The Project Gutenberg EBook of The Systematics of the Frogs of the Hyla Rubra Group in Middle America, by Juan R. León This eBook is for the use of anyone anywhere at no cost and with almost no restrictions whatsoever. You may copy it, give it away or re-use it under the terms of the Project Gutenberg License included with this eBook or online at www.gutenberg.org Title: The Systematics of the Frogs of the Hyla Rubra Group in Middle America Author: Juan R. León Release Date: May 24, 2010 [EBook #32505] Language: English Character set encoding: ISO-8859-1 *** START OF THIS PROJECT GUTENBERG EBOOK FROGS OF HYLA RUBRA GROUP *** Produced by Chris Curnow, Joseph Cooper and the Online Distributed Proofreading Team at https://www.pgdp.net
University of Kansas Publications, Museum of Natural History
Editors: Frank B. Cross, Philip S. Humphrey, Robert M. Mengel.
Volume 18, No. 6, pp. 505-545, 7 figs., 4 pls.
Published December 2, 1969
University of Kansas
Lawrence, Kansas
PRINTED BY
ROBERT R. (BOB) SANDERS, STATE PRINTER
TOPEKA, KANSAS
1969
The Systematics of the Frogs of the
Hyla rubra
Group in Middle America
BY
JUAN R. LEÓN
CONTENTS
| page | ||
| Introduction | 508 | |
| Acknowledgments | 508 | |
| Materials and Methods | 509 | |
| The Hyla rubra Group | 509 | |
| Key to Species and Subspecies | 510 | |
| Key to Known Tadpoles | 511 | |
| Accounts of Species and Subspecies | 511 | |
| Hyla boulengeri (Cope) | 511 | |
| Hyla foliamorta Fouquette | 520 | |
| Hyla rubra Laurenti | 524 | |
| Hyla elaeochroa Cope | 525 | |
| Hyla staufferi Cope | 532 | |
| Hyla staufferi staufferi Cope | 537 | |
| Hyla staufferi altae Dunn | 540 | |
| Evolutionary History | 540 | |
| Literature Cited | 543 | |
Introduction
The tree frogs of the Hyla rubra group are abundant and form a conspicuous element of the Neotropical frog fauna. Representatives of the group occur from lowland México to Argentina; the greatest diversity is reached in the lowlands of southeastern Brazil (Cochran, 1955). The group apparently originated in South America; the endemic Central American species evolved from stocks that invaded Middle America after the closure of the Colombian Portal in the late Pliocene.
Dunn (1933) partially defined the rubra group as it occurs in Central America. Cope (1865, 1876, 1887), Brocchi (1881), Boulenger (1882), Günther (1901), Noble (1918), Kellogg (1932), Dunn and Emlen (1932), Stuart (1935), and Gaige (1936) dealt with the Middle American species now considered to make up the rubra group. More recently, Taylor (1952, 1958), Fouquette (1958), Starrett (1960), and Duellman (1960, 1963, 1966a) studied aspects of the taxonomy and biology of the species of this group. The five species of the rubra group in Central America have received ten different names. One species, Hyla staufferi, has received five names (two subspecies are recognized herein). Hyla boulengeri was named in the genus Scytopis, but the type species of Scytopis is a member of the genus Phrynohyas Fitzinger, 1843 (Duellman, 1956.)
Little has been published concerning the ecology, life history, osteology, and mating calls of the Middle American species of this group. The purpose of the present report is to describe the species occurring in Middle America and to comment on their distributions, ecology, cranial osteology, and mating calls, and in so doing provide evidence for the evolutionary history of the species inhabiting Middle America.
Acknowledgments
For permission to examine specimens in their care, I am grateful to Drs. Richard G. Zweifel, American Museum of Natural History (AMNH); Robert F. Inger, Field Museum of Natural History (FMNH); Ernest E. Williams, Museum of Comparative Zoology (MCZ); Hobart M. Smith, University of Illinois Museum of Natural History (UIMNH); Charles F. Walker, University of Michigan Museum of Zoology (UMMZ); Jay M. Savage, University of Southern California (USC); James A. Peters, United States National Museum (USNM); Richard J. Baldauf, Texas Cooperative Wildlife Collection (TCWC); and W. Frank Blair, Texas Natural History Collection (TNHC). KU refers to specimens in the Museum of Natural History, University of Kansas. For the loan of tape-recordings of mating calls I thank Drs. W. Frank Blair, University of Texas, and Richard G. Zweifel, American Museum of Natural History.
I am indebted to the Ford Foundation-Universidad de Oriente (Venezuela) Science Project for a scholarship which enabled me to study for two years at The University of Kansas, foster institution of the project. I have benefited by being able to work in the Museum of Natural History at The University of Kansas and I am grateful to Dr. E. Raymond Hall, Director, for providing space and equipment.
I gratefully acknowledge the assistance and advice of Dr. William E. Duellman, who suggested and directed this work, made available specimens under his care and gave much of his time in reading the manuscript and suggesting improvements. I am grateful to Dr. Frank B. Cross who critically read the manuscript and made many editorial suggestions. I am indebted to Linda Trueb for assistance with the osteological aspects of this study; she helped to clarify many confusing points. I am grateful to Charles W. Myers for making available his field notes on these frogs in Panamá, to Arthur C. Echternacht for reading part of the manuscript, and to John D. Lynch for many suggestions and helpful criticisms. The illustrations were executed by David M. Dennis.
Materials and Methods
For the purposes of the present study I examined 1383 preserved specimens, 50 skeletons, and 9 lots of tadpoles. External characteristics used in the analysis of variation are those currently employed in the study of anuran systematics. Twelve measurements and six proportions were taken in the manner described by Duellman (1956). Only the most important references are given in the synonymies, except those of the two subspecies of Hyla staufferi, which are more nearly complete. The taxonomic history of each frog is discussed under Remarks in each account. The cranial osteology was studied by using skeletons and cleared and stained specimens of all species. Developmental stages of tadpoles were determined from Gosner's (1960) table. Personal field work in Central America in the summer of 1966 provided an opportunity to make observations on the ecology, calling sites, and color in life; these data were supplemented by field notes from, and discussions with, Dr. William E. Duellman and Charles W. Myers.
The mating calls of the frogs were recorded in the field on Magnemite and Uher Tape Recorders by Dr. Duellman in the course of his work on the hylid frogs of Middle America—supported by grants from the National Science Foundation (G-9827 and GB-1441). These recordings, plus those borrowed from other institutions, provided 50 tapes for analysis of the mating calls. The calls were analyzed on a Vibralyzer (Kay Electric Company).
The Hyla rubra Group
Definition.—The species forming the group are small to moderate-sized tree frogs (maximum snout-vent length of males of various species 20-49 mm.), distinguished from other groups in the genus Hyla as follows: Brown, grayish brown, or yellowish tan above; thighs plain, marbled with dark brown, or having vertical bands; vocal sac single, median, subgular; snout flat, protruding, rounded or pointed; webbing between fingers reduced or absent; web between first and second toes reduced to fringe on second toe, rest of toes about half webbed; tarsal fold reduced or absent; shanks robust; inner metatarsal tubercle larger than outer; prevomerine teeth on transverse ridges between small to large sized choanae; skull generally longer than wide; nasals large (length more than 40 per cent total length of skull) and having pointed maxillary processes; maxillary bearing small ventromedial palatine process; quadratojugal slender, always joined to maxillary by bony suture; auditory region of proötic slender and short; delicate spatulate columella ventral to crista parotica, broad basally, compressed anterolaterally, slightly rounded distally; anterior arm of squamosal extending about half distance to maxillary; sphenethmoid wider than long; frontoparietal fontanelle present or absent; prevomerine, premaxillary, and maxillary teeth present; prevomer with two lateral processes forming incomplete bony margin to internal nares; tadpoles having pointed xiphicercal tail, snout short, rounded; 2/3 tooth rows; dorsal fin deeper than ventral fin; sinistral spiracle; short dextral anal tube not reaching edge of ventral fin; mating calls consisting of single long note or series of short notes.
Composition.—This group contains about 24 currently recognized species, most of which occur in Brazil. Only five species—boulengeri, elaeochroa, foliamorta, rubra, and staufferi with two subspecies—occur in Central America. Hyla boulengeri and rubra are widespread in South America, and foliamorta occurs in Colombia, whereas the other species are known only from Middle America.
Distribution.—The species of the Hyla rubra group range from the lowlands of northern Argentina and Bolivia to southern Tamaulipas and Guerrero, México.
Comments.—In Central America two subgroups of species can be recognized. Hyla boulengeri and H. foliamorta are distinctive in the large size of adults (snout-vent lengths 41-49 mm.); both have prominent bars on the thighs, a well-defined interorbital triangular mark, blotches or spots dorsally, and large choanae. Hyla elaeochroa, H. rubra, and H. staufferi are smaller (snout-vent lengths 29-40 mm.); they have the thighs weakly barred or vermiculate anteriorly and posteriorly or unmarked, an ill-defined interorbital triangular mark, linear markings dorsally, and small choanae.
Key to Species and Subspecies
1. Larger frogs (males to 49 mm. snout-vent length); thighs strongly barred; supratympanic fold black; dorsum blotched or spotted2
Smaller frogs (males to 40 mm. snout-vent length); thighs weakly barred or plain; supratympanic fold pale brown; dorsum usually having linear pattern3
2. Dorsum tuberculate; snout subacuminate; vocal sac flecked with brown; tarsal fold rudimentary; web absent between fingers; black spots absent in scapular regionH. boulengeri
Dorsum smooth; snout pointed; vocal sac dark gray; tarsal fold absent; trace of web between fingers; two or more elongate dark spots in scapular regionH. foliamorta
3. Snout-vent length more than 30 mm.; tympanum 2/3 to ¾ diameter of eye; prevomerine elevations about size of choanae4
Snout-vent length less than 30 mm.; tympanum less than ½ diameter of eye; prevomerine elevations smaller than choanae5
4. Thighs mottled posteriorly; discs on fingers about ½ size of tympanum; faint dark line from nostril to eyeH. rubra
Thighs faintly barred or plain posteriorly; discs on fingers about size of tympanum; distinct dark line from nostril to eye H. elaeochroa
5. Dorsum brown with irregular dorsolateral stripes and interrupted paravertebral stripes; two transverse bars on shanks; interorbital bar presentH. staufferi staufferi
Dorsum gray with complete dorsolateral and paravertebral stripes; longitudinal stripe on shank; interorbital bar absent H. staufferi altae
Key to Known Tadpoles
1. Entire lower beak black; beaks moderate-sized, serrate; dorsal fin high, extending to middle of back2
No more than half of lower beak black; beaks small, finely serrate; dorsal fin lower, barely extending onto body3
2. Papillae present only laterally H. boulengeri Papillae present laterally and ventrallyH. foliamorta
3. Distinct brown stripe from nostril to eye; two stripes below eye,H. elaeochroa
Faint stripe from nostril to eye; no stripe below eye H. staufferi
Accounts of Species and Subspecies
Hyla boulengeri (Cope)
Scytopis boulengeri Cope, Bull. U.S. Natl. Mus., 32:12, December 1, 1887 [Holotype.—USNM 13974, from "Nicaragua"; J. A. McNiel, collector].
Hyla boulengeri: Günther, Biologia Centrali-Americana, Reptilia and Batrachia, p. 267, June 1901. Noble, Bull. Amer. Mus. Nat. Hist, 38:339, June 1918. Taylor, Univ. Kansas Sci. Bull., 35:856, July 1, 1952.
Diagnosis.—Size large (♂ to 49 mm., ♀ to 53 mm.); skull as long as wide; frontoparietal fontanelle present; snout subacuminate; canthus not pronounced; choanae large; tongue cordiform, slightly longer than broad; interorbital triangle tubercular; skin on dorsum tuberculate; tarsal fold reduced or absent; thighs, shanks, and tarsi boldly barred with dark brown and pale yellow-green in life.
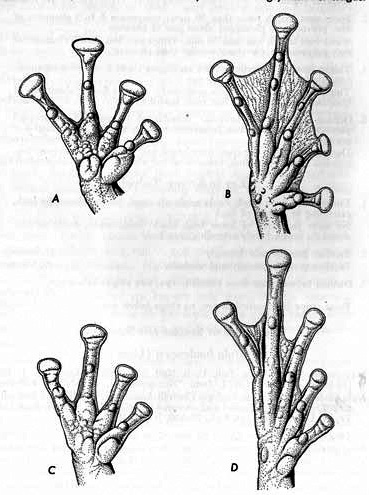
Description.—Head flattened, longer than wide; snout projecting beyond lower lip; loreal region oblique; canthus not pronounced; length of eye less than interorbital distance; tympanum large, about 70 per cent of diameter of eye; interorbital triangle distinct; arms short; fingers lacking web; palmar tubercle tripartite; subarticular tubercles distinct; long tubercle on base of first finger; discs truncate; legs long; tarsal fold reduced or absent; inner metatarsal tubercle rounded, larger than outer, both elevated; subarticular tubercles distinct; one phalanx free of web on second, third, and fifth toes, three free on fourth toe (Fig. 1A and B); skin tuberculate on dorsum, less so on flanks; skin of belly granular, that on chest and throat weakly granular; tongue cordiform, longer than wide, free and notched behind; vocal slits large, lateral to tongue.
Fig. 1. A and B.—Hand and foot of Hyla boulengeri (KU 102173), × 3. C and D.—Hand and foot of Hyla s. staufferi× (KU 57790), × 6
In life, dorsum tan or brown with dark spots on snout, head, and scapular region; interorbital triangle and blotch posteriorly on dorsum dark brown; flanks pale green; groin pale green or orange, mottled with dark brown; thighs tan or brown above with dark transverse bars on anterior and posterior surfaces; spaces between bars green or orange; inner surfaces of shanks and outer surfaces of tarsi brown and orange; foot brown above; forelimbs brown and pale green above, weakly barred; belly creamy white with scattered brown spots; vocal sac creamy white flecked with brown; lower jaw brown with white spots on lips (Pl. 1A).
In preservative, head and dorsum dark brown with triangular spot between eyes; dark spots on head and scapular region and dark brown blotch posteriorly on dorsum; flanks creamy white with brown spots; groin creamy white mottled with dark brown; thighs brown above with dark brown transverse bars on anterior and posterior surfaces; inner surfaces of shanks and outer surfaces of tarsi barred with pale brown; dorsal surface of foot mottled brown and creamy white; ventral surface of foot and toes pale brown; forelimbs faintly barred with pale brown; belly white with a few pale brown spots; vocal sac flecked with pale brown; lower jaw marked with small white spots on lips.
Variation.—Geographic variation is evident in the snout-vent length, tibia length, and foot length, all in relation to snout-vent length, and the relative size of the tympanum to the eye (Table 1). The largest specimens are from the humid Pacific lowlands of Costa Rica; individuals from the Caribbean lowlands of Costa Rica, Canal Zone, and South America are smaller. A general trend for increase in size extends from South America to the Pacific lowlands of Costa Rica.
Most variation in color does not seem to be correlated with geography; color variation is nearly as great within a given population as between separated populations. However, most specimens from Rincón de Osa, Puntarenas Province, Costa Rica, are dusky brown, but a few are darker. In comparison with specimens from the Caribbean lowlands of Central America, specimens from the Pacific slopes in Costa Rica have a darker interorbital triangle. In some specimens from the latter area rugosities are present on the borders of the interorbital triangle, on the snout, on the upper eyelid, and on the heel. Specimens from the Caribbean lowlands are less tuberculate, and most individuals from there lack rugosities on the tarsus. Living individuals from Puerto Viejo, Heredia Province, Costa Rica, and from the Canal Zone, Panamá, are brown above with a metallic green tint. Rugosities are present on the posterior edges of the forelimbs in some specimens from throughout the range. In most respects, specimens from the Canal Zone resemble those from the Caribbean lowlands of Costa Rica more than they resemble those from the Pacific lowlands of Costa Rica, but some individuals from the Canal Zone are less metallic above and have small white spots dorsally on the body, head, and limbs.
A moderate amount of color change from night to day has been noted. At night, a male from Puerto Viejo, Heredia Province, Costa Rica, was grayish tan above with slightly darker markings; the flanks were pale yellowish green. By day, the dorsum was brown with darker markings, and the throat was pale gray with white flecks; the rest of the venter was creamy white. The groin was pale green with black mottling; the anterior and posterior surfaces of the thighs and inner edges of the tarsi were greenish yellow with black bars.
Table 1.—Geographic Variation in Size and Proportions in Males of
Hyla boulengeri.
(Means in parentheses below observed ranges.)
| Locality | N |
Snout-vent
length (mm.) |
Tibia length/
snout-vent |
Tympanum/
eye |
Foot length/
snout-vent |
| Costa Rica: Tilarán | 23 |
37.4-48.7
(43.8) |
0.52-0.58
(0.55) |
0.62-0.76
(0.71) |
0.39-0.45
(0.42) |
| Costa Rica: Rincón de Osa | 10 |
41.4-46.1
(44.0) |
0.54-0.60
(0.57) |
0.68-0.80
(0.74) |
0.40-0.45
(0.43) |
| Costa Rica: Alajuela Province | 13 |
35.6-43.1
(39.8) |
0.55-0.60
(0.57) |
0.63-0.78
(0.69) |
0.41-0.46
(0.44) |
| Costa Rica: Puerto Viejo | 25 |
37.5-42.9
(41.6) |
0.51-0.62
(0.55) |
0.63-0.79
(0.71) |
0.38-0.46
(0.43) |
| Costa Rica: Suretka | 9 |
38.7-42.0
(41.0) |
0.56-0.58
(0.56) |
0.53-0.72
(0.62) |
0.35-0.45
(0.42) |
| Panamá: Canal Zone | 16 |
36.7-42.9
(39.0) |
0.52-0.58
(0.54) |
0.70-0.78
(0.74) |
0.40-0.44
(0.42) |
| Venezuela: Santomé | 4 |
35.5-40.9
(38.5) |
0.45-0.48
(0.46) |
0.63-0.67
(0.65) |
0.36-0.40
(0.38) |
Table 2.—Comparison of Mating Calls in the
Hyla rubra
Group.
(Means in parentheses below observed ranges.)
| Species | N |
Notes per
call group |
Duration
of note (sec.) |
Pulses
per second |
Fundamental
frequency (cps) |
Major frequencies (cps) | |
| Lower | Upper | ||||||
| H. boulengeri | 8 | 1 |
0.24-0.47
(0.35) |
80-120
(101) |
70-74
(71) |
1400-1820
(1611) |
2520-3182
(2840) |
| H. foliamorta | 7 | 1 |
0.23-1.86
(0.69) |
50-60
(51) |
52-61
(56) |
912-1026
(918) |
2736-3477
(3055) |
| H. elaeochroa | 15 |
2-95
(19) |
0.12-0.24
(0.17) |
40-50
(42) |
48-65
(57) |
1254-1586
(1499) |
2562-3477
(2911) |
| H. s. staufferi | 18 |
2-77
(23) |
0.13-0.23
(0.18) |
100-130
(120) |
96-130
(106) |
1582-1872
(1743) |
1962-3744
(3056) |
| H. s. altae | 7 |
2-22
(11) |
0.14-0.18
(0.15) |
110-130
(120) |
104-117
(112) |
1853-2106
(2008) |
3379-4056
(3775) |
Cranial Osteology.—The skull of Hyla boulengeri is as long as it is wide, and is flat; the premaxillary is small and bears 13 to 17 teeth (mean for 6 specimens, 14.9). The alary processes of the premaxillaries are widely separated, concave posteriorly, and vertical. Ventrally, the premaxillary is connected to the prevomer by bony tissues. The maxillary is slender and bears 70 to 91 teeth (mean for 6 specimens 78.1). The pars facialis of the maxillary is laterally convex and about four times as high as the pars dentalis.
The nasal is large (its length about 40 per cent of total length of skull), and pointed anteriorly and posteriorly in dorsal view. The nasals are separated anteromedially by the cartilaginous septum nasi. One or two protuberances are present on the midlateral concavity of the nasal. Posteriorly, the nasal overlaps the sphenethmoid and articulates with the palatine. Dorsally the sphenethmoid is large, pentagonal, and completely ossified. The frontoparietal is elongate, smooth, and bears a small supraorbital process on the anterior edge of the orbit. A keyhole-shaped frontoparietal fontanelle is present; the fontanelle is narrow anteriorly and wide posteriorly.
The bony part of the proötic is separated dorsally from the squamosal by the cartilaginous crista parotica. The squamosal is small, its anterior arm slender and pointed. The posterior arm of the squamosal is pointed terminally and articulates with the proötic medially.
The prevomer is large and elongate. Anteriorly the prevomer is connected to the maxillary-premaxillary articulation; posteriorly, the prevomer is separated from the sphenethmoid by cartilage. Each prevomer bears six to nine teeth. The palatine is present and edentate. The anterior end of the parasphenoid is broad (less pointed than in Hyla foliamorta). The pterygoid is slender and well developed.
Natural History.—Hyla boulengeri inhabits humid lowland tropical forests and breeds in temporary ponds. Clasping pairs and gravid females were observed at Puerto Viejo, Heredia Province, Costa Rica, on June 21, 1966. Males were calling from depressions in decaying logs and stumps, in forked stems, and from leaves of broad-leafed plants near the pond. Males were observed in late July and early August calling from Calathea and Heliconia leaves at the edge of a pond in the wet forest of the Osa Peninsula. William E. Duellman informed me that he collected calling males in January at El Real, Darién, and in March at Almirante, Bocas del Toro, Panamá. Taylor (1952) found calling males in June at Turrialba, Cartago Province, Costa Rica, and Dunn (1931a) observed males calling in July, November, and December in Panamá. Gravid females have been found from April to August. Breeding activities of Hyla boulengeri always seem to be associated with temporary ponds; in Central America breeding apparently takes place throughout most of the year.
The mating call of Hyla boulengeri consists of one short, moderately low-pitched note. Each note has a duration of 0.24 to 0.47 second and is repeated at intervals of one second to several minutes. The notes have 80 to 120 pulses per second, a fundamental frequency of about 70 cycles per second, and a dominant frequency of 2,840 cycles per second (Table 2, Pl. 3A).
The eggs are deposited in a mass in the water. No information is available concerning early development. Tadpoles in advanced stages of development were found in a temporary pond at Rincón de Osa, Puntarenas Province, Costa Rica. The pond was about 10 cm. deep, had a muddy bottom and lacked vegetation. Three recently metamorphosed young were found in mid-August, 1966, on grass at the edge of another temporary pond in the forest.
Tadpoles—Twelve tadpoles are available. These were collected at Rincón de Osa, Puntarenas Province, Costa Rica. The maximum size represented is 34.0 mm., total length (stage 42 of development).
A typical tadpole in stage 36 of development (KU 104295) has a body length of 12.0 mm., tail length of 20.0 mm., and total length of 32.0 mm. Other characters are as follows: depth of tail equal to length of body; body deeper than wide; distance between eye and nostril equal to that between nostril and tip of snout; mouth anteroventral, upper and lower lips bare; papillae present laterally; tooth rows 2/3; upper rows about equal in length; first upper row slightly, and second upper row widely, interrupted medially; lower rows about equal in length, shorter than upper rows; third lower row containing 5-10 large teeth; beak strong, serrate; spiracle nearer anus than eye; anal aperture not extending to border of ventral fin; caudal musculature slender posteriorly, extending to tip of pointed tail; dorsal fin extending to middle of body, slightly deeper than ventral fin; posterior three fourths of tail spotted; rest of tail and body gray-brown or transparent; hindlimbs flecked or spotted with brown (Table 3, Fig. 2A and 3A).
Table 3.—Sizes of Tadpoles of
Hyla boulengeri
in Relation to Developmental Stages.
(Means in parentheses below observed ranges; measurements in mm.)
| Stage | N | Body length | Tail length | Total length |
| 30 | 1 | 11.0 | 22.2 | 33.2 |
| 35 | 1 | 11.0 | 12.0 | 23.0 |
| 36 | 3 |
9.5-12.0
(11.2) |
20.0-21.5
(20.5) |
31.0-32.0
(31.7) |
| 38 | 2 | 11.5 | 22.0 | 33.5 |
| 42 | 2 |
10.5-13.0
(11.8) |
21.0-22.0
(21.5) |
32.5-34.0
(33.3) |
| 44 | 2 |
14.0-15.0
(14.5) |
8.0-15.0
(12.5) |
22.0-30.0
(26.0) |
| 46 | 1 | 15.0 | — | 15.0 |
A recently metamorphosed young has a snout-vent length of 15 mm.; the head is as long as wide, the eyes are prominent; the limbs are weakly barred; the skin is rugose above and granular below. The venter is immaculate; the dorsum and limbs are gray-brown in preservative (pale green in life). The interorbital space, supratympanic fold, and scapular region are darker than the rest of the body; the fingers lack webbing; the webbing on the foot is the same as in adults; small metatarsal tubercles are present, but the tarsal fold is absent.
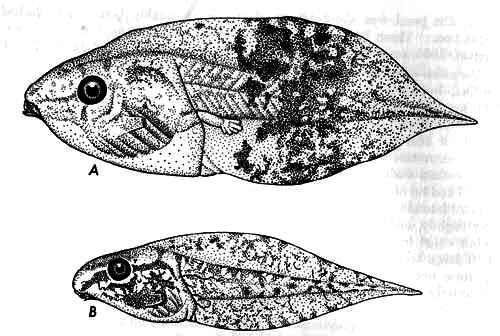
Fig. 2. Tadpoles of (A) Hyla boulengeri (KU 104295) and (B) Hyla elaeochroa (KU 104134), × 3.
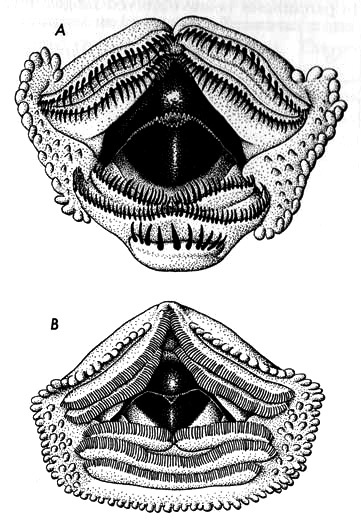
Fig. 3. Mouthparts of tadpoles of (A) Hyla boulengeri (KU 104295) and (B) Hyla elaeochroa (KU 104134), × 25.
Remarks.—Cope (1887:12) described Scytopis boulengeri from Nicaragua. Günther (1901:267) placed boulengeri in the genus Hyla, and stated that Cope possibly placed boulengeri in the genus Scytopis on the supposition that it had an accumulation of "sebaceous glands" above the tympanum. Noble (1918:339) redescribed Hyla boulengeri on the basis of three specimens from Zelaya Province, Nicaragua, and noted that the glands were not prominent in any of the specimens. Duellman (1956:8) showed that Scytopis hebes (generotype of Scytopis by monotypy) is a Phrynohyas, and thus placed Scytopis Cope, 1862, in the synonymy of Phrynohyas Fitzinger, 1843.
Dunn and Emlen (1932:25) placed Hyla lancasteri Barbour in the synonymy of Hyla boulengeri; the former was known solely from one juvenile. They made no qualifying statements, but probably they were impressed by the strongly barred thighs, a coloration known among Central American hylids at that time only in Hyla boulengeri (Duellman, 1966a:271). Taylor (1952:856) followed Dunn and Emlen with reservation and noted some differences. Duellman (1966a:271) showed that the holotype of lancasteri was a juvenile of a species subsequently named as Hyla moraviaensis by Taylor (1952:865).
In Central America, Hyla boulengeri can be confused only with Hyla foliamorta; the latter is restricted to central and eastern Panamá and northern Colombia. The snout of foliamorta is more pointed and protruding, and the vocal sac is darker than in boulengeri; the groin of foliamorta usually is creamy white, whereas boulengeri usually has a dark spot. The skulls differ in that boulengeri has a frontoparietal fontanelle, the prevomer is larger and elongate, anteriorly connected to the premaxillary, and posteriorly separated from the sphenethmoid by cartilage; foliamorta lacks a fontanelle, the prevomer is smaller, anteriorly separated from the premaxillary by cartilage, but connected by a bony suture to the sphenethmoid. The mating call of boulengeri differs by having shorter notes, twice as many pulses per second, a higher fundamental frequency, and more closely approximated major frequencies than does that of foliamorta.
Hyla boulengeri need not be compared in detail with the other Central American members of the Hyla rubra group, because all of them are smaller and have shorter snouts, smoother skin, and dissimilar color patterns.
Distribution.—In Central America Hyla boulengeri inhabits the forested lowlands in locally humid areas in Guanacaste Province, Costa Rica, and in the humid Golfo Dulce region of Costa Rica; it occurs on the Carribbean lowlands from central Nicaragua to South America, where it ranges to Guyana and Ecuador. The highest elevations where H. boulengeri has been found are 620 meters at Turrialba, Cartago Province, and 700 meters at Tilarán, Guanacaste Province, Costa Rica (Fig. 4).
Specimens Examined.—Costa Rica: Alajuela: 9 km N Ciudad Quesada, near La Florencia, USC 8059 (4); 18 km N Florencia, USC 2624; Laguna Monte Alegre, KU 64334; Las Playuelas, 11 km S Los Chiles, USC 7216, 7217 (2), 7219; 3 km NE Muelle del Arenal, USC 2644 (5). Cartago: Turrialba, KU 24741. Guanacaste: 7 km N Liberia, USC 8096 (2), 8138 (6); 13.6 km N Liberia, USC 8151, 8171 (2); 20.5 km S Liberia, USC 8205; Taboga, 20 km SE Las Cañas, KU 102170, USC 7166; 4 km NE Tilarán, USC 8023; 6 km NE Tilarán, USC 523 (3), 6262, 7019. Heredia: Puerto Viejo, KU 64323-7 (skeletons), 104351-3 (skeletons), 64330-3, 103592-620; 1 km NE Puerto Viejo, UMMZ 126042; 1 km S Puerto Viejo, KU 84983-4 (skeletons), 86317-22, 87774 (skeleton); 4.2 km W Puerto Viejo, KU 64329, 64328 (skeleton). Limón: Mountain Cow Creek, near Banano, KU 37031, 41067 (skeleton); 3 km S Río Tortuguero, AMNH 69057; Suretka, KU 36482-8, 36699. Puntarenas: 4.8 km S Bahía Rincón on NW side Río Rincón, USC 705; Parrita, USC 6163; 4.5 km W Rincón de Osa, KU 102177-9, 104295-6 (tadpoles); 6 km SW Rincón de Osa, KU 102171-6; 4.4 km NW Villa Neilly, USC 8003; 10.5 km WNW Villa Neilly, KU 64321. San José: 21 km WSW San Isidro el General, KU 34104-6.
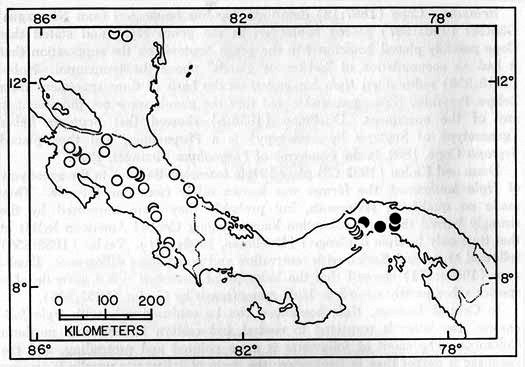
Fig. 4. Map showing locality records for Hyla boulengeri (circles) and H. foliamorta (dots).
Panamá: Bocas del Toro: 3.2 km W Almirante, KU 95978. Canal Zone: Barro Colorado Island, FMNH 13379; near Clayton Reservation, UIMNH 42000; 2.6 km SW Fort Kobbe, KU 95977; Miraflores Locks, AMNH 69764-5; Summit, AMNH 73445, KU 97777, 101540-9, 104350 (skeleton). Colón: Río Gatuncillo, near Nuevo San Juan, KU 95976. Darién: El Real, KU 80451-3.
Hyla foliamorta Fouquette
Hyla foliamorta Fouquette, Herpetologica, 14:125, April 25, 1958 [Holotype.—TNHC 23109, 11 km. NW Miraflores Locks, Canal Zone, Panamá; M. J. Fouquette, Jr. collector].
Diagnosis.—Size medium (♂ to 43 mm., ♀ to 41 mm.); skull longer than wide; frontoparietal fontanelle absent; snout acuminate, projecting; interorbital triangle bordered by white lines; scapular region having two or more elongate spots; dorsum smooth; vocal sac dark gray; groin creamy white; traces of web between fingers.
Description.—Head flattened, longer than wide; snout flat, pointed, protruding beyond lower lip; loreal region slightly concave; canthus moderately prominent; eyes smaller than interorbital space; tympanum distinct, 55 to 75 per cent of diameter of eye, smaller than internarial space; arms short; fingers having rudimentary webs; median palmar tubercle tripartite; inner palmar tubercle on base of first finger flat; subarticular tubercles distinct; discs of fingers smaller than diameter of tympanum; legs long; tarsal fold lacking; inner metatarsal tubercle larger than outer; one phalanx free on second, third, and fifth toes, two and one half phalanges free on fourth toe; narrow fringe continuing from web to discs of toes; discs of toes about the size of those on fingers; skin smooth on dorsum and flanks, that on belly and posterior part of thighs granular; tongue oval, longer than wide; vocal slits oblique, about one half length of tongue.
In life, dorsum pale tan to pale reddish brown with irregular reddish brown markings; small dark spots on head; distinct dark brown triangular mark between eyes, bordered by thin white lines; apex of triangle always directed backward; supratympanic fold with black edge; scapular region having two to five small, elongate black spots; belly creamy tan with small brown spots; vocal sac uniformly dark brown with scattered creamy tan flecks; upper jaw dark brown; limbs creamy white below with scattered brown spots; groin marked with small brown spots in some specimens; anterior and posterior surfaces of thighs yellow-orange with three distinct black blotches; two dark bands on upper surface of shanks; webbing of feet yellowish tan with brown mottlings (Pl. 1B).
In preservative, dorsum brown or gray with darker markings; interorbital triangle distinct, bordered by white lines; supratympanic fold with black edge; two or more small elongate black spots in scapular region; belly white with numerous brown flecks; edge of upper lip dark brown; vocal sac dark gray; undersides of limbs creamy white; groin creamy white with or without brown spots; anterior and posterior surfaces of thighs having three black blotches separated by creamy white spaces; shanks having two brown bands; webbing of feet mottled with brown.
Variation.—Twenty-eight breeding males from the area between Chepo and Tocumen, Panamá, have snout-vent lengths of 39.0 mm. to 46.0 mm. (mean 42.5 mm.). In these specimens, the ratio of the tibia length to the snout-vent length is 0.54 to 0.61 (mean, 0.57); the ratio of the diameter of the tympanum to that of the eye is 0.55 to 0.75 (mean, 0.67). One female has a snout-vent length of 41.0 mm., tibia/snout-vent length ratio of 0.57, and tympanum/eye ratio of 0.76. Two to five (usually three) elongate black spots are present in the scapular region in different individuals. The flanks in some are spotted with brown; in others they are creamy white. A small black spot is present in the groin of some specimens. Usually two to four blotches are present on the anterior and posterior surfaces of the thighs; in some specimens the blotches are reduced to small spots. One or two brown spots are present proximally on the shanks in most specimens. In some individuals tuberculations are scattered on the head and in the tympanic and scapular regions, but the dorsum is smooth in most specimens; the belly is creamy white flecked with brown.
Cranial Osteology.—The skull of Hyla foliamorta is flat and longer than it is wide. The premaxillary is small and bears 13 to 16 teeth (mean for 2 specimens, 14.8). The alary process of the premaxillary is vertical and concave posteriorly. Ventrally, the premaxillary is completely separated from the prevomer by cartilage. The maxillary is slender; each bears 77 to 84 teeth (mean for 2 specimens, 81). The pars facialis of the maxillary is laterally convex and less than three times the height of the pars dentalis.
The nasal is large and pointed anteriorly and posteriorly in dorsal view. The length of the nasal comprises about 40 per cent of the total length of the skull. The nasals are separated anteromedially by the cartilaginous septum nasi. One protuberance is present on the midlateral concavity of the nasal. Posteriorly, the nasal overlaps the sphenethmoid; posterolaterally the nasal articulates with the palatine. The sphenethmoid is completely ossified and pentagonal in dorsal view. The frontoparietal is elongate, without a pronounced anterior supraorbital process. The frontoparientals are sutured medially throughout their lengths; the frontoparietal fontanelle is absent.
The bony part of the proötic is narrowly separated dorsolaterally from the squamosal by the cartilaginous crista parotica. The squamosal is large; the anterior arm is pointed. The posterior arm of the squamosal is broad, rounded terminally, and articulates with the proötic medially.
The prevomer is short and separated anteriorly from the premaxillary and maxillary by cartilage. The posterior margin of the prevomer has a bony articulation with the sphenethmoid. Each prevomer bears five to seven teeth. The palatine is small and edentate. The anterior end of the parasphenoid is narrow (more pointed than in Hyla boulengeri). The pterygoid is slender and well developed (Fig. 5A).
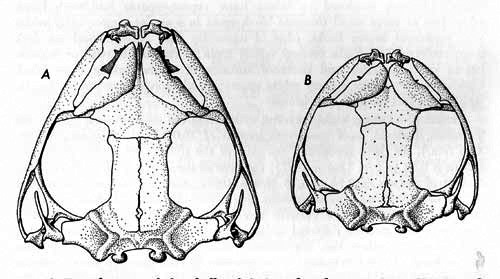
Fig. 5. Dorsal views of the skulls of (A) Hyla foliamorta (KU 77687) and (B) H. elaeochroa (KU 68289), × 3.
Natural History.—Hyla foliamorta inhabits lowland forests in eastern Panamá and breeds in temporary ponds. Males have been observed calling from grasses, bushes, and emergent vegetation near temporary ponds and ditches along roads. William E. Duellman informed me that he found a breeding congregation of this species in June near Chepo, Panamá, where males were calling from spiny palms at the edge of a woodland pond. Fouquette (1958) found calling males in May, August, and September near Miraflores Locks, Canal Zone. Calling stations vary from one to two meters above ground. No clasping pairs have been found; only one female is known (KU 101589, from 8 km NE Tocumen, Panamá); this gravid individual was collected in early June.
The mating call of Hyla foliamorta consists of one pulsed, low-pitched, moderate trill of about O.5 second duration. Each note is repeated at intervals of 5 seconds to a few minutes. The notes have about 50 pulses per second, a fundamental frequency of 56 cycles per second and a dominant frequency of about 3000 cycles per second (Table 2, Pl. 3B).
Egg deposition sites are unknown. No information is available concerning early development, and little is known about the breeding season of Hyla foliamorta. Probably its breeding activities are restricted to the rainy months.
Tadpoles.—Eight tadpoles were collected from a weedy temporary pond near Chepo, Panamá, in early June.
A typical tadpole in stage 35 of development (KU 104244) has a body length of 9.5 mm., tail length of 25.0 mm., and a total length of 34.5 mm. Other characters are as follows: depth of tail equal to length of body; body deeper than wide; distance between eye and nostril equal to distance between eye and spiracle; mouth anteroventral; median part of upper lip bare; rest of lip having one row of papillae; a few other rows of small papillae at corners of mouth; tooth rows 2/3; first upper row entire, second upper row interrupted medially, shorter than first; lower rows shorter than upper rows, third shortest; beak moderately robust; spiracle nearer eye than anus; anal tube short, aperture not extending to border of ventral fin; caudal musculature slender, extending to tip of pointed tail; dorsal fin extending onto body (Table 4).
Table
4.—Sizes of Tadpoles of
Hyla foliamorta
in Relation to Developmental Stages.
(Means in parentheses below observed ranges; measurements in mm.)
| Stage | N | Body length | Tail length | Total length |
| 25 | 2 |
5.0-5.2
(5.1) |
8.0-8.5
(8.3) |
13.0-13.7
(13.4) |
| 26 | 3 |
7.0-7.5
(7.2) |
12.0-12.4
(12.1) |
17.0-19.5
(18.6) |
| 28 | 2 |
6.5-7.0
(6.8) |
18.0 | 25.0 |
| 35 | 1 | 9.5 | 25.0 | 34.5 |
In life, yellow above, white below; caudal fin greenish yellow with black or gray reticulations; dark line from snout to eye; dark spot behind eye; tail unpigmented except for fine dark reticulations. In preservative body creamy white, transparent below with dark pigment above in some specimens.
Remarks.—Hyla foliamorta can be confused only with Hyla boulengeri. The differences between adults of these species were discussed in Remarks on H. boulengeri. The tadpoles of foliamorta have labial papillae on the lower lip and a stripe between the eye and the tip of the snout. By comparison the tadpoles of boulengeri have a bare lower lip and no stripe between the eye and the tip of the snout.
Distribution.—Hyla foliamorta inhabits the subhumid Pacific lowlands (elevations of less than 100 meters) of Central Panamá and Caribbean lowlands of northern Colombia (Fig. 4).
Specimens Examined.—Panamá: Panamá: 3 km WSW Chepo, KU 77164-9, 101573-5, 104243-4 (tadpoles); 6 km WSW Chepo, KU 77170, 101576-8; 1.5 km SW Naranjal, KU 77171, 77687 (skeleton); 2 km N Tocumen, KU 101579-83, 104349 (skeleton); 8 km NE Tocumen, KU 101584-92.
No specific locality: TNHC 24401.
Hyla rubra Laurenti
Hyla rubra Laurenti, Synopsis Reptilium Emendatum, p. 35, 1768. Daudin, Hist. Nat. Rainettes Grenouilles Crapauds, II:26, 1802. Daudin, Hist. Nat. Particuliere Reptiles, 8:53, 1803. Günther, Catalogue Batrachia Salientia Brit. Mus., p. 110, 1859. Boulenger, Catalogue Batrachia Salientia s. Ecaudata, p. 403, February 1, 1882. Dunn, Occas. Papers, Boston Soc. Nat. Hist., 5:413, October 10, 1931.
Hyla elaeochroa (part): Dunn and Emlen, Proc. Acad. Nat. Sci. Philadelphia, 84:25, March 22, 1932.
Diagnosis.—Size medium; skull longer than wide; frontoparietal fontanelle absent in adults; snout subovoid; choanae rounded; dorsal stripes present; black vermiculations on posterior surfaces of thighs.
Description.—Head flat, longer than wide; snout long, subovoid, slightly protruding beyond lower lip; loreal oblique, concave; canthus rounded, indistinct; diameter of eye about equal to interorbital space; tympanum large, about three fifths diameter of eye, smaller than internarial distance; supratympanic fold indistinct; arms short; fingers free of webs; subarticular tubercles distinct; median palmar tubercle large, bifid; inner palmar tubercle on base of first finger flat, elongate; disc of third finger about one half diameter of tympanum; legs moderately long; tarsal fold absent; inner metatarsal tubercle distinct, oval; toes about half webbed; web on fourth toe extending to disc; discs of toes about size of those on fingers; skin smooth above with small granules on head and in scapular region in some specimens; skin on flanks, throat, belly, and lower surfaces of thighs granular; tongue oval, longer than wide, not free behind; choanae small, oval; vocal slits long, lateral to tongue.
In preservative, dorsum pale brown with darker dorsolateral stripes; narrow dark brown line from nostril to eye; groin, anterior surface of thighs, and posteroventral surfaces of shanks creamy tan with dark brown vermiculations; white spots present on thighs in some specimens; throat flecked with brown; belly creamy white or gray.
Remarks.—The taxonomic history of Hyla rubra Laurenti is confused. Seba (1734:70) illustrated and diagnosed a frog for which he used the name Ranula, Americana, Rubra. Linnaeus (1758:213) considered Seba's frog to be a variety of Hyla arborea. Laurenti (1768:35) apparently examined the same individual that Seba called Ranula, Americana, Rubra. For this specimen, Laurenti used the binominal Hyla rubra and provided a brief diagnosis. The type locality was given as America.
Daudin (1802:26) redescribed the same specimen(s?) treated by Seba and Laurenti and provided a fairly good description and figures. Daudin restricted the type locality to Surinam and indicated that Marin de Baize was the probable collector. Daudin (1802:26 and 1803:53) neglected to consider Laurenti's work, but he applied the same name used by Laurenti. Most authors have credited Hyla rubra to Daudin, but Rivero (1961:120) noted that Hyla rubra Laurenti, 1768, has priority over Hyla rubra Daudin, 1802. Since both Laurenti and Daudin worked on Seba's material, it is reasonable to assume that Daudin redescribed the same frog that was named by Laurenti; this was not an uncommon practice in the early nineteenth century. Thus I conclude that Hyla rubra Daudin, 1802, is a junior synonym of Hyla rubra Laurenti, 1768.
Dunn (1931a:413) first reported Hyla rubra from Central America; he recorded the species from the Canal Zone and San Pablo, Panamá. I have examined the material of Hyla rubra from Panamá deposited in various museums. Most of the specimens are faded, discolored, and do not have distinct brown vermiculations on the thighs. The specimens seem to be more like Hyla rubra than any of the other species in the rubra group. The presence of oval choanae and a tympanum larger than the largest finger disc separate these specimens from Hyla elaeochroa, a species with which rubra has been confused. Hyla elaeochroa does not occur in the Canal Zone or eastern Panamá. All museum specimens from Nicaragua, Costa Rica, and western Panamá that have been called Hyla rubra, plus those mentioned by Dunn and Emlen (1932:25) and Dunn (1933:61) are Hyla elaeochroa.
The taxonomic status of the many South American populations referred to Hyla rubra and of other populations now recognized as different species is not clear at the present time. Considerable variation in external characters and in cranial features has been observed in South American rubra. A review of the taxonomy of these populations is beyond the scope of this paper. Possibly the Central American specimens herein referred to rubra will ultimately be found to be specifically distinct from those in Surinam. Since I have no osteological material from Central America, I have been unable to describe the cranium in this account. Furthermore, I have no data on the ecology and life history of rubra in Central America.
Distribution.—Hyla rubra inhabits lowland tropical forests from central-eastern Panamá to northern South America and thence through lowlands east of the Andes to northern Argentina (Fig. 6).
Specimens Examined.—Panamá: Canal Zone: Gatun, UMMZ 52720 (2); Madden Dam, FMNH 67820; no specific locality, UMMZ 56517 (3), USNM 37863. Colón: Cerro Bruja, MCZ 13248. Darién: El Real, USNM 140569-70, 140573. Panamá: Juan Díaz, MCZ 17973; Las Sabanas, MCZ 17581; Río Trinidad, UMMZ 64003; San Pablo, MCZ 1398-9.
Hyla elaeochroa Cope
Hyla elaeochroa Cope, Jour. Acad. Nat. Sci. Philadelphia, 8:105, 1876 [Holotype.—USNM 30689, Sipurio, Limón Province, Costa Rica; William M. Gabb collector]. Günther, Biologia Centrali-Americana, Reptilia and Batrachia, p. 265, June 1901. Taylor, Univ. Kansas Sci. Bull., 35:859, July 1, 1952. Duellman, Univ. Kansas Publ., Mus. Nat. Hist., 17:270, June 17, 1966.
Hyla quinquevittata Cope, Proc. Amer. Philos. Soc., 23:273, April 1887 [Holotype.—USNM 14187, Nicaragua; J. F. Bransford collector]. Günther, Biologia Centrali-Americana, Reptilia and Batrachia, p. 268, June 1901. Noble, Bull. Amer. Mus. Nat. Hist., 38:340, June 1918.
Hyla rubra (part): Dunn and Emlen, Proc. Acad. Nat. Sci. Philadelphia, 84:25, March 22, 1932.
Hyla dulcensis Taylor, Univ. Kansas Sci. Bull., 39:37, November 18, 1958 [Holotype.—KU 32168, Golfito, Puntarenas Province, Costa Rica; Edward H. Taylor collector].
Diagnosis.—Size medium (♂ to 38 mm., ♀ to 40 mm.); skull wider than long; nasals truncate anteriorly; frontoparietal fontanelle moderate in size; snout slightly protruding; tympanum about size of largest discs on fingers; dorsum marked by longitudinal stripes; dark stripe between eye and nostril; in life tan to olive-green with or without dark mark between eyes; bones greenish blue.
Description.—Head flat, longer than wide; snout long, rounded, protruding beyond mouth; canthus indistinct; length of eye equal to interorbital distance; loreal region not pronounced; tympanum distinct and about two-fifths diameter of eye; interorbital triangle present or absent; arms short; trace of web between fingers, extending as fringe along sides of fingers; first finger very short with small disc; other discs about size of those on toes; discs on third finger and fourth toe as large as tympanum; outer palmar tubercle moderate in size, partly bifid; inner palmar tubercle large, elongate, flat; subarticular tubercles distinct; legs moderately long; tarsal fold absent; inner metatarsal tubercle flat; outer metatarsal tubercle smaller, indistinct; subarticular tubercles moderate in size; fringe on toes to tip of disc of second toe; rest of toes about two-thirds webbed; foot length about two fifths snout-vent length; tibia length about one half snout-vent length; skin above smooth or with minute pustules; belly finely granular; ventral surfaces of thighs and areas below anus granular; skin on ventral surfaces of limbs smooth; tongue relatively large, longer than wide, barely notched behind; vocal slits elongate, lateral to tongue; choanae medium in size. In life, dorsum yellowish brown, olive green, or grayish brown with dark brown spots on snout, dark brown stripe from nostril to eye, dark yellow-brown interorbital triangle, and dark supratympanic region; generally five interrupted longitudinal dark brown stripes on dorsum (one on each flank, pair of paravertebral and one vertebral); flanks pale yellow; groin yellowish brown; thighs marked with one or two transverse yellow-brown blotches; shanks with two or three yellow-brown blotches above; spaces between blotches on thighs, shanks, tarsi, and feet yellow; brown spots on tarsi and in some specimens on feet; arm pale yellow with pale brown spots; belly creamy white having slight blue-green tint; vocal sac and chin yellow; axillary region yellow, blue-green in some specimens (Pl. 2A).
In preservative, head and dorsum yellowish brown; dark brown stripe from nostril to eye; dark brown spots on snout; a dark brown interorbital triangle with apex directed backward; dark brown supratympanic region; dorsal stripes same as in living individuals; flanks pale yellow with brown spots in some specimens; groin creamy white; thighs and shanks having or lacking transverse dark brown blotches; spaces between blotches creamy white or yellow-brown; arms pale yellowish brown; belly and vocal sac creamy white.
Variation.—Geographic variation in size and some proportions, such as the ratio of tibia length to snout-vent length and the ratio of the diameter of the tympanum to that of the eye, have been observed in this species. The largest individuals are from the Golfo Dulce region (samples from Piedras Blancas and Rincón de Osa), Puntarenas Province, Costa Rica. The smallest individuals are from El Recreo, Zelaya Province, Nicaragua, and from the Caribbean lowlands of Costa Rica.
The diameter of the tympanum is proportionately larger (relative to the size of the eye) in males from Tilarán, Guanacaste Province; the tympanum is nearly as large in males from Piedras Blancas, Puntarenas Province, and Puerto Viejo, Heredia Province, Costa Rica. The lowest ratios occur in individuals from Almirante, Bocas del Toro, Panamá, in specimens from the Caribbean lowlands of Costa Rica (except Puerto Viejo), and in those from El Recreo, Zelaya Province, Nicaragua. In general, the tympanum is proportionately larger in females than in males; the tympanum is largest in females from the Pacific lowlands of Costa Rica (Table 5).
Color variation has been observed in individuals from the same population, as well as in individuals from different localities, between males and females, and from night to day. In life, most individuals from the Pacific lowlands of Costa Rica are dark tan to greenish gray above with dark brown longitudinal stripes that are entire or broken, but some specimens (mostly males) are dusky brown and lack longitudinal stripes or an interorbital triangle; females usually have the dark interorbital triangle and the stripes on the dorsum. Individuals from Turrialba, Cartago Province, Costa Rica, are pale olive-tan with olive-brown markings. Individuals from Puerto Viejo, Heredia Province, Costa Rica, are uniformly yellowish brown with or without dark longitudinal stripes. Specimens from El Recreo, Zelaya Province, Nicaragua, are like those from Puerto Viejo. Males from Almirante, Bocas del Toro, Panamá, are pale brown with dark brown longitudinal stripes and an indistinct interorbital triangle. Females have a distinct interorbital triangle and dark brown blotches on the thighs and shanks.
By night, the dorsum usually is pale yellow, and the belly is creamy white. By day, the dorsum is dark tan; the stripes and spots are darker, and the belly is yellowish white. Taylor (1952) noticed that considerable variation in color pattern occurred from night to day in individuals from Turrialba, Cartago Province, Costa Rica. At night some individuals lacked a dorsal pattern, but by day many of these individuals developed dorsal stripes.
Cranial Osteology.—The skull of Hyla elaeochroa is slightly wider than it is long, and flat. The premaxillary is small and bears 10 to 15 teeth (mean for 9 specimens, 12.3). The alary process of the premaxillary is small, vertical, and slightly concave posteriorly. Ventrally, the premaxillary is partially united to the prevomer by ossification. The maxillary is slender and bears 70 to 82 teeth (mean for 9 specimens, 74.3). The pars facialis of the maxillary is laterally convex and is about twice as high as the pars dentalis.
The nasal is large, robust, anteriorly truncate, but pointed posteriorly in dorsal view. The nasal comprises about 45 per cent of the total length of the skull. There is an anterior cartilaginous septum nasi separating the two nasals; the latter overlap the sphenethmoid posteriorly. Each nasal bears a shallow concavity in the midlateral side and lacks a maxillary process. Dorsally, the sphenethmoid is wider than long, roughly pentagonal in shape; the frontoparietal is elongate, smooth, and bears a small anterior supraorbital process. The sphenethmoid and frontoparietal form the anterior margin of the frontoparietal fontanelle; the fontanelle is narrow anteriorly and wider posteriorly (Fig. 5B).
The entire distal surface of the proötic is in contact with the posterior arm of the squamosal. A narrow cartilaginous crista parotica is visible dorsally in some specimens. The squamosal is broad posteriorly but its anterior arm is slender and not in contact with the maxillary.
Table 5.—Geographic Variation in Size and Proportions in Males of
Hyla elaeochroa.
(Means in parentheses below observed ranges.)
| Locality | N |
Snout-vent
length (mm.) |
Tibia length/
snout-vent |
Tympanum/
eye |
Foot length/
snout-vent |
| Nicaragua: El Recreo | 9 |
28.0-30.3
(29.3) |
0.51-0.57
(0.55) |
0.47-0.59
(0.51) |
0.39-0.54
(0.41) |
| Costa Rica: Tilarán | 21 |
28.8-33.6
(30.6) |
0.47-0.57
(0.52) |
0.48-0.65
(0.59) |
0.40-0.46
(0.41) |
| Costa Rica: Puerto Viejo | 22 |
26.3-32.4
(29.7) |
0.49-0.54
(0.52) |
0.48-0.65
(0.57) |
0.38-0.45
(0.42) |
| Costa Rica: Turrialba | 95 |
28.1-35.0
(30.6) |
0.47-0.56
(0.51) |
0.47-0.68
(0.56) |
0.37-0.46
(0.41) |
| Costa Rica: Bataán, Limón, and Suretka | 26 |
26.3-32.7
(30.0) |
0.47-0.54
(0.51) |
0.45-0.66
(0.50) |
0.36-0.44
(0.41) |
| Costa Rica: Piedras Blancas | 21 |
33.3-37.7
(35.2) |
0.50-0.54
(0.51) |
0.48-0.64
(0.57) |
0.40-0.46
(0.43) |
| Costa Rica: Rincón de Osa | 24 |
31.4-35.9
(34.1) |
0.50-0.56
(0.53) |
0.45-0.61
(0.54) |
0.40-0.46
(0.43) |
| Panamá: Bocas del Toro | 6 |
31.0-33.5
(32.1) |
0.49-0.54
(0.51) |
0.47-0.50
(0.48) |
0.41-0.43
(0.42) |

PLATE 1
Living
Hyla: (A)
H. boulengeri
(KU 86322) and (B)
H. foliamorta
(KU 101576), × 2.

PLATE 2
Living
Hyla:
(A)
H. elaeochroa
(KU 91688), (B)
H. staufferi staufferi
(KU 57791), and (C)
H. staufferi altae
(KU 101688), × 2.
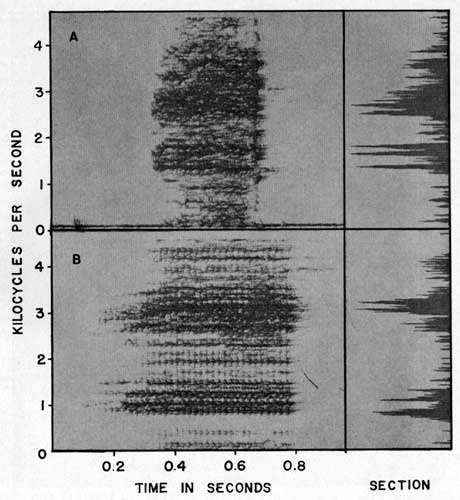
PLATE 3
Audiospectrograms and sections of mating calls of (A)
Hyla boulengeri
(KU Tape No. 511) and (B)
H. foliamorta
(KU Tape No. 288).
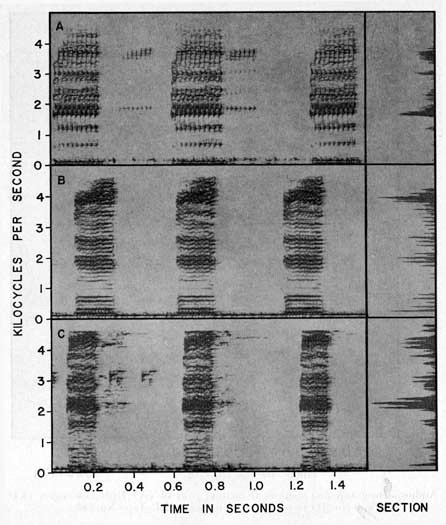
PLATE 4
Audiospectrograms and sections of mating calls of (A)
Hyla elaeochroa
(KU Tape No. 97), (B)
H. s. staufferi
(KU Tape No. 93), and (C)
H. staufferi altae
(KU Tape No. 502).
The prevomer is short, and broadest anteriorly. The prevomer is joined to the premaxillary by ossification. The posterior margin of the prevomer bears a narrow cartilaginous articulation with the sphenethmoid. The anterolateral and posterolateral processes of the prevomer form an incomplete bony margin to the small choanae; each prevomer bears four to seven teeth. The palatine is small, curved anteriorly and edentate. The anterior part of the parasphenoid is robust and ends in a point. The pterygoid is slender and weakly developed.
Natural History.—Hyla elaeochroa inhabits humid lowland tropical forests in lower Central America and breeds in temporary ponds. Clasping pairs, gravid females, and calling males have been found mostly in June, July, and August. William E. Duellman informed me that he also found males calling in mid-February, late April, and May. Duellman (1967) reported detailed observations of the social organization in the mating call of Hyla elaeochroa. The choruses in this species are initially organized, but when many individuals call, the chorus loses organization. I observed this species breeding in a temporary pond at Puerto Viejo, Heredia Province, Cost Rica, in late June. Calling males and clasping pairs were extremely abundant within a few hours after a heavy rain. Males were mostly found calling from low emergent herbs in the pond and less commonly from bushes and trees to heights of six meters above the water. Calling males were also observed at Ricón de Osa, Puntarenas Province, Costa Rica, in late July. These breeding individuals were found in a shallow pond at the edge of a wet forest. Calling stations were less than two meters in height. John D. Lynch informed me that after a heavy rain in early August, he found several hundred individuals congregated in a small grassy pond less than a foot deep, at Rincón de Osa. Males were calling from sites on grass stems a few centimeters above the water.
The mating call of Hyla elaeochroa consists of short notes, repeated at intervals of about 0.40 second. Each note has a duration of 0.12 to 0.24 second. The fundamental frequency varies from 48 to 65 cycles per second, and the notes have 40-50 pulses per second; the dominant frequency is at about 2,900 cycles per second (Table 2, Pl. 4A).
The eggs are deposited in a mass in the water near floating vegetation. William E. Duellman informed me that he observed hatchlings oriented vertically with the tip of the mouth at the surface of the water. They gradually sank to bottom, but swam back to surface again. No additional information is available concerning early development. Tadpoles have been found in shallow grassy ponds in clearings and in temporary woodland ponds.
Tadpoles.—Three hundred and thirty-one tadpoles in various stages of development are available. Thirty-five tadpoles in stage 35 have a mean body length of 8.1 mm. (8.0-9.0 mm.), tail length of 17.7 mm. (15.0-19.5 mm.), and total length of 25.9 mm. (23.0-27.5 mm.). The largest tadpole examined is in stage 40 and has a total length of 34.5 mm. (Table 6).
A typical tadpole, stage 35 of development (KU 104134, from Puerto Viejo, Heredia Province, Costa Rica), has a body length of 9.1 mm., tail length of 17.7 mm., and a total length of 26.8 mm. Other characters are as follows: body depressed anteriorly; body length greater than depth of tail; internarial space as broad as interorbital distance; nostril equidistant between eye and tip of snout; eyes moderately large; mouth anteroventral and triangular; median fourth of upper lip bare; rest of lip bordered by one row of papillae; clumps of small papillae at corners of mouth; tooth rows 2/3; upper rows equal in length; second row interrupted medially; lower rows shorter than upper rows, diminishing in length; beak rather weak with small serrations; spiracle short and nearer eyes than anus; anal opening not reaching edge of ventral fin; caudal musculature attenuated distally (Figs. 2B and 3B).
Table
6.—Sizes of Tadpoles of
Hyla elaeochroa
in Relation to Developmental Stages.
(Means in parentheses below observed ranges; measurements in mm.)
| Stage | N | Body length | Tail length | Total length |
| 24 | 2 |
4.0-4.0
(4.0) |
8.5-9.0
(8.8) |
12.5-13.0
(12.8) |
| 25 | 64 |
5.0-6.5
(5.7) |
8.5-15.0
(11.8) |
13.5-21.5
(17.6) |
| 27 | 30 |
7.0-7.5
(7.1) |
13.0-16.0
(14.2) |
20.0-23.0
(21.3) |
| 30 | 15 |
7.0-8.0
(7.3) |
13.0-16.5
(15.0) |
20.0-24.0
(22.4) |
| 32 | 30 |
7.5-8.5
(7.8) |
15.0-17.0
(16.1) |
22.5-25.0
(23.8) |
| 35 | 35 |
8.0-9.0
(8.1) |
15.0-19.5
(17.7) |
23.0-27.5
(25.9) |
| 37 | 22 |
8.5-9.5
(9.0) |
16.0-22.0
(18.8) |
25.0-31.0
(27.8) |
| 39 | 14 |
9.5-10.5
(9.9) |
19.0-24.9
(21.1) |
28.5-33.5
(31.0) |
| 40 | 27 |
7.0-11.5
(9.1) |
15.0-23.0
(22.0) |
23.0-34.5
(31.2) |
| 43 | 10 |
8.0-12.0
(10.2) |
11.0-17.0
(13.5) |
20.0-26.0
(23.7) |
| 45 | 16 |
10.0-12.0
(11.2) |
1.0-7.0
(3.4) |
12.0-17.0
(14.6) |
| 46 | 45 |
11.0-13.0
(11.8) |
— | — |
In life, dorsum yellowish tan with gray-brown mottling; belly and ventrolateral surfaces silvery-gold or white; black stripe from tip of snout to eye; two black blotches below eye, another blotch extending from eye to base of caudal musculature; caudal musculature and fins gray-brown. In preservative, yellowish tan and silvery-gold colors lost; black reticulations present on tail.
Remarks.—Cope (1876:105) described Hyla elaeochroa from Sipurio, Limón Province, Costa Rica. He based his description on a small specimen, 26.0 mm. in snout-vent length, having a dorsum uniformly colored and lacking an interorbital triangle and blotches on the thighs. Cope (1887) described pigmented specimens from Nicaragua as Hyla quinquevittata, which he diagnosed as having dark brown bars on the hind limbs and five dark brown longitudinal stripes on the dorsum, the median one of which was expanded anteriorly so as to form a large triangular spot between the eyes. He thought this species was related to Hyla eximia Baird and noted that "the hinder legs are much larger; the muzzle is more acuminate and the color bands are much wider" than in eximia. Cope did not compare quinquevittata with elaeochroa, which he had described ten years before. Günther (1901:268), Noble (1918:340), and Nieden (1923:251) regarded both elaeochroa and quinquevittata as valid species. Dunn and Emlen (1932:25) regarded both as synonyms of Hyla rubra, but they made no qualifying statements. Taylor (1952:859) placed quinquevittata as a synonym of elaeochroa and indicated that rubra was another species.
Taylor (1958:37) described Hyla dulcensis from the humid tropical forests of Golfo Dulce, Puntarenas Province, Costa Rica. He thought this species was "related to H. elaeochroa but differs in its somewhat larger size, smaller finger and toe discs, the obsolete canthus rostralis; the loreal region concave and the choanae larger." Duellman (1966a:270) compared adults, tadpoles, and mating calls of dulcensis and elaeochroa and concluded that a single species was involved.
Hyla elaeochroa can be easily confused with the closely related Hyla staufferi. Although the durations of the calls are similar, the call of elaeochroa has only about one third the number of pulses per second, a much lower fundamental frequency, and a lower dominant frequency than that of staufferi. Hyla elaeochroa is larger and has a less pointed snout than does staufferi. Although the skulls of the two species are similar, that of elaeochroa differs in having broad palatines and comparatively larger nasals that are truncate anteriorly. In staufferi the nasal is rounded anteriorly and the palatine is absent.
Distribution.—Hyla elaeochroa occurs on the Caribbean lowlands from western Panamá through Costa Rica to eastern Nicaragua, and on the Pacific lowlands of southeastern Costa Rica and extreme western Panamá. Most localities where it has been collected are below 800 meters, but the species has been found at two localities above 1000 meters (El Silencio and Pacuare, Cartago Province) on the Caribbean slopes of the Cordillera de Talamanca, Costa Rica (Fig. 6).
Specimens Examined.—Nicaragua: Zelaya: El Recreo, UMMZ 79721 (9).
Costa Rica: Alajuela: Laguna Monte Alegre, KU 64499. Cartago: 2 km E Chitaría, KU 107058; El Silencio, 14.4 km NE Turrialba, KU 107059-60; 4.6 km ENE Pacuare, KU 64451-75, 64628-37; 4 km S Pavones, KU 64500; Turrialba (Instituto Interamericano de Ciéncias Agrícolas), KU 30305-26, 24616-57, 30337-54, 31776-91, 31803, 31807-15, 64413-50, 68283-87 (skeletons), 68390-1 (young), 35042 (eggs), 25207-8 (skeletons), 25221 (skeleton), 41073-83 (skeletons). Guanacaste: 2 km E Tilarán, KU 86356-77, 87667-8 (young). Heredia: Puerto Viejo, KU 36696, 46466, 64501-17, 68288-91, 68387, 68388-9 (young), 91803 (young), 91688-9, 104134 (tadpoles), 104135 (young), 104354-6 (skeletons); 1.5 km N Puerto Viejo, KU 64518-23, 68386 (tadpoles); 1 km S Puerto Viejo, KU 84985-6 (skeletons), 87669 (young), 87772-3 (skeletons). Limón: Bataán, KU 30327-36; La Lola, KU 64478-98, 68281-2 (skeletons); Los Diamantes, KU 31800-02, 64476-7; Peralta, KU 31816-21; Puerto Limón, KU 31792-99; Suretka, KU 36467-79, 36697, 41084. Puntarenas: 5 km NW Buenos Aires, KU 107057; 10 km E Esparta, KU 87666 (tadpoles); Golfito, KU 32166-8; 8 km E Palmer Norte KU 93939; 10.7 km SE Palmar Sur, KU 93938 (skeleton), 93940-51, 93952 (eggs), 93953-6 (tadpoles); Piedras Blancas, KU 103646-59; 4.5 km W Rincón de Osa, KU 102208-41, 104298 (tadpoles).
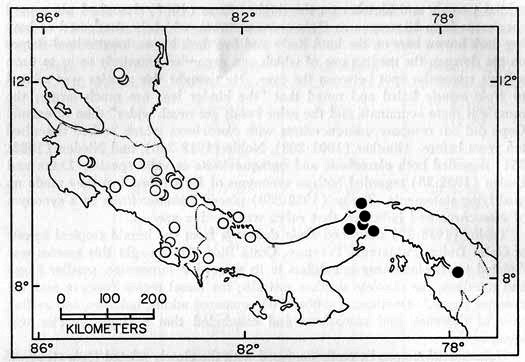
Fig. 6. Map showing locality records for Hyla elaeochroa (circles) and H. rubra (dots).
Panamá: Bocas del Toro: Almirante, KU 80079; Isla Bastimentos, KU 96008-11; Río Cricamola, 3.7 km from coast, KU 96012. Chiriquí: Río Gariché, 8.3 km ESE Paso de Canoas, KU 101571-2.
Hyla staufferi Cope
Hyla staufferi Cope, Proc. Acad. Nat. Sci. Philadelphia, 17:165, October 1, 1865 [Holotype.—USNM 15317, Orizaba, Veracruz, México; Francis Sumichrast collector].
Diagnosis.—Small frogs (♂ to 29 mm., ♀ to 31.6 mm.); skull longer than wide; palatine absent; large cartilaginous crista parotica present; snout flat, elongate and protruding; dark interorbital bar and dorsal stripes usually present.
Description.—Head flat, especially in females, longer than wide; snout long, protruding beyond mouth; loreal region concave; canthus ill-defined; length of eye greater than internarial distance or width of eyelid; length of eye less than interorbital space; tympanum distinct; interorbital spot irregular; supratympanic fold faint; arms short; fingers free of webs; discs on third and fourth fingers equal to diameter of tympanum; inner metatarsal tubercle on base of first finger distinct; first finger shorter than second; palmar tubercle distinct (Fig. 1C); legs short (usually less than 50 per cent of snout-vent length); tarsal fold absent; metatarsal tubercles small, outer tubercle smaller than inner; subarticular tubercles small, simple, distinct; toes less than half webbed (Fig. 1D); skin smooth above with a few small pustules on head, scapular region, flanks, and supratympanic region; arms and legs smooth; skin of belly coarsely granular; posteroventral surfaces of thighs finely granular; tongue small, rounded, longer than wide, slightly free and notched posteriorly; vocal slits small, lateral to tongue; choanae moderate in size.
Variation.—The largest males of Hyla staufferi are from Jalapa, Guatemala, and from San Salvador, El Salvador. In these samples the average snout-vent length is 27 mm. In Panamanian specimens the average snout-vent length is 23.6 mm. Slight variation in the ratio of tibia length to snout-vent length exists throughout the range; more variation exists in the ratio of the diameter of the tympanum to that of the eye; the tympanum is proportionately larger in northern populations (Table 7). The primary differences between Panamanian and more northern populations are in size, color pattern on the dorsum and shanks, amount of webbing between the toes, and duration of notes in the mating call (Table 2, Pl. 4).
The color in Panamanian staufferi is gray or gray-brown with a pair of distinct, complete, dark brown dorsolateral stripes, a pair of entire paravertebral stripes, and in some specimens a vertebral stripe. About five per cent of the individuals have interrupted stripes on the dorsum, whereas in the more northern populations complete paravertebral stripes are present in less ten per cent of the specimens; when complete stripes are present, they are irregular. The dorsal ground color in non-Panamanian specimens is brown, olive-brown, or dark brown.
Transverse bars are present on the shanks in Hyla staufferi from Costa Rica northward to México, whereas in Panamá all the individuals have a longitudinal stripe on the shank (Table 7, Pl. 2). The interorbital spot or bar is more noticeable in northern populations than in specimens from Panamá. Frogs from Costa Rica and northward have the toes about three fourths webbed, whereas in Panamá the toes are about two fifths webbed. The mating calls of the northern and Panamanian populations are similar, but the notes have a longer duration in the northern populations and a higher dominant frequency in Panamanian populations.
Hyla staufferi is the most variable member of the Hyla rubra group in Central America. The Panamanian populations are geographically separated from the Costa Rican and more northern populations by an area of tropical rainforest in the Golfo Dulce region in southeastern Costa Rica and adjacent Panamá. Hyla staufferi does not occur on the Caribbean versant of Costa Rica and Panamá. The Golfo Dulce region and the Caribbean versant are humid and inhabited by Hyla elaeochroa. Hyla staufferi is an inhabitant of subhumid and xeric areas.
On the basis of the discontinuous variation in several characters which correlate with the disjunct distribution of the two populations, two subspecies of Hyla staufferi are recognized. The accounts that follow apply equally to each.
Cranial Osteology.—The skull of Hyla staufferi is flat and longer than wide. The premaxillary is small and bears 9 to 13 teeth (mean for 5 specimens, 11.3). The alary process of the premaxillary is small, concave posteriorly and vertical. Ventrally, the premaxillary is united to the prevomers by partially ossified cartilage. The maxillary is slender and usually bears 49 to 70 teeth (mean for 5 specimens, 60.7). The pars facialis of the maxillary is convex and less than twice the height of the pars dentalis.
The nasal is large, rounded anteriorly, and pointed posteriorly in dorsal view. The nasal comprises about 40 per cent of the total length of the skull. Anteromedially the two nasals converge; posteriorly they overlap the sphenethmoid. The nasals lack a concavity in the midlateral surface. Dorsally, the sphenethmoid is wider than long, roughly pentagonal in shape; the frontoparietal is elongate, narrow, and smooth, with a small supraorbital process anteriorly. The frontoparietal fontanelle is narrow anteriorly and wide posteriorly.
Table 7.—Geographic Variation in Size and Color in Males of
Hyla staufferi.
(Means in parentheses below observed ranges.)
| Locality | N |
Snout-vent
length (mm.) |
Complete
dorsal stripes (per cent) |
Barred shanks
(per cent) |
| Veracruz | 47 |
23.0-27.3
(25.4) |
0.0 | 100 |
| Campeche | 20 |
24.6-27.5
(25.5) |
0.0 | 100 |
| Oaxaca | 75 |
24.0-28.7
(26.4) |
9.3 | 100 |
| Chiapas | 20 |
23.2-27.8
(25.5) |
10.0 | 100 |
| Guatemala | 22 |
25.0-29.0
(26.9) |
10.9 | 100 |
| El Salvador | 21 |
24.7-28.6
(27.0) |
0.0 | 100 |
| Honduras | 34 |
20.6-27.0
(24.9) |
3.3 | 100 |
| Nicaragua | 67 |
21.5-26.8
(24.9) |
3.0 | 92.7 |
| Costa Rica | 54 |
20.7-26.6
(24.2) |
5.5 | 98.1 |
| Total Non-Panamanian | 360 |
20.7-29.0
(25.9) |
5.4 | 98.3 |
| Panamá | 72 |
21.7-26.0
(23.6) |
94.5 | 0.0 |
Only a narrow connection exists between the posterior, pointed arm of the squamosal and the lateral edge of the proötic. The crista parotica is visible dorsally along the lateral edge of the bony proötic. The squamosal is narrow anteriorly and posteriorly.
The prevomers are short and separated anteriorly by partly ossified cartilage of the overlying solum nasi. The prevomer is joined to the premaxillary by cartilage. The posterior margin of the prevomer articulates directly with the sphenethmoid. The anterolateral and posterolateral processes of the prevomers form the incomplete bony internal margin of the choanae. Each prevomer bears three to six teeth. The palatine is absent. The anterior part of the parasphenoid is narrow and ends in a point. The pterygoid is slender and weakly developed.
Natural History.—Throughout its range Hyla staufferi occurs in subhumid forests and savannas; consequently, the breeding activities are limited by the seasonal occurrence of rainfall, which accumulates in temporary ponds where this species breeds. Clasping pairs and gravid females have been found mostly from June to August throughout its range. This species was observed calling at Finca Taboga, Guanacaste Province, Costa Rica, in mid-July. The males were calling from temporary grassy and weedy ponds in which Hyla microcephala also was calling, but the two species had different calling sites. Hyla staufferi called at stations at heights of five to 80 cm. near the edge of the pond, whereas Hyla microcephala called from emergent vegetation in the middle of the pond. Charles W. Myers informed me that at Penonomé, Coclé, Panamá, he found staufferi calling from grass in puddles where microcephala was absent, and at El Caño, Coclé, Panamá, staufferi was calling from higher sites ("several inches to a few feet above water") than microcephala.
Stuart (1948:34) reported breeding individuals from La Libertad, Guatemala, after rainfall in late May, and Schmidt and Stuart (1941:239) reported staufferi breeding in July in the Salamá basin, Alta Verapaz, Guatemala. Stuart (1935:38) and Duellman (1960:63 and 1963:226) agreed that this species breeds early in the rainy season. However, Rand (1957:519) stated that in El Salvador "these frogs did not begin to call until almost a month and a half after the beginning of the rains." Blair (1960:133) reported that males call in June and July in Chiapas, Oaxaca, Veracruz, and Tamaulipas, México.
The mating call of this species is a series of closely spaced notes having a fundamental frequency of about 100 cycles per second. Each note has a duration of 0.13 to 0.23 second, repeated at intervals that are longer than the duration of the call. The notes are moderately low-pitched and have a dominant frequency of more than 3,000 cycles per second and about 120 pulses per second (Table 2).
Tadpoles.—Measurements of the 33 tadpoles that are available are given in Table 8. The largest tadpole examined is in stage 38 and has a total length of 29.5 mm.
A typical tadpole in stage 38 of development (KU 104162, 5 km ESE Córdoba, Veracruz, México) has a body length of 10 mm., tail length of 19.5 mm., and a total length of 29.5 mm. Other characters are as follows: body as deep as wide, depressed anteriorly; body as long as depth of tail; interorbital space greater than distance between eye and snout but equal to internarial space; nostril equidistant between eye and tip of snout; distance between spiracle and eye less than distance between eye and snout; eyes large, situated dorsolaterally; mouth anteroventral, approximately triangular in outline; one row of papillae covering lower lip and all except median fourth of upper lip; scattered papillae at corners of mouth; tooth rows 2/3; first upper row entire, second row interrupted medially, shorter than first; lower rows shorter than upper rows; beak weak; spiracle short and nearer eyes than anus; anal opening not reaching edge of ventral fin; dorsal fin barely extending onto body; caudal musculature pointed distally.
Table 8.—Sizes of Tadpoles of
Hyla s. staufferi
in Relation to Developmental Stages.
(Means in parentheses below observed ranges; measurements in mm.)
| Stage | N | Body length | Tail length | Total length |
| 25 | 3 |
6.0-7.0
(6.7) |
12.0-13.0
(12.5) |
18.0-20.0
(19.2) |
| 26 | 2 |
7.0-7.5
(7.3) |
14.0-15.0
(14.5) |
21.5-22.0
(21.8) |
| 27 | 9 |
7.0-8.0
(7.6) |
13.0-17.0
(14.5) |
21.0-25.0
(22.0) |
| 32 | 1 | 8.5 | 15.5 | 24.0 |
| 36 | 2 |
8.0-10.0
(9.0) |
16.5-17.0
(16.8) |
25.0-26.5
(25.8) |
| 38 | 6 |
9.0-10.0
(9.6) |
19.0-20.5
(19.5) |
28.0-29.5
(29.1) |
| 41 | 1 | 10.0 | 14.0 | 24.0 |
| 42 | 6 |
11.0-14.0
(11.8) |
10.0-13.0
(11.9) |
20.0-29.0
(24.8) |
| 45 | 1 | 12.5 | 0.5 | 13.0 |
| 46 | 1 | 13.0 | — | — |
In life, body pale olive-tan, belly silvery white with pinkish-orange reticulations in some specimens; tail creamy white with silvery flecks and black or brown reticulations. In preservative, tan and pinkish-orange coloration lost; body transparent, reticulations on tail present.
Remarks.—Hyla staufferi was described by Cope (1865:195) on the basis of specimens from Orizaba, Veracruz, México. He described the color pattern as "color above dark olive, with a short black bar over each scapula, and one from eye to eye, with a trace along the coccyx." Cope (1887:14) placed staufferi as a subspecies of Hyla eximia, but he did not justify his action. Günther (1901:262) also considered staufferi to be conspecific with eximia without making any qualifying statement. Dunn and Emlen (1932:24) named Hyla culex from Tela, Honduras, on the basis of a male (MCZ 16098) having a snout-vent length of 25.1 mm., and a female (USNM 20267) from Patuca, Honduras. They diagnosed the species as having "discs larger than tympanum ... black interorbital triangle, traces of black dorsal marking; three black bars on anterior and posterior face of thighs, two black bars on tibia, on tarsus and on forearm." The holotype now is faded but has some of the pattern described. Dunn and Emlen did not compare culex with staufferi but did compare it with boulengeri and rubra.
Dunn (1933:61) named Hyla altae from Summit, Canal Zone. His description was based on a male (MCZ 17972) having a snout-vent length of 25.1 mm., the color pattern was described as "gray with four darker dorsal stripes ... a faint trace of mid-dorsal striping...." Dunn defined the Hyla rubra group and recognized boulengeri, altae, culex, and rubra as members. Hyla elaeochroa and staufferi were omitted from his key to the group in Central America.
Kellogg (1932:174) compared staufferi with eximia and concluded that the two were probably distinct species. Stuart (1935:38) considered altae to be a synonym of culex. Gaige (1936:293) considered altae and culex to be conspecific but regarded staufferi as a different species. She also suggested that staufferi was not related to eximia but belonged to the rubra group. Taylor (1952:865) and Duellman (1966a:274) considered altae and culex to be synonyms of staufferi.
The only other worker besides Cope and Günther to consider Hyla staufferi as a member of the eximia group was Blair (1960:129), who suggested the relationship on the basis of similarities in the structure of the calls of eximia and staufferi. Taylor (1938:421) and Smith and Taylor (1948:78) excluded staufferi from the eximia group on the basis of morphological characteristics. I consider culex to be inseparable from staufferi, whereas altae is recognizable as a Panamanian subspecies of staufferi.
Hyla staufferi staufferi Cope, New Combination
Hyla staufferi Cope, Proc. Acad. Nat. Sci. Philadelphia, 17:195, October 1865 [Holotype.—USNM 15317, Orizaba, Veracruz, México; Francis Sumichrast collector], Brocchi, Mission Scientifique au Mexique et dans L'Amerique Centrale, 1881, p. 36. Boulenger, Catalogue, of the Bratrachia Salientia s. Ecaudata, p. 400, February 1, 1882. Kellogg, Bull. U.S. Natl. Mus., 160:173, March 31, 1932. Smith and Taylor, Bull. U.S. Natl. Mus., 194:88, 1948. Taylor, Univ. Kansas Sci. Bull., 35:862, July 1, 1952. Rand, Fieldiana Zool. Chicago Nat. Hist. Mus., 34:518, April 18, 1957. Duellman, Univ. Kansas Publ, Mus. Nat. Hist., 17:274, June 17, 1966.
Hyla eximia staufferi Cope, Bull. U.S. Natl. Mus., 32:14, January 16, 1887.
Hyla eximia (part): Günther, Biologia Centrali-Americana, Reptilia and Batrachia, p. 261, June 1901. Nieden, Das Tierreich, Anura I, p. 245, June 1923.
Hyla culex Dunn and Emlen, Proc. Acad. Nat. Sci. Philadelphia, 84:24, March 22, 1932 [Holotype.—MCZ 16098, Tela Honduras; Raymond A. Stadelman collector]. Stuart, Misc. Publ., Univ. Michigan Mus. Zool., 29:38, October 1935. Gaige, Carnegie Inst. Washington Publ., 457:293, 1936.
Diagnosis.—Small frogs (♂ to 29 mm., ♀ to 31.6 mm.); dorsolateral stripes irregular; paravertebral stripes usually broken; two or three transverse bars on shanks; thighs spotted or not; arms usually barred; interorbital bar usually present; toes about three fourths webbed; color brown, tan, or olive-green.
Variation.—Three hundred and sixty males chosen at random from throughout the range have snout-vent lengths of 20.7 to 29 mm. (25.9 mm.). The smallest individuals are from Costa Rica and Nicaragua (means 24.2 and 24.4 mm., respectively). The largest individuals are from Guatemala and El Salvador (mean of each 27.0 mm.). The ratio of the diameter of the tympanum to that of the eye is more than 60 per cent in most samples, but in those from Costa Rica and British Honduras it is smaller. The color pattern is highly variable. Some specimens are dark brown or pale brown in color. Incomplete dorsal stripes are present in 94.6 per cent of the specimens, and transverse bars are present on the shanks in 98.3 per cent of the specimens. The interorbital spot varies from transverse to longitudinal in position, and an irregular white line extends from the upper jaw to the arm in some specimens (Table 7).
Distribution.—Hyla staufferi staufferi inhabits savanna and subhumid and xeric forests in the lowlands and moderate elevations from southern Tamaulipas southward to Nicaragua on the Caribbean versant and from Guerrero, México to northwestern Costa Rica on the Pacific lowlands (Fig. 7). Duellman (1963:226) commented that a specimen from Chinajá, Guatemala, possibly was transported there in the cargo from Toocog, because with this one exception the species is unknown in tropical rainforest in Guatemala.
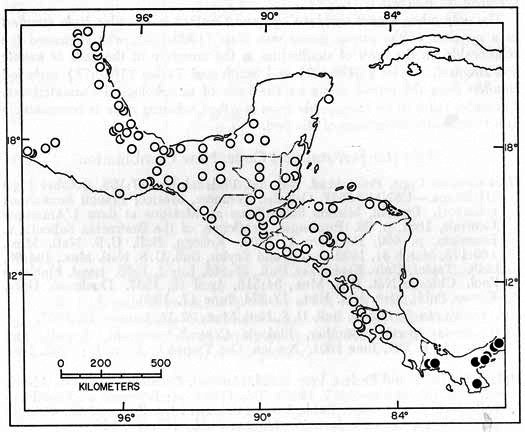
Fig. 7. Map showing locality records for Hyla staufferi staufferi (circles) and H. staufferi altae (dots).
Specimens Examined.—México: Campeche: 5 km S Champotón KU 71296-7; 7 km W Escárcega, KU 71298-308; 13 km W, 1 km N Escárcega, KU 71309-10, 75090-4. Chiapas: 32 km S Arriaga, KU 57789-92; 4 km N Ixtapa, KU 5776-81; 3.6 km SW Las Cruces, KU 37740; 17 km S Las Cruces, KU 57793-4; 24 km S Las Cruces, KU 104160 (tadpoles); 11 km S Tapachula, KU 57782-8, 60000 (young). Guerrero: El Limoncito, near La Venta, KU 31392-401; Mexcala, near Balsa River, KU 31391; Organos, S El Trienta, KU 31390. Oaxaca: 26 km N Matías Romero, KU 33878-82; 2.5 km S Pochutla, KU 59924-7 (skeletons); 5 km S Pochutla, KU 57795-801; 3.2 km E Tapanatepec, KU 37877-902; 17.6 km WNW Tapanatepec, KU 65033-4; Temascal, USC 8243 (8); 3.2 km S Tolocita, KU 39657-8; 0.5 km Tuxtepec, KU 87073-81, 87610 (tadpoles); 17 km S Tuxtepec, KU 65035-7; 1 km W Zanatepec, KU 104161 (tadpoles). Quintana Roo: Isla Cozumel, 3.5 km N San Miguel, KU 71710-11 (young). San Luis Potosí: Valles, KU 31490. Tabasco: Teapa, UMMZ 118887 (3), 119203 (13); 9.6 km N Teapa, UMMZ 119202; 24 km N Teapa, UMMZ 119961 (5); 29 km N Teapa, UMMZ 119960; 3.5 km S Villahermosa, UMMZ 119201 (2); 17.6 km S Villahermosa, UMMZ 119200 (8). Tamaulipas: 1 km E Chamal, UMMZ 110706; Gómez Farías, UMMZ 110701 (3); 5 km SE Gómez Farías, UMMZ 110705; 8 km NE Gómez Farías, UMMZ 11282 (2), 11283 (3); Kilometer 615 between Río Limón and Llera, UMMZ 80455 (2); 5 km W San Geraldo, UMMZ 110702 (4), 110703 (3); 8 km W San Geraldo, near Río Frío, UMMZ 110704 (5). Veracruz: 3 km SW Boca del Río, KU 10494-8; 5 km SW Boca del Río, KU 23701; 5 km ESE Córdoba, KU 104162 (tadpoles); Cuautlapán, KU 57098-102, 26787; Hacienda Tamiahua, Cabo Rojo, KU 62871; 2 km ENE Mata Oscura, KU 105627; 5 km SE Paso del Toro, KU 40144; Portrero Viejo, KU 23911-2, 26786, 27413, 57094-7.
Guatemala: Alta Verapaz: Chinajá, KU 57769; Finca La Cubilquitz, UMMZ 90871, 90872 (5), 91379 (2). Baja Verapaz: 1 km S San Jerónimo, UMMZ 84077 (7), 84078 (14). Chiquimula: 1.6 km SE Chiquimula, UMMZ 98114 (2); Esquipulas, UMMZ 106784 (4), 106785 (14). El Petén: No specific locality, USNM 25143, 24825-6; La Libertad, FMNH 27096-7, KU 57770, UMMZ 75339 (15), 75340 (15), UMMZ 94341-2. Esquintla: 20 km N San José, AMNH 74369-76. Guatamala: 16 km NE Guatamala, KU 43539. Izábal: Puerto Barrios, TCWC 16671-73, 16646-56; 2.5 km NE Río Blanco, KU 57774-5. Jalapa: Jalapa, UMMZ 106788 (44). Jutiapa: Finca La Trinidad, UMMZ 107730 (12), 107731 (16); Jutiapa, UMMZ 106786 (2). Zacapa: 14 km ENE Mayuelas, KU 57773; 7 km ENE Río Hondo, KU 57771-2, 59999 (young).
British Honduras: Belize: Belize, FMNH 4406. El Cayo: San Agustín, UMMZ 80741 (8). Stann Creek: 10 km S Stann Creek on Hummingbird Highway, UMMZ 125720-1.
El Salvador: Cuscatlán: 7 km WNW Cojutepeque, TNHC 32004-10. La Libertad: 16 km NW Santa Tecla, KU 43540-1. La Union: 2.5 km Santa Rosa, TCWC 16669-70. Morazán: Dividendero, USNM 73288-92. San Salvador: San Salvador, FMNH 65101-06, KU 61932-44, 61989-92, 62152 (eggs), USNM 117588, 118391 (3), 118394; 1.6 km NW San Salvador, KU 43162-3.
Honduras: Atlantidad: Ceiba, USNM 117592. Choluteca: Choluteca, KU 85361-6; 2 km E Choluteca, UMMZ 118395 (7); 3.2 km NE Choluteca, KU 100500-01; 6.2 km E Choluteca, KU 65046-56; 10 km E Choluteca, KU 65045: 5 km S Choluteca, USC 2700 (4). Colón: Isla Guanaja (Islas de la Bahía), TCWC 21551, TNHC 32011. Cortés: Agua Azul, TCWC 19178-9; East side Lago Yojoa, KU 65038-44. El Paraiso: Valle de Jamastrán, AMNH 54800-04. Francisco Morazán: Escuela Agrícola Panamericana, AMNH 54963-73; 14.5 km NW Comayaguela, KU 100499; El Zamorano, KU 103224; 29 km N Tegucigalpa, TNHC 32003, 32012.
Nicaragua: Chinandega: Finca San Isidro, 10 km S Chinandega, KU 85311-33. Managua: 13 km E Managua, KU 85339; 2 km S Tipitapa, KU 85334-8. Rivas: 9.5 km SE Rivas, KU 85355-6; 18 km SE Rivas, KU 85354; 7.7 km NE San Juan del Sur, KU 85346-53; 16.5 km NE San Juan del Sur, KU 85340-5; 5 km SE San Pablo, KU 43151-61. Zelaya: Isla Grande del Maiz, KU 85357-60.
Costa Rica: Alajuela: Los Chiles, USC 7215 (2), 7217. Guanacaste: 4 km W Bagaces, USC 7019 (5); Finca Taboga, KU 102265-5; 12 km S La Cruz, USC 8091; Las Cañas, KU 41113 (skeleton); 27 km N Las Cañas, USC 8171 (5); Guardia, Río Tempisque, USC 8214; 10 km N Guardia, KU 102266-7; 1.6 km N Guayabo de Bagaces, USC 7023 (3); Liberia, KU 36510-22; 4 km W Liberia, KU 36449-64, USC 102 (10), 103 (9), 104 (7), 105; 6 km N Liberia. USC 8096; 8 km NNW Liberia, KU 65032; 14.5 km N Liberia, USC 8079, 8138 (2); 14.5 km S Liberia, USC 8238 (5); 6 km N Nicoya, USC 8229 (11); 4 km S Nicoya, USC 8230, 8231; Peñas Blancas, KU 102263; 8.6 km ESE Playa del Coco, USC 8137 (14); 21 km E Playa del Coco, USC 8138 (2); Santa Cruz, USC 8232 (2); 3 km E Santa Rosa, TCWC 16663-68; Tenorio, KU 32159; Tilarán, KU 36509. Puntarenas: 10 km WNW Esparta, KU 65022-9, 68614 (skeleton); 4.5 km WNW Esparta, KU 65030; 12 km WNW Esparta, KU 65031; 6 km E Esparta, KU 86477; Hotel Maribella, KU 32157-8; 3 km W Puntarenas, TCWC 16657-62.
Hyla staufferi altae Dunn, New Combination
Hyla altae Dunn, Occas. Papers Boston Soc. Nat. Hist., 8:61, June 7, 1933 [Holotype.—MCZ 17972, Summit, Canal Zone, Panamá; Emmett R. Dunn collector].
Hyla culex: Stuart, Misc. Publ. Univ. Michigan Mus. Zool., 29:38, October 1, 1935. Gaige, Carnegie Inst. Washington Publ., 457:293, 1936.
Hyla staufferi: Taylor, Univ. Kansas Sci. Bull., 35:862, July 1, 1952. Duellman, Univ. Kansas Publ., Mus. Nat. Hist., 17:274, June 17, 1966.
Diagnosis.—Small frogs (♂ to 26 mm., ♀ to 27 mm.); dorsolateral and paravertebral stripes complete; longitudinal dark gray stripe on shank; thighs unmarked; interorbital bar usually absent; toes about three fifths webbed; gray to brownish gray above.
Variation.—Hyla staufferi altae is less variable in size, proportions, and color pattern than is H. s. staufferi. The size varies from 21.7 to 26 mm. (23.6) in 72 males. The ratio of tibia to snout-vent length is 0.42 to 0.50 (0.45), slightly less than in the northern subspecies. In color pattern 94.5 per cent of the individuals have complete dorsal stripes, and all have a longitudinal stripe on the shank (Table 7).
Distribution.—This subspecies is restricted to subhumid forests and savannas on the Pacific lowlands of Panamá. Hyla s. altae is presently known to occur from Chepo in east-central Panamá through the Azuero Peninsula to Concepción, Chiriquí, in western Panamá (Fig. 7).
Specimens Examined.—Panamá: Canal Zone: No specific locality, TNHC 24406; 2.8 km SW Fort Kobbe, KU 101679. Chiriquí: 14.4 km E Concepción, AMNH 69799-801; 6.6 km N David, TNHC 32013-4; 2 km S David, AMNH 68802. Coclé: 1 km NE El Caño, KU 101662-75; El Valle de Antón, AMNH 59601-5, KU 77333-47; 7 km SSW Penonomé, KU 101654-61. Los Santos: Tonosí, KU 101246 (tadpoles), 101697-701. Panamá: 2 km WSW Chepo, KU 101680-8; 6 km WSW Chepo, KU 77324-27; El Cangrejo (Panamá), KU 101676-8; Nueva Gorgona, AMNH 69991, 69798; 1.5 km W Pacora, KU 77328-32; 2 km N Tocumen, KU 101689-95; 8 km NE Tocumen, KU 101696.
Evolutionary History
My assumptions regarding the evolutionary history of the Hyla rubra group in Central America were derived partly from interpretations of the evolutionary history of other animal groups (Simpson, 1943, 1965; Dunn, 1931b; Stuart, 1950; Duellman, 1958, 1960, 1963, 1965; and Duellman and Trueb, 1966). The origin and early evolution of the group probably occurred prior to the Mid-Pliocene in the lowlands of South America, because the greatest diversity of the group is in Brazil. Differentiation into two or more subgroups took place in South America prior to the late Pliocene. At the end of the Pliocene, shortly after the closure of the Colombian Portal, many South American animals migrated into Central America (Simpson, 1943, Maldonado-Koerdell, 1964, and Savage, 1966). It is likely that the Hyla rubra group entered Central America at that time; apparently two stocks (rubra-elaeochroa-staufferi stock and boulengeri-foliamorta stock) migrated into Central America.
Hyla elaeochroa is closely related to rubra and probably differentiated from rubra through spatial isolation. Thus, we have elaeochroa in Central America and rubra in South America; most likely only in relatively recent times has rubra migrated into eastern Panamá from northern South America. The differentiation and dispersal of elaeochroa and staufferi took place in Central America after the Pliocene. Probably the events of the Pleistocene resulted in the isolation of populations. One of these (Hyla staufferi stock) was restricted in the subhumid Pacific lowlands, whereas the Hyla elaeochroa stock occupied the tropical wet forests of the Caribbean lowlands. Hyla elaeochroa apparently more closely resembled the parental stock by being restricted to the tropical rain forests, whereas staufferi adapted to subhumid environments and thereby was able to disperse throughout most of the subhumid regions of Central America.
After geographical separation took place the initial genetic divergence between the two populations was maintained by means of ecological and ethological isolating mechanisms. Under these circumstances it can be supposed that the different ecological preferences of elaeochroa and staufferi depend on the climatic changes that took place during the Pleistocene. On this basis it may be proposed that when the original prototype broke up into the two incipient species, the staufferi stock became physiologically and behaviorally adapted to subhumid conditions and dispersed into dry areas of the lowlands of Middle America. The tropical evergreen forests on the Caribbean side of lower Central America and the uplift of the Talamanca range in the Pliocene were barriers to the dispersal of staufferi. Consequently, this frog dispersed along the Pacific lowlands.
At the present time staufferi occupies the length of the Pacific lowlands in Central America, except in the rainforest of the Golfo Duce region, which apparently is a relict stand and now separates the ranges of two subspecies of Hyla staufferi. This species crossed the central Nicaraguan lowlands and reached the Caribbean lowlands of Nicaragua and nuclear Central America. The species migrated through the subhumid corridor in northern Honduras and eastern Guatemala (Comayagua Valley in Honduras and the Motagua Valley of Guatemala) to the Isthmus of Tehuantepec. Duellman (1960) hypothesized "that during the times of glacial advances (Pleistocene) the lowlands of the Isthmus probably were more extensive and had more semiarid tropical environments than at the present" and that when semiarid environments were continuous from the Pacific slope across the isthmus to the Gulf lowlands staufferi and other amphibians migrated northward to southeastern Tamaulipas, México.
Hyla elaeochroa dispersed along Caribbean lowland routes. This species not only occurs in the wet forests of the Golfo Dulce region but also in Guanacaste. It is possible that elaeochroa entered Guanacaste and moved to the Golfo Dulce region when the intervening area was less xeric than now (Duellman, 1966b). Hyla elaeochroa extended its range to eastern Nicaragua, but even though northeastern Nicaragua has over 2,000 mm. of precipitation annually (Vivo Escoto, 1964), this species has not spread into Honduras and Guatemala.
Hyla boulengeri is widespread in Amazonian and northern South America, whereas foliamorta occurs only in eastern Panamá and in north-central Colombia. The ancestral boulengeri-foliamorta stock probably invaded Central America in the late Pliocene and dispersed through humid forested environments to Nicaragua. Apparently a peripheral population established itself in the dry Pacific lowlands of Panamá. This population differentiated from boulengeri of the humid Caribbean lowlands and evolved into foliamorta, which subsequently expanded its range into Colombia.
LITERATURE CITED
Blair, W. F.
| 1960. | Mating call as evidence of relations in the Hyla eximia group. Southwestern Nat., 5(3):129-135. November 1. |
Boulenger, G. A.
| 1882. | Catalogue of the Batrachia Salientia s. Ecaudata, 2nd. ed., pp. 1-503, pls. 1-30. February. |
Brocchi, P.
| 1881-1883. | Etude des batraciens de l'Amerique Centrale. Mission Scientifique au Mexique et dans l'Amerique Centrale, Liv. 1, pp. 1-122, pls. 1-21. |
Cochran, D. M.
| 1955. | Frogs of southeastern Brazil. Bull. U.S. Natl. Mus., 206:1-423. |
Cope, E. D.
| 1865. | Third contribution to the Herpetology of Tropical America. Proc. Acad. Nat. Sci. Philadelphia, 17:195. |
| 1876. | On the batrachia and reptilia of Costa Rica. Jour. Acad. Nat. Sci. Philadelphia, new series, 8:93-154, pls. 23-28. 1887. Thirteenth contribution to the Herpetology of Tropical America. Proc. Amer. Philos. Society, 23:273. April. |
Daudin, F. M.
| 1802. | Histoire naturelle des rainettes, des grenouilles et des crapauds. Paris, pp. 1-71, pls. 1-33. |
| 1803. | Histoire naturelle générale et particulière des reptiles. Paris, 8:1-439. |
Duellman, W. E.
| 1956. | The frogs of the hylid Genus Phrynohyas Fitzinger, 1843. Misc. Publ. Mus. Zool., Univ. Mich., 96:1-47, pls. 1-6. February 21. |
| 1958. | A monographic study of the colubrid snakes Genus Leptodeira. Bull. Amer. Mus. Nat. Hist., 114:1-152, pls. 1-31. February 24. |
| 1960. | A distributional study of the amphibians of the Isthmus of Tehuantepec, México. Univ. Kansas Publ., Mus. Nat. Hist, 13:19-72, pls. 1-8. August 16. |
| 1963. | Amphibians and reptiles of the rainforests of southern El Petén, Guatemala. Univ. Kansas Publ., Mus. Nat. Hist., 15:205-249, pls. 7-10. October 4. |
| 1965. | A biogeographic account of the herpetofauna of Michoacán, México. Univ. Kansas Publ., Mus. Nat. Hist., 15:627-709, pls. 29-36. December 30. |
| 1966a. | Taxonomic notes on some Mexican and Central American hylid frogs. Univ. Kansas Publ., Mus. Nat. Hist., 17:263-279. June 17. |
| 1966b. | The Central American herpetofauna: An ecological perspective. Copeia, 4:700-719. December 23. |
| 1967. | Social organization in the mating calls of some Neotropical Anurans. Amer. Midl. Nat., 77(1):156-163. January. |
Duellman, W. E., and L. Trueb
| 1966. | Neotropical hylid frogs, Genus Smilisca. Univ. Kansas Publ., Mus. Nat. Hist., 17:281-375, pls. 1-12. July 14. |
Dunn, E. R.
| 1931a. | The amphibians of Barro Colorado Island. Occas. Papers Boston Soc. Nat. Hist., 5:403-421. October 10. |
| 1931b. | The herpetological fauna of the Americas. Copeia, 3:106-119. October 30. |
| 1933. | A new Hyla from Panamá Canal Zone. Occas. Papers Boston Soc. Nat. Hist., 8:61-64. June 7. |
Dunn, E. R., and J. Emlen
| 1932. | Reptiles and amphibians from Honduras. Proc. Acad. Nat. Sci. Philadelphia, 84:24-25. March 22. |
Fouquette, M. J.
| 1958. | A new tree frog, Genus Hyla, from the Canal Zone. Herpetologica, 14:125-128. April 25. |
Gaige, H. T.
| 1936. | Some reptiles and amphibians from Yucatan and Campeche, México. Carnegie Inst. Washington Publ., 457:289-304. |
Gosner, K. L.
| 1960. | A simplified table for staging anuran embryos and larvae with notes on identification. Herpetologica, 16:183-190. June 17. |
Günther, A. C. L.
| 1859. | Catalogue of the Batrachia Salientia in the British Museum. pp. 1-160. |
| 1885-1902. | Biologia Centrali Americana. Reptilia and Batrachia. London, xx + 326 pp., pls. 1-76. |
Kellogg, R.
| 1932. | Mexican tailless amphibians in the United States National Museum, Bull. U.S. Natl. Mus., 160:1-224. |
Laurenti, J. N.
| 1768. | Specimen medicum exhibens synopsin reptilium emendatum cum experimentis circa venema et antidota reptilium austriacorum. Vienna, pp. 1-216. |
Linnaeus, C.
| 1758. | Systema naturae. Holmiae, ed. 10 reformata, I:1-532. |
Maldonado-Koerdell, M.
| 1964. | Geohistory and paleography of Middle America. In R. Wauchope and R. C. West (Eds.). Handbook of Middle American Indians, Vol. I, Univ. Texas Press, Austin, 570 pp. |
Nieden, F.
| 1923. | Amphibia:Anura I. Das Tierreich. Berlin, 584 pp. |
Noble, G. K.
| 1918. | The amphibians collected by the American Museum expedition to Nicaragua in 1916. Bull. Amer. Mus. Nat. Hist., 38:311-347. |
Rand, A. S.
| 1957. | Notes on amphibians and reptiles from El Salvador. Fieldiana, Zool., Chicago Nat. Hist. Mus., 43:505-534. April 18. |
Rivero, J. A.
| 1961. | Salientia of Venezuela. Bull. Mus. Comp. Zool., 126:1-207. November. |
Savage, J. M.
| 1966. | The origins and history of the Central American herpetofauna. Copeia, 4:719-756. December 23. |
Schmidt, K. P., and L. C. Stuart
| 1941. | The herpetological fauna of the Salamá Basin, Baja Verapaz, Guatemala. Field Mus. Nat. Hist., Zool. Ser., 24:233-247. |
Seba, A.
| 1734. | Locupletissimi rerum naturalium thesauri accurata descriptio, et iconibus artificissimis expressio, per universam physis historiam. Amsterdam, I xxxiv + 178 pp., pls. 1-111. |
Simpson, G. G.
| 1943. | Turtles and the origin of the fauna of Latin America. Amer. Jour. Sci., 24:413-429. |
| 1965. | The geography of evolution. Chilton Books Publishers. Philadelphia, pp. 1-349. July. |
Smith, H. M. and E. H. Taylor
| 1948. | An annotated checklist and key to the amphibians of México. Bull. U.S. Natl. Mus., 194:1-118. |
Starrett, P.
| 1960. | Description of tadpoles of Middle American frogs. Misc. Publ. Mus. Zool., Univ. Michigan, 110:1-37, pl. 1. |
Stuart, L. C.
| 1935. | A contribution to a knowledge of the herpetology of a portion of the savanna region of Central Petén, Guatemala. Misc. Publ. Mus. Zool., Univ. Michigan, 29:1-56, pls. 1-4. October 1. |
| 1948. | The amphibians and reptiles of Alta Verapaz, Guatemala. Misc. Publ. Mus. Zool., Univ. Michigan, 69:1-109. June 12. |
| 1950. | A geographic study of the herpetofauna of Alta Verapaz, Guatemala. Contr. Lab. Vert. Biol., Univ. Michigan, 45:1-77, pls. 1-9. May. |
Taylor, E. H.
| 1938. | Frogs of the Hyla eximia group, with description of two new species. Univ. Kansas Sci. Bull., 25:421-445. June 1. |
| 1952. | The frogs and toads of Costa Rica. Univ. Kansas Sci. Bull., 35:577-942. July 1. |
| 1958. | Additions to the known herpetological fauna of Costa Rica, with comments on other species. No. III. Univ. Kansas Sci. Bull., 34:3-40. November 18. |
Vivo Escoto, J. A.
| 1964. | Weather and climate of México and Central America. In R. Wauchope and R. C. West (Eds.), Handbook of Middle American Indians. Vol. 1, Univ. Texas Press, Austin, 570 pp. |
Transmitted February 7, 1969.
End of the Project Gutenberg EBook of The Systematics of the Frogs of the
Hyla Rubra Group in Middle America, by Juan R. León
*** END OF THIS PROJECT GUTENBERG EBOOK FROGS OF HYLA RUBRA GROUP ***
***** This file should be named 32505-h.htm or 32505-h.zip *****
This and all associated files of various formats will be found in:
https://www.gutenberg.org/3/2/5/0/32505/
Produced by Chris Curnow, Joseph Cooper and the Online
Distributed Proofreading Team at https://www.pgdp.net
Updated editions will replace the previous one--the old editions
will be renamed.
Creating the works from public domain print editions means that no
one owns a United States copyright in these works, so the Foundation
(and you!) can copy and distribute it in the United States without
permission and without paying copyright royalties. Special rules,
set forth in the General Terms of Use part of this license, apply to
copying and distributing Project Gutenberg-tm electronic works to
protect the PROJECT GUTENBERG-tm concept and trademark. Project
Gutenberg is a registered trademark, and may not be used if you
charge for the eBooks, unless you receive specific permission. If you
do not charge anything for copies of this eBook, complying with the
rules is very easy. You may use this eBook for nearly any purpose
such as creation of derivative works, reports, performances and
research. They may be modified and printed and given away--you may do
practically ANYTHING with public domain eBooks. Redistribution is
subject to the trademark license, especially commercial
redistribution.
*** START: FULL LICENSE ***
THE FULL PROJECT GUTENBERG LICENSE
PLEASE READ THIS BEFORE YOU DISTRIBUTE OR USE THIS WORK
To protect the Project Gutenberg-tm mission of promoting the free
distribution of electronic works, by using or distributing this work
(or any other work associated in any way with the phrase "Project
Gutenberg"), you agree to comply with all the terms of the Full Project
Gutenberg-tm License (available with this file or online at
https://gutenberg.org/license).
Section 1. General Terms of Use and Redistributing Project Gutenberg-tm
electronic works
1.A. By reading or using any part of this Project Gutenberg-tm
electronic work, you indicate that you have read, understand, agree to
and accept all the terms of this license and intellectual property
(trademark/copyright) agreement. If you do not agree to abide by all
the terms of this agreement, you must cease using and return or destroy
all copies of Project Gutenberg-tm electronic works in your possession.
If you paid a fee for obtaining a copy of or access to a Project
Gutenberg-tm electronic work and you do not agree to be bound by the
terms of this agreement, you may obtain a refund from the person or
entity to whom you paid the fee as set forth in paragraph 1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be
used on or associated in any way with an electronic work by people who
agree to be bound by the terms of this agreement. There are a few
things that you can do with most Project Gutenberg-tm electronic works
even without complying with the full terms of this agreement. See
paragraph 1.C below. There are a lot of things you can do with Project
Gutenberg-tm electronic works if you follow the terms of this agreement
and help preserve free future access to Project Gutenberg-tm electronic
works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the Foundation"
or PGLAF), owns a compilation copyright in the collection of Project
Gutenberg-tm electronic works. Nearly all the individual works in the
collection are in the public domain in the United States. If an
individual work is in the public domain in the United States and you are
located in the United States, we do not claim a right to prevent you from
copying, distributing, performing, displaying or creating derivative
works based on the work as long as all references to Project Gutenberg
are removed. Of course, we hope that you will support the Project
Gutenberg-tm mission of promoting free access to electronic works by
freely sharing Project Gutenberg-tm works in compliance with the terms of
this agreement for keeping the Project Gutenberg-tm name associated with
the work. You can easily comply with the terms of this agreement by
keeping this work in the same format with its attached full Project
Gutenberg-tm License when you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern
what you can do with this work. Copyright laws in most countries are in
a constant state of change. If you are outside the United States, check
the laws of your country in addition to the terms of this agreement
before downloading, copying, displaying, performing, distributing or
creating derivative works based on this work or any other Project
Gutenberg-tm work. The Foundation makes no representations concerning
the copyright status of any work in any country outside the United
States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other immediate
access to, the full Project Gutenberg-tm License must appear prominently
whenever any copy of a Project Gutenberg-tm work (any work on which the
phrase "Project Gutenberg" appears, or with which the phrase "Project
Gutenberg" is associated) is accessed, displayed, performed, viewed,
copied or distributed:
This eBook is for the use of anyone anywhere at no cost and with
almost no restrictions whatsoever. You may copy it, give it away or
re-use it under the terms of the Project Gutenberg License included
with this eBook or online at www.gutenberg.org
1.E.2. If an individual Project Gutenberg-tm electronic work is derived
from the public domain (does not contain a notice indicating that it is
posted with permission of the copyright holder), the work can be copied
and distributed to anyone in the United States without paying any fees
or charges. If you are redistributing or providing access to a work
with the phrase "Project Gutenberg" associated with or appearing on the
work, you must comply either with the requirements of paragraphs 1.E.1
through 1.E.7 or obtain permission for the use of the work and the
Project Gutenberg-tm trademark as set forth in paragraphs 1.E.8 or
1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted
with the permission of the copyright holder, your use and distribution
must comply with both paragraphs 1.E.1 through 1.E.7 and any additional
terms imposed by the copyright holder. Additional terms will be linked
to the Project Gutenberg-tm License for all works posted with the
permission of the copyright holder found at the beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm
License terms from this work, or any files containing a part of this
work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this
electronic work, or any part of this electronic work, without
prominently displaying the sentence set forth in paragraph 1.E.1 with
active links or immediate access to the full terms of the Project
Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary,
compressed, marked up, nonproprietary or proprietary form, including any
word processing or hypertext form. However, if you provide access to or
distribute copies of a Project Gutenberg-tm work in a format other than
"Plain Vanilla ASCII" or other format used in the official version
posted on the official Project Gutenberg-tm web site (www.gutenberg.org),
you must, at no additional cost, fee or expense to the user, provide a
copy, a means of exporting a copy, or a means of obtaining a copy upon
request, of the work in its original "Plain Vanilla ASCII" or other
form. Any alternate format must include the full Project Gutenberg-tm
License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying,
performing, copying or distributing any Project Gutenberg-tm works
unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing
access to or distributing Project Gutenberg-tm electronic works provided
that
- You pay a royalty fee of 20% of the gross profits you derive from
the use of Project Gutenberg-tm works calculated using the method
you already use to calculate your applicable taxes. The fee is
owed to the owner of the Project Gutenberg-tm trademark, but he
has agreed to donate royalties under this paragraph to the
Project Gutenberg Literary Archive Foundation. Royalty payments
must be paid within 60 days following each date on which you
prepare (or are legally required to prepare) your periodic tax
returns. Royalty payments should be clearly marked as such and
sent to the Project Gutenberg Literary Archive Foundation at the
address specified in Section 4, "Information about donations to
the Project Gutenberg Literary Archive Foundation."
- You provide a full refund of any money paid by a user who notifies
you in writing (or by e-mail) within 30 days of receipt that s/he
does not agree to the terms of the full Project Gutenberg-tm
License. You must require such a user to return or
destroy all copies of the works possessed in a physical medium
and discontinue all use of and all access to other copies of
Project Gutenberg-tm works.
- You provide, in accordance with paragraph 1.F.3, a full refund of any
money paid for a work or a replacement copy, if a defect in the
electronic work is discovered and reported to you within 90 days
of receipt of the work.
- You comply with all other terms of this agreement for free
distribution of Project Gutenberg-tm works.
1.E.9. If you wish to charge a fee or distribute a Project Gutenberg-tm
electronic work or group of works on different terms than are set
forth in this agreement, you must obtain permission in writing from
both the Project Gutenberg Literary Archive Foundation and Michael
Hart, the owner of the Project Gutenberg-tm trademark. Contact the
Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable
effort to identify, do copyright research on, transcribe and proofread
public domain works in creating the Project Gutenberg-tm
collection. Despite these efforts, Project Gutenberg-tm electronic
works, and the medium on which they may be stored, may contain
"Defects," such as, but not limited to, incomplete, inaccurate or
corrupt data, transcription errors, a copyright or other intellectual
property infringement, a defective or damaged disk or other medium, a
computer virus, or computer codes that damage or cannot be read by
your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right
of Replacement or Refund" described in paragraph 1.F.3, the Project
Gutenberg Literary Archive Foundation, the owner of the Project
Gutenberg-tm trademark, and any other party distributing a Project
Gutenberg-tm electronic work under this agreement, disclaim all
liability to you for damages, costs and expenses, including legal
fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT
LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE
PROVIDED IN PARAGRAPH F3. YOU AGREE THAT THE FOUNDATION, THE
TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE
LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR
INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH
DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a
defect in this electronic work within 90 days of receiving it, you can
receive a refund of the money (if any) you paid for it by sending a
written explanation to the person you received the work from. If you
received the work on a physical medium, you must return the medium with
your written explanation. The person or entity that provided you with
the defective work may elect to provide a replacement copy in lieu of a
refund. If you received the work electronically, the person or entity
providing it to you may choose to give you a second opportunity to
receive the work electronically in lieu of a refund. If the second copy
is also defective, you may demand a refund in writing without further
opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth
in paragraph 1.F.3, this work is provided to you 'AS-IS' WITH NO OTHER
WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT LIMITED TO
WARRANTIES OF MERCHANTIBILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied
warranties or the exclusion or limitation of certain types of damages.
If any disclaimer or limitation set forth in this agreement violates the
law of the state applicable to this agreement, the agreement shall be
interpreted to make the maximum disclaimer or limitation permitted by
the applicable state law. The invalidity or unenforceability of any
provision of this agreement shall not void the remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the
trademark owner, any agent or employee of the Foundation, anyone
providing copies of Project Gutenberg-tm electronic works in accordance
with this agreement, and any volunteers associated with the production,
promotion and distribution of Project Gutenberg-tm electronic works,
harmless from all liability, costs and expenses, including legal fees,
that arise directly or indirectly from any of the following which you do
or cause to occur: (a) distribution of this or any Project Gutenberg-tm
work, (b) alteration, modification, or additions or deletions to any
Project Gutenberg-tm work, and (c) any Defect you cause.
Section 2. Information about the Mission of Project Gutenberg-tm
Project Gutenberg-tm is synonymous with the free distribution of
electronic works in formats readable by the widest variety of computers
including obsolete, old, middle-aged and new computers. It exists
because of the efforts of hundreds of volunteers and donations from
people in all walks of life.
Volunteers and financial support to provide volunteers with the
assistance they need are critical to reaching Project Gutenberg-tm's
goals and ensuring that the Project Gutenberg-tm collection will
remain freely available for generations to come. In 2001, the Project
Gutenberg Literary Archive Foundation was created to provide a secure
and permanent future for Project Gutenberg-tm and future generations.
To learn more about the Project Gutenberg Literary Archive Foundation
and how your efforts and donations can help, see Sections 3 and 4
and the Foundation web page at https://www.pglaf.org.
Section 3. Information about the Project Gutenberg Literary Archive
Foundation
The Project Gutenberg Literary Archive Foundation is a non profit
501(c)(3) educational corporation organized under the laws of the
state of Mississippi and granted tax exempt status by the Internal
Revenue Service. The Foundation's EIN or federal tax identification
number is 64-6221541. Its 501(c)(3) letter is posted at
https://pglaf.org/fundraising. Contributions to the Project Gutenberg
Literary Archive Foundation are tax deductible to the full extent
permitted by U.S. federal laws and your state's laws.
The Foundation's principal office is located at 4557 Melan Dr. S.
Fairbanks, AK, 99712., but its volunteers and employees are scattered
throughout numerous locations. Its business office is located at
809 North 1500 West, Salt Lake City, UT 84116, (801) 596-1887, email
business@pglaf.org. Email contact links and up to date contact
information can be found at the Foundation's web site and official
page at https://pglaf.org
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
gbnewby@pglaf.org
Section 4. Information about Donations to the Project Gutenberg
Literary Archive Foundation
Project Gutenberg-tm depends upon and cannot survive without wide
spread public support and donations to carry out its mission of
increasing the number of public domain and licensed works that can be
freely distributed in machine readable form accessible by the widest
array of equipment including outdated equipment. Many small donations
($1 to $5,000) are particularly important to maintaining tax exempt
status with the IRS.
The Foundation is committed to complying with the laws regulating
charities and charitable donations in all 50 states of the United
States. Compliance requirements are not uniform and it takes a
considerable effort, much paperwork and many fees to meet and keep up
with these requirements. We do not solicit donations in locations
where we have not received written confirmation of compliance. To
SEND DONATIONS or determine the status of compliance for any
particular state visit https://pglaf.org
While we cannot and do not solicit contributions from states where we
have not met the solicitation requirements, we know of no prohibition
against accepting unsolicited donations from donors in such states who
approach us with offers to donate.
International donations are gratefully accepted, but we cannot make
any statements concerning tax treatment of donations received from
outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation
methods and addresses. Donations are accepted in a number of other
ways including including checks, online payments and credit card
donations. To donate, please visit: https://pglaf.org/donate
Section 5. General Information About Project Gutenberg-tm electronic
works.
Professor Michael S. Hart was the originator of the Project Gutenberg-tm
concept of a library of electronic works that could be freely shared
with anyone. For thirty years, he produced and distributed Project
Gutenberg-tm eBooks with only a loose network of volunteer support.
Project Gutenberg-tm eBooks are often created from several printed
editions, all of which are confirmed as Public Domain in the U.S.
unless a copyright notice is included. Thus, we do not necessarily
keep eBooks in compliance with any particular paper edition.
Most people start at our Web site which has the main PG search facility:
https://www.gutenberg.org
This Web site includes information about Project Gutenberg-tm,
including how to make donations to the Project Gutenberg Literary
Archive Foundation, how to help produce our new eBooks, and how to
subscribe to our email newsletter to hear about new eBooks.