UMMZ 115182; Coyul, USNM 114512; Garza Mora, UIMNH 40967-8; Juchatengo, KU 87193; 17 km. NE Juchatengo, KU 87645 (tadpoles), 87646 (young); Juchitán, USNM 70400; Lagartero, UIMNH 9514; Matías Romero, AMNH 52143-5; 25 km. N Matías Romero, KU 33822-8; 7 km. S Matías Romero, UIMNH 42703; Mirador, AMNH 6277, 13832-9, 13842-55; Mira León, 1.6 km. N Huatulco, UIMNH 9503-4; Mixtequillo, AMNH 13924; Pochutla, KU 57167-81, UIMNH 9505-13; Quiengola, AMNH 51817, 52146; Río del Corte, UIMNH 48677; Río Mono Blanco, UIMNH 36831; Río Sarabia, 5 km. N Sarabia, UMMZ 115180 (4); 2.5 km. N Salina Cruz, KU 57165-6; San Antonio, UIMNH 37286; 5 km. NNW San Gabriel Mixtepec, KU 87192; San Pedro del Istmo, UIMNH 37197; Santo Domingo, USNM 47120-2; 3.7 km. N Sarabia, UMMZ 115181 (3); Tapanatepec, KU 37793 (skeleton), 37794, UIMNH 9542, UMMZ 115183; between Tapanatepec [Pg 300] and Zanatepec, UIMNH 42704-25; Tecuane, UMMZ 82163 (3); Tehuantepec, AMNH 52625, 52639, 53470, UMMZ 82157-8, 82159 (9), 82160 (4), 82161 (8), 82162 (12), 112844-5, 118703, USNM 10016, 30171-4, 30188; 4.5 km. W Tehuantepec, KU 59801-12 (skeletons), 69102-3 (skeletons); 10 km. S Tehuantepec, KU 57163-4; Temazcal, USC 8243 (3); 3 km. S Tolocita, KU 39666-9; Tolosa, AMNH 53605; Tuxtepec, UMMZ 122098 (2); 2 km. S Valle Nacional, KU 87194-5; 11 km. N Vista Hermosa, KU 87196, 87639-41 (tadpoles), 87642-3 (young), 87644 (tadpoles); Yetla, KU 87197.
Puebla: 16 km. SW Mecatepec (Veracruz), UIMNH 3657-8; San Diego, AMNH 57714, USNM 114511; Vegas de Suchil, AMNH 57712; Villa Juárez, UF 11205.
Quintana Roo: Cóba, CNHM 26937; Esmeralda, UMMZ 113551; 4 km. NNE Felipe Carrillo Puerto, KU 71417-8; Pueblo Nuevo X-Can, KU 71405; 10 km. ENE Pueblo Nuevo X-Can, KU 71406; 4 km. WSW Puerto Juárez, KU 71407-11, 71721 (tadpoles); 12 km. W Puerto Juárez, KU 71412-6; San Miguel, Isla de Cozumel, UMMZ 78542 (6), 78543 (10), 78544 (2); 3.5 km. N San Miguel, Isla de Cozumel, KU 71419-22; 10 km. E San Miguel, Isla de Cozumel, UMMZ 78541; Telantunich, CNHM 26950.
San Luis Potosí: Ciudad Valles, AMNH 57776-81 (12), CNHM 37193, 102297, KU 23705; 21 km. N Ciudad Valles, UMMZ 118377; 6 km. E Ciudad Valles, UF 3524; 24 km. E Ciudad Valles, UF 7340 (2); 5 km. S Ciudad Valles, UIMNH 30751; 16 km. S Ciudad Valles, AMNH 52953; 30 km. S Ciudad Valles, CNHM 102394, 102402, 102412, UIMNH 30749-50; 63 km. S Ciudad Valles, UIMNH 19247-58; Pujal, UMMZ 99872 (2); Río Axtla, near Axtla, AMNH 53211-5, 59516, KU 23706; Tamazunchale, AMNH 52675, CNHM 39621-2, 102226, 102281, UF 7615 (2), UIMNH 26596, UMMZ 99506 (9), 118701 (2), USNM 114468; 17 km. N Tamazunchale, UIMNH 3659; 2.4 km. S Tamazunchale, AMNH 57743; 17 km. E Tamuin, UF 11202 (2); Xilitla, UIMNH 19259-60.
Sinaloa: 8 km. N. Carrizalejo, KU 78133; 4 km. NE Concordia, KU 73914; 5 km. SW Concordia, KU 75438-9; 6 km. E Cosalá, KU 73910; Costa Rica, 16 km. S. Culiacán, UIMNH 34887-9; 51 km. SSE Culiacán, KU 37792; El Dorado, KU 60392; 1.6 km. NE El Fuerte, CNHM 71468; Isla Palmito del Verde, middle, KU 73916-7; 21 km. NNE Los Mochis, UIMNH 40536-7; Matatán, KU 73913; 7.3 km. SW Matatán, KU 78464, 78466-70; Mazatlán, AMNH 12562, UMMZ 115197 (3); 57 km. N Mazatlán, UIMNH 38364; Plomosas, USNM 47439-40; Presidio, UIMNH 30811, USNM 14082; Rosario, KU 73911-2; 5 km. E Rosario, UIMNH 7360-76; 8 km. SSE Rosario, KU 37625; 5 km. SW San Ignacio, KU 78465; 1.6 km. ENE San Lorenzo, KU 47917-24; Teacapán, Isla Palmito del Verde, KU 73915; 9.6 km. NNW Teacapán, KU 91410; Villa Unión, KU 78471; 9 km. NE Villa Unión, KU 75434-7; 1 km. W Villa Unión, AMNH 59284.
Sonora: Guiracoba, AMNH 51225-38 (25).
Tabasco: 4 km. NE Comalcalco, AMNH 60313; Teapa, UMMZ 119943; 5 km. N Teapa, UMMZ 119940, 119944, 122997 (2); 10 km. N Teapa, UMMZ 119187, 119188 (2); 13 km. N Teapa, UMMZ 119941 (2), 119945 (3), 120254 (2); 21 km. N Teapa, UMMZ 119942, 119947; 29 km. N Teapa, UMMZ 119946 (11); Tenosique, USNM 114505-7.
Tamaulipas: Acuña, UMMZ 99864; 5 km. S Acuña, UMMZ 101180; 13 km. N Antiguo Morelos, UIMNH 40532-5; 3 km. S Antiguo Morelos, UF 11204; 3 km. NE Chamal, UMMZ 102867; 1.6 km. E Chamal, UMMZ 110734; Ciudad Mante, UMMZ 80957, 80958 (3), 106400 (3); 16 km. N Ciudad Victoria, CNHM 102408; 34 km. NE Ciudad Victoria, KU 60395-411; 8.8 km. S Ciudad Victoria, UIMNH 19261-3; 11 km. W Ciudad Victoria, UIMNH 30924; 16 km. W Ciudad Victoria, UIMNH 30825; 3 km. W El Carizo, UMMZ 111279; Gómez Farías, UMMZ 110837-8; 8 km. NE Gómez Farías, UMMZ 102265, 102916 (4), 102917, 104110 (5), 105493, 110836 (2), 111274-7; 8 km. NW Gómez Farías, UMMZ 101178 (7), 101179 (3), 101362-3, 101364 (2), 108799 (2), 110129, 111278, 111280; 8 km. W Gómez Farías, UMMZ 102859 (2); 16 km. W Gonzales, KU 37795-6; Jiménez, KU 60412; La Clementina, 6 km. [Pg 301] W Forlan, USNM 106244; Limón, UIMNH 30831; Llera, USNM 140137-40; 3 km. E Llera, UIMNH 16858; 21 km. S Llera, UIMNH 30828-9; 23 km. S Llera, UIMNH 30830; 11 km. SW Ocampo, UMMZ 118956; 22 km. W, 5 km. S Piedra, KU 37568-71; Rio Sabinas, UMMZ 97976; 5 km. W San Gerardo, UMMZ 110733 (2); Santa Barbara, UMMZ 111272-3; Villagrán, CNHM 102280, 102287, 102299, 102309, UIMNH 30826-7; 1.7 km. W Xicotencatl, UMMZ 115179.
Veracruz: 1.6 km. NW Acayucan, UMMZ 115189; 28.5 km. SE Alvarado, UMMZ 119933; 2.4 km. SSW Amatitlán, UMMZ 115195; Barranca Metlac, UIMNH 38365; Boca del Río, UIMNH 26619-30, UMMZ 74954 (9); 16 km. S Boca del Río, UIMNH 26631; between Boca del Río and Anton Lizardo, UIMNH 42701; Canadá, CNHM 102397; Catemaco, UMMZ 118702 (4); Ciudad Alemán, UMMZ 119608 (3); Córdoba, CNHM 38665-7, USNM 30410-3; 5.2 km. ESE Córdoba, KU 71423-35, 89924 (skeleton); 7 km. ESE Córdoba, UMMZ 115176 (4); Cosamaloapan, UMMZ 115193 (2); Coyame, UIMNH 36853-6, 38366, UMMZ 111461 (3), 111462-3; 1 km. SE Coyame, UMMZ 121202 (3); Cuatotolapam, UMMZ 41625-39; Cuautlapan, CNHM 38664, 70591-600, 102218, 102398, KU 26300, 26302, 26309, 26312-3, 26315-6, 26321, 26336, 26339, 26347 (skeleton), 26354, 55614-21 (skeletons), UIMNH 11236-67, 11269-71, 11273, 26611-8, 30792-5, UMMZ 85466 (6), 115173 (25), 115175 (7), USNM 114433-57; Dos Ríos, CNHM 39623; 5 km. ENE El Jobo, KU 23843, 23845, 23847; 6.2 km. E Encero, UIMNH 30835; Escamilo, CNHM 102298, UIMNH 30788; 1 km. N Fortín, UF 11201; 4 km. SW Huatusco, UMMZ 115177; 1 km. SW Huatusco, UMMZ 123119; 10 km. SE Hueyapan, UMMZ 115190; 20 km. S Jesús Carranza, KU 23844, 23846, 27399; 38 km. SE Jesús Carranza, KU 23417; Laguna Catemaco, UMMZ 119932 (62); 1.6 km. N La Laja, UIMNH 3651; La Oaxaqueña, AMNH 43930-1; 17 km. E Martínez de la Torre, UIMNH 36630-2; 6.2 km. W Martínez de la Torre, UIMNH 3652-4; Minatitlán, AMNH 52141-2; Mirador, USNM 25097-8, 115178; 6 km. S Monte Blanco, UF 11200 (4); 21 km. E Nanchital, UMMZ 123004; 2 km. S Naranja, UMMZ 115188 (3); 1.6 km. NE Novillero, UMMZ 115194 (2); 3 km. NE Novillero, UMMZ 115196; 5.2 km. NE Novillero, UMMZ 115192 (4); 6 km. NE Novillero, UMMZ 115191; 5 km. N Nuevo Colonia, UMMZ 105066; Orizaba, USNM 16563-6; 4 km. NE Orizaba, UMMZ 120251 (2); Panuco, UMMZ 118922; Paraje Nuevo, UMMZ 85465 (2), 85467 (35), 85468 (36); Paso del Macho, UIMNH 49281; Paso de Talaya, Jicaltepec, USNM 32365, 84420; Pérez, CNHM 1686 (5); 20 km. N Piedras Negras, Río Blanco, KU 23708; Plan del Río, KU 26310, 26333-5, 26345, 26354, UMMZ 102069, 102070 (5); Potrero, UIMNH 49282-5, UMMZ 88799, 88805, 88806 (2), USNM 32391-5; Potrero Viejo, CNHM 102296, KU 26301, 26304-5, 26307-8, 26311, 26317-20, 26323-25, 26326-8 (skeletons), 26329-31, 26332 (skeleton), 26337-8, 26340-4, 26346, 26348, 26351, 26353, 27400-12, UIMNH 30800, UMMZ 88800 (2), 88802 (15), 88803 (9), 88804, USNM 114458-67; 5 km. S Potrero Viejo, KU 26303, 26314, 26322; Puente Nacional, UIMNH 21783-8; 3 km. N Rinconada, UMMZ 122099 (5); Río de las Playas, USNM 118635-6; Río Seco, UMMZ 88801 (9); Rodriguez Clara, CNHM 102225; San Andrés Tuxtla, CNHM 102213, 102222, 102227, 102247, UIMNH 30789-91; 10 km. NW San Andrés Tuxtla, UMMZ 119935; 13.4 km. NW San Andrés Tuxtla, UMMZ 119939 (2); 19.8 km. NW San Andrés Tuxtla, UMMZ 119938; 27.2 km. NW San Andrés Tuxtla, UMMZ 119936 (6); 48 km. NW San Andrés Tuxtla, UMMZ 119937; 4 km. W San Andrés Tuxtla, UMMZ 115187; 37.4 km. S San Andrés Tuxtla, UMMZ 119934 (12); 15 km. ESE San Juan de la Punta, KU 23707; San Lorenzo, USNM 123508-12; 3 km. SW San Marcias KU 23841; 1.5 km. S Santa Rosa, UIMNH 42702; 2 km. S Santiago Tuxtla, UMMZ 121201 (4); Sauzel, UMMZ 121239; 14 km. E Suchil, UIMNH 46880; 15 km. S Tampico (Tamaulipas), UMMZ 103322 (4); 4 km. N Tapalapan, UMMZ 115186 (2); Tecolutla, UIMNH 42677-700; 16 km. NW Tehuatlán, UIMNH 3660-3; 5 km. S Tehuatlán, KU 23842; Teocelo, KU 26306; Tierra Colorado, CNHM 102393, 102395-6, UIMNH 30789-91; Veracruz, AMNH 6301-4, 59398-402, UIMNH 30801, UMMZ 115174, 122060 (2); 24 km. W Veracruz, CNHM 104570-2.
[Pg 302] Yucatán: No specific locality, CNHM 548, 49067, USNM 32298; Chichén-Itzá, CNHM 20636, 26938-49, 36559-62, UIMNH 30742-6, UMMZ 73173 (6), 73174 (14), 73175 (14), 73178-9, 76171, 83107 (2), 83108, 83109 (2), 83915 (30), USNM 72744; 9 km. E Chichén-Itzá, KU 71438-9; 12 km. E Chichén-Itzá, KU 71440; Mérida, CNHM 40659-66, UIMNH 30747-8, UMMZ 73182; 6 km. S Mérida, KU 75194; 8.8 km. SE Ticul, UMMZ 114296; Valladolid, CNHM 26934-6; Xcalah-op, CNHM 53906-14; 3.5 km. E Yokdzonot, KU 71441-3, 71720 (tadpoles).
British Honduras: Belize, CNHM 4153, 4384-5, 4387, UMMZ 75310, USNM 26065; Bokowina, CNHM 49064-5; Cocquercote, UMMZ 75331 (2); Cohune Ridge, UMMZ 80738 (15); Double Falls, CNHM 49066; El Cayo, UMMZ 75311; 6 km. S El Cayo, MCZ 37856; Gallon Jug, MCZ 37848-55; Manatee, CNHM 4264-7; Mountain Pine Ridge, MCZ 37857-8; San Augustin, UMMZ 80739; San Pedro, Columbia, MCZ 37860-2; Valentin, UMMZ 80735 (4), 80736 (2), 80737 (2); 5 km. S Waha Loaf Creek, MCZ 37859.
Guatemala: Alta Verapaz: 5.1 km. NE Campur, KU 68464 (tadpoles), 67465 (young); 28.3 km. NE Campur, KU 64203-22, 68183-4 (skeletons); Chamá, MCZ 15792-3, UMMZ 90895 (7), 90896 (5), 90897 (29), 90898 (12), 90899; Chinajá, KU 55939-41, 57193-8, 60018-20 (tadpoles), 60021 (eggs), 60022 (tadpoles); Cobán, CNHM 21006; Cubilquitz, UMMZ 90902 (10); Finca Canihor, UMMZ 90908; Finca Chicoyou, KU 57246-8, 60026 (young), 64202, 68466-7 (tadpoles); Finca Los Alpes, KU 64197-201, 68463 (tadpoles); Finca Los Pinales, UMMZ 90903 (2); Finca Tinajas, BYU 16031; Finca Volcán, UMMZ 90905 (4), 90906-7; Panzos, MNHN 6313, UMMZ 90904; Samac, UMMZ 90900; Samanzana, UMMZ 90901 (6).
Baja Verapaz: Chejel, UMMZ 90909 (7), 90910 (3); San Gerónimo, UMMZ 84076 (16).
Chiquimula: 1.6 km. SE Chiquimula, UMMZ 98112; Esquipulas, UMMZ 106793 (28).
El Petén: 20 km. NNW Chinajá (Alta Verapaz), KU 57199-240; Flores, UMMZ 117985; La Libertad, KU 60024 (young), UMMZ 75313-20, 75323 (2), 75324 (7), 75325 (13), 75326 (2), 75327 (11), 75328 (12), 75329 (2); 3 km. SE La Libertad, KU 57243-4; 13 km. S La Libertad, MCZ 21458 (2); Pacomon, USNM 71334; Piedras Negras, USNM 114469-71; Poptún, UMMZ 120475; Poza de la Jicotea, USNM 114672; Ramate-Yaxha trail, UMMZ 75321; Río de la Pasión between Sayaxché and Subín, KU 57151; Río San Román, 16 km. NNW Chinajá (Alta Verapaz), KU 55942-6; Sacluc, USNM 25131; Sayaxché, KU 57144-5; Tikal, UMMZ 117983 (7), 117984 (5), 117993 (5), 120474 (5); Toocog, KU 57241-2, 60023 (young), 60025 (young); Uaxactún, UMMZ 70401-3; Yaxha, UMMZ 75322; 19 km. E Yaxha, UMMZ 75330 (4).
El Quiché: Finca Tesoro, UMMZ 89166 (3), 90549 (tadpoles).
Escuintla: Río Guacalate, Masagua, USNM 125239; Tiquisate, UMMZ 98262 (7).
Guatemala: 16 km. NE Guatemala, KU 43545-53.
Huehuetenango: Finca San Rafael, 16 km. SE Barillas, CNHM 40912-6; 45 km. WNW Huehuetenango, KU 64223-4; Jacaltenango, UMMZ 120080 (6), 120081 (14), 120082 (13).
Izabál: 2 km. SW Puerto Matías de Gálvez, KU 60027-8 (tadpoles); Quiriguá, CNHM 20587, UMMZ 70060.
Jalapa: Jalapa, UMMZ 98109, 106792 (11).
Jutiapa: Finca La Trinidad, UMMZ 107728 (10); Jutiapa, UMMZ 106789; 1.6 km. SE Mongoy, KU 43069; Santa Catarina Mita, UMMZ 106790.
Progreso: Finca Los Leones, UMMZ 106791.
Quetzaltenango: Coatepeque, AMNH 62204.
Retalhueleu: Casa Blanca, UMMZ 107725 (18); Champerico, UMMZ 107726 (3).
San Marcos: Talisman Bridge, USNM 128056-7.
[Pg 303] Santa Rosa: Finca La Guardiana, UMMZ 107727 (6); Finca La Gloria, UMMZ 107724 (6); 1.6 km. WSW El Molino, KU 43065-8.
El Salvador: La Libertad: 16 km. NW Santa Tecla, KU 43542-4. Morazán: Divisadero, USNM 73284. San Salvador: San Salvador, CNHM 65087-99, KU 61955-88, 62138-9 (skeletons), 62154 (eggs), 62155-60 (tadpoles), 68462 (tadpoles), UMMZ 117586 (3), 118380 (3), USNM 140278.
Honduras: State unknown: Guaimas, UMMZ 58391. Atlantidad: Isla de Roatán, CNHM 34551-4; La Ceiba, USNM 64985, 117589-91; Lancetilla, MCZ 16207-11; Tela, MCZ 15774-5, 28080, UMMZ 58418, USNM 82173-4. Choluteca: 1.5 km. NW Choluteca, KU 64228-32; 10 km. NW Choluteca, KU 64233; 10 km. E Choluteca, KU 64226-7; 12 km. E Choluteca, KU 64225; 5 km. S Choluteca, USC 2700 (2). Colón: Bambú, UF 320; Belfate, AMNH 45692-5; Patuca, USNM 20261. Comayagua: La Misión, 3.5 leagues N Siguatepeque, MCZ 26424-5. Copán: Copán, UMMZ 83026 (2). Cortés: Cofradía, AMNH 45345-6; Hacienda Santa Ana, CNHM 4724-31; Lago de Yojoa, MCZ 26410-1; Río Lindo, AMNH 54972. El Paraiso: El Volcán, MCZ 26436. Francisco Morazán: Tegucigalapa, BYU 18301-4, 18837-41, MCZ 26395-7, USNM 60499. Gracias A Dios: Río Segovia, MCZ 24543. Santa Barbara: Santa Barbara, USNM 128062-5.
Nicaragua: Chinandega: 4 km. N, 2 km. W Chichigalpa, KU 85385; Chinandega, MCZ 2632; Río Tama, USNM 40022; San Antonio, KU 84944-9 (skeletons), 85386-403. Chontales: 1 km. NE Acoyapa, KU 64234. Estelí: Finca Daraili, 5 km. N, 15 km. E Condega, KU 85404-8; Finca Venecia, 7 km. N, 16 km. E Condega, KU 85409. León: 1.6 km. ENE Poneloya, KU 43084-5. Managua: Managua, USNM 79989-90; 8 km. NW Managua, KU 43094-110; 20 km. NE Managua, KU 85412; 21 km. NE Managua, KU 85413-4; 5 km. SW Managua, KU 43086-93; 2 km. N Sabana Grande, KU 85411; 3 km. N Sabana Grande, KU 43070-8; 20 km. S, 0.5 km. W Tipitapa, KU 85410. Matagalpa: Guasqualie, UMMZ 116493; Matagalpa, UMMZ 116492; 19 km. N Matagalpa, UMMZ 116494. Río San Juan: Greytown, USNM 19585-6, 19767-8. Rivas: Javillo, UMMZ 123001; Moyogalpa, Isla Ometepe, KU 85428-37, 87706 (tadpoles); Peñas Blancas, KU 85417; Río Javillo, 3 km. N, 4 km. W Sapoá, KU 85418-20, 85438 (skeleton); 13.1 km. SE Rivas, KU 85415; 14.8 km. SE Rivas, KU 85421-3; 11 km. S, 3 km. E Rivas, KU 85416; 16 km. S Rivas, MCZ 29009-10; 7.7 km. NE San Juan del Sur, KU 85426-7; 16.5 km. NE San Juan del Sur, KU 85424-5, 87705 (young); 5 km. SE San Pablo, KU 43079-83. Zelaya: Cooley, AMNM 7063-8, 8019-20, 8022, 8034-5; Cukra, AMNH 8016-7; Musahuas, Río Huaspuc, AMNH 58428-31; 11 km. NW Rama, Río Siquia, UMMZ 79708, 79709 (5), 79710 (2); Río Escondido, USNM 19766, 20701; Río Siquia at Río Mico, UMMZ 79707 (10); Sioux Plantation, AMNH 7058-61, 8023-33.
Costa Rica: Alajuela: Los Chiles, AMNH 54639; Orotina, MCZ 7960-1; San Carlos, USNM 29991. Guanacaste: La Cruz, USC 8232 (3); 4.3 km. NE La Cruz, UMMZ 123002; 18.4 km. S La Cruz, USC 8136; 23.5 km. S La Cruz, USC 8094 (4); 3 km. W La Cruz, USC 8233 (4); 2 km. NE Las Cañas, KU 64235-7; Las Huecas, UMMZ 71212-3; Liberia, KU 36787, USC 8161; 11.5 km. N Liberia, USC 8149; 13 km. N Liberia, USC 8139; 22.4 km. N Liberia, USC 8126; 8 km. NNW Liberia, KU 64238; 8.6 km. ESE Playa del Coco, USC 8137; 21.8 km. ESE Playa del Coco, USC 8138; Río Piedra, 1.6 km. W Bagaces, USC 7027; Río Bebedero, 5 km. S Bebedero, KU 64158; 5 km. NE Tilarán, KU 36782-6. Heredia: 13 km. SW Puerto Viejo, KU 64142-6. Limón: Batán, KU 34927; Guacimo, USC 621; Pandora, USC 505 (3); Suretka, KU 36788-9; Tortugero, UF 7697, 10540-2. Puntarenas: Barranca, CNHM 35254-6; 15 km. WNW Barranca, KU 64155-7, UMMZ 118381; 18 km. WNW Barranca, UMMZ 118382 (4); 4 km. WNW Esparta, KU 64159-96, 68178-82 (skeletons); 19 km. NW Esparta, KU 64147-54.
Diagnosis.—Size moderately large ([M] 56.0 mm., [F] 70.0 mm.); skull as long as wide; frontoparietal fontanelle large; narrow supraorbital flanges having irregular margins anteriorly; large squamosal not in contact with maxillary; tarsal fold moderately wide, full length of tarsus; inner metatarsal tubercle moderately large, low, flat, elliptical; hind limbs relatively long; tibia usually more than 52 per cent of snout-vent length; labial stripe silvery-white; lips lacking vertical bars; loreal region pale green; pale bronze-colored stripe from nostril along edge of eyelid to point above tympanum narrow, bordered below by narrow dark brown stripe from nostril to eye, and broad dark brown postorbital mark encompassing tympanum and terminating above insertion of arm; flanks, dark brown with large pale blue spots; anterior and posterior surfaces of thighs dark brown with small pale blue spots on thighs. (Foregoing combination of characters distinguishing S. cyanosticta from any other species in genus.)
Description and Variation.—The largest males are from Piedras Negras, El Petén, Guatemala, and they average 52.5 mm. in snout-vent length whereas males from Los Tuxtlas, Veracruz, average 50.6 mm. and those from northern Oaxaca 50.3 mm. The smallest breeding male has a snout-vent length of 44.6 mm. The average ratio of tibia length to snout-vent length is 54.8 per cent in males from Piedras Negras, and 56.4 and 56.3 per cent in males from Los Tuxtlas and Oaxaca, respectively. The only other character showing noticeable geographic variation is the size of the tympanum. The average ratio of the diameter of the tympanum to the diameter of the eye is 76.3 per cent in males from Piedras Negras, 71.8 from Oaxaca, and 66.9 from Los Tuxtlas.
The dorsal ground color of Smilisca cyanosticta is pale green to tan and the venter is creamy white. The dorsum is variously marked with dark olive-green or dark brown spots or blotches (Pl. 6B). An interorbital dark bar usually is present. The most extensive dark area is a V-shaped mark in the occipital region with the anterior branches not reaching the eyelids; this mark is continuous, by means of a narrow mid-dorsal mark, with an inverted V-shaped mark in the sacral region. In many specimens this dorsal marking is interrupted, resulting in irregular spots. In some specimens the dorsum is nearly uniform pale green or tan with a few small, dark spots. The hind limbs are marked by dark transverse bands, usually three or four each on the thigh and shank, and two or three on the tarsus. The webbing on the feet is brown. The loreal region is pale green, bordered above by a narrow, dark brown canthal stripe extending from the nostril to the orbit, which is bordered above by a narrow, bronze-colored stripe extending from the nostril along the edge of the eyelid to a point above the tympanum. The upper lip is white. A broad dark brown mark extends posteriorly from the orbit and encompasses the tympanum to a point above the insertion of the forelimb. The flanks are dark brown with many pale blue, rounded spots, giving the impression of a pale [Pg 305] blue ground color with dark brown mottling enclosing spots. The anterior and posterior surfaces of the thighs are dark brown with many small pale blue spots. The inner surfaces of the shank and tarsus are colored like the posterior surfaces of the thighs. Pale blue spots are usually present on the proximal segments of the second and third toes. A distinct white stripe is present on the outer edge of the tarsus and fifth toe; on the tarsus the white stripe is bordered below by dark brown. A white stripe also is present on the outer edge of the forearm and fourth finger. The anal region is dark brown, bordered above by a narrow transverse white stripe. The throat in breeding males is dark, grayish brown with white flecks.
No geographic variation in the dorsal coloration is evident. Specimens from the eastern part of the range (Piedras Negras and Chinajá, Guatemala) have bold, dark reticulations on the flanks enclosing large pale blue or pale green spots, which fade to tan in preservative. Specimens from Oaxaca and Veracruz characteristically have finer dark reticulations on the flanks enclosing smaller blue spots; in many of these specimens the ventrolateral spots are smallest and are white.
All living adults are easily recognized by the presence of pale, usually blue, spots on the flanks and thighs. Individuals under cover by day have a tan dorsum with dark brown markings. A hiding individual at Chinajá, Alta Verapaz, Guatemala (KU 55936), had a pale tan dorsum when found; later the dorsal color changed to chocolate brown. A pale green patch was present below the eye; the spots on the posterior surfaces of the thighs were pale blue, and those on the flanks were yellowish green. A calling male obtained 10 kilometers north-northwest of Chinajá (KU 55934) was reddish brown when found at night; later the dorsal color changed to pale tan. A green patch below the eye was persistent. Usually these frogs are green at night. The coloration of an adult male (KU 87201) from 11 kilometers north of Vista Hermosa, Oaxaca, México, was typical: "When calling dorsum pale green; later changed to dull olive-green. Flanks dark brown with pale blue spots in axilla and groin and bluish white flecks on mid-flank. Anterior and posterior surfaces of thighs, inner surfaces of shanks, and median dorsal surfaces of tarsi dark brown with blue spots. Canthal and postorbital stripes dark chocolate brown; labial stripe creamy white. Forearm, tarsal, and anal stripes pale cream-color. Throat dark brown with yellow flecks; belly and ventral surfaces of limbs creamy buff; webs pinkish tan; iris deep bronze, brown below pupil." (Duellman, field notes, June 24, 1964.)
Some individuals have both green and brown coloration in life. An individual obtained at night on the south slope of Volcán San Martin, Veracruz, México, had a pale tan dorsum changing peripherally to pale green. The dorsal markings were dark brown and dark olive-green.
In contrast to the color-changes noted above, the labial region below the eye is always pale green, and pale spots are always present on the flanks and thighs in adults. The iris is invariably golden or bronze above and darker, usually brown, below. Minute black flecks are present on the iris, and in some individuals these flecks are so numerous that the eye appears gray.
Recently metamorphosed young have pale tan flanks, and the posterior surfaces of the thighs are orange-yellow; pale spots are absent. A juvenile (KU 55935) from Chinajá, Alta Verapaz, Guatemala, having a snout-vent length of 35.0 mm. was pale yellowish tan above with olive-green markings; the flanks were dark brown with pale blue spots, and the anterior and posterior [Pg 306] surfaces of the thighs were uniform bright tomato red. A juvenile (UMMZ 121298), 18.6 mm. in snout-vent length, from the southeast slope of Volcán San Martín, Veracruz, México, had pale tan flanks lacking blue spots, but had red thighs. Apparently the ontogenetic changes in coloration proceed as follows: (1) flanks pale tan and thighs orange-yellow, both lacking spots, (2) flanks pale tan and thighs red, both lacking spots, (3) flanks dark brown with blue spots and thighs red, lacking spots, and (4) flanks and thighs dark brown, both having pale blue spots.
Natural History.—Smilisca cyanosticta inhabits humid tropical forest and cloud forest from the lowlands to elevations of about 1200 meters in Los Tuxtlas and to about 900 meters in northern Oaxaca. In these moist environments the frogs apparently are active throughout the year. Active individuals have been obtained in January, July, and August in Los Tuxtlas, Veracruz, in June and July in northern Oaxaca, in February and March at Chinajá, Guatemala, and Taylor and Smith reported (1945:589) activity in May at Piedras Negras, Guatemala. Calling males were observed as follows; in a rain barrel 11 kilometers north of Vista Hermosa, Oaxaca, México, on June 23-28, 1964; in a quiet pool in a stream 8 kilometers south of Yetla, Oaxaca, México, in July, 1963 (Dale L. Hoyt, personal communication); in and near springs flowing into a stream at Dos Amates, Veracruz, México, on August 4, 1959 (Douglas Robinson, personal communication); and in a water-filled depression in a log 10 kilometers west-northwest of Chinajá, Guatemala, on March 13, 1960. Taylor and Smith (1945:589) reported that individuals were found at night on the ground at the edge of temporary pools at Piedras Negras, Guatemala, on May 28-29, 1939. A clasping pair was found on a rock at the edge of a small stream on the south slope of Volcán San Martín, Veracruz, México, on July 11, 1959 (Douglas Robinson, personal communication).
Only one individual has been observed in a tree at night. In the daytime, individuals were found in elephant ear plants (Xanthosoma) at Chinajá, Guatemala.
The breeding call consists of one or two moderately short notes that are lower pitched than those of S. baudini, but higher pitched than those of S. phaeota. Each note has a duration of 0.25 to 0.45 seconds and is repeated at intervals of one-half minute to several minutes. Each note is a vibrant "waunk," having 110 to 180 pulses per second and dominant frequency of 1600 to 2100 cycles per second (Pl. 10B).
Apparently the eggs are deposited as loose clumps in the water. In eggs in the yolk plug stage of development, the diameter of the embryo is about 2.3 mm.; that of the outer envelope is 4.0 mm. Hatchling tadpoles have total lengths of 5.8 to 6.5 mm. and body lengths of 2.8 to 3.1 mm. The external gills are moderately long, slender, and filamentous; the yolk sac is still moderately large. The body and anterior part of the caudal musculature are dark brown; posteriorly the caudal musculature is pale brown. The caudal fins are creamy tan. The oral discs are large and ovoid. The growth of the tadpole is summarized in Table 10.
A typical tadpole in stage 30 of development (KU 87652 from 11 km. N Vista Hermosa, Oaxaca, México) can be described as follows:
Body length 9.5 mm.; tail length 15.5 mm.; total length 25.0 mm.; body slightly wider than deep; snout rounded laterally, broadly ovoid dorsally; eyes widely separated, directed dorsolaterally; nostril about midway between eye [Pg 307] and tip of snout; mouth anteroventral; spiracle sinistral, slightly posterior to midpoint of body and slightly below midline; anal tube dextral; caudal musculature slender, barely curved upward distally; dorsal fin not extending onto body, depth of dorsal fin slightly more than that of ventral fin on mid-length of tail; dorsal part of body dark brown; ventral surfaces transparent, lacking pigment; posterior edge of body pale cream-color; caudal musculature creamy white with interconnected brown spots; caudal fins transparent with small brown blotches on dorsal fin and posterior half of ventral fin; iris coppery bronze in life (Fig. 12). Mouth small, median part of upper lip bare; rest of mouth bordered by single row of bluntly rounded papillae; lateral fold present; tooth rows 2/3; all tooth-rows approximately equal in length; second upper row broadly interrupted medially; other rows complete; upper beak moderately deep, forming broad arch with slender lateral processes; lower beak slender, broadly V-shaped; both beaks finely serrate (Fig. 15C).
All tadpoles having fully developed mouth parts have 2/3 tooth rows. Little variation is noticeable in coloration. In many specimens the posterior edge of the body is dark brown instead of pale cream-color. Mottling is rather dense on the caudal fins in all specimens; in some individuals pigment is concentrated along the anterior one-third of the lateral groove. In life the body is dark brown with greenish gold flecks ventrally; the caudal musculature is gray.
In each of two recently metamorphosed young the snout-vent length is 14.0 mm. Coloration of young in life (KU 87653 from 11 km. N Vista Hermosa, Oaxaca, México): "Dorsum pale tan with dark brown markings. Thighs orange-yellow; labial stripe white; iris bronze" (Duellman, field notes, July 10, 1964.)
Remarks.—Smith (1953:150) named cyanosticta as a subspecies of Hyla phaeota. The differences in cranial characters and certain external characters between phaeota and cyanosticta indicate that they are distinct species. Furthermore, a gap of about 350 kilometers exists between the known geographic ranges of the two kinds.
Distribution.—Smilisca cyanosticta inhabits wet forests on the Atlantic slope of southern México and northern Central America from northern Oaxaca and southern Veracruz through northern Chiapas in México and into El Petén and northern Alta Verapaz in Guatemala (Fig. 2). Apparently the range is discontinuous, for in southern México the species is found in cloud forest at elevations of 830 to 900 meters on the northern slopes of the Sierra de Juárez. In the Sierra de Los Tuxtlas in southern Veracruz the species is found in wet forest at elevations of 300 to 1200 meters, but is absent in the intervening lowlands characterized by drier forest. In the west forests of northern Alta Verapaz and El Petén, Guatemala, the species is found at low elevations.
Specimens examined.—78, as follows: Mexico: Chiapas: Monte Libano, MCZ 28271-9; 8 km. N San Fernando, 24 km. NE Tuxtla Gutierrez, UIMNH 41588. Oaxaca: 11 km. N Vista Hermosa, KU 84918-20 (skeletons), 87198-212, 87647 (eggs), 87648-52 (tadpoles), 87653 (young), UIMNH 57199-201; 8 km. S Yetla, KU 87213, UMMZ 124838 (8). Veracruz: Coyame, UMMZ 111459-60; between Coyame and Tebanco, UMMZ 121198; Dos Amates, UMMZ 121297; between Laguna de Catemaco and Volcán San Martín, UMMZ 121200; Volcán San Martín, UIMNH 35403-4, 35408-12, UMMZ 118163; SE slope Volcán San Martín, UMMZ 121199, 121295 (2), 121296, 121298.
Guatemala: Alta Verapaz: Chinajá, KU 55935-7, 55938 (skeleton). El
[Pg 308] Petén: 10 km. NNW Chinajá (Alta Verapaz), KU 55934; Piedras Negras, CNHM 99006-7, 99227, UIMNH 28853, USNM 111139-41, 111143-7; 8 km. S Piedras Negras, CNHM 99008; Semicoch, USNM 35907.

Diagnosis.—Size large ([M] 65 mm., [F] 78 mm.); skull as long as wide, lacking frontoparietal fontanelle; large supraorbital flanges having straight edges and extending posterolaterally; large squamosal not in contact with maxillary; tarsal fold moderately wide, full length of tarsus; inner metatarsal tubercle moderately large, low, flat, elliptical; hind limbs relatively long, tibia averaging more than 53 per cent of snout-vent length; labial stripe silvery white; lips lacking vertical bars; loreal region pale green; dark brown or black tympanic mark dispersing into brown venated pattern on flanks; posterior surfaces of thighs pale brown, with or without darker flecks or small cream-colored flecks. (Foregoing combination of characters distinguishing S. phaeota from any other species in genus.)
| Table 2.—Geographic Variation in Size and Proportions in Males of Smilisca phaeota. (Means in Parentheses Below Observed Ranges; Data Based of Ten Specimens From Each Locality.) | |||
| Locality | Snout-vent length |
Head width/ snout-vent length |
Interorbital distance/ head width |
| Bonanza, Nicaragua | 40.8-47.7 | 34.1-38.0 | 31.0-36.1 |
| (43.7) | (36.3) | (35.4) | |
| Heredia Prov., Costa Rica | 46.3-59.0 | 32.5-36.0 | 30.5-39.6 |
| (51.7) | (35.0) | (34.7) | |
| Puntarenas Prov., Costa Rica | 53.6-64.9 | 32.6-36.1 | 31.0-38.0 |
| (61.4) | (34.5) | (34.4) | |
| Canal Zone, Panamá | 52.4-65.5 | 33.5-37.6 | 31.3-37.2 |
| (56.5) | (35.6) | (34.7) | |
| Río Quesada, Colombia | 52.6-61.0 | 33.1-37.1 | 30.1-33.9 |
| (56.0) | (35.0) | (32.1) | |
Description and Variation.—For the purposes of analyzing geographic variation in size and proportions, measurements were taken on ten adult males from each of five samples throughout the range of the species. Aside from the data summarized in Table 2, the average ratio of tibia length to snout-vent length is noticeably less in Colombian specimens (53.4 per cent, as compared with 54.8 to 57.8 per cent in the other samples) and the ratio of head length to [Pg 310] snout-vent length is noticeably less in Costa Rican specimens (33.5 per cent as compared with 34.9 to 35.1 per cent in the other samples). Also, specimens from Heredia Province, Costa Rica, have a relatively smaller tympanum (62.7 to 80.4 [mean 68.4] per cent of the diameter of the eye, as compared with means of 74.0 to 77.9 per cent in the other samples).
Two populations are distinctive as regards the size of adult males. Specimens from the northern Caribbean lowlands of Nicaragua (Bonanza, the northernmost locality for the species) are remarkably small. Males having snout-vent lengths of between 40 and 43 mm. were breeding; the largest male found had a snout-vent length of 47.7 mm. The other extreme in size is attained in specimens from the Pacific lowlands of eastern Costa Rica and western Panamá, where most breeding males have snout-vent lengths of more than 55 mm.; the largest male had a snout-vent length of 64.9 mm.
The rather striking differences in size among certain samples and the minor differences in proportions among other samples show no geographic trends. Instead, the variations apparently are random among the samples. The data presented here possibly are the results of inadequate sampling, but more likely reflect actual differences in the populations.
The dorsal ground color of Smilisca phaeota is pale green to tan; the venter is creamy white. The dorsum is variously marked with dark olive-green or dark brown spots or blotches (Pl. 6C). A dark interorbital bar is usually present. Usually a large dark dorsal mark extends from the occiput to the sacral region, but in many individuals this blotch is replaced by two or three dark marks. The dorsal markings are irregular in shape and do not tend to form transverse bands or longitudinal bars. The hind limbs are marked by dark transverse bands, usually four or five on the thigh, five or six on the shank, and four on the tarsus. Two or three narrow bands are usually present on the proximal part of the fourth toe. The webbing on the feet is brown. The loreal region is pale green, bordered above by a narrow dark brown canthal stripe extending from the nostril to the orbit. The upper lip is silvery white. A broad dark brown or black mark extends posteriorly from the orbit, encompassing the tympanum, to a point above the insertion of the forelimb. The flanks are pale green or pale tan and marked with a fine dark brown or black venation. The anterior surfaces of the thighs usually are pale brown or grayish tan, sometimes having small, indistinct darker flecks. The posterior surfaces of the thighs are similarly colored, but in most specimens small but distinct dark flecks are present; in some specimens small cream-colored spots are also present on the posterior surfaces of the thighs. A distinct, narrow creamy white anal stripe usually is present. A distinct white stripe is present on the outer edge of the tarsus and fifth toe; on the tarsus the white stripe is bordered below by dark brown. A white stripe also is present on the outer edge of the forearm and fourth finger. In breeding males the throat is dark gray.
Little geographic variation in color or pattern is evident. Few, if any, specimens from the Pacific lowlands of South America are green in life. (We have seen no living individuals from South America.) Some living individuals from Costa Rica and all those seen alive from Nicaragua have a tint of pale blue on the flanks. In some specimens the dorsal pattern is so faint as to be barely discernible, whereas in most specimens the pattern is bold.
The coloration in the living frogs is highly variable due to extreme metachrosis. Individuals of this species are capable of changing the dorsal coloration [Pg 311] from green to brown in a short period of time. Both green and brown individuals have been found active at night. Usually those individuals found hiding by day are brown. One individual from Finca La Sumbadora, Panamá (now KU 91914), was kept alive in the laboratory for nearly one month. This individual usually was pale green with tan dorsal markings at night and tan with pale green markings by day. On occasion the pale green dorsal markings were boldly outlined by bright dark green.
In living individuals from throughout the range of the species the iris is a bronze color, darkest medially with fine black reticulations.
Natural History.—Smilisca phaeota inhabits humid lowland tropical forest and seldom ascends the foothills to more than 1,000 meters. The rather equable climatic conditions, especially more or less evenly distributed rainfall throughout the year, permit this frog to be active most of the year. Dunn (1931:413) reported males calling on Barro Colorado Island, Panamá, in February and in July, and Breder (1946:416) noted calling individuals in the Chucanaque drainage of Darién, Panamá in January, March, July, August and October and in Costa Rica in April through August inclusively. Calling males were found at Bonanza, Nicaragua in March and in July.
At all times of year the usual daytime retreats for these frogs are near water; the frogs have been found in elephant ear plants (Xanthosoma) and in bromeliads; occasional individuals have been found sitting on shaded branches of bushes and trees. None has been observed on the ground or beneath ground-cover by day.
The length of the breeding season cannot be determined definitely. The earliest date on which eggs have been found is May 23; Gaige, Hartweg, and Stuart (1937:5) reported a gravid female taken at El Recreo, Nicaragua, in September, and we have a gravid female taken at Almirante, Panamá, in March.
Males usually call from secluded spots at the edge of water. All calling males that we observed were on the ground within a few centimeters of the water. The males usually are hidden beneath an overhanging leaf or some other cover; they definitely do not sit in the open like Smilisca baudini. Most calling males and clasping pairs have been found at the edges of small pools or shallow ditches, although occasional individuals are found at the edges of large ponds or streams.
The breeding call consists of one or two moderately short, low-pitched notes (duration 0.33 to 0.42 seconds), repeated at intervals of about 20 seconds to several minutes. Each note is a low, vibrant "wauk," having 100 to 130 pulses per second and a dominant frequency of 330 to 420 cycles per second (Pl. 10C).
The eggs are deposited in loose clumps amidst vegetation in the water. Hatchling tadpoles have total lengths of 8.7 to 10.6 mm., and body lengths of 4.1 to 4.5 mm. The external gills are long and filamentous, and the yolk sac is large. The head and caudal musculature are dark brownish black, and the caudal fins are gray. The oral discs are large and roughly circular. The growth and development of the tadpoles are summarized in table 11 and figure 16.
A typical tadpole in stage 30 of development (KU 68482 from the Río Chitaría, Cartago Province, Costa Rica) may be described as follows: body length 9.7 mm.; tail length 14.6 mm.; total length 24.3 mm.; body as wide as deep; snout rounded dorsally and laterally; eyes widely separated, directed [Pg 312] dorsolaterally; nostril about midway between eye and tip of snout; mouth anteroventral; spiracle sinistral, about midway on length of body and slightly below midline; anal tube dextral; caudal musculature slender, curved upward distally; dorsal fin extending onto body; depth of dorsal fin slightly less than that of ventral fin at mid-length of tail; dorsal part of body pale brown; ventral surfaces transparent with scattered pigment; pale cream-colored, crescent-shaped mark on posterior edge of body; caudal musculature pale creamy tan with scattered pale brown spots; caudal fins transparent with scattered small brown blotches on dorsal and ventral fins; iris pale bronze in life (Fig. 13); mouth small; median part of upper lip bare; rest of mouth bordered by one row of pointed papillae; lateral fold present; tooth-rows 2/3, first upper row longest; second upper row slightly shorter, broadly interrupted medially; three lower rows complete, equal in length, slightly shorter than second upper row; upper beak moderately deep, forming broad arch with slender lateral processes; lower beak slender, broadly V-shaped; both beaks serrate (Fig. 15E).
In tadpoles having fully developed mouthparts the tooth-row formula of 2/3 is invariable. The pale crescent-shaped mark on the posterior part of the body curves anterodorsally on the dorsal surface of the body. These marks in dorsal view give the appearance of a pair of short, pale stripes on the posterior part of the body. Most specimens from Costa Rica have the pale coloration like that described above, but some individuals (notable KU 87683 from Guápiles, Costa Rica, KU 87707 from Finca Tepeyac, Nicaragua, and KU 87708 from Bonanza, Nicaragua) have much more pigment. In these specimens the same color pattern obtains as in the pallid individuals, but the pigmentation is dense. This is especially noticeable on the tail.
Recently metamorphosed young have snout-vent lengths of 12.7 to 16.7 mm. (average, 14.3 mm. in eleven specimens). Coloration of young in life (KU 68484 from Río Chitaría, Cartago Province, Costa Rica): "Dorsum pale tan; side of head and flanks darker brown, separated from tan dorsum by an indistinct cream stripe. Limbs pale yellow; thighs flecked with brown; shank and tarsus yellowish tan with indistinct brown bars. Soles of feet brown. Belly white; throat dusty cream flecked with silvery white. Upper lip silvery white. Iris bright gold with black flecks. Heels, tarsal and anal stripes white" (Duellman, field notes, May 23, 1961).
Remarks.—Peters (1863:463) named Hyla labialis from the "region of Bogotá, Colombia", but in 1873 regarded his new species as identical with Hyla phaeota Cope, 1862, from Turbo, Colombia. The name Hyla labialis has been used for frogs from the northern Andes in Colombia (see Dunn, 1944:72, and Stebbins and Hendrickson, 1959:522, for discussion of nomenclature). Rivero (1961:131) used the name Hyla vilsoniana Cope, 1899, for the frogs from the northern Andes previously referred to Hyla labialis. A review of the nomenclature and taxonomy of these frogs, which superficially resemble Smilisca but are unrelated, is beyond the scope of the present study.
Hyla baudini dolomedes Barbour, 1923, is based on a small Smilisca phaeota (MCZ 8539) having a snout-vent length of 45.5 mm. Dunn (1931a:413) placed dolomedes in the synonymy of Smilisca phaeota. We have examined the holotype of dolomedes and agree with Dunn's assignment.
Smith (1953:150) described Hyla phaeota cyanosticta from Guatemala. Our studies on the external morphology, coloration, and especially the cranial osteology provide evidence that cyanosticta is a species distinct from phaeota.
[Pg 313] Distribution.—Smilisca phaeota inhabits humid tropical forests from northeastern Nicaragua southward on the Caribbean lowlands to elevations of about 1000 meters and on the Pacific lowlands of Costa Rica, exclusive of the arid regions of Guanacaste, throughout the lowlands of Panamá, exclusive of the savannas of the Pacific lowland and the Azuero Peninsula, and southward on the Pacific slopes of South America through Colombia to west-central Ecuador; also the valleys of the Río Cauca and Río Magdalena in Colombia (Fig. 2).
Specimens examined.—528, as follows: Nicaragua: Matagalpa: Finca Tepeyac, 10 km. N, 9 km. E Matagalpa, KU 85439, 87707 (tadpoles); Matagalpa, MCZ 3546-7, UMMZ 92367; 19 km. N Matagalpa, UMMZ 116495-6. Zelaya: Bonanza, KU 84854-62, 84950-2 (skeletons), 85440-50, 87708-9 (tadpoles); Cukra, AMNH 80618; Río Mico, 16 km. E Recreo, UMMZ 79711 (6), 79712 (4); junction Río Mico and Río Siguia, UMMZ 79713 (10); Río Siguia, 11 km. NW Rama, UMMZ 79714 (14), 79715 (11), 79716 (21), 79717, 79718 (3).
Costa Rica: Alajuela: Cinchona, KU 32255, 64286-8; 5 km. S Ciudad Quesada, USC 8077; Laguna Monte Alegre, KU 64289-90; Las Playuelas, 11 km. S Los Chiles, USC 7216; San Carlos, USNM 29961.
Cartago: Moravia de Turrialba, KU 32212-47, 37133-5, 41093 (skeleton), 64280-1, USC 7243 (3); Peralta, KU 32271-2; Río Chitaría, 3 km. NNE Pavones, KU 64273-9, 68477 (eggs), 68478-83 (tadpoles), 68484 (young); Río Reventazón, MCZ 29196-203, UMMZ 117677 (9); Turrialba, KU 25720-2, 32209-11, 32266-8, 32273-4, 37136-67, 41090-2 (skeletons), 64270-2, MCZ 29221, 29222 (tadpoles), 29269-70, USNM 29934.
Guanacaste: Tilarán, KU 36805-7; 8 km. NE Tilarán, KU 36803-4.
Heredia: Barranca del Río Sarapiquí below Isla Bonita, KU 64282-3; Cariblanco, KU 32256-60, 41094 (skeleton), 64284, MCZ 7967; Isla Bonita, KU 32250-4; 4.2 km. W Puerto Viejo, KU 64285, 68485; 7.5 km. W Puerto Viejo, KU 68486; 1 km. S Puerto Viejo, KU 86518.
Limón: Bambú, USC 7182 (4); Batán, UMMZ 118582; Coén, MCZ 9825; La Lola, KU 32262-4, UF 4029, UMMZ 117678 (3); Los Diamantes, CNHM 101295-8, KU 25723-4, 32265, 64267-9; Pandora, UMMZ 122650 (2), USC 7188 (3), 7190; Puerto Limón, KU 32261; Río Larí at Río Dipari, 21 km. SW Amubre, USC 7177; Río Toro Amarillo, 7 km. W Guápiles, KU 86519, 87683 (tadpoles); Suretka, KU 36808-10, 37168.
Puntarenas: Agua Buena, KU 36790; 1.6 km. E Buenos Aires, UMMZ 117578; 3 km. NW Buenos Aires, KU 64304; 4 km. N, 15 km. W Dominical, KU 68491-2 (tadpoles); Esparta, MCZ 8029-30, 8032; Golfito, KU 32270; 6 km. E Golfito, KU 84999-500 (skeletons); Gromaco, UMMZ 123677 (4); Palmar, KU 32269; 4 km. ESE Palmar Sur, KU 64305-6; 5.6 km. SE Palmar Sur, KU 68489 (tadpoles); 7.0 km. SE Palmar Sur, KU 68490 (young); 8.5 km. SE Piedras Blancas, KU 64292-303; Quebrada Boruca, 22 km. E Palmar Norte, KU 64291; Rincón, "Camp Seattle," Peninsula de Osa, UMMZ 123676 (3), USC 7254; Río Ferruviosa, 7 km. S Rincón, USC 7235; 1.6 km. WNW Villa Neily, KU 68493 (young), 68494 (tadpoles).
San José: San Isidro el General, KU 32249, UMMZ 75025; 10 km. N San Isidro el General, MCZ 29099-103; 13 km. WSW San Isidro el General, KU 86517; 15 km. WSW San Isidro el General, KU 68487 (tadpoles), 68488 (young), 68495 (young); 20 km. WSW San Isidro el General, KU 32248.
Panama: No province: Cano Saddle, USNM 69588; Punta de Pena, USNM 38733; Quipo, AMNH 18925-6. Bocas del Toro: Almirante, KU 80080, 91835-6; 1.6 km. W Almirante, KU 91837; 3 km. W Almirante, KU 91824 (skeleton), 91838-43, 91906-7; 11 km. NW Almirante, CNHM 67853-61; 13 km. W Almirante, KU 91825-7 (skeletons), 91844-9; Fish Creek, KU 92329; Isla Popa, KU 91850-1. Canal Zone: Barro Colorado Island, CNHM 6007, 13316, 13325, 13331, 13360-2, 13377-8, MCZ 24191-5, UF 7523, UMMZ 63547-60, 64457, 69497 (3); 3.7 km. W Cocoli, KU 67916; Fort Sherman, MCZ 10139; Gatun, MCZ 35644; Junction roads C25B and C16, TNHC 23839; [Pg 314] Madden Forest Preserve, TNHC 23837-8. Coclé: El Valle, KU 77521-4, 77649 (tadpole), TNHC 23369. Comarca del Barú: Progreso, UMMZ 61085-9. Colón: Achiote, KU 77516-20, 77648 (young); Río Candelaria, CNHM 67851-2. Darién: Río Esnápe, Sambú Valley, MCZ 8539; Río Sucubti, Chalichiman's Creek, AMNH 40512; Camp Creek, AMNH 40758-9; Camp Creek, Camp Townsend, AMNH 40988. Panamá: NW slope Cerro Prominente, KU 80459; Finca La Sumbadora, KU 91914 (skeleton). Chiriquí: 2 km. W Concepción, AMNH 68910.
Columbia: Antioquia: Puerto Berrio, CNHM 30805 (Goin); Turbo, USNM 39899. Caldas: Pueblorrica, Santa Cecilia, CNHM 54768-71 (Goin). Choco: No specific locality, AMNH 3984-6; Andagoya, BMNH 1915.10.21. 69-70, CNHM 81857 (Goin); Golfo de Urabá, CNHM 63881 (Goin); Peña Lisa, Condoto, BMNH 1913.11.12. 118-125, 1913.11.12. 137-146 (Goin); Pizarro, CNHM 4451-3, 4455-61 (Goin); Río San Juan, Playa del Oro, CNHM 54772 (Goin); Río Quesada, AMNH 13615-77; 37 km. up Río Puné, AMNH 13688; 48 km. up Río Puné, AMNH 13689. Narino: Tumaco, Río Rosario, CJG 2310-13 (Goin). Valle: Buenaventura, BMNH 1895.11.16.82 (Goin); Raposa, WAT 166, 346-47, 388 (Goin); Río Calima above Córdoba, CJG 2249-57 (Goin).
Ecuador: No province: Bulun, AMNH 10620. Esmeraldas: Cachabé, AMNH 10625-8; Río Capayas, CNHM 35712; Río Sapaya, UMMZ 58910 (5); Salidero, AMNH 10623-4; San Javier, AMNH 10618. Guayas: Hacienda Balao Chico, UMMZ 123904. Imbabura: Pambelar, AMNH 10629, 10631. Pichincha: Hacienda Espinosa, 9 km. W Santo Domingo de los Colorados, KU 40220.
Diagnosis.—Size small ([M] 38.0 mm., [F] 46.0 mm.), differing from other species in the genus by the following combination of characters: skull about as long as broad; frontoparietal fontanelle keyhole-shaped; supraorbital flanges absent; squamosal small, not in contact with maxillary; bony portion of ethmoid terminating at anterior edge of orbit; tarsal fold weak, two-thirds length of tarsus; inner metatarsal tubercle small, low, flat, elliptical; snout rounded in dorsal profile; lips thin and flaring; fingers having only vestige of web; toes one-half webbed; diameter of tympanum about two-thirds that of eye; narrow labial stripe white; pair of dark brown (sometimes interconnected) stripes on tan dorsum; no blue spots on flanks or thighs; vocal sac in breeding males pale brown. (Foregoing combination of characters distinguishing S. puma from other species in genus.)
Description and variation.—Ten breeding males from the vicinity of Puerto Viejo, Heredia Province, Costa Rica, have snout-vent lengths of 32.5 to 37.9 mm. (34.8 mm.). In these specimens, the length of the tibia to the snout-vent length is 0.48 to 0.53 (0.51), and the tympanum/eye ratio is 0.52 to 0.72 (0.65). Seven females have snout-vent lengths of 40.8 to 45.8 mm. (43.9 mm.). [Pg 315] No individual has more than a vestige of a web between the second and third and fourth fingers. None has a web between the first and second fingers. Breeding males lack nuptial excrescences on the thumbs. The vocal sac is moderately large and bilobed.
In preserved specimens the dorsal ground color varies from yellowish tan to grayish brown. All specimens have dark brown dorsal markings in the form of a pair of dorsal stripes, variously modified (Pl. 7A). In some specimens, such as KU 91716, the stripes are discrete and extend from the postorbital region nearly to the vent. In most specimens the stripes are connected by a transverse mark in the scapular region and in many others also by a transverse mark in the sacral region. In some specimens the stripes are fragmented posteriorly; fragmentation is extreme in KU 30300, in which the dorsal pattern consists of two series of dark longitudinal dashes. The other extreme is a nearly complete fusion of the stripes, as in KU 91714. A dark brown interorbital bar usually extends onto the eyelids, but in some specimens this is reduced to a short V-shaped mark or small spot between the eyes. There is no dark post-tympanic mark, but dark brown pigment forms a venated pattern from the axilla to the mid-flank; the inguinal region is white, finely mottled with dark brown. The dorsal surfaces of the hind limbs are colored like the body and have two or three dark brown transverse marks on the thighs, three to five marks on the shanks, and one or two marks or irregularly arranged dark flecks on the tarsi. The anterior and posterior surfaces of the thighs are pale tan to brown. The webbing of the feet is tan to grayish brown. A narrow white labial stripe, white anal stripe, and narrow white stripes on the tarsi and outer edges of the forelimbs are invariably present. The ventral surfaces are creamy white.
In life the dorsum is tan or pale brown with dark brown markings. Some individuals have scattered metallic green flecks on the dorsum. The flanks are mottled dark brown and creamy white. The posterior surfaces of the thighs are dark brown. The vocal sacs are grayish brown, and the iris is a deep bronze color.
Natural History.—Smilisca puma inhabits humid lowland tropical forests having more or less evenly distributed rainfall throughout the year. The equable climatic conditions seemingly permit these frogs to be active throughout most of the year. Taylor (1952:846) found calling males at Batán, Costa Rica, on July 20, 1951. We found the species breeding near Puerto Viejo, Costa Rica, on February 19, June 18, July 13, and July 31. Specimens of calling males from Costa Rica in the collection at the University of Southern California were obtained in February at La Fortuna, on August 22 at Los Diamantes, on August 30 at Jabillos, and on September 5 at La Lola. Gravid females were collected in June, July and August.
Males call from shallow water. All breeding congregations of this species that we have found were in a grassy marsh, 7.5 kilometers west of Puerto Viejo, Costa Rica. Tadpoles were found in water-filled depressions in the marsh at night. When first observed, tadpoles were near the surface of the water; they responded to light by quickly taking refuge in the dense grass. No tadpoles were observed by day.
The breeding call consists of a low squawk, usually followed by a series of one or more rattling secondary notes (duration of primary notes, 0.06-0.35 [Pg 316] seconds; of secondary notes, 0.10 to 0.47 seconds), repeated at intervals of 5 to 55 seconds. The primary notes have 187 to 240 pulses per second and major frequencies of about 740 to 1870 cycles per second (Pl. 11A).
Only six tadpoles are available for study. Four of them in stage 34 of development have body lengths of 9.0 to 9.5 mm., tail lengths of 14.0 to 15.0 mm., and total lengths of 23.0 to 24.5 mm. One tadpole in stage 38 and one in stage 40 have total lengths of 31.0 mm. A typical tadpole in stage 34 of development (KU 91807 from 7.5 km. W Puerto Viejo, Heredia Province, Costa Rica) has a body length of 9.5 mm., tail length of 15.0 mm., and total length of 24.5 mm.; body about three-fourths as deep as wide; snout rounded dorsally and laterally; eyes widely separated, directed dorsolaterally; nostril about midway between eye and tip of snout; mouth anteroventral; spiracle sinistral, about two-thirds distance from snout to posterior end of body and slightly below midline; anal tube dextral; caudal musculature slender, barely curved upward distally; dorsal fin extending onto body; at mid-length of tail, depth of caudal musculature equal to that of dorsal fin and ventral fin; body grayish brown, palest ventrally; caudal musculature pale creamy yellow with bold gray reticulations; caudal fins transparent with gray reticulations anteriorly and black flecks posteriorly on both fins (Fig. 14A). Median part of upper lip bare; rest of mouth bordered by two rows of short blunt papillae; lateral fold present; tooth-rows 2/3; upper rows equal in length; second upper row broadly interrupted medially; three lower rows complete, first and second rows equal in length, slightly shorter than upper rows; third lower row noticeably shorter; upper beak shallow, forming broad, continuous arch with slender lateral processes; lower beak slender, broadly V-shaped, both beaks finely serrate (Fig. 15B).
All six tadpoles are colored alike, except that in the larger specimens scattered white flecks are present on the ventral surface of the body, and the dark reticulations continue farther posteriorly on the caudal fins than in the smaller tadpoles. In two specimens the third lower tooth-row is only about one-half the length of the other lower rows, and in one specimen the second lower tooth-row is shorter than the first. Coloration of tadpoles in life: "Body olive-brown with silvery green flecks laterally. Caudal musculature olive-brown with greenish tan flecks. Fins brown with greenish gold flecks. Iris deep bronze." (Duellman, field notes, February 19, 1965).
One recently metamorphosed young (KU 91808) has a snout-vent length of 12.4 mm. In life this frog had a pale tan dorsum with dark brown markings, yellowish tan posterior surfaces of thighs, grayish brown throat, and bronze iris.
Remarks.—The identity of Cope's Hyla puma has not been known. The name has appeared in various compilations, but no workers have referred any of their specimens to that species. Examination of the holotype (USNM 13735), an adult female, revealed the presence of the following combination of characters: snout-vent length 45.8 mm., snout blunt above and rounded laterally, nostrils close to tip of snout, lips thin and flaring, a vestige of a web on the hands, feet about one-half webbed, tarsal fold weak and extending about two-thirds length of tarsus, dorsal markings consisting of a faded dark interorbital bar and a pair of faded longitudinal brown marks connected by a transverse band in the scapular region. The type agrees well with specimens of Smilisca wellmanorum (Taylor, 1952); the vestigial webbing on the [Pg 317] hands and the dorsal coloration are especially significant. Consequently, we consider Hyla wellmanorum Taylor, 1952, to be a synonym of Hyla puma Cope, 1885. Cope gave only "Nicaragua" as the locality for Hyla puma. The specimen was part of a collection received at the United States National Museum from Lt. J. F. Moser. Among the species in the collection are Dentrobates pumilio, Phyllomedusa helenae, Corythophanes cristatus, Pliocercus dimidatus, Tretanorhinus nigroluteus, and others characteristically found on the Caribbean lowlands of Central America. Thus, it seems reasonable to assume that the type specimen of Hyla puma came from the Caribbean lowlands. Though no other Nicaraguan specimens have been found by us, numerous specimens are known from the Caribbean lowlands of Costa Rica.
Cochran (1961:58), in her catalogue of type specimens in the United States National Museum, listed Hyla puma Cope, 1885, as a synonym of Hyla molitor Schmidt, 1857. She made no qualifying statements. Schmidt (1858:246), in his descriptions of the species in the year following his publication of the names and Latin diagnoses, stated: "Dorsum uniformly gray, more intensive on back, fading away laterally and on extremities; in every-day-life this blue would be called Mueller's Blau. A delicately dotted black line runs on the canthus rostralis from the opening of the nose to the corner of the eye. In the armpits, on the flanks and the thighs two of our three specimens have black marblings." [Free translation] Certainly on the basis of coloration Hyla puma is distinctly different from Hyla molitor.
Distribution.—This species lives in the wet, forested regions of the Caribbean lowlands of Costa Rica and presumably southern Nicaragua (Fig. 3). All specimens are from low elevations; the highest known elevation for the occurrence of this frog is 285 meters at Laguna Bonilla.

Specimens examined.—62, as follows: Nicaragua: No specific locality, USNM 13735.
Costa Rica: Alajuela: Jabillos, 5 km. N Santa Clara, USC 8058 (6); 5 km. W La Fortuna, USC 8078 (2); Río La Fortuna at La Fortuna, USC 7151 (3). Cartago: Laguna Bonilla, tunnel camp near Peralta, KU 32171. Heredia: Puerto Viejo, KU 86521; 5.9 km. W Puerto Viejo, KU 64307; 7.5 km. W Puerto [Pg 318] Viejo, KU 64308-10, 64311 (skeleton), 64312-15, 68635-6 (skeletons), 85001-2 (skeletons), 86520, 87770-1 (skeletons), 91709-16, 91791-2, 91807 (tadpoles), 91808 (young). Limon: Batán, KU 30300-2; La Lola, KU 32169, USC 141, 201, 8067; Los Diamantes, KU 32170, UMMZ 118470 (6), USC 212; 2.4 km. E Los Diamantes, USC 8049 (5).
Holotype.—Adult male, KU 91852 from a small stream at the north edge of the village of El Volcán, Chiriquí Province, Panamá, elevation 1280 meters; obtained on Feb. 5, 1965, by William E. Duellman.
Paratypes.—KU 91853-74, collected with the holotype.
Diagnosis.—Size moderate ([M] 45.0 mm., [F] 62.2 mm.); skull wider than long, having large, ovoid frontoparietal fontanelle; supraorbital flanges absent; squamosal small, not contacting maxillary; bony section of ethmoid extending anteriorly between nasals; tarsal fold weak, full length of tarsus; inner metatarsal tubercle low, flat, elliptical; lips thick, rounded, not flaring; fingers one-third webbed; toes three-fourths webbed; diameter of tympanum about one-half that of eye; margin of upper lip faintly marked by interrupted creamy white stripe; dark spots on dorsum; pale flecks on flanks and posterior surfaces of thighs; vocal sacs in breeding males dark brown. (Foregoing combination of characters distinguishing S. sila from any other species in genus.)
Description of holotype.—Snout-vent length 36.6 mm.; tibia length 19.8 mm., 54.1 per cent of snout-vent length; foot length 15.5 mm., 42.3 per cent of snout-vent length; head length 12.7 mm., 34.7 per cent of snout-vent length; head width 13.3 mm., 36.8 per cent of snout-vent length; snout short, in lateral profile truncate, only slightly rounded above, in dorsal profile rounded; canthus rounded; loreal region noticeably concave; lips thick, rounded, not flaring; nostrils not protuberant, directed laterally; internarial distance 3.0 mm.; internarial area flat; top of head flat; interorbital distance 3.5 mm., 26.3 per cent of head width; diameter of eye 4.2 mm., thrice distance (1.4 mm.) from tympanum to eye, and half again distance (2.8 mm.) from orbit to nostril; pupil horizontally ovoid; width of eyelid 2.8 mm., 21.1 per cent of head width; dermal fold from posterior corner of orbit covering upper edge of tympanum to point above insertion of forelimb; diameter of tympanum 2.3 mm., 54.7 per cent of diameter of eye; no axillary membrane; arms moderately robust; weak fold on wrist; faintly scalloped fold along ventrolateral margin of forearm; fingers short, slender; fingers from shortest to longest, 1-2-4-3; vestige of web between first and second fingers; others about two-fifths webbed; discs moderate, diameter of that on third finger about one-third diameter of eye; triangular outer palmar tubercle; elliptical inner palmar tubercle on base of pollex; subarticular tubercles large, conical, none bifid; supernumerary tubercles few, large, conical; brown nuptial excrescence on prepollex; heels overlap by about one-fifth length of shank when hind limbs adpressed; tibiotarsal articulation extending to nostril; tarsal fold weak, extending nearly full length of tarsus; inner metatarsal tubercle elliptical, flat; outer metatarsal tubercle absent; toes moderately long; toes from shortest to longest, 1-2-3-5-4, third and fifth about equal in length; discs about same size as those on fingers; webbing [Pg 319] extending to middle of penultimate phalanx on all toes, except only to distal end of antepenultimate phalanx of fourth toe; subarticular tubercles round; supernumerary tubercles large, round, present only on proximal digits. Anal opening directed posteriorly at level of upper edge of thighs; no noticeable anal sheath; flat tubercles ventrolateral to anal opening large; skin of chest, belly, and posterior surfaces of thighs granular; other surfaces smooth; tongue broadly cordiform, shallowly notched posteriorly, and barely free behind; vomerine teeth 4-4, situated on ventral surfaces of separated rounded prominences between posterior margins of small, ovoid inner nares; vocal slits long, each situated along inner margin of ramus; color (in preservative) pinkish tan above with irregular olive-brown markings forming interconnected spots on back; four bars on dorsal surface of each thigh; five bars on shank, and three on tarsus; inguinal region white with black mottling; posterior surfaces of thighs yellowish tan proximally, dark brown distally; margins of lips grayish white with brown markings; ventral surfaces of hands and feet grayish brown; belly and posterior part of throat creamy white; anterior part of throat brown.
Description and variation.—Ten breeding males from Finca La Sumbadora, Panamá, have snout-vent lengths of 40.0 to 44.8 mm. (42.3 mm.). In these specimens the tibia/snout-vent length ratio is 0.50 to 0.57 (0.54), and the tympanum/eye ratio is 0.48 to 0.58 (0.53). There is a geographic gradient in size; specimens from the western part of the range (southern Costa Rica) are smaller than those from the eastern part of the range (eastern Panamá). Five males from the Pacific lowlands of southern Costa Rica have snout-vent lengths of 31.6 to 38.2 mm. (34.7 mm.); ten males from El Volcán, Chiriquí, Panamá, 32.6 to 37.9 mm. (36.4 mm.), and eight males from Barro Colorado Island, Canal Zone, 38.2 to 42.0 mm. (35.6 mm.). These are smaller than the males from Finca La Sumbadora, which is east of the Canal Zone. Ten females from El Volcán have snout-vent lengths of 44.2 to 55.6 mm. (49.2 mm.), as compared 56.1 to 62.2 mm. (58.2 mm.) in three females from Finca La Sumbadora.
Large females have scattered small tubercles on the head and back; tubercles occur in males from Costa Rica and in some males from western Panamá. The truncate snout is characteristic of both sexes.
The coloration of Smilisca sila consists of a gray, tan, or pale reddish brown dorsal ground color and a creamy white venter. The dorsum is marked by dark brown, olive-brown, or dark reddish brown spots or blotches (Pl. 7B). Usually the blotches are discrete, but in some individuals they are interconnected and form an irregular dark mark on the dorsum. There is no tendency for the blotches to form transverse bars as in Smilisca sordida. In one specimen (KU 80467) the blotches are fused and form two wide irregular longitudinal stripes, as in Smilisca puma. In some females the dorsal markings are reduced to a few small spots or are nearly absent (KU 92332), whereas in other females the dorsal markings are bold. In one female (KU 91894) the dorsal markings are narrowly bordered by pale blue, and numerous pale blue flecks are present on the pale brown dorsum. In many individuals of both sexes small white flecks are present on the dorsal surfaces.
Usually the flanks and posterior surfaces of the thighs have black mottling enclosing pale blue spots and flecks, respectively. The dorsal surfaces of the limbs are marked by dark brown transverse bars; usually three or four bars are present on each forearm, thigh, and shank. The coloration of the flanks and limbs varies geographically. Specimens from southern Costa Rica and western Panamá have distinct bars on the limbs; the posterior surfaces of the thighs have brown reticulations enclosing small blue flecks in specimens from [Pg 320] Costa Rica and bolder, black reticulations enclosing large pale blue spots in specimens from western Panamá. In specimens from Costa Rica the flanks are brown with pale blue flecks, whereas in those from Chiriquí, Panamá, the flanks are pale blue with dark brown mottling in the inguinal region. Frogs from El Valle and Cerro la Campana usually have distinct bars on the limbs; the posterior surfaces of the thighs are colored as in frogs from Chiriquí, and the inguinal region is pale blue with coarse brown mottling. Specimens from Barro Colorado Island are marked like those from El Valle and Cerro la Campana, except that on the posterior surfaces of the thighs fine black reticulations enclose many dark blue spots. In specimens from Darién and from Panamá Province east of the Canal Zone (Altos de Pacora, Cerro Jefe, Finca La Sumbadora, and Río Pacora), the markings on the dorsal surfaces of the limbs are indistinct or absent in males, but distinct in some females. Intense brown and black pigment forms fine reticulations delimiting bold blue spots on the flanks; this coloration extends to the axilla in many specimens. Fine black reticulations enclose many dark blue spots on the posterior surfaces of the thighs.
In females, the throat is creamy white; in some specimens scattered brown flecks are present on the chin and throat. In breeding males the anterior part of the throat is dark gray or dark brown.
The coloration in life is as variable as it is in preservative. In life the holotype had a tan dorsum with dark olive-green irregular markings and small green flecks. The limbs were tan with dark brown transverse bars. The flanks were grayish tan anteriorly; the inguinal region and posterior surfaces of thighs were blue with black mottling. The belly was creamy white, and the throat was brown with creamy yellow flecks. The iris was a dull bronze color. Among the paratypes, some individuals had green flecks, others did not. The inguinal region and posterior surfaces of the thighs were pale blue, pale yellowish green, or grayish tan with black mottling. The blue was most noticeable in females.
Colors of a male from Finca La Sumbadora, Panamá, were described as follows: "Dorsum olive-brown; irregular dark brown blotches, pale green flecks, and raised creamy yellow spots on dorsal surfaces; belly creamy white; throat grayish brown; undersides of limbs grayish tan; groin, anterior and posterior surface of thigh, inner surface of shank, anterior edge of tarsus, and proximal parts of third and fourth toes pale blue marbled with dark brown and black; webbing brown; iris pale bronze, finely reticulated with black." (Duellman, field notes, January 28, 1964.)
A female (now KU 91894) from Altos de Pacora, Panamá, was described as follows: "An irregular dark brown, green-bordered figure on head and back; dark brown, green-bordered bands on limbs—all on a lighter brown and heavily green-spotted background. These markings are more vivid at night than during the day. Lower sides, from midbody onto front of thighs and rear of thighs onto venter of shanks to heels and thence dorsally onto basal portions of toes heavily blue spotted on a light brown (front of thighs and venter of shanks) to blackish brown background. Venter cream. Iris gray-brown, finely veined with dark brown." (Charles W. Myers, field notes, December 14, 1964.) Note that in the earlier discussion of coloration of preserved specimens, the green spots and borders have changed to pale blue after six months in alcohol.
[Pg 321] In living individuals from Costa Rica and Panamá west of the Canal Zone, the blue coloration on the flanks and thighs is much less conspicuous than in specimens from eastern Panamá. The color of the iris is variable, even in frogs from one locality. The coloration of the iris in 13 living frogs (now KU 92333-45) from Valle Hornito, Chiriquí, Panamá, was described as follows: "Iris variable—from pale to dark brown; in a few the iris has a golden cast to the brown; in a few others the lower half of the iris is pale gray with the upper half being light brown." (Charles W. Myers, field notes, April 24, 1965).
Natural history.—Smilisca sila inhabits the Pacific slopes of lower Central America where a pronounced dry season occurs. We have records of males calling in December through May and also in August (latter date from El Volcán, Chiriquí, Panamá). The breeding season seems to be correlated with the time of the year when the water is clear and at a low level in the streams where these frogs breed.
Males call from the edges of small, shallow streams, from rocks in the streams, or less frequently from vegetation overhanging the streams. Females are most frequently found on the banks of streams, and clasping pairs usually are in shallow pools in streams. One individual was found in a bromeliad about three meters above the ground in the daytime.
The breeding call consists of a low squawk, usually followed by a series of one or more rattling secondary notes (duration of primary notes, 0.06 to 0.28 seconds; of secondary notes, 0.14 to 0.48 seconds), repeated at intervals of 4 to 20 seconds. The primary notes have 97 to 120 pulses per second and major frequencies of about 900 to 2220 cycles per second (Pl. 11B).
Eggs were obtained artificially in the field; the average length of ten embryos in the neural groove stage is 2.4 mm., and the average diameter of the outer envelope is 4.9 mm. Hatchlings have large, conical oral discs, heavy gills, and a large amount of yolk; their average total length is 6.3 mm.
Tadpoles have been found in pools in clear streams; some tadpoles have been observed to cling by their mouths to rocks in the stream; others were found on the bottom where they seek refuge among pebbles or under rocks and leaves. A complete developmental series of tadpoles is not available. Eleven tadpoles in stage 25 of development have body lengths of 8.3 to 10.2 mm. (9.3 mm.), tail lengths of 17.3 to 21.0 mm. (18.8 mm.), and total lengths of 25.9 to 31.0 mm. (28.1 mm.). One tadpole in stage 41 and one in stage 42 have body lengths of 11.5 and 12.5 mm., tail lengths of 27.2 and 29.5 mm., and total lengths of 38.7 and 42.0 mm., respectively. The snout-vent lengths of two specimens in stage 43 and one in stage 45 are 12.7, 13.0, and 13.6 mm., respectively.
A typical tadpole in stage 25 of development (KU 80620 from Finca La Sumbadora, Panamá) has a body length of 9.5 mm., tail length of 19.0 mm., and a total length of 28.5 mm.; body only slightly wider than deep, nearly flat dorsally; snout broadly rounded in dorsal view, bluntly rounded in lateral view; eyes widely separated, directed dorsolaterally; nostril slightly closer to eye than to tip of snout; mouth ventral; spiracle sinistral, located about two-thirds distance from snout to posterior edge of body; anal tube dextral; caudal musculature moderately heavy, straight; dorsal fin not extending onto body; fins deepest at about two-fifths length of tail, where depth of caudal musculature about equal to depth of dorsal and depth of ventral fin; musculature [Pg 322] extending nearly to tip of tail; body dark grayish brown above and pale grayish tan below with small dark brown spots dorsally and white flecks laterally; caudal musculature pale tan with dark brown flecks over entire surface and dark brown streaks on posterior one-half of ventral fin and on all of dorsal fin (Fig. 14B). Median one-third of upper lip bare; rest of mouth bordered by a single row of conical papillae; lateral fold present; tooth rows 2/3; upper rows cone-shaped, about equal in length, broadly ∧-shaped; second upper row narrowly interrupted medially; lower rows complete, about equal in length, but slightly shorter than upper rows; upper beak moderately massive, its inner surface forming a continuous arch with short lateral processes; lower beak broadly ∨-shaped; both beaks finely serrate (Fig. 15D).
Tadpoles from El Volcán, Chiriquí (KU 91833), are more heavily pigmented than those from Finca La Sombadora; the spots on the tail are larger. In life these tadpoles had dark brownish black bodies with golden and green lichenous flecks; the tail was tan with dark brown markings, and the iris was a grayish bronze color. In life tadpoles from Finca La Sumbadora were olive-tan above and dark gray with pale bluish gray irridescent spots ventrally. The caudal musculature was creamy tan with brown flecks and streaks, and the iris was pale bronze.
Metamorphosing young have been found on vegetation at the edge of streams and have been raised in the laboratory. Seven recently metamorphosed young have snout-vent lengths of 13.6 to 15.6 mm. (14.6 mm.). A living juvenile (KU 91913) raised in the laboratory from a tadpole obtained at Finca La Sumbadora had a brown dorsum with darker brown markings, a white spot below the eye, and a narrow white labial stripe. The belly was white; the flanks were brown with white spots, and the posterior surfaces of the thighs were yellowish tan. The iris was a golden bronze color with much black reticulation.
Remarks.—This species has been confused with Smilisca sordida; most authors have referred both species to Hyla (Smilisca) gabbi. Examination of the types of Hyla sordida, gabbi, salvini, and nigripes revealed that all of the names were referable to a single species (S. sordida), and that the small, blunt-snouted species in Panamá and southern Costa Rica probably was without a name. Possibly Hyla molitor Schmidt (1857) is based on the species that we have named S. sila, but several discrepancies in his description, plus the unknown provenance of the type, have led us to discount the applicability of that name to the species under consideration.
Distribution.—Smilisca sila ranges along the Pacific slopes and lowlands of southern Costa Rica and Panamá at elevations from sea level to about 1300 meters; in northern South America the species occurs in the Caribbean lowlands and in the valleys of the northward draining rivers of Colombia (Fig. 3).
Specimens examined, 234, as follows: Costa Rica: Puntarenas: 6 km. E Golfito, KU 91717; Quebrada Boruca, 22 km. E Palmar Norte, KU 64265-6; Río Zapote, 7 km. E Palmar Norte, USC 7100 (2). San José: San Isidro el General, KU 28200; 14 km. NW San Isidro el General, USC 7098 (2); 15 km. WSW San Isidro el General, USC 7097.
Panama: Canal Zone: Barro Colorado Island, AMNH 62320-3, CNHM 13324, 13326-8, 13330, 13338, 13359, 13423-5, KU 80460-6, 80619 (young), 80625 (skeleton), UMMZ 63542-6, USC 7051. Chiriquí: Boquete, AMNH 69815, UMMZ 58441-5; El Volcán, KU 77413, 91828-31 (skeletons), 91852-74, 91832 (eggs), 91833 (tadpoles); 6 km. S El Volcán, CNHM 60442; 16 km. NNW El Volcán, KU 91879-90; Finca Palosanto, 6 km. WNW El Volcán, [Pg 323] KU 77406-12, 77692 (skeleton), 91875-7, 92330-1; Río Colorado, 17 km. NNW El Volcán, KU 91878, 92332; Valle Hornito, 19 km. NE Gualaca, KU 92333-45. Coclé: El Valle, AMNH 55440-5 (13), 59607-14, CNHM 48140, 60349-2, 60387-92, 60401-4, 60443, 67842-5, KU 91834 (young), 91902-4, TNHC 23751-2, USNM 140653. Colón: Río Candelaria, AMNH 53708-15, CNHM 67826-36. Darién: Camp Creek, Camp Townsend, AMNH 40756-7, 40936-9, 40992; Río Chico, AMNH 39784, 40986-7; Río Pita, CNHM 67823-5; Tacarcuna, USNM 141796-802; Three Falls Creek, AMNH 41684, 51788. Los Santos: Cerro Hoya, USNM 148213-4; Lajamina, Río Puria, KU 67915. Panamá: Altos de Pacora, KU 91894; Cerro Jefe, KU 91895-6; Cerro La Campana, CNHM 67846, KU 91897-900, USNM 139689; Finca La Sumbadora, KU 80467-81, 80620 (tadpoles), 91910 (eggs), 91911-2 (tadpoles), 91913 (young), 91908-9 (skeletons); Río Calobra, USNM 53722, Río Pacora, 9 km. NNE Pacora, KU 91901. Veraguas: Cerro Carbunco, USNM 129066; Cerro Tute, CNHM 67837-41; Isla Cebaco, Río Platanal, KU 91891-3.
Colombia: Antioquia: Urabá, Villa Arteaga, CNHM 63893 (Goin). Atlantico: Sabanalarga, Río Causa, AMNH 14506.
Diagnosis.—Size moderate ([M] 45 mm., [F] 64 mm.); skull slightly wider than long, having large and elongate frontoparietal fontanelle; supraorbital flanges absent; squamosal small, not contacting maxillary; bony section of ethmoid terminating just anterior to anterior edge of orbit; tarsal fold weak, full length of tarsus; inner metatarsal tubercle long, low, flat, elliptical; lips thin and flaring; [Pg 324] fingers one-half webbed; toes four-fifths webbed; diameter of tympanum about one-half that of eye; no white labial stripe; dorsal dark markings irregular, sometimes forming broad transverse bars; pale flecks on flanks and usually on posterior surfaces of thighs; vocal sacs in breeding males white. (Foregoing combination of characters distinguishing S. sordida from any other species in genus.)
Description and variation.—Ten breeding males from 15 to 20 kilometers west-southwest of San Isidro el General, San José, Costa Rica, have snout-vent lengths of 38.1 to 42.6 mm. (40.5 mm.). In these specimens, the tibia/snout-vent length ratio is 0.50 to 0.54 (0.52), and the tympanum/eye ratio is 0.45 to 0.57 (0.49). Specimens from the Pacific slopes of Costa Rica are larger than those from the Meseta Central and the Caribbean lowlands. Ten males from 6 kilometers east of Golfito, Puntarenas, have snout-vent lengths of 38.4 to 44.6 mm. (41.8 mm.), and five males from Rincón, Peninsula de Osa, have snout-vent lengths of 38.8 to 41.6 mm. (40.3 mm.). Snout-vent lengths of ten males from La Fortuna, Alajuela, are 31.9 to 36.0 mm. (34.4 mm.), of ten males from Pandora, Limón, 33.8 to 37.6 mm. (35.9 mm.), and of ten males from Escazú and Río Jorco on the Meseta Central, 34.3 to 37.6 mm. (36.0 mm.). Eight females from the Río Jorco on the Meseta Central have snout-vent lengths of 48.8 to 53.8 mm. (50.4 mm.), and six females from various localities on the Pacific slopes of Costa Rica have snout-vent lengths of 56.5 to 64.0 mm. (59.8 mm.). The only noticeable differences in proportions between males and females is in the tympanum/eye ratio; for example, this ratio is 0.47 to 0.53 (0.49) and 0.54 to 0.68 (0.61) in ten males and eight females, respectively, from the Meseta Central.
The shape of the snout and the associated cranial elements of S. sordida vary geographically and ontogenetically. Specimens from the Caribbean lowlands have blunt snouts in lateral view; those from the Pacific lowlands have longer, more slender snouts that are pointed in lateral view, and those from the Meseta Central are intermediate in snout shape between the two lowland populations (Fig. 4). These differences in shape of the snout are dependent on the nature of the underlying cranial bones, principally the maxillaries and nasals. In specimens from the Caribbean lowlands the nasals are long, wide, and narrowly separated from the ethmoid; the anterior edge is just posterior to the nostril. The maxillary flanges are nearly vertical. In specimens from the Pacific lowlands the nasals are relatively shorter, narrower, and rather widely separated from the ethmoid; the anterior edges of the nasals do not extend so far forward as in specimens from the Caribbean lowlands. The maxillary flanges slant medially. In these cranial characters, specimens from the Meseta Central are intermediate between the two lowland populations.
Superimposed on this geographic variation are ontogenetic changes, which are most noticeable in males. In smaller, and presumably younger, specimens the snouts are more pointed than in larger specimens; consequently some small males from the Caribbean lowlands resemble larger males from the Pacific lowlands, since the nasals and maxillaries of the former are not fully ossified. In addition, in small breeding males the ethmoid is only about one-half ossified, a large frontoparietal foramen is present, the anterior arm of the squamosal extends only about one-fourth the distance to the maxillary (two-thirds the distance in larger specimens), and the tegmen tympani are short, as compared with the long, thin elements in larger specimens.

The dorsal ground-color of Smilisca sordida is gray to pale tan or reddish brown; the venter is white. The dorsum is variously marked with dark gray, dark brown, reddish brown, or olive-green spots or blotches (Pl. 7C). A dark interorbital bar usually is present. The dorsal markings on the body usually consist of a blotch, or two or more spots, on the occiput, in the scapular region, and in the sacral region. In many specimens, especially females, these markings are in the form of broad transverse bars. A female (USC 7164) from [Pg 326] Las Cañas, Guanacaste, Costa Rica, has a tan dorsum with many black flecks and round brown spots bordered by darker brown. One female (KU 91763) from the Río Jorco, San José, Costa Rica, has a unicolor tan dorsum. Some individuals have scattered, small white spots on the dorsum; these are most evident in a male (USC 7153) from La Fortuna, Alajuela. White labial stripes and anal stripes are absent in all specimens.
The limbs are marked by dark brown transverse bars; these are indistinct in some specimens from the Meseta Central and Caribbean lowlands, whereas they are distinct in all specimens from the Pacific lowlands. Specimens from the Caribbean lowlands have two to six bars on each shank, whereas specimens from the Pacific slopes have four to six bars on each shank, and specimens from the Meseta Central have as many as eight bars on each shank. A narrow, sometimes broken white line is present on the ventrolateral edge of the forearm. The webbing on the hand is tan or pale gray, and the ventral surfaces of the tarsi and the webbing on the feet are dark gray or brown. Breeding males have dark brown nuptial excrescences on the prepollex.
The flanks and posterior surfaces of the thighs usually are marked by bluish white and creamy tan flecks, respectively, but vary considerably. In specimens from the Caribbean lowlands a small amount of flecking is present in the inguinal region, and on the posterior surfaces of the thighs flecks are few or absent. In specimens from the Meseta Central, numerous large flecks or small, round spots (pale bluish white in life) are on the posterior half of the flanks; small flecks are on the posterior surfaces of the thighs. Specimens from the Pacific slopes and lowlands of southern Costa Rica (Puntarenas and San José Provinces) have bold mottling of black and bluish white on the flanks and many bluish white flecks on the posterior surfaces of the thighs. The flanks are reticulated from the axilla to the groin in two females (UMMZ 123684 and USC 7236) from Rincón, Peninsula de Osa. In specimens from the Pacific slopes of Guanacaste in northwestern Costa Rica, flecks are present in the inguinal region; indistinct flecks are on the posterior surfaces of the thighs.
The throat is immaculate in specimens from the Caribbean lowlands in Limón Province; the throats are dusky laterally in most other specimens except some from the Meseta Central, in which the throats are heavily flecked with black. This variation occurs in males and females.
The color and pattern in life are highly variable. A composite description of living individuals (now KU 91718-41) from 6 kilometers east of Golfito, Puntarenas, Costa Rica, illustrates the variability: "Dorsum pale olive-green, fading to tan posteriorly, or tan all over with dark olive-green or dark brown spots on back and bars on limbs. Flanks dark brown with cream, greenish gray, or bluish gray mottling. Posterior surfaces of thighs dark brown with pale blue, pale green, or tan flecks. Iris creamy silver. Throats white with some brown flecks peripherally." (Duellman, Field notes, February 15, 1965.) A male from the Río Jorco, San José, Costa Rica, was dull olive-tan above with olive-green marks; the flanks were brown with pale tan flecks, and the posterior surfaces of the thighs were pale brown with cream-colored flecks. Six females from the same locality were reddish brown above with olive-brown or dark brown markings; one was uniform orange-tan, and another was dull olive-green with darker markings.
The color of the iris in living frogs varies from creamy silver to grayish yellow or bronze with a variable amount of black reticulation.
[Pg 327] Natural History.—Smilisca sordida is not associated with any one type of vegetation; instead it lives in the vicinity of rocky streams having low gradients. Breeding takes place primarily in the dry season, when the water in the streams is clear and at a low level. Through most of the range of S. sordida showers, or even short heavy rains, occur in the dry season. After such rains the breeding activity is maximal. Breeding congregations have been found from December through April, but a few calling males and gravid females have been taken in June, July, and August. In the rainy season non-breeding individuals are found sitting on bushes near streams at night. Taylor (1952:843) found specimens in bromeliads by day.
Males usually call from rocks or gravel bars in, or at the edge of, streams. Some individuals perch in low bushes overhanging the streams, and some sit in shallows in the streams. Clasping pairs have been found on the banks of streams and in shallow water in streams.
The breeding call consists of one to six moderately short, rather high-pitched notes (duration 0.18 to 0.45 seconds) repeated at intervals of 12 seconds to several minutes. Each note is a vibrant rattle having 78 to 135 pulses per second and major frequences of about 1200 to 2600 cycles per second (Pl. 11C).
The tadpoles live in shallow parts of the streams, where they cling to the surfaces of small rocks and hide beneath leaves and rocks. A complete developmental series of tadpoles is not available; measurements of those stages examined are summarized in Table 12.
A typical tadpole in stage 36 of development (KU 68475 from 15 km. WSW of San Isidro el General, Costa Rica) has a body length of 11.7 mm., tail length of 22.8 mm., and a total length of 34.5 mm.; body about three-fourths as deep as wide; snout broadly rounded in dorsal view, sloping and rounded in lateral view; eyes widely separated, directed dorsolaterally; nostril slightly closer to eye than to tip of snout; mouth ventral; spiracle sinistral, about two-thirds distance from snout to posterior end of body and slightly below midline; anal tube dextral; caudal musculature heavy, straight; dorsal fin not extending onto body; fins deepest at about mid-length of tail; there depth of caudal musculature equal to depth of dorsal fin and half again as deep as ventral fin; musculature extending nearly to tip of tail; body reddish brown above and pale grayish brown with white flecks below; caudal musculature pale tan with brown flecks; a series of reddish brown dashes at base of caudal fin separated from others in series and from dashes on other side by creamy white; fins transparent with reddish brown flecks on posterior one-half of ventral fin and on all of dorsal fin (Fig. 14C). Mouth bordered by two rows of short, pointed papillae; lateral fold present; tooth-rows 2/3; upper rows equal in length; second upper row narrowly interrupted medially; three lower rows complete, nearly as long as upper rows, deeply indented medially; upper beak robust, inner surface not forming continuous arch with short lateral processes; lower beak deep, V-shaped; both beaks bearing short serrations (Fig. 15F).
Little variation occurs in structure. In some specimens the second upper tooth-row is complete; no individuals were found to have the row broadly interrupted medially.
The series of dark dashes on the dorsal edge of the caudal musculature is diagnostic of all stages studied. In life, tadpoles from 15 and 20 kilometers west-southwest of San Isidro el General, Costa Rica, had a tan body, often [Pg 328] with an olive-tan tinge; the caudal musculature was tan; the flecks and dashes were dull red or reddish brown. Tadpoles from 6 kilometers east of Golfito, Costa Rica, had bodies with olive-green flecks. The caudal musculature was brown with bluish green flecks; the fins were transparent with reddish brown flecks. The belly was a silvery golden color. Tadpoles from Bajos de Jorco, Costa Rica, had brown bodies with bluish green flecks; the tail and fins had reddish brown flecks and dashes. The iris was a bronze color in specimens from all three localities, as well as in the young mentioned in the following paragraph.
Nine recently metamorphosed young were found on vegetation at the edges of streams in April. These specimens have snout-vent lengths of 13.1 to 15.7 mm. (14.9 mm.) and in life were pale greenish tan or olive-tan above and white below. The hands, feet, and thighs were pale yellowish tan.
Remarks.—The foregoing synonymies indicate that confusion has existed in the application of various names, to this species, as well as in use of the names sordida and gabbi to include the species that we describe and name Smilisca sila. Correct allocation of the names involved was possible only after studying and comparing the type specimens, for the descriptions given by the various authors are not sufficiently explicit to determine the nature of many essential features.
The presence of a rounded snout and a long white throat in males distinguishes S. sordida from S. sila, which has a high truncate snout and short dark throat in males. The two syntypes of Hyla sordida Peters, 1863, (ZMB 3141) are males having snout-vent lengths of 36.9 and 37.0 mm. The two syntypes of Hyla gabbi Cope, 1876 (USNM 30658-9), are females having snout-vent lengths of 52.8 and 53.7 mm., respectively. Also included in the collections made by Gabb is eastern Costa Rica are two males (USNM 30685-6), which Cope (1876) named and described as Hyla nigripes. These specimens are soft and faded, but are recognizable as the same as Hyla sordida Peters; the syntypes of Hyla nigripes have snout-vent lengths of 37.6 and 37.7 mm. We have examined one of the syntypes of Hyla salvini Boulenger, 1882 (BMNH 1947.2.24.13), a female having a snout-vent length of 54.6 mm. We are convinced that all of these type specimens are representatives of one species, the earliest name for which is Hyla sordida Peters, 1863. The type localities for three of the named species are in Costa Rica—H. gabbi from Sipurio on the Caribbean lowlands, H. nigripes from the Caribbean slopes of Pico Blanco, and H. salvini from Cartago on the Meseta Central. The type locality of H. sordida was given as "Veraguas" by Peters (1863). At that time Veraguas was often considered to be most of western Panamá. Though we have not seen Panamanian specimens other than the types of S. sordida and one specimen from the Pacific lowlands of western Panamá, the species probably occurs on the Caribbean slopes of western Panamá. The species has been taken on the Caribbean lowlands of Costa Rica within a few kilometers of Panamá; collecting on the Caribbean slopes in the provinces of Bocas del Toro and Veraguas should reveal the presence of Smilisca sordida there.
Distribution.—Smilisca sordida is found along the Pacific slopes and lowlands from Guanacaste, Costa Rica, southeastward to extreme western Panamá, to elevations of about 1200 meters on the Meseta Central in Costa Rica, and on the Caribbean slopes and lowlands of Costa Rica and probably adjacent Panamá (Fig. 5). One specimen purportedly comes from "Río Grande, Nicaragua."
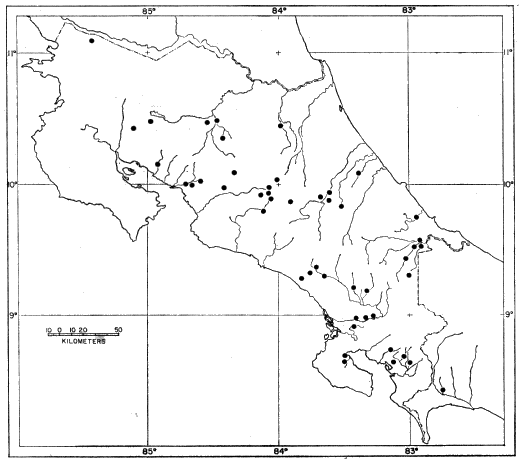
Specimens examined.—412, as follows: Nicaragua: "Río Grande" (? Depto. Zelaya), MCZ 2634.
Costa Rica: Alajuela: Between Atena and Salto de San Mateo, USC 6185; 8 km. N Ciudad Quesada, USC 7155 (4); La Fortuna, USC 7153 (20); 3 km. E La Fortuna, USC 7150; San Carlos, USNM 29969; Sarchi, KU 32990-9, 36792-3.
Cartago: Cartago, BMNH 1947.2.24.13; headwaters of Río Pacuare, USC 119; Instituto Interamericano de Ciéncias Agricolas, Turrialba, KU 37012, USC 420, 437; Río Reventazón, Turrialba, MCZ 29268: 10 km. N Río Reventazón bridge, USC 7073; 5 km. SW Río Reventazón bridge on Paraiso-Orosi road, USC 669; Turrialba, UMMZ 118405, USC 455, USNM 29936-9.
Heredia: Puerto Viejo, KU 36791.
Guanacaste: Las Cañas, USC 7164; Santa Cecilia, MCZ 7924-5; Tilarán, USC 7161 (5).
Limón: Bambú, USC 7171 (2), 7183 (13); La Lola, USC 820 (6), 6083-94, 8064, 8071; Pandora, USC 7188 (7), 7189, 7190 (3), 7191 (5); Pico Blanco, USNM 30685-6; Río Larí, 14-16 km. SW Amubre, USC 7179, 7180 (10); Sipurio, USNM 30658-9; Suretka, KU 36764, 36765 (skeleton), 36766-78.
Puntarenas: 6 km. N Dominical, KU 91749-50, 91811 (young), 91812 (tadpoles); Esparta, MCZ 8028; 6 km. E Golfito, KU 91718-41, 91809 (young), 91810 (tadpoles), 91816-9 (skeletons), USC 7103 (23); Quebrada Agua Buena, 3 km. SW Rincón de Osa, USC 7236 (6); Quebrada Boruca, 22 km. E Palmar Norte, KU 64264; Rincón de Osa, Camp Seattle, UMMZ 123680-5, S-2792 [Pg 330] (skeleton), USC 705 (5), 6023, 7254; Río Barranca, USC 7119 (2); Río Ceiba, 6 km. NW Buenos Aires, KU 91747-8, USC 7112 (7); Río Ciruelitas, 16 km. NW Esparta, USC 7121 (3); Río Claro, 14.2 km. NW Villa Neily, USC 7110 (4); Río Ferruviosa, 7 km. S Rincón de Osa, USC 7235 (4); Río Lagarto at Pan-American Hwy. (Guanacaste Border), USC 7122 (4); Río La Vieja, 30 km. E Palmar Norte, KU 87684 (tadpoles), 91743-6, USC 7083 (2); Río Oro, 28.5 km. NW Villa Neily, KU 91742; Río Volcán, 10 km. W Buenos Aires, USC 7113; Río Zapote, 7 km. E Palmar, USC 7100 (4); 3-5 km. W Palmar, USC 7101 (18); 7 km. SE Palmar, KU 64261-3; 1.2 km. NW Villa Neily, USC 8032; 3 km. NW Villa Neily, USC 7109 (20); 5 km. NW Villa Neily, USC 6176, 8035.
San José: Bajos de Jorco, KU 91813 (tadpoles); Escazú, KU 34863, 34869-75, USC 813; between Monrovia and La Hondura, ± 0.5 km. N Santa Rosa, USC 302 (2); Paso Ancho, Río Jorco, UMMZ 122649 (6), USC 530 (3); Río Jorco, near Desamparados, KU 91757-65, 91796-7, 91820-3 (skeletons), USC 228, 513, 7117 (7); Río Peje, 10 km. SSE San Isidro el General, USC 7115 (3); Río Tiriví, MCZ 7972; San Isidro el General, CNHM 101096, KU 28201, 32989, UMMZ 72024; 15 km. WSW San Isidro el General, KU 64245-56, 68473 (tadpoles), 68474 (young), 68475 (tadpoles), 86516, 91754-6, 91793-5, USC 7097 (6); 17.1 km. WSW San Isidro el General, USC 6047; 18 km. WSW San Isidro el General, USC 689; 20 km. WSW San Isidro el General, KU 64257-9, 64260 (skeleton), 68468 (young), 68469 (tadpoles), 68470 (young), 68471-2 (tadpoles), 68476 (young), 68633-4 (skeletons), 91751-3; San José, AMNH 7501-4, USC 298; Santa Rosa, Río Virilla, USC 7145.
Panama: Chiriquí: Río Jacu, 5.8 km. ESE Paso Canoas, KU 91905. "Veraguas," ZMB 3141 (2).
In attempting to assay the taxonomic significance of skeletal differences we are faced with a dearth of data on the skeletons of frogs in general and hylids in particular. Recent reviews by Brattstrom (1957) and Hecht (1962, 1963) have been concerned with general salientian classification and phylogeny, principally at the family level. Savage and Carvalho (1953), Griffiths (1959), and Baldauf (1959) used osteological characters in determining the taxonomic status of the families Pseudidae, Brachycephalidae, and Bufonidae, respectively. Carvalho (1954) presented osteological evidence for the generic separation of New World microhylids. Zweifel (1956) and Tihen (1962) used osteological characters at the levels of the species-group and species in their respective studies on Scaphiopus and Bufo. Little has been recorded about the skeletons of the hylids. Goin (1961) mentioned dentigerous elements and cranial co-ossification in his synopsis of the genera of hylids. Copland (1957) in his review of the Hyla of Australia, Funkhouser (1957) in her revision of Phyllomedusa, and Zweifel (1958) in his review of Nyctimystes did not consider skeletal characters.
Some osteological studies on hylids have yielded worthwhile information. Mittleman and List (1953) used osteological characters in defining the genus Limnaoedus: Starrett (1960) used cranial characters in combination with jaw musculature in defining the genus Smilisca, and Duellman (1964) used cranial characters in delimiting the Hyla bistincta group. Brief descriptions of cranial structure were given for Phrynohyas (Duellman, 1956) and Ptychohyla (Duellman, 1963a); specific and sexual differences in the skulls of Hyla chaneque and Hyla taeniopus were pointed out by Duellman (1965). Stokely [Pg 331] and List (1954) described early cranial development in the hylid Pseudacris triseriata triseriata.
Because our knowledge of the skeleton in hylids is so incomplete, we are not attempting to place Smilisca in the general scheme of hylid phylogeny on the basis of skeletal characters. Instead, our purposes are to describe the skeleton and its ontogenetic development in one member of the genus (S. baudini), and to make comparisons that show taxonomic differences in osteological characters among species of Smilisca.
The study of 68 dried skeletons and 25 cleared and stained preparations, including an ontogenetic series of S. baudini, has resulted in an understanding of the progressive development of skeletal elements and a knowledge of interspecific and intraspecific variation in these elements. Furthermore, investigations of the osteology have provided correlations between some cranial characters and certain aspects of external morphology.
The following description is based primarily on an adult female (KU 68184):
Skull.—The skull is large, solid, and broader than long; the greatest width is between the sutures of quadratojugal and maxillary on either side of the skull (Pls. 2-3). The maxillaries bear well-developed dorsal flanges, curve gently, join the moderately convex premaxillaries anteriorly and form a slightly truncate snout. The combined premaxillary width is about one-fourth the width of the skull. The premaxillaries are separated medially, and laterally from the maxillaries by sutures. Each premaxillary bears a dorsomedial alary process, which is anteriorly convex and four times as high as the depth of the lateral wing of premaxillary; each premaxillary also has a ventromedial palatine process that projects dorsally from the lingual edge of the premaxillary. The septomaxillaries are closely associated dorsally with the premaxillaries immediately lateral to the prenasal processes.
The nasals are large, widest anteriorly and narrowing posteriorly, parallel to maxillaries, and not separated from the ethmoid by cartilage. The nasals bear long, delicate maxillary processes extending nearly to the maxillaries. Anteriorly, the nasals are widely separated by the partially ossified internasal septum, which is in contact with the premaxillaries between the prenasal processes; the anterior points of the nasals lie approximately one-half the distance between the anterior ends of the ethmoid and the premaxillaries. The ethmoid is large and completely ossified; the margins are smooth. The trunate anterior edge lies between the nasals and is in contact with the internasal septum. The frontoparietals are large, smooth-margined, and bear large supraorbital flanges curving posterolaterally at the rear of the orbit. A small, oval foramen involves the posterior part of the ethmoid and anterior portion of frontoparietals; continued ossification in older specimens fills in the foramen, thereby resulting in a solidly roofed cranium. The auditory regions are relatively massive and bear narrow tegmen tympani; the distal ends of the tegmen tympani are medial to the lateral edge of the pterygoids in dorsal view. The squamosals are large; the long anterior arm is separated from the maxillary by a suture. The delicate, spindle-shaped columellae lie ventral to the tegmen tympani and squamosals, are spatulate distally, and have a broad basal attachment to the auditory region.
The vomers are moderately large and are in contact anteriorly with the premaxillaries and posteriorly with the ethmoid. Each vomer has two wide serrated flanges laterally. The tooth-bearing parts of the vomers are widely separated and at a slight angle to one another; the vomers terminate medially in two pointed processes on the ethmoid. The palatines are edentate, but bear strong ridges throughout their lengths. They are broadly in contact with the maxillary, are narrow medially, and are attached by pointed processes to [Pg 332] the medial part of the ethmoid. The pterygoids are large, attached to the maxillaries immediately anterior and medial to the squamosal-maxillary connection, bear well-developed pedicles, which are broadly attached to the proötic, and a wide wing is in contact posteriorly with the distal two-thirds of the quadrate.
The angular makes up most of the lower jaw, bears a broad articular surface posteriorly, and has a small coronoid process on the lingual edge; anteriorly the angular is separated from the dentary and mentomecklian by Meckel's cartilage. The dentary lies external to the angular and extends from the mentomecklian to approximately the mid-length of the angular. The mentomecklians are ossified, but separated by cartilage medially.
Hyoid.—The hyoid plate is curved, thin, and mostly cartilaginous, but calcined posteriorly (Fig. 6). The anterior cornua are slender, cartilaginous, and curve anteromedially from the hyoid plate and thence laterally and posteriorly, to attach to the posterior surface of the proötics. The lateral cornua are broad, flat, cartilaginous lateral extensions from the bases of the anterior cornua. The posterior cornua are bony, except distally.

Vertebral Column.—The atlas lacks transverse processes and a neural crest, whereas transverse processes are present on the other seven presacral vertebrae, and knoblike neural crests are present on the second, third, and fourth vertebrae; a faint neural ridge is visible on the fifth vertebra. The transverse processes are directed laterally on the second and sixth vertebrae, ventrolaterally on the third, posterolaterally on the fourth and fifth, and anterolaterally on the seventh and eighth. The processes are slightly expanded on the fourth, and more so on the fifth, vertebra. The sacral diapophyses are expanded and have a border of calcified cartilage laterally. There are two sacral condyles. The slender coccyx has a thin dorsal ridge on the anterior three-fourths of its length.
Pectoral Girdle.—The omosternum is large, ovoid, and cartilaginous; the sternum is a thin cartilaginous sheet deeply notched posteriorly and is not differentiated into episternal and xiphisternal elements. The coracoids are robust, twice as stout as the clavicles. The epicoracoidal cartilages overlap in the usual arciferal manner, except that they are fused anteriorly between the slender clavicles. The clavicles are strongly arched. The clavicle, coracoid, and scapula on each side form a bony articulation at the glenoid fossa. A bifurcation of the ventral end of the scapula results in a large glenoid foramen. The scapula is flat and expanded dorsally; the suprascapula is broad, flat, and calcified in large adults. In young specimens no distinct ossification of the cleithrum or ossification of endochondral centers are evident.
Arm and Hand.—The humerus is equally well-developed in both sexes and has a prominent lateral crest. The radius and ulna are completely fused. A bony prepollex is present in both sexes. The metacarpals are about equal in length. The phalangeal formula is 2-2-3-3; the terminal phalanges are claw-shaped.
Pelvic Girdle.—The ilia are long, slender, and slightly curved. A thin ridge projects laterally from the dorsal edge of the posterior one-half of each ilium. The ilial prominence is large and knoblike when viewed from above. The anterior edge of the ilial prominence is at the level of the anterior edge of the acetabular border. The dorsal acetabular expansion is small. The pubis is slender, and the ischium is elevated and robust.
Leg and Foot.—The slightly curved femur has a distinct crest proximally on the posterior surface. The nearly straight tibio-fibula is slightly longer than the femur. The tibial and fibial elements are completely fused but have a distinct cleft between them. A small foramen exists at the mid-length of the tibio-fibula. The fibulare (calcaneum) is much more robust than the tibiale (astragalus). The prehallux is large and flat. The metatarsals of the third, fourth, and fifth digits are equal in length; the metatarsal of the second is somewhat shorter, and that of the first is much shorter. The phalangeal formula is 2-2-3-4-3; the terminal phalanges are claw-shaped.
The following description of development of the skull of Smilisca baudini is based on the examination of 12 cleared and stained specimens. In table 3 the cranial bones are listed in the left hand column in the approximate order of their appearance in the young frogs. Across the top of the table selected specimens designated by developmental stage or snout-vent length are listed. It should be noted that although each individual, from left to right, has an increasing number of ossified bones, the correlation with increasing size is imperfect; the precise ages of the individuals are unknown.
The first bones to appear are the septomaxillaries, frontoparietals, part of the exoccipital, and the parasphenoid in developmental stage 40. The frontoparietals are represented by two slender ossifications dorsomedial to the orbits; the septomaxillaries are present as small ossifications anterior to the nasal capsules (Pl. 1A). The parasphenoid is present as a faint median ossification, and the exoccipital shows some ossification.
| Table 3.—The Order of Occurrence of Cranial Ossifications in the Skull of Smilisca baudini. Where Numbers Are Divided by a Slash Mark, the Left and Right Symbols Correspond to the Left and Right Sides of the Skull, Respectively. | |||||||
| Bone | Stage 40 | Stage 44 | 12.6 mm. | 13.9 mm. | 32.0 mm. | 27.0 mm. | 20.1 mm. |
| Frontoparietal | X | X | X | X | X | X | X |
| Parasphenoid | X | X | X | X | X | X | X |
| Septomaxillaries | X | X | X | X | X | X | X |
| Exoccipitals | X | X | X | X | X | X | X |
| Squamosals | — | X | X | X | X | X | X |
| Premaxillaries | — | X | X | X | X | X | X |
| Maxillaries | — | X | X | X | X | X | X |
| Nasals | — | — | X | X | X | X | X |
| Pterygoids | — | — | X | X | X | X | X |
| Vomers | — | — | — | X | X | X | X |
| Palatines | — | — | — | X | X | X | X |
| Quadratojugals | — | — | — | X | X | X | X |
| Ethmoid | — | — | — | — | X | X | X |
| Columellas | — | — | — | — | X | X | X |
| Supraorbital Flanges | — | — | — | — | — | X | X |
| Proötics | — | — | — | — | — | — | X |
| Vomerine Teeth | — | — | 1/1 | 4/3 | 5/5 | 3/3 | 5/4 |
| Maxillary Teeth | — | 0/7 | 3/5 | 6/5 | 30/31 | 30/26 | 37/36 |
| Premaxillary Teeth | — | 2/4 | 3/3 | 5/5 | 7/6 | 8/6 | 8/7 |
The dentigerous bones are among the most rapidly developed, although not the first to appear. They are present in developmental stage 44 before metamorphosis is completed. The maxillaries bear a few teeth anteriorly and are ossified posteriorly to a point one-third of the distance from the anterior to the posterior edge of the orbit. Ossification lengthens the posterior termini of the maxillaries to the posterior edge of the orbit. In front of the anterior margin of the orbit, bone is proliferated dorsal to the main axes of the maxillaries and forms moderate dorsal maxillary flanges. The premaxillaries appear simultaneously with the maxillaries. Initially they are widely separated medially from each other, and laterally from the developing maxillaries; each bears two or three teeth, large dorsally blunt alary processes, and small [Pg 335] palatine processes. The median and lateral edges of the prenasal processes lengthen heterochronously, causing the median edges to be longest and to lie slightly dorsal to the level of the septomaxillaries. After the maxillaries and premaxillaries develop, the vomers appear as small horizontal ossifications anterior to the parasphenoid. Ossification begins in the lateral flanges, then in the prevomerine processes, and lastly in the posterior dentigerous parts of the bones; the prevomerine processes are the last parts of the vomers to ossify completely.
Initially the frontoparietals are present as thin rods of ossification dorsomedial to the orbits; the frontoparietals extend from the anterior to the posterior end of the orbit by developmental stage 44. The anterior ends of the bones remain thin and pointed; ossification progresses medially from the midpoint of the length of the orbit and posteriorly to the level of the exoccipital; a median center of ossification joins the frontoparietals posteriorly, thereby forming the posterior border of the frontoparietal fontanelle. The supraorbital flanges of the frontoparietals do not appear until all other cranial bones are ossified, or nearly so. The most rapid ossification begins laterally at the posterior edge of the orbit and decreases anteriorly over the posterior half of the orbit. This differential rate of proliferation of bone results in the pattern of development of the supraorbital flanges shown in figure 7. The nasals appear as thin slivers of bone half way between the anterior ends of the frontoparietals and the end of the snout. As ossification proceeds the nasals assume a triangular shape in dorsal view. The anterior ends are pointed; the lateral margins are parallel to the maxillaries. The posteromedial points do not reach the lateral margins of the ethmoid, and the maxillary processes extend about three-fourths the distance from the bodies of the nasals to the maxillaries. Following the union of the frontoparietals posteriorly, the nasals widen anteriorly and are narrower at the midpoints of their long axes than anteriorly or posteriorly. With further ossification the maxillary processes extend to the maxillaries and form complete bony anterior margins to the orbits; the mid-parts of the nasals widen (Pl. 1B).
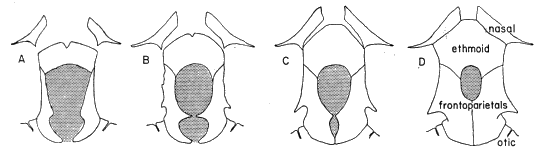
The parasphenoid is the first of the palatal bones to appear. At metamorphosis the bone is well developed; the anterior tip is situated just in front of the anterior edge of the orbit, and posteriorly the lateral processes extend laterally beyond the ossified parts of the auditory region. The pterygoids do not appear until metamorphosis, when ossification is evident in only the mid-parts of the posterolateral arms. Ossification follows in the mid-parts of the [Pg 336] anterolateral arms and occurs last in the pterygoid pedicles. The palatines do not appear until all three arms of the pterygoids are at least partly ossified. Ossification proceeds rapidly from the maxillaries medially to the unossified ethmoid, which is the last of the cranial bones to appear. Initially it is extremely shallow; dorsally it is widely separated from the nasals, and ventrally the posterior margin meets the anterior point of the parasphenoid. In dorsal view, ossification proceeds anteriorly between the nasals and posteriorly, ventral to the frontoparietals; ventrally, ossification proceeds posteriorly dorsal to the parasphenoid.
The ventral arms of the squamosal and the supraoccipital region of the exoccipital are the first occipital bones to appear. Ossification follows in the regions of the semicircular canals and occipital condyles. The dorsal end of the ventral arm of the squamosal and the posterior arm of the squamosal ossify as a unit at the same time the quadratojugal appears. Shortly thereafter the anterior arm of the squamosal ossifies, the distal part of the columella appears, and the anterior and lateral parts of the auditory region ossify.
The angular and dentary of the lower jaw appear concurrently with the dentigerous bones. Initially, the angular is short and broad; the articular surface is absent, and the anterior end is slightly overlapped by the dentary. The mentomecklians do not ossify until approximately the same time that the quadratojugal appears in the upper jaw.
The genus Smilisca is characterized by the following combination of cranial osteological characters: (1) A large amount of bone is involved in the skull and a minimal amount of cartilage and/or secondarily ossified cartilage; co-ossification is absent. (2) The skulls are uniformly broad with angular lateral margins, and truncate anteriorly. (3) An internasal septum and quadratojugals are present. (4) A well-developed squamosal minimally extends one-fourth the distance from the dorsal end of the quadrate to the maxillary, and maximally is separated from the maxillary by a suture. (5) The ethmoid is large; the distance between the anterior end of the ethmoid and the anterior edge of the premaxillary varies between 15 and 20 per cent of the total length of the skull.
On the basis of cranial osteology two species-groups can be recognized within the genus Smilisca. The sordida group, comprising S. sordida and puma, is characterized by a broad skull in which the lateral margins of the maxillaries are relatively straight anterior to the orbit. The moderate-sized nasals are rounded anteriorly, and bear relatively short, sometimes blunt, maxillary processes. The long axes of the nasals are not parallel to the maxillaries. The ethmoid is proportionately small in the sordida group. The bony part of the ethmoid terminates near the anterior edge of the orbits and does not extend anteriorly between the nasals; the entire anterior margin of the ethmoid is separated from the nasals by cartilage. The squamosals are generally small. They are narrow in dorsal view, and minimally extend one-fourth the distance from the dorsal end of the quadrate to the maxillary, and maximally, two-thirds the distance. The tegmen tympani are relatively small (Fig. 8).
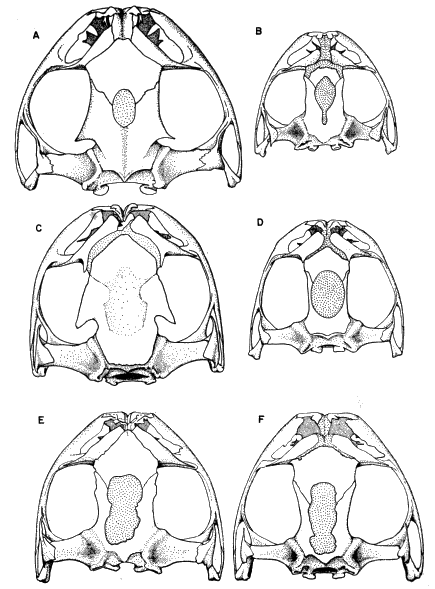
In contrast to the tendency for reduction of cranial parts in the sordida group, the baudini group, constituted by S. cyanosticta, phaeota, and baudini, is characterized by more ossification of the cranial elements. The skull is broad; the lateral margins are less angular and are gently curved, rather than straight as in the sordida group. The nasals tend to be larger with the long axes parallel to the maxillary. Anteriorly the nasals are pointed, and posteriorly [Pg 338] they bear long, delicate palatine processes extending to the maxillary. The ethmoid is fully ossified, extends anteriorly between the nasals, and laterally is separated by a suture from the nasals if the latter are fully ossified. The squamosals are large, and wide in dorsal view. They minimally extend one-fourth the distance from the dorsal end of the quadrate to the maxillary, and maximally are sutured to the maxillary. The tegmen tympani are massive.
Smilisca sila is intermediate between the two species-groups described. The skull is broad; the lateral margins are gently curved, and have a pronounced angularity just anterior to the palatines which results in a broad, truncate snout. The nasals are moderate in size; because of the anterior angularity of the lateral margins, the long axes of the nasals lie parallel to the maxillary. The nasals are only slightly pointed anteriorly, and posteriorly they bear short, blunt palatine processes and medial processes in contact with the lateral corners of the ethmoid. The ethmoid is fully ossified, but does not extend anteriorly between the nasals. The squamosals are moderate in size and extend one-fourth the distance from the dorsal end of the quadrate to the maxillary. The tegmen tympani are relatively large, but proportionately short.
The cranial characters utilized in the analysis of species groups (general shape, nature of the nasals, ethmoid, squamosals, and tegmen tympani), together with other characters, such as the relative height and shape of the prenasal processes, the extent of the internasal septum, and the nature of the vomers, frontoparietals, maxillaries and pterygoids are useful in distinguishing the various species (Table 4, Fig. 8), as well as in establishing relationships within the species-groups.
Within the sordida group, S. sordida and S. puma can be distinguished by the following characters: The bony part of the ethmoid terminates posterior to the anterior edge of the orbit and is thus widely separated from the nasals by cartilage in S. puma. In S. sordida the bony part of the ethmoid always terminates at a level equal to, or slightly in front of the anterior edge of the orbit; therefore, less cartilage exists between the ethmoid and nasals in S. sordida than in S. puma. The width of the premaxillary comprises about 30 per cent of the width of the skull in S. sordida and 20 per cent in S. puma. The proportion of the length of the skull anterior to the bony part of the ethmoid in S. sordida is approximately 21 per cent, as compared with about 29 per cent in S. puma. The prenasal processes are convex in S. sordida and straight in S. puma.
The marked ontogenetic variation in S. sordida is considered in more detail in the account of that species, but it is pertinent to the present discussion to note that with respect to some features of the skull some young breeding specimens of S. sordida are intermediate in appearance between large females of S. sordida and adults of S. puma. In some breeding males (usually the smaller individuals) of S. sordida the bony part of the ethmoid terminates at the anterior edge of the orbit and is widely separated from the nasals by cartilage. In small individuals S. sordida, especially in males, and in adults of S. puma the tegmen tympani are relatively short, whereas in adult females of S. sordida these elements are long and slender. In the smaller specimens of S. sordida and in S. puma the squamosal is small; it extends only about one-fourth of the distance to the maxillary in the smaller S. sordida and about one-half the distance in S. puma. The more massive squamosal in large adult females of S. sordida extends at least two-thirds of the distance to the maxillary.
| Table 4.—Comparative Cranial Osteology of Smilisca. | ||||||
| Character | S. baudini | S. cyanosticta | S. phaeota | S. puma | S. sila | S. sordida |
| Alary Processes | Four times as high as lateral wing of premaxillary; anteriorly convex. | Three times as high as lateral wing of premaxillary; anteriorly convex. | Two and one-half times as high as lateral wing of premaxillary; anteriorly convex. | Two times as high as lateral wing of premaxillary; straight. | One and one-half times as high as lateral wing of premaxillary; straight. | Two and one-half times as high as lateral wing of premaxillary; slightly convex anteriorly. |
| Nasals | Long, wide anteriorly, narrowing posteriorly; attached to ethmoid. | Long, widest posteriorly; attached to ethmoid. | Long, widest anteriorly and posteriorly, bearing posteromedial process; not attached to ethmoid. | Short, narrow, not attached to ethmoid. | Short, wide, bearing small posteromedial processes; not attached to ethmoid. | Moderately long narrowest anteriorly and posteriorly; not attached to ethmoid. |
| Ethmoid | Long; entirely ossified; smooth margins. | Long, entirely ossified; smooth margins. | Long, entirely ossified; smooth margins. | Short, about two-thirds ossified; irregular margins. | Moderately long; entirely ossified; smooth margins | Short; one-half to entirely ossified; irregular margins. |
| Frontoparietal | Small, ovid fontanelle present or absent; long, pointed postorbital processes curving along posterior border of orbit. | Large fontanelle, two and one-half times as long as wide; narrow supraorbital flanges with irregular margins. | Fontanelle absent; large supraorbital flanges having straight edges and extending posterolaterally. | Keyhole-shaped fontanelle; smooth margins; flanges absent. | Large, ovoid fontanelle; smooth margins; flanges absent. | Large, elongate fontanelle; smooth margins; flanges absent. |
| Squamosal | Large: anterior arm in contact with maxillary. | Large; anterior arm in contact with maxillary. | Large; anterior arm extending 1/2-2/3 way to maxillary. | Small; anterior arm extending 1/2 way to maxillary. | Moderately large; anteriorarm extending 1/4 way to maxillary. | Moderately small; anterior arm extending 1/4-2/3 way to maxillary. |
Within the baudini group, the skull of S. cyanosticta is the most generalized of the three species; the cranial characters are intermediate between S. phaeota and S. baudini. The lateral margins of the skull in S. cyanosticta are gently curved, and have an angularity anterior to the palatine-maxillary suture; the anterior margins are less angular in S. phaeota, which has a broader snout. Posteriorly in S. baudini the margins are slightly curved medially, and the greatest width of the skull is between the quadratojugal-maxillary sutures on either side of the skull. The frontoparietals of S. cyanosticta bear slightly irregular lateral margins and a large fontanelle. There is a tendency for obliteration of the fontanelle with increasing age in both S. baudini and S. cyanosticta; the lateral margins of the frontoparietals bear large supraorbital flanges in both of these species. In S. phaeota the flanges are most prominent; they extend posterolaterally with straight margins along two-thirds of the length of the orbit and terminate in rather blunt points. The broad interorbital flanges result in a relatively broad external interorbital distance. In S. baudini the flanges are curved posterolaterally around the orbit and terminate in sharp, thin points. The tegmen tympani of all three species are massive. In S. cyanosticta the proötics slope posteriorly, whereas they slope anteriorly in S. baudini and S. phaeota.
The skulls of S. cyanosticta and S. baudini are alike in certain respects. The squamosals of both species are large and connected to the maxillary by a bony connection; the squamosals of S. phaeota are large, but extend only two-thirds of the distance from the dorsal end of the quadrate to the maxillary. In S. baudini and S. cyanosticta the nasals are separated throughout their lengths from the ethmoid, whereas the nasals of S. phaeota are separated from the ethmoid by cartilage. The latter separation is due to an incomplete ossification of the nasals in S. phaeota. The bony part of each nasal is constricted in the middle of the long axis of the bone, and the nasals are widest anteriorly; posteriorly each nasal bears a medial process, which is narrowly separated from the lateral edge of the ethmoid.
| Table 5.—Variation in the Number of Teeth in the Species of Smilisca. (All Are Males; N = Number of Jaws, or Twice the Number of Individuals; Means Are Given in Parentheses After the Observed Ranges.) | ||||||
| Species | N | Maxillary | Premaxillary | Vomerine | ||
| S. baudini | 20 | 49-65 (56.0) | 9-16 (13.6) | 5-9 (7.2)) | ||
| S. cyanosticta | 8 | 50-64 (57.9) | 10-12 (10.8) | 4-11 (7.1)) | ||
| S. phaeota | 20 | 50-68 (58.1) | 10-15 (12.1) | 5-9 (7.3)) | ||
| S. puma | 6 | 60-67 (63.6) | 11-13 (12.0) | 4-7 (5.3)) | ||
| S. sila | 8 | 48-60 (52.9) | 10-14 (11.3) | 5-7 (5.7)) | ||
| S. sordida | 12 | 39-55 (44.2) | 7-11 (9.3) | 4-6 (5.2) | ||
The teeth of all species of Smilisca are spatulate and bifid. The numbers of maxillary, premaxillary, and vomerine teeth are summarized in Table 5. Smaller and presumably younger specimens of all species of Smilisca have fewer teeth than do larger specimens of the same species. This correlation [Pg 341] between size and number of teeth does not exist as an interspecific trend within the genus; for example, the smallest species in the genus, S. puma, has the highest number of maxillary teeth. In small specimens of a given species wide gaps are present between the maxillary teeth posteriorly; in large specimens the gaps are filled by teeth, beginning anteriorly and progressing posteriorly, until the maxillary dentition is continuous.
No extensive study of the muscular system was undertaken, but certain muscles know to be of taxonomic importance were studied.
Jaw Musculature.—Starrett (1960) pointed out the unique jaw musculature in Smilisca. In this genus M. depressor mandibulae consists of two parts, one arising from the dorsal fascia and one from the posterior arm of the squamosal. Two muscles arise from the anterior arm of the squamosal and insert on the lateral face of the mandible. Of these muscles, M. adductor mandibulae posterior subexternus lies medial to the mandibular branch of the trigeminal nerve; the other, M. adductor mandibulae externus superficialis, lies lateral to the same nerve (Fig. 9). In most other hylids the latter muscle is absent. No significant variation in the position of the muscles was noted in the various species of Smilisca, though M. adductor mandibulae originate somewhat more anteriorly in S. baudini and S. cyanosticta than in the other members of the genus, all of which have a shorter anterior arm of the squamosal that does not reach the maxillary. The two separate parts of M. depressor mandibulae are not so widely separated in members of the sordida group as in the baudini group.


Throat Musculature.—The frogs that comprise the genus Smilisca are characterized by paired subgular vocal sacs, essentially the same as those in Triprion (Duellman and Klaas, 1964). The following description is based on Smilisca baudini (Fig. 10).
M. submentalis lies in the anterior angle of the lower jaw, is thick, and consists of transverse fibers extending between the dentaries. M. submaxillaris is thin and arises from the whole of the inner surface of the lower jaw, except for the anterior angle occupied by M. submentalis. Anteriorly M. submaxillaris is broadly attached by fascia to M. hyoglossus and M. geniohyoideus, which lie dorsal to M. submaxillaris. Medially this attachment continues posteriorly for about one-half the length of the hyoglossus. Posteriorly M. submaxillaris is folded and attached to M. sternoradialis of the pectoral girdle. The vocal sacs are formed by a pair of posterolateral evaginations of M. submaxillaris; a broad connection between the pouches allows free passage of air between the pouches.
The deeper throat musculature is essentially the same as that described for Phrynohyas spilomma by Duellman (1956), except for slight differences in the place of attachment on the hyoid.
The skin of Smilisca is typical of that of most hylids in organization and structure. Smilisca sila is distinguished from other members of the genus by the presence of small wartlike protrusions and peculiar white, pustular spots on the dorsum. The wartlike structures are composed of three or four epidermal cells, which protrude from the surface of the epidermis; the structures are covered by a slightly thickened layer of keratin. The white pustules are slightly elevated above the surrounding skin. Internally they consist of aggregations [Pg 343] of swollen, granular, pigment-cells (perhaps lipophores) lying between the epidermis and the melanophores.
Dried skins of all species of Smilisca were sent to José M. Cei, Instituto Nacional de Cuyo, Mendoza, Argentina, for biochemical screening by means of the chromatographic techniques described by Erspamer and Cei (1963). The species in the baudini group have detectable amounts of penta-hydroxi-trypatamine, whereas only a trace is present in the other species. Furthermore, species in the baudini group differ from S. sila and the sordida group in lacking, or having only a trace of, tryptophan-containing polypeptides. These superficial biochemical tests support the arrangement of species as ascertained by conventional taxonomic characters.
The features of external morphology that were studied in connection with the taxonomy of the genus Smilisca are discussed below.
The frogs of the genus Smilisca are medium to large tree frogs. The three species comprising the baudini group (S. baudini, cyanosticta, and phaeota) are notably larger than S. puma, sila, and sordida (Table 6). The largest specimen that we examined is a female of S. baudini having a snout-vent length of 90 mm. Smilisca puma is the smallest species; the largest male has a snout-vent length of 38 mm. and the largest female, 46 mm.
| Table 6.—Comparison of Sizes and Certain Proportions of the Species of Smilisca. (Means in Parentheses Below Observed Ranges; Data for Males Only.) | ||||
| Species | N | Snout-vent length |
Tibia length/ snout-vent |
Tympanum/ eye |
| S. baudini | 140 | 47.3-75.9 | 42.1-53.6 | 56.1-94.4 |
| (58.7) | (47.8) | (73.5) | ||
| S. cyanosticta | 40 | 44.6-56.8 | 51.9-59.7 | 62.7-88.4 |
| (50.7) | (56.0) | (71.4) | ||
| S. phaeota | 50 | 40.8-65.5 | 50.9-60.2 | 62.7-85.5 |
| (53.9) | (55.5) | (76.6) | ||
| S. puma | 20 | 31.9-38.1 | 48.2-53.1 | 52.1-72.2 |
| (34.7) | (51.3) | (64.9) | ||
| S. sila | 33 | 31.6-44.8 | 49.7-58.1 | 47.6-58.3 |
| (37.7) | (54.8) | (53.2) | ||
| S. sordida | 55 | 31.9-44.6 | 50.5-57.1 | 46.5-57.1 |
| (37.9) | (53.4) | (49.1) | ||
No outstanding differences in proportions exist between species, although [Pg 344] certain proportions are sufficiently different in some species to warrant mention. Smilisca baudini is a more squat and stocky frog than other members of the genus; this is reflected in the somewhat shorter hind legs (Table 6). The size of the tympanum relative to that of the eye is highly variable within samples of a given species. Even so, noticeable differences in the tympanum/eye ratio are apparent. Members of the baudini group have the largest tympani, whereas S. sila and sordida have the smallest, and S. puma is intermediate (Table 6).
Although all members of the genus have rather truncate snouts, subtle differences exist among the species (Pl. 12). Smilisca sila has the shortest snout; that of S. baudini is only slightly longer. The snouts of S. cyanosticta and puma are nearly square in lateral profile, whereas those of S. phaeota and sordida are slightly inclined. The shape of the snout is relatively uniform within each species and displays no noticeable sexual dimorphism, except in S. sordida, in which there are sexual differences and geographic variation (see p. 324).
The characters of the hands and feet are among the most taxonomically important external features in Smilisca. Consistent differences exist in relative lengths of the digits, size of subarticular tubercles, size and number of supernumerary tubercles, size and shape of the inner metatarsal tubercle, and amount of webbing (Pls. 4 and 5). In the baudini group the series of species (baudini-phaeota-cyanosticta) show a progressive increase in amount of webbing in the hand and a decrease in number, and corresponding increase in size, of supernumerary tubercles. The amount of webbing in the feet of S. baudini and phaeota is about the same, but the webbing is slightly more extensive in S. cyanosticta. Smilisca puma is unique in the genus in lacking webbing in the hand; furthermore, this species is distinctive in having many large subarticular tubercles on the hand and a relatively small inner metatarsal tubercle. The two stream-inhabitants, S. sila and sordida, have shorter and stouter fingers than the other species. The webbing is most extensive in both the hands and feet of these species, which also are distinctive in having many small supernumerary tubercles on the feet.
Minor ontogenetic changes in structure involve the shape of the snout, relative size of the eye, development of the tympanum, and amount of webbing in the hand. In recently metamorphosed young the snout is more rounded than in adults; the canthus and loreal concavity are not evident. Usually the tympanum is not differentiated in recently metamorphosed young, and the eye is proportionately large. The webbing in the feet is completely developed at metamorphosis, but young individuals have noticeably less webbing in the hand than do adults of the same species.
Some of the most distinctive characters of the species of Smilisca are color and pattern of the living frogs. Although many chromatic features are lost or subdued in preserved specimens, the patterns usually persist.
Change in color, well known in frogs, is common in hylids, especially in species having green dorsal surfaces (Phyllomedusa is a notable exception). The non-green Smilisca (puma, sila, and sordida) changes color, but this mostly is a change in intensity of color. In these species the markings usually are most distinct at night; frequently by day the frogs become pallid. The most striking examples of metachrosis in Smilisca are found in the baudini group, in which the dorsal ground-color changes from green to tan; correlated with the change in ground-color may be a corresponding change in the dorsal markings, but the dorsal markings may change to the opposite color.
Chromosomes of all six species of Smilisca were studied by means of the propriono-orcein squash technique described by Duellman and Cole (1965). Karyotype analysis was attempted for several species by means of intraperitoneal injections of colchicine, which affected the mitotic cells as desired, but the testes examined contained too few mitotic cells to allow accurate determination of karyotypes.
Haploid (n) chromosome numbers were determined from cells in diakinesis, metaphase I, and metaphase II of meiosis. Diploid (2n) chromosome numbers were determined from cells in late prophase and metaphase of mitosis. Chromosome counts from as few as 23 meiotic cells of S. phaeota and as many as 80 cells of S. sordida reveal a constant haploid (n) number of 12; counts of chromosomes in one to five mitotic cells in all species, except S. sila, reveal that the diploid (2n) number is 24.
Like most hylid frogs Smilisca is most readily collected and observed when individuals congregate for breeding.
Smilisca breeds primarily in quiet water and reaches its height of breeding activity at times of plentiful rainfall,—usually from May through October. Through most of its range Smilisca baudini breeds in those months, but in some places where abundant rain falls in other seasons, the species breeds at those times. For example, in southern El Petén and northern Alta Verapaz, Guatemala, Smilisca baudini has been found breeding in February and March. The other pond-breeding species (S. cyanosticta, phaeota, and puma) live in regions lacking a prolonged dry season, and possibly they breed throughout the year, but breeding activity seems to be greatest in the rainiest months.
The two stream-breeding species (S. sila and sordida) breed in the dry season when the streams are low and clear, principally in December through April. At high elevations the species sometimes breed in the rainy season; also, individuals sometimes breed in the short dry season (summer canicula) in July and August.
[Pg 346] At several localities species have been found breeding at different times of the year: S. baudini in March and July at Chinajá, Guatemala; S. phaeota in April and August at Palmar Sur, Costa Rica; S. puma in February and July at Puerto Viejo, Costa Rica; and S. sila in February, April, and August at El Volcan, Panamá. These observations indicate only that the population breeds at more than one time in the year, but do not provide any evidence on the breeding cycles of the individual frogs. This is one important aspect of the natural history of Smilisca for which we lack data.
All members of the genus Smilisca presumably deposit their eggs in water.
Smilisca baudini usually breeds in temporary rain pools; often these are nothing more than shallow, muddy puddles. In other instances the sites are extensive ditches or large flooded areas (Pl. 8, Fig. 1). This species is an opportunistic breeder, and males gather at any of a wide variety of suitable breeding sites that are formed by torrential rains in the early part of the rainy season. Smilisca baudini nearly always breeds in open pools having bare earthen edges. Frequently congregations of S. baudini are found at such small pools, but are absent from nearby large ponds surrounded by vegetation.
Little is known of the breeding habits of S. cyanosticta, which inhabits humid forests on foothills and lowlands. Apparently its breeding sites are not unlike those of S. phaeota, which usually are pools surrounded by vegetation (Pl. 8, Fig. 2), although sometimes males of S. cyanosticta call from open muddy puddles. In uplands, where standing water is uncommon, this species breeds in quiet pools in streams.
Smilisca puma breeds in grass-choked ponds and marshes, where the males call from bases of dense clumps of grass in the water (Pl. 9, Fig. 1).
Smilisca sila and S. sordida differ noticeably from other species in the genus by breeding exclusively in streams, where males usually call from rocks or gravel bars in or at the edges of streams (Pl. 9, Fig. 2); sometimes individuals perch on bushes overhanging streams. In the streams, or parts of streams, utilized by these frogs the water is clear, shallow, and has a slow gradient; occasional males have been found calling along cascading mountain streams.
Breeding choruses composed of ten or more species of frogs are not uncommon in Middle America, but Smilisca usually breeds alone or with one or two other species and at the most five others. This tendency towards solitary breeding possibly is the result of selection of breeding sites that are unsuitable to many other species of frogs. Nevertheless, many other species of frogs have been found at the breeding sites with the various species of Smilisca; these breeding associates (Table 7) are most numerous for S. baudini, which has a broad geographic range, including a variety of habitats.
Calling sites.—All species of Smilisca usually call from the ground, including rocks and gravel bars; some individuals sit in shallow water near the edge of the pool or stream. Sometimes males of S. baudini, sila, and sordida call from low bushes or trees near the breeding site. One S. baudini was observed calling while it was floating on the surface of a pond. Smilisca cyanosticta, phaeota, and puma call from secluded places at the edge of the water or in the water, whereas S. baudini, sila and sordida call from open situations.
| Table 7.—Breeding Associates of the Various Species of Smilisca. | ||||||
| Associate | S. baudini | S. cyanosticta | S. phaeota | S. puma | S. sila | S. sordida |
| Rhinophrynus dorsalis | X | — | — | — | — | — |
| Leptodactylus bolivianus | — | — | X | — | — | — |
| Leptodactylus labialis | X | — | X | — | — | — |
| Leptodactylus melanonotus | X | — | X | X | X | — |
| Leptodactylus occidentalis | X | — | — | — | — | — |
| Leptodactylus quadrivittatus | — | — | X | — | — | — |
| Leptodactylus pentadactylus | — | — | X | X | — | X |
| Engystomops pustulosus | X | — | X | — | — | — |
| Bufo canaliferus | X | — | — | — | — | — |
| Bufo cavifrons | — | X | — | — | — | — |
| Bufo coccifer | X | — | — | — | — | — |
| Bufo coniferus | — | — | X | — | — | — |
| Bufo cristatus | — | X | — | — | — | — |
| Bufo gemmifer | X | — | — | — | — | — |
| Bufo haematiticus | — | — | X | — | X | X |
| Bufo kellogi | X | — | — | — | — | — |
| Bufo luetkeni | X | — | — | — | — | — |
| Bufo marinus | X | — | X | X | X | X |
| Bufo marmoreus | X | — | — | — | — | — |
| Bufo mazatlanensis | X | — | — | — | — | — |
| Bufo melanochloris | — | — | X | — | X | X |
| Bufo perplexus | X | — | — | — | — | — |
| Bufo typhonius | — | — | X | — | X | — |
| Atelopus varius | — | — | — | — | X | X |
| Diaglena reticulata | X | — | — | — | — | — |
| Diaglena spatulata | X | — | — | — | — | — |
| Table 7.—Continued | ||||||
| Associate | S. baudini |
S. cyanosticta |
S. phaeota |
S. puma |
S. sila |
S. sordida |
| Hyla boulengeri | — | — | X | — | — | — |
| Hyla colymba | — | — | — | — | X | — |
| Hyla ebraccata | X | — | X | — | — | — |
| Hyla elaeochroa | — | — | X | X | — | — |
| Hyla eximia | X | — | — | — | — | — |
| Hyla legleri | — | — | — | — | — | X |
| Hyla microcephala | X | — | X | — | — | — |
| Hyla phlebodes | — | — | X | X | — | — |
| Hyla picta | X | — | — | — | — | — |
| Hyla robertmertensi | X | — | — | — | — | — |
| Hyla rosenbergi | — | — | X | — | — | — |
| Hyla rufioculis | — | — | — | — | — | X |
| Hyla smithi | X | — | — | — | — | — |
| Hyla staufferi | X | — | — | — | — | — |
| Hyla walkeri | X | — | — | — | — | — |
| Phrynohyas inflata | X | — | — | — | — | — |
| Phrynohyas spilomma | X | — | — | — | — | — |
| Phrynohyas venulosa | X | — | — | — | — | — |
| Phyllomedusa callidryas | X | — | X | — | — | — |
| Phyllomedusa dacnicolor | X | — | — | — | — | — |
| Phyllomedusa moreleti | X | X | — | — | — | — |
| Pternohyla fodiens | X | — | — | — | — | — |
| Smilisca baudini | X | X | — | — | — | — |
| Smilisca cyanosticta | X | X | — | — | — | — |
| Smilisca phaeota | — | — | X | — | — | — |
| Smilisca puma | — | — | — | X | — | — |
| Table 7.—Concluded | ||||||
| Associate | S. baudini |
S. cyanosticta |
S. phaeota |
S. puma |
S. sila |
S. sordida |
| Smilisca sila | — | — | — | — | X | X |
| Smilisca sordida | — | — | X | — | X | X |
| Triprion petasatus | X | — | — | — | — | — |
| Cochranella fleischmanni | — | — | — | — | X | X |
| Centrolene prosoblepon | — | — | — | — | X | — |
| Gastrophryne elegans | X | — | — | — | — | — |
| Gastrophryne olivacea | X | — | — | — | — | — |
| Gastrophryne usta | X | — | — | — | — | — |
| Hypopachus alboventer | X | — | — | — | — | — |
| Hypopachus caprimimus | X | — | — | — | — | — |
| Hypopachus inguinalis | X | — | — | — | — | — |
| Hypopachus maculatus | X | — | — | — | — | — |
| Hypopachus oxyrrhinus | X | — | — | — | — | — |
| Hypopachus variolosus | X | — | — | — | — | — |
| Rana palmipes | X | — | X | X | — | — |
| Rana pipiens | X | — | — | — | — | — |
| Rana warschewitschi | — | — | X | — | X | X |
Chorus structure.—Limited observations on some of the species of Smilisca show a definite organization of the calling behavior of individuals. Smilisca baudini and S. phaeota call in duets. This is especially noticeable in S. baudini, in which the members of a duet often call from sites separated by only a few centimeters. The call of S. baudini consists of a series of like notes (see description of call in following section); the duration of each note is about equal to the interval between notes. Normally one individual utters one note, pauses, and utters a single note again, or series of two or three notes. If there is no response, the first individual often waits several seconds or even several minutes and then repeats the call. The second individual usually responds after the first or second note of the sequence. The notes of the second individual usually are spaced so that they are emitted in the intervals between the notes of the first individual. This can be shown diagrammatically by having [Pg 350] the figure "1" represent notes of the first individual and figure "2," the notes of the second; an empty interval is represented by "0":
Usually a chorus is initiated by one duet and is quickly picked up by other individuals also calling in duets. A numerical representation of a chorus of eight frogs would approximate the following organization:
3-0-3-4-3-4-3-4-3-4-3-4-3
5-6-5-6-5-6-5-6-5-6-5-6
7-8-7-8-7-8-7-8-7-8-7-8
After the first one or two duets are initiated, the second individuals in the following duets usually call immediately after their respective partners have given the first notes. The other noteworthy aspect about the organization is that the entire chorus usually stops abruptly. Normally the first duet stops calling shortly before the others, but this is not invariable. Often one duet or one individual will emit several notes after the rest of the frogs have become silent. An interval of several minutes sometimes elapses before the chorus begins again. Successive choruses apparently are initiated by the same duet. Responses can be initiated artificially by imitating the call, and sometimes any loud noise will start a chorus.
Similar duets have been observed in S. phaeota. In this species the intervals are often much longer than the notes, and if two males are calling in close proximity, their calls can be mistaken for those of one individual. Smilisca phaeota does not congregate in large numbers; usually only two males call from one restricted site.
Smilisca sila has a call consisting of a primary note followed by one or more secondary notes. Males often call in duets, but not necessarily so. In a duet, the first male usually utters only primary notes until the second individual responds; then each individual produces a rapid series of secondary notes.
Smilisca puma also produces primary and secondary notes. Although individuals sometimes call alone, duets, trios, or quartets were more common. The chorus is initiated by one individual uttering primary notes until joined by the second, third, and fourth frogs. In one quartet in a marsh 7.5 kilometers west of Puerto Viejo, Costa Rica, on February 19, 1965, the same individual initiated four consecutive choruses. Each time the second member of the chorus was the same; the third and fourth frogs joined the chorus nearly simultaneously.
Individuals of S. sordida are usually irregularly situated along a stream. No duets or other combinations of individuals are apparent in the chorus structure, but once an individual calls, a frog nearby calls almost immediately; then a frog near the second individual calls, and so on. The resulting series of calls gives the impression that the sound is moving along the stream as successive individuals join the chorus and the first callers become quiet. It is not known if the same individual initiates successive choruses or if the order of calling is the same in subsequent choruses.
These limited observations on chorus structure in Smilisca show the presence of behavioral organization. The methods of establishing the organization and the significance of the call-order in breeding have yet to be discovered.
Calling males of S. baudini are often close together; some individuals have been observed almost touching one another, but no indication of territoriality [Pg 351] or aggressive behavior has been witnessed. The more distant spacing of the stream-breeding species S. sila and S. sordida may be a function of calling-territories, but no direct evidence is available to substantiate this supposition.
Sex recognition and amplexus.—Observations on Smilisca baudini indicate that the calls of males attract females. At Tehuantepec, Oaxaca, México, a female was first observed about two meters away from a male calling at the edge of a rain pool; in a series of short hops she progressed directly towards the male, although vegetation obscured him until she was less than a meter away. When she approached to within about 20 centimeters of the male, he took notice of her, moved to her, and clasped her. At Chinajá, Alta Verapaz, Guatemala, a female swam directly across a pool about three meters wide to a calling male. Her line of movement took her within a few centimeters of a silent male, to whom she paid no attention. She stopped just in front of the calling male, which immediately clasped her. At a large muddy pond 4 kilometers west-northwest of Esparta, Puntarenas, Costa Rica, a female was observed swimming toward a small submerged tree; a male was calling from a branch about one meter above the water. The female climbed to a branch about 20 centimeters below the male, which upon seeing her there immediately jumped down and clasped her. These few observations of S. baudini show that in this species females are capable of locating calling males by means of phono-orientation; visual reception on the part of females seems to be secondary. Contrariwise, males apparently become aware of the proximity of females by seeing them; once a male sees a female he usually tries to clasp her. Possibly the males receive stimuli by means of chemo-reception, but in each observed instance the male obviously looked at the female.
Amplexus is axillary in all members of the genus. Normally amplexing males hunch their backs and press their chins to the females' backs. Clasping pairs are usually found at the edge of the water, but sometimes amplexus takes place in trees or bushes.
Egg deposition.—Oviposition has been observed only in Smilisca baudini. On the night of June 28, 1961, at Chinajá, Alta Verapaz, Guatemala, a clasping pair was observed at the edge of a shallow rain pool. After sitting for several minutes in shallow water, the female (with male on her back) swam part way across the pool and grasped an emergent stick with one hand. The female's body was nearly level with the surface of the water, and her hind legs were outstretched as deposition commenced; eggs were extruded rapidly. After a few seconds the female moved slowly to another twig a few centimeters away and deposited more eggs. This process was repeated until the female was spent. The spawn resulted in a surface film covering roughly one square meter. It is doubtful if this type of egg deposition occurs in any other species in the genus, especially those that lay their eggs in streams.
The breeding calls of the six species of Smilisca are alike in their explosive nature. Calls are emitted quickly with a short burst of air filling the vocal sac, which immediately deflates. Phonetically the calls can be described as a single "wonk" or series of such notes in S. baudini and S. cyanosticta, a low growl in S. phaeota, a relatively high pitched rattle in S. sordida, and a low [Pg 352] squawk usually followed by one or more rattling secondary notes in S. puma and S. sila. Quantitatively, the calls of the six species differ in number of notes, duration of notes, and in pitch (Table 8, Pls. 10 and 11). Although no measurements were taken on the intensity of the calls, we observed in the field that each of the species has a loud voice. The call of S. baudini seems to carry farther than any of the others.
| Table 8.—Comparison of Breeding Calls in Smilisca. (Observed Range Given in Parentheses Below Mean. In Species Having Primary and Secondary Notes, Only the Primary Notes Are Analyzed Here.) | |||||||
| Species | N | Notes per call group |
Duration of note (seconds) |
Pulses per second |
Fundamental frequency (cps) |
Major frequencies (cps) | |
| Lower | Upper | ||||||
| S. baudini | 20 | 8.0 | 0.11 | 174.7 | 166.2 | 351 | 2507 |
| (2-15) | (0.09-0.13) | (140-195) | (135-190) | (175-495) | (2400-2725) | ||
| S. cyanosticta | 10 | 1.2 | 0.38 | 147.0 | 145.1 | 841 | 1894 |
| (1-2) | (0.25-0.45) | (110-180) | (135-160) | (480-975) | (1600-2100) | ||
| S. phaeota | 10 | 1.6 | 0.31 | 116.0 | 143.0 | 372 | — |
| (1-2) | (0.10-0.45) | (100-130) | (110-165) | (330-495) | |||
| S. puma | 28 | 3.7 | 0.13 | 208.2 | 145.6 | 743 | 1868 |
| (2-10) | (0.06-0.35) | (187-240) | (125-200) | (495-980) | (1456-2240) | ||
| S. sila | 15 | 2.4 | 0.16 | 108.5 | 103.0 | 899 | 2218 |
| (1-6) | (0.06-0.28) | (97-120) | (90-115) | (665-1180) | (1980-2700) | ||
| S. sordida | 19 | 1.7 | 0.29 | 104.7 | 123.1 | 1216 | 2694 |
| (1-6) | (0.18-0.45) | (78-135) | (90-140) | (1150-1540) | (2340-2990) | ||
[Pg 353] Call rate.—The rate at which call-groups are produced varies from one every few seconds to one in several minutes. In S. baudini, cyanosticta, phaeota, and sordida, call-groups are produced as frequently as every 12 seconds, but usually more time elapses between call groups. In S. sordida, five or more minutes sometimes elapse between call-groups. The interval is somewhat less in S. phaeota. Calls are repeated at much shorter intervals in S. puma (5-55 seconds) and S. sila (4-20 seconds).
Notes per call-group.—Except for S. puma and S. sila, the series of notes produced in any given call of a species of Smilisca is essentially the same; there is no differentiation into primary and secondary notes. Smilisca cyanosticta and S. phaeota emit only one or two relatively long notes per call-group, whereas S. baudini and S. sordida produce as many as 15 and 6 notes, respectively. Males of S. puma and S. sila often produce only the primary note; sometimes this is done several times before the secondary notes are produced. For example, one S. puma (KU 91711; tape No. 379) produced the following number of notes in consecutive call-groups: 1, 1, 1, 1, 2, 2, 3, 1, 4; secondary notes are present in only four of the nine call-groups. A typical series of consecutive call-groups in S. sila (KU 91852; Tape No. 385) has 1, 1, 1, 2, 4, 2 notes per call-group; secondary notes are present in only half of the call-groups. Smilisca puma apparently always produces at least two primary notes before emitting secondary notes; sometimes only primary notes are produced in one series of calls. The number of secondary notes following a given primary varies from one to nine; the modal number is one, and the mean is three in 27 call-groups. Smilisca sila frequently begins a series of calls with two or more primary notes, but sometimes the first primary note is followed immediately by two or more secondary notes. The number of secondary notes following a given primary varies from one to five; the modal number is one, and the average is two in 13 call-groups.
Duration.—The average duration of call-groups consisting of two or more notes is 1.18 seconds in S. baudini; 1.02 in cyanosticta, 0.91 in phaeota, 1.32 in puma, 1.48 in sila, and 1.29 in sordida. Although there is considerable variation in the lengths of the notes (only primary notes in S. puma and sila are considered here), S. cyanosticta, phaeota, and sordida have noticeably longer notes than do the other species (Table 8). The secondary notes are longer than the primary notes in S. puma (average 0.27 secs. as compared with 0.13 secs.) and in S. sila (average 0.25 secs., as compared with 0.16 secs.).
Note repetition rate.—The rate at which notes in call-groups containing two or more notes are produced varies in S. baudini from 2.5 to 7.1 (average, 3.7) calls per second; cyanosticta, 1.8-2.1 (1.9); phaeota, 2.0-2.4 (2.2); puma, 1.9-2.9 (2.2); sila, 1.3-2.4 (1.8); and sordida, 1.5-2.6 (2.1). Smilisca baudini, which has notes of short duration (0.09 to 0.13 seconds), has the fastest note-repetition rate. Although the individual notes of S. cyanosticta and S. phaeota are relatively long (average, 0.38 and 0.31 seconds, respectively), the intervals [Pg 354] between the notes is short; consequently, their note-repetition rates do not differ greatly from those of S. puma and S. sila, which have shorter notes (average, 0.13 and 0.16 seconds, respectively) but longer intervals between notes.
Pulse rate.—Pulses vary in frequency from 78 to 240 per second in the calls analyzed (only primary notes in S. puma and S. sila), but the variation in any given species is much less than that in the entire genus (Table 8). Smilisca puma is outstanding in having a high pulse rate, which is approached only by that of S. baudini. Even in the species having the lowest pulse rates, the pulsations are not audible. The secondary notes produced by S. puma and S. sila have a slower pulse rate than the primary notes; often the pulses are audible. In S. puma the pulse rate of secondary notes is sometimes as low as 48 pulses per second, and in S. sila still lower (as low as 40 pulses per second). The upper limits of pulse rate in the secondary notes in these species merge imperceptibly with the rates of the primary note; consequently, on the basis of pulse rate alone it is not always possible to distinguish primary from secondary notes.
Frequency.—Smilisca produces noisy (as opposed to more musical) calls, and the energy is distributed throughout the frequency spectrum; the calls are poorly modulated, except in S. sordida, in which two usually discrete bands of frequency are present (Pl. 11C). For the most part the calls of Smilisca consist of little modified energy of the fundamental frequency and of its harmonics, some of which are emphasized.
The upper frequency range varies within each species and even within the calls of one individual. Smilisca phaeota has the lowest upper frequencies; no calls ranged above 4400 cycles per second (cps.), and half of the calls never exceeded 3000 cps. Smilisca cyanosticta produces calls in which the upper frequency is below 7000 cps. and usually below 6000 cps. Likewise, S. puma produces calls that are below 7000 cps., whereas S. sila has frequencies of up to 8400 cps. In both S. baudini and S. sordida, the highest frequencies attained are about 9100 cps. Variation in the highest frequencies in a series of consecutive calls by one individual frog was noted in all species. Such variation is especially prevalent in S. puma; for example one individual (KU 87771; Tape No. 376) recorded at a temperature of 24° C. at 7.5 kilometers west of Puerto Viejo, Heredia Province, Costa Rica, on July 31, 1964, produced three consecutive primary notes having upper frequencies of about 6000, 4000, and 4000 cps., respectively. Apparently in a given species the production of the higher frequencies in some notes and not in others is correlated with the amount of distention of the vocal sac and is not dependent upon the structure or tension of the vocal cords.
Although the dominant frequency in S. sordida is lower than that in S. baudini and S. cyanosticta, the call of the former is audibly higher-pitched. This is due primarily to the emphasis on certain harmonics at a high frequency (sometimes as high as 9000 cps.) in S. sordida, whereas in S. baudini and other species, if harmonics are present at those frequencies, they are not emphasized.
The fundamental frequencies are as low as 90 cps. in S. sila and S. sordida and as high as 200 cps. in S. puma (Table 8). The fundamental frequency seemingly is relatively unimportant in determining the general pitch of the call, a characteristic most dependent on the dominant frequency and emphasized harmonics in the higher-frequency spectrum. In none of the species is the [Pg 355] fundamental the dominant frequency. In the low-pitched call of S. phaeota the dominant frequency is the third harmonic (the second harmonic above the fundamental frequency, which is the first harmonic). In all other species a much higher harmonic is dominant; for examples, in S. cyanosticta harmonics from 10 to 15 are dominant; in S. baudini, 15-19; and S. sila, 20-30.
A glance at the audiospectrographs and their accompanying sections (Pls. 10 and 11) reveals the presence of two emphasized bands of frequency in all species except S. phaeota, in which only the lower band is present. These two bands of emphasized harmonics are part of a continuous, or nearly continuous, spread of energy throughout the frequency spectrum, except in S. sordida in which the bands are usually distinct. As shown in the sections, certain harmonics in each of the bands are emphasized with nearly equal intensity. Therefore, with the exception of S. phaeota, the calls of Smilisca are characterized by two major frequencies, one of which is the dominant frequency and the other is a subdominant frequency (Table 8). The upper major frequency is dominant in all calls in S. baudini and S. cyanosticta, but either major frequency may be dominant in other species. The upper major frequency is dominant in 65 per cent of calls by S. puma, 87 per cent in S. sila, and 68 per cent in S. sordida. Individuals of these three species sometimes produce a series of calls in which the dominant frequency changes from one of the major frequencies to the other. Four consecutive notes emitted by an individual of S. sordida recorded 13 kilometers east-northeast of Golfito, Puntarenas Province, Costa Rica, had dominant frequencies of 910, 1950, and 750 cps., respectively. In each case, an alternation of major frequencies took place in respect to dominance. An individual of S. puma from 7.5 kilometers west of Puerto Viejo, Costa Rica, produced a primary note followed by one secondary note; each note had major frequencies at 600 and 1800 cps.; the dominant frequency of the primary note was at 1800 cps., whereas in the secondary note the dominant frequency was at 600 cps. The difference in emphasis on the major frequencies is so slight that shift in dominance is not audible.
Effect of temperature on calls.—The present data are insufficient to test statistically the correlation between temperature and variation within certain components of the calls in Smilisca, but even a crude graph shows some general correlations. The widest range of temperatures is associated with the recordings of S. baudini. Three individuals recorded at a temperature of 30° C. at Tehuantepec, Oaxaca, had pulse rates of 180 pulses per second and fundamental frequencies of 160-180 cps., as compared with an individual recorded at a temperature of 17° C., which had a pulse rate of 140 and a fundamental frequency of 135 cps. All individuals of S. baudini recorded at higher temperatures had faster pulse rates and higher fundamental frequencies. Pulse rates differ in the other species in the genus but less strikingly (probably owing to narrower ranges of temperatures at which recordings were made). In five recordings of S. sordida made at 20° C. the pulse rate is 80-90, as compared with four recordings made at 25° C. having pulse rates of 120-135. Thirteen recordings of S. sila made at 17° C. have pulse rates of 97-112 (average 105); one individual recorded at 26° C. has 120 pulses per second. Seemingly no correlation exists between temperature and other characteristics of the calls, such as duration and rate of note-repetition.
The breeding call as an isolating mechanism.—Blair (1958), Bogert (1960), Duellman (1963a), Fouquette (1960), Johnson (1959), and others have provided [Pg 356] evidence that the breeding calls of male hylids (and other anurans) serve as isolating mechanisms in sympatric species. In summarizing this discussion of the breeding calls of Smilisca we want to point out what seem to be important differences in the calls that may prevent interspecific hybridization in sympatric species of Smilisca.
The genus is readily divided into two species-groups on morphological characters; this division is supported by the breeding calls. In the species of the baudini group the calls are unmodulated and lack secondary notes. In the sordida group the calls either have secondary notes or are modulated.
Smilisca baudini occurs sympatrically with S. cyanosticta and S. phaeota; where they occur together, both species sometimes breed in like places at the same time. We are not aware of these species breeding synchronously at exactly the same site, although S. baudini and S. cyanosticta were calling on the same nights and less than 100 meters apart in Oaxaca in June, 1964. Regardless of their respective breeding habits, sympatric species have calls that differ notably. Except for the higher fundamental and dominant frequencies, the calls of S. cyanosticta and S. phaeota closely resemble one another, but the calls of both species differ markedly from that of S. baudini. The geographic ranges of S. cyanosticta and S. phaeota are widely separated.
The calls of the allopatric species S. puma and S. sila are not greatly different. Smilisca sordida has a distinctive call and occurs sympatrically with S. puma and S. sila. In the streams in southern Costa Rica S. sordida and S. sila breed synchronously, but the high-pitched modulated call of the former is notably different from the lower, unmodulated call of S. sila.
The data indicate that the calls of related sympatric species differ more than the calls of related allopatric species. We postulate that these differences evolved to support the reproductive isolation of the sympatric species. The data are insufficient to determine geographic variation in the calls and to determine if differences in the calls are enhanced in areas of sympatry as compared with the allopatric parts of the ranges.
Other calls.—As stated previously, there is no direct evidence of territoriality in Smilisca; we have heard no calls that can be definitely identified as territorial. Single notes of S. baudini, phaeota, and sila have been heard by day, just prior to rains, or during, or immediately after rains. Such calls can be interpreted as "rain calls," which are well known in Hyla eximia and Hyla squirella. Distress calls are known in several species of Rana and in Leptodactylus pentadactylus; such calls result from the rapid expulsion of air over the vocal cords and with the mouth open. Distress calls have been heard from S. baudini. At Charapendo, Michoacán, México, a male that had one hind limb engulfed by a Leptodeira maculata emitted several long, high-pitched cries. A clasping pair of S. baudini was found in a bush at the edge of a marshy stream 2 kilometers northeast of Las Cañas, Guanacaste Province, Costa Rica. When the pair was grasped, the female emitted a distress call.
Eggs of S. baudini, cyanosticta, and phaeota have been found in the field, and eggs of S. sila have been observed in the laboratory. The eggs of S. puma and sordida are unknown. Insofar as known, Smilisca baudini is unique in the genus in depositing the eggs in a surface film. Each egg is encased in a [Pg 357] vitelline membrane, but individual outer envelopes are lacking. The eggs are small; the diameter of recently-deposited eggs is about 1.3 mm. and that of the vitelline membrane is about 1.5 mm. The eggs of S. cyanosticta and phaeota are deposited in clumps, and the eggs are larger than those of S. baudini. Diameters of eggs of S. cyanosticta are about 2.3 mm., and those of the outer envelopes are about 4.0 mm. Artificially fertilized eggs of S. sila raised in the laboratory have diameters of about 2.4 mm.; the diameter of the outer envelopes is about 4.9 mm.
In order to determine the reproductive potential of the six species, ovulated eggs were removed from females and counted. The numbers of eggs recorded are: 3 S. baudini—2620, 2940, 3320; 1 S. cyanosticta—910; 3 S. phaeota—1665, 1870, 2010; 1 S. puma—518; 3 S. sila—369, 390, 473; 3 S. sordida—524, 702, 856. These limited data indicate that the large species (S. baudini, cyanosticta, and phaeota) have more eggs than do the smaller species. The stream-breeding species (S. sila and sordida) have relatively few eggs by comparison with the pond-breeders. Possibly this is a function of size of eggs rather than a correlation with the site of egg-deposition.
The acquisition of tadpoles of all of the species of Smilisca has made possible the use of larval characters in erecting a classification and in estimating the phylogenetic relations of the several species. Furthermore, developmental series of tadpoles of four species allow a comparison of the growth and development in these species. Throughout the discussion of tadpoles we have referred to the various developmental stages by the Stage Numbers proposed by Gosner (1960).
Tadpoles of the genus Smilisca are of a generalized hylid type, having 2/3 tooth-rows, unspecialized beaks, mouth partly or completely bordered by papillae, lateral fold present in the lips, spiracle sinistral, anal tube dextral, and caudal musculature extending nearly to tip of caudal fin. Although minor differences exist in coloration, proportions, and mouthparts, no great modifications of the basic structure are present.
The larval characters of the species of Smilisca are compared below and illustrated in Figures 11-15.
Shape and Proportions.—The bodies of S. baudini, cyanosticta, phaeota, and puma are rounded and about as wide as deep; the eyes are moderately large and directed dorsolaterally, and the nostrils are about midway between the bluntly rounded snout and the eyes. The mouths are medium-sized and directed anteroventrally. The bodies of tadpoles of S. sila and sordida are slightly compressed dorso-ventrally. The snout is moderately long and sloping; the eyes are larger and directed more dorsally than in the other species, and the nostrils are closer to the eyes than the snout. The mouths are moderately large and directed ventrally.
The tail is about half again as long as the body in S. baudini, cyanosticta, [Pg 358] phaeota, and puma; in these species the caudal musculature is moderately heavy, and the caudal fins are deep. The caudal musculature is upturned distally in S. baudini and phaeota, and the dorsal fin extends anteriorly onto the body in these two species and in S. puma. The tail is about twice as long as the body in S. sila and sordida. In both species the caudal fins are shallow in comparison with the depth of the caudal musculature, especially in S. sordida (Fig. 14); in neither species does the dorsal fin extend anteriorly onto the body.

Mouthparts.—The mouth of S. sordida is completely bordered by two rows of papillae, whereas in the other species the median part of the upper lip is devoid of papillae. Smilisca baudini and puma have two rows of papillae; S. sila has one complete row (except medially on the upper lip) and one incomplete row, and S. cyanosticta and phaeota have only one row (Fig. 15). All species have numerous papillae in the lateral fold; the fewest lateral papillae [Pg 359] are found in S. cyanosticta and phaeota. Although all species have two rows of teeth in the upper jaw and three rows in the lower jaw, specific differences in the nature of the rows exist between certain species. The second upper tooth-row is narrowly interrupted medially in S. sila and sordida and broadly interrupted in the other species. The first upper row is strongly arched in S. puma, moderately arched in S. baudini and sila, and weakly arched in the other species. In all species the third lower tooth-row is the shortest, only slightly so in S. sila and sordida, but only about half the length of the second lower row in S. puma.

The beaks are well developed and finely serrate in all species. The lower, broadly V-shaped, beak is slender in S. puma, rather robust in S. baudini and [Pg 360] sila, and moderately heavy in the other species. The lateral processes of the upper beak are shortest in S. puma and longest in S. baudini and sordida. In the latter the inner margin of the upper beak and lateral process have the form of a shallow S, whereas in the other species the inner margin of the upper beak forms a continuous arch with the lateral processes (Fig. 15).

Coloration.—The tadpoles of Smilisca lack the bright colors or bold markings characteristic of some hylid tadpoles; even so, the subdued colors and arrangement of pigments provide some distinctive markings by which the species can be distinguished from one another. The species comprising the baudini group (S. baudini, cyanosticta, and phaeota) are alike in having the body brown or grayish brown dorsally and transparent with scattered brown pigment ventrally. A cream-colored, crescent-shaped mark is present on the posterior edge of the body; this mark is usually most noticeable in S. baudini and least so in S. cyanosticta. Other differences in coloration in members of the baudini group are relative and subtle. Smilisca phaeota usually is more [Pg 361] pallid than baudini, and cyanosticta usually is darker than baudini; both species have larger dark markings on the tail than does S. phaeota. Smilisca baudini has a dark streak on the middle of the anterior one-fourth of the tail (Figs. 11-13).
Smilisca puma is distinctive in having a grayish brown body and dark gray reticulations on the tail. Smilisca sila and sordida are distinctive in having pairs (sometimes interconnected) of dark marks on the dorsal surfaces of the caudal musculature, and in dorsal view the tail appears to be marked with dark and pale creamy tan transverse bars. These dark marks, as well as the small flecks on the tail, are brown in S. sila and red in sordida. Smilisca sila has dark brown flecks on the dorsal surface of the body and small white flecks laterally; these markings are absent in S. sordida (Fig. 14).
Descriptions of the coloration of living tadpoles are given in the accounts of the species.

Information on the growth and development of Middle American hylids is scanty. Adequate descriptions have been published for Phyllomedusa annae (Duellman, 1963b), Phrynohyas venulosa (Zweifel, 1964), and Triprion petasatus (Duellman and Klaas, 1964). Material is available for adequate descriptions of the developmental stages of four species of Smilisca (Tables 9-12, Figs. 11-13). Because none of the tadpoles was raised from hatching to metamorphosis, the rate of growth and duration of the larval stages are unknown.
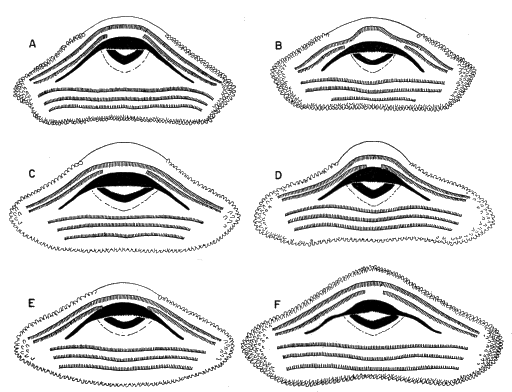
| Table 9.—Growth and Development of Tadpoles of Smilisca baudini. (Means Are Given in Parentheses After the Observed Ranges.) | ||||
| Stage | N | Total length | Body length | Tail length |
| 21 | 10 | 5.1-5.4 (5.22) | 2.6-2.7 (2.54) | 2.5-2.7 (2.58) |
| 24 | 10 | 6.0-6.5 (6.20) | 2.3-2.6 (2.45) | 3.5-3.9 (3.69) |
| 25 | 10 | 7.2-8.3 (7.78) | 3.0-3.3 (3.14) | 4.2-5.0 (4.64) |
| 27 | 10 | 18.5-21.5 (20.22) | 8.0-9.0 (8.38) | 10.4-13.0 (11.84) |
| 29 | 10 | 21.5-24.5 (22.60) | 8.5-10.0 (9.25) | 12.5-14.5 (13.35) |
| 37 | 3 | 28.5-31.0 (30.00) | 11.0-12.5 (11.67) | 17.5-19.0 (18.00) |
| 38 | 10 | 35.0-37.5 (35.50) | 12.0-13.5 (12.80) | 21.5-24.0 (22.70) |
| 40 | 2 | 34.0-37.0 (35.50) | 12.5-13.5 (13.00) | 21.5-23.5 (22.50) |
| 41 | 10 | 34.0-37.0 (35.50) | 12.5-13.5 (13.00) | 21.5-23.5 (22.50) |
| 42 | 3 | 24.0-30.0 (27.00) | 12.5-13.0 (12.67) | 11.5-17.0 (14.33) |
| 45 | 6 | 14.0-24.0 (17.58) | 12.5-14.0 (13.37) | 1.5-10.0 (4.17) |
| 46 | 23 | —— | 12.0-15.5 (13.34) | —— |
| Table 10.—Growth and Development of Tadpoles of Smilisca cyanosticta. (Means Are Given in Parentheses After the Observed Ranges.) | ||||
| Stage | N | Total length | Body length | Tail length |
| 21 | 10 | 5.8-6.5 (6.28) | 2.8-3.1 (3.00) | 3.0-3.5 (3.28) |
| 25 | 10 | 7.9-9.2 (8.44) | 2.7-3.2 (2.96) | 4.8-6.0 (5.48) |
| 30 | 7 | 22.5-25.0 (23.50) | 8.5-9.5 (9.00) | 14.0-15.5 (14.57) |
| 36 | 10 | 27.0-30.0 (28.75) | 9.5-11.5 (10.80) | 17.0-18.5 (17.95) |
| 42 | 2 | 26.0-27.0 (26.50) | 10.00 | 16.0-17.0 (16.50) |
| 46 | 2 | — | 14.00 | — |
Hatchlings of three species (S. baudini, cyanosticta, and phaeota) are available. These larvae have non-functional eyes and large oral suckers. By the time the larvae have developed to stage 21, external gills are present, the caudal musculature and caudal fin have been differentiated, and the head is distinguishable from the body. In stage 21 oral suckers and a large amount of yolk are still present.
The developmental data on the four species show no significant variations; consequently, we will describe the development of only one species, Smilisca phaeota (Table 11, Figs. 13 and 16).
Stage 21.—Bulging cream-colored yolk mass, transparent cornea, and moderately long, unbranched filamentous gills, and oral suckers present; mouth having irregular papillae on lower lip; teeth and beaks absent; caudal myomeres distinct; pigmentation uniform over body and caudal musculature; caudal fin transparent with scattered small flecks.

Stage 25.—Operculum complete; gills absent; sinistral spiracle apparently functional; cloacal tail-piece, nasal capsules, and external nares present; gut [Pg 364] partly formed; mouth bordered by single row of papillae, except medially; small papillae present in lateral fold of lips; two upper and three lower tooth-rows present, but not fully developed; beaks apparently fully developed; depth of dorsal and ventral fins less than depth of caudal musculature: tip of tail upturned; pigment on body most dense on dorsum and sides; faint, nearly pigmentless crescent-shaped mark on posterior edge of body; concentrations of pigment forming small spots on tail.
Stage 28.—Mouthparts complete; limb bud about half as long as thick; other structural features and coloration closely resemble those in stage 25.
Stage 30.—Limb bud approximately twice as long as thick; body as deep as wide; dorsal fin deepest just posterior to body; ventral fin deeper than caudal musculature; tail sharply upturned distally; anal tube dextral; brown pigment sparse on flanks.
| Table 11.—Growth and Development of Tadpoles of Smilisca phaeota. (Means Are Given in Parentheses After the Observed Ranges.) | ||||
| Stage | N | Total length | Body length | Tail length |
| 15 | 10 | — | 1.9-2.1 (1.97) | — |
| 16 | 8 | — | 2.0-2.2 (2.07) | — |
| 18 | 4 | — | 2.2-2.6 (2.31) | — |
| 21 | 3 | 7.9-8.6 (8.21) | 4.1-4.5 (4.31) | 3.8-4.1 (3.92) |
| 25 | 10 | 8.7-10.6 (9.69) | 4.5-4.8 (4.64) | 4.3-6.0 (5.05) |
| 26 | 11 | 12.3-16.1 (14.01) | 4.2-6.3 (5.60) | 6.7-9.8 (8.41) |
| 27 | 10 | 13.0-15.7 (14.28) | 4.9-6.2 (5.40) | 7.7-10.5 (8.88) |
| 28 | 13 | 13.9-20.9 (15.62) | 5.2-8.3 (5.75) | 8.5-12.6 (9.85) |
| 29 | 8 | 17.8-22.3 (19.79) | 6.3-8.4 (7.19) | 11.5-14.0 (12.60) |
| 30 | 9 | 20.3-24.8 (22.85) | 8.1-10.5 (9.32) | 10.5-15.5 (13.53) |
| 31 | 5 | 24.1-28.5 (26.61) | 9.4-11.2 (10.59) | 14.7-17.3 (16.02) |
| 34 | 5 | 24.8-29.4 (27.31) | 9.2-11.6 (10.73) | 15.6-18.5 (16.80) |
| 36 | 3 | 30.0-30.1 (30.07) | 10.1-12.2 (11.15) | 18.9-20.0 (19.44) |
| 37 | 4 | 28.9-34.1 (31.75) | 11.5-12.4 (11.88) | 17.4-22.5 (19.88) |
| 38 | 1 | 28.98 | 12.88 | 16.10 |
| 39 | 2 | 35.6-36.9 (36.25) | 14.00 | 21.6-22.9 (22.25) |
| 40 | 2 | 32.3-39.8 (36.05) | 14.00 | 18.3-21.8 (20.05) |
| 43 | 2 | 21.5-23.0 (22.25) | 14.2-14.8 (14.45) | 6.8-8.8 (7.80) |
| 44 | 4 | — | 14.5-15.6 (15.08) | — |
| 46 | 11 | — | 12.7-16.7 (14.26) | — |
| Table 12.—Growth and Development of Tadpoles of Smilisca sordida. (Means Are Given in Parentheses After the Observed Ranges.) | ||||
| Stage | N | Total length | Body length | Tail length |
| 25 | 6 | 25.5-28.0 (26.1) | 9.0-9.5 (9.3) | 16.2-18.5 (16.7) |
| 33 | 2 | 28.5-30.0 (29.3) | 10.2-10.5 (10.4) | 18.0-19.8 (18.9) |
| 36 | 8 | 29.5-34.5 (32.3) | 10.2-11.7 (10.8) | 19.3-23.0 (21.5) |
| 37 | 7 | 31.6-37.5 (34.6) | 11.0-12.5 (11.5) | 21.6-25.0 (23.2) |
| 41 | 3 | 33.0-37.2 (35.2) | 11.6-12.2 (11.9) | 21.4-25.2 (23.2) |
| 43 | 1 | —— | 12.4 | —— |
| 46 | 9 | —— | 13.1-15.7 (14.9) | —— |
Stages 34, 36, 37, and 38.—Stage 34, foot paddle-shaped with four toe buds; stage 36, five toe buds; stages 37 and 38, lengthening of toes. In all four stages, spiracle persistent, and pigmentation resembling that of early stages.
Stage 39.—Metatarsal tubercle present; greatest total length (36.9 mm.) attained.
Stage 40.—Subarticular tubercles prominent; skin over forelimbs transparent; cloacal tail-piece and spiracle absent; outer tooth-rows degenerating; caudal fins shallower than in preceding stages; distal part of tail nearly straight; size of dark markings on tail decreased; pigment present on hind limb.
Stage 43.—Forelimbs erupted; larval mouthparts absent; corner of mouth between nostril and eye; transverse bands present on hind limbs; tail greatly reduced (about 8 mm. in length).
Stage 44.—Sacral hump barely noticeable; tail reduced to a stub; corner of mouth at level of pupil of eye; dorsal surfaces pale olive-green; venter white.
Changes proceed in a definite pattern during the growth and development of tadpoles. Larval teeth are absent in hatchlings; the inner tooth-rows develop first, and the third lower row last. At metamorphosis the third lower row is the first to be lost. The tail increases gradually in length relative to the body. In stage 25 the tail is 52.1 per cent of the total length, and in stage 36, 64.6 per cent. In later stages the tail becomes relatively shorter through resorption. Duellman and Klaas (1964:320) noted a great size-variation in Triprion tadpoles in stage 25. No such variation is apparent in any stage of any of the species of Smilisca studied.
The growth and development of the other species of Smilisca do not differ significantly from that of S. phaeota. The tadpoles of S. sila and sordida from streams have relatively longer tails at hatching. For example, in tadpoles of S. sordida the average length of tail is 64.0 per cent of the body-length in stage 25, and in stage 37, 67.0 per cent.
The tadpoles of S. baudini, cyanosticta, phaeota, and puma are pelagic inhabitants of shallow ponds. Early stages of S. baudini in which external gills [Pg 366] are present have been observed to hang vertically with the gills spread out at the surface of the water, a behavior noted by Zweifel (1964:206) in tadpoles of Phrynohyas venulosa, which also develop in warm, standing water having a relatively low oxygen-tension. When disturbed the pelagic tadpoles usually dive and seek shelter amidst vegetation or in mud on the bottom. This behavior was observed in S. baudini, cyanosticta, and phaeota by day and at night. No tadpoles of S. puma were observed by day; those seen at night were near the surface of small water-filled depressions in a grassy marsh; they responded to light by taking refuge in the dense grass. Perhaps tadpoles of this species are negatively phototactic and remain hidden by day.
The stream-inhabiting tadpoles of S. sila and sordida live in clear pools in rocky streams, where they were observed to cling by their mouths to rocks in the stream and to seek shelter amidst pebbles or beneath rocks and leaves on the bottom. These tadpoles are not found in shallow riffles.
We have not found tadpoles of two species of Smilisca in the same body of water and therefore cannot offer observations on ecological relationships in sympatric situations.
Identifiable hylid remains are known from the Miocene to the Recent, but these fossils are mostly fragmentary and provide little useful information regarding the phylogenetic relationships of living genera. Frogs of the genus Smilisca are generalized and show no striking adaptations, either in their structure or in their modes of life history.
In attempting to understand the relationships of the species of Smilisca we have emphasized osteological characters. The phylogeny suggested by these characters is supported by other lines of evidence, including external morphology, tadpoles, and breeding calls.
Our concept of the prototype of the genus Smilisca is a moderate-sized hylid having: (1) a well-developed frontoparietal fontanelle, (2) frontoparietal lacking lateral processes, (3) no bony squamosal-maxillary arch, (4) a fully ossified ethmoid, (5) paired subgular vocal sac, (6) moderately webbed fingers and toes, (7) relatively few supernumerary tubercles on the digits, (8) eggs deposited in clumps in ponds, (9) anteroventral mouth in tadpoles bordered by one row of labial papillae, but median part of upper lip bare, (10) tail relatively short and deep in tadpoles, and (11) a breeding call consisting of a series of like notes.
Two phyletic lines evolved from this prototype. The first of these was the stock that gave rise to the baudini group. The evolutionary changes that took place in this line included increase in size, development of a lateral curvature of the maxillary, and an increased amount of cranial ossification, especially in the dermal roofing bones. This phyletic line retained the larval characters and breeding call of the prototype. The second phyletic line gave rise to the sordida group and diverged from the prototype in the development of an angular maxillary and a breeding call consisting of a primary note followed by secondary notes. The frogs in this phyletic line retained the moderate [Pg 367] size of the prototype and did not develop additional dermal bone. Our concept of the phylogenetic relationships is shown graphically in Figure 17.
Within the baudini group one stock retained separate nasals and did not develop a bony squamosal-maxillary arch, but broad lateral processes developed on the frontoparietals. The tadpoles remained unchanged from the primitive type. This stock evolved into S. phaeota. In the other stock the nasals became fully ossified and a bony squamosal-maxillary arch developed. One branch of this second stock retained tadpoles having only one row of labial papillae and did not develop lateral processes on the frontoparietals; this branch evolved into S. cyanosticta. The other branch diverged and gave rise to S. baudini by developing relatively shorter hind legs, large lateral processes on the frontoparietals, and tadpoles having two rows of labial papillae.
Within the sordida group the cranial features remained unchanged in one line, which gave rise to S. sila, whereas in a second line the nasals were reduced, and their long axes shifted with the result that they are not parallel to the maxillaries; the amount of ossification of the ethmoid was reduced, and the tadpoles developed two rows of labial papillae. In this second line one branch retained the pond-breeding habits and gave rise to S. puma, whereas a second branch became adapted to stream-breeding and gave rise to S. sordida.

Certain aspects of this proposed phylogeny warrant further comment. Features such as the deposition of additional bone that roofs the skull or that forms lateral projections from the frontoparietals, like those in S. baudini and phaeota, are minor alterations of dermal elements and not basic modifications [Pg 368] of the architecture of the skull. Consequently, we hypothesize the independent development of these dermal changes in S. baudini and phaeota. Similar kinds of dermal modifications have evolved independently in many diverse groups of frogs.
Likewise, we propose the parallel development of stream-adapted tadpoles in S. sordida and sila; in both cases the tadpoles adapted to changing environmental conditions (see following section on evolutionary history). Tadpoles of S. sordida already had two rows of labial papillae before entering the streams; subsequently the tadpoles developed complete rows of papillae, ventral mouths and long tails having low fins. Possibly the tadpoles of S. sila had two rows of labial papillae prior to their adapting to stream conditions; in the process of adapting they developed ventral mouths and long tails having low fins. Similar modifications in tadpoles have occurred in many diverse groups of Middle American hylids, such as Plectrohyla, Ptychohyla, the Hyla uranochroa group, and the Hyla taeniopus group.
Our lack of concern about coloration is due to the fact that, with the exception of the blue spots on the flanks and posterior surfaces of the thighs in some species, the coloration of Smilisca, consisting of a pattern of irregular dark marks on a paler dorsum and dark transverse bars on the limbs, is not much different from that of many other Neotropical hylids. Blue is a structural color, rare among Amphibia, which is achieved by the absence of lipophores above the guanophores. Thus, the incident light rays at the blue end of the spectrum are reflected by the guanophores without interference by an overlying yellow lipophore screen. According to Noble (1931), lipophores are capable of amoeboid movement that permits shifts in their positions, between or beneath the guanophores. We do not know whether this behavior of lipophores is widespread and is effected in response to environmental changes, or whether it is a genetically controlled attribute that is restricted in appearance. If the latter is the case we must assume that the prototype of Smilisca possessed such an attribute which was lost in S. baudini, phaeota, and puma. The development of blue spots is not constant in S. sordida and S. sila; in S. cyanosticta the spots range in color from blue to pale green.
The coloration of the tadpoles is not distinctive, except for the presence of dorsal blotches on the tails of S. sila and sordida. However, the similarity in pattern cannot be interpreted as indicating close relationships because nearly identical patterns are present in Hyla legleri and some species of Prostherapis. This disruptive coloration seems to be directly associated with the pebble-bottom, stream-inhabiting tadpoles.
In the baudini group, S. phaeota and cyanosticta are allopatric, whereas S. baudini occurs sympatrically with both of those species. The call of S. baudini differs notably from the calls of S. phaeota and cyanosticta, which are more nearly alike. Although in the phylogenetic scheme proposed here S. sila is considered to be more distantly related to S. puma than is S. sordida, the calls of S. sila and puma more closely resemble one another than either resembles that of S. sordida. Smilisca sila and puma are allopatric, whereas S. sordida is broadly sympatric with both of those species. We assume that in their respective phyletic lines the differentiation of both S. baudini and sordida was the result of genetic changes in geographically isolated populations. Subsequently, each species dispersed into areas inhabited by other members of their respective groups. Selection for differences in the breeding calls helped to [Pg 369] reinforce other differences in the populations and thereby aided in maintaining specificity.
With respect to temporal and spatial aspects of evolution in Smilisca, we have tried to correlate the phylogenetic evidence on Smilisca with the geologic data on Middle America presented by Lloyd (1963), Vinson and Brineman (1963), Guzmán and Cserna (1963), Maldonado-Koerdell (1964), and Whitmore and Stewart (1965). Likewise, we have borne in mind the evidence for, and ideas about, the evolution of the Middle American herpetofauna given by Dunn (1931b), Schmidt (1943), Stuart (1950, 1964) Duellman (1958, MS), and Savage (MS).
According to Stuart's (1950) historical arrangement of the herpetofauna, Smilisca is a member of the Autochthonous Middle American Faunal Element, and according to Savage's (MS) arrangement the genus belongs to the Middle American Element, a fauna which was derived from a generalized tropical American unit that was isolated in tropical North America by the inundation of the Isthmian Link in early Tertiary, that developed in situ in tropical North America, and that was restricted to Middle America by climatic change in the late Cenozoic.
Savage (MS) relied on the paleogeographic maps of Lloyd (1963) to hypothesize the extent and centers of differentiation of the Middle American Faunal Element. According to Lloyd's concept, Middle America in the Miocene consisted of a broad peninsula extending southeastward to about central Nicaragua, separated from the Panamanian Spur of continental South America by shallow seas. A large island, the Talamanca Range, and remnants of the Guanarivas Ridge formed an archipelago in the shallow sea. The recent discovery of remains of mammals having definite North American affinities in the Miocene of the Canal Zone (Whitmore and Stewart, 1965) provides substantial evidence that at least a peninsula was continuous southeastward from Nuclear Central America to the area of the present Canal Zone in early mid-Miocene time. South America was isolated from Central America by the Bolivar Trough until late mid-Pliocene.
Thus, in the mid-Tertiary the broad peninsula of Nuclear Central America, which consisted of low and moderately uplifted regions having a tropical mesic climate, provided the site for the evolution of Smilisca. It is not possible to determine when the genus evolved, but to explain the differentiation of the species it is unnecessary to have the ancestral Smilisca present prior to the Miocene.
We view the Miocene Smilisca as the prototype described in the preceding section, and suppose that it lived in the mesic tropical environment of the eastern part of the Central American Peninsula (in what is now Costa Rica and western Panamá). Two stocks differentiated, probably in middle Miocene times; one of these, the ancestral stock of the baudini group, was widespread on the Caribbean lowlands from the Nicaraguan Depression to the Bolivar Trough, and the other, the ancestral stock of the sordida group, was restricted to the Pacific lowlands of the same region. In late Miocene time the ancestral stock of the baudini group dispersed northwestward around the deep embayment in the Nicaraguan depression into upper Central America (in what is now Honduras and Guatemala) and thence into southern México. Apparently [Pg 370] differentiation took place on each side of the Nicaraguan Depression; the frogs to the south of the depression evolved into S. phaeota, whereas those to the north of the depression represented the stock from which S. baudini and cyanosticta arose. Prior to the uplift of the mountains in the late Miocene and the Pliocene the baudini-cyanosticta stock probably was widespread in northwestern Central America. The elevation of the mountains resulted in notable climatic changes, principally the development of sub-humid environments on the Pacific lowlands. The frogs living on the Pacific lowlands became adapted to sub-humid conditions and developed into S. baudini. The stock on the Caribbean lowlands remained in mesic environments and evolved into S. cyanosticta.
Possibly in the middle Miocene before the Talamanca Range in Costa Rica and western Panamá was greatly uplifted, the ancestral stock of the sordida group invaded the Caribbean lowlands of what is now Costa Rica. The subsequent elevation of the Talamanca Range in the Pliocene effectively isolated the ancestral stock of S. sila on the Pacific lowlands from the puma-sordida stock on the Caribbean lowlands. The former was subjected to the sub-humid conditions which developed on the Pacific lowlands when the Talamanca Range was uplifted. It adapted to the sub-humid environment by living along streams and evolving stream-adapted tadpoles. On the Caribbean side of the Talamanca Range the puma-sordida stock inhabited mesic environments. The stock that evolved into S. puma remained in the lowlands as a pond-breeding frog, whereas those frogs living on the slopes of the newly elevated mountains became adapted for their montane existence by developing stream-adapted tadpoles and thus differentiated into S. sordida.
Probably the six species of Smilisca were in existence by the end of the Pliocene; at that time a continuous land connection existed from Central America to South America. The climatic fluctuations in the Pleistocene, and the post-Wisconsin development of present climatic and vegetational patterns in Middle America, brought about the present patterns of distribution of the species. From its place of origin on the Caribbean lowlands of lower Central America, S. phaeota dispersed northward into Nicaragua and southward along the Pacific slopes of northwestern South America. Perhaps in the late Pleistocene or in post-Wisconsin time when mesic conditions were more widespread than now, S. phaeota moved onto the Pacific lowlands of Costa Rica. Its route could have been through the Arenal Depression. Subsequent aridity restricted its range on the Pacific lowlands to the Golfo Dulce region. Climatic fluctuation in northern Central America restricted the distribution of S. cyanosticta to mesic habitats on the slopes of the Mexican and Guatemalan highlands and to certain humid areas on the lowlands. Smilisca baudini was well adapted to sub-humid conditions, and the species dispersed northward to the Rio Grande Embayment and to the edge of the Sonoran Desert and southward into Costa Rica. In southern México and Central America the species invaded mesic habitats. Consequently, in some areas it is sympatric with S. cyanosticta and phaeota.
Smilisca puma dispersed northward onto the Caribbean lowlands of southern Nicaragua. Its southward movements probably were limited by the ridges of the Talamanca Range that extend to the Caribbean coast in the area of Punta Cahuita in Costa Rica. Smilisca sila dispersed along the Pacific lowlands and slopes of the mountains from eastern Costa Rica and western Panamá through [Pg 371] eastern Panamá to northern Colombia. Climatic fluctuation in the Pleistocene evidently provided sufficient altitudinal shifts in environments in the Talamanca Range to permit S. sordida to move onto the Pacific slopes. From its upland distribution the species followed streams down to both the Caribbean and Pacific lowlands, where it is sympatric with S. puma on the Caribbean lowlands and S. sila on the Pacific lowlands.
The evolution of the species-groups of Smilisca was effected through isolation by physical barriers in the Cenozoic; the differentiation of the species was initiated by further isolation of populations by changes in physiography and climate. Present patterns of distribution resulted from Pleistocene and post-Wisconsin climatic changes. Today, sympatric species have different breeding habits and breeding calls which reinforce the differences in morphology.
The genus Smilisca is composed of six species of tree frogs; each species is defined on the basis of adult morphology, larval characters, and breeding behavior. Keys are provided to aid in the identification of adults and of tadpoles.
Analysis of the characters and examination of type specimens indicates that several currently-recognized taxa are synonymous, as follows:
2. Hyla gabbi Cope, 1876 = Smilisca sordida.
3. Hyla manisorum Taylor, 1954 = Smilisca baudini.
4. Hyla nigripes Cope, 1876 = Smilisca sordida.
5. Hyla wellmanorum Taylor, 1952 = Smilisca puma.
Smilisca phaeota cyanosticta Smith, 1953 is elevated to specific rank, and one new species, Smilisca sila, is named and described.
The skeletal system of developmental stages and the adult of Smilisca baudini is described, and the skull is compared with that of other members of the genus.
The tadpoles are described, compared, and illustrated; the larval development of Smilisca phaeota is described.
Breeding behavior and breeding calls are described and compared. Some species of Smilisca have breeding choruses. Two species, S. sila and sordida, breed in streams, whereas the others breed in ponds.
The genus is considered to be part of the Middle American Faunal Element; the species are thought to have differentiated in response to ecological diversity and historical opportunities provided by Cenozoic changes in physiography and climate.
Transmitted March 14, 1966.
31-3430
MUSEUM OF NATURAL HISTORY
Institutional libraries interested in publications exchange may obtain this series by addressing the Exchange Librarian, University of Kansas Library, Lawrence, Kansas. Copies for individuals, persons working in a particular field of study, may be obtained by addressing instead the Museum of Natural History, University of Kansas, Lawrence, Kansas. When individuals request copies from the Museum, 25 cents should be included, for each 100 pages or part thereof, for the purpose of defraying the costs of wrapping and mailing. For certain longer papers an additional amount, indicated below, toward some of the costs of production, is to be included.
* An asterisk designates those numbers of which the Museum's supply is exhausted.
| Vol. 1. | Nos. 1-26 and index. Pp. 1-638, 1946-1950. | |
| *Vol. 2. | (Complete) Mammals of Washington. By Walter W. Dalquest. Pp. 1-444, 140 figures in text. April 9, 1948. | |
| Vol. 3. | *1. | The avifauna of Micronesia, its origin, evolution, and distribution. By Rollin H. Baker. Pp. 1-359, 16 figures in text. June 12, 1951. |
| *2. | A quantitative study of the nocturnal migration of birds. By George H. Lowery, Jr. Pp. 361-472, 47 figures in text. June 29, 1951. | |
| 3. | Phylogeny of the waxwings and allied birds. By M. Dale Arvey. Pp. 473-530, 49 figures in text, 13 tables. October 10, 1951. | |
| *4. | Birds from the state of Veracruz, Mexico. By George H. Lowery, Jr., and Walter W. Dalquest. Pp. 531-649, 7 figures in text, 2 tables. October 10, 1951. | |
| Index. Pp. 651-681. | ||
| *Vol. 4. | (Complete) American weasels. By E. Raymond Hall. Pp. 1-466, 41 plates, 31 figures in text. December 27, 1951. | |
| Vol. 5. | Nos. 1-37 and index. Pp. 1-676, 1951-1953. | |
| *Vol. 6. | (Complete) Mammals of Utah, taxonomy and distribution. By Stephen D. Durrant. Pp. 1-549, 91 figures in text, 30 tables. August 10, 1952. | |
| Vol. 7. | Nos. 1-15 and index. Pp. 1-651, 1952-1955. | |
| Vol. 8. | Nos. 1-10 and index. Pp. 1-675, 1954-1956. | |
| Vol. 9. | Nos. 1-23 and index. Pp. 1-690, 1955-1960. | |
| Vol. 10. | Nos. 1-10 and index. Pp. 1-626, 1956-1960. | |
| Vol. 11. | Nos. 1-10 and index. Pp. 1-703, 1958-1960. | |
| Vol. 12. | *1. | Functional morphology of three bats: Eumops, Myotis, Macrotus. By Terry A. Vaughan. Pp. 1-153, pls. 1-4, 24 figures in text. July 8, 1959. |
| *2. | The ancestry of modern Amphibia: a review of the evidence. By Theodore H. Eaton, Jr. Pp. 155-180, 10 figures in text. July 10, 1959. | |
| 3. | The baculum in microtine rodents. By Sidney Anderson. Pp. 181-216, 49 figures in text. February 19, 1960. | |
| *4. | A new order of fishlike Amphibia from the Pennsylvanian of Kansas. By Theodore H. Eaton, Jr., and Peggy Lou Stewart. Pp. 217-240, 12 figures in text. May 2, 1960. | |
| 5. | Natural history of the Bell Vireo, Vireo bellii Audubon. By Jon C. Barlow. Pp. 241-296, 6 figures in text. March 7, 1962. | |
| 6. | Two new pelycosaurs from the lower Permian of Oklahoma. By Richard C. Fox. Pp. 297-307, 6 figures in text. May 21, 1962. | |
| 7. | Vertebrates from the barrier island of Tamaulipas, México. By Robert K. Selander, Richard F. Johnston, B. J. Wilks, and Gerald G. Raun. Pp. 309-345, pls. 5-8. June 18, 1962. | |
| 8. | Teeth of edestid sharks. By Theodore H. Eaton, Jr. Pp. 347-362, 10 figures in text. October 1, 1962. | |
| 9. | Variation in the muscles and nerves of the leg in two genera of grouse (Tympanuchus and Pedioecetes). By E. Bruce Holmes. Pp. 363-474, 20 figures in text. October 25, 1963. $1.00. | |
| 10. | A new genus of Pennsylvanian fish (Crossopterygii, Coelacanthiformes) from Kansas. By Joan Echols. Pp. 475-501, 7 figures in text. October 25, 1963. | |
| 11. | Observations on the Mississippi kite in southwestern Kansas. By Henry S. Fitch. Pp. 503-519. October 25, 1963. | |
| 12. | Jaw musculature of the Mourning and White-winged doves. By Robert L. Merz. Pp. 521-551, 22 figures in text. October 25, 1963. | |
| 13. | Thoracic and coracoid arteries in two families of birds, Columbidae and Hirundinidae. By Marion Anne Jenkinson. Pp. 553-573, 7 figures in text. March 2, 1964. | |
| 14. | The breeding birds of Kansas. By Richard F. Johnston. Pp. 575-655, 10 figures in text. May 18, 1964. 75 cents. | |
| 15. | The adductor muscles of the jaw in some primitive reptiles. By Richard C. Fox. Pp. 657-680, 11 figures in text. May 18, 1964. | |
| Index. Pp. 681-694. | ||
| Vol. 13.[Pg ii] | 1. | Five natural hybrid combinations in minnows (Cyprinidae). By Frank B. Cross and W. L. Minckley. Pp. 1-18. June 1, 1960. |
| 2. | A distributional study of the amphibians of the Isthmus of Tehuantepec, México. By William E. Duellman. Pp. 19-72, pls. 1-8, 3 figures in text. August 16, 1960. 50 cents. | |
| 3. | A new subspecies of the slider turtle (Pseudemys scripta) from Coahulia, México. By John M. Legler. Pp. 73-84, pls. 9-12, 3 figures in text. August 16, 1960. | |
| *4. | Autecology of the copperhead. By Henry S. Fitch. Pp. 85-288, pls. 13-20, 26 figures in text. November 30, 1960. | |
| 5. | Occurrence of the garter snake, Thamnophis sirtalis, in the Great Plains and Rocky Mountains. By Henry S. Fitch and T. Paul Maslin. Pp. 289-308, 4 figures in text. February 10, 1961. | |
| 6. | Fishes of the Wakarusa river in Kansas. By James E. Deacon and Artie L. Metcalf. Pp. 309-322, 1 figure in text. February 10, 1961. | |
| 7. | Geographic variation in the North American cyprinid fish, Hybopsis gracilis. By Leonard J. Olund and Frank B. Cross. Pp. 323-348, pls. 21-24, 2 figures in text. February 10, 1961. | |
| 8. | Descriptions of two species of frogs, genus Ptychohyla; studies of American hylid frogs, V. By William E. Duellman. Pp. 349-357, pl. 25, 2 figures in text. April 27, 1961. | |
| 9. | Fish populations, following a drought, in the Neosho and Marais des Cygnes rivers of Kansas. By James Everett Deacon. Pp. 359-427, pls. 26-30, 3 figures in text. August 11, 1961. 75 cents. | |
| 10. | Recent soft-shelled turtles of North America (family Trionychidae). By Robert G. Webb. Pp. 429-611, pls. 31-54, 24 figures in text, February 16, 1962. $2.00. | |
| Index. Pp. 613-624. | ||
| Vol. 14. | 1. | Neotropical bats from western México. By Sydney Anderson. Pp. 1-8. October 24, 1960. |
| 2. | Geographic variation in the harvest mouse. Reithrodontomys megalotis, on the central Great Plains and in adjacent regions. By J. Knox Jones, Jr., and B. Mursaloglu. Pp. 9-27, 1 figure in text. July 24, 1961. | |
| 3. | Mammals of Mesa Verde National Park, Colorado. By Sydney Anderson. Pp. 29-67, pls. 1 and 2, 3 figures in text. July 24, 1961. | |
| 4. | A new subspecies of the black myotis (bat) from eastern Mexico. By E. Raymond Hall and Ticul Alvarez. Pp. 69-72, 1 figure in text. December 29, 1961. | |
| 5. | North American yellow bats, "Dasypterus," and a list of the named kinds of the genus Lasiurus Gray. By E. Raymond Hall and J. Knox Jones, Jr. Pp. 73-98, 4 figures in text. December 29, 1961. | |
| 6. | Natural history of the brush mouse (Peromyscus boylii) in Kansas with description of a new subspecies. By Charles A. Long. Pp. 99-110, 1 figure in text. December 29, 1961. | |
| 7. | Taxonomic status of some mice of the Peromyscus boylii group in eastern Mexico, with description of a new subspecies. By Ticul Alvarez. Pp. 111-120, 1 figure in text. December 29, 1961. | |
| 8. | A new subspecies of ground squirrel (Spermophilus spilosoma) from Tamaulipas, Mexico. By Ticul Alvarez. Pp. 121-124. March 7, 1962. | |
| 9. | Taxonomic status of the free-tailed bat, Tadarida yucatanica Miller. By J. Knox Jones, Jr., and Ticul Alvarez. Pp. 125-133, 1 figure in text. March 7, 1962. | |
| 10. | A new doglike carnivore, genus Cynaretus, from the Clarendonian Pliocene, of Texas. By E. Raymond Hall and Walter W. Dalquest. Pp. 135-138, 2 figures in text. April 30, 1962. | |
| 11. | A new subspecies of wood rat (Neotoma) from northeastern Mexico. By Ticul Alvarez. Pp. 139-143. April 30, 1962. | |
| 12. | Noteworthy mammals from Sinaloa, Mexico. By J. Knox Jones, Jr., Ticul Alvarez, and M. Raymond Lee. Pp. 145-159, 1 figure in text. May 18, 1962. | |
| 13. | A new bat (Myotis) from Mexico. By E. Raymond Hall. Pp. 161-164, 1 figure in text. May 21, 1962. | |
| *14. | The mammals of Veracruz. By E. Raymond Hall and Walter W. Dalquest. Pp. 165-362, 2 figures. May 20, 1963. $2.00. | |
| 15. | The recent mammals of Tamaulipas, México. By Ticul Alvarez. Pp. 363-473, 5 figures in text. May 20, 1963. $1.00. | |
| 16. | A new subspecies of the fruit-eating bat, Sturnira ludovici, from western Mexico. By J. Knox Jones, Jr., and Gary L. Phillips. Pp. 475-481, 1 figure in text. March 2, 1964. | |
| 17. | Records of the fossil mammal Sinclairella, Family Apatemyidae, from the Chadronian and Orellan. By William A. Clemens, Jr. Pp. 483-491, 2 figures in text. March 2, 1964. | |
| 18. | The mammals of Wyoming. By Charles A. Long. Pp. 493-758, 82 figures in text. July 6, 1965. $3.00. | |
| Index. Pp. 759-784. | ||
| Vol. 15. [Pg iii] | 1. | The amphibians and reptiles of Michoacán, México. By William E. Duellman. Pp. 1-148, pls. 1-6, 11 figures in text. December 20, 1961. $1.50. |
| 2. | Some reptiles and amphibians from Korea. By Robert G. Webb, J. Knox Jones, Jr., and George W. Byers. Pp. 149-173. January 31, 1962. | |
| 3. | A new species of frog (Genus Tomodactylus) from western México. By Robert G. Webb. Pp. 175-181, 1 figure in text. March 7, 1962. | |
| 4. | Type specimens of amphibians and reptiles in the Museum of Natural History,
the University of Kansas. By William E. Duellman and Barbara Berg. Pp. 183-204. October 26, 1962. |
|
| 5. | Amphibians and Reptiles of the Rainforests of Southern El Petén, Guatemala. By William E. Duellman. Pp. 205-249, pls. 7-10, 6 figures in text. October 4, 1963. | |
| 6. | A revision of snakes of the genus Conophis (Family Colubridae, from Middle America). By John Wellman. Pp. 251-295, 9 figures in text. October 4, 1963. | |
| 7. | A review of the Middle American tree frogs of the genus Ptychohyla. By William E. Duellman. Pp. 297-349, pls. 11-18, 7 figures in text. October 18, 1963. 50 cents. | |
| 8. | Natural history of the racer Coluber constrictor. By Henry S. Fitch. Pp. 351-468, pls. 19-22, 20 figures in text. December 30, 1963. $1.00. | |
| 9. | A review of the frogs of the Hyla bistincta group. By William E. Duellman. Pp. 469-491, 4 figures in text. March 2, 1964. | |
| 10. | An ecological study of the garter snake, Thamnophis sirtalis. By Henry S. Fitch. Pp. 493-564, pls. 23-25, 14 figures in text. May 17, 1965. | |
| 11. | Breeding cycle in the ground skink, Lygosoma laterale. By Henry S. Fitch and Harry W. Greene. Pp. 565-575, 3 figures in text. May 17, 1965. | |
| 12. | Amphibians and reptiles from the Yucatan Peninsula, México. By William E. Duellman. Pp. 577-614, 1 figure in text. June 22, 1965. | |
| 13. | A new species of turtle, genus Kinosternon, from Central America. By John M. Legler. Pp. 615-625, pls. 26-28, 2 figures in text. June 20, 1965. | |
| 14. | A biogeographic account of the herpetofauna of Michoacán, México. By William E. Duellman. Pp. 627-709, pls. 29-36, 5 figures in text. December 30, 1965. | |
| 15. | Amphibians and reptiles of Mesa Verde National Park, Colorado. By Charles L. Douglas. Pp. 711-744, pls. 37 and 38, 6 figures in text. March 7, 1966. | |
| Index in preparation. | ||
| Vol. 16. | 1. | Distribution and taxonomy of mammals of Nebraska. By J. Knox Jones, Jr. Pp. 1-356, plates 1-4, 82 figures in text. October 1, 1964. $3.50. |
| 2. | Synopsis of the lagomorphs and rodents of Korea. By J. Knox Jones, Jr., and David H. Johnson. Pp. 357-407. February 12, 1965. | |
| 3. | Mammals from Isla Cozumel, Mexico, with description of a new species of harvest mouse. By J. Knox Jones, Jr. and Timothy E. Lawlor. Pp. 409-419, 1 figure in text. April 13, 1965. | |
| 4. | The Yucatan deer mouse, Peromyscus yucatanicus. By Timothy E. Lawlor. Pp. 421-438, 2 figures in text. July 20, 1965. | |
| 5. | Bats from Gautemala. By J. Knox Jones, Jr. Pp. 439-472. April 18, 1966. | |
| More numbers will appear in volume 16. | ||
| Vol. 17. | 1. | Localities of fossil vertebrates obtained from the Niobrara Formation (Cretaceous) of Kansas. By David Bardack. Pp. 1-14. January 22, 1965. |
| 2. | Chorda tympani branch of the facial nerve in the middle ear of tetrapods. By Richard C. Fox. Pp. 15-21. June 22, 1965. | |
| 3. | Fishes of the Kansas River System in relation to zoogeography of the Great Plains. By Artie L. Metcalf. Pp. 23-189, 4 figures in text, 51 maps. March 24, 1966. | |
| 4. | Factors affecting growth and production of channel catfish, Ictalurus punctatus. By Bill A. Simco and Frank B. Cross. Pp. 191-256, 13 figures in text. June 6, 1966. | |
| 5. | A new species of fringe-limbed tree frog, genus Hyla, from Darién, Panamá. By William E. Duellman. Pp. 257-262, 1 figure in text. June 17, 1966. | |
| 6. | Taxonomic notes on some Mexican and Central American hylid frogs. By William E. Duellman. Pp. 263-279. June 17, 1966. | |
| 7. | Neotropical hylid frogs, genus Smilisca. By William E. Duellman and Linda Trueb. Pp. 281-375, pls. 1-12, 17 figures in text. July 14, 1966. | |
| More numbers will appear in volume 17. | ||
| Page | Correction | |
| 287 | cleared ⇒ cleaned | |
| 292 | Data based of => Data based on | |
| 298 | CNMH ⇒ CNHM | |
| 299 | Acahuitzotla ⇒ Acahuizotla | |
| 304 | cyanostica ⇒ cyanosticta | |
| 305 | Quatemala ⇒ Guatemala | |
| 307 | cyanostica ⇒ cyanosticta | |
| 313 | Matagalapa ⇒ Matagalpa | |
| 322 | Carribean ⇒ Caribbean | |
| 323 | Centralia ⇒ Centrali | |
| 336 | proportionaely ⇒ proportionately | |
| 346 | noticably ⇒ noticeably | |
| 372 | Carvaljo ⇒ Carvalho | |
| 375 | Dumeril ⇒ Duméril | |
| ii | trutles ⇒ turtles |