Project Gutenberg's Form and Function, by E. S. (Edward Stuart) Russell
This eBook is for the use of anyone anywhere at no cost and with
almost no restrictions whatsoever. You may copy it, give it away or
re-use it under the terms of the Project Gutenberg License included
with this eBook or online at www.gutenberg.org
Title: Form and Function
A Contribution to the History of Animal Morphology
Author: E. S. (Edward Stuart) Russell
Release Date: January 23, 2007 [EBook #20426]
Language: English
Character set encoding: UTF-8
*** START OF THIS PROJECT GUTENBERG EBOOK FORM AND FUNCTION ***
Produced by Suzanne Lybarger, Turgut Dincer and the Online
Distributed Proofreading Team at http://www.pgdp.net (This
file was produced from images generously made available
by The Internet Archive/Canadian Libraries)
FORM AND FUNCTION
A CONTRIBUTION TO THE
HISTORY OF ANIMAL MORPHOLOGY
By E. S. RUSSELL,
M.A., B.Sc., F.Z.S.
ILLUSTRATED
LONDON
JOHN MURRAY, ALBEMARLE STREET, W.
1916
All rights reserved
Transcriber's Note: Obvious printer errors have been corrected, all other inconsistencies
in spelling and punctuation are as in the original.
PREFACE
This book is not intended to be a
full or detailed history of animal morphology: a complete account
is given neither of morphological discoveries nor of morphological
theories. My aim has been rather to call attention to the existence
of diverse typical attitudes to the problems of form, and to trace
the interplay of the theories that have arisen out of them.
The main currents of morphological thought are to my mind
three—the functional or synthetic, the formal or
transcendental, and the materialistic or disintegrative.
The first is associated with the great names of Aristotle,
Cuvier, and von Baer, and leads easily to the more open vitalism of
Lamarck and Samuel Butler. The typical representative of the second
attitude is E. Geoffroy St. Hilaire, and this habit of thought has
greatly influenced the development of evolutionary morphology.
The main battle-ground of these two opposing tendencies is the
problem of the relation of function to form. Is function the
mechanical result of form, or is form merely the manifestation of
function or activity? What is the essence of
life—organisation or activity?
The materialistic attitude is not distinctively biological, but
is common to practically all fields of thought. It dates back to
the Greek atomists, and the triumph of mechanical science in the
19th century has induced many to accept materialism as the only
possible scientific method. In biology it is more akin to the
formal than to the functional attitude.
In the course of this book I have not hidden my own sympathy
with the functional attitude. It appears to me probable that more
insight will be gained into the real nature of life and
organisation by concentrating on the active response of the animal,
as manifested both in behaviour and in morphogenesis, particularly
in the post-embryonic stages, than by giving attention exclusively
to the historical aspect of structure, as is the custom of "pure
morphology." I believe we shall only make progress in this
direction if we frankly adopt the simple everyday conception of
living things—which many of us have had drilled out of
us—that they are active, purposeful agents, not mere
complicated aggregations of protein and other substances. Such an
attitude is probably quite as sound philosophically as the opposing
one, but I have not in this place attempted any justification of
it. I have touched very lightly upon the controversy between
vitalism and materialism which has been revived with the early
years of the present century. It hardly lends itself as yet to
historical treatment, and I could hardly hope to maintain with
regard to it that objective attitude which should characterise the
historian.
The main result I hope to have achieved with this book is the
demonstration, tentative and incomplete as it is, of the essential
continuity of animal morphology from the days of Aristotle down to
our own time. It is unfortunately true that modern biology, perhaps
in consequence of the great advances it has made in certain
directions, has to a considerable extent lost its historical
consciousness, and if this book helps in any degree to counteract
this tendency so far as animal morphology is concerned, it will
have served its purpose.
I owe a debt of gratitude to my friends Dr James F. Gemmill and
Prof. J. Arthur Thomson for much kindly encouragement and helpful
criticism. The credit for the illustrations is due to my wife, Mrs
Jehanne A. Russell. One is from Nature; the others are drawn from
the original figures.
E. S. R.
Chelsea,
1916.
CONTENTS
| CHAP. |
|
Page |
| I. |
The Beginnings of
Comparative Anatomy |
1 |
| II. |
Comparative Anatomy
before Cuvier |
17 |
| III. |
Cuvier |
31 |
| IV. |
Goethe |
45 |
| V. |
Etienne Geoffroy St
Hilaire |
52 |
| VI. |
The Followers of Etienne
Geoffroy St Hilaire |
79 |
| VII. |
The German
Transcendentalists |
89 |
| VIII. |
Transcendental Anatomy in
England—Richard Oven |
102 |
| IX. |
Karl Ernst von
Baer |
113 |
| X. |
The Embryological
Criterion |
133 |
| XI. |
The
Cell-Theory |
169 |
| XII. |
The Close of the
Pre-evolutionary Period |
190 |
| XIII. |
The Relation of Lamarck
and Darwin to Morphology |
213 |
| XIV. |
Ernst Haeckel and Carl
Gegenbaur |
246 |
| XV. |
Early Theories on the
Origin of Vertebrates |
268 |
| XVI. |
The Germ-layers and
Evolution |
288 |
| XVII. |
The Organism as an
Historical Being |
302 |
| XVIII. |
The Beginnings of Causal
Morphology |
314 |
| XIX. |
Samuel Butler and the
Memory Theories of Heredity |
335 |
| XX. |
The Classical Tradition
in Modern Morphology |
345 |
|
Index |
365 |
ILLUSTRATIONS
| FIG. |
|
Page |
| 1. |
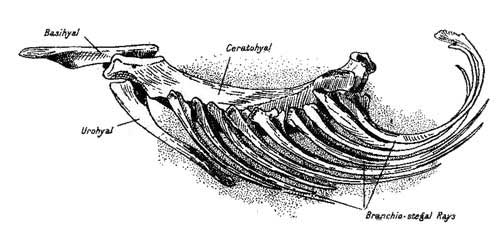
Hyoid Arch of the Conger.
(Original.) |
58 |
| 2. |
"Vertebra" of a
Pleuronectid. (Geoffroy.) |
61 |
| 3. |
Abdominal Segment of the
Lobster. (Geoffroy.) |
63 |
| 4. |
Ideal Typical Vertebra.
(Owen.) |
102 |
| 5. |
Natural Typical Vertebra.
(Owen.) |
103 |
| 6. |
The Archetype of the
Vertebrate Skeleton. (Owen.) |
105 |
| 7. |
Ideal Transverse Section
of a Vertebrate Embryo. (Von Baer.) |
119 |
| 8. |
Gill-slits of the Pig
Embryo. (Rathke.) |
134 |
| 9. |
Meckel's Cartilage and
Ear-ossicles in Embryo of Pig. (Reichert.) |
145 |
| 10. |
Cranial Vertebræ
and Visceral Arches in Embryo of Pig. (Reichert.) |
148 |
| 11. |
Embryonic Cranium of the
Adder. (Rathke.) |
152 |
| 12. |
Transverse Section of
Chick Embryo. (Remak.) |
211 |
| 13. |
Development of the
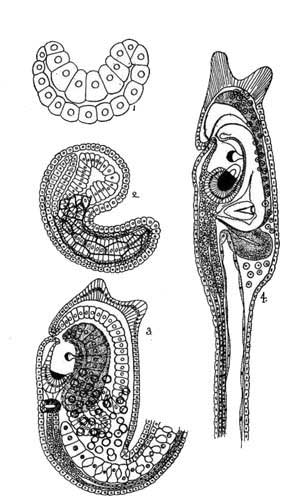
Ascidian Larva (Kowalevsky.) |
272 |
| 14. |
Transverse Section of the
Worm Nais. (Semper.) |
280 |
| 15. |
The Five Primary Stages
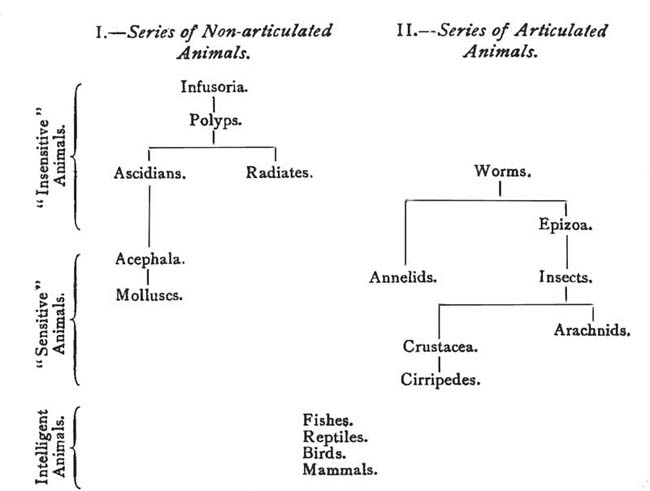
of Ontogeny. (Haeckel.) |
292 |
001
FORM AND FUNCTION
CHAPTER I
THE BEGINNINGS OF COMPARATIVE ANATOMY
The first name of which the history
of anatomy keeps record is that of Alcmaeon, a contemporary of
Pythagoras (6th century B.C.). His interests appear to have been
rather physiological than anatomical. He traced the chief nerves of
sense to the brain, which he considered to be the seat of the soul,
and he made some good guesses at the mechanism of the organs of
special sense. He showed that, contrary to the received opinion,
the seminal fluid did not originate in the spinal cord. Two
comparisons are recorded of his, one that puberty is the equivalent
of the flowering time in plants, the other that milk is the
equivalent of white of egg.[1]
Both show his bias towards looking at the functional side of living
things. The latter comparison reappears in Aristotle.
A century later Diogenes of Apollonia gave a description of the
venous system. He too placed the seat of sensation in the brain. He
assumed a vital air in all living things, being in this influenced
by Anaximenes whose primitive matter was infinite air. In following
out this thought he tried to prove that both fishes and oysters
have the power of breathing.[2]
A more strictly morphological note is struck by a curious saying
of Empedocles (4th century B.C.), that "hair and foliage and the
thick plumage of birds are one."[3]
002In the collected writings of Hippocrates and
his school, the Corpus Hippocraticum, of which no part is
later than the end of the 5th century, there are recorded many
anatomical facts. The author of the treatise "On the Muscles" knew,
for instance, that the spinal marrow is different from ordinary
marrow and has membranes continuous with those of the brain.
Embryos of seven days (!) have all the parts of the body plainly
visible. Work on comparative embryology is contained in the
treatise "On the Development of the Child."[4]
The author of the treatise "On the Joints," which Littré
calls "the great surgical monument of antiquity," is to be credited
with the first systematic attempt at comparative anatomy, for he
compared the human skeleton with that of other Vertebrates.
Aristotle (384-322 B.C.)[5]
may fairly be said to be the founder of comparative anatomy, not
because he was specially interested in problems of "pure
morphology," but because he described the structure of many animals
and classified them in a scientific way. We shall discuss here the
morphological ideas which occur in his writings upon
animals—in the Historia Animalium, the De Partibus
Animalium, and the De Generatione Animalium.
The Historia Animalium is a most comprehensive work, in
some ways the finest text-book of Zoology ever written. Certainly
few modern text-books take such a broad and sane view of living
creatures. Aristotle never forgets that form and structure are but
one of the many properties of living things; he takes quite as much
interest in their behaviour, their ecology, distribution,
comparative physiology. He takes a special interest in the
comparative physiology of reproduction. The Historia
Animalium contains a description of the form and structure of
man and of as many animals as Aristotle was acquainted
with—and he was acquainted with an astonishingly large
number. The later De Partibus Animalium is a treatise on
the causes of the form and 003structure of animals. Owing to the
importance which Aristotle ascribed to the final cause this work
became really a treatise on the functions of the parts, a
discussion of the problems of the relation of form to function, and
the adaptedness of structure.
Aristotle was quite well aware that each of the big groups of
animals was built upon one plan of structure, which showed endless
variations "in excess and defect" in the different members of the
group. But he did not realise that this fact of community of plan
constituted a problem in itself. His interest was turned towards
the functional side of living things, form was for him a secondary
result of function.
Yet he was not unaware of facts of form for which he could not
quite find a place in his theory of organic form, facts of form
which were not, at first sight at least, facts of function. Thus he
was aware of certain facts of "correlation," which could not be
explained off-hand as due to correlation of the functions of the
parts. He knew, for instance, that all animals without front teeth
in the upper jaw have cotyledons, while most that have front teeth
on both jaws and no horns have no cotyledons (De Gen., ii.
7).
Speaking generally, however, we find in Aristotle no purely
morphological concepts. What then does morphology owe to Aristotle?
It owes to him, first, a great mass of facts about the
structure of animals; second, the first scientific
classification of animals;[6]
third, a clear enunciation of the fact of community of plan
within each of the big groups; fourth, an attempt to explain
certain instances of the correlation of parts; fifth, a
pregnant distinction between homogeneous and heterogeneous parts;
sixth, a generalisation on the succession of forms in
development; and seventh, the first enunciation of the idea
of the Échelle des êtres.
(1) What surprises the modern reader of the Historia
Animalium perhaps more than anything else is the extent and
variety of Aristotle's knowledge of animals. He 004describes more
than 500 kinds.[7] Not only does he know the
ordinary beasts, birds, and fishes with which everyone is
acquainted, but he knows a great deal about cuttlefish, snails and
oysters, about crabs, crawfish (Palinurus), lobsters,
shrimps, and hermit crabs, about sea-urchins and starfish,
sea-anemones and sponges, about ascidians (which seem to have
puzzled him not a little!). He has noticed even fish-lice and
intestinal worms, both flat and round. Of the smaller land animals,
he knows a great many insects and their larvæ. The extent of
his anatomical knowledge is equally surprising, and much of it is
clearly the result of personal observation. No one can read his
account of the internal anatomy of the chameleon (Hist.
Anim., ii.), or his description of the structure of cuttlefish
(Hist. Anim., iv), or that touch in the description of the
hermit crab (Hist. Anim., iv.)—"Two large eyes ...
not ... turned on one side like those of crabs, but straight
forward"—without being convinced that Aristotle is speaking
of what he has seen. Naturally he could not make much of the
anatomy of small insects and snails, and, to tell the truth, he
does not seem to have cared greatly about the minutiæ of
structure. He was too much of a Greek and an aristocrat to care
about laborious detail.
Not only did he lay a foundation for comparative anatomy, but he
made a real start with comparative embryology. Medical men before
him had known many facts about human development; Aristotle seems
to have been the first to study in any detail the development of
the chick. He describes this as it appears to the naked eye, the
position of the embryo on the yolk, the palpitating spot at the
third day, the formation of the body and of the large sightless
eyes, the veins on the yolk, the embryonic membranes, of which he
distinguished two.
(2) Aristotle had various systems of classifying animals. They
could be classified, he thought, according to their structure,
their manner of reproduction, their manner of life, their mode of
locomotion, their food, and so on. Thus you 005might, in
addition to structural classifications, divide animals into
gregarious, solitary and social, or land animals into troglodytes,
surface-dwellers, and burrowers (Hist. Anim., i.).
He knew that dichotomous classifications were of little use for
animals (De Partibus, i. 3) and he explicitly and in so many
words accepted the principle of all "natural" classification, that
affinities must be judged by comparing not one but the sum total of
characters. As everyone knows, he was the first to distinguish the
big groups of animals, many of which were already distinguished
roughly by the common usages of speech. Among his Sanguinea he did
little more than define with greater exactitude the limits of the
groups established by the popular classification. Among the
"exsanguineous" animals, however, corresponding to our
Invertebrates, he established a much more definite classification
than the popular, which is apt to call them indiscriminately
"shellfish," "insects," or "creeping things." He went beyond the
superficialities of popular classification, too, in clearly
separating Cetacea from fishes. He had some notion of species and
genera in our sense. He distinguished many species of
cuttlefish—Octopus (Polypus) of which there were many
kinds, Eledone (Moschites) which he knew to have only one
row of suckers while Octopus has two, Argonauta,
Nautilus, Sepia, and apparently Loligo media (= his
Teuthis) and L. vulgaris (or forbesii) which seems to
be his Teuthos. He had a grasp of the principles which should be
followed in judging of the natural affinities of species. For
example, he knew that the cuckoo resembles a hawk. "But," he says,
"the hawk has crooked talons, which the cuckoo has not, nor does it
resemble the hawk in the form of its head, but in these respects is
more like the pigeon than the hawk, which it resembles in nothing
but its colour; the markings, however, upon the hawk are like
lines, while the cuckoo is spotted" (Hist. Anim.,
Cresswell's trans., p. 147, London, 1862).
The groups he distinguished were—man, viviparous
quadrupeds, oviparous quadrupeds, birds, fishes, Cetacea,
Cephalopoda, Malacostraca (= higher Crustacea), Insecta (= annulose
animals), Testacea (= molluscs, echinoderms, ascidians). A class of
Acalephæ, including sea-anemones and 006sponges, was grouped with the
Testacea. The first five groups were classed together as
sanguineous, the others as exsanguineous, from the presence or
absence of red blood.
Besides these classes "there are," he says, "many other
creatures in the sea which it is not possible to arrange in any
class from their scarcity" (Creswell, loc. cit., p. 90).
(3) Aristotle's greatest service to morphology is his clear
recognition of the unity of plan holding throughout each of the
great groups.
He recognises this most clearly in the case of man and the
viviparous quadrupeds, with whose structure he was best acquainted.
In the Historia Animalium he takes man as a standard, and
describes his external and internal parts in detail, then considers
viviparous quadrupeds and compares them with man. "Whatever parts a
man has before, a quadruped has beneath; those that are behind in
man form the quadruped's back" (Cresswell, loc. cit., p.
26). Apes, monkeys, and Cynocephali combine the characteristics of
man and quadrupeds. He notices that all viviparous quadrupeds have
hair. Oviparous quadrupeds resemble the viviparous, but they lack
some organs, such as ears with an external pinna, mammæ,
hair. Oviparous bipeds, or birds, also "have many parts like the
animals described above." He does not, however, seem to realise
that a bird's wings are the equivalent of a mammal's arms or
fore-legs. Fishes are much more divergent; they possess no neck,
nor limbs, nor testicles (meaning a solid ovoid body such as the
testis in mammals), nor mammæ. Instead of hair they have
scales.
Speaking generally, the Sanguinea differ from man and from one
another in their parts, which may be present or absent, or exhibit
differences in "excess and defect," or in form. Unity of plan
extends to all the principal systems of organs. "All sanguineous
animals have either a bony or a spinous column. The remainder of
the bones exist in some animals; but not in others, for if they
have the limbs they have the bones belonging to them" (Cresswell,
loc. cit., p. 60). "Viviparous animals with blood and feet
do not differ much in their bones, but rather by analogy, in
hardness, softness, and size" (Cresswell, loc. cit., p. 59).
007The
venous system, too, is built upon the same general plan throughout
the Sanguinea. "In all sanguineous animals, the nature and origin
of the principal veins are the same, but the multitude of smaller
veins is not alike in all, for neither are the parts of the same
nature, nor do all possess the same parts" (Cresswell, loc.
cit., p. 56). It will be noticed in the first and last of these
three quotations that Aristotle recognises the fact of correlation
between systems of organs—between limbs and bones, and
between blood-vessels and the parts to which they go.
Sanguineous animals all possess certain organs—heart,
liver, spleen, kidneys, and so on. Other organs occur in most of
the classes—the œsophagus and the lungs. "The position
which these parts occupy is the same in all animals [sc.
Sanguinea]" (Cresswell, loc. cit., p. 39).
Unity of plan is observable not only in the Sanguinea, but also
within each of the other large groups. Aristotle recognises that
all his cuttlefish are alike in structure. Among his Malacostraca
he compares point by point the external parts of the carabus
(Palinurus), and the astacus (Homarus), and he
compares also the general internal anatomy of the various "genera"
he distinguishes. As regards Testacea, he writes, "The nature of
their internal structure is similar in all, especially in the
turbinated animals, for they differ in size and in the relations of
excess; the univalves and bivalves do not exhibit many differences"
(Cresswell, loc. cit., p. 83). There is an interesting
remark about "the creature called carcinium" (hermit-crab), that it
"resembles both the Malacostraca and the Testacea, for this in its
nature is similar to the animals that are like carabi, and it is
born naked" (Cresswell, loc. cit., p. 85). In the last
phrase we may perhaps read the first recognition of the
embryological criterion.
With the recognition of unity of plan within each group
necessarily goes the recognition of what later morphology calls the
homology of parts. The parts of a horse can be compared one by one
with the parts of another viviparous quadruped; in all the animals
belonging to the same class the parts are the same, only they
differ in excess or defect—these remarks are placed in the
forefront of the 008Historia Animalium. Generally
speaking, parts which bear the same name are for Aristotle
homologous throughout the class. But he goes further and notes the
essential resemblance underlying the differences of certain parts.
He classes together nails and claws, the spines of the hedgehog,
and hair, as being homologous structures. He says that teeth are
allied to bones, whereas horns are more nearly allied to skin
(Hist. Anim., iii.). This is an astonishingly happy guess,
considering that all he had to go upon was the observation that in
black animals the horns are black but the teeth white. One cannot
but admire the way in which Aristotle fixes upon apparently trivial
and commonplace facts, and draws from them far-reaching
consequences. He often goes wrong, it is true, but he always errs
in the grand manner.
While Aristotle certainly recognised the existence of
homologies, and even had a feeling for them, he did not clearly
distinguish homology from analogy. He comes pretty near the
distinction in the following passage. After explaining that in
animals belonging to the same class the parts are the same,
differing only in excess or defect, he says, "But some animals
agree with each other in their parts neither in form nor in excess
and defect, but have only an analogous likeness, such as a bone
bears to a spine, a nail to a hoof, a hand to a crab's claw, the
scale of a fish to the feather of a bird, for that which is a
feather in the bird is a scale in the fish" (Cresswell, loc.
cit., p. 2). One of these comparisons is, however, a homology
not an analogy, and the last phrase throws a little doubt upon the
whole question, for it is not made clear whether it is position or
function that determines what are equivalent organs.
In the De Partibus Animalium there occurs the following
passage:—"Groups that only differ in degree, and in the more
or less of an identical element that they possess, are aggregated
under a single class; groups whose attributes are not identical but
analogous are separated. For instance, bird differs from bird by
gradation, or by excess and defect; some birds have long feathers,
others short ones, but all are feathered. Bird and Fish are more
remote and only agree in having analogous organs; for what in the
bird is feather, 009in the fish is scale. Such analogies can
scarcely, however, serve universally as indications for the
formation of groups, for almost all animals present analogies in
their corresponding parts."[8]
It is thus similarity in form and structure which determines the
formation of the main groups. Within each group the parts differ
only in degree, in largeness or smallness, softness and hardness,
smoothness or roughness, and the like (loc. cit., i., 4,
644b). These passages show that Aristotle had some
conception of homology as distinct from analogy. He did not,
however, develop the idea. What Aristotle sought in the variety of
animal structure, and what he found, were not homologies, but
rather communities of function, parts with the same attributes. His
interest was all in organs, in functioning parts, not in the
mere spatial relationship of parts.
This comes out clearly in his treatise On the Parts of
Animals, which is subsequent to, and the complement of, his
History of Animals. The latter is a description of the
variety of animal form, the former is a treatise on the functions
of the parts. He describes the plan of the De Partibus
Animalium as follows:—"We have, then, first to describe
the common functions, common, that is, to the whole animal kingdom,
or to certain large groups, or to members of a species. In other
words, we have to describe the attributes common to all animals, or
to assemblages, like the class of Birds, of closely allied groups
differentiated by gradation, or to groups like Man not
differentiated into subordinate groups. In the first case the
common attributes may be called analogous, in the second generic,
in the third specific" (i, 5, 645b, trans. Ogle). The
alimentary canal is a good example of a part which is "analogous"
throughout the animal kingdom, for "all animals possess in common
those parts by which they take in food, and into which they receive
it" (Cresswell, loc. cit., p. 6).
The De Partibus Animalium becomes in form a comparative
organography, but the emphasis is always on function and community
of function. Thus he treats of bone, "fish-spine," and cartilage
together (De Partibus, ii., 9, 655a), because
they have the same function, though he says 010elsewhere that
they are only analogous structures (ii., 8, 653b). In
the same connection he describes also the supporting tissues of
Invertebrates—the hard exoskeleton of Crustacea and Insects,
the shell of Testacea, the "bone" of Sepia (ii., 8,
654a). Aristotle took much more interest in analogies,
in organs of similar function, than in homologies. He did recognise
the existence of homologies, but rather malgré lui,
because the facts forced it upon him.
His only excursion into the realm of "transcendental anatomy" is
his comparison of a Cephalopod to a doubled-up Vertebrate whose
legs have become adherent to its head, whose alimentary canal has
doubled upon itself in such a way as to bring the anus near the
mouth (De Partibus, iv., 9, 684b). It is clear,
however, that Aristotle did not seek to establish by this
comparison any true homologies of parts, but merely analogies, thus
avoiding the error into which Meyranx and Laurencet fell more than
two thousand years later in their paper communicated to the
Académie des Sciences, which formed the starting-point of
the famous controversy between Cuvier and E. Geoffroy St Hilaire
(see Chap. V., below).
Moreover, Aristotle did not so much compare a Cephalopod with a
doubled-up Vertebrate as contrast Cephalopods (and also Testacea)
with all other animals. Other animals have their organs in a
straight line; Cephalopods and Testacea alone show this peculiar
doubling up of the body.
(4) Aristotle was much struck with certain facts of correlation,
of the interdependence of two organs which are not apparently in
functional dependence on one another. Such correlation may be
positive or negative; the presence of one organ may either entail
the presence of the other, or it may entail its absence. Aristotle
has various ways of explaining facts of correlation. He observed
that no animal has both tusks and horns, but this fact could easily
be explained on the principle that Nature never makes anything
superfluous or in vain. If an animal is protected by the possession
of tusks it does not require horns, and vice versa. The
correlation of a multiple stomach with deficient 011development of
the teeth (as in Ruminants) is accounted for by saying that the
animal needs its complex stomach to make up for the shortcomings of
its teeth! (De Partibus, iii., 14, 674b.) Other
examples of correlation were not susceptible of this explanation in
terms of final causes. He lays stress on the fact, in the main
true, of the inverse development of horns and front teeth in the
upper jaw, exemplified in Ruminants. He explains the fact in this
way. Teeth and horns are formed from earthy matter in the body and
there is not enough to form both teeth and horns, so "Nature by
subtracting from the teeth adds to the horns; the nutriment which
in most animals goes to the former being here spent on the
augmentation of the latter" (De Partibus, iii., 2,
664a, trans. Ogle). A similar kind of explanation is
offered of the fact that Selachia have cartilage instead of bone,
"in these Selachia Nature has used all the earthy matter on the
skin [i.e., on the placoid scales]; and she is unable to
allot to many different parts one and the same superfluity of
material" (De Partibus, ii., 9, 655a, trans.
Ogle). Speaking generally, "Nature invariably gives to one part
what she subtracts from another" (loc. cit., ii., 14,
658a).
This thought reappears again in the 19th century in E. Geoffroy
St Hilaire's loi de balancement and also in Goethe's
writings on morphology. For Aristotle it meant that Nature was
limited by the nature of her means, that finality was limited by
necessity. Thus in the larger animals there is an excess of earthy
matter, as a necessary result of the material nature of the animal;
this excess is turned by Nature to good account, but there is not
enough to serve both for teeth and for horns (loc. cit.,
iii., 2, 663b).
But there are other instances of correlation which seem to have
taxed even Aristotle's ingenuity beyond its powers. Thus he knew
that all animals (meaning viviparous quadrupeds) with no front
teeth in the upper jaw have cotyledons on their fœtal membranes,
and that most animals which have front teeth in both jaws and no
horns have no cotyledons (De Generatione, ii., 7). He offers
no explanation of this, but accepts it as a fact.
We may conveniently refer here to one or two other ideas of
Aristotle regarding the causes of form. He makes the 012profound remark
that the possible range of form of an organ is limited to some
extent by its existing differentiation. Thus he explains the
absence of external (projecting) ears in birds and reptiles by the
fact that their skin is hard and does not easily take on the form
of an external ear (De Partibus, ii, 12). The fact of the
inverse correlation is certain; the explanation is, though very
vague, probably correct.
In one passage of the De Partibus Aristotle clearly
enunciates the principle of the division of labour, afterwards
emphasised by H. Milne-Edwards. In some insects, he says, the
proboscis combines the functions of a tongue and a sting, in others
the tongue and the sting are quite separate. "Now it is better," he
goes on, "that one and the same instrument shall not be made to
serve several dissimilar ends; but that there shall be one organ to
serve as a weapon, which can then be very sharp, and a distinct one
to serve as a tongue, which can then be of spongy texture and fit
to absorb nutriment. Whenever, therefore, Nature is able to provide
two separate instruments for two separate uses, without the one
hampering the other, she does so, instead of acting like a
coppersmith who for cheapness makes a spit and lampholder in one"
(iv., 6, 683a).
(5) The first sentence of the Historia Animalium
formulates, with that simplicity and directness which is so
characteristic of Aristotle, the distinction between homogeneous
and heterogeneous parts, in the mass the distinction between
tissues and organs. "Some parts of animals are simple, and these
can be divided into like parts, as flesh into pieces of flesh;
others are compound, and cannot be divided into like parts, as the
hand cannot be divided into hands, nor the face into faces. All the
compound parts also are made up of simple parts—the hand, for
example, of flesh and sinew and bone" (Cresswell, loc. cit.,
p. 1).
In the De Partibus Animalium he broadens the conception
by adding another form of composition. "Now there are," he says,
"three degrees of composition; and of these the first in order, as
all will allow, is composition out of what some call the elements,
such as earth, air, water, fire.... 013The second degree of composition
is that by which the homogeneous parts of animals, such as bone,
flesh, and the like, are constituted out of the primary substances.
The third and last stage is the composition which forms the
heterogeneous parts, such as face, hand, and the rest" (ii., 1,
646a, trans. Ogle).
In the Historia Animalium the homogeneous parts are
divided into (1) the soft and moist (or fluid), such as blood,
serum, flesh, fat, suet, marrow, semen, gall, milk, phlegm,
fæces and urine, and (2) the hard and dry (or solid), such as
sinew, vein, hair, bone, cartilage, nail, and horn. It would appear
from this enumeration that Aristotle's distinction of simple and
complex parts does not altogether coincide with our distinction of
tissues and organs. We should not call vein a tissue, nor do we
include under this heading non-living secretions. But in the De
Partibus Animalium Aristotle, while still holding to the
distinction set forth above, is alive to the fact that his simple
parts include several different sorts of substances. He
distinguishes among the homogeneous parts three sets. The first of
these comprises the tissues out of which the heterogeneous parts
are constructed, e.g., flesh and bone; the second set form
the nutriment of the parts, and are invariably fluid; while the
third set are the residue of the second and constitute the residual
excretions of the body (ii., 2, 647b). He sees clearly
the difficulty of calling vein or blood-vessel a simple part, for
while a blood-vessel and a part of it are both blood-vessel, as we
should say vascular tissue, yet a part of a blood-vessel is not a
blood-vessel. There is form superadded to homogeneity of structure
(ii., 2, 647b). Similarly for the heart and the other
viscera. "The heart, like the other viscera, is one of the
homogeneous parts; for, if cut up, its pieces are homogeneous in
substance with each other. But it is at the same time heterogeneous
in virtue of its definite configuration" (ii., 1, 647a,
trans. Ogle).
Aristotle, therefore, came very near our conception of tissue.
He was of course not a histologist; he describes not the structure
of tissues, which he could not know, but rather their distribution
within the organism; his section on the homogeneous parts of
Sanguinea (Historia Animalium, iii., 014second half) is
largely a comparative topographical anatomy; in it, for instance,
he describes the venous and skeletal systems.
This distinction which Aristotle drew plays an important part in
all his writings on animals, particularly in his theory of
development. It was a distinction of immense value, and is full of
meaning even at the present day. No one has ever given a better
definition of organ than is implied in Aristotle's description of
the heterogeneous parts—"The capacity of action resides in
the compound parts" (Cresswell, loc. cit., p. 7). The
heterogeneous parts were distinguished by the faculty of doing
something, they were the active or executive parts. The homogeneous
parts were distinguished mainly by physical characters (De
Generatione, i., 18), but certain of them had other than purely
physical properties, they were the organs of touch (De
Partibus, ii., 1, 647a).
(6) In a passage in the De Generatione (ii, 3) Aristotle
says that the embryo is an animal before it is a particular animal,
that the general characters appear before the special. This is a
foreshadowing of the essential point in von Baer's law (see Chap. IX. below).
He considers also that tissues arise before organs. The
homogeneous parts are anterior genetically to the heterogeneous
parts and posterior to the elementary material (De Partibus,
ii., 1, 646b).
(7) We meet in Aristotle an idea which later acquired
considerable vogue, that of the Échelle des
êtres(or "scale of beings"), that organisms, or even all
objects organic or inorganic, can be arranged in a single ascending
series. The idea is a common one; its first literary expression is
found perhaps in primitive creation-myths, in which inorganic
things are created before organic, and plants before animals. It
may be recognised also in Anaximander's theory that land animals
arose from aquatic animals, more clearly still in Anaxagoras'
theory that life took its origin on this globe from vegetable germs
which fell to earth with the rain. Anaxagoras considered animals
higher in the scale than plants, for while the latter participated
in pleasure (when they 015grew) and pain (when they lost their leaves),
animals had in addition "Nous." In Empedocles' theory of evolution,
the vegetable world preceded the animal. Plato, in the
Timaeus, describes the whole organic world as being formed
by degradation from man, who is created first. Man sinks first into
woman, then into brute form, traversing all the stages from the
higher to the lower animals, and becoming finally a plant. This is
a reversal of the more usual notion, but the idea of gradation is
equally present.
Aristotle seems not to have believed in any transformation of
species, but he saw that Nature passes gradually from inanimate to
animate things without a clear dividing line. "The race of plants
succeeds immediately that of inanimate objects" (Cresswell, loc.
cit., p. 94). Within the organic realm the passage from plants
to animals is gradual. Some creatures, for example, the
sea-anemones and sponges, might belong to either class.
Aristotle recognised also a natural series among the groups of
animals, a series of increasing complexity of structure. He begins
his study of structure with man, who is the most intricate, and
then takes up in turn viviparous and oviparous quadrupeds, then
birds, then fishes. After the Sanguinea he considers the
Exsanguinea, and of the latter first the most highly organised, the
Cephalopods, and last the simplest, the lower members of his class
of the Testacea. In treating of generation (in Hist.
Animalium, v.) he reverses this order. In the De
Generatione (Book ii., 1) there is given another serial
arrangement of animals, this time in relation to their manner of
reproduction. There is a gradation, he says, of the following
kind:—
1. Internally viviparous Sanguinea |
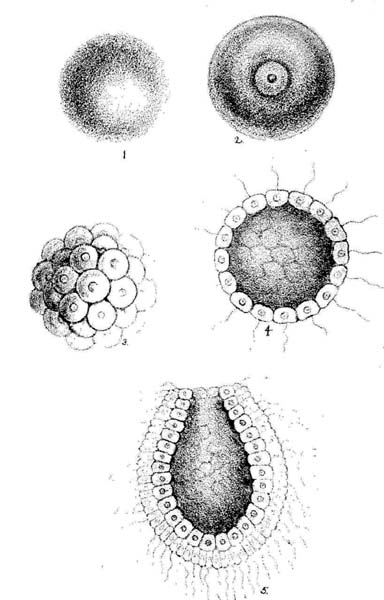
 |
producing a perfect animal |
2. Externally viviparous Sanguinea |
3. Oviparous
Sanguinea—producing a perfect egg. |
4. Animals producing an imperfect
egg (one which increases in size after being laid). |
5. Insects, producing a scolex (or
grub). |
In Aristotle's view the gradation of organic forms is the
consequence, not the cause, of the gradation observable in their
activities. Plants have no work to do beside nutrition, 016growth,
and reproduction; they possess only the nutritive soul. Animals
possess in addition sensation and the sensitive or perceptive
soul—"their manner of life differs in their having pleasure
in sexual intercourse, in their mode of parturition and rearing
their young" (Hist. Anim., viii., trans. Cresswell, p. 195).
Man alone has the rational soul in addition to the two lower
kinds.
As it is put in the De Partibus (ii., 10,
656a, trans. Ogle), "Plants, again, inasmuch as they are
without locomotion, present no great variety in their heterogeneous
parts. For, where the functions are but few, few also are the
organs required to effect them.... Animals, however, that not only
live but feel, present a greater multiformity of parts, and this
diversity is greater in some animals than in others, being most
varied in those to whose share has fallen not mere life but life of
high degree. Now such an animal is man."
With the great exception of Aristotle, the philosophers of
Greece and Rome made little contribution to morphological theory.
Passing mention may be made of the Atomists—Leucippus,
Democritus, and their great disciple Lucretius, who in his
magnificent poem "De Natura Rerum" gave impassioned expression to
the materialistic conception of the universe. But the full effect
of materialism upon morphology does not become apparent till the
rise of physiology in the 17th and 18th centuries, and reaches its
culmination in the 19th century. The evolutionary ideas of
Lucretius exercised no immediate influence upon the development of
morphology.
017
CHAPTER II
COMPARATIVE ANATOMY BEFORE CUVIER
For two thousand years after Aristotle little advance was made
upon his comparative anatomy. Knowledge of the human body was
increased not long after his death by Herophilus and Erasistratus,
but not even Galen more than four centuries later made any
essential additions to Aristotle's anatomy.
During the Middle Ages, particularly after the introduction to
Europe in the 13th century of the Arab texts and commentaries,
Aristotle dominated men's thoughts of Nature. The commentary of
Albertus Magnus, based upon that of Avicenna, did much to impose
Aristotle upon the learned world. Albertus seems to have contented
himself with following closely in the footsteps of his master.
There are noted, however, by Bonnier certain improvements made by
Albertus on Aristotle's view of the seriation of living things. "He
is the first," writes Bonnier, "to take the correct view that fungi
are lower plants allied to the most lowly organised animals. From
this point there start, for Albertus Magnus, two series of living
creatures, and he regards the plant series as culminating in the
trees which have well-developed flowers."[9]
Aristotle's influence is predominant also in the work of Edward
Wotton (1492-1555), who in his book De differentiis
animalium adopted a classification similar to that proposed by
Aristotle. He too laid stress upon the gradation shown from the
lower to the higher forms.
In the 16th century, two groups of men helped to lay foundations
for a future science of comparative anatomy—the 018great Italian
anatomists Vesalius, Fallopius and Fabricius, and the first
systematists (though their "systems" were little more than
catalogues) Rondeletius, Aldrovandus and Gesner.
The anatomists, however, took little interest in problems of
pure morphology; the anatomy of the human body was for them simply
the necessary preliminary of the discovery of the functions of the
parts—they were quite as much physiologists as
anatomists.
One of them, Fabricius, made observations on the development of
the chick (1615). Harvey, who was a pupil of Fabricius, likewise
published an account of the embryology of the chick.[10] In his philosophy and habit of thought
Harvey was a follower of Aristotle. It is worth noting that in his
Exercitationes anatomicae de motu cordis (1628) there is a
passage which dimly foreshadows the law of recapitulation in
development which later had so much vogue.[11]
A stimulating contribution to comparative anatomy was made by
Belon,[12] who published in 1555 a
Histoire de la nature des Oyseaux, in which he showed
opposite one another a skeleton of a bird and of a mammal, giving
the same names to homologous bones. The anatomy of animals other
than man was indeed not altogether neglected at this time. Coiter
(1535-1600) studied the anatomy of Vertebrates, discovering among
other things the fibrous structure of the brain. Carlo Ruini of
Bologna wrote in 1598 a book on the anatomy of the horse.[13] Somewhat later Severino, professor at
Naples, dissected many animals and came to the conclusion 019that they
were built upon the same plan as man.[14]
Willis, of Oxford and London, in his Cerebri Anatome (1659)
recognised the necessity for comparative study of the structure of
the brain. He found out that the brain of man is very like that of
other mammals, the brain of birds, on the contrary, like that of
fishes![15] He described the anatomy
of the oyster and the crayfish. He had, however, not much feeling
for morphology.
The foundation of the Jardin des Plantes at Paris in 1626 and
the subsequent addition to it of a Museum of Natural History and a
menagerie gave a great impulse to the study of comparative anatomy
by supplying a rich material for dissection. Advantage was taken of
these facilities, particularly by Claude Perrault and Duverney.[16] In a volume entitled De la
Mécanique des Animaux, Perrault recognises clearly the
idea of unity of type, and even pushes it too far, seeking to prove
that in plants there exists an arterial system and veins provided
with valves.[17]
The beginning of the 17th century saw the invention of the
microscope, which was to have such an enormous influence upon the
development of biological studies. It did not come into scientific
use until well on in the middle of the century. Just before it came
into use Francis Glisson (1597-1677), an Englishman, gave in the
introduction to his treatise on the liver an account of the notions
then current on the structure of organic bodies. He classifies the
parts as "similar" and "organic," the former determined by their
material, the latter by the form which they assume. The similar
parts are divided into the sanguineous or rich in blood and the
spermatic. Both sets are further subdivided according to their
physical characters,[18] the latter, for
instance, into the hard, soft, and tensile tissues. The
classification resembles greatly that propounded by Aristotle,
though it is notably inferior in the details of its working
out.
020For Aristotle, as for all anatomists before
the days of the microscope, the tissues were not much more than
inorganic substances, differing from one another in texture, in
hardness, and other physical properties. They possessed indeed
properties, such as contractility, which were not inorganic, but as
far as their visible structure was concerned there was little to
raise them above the inorganic level. The application of the
microscope changed all that, for it revealed in the tissues an
organic structure as complex in its grade as the gross and visible
structure of the whole organism. Of the four men who first made
adequate use of the new aid, Malpighi, Hooke, Leeuenhoek, and
Swammerdam, the first-named contributed the most to make current
the new conceptions of organic structure. He studied in some detail
the development of the chick. He described the minute structure of
the lungs (1661), demonstrating for the first time, by his
discovery of the capillaries, the connection of the arteries with
the veins. In his work, De viscerum structura (1666), he
describes the histology of the spleen, the kidney, the liver, and
the cortex of the brain, establishing among other things the fact
that the liver was really a conglomerate gland, and discovering the
Malpighian bodies in the kidney. This work was done on a broad
comparative basis. "Since in the higher, more perfect, red-blooded
animals, the simplicity of their structure is wont to be involved
by many obscurities, it is necessary that we should approach the
subject by the observation of the lower, imperfect animals."[19] So he wrote in the De viscerum
structura, and accordingly he studied the liver first in the
snail, then in fishes, reptiles, mammals, and finally man. In the
introduction to his Anatome plantarum (1675), in which he
laid the foundations of plant histology, he vindicates the
comparative method in the following words:—"In the enthusiasm
of youth I applied myself to Anatomy, and although I was interested
in particular problems, yet I dared to pry into them in the higher
animals. But since these matters enveloped in peculiar mystery
still lie in obscurity, they require the comparison of simpler
conditions, and so the investigation of insects[20] 021at once attracted me; finally, since this
also has its own difficulties I applied my mind to the study of
plants, intending after prolonged occupation with this domain, to
retrace my steps by way of the vegetable kingdom, and get back to
my former studies. But perhaps not even this will be sufficient;
since the simpler world of minerals and the elements should have
been taken first. In this case, however, the undertaking becomes
enormous and far beyond my powers."[21]
There is something fine in this life of broad outlines, devoted
whole-heartedly to an idea, to a plan of research, which required a
lifetime to carry out.
An important histological discovery dating from this time is
that of the finer structure of muscle, made by Stensen (or Steno)
in 1664. He described the structure of muscle-fibres, resolving
them into their constituent fibrils.
To the microscope we owe not only histology but the comparative
anatomy of the lower animals. Throughout the 17th and 18th
centuries the discovery of structure in the lower animals went on
continuously, as may be read in any history of Zoology.[22] We content ourselves here with mentioning
only some representative names.
In the 17th century Leeuenhoek, applying the microscope almost
at random, discovered fact after fact, his most famous discovery
being that of the "spermatic animalcules."
Swammerdam studied the metamorphoses of insects and made
wonderfully minute dissections of all sorts of animals, snails and
insects particularly. He described also the development of the
frog. It is curious to see what a grip his conception of
metamorphosis had upon him when he 022homologises the stages of the
frog's development with the Egg, the Worm, and the Nymph of insects
(Book of Nature, p. 104, Eng. trans., 1785). He even speaks
of the human embryo as being at a certain stage a Man-Vermicle.
In the 18th century, Réaumur and Bonnet continued the
minute study of insects, laying more stress, however, on their
habits and physiology than upon their anatomy. Lyonnet made a most
laborious investigation of the anatomy of the willow-caterpillar
(1762). John Hunter (1728-93) dissected all kinds of animals, from
holothurians to whales. His interest was, however, that of the
physiologist, and he was not specially interested in problems of
form. It is interesting to note a formulation in somewhat confused
language of the recapitulation theory. The passage occurs in his
description of the drawings he made to illustrate the development
of the chick. It is quoted in full by Owen (J. Hunter,
Observations on certain Parts of the Animal Œconomy, with
Notes by Richard Owen. London, 1837. Preface, p. xxvi). We give
here the last and clearest sentence—"If we were to take a
series of animals from the more imperfect to the perfect, we should
probably find an imperfect animal corresponding with some stage of
the most perfect."
The tendency of the time was not towards morphology, but rather
to general natural history and to systematics, the latter under the
powerful influence of Linnæus (1707-1778). The former tendency is
well represented by Réaumur (1683-1757) with his
observations on insects, the digestion of birds, the regeneration
of the crayfish's legs, and a hundred other matters. To this
tendency belong also Trembley's famous experiments on Hydra (1744),
and Rösel von Rosenhof's Insektenbelustigungen
(1746-1761).
Bonnet (1720-1793) deserves special mention here, since in his
Traité d'Insectologie (1745), and more fully in his
Contemplation de la Nature (1764), he gives the most
complete expression to the idea of the Échelle des
êtres.
This idea seems to have taken complete possession of his
imagination. He extends it to the universe. Every world has its own
scale of beings, and all the scales when joined together form but
one, which then contains all the possible orders of perfection. At
the end of the Preface to his Traité 023d'Insectologie (Œuvres, i.,
1779) he gives a long table, headed "Idée d'une
Échelle des êtres naturels," and rather resembling a
ladder, on the rungs of which the following names
appear:—
Man. |
Shell Fish. |
Stones. |
Orang-utan. |
Tube-worms. |
Figured stones. |
Ape. |
Clothes-worms. |
Crystals. |
| |
|
|
Quadrupeds. |
Insectes. |
Salts. |
Flying squirrel. |
Gall insectes. |
Vitriols. |
Bat. |
Taenia. |
|
Ostrich. |
Polyps. |
Metals. |
| |
Sea Nettles. |
|
Birds. |
Sensitive plant. |
Half-metals. |
Aquatic birds. |
|
|
Amphibious birds. |
Plants. |
Sulphurs. |
Flying Fish. |
Lichens. |
Bitumens. |
| |
Moulds. |
|
Fish. |
Fungi, Agarics. |
Earths. |
Creeping fish. |
Truffles. |
Pure earth. |
Eels. |
Corals, and Coralloids. |
|
Water sepents. |
Lithophytes. |
Water. |
| |
Asbestos. |
|
Serpents. |
Talc, Gypsums. |
Air. |
Slugs. |
Selenites, Slates. |
|
Snails. |
|
Fire. |
| |
|
|
| |
|
More subtile matter. |
The nature of the transitional forms which he inserts between
his principal classes show very clearly his entire lack of
morphological insight—the transitions are functional. The
positions assigned to clothes-moths and corals are very curious!
The whole scheme, so fantastic in its details, was largely
influenced by Leibniz's continuity philosophy, and is in no way an
improvement on the older and saner Aristotelian scheme.
Robinet, in the fifth volume of his book De la nature
(1761-6), foreshadows the somewhat similar views of the German
transcendentalists. "All beings," he writes, "have been conceived
and formed on one single plan, of which they are the endlessly
graduated variations: this prototype is the human form, the
metamorphoses of which are to be considered as so many steps
towards the most excellent form of being."[23]
024The idea of a gradation of beings appears
also in Buffon (1707-1788), but here it takes more definitely its
true character as a functional gradation.[24] "Since everything in Nature shades into
everything else," he says, "it is possible to establish a scale for
judging of the degrees of the intrinsic qualities of every
animal."[25]
He is quite well aware that the groups of Invertebrates are
different in structural plan from the Vertebrates—"The animal
kingdom includes various animated beings, whose organisation is
very different from our own and from that of the animals whose body
is similarly constructed to ours."[26]
He limits himself to a consideration of the Vertebrates, deeming
that the economy of an oyster ought not to form part of his subject
matter! He has a clear perception of the unity of plan which reigns
throughout the vertebrate series.[27]
What is new in Buffon is his interpretation of the unity of plan.
For the first time we find clearly expressed the thought that unity
of plan is to be explained by community of origin.
Buffon's utterances on this point are, as is well known,
somewhat vacillating. The famous passage, however, which occurs in
his account of the Ass shows pretty clearly that Buffon saw no
theoretical objection to the descent of all the varied species of
animals from one single form. Once admit, he argues, that within
the bounds of a single family one species may originate from the
type species by "degeneration," then one might reasonably suppose
that from a single being Nature could in time produce all the other
organised beings.[28] Elsewhere,
e.g., in the discourse De la Dégéneration
des Animaux,[29] Buffon expresses
himself with more caution. He finds that it is possible to reduce
the two hundred species of quadrupeds which he has described to
025quite a small number of families "from which
it is not impossible that all the rest are derived."[30] Within each of the families the species
branch off from a parent or type species. This we may note is a
great advance on the linear arrangement implied in the idea of an
Échelle des êtres.[31]
It is a mistake to suppose that Buffon was par excellence a
maker of hypotheses. On the contrary he saw things very sanely and
with a very open mind. He expressly mentions the great difficulties
which one encounters in supposing that one species may arise from
another by "degeneration." How does it happen that two individuals
"degenerate" just in the right direction and to the right stage so
as to be capable of breeding together? How is it that one does not
find intermediate links between species? One is reminded of the
objections, not altogether without validity, which were made to the
Darwinian theory in its early days. I cannot agree with those who
think that Buffon was an out-and-out evolutionist, who concealed
his opinions for fear of the Church. No doubt he did trim his
sails—the palpably insincere "Mais non, il est certain, par
la révélation, que tous les animaux ont
également participé à la grace de la
création,"[32] following hard upon
the too bold hypothesis of the origin of all species from a single
one, is proof of it. But he was too sane and matter-of-fact a
thinker to go much beyond his facts, and his evolution doctrine
remained always tentative. One thing, however, he was sure of, that
evolution would give a rational foundation to the classification
which, almost in spite of himself, he recognised in Nature. If, and
only if, the species of one family originated from a single type
species, could families, be founded rationally, avec
raison.
Buffon was, curiously enough, rather unwilling to recognise any
systematic unit higher than the species. Strictly speaking there
are only individuals in Nature; but there 026are also groups of
individuals which resemble one another from generation to
generation and are able to breed together. These are
species—Buffon adheres to the genetic definition of
species—and the species is a much more definite unit than the
genus, the order, the class, which are not divisions imposed by us
upon Nature. Species are definitely discontinuous,[33] and this is the only discontinuity which
Nature shows us. Buffon put his views into practice in his
Histoire Naturelle, where he describes species after
species, never uniting them into larger groups. We have seen,
however, how the facts forced upon him the conception of the
"family."
Buffon was no morphologist. He left to Daubenton what one might
call the "dirty work" of his book, the dissection and minute
description of the animals treated.
But Buffon was a man of genius, and accordingly his ideas on
morphology are fresh and illuminating. Few naturalists have been so
free from the prejudices and traditions of their trade. He makes in
the Discours sur la Nature des Animaux[34] a distinction, which Bichat and Cuvier
later developed with much profit, between the "animal" and the
"vegetative" part of animals.[35]
The vegetative or organic functions go on continuously, even in
sleep, and are performed by the internal organs, of which the heart
is the central one. The active waking life of the animal, that part
of its life which distinguishes it from the plant, involves the
external parts—the sense-organs and the extremities. An
animal is, as it were, made up of a complex of organs performing
the vegetative functions, assimilation, growth, and reproduction,
surrounded by an envelope formed by the limbs, the sense-organs,
the nerves and the brain, which is the centre of this "envelope."[36] Animals may differ from one another
enormously in the external parts, particularly in the appendicular
skeleton, without showing any great difference in the plan and
arrangement of their internal organs. 027Quadrupeds, Cetacea, birds,
amphibians and fish are as unlike as possible in external form and
in the shape of their limbs; but they all resemble one another in
their internal organs. Let the internal organs change,
however—the external parts will change infinitely more, and
you will get another animal, an animal of a totally different
nature. Thus an insect has a most singular internal economy, and,
in consequence, you find it is in every point different from any
vertebrate animal.
In this contrast, on the whole justified, between the importance
of variations in the "vegetative" and variations in the "animal"
parts, one may see without doing violence to Buffon's thought, an
indication of the difference between homology and analogy. It is
usually in the external parts, in the organs by which the animal
adapts itself to its environment, that one meets with the greatest
number of analogical resemblances. This contrast of vegetative and
animal parts and their relative importance for the discovery of
affinities was at any rate a considerable step towards an analysis
of the concept of unity of plan.
To Xavier Bichat (1771-1802) belongs the credit of working out
in detail the distinction drawn by Aristotle and Buffon between the
animal and the vegetative functions. Bichat was not a comparative
anatomist; his interest lay in human anatomy, normal and
pathological. So his views are drawn chiefly from the consideration
of human structure.
He classifies functions into those relating to the individual
and those relating to the species. The functions pertaining to the
individual may be divided into those of the animal and those of the
organic life.[37] "I call animal
life that order of functions which connects us with surrounding
bodies; signifying thereby that this order belongs only to animals"
(p. lxxviii.). Its organs are the afferent and efferent nerves, the
brain, the sense-organs and the voluntary muscles; the brain is its
central organ. "Digestion, circulation, respiration, exhalation,
absorption, secretion, nutrition, calorification, or production of
animal heat, compose organic life, whose principal and central
organ is the heart" (p. lxxix.).
The contrast of the animal and the organic life runs 028through
all Bichat's work; it receives classical expression in his
Recherches Physiologiques sur la Vie et la Mort (1800). The
plant and the animal stand for two different modes of living. The
plant lives within itself, and has with the external world only
relations of nutrition; the animal adds to this organic life a life
of active relation with surrounding things (3rd ed., 1805, p. 2).
"One might almost say that the plant is the framework, the
foundation of the animal, and that to form the animal it sufficed
to cover this foundation with a system of organs fitted to
establish relations with the world outside. It follows that the
functions of the animal form two quite distinct classes. One class
consists in a continual succession of assimilation and excretion;
through these functions the animal incessantly transforms into its
own substance the molecules of surrounding bodies, later to reject
these molecules when they have become heterogeneous to it. Through
this first class of functions the animal exists only within itself;
through the other class it exists outside; it is an inhabitant of
the world, and not, like the plant, of the place which saw its
birth. The animal feels and perceives its surroundings, reflects
its sensations, moves of its own will under their influence, and,
as a rule, can communicate by its voice its desires and its fears,
its pleasures or its pains. I call organic life the sum of the
functions of the former class, for all organised creatures, plants
or animals, possess them to a more or less marked degree, and
organised structure is the sole condition necessary to their
exercise. The combined functions of the second class form the
'animal' life, so named because it is the exclusive attribute of
the animal kingdom" (pp. 2-3).
In both lives there is a double movement, in the animal life
from the periphery to the centre and from the centre to the
periphery, in the organic life also from the exterior to the
interior and back again, but here a movement of composition and
decomposition. As the brain mediates between sensation and motion,
so the vascular system is the go-between of the organs of
assimilation and the organs of dissimilation.
The most essential structural difference between the organs of
animal life and the organs of organic life is, in man and the higher
animals at least, the symmetry of 029the one set and the irregularity
of the other—compare the symmetry of the nerves and muscles
of the animal life with the asymmetrical disposition of the
visceral muscles and the sympathetic nerves, which belong to the
organic life.
Noteworthy differences exist between the two lives with respect
to the influence of habit. Everything in the animal life is under
the dominion of habit. Habit dulls sensation, habit strengthens the
judgment. In the organic life, on the contrary, habit exercises no
influence. The difference comes out clearly in the development of
the individual. The organs of the organic life attain their full
perfection independently of use; the organs of the animal life
require an education, and without education they do not reach
perfection (loc. cit., p. 127).
Bichat was the founder of what was known for a time as General
Anatomy—the study of the constituent tissues of the body in
health and disease. His classification of tissues was macroscopical
and physiological; he relied upon texture and function in
distinguishing them rather than upon microscopical structure. The
tissues he distinguished are as follows:—[38]
| 1. |
The cellular membrane. |
12. |
Fibro-cartilage. |
| 2. |
Nerves of animal life. |
13. |
Muscles of organic life. |
| 3. |
Nerves of organic life. |
14. |
Muscles of animal life. |
| 4. |
Arteries. |
15. |
Mucous membrane. |
| 5. |
Veins. |
16. |
Serous membrane. |
| 6. |
Exhalants. |
17. |
Synovial membrane. |
| 7. |
Absorbents and glands. |
18. |
The Glands. |
| 8. |
Bones. |
19. |
The Dermis. |
| 9. |
Medulla. |
20. |
Epidermis. |
| 10. |
Cartilage. |
21. |
Cutis. |
| 11. |
Fibrous tissue. |
|
|
The "cellular membrane" seems to mean undifferentiated
connective tissue; "exhalants" are imperceptible tubes arising from
the capillaries and secreting fat, serum, marrow, etc.; the
"absorbents and glands" are the lymphatics and the lymphatic
glands.
In Bichat's eyes this resolution of the organism into 030tissues
had a deeper significance than any separation into organs, for to
each tissue must be attributed a vie propre, an individual
and peculiar life. "When we study a function we must consider the
complicated organ which performs it in a general way; but if we
would be instructed in the properties and life of that organ we
must absolutely resolve it into its constituent parts."[39] The tissues have, too, a great importance
for pathology, for diseases are often diseases of tissues rather
than of organs.[40]
031
CHAPTER III
CUVIER
Cuvier was perhaps the greatest of
comparative anatomists; his work is, in the best sense of the word,
classical.
Like all his predecessors, like Aristotle, like the Italian
anatomists, Cuvier studied structure and function together, even
gave function the primacy.
Some functions, he says,[41]
are common to all organised bodies—origin by generation,
growth by nutrition, end by death. There are also secondary
functions. Of these the most important, in animals at least, are
the faculties of feeling and moving. These two faculties are
necessarily bound up together; if Nature has given animals
sensation she must also have given them the power of movement, the
power to flee from what is harmful and draw near to what is good.
These two faculties determine all the others. A creature that feels
and moves requires a stomach to carry food in. Food requires
instruments to divide it, liquids to digest it. Plants, which do
not feel and do not move, have no need of a stomach, but have roots
instead. Thus the "Animal Functions" of feeling and moving
determine the character of the organs of the second order, the
organs of digestion. These in their turn are prior to the organs of
circulation, which are a means to the end of distributing the
nutrient fluid or blood to all parts of the body. These organs of
the third order are not only dependent on those of the second
order, but are also not even necessary, for many animals are
without them. Only animals with a circulatory system can have
definite breathing organs—lungs or gills. 032Plants, and
animals without a circulation, breathe by their whole surface.
There is accordingly a rational order of functions, and
therefore of the systems of organs which perform them. The most
important are the Animal Functions, with their great organ-system,
the neuro-muscular mechanism. Then come the digestive functions,
and after them, and in a sense accessory to them, the functions and
organs of circulation and respiration. The last three may be
grouped as the Vital Functions.
The Animal Functions not only determine the character of the
Vital Functions, but influence also the primary faculty of
generation, for animals' power of movement has rendered their mode
of fecundation more simple, has therefore had an effect on their
organs of generation.
This division into "Animal" and "Vital" functions recalls
Buffon's and Bichat's distinction of the "animal" and the
"vegetative" lives. Cuvier apparently took this idea from Buffon,
for he says that a plant is an animal that sleeps.[42] But the idea is as old as Aristotle, who
discusses the "sleep" of embryos and of plants in the last book of
the De Generatione animalium. The distinction between animal
and vegetative life is, of course, based for Aristotle in the
difference between the ψυχή
ἀισθητική and the
ψυχή
θρέπτική. Cuvier, like
Aristotle, Buffon, and Bichat, makes the heart the centre of
the "vegetative" organs.
It is important to note that Cuvier puts function before
structure, and infers from function what the organ will be.
"Plants," he writes, "having few faculties, have a very simple
organisation."[43] It is only after having
discussed and classified functions that Cuvier goes on to examine
organs.
First his views on the composition of the animal body. Aristotle
distinguished three degrees of composition—the "elements,"
the homogeneous parts, and the heterogeneous parts or organs.
Cuvier does the same. Some small advance has been made in the two
thousand years' interval, due in the first place to the progress of
chemistry, and in the second to the invention of the microscope. To
the first circumstance Cuvier owes his knowledge that the inorganic
033substances forming the first degree of
composition are principally C, N, H, O, and P, combined to form
albumen, fibrine, and the like, which are in their turn combined to
form the solids and fluids of the body. To the latter circumstance
Cuvier owes the statement that the finest fragments into which
mechanical division can resolve the organism are little flakes and
filaments, which, joined up loosely together, form a "cellulosity."
The discovery of the true cellular nature of animal tissues did not
come till much later, till some years after Cuvier's death in 1832.
Knowledge of histological detail was, however, considerable by the
beginning of the 19th century. Cuvier knew, for example, that each
muscle fibre has its own nerve fibre. But he gives no elaborate
account of the homogeneous parts, no detailed histology. On the
other hand his treatment of the heterogeneous parts or organs is
detailed and masterly.[44]
The main systems of organs are, in order of importance, the
nervous and muscular, the digestive, the circulatory, and the
respiratory. Each organ or system of organs may have many forms. If
any form of any organ could exist in combination with any form of
all the others there would be an enormous number of combinations
theoretically possible. But these combinations do not all exist in
Nature, for organs are not merely assembled
(rapprochés), but act upon one another, and act all
together for a common end. Accordingly only the combinations that
fulfil these conditions exist in Nature. Cuvier thus dismisses the
question of a science of possible organic forms and considers only
the forms or combinations actually existing. This question of the
possibility of a "theoretical" morphology of living things, after
the fashion of the morphology of crystals with their sixteen
possible types, was raised in later years by K. G. Carus, Bronn, and
Haeckel.
Organisms, then, are harmonious combinations of organs, and the
harmony is primarily a harmony of functions. Every function depends
upon every other, and all are necessary. The harmony of organs and
their mutual dependence are the results of the interdependence of
function. This thought, the recognition of the functional unity of
the 034organism, is the fundamental one at the base
of all Cuvier's work. Before him men had recognised more or less
clearly the harmony of structure and function, and had based much
of their work upon this unanalysed assumption. Cuvier was the first
naturalist to raise this thought to the level of a principle
peculiar to natural history. "It is on this mutual dependence of
the functions and the assistance which they lend one to another
that are founded the laws that determine the relations of their
organs; these laws are as inevitable as the laws of metaphysics and
mathematics, for it is evident that a proper harmony between organs
that act one upon another is a necessary condition of the existence
of the being to which they belong."[45]
This rational principle, peculiar to natural history, Cuvier
calls the principle of the conditions of existence, for the
following reason:—"Since nothing can exist that does not
fulfil the conditions which render its existence possible, the
different parts of each being must be co-ordinated in such a way as
to render possible the existence of the being as a whole, not only
in itself, but also in its relations with other beings, and the
analysis of these conditions often leads to general laws which are
as certain as those which are derived from calculation or from
experiment."[46]
By "conditions of existence" he means something quite different
from what is now commonly understood. The idea of the external
conditions of existence, the environment, enters very little into
his thought. He is intent on the adaptations of function and organ
within the living creature—a point of view rather neglected
nowadays, but essential for the understanding of living things. The
very condition of existence of a living thing, and part of the
essential definition of it, is that its parts work together for the
good of the whole.
The principle of the adaptedness of parts may be used as an
explanatory principle, enabling the naturalist to trace out in
detail the interdependence of functions and their organs. When you
have discovered how one organ is adapted to another and to the
whole, you have gone a certain way towards understanding it. That
is 035using teleology as a regulative principle, in
Kant's sense of the word. Cuvier was indeed a teleologist after the
fashion of Kant, and there can be no doubt that he was influenced,
at least in the exposition of his ideas, by Kant's Kritik der
Urtheilskraft, which appeared ten years before the publication
of the Leçons d'Anatomie Comparée. Teleology
in Kant's sense is and will always be a necessary postulate of
biology. It does not supply an explanation of organic forms and
activities, but without it one cannot even begin to understand
living things. Adaptedness is the most general fact of life, and
innumerable lesser facts can be grouped as particular cases of it,
can be, so far, understood.
Cuvier's famous principle of correlation, the corner-stone of
his work, is simply the practical application to the facts of
structure of the principle of functional adaptedness. By the
principle of correlation, from one part of an animal, given
sufficient knowledge of the structure of its like, you can in a
general way construct the whole. "This must necessarily be so: for
all the organs of an animal form a single system, the parts of
which hang together, and act and re-act upon one another; and no
modifications can appear in one part without bringing about
corresponding modifications in all the rest."[47] The logical basis of the principle is
sound. The functions of the parts are all intimately bound up with
one another, and one function cannot vary without bringing in its
train corresponding modifications in the others. Structure and
function are bound up together; every modification of a function
entails therefore the modification of an organ. Hence from the
shape of one organ you can infer the shape of the other
organs—if you have sufficiently extensive empirical knowledge
of functions, and of the relation of structure to function in each
kind of organ. Given an alimentary canal capable of digesting only
flesh, and possessing therefore a certain form, you know that the
other functions must be adapted to this particular function of the
alimentary canal. The animal must have keen sight, fine smell,
speed, agility, and strength in paws and jaws. These particular
functions must have correspondingly modified organs, 036well-developed
eyes and ears, claws and teeth. Further, you know from experience
that such and such definitely modified organs are invariably found
with the carnivorous habit, carnassial teeth, for example, and
reduced clavicles. From a "carnivorous" alimentary canal, then, you
can infer with certainty that the animal possessed carnassial teeth
and the other structural peculiarities of carnivorous animals,
e.g., the peculiar coronoid process of the mandible. From
the carnassial tooth you can infer the reduced clavicle, and so on.
"In a word, the form of the tooth implies the form of the condyle;
that of the shoulder blade that of the claws, just as the equation
of a curve implies all its properties."[48]
Similarly the great respiratory power of birds is correlated
with their great muscular strength, and renders necessary great
digestive powers. Hence the correlated structure of lungs, muscles
and their attachments, and alimentary canal, in birds.
Not only do systems of organs, by being adjusted to special
modifications of function, influence one another, but so also do
parts of the same organ. This is noticeably the case with the
skeleton, where hardly a facet can vary without the others varying
proportionately, so that from one bone you can up to a certain
point deduce all the rest.
We deduce the necessity, the constancy, of these co-existences
of organs from the observed reciprocal influence of their
functions. That being established, we can argue from observed
constancy of relation between two organs an action of one upon the
other, and so be led to a discovery of their functions. But even if
we do not discover the functional interdependencies of the parts,
we can use the established fact of the constant co-existence of two
parts as proof of a functional correlation between them.
Correlation is either a rational or an empirical principle,
according as we know or do not know the interdependence of function
of which it is the expression. Even when we apply the rational
principle of correlation it would be useless in our hands if we had
not extensive empirical knowledge; when we use an empirical rule of
correlation we depend entirely upon observation. "There are a great
many cases," writes 037Cuvier,[49]
"where our theoretical knowledge of the relations of forms would
not suffice, if it were not filled out by observation," that is to
say, there are many cases of correlation not yet explicable in
terms of function. From a hoof you can deduce the main characters
of herbivores (with a certain amount of assistance from your
empirical knowledge of herbivores), but could you from a cloven
hoof deduce that the animal is a ruminant, unless you had observed
the constancy of relation, not directly explicable in terms of
function, between cloven hoofs and chewing the cud? Or could you
deduce from the existence of frontal horns that the animal
ruminates? "Nevertheless, since these relations are constant, they
must necessarily have a sufficient cause; but as we are ignorant of
this cause, observation must supplement theory; observation
establishes empirical laws which become almost as certain as the
rational laws, when they are based upon a sufficient number of
observations.... But that there exist all the same hidden reasons
for all these relations is partly revealed by observation itself,
independently of general philosophy."[50]
That is to say, even correlations for which no explanation in terms
of function can be supplied are probably in reality functional
correlations. This may, in some cases, be inferred from the graded
correspondence of two sets of organs. For example, ungulates which
do not ruminate, and have not a cloven hoof, have a more perfect
dentition and more bones in the foot than the true cloven-hoofed
ruminants. There is a correlation between the state of development
of the teeth and of the foot. This correlation is a graded one, for
camels, which have a more perfect dentition than other ruminants,
have also a bone more in their tarsus. It seems probable,
therefore, that there is some reason, that is, some explanation in
terms of function, for this case of correlation.
Nevertheless, the fact remains that many correlations are not
explicable in terms of function, and the substitution of
correlation as an empirical principle for correlation as a rational
principle marks for Cuvier a step away from his functional
comparative anatomy towards a pure morphology. It is significant
that in later times the term correlation 038has come to be applied more
especially to the purely empirical constancies of relation, and has
lost most of its functional significance. But the correlation of
the parts of an organism is no mere mathematical concept, to be
expressed by a coefficient, but something deeper and more
vital.
Cuvier interpreted the functional dependence of the parts in
terms of what we now call the general metabolism. He had a clear
vision of the constant movement of molecules in the living tissue,
combining and recombining, of the organism taking in and
intercalating molecules from outside from the food and rejecting
molecules in the excretions, a ceaseless tourbillon vital.
"This general movement, universal in every part, is so unmistakably
the very essence of life that parts separated from a living body
straightway die."[51] The organisation of
the body, the arrangement of its solids and liquids, is adapted to
further the tourbillon vital. "Each part contributes to this
general movement its own particular action and is affected by it in
particular ways, with the result that, in every being, life is a
unity which results from the mutual action and reaction of all its
parts."[52]
Cuvier, however, did not resolve life into metabolism, nor
reduce vital happenings to the chemical level. The form of
organised bodies is more essential than the matter of which they
are composed, for the matter changes ceaselessly while the form
remains unchanged. It is in form that we must seek the differences
between species, and not in the combinations of matter, which are
almost the same in all.[53]
The differences are to be sought at the level of the second and
third degrees of composition.
The existence of differences of form introduces a new problem,
the problem of diversity. There are only a few possible
combinations of the principal organs, but as you get down to less
important parts the possible scope of variation is greatly
increased, and most of the possible variations do exist. Nature
seems prodigal of form, of form which needs not to be useful in
order to exist. "It needs only to be possible, i.e., of
039such
a character that it does not destroy the harmony of the whole."[54] We seize here the relation of the
principle of the adaptedness of parts to the problem of the variety
of form. The former is in a sense a regulative and conservative
principle which lays down limits beyond which variation may not
stray. In itself it is not a fountain of change; there must be
another cause of change. This thought is of great importance for
theories of descent.
Cuvier has no theory to account for the variety of form: he
contents himself with a classification. There are two main ways of
classifying forms; you may classify according to single organs or
according to the totality of organs. By the first method you can
have as many classifications as you have organs, and the
classifications will not necessarily coincide. Thus you can divide
animals according to their organs of digestion into two classes,
those in which the alimentary canal is a sac with one opening
(zoophytes) and those in which the canal has two openings,[55] a curious forestalment, in the rough, of
the modern division of Metazoa into Cœlentera and
Cœlomata.
It is only by taking single organs that you can arrange animals
into long series, and you will have as many series as you take
organs. Only in this way can you form any Échelle des
êtres or graded series; and you can get even this kind
of gradation only within each of the big groups formed on a common
plan of structure; you can never grade, for example, from
Invertebrates to Vertebrates through intermediate forms[56] (which is perfectly true, in spite of
Amphioxus and Balanoglossus!).
In the Règne Animal Cuvier restricts the
application of the idea of the Échelle within even
narrower limits, refusing to admit its validity within the bounds
of the vertebrate phylum, or even within the vertebrate classes.
This seems, however, to refer to a seriation of whole organisms and
not of organs, so that the possibility of a seriation of organs
within a class is not denied. Cuvier was, above all, a positive
spirit, and he looked askance at all speculation which went beyond
the facts. "The pretended scale of beings," he wrote, "is only
040an
erroneous application to the totality of creation of partial
observations, which have validity only when confined to the sphere
within which they were made."[57]
This remark, which is after all only just, perfectly expresses
Cuvier's attitude to the transcendental theories, and was probably
a protest against the sweeping generalisations of his colleague,
Etienne Geoffroy St Hilaire.
A true classification should be based upon the comparison of all
organs, but all organs are not of equal value for classification,
nor are all the variations of each organ equally important. In
estimating the value of variations more stress should be laid on
function than on form, for only those variations are important
which affect the mode of functioning. These are the principles on
which Cuvier bases the classification of animals given in the
Leçons, Article V., "Division des animaux
d'après l'ensemble de leur organisation." The scheme of
classification actually given in the Leçons recalls
curiously that of Aristotle, for there is the same broad division
into Vertebrates, with red blood, and Invertebrates, almost all
with white blood. Nine classes altogether are
distinguished—Mammals, Birds, Reptiles, Fishes, Molluscs,
Crustacea, Insects, Worms, Zoophytes (including Echinoderms and
Cœlenterates).
A maturer theory and practice of classification is given in the
Règne Animal of seventeen years later. Here the
principle of the subordination of characters (which seems to have
been first explicitly stated by the younger de Jussieu in his
Genera Plantarum, 1789,[58]) is more clearly recognised. The properties or
peculiarities of structure which have the greatest number of
relations of incompatibility and coexistence, and therefore
influence the whole in the greatest degree, are the important or
dominating characters, to which the others must be subordinated in
classification. These dominant characters are also the most
constant.[59] In deciding which
characters are the most important Cuvier makes use of his
fundamental classification of functions and organs into two main
sets. "The heart and the organs of circulation are 041a kind of centre
for the vegetative functions, as the brain and the spinal cord are
for the animal functions."[60]
These two organ-systems vary in harmony, and their characters must
form the basis for the delimitation of the great groups. Judged by
this standard there are four principal types of form,[61] of which all the others are but
modifications. These four types are Vertebrates, Molluscs,
Articulates, and Radiates. The first three have bilateral, the last
has radial symmetry. Vertebrates and Molluscs have blood-vessels,
but Articulates show a functional transition from the blood-vessel
to the tracheal system. Radiates approach the homogeneity of
plants; they appear to lack a distinct nervous system and sense
organs, and the lowest of them show only a homogeneous pulp which
is mobile and sensitive. All four classes are principally
distinguished from one another by the broad structural relations of
their neuromuscular system, of the organs of the animal functions.
Vertebrates have a spinal cord and brain, an internal skeleton
built on a definite plan, with an axis and appendages; in Molluscs
the muscles are attached to the skin and the shell, and the nervous
system consists of separate masses; Articulates have a hard
external skeleton and jointed limbs, and their nervous system
consists of two long ventral cords; Radiates have ill-defined
nervous and muscular systems, and in their lowest forms possess the
animal functions without the animal organs.
This well-rounded classification of animal forms is in a sense
the crown of Cuvier's work, for the principle of the subordination
of characters, in the interpretation which he gives to it, is a
direct application of his principle of functional correlation. Each
of the great groups is built upon one plan. The idea of the unity
of plan has become for Cuvier a commonplace of his thought, and it
is tacitly recognised in all his anatomical work. But he never
takes it as a hard-and-fast principle which must at all costs be
imposed upon the facts.
Cuvier has become known as the greatest champion of the fixity
of species, but it is not often recognised that his 042attitude to this
problem is at least as scientific as that of the evolutionists of
his own and later times. No doubt he became dogmatic in his
rejection of evolution-theory, but he was on sure ground in
maintaining that the evolutionists of his day went beyond their
facts. He considered that certain forms (species) have reproduced
themselves from the origin of things without exceeding the limits
of variation. His definition of a species was, "the individuals
descended from one another or from common parents, together with
those that resemble them as much as they resemble one another."[62] "These forms are neither produced nor do
they change of themselves; life presupposes their existence, for it
cannot arise save in organisations ready prepared for it."[63]
He based his rejection of all theories of descent upon the
absence of definite evidence for evolution. If species have
gradually changed, he argued, one ought to find traces of these
gradual modifications.[64]
Palæontology does not furnish such traces. Again, the limits
of variation, even under domestication, are narrow, and the most
extreme variation does not fundamentally alter the specific type.
Thus the dog has varied perhaps most of all, in size, in shape, in
colour. "But throughout all these variations the relations of the
bones remain the same, and the form of the teeth never changes to
an appreciable extent; at most there are some individuals in which
an additional false molar develops on one side or the other."[65] This second objection is the objection of
the morphologist. It would be an interesting study to compare
Cuvier's views on variation with those of Darwin, who was
essentially a systematist.
Cuvier's first objection was of course determined to some extent
by the imperfection of the palæontological knowledge of his
time. But even at the present day the objection has a certain
force, for although we have definite evidence of many serial
transformations of one species into another along a single line,
for example, Neumayr's Paludina series, 043yet at any one
geological level the species, the lines of descent, are all
distinct from one another.[66]
Cuvier recognised very clearly that there is a succession of
forms in time, and that on the whole the most primitive forms are
the earliest to appear. Mammals are later than reptiles, and fishes
appear earlier than either. As Depéret puts it, "Cuvier not
only demonstrated the presence in the sedimentary strata of a
series of terrestrial faunas superimposed and distinct, but he was
the first to express, and that very clearly, the idea of the
gradual increase in complexity of these faunas from the oldest to
the most recent" (p. 10).
He did not believe that the fauna of one epoch was transformed
into the fauna of the next. He explained the disappearance of the
one by the hypothesis of sudden catastrophes, and the appearance of
the next by the hypothesis of immigration. He nowhere advanced the
hypothesis of successive new creations. "For the rest, when I
maintain that the stony layers contain the bones of several genera
and the earthy layers those of several species which no longer
exist, I do not mean that a new creation has been necessary to
produce the existing species, I merely say that they did not exist
in the same localities and must have come thither from
elsewhere."[67] It was left to d'Orbigny
to teach the doctrine of successive creations, of which he
distinguished twenty-seven (Cours élémentaire de
palaeontologie stratigraphique, 1849).
Cuvier, however, can hardly have believed that all species were
present at the beginning, since he does admit a progression of
forms. Probably he had no theory on the subject, for theories
without facts had little interest for him. At any rate it is a
mistake to think that Cuvier was a supporter of the theological
doctrine of special creation. His philosophy of Nature was
mechanistic, and he dedicated his Recherches sur les Ossemens
Fossiles to his friend Laplace. He admitted the idea of
evolution at least so far as to conceive of a development of man
from a savage 044to a civilised state.[68] He refused to accept the extravagant
evolutionary theory of Demaillet and the somewhat confused theory
of Lamarck (whom he joins with Demaillet),[69] just as he rejected the transcendental theories
of Geoffroy St Hilaire, because they seemed to him not based upon
facts.
045
CHAPTER IV
GOETHE
Science, in so far as it rises above
the mere accumulation of facts, is a product of the mind's creative
activity. Scientific theories are not so much formulæ
extracted from experience as intuitions imposed upon experience. So
it was that Goethe, who was little more than a dilettante,[70] seized upon the essential principles of a
morphology some years before that morphology was accepted by the
workers.
Goethe is important in the history of morphological method
because he was the first to bring to clear consciousness and to
express in definite terms the idea on which comparative anatomy
before him was based, the idea of the unity of plan. We have seen
that this idea was familiar to Aristotle and that it was recognised
implicitly by all who after him studied structure comparatively. In
Goethe's time the idea had become ripe for expression. It was used
as a guiding principle in Goethe's youth particularly by Vicq
d'Azyr and by Camper. The former (1748-1794), who discovered[71] in the same year as Goethe (1784) the
intermaxillary bone in man, pointed out the homology in structure
between the fore limb and the hind limb, and interpreted certain
rudimentary bones, the intermaxillaries and rudimentary clavicles,
in the light of the theory that Vertebrates are built upon one
single plan of structure.
"Nature seems to operate always according to an original and
general plan, from which she departs with regret and 046whose traces we
come across everywhere" (Vicq d'Azyr, quoted by Flourens,
Mém. Acad. Sci., XXIII., p. xxxvi.).
Peter Camper (1722-1789), we are told by Goethe himself in his
Ostéologie, was convinced of the unity of plan holding
throughout Vertebrates; he compared in particular the brain of
fishes with the brain of man.
The idea of the unity of plan had not yet become limited and
defined as a strictly scientific theory; it was an idea common to
philosophy, to ordinary thought, and to anatomical science. We find
it expressed by Herder (who perhaps got it from Kant) in his
Ideen zur Philosophie der Geschichte der Menschheit (1784),
and it is possible that Goethe became impressed with the importance
of the idea through his conversations with Herder. Be that as it
may, it is certain that Goethe sought for the intermaxillaries in
man only because he was firmly convinced that the skeleton in all
the higher animals was built upon one common plan and that
accordingly bones such as the intermaxillaries, found well
developed in some animals, must also be found in man. The idea was
not drawn from the facts, but the facts were interpreted and even
sought for in the light of the idea. "I eagerly worked upon a
general osteological scheme, and had accordingly to assume that all
the separate parts of the structure, in detail as in the whole,
must be discoverable in all animals, because on this supposition is
built the already long begun science of comparative anatomy."[72]
The principle comes to clear expression in his Erster Entwurf
einer allgemeinen Einleitung in die vergleichende Anatomie
(1795).[73] He writes:—"On this
account an attempt is here made to arrive at an anatomical type, a
general picture in which the forms of all animals are contained in
potentia, and by means of which we can describe each animal in an
invariable order."[74] His aim is to
discover a general scheme of the constant in organic parts, a
scheme into which all animals will fit equally well, and no animal
better than the rest. When we remember that the type to which
anatomists before him had, consciously or unconsciously, referred
all 047other structure was man himself, we see that
in seeking after an abstract generalised type Goethe was reaching
out to a new conception. The fact that only the structure of man
and the higher animals was at all well-known in his time led Goethe
to think that his general Typus would hold for the lower animals as
well, though it was to be arrived at primarily from a study of the
higher animals. All he could assert of the entire animal kingdom
was that all animals agreed in having a head, a middle part, and an
end part, with their characteristic organs, and that accordingly
they might, in this respect at least, be reduced to one common
Typus. Goethe's knowledge of the lower animals was not
extensive.
Though Goethe did not work out a criterion of the homology of
parts with any great clearness, he had an inkling of the principle
later developed by E. Geoffroy St Hilaire, and called by him the
"Principle of Connections." According to this principle, the
homology of a part is determined by its position relative to other
parts. Goethe expresses it thus:—"On the other hand the most
constant factor is the position in which the bone is invariably
found, and the function to which it is adapted in the organic
edifice."[75] But from this sentence it
is not clear that Goethe understood the principle as one of form
independent of function, for he seems to consider that the homology
of an organ is partly determined by the function which it performs
for the whole. He wavers between the purely formal or morphological
interpretation of the principle of connections and the functional.
We find him in the additions to the Entwurf (1796),
saying:—"We must take into consideration not merely the
spatial relations of the parts, but also their living reciprocal
influence, their dependence upon and action on one another."[76] But in seeking for the intermaxillary bone
in man he was guided by its position relative to the
maxillaries—it must be the bone between the anterior ends of
the maxillaries, a bone whose limits are indicated in the adult
only by surface grooves.
As a matter of fact Goethe's morphological views are neither
very clearly expressed nor very consistent. This 048comes out in his
treatment of the relation between structure and function. Sometimes
he takes the view that structure determines function. "The parts of
the animal," he writes, "their reciprocal forms, their relations,
their particular properties determine the life and habits of the
creature."[77] We are not to explain, he
says, the tusks of the Babirussa by their possible use, but
we must ask how it comes to have tusks. In the same way we must not
suppose that a bull has horns in order to gore, but we must
investigate the process by which it comes to have horns to gore
with. This is the rigorous morphological view. On the other hand he
admits elsewhere that function may influence form. Apparently he
did not work out his ideas on this point to logical clearness, and
Rádl[78] is probably correct in
saying that the following quotation with its double assertion
represents most nearly Goethe's position:—
"Also bestimmt die Gestalt die Lebensweise des Thieres, Und die
Weise zu leben, sie wirkt auf alle Gestalten Mächtig
zurück."[79]
His best piece of purely morphological work was his theory of
the metamorphosis of plants. Stripped of its vaguer elements, and
of the crude attempt to explain differences in the character of
plant organs by differences in the degree of "refinement" of the
sap supplied to them, the theory is that stem-leaves, sepals,
petals, and stamens are all identical members or appendages. These
appendages differ from one another only in shape and in degree of
expansion, stem-leaves being expanded, sepals contracted, petals
expanded, and so on alternately. It is equally correct to call a
stamen a contracted petal, and a petal an expanded stamen, for no
one of the organs is the type of the others, but all equally are
varieties of a single abstract plant-appendage.
What Goethe considered he had proved for the appendages of
plants he extended to all living things. Every living thing is a
complex of living independent beings, which "der 049Idee, der Anlage
nach," are the same, but in appearance may be the same or similar,
different or unlike.[80] Not only is there a
primordial animal and a primordial plant, schematic forms to which
all separate species are referable, but the parts of each are
themselves units, which "der Idee nach," are identical inter
se. This fantasy can hardly be taken seriously as a scientific
theory; it seems, however, to have been what guided Goethe in his
"discovery" of the vertebral nature of the skull. Just as the fore
limb can be homologised with the hind limb, so, reasoning by
analogy, the skull should be capable of being homologised with the
vertebræ. To what ludicrous extremes this doctrine of the
repetition of parts within the organism was pushed we shall see
when we consider the theories of the German transcendentalists of
the early nineteenth century.
Though Goethe's morphological views were lacking in definiteness
he hit upon one or two ideas which proved useful. Thus he
enunciated the "law of balance" long before Etienne Geoffroy St
Hilaire, the law "that to no part can anything be added, without
something being taken away from another part, and vice
versa."[81] He saw, too, what a help
to the interpretation of adult structure the study of the embryo
would be, for many bones which are fused in the adult are separate
in the embryo.[82] This also was a point to
which the later transcendentalists gave considerable attention.
So far we have spoken of Goethe as if he were merely the prophet
of formal morphology; we have pointed out how he brought to clear
expression the morphological principle implicit in the idea of
unity of type, and how he seized upon some important guiding ideas,
such as the principle of connections. But Goethe was not a
formalist, and he was very far from the static conception of life
which is at the base of pure morphology. His interest was not in
Gestalt or fixed form, Bildung or form change. He saw
that Gestalt was but a momentary phase of Bildung,
and could be considered apart and in itself only by an abstraction
fatal to all understanding of the living thing. Mephistopheles
050scoffs at the scholars who would explain a
living creature by anatomising it:
"Dann hat er die Theile in seiner Hand,
Fehlt leider! nur das geistige Band."
[83]
Goethe kept clear of this mistake; he knew that the artist comes
nearer to the truth than the analyst.
In the fragment entitled Bildung und Umbildung organischer
Naturen (1807), introductory to a reprint of his paper on the
"Metamorphosis of Plants," we get an exposition of his general
views on living things. He points out there how we try to
understand things by separating them into their parts. We can, it
is true, resolve the organism into its structural elements, but we
cannot recompose it or endow it with life by joining up the parts.
Hence we require some other means of understanding it. "In all ages
even among scientific men there can be discerned a yearning to
apprehend the living form as such, to grasp the connection of their
external visible parts, to interpret them as indications of the
inner activity, and so, in a certain measure, to master the whole
conceptually." This science which should discover the inner meaning
of organic Bildung is called Morphology.[84] In Morphology we should not speak of
Gestalt or fixed form, or if we do we should understand by
it only a momentary phase of Bildung. Form is of interest
not in itself but only as the manifestation of the inner activity
of the living being. Over development, he says elsewhere, there
presides a formative force, a bildende Kraft or
Bildungstrieb, which works out the idea of the organism.
Living things, in his view of them, strive to manifest an idea.
They are Nature's works of art—and so, incidentally, they
require an artist to interpret them.
This profound conception of the nature of life is applied not
only to the growing changing individual but also to the whole
changing world of organisms. They are all manifestations of a
living shaping power which moulds them. This shaping power,
immanent in all life, is conceived to work according to a general
plan, and so we get an explanation of 051the fact that living things
seem simply varieties of one common type.
"If we once recognise," says Goethe, "that the creative spirit
brings into being and shapes the evolution of the more perfect
organic creatures according to a general scheme, is it altogether
impossible to represent this original plan if not to the senses at
least to the mind...?"[85]
Such an interpretation of the unity of plan reaches perhaps
beyond the bounds of science.
052
CHAPTER V
ETIENNE GEOFFROY SAINT-HILAIRE
E. Geoffrey made an experiment, unsuccessful but instructive. He
tried to found a science of pure morphology; he failed: his failure
showed, once and for all, that a pure morphology of organic forms
is impracticable.
Already, in 1796, in one of his earliest memoirs,[86] Geoffroy was guided by the idea that
Nature has formed all living things upon one plan. Organs which
seem anomalous are merely modifications of the normal; the trunk of
an elephant is formed by the excessively prolonged nostrils, the
horn of a rhinoceros is simply a mass of adhering hairs. In
general, however varied their form, all organs are simply
variations of a common scheme; Nature employs no new organs. Organs
which are rudimentary, such as the clavicles in the ostrich and the
nictitating membrane in man, bear witness to the unity of plan. In
this Geoffroy goes no further than his predecessors. They too had
recognised homologies of organs; they too had interpreted
rudimentary organs as vestiges of an original plan.
In a series of papers published in 1807, Geoffroy took a further
step, and sought to establish homologies which were not
obvious—homologies, too, not so much of organs as of
parts.
These memoirs (published in the Annales du Muséum
d'Histoire naturelle, vols. ix. and x., 1807) dealt with the
homology between the bones of the pectoral fin and girdle in fish
and the bones of the arm and shoulder-girdle in higher 053Vertebrates,
with the homologies of the bones of the sternum, and with the
determination of the pieces of the skull, particularly in the
crocodile. All Geoffroy's morphological doctrine is found in them,
but for the full expression of his views we must take his chief
work, the Philosophie anatomique, particularly the first
volume (1818). This volume contains, beside the important "Discours
préliminaire" and "Introduction" which we shall presently
consider in detail, five memoirs, which deal with the various bones
connected with the respiratory organs in fishes (the bones of the
operculum, of the hyoid, of the branchial arches, of the pectoral
girdle), and seek to discover their homologies with corresponding
bones in air-breathing Vertebrates.
"Can the organisation of vertebrated animals be referred to one
uniform type?" This is the question with which the Philosophie
anatomique opens, the question to which the whole book is an
answer. But is it not generally acknowledged by naturalists that
Vertebrates are built upon one uniform plan, that, for instance,
the fore limb may be modified for running, climbing, swimming, or
flying, yet the arrangement of the bones remain the same? How else
could there be a "natural method" of classification?[87]
But the homologies so drawn repose upon a vague and confused
feeling for likenesses; they are not based upon an explicit
principle. What general principle can be applied? "Now it is
evident that the sole general principle one can apply is given by
the position, the relations, and the dependencies of the parts,
that is to say, by what I name and include under the term of
connections." For instance, the part known as the hand in
man and generally as the fore foot in other Vertebrates, is the
fourth part in order in the anterior member, and its homologue can
always be recognised by this fact of its connections (p. xxvi.).
The principle of connections serves as a guide in tracing an organ
through all its functional transformations, for "an organ can be
deteriorated, atrophied, annihilated, but not transposed" (p.
xxx.).
It is this principle which enables one to follow out in detail
the further fundamental conception that in every 054Vertebrate there
are found the same "organic materials," or units of construction.
This conception, which Geoffroy calls the Théorie des
analogues (p. xxxii.), is clearly one part of the old idea of
the unity of type; it teaches the unity of composition of
organic beings, while the Principe des connexions adds the
unity of plan.
Both conceptions are logically implicit in the vague notion of
unity of type; Geoffroy disengaged them, and pushed each to its
logical extreme.
Most of the ordinary homologies of structure in air-breathing
Vertebrates have already been seized, he continues, for they are
more or less obvious, and many intermediate states exist (p.
xxxiv.). But ordinary methods of comparison fail when the attempt
is made to homologise the structure of fishes with that of
air-breathing Vertebrates, for the homologies are anything but
obvious and no intermediate organs are found.
Most air-breathing Vertebrates have a larynx, a trachea, and
bronchi, which are absent in fish; and fish have many parts which
seem to be absent in higher Vertebrates. But apply the "Theory of
Analogues"; it teaches that there can be no organ peculiar to fish
and not found in other Vertebrates; apply the "Principle of
Connections," it will show which organs are homologous in the two
types (p. xxxv.).
Comparative anatomists, with few exceptions, had hitherto taken
man as the type, and referred all structure to his; Geoffroy's
principles led him to give preference to no one animal in
particular, but to seize upon each part in the species in which it
reaches the maximum of its development (p. xxxvi.). He is thus led
to refer all structures to a generalised abstract type. In this
abstract type each organ exists at the maximum of its development,
each organ shows all its potentialities realised. In a way,
therefore, this type, this abstraction, gives the scheme of the
possible transformations of each organ.
It is true Geoffroy does not refer to this "Archetype" in so
many words, but it must always have been vaguely present in his
mind. He has this idea in his head when he says in one of his later
works, "There is, philosophically 055speaking, only a single animal."[88] The "single animal" is simply the
generalised type.
Having laid down his two principles Geoffroy goes on to apply
them to the difficult case of the comparison of the skeleton of
fish with the skeleton of the higher Vertebrates. "My present task
is to demonstrate that there is no part of the bony framework of
fishes that cannot find its analogue in the other vertebrated
animals."[89] It seems at first sight
that many bones are peculiar to fish, formed expressly for
performing the functions which fish do not share with higher
animals. These are the bones connected with respiration—the
operculum, the branchiostegal rays, the branchial arches, and
others. That the peculiar bones should be connected with the
respiratory functions is only natural, for the contrast between
fish and higher Vertebrates is essentially a contrast between
water-breathing and air-breathing animals. Considering first the
general form of the skeleton in fish, we are met at once with a
difficulty; there is no obvious homologue in fishes of the neck,
the trunk, and the abdomen of higher animals. What apparently
corresponds to the trunk is in fishes crowded close up under the
head. But, after all, it is not of the essence of the vertebrate
type to have the trunk and the abdomen attached at definite and
invariable distances along the vertebral column—that is a
notion surviving from the anatomy which made man its type. The
"trunk" differs in position according to the class, in quadrupeds,
birds, and fishes (p. 9). Now, says Geoffroy, allow me this one
hypothesis, that the trunk with its organs can, as it were, move
bodily along the vertebral column, so as to be found in one class
near the front end of the vertebral column, in another about the
middle, and in a third near the end, then I can show you in detail
that the constituent parts of this trunk are found in all classes
to be invariably in the same positions relatively to one another
(p. 10). It is important to note this hypothesis of a "metastasis"
which Geoffroy makes, for it is the key to the understanding of
many of the far-fetched homologies which he tries to establish. It
is, of course, clear that this hypothesis is in formal
contradiction 056with his principal hypothesis of the
invariability of connections, and that he, so to speak, gets a hold
on his fish to apply his principle of connections only by admitting
at the very outset an exception to his primary principle. A further
application of the hypothesis of metastasis will be noticed below
in connection with the determination of the sternum of fishes. We
note here an interpretation of the first metastasis in terms of
functional adaptation. "The constant and violent action of the
tail, if it does not go so far as actually to displace and move
forward the internal organs, at least fits in well with an
arrangement in which the organs are so disposed" (p. 99).
The first memoir deals with the homologies of the opercular
bones. Geoffroy considers that the external opening of the ear
corresponds to the external opening of the gill-chamber, which lies
between the operculum and the pectoral girdle. The ear communicates
with the buccal cavity by the Eustachian tube, so does the
branchial chamber by means of the gill-slits. The auditory chamber
of higher Vertebrates is, therefore, the homologue of the branchial
chamber in fish; the opercular bones in fish and the ossicles of
the ear in other Vertebrates stand in close relation to this
chamber; therefore the opercular bones are the homologues of the
ossicles of the ear, the interoperculum corresponding to the
malleus, the suboperculum to the lenticular, the minute lower part
of the suboperculum to the incus, the operculum to the stapes, and
the pre-operculum to the tympanic ring. In making these particular
determinations Geoffroy professes to be led by his principle of
connections. The pre-operculum has, he says, the same connections
with neighbouring bones as the tympanic bone in other Vertebrates,
and the other pieces of the gill-cover are homologised with
particular ear-ossicles according to the order in which they stand
to one another. The second memoir in the book deals with the
sternum, and affords a very good example of Geoffroy's method of
dealing with the facts of structure. We shall omit here any
detailed reference to the other three memoirs, which deal with the
hyoid, with the branchial arches and the structures which
correspond in air-breathing Vertebrates, and with the bones of the
shoulder-girdle.
057In the memoir on the sternum Geoffroy's first
care is to arrive at a definition of what a sternum is. He defines
it partly by its functions, partly by its connections, as the
system of bones which covers and protects the thorax, and gives
attachment to certain groups of muscles.
The most highly developed sternum (according to this definition)
is the plastron of the tortoise, whose structure it dominates (p.
103). It is important, therefore, to determine of how many bones
the plastron is composed, since the full number of elementary parts
of which an organ is composed is best seen when the organ is at the
maximum of its development. There are nine bones in the plastron of
the tortoise. "The conclusion to be drawn from this is that every
sternum, provided that it is not inhibited in its development by
some obstacle, is composed of nine elementary parts" (p.
105). These nine bones are in Geoffroy's nomenclature, the
episternals, the hyosternals, the hyposternals, the xiphisternals,
which are all paired bones, and the entosternal, which is unpaired.
The arrangement of them is in the tortoise:—
The articulations in the tortoise are indicated by the
connecting lines. Geoffroy tries to show that the sternum in other
animals is composed of these nine bones, or at least of a certain
number of them, always in the same invariable relative positions.
Thus in birds the sternum consists of five pieces, of a huge keeled
entosternal, and of two "annexes" on either side, which are the
hyo-and hyposternals. These are separate only in young birds.
Occasionally, especially in 058young birds, rudiments of episternals and
xiphisternals also occur. The minuteness of the episternals and the
xiphisternals
Fig 1.—Hyoid Arch of the Conger.(Original)
may be attributed to the gigantic size of the entosternal, in
accordance with the Loi de balancement. In the other
air-breathing Vertebrates the nine sternal elements can according
to Geoffroy be discovered without 059great difficulty. But when we come
to the determination of the sternum in fishes, difficulties abound,
which Geoffroy solves in the following way. He points out that
between the clavicles (cleithra) and the hyoid bone
(basihyal) in fishes there is a long median bone
(urohyal) which is attached in front by two strong tendons
to the horns of the hyoid and is free behind (see Fig. 1). Gouan (1720) had seen in this bone the
homologue of the sternum. Geoffroy adopts this view, but considers
that this bone alone cannot represent the whole sternum. He finds
the representatives of other bones of the sternum in the large
bones (epihyal and ceratohyal, or the two pieces of
the ceratohyal) which are comprised in the hyoid arch. But
he is immediately met by the difficulty that this complex of bones
is situated in front of the pectoral girdle, whereas the sternum in
higher Vertebrates lies behind the pectoral girdle. He reflects,
however, that the gills of fish, situated in front of the
clavicles, are merely the lungs under another name. The gills have
become shifted forward by a metastasis similar to that which
brought the whole thoracic organs far forward in fish. This being
so, their supporting elements, the sternum and the ribs, must have
moved with them, and are hence to be found in front of the pectoral
girdle.
Geoffroy's next step is to point out that the only possible
homologues of sternal ribs are the branchiostegal rays, which arise
from the large bones of the hyoid arch. If these are sternal ribs,
the bones to which they are attached must be the hyo- and
hyposternals or "annexes," the bones from which in birds the ribs
take their origin.
The unpaired sternal bone (urohyal) cannot be homologous
with the entosternal, for it has no connections with the annexes.
He decides that it must represent the episternals, for in some
young birds there is a two-headed episternal to which two strong
tendons are attached, just in the same way as the unpaired piece in
fish is bound to the bones of the hyoid by two tendons. "Thus it is
not the sternum as a whole that has shifted in front of the
clavicles and covered with its side pieces the gills placed there;
it is a piece exclusively piscine, in the sense that it is only in
the class of 060fishes that it reaches the maximum of
its development" (p. 83).
To sum up, the sternum in all four vertebrate classes is
composed of the same elements, arranged always in the same way.
"One is ... led to the conception of an ideal type of sternum for
all Vertebrates, which then, considered from a lower standpoint,
resolves itself into several secondary forms according as the whole
or the majority of the constituent materials are employed, or even
as these elements come to change their respective dimensions or
proportions" (p. 134). As to the elementary constituents, "they
give proof of individuality, and sometimes even, in certain
abnormalities, of independence, and rise to the level of primary
organisatory materials" (p. 132). What holds good for the sternum
holds good for other organs—and accordingly the unity of plan
and composition can be demonstrated for all the organs of
Vertebrates.
Soon after the publication of the Philosophie anatomique
(1818) Geoffroy went further in his search for unity, and
maintained that the structure of insects and Crustacea could be
reduced to the vertebrate type.
He proposed to replace Cuvier's classification of the animal
kingdom into the four large groups, Vertebrata, Mollusca,
Articulata, and Radiata by the following classification:—[90]
| Vertébrés |
 |
Hauts-Vertébrés (Vertebrata, Cuv.). |
| Dermo-Vertébrés (Articulata, Cuv.). |
| .Invertébrés. |
|
Mollusques (Mollusca, Cuv.) |
| Rayonnés (Radiata, Cuv.). |
The idea upon which is based the comparison of Articulates with
Vertebrates is that each skeletal segment of Articulates is a
vertebra. In the Hauts-vertébrés the vertebræ are
internal; in the Dermo-vertébrés they are external.
"Every animal lives either outside or inside its vertebral
column."[91] The essence of a vertebra
is not its form, nor its function, but its composition from four
061elementary pieces which unite round a central
space (Isis, loc. cit., p. 532). Serres had shown that in
the higher animals every vertebra is formed from four centres of
ossification, that the body of the vertebra is at first tubular,
and that afterwards it becomes filled up. In lobsters and crabs
each segment is composed of four elementary pieces, as may be seen
most easily in young ones. "Accordingly each segment corresponds to
a true vertebra in composition: there is the same number of
'materials,' the same order in the course of ossification, the same
kind of articulation, the same annular arrangement, the same empty
space in the middle" (p. 534). The only difference is that in
Articulates the central space is very great and contains all the
organs of the body, whereas in the higher Vertebrates the body of
the vertebra becomes completely filled up. In the thoracic region
of Crustacea it is not the whole segment with part of the carapace
which corresponds to a vertebra, but merely the part round the
ventral nerve-cord (endophragmal skeleton).

Fig. 2.
"Vertebra" of a Pleuronectid.
(After Geoffroy.)
If the skeleton of the segment in Articulates corresponds to the
body of a vertebra and is here external, then the appendages of the
Articulate must correspond to ribs (p. 538). The full development
of this thought is found in a Memoir of 1822, "Sur la
vertèbre."[92] He takes as the
typical vertebra that of a Pleuronectid, probably the turbot. His
original figure is reproduced (Fig. 2).
He includes as part of the vertebra not only the neural (e′, e″)
and hæmal (o′, o″) arches, but also, above and below these,
the radialia (a″, u′) and the fin-rays (a′, u″). (Neither the
radialia nor the fin-rays are, 062by the way, in the same transverse
plane as the body of the vertebra). Every vertebra, he considers,
contains these nine pieces—the cycleal (or body), the two
perials (e′, e″) and the two epials (a′, a″) above, the two paraals
(o′, o″) and the two cataals (u′, u″) below. The epials and the
cataals are in reality paired bones which in fish mount one on top
of the other to support the median fins. In the cranial
region—the skull is formed of modified vertebræ—the
epials and perials open out so as to form the walls and roof of the
brain; in the thoracic region the paraals and cataals reach their
maximum of development and perform the same service for the
thoracic organs, the paraals becoming vertebral, and the cataals
sternal, ribs.
We have seen that in Arthropods the body of the vertebra
(cycleal) forms the open ring of the segment, which lies
immediately under the skin, the vertebral tube coinciding with the
epidermal tube. The homologues of the other eight pieces of the
vertebra must accordingly be sought in the external appendages. At
first sight there seems here a contradiction of the principle of
connections, for the appendages in Arthropods are lateral, whereas
the paired bones of the vertebra are dorsal and ventral. But there
is in reality no contradiction, for "what our law of connections
absolutely requires is that all organs, whether internal or
external, should stand to one another in the same relations; but it
is all one whether the box (coffre) that encloses them lies
with this or that side on the ground. What similarities in the
organisation of man and the digitate mammals, and yet what
differences between their attitudes when standing! The same holds
true as regards the normal attitudes of the pleuronectids and the
other fishes" (p. 107).
The exact way in which Geoffroy homologised the parts of
the appendages in Arthropods with the paired pieces of the typical
vertebra is best shown by the reproduction of his figure of an
abdominal segment of the lobster (Fig. 3), in
which the parts homologous with those represented in the figure of
the typical vertebra (Fig. 2) are indicated by
the same letters. The ingenuity of the comparison is
astonishing.
063The comparison of the Arthropod with the
Vertebrate is extended also to the internal organs. The internal
organs of the Arthropod are shown to stand in the same order to one
another as in the Vertebrate, only the organs are inverted. Thus
the nervous system is dorsal in the Vertebrate, ventral in the
Arthropod. Turn the Arthropod on its back and the relative
positions of the systems of organs are the same as in the
Vertebrate. The relation of the organs to the external tube is of
course different in Arthropods and Vertebrates, but this is no
contradiction of the principle of connections. "Such a tube,
although it is the organs essential to life that it contains, can
yet behave in different ways with regard to the mass of these
organs: the principle of connections demands only that all the
organs maintain with one another fixed and definite relations; but
the principle would be in no way invalidated if the whole mass had
rotated inside the tube" (p. 112).
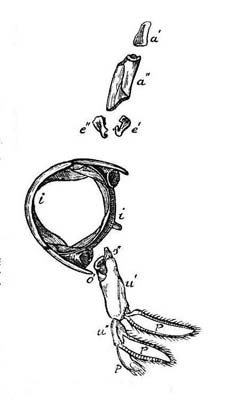 Fig.
Fig. 3.
Abdominal Segment of the Lobster.
(After Geoffroy.)
Geoffroy pushed the analogy between Arthropods and Vertebrates
very far, for he asserted that every piece in the skeleton of an
insect was homologous with some bone in Vertebrates, that it stood
always in its proper place, and remained faithful to at least one
of its connections.[93] It does not appear
that he attempted to prove in detail this very big assumption, but
the beginnings of a detailed comparison are found in the paper of
1820, Sur l'organisation des insectes. Six segments are
distinguished in an insect—the head, the three divisions
064of
the thorax, the abdomen, and the terminal segment of the abdomen
(p. 455).
The skeleton of the insect's head is said to correspond to the
bones of the face, to the bones of the cerebrum and to the hyoid of
higher Vertebrates, the skeleton of the prothorax to the bones of
the cerebellum, of the palate, and the pieces of the larynx, the
skeleton of the mesothorax to the parietals, interparietals, and
opercular bones, and that of the metathorax to the skeleton of the
thorax of Vertebrates. The pieces of the abdomen and of the
terminal segment correspond to the bones of the abdomen and coccyx
(p. 458). It does not need the subsequent likening of the hind
wings of insects to the air bladder of fish, and of the stigmata to
the pores of the lateral line, to convince one finally of the
fancifulness of the whole comparison.
In 1830 two young naturalists, Meyranx and Laurencet, presented
to the Académie des Sciences a memoir in which they likened
a Cephalopod to a Vertebrate bent back at the level of the
umbilicus, saying that the Vertebrate in this position had all its
organs in the same order as in the Cephalopod. Geoffroy took up
this idea with enthusiasm, seeing in it a further application of
his master-idea of the unity of plan and composition. By means of
this comparison Mollusca definitely took their place in the
Échelle des êtres, after the Articulata, just
as Geoffroy had maintained in 1820, saying that crabs formed a link
between the other Crustacea and the molluscs.[94] The comparison brought him nearer to the
end he had in view, the reference of all animal structure to one
single type.
But in championing the memoir of Meyranx and Laurencet, Geoffroy
found himself in direct antagonism with Cuvier, who held that his
four "Embranchements" had each a separate and distinct plan of
structure. In a paper read to the Academy in February 1830,[95] Cuvier easily demolished the crude
comparison of the Cephalopod to the Vertebrate. He gave diagrams of
the internal organs of a Cephalopod and of a Vertebrate bent back
in the manner indicated by Meyranx and Laurencet, and he showed in
065detail that the arrangement of the main
organs was quite different, that the likeness would have been much
greater if the Cephalopod had been likened to a Vertebrate doubled
up the other way,[96] but that even then
the arrangement of the organs would not be the same. The organs,
too, of the Cephalopod are differently constructed. He sums up his
criticism by saying:—"I give true and summary expression to
all these facts when I say that Cephalopods have several organs in
common with Vertebrates, which fulfil in either case similar
functions, but that these organs are differently arranged with
respect to one another, and often constructed in a different way;
that they are in Cephalopods accompanied by several other organs
which Vertebrates do not possess, whilst the latter on their side
have many organs which Cephalopods lack" (p. 257). Geoffroy could
not accept this commonsense view of the matter, but made a fight
for his transcendental theories. This was the beginning of the
famous controversy between Geoffroy and Cuvier which so excited the
interest of Goethe. It was a struggle between "comparative anatomy"
and "morphology," between the commonsense teleological view of
structure and the abstract, transcendental. Geoffroy brought
forward all his theories on the homology of the skeleton of fish
with the skeleton of higher Vertebrates, and tried to prove by them
his great principle of the unity of plan and composition; Cuvier
took Geoffroy's homologies one by one, and showed how very slight
was their foundation. Cuvier was on sure ground in insisting upon
the observable diversities of structural type, and his vast
knowledge enabled him to score a decisive victory.[97]
The controversy was not, as we are sometimes told, a controversy
between a believer in evolution and an upholder of the fixity of
species, although it raised a question upon which evolution theory
was to throw some light.
066In these Darwinian days Geoffroy has reaped a
little posthumous glory as an early believer in evolution. That he
did believe in evolution to a limited extent is certain; that his
theory of evolution was, as it were, a by-product of his life-work,
is also certain. Geoffroy was primarily a morphologist and a seeker
after the unity hidden under the diversity of organic form. His
theory of evolution had as good as no influence upon his
morphology, for he did not to any extent interpret unity of plan as
being due to community of descent. His morphological,
non-evolutionary standpoint comes out quite clearly in several
places in the Philosophie anatomique. He does not derive the
structure of the higher Vertebrates from the simpler structure of
the lower, but when he finds in fish a part at the maximum of its
development, he speaks of the same part, rudimentary in the higher
forms, as being, as it were, held in reserve for use in the fish.
Thus, speaking of the episternal in fish which forms the central
piece of its sternum, he says, "it is a bone that is rudimentary in
birds (one might almost add a bone that is held in reserve in birds
for this fate) which is destined to form in the centre the
principal keel of this new machine" (p. 84). Again, with reference
to the homology of the ossicles of the ear with the opercular bones
in fish, "employing other resources equally hidden and rudimentary,
Nature makes profitable use of the four tiny ossicles lodged in the
auditory passage, and, raising them in fish to the greatest
possible dimensions, forms from them these broad opercula...." (p.
85). Or you may take it the other way about, and start from the
organisation of fishes; opercular bones are of no use to
air-breathing animals, so they dwindle away, and are pressed into
the service of the ear, although they are of little use in hearing
(p. 46).
There is here no thought of evolution; in later years, however,
his researches upon fossil crocodilians led him to consider the
possibility that the living species were descended from the
antediluvian. For the factors of the transformation he refers to
Lamarck's hypotheses.[98]
In a memoir of 1828,[99] 067dealing with the
possible genetic relation of living to fossil species, he still
regards the question as more or less open. Although fossil species
are mostly different from living species are we therefore to
conclude, he asks, that they are not the ancestors of the present
day forms? "The contrary idea arises more naturally in the mind;
for otherwise the six-days' creation would have had to be repeated
and new beings produced by a fresh creation. Now this proposition,
contrary as it is to the most ancient historical traditions, is
inadmissible" (p. 210). It is sufficiently clear from this
quotation that Geoffroy was thinking only of a transformation of
the antediluvian species created by God, and by no means of an
evolution of all species from one primitive type. In matters of
religion Geoffroy was orthodox. He goes on to point out how great a
resemblance there is in essential structure between fossil and
living species. All find their place in one scheme of
classification; does it not seem that all are modifications "of one
single being, of that abstract being or common type, which it is
always possible to denote by the same name?" (p. 211). This type is
abstract, not actual, and it is certainly not conceived as an
original ancestor of all animals.
The fullest development of Geoffroy's views on evolution is
found in his memoir "Le degré d'influence du monde ambiant
pour modifier les formes animales."[100] Here the relation of his evolution-theory to his
morphology is pointed out. The principle of unity of plan and
composition cannot be the final goal of zoology; there must follow
on it a philosophical study of the differences between
organic forms. The causes of these differences are to be found in
the environment (pp. 66-7). Geoffroy seems here to be moving from
a pure to a causal morphology. It is probable, he continues, that
living species have descended by uninterrupted generation from the
antediluvian species (p. 74), and that they have in the process
become modified through external influences.
Now of all functions respiration is the most important, and upon
respiration everything is regulated. "If it be admitted that the
slow progression of the centuries has 068brought in its train
successive changes in the proportion of the different elements of
the atmosphere, it follows as a rigorously necessary consequence
that the organisation has been proportionately influenced by them"
(p. 76). The respiratory milieu changes, the species change with
it, or are eliminated (p. 79). We may see, perhaps, in the stress
which Geoffroy lays upon respiration and the respiratory milieu a
result of his constant obsession with the comparison of fish with
air-breathing Vertebrates.
In the first geological period, we read in another Memoir of the
same year,[101] when ammonites and
Gryphæa flourished, hot-blooded animals with lungs
could not exist. "A lung constructed like that of mammals and birds
would not have been adapted to the essence of the respiratory
element such as in my conception of it the system of the environing
air used to be"[102] (p. 58).
Geoffroy does not tell us exactly how the milieu is to act upon
the organism; the whole theory is little more than a sketch and a
pointing out of the way for future research—and in this
prophetic enough. The action of external agents was apparently
considered as physical, and no power of active adaptation was
ascribed to the organism.
From a passage in the memoir "Sur la Vertèbre" we may
perhaps infer that he believed increasing complexity of structure
to be due to a realisation of potentialities, to the development of
parts present in the lower animals only in potency—"the
organisation ... only awaits favourable conditions to rise, by
addition of parts, from the simplicity of the first formations to
the complication of the creatures at the head of the scale" (p.
112). Evolution takes place as the environment allows, and in a
sense in opposition to the environment.
He believed in saltatory evolution, for he considered that the
lower oviparous Vertebrates could not be transformed into birds by
slow modification, but only by a sudden transformation of their
lungs, which would bring about the other characteristics of birds
(p. 80). He considered, too, 069that transformations could arise by means
of monstrous development (p. 86). In this connection the
experiments which he made on the hen's egg[103] in order to produce artificial monstrosities are
significant, though his purpose was rather to obtain proof of the
inadequacy of the preformation hypothesis.[104]
It seems probable enough that if Geoffroy had developed his
views on evolution he would finally have been led to interpret
unity of plan in terms of genetic relationship. But as it was he
remained at his morphological standpoint. He did not interpret
rudimentary organs as useless heritages of the past; he preferred
to think that Nature had prepared double means for the same
function, one or other being predominant according as the animal
lived in the water or on the land. "To the animal that lives
exclusively in the air Nature has granted an organisation suited to
this mode of respiration, without however suppressing the other
corresponding means, that is to say, without depriving it of a
second system which is applicable only to the mode of respiration
by the intermediary of water, and vice versa."[105]
He seems, in one instance at least, to have hit upon the
root-idea of the biogenetic law, but he was far from appreciating
its significance. He recognised that an amphibian in its
development passed through a stage when it was in all essentials
similar to a fish, and he saw in this visible transformation a
picture of the evolutionary transformation. "An amphibian," he
writes,[106] "is at first a fish
under the name of tadpole, and then a reptile [sic] under
that of frog.... In this observed fact is realised what we have
above represented as an hypothesis, the transformation of one
organic stage into the stage immediately superior." But it is not
clear that he considered the development of the amphibian to be a
repetition of its ancestral history.
He went, however, a certain length towards recognising the main
principle of a law which was a commonplace of 070German
transcendental thought, and was developed later by his disciple E.
Serres, the law that the higher animals repeat during their
development the main features of the adult organisation of animals
lower in the scale. Thus he compared fish as regards certain parts
of their structure with the fœtus of mammals. He compared
also Articulates with embryonic Vertebrates in respect of their
vertebræ, for in the higher Vertebrates the body of the
vertebra is tubular at an early stage of development, and in
Articulates the body of the vertebra remains tubular permanently
(supra, p. 61). As regards their vertebræ, "insects
occupy a place in the series of the ages and developments of the
vertebrate animals, that is to say, they realise one of the states
of their embryo, as fishes do one of the states of their
fœtal condition."[107]
This idea was destined to exercise a great influence upon the
development of morphology. A further development of the thought is
that certain abnormalities in the higher animals, resulting from
arrest of development, represent states of organisation which are
permanent in the lower animals.[108]
So far we have considered Geoffroy's theories in their
application to the facts. We go on to discuss the theories
themselves, and the general conception of living things which
underlies them.
The principle of unity of plan and composition is the keynote of
Geoffroy's work. It states that the same materials of organisation
are to be found in all animals, and that these materials stand
always in the same general spatial relations to one another. The
"materials of organisation" are not necessarily organs in the
physiological sense, and indeed the principle of the unity of plan
cannot be upheld if the unity has reference to organs only. This
became clear to Geoffroy, especially in his later years. In 1835 he
wrote, speaking of the principle of the unity of plan, "I have,
moreover, regenerated this principle, and obtained for it
universality of application, by showing that it is not always the
organs as a whole, but merely the materials composing each 071organ,
that can be reduced to unity."[109] Even in the Philosophie anatomique he
deals rather with parts than with organs; he deals, for instance,
with the elementary parts of the sternum, not with the organ
"sternum" in its totality. The functions of the sternum vary, and
the primary protective function of the sternum may be assumed by
quite other parts, e.g., by the clavicles in fish, which
protect the heart.[110]
True homologies can be established between materials of
organisation but not always between organs, which may be composed
of different "materials."
Almost as a corollary to this comes the further view that form
is of little importance in determining homologies. An organ is
essentially an instrument for doing a particular kind of work, and
its form is determined by its function. Organs which perform the
same function are usually similar in form though the elementary
materials composing them may be different. This is seen in many
cases of convergence. Organs, therefore, which perform the same
function and are similar in external form are not necessary
homologous. Conversely, the same complex of materials, say a fore
limb, may take on the most varied shapes according as the function
of the organ changes—but homology remains though form
changes. Accordingly, form is one of the least important elements
to be considered in determining a homology. "Nature," he wrote in
one of his early papers, "tends to repeat the same organs in the
same number and in the same relations, and varies to infinity only
their form. In accordance with this principle I shall have to draw
my conclusions, in the determining the bones of the fish's skull,
not from a consideration of their form, but from a consideration of
their connections."[111]
Again, after comparing a vertebra of the Aurochs with an
abdominal segment of the crab, he says, "I have insisted upon an
identity which has extended to the least important relation of all,
that of form."[112]
072Geoffroy's morphological units or materials
of organisation were in the case of the skeleton—with which
his researches principally deal—the single bones. But the
interesting point is that he sought his skeleton-units in the
embryo, and considered each separate centre of ossification as a
separate bone. Coalescence of bones originally separate is one of
the most usual events in development, and it is an occurrence
which, more than any other, tends to obscure homologies. Because of
its coalescence with the maxillaries, the intermaxillary in man was
not discovered until Vicq d'Azyr and Goethe found it separate in
the embryo. Apparently quite independently of Goethe, Geoffroy hit
upon this plan of seeking in the embryo the primary elements or
materials of organisation. In an early paper on the skull of
Vertebrates,[113] where he is concerned
with showing that each bone of the fish's skull has its homologue
in the skull of higher Vertebrates, he is faced with the difficulty
that the skull of the fish has more bones than the skull of higher
Vertebrates. "Having had the inspiration," he writes, "to reckon as
many bones as there are distinct centres of ossification, and
having made a consistent trial of this method, I have been able to
appreciate the correctness of the idea: fish, in their earliest
stages, are in the same conditions relatively to their development
as the fœtuses of mammals, and hence bear out the theory" (p.
344). So, too, in dealing with the homologies of the sternal
elements (supra, p. 57) he treats as separate bones the
"annexes" of the sternum in birds, though these are separate only
in the young.
If the same materials of organisation are present in all
animals, and if they are arranged always in the same positions
relatively to one another, how does it come about that animal forms
are so varied, what explanation can be offered of the diversities
of organic structure? Geoffroy's main answer to this question is
his Loi de balancement. The law was enunciated by him
already in 1807.[114] We take the
following quotation, which represents his thought most nearly, from
the Cours de l'histoire naturelle des Mammifères
(1829). "According to our manner of regarding the 073organisation of
mammals, there is only a single animal modified by the inverse
reciprocal variation of all or some of its parts. Now, from the
fact that there is only one single general animal, it follows that
for each section of its components or for each of its organs there
is available only a given quantity of formative materials. Now
suppose that the distribution of these materials has not been made
in such a way as to ensure an exact equilibrium between all the
parts concerned, one organ will get more than its share, another
less. My law of the compensation of organs is founded on these
principles" (i., Leçon 16, p. 12). "The atrophy of
one organ turns to the profit of another; and the reason why this
cannot be otherwise is simple, it is because there is not an
unlimited supply of the substance required for each special
purpose."[115] The nutritive material
available is limited for each species; if one part gets more than
its share the other parts must get less—that is all the law
means. As an example, take the minuteness of the episternals and
xiphisternals in birds, as contrasted with the huge size of the
entosternal. "The minuteness of the episternals and xiphisternals
might be imputed to this gigantic piece diverting to its own profit
the nutritive fluid, since the bigger it is the smaller these
are."[116]
One has constantly to remember in dealing with Geoffroy's
theories that he was not an evolutionist, but purely a
morphologist. It is therefore, perhaps, to ask too much to require
of him an explanation of the causes of diversity. The morphologist
describes, classifies, generalises; he does not seek for causes.
But we must leave this question aside in order to discuss how far
Geoffroy's theory of the unity of plan and composition fits the
facts. As Geoffroy himself admitted on several occasions, his
theory was an à priori one, a theory hit upon by
hasty induction, then erected into a principle and imposed upon the
facts. No more than Goethe did he extract his principle from a
sufficient mass of data.
Now he found his theory to be in its pure form unworkable; he
found, for example, that the skeleton of fishes 074could not be
compared directly, bone for bone, with the skeleton of higher
Vertebrates; he had to admit differences of position of whole sets
of organs in the two groups, he had to admit various
metastases, before he could bring the skeleton of fish into
line. And these metastases are due to functional
requirements—for example, the forward position of sternum and
thoracic organs in fish is an adaptation to swimming.
So he does not so much demonstrate the unity of plan of whole
organisms as the unity of plan of particular corresponding parts of
them. Thus he does not prove or attempt to prove that Articulates
are in all points like Vertebrates, but simply that their skeleton
is built upon the same plan as that of Vertebrates. The rest of the
organs, while still comparable with the organs of Vertebrates,
stand in different relations to the skeleton. An Articulate
therefore, on his own showing, is not, as a whole, built
upon the same general structural plan as a Vertebrate.
Further, he does not always remain true to his principles, for
he does not establish homologies of parts entirely by their
connections but sometimes by their functions as well. Thus the
sternum, or rather the complex of sternal elements, is defined and
discovered in particular cases not by its connections only but also
by its functions. The framework of the gills is homologised part by
part with the framework of the lungs, not because the relations of
the framework to the rest of the skeleton are the same in fish and
air-breathing Vertebrates, but simply because gills are considered
the equivalents of lungs—a comparison which is purely
physiological.
Even with these concessions to the functional view of living
things, Geoffroy was unable to make good his contention that all
animals are built upon the same plan. His arguments failed to carry
conviction to his contemporaries, and Cuvier in particular
subjected them to destructive, and indeed final, criticism.
The paper, already referred to, in which Cuvier disposed of the
transcendentalists' comparison of Cephalopods and Vertebrates is of
great significance, for it states in the 075clearest way the radical
opposition between the functional and the formal attitudes to
living things.
Cuvier points out that if by unity of composition is meant
identity, then the statement that all animals show the same
composition is simply not true—compare a polyp with a
man!—on the other hand, if by unity is meant simply
resemblance or homology, the statement is true within certain
limits, but it has been employed as a principle since the days of
Aristotle, and the theory of unity of composition is original only
in so far as it is false. He admits, however, that Geoffroy has
seized upon many hidden homologies, especially by his valuable
discovery of the importance of fœtal structure. In all this
Cuvier is undoubtedly right. Unity of plan and composition, as
Geoffroy conceived it, simply does not exist. Cuvier goes on to say
that this principle of Geoffroy's, in the greatly modified form in
which it can be accepted, and has been accepted from the dawn of
zoology, is not the sole and unique principle of the science. On
the contrary, it is merely a subordinate principle, subordinate to
a higher and more fruitful principle, that, namely, of the
conditions of existence, of the adaptation (convenance) of
the parts, of the co-ordination of the parts for the rôle
which the animal is to play in Nature. "That is the true
philosophical principle," he says, "whence may be deduced the
possibility of certain resemblances, the impossibility of certain
others; it is the rational principle from which follows the
principle of the unity of plan and composition, and in which at the
same time it finds those limits, which some would like to
disregard" (p. 248).
Geoffroy's position is the direct contrary. He holds that the
principle of the unity of plan and composition is the true base of
natural history,[117]and that this unity
limits the possible transformations of the organism. Thus, speaking
of the influence of the respiratory medium, he says, "All the same
this influence of the external world, if it has ever become a cause
which disturbed organisation, must necessarily have been confined
within fairly narrow limits; animals must have opposed to it
certain conditions inherent to their nature, the existence of the
same materials composing them, and a 076manifest tendency to resemble one
another, and to reproduce invariably the same primordial type."[118] Unity of plan and
composition is, on this view, prior to adaptation and limits
adaptation. Cuvier's view, on the contrary, is that the necessity
of functional and ecological adaptation accounts for the repetition
of the same types of structure. There are, of all the possible
combinations of organs, only a few viable types—those whose
structure is adapted to their life. Therefore it is reasonable that
these few types should be repeated in innumerable exemplars. One
must remember, in order to appreciate Cuvier's view, that he was
not obsessed, as we are, by the idea of evolution.
Cuvier thought in terms of organs, not in terms of "materials of
organisation." He held that the resemblances between the organs of
one class of animals and the organs of another were due to the
similarity of their functions. "Let us conclude, then, that if
there are resemblances between the organs of fish and those of
other classes, it is only in the measure that there is a
resemblance between their functions."[119] There are only a few kinds of organs, each
adapted for a particular function, and these organs are necessarily
repeated from class to class.—"As the animal kingdom has
received only a limited number of organs, it is inevitable that
some at least of these organs should be common to several
classes."[120]
Geoffroy thought in terms of "materials," of parts of indefinite
function, parts which might take on any function. He insists upon
the necessity of disregarding function when tracing out the unity
of composition. He considers, in direct opposition to Cuvier's
interpretation of structural resemblance as due to similarity of
function, that unity of composition is the primary fact, and
similarity of function subsidiary. In his reply in the
Mammifères (1829) to Cuvier's criticisms in the
Histoire naturelle des Poissons (1828), he insists on the
necessity of excluding function from consideration in any truly
philosophical treatment of comparative anatomy (Discours
prél., p. 25). Cuvier held that function determined
structure, or at least that the necessity 077of adaptation ruled the
transformations of form. Geoffroy considered that structure
determined function, that changes of structure, however they might
arise, caused changes of function. "Animals," he writes, "have no
habits but those that result from the structure of their organs; if
the latter varies, there vary in the same manner all their springs
of action, all their faculties and all their actions."[121]
Again, "a vegetarian régime is imposed upon the
Quadrumana by their possession of a somewhat ample stomach, and
intestines of moderate length."[122] The hand of the bat has become so modified as to
constrain the bat to live in the air.[123]
The best example of Geoffroy's insistence upon the priority of
structure to function, and so of his purely morphological attitude,
is perhaps his interpretation, already alluded to, of the
appendages of Articulates. The segments of the Articulate are, he
says, the equivalents of the bodies of the vertebræ of higher
forms. Now "from the circumstance that the vertebra is external, it
results that the ribs must be so too; and, as it is impossible that
organs of such a size can remain passive and absolutely
functionless, these great arms, hanging there continually at the
disposition of the animal, are pressed into the service of
progression, and become its efficient instruments."[124] The ribs become locomotory appendages.
We may compare the similar thought that the ear ossicles are
simply opercular bones reduced and turned to other uses.
Geoffroy could not but recognise the correlation of structure to
function, for this is a fact which imposes itself upon every
observer. He recognised also correlation between functions, as when
he pointed out the connection between increased respiration and
enhanced muscular activity in birds.[125] He interpreted structure at times in terms of
function, the short, strong clavicle of the mole as an adaptation
to digging, the keeled sternum of birds as an adaptation to flying,
and so on. But we may say that his whole tendency was to disregard
function, to look upon it as subsidiary. He protests against
arguing from function and habits to structure, as an "abuse 078of final
causes."[126] He was not so convinced
as Cuvier was of the all-importance of functional correlation; in
this view he was probably confirmed by his work on teratology. It
did not surprise him that Insects, in which lungs, heart and
circulation have disappeared(!), should yet have a skeleton built
upon the same plan as the skeleton of Vertebrates, which possess
these organs; the correlation of organ-systems is not so close as
to prevent this.[127] So too, although
the other organs of the insect are all inside the body of the
vertebræ, they are yet comparable with the organs of
Vertebrates.[128] The existence of
rudimentary organs also seemed to him an argument against too
strict a correlation of parts.
The contrast between the teleological attitude, with its
insistence upon the priority of function to structure, and the
morphological attitude, with its conviction of the priority of
structure to function, is one of the most fundamental in
biology.
Cuvier and Geoffroy are the greatest representatives of these
opposing views. Which of them is right? Is there nothing more in
the unity and diversity of organic forms than the results of
functional adaptation, or is Geoffroy right in insisting upon an
element of unity which cannot be explained in terms of adaptation?
If there be an irreducible element of unity, is there any truth in
Geoffroy's suggestion that this unity results from a power which is
exercised in the world of atoms where are elements of inalterable
character?[129]
The problem as Geoffroy and Cuvier understood it was not an
evolutionary one. But the problem exists unchanged for the
evolutionist, and evolution-theory is essentially an attempt to
solve it in the one direction or the other. Theories such as
Darwin's, which assume a random variation which is not primarily a
response to environmental changes, answer the problem in Geoffroy's
sense. Theories such as Lamarck's, which postulate an active
responsive self-adaptation of the organism, are essentially a
continuation and completing of Cuvier's thought.
079
CHAPTER VI
THE FOLLOWERS OF ETIENNE GEOFFROY SAINT-HILAIRE
Geoffroy's theories were not
generally accepted by his contemporaries, but his methods had
considerable influence, especially in France, where many made
essays in pure morphology.
His chief follower was Serres, who is mentioned indeed in the
Philosophie anatomique as a fellow-worker. Serres was
primarily a medical anatomist; his interest lay in human anatomy
and embryology, normal and pathological.
His best early work was an Anatomie comparée du
cerveau (1824-26), which met with a flattering reception from
Cuvier.[130] He laid great stress
upon the development of the brain and spinal cord in the different
classes, and was quick to point out analogies not only between
adult but also between embryonic structures. He paid much attention
to cases of correlation, and noted a great many; he observed, for
instance, a constant relation between the development of the spinal
cord and of the corpora quadrigemina, and between the size of the
corpora quadrigemina and the volume of the optic nerves and eyes.
In this the influence of Cuvier is unmistakable.
Serres' early theoretical views are to be found in a series of
papers in the Annales des Sciences naturelles,[131] under the general title Recherches
d'Anatomie transcendante, sur les Lois de l'Organogénie
appliquées à l'anatomie pathologique, also
published separately. We follow these papers in our exposé
of Serres' doctrine, reserving for a future chapter (Chap. XII.) the consideration of his matured views of
thirty years later.
080In the first of them he points out how
neither position nor function has proved altogether sufficient to
establish homologies. In the early days anatomists were guided by
form; when form failed them, they traced an organ in its changes
throughout the series of animals by considering its function. This
method was satisfactory enough as regards the organs of the
nutritive life. But in the organs of the life of relation, in the
nervous system, the functions of the parts were difficult to
discover, and their form very changeful. Hence a new principle was
required, and Serres found it in the thought which he probably owed
to the German transcendentalists (see Chap.
VII.), that the permanent structure of the lower animals could
be compared with phases in the development of the higher, and
particularly of man, or, as he put it, that comparative anatomy was
often only a fixed and permanent anthropogeny, and anthropogeny a
fugitive and transitory comparative anatomy (xi., p. 106).
"In rising towards the first formations," he writes,
"transcendental anatomy recognised that one and the same organ,
however complicated its definitive form might be, repeated in its
transitory states the organic simplicities of the lower classes.
Thus the primitive heart of birds was first of all a canal, then a
pocket or single cavity, then finally the complex organ of the
class. Comparative anatomy was thus seen to be repeated and
reproduced by embryogeny" (xii., p. 85).
His explanation of the fact of repetition is that, "in animals
belonging to the lower classes the formative force, whatever
it may be, has a less energetic impulsion than in the higher
animals, and hence the organs pass through only a part of the
transformations which those of the higher forms undergo; and it is
for this reason that they show permanently the organic dispositions
which are only transitory in the embryo of man and the higher
Vertebrates. Hence these double aortas, these double venæ
cavæ which one observes more or less constantly among
reptiles" (xxi., p. 48).
The number of stages in embryogeny is proportionate to the
complexity of the adult; the younger the embryo the simpler its
organs—such is the general formula of the relation between
the embryo and the adult. But here in Serres' 081doctrine of
parallelism a complication enters. He observed that embryonic
organs did not always develop in a piece, by simple growth, but
often were formed by the union of separately formed parts or
layers. Thus the kidney in man is formed by the fusion of a number
of "little kidneys," and the spinal cord reaches its full
development by the laying down of successive layers within it. He
was greatly impressed with this fact, which, as a convinced
believer in epigenesis, he used with great effect against the
preformistic theories. "This method of isolated formation," he
wrote, "is noticed in early stages in the thyroid, the liver, the
heart, the aorta, the intestinal canal, the womb, the prostate, the
clitoris, and the penis" (xi., p. 69). So, too, in the development
of the skeleton, ossification proceeds from separate centres,
foramina are formed by the fusion of separate bones round them. In
his memoir, Lois d'Osteogénie (1819), Serres
established several laws of ossification based upon this principle
of separate formation.[132]
How is the fact of multiple formation to be reconciled with the
principle of repetition, according to which organs are simplest in
the early embryo and in the lower animals? But observation shows
that, as a rule, the further down the scale you go the more divided
organs become—the more numerous the bones of the skull, for
example. There is thus a parallel between multiple formation of
organs in the embryos of the higher Vertebrates and their
subdivided state in the lower. Take, for example, the kidney. In
the genus Felis, and in birds, each kidney has two lobes, in
the elephant four, in the otter ten, in the ox twelve to fourteen.
The human kidney in its development starts with about a dozen
lobes, and the number diminishes as the kidney grows. Thus the
permanent state of the kidney in the animals mentioned is
reproduced by the stages of its development in man (xii., p.
126).
So, too, at the second or third month the uterus of the human
embryo is bicornuate, and afterwards passes through stages
comparable to the adult and permanent uterus of rodents, ruminants,
and carnivores. There is indeed a time in the development of the
human embryo when it resembles 082in many of its organs the adult
stage of various lower animals. It is about this time that it
possesses a tail.
We note that Serres' theory of parallelism applies, strictly
speaking, only to organs, not to organisms, although he, too,
readily fell into the error of supposing that the organisation of
an embryo could be compared as a whole with the adult organisation
of an animal lower in the scale. Thus he wrote in one of his later
papers[133]—"As our researches
have made clear, an animal high in the organic scale only reaches
this rank by passing through all the intermediate states which
separate it from the animals placed below it. Man only becomes man
after traversing transitional organisatory states which assimilate
him first to fish, then to reptiles, then to birds and mammals."
Serres was not altogether free from the besetting sin of the
transcendentalists—hasty generalisation.
The law of parallelism applied not only to Vertebrates but also
to Invertebrates. In a short paper[134] of 1824 Serres attempted an explanation of the
nervous system of Invertebrates. Invertebrates, he considered,
lacked the cerebrospinal axis of Vertebrates, and their nervous
system was the homologue of the sympathetic system of Vertebrates.
The relation of the invertebrate to the vertebrate nervous system
being thus fixed, can the nervous system of Invertebrates be
reduced to one plan? It does not seem possible to establish a
common plan for the adult nervous systems. But apply the principle
of parallelism, which has proved so valuable within the limits of
the vertebrate series. Taking insects as the highest class, we find
that there are three stages in the development of their nervous
system; in the first the nervous system is composed of two separate
strands, in the second the strands unite round the œsophagus, in
the third they unite also behind. Now in Bulla aperta, stage
(1) is permanent; in Clio, Doris, Aplysia,
Tritonia, Sepia, Helix, stage (2) is
permanent, and in Unio stage (3). In fact, all the varieties
of the nervous system of molluscs fall into one or other of these
three classes. "It follows, then, that as regards their nervous
system, the Mollusca are more or less advanced larvæ of
insects" (p. 380). The law of parallelism 083is here applied to single
organ-systems, but in later years Serres applied it to whole
organisations also, saying that the lower Invertebrates were
permanent embryos of the higher.
In the paper of 1834, already referred to, Serres pushed his
speculations further and attempted to establish the unity of type
of all animals, Vertebrates and Invertebrates alike—a
favourite pastime of the transcendentalists. It is incontestable,
he admits, that adult Invertebrates are quite different in
structure from adult Vertebrates, "but if one regards them as what
I take them to be, namely, permanent embryos, and if one
compares their organisation with the embryogeny of Vertebrates, one
sees the differences disappear, and from their analogies arise a
crowd of unsuspected resemblances" (loc. cit., p. 247).
The last point of Serres' doctrine which calls for remark is his
interpretation of abnormalities as being often comparable to grades
of structure permanent in the lower animals. Thus the double aorta
which may occur as an abnormality in man is the normal and
permanent state in reptiles. This idea, of course, he got from
Etienne Geoffroy St Hilaire. It is further developed in his
"Théorie des formations et des déformations
organiques appliquée à l'anatomie comparée des
monstruosités (1832), and in his final large memoir of
1860 (see below, p. 205).
In 1816 appeared a fine piece of work by J. C. Savigny on the
homologies of the appendages in Articulates. The standpoint was
that of pure morphology. "I am convinced," he wrote, "that when a
more complete examination has been made of the mouth of insects,
properly so called, that is to say, having six legs and two
antennæ, it will be found that whatever form it affects it is
always essentially composed of the same elements.... The organ
remains the same, only the function is modified or
changed—such is Nature's constant plan."[135] In this the influence of Geoffroy can be
traced; but the work was very free from the exaggerations of the
transcendentalists, and many of Savigny's homologies are accepted
even to-day. The first memoir dealt with the mouth-parts of
insects; the 084second with the anterior appendages of
Articulates generally. Savigny shows that the mouth-parts of
insects can be reduced to the type shown in Orthoptera, where there
are clearly two mandibles, two maxillæ, and a lower lip
formed by the fusion of two second maxillæ. All other insects
have these same mouth-parts, disposed in the same order, however
much their form may have been modified in response to new
functions. He goes on to compare the anterior set of appendages in
a long series of Articulates, in Julus, Scolopendra,
Cancer, Gammarus, Cyamus, Nymphon,
Phalangium, Apus, Caligus, Limulus, and
a few others. For Crustacea he established the homologies now
accepted, of the mandibles with the mandibles of insects, of the
first and second pairs of maxillæ with the parts so named in
insects, and so on. He is quite clear that the maxillipedes of
Crustacea are the homologues of the feet of Hexapoda. "Their
disposition must lead one to think that the six anterior feet of
Julus, that is to say, all the feet of the Hexapoda, are
here transformed into jaws" (loc. cit., p. 48). In
Scolopendra also there is a similar transformation of two
pairs of legs into auxiliary jaws. In Gammarus, where there
is only the first pair of maxillipedes, the other two pairs have
become "retransformed" into feet. We find him supporting his
comparison of the three anterior pairs of legs in Julus to
the three pairs of legs in insects by an argument drawn from
embryology; for only the first three pairs of feet are present in
Julus at birth (Degeer), "an observation, which, together
with their position, should cause them to be considered as the
representatives of the six thoracic feet of Hexapoda" (p. 44).
His comparison of the Arachnid appendages with those of insects
and Crustacea is very curious. As his starting-point he takes
Cyamus, which has antennæ (two pairs) and mouth parts
(four pairs) as in many Crustacea, and then seven pairs of legs; he
compares with it Nymphon, which has in all seven pairs of
appendages. These appendages he homologises with the seven pairs of
legs of Cyamus, so that the first appendage in
Nymphon corresponds to the seventh appendage of
Cyamus. This homology is extended to all Arachnids; their
first two pairs of appendages, however 085they may be modified as
"false" mandibles and "false" maxillæ, really correspond to
the second and third maxillipedes in Crustacea, and to the second
and third pairs of feet in insects. It is interesting to note that
he treats Limulus as an Arachnid, pointing out that there is
as much difference between Apus and Limulus as
between Cancer and Phalangium. He describes the
"gnathobases" in Phalangium and Limulus. We may note
that he had just an inkling of the modern doctrine that all the
appendages of Articulates consist of a basal joint bearing an inner
and an outer terminal piece, for he observes that the "cirri" of
the maxillipedes of Crustacea give the appendage the same bifid
appearance as the appendages of the abdomen and the thoracic legs
of Mysis (p. 50).
V. Audouin, in his memoir, Recherches anatomiques sur le
thorax des animaux articulés,[136] applied the principle of the unity of plan and
composition to the exoskeleton of insects, Crustaceans, and
Arachnids. His guiding ideas were, "(1) that the skeleton of
articulated animals is formed of a definite number of pieces, which
are either distinct or intimately fused with one another; (2) that
in many cases, some pieces diminish or altogether disappear, while
others reach an excessive development; (3) that the increase of one
piece seems to exert on the neighbouring pieces a kind of influence
which explains all the differences one finds between the
individuals of each order, family and genus" (Sep. copy, p. 16p.
Geoffroy had already stated, without proof, that the parts of the
Arthropod's skeleton, however they might change in shape and size,
remained faithful to the principle of connections, at least at
their points of insertion.[137] Audouin gave the detailed demonstration of this
by his accurate and minute determination of the pieces of the
arthropod skeleton. He recognised that the body of Arthropods was
made up of a series of similar rings, and that even the compact
head of insects consisted of fused segments. In each segment
Audouin distinguished a fixed number of hard chitinous parts, the
dorsal tergum, the ventral sternum, the lateral "flanc" of three
pieces, all to be recognised by their positions 086relative to one
another. Many of the names which he proposed are still in use; it
was he who introduced the terms prothorax, mesothorax, and
metathorax, for the three segments of the insect's thorax. He used
Geoffroy's Loi de balancement to explain cases of
correlative development, such as the relation between the size of
the front wings and the development of the mesothorax. In another
paper Audouin compared the three pieces of the dorsal skeleton of
Trilobites to the tergum and the upper part of the "flanc."[138] In a third paper of about the same time he
tried to establish the homologies of the segments throughout the
Articulate series—with less success than Savigny.
Later on, in conjunction with Milne-Edwards, he demonstrated the
unity of composition of the nervous system in Crustacea, showing
how the concentrated system of the crab was formed by the same
series of ganglia as in the Macrura.
The entomologist Latreille also tackled the problem of the
homologies of the segments in the different classes of Arthropods
(Cuvier, loc. cit., p. cclxxii.). He thought he could find
fifteen segments in all Arthropods. He made the retrograde step of
likening the head of insects to a single segment. But some of his
homologies showed morphological insight, e.g., his
comparison of the "first jaws" of Arachnids to antennæ,
because they were placed above the upper lip. It was he who first
pointed out the resemblance of the leaf-like gills of Ephemerid
larvæ to wings, and suggested that wings were "a sort of
tracheal feet."
He made also a rather hazy and speculative contribution on
Okenian lines to the problem of the relation of Arthropods to
Vertebrates, likening the carapace of Crustacea to an enormously
developed hyoid, the appendages of the tail to the ventral and anal
fins of fish. The masticatory organs of Arthropods were jaws
disjointed at their symphysis; antennæ, nostrils turned
outside in.
Dugès also made a comparison of Articulates with
Vertebrates.[139] He did not accept
Geoffroy's vertebral theory 087of the Arthropod skeleton, though he
admitted that in Arthropods the dorsal surface was turned towards
the ground, basing this assumption on the position of the nervous
system, and also, curiously enough, on the inverted position of the
embryo on the lower surface of the yolk. He considered that the
mandibles and first maxillæ of Arthropods were the homologues
of the upper and lower jaws of Vertebrates, adducing as
confirmatory evidence the fact that in snakes the rami are
separate. The labium was the equivalent of the hyoid, the labial
palps and maxillipedes the equivalent of the "hyoid" elements which
form the branchial arches.
But Dugès' main contribution to morphological method was
his conception of the living organism as a colony of lesser units,
which were themselves real "organisms." "By organism the
author means a complex of organs which taken together suffice to
constitute, ideally or actually, a complete animal. An 'organism'
is, as it were, an elementary or simple animal; several organisms
combined form a complex animal" (p. 255). Dugès hit upon
this principle, which was first suggested to him by A.
Moquin-Tandon's work on the leech (1827), as a great aid in
demonstrating the unity of plan and composition throughout the
animal kingdom.[140] According to his
view there are three main types of animals—(1) Biserials,
including bilaterally symmetrical animals, composed of two parallel
series of "organisms"; (2) Radiates, composed of "organisms"
arranged like the spokes of a wheel; and (3) Raceme-animals, in
which the separate "organisms" were disposed more or less
irregularly, in bunches (p. 257). The unitary "organism" is
supposed to be the same in all, only the arrangement differing.
Dugès of course admitted that the centralisation of the
complete organism became greater the higher it stood in the scale,
and that this held good also in individual development. The
appendages of Articulates and Vertebrates were thought of as the
members of as many separate organisms. He went so far as to suggest
that the 088fingers of a man's hand were the free
extremities of as many thoracic members.
Dugès' conception of the organism has often been revived
since in a saner form, e.g., by E. Perrier, and it has a
certain validity. It has much affinity with the similar conceptions
of Goethe and the German transcendentalists.
089
CHAPTER VII
THE GERMAN TRANSCENDENTALISTS
To complete our historical survey of
the morphology of the early 19th century we have now to turn back
some way and consider the curious development of morphological
thought in Germany under the influence of the Philosophy of
Nature. We have already seen many of these notions foreshadowed
by Goethe, who had considerable affinity with the
transcendentalists, but the full development of transcendental
habits of thought comes a little later than the bulk of Goethe's
scientific work, and owes more to Kielmeyer and Oken than to Goethe
himself.
A great wave of transcendentalism seems to have passed over
biological thought in the early 19th century, arising mainly in
Germany, but powerfully affecting, as we have seen, the thought of
Geoffroy and his followers. Many ideas were common to the French
and German schools of transcendental anatomy, the fundamental
conception that there exists a unique plan of structure, the idea
of the scale of beings, the notion of the parallelism between the
development of the individual and the evolution of the race. It is
difficult to disentangle the part played by each school and to
determine which should have the credit for particular theories and
discoveries. The philosophy seems to have come chiefly from
Germany, the science from France. It must be borne in mind that
German comparative anatomy was largely derivative from French, that
the Paris Museum was the acknowledged anatomical centre, and that
Cuvier was its acknowledged head.
It is probably correct to say that the credit mainly belongs to
the German transcendental school for the law 090of the
parallelism between the stages of individual development and the
stages of the scale of beings, and the theory of the repetition or
multiplication of parts within the individual. The vertebral theory
of the skull is a particular application of the second of these
generalisations.
The law of parallelism[141] seems to have been expressed first by Kielmeyer
(1793),[142] who gave to it a
physiological form, saying that the human embryo shows at first a
purely vegetative life, then becomes like the lower animals, which
move but have no sensation, and finally reaches the level of the
animals that both feel and move.
The idea was next taught by Autenrieth in 1797.[143]
Oken (1779-1851) in his early tract Die Zeugung (1805),
and in his Lehrbuch der Naturphilosophie (1809-11)
elaborated the thought, and taught that every animal in its
development passes through the classes immediately below it.
"During its development the animal passes through all stages of the
animal kingdom. The fœtus is a representation of all animal
classes in time."[144] The Insect, for
example, is at first Worm, next Crab, then a perfect volant animal
with limbs, a Fly (ibid., p. 542).
As Nature is "the representation of the individual activities of
the spirit," so the animal kingdom is the representation of the
activities or organs of man. The animal kingdom is therefore "a
dismemberment of the highest animal, i.e., of Man" (p. 494).
Now "animals are gradually perfected, entirely like the single
animal body, by adding organ unto organ"—the way of evolution
is the way of development. Hence "animals are only the persistent
fœtal stages or conditions of Man," who is the microcosm, and
contains within himself all the animal kingdom.
Oken was himself a careful student of embryology; von Baer[145] speaks of his work
(published in Oken and Kieser, Beiträge zur vergleichenden
Zoologie, Anatomie und Physiologie, 0912 pts., 1806-7) as forming
the turning-point in our understanding of the
mammalian ovum. He had accordingly actually observed a resemblance
in certain details of structure between the human fœtus and
the lower animals; but the peculiar form which the law took in his
hands was a consequence of his hazy philosophy. He saw the relation
of teratological to fœtal structure, for he affirmed that
"malformations are only persistent fœtal conditions" (p.
492).
The idea of comparing the embryo of higher animals with the
adult of lower was widely spread at this time among German
zoologists. We find, for example, in Tiedemann's brilliant little
textbook[146] the statement that
"Every animal, before reaching its full development, passes through
the stage of organisation of one or more classes lower in the
scale, or, every animal begins its metamorphosis with the simplest
organisation" (p. 57).
Thus the higher animals begin life as a kind of fluid animal
jelly which resembles the substance of a polyp; the young mammal,
like the lower Vertebrates, has only a simple circulation, and,
like them, lives in water (the amniotic fluid); the frog is first
like a worm, then develops gills and becomes like a fish (p. 57).
In his work on the anatomy of the brain,[147] Tiedemann established the homology of the optic
lobes in birds by comparing them with fœtal corpora
quadrigemina in man (see Serres, Ann. Sci. nat., xii., p.
112).
J. F. Meckel, in 1811, devoted a long essay to a detailed proof
of the parallelism between the embryonic states of the higher
animals and the permanent states of the lower animals. In a
previous memoir in the same collection[148] (i., 1, 1808) he had made some comparisons of
this kind in dealing with the development of the human fœtus;
in this memoir (ii., 1, 1811) he brings together all the facts
which seem to prove the parallelism.
His collection of facts is a very heterogeneous one; he mingles
morphological with physiological analogies, and makes the most
far-fetched comparisons between organs 092belonging to animals of the
most diverse groups. He compares, for instance, the placenta with
the gills of fish, of molluscs and of worms, homologising the
cotyledons with the separate tufts of gills in Tethys,
Scyllæa and Arenicola(p. 26). This is purely a
physiological analogy. He compares the closed anus of the early
human embryo with the permanent absence of an anus in
Cœlentera, and the embryo's lack of teeth with the absence of
teeth in many reptiles and fish, in birds, and in many Cetacea (p.
46).[149] These are merely chance
resemblances of no morphological importance. He considers
bladderworms as animals which have never escaped from their amnion,
and Volvox as not having developed beyond the level of an
egg (p. 7). He lays much stress upon likeness of shape and of
relative size, comparing, for instance, the large multilobate liver
of the human fœtus with the many-lobed liver of lower
Vertebrates and of Invertebrates. In general he shows himself, in
his comparisons, lacking in morphological insight.
His treatment of the vascular system affords perhaps the best
example of his method (pp. 8-25). The simplest form of heart is the
simple tubular organ in insects, and it is under this form that the
heart first appears in the developing chick. The bent form of the
embryonic heart recalls the heart of spiders; it lies at first
free, as in the mollusc Anomia. The heart consists at first
of one chamber only, recalling the one-chambered heart of
Crustacea. A little later three chambers are developed, the
auricle, ventricle, and aortic bulb; at this stage there is a
resemblance to the heart of fish and amphibia. At the end of the
fourth day the auricle becomes divided into two, affording a
parallel with the adult heart of many reptiles.
In his large text-book of a somewhat later date, the System
der vergleichenden Anatomie (i., 1821), he works out the idea
again and gives to it a much wider theoretic sweep, hinting that
the development of the individual is a repetition of the
evolutionary history of the race. Meckel was a timid believer in
evolution. He thought it quite possible that much of the variety of
animal form was due to a process of 093evolution caused by forces
inherent in the organism. "The transformations," he writes, "which
have determined the most remarkable changes in the number and
development of the instruments of organisation are incontestably
much more the consequence of the tendency, inherent in organic
matter, which leads it insensibly to rise to higher states of
organisation, passing through a series of intermediate states."[150]
His final enunciation of the law of parallelism in this same
volume shows that he considered the development of the individual
to be due to the same forces that rule evolution. "The development
of the individual organism obeys the same laws as the development
of the whole animal series; that is to say, the higher animal, in
its gradual evolution, essentially passes through the permanent
organic stages which lie below it; a circumstance which allows us
to assume a close analogy between the differences which exist
between the diverse stages of development, and between each of the
animal classes" (p. 514).
He was not, of course, able fully to prove his contention that
the lower animals are the embryos of the higher, and we gather from
the following passage that he could maintain it only in a somewhat
modified form. "It is certain," he writes, "that if a given organ
shows in the embryo of a higher animal a given form, identical with
that shown throughout life by an animal belonging to a lower class,
the embryo, in respect of this portion of its economy, belongs to
the class in question" (p. 535). The embryo of a Vertebrate might
at a certain stage of development, be called a mollusc, if for
instance, it had the heart of a mollusc.
He admits, too, that the highest animal of all does not pass
through in his development the entire animal series. But the embryo
of man always and necessarily passes through many animal stages, at
least as regards its single organs and organ-systems, and this is
enough in Meckel's eyes to justify the law of parallelism (p.
535).
In his excellent discussion of teratology Meckel points out how
the idea of parallelism throws light upon certain 094abnormalities
which are found to be normal in other (lower) forms (p. 556).[151]
We may refer to one other statement of the law of
parallelism—by K. G. Carus in his Lehrbuch der
vergleichenden Anatomie (Leipzig, 1834). The standpoint is
again that of Naturphilosophie. It is a general law of
Nature, Carus thinks, that the higher formations include the lower;
thus the animal includes the vegetable, for it possesses the
"vegetative" as well as the "animal" organs. So it is, too, by a
rational necessity that the development of a perfect animal repeats
the series of antecedent formations.
As we have said, the main credit for the enunciation of the law
of parallelism belongs to the German transcendental school; but the
law owes much also to Serres, who, with Meckel, worked out its
implications. It might for convenience, and in order to distinguish
it from the laws later enunciated by von Baer and Haeckel, be called
the law of Meckel-Serres.
Under the "theory of the repetition or multiplication of parts
within the organism" may be included, first, generalisations on the
serial homology of parts, and second, more or less confused
attempts to demonstrate that the whole organisation is repeated in
certain of the parts. The recognition of serial homologies
constituted a real advance in morphology; the "philosophical" idea
of the repetition of the whole in the parts led to many
absurdities. It led Oken to assert that in the head the whole trunk
is repeated, that the upper jaw corresponds to the arms, the lower
to the legs, that in each jaw the same bony divisions exist as in
the limbs, the teeth, for instance, corresponding to the claws
(loc. cit., p. 408). It led him to distinguish "two animals"
in every body—the cephalic and the sexual animal. Each of
these has its own organs; thus "in the perfect animal there are two
intestinal systems thoroughly distinct from each other, two
intestines which belong to two different animals, the sexual and
cephalic animal, or the plant and the animal" (p. 382). The
intestine of the sexual animal is the large intestine; the lungs of
the sexual animal are the kidneys, its glottis is the urethra, its
mouth the anus. So, too, the mouth is the stomach of the head. On
another line of thought the 095sternum is a ventral vertebral column.
Limbs are connate ribs, the digits indicating the number of ribs
included (cf. Dugès, supra, p. 88).
J. F. Meckel[152] discusses
"homologies" of this kind in the thorough and pedestrian way so
characteristic of him. Not only, he says, are the right and left
halves of the body comparable with one another, but also the upper
and the lower, the dividing line being drawn at the level of the
diaphragm. The lumbar complex corresponds to the skull, the anus to
the mouth, the urino-genital opening to the nasal opening; in
general, the urino-genital system corresponds to the respiratory,
the kidneys to the lungs, the ureters to bronchi, the testes and
ovaries to the thymus (he had observed the physiological relation
between the development of the thymus and the state of the genital
organs), the prostate and the uterus to the thyroid gland, and the
penis and clitoris to the tongue. The fore-limbs and girdle
correspond in detail with the hind limbs and the pelvis—a
point already worked out by Vicq d'Azyr; the dorsal and ventral
halves of the body are likewise comparable in some respects, the
sternum, for example, answering in the arrangement of its bones,
muscles and arteries to the vertebral column. The skeleton of each
member is in some respects a repetition of the vertebral
column.
His brother, D. A. Meckel,[153] worked out an elaborate comparison between the
alimentary canal and the genital organs, basing the legitimacy of
the comparison upon early embryological relations and upon the
state of things in Cœlentera, where genital and digestive
organs occupy the same cavity. In his view the uterus corresponded
to the stomach, the vagina to the œsophagus, the fallopian
tubes to the intestine, and so on.
The vertebral theory of the skull took its origin from the same
habit of thought. As part of the wider idea of the metameric
repetition of parts it had some scientific worth, but the theory
was pushed too far, and the facts were twisted to suit it. Among
annulate animals the theory of repetition found ample scope; Oken
was able to compare with justice 096the jaws of crabs and insects with
their other limbs, as Savigny did later in a more scientific way.
Among Vertebrates the application of the theory of serial
repetition was not so obvious, except in the case of the
vertebræ. Goethe seems to have been the first to hit upon the
idea that the skull is composed of a number of vertebræ,
serially homologous with those of the vertebral column. He tells us
that the idea flashed into his mind when contemplating in the
Jewish cemetery at Venice a dried sheep's skull. The discovery was
made in 1790, but not published till 1820.[154]
The idea seems to have been taught by Kielmeyer, one of the
earliest of the "philosophers of nature," but it was not published
by him.
In a book (Cours d'Études médicales),
published in 1803, Burdin assimilated the skull to the vertebral
column.
Oken, in an inaugural dissertation (Programm) Ueber die
Bedeutung der Schädelknochen,[155] published in 1807, gave to the theory its
necessary development. Autenrieth, also in 1807,[156] distinguishing separate ganglia in the
brain, was not far from the hypothesis that each of these ganglia
must have its separate vertebra.
In 1808 Duméril read a paper to the Académie des
Sciences in which he compared the skull to a gigantic vertebra,
basing his hypothesis on the similarity existing between the crests
and depressions on the hinder part of the skull and those on the
posterior surfaces of the vertebræ.
After Oken's work the vertebral theory was taken up generally by
both the German and the French anatomists. Spix published in 1815 a
large volume on the skull, entitled Cephalogenesis,
distinguishing (as Oken did at first) three cranial vertebræ.
Bojanus in his Anatome testudinis europæae (1819), and
in a series of papers in Isis (1817-1819, and 1821)
established the existence of a fourth cranial vertebra, and this
was accepted by Oken in the later editions of his Lehrbuch.
Meckel and Carus among the Germans, de Blainville and E. Geoffroy
among the French, contributed to 097the development of the theory. In
England the theory was championed particularly by Richard Owen.
It was one thing to assert in a moment of inspiration that the
skull was composed of modified vertebræ; it was quite another
to demonstrate the relation of the separate bones of the skull to
the supposed vertebræ. Upon this much uncertainty reigned;
there was not even unanimity as to the number of vertebræ to
be distinguished. Goethe found six vertebræ in the skull;
Spix, and at first Oken, three only, Geoffroy seven; the accepted
orthodox number seems to have been four (Bojanus, Oken, Owen).
As an example of the method of treatment adopted we may take
Oken's matured account of the composition of the cranial
vertebræ, as given in the English translation of his
Lehrbuch. "To a perfect vertebra," he says, "belong at least
five pieces, namely, the body, in front the two ribs, behind the
two arches or spinous processes" (p. 370). In the cervical
vertebræ the transverse processes represent the ribs. The
skull consists of four vertebræ, the occipital, the parietal,
the frontal and the nasal, or, named after the sense with which
each is associated, the auditory, the lingual, the ocular and the
olfactory. The "bodies" of these vertebræ are the body of the
occipital (basioccipital), the two bodies of the sphenoid (basi- and
pre-sphenoid), and the vomer. The transverse processes of each are
the condyles of the occipitals (exoccipitals), the alæ of the
two sphenoids (alisphenoids and orbitosphenoids) and the lateral
surfaces of the vomer. The arches or spinous processes are the
occipital crest, the parietals, the frontals, and the nasals.
The cranium is thus composed of four rings of bone, each
composed of the typical elements of a vertebra.
The arbitrary nature of the comparison is obvious enough. As
Cuvier pointed out in the posthumous edition of his
Leçons, it is only the occipital segment that shows
any real analogy with a vertebra—an analogy which Cuvier
ascribed to similarity of function. He admitted a faint resemblance
of the parietal segment to a vertebra:—"The body of the
sphenoid does indeed look like a repetition of the basioccipital,
but having a different function it takes on another form,
especially above, by reason of its posterior 098clinoid
apophyses."[157] He denied the
resemblance of the frontal and nasal "vertebræ" to true
vertebræ, pointing out that both parietals and frontals are
bones specially developed for the purpose of roofing over and
protecting the cerebrum.
A very curious development was given to the vertebral theory by
K. G. Carus, who seems to have taken as his text a saying of Oken's,
that the whole skeleton is only a repeated vertebra.[158] His system is worthy of some
consideration, for he tries to work out a geometry of the
skeleton.[159]
His method of deduction is a good example of pure
Naturphilosophie. Life, he says, is the development of
something determinate from something indeterminate. A finite
indeterminate thing, that is, a liquid, must take a spherical form
if it is to exist as an individual. Hence the sphere is the
prototype of every organic body. Development takes place by
antagonism, by polarity, typically by the division and
multiplication of the sphere. In the course of development the
sphere may change, by expansion into an egg-shaped body, or by
contraction into a crystalline form, the changes due to expansion
being typical of living things, those due to contraction being
typical of dead. At the surface of the primitive living sphere is
developed the protective dermatoskeleton, which naturally
takes the shape of a hollow sphere; round the digestive cavity
which is formed in the living sphere is developed the
splanchnoskeleton; round the nervous system (which is, as it
were, the animal within the animal) is developed the
neuroskeleton. All skeletal formations belong to one or
other of these systems.
Carus defines his aim to be the discovery of the inner law which
presides over the formation of the skeleton throughout the animal
kingdom; he desires to know "how such and such a formation is
realised in virtue of the eternal laws of reason" (iii., p. 93).
Here we touch the kernel of Naturphilosophie—the
search for rational laws which are active in Nature; the discontent
with merely empirical laws.
099The thesis which Carus sustains is that all
forms of skeleton, whether of dermatoskeleton, splanchnoskeleton,
or neuroskeleton, can be deduced from the hollow sphere, which is
the primary form of any skeleton whatsoever (p. 95). That means, put
empirically, that every skeleton can be represented schematically
by a number of hollow spheres, suitably modified in shape, and
suitably arranged. The chief modification in shape exhibited by
bones is one which is intermediate between the organic and the
crystalline series of modifications of the sphere. The organic
modifications are bounded by curved lines, the crystalline by
straight; the intermediate partly by curved and partly by straight
lines. They are the dicone (the shape of a diabolo) and the
cylinder. These forms must necessarily be of importance for the
skeleton, which is intermediate between the organic and the
inorganic. "The dicone embodies the real significance of the bone,"
writes Carus. Each dicone and cylinder composing the skeleton is
called by Carus a vertebra.
We may expect then all skeletons to be composed of spheres,
cylinders and dicones in diverse arrangements. Nature being
infinite, all the possible types of arrangement of these elements
must exist in the test or skeleton of some animal, living, fossil,
or to come (p. 127). One conceives easily what the main types of
skeleton must be. In some animals, e.g., sea-urchins, the
skeleton is a simple sphere; in others, e.g., starfish,
secondary rows of spheres radiate out from a central sphere or
ring; in annulate animals the skeleton consists of a row of
partially fused spheres.
In Vertebrates the arrangement is more complex. There are first
the protovertebral rings of the dermatoskeleton, these being
principally the ribs, limb-girdles, and jaws. Round the central
nervous system are developed the deutovertebral rings of the
neuroskeleton (vertebræ in the ordinary sense). The apophyses
and bodies of the vertebræ, and the bones of the members[160] are composed of columns
of tritovertebræ, or vertebræ of the third order. Thus
the whole vertebrate skeleton is a particular arrangement of
vertebræ, which 100in their turn are modifications of the
primary hollow sphere.
The German transcendentalists were more or less contemporary
with E. Geoffroy, and no doubt influenced him, especially in his
later years, as they certainly did his follower Serres. Oken indeed
wrote, in a note[161] appended to
Geoffroy's paper on the vertebral column of insects, that "Mr
Geoffroy [sic] is without a doubt the first to introduce in
France Naturphilosophie into comparative anatomy, that is to
say, that philosophy one of whose doctrines it is to seek after the
signification of organs in the scale of organised beings."
This is, however, an exaggeration, for Geoffroy was primarily a
morphologist, whereas the morphology of the German
transcendentalists was only a side-issue of their
Naturphilosophie.
Geoffroy, on his part, exercised some influence on the
transcendentalists. He asserts[162] indeed that Spix got some of the ideas published
in the Cephalogenesis (1815) from attending his course of
lectures in 1809. It is certainly the case that Spix published
before Geoffroy the view that the opercular bones are homologous
with the ear-ossicles, adopting, however, a different homology for
the separate bones.[163]
Some speculations seem to have been common to both
schools—for instance, the law of Meckel-Serres, the vertebral
theory of the skull, and the recognition of serial homology in the
appendages of Arthropods (Savigny, Oken). Latreille and
Dugès, as well as Serres, clearly show in their theoretical
views the influence of Oken and the other transcendentalists.
Geoffroy's principle of connections and law of compensation were
recognised by some at least of the Germans.
But whatever his actual historical relations may have been with
the German school, Geoffroy was vastly their superior in the matter
of pure morphology. He alone brought to clear consciousness the
principles on which a pure morphology could be based: the Germans
were transcendental philosophers first, and morphologists
after.
101One understands from this how J. F. Meckel,
who was in some ways the leading comparative anatomist in Germany
at this time, could be at once a transcendentalist and an opponent
of Geoffroy. Meckel had a curiously eclectic mind. A disciple of
Cuvier, having studied in 1804-6 the rich collections at the Museum
in Paris, the translator of Cuvier's Leçons d'anatomie
comparée, he earned for himself the title of the "German
Cuvier," partly through the publication of his comprehensive
textbook (System der vergl. Anatomie, 5 vols.), partly by
his extensive and many-sided research work, partly by his
authoritative teaching. His System shows in almost every
page of its theoretical part the influence of Cuvier; and it is
through having assimilated Cuvier's teaching as to the importance
of function that Meckel combats Geoffroy's law of connections, at
least in its rigorous form. He submits that the connections of
bones and muscles must change in relation to functional
requirements. He rejects Geoffroy's theory of the vertebrate nature
of Articulates. Generally throughout his work the functional point
of view is well to the fore.
Yet at heart Meckel was a transcendentalist of the German
school. His vagaries on the subject of "homologues" leave no doubt
about that, and, in spite of Cuvier, he believed, though not very
firmly, in the existence of one single type of structure.
A Cuverian by training, his lack of morphological sense threw
him into the ranks of the transcendentalists, to whom perhaps he
belonged by nature.
102
CHAPTER VIII
TRANSCENDENTAL ANATOMY IN ENGLAND—RICHARD OWEN
Richard Owen is the epigonos of
transcendental morphology; in him its guiding ideas find clear
expression, and in his writings are no half-truths struggling for
utterance.
Fig. 4.—Ideal
Typical Vertebra. (After Owen.)
But he was, though a staunch transcendentalist, an
eclectic of the older ideas current in his time; for he picked out what was
best in the older systems—Cuvier's teleology, Geoffroy's
principle of connections, Oken's idea of the serial repetition of
parts. In particular, he assimilated the teaching of Cuvier, the
great opponent of the transcendentalists, and reconciled it 103in part
with his own transcendentalism. His main theoretical views are to
be found in his volume On the Archetype and Homologies of the
Vertebrate Skeleton (London, 1848). The master-idea of the book
is that the vertebrate skeleton consists of a series of comparable
segments, each of which Owen calls a vertebra.
Fig 5.—Natural
Typical Vertebra; Thorax of a Bird. (After Owen.)
His definition of a vertebra is,
"one of those segments of the endo-skeleton which constitute the
axis of the body, and the protecting canals of the nervous and
vascular trunks" (p. 81). The parts of a typical vertebra are shown
in Fig. 4, which is copied from Owen's Fig.
14.
104In Fig. 5 (page 103) is
shown an actual vertebra, as Owen conceives it, the "vertebra"
being that of a bird.
A segment of sternum is included as the "hæmal spine" of
the vertebra (hs); the vertebral rib is the "pleurapophysis"
(pl); the sternal rib the "hæmapophysis" (h);
the uncinate process of the vertebral rib is known as the
"diverging appendage" (a). The whole vertebrate skeleton is
composed of a series of vertebræ which show these typical parts.
We arrive thus at the conception of an "Archetype" of the
vertebrate skeleton, such as is represented in Fig. 6.
The archetype is only a scheme of what is usually constant in
the vertebrate skeleton, and both the number and the arrangement of
the bones in any real Vertebrate are subject to variation. "It has
been abundantly proved," Owen writes, towards the end of his
volume, "that the idea of a natural segment (vertebra) of the
endoskeleton does not necessarily involve the presence of a
particular number of pieces, or even a determinate and unchangeable
arrangement of them. The great object of my present labour has been
to deduce ... the relative value and constancy of the different
vertebral elements, and to trace the kind and extent of their
variations within the limits of a plain and obvious maintenance of
a typical character" (p. 146).
It goes without saying that Owen considered the skull to be
formed of vertebræ—the vertebral theory of the skull
was, in his system, a deduction from the vertebral theory of the
skeleton. He recognised four cranial vertebræ; the arrangement of
them, and the relation of their constituent bones to the parts of
the typical vertebra are shown in the table appearing on page 106.
So far as their first three elements are concerned, these
vertebræ are practically identical with the vertebræ
distinguished in the classical vertebral theory of the skull, as
enunciated by Oken. A divergence appears with the determination of
the other elements of the vertebræ. The upper and lower jaws
are associated with the nasal and frontal vertebræ
respectively, not however as limbs of the head, but as constituent
elements of these vertebræ. In the same way the hyoid
apparatus is part and parcel of the parietal vertebra, and the
pectoral girdle and fore-limbs part of the occipital vertebra.
105
Fig. 6.—The
Archetype of the Vertebrate Skeleton. (After Owen.)
106
Cranial Vertebræ.[164] (After Owen, 1848, p. 165.)
| Vertebræ. |
Occipital. |
Parietal. |
Frontal. |
Nasal. |
| Centra. |
Basioccipital. |
Basisphenoid. |
Presphenoid. |
Vomer. |
| Neurapopbyses. |
Exoccipital. |
Alisphenoid. |
Orbitosphenoid. |
Prefrontal. |
| Neural Spines. |
Supraoccipital. |
Parietal. |
Frontal. |
Nasal. |
| Parapopbyses. |
Paroccipital. |
Mastoid. |
Postfrontal. |
None. |
| Pleurapophyses. |
Scapular. |
Stylohyal. |
Tympanic. |
Palatal. |
| Hæmapophyses. |
Coracoid. |
Ceratohyal. |
Articular. |
Maxillary. |
| Hæmal Spines. |
Episternum. |
Basihyal. |
Dentary. |
Premaxillary. |
| Diverging Appendage. |
Fore-limb or Fin. |
Branchio-stegals. |
Operculum. |
Pterygoid and Zygoma. |
Owen's reasons for considering the pectoral girdle and the
fore-limb part of the occipital vertebra are as follows. In fish
the pectoral girdle is slung to the skull by means of the
post-temporal bone (supra-scapula, according to Owen) which abuts
on the occipital arch. In Lepidosiren, whose skeleton
resembles the archetype in many ways, the pectoral girdle is
likewise attached to the occipital segment.
In most other Vertebrates the pectoral girdle has shifted
backwards along the vertebral column, by a "metastasis" (Geoffroy)
similar to that by which the pelvic fins in many fish have shifted
up close to the pectoral girdle. The scapula (with supra-scapula)
is the pleurapophysis, the coracoid the hæmapophysis, of the
occipital vertebra. The clavicle is homologised with the slender
bone in fish now known as the post-clavicle, which shows a
connection with the first or atlas vertebra of the vertebral
column, forming, according to Owen, the hæmapophysis of the
atlas. Owen considers it no objection to this view that in other
Vertebrates the 107clavicle is anterior to the
coracoid—"its anterior position to the coracoid in the
air-breathing Vertebrata is no valid argument against the
determination, since in these we have shown that the true scapular
arch is displaced backwards" (On the Nature of Limbs, p. 63,
London, 1849). In the pelvic girdle the ilium corresponds to the
scapula, the ischium to the coracoid, the pubis to the clavicle.
Hence the ilium is a pleurapophysis, the ischium and pubis are both
hæmapophyses. The fore-limb is the developed "appendage" of
the occipital vertebra, the hind-limb the developed "appendage" of
the pelvic vertebra. They are serially homologous with, for
example, the uncinate processes of the ribs in birds (see Figs. 5
and 6). The fore-limb is a simple filament in Lepidosiren,
and presents few joints in Proteus and Amphiuma; in
other air-breathing Vertebrates it shows a more complete
development, the humerus, radius and ulna, and the bones of the
wrist and hand becoming differentiated out.
As the fore-limb is equivalent to a single bone of the
archetype, it is said to be, in its developed state,
"teleologically compound" (p. 103).
Since in the archetype every vertebra has its appendage, more
than two pairs of locomotory limbs might have been developed. "Any
given appendage might have been the seat of such developments as
convert that of the pelvic arch into a locomotive limb; and the
true insight into the general homology of limbs leads us to
recognise many potential pairs in the typical endoskeleton. The
possible and conceivable modifications of the vertebrate archetype
are far from having been exhausted in the forms which have hitherto
been recognised, from the primæval fishes of the
palæozoic ocean of this planet up to the present time" (p.
102). It is not of the essence of the vertebrate type to be
tetrapodal.
In determining homologies Owen remained true to Geoffroy's
principle of connections. Speaking of an attempt which had been
made to determine homologies by the mode of development, he writes,
"There exists doubtless a close general resemblance in the mode of
development of homologous parts; but this is subject to
modification, like the forms, proportions, functions, and very
substance of such 108parts, without their essential homological
relationships being thereby obliterated. These relationships are
mainly, if not wholly, determined by the relative position and
connection of the parts, and may exist independently of form,
proportions, substance, function and similarity of development. But
the connections must be sought for at every period of development,
and the changes of relative position, if any, during growth, must
be compared with the connections which the part presents in the
classes where vegetative repetition is greatest and adaptive
modification least" (p. 6). It is interesting to note that in
Owen's opinion comparative anatomy explains embryology. Thus the
scapula, which is the pleurapophysis of the occipital vertebra, is
vertical on its first appearance in the embryo of tetrapoda, and
lies close up to the head (On the Nature of Limbs, p.
49)—the embryo shows a greater resemblance to the archetype
than the adult. "We perceive a return to it, as it were, in the
early phases of development of the highest organised of the
actually existing species, or we ought rather to say that
development starts from the old point; and thus, in regard to the
scapula, we can explain the constancy of its first appearance close
to the head, whether in the human embryo or in that of the swan,
also its vertical position to the axis of the spinal column, by its
general homology as the rib or 'pleurapophysis' of the occipital
vertebra" (Limbs, p. 56).
We owe to Owen the first clear distinction between "homologous"
and "analogous" organs; it was he who first proposed the terms
"homologue" and "analogue," which he defined as
follows:—"Analogue. A part or organ in one animal
which has the same function as another part or organ in a different
animal." "Homologue. The same organ in different animals
under every variety of form and function."[165]
He introduced also useful distinctions between Special, General,
and Serial Homology. "The relations of homology," he writes, "are
of three kinds: the first is that above defined, viz., the
correspondency of a part or organ, determined by its relative
position and connections, with a part or organ in a different
animal; the determination of which homology indicates that such
animals are constructed on a common 109type; when, for example, the
correspondence of the basilar process of the human occipital bone
with the distinct bone called 'basi-occipital' in a fish or
crocodile is shown, the special homology of that process is
determined. A higher relation of homology is that in which a part
or series of parts stands to the fundamental or general type, and
its enunciation involves and implies a knowledge of the type on
which a natural group of animals, the Vertebrate, for example, is
constructed. Thus when the basilar process of the human occipital
bone is determined to be the 'centrum' or 'body' of the last
cranial vertebra, its general homology is enunciated.
"If it be admitted that the general type of the vertebrate
endoskeleton is rightly represented by the idea of a series of
essentially similar segments succeeding each other longitudinally
from one end of the body to the other, such segments being for the
most part composed of pieces similar in number and arrangement, and
though sometimes extremely modified for special functions, yet
never so as to wholly mask their typical character—then any
given part of one segment may be repeated in the rest of the
series, just as one bone may be reproduced in the skeletons of
different species, and this kind of repetition or representative
relation in the segments of the same skeleton I call 'serial
homology'" (p. 7). As an example of serial homology we might take
the centra of the vertebræ—the vomer, the presphenoid,
the basisphenoid, the basioccipital and the series of centra in the
spinal column. Such serially repeated parts are called
homotypes (p. 8).
Not all the bones of the vertebrate skeleton are included in the
archetype as constituents of the vertebræ. Thus the branchial
and pharyngeal arches are accounted part of the splanchnoskeleton,
as belonging to the same category as the heart bone of some
ruminants, and the ossicles of the stomach in the lobster (p. 70).
The ossicles of the ear in mammals are "peculiar mammalian
productions in relation to the exalted functions of a special organ
of sense" (p. 140, f.n.). This recognition of a possible
development of new organs to meet new functions shows unmistakably
the influence of Cuvier. Owen was indeed well aware of the
importance of the functional aspect of living things, and he often
adopted 110the teleological point of view. As a true
morphologist, however, he held that the principle of adaptation
does not suffice to explain the existence of special homologies.
The ossification of the bones of the skull from separate centres
may be purposive in Eutheria, in that it prevents injury to the
skull at birth; but how explain on teleological principles the
similar ossification from separate centres in marsupials, birds and
reptiles? How explain above all the fact that the centres are the
same in number and relative position in all these groups? Surely we
must accept the idea of an archetype "on which it has pleased the
divine Architect to build up certain of his diversified living
works" (p. 73).
In his study of centres of ossification, Owen made in point of
theory a distinct advance on his predecessors. We saw that Geoffroy
recognised the importance of studying the ossification of the
skeleton, and that Cuvier accepted such embryological evidence as
an aid in determining homologies. Owen pointed out that it was
necessary to distinguish between centres of ossification which were
teleological in import and such as were purely indicative of
homological relationships. Many bones, single in the adult, arise
from separate centres of ossification, but we must distinguish
between "those centres of ossification that have homological
relations, and those that have only teleological ones; i.e.,
between the separate points of ossification of a human bone which
typify vertebral elements, often permanently distinct bones in the
lower animals; and the separate points which, without such
signification, facilitate the progress of osteogeny, and have for
their obvious final cause the well-being of the growing animal" (p.
105). There is, for example, a teleological reason why in mammals
and leaping Amphibia (e.g., frogs), the long bones should
ossify first at their ends, for the brain is thus protected from
concussion; in reptiles that creep there is less danger of
concussion, and the long bones ossify in the middle (p. 105). But
there is no teleological reason why the coracoid process of the
scapula should in all mammals develop from a separate centre. The
coracoid is however a real vertebral element (hæmapophysis),
and in monotremes, birds and reptiles it is in the adult a large
and separate bone. Its ossification from a separate 111centre in
mammals has therefore a homological significance. The scapula in
mammals is an example of what Owen calls a "homologically compound"
bone. All those bones which are formed by a coalescence of parts
answering to distinct elements of the typical vertebra are
"homologically compound" (p. 105). On the other hand, "All those
bones which represent single vertebral elements are 'teleologically
compound' when developed from more than one centre, whether such
centres subsequently coalesce, or remain distinct, or even become
the subject of individual adaptive modifications, with special
joints, muscles, etc., for particular offices" (p. 106). The
limb-skeleton, corresponding as it does to a single bone of the
archetype, is the typical example of a teleologically compound
bone. Owen in his definition of teleological compoundness has
combined two kinds of adaptation—(1) temporary adaptation of
bones to the exigencies of development, birth and growth
(e.g., development of long bones from separate centres); (2)
definitive adaptation of a skeletal part to the functions which it
has to perform (e.g., teleological structure of limbs). Such
adaptations are, so to speak, grafted on the archetype.
Owen's general views on the nature of living things merit some
attention. Organic forms, according to Owen, result from the
antagonistic working of two principles, of which one brings about a
vegetative repetition of structure, while the other, a teleological
principle, shapes the living thing to its functions. The former
principle is illustrated in the archetype of the vertebrate
skeleton, in the segmentation of the Articulates, in the almost
mathematical symmetry of Echinoderms, and the actually crystalline
spicules of sponges. It is the same principle which causes
repetition of the forms of crystals in the inorganic world. "The
repetition of similar segments in a vertebral column, and of
similar elements in a vertebral segment, is analogous to the
repetition of similar crystals as the result of polarising force in
the growth of an inorganic body" (p. 171). This "general polarising
force" it is which mainly produces the similarity of forms, the
repetition of parts, and generally the signs of the unity of
organisation. The adaptive or "special organising force" or
ἰδέα, on the other hand, produces the
diversity of organic 112beings. In every species these two forces are
at work, and the extent to which the general polarising or
"vegetative-repetition-force" is subdued by the teleological is an
index of the grade of the species.
This view is analogous to the Geoffroyan conception that the
diversity of form is limited by the unity of plan. Owen thus ranges
himself with Geoffroy against Cuvier, who considered that diversity
of form is limited only by the principle of the adaptation of
parts.
113
CHAPTER IX
KARL ERNST VON BAER
Von Baer was recognised as the
founder of embryology even by his contemporaries. His predecessors,
Aristotle,[166] Fabricius,[167] Harvey,[168] Malpighi,[169] Haller,[170] Wolff,[171] had made a beginning with the study of
development; von Baer, by the thoroughness of his observation and
the strength of his analysis, made embryology a science.
It was to one of the German transcendentalists that von Baer
owed the impulse to study development. Ignatius Döllinger,
Professor in Würzburg, induced three of his pupils, Pander,
d'Alton and von Baer, to devote themselves to embryological
research. The development of animals was at this time little known,
in spite of recent work by Meckel (1815 and 1817), Tiedemann
(Anatomie u. Bildungsgeschichte des Gehirns, 1816), by Oken
(loc. cit., supra, p. 90), and some others.
Pander, with whom apparently Döllinger and d'Alton
collaborated, was the first to publish his results;[172] von Baer, who through absence from
Würzburg had for a time dropped his embryological studies,
started to work in 1819, after the publication of Pander's
treatise, and produced in 1828 the first volume of his master-work,
Ueber Entwickelungsgeschichte 114der Thiere. Beobachtung und
Reflexion (Königsberg, 1828). The second volume followed
in 1837, but dates really from 1834, and was published in an
incomplete form. This second volume is intended as an introduction
to embryology for the use of doctors and science students. In it
von Baer describes in full detail the development of many
vertebrate types—chick, tortoise, snake, lizard, frog, fish,
several mammals and man, basing his remarks largely upon his
personal observations, but taking account also of all contemporary
work. A separate account of the development of a fish (Cyprinus
blicca) appeared in 1835.[173]
We shall concentrate attention on the first volume. This volume
contains the first full and adequate account of the development of
the chick, followed by a masterly discussion of the laws of
development in general.
When we consider that von Baer worked chiefly with a simple
microscope and dissecting needles, the minuteness and accuracy of
his observations are astonishing. He described the main facts
respecting the development of all the principal organs, and if,
through lack of the proper means of observation, he erred in
detail, he made up for it by his masterly understanding and
profound analysis of the essential nature of development. His
account of the development of the chick is a model of what a
scientific memoir ought to be; the series of "Scholia" which follow
contain the deductions he made from the data, and, in so far as
they are direct generalisations from experience, they are valid for
all time.
The first Scholion is directed against the theory of
preformation, and succeeds in refuting it on the ground of simple
observation. The theme of the second Scholion is that the essential
nature (die Wesenheit) of the animal determines its
differentiation, that no stage of development is solely determined
by the antecedent stage, but that throughout all stages the
Wesenheit or idea of the definitive whole exercises
guidance. This guidance is shown most clearly in the regulatory
processes of the germ, whereby the large individual variations
commonly presented by the 115early embryo are compensated for or
neutralised in the course of further development. Baer in this
shows himself a vitalist.
It is, however, the third and subsequent Scholia which must here
particularly occupy our attention, for it is in these that von Baer
comes to grips with morphological problems. Already in the second
Scholion he had definitely enunciated the law which runs as a theme
throughout the volume, the observational and the theoretical part
alike, the law that development is essentially a process of
differentiation by which the germ becomes ever more and more
individualised. "The essential result of development," he writes,
"when we consider it as a whole, is the increasing independence
(Selbständigkeit) of the developing animal" (p. 148).
In the third Scholion he elaborates this thought and shows that
differentiation takes place in triple wise. The three processes of
differentiation are "primary differentiation" or layer-formation,
"histological differentiation" within the layers, and the
"morphological differentiation" of primitive organs.
The first of these differentiations in time is the formation of
the germ-layers, which takes place by a splitting or separation of
the blastoderm into a series of superimposed lamellæ. Baer's
account of the process in the chick is as follows:—
"First of all, the germ separates out into heterogeneous layers,
which with advancing development acquire ever greater
individuality, but even on their first appearance show rudiments of
the structures which will characterise them later. Thus in the germ
of the bird, so soon as it acquires consistency at the beginning of
incubation, we can distinguish an upper smooth continuous surface
and a lower more granular surface. The blastoderm separates
thereupon into two distinct layers, of which the lower develops
into the plastic body-parts of the embryo, the upper into the
animal parts; the lower shows clearly a further division into two
closely connected subsidiary layers—the mucous layer and the
vessel-layer; the original upper layer also shows a division into
two, which form respectively the skin and the parts which I have
called the true ventral and dorsal 116plates—parts which contain
in an undifferentiated state the skeletal and muscular systems, the
connective tissues, and the nerves belonging to these. In order to
have a convenient term for future use, I have named this layer the
muscle-layer" (p. 153).
The process of delamination results then in the formation of
four layers, of which the upper two (composing the "animal" or
"serous" layer) will give origin to the animal (neuromuscular) part
of the body, the lower pair to the plastic or vegetative organs.
The uppermost layer will form the external covering of the embryo,
and also the amniotic folds; from it there differentiates out at a
very early stage the rudiment of the central nervous system,
forming a more or less independent layer. Below the outermost layer
lies the layer from which are formed the muscular and skeletal
systems, and beneath this "muscle-layer" comes the "vessel-layer,"
which gives origin to the main blood-vessels. The innermost layer
of the four will form the mucous membrane of the alimentary canal
and its dependencies; at the present stage, however, it is, like
the other layers, a flat plate.
From all these layers tubes are developed by the simple bending
round of their edges. The outermost layer becomes the investing
skin-tube of the embryo; the layer for the nervous system forms the
tubular rudiment of the brain and spinal cord; the mucous layer
curls round to form the alimentary tube; the muscle layer grows
upwards and downwards to form the fleshy and osseous tube of the
body wall; even the vessel layer forms a tube investing the
alimentary canal, but a part of it goes to form the medial
"Gekröse," or mesenterial complex, which departs considerably
from the tubular form.
When these tubes or "fundamental organs" are formed the process
of primary differentiation is complete. The fundamental organs,
however, have at no time actually the form of tubes; they exist as
tubes only ideally, for morphological and histological
differentiation go on concurrently with the process of primary
differentiation.
Through morphological differentiation the various parts of the
fundamental organs become specialised, through 117unequal growth,
first into the primitive organs and then into the functional organs
of the body. "Single sections of the tubes originally formed from
the layers develop individual forms, which later acquire special
functions: these functions are in the most general way subordinate
elements of the function of the whole tube, but yet differ from the
functions of other sections. Thus the nerve-tube differentiates
into sense-organs, brain and spinal cord, the alimentary tube into
mouth cavity, œsophagus, stomach, intestine, respiratory
apparatus, liver, bladder, etc. This specialisation in development
is bound up with increased or diminished growth" (p. 155). Rapid
growth concentrated at one point brings about an evagination; in
this manner are formed the sense-organs from the nerve-tube, the
liver and lungs from the alimentary tube. Or increased growth over
a section of a tube causes it to swell out; in this wise the brain
develops from the nerve-tube, the stomach from the alimentary tube.
The segmentation which soon becomes so marked, particularly in the
muscle layer, is also due to a process of morphological
differentiation.
At the same time that the organs of the body are being thus
roughly blocked out and moulded from the germ-layers the third
process of differentiation is actively going on. "In addition to
the differentiation of the layers, there follows later another
differentiation in the substance of the layers, whereby cartilage,
muscle and nerve separate out, a part also of the mass becoming
fluid and entering the bloodstream" (p. 154). Through histological
differentiation the texture of the layers and incipient organs
becomes individualised. In its earliest appearance the germ
consists of an almost homogeneous mass, containing clear or dark
globules suspended in its substance (ii., p. 92). This homogeneity
gives place to heterogeneity; the structureless mass becomes
fibrous to form muscles, hardens to form cartilage or bone, becomes
liquid to form the blood, differentiates in a hundred other
ways—into absorbing and secreting tissues, into nerves and
ganglia, and so forth. It will be noticed that the concept of
histological differentiation is independent of the cell-theory; it
signifies that textural differentiation which leads to the
formation of tissues in 118Bichat's sense. The tissues and the
germ-layers stand in fairly close relation with one another, for
while certain tissues are formed chiefly but not exclusively in one
layer, others are formed only in one layer and never elsewhere. For
example, peripheral nerves are for the most part formed in the
muscle layer, though the bulk of the nervous tissue is formed in
the walls of the nerve tube; similarly blood and blood-vessels may
arise from almost any layer, though their chief seat of origin is
the vessel-layer; on the other hand, bone is formed only in the
muscle-layer (i., p. 155, ii., pp. 92-3).
This relation of tissue to germ-layer was more fully discussed
and brought into greater prominence by Remak, from the standpoint
of the cell-theory, and it will occupy us in a later chapter (Chap. XII.).
The fourth Scholion elaborates the analysis of developmental
processes still further, and discusses in particular the scheme of
development which is shown by the Vertebrata. The characteristic
structure of the vertebrate body is brought about by a "double
symmetrical" rolling together of the germ-layers, whereby two main
tubes are formed, one above and one below the axis of the body,
which is the chorda. The dorsal tube is formed by the two animal
layers, the ventral tube by all the layers combined (see Fig. 7).
The process is indicated with sufficient clearness in the
diagram. It will be seen that the real foundation and framework of
the arrangement is the muscle-layer, with its two tubes, one
surrounding the central nervous system and forming the "dorsal
plates," the other surrounding the body cavity and forming the
"ventral plates." In the dorsal plates, which early show metameric
segmentation, the investing skeleton of the neural axis develops;
in the ventral plates are formed the ribs, the ventral arches of
the vertebræ, the hyoid, the lower jaw and other skeletal
structures.
The alimentary or "mucous" tube and the part of the vessel layer
which invests it become so closely bound up with one another as to
form a single primitive organ—the alimentary canal. The
muscles of the alimentary canal are accordingly in all probability
developed in the investing part of the vessel layer. From the
"Gekröse," or remaining part of the vessel layer develop the
Wolffian bodies (Urnieren, 119Pronephros), the kidneys, the sex
glands, and the series of "blood-glands"—suprarenals,
thyroid, thymus and spleen. Baer did not attach any special
morphological significance to the peritoneal lining of the body
cavity, as is done in more modern forms of the germ-layer theory.
The gill-slits were largely formed by outgrowths from the
alimentary canal.
Fig. 7.—Ideal
Transverse Section of a Vertebrate Embryo. (After von Baer.)
| a. Chorda. |
e. Vessel-layer. |
i. Amnion. |
| b. Dorsal plates. |
f. Alimentary tube. |
k. Serous membrane. |
| c. Ventral plates. |
g. Pronephros. |
l. Tolk sac. |
| d. Spinal cord. |
h. Skin. |
|
In his germ-layer theory von Baer was influenced a good deal by
Pander, to whom the actual discovery of the process of
layer-formation is due. Pander, however, had distinguished only
three germ-layers, an upper "serous" layer, a lower "mucous" layer
and a middle "vessel-layer." He it was who introduced the terms
"Keimhaut" (blastoderm) and "Keimblatt" (germ-layer).
The honour of being the founder of the germ-layer theory is
sometimes attributed to C. F. Wolff, notably by Kölliker and O.
Hertwig. Wolff, it is true, in his memoir De formatione
intestinorum (1768-9) showed that the alimentary canal was
120first formed as a flat plate which folded
round to form a tube, and in a somewhat vaguely worded passage he
hinted that a similar mode of origin might be found to hold good
for the other organ-systems. But it seems clear that Wolff had no
definite conception of the process of layer-formation as the first
and necessary step in all differentiation. This, at any rate, was
von Baer's opinion, who assigns to Pander the glory of the
discovery of the germ-layers. "You," he writes, "through your
clearer recognition of the splitting of the germ—a process
which remained dark to Wolff—have shed a light upon all forms
of development" (p. xxi.).
We have now seen, following von Baer's exposition, how
development is essentially a process of differentiation, a progress
from the general to the special, from the homogeneous to the
heterogeneous; we have analysed the process into its three
subordinate processes—primary, histological and morphological
differentiation. So far we have considered development in general
and the laws which govern it; we have now to consider the varieties
of development which the animal kingdom offers in such profusion,
in order to discover what relations exist between them. This is the
problem set in the fifth Scholion. Baer at once brings us face to
face with the solution of the problem attempted in the
Meckel-Serres law. It is a generally received opinion, he writes,
that the higher animals repeat in their development the adult
stages of the lower, and this is held to be the essential law
governing the relation of the variety of development to the variety
of adult form. This opinion arose when there was little real
knowledge of embryology; it threw light indeed upon certain cases
of monstrous development, but it was pushed altogether too far. It
complicated itself with a belief in a historical
evolution;—"People gradually learnt to think of the different
animal forms as developed one from another—and seemed, in
some circles at least, determined to forget that this metamorphosis
could only be conceptual" (p. 200). At the same time the theory of
parallelism led men to rehabilitate the outworn conception of the
scale of beings, to maintain that animals form one single series of
increasing complexity, a scale which the higher members must mount
step by step in their 121development—from which it followed that
evolution, whether conceived as an ideal or as an historical
process, could take place only along one line, could be only
progressive or regressive. Not all the supporters of the theory of
parallelism held these extreme views, but conclusions of this kind
were natural and logical enough.
Von Baer had soon found in the course of his embryological
studies that the facts did not at all fit in with the doctrine of
parallelism; the developing chick, for example, was at a very early
stage demonstrably a Vertebrate, and did not recapitulate in its
early stages the organisation of a polyp, a worm or a mollusc. He
had published his doubts in 1823, but his final confutation of the
theory of parallelism is found in this Scholion.
If it were true, he says, that the essential thing in the
development of an animal is this repetition of lower organisations,
then certain deductions could be drawn, which one would expect to
find confirmed in Nature. The first deduction would be that no
structures should appear in the embryo of the higher animals that
are not found in the lower animals. But this is not confirmed by
the facts—no adult among the lower animals, for instance, has
a yolk-sac like that of the chick embryo. Again, if the law of
parallelism were true, the mammalian embryo would have to repeat
the organisation of, among other groups, insects and birds. But the
embryo in utero is surrounded by fluid and cannot possibly
breathe free air, so it cannot possibly repeat the structure of
either insects or birds, which are pre-eminently air-organisms.
Generally speaking, indeed, we find in all the higher embryos
special structures which adapt them to the very special conditions
of their development, and these we never find as permanent
structures in the lower animals. The supporters of the theory of
parallelism might, however, admit the existence of such special
embryonic organs without greatly prejudicing their case, for these
temporary organs stand to some extent outside the scope of the
theory.
But they would have to face a second and more important
deduction from their views, namely, that the higher animals should
repeat at every stage of their development the whole organisation
of some lower animal, and not merely agree 122with them in isolated details
of structure. The deduction is, however, not borne out by the
facts. The embryo of a mammal resembles in many points, at
different stages of its development, the adult state of a fish; it
has gill-slits and complete aortic arches, a two-chambered heart,
and so on. But at no time does it combine all the essential
characters of a fish; nor has it ever the tail of a fish, nor the
fins, nor the shape. Any recapitulation there may be is a
recapitulation of single organs, there is never a repetition of the
complete organisation of a fish. This is indeed the fundamental
criticism of the theory of parallelism; and if it applies even
within the limits of the vertebrate phylum, so much the more does
it apply to comparisons between embryonic Vertebrates and adult
Invertebrates.
There are also some lesser arguments which might be urged
against the theory of parallelism. If the theory were strictly
true, no state which is permanent in a higher animal could be
passed through by an animal lower in the scale. But birds, which
are lower in the scale than mammals, pass through a stage in which
they resemble mammals in certain respects much more than they do
when adult, for in an embryonic condition they agree with mammals
in having no feathers, no air sacs, no pneumatic sacs in the bones,
no beak. Their brain also resembles that of mammals more in an
earlier stage than it does later. So, too, myriapods and
hydrachnids have at birth three pairs of feet, and resemble at this
stage adult insects, which form a higher class.
Again, were the analogy between the development of the
individual and the evolution of the Échelle des
êtres complete, organs and organ-systems ought to
develop in the individual in the order in which they appear in the
scale of beings. But this is not always the case. In fish the
hinder extremity develops only its terminal joint, while in the
embryos of higher animals the basal joint is the first to
appear.
Another consequence one would expect to find realised, were the
theory of parallelism correct, is the late appearance in
development of parts which are confined to the higher animals. In
the development of a Vertebrate accordingly 123one would not
expect the vertebræ to appear before the embryo had passed
through many Invertebrate stages. But experience shows the direct
contrary, for in the chick the rudiments of the vertebral axis
appear sooner than any other part.
The theory of parallelism or recapitulation then is not borne
out by the facts, and clearly cannot be the law which we are
seeking. But what then is the true relation between the variety of
development and the variety of adult structure? Before answering
this question we must review the varied forms of adult organisation
and consider in what relations they stand to one another. In
particular we must enquire whether they belong to one type or to
many. One point is here cardinal—we must distinguish between
the type of organisation and the grade of
differentiation. By "type" von Baer means the structural plan of
the organism. "I call the type the spatial relationship of
the organic elements and organs" (p. 208). Each type of
organisation characterises one of the big groups of animals; the
lesser groups represent "grade" modifications of the type. "The
product of the degree of differentiation and the type gives the
several great groups of animals which are called classes" (p. 208).
Ausbildung (differentiation) takes place in one or other of
several directions, in adaptation, for instance, to life in the
water or to life in the air.
There are, von Baer considers, four main types—(1) the
peripheral or radiate type, (2) the longitudinal type, (3) the
massive or molluscan type, (4) the vertebrate type. The radiate
type is shown by discoid infusoria, by medusæ, by starfish
and their allies. The longitudinal type characterises such genera
as Vibrio, Filaria, Gordius, and all the
annulate animals. Mollusca, rotifers, polyzoa, and such infusoria
as are not included in types (1) and (2) belong to the massive
type, in which the body and its parts form rounded masses. The
longitudinal type is predominantly "animal," the massive type
predominantly "plastic" (vegetative). The vertebrate type has both
the "animal" and the "plastic" organs highly developed. In the
symmetrical arrangement of the animal parts it resembles the
longitudinal type; its 124plastic parts with their asymmetrical
arrangement and rounded shape belong to the massive type.
These types of von Baer inevitably recall the "Embranchements"
of Cuvier, with which they more or less coincide. It seems that von
Baer arrived at his types (from the study of adult structure)
independently of Cuvier, though the priority of publication rests
with Cuvier.[174]
Now it is clear that the development of the individual, which is
essentially an Ausbildung, a differentiation, is directly
comparable with the grade-differentiation of forms within the type.
And just as the type rules all its varied modifications, so does
the development of the individual take place always within the
bounds imposed by type. This is von Baer's chief contribution to
the theory of embryonic relationships—the law that "the type
of organisation determines the manner of development" (p. xxii.).
Development is not merely from the general to the
special—there are at least four distinct "general" types,
from which the special is developed. The type is fixed in the very
earliest stages of development—the embryo of a Vertebrate is
from the very beginning a Vertebrate (p. 220), and it shows at no
time any agreement in total organisation with any Invertebrate. The
types are independent of one another; differentiation and
development follow a different course in each of them. Not but what
some analogies can be found between the very earliest stages of
embryos of different type. Thus vertebrate and annulate embryos
agree in certain points at the time of the formation of the
primitive streak. And in the earliest stage of all, the egg-stage,
there is probably agreement between all the types. In eggs with
yolk, whether vertebrate or annulate, there is always a separation
into an animal and a plastic layer. It seems, too, as if a hollow
sphere were a constant stage in the development of all animals (pp.
224, 258). Apart from these analogies, development takes an
entirely independent course in each of the four main types, and no
embryo of one of the higher types repeats in its development the
peculiar organisation of any adult of the lower types.
125If we consider now development within the
type, which is the only legitimate thing to do, we arrive at
certain laws governing the relation of embryos to one another. For
instance, at a certain stage vertebrate embryos are uncommonly
alike. Von Baer had two in spirit which he was unable to assign to
their class among amniotes; they might have been lizard, bird, or
mammal, he could not say definitely which.[175] Generally the farther back we go in the
development of Vertebrates the more alike we find the embryos. The
type-characters are first to appear, then the class characters,
then the characters distinguishing the lesser classificatory
groups. "From a more general type the special gradually emerges"
(p. 221). The chick is first a Vertebrate, then a land-vertebrate,
then a bird, then a land-bird, then a gallinaceous bird, and
finally Gallus domesticus. Development within the type is a
progress from the general to the special, a real evolution. The
more divergent two adults are, the farther back we must go in their
development to find an agreement between their embryos. We can sum
up the case in the following laws:—
"(1) That the general characters of the big group to which
the embryo belongs appear in development earlier than the special
characters. In agreement with this is the fact that the
vesicular form is the most general form of all; for what is common
in a greater degree to all animals than the opposition of an
internal and an external surface?
"(2) The less general structural relations are formed after
the more general, and so on until the most special appear.
"(3) The embryo of any given form, instead of passing through
the state of other definite forms, on the contrary separates itself
from them.
"(4) Fundamentally the embryo of a higher animal form
126never resembles the adult of another
animal form, but only its embryo" (p. 224).
These laws relating to development within the limits of type are
destructive of even a limited application of the theory of
parallelism, for not even within the limits of the type is there a
real scale which the higher forms must mount; each embryo develops
for itself, and diverges sooner or later from the embryos of other
species, the divergence coming earlier the greater the difference
between the adult forms. It is only because the lower
less-differentiated adult forms happen to be little divergent from
the generalised or embryonic type, that they show a certain
similarity with the embryos of the higher more differentiated
members of the group. Such similarity, however, is due to no
necessary law governing the development of the higher animals; it
is, on the contrary, merely a consequence of the organisation of
these lower animals (p. 224).
Von Baer goes on to show what are the distinguishing
embryological characters of the types and classes, working out a
dichotomous schema of development, which each embryo must follow,
branching off early or late to its terminal point, according to the
lower or higher goal it has to reach.
One important consequence for morphology results from von Baer's
laws of differentiation within the type. If the embryo develops
from the general to the special, then the state in which each organ
or organ-system first appears must represent the general or typical
state of that organ within the group. Embryology will therefore be
of great assistance to comparative anatomy, whose chief aim it is
to discover the generalised type, the common plan of structure,
upon which the animals of each big group are built. And the surest
way to determine the true homologies of parts will be to study
their early development. "For since each organ becomes what it is
only through the manner of its development, its true value can be
recognised only from its method of formation. At present, we form
our judgments by an undefined intuition, instead of regarding each
organ merely as an isolated product of its fundamental organ, and
discerning from this standpoint the correspondences and
dissimilarities in the different types" (p. 233). Parts, therefore,
which 127develop from the same "fundamental organ,"
and in the last resort from the same germ-layer, have a certain
kinship, which may even reach the degree of exact homology.
Now since the mode of development in each type is peculiar to
that type, organs of the same name in different types must not
necessarily be accounted homologous, even if they correspond
exactly with one another in their general functional
relations to the rest of the organs. Thus the central nervous
system of Arthropods must not be homologised with the central
nervous system of Vertebrates, for it develops in a different
manner. So, too, the brain of Arthropods or of Mollusca is not
strictly comparable with the brain of Vertebrates. Again, the
air-tubes or tracheæ of insects are, like the trachea and
bronchi of many Vertebrates, air-breathing organs. But the two
organs are not homologous, for the air-tubes of Vertebrates are
developed from the alimentary tube ("fundamental organ" of the
alimentary system, developed from the vegetative layer), while the
air-tubes of insects arise either by histological differentiation,
or by invagination of the skin (p. 236). Organs can be homologous
only within the limits of the big groups; there can be no question
of homology between members of different types.
The development of plants, like the development of animals, is
essentially a progress from the general to the special (p. 242).
Botanists have not been troubled by any recapitulation theory, and
in founding their big groups, Acotyledons, Monocotyledons, and
Dicotyledons, upon embryological characters, they were guided by
true principles, which ought indeed to be followed in zoology. If
we knew the development of all kinds of animals sufficiently well,
then the best way to classify them would be according to the
characters they show in their early development, for it is in early
development that they show the characters of the type in their most
generalised form. As it is, we have in our ignorance to establish
the big groups by the study of adult structure, but we find, on
putting together all we know of comparative embryology, that a
classification of animals according to the mode of their
development gives, as is only natural, the same four 128groups as does
the study of adult structure. The four types of development are
thus:—
(1) The double-symmetrical, which is found in Vertebrates. It is
called the double-symmetrical, because in Vertebrates development
takes place from a central axis (notochord) in two directions,
upwards and downwards, in such a way that two tubes are formed, one
above and one below the axis. (2) The second type is the
symmetrical, which is shown by Annulates. A primitive streak is
formed on the ventral surface of the yolk; development proceeds
symmetrically on both sides of the streak. (3) Radiate development
is probably typical of the radiate structural type. (4) In the
massive type, the development seems to be a spiral one.
Common to most modes is a separation of the germ into animal and
plastic layers, a separation which seems to be conditioned largely
by the presence of yolk. A classification based upon embryological
characters ought to be applied even to the lesser groups and would
here prove itself of service. Embryology, for instance, fully
supports de Blainville's separation of Batrachia from true
reptiles,[176] for reptiles develop an
amnion and Batrachia do not.
We come now to the sixth and last Scholion. Development is a
true evolution of the special from the general, so runs von Baer's
most general law of all. This can be expressed in a slightly
different way, and the words which he chooses in the sixth Scholion
to express this final and most general result are these:—"The
developmental history of the individual is the history of the
growing individuality in every respect" (p. 263). The greatest
modern treatise on embryology ends on a splendid note. One creative
thought rules all the forms of life. And more—"It is this
same thought that in cosmic space gathered the scattered masses
into spheres and bound them together in the solar system, the same
that from the weathered dust on the surface of the metallic planets
brought forth the forms of life. And this thought is nought else
but life itself, and the words and syllables in which life
expresses itself are the varied forms of the living" (p. 264).
Von Baer reminds one greatly of Cuvier. There is 129the same sheer
intellectual power, the same sanity of mind, the same synthetic
grip. Von Baer, like Cuvier, never forgot that he was working with
living things; he was saturated, like Cuvier, with the sense of
their functional adaptedness. In his paper on the external and
internal skeleton[177] he gives a masterly
analysis of the functional modifications of the limbs in
Vertebrates, and the whole paper indeed, with its sober attack on
transcendentalism, is a vindication as much of the functional point
of view as of the importance of embryology.
Both Cuvier and von Baer, by the very sanity of their views,
found themselves in partial opposition to the theories current in
their time. Cuvier was the critic of Geoffroy and the
transcendentalists, of Lamarck and the believers in the
Échelle des êtres, evolutionary or ideal. Von
Baer also, though influenced greatly by Naturphilosophie,
turned against the exaggerations of the transcendental school, and
by his unanswerable criticism of the theory of parallelism took
away the ground from those who too easily believed in an historical
evolution.[178]
We have seen what were von Baer's criticisms of the theory of
parallelism. If we turn to the later writings of Cuvier we find the
essential criticism expressed in similar terms. Speaking of an
attempt which had been made to show that fish were molluscs
developed to a higher degree, he wrote in 1828,[179] "Let us draw the conclusion that even if
these animals can be spoken of as ennobled molluscs, as molluscs
raised to a higher power, or if they are embryos of reptiles, the
beginnings of reptiles, this can be true of them only in an
abstract and metaphysical sense, and that even this abstract
statement would be very far from giving an accurate idea of their
organisation." From the fact that the respiratory and circulatory
organs of fish greatly resemble those of tadpoles the conclusion
has been drawn that fish are 130in a sense embryos of Amphibia (p. 547).
But this manner of viewing things is none the less vicious, "for
this reason ... that it considers only one or two points and
neglects all the others" (p. 548), and is directly contrary to
common sense. There is never a recapitulation of total
organisations, only at the most of single organs.
It will be remembered that Cuvier opposed and demolished the
theory of the Échelle des êtres, not only by
showing that there were in Nature four entirely different plans of
animal structure, but also by demonstrating that even the animals
of each single Embranchement could not readily be arranged
in one series, that a serial arrangement was really valid only for
their separate organs. Von Baer also held that there are four
distinct types of structure; he, too, combated the idea of
gradation within the limits of the type. In so far as species
represent successive stages in the development, the
Ausbildung, of the type, so far can the idea of a scale of
beings be applied. But the members of a type follow not one line of
evolution but several diverging lines, in direct adaptation to
different environmental conditions, so that a serial arrangement of
them is not as a rule possible. It may be possible to establish a
serial arrangement of single organs from the simplest to the most
complex. But each organ or organ-system will require a different
serial arrangement, for the different systems vary on different
lines and an animal may be highly developed in respect of one
system and little developed in respect of all the others. Man, for
instance, is the highest animal only in respect of his nervous
system. The idea of the scale of beings has therefore only a very
limited application even within the limits of the type. Applied to
the whole animal kingdom it becomes merely absurd.
Another point of resemblance between Cuvier and von Baer was
that Cuvier, though essentially a student of adult structure, did
recognise the importance of embryology; following up some
observations of Dutrochet he studied the fœtal membrane of
mammals and tried to establish their homologies.[180] And in his criticism of the vertebral
theory of the skull he advanced as an argument against the
basisphenoid 131being a vertebral centrum the fact
(established by Kerkring, 1670), that it develops from two
centres.[181] Von Baer's relation to
transcendental anatomy was in some ways a close one, though he was
a trenchant critic of the extreme views of the school.[182] He took from Oken the idea that a simple
fundamental plan rules the organisation of all Vertebrates; "That
jaws and limbs are modifications of one fundamental form is readily
apparent, and, after Oken, the fact ought to be accepted by the
majority of those naturalists who do not refuse to admit the
existence of a general type from which the diversity of structure
is developed" (i., p. 192). He accepted the vertebral theory of the
skull in its main lines, and used his embryological knowledge to
support the idea that jaws correspond to limbs—the latter
point as part of the transcendental idea that the hind end of the
body repeats the organisation of the anterior part (i., p. 192).
The particular form which his theory of the relation of jaws to
limbs took is shown in the following passage:—"The maxillary
bone has ... the significance of an extremity and at the same time
that of a rib or lower arch of a vertebra, just as the pelvic bones
unite in themselves the signification of ribs and proximal members
of the hinder extremity" (Meckel's Archiv, p. 367,
1826).
He appreciated the morphological idea of the serial repetition
of parts, and gave it accurate formulation. The whole vertebrate
body, he considered, was composed of a longitudinal series of
morphological elements, each of which was made up a section
from each of the fundamental organs—a vertebra, a section of
the nerve-cord, and so on (Entwickelungsgeschichte, ii., p.
53). Groups of these morphological elements formed morphological
divisions, such as the vertebral segments of the head with
their highly developed neural arches, or the segments of the neck
with their undeveloped hæmal arches. The morphological
elements are clearly shown only in the animal parts, but there are
indications in the embryo of a segmentation also of the vegetative
parts,—the gill-slits, for instance, and the vascular arches.
132The
vegetative parts, however, develop on the whole unsymmetrically
(cf. Bichat). These elements which von Baer distinguishes
are morphological units, as he himself points out, contrasting them
with organs which are not usually units in a morphological sense.
"We call organ," he writes, "each part that has by reason of its
form or its function a certain distinctiveness, but this concept is
very indefinite, and possesses, from a morphological point of view,
little value. For this reason it seems necessary to introduce into
scientific morphology the concepts of morphological elements and
divisions" (ii., p. 84).
Von Baer exercised a very considerable influence upon the
subsequent trend of morphological theory. By his criticism of the
Meckel-Serres theory, he rid morphology for a time of an idea which
was leading it astray; by his substitution of the law that
development is always from the general to the special, he set
morphologists looking for the archetype in the embryo, not in the
adult alone, and made them realise that homologies could often best
be sought in the earliest stages of development; by formulating the
germ-layer theory he supplied morphologists with a new criterion of
homology, based upon the special relations of the parts
(germ-layers) which are first differentiated in all development. He
made the study of development an essential part of morphology.
133
CHAPTER X
THE EMBRYOLOGICAL CRITERION
Pander's work of 1817 was the
forerunner of an embryological period in which men's hopes and
interest centred round the study of development. "With bewilderment
we saw ourselves transported to the strange soil of a new world,"
wrote Pander, and many shared his hopeful enthusiasm. K. E. von
Baer's Entwickelungsgeschichte was by far the greatest
product of this time, but it stands in a measure apart; we have in
this chapter to consider the lesser men who were Baer's
contemporaries, friends, followers or critics.
It was largely a German science, this new embryology, and its
leaders were all personally acquainted. Pander, von Baer and Rathke
were on friendly terms with one another; von Baer dedicated his
master-work to Pander; Rathke dedicated the second volume of his
Abhandlungen to von Baer. Interest in the new science was,
however, not confined to Germany. In Italy, Rusconi commenced in
1817 his pioneer researches on the development of the Amphibia with
a Descrizione anatomica degli organi della circolazione delle
larve delle Salamandre aquatiche (Pavia), in which he traced
the metamorphoses of the aortic arches. This was followed in 1822
by his Amours des Salamandres aquatiques (Milan), and in
1826 by his memoir Du développement de la grenouille
(Milan). In this last paper he described how the dark upper
hemisphere of the frog's egg grows down over the lower white
hemisphere and leaves free only the yolk plug; he observed the
segmentation cavity and the archenteron, but thought that the
former became 134the alimentary canal; he observed and
interpreted rightly the formation of the medullary folds. The
circular blastopore in the frog in later years often went by the
name of the anus of Rusconi.
In France Dutrochet[183] investigated the fœtal membranes in
various vertebrate classes; Prévost and Dumas studied the
very earliest stages of development in birds, mammals and amphibia
(Ann. Sci. nat., ii., iii., 1824, xii., 1827).
 Fig.
Fig. 8.
Gill-slits of the Pig Embryo.
(After Rathke.)
A little later came Dugès' studies of the osteology and
myology of developing amphibia (1834),[184] and Coste's careful
researches into the early developmental history of mammals.[185]
It was in 1825 that Heinrich Rathke (1793-1860), published his
famous discovery of gill-slits in the embryo of a mammal,[186] a discovery which aroused considerable
interest, and greatly stimulated embryological research. He
describes how in a young embryo of a pig he saw four slits in the
region of the neck, going right through into the œsophagus.
They were separated by partitions which he called
Kiemenbogen (gill-arches), and immediately in front of the
first gill-slit lay the developing lower jaw. He compared these
gill-slits with those of a dogfish. We reproduce his drawing of the
pig-embryo (Isis, Pl. IV., fig. 1).
Later in the same year Rathke discovered gill-slits in the
chick,[187] in this case finding
only three. He described growing out from in front of the first
slit a structure which he compared to the operculum or gill-cover
of a fish.
These discoveries were confirmed and extended for the 135chick[188] by the embryologist
Huschke, a pupil of Oken. Like Rathke, he found only three
indubitable gill-slits, but he noticed that the body-wall in front
of the first gill-slit was really composed of two arches, which
were on the whole similar to the gill-arches. The hinder of these
two seemed to him to be a horn of the hyoid, the front one, which
was bent at an angle, to be the rudiment of the upper and lower
jaws (p. 401). Between these two arches he found an opening, just
as between two gill-arches a gill-slit. This opening led into the
mouth-cavity, and according to Huschke it became the external
ear-passage. He discovered also three pairs of aortic arches in
close relation with the gill-arches, so close indeed, that he did
not hesitate to call them gill-arteries, and to recognise their
resemblance with the aortic arches of fish. He traced, in part at
least, the metamorphosis which these aortic arches undergo. This
part of his discovery he developed in fuller detail in a paper of
1828,[189] in which he gave some
excellent figures.
Shortly after Huschke's first paper, von Baer published his
views and observations on this subject in a short memoir in
Meckel's Archiv.[190] In this paper he confirmed Rathke's discovery,
and described the slits and arches in the dog and the chick. Both
Rathke and he found gill-slits in the human embryo about this time
(p. 557). There were generally present, he found, four gill-slits,
and, as Rathke had suggested, the first gill-arch became the lower
jaw. Von Baer also confirmed Rathke's discovery of the operculum,
assigning it, however, to the second gill-arch. He refused to
accept Huschke's derivation of the auditory meatus from the first
gill-slit. Von Baer saw what had escaped Rathke and Huschke, that
there were, not three nor four, but as many as five aortic
arches.
136In his view of the metamorphosis of the
aortic arches in the chick the first two pairs disappeared
completely, the third pair gave rise to the arteries of the head
and the fore-limbs, the right side of the fourth arch became the
aorta, the left half of the fourth and the right half of the fifth
arch became the pulmonary arteries, while the left half of the
fifth arch disappeared. This schema, which for the last three
arches was the same as Huschke's, von Baer upheld for the chick
even in the second volume of his Entwickelungsgeschichte (p.
116); he rectified it, however, for mammals in the same volume (p.
212), deriving both pulmonary arteries from the fifth arch, and the
aorta from the fourth left. He fully recognised the great analogy
of the embryonic arrangement of gill-arches and gill-arteries in
Tetrapoda with their arrangement in fish (i., pp. 53, 73).
Huschke, in a paper of 1832,[191] chiefly devoted to the development of the eye,
figured and described the developing upper and lower jaws, and
maintained against von Baer that the first slit turns into the
auditory meatus and the Eustachian tube.
These were the first papers of the embryological period. Before
going on to discuss the principles which guided embryological
research during the next ten or twenty years it is convenient to
note what were the main lines of work characterising the
period.
The typical figure of the period is Rathke, who produced a great
deal of first-class embryological work. He was, even more than von
Baer, a comparative embryologist, and there were few groups of
animals that he did not study. His first large publication, the
Beiträge zur Geschichte der Thierwelt (i.-iv., Halle,
1820-27), contained much anatomical work in addition to the purely
embryological; he commenced here his series of papers on the
development of the genital and urinary organs, continued in the
Abhandlungen zur Bildungsund Entwickelungs-Geschichte des
Menschen und der Thiere (i., ii., Leipzig, 1832-3). A
fellow-worker in this line was Johannes Müller, whose
Bildungsgeschichte der Genitalien (Düsseldorf) appeared
in 1830.
In a memoir on the development of the crayfish which 137appeared
in 1829,[192] Rathke found in an
Invertebrate confirmation of the germ-layer theory propounded by
Pander and von Baer. He was greatly struck by the inverted position
of the embryo with respect to the yolk. In following out the
development of the appendages he noticed how much alike were jaws
and legs in their earliest stage, and how this supported Savigny's
contention that the limbs of Arthropods belonged to one single type
of structure. In his paper (1832) on the development of the
fresh-water Isopod, Asellus,[193] Rathke returns to this point. Commenting on the
original similarity in development of antennæ, jaws and legs,
he writes, "Whatever the doubts one may have reserved as to the
intimate relation existing between the jaws and feet of articulate
animals after the researches of Savigny on this subject and mine on
developing crayfish, they must all fall to the ground when one
examines with care the development of the fresh-water Asellus" (p.
147 of French translation).
Further comparative work by Rathke is found in the two volumes
of Abhandlungen and in a book, Zur Morphologie,
Reisebemerkungen aus Taurien (1837), which contains
embryological studies of many different types, including a study of
the uniform plan of arthropod limbs. Later on Rathke devoted
himself more to vertebrate embryology, producing among other works
his classical papers on the development of the adder (1839), of the
tortoise (1848), and of the crocodile (1866). He laid the
foundations of all subsequent knowledge of the development of the
blood-vascular system in a series of papers of various dates from
1838 to 1856. The diagrams in his paper on the aortic arches of
reptiles (1856) were for long copied in every text-book.
Rathke was a foremost worker in another important line of
embryological work, the study of the development of the skeleton
and particularly of the skull. We shall discuss the 138history of the
embryological study of the skull in some detail below; meantime, we
note the two other important lines of research which characterise
this period. One is the intensive study of the development of the
human embryo, a study pursued by, among others, Pockels, Seiler,
Breschet, Velpeau, Bischoff, Weber, Müller, and Wharton
Jones.[194] The other important
line—the early development of the Mammalia—was worked
chiefly by Valentin,[195] Coste,[196] and, above all, by Bischoff, whose series of
papers[197] was justly recognised as
classical.
What interests us chiefly in the work of this embryological
period is, of course, the relation of embryology to comparative
anatomy and to pure morphology. The embryologists were not slow to
see that their work threw much light upon questions of homology,
and upon the problem of the unity of plan. Von Baer, we have seen,
recognised this clearly in 1828; Rathke, in one of his most
brilliant papers, the Anatomische-philosophische Untersuchungen
über den Kiemenapparat und das Zungenbein (Riga and
Dorpat, 1832), used the facts of development with great effect to
show the homology of the gill-arches and hyoid throughout the
vertebrate series; Johannes Müller made great use of
embryology in his classical Vergleichende Anatomie der
Myxinoiden (i. Theil, 1836), and, according to his pupil
Reichert, firmly held the opinion that embryology was the final
court of appeal in disputed points of comparative anatomy;[198] Reichert himself in a book of 1838
(Vergleichende 139Entwickelungsgeschichte des Kopfes der
nackten Amphibien) discussed the two different methods of
arriving at the "Type"—the anatomical method of comparing
adults, and the embryological method of comparing embryogenies. Of
the embryological method, he says, "Its aim is to distinguish
during the formation of the organism the originally given, the
essence of the type, and to classify and interpret what is added or
altered in the further course of development. Embryologists watch
the gradual building up of the organism from its foundations, and
distinguish the fundament, the primordial form, the type, from the
individual developments; they reach thus, following Nature in a
certain measure, the essential structure of the organism, and
demonstrate the laws that manifest themselves during embryogeny"
(p. vi.). The embryologists, influenced in this greatly by von
Baer, gradually felt their way to substituting for the "Archetype"
of pure morphology what one may perhaps best call the
embryological archetype. How the transition was made we can
best see by following out the course of discovery in one particular
line. We choose for this purpose the development of the skull, a
subject which excited much interest at this time and upon which
much quite fundamental work was done, particularly by Rathke and
Reichert.
Following up his discovery of gill-slits and arches in the
embryos of birds and mammals, Rathke in two papers of 1832[199] and 1833[200] worked out the detailed homologies of the
gill-arches in the higher Vertebrates. He describes how in the
embryo of the Blenny there is a short, thick arch between the first
gill-slit and the mouth. A furrow appears down the middle of the
arch dividing it incompletely into two. In the anterior halves a
cartilaginous rod is developed which is connected with the skull;
these rods become on either side the lower jaw and "quadrate." In
the posterior halves two similar rods are formed which develop into
the hyoid. The hyoid is at first connected with the skull, 140but
afterwards frees itself and becomes slung to the "quadrate." From
the hinder edge of the hyoid arch grows out the membranous
operculum, in which develop later the opercular bones and
branchiostegal rays. The upper jaw is an independent outgrowth of
the serous layer.
The serial homology of the lower jaw and quadrate with the hyoid
and with the true gill-arches was thus established in fish, and
Rathke had little difficulty in demonstrating a similar origin of
lower jaw and hyoid in the embryos of higher Vertebrates. He could
even, as we have noted before, find the homologue of the operculum
in a flap which grows out from the hyoid arch in the embryo of
birds.
But Rathke could not altogether shake himself free from the
transcendental notion of the homology of jaws with ribs, and this
led him to draw a certain distinction between the first two and the
remaining gill-arches, by which the homology of the former with the
ribs was asserted and the homology of the latter denied. He thought
he could show that the skeletal structures (lower jaw, "quadrate,"
and hyoid) of the first two arches were formed in the serous layer,
just like true ribs, and like them in close connection with the
vertebral skeletal axis. The other, "true," gill-arches appeared to
him to be formed in the mucous layer, in the lining of the
alimentary canal. They had no direct connection with the vertebral
column, and seemed therefore to belong to what Carus[201] had called the visceral or
splanchno-skeleton. He did not, however, let this distinction
hinder him from asserting the substantial homology of all the
gill-arches inter se, the first two included.
Rathke's discoveries relative to the development of the jaws,
the hyoid and the operculum, enabled him to make short work of the
homologies proposed for them by the transcendentalists. He could
prove from embryology that the jaws were not the equivalent of
limbs, as so many Okenians believed. He could reject, with a mere
reference to the facts of development, Geoffroy's comparison of the
hyoid and the branchiostegal rays in fish with sternum and ribs. He
could show the emptiness of the attempts made 141by Carus,
Treviranus, de Blainville and Geoffroy, to establish by anatomical
comparison the homologies of the opercular bones, for he could show
that these bones were peculiar to fish, and were scarcely
indicated, and that only temporarily, in the development of other
Vertebrates.[202] He did not, however,
himself realise the relation of the ear-ossicles to the
gill-arches, though he knew that Spix and Geoffroy were quite wrong
in homologising them with the opercular bones in fish. He
described, it is true, the development of the external meatus of
the ear and the Eustachian tube from the slit which appears between
the first and the second arch, as Huschke had done before him; he
described, in confirmation of Meckel, the "Meckelian process" of
the hammer running down inside the lower jaw; but the discovery of
the true homologies of the ear-ossicles was not made until a year
or two later by Reichert.
In his further study of the development of Blennius
viviparus, Rathke observed some important facts about the
development of the vertebral column and skull. He found that the
vertebral centra were first formed as rings in the chorda-sheath,
which give off neural and hæmal processes. The vertebra later
ossifies from four centres. The chorda (notochord) is prolonged
some little way into the head, and the base of the cranium is
formed by the expanded sheath, which reaches forward in front of
the end of the notochord. This cranial basis shows a division into
three segments, in which Rathke was inclined to see an indication
of three cranial vertebræ. (It turned out that this division
into three segments did not really exist, and Rathke later
acknowledged that he had made an error of observation.) The side
walls of the skull grow out from this base and form a fibrous
capsule for the brain. The cranial section of the chorda itself
shows no sign of segmentation; but later on the cranial portion of
the chorda-sheath ossifies, like the vertebræ, from several
centres. The vomer, which, in the classical form of the vertebral
theory of the skull, was the centrum of the fourth, or foremost,
cranial vertebra, does not, according to Rathke, develop in
continuity with the cranial basis and the chorda sheath, but
develops separately in the facial region.
142Von Baer, like Rathke at this time, was also
to some extent a believer in the vertebral theory of the skull. In
his second volume (1834, pub. 1837) he holds that the development
of the skull, as the sum of the anterior vertebral arches, is in
general the same as that of the other neural arches, and is
modified only by the great bulk of the brain
(Entwickelungsgeschichte, ii., p. 99). He had, however, some
doubts as to the entire correctness of the vertebral theory, doubts
suggested by a study of the developing skull. "In the course of the
formation of the head in the higher animals, something additional
is introduced which does not originally belong to the cranial
vertebræ. At first we see the vertebration in the hinder
region of the skull very clearly. Afterwards it becomes suddenly
indistinct, as if some new formation overlaid it" (i., p. 194).
Even more clearly is his doubt expressed in his paper on
Cyprinus. "Upon the formation of the vertebral column only
this need be said, that at this stage the notochord is very clearly
seen, and the upper and lower arches and spinous processes are
visible right to the end of the tail, but the separation into
vertebræ ceases abruptly where the back passes into the head.
I do not hesitate to assert that bony fish, too, have at this
stage an unsegmented cartilaginous cranium (as cartilaginous
fish have all their life), the prominences and hollows of which
constitute its only resemblance with the vertebral type" (1835, p.
19).
A convinced supporter of the vertebral theory was Johannes
Müller, who, in his classical memoir on the Myxinoids,[203] discussed at some length the relation
between the development of the vertebræ and the development
of the skull. His memoir is principally devoted to comparative
anatomy, but in treating of the skeleton he pays much attention to
development. He describes the formation of the vertebræ in
elasmobranch embryos; for the facts regarding other Vertebrates he
relies largely on work by Rathke (Blennius, 1833) and
Dugès (1834). He recognises as the basis of his comparisons
the homology of the notochord 143in all vertebrate embryos with the
persistent notochord which forms the chief part or the whole of the
vertebral column in the Cyclostomes. The notochord possesses an
inner and an outer sheath and the outer sheath is continuous with
the basis cranii (p. 92). It is in the outer sheath that the
vertebræ develop—from four separate pieces, in fish at
least, plus an additional element which helps to form the centrum.
The skull of Vertebrates consists, according to Müller, of
three vertebræ, whose centra are the basioccipital, the
basisphenoid and the presphenoid. Other bones besides those
belonging to the vertebræ are present, but this formation out
of three vertebræ gives the essential schema for the skull.
Now the brain capsule, like the sheath of the spinal cord, is a
development from the outer sheath of the notochord. If the skull
consists of vertebræ we should expect the centra of the
skull-vertebræ to develop in the outer sheath at the sides of
the cranial section of the notochord as two separate halves, just
as do the bodies of the vertebræ; we should expect further
the cartilaginous side-walls of the cranium to develop in the
membranous brain-sheath just as the neural arches develop in the
membranous sheath of the spinal column. In Rathke's discovery (!)
of a segmentation of the basis cranii into three parts, and
of the isolated formation of the vomer, Müller sees a
confirmation of his view that the skull is composed of three and
not four vertebræ. But there is nothing in Rathke's
observations to support the idea that the centra of the cranial
vertebræ are formed from separate halves. Müller has to
be content with a reference to the state of things in
Ammocoetes (which, by the way, he did not know to be the
young of Petromyzon). In the simple skull of
Ammocoetes the base is formed chiefly by two cartilaginous
bars lying more or less parallel with the longitudinal axis of the
skull and embracing with their hinder ends the cranial portion of
the notochord.
These bars, declares Müller, are clearly the still separate
halves of the pars basilaris cranii, and represent the
divided centra of the two hinder cranial vertebræ. To
complete the parallel between the development of the skull and of
the vertebræ, it would have been necessary to show that the
side walls of the cranium developed in a similar manner from 144separate
pieces. Müller could not prove this point from the available
embryological data, and indeed the facts which he did use had to be
twisted to suit his theory. A curious apparent confirmation of his
idea that the centra of the cranial vertebræ are formed from
separate halves was supplied in 1839 by Rathke's discovery of the
trabeculæ in the embryonic skull of the adder.
The next big step in the study of the development of the skull
was taken by a pupil of Müller, C. B. Reichert, who showed in
his work very distinct traces of his master's influence. Reichert's
first and most important contribution to the subject was his paper
on the metamorphosis of the gill, or, as he called them, the
visceral arches in Vertebrates,[204] particularly in the two higher classes. Reichert
describes the similar origin in embryo of bird and mammal (pig) of
three "visceral" arches. These arches stand in close relation to
the three cranial vertebræ which Reichert, like Müller,
distinguishes. He makes the retrograde step of admitting only three
aortic arches, and he is not inclined to consider the three
visceral arches as equivalent to the gill-arches of fish—in
his opinion they have more analogy with ribs, though differing
somewhat from ribs in their later modifications. The visceral
arches are processes of the visceral plates (von Baer), which grow
downwards and meet in the middle line, leaving between one another
and the undivided body wall three visceral slits opening into the
pharynx. The first visceral process is different in shape from the
others, for it sends forward, parallel with the head and at right
angles to its downward portion, an upper portion in which later the
upper jaw is formed. The other two processes are straight. From the
hinder edge of the second visceral arch there develops, as Rathke
had seen, a fold which is comparable with the operculum of fish.
The first slit develops externally into the ear-passage, internally
into the Eustachian tube, and in the middle a partition forms the
tympanic ring and tympanum. Inside each of the visceral processes
on either side a cartilaginous rod develops. In 145the first
process this rod shows three segments, of which the first lies
inside that portion of the process which is parallel with the head.
This upper segment forms the foundation for the bones of the upper
jaw. The lowest segment of the cartilaginous rod becomes Meckel's
cartilage, and on the outer side of this the bones of the lower jaw
are formed. The middle segment becomes in mammals the incus (one of
the ear-ossicles), and in birds the quadrate. Meckel's cartilage,
which was discovered by Meckel[205] in fish, amphibians and birds, is a long strip
of cartilage which runs from the ear-ossicle known as the hammer in
mammals,[206] to the inside of the
mandible. Reichert shows how this relation comes about.
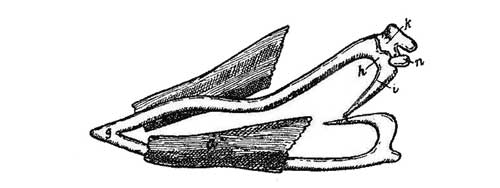
Fig.
9.—Meckel's Cartilage and Ear-ossicles in Embryo of Pig.
(After Reichert.)
| a. Mandible. |
h. Hammer. |
k. Incus. |
| g. Meckel's cartilage. |
i. Handle of Hammer. |
n. Stapes. |
The hammer, according to his observations on the embryo
of the pig, is simply the proximal end of Meckel's cartilage, which
later becomes separated off from the long distal portion (see Fig. 9).
The third ear-ossicle of mammals, the
stapes, comes not from the first arch but from the second. The
cartilaginous rod of the second arch segments like the first into
three pieces. Of these the uppermost disappears, the middle one,
which lies close up to the labyrinth of the ear, becomes the
stapes, and the lowest becomes the anterior 146horn of the
hyoid. The stapes forms a close connection with the hammer and the
incus. In birds, where there is a single ear-ossicle, the
columella, the middle piece of arch I forms, as we have seen, the
quadrate, by means of which the lower jaw is joined to the skull.
The proximal end of Meckel's cartilage, which in mammals forms the
hammer, here gives the articular surface between the lower jaw and
the quadrate. The columella is formed from the middle piece of the
three into which the cartilage of the second arch segments. It is,
therefore, the homologue of the stapes in mammals. The third arch
takes a varying share, together with the second, in the formation
of the hyoid apparatus.
In this paper Reichert made a distinct advance on the previous
workers in the same field—Rathke, Huschke, von Baer, Martin
St Ange, Dugès. Huschke was indeed the first to suggest that
both upper and lower jaws were formed in the first gill-arch. But
both von Baer and Rathke[207] held that the upper jaw developed as a special
process independent of the lower jaw rudiment, and the actual proof
that the upper jaw is a derivative of the first visceral arch seems
to have been first supplied by Reichert. His brilliant work on the
development of the ear-ossicles founded what we may justly call the
classical theory of their homologies. His views were attacked and
in some points rectified, but the main homologies he established
are even now accepted by many, perhaps the majority of
morphologists.
In a paper of 1838 on the comparative embryology of the skull in
Amphibia,[208] Reichert added to his
results for mammals and birds an account of the fate of the first
and second visceral arches in Anura and Urodela.
The first visceral arch, he found, gave in Amphibia practically
the same structures as in the higher Vertebrates. Its skeleton
segmented, as in mammals and birds, into three parts; the upper
part gave rise to the palatine and pterygoid in Anura, but seemed
to disappear in Urodeles, where the so-called palatine and
pterygoid developed in the mucous membrane of the mouth; the middle
part gave, as in birds, 147the quadrate, which formed a suspensorium for
both arches; the lower part, as Meckel's cartilage, formed a
foundation for the bones of the lower jaw. Of arch II., the lower
part became the horn of the hyoid, the upper part had a varying
fate. In some Anura it formed the ossicle of the ear (homologue of
the columella of birds and the stapes of mammals), in others it
disappeared. In reptiles the upper segment of the second arch
formed, as in birds, the columella.
The account of the metamorphoses of the visceral arches in
Amphibia forms only a small part of Reichert's memoir of 1838, the
chief object of which was to discover the general "typus" of the
vertebrate skull, and to follow out its modifications in the
different classes. Von Baer had shown that the generalised type
appeared most clearly in the early embryo; Reichert therefore
sought the archetype of the skull in the developing embryo. He
brought to his task the preconceived notion that the skull could be
reduced to an assemblage of vertebræ, but he saw that
comparative anatomy alone could not effect this reduction; he had
recourse, therefore, to embryology, hoping to find in the
simplified structure of the embryo clear indications of three
primitive cranial vertebræ (p. 121, 1837).
 Fig.
Fig. 10.
Cranial Vertebræ and Visceral Arches
in Embryo of Pig. Ventral Aspect.
(After Reichert.)
In the head he distinguished two tubes, the upper formed by the
dorsal plates, the lower by the ventral or visceral plates. Both of
these tubes were derived from the serous or animal layer
(cf. von Baer, supra, p. 118). The walls of the lower
tube were formed by the visceral processes, within which later the
skeleton of the visceral arches developed. The walls of the upper
tube formed the bones and muscles of the cranium proper. The facial
part of the head was formed by elements from both upper and lower
tubes. The dorsal tube showed signs of a division into three
cranial vertebræ (Urwirbeln, primitive
vertebræ). In mammals and birds, as Reichert had shown in his
1837 paper, the three cranial vertebræ were indicated by
transverse furrows on the ventral surface of the still membranous
skull (see Fig. 10, p. 148).
Even in mammals and birds, however, the positions of the eye,
the ear-labyrinth, and the three visceral arches were the safest
guides to the delimitation of the cranial vertebræ 148(pp.
134-138, 1837). In Amphibia generally there were no definite lines
of separation on the skull itself. "At this stage," he writes of
the cartilaginous cranium of the frog, "we find no trace of a
veritable division into vertebræ in the cartilaginous trough
formed by the basis cranii and the side parts. On the
contrary, it is quite continuous, as it is also in the higher
Vertebrates during the process of chondrification" (p. 44, 1838).
The vertebræ in the membranous or cartilaginous skull could
be delimited in Amphibia by the help of the eye and the
ear-labyrinth, which lie more or less between the first and second,
and the second and third vertebræ, but, above all, by the
vesicles of the brain.
As in the higher Vertebrates, the visceral arches are associated
with the cranial vertebræ as their ventral extensions, being
equivalent to the visceral plates which form the ventral portion of
the "primitive vertebræ" or primitive segments of the
trunk.
If the three cranial vertebræ are not very distinct in the
early stages of development when the skull is still membranous or
cartilaginous, they become clearly delimited when ossification sets
in. Three rings of bone forming three more or less complete
vertebræ are the final result of ossification. The
composition of these rings is as follows:—
|
Base. |
Sides. |
Top. |
| First vertebra |
Presphenoid |
Orbitosphenoids |
Frontals |
| Second vertebra |
Basisphenoid |
Alisphenoids |
Parietals |
| Third vertebra |
Basioccipital |
Exoccipitals |
Supraoccipital |
149The other bones of the skull are not included
in the vertebræ, and this is in large part due to the fact
that the sense capsules are formed separately from the cranium (p.
29, 1838). The ear-labyrinth, it is true, fuses indissolubly with
the cranium at a later period, but the bones which develop in its
capsule are not for all that integral parts of the primitive
cranial vertebræ. This point, it is interesting to note, had
already been made by Oken in his Programm (1807). But many
of the bones developed in relation to the sense organs can find
their place in the generalised embryonic schema or archetype of the
vertebrate skull, for they are of very constant occurrence during
early development.
Having arrived at a generalised embryonic type for the
vertebrate skull, of which the fundamental elements are the three
cranial vertebræ and their arches, Reichert goes on to
discuss the particular forms under which the skull appears in adult
Vertebrates. He accepts in general von Baer's law that the
characters of the large groups appear earlier in embryogeny than
the characters of the lesser classificatory divisions. "When we
observe new and not originally present rudiments in very early
embryonic stages, as, for instance, that for the lacrymals, the
probability is that they belong to the distinctive development of
one of the larger vertebrate groups. From these are to be
carefully distinguished such rudiments as arise later during
ossification, mostly as ossa intercalaria, in order to give
greater strength to the skull in view of the greater development of
the brain, etc.; the latter give their individual character to the
smaller vertebrate groups, and comprise such bones as the
vomer, the Wormian bones, the lowermost turbinal,
etc." (p. 63, 1838).
He did not accept the Meckel-Serres law of parallelism. He
recognised the great similarity between the unsegmented
cartilaginous cranium of Elasmobranchs, and the primordial cranium
of the embryos of the higher Vertebrates, but he did not think that
the cranium of Elasmobranchs was simply an undeveloped or embryonic
stage of the skulls of the higher forms. Rather "do the
Holocephala, Plagiostomata, and Cyclostomata
appear to us to be lower developmental stages individually
differentiated, so that the other fully differentiated Vertebrates
cannot easily be referred directly 150to their type" (p. 152, 1838). The
skull of these lower fishes is itself a specialised one; it is an
individualised modification of a simple type of skull. And this
holds good in general of the skulls of the lower
Vertebrates—they are individualised exemplars of a simple
general type, not merely unmodified embryonic stages of the greatly
differentiated skulls of the higher Vertebrates (p. 250, 1838).
Differentiation within the vertebrate phylum is therefore not
uniserial, but takes place in several directions. Reichert
describes two sorts of modifications of the typical
skull—class modifications and functional modifications. The
causes of the modifications which characterise classificatory
groups are unknown; the second class of modifications occur in
response to adaptational requirements.
Reichert's two papers are of considerable importance, and
Müller's remark in his review[209] of them is on the whole justified. "These
praiseworthy investigations supply from the realm of embryology new
and welcome foundations for comparative anatomy" (p.
clxxxvii.).
The development of the skull was, however, more thoroughly
worked out by Rathke, and with less theoretical bias, in his
classical paper on the adder.[210] This memoir of Rathke's is an exhaustive one and
deals with the development of all the principal organ-systems, but
particularly of the skeletal and vascular. He confirmed in its
essentials Reichert's account of the metamorphoses of the first two
visceral arches, describing how the rudiment of the skeleton of the
first arch appears as a forked process of the cranial basis, the
upper prong developing into the palatine and pterygoid, the lower
forming Meckel's cartilage, while the quadrate develops from the
angle of the fork. The actual bone of the upper jaw (maxillary)
develops outside and separate from the palato-pterygoid bar. The
cartilaginous rod supporting the second visceral arch divides into
three pieces on each side, of which the lower two form the hyoid,
the uppermost the columella. Like Reichert he held the visceral
arches to be parts of the visceral plates, containing, however,
elements from all three germ-layers—the serous, mucous, and
vessel layers.
151The first gill-slit, or, as Rathke here
prefers to call it, pharyngeal slit, closes completely in snakes
and in Urodeles. It forms the Eustachian tube in all other
Tetrapoda. As regards the vertebræ, Rathke describes them as
being formed in the sheath of the chorda from paired rudiments,
each of which sends two branches upwards, and two branches
downwards. The two inner pairs of processes coalesce round the
chorda, and later form the centrum; the upper outer pair meet above
the spinal column; the lower outer pair form ribs. The odontoid
process of the axis vertebra is the centrum of the atlas (p. 120).
The formation of vertebral rudiments begins close behind the
ear-labyrinth, but in front of this the chorda-sheath gives origin
to a flat membranous plate which afterwards becomes cartilaginous.
This plate reaches forward below the third cerebral vesicle as far
as the infundibulum. The notochord ends in this plate, which is the
basis cranii, just at the level of the ear-labyrinth. In no
Vertebrate does the notochord extend farther forward (p. 122). The
basis cranii gives off three trabeculæ. The middle one
is small and sticks up behind the infundibulum; it is absent in
fish and Amphibia, and soon disappears during the development of
the higher forms. The lateral trabeculæ are long bars which
curve round the infundibulum and reach nearly to the front end of
the head. Together they are lyre-shaped. The cranial basis and the
trabeculæ are formed, like the vertebræ, in the sheath
of the notochord, and the only differences between the two in the
early stage of their development are that the formative mass for
the cranial basis is much greater in amount than that for the
vertebræ, and that the cranial basis by means of its
processes, the trabeculæ, reaches well in front of the
terminal portion of the notochord (p. 36). The capsule for the
ear-labyrinth develops quite independently of the cranial basis and
the notochord. It resembles on its first appearance, in form,
position, composition, and connections, the ear-capsule of
Cyclostomes, and so do the ear-capsules of all embryonic
Vertebrates (p. 39). It manifests clearly the embryonic archetype,
... "there exists one single and original plan of formation, as we
may suppose, upon which is built the labyrinth of Vertebrates in
general" 152(p. 40). When ossification sets in, the
ear-capsule forms three bones, of which two fuse with the
supraoccipital and exoccipitals.
During the formation of the ear-capsule the cranial basis
develops from a plate to a trench, for in its hinder section the
side parts grow up to form the side walls of the brain, in exactly
the same way as the processes of the vertebral rudiments grow up to
enclose the spinal column (pp. 122, 192). The foundations of the
skull are now complete, and ossification gradually sets in.
Fig.
11.—Embrionic Cranium of the Adder. Ventral Aspect.
(After Rathke.)
| a. Basioccipital. |
d. Basisphenoid. |
g. Trabeculæ. |
| b. Exoccipital. |
c. Alisphenoid. |
h. Foramen. |
| c. Ear capsule. |
f. Orbitosphenoid. |
i. Ptuitary space. |
The basioccipital is formed in the posterior part of the basis cranii, and the
exoccipitals in the side walls of the trench in continuity with the
fundament of the basioccipital (see Fig. 11).
The supraoccipital is formed in cartilage above the exoccipitals.
The basisphenoid develops, like the basioccipital, in the flat
basis cranii, but towards its anterior edge, between the
large foramen (h) and the pituitary space (i). It is
formed from two centres, each of which is originally a ring round
the carotid foramen. The presphenoid develops in isolation between
the lateral trabeculæ, just behind the point where 153they fuse.
The side parts of the basisphenoid and presphenoid (forming the
alisphenoids and the orbitosphenoids respectively) develop in
cartilage separately from the cranial basis, not like the
exoccipitals in continuity with it. The hinder parts of the
trabeculæ become enclosed by two processes of the
basisphenoid; their front parts remain in a vestigial and
cartilaginous state alongside the presphenoid. The frontals and
parietals show a peculiar mode of origin in the adder, differing
from their origin in other Vertebrates. The frontals develop in
continuity with the orbitosphenoids, the parietals in continuity
with the alisphenoids, and so have much resemblance with the
vertebral neural arches which surround the spinal column (p.
195).
Through Rathke's work the real embryonic archetype of the
vertebrate skull was for the first time disclosed. Rathke discussed
this archetype and its relation to the vertebral theory of the
skull in another paper of the same year (1839), but before going on
to this paper, we shall quote from the paper on the adder the
following passage, remarkable for the clear way in which the idea
of the embryological archetype is expressed. "Whatever differences
may appear in the development of Vertebrates, there yet exists for
the different classes and orders a universally valid idea (plan,
schema, or type) ruling the first formation of their separate
parts. This idea must first be worked out, though possibly with
modifications, before more special ideas can find play. The result
of the latter process, however, is that what was formed by the
first idea is not so much hidden as partially or wholly destroyed"
(p. 135).
Rathke's general paper on the development of the skull in
Vertebrates[211] treats the matter on a
broader comparative basis than his paper on the adder, and takes
into account all the vertebrate classes, in so far as their
development was then known. He here makes the interesting
suggestion, later entirely confirmed, that the basis cranii
or basilar plate is first laid down as two strips, one on each side
of the chorda—the structures now known as parachordals (pp.
6, 27). For this supposition, he thinks, speaks the structure of the
154skull in Ammocoetes, which in this
respect is the simplest of all Vertebrates (pp. 6, 22). In
Ammocoetes, as Johannes Müller had shown, the
foundation of the skull is formed by two long cartilaginous bars,
between the hinder portions of which the notochord ends. In these
Rathke was inclined to see the homologues of his trabeculæ,
and of the parachordals which he was ready to assume from his
embryological observations.
Müller was, of course, very ready to accept Rathke's
opinions on this subject, for he considered that they supported his
own theory of the vertebral nature of the skull. After describing
in his Handbuch der Physiologie the cartilaginous bands in
Ammocoetes and their highly differentiated homologues in the
Myxinoids, he writes in the later editions, "Hence we see that in
the cranium, as in the spinal column, there are at first developed
at the sides of the chorda dorsalis two symmetrical elements, which
subsequently coalesce, and may wholly enclose the chorda. Rathke
has recently observed, in the embryos of serpents and other
animals, before the formation of the proper cranial vertebræ, two
symmetrical bands of cartilage, similar to those which I discovered
as a persistent structure in Ammocoetes.... At a later
period the basis cranii of vertebrate animals contains three
parts analogous to the bodies of vertebræ, the most anterior
of which, in the majority of animals, is generally small, and its
development frequently abortive, whilst in man and mammiferous
animals the three are very distinct. These parts are developed by
the formation of three distinct points of ossification, one behind
the other, in the basilar cartilage."[212]
Rathke was very cautious about accepting the vertebral theory of
the skull; he saw that the facts of development were not altogether
favourable to the theory, and he gave his adherence with many
reservations and saving clauses. His general attitude may be summed
up as follows.[213]
155The chorda sheath is the common matrix of the
vertebræ and of a large part of the skull. The basilar plate
and the trabeculæ, which are developed from the chorda
sheath, give origin to three bones, which might possibly be
considered equivalent to vertebral centra—the basioccipital,
the basisphenoid, and the Riechbein (ethmoid). The
Riechbein develops from the fused ends of the
trabeculæ. The presphenoid might also be considered as a
vertebral body, but it develops independently of the basilar plate
and trabeculæ.
Now of these bones, the basioccipital is in every way equivalent
to a vertebral centrum, for it develops in the basilar plate round
the notochord. With the exoccipitals, which arise just like neural
arches, it forms a true vertebra. The supraoccipital is an
accessory bone developed in relation to bigger brains. The
basisphenoid appears in the basilar plate, but in front of the
notochord, nor does it arise in exactly the same way as the centrum
of a vertebra. The basisphenoid with the alisphenoids, which
develop independently in the side walls of the brain, may, however,
still be considered as forming a vertebra, though the resemblance
is not so great as in the case of the occipital ring. The
presphenoid, being long and pointed, is very unlike a vertebral
body. The orbitosphenoids develop separately from it. The ethmoid
also differs from a vertebra, for it surrounds not the whole
nervous axis as the two hinder "vertebræ" do, but only two
prolongations of it, the olfactory lobes. In its development and
final form it shows no particular resemblance to a vertebra. Its
body, the pars perpendicularis (mesethmoid) shows no
similarity with a vertebral centrum. Completing the three hinder
cranial "vertebræ" and roofing in the brain are the
supraoccipital, the parietals and the frontals. The premaxillaries,
vomer, and nasals do not belong to the cranial scheme; they are
covering bones connected with the ethmoid. So, too, the ear-capsule
is not part of the cranial vertebræ, but is rather to be
compared to the intercalary bones in the vertebral column of
certain fish. Summing up as regards the cranial vertebræ
Rathke writes, "We find that the four different groups of bones,
consisting of the basioccipital with its intercalary (the
supraoccipital), the basisphenoid with its intercalaries 156(parietals), the presphenoid with its
intercalaries (frontals), and the ethmoid with its outgrowths
(turbinals and cribriform plate), taking them in order from behind
forwards, show an increasing divergence from the plan according to
which vertebræ as commonly understood develop, so that the
basioccipital shows the greatest resemblance to a vertebra, the
ethmoid the least" (p. 30).
In a posthumous volume published in 1861 the same opinion is put
forward. "In the head, too," he writes, "some vertebræ can be
recognised, although in a more or less modified form. Yet at most
only four cranial vertebræ can be assumed, and these differ
from ordinary well-developed vertebræ in their manner of
formation the more the farther forward they lie."[214]
Rathke was an able and careful critic of the vertebral theory of
the skull, but he accepted it in the main. Actual attack on the
theory upon embryological grounds was begun by C. Vogt, in his work
on the development of Coregonus,[215] and in his paper on the development of
Alytes.[216] He described for
Coregonus an origin of the skull in the main similar to that
established by Rathke for the adder. There was a "nuchal plate" in
which the front end of the notochord was imbedded; the notochord
ended at the level of the labyrinth; there were two lateral bands,
comparable to Rathke's lateral trabeculæ; a "facial plate"
was also formed, which seems on the whole equivalent to the plate
formed by the fused anterior ends of the trabeculæ. A little
later the cranium formed a complete cartilaginous box surrounding
the brain, very similar to the adult cranium of a shark.
In his criticism of the vertebral theory of the skull, Vogt
started by defining the vertebra as a ring formed round the chorda.
Now since only the occipital segment of the skull is formed
actually round the notochord, the parts of the skull 157lying in front
of this cannot themselves be vertebræ, though they may be
considered as prolongations of the occipital or nuchal vertebra.
"We must regard the nuchal plate as a true vertebra, modified, it
is true, in its formation and development by its particular
functions. Now, since the notochord ends with the nuchal plate we
can no longer regard as vertebræ the parts of the skull that
lie beyond, such as the lateral processes of the cranium and the
facial plate, for they have no relation with the notochord" (p.
123).
To support this view he adduced the fact that the vertebral
divisions (primitive vertebræ) visible in the trunk do not
extend into the head. He used precisely the same arguments in his
paper on Alytes to destroy the vertebral theory of the
skull. We quote the following passage translated by Huxley (1864,
p. 295) from this paper. "It has therefore become my distinct
persuasion that the occipital vertebra is indeed a true vertebra,
but that everything which lies before it is not fashioned upon the
vertebrate type at all, and that efforts to interpret it in such a
way are vain; that, therefore, if we except that vertebra
(occipital) which ends the spinal column anteriorly, there are no
cranial vertebræ at all."
L. Agassiz, himself a pupil of Döllinger, in the general
part (1844) of his Recherches sur les Poissons fossiles
(Neuchâtel, 1833-43), repeats in the main his pupil Vogt's
criticism of the vertebral theory (vol. i., pp. 125-9).
These arguments of Vogt and Agassiz were not considered by
Müller to dispose of the theory,[217] which maintained a firm hold even upon
embryologists. It was still upheld by Reichert, and Kölliker
in 1849 showed himself convinced of its general validity.
A useful step in the analysis of the concept "vertebra" was
taken by Remak,[218] who showed what a
complex affair the formation of vertebræ really is, involving
as it does a complete resegmentation (Neugliederung) of the
vertebral column, whereby the original vertebral bodies were
replaced by the secondary definitive bodies (p. 143). Remak showed,
as he thought, that the protovertebral segmentation of the dorsal
158muscle-plates did not extend into the head,
and he denied Reichert's assertion (1837) that the cranial basis in
mammals showed transverse grooves delimiting three cranial
vertebræ (p. 36). The gill-slits, he considered, could not
possibly be regarded as marking the limits of head
vertebræ.
In 1858 appeared Huxley's well-known Croonian Lecture, On the
Theory of the Vertebrate Skull,[219] in which he stated with great clearness and
force the case for the embryological method of determining
homologies, and criticised with vigour the vertebral theory of the
skull. By this time the two rival methods in morphology had become
clearly differentiated, and Huxley was able to contrast them, or at
least to show how necessary the new embryological method was as a
corrective and a supplement to the older anatomical, or, as he
calls it, "gradation" method. Applied to the "Theory of the Skull,"
the gradation method consists in comparing the parts of the skull
and vertebral column in adult animals with respect to their form
and connections. "Using the other method, the investigator traces
back skull and vertebral column to their earliest embryonic states
and determines the identity of parts by their developmental
relations" (p. 541). This second method is the final and ultimate.
"The study of the gradations of structure presented by a series of
living beings may have the utmost value in suggesting homologies,
but the study of development alone can finally demonstrate them"
(p. 541). As an example of the utility and, indeed, the necessity
of applying the embryological method Huxley takes the case of the
quadrate bone in birds. This bone had been generally regarded by
anatomists as the equivalent of the tympanic of mammals, on account
of its connection with the tympanum; but Reichert showed (1837)
that the same segment of the first visceral arch developed into the
incus in mammals, and into the quadrate in birds, and that
therefore the quadrate was homologous with the incus. Similarly, on
developmental grounds, the malleus or hammer of mammals is the
homologue of the articular of birds, since both are developed from
a portion 159of Meckel's cartilage identical in form and
connections in the two groups. The homologies of the bones
connected with the jaws in bony fishes had long been a subject of
contention among comparative anatomists; Huxley shows from his
personal observations how the development of the visceral arches
throws light upon these difficulties. The mandibular arch in the
developing fish is abruptly angled, as in the embryo of Tetrapoda;
the upper prong of it ossifies into the palatine and pterygoid; at
the angle is formed the quadrate (jugal, Cuvier), and to the
quadrate is articulated the lower jaw, which ossifies round the
lower prong or Meckel's cartilage. The scheme of development of the
jaws is accordingly similar in fish to what it is in other
Vertebrates, and this similarity of development enables Huxley to
recognise what are the true homologues of the quadrate, the
palatine and the pterygoid in adult bony fish, and to prove that
the symplectic and the metapterygoid (tympanal, Cuvier) are bones
peculiar to fish. In developing Amphibia Huxley found a
suspensorium of hyoid and mandibular arches similar to the
hyomandibular of fish.
Tackling his main problem of the unity of plan of the vertebrate
skull, Huxley shows, by a careful discussion of the anatomical
relationships of the chief bones in typical examples of all
vertebrate classes, that there is on the whole unity of plan as
regards the osseous skull. This unity of composition can be
established, on the gradation method, by considering the
connections of the bones of the skull with one another, their
relations to the parts of the brain and to the foramina of the
principal cranial nerves. The assistance of the embryological
method is, however, necessary in determining many points with
regard to the bones developed in relation to the visceral arches.
But there is a further step to be taken. "Admitting ... that a
general unity of plan pervades the organisation of the ossified
skull, the important fact remains that many vertebrated
animals—all those fishes, in fact, which are known as
Elasmobranchii, Marsipobranchii, Pharyngobranchii and
Dipnoi have no bony skull at all, at least in the sense in
which the words have hitherto been used" (p. 571). The membranous
or cartilaginous skull of these fishes shows a general resemblance
in its main 160features to the ossified skull of other
Vertebrates; the relations of the ear to the vagus and trigeminal
nerves are, for instance, the same in both; the main regions of the
cartilaginous skull can be homologised with definite bones or
groups of bones in the bony skull; but discrepancies occur. It is
again to development that we must turn to discover the true
relationship of the cartilaginous to the ossified skull. "The study
of the development of the ossified vertebrate skull ...
satisfactorily proves that the adult crania of the lower
Vertebrata are but special developments[220] of conditions through which the embryonic
crania of the highest members of the sub-kingdom pass" (p. 573). It
is with the embryonic cranium of higher Vertebrates that the adult
skull of the lower fishes must be compared, and the comparison will
show a substantial though not a complete agreement between them.
Thus, speaking of the development of the frog's skull, Huxley
writes:—"If, bearing in mind the changes which are undergone
by the palatosuspensorial apparatus, ... we now compare the stages
of development of the frog's skull with the persistent conditions
of the skull in the Amphioxus, the lamprey, and the shark,
we shall discover the model and type of the latter in the former.
The skull of the Amphioxus presents a modification of that
plan which is exhibited by the frog's skull when its walls are
still membranous and the notochord is not yet embedded in
cartilage. The skull of the lamprey is readily reducible to the
same plan of structure as that which is exhibited by the tadpole
when its gills are still external and its blood colourless. And
finally, the skull of the shark is at once intelligible when we
have studied the cranium in further advanced larvæ, or its
cartilaginous basis in the adult frog" (p. 577). Development,
therefore, proves what comparative anatomy could only
foreshadow—the unity of plan of all vertebrate skulls,
ossified and unossified alike. "We have thus attained to a theory
or general expression of the laws of structure of the skull. All
vertebrate skulls are originally alike; in all (save
Amphioxus?) the base of the primitive cranium undergoes the
mesocephalic flexure, behind which the notochord terminates, while
immediately in front of it 161the pituitary body is developed;[221] in all, the cartilaginous cranium has
primarily the same structure—a basal plate enveloping the end
of the notochord and sending forth three processes, of which one is
short and median, while the other two, the lateral trabeculæ,
pass on each side of the space on which the pituitary body rests,
and unite in front of it; in all, the mandibular arch is primarily
attached behind the level of the pituitary space, and the auditory
capsules are enveloped by a cartilaginous mass, continuous with the
basal plate between them. The amount of further development to
which the primary skull may attain varies, and no distinct
ossifications at all may take place in it; but when such
ossification does occur, the same bones are developed in similar
relations to the primitive cartilaginous skull" (p. 578).
In a word, there is a general plan or primordial type which is
manifested in the higher forms most clearly in their earliest
development—an embryological archetype therefore.
Huxley now goes on to consider the relation of this general plan
or type of the skull to the structure and development of the
vertebral column. Does the skull in its development show any signs
of a composition out of several vertebræ? The vertebral
column develops as a segmented structure round the notochord; the
skull develops first as an unsegmented plate extending far beyond
the notochord. The processes of this basilar plate, the
trabeculæ, are quite unlike anything in the vertebral column.
It is true that when the process of ossification begins, separate
bones are differentiated in the basilar plate one in front of the
other, giving an appearance of segmentation. The hindmost of these
bones, the basioccipital, ossifies round the notochord, quite like
a vertebral centrum, and its side parts which form the occipital
arch develop in a "remotely similar" way to the neural arches of
the vertebræ. The next bone, however, the basisphenoid,
develops in front of the notochord, and shows very little analogy
with a vertebral body. The analogy is even more far-fetched when
applied to the axial 162bones in front of the basisphenoid. The
cranium might indeed be divided upon ossification into a series of
segments bearing a more or less remote analogy with vertebræ.
"In the process of ossification there is a certain analogy between
the spinal column and the cranium, but that analogy becomes weaker
and weaker as we proceed towards the anterior end of the skull" (p.
585). The best way to state the facts is to say that both skull and
vertebral column start in their development from the same point,
but immediately begin to diverge. The clear indications of
segmentation which fully ossified adult skulls undoubtedly show
are, therefore, secondary, and the vertebral theory of the skull,
which was originally based upon the appearance of such fully
ossified crania, is on the whole negatived by embryology.
We have now to turn back a few years in order to follow up
another line of discovery which had an important bearing upon the
theory of the vertebrate skull—the working out of the
distinction between membrane and cartilage bones.
As early as 1731, R. Nesbitt,[222] in two lectures delivered to the Royal College
of Surgeons, demonstrated that in the human fœtus some bones
were formed not in cartilage but directly in fibrous tissue, and
this observation was confirmed by other human anatomists,
particularly by Sharpey at a considerably later date. In 1822
Arendt[223] focussed attention upon
the remarkable structure of the skull of the Pike, with its
cartilaginous brain-box studded all over with bony plaques, an
arrangement which had already attracted the interest of Cuvier and
Meckel. K. E. von Baer[224] in 1826 discussed at some length the relation
between the bony and the cartilaginous skull in fishes, with
particular reference to the sturgeon, coming to the following just
conclusion:—"If we consider the fibrous skeleton of
Ammocoetes as the first foundation of the skeleton of
Vertebrates, we can form a 163series among the cartilaginous fishes,
according as a cartilaginous skeleton penetrates more and more into
this fibrous foundation. In the same way the process of
ossification supplants the cartilaginous skeleton. So long as the
ossifications lie in the skin, as in the sturgeon, they form
corneous bones (Hornknochen), but when they lie under the
skin, they form true bones, e.g., the bones of the skull in
the pike" (p. 374).
Embryologists soon become aware that a similar distinction
between a primitive cartilaginous foundation and a secondary
overlying ossification of the skull showed itself in the
development of all Vertebrates. Dugès, in his Recherches
sur l'ostéologie et la myologie des Batraciens (1834),
distinguished between such bones as are formed by direct
ossification of the cartilaginous groundwork of the skull, and such
as are developed in the periosteal fibrous tissue.
Reichert in 1838[225] noted that several of the skull bones in
Amphibia are formed without the intermediary of cartilage, such as
the nasals, the maxillaries and the lacrymals. So, too, the
frontals and parietals of Teleosts developed independently of the
cartilaginous skull, and belonged to the skeletal system of the
skin, not to the true vertebral axial skeleton (pp. 215-6). Even
more interesting was his discovery, afterwards confirmed by
Hertwig,[226] that in the newt several
bones connected with the palate were formed in the mucous membrane
of the mouth by the fusion of a number of little conical teeth (p.
97). Certain of these bones he considered to be the substitutes,
not the equivalents, of the palatine and pterygoid of other
Vertebrates, which are formed from the upper part of the first
visceral arch, a part missing in the newt (p. 100). Owing to the
difference of development he would not homologise these bones in
the newt with the palatine and pterygoid of other Vertebrates. He
recognised also that the bone now known as the parasphenoid was
developed in the frog in the mucous membrane of the mouth, and had
originally no connection with the cranial basis (p. 34). Rathke in
1839 also allowed the distinction between cartilage and membrane
bone, but laid no stress upon it (Entw. d. Natter., p.
197).
164Jacobson in 1842[227] introduced the useful term, "primordial
cranium," for the primitive cartilaginous foundation of the skull,
and drew a sharp distinction between cartilage bones and membrane
bones.
In his Recherches sur les Poissons fossiles,[228] L. Agassiz used Vogt's work on the
development of Coregonus to establish a classification of
the bones of the skull in fish, a classification which had the
merit of drawing a sharp distinction between the cartilaginous
groundwork and the "protective plates" of the fish's skull. He
recognised that the protective plates developed in a different way
from the other bones of the skull. "We must distinguish," he
writes, "two kinds of ossification; one which tends to transform
the primitive parts of the embryonic cranium directly into bone,
and another which leads to the deposition of protective plates
round this core, which develop not only upon the upper surface, as
has hitherto been supposed, but also on the lateral walls and on
the lower surface of the cranium" (p. 112). In the skull of all fish
there are three elements—(1) the cartilaginous base,
including the nuchal plate, the trabeculæ and the facial
plate, together with the auditory capsules; (2) the cartilaginous
cerebral envelope; (3) the bony protective plates (absent in
Elasmobranchs). The bones developed in relation to these cranial
elements can be classified as follows:—(1) the basioccipital,
exoccipitals (paroccipitals?), supraoccipital and "petrous"
(rocher), developed from the nuchal plate; the ali- and
orbito-sphenoids developed from the trabeculæ; the "cranial
ethmoid"[229] developed from the
facial plate; (2) the parietals, frontals and nasals formed from
the "superior" protective plate; the "anterior" and "posterior"
frontals and the temporal, from the "lateral" plates; the body of
the sphenoid and the vomer from the "inferior" plates. The other
element, the cartilaginous brain-box, does not ossify, and tends to
become absorbed (p. 124).
In 1849 Kölliker published a paper[230] dealing with the 165morphological significance of the
distinction between membrane and cartilage bones, and in 1850[231] he defended his views
against the criticisms of Reichert[232] in a further note entitled Die Theorie des
Primordialschädels festgehalten. It is convenient to
consider these papers together. Kölliker held that there was
(1) a histological and (2) a morphological difference between the
two categories of bones. The histological development of the two
kinds was different, but this difference was not sufficient to
establish a morphological distinction between them, a distinction
in their anatomical Bedeutung. The true morphological
distinction between them was their development in different
skeleton-forming layers. Membrane bones were developed in fibrous
tissue lying between the skin and the deep layer which formed the
primordial cranium, and it was this formation in a separate layer
that gave them a different morphological significance from the
bones formed directly in the deep layer. Kölliker's
distinction, therefore, was between the bones formed in the
primordial cartilaginous cranium on the one hand, and the
superficial ossifications in fibrous tissue on the other hand. The
cartilaginous cranium in Kölliker's opinion was formed upon
the vertebral type, and the membrane bones were accessory. This, at
least, was his opinion in 1849. In 1850, after Stannius had shown
that membrane bones occurred as integral parts of the
vertebræ in certain fish, he modified his view of the
membrane bones, and admitted them, at least in some cases, as
constituents of the cranial vertebræ.
On this morphological distinction of membrane and cartilage
bones future comparative osteology was to be based:—
"My sole aim is to state again the principle upon which
comparative osteology is to be based and extended, and this is that
first place should be assigned to anatomical considerations, and
among these to the manner of origin of the whole bone in relation
to the skeleton-forming layers" (1850, p. 290).
The homologies established by this new principle might 166run
counter to the homologies indicated by the study of adult
structure. "Thus, for instance, although the lower jaw in position,
function, form and shape, appears to be the same bone throughout,
yet it must be admitted that it shows a difference in the different
classes. In Mammals and Man it is an entirely secondary bone (an
extremity according to Reichert), in Birds, Amphibia and Fishes
only partially so, for its articular belongs to Meckel's cartilage
and is accordingly analogous to a rib; indeed, in the Plagiostomes,
etc., the whole lower jaw along with the articular is a persistent
Meckel's cartilage" (p. 290, 1850).
So, too, the supraoccipital in man cannot be fully homologised
with the supraoccipital of many mammals, for its upper half arises
at first in isolation as a secondary bone (p. 290).
Reichert objected to the distinction drawn by Kölliker, and
denied that there was either a histological or a morphological
difference between membrane and cartilage bones. It was shown a few
years later by H. Müller[233] that there was in truth no essential difference
in histological development between the two categories of bone,
that the cartilage cells were replaced by bone cells identical with
those taking part in the formation of membrane bones. The
morphological distinction continued however to be recognised,
particularly by the embryologists. Rathke in his volume of 1861[234] classified the bones of
the skull according to their origin from the primordial cranium or
from the overlying fibrous layer, distinguishing as membrane bones,
the parietals, frontals, nasals, lachrymals, maxillaries and
premaxillaries, jugals, tympanic, parts of the "temporal," vomer,
part of the supraoccipitals in some mammals, and the mandible (with
the exception of the articular in such as have a quadrate bone).
Huxley was also inclined in 1864[235] to recognise the distinction, but he writes with
some reserve:—"Is there a clear line of demarcation between
membrane bones and cartilage bones? Are certain bones always
developed primarily from cartilage, while certain others as
constantly originate in membrane? And further, 167if a membrane
bone is found in the position ordinarily occupied by a cartilage
bone, is it to be regarded merely as the analogue and not as the
homologue of the latter?" (p. 296).
We may note here that many comparative anatomists of the period
were quite ready to decide Huxley's last question in a sense
favourable to the older, purely anatomical, view of homology. Owen,
for instance, held that difference of development did not disturb
homologies established by form and connections. "Parts are
homologous," he writes, "in the sense in which the term is used in
this work, which are not always similarly developed: thus the 'pars
occipitalis stricte dicta,' etc., of Soemmering is the special
homologue of the supraoccipital bone of the cod, although it is
developed out of pre-existing cartilage in the fish and out of
aponeurotic membrane in the human subject."[236] Similarly he pointed to the diversities of
development of the vertebral centrum in the different vertebrate
classes as proof that development could not always be relied upon
in deciding homologies (p. 89). But he could not deny that the
archetype was better shown in the embryo than in the adult
(supra, p. 108).
J. V. Carus[237] likewise stood firm
for the older method of determining homologies by comparison of
adult structure. "We can regard as homologous," he writes, "only
those parts which in the fully formed animal possess a like
position and show the same topographical relations to the
neighbouring parts" (p. 389). Parts homologous in this sense might
develop in different ways, but no great importance was to be
attached to such a circumstance. Membrane and cartilage bones
developed in practically the same way, from the same
skeleton-forming layer, and no morphological significance attached
to their distinction (pp. 227, 457). Embryology was of considerable
value in helping to determine homologies, but the evidence that it
supplied was contributory, not conclusive. Perhaps the greatest
service which the study of development rendered was to disentangle,
by a comparison of the earliest embryos, the generalised type (p.
389).
168We have now traced, by our historical study
of the theory of the skull, the gradual evolution of the tendency
to find in development the surest guide to determining homologies.
We have seen how the embryological "type" came to be substituted,
in whole or in part, for the anatomical "type" derived from the
study of adult structure. But we have had to do only with a
modification, not with a transformation, of the criterion of
homology recognised by the anatomists. Homology is still determined
by position, by connections, in the embryo as in the adult.
"Similarity of development" has become the criterion of homology in
the eyes of the embryologist, but "similarity of development"
means, not identity of histological differentiation, but similarity
of connections throughout the course of development. For the
purposes of morphology, development has to be considered as an
orderly sequence of successive forms, not in its real nature as a
process essentially continuous. Morphology has to replace the
living continuity by a kinematographic succession of stages. Since
it is the earliest of these stages that manifest the simplest and
most generalised structural relations of the parts, it is in the
earlier stages that homologies can be most easily determined. But
these homologies are still determined solely by the relative
positions and connections of the parts, just as homologies are
determined in the last of all the stages of development, the adult
state. And since the generalised type is shown most clearly in the
earliest stages and tends to become obscured by later
differentiation, homologies observed in embryonic life are to be
upheld even if the relations in adult life seem to indicate
different interpretations.
169
CHAPTER XI
THE CELL-THEORY.
With the founding of the cell-theory by Schwann in 1839 an
important step was taken in the analysis of the degrees of
composition of the animal body. Aristotle had distinguished
three—the unorganised material, itself compounded of the four
primitive elements, earth and water, air and fire, the homogeneous
parts or tissues and the heterogeneous parts or organs, and this
conception was retained with little change even to the days of
Cuvier and von Baer. Those of the old anatomists who speculated on
the relations of organic elements to one another were dominated by
Aristotle's simple and profound classification, and proposed
schemes which differed from his only in detail. Bichat enlarged and
deepened the concept of tissue, but the degree of composition below
this was for him, as for all anatomists of his time, a fibrous or
pulpy "cellulosity," living, indeed, but showing no uniform and
elemental structure. It was Schwann's merit to interpose between
the tissue and the mere unorganised material a new element of
structure, the cell. And, as it happened, a few years before
Schwann published his cell-theory, Dujardin hinted at another
degree of composition which was later to take its place between the
cell and the chemical elements—sarcode or protoplasm.
As is well known, the concept of the cell arose first in botany.
Robert Hooke discovered cells in cork and pith in 1667, and his
discovery was followed up by Grew and Malpighi in 1671, and by
Leeuenhoek in 1695. But they did not conceive the cell as a living,
independent, structural unit. They were interested in the
physiology of the plant 170as a whole, how it lived and nourished
itself, and they studied cells and sieve-tubes, wood fibres and
tracheæ with a view rather to finding out their functions and
their significance for the life of the plant than to discovering
the minutiæ of their structure. The same attitude was taken
up by the few botanists who in the 18th century paid any heed to
the microscopical anatomy of plants. For C. F. Wolff,[238] the formation of cells was a result of the
secretion of drops of sap in the fundamental substance of the
plant, this substance remaining as cell-walls when cell-formation
was completed—no idea here of cells as units of
structure.
In the early 19th century, interest in plant anatomy revived
somewhat, and much work was done by Treviranus, Mirbel,
Moldenhawer, Meyen and von Mohl.[239] As a result of their work the fact was
established that the tissues of plants are composed of elements
which can, with few exceptions, be reduced to one simple
fundamental form—the spherical closed cell. Thus the vessels
of plants are formed by coalescence of cells, fibres by the
elongation of cells and the thickening and toughening of their
walls. At this time, interest was concentrated on the cell-wall, to
the almost total neglect of the cell-contents; the "matured
framework" of plant cells, to use Sach's convenient phrase, was the
chief, almost the sole, object of study. And it was natural enough
that the mere architecture of the plant should monopolise interest,
that the composition of the tissues out of the cells, and the
fitting together of the tissues to form the plant should awaken and
hold the curiosity of the investigator; even the modifications of
the cell-walls themselves, their rings and spiral thickenings and
pits, offered a fascinating field of enquiry.
The idea that the cell-contents might show a characteristic and
individual structure had hardly dawned upon botanists when
Schleiden published his famous paper, Beiträge zur
Phytogenesis.[240] Schleiden's theme
in this paper is the origin 171and development of the plant cell, a
subject then very obscure, in spite of pioneer work by Mirbel. A
few years before, Robert Brown had called attention to the presence
in the epidermal cells of orchids and other plants of a
characteristic spot which he called the areola or nucleus.[241] Schleiden saw the importance of this
discovery, confirmed the constant presence of the nucleus in young
cells, and held it to be an elementary organ of the cell. He named
it the cytoblast because, in his opinion, it formed the cell. It
was embedded in a peculiar gummy substance, the cytoblastem, which
formed a lining to the cellulose cell-wall. Within the nucleus
there was often a small dark spot or sphere—the nucleolus.
The nucleus, Schleiden thought, originated as a minute granule in
the cytoblastem which gradually increased in size, becoming first a
nucleolus (Kernchen), and then, by further condensation of
matter round it, a nucleus. Several nuclei might be formed in this
way in a single cell. New cells took their origin directly from a
full-grown nucleus, in a peculiar way which Schleiden describes as
follows:—"As soon as the cytoblasts have reached their full
size a delicate transparent vesicle arises on their surface; this
is the young cell, which at first takes the shape of a very flat
segment of a sphere, of which the plane surface is formed by the
cytoblast, the convex side by the young cell itself, which lies
upon the cytoblast like a watch-glass on a watch" (p. 145). The
young cells increase in size and fill up the cavity of the old
cell, which is in time resorbed. Cell-development always takes
place within existing cells, and either one or many new cells may
be formed within the mother-cell. Schleiden's views on
cell-formation were drawn from some rather imperfect observations
on the embryo-sac and pollen-tube, but he extended his theory to
cell-formation in general. Though wrong in almost all respects the
theory had at least the merit of fixing attention upon the really
important constituents of the cell, the nucleus and the
cell-plasma. To Schleiden, too, we owe the conception of the cell
as a more or less independent living unity, whose life is not
entirely identified with the life of the plant as a whole. "Each
cell," he writes, "carries on a double life; one a quite
independent and self-contained 172life, the other a dependent life
in so far as the cell has become an integral part of the plant" (p.
138).
So long as the definition of the plant cell embraced little more
than the hardened cell-wall it was little wonder that "cells" in
this sense were not recognised in animal tissues, except in a few
exceptional cases—as in the notochord by Johannes
Müller.[242] Careful observation of
animal tissues discovered in some cases the existence of
discontinuous units of structure, but these were not, as a rule,
recognised before 1838 as analogous to plant cells. Von Baer, for
example, observed that the young chick embryo was composed partly
of an albuminous mass and partly of Kügelchen or little
globules suspended in it (Entwickelungsgeschichte, i., pp.
19, 144). Since such Kügelchen disposed in a row formed
the notochord (i., p. 145) it seems probable that his
Kügelchen were really cells. Similarly A. de
Quatrefages[243] in 1834 saw and figured
segmentation spheres in the developing egg of Limnæa,
but he called them globules and did not recognise their analogy
with the cells of plants. According to M'Kendrick,[244] Fontana, so far back as 1781,[245] described cells with nuclei in various
tissues, and used acids and alkalis to bring out their structure
more clearly. But it was not till 1836-7-8 that a fairly widespread
occurrence of cells in animal tissues was recognised. The pioneer
in this seems to have been Purkinje, who described cells in the
choroidal plexus in 1836,[246] and compared gland cells with the cells of
plants in 1837.[247] Henle in 1837[248] and 1838[249] described various kinds of epithelial
tissue, distinguishing them according to the kind of cell composing
them; he also discovered the mode of growth of stratified
epithelium. 173Valentin[250] appears to have seen cells in cartilage and
epithelium even before Henle, and to have observed cells in the
blastoderm of the chick. In his report on the progress of anatomy
during 1838 Johannes Müller was able to refer to quite a
number of papers dealing with the occurrence of cells in animal
tissues. In addition to those already noted, he mentions work by
Breschet and Gluge on the cells of the umbilical cord, by Dumortier
on the cells in the liver of molluscs, by Remak and by Purkinje on
nerve cells, by Donné on the cells of the conjuctiva, cornea
and lens. He reports, too, that Turpin had compared the epithelial
cells of the vagina with the cell-tissue of plants. Müller
himself had not only recognised the cellular nature of the
notochord, but had observed the cells of the vitreous humour, fat
cells and pigment cells, and even the nuclei of cartilage cells.
From Schwann (1839) we learn that C. H. Schults had followed back
the corpuscles of the blood to their original state of nucleated
cells, and that Werneck had recognised cells in the embryonic lens.
A preliminary notice of Schwann's own work appeared in 1838
(Froriep's Notizen, No. 91, 1838), the full memoir in 1839,
under the title Mikroskopische Untersuchungen über die
Uebereinstimmung in der Struktur und dem Wachstume der Tiere und
Pflanzen.[251]
Theodor Schwann was a pupil of Johannes Müller, and we know
that Müller took much interest in the new histology. It is
probably to his influence that we owe Schwann's brilliant work on
the cell, which appeared just after Schwann left Berlin for
Löwen. Schwann was himself, as his later work showed, more a
physiologist than a morphologist; he did quite fundamental work on
enzymes, discovering and isolating the pepsin of the gastric juice;
he proved that yeast was not an inorganic precipitate but a mass of
living cells; he carried out experiments directed to show that
spontaneous generation does not occur. We shall see in his
treatment of the cell-theory clear indications of his physiological
174turn
of mind. Schwann was only twenty-nine when his master-work
appeared, and the book is clearly the work of a young man. It has
the clear structure, the logical finish, which the energy of youth
imparts to its chosen work. So the work of Rathke's prime, the
Anatomische-philosophische Untersuchungen of 1832 shows more
vigour and a more reasoned structure than his later papers.
Schwann's book is indeed a model of construction and cumulative
argument, and even for this reason alone justly deserves to rank as
a classic.
The first section of his book is devoted to a detailed study of
the structure and development of cartilage cells and of the cells
of the notochord, and to a comparison of these with plant cells. He
accepts Schleiden's account of the origin and development of nuclei
and cells as a standard of comparison; and he seeks to show that
nucleus and nucleolus, cell-wall and cell-contents, show the same
relations and behave in the same manner in these two types of
animal cells as in the plant-cells studied by Schleiden. The types
of cell which he chose for this comparison are the most plant-like
of all animal cells, and he was even able to point to a thickening
of the cell-wall in certain cartilage cells, analogous to the
thickening which plays so important a part in the outward
modification of plant-cells. The analogy indeed in structure and
development between chorda and cartilage cells and the cells of
plants seemed to him complete. The substance of the notochord
consisted of polyhedral cells having attached to their wall an oval
disc similar in all respects to the nucleus of the plant-cell, and
like it containing one or more nucleoli. Inside the mother-cell
were to be found young developing cells of spherical shape, lacking
however a nucleus. Cartilage was even more like plant tissue. It
was composed of cells, each with its cell membrane. The cells lay
close to one another, separated only by their thickened cell-wall
and the intercellular matrix, showing thus even the general
appearance of the cellular tissue of plants. They contained a
nucleus with one or two nucleoli, and the nucleus was often
resorbed, as in plants, when the cell reached its full development.
Other nuclei were in many cases present in the cell, round which
young cells could be 175seen to develop, in exactly the same manner
as in plants. These nuclei had accordingly the same significance as
the nuclei of plants, and deserved the same name of cytoblasts or
cell-generators. The true nucleus of the cartilage cell was
probably in the same way the original generator of the
mother-cell.
Having proved the identity in structure and function of the
cells of these selected tissues with the cells of plants, as
conceived by Schleiden, Schwann had still to show that the
generality of animal tissues consisted either in their adult or in
their embryonic state of similar cells. This demonstration occupies
the second and longest section of his book.
His method is throughout genetic; he seeks to show, not so much
that all animal tissues are actually in their finished state
composed of cells and modifications of cells, as that all tissues,
even the most complex, are developed from cells analogous in
structure and growth with the cells of plants.
All animals develop from an ovum; it was his first task to
discover whether the ovum was or was not a cell. It happened that,
some years before Schwann wrote, a good deal of work had been done
on the minute structure of the ovum, particularly by Purkinje and
von Baer. Purkinje in 1825[252] discovered and described in the unfertilised egg
of the fowl a small vesicle containing granular matter, which he
named the Keimbläschen or germinal vesicle. It
disappeared in the fertilised egg. As early as 1791 Poli had seen
the germinal vesicle in the eggs of molluscs, but the first
adequate account was given by Purkinje. In 1827[253] von Baer discovered the true ova of
mammals and cleared up a point which had been a stumbling block
ever since the days of von Graaf, who had described as the ova the
follicles now bearing his name.[254] Even von Graaf had noticed that the early
uterine eggs were smaller than the supposed ovarian eggs;
Prévost and Dumas[255] had observed the presence in the Graafian
follicle of a minute spherical body, which, however, they hesitated
to call the ovum; it was left to von Baer to elucidate the
structure of the follicle and to prove 176that this small sphere was
indeed the mammalian ovum. His discovery was confirmed by Sharpey
and by Allen Thomson. Von Baer found the germinal vesicle in the
eggs of frogs, snakes, molluscs, and worms, but not in the
mammalian ovum; he considered the whole mammalian ovum to be the
equivalent of the germinal vesicle of birds—a comparison
rightly questioned by Purkinje (1834). In 1834 Coste[256] discovered in the ovum of the rabbit a
vesicle which he considered to be the germinal vesicle of Purkinje;
he observed that it disappeared after fertilisation. Independently
of Coste, and very little time after him, Wharton Jones[257] found the germinal vesicle in the
mammalian ovum. Valentin in 1835,[258] Wagner in 1836,[259] and Krause in 1837,[260] added considerably to the existing knowledge of
the structure of the ovum. Wagner in his Prodromus called
attention to the widespread occurrence, within the germinal vesicle
of a darker speck which he called the Keimfleck or germinal
spot, known sometimes as Wagner's spot. He recognised the
Keimfleck in the ova of many classes of animals from mammals
to polyps. Frequently more than one Keimfleck occurred.
Schwann had therefore a good deal of exact knowledge to go upon
in discussing the significance of the ovum for the cell-theory.
There were two possible interpretations. Either the ovum was a cell
and the germinal vesicle its nucleus, or else the germinal vesicle
was itself a cell within the larger cell of the ovum and the
germinal spot was its nucleus. Schwann had some difficulty in
deciding which of these views to adopt, but he finally inclined to
the view that the ovum is a cell and the germinal vesicle its
nucleus, basing his opinion largely upon observations by Wagner
which tended to prove that the germinal vesicle was formed 177first and
the ovum subsequently formed round it. But the ovum was not, in
Schwann's view, a simple cell, for within it were contained
yolk-granules, one set apparently containing a nucleus, the others
not. Even the second set, those composing the yellow yolk, were
considered by Schwann to deserve the name of cells, because,
although a nucleus could not be observed in them, they had a
definite membrane, distinct from their contents—a conception
of the cell obviously dating from the earliest botanical notions of
cells as little sacs. The yolk cells were not mere dead food
material but living units which took part in the subsequent
development of the egg. The relation between the unfertilised egg
and the blastoderm which arises from it is not made altogether
clear by Schwann. According to his account the cells of the
blastoderm are formed actually in the ovum. Round the nucleus of
the egg appears a Niederschlag or precipitate which is the
rudiment of the blastoderm (p. 68). When the egg leaves the ovary
the nucleus disappears, leaving behind it this rudiment of the
blastoderm, which rapidly grows and increases in size. The
blastoderm of the chick before incubation is found to be composed
of spherical anucleate bodies which Schwann considers to be cells,
because they almost certainly develop into the cells of the
incubated blastoderm, which are clearly recognisable as such after
eight hours' incubation. The serous and mucous layers can be
distinguished after sixteen hours' incubation, and it is found that
the cells of the serous layer contain definite nuclei, though such
seem to be absent in the cells of the mucous layer. Between the two
layers other cells are formed belonging to the vessel layer, which
is, however, in Schwann's opinion not a very definitely
individualised layer.
Schwann's next step is a detailed demonstration of the origin of
each tissue from simple cells such as those composing the incubated
blastoderm.
"The foregoing investigation has taught us that the whole ovum
shows nothing but a continual formation and differentiation of
cells, from the moment of its appearance up to the time when,
through the development of the serous and mucous layers of the
blastoderm, the foundation is given for all the tissues
subsequently appearing: we have 178found this common parent of all
tissues itself to consist of cells; our next task must be to
demonstrate not only in this general way that tissues originate
from cells, but also that the special formative mass of each tissue
is composed of cells, and that all tissues are either constituted
by simple cells or by one or other of the manifold kinds of
modified cells" (p. 71). Five classes of tissue can be
distinguished, according to the extent and manner of the
modifications which the cells composing them have undergone. There
are first of all independent and isolated cells, such as the
corpuscles of the blood and lymph, not forming a coherent tissue in
the ordinary sense. Next there are the assemblages of cells lying
in contiguity with one another, but not in any way fused; examples
of this class are the epidermal tissues and the lens of the eye. In
the third class come tissues the cells of which have fused by their
walls, but whose cell-cavities are not in continuity, such as
osseous tissue and cartilage. In the tissues of the fourth class,
comprising the most highly specialised of all, not only are the
cell-walls continuous but also the cell-cavities; to this class
belong muscle, nerve and capillary vessels. A fifth class, of
rather a special nature, includes the fibrous tissues of all kinds.
This is the first classification of tissues upon a cellular basis,
and it marks the foundation of a new histology which took the place
of the "general anatomy" of Bichat. The exhaustive account which
Schwann gives of the structure and development of the tissues in
this section of his book constitutes the first systematic treatise
on histology in the modern sense, and it is still worth reading, in
spite of many errors in detail.
Schwann found it easy to demonstrate the cellular nature of the
tissues of his first three classes. With the other two classes he
had more difficulty. Fibres of all kinds, he considered, arose by
an elongation of cells, which afterwards split longitudinally into
long strips, forming as the case might be white or elastic fibrous
tissue. Muscle-fibres and nerve-fibres were formed in a totally
different way, by coalescence of cells; each separate muscle-fibre
and nerve-fibre was thus a compound cell. Capillaries, Schwann
held, were formed by cells hollowed out like drain-pipes, and 179set end to
end—a mistaken view soon corrected by Vogt (Embryologie
des Salmones, p. 206, 1842).
In this detail part of his book Schwann accumulates material for
a general theory of the cell which he develops in the third and
last section. Taking up the physiological or dynamical standpoint,
he points out that one process is common to all growth and
development of tissues both in animals and plants, namely, the
formation of cells, a process which he conceives to take place in
the following manner. There is, first of all, a structureless
substance, the cytoblastem, the matrix in which all cells
originate. The cytoblastem may be either inside the cells, or, more
usually, in the spaces between them. It is not a substance of
definite chemical and physical properties, for the matrix of
cartilage and the plasma of the blood alike come within the
definition. It has largely the significance of food material for
the developing cells. In plants, according to Schleiden, cells are
never formed in the intercellular substance—the cytoblastem
is within the cells; but extracellular cell formation seems to be
the general rule in animals. An intracellular formation of cells
occurs only in the ovum, in cartilage cells and chorda cells and in
a few others, and even there it is not the exclusive method of
formation; a formation of cells within cells never occurs in
muscles and nerves, nor in fibrous tissue (p. 204). In the
cytoblastem granules appear, which gradually increase in size and
take on the characteristic shape of nuclei; round each of these a
young cell is formed. Sometimes the young cells appear to have no
nuclei, as in the intracellular brood of chorda cells, but, as a
rule, a nucleus is clearly visible. The nucleus is indeed the most
characteristic constituent of the cell. "The most important and
most constant criterion of the existence of a cell is the presence
or absence of the nucleus," writes Schwann near the beginning of
his book (p. 43).
As a general rule the nucleolus is formed first, and round it by
a sort of condensation or concretion the nucleus, which is
frequently hollow, and round this again, by a somewhat similar
process, the cell. "The whole process of the formation of a cell
consists in the precipitation round a small previously formed
corpuscle (the nucleolus) of first one layer 180(the nucleus)
and then later round this a second layer (the cell substance)" (p.
213). The outermost layer of the cell usually thickens to form the
membrane, but this membrane formation does not always occur, and
the membrane is not present in all cells. The nucleus is formed in
exactly the same manner as the cell, and it might with much truth
itself be called a cell—a cell of the first order, while
ordinary nucleated cells might be designated cells of the second
order (p. 212). In anucleate cells there is probably only a single
process of layer formation round an infinitely small nucleolus. In
almost all nucleate cells the nucleus is resorbed when the cell
reaches its full development, and it is larger and more important
the younger the cell is.
The cell was for Schwann not a morphological concept at all, but
a physiological; the cell was a dynamical, not a statical unit.
Cell-formation was the process at the back of all production of
life, and cells were the centres of all vital activity. Each cell
was itself an organism, and its life and activities were to some
extent independent of the lives and activities of all the other
cells. The multicellular organism was a colony of unicellular
organisms, and its life was a sum of the lives of its constituent
elements. This "theory of the organism," which holds so important a
place in biology even at the present day, is developed by Schwann
in the concluding pages of his book.
He begins by contrasting the teleological with the materialistic
conception of living things. In the teleological view, a special
force works in the living organism, guiding and directing its
activities towards a purposeful end. According to the materialistic
view there are no other forces at work in the living organism than
those which act in the inorganic realm, or at least there are none
but forces at one with these in their blindness and necessity.
True, the purposiveness of living processes cannot be denied; but
its ground lies, according to this view, not in a vital force which
guides and rules the individual life, but in the original creation
and collocation of matter according to a rational plan. The
purposiveness of life is part of the purposiveness of the universe;
just as the stars circle for ever in harmoniously adjusted paths,
so do the processes of life work together 181towards a common end. Both
are the inevitable result of the original distribution of matter in
the primitive chaos, a distribution fixed by a rational and
foreknowing Being (p. 222).
Which of the two conceptions is to be adopted in biology?
Teleological explanations have long been banished from the physical
sciences, and in biology they are only a last resort when physical
explanations have proved incomplete (p. 223). And if the ground of
the purposiveness of living Nature is the same as the ground of the
purposiveness of the universe, is it not reasonable to suppose that
explanations which have proved satisfactory for inorganic things
will in time with sufficient knowledge prove adequate also for
organic things?
The teleological conception, again, leads to difficulties
particularly when it is applied to the facts of reproduction. If we
suppose that a vital force unifies and coordinates the organism and
is its very essence, we must also suppose that this force is
divisible and that a part of it—separated in
reproduction—can bring about the same results as the whole.
If on the contrary the forces having play in the organism are the
mere result of the particular combination of the matter composing
it, the reconstruction of a particular combination of molecules in
the ovum is all that is necessary to set development a-going along
exactly the course taken by the ovum of the parent. Another
argument against the teleological view is derived from the facts of
the cell-theory. The cell-theory tells us that the molecules of the
living body are not immediately built up in manifold combinations
to form the organism, but are formed first into unit-constructions
or cells, and that these units of composition are invariably formed
in all development, of plants and animals alike, however diverse
the goal of development may be. If there were a vital principle
would we not expect to find that, scorning this roundabout way of
reaching its goal, it went straight to the mark, taking a different
and distinctive course for each individual development, building up
the organism direct without the intermediary of cells? But since
there is a universal principle of development, namely, the
formation of cells, does it not seem that the cells must be the
true 182organisms, that the whole "individual"
organism must be an aggregate of cells, and that the concept of
individuality applied to the organism is accordingly a logical
fiction? And it is just upon this notion of the individuality of
the organism that the teleological concept is based. The
teleological view can perhaps not be completely refuted until the
adequacy of materialistic explanations has been finally shown; but
it is certain that the most promising method for research is the
materialistic (p. 226).
"We start out then from the assumption that the basis of the
organism is not a force acting according to a definite plan; on the
contrary, the organism arises through the action of blind and
necessary laws, of forces which are as much implicit in matter as
those of the inorganic world. Since the chemical elements in
organic Nature differ in no way from those of inorganic Nature, the
ground or cause of organic phenomena can consist only in a
different mode of combination of matter, either in a peculiar mode
of combination of the elementary atoms to form atoms of the second
order, or in the particular arrangement of these compound molecules
to form the separate morphological units of the organism or the
whole organism itself" (p. 226). Accepting then the materialistic
conception of the organism, we have to consider this further
problem. Does the ground of organic processes lie in the whole
organism or in its elementary parts? Translated into terms of
metabolism—note the physiological point of view—the
question runs, are metabolic processes the result of the molecular
construction of the organism as a whole, or does the centre of
metabolic activity lie in the cell? Is it the cell rather than the
organism that is the immediate agent of assimilatory processes? In
the first alternative the cause of the growth of the constituent
parts lies in the totality of the organism; in the other
alternative:—"Growth is not the result of a force having its
ground in the organism as a whole, but each of the elementary parts
possesses a force of its own, a life of its own, if you will; that
is to say, in each elementary part the molecules are so combined as
to set free a force whereby the cell is enabled to attract new
183molecules and so to grow, and the whole
organism exists only through the reciprocal action of the single
elementary parts.... In this eventuality it is the elementary parts
that form the active element in nutrition, and the totality of the
organism can be indeed a condition, but on this view it cannot be a
cause" (p. 227).
To help in the decision of this question, appeal must be made to
the facts established as to the cellular nature of the organism and
of its reproductive elements. We know that every organism is
composed of cells, which are formed and grow according to the same
laws wherever they are found, whose formation therefore is
everywhere due to the same forces. If we find that certain of these
cells—all of which we know to be essentially identical one
with another—have the power when separated from the others of
growing and developing into new organisms, we can infer that not
only such cells but also all other cells have this assimilatory
power. The ova of animals, the spores of plants, the isolated cells
of lower organisms in general, all show the power of separate
assimilation and development. "We must therefore, in general,
ascribe to the cell an individual life, that is to say, the
combination of the molecules in the single cell does suffice to
produce the force whereby the cell is enabled to draw to itself new
molecules. The ground of nutrition and growth lies not in the
organism as a whole, but in the separate elementary parts, the
cells. The fact that it is not every cell that can continue to grow
when separated from the organism is not in itself an objection to
this theory, any more than it is an objection to the individual
life of a bee that it cannot continue to exist apart from the
swarm. The activation of the forces existing within the cell
depends on conditions which the cell encounters only in connection
with the whole" (pp. 228-9).
Schwann's next step is to discover what are the essential forces
active in the cell, and here he enters the realm of hypothesis. He
finds they can be reduced to two—an attractive force and a
metabolic force. The attractive force is seen in the process of
cell-formation, where first of all the nucleolus is formed by a
concentration and precipitation of substances found free in the
cytoblastem, and in the same 184way the nucleus and later the cell are
laid down as concentric precipitates from the cytoblastem.
Cell-formation also involves the second or metabolic force, by
means of which the cell alters the chemical composition of the
medium surrounding it so as to prepare it for assimilation.
Schwann's attractive force brings about the actual taking up of the
prepared substance; his metabolic force is the cause of the
digestion of food substances, and is nearly identical with enzyme
action. With what inorganic process, he now asks (p. 239), can the
process of cell-formation be most nearly compared, and the answer
obviously is, with the process of crystallisation. Cells are, it is
true, quite different in shape and consistency from crystals, and
they grow by intussusception, not by apposition—their plastic
or attractive forces seem therefore to be different. A still more
important difference is that the metabolic force is peculiar to the
cell. Yet there are important analogies between crystals and cells.
They agree in the important respect that they both grow in
solutions at the cost of the dissolved substance, according to
definite laws, and develop a definite and characteristic shape. It
might even be maintained, Schwann thinks, that the attractive force
of crystals is really identical with that of cells, and that the
difference in result is due merely to the difference between the
substance of the cell and the substance of the crystal. He points
out how organic bodies are remarkable for their powers of
imbibition, and he seeks to show that the cell is the form under
which a body capable of imbibition must necessarily crystallise,
and that the organism is an aggregate of such imbibition-crystals.
The analogy between crystallisation and cell-formation he works out
in the following manner:—"The substance of which cells are
composed possesses the power of chemically transforming the
substance with which it is in immediate contact, in somewhat the
same way as the well-known preparation of platinum changes alcohol
into acetic acid. Each part of the cell possesses this property. If
now the cytoblastem is altered by an already formed cell in such a
way that a substance is formed that cannot become part of the cell,
it crystallises out first as the nucleolus of a new cell. This in
its turn alters the composition of the cytoblastem. A part of the
transfomed 185substance may remain in solution in the
cytoblastem or may crystallise out as the beginning of a new cell;
another part, the cell-substance, crystallises round the nucleolus.
The cell-substance is either soluble in the cytoblastem and
crystallises out only when the latter is saturated with it, or it
is insoluble and crystallises as soon as it is formed, according to
the aforementioned laws of the crystallisation of
imbibition-bodies; it forms thus one or more layers round the
nucleolus, etc. If one imagines cell-formation to take place in
this way, one is led to think of the plastic force of the cell as
identical with the force by means of which a crystal grows" (pp.
249-50).
Two difficulties have to be faced by this theory—(1) the
origin of the metabolic power of the cells, (2) the reason why the
cells arrange themselves so as to form an organism of complex and
definite structure. Schwann tries to explain the origin of the
"metabolic" action, the analogy of which with the contact-action of
colloidal platinum he recognises, by attributing it to the peculiar
structural arrangements of molecules. In attempting to account for
the harmonious structure of the organism he points to the analogy
of ordinary crystals, which often form complex and regular
tree-like arrangements; plants in particular resemble these
regularly shaped crystal-aggregates.
The whole ingenious theory is offered merely as an hypothesis
and a guide to research. It is interesting as one of the most
carefully thought-out attempts ever made to give a thorough-going
materialistic account of the origin and development of organic
form, and it arose directly out of the cell-theory.
Schleiden and Schwann started out from an erroneous theory of
the origin and development of cells, which impaired to some extent
the value of their results. It was not long, however, before their
theory of the origin of cells by "crystallisation" from an intra- or
extra-cellular cytoblastem was challenged and overthrown, and the
generalisation that cells originate by division from pre-existing
cells put in its place.
This was established for plant cells by Meyen, Unger, von Mohl,
Naegeli and Hofmeister in or about the 186forties.[261] Criticism of the Schwann-Schleiden theory
from the zoological side was suggested by the study of the
segmentation of the ovum—the developmental process in which
the multiplication of cells is most easily observed. The
segmentation of the ovum was well known to Schwann, for the process
had been described in the frog by Prévost and Dumas in 1824,[262] in the frog and newt by
Rusconi,[263] and an elaborate study
of the process in the frog had been made by von Baer.[264] Schwann indeed suspected that there must
be some connection between the segmentation of the ovum and the
formation of cells, but he did not realise that the cellular
blastoderm of the chick was formed by the division or segmentation
of the egg-cell.
Segmentation was soon found to be of widespread occurrence. Von
Siebold in 1837 described the process in Entozoa,[265] and in the same year Lovén saw
segmentation in Campanularia,[266] and Sars in the starfish and in Nudibranchs.[267]
In 1838 Bischoff[268] observed segmentation in the mammalian ovum, and
the whole course of segmentation in the ovum of the rabbit from the
2-celled to the morula stage was carefully described and figured by
Barry[269] in 1839. C. Vogt[270] in 1842 described segmentation in
Coregonus and Alytes. The discovery of segmentation
in the ovum of 187birds was not made until 1847, by Bergmann,[271] confirmed
independently by Coste[272] in 1850. By 1848 segmentation had been noted in
Hydra and various hydroids, in acalephs, in starfish,
polyzoa, nematodes, rotifers, leeches, oligochætes,
polychætes, in most groups of molluscs and arthropods, and in
all the vertebrate classes.[273]
The process was at first held to be merely one of yolk-division,
or Dotterfurchung, and its details were by most interpreted
in the light of the Schleiden-Schwann theory of cell-formation.
The first steps towards a truer conception of the process seem
to have been taken by Bergmann, who in 1841[274] called attention to the presence of nuclei in
the segmentation-spheres of the frog's egg, and by Bagge in the
same year, who observed that division of the nuclei preceded the
multiplication of the segmentation spheres.[275] He considered the nuclei to be anucleate cells,
and the same view was taken by Kölliker in 1843.[276] Next year, however, in his classical paper
on Cephalopod development[277] Kölliker came to the opinion that they were
really nuclei. He showed that segmentation was brought about by
cell-division, that between "total" and "partial" segmentation
there was a difference of degree and not of kind, and that the
cells of the body were formed by division of the segmentation
spheres. He held, however, that the nuclei multiplied endogenously
and not by division. The division of nuclei was observed by Coste
in 1846.[278] Leydig in 1848[279] took the necessary step in advance and
maintained that the nuclei as well as the cells increased always by
division. He was supported by Remak, who in a paper of 1852,[280] and more fully in his
monumental 188Untersuchungen über die Entwickelung
der Wirbelthiere (Berlin, 1850-55), proved that in the frog's
egg at least segmentation was a simple process of cell-division,
initiated always by division of the nucleus.[281]
One point Remak left undecided—the fate of the
Keimbläschen or egg-nucleus. It was generally held,
even so late as the 'fifties, that the egg-nucleus disappeared just
before segmentation began—Bischoff clung to this belief even
in 1877.[282] Though Barry had held in
1839 that the egg-nucleus does not disappear in segmentation, J.
Müller seems to have been the first actually to prove that it
forms by division the nuclei of the first two segmentation spheres.
He furnished the demonstration in the egg of Entoconcha
mirabilis,[283] and his paper was
known to Remak, who could not, however, observe a similar division
of the egg-nucleus in the frog. Müller's discovery was
confirmed for Oceania armata by Gegenbaur,[284] and for Notommata sieboldii by
Leydig.[285]
In 1854 Virchow,[286] previously a supporter of Schwann, crystallised
the new views in the famous phrase—Omnis cellula e
cellula—and gave wide publicity to them in his classical
lectures on Cellular Pathology, delivered in 1858.[287] The new doctrine of cell-formation was
also taught by Leydig[288] in his text-book of histology, published in
1857.
The Schleiden-Schwann theory of the origin of cells by
generation in a cytoblastem was now definitely overthrown.
The importance of the protoplasmic content of the cell was
brought into prominence through the work of Dujardin,[289] 189Purkinje,[290] Cohen[291] and Max Schultze.[292] The last-named in 1861 proposed a definition of
the cell which might be accepted at the present day. "A cell," he
wrote, "is a little blob of protoplasm containing a nucleus" (p.
11).
190
CHAPTER XII
THE CLOSE OF THE PRE-EVOLUTIONARY PERIOD
The influence of the cell-theory on morphology was not
altogether happy. The cell-theory was from the first physiological;
cells were looked upon as centres of force rather than elements of
form, and the explanation of all the activities of the organism was
sought in the action of these separate dynamic centres. There
resulted a certain loss of feeling for the problems of form. The
organism was seen no longer as a cunningly constructed complex of
organs, tissues and cells; it had become a mere cell-aggregate; the
higher elements of form were disregarded and ignored.
We have seen this physiological attitude expressed with the
utmost clearness by the founder of the cell-theory himself; we
shall see the same attitude taken up by most of his successors.
Thus Vogt, who was later to become one of the protagonists of
materialism in Germany, developed in his memoir on the embryology
of Coregonus[293] the theory of the independent or individual life
of the cell. "Each cell," he wrote, "represents in some measure a
separate organism, and while their development necessarily conforms
to the general plan and the particular tendencies of the parent
organism, they nevertheless each follow their own particular
tendency and do not lose their independence until, by reason of the
metamorphoses which they undergo, they lose their cellular nature"
(p. 275).
And again, "... we are obliged to admit the existence in the
cell of an independent life, which makes its development
self-sufficient.... Each cell consequently represents a little
independent organism, which assimilates foreign substances, builds
them up, and rejects those that are useless; 191from this point
of view the embryo can be compared up to a certain point with a
zoophyte stock, of which each polyp, while living its own
independent life, is yet incorporated in the common corm, which
impresses its distinctive character upon every polyp" (p. 293).
Classical expression was given to the "colonial theory" of the
organism by Virchow in his lectures on "Cellular Pathology."[294] For Virchow the organism
resolves itself into an assemblage of living centres, the cells;
the organism has no real existence as a unity, for there is no one
single centre from which its activities are ruled. Even the nervous
system, which appears to act as a co-ordinating centre, is itself
an aggregate of discrete cells. "A tree is a body of definite and
orderly composition, the ultimate elements of which, in every part
of it, in leaf and root, in stem and flower, are cellular
elements—so also are animal forms. Every animal is a sum
of vital units, each of which possesses the full
characteristics of life. The character and the unity of life cannot
be found in one definite point of a higher organisation, for
example in the brain of man, but only in the definite, constantly
recurring disposition shown individually by each single element. It
follows that the composition of the major organism, the so-called
individual, must be likened to a kind of social arrangement or
society, in which a number of separate existences are dependent
upon one another, in such a way, however, that each element
possesses its own particular activity, and, although receiving the
stimulus to activity from the other elements, carries out its own
task by its own powers" (2nd ed., pp. 12-13).
Analysis, decomposition, or disintegration of the organism is
here pushed to its extreme point, and the problem of recomposition,
synthesis and co-ordination shirked or forgotten.
The harmful influence of the cell-theory upon morphology did not
pass unnoticed by the broader-minded zoologists of the day.
Virchow's earlier paper[295] on the application of the 192cell-theory to
physiology and pathology called forth a vigorous protest from
Reichert,[286] who discussed in a very
instructive way the contrast between the older "systematic" and the
newer "atomistic" attitude to living Nature.
Is it really true, he asks, that the cell is the dominant
element in all organisation; is the cell comparable in importance
to the atom of the chemists; or is it not rather the servant of a
higher regulatory power? Johannes Müller, who was Reichert's
master, had in his Physiology[297] argued splendidly for the existence of a
creative force which guides and rules development, and brings to
pass that unity and harmony of composition which distinguish living
things from inorganic products. Reichert sought in vain in the
writings of the biological "atomists" for any smallest recognition
of these broader characteristics of living things upon which
Müller had rightly laid stress. For the atomists the cell was
the only element of form; they ignored the combination of cells to
form tissues, of tissues to form organs, of organs to form an
organism. For the morphologists the cell was one element among
many, and the lowest of all.
The difference of attitude is clearly shown if we consider from
the two points of view a complicated organ-system such as the
central nervous system. The atomist sees in this a mere aggregate
of cells or at the most of groups of cells. "The morphologist," on
the other hand, "sees in the central nervous system a
proximate element in the composition of the body—a
primitive organ. From this point of view he apprehends and judges
its morphological relations with, in the first place, the other
co-ordinated primitive organs in the system as a whole; in all this
the cells remain in the background, and have nothing to do directly
with the determination of these morphological relations" (p. 6).
Within the nervous system there are separate organs which stand to
one another in a definite morphological and functional
relationship. These organs are, it is true, composed of cells; but
between the form and connections of 193these organs and the cells which
compose them there is no direct and necessary relation (p. 6). It
is true that the cell is the ultimate element of organic form, and
that all development takes place by multiplication and form-change
of cells. Yet is the cell in all this not independent of the unity
of the developing embryo, and what the cells produce, they produce,
so to speak, not of their own free will, nor by chance, but under
the guiding influence of the unity of the whole, and in a certain
measure as its agents (p. 7). The atomists will not admit the truth
of this; they see in development nothing more than a process of the
form-change and multiplication of cells. The full meaning of
development escapes them, for they take no cognisance of the
increasing complexity of the embryo, of the separating-out of
tissues, of the moulding of organs, of the harmonious adaptation
and adjustment of the parts to form a working whole.
In general, the fault of the atomists is that they do not
respect the limits which Nature herself has prescribed to the
process of logical analysis and disintegration of the organism;
they do not recognise the existence of natural and rational units
or unities; they forget the one great principle of rational
analysis, "that, by universally valid, inductive, logical method,
natural objects must in all cases be accepted and dealt with in the
combination and concatenation in which they are given" (p. 10).
The atomists at least recognised one natural organic element,
the cell; the materialistic physiologists of the time resolved even
this unity into an aggregate of inorganic compounds, and regarded
the organism itself as nothing but a vastly complicated
physico-chemical mechanism. From this point of view morphology had
no right of existence, and we find Ludwig, one of the foremost of
the materialistic school, maintaining that morphology was of no
scientific importance, that it was nothing more than an artistic
game, interesting enough, but completely superseded and robbed of
all value by the advance of materialistic physiology.[298]
Naturally enough, morphologists did not accept this rather
contemptuous estimate of their science, but held 194firmly to the
morphological attitude. So Leuckart in his reply to Ludwig, so
Rathke in a letter to Leuckart published in that reply, so Reichert
in his Bericht, so J. V. Carus in his System der
thierischen Morphologie,[299] upheld the validity, the independence, of
morphological methods. Leuckart and Rathke called attention to the
absolute impossibility of explaining by materialistic physiology
the unity of plan underlying the diversity of animal form. J. V..
Carus, who was convinced of the validity of physiological methods
within their proper sphere, drew a sharp distinction between
systematics and morphology on the one hand, and physiology on the
other. Physiology had nothing to do with the problems of form at
all; its business was to study the physical and chemical processes
which lay at the base of all vital activities. Morphology, on its
part, had to accept form as something given, and to study the
abstract relations of forms to one another. "On this point," he
writes, "stress is to be laid, that morphology has to do with
animal form as something given by Nature, that though it
follows out the changes taking place during the development of an
animal and tries to explain them, it does not enquire after the
conditions whose necessary and physical consequence this form
actually is" (p. 24). He expressed indeed a pious hope (p. 25) that
physiology might one day be so far advanced that it could attempt
with some hope of success to discover the physico-chemical
determinism of form, but this remained with him merely a pious
hope. Reichert, in his Bericht, applied to the rather wild
theorisings of the physiologist Ludwig the same clear commonsense
criticism that he bestowed on the other "atomists."
It would take too long to describe the great development that
materialistic physiology took at this time, and to show how the
separation of morphology from physiology, which originally took
place away back in the 17th century, had by this time become almost
absolute. The years towards the end of the first half of the
century marked indeed the beginning of the classical period as well
of physiology as of dogmatic materialism. Moleschott and Buchner
popularised materialism in Germany in the 'fifties, while Ludwig,
du Bois 195Reymond and von Helmholtz began to apply the
methods of physics to physiology. In France, Claude Bernard was at
the height of his activity, rivalled by workers almost as great.
The doctrine of the conservation of energy was established about
this same time.
Between the cell-theory on the one side, and physiology on the
other, it was a wonder that morphology kept alive at all. The only
thing that preserved it was the return to the sound Cuvierian
tradition which had been made by many zoologists in the 'thirties
and 'forties. It is a significant fact that this return to the
functional attitude coincided in the main with the rise of marine
zoology, and that the man who most typically preserved the
Cuvierian attitude, H. Milne-Edwards, was also one of the first and
most consistent of marine biologists. Milne-Edwards describes in
his interesting Rapport sur les Progrès récents
des Sciences zoologiques en France (Paris) 1867, how "About the
year 1826, two young naturalists, formed in the schools of Cuvier,
Geoffroy and Majendie, considered that zoology, after having been
purely descriptive or systematic and then anatomical, ought to take
on a more physiological character; they considered that it was not
enough to observe living objects in the repose of death, and that
it was desirable to get to understand the organism in action,
especially when the structure of these animals was so different
from that of man that the notions acquired as to the special
physiology of man could not properly be applied to them" (p. 17).
The two young naturalists were H. Milne-Edwards and V. Audouin. In
pursuance of these excellent ideas they set to work to study the
animals of the seashore, producing in 1832-4 two volumes of
Recherches pour servir à l'histoire naturelle du littoral
de la France. After Audouin's early death A. de Quatrefages was
associated with Milne-Edwards in this pioneer work, and their
valiant struggles with insufficient equipment and lack of all
laboratory accommodation, and the rich harvest they reaped, may be
read of in Quatrefage's fascinating account of their journeyings.[300] Note that though they
called themselves 196physiologists they meant by physiology
something very different from the mere physical and chemical study
of living things. They were interested, as Cuvier was, primarily in
the problems of form; they sought to penetrate the relation between
form and function; their chief aim was, therefore, the study not of
physiology[301] in the restricted sense,
but physiological morphology. As a matter of fact they produced
more taxanomic and anatomical work than work on physiological
morphology, but this was only natural, since such a wealth of new
forms was disclosed to their gaze. Milne-Edwards' masterly
Histoire Naturelle des Crustacés[302] and A. de Quatrefage's Histoire
Naturelle des Annelés marins et d'eau douce[303] were typical products of their
activity.
In the North, men like Sars and Lovén were starting to
work on the littoral fauna of the fjords; in Britain, Edward Forbes
was opening up new worlds by the use of the dredge; Johannes
Müller was using the tow-net to gather material for his
masterly papers on the metamorphoses of Echinoderms.[304] Work on the taxonomy and anatomy of marine
animals was in general in full swing by the 'fifties and
'sixties.
This return to Nature and to the sea had a very beneficial
effect upon morphology, bringing it out from the laboratory to the
open air and the seashore. It saved morphology from formalism and
aridity, and in particular from a certain narrowness of outlook
born of too close attention paid to the details of microscopical
anatomy. It brought morphologists face to face again with the
wonderful diversity of organic forms, with the unity of plan
underlying that diversity, with the admirable adjustment of organ
to function and of both to the life of the whole.
Milne-Edwards' theoretical views, as expounded in his
Introduction à la zoologie générale
(1851), well reflect this Cuvierian attitude.[305] He acknowledges himself the debt he 197owes to
Cuvier; "the further I advance in the study of the sciences which
he cultivated with so sure a hand," he writes in 1867, "the more I
venerate him."
Milne-Edwards frankly takes up the teleological standpoint, and
interprets organic forms on the assumption that they are purposive
and rationally constructed. "To arrive at an understanding of the
harmony of the organic creation," he writes, "it seemed to me that
it would be well to accept the hypothesis that Nature has gone
about her work as we would do ourselves according to the light of
our own intelligence, if it were given us to produce a similar
result. Comparing and studying living things as if they were
machines created by the industry of man, I have tried to grasp the
manner in which they might have been invented, and the principles
whose application would have led to the production of such an
assemblage of diversified instruments" (p. 435). The problem is to
discover the laws which rule the diversity of organic forms. The
first and most obvious of these laws is the "law of economy," or
the law of unity of type. Nature, as Cuvier pointed out, has not
had recourse to all the possible forms and combinations of organs;
she appears to work with a limited number of types and to get the
greatest possible diversity out of these by varying the proportions
of the constitutive materials of structure. Within the limits of
each type Nature has brought about diversity by raising her
creatures to different degrees of perfection. This is the second
law of organic form, and it is this law that Milne-Edwards chiefly
elaborates. Degrees of perfection mean for him, as for Aristotle,
primarily degrees of perfection of function, but since structure is
necessarily in close relation with function, perfection of function
brings in its train increased perfection of organisation. This can
only be attained by a division of labour[306] 198among the organs and by their consequent
differentiation. An animal is like a workshop where some
complicated product is manufactured, and the organs are like the
workmen. Each workman has his own special piece of work to do, at
which he becomes thoroughly expert; and the finished product is
manufactured more rapidly and efficiently by the co-operation of
workers each skilled in one department than it would be if each
workman had to produce the whole. Applied to the organism this
principle of the division of labour means the differentiating out
of the separate functions, their localisation in different parts of
the organism, and their co-ordination to produce a combined
result.
This differentiation of functions implies a corresponding
differentiation of organs, but it is functional differentiation
which always takes the lead. "Where division of labour has not been
introduced into the organism there must exist a great simplicity of
structure. But just as uniformity in the functions of the different
parts of the body implies a uniformity in their mode of
constitution, so diversity in function must be accompanied by
particularities in structure; and, in consequence also, the number
of dissimilar parts must be augmented and the complication of the
machine increased" (p. 463). Since function comes before form there
is not always a special organ for every function. "It is a grave
error to believe that a particular function can be performed only
by one and the same organ. Nature can arrive at the desired result
by various ways, and when we look down through the animal kingdom
from the highest to the lowest forms we see that the function does
not disappear even when the special instrument provided for the
purpose in the higher types ceases to exist" (p 470).
Nature, holding fast to the law of economy, does not even always
create a new organ for a new function; she may simply adapt an
undifferentiated part to special functions, or she may even convert
to other uses an organ already specialised (p. 464). So, for
example, the function of respiration is in the lowest animals
diffused indifferently over the whole surface of the body, and only
as organisation advances is it localised in special organs, such as
gills. Now 199suppose that Nature wishes to adapt a fish,
which breathes by gills, to life in the air; she does not create an
organ specially for this purpose, but utilises the moist
gill-chamber (e.g., in Anabas scandens), modifying it
in certain ways so that the fish can take advantage of the oxygen
it contains. But this gill-chamber lung is at best a makeshift, and
when she comes to the more definitely terrestrial Amphibia Nature
gives up the attempt to use the gill-chamber as a lung, and creates
a new organ, the true vertebrate lung, specially adapted for
breathing air (p. 475).
But whatever means Nature adopts, her aim is always the
same—to specialise, to differentiate, to produce diversity
from uniformity.
Differentiation not only raises the level of organisation; it
usually also takes the direction of adaptation to particular habits
of life, and this is perhaps the most fruitful cause of diversity.
Everywhere we find animals specialised in adaptation to their
environment—to life in air or water, or on land—and
many of their most striking differences are due to this cause. But
adaptation may also act in reducing diversity, for there
necessarily occur many instances of parallel adaptation or
convergence. So we get the extraordinary parallelism between the
families of marsupials and the orders of placentals,[307] the remarkable similarity between the
respiratory organs of land-crabs and air-breathing fish—to
mention only two out of an immense range of analogous facts.
The last cause of diversity that Milne-Edwards adduces is what
he calls a "borrowing" of peculiarities of structure from another
systematic group. Thus, "among reptiles, the tortoises seem to have
borrowed from birds some of their characteristic features of
organisation; and among the sauroid fishes the piscine type seems
to have been influenced by the type from which reptiles are
derived" (p. 479). So many riddles that, a little later on,
stimulated the ingenuity of the evolutionists!
Such, then, were the factors which Milne-Edwards 200considered
adequate to explain the rich variety of animal forms. We cannot do
better than quote his own summary of his doctrine:—"To sum
up, then, the great differences introduced by Nature into the
constitution of animals seem to depend essentially upon the
existence of a certain number of general plans or distinct types,
upon the perfecting in various degrees either of the whole or of
parts of each of these structural plans, upon the adaptation of
each type to varied conditions of existence, and upon the secondary
imitation of foreign types by certain derivatives of each
particular type" (p. 480).
We have laid stress on the fact that Milne-Edwards put function
before form, for this is the mark of the true Cuvierian. With it
goes the belief that Nature forms new parts to meet new
requirements, that she is not limited, as Geoffroy thought, to a
definite number of "materials of organisation," but can produce
others at need. Cuvier held, for example, that many of the muscles
and even the bones of fish were peculiar to them, and without
homologues in the other Vertebrates, having been created by Nature
for special ends.[308] So, too, Johannes
Müller, who in many ways and not least in his sane vitalism
was a follower of the Cuvierian tradition, recognised that many of
the complicated cartilages in the skull of Cyclostomes were
specially formed for the important function of sucking, and had no
equivalent in other fish.[309]
So, too, the embryologists after Cuvier often came across
instances of the special formation of parts to meet temporary
needs. Thus Reichert interpreted the "palatine" and "pterygoid,"
which are formed in the mouth of the newt larva by a fusion of
conical teeth, as special adaptations to enable the little larva to
lead a carnivorous life.[310]
Not many years after the publication of Milne-Edwards'
Introduction à la zoologie générale
(1851) there appeared a book by H. G. Bronn in which was offered a
very similar analysis of organic diversity. The curious thing was
that 201Bronn approached the problem from quite a
different standpoint, from the standpoint, indeed, of
Naturphilosophie. Of this the title of the book is itself
sufficient proof—Morphologische Studien über die
Gestaltungs-gesetze der Naturkörper überhaupt und der
organischen insbesondere (Leipzig and Heidelberg, 1858).[311] The linking up of
organic with inorganic form is characteristic; there is much talk,
too, in the book of Urstoffe and Urkräfte, but
underlying the Naturphilosophie we can trace the same
Cuvierian treatment of form, and see crystallise out laws of
progressive development that bear no small analogy with the laws
established by Milne-Edwards.
According to Bronn, the ideal fundamental form of the plant is
an ovoid or strobiloid[312] body, for a plant reaches out in two directions
in search of food—towards the sun and towards the earth.
Animals differ from plants in being endowed with sensation and
mobility (cf. Aristotle and Cuvier), and it is this
characteristic that gives them their distinctive form. The main
types of animal form—the Amorphozoa, Actinozoa, and
Hemisphenozoa—are essentially adaptations to particular modes
of locomotion. Animals either are fixed, or they move in all
directions without reference to any definite axis, or they move in
one main direction.
The Amorphozoa or shapeless animals include many of the Protozoa
and sponges; they have no typical form, and most of them are
sessile. The Actinozoa include such animals as the Cœlentera,
which are fixed, and the Echinoderms, which have a central point
and move indifferently along any radial axis; their form differs
from the strobiloid mainly in having radiate rather than spiral
symmetry. The Hemisphenozoa, or bilaterally symmetrical animals,
include all those that habitually move forward; they have a front
end and a hind end, a dorsal surface and a ventral, and the mouth,
sense-organs and "brain" are concentrated 202in the front end to form a
head—all in direct adaptation to this forward movement; they
make up the vast majority of animals.
The fundamental forms of living things are, however, merely so
many themes on which a multitude of further variations are woven,
through the action of the laws which rule the detail of organic
diversities. These further laws may be set down under four main
heads. Under the first comes the law of the existence of certain
fundamentally distinct structural types, which are distinguished
from one another by their ground-form, by the number of
organ-systems, and by the number of homotypic organs they possess,
but principally by the relative position of the organs to one
another (principle of connections). The form and connections of the
nervous system are of particular importance in distinguishing the
types (cf. Cuvier). The second factor in the diversity of
organic form is the action of certain laws of progressive
development[313]
(Entwickelungsgesetze), which bear the same relation to the
development of the animal kingdom as the laws of individual
development bear to the development of the embryo, for organs
appear in the different animal series in much the same order and
manner as they develop in the individual. These laws are (1)
progressive differentiation of functions and organs; (2) numerical
reduction of serially repeated parts; (3) concentration of
functions and their organs in particular parts of the body; (4)
centralisation of organ-systems and parts of such, so that they
come to depend upon one central organ; (5) internalisation of the
"noblest" organs, unless these are necessarily external, and (6)
increase in size of the whole or of parts. Of these the law of
differentiation is by far the most important, and most of the
others are in a sense merely special cases of this fundamental law.
To this law of differentiation is due the increase in complexity or
perfection of organisation which is shown by all the animal series.
Bronn himself recognised the great similarity of this law of
progressive differentiation to Milne-Edwards' principle of the
division of labour; he seems, however, to have arrived at it
independently.
203Bronn's third factor in the production of
variety of form is adaptation to environment, or better, functional
response to environment. Bronn gives an excellent account of
adaptational modifications and calls attention, just as
Milne-Edwards did, to the numerous analogies of structure which
adaptation brings about. He works out the interesting view that
there is some connection between classificatory groups and
adaptational forms, especially such as are connected with the
function of locomotion:—"Based upon a common characteristic
method of locomotion are whole or nearly whole sub-phyla
(Hexapoda), classes (mammals and reptiles, birds, fishes,
gastropods, pteropods, brachiopods, Bryozoa, Rotifera, jelly-fish,
polypes, sponges), sub-classes (mobile and immobile lamellibranchs,
echinoderms, walking and swimming Crustacea, parasitic and
free-living worms, and so on), often, however, only orders and
quite small groups (snakes, eels, bats, sepias, medusæ,
etc.)" (p. 141).
It was characteristic of the 'forties and 'fifties that
transcendental anatomy, along with Nature-philosophy, went rather
out of fashion, its false simplicities and premature
generalisations being overwhelmed by the flood of new discoveries.
A few stalwarts indeed upheld transcendental views. We have already
discussed the morphological system built up by Richard Owen in the
late 'forties, a system transcendental in its main lines. We have
seen the vertebral theory of the skull still maintained in the
'fifties by such men as Reichert and Kölliker, and we find
J. V.. Carus in 1853[314] taking it as almost
conclusively proved.[315]
We may mention, too, as showing clear marks of the influence of
transcendental ideas, L. Agassiz's work on the principles of
classification.[316] And Serres, who was
Geoffroy's 204chief disciple, recanted not a whit of his
doctrine of recapitulation, but re-affirmed and expanded it from
time to time, and particularly in a lengthy memoir published in
1860.[317] But in general we may
say that pure morphology in the Geoffroyan or Okenian sense was
becoming gradually discredited. A curious indication of this is
seen in the fact that not only the idea but the very word
"Archetype" came to be regarded with suspicion. Thus even J. V..
Carus, who had much affinity with the transcendentalists, wrote of
the vertebrate archetype (which he took over almost bodily from
Owen)—"It may here be observed that this schema may be used
as a methodological help, but it is not to be placed in the
foreground" (loc. cit., p. 395). Huxley, who was definitely
a follower of von Baer, was much more outspoken with regard to
ideal types. In an important memoir on the general anatomy of the
Gastropoda and Cephalopoda,[318] he set himself the task of reducing all their
complex forms to one type. In summing up, he writes:—"From
all that has been stated, I think that it is now possible to form a
notion of the archetype of the Cephalous Mollusca, and I beg it to
be understood that in using this term, I make no reference to any
real or imaginary 'ideas' upon which animal forms are modelled. All
that I mean is the conception of a form embodying the most general
propositions that can be affirmed respecting the Cephalous
Mollusca, standing in the same relation to them as the diagram to a
geometrical theorem, and like it, at once imaginary and true" (i.,
p. 176). Again, in his Croonian lecture on the theory of the
vertebrate skull, he remarks that a general diagram of the skull
could easily be given. "There is no harm," he continues, "in
calling such a convenient diagram the 'Archetype' of the skull, but
I prefer to avoid a word whose connotation is so fundamentally
opposed to the spirit of modern science" (Sci. Memoirs, vol.
i., p. 571).
It is instructive to find that between Serres and Milne-Edwards
there existed the same antagonism as between von 205Baer and the
German transcendentalists. Milne-Edwards was a constant critic of
the law of parallelism which Serres continued to uphold with little
modification for over thirty years, just as von Baer was a critic
of that form of the doctrine which was current in the early part of
the century. As early as 1833, Milne-Edwards, through his studies
of crustacean development,[319] had come to the conclusion, independently of von
Baer, that development always proceeded from the general to the
special; that class characters appeared before family characters,
generic characters before specific. In an interesting paper
published in 1844,[320] he discussed the
relation of this law of development to the problems of
classification, and arrived at results almost identical with those
set forth by von Baer in his Fifth Scholion.
Like von Baer he rejected completely the theory of parallelism
and the doctrine of the scale of beings; like von Baer he held that
the type of organisation—of which there are several—is
manifested in the very earliest stages and becomes increasingly
specialised throughout the course of further development; like von
Baer, too, he sketched a classification based upon embryological
characters.
These views were further developed in his volume of 1851, and
also in his Rapport of 1867.
They brought him into conflict with his confrere in the Academy
of Sciences, Étienne Serres, who in a number of papers
published in the 'thirties and 'forties,[321] and particularly in his comprehensive memoir of
1860, still maintained the theory of parallelism and the doctrine
of the absolute unity of type. His memoir of 1860 shows how
completely Serres was under the domination of transcendental ideas.
Much of it indeed goes back to Oken. "The animal kingdom," he
writes, "may be considered in its entirety as a single ideal and
complex being" (p. 141). His views have become a little more
complicated since his first exposition of them in 1827, 206and he has
been forced to modify in some respects the rigour of his doctrine.
But he still holds fast to the main thesis of
transcendentalism—the absolute unity of plan of all animals,
vertebrate and invertebrate alike,[322] the gradual perfecting of organisation from
monad to man, the repetition in the embryogeny of the higher
animals of the "zoogeny" of the lower.
He recognised, however, that the idea of a simple scale of
beings is only an abstraction, and that the true repetition is of
organs rather than of organisms. He was willing even to admit, at
least in the later pages of his memoir, that there might be not one
animal series but several parallel series, as had been suggested by
Isidore Geoffroy St Hilaire (p. 749). In general, his views are now
less dogmatic than they were in his earlier writings, but they are
not for all that changed in any essential. For, in summing up his
main results, he writes, "The whole animal kingdom can in some
measure be regarded ideally as a single animal, which, in the
course of formation and metamorphosis in its diverse
manifestations, here and there arrests its own development, and
thus determines at each point of interruption, by the very state it
has reached, the distinctive characters of the phyla, the classes,
families, genera, and species" (p. 833).[323]
To settle the dispute pending between two of its most
illustrious members, the Academy proposed in 1853, as the subject
of one of its prizes, "the positive determination of the
resemblances and differences in the comparative development of
Vertebrates and Invertebrates." A memoir was presented the next
year by Lereboullet[324] which met with the approval of the Academy in so
far as its statements of fact were concerned, but seemed to them to
require amplification 207in its theoretical part. But even in this
memoir Lereboullet was able to show that the balance of evidence
was greatly in favour of Milne-Edwards' views, and his general
conclusions in 1854 were that "in the presence of such fundamental
differences, one is obliged to give up the idea of one single plan
in the formation of animals; while, on the contrary, the existence
of diverse plans or types is clearly demonstrated by all the facts"
(p. 79). To fulfil the Academy's requirements, Lereboullet
continued his work, and in 1861-63 he published a series of
elaborate monographs[325] on the embryology of the trout, the lizard and
the pond-snail Lymnæa, and rounded off his work with a
full discussion[326] of the theoretical
questions involved. In this considered and authoritative judgment
he completely disposed of Serres' theories of the unity of plan and
the unity of genetic formation. Except in the very earliest stages
of oogenesis there is no real similarity between the development of
a Zoophyte, a Mollusc, an Articulate and a Vertebrate, but each is
stamped from the beginning with the characteristics of its type.
The lower animals are not, and cannot possibly be the permanent
embryos of the higher animals. "The results which I have obtained,"
he writes, "are diametrically opposed to the theory of the
zoological series constituted by stages of increasing perfection, a
theory which tries to demonstrate in the embryonic phases of the
higher animals a repetition of the forms which characterise the
lower animals, and which has led to the assertion that the latter
are permanent embryos of the former. The embryo of a Vertebrate
shows the vertebrate type from the very beginning, and retains this
type throughout the whole course of its development; it never is,
and never can be, either a Mollusc or an Articulate" (xx., p.
54).
"We are led to establish ... as the general result of our
researches, the existence of several types, and, consequently, of
different plans, in the development of animals. These different
types are manifested from the very beginning of embryonic life; the
characters distinguishing them are therefore 208primordial, and
we can say with M. Milne-Edwards that everything goes to prove
that the distinction established by Nature between animals
belonging to different phyla is a primordial distinction" (p.
58).
In other directions also von Baer's work was confirmed and
extended by later observers—those parts of it particularly
that had reference to the germ-layer theory, and to the concept of
histological differentiation. His germ-layer theory was accepted in
its main lines by Rathke, Bischoff and Lereboullet, and applied by
them to the multitude of new facts they discovered. Rathke, in
particular, was a firm upholder of the doctrine, and made
considerable use of it in his writings.[327] Even before the publication of von Baer's book
he had interpreted in terms of the germ-layer theory sketched by
his friend Pander the splitting of the blastoderm which occurs in
the early development of Astacus, whereby there are formed a
serous and a mucous layer, one inside the other—like the
coats of an onion, to use his own expressive phrase.[328]
An ingenious application of the Pander-Baer theory was made by
Huxley, who compared the outer and inner cell-layers which form the
groundwork of the Cœlentera with the serous and mucous layers of
the vertebrate germ.[329] He laid stress, it is true, rather on the
physiological than on the morphological resemblance. "A complete
identity of structure," he writes, "connects the 'foundation
membranes' of the Medusæ with the corresponding organs in the
rest of the series; and it is curious to remark, that throughout,
the outer and inner membranes appear to bear the same physiological
relation to one another as do the serous and mucous layers of the
germ; the outer becoming developed into the muscular system, and
giving rise to the organs of offence and defence; the inner, on the
other hand, appearing to be more closely subservient to the
purposes of nutrition and generation" (p. 24). Von Baer had already
hinted at this homology 209in the second volume of his
Entwickelungsgeschichte (1837), where he says with reference
to the separation of the blastoderm of the chick into two layers.
"Yet originally there are not two distinct or even separable
layers, it is rather the two surfaces of the germ which show this
differentiation, just as polyps show the same contrast of an
external surface and an internal digestive surface. In between the
two layers there is in our germ as in the polyp an indifferent
mass" (p. 67). The terms ectoderm and entoderm were introduced by
Allman[330] in 1853 for the two
cell-layers in the Hydrozoa.
Remak is the second great name in the history of the germ-layer
theory. He had the great advantage over von Baer of being able to
make use of the cell-theory in interpreting the formation of the
germ-layers. Microscopical technique also had been greatly improved
since 1828.[331]
Remak's greatest service was that he put the germ-layer theory
in direct relation with the cell-theory by demonstrating the
cellular continuity from egg-cell to tissue, and by showing that
each germ-layer possessed distinctive histological characteristics.
Hardly less important was his clear marking-off of the "middle
layer" as a separate and distinct layer of the germ. He it was who
introduced the modern conception of the mesoderm, and cleared up
the confusion in which Pander and von Baer had left the organs
formed between the serous and the mucous layer. Remak's middle
layer was a different thing from Pander's ill-defined
"vessel-layer"; it included and unified from a new point of view
the "vessel" and "muscle" layers of von Baer.
There are in the unincubated blastoderm of the chick, according
to Remak,[332] two cell-layers, of
which the undermost 210subsequently splits into two. Three layers
are thus formed—the upper, middle and lower. The upper layer
differentiates into a medullary plate and an epidermic plate
(Remak's Hornblatt), and gives origin to the medullary tube
with all its evaginations, and to the skin with all its derivatives
and pockets. It forms such diverse structures as the brain, the
spinal cord, the eye, the ear, the mouth, hairs, feathers, nails,
sweat-glands, lacrymal glands, and so forth. All these parts are
connected directly or indirectly with sensation, and the upper
germ-layer may accordingly be called the sensory layer. The
lower layer gives rise to the epithelium and the proper tissue of
the alimentary canal and its derivatives, as the liver, lungs,
pancreas, kidneys, thyroid, thymus, etc. These parts are all
concerned in the processes of assimilation and dissimilation, and
the lower layer may accordingly be called the trophic layer.
Now between the upper or sensory layer and the lower or trophic
layer there exists, in spite of their very different functions, a
close histological likeness, for both are essentially epithelial
layers. The resemblance is particularly strong if we compare the
lower layer with the Hornblatt of the upper layer—both
consist of epithelial tissue, and of its derivative, glandular
tissue, and form neither vessels nor nerves. The middle layer, on
the contrary, forms nerves and muscles, vessels and connective
tissue, and little or no epithelium. It does not form all the
blood-vessels without exception (and so cannot be called the
vessel-layer), for the blood-vessels of the central nervous system
are in all probability formed from the upper layer. So, too, it
does not form all the nerves and muscles—the optic and
auditory nerves and the nerves and muscles of the iris probably
arise in the upper layer. But, in spite of these exceptions, its
general histological character is so well defined that it may be
contrasted with the other two as preeminently the layer that forms
muscular, nervous, vascular and connective tissue. In view of its
functional significance, it may be called the motory layer,
or better, since it forms also the sexual glands, the
motor-germinative layer. The middle layer, early in its
history, shows a division into dorsal plates
(Urwirbelplatten) and ventral plates (Seitenplatten).
The former exhibit almost as soon as they are 211formed the
characteristic proto-vertebral segmentation, the latter split to
form the pleuro-peritoneal or body-cavity. Remak describes the
latter process as follows:—"In the region of the trunk, where
a greater independence of the fate of the alimentary canal and its
annexes becomes necessary for the voluntary executive organs, the
ventral plates undergo a process of splitting, leading to the
formation of the sensitive part of the integument (the
Hautplatten), the muscular part of the alimentary tube (the
Darmfaserplatten), and the mother-tissue of the generative
organs (the Mittelplatten). From the Hautplatten
there develops, without the dorsal plates seeming to take any part
in the process, the rudiment of the extremities" (p. 79).
His Darmfaserplatten form the nervous and muscular tissue
of the alimentary canal and its dependencies, and also the heart;
the Hautplatten form the general body-wall (exclusive of the
skin) and the appendages. In the embryo they line the amniotic
cavity. The skeleton and peripheral nerves originate wholly within
the middle layer.
Remak's conception of the relations of the three germ-layers to
one another and to the body-cavity is well illustrated in Fig. 12.
Fig.
12.—Transverse Section of Chick Embryo. (After Remak.)
| h. |
Epidermis. |
hp. |
 |
"Hautplatte" |
x. |
Edge of the smniotic fold. |
| m. |
Spinal cord. |
and |
ph. |
Pleuro-Peritonial cavity. |
| mu. |
Dorsal plate. |
um. |
|
|
| ug. |
Pronephric duct. |
mp. |
"Mittelplatte" |
d. |
Epithelium of alimentary canal. |
| pa. |
Aortic root. |
df. |
"Darmfaser platte." |
|
|
In his germ-layer theory Remak's standpoint is histological
212rather than morphological. The distinction
which he draws between the sensory and trophic layers on the one
hand, and the motor-germinative layer on the other, is entirely a
histological one. The greater part of his book, indeed, is devoted
to a study of the histogenesis of the different organs of the body;
he is bent chiefly upon unravelling the part which each germ-layer
takes in the formation of each tissue and organ.
His generalisation that two of the germ-layers give rise
exclusively or almost exclusively to one kind of tissue excited
great interest at the time, and gave the direction to histogenetic
research for quite a number of years, though in the end it turned
out to be insufficiently founded.
Though Remak's germ-layer theory had thus principally a
histological orientation, it laid down the main lines of the modern
morphological treatment of the germ-layers.
213
CHAPTER XIII
THE RELATION OF LAMARCK AND DARWIN TO MORPHOLOGY.
It is a remarkable fact that morphology took but a very little
part in the formation of evolution-theory. When one remembers what
powerful arguments for evolution can be drawn from such facts as
the unity of plan and composition and the law of parallelism, one
is astonished to find that it was not the morphologists at all who
founded the theory of evolution.
It is true that the noticeable resemblances of animals to one
another, the possibility of arranging them in a system, the vague
perception of an all-pervading plan of structure, did suggest to
many minds the thought that systematic affinities might be due to
blood-relationship. Thus Leibniz considered that the cat tribe
might possibly be descended from a common ancestor,[333] and another great philosopher, Immanuel
Kant, was led by his perception of the unity of type to suggest as
possible the derivation of the whole organic realm from one parent
form, or even ultimately from inorganic matter. In the course of
his masterly discussion of mechanism and teleology,[334] he writes, "The agreement of so many
genera of animals in a certain common schema, which appears to be
fundamental not only in the structure of their bones, but also in
the disposition of their remaining parts—so that with an
admirable simplicity of original outline, a great variety of
species has been produced by the shortening of one member and the
lengthening of another, the involution of this part and the
evolution of that—allows a ray of hope, however faint, to
penetrate into our minds, that here 214something may be accomplished by
the aid of the principle of the mechanism of Nature (without which
there can be no natural science in general). This analogy of forms,
which with all their differences seem to have been produced
according to a common original type, strengthens our suspicions of
an actual relationship between them in their production from a
common parent, through the gradual approximation of one
animal-genus to another—from those in which the principle of
purposes seems to be best authenticated, i.e., from man down
to the polype, and again from this down to mosses and lichens, and
finally to the lowest stage of Nature noticeable by us, viz., to
crude matter."[335]
So, too, Buffon's evolutionism was suggested by his study of the
structural affinities of animals, and Erasmus Darwin in his
Zoonomia (1794) brought forward as one of the strongest
proofs of evolution, "the essential unity of plan in all
warm-blooded animals."[336]
But, as a matter of historical fact, no morphologist, not even
Geoffroy, deduced from the facts of his science any comprehensive
theory of evolution. The pre-Darwinian morphologists were
comparatively little influenced by the evolution-theories current
in their day, and it was in the anatomist Cuvier and the
embryologist von Baer that the early evolutionists found their most
uncompromising opponents.
Speaking generally, and excepting for the moment the theory of
Lamarck, we may say that the evolution-theories of the 18th and
19th centuries arose in connection with the transcendental notion
of the Échelle des êtres, or scale of
perfection. This notion, which plays so great a part in the
philosophy of Leibniz, was very generally accepted about the middle
of the 18th century, and received complete and even exaggerated
expression from Bonnet and Robinet. Buffon also was influenced by
it. Towards the beginning of the 19th century the idea was taken up
eagerly by the transcendental school and by them given, in their
theories of the 215"one animal," a more morphological turn.
Their recapitulation theory was part and parcel of the same general
idea.
One understands how easily the notion of evolution could arise
in minds filled with the thought of the ideal progression of the
whole organic kingdom towards its crown and microcosm, man. Their
theory of recapitulation led them to conceive evolution as the
developmental history of the one great organism.[337] Many of them wavered between the
conception of evolution as an ideal process, as a
Vorstellungsart, and the conception of it as an historical
process. Bonnet, Oken, and the majority of the transcendentalists
seem to have chosen the former alternative; Robinet, Treviranus,
Tiedemann, Meckel, and a few others held evolution to be a real
process.
We have already in previous chapters[338] briefly noticed the relation of one or two of
the transcendental evolution-theories to morphology, and there is
little more to be said about them here. They had as good as no
influence upon morphological theory, nor indeed upon biology in
general.[339] It is different with the
theory of Lamarck, which, although it had little influence upon
biological thought during and for long after the lifetime of its
author, is still at the present day a living and developing
doctrine.
Lamarck's affinity with the transcendentalists was in many ways
a close one, but he differed essentially in being before all a
systematist. Nor is the direct influence of the German
transcendentalists traceable in his work—his spiritual
ancestors are the men of his own race, the materialists Condillac
and Cabanis, and Buffon, whose friend he was. The idea of a
gradation of all animals from the lowest to the highest was always
present in Lamarck's mind, and links him up, perhaps through
Buffon, with the school of Bonnet. The idea of the
Échelle des êtres had for him much less a 216morphological orientation than it had even
for the transcendentalists, for he was lacking almost completely in
the sense for morphology. Lamarck's scientific, as distinguished
from his speculative work, was exclusively systematic, and it was
systematics of a very high order. He introduced many reforms into
the general classification of animals. He was the first clearly to
separate Crustacea (1799), and a little later (1800) Arachnids,
from insects. He reduced to a certain orderliness the neglected
tribes of the Invertebrates, and wrote what was for long the
standard work on their systematics—the Histoire naturelle
des Animaux sans Vertèbres (1816-22). His speculative
work on biology is contained in three publications, the small book
entitled Considérations sur l'organisation des corps
vivants (1802), the larger work of 1809, the Philosophie
zoologique, and the introductory matter to his Animaux sans
Vertèbres (vol. i., 1816).
It is no easy matter to give in short compass an account of
Lamarck's biological philosophy. He is an obscure writer, and often
self-contradictory.
In the first part of the Philosophie zoologique Lamarck
is largely pre-occupied with the problem of whether species are
really distinct, or do not rather grade insensibly into one
another. As a systematist of vast experience Lamarck knew how
difficult it is in practice to distinguish species from varieties.
"The more," he writes, "we collect the productions of Nature, the
richer our collections become, the more do we see almost all the
gaps filled up and the lines of separation effaced. We find
ourselves reduced to an arbitrary determination, which sometimes
leads us to seize upon the slightest differences of varieties, and
form from them the distinctive character of what we call a species,
and at other times leads us to consider as a variety of a certain
species individuals a little bit different, which others regard as
forming a separate species."[340]
For Lamarck, as for Darwin later, the chief problem was not the
evolution and differentiation of types of structure, but the mode
of origin of species.
Lamarck is at great pains to show how arbitrary are our 217determinations of species, and how artificial
the classificatory groups which we distinguish in Nature. Strictly
speaking, there are in Nature only individuals, "... this is
certain, that among her products Nature has in reality formed
neither classes, nor orders, nor families, nor genera, nor constant
species, but only individuals which succeed one another and
resemble those that produced them. Now, these individuals belong to
infinitely diversified races, which shade into one another under
all the forms and in all the degrees of organisation, and each of
which maintains itself without change, so long as no cause of
change acts upon it" (p. 41).
But there is a natural order in the animal kingdom, a
progression from the simpler to the more complex organisations, a
natural Échelle des êtres.
This order is shown by the relation to one another of the large
classificatory groups, for they can be arranged in series from the
simplest to the most complex, somewhat as follows:—
| 1. Infusoria. |
6. Arachnids. |
11. Fishes. |
| 2. Polyps. |
7. Crustacea. |
12. Reptiles. |
| 3. Radiates. |
8. Annelids. |
13. Birds. |
| 4. Worms. |
9. Cirripedes. |
14. Mammals. |
| 5. Insects. |
10. Molluscs. |
|
But the order of Nature is essentially continuous, and the
limits of even the best defined of these classes are in reality
artificial—"if the order of Nature were perfectly known in a
kingdom, the classes which we should be forced to establish in it
would always constitute entirely artificial sections" (p. 45).
In the same way the lesser classificatory groups represent
smaller sections of the one unique order of Nature. Note that
Lamarck's Échelle is in no way a morphological one,
and was not intended to be such. It is a scale of increasing
physiological differentiation, and the stages of it are marked by
the acquirement of this or that new organ (cf. Oken).
"Observation of their state convinces one that in order to produce
them successively Nature has proceeded gradually from the simpler
to the more complex. Now Nature, having had in mind the realisation
of a plan of organisation 218which would permit of the greatest perfecting
(that of the Vertebrates), a plan very different from those which
she has been obliged to form as a preliminary to reaching it, one
understands that, among the multitude of animals, one must
necessarily come across not a single system of organisation which
has become progressively perfected, but diverse very distinct
systems, each of which has come into existence at the moment when
each primary organ first put in its appearance" (p. 171).
For Lamarck this order of Nature was not merely
ideal—Nature had actually formed the classes successively,
proceeding from the simpler to the more complex; she had brought
about this evolution by transforming the primitive species of
animals, raising them to higher degrees of organisation, and
modifying them in relation to the environment in which they found
themselves.
Lamarck's theory of evolution is worked out in great detail in
his Philosophie zoologique, but the exposition is diffuse
and disconnected; it is better in giving an account of it to follow
the more concise, mature and general exposition which he gives in
the Introduction to his Histoire naturelle des Animaux sans
Vertèbres.[341] Near the beginning of the Introduction Lamarck
gives us in a few short "Fundamental Principles" the main lines of
his general philosophy. He is a confirmed materialist. Every fact
and phenomenon is essentially physical and owes its existence or
production entirely to material bodies or to relations between
them. All change and all movement is in the last resort due to
mechanical causes. Every fact or phenomenon observed in a living
body is at once a physical fact or phenomenon and a product of
organisation (p. 19). Life, thought and sensation are not
properties of matter, but result from particular material
combinations.
His thorough-going materialism is most clearly shown in its
relation to living things in the first three of the "Zoological
Principles and Axioms," which are developed further on in the
book.
These are as follows:—"1. No kind or particle of matter
219can
have in itself the power of moving, living, feeling, thinking, nor
of having ideas; and if, outside of man, we observe bodies endowed
with all or one of these faculties, we ought to consider these
faculties as physical phenomena which Nature has been able to
produce, not by employing some particular kind of matter which
itself possesses one or other of these faculties, but by the order
and state of things which she has constituted in each organisation
and in each particular system of organs.
"2. Every animal faculty, of whatever nature it may be, is an
organic phenomenon, and results from a system of organs or an
organ-apparatus which gives rise to it and upon which it is
necessarily dependent.
"3. The more highly a faculty is developed the more complex is
the system of organs which produces it, and the higher the general
organisation; the more difficult also does it become to grasp its
mechanism. But the faculty is none the less a phenomenon of
organisation, and for that reason purely physical" (p. 104).
According to these "axioms" function is a direct and mechanical
effect of structure.
The curious thing is that in spite of his avowed materialism,
Lamarck's conception of life and evolution is profoundly
psychological, and from the conflict of his materialism and his
vitalism (of which he was himself hardly conscious), arise most of
the obscurities and the irreductible self-contradiction of his
theory.
Lamarck divided animals (psychologically!) into three great
groups—apathetic or insensitive animals, animals endowed with
sensation, and intelligent animals. The first group, which comprise
all the lower Invertebrates, are distinguished from other animals
by the fact that their actions are directly and mechanically due to
the excitations of the environment; they have no principle of
reaction to external influences, but passively prolong into action
the excitations they receive from without. They are
irritable merely. The second group are distinguished from
the first by their possessing, in addition to irritability, a power
which Lamarck calls the sentiment intérieur. He has
some difficulty in defining exactly what he means by it:—"I
220have
no term to express this internal power possessed not only by
intelligent animals but also by those that are endowed merely with
the faculty of sensation; it is a power which, when set in action
by the feeling of a need, causes the individual to act at once,
i.e., in the very moment of the sensation it experiences;
and if the individual is of those that are endowed with
intelligence it nevertheless acts in such a case entirely without
premeditation and before any mental operation has brought its
will into play" (p. 24).
It is the power we call instinct in animals (p. 25), and it
implies neither consciousness nor will. It acts by transforming
external into internal excitations.
To this second group of animals, possessing the sentiment
intérieur, belong the higher Invertebrates, notably
insects and molluscs. Only animals possessed of a more or less
centralised nervous system can manifest this sentiment, or
principle of (unconscious) reaction to external stimuli.
The higher animals, or the four Vertebrate classes, form the
group of "intelligent animals." In virtue of their more complex
organisation they possess in addition to the sentiment
intérieur the faculties of intelligence and will.
Now, broadly put, Lamarck's theory of evolution is that new
organs are formed in direct reaction to needs (besoins)
experienced by the sentiment intérieur. The
sentiment intérieur is therefore the cause not only
of instinctive action but also of all morphogenetic processes. Will
and intelligence (which are confined to a relatively small number
of animals) have little or nothing to do directly with
evolution.
To understand the working-out of Lamarck's evolution-theory we
must revert to his conception of the Échelle des
êtres. What he wrote in the Philosophie zoologique
is here repeated in the work of 1816 with little modification.
There is a real progression from the simpler to the more complex
organisations; Nature has gradually complicated her creatures by
giving them new organs and therefore new faculties.
It is interesting to note that Lamarck expressly refers to
Bonnet (p. 110), but refuses to accept his view of an
Échelle extending down into the inorganic. Like
Bonnet, however, 221and like the German transcendentalists,
Lamarck makes man the goal of evolution (p. 116). He makes it quite
clear that his Échelle is a functional one, for he
links Vertebrates to molluscs even while expressly admitting that
they are not connected by any structural intermediates (p. 123). He
does not fall into the error of the transcendentalists and assume
that Vertebrates and Invertebrates alike are formed upon one common
plan of structure.
The progression of organisation shown by the animal kingdom has
not been altogether regular and uninterrupted:—"The
progression in complexity of organisation shows here and there, in
the general animal series, anomalies induced by the influence of
environment and by the influence of the habits contracted"
(Phil. zool., i., p. 145).
There are thus really two causes at work to produce the variety
of organisation as it appears to us, one which tends to produce a
regular increase in complexity, and one which disturbs and
diversifies this regular advance.
The first cause Lamarck calls the vital power (pouvoir de la
vie); the other may be called the influence of circumstance
(Anim. s. Vert., p. 134). To the latter cause are due the
lacunæ, the blind alleys, and the complications which the
otherwise simple scale of perfection shows.
To explain both these aspects of evolution Lamarck propounded in
his volume of 1816 four laws, which read as follows:—
"First Law.—Life, by its own forces, tends
continually to increase the volume of every body possessing it, and
to extend the dimensions of its parts, up to a limit which it
brings about itself.
"Second Law.—The production of a new organ in an
animal body results from the arisal and continuance of a new need,
and from the new movement which this need brings into being and
sustains.
"Third Law.—The degree of development of organs and
their force of action are always proportionate to the use made of
these organs.
"Fourth Law.—All that has been acquired, imprinted
or changed in the organisation of the individual during the course
of its life is preserved by generation and transmitted 222to the new
individuals that descend from the individual so modified" (pp.
151-2).
It is mainly but not entirely by reason of the first of these
laws that organisation tends to progress, and mainly by reason of
the second and third that difference of environment brings about
diversity of organisation. In virtue of the fourth law the
acquirements of the individual become the property of the race.
Lamarck's exposition of his first law, that life tends by its
own powers to enlarge and extend its bodily instrument, is vague
and difficult to understand. He has already explained some pages
back how the first organisms arose by spontaneous generation in the
form of minute gelatinous utricles (cf. Oken). He conceives
that it is in the movements of the fluids proper to the organism
that the power resides to enlarge and extend the body. Nutrition
alone is not sufficient to bring about extension; a special force
is required, acting from within outwards (p. 153). In the most
primitive organisms the movements of the vital fluids are weak and
slow, but in the course of evolution they gradually accelerate,
and, becoming more rapid, trace out canals in the delicate tissue
which contains them, and finally form organs.
Subtle fluids play a great part in Lamarck's biology: they take
the place of the soul or entelechy which the vitalists would
postulate to explain organic happenings. Lamarck seems in this to
follow certain of the old materialists, who conceived the soul to
be formed of a matter more subtle than the ordinary.[342]
In his second law Lamarck's essentially vitalistic attitude
comes out very clearly, for it states that a psychological moment
enters into all new production of form, that the ultimate cause of
the development of new form is the need felt by the organism. This
need is of course not a conscious one, it is a need perceived by
the sentiment intérieur.
223In the large group of apathetic or
insensitive animals, which do not possess this faculty, needs
cannot be experienced; accordingly new organs are here formed
directly and mechanically, by the movements of the vital fluids set
in action by excitations from without—the evolution, like the
behaviour, of these animals is due to the direct and physical
action of the environment. "But this is not the case with the more
highly organised animals which possess feeling. They
experience needs, and each need felt, acting upon their 'inner
feeling,' immediately directs the fluids and the forces to the part
of the body where action can satisfy the need. Now, if there exists
at this point an organ capable of performing the required action,
it is quickly stimulated to act; and if the organ does not exist
and the need is pressing and sustained, bit by bit the organ is
produced and developed in proportion to the continuity and the
energy of its use" (p. 155).
In intelligent animals the sentiment intérieur may
be moved by thought or will.
As an example of the way in which the law works Lamarck takes
the hypothetical case of a gastropod mollusc, which as it creeps
along experiences dimly the need to feel the objects in front of
it. It makes an effort (unconscious, be it noted) to touch these
objects with the anterior portions of its head, and sends forward
continually to these parts a great volume of nervous and other
fluids. From these efforts and the repeated afflux of fluids there
must result a development of the nerves supplying these parts. And
as, along with the nervous fluids, nutritive juices constantly flow
to the parts, there must result the formation of two or four
tentacles in the places to which these fluids are directed. A
curious mixture of mechanistic "explanations" and vitalistic
hypothesis!
In his third law, that use and disuse are powerful to modify
organs, Lamarck is upon more solid ground, and can point to many
instances of the visible effect of these factors of change. It is
of course rather closely bound up with his second law and may even
be regarded as an extension of it.
The law has reference to one of the most powerful means 224employed
by Nature to diversify species, a means which comes into play
whenever the environment changes. The cause of the great diversity
shown by animal species is indeed ultimately to be sought in the
environment. As the imperfect and earliest forms developed they
spread over the earth and invaded the utmost corners of
it:—"One can imagine what an enormous variety of habitats,
stations, climates, available foods, environing media, etc.,
animals and plants have had to endure, as the existing species were
forced to change their place of abode. And although these changes
have taken place with extreme slowness ... their reality,
necessitated by various causes, has none the less induced the
species affected by them slowly to change their manner of life and
their habitual actions. Through the effects of the second and third
of the laws cited above, these induced activity-changes must have
brought into being new organs, and must have been able to develop
them further if more frequent use was made of them; they must in
the same way have been capable of bringing about the degeneration
and finally the complete disappearance of existing organs which had
become useless" (p. 161).
On the other hand, if the environment does not change, species
remain constant.
It is to be noted that change in environment is rather the
occasion than the cause of modification; the environment induces
the organism to change its habitual way of life; it sets up new
needs, to satisfy which the organism must modify its structure. It
is the organism that takes the active part in all this, the action
of the environment is indirect.
Of Lamarck's fourth law, which asserts the transmission of
acquired characters, little need here be said in the way of
exposition. Upon the truth of it depends of course Lamarck's whole
theory. He himself never dreamed that anyone would ever dispute
it.
Lamarck sums up as follows:—"By the four laws which I have
just enunciated all the facts of organisation seem to me to be
easily explained; the progression in the complexity of organisation
of animals, and in their faculties, seems to me easy to conceive;
so, too, the means which Nature has employed to diversify animals,
and bring them to the 225state in which we now see them, become easily
determinable" (p. 168).
It is never made quite clear, we may note in passing, how far
his second and third laws tend to bring about an increase in
complexity, in addition to diversifying animals.[343]
"The function creates the organ," this would seem to be the
kernel of Lamarck's doctrine. But how does he reconcile this
essentially vitalistic conception with his strictly materialistic
philosophy?
We have seen that irritability, the sentiment
intérieur, and intelligence itself, are the effects of
organisation. We are told farther on that both the sentiment
and intelligence are caused by nervous fluids. A great part of both
the Philosophie zoologique and the introduction to the
Animaux sans Vertèbres is given up to the exposition
of a materialistic psychology of animals and man, based entirely
upon this hypothesis of nervous fluids. Thus habits are due to the
fluids hollowing out definite paths for themselves.
The sentiment intérieur acts by directing the
movements of the subtle fluids of the body (which are themselves
modifications of the nervous fluids) upon the parts where a new
organ is needed. But if it is itself only a result of the movement
of nervous fluids? Again, how can a need be "felt" by a nervous
fluid? This is an entirely psychological notion and cannot be
applied to a purely material system. Whence arises the power of the
sentiment intérieur to canalise the energies of the
organism, so to direct and co-ordinate them that they build up
purposive structures, or effect purposive actions (as in all
instinctive behaviour)? Either the sentiment
intérieur is a psychological faculty, or it is
nothing.
There is no doubt that, as expressed by Lamarck, the conception
conceals a radical confusion of thought. It is not possible to be a
thorough-going materialist, and at the 226same time to believe that new
organs are formed in direct response to needs felt by the organism.
Lamarck could never resolve this antinomy, and his speculations
were thrown into confusion by it. To this cause is due the frequent
obscurity of his writings.
Should we be right in laying stress upon the psychological side
of Lamarck's theory, and disregarding the materialistic dress in
which, perhaps under the influence of the materialism current in
his youth, he clothed his essentially vitalistic thought?
Everything goes to prove it—his constant preoccupation with
psychological questions, his tacit assimilation of organ-formation
to instinctive behaviour, his constant insistence on the importance
of besoin and habitude.
Let us not forget the profundity of his main idea, that,
exception made for the lower forms, the animal is essentially
active, that it always reacts to the external world, is
never passively acted upon. Let us not forget that he pointed out
the essentially psychological moment implied in all processes of
individual adaptation. With keen insight he realised that conscious
intelligence counts for little in evolution, and focussed attention
upon the unconscious but obscurely psychical processes of instinct
and morphogenesis.
Not without reason have the later schools of evolutionary
thought, who developed the psychological and vitalistic side of his
doctrine, called themselves Neo-Lamarckians.
We shall say then that Lamarck, in spite of his materialism, was
the founder of the "psychological" theory of evolution.
Lamarck stood curiously aloof and apart from the scientific
thought of his day.[344] He took no interest in the morphological
problems that filled the minds of Cuvier and Geoffroy; he had
indeed no feeling at all for morphology. He did not realise, like
Cuvier, the convenance des parties, the marvellous
co-ordination of parts to form a whole; he had little conception of
what is really implied in the word "organism." He was not, like
Geoffroy, imbued with a lively sense of the unity of plan and
composition, and of the significance of vestigial 227organs as
witnesses to that unity. He seems not to have known of the
recapitulation theory, of which he might have made such good use as
powerful evidence for evolution. Even with the German
transcendentalists, with whom in the looseness of his
generalisations he shows some affinity, he seems not to have been
specially acquainted.
He was interested more in the problems suggested to him by his
daily work in the museum. He wanted to know why species graded so
annoyingly into one another; he wanted to examine critically his
haunting suspicion that species were really not distinct, and that
classification was purely conventional. The question, too, of the
adaptation of species to their environment, the problem of
ecological adaptation, in distinction to that of functional
adaptation which interested Cuvier so greatly, came vividly before
him as he worked through the vast collections of the museum. He was
the first systematist to occupy himself in a philosophical manner
with the problems of general biology. He introduced new problems
and a new way of looking at old. With Lamarck the problem of
species and the problem of ecological adaptation enter into general
biology.
The one point in which he does definitely carry on the thought
of his predecessors is his conception of the animal kingdom as
forming a scale of (functional) perfection. He did not go to the
same extreme as Bonnet; he did not even consider that the animal
series was a continuation of the vegetable series; in his opinion
they formed two diverging scales. He recognised, too, that among
animals there was no simple and regular gradation from the lowest
to the highest, but that the orderly progression was disturbed and
diverted by the necessity of adaptation to different environments.
It is interesting to note that in developing this idea he arrived
at a roughly accurate distinction between homologous and analogous
structures. More importance, he thought, was to be attributed in
classifying animals to characters which appeared due to the "plan
of Nature" than to such as were produced by an external modifying
cause (p. 299). But he did not formulate the distinction in any
strictly morphological way.
As his ideas developed he laid less stress upon the simplicity
228and
continuity of the scale; in his supplementary remarks to the
Introduction of 1816 he admits that the series is really very much
branched, and even that there may be two distinct series among
animals instead of one. His last schema of the course of evolution
shows no little analogy with the genealogical trees of Darwinian
speculation. It is headed "The presumed Order of the
formation of Animals, showing two separate partly-branching
series," and it reads as follows:—
It is interesting to note that Vertebrates are placed between
the two series, and are now not linked on directly to any
Invertebrate group.
Lamarck's theory had little success. There is evidence, however,
that both Meckel and Geoffroy owed a good many of their
evolutionary ideas to Lamarck, and Cuvier paid him at least the
compliment of criticising his theory,[345] not distinguishing it, however, very clearly
from the evolutionary theories of the transcendentalists. But,
speaking generally, Lamarck's theory of evolution exercised very
little influence upon his 229contemporaries. This was probably due partly
to the obscurity and confusion of his thought, partly to his lack
of sympathy with the biological thought of his day, which was
preponderatingly morphological.
It was not that men's minds were not ripe for evolution, for in
the early decades of the 19th century evolution was in the air.
There were few of von Baer's contemporaries who had not read
Lamarck;[346] Erasmus Darwin's
Zoonomia ran through three editions, and was translated into
German, French and Italian;[347] German philosophy was full of the idea of
evolution.
There was no unreadiness to accept the derivation of present-day
species from a primordial form—if only some solid evidence
for such derivation were forthcoming. Cuvier and von Baer, as we
have seen, combated the current evolution theories on the ground
that the evidence was insufficient, but von Baer at least had no
rooted objection to evolution. In an essay of 1834, entitled The
Most General Law of Nature in all Development,[348] von Baer expressed belief in a limited
amount of evolution. In this paper he did not admit that all
animals have developed from one parent form, and he refused to
believe that man has descended from an ape; but, basing his
supposition upon the facts of variability and upon the evidence of
palæontology, he went so far as to maintain that many species have
evolved from parent stocks. In the absence of conclusive proofs he
did not commit himself to a belief in any extended or comprehensive
process of evolution.
Imbued as he was with the idea of development von Baer saw in
evolution a process essentially of the same nature as the
development of the individual. Evolution, like development, was due
to a Bildungskraft or formative force. The ultimate law of
all becoming was that "the history of Nature is nothing but the
history of the ever-advancing victory of spirit over matter" (p.
71). In a later essay (1835) in the same volume he says that all
natural science is nothing but a long commentary on the single
phrase Es werde!. (p. 86).
As we shall see, von Baer adopted in later years the same 230attitude
to Darwinism as he did to the evolution theories in vogue in his
youth.
Although in the twenty or thirty years before the publication of
the Origin of Species (1859) no evolution theory of any
importance was published, and although the great majority of
biologists believed in the constancy of species, there were not
wanting some who, like von Baer, had an open mind on the subject,
or even believed in the occurrence of evolutionary processes of
small scope. Isidore Geoffroy St Hilaire, the son of the great
Etienne Geoffroy St Hilaire, seems to have held that species might
be formed from varieties. The law which L. Agassiz thought he could
establish,[349] of the parallelism
between palæontological succession, systematic rank, and
embryological development, tended to help the progress of
evolutionary ideas. J. V.. Carus, who afterwards became a supporter
of Darwin, seems already, in 1853, to have inferred from Agassiz's
law the probability of evolution.[350]
But no evolution theory was taken very seriously before 1859,
when the Origin of Species was published.
Like Lamarck, Charles Darwin was, neither by inclination nor by
training, a morphologist. In his youth he was a collector, a
sportsman and a field geologist. His voyage round the world on the
Beagle aroused in him keen interest in the problem of
species—their variety, their variation according to place and
time, their adaptedness to environment. The conviction gradually
took possession of his mind that the puzzling facts of geographical
range and geological succession which he observed wherever he went
were explicable only on the hypothesis that species change. He was
not satisfied with the theories of evolution that had been proposed
by his grandfather, by Lamarck, and by E. Geoffroy St
Hilaire—he did not indeed understand these theories any too
well. He resolved to work out the problem in his own way, for his
own satisfaction. He tells us all this very clearly in his
autobiography. "During the voyage 231of the Beagle I had been
deeply impressed by discovering in the Pampean formation great
fossil animals covered with armour like that on the existing
armadillos; secondly, by the manner in which closely allied animals
replace one another in proceeding southwards over the continent;
and thirdly, by the South American character of most of the
productions of the Galapagos archipelago, and more especially by
the manner in which they differ slightly on each island of the
group; some of the islands appearing to be very ancient in a
geological sense.
"It was evident that such facts as these, as well as many
others, could only be explained on the supposition that species
gradually become modified; and the subject haunted me. But it was
equally evident that neither the action of the surrounding
conditions, nor the will of the organisms (especially in the case
of plants) could account for the innumerable cases in which
organisms of every kind are beautifully adapted to their habits of
life—for instance, a woodpecker or a tree-frog to climb
trees, or a seed for dispersal by hooks or plumes. I had always
been much struck by such adaptations, and until these could be
explained it seemed to me almost useless to endeavour to prove by
indirect evidence that species have been modified."[351]
All Darwin's varied subsequent work revolved round these, for
him, essential problems—How do species change, and how do
they become adapted to their environment? He never ceased to be
essentially a field naturalist, and his theory of natural selection
would have been an empty and abstract thing if his vast knowledge
and understanding of the "web of life" had not given it colour and
form. He never lost touch with the living thing in its living,
breathing reality—even plants he rightly regarded as active
things, full of tricks and contrivances for making their way in the
world. No one ever realised more vividly than he the delicacy and
complexity of the adaptations to environment which are the
necessary condition of success in the struggle for existence.
Almost his greatest service to biology was that he made 232biologists
realise as they never did before the vast importance of
environment. He took biology into the open air, away from the
museum and the dissecting-room.
Naturally this attitude was not without its drawbacks. It led
him to take only a lukewarm interest in the problems of morphology.
It is true he used the facts of morphology with great effect as
powerful arguments for evolution, but it was not from such facts
that he deduced his theory to account for evolution. It is
questionable indeed whether the theory of natural selection is
properly applicable to the problems of form. It was invented to
account for the evolution of specific differences and of ecological
adaptations; it was not primarily intended as an explanation of the
more wonderful and more mysterious facts of the convenance des
parties and the interaction of structure and function. Perhaps
Darwin did not realise this inner aspect of adaptation quite so
vividly as he did the more superficial adaptation of organisms to
their environment. It was, perhaps, his lack of morphological
training and experience that led him to disregard the problems of
form, or at least to realise very insufficiently their
difficulty.
It is in any case very significant that only a small part of his
Origin of Species is devoted to the discussion of
morphological questions—only one chapter out of the fourteen
contained in the first edition.
Though the theory of natural selection took little account of
the problems of form, Darwin's masterly vindication of the theory
of evolution was of immense service to morphology, and Darwin
himself was the first to point out what a great light evolution
threw upon all morphological problems. In a few pages of the
Origin he laid the foundations of evolutionary
morphology.
We have here to consider his interpretation of morphological
facts and its relation to the current morphology of his time.
The sketch of his theory, written in 1842,[352] shows a very significant division into two
parts—the first dealing with the positive facts of
variability and the theory of natural selection, 233the second with
the general evidence for evolution. It is in the second part that
the paragraphs on morphological matters occur. In paragraph 7, on
affinities and classification, Darwin points out that on the theory
of evolution homological relationship would be real relationship,
and the natural system would really be genealogical. In the next
paragraph he notes that evolution would account for the unity of
type in the great classes, for the metamorphosis of organs, and for
the close resemblance which early embryos show to one another. It
is of special interest to note that he definitely rejects the
Meckel-Serres theory of recapitulation. "It is not true," he
writes, "that one passes through the form of a lower group, though
no doubt fish more nearly related to fœtal state" (p. 42).
The greater divergence which adults show seems to him to be due to
the fact that selection acts more on the later than on the
embryonic stages. He realises very clearly how illuminative the
theory of evolution is when applied to the puzzling facts of
embryonic development. "The less differences of
fœtus—this has obvious meaning on this view: otherwise
how strange that a horse, a man, a bat should at one time of life
have arteries, running in a manner which is only intelligibly
useful in a fish! The natural system being on theory genealogical,
we can at once see why fœtus, retaining traces of the
ancestral form, is of the highest value in classification" (p.
45).
Abortive organs, too, gain significance on the evolutionary
hypothesis. "The affinity of different groups, the unity of types
of structure, the representative forms through which fœtus
passes, the metamorphosis of organs, the abortion of others, cease
to be metaphorical expressions and become intelligible facts" (p.
50).
In general, organisms can be understood only if we take into
account the cardinal fact that they are historical beings. "We must
look at every complicated mechanism and instinct as the summary of
a long history of useful contrivances much like a work of art" (p.
51).[353]
Already in 1842 Darwin had seized upon the main principles of
evolutionary morphology: the indications then given are elaborated
in the thirteenth chapter of the234 Origin of Species (1st ed.,
1859). A good part of this chapter is given up to a discussion of
the principles of classification, only a few pages dealing with
morphology proper. But, as Darwin rightly saw, the two things are
inseparable.
We note first that there is no hint of the "scale of
beings"—Darwin conceives the genealogical tree as many
branched. Animals can be classed in "groups under groups," and
cannot be arranged in one single series.
He discusses first what kind of characters have the greatest
classificatory value. Certain empirical rules have been recognised,
more or less consciously, by systematists—that analogical
characters are less valuable than homological, that characters of
great physiological importance are not always valuable for
classificatory purposes, that rudimentary organs are often very
useful, and so on. He finds that as a general rule "the less any
part of the organisation is concerned with special habits, the more
important it becomes for classification" (p. 414), and adduces in
support Owen's remark that the generative organs afford very clear
indications of affinities, since they are unlikely to be modified
by special habits. These rules of classification can be explained
"on the view that the natural system is founded on descent with
modification; that the characters which naturalists consider as
showing true affinity ... are those which have been inherited from
a common parent, and, in so far, all true classification is
genealogical; that community of descent is the hidden bond which
naturalists have been unconsciously seeking, and not some unknown
plan of creation, or the enunciation of general propositions, and
the mere putting together and separating objects more or less
alike" (p. 420).
In general, then, homological characters are more valuable for
classificatory purposes because they have a longer pedigree than
analogical characters, which represent recent acquirements of the
race.
Coming to morphology proper, Darwin takes up the question of the
unity of type, and the homology of parts, for which the unity of
type is but a general expression.
He treats this on the same lines as E. Geoffroy St Hilaire, and
Owen, referring indeed specifically to Geoffroy's law of
connections. "What can be more curious," he asks, 235"than that the
hand of a man, formed for grasping, that of a mole for digging, the
leg of a horse, the paddle of the porpoise, and the wing of the
bat, should all be constructed on the same pattern, and should
include similar bones, in the same relative positions? Geoffroy St
Hilaire has strongly insisted on the high importance of relative
position or connection in homologous parts; they may differ to
almost any extent in form and size, and yet remain connected
together in the same invariable order" (p. 434).
The unity of plan cannot be explained on teleological grounds,
as Owen has admitted in his Nature of Limbs, nor is it
explicable on the hypothesis of special creation (p. 435). It can
be understood only on the theory that animals are descended from
one another and retain for innumerable generations the essential
organisation of their ancestors. "The explanation is to a large
extent simple on the theory of the selection of successive slight
modifications—each modification being profitable in some way
to the modified form, but often affecting by correlation other
parts of the organisation. In changes of this nature, there will be
little or no tendency to alter the original pattern or to transpose
the parts.... If we suppose that the ancient progenitor, the
archetype as it may be called, of all animals, had its limbs
constructed on the existing general pattern, for whatever purpose
they served, we can at once perceive the plain significance of the
homologous construction of the limbs throughout the whole class"
(p. 435).
We may note three important points in this passage—first,
the identification of the archetype with the common progenitor;
second, the view that progressive evolution is essentially
adaptive, and dominated by natural selection; and third, the
petitio principii involved in the assumption that adaptive
modification brings inevitably in its train the necessary
correlative changes.
In his section on morphology Darwin shows clearly the influence
of Owen, and through him of the transcendental anatomists. He
refers to the transcendental idea of "metamorphosis," as
exemplified in the vertebral theory of the skull and the theory of
the plant appendage, and shows how, on the hypothesis of descent
with modification, "metamorphosis" 236may now be interpreted literally,
and no longer figuratively merely (p. 439).
Very great interest attaches to Darwin's treatment of
development, for post-Darwinian morphology was based to a very
large extent on the presumed relation between the development of
the individual and the evolution of the race. Just as he kept clear
of the notion of the scale of beings, so he avoided the snare of
the Meckel-Serres theory of recapitulation, according to which the
embryo of the highest animal, man, during its development climbs
the ladder upon the rungs of which the whole animal series is
distributed, in its gradual progression from simplicity to
complexity. The law of development which he adopts is that of von
Baer, which states that development is essentially differentiation,
and that as a result embryos belonging to the same group resemble
one another the more the less advanced they are in development.
There can be little doubt that he was indebted to von Baer for the
idea, and in the later editions of the Origin he
acknowledges this by quoting the well-known passage in which von
Baer tells how he had two embryos in spirit which he was unable to
refer definitely to their proper class among Vertebrates.[354]
Not only are embryos more alike than adults, because less
differentiated, but it is in points not directly connected with the
conditions of existence, not strictly adaptive, that their
resemblance is strongest (p. 440)—think, for instance, of the
arrangement of aortic arches common to all vertebrate embryos.
Larval forms are to some extent exceptions to this rule, for they
are often specially adapted to their particular mode of life, and
convergence of structure may accordingly result. All these facts
require an explanation. "How, then, can we explain these several
facts in embryology—namely, the very general, but not
universal, difference in structure between the embryo and the
adult—of parts in the same individual embryo, which
ultimately become very unlike and serve for different purposes,
being at this early period of growth alike—of embryos of
different species within the same class, generally but not
universally, 237resembling each other—of the structure
of the embryo not being closely related to its conditions of
existence, except when the embryo becomes at any period of life
active and has to provide for itself—of the embryo apparently
having sometimes a higher organisation than the mature animal, into
which it is developed" (pp. 442-3). Obviously all these facts are
formally explained by the doctrine of descent. But Darwin goes
further, he tries to show exactly how it is that the embryos
resemble one another more than the adults. He thinks that the
phenomenon results from two principles—first, that
modifications usually supervene late in the life of the individual;
and second, that such modifications tend to be inherited by the
offspring at a corresponding, not early, age (p. 444).
Thus, applying these principles to a hypothetical case of the
origin of new species of birds from a common stock, he
writes:—"... from the many slight successive steps of
variation having supervened at a rather late age and having been
inherited at a corresponding age, the young of the new species of
our supposed genus will manifestly tend to resemble each other much
more closely than do the adults, just as we have seen in the case
of pigeons"[355] (pp. 446-7).
Since the embryo shows the generalised type, the structure of
the embryo is useful for classificatory purposes. "For the embryo
is the animal in its less modified state; and in so far it reveals
the structure of its progenitor" (p. 449)—the embryological
archetype reveals the ancestral form. "Embryology rises greatly in
interest, when we thus look at the embryo as a picture, more or
less complete, of the parent form of each great class of animals"
(p. 450)—a prophetic remark, in view of the enormous
subsequent development of phylogenetic speculation.
We may sum up by saying that Darwin interpreted von Baer's law
phylogenetically.
The rest of the chapter is devoted to a discussion of abortive
and vestigial organs, whose existence Darwin 238naturally turns
to great advantage in his argument for evolution. Throughout the
whole chapter Darwin's preoccupation with the problems of
classification is clearly manifest.
On the question as to whether descent was monophyletic or
polyphyletic Darwin expressed no dogmatic opinion. "I believe that
animals have descended from at most only four or five progenitors,
and plants from an equal or lesser number.... I should infer from
analogy that probably all the organic beings which have ever lived
on this earth have descended from one primordial form, into which
life was first breathed" (p. 484).
Darwin rightly laid much stress upon the morphological evidence
for evolution,[356] which he considered
to be weighty. It probably contributed greatly to the success of
his theory. Though he himself did little or no work in pure
morphology, he was alive to the importance of such work,[357] and followed with interest the progress of
evolutionary morphology, incorporating some of its results in later
editions of the Origin, and in his Descent of Man
(1871).
In his morphology Darwin was hardly up to date. He does not seem
to have known at first hand the splendid work of the German
morphologists, such as Rathke and Reichert; he pays no attention to
the cell-theory, nor to the germ-layer theory. His sources are, in
the main, Geoffroy St Hilaire, Owen, von Baer, Agassiz,
Milne-Edwards, and Huxley.
Perhaps his greatest omission was that he did not give any
adequate treatment of the problem of functional adaptation and the
correlation of parts. It is not too much to say that Darwin not
only disregarded these problems almost entirely, but by his
insistence upon ecological adaptation and upon certain superficial
aspects of correlation, succeeded in giving to the words
"adaptation" and "correlation" 239a new signification, whereby they
lost to a large extent their true and original functional
meaning.
It is true that Darwin himself, as well as his successors,
believed that natural selection was all-powerful to account for the
evolution of the most complicated organs, but it may be questioned
whether he realised all the conditions of the problem of which he
thus easily disposed. He says, rightly, in an important passage,
that "It is generally acknowledged that all organic beings have
been formed on two great laws—Unity of Type, and the
Conditions of Existence. By unity of type is meant that fundamental
agreement in structure which we see in organic beings of the same
class, and which is quite independent of their habits of life. On
my theory, unity of type is explained by unity of descent. The
expression of conditions of existence, so often insisted upon by
the illustrious Cuvier, is fully embraced by the principle of
natural selection. For natural selection acts by either now
adapting the varying parts of each being to its organic and
inorganic conditions of life:[358] or by having adapted them during past periods of
time: the adaptations being aided in many cases by the increased
use or disuse of parts, being affected by the direct action of the
external conditions of life, and subjected in all cases to the
several laws of growth and variation. Hence, in fact, the law of
the Conditions of Existence is the higher law; as it includes,
through the inheritance of former variations and adaptations, that
of Unity of Type" (Origin, 6th ed., Pop. Impression, pp.
260-1). It is clear that Darwin took the phrase "Conditions of
Existence" to mean the environmental conditions, and the law of the
Conditions of Existence to mean the law of adaptation to
environment. But that is not what Cuvier meant by the phrase: he
understood by it the principle of the co-ordination of the parts to
form the whole, the essential condition for the existence of any
organism whatsoever (see above, Chap. III., p.
34).
Of this thought there is in Darwin little trace, and that is why
he did not sufficiently appreciate the weight of the argument
brought against his theory that it did not account for the
correlation of variations.
240Darwin's conception of correlation was
singularly incomplete. As examples of correlation he advanced such
trivial cases as the relation between albinism, deafness and blue
eyes in cats, or between the tortoise-shell colour and the female
sex. He used the word only in connection with what he called
"correlated variation," meaning by this expression "that the whole
organisation is so tied together during its growth and development,
that when slight variations in any one part occur, and are
accumulated through natural selection, other parts become modified"
(6th ed., p. 177). He took it for granted that the "correlated
variations" would be adapted to the original variation which was
acted upon by natural selection, and he saw no difficulty in the
gradual evolution of a complicated organ like the eye if only the
steps were small enough. "It has been objected," he writes, "that
in order to modify the eye and still preserve it as a perfect
instrument, many changes would have to be effected simultaneously,
which, it is assumed, could not be done through natural selection;
but as I have attempted to show in my work on the variation of
domestic animals, it is not necessary to suppose that the
modifications were all simultaneous, if they were extremely slight
and gradual" (6th ed., p. 226).
In post-Darwinian speculation the difficulty of explaining
correlated variation by natural selection alone became more acutely
realised, and it was chiefly this difficulty that led Weismann to
formulate his hypothesis of germinal selection as a necessary
supplement to the general selection theory.
The change in the conception of correlation which Darwin's
influence brought about has been very clearly stated by E. von
Hartmann,[359] from whom the following
is taken:—"While the correlation of parts in the organism was
before Darwin regarded exclusively from the standpoint of
morphological systematics, Darwin tried to look at it from the
standpoint of physiological and genealogical development, and in so
doing he put the standpoint of morphological systematics in the
shade. But the more we are now beginning to realise that systematic
relationship does not necessarily 241imply genetic affinity the more
must the correlation of parts come back into favour as a systematic
principle. While Darwin only, as it were, against his will, relied
on the law of correlation as a last resort when all other help
failed, this law must be regarded, from the standpoint of the
orderly inner determination of all organic form-change, as having
the rank of the highest principle of all, a principle which rules
parallel, divergent and convergent evolution" (pp. 47-8).
Further on, following Rádl, he characterises Darwin's
attitude to the law of correlation in these terms:—"Darwin's
interest is entirely focussed on the variation, the function, the
causes of form-production, in short, upon evolution. Accordingly he
regards correlation essentially as correlative variation in the
sense of a departure from the given type. With morphological
correlation in different types Darwin troubles himself not
at all, nor with correlation in the normal development of a type"
(p. 49).
Cuvier's conception of the convenance des parties,
essential to all biology, remained on the whole foreign to Darwin's
thought, and to the thought of his successors.
It was indeed one of their boasts that they had finally
eliminated all teleology from Nature. The great and immediate
success which Darwinism had among the younger generation of
biologists and among scientific men in general was due in large
part to the fact that it fitted in well with the prevailing
materialism of the day, and gave solid ground for the hope that in
time a complete mechanistic explanation of life would be
forthcoming. "Darwinismus" became the battle-cry of the militant
spirits of that time.
It was precisely this element in Darwinism that was repugnant to
most of Darwin's opponents, in whose ranks were found the majority
of the morphologists of the old school. They found it impossible to
believe that evolution could have come about by fortuitous
variation and fortuitous selection; they objected to Darwin that he
had enunciated no real Entwickelungsgesetz, or law governing
evolution. They were not unwilling to believe that evolution was a
real process, though many drew the line at the derivation of man
from apes, but they felt that if evolution had really taken place,
it must have been under the guidance of some principle 242of development,
that there must have been manifested in evolution some definite and
orderly tendency towards perfection.[360]
No one expressed this objection with greater force than did von
Baer, in a series of masterly essays[361] which the Darwinians, through sheer inability to
grasp his point of view, dismissed as the maunderings of old age.
In these essays von Baer pointed out the necessity for the
teleological point of view, at least as complementary to the
mechanistic. His general position is that of the "statical"
teleology—to use Driesch's term—of Kant and Cuvier. His
attitude to Darwinism is determined by his teleology. He admits,
just as in 1834, a limited amount of evolution; he criticises the
evolution theory of Darwin on the same lines exactly as forty or
fifty years previously he had criticised the recapitulation and
evolution-theories of the transcendentalists—principally on
the ground that their deductions far outrun the positive facts at
their disposal. He rejects the theory of natural selection
entirely, on the ground that evolution, like development, must have
an end or purpose (Ziel)—"A becoming without a purpose
is in general unthinkable" (p. 231); he points out, too, the
difficulty of explaining the correlation of parts upon the
Darwinian hypothesis. His own conception of the evolutionary
process is that it is essentially zielstrebig or guided by
final causes, that it is a true evolutio or differentiation,
just as individual development is an orderly progress from the
general to the special. He believed in saltatory evolution, in
polyphyletic descent, and in the greater plasticity of the organism
in earlier times.
The idea of saltatory evolution he took from Kölliker, who
shortly after the publication of the Origin promulgated
243in a
critical note on Darwinism a sketch of his theory of "heterogeneous
generation."[362]
Kölliker's attitude is typical of that taken up by many of
the morphologists of the day.[363] He accepts evolution completely, but rejects
Darwinism because it recognises no Entwickelungsgesetz, or
principle of evolution. For the Darwinian theory of evolution
through the selection of small fortuitous variations he would
substitute the theory of evolution through sudden, large
variations, brought about by the influence of a general law of
evolution. This is his theory of heterogeneous generation. "The
fundamental idea of this hypothesis is that under the influence of
a general law of evolution creatures produce from their germs
others which differ from them" (p. 181). It is to be noticed that
Kölliker laid more stress upon the Entwickelungsgesetz
than upon the saltatory nature of variation, for he says a few
pages further on—"the notion at the base of my theory is that
a great evolutionary plan underlies the development of the whole
organised world, and urges on the simpler forms towards ever higher
stages of complexity" (p. 184). Saltatory evolution was not the
essential point of the theory:—"Another difference between
the Darwinian hypothesis and mine is that I postulate many
saltatory changes, but I will not and indeed cannot lay the chief
stress upon this point, for I have not intended to maintain that
the general law of evolution which I hold to be the cause of the
creation of organisms, and which alone manifests itself in the
activity of generation, cannot also so act that from one form
others quite gradually arise" (p. 185). He put forward the
hypothesis of saltatory variation because it seemed to him to
lighten many of the difficulties of Darwinism—the lack of
transition forms, the enormous time required for evolution, and so
on. It should be noted that Kölliker regarded his principle of
evolution as mechanical.
244It would take too long to show in detail how
a belief in innate laws of evolution was held by the majority of
Darwin's critics. A few further examples must suffice.
Richard Owen, who in 1868[364] admitted the possibility of evolution, held that
"a purposive route of development and change, of correlation and
interdependence, manifesting intelligent Will, is as determinable
in the succession of races as in the development and organisation
of the individual. Generations do not vary accidentally, in any and
every direction; but in pre-ordained, definite, and correlated
courses" (p. 808).
He conceived change to have taken place by abrupt variation,
independent of environment and habit, by "departures from parental
type, probably sudden and seemingly monstrous, but adapting the
progeny inheriting such modifications to higher purposes" (p. 797).
He believed spontaneous generation to be a phenomenon constantly
taking place, and constantly giving the possibility of new lines of
evolution.
E. von Hartmann in his Philosophie des Unbewussten (1868)
and in his valuable essay on Wahrheit und Irrtum im
Darwinismus (1874) criticised Darwinism in a most suggestive
manner from the vitalistic standpoint. He drew attention to the
importance of active adaptation, the necessity for assuming
definite and correlated variability, and to the evidence for the
existence of an immanent, purposive, but unconscious principle of
evolution, active as well in phylogenetic as in individual
development.
In France H. Milne-Edwards[365] stated the problem thus:—"In the present
state of science, ought we to attribute to modifications dependent
on the action of known external agents the differences in the
organic types manifested by the animals distributed over the
surface of the globe either at the present day, or in past
geological ages? Or must the origin of types transmissible by
heredity be attributed to causes of another order, to forces whose
effects are not apparent in the present state of things, to a
creative power 245independent of the general properties of
organisable matter such as we know them to-day?" (p. 426)
He concluded that the action of environment, direct or indirect,
was insufficient to account for the diversity of organic forms, and
rejected Darwin's theory completely. He thought it likely that the
successive faunas which palæontology discloses have originated
from one another by descent. But he thought that the process by
which they evolved should rightly be called "creation." The word
was of course not to be taken in a crude sense. When the zoologist
speaks of the "creation" of a new species, "he in no way means that
the latter has arisen from the dust, rather than from a
pre-existing animal whose mode of organisation was different; he
merely means that the known properties of matter, whether inert or
organic, are insufficient to bring about such a result, and that
the intervention of a hidden cause, of a power of some higher
order, seems to him necessary" (p. 429).
The criticism of Darwinism exercised by the older currents of
thought remained on the whole without influence. It was under the
direct inspiration of the Darwinian theory that morphology
developed during the next quarter of a century.
246
CHAPTER XIV
ERNST HAECKEL AND CARL GEGENBAUR
At the time when Darwin's work
appeared there already existed, as we have seen, a fully formed
morphology with set and definite principles. The aim of this
pre-evolutionary morphology had been to discover and work out in
detail the unity of plan underlying the diversity of forms, to
disentangle the constant in animal form and distinguish from it the
accessory and adaptive. The main principle upon which this work was
based was the principle of connections, so clearly stated by
Geoffroy. The principle of connections served as a guide in the
search for the archetype, and this search was prosecuted in two
directions—first, by the comparison of adult structure; and
second, by the comparative study of developing embryos. It was
found that the archetype was shown most clearly by the early
embryo, and this embryological archetype came to be preferred
before the archetype of comparative anatomy. It became apparent
also that the parts first formed (germ-layers) were of primary
importance for the establishing of homologies.
While practically all morphologists were agreed as to the main
principles of their science, they yet showed, as regards their
general attitude to the problems of form, a fairly definite
division into two groups, of which one laid stress upon the
intimate relation existing between form and function, while the
other disregarded function completely, and sought to build up a
"pure" or abstract morphology. In opposition to both groups, in
opposition really to morphology altogether, a movement had gained
strength which tended towards the analysis and disintegration of
the organism. This movement took its origin in the current 247materialism of the day, and found expression
particularly in the cell-theory and in materialistic
physiology.
The separation between morphology as the science of form and
physiology as the science of the physics and chemistry of the
living body had by Darwin's day become well-nigh absolute.
The morphology of the 'fifties lent itself readily to
evolutionary interpretation. Darwin found it easy to give a formal
solution of all the main problems which pre-evolutionary morphology
had set—he was able to interpret the natural system of
classification as being in reality genealogical, systematic
relationship as being really blood-relationship; he was able to
interpret homology and analogy in terms of heredity and adaptation;
he was able to explain the unity of plan by descent from a common
ancestor, and for the concept of "archetype" to substitute that of
"ancestral form."
The current morphology, Darwin found, could be taken over, lock,
stock and barrel, to the evolutionary camp.
In what follows we shall see that the coming of evolution made
surprisingly little difference to morphology, that the same methods
were consciously or unconsciously followed, the same mental
attitudes taken up, after as before the publication of the
Origin of Species.
Darwin himself was not a professional morphologist; the
conversion of morphology to evolutionary ideas was carried out
principally by his followers, Ernst Haeckel and Carl Gegenbaur in
Germany, Huxley, Lankester, and F. M. Balfour in England.
It was in 1866 that Haeckel's chief work appeared, a General
Morphology of Organisms,[366] which was intended by its author to bring all
morphology under the sway and domination of evolution.
It was a curious production, this first book of Haeckel's, and
representative not so much of Darwinian as of pre-Darwinian 248thought.
It was a medley of dogmatic materialism, idealistic morphology, and
evolution theory; its sources were, approximately, Büchner,
Theodor Schwann, Virchow, H. G. Bronn, and, of course, Charles
Darwin.
It was scarcely modern even on its first appearance, and many
regarded it, not without reason, as a belated offshoot of
Naturphilosophie.
Its materialism is of the most intransigent character. The form
and activities of living things are held to be merely the
mechanical result of the physical and chemical composition of their
bodies. The simplest living things, the Monera, are nothing more
than homogeneous masses of protein substance. "They live, but
without organs of life; all the phenomena of their life, nutrition
and reproduction, movement and irritability, appear here as merely
the immediate outcome of formless organic matter, itself an albumen
compound" (p. 63, 1906).
Teleology, the Achilles' heel of Kant's (otherwise sound!)
philosophy, is to be regarded as a totally refuted and antiquated
doctrine, definitely put out of court by Darwinism.
Haeckel works out his materialistic philosophy of living things
very much after the fashion of Schwann. There is the same talk of
cells as organic crystals, of crystal trees, of the analogy between
assimilation by the cell and the growth of crystals in a mother
liquid. Heredity and adaptation are shown equally as well by
crystals as by organisms; for heredity, or the internal
Bildungstrieb (!), is the mechanical effect of the material
structure of the crystal or the germ, and adaptation, or the
external Bildungstrieb, is a name for the modifications
induced by the environment. Adaptation so defined comes to be
synonymous with the fortuitous variation which plays so great a
part in Darwin's theory of natural selection.
It goes without saying that Haeckel allowed to the organism no
other nor higher individuality than belongs to the crystal, and
took no account at all of that harmonious interaction of the organs
which Cuvier called the principle of the "conditions of existence."
The concept of correlation had simply no meaning for Haeckel. The
analysis and 249disintegration of the organism was pushed by
him to its logical extreme, and in this also he was a child of his
time.
A no less important influence clearly visible in the General
Morphology is the idealistic morphology of men like K. G. Carus
and H. G. Bronn. In previous chapters we have seen how K. G. Carus
attempted to work out a geometry of the organism, and how Bronn
tried in a modest way to found a stereometrical morphology, but had
the grace not to push his stereometry à l'outrance,
recognising very wisely that the greater part of organic form is
functionally determined. Haeckel took over this idea[367] and pushed it to wild extremes, founding a
new science of "Promorphology" of which he was the
greatest—and only—exponent.[368]
This "science" dealt with axes and planes, poles and angles, in
a veritable orgy of barbarous technical terms. It was intended to
be a "crystallography of the organic," and to lay the foundations
of a mechanistic morphology, or morphography at least.
How it was to be linked up with the physics and chemistry of
living matter on the one hand and with the ordinary morphology of
real animals on the other, was never made quite clear.
The science of Promorphology has no historical significance; it
is interesting only because it illustrates Haeckel's close affinity
with the idealistic morphologists.
Another abortive science of Haeckel's, the science of Tectology,
was equally a heritage from idealistic morphology. Tectology is the
science of the composition of organisms from individuals of
different orders. There were six orders of individuals:—(1)
Plastids (Cytodes and cells); (2) Organs (including cell-fusions,
tissues, organs, organ-systems); (3) Antimeres (homotypic parts,
i.e., halves or rays); (4) Metameres (homodynamic parts,
i.e., segments); (5) Persons (individuals in the ordinary
sense); (6) Corms (colonial animals).
The thought is essentially transcendental, and recalls 250the
"theory of the repetition of parts," of which so much use was made
by the German transcendentalists, such as Goethe,[369] Oken, Meckel and K. G. Carus, as well as by
Dugès.
The third, and naturally the most important, ingredient in the
General Morphology was the doctrine of evolution, in the
form given to it by Darwin. We have here no concern with Haeckel's
evolutionary philosophy, with the way in which he combined his
evolutionism and his materialism to form a queer Monism of his own.
We are interested only in the way he applied evolution to
morphology, what modifications he introduced into the principles of
the science, and in general in what way he interpreted the facts
and theories of morphology in the light of the new knowledge.
We find that he repeats very much what Darwin said, giving, of
course, more detail to the exposition, and elaborating,
particularly in his recapitulation theory or "biogenetic law,"
certain doctrines not explicitly stated by Darwin.
Like Darwin he held that the natural system is in reality
genealogical. "There exists," he writes, "one single connected
natural system of organisms, and this single natural system is the
expression of real relations which actually exist between all
organisms, alike those now in being on the earth and those that
have existed there in some past time. The real relations which
unite all living and extinct organisms in one or other of the
principal groups of the natural system, are genealogical: their
relationship in form is blood-relationship; the natural system is
accordingly the genealogical tree of organisms, or their
genealogema.... All organisms are in the last resort descendants of
autogenous Monera, evolved as a consequence of the divergence of
characters through natural selection. The different subordinate
groups of the natural system, the categories of the class, order,
family, genus, etc., are larger or smaller branches of the
genealogical tree, and the degree of their divergence indicates the
degree of genealogical affinity of the 251related organisms with one
another and with the common ancestral form" (ii., p. 420).
The degree of systematic relationship is thus the degree of
genealogical affinity. It follows that the natural system of
classification may be converted straightway into a genealogical
tree, and this is actually what Haeckel does in the General
Morphology. The genealogical trees depicted in the second
volume (plates i.-viii.) are nothing more than graphic
representations of the ordinary systematic relationships of
organisms, with a few hypothetical ancestral groups or forms thrown
in to give the whole a genealogical turn.
If the genealogical tree is truly represented by the natural
system, it would seem that for each genus a single ancestral form
must be postulated, for each group of genera a single more
primitive form, and so in general for each of the higher
classificatory categories, right up to the phylum. Species of one
genus must be descended from a generic ancestral form, genera of
one family from a single family Urform, and so on for the
higher categories.
This consequence was explicitly recognised by Haeckel. "Genera
and families," he writes, "as the next highest systematic grades,
are extinct species which have resolved themselves into a divergent
bunch of forms (Formenbüschel)" (ii., p. 420).
The archetype of the genus, family, order, class and phylum was
thus conceived to have had at some past time a real existence.
The natural system of classification is based upon a proper
appreciation of the distinction between homological and analogical
characters. Haeckel, following Darwin, naturally interprets the
former as due to inheritance, the latter as due to adaptation,
using these words, we may note, in their accepted meaning and not
in the abstract empty sense he had previously attributed to them.[370] Similarly the "type of
organisation," in von Baer's sense, was due to heredity, the "grade
of differentiation" to adaptation.
So far Haeckel merely emphasised what Darwin had already said in
the Origin of Species. But by his statement of the
"biogenetic law," and particularly by the clever use 252he made of it,
Haeckel went a step beyond Darwin, and exercised perhaps a more
direct influence upon evolutionary morphology than Darwin
himself.
Haeckel was not the original discoverer of the law of
recapitulation. It happened that a few years before the publication
of Haeckel's General Morphology, a German doctor, Fritz
Müller by name, stationed in Brazil, had been working on the
development of Crustacea under the direct inspiration of Darwin's
theory, and had published in 1864 a book[371] in which he showed that individual development
gave a clue to ancestral history.
He conceived that progressive evolution might take place in two
different ways. "Descendants ... reach a new goal, either by
deviating sooner or later whilst still on the way towards the form
of their parents, or by passing along this course without
deviation, but then instead of standing still advancing still
farther" (Eng. trans., p. 111). In the former case the
developmental history of descendants agrees with that of the
ancestors only up to a certain point and then diverges. "In the
second case the entire development of the progenitors is also
passed through by the descendants, and, therefore, so far as the
production of a species depends upon this second mode of progress,
the historical development of the species will be mirrored in its
developmental history" (p. 112).
Of course the recapitulation of ancestral history will be
neither literal nor extended. "The historical record preserved in
developmental history is gradually effaced as the
development strikes into a constantly straighter course from the
egg to the perfect animal, and it is frequently
sophisticated by the struggle for existence which the
free-living larvæ have to undergo" (p. 114).
It follows that "the primitive history of a species will be
preserved in its developmental history the more perfectly the
longer the series of young stages through which it passes by
uniform steps; and the more truly, the less the mode of life of the
young departs from that of the adults, and the less the
peculiarities of the individual young states can be conceived 253as
transferred back from later ones in previous periods of life, or as
independently acquired" (p. 121).
Applying these principles to Crustacea, he concluded that the
shrimp Peneus with its long direct development gave the best
and truest picture of the ancestral history of the Malacostraca,
and that accordingly the nauplius and the zoaea larvæ
represented important ancestral stages. He conceived it possible so
to link up the various larval forms of Crustacea as to weave a
picture of the primeval history of the class, and he made a plucky
attempt to work out the phylogeny of the various groups.
The thought that development repeats evolution was already
implicit in the first edition of the Origin, but the credit
for the first clear and detailed exposition of it belongs to F.
Müller.
In much the same form as it was propounded by Müller it was
adopted by Haeckel, and made the corner-stone of his evolutionary
embryology. Haeckel gave it more precise and more technical
formulation, but added nothing essentially new to the idea.
It is convenient to use his term for it—the biogenetic law
(Biogenetische Grundgesetz)—to distinguish it from the
laws of Meckel-Serres and von Baer, with which it is so often
confused.
Haeckel's statement of it may best be summarised in his own
words, "Ontogeny, or the development of the organic individual,
being the series of form-changes which each individual organism
traverses during the whole time of its individual existence, is
immediately conditioned by phylogeny, or the development of the
organic stock (phylon) to which it belongs.
"Ontogeny is the short and rapid recapitulation of phylogeny,
conditioned by the physiological functions of heredity
(reproduction) and adaptation (nutrition). The organic individual
(as a morphological individual of the first to the sixth order)
repeats during the rapid and short course of its individual
development the most important of the form-changes which its
ancestors traversed during the long and slow course of their
palæontological evolution according to the laws of heredity
and adaptation.
254"The complete and accurate repetition of
phyletic by biontic development is obliterated and abbreviated by
secondary contraction, as ontogeny strikes out for itself an ever
straighter course; accordingly, the repetition is the more complete
the longer the series of young stages successively passed
through.
"The complete and accurate repetition of phyletic by biontic
development is falsified and altered by secondary adaptation, in
that the bion[372] during its
individual development adapts itself to new conditions: accordingly
the repetition is the more accurate the greater the resemblance
between the conditions of existence under which respectively the
bion and its ancestors developed" (ii., p. 300).
The last two propositions, it will be observed, are taken over
almost verbally from F. Müller.
Now we have seen that the natural system of classification gives
a true picture of the genealogical relationships of organisms, that
the smaller and larger classificatory groups correspond to greater
or lesser branches of the genealogical tree. If ontogeny is a
recapitulation of phylogeny, we must expect to find the embryo
repeating the organisation first of the ancestor of the phylum,
then of the ancestor of the class, the order, the family and the
genus to which it belongs. There must be a threefold parallelism
between the natural system, ontogeny and phylogeny (ii., pp.
421-2).
It will be observed that there is here implied an analogy
between the biogenetic law and the law of von Baer, for both assert
that development proceeds from the general to the special, that the
farther back in development you go the more generalised do you find
the structure of the embryo; both assert, too, that differentiation
of structure takes place not in one progressive or regressive line,
but in several diverging directions.
But the analogy between the biogenetic law and the Meckel-Serres
law is even more obvious, and the resemblance between the two is
much more fundamental. It is a significant fact that in his theory
of the threefold 255parallelism Haeckel merely resuscitated in an
evolutionary form a doctrine widely discussed in the 'forties and
'fifties,[373] and championed
particularly by L. Agassiz,[374] a doctrine which must be regarded as a
development or expansion of the Meckel-Serres law.[375] It is the view that a parallelism exists
between the natural system, embryonic development, and
palæontological succession. Actually, as Agassiz stated it,
the doctrine applied neither to types, nor as a general rule to
classes, but merely to orders. It was well exemplified, he thought,
in Crinoids:—"The successive stages of the embryonic growth
of Crinoids typify, as it were, the principal forms of Crinoids
which characterise the successive geological formations. First, it
recalls the Cistoids of the palæozoic rocks, which are
represented in its simple spheroidal head; next the few-plated
Platycrinoids of the Carboniferous period; next the Pentacrinoids
of the Lias and Oolite with their whorls of cirrhi; and finally,
when freed from its stem, it stands as the highest Crinoid, as the
prominent type of the family in the present period" (p. 171).
The Meckel-Serres law, it will be remembered, expressed the idea
that the higher animals repeat in their ontogeny the adult
organisation of animals lower in the scale. Since Haeckel
recognised clearly that a linear arrangement of the animal kingdom
was a mere perversion of reality, and that a branching arrangement
of groups more truly represented the real relations of animals to
one another, he could not of course entertain the Meckel-Serres
theory in its original form. But he accepted the main tenet of it
when he asserted that each stage of ontogeny had its counterpart in
an adult ancestral form. Such ancestral forms might or might not be
in existence as 256real species at the present day; they might
or might not be discoverable as fossils. That they had real
existence either now or at some past epoch Haeckel never doubted.
In his construction of phylogenetic trees he was so confident in
the truth of his biogenetic law that he largely disregarded and
consistently minimised the importance of the evidence from
palæontology.
The biogenetic law differed from the Meckel-Serres law chiefly
in the circumstance that many of the adult lower forms whose
organisation was supposed to be repeated in the development of the
higher animals were purely hypothetical, being deduced directly
from a study of ontogeny and systematic relationships. The
hypothetical ancestral forms which the theory thus postulated
naturally took their place in the natural system, for they were
merely the concrete projections or archetypes of the classificatory
groups.
The transcendentalists, of course, conceived evolution, whether
real or ideal, as a uniserial process, whereas Haeckel conceived it
as multiserial and divergent. It is here that the superficial
agreement of the biogenetic law with the law of von Baer comes
in.
We might almost sum up the relation of the biogenetic law to the
laws of von Baer and Meckel-Serres by saying that it was the
Meckel-Serres law applied to the divergent differentiation upheld
by von Baer instead of to the uniserial progression believed in by
the transcendentalists.
How near in practice Haeckel's law came to the recapitulation
theory of the transcendentalists may be seen in passages like the
following, with its partial recognition of the Échelle
idea:[376]—"As so high and
complicated an organism as that of man ... rises upwards from a
simple cellular state, and as it progresses in its differentiating
and perfecting, it passes through the same series of
transformations which its animal progenitors have passed through,
during immense spaces of time, inconceivable ages ago.... Certain
very early and low stages in the development of man, and other
vertebrate animals in general, correspond completely in many points
of structure with conditions which 257last for life in the lower fishes.
The next phase which follows on this presents us with a change of
the fish-like being into a kind of amphibious animal. At a later
period the mammal, with its special characteristics, develops out
of the amphibian, and we can clearly see, in the successive stages
of its later development, a series of steps of progressive
transformation which evidently correspond with the differences of
different mammalian orders and families."[377]
The biogenetic law went beyond both the Meckel-Serres law and
the law of von Baer in that it recognised that the ancestral
history of the species accounts in part for the course which the
development of the individual takes, that in a certain sense,
though not in the crude way supposed by Haeckel, phylogeny is the
cause of ontogeny. This thought, that the organism is before all an
historical being, is of course implied in the evolution idea, is
indeed the essential core of it. Take away this element from the
biogenetic law—not a difficult matter—and it becomes
merely a law of idealistic morphology, applicable to evolution
considered as an ideal process, as the progressive development in
the Divine thought of archetypal models.
As a book, the General Morphology suffers a good deal
from the arid, schematic, almost scholastic manner of exposition
adopted. Haeckel's Prussian mania for organisation, for absolute
distinctions, for iron-bound formalism, is here given full scope. A
treatment less adequate to the variety, fluidity and changeableness
of living things could hardly be imagined.
His doctrine, though it remains essentially unchanged, receives
in his later works a less formal and more concrete expression, and,
in particular, his views on the biogenetic law undergo some small
modification.
Even in the General Morphology Haeckel had recognised
that ontogeny is neither a complete nor an entirely accurate
recapitulation of phylogeny; he had admitted, following F.
Müller, that the true course of recapitulation was frequently
modified by larval and fœtal adaptations. As time went on, he
was forced to hedge more and more on this point, and finally in his
Anthropogenie (1874) and his second 258paper on the Gastræa
theory (1875),[378] he had to work out
a distinction between palingenetic and cenogenetic characters, of
which much use was made by subsequent writers.
The distinction may be given in Haeckel's own
words:—"Those ontogenetic processes," he writes, "which are
to be referred immediately, in accordance with the biogenetic law,
to an earlier completely developed independent ancestral
form, and are transmitted from this by heredity,
obviously possess primary importance for the understanding
of the casual-physiological relations; on the other hand, those
developmental processes which appear subsequently through
adaptation to the needs of embryonic or larval life, and
accordingly can not be regarded as repeating the
organisation of an earlier independent ancestral form, can clearly
have for the understanding of the ancestral history only a quite
subordinate and secondary importance.
"The first I have named palingenetic, the second
cenogenetic. Considered from this critical standpoint, the
whole of ontogeny falls into two main parts:—First,
palingenesis, or 'epitomised history'
(Auszugsgeschichte), and second, cenogenesis, or
'counterfeit history' (Fälschungsgeschichte). The first
is the true ontogenetic epitome or short recapitulation of past
evolutionary history; the second is the exact contrary, a new
foreign ingredient, a falsifying or concealing of the epitome of
phylogeny."[379]
As examples of palingenetic processes in the development of
Amniotes, for instance, may be quoted the separation of two primary
germ-layers, the formation of a simple notochord between medullary
tube and alimentary canal, the appearance of a simple cartilaginous
cranium, of the gill-arches and their vessels, of the primitive
kidneys, the primitive tubular heart, the paired aortæ and
the cardinal veins, the hermaphroditic rudiment of the gonads, and
so on. Cenogenetic processes, on the other hand, include such
phenomena as the formation of yolk and the embryonic membranes, the
temporary allantoic circulation, the navel, the curved and
contracted shape of the embryo, and the like.
The most important phenomena to be included under 259the general
heading of cenogenesis are, first, the occurrence of food-yolk, and
second, those anomalies of development which are classed by Haeckel
as heterochronies and heterotopies.
It is to the influence of the different amounts of yolk present
in the egg that are due the great differences in the segmentation
and gastrulation processes, which almost mask their true
significance.
Heterochronic processes are such as arise through the
dislocation of the proper phylogenetic order of succession:
heterotopic processes in the same way are caused by a wandering of
cells from one germ-layer to another. The two classes of phenomena
are disturbances either of the proper spatial or of the proper
temporal relation of the parts during development.
Heterochrony shows itself, as a rule, either as an acceleration
or as a retardation of developmental events, as compared with their
relative time of occurrence during phylogeny. Thus the notochord,
the brain, the eyes, the heart, appear earlier in the ontogenetic
than in the phylogenetic series, while, on the other hand, the
septum of the auricles appears in the development of the higher
Vertebrates before the ventricular septum, which is undoubtedly a
reversal of the phylogenetic order.
Cases of heterotopy, or of organs being developed in a position
or a germ-layer other than that in which they originally arose in
phylogeny, are not so easy to find. According to Haeckel, the
origin of the generative products in the mesoderm is a heterotopic
phenomenon, for he considers that they must have originated
phylogenetically in one of the two primary layers, ectoderm or
endoderm.
It is worthy of note that the help of comparative anatomy is
admittedly required in deciding what processes are palingenetic and
what cenogenetic (p. 412).
Haeckel's morphological notions, and particularly his biogenetic
law, excited a good deal of adverse criticism from men like His,
Claus, Salensky, Semper and Goette. Nor was his principal work, the
General Morphology, received with much favour. Nevertheless,
since he did express, 260though in a crude, dogmatic and extreme
manner, the main hypotheses upon which evolutionary morphology is
founded, his historical importance is considerable. He cannot
perhaps be regarded as typical of the morphologists of his
time—he was too trenchantly materialistic, too much the
populariser of a crude and commonplace philosophy of Nature. In
point of concrete achievement in the field of pure research he fell
notably behind many of his contemporaries.
His friend, Carl Gegenbaur, who gained a great and well-deserved
reputation by his masterly studies on vertebrate morphology,[380] was a sounder man, and
probably exercised a wider and certainly a more wholesome influence
upon the younger generation of professional morphologists than the
more brilliant Haeckel. It is true that in his famous
Grundzüge der vergleichenden Anatomie, the second
edition of which, published in 1870, soon came to be regarded as
the classical text-book of evolutionary morphology, Gegenbaur
enunciated very much the same general principles as Haeckel, and
referred to the Generelle Morphologie as the chief and
fundamental work on animal morphology. But in Gegenbaur's pages the
Haeckelian doctrines are modified and subdued by the strong
commonsense and thorough appreciation of the older classical or
Cuvierian morphology that characterise Gegenbaur's work. According
to Haeckel,[381] Gegenbaur was greatly
influenced by J. Müller, who, as we know, laid as much stress
on function as on form.
The "General Part" of Gegenbaur's text-book is in many ways a
significant document and deserves close attention.
We note first of all that physiology and morphology are
considered by Gegenbaur to be entirely distinct sciences, with
different subject-matter and different methods. "The task of
physiology is the investigation of the functions of the animal body
or of its parts, the referring back of these functions to
elementary processes and their explanation by 261general laws.
The investigation of the material substratum of these functions, of
the form of the body and its parts, and the explanation of this
form, constitute the task of Morphology" (2nd ed., p. 3).
Morphology falls naturally into two divisions—comparative
anatomy and embryology. The method of comparative anatomy is
comparison (p. 6), and in employing this method account is
to be taken of "the spatial relations of the parts to one another,
their number, extent, structure, and texture." Through comparison
one is enabled to arrange organs in continuous series, and it comes
out very clearly during this proceeding "that the physiological
value of an organ is by no means constant throughout the different
form-states of the organ, that an organ, through the mere
modification of its anatomical relations, can subserve very
different functions. Exclusive regard for their physiological
functions would place morphologically related organs in different
categories. From this it follows that in comparative anatomy we
should never in the first place consider the function of an organ.
The physiological value comes only in the second place into
consideration, when we have to reconstruct the relations to the
organism as a whole of the modification which an organ has
undergone as compared with another state of it. In this way
comparative anatomy shows us how to arrange organs in series;
within these series we meet with variations which sometimes are
insignificant and sometimes greater in extent; they affect the
extent, number, shape, and texture of the parts of an organ, and
can even, though only in a slight degree, lead to alterations of
position" (p. 6).
Geoffroy St Hilaire would have subscribed to every word of this
vindication of his "principle of connections."
Between comparative anatomy and embryology there exists a close
connection, for the one throws light on the other. "While in some
cases the same organ shows only slight modifications in its
development from its early beginnings to its perfect state, in
other cases the organ is subjected to manifold modifications before
it reaches its definitive form; we see parts appear in it which
later disappear, we observe alterations in it in all its anatomical
relations, alterations which may even affect its texture. This
262fact
is of great importance, for those changes which an organ undergoes
during its individual development lead through states which the
organ in other cases permanently shows, or at the least the first
appearance of the organ is the equivalent of a permanent state in
another organism. If then the fully developed organ is in any
special case so greatly modified that its proper relation to some
organ-series is obscured, this relation may be cleared up by a
knowledge of the organ's development. The earlier state indicated
in this way enables one to find with ease the proper place for the
organ and so insert it into an already known series. The relations
which we observe in an organ-seriation are then the equivalent of
processes which in certain cases take place in a similar manner
during the individual development of an organ. Embryology enters
therefore into the closest connection with comparative anatomy....
It teaches us to know organs in their earliest states, and connects
them up with the permanent states of others, whereby they fill up
the gaps which we meet with in the various series formed by the
fully developed organs of the body" (pp. 6-7).
This recognition of the parallelism between comparative anatomy
and embryology is, of course, the kernel of the Meckel-Serres law.
For Gegenbaur it had a very definite evolutionary meaning—he
subscribed to the evolutionary form of it, the biogenetic law. How
near his conception of the relation between ontogeny and phylogeny
came to the old Meckel-Serres law may be gauged from the following
passage, taken from a later work:—"Ontogeny thus represents,
to a certain degree, palæontological development abbreviated or
epitomised. The stages which are passed through by higher organisms
in their ontogeny correspond to stages which are maintained in
others as the definitive organisation. These embryonic stages may
accordingly be explained by comparing them with the mature stages
of lower organisms, since we regard them as forms inherited from
ancestors belonging to such lower stages"[382] (p. 6).
It is worth noting that in Gegenbaur's opinion comparative 263anatomy
was prior in importance to embryology, that embryology could hardly
exist as an independent science, since it must seek the
interpretation of its facts always in the facts of comparative
anatomy (Grundzüge, pp. 7-8).
While Gegenbaur was at one with all "pure" morphologists,
whether evolutionary or pre-evolutionary, in minimising as far as
possible the importance of function in the study of form, he was
too cautious and sober a thinker not to recognise the immense part
which function really plays. Thus he classified organs, according
to their function, into those that established relations with the
external world and those that had to do with nutrition and
reproduction, very much as Bichat had done before him.
Like Darwin, Haeckel and most evolutionists, he interpreted the
homological resemblances of animals as being due to heredity, their
differences as due to adaptation,[383] but he did not adopt Haeckel's crude and shallow
definition of these terms. For Gegenbaur heredity was a convenient
expression for the fact of transmission, and was not explained
offhand as the mere mechanical result of a certain material
structure handed down from germ to germ. Adaptation he defined in a
way which took the fullest account of function, and was as far as
possible removed from Haeckel's definition of it as the direct
mechanical effect of the environment upon the organism. "The
organism is altered," writes Gegenbaur, "according to the
conditions which influence it. The consequent Adaptations
are to be regarded as gradual, but steadily progressive, changes in
the organisation, which are striven after during the individual
life of the organism, preserved by transmission in a series of
generations, and further developed by means of natural selection.
What has been gained by the ancestor becomes the heritage of the
descendant. Adaptation and Transmission are thus alternately
effective, the former representing the modifying, the latter the
conservative 264principle.... Adaptation is commenced by a
change in the function of organs, so that the physiological
relations of organs play the most important part in it. Since
adaptation is merely the material expression of this change of
function, the modification of the function as much as its
expression is to be regarded as a gradual process. In Adaptation,
the closest connection between the function and the structure of an
organ is thus indicated. Physiological functions govern, in a
certain sense, structure; and so far what is morphological is
subordinated to what is physiological" (Elements, pp. 8-9).
Gegenbaur recognised also that morphological differentiation
depended largely on the physiological division of labour
(Grundzüge, p. 49).
It is clear that Gegenbaur realised vividly the importance of
function, and in this respect, as in others, he is far beyond
Haeckel. The same thing comes out markedly in his treatment of
correlation. Haeckel had no slightest feeling for the true meaning
of correlation. For him, as for Darwin, it reduced itself to a law
of correlative variation, according to which "actual adaptation not
only changes those parts of the organism which are directly
affected by its influence, but other parts also, not directly
affected by it."[384] Such "correlative
adaptation" was due to nutrition being a "connected, centralised
activity."
Gegenbaur, on the contrary, had a firm grasp of the Cuvierian
conception, and expressed it in unmistakable terms. "As indeed
follows from the conception of life as the harmonious expression of
a sum of phenomena rigorously determining one another, no activity
of an organ can in reality be thought of as existing for itself.
Each kind of function (Verrichtung) presupposes a series of
other functions, and accordingly every organ must possess close
relations with, and be dependent on, all the others"
(Grundzüge, p. 71). The organism must be regarded as an
individual whole which is as much conditioned by its parts as one
part is conditioned by the others. For an understanding of
correlation a knowledge of functions, and of the functional
relations of the organism to its environment, is clearly
indispensable.
265Gegenbaur's morphological system was
out-and-out evolutionary. "The most important part of the business
of comparative anatomy," in Gegenbaur's eyes, "is to find
indications of genetic connection in the organisation of the animal
body" (Elements, p. 67).
The most important clue to discovering this genetic connection
is of course that given by homology; it is indeed the main
principle of evolutionary morphology that what is common in
organisation is due to common descent, what is divergent is due to
adaptation. "Homology ... corresponds to the hypothetical genetic
relationship. In the more or the less clear homology, we have the
expression of the more or less intimate degree of relationship.
Blood-relationship becomes dubious exactly in proportion as the
proof of homologies is uncertain" (Elements, p. 63).
It is worth noting that while Gegenbaur agrees with Haeckel
generally that morphological relationships are really genealogical,
that, for instance, each phylum has its ancestral form, he enters a
caution against too hastily assuming the existence of a genetic
relation between two forms on the basis of the comparison of one or
two organs. "In treating comparative anatomy from the genealogical
standpoint required by the evolution-theory," he writes, "we have
to take into consideration the fact that the connections can almost
never be discovered in the real genealogically related objects, for
we have almost always to do with the divergent members of an
evolutionary series. We derive, for instance, the circulatory
system of insects from that of Crustacea ... but there exists
neither a form that leads directly from Crustacea to insects nor
any organisatory state (Organisationszustand), which as such
shows the transition. Even when one point of organisation can be
denoted as transitional, numerous other points prevent us from
regarding the whole organism strictly in the same light"
(Grundzüge, p. 75). The real ancestral forms cannot, as
a rule, be discovered among living species, nor often as extinct.
"When we arrange allied forms in series by means of comparison, and
seek to derive the more complex from the simpler, we recognise in
the lower and simpler forms only similarities 266with the
ancestral form, which remains essentially hypothetical" (p.
75).
The facts of development, Gegenbaur goes on to say, help us out
greatly in our search for ancestral forms, for the early stages in
the ontogeny of a highly organised animal give us some idea of the
organisation of its original ancestor. Characters common to the
early ontogeny of all the members of a large group are particularly
important in this respect (cf. von Baer's law).
Gegenbaur distinguishes homologous or morphologically equivalent
structures from such as are analogous or physiologically
equivalent, just as did Owen and the older anatomists. Like von
Baer he recognises homologies, as a rule, only within the type.
He contributed, however, to the common stock a useful analysis
of the concept of homology, and established certain classes and
degrees of it. He distinguished first between general and special
homology, in quite a different sense from Owen.
General homology, in Gegenbaur's sense, relates to resemblances
of organs within the organism, and includes four kinds of
resemblance, homotypy, homodynamy, homonomy and homonymy. Right and
left organs are homotypic, metameric organs are homodynamic;
homonomy is the relation exemplified by fin-rays or fingers, which
are arranged with reference to a transverse axis of the body;
homonymy is a sort of metamerism in secondary parts (not the main
axis) of the body, and is shown by the various divisions of the
appendages (Grundzüge, p. 80).
Special homology, on the other hand, relates to resemblances
between organs in different animals. The interesting thing is that
Gegenbaur defines it genetically. Special homology is the name we
give "to the relations which obtain between two organs which have
had a common origin, and which have also a common embryonic
history" (Elements, p. 64). This is his definition; but, in
practice, Gegenbaur establishes homologies by comparison just as
the older anatomists did, and infers common descent from homology,
not homology from common descent.
"Special homology," he continues, "must be again separated 267into
sub-divisions, according as the organs dealt with are essentially
unchanged in their morphological characters, or are altered by the
addition or removal of parts" (p. 65). In the former case the
homology is said to be "complete," in the latter "incomplete." Thus
the bones of the upper arm are completely homologous throughout all
vertebrate classes from Amphibia upwards, while the heart of a fish
is incompletely homologous with the heart of a mammal.
Independently of Gegenbaur, Sir E. Ray Lankester proposed in
1870 a genetic definition of homology.[385] He proposed, indeed, to do away with the term
homology altogether, on the ground that it included many
resemblances which were obviously not due to common
descent—as, for instance, the resemblance of metameres. So,
too, organs which were homologous in the ordinary sense, as the
heart of birds and mammals, might have arisen separately in
evolution. He proposed, therefore, that "structures which are
genetically related, in so far as they have a single representative
in a common ancestor," should be called homogenous(p. 36).
All other resemblances were to be called homoplastic.
"Homoplasy includes all cases of close resemblance of form which
are not traceable to homogeny, all details of agreement not
homogenous, in structures which are broadly homogenous, as well as
in structures having no genetic affinity" (p. 41). Serial homology,
for instance, was a case of homoplasy.
The term "analogy" was to be retained for cases of functional
resemblance, whether homogenetic or not.
The attempt was an interesting one, but most morphologists
wisely adhered to the old concept of homology, in spite of
Lankester's declaration that this belonged to an older "Platonic"
philosophy, and ought to be superseded by a term more consonant
with the new philosophy of evolution.
268
CHAPTER XV
EARLY THEORIES ON THE ORIGIN OF VERTEBRATES
Haeckel and Gegenbaur set the fashion
for phylogenetic speculation, and up to the middle 'eighties, when
the voice of the sceptics began to make itself heard, the chief
concern of the younger morphologists was the construction of
genealogical trees. The period from about 1865 to 1885 might well
be called the second speculative or transcendental period of
morphology, differing only from the first period of
transcendentalism by the greater bulk of its positive achievement.
It must be remembered that the later workers (at least towards the
end of this period) had immense advantages over their predecessors
in the matter of equipment and technique; they possessed
well-fitted laboratories in the university towns and by the sea;
they had at their command perfected microscopes and microtomes;
while the whole new technique of microscopical anatomy with its
endless variety of stains and reagents made it possible for the
tyro to confirm in a day what von Baer and Müller had taken
weeks of painful endeavour to discover.[386] But the democratisation of morphology which
followed upon the facilitation of its means of research left an
evil heritage of detailed and unintelligent work to counterbalance
the very great and real advances which technical improvements alone
rendered possible.
This period of rapid development, which set in soon after the
coming of evolution and multiplied the concrete 269facts of
morphology an hundredfold, may for our present purpose be
conveniently divided into two somewhat overlapping periods, of
which the second may be said to begin with the enunciation by
Haeckel of his Gastræa theory. Within the first period fall
the evolutionary speculations associated with the names of
Kowalevsky, Dohrn, Semper, and others; the characteristic of the
second period is the preponderating influence exercised upon
phylogenetic speculations by the germ-layer doctrine in its two
main evolutionary developments, the Gastræa and Cœlom
theories.
In the first period we might again distinguish two main
tendencies, according as speculations were based mainly upon
anatomical or mainly upon embryological considerations, and it so
happens that these two tendencies are very well illustrated by the
various theories as to the origin of Vertebrates which began to
appear towards the 'seventies. We shall accordingly, in this
chapter, consider very briefly the history of the earlier views on
the phylogeny of the vertebrate stock.
In the early days, before the other claimants to the dignity of
ancestral form to the Vertebrates—Balanoglossus,
Nemertines and the rest—had put in an appearance, there were
two main views on the subject, one upheld by Haeckel, Kowalevsky
and others, to the effect that the proximate ancestor of
Vertebrates was a form somewhat resembling the ascidian tadpole,
the other supported principally by Dohrn and Semper that
Vertebrates and Arthropods traced their descent to a common
segmented annelid or pro-annelid ancestor. The former view is
historically prior, and arose directly out of the brilliant
embryological investigations of A. Kowalevsky, who proved himself
to be a worthy successor of the great comparative embryologist
Rathke. His work was indeed a true continuation of Rathke's. It was
not directly inspired by evolution, though it supplied much useful
confirmation of the theory—you may read Kowalevsky's earlier
memoirs and not realise that they were written several years after
the publication of the Origin of Species.
His first paper of evolutionary importance was a note in 270Russian on
the development of Amphioxus, published in 1865. This subject was
followed up in two papers which appeared in 1867[387] and 1877.[388] In his papers on Amphioxus Kowalevsky made out
the main features in the development of this primitive form, and
showed that the chief organs were formed in essentially the same
way as in Vertebrates; he described the formation of the
archenteron by invagination, the appearance of the medullary folds,
which coalesced to form the neural canal, the formation of the
notochord and of the gill-slits. At first he made the mistake of
supposing that the body-cavity arose from the segmentation-cavity,
but in his later paper he rightly surmised that it was formed from
the cavities of the "primitive vertebræ," or mesodermal
segments. The origin of the notochord from the endoderm was also
not made out by Kowalevsky in his paper of 1867.
Although many important details remained to be discovered by
later investigators,[389] Kowalevsky's work at once made the development
of Amphioxus the key to vertebrate embryology, the typical ontogeny
with which all others could be compared.
Meanwhile, in 1866 and 1871, Kowalevsky had communicated memoirs
of even greater interest,[390] in which he showed that the simple Ascidians
developed in an extraordinarily similar way to Amphioxus and hence
to Vertebrates in general. His proof that Ascidians also develop on
the vertebrate type aroused great interest at the time, and was
naturally acclaimed by the evolutionists as a striking piece of
evidence in favour of their doctrine. The systematic position of
the Ascidians was at that time quite uncertain; they were grouped,
as a rule, with the Mollusca, and certainly no one suspected that
their well-known 271tailed larvæ, first seen by Savigny,
showed any but the most superficial analogy with the tadpoles of
Amphibia. Kowalevsky's papers put a different complexion on the
matter. In the first of them he showed how the nervous system of
the simple Ascidian developed from ectodermal folds just as it did
in Amphioxus and Vertebrates, how gill-slits were formed in the
walls of the pharynx, and how there existed in the ascidian larva a
structure which in position and mode of development was the strict
homologue of the vertebrate notochord. In his second paper he
entered into much more detail, and published some excellent
figures, often reproduced since (see Fig. 13),
but the proof of the affinity between Vertebrates and Ascidians was
in all essentials complete in his paper of 1866.
272
Fig.
13.—Development of the Ascidian Larva. (After
Kowalevsky.)
Kowalevsky's results were accepted by Haeckel, Gegenbaur,
Darwin,[391] and many others as
conclusive evidence of the origin of Vertebrates from a form
resembling the ascidian tadpole; they were extended and amplified
by Kupffer[392] in 1870, later by van
Beneden and Julin[393] and numerous other
workers; they were adversely criticised by Metschnikoff[394] and von Baer,[395] as well as by H. de Lacaze-Duthiers and A.
Giard.[396] Lacaze-Duthiers and von
Baer both held fast to the old view that Ascidians were directly
comparable with Lamellibranch molluscs; they denied the homology of
the ascidian nervous system with that of Vertebrates, von Baer
being at great pains to show that the ascidian nerve-centre was
really ventral in position. He pointed out also that the
"notochord" was confined to the tail of the ascidian larva. Giard's
attitude was by no means so uncompromising, and the criticisms he
passed on the Kowalevsky theory are both subtle and instructive. He
admits that there exists a real homology between, for instance, the
notochord of Vertebrates and that of Ascidians. "But," he adds, "it
is too often forgotten 273that homology does not necessarily mean an
immediate common origin or close relationship. There exist,
doubtless, homologies of great atavistic importance—I
consider as such, for example, the formation of the cavity of
Rusconi [the archenteron] in Ascidians and lower Vertebrates. But
there are also adaptive and purely analogical homologies, such as
the interdigital palmation of aquatic birds, amphibians and
mammals. These are not purely analogous organs, for they can be
superposed one on another, which is not the case with simply
analogous structures (the bat's wing, for example, cannot be
superposed on the bird's wing); they are homologous formations,
resulting from the adaptation of the same fundamental organs to
identical functions. Such is, in my opinion, the nature of the
homology existing between the tail of the ascidian tadpole and that
of Amphioxus or of young amphibians. The ascidian larva, having no
cilia and being necessarily motile, requires for the insertion of
its muscles or contractile organs ... a central flexible axis, a
true chorda dorsalis analogous to that of Vertebrates" (pp. 278-9).
This point of view is strengthened by the fact that in
Molgula, studied by Lacaze-Duthiers, the embryo is
practically stationary, and forms no notochord, nor ever develops
sense-organs in the cerebral vesicle.
Giard's general conclusion is that "the true homology with
Vertebrates ceases after the formation of the cavity of Rusconi and
the medullary groove: the homologies established by Kowalevsky for
the notochord and the relations of the digestive tube and nervous
systems are not atavistic, but adaptive, homologies" (p. 282).
There is accordingly no close genetic relationship between
Ascidians and Vertebrates.
Giard's criticisms did not avail to check the vogue of the new
theory, which soon became an accepted article of faith in most
morphological circles.[397] The fall of the Ascidians from their larval high
estate provided the text for many a Darwinian sermon.
274Some years after the genetic relationship of
Ascidians and Vertebrates had been established, a rival theory of
the origin of Vertebrates made its appearance—a theory which
was practically a rehabilitation in a somewhat altered form of the
old Geoffroyan conception that Vertebrates are Arthropods walking
on their backs. This was the so-called Annelid theory of Dohrn and
Semper. Both Dohrn and Semper started out from the fact that
Annelids and Vertebrates are alike segmented animals, and it was an
essential part of their theory that this resemblance was due to
descent from a common segmented ancestor. Both laid great stress on
the fact that the main organs in Vertebrates are arranged in the
same way as in an Annelid lying on its back, the nervous system
being uppermost, the alimentary system coming next, and below this
the vascular.
Dohrn's earlier views are contained in the fascinating little
book published in 1875, which bears the title Der Ursprung der
Wirbelthiere und das Princip des Functionswechsel (Leipzig). He
followed this up by a long series of studies on vertebrate anatomy
and embryology,[398] in which he
modified his views in certain details. We shall confine our
attention to the first sketch of his theory.
If the Vertebrate is conceived to have evolved from a primitive
Annelid which took to creeping or swimming ventral surface
uppermost, a difficulty at once arises with regard to the relative
positions of the "brain" and the mouth. In Vertebrates the brain,
like the rest of the nervous system, is dorsal to the mouth and the
alimentary canal; in an inverted Annelid, however, the brain is
ventral to the mouth and is connected with the dorsal nerve cord by
commissures passing round the œsophagus. It would seem,
therefore, that the primitive Vertebrate must have acquired either
a new brain or a new mouth. Dohrn took the latter view. He supposed
that the original mouth of the primitive ancestor lay between the
crura cerebelli in the fossa rhomboidea, and that in
Vertebrates this mouth has been replaced functionally by a new
ventrally placed mouth, formed by the 275medial coalescence of a pair
of gill-slits.[399] Probably the two
mouths at one period co-existed, and the older one was ousted by
the growing functional importance of the newer mouth.
The gill-slits were considered by Dohrn to be derived from the
segmental organs of Annelids, which were present originally in
every segment of the primitive ancestor. The gills were at first
external, like the gills of many Chætopods at the present
day. For their support cartilaginous gill-arches naturally arose in
the body-wall, and the superficial musculature became attached to
these bars. "There existed in all the segments of the
Annelid-ancestors of Vertebrates gills with cartilaginous skeleton
and gill-arches in the body wall. Each gill had its veins and
arteries, each had its branch of the ventral nerve-cord, and
between each successive pair of gills a segmental organ opened to
the exterior" (p. 14, 1875). The paired fins and limbs of the
Vertebrate arose by the functional transformation of two pairs of
these gills. The anterior gills became the definitive internal
gills of the Vertebrate, for they gradually shifted into the mouths
of the anterior segmental organs, which had already acquired an
opening into the pharynx and had been transformed into true
gill-slits. The posterior gills degenerated and disappeared, but
their arches remained as ribs. Gill-arches and ribs were
accordingly homologous structures and formed a parietal
skeleton. The vertebrate anus, like the mouth, was probably
secondary and formed from a pair of gill-slits, the post-anal gut
of vertebrate embryos hinting that the original anus was terminal
as in Annelids. The unpaired fins of fish were originally paired
and possibly arose from the coalescence of rows of parapodia. Dohrn
assumed also that the primitive Annelid ancestor must have
possessed a notochord to give support in swimming.
If Vertebrates arose from primitive Annelid ancestors, how
account for Amphioxus and the Ascidians, which seem to 276be the most
primitive living Vertebrates and yet show no particular annelidan
affinities? Dohrn tries to answer this awkward question by showing
that these forms are not primitive but degenerate. He points out
first that Cyclostomes are degenerate fish, half specialised and
half degraded in adaptation to a parasitic mode of life. He thinks
that if an Ammocoetes were to become sexually mature and
degenerate still further, forms would result which would resemble
Amphioxus, and ultimately, if the process of degeneration went far
enough, larval Ascidians. Amphioxus therefore might well be
considered an extremely simplified and degenerate Cyclostome, and
the ascidian larva the last term of this degeneration-series. Both
Amphioxus and the Ascidians would accordingly be descended from
fish, instead of fish being evolved from them.
Dohrn conceived that the transformation of the Annelid into the
Vertebrate took place mainly by reason of an important transforming
principle, which he calls the principle of function-change. Each
organ, Dohrn thinks, has besides its principal function a number of
subsidiary functions which only await an opportunity to become
active. "The transformation of an organ takes place by reason of
the succession of the functions which one and the same organ
possesses. Each function is a resultant of several components, of
which one is the principal or primary function, while the others
are the subsidiary or secondary functions. The weakening of the
principal function and the strengthening of a subsidiary function
alters the total function; the subsidiary function gradually
becomes the chief function, the total function becomes quite
different, and the consequence of the whole process is the
transformation of the organ" (p. 60). Examples of function-change
are not difficult to find. Thus the stomach in most Vertebrates
performs both a chemical and a mechanical function, but in some
forms a part of it specialises in the mechanical side of the work
and becomes a gizzard, while the remaining part confines its
energies to the secretion of the gastric juice. So, too, it is
through function-change that certain of the ambulatory appendages
of Arthropods have become transformed into jaws—their
function as graspers of food has gradually prevailed over their
main 277function as walking limbs. In the evolution
of Vertebrates from Annelids the principle came into action in many
connections—in the formation of a new mouth from gill-slits,
in the transformation of gills into fins and limbs, of segmental
organs into gill-slits, and so on. Dohrn tells us that the
principle of function-change was suggested to him by Mivart's
Genesis of Species (1870), and he points out how it enables
a partial reply to be made to the dangerous objection raised
against the theory of natural selection that the first beginnings
of new organs are necessarily useless in the struggle for
existence.
We may note in passing that a somewhat similar idea was later
applied by Kleinenberg to the explanation of some of the ancestral
features of development. He pointed out in his classical memoir on
the embryology of the Annelid Lopadorhynchus[400] that many embryonic organs seem to be
formed for the sole purpose of providing the necessary stimulus for
the development of the definitive organs. Thus the notochord is the
necessary forerunner of the vertebral column, cartilage the
precursor of bone. "From this point of view," he writes, "many
rudimentary organs appear in a different light. Their obstinate
reappearance throughout long phylogenetic series would be hard to
understand were they really no more than reminiscences of bygone
and forgotten stages. Their significance in the processes of
individual development may in truth be far greater than is
generally recognised. When in the course of the phylogeny they have
played their part as intermediary organs
(Vermittelungsorgane) they assume the same function in the
ontogeny. Through the stimulus or by the aid of these organs, now
become rudimentary, the permanent parts of the embryo appear and
are guided in their development; when these have attained a certain
degree of independence, the intermediary organ, having played its
part, may be placed upon the retired list."[401]
Dohrn was well aware of the functional, or as he calls 278it, the
physiological, orientation of his principle, and he rightly
regarded this as one of its chief merits. He held that morphology
became too abstract and one-sided if it disregarded physiology
completely; he saw clearly that the evolution of function was quite
as important a problem as the evolution of form, and that neither
could be solved in isolation from the other. "The concept of
function-change is purely physiological;" he writes, "it contains
the elements out of which perhaps a history of the evolution of
function may gradually arise, and for this very reason it will be
of great utility in morphology, for the evolutionary history of
structure is only the concrete projection of the content and course
of the evolution of function, and cannot be comprehended apart from
it" (p. 70).[402]
It is very instructive in this connection to note that Dohrn was
not, like so many of his contemporaries, a dogmatic materialist,
but upheld the commonsense view that vital phenomena must, in the
first instance at least, be accepted as they are. "It is for the
time being irrelevant," he writes, "to squabble over the question
as to whether life is a result of physico-chemical processes or an
original property (Urqualität) of all being.... Let us
take it as given" (p. 75).
Semper's speculations on the genetic affinity of Articulates and
Vertebrates are contained in two papers[403] which appeared about the same time as Dohrn's.
He openly acknowledges that his work is essentially a continuation
of Geoffroy's transcendental speculations, and gives in his second
paper a good historical account of the views of his great
predecessor. It is a significant fact that evolutionary
morphologists very generally held that Geoffroy was right in
maintaining against Cuvier[404] the unity of plan of the whole 279animal kingdom,
for they saw in this a strong argument for the monophyletic descent
of all animals from one common ancestral form.
In his first paper Semper does little more than break ground; he
insists on the fact that both Annelids and Vertebrates are
segmented animals, and he points out how close is the analogy
between the nephridia or "segmental organs" of the former and the
excretory (mesonephric) tubules of the latter, upon which he
published in the same volume an extensive memoir. At this time he
considered Balanoglossus—by reason of its gill-slits
(its notochord he did not know)—to be the nearest living
representative of the ancestral form of Vertebrates and
Annelida.
His second paper is a more exhaustive piece of work and deals
with every aspect of the problem, both from an anatomical and from
an embryological standpoint. It is consciously and admittedly an
attempt to apply Geoffroy's principle of the unity of plan and
composition to the three great metameric groups, the Annelida,
Arthropoda, and Vertebrata. Semper follows Geoffroy's lead very
closely in maintaining that it is not the position of the organs
relative to the ground that must be taken into account in
establishing their homologies, but solely their spatial relations
one to another. He holds that dorsum and venter are terms of purely
physiological import, and he proposes to substitute for them the
terms neural and cardial (better, hæmal) surfaces, either of
which may be either dorsal or ventral in position.
Having established this primary principle, Semper has little
difficulty in showing that the main organs of the body lie to one
another in the same relative positions in Annelida, Arthropoda, and
Vertebrata; and this, together with the metameric segmentation
common to them all, constitutes his first great argument in favour
of their genetic relationship. But he has still to show that
Annelids possess at least the rudiments of certain organs which
seem to be peculiar to Vertebrates, as the gill-slits, the
notochord, and a nervous system developed from the ectoderm of the
"dorsal" surface. He takes particular cognisance also of the old
distinction drawn by von Baer, that Vertebrates show a
"double-symmetrical" mode of development (evolutio
bigemina), the 280dorsal muscle-plates forming a tube above the
notochord, the ventral plates a tube below the notochord, whereas
Articulates do not possess this axis, and form only one tube,
namely, that round the "vegetative" organs (evolutio
gemina). Semper is at pains to prove that evolutio
bigemina is characteristic also of Annelidan development.
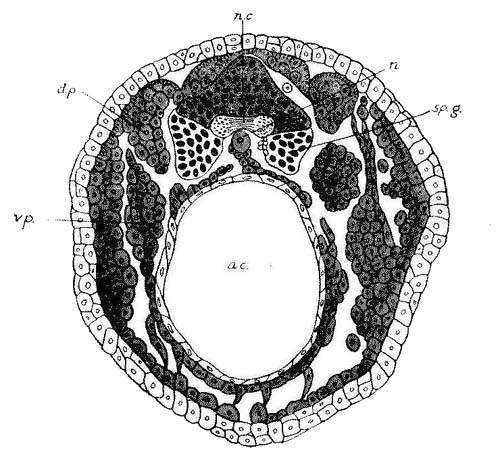
Fig.
14.—Transverse Section (Inverted) of the Worm Nais. (After
Semper.)
| a.c. Alimentary canal. |
sp.g. Spinal ganglion. |
d.p. Neural muscle-plate. |
| n.c. Nerve cord. |
n. Notechord. |
v.p. Haemal muscle-plate. |
He gets his facts from an elaborate study of the process of
budding in the Naidæ, making the somewhat risky
assumption that regeneration takes essentially the same course as
embryonic development.
He succeeds in showing—to his own satisfaction at
least—that in the formation of new segments in Nais
and Chætogaster a strand of cells appears between the
alimentary canal and the nerve-cord, and that from this axial
strand the 281hæmal muscle-plates grow out dorsally
round the alimentary canal and the neural muscle-plates ventrally
round the nerve-cord (see Fig. 14).
This strand of cells, he concludes, must clearly be the
notochord, and the type of development is obviously the
double-symmetrical met with in Vertebrates.
The nervous system Semper found to develop in the buds of
Nais and Chætogaster by an ectodermal
thickening, just as in some Vertebrates. The cerebral ganglion was
formed by the ends of the nerve-cord growing up round the
œsophagus and fusing with the paired "sense-plates" which
develop from the ectoderm of the head. The cerebral ganglion is
accordingly only secondarily hæmal in position, and there is
no need therefore to seek in Vertebrates for the homologue of the
œsophageal commissures of Annelids, as, for instance,
Schneider did.
Since the mouth opens on the neural surface in Annelids and on
the hæmal surface in Vertebrates, Semper considers that they
cannot be equivalent structures, and he finds the homologue of the
Vertebrate mouth in a little pit on the hæmal surface of the
head in the leech Clepsine (also in the true mouth of
Turbellaria and the proboscis-opening in Nemertines). The primitive
Annelid mouth, however, does not appear in the embryogeny of
Vertebrates, for the great development of the brain crowds it out
of existence.
The homologues of the gill-slits Semper finds in two little
canals in the head of Chætogaster, which open from the
pharynx to the exterior. In Sabellids he describes an elaborate
system of gill-canals, with a supporting cartilaginous framework
which forms a real Kiemenkorb or gill-basket, comparable
with that of Amphioxus.
Gill-slits, notochord, relation of nervous system, mesonephric
tubules, are thus common to Annelids and Vertebrates—what
further proof could one desire of the close relationship of these
groups? Yet Semper enters into refinements of comparison, seeing,
for instance, in the lateral portions of the ventral ganglia (Fig. 14, sp. g.) the homologues of the
spinal ganglia of Vertebrates, and comparing the lateral line of
sense organs in Annelids with the lateral line in Anamnia.
He will not admit that Amphioxus and the Ascidians 282show a closer
resemblance to Vertebrates than his beloved Annelids. Amphioxus, he
thinks, is not a Vertebrate, and Ascidians, though sharing with
Annelids the possession of a notochord, gill-slits, and a "dorsal"
nervous system, yet are further removed from Vertebrates than the
latter by reason of their lacking that essential characteristic of
Vertebrates, metameric segmentation.
Not content with establishing the unity of plan of Annelids,
Arthropods, and Vertebrates, Semper tries to link on the Annelids,
as the most primitive group of the three, to the unsegmented worms,
and particularly to the Turbellaria. His speculations on this
matter may be summed up somewhat as follows:—The common
ancestor of all segmented animals is a segmented worm-like form,
not quite like any existing type, resembling the Turbellaria in
having two nerve strands on the dorsal side and no œsophageal
ring, potentially able to develop either the Vertebrate or the
Annelid mouth, and so to give origin both to the Articulate and to
the Vertebrate series. The common ancestor alike of unsegmented
worms and of all segmented types is probably the trochosphere
larva, which in the Vertebrates is represented by the simple
Keimblase or blastula.
The Annelid theory of Dohrn and Semper was perhaps not so widely
accepted as the rival Ascidian theory, but it counted not a few
adherents and gave a certain stimulus to comparative morphology.
F. M. Balfour, who pointed out about the same time as Semper the
analogy between the nephridia of Annelids and the mesonephric
tubules of Vertebrates,[405] while not accepting the actual theories of Dohrn
and Semper, took up a distinctly favourable attitude to the general
idea that Annelids and Vertebrates were descended from a common
segmented ancestor. Discussing this question in his classical work
on the development of Elasmobranch fishes,[406] Balfour came to the conclusion "that we must
look for the ancestors of the Chordata, not in allies 283of the present
Chætopoda, but in a stock of segmented forms descended from
the same unsegmented types as the Chætopoda, but in which two
lateral nerve-cords, like those of Nemertines, coalesced dorsally
instead of ventrally to form a median nervous cord. This group of
forms, if my suggestion as to their existence is well founded,
appears now to have perished."[407]
He held that while there was much to be said for the interchange
of dorsal and ventral surfaces postulated by Dohrn and Semper, the
difficulties involved in the supposition were too great; he
preferred, therefore, to assume that the present Vertebrate mouth
was primitive, and not a secondary formation.
His views as to the phylogeny of the Chordata and the genetic
relation of the various classes to one another are exhibited in the
following schema,[408] names of
hypothetical groups being printed in capitals, names of degenerate
groups in italics:—
284The hypothetical ancestral forms
(Protochordata) possessed a notochord, a ventral suctorial mouth
and numerous gill-slits, and were presumably descended from the
common ancestor of Annelids and Vertebrates. Amphioxus and the
Ascidians found their place in this schema as degenerate offshoots
of the ancestral Protochordates, while the Cyclostomes were in the
same way the degenerate modern representatives of the ancestral
Protovertebrates.
Balfour's suggestion, that the nervous system in Annelids and
Vertebrates might have arisen by the dorsal or ventral coalescence
of the lateral nerve cords found in their common ancestor, bore
fruit in the speculations of Hubrecht,[409] on the relation of Nemertines to
Vertebrates.
The Annelid theory was firmly supported by Eisig, who in his
elaborate monograph on the Capitellidæ[410] maintained against Fürbringer the
genetic identity of the Annelidan nephridia with the kidney tubules
of Vertebrates. The independent discovery by E. Meyer[411] and J. T. Cunningham,[412] of an internal segmental duct in
Lanice, into which several nephridia opened, seemed to
strengthen this view.
Following Ehlers,[413] Eisig found the homologue of the notochord in
the accessory intestine of the Capitellidæ and
Eunicidæ, which he supposed might easily be
transformed, according to the principle of function-change, from a
respiratory to a supporting organ. He finally disposed of the
alternative notion that the notochord was represented in Annelids
by the "giant-fibres" or neurochordal strands which lie close above
the nerve-cord, a view held by Kowalevsky,[414] and for a time by Semper. These strands were
285shown by Eisig, and by Spengel, to be the
neurilemmar sheaths of thick nerve fibres which had in many cases
degenerated. The view that the content of the neurochordal tubes
was nervous in nature was first promulgated by Leydig in 1864.
Much difference of opinion reigned as to the true homologies of
the brain and mouth of Annelids and Vertebrates. Beard[415] and others got over the difficulty of the
hæmal position of the cerebral ganglion in Annelids by
supposing that it degenerated and disappeared altogether in the
Annelidan ancestor of Vertebrates, and that accordingly it had no
homologue in the Vertebrate nervous system. Beard put forward also
the ingenious theory that the hypophysis represents the old
Annelidan mouth.
Van Beneden and Julin[416] assumed that in the ancestors of Vertebrates the
œsophagus shifted forward between the still unconnected lobes
of the brain to open on the hæmal surface.
The fundamental assumption of the Annelid theory, that dorsal
and ventral surfaces are morphologically interchangeable, seemed
rather bold to many zoologists, and Gegenbaur[417] voiced a common opinion when he rejected
as unscientific the comparison of the ventral nerve cord of
Articulates with the dorsal nervous system of Vertebrates.
The Balanoglossus theory of Vertebrate descent also
belongs, at least in its first form, to the earlier group of
evolutionary speculations. The gill-slits of Balanoglossus
were discovered by Kowalevsky as early as 1866.[418] Tornaria was discovered by J.
Müller in 1850, but by him considered an Asterid larva; its
true nature as the larva of Balanoglossus was made out by
Metschnikoff in 1870, who also remarked upon its extraordinary
likeness to the larvæ of Echinoderms.[419] 286That it had some relationship with
Vertebrates was recognised by Semper, Gegenbaur and others, but the
full working-out of its Vertebrate affinities is due to Bateson.[420]
Bateson broke completely with the Dohrn-Semper view that the
metamerism of Articulates and Vertebrates must be put down to
inheritance from a common ancestor. He held that metamerism was
merely a special manifestation of the general property of
repetition, common to all living things (cf. Owen's
"vegetative force"), and that accordingly "however far back a
segmented ancestor of a segmented descendant may possibly be found,
yet ultimately the form has still to be sought for in which these
repetitions had their origin" (p. 549). The meaning of the
phenomenon was obscure, but he was convinced that the explanation
was not to be found in ancestry. "This much alone is clear," he
wrote, "that the meaning of cases of complex repetition will not be
found in the search for an ancestral form, which, itself presenting
this same character, may be twisted into a representation of its
supposed descendant. Such forms there may be, but in finding them
the real problem is not even resolved a single stage; for from
whence was their repetition derived? The answer to this question
can only come in a fuller understanding of the laws of growth and
of variation, which are as yet merely terms" (pp. 548-9). It was in
following up this line of thought that Bateson produced his
monumental Materials for the Study of Variation (1894).
He found a strong positive argument for his theory that
Vertebrates are descended from unsegmented forms in the fact that
the notochord arises as an unsegmented structure. With the
notochord he homologised the supporting rod in the proboscis of
Balanoglossus, which like the notochord arises from the
dorsal wall of the archenteron, and has a vacuolated structure. The
gill-slits of Balanoglossus, with their close resemblance in
detail to those of Amphioxus, Bateson also used as an argument in
favour of the phylogenetic relationship of the Enteropneusta and
Vertebrata, 287together with the formation from the ectoderm
of a dorsal nerve tube.
Bateson's views attracted considerable attention, and were
thought by many to lighten appreciably the obscurity in which the
origin of Vertebrates was wrapped. Thus Lankester wrote in his
article on Vertebrates[421] in the Encyclopedia Britannica:—"It
seems that in Balanoglossus we at last find a form which,
though no doubt specialised for its burrowing sand-life, and
possibly to some extent degenerate, yet has not to any large extent
fallen from an ancestral eminence. The ciliated epidermis, the long
worm-like form, and the complete absence of segmentation of the
body-muscles lead us to forms like the Nemertines. The great
proboscis of Balanoglossus may well be compared to the
invaginable organ similarly placed in the Nemertines. The collar is
the first commencement of a structure destined to assume great
importance in Cephalochorda and Craniata, and perhaps
protective of a single gill-slit in Balanoglossus before the
number of those apertures had been extended. Borrowing, as we may,
the nephridia from the Nemertines, and the lateral in addition to
the dorsal nerve, we find that Balanoglossus gives the most
hopeful hypothetical solution of the pedigree of Vertebrates."
Much doubt was cast upon the Chordate affinities of the
Enteropneusta by Spengel in his monograph of the group,[422] but when the development of the
cœlom came to be more thoroughly worked out in
Balanoglossus and Amphioxus, the striking resemblance in
this respect between the two forms gave additional support to the
Batesonian view.[423]
288
CHAPTER XVI
THE GERM-LAYERS AND EVOLUTION
In his papers of 1866 and 1867
Kowalevsky had remarked upon the widespread occurrence of a certain
type or fundamental plan of early embryonic development,
characterised by the formation, through invagination, of a
two-layered sac, whose cavity became the alimentary canal. This
developmental archetype was manifested in, for instance,
Sagitta,[424] Rana,[425] Lymnæa,[426] Astacus,[427] Phoronis,[428] Asterias,[429] Ascidia,[428] the Ctenophora,[428] and Amphioxus.[428] He noticed also that the
invagination-opening often became the definitive anus. Further
instances of this mode of development were later observed by
Metschnikoff[430] and by Kowalevsky[431] himself, but it was left
to Haeckel to generalise these observations and build up from them
his famous Gastræa theory. This was first enunciated in his
monograph of the calcareous sponges,[432] and worked out in detail in a series of papers
published in 1874-76.[433]
289Haeckel maintained that the "gastrula" stage
occurred in the development of all Metazoa, and that it was
typically formed, by invagination, from a hollow sphere of cells or
"blastula." This typical formation might be masked by cenogenetic
modifications caused chiefly by the presence of yolk. The gastrula
stage was the palingenetic repetition of the ancestral form of all
Metazoa, the Gastræa.
From the Gastræa theory there followed at once two
consequences, (1) that ectoderm and endoderm, invagination-cavity
(Urdarm) and gastrula-mouth (Urmund or
Protostoma), were, with all their derivatives, homologous,
because homogenous, throughout the Metazoa, and (2) that the
descent of the Metazoa had been monophyletic, since all were
derived from the ancestral Gastræa. Huxley's suggestion
(supra, p. 208) that the outer and inner layers in
Cœlentera were homologous with the ectoderm and endoderm of
the germ was thus fully confirmed and greatly extended.
The great importance of the Gastræa theory lay in the fact
that it linked up, by means of the biogenetic law, the germ-layer
theory with the doctrine of evolution. It supplied an evolutionary
interpretation of the earliest and most important of embryogenetic
events, the process of layer-formation. Upon the Gastræa
theory or its implications were founded most of the phylogenetic
speculations which subsequently appeared.
Upon the Gastræa theory Haeckel based a system of
phylogenetic classification which was intended to replace Cuvier's
and von Baer's doctrine of Types. This took the form of a
monophyletic ancestral tree. Its main outlines are given on p. 290
in graphic form, combined and modified from the table on p. 53 of
the 1874 paper and the genealogical tree given in the
Kalkschwämme.[434]
290
Monophyletic Genealogical Tree of the Animal
Kingdom, based upon the Gastræa Theory and the Homology of
the Germ Layers.
The scheme is in many respects an interesting and important one.
The great contrast between the Protozoa, or animals with neither
gut nor germ-layers, and the Metazoa, 291which possess both
structures, is for the first time clearly brought out. The
derivation of all the Metazoa from a single ancestral form, the
Gastræa, leads to the conclusion that the types are not
distinct from one another as Cuvier and von Baer supposed, but
agree in the one essential point, in the possession of an
archenteron (Lankester, 1875), and an ectoderm and endoderm
which are homologous throughout all the Metazoan phyla. Finally, in
the separation of the sponges, Cœlenterata and Acœlomi
as animals lacking a body cavity or cœlom[435] from the four higher
phyla, which are essentially Cœlomati, there is contained the
germ of a conception which later became of importance.
Somewhat similar views as to the importance of the germ-layer
theory for the phylogenetic classification of animals were
published by Sir E. Ray Lankester in 1873.[436] He distinguished three
grades of animals—the Homoblastica, Diploblastica, and
Triploblastica. The first included the Protozoa, the second the
Cœlenterata, the third the other five phyla, distinguished by the
possession of a third layer, the mesoderm, and a "blood-lymph"
cavity enclosed therein. He used the germ-layer theory to prove the
essential unity of type of all the Triploblastica.
The Gastræa theory gave point and substance to the
biogenetic law, and enabled Haeckel to state much more concretely
the parallelism existing between ontogeny and phylogeny. He was
able to assert that five primordial stages, each representing a
primitive ancestral form, recurred with regularity in the very
earliest development of all Metazoa.[437] These were the monerula, cytula, morula,
blastula, and gastrula (see Fig. 15). The
monerula was the fertilised ovum after the disappearance of the
germinal vesicle;[438] it was the equivalent of the primordial
anucleate Monera which are the ancestors of all animals.
292
Fig. 15.—The
Five Primary Stages of Ontogeny. (After Haeckel.)
| 1. Monerula. |
2. Cytula. |
3. Morula. |
4. Blastula. |
5. Gastrula. |
293The ovum after the nucleus had been re-formed became the cytula, which
was the ontogenetic counterpart of the amœba. The morula, a
compact mulberry-like congeries of segmentation-cells, corresponded
to the synamœba, or earliest association of undifferentiated
amœboid cells to form the first multicellular organism. The
blastula, or hollow sphere of segmentation cells, usually ciliated,
was reminiscent of the planæa, an ancestral free-swimming
form whose nearest living relation is the spherical
Magosphæra. The gastrula, finally, is the two-layered
sac formed from the blastula, typically by invagination of its
wall. It repeats the organisation of the gastræa, which is
the common ancestor of all Metazoa, and finds its nearest living
counterpart in the simple "sponges" Haliphysema and
Gastrophysema.[439] The ancestral line of all the higher animals
begins with the five hypothetical forms of the moneron,
amœba, synamœba, planæa, and gastræa.
We may take the following account[440] of the phylogeny of the human species, from the
gastræa stage onwards, as typical of Haeckel's speculations
on the evolution of the higher forms. The progenitors of man are,
after the Gastræada:—
1. Turbellaria.
*2. Scolecida. (Worms with a
cœlom, probably represented at the present day by
Balanoglossus.)
*3. Himatega. (Evolved from Scolecida by
formation of dorsal nerve-tube and chorda, and resembling tailed
larvæ of Ascidians.)
4. Acrania. (With metameric
segmentation. Including Amphioxus.)
5. Monorrhina.
(Cyclostomes.)
6. Selachia.
7. Dipneusta.
8. Sozobranchia. (Amphibia
with permanent gills.)
2949. Sozura. (Tailed
Amphibia.)
*10. Protamnia.
*11. Promammalia.
12. Marsupialia.
13. Prosimiæ.
14. Menocerca. (Tailed apes.)
15. Anthropoides.
16. Pithecanthropi.
17. Homines.
It will be noticed that except for the hypothetical forms
(marked with an asterisk), which are themselves generalised
classificatory groups, the ancestral forms belong to
long-recognised classes. The whole course of the evolution follows
well-worn systematic lines. This is typical of Haeckel's
phylogenetic speculations.
A more abstractly morphological scheme of the evolution of
Vertebrates is given in the Systematic Phylogeny of 1895.[441] The ontogenetic and
ancestral stages are arranged in parallel columns thus:—
| Cytula. |
|
Cytæa (Protozoa). |
| Morula. |
|
Moræa (Cœnobium of
Protozoa). |
| Blastula. |
|
Blastæa (Volvocina,
etc.). |
| Depula (invaginated blastula). |
|
Depæa. |
| Gastrula. |
|
Gastræa (cf. Olynthus,
Hydra, and primitive Coelentera). |
| Cœlomula (with one pair of
cœlom-pockets). |
|
Cœlomæa (cf. Sagitta,
Ascidia, and primitive Helminthes). |
| Chordula (with medullary tube and
chorda). |
|
Chordæa (cf. Ascidian larva
and larva of Amphioxus). |
| Spondula (with segmented mesoderm). |
|
Prospondylus (Primitive Vertebrate). |
This scheme differs from the earlier one chiefly in taking into
account certain advances, notably as regards the cytology of the
fertilised ovum and the true nature of the cœlom, which had
been made in the interval of some twenty years.
Haeckel's Gastræa theory, though it exercised a great
influence upon the subsequent trend of phylogenetic speculation,
was by no means universally accepted telle quelle. Opinions
differed considerably as to the primitive mode of 295origin of the
two-layered sac which was very generally admitted to be of constant
occurrence in early embryogeny. Ray Lankester, in his paper of
1873, and more fully in 1877,[442] propounded a "Planula" theory, according to
which the ancestral form of the Metazoa was a two-layered closed
sac formed typically by delamination, less often by invagination.
He denied that the invagination opening (which he named the
blastopore) represented the primitive mouth,[443] holding that this was typically formed by
an "inruptive" process at the anterior end of the planula, which
led to the formation of a "stomodæum." A similar process at
the posterior end gave rise to the anus and the
"proctodæum."
The question as to whether delamination or invagination was to
be considered the more primitive process was discussed in detail by
Balfour,[444] without, however, any
very definite conclusion being reached. He held that both processes
could be proved in certain cases to be purely secondary or
adaptive, and that accordingly there was nothing to show that
either of them reproduced the original mode of transition from the
Protozoa to the ancestral two-layered Metazoa (p. 342). He by no
means rejected the theory that the Gastræa, "however evolved,
was a primitive form of the Metazoa," but, having regard to the
great variations shown in the relation of the blastopore to mouth
and anus (pp. 340-1), he was inclined to think that if the gastrula
had any ancestral characters at all, these could only be of the
most general kind. Balfour's attitude perhaps best represents the
general consensus of opinion with regard to the Gastræa
theory.
From the same origins as the Gastræa theory arose the
theory of the cœlom. The term dates back to Haeckel in 1872,
and the observations which first led up to the theory were made by
the men who supplied the foundations of the Gastræa
theory—A. Agassiz, Metschnikoff and Kowalevsky. 296But it was not
Haeckel himself who enunciated the cœlom theory.
It will be remembered that Remak introduced in 1855 the
conception of the mesoderm as an independent layer derived from the
endoderm. The pleuro-peritoneal or body-cavity was formed as a
split in the "ventral plates" of the mesoderm. Haeckel's
"cœlom" corresponded to the "pleuro-peritoneal cavity" of
Remak, but his view of the origin of the mesoderm brought him much
closer to von Baer's conception of the origin of two
secondary layers from ectoderm and endoderm respectively than to
Remak's conception of the mesoderm as a single independent
layer.
Much uncertainty reigned at the time as to the exact manner of
origin of the mesoderm;[445] some held that it developed from the ectoderm,
others that it originated in the endoderm, while still others, and
among them Haeckel, considered that part of it came from the
ectoderm and part from the endoderm (pp. 23-4, 1874).
The solution of the problem came from those observations on the
development of the lower forms to which we have just alluded.
The early history of these discoveries and of the theory which
grew out of them has been well summarised by Lankester,[446] and may conveniently be given in his own
words:—
"As far back as 1864 Alexander Agassiz ("Embryology of the
Star-fish," in Contributions to the Natural History of the
United States, vol. v., 1864) showed in his account of the
development of Echinoderma that the great body-cavity of those
animals developed as a pouch-like outgrowth of the archenteron of
the embryo, whilst a second outgrowth gave rise to their ambulacral
system; and in 1869 Metschnikoff (Mém. de l'Acad.
impériale des Sciences de St Pétersbourg, series
vii., vol. xiv., 1869), confirmed the observations of Agassiz, and
showed that in Tornaria (the larva of Balanoglossus) a similar
formation of body-cavities by pouch-like outgrowths of the
archenteron took place. 297Metschnikoff has further the credit of
having, in 1874 (Zeitsch. wiss. Zoologie, vol. xxiv., p. 15,
1874), revived Leuckart's theory of the relationship of the
cœlenteric apparatus of the Enterocœla to the digestive
canal and body-cavities of the higher animals. Leuckart had in 1848
maintained that the alimentary canal and the body-cavity of higher
animals were united in one system of cavities in the
Enterocœla (Verwandschaftsverhältnisse der
wirbellosen Thiere, Brunswick, 1848). Metschnikoff insisted
upon such a correspondence when comparing the Echinoderm larva,
with its still continuous enteron and cœlom, to a Ctenophor,
with its permanently continuous system of cavities and canals.
Kowalevsky, in 1871, showed that the body-cavity of Sagitta was
formed by a division of the archenteron into three parallel
cavities, and in 1874 demonstrated the same fact for the
Brachiopoda. In 1875 (Quart. Journ. Micr. Sci., vol. xv., p.
52) Huxley proposed to distinguish three kinds of body-cavity: the
schizocœl, formed by the splitting of the mesoblast, as in
the chick's blastoderm; the enterocœl, formed by pouching of
the archenteron, as in Echinoderms, Sagitta and Brachiopoda; and
the epicœl.... Immediately after this I put forward the
theory of the uniformity of origin of the cœlom as an
enterocœl (Quart. Journ. Micr. Sci., April, 1875)....
My theory of the cœlom as an enterocœl was accepted by
Balfour and was greatly strengthened by his observations on the
derivation of both notochord and mesoblastic somites from
archenteron in the Elasmobranchs, and by the publication in 1877 by
Kowalevsky of his second paper on the development of
Amphioxus—in which the actual condition which I had supposed
to exist in the Vertebrata was shown to occur, namely, the
formation of the mesoblast as paired pouches in which a narrow
lumen exists, but is practically obliterated on the nipping-off of
the pouch from the archenteron, after which process it opens out
again as cœlom" (pp. 16-18).
The enterocœlic theory was taken up by O. and R. Hertwig
as an essential part of their Cœlomtheorie.[447] In 298a lengthy series of monographs these
workers made a comparative study of the mode of formation of the
middle layer, and arrived at a coherent theory of its origin. They
distinguished in the middle layer two quite distinct elements, the
mesoblast proper, formed by the evagination of the walls of the
archenteron, and the mesenchyme, formed by free cells budded off
from the germ-layers. The following passage gives a good idea of
their views and of the phylogenetic implications
involved:—"Ectoblast and entoblast are the two primary
germ-layers which arise from the invagination of the blastula; they
are always the first to be laid down, and they can be directly
referred back to a simple ancestral form, the Gastræa; they
form the limits of the organism towards the exterior and towards
the archenteron. The parietal and visceral mesoblast, or the two
middle layers, are always of later origin, and arise through
evagination or plaiting of the entoblast, the remainder of which
can now be distinguished as secondary entoblast from the primary.
They form the walls of a new cavity, the enterocœl, which is
to be regarded as a nipped-off diverticulum of the archenteron.
Just as the two-layered animals can be derived from the
Gastræa, so can the four-layered animals be derived from a
Cœlom form. Embryonic cells, which become singly detached
from their epitheliar connections we consider to be something quite
different from the germ-layers, and accordingly we call them by the
special name of mesenchyme germs or primary cells of the
mesenchyme. They may develop both in two-layered and in
four-layered animals. Their function is to form between the
epithelial limiting layers a secreted tissue (Secretgewebe)
or connective tissue with scattered cells, which cells can undergo,
like the epithelial elements, the most varied modifications....
This secreted tissue in its simple or in its differentiated state,
with all its derivatives, we call the mesenchyme" (p. 122).
The important point for us is that, just as all Metazoa were
considered by Haeckel to be descended from the Gastræa, so
all Cœlomati were held by the Hertwigs to be derived from an
original cœlomate Urform. In both cases an
embryological archetype becomes a hypothetical ancestral form.
The Cœlom theory was considerably modified, extended 299and
developed by later workers, particularly as regards the relations
to the cœlom of the genital organs and ducts and the nephridia,
but no special methodological interest attaches to these further
developments.[448] We shall here focus
attention upon one interesting line of speculation followed out in
this country particularly by Sedgwick—the theory of the
Actinozoan ancestry of segmented animals. Its relation to the
Cœlom theory lies in the fact that Sedgwick regarded the
segmentation of the body as moulded upon the segmentation of the
mesoblast, which in its turn, as Kowalevsky and Hatschek had shown,
was a consequence of its mode of origin as a series of pouches of
the archenteron. In other respects Sedgwick's speculations link on
more closely to the Gastræa theory, for one of his main
contentions is that the blastopore or Urmund is homologous
throughout at least the three metameric phyla. In following up
Balfour's observations on the development of Peripatus,[449] Sedgwick was struck with
the close resemblance existing between the elongated slit-like
blastopore of this form (giving rise to both mouth and anus), with
its border of nervous tissue, and the slit-like mouth of the
Actinozoan (functioning both as mouth and anus), round which, as
the Hertwigs had shown, there lies a special concentration of nerve
cells and nerve fibres. He found another point of resemblance in
the gastric pouches of the Actinozoa, which he homologised directly
with the enterocœlic pouches of the Cœlomati. He was
led to enunciate the following theses:—[450] (1) that the mouth and anus of Vermes,
Mollusca, Arthopoda, and probably Vertebrata, is derived from the
elongated mouth of an ancestor resembling the Actinozoa; (2) that
somites are derived from a series of archenteric pouches, like
those of Actinozoa and Medusæ; (3) that excretory organs
(nephridia, segmental organs) are derived from parts of these
pouches which in the ancestral form, as in many polyps, were
connected by a circular or longitudinal canal, and opened 300to the
exterior by pores. This longitudinal canal was lost in
Invertebrates, but persisted in Vertebrates as the pronephric duct,
while the pores remained in Invertebrates and disappeared in
Vertebrates; (4) that the tracheæ of Arthropods, as well as
the canal of the central nervous system in Vertebrates, are to be
traced back to certain ectodermal pits in the diploblastic ancestor
comparable to the sub-genital pits of the Scyphomedusæ. These
ectodermal pits were all originally respiratory organs. "The
essence of all these propositions," he writes, "lies in the fact
that the segmented animals are traced back not to a triploblastic
unsegmented ancestor, but to a two-layered Cœlenterate-like
animal with a pouched gut, the pouching having arisen as a result
of the necessity for an increase in the extent of the vegetative
surfaces in a rapidly enlarging animal (for circulation and
respiration)" (p. 47). "I have attempted to show," he writes
further on, "that the majority of the Triploblastica ... are built
upon a common plan, and that that plan is revealed by a careful
examination of the anatomy of Cœlenterata; that all the most
important organ-systems of these Triploblastica are found in a
rudimentary condition in the Cœlenterata; and that all the
Triploblastica referred to must be traced back to a diploblastic
ancestor common to them and the Cœlenterata" (p. 68). The main
assumption was that the neural or blastoporal surface must be
homologous throughout the Metazoa, though it was dorsal in the
Chordata, ventral in the Annelida and Arthropoda. He derived the
central nervous system of the Chordata from the circumoral ring of
the common ancestor by means of the hypothesis that both the
pre-blastoporal and the post-blastoporal parts of it disappeared.[451]
The characteristic relation of the central nervous system to the
blastopore in Annelida and Vertebrates had already been pointed out
by Kowalevsky,[452] who had also
sketched a theory of the common descent of these two phyla from an
ancestral form in which the nervous system encircled the
blastopore.
301In 1882, before the publication of Sedgwick's
papers, A. Lang[453] had put forward the
somewhat similar view that the stomach-diverticula of the
Turbellaria, which he had found to be segmentally arranged in
certain Triclads, were the morphological equivalents of the
enterocœlic pouches of higher animals. This view, however, he
soon gave up.[454] Sedgwick's views
found a supporter in A. A. W. Hubrecht,[455] who utilised them in connection both with his
speculations on the relation of Nemertines to Vertebrates, and with
his exhaustive work on the early development of the Mammalia. He
postulated as the far-back ancestor of Vertebrates, "an
actinia-like, vermiform being, elongated in the direction of the
mouth-slit" (p. 410, 1906), and derived the central nervous system
from the circum-oral ring of this primitive form, the notochord
from its stomodæum, and the cœlom from the peripheral
parts of the gastric cavity (p. 169, 1909).
302
CHAPTER XVII
THE ORGANISM AS AN HISTORICAL BEING
"Of late the attempt to arrange
genealogical trees involving hypothetical groups has come to be the
subject of some ridicule, perhaps deserved. But since this is what
modern morphological criticism in great measure aims at doing, it
cannot be altogether profitless to follow this method to its
logical conclusions. That the results of such criticism must be
highly speculative, and often liable to grave error, is
evident."
The quotation is from Bateson's paper of 1886, and it is
symptomatic of the change which was soon to come over morphological
thought. New interests, new lines of work, began to usurp the place
which pure morphology had held so long.
This is accordingly a convenient stage at which to take stock of
what has gone before, to consider the relation of evolutionary
morphology to the transcendental and the Cuvierian schools of
thought which preceded it, and to make clear what new element
evolution-theory added to morphology.
The close analogy between evolutionary and transcendental
morphology has already been remarked upon and illustrated in the
last three chapters. We have seen that the coming of evolution made
comparatively little difference to pure morphology, that no new
criteria of homology were introduced, and that so far as pure
morphology was concerned, evolution might still have been conceived
as an ideal process precisely as it was by the transcendentalists.
The principle of connections still remained the guiding thread of
morphological work; the search for archetypes, whether anatomical
303or
embryological, still continued in the same way as before, and it
was a point of subordinate importance that, under the influence of
the evolution-theory, these were considered to represent real
ancestral forms rather than purely abstract figments of the
intelligence. The law of Meckel-Serres was revived in an altered
shape as the law of the recapitulation of phylogeny by ontogeny;
the natural system of classification was passively inherited, and,
by a petitio principii, taken to represent the true course
of evolution. It is true that the attempt was made to substitute
for the concept of homology the purely genetic concept of homogeny,
but no inkling was given of any possible method of recognising
homogeny other than the well-worn methods generally employed in the
search after homologies.
There was a close spiritual affinity between the speculative
evolutionists and the transcendentalists. Both showed the same
subconscious craving for simplicist conceptions—the
transcendentalists clung fast to the notion of the absolute unity
of type, of the ideal existence of the "one animal," and the
evolutionists did precisely the same thing when they blindly and
instinctively accepted the doctrine of the monophyletic descent of
all animals from one primeval form. Geoffroy persisted in regarding
Arthropods as being built on the same plan as Vertebrates: Dohrn
and Semper did nothing different when they derived both groups from
an ancestor combining the main characters of both. The
determination to link together all the main phyla of the animal
kingdom and to force them all into a single mould was common to
evolutionary and pre-evolutionary transcendentalists alike.
From the fact that all Metazoa develop from an ovum which is a
simple cell, the evolutionists inferred that all must have arisen
from one primordial cell. From the fact that the next step in
development is the segmentation of the ovum, they argued that the
ancestral Metazoa came into being through the division of the
primal Protozoon with aggregation of the division-products. From
the fact that a gastrula stage is very commonly formed when
segmentation has been completed, they assumed that all germ-layered
animals were descended from an ancestral Gastræa.
304They quite ignored the possibility that a
different explanation of the facts might be given; they seized upon
the simplest and most obvious solution because it satisfied their
overwhelming desire for simplification. But is the simplest
explanation always the truest—especially when dealing with
living things? One may be permitted to doubt it. It is easy to
account for the structural resemblance of the members of a
classificatory group, by the assumption that they are all descended
from a common ancestral form; it is easy to postulate any number of
hypothetical generalised types; but in the absence of positive
evidence, such simplicist explanations must always remain doubtful.
The evolutionists, however, had no such scruples.
Phylogenetic method differed in no way from
transcendental—except perhaps that it had learnt from von
Baer and from Darwin to give more weight to embryology. The
criticisms passed by Cuvier and von Baer upon the
transcendentalists and their recapitulation theory might with equal
justice be applied to the phylogenetic speculations which were
based on the biogenetic law. There was the same tendency to fix
upon isolated points of resemblance and disregard the rest of the
organisation. Thus, on the ground of a presumed analogy of certain
structures to the vertebrate notochord, several invertebrate
groups, as the Enteropneusta, the Rhabdopleura, the Nemertea, were
supposed to be, if not ancestral, at least offshoots from the
direct line of vertebrate descent. And if other points of
resemblance could in some of these cases be discovered, yet no
successful attempt was made to show that the total organisation of
any of these forms corresponded with that of the Vertebrate type.
With the possible exception of the Ascidian theory, all the
numerous theories of vertebrate descent suffered from this
irremediable defect, and none carried complete conviction.
In spite of the efforts of the evolutionists, as of those of the
transcendentalists, the phyla or "types" remained distinct, or at
best connected by the most general of bonds.
The close affinity of transcendentalists and evolutionists is
shown very clearly in their common contrast in habits of 305thought
with the Cuvierian school. It is the cardinal principle of pure
morphology that function must be excluded from consideration. This
is a necessary and unavoidable simplification which must be carried
out if there is to be a science of pure form at all. But this
limitation of outlook, if carried over from morphology to general
biology becomes harmful, since it wilfully ignores one whole side
of life—and that the most important. The functional point of
view is clearly indispensable for any general understanding of
living things, and this is where the Cuvierian school has the
advantage over the transcendental—its principles are
applicable to biology in general.
Geoffroy and Cuvier in pre-evolutionary times well typified the
contrast between the formal and the functional standpoints. For
Geoffroy form determined function, while for Cuvier function
determined form. Geoffroy held that Nature formed nothing new, but
adapted existing "materials of organisation" to meet new needs.
Cuvier, on the other hand, was always ready to admit Nature's power
to form entirely new organs in response to new functional
requirements.
The evolutionists followed Geoffroy rather than Cuvier. They
laid great store by homological resemblances, and dismissed
analogies of structure as of little interest. They were singularly
unwilling to admit the existence of convergence or of parallel
evolution, and they held very firmly the distinctively Geoffroyan
view that Nature is so limited by the unity of composition that she
can and does form no new organs.
By no one has this underlying principle of evolutionary
morphology been more explicitly recognised than by Hubrecht, who in
his paper of 1887, after summarising the points of resemblance
between Nemertines and Vertebrates which led him to assume a
genetic connection between them, writes as follows:—"At the
base of all the speculations contained in this chapter lies the
conviction, so strongly insisted upon by Darwin, that new
combinations or organs do not appear by the action of natural
selection unless others have preceded, from which they are
gradually derived by a slow change and differentiation.
306"That a notochord should develop out of the
archenteric wall because a supporting axis would be beneficial to
the animal may be a teleological assumption, but it is at the same
time an evolutional heresy. It would never be fruitful to try to
connect the different variations offered, e.g., by the
nervous system throughout the animal kingdom, if similar
assumptions were admitted, for there would be then quite as much to
say for a repeated and independent origin of central nervous
systems out of indifferent epiblast just as required in each
special case. These would be steps that might bring us back a good
way towards the doctrine of independent creations. The remembrance
of Darwin's, Huxley's, and Gegenbaur's classical foundations, and
of Balfour's and Weismann's brilliant superstructures, ought to
warn us away from these dangerous regions" (p. 644).
This same prejudice lies at the root of the idea of
Functionswechsel, in spite of the general functional
orientation of that idea.
Dohrn's constant assumption is that Nature makes shift with old
organs wherever possible, instead of forming new ones. He derives
gill-slits from segmental organs, fins and limbs from gills, ribs
from gill-arches, and so on, instead of admitting that these organs
might quite as well have arisen independently. He objects on
principle to the origin of organs de novo. Thus, rebutting
the suggestion that certain organs which are not found in the lower
Vertebrates might have arisen as new formations, he
writes:—"Against this supposition the whole weight of all
those objections can be directed that are to be brought in general
against the method of explanation which consists in appealing
without imperative necessity to the Deus ex machina, 'New
formation,' which is neither better nor worse than Generatio
equivoca" (p. 21).
Of a similar nature was the objection to convergence.[456]
Why, we may ask, were morphologists so unwilling to 307admit the
creative power of life? Dohrn, for instance, was fully aware of the
great transforming influence exerted by function upon
form—his theory of Functionswechsel regards as the
most powerful agent of change the activity of the animal, its
effort to make the best use of its organs, to apply them at need in
new ways to meet new demands. Why then did he not go a step further
and admit that the animal could by its own subconscious efforts
form entirely new organs? Why did most morphologists join with him
in belittling the organism's power of self-transformation?
The reasons seem to have been several. There is first the
fundamental reason, that the idea of an active creative organism is
repugnant to the intelligence, and that we try by all means in our
power to substitute for this some other conception. In so doing we
instinctively fasten upon the relatively less living side of
organisms—their routine habits and reflexes, their routine
structure—and ignore the essential activity which they
manifest both in behaviour and in form-change.
We tend also to lay the causes of form-change, of evolution, as
far as possible outside the living organism. With Darwin we seek
the transforming factors in the environment rather than within the
organism itself. We fight shy of the Lamarckian conception that the
living thing obscurely works out its own salvation by blind and
instinctive effort. We like to think of organisms as machines, as
passive inventions[457] gradually perfected
from generation to generation by some external agency, by
environment or by natural selection, or what you will. All this
makes us chary of believing that Nature is prodigal of new
organs.
Other causes of the unwillingness of morphologists to admit the
new formation of organs are to be sought in the main principle of
pure morphology itself, that the unity of plan imposes an iron
limit upon adaptation, and in the 308powerful influence exercised at
the time by materialistic habits of thought. Teleology had become a
bugbear to the vast majority of biologists, and all real
understanding of the Cuvierian attitude seems, in most cases, to
have been lost, although, curiously enough, teleological
conceptions were often unconsciously introduced in the course of
discussions on the "utility" of organs in the struggle for
existence.
Evolutionary morphology, being for the most part a form of pure
or non-functional morphology, agreed then in all essential respects
with pre-evolutionary or transcendental morphology.
But it contained the germ of a new conception which threw a new
light upon the whole science of morphology. This was the conception
of the organism as an historical being.
We have seen this thought expressed with the utmost clearness by
Darwin himself (supra, p. 233). In his eyes the structure
and activities of the living thing were a heritage from a remote
past, the organism was a living record of the achievements of its
whole ancestral line. What a light this conception threw upon all
biology! "When we no longer look at an organic being as a savage
looks at a ship as something wholly beyond his comprehension; when
we regard every production of Nature as one which has had a long
history; when we contemplate every complex structure and instinct
as the summing-up of many contrivances, each useful to the
possessor, in the same way as any great mechanical invention is the
summing-up of the labour, the experience, the reason, and even the
blunders of numerous workmen; when we thus view each organic being,
how far more interesting—I speak from experience—does
the study of natural history become!" (Origin, 6th ed., pp.
665-6).
Sedgwick expressed the same thing from the morphological point
of view when he wrote, with reference to the ancestral significance
of the blastopore:—"If there is anything in the theory of
evolution, every change in the embryo must have had a counterpart
in the history of the race, and it is our business as morphologists
to find it out" (p. 49, 1884).
By the evolution-theory the problems of form were linked
indissolubly with the problem of heredity. Unity of plan 309could no
longer be explained idealistically as the manifestation of Divine
archetypal ideas; it had a real historical basis, and was due to
inheritance from a common ancestor. The evolution-theory gave
meaning and intelligibility to the transcendental conception of the
unity of plan; in particular it supplied a simple and satisfying
explanation of those puzzling vestigial organs, whose existence was
such a stumbling-block to the teleologists. It enabled the
biogenetic law to be substituted for the laws of Meckel-Serres and
von Baer, as being in some measure a combination and interpretation
of both.
Where the concept of evolution proved itself particularly useful
was in the interpretation of structures which were not immediately
conditioned by adaptation to present requirements, such as, for
instance, the arrangement of gill-slits and aortic arches in the
fœtus of land Vertebrates. Such "heritage characters" could
only be explained on the hypothesis that they had once had
functional or adaptational meaning. Why, for instance, should the
blastopore so often appear as a long slit, closing by concrescence,
unless this had been the original method of its formation in remote
Cœlenterate ancestors?
The point hardly requires elaboration, since it has become an
integral part of all our thinking on biological problems. It may be
as well, however, for the sake of continuity, to give one or two
examples of the historical interpretation of animal structures. The
first may conveniently be the phylogenetic interpretation of the
contrast between "membrane" and "cartilage" bones.
In his Grundzüge of 1870, Gegenbaur made the
suggestion that the investing or membrane bones were derived
phylogenetically from integumentary ossifications, and this was
worked out in detail a few years later by O. Hertwig.[458]
Many years before, several observers—J. Müller,
Williamson, and Steenstrup—had been struck with the
resemblance existing between the placoid scales and the teeth of
Elasmobranch fishes. Hertwig followed up this clue, and came to the
conclusion not only that placoid scales and teeth were 310strictly
homologous, but also that all membrane bones were derived
phylogenetically from ossifications present in the skin or in the
mucous membrane of the mouth, just as cartilage bones were derived
from the cartilaginous skeletons of the primitive Vertebrates. In
some cases this manner of derivation could even be observed in
ontogeny, as Reichert had seen in the Newt, where certain bones in
the roof of the mouth are actually formed by the concrescence of
little teeth, (supra, p. 163). Hertwig considered that the
following bones were originally formed by coalescence of
teeth—parasphenoid, vomer, palatine, pterygoid, the
tooth-bearing part of the pre-maxillary, the maxillary, the dentary
and certain bones of the hyo-mandibular skeleton of Teleosts. All
the investing bones (Deckknochen) of the skull were of
common origin, and could be traced back to integumentary skeletal
plates, which in the ancestral fish formed a dense carapace.
These conclusions were accepted by Kölliker himself, who
wrote in his Entwickelungsgeschichte (1879)—"The
distinction between the primary or primordial, and the investing or
secondary bones is from the morphological standpoint sharp and
definite. The former are ossifications of the (cartilaginous)
primordial skeleton, the latter are formed outside this skeleton,
and are probably all ossifications of the skin or the mucous
membrane" (p. 464).
Gegenbaur[459] consistently upheld the
phylogenetic derivation of investing bones from dermal
ossifications, and even went further and derived substitutionary
bones as well from the integument, thus establishing a direct
comparison between the skeletal formations of Vertebrates and
Invertebrates. Investing bones were actual integumentary
ossifications which had gradually sunk beneath the skin to become
part of the internal skeleton; substitutionary bones were produced
by cells (osteoblasts) which were ultimately derived from the
integument.[460]
311A
further instance of the historical interpretation of animal
structure, taken from quite a different field, is afforded by the
speculations of Dollo[461] on the ancestral history of the Marsupials. In a
brilliant paper of 1880[462] Huxley made the suggestion that the ancestors of
Marsupials were arboreal forms. "I think it probable," he wrote,
"from the character of the pes, that the primitive forms, whence
the existing Marsupialia have been derived, were arboreal animals;
and it is not difficult, I conceive, to see that, with such habits,
it may have been highly advantageous to an animal to get rid of its
young from the interior of its body at as early a period of
development as possible, and to supply it with nourishment during
the later periods through the lacteal glands, rather than through
an imperfect form of placenta" (p. 655). Dollo followed up this
suggestion, which had in the meantime been strengthened by Hill's
discovery of a true allantoic placenta in Perameles, by
demonstrating in the foot of present-day Marsupials certain
features which could only be interpreted as inherited from a time
when the ancestors of Marsupials were tree-living animals. These
were the occurrence of an opposable big toe (when this was present
at all), the great development of the fourth toe, the reduction and
partial syndactylism of the second and third toes, and in some
cases the regression of the nails. These characters were shown to
be typical of arboreal Vertebrates, and their occurrence in forms
not arboreal indicated that these were descended from tree-living
ancestors. Traces of an arboreal ancestry could be demonstrated
even in the marsupial mole Notoryctes.
These are only two examples out of hundreds that might be given.
Present day structure was interpreted in the light of past history;
the common element in organic form was seen to be due to common
descent; the existence of vestigial and non-functional organs was
no longer a riddle.
There was even a tendency to concentrate attention upon the
historical side of structure, upon what the animal passively
inherited rather than upon what it personally 312achieved.
Homologies were considered more interesting than analogies,
vestigial organs more interesting than fœtal and larval
adaptations. Convergence was anathema. The dead-weight of the past
was appreciated at its full and more than its full value; and the
essential vital activity of the living thing, so clearly shown in
development and regeneration, was ignored or forgotten.
But evolutionary morphology for all practical purposes was a
development of pure or idealistic morphology, and was powerless to
bring to fruit the new conception with which evolution-theory had
enriched it. The reason is not far to seek. Pure morphology is
essentially a science of comparison which seeks to disentangle the
unity hidden beneath the diversity of organic form. It is not
immediately concerned with the causes of organic
diversity—that is rather the task of the sciences of the
individual, heredity and development. To take an example—the
recapitulation theory may legitimately be used as a law of pure
morphology, as stating the abstract relation of ontogeny to
phylogeny, and the probable line of descent of any organism may be
deduced from it, as a mere matter of the ideal derivation of one
form from another; but an explanation of the reason for the
recapitulation of ancestral history during development can clearly
not be given by pure morphology unaided. From the fact that the
common starfish shows in the course of its development distinct
traces of a stalk[463] it is possible to
infer, taking other evidence also into consideration, that the
ancestors of the starfish were at one stage of their existence
stalked and sessile organisms. But this leaves unanswered the
question as to how and why the starfish does still repeat after so
many millions of years part of the organisation of one of its
remote ancestors. Why is this feature retained, and by what means
has it been conserved through countless generations? It is clear
that the answer can be given only by a science of the causes of the
production and retention of form, by a causal morphology, based
upon a study of heredity and development.
From the point of view of the pure morphologist the
recapitulation theory is an instrument of research enabling 313him to
reconstruct probable lines of descent; from the standpoint of the
student of development and heredity the fact of recapitulation is a
difficult problem whose solution would perhaps give the key to a
true understanding of the real nature of heredity.
To make full use of the conception of the organism as an
historical being it is necessary then to understand the causal
nexus between ontogeny and phylogeny.
We shall see in the next chapter that the transformation of
morphology from a comparative to a causal science did take place
towards the end of the century, and that some progress was made
towards an understanding of the relation between individual
development and ancestral history, particularly by Roux and Samuel
Butler, working with the fruitful Lamarckian conception of the
transforming power of function.
314
CHAPTER XVIII
THE BEGINNINGS OF CAUSAL MORPHOLOGY
Until well into the 'eighties animal
morphology remained a purely descriptive science, content to state
and summarise the relations between the coexistent and successive
form-states of the same and of different animals. No serious
attempt had been made to discover the causes which led to the
production of form in the individual and in the race.
It is true that evolution-theory had offered a simple solution
of the great problem of the unity in diversity of animal forms, but
this solution was formal merely, and went little beyond that
abstract deduction of more complex from simpler forms, which had
been the main operation of pre-evolutionary morphology. Little was
known of the actual causes of ontogeny, and nothing at all of the
causes of phylogeny; it was, for instance, mere rhetoric on
Haeckel's part to proclaim that phylogeny was the mechanical cause
of ontogeny.
Animal physiology, on its side, had developed in complete
isolation from morphology into a science of the functioning of the
adult and finished animal, considered as a more or less stable
physico-chemical mechanism. Since the days of Ludwig, Claude
Bernard and E. du Bois Reymond, the physiologists' chief care had
been to analyse vital activities into their component physical and
chemical processes, and to trace out the interchange of matter and
energy between the organism and its environment. Physiologists had
left untouched, perhaps wisely, the much more difficult problem of
the causes of the development of form. For all practical purposes
they took the animal-machine as given, and did not trouble about
its mode of origin. They held indeed 315that form-production was due to a
complex of physico-chemical causes, which they hoped some day to
unravel;[464] but this future
physiology of development remained quite embryonic.
Physiology then had not really come into contact with the
problems of form, and it could give the morphologist no direct help
when he turned to investigate the causes of form-production. It
had, however, a determining influence upon the methods of those who
first broke ground in this No Man's Land between morphology proper
and physiology. But it is significant that it was a morphologist
and not a physiologist that did the first spade-work.
The pioneer in this field, both as investigator and as thinker,
was W. Roux, who sketched in the 'eighties the main outlines of a
new science of causal morphology, to which he gave the name of
Entwicklungsmechanik. The choice of name was deliberate, and
the word implied, first, that the new science was essentially an
investigation of the development of form, not of the mode of action
of a formed mechanism, and second, that the methods to be adopted
were mechanistic.[465]
Though Roux was the only begetter of the science of
Entwicklungsmechanik, he was, of course, not the first to
investigate experimentally the formative processes of animal life.
Study of regeneration dates back to Trembley (1740-44), Réaumur
(1742), Bonnet (1745), and Spallanzani (1768-82),[466] and in the years preceding Roux's activity
good work was done by Philipeaux. A beginning had been made with
experimental teratology by E. Geoffroy St Hilaire and others, and
the work of C. Dareste[467] remains classical. Back in the 18th century,
some of John Hunter's experiments had a bearing upon the problems
of form; his work on transplantation was followed up in the 19th
century by Flourens, p. Bert, Ollier and many others. In founding
in 1872 the Archives de Zoologie expérimentale et
générale H. de Lacaze-Duthiers 316put forward in
his introduction a powerful plea for the use of the experimental
method in zoology.
In some ways more directly connected with
Entwicklungsmechanik was His's attempt in 1874[468] to explain on mechanical principles the
formation of certain of the embryonic organs by the bendings and
foldings of tubes or plates of cells. "His compared the various
layers of the chick embryo to elastic plates and tubes; out of
these he suggested that some of the principal organs might be
moulded by mere local inequalities of growth—the ventricles
of the brain, for instance, the alimentary canal, the
heart—and he further succeeded in imitating the formation of
these organs by folding, pinching, and cutting india-rubber tubes
and plates in various ways."[469]
But Roux was undoubtedly the first to make a systematic survey
of the problems to be solved and to work out an organised method of
attack. His earliest work deals with the important problem of
functional adaptation—its importance to the organism, and its
possible mechanistic explanation. The first paper[470] was a study of the branching and
distribution of the arteries in the human body (1878), and a second
paper on the same subject followed in 1879.[471]
In these papers Roux showed how the development of the
blood-vascular system was largely determined by direct adaptation
to functional requirements, and he inferred the existence in the
vascular tissues of certain vital properties, in virtue of which
the functional adaptation of the blood-vessels came about. Thus the
intima or inner lining must possess the faculty of so reacting to
the friction set up by the blood-current as to oppose the least
possible resistance to its flow; the muscular coats must react to
increased pressure by growing thicker, and so on.
These papers were followed in 1881 by his well-known 317book,
Der Kampf der Theile im Organismus, which contained the
working-out of his mechanistic explanation of functional
adaptation, and most of the elements of his general
"causal-analytical" theory of form production. The significance of
the book was popularly considered at the time to lie in its
supposed application of the selection idea to the explanation of
the internal adaptedness of animal structure—in the theory of
"cellular selection," and the book owed its success to its fitting
in so well with the prevalent Darwinism of the day. But its real
importance, as a big step towards causal morphology, was naturally
not so fully appreciated.
During the next few years Roux continued his studies on
functional adaptation,[472] and at the same time made a new departure by
inaugurating, almost contemporaneously with the physiologist
Pflüger, the study of experimental embryology. Isolated
observations had previously been made upon the development of
single blastomeres or parts of blastulæ, by Haeckel and Chun
for instance,[473] but Roux[474] and Pflüger[475] were the first to investigate the subject
systematically, choosing for their work the egg of the frog.[476] Roux continued for many
years to follow up this line of work.[477]
In 1890 he drew up a programme and manifesto[478] of Entwicklungsmechanik as "an
anatomical science of the 318future," and in 1895 he founded the famous
Archiv für Entwicklungsmechanik,[479] publishing in the same year the two large
volumes of his collected papers,[480] of which the first volume dealt with functional
adaptation, the second with experimental embryology.
His subsequent work includes several important general papers;[481] besides a number of
special memoirs dealing with the factors of development, and with
his original subject, functional adaptation.[482]
In our sketch of his views we shall have occasion to refer
particularly to his publications of 1881, 1895 (the
Einleitung), 1902, 1905, and 1910.
Although Roux's biological philosophy is out-and-out
mechanistic, he yet recognises the difficulty, even the
impossibility, of straightway reducing development to the
physico-chemical level. He tries to steer a course midway between
the simplicist conceptions of the materialists and the
"metaphysics" of the neo-vitalist school, which the experimental
study of development and regeneration soon brought into being. In
1895 he writes:—"The too simple mechanistic conception on the
one hand, and the metaphysical conception on the other represent
the Scylla and Charybdis, between which to sail is indeed
difficult, and so far by few satisfactorily accomplished; it cannot
be denied that with the increase of knowledge the seduction of the
second has lately notably increased" (p. 23).
The via media adopted by Roux is the analysis of
development, not directly into simple physico-chemical processes,
but into more complex organic processes dependent 319upon the
fundamental properties of living matter. The aim of
Entwicklungsmechanik is defined by Roux to be the reduction
of developmental events to the fewest and simplest
Wirkungsweisen, or causal processes.[483] Two classes of causal processes may be
distinguished, as "complex components" and "simple components" of
development. The latter are directly explicable by the laws of
physics and chemistry; the former, while in essence
physico-chemical, are yet so very complicated that they cannot at
present be reduced to physico-chemical terms. The ultimate aim of
Entwicklungsmechanik is to reduce development to its "simple
components," but its main task at the present day and for many
years to come is the analysis of development into its "complex
components."
These complex components must be accepted as having much of the
validity of physical and chemical laws. They are mysterious in the
sense that they cannot yet be explained mechanistically, but they
are constant in their action, and under the same conditions produce
always the same effect—hence they may be made the subject of
strictly scientific study. They represent biological
generalisations, in their way of equal validity with the
generalisations of physics and chemistry.
The principal "complex components" which Roux recognises are
somewhat as follows:—First come the elementary cell-functions
of assimilation and dissimilation, growth, reproduction and
heredity, movement and self-division (as a special co-ordination of
cell-movements). Then at a somewhat higher level,
self-differentiation, and the trophic reaction to functional
stimuli. Components of even greater complexity may also be
distinguished, as, for instance, the biogenetic law. The various
tropisms exhibited in development may be regarded as "directive"
complex components. There must be added, not as being itself a
component, but rather as a mode or peculiar property of all
functioning, the omnipresent faculty of self-regulation.
It will be noticed that Roux's "complex components" are 320simply the
general properties or functions of organised matter.
Expressing Roux's thought in another way, we might say that life
can only be defined functionally, i.e., by an enumeration of
the "complex components" or elementary functions which all living
beings manifest, even down to the very simplest. "Living beings,"
writes Roux, "can at present be defined with any approach to
completeness only functionally, that is to say, through
characterisation of their activities, for we have an adequate
acquaintance with their functions in a general way, though our
knowledge of particulars is by no means complete" (p. 105, 1905).
Defined in the most general and abstract way, living things are
material objects which persist in spite of their metabolism, and,
by reason of their power of self-regulation, in spite also of the
changes of the environment. This is the "functional
minimum-definition of life" (pp. 106-7, 1905).
We may now go on to consider the relation of function to form
throughout the course of development. Roux distinguishes in all
development two periods, in the first of which the organ is formed
prior to and independent of its function, while in the second the
differentiation and growth of the organ are dependent on its
functioning. Latterly (1906 and 1910) Roux has distinguished three
periods, counting as the second the transition period when form is
partly self-determined, partly determined by functioning. As this
conception of Roux's is of the greatest importance we shall follow
it out in some detail.
The idea was first elaborated in the Kampf der Theile
(1881), where he wrote:—"There must be distinguished in the
life of all the parts two periods, an embryonic in the broad sense,
during which the parts develop, differentiate and grow of
themselves, and a period of completer development, during which
growth, and in many cases also the balance of assimilation over
dissimilation, can come about only under the influence of stimuli"
(p. 180). There is thus a period of self-differentiation in which
the organs are roughly formed in anticipation of functioning, and a
period of functional development in which the organs are perfected
through functioning and only through functioning. The two 321 periods
cannot be sharply separated from one another, nor does the
transition from the one to the other occur at the same time in the
different tissues and organs.
The conception is more fully expressed in 1905 as
follows:—"This separation (of development into two periods)
is intended only as a first beginning. The first period I called
the embryonic period κατ'
ἐξοχήν, or the period of
organ-rudiments. It includes the 'directly inherited' structures,
i.e., the structures which are directly predetermined in the
structure of the germ-plasm, as, for instance, the first
differentiation of the germ, segmentation, the formation of the
germ-layers and the organ-rudiments, as well as the next stage of
'further differentiation,' and of independent growth and
maintenance, that is, of growth and maintenance which take place
without the functioning of the organs.
"This is accordingly the period of direct fashioning through the
activity of the formative mechanism implicit in the germ-plasm,
also the period of the self-conservation of the formed parts
without active functioning.
"The second period is the period of 'functional
form-development.' It includes the further differentiation and the
maintenance in their typical form of the organs laid down in the
first period; and this is brought about by the exercise of the
specific functions of the organs. This period adds the finishing
touches to the finer functional differentiation of the organs, and
so brings to pass the 'finer functional harmony' of all organs with
the whole. The formative activity displayed during this period
depends upon the circumstance that the functional stimulus, or
rather the exercise by the organs of their specific functions, is
accompanied by a subsidiary formative activity, which acts partly
by producing new form and partly by maintaining that which is
already formed.... Between the two periods lies presumably a
transition period, an intermediary stage of varying duration in the
different organs, in which both classes of causes are concerned in
the further building-up of the already formed, those of the first
period in gradually decreasing measure, those of the second in an
increasing degree" (pp. 94-6, 1905).
In the first period the organ forms or determines the 322function,
in the second period the function forms the organ, or at least
completes its differentiation. It is characteristic that in the
first period functionally adapted structure appears in the complete
absence of the functional stimulus.
The explanation of the difference between the two periods is to
be found in the different evolutionary history of the characters
formed during each. First-period characters are inherited
characters, and taken together constitute the historical basis of
the organism's form and activity; second-period characters are
those of later acquirement which have not yet become incorporated
in the racial heritage.
Inherited characters appear in development in the absence of the
stimulus that originally called them forth; acquired characters are
those that have not yet freed themselves from this dependence upon
the functional stimulus. First-period characters were originally,
like second-period characters, entirely dependent for their
development upon the functional stimuli in response to which they
arose, and only gradually in the course of generations did they
gain that independence of the functional stimulus which stamps them
as true inherited characters. Speaking of the formative stimuli
which are active in second-period development, Roux
writes:—"These stimuli can also produce new structure, which
if it is constantly formed throughout many generations finally
becomes hereditary, i.e., develops in the descendants in the
absence of the stimuli, becomes in our sense embryonic" (p. 180,
1881). Again, "form-characteristics which were originally acquired
in post-embryonic life through functional adaptation may be
developed in the embryo without the functional stimulus, and may in
later development become more or less completely differentiated,
and retain this differentiation without functional activity or with
a minimum of it. But in the continued absence of functional
activity they become atrophied ... and in the end disappear" (p.
201, 1881).
This conception of the nature of hereditary transmission is an
important one, and constitutes the first big step towards a real
understanding of the historical element in organic form and
activity. It supplies a practical criterion for the distinguishing
of "heritage" characters from acquired 323characters, of palingenetic
from cenogenetic—a criterion which descriptive morphology was
unable to find.[484] The introduction of
a functional moment into the concept of heredity was a
methodological advance of the first importance, for it linked up in
an understandable way the problems of embryology, and indirectly of
all morphology, with the problem of hereditary transmission, and
gave form and substance to the conception of the organism as an
historical being.
It is this element in Roux's theories that puts them so far in
advance of those of Weismann. Weismann did not really tackle the
big problem of the relation of form to function, and he left no
place in his mechanical system of preformation for functional or
second-period development; he conceived all development to be in
Roux's sense embryonic, and due to the automatic unpacking of a
complex germinal organisation. Roux himself was to a certain extent
a preformationist, for the development of his first-period
characters is conditioned by the inherited organisation of the
germ-plasm, and is purely automatic. It was indeed his experiments
on the frog's egg (1888) that supplied some of the strongest
evidence in favour of the mosaic theory of development. The number
of Anlagen which he postulates in the germ is however small,
and the germ-plasm in his conception of it has a relatively simple
structure (p. 103, 1905).
The transmission of acquired characters forms, of course, an
integral part of Roux's conception of heredity and development, for
without this transmission second-stage characters could not be
transformed into first-stage characters. He discusses this
difficult question at some length in the Kampf der Theile,
coming to the conclusion that such transmission takes place in
small degree and gradually, and that many generations are required
before a new character can become hereditary. He thinks that
acquired characters are probably transmitted at the chemical level.
It is conceivable that acquired form-changes are dependent on 324chemical
changes, or are correlative with such, and that, since the
germ-cells stand in close metabolic relations with the soma, these
chemical changes may soak through to the germ-cells and so modify
them that a predisposition will appear in the descendants towards
similar form-changes.[485] From this point of view the problem of
transmission might be merged in the broader problem of the
production of form through chemical processes—the central
problem of all development.
Inherited characters develop by an automatic process of
self-differentiation, and the separate parts of the embryo show
during this first period a surprising functional independence of
one another. But this state of things changes progressively as the
second period is reached, until finally all form-production and
maintenance and all correlation depend upon functioning. It is in
the first period of automatic development through internal
"determining" factors that the "developmental" functions in the
strict sense, e.g. automatic growth, division and
self-differentiation, are most clearly shown. In the second or
"functional" period the formative influence of function upon
structure comes into play, and development becomes largely a matter
of "functional adaptation" to functional requirements.
All structure, according to Roux, is either functional or
non-functional. The former includes all structure that is adapted
to subserve some function. "Such 'functional structures' are, for
example, the composition of striated muscle fibres out of
fibrillæ and these out of muscle-prisms, or again the length
and thickness of the muscles, the static structure of the bones,
the composition of the stomach and the blood-vessels out of
longitudinal and circular fibres, the external shape of the
vertebral centra and of the cuneiform bones of the foot" (p. 73,
1910). Indeed, as Cuvier had already pointed out, practically every
organ in the body shows a functional structure which is accurately
and minutely adjusted to the function it is intended to perform.
Thus, to take some further examples, the arteries are admirably
adapted as regards size of lumen, elasticity of wall, direction of
branching, to conduct the blood to all parts of the body 325with the
least possible waste of the propelling power through frictional
resistance. So, too, the spongy substance of the long bones is
arranged in lamellæ which take the direction of the principal
stresses and strains which fall upon the bones in action.
Functional structure may be formed either in the first or in the
second period of development, may be either inherited or acquired,
but it reaches its full differentiation only in the second period,
i.e., under the influence of functioning. Practically
speaking, functional structure is directly dependent for its full
development and for its continued conservation upon the exercise of
the particular function which it serves. In the second period, but
not in the first, increased use leads to hypertrophy of the
functional structure, disuse to atrophy.
From functional structure is to be distinguished nonfunctional
structure, which has no relation to the bodily functions—is
neither adapted to perform any of these, nor has arisen as a
by-product of functional activity. "To this category belong, for
example, among typical structures, the triangular form of the
cross-section of the tibia, the dolicocephalic or brachycephalic
shape of the skull, most of the external characters distinguishing
genera and species, many of the external features of the embryo
which change in the course of development, besides most of the
abnormal forms shown by monstrosities, tumours, etc." (p. 74,
1910). Non-functional structure is not affected by functional
adaptation, and may accordingly be left out of consideration
here.
Now the influence of functioning upon the form and structure of
an organ is twofold. There is first the immediate change brought
about by the very act of functioning—for example, the
shortening and thickening of skeletal muscles when they act. This
is a purely temporary change, for the organ at once returns to its
normal quiescent state as soon as it ceases to function. Such
temporary functional change, brought about in the moment of
functioning, is usually dependent for its initiation upon some
neuro-muscular mechanism, though it may be elicited also by a
chemical stimulus. It is thus always a phenomenon of "behaviour."
326"From such temporary changes are sharply to
be distinguished all permanent alterations which first appear in
perceptible fashion through oft-repeated or long-continued,
enhanced functional activity. These produce a new and lasting
internal equilibrium of the organ, consisting in an insertion of
new molecules or a rearrangement of old. For this reason they
outlast the periods of functional form-change, or, if as in the
case of the muscles they themselves alter during functional
activity, they regain their state when the organ ceases to
function" (p. 72, 1910). "Oft-repeated exercise or heightened
exercise of the specific functions, or repeated action of the
functional stimuli which determine them, produces, as we have said
before, true form-changes as a by-product. These are of two kinds.
In so far as these form-changes facilitate the repetition of the
specific functions, I have called them functional
adaptations.... Such as do not improve the functioning of the
organ are indeed by-products of functioning, but without adaptive
character; they do not belong to the class of functional
adaptations at all" (p. 75, 1910).
We may now enquire in what way functional adaptations can arise
as by-products of functioning.
It is clear that natural selection in the sense of individual or
"personal" selection cannot adequately explain the origin of
functional structure and the functional harmony of structure, for
thousands of cells would have to vary together in a purposive way
before any real advantage could be gained in the struggle for
existence, and it is in the highest degree unlikely that this
should come about by chance variation.[486] The development of purposive internal structure
is only to be explained by the properties of the tissues
concerned.
In illustration and proof of the statement that functional
adaptation is due to the properties of the tissues we may adduce
the development and regulation of the blood-vascular 327system, which
has been thoroughly studied from this point of view by Roux and
Oppel (1910).
It appears that only the very first rudiments of the vascular
system are laid down in the short first period of automatic
non-functional development. All the subsequent growth and
differentiation of the blood-vessels falls into the second period,
and is due wholly or in great part to direct functional adaptation
to the requirements of the tissues. Thus from the rudiments formed
in the first period there sprout out the definitive vessels in
direct adaptation to the food-consumption of the tissues they are
to supply. The size, direction and intimate structure of these
vessels are accurately adjusted to the part they play in the
economy of the whole, and this adjustment is brought about in
virtue of the peculiar properties or reaction-capabilities of the
different tissues of which the blood-vessels are composed.
The properties which Roux finds himself compelled to postulate
in the vascular tissues, after a thorough-going analysis of the
different kinds of functional adaptation shown by the
blood-vessels, are summarised by him as follows:—
"(1) The faculty—depending on a direct sensibility
possessed by the endothelium and perhaps also by the other layers
of the intima—of yielding to the impact of the blood, so far
as the external relations of the vessel permit. In this way the
wall adapts itself to the hæmodynamically conditioned
'natural' shape of the blood-stream, and reaches this shape as
nearly as possible." Through this faculty of the lining tissue of
the blood-vessels, the size of the lumen and the direction of
branching are so regulated as to oppose the least possible
resistance to the flow of the blood.
"(2) The faculty possessed by the endothelium of the capillaries
of each organ of adapting itself qualitatively to the particular
metabolism of the organ." This adaptedness of the capillaries is,
however, more usually an inherited state, i.e., brought
about in the first period of development.
"(3) The faculty possessed by the capillary walls of being
stimulated to sprout out and branch by increased functioning, 328i.e., by increased diffusion, and
their power to exhibit a chemically conditioned cytotropism, which
causes the sprouts to find one another and unite. A similar process
can be directly observed in isolated segmentation-cells, which tend
to unite in consequence of a power of mutual attraction.
"(4) The faculty of developing normal arterial walls in response
to strong intermittent pressure, and normal venous walls in
response to continuous lesser pressure." It has been shown, for
instance, by Fischer and Schmieden that in dogs a section of vein
transplanted into an artery takes on an arterial structure, at
least as regards the circular musculature, which doubles in
thickness.
"(5) The power to regulate the normal[487] length of the arteries and veins, in adaptation
to the growth of the surrounding tissues, in such a way that the
stretching action of the blood-stream brings the vessel to its
proper functional length.
"(6) The power to form, in response to slight increases in
longitudinal tension, new structural parts which take their place
alongside the existing longitudinal fibres.
"(7) The power to regulate the width of the circular musculature
according to the degree of food-consumption by the tissues, in
response to nerve impulses initiated in these tissues.
"(8) The power possessed by the circular musculature of
responding to such continuous functional widening, by the formation
of new structural parts in the circular musculature, and so of
widening the vessel permanently or by this new formation of
muscular fibres thickening the circular musculature.
"(9) The faculty of being stimulated by increased blood-pressure
to produce the same structural changes as mentioned in par. 8,
though here the response is otherwise conditioned" (pp. 126-7,
1910).
It is by virtue of the tissue-properties detailed above that the
complex functional adaptations of the blood-vessels come about.
The development of the vascular system is no mere automatic and
mechanical production of form, apart from 329and independent of
functioning; it implies a living and co-ordinated activity of the
tissues and organs concerned, a power of active response to
foreseen and unforeseen contingencies. Form is then not something
fixed and congealed—it is the ever-changing manifestation of
functional activity. "Since most of the structure and form of the
blood-vessels arises in direct adaptation to function, the vessels
of adult men and animals are no fixed structures, which, once
formed, retain their form and structural build unchanged throughout
life; on the contrary, they require even for their continued
existence the stimulus of functional activity.... The fully formed
blood-vessels are no static structures, such as they appear to be
according to the teaching of normal histology, and such as they
have long been taken to be. Observation and description of normal
development never shows us anything but the visible side of organic
happenings, the products of activity, and leaves us ignorant
of the real processes of form-development and form-conservation,
and of their causes" (p. 125, 1910).
The real thing in organisation is not form but activity. It is
in this return to the Cuvierian or functional attitude to the
problems of form that we hold Roux's greatest service to biology to
consist. The attitude, however, seems to smack of vitalism, and
Roux, as we have seen, is no vitalist. He holds that the marvellous
and apparently purposive tissue-qualities which underlie all
processes of functional adaptation have arisen "naturally," in the
course of evolution, by the action of natural selection upon the
various properties, useful and useless, which appeared fortuitously
in the primary living organisms. He is, moreover, deeply imbued
with the materialistic philosophy of his youth, and it is indeed
one of the chief characteristics of his system that he states the
fundamental properties or qualities of life in terms of metabolism.
A vital quality is for Roux a special process or mode of
assimilation. The faculty of "morphological assimilation" whereby
form is imposed upon formless chemical processes is the ultimate
term of Roux's analysis—"the most general, most essential,
and most characteristic formative activity of life" (p. 631,
1902).
We have now to consider very briefly the early results 330achieved
by Roux's fellow-workers in the field of causal morphology. As D.
Barfurth points out,[488] the years 1880-90 saw a general awakening of
interest in experimental morphology, and it is hard to say whether
Roux's work was cause or consequence. "There fall into this
period," writes Barfurth, "the experimental investigations by Born
and Pflüger on the sexual difference in frogs (1881), by
Pflüger on the parthenogenetic segmentation of Amphibian ova,
on crossing among the Amphibia, and on other important subjects
(1882). In the following year (1883) appeared two papers of
fundamental importance, by E. Pflüger and W. Roux:
Pflüger publishing his researches on 'the influence of gravity
on cell-division,' Roux his experimental investigations on 'the
time of the determination of the chief planes in the
frog-embryo.'... In the same year appeared A. Rauber's experimental
studies 'on the influence of temperature, atmospheric pressure, and
various substances on the development of animal ova,' which have
brought many similar works in their train. The following year
(1884) saw a lively controversy on Pflüger's
gravity-experiments with animal eggs, in which took part
Pflüger, Born, Roux, O. Hertwig and others, and in this year
appeared work by Roux dealing with the experimental study of
development, and in particular giving the results of the first
definitely localised pricking-experiments on the frog's egg (in the
Schles. Gesell. f. vaterl. Kultur, 15th Feb. 1884), also the
important researches of M. Nussbaum and Gruber (followed up later
by Verworn, Hofer and Balbiani) on Protozoa, and other experimental
work" (pp. xi.-xii.).
In 1888 appeared a famous paper by W. Roux,[489] in which he described how he had succeeded
in killing by means of a hot needle one of the two first
blastomeres of the frog's egg, and how a half-embryo had developed
from the uninjured cell. Some years before[490] he had enunciated, at about the same time as
Weismann, the view that development 331was brought about by a qualitative
division of the germ-plasm contained in the nucleus, and that the
complicated process of karyokinetic or mitotic division of the
nucleus was essentially adapted to this end. He conceived that
development proceeded by a mosaic-like distribution of potencies to
the segmentation-cells, that, for instance, the first segmentation
furrow separated off the material and potencies for the right half
of the embryo from those for the left half. He had tried to show
experimentally that the first furrow in the frog's egg coincided
with the sagittal plane of the embryo,[491] and his later success in obtaining a half-embryo
from one of the first two blastomeres seemed to establish the
"mosaic theory" conclusively.
Roux's needle-experiment aroused much interest, especially as
Weismann's theory of heredity was then being keenly discussed.
Chabry had published in 1887 some interesting results on the
Ascidian egg,[492] which strongly
supported the Roux-Weismann theory. Considerable astonishment was
therefore caused by Driesch's announcement in 1891[493] that he had
obtained complete larvæ from single blastomeres of the
sea-urchin's egg isolated at the two-celled stage. He followed this
up in the next year[493] by showing that whole embryos could be produced
from one or more blastomeres isolated at the four-cell stage.
Similar or even more striking results were obtained by E. B. Wilson
on Amphioxus,[494] and Zoja on medusæ.[495] Driesch succeeded also in disturbing the
normal course and order of segmentation by compressing the eggs of
the sea-urchin between glass plates, and yet obtained normal
embryos. Similar pressure-experiments were carried out on the frog
by O. Hertwig,[496] and on
Nereis by E. B. Wilson,[497] with analogous results.
In 1895 O. Schultze[498] showed that if the frog's egg is held between
two plates and inverted at the two-celled stage 332there are formed
two embryos instead of one. In the same year T. H. Morgan[499] repeated Roux's fundamental experiment of
destroying one of the two blastomeres, but inverted the egg
immediately after the operation—a whole embryo of half size
resulted. A year or two later Herlitzka[500] found that if the first two blastomeres of the
newt's egg were separated by constriction, two normal embryos of
rather more than half normal size were formed.
The main result of the first few years' work on the development
of isolated blastomeres was to show that the mosaic theory was not
strictly true, and that the hypothesis of a qualitative division of
the nucleus was on the whole negatived by the facts.
Evidence soon accumulated that the cytoplasm of the egg stood
for much in the differentiation of the embryo. A number of years
previously Chun had made the discovery that single blastomeres of
the Ctenophore egg, isolated at the two-celled stage, gave
half-embryos. This was in the main confirmed by Driesch and Morgan
in 1896,[501] and they made the
further interesting discovery that the same defective larvæ
could be obtained by removing from the unsegmented egg a large
amount of cytoplasm. Conclusive proof of the importance of the
cytoplasm was obtained soon after by Crampton,[502] who removed the anucleate "yolk-lobe" from
the egg of the mollusc Ilyanassa at the two-celled stage,
and obtained larvæ which lacked a mesoblast. This result was
brilliantly confirmed and extended some years later by E. B.
Wilson,[503] working on the egg of
Dentalium. He found that if the similar anucleate "polar
lobe" of this form is removed at the two-celled stage, deficient
larvæ are formed, in which the post-trochal region and the
apical organ are absent. He further showed that in the unsegmented
but mature egg prelocalised cytoplasmic regions can be
distinguished, which later become separated from one another
through the segmentation of the egg. The segmentation-cells into
which these cytoplasmic substances are thus segregated show a
marked specificity of development, 333giving rise, even when isolated,
to definite organs of the embryo. Wilson concluded that the
cytoplasm of the egg contains a number of specific organ-forming
stuffs, which have a definite topographical arrangement in the egg.
Development is thus due in part to a qualitative division not of
the nucleus but of the cytoplasm. Corroborative evidence of the
existence of cytoplasmic organ-forming stuffs has been supplied for
several other species, e.g., Patella (Wilson),
Cynthia (Conklin), Cerebratulus (Zeleny), and
Echinus (Boveri).
It is interesting to recall that so long ago as 1874 W. His[504] put forward the theory
that there exist in the blastoderm and even in the egg prelocalised
areas, which contain the formative material for each organ of the
embryo, and from which the embryo is developed by a simple process
of unequal growth.
The experimental study of form was prosecuted in many other
directions besides that of experimental embryology. The study of
regeneration and of regulatory processes attracted many workers,
among whom may be mentioned T. H. Morgan, C. M. Child, and H.
Driesch. In an interesting series of papers C. Herbst applied the
principles of the physiology of stimulus to the interpretation of
development.[505] The formative power of
function was studied in Germany by Roux and his pupils, Fuld, O.
Levy, Schepelmann and others, particularly by E. Babák. In
France, F. Houssay inaugurated[506] an important series of memoirs by himself and
his pupils on "dynamical morphology," the most important memoir
being his own valuable discussion of the functional significance of
form in fishes.[507] The principles of
his dynamical morphology were first laid down in his book La
Forme et la Vie (1900).
The famous experiments of Loeb, Delage and others on artificial
parthenogenesis may also be mentioned, though their connection with
morphology is somewhat remote.
334The period was characterised also by the
lively discussion of first principles, in which Driesch took a
leading part. Materialistic methods of interpretation were upheld
by perhaps the majority of biologists, but vitalism found powerful
support.
335
CHAPTER XIX
SAMUEL BUTLER AND THE MEMORY THEORIES OF HEREDITY
We have laid stress upon the
distinction established by Roux between the two stages of
development—the automatic and the functional—because of
the light which it seems to throw upon the phylogenetic relation of
form to function. We have pointed out, too, the paramount rôle that
function plays in Roux's theories of development and heredity, and
we have brought out the close kinship existing between his theory
and that of Lamarck. For Roux, as for Lamarck, the function creates
the organ, and it is only after long generations that the organ
appears before the function.
It so happened that just about the time when Roux's papers were
beginning to appear a brilliant attempt was made by Samuel Butler
to revive and complete the Lamarckian doctrine.
A man of singular freshness and openness of mind, combining in
an extraordinary degree extreme intellectual subtlety with a
childlike simplicity of outlook, Butler was one of the most
fascinating figures of the 19th century. He was not a professional
biologist, and much of his biological work is, for that reason,
imperfect. But he brought to bear upon the central problems of
biology an unbiassed and powerful intelligence, and his attitude to
these problems, just because it is that of a cultivated layman, is
singularly illuminating.
He was not well acquainted with biological literature; he seems
to have hit upon the main ideas of his theory of life and habit in
complete independence of Lamarck, and 336only later to have become
aware that Lamarck had in a measure forestalled him. He puts this
very beautifully in the following passage from his chief biological
work Life and Habit (1877[508]):—"I admit that when I began to write upon
my subject I did not seriously believe in it. I saw, as it were, a
pebble upon the ground, with a sheen that pleased me; taking it up,
I turned it over and over for my amusement, and found it always
grow brighter and brighter the more I examined it. At length I
became fascinated, and gave loose rein to self-illusion. The aspect
of the world changed; the trifle which I had picked up idly had
proved to be a talisman of inestimable value, and had opened a door
through which I caught glimpses of a strange and interesting
transformation. Then came one who told me that the stone was not
mine, but that it had been dropped by Lamarck, to whom it belonged
rightfully, but who had lost it; whereon I said I cared not who was
the owner, if only I might use it and enjoy it. Now, therefore,
having polished it with what art and care one who is no jeweller
could bestow upon it, I return it, as best I may, to its possessor"
(p. 306). In one of his later works, however, Butler made up for
his first neglect of his predecessors by giving what is undeniably
the best account in English literature of the work of Buffon,
Lamarck, and Erasmus Darwin—in his Evolution, Old and
New (1879). Many of his facts he took from Charles Darwin,
whose theory of natural selection he bitterly opposed, in the two
books just mentioned and in Unconscious Memory (1880) and
Luck or Cunning (1887).
Butler's main thesis is that living things are active,
intelligent agents, personally continuous with all their ancestors,
possessing an intense but unconscious memory of all that their
ancestors did and suffered, and moving through habit from the
spontaneity of striving to the automatism of remembrance.
The primary cause of all variation in structure is the active
response of the organism to needs experienced by it, and the
indispensable link between the outer world and the creature itself
is that same "sense of need" upon which 337Lamarck insisted. "According
to Lamarck, genera and species have been evolved, in the main, by
exactly the same process as that by which human inventions and
civilisations are now progressing; and this involves that
intelligence, ingenuity, heroism, and all the elements of romance,
should have had the main share in the development of every herb and
living creature around us" (Life and Habit, p. 253).
Variations are indubitably the raw material of evolution—"The
question is as to the origin and character of these variations. We
say they mainly originate in a creature through a sense of its
needs, and vary through the varying surroundings which will cause
those needs to vary, and through the opening-up of new desires in
many creatures, as the consequence of the gratification of old
ones; they depend greatly on differences of individual capacity and
temperament; they are communicated, and in the course of time
transmitted, as what we call hereditary habits or structures,
though these are only, in truth, intense and epitomised memories of
how certain creatures liked to deal with protoplasm" (p. 267).
Butler's theory then is essentially a bold and enlightened
Lamarckism, completed and rounded off by the conception that
heredity too is a psychological process, of the same nature as
memory.
In seeking to establish a close analogy between memory and
heredity Butler starts out from the fact of common experience, that
actions which on their first performance require the conscious
exercise of will and intelligence, and are then carried out with
difficulty and hesitation, gradually through long-continued
practice come to be performed easily and automatically, without the
conscious exercise of intelligence or will.
He tries to show that this is a general law—that knowledge
and will become intense and perfect only when through
long-continued exercise they become automatic and
unconscious—and he applies this conception to the elucidation
of development.
Developmental processes, especially the early ones (of Roux's
first stage) are automatic and unconscious, and yet imply the
possession by the embryo of a wonderfully perfect 338knowledge of the
processes to be gone through, and an assured power of will and
judgment. Is it conceivable, says Butler, that the embryo can do
all these things without knowing how to do them, and without having
done them before? "Shall we say ... that a baby of a day old sucks
(which involves the whole principle of the pump, and hence a
profound practical knowledge of the laws of pneumatics and
hydrostatics), digests, oxygenises its blood (millions of years
before Sir Humphrey Davy discovered oxygen), sees and
hears—all most difficult and complicated operations,
involving a knowledge of the facts concerning optics and acoustics,
compared with which the discoveries of Newton sink into utter
insignificance? Shall we say that a baby can do all these things at
once, doing them so well and so regularly, without being even able
to direct its attention to them, and without mistake, and at the
same time not know how to do them, and never have done them
before?" (p. 54). Assuredly not.
The only possible explanation is that the embryo's ancestors
have done these things so often, throughout so many millions of
generations, that the embryo's knowledge of how to do them has
become unconscious and automatic by reason of this age-long
practice. This implies that there is in a very real sense actual
personal continuity between the embryo and all its ancestors, so
that their experiences are his, their memory also his. "We must
suppose the continuity of life and sameness between living beings,
whether plants or animals, to be far closer than we have hitherto
believed; so that the experience of one person is not enjoyed by
his successor, so much as that the successor is bona fide
but a part of the life of his progenitor, imbued with all his
memories, profiting by all his experiences—which are, in
fact, his own—and only unconscious of the extent of his own
memories and experiences owing to their vastness and already
infinite repetitions" (p. 50). It is very suggestive in this
connection, he continues—"I. That we are most conscious
of, and have most control over, such habits as speech, the
upright position, the arts and sciences, which are acquisitions
peculiar to the human race, always acquired after birth, and not
common 339to ourselves and any ancestor who had not
become entirely human.
"II. That we are less conscious of, and have less control
over, eating and drinking, swallowing, breathing, seeing and
hearing, which were acquisitions of our prehuman ancestry, and for
which we had provided ourselves with all the necessary apparatus
before we saw light, but which are, geologically speaking, recent,
or comparatively recent.
"III. That we are most unconscious of, and have least control
over, our digestion and circulation, which belonged even to our
invertebrate ancestry, and which are habits, geologically speaking,
of extreme antiquity.... Does it not seem as though the older and
more confirmed the habit, the more unquestioning the act of
volition, till, in the case of the oldest habits, the practice of
succeeding existences has so formulated the procedure, that, on
being once committed to such and such a line beyond a certain
point, the subsequent course is so clear as to be open to no
further doubt, to admit of no alternative, till the very power of
questioning is gone, and even the consciousness of volition" (pp.
51-2).
The hypothesis then, that heredity and development are due to
unconscious memory, finds much to support it—"the
self-development of each new life in succeeding
generations—the various stages through which it passes (as it
would appear, at first sight, without rhyme or reason), the manner
in which it prepares structures of the most surpassing intricacy
and delicacy, for which it has no use at the time when it prepares
them, and the many elaborate instincts which it exhibits
immediately on, and indeed before, birth—all point in the
direction of habit and memory, as the only causes which could
produce them" (p. 125). The hypothesis explains, for instance, the
fact of recapitulation:—"Why should the embryo of any animal
go through so many stages—embryological allusions to
forefathers of a widely different type? And why, again, should the
germs of the same kind of creature always go through the same
stages? If the germ of any animal now living is, in its simplest
state, but part of the personal identity of one of the original
germs of all life whatsoever, and hence, if any now living organism
must be considered without quibble as being itself millions of
years 340old, and as imbued with an intense though
unconscious memory of all that it has done sufficiently often to
have made a permanent impression; if this be so, we can answer the
above questions perfectly well. The creature goes through so many
intermediate stages between its earliest state as life at all, and
its latest development, for the simplest of all reasons, namely,
because this is the road by which it has always hitherto travelled
to its present differentiation; this is the road it knows, and into
every turn and up or down of which it has been guided by the force
of circumstances and the balance of considerations" (pp.
125-6).
The hypothesis explains also the way in which the orderly
succession of stages in embryogeny is brought about, for we can
readily understand that the embryo will not remember any stage
until it has passed through the stage immediately preceding it.
"Each step of normal development will lead the impregnated ovum up
to, and remind it of, its next ordinary course of action, in the
same way as we, when we recite a well-known passage, are led up to
each successive sentence by the sentence which has immediately
preceded it.... Though the ovum immediately after impregnation is
instinct with all the memories of both parents, not one of these
memories can normally become active till both the ovum itself and
its surroundings are sufficiently like what they respectively were,
when the occurrence now to be remembered last took place. The
memory will then immediately return, and the creature will do as it
did on the last occasion that it was in like case as now. This
ensures that similarity of order shall be preserved in all the
stages of development in successive generations" (pp. 297-8).
Abnormal conditions of development will cause the embryo to
pause and hesitate, as if at a loss what to do, having no ancestral
experience to guide it. Abnormalities of development represent the
embryo's attempt to make the best of an unexpected situation. Or,
as Butler puts it, "When ... events are happening to it which, if
it has the kind of memory we are attributing to it, would baffle
that memory, or which have rarely or never been included in the
category of its recollections, it acts precisely as a creature
acts 341when its recollection is disturbed, or
when it is required to do something which it has never done
before" (p. 132). "It is certainly noteworthy that the embryo
is never at a loss, unless something happens to it which has not
usually happened to its forefathers, and which in the nature of
things it cannot remember" (p. 132).
Butler's teleological conception of organic evolution was of
course completely antagonistic to the naturalistic conceptions
current in his time. In one of his later books he repeats Paley's
arguments in favour of design, and to the question, "Where, then,
is your designer of beasts and birds, of fishes, and of plants?" he
replies: "Our answer is simple enough; it is that we can and do
point to a living tangible person with flesh, blood, eyes, nose,
ears, organs, senses, dimensions, who did of his own cunning, after
infinite proof of every kind of hazard and experiment, scheme out
and fashion each organ of the human body. This is the person whom
we claim as the designer and artificer of that body, and he is the
one of all others the best fitted for the task by his antecedents,
and his practical knowledge of the requirements of the
case—for he is man himself. Not man, the individual of any
given generation, but man in the entirety of his existence from the
dawn of life onwards to the present moment" (Evolution, Old and
New, p. 30, 1879).
Butler's theory of life and habit remained only a sketch, and he
was perhaps not fully aware of its philosophical implications.
Since Butler's time, a new complexion has been put upon biological
philosophy by the profound speculations of Bergson.
But it is not impossible that the future development of
biological thought will follow some such lines as those which he
tentatively laid down.
Butler was not the first to suggest that there is a close
connection between heredity and memory—it is a thought likely
to occur to any unprejudiced thinker. The first enunciation of it
which attracted general attention was that contained in Hering's
famous lecture "On Memory as a general Function of organised
Matter."[509] Butler was not 342aware of
Hering's work when he published his Life and Habit, but in
Unconscious Memory (1880) he gave full credit to Hering as
the first discoverer, and supplied an admirable translation of
Hering's lecture. As far as the assimilation of heredity to memory
is concerned Hering and Butler have much in common, but Hering did
not share Butler's Lamarckian and vitalistic views, preferring to
hold fast, for the practical purposes of physiology at all events,
to the general accepted theory of the parallelism between psychical
and physical processes. He was inclined to regard memory in the
ordinary sense as a function of the brain, and memory in general as
a function of all organised matter. Speaking of the psychical life,
he says, "Thus the cause which produces the unity of all single
phenomena of consciousness must be looked for in unconscious life.
As we know nothing of this except what we learn from our
investigations of matter, and since in a purely empirical
consideration, matter and the unconscious must be regarded as
identical, the physiologist may justly define memory in a wider
sense to be a faculty of the brain, the results of which to a great
extent belong to both consciousness and unconsciousness."[510] Hering's views were supported by
Haeckel.[511]
In 1893 an American, H. F. Orr,[512] tried to work out a theory of development and
heredity based upon the fundamental idea "that the property which
is the basis of bodily development in organisms is the same
property which we recognise as the basis of psychic activity and
psychic development." He tried also to explain the recapitulation
of phylogeny by ontogeny as due to habit.
The neo-Lamarckian school of American palæontologists were
also in sympathy with the memory idea, and this was expressed most
clearly perhaps by Cope.[513]
In 1904 appeared the work on this subject which has attracted
the most attention—R. Semon's Die Mneme.[514] 343This was an elaborate treatment of the
question from the materialistic point of view, the main assumption
of Semon's theory being that the action of a stimulus upon the
organism leaves a more or less permanent material trace or
"engramm," of such a nature as to modify the subsequent action of
the organism.
Applied to the explanation of heredity and development, Semon's
theory comes to very much the same as Weismann's, with engramms
substituted for determinants, but it has the great advantage of
allowing for the transmission of acquired characters. The
application of the concept of stimulus is valuable and suggestive,
but it seems to us that the memory theory of heredity can be
properly utilised only by adopting a frankly Lamarckian and
vitalistic standpoint, and this standpoint Semon expressly combats.
As Ward[515] points out in his
illuminating lecture on heredity and memory—"Records or
memoranda alone are not memory, for they presuppose it. They
may consist of physical traces, but memory, even when called
'unconscious,' suggests mind; for, as we have seen, the automatic
character implied by this term 'unconscious' presupposes foregone
experience.... The mnemic theory then, if it is to be worth
anything, seems to me clearly to require not merely physical
records or 'engrams,' but living experience or tradition. The
mnemic theory will work for those who can accept a monadistic or
pampsychist interpretation of the beings that make up the world,
who believe with Spinoza and Leibniz that 'all individual things
are animated albeit in divers degree'" (pp. 55-6).
Perhaps the best and most ingenious treatment of memory and
heredity from a physical standpoint is that offered by E. Rignano
in his book, Sur la transmissibilité des
caractères acquis.[516] Rignano seeks to construct a physico-chemical
"model" which will explain both heredity and memory.
His system, which is based more firmly upon the facts of
experimental embryology than Semon's, postulates the existence of
"specific nervous accumulators." The essential 344hypothesis set
up is that every functional stimulus is transformed into specific
vital energy, and deposits in the nucleus of the cell a specific
substance which is capable of discharging, in an inverse direction,
the nervous current which has formed it, as soon as the dynamical
equilibrium of the organism is restored to the state in which it
was when the original stimulus acted upon it. These specific
nuclear substances, different for each cell, are accumulated also
in the nuclei of the germinal substance, constituting what Rignano
calls the central zone of development. That is to say, each
functional adaptation changes slightly the dynamical equilibrium of
the organism, and this change in the system of distribution of the
nervous currents leads to the deposit in the central zone of
development of a new specific substance. In the development of the
next individual this new specific element enters into activity, and
reproduces the nervous current which has formed it, as soon as the
organism reaches the same conditions of dynamical equilibrium as
those obtaining when the stimulus acted on the parent.
Development can thus be regarded as consisting of a number of
stages, at each of which new specific elements enter automatically
into play and lead the embryo from that stage to the stage
succeeding. The germinal substance on this theory of Rignano's is
to be regarded as being composed of a large number of specific
elements, originally formed as a result of each new functional
adaptation, but now forming part of the hereditary equipment.
The theory represents an advance upon the more static
conceptions of Semon. It owes much to Roux's influence.
In this country, the mnemic theories have been championed
particularly by M. Hartog[517] and Sir Francis Darwin.[518]
345
CHAPTER XX
THE CLASSICAL TRADITION IN MODERN MORPHOLOGY
To write a history of contemporary
movements from a purely objective standpoint is well recognised to
be an impossible task. It is difficult for those in the stream to
see where the current is carrying them: the tendencies of the
present will only become clear some twenty years in the future.
I propose, therefore, in this concluding chapter to deal only
with certain characteristics of modern work on the problems of form
which seem to me to be derived directly from the older classical
tradition of Cuvier and von Baer.
The present time is essentially one of transition. Complete
uncertainty reigns as to the main principles of biology. Many of us
think that the materialistic and simplicist method has proved a
complete failure, and that the time has come to strike out on
entirely different lines. Just in what direction the new biology
will grow out is hard to see at present, so many divergent
beginnings have been made—the materialistic vitalism of
Driesch, the profound intuitionalism of Bergson, the psychological
biology of Delpino, Francé, Pauly, A. Wagner and W.
Mackenzie. But if any of these are destined to give the future
direction to biology, they will in a measure only be bringing
biology back to its pre-materialistic tradition, the tradition of
Aristotle, Cuvier, von Baer and J. Müller. It may well be that
the intransigent materialism of the 19th century is merely an
episode, an aberration rather, in the history of biology—an
aberration brought about by the over-rapid development of a
materialistic and luxurious civilisation, in which man's material
means have outrun his mental and moral growth.
346Two movements seem significant in the
morphology of the last decade or so of the 19th
century—first, the experimental study of form, and second,
the criticism of the concepts or prejudices of evolutionary
morphology.
The period was characterised also by the great interest taken in
cytology, following upon the pioneer work of Hertwig, van Beneden
and others on the behaviour of the nuclei in fertilisation and
maturation.[519] This line of work gained
added importance in connection with contemporary research and
speculation on the nature of hereditary transmission, and it has in
quite recent years received an additional stimulus from the
re-discovery of Mendelian inheritance. Its importance, however,
seems to lie rather in its possible relation to the problems of
heredity than in any meaning it may have for the problems of form.
More significant is the revolt against the cell-theory started by
Sedgwick[520] and Whitman,[521] on the ground that the organism is
something more than an aggregation of discrete, self-centred
cells.
The experimental work on the causes of the production and
restoration of form infused new life into morphology. It opened
men's eyes to the fact that the developing organism is very much a
living, active, responsive thing, quite capable of relinquishing at
need the beaten track of normal development which its ancestors
have followed for countless generations, in order to meet
emergencies with an immediate and purposive reaction. It was cases
of this kind, cases of active regulation in development and
regeneration, that led men like G. Wolff and H. Driesch to cast off
the bonds of dogmatic Darwinism and declare boldly for vitalism and
teleology.
There was the famous case of the regeneration of the lens in
Amphibia from the edge of the iris—an entirely novel mode of
origin, not occurring in ontogeny. The fact seems to have been
discovered first by Colucci in 1891, and independently by G. Wolff
in 1895.[522] The experiment was later
repeated and confirmed by Fischel and other workers. 347Wolff drew from
this and other facts the conclusion that the organism possesses a
faculty of "primary purposiveness" which cannot have arisen through
natural selection.[523] And, as is well
known, Driesch derived one of his most powerful arguments in favour
of vitalism from the extraordinary regenerative processes shown by
Tubularia and Clavellina in the course of which the
organism actually demolishes and rebuilds a part or the whole of
its structure. But under the influence of physiologists like Loeb
many workers held fast to materialistic methods and
conceptions.
The great variety of regulative response of which the organism
showed itself capable made it very difficult for the morphologist
to uphold the generalisations which he had drawn from the facts of
normal undisturbed development. The germ-layer theory was found
inadequate to the new facts, and many reverted to the older
criterion of homology based on destiny rather than origin. The
trend of opinion was to reject the ontogenetic criterion of
homology, and to refuse any morphological or phylogenetic value to
the germ-layers.[524]
The biogenetic law came more and more into disfavour, as the
developing organism more and more showed itself to be capable of
throwing off the dead-weight of the past, and working out its own
salvation upon original and individual lines.[525] A. Giard in particular called attention to
a remarkable group of facts which went to show that embryos or
larvæ of the same or closely allied species might develop in
most dissimilar ways according to the conditions in which they
found themselves.[526] His classical case
of 348"pœcilogeny" was that of the shrimp
Palæmonetes varians, the fresh-water form of which
develops in an entirely different way from the salt-water form.
Experimental workers indeed were inclined to rule the law out of
account, to disregard completely the historical element in
development, and this was perhaps the chief weakness of the
neo-vitalist systems which took their origin in this experimental
work.
From the side also of descriptive morphology the biogenetic law
underwent a critical revision. It was studied as a fact of
embryology and without phylogenetic bias by men like Oppel, Keibel,
Mehnert, O. Hertwig and Vialleton,[527] and they arrived at a critical estimate of it
very similar to that of von Baer.
Theoretical objections to the biogenetic law had been raised
from time to time by many embryologists, but the positive testing
of it by the comparison of embryos in respect of the degree of
development of their different organs starts with Oppel's work of
1891.[528] He studied a large
number of embryos of different species at different stages of their
development, and determined the relative time of appearance of the
principal organs and their relative size. His results are
summarised in tabular form and have reference to all the more
important organs. He was led to ascribe a certain validity to the
biogenetic law, but he drew particular attention to the very
considerable anomalies in the time of appearance which are shown by
many organs, anomalies which had been classed by Haeckel under the
name of heterochronies.
Oppel's main conclusions were as follows:—"There are found
in the developmental stages of different Vertebrates 'similar
ontogenetic series,' that is to say, Vertebrates show at definite
stages similarities with one another in the degree of development
of the different organs. Early stages resemble one another, so also
do later stages; equivalent stages of closely allied species
resemble one another, and older stages of lower animals resemble
younger stages of 349higher animals; young stages are more alike
than old stages.... The differences which these similar series show
(for which reason they cannot be regarded as identical) may be
designated as temporal disturbances in the degree of development of
the separate organs or organ-systems. Some organs show very
considerable temporal dislocations, others a moderate amount,
others again an inconsiderable amount. Among the developmental
stages of various higher animals can be found some which correspond
to the ancestral forms and also to the lower types which resemble
these ancestral forms. On the basis of the tabulated data here
given there can be distinguished with certainty in the ontogeny of
Amniotes a pro-fish stage, a fish-stage, a land-animal stage, a
pro-amniote stage, and following on these a fully developed
reptile, bird or mammal stage."[529]
Oppel's methods were employed by Keibel[530] in his investigations on the development of the
pig, which formed the model for the well-known series of
Normentafeln of the ontogeny of Vertebrates which were
issued in later years under Keibel's editorship. Keibel was more
critical of the biogenetic law than Oppel, and he held that the
ancestral stages distinguished by Oppel could not be satisfactorily
established. He suggested an interesting explanation of
heterochrony in development, according to which the premature or
retarded appearance of organs in ontogeny stands in close relation
with the time of their entering upon functional activity. Thus in
many mammals the mesodermal part of the allantois often appears
long before the endodermal part, though this is phylogenetically
older. This Keibel ascribes to the fact that the endodermal part is
almost functionless. "One can directly affirm," he writes, "that
the time of appearance of an organ depends in an eminent degree
upon the time when it has to enter upon functional 350activity. This
moment is naturally dependent upon the external conditions. Among
the highest Vertebrates, the mammals, the traces of phylogeny shown
in ontogeny are to a great extent obliterated through the
adaptation of ontogeny to the external conditions, and through the
modifications which the germs of more highly organised animals
necessarily exhibit from the very beginning as compared with germs
which do not reach such a high level of development" (p. 754,
1897).
Study of individual variation in the time of appearance of the
organs in embryos of the same species was prosecuted with
interesting results by Bonnet,[531] Mehnert,[532] and Fischel.[533] Fischel found that variability was greatest
among the younger embryos, and became progressively less in later
stages. Like von Baer (supra, p. 114) he inferred that
regulatory processes were at work during development which brought
divergent organs back to the normal and enabled them to play their
part as correlated members of a functional whole.
Important theoretical views were developed by Mehnert[534] in a series of publications appearing from
1891 to 1898. Like Keibel, Mehnert emphasised the importance of
function in determining the late or early appearance of organs, but
he conceived the influence of function to be exerted not only in
ontogeny, but also throughout the whole course of phylogeny, by
reason of the transmission to descendants of the effects of
functioning in the individual life.
In his paper of 1897 Mehnert details the results of an extensive
examination of the development of the extremities throughout the
Amniote series. He finds that in all cases a pentadactylate
rudiment is formed, even in those forms in 351which only a few of the
elements of the hand or foot come to full development. But whereas
in forms with a normally developed hand, e.g. the tortoise
and man, all the digits develop and differentiate at about the same
rate, in forms which have in the adult reduced digits, e.g.
the ostrich and the pig, these vestigial digits undergo a very slow
and incomplete differentiation, while the others develop rapidly
and completely. He draws a general distinction between organs that
are phylogenetically progressive and such as are phylogenetically
regressive, and seeks to prove that progressive organs show an
ontogenetic acceleration and regressive organs a retardation.[535] The acceleration or
retardation affects not only the mass-growth of the organs, but
also their histological differentiation.
Now between progression and functioning and between regression
and functional atrophy there is obviously a close connection. Loss
of function is well known to be one of the chief causes of the
degeneration of organs in the individual life, and on the other
hand, as Roux has pointed out, all post-embryonic development is
ruled and guided by functioning. It is thus in the long run
functioning that brings about phylogenetic progression, absence of
functional activity that causes phylogenetic regression. This comes
about through the transmission of acquired functional characters, a
transmission which Mehnert conceives to be extraordinarily accurate
and complete.
In general Mehnert adopts the functional standpoint of Cuvier,
von Baer, and Roux. His considered judgment as to the phylogenetic
value of the biogenetic law closely resembles that formed by von
Baer, for he admits recapitulation only as regards the single
organs, not as regards the organism as a whole. He has, however,
much more sympathy with the 352law than either Keibel or Oppel, though he
agrees that it cannot be used for the construction of ancestral
trees. But he ascribes to it as a fact of development considerable
importance. The following passage gives a good summary of his view
as to the scope and validity of the law. "The biogenetic law has
not been shaken by the attacks of its opponents. The assertion is
still true that individual organogenesis is exclusively dependent
on phylogeny. But we must not expect to find that all the stages in
the development of the separate organs, which coexisted in any
member of the phylogenetic series, appear at the same time
in the individual ontogeny of the descendants, because each organ
possesses its own specific rate of development. In this way it
comes about naturally that organs which become differentiated
rapidly, as, for example, the medullary tube, as a rule dominate
earlier periods of ontogeny than do the organs of locomotion. For
the same reason the cerebral hemispheres of man are almost as large
in youth as in maturity. The picture which an embryo gives is not a
repetition in detail of one and the same phylogenetic stage; it
consists rather of an assemblage of organs, some of which are at a
phyletically early stage of development, while others are at a
phyletically older stage."[536]
A different line of attack was that adopted by O. Hertwig in a
series of papers, which contain also what is perhaps the best
critical estimate of the present position and value of descriptive
morphology.[537]
It had not escaped the notice of many previous observers that
quite early embryos not infrequently show specific characters even
before the characters proper to their class, order and genus are
developed—in direct contradiction of the law of von Baer.
Thus L. Agassiz[538] had remarked 353in 1859
that specific characteristics were often developed precociously.
"The Snapping Turtle, for instance, exhibits its small crosslike
sternum, its long tail, its ferocious habits, even before it leaves
the egg, before it breathes through lungs, before its derm is
ossified to form a bony shield, etc.; nay, it snaps with its gaping
jaws at anything brought near, when it is still surrounded by its
amnion and allantois, and its yolk still exceeds in bulk its whole
body" (p. 269).
Wilhelm His,[539] in the course of an
acute and damaging criticism of the biogenetic law as enunciated by
Haeckel, showed clearly that by careful examination the very
earliest embryos of a whole series of Vertebrates could be
distinguished with certainty from one another. "An identity in
external form of different animal embryos, despite the common
affirmation to the contrary, does not exist. Even at early stages
in their development embryos possess the characters of their class
and order, nay, we can hardly doubt, of their species and sex, and
even their individual characteristics" (201).
This specificity of embryos was affirmed with even greater
confidence by Sedgwick in a paper critical of von Baer's law.[540] He wrote:—"If v.
Baer's law has any meaning at all, surely it must imply that
animals so closely allied as the fowl and duck would be
indistinguishable in the early stages of development; and that in
two species so closely similar that I was long in doubt whether
they were distinct species, viz., Peripatus capensis and
Balfouri, it would be useless to look for embryonic
differences; yet I can distinguish a fowl and a duck embryo on the
second day by the inspection of a single transverse section through
the trunk, and it was the embryonic differences between the
Peripatuses which led me to establish without hesitation the two
separate species.... I need only say ... that a species is distinct
and distinguishable from its allies from the very earliest stages
all through the development, although these embryonic differences
do not necessarily implicate the same organs as do the adult
differences" (p. 39).
354Hertwig interprets this fact of the specific
distinctness of closely allied embryos in the light of the
preformistic conception of heredity. According to this view the
whole adult organisation is represented in the structure of the
germ-plasm contained in the fertilised ovum, from which it follows
that the ova of two different species, and also their embryos at
every stage of development, must be as distinct from one another as
are the adults themselves, even though the differences may not be
so obvious. If this be the case there can be no real recapitulation
in ontogeny of the phylogeny of the race, for the egg-cell
represents not the first term in phylogeny, but the last. The
egg-cell is the organism in an undeveloped state; it has a
vastly more complicated structure than was possessed by the
primordial cell from which its race has sprung, and it can in no
way be considered the equivalent of this ancestral cell.
Hertwig puts this vividly when he says that "the hen's egg is no
more the equivalent of the first link in the phylogenetic chain
than is the hen itself" (p. 160, 1906, b).
If ontogeny is not a recapitulation of phylogeny, how is it that
the early embryonic stages are so alike, even in animals of widely
different organisation? Hertwig's answer to this is very
interesting. He takes the view that many of the processes
characterising early embryonic development are the means
necessarily adopted for attaining certain ends. Such are the
processes of segmentation, the formation of a blastula, of
cell-layers, of medullary folds where the nervous system is a
closed tube, the formation of the notochord as a necessary
condition of the development of the vertebral column, and so on.
"Looked at from this standpoint it cannot surprise us that in all
animal phyla the earliest embryonic processes take place in similar
fashion, so that we observe the occurrence both in Vertebrates and
Invertebrates of a segmentation-process, a morula-stage, a blastula
and a gastrula. If now these developmental processes do not depend
on chance, but, on the contrary, are rooted in the nature of the
animal cell itself, we have no reason for inferring from the
recurrence of a similar segmentation-process, morula, blastula, and
gastrula in all classes of the animal kingdom the common descent of
all animals from one 355blastula-like or gastrula-like ancestral
form. We recognise rather in the successive early stages of animal
development only the manifestation of special laws, by which the
shaping of animal forms (as distinct from plant forms) is brought
about" (p. 178, 1906, b).
"The principal reason why certain stages recur in ontogeny with
such constancy and always in essentially the same manner is that
they provide under all circumstances the necessary pre-conditions
through which alone the later and higher stages of ontogeny can be
realised. The unicellular organism can by its very nature transform
itself into a multicellular organism only by the method of
cell-division. Hence, in all Metazoa, ontogeny must start with a
segmentation-process, and a similar statement could be made with
regard to all the later stages" (p. 57, 1906, a).
Similarities in early development are therefore no evidence of
common descent, and in the same way the resemblances of adult
animals, subsumed under the concepts of homology and the unity of
plan, are not necessarily due to community of descent, but may also
be brought about by the similarity or identity of the laws which
govern the evolution of these animals. In the absence, therefore,
of positive evidence as to the actual lines of descent (to be
obtained only from palæontology), homological resemblance
cannot be taken as proof of blood relationship, for homology is a
wider concept than homogeny. The only valid definition of homology
is that adopted in pre-evolutionary days, when those organs were
considered homologous "which agree up to a certain point in
structure and composition, in position, arrangement, and relation
to the neighbouring organs, and accordingly possess identical
functions and uses in the organism" (p. 151, 1906, b).
The concept of homology has thus a value quite independent of
any evolutionary interpretation which may be superadded to it.
"Homology is a mental concept obtained by comparison, which under
all circumstances retains its validity, whether the homology finds
its explanation in common descent or in the common laws that rule
organic development" (p. 151, 1906, b). As A. Braun long ago
pointed out, "It is not descent which decides in matters 356of
morphology, but, on the contrary, morphology which has to decide as
to the possibility of descent."[541]
Hertwig, in a word, reverts to the pre-evolutionary conception
of homology. "We see in homology," he writes, "only the expression
of regularities (Gesetzmässigkeiten) in the
organisation of the animals showing it, and we regard the question,
how far this homology can be explained by common descent and how
far by other principles, as for the present an open one, requiring
for its solution investigations specially directed towards its
elucidation" (p. 179, 1906, b).
Holding, as he does, that no definite conclusions can be drawn
from the facts of comparative anatomy and embryology as to the
probable lines of descent of the animal kingdom, Hertwig accords
very little value to phylogenetic speculation. It is, he admits,
quite probable that the archetype of a class represents in a
general sort of way the ancestral form, but this does not, in his
opinion, justify us in assuming that such generalised types ever
existed and gave origin to the present-day forms. "It is not
legitimate to picture to ourselves the ancestral forms of the more
highly organised animals in the guise of the lower animals of the
present day—and that is just what we do when we speak of
Proselachia, Proamphibia and Proreptilia" (p. 155, 1906, b).
He rejects on the same general grounds the evolutionary dogma of
monophyletic or almost monophyletic descent, and admits with
Kölliker, von Baer, Wigand, Naegeli and others that evolution
may quite well have started many times and from many different
primordial cells.
There is indeed a great similarity between the views developed
by O. Hertwig and those held by the older critics of
Darwinism—von Baer, Kölliker, Wigand, E. von Hartmann
and others. It is true the philosophical standpoint is on the whole
different, for while many of that older generation were vitalists
Hertwig belongs to the mechanistic school.
But both Hertwig and the older school agree in pointing out the
petitio principii involved in the assumption that the 357archetype
represents the ancestral form; both reject the simplicist
conception of a monophyletic evolution (which may be likened to the
"one animal" idea of the transcendentalists); both admit the
possibility that evolution has taken place along many separate and
parallel lines, and explain the correspondences shown by these
separate lines by the similarity of the intrinsic laws of
evolution; finally, both emphasise the fact that we know nothing of
the actual course of evolution save the few indications that are
furnished by palæontology, and both insist upon the unique
importance of the palæontological evidence.[542]
It was a curious but very typical characteristic of evolutionary
morphology that its devotees paid very little attention to the
positive evidence accumulated by the palæontologists,[543] but shut themselves up in their tower of
ivory and went on with their work of constructing ideal
genealogies. It was perhaps fortunate for their peace of mind that
they knew little of the advances made by palæontology, for
the evidence acquired through the study of fossil remains was
distinctly unfavourable to the pretty schemes they evolved.
As Neumayr, Zittel, Depéret, Steinmann and others have
pointed out, the palæontological record gives remarkably
little support to the ideal genealogies worked out by
morphologists. There is, for instance, a striking absence of
transition forms between the great classificatory groups. A few
types are known which go a little way towards bridging over the
gaps—the famous Archæopteryx, for
example—but these do not always represent the actual
phylogenetic links. There is an almost complete absence of the
archetypal ancestral forms which are postulated by evolutionary
morphology. Amphibia do not demonstrably evolve from an archetypal
Proamphibian, nor do mammals derive from a single generalised
Promammalian type. Few of the hypothetical ancestral types imagined
by Haeckel have ever 358been found as fossils. The great
classificatory groups are almost as distinct in early fossiliferous
strata as they are at the present day. As Depéret says in
his admirable book,[544] in the course of a presentation of the matured
views of the great Karl von Zittel, "We cannot forget that there
exist a vast number of organisms which are not connected by any
intermediate links, and that the relations between the great
divisions of the animal and vegetable kingdoms are much less close
than the theory [of evolution] demands. Even the
Archæopteryx, the discovery of which made so much stir and
appeared to establish a genetic relation between classes so
distinct as Birds and Reptiles, fills up the gap only imperfectly,
and does not indicate the point of bifurcation of these two
classes. Intermediate links are lacking between Amphibia and
Reptiles. Mammals, too, occupy an isolated position, and no
zoologist can deny that they are clearly demarcated from other
Vertebrates; indeed, no fossil mammal is certainly known which
comes nearer to the lower Vertebrates than does Ornithorhynchus at
the present day" (p. 115).
To take a parallel from the Invertebrata, B. B. Woodward,[545] after discussing the phylogeny of the
Mollusca as worked out by the morphologists and comparing it with
the probable actual course of the evolution of the group, as
evidenced by fossil shells, sums up as follows:—"The
lacunæ in our knowledge of the interrelationships of the
members of the various families and orders of Mollusca are slight
however, compared with the blank caused by the total absence from
palæontological history of any hint of passage forms between
the classes themselves, or between the Mollusca and their nearest
allies. Nor is this hiatus confined to the Molluscan phylum; it is
the same for all branches of the animal kingdom. There is
circumstantial evidence that transitional forms must have existed,
but of actual proof none whatever. All the classes of Mollusca
appear fully fledged, as it were. No form has as yet been
discovered of which it could be said that it in any way approached
the 359hypothecated prorhipidoglossate mollusc,
still less one linking all the classes" (p. 79).
Pointing in the same direction as the absence of transitional
forms is the undeniable fact that all the great groups of animals
appear with all their typical characters at a very early geological
epoch. Thus, in the Silurian age a very rich fauna has already
developed, and representatives are found of all the main
Invertebrate groups—sponges, corals, hydroid colonies, five
types of Echinoderms, Bryozoa, Brachiopods, Worms, many types of
Mollusca and Arthropoda. Of Vertebrates, at least two types of fish
are present—Ganoids and Elasmobranchs. In the very earliest
fossiliferous rocks of all, the Precambrian formation, there are
remains of Molluscs, Trilobites and Gigantostraca, similar to those
which flourished in Cambrian and Silurian times.
The contributions of palæontology to the solution of the
problems of descent posed by morphology are, however, not all of
this negative character. The law of recapitulation is in some
well-controlled cases triumphantly vindicated by
palæontology. Thus Hyatt and others found that in Ammonites
the first formed coils of the shell often reproduce the characters
belonging to types known to be ancestral, and what is more they
have demonstrated the actual occurrence of the phenomenon known as
acceleration or tachygenesis, often postulated by speculative
morphologists.[546] This is the
tendency universally shown by embryos to reproduce the characters
of their ancestors at earlier and earlier stages in their
development.
The most valuable contribution made by palæontologists to
morphology and to the theory of evolution arose out of the careful
and methodical study of the actual succession of fossil forms as
exemplified in limited but richly represented groups. Classical
examples were the researches of Hilgendorf[547] on the evolution of Planorbis multiformis
in the lacustrine deposits of Steinheim, those of Waagen[548] on the 360phylogeny of Ammonites
subradiatus, and the work of Neumayr and Paul[549] on Paludina (Vivipara).
These investigations demonstrated that it was possible to follow
out step by step in superjacent strata the actual evolution of
fossil species and to establish the actual "phyletic series."
To take an example from among the Vertebrates, Depéret
has shown (loc. cit., pp. 184-9), that the European
Proboscidea, belonging to the three different types of the
Elephants, Mastodons and Dinotheria, have evolved since the
Oligocene epoch along five distinct but continuous lines. The
Dinotherian stock is represented at the beginning of the Miocene by
the relatively small form D. cuvieri; this changes
progressively throughout Miocene times into D. laevius,
D. giganteum, and D. gigantissimum. Among the
Mastodons two quite distinct phyletic series can be distinguished,
the first commencing with Palæomastodon beadnelli of
the Oligocene, and evolving between the Miocene and Pliocene into
Mastodon arvernensis, after traversing the forms M.
angustidens and M. longirostris, the second starting
with the M. turicensis of the Lower Miocene and evolving
through M. borsoni into the M. americanus of the
Quaternary. The phyletic series of the true elephants in Europe are
relatively short, and go back only to the Quaternary, Elephas
antiquus giving origin to the Indian elephant, E.
priscus to the African.
The careful study of phyletic series brought to light the
significant fact that these lines of filiation tend to run for long
stretches of time parallel to, and distinct from one another,
without connecting forms. This is clearly exemplified in the case
of the Proboscidea, and many other examples could be quoted. Almost
all rich genera are polyphyletic in the sense that their component
species evolve along separate and parallel lines of descent.[550] "Such great genera as
the genus Hoplites among the Ammonites, the genus
Cerithium among the Gastropoda, the genus Pecten or
the genus 361Trigonia among the Lamellibranchs,
each comprise perhaps more than twenty independent phyletic series"
(Depéret, p. 200).
Variation along the phyletic lines is gradual[551] and determinate, and appears to obey
definite laws. The earliest members of a phyletic series are
usually small in size and undifferentiated in structure, while the
later members show a progressive increase in size and complexity.
Rapid extinction often supervenes soon after the line has reached
the maximum of its differentiation.
The general picture which palæontology gives us of the
evolution of the animal kingdom is accordingly that of an immense
number of phyletic lines which evolve parallel to one another, and
without coalescing, throughout longer or shorter periods of
geological times. "Each of these lines culminates sooner or later
in mutations of great size and highly specialised characters, which
become extinct and leave no descendants. When one line disappears
by extinction it hands the torch, so to speak, to another line
which has hitherto evolved more slowly, and this line in its turn
traverses the phases of maturity and old age which lead it
inevitably to its doom. The species and genera of the present day
belong to lines that have not reached the senile phase; but it may
be surmised that some of them, e.g. elephants, whales, and
ostriches, are approaching this final phase of their existence"
(Depéret, p. 249).
It is one of the paradoxes of biological history that the
palæontologists have always laid more stress upon the
functional side of living things than the morphologists, and have,
as a consequence, shown much more sympathy for the Lamarckian
theory of evolution. The American palæontologists in
particular—Cope, Hyatt, Ryder, Dall, Packard,
Osborn—have worked out a complete neo-Lamarckian theory based
upon the fossil record.
The functional point of view was well to the fore in the works
of those great palæontologists, L. Rütimeyer (1825-1895)
and V. O. Kowalevsky (1842-83), who seem to have carried on the
splendid tradition of Cuvier. Speaking of 362Kowalevsky's classical
memoir, Versuch einer natürlichen Classification der
fossilen Hufthiere, Osborn[552] writes:—"This work is a model union of the
detailed study of form and function with theory and the working
hypothesis. It regards the fossil not as a petrified skeleton, but
as having belonged to a moving and feeding animal; every joint and
facet has a meaning, each cusp a certain significance. Rising to
the philosophy of the matter, it brings the mechanical perfection
and adaptiveness of different types into relation with environment,
with changes of herbage, with the introduction of grass. In this
survey of competition it speculates upon the causes of the rise,
spread, and extinction of each animal group. In other words, the
fossil quadrupeds are treated biologically—so far as
is possible in the obscurity of the past" (p. 8). The same high
praise might with justice be accorded to the work of Cope on the
functional evolution of the various types of limb-skeleton in
Vertebrates, and on the evolution of the teeth as well as to the
work of other American palæontologists, including Osborn
himself.
Osborn's law of "adaptive radiation," which links on to Darwin's
law of divergence,[553] constitutes a
brilliant vindication of the functional point of view. "According
to this law each isolated region, if large and sufficiently varied
in its topography, soil, climate, and vegetation, will give rise to
a diversified mammalian fauna. From primitive central types
branches will spring off in all directions, with teeth and
prehensile organs modified to take advantage of every possible
opportunity of securing food, and in adaptation of the body, limbs
and feet to habitats of every kind, as shown in the diagram [on p.
363]. The larger the region and the more diverse the conditions,
the greater the variety of mammals which will result.
"The most primitive mammals were probably small insectivorous or
omnivorous forms, therefore with simple, short-crowned teeth, of
slow-moving, ambulatory, terrestrial, or arboreal habit, and with
short feet provided with claws. In seeking food and avoiding
enemies in different habitats 363the limbs and feet radiate in four diverse
directions; they either become fossorial or adapted to
digging habits, natatorial or adapted to amphibious
and finally to aquatic habits, cursorial or adapted
to swift-moving, terrestrial progression, arboreal or
adapted to tree life. Tree life leads, as its final stage, into
the parachute types of the flying squirrels and phalangers, or
into the true flying types of the bats.... Similarly in the case of
the teeth, insectivorous and omnivorous types appear to be more
central and ancient than either the exclusively carnivorous or
herbivorous types. Thus the 364extremes of carnivorous adaptation, as in
the case of the cats, of omnivorous adaptation, as in the case of
the bears, of herbivorous adaptation, as in the case of the horses,
or myrmecophagous adaptation, as in the case of the anteaters, are
all secondary" (loc. cit., pp. 23-4).
We have now reached the end of our historical survey of the
problems of form. What the future course of morphology will be no
one can say. But one may hazard the opinion that the present
century will see a return to a simpler and more humble attitude
towards the great and unsolved problems of animal form. Dogmatic
materialism and dogmatic theories of evolution have in the past
tended to blind us to the complexity and mysteriousness of vital
phenomena. We need to look at living things with new eyes and a
truer sympathy. We shall then see them as active, living,
passionate beings like ourselves, and we shall seek in our
morphology to interpret as far as may be their form in terms of
their activity.
This is what Aristotle tried to do, and a succession of
master-minds after him. We shall do well to get all the help from
them we can.
365
INDEX
- Actinozoan Theory of Vertebrate
Descent, 299-300
- Adaptation as Conservative Principle—
- Adaptation, Ecological—
- Von Baer, 123
- H. Milne-Edwards, 199
- Lamarck, 221, 222, 223, 224, 227
- Treviranus, 225 f.n.
- C. Darwin, 231-2, 235, 239
- Haeckel, 248, 263
- Gegenbaur, 263
- V.O. Kowalevsky, 362
- Osborn, 362-4
- Adaptation, Ecological, and Classification—
- Adaptation of Parts. See "Correlation, Functional," and
"Conditions of Existence"
- Adaptive Radiation (Osborn), 362-4
- Agassiz, A., 288 f.n., 295
- Agassiz, L.—
- Criticism of Vertebral Theory of Skull, 157
- Membrane and Cartilage Bones, 164
- Transcendentalism, 203
- Classification, 203 f.n.
- Three-fold Parallelism, 203, 255
- Influence on Darwin, 238
- Specific Distinctness of Embryos, 353
- Albertus Magnus, 17
- Alcmæon, 1
- Aldrovandus, 18
- Allman, 209
- Analogy. _See also_ Homology.
- Aristotle, 8-10
- Owen, 108
- Haeckel, 251
- Gegenbaur, 266
- Lankester, 267
- Anaxagoras, 14
- Anaximander, 14
- Anaximenes, 1
- Animal and Vegetative Lives—
- Aristotle, 16, 32
- Buffon, 26-7
- Bergson, 26 f.n.
- Cuvier, 2626, 32
- Bichat, 27-9
- Oken, 29694
- K. G. Carus, 94
- Von Baer, 116, 123, 131
- Remak (Sensory and trophic layers), 210
- Gegenbaur, 263
- Annelid Theory of Vertebrate Descent, 274-85, 301
- Archetype, Anatomical, 246, 302-3
- E. Geoffroy, 54, 67
- Owen, 104-7, 110
- J. V. Carus, Huxley, 204
- C. Darwin, 238 f.n.
- Archetype, Anatomical, as Ancestral—
- C. Darwin, 235, 247
- Haeckel, 251
- Gegenbaur, 265
- Sedgwick, 300
- Criticism of this idea—
- Archetype, Embryological, 168, 246, 302-3
- Archetype, Embryological, as Ancestral—
- Arendt, 162
- Aristotle, 2-16, 17, 345, 364
- _Historia Animalium_, 2
- _De Partibus Animalium_, 2, 9
- Knowledge of Animals, 3, 4
- Comparative Embryology, 4
- Classification of Animals, 4-6
- Unity of Plan, 6-7, 10
- Homology and Analogy, 7-10
- Teleology and Correlation, 10-12
- Law of Compensation, 11
- Division of Labour, 12
- Degrees of Composition--homogeneous and heterogeneous parts, 12-14, 169
- Law of Development (Von Baer), 14
- Scale of Beings, 14-16
- Functional attitude, 15-16, 197
- Animal and Vegetative Lives, 16, 32
- Ascidian Theory of Vertebrate Descent, 269-73, 304
- Atomists, 16
- Atomists, "Biological," 192-4
- Audouin, V.—
- Unity of plan in Arthropods, 85-6
- Law of Compensation, 86
- Marine Zoology, 195
- Autenrieth, 90, 96
- Avicenna, 17
-
- Babák, E., 333
- Baer, K. E. von, 113-32, 133, 251, 304, 345, 356
- Founder of Embryology, 113
- Entwickelungsgeschichte der Thiere, 114
- Regulation of Development, 114, 350
- Development as Differentiation, 115, 128
- Germ-Layer Theory, 115-6, 118-119, 208-9, 296
- Morphological Differentiation, 116-7
- Histological Differentiation, 117-8
- Tissues and Germ-Layers, 118
- Double symmetrical Development, 118, 279
- Criticism of Meckel-Serres Law, 120-3, 304
- Theory of Types, 123-4, 289, 291
- Law of Development, 124-6
- Embryological Criterion, 126-8, 132, 138
- Embryological Archetype, 126, 132
- Types of Development, 127-8
- Von Baer and Cuvier, 128-30
- Functional attitude, 129
- Relation to Transcendentalists, 129, 131
- Criticism of Scale of Beings, 130
- Vertebral Theory of Skull, 131, 142
- Serial Homology, 131-2
- Gill-slits, Gill-arches and Aortic arches, 135-6, 146
- Membrane and Cartilage Bones, 162-3
- Degrees of Composition, 172
- Ova of Mammals, 175-6
- Segmentation of Ovum, 186
- Criticism of Evolution Theory, 229, 242
- Influence on Darwin, 236, 238
- Criticism of Darwinism, 242
- 367Teleology and Correlation, 242
- On Ascidians, 271
- Baer's Law. See "Development, Von Baer's Law"
- Bagge, 187
- Balanoglossus Theory of Vertebrate Descent, 285-7
- Balbiani, 330
- Balfour, F. M., 247, 299
- Annelid Theory, 282-4
- Gastrulation and Gastræa Theory, 295
- Mesoderm, 296 f.n.
- Cœlom, 297
- Barfurth, D., 330
- Barry, M., 186, 188
- Bateson, W.—
- Metamerism, Vegetative Repetition, 286
- Balanoglossus Theory, 286-7
- On Phylogenetic Speculation, 302
- Beard, J., 285
- Belon, 18
- Beneden van, and Julin, 271, 285, 346
- Bensley, A. B., 311 f.n.
- Bergmann, 187
- Bergson, H., 26 f.n., 341, 345
- Bernard, Claude, 195, 314
- Bert, P., 315
- Bichat, X., 27-30, 118, 132, 169, 178, 263
- Animal and Vegetative Lives, 27-9
- "General Anatomy," 29-30
- Vie propre of Tissues, 30
- Biogenetic Law. See "Development, Haeckel's Law"
- Bischoff, 138
- Blainville, de, 96, 128, 141, 199 f.n.
- Bojanus, 96, 97
- Bonnet, C.—
- Bonnet, R., 350
- Bonnier, G., on Albertus Magnus, 17
- Born, G., 330
- Boveri, T., 270 f.n., 333
- Braem, 347 f.n.
- Braun, A., 355
- Breschet, 138, 173
- Bronn, H. G., 200-3, 248
- Naturphilosophie, 201
- Functional attitude, 201-3
- Geometry of Organism, 201, 249
- Theory of Types, 202
- Principle of Connections, 202
- Intrinsic Laws of Evolution, 202
- Division of Labour, 202
- Ecological Adaptation and Classification, 203
- Brown, R., 171
- Bruch, C., 203 f.n.
- Büchner, 194, 248
- Buffon, 24-7, 336
- Scale of Beings, 24, 215
- Unity of Plan, 24
- Evolution, 24-5, 214
- Classification, 25-6
- Animal and Vegetative Lives, 26-7
- Homology and Analogy, 27
- Burckhardt, R., 3 f.n., 268 f.n.
- Burdin, 96
- Burmeister, 249 f.n.
- Butler, S., 226 f.n., 313, 335-42
- Relation to Lamarck, 335-7
- Psychological Vitalism, 336-41
- Heredity and Memory, 337-41
- The Two Stages of Development, 337-9
- Consciousness and Habit, 337-9
- Recapitulation Theory, 339-40
- Teleology, 341
-
- Cabanis, 215
- Camper, P., 45, 46
- Carter, 293 f.n.
- 368Carus, J. V..—
- Criticism of Embryological Criterion, 167
- Morphology and Physiology, 194
- Vertebral Theory of Skull, 203
- On Archetype, 204
- Evolution, 230
- Carus, K. G.—
- Law of Parallelism, 94, 249
- Vertebral Theory, 96
- Geometry of Skeleton, 98-100
- Splanchnoskeleton, 98, 140
- Causal Morphology, 312-3, 315-34
- Cell-Theory—
- Schwann, 169, 173-86,
188
- C. F. Wolff, 170
- Schleiden, 170-2
- Criticism of Schwann-Schleiden Theory, 185-8
- Cell-Theory and Germ-Layer Theory—
- Cell-Theory as Disintegrative—
- Schwann, 180-5, 248
- Vogt, 190-1
- Virchow, 191
- Haeckel, 248
- Criticism of this idea—
- Reichert, 192-3, 194
- J. V.. Carus, 194
- Sedgwick, Whitman, 346
- Cell-Theory, Influence on Morphology, 190
- Cenogenesis, 258-9, 323
- Chabry, 331
- Child, C. M., 333
- Chun, C, 317, 332
- Classification of Animals—
- Aristotle, 4-6
- Rondeletius, Aldrovandus, Gesner, 18
- Linnæus, 22
- Buffon, 25-6
- Cuvier, 39-41
- E. Geoffroy, 60
- L. Agassiz, 203 f.n.
- Lamarck, 216-7, 227,
228
- Classification and Ecological Adaptation (Bronn), 203
- Classification as Genealogical—
- Buffon, 24-5
- Lamarck, 218, 228
- C. Darwin, 233, 234,
247
- Haeckel, 250-1, 254
- Criticism of this idea, 303, 304,
- Classification, Phylogenetic—
- Claus, 259
- Co-adaptation, 326 f.n.
- Cœlom—
- Cœlom, Theory of, 295-301
- Cohen, 189
- Coiter, 18
- Colucci, 346
- Compensation, Law of—
- Aristotle, 11
- Goethe, 49
- E. Geoffroy, 72-3
- Audouin, 86
- German Transcendentalists, 100
- Condillac, 215
- Conditions of Existence, Principle of—
- Cuvier, 34, 75-6, 239
- Gegenbaur, 263-4
- Roux, 324, 326
- Spencer, Weismann, 326 f.n.
- Disregard for—
- Conklin, 333
- Connections, Principle of—
- Goethe, 47
- E. Geoffroy, 53-4, 62-3, 71, 74, 261
- Audouin, 85
- 369German Transcendentalists, 100
- J. F. Meckel, 101
- Owen, 107-8
- Bronn, 202
- C. Darwin, 234-5
- Gegenbaur, 261
- Semper, 279
- In Embryology, 168
- Main Principle of Morphology, 246, 302
- Convergence—
- Milne-Edwards, 199
- I. Geoffroy St Hilaire, 199 f.n., 206
- C. Darwin, 236
- Friedmann, Willey, Vialleton, 306
f.n.
- Convergence, Rejected by Evolutionary Morphologists, 305, 312
- Cope, E. D., 342, 357
f.n., 361, 362
- Correlation, Functional—
- Aristotle, 10-12
- Cuvier, 35-8, 239, 241
- E. Geoffroy, 77
- Von Hartmann, 240-1
- Rádl, 240 f.n., 241
- Von Baer, 242
- Gegenbaur, 264
- Disregarded by—
- Coste, 138, 176, 187
- Crampton, 332
- Cunningham, J. T., 284
- Cuvier, 26, 31-44, 89, 196, 197, 199 f.n., 278, 345, 361
- Functional attitude, 31-6, 65, 75-8, 200, 305
- Animal and Vegetative Lives, 32
- Degrees of Composition, 32-3
- Teleology, 33-5
- Functional Adaptedness, 33-5, 324
- Principle of Conditions of Existence, 34,
75-6, 239
- Correlation, 35-8, 239, 241
- Metabolism, 38
- Adaptation as Conservative Principle, 39, 76
- Classification, 39-41
- Principle of Subordination of Characters, 40
- Criticism of Scale of Beings, 39-40, 130
- Type Theory, 41, 124,
289, 291
- Criticism of Evolution-Theory, 41-4, 129, 304
- Variation, Limits of, 42
- Palæontological Succession, 43
- Polemic with Geoffroy, 64-5, 74-8
- Criticism of Vertebral Theory of Skull, 97-8
- Influence on J. F. Meckel, 101
- Criticism of Meckel-Serres Law, 129-30, 304
- As Embryologist, 130
- Criticism of Lamarck, 228
- Cytology, 346
- Cytoplasm of Egg, Organ-forming Stuffs, 332-3
-
- Dall, 361
- D'Alton, 113
- Dareste, C., 315
- Darwin, Charles, 78, 230-41, 271, 304, 307, 336, 362
- Systematist and Field Naturalist, 230, 231
- Palæontological Succession, 231
- Ecological Adaptation, 231-2, 235, 239
- Species Problem, 231
- Functional Adaptation, Disregard for, 232,
238-41
- Classification as genealogical, 233, 234, 247
- Unity of Plan due to Community of Descent, 233, 234-5, 239, 247
- Embryological Archetype as ancestral, 233,
236-7
- 370
Rejects Meckel-Serres Law, 233, 236
- Interpretation of Vestigial Organs, 233, 237
- Organism as Historical Being, 233, 308
- Rejects Scale of Beings, 234
- Homology, 234-5, 247
- Principle of Connections, 234-5
- Anatomical Archetype as ancestral, 235, 247
- Von Baer's Law interpreted phylogenetically, 236-7
- Modifications inherited at corresponding age, 237
- Monophyletism and Polyphyletism, 238
- Causes of Success, 238, 241
- Darwin, Erasmus, 214, 226 f.n., 229, 336
- Darwin, Sir Francis, 344
- Daubenton, 26
- Degrees of Composition—
- Aristotle, 12-14, 169
- Glisson, 19
- Malpighi, 20
- Bichat, 29-30
- Cuvier, 32-3,
- Dujardin, 169, 188
- Von Baer, 172
- Effect of Invention of Microscope, 20
- Relation to Cell-Theory, 169
- Delage, 333
- Delage and Hérouard, 273 f.n.
- Delpino, 345
- Demaillet, 44
- Democritus, 16
- Depéret, C., 357
- On Cuvier, 43
- Absence of intermediary forms in Palæontology, 358
- Phyletic series and Polyphyletism, 360-1
- Development, Von Baer's Law—
- Aristotle, 14
- Von Baer, 124-6
- Prévost and Dumas, 125 f.n.
- Reichert, 149-50, 351
f.n.
- Milne-Edwards, 205-8
- Lereboullet, 206-8
- Criticised by—
- Phylogenetic Interpretation of—
- Development, Biogenetic Law (Haeckel)—
- Haeckel, 251, 253-9,
291-4
- F. Müller, 252-3, 254, 257
- Gegenbaur, 262
- Roux, 319
- Butler, 339-40
- Orr, 342
- Criticism of—
- Relation to Laws of Meckel-Serres and Von Baer, 254, 256, 257, 303, 309
- Relation to Heredity and Development, 312-3
- Influence of Causal Morphology, 347-8
- Palæontological Evidence for, 359
- Development, Meckel-Serres Law—
- Harvey, 18
- Hunter, 22
- E. Geoffroy, 69-70, 72
- Serres, 80-3, 94, 203-4, 205-6
- Kielmeyer, Autenrieth, Oken, 90
- 371
Tiedemann, 91
- J. F. Meckel, 91-3
- K. G. Carus, 94
- Criticism of—
- Analogy with Biogenetic Law, 254-7, 262, 303, 304, 309
- Development, Meckel-Serres Law, Theory of Three-fold
Parallelism—
- L. Agassiz, 230, 255
- Tiedemann, Vogt, 255 f.n.
- Haeckel, 254-5
- Development, The two periods of—
- Diogenes of Apollonia, 1
- Disintegration. See "Materialistic
Attitude"
- Division of Labour, Principle of—
- Aristotle, 12
- Milne-Edwards, 197-8
- Bronn, 202
- Gegenbaur, 264
- Dohrn, A., 269, 274-8
- Annelid Theory of Vertebrate Descent, 274-7, 303
- Principle of Function-Change, 276-8, 307
- Functional Attitude, 277-8, 307
- Formal Attitude, 306
- Döllinger, I., 113, 157
- Dollo, 311
- Donné, 173
- D'Orbigny, 43
- Driesch, H., 242, 331, 332, 333, 334, 345, 346-7
- Dugès, A., 86-8, 100, 134, 142, 146
- Unity of Plan, 87
- Polyzoic conception of Organism, 87-8
- Membrane and Cartilage Bones, 163
- Dujardin, 169, 188
- Dumas. See Prévost and Dumas
- Duméril, 96
- Dumortier, 173
- Dutrochet, 99 f.n., 130, 134
- Duverney, 19
-
- Ear-Ossicles, Homology of—
- Échelle des êtres. See "Scale of
Beings."
- Ehlers, 284
- Eisig, H., 284, 285
- Embryology, Comparative, Early Workers—
- Aristotle, 4, 113
- Fabricius, Harvey, 18, 113
- Malpighi, 20, 113
- Oken and Kieser, 90, 113
- Haller, C. F. Wolff, J. F. Meckel, Tiedemann, 113
- Embryology, Experimental, 317, 318, 330-3
- Embryological Archetype. See "Archetype,
Embryological"
- Embryological Criterion of Homology, 133-168, 347
- Goethe, 49
- E. Geoffroy, 72, 110
- Cuvier, 75, 110, 130
- Owen, 110-1
- Von Baer, 126-8, 132,
138
- Rathke, 138, 140-1
- J. Müller, 138
- Reichert, 138-9, 144-7, 163
- Vogt, 156-7
- Huxley, 158-9, 166
- Kölliker, 165-6
- Criticised by—
- Empedocles, 1, 15
- 372Engramm (Semon), 343
- Entwicklungsgesetz. See "Evolution, Intrinsic Laws
of"
- Entwicklungsmechanik, 315
- Erasistratus, 17
- Evolution Theory—
- Lucretius, 16
- Buffon, 24-5, 214
- Cuvier's criticism, 41-4, 129, 304
- E. Geoffroy, 66-9, 73, 228
- J. F. Meckel, 92-3, 215, 228
- Leibniz, 213
- Kant, 213-4
- Erasmus Darwin, 214, 229
- C. Bonnet, Oken, Robinet, Treviranus, 215
- Tiedemann, 215, 255
f.n.
- Lamarck, 215-29
- Von Baer, 229, 242
- I. Geoffroy St Hilaire, J. V.. Carus, 230
- Charles Darwin, 230-41
- Von Hartmann, 240-1, 244, 356
- Kölliker, 243
- Owen, 244
- Milne-Edwards, 244-5
- Haeckel, 250-9
- Gegenbaur, 265
- The Organism as an Historical Being, 308-13
- C. Darwin, 233, 308
- Haeckel, 252, 257
- Sedgwick, 308
- Roux, 313, 322-4
- Butler, 313, 336-41
- Evolution-Theory, Influence on Morphology, 302-13
- Evolution, Intrinsic Laws of, 241
- J. F. Meckel, 93
- Bronn, 202
- Von Baer, 229, 242,
356
- Kölliker, Naegeei, 243, 356
- Owen, 244
- Von Hartmann, 244, 356
- Milne-Edwards, 244-5
- O. Hertwig, 354-5, 356-7
- Wigand, 356
- Depéret, 361
-
- Fabricius, 18,
113
- Fallopius, 18
- Fischel, 346, 350
- Fischer, 328
- Fleischmann, 357 f.n.
- Flourens, 46, 315
- Fontana, 172
- Forbes, E., 196
- Formal Attitude, 246, 305
- Goethe, 49
- E. Geoffroy, 62-3, 71, 75-8, 305
- Haeckel, 249, 257, 260
- Gegenbaur, 261, 263
- Semper, 279
- Adopted by Evolutionary Morphologists, 302-8, 311-2, 314
- Hubrecht, 305-6
- Dohrn, 306
- Francé, R., 345
- Friedmann, 306 f.n.
- Fuld, 333
- Functional Adaptation, 316-7, 318, 320-9, 333, 344, 351
- Functional Attitude—
- Aristotle, 15-6, 197
- Bichat, 27-9
- Cuvier, 31-6, 65, 75-8, 200, 305
- Goethe, 49-50
- J. F. Meckel, 101
- Owen, 109, 110, 111
- Von Baer, 129
- Milne-Edwards, 195, 197-200
- J. Müller, Reichert, 200
- Bronn, 201-3
- Lamarck, 222-6, 307,
335
- Gegenbaur, 260, 263-4
- Dohrn, 277-8, 307
- Roux, 320-9, 335
- Houssay, 333
- Butler, 336-41
- G. Wolff, 346
- Driesch, 346-7
- Giard, 347
- 373
E. Schulz, 347 f.n.
- Keibel, 349-50
- Mehnert, 350-1
- American Palæontologists, 361, 362
- Rütimeyer, 361
- V. O. Kowalevsky, 361-2
- Osborn, 362-4
- Function-Change, Principle of—
- Fürbringer, M., 282 f.n., 284, 323 f.n.
-
- Galen, 17
- Gastræa Theory, 269, 288-95, 298, 299-301, 303
- Gastrula, Discovery of, 288
- Gaupp, E., 310 f.n.
- Gegenbaur, C., 247, 260-7, 271, 285, 286, 288 f.n.
- Division of Egg-nucleus, 188
- Functional Attitude, 260, 263-4
- Formal Attitude, 261, 263
- Principle of Connections, 261
- Embryology and Comparative Anatomy, 261-2,
263
- Biogenetic and Meckel-Serres Laws, 262
- Homology, 261, 263,
265, 266-7
- Adaptation and Correlation, 263-4
- Archetype as ancestral, 263 f.n, 265
- On Phylogenetic Speculation, 265-6
- Embryological Archetype, 266
- Membrane and Cartilage Bones, 309, 310
- Gemmill, J. F., 285 f.n., 312 f.n.
- Geoffroy, Etienne, St Hilaire, 40, 52-78, 141
- Unity of Plan, 52-65, 70 ff., as conservative, 75, 78
- Principle of Connections, 53-4, 62-3, 71, 74, 261
- Unity of Composition, 54, 70-1, 75-6, 200, 305
- Archetype, 54, 67
- Metastasis, 55-6, 59,
74
- Opercular Bones, 56
- Unity of Composition of Sternum, 57-60
- Classification, 60
- Vertebrates and Articulates, 60-4,
274, 278-9, 303
- Formal Attitude, 62-3, 65, 71, 75-8, 305
- Cephalopods and Vertebrates, 64-5
- Scale of Beings, 64
- Polemic with Cuvier, 64-5, 74-8
- Evolution, 66-9, 73,
228
- Biogenetic Law, 69
- Teratology, 69, 315
- Meckel-Serres Law, 70, 72
- Criteria of Homology, 71, 72, 110
- Law of Compensation, 72-3
- Criticism of his Principles, 74
- Relation to German Transcendentalists, 89,
100-1
- Vertebral Theory of Skull, 96, 97
- Influence on Darwin, 234-5, 238
- Geoffroy, Isidore, St Hilaire, 65 f.n., 199 f.n., 230
- Geometry of the Organism, 33
- K. G. Carus, 98-100, 249
- Bronn, 201, 249
- Haeckel, J. Müller, Burmeister, G. Jäger, 249
- Germinal Vesicle (Egg-nucleus), 175-7, 188, 291 f.n.
- Germ-Layer Theory—
- Von Baer, 115-6, 118-9, 208-9, 296
- Pander, 119-20, 209
- C. F. Wolff, 119-20
- Rathke, 136, 208
- Lereboullet, Bischoff, 208
- Huxley, 208, 289
- Remak, 209-12, 296
- 374Germ-Layers and Gastræa Theory—
- Haeckel, 289-95
- Lankester, Balfour, 295
- Germ-Layer Theory, Influence of Causal Morphology on, 347
- Gesner, 18
- Giard, A.—
- On Ascidian Theory, 271-3
- Adaptive Homology, 273
- Pœcilogeny, 347-8
- Glisson, F., 19
- Gluge, 173
- Goebel, K., 356 f.n.
- Goethe, 45-51, 65, 89, 250
- Unity of Plan, 45-7, 51
- Homology, 47
- Principle of Connections, 47
- Formal and Functional Attitudes, 48-50
- Teleology, 48
- Metamorphosis of Plants, 48
- Repetition of parts, 48-9
- Vertebral Theory of Skull, 49, 96, 97
- Law of Compensation, 49
- Embryological Criterion, 49
- Organisms as Nature's Works of Art, 50
- Goette, 259
- Graaf, von, 175
- Grew, N., 169
- Gruber, 330
-
- Haeckel, Ernst, 247-60, 271, 314, 342, 353, 357
- His sources, 248-50
- Materialism, 248, 250
- On Teleology, Heredity and Adaptation, 248, 263
- Correlation, Disregard for, 248, 264
- Geometry of the Organism (Promorphology), 249
- Repetition of Parts (Tectology), 249-50
- Classification as Genealogical, 250-1, 254
- Archetype as ancestral, 251
- Homology and Analogy, 251
- Biogenetic Law, 251, 253-9, 291-4
- Three-fold parallelism, 254-5
- Scale of Beings, 255, 256-7
- Organism as an Historical Being, 257
- Prussianism, 257
- Palingenesis, 258
- Cenogenesis, 258-9
- Heterotopy, Heterochrony, 259
- Gastræa Theory, 269, 288-95
- Phylogenetic Classification, 289-94
- Criticism of Theory of Types, Monophyletism, 289, 291
- Gastræa Theory and Biogenetic Law, 291-4
- Primary stages of Ontogeny and Phylogeny, 291-3
- Cœlom, 291, 295, 296
- Experimental Embryology, 317
- Haller, 113
- Harting, 284 f.n.
- Hartmann, E. von—
- On Darwin's conception of correlation, 240-1
- Evolution, 244, 356
- Hartog, M., 344
- Harvey, 18, 113
- Hatschek, 270 f.n., 299
- Helmholtz, H. von, 195
- Henle, 172
- Hensen, V., 209 f.n.
- Herbst, C., 333
- Herder, 46
- Heredity and Memory, 336-44
- Hering, E., 341-2
- "Heritage" Characters, 309, 322
- Herlitzka, 332
- Herophilus, 17
- Hertwig, O., 163, 330, 331, 346
- On C. F. Wolff, 119
- 375
Fertilisation, 291 f.n.
- Membrane and Cartilage Bones, 309-10
- Biogenetic Law, 352, 354-5
- Von Baer's Law, 354
- Intrinsic Laws of Evolution, 354-5, 356-7
- Homology not necessarily Homogeny, 355-7
- Unity of Plan not necessarily due to Community of Descent, 355-7
- On Phylogenetic Speculation, 356
- Hertwig, O. and R.—
- Cœlom Theory, 297-8
- Nervous System of Cœlentera, 299
- Heterochrony, 259, 348, 349-52
- Heterogeneous Generation (Kölliker), 243
- Heterotopy, 259
- Hilgendorf, 359
- Hill, 311
- Hippocratic Treatises, 2
- His, W., 206 f.n., 209 f.n.
- Causal Morphology, 316
- Cytoplasm of Egg, Organ-forming Stuffs, 333
- Specific Distinctness of Embryos, 353
- Histological Differentiation (von Baer), 117-8
- Histology. See also "Cell-Theory"
- Hofer, B., 330
- Hofmeister, 185
- Homogeny, 267, 303,
355
- Homology, 168, 303,
355-7. See also "Connections, Principle
of," and "Embryological Criterion"
- Aristotle, 7-10
- Belon, 18
- Buffon, 27
- Goethe, 47
- E. Geoffroy, 53, 71
- Serres, 80
- Owen, 107-9
- Lamarck, 227
- C. Darwin, 234-5, 247
- Haeckel, 251
- Gegenbaur, 261, 263,
265, 266-7
- Giard, 273
- Semper, 279
- O. Hertwig, 355-7
- Braun, 355
- Homology, Genetic Definition of—
- Gegenbaur, 266
- Lankester, 267
- O. Hertwig's criticism, 355-7
- Homoplasy, 267
- Hooke, R., 20, 169
- Houssay, F., 19 f.n., 333
- Hubrecht, A. A. W., 284, 295 f.n., 301, 305-6
- Hunter, J., 22, 315
- Huschke, 134-5, 136,
141, 146
- Huxley, T. H., 157, 238, 247
- On Rathke, 154 f.n.
- Embryological Criterion, 158-9, 166
- Embryological Archetype, 159-61
- Criticism of Vertebral Theory of Skull, 161-2
- Membrane and Cartilage Bones, 166-7
- On Archetype, 204
- Germ-Layer Theory, 208, 289
- Criticism of Three-fold Parallelism, 230
f.n.
- Cœlom, 297
- Ancestry of Marsupials, 311
- Hyatt, A., 359, 361
-
- Instinct and Morphogenesis, Analogy
of, vi., 307, 312
-
- 376Jacobson , 164
- Jäger, G., 249 f.n.
- Jardin des Plantes, Paris, 19
- Jenkinson, J. W., 347 f.n.
- Jones, Wharton, 138, 176
- Julin, C., 271, 285
- Jussieu, de, 40
-
- Kant, I.—
- Keibel, F., 348, 349-50
- Kerkring, 131
- Kielmeyer, 89, 90, 96
- Kieser, 90
- Kleinenberg, N., 277
- Kohlbrugge, J., 44 f.n., 65 f.n.
- Kölliker, A.—
- On C. F. Wolff, 119
- Vertebral Theory of Skull, 157
- Membrane and Cartilage Bones, 164-6, 310
- Embryological Criterion, 165-6
- Cell-division, 187
- Intrinsic Laws of Evolution, 243, 356
- Saltatory Variation, 243
- Kowalevsky, A., 269-71, 284, 285, 299, 300
- Kowalevsky, V. O., 361-2
- Krause, 176
- Kupffer, 271
-
- Lacaze-Duthiers, H. de, 203 f.n., 315-6
- Lamarck, 44, 66, 78, 215-29
- Relation to Buffon, 215
- Scale of Beings, 215-8, 220-1, 227-8
- As Evolutionary, 218, 220
- Classification, 216-7, 227, 228
- Species Problem, 216, 227
- Materialism, 218-9, 222-3, 225-6
- Psychological Vitalism, 219, 220-6, 307, 335
- Sentiment intérieur, 219-20,
222-3, 225
- Ecological Adaptation, 221, 222, 223, 224, 227
- Laws of Evolution, 221-5
- Transmission of Acquired Characters, 221-2, 224
- Subtle Fluids, 222
- Use and Disuse, 223-4
- Independence of Current Thought, 226-7
- Homology and Analogy, 227
- Reception of his Theory, 228-9
- Lamarck and Butler, 335-7
- Lang, A., 301
- Lankester, Sir E. Ray, 247
- Homology, Homogeny, Homoplasy, and Analogy, 267
- Balanoglossus Theory of Vertebrate Descent, 287
- Germ-Layer Theory and Phylogenetic Classification, 291
- Planula Theory, 295
- On Cœlom Theory, 296-7, 299 f.n.
- Latreille, 86, 100
- Laurencet, 64
- Lavocat, 203 f.n.
- Leeuenhoek, 20, 21,
169
- Leibniz, 23, 213, 343
- Lereboullet—
- Von Baer's Law, 206-8
- Germ-layer Theory, 208
- Gastrula, 288 f.n.
- Leucippus, 16
- Leuckart, 193 f.n., 194, 297
- Levy, O., 333
- Leydig, 187, 188, 275 f.n., 285
- Linnæus, 22
- Loeb, J., 333, 347
- 377Loi de Balancement. See
"Compensation, Law of"
- Lovén, 186, 196
- Lucretius, 16
- Ludwig, 193, 194, 314
- Lyell, Sir C., 228 f.n.
- Lyonnet, 22
-
- Macbride, E. W., 287 f.n.
- M'Kendrick, J.—
- Mackenzie, W., 345
- Malpighi, M., 20-1, 113, 169
- Marine Zoology, Rise of, 195-6
- Materialistic Attitude, 246-7, 345, 364
- Schwann, 180-5
- Vogt, 190-1
- Virchow, 191
- Ludwig, 193
- Materialistic Physiology, 193-4, 314-5, 347
- Lamarck, 218-9, 222-3, 225-6
- The Darwinians, 241, 308
- Haeckel, 248, 250
- Roux, 315, 317, 318-9, 329
- Semon, 343
- Rignano, 344
- Loeb, 347
- Criticism of this attitude—
- Meckel, D. A., 95
- Meckel, J. F., 113
- Meckel-Serres Law, 91-3
- Evolution, 92-3, 215,
228
- Teratology, 93-4
- Repetition of Parts, 95
- Vertebral Theory of Skull, 96
- Eclecticism, 101
- Meckel's Cartilage, 141, 145
- Meckel-Serres Law. See "Development, Meckel-Serres
Law"
- Mehnert, E., 348, 350-2
- Membrane and Cartilage Bones, 162-7, 309-10
- Memory and Heredity, 336-44
- Mendelism, 346
- Mesenchyme, 298
- Mesoderm, 209-11, 296, 297, 298
- Metabolism—
- Metamerism, 94, 95,
100, 109, 131-2, 266-7, 274-5, 279, 282, 286, 299, 301
- Metamorphosis of Plants, 48, 235
- Metastasis, Principle of—
- Metschnikoff, E., 278 f.n., 285, 288
- Meyen, 170, 185
- Meyer, E., 284
- Meyranx, 64
- Microscope, Invention of, 19
- Milne-Edwards, H., 12, 86, 238
- Marine Zoology, 195
- Functional Attitude, 195, 197-200
- Unity of Plan, 197
- Division of Labour, 197-8
- Ecological Adaptation, Convergence, 199
- Von Baer's Law, Polemic with Serres, 204-8
- Evolution, 244-5
- Mirbel, 170, 171
- Mivart, St G., 277
- Mohl, von, 170, 185
- Moldenhawer, 170
- Moleschott, 194
- Moquin-Tandon, A., 87
- Morgan, T. H., 317 f.n., 332, 333, 347 f.n.
- Mosaic Theory of Development, 330-3
- Müller, F., Biogenetic Law, 252-3, 254, 257
- Müller, H., 166
- 378Müller, J., 136, 209 f.n., 260, 285, 309, 345
- Embryological Criterion, 138
- Vertebral Theory of Skull, 142-4, 154, 157
- On Reichert, 150
- Cell Theory, 172-3
- Division of Egg-nucleus, 188
- Vitalism, 192
- Marine Zoology, 196
- Functional Attitude, 200
- Mutations (Waagen), 361 f.n.
-
- Naegeli, 185,
243 f.n., 356
- Naturphilosophie. See "Philosophy of Nature"
- Nesbitt, R., 162
- Neumayr, 357, 360
- Nussbaum, M., 330
-
- Oken, L., 89,
113, 131, 134, 149
- Meckel-Serres Law, 90-1
- Teratology, 91
- Repetition of Parts, 94-5
- Serial Homology, 95-6, 100
- Vertebral Theory, 96, 97, 98
- On Geoffroy, 100
- Influence on Serres, 205
- Evolution, 215
- Ollier, 315
- Oppel, A., 318 f.n., 324 f.n., 327, 348-9
- Orr, H. F., 342
- Osborn, H. F., 214 f.n., 361
- On V. O. Kowalevsky, 362
- Functional Attitude, 362-4
- Law of Adaptive Radiation, 362-4
- Owen, R., 97, 102-12,
204
- Eclecticism, 102
- Vertebral Theory of Skeleton, 103-7
- Archetype of Vertebrate Skeleton, 104-7,
110
- Vertebral Theory of Skull, 104-6
- Metastasis, 106
- Principle of Connections, 107-8
- Anatomy and Embryology, 108
- Homology and Analogy, 108
- Classes of Homology, 108-9, 266
- Functional Attitude, 109, 110, 111
- Embryological Criterion, 110, 167
- Homological and Teleological Compoundedness, 110-1
- Vegetative Repetition of Parts, 111, 286
- Unity of Plan as Conservative Principle, 112
- Influence on Darwin, 234, 235, 238
- Evolution, 244
-
- Packard, 361
- Palæontological Record, 357-61
- Absence of connecting forms, 357-9
- Biogenetic Law, 359
- Phyletic Series, 359-61
- Palæontological Succession—
- Cuvier, 43
- E. Geoffroy, 67
- L. Agassiz, 230, 255
- C. Darwin, 231
- Milne-Edwards, 245
- Tiedemann, 255 f.n.
- Paley, W., 341
- Palingenesis (Haeckel), 258, 323
- Pander, 113, 119-20,
133, 208, 209
- Parallelism, Theory of. See "Development, Meckel-Serres
Law"
- Three-fold. See "Development, Meckel-Serres Law"
- Paris Museum of Natural History, 19, 89, 101
- Paul, 360
- Pauly, A., 345
- Perrault, C., 19
- Perrier, E., 88, 359
f.n.
- Pflüger, E., 317, 330
- Philipeaux, 315
- 379"Philosophy of Nature," 89, 94, 98,
203, 248
- Phyletic Series, 359-61
- Physiology, Separation from Morphology, 194, 247, 260, 314
- Physiology of Development, 315
- Planula Theory (Lankester), 295
- Plato, 15
- Pockels, 138
- Pœcilogeny (Giard), 347-8
- Poli, 175
- Polyphyletism—
- Darwin, 238
- Von Baer, 242, 356
- Kölliker, Wigand, Naegeli, 356
- Depéret, 360-1
- Steinmann, 360 f.n.
- Polyzoic Conception of Organism—
- Prévost and Dumas, 125 f.n., 134, 175, 186
- Promorphology (Haeckel), 249
- Protoplasm, 169, 188-9
- Purkinje, 172, 173,
175, 176, 189
-
- Quatrefages, A. de, 172, 195-6
-
- Rádl, E., on Goethe, 48
- Correlation, 240 f.n., 241
- On Darwin's Critics, 242 f.n.
- On Cuvier's Critics, 278 f.n.
- Rathke, H., 133, 136-7, 174, 194, 269, 351 f.n.
- Discovery of Gill-slits in Pig and Chick, 134
- Discovery of Gill-slits in Man, 135
- Germ-Layer Theory, 136, 208
- Embryological Criterion, 138, 140-1
- Homologies of Gill-arches, 139-41, 146, 150
- Development of Skull, 141, 150-4
- Vertebral Theory of Skull, 141, 154-6
- Embryological Archetype, 151, 153
- Membrane and Cartilage Bones, 163, 166
- Rauber, A., 330
- Réaumur, 22, 315
- Recapitulation Theory. See "Development, Biogenetic
Law"
- Regeneration, 315, 318, 333, 346
- Regulatory Processes in Development, 114,
319, 333, 346-7, 350
- Reichert, C. B., Embryological Criterion, 138-9, 144-7, 163 Archetype, 139, 147, 149
- Homologies of Gill-arches and Ear-ossicles, 144-7
- Vertebral Theory of Skull, 147-9, 157
- Von Baer's Law, 149-50, 351 f.n.
- Membrane and Cartilage Bones, 163, 165, 166, 310
- Criticism of "Biological Atomists," 192-3,
194
- Functional Attitude, 193, 200
- Remak, R., 118, 288
f.n.
- On Vertebræ, 157
- Cell Theory, 173, 187-8, 209
- Microscopical Technique, 209 f.n.
- Germ-Layer Theory, 209-12, 296
- Cells, Tissues and Germ-Layers, 209-12
- Mesoderm, 209-11
- Cœlom, 211, 296
- Repetition of Parts within the Organism, Theory of. See
also "Vertebral Theory of Skull"
- Goethe, 48-9
- Dugès, 87-8
- Oken, 94-5
- J. F. Meckel, D. A. Meckel, 95
- Haeckel (Tectology), 249-50
- Reymond, E. du Bois, 194, 314
- Rignano, E., 343-4
- Robinet, 23, 215
- Rondeletius, 18
- Rosenhof, Rösel von, 22
- 380Roux, W., 313, 315-29, 344, 351
- Entwicklungsmechanik, 315, 317-8
- Materialistic Attitude, 315, 317, 318-9, 329
- Functional Adaptation, 316-7, 318, 320-9, 333
- Experimental Embryology, 317, 318, 330-1
- Simple and Complex Components, 318-20
- Functional Definition of Life, 320
- Functional Attitude, 320-9, 335
- The Two Periods of Development, 320-4, 325, 327, 335
- Mosaic Theory of Development, 323, 330-1
- Metabolism, 324, 329
- Structure, Functional and Non-functional, 324-6
- Functional Unity of Organism, 326
- Functional Adaptation of Blood-vessels, 326-9
- Form as manifestation of Activity, 329
- Ruini, C., 18
- Rusconi, 133-4, 186
- Rütimeyer, L., 361
- Ryder, 361
-
- Sachs, J. von, 170
- St Ange, M., 146
- Salensky, 259
- Saltatory Variation—
- Sarcode, 169
- Sars, M., 186, 196
- Savigny, J. C., 83-5, 100, 137, 271
- Scale of Beings, 89, 206, 214-5
- Aristotle, 14-6
- Anaximander, Anaxagoras, 14
- Empedocles, Plato, 15
- Albertus Magnus, 17
- C. Bonnet, 22-3
- Robinet, 23
- Buffon, 24
- E. Geoffroy, 64
- Lamarck, 215-8, 220-1, 227-8
- As Evolutionary, 218, 220
- Haeckel, 256-7
- Criticism of this idea—
- Cuvier, 39-40, 130
- Von Baer, 130
- Milne-Edwards, 205
- Lereboullet, 207
- Darwin, 234
- Haeckel, 255
- Relation to Evolution-Theory, 214-5
- Schepelmann, 333
- Schleiden, 170-2
- Schmieden, 328
- Schults, C. H., 173
- Schultze, Max, 189
- Schultze, O., 331
- Schulz, E., 347 f.n.
- Schwann, Theodor, 169, 173-86, 248
- Physiological Standpoint, 173, 179, 180, 182
- Development of Cells, 174-5, 179-80
- Cellular Nature of Ovum, 175-7
- Development of Tissues from Cells, 177-8
- Histology, 178
- Materialism and Teleology, 180-3, 185
- Cell-metabolism, 182-5
- Cells as organic Crystals, 184-5
- Sedgwick, A., 347 f.n.
- Actinozoan Theory of Vertebrate Descent, 299-300
- Metamerism, 299
- Embryological Archetype, 300
- Organism as Historical Being, 308
- Cell-Theory, 346
- Von Baer's Law, 353
- 381Segmentation of Ovum, 186-8
- Seiler, 138
- Selection, Natural and Artificial, 307
f.n.
- Self-Differentiation (Roux), 319, 320-1, 322, 323, 324, 327
- Self-Regulation (Roux), 319
- Semon, R., 342-3
- Semper, C., 259, 269,
278-82, 284, 286
- Annelid Theory, 274, 278-82
- Metamerism, 274, 279,
282
- Follower of Geoffroy, 278
- Unity of Plan and Composition, 279, 303
- Principle of Connections, 279
- Formal Attitude, 279
- Sentiment intérieur (Lamarck), 219-20, 222-3, 225
- Serial Homology. See "Metamerism"
- Serres, E., 79-83, 91, 100, 205-6, 257 f.n.
- Criteria of Homology, 80
- Law of parallelism, 80-3, 94, 203-4, 205-6
- Law of Multiple Formation, 80-1
- Unity of Plan, 83, 205, 206
- Teratology, 83
- Meckel's Cartilage, 145 f.n.
- Transcendentalism, 205-6
- Concrescence Theory, 206 f.n.
- Severino, 18
- Sharpey, 162, 176
- Siebold, von, 186
- Skull, Development of, 139-62. See
also "Vertebral Theory"
- Spallanzani, 315
- Species-Problem—
- Spencer, H., 326 f.n.
- Spengel, 285, 287
- Spinoza, 343
- Spix, 96, 97, 100, 141
- Stannius, 165
- Steenstrup, 309
- Steinmann, G., 357, 360 f.n.
- Stensen (Steno), 21
- Swammerdam, 20, 21-2
-
- Tachygenesis, 359
- Technique, Microscopical, 209 f.n., 268
- Tectology (Haeckel), 249
- Teleology—
- Aristotle, 10
- Cuvier, 33-5
- Kant, 35, 213, 242
- Von Baer, 242
- Owen, Von Hartmann, 244
- Butler, 341
- G. Wolff, Driesch, 346
- Criticism of—
- Goethe, 48
- Schwann, 180-2
- The Darwinians, 241
- Haeckel, 248
- Evolutionary Morphologists, 308
- Teratology, 69, 83,
91, 93, 315
- Thienemann, 23 f.n.
- Thompson, D'Arcy W., 2 f.n.
- Thomson, A., 176
- Thomson, J. Arthur, 215 f.n.
- Tiedemann, 91, 113,
215, 255 f.n.
- Tissues and Germ-Layers, 118, 209-12
- Transcendental Anatomy, Relation to Evolutionary Morphology, 302-8, 312
- Transcendentalism, French and German Schools, 89, 100
- Trembley, 22, 315
- Treviranus, 141, 170,
215, 225 f.n.
- Turpin, 173
- Types, Theory of (Cuvier and Von Baer)—
- Type-Theory and Evolution, 304
-
- Unger, 185
- Unity of Composition, Principle of, Geoffroy, 54, 70-2, 75-6, 200, 305
- Unity of Plan, 88, 241, 278-9, 303, 312. See also
"Archetype"
- Aristotle, 6-7, 10
- Belon, Severino, 18
- Perrault, 19
- Robinet, 23
- Buffon, 24
- Cuvier, 41
- Goethe, 45-7, 51
- Vicq D'Azyr, 45
- Camper, 45, 46
- Herder, 46
- Kant, 46, 213-4
- E. Geoffroy, 52-65, 70 ff.
- Serres, 83, 205, 206
- Savigny, 83
- Audouin, 85-6
- Latreille, 86
- Dugès, 86-7
- J. F. Meckel, 101
- Milne-Edwards, 197
- Semper, 279
- Haeckel, 289, 291
- Lankester, 291
- Unity of Plan as due to Community of Descent—
- Unity of Plan as Conservative Principle—
- E. Geoffroy, 75, 78
- Owen, 112
- Gegenbaur, 263-4
- Evolutionary Morphologists, 307
-
- Valentin, 138,
173, 176
- Variation, Limits of, Cuvier, 42
- Vegetative Repetition of Parts—
- Velpeau, 138
- Vertebral Theory of Skull, 49, 96-9, 104-6, 131, 141-4, 147-9, 154-7, 161-2, 165, 203, 235, 310 f.n.
- Vertebrate Descent, 269-87, 299-301, 304
- Verworn, M., 330
- Vesalius, 18
- Vestigial Organs, 233, 237, 309, 312
- Vialleton, L., 306 f.n., 348
- Vicq d'Azyr, 45, 95
- Virchow, R., 188, 191
- Vitalism, Psychological—
- Lamarck, 219, 220-6,
307, 335
- Butler, 336-41
- Orr, Cope, 342
- Ward, 343
- Delpino, Francé, Pauly, A. Wagner, Mackenzie, 345
- Vogt, C.—
- Criticism of Vertebral Theory, 156-7
- Capillaries, 179
- Segmentation, 186
- Materialistic Attitude, 190-1
- Threefold Parallelism, 255 f.n.
-
- Waagen, 359, 361 f.n.
- Wagner, A., 345
- Wagner, R., 176
- Ward, J., 343
- Weber, 138
- Weismann, A., 240, 323, 326 f.n., 330-1, 343
- Werneck, 173
- Whitman, C. O., 346
- Wigand, A., 242 f.n., 356
- 383Willey, A., 273 f.n., 306 f.n.
- Williamson, 309
- Willis, 19
- Wilson, E. B., 331, 332-3, 346 f.n., 347 f.n.
- Wolff, C. F., 113
- Germ-layer Theory, 119-20
- Cells, 170
- Wolff, G., 346-7
- Woodward, B. B., 358
- Wotton, E., 17
-
- Zeleny, 333
- Zittel, K. von, 357, 358
- Zoja, 331
384
PRINTED BY
OLIVER AND BOYD,
EDINBURGH, SCOTLAND
HEREDITY. By J. Arthur Thompson, M.A., LL.D., Regius
Professor of Natural History in the University of Aberdeen. With
numerous Illustrations. 9s. net.
Heredity and Inheritance: Defined and Illustrated--The Physical Basis
of Inheritance--Heredity and Variation--Common Modes of
Inheritance--Reversion and allied Phenomena--Telegony and other
Dispute Questions--The Transmission of Acquired Characters--Heredity
and Disease--Statistical Study of Inheritance--Experimental Study of
Inheritance--History of Theories of Heredity and Inheritance--Heredity
and Development--Heredity and Sex--Social Aspects of Biological
Results--Bibliography--Subject-Index to Bibliography--Index.
"We all know books of science which we ought to read with pleasure,
but to which we turn with shrinking. Full, perhaps, of new facts and
ideas, they are so expressed as to bore consumedly. 'Heredity' belongs
to another category. He who runs may read, even if he be a beginner,
and he who reads will probably not cease to run until he has traversed
the last page."--Nature.
"This is certainly the best modern book on heredity to recommend to
the student and the intelligently curious."--Science Progress.
"May be regarded as the standard work of reference on this subject. As
a judicial summary of an exceedingly difficult and controversial
subject it is masterly, while in the matter of clearness of exposition
it has no rival."--Knowledge.
RECENT ADVANCES IN THE STUDY OF VARIATION, HEREDITY AND EVOLUTION. By
Robert H. Lock, M.A., sometime Fellow of Gonville and Caius College,
Cambridge, late Assistant-Director of the Royal Botanic Gardens,
Ceylon. With Portraits and other Illustrations. Revised by L.
Doncaster, D.Sc., F.R.S. With a Biographical Note by Bella Sidnay
Woolf (Mrs R. H. Lock). Crown 8vo.
MICROSCOPY. The Construction, Theory, and Use of the Microscope. By
Edmund J. Spitta, F.R.A.S., F.R.M.S., etc. With numerous Diagrams and
Illustrations. Second Edition. 12s. 6d. net.
"Let us hasten to urge every student of the microscope who wishes to
gain a thorough understanding of its principles and possibilities and
its defects, and every user of the instrument who desires a work of
reference to which he may turn for an explanation of some unexplained
optical phenomenon, or for particulars of up-to-date apparatus, to
procure a copy of Mr Spitta's book without delay."--Nature.
CONVERGENCE IN EVOLUTION. By Arthur
Willey, D.Sc. (Lond.); Hon. M.A. (Cantab.); F.R.S. With
Diagrams. Demy 8vo. 7s. 6d. net.
This work brings together some scattered facts of parallel development
of outward form and internal structure in the Animal kingdom, introducing
new cases and fresh interpretations. It is, taken as a whole, an
original contribution to the theory of organic evolution, with special reference
to the forms of Animal life.
THE HEREDITY OF ACQUIRED CHARACTERS IN PLANTS. An aspect of the true
Darwinism based on Personal Observations and Experiments. By the Rev.
Prof. George Henslow. With Illustrations. Demy 8vo. 6s. net.
A HANDBOOK OF PHYSIOLOGY. By W.D.
Halliburton, M.D., F.R.S., Professor of Physiology, King's
College, London. Eleventh Edition, being the Twenty-fourth
of Kirkes'. With nearly Seven Hundred Illustrations, including
some Coloured Plates. Large Crown 8vo. 15s. net.
"One of the best manuals for the student which we possess ... the book is
an eminently trustworthy one, and will prove a valuable foundation for, and
introduction to, the large treatises on physiology."—Lancet.
THE BACTERIOLOGY OF MILK. By Harold
Swithinbank, of the Bacteriological Research Laboratory,
Durham, and Sir George Newman, M.D., F.R.S.E., D.P.H.,
Chief Medical Officer, Board of Education. With Special
Chapters also by Dr Newman on Spread of Disease by Milk
and the Control of the Milk Supply. With numerous Illustrations.
Royal 8vo. 25s. net.
"Ought to find a place in the library of every medical officer of health and
of every milk-producer. Scientific in method and lucid in exposition, the authors
have given us a really invaluable text-book."—Spectator.
BACTERIOLOGY AND THE PUBLIC HEALTH. By Sir George Newman, M.D., F.R.S.
(Edin.), D.P.H., Chief Medical Officer, Board of Education.
With Illustrations. Medium 8vo. 21s. net.
"The present work, though nominally a third edition of 'Bacteria in
Relation to the Economy of Nature, Industrial Processes, and the Public
Health,' is virtually a new book, written with the object of supplying all
that is necessary for the student of hygiene and the officer of health to know,
so far as every-day problems of sanitation and preventive medicine demand....
Dr Newman has done a good work in producing a treatise which places at
the service of the community what is known about all these topics."—Daily
Telegraph.
THE RECENT DEVELOPMENT OF PHYSICAL
SCIENCE. By W.C.D. Whetham, M.A.,
F.R.S. Illustrated. Large Crown 8vo. 5s. net.
The Philosophical Basis of Physical Science—The Liquefaction
of Gases and the Absolute Zero of Temperature—Fusion and Solidification—The
Problems of Solution—The Conduction of Electricity
Through Gases—Radio-Activity—Atoms and Æther—Astro-Physics—Index.
THE REALM OF NATURE. An Outline of
Physiography. By H.R. Mill, D.SC, LL.D., Director of
the British Rainfall Organisation. Second Edition. Revised
and entirely reset. With 19 coloured Maps and 73 Illustrations
in the text. Crown 8vo. 5s. net.
"Dr Mill is to be congratulated on having now brought his information, so
far as space permitted, well up to date. The most striking features of the work
are its comprehensiveness and conciseness.... It would, indeed, be difficult to
point to any other English work on physiography giving so much trustworthy
matter in equally condensed form, yet so readable."—Athenæum.
NATURE AND ORIGIN OF FIORDS. By
J.W. Gregory, D.Sc., F.R.S., Author of "The Dead
Heart of Australia." With Illustrations. Demy 8vo. 16s. net.
Professor T.G. Bonney says in Nature, 12th Feb. 1914:—"But we must
conclude, and do this by expressing our hearty thanks to him for this admirable
history of fiords and other forms of inlets of the sea. It will be a great boon to
students, for it is a veritable encyclopædia, full of important facts."
MECHANISM, LIFE AND PERSONALITY.
An Examination of the Mechanistic Theory of Life and Mind.
By J.S. Haldane, M.D., LL.D., F.R.S., Fellow of New
College and Reader in Physiology, University of Oxford.
Crown 8vo. 2s. 6d. net.
"Dr Haldane has succeeded in packing an immense amount of knowledge and
thought into the compass of a small volume. The complexity of his themes has
never for a moment betrayed him into ambiguity either of thought or expression,
and the pervading temptation to stray into bypaths, the failure to resist which
makes the weakness of so much otherwise fine work of this class, has been most
successfully resisted. The clarity of the book may fairly be described as
remarkable."—Sunday Times.
THE
PROGRESSIVE SCIENCE SERIES
THE INTERPRETATION OF RADIUM. By Frederick
Soddy, M.A., Independent Lecturer in Physical Chemistry and
Radio-activity in the University of Glasgow. With Illustrations. 6s. net.
HEREDITY. By J. Arthur Thomson, M.A, Regius
Professor of Natural History in the University of Aberdeen. Illustrated.
9s. net.
THE PROBLEM OF AGE, GROWTH, & DEATH.
A Study of Cytomorphosis. By Charles S. Minot, LL.D. (Yale,
Toronto), D.Sc. (Oxford). Illustrated. 6s. net.
THE SOLAR SYSTEM. A Study of Recent Observations.
By Charles Lane Poor, Professor of Astronomy in Columbia
University. Illustrated. 6s. net.
PROBLEMS OF LIFE AND REPRODUCTION.
By Marcus Hartog, M.A., D.Sc, Professor of Biology in the University,
Cork. Illustrated. 7s.* 6d. net.
CLIMATE. Considered Especially in Relation to Man.
By Robert de Courcy Ward, Assistant Professor of Climatology in
Harvard University. Illustrated. 6s. net.
HYGIENE OF NERVES AND MIND IN HEALTH
AND DISEASE. By August Forel, M.D. Translated from the German
by A. Atkins. Illustrated. 6s. net.
INFECTION AND IMMUNITY. By George S.
Sternberg, M.D., LL.D. 6s. net.
THE STARS. A Study of the Universe. By Professor
Simon Newcomb. Illustrated. 6s. net.
A BOOK OF WHALES. By F.E. Beddard, M.A.,
F.R.S. (The Editor). Illustrated. 6s. net.
THE STUDY OF MAN: An Introduction to Ethnology.
By Professor A.C. Haddon, D.SC, M.A., M.R.I.A. Illustrated.
6s. net.
THE GROUNDWORK OF SCIENCE. A Study of
Epistemology. By St. George Mivart, M.D., PH.D., F.R.S. 6s. net.
EARTH SCULPTURE; or, The Origin of Land Forms.
By Professor Geikie, LL.D., F.R.S. Illustrated. 6s. net.
RIVER DEVELOPMENT. As Illustrated by the Rivers
of North America. By Professor I.C. Russell. Illustrated. 6s. net.
VOLCANOES: Their Structure and Significance. By
Professor Bonney, D.SC, F.R.S. Illustrated. 6s. net.
EARTHQUAKES, In the Light of the New Seismology.
By Clarence E. Dutton, Major U.S.A. Illustrated. 6s. net.
Life and Works of
CHARLES DARWIN
THE ORIGIN OF SPECIES BY MEANS OF
NATURAL SELECTION. 6s. net. Popular Edition. 2s. 6d. net. Also in
Paper Covers, 1s. net.
DESCENT OF MAN, AND SELECTION IN
RELATION TO SEX. With Illustrations. 3s. 6d. net.
VARIATION OF ANIMALS AND PLANTS
UNDER DOMESTICATION. Woodcuts. 2 vols. 15s. net. Popular
Edition. 7s. 6d. net.
EXPRESSION OF THE EMOTIONS IN MAN
AND ANIMALS. With Illustrations. 9s. net. Popular Edition. 3s. 6d. net.
VARIOUS CONTRIVANCES BY WHICH ORCHIDS
ARE FERTILIZED BY INSECTS. Woodcuts. 7s. 6d. net.
Popular Edition. 3s. 6d. net.
MOVEMENTS AND HABITS OF CLIMBING
PLANTS. Popular Edition. 3s. 6d. net.
INSECTIVOROUS PLANTS. Popular Edition. 3s. 6d. net.
CROSS AND SELF-FERTILIZATION IN THE
VEGETABLE KINGDOM. 9s. net.
DIFFERENT FORMS OF FLOWERS ON
PLANTS OF THE SAME SPECIES. 7s. 6d. net.
FORMATION OF VEGETABLE MOULD
THROUGH THE ACTION OF WORMS. Illustrations. 6s. net. Popular
Edition. 3s. 6d. net.
JOURNAL OF A NATURALIST DURING A
VOYAGE ROUND THE WORLD IN H.M.S. "BEAGLE." With 100
Illustrations. Medium 8vo. 21s. net. Popular Edition. With 16 full-page
Plates. 2s. 6d. net.
LIFE and LETTERS of CHARLES DARWIN.
With an Autobiographical Chapter. Edited by FRANCIS DARWIN, F.R.S.
With 3 Portraits and Illustrations. 3 vols. 8vo. 36s.
CHARLES DARWIN: An Autobiography. With
Selections from his Letters by FRANCIS DARWIN. Portrait, 7s. 6d. net. Popular
Edition. 2s. 6d. net.
MORE LETTERS OF CHARLES DARWIN.
A Record of his Work in a Series of hitherto Unpublished Letters. Edited by
FRANCIS DARWIN and A.C. SEWARD. With Portraits. 2 vols. Demy 8vo.
32s. net.
THERAPEUTICS OF THE CIRCULATION.
By Sir T. Lauder Brunton, Bart., M.D., LL.D., F.R.C.P.,
F.R.S., Consulting Physician to St Bartholomew's Hospital.
New and Revised Edition. With Illustrations. 5s. net.
In this new edition so much matter has been added that the book has been
practically re-written. It is intended to supplement, and not to replace, the usual
text books dealing with circulation.
THE PREVENTION OF MALARIA. By Sir
Ronald Ross, K.C.B., F.R.S., etc. With Contributions
by twenty of the Leading Experts. With Illustrations.
Demy 8vo. 21s. net.
"A thoroughly sound and comprehensive treatise; Sir Ronald Ross and
his colleagues have turned out work worthy of their high reputations. The
student of malaria in all respects will find in this work the most complete
exposition of the subject in medical literature."—Lancet.
THE HOUSE-FLY: Disease Carrier. An Account of
its dangerous activities and of the means of destroying it.
By Leland O. Howard, Ph.D. With numerous Illustrations.
6s. net.
In this book the Chief of the U.S. Bureau of Entomology sets forth complete
information about the fly.
After describing the nature of the common house-fly, its habits and methods
of breeding, he proves his case against it as a carrier of disease, and goes on to
what will be the most interesting section to most readers—that on remedies and
preventive measures. A special point is made or the possibilities of action by
communities, with suggestions as to organisation, publicity, interesting the
children, and the work of Boards of Health.
WORKS BY EDWARD HALFORD ROSS,
M.R.C.S. (Eng.), L.R.C.P. (Lond.).
Of the John Howard McFadden Researches; the Lister Institute
of Preventive Medicine, London; and sometime Health Officer,
Port Said, the Suez Canal District and Cairo.
THE REDUCTION OF DOMESTIC MOSQUITOS.
Instructions for the use of Municipalities,
Town Councils, Health Officers, Sanitary Inspectors, and
Residents in Warm Climates. Demy 8vo. 5s. net.
THE REDUCTION OF DOMESTIC FLIES.
With Illustrations. Demy 8vo. 5s. net.
LONDON: JOHN MURRAY, ALBEMARLE STREET, W.
End of Project Gutenberg's Form and Function, by E. S. (Edward Stuart) Russell
*** END OF THIS PROJECT GUTENBERG EBOOK FORM AND FUNCTION ***
***** This file should be named 20426-h.htm or 20426-h.zip *****
This and all associated files of various formats will be found in:
http://www.gutenberg.org/2/0/4/2/20426/
Produced by Suzanne Lybarger, Turgut Dincer and the Online
Distributed Proofreading Team at http://www.pgdp.net (This
file was produced from images generously made available
by The Internet Archive/Canadian Libraries)
Updated editions will replace the previous one--the old editions
will be renamed.
Creating the works from public domain print editions means that no
one owns a United States copyright in these works, so the Foundation
(and you!) can copy and distribute it in the United States without
permission and without paying copyright royalties. Special rules,
set forth in the General Terms of Use part of this license, apply to
copying and distributing Project Gutenberg-tm electronic works to
protect the PROJECT GUTENBERG-tm concept and trademark. Project
Gutenberg is a registered trademark, and may not be used if you
charge for the eBooks, unless you receive specific permission. If you
do not charge anything for copies of this eBook, complying with the
rules is very easy. You may use this eBook for nearly any purpose
such as creation of derivative works, reports, performances and
research. They may be modified and printed and given away--you may do
practically ANYTHING with public domain eBooks. Redistribution is
subject to the trademark license, especially commercial
redistribution.
*** START: FULL LICENSE ***
THE FULL PROJECT GUTENBERG LICENSE
PLEASE READ THIS BEFORE YOU DISTRIBUTE OR USE THIS WORK
To protect the Project Gutenberg-tm mission of promoting the free
distribution of electronic works, by using or distributing this work
(or any other work associated in any way with the phrase "Project
Gutenberg"), you agree to comply with all the terms of the Full Project
Gutenberg-tm License (available with this file or online at
http://gutenberg.org/license).
Section 1. General Terms of Use and Redistributing Project Gutenberg-tm
electronic works
1.A. By reading or using any part of this Project Gutenberg-tm
electronic work, you indicate that you have read, understand, agree to
and accept all the terms of this license and intellectual property
(trademark/copyright) agreement. If you do not agree to abide by all
the terms of this agreement, you must cease using and return or destroy
all copies of Project Gutenberg-tm electronic works in your possession.
If you paid a fee for obtaining a copy of or access to a Project
Gutenberg-tm electronic work and you do not agree to be bound by the
terms of this agreement, you may obtain a refund from the person or
entity to whom you paid the fee as set forth in paragraph 1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be
used on or associated in any way with an electronic work by people who
agree to be bound by the terms of this agreement. There are a few
things that you can do with most Project Gutenberg-tm electronic works
even without complying with the full terms of this agreement. See
paragraph 1.C below. There are a lot of things you can do with Project
Gutenberg-tm electronic works if you follow the terms of this agreement
and help preserve free future access to Project Gutenberg-tm electronic
works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the Foundation"
or PGLAF), owns a compilation copyright in the collection of Project
Gutenberg-tm electronic works. Nearly all the individual works in the
collection are in the public domain in the United States. If an
individual work is in the public domain in the United States and you are
located in the United States, we do not claim a right to prevent you from
copying, distributing, performing, displaying or creating derivative
works based on the work as long as all references to Project Gutenberg
are removed. Of course, we hope that you will support the Project
Gutenberg-tm mission of promoting free access to electronic works by
freely sharing Project Gutenberg-tm works in compliance with the terms of
this agreement for keeping the Project Gutenberg-tm name associated with
the work. You can easily comply with the terms of this agreement by
keeping this work in the same format with its attached full Project
Gutenberg-tm License when you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern
what you can do with this work. Copyright laws in most countries are in
a constant state of change. If you are outside the United States, check
the laws of your country in addition to the terms of this agreement
before downloading, copying, displaying, performing, distributing or
creating derivative works based on this work or any other Project
Gutenberg-tm work. The Foundation makes no representations concerning
the copyright status of any work in any country outside the United
States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other immediate
access to, the full Project Gutenberg-tm License must appear prominently
whenever any copy of a Project Gutenberg-tm work (any work on which the
phrase "Project Gutenberg" appears, or with which the phrase "Project
Gutenberg" is associated) is accessed, displayed, performed, viewed,
copied or distributed:
This eBook is for the use of anyone anywhere at no cost and with
almost no restrictions whatsoever. You may copy it, give it away or
re-use it under the terms of the Project Gutenberg License included
with this eBook or online at www.gutenberg.org
1.E.2. If an individual Project Gutenberg-tm electronic work is derived
from the public domain (does not contain a notice indicating that it is
posted with permission of the copyright holder), the work can be copied
and distributed to anyone in the United States without paying any fees
or charges. If you are redistributing or providing access to a work
with the phrase "Project Gutenberg" associated with or appearing on the
work, you must comply either with the requirements of paragraphs 1.E.1
through 1.E.7 or obtain permission for the use of the work and the
Project Gutenberg-tm trademark as set forth in paragraphs 1.E.8 or
1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted
with the permission of the copyright holder, your use and distribution
must comply with both paragraphs 1.E.1 through 1.E.7 and any additional
terms imposed by the copyright holder. Additional terms will be linked
to the Project Gutenberg-tm License for all works posted with the
permission of the copyright holder found at the beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm
License terms from this work, or any files containing a part of this
work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this
electronic work, or any part of this electronic work, without
prominently displaying the sentence set forth in paragraph 1.E.1 with
active links or immediate access to the full terms of the Project
Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary,
compressed, marked up, nonproprietary or proprietary form, including any
word processing or hypertext form. However, if you provide access to or
distribute copies of a Project Gutenberg-tm work in a format other than
"Plain Vanilla ASCII" or other format used in the official version
posted on the official Project Gutenberg-tm web site (www.gutenberg.org),
you must, at no additional cost, fee or expense to the user, provide a
copy, a means of exporting a copy, or a means of obtaining a copy upon
request, of the work in its original "Plain Vanilla ASCII" or other
form. Any alternate format must include the full Project Gutenberg-tm
License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying,
performing, copying or distributing any Project Gutenberg-tm works
unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing
access to or distributing Project Gutenberg-tm electronic works provided
that
- You pay a royalty fee of 20% of the gross profits you derive from
the use of Project Gutenberg-tm works calculated using the method
you already use to calculate your applicable taxes. The fee is
owed to the owner of the Project Gutenberg-tm trademark, but he
has agreed to donate royalties under this paragraph to the
Project Gutenberg Literary Archive Foundation. Royalty payments
must be paid within 60 days following each date on which you
prepare (or are legally required to prepare) your periodic tax
returns. Royalty payments should be clearly marked as such and
sent to the Project Gutenberg Literary Archive Foundation at the
address specified in Section 4, "Information about donations to
the Project Gutenberg Literary Archive Foundation."
- You provide a full refund of any money paid by a user who notifies
you in writing (or by e-mail) within 30 days of receipt that s/he
does not agree to the terms of the full Project Gutenberg-tm
License. You must require such a user to return or
destroy all copies of the works possessed in a physical medium
and discontinue all use of and all access to other copies of
Project Gutenberg-tm works.
- You provide, in accordance with paragraph 1.F.3, a full refund of any
money paid for a work or a replacement copy, if a defect in the
electronic work is discovered and reported to you within 90 days
of receipt of the work.
- You comply with all other terms of this agreement for free
distribution of Project Gutenberg-tm works.
1.E.9. If you wish to charge a fee or distribute a Project Gutenberg-tm
electronic work or group of works on different terms than are set
forth in this agreement, you must obtain permission in writing from
both the Project Gutenberg Literary Archive Foundation and Michael
Hart, the owner of the Project Gutenberg-tm trademark. Contact the
Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable
effort to identify, do copyright research on, transcribe and proofread
public domain works in creating the Project Gutenberg-tm
collection. Despite these efforts, Project Gutenberg-tm electronic
works, and the medium on which they may be stored, may contain
"Defects," such as, but not limited to, incomplete, inaccurate or
corrupt data, transcription errors, a copyright or other intellectual
property infringement, a defective or damaged disk or other medium, a
computer virus, or computer codes that damage or cannot be read by
your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right
of Replacement or Refund" described in paragraph 1.F.3, the Project
Gutenberg Literary Archive Foundation, the owner of the Project
Gutenberg-tm trademark, and any other party distributing a Project
Gutenberg-tm electronic work under this agreement, disclaim all
liability to you for damages, costs and expenses, including legal
fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT
LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE
PROVIDED IN PARAGRAPH F3. YOU AGREE THAT THE FOUNDATION, THE
TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE
LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR
INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH
DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a
defect in this electronic work within 90 days of receiving it, you can
receive a refund of the money (if any) you paid for it by sending a
written explanation to the person you received the work from. If you
received the work on a physical medium, you must return the medium with
your written explanation. The person or entity that provided you with
the defective work may elect to provide a replacement copy in lieu of a
refund. If you received the work electronically, the person or entity
providing it to you may choose to give you a second opportunity to
receive the work electronically in lieu of a refund. If the second copy
is also defective, you may demand a refund in writing without further
opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth
in paragraph 1.F.3, this work is provided to you 'AS-IS' WITH NO OTHER
WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT LIMITED TO
WARRANTIES OF MERCHANTIBILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied
warranties or the exclusion or limitation of certain types of damages.
If any disclaimer or limitation set forth in this agreement violates the
law of the state applicable to this agreement, the agreement shall be
interpreted to make the maximum disclaimer or limitation permitted by
the applicable state law. The invalidity or unenforceability of any
provision of this agreement shall not void the remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the
trademark owner, any agent or employee of the Foundation, anyone
providing copies of Project Gutenberg-tm electronic works in accordance
with this agreement, and any volunteers associated with the production,
promotion and distribution of Project Gutenberg-tm electronic works,
harmless from all liability, costs and expenses, including legal fees,
that arise directly or indirectly from any of the following which you do
or cause to occur: (a) distribution of this or any Project Gutenberg-tm
work, (b) alteration, modification, or additions or deletions to any
Project Gutenberg-tm work, and (c) any Defect you cause.
Section 2. Information about the Mission of Project Gutenberg-tm
Project Gutenberg-tm is synonymous with the free distribution of
electronic works in formats readable by the widest variety of computers
including obsolete, old, middle-aged and new computers. It exists
because of the efforts of hundreds of volunteers and donations from
people in all walks of life.
Volunteers and financial support to provide volunteers with the
assistance they need, is critical to reaching Project Gutenberg-tm's
goals and ensuring that the Project Gutenberg-tm collection will
remain freely available for generations to come. In 2001, the Project
Gutenberg Literary Archive Foundation was created to provide a secure
and permanent future for Project Gutenberg-tm and future generations.
To learn more about the Project Gutenberg Literary Archive Foundation
and how your efforts and donations can help, see Sections 3 and 4
and the Foundation web page at http://www.pglaf.org.
Section 3. Information about the Project Gutenberg Literary Archive
Foundation
The Project Gutenberg Literary Archive Foundation is a non profit
501(c)(3) educational corporation organized under the laws of the
state of Mississippi and granted tax exempt status by the Internal
Revenue Service. The Foundation's EIN or federal tax identification
number is 64-6221541. Its 501(c)(3) letter is posted at
http://pglaf.org/fundraising. Contributions to the Project Gutenberg
Literary Archive Foundation are tax deductible to the full extent
permitted by U.S. federal laws and your state's laws.
The Foundation's principal office is located at 4557 Melan Dr. S.
Fairbanks, AK, 99712., but its volunteers and employees are scattered
throughout numerous locations. Its business office is located at
809 North 1500 West, Salt Lake City, UT 84116, (801) 596-1887, email
business@pglaf.org. Email contact links and up to date contact
information can be found at the Foundation's web site and official
page at http://pglaf.org
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
gbnewby@pglaf.org
Section 4. Information about Donations to the Project Gutenberg
Literary Archive Foundation
Project Gutenberg-tm depends upon and cannot survive without wide
spread public support and donations to carry out its mission of
increasing the number of public domain and licensed works that can be
freely distributed in machine readable form accessible by the widest
array of equipment including outdated equipment. Many small donations
($1 to $5,000) are particularly important to maintaining tax exempt
status with the IRS.
The Foundation is committed to complying with the laws regulating
charities and charitable donations in all 50 states of the United
States. Compliance requirements are not uniform and it takes a
considerable effort, much paperwork and many fees to meet and keep up
with these requirements. We do not solicit donations in locations
where we have not received written confirmation of compliance. To
SEND DONATIONS or determine the status of compliance for any
particular state visit http://pglaf.org
While we cannot and do not solicit contributions from states where we
have not met the solicitation requirements, we know of no prohibition
against accepting unsolicited donations from donors in such states who
approach us with offers to donate.
International donations are gratefully accepted, but we cannot make
any statements concerning tax treatment of donations received from
outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation
methods and addresses. Donations are accepted in a number of other
ways including checks, online payments and credit card donations.
To donate, please visit: http://pglaf.org/donate
Section 5. General Information About Project Gutenberg-tm electronic
works.
Professor Michael S. Hart is the originator of the Project Gutenberg-tm
concept of a library of electronic works that could be freely shared
with anyone. For thirty years, he produced and distributed Project
Gutenberg-tm eBooks with only a loose network of volunteer support.
Project Gutenberg-tm eBooks are often created from several printed
editions, all of which are confirmed as Public Domain in the U.S.
unless a copyright notice is included. Thus, we do not necessarily
keep eBooks in compliance with any particular paper edition.
Most people start at our Web site which has the main PG search facility:
http://www.gutenberg.org
This Web site includes information about Project Gutenberg-tm,
including how to make donations to the Project Gutenberg Literary
Archive Foundation, how to help produce our new eBooks, and how to
subscribe to our email newsletter to hear about new eBooks.