The Project Gutenberg eBook, The Descent of Man and Selection in Relation
to Sex, by Charles Darwin
This eBook is for the use of anyone anywhere at no cost and with
almost no restrictions whatsoever. You may copy it, give it away or
re-use it under the terms of the Project Gutenberg License included
with this eBook or online at www.gutenberg.org
Title: The Descent of Man and Selection in Relation to Sex
Volume II (1st Edition)
Author: Charles Darwin
Release Date: June 25, 2011 [eBook #36520]
Language: English
Character set encoding: UTF-8
***START OF THE PROJECT GUTENBERG EBOOK THE DESCENT OF MAN AND SELECTION IN RELATION TO SEX***
E-text prepared by Steven Gibbs, Turgut Dincer,
and the Online Distributed Proofreading Team
(http://www.pgdp.net)
THE
DESCENT OF MAN,
AND
SELECTION IN RELATION TO SEX.
By CHARLES DARWIN, M.A., F.R.S., &c.
IN TWO VOLUMES.—Vol. II.
WITH ILLUSTRATIONS.
LONDON:
JOHN MURRAY, ALBEMARLE STREET.
1871.
[The right of Translation is reserved.]
ERRATA.
VOL. I.
| Page |
line |
For |
read |
| 27 |
13 |
kaolo |
koala. |
| 31 |
6 |
prostratica |
prostatica. |
| 59, note86 |
2 |
speech |
species. |
| 74, note107 |
— |
Browne |
Brown. |
| 118, note167 |
— |
Vol. I. |
Vol. II. |
| 128, note184 |
4 |
Before vol. xiv. |
insert ‘Proc. Royal Soc. |
| 208 |
2 |
prostratica |
prostatica. |
| 322 |
5 |
Actineæ |
Actiniæ. |
| 324 |
30 |
land-shells |
land-snails. |
| 330 |
16 |
figs. 4 and 5 |
figs. 4, 5, and 6. |
| 334 |
17 |
Birgos |
Birgus. |
| 339 |
8 |
attractions |
attentions. |
| 341 |
3 |
dragon-flys |
dragon-flies. |
| 378 |
17 |
Typhæus |
Typhœus. |
| 384 |
31 |
tesselatum |
tessellatum. |
| 397 |
9 |
Hypopira |
Hypopyra. |
| 405 |
21 |
Acrœidæ |
Acræidæ. |
VOL. II.
| 32 |
30 |
chamelion |
chameleon. |
| 115 |
4 |
mail |
male. |
| 178 |
23 |
Chloehaga |
Chloephaga. |
| 227, note281 |
— |
Ramphaston |
Ramphastos. |
| 240, note289 |
— |
Mr. H. Brown |
Mr. R. Brown. |
| 240, note290 |
2 |
elephus |
elaphus. |
| 242 |
14 |
walruses |
narwhals. |
| 339 |
27 |
Durfur |
Darfur. |
v
CONTENTS. |
|
PART II. |
SEXUAL SELECTION—continued. |
| CHAPTER XII. |
| Secondary Sexual Characters of Fishes, Amphibians,
and Reptiles. |
| Fishes: Courtship and battles of the males—Larger size of the
females—Males, bright colours and ornamental appendages;
other strange characters—Colours and appendages acquired by
the males during the breeding-season alone—Fishes with both
sexes brilliantly coloured—Protective colours—The less conspicuous
colours of the female cannot be accounted for on the
principle of protection—Male fishes building nests, and taking
charge of the ova and young. Amphibians: Differences in
structure and colour between the sexes—Vocal organs. Reptiles:
Chelonians—Crocodiles—Snakes, colours in some cases
protective—Lizards, battles of—Ornamental appendages—Strange
differences in structure between the sexes—Colours—Sexual
differences almost as great as with birds |
1-37 |
| CHAPTER XIII. |
| Secondary Sexual Characters of Birds. |
| Sexual differences—Law of battle—Special weapons—Vocal
organs—Instrumental music—Love-antics and dances—Decorations,
permanent and seasonal—Double and single annual
moults—Display of ornaments by the males |
38-98 |
| CHAPTER XIV. |
| viBirds—continued |
| Choice exerted by the female—Length of courtship—Unpaired
birds—Mental qualities and taste for the beautiful—Preference
or antipathy shewn by the female for particular males—Variability
of birds—Variations sometimes abrupt—Laws of variation—Formation
of ocelli—Gradations of character—Case of
Peacock, Argus pheasant, and Urosticte |
99-153 |
| CHAPTER XV. |
| Birds—continued. |
| Discussion why the males alone of some species, and both sexes
of other species, are brightly coloured—On sexually-limited
inheritance, as applied to various structures and to brightly-coloured
plumage—Nidification in relation to colour—Loss of
nuptial plumage during the winter |
154-182 |
| CHAPTER XVI. |
| Birds—continued. |
| The immature plumage in relation to the character of the plumage
in both sexes when adult—Six classes of cases—Sexual differences
between the males of closely-allied or representative species—The
female assuming the characters of the male—Plumage of
the young in relation to the summer and winter plumage of the
adults—On the increase of beauty in the Birds of the World—Protective
colouring—Conspicuously-coloured birds—Novelty
appreciated—Summary of the four chapters on birds |
183-238 |
| CHAPTER XVII. |
| vii
Secondary Sexual Characters of Mammals. |
| The law of battle—Special weapons, confined to the males—Cause
of absence of weapons in the female—Weapons common to both
sexes, yet primarily acquired by the male—Other uses of such
weapons—Their high importance—Greater size of the male—Means
of defence—On the preference shewn by either sex in the
pairing of quadrupeds |
239-273 |
| CHAPTER XVIII. |
| Secondary Sexual Characters of Mammals.—continued. |
| Voice—Remarkable sexual peculiarities in seals—Odour—Development
of the hair—Colour of the hair and skin—Anomalous
case of the female being more ornamented than the male—Colour
and ornaments due to sexual selection—Colour acquired
for the sake of protection—Colour, though common to both
sexes, often due to sexual selection—On the disappearance of
spots and stripes in adult quadrupeds—On the colours and
ornaments of the Quadrumana—Summary |
274-315 |
| CHAPTER XIX. |
| Secondary Sexual Characters of Mammals.—continued. |
| Differences between man and woman—Causes of such differences
and of certain characters common to both sexes—Law of battle—Differences
in mental powers—and voice—On the influence
of beauty in determining the marriages of mankind—Attention
paid by savages to ornaments—Their ideas of beauty in woman—The
tendency to exaggerate each natural peculiarity |
316-354 |
| CHAPTER XX. |
| Secondary Sexual Characters of Man—continued. |
| On the effects of the continued selection of women according to a
different standard of beauty in each race—On the causes which
interfere with sexual selection in civilised and savage nations—Conditions
favourable to sexual selection during primeval
times—On the manner of action of sexual selection with mankind—On
the women in savage tribes having some power to
choose their husbands—Absence of hair on the body, and development
of the beard—Colour of the skin—Summary |
355-384 |
| CHAPTER XXI. |
| General Summary and Conclusion. |
| Main conclusion that man is descended from some lower form—Manner
of development—Genealogy of man—Intellectual and
moral faculties—Sexual selection—Concluding remarks |
385-405 |
| Index |
406 |
ix
POSTSCRIPT.
Vol. I. pp. 297-299.—I have fallen into a serious and
unfortunate error, in relation to the sexual differences
of animals, in attempting to explain what seemed to
me a singular coincidence in the late period of life
at which the necessary variations have arisen in many
cases, and the late period at which sexual selection
acts. The explanation given is wholly erroneous, as
I have discovered by working out an illustration in
figures. Moreover, the supposed coincidence of period
is far from general, and is not remarkable; for, as I
have elsewhere attempted to show, variations arising
early in life have often been accumulated through
sexual selection, being then commonly transmitted to
both sexes. On the other hand, variations arising late
in life cannot fail to coincide approximately in period
with that of the process of sexual selection. Allusions
to these erroneous views reappear in Vol. II. pp. 161
and 237.
1
SEXUAL SELECTION.
CHAPTER XII.
Secondary Sexual Characters of Fishes, Amphibians,
and Reptiles.
Fishes: Courtship and battles of the males—Larger size of the
females—Males, bright colours and ornamental appendages;
other strange characters—Colours and appendages acquired by
the males during the breeding-season alone—Fishes with both
sexes brilliantly coloured—Protective colours—The less conspicuous
colours of the female cannot be accounted for on the
principle of protection—Male fishes building nests, and taking
charge of the ova and young. Amphibians: Differences in
structure and colour between the sexes—Vocal organs. Reptiles:
Chelonians—Crocodiles—Snakes, colours in some cases
protective—Lizards, battles of—Ornamental appendages—
Strange differences in structure between the sexes—Colours—Sexual
differences almost as great as with birds.
We have now arrived at the great sub-kingdom of the
Vertebrata, and will commence with the lowest class,
namely Fishes. The males of Plagiostomous fishes
(sharks, rays) and of Chimæroid fishes are provided
with claspers which serve to retain the female, like the
various structures possessed by so many of the lower
animals. Besides the claspers, the males of many rays
have clusters of strong sharp spines on their heads,
and several rows along “the upper outer surface of their
pectoral fins.” These are present in the males of some
species, which have the other parts of their bodies2
smooth. They are only temporarily developed during
the breeding-season; and Dr. Günther suspects that they
are brought into action as prehensile organs by the
doubling inwards and downwards of the two sides of the
body. It is a remarkable fact that the females and not
the males of some species, as of Raia clavata, have their
backs studded with large hook-formed spines.1
Owing to the element which fishes inhabit, little
is known about their courtship, and not much about
their battles. The male stickleback (Gasterosteus leiurus)
has been described as “mad with delight” when
the female comes out of her hiding-place and surveys
the nest which he has made for her. “He darts round
her in every direction, then to his accumulated materials
for the nest, then back again in an instant;
and as she does not advance he endeavours to push
her with his snout, and then tries to pull her by the
tail and side-spine to the nest.”2 The males are said
to be polygamists;3 they are extraordinarily bold and
pugnacious, whilst “the females are quite pacific.”
Their battles are at times desperate; “for these puny
combatants fasten tight on each other for several
seconds, tumbling over and over again, until their
strength appears completely exhausted.” With the
rough-tailed stickleback (G. trachurus) the males whilst
fighting swim round and round each other, biting and
endeavouring to pierce each other with their raised lateral
spines. The same writer adds,4 “the bite of these little
3furies is very severe. They also use their lateral spines
with such fatal effect, that I have seen one during a
battle absolutely rip his opponent quite open, so that
he sank to the bottom and died.” When a fish is
conquered, “his gallant bearing forsakes him; his gay
colours fade away; and he hides his disgrace among
his peaceable companions, but is for some time the
constant object of his conqueror’s persecution.”
The male salmon is as pugnacious as the little stickleback;
and so is the male trout, as I hear from Dr.
Günther. Mr. Shaw saw a violent contest between two
male salmons which lasted the whole day; and Mr. R.
Buist, Superintendent of Fisheries, informs me that he
has often watched from the bridge at Perth the males
driving away their rivals whilst the females were spawning.
The males “are constantly fighting and tearing
each other on the spawning-beds, and many so injure
each other as to cause the death of numbers, many
being seen swimming near the banks of the river in
a state of exhaustion, and apparently in a dying
state.”5 The keeper of the Stormontfield breeding-ponds
visited, as Mr. Buist informs me, in June, 1868,
the northern Tyne, and found about 300 dead salmon,
all of which with one exception were males; and he was
convinced that they had lost their lives by fighting.
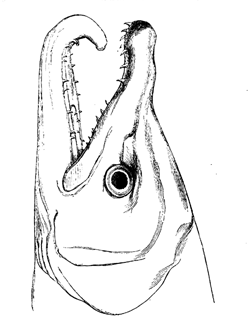
Fig. 26. Head of male of common salmon (Salmo salar) during the breeding-season.
[This drawing, as well as all the others in the present chapter, have been executed by
the well-known artist, Mr. G. Ford, under the kind superintendence of Dr. Günther, from
specimens in the British Museum.]
The most curious point about the male salmon is
that during the breeding-season, besides a slight change
in colour, “the lower jaw elongates, and a cartilaginous
projection turns upwards from the point, which, when
the jaws are closed, occupies a deep cavity between
4the intermaxillary bones of the upper jaw.”6 (Figs.
26 and 27.) In our salmon this change of structure
lasts only during the breeding-season; but in the Salmo
lycaodon of N.W. America the change, as Mr. J. K.
5Lord7 believes, is permanent and best marked in the
older males which have previously ascended the rivers.
In these old males the jaws become developed into immense
hook-like projections, and the teeth grow into
regular fangs, often more than half an inch in length.
With the European salmon, according to Mr. Lloyd,8
6the temporary hook-like structure serves to strengthen
and protect the jaws, when one male charges another
with wonderful violence; but the greatly developed
teeth of the male American salmon may be compared
with the tusks of many male mammals, and they
indicate an offensive rather than a protective purpose.
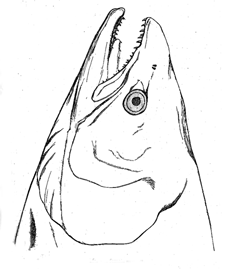
Fig. 27. Head of female salmon.
The salmon is not the only fish in which the teeth
differ in the two sexes. This is the case with many
rays. In the thornback (Raia clavata) the adult male
has sharp, pointed teeth, directed backwards, whilst
those of the female are broad and flat, forming a pavement;
so that these teeth differ in the two sexes of the
same species more than is usual in distinct genera of
the same family. The teeth of the male become sharp
only when he is adult: whilst young they are broad and
flat like those of the female. As so frequently occurs
with secondary sexual characters, both sexes of some
species of rays, for instance R. batis, possess, when adult,
sharp, pointed teeth; and here a character, proper to
and primarily gained by the male, appears to have been
transmitted to the offspring of both sexes. The teeth
are likewise pointed in both sexes of R. maculata, but
only when completely adult; the males acquiring them
at an earlier age than the females. We shall hereafter
meet with analogous cases with certain birds, in which
the male acquires the plumage common to both adult
sexes, at a somewhat earlier age than the female.
With other species of rays the males even when old
never possess sharp teeth, and consequently both sexes
when adult are provided with broad, flat teeth like
those of the young, and of the mature females of
the above-mentioned species.9 As the rays are bold,
7strong and voracious fishes, we may suspect that the
males require their sharp teeth for fighting with their
rivals; but as they possess many parts modified and
adapted for the prehension of the female, it is possible
that their teeth may be used for this purpose.
In regard to size, M. Carbonnier10 maintains that
with almost all fishes the female is larger than the male;
and Dr. Günther does not know of a single instance
in which the male is actually larger than the female.
With some Cyprinodonts the male is not even half as
large as the female. As with many kinds of fishes the
males habitually fight together; it is surprising that
they have not generally become through the effects of
sexual selection larger and stronger than the females.
The males suffer from their small size, for according to
M. Carbonnier they are liable to be devoured by the
females of their own species when carnivorous, and no
doubt by other species. Increased size must be in
some manner of more importance to the females, than
strength and size are to the males for fighting with
other males; and this perhaps is to allow of the production
of a vast number of ova.
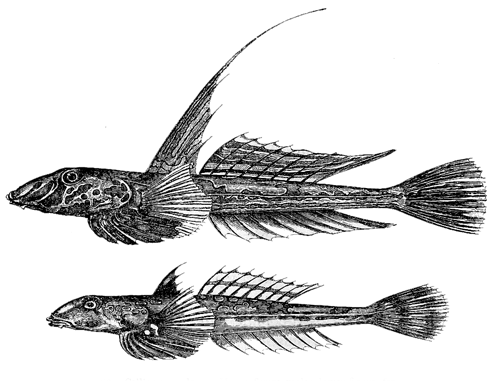
Fig. 28. Callionymus lyra. Upper figure, male; lower figure, female.
In many species the male alone is ornamented with
bright colours; or these are much brighter in the male
than the female. The male, also, is sometimes provided
with appendages which appear to be of no more use to
him for the ordinary purposes of life than are the tail-feathers
to the peacock. I am indebted for most of the
following facts to the great kindness of Dr. Günther.
There is reason to suspect that many tropical fishes
differ sexually in colour and structure; and there are
some striking cases with our British fishes. The male
Callionymus lyra has been called the gemmeous dragonet
8“from its brilliant gem-like colours.” When freshly
taken from the sea the body is yellow of various shades,
striped and spotted with vivid blue on the head; the
dorsal fins are pale brown with dark longitudinal bands;
the ventral, caudal and anal fins being bluish-black.
The female, or sordid dragonet, was considered by Linnæus
and by many subsequent naturalists as a distinct
species; it is of a dingy reddish-brown, with the dorsal
fin brown and the other fins white. The sexes differ also
in the proportional size of the head and mouth, and in
the position of the eyes;11 but the most striking difference
is the extraordinary elongation in the male (fig. 28)
9of the dorsal fin. The young males resemble in structure
and colour the adult females. Throughout the
genus Callionymus,12 the male is generally much more
brightly spotted than the female, and in several species,
not only the dorsal, but the anal fin of the male is much
elongated.
The male of the Cottus scorpius, or sea-scorpion, is more
slender and smaller than the female. There is also a
great difference in colour between them. It is difficult,
as Mr. Lloyd13 remarks, “for any one, who has not seen
this fish during the spawning-season, when its hues are
brightest, to conceive the admixture of brilliant colours
with which it, in other respects so ill-favoured, is at
that time adorned.” Both sexes of the Labrus mixtus,
although very different in colour, are beautiful; the male
being orange with bright-blue stripes, and the female
bright-red with some black spots on the back.
In the very distinct family of the Cyprinodontidæ—inhabitants
of the fresh waters of foreign lands—the
sexes sometimes differ much in various characters. In
the male of the Mollienesia petenensis,14 the dorsal fin is
greatly developed and is marked with a row of large,
round, ocellated, bright-coloured spots; whilst the same
fin in the female is smaller, of a different shape, and
marked only with irregularly-curved brown spots. In
the male the basal margin of the anal fin is also a little
produced and dark-coloured. In the male of an allied
form, the Xiphophorus Hellerii (fig. 29), the inferior
margin of the anal fin is developed into a long filament,
10which is striped, as I hear from Dr. Günther, with bright
colours. This filament does not contain any muscles,
and apparently cannot be of any direct use to the fish.
As in the case of the Callionymus, the males whilst
young resemble in colour and structure the adult
females. Sexual differences such as these may be
strictly compared with those which are so frequent
with gallinaceous birds.15

Fig. 29. Xiphophorus Hellerii. Upper figure, male; lower figure, female.
11
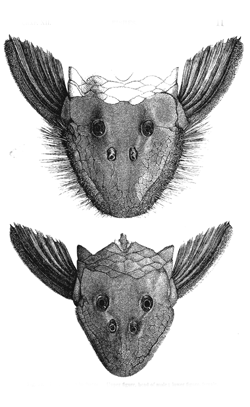
Fig. 30. Plecostomus barbatus. Upper figure, head of male; lower figure, female.
In a siluroid fish, inhabiting the fresh waters of South
America, namely the Plecostomus barbatus16 (fig. 30),
the male has its mouth and interoperculum fringed with
a beard of stiff hairs, of which the female shews hardly
a trace. These hairs are of the nature of scales. In
another species of the same genus, soft flexible tentacles
project from the front part of the head of the
12male, which are absent in the female. These tentacles
are prolongations of the true skin, and therefore are not
homologous with the stiff hairs of the former species; but
it can hardly be doubted that both serve the same
purpose. What this purpose may be it is difficult to
conjecture; ornament does not here seem probable, but
we can hardly suppose that stiff hairs and flexible filaments
can be useful in any ordinary way to the males
alone. The Monacanthus scopas, which was shewn to me
in the British Museum by Dr. Günther, presents a nearly
analogous case. The male has a cluster of stiff, straight
spines, like those of a comb, on the sides of the tail; and
these in a specimen six inches long were nearly an inch
and a half in length; the female has on the same place
a cluster of bristles, which may be compared with those
of a tooth-brush. In another species, the M. peronii, the
male has a brush like that possessed by the female of
the last species, whilst the sides of the tail in the female
are smooth. In some other species the same part of the
tail can be perceived to be a little roughened in the
male and perfectly smooth in the female; and lastly in
others, both sexes have smooth sides. In that strange
monster, the Chimæra monstrosa, the male has a hook-shaped
bone on the top of the head, directed forwards,
with its rounded end covered with sharp spines; in the
female “this crown is altogether absent,” but what its
use may be is utterly unknown.17
The structures as yet referred to are permanent in the
male after he has arrived at maturity; but with some
Blennies and in another allied genus18 a crest is developed
on the head of the male only during the breed13ing-season,
and their bodies at the same time become
more brightly-coloured. There can be little doubt that
this crest serves as a temporary sexual ornament, for the
female does not exhibit a trace of it. In other species
of the same genus both sexes possess a crest, and in at
least one species neither sex is thus provided. In this
case and in that of the Monacanthus, we have good instances
to how great an extent the sexual characters of
closely-allied forms may differ. In many of the Chromidæ,
for instance in Geophagus and especially in Cichla,
the males, as I hear from Professor Agassiz,19 have a conspicuous
protuberance on the forehead, which is wholly
wanting in the females and in the young males. Professor
Agassiz adds, “I have often observed these fishes
at the time of spawning when the protuberance is
largest, and at other seasons when it is totally wanting
and the two sexes shew no difference whatever in the
outline of the profile of the head. I never could
ascertain that it subserves any special function, and
the Indians on the Amazon know nothing about its
use.” These protuberances in their periodical appearance
resemble the fleshy caruncles on the heads of certain
birds; but whether they serve as ornaments must
remain at present doubtful.
The males of those fishes, which differ permanently in
colour from the females, often become more brilliant, as
I hear from Professor Agassiz and Dr. Günther, during
the breeding-season. This is likewise the case with a
multitude of fishes, the sexes of which at all other
seasons of the year are identical in colour. The tench,
roach, and perch may be given as instances. The male
salmon at this season is “marked on the cheeks with
14orange-coloured stripes, which give it the appearance
of a Labrus, and the body partakes of a golden-orange
tinge. The females are dark in colour, and are commonly
called black-fish.”20 An analogous and even
greater change takes place with the Salmo eriox or bull-trout;
the males of the char (S. umbla) are likewise at
this season rather brighter in colour than the females.21
The colours of the pike (Esox reticulatus) of the United
States, especially of the male, become, during the
breeding-season, exceedingly intense, brilliant, and iridescent.22
Another striking instance out of many is
afforded by the male stickleback (Gasterosteus leiurus),
which is described by Mr. Warington,23 as being then
“beautiful beyond description.” The back and eyes of
the female are simply brown, and the belly white. The
eyes of the male, on the other hand, are “of the most
splendid green, having a metallic lustre like the
green feathers of some humming-birds. The throat
and belly are of a bright crimson, the back of an
ashy-green, and the whole fish appears as though it
were somewhat translucent and glowed with an internal
incandescence.” After the breeding-season
these colours all change, the throat and belly become
of a paler red, the back more green, and the glowing
tints subside.
That with fishes there exists some close relation
between their colours and their sexual functions we can
clearly see;—firstly, from the adult males of certain
species being differently coloured from the females, and
often much more brilliantly;—secondly, from these same
15
males, whilst immature, resembling the mature females;—and,
lastly, from the males, even of those species
which at all other times of the year are identical in
colour with the females, often acquiring brilliant tints
during the spawning-season. We know that the males
are ardent in their courtship and sometimes fight desperately
together. If we may assume that the females
have the power of exerting a choice and of selecting the
more highly-ornamented males, all the above facts
become intelligible through the principle of sexual
selection. On the other hand, if the females habitually
deposited and left their ova to be fertilised by
the first male which chanced to approach, this fact
would be fatal to the efficiency of sexual selection; for
there could be no choice of a partner. But, as far
as is known, the female never willingly spawns except
in the close presence of a male, and the male never
fertilises the ova except in the close presence of
a female. It is obviously difficult to obtain direct
evidence with respect to female fishes selecting
their partners. An excellent observer,24 who carefully
watched the spawning of minnows (Cyprinus phoxinus),
remarks that owing to the males, which were ten times
as numerous as the females, crowding closely round
them, he could “speak only doubtfully on their operations.
When a female came among a number of
males they immediately pursued her; if she was not
ready for shedding her spawn, she made a precipitate
retreat; but if she was ready, she came boldly in
among them, and was immediately pressed closely by
a male on each side; and when they had been in that
situation a short time, were superseded by other two,
who wedged themselves in between them and the
16female, who appeared to treat all her lovers with
the same kindness.” Notwithstanding this last statement,
I cannot, from the several previous considerations,
give up the belief that the males which are
the most attractive to the females, from their brighter
colours or other ornaments, are commonly preferred by
them; and that the males have thus been rendered
more beautiful in the course of ages.
We have next to inquire whether this view can be
extended, through the law of the equal transmission of
characters to both sexes, to those groups in which the
males and females are brilliant in the same or nearly
the same degree and manner. In such a genus as
Labrus, which includes some of the most splendid
fishes in the world, for instance, the Peacock Labrus
(L. pavo), described,25 with pardonable exaggeration, as
formed of polished scales of gold encrusting lapis-lazuli,
rubies, sapphires, emeralds and amethysts, we
may, with much probability, accept this belief; for we
have seen that the sexes in at least one species differ
greatly in colour. With some fishes, as with many of
the lowest animals, splendid colours may be the direct
result of the nature of their tissues and of the surrounding
conditions, without any aid from selection. The
goldfish (Cyprinus auratus), judging from the analogy
of the golden variety of the common carp, is, perhaps,
a case in point, as it may owe its splendid colours to
a single abrupt variation, due to the conditions to
which this fish has been subjected under confinement.
It is, however, more probable that these colours have
been intensified through artificial selection, as this species
has been carefully bred in China from a remote
17period.26 Under natural conditions it does not seem
probable that beings so highly organised as fishes, and
which live under such complex relations, should become
brilliantly coloured without suffering some evil or receiving
some benefit from so great a change, and consequently
without the intervention of natural selection.
What, then, must we conclude in regard to the many
fishes, both sexes of which are splendidly coloured?
Mr. Wallace27 believes that the species which frequent
reefs, where corals and other brightly-coloured organisms
abound, are brightly coloured in order to escape detection
by their enemies; but according to my recollection
they were thus rendered highly conspicuous. In the
freshwaters of the Tropics there are no brilliantly-coloured
corals or other organisms for the fishes to
resemble; yet many species in the Amazons are beautifully
coloured, and many of the carnivorous Cyprinidæ
in India are ornamented with “bright longitudinal
lines of various tints.”28 Mr. M’Clelland, in
describing these fishes goes so far as to suppose that
“the peculiar brilliancy of their colours” serves as “a
better mark for kingfishers, terns, and other birds
which are destined to keep the number of these fishes
in check;” but at the present day few naturalists will
18admit that any animal has been made conspicuous as an
aid to its own destruction. It is possible that certain
fishes may have been rendered conspicuous in order to
warn birds and beasts of prey (as explained when treating
of caterpillars) that they were unpalatable; but it
is not, I believe, known that any fish, at least any freshwater
fish, is rejected from being distasteful to fish-devouring
animals. On the whole, the most probable
view in regard to the fishes, of which both sexes are
brilliantly coloured, is that their colours have been
acquired by the males as a sexual ornament, and have
been transferred in an equal or nearly equal degree to
the other sex.
We have now to consider whether, when the male
differs in a marked manner from the female in colour
or in other ornaments, he alone has been modified,
with the variations inherited only by his male offspring;
or whether the female has been specially modified and
rendered inconspicuous for the sake of protection, such
modifications being inherited only by the females. It is
impossible to doubt that colour has been acquired by
many fishes as a protection: no one can behold the
speckled upper surface of a flounder, and overlook its
resemblance to the sandy bed of the sea on which it
lives. One of the most striking instances ever recorded
of an animal gaining protection by its colour (as far
as can be judged in preserved specimens) and by
its form, is that given by Dr. Günther29 of a pipefish,
which, with its reddish streaming filaments, is
hardly distinguishable from the sea-weed to which it
clings with its prehensile tail. But the question now
under consideration is whether the females alone have
been modified for this object. Fishes offer valuable
19evidence on this head. We can see that one sex will
not be modified through natural selection for the sake
of protection more than the other, supposing both to
vary, unless one sex is exposed for a longer period
to danger, or has less power of escaping from such
danger than the other sex; and it does not appear that
with fishes the sexes differ in these respects. As far as
there is any difference, the males, from being generally
of smaller size, and from wandering more about, are
exposed to greater danger than the females; and yet,
when the sexes differ, the males are almost always the
most conspicuously coloured. The ova are fertilised
immediately after being deposited, and when this process
lasts for several days, as in the case of the salmon,30
the female, during the whole time, is attended by the
male. After the ova are fertilised they are, in most
cases, left unprotected by both parents, so that the
males and females, as far as oviposition is concerned,
are equally exposed to danger, and both are equally
important for the production of fertile ova; consequently
the more or less brightly-coloured individuals of either
sex would be equally liable to be destroyed or preserved,
and both would have an equal influence on the
colours of their offspring or the race.
Certain fishes, belonging to several families, make
nests; and some of these fishes take care of their
young when hatched. Both sexes of the brightly-coloured
Crenilabrus massa and melops work together in
building their nests with sea-weed, shells, &c.31 But the
males of certain fishes do all the work, and afterwards
take exclusive charge of the young. This is the case
20with the dull-coloured gobies,32 in which the sexes are
not known to differ in colour, and likewise with the
sticklebacks (Gasterosteus), in which the males become
brilliantly coloured during the spawning-season. The
male of the smooth-tailed stickleback (G. leiurus) performs
during a long time the duties of a nurse with
exemplary care and vigilance, and is continually
employed in gently leading back the young to the
nest when they stray too far. He courageously
drives away all enemies, including the females of his
own species. It would indeed be no small relief to the
male if the female, after depositing her eggs, were
immediately devoured by some enemy, for he is forced
incessantly to drive her from the nest.33
The males of certain other fishes inhabiting South
America and Ceylon, and belonging to two distinct
orders, have the extraordinary habit of hatching the
eggs laid by the females within their mouths or branchial
cavities.34 With the Amazonian species which follow
this habit, the males, as I am informed by the kindness
of Professor Agassiz, “not only are generally brighter
than the females, but the difference is greater at
the spawning-season than at any other time.” The
species of Geophagus act in the same manner; and in
this genus, a conspicuous protuberance becomes developed
on the forehead of the males during the breeding-season.
With the various species of Chromids, as Professor
Agassiz likewise informs me, sexual differences
21in colour may be observed, “whether they lay their
eggs in the water among aquatic plants, or deposit
them in holes, leaving them to come out without
further care, or build shallow nests in the river-mud,
over which they sit, as our Promotis does. It ought
also to be observed that these sitters are among the
brightest species in their respective families; for
instance, Hygrogonus is bright green, with large
black ocelli, encircled with the most brilliant red.”
Whether with all the species of Chromids it is the male
alone which sits on the eggs is not known. It is,
however, manifest that the fact of the eggs being protected
or unprotected, has had little or no influence on
the differences in colour between the sexes. It is further
manifest, in all the cases in which the males take
exclusive charge of the nests and young, that the
destruction of the brighter-coloured males would be far
more influential on the character of the race, than the
destruction of the brighter-coloured females; for the
death of the male during the period of incubation or
nursing would entail the death of the young, so that
these could not inherit his peculiarities; yet, in many
of these very cases the males are more conspicuously
coloured than the females.
In most of the Lophobranchii (Pipe-fish, Hippocampi,
&c.) the males have either marsupial sacks or
hemispherical depressions on the abdomen, in which
the ova laid by the female are hatched. The males
also shew great attachment to their young.35 The
sexes do not commonly differ much in colour; but Dr.
Günther believes that the male Hippocampi are rather
brighter than the females. The genus Solenostoma,
22however, offers a very curious exceptional case,36 for the
female is much more vividly coloured and spotted than
the male, and she alone has a marsupial sack and
hatches the eggs; so that the female of Solenostoma
differs from all the other Lophobranchii in this latter
respect, and from almost all other fishes, in being more
brightly coloured than the male. It is improbable that
this remarkable double inversion of character in the
female should be an accidental coincidence. As the
males of several fishes which take exclusive charge of
the eggs and young are more brightly coloured than
the females, and as here the female Solenostoma takes
the same charge and is brighter than the male, it might
be argued that the conspicuous colours of the sex which
is the most important of the two for the welfare of the
offspring must serve, in some manner, as a protection.
But from the multitude of fishes, the males of which
are either permanently or periodically brighter than
the females, but whose life is not at all more important
than that of the female for the welfare of the species,
this view can hardly be maintained. When we treat of
birds we shall meet with analogous cases in which
there has been a complete inversion of the usual attributes
of the two sexes, and we shall then give what
appears to be the probable explanation, namely, that
the males have selected the more attractive females,
instead of the latter having selected, in accordance with
the usual rule throughout the animal kingdom, the more
attractive males.
On the whole we may conclude, that with most fishes,
in which the sexes differ in colour or in other orna23mental
characters, the males originally varied, with their
variations transmitted to the same sex, and accumulated
through sexual selection by attracting or exciting the
females. In many cases, however, such characters have
been transferred, either partially or completely, to the
females. In other cases, again, both sexes have been
coloured alike for the sake of protection; but in no
instance does it appear that the female alone has had
her colours or other characters specially modified for
this purpose.
The last point which need be noticed is that in many
parts of the world fishes are known to make peculiar
noises, which are described in some cases as being
musical. Very little has been ascertained with respect
to the means by which such sounds are produced, and
even less about their purpose. The drumming of the
Umbrinas in the European seas is said to be audible
from a depth of twenty fathoms. The fishermen of
Rochelle assert “that the males alone make the noise
during the spawning-time; and that it is possible by
imitating it, to take them without bait.”37 If this
statement is trustworthy, we have an instance in this,
the lowest class of the Vertebrata, of what we shall
find prevailing throughout the other vertebrate classes,
and which prevails, as we have already seen, with
insects and spiders; namely, that vocal and instrumental
sounds so commonly serve as a love-call or as
a love-charm, that the power of producing them was
probably first developed in connection with the propagation
of the species.
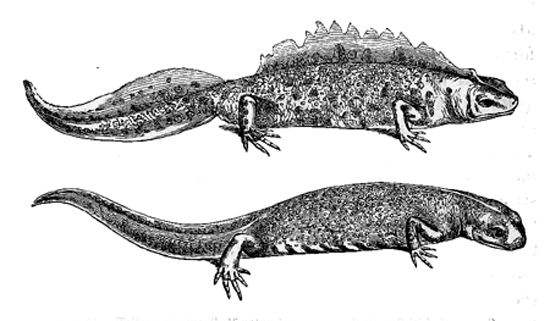
Fig. 31. Triton cristatus (half natural size, from Bell’s ‘British Reptiles’).
Upper figure, male during the breeding-season; lower figure, female.
24
Amphibians.
Urodela.—First for the tailed amphibians. The
sexes of salamanders or newts often differ much both
in colour and structure. In some species prehensile
claws are developed on the forelegs of the males
during the breeding-season; and at this season in
the male Triton palmipes the hind-feet are provided
with a swimming web, which is almost completely
absorbed during the winter; so that their feet then
resemble those of the female.38 This structure no doubt
aids the male in his eager search and pursuit of the
female. With our common newts (Triton punctatus
and cristatus) a deep, much-indented crest is developed
along the back and tail of the male during the breeding-season,
being absorbed during the winter. It is
not furnished, as Mr. St. George Mivart informs me,
25with muscles, and therefore cannot be used for locomotion.
As during the season of courtship it becomes
edged with bright colours, it serves, there can hardly
be a doubt, as a masculine ornament. In many species
the body presents strongly contrasted, though lurid
tints; and these become more vivid during the
breeding-season. The male, for instance, of our common
little newt (Triton punctatus) is “brownish-grey
above, passing into yellow beneath, which in the
spring becomes a rich bright orange, marked everywhere
with round dark spots.” The edge of the crest
also is then tipped with bright red or violet. The
female is usually of a yellowish-brown colour with
scattered brown dots; and the lower surface is often
quite plain.39 The young are obscurely tinted. The
ova are fertilised during the act of deposition and
are not subsequently tended by either parent. We
may therefore conclude that the males acquired their
strongly-marked colours and ornamental appendages
through sexual selection; these being transmitted either
to the male offspring alone or to both sexes.
Anura or Batrachia.—With many frogs and toads
the colours evidently serve as a protection, such as
the bright green tints of tree-frogs and the obscure
mottled shades of many terrestrial species. The most
conspicuously coloured toad which I ever saw, namely
the Phryniscus nigricans40 had the whole upper surface
of the body as black as ink, with the soles of the feet
and parts of the abdomen spotted with the brightest
vermilion. It crawled about the bare sandy or open
grassy plains of La Plata under a scorching sun, and
26could not fail to catch the eye of every passing creature.
These colours may be beneficial by making this toad
known to all birds of prey as a nauseous mouthful;
for it is familiar to every one that these animals
emit a poisonous secretion, which causes the mouth
of a dog to froth, as if attacked by hydrophobia. I
was the more struck with the conspicuous colours of
this toad, as close by I found a lizard (Proctotretus
multimaculatus) which, when frightened, flattened its
body, closed its eyes, and then from its mottled tints
could hardly be distinguishable from the surrounding
sand.
With respect to sexual differences of colour, Dr. Günther
knows of no striking instance with frogs or toads;
yet he can often distinguish the male from the female,
by the tints of the former being a little more intense.
Nor does Dr. Günther know of any striking difference
in external structure between the sexes, excepting the
prominences which become developed during the breeding-season
on the front-legs of the male, by which he
is enabled to hold the female. The Megalophrys montana41
(fig. 32) offers the best case of a certain amount of
structural difference between the sexes; for in the male
the tip of the nose and the eyelids are produced into
triangular flaps of skin, and there is a little black
tubercle on the back—characters which are absent, or
only feebly developed, in the females. It is surprising
that frogs and toads should not have acquired more
strongly-marked sexual differences; for though cold-blooded,
their passions are strong. Dr. Günther informs
me that he has several times found an unfortunate
female toad dead and smothered from having been
so closely embraced by three or four males.
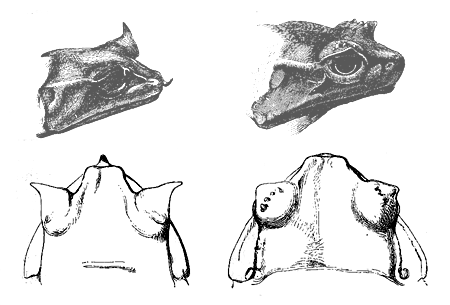
Fig. 32. Megalophrys montana. The two left-hand figures, the male; the two
right-hand figures, the female.
27
These animals, however, offer one interesting sexual
difference, namely in the musical powers possessed by
the males; but to speak of music, when applied to the
discordant and overwhelming sounds emitted by male
bull-frogs and some other species, seems, according to
our taste, a singularly inappropriate expression. Nevertheless
certain frogs sing in a decidedly pleasing manner.
Near Rio de Janeiro I used often to sit in the
evening to listen to a number of little Hylæ, which,
perched on blades of grass close to the water, sent
forth sweet chirping notes in harmony. The various
sounds are emitted chiefly by the males during the
breeding-season, as in the case of the croaking of our
common frog.42 In accordance with this fact the vocal
organs of the males are more highly developed than
those of the females. In some genera the males alone
28are provided with sacs which open into the larynx.43
For instance, in the edible frog (Rana esculenta) “the
sacs are peculiar to the males, and become, when filled
with air in the act of croaking, large globular bladders,
standing out one on each side of the head, near
the corners of the mouth.” The croak of the male is
thus rendered exceedingly powerful; whilst that of the
female is only a slight groaning noise.44 The vocal
organs differ considerably in structure in the several
genera of the family; and their development in all
cases may be attributed to sexual selection.
Reptiles.
Chelonia.—Tortoises and turtles do not offer well-marked
sexual differences. In some species, the tail
of the male is longer than that of the female. In
some, the plastron or lower surface of the shell of the
male is slightly concave in relation to the back of the
female. The male of the mud-turtle of the United
States (Chrysemys picta) has claws on its front-feet twice
as long as those of the female; and these are used when
the sexes unite.45 With the huge tortoise of the Galapagos
Islands (Testudo nigra) the males are said to
grow to a larger size than the females: during the
pairing-season, and at no other time, the male utters a
hoarse, bellowing noise, which can be heard at the distance
of more than a hundred yards; the female, on
the other hand, never uses her voice.46
Crocodilia.—The sexes apparently do not differ in
29colour; nor do I know that the males fight together,
though this is probable, for some kinds make a prodigious
display before the females. Bartram47 describes
the male alligator as striving to win the female by
splashing and roaring in the midst of a lagoon, “swollen
to an extent ready to burst, with his head and tail
lifted up, he spins or twirls round on the surface of
the water, like an Indian chief rehearsing his feats
of war.” During the season of love, a musky odour
is emitted by the submaxillary glands of the crocodile,
and pervades their haunts.48
Ophidia.—I have little to say about Snakes. Dr.
Günther informs me that the males are always smaller
than the females, and generally have longer and slenderer
tails; but he knows of no other difference in
external structure. In regard to colour, Dr. Günther
can almost always distinguish the male from the female
by his more strongly-pronounced tints; thus the black
zigzag band on the back of the male English viper is
more distinctly defined than in the female. The difference
is much plainer in the Rattle-snakes of N. America,
the male of which, as the keeper in the Zoological
Gardens shewed me, can instantly be distinguished from
the female by having more lurid yellow about its whole
body. In S. Africa the Bucephalus capensis presents an
analogous difference, for the female “is never so fully
variegated with yellow on the sides, as the male.”49
The male of the Indian Dipsas cynodon, on the other
hand, is blackish-brown, with the belly partly black,
whilst the female is reddish or yellowish-olive with the
belly either uniform yellowish or marbled with black.
30
In the Tragops dispar of the same country, the male is
bright green, and the female bronze-coloured.50 No
doubt the colours of some snakes serve as a protection,
as the green tints of tree-snakes and the various mottled
shades of the species which live in sandy places; but
it is doubtful whether the colours of many kinds, for
instance of the common English snake or viper, serve
to conceal them; and this is still more doubtful with
the many foreign species which are coloured with extreme
elegance.
During the breeding-season their anal scent-glands
are in active function;51 and so it is with the same
glands in lizards, and as we have seen with the submaxillary
glands of crocodiles. As the males of most
animals search for the females, these odoriferous glands
probably serve to excite or charm the female, rather
than to guide her to the spot where the male may be
found.52 Male snakes, though appearing so sluggish,
are amorous; for many have been observed crowding
round the same female, and even round the dead body
of a female. They are not known to fight together
from rivalry. Their intellectual powers are higher than
might have been anticipated. An excellent observer
in Ceylon, Mr. E. Layard,53 saw a Cobra thrust its head
through a narrow hole and swallow a toad. “With
31this incumbrance he could not withdraw himself;
finding this, he reluctantly disgorged the precious
morsel, which began to move off; this was too much
for snake philosophy to bear, and the toad was again
seized, and again was the snake, after violent efforts
to escape, compelled to part with its prey. This time,
however, a lesson had been learnt, and the toad was
seized by one leg, withdrawn, and then swallowed in
triumph.”
It does not, however, follow because snakes have
some reasoning power and strong passions, that they
should likewise be endowed with sufficient taste to
admire brilliant colours in their partners, so as to
lead to the adornment of the species through sexual
selection. Nevertheless it is difficult to account in
any other manner for the extreme beauty of certain
species; for instance, of the coral-snakes of S. America,
which are of a rich red with black and yellow transverse
bands. I well remember how much surprise I felt at
the beauty of the first coral-snake which I saw gliding
across a path in Brazil. Snakes coloured in this peculiar
manner, as Mr. Wallace states on the authority of Dr.
Günther,54 are found nowhere else in the world except
in S. America, and here no less than four genera occur.
One of these, Elaps, is venomous; a second and widely-distinct
genus is doubtfully venomous, and the two others
are quite harmless. The species belonging to these distinct
genera inhabit the same districts, and are so like
each other, that no one “but a naturalist would distinguish
the harmless from the poisonous kinds.” Hence,
as Mr. Wallace believes, the innocuous kinds have probably
acquired their colours as a protection, on the
principle of imitation; for they would naturally be
32thought dangerous by their enemies. The cause, however,
of the bright colours of the venomous Elaps
remains to be explained, and this may perhaps be
sexual selection.
Lacertilia.—The males of some, probably of many
kinds of lizards fight together from rivalry. Thus the
arboreal Anolis cristatellus of S. America is extremely
pugnacious: “During the spring and early part of the
summer, two adult males rarely meet without a contest.
On first seeing one another, they nod their heads
up and down three or four times, at the same time
expanding the frill or pouch beneath the throat; their
eyes glisten with rage, and after waving their tails
from side to side for a few seconds, as if to gather
energy, they dart at each other furiously, rolling over
and over, and holding firmly with their teeth. The
conflict generally ends in one of the combatants losing
his tail, which is often devoured by the victor.” The
male of this species is considerably larger than the female;55
and this, as far as Dr. Günther has been able to
ascertain, is the general rule with lizards of all kinds.
The sexes often differ greatly in various external
characters. The male of the above-mentioned Anolis
is furnished with a crest, which runs along the back and
tail, and can be erected at pleasure; but of this crest
the female does not exhibit a trace. In the Indian
Cophotis ceylanica, the female possesses a dorsal crest,
though much less developed than in the male; and
so it is, as Dr. Günther informs me, with the females
of many Iguanas, Chameleons and other lizards. In
some species, however, the crest is equally developed in
both sexes, as in the Iguana tuberculata. In the genus
33Sitana, the males alone are furnished with a large
throat-pouch (fig. 33), which can be folded up like a
fan, and is coloured blue, black, and red; but these
splendid colours are exhibited only during the pairing-season.
The female does not possess even a rudiment
of this appendage. In the Anolis cristatellus, according
to Mr. Austen, the throat-pouch, which is bright
red marbled with yellow, is present, though in a rudimental
condition, in the female. Again, in certain
other lizards, both sexes are equally well provided with
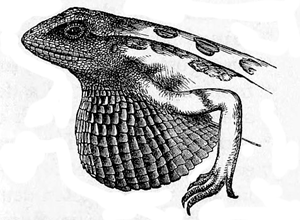 Fig. 33. Sitana minor. Male, with the gular
pouch expanded (from Günther’s ‘Reptiles
of India’).
throat-pouches. Here, as
in so many previous cases,
we see with species belonging
to the same group,
the same character confined
to the males, or more
largely developed in the
males than in the females,
or equally developed in
both sexes. The little lizards
of the genus Draco,
which glide through the air on their rib-supported parachutes,
and which in the beauty of their colours baffle
description, are furnished with skinny appendages to the
throat, “like the wattles of gallinaceous birds.” These
become erected when the animal is excited. They occur
in both sexes, but are best developed in the male when
arrived at maturity, at which age the middle appendage
is sometimes twice as long as the head. Most of the
species likewise have a low crest running along the
neck; and this is much more developed in the full-grown
males, than in the females or young males.56
Fig. 33. Sitana minor. Male, with the gular
pouch expanded (from Günther’s ‘Reptiles
of India’).
throat-pouches. Here, as
in so many previous cases,
we see with species belonging
to the same group,
the same character confined
to the males, or more
largely developed in the
males than in the females,
or equally developed in
both sexes. The little lizards
of the genus Draco,
which glide through the air on their rib-supported parachutes,
and which in the beauty of their colours baffle
description, are furnished with skinny appendages to the
throat, “like the wattles of gallinaceous birds.” These
become erected when the animal is excited. They occur
in both sexes, but are best developed in the male when
arrived at maturity, at which age the middle appendage
is sometimes twice as long as the head. Most of the
species likewise have a low crest running along the
neck; and this is much more developed in the full-grown
males, than in the females or young males.56
34
There are other and much more remarkable differences
between the sexes of certain lizards. The
male of Ceratophora aspera bears on the extremity of
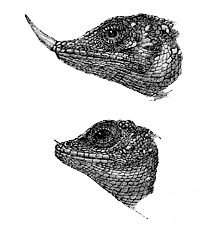 Fig. 34. Ceratophora Stoddartii. Upper figure, male; lower figure, female.
his snout an appendage half as long as the head. It
is cylindrical, covered with scales, flexible, and apparently
capable of erection: in the female it is quite
rudimental. In a second species of the same genus a
terminal scale forms a minute horn on the summit
of the flexible appendage; and in a third species (C.
Stoddartii, fig. 34) the whole appendage is converted
into a horn, which is usually
of a white colour, but assumes
a purplish tint when
the animal is excited. In
the adult male of this latter
species the horn is half an
inch in length, but is of quite
minute size in the female and
in the young. These appendages,
as Dr. Günther has
remarked to me, may be
compared with the combs of
gallinaceous birds, and apparently
serve as ornaments.
Fig. 34. Ceratophora Stoddartii. Upper figure, male; lower figure, female.
his snout an appendage half as long as the head. It
is cylindrical, covered with scales, flexible, and apparently
capable of erection: in the female it is quite
rudimental. In a second species of the same genus a
terminal scale forms a minute horn on the summit
of the flexible appendage; and in a third species (C.
Stoddartii, fig. 34) the whole appendage is converted
into a horn, which is usually
of a white colour, but assumes
a purplish tint when
the animal is excited. In
the adult male of this latter
species the horn is half an
inch in length, but is of quite
minute size in the female and
in the young. These appendages,
as Dr. Günther has
remarked to me, may be
compared with the combs of
gallinaceous birds, and apparently
serve as ornaments.
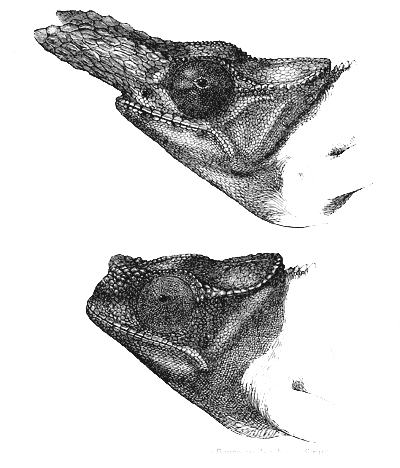
Fig. 35. Chamæleon bifurcus. Upper figure, male; lower figure, female.
In the genus Chamæleon we come to the climax of
difference between the sexes. The upper part of the
skull of the male C. bifurcus (fig. 35), an inhabitant of
Madagascar, is produced into two great, solid, bony projections,
covered with scales like the rest of the head;
and of this wonderful modification of structure the female
exhibits only a rudiment. Again, in Chamæleon Owenii
(fig. 36), from the West Coast of Africa, the male bears
35on his snout and forehead three curious horns, of which
the female has not a trace. These horns consist of
an excrescence of bone covered with a smooth sheath,
forming part of the general integuments of the body,
so that they are identical in structure with those of a
bull, goat, or other sheath-horned ruminant. Although
the three horns differ so much in appearance from
the two great prolongations of the skull in C. bifurcus,
we can hardly doubt that they serve the same general
purpose in the economy of these two animals. The36
first conjecture which will occur to every one is that
they are used by the males for fighting together; but
Dr. Günther, to whom I am indebted for the foregoing
details, does not believe that such peaceable creatures
would ever become pugnacious. Hence we are
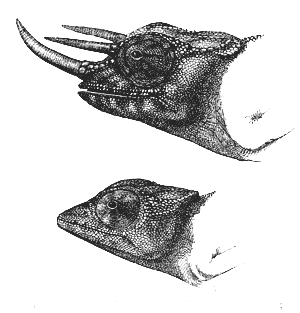 Fig. 36. Chamæleon Owenii. Upper figure, male;
lower figure, female.
driven to infer that
these almost monstrous
deviations
of structure serve
as masculine ornaments.
Fig. 36. Chamæleon Owenii. Upper figure, male;
lower figure, female.
driven to infer that
these almost monstrous
deviations
of structure serve
as masculine ornaments.
With many kinds
of lizards, the sexes
differ slightly in colour,
the tints and
stripes of the males
being brighter and
more distinctly defined
than in the
females. This, for
instance, is the case
with the previously-mentioned Cophotis and with the
Acanthodactylus capensis of S. Africa. In a Cordylus
of the latter country, the male is either much redder or
greener than the female. In the Indian Calotes nigrilabris
there is a greater difference in colour between
the sexes; the lips also of the male are black, whilst
those of the female are green. In our common
little viviparous lizard (Zootoca vivipara) “the under
side of the body and base of the tail in the male are
bright orange, spotted with black; in the female
these parts are pale greyish-green without spots.”57
We have seen that the males alone of Sitana possess a
37throat-pouch; and this is splendidly tinted with blue,
black, and red. In the Proctotretus tenuis of Chile the
male alone is marked with spots of blue, green, and
coppery-red.58 I collected in S. America fourteen
species of this genus, and though I neglected to record
the sexes, I observed that certain individuals alone were
marked with emerald-like green spots, whilst others
had orange-coloured gorges; and these in both cases
no doubt were the males.
In the foregoing species, the males are more brightly
coloured than the females, but with many lizards both
sexes are coloured in the same elegant or even magnificent
manner; and there is no reason to suppose that
such conspicuous colours are protective. With some
lizards, however, the green tints no doubt serve for
concealment; and an instance has already been incidently
given of one species of Proctotretus which
closely resembles the sand on which it lives. On the
whole we may conclude with tolerable safety that the
beautiful colours of many lizards, as well as various
appendages and other strange modifications of structure,
have been gained by the males through sexual selection
for the sake of ornament, and have been transmitted
either to their male offspring alone or to both sexes.
Sexual selection, indeed, seems to have played almost as
important a part with reptiles as with birds. But the
less conspicuous colours of the females in comparison
with those of the males cannot be accounted for, as
Mr. Wallace believes to be the case with birds, by the
exposure of the females to danger during incubation.
38
CHAPTER XIII.
Secondary Sexual Characters of Birds.
Sexual differences—Law of battle—Special weapons—Vocal
organs—Instrumental music—Love-antics and dances—Decorations,
permanent and seasonal—Double and single
annual moults—Display of ornaments by the males.
Secondary sexual characters are more diversified and
conspicuous in birds, though not perhaps entailing more
important changes of structure, than in any other class
of animals. I shall, therefore, treat the subject at considerable
length. Male birds sometimes, though rarely,
possess special weapons for fighting with each other.
They charm the females by vocal or instrumental music
of the most varied kinds. They are ornamented by
all sorts of combs, wattles, protuberances, horns, air-distended
sacs, top-knots, naked shafts, plumes and
lengthened feathers gracefully springing from all parts
of the body. The beak and naked skin about the head,
and the feathers are often gorgeously coloured. The
males sometimes pay their court by dancing, or by fantastic
antics performed either on the ground or in the air.
In one instance, at least, the male emits a musky odour
which we may suppose serves to charm or excite the
female; for that excellent observer, Mr. Ramsay,59 says
of the Australian musk-duck (Biziura lobata) that “the
smell which the male emits during the summer
months is confined to that sex, and in some individuals
is retained throughout the year; I have never
39even in the breeding-season, shot a female which had
any smell of musk.” So powerful is this odour during
the pairing-season, that it can be detected long before
the bird can be seen.60 On the whole, birds appear to
be the most æsthetic of all animals, excepting of course
man, and they have nearly the same taste for the beautiful
as we have. This is shewn by our enjoyment of
the singing of birds, and by our women, both civilised
and savage, decking their heads with borrowed plumes,
and using gems which are hardly more brilliantly
coloured than the naked skin and wattles of certain
birds.
Before treating of the characters with which we are
here more particularly concerned, I may just allude to
certain differences between the sexes which apparently
depend on differences in their habits of life; for such
cases, though common in the lower, are rare in the
higher classes. Two humming-birds belonging to the
genus Eustephanus, which inhabit the island of Juan
Fernandez, were long thought to be specifically distinct,
but are now known, as Mr. Gould informs me, to be the
sexes of the same species, and they differ slightly in the
form of the beak. In another genus of humming-birds
(Grypus), the beak of the male is serrated along the
margin and hooked at the extremity, thus differing
much from that of the female. In the curious Neomorpha
of New Zealand, there is a still wider difference in
the form of the beak; and Mr. Gould has been informed
that the male with his “straight and stout beak” tears
off the bark of trees, in order that the female may
feed on the uncovered larvæ with her weaker and more
curved beak. Something of the same kind may be
observed with our goldfinch (Carduelis elegans), for I
40am assured by Mr. J. Jenner Weir that the bird-catchers
can distinguish the males by their slightly
longer beaks. The flocks of males, as an old and trustworthy
bird-catcher asserted, are commonly found feeding
on the seeds of the teazle (Dipsacus) which they
can reach with their elongated beaks, whilst the females
more commonly feed on the seeds of the betony or
Scrophularia. With a slight difference of this nature
as a foundation, we can see how the beaks of the two
sexes might be made to differ greatly through natural
selection. In all these cases, however, especially in
that of the quarrelsome humming-birds, it is possible
that the differences in the beaks may have been first
acquired by the males in relation to their battles, and
afterwards led to slightly changed habits of life.
Law of Battle.—Almost all male birds are extremely
pugnacious, using their beaks, wings, and legs for fighting
together. We see this every spring with our robins and
sparrows. The smallest of all birds, namely the humming-bird,
is one of the most quarrelsome. Mr. Gosse61
describes a battle, in which a pair of humming-birds
seized hold of each other’s beaks, and whirled round
and round, till they almost fell to the ground; and M.
Montes de Oca, in speaking of another genus, says that
two males rarely meet without a fierce aerial encounter:
when kept in cages “their fighting has mostly ended
in the splitting of the tongue of one of the two, which
then surely dies from being unable to feed.”62 With
Waders, the males of the common water-hen (Gallinula
chloropus) “when pairing, fight violently for the females:
they stand nearly upright in the water and strike
with their feet.” Two were seen to be thus engaged
41for half an hour, until one got hold of the head of the
other which would have been killed, had not the observer
interfered; the female all the time looking on as
a quiet spectator.63 The males of an allied bird (Gallicrex
cristatus), as Mr. Blyth informs me, are one third
larger than the females, and are so pugnacious during
the breeding-season, that they are kept by the natives
of Eastern Bengal for the sake of fighting. Various
other birds are kept in India for the same purpose, for
instance the Bulbuls (Pycnonotus hæmorrhous) which
“fight with great spirit.”64
The polygamous Ruff (Machetes pugnax, fig. 37) is
notorious for his extreme pugnacity; and in the spring,
the males, which are considerably larger than the
females, congregate day after day at a particular spot,
where the females propose to lay their eggs. The
fowlers discover these spots by the turf being trampled
somewhat bare. Here they fight very much like game-cocks,
seizing each other with their beaks and striking
with their wings. The great ruff of feathers round the
neck is then erected, and according to Col. Montagu
“sweeps the ground as a shield to defend the more
tender parts;” and this is the only instance known
to me in the case of birds, of any structure serving as a
shield. The ruff of feathers, however, from its varied
and rich colours probably serves in chief part as an
ornament. Like most pugnacious birds, they seem
always ready to fight, and when closely confined often
kill each other; but Montagu observed that their
pugnacity becomes greater during the spring, when the
long feathers on their necks are fully developed; and
at this period the least movement by any one bird
42provokes a general battle.65 Of the pugnacity of web-footed
birds, two instances will suffice: in Guiana “bloody
fights occur during the breeding-season between the
43males of the wild musk-duck (Cairina moschata);
and where these fights have occurred the river
is covered for some distance with feathers.”66 Birds
which seem ill-adapted for fighting engage in fierce
conflicts; thus with the pelican the stronger males
drive away the weaker ones, snapping with their
huge beaks and giving heavy blows with their wings.
Male snipes fight together, “tugging and pushing each
other with their bills in the most curious manner
imaginable.” Some few species are believed never to
fight; this is the case, according to Audubon, with one
of the woodpeckers of the United States (Picus auratus),
although “the hens are followed by even half a dozen
of their gay suitors.”67

Fig. 37. The Ruff or Machetes pugnax (from Brehm’s ‘Thierteben’).
The males of many birds are larger than the females,
and this no doubt is an advantage to them in their
battles with their rivals, and has been gained through
sexual selection. The difference in size between the
two sexes is carried to an extreme point in several
Australian species; thus the male musk-duck (Biziura)
and the male Cincloramphus cruralis (allied to our
pipits) are by measurement actually twice as large as
their respective females.68 With many other birds the
females are larger than the males; and as formerly
remarked, the explanation often given, namely that the
females have most of the work in feeding their young,
will not suffice. In some few cases, as we shall hereafter
see, the females apparently have acquired their
greater size and strength for the sake of conquering
other females and obtaining possession of the males.
44
The males of many gallinaceous birds, especially
of the polygamous kinds, are furnished with special
weapons for fighting with their rivals, namely spurs,
which can be used with fearful effect. It has been
recorded by a trustworthy writer69 that in Derbyshire
a kite struck at a game-hen accompanied by her
chickens, when the cock rushed to the rescue and drove
his spur right through the eye and skull of the
aggressor. The spur was with difficulty drawn from
the skull, and as the kite though dead retained his
grasp, the two birds were firmly locked together; but
the cock when disentangled was very little injured.
The invincible courage of the game-cock is notorious:
a gentleman who long ago witnessed the following
brutal scene, told me that a bird had both its legs
broken by some accident in the cock-pit, and the owner
laid a wager that if the legs could be spliced so that
the bird could stand upright, he would continue fighting.
This was effected on the spot, and the bird fought
with undaunted courage until he received his death-stroke.
In Ceylon a closely-allied and wild species,
the Gallus Stanleyi, is known to fight desperately “in
defence of his seraglio,” so that one of the combatants
is frequently found dead.70 An Indian partridge (Ortygornis
gularis), the male of which is furnished with
strong and sharp spurs, is so quarrelsome, “that the
scars of former fights disfigure the breast of almost
every bird you kill.”71
The males of almost all gallinaceous birds, even those
which are not furnished with spurs, engage during the
breeding-season in fierce conflicts. The Capercailzie and
45Black-cock (Tetrao urogallus and T. tetrix), which are
both polygamists, have regular appointed places, where
during many weeks they congregate in numbers to
fight together and to display their charms before the
females. M. W. Kowalevsky informs me that in Russia
he has seen the snow all bloody on the arenas where
the Capercailzie have fought; and the Black-cocks
“make the feathers fly in every direction,” when
several “engage in a battle royal.” The elder Brehm
gives a curious account of the Balz, as the love-dance
and love-song of the Black-cock is called in Germany.
The bird utters almost continuously the most strange
noises: “he holds his tail up and spreads it out like a
fan, he lifts up his head and neck with all the feathers
erect, and stretches his wings from the body. Then
he takes a few jumps in different directions, sometimes
in a circle, and presses the under part of his
beak so hard against the ground that the chin-feathers
are rubbed off. During these movements he beats
his wings and turns round and round. The more
ardent he grows the more lively he becomes, until at
last the bird appears like a frantic creature.” At
such times the black-cocks are so absorbed that they
become almost blind and deaf, but less so than the
capercailzie: hence bird after bird may be shot on
the same spot, or even caught by the hand. After
performing these antics the males begin to fight: and
the same black-cock, in order to prove his strength over
several antagonists, will visit in the course of one morning
several Balz-places, which remain the same during
successive years.72
46
The peacock with his long train appears more like a
dandy than a warrior, but he sometimes engages in
fierce contests: the Rev. W. Darwin Fox informs me
that two peacocks became so excited whilst fighting at
some little distance from Chester that they flew over
the whole city, still fighting, until they alighted on the
top of St. John’s tower.
The spur, in those gallinaceous birds which are thus
provided, is generally single; but Polyplectron (see
fig. 51, p. 90) has two or more on each leg; and one of
the Blood-pheasants (Ithaginis cruentus) has been seen
with five spurs. The spurs are generally confined to the
male, being represented by mere knobs or rudiments in
the female; but the females of the Java peacock (Pavo
muticus) and, as I am informed by Mr. Blyth, of the small
fire-backed pheasant (Euplocamus erythropthalmus) possess
spurs. In Galloperdix it is usual for the males to
have two spurs, and for the females to have only one
on each leg.73 Hence spurs may safely be considered as
a masculine character, though occasionally transferred
in a greater or less degree to the females. Like most
other secondary sexual characters, the spurs are highly
variable both in number and development in the same
species.

Fig. 38. Palamedea cornuta (from Brehm), shewing the double-wing-spurs, and the
filament on the head.
Various birds have spurs on their wings. But the
Egyptian goose (Chenalopex ægyptiacus) has only “bare
obtuse knobs,” and these probably shew us the first
steps by which true spurs have been developed in other
allied birds. In the spur-winged goose, Plectropterus
gambensis, the males have much larger spurs than the
females; and they use them, as I am informed by Mr.
Bartlett, in fighting together, so that, in this case, the
47wing-spurs serve as sexual weapons; but according to
Livingstone, they are chiefly used in the defence of the
young. The Palamedea (fig. 38) is armed with a pair of48
spurs on each wing; and these are such formidable weapons
that a single blow has driven a dog howling away.
But it does not appear that the spurs in this case, or in
that of some of the spur-winged rails, are larger in the
male than in the female.74 In certain plovers, however,
the wing-spurs must be considered as a sexual character.
Thus in the male of our common peewit (Vanellus cristatus)
the tubercle on the shoulder of the wing becomes
more prominent during the breeding-season, and the
males are known to fight together. In some species
of Lobivanellus a similar tubercle becomes developed
during the breeding-season “into a short horny spur.”
In the Australian L. lobatus both sexes have spurs, but
these are much larger in the males than in the females.
In an allied bird, the Hoplopterus armatus, the spurs
do not increase in size during the breeding-season; but
these birds have been seen in Egypt to fight together,
in the same manner as our peewits, by turning suddenly
in the air and striking sideways at each other, sometimes
with a fatal result. Thus also they drive away
other enemies.75
The season of love is that of battle; but the males
of some birds, as of the game-fowl and ruff, and even
the young males of the wild turkey and grouse,76 are
ready to fight whenever they meet. The presence of
the female is the teterrima belli causa. The Bengali
49baboos make the pretty little males of the amadavat
(Estrelda amandava) fight together by placing three
small cages in a row, with a female in the middle;
after a little time the two males are turned loose, and
immediately a desperate battle ensues.77 When many
males congregate at the same appointed spot and fight
together, as in the case of grouse and various other
birds, they are generally attended by the females,78
which afterwards pair with the victorious combatants.
But in some cases the pairing precedes instead of succeeding
the combat: thus, according to Audubon,79
several males of the Virginian goat-sucker (Caprimulgus
Virginianus) “court, in a highly entertaining
manner, the female, and no sooner has she made her
choice, than her approved gives chase to all intruders,
and drives them beyond his dominions.” Generally
the males try with all their power to drive away or kill
their rivals before they pair. It does not, however,
appear that the females invariably prefer the victorious
males. I have indeed been assured by M. W. Kowalevsky
that the female capercailzie sometimes steals
away with a young male who has not dared to enter
the arena with the older cocks; in the same manner as
occasionally happens with the does of the red-deer in
Scotland. When two males contend in presence of a
single female, the victor, no doubt, commonly gains his
50desire; but some of these battles are caused by wandering
males trying to distract the peace of an already
mated pair.80
Even with the most pugnacious species it is probable
that the pairing does not depend exclusively on the
mere strength and courage of the male: for such
males are generally decorated with various ornaments,
which often become more brilliant during the breeding-season,
and which are sedulously displayed before the
females. The males also endeavour to charm or excite
their mates by love-notes, songs, and antics; and
the courtship is, in many instances, a prolonged affair.
Hence it is not probable that the females are indifferent
to the charms of the opposite sex, or that they are
invariably compelled to yield to the victorious males.
It is more probable that the females are excited, either
before or after the conflict, by certain males, and thus
unconsciously prefer them. In the case of Tetrao umbellus,
a good observer81 goes so far as to believe that
the battles of the males “are all a sham, performed
to show themselves to the greatest advantage before
the admiring females who assemble around; for I
have never been able to find a maimed hero, and
seldom more than a broken feather.” I shall have
to recur to this subject, but I may here add that with
the Tetrao cupido of the United States, about a score of
males assemble at a particular spot, and strutting about
make the whole air resound with their extraordinary
noises. At the first answer from a female the males
begin to fight furiously, and the weaker give way; but
then, according to Audubon, both the victors and vanquished
search for the female, so that the females must
51either then exert a choice, or the battle must be renewed.
So, again, with one of the Field-starlings of
the United States (Sturnella ludoviciana) the males
engage in fierce conflicts, “but at the sight of a female
they all fly after her, as if mad.”82
Vocal and instrumental Music.—With birds the voice
serves to express various emotions, such as distress, fear,
anger, triumph, or mere happiness. It is apparently
sometimes used to excite terror, as with the hissing
noise made by some nestling-birds. Audubon83 relates
that a night-heron (Ardea nycticorax, Linn.) which he
kept tame, used to hide itself when a cat approached,
and then “suddenly start up uttering one of the most
frightful cries, apparently enjoying the cat’s alarm
and flight.” The common domestic cock clucks to
the hen, and the hen to her chickens, when a dainty
morsel is found. The hen, when she has laid an egg,
“repeats the same note very often, and concludes with
the sixth above, which she holds for a longer time;”84
and thus she expresses her joy. Some social birds
apparently call to each other for aid; and as they flit
from tree to tree, the flock is kept together by chirp
answering chirp. During the nocturnal migrations of
geese and other water-fowl, sonorous clangs from the
van may be heard in the darkness overhead, answered
by clangs in the rear. Certain cries serve as danger-signals,
which, as the sportsman knows to his cost, are
well understood by the same species and by others.
The domestic cock crows, and the humming-bird chirps,
in triumph over a defeated rival. The true song, how52ever,
of most birds and various strange cries are chiefly
uttered during the breeding-season, and serve as a
charm, or merely as a call-note, to the other sex.
Naturalists are much divided with respect to the object
of the singing of birds. Few more careful observers ever
lived than Montagu, and he maintained that the “males
of song-birds and of many others do not in general
search for the female, but, on the contrary, their
business in the spring is to perch on some conspicuous
spot breathing out their full and amorous notes, which,
by instinct, the female knows, and repairs to the spot to
choose her mate.”85 Mr. Jenner Weir informs me that
this is certainly the case with the nightingale. Bechstein,
who kept birds during his whole life, asserts, “that
the female canary always chooses the best singer, and
that in a state of nature the female finch selects that
male out of a hundred whose notes please her most.”86
There can be no doubt that birds closely attend to
each other’s song. Mr. Weir has told me of the case of
a bullfinch which had been taught to pipe a German
waltz, and who was so good a performer that he cost
ten guineas; when this bird was first introduced into
a room where other birds were kept and he began to
sing, all the others, consisting of about twenty linnets
and canaries, ranged themselves on the nearest side of
their cages, and listened with the greatest interest to
the new performer. Many naturalists believe that the
singing of birds is almost exclusively “the effect of rivalry
and emulation,” and not for the sake of charming
their mates. This was the opinion of Daines Barrington
and White of Selborne, who both especially attended to
53this subject.87 Barrington, however, admits that “superiority
in song gives to birds an amazing ascendancy
over others, as is well known to bird-catchers.”
It is certain that there is an intense degree of rivalry
between the males in their singing. Bird-fanciers
match their birds to see which will sing longest; and
I was told by Mr. Yarrell that a first-rate bird will
sometimes sing till he drops down almost dead, or,
according to Bechstein,88 quite dead from rupturing a
vessel in the lungs. Whatever the cause may be,
male birds, as I hear from Mr. Weir, often die suddenly
during the season of song. That the habit of
singing is sometimes quite independent of love is clear,
for a sterile hybrid canary-bird has been described89
as singing whilst viewing itself in a mirror, and then
dashing at its own image; it likewise attacked with
fury a female canary when put into the same cage.
The jealousy excited by the act of singing is constantly
taken advantage of by bird-catchers; a male, in good
song, is hidden and protected, whilst a stuffed bird, surrounded
by limed twigs, is exposed to view. In this
manner a man, as Mr. Weir informs me, has caught, in
the course of a single day, fifty, and in one instance
seventy, male chaffinches. The power and inclination
to sing differ so greatly with birds that although the
price of an ordinary male chaffinch is only sixpence,
Mr. Weir saw one bird for which the bird-catcher asked
three pounds; the test of a really good singer being
that it will continue to sing whilst the cage is swung
round the owner’s head.
That birds should sing from emulation as well as for
54the sake of charming the female, is not at all incompatible;
and, indeed, might have been expected to go
together, like decoration and pugnacity. Some authors,
however, argue that the song of the male cannot serve
to charm the female, because the females of some few
species, such as the canary, robin, lark, and bullfinch,
especially, as Bechstein remarks, when in a state of
widowhood, pour forth fairly melodious strains. In
some of these cases the habit of singing may be in part
attributed to the females having been highly fed and
confined,90 for this disturbs all the usual functions connected
with the reproduction of the species. Many instances
have already been given of the partial transference
of secondary masculine characters to the female,
so that it is not at all surprising that the females of some
species should possess the power of song. It has also
been argued, that the song of the male cannot serve as
a charm, because the males of certain species, for instance,
of the robin, sing during the autumn.91 But
nothing is more common than for animals to take pleasure
in practising whatever instinct they follow at other
times for some real good. How often do we see birds
which fly easily, gliding and sailing through the air
obviously for pleasure. The cat plays with the captured
mouse, and the cormorant with the captured fish.
The weaver-bird (Ploceus), when confined in a cage,
amuses itself by neatly weaving blades of grass between
the wires of its cage. Birds which habitually fight
during the breeding-season are generally ready to fight
at all times; and the males of the capercailzie sometimes
hold their balzens or leks at the usual place of
55assemblage during the autumn.92 Hence it is not at all
surprising that male birds should continue singing for
their own amusement after the season for courtship is
over.
Singing is to a certain extent, as shewn in a previous
chapter, an art, and is much improved by practice.
Birds can be taught various tunes, and even the unmelodious
sparrow has learnt to sing like a linnet.
They acquire the song of their foster-parents,93 and
sometimes that of their neighbours.94 All the common
songsters belong to the Order of Insessores, and their
vocal organs are much more complex than those of
most other birds; yet it is a singular fact that some
of the Insessores, such as ravens, crows, and magpies,
possess the proper apparatus,95 though they never sing,
and do not naturally modulate their voices to any great
extent. Hunter asserts96 that with the true songsters
the muscles of the larynx are stronger in the males
than in the females; but with this slight exception there
is no difference in the vocal organs of the two sexes,
although the males of most species sing so much better
and more continuously than the females.
It is remarkable that only small birds properly sing.
The Australian genus Menura, however, must be excepted;
for the Menura Alberti, which is about the size
of a half-grown turkey, not only mocks other birds, but
“its own whistle is exceedingly beautiful and varied.”
The males congregate and form “corroborying places,”
where they sing, raising and spreading their tails like
56peacocks and drooping their wings.97 It is also remarkable
that the birds which sing are rarely decorated
with brilliant colours or other ornaments. Of our British
birds, excepting the bullfinch and goldfinch, the best
songsters are plain-coloured. The kingfisher, bee-eater,
roller, hoopoe, woodpeckers, &c., utter harsh cries; and
the brilliant birds of the tropics are hardly ever songsters.98
Hence bright colours and the power of song
seem to replace each other. We can perceive that if the
plumage did not vary in brightness, or if bright colours
were dangerous to the species, other means would have
to be employed to charm the females; and the voice
being rendered melodious would offer one such means.

Fig. 39. Tetrao cupido; male. (From Brehm.)
In some birds the vocal organs differ greatly in the
two sexes. In the Tetrao cupido (fig. 39) the male has
two bare, orange-coloured sacks, one on each side of the
neck; and these are largely inflated when the male,
during the breeding-season, makes a curious hollow
sound, audible at a great distance. Audubon proved
that the sound was intimately connected with this apparatus,
which reminds us of the air-sacks on each side
of the mouth of certain male frogs, for he found that
the sound was much diminished when one of the sacks
of a tame bird was pricked, and when both were pricked
it was altogether stopped. The female has “a somewhat
similar, though smaller, naked space of skin on
the neck; but this is not capable of inflation.”99 The
57male of another kind of grouse (Tetrao urophasianus),
whilst courting the female, has his “bare yellow œsophagus
inflated to a prodigious size, fully half as large
as the body;” and he then utters various grating,58
deep hollow tones. With his neck-feathers erect, his
wings lowered and buzzing on the ground, and his long
pointed tail spread out like a fan, he displays a variety
of grotesque attitudes. The œsophagus of the female
is not in any way remarkable.100
It seems now well made out that the great throat-pouch
of the European male bustard (Otis tarda), and
of at least four other species, does not serve, as was
formerly supposed, to hold water, but is connected with
the utterance during the breeding-season of a peculiar
sound resembling “ock.” The bird whilst uttering this
sound throws himself into the most extraordinary attitudes.
It is a singular fact that with the males of the
same species the sack is not developed in all the individuals.101
A crow-like bird inhabiting South America
(Cephalopterus ornatus, fig. 40) is called the umbrella-bird,
from its immense top-knot, formed of bare white
quills surmounted by dark-blue plumes, which it can
elevate into a great dome no less than five inches in
diameter, covering the whole head. This bird has on
its neck a long, thin, cylindrical, fleshy appendage, which
is thickly clothed with scale-like blue feathers. It probably
serves in part as an ornament, but likewise as a
resounding apparatus, for Mr. Bates found that it is
connected “with an unusual development of the trachea
and vocal organs.” It is dilated when the bird utters
its singularly deep, loud, and long-sustained fluty note.
59The head-crest and neck-appendage are rudimentary
in the female.102

Fig. 40. The Umbrella-bird or Cephalopterus ornatus (male, from Brehm).
The vocal organs of various web-footed and wading
birds are extraordinarily complex, and differ to a certain
extent in the two sexes. In some cases the trachea is
convoluted, like a French horn, and is deeply embedded
in the sternum. In the wild swan (Cygnus ferus) it is
60more deeply embedded in the adult male than in the
female or young male. In the male Merganser the
enlarged portion of the trachea is furnished with an
additional pair of muscles.103 But the meaning of these
differences between the sexes of many Anatidæ is not
at all understood; for the male is not always the more
vociferous; thus with the common duck, the male hisses,
whilst the female utters a loud quack.104 In both sexes of
one of the cranes (Grus virgo) the trachea penetrates
the sternum, but presents “certain sexual modifications.”
In the male of the black stork there is also a well-marked
sexual difference in the length and curvature of
the bronchi.105 So that highly important structures have
in these cases been modified according to sex.
It is often difficult to conjecture whether the many
strange cries and notes, uttered by male birds during
the breeding-season, serve as a charm or merely as a
call to the female. The soft cooing of the turtle-dove
and of many pigeons, it may be presumed, pleases the
female. When the female of the wild turkey utters her
call in the morning, the male answers by a different
note from the gobbling noise which he makes, when
with erected feathers, rustling wings and distended
wattles, he puffs and struts before her.106 The spel of
the black-cock certainly serves as a call to the female,
for it has been known to bring four or five females
61from a distance to a male under confinement; but as
the black-cock continues his spel for hours during
successive days, and in the case of the capercailzie
“with an agony of passion,” we are led to suppose
that the females which are already present are thus
charmed.107 The voice of the common rook is known
to alter during the breeding-season, and is therefore in
some way sexual.108 But what shall we say about the
harsh screams of, for instance, some kinds of macaws;
have these birds as bad taste for musical sounds as
they apparently have for colour, judging by the inharmonious
contrast of their bright yellow and blue
plumage? It is indeed possible that the loud voices
of many male birds may be the result, without any
advantage being thus gained, of the inherited effects of
the continued use of their vocal organs, when they are
excited by the strong passions of love, jealousy, and
rage; but to this point we shall recur when we treat of
quadrupeds.
We have as yet spoken only of the voice, but the
males of various birds practise, during their courtship,
what may be called instrumental music. Peacocks
and Birds of Paradise rattle their quills together,
and the vibratory movement apparently serves merely
to make a noise, for it can hardly add to the beauty
of their plumage. Turkey-cocks scrape their wings
against the ground, and some kinds of grouse thus
produce a buzzing sound. Another North American
grouse, the Tetrao umbellus, when with his tail erect,
his ruffs displayed, “he shows off his finery to the
females, who lie hid in the neighbourhood,” drums
rapidly with his “lowered wings on the trunk of a
62fallen tree,” or, according to Audubon, against his own
body; the sound thus produced is compared by some to
distant thunder, and by others to the quick roll of a
drum. The female never drums, “but flies directly to
the place where the male is thus engaged.” In the
Himalayas the male of the Kalij pheasant “often makes
a singular drumming noise with his wings, not unlike
the sound produced by shaking a stiff piece of cloth.”
On the west coast of Africa the little black-weavers
(Ploceus?) congregate in a small party on the bushes
round a small open space, and sing and glide through
the air with quivering wings, “which make a rapid
whirring sound like a child’s rattle.” One bird after
another thus performs for hours together, but only during
the courting-season. At this same season the males of
certain nightjars (Caprimulgus) make a most strange
noise with their wings. The various species of woodpeckers
strike a sonorous branch with their beaks, with
so rapid a vibratory movement that “the head appears
to be in two places at once.” The sound thus produced
is audible at a considerable distance, but cannot
be described; and I feel sure that its cause would
never be conjectured by any one who heard it for the
first time. As this jarring sound is made chiefly during
the breeding-season, it has been considered as a love-song;
but it is perhaps more strictly a love-call. The
female, when driven from her nest, has been observed
thus to call her mate, who answered in the same
manner and soon appeared. Lastly the male Hoopoe
(Upupa epops) combines vocal and instrumental music;
for during the breeding-season this bird, as Mr. Swinhoe
saw, first draws in air and then taps the end of its
beak perpendicularly down against a stone or the trunk
of a tree, “when the breath being forced down the
tubular bill produces the correct sound.” When the63
male utters its cry without striking his beak the sound
is quite different.109

Fig. 41. Outer tail-feather of Scolopax gallinago (from Proc. Zool. Soc. 1858).
In the foregoing cases sounds are made by the aid
of structures already present and otherwise necessary;
but in the following cases certain feathers have been
specially modified for the express purpose of producing
the sounds. The drumming, or bleating, or neighing, or
thundering noise, as expressed by different observers,
which is made by the common snipe (Scolopax gallinago)
must have surprised every one who has ever heard it.
This bird, during the pairing-season, flies to “perhaps a
thousand feet in height,” and after zig-zagging about
for a time descends in a curved line, with outspread tail
and quivering pinions, with surprising velocity to the
earth. The sound is emitted only during this rapid
descent. No one was able to explain the cause, until
M. Meves observed that on each side of the tail the outer
feathers are peculiarly formed (fig. 41), having a stiff
sabre-shaped shaft, with the oblique barbs of unusual
length, the outer webs being strongly bound together.
64
He found that by blowing on these feathers, or by fastening
them to a long thin stick and waving them rapidly
through the air, he could exactly reproduce the drumming
noise made by the living bird. Both sexes are
furnished with these feathers, but they are generally
 Fig. 42. Outer tail-feather of Scolopax frenata.
Fig. 42. Outer tail-feather of Scolopax frenata.
 Fig. 43. Outer tail-feather of Scolopax javensis.
larger in the male than
in the female, and emit
a deeper note. In some
species, as in S. frenata
(fig. 42), four feathers,
and in S. javensis (fig.
43), no less than eight
on each side of the tail
are greatly modified. Different tones are emitted by
the feathers of the different species when waved through
the air; and the Scolopax Wilsonii of the United States
makes a switching noise whilst descending rapidly to
the earth.110
Fig. 43. Outer tail-feather of Scolopax javensis.
larger in the male than
in the female, and emit
a deeper note. In some
species, as in S. frenata
(fig. 42), four feathers,
and in S. javensis (fig.
43), no less than eight
on each side of the tail
are greatly modified. Different tones are emitted by
the feathers of the different species when waved through
the air; and the Scolopax Wilsonii of the United States
makes a switching noise whilst descending rapidly to
the earth.110
In the male of the Chamæpetes unicolor (a large gallinaceous
bird of America) the first primary wing-feather
is arched towards the tip and is much more attenuated
than in the female. In an allied bird, the Penelope
nigra, Mr. Salvin observed a male, which, whilst it
flew downwards “with outstretched wings, gave forth
a kind of crashing, rushing noise,” like the falling
of a tree.111 The male alone of one of the Indian
bustards (Sypheotides auritus) has its primary wing-feathers
greatly acuminated; and the male of an allied
65species is known to make a humming noise whilst
courting the female.112 In a widely different group of
birds, namely the Humming-birds, the males alone
 Fig. 44. Primary wing-feather of a
Humming-bird, the Selasphorus platycercus
(from a sketch by Mr. Salvin).
Upper figure, that of male; lower
figure, corresponding feather of female.
of certain kinds have either
the shafts of their primary
wing-feathers broadly dilated,
or the webs abruptly excised
towards the extremity. The
male, for instance, of Selasphorus
platycercus, when adult,
has the first primary wing-feather
(fig. 44), excised in
this manner. Whilst flying
from flower to flower he
makes “a shrill, almost whistling, noise;”113 but it
did not appear to Mr. Salvin that the noise was intentionally
made.
Fig. 44. Primary wing-feather of a
Humming-bird, the Selasphorus platycercus
(from a sketch by Mr. Salvin).
Upper figure, that of male; lower
figure, corresponding feather of female.
of certain kinds have either
the shafts of their primary
wing-feathers broadly dilated,
or the webs abruptly excised
towards the extremity. The
male, for instance, of Selasphorus
platycercus, when adult,
has the first primary wing-feather
(fig. 44), excised in
this manner. Whilst flying
from flower to flower he
makes “a shrill, almost whistling, noise;”113 but it
did not appear to Mr. Salvin that the noise was intentionally
made.
Lastly, in several species of a sub-genus of Pipra or
Manakin, the males have their secondary wing-feathers
modified, as described by Mr. Sclater, in a still more
remarkable manner. In the brilliantly-coloured P.
deliciosa the first three secondaries are thick-stemmed
and curved towards the body; in the fourth and fifth
(fig. 45, a) the change is greater; and in the sixth
and seventh (b, c) the shaft “is thickened to an
extraordinary degree, forming a solid horny lump.”
The barbs also are greatly changed in shape, in comparison
with the corresponding feathers (d, e, f) in the
female. Even the bones of the wing which support
these singular feathers in the male are said by Mr.
Fraser to be much thickened. These little birds make
66an extraordinary noise, the first “sharp note being not
unlike the crack of a whip.”114

Fig. 45. Secondary wing-feathers of Pipra deliciosa(from Mr. Sclater, in Proc, Zool. Soc.
1860). The three upper feathers, a, b, c, from the male; the three lower corresponding
feathers, d, e, f, from the female.
a. and d. Fifth secondary wing-feather of male and female, upper surface. b and e.
Sixth secondary, upper surface. c and f. Seventh secondary, lower surface.
The diversity of the sounds, both vocal and instrumental,
made by the males of many species during the
67breeding-season, and the diversity of the means for
producing such sounds, are highly remarkable. We
thus gain a high idea of their importance for sexual
purposes, and are reminded of the same conclusion with
respect to insects. It is not difficult to imagine the steps
by which the notes of a bird, primarily used as a mere
call or for some other purpose, might have been improved
into a melodious love-song. This is somewhat
more difficult in the case of the modified feathers, by
which the drumming, whistling, or roaring noises are
produced. But we have seen that some birds during
their courtship flutter, shake, or rattle their unmodified
feathers together; and if the females were led to select
the best performers, the males which possessed the
strongest or thickest, or most attenuated feathers, situated
on any part of the body, would be the most
successful; and thus by slow degrees the feathers might
be modified to almost any extent. The females, of
course, would not notice each slight successive alteration
in shape, but only the sounds thus produced. It is a
curious fact that in the same class of animals, sounds
so different as the drumming of the snipe’s tail, the
tapping of the woodpecker’s beak, the harsh trumpet-like
cry of certain water-fowl, the cooing of the turtle-dove,
and the song of the nightingale, should all be
pleasing to the females of the several species. But
we must not judge the tastes of distinct species by a
uniform standard; nor must we judge by the standard
of man’s taste. Even with man, we should remember
what discordant noises, the beating of tom-toms and
the shrill notes of reeds, please the ears of savages.
Sir S. Baker remarks,115 that “as the stomach of the
Arab prefers the raw meat and reeking liver taken
68hot from the animal, so does his ear prefer his equally
coarse and discordant music to all other.”
Love-Antics and Dances.—The curious love-gestures
of various birds, especially of the Gallinaceæ, have
already been incidentally noticed; so that little need
here be added. In Northern America, large numbers
of a grouse, the Tetrao phasianellus, meet every morning
during the breeding-season on a selected level spot,
and here they run round and round in a circle of about
fifteen or twenty feet in diameter, so that the ground
is worn quite bare, like a fairy-ring. In these Partridge-dances,
as they are called by the hunters, the
birds assume the strangest attitudes, and run round, some
to the left and some to the right. Audubon describes
the males of a heron (Ardea herodias) as walking
about on their long legs with great dignity before
the females, bidding defiance to their rivals. With
one of the disgusting carrion-vultures (Cathartes
jota) the same naturalist states that “the gesticulations
and parade of the males at the beginning of the
love-season are extremely ludicrous.” Certain birds
perform their love-antics on the wing, as we have seen
with the black African weaver, instead of on the
ground. During the spring our little whitethroat
(Sylvia cinerea) often rises a few feet or yards in the
air above some bush, and “flutters with a fitful and
fantastic motion, singing all the while, and then drops
to its perch.” The great English bustard throws
himself into indescribably odd attitudes whilst courting
the female, as has been figured by Wolf. An allied
Indian bustard (Otis bengalensis) at such times “rises
perpendicularly into the air with a hurried flapping
of his wings, raising his crest and puffing out the
feathers of his neck and breast, and then drops to the69
ground;” he repeats this manœuvre several times
successively, at the same time humming in a peculiar
tone. Such females as happen to be near “obey this
saltatory summons,” and when they approach he
trails his wings and spreads his tail like a turkey-cock.116
But the most curious case is afforded by three allied
genera of Australian birds, the famous Bower-birds,—no
doubt the co-descendants of some ancient species
which first acquired the strange instinct of constructing
bowers for performing their love-antics. The bowers
(fig. 46), which, as we shall hereafter see, are highly
decorated with feathers, shells, bones and leaves, are
built on the ground for the sole purpose of courtship,
for their nests are formed in trees. Both sexes assist
in the erection of the bowers, but the male is the principal
workman. So strong is this instinct that it is
practised under confinement, and Mr. Strange has
described117 the habits of some Satin Bower-birds,
which he kept in his aviary in New South Wales. “At
times the male will chase the female all over the
aviary, then go to the bower, pick up a gay feather
or a large leaf, utter a curious kind of note, set all
his feathers erect, run round the bower and become
so excited that his eyes appear ready to start from
his head; he continues opening first one wing, and
then the other, uttering a low, whistling note, and,
70like the domestic cock, seems to be picking up something
from the ground, until at last the female goes
gently towards him.” Captain Stokes has described
the habits and “play-houses” of another species, the
71Great Bower-bird, which was seen “amusing itself by
flying backwards and forwards, taking a shell alternately
from each side, and carrying it through the
archway in its mouth.” These curious structures,
formed solely as halls of assemblages, where both sexes
amuse themselves and pay their court, must cost the
birds much labour. The bower, for instance, of the
fawn-breasted species, is nearly four feet in length,
eighteen inches in height, and is raised on a thick
platform of sticks.

Fig. 46. Bower-bird, Chlamydera maculata, with bower (from
Brehm).
Decoration.—I will first discuss the cases in which the
males are ornamented either exclusively or in a much
higher degree than the females; and in a succeeding
chapter those in which both sexes are equally ornamented,
and finally the rare cases in which the female is
somewhat more brightly-coloured than the male. As with
the artificial ornaments used by savage and civilised men,
so with the natural ornaments of birds, the head is the
chief seat of decoration.118 The ornaments, as mentioned
at the commencement of this chapter, are wonderfully
diversified. The plumes on the front or back of the
head consist of variously-shaped feathers, sometimes
capable of erection or expansion, by which their beautiful
colours are fully displayed. Elegant ear-tufts (see
fig. 39 ante) are occasionally present. The head is
sometimes covered with velvety down like that of the
pheasant; or is naked and vividly coloured; or supports
fleshy appendages, filaments, and solid protuberances.
The throat, also, is sometimes ornamented with a beard,
or with wattles or caruncles. Such appendages are
generally brightly coloured, and no doubt serve as
72ornaments, though not always ornamental in our eyes;
for whilst the male is in the act of courting the female,
they often swell and assume more vivid tints, as in the
case of the male turkey. At such times the fleshy appendages
about the head of the male Tragopan pheasant
(Ceriornis temminckii) swell into a large lappet
on the throat and into two horns, one on each side of
the splendid top-knot; and these are then coloured of
the most intense blue which I have ever beheld. The
African hornbill (Bucorax abyssinicus) inflates the
scarlet bladder-like wattle on its neck, and with its
wings drooping and tail expanded “makes quite a grand
appearance.”119 Even the iris of the eye is sometimes
more brightly coloured in the male than in the female;
and this is frequently the case with the beak, for
instance, in our common blackbird. In Buceros corrugatus,
the whole beak and immense casque are
coloured more conspicuously in the male than in the
female; and “the oblique grooves upon the sides of
the lower mandible are peculiar to the male sex.”120
The males are often ornamented with elongated feathers
or plumes springing from almost every part of the
body. The feathers on the throat and breast are sometimes
developed into beautiful ruffs and collars. The
tail-feathers are frequently increased in length; as we
see in the tail-coverts of the peacock, and in the tail of
the Argus pheasant. The body of this latter bird is not
larger than that of a fowl; yet the length from the end
of the beak to the extremity of the tail is no less than
five feet three inches.121 The wing-feathers are not
elongated nearly so often as the tail-feathers; for their
73elongation would impede the act of flight. Yet the
beautifully ocellated secondary wing-feathers of the
male Argus pheasant are nearly three feet in length;
and in a small African nightjar (Cosmetornis vexillarius)
one of the primary wing-feathers, during the
breeding-season, attains a length of twenty-six inches,
whilst the bird itself is only ten inches in length.
In another closely-allied genus of nightjars, the shafts
of the elongated wing-feathers are naked, except at
the extremity, where there is a disc.122 Again, in
another genus of nightjars, the tail-feathers are even
still more prodigiously developed; so that we see the
same kind of ornament gained by the males of closely-allied
birds, through the development of widely different
feathers.
It is a curious fact that the feathers of birds belonging
to distinct groups have been modified in almost exactly
the same peculiar manner. Thus the wing-feathers
in one of the above-mentioned nightjars are bare
along the shaft and terminate in a disc; or are, as
they are sometimes called, spoon or racket-shaped.
Feathers of this kind occur in the tail of a motmot
(Eumomota superciliaris), of a kingfisher, finch, humming-bird,
parrot, several Indian drongos (Dicrurus
and Edolius, in one of which the disc stands vertically),
and in the tail of certain Birds of Paradise. In these
latter birds, similar feathers, beautifully ocellated,
ornament the head, as is likewise the case with some
gallinaceous birds. In an Indian bustard (Sypheotides
auritus) the feathers forming the ear-tufts, which are
about four inches in length, also terminate in discs.123
74The barbs of the feathers in various widely-distinct
birds are filamentous or plumose, as with some Herons,
Ibises, Birds of Paradise and Gallinaceæ. In other
cases the barbs disappear, leaving the shafts bare; and
these in the tail of the Paradisea apoda attain a length
of thirty-four inches.124 Smaller feathers when thus
denuded appear like bristles, as on the breast of the
turkey-cock. As any fleeting fashion in dress comes
to be admired by man, so with birds a change of
almost any kind in the structure or colouring of the
feathers in the male appears to have been admired by
the female. The fact of the feathers in widely distinct
groups, having been modified in an analogous
manner, no doubt depends primarily on all the feathers
having nearly the same structure and manner of development,
and consequently tending to vary in the same
manner. We often see a tendency to analogous variability
in the plumage of our domestic breeds belonging
to distinct species. Thus top-knots have appeared in
several species. In an extinct variety of the turkey,
the top-knot consisted of bare quills surmounted with
plumes of down, so that they resembled, to a certain
extent, the racket-shaped feathers above described. In
certain breeds of the pigeon and fowl the feathers are
plumose, with some tendency in the shafts to be naked.
In the Sebastopol goose the scapular feathers are greatly
elongated, curled, or even spirally twisted, with the
margins plumose.125

Fig. 47. Paradisea rubra, male (from Brehm).

Fig. 48. Lophornis ornatus, male and female (from Brehm).
In regard to colour hardly anything need here be
said; for every one knows how splendid are the tints
75of birds, and how harmoniously they are combined.
The colours are often metallic and iridescent. Circular
spots are sometimes surrounded by one or more differently
shaded zones, and are thus converted into ocelli.
76
Nor need much be said on the wonderful differences
between the sexes, or of the extreme beauty of the
males of many birds. The common peacock offers a
striking instance. Female Birds of Paradise are
77obscurely coloured and destitute of all ornaments, whilst
the males are probably the most highly decorated of
all birds, and in so many ways, that they must be
seen to be appreciated. The elongated and golden-78orange
plumes which spring from beneath the wings
of the Paradisea apoda (see fig. 47 of P. rubra, a much
less beautiful species), when vertically erected and made
to vibrate, are described as forming a sort of halo, in
the centre of which the head “looks like a little
emerald sun with its rays formed by the two plumes.”126
In another most beautiful species the head is bald,
“and of a rich cobalt blue, crossed by several lines of
black velvety feathers.”127

Fig. 49. Spathura underwoodi, male and female (from Brehm).
Male humming-birds (figs. 48 and 49) almost vie
with Birds of Paradise in their beauty, as every one will
admit who has seen Mr. Gould’s splendid volumes or his
rich collection. It is very remarkable in how many
different ways these birds are ornamented. Almost every
part of the plumage has been taken advantage of and
modified; and the modifications have been carried, as
Mr. Gould shewed me, to a wonderful extreme in some
species belonging to nearly every subgroup. Such cases
are curiously like those which we see in our fancy
breeds, reared by man for the sake of ornament: certain
individuals originally varied in one character, and other
individuals belonging to the same species in other
characters; and these have been seized on by man and
augmented to an extreme point—as the tail of the
fantail-pigeon, the hood of the jacobin, the beak and
wattle of the carrier, and so forth. The sole difference
between these cases is that in the one the result is due
to man’s selection, whilst in the other, as with Humming-birds,
Birds of Paradise, &c., it is due to sexual
selection,—that is to the selection by the females of the
more beautiful males.
79
I will mention only one other bird, remarkable from
the extreme contrast in colour between the sexes,
namely the famous Bell-bird (Chasmorhynchus niveus) of
S. America, the note of which can be distinguished at
the distance of nearly three miles, and astonishes every
one who first hears it. The male is pure white, whilst
the female is dusky-green; and the former colour with
terrestrial species of moderate size and inoffensive
habits is very rare. The male, also, as described by
Waterton, has a spiral tube, nearly three inches in
length, which rises from the base of the beak. It is jet-black,
dotted over with minute downy feathers. This
tube can be inflated with air, through a communication
with the palate; and when not inflated hangs down on
one side. The genus consists of four species, the males
of which are very distinct, whilst the females, as described
by Mr. Sclater in a most interesting paper,
closely resemble each other, thus offering an excellent
instance of the common rule that within the same
group the males differ much more from each other than
do the females. In a second species (C. nudicollis) the
male is likewise snow-white, with the exception of a
large space of naked skin on the throat and round the
eyes, which during the breeding-season is of a fine green
colour. In a third species (C. tricarunculatus) the head
and neck alone of the male are white, the rest of the
body being chesnut-brown, and the male of this species
is provided with three filamentous projections half as
long as the body—one rising from the base of the beak
and the two others from the corners of the mouth.128
The coloured plumage and certain other ornaments of
80the males when adult are either retained for life or are
periodically renewed during the summer and breeding-season.
At this season the beak and naked skin about
the head frequently change colour, as with some herons,
ibises, gulls, one of the bell-birds just noticed, &c. In
the white ibis, the cheeks, the inflatable skin of the
throat, and the basal portion of the beak, then become
crimson.129 In one of the rails, Gallicrex cristatus a large
red caruncle is developed during this same period on
the head of the male. So it is with a thin horny crest
on the beak of one of the pelicans, P. erythrorhynchus;
for after the breeding-season, these horny crests are
shed, like horns from the heads of stags, and the shore
of an island in a lake in Nevada was found covered
with these curious exuviæ.130
Changes of colour in the plumage according to the
season depend firstly on a double annual moult, secondly
on an actual change of colour in the feathers themselves,
and thirdly on their dull-coloured margins being periodically
shed, or on these three processes more or less
combined. The shedding of the deciduary margins may
be compared with the shedding by very young birds
of their down; for the down in most cases arises from
the summits of the first true feathers.131
With respect to the birds which annually undergo a
double moult, there are, firstly, some kinds, for instance
snipes, swallow-plovers (Glareolæ), and curlews, in
which the two sexes resemble each other and do not
change colour at any season. I do not know whether
the winter-plumage is thicker and warmer than the
81summer-plumage, which seems, when there is no change
of colour, the most probable cause of a double moult.
Secondly, there are birds, for instance certain species of
Totanus and other grallatores, the sexes of which resemble
each other, but have a slightly different summer
and winter plumage. The difference, however, in colour
in these cases is so slight that it can hardly be an
advantage to them; and it may, perhaps, be attributed
to the direct action of the different conditions to which
the birds are exposed during the two seasons. Thirdly,
there are many other birds the sexes of which are
alike, but which are widely different in their summer
and winter plumage. Fourthly, there are birds, the
sexes of which differ from each other in colour; but
the females, though moulting twice, retain the same
colours throughout the year, whilst the males undergo
a change, sometimes, as with certain bustards, a great
change of colour. Fifthly and lastly, there are birds
the sexes of which differ from each other in both
their summer and winter plumage, but the male undergoes
a greater amount of change at each recurrent
season than the female—of which the Ruff (Machetes
pugnax) offers a good instance.
With respect to the cause or purpose of the differences
in colour between the summer and winter plumage, this
may in some instances, as with the ptarmigan,132 serve
during both seasons as a protection. When the difference
between the two plumages is slight it may
perhaps be attributed, as already remarked, to the
82direct action of the conditions of life. But with many
birds there can hardly be a doubt that the summer
plumage is ornamental, even when both sexes are alike.
We may conclude that this is the case with many
herons, egrets, &c., for they acquire their beautiful
plumes only during the breeding-season. Moreover,
such plumes, top-knots, &c., though possessed by both
sexes, are occasionally a little more highly developed in
the male than in the female; and they resemble the
plumes and ornaments possessed by the males alone
of other birds. It is also known that confinement, by
affecting the reproductive system of male birds, frequently
checks the development of their secondary
sexual characters, but has no immediate influence on
any other characters; and I am informed by Mr.
Bartlett that eight or nine specimens of the Knot
(Tringa canutus) retained their unadorned winter plumage
in the Zoological Gardens throughout the year,
from which fact we may infer that the summer plumage
though common to both sexes partakes of the nature
of the exclusively masculine plumage of many other
birds.133
From the foregoing facts, more especially from
neither sex of certain birds changing colour during
either annual moult, or changing so slightly that the
change can hardly be of any service to them, and from
the females of other species moulting twice yet retaining
the same colours throughout the year, we may conclude
that the habit of moulting twice in the year has
83not been acquired in order that the male should assume
during the breeding-season an ornamental character;
but that the double moult, having been originally
acquired for some distinct purpose, has subsequently
been taken advantage of in certain cases for gaining a
nuptial plumage.
It appears at first sight a surprising circumstance
that with closely-allied birds, some species should regularly
undergo a double annual moult, and others only a
single one. The ptarmigan, for instance, moults twice
or even thrice in the year, and the black-cock only
once: some of the splendidly-coloured honey-suckers
(Nectariniæ) of India and some sub-genera of obscurely-coloured
pipits (Anthus) have a double, whilst others
have only a single annual moult.134 But the gradations
in the manner of moulting, which are known to
occur with various birds, shew us how species, or whole
groups of species, might have originally acquired their
double annual moult, or having once gained the habit,
have again lost it. With certain bustards and plovers
the vernal moult is far from complete, some feathers
being renewed, and some changed in colour. There is
also reason to believe that with certain bustards and
rail-like birds, which properly undergo a double moult,
some of the older males retain their nuptial plumage
throughout the year. A few highly modified feathers
may alone be added during the spring to the plumage,
as occurs with the disc-formed tail-feathers of certain
drongos (Bhringa) in India, and with the elongated
feathers on the back, neck, and crest of certain herons.
By such steps as these, the vernal moult might be ren84dered
more and more complete, until a perfect double
moult was acquired. A gradation can also be shewn to
exist in the length of time during which either
annual plumage is retained; so that the one might
come to be retained for the whole year, the other being
completely lost. Thus the Machetes pugnax retains
his ruff in the spring for barely two months. The
male widow-bird (Chera progne) acquires in Natal his
fine plumage and long tail-feathers in December or
January and loses them in March; so that they are
retained during only about three months. Most species
which undergo a double moult keep their ornamental
feathers for about six months. The male, however, of
the wild Gallus bankiva retains his neck-hackles for
nine or ten months; and when these are cast off, the
underlying black feathers on the neck are fully exposed
to view. But with the domesticated descendant of this
species, the neck-hackles of the male are immediately
replaced by new ones; so that we here see, with respect
to part of the plumage, a double moult changed under
domestication into a single moult.135
The common drake (Anas boschas) is well known after
the breeding-season to lose his male plumage for a
period of three months, during which time he assumes
that of the female. The male pintail-duck (Anas
acuta) loses his plumage for the shorter period of
six weeks or two months; and Montagu remarks that
85“this double moult within so short a time is a most
extraordinary circumstance, that seems to bid defiance
to all human reasoning.” But he who believes in the
gradual modification of species will be far from feeling
surprise at finding gradations of all kinds. If the male
pintail were to acquire his new plumage within a still
shorter period, the new male feathers would almost
necessarily be mingled with the old, and both with
some proper to the female; and this apparently is the
case with the male of a not distantly-allied bird, namely
the Merganser serrator, for the males are said to
“undergo a change of plumage, which assimilates them
in some measure to the female.” By a little further
acceleration in the process, the double moult would be
completely lost.136
Some male birds, as before stated, become more
brightly coloured in the spring, not by a vernal moult,
but either by an actual change of colour in the feathers,
or by their obscurely-coloured deciduary margins being
shed. Changes of colour thus caused may last for a
longer or shorter time. With the Pelecanus onocrotalus
a beautiful rosy tint, with lemon-coloured marks on the
breast, overspreads the whole plumage in the spring; but
these tints, as Mr. Sclater states, “do not last long, disappearing
generally in about six weeks or two months
after they have been attained.” Certain finches shed
the margins of their feathers in the spring, and then become
brighter-coloured, while other finches undergo no
such change. Thus the Fringilla tristis of the United
States (as well as many other American species), exhibits
its bright colours only when the winter is past,
whilst our goldfinch, which exactly represents this bird
86in habits, and our siskin, which represents it still more
closely in structure, undergo no such annual change.
But a difference of this kind in the plumage of allied
species is not surprising, for with the common linnet,
which belongs to the same family, the crimson forehead
and breast are displayed only during the summer in
England, whilst in Madeira these colours are retained
throughout the year.137
Display by Male Birds of their Plumage.—Ornaments
of all kinds, whether permanently or temporarily gained,
are sedulously displayed by the males, and apparently
serve to excite, or attract, or charm the females. But
the males will sometimes display their ornaments, when
not in the presence of the females, as occasionally occurs
with grouse at their balz-places, and as may be noticed
with the peacock; this latter bird, however, evidently
wishes for a spectator of some kind, and will shew off
his finery, as I have often seen, before poultry or even
pigs.138 All naturalists who have closely attended to the
habits of birds, whether in a state of nature or under
confinement, are unanimously of opinion that the males
delight to display their beauty. Audubon frequently
speaks of the male as endeavouring in various ways to
charm the female. Mr. Gould, after describing some
peculiarities in a male humming-bird, says he has no
doubt that it has the power of displaying them to the
greatest advantage before the female. Dr. Jerdon139
87insists that the beautiful plumage of the male serves
“to fascinate and attract the female.” Mr. Bartlett, at
the Zoological Gardens, expressed himself to me in the
strongest terms to the same effect.
It must be a grand sight in the forests of India “to
come suddenly on twenty or thirty peafowl, the males
displaying their gorgeous trains, and strutting about
in all the pomp of pride before the gratified females.”
The wild turkey-cock erects his glittering plumage,
expands his finely-zoned tail and barred wing-feathers,
and altogether, with his gorged crimson and blue wattles,
makes a superb, though, to our eyes, grotesque
appearance. Similar facts have already been given
with respect to grouse of various kinds. Turning to
another Order. The male Rupicola crocea (fig. 50) is
one of the most beautiful birds in the world, being of
a splendid orange, with some of the feathers curiously
truncated and plumose. The female is brownish-green,
shaded with red, and has a much smaller
crest. Sir R. Schomburgk has described their courtship;
he found one of their meeting-places where ten
males and two females were present. The space was
from four to five feet in diameter, and appeared to have
been cleared of every blade of grass and smoothed as
if by human hands. A male “was capering to the
apparent delight of several others. Now spreading
its wings, throwing up its head, or opening its tail
like a fan; now strutting about with a hopping gait
until tired, when it gabbled some kind of note, and
was relieved by another. Thus three of them successively
took the field, and then, with self-approbation,
withdrew to rest.” The Indians, in order to
obtain their skins, wait at one of the meeting-places
till the birds are eagerly engaged in dancing, and then
are able to kill, with their poisoned arrows, four or five88
males, one after the other.140 With Birds of Paradise
a dozen or more full-plumaged males congregate in a
tree to hold a dancing-party, as it is called by the
natives; and here flying about, raising their wings,
elevating their exquisite plumes, and making them
vibrate, the whole tree seems, as Mr. Wallace re89marks,
to be filled with waving plumes. When thus
engaged, they become so absorbed that a skilful archer
may shoot nearly the whole party. These birds, when
kept in confinement in the Malay Archipelago, are said
to take much care in keeping their feathers clean; often
spreading them out, examining them, and removing
every speck of dirt. One observer, who kept several
pairs alive, did not doubt that the display of the male
was intended to please the female.141

Fig. 50. Rupicola crocea, male (from Brehm).
The gold pheasant (Thaumalea picta) during his courtship
not only expands and raises his splendid frill, but
turns it, as I have myself seen, obliquely towards the
female on whichever side she may be standing, obviously
in order that a large surface may be displayed before
her.142 Mr. Bartlett has observed a male Polyplectron
(fig. 51) in the act of courtship, and has shewn me a
specimen stuffed in the attitude then assumed. The
tail and wing-feathers of this bird are ornamented with
beautiful ocelli, like those on the peacock’s train. Now
when the peacock displays himself, he expands and
erects his tail transversely to his body, for he stands in
front of the female, and has to shew off, at the same
time, his rich blue throat and breast. But the breast
of the Polyplectron is obscurely coloured, and the ocelli
are not confined to the tail-feathers. Consequently the
Polyplectron does not stand in front of the female; but
he erects and expands his tail-feathers a little obliquely,
90lowering the expanded wing on the same side, and
raising that on the opposite side. In this attitude the
ocelli over the whole body are exposed before the eyes
of the admiring female in one grand bespangled ex91panse.
To whichever side she may turn, the expanded
wings and the obliquely-held tail are turned towards
her. The male Tragopan pheasant acts in nearly the
same manner, for he raises the feathers of the body,
though not the wing itself, on the side which is opposite
to the female, and which would otherwise be concealed,
so that nearly all the beautifully-spotted feathers are
exhibited at the same time.

Fig. 51. Polyplectron chinquis. male (from Brehm.)
The case of the Argus pheasant is still more striking.
The immensely developed secondary wing-feathers, which
are confined to the male, are ornamented with a row of
from twenty to twenty-three ocelli, each above an inch in
diameter. The feathers are also elegantly marked with
oblique dark stripes and rows of spots, like those on the
skin of a tiger and leopard combined. The ocelli are
so beautifully shaded that, as the Duke of Argyll remarks,143
they stand out like a ball lying loosely within
a socket. But when I looked at the specimen in the
British Museum, which is mounted with the wings expanded
and trailing downwards, I was greatly disappointed,
for the ocelli appeared flat or even concave.
Mr. Gould, however, soon made the case clear to me,
for he had made a drawing of a male whilst he was displaying
himself. At such times the long secondary
feathers in both wings are vertically erected and expanded;
and these, together with the enormously elongated
tail-feathers, make a grand semicircular upright
fan. Now as soon as the wing-feathers are held in this
position, and the light shines on them from above, the
full effect of the shading comes out, and each ocellus at
once resembles the ornament called a ball and socket.
These feathers have been shewn to several artists, and all
have expressed their admiration at the perfect shading.
92
It may well be asked, could such artistically-shaded
ornaments have been formed by means of sexual selection?
But it will be convenient to defer giving an
answer to this question until we treat in the next
chapter of the principle of gradation.
The primary wing-feathers, which in most gallinaceous
birds are uniformly coloured, are in the Argus
pheasant not less wonderful objects than the secondary
wing-feathers. They are of a soft brown tint with
numerous dark spots, each of which consists of two or
three black dots with a surrounding dark zone. But
the chief ornament is a space parallel to the dark-blue
shaft, which in outline forms a perfect second feather
lying within the true feather. This inner part is
coloured of a lighter chesnut, and is thickly dotted
with minute white points. I have shewn this feather to
several persons, and many have admired it even more
than the ball-and-socket feathers, and have declared
that it was more like a work of art than of nature.
Now these feathers are quite hidden on all ordinary
occasions, but are fully displayed when the long secondary
feathers are erected, though in a widely different
manner; for they are expanded in front like two little
fans or shields, one on each side of the breast near the
ground.
The case of the male Argus pheasant is eminently
interesting, because it affords good evidence that the
most refined beauty may serve as a charm for the
female, and for no other purpose. We must conclude
that this is the case, as the primary wing-feathers are
never displayed, and the ball-and-socket ornaments are
not exhibited in full perfection, except when the male
assumes the attitude of courtship. The Argus pheasant
does not possess brilliant colours, so that his success in
courtship appears to have depended on the great size of93
his plumes, and on the elaboration of the most elegant
patterns. Many will declare that it is utterly incredible
that a female bird should be able to appreciate fine
shading and exquisite patterns. It is undoubtedly a
marvellous fact that she should possess this almost
human degree of taste, though perhaps she admires
the general effect rather than each separate detail.
He who thinks that he can safely gauge the discrimination
and taste of the lower animals, may deny that
the female Argus pheasant can appreciate such refined
beauty; but he will then be compelled to admit that
the extraordinary attitudes assumed by the male during
the act of courtship, by which the wonderful beauty of
his plumage is fully displayed, are purposeless; and
this is a conclusion which I for one will never admit.
Although so many pheasants and allied gallinaceous
birds carefully display their beautiful plumage before
the females, it is remarkable, as Mr. Bartlett informs
me, that this is not the case with the dull-coloured
Eared and Cheer pheasants (Crossoptilon auritum and
Phasianus Wallichii); so that these birds seem conscious
that they have little beauty to display. Mr.
Bartlett has never seen the males of either of these
species fighting together, though he has not had such
good opportunities for observing the Cheer as the
Eared pheasant. Mr. Jenner Weir, also, finds that
all male birds with rich or strongly-characterised
plumage are more quarrelsome than the dull-coloured
species belonging to the same groups. The goldfinch,
for instance, is far more pugnacious than the
linnet, and the blackbird than the thrush. Those birds
which undergo a seasonal change of plumage likewise
become much more pugnacious at the period when
they are most gaily ornamented. No doubt the males
of some obscurely-coloured birds fight desperately94
together, but it appears that when sexual selection
has been highly influential, and has given bright
colours to the males of any species, it has also very
often given a strong tendency to pugnacity. We
shall meet with nearly analogous cases when we treat
of mammals. On the other hand, with birds the power
of song and brilliant colours have rarely been both
acquired by the males of the same species; but in this
case, the advantage gained would have been identically
the same, namely success in charming the female.
Nevertheless it must be owned that the males of several
brilliantly-coloured birds have had their feathers specially
modified for the sake of producing instrumental
music, though the beauty of this cannot be compared,
at least according to our taste, with that of the vocal
music of many songsters.
We will now turn to male birds which are not
ornamented in any very high degree, but which
nevertheless display, during their courtship, whatever
attractions they may possess. These cases are in some
respects more curious than the foregoing, and have been
but little noticed. I owe the following facts, selected
from a large body of valuable notes, sent to me by Mr.
Jenner Weir, who has long kept birds of many kinds, including
all the British Fringillidæ and Emberizidæ. The
bullfinch makes his advances in front of the female,
and then puffs out his breast, so that many more of the
crimson feathers are seen at once than otherwise would
be the case. At the same time he twists and bows his
black tail from side to side in a ludicrous manner. The
male chaffinch also stands in front of the female, thus
shewing his red breast, and “blue bell,” as the fanciers
call his head; the wings at the same time being slightly
expanded, with the pure white bands on the shoulders
thus rendered conspicuous. The common linnet distends95
his rosy breast, slightly expands his brown wings and
tail, so as to make the best of them by exhibiting their
white edgings. We must, however, be cautious in concluding
that the wings are spread out solely for display,
as some birds act thus whose wings are not beautiful.
This is the case with the domestic cock, but it is always
the wing on the side opposite to the female which is
expanded, and at the same time scraped on the ground.
The male goldfinch behaves differently from all other
finches: his wings are beautiful, the shoulders being
black, with the dark-tipped wing-feathers spotted with
white and edged with golden yellow. When he courts
the female, he sways his body from side to side, and
quickly turns his slightly expanded wings first to
one side then to the other, with a golden flashing effect.
No other British finch, as Mr. Weir informs me, turns
during his courtship from side to side in this manner;
not even the closely-allied male siskin, for he would not
thus add to his beauty.
Most of the British Buntings are plain-coloured birds;
but in the spring the feathers on the head of the male
reed-bunting (Emberiza schœniculus) acquire a fine black
colour by the abrasion of the dusky tips; and these are
erected during the act of courtship. Mr. Weir has kept
two species of Amadina from Australia: the A. castanotis
is a very small and chastely-coloured finch, with
a dark tail, white rump, and jet-black upper tail-coverts,
each of the latter being marked with three large
conspicuous oval spots of white.144 This species, when
courting the female, slightly spreads out and vibrates
these parti-coloured tail-coverts in a very peculiar
manner. The male Amadina Lathami behaves very
96differently, exhibiting before the female his brilliantly-spotted
breast and scarlet rump and scarlet upper tail-coverts.
I may here add from Dr. Jerdon, that the Indian
Bulbul (Pycnonotus hæmorrhous) has crimson under
tail-coverts, and the beauty of these feathers, it might
be thought, could never be well exhibited; but the
bird “when excited often spreads them out laterally,
so that they can be seen even from above.”145 The
common pigeon has iridescent feathers on the breast,
and every one must have seen how the male inflates his
breast whilst courting the female, thus showing off these
feathers to the best advantage. One of the beautiful
bronze-winged pigeons of Australia (Ocyphaps lophotes)
behaves, as described to me by Mr. Weir, very differently:
the male, whilst standing before the female,
lowers his head almost to the ground, spreads out and
raises perpendicularly his tail, and half expands his
wings. He then alternately and slowly raises and depresses
his body, so that the iridescent metallic feathers
are all seen at once, and glitter in the sun.
Sufficient facts have now been given to shew with
what care male birds display their various charms, and
this they do with the utmost skill. Whilst preening
their feathers, they have frequent opportunities for admiring
themselves and of studying how best to exhibit
their beauty. But as all the males of the same species
display themselves in exactly the same manner, it
appears that actions, at first perhaps intentional, have
become instinctive. If so, we ought not to accuse birds
of conscious vanity; yet when we see a peacock strutting
about, with expanded and quivering tail-feathers, he
seems the very emblem of pride and vanity.
The various ornaments possessed by the males are
97certainly of the highest importance to them, for they
have been acquired in some cases at the expense of
greatly impeded powers of flight or of running. The
African nightjar (Cosmetornis), which during the pairing-season
has one of its primary wing-feathers developed
into a streamer of extreme length, is thus much
retarded in its flight, although at other times remarkable
for its swiftness. The “unwieldy size” of the
secondary wing-feathers of the male Argus pheasant
are said “almost entirely to deprive the bird of flight.”
The fine plumes of male Birds of Paradise trouble them
during a high wind. The extremely long tail-feathers
of the male widow-birds (Vidua) of Southern Africa
render “their flight heavy;” but as soon as these are
cast off they fly as well as the females. As birds always
breed when food is abundant, the males probably do
not suffer much inconvenience in searching for food
from their impeded powers of movement; but there can
hardly be a doubt that they must be much more liable
to be struck down by birds of prey. Nor can we doubt
that the long train of the peacock and the long tail
and wing-feathers of the Argus pheasant must render
them a more easy prey to any prowling tiger-cat than
would otherwise be the case. Even the bright colours
of many male birds cannot fail to make them conspicuous
to their enemies of all kinds. Hence it probably
is, as Mr. Gould has remarked, that such birds are
generally of a shy disposition, as if conscious that their
beauty was a source of danger, and are much more difficult
to discover or approach, than the sombre-coloured
and comparatively tame females, or than the young
and as yet unadorned males.146
98
It is a more curious fact that the males of some birds
which are provided with special weapons for battle, and
which in a state of nature are so pugnacious that they
often kill each other, suffer from possessing certain
ornaments. Cock-fighters trim the hackles and cut off
the comb and gills of their cocks; and the birds are
then said to be dubbed. An undubbed bird, as Mr.
Tegetmeier insists, “is at a fearful disadvantage: the
comb and gills offer an easy hold to his adversary’s
beak, and as a cock always strikes where he holds,
when once he has seized his foe, he has him entirely
in his power. Even supposing that the bird is not
killed, the loss of blood suffered by an undubbed cock
is much greater than that sustained by one that has
been trimmed.”147 Young turkey-cocks in fighting
always seize hold of each other’s wattles; and I presume
that the old birds fight in the same manner. It
may perhaps be objected that the comb and wattles are
not ornamental, and cannot be of service to the birds
in this way; but even to our eyes, the beauty of
the glossy black Spanish cock is much enhanced by his
white face and crimson comb; and no one who has ever
seen the splendid blue wattles of the male Tragopan
pheasant, when distended during the act of courtship,
can for a moment doubt that beauty is the object gained.
From the foregoing facts we clearly see that the plumes
and other ornaments of the male must be of the highest
importance to him; and we further see that beauty in
some cases is even more important than success in battle.
99
CHAPTER XIV.
Birds—continued.
Choice exerted by the female—Length of courtship—Unpaired
birds—Mental qualities and taste for the beautiful—Preference
or antipathy shewn by the female for particular males—Variability
of birds—Variations sometimes abrupt—Laws of variation—Formation
of ocelli—Gradations of character—Case of
Peacock, Argus pheasant, and Urosticte.
When the sexes differ in beauty, in the power of
singing, or in producing what I have called instrumental
music, it is almost invariably the male which
excels the female. These qualities, as we have just
seen, are evidently of high importance to the male.
When they are gained for only a part of the year, this
is always shortly before the breeding-season. It is the
male alone who elaborately displays his varied attractions,
and often performs strange antics on the ground
or in the air, in the presence of the female. Each
male drives away or, if he can, kills all his rivals.
Hence we may conclude, that it is the object of the
male to induce the female to pair with him, and for
this purpose he tries to excite or charm her in various
ways; and this is the opinion of all those who have
carefully studied the habits of living birds. But there
remains a question which has an all important bearing
on sexual selection, namely, does every male of the
same species equally excite and attract the female? or
does she exert a choice, and prefer certain males? This
question can be answered in the affirmative by much100
direct and indirect evidence. It is much more difficult
to decide what qualities determine the choice of the
females; but here again we have some direct and indirect
evidence that it is to a large extent the external
attractions of the male, though no doubt his vigour,
courage, and other mental qualities come into play.
We will begin with the indirect evidence.
Length of Courtship.—The lengthened period during
which both sexes of certain birds meet day after day
at an appointed place, probably depends partly on the
courtship being a prolonged affair, and partly on the reiteration
of the act of pairing. Thus in Germany and
Scandinavia the balzens or leks of the Black-cocks,
last from the middle of March, all through April into
May. As many as forty or fifty, or even more birds
congregate at the leks; and the same place is often frequented
during successive years. The lek of the Capercailzie
lasts from the end of March to the middle or
even end of May. In North America “the partridge
dances” of the Tetrao phasianellus “last for a month
or more.” Other kinds of grouse both in North America
and Eastern Siberia148 follow nearly the same habits.
The fowlers discover the hillocks where the Ruffs congregate
by the grass being trampled bare, and this shews
that the same spot is long frequented. The Indians of
Guiana are well acquainted with the cleared arenas,
where they expect to find the beautiful Cocks of the
Rock; and the natives of New Guinea know the trees
where from ten to twenty full-plumaged male Birds of
101
Paradise congregate. In this latter case it is not expressly
stated that the females meet on the same trees,
but the hunters, if not specially asked, would not probably
mention their presence, as their skins are valueless.
Small parties of an African weaver (Ploceus) congregate,
during the breeding-season, and perform for
hours their graceful evolutions. Large numbers of the
Solitary snipe (Scolopax major) assemble during the
dusk in a morass; and the same place is frequented for
the same purpose during successive years; here they
may be seen running about “like so many large rats,”
puffing out their feathers, flapping their wings, and
uttering the strangest cries.149
Some of the above-mentioned birds, namely, the
black-cock, capercailzie, pheasant-grouse, the ruff, the
Solitary snipe, and perhaps some others, are, as it is
believed, polygamists. With such birds it might have
been thought that the stronger males would simply
have driven away the weaker, and then at once have
taken possession of as many females as possible; but if
it be indispensable for the male to excite or please the
female, we can understand the length of the courtship
and the congregation of so many individuals of both
sexes at the same spot. Certain species which are
strictly monogamous likewise hold nuptial assemblages;
this seems to be the case in Scandinavia with one of
the ptarmigans, and their leks last from the middle
of March to the middle of May. In Australia the lyre-bird
or Menura superba forms “small round hillocks,”
102and the M. Alberti scratches for itself shallow holes, or,
as they are called by the natives, corroborying places,
where it is believed both sexes assemble. The meetings
of the M. superba are sometimes very large; and
an account has lately been published150 by a traveller,
who heard in a valley beneath him, thickly covered
with scrub, “a din which completely astonished” him;
on crawling onwards he beheld to his amazement about
one hundred and fifty of the magnificent lyre-cocks,
“ranged in order of battle, and fighting with indescribable
fury.” The bowers of the Bower-birds are
the resort of both sexes during the breeding-season;
and “here the males meet and contend with each other
“for the favours of the female, and here the latter
assemble and coquet with the males.” With two of
the genera, the same bower is resorted to during many
years.151
The common magpie (Corvus pica, Linn.), as I have
been informed by the Rev. W. Darwin Fox, used to
assemble from all parts of Delamere Forest, in order
to celebrate the “great magpie marriage.” Some
years ago these birds abounded in extraordinary numbers,
so that a gamekeeper killed in one morning
nineteen males, and another killed by a single shot
seven birds at roost together. Whilst they were so
numerous, they had the habit very early in the spring
of assembling at particular spots, where they could be
seen in flocks, chattering, sometimes fighting, bustling
and flying about the trees. The whole affair was
evidently considered by the birds as of the highest
importance. Shortly after the meeting they all separated,
and were then observed by Mr. Fox and others
103to be paired for the season. In any district in which
a species does not exist in large numbers, great assemblages
cannot, of course, be held, and the same species
may have different habits in different countries. For
instance, I have never met with any account of regular
assemblages of black game in Scotland, yet these assemblages
are so well known in Germany and Scandinavia
that they have special names.
Unpaired Birds.—From the facts now given, we
may conclude that with birds belonging to widely-different
groups their courtship is often a prolonged, delicate,
and troublesome affair. There is even reason to
suspect, improbable as this will at first appear, that
some males and females of the same species, inhabiting
the same district, do not always please each other and
in consequence do not pair. Many accounts have been
published of either the male or female of a pair having
been shot, and quickly replaced by another. This has
been observed more frequently with the magpie than
with any other bird, owing perhaps to its conspicuous
appearance and nest. The illustrious Jenner states
that in Wiltshire one of a pair was daily shot no less
than seven times successively, “but all to no purpose,
for the remaining magpie soon found another mate;”
and the last pair reared their young. A new partner
is generally found on the succeeding day; but Mr.
Thompson gives the case of one being replaced on the
evening of the same day. Even after the eggs are
hatched, if one of the old birds is destroyed a mate
will often be found; this occurred after an interval
of two days, in a case recently observed by one of
Sir J. Lubbock’s keepers.152 The first and most obvious
104conjecture is that male magpies must be much more
numerous than the females; and that in the above cases,
as well in many others which could be given, the males
alone had been killed. This apparently holds good in
some instances, for the gamekeepers in Delamere Forest
assured Mr. Fox that the magpies and carrion-crows
which they formerly killed in succession in large numbers
near their nests were all males; and they accounted
for this fact by the males being easily killed
whilst bringing food to the sitting females. Macgillivray,
however, gives, on the authority of an excellent
observer, an instance of three magpies successively
killed on the same nest which were all females; and
another case of six magpies successively killed whilst
sitting on the same eggs, which renders it probable
that most of them were females, though the male will
sit on the eggs, as I hear from Mr. Fox, when the
female is killed.
Sir J. Lubbock’s gamekeeper has repeatedly shot, but
how many times he could not say, one of a pair of jays
(Garrulus glandarius), and has never failed shortly
afterwards to find the survivor rematched. The Rev.
W. D. Fox, Mr. F. Bond, and others, have shot one of a
pair of carrion-crows (Corvus corone), but the nest was
soon again tenanted by a pair. These birds are rather
common; but the peregrine falcon (Falco peregrinus)
is rare, yet Mr. Thompson states that in Ireland “if
either an old male or female be killed in the breeding-season
(not an uncommon circumstance), another
mate is found within a very few days, so that the
eyries, notwithstanding such casualties, are sure to
turn out their complement of young.” Mr. Jenner
Weir has known the same thing to occur with the peregrine
falcons at Beachy Head. The same observer
informs me that three kestrels, all males (Falco tinnunculus),105
were killed one after the other whilst attending
the same nest; two of these were in mature plumage,
and the third in the plumage of the previous year.
Even with the rare golden eagle (Aquila chrysaëtos),
Mr. Birkbeck was assured by a trustworthy gamekeeper
in Scotland, that if one is killed, another is soon found.
So with the white owl (Strix flammea), it has been
observed that “the survivor readily found a mate, and
the mischief went on.”
White of Selborne, who gives the case of the owl,
adds that he knew a man, who from believing that
partridges when paired were disturbed by the males
fighting, used to shoot them; and though he had
widowed the same female several times she was always
soon provided with a fresh partner. This same naturalist
ordered the sparrows, which deprived the house-martins
of their nests, to be shot: but the one which
was left, “be it cock or hen, presently procured a mate,
and so for several times following.” I could add analogous
cases relating to the chaffinch, nightingale, and
redstart. With respect to the latter bird (Phœnicura
ruticilla), the writer remarks that it was by no means
common in the neighbourhood, and he expresses much
surprise how the sitting female could so soon give effectual
notice that she was a widow. Mr. Jenner Weir has
mentioned to me a nearly similar case: at Blackheath
he never sees or hears the note of the wild bullfinch, yet
when one of his caged males has died, a wild one in the
course of a few days has generally come and perched
near the widowed female, whose call-note is far from
loud. I will give only one other fact, on the authority
of this same observer; one of a pair of starlings (Sturnus
vulgaris) was shot in the morning; by noon a new mate
was found; this was again shot, but before night the
pair was complete; so that the disconsolate widow or106
widower was thrice consoled during the same day. Mr.
Engleheart also informs me that he used during several
years to shoot one of a pair of starlings which built in a
hole in a house at Blackheath; but the loss was always
immediately repaired. During one season he kept an
account and found that he had shot thirty-five birds from
the same nest; these consisted of both males and females,
but in what proportion he could not say: nevertheless
after all this destruction, a brood was reared.153
These facts are certainly remarkable. How is it that
so many birds are ready immediately to replace a lost
mate? Magpies, jays, carrion-crows, partridges, and
some other birds, are never seen during the spring by
themselves, and these offer at first sight the most
perplexing case. But birds of the same sex, although
of course not truly paired, sometimes live in pairs or in
small parties, as is known to be the case with pigeons
and partridges. Birds also sometimes live in triplets,
as has been observed with starlings, carrion-crows, parrots,
and partridges. With partridges two females have
been known to live with one male, and two males with
one female. In all such cases it is probable that the
union would be easily broken. The males of certain
birds may occasionally be heard pouring forth their
love-song long after the proper time, shewing that they
have either lost or never gained a mate. Death from
accident or disease of either one of a pair, would leave
the other bird free and single; and there is reason to
believe that female birds during the breeding-season
107are especially liable to premature death. Again, birds
which have had their nests destroyed, or barren pairs, or
retarded individuals, would easily be induced to desert
their mates, and would probably be glad to take what
share they could of the pleasures and duties of rearing
offspring, although not their own.154 Such contingencies
as these probably explain most of the foregoing cases.155
Nevertheless it is a strange fact that within the same
district, during the height of the breeding-season,
there should be so many males and females always
ready to repair the loss of a mated bird. Why do not
such spare birds immediately pair together? Have
we not some reason to suspect, and the suspicion has
occurred to Mr. Jenner Weir, that inasmuch as the
act of courtship appears to be with many birds a prolonged
and tedious affair, so it occasionally happens
that certain males and females do not succeed during
the proper season, in exciting each other’s love, and
consequently do not pair? This suspicion will appear
somewhat less improbable after we have seen what
108strong antipathies and preferences female birds occasionally
evince towards particular males.
Mental Qualities of Birds, and their taste for the
beautiful.—Before we discuss any further the question
whether the females select the more attractive males
or accept the first whom they may encounter, it will be
advisable briefly to consider the mental powers of birds.
Their reason is generally, and perhaps justly, ranked
as low; yet some facts could be given156 leading to an
opposite conclusion. Low powers of reasoning, however,
are compatible, as we see with mankind, with
strong affections, acute perception, and a taste for the
beautiful; and it is with these latter qualities that we
are here concerned. It has often been said that parrots
become so deeply attached to each other that when
one dies the other for a long time pines; but Mr.
Jenner Weir thinks that with most birds the strength
of their affection has been much exaggerated. Nevertheless
when one of a pair in a state of nature has
been shot, the survivor has been heard for days afterwards
uttering a plaintive call; and Mr. St. John
gives157 various facts proving the attachment of mated
birds. Starlings, however, as we have seen, may be
consoled thrice in the same day for the loss of their
mates. In the Zoological Gardens parrots have clearly
109recognised their former masters after an interval of
some months. Pigeons have such excellent local memories
that they have been known to return to their
former homes after an interval of nine months, yet, as
I hear from Mr. Harrison Weir, if a pair which would
naturally remain mated for life be separated for a few
weeks during the winter and matched with other birds,
the two, when brought together again, rarely, if ever,
recognise each other.
Birds sometimes exhibit benevolent feelings; they
will feed the deserted young even of distinct species,
but this perhaps ought to be considered as a mistaken
instinct. They will also feed, as shewn in an earlier
part of this work, adult birds of their own species
which have become blind. Mr. Buxton gives a curious
account of a parrot which took care of a frost-bitten and
crippled bird of a distinct species, cleansed her feathers
and defended her from the attacks of the other parrots
which roamed freely about his garden. It is a still
more curious fact that these birds apparently evince
some sympathy for the pleasures of their fellows. When
a pair of cockatoos made a nest in an acacia tree, “it
was ridiculous to see the extravagant interest taken
in the matter by the others of the same species.”
These parrots, also, evinced unbounded curiosity, and
clearly had “the idea of property and possession.”158
Birds possess acute powers of observation. Every
mated bird, of course, recognises its fellow. Audubon
states that with the mocking-thrushes of the United
States (Mimus polyglottus) a certain number remain all
the year round in Louisiana, whilst the others migrate
to the Eastern States; these latter, on their return,
110are instantly recognised, and always attacked, by their
Southern brethren. Birds under confinement distinguish
different persons, as is proved by the strong and
permanent antipathy or affection which they shew,
without any apparent cause, towards certain individuals.
I have heard of numerous instances with jays,
partridges, canaries, and especially bullfinches. Mr.
Hussey has described in how extraordinary a manner
a tamed partridge recognised everybody; and its likes
and dislikes were very strong. This bird seemed “fond
of gay colours, and no new gown or cap could be put
on without catching his attention.”159 Mr. Hewitt
has carefully described the habits of some ducks (recently
descended from wild birds), which, at the approach
of a strange dog or cat, would rush headlong into
the water, and exhaust themselves in their attempts
to escape; but they knew so well Mr. Hewitt’s own
dogs and cats that they would lie down and bask in the
sun close to them. They always moved away from a
strange man, and so they would from the lady who
attended them, if she made any great change in her
dress. Audubon relates that he reared and tamed a
wild turkey which always ran away from any strange
dog; this bird escaped into the woods, and some days
afterwards Audubon saw, as he thought, a wild turkey,
and made his dog chase it; but to his astonishment,
the bird did not run away, and the dog, when he came
up, did not attack the bird, for they mutually recognised
each other as old friends.160
Mr. Jenner Weir is convinced that birds pay particular
attention to the colours of other birds, sometimes
111out of jealousy, and sometimes as a sign of kinship.
Thus he turned a reed-bunting (Emberiza schœniculus),
which had acquired its black head, into his aviary,
and the new-comer was not noticed by any bird, except
by a bullfinch, which is likewise black-headed. This
bullfinch was a very quiet bird, and had never before
quarrelled with any of its comrades, including another
reed-bunting, which had not as yet become black-headed:
but the reed-bunting with a black head was
so unmercifully treated, that it had to be removed.
Mr. Weir was also obliged to turn out a robin, as it
fiercely attacked all birds with any red in their plumage,
but no other kinds; it actually killed a red-breasted
crossbill, and nearly killed a goldfinch. On
the other hand, he has observed that some birds, when
first introduced into his aviary, fly towards the species
which resemble them most in colour, and settle by
their sides.
As male birds display with so much care their fine
plumage and other ornaments in the presence of the
females, it is obviously probable that these appreciate
the beauty of their suitors. It is, however, difficult to
obtain direct evidence of their capacity to appreciate
beauty. When birds gaze at themselves in a looking-glass
(of which many instances have been recorded) we
cannot feel sure that it is not from jealousy at a supposed
rival, though this is not the conclusion of some
observers. In other cases it is difficult to distinguish
between mere curiosity and admiration. It is perhaps
the former feeling which, as stated by Lord Lilford,161
attracts the Ruff strongly towards any bright object,
so that, in the Ionian Islands, it “will dart down to a
bright-coloured handkerchief, regardless of repeated
112shots.” The common lark is drawn down from the
sky, and is caught in large numbers, by a small mirror
made to move and glitter in the sun. Is it admiration
or curiosity which leads the magpie, raven, and some
other birds to steal and secrete bright objects, such as
silver articles or jewels?
Mr. Gould states that certain humming-birds decorate
the outside of their nests, “with the utmost taste;
they instinctively fasten thereon beautiful pieces of
flat lichen, the larger pieces in the middle, and the
smaller on the part attached to the branch. Now
and then a pretty feather is intertwined or fastened
to the outer sides, the stem being always so placed,
that the feather stands out beyond the surface.” The
best evidence, however, of a taste for the beautiful is
afforded by the three genera of Australian bower-birds
already mentioned. Their bowers (see fig. 46, p. 70),
where the sexes congregate and play strange antics, are
differently constructed, but what most concerns us is, that
they are decorated in a different manner by the several
species. The Satin bower-bird collects gaily-coloured
articles, such as the blue tail-feathers of parrakeets,
bleached bones and shells, which it sticks between the
twigs, or arranges at the entrance. Mr. Gould found
in one bower a neatly-worked stone tomahawk and a
slip of blue cotton, evidently procured from a native
encampment. These objects are continually rearranged,
and carried about by the birds whilst at play. The
bower of the Spotted bower-bird “is beautifully lined
with tall grasses, so disposed that the heads nearly
meet, and the decorations are very profuse.” Round
stones are used to keep the grass-stems in their proper
places, and to make divergent paths leading to the
bower. The stones and shells are often brought from
a great distance. The Regent bird, as described by113
Mr. Ramsay, ornaments its short bower with bleached
land-shells belonging to five or six species, and with
“berries of various colours, blue, red, and black, which
give it when fresh a very pretty appearance. Besides
these there were several newly-picked leaves and
young shoots of a pinkish colour, the whole shewing a
decided taste for the beautiful.” Well may Mr. Gould
say “these highly decorated halls of assembly must be
regarded as the most wonderful instances of bird-architecture
yet discovered;” and the taste, as we see, of
the several species certainly differs.162
Preference for particular Males by the Females.—Having
made these preliminary remarks on the discrimination
and taste of birds, I will give all the facts known
to me, which bear on the preference shewn by the female
for particular males. It is certain that distinct species of
birds occasionally pair in a state of nature and produce
hybrids. Many instances could be given: thus Macgillivray
relates how a male blackbird and female thrush
“fell in love with each other,” and produced offspring.163
Several years ago eighteen cases had been recorded of
the occurrence in Great Britain of hybrids between the
black grouse and pheasant;164 but most of these cases
may perhaps be accounted for by solitary birds not
finding one of their own species to pair with. With
other birds, as Mr. Jenner Weir has reason to believe,
hybrids are sometimes the result of the casual intercourse
of birds building in close proximity. But these
114remarks do not apply to the many recorded instances of
tamed or domestic birds, belonging to distinct species,
which have become absolutely fascinated with each
other, although living with their own species. Thus
Waterton165 states that out of a flock of twenty-three
Canada geese, a female paired with a solitary Bernicle
gander, although so different in appearance and size;
and they produced hybrid offspring. A male Wigeon
(Mareca penelope), living with females of the same
species, has been known to pair with a Pintail duck,
Querquedula acuta. Lloyd describes the remarkable
attachment between a shield-drake (Tadorna vulpanser)
and a common duck. Many additional instances could
be given; and the Rev. E. S. Dixon remarks that “Those
who have kept many different species of geese together,
well know what unaccountable attachments
they are frequently forming, and that they are quite
as likely to pair and rear young with individuals of a
race (species) apparently the most alien to themselves,
as with their own stock.”
The Rev. W. D. Fox informs me that he possessed at
the same time a pair of Chinese geese (Anser cygnoides),
and a common gander with three geese. The two lots
kept quite separate, until the Chinese gander seduced
one of the common geese to live with him. Moreover,
of the young birds hatched from the eggs of the common
geese, only four were pure, the other eighteen proving
hybrids; so that the Chinese gander seems to have
had prepotent charms over the common gander. I will
115give only one other case; Mr. Hewitt states that a wild
duck, reared in captivity, “after breeding a couple of
seasons with her own mallard, at once shook him off
on my placing a male Pintail on the water. It was
evidently a case of love at first sight, for she swam
about the new-comer caressingly, though he appeared
evidently alarmed and averse to her overtures of
affection. From that hour she forgot her old partner.
Winter passed by, and the next spring the Pintail
seemed to have become a convert to her blandishments,
for they nested and produced seven or eight
young ones.”
What the charm may have been in these several
cases, beyond mere novelty, we cannot even conjecture.
Colour, however, sometimes comes into play; for in
order to raise hybrids from the siskin (Fringilla spinus)
and the canary, it is much the best plan, according to
Bechstein, to place birds of the same tint together.
Mr. Jenner Weir turned a female canary into his aviary,
where there were male linnets, goldfinches, siskins,
greenfinches, chaffinches, and other birds, in order to
see which she would choose; but there never was any
doubt, and the greenfinch carried the day. They paired
and produced hybrid offspring.
With the members of the same species the fact of the
female preferring to pair with one male rather than
with another is not so likely to excite attention, as
when this occurs between distinct species. Such cases
can best be observed with domesticated or confined
birds; but these are often pampered by high feeding,
and sometimes have their instincts vitiated to an extreme
degree. Of this latter fact I could give sufficient
proofs with pigeons, and especially with fowls, but they
cannot be here related. Vitiated instincts may also
account for some of the hybrid unions above referred116
to; but in many of these cases the birds were allowed
to range freely over large ponds, and there is no reason
to suppose that they were unnaturally stimulated by
high feeding.
With respect to birds in a state of nature, the first
and most obvious supposition which will occur to everyone
is that the female at the proper season accepts the
first male whom she may encounter; but she has at
least the opportunity for exerting a choice, as she is
almost invariably pursued by many males. Audubon—and
we must remember that he spent a long life in
prowling about the forests of the United States and
observing the birds—does not doubt that the female
deliberately chooses her mate; thus, speaking of a woodpecker,
he says the hen is followed by half-a-dozen gay
suitors, who continue performing strange antics, “until
a marked preference is shewn for one.” The female of
the red-winged starling (Agelæus phœniceus) is likewise
pursued by several males, “until, becoming fatigued,
she alights, receives their addresses, and soon makes
a choice.” He describes also how several male nightjars
repeatedly plunge through the air with astonishing
rapidity, suddenly turning, and thus making a
singular noise; “but no sooner has the female made
her choice, than the other males are driven away.”
With one of the vultures (Cathartes aura) of the United
States, parties of eight or ten or more males and females
assemble on fallen logs, “exhibiting the strongest desire
to please mutually,” and after many caresses, each male
leads off his partner on the wing. Audubon likewise
carefully observed the wild flocks of Canada geese (Anser
Canadensis), and gives a graphic description of their
love-antics; he says that the birds which had been previously
mated “renewed their courtship as early as the
month of January, while the others would be contend117ing
or coquetting for hours every day, until all seemed
satisfied with the choice they had made, after which,
although they remained together, any person could
easily perceive that they were careful to keep in pairs.
I have observed also that the older the birds, the
shorter were the preliminaries of their courtship.
The bachelors and old maids, whether in regret, or
not caring to be disturbed by the bustle, quietly
moved aside and lay down at some distance from the
rest.”166 Many similar statements with respect to other
birds could be cited from this same observer.
Turning now to domesticated and confined birds, I
will commence by giving what little I have learnt respecting
the courtship of fowls. I have received long
letters on this subject from Messrs. Hewitt and Tegetmeier,
and almost an essay from the late Mr. Brent.
It will be admitted by every one that these gentlemen,
so well known from their published works, are careful
and experienced observers. They do not believe that the
females prefer certain males on account of the beauty of
their plumage; but some allowance must be made for
the artificial state under which they have long been
kept. Mr. Tegetmeier is convinced that a game-cock,
though disfigured by being dubbed with his hackles
trimmed, would be accepted as readily as a male retaining
all his natural ornaments. Mr. Brent, however,
admits that the beauty of the male probably aids in
exciting the female; and her acquiescence is necessary.
Mr. Hewitt is convinced that the union is by no means
left to mere chance, for the female almost invariably
prefers the most vigorous, defiant, and mettlesome male;
hence it is almost useless, as he remarks, “to attempt
118true breeding if a game-cock in good health and condition
runs the locality, for almost every hen on leaving
the roosting-place will resort to the game-cock, even
though that bird may not actually drive away the male
of her own variety.” Under ordinary circumstances the
males and females of the fowl seem to come to a mutual
understanding by means of certain gestures, described
to me by Mr. Brent. But hens will often avoid the
officious attentions of young males. Old hens, and
hens of a pugnacious disposition, as the same writer
informs me, dislike strange males, and will not yield
until well beaten into compliance. Ferguson, however,
describes how a quarrelsome hen was subdued by the
gentle courtship of a Shanghai cock.167
There is reason to believe that pigeons of both sexes
prefer pairing with birds of the same breed; and dovecot-pigeons
dislike all the highly improved breeds.168 Mr.
Harrison Weir has lately heard from a trustworthy
observer, who keeps blue pigeons, that these drive
away all other coloured varieties, such as white, red,
and yellow; and from another observer, that a female
dun carrier could not be matched, after repeated trials,
with a black male, but immediately paired with a dun.
Generally colour alone appears to have little influence
on the pairing of pigeons. Mr. Tegetmeier, at my request,
stained some of his birds with magenta, but they
were not much noticed by the others.
Female pigeons occasionally feel a strong antipathy
towards certain males, without any assignable cause.
Thus MM. Boitard and Corbié, whose experience extended
over forty-five years, state: “Quand une femelle
119éprouve de l’antipathie pour un mâle avec lequel on
veut l’accoupler, malgré tous les feux de l’amour,
malgré l’alpiste et le chènevis dont on la nourrit
pour augmenter son ardeur, malgré un emprisonnement
de six mois et même d’un an, elle refuse constamment
ses caresses; les avances empressées, les
agaceries, les tournoiemens, les tendres roucoulemens,
rien ne peut lui plaire ni l’émouvoir; gonflée, boudeuse,
blottie dans un coin de sa prison, elle n’en sort
que pour boire et manger, ou pour repousser avec une
espèce de rage des caresses devenues trop pressantes.”169
On the other hand, Mr. Harrison Weir has himself
observed, and has heard from, several breeders, that a
female pigeon will occasionally take a strong fancy for
a particular male, and will desert her own mate for
him. Some females, according to another experienced
observer, Riedel,170 are of a profligate disposition, and
prefer almost any stranger to their own mate. Some
amorous males, called by our English fanciers “gay
birds,” are so successful in their gallantries, that, as
Mr. H. Weir informs me, they must be shut up, on
account of the mischief which they cause.
Wild turkeys in the United States, according to
Audubon, “sometimes pay their addresses to the domesticated
females, and are generally received by them
with great pleasure.” So that these females apparently
prefer the wild to their own males.171
Here is a more curious case. Sir R. Heron during
many years kept an account of the habits of the peafowl,
which he bred in large numbers. He states that
120“the hens have frequently great preference to a particular
peacock. They were all so fond of an old pied
cock, that one year, when he was confined though
still in view, they were constantly assembled close
to the trellice-walls of his prison, and would not suffer
a japanned peacock to touch them. On his being let
out in the autumn, the oldest of the hens instantly
courted him, and was successful in her courtship.
The next year he was shut up in a stable, and then
the hens all courted his rival.”172 This rival was a
japanned or black-winged peacock, which to our eyes
is a more beautiful bird than the common kind.
Lichtenstein, who was a good observer and had
excellent opportunities of observation at the Cape of
Good Hope, assured Rudolphi that the female widow-bird
(Chera progne) disowns the male, when robbed of
the long tail-feathers with which he is ornamented
during the breeding-season. I presume that this observation
must have been made on birds under confinement.173
Here is another striking case; Dr. Jaeger,174
director of the Zoological Gardens of Vienna, states
that a male silver pheasant, who had been triumphant
over the other males and was the accepted lover of the
females, had his ornamental plumage spoiled. He
was then immediately superseded by a rival, who got
the upper hand and afterwards led the flock.
Not only does the female exert a choice, but in some
few cases she courts the male, or even fights for his
possession. Sir R. Heron states that with peafowl, the
121first advances are always made by the female; something
of the same kind takes place, according to
Audubon, with the older females of the wild turkey.
With the capercailzie, the females flit round the male,
whilst he is parading at one of the places of assemblage,
and solicit his attention.175 We have seen that a tame
wild-duck seduced after a long courtship an unwilling
Pintail drake. Mr. Bartlett believes that the Lophophorus,
like many other gallinaceous birds, is naturally
polygamous, but two females cannot be placed in the
same cage with a male, as they fight so much together.
The following instance of rivalry is more surprising as
it relates to bullfinches, which usually pair for life.
Mr. Jenner Weir introduced a dull-coloured and ugly
female into his aviary, and she immediately attacked
another mated female so unmercifully that the latter
had to be separated. The new female did all the courtship,
and was at last successful, for she paired with the
male; but after a time she met with a just retribution,
for, ceasing to be pugnacious, Mr. Weir replaced the
old female, and the male then deserted his new and
returned to his old love.
In all ordinary cases the male is so eager that he will
accept any female, and does not, as far as we can judge,
prefer one to the other; but exceptions to this rule, as
we shall hereafter see, apparently occur in some few
groups. With domesticated birds, I have heard of only
one case in which the males shew any preference for
particular females, namely, that of the domestic cock,
who, according to the high authority of Mr. Hewitt,
prefers the younger to the older hens. On the other
122hand, in effecting hybrid unions between the male
pheasant and common hens, Mr. Hewitt is convinced
that the pheasant invariably prefers the older birds.
He does not appear to be in the least influenced by
their colour, but “is most capricious in his attachments.”176
From some inexplicable cause he shews the
most determined aversion to certain hens, which no
care on the part of the breeder can overcome. Some
hens, as Mr. Hewitt informs me, are quite unattractive
even to the males of their own species, so that they
may be kept with several cocks during a whole season,
and not one egg out of forty or fifty will prove
fertile. On the other hand with the Long-tailed duck
(Harelda glacialis), “it has been remarked,” says
M. Ekström, “that certain females are much more
courted than the rest. Frequently, indeed, one sees
an individual surrounded by six or eight amorous
males.” Whether this statement is credible, I know
not; but the native sportsmen shoot these females in
order to stuff them as decoys.177
With respect to female birds feeling a preference for
particular males, we must bear in mind that we can
judge of choice being exerted, only by placing ourselves
in imagination in the same position. If an
inhabitant of another planet were to behold a number
of young rustics at a fair, courting and quarrelling
over a pretty girl, like birds at one of their places of
assemblage, he would be able to infer that she had the
power of choice only by observing the eagerness of the
wooers to please her, and to display their finery. Now
with birds, the evidence stands thus; they have acute
powers of observation, and they seem to have some
123taste for the beautiful both in colour and sound. It
is certain that the females occasionally exhibit, from
unknown causes, the strongest antipathies and preferences
for particular males. When the sexes differ in
colour or in other ornaments, the males with rare exceptions
are the most highly decorated, either permanently
or temporarily during the breeding-season. They
sedulously display their various ornaments, exert their
voices, and perform strange antics in the presence of the
females. Even well-armed males, who, it might have
been thought, would have altogether depended for
success on the law of battle, are in most cases highly
ornamented; and their ornaments have been acquired
at the expense of some loss of power. In other cases
ornaments have been acquired, at the cost of increased
risk from birds and beasts of prey. With various species
many individuals of both sexes congregate at the
same spot, and their courtship is a prolonged affair.
There is even reason to suspect that the males and
females within the same district do not always succeed
in pleasing each other and pairing.
What then are we to conclude from these facts and
considerations? Does the male parade his charms with
so much pomp and rivalry for no purpose? Are we not
justified in believing that the female exerts a choice,
and that she receives the addresses of the male who
pleases her most? It is not probable that she consciously
deliberates; but she is most excited or attracted
by the most beautiful, or melodious, or gallant
males. Nor need it be supposed that the female
studies each stripe or spot of colour; that the peahen,
for instance, admires each detail in the gorgeous train of
the peacock—she is probably struck only by the general
effect. Nevertheless after hearing how carefully the
male Argus pheasant displays his elegant primary124
wing-feathers, and erects his ocellated plumes in the
right position for their full effect; or again, how the
male goldfinch alternately displays his gold-bespangled
wings, we ought not to feel too sure that the female
does not attend to each detail of beauty. We can
judge, as already remarked, of choice being exerted,
only from the analogy of our own minds; and the
mental powers of birds, if reason be excluded, do not
fundamentally differ from ours. From these various
considerations we may conclude that the pairing of
birds is not left to chance; but that those males, which
are best able by their various charms to please or excite
the female, are under ordinary circumstances accepted.
If this be admitted, there is not much difficulty in
understanding how male birds have gradually acquired
their ornamental characters. All animals present individual
differences, and as man can modify his domesticated
birds by selecting the individuals which appear
to him the most beautiful, so the habitual or even occasional
preference by the female of the more attractive
males would almost certainly lead to their modification;
and such modifications might in the course of time be
augmented to almost any extent, compatible with the
existence of the species.
Variability of Birds, and especially of their secondary
Sexual Characters.—Variability and inheritance are the
foundations for the work of selection. That domesticated
birds have varied greatly, their variations being
inherited, is certain. That birds in a state of nature
present individual differences is admitted by every
one; and that they have sometimes been modified into
distinct races, is generally admitted.178 Variations are
125of two kinds, which insensibly graduate into each other,
namely, slight differences between all the members of
the same species, and more strongly-marked deviations
which occur only occasionally. These latter are rare
with birds in a state of nature, and it is very doubtful
whether they have often been preserved through selection,
and then transmitted to succeeding generations.179
Nevertheless, it may be worth while to give the few
cases relating chiefly to colour (simple albinism and
melanism being excluded), which I have been able to
collect.
Mr. Gould is well known rarely to admit the existence
of varieties, for he esteems very slight differences as
specific; now he states180 that near Bogota certain humming-birds
belonging to the genus Cynanthus are
divided into two or three races or varieties, which differ
from each other in the colouring of the tail,—“some
126having the whole of the feathers blue, while others
have the eight central ones tipped with beautiful
green.” It does not appear that intermediate gradations
have been observed in this or the following
cases. In the males alone of one of the Australian
parrakeets “the thighs in some are scarlet, in others
grass-green.” In another parrakeet of the same
country “some individuals have the band across the
wing-coverts bright-yellow, while in others the same
part is tinged with red.”181 In the United States
some few of the males of the Scarlet Tanager (Tanagra
rubra) have “a beautiful transverse band of glowing
red on the smaller wing-coverts;”182 but this variation
seems to be somewhat rare, so that its preservation
through sexual selection would follow only under
unusually favourable circumstances. In Bengal the
Honey buzzard (Pernis cristata) has either a small
rudimental crest on its head, or none at all; so slight a
difference however would not have been worth notice,
had not this same species possessed in Southern India
“a well-marked occipital crest formed of several graduated
feathers.”183
The following case is in some respects more interesting.
A pied variety of the raven, with the head, breast,
abdomen, and parts of the wings and tail-feathers white,
is confined to the Feroe Islands. It is not very rare
there, for Graba saw during his visit from eight to
ten living specimens. Although the characters of this
variety are not quite constant, yet it has been named
by several distinguished ornithologists as a distinct
species. The fact of the pied birds being pursued and
127persecuted with much clamour by the other ravens of
the island was the chief cause which led Brünnich to
conclude that it was specifically distinct; but this is
now known to be an error.184
In various parts of the northern seas a remarkable
variety of the common Guillemot (Uria troile) is found;
and in Feroe, one out of every five birds, according to
Graba’s estimation, consists of this variety. It is characterised185
by a pure white ring round the eye, with
a curved narrow white line, an inch and a half in
length, extending back from the ring. This conspicuous
character has caused the bird to be ranked by
several ornithologists as a distinct species under the
name of U. lacrymans, but it is now known to be merely
a variety. It often pairs with the common kind, yet
intermediate gradations have never been seen; nor is
this surprising, for variations which appear suddenly
are often, as I have elsewhere shewn,186 transmitted
either unaltered or not at all. We thus see that two
distinct forms of the same species may co-exist in the
same district, and we cannot doubt that if the one had
possessed any great advantage over the other, it would
soon have been multiplied to the exclusion of the latter.
If, for instance, the male pied ravens, instead of being
persecuted and driven away by their comrades, had
been highly attractive, like the pied peacock before
mentioned, to the common black females, their numbers
would have rapidly increased. And this would have
been a case of sexual selection.
128
With respect to the slight individual differences which
are common, in a greater or less degree, to all the
members of the same species, we have every reason
to believe that they are by far the most important
for the work of selection. Secondary sexual characters
are eminently liable to vary, both with animals in a
state of nature and under domestication.187 There is
also reason to believe, as we have seen in our eighth
chapter, that variations are more apt to occur in the
male than in the female sex. All these contingencies
are highly favourable for sexual selection. Whether
characters thus acquired are transmitted to one sex
or to both sexes, depends exclusively in most cases,
as I hope to shew in the following chapter, on the
form of inheritance which prevails in the groups in
question.
It is sometimes difficult to form any opinion whether
certain slight differences between the sexes of birds
are simply the result of variability with sexually-limited
inheritance, without the aid of sexual selection,
or whether they have been augmented through this
latter process. I do not here refer to the innumerable
instances in which the male displays splendid colours
or other ornaments, of which the female partakes only
to a slight degree; for these cases are almost certainly
due to characters primarily acquired by the male,
having been transferred, in a greater or less degree, to
the female. But what are we to conclude with respect
to certain birds in which, for instance, the eyes differ
slightly in colour in the two sexes?188 In some cases
the eyes differ conspicuously; thus with the storks
129of the genus Xenorhynchus those of the male are
blackish-hazel, whilst those of the females are gamboge-yellow;
with many hornbills (Buceros), as I hear
from Mr. Blyth,189 the males have intense crimson, and
the females white eyes. In the Buceros bicornis, the
hind margin of the casque and a stripe on the crest of
the beak are black in the male, but not so in the female.
Are we to suppose that these black marks and the
crimson colour of the eyes have been preserved or augmented
through sexual selection in the males? This
is very doubtful; for Mr. Bartlett shewed me in the
Zoological Gardens that the inside of the mouth of this
Buceros is black in the male and flesh-coloured in the
female; and their external appearance or beauty would
not be thus affected. I observed in Chili190 that the
iris in the condor, when about a year old, is dark-brown,
but changes at maturity into yellowish-brown in the
male, and into bright red in the female. The male
has also a small, longitudinal, leaden-coloured, fleshy
crest or comb. With many gallinaceous birds the
comb is highly ornamental, and assumes vivid colours
during the act of courtship; but what are we to think
of the dull-coloured comb of the condor, which does
not appear to us in the least ornamental? The same
question may be asked in regard to various other
characters, such as the knob on the base of the beak of
the Chinese goose (Anser cygnoides), which is much
larger in the male than in the female. No certain
answer can be given to these questions; but we ought
to be cautious in assuming that knobs and various
fleshy appendages cannot be attractive to the female,
when we remember that with savage races of man
130various hideous deformities=-deep scars on the face
with the flesh raised into protuberances, the septum
of the nose pierced by sticks or bones, holes in the ears
and lips stretched widely open—are all admired as
ornamental.
Whether or not unimportant differences between the
sexes, such as those just specified, have been preserved
through sexual selection, these differences, as well as
all others, must primarily depend on the laws of variation.
On the principle of correlated development, the
plumage often varies on different parts of the body, or
over the whole body, in the same manner. We see
this well illustrated in certain breeds of the fowl. In
all the breeds the feathers on the neck and loins of
the males are elongated, and are called hackles; now
when both sexes acquire a top-knot, which is a new
character in the genus, the feathers on the head of the
male become hackle-shaped, evidently on the principle
of correlation; whilst those on the head of the female
are of the ordinary shape. The colour also of the
hackles forming the top-knot of the male, is often correlated
with that of the hackles on the neck and loins,
as may be seen by comparing these feathers in the
Golden and Silver-spangled Polish, the Houdans, and
Crève-cœur breeds. In some natural species we may
observe exactly the same correlation in the colours of
these same feathers, as in the males of the splendid
Golden and Amherst pheasants.
The structure of each individual feather generally
causes any change in its colouring to be symmetrical;
we see this in the various laced, spangled, and pencilled
breeds of the fowl; and on the principle of
correlation the feathers over the whole body are often
modified in the same manner. We are thus enabled
without much trouble to rear breeds with their plum131age
marked and coloured almost as symmetrically
as in natural species. In laced and spangled fowls
the coloured margins of the feathers are abruptly
defined; but in a mongrel raised by me from a black
Spanish cock glossed with green and a white game
hen, all the feathers were greenish-black, excepting
towards their extremities, which were yellowish-white;
but between the white extremities and the black
bases, there was on each feather a symmetrical, curved
zone of dark-brown. In some instances the shaft of
the feather determines the distribution of the tints;
thus with the body-feathers of a mongrel from the
same black Spanish cock and a silver-spangled Polish
hen, the shaft, together with a narrow space on each
side, was greenish-black, and this was surrounded by
a regular zone of dark-brown, edged with brownish-white.
In these cases we see feathers becoming symmetrically
shaded, like those which give so much
elegance to the plumage of many natural species. I
have also noticed a variety of the common pigeon
with the wing-bars symmetrically zoned with three
bright shades, instead of being simply black on a slaty-blue
ground, as in the parent-species.
In many large groups of birds it may be observed
that the plumage is differently coloured in each species,
yet that certain spots, marks, or stripes, though likewise
differently coloured, are retained by all the species.
Analogous cases occur with the breeds of the pigeon,
which usually retain the two wing-bars, though they
may be coloured red, yellow, white, black, or blue, the
rest of the plumage being of some wholly different tint.
Here is a more curious case, in which certain marks
are retained, though coloured in almost an exactly
reversed manner to what is natural; the aboriginal
pigeon has a blue tail, with the terminal halves of the132
outer webs of the two outer tail-feathers white; now
there is a sub-variety having a white instead of a blue
tail, with precisely that small part black which is white
in the parent-species.191
Formation and variability of the Ocelli or eye-like
Spots on the Plumage of Birds.—As no ornaments are
more beautiful than the ocelli on the feathers of various
birds, on the hairy coats of some mammals, on the
scales of reptiles and fishes, on the skin of amphibians,
on the wings of many Lepidoptera and other insects,
they deserve to be especially noticed. An ocellus consists
of a spot within a ring of another colour, like the
pupil within the iris, but the central spot is often surrounded
by additional concentric zones. The ocelli on
the tail-coverts of the peacock offer a familiar example,
as well as those on the wings of the peacock-butterfly
(Vanessa). Mr. Trimen has given me a description of
a S. African moth (Gynanisa Isis), allied to our Emperor
moth, in which a magnificent ocellus occupies nearly the
whole surface of each hinder wing; it consists of a black
centre, including a semi-transparent crescent-shaped
mark, surrounded by successive ochre-yellow, black,
ochre-yellow, pink, white, pink, brown, and whitish zones.
Although we do not know the steps by which these
wonderfully beautiful and complex ornaments have been
developed, the process at least with insects has probably
been a simple one; for, as Mr. Trimen writes to me,
“no characters of mere marking or coloration are so
unstable in the Lepidoptera as the ocelli, both in
number and size.” Mr. Wallace, who first called my
attention to this subject, shewed me a series of specimens
of our common meadow-brown butterfly (Hip133parchia Janira) exhibiting numerous gradations from
a simple minute black spot to an elegantly-shaded
ocellus. In a S. African butterfly (Cyllo Leda
belonging to the same family, the ocelli are even still
more variable. In some specimens (A, fig. 52) large
spaces on the upper surface of the wings are coloured
black, and include irregular white marks; and from
this state a complete gradation can be traced into a
tolerably perfect (A1) ocellus, and this results from the
contraction of the irregular blotches of colour. In
another series of specimens a gradation can be followed
from excessively minute white dots, surrounded by a
scarcely visible black line (B), into perfectly symmetrical
and large ocelli (B1).192 In cases like these, the
134development of a perfect ocellus does not require a
long course of variation and selection.
Fig. 52. Cyllo leda, Linn., from a drawing by Mr. Trimen, shewing the extreme range
of variation in the ocelli.
| A. Specimen, from Mauritius, upper surface of fore-wing |
B. Specimen, from Java, upper surface of hind-wing. |
| A1. Specimen, from Natal, ditto. |
B1. Specimen, from Mauritius, ditto. |
With birds and many other animals it seems, from
the comparison of allied species, to follow, that circular
spots are often generated by the breaking up
and contraction of stripes. In the Tragopan pheasant
faint white lines in the female represent the beautiful
white spots in the male;193 and something of the
same kind may be observed in the two sexes of the
Argus pheasant. However this may be, appearances
strongly favour the belief that, on the one hand, a dark
spot is often formed by the colouring-matter being
drawn towards a central point from a surrounding
zone, which is thus rendered lighter. And, on the other
hand, that a white spot is often formed by the colour
being driven away from a central point, so that it accumulates
in a surrounding darker zone. In either case
an ocellus is the result. The colouring matter seems
to be a nearly constant quantity, but is redistributed,
either centripetally or centrifugally. The feathers of
the common guinea-fowl offer a good instance of white
spots surrounded by darker zones; and wherever the
white spots are large and stand near each other, the
surrounding dark zones become confluent. In the same
wing-feather of the Argus pheasant dark spots may
be seen surrounded by a pale zone, and white spots
by a dark zone. Thus the formation of an ocellus
in its simplest state appears to be a simple affair.
By what further steps the more complex ocelli, which
135are surrounded by many successive zones of colour,
have been generated, I will not pretend to say. But
bearing in mind the zoned feathers of the mongrel
offspring from differently-coloured fowls, and the extraordinary
variability of the ocelli in many Lepidoptera,
the formation of these beautiful ornaments can hardly
be a highly complex process, and probably depends on
some slight and graduated change in the nature of the
tissues.
Gradation of Secondary Sexual Characters.—Cases
of gradation are important for us, as they shew that
it is at least possible that highly complex ornaments
may have been acquired by small successive steps.
In order to discover the actual steps by which the
male of any existing bird has acquired his magnificent
colours or other ornaments, we ought to behold
the long line of his ancient and extinct progenitors;
but this is obviously impossible. We may, however,
generally gain a clue by comparing all the species of
a group, if it be a large one; for some of them will
probably retain, at least in a partial manner, traces of
their former characters. Instead of entering on tedious
details respecting various groups, in which striking
instances of gradation could be given, it seems the best
plan to take some one or two strongly-characterised
cases, for instance that of the peacock, in order to discover
if any light can thus be thrown on the steps by
which this bird has become so splendidly decorated.
The peacock is chiefly remarkable from the extraordinary
length of his tail-coverts; the tail itself not
being much elongated. The barbs along nearly the
whole length of these feathers stand separate or are
decomposed; but this is the case with the feathers
of many species, and with some varieties of the136
domestic fowl and pigeon. The barbs coalesce towards
the extremity of the shaft to form the oval disc or
ocellus, which is certainly one of the most beautiful
objects in the world. This consists of an iridescent,
intensely blue, indented centre, surrounded by a rich
green zone, and this by a broad coppery-brown zone,
and this by five other narrow zones of slightly-different
iridescent shades. A trifling character in the disc perhaps
deserves notice; the barbs, for a space along one
of the concentric zones are destitute, to a greater or
less degree, of their barbules, so that a part of the disc
is surrounded by an almost transparent zone, which
gives to it a highly-finished aspect. But I have elsewhere
described194 an exactly analogous variation in the
hackles of a sub-variety of the game-cock, in which
the tips, having a metallic lustre, “are separated from
the lower part of the feather by a symmetrically-shaped
transparent zone, composed of the naked portions
of the barbs.” The lower margin or base of
the dark-blue centre of the ocellus is deeply indented
on the line of the shaft. The surrounding zones likewise
shew traces, as may be seen in the drawing
(fig. 53), of indentations, or rather breaks. These indentations
are common to the Indian and Javan peacocks
(Pavo cristatus and P. muticus); and they seemed
to me to deserve particular attention, as probably connected
with the development of the ocellus; but for a
long time I could not conjecture their meaning.
If we admit the principle of gradual evolution, there
must formerly have existed many species which presented
every successive step between the wonderfully
elongated tail-coverts of the peacock and the short tail-coverts
137of all ordinary birds; and again between the
magnificent ocelli of the former, and the simpler ocelli
or mere coloured spots of other birds; and so with
all the other characters of the peacock. Let us look
to the allied Gallinaceæ for any still-existing gradations.
The species and sub-species of Polyplectron
inhabit countries adjacent to the native land of the
peacock; and they so far resemble this bird that they
are sometimes called peacock-pheasants. I am also
informed by Mr. Bartlett that they resemble the peacock
in their voice and in some of their habits. During
138the spring the males, as previously described, strut
about before the comparatively plain-coloured females,
expanding and erecting their tail and wing-feathers,
which are ornamented with numerous ocelli. I request
the reader to turn back to the drawing (fig. 51, p. 90)
of a Polyplectron. In P. Napoleonis the ocelli are
confined to the tail, and the back is of a rich metallic
blue, in which respects this species approaches
the Java peacock. P. Hardwickii possesses a peculiar
top-knot, somewhat like that of this same kind
of peacock. The ocelli on the wings and tail of the
several species of Polyplectron are either circular
or oval, and consist of a beautiful, iridescent, greenish-blue
or greenish-purple disc, with a black border.
This border in P. chinquis shades into brown which
is edged with cream-colour, so that the ocellus is
here surrounded with differently, though not brightly,
shaded concentric zones. The unusual length of the
tail-coverts is another highly remarkable character
in Polyplectron; for in some of the species they are
half as long, and in others two-thirds of the length
of the true tail-feathers. The tail-coverts are ocellated,
as in the peacock. Thus the several species
of Polyplectron manifestly make a graduated approach
in the length of their tail-coverts, in the zoning
of the ocelli, and in some other characters, to the
peacock.
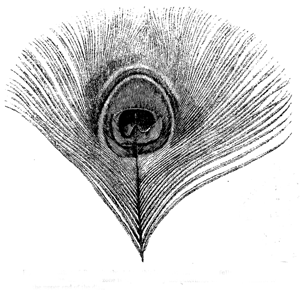
Fig. 53. Feather of Peacock, about two-thirds of natural size, carefully drawn by Mr.
Ford. The transparent zone is represented by the outermost white zone, confined to
the upper end of the disc.
Notwithstanding this approach, the first species of
Polyplectron which I happened to examine almost made
me give up the search; for I found not only that the
true tail-feathers, which in the peacock are quite plain,
were ornamented with ocelli, but that the ocelli on
all the feathers differed fundamentally from those of
the peacock, in there being two on the same feather,
(fig. 54), one on each side of the shaft. Hence I139
concluded that the early progenitors of the peacock
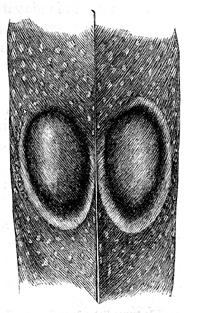 Fig. 54. Part of a tail-covert of Polyplectron
chinquis, with two oval ocelli
of nat. size.
Fig. 54. Part of a tail-covert of Polyplectron
chinquis, with two oval ocelli
of nat. size.
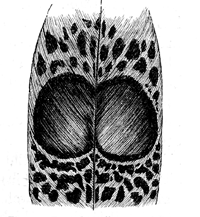 Fig. 55. Part of a tail-covert of Polyplectron
malaccense, with the two
oval ocelli, partially confluent, of nat. size.
could not have resembled in
any degree a Polyplectron.
But on continuing my search,
I observed that in some of
the species the two ocelli
stood very near each other;
that in the tail-feathers of
P. Hardwickii they touched
each other; and, finally, that
in the tail-coverts of this same
species as well as of P. malaccense
(fig. 55) they were
actually confluent. As the
central part alone is confluent,
an indentation is left at both
the upper and lower ends;
and the surrounding coloured zones
zones are likewise indented.
Fig. 55. Part of a tail-covert of Polyplectron
malaccense, with the two
oval ocelli, partially confluent, of nat. size.
could not have resembled in
any degree a Polyplectron.
But on continuing my search,
I observed that in some of
the species the two ocelli
stood very near each other;
that in the tail-feathers of
P. Hardwickii they touched
each other; and, finally, that
in the tail-coverts of this same
species as well as of P. malaccense
(fig. 55) they were
actually confluent. As the
central part alone is confluent,
an indentation is left at both
the upper and lower ends;
and the surrounding coloured zones
zones are likewise indented.
A single ocellus is thus
formed on each tail-covert,
though still plainly betraying
its double origin. These confluent
ocelli differ from the
single ocelli of the peacock
in having an indentation at
both ends, instead of at the
lower or basal end alone. The
explanation, however, of this
difference is not difficult; in
some species of Polyplectron
the two oval ocelli on the
same feather stand parallel
to each other; in other species (as in P. chinquis) they
converge towards one end; now the partial confluence
of two convergent ocelli would manifestly leave a much140
deeper indentation at the divergent than at the convergent
end. It is also manifest that if the convergence
were strongly pronounced and the confluence complete,
the indentation at the convergent end would tend to be
quite obliterated.
The tail-feathers in both species of peacock are entirely
destitute of ocelli, and this apparently is related
to their being covered up and concealed by the long
tail-coverts. In this respect they differ remarkably from
the tail-feathers of Polyplectron, which in most of the
species are ornamented with larger ocelli than those on
the tail-coverts. Hence I was led carefully to examine
the tail-feathers of the several species of Polyplectron
in order to discover whether the ocelli in any of them
shewed any tendency to disappear, and, to my great
satisfaction, I was successful. The central tail-feathers
of P. Napoleonis have the two ocelli on each side of the
shaft perfectly developed; but the inner ocellus becomes
less and less conspicuous on the more exterior tail-feathers,
until a mere shadow or rudimentary vestige is
left on the inner side of the outermost feather. Again,
in P. malaccense, the ocelli on the tail-coverts are, as we
have seen, confluent; and these feathers are of unusual
length, being two-thirds of the length of the tail-feathers,
so that in both these respects they resemble the
tail-coverts of the peacock. Now in this species the two
central tail-feathers alone are ornamented, each with two
brightly-coloured ocelli, the ocelli having completely
disappeared from the inner sides of all the other tail-feathers.
Consequently the tail-coverts and tail-feathers
of this species of Polyplectron make a near approach
in structure and ornamentation to the corresponding
feathers of the peacock.
As far, then, as the principle of gradation throws
light on the steps by which the magnificent train of
the peacock has been acquired, hardly anything more141
is needed. We may picture to ourselves a progenitor
of the peacock in an almost exactly intermediate condition
between the existing peacock, with his enormously
elongated tail-coverts, ornamented with single
ocelli, and an ordinary gallinaceous bird with short
tail-coverts, merely spotted with some colour; and we
shall then see in our mind’s eye, a bird possessing
tail-coverts, capable of erection and expansion, ornamented
with two partially confluent ocelli, and long
enough almost to conceal the tail-feathers,—the latter
having already partially lost their ocelli; we shall
see in short, a Polyplectron. The indentation of the
central disc and surrounding zones of the ocellus in both
species of peacock, seems to me to speak plainly in
favour of this view; and this structure is otherwise inexplicable.
The males of Polyplectron are no doubt very
beautiful birds, but their beauty, when viewed from a
little distance, cannot be compared, as I formerly saw
in the Zoological Gardens, with that of the peacock.
Many female progenitors of the peacock must, during
a long line of descent, have appreciated this superiority;
for they have unconsciously, by the continued preference
of the most beautiful males, rendered the peacock
the most splendid of living birds.
Argus pheasant.—Another excellent case for investigation
is offered by the ocelli on the wing-feathers of
the Argus pheasant, which are shaded in so wonderful a
manner as to resemble balls lying within sockets, and
which consequently differ from ordinary ocelli. No one,
I presume, will attribute the shading, which has excited
the admiration of many experienced artists, to chance—to
the fortuitous concourse of atoms of colouring
matter. That these ornaments should have been formed
through the selection of many successive variations, not
one of which was originally intended to produce the142
ball-and-socket effect, seems as incredible, as that one
of Raphael’s Madonnas should have been formed by
the selection of chance daubs of paint made by a
long succession of young artists, not one of whom intended
at first to draw the human figure. In order to
discover how the ocelli have been developed, we cannot
look to a long line of progenitors, nor to various
closely-allied forms, for such do not now exist. But
fortunately the several feathers on the wing suffice
to give us a clue to the problem, and they prove to
demonstration that a gradation is at least possible from
a mere spot to a finished ball-and-socket ocellus.
The wing-feathers, bearing the ocelli, are covered with
dark stripes or rows of dark spots, each stripe or row
running obliquely down the outer side of the shaft to
an ocellus. The spots are generally elongated in a
transverse line to the row in which they stand. They
often become confluent, either in the line of the row—and
then they form a longitudinal stripe—or transversely,
that is, with the spots in the adjoining rows,
and then they form transverse stripes. A spot sometimes
breaks up into smaller spots, which still stand in
their proper places.
It will be convenient first to describe a perfect ball-and-socket
ocellus. This consists of an intensely black
circular ring, surrounding a space shaded so as exactly
to resemble a ball. The figure here given has been
admirably drawn by Mr. Ford, and engraved, but a woodcut
cannot exhibit the exquisite shading of the original.
The ring is almost always slightly broken or interrupted
(see fig. 56) at a point in the upper half, a little to the
right of and above the white shade on the enclosed
ball; it is also sometimes broken towards the base on
the right hand. These little breaks have an important
meaning. The ring is always much thickened, with the
edges ill-defined towards the left-hand upper corner143
the feather being held erect, in the position in which it
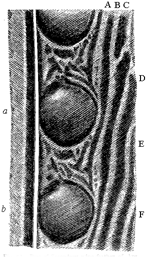 Fig. 56. Part of Secondary wing-feather of Argus
pheasant, shewing two, a and b, perfect ocelli.
A, B, C, &c., dark stripes running obliquely down,
each to an ocellus.
Fig. 56. Part of Secondary wing-feather of Argus
pheasant, shewing two, a and b, perfect ocelli.
A, B, C, &c., dark stripes running obliquely down,
each to an ocellus.
[Much of the web on both sides, especially to the
left of the shaft, has been cut off.]
is here drawn. Beneath
this thickened
part there is on the
surface of the ball an
oblique almost pure-white
mark, which
shades off downwards
into a pale-leaden hue,
and this into yellowish
and brown tints,
which insensibly become
darker and darker
towards the lower
part of the ball. It
is this shading which
gives so admirably the
effect of light shining
on a convex surface.
If one of the balls be
examined, it will be
seen that the lower
part is of a browner
tint and is indistinctly
separated by a curved
oblique line from the
upper part, which is
yellower and more leaden; this oblique line runs at
right angles to the longer axis of the white patch of
light, and indeed of all the shading; but this difference
in the tints, which cannot of course be shewn in the
woodcut, does not in the least interfere with the perfect
shading of the ball.195 It should be particularly ob144served
that each ocellus stands in obvious connection
with a dark stripe, or row of dark spots, for both occur
indifferently on the same feather. Thus in fig. 56 stripe
A runs to ocellus a; B runs to ocellus b; stripe C is
broken in the upper part, and runs down to the next
succeeding ocellus, not represented in the woodcut; D
to the next lower one, and so with the stripes E and F.
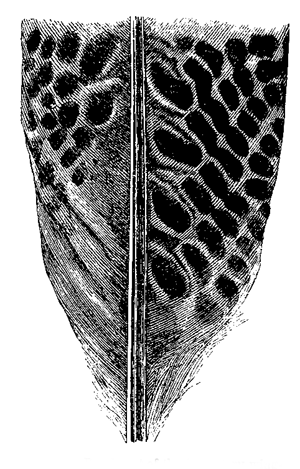 Fig. 57. Basal part of the Secondary wing-feather,
nearest to the body.
Lastly, the several ocelli
are separated from each
other by a pale surface
bearing irregular black
marks.
Fig. 57. Basal part of the Secondary wing-feather,
nearest to the body.
Lastly, the several ocelli
are separated from each
other by a pale surface
bearing irregular black
marks.
I will next describe
the other extreme of the
series, namely the first
trace of an ocellus. The
short secondary wing-feather
(fig. 57), nearest
to the body, is marked
like the other feathers,
with oblique, longitudinal,
rather irregular, rows
of spots. The lowest spot,
or that nearest the shaft,
in the five lower rows (excluding
the basal row) is
a little larger than the
other spots in the same row, and a little more elon145gated
in a transverse direction. It differs also from
the other spots by being bordered on its upper side
with some dull fulvous shading. But this spot is not in
any way more remarkable than those on the plumage
of many birds, and might easily be quite overlooked.
The next higher spot in each row does not differ at all
from the upper ones in the same row, although in
the following series it becomes, as we shall see, greatly
modified. The larger spots occupy exactly the same
relative position on this feather as those occupied by the
perfect ocelli on the longer wing-feathers.
By looking to the next two or three succeeding
secondary wing-feathers, an absolutely insensible gradation
can be traced from one of the above-described
lower spots, together with the next higher one in the
same row, to a curious ornament, which cannot be called
an ocellus, and which I will name, from the want of a
better term, an “elliptic ornament.” These are shewn
in the accompanying figure (fig. 58). We here see
several oblique rows, A, B, C, D (see the lettered diagram),
&c., of dark spots of the usual character. Each
row of spots runs down to and is connected with one of
the elliptic ornaments, in exactly the same manner as
each stripe in fig. 56 runs down to, and is connected with,
one of the ball-and-socket ocelli. Looking to any one
row, for instance, B, the lowest spot or mark (b) is
thicker and considerably longer than the upper spots,
and has its left extremity pointed and curved upwards.
This black mark is abruptly bordered on its upper side
by a rather broad space of richly-shaded tints, beginning
with a narrow brown zone, which passes into orange,
and this into a pale leaden tint, with the end towards
the shaft much paler. This mark corresponds in every
respect with the larger, shaded spot, described in the
last paragraph (fig. 57), but is more highly deve146loped
and more brightly coloured. To the right and
above this spot (b), with its bright shading, there is a
long, narrow, black mark (c), belonging to the same
row, and which is arched a little downwards so as to
face (b). It is also narrowly edged on the lower side
with a fulvous tint. To the left of and above c, in the
same oblique direction, but always more or less distinct
from it, there is another black mark (d). This mark is
generally sub-triangular and irregular in shape, but in
the one lettered in the diagram is unusually narrow,
elongated, and regular. It apparently consists of a
lateral and broken prolongation of the mark (c), as I147
infer from traces of similar prolongations from the
succeeding upper spots; but I do not feel sure of this.
These three marks, b, c, and d, with the intervening
bright shades, form together the so-called elliptic ornament.
These ornaments stand in a line parallel to
the shaft, and manifestly correspond in position with the
ball-and-socket ocelli. Their extremely elegant appearance
cannot be appreciated in the drawing, as the orange
and leaden tints, contrasting so well with the black
marks, cannot be shewn.
Fig. 58. Portion of one of the Secondary wing-feathers near to the body; shewing the
so-called elliptic ornaments. The right-hand figure is given merely as a diagram
for the sake of the letters of reference.
| A, B, C, &c. Rows of spots running down
to and forming the elliptic ornaments. |
c. The next succeeding spot or mark in
the same row. |
| b. Lowest spot or mark in row B. |
d. Apparently a broken prolongation of
the spot c in the same row B. |
Between one of the elliptic ornaments and a perfect
ball-and-socket ocellus, the gradation is so perfect that
it is scarcely possible to decide when the latter term
ought to be used. I regret that I have not given an
additional drawing, besides fig. 58, which stands about
half-way in the series between one of the simple spots
and a perfect ocellus. The passage from the elliptic
ornament into an ocellus is effected by the elongation
and greater curvature in opposed directions of the lower
black mark (b), and more especially of the upper one
(c), together with the contraction of the irregular sub-triangular
or narrow mark (d), so that at last these
three marks become confluent, forming an irregular
elliptic ring. This ring is gradually rendered more
and more circular and regular, at the same time increasing
in diameter. Traces of the junction of all
three elongated spots or marks, especially of the two
upper ones, can still be observed in many of the most
perfect ocelli. The broken state of the black ring on
the upper side of the ocellus in fig. 56 was pointed
out. The irregular sub-triangular or narrow mark
(d) manifestly forms, by its contraction and equalisation,
the thickened portion of the ring on the left
upper side of the perfect ball-and-socket ocellus. The
lower part of the ring is invariably a little thicker than148
the other parts (see fig. 56), and this follows from the
lower black mark of the elliptic ornament (b) having
been originally thicker than the upper mark (c). Every
step can be followed in the process of confluence and
modification; and the black ring which surrounds the ball
of the ocellus is unquestionably formed by the union
and modification of the three black marks, b, c, d, of the
elliptic ornament. The irregular zigzag black marks
between the successive ocelli (see again fig. 56) are
plainly due to the breaking up of the somewhat more
regular but similar marks between the elliptic ornaments.
The successive steps in the shading of the ball-and-socket
ocelli can be followed out with equal clearness.
The brown, orange, and pale-leaden narrow zones which
border the lower black mark of the elliptic ornament
can be seen gradually to become more and more softened
and shaded into each other, with the upper lighter part
towards the left-hand corner rendered still lighter, so as
to become almost white. But even in the most perfect
ball-and-socket ocelli a slight difference in the tints,
though not in the shading, between the upper and lower
parts of the ball can be perceived (as was before especially
noticed), the line of separation being oblique,
in the same direction with the bright coloured shades
of the elliptic ornaments. Thus almost every minute
detail in the shape and colouring of the ball-and-socket
ocelli can be shewn to follow from gradual changes in
the elliptic ornaments; and the development of the
latter can be traced by equally small steps from the
union of two almost simple spots, the lower one (fig. 57)
having some dull fulvous shading on the upper side.
The extremities of the longer secondary feathers
which bear the perfect ball-and-socket ocelli are peculiarly
ornamented. (Fig. 59.) The oblique longitudinal149
stripes suddenly cease upwards and become confused,
and above this limit the whole upper end of the feather
(a) is covered with white dots, surrounded by little
black rings, standing on a dark ground. Even the
 Fig. 59. Portion near summit of one of
the Secondary wing-feathers, bearing
perfect ball-and-socket ocelli.
Fig. 59. Portion near summit of one of
the Secondary wing-feathers, bearing
perfect ball-and-socket ocelli.
a. Ornamented upper part.
b. Uppermost, imperfect ball-and-socket
ocellus. (The shading above the
white mark on the summit of the
ocellus is here a little too dark.)
c. Perfect ocellus.
oblique stripe belonging to
the uppermost ocellus (b)
is represented only by a
very short irregular black
mark with the usual, curved,
transverse base. As this
stripe is thus abruptly cut
off above, we can understand,
from what has gone before,
how it is that the upper
thickened part of the ring is
absent in the uppermost ocellus;
for, as before stated, this
thickened part is apparently
formed by a broken prolongation
of the next higher
spot in the same row. From
the absence of the upper
and thickened part of the
ring, the uppermost ocellus,
though perfect in all other
respects, appears as if its top
had been obliquely sliced off.
It would, I think, perplex
any one, who believes that
the plumage of the Argus pheasant
was created as we
now see it, to account for the
imperfect condition of the
uppermost ocelli. I should add that in the secondary
wing-feather farthest from the body all the ocelli are150
smaller and less perfect than on the other feathers,
with the upper parts of the external black rings deficient,
as in the case just mentioned. The imperfection
here seems to be connected with the fact that the
spots on this feather shew less tendency than usual to
become confluent into stripes; on the contrary, they are
often broken up into smaller spots, so that two or three
rows run down to each ocellus.
We have now seen that a perfect series can be followed,
from two almost simple spots, at first quite
distinct from each other, to one of the wonderful ball-and-socket
ornaments. Mr. Gould, who kindly gave me
some of these feathers, fully agrees with me in the completeness
of the gradation. It is obvious that the stages
in development exhibited by the feathers on the same
bird do not at all necessarily shew us the steps which
have been passed through by the extinct progenitors of
the species; but they probably give us the clue to the
actual steps, and they at least prove to demonstration
that a gradation is possible. Bearing in mind
how carefully the male Argus pheasant displays his
plumes before the female, as well as the many facts
rendering it probable that female birds prefer the more
attractive males, no one who admits the agency of
sexual selection, will deny that a simple dark spot with
some fulvous shading might be converted, through
the approximation and modification of the adjoining
spots, together with some slight increase of colour,
into one of the so-called elliptic ornaments. These
latter ornaments have been shewn to many persons,
and all have admitted that they are extremely pretty,
some thinking them even more beautiful than the
ball-and-socket ocelli. As the secondary plumes became
lengthened through sexual selection, and as
the elliptic ornaments increased in diameter, their151
colours apparently became less bright; and then the
ornamentation of the plumes had to be gained by
improvements in the pattern and shading; and this
process has been carried on until the wonderful ball-and-socket
ocelli have been finally developed. Thus we
can understand—and in no other way as it seems to
me—the present condition and origin of the ornaments
on the wing-feathers of the Argus pheasant.
From the light reflected by the principle of gradation;
from what we know of the laws of variation;
from the changes which have taken place in many
of our domesticated birds; and, lastly, from the character
(as we shall hereafter more clearly see) of the
immature plumage of young birds—we can sometimes
indicate with a certain amount of confidence, the probable
steps by which the males have acquired their
brilliant plumage and various ornaments; yet in many
cases we are involved in darkness. Mr. Gould several
years ago pointed out to me a humming-bird, the
Urosticte benjamini, remarkable from the curious differences
presented by the two sexes. The male, besides
a splendid gorget, has greenish-black tail-feathers, with
the four central ones tipped with white; in the female,
as with most of the allied species, the three outer tail-feathers
on each side are tipped with white, so that the
male has the four central, whilst the female has the six
exterior feathers ornamented with white tips. What
makes the case curious is that, although the colouring
of the tail differs remarkably in both sexes of many
kinds of humming-birds, Mr. Gould does not know a
single species, besides the Urosticte, in which the male
has the four central feathers tipped with white.
The Duke of Argyll, in commenting on this case,196
152passes over sexual selection, and asks, “What explanation
does the law of natural selection give of such
specific varieties as these?” He answers “none
whatever;” and I quite agree with him. But can
this be so confidently said of sexual selection? Seeing
in how many ways the tail-feathers of humming-birds
differ, why should not the four central feathers have
varied in this one species alone, so as to have acquired
white tips? The variations may have been gradual, or
somewhat abrupt as in the case recently given of the
humming-birds near Bogota, in which certain individuals
alone have the “central tail-feathers tipped
with beautiful green.” In the female of the Urosticte
I noticed extremely minute or rudimental white
tips to the two outer of the four central black tail-feathers;
so that here we have an indication of change
of some kind in the plumage of this species. If we grant
the possibility of the central tail-feathers of the male
varying in whiteness, there is nothing strange in such
variations having been sexually selected. The white
tips, together with the small white ear-tufts, certainly
add, as the Duke of Argyll admits, to the beauty of the
male; and whiteness is apparently appreciated by other
birds, as may be inferred from such cases as the snow-white
male of the Bell-bird. The statement made by
Sir E. Heron should not be forgotten, namely that his
peahens, when debarred from access to the pied peacock,
would not unite with any other male, and during that
season produced no offspring. Nor is it strange that
variations in the tail-feathers of the Urosticte should
have been specially selected for the sake of ornament,
for the next succeeding genus in the family takes its
name of Metallura from the splendour of these feathers.
Mr. Gould, after describing the peculiar plumage of the
Urosticte, adds, “that ornament and variety is the sole153
object, I have myself but little doubt.”197 If this be
admitted, we can perceive that the males which were
decked in the most elegant and novel manner would
have gained an advantage, not in the ordinary struggle
for life, but in rivalry with other males, and would
consequently have left a larger number of offspring to
inherit their newly-acquired beauty.
154
CHAPTER XV.
Birds—continued.
Discussion why the males alone of some species, and both sexes of
other species, are brightly coloured—On sexually-limited inheritance,
as applied to various structures and to brightly-coloured
plumage—Nidification in relation to colour—Loss of
nuptial plumage during the winter.
We have in this chapter to consider, why with many
kinds of birds the female has not received the same
ornaments as the male; and why with many others,
both sexes are equally, or almost equally, ornamented?
In the following chapter we shall consider why in some
few rare cases the female is more conspicuously coloured
than the male.
In my ‘Origin of Species’198 I briefly suggested that
the long tail of the peacock would be inconvenient, and
the conspicuous black colour of the male capercailzie
dangerous, to the female during the period of incubation;
and consequently that the transmission of these characters
from the male to the female offspring had been
checked through natural selection. I still think that
this may have occurred in some few instances: but after
mature reflection on all the facts which I have been
able to collect, I am now inclined to believe that
when the sexes differ, the successive variations have
generally been from the first limited in their transmission
to the same sex in which they first appeared. Since
my remarks appeared, the subject of sexual coloration
155has been discussed in some very interesting papers by
Mr. Wallace,199 who believes that in almost all cases the
successive variations tended at first to be transmitted
equally to both sexes; but that the female was saved,
through natural selection, from acquiring the conspicuous
colours of the male, owing to the danger which she
would thus have incurred during incubation.
This view necessitates a tedious discussion on a
difficult point, namely whether the transmission of a
character, which is at first inherited by both sexes, can
be subsequently limited in its transmission, by means
of selection, to one sex alone. We must bear in mind,
as shewn in the preliminary chapter on sexual selection,
that characters which are limited in their development
to one sex are always latent in the other.
An imaginary illustration will best aid us in seeing
the difficulty of the case: we may suppose that a
fancier wished to make a breed of pigeons, in which
the males alone should be coloured of a pale blue,
whilst the females retained their former slaty tint. As
with pigeons characters of all kinds are usually transmitted
to both sexes equally, the fancier would have
to try to convert this latter form of inheritance into
sexually-limited transmission. All that he could do
would be to persevere in selecting every male pigeon
which was in the least degree of a paler blue; and the
natural result of this process, if steadily carried on for
a long time, and if the pale variations were strongly
inherited or often recurred, would be to make his whole
stock of a lighter blue. But our fancier would be compelled
to match, generation after generation, his pale
blue males with slaty females, for he wishes to keep the
156latter of this colour. The result would generally be
the production either of a mongrel piebald lot, or more
probably the speedy and complete loss of the pale-blue
colour, for the primordial slaty tint would be transmitted
with prepotent force. Supposing, however, that
some pale-blue males and slaty females were produced
during each successive generation, and were always
crossed together; then the slaty females would have,
if I may use the expression, much blue blood in their
veins, for their fathers, grandfathers, etc., will all have
been blue birds. Under these circumstances it is conceivable
(though I know of no distinct facts rendering
it probable) that the slaty females might acquire so
strong a latent tendency to pale-blueness, that they
would not destroy this colour in their male offspring,
their female offspring still inheriting the slaty tint. If
so, the desired end of making a breed with the two
sexes permanently different in colour might be gained.
The extreme importance, or rather necessity, of the
desired character in the above case, namely, pale-blueness,
being present though in a latent state in the
female, so that the male offspring should not be deteriorated,
will be best appreciated as follows: the male
of Sœmmerring’s pheasant has a tail thirty-seven inches
in length, whilst that of the female is only eight inches;
the tail of the male common pheasant is about twenty
inches, and that of the female twelve inches long. Now
if the female Sœmmerring pheasant with her short tail
were crossed with the male common pheasant, there
can be no doubt that the male hybrid offspring would
have a much longer tail than that of the pure offspring of
the common pheasant. On the other hand, if the female
common pheasant, with her tail nearly twice as long as
that of the female Sœmmerring pheasant, were crossed
with the male of the latter, the male hybrid offspring157
would have a much shorter tail than that of the pure
offspring of Sœmmerring’s pheasant.200
Our fancier, in order to make his new breed with the
males of a decided pale-blue tint, and the females unchanged,
would have to continue selecting the males
during many generations; and each stage of paleness
would have to be fixed in the males, and rendered
latent in the females. The task would be an extremely
difficult one, and has never been tried, but might possibly
succeed. The chief obstacle would be the early
and complete loss of the pale-blue tint, from the necessity
of reiterated crosses with the slaty female, the
latter not having at first any latent tendency to produce
pale-blue offspring.
On the other hand, if one or two males were to vary
ever so slightly in paleness, and the variations were
from the first limited in their transmission to the male
sex, the task of making a new breed of the desired
kind would be easy, for such males would simply have
to be selected and matched with ordinary females. An
analogous case has actually occurred, for there are
breeds of the pigeon in Belgium201 in which the males
alone are marked with black striæ. In the case of the
fowl, variations of colour limited in their transmission to
the male sex habitually occur. Even when this form of
inheritance prevails, it might well happen that some
of the successive steps in the process of variation might
be transferred to the female, who would then come to
resemble in a slight degree the male, as occurs in some
breeds of the fowl. Or again, the greater number, but
158not all, of the successive steps might be transferred to
both sexes, and the female would then closely resemble
the male. There can hardly be a doubt that this is
the cause of the male pouter pigeon having a somewhat
larger crop, and of the male carrier pigeon having somewhat
larger wattles, than their respective females; for
fanciers have not selected one sex more than the other,
and have had no wish that these characters should be
more strongly displayed in the male than in the female,
yet this is the case with both breeds.
The same process would have to be followed, and the
same difficulties would be encountered, if it were desired
to make a breed with the females alone of some new
colour.
Lastly, our fancier might wish to make a breed with
the two sexes differing from each other, and both from
the parent-species. Here the difficulty would be extreme,
unless the successive variations were from the
first sexually limited on both sides, and then there would
be no difficulty. We see this with the fowl; thus the
two sexes of the pencilled Hamburghs differ greatly
from each other, and from the two sexes of the aboriginal
Gallus bankiva; and both are now kept constant
to their standard of excellence by continued selection,
which would be impossible unless the distinctive characters
of both were limited in their transmission. The
Spanish fowl offers a more curious case; the male has
an immense comb, but some of the successive variations,
by the accumulation of which it was acquired, appear
to have been transferred to the female; for she has a
comb many times larger than that of the females of the
parent-species. But the comb of the female differs in
one respect from that of the male, for it is apt to lop
over; and within a recent period it has been ordered
by the fancy that this should always be the case, and159
success has quickly followed the order. Now the lopping
of the comb must be sexually limited in its transmission,
otherwise it would prevent the comb of the
male from being perfectly upright, which would be
abhorrent to every fancier. On the other hand the
uprightness of the comb in the male must likewise be
a sexually-limited character, otherwise it would prevent
the comb of the female from lopping over.
From the foregoing illustrations, we see that even
with almost unlimited time at command, it would be
an extremely difficult and complex process, though
perhaps not impossible, to change through selection
one form of transmission into the other. Therefore,
without distinct evidence in each case, I am unwilling
to admit that this has often been effected with natural
species. On the other hand by means of successive
variations, which were from the first sexually limited
in their transmission, there would not be the least
difficulty in rendering a male bird widely different in
colour or in any other character from the female; the
latter being left unaltered, or slightly altered, or specially
modified for the sake of protection.
As bright colours are of service to the males in their
rivalry with other males, such colours would be selected,
whether or not they were transmitted exclusively to the
same sex. Consequently the females might be expected
often to partake of the brightness of the males to a
greater or less degree; and this occurs with a host of
species. If all the successive variations were transmitted
equally to both sexes, the females would be
undistinguishable from the males; and this likewise
occurs with many birds. If, however, dull colours were
of high importance for the safety of the female during
incubation, as with many ground birds, the females
which varied in brightness, or which received through160
inheritance from the males any marked accession of
brightness, would sooner or later be destroyed. But
the tendency in the males to continue for an indefinite
period transmitting to their female offspring their own
brightness, would have to be eliminated by a change in
the form of inheritance; and this, as shewn by our
previous illustration, would be extremely difficult. The
more probable result of the long-continued destruction
of the more brightly-coloured females, supposing the
equal form of transmission to prevail, would be the lessening
or annihilation of the bright colours of the males,
owing to their continually crossing with the duller
females. It would be tedious to follow out all the
other possible results; but I may remind the reader, as
shewn in the eighth chapter, that if sexually-limited
variations in brightness occurred in the females, even if
they were not in the least injurious to them and consequently
were not eliminated, yet they would not be
favoured or selected, for the male usually accepts any
female, and does not select the more attractive individuals;
consequently these variations would be liable
to be lost, and would have little influence on the
character of the race; and this will aid in accounting
for the females being commonly less brightly-coloured
than the males.
In the chapter just referred to, instances were given,
and any number might have been added, of variations
occurring at different ages, and inherited at the same
age. It was also shewn that variations which occur late
in life are commonly transmitted to the same sex in
which they first appeared; whilst variations occurring
early in life are apt to be transmitted to both sexes;
not that all the cases of sexually-limited transmission
can thus be accounted for. It was further shewn that
if a male bird varied by becoming brighter whilst161
young, such variations would be of no service until
the age for reproduction had arrived, and there was
competition between rival males. If we suppose that
three-fourths of the young males of any species are
on an average destroyed by various enemies; then the
chances would be as three to one against any one individual
more brightly-coloured than usual surviving to
propagate its kind. But in the case of birds which
live on the ground and which commonly need the
protection of dull colours, bright tints would be far
more dangerous to the young and inexperienced than
to the adult males. Consequently the males which
varied in brightness whilst young would suffer much
destruction and be eliminated through natural selection;
on the other hand the males which varied in
this manner when nearly mature, notwithstanding that
they were exposed to some additional danger, might survive,
and from being favoured through sexual selection,
would procreate their kind. The brightly-coloured young
males being destroyed and the mature ones being successful
in their courtship, may account, on the principle
of a relation existing between the period of variation
and the form of transmission, for the males alone of
many birds, having acquired and transmitted brilliant
colours to their male offspring alone. But I by no means
wish to maintain that the influence of age on the form
of transmission is indirectly the sole cause of the great
difference in brilliancy between the sexes of many birds.
As with all birds in which the sexes differ in colour, it
is an interesting question whether the males alone have
been modified through sexual selection, the females
being left, as far as this agency is concerned, unchanged
or only partially changed; or whether the females have
been specially modified through natural selection for the
sake of protection, I will discuss this question at con162siderable
length, even at greater length than its intrinsic
importance deserves; for various curious collateral points
may thus be conveniently considered.
Before we enter on the subject of colour, more
especially in reference to Mr. Wallace’s conclusions,
it may be useful to discuss under a similar point of
view some other differences between the sexes. A
breed of fowls formerly existed in Germany202 in which
the hens were furnished with spurs; they were good
layers, but they so greatly disturbed their nests with
their spurs that they could not be allowed to sit on their
own eggs. Hence at one time it appeared to me probable
that with the females of the wild Gallinaceæ
the development of spurs had been checked through
natural selection, from the injury thus caused to their
nests. This seemed all the more probable as the wing-spurs,
which could not be injurious during nidification,
are often as well developed in the female as in the
male; though in not a few cases they are rather larger
in the male. When the male is furnished with leg-spurs
the female almost always exhibits rudiments of
them,—the rudiment sometimes consisting of a mere
scale, as with the species of Gallus. Hence it might
be argued that the females had aboriginally been furnished
with well-developed spurs, but that these had
subsequently been lost either through disuse or natural
selection. But if this view be admitted, it would have
to be extended to innumerable other cases; and it implies
that the female progenitors of the existing spur-bearing
species were once encumbered with an injurious
appendage.
In some few genera and species, as in Galloperdix,
Acomus, and the Javan peacock (Pavo muticus), the
163females, as well as the males, possess well-developed
spurs. Are we to infer from this fact that they construct
a different sort of nest, not liable to be injured
by their spurs, from that made by their nearest allies,
so that there has been no need for the removal of their
spurs? Or are we to suppose that these females especially
require spurs for their defence? It is a more probable
conclusion that both the presence and absence of
spurs in the females result from different laws of inheritance
having prevailed, independently of natural selection.
With the many females in which spurs appear
as rudiments, we may conclude that some few of the
successive variations, through which they were developed
in the males, occurred very early in life, and were
as a consequence transferred to the females. In the
other and much rarer cases, in which the females possess
fully developed spurs, we may conclude that all
the successive variations were transferred to them; and
that they gradually acquired the inherited habit of not
disturbing their nests.
The vocal organs and the variously-modified feathers
for producing sound, as well as the proper instincts
for using them, often differ in the two sexes, but are
sometimes the same in both. Can such differences be
accounted for by the males having acquired these organs
and instincts, whilst the females have been saved from
inheriting them, on account of the danger to which
they would have been exposed by attracting the attention
of birds or beasts of prey? This does not
seem to me probable, when we think of the multitude
of birds which with impunity gladden the country with
their voices during the spring.203 It is a safer conclu164sion
that as vocal and instrumental organs are of special
service only to the males during their courtship, these
organs were developed through sexual selection and
continued use in this sex alone—the successive variations
and the effects of use having been from the first
limited in their transmission in a greater or less degree
to the male offspring.
Many analogous cases could be advanced; for instance
the plumes on the head, which are generally
longer in the male than in the female, sometimes of
equal length in both sexes, and occasionally absent in
the female,—these several cases sometimes occurring
in the same group of birds. It would be difficult to
account for a difference of this kind between the sexes
on the principle of the female having been benefited by
possessing a slightly shorter crest than the male, and its
consequent diminution or complete suppression through
natural selection. But I will take a more favourable
case, namely, the length of the tail. The long train
of the peacock would have been not only inconvenient
but dangerous to the peahen during the period of incubation
and whilst accompanying her young. Hence
there is not the least à priori improbability in the
development of her tail having been checked through
natural selection. But the females of various pheasants,
which apparently are exposed on their open nests
to as much danger as the peahen, have tails of considerable
length. The females as well as the males
of the Menura superba have long tails, and they build
a domed nest, which is a great anomaly in so large a
bird. Naturalists have wondered how the female Menura
could manage her tail during incubation; but it
165is now known204 that she “enters the nest head first,
and then turns round with her tail sometimes over
her back, but more often bent round by her side.
Thus in time the tail becomes quite askew, and is a
tolerable guide to the length of time the bird has
been sitting.” Both sexes of an Australian kingfisher
(Tanysiptera sylvia) have the middle tail-feathers greatly
lengthened; and as the female makes her nest in a
hole, these feathers become, as I am informed by Mr.
R. B. Sharpe, much crumpled during nidification.
In these two cases the great length of the tail-feathers
must be in some degree inconvenient to the female;
and as in both species the tail-feathers of the female
are somewhat shorter than those of the male, it might
be argued that their full development had been prevented
through natural selection. Judging from these
cases, if with the peahen, the development of the tail
had been checked only when it became inconveniently
or dangerously long, she would have acquired a much
longer tail than she actually possesses; for her tail is
not nearly so long, relatively to the size of her body,
as that of many female pheasants, nor longer than that
of the female turkey. It must also be borne in mind,
that in accordance with this view as soon as the tail of
the peahen became dangerously long, and its development
was consequently checked, she would have continually
reacted on her male progeny, and thus have
prevented the peacock from acquiring his present magnificent
train. We may therefore infer that the length
of the tail in the peacock and its shortness in the peahen
are the result of the requisite variations in the
male having been from the first transmitted to the male
offspring alone.
166
We are led to a nearly similar conclusion with
respect to the length of the tail in the various species
of pheasants. In the Eared pheasant (Crossoptilon
auritum) the tail is of equal length in both sexes,
namely, sixteen or seventeen inches; in the common
pheasant it is about twenty inches long in the male,
and twelve in the female; in Sœmmerring’s pheasant,
thirty-seven inches in the male, and only eight in the
female; and lastly in Reeve’s pheasant it is sometimes
actually seventy-two inches long in the male and sixteen
in the female. Thus in the several species, the
tail of the female differs much in length, irrespectively
of that of the male; and this can be accounted for
as it seems to me, with much more probability, by the
laws of inheritance,—that is by the successive variations
having been from the first more or less closely
limited in their transmission to the male sex,—than by
the agency of natural selection, owing to the length of
tail having been injurious in a greater or less degree
to the females of the several species.
We may now consider Mr. Wallace’s arguments, in
regard to the sexual coloration of birds. He believes
that the bright tints originally acquired through sexual
selection by the males, would in all or almost all cases
have been transmitted to the females, unless the transference
had been checked through natural selection.
I may here remind the reader that various facts
bearing on this view have already been given under
reptiles, amphibians, fishes, and lepidoptera. Mr.
Wallace rests his belief chiefly, but not exclusively, as
we shall see in the next chapter, on the following statement,205
that when both sexes are coloured in a strikingly-167conspicuous
manner the nest is of such a nature as to
conceal the sitting bird; but when there is a marked
contrast of colour between the sexes, the male being
gay and the female dull-coloured, the nest is open and
exposes the sitting bird to view. This coincidence,
as far as it goes, certainly supports the belief that the
females which sit on open nests have been specially
modified for the sake of protection. Mr. Wallace
admits that there are, as might have been expected,
some exceptions to his two rules, but it is a question
whether the exceptions are not so numerous as seriously
to invalidate them.
There is in the first place much truth in the Duke
of Argyll’s remark206 that a large domed nest is more
conspicuous to an enemy, especially to all tree-haunting
carnivorous animals, than a smaller open nest. Nor
must we forget that with many birds which build open
nests the males sit on the eggs and aid in feeding the
young as well as the females: this is the case, for instance,
with Pyranga æstiva,207 one of the most splendid
birds in the United States, the male being vermilion,
and the female light brownish-green. Now if brilliant
colours had been extremely dangerous to birds whilst
sitting on their open nests, the males in these cases
would have suffered greatly. It might, however, be of
such paramount importance to the male to be brilliantly
coloured, in order to beat his rivals, that this
would more than compensate for some additional danger.
Mr. Wallace admits that with the King-crows (Dicrurus),
Orioles, and Pittidæ, the females are conspicuously
coloured, yet they build open nests; but he
urges that the birds of the first group are highly pug168nacious
and could defend themselves; that those of the
second group take extreme care in concealing their
open nests, but this does not invariably hold good;208
and that with the birds of the third group the females
are brightly coloured chiefly on the under surface. Besides
these cases the whole great family of pigeons,
which are sometimes brightly, and almost always conspicuously
coloured, and which are notoriously liable to
the attacks of birds of prey, offers a serious exception
to the rule, for pigeons almost always build open and
exposed nests. In another large family, that of the
Humming-birds, all the species build open nests, yet
with some of the most gorgeous species the sexes are
alike; and in the majority, the females, though less
brilliant than the males, are very brightly coloured.
Nor can it be maintained that all female humming-birds,
which are brightly coloured, escape detection by
their tints being green, for some display on their upper
surfaces red, blue, and other colours.209
In regard to birds which build in holes or construct
domed nests, other advantages, as Mr. Wallace remarks,
besides concealment are gained, such as shelter from
the rain, greater warmth, and in hot countries protection
from the rays of the sun;210 so that it is no valid
169objection to his view that many birds having both sexes
obscurely coloured build concealed nests.211 The female
Hornbills (Buceros), for instance, of India and Africa
are protected, during nidification, with extraordinary
care, for the male plaisters up the hole in which the
female sits on her eggs, and leaves only a small orifice
through which he feeds her; she is thus kept a close
prisoner during the whole period of incubation;212 yet
female hornbills are not more conspicuously coloured
than many other birds of equal size which build open
nests. It is a more serious objection to Mr. Wallace’s
view, as is admitted by him, that in some few groups the
males are brilliantly coloured and the females obscure,
and yet the latter hatch their eggs in domed nests.
This is the case with the Grallinæ of Australia, the
Superb Warblers (Maluridæ) of the same country,
the Sun-birds (Nectariniæ), and with several of the
Australian Honey-suckers or Meliphagidæ.213
If we look to the birds of England we shall see that
there is no close and general relation between the
colours of the female and the nature of the nest constructed
by her. About forty of our British birds (excluding
those of large size which could defend themselves)
build in holes in banks, rocks, or trees, or construct
domed nests. If we take the colours of the
female goldfinch, bullfinch, or blackbird, as a standard
of the degree of conspicuousness, which is not highly
dangerous to the sitting female, then out of the above
forty birds, the females of only twelve can be considered
170as conspicuous to a dangerous degree, the remaining
twenty-eight being inconspicuous.214 Nor is there any
close relation between a well-pronounced difference in
colour between the two sexes, and the nature of the
nest constructed. Thus the male house-sparrow (Passer
domesticus) differs much from the female, the male
tree-sparrow (P. montanus) differs hardly at all, and yet
both build well-concealed nests. The two sexes of the
common fly-catcher (Muscicapa grisola) can hardly be
distinguished, whilst the sexes of the pied fly-catcher
(M. luctuosa) differ considerably, and both build in holes.
The female blackbird (Turdus merula) differs much,
the female ring-ouzel (T. torquatus) differs less,
and the female common thrush (T. musicus) hardly
at all from their respective males; yet all build open
nests. On the other hand, the not very distantly-allied
water-ouzel (Cinclus aquaticus) builds a domed
nest, and the sexes differ about as much as in the case
of the ring-ouzel. The black and red grouse (Tetrao
tetrix and T. Scoticus) build open nests, in equally well-concealed
spots, but in the one species the sexes differ
greatly, and in the other very little.
Notwithstanding the foregoing objections, I cannot
doubt, after reading Mr. Wallace’s excellent essay,
171that looking to the birds of the world, a large majority
of the species in which the females are conspicuously
coloured (and in this case the males with rare exceptions
are equally conspicuous), build concealed nests for the
sake of protection. Mr. Wallace enumerates215 a long
series of groups in which this rule holds good; but it
will suffice here to give, as instances, the more familiar
groups of kingfishers, toucans, trogons, puff-birds (Capitonidæ),
plaintain-eaters (Musophagæ), woodpeckers, and
parrots. Mr. Wallace believes that in these groups,
as the males gradually acquired through sexual selection
their brilliant colours, these were transferred to
the females and were not eliminated by natural selection,
owing to the protection which they already enjoyed
from their manner of nidification. According to this
view, their present manner of nesting was acquired
before their present colours. But it seems to me
much more probable that in most cases as the females
were gradually rendered more and more brilliant from
partaking of the colours of the male, they were gradually
led to change their instincts (supposing that they
originally built open nests), and to seek protection by
building domed or concealed nests. No one who studies,
for instance, Audubon’s account of the differences in the
nests of the same species in the Northern and Southern
United States,216 will feel any great difficulty in admitting
that birds, either by a change (in the strict sense
of the word) of their habits, or through the natural
selection of so-called spontaneous variations of instinct,
might readily be led to modify their manner of
nesting.
172
This way of viewing the relation, as far as it holds
good, between the bright colours of female birds and
their manner of nesting, receives some support from
certain analogous cases occurring in the Sahara Desert.
Here, as in most other deserts, various birds, and many
other animals, have had their colours adapted in a wonderful
manner to the tints of the surrounding surface.
Nevertheless there are, as I am informed by the Rev.
Mr. Tristram, some curious exceptions to the rule; thus
the male of the Monticola cyanea is conspicuous from
his bright blue colour, and the female almost equally
conspicuous from her mottled brown and white plumage;
both sexes of two species of Dromolæa are of a lustrous
black; so that these three birds are far from receiving
protection from their colours, yet they are able to survive,
for they have acquired the habit, when in danger,
of taking refuge in holes or crevices in the rocks.
With respect to the above-specified groups of birds,
in which the females are conspicuously coloured and
build concealed nests, it is not necessary to suppose
that each separate species had its nidifying instinct
specially modified; but only that the early progenitors
of each group were gradually led to build domed or
concealed nests; and afterwards transmitted this instinct,
together with their bright colours, to their modified
descendants. This conclusion, as far as it can be
trusted, is interesting, namely, that sexual selection,
together with equal or nearly equal inheritance by both
sexes, have indirectly determined the manner of nidification
of whole groups of birds.
Even in the groups in which, according to Mr. Wallace,
the females from being protected during nidification,
have not had their bright colours eliminated
through natural selection, the males often differ in a
slight, and occasionally in a considerable degree, from173
the females. This is a significant fact, for such differences
in colour must be accounted for on the principle
of some of the variations in the males having been from
the first limited in their transmission to the same sex;
as it can hardly be maintained that these differences,
especially when very slight, serve as a protection to
the female. Thus all the species in the splendid group
of the Trogons build in holes; and Mr. Gould gives
figures217 of both sexes of twenty-five species, in all of
which, with one partial exception, the sexes differ sometimes
slightly, sometimes conspicuously, in colour,—the
males being always more beautiful than the females,
though the latter are likewise beautiful. All the
species of kingfisher build in holes, and with most of
the species the sexes are equally brilliant, and thus far
Mr. Wallace’s rule holds good; but in some of the
Australian species the colours of the females are rather
less vivid than those of the male; and in one splendidly-coloured
species, the sexes differ so much that
they were at first thought to be specifically distinct.218
Mr. R. B. Sharpe, who has especially studied this
group, has shewn me some American species (Ceryle)
in which the breast of the male is belted with
black. Again, in Carcineutes, the difference between
the sexes is conspicuous: in the male the upper surface
is dull-blue banded with black, the lower surface
being partly fawn-coloured, and there is much red
about the head; in the female the upper surface is
reddish-brown banded with black, and the lower surface
white with black markings. It is an interesting fact,
as shewing how the same peculiar style of sexual
174colouring often characterises allied forms, that in three
species of Dacelo the male differs from the female only
in the tail being dull-blue banded with black, whilst
that of the female is brown with blackish bars; so that
here the tail differs in colour in the two sexes in exactly
the same manner as the whole upper surface in the
sexes of Carcineutes.
With parrots, which likewise build in holes, we find
analogous cases: in most of the species both sexes are
brilliantly coloured and undistinguishable, but in not a
few species the males are coloured rather more vividly
than the females, or even very differently from them.
Thus, besides other strongly-marked differences, the
whole under surface of the male King Lory (Aprosmictus
scapulatus) is scarlet, whilst the throat and chest of the
female is green tinged with red: in the Euphema splendida
there is a similar difference, the face and wing-coverts
moreover of the female being of a paler blue
than in the male.219 In the family of the tits (PARINÆ),
which build concealed nests, the female of our common
blue tomtit (Parus cæruleus) is “much less brightly
coloured” than the male; and in the magnificent Sultan
yellow tit of India the difference is greater.220
Again in the great group of the woodpeckers,221 the
sexes are generally nearly alike, but in the Megapicus
validus all those parts of the head, neck, and
breast, which are crimson in the male are pale brown
in the female. As in several woodpeckers the head of
the male is bright crimson, whilst that of the female is
175plain, it occurred to me that this colour might possibly
make the female dangerously conspicuous, whenever
she put her head out of the hole containing her nest,
and consequently that this colour, in accordance with
Mr. Wallace’s belief, had been eliminated. This view is
strengthened by what Malherbe states with respect to
Indopicus carlotta; namely, that the young females,
like the young males, have some crimson about their
heads, but that this colour disappears in the adult
female, whilst it is intensified in the adult male. Nevertheless
the following considerations render this view
extremely doubtful: the male takes a fair share in
incubation,222 and would be thus far almost equally
exposed to danger; both sexes of many species have
their heads of an equally bright crimson; in other
species the difference between the sexes in the amount
of scarlet is so slight that it can hardly make any
appreciable difference in the danger incurred; and
lastly, the colouring of the head in the two sexes
often differs slightly in other ways.
The cases, as yet given, of slight and graduated
differences in colour between the males and females
in the groups, in which as a general rule the sexes
resemble each other, all relate to species which build
domed or concealed nests. But similar gradations may
likewise be observed in groups in which the sexes
as a general rule resemble each other, but which build
open nests. As I have before instanced the Australian
parrots, so I may here instance, without giving any
details, the Australian pigeons.223 It deserves especial
notice that in all these cases the slight differences in
176plumage between the sexes are of the same general
nature as the occasionally greater differences. A good
illustration of this fact has already been afforded by
those kingfishers in which either the tail alone or
the whole upper surface of the plumage differs in the
same manner in the two sexes. Similar cases may be
observed with parrots and pigeons. The differences in
colour between the sexes of the same species are, also,
of the same general nature as the differences in colour
between the distinct species of the same group. For
when in a group in which the sexes are usually alike,
the male differs considerably from the female, he is
not coloured in a quite new style. Hence we may
infer that within the same group the special colours of
both sexes when they are alike, and the colours of the
male, when he differs slightly or even considerably from
the female, have in most cases been determined by the
same general cause; this being sexual selection.
It is not probable, as has already been remarked,
that differences in colour between the sexes, when very
slight, can be of service to the female as a protection.
Assuming, however, that they are of service, they might
be thought to be cases of transition; but we have no
reason to believe that many species at any one time
are undergoing change. Therefore we can hardly
admit that the numerous females which differ very
slightly in colour from their males are now all commencing
to become obscure for the sake of protection.
Even if we consider somewhat more marked sexual differences,
is it probable, for instance, that the head of the
female chaffinch, the crimson on the breast of the female
bullfinch,—the green of the female greenfinch,—the
crest of the female golden-crested wren, have all been
rendered less bright by the slow process of selection for
the sake of protection? I cannot think so; and still less177
with the slight differences between the sexes of those
birds which build concealed nests. On the other hand,
the differences in colour between the sexes, whether
great or small, may to a large extent be explained
on the principle of the successive variations, acquired
by the males through sexual selection, having been
from the first more or less limited in their transmission
to the females. That the degree of limitation should
differ in different species of the same group will not
surprise any one who has studied the laws of inheritance,
for they are so complex that they appear to us in our
ignorance to be capricious in their action.224
As far as I can discover there are very few groups
of birds containing a considerable number of species,
in which all have both sexes brilliantly coloured
and alike; but this appears to be the case, as I hear
from Mr. Sclater, with the Musophagæ or plaintain-eaters.
Nor do I believe that any large group
exists in which the sexes of all the species are widely
dissimilar in colour: Mr. Wallace informs me that
the chatterers of S. America (COTINGIDÆ) offer one of
the best instances; but with some of the species, in
which the male has a splendid red breast, the female
exhibits some red on her breast; and the females of
other species shew traces of the green and other colours
of the males. Nevertheless we have a near approach
to close sexual similarity or dissimilarity throughout
several groups: and this, from what has just been said
of the fluctuating nature of inheritance, is a somewhat
surprising circumstance. But that the same
laws should largely prevail with allied animals is
not surprising. The domestic fowl has produced a
178great number of breeds and sub-breeds, and in these
the sexes generally differ in plumage; so that it has
been noticed as a remarkable circumstance when in
certain sub-breeds they resemble each other. On the
other hand, the domestic pigeon has likewise produced
a vast number of distinct breeds and sub-breeds, and
in these, with rare exceptions, the two sexes are identically
alike. Therefore if other species of Gallus and
Columba were domesticated and varied, it would not be
rash to predict that the same general rules of sexual
similarity and dissimilarity, depending on the form of
transmission, would, in both cases, hold good. In a
similar manner the same form of transmission has generally
prevailed throughout the same natural groups,
although marked exceptions to this rule occur. Within
the same family or even genus, the sexes may be
identically alike or very different in colour. Instances
have already been given relating to the same genus,
as with sparrows, fly-catchers, thrushes and grouse. In
the family of pheasants the males and females of almost
all the species are wonderfully dissimilar, but are quite
similar in the eared pheasant or Crossoptilon auritum.
In two species of Chloephaga, a genus of geese, the
males cannot be distinguished from the females, except
by size; whilst in two others, the sexes are so unlike
that they might easily be mistaken for distinct species.225
The laws of inheritance can alone account for the
following cases, in which the female by acquiring at
a late period of life certain characters proper to the
male, ultimately comes to resemble him in a more or
less complete manner. Here protection can hardly
have come into play. Mr. Blyth informs me that
the females of Oriolus melanocephalus and of some
179allied species, when sufficiently mature to breed, differ
considerably in plumage from the adult males; but
after the second or third moults they differ only in
their beaks having a slight greenish tinge. In the
dwarf bitterns (Ardetta), according to the same authority,
“the male acquires his final livery at the
first moult, the female not before the third or fourth
moult; in the meanwhile she presents an intermediate
garb, which is ultimately exchanged for the
same livery as that of the male.” So again the
female Falco peregrinus acquires her blue plumage
more slowly than the male. Mr. Swinhoe states that
with one of the Drongo shrikes (Dicrurus macrocercus)
the male whilst almost a nestling, moults his soft
brown plumage and becomes of a uniform glossy
greenish-black; but the female retains for a long time
the white striæ and spots on the axillary feathers;
and does not completely assume the uniform black
colour of the male for the first three years. The same
excellent observer remarks that in the spring of the
second year the female spoonbill (Platalea) of China resembles
the male of the first year, and that apparently
it is not until the third spring that she acquires the
same adult plumage as that possessed by the male at a
much earlier age. The female Bombycilla carolinensis
differs very little from the male, but the appendages,
which like beads of red sealing-wax ornament the wing-feathers,
are not developed in her so early in life as in
the male. The upper mandible in the male of an Indian
parrakeet (Palæornis Javanicus) is coral-red from his
earliest youth, but in the female, as Mr. Blyth has
observed with caged and wild birds, it is at first black
and does not become red until the bird is at least a year
old, at which age the sexes resemble each other in all
respects. Both sexes of the wild turkey are ultimately180
furnished with a tuft of bristles on the breast, but in
two-year-old birds the tuft is about four inches long in
the male and hardly apparent in the female; when,
however, the latter has reached her fourth year, it is
from four to five inches in length.226
In these cases, the females follow a normal course of
development in ultimately becoming like the males; and
such cases must not be confounded with those in which
diseased or old females assume masculine characters,
or with those in which perfectly fertile females, whilst
young, acquire through variation or some unknown cause
the characters of the male.227 But all these cases have
so much in common that they depend, according to the
hypothesis of pangenesis, on gemmules derived from each
part of the male being present, though latent, in the female;
their development following on some slight change
in the elective affinities of her constituent tissues.
A few words must be added on changes of plumage
in relation to the season of the year. From reasons
formerly assigned there can be little doubt that the
elegant plumes, long pendant feathers, crests, &c., of
egrets, herons, and many other birds, which are developed
and retained only during the summer, serve
exclusively for ornamental or nuptial purposes, though
181common to both sexes. The female is thus rendered
more conspicuous during the period of incubation than
during the winter; but such birds as herons and egrets
would be able to defend themselves. As, however,
plumes would probably be inconvenient and certainly
of no use during the winter, it is possible that the
habit of moulting twice in the year may have been
gradually acquired through natural selection for the
sake of casting off inconvenient ornaments during the
winter. But this view cannot be extended to the many
waders, in which the summer and winter plumages
differ very little in colour. With defenceless species,
in which either both sexes or the males alone become
extremely conspicuous during the breeding-season,—or
when the males acquire at this season such long
wing or tail-feathers as to impede their flight, as with
Cosmetornis and Vidua,—it certainly at first appears
highly probable that the second moult has been gained
for the special purpose of throwing off these ornaments.
We must, however, remember that many birds, such as
Birds of Paradise, the Argus pheasant and peacock, do
not cast their plumes during the winter; and it can
hardly be maintained that there is something in the
constitution of these birds, at least of the Gallinaceæ,
rendering a double moult impossible, for the ptarmigan
moults thrice in the year.228 Hence it must be considered
as doubtful whether the many species which
moult their ornamental plumes or lose their bright
colours during the winter, have acquired this habit on
account of the inconvenience or danger which they would
otherwise have suffered.
I conclude, therefore, that the habit of moulting
twice in the year was in most or all cases first acquired
182for some distinct purpose, perhaps for gaining a warmer
winter covering; and that variations in the plumage
occurring during the summer were accumulated through
sexual selection, and transmitted to the offspring at the
same season of the year. Such variations being inherited
either by both sexes or by the males alone, according
to the form of inheritance which prevailed. This
appears more probable than that these species in all
cases originally tended to retain their ornamental
plumage during the winter, but were saved from this
through natural selection, owing to the inconvenience
or danger thus caused.
I have endeavoured in this chapter to shew that the
arguments are not trustworthy in favour of the view
that weapons, bright colours, and various ornaments,
are now confined to the males owing to the conversion,
by means of natural selection, of a tendency to the equal
transmission of characters to both sexes into transmission
to the male sex alone. It is also doubtful whether
the colours of many female birds are due to the preservation,
for the sake of protection, of variations which
were from the first limited in their transmission to the
female sex. But it will be convenient to defer any
further discussion on this subject until I treat, in the
following chapter, on the differences in plumage between
the young and old.
183
CHAPTER XVI.
Birds—concluded.
The immature plumage in relation to the character of the plumage
in both sexes when adult—Six classes of cases—Sexual differences
between the males of closely-allied or representative species—The
female assuming the characters of the male—Plumage of
the young in relation to the summer and winter plumage of the
adults—On the increase of beauty in the Birds of the World—Protective
colouring—Conspicuously-coloured birds—Novelty
appreciated—Summary of the four chapters on Birds.
We must now consider the transmission of characters
as limited by age in reference to sexual selection.
The truth and importance of the principle of inheritance
at corresponding ages need not here be discussed,
as enough has already been said on the subject. Before
giving the several rather complex rules or classes of
cases, under which all the differences in plumage
between the young and the old, as far as known to
me, may be included, it will be well to make a few
preliminary remarks.
With animals of all kinds when the young differ in
colour from the adults, and the colours of the former are
not, as far as we can see, of any special service, they
may generally be attributed, like various embryological
structures, to the retention by the young of the character
of an early progenitor. But this view can be maintained
with confidence, only when the young of several species
closely resemble each other, and likewise resemble
other adult species belonging to the same group; for
the latter are the living proofs that such a state of
things was formerly possible. Young lions and pumas184
are marked with feeble stripes or rows of spots, and as
many allied species both young and old are similarly
marked, no naturalist, who believes in the gradual
evolution of species, will doubt that the progenitor of
the lion and puma was a striped animal, the young
having retained vestiges of the stripes, like the kittens
of black cats, which when grown up are not in the least
striped. Many species of deer, which when mature are
not spotted, are whilst young covered with white spots,
as are likewise some few species in their adult state.
So again the young in the whole family of pigs (Suidæ),
and in certain rather distantly-allied animals, such as
the tapir, are marked with dark longitudinal stripes;
but here we have a character apparently derived from
an extinct progenitor, and now preserved by the young
alone. In all such cases the old have had their colours
changed in the course of time, whilst the young have
remained but little altered, and this has been effected
through the principle of inheritance at corresponding
ages.
This same principle applies to many birds belonging
to various groups, in which the young closely resemble
each other, and differ much from their respective adult
parents. The young of almost all the Gallinaceæ, and
of some distantly-allied birds such as ostriches, are
whilst covered with down longitudinally striped; but
this character points back to a state of things so remote
that it hardly concerns us. Young crossbills
(Loxia) have at first straight beaks like those of other
finches, and in their immature striated plumage they
resemble the mature redpole and female siskin, as well
as the young of the goldfinch, greenfinch, and some
other allied species. The young of many kinds of
buntings (Emberiza) resemble each other, and likewise
the adult state of the common bunting, E. miliaria.185
In almost the whole large group of thrushes
the young have their breasts spotted—a character
which is retained by many species throughout life,
but is quite lost by others, as by the Turdus migratorius.
So again with many thrushes, the feathers on
the back are mottled before they are moulted for the
first time, and this character is retained for life by
certain eastern species. The young of many species of
shrikes (Lanius), of some woodpeckers, and of an Indian
pigeon (Chalcophaps indicus), are transversely striped
on the under surface; and certain allied species or
genera when adult are similarly marked. In some
closely-allied and resplendent Indian cuckoos (Chrysococcyx),
the species when mature differ considerably
from each other in colour, but the young cannot be distinguished.
The young of an Indian goose (Sarkidiornis
melanonotus) closely resemble in plumage an allied
genus, Dendrocygna, when mature.229 Similar facts will
hereafter be given in regard to certain herons. Young
black grouse (Tetrao tetrix) resemble the young as well
as the old of certain other species, for instance the red
grouse or T. scoticus. Finally, as Mr. Blyth, who has
attended closely to this subject, has well remarked, the
natural affinities of many species are best exhibited in
their immature plumage; and as the true affinities of
all organic beings depend on their descent from a
common progenitor, this remark strongly confirms the
belief that the immature plumage approximately shews
us the former or ancestral condition of the species.
186
Although many young birds belonging to various
orders thus give us a glimpse of the plumage of their
remote progenitors, yet there are many other birds, both
dull-coloured and bright-coloured, in which the young
closely resemble their parents. With such species the
young of the different species cannot resemble each other
more closely than do the parents; nor can they present
striking resemblances to allied forms in their adult
state. They give us but little insight into the plumage
of their progenitors, excepting in so far that when the
young and the old are coloured in the same general
manner throughout a whole group of species, it is probable
that their progenitors were similarly coloured.
We may now consider the classes of cases or rules
under which the differences and resemblances, between
the plumage of the young and the old, of both sexes or
of one sex alone, may be grouped. Rules of this kind
were first enounced by Cuvier; but with the progress
of knowledge they require some modification and amplification.
This I have attempted to do, as far as the
extreme complexity of the subject permits, from information
derived from various sources; but a full essay
on this subject by some competent ornithologist is
much needed. In order to ascertain to what extent
each rule prevails, I have tabulated the facts given in
four great works, namely, by Macgillivray on the birds
of Britain, Audubon on those of North America, Jerdon
on those of India, and Gould on those of Australia. I
may here premise, firstly, that the several cases or rules
graduate into each other; and secondly, that when the
young are said to resemble their parents, it is not
meant that they are identically alike, for their colours
are almost always rather less vivid, and the feathers
are softer and often of a different shape.
187
RULES OR CLASSES OF CASES.
I. When the adult male is more beautiful or conspicuous
than the adult female, the young of both sexes
in their first plumage closely resemble the adult female,
as with the common fowl and peacock; or, as occasionally
occurs, they resemble her much more closely than
they do the adult male.
II. When the adult female is more conspicuous than
the adult male, as sometimes though rarely occurs, the
young of both sexes in their first plumage resemble
the adult male.
III. When the adult male resembles the adult female,
the young of both sexes have a peculiar first plumage
of their own, as with the robin.
IV. When the adult male resembles the adult female,
the young of both sexes in their first plumage resemble
the adults, as with the kingfisher, many parrots, crows,
hedge-warblers.
V. When the adults of both sexes have a distinct
winter and summer plumage, whether or not the male
differs from the female, the young resemble the adults
of both sexes in their winter dress, or much more rarely
in their summer dress, or they resemble the females
alone; or the young may have an intermediate character;
or again they may differ greatly from the adults
in both their seasonal plumages.
VI. In some few cases the young in their first plumage
differ from each other according to sex; the young
males resembling more or less closely the adult males,
and the young females more or less closely the adult
females.
Class I.—In this class, the young of both sexes
resemble, more or less closely, the adult female, whilst
the adult male differs, often in the most conspicuous188
manner, from the adult female. Innumerable instances
in all Orders could be given; it will suffice to call to
mind the common pheasant, duck, and house-sparrow.
The cases under this class graduate into others. Thus
the two sexes when adult may differ so slightly, and the
young so slightly from the adults, that it is doubtful
whether such cases ought to come under the present, or
under the third or fourth classes. So again the young
of both sexes, instead of being quite alike, may differ
in a slight degree from each other, as in our sixth class.
These transitional cases, however, are few in number,
or at least are not strongly pronounced, in comparison
with those which come strictly under the present class.
The force of the present law is well shewn in those
groups, in which, as a general rule, the two sexes and
the young are all alike; for when the male in these
groups does differ from the female, as with certain parrots,
kingfishers, pigeons, &c., the young of both sexes
resemble the adult female.230 We see the same fact exhibited
still more clearly in certain anomalous cases;
thus the male of Heliothrix auriculata (one of the humming-birds)
differs conspicuously from the female in
having a splendid gorget and fine ear-tufts, but the
female is remarkable from having a much longer tail
than that of the male; now the young of both sexes
189resemble (with the exception of the breast being spotted
with bronze) the adult female in all respects including
the length of her tail, so that the tail of the male
actually becomes shorter as he reaches maturity, which
is a most unusual circumstance.231 Again, the plumage
of the male goosander (Mergus merganser) is more conspicuously
coloured, with the scapular and secondary
wing-feathers much longer than in the female, but differently
from what occurs, as far as I know, in any other
bird, the crest of the adult male, though broader than
that of the female, is considerably shorter, being only a
little above an inch in length; the crest of the female
being two and a half inches long. Now the young of
both sexes resemble in all respects the adult female,
so that their crests are actually of greater length though
narrower than in the adult male.232
When the young and the females closely resemble
each other and both differ from the male, the most obvious
conclusion is that the male alone has been modified.
Even in the anomalous cases of the Heliothrix
and Mergus, it is probable that originally both adult
sexes were furnished, the one species with a much elongated
tail, and the other with a much elongated crest,
these characters having since been partially lost by the
adult males from some unexplained cause, and transmitted
in their diminished state to their male offspring
alone, when arrived at the corresponding age of maturity.
The belief that in the present class the male
alone has been modified, as far as the differences between
the male and the female together with her
young are concerned, is strongly supported by some
190remarkable facts recorded by Mr. Blyth,233 with respect
to closely-allied species which represent each other in
distinct countries. For with several of these representative
species the adult males have undergone a certain
amount of change and can be distinguished; the
females and the young being undistinguishable, and
therefore absolutely unchanged. This is the case with
certain Indian chats (Thamnobia), with certain honey-suckers
(Nectarinia), shrikes (Tephrodornis), certain
kingfishers (Tanysiptera), Kallij pheasants (Gallophasis),
and tree-partridges (Arboricola).
In some analogous cases, namely with birds having
a distinct summer and winter plumage, but with the
two sexes nearly alike, certain closely-allied species
can easily be distinguished in their summer or nuptial
plumage, yet are undistinguishable in their winter as
well as in their immature plumage. This is the case
with some of the closely-allied Indian wagtails or Motacillæ.
Mr. Swinhoe234 informs me that three species of
Ardeola, a genus of herons, which represent each other
on separate continents, are “most strikingly different”
when ornamented with their summer plumes, but are
hardly, if at all, distinguishable during the winter. The
young also of these three species in their immature
plumage closely resemble the adults in their winter
dress. This case is all the more interesting because
with two other species of Ardeola both sexes retain,
during the winter and summer, nearly the same plum191age
as that possessed by the three first species during
the winter and in their immature state; and this plumage,
which is common to several distinct species at
different ages and seasons, probably shews us how the
progenitor of the genus was coloured. In all these
cases, the nuptial plumage which we may assume was
originally acquired by the adult males during the breeding-season,
and transmitted to the adults of both sexes
at the corresponding season, has been modified, whilst
the winter and immature plumages have been left unchanged.
The question naturally arises, how is it that in these
latter cases the winter plumage of both sexes, and in
the former cases the plumage of the adult females, as
well as the immature plumage of the young, have not
been at all affected? The species which represent each
other in distinct countries will almost always have
been exposed to somewhat different conditions, but we
can hardly attribute the modification of the plumage
in the males alone to this action, seeing that the
females and the young, though similarly exposed, have
not been affected. Hardly any fact in nature shews
us more clearly how subordinate in importance is the
direct action of the conditions of life, in comparison
with the accumulation through selection of indefinite
variations, than the surprising difference between the
sexes of many birds; for both sexes must have consumed
the same food and have been exposed to the
same climate. Nevertheless we are not precluded from
believing that in the course of time new conditions
may produce some direct effect; we see only that this
is subordinate in importance to the accumulated results
of selection. When, however, a species migrates into
a new country, and this must precede the formation of
representative species, the changed conditions to which192
they will almost always have been exposed will cause
them to undergo, judging from a widely-spread analogy,
a certain amount of fluctuating variability. In this
case sexual selection, which depends on an element
eminently liable to change—namely the taste or admiration
of the female—will have had new shades of colour
or other differences to act on and accumulate; and as
sexual selection is always at work, it would (judging
from what we know of the results on domestic animals
of man’s unintentional selection), be a surprising fact if
animals inhabiting separate districts, which can never
cross and thus blend their newly-acquired characters,
were not, after a sufficient lapse of time, differently
modified. These remarks likewise apply to the nuptial
or summer plumage, whether confined to the males or
common to both sexes.
Although the females of the above closely-allied
species, together with their young, differ hardly at all
from each other, so that the males alone can be distinguished,
yet in most cases the females of the species
within the same genus obviously differ from each other.
The differences, however, are rarely as great as between
the males. We see this clearly in the whole family of
the Gallinaceæ: the females, for instance, of the common
and Japan pheasant, and especially of the gold and
Amherst pheasant—of the silver pheasant and the wild
fowl—resemble each other very closely in colour, whilst
the males differ to an extraordinary degree. So it is
with the females of most of the Cotingidæ, Fringillidæ,
and many other families. There can indeed be no doubt
that, as a general rule, the females have been modified
to a less extent than the males. Some few birds,
however, offer a singular and inexplicable exception;
thus the females of Paradisea apoda and P. papuana
differ from each other more than do their respective193
males;235 the female of the latter species having the
under surface pure white, whilst the female P. apoda is
deep brown beneath. So, again, as I hear from Professor
Newton, the males of two species of Oxynotus (shrikes),
which represent each other in the islands of Mauritius
and Bourbon,236 differ but little in colour, whilst the
females differ much. In the Bourbon species the female
appears to have partially retained an immature condition
of plumage, for at first sight she “might be taken for
the young of the Mauritian species.” These differences
may be compared with those which occur, independently
of selection by man, and which we cannot explain, in
certain sub-breeds of the game-fowl, in which the females
are very different, whilst the males can hardly be distinguished.237
As I account so largely by sexual selection for the
differences between the males of allied species, how can
the differences between the females be accounted for
in all ordinary cases? We need not here consider the
species which belong to distinct genera; for with these,
adaptation to different habits of life, and other agencies,
will have come into play. In regard to the differences
between the females within the same genus, it appears to
me almost certain, after looking through various large
groups, that the chief agent has been the transference,
in a greater or less degree, to the female of the characters
acquired by the males through sexual selection.
In the several British finches, the two sexes differ either
very slightly or considerably; and if we compare the
females of the greenfinch, chaffinch, goldfinch, bullfinch,
crossbill, sparrow, &c., we shall see that they
194differ from each other chiefly in the points in which
they partially resemble their respective males; and the
colours of the males may safely be attributed to sexual
selection. With many gallinaceous species the sexes
differ to an extreme degree, as with the peacock, pheasant,
and fowl, whilst with other species there has been a
partial or even complete transference of character from
the male to the female. The females of the several
species of Polyplectron exhibit in a dim condition, and
chiefly on the tail, the splendid ocelli of their males.
The female partridge differs from the male only in the
red mark on her breast being smaller; and the female
wild turkey only in her colours being much duller. In
the guinea-fowl the two sexes are undistinguishable.
There is no improbability in the plain, though peculiar
spotted plumage of this latter bird having been
acquired through sexual selection by the males, and
then transmitted to both sexes; for it is not essentially
different from the much more beautifully-spotted
plumage, characteristic of the males alone of the Tragopan
pheasants.
It should be observed that, in some instances, the
transference of characters from the male to the female
has been effected apparently at a remote period, the
male having subsequently undergone great changes,
without transferring to the female any of his later-gained
characters. For instance, the female and the
young of the black-grouse (Tetrao tetrix) resemble
pretty closely both sexes and the young of the red-grouse
T. Scoticus; and we may consequently infer
that the black-grouse is descended from some ancient
species, of which both sexes were coloured in nearly
the same manner as the red-grouse. As both sexes of
this latter species are more plainly barred during the
breeding-season than at any other time, and as the male195
differs slightly from the female in his more strongly-pronounced
red and brown tints,238 we may conclude
that his plumage has been, at least to a certain extent,
influenced by sexual selection. If so, we may further
infer that the nearly similar plumage of the female
black-grouse was similarly produced at some former
period. But since this period the male black-grouse
has acquired his fine black plumage, with his forked and
outwardly-curled tail-feathers; but of these characters
there has hardly been any transference to the female,
excepting that she shews in her tail a trace of the curved
fork.
We may therefore conclude that the females of distinct
though allied species have often had their plumage
rendered more or less different by the transference in
various degrees, of characters acquired, both during
former and recent times, by the males through sexual
selection. But it deserves especial attention that
brilliant colours have been transferred much more
rarely than other tints. For instance, the male of
the red-throated bluebreast (Cyanecula suecica) has
a rich blue breast, including a sub-triangular red
mark; now marks of approximately the same shape
have been transferred to the female, but the central
space is fulvous instead of red, and is surrounded by
mottled instead of blue feathers. The Gallinaceæ offer
many analogous cases; for none of the species, such as
partridges, quails, guinea-fowls, &c., in which the colours
of the plumage have been largely transferred from the
male to the female, are brilliantly coloured. This is
well exemplified with the pheasants, in which the male
is generally so much more brilliant than the female;
but with the Eared and Cheer pheasants (Crossoptilon
196
auritum and Phasianus Wallichii) the two sexes closely
resemble each other and their colours are dull. We
may go so far as to believe that if any part of the
plumage in the males of these two pheasants had been
brilliantly coloured, this would not have been transferred
to the females. These facts strongly support Mr.
Wallace’s view that with birds which are exposed to
much danger during nidification, the transference of
bright colours from the male to the female has been
checked through natural selection. We must not,
however, forget that another explanation, before given,
is possible; namely, that the males which varied and
became bright, whilst they were young and inexperienced,
would have been exposed to much danger,
and would generally have been destroyed; the older
and more cautious males, on the other hand, if they
varied in a like manner, would not only have been able
to survive, but would have been favoured in their
rivalry with other males. Now variations occurring
late in life tend to be transmitted exclusively to the
same sex, so that in this case extremely bright tints
would not have been transmitted to the females. On
the other hand, ornaments of a less conspicuous kind,
such as those possessed by the Eared and Cheer pheasants,
would not have been dangerous, and if they appeared
during early youth, would generally have been
transmitted to both sexes.
In addition to the effects of the partial transference
of characters from the males to the females, some of the
differences between the females of closely-allied species
may be attributed to the direct or definite action of
the conditions of life.239 With the males any such
197action would generally have been masked by the
brilliant colours gained through sexual selection; but
not so with the females. Each of the endless diversities
in plumage, which we see in our domesticated
birds is, of course, the result of some definite cause;
and under natural and more uniform conditions, some
one tint, assuming that it was in no way injurious, would
almost certainly sooner or later prevail. The free
intercrossing of the many individuals belonging to the
same species would ultimately tend to make any change
of colour, thus induced, uniform in character.
No one doubts that both sexes of many birds have
had their colours adapted for the sake of protection;
and it is possible that the females alone of some species
may have been thus modified. Although it would be a
difficult, perhaps an impossible process, as shewn in the
last chapter, to convert through selection one form of
transmission into another, there would not be the least
difficulty in adapting the colours of the female, independently
of those of the male, to surrounding objects,
through the accumulation of variations which were from
the first limited in their transmission to the female sex.
If the variations were not thus limited, the bright tints of
the male would be deteriorated or destroyed. Whether
the females alone of many species have been thus
specially modified, is at present very doubtful. I wish
I could follow Mr. Wallace to the full extent; for the
admission would remove some difficulties. Any variations
which were of no service to the female as a protection
would be at once obliterated, instead of being lost
simply by not being selected, or from free intercrossing,
or from being eliminated when transferred to the male
and in any way injurious to him. Thus the plumage of
the female would be kept constant in character. It
would also be a relief if we could admit that the obscure198
tints of both sexes of many birds had been acquired and
preserved for the sake of protection,—for example, of
the hedge-warbler or kitty-wren (Accentor modularis and
Troglodytes vulgaris), with respect to which we have no
sufficient evidence of the action of sexual selection.
We ought, however, to be cautious in concluding that
colours which appear to us dull, are not attractive to the
females of certain species; we should bear in mind such
cases as that of the common house-sparrow, in which
the male differs much from the female, but does not
exhibit any bright tints. No one probably will dispute
that many gallinaceous birds which live on the open
ground have acquired their present colours, at least in
part, for the sake of protection. We know how well they
are thus concealed; we know that ptarmigans, whilst
changing from their winter to their summer plumage,
both of which are protective, suffer greatly from birds
of prey. But can we believe that the very slight differences
in tints and markings between, for instance,
the female black and red-grouse serve as a protection?
Are partridges, as they are now coloured, better protected
than if they had resembled quails? Do the
slight differences between the females of the common
pheasant, the Japan and golden pheasants, serve as a
protection, or might not their plumages have been
interchanged with impunity? From what Mr. Wallace
has observed of the habits of certain gallinaceous
birds in the East he thinks that such slight differences
are beneficial. For myself, I will only say that I am
not convinced.
Formerly when I was inclined to lay much stress on
the principle of protection, as accounting for the less
bright colours of female birds, it occurred to me that
possibly both sexes and the young might aboriginally
have been brightly coloured in an equal degree; but199
that subsequently, the females from the danger incurred
during incubation, and the young from being
inexperienced, had been rendered dull as a protection.
But this view is not supported by any evidence, and is
not probable; for we thus in imagination expose during
past times the females and the young to danger, from
which it has subsequently been necessary to shield their
modified descendants. We have, also, to reduce, through
a gradual process of selection, the females and the young
to almost exactly the same tints and markings, and to
transmit them to the corresponding sex and period of
life. It is also a somewhat strange fact, on the supposition
that the females and the young have partaken
during each stage of the process of modification of a
tendency to be as brightly coloured as the males, that the
females have never been rendered dull-coloured without
the young participating in the same change; for there
are no instances, as far as I can discover, of species with the
females dull-coloured and the young bright-coloured. A
partial exception, however, is offered by the young of certain
woodpeckers, for they have “the whole upper part
of the head tinged with red,” which afterwards either
decreases into a mere circular red line in the adults of
both sexes, or quite disappears in the adult females.240
Finally, with respect to our present class of cases,
the most probable view appears to be that successive
variations in brightness or in other ornamental characters,
occurring in the males at a rather late period of
life have alone been preserved; and that most or all
of these variations owing to the late period of life at
which they appeared, have been from the first transmitted
only to the adult male offspring. Any varia200tions
in brightness which occurred in the females or in
the young would have been of no service to them, and
would not have been selected; moreover, if dangerous,
would have been eliminated. Thus the females and the
young will either have been left unmodified, or, and
this has much more commonly occurred, will have been
partially modified by receiving through transference from
the males some of the successive variations. Both sexes
have perhaps been directly acted on by the conditions
of life to which they have long been exposed; but the
females from not being otherwise much modified will
best exhibit any such effects. These changes and all
others will have been kept uniform by the free intercrossing
of many individuals. In some cases, especially
with ground birds, the females and the young may possibly
have been modified, independently of the males,
for the sake of protection, so as to have acquired the
same dull-coloured plumage.
Class II. When the adult female is more conspicuous
than the adult male, the young of both sexes in their first
plumage resemble the adult male.—This class is exactly
the reverse of the last, for the females are here more
brightly coloured or more conspicuous than the males;
and the young, as far as they are known, resemble
the adult males instead of the adult females. But the
difference between the sexes is never nearly so great
as occurs with many birds in the first class, and the
cases are comparatively rare. Mr. Wallace who first
called attention to the singular relation which exists
between the less bright colours of the males and their
performing the duties of incubation, lays great stress on
this point,241 as a crucial test that obscure colours have
201been acquired for the sake of protection during the
period of nesting. A different view seems to me more
probable. As the cases are curious and not numerous,
I will briefly give all that I have been able to find.
In one section of the genus Turnix, quail-like birds,
the female is invariably larger than the male (being
nearly twice as large in one of the Australian species)
and this is an unusual circumstance with the Gallinaceæ.
In most of the species the female is more distinctly
coloured and brighter than the male,242 but in some
few species the sexes are alike. In Turnix taigoor of
India the male “wants the black on the throat and neck,
and the whole tone of the plumage is lighter and less
pronounced than that of the female.” The female
appears to be more vociferous, and is certainly much
more pugnacious than the male; so that the females
and not the males are often kept by the natives for
fighting, like game-cocks. As male birds are exposed
by the English bird-catchers for a decoy near a trap,
in order to catch other males by exciting their rivalry,
so the females of this Turnix are employed in India.
When thus exposed the females soon begin their “loud
purring call, which can be heard a long way off,
and any females within ear-shot run rapidly to the
spot, and commence fighting with the caged bird.”
In this way from twelve to twenty birds, all breeding-females,
may be caught in the course of a single
day. The natives assert that the females after laying
their eggs associate in flocks, and leave the males to
sit on them. There is no reason to doubt the truth
of this assertion, which is supported by some observa202tions
made in China by Mr. Swinhoe.243 Mr. Blyth
believes, that the young of both sexes resemble the
adult male.

Fig. 60. Rhynchæa capensis (from Brehm).
The females of the three species of Painted Snipes
(Rhynchæa) “are not only larger, but much more richly
coloured than the males.”244 With all other birds, in
which the trachea differs in structure in the two sexes
203it is more developed and complex in the male than in
the female; but in the Rhynchæa Australis it is simple
in the male, whilst in the female it makes four distinct
convolutions before entering the lungs.245 The female
therefore of this species has acquired an eminently
masculine character. Mr. Blyth ascertained, by examining
many specimens, that the trachea is not convoluted
in either sex of R. Bengalensis, which species
so closely resembles R. Australis that it can hardly be
distinguished except by its shorter toes. This fact is
another striking instance of the law that secondary
sexual characters are often widely different in closely-allied
forms; though it is a very rare circumstance
when such differences relate to the female sex. The
young of both sexes of R. Bengalensis in their first
plumage are said to resemble the mature male.246
There is also reason to believe that the male undertakes
the duty of incubation, for Mr. Swinhoe247 found the
females before the close of the summer associated in
flocks, as occurs with the females of the Turnix.
The females of Phalaropus fulicarius and P. hyperboreus
are larger, and in their summer plumage “more gaily
attired than the males.” But the difference in colour
between the sexes is far from conspicuous. The male
alone of P. fulicarius undertakes, according to Professor
Steenstrup, the duty of incubation, as is likewise shewn
by the state of his breast-feathers during the breeding-season.
The female of the dotterel plover (Eudromias
morinellus) is larger than the male, and has the red
and black tints on the lower surface, the white crescent
on the breast, and the stripes over the eyes, more
strongly pronounced. The male also takes at least a
204share in hatching the eggs; but the female likewise
attends to the young.248 I have not been able to discover
whether with these species the young resemble the
adult males more closely than the adult females; for
the comparison is somewhat difficult to make on account
of the double moult.
Turning now to the Ostrich order: the male of the common
cassowary (Casuarius galeatus) would be thought
by any one to be the female, from his smaller size and
from the appendages and naked skin about his head
being much less brightly coloured; and I am informed
by Mr. Bartlett that in the Zoological Gardens it is
certainly the male alone who sits on the eggs and takes
care of the young.249 The female is said by Mr. T. W.
Wood250 to exhibit during the breeding-season a most
pugnacious disposition; and her wattles then become
enlarged and more brilliantly coloured. So again the
female of one of the emus (Dromœus irroratus) is considerably
larger than the male, and she possesses a
slight top-knot, but is otherwise undistinguishable in
plumage. She appears, however, “to have greater
power, when angry or otherwise excited, of erecting,
like a turkey-cock, the feathers of her neck and
205breast. She is usually the more courageous and
pugilistic. She makes a deep hollow guttural boom,
especially at night, sounding like a small gong. The
male has a slenderer frame and is more docile, with
no voice beyond a suppressed hiss when angry, or a
croak.” He not only performs the whole duty of
incubation, but has to defend the young from their
mother; “for as soon as she catches sight of her progeny
she becomes violently agitated, and notwithstanding
the resistance of the father appears to use
her utmost endeavours to destroy them. For months
afterwards it is unsafe to put the parents together,
violent quarrels being the inevitable result, in which
the female generally comes off conqueror.”251 So
that with this emu we have a complete reversal not
only of the parental and incubating instincts, but of
the usual moral qualities of the two sexes; the females
being savage, quarrelsome and noisy, the males gentle
and good. The case is very different with the African
ostrich, for the male is somewhat larger than the female
and has finer plumes with more strongly contrasted
colours; nevertheless he undertakes the whole
duty of incubation.252
I will specify the few other cases known to me, in
which the female is more conspicuously coloured than
the male, although nothing is known about their manner
of incubation. With the carrion-hawk of the Falkland
Islands (Milvago leucurus) I was much surprised
to find by dissection that the individuals, which had
all their tints strongly pronounced, with the cere and
legs orange-coloured, were the adult females; whilst
206those with duller plumage and grey legs were the males
or the young. In an Australian tree-creeper (Climacteris
erythrops) the female differs from the male in
“being adorned with beautiful, radiated, rufous markings
on the throat, the male having this part quite
plain.” Lastly in an Australian nightjar “the female
always exceeds the male in size and in the brilliance
of her tints; the males, on the other hand, have two
white spots on the primaries more conspicuous than
in the female.”253
We thus see that the cases in which female birds are
more conspicuously coloured than the males, with the
young in their immature plumage resembling the adult
males instead of the adult females, as in the previous
class, are not numerous, though they are distributed in
various Orders. The amount of difference, also, between
the sexes is incomparably less than that which frequently
occurs in the last class; so that the cause of the difference,
whatever it may have been, has acted on the females
in the present class either less energetically or less
persistently than on the males in the last class. Mr.
Wallace believes that the males have had their colours
207rendered less conspicuous for the sake of protection
during the period of incubation; but the difference
between the sexes in hardly any of the foregoing cases
appears sufficiently great for this view to be safely accepted.
In some of the cases the brighter tints of the
female are almost confined to the lower surface, and the
males, if thus coloured, would not have been exposed to
danger whilst sitting on the eggs. It should also be
borne in mind that the males are not only in a slight
degree less conspicuously coloured than the females, but
are of less size, and have less strength. They have, moreover,
not only acquired the maternal instinct of incubation,
but are less pugnacious and vociferous than the
females, and in one instance have simpler vocal organs.
Thus an almost complete transposition of the instincts,
habits, disposition, colour, size, and of some points of
structure, has been effected between the two sexes.
Now if we might assume that the males in the present
class have lost some of that ardour which is usual to
their sex, so that they no longer search eagerly for the
females; or, if we might assume that the females have
become much more numerous than the males—and in
the case of one Indian Turnix the females are said to be
“much more commonly met with than the males”254—then
it is not improbable that the females would have
been led to court the males, instead of being courted by
them. This indeed is the case to a certain extent, with
some birds, as we have seen with the peahen, wild turkey,
and certain kinds of grouse. Taking as our guide the
habits of most male birds, the greater size and strength
and the extraordinary pugnacity of the females of the
Turnix and Emu, must mean that they endeavour to
drive away rival females, in order to gain possession of
208the male; and on this view, all the facts become clear;
for the males would probably be most charmed or excited
by the females which were the most attractive to
them by their brighter colours, other ornaments, or
vocal powers. Sexual selection would then soon do its
work, steadily adding to the attractions of the females;
the males and the young being left not at all, or but
little modified.
Class III. When the adult male resembles the adult
female, the young of both sexes have a peculiar first plumage
of their own.—In this class both sexes when adult
resemble each other, and differ from the young. This
occurs with many birds of many kinds. The male robin
can hardly be distinguished from the female, but the
young are widely different with their mottled dusky-olive
and brown plumage. The male and female of the
splendid scarlet Ibis are alike, whilst the young are
brown; and the scarlet-colour, though common to both
sexes, is apparently a sexual character, for it is not well
developed with birds under confinement, in the same
manner as often occurs in the case of brilliantly coloured
male birds. With many species of herons the
young differ greatly from the adults, and their summer
plumage, though common to both sexes, clearly has
a nuptial character. Young swans are slate-coloured,
whilst the mature birds are pure white; but it would be
superfluous to give additional instances. These differences
between the young and the old apparently depend,
as in the two last classes, on the young having
retained a former or ancient state of plumage, which has
been exchanged for a new plumage by the old of both
sexes. When the adults are brightly coloured, we may
conclude from the remarks just made in relation to the
scarlet ibis and to many herons, and from the analogy of
the species in the first class, that such colours have been209
acquired through sexual selection by the nearly mature
males; but that, differently from what occurs in the
two first classes, the transmission, though limited to the
same age, has not been limited to the same sex. Consequently
both sexes when mature resemble each other
and differ from the young.
Class IV. When the adult male resembles the adult
female, the young of both sexes in their first plumage
resemble the adults.—In this class the young and the
adults of both sexes, whether brilliantly or obscurely
coloured, resemble each other. Such cases are, I think,
more common than those in the last class. We have
in England instances in the kingfisher, some woodpeckers,
the jay, magpie, crow, and many small dull-coloured
birds, such as the hedge-warbler or kitty-wren.
But the similarity in plumage between the young and
the old is never absolutely complete, and graduates away
into dissimilarity. Thus the young of some members of
the kingfisher family are not only less vividly coloured
than the adults, but many of the feathers on the lower
surface are edged with brown,255—a vestige probably of
a former state of the plumage. Frequently in the same
group of birds, even within the same genus, for instance
in an Australian genus of parrakeets (Platycercus), the
young of some species closely resemble, whilst the
young of other species differ considerably from their
parents of both sexes, which are alike.256 Both sexes
and the young of the common jay are closely similar;
but in the Canada jay (Perisoreus canadensis) the young
differ so much from their parents that they were formerly
described as distinct species.257
210
Before proceeding, I may remark that under the
present and two next classes of cases the facts are so
complex, and the conclusions so doubtful, that any one
who feels no especial interest in the subject had better
pass them over.
The brilliant or conspicuous colours which characterise
many birds in the present class, can rarely
or never be of service to them as a protection;
so that they have probably been gained by the males
through sexual selection, and then transferred to the
females and the young. It is, however, possible that
the males may have selected the more attractive females;
and if these transmitted their characters to
their offspring of both sexes, the same results would
follow as from the selection of the more attractive
males by the females. But there is some evidence that
this contingency has rarely, if ever, occurred in any of
those groups of birds, in which the sexes are generally
alike; for if even a few of the successive variations had
failed to be transmitted to both sexes, the females
would have exceeded to a slight degree the males
in beauty. Exactly the reverse occurs under nature;
for in almost every large group, in which the sexes
generally resemble each other, the males of some few
species are in a slight degree more brightly coloured
than the females. It is again possible that the females
may have selected the more beautiful males, these males
having reciprocally selected the more beautiful females;
but it is doubtful whether this double process of selection
would be likely to occur, owing to the greater
eagerness of one sex than the other, and whether it
would be more efficient than selection on one side
alone. It is, therefore, the most probable view that
sexual selection has acted, in the present class, as far
as ornamental characters are concerned, in accordance211
with the general rule throughout the animal kingdom,
that is, on the males; and that these have
transmitted their gradually-acquired colours, either
equally or almost equally, to their offspring of both
sexes.
Another point is more doubtful, namely, whether the
successive variations first appeared in the males after
they had become nearly mature, or whilst quite young.
In either case sexual selection must have acted on
the male when he had to compete with rivals for
the possession of the female; and in both cases the
characters thus acquired have been transmitted to both
sexes and all ages. But these characters, if acquired
by the males when adult, may have been transmitted
at first to the adults alone, and at some subsequent
period transferred to the young. For it is known that
when the law of inheritance at corresponding ages
fails, the offspring often inherit characters at an
earlier age than that at which they first appeared
in their parents.258 Cases apparently of this kind have
been observed with birds in a state of nature. For
instance Mr. Blyth has seen specimens of Lanius
rufus and of Colymbus glacialis which had assumed
whilst young, in a quite anomalous manner, the adult
plumage of their parents.259 Again, the young of the
common swan (Cygnus olor) do not cast off their dark
feathers and become white until eighteen months or
two years old; but Dr. F. Forel has described the case
of three vigorous young birds, out of a brood of four,
which were born pure white. These young birds were
not albinoes, as shewn by the colour of their beaks
212and legs, which nearly resembled the same parts in
the adults.260
It may be worth while to illustrate the above three
modes by which, in the present class, the two sexes
and the young may have come to resemble each other,
by the curious case of the genus Passer.261 In the
house-sparrow (P. domesticus) the male differs much
from the female and from the young. These resemble
each other, and likewise to a large extent both sexes
and the young of the sparrow of Palestine (P. brachydactylus),
as well as of some allied species. We may
therefore assume that the female and young of the
house-sparrow approximately shew us the plumage of
the progenitor of the genus. Now with the tree-sparrow
(P. montanus) both sexes and the young closely resemble
the male of the house-sparrow; so that they have all
been modified in the same manner, and all depart from
the typical colouring of their early progenitor. This
may have been effected by a male ancestor of the tree-sparrow
having varied, firstly, when nearly mature, or,
secondly, whilst quite young, having in either case transmitted
his modified plumage to the females and the
young; or, thirdly, he may have varied when adult and
transmitted his plumage to both adult sexes, and, owing
to the failure of the law of inheritance at corresponding
ages, at some subsequent period to his young.
It is impossible to decide which of these three modes
has generally prevailed throughout the present class of
cases. The belief that the males varied whilst young,
and transmitted their variations to their offspring of
213both sexes is perhaps the most probable. I may here
add that I have endeavoured, with little success, by
consulting various works, to decide how far with birds
the period of variation has generally determined the
transmission of characters to one sex or to both. The
two rules, often referred to (namely, that variations
occurring late in life are transmitted to one and the
same sex, whilst those which occur early in life are
transmitted to both sexes), apparently hold good in
the first,262 second, and fourth classes of cases; but
they fail in an equal number, namely, in the third,
often in the fifth,263 and in the sixth small class.
They hold good, however, as far as I can judge, with a
considerable majority of the species of birds. Whether
or not this be so, we may conclude from the facts
given in the eighth chapter that the period of variation
has been one important element in determining the
form of transmission.
With birds it is difficult to decide by what standard
we ought to judge of the earliness or lateness of the
period of variation, whether by the age in reference to
the duration of life, or to the power of reproduction,
or to the number of moults through which the species
passes. The moulting of birds, even within the same
family, sometimes differs much without any assignable
214cause. Some birds moult so early, that nearly all
the body-feathers are cast off before the first wing-feathers
are fully grown; and we cannot believe that
this was the primordial state of things. When the period
of moulting has been accelerated, the age at which
the colours of the adult plumage were first developed
would falsely appear to us to have been earlier than
it really was. This may be illustrated by the practice
followed by some bird-fanciers, who pull out a few
feathers from the breast of nestling bullfinches, and
from the head or neck of young gold pheasants, in
order to ascertain their sex; for in the males these
feathers are immediately replaced by coloured ones.264
The actual duration of life is known in but few birds, so
that we can hardly judge by this standard. And with
reference to the period at which the powers of reproduction
are gained, it is a remarkable fact that various
birds occasionally breed whilst retaining their immature
plumage.265
The fact of birds breeding in their immature plumage
seems opposed to the belief that sexual selection has
215played as important a part, as I believe it has, in
giving ornamental colours, plumes, &c., to the males,
and, by means of equal transmission, to the females of
many species. The objection would be a valid one, if
the younger and less ornamented males were as successful
in winning females and propagating their kind,
as the older and more beautiful males. But we have
no reason to suppose that this is the case. Audubon
speaks of the breeding of the immature males of Ibis
tantalus as a rare event, as does Mr. Swinhoe, in regard
to the immature males of Oriolus.266 If the young
of any species in their immature plumage were more
successful in winning partners than the adults, the
adult plumage would probably soon be lost, as the
males which retained their immature dress for the
longest period would prevail, and thus the character of
the species would ultimately be modified.267 If, on the
other hand, the young never succeeded in obtaining a
female, the habit of early reproduction would perhaps
be sooner or later quite eliminated, from being superfluous
and entailing waste of power.
The plumage of certain birds goes on increasing in
216beauty during many years after they are fully mature;
this is the case with the train of the peacock, and with
the crest and plumes of certain herons; for instance, the
Ardea Ludovicana;268 but it is very doubtful whether
the continued development of such feathers is the
result of the selection of successive beneficial variations,
or merely of continuous growth. Most fishes continue
increasing in size, as long as they are in good health
and have plenty of food; and a somewhat similar law
may prevail with the plumes of birds.
Class V. When the adults of both sexes have a distinct
winter and summer plumage, whether or not the male
differs from the female, the young resemble the adults of
both sexes in their winter dress, or much more rarely in
their summer dress, or they resemble the females alone;
or the young may have an intermediate character; or
again, they may differ greatly from the adults in both
their seasonal plumages.—The cases in this class are
singularly complex; nor is this surprising, as they
depend on inheritance, limited in a greater or less
degree in three different ways, namely by sex, age,
and the season of the year. In some cases the individuals
of the same species pass through at least five
distinct states of plumage. With the species, in which
the male differs from the female during the summer
season alone, or, which is rarer, during both seasons,269
the young generally resemble the females,—as with
the so-called goldfinch of North America, and apparently
with the splendid Maluri of Australia.270 With
217the species, the sexes of which are alike during both
the summer and winter, the young may resemble
the adults, firstly, in their winter dress; secondly,
which occurs much more rarely, in their summer
dress; thirdly, they may be intermediate between
these two states; and, fourthly, they may differ greatly
from the adults at all seasons. We have an instance
of the first of these four cases in one of the egrets
of India (Buphus coromandus), in which the young and
the adults of both sexes are white during the winter,
the adults becoming golden-buff during the summer.
With the Gaper (Anastomus oscitans) of India we
have a similar case, but the colours are reversed;
for the young and the adults of both sexes are grey
and black during the winter, the adults becoming white
during the summer.271 As an instance of the second
case, the young of the razor-bill (Alca torda, Linn.),
in an early state of plumage, are coloured like the
adults during the summer; and the young of the
white-crowned sparrow of North America (Fringilla
leucophrys), as soon as fledged, have elegant white
stripes on their heads, which are lost by the young and
the old during the winter.272 With respect to the third
case, namely, that of the young having an intermediate
character between the summer and winter adult plumages,
Yarrell273 insists that this occurs with many
218waders. Lastly, in regard to the young differing
greatly from both sexes in their adult summer and
winter plumages, this occurs with some herons and
egrets of North America and India,—the young alone
being white.
I will make only a few remarks on these complicated
cases. When the young resemble the female in her
summer dress, or the adults of both sexes in their winter
dress, the cases differ from those given under Classes I.
and III. only in the characters originally acquired by
the males during the breeding-season, having been
limited in their transmission to the corresponding season.
When the adults have a distinct summer and winter
plumage, and the young differ from both, the case is
more difficult to understand. We may admit as probable
that the young have retained an ancient state
of plumage; we can account through sexual selection
for the summer or nuptial plumage of the adults, but
how are we to account for their distinct winter plumage?
If we could admit that this plumage serves in all cases
as a protection, its acquirement would be a simple
affair; but there seems no good reason for this admission.
It may be suggested that the widely different
conditions of life during the winter and summer have
acted in a direct manner on the plumage; this may
have had some effect, but I have not much confidence
in so great a difference, as we sometimes see, between
the two plumages having been thus caused. A more
probable explanation is, that an ancient style of plumage,
partially modified through the transference of some
characters from the summer plumage, has been retained
by the adults during the winter. Finally, all the cases
in our present class apparently depend on characters
acquired by the adult males, having been variously
limited in their transmission according to age, season,219
and sex; but it would not be worth while to attempt to
follow out these complex relations.
Class VI. The young in their first plumage differ
from each other according to sex; the young males
resembling more or less closely the adult males, and the
young females more or less closely the adult females.—The
cases in the present class, though occurring in
various groups, are not numerous; yet, if experience
had not taught us to the contrary, it seems the most
natural thing that the young should at first always
resemble to a certain extent, and gradually become
more and more like, the adults of the same sex. The
adult male blackcap (Sylvia atricapilla) has a black
head, that of the female being reddish-brown; and I
am informed by Mr. Blyth, that the young of both sexes
can be distinguished by this character even as nestlings.
In the family of thrushes an unusual number of similar
cases have been noticed; the male blackbird (Turdus
merula) can be distinguished in the nest from the female,
as the main wing-feathers, which are not moulted so
soon as the body-feathers, retain a brownish tint until the
second general moult.274 The two sexes of the mocking
bird (Turdus polyglottus, Linn.) differ very little
from each other, yet the males can easily be distinguished
at a very early age from the females by shewing
more pure white.275 The males of a forest-thrush
and of a rock-thrush (viz. Orocetes erythrogastra and
Petrocincla cyanea) have much of their plumage of a
fine blue, whilst the females are brown; and the nestling
males of both species have their main wing and tail-feathers
edged with blue, whilst those of the female are
220edged with brown.276 So that the very same feathers
which in the young blackbird assume their mature character
and become black after the others, in these two
species assume this character and become blue before
the others. The most probable view with reference to
these cases is that the males, differently from what
occurs in Class I., have transmitted their colours to
their male offspring at an earlier age than that at
which they themselves first acquired them; for if they
had varied whilst quite young, they would probably
have transmitted all their characters to their offspring
of both sexes.277
In Aïthurus polytmus (one of the humming-birds)
the male is splendidly coloured black and green, and
two of the tail-feathers are immensely lengthened; the
female has an ordinary tail and inconspicuous colours;
now the young males, instead of resembling the adult
female, in accordance with the common rule, begin
from the first to assume the colours proper to their
sex, and their tail-feathers soon become elongated.
I owe this information to Mr. Gould, who has given
me the following more striking and as yet unpublished
case. Two humming-birds belonging to the
genus Eustephanus, both beautifully coloured, inhabit
the small island of Juan Fernandez, and have always
been ranked as specifically distinct. But it has lately
been ascertained that the one, which is of a rich ches221nut-brown
colour with a golden-red head, is the male,
whilst the other, which is elegantly variegated with
green and white with a metallic-green head, is the female.
Now the young from the first resemble to a
certain extent the adults of the corresponding sex, the
resemblance gradually becoming more and more complete.
In considering this last case, if as before we take the
plumage of the young as our guide, it would appear
that both sexes have been independently rendered
beautiful; and not that the one sex has partially transferred
its beauty to the other. The male apparently
has acquired his bright colours through sexual selection
in the same manner as, for instance, the peacock or
pheasant in our first class of cases; and the female in
the same manner as the female Rhynchæa or Turnix
in our second class of cases. But there is much difficulty
in understanding how this could have been
effected at the same time with the two sexes of the
same species. Mr. Salvin states, as we have seen in
the eighth chapter, that with certain humming-birds
the males greatly exceed in number the females, whilst
with other species inhabiting the same country the
females greatly exceed the males. If, then, we might
assume that during some former lengthened period the
males of the Juan Fernandez species had greatly exceeded
the females in number, but that during another
lengthened period the females had greatly exceeded
the males, we could understand how the males at one
time, and the females at another time, might have been
rendered beautiful by the selection of the brighter-coloured
individuals of either sex; both sexes transmitting
their characters to their young at a rather earlier
age than usual. Whether this is the true explanation I222
will not pretend to say; but the case is too remarkable
to be passed over without notice.
We have now seen in numerous instances under all
six classes, that an intimate relation exists between the
plumage of the young and that of the adults, either of
one sex or both sexes. These relations are fairly well
explained on the principle that one sex—this being in
the great majority of cases the male—first acquired
through variation and sexual selection bright colours
or other ornaments, and transmitted them in various
ways, in accordance with the recognised laws of inheritance.
Why variations have occurred at different
periods of life, even sometimes with the species of the
same group, we do not know; but with respect to
the form of transmission, one important determining
cause seems to have been the age at which the variations
first appeared.
From the principle of inheritance at corresponding
ages, and from any variations in colour which occurred
in the males at an early age not being then selected, on
the contrary being often eliminated as dangerous, whilst
similar variations occurring at or near the period of
reproduction have been preserved, it follows that the
plumage of the young will often have been left unmodified,
or but little modified. We thus get some insight
into the colouring of the progenitors of our existing
species. In a vast number of species in five out of our
six classes of cases, the adults of one sex or both are
brightly coloured, at least during the breeding-season,
whilst the young are invariably less brightly coloured
than the adults, or are quite dull-coloured; for no instance
is known, as far as I can discover, of the young
of dull-coloured species displaying bright colours, or223
of the young of brightly-coloured species being more
brilliantly coloured than their parents. In the fourth
class, however, in which the young and the old resemble
each other, there are many species (though by no means
all) brightly-coloured, and as these form whole groups,
we may infer that their early progenitors were likewise
brightly-coloured. With this exception, if we look to
the birds of the world, it appears that their beauty
has been greatly increased since that period, of which
we have a partial record in their immature plumage.
On the Colour of the Plumage in relation to Protection.—It
will have been seen that I cannot follow
Mr. Wallace in the belief that dull colours when confined
to the females have been in most cases specially
gained for the sake of protection. There can, however,
be no doubt, as formerly remarked, that both sexes of
many birds have had their colours modified for this
purpose, so as to escape the notice of their enemies; or,
in some instances, so as to approach their prey unobserved,
in the same manner as owls have had their
plumage rendered soft, that their flight may not be
overheard. Mr. Wallace remarks278 that “it is only
in the tropics, among forests which never lose their
foliage, that we find whole groups of birds, whose
chief colour is green.” It will be admitted by every
one, who has ever tried, how difficult it is to distinguish
parrots in a leaf-covered tree. Nevertheless, we must remember
that many parrots are ornamented with crimson,
blue, and orange tints, which can hardly be protective.
Woodpeckers are eminently arboreal, but, besides green
species, there are many black, and black-and-white
kinds—all the species being apparently exposed to
224nearly the same dangers. It is therefore probable
that strongly-pronounced colours have been acquired
by tree-haunting birds through sexual selection, but
that green tints have had an advantage through
natural selection over other colours for the sake of
protection.
In regard to birds which live on the ground, everyone
admits that they are coloured so as to imitate the
surrounding surface. How difficult it is to see a partridge,
snipe, woodcock, certain plovers, larks, and
nightjars when crouched on the ground. Animals inhabiting
deserts offer the most striking instances, for the
bare surface affords no concealment, and all the smaller
quadrupeds, reptiles, and birds depend for safety on
their colours. As Mr. Tristram has remarked,279 in
regard to the inhabitants of the Sahara, all are protected
by their “isabelline or sand-colour.” Calling to
my recollection the desert-birds which I had seen in
South America, as well as most of the ground-birds
in Great Britain, it appeared to me that both sexes
in such cases are generally coloured nearly alike. Accordingly
I applied to Mr. Tristram, with respect to the
birds of the Sahara, and he has kindly given me the
following information. There are twenty-six species,
belonging to fifteen genera, which manifestly have had
their plumage coloured in a protective manner; and
this colouring is all the more striking, as with most
of these birds it is different from that of their congeners.
Both sexes of thirteen out of the twenty-six
species are coloured in the same manner; but these
belong to genera in which this rule commonly prevails,
so that they tell us nothing about the protective
colours being the same in both sexes of desert-birds. Of
225the other thirteen species, three belong to genera in
which the sexes usually differ from each other, yet they
have the sexes alike. In the remaining ten species,
the male differs from the female; but the difference is
confined chiefly to the under surface of the plumage,
which is concealed when the bird crouches on the
ground; the head and back being of the same sand-coloured
hue in both sexes. So that in these ten
species the upper surfaces of both sexes have been
acted on and rendered alike, through natural selection,
for the sake of protection; whilst the lower surfaces of
the males alone have been diversified through sexual
selection, for the sake of ornament. Here, as both
sexes are equally well protected, we clearly see that the
females have not been prevented through natural selection
from inheriting the colours of their male parents:
we must look to the law of sexually limited transmission,
as before explained.
In all parts of the world both sexes of many soft-billed
birds, especially those which frequent reeds or
sedges, are obscurely coloured. No doubt if their
colours had been brilliant, they would have been
much more conspicuous to their enemies; but whether
their dull tints have been specially gained for the
sake of protection seems, as far as I can judge, rather
doubtful. It is still more doubtful whether such
dull tints can have been gained for the sake of ornament.
We must, however, bear in mind that male
birds, though dull-coloured, often differ much from
their females, as with the common sparrow, and this
leads to the belief that such colours have been gained
through sexual selection, from being attractive. Many
of the soft-billed birds are songsters; and a discussion
in a former chapter should not be forgotten, in which
it was shewn that the best songsters are rarely orna226mented
with bright tints. It would appear that female
birds, as a general rule, have selected their mates
either for their sweet voices or gay colours, but not
for both charms combined. Some species which are
manifestly coloured for the sake of protection, such
as the jack-snipe, woodcock, and nightjar, are likewise
marked and shaded, according to our standard
of taste, with extreme elegance. In such cases we
may conclude that both natural and sexual selection
have acted conjointly for protection and ornament.
Whether any bird exists which does not possess some
special attraction, by which to charm the opposite sex,
may be doubted. When both sexes are so obscurely
coloured, that it would be rash to assume the agency
of sexual selection, and when no direct evidence can
be advanced shewing that such colours serve as a protection,
it is best to own complete ignorance of the
cause, or, which comes to nearly the same thing, to
attribute the result to the direct action of the conditions
of life.
There are many birds both sexes of which are conspicuously,
though not brilliantly coloured, such as
the numerous black, white, or piebald species; and
these colours, are probably the result of sexual selection.
With the common blackbird, capercailzie, black-cock,
black Scoter-duck (Oidemia), and even with one
of the Birds of Paradise (Lophorina atra), the males
alone are black, whilst the females are brown or mottled;
and there can hardly be a doubt that blackness
in these cases has been a sexually selected character.
Therefore it is in some degree probable that the complete
or partial blackness of both sexes in such birds
as crows, certain cockatoos, storks, and swans, and many
marine birds, is likewise the result of sexual selection,
accompanied by equal transmission to both sexes;227
for blackness can hardly serve in any case as a protection.
With several birds, in which the male alone
is black, and in others in which both sexes are black,
the beak or skin about the head is brightly coloured,
and the contrast thus afforded adds greatly to their
beauty; we see this in the bright yellow beak of the
male blackbird, in the crimson skin over the eyes of
the black-cock and capercailzie, in the variously and
brightly-coloured beak of the Scoter-drake (Oidemia),
in the red beak of the chough (Corvus graculus, Linn.),
of the black swan, and black stork. This leads me to
remark that it is not at all incredible that toucans may
owe the enormous size of their beaks to sexual selection,
for the sake of displaying the diversified and vivid
stripes of colour, with which these organs are ornamented.280
The naked skin at the base of the beak and
round the eyes is likewise often brilliantly coloured;
and Mr. Gould, in speaking of one species,281 says that
the colours of the beak “are doubtless in the finest
and most brilliant state during the time of pairing.”
There is no greater improbability in toucans being
encumbered with immense beaks, though rendered as
light as possible by their cancellated structure, for
an object falsely appearing to us unimportant, namely,
the display of fine colours, than that the male Argus
228pheasant and some other birds should be encumbered
with plumes so long as to impede their flight.
In the same manner, as the males alone of various
species are black, the females being dull-coloured;
so in a few cases the males alone are either wholly
or partially white, as with the several Bell-birds
of South America (Chasmorhynchus), the Antarctic
goose (Bernicla antarctica), the silver pheasant, &c.,
whilst the females are brown or obscurely mottled.
Therefore, on the same principle as before, it is probable
that both sexes of many birds, such as white
cockatoos, several egrets with their beautiful plumes,
certain ibises, gulls, terns, &c., have acquired their
more or less completely white plumage through sexual
selection. The species which inhabit snowy regions of
course come under a different head. The white plumage
of some of the above-named birds appears in
both sexes only when they are mature. This is
likewise the case with certain gannets, tropic-birds,
&c., and with the snow-goose (Anser hyperboreus). As
the latter breeds on the “barren grounds,” when not
covered with snow, and as it migrates southward during
the winter, there is no reason to suppose that its snow-white
adult plumage serves as a protection. In the
case of the Anastomus oscitans previously alluded to,
we have still better evidence that the white plumage
is a nuptial character, for it is developed only during
the summer; the young in their immature state, and
the adults in their winter dress, being grey and black.
With many kinds of gulls (Larus), the head and neck
become pure white during the summer, being grey
or mottled during the winter and in the young state.
On the other hand, with the smaller gulls, or sea-mews
(Gavia), and with some terns (Sterna), exactly the reverse
occurs; for the heads of the young birds during229
the first year, and of the adults during the winter, are
either pure white, or much paler-coloured than during
the breeding-season. These latter cases offer another
instance of the capricious manner in which sexual selection
appears often to have acted.282
The cause of aquatic birds having acquired a white
plumage so much more frequently than terrestrial birds,
probably depends on their large size and strong powers
of flight, so that they can easily defend themselves or
escape from birds of prey, to which moreover they are
not much exposed. Consequently sexual selection has
not here been interfered with or guided for the sake of
protection. No doubt, with birds which roam over the
open ocean, the males and females could find each
other much more easily when made conspicuous either
by being perfectly white, or intensely black; so that
these colours may possibly serve the same end as the
call-notes of many land-birds. A white or black bird,
when it discovers and flies down to a carcase floating
on the sea or cast up on the beach, will be seen from
a great distance, and will guide other birds of the same
and of distinct species, to the prey; but as this would
be a disadvantage to the first finders, the individuals
which were the whitest or blackest would not thus have
procured more food than the less strongly coloured
individuals. Hence conspicuous colours cannot have
been gradually acquired for this purpose through natural
selection.283
230
As sexual selection depends on so fluctuating an
element as taste, we can understand how it is that within
the same group of birds, with habits of life nearly
the same, there should exist white or nearly white,
as well as black, or nearly black species,—for instance,
white and black cockatoos, storks, ibises, swans, terns,
and petrels. Piebald birds likewise sometimes occur
in the same groups, for instance, the black-necked
swan, certain terns, and the common magpie. That
a strong contrast in colour is agreeable to birds, we
may conclude, by looking through any large collection
of specimens or series of coloured plates, for the sexes
frequently differ from each other in the male having
the pale parts of a purer white, and the variously
coloured dark parts of still darker tints than in the
female.
It would even appear that mere novelty, or change
for the sake of change, has sometimes acted like a
charm on female birds, in the same manner as changes
of fashion with us. The Duke of Argyll says,284—and I
am glad to have the unusual satisfaction of following
for even a short distance in his footsteps—“I am more
and more convinced that variety, mere variety, must
be admitted to be an object and an aim in Nature.”
I wish the Duke had explained what he here means by
Nature. Is it meant that the Creator of the universe
ordained diversified results for His own satisfaction, or for
that of man? The former notion seems to me as much
wanting in due reverence as the latter in probability.
Capriciousness of taste in the birds themselves appears
a more fitting explanation. For example; the males
231of some parrots can hardly be said to be more beautiful,
at least according to our taste, than the females, but
they differ from them in such points, as the male
having a rose-coloured collar instead of, as in the
female, “a bright emeraldine narrow green collar;” or
in the male having a black collar instead of “a yellow
demi-collar in front,” with a pale roseate instead of a
plum-blue head.285 As so many male birds have for
their chief ornament elongated tail-feathers or elongated
crests, the shortened tail, formerly described in the
male of a humming-bird, and the shortened crest of
the male goosander almost seem like one of the many
opposite changes of fashion which we admire in our
own dresses.
Some members of the heron family offer a still more
curious case of novelty in colouring having apparently
been appreciated for the sake of novelty. The
young of the Ardea asha are white, the adults being
dark slate-coloured; and not only the young, but the
adults of the allied Buphus coromandus in their winter
plumage are white, this colour changing into a rich
golden-buff during the breeding-season. It is incredible
that the young of these two species, as well as of some
other members of the same family,286 should have been
specially rendered pure white and thus made conspicuous
to their enemies; or that the adults of one of
these two species should have been specially rendered
white during the winter in a country which is never
232covered with snow. On the other hand we have reason
to believe that whiteness has been gained by many birds
as a sexual ornament. We may therefore conclude that
an early progenitor of the Ardea asha and the Buphus
acquired a white plumage for nuptial purposes, and
transmitted this colour to their young; so that the
young and the old became white like certain existing
egrets; the whiteness having afterwards been retained
by the young whilst exchanged by the adults for more
strongly pronounced tints. But if we could look still
further backwards in time to the still earlier progenitors
of these two species, we should probably see the adults
dark-coloured. I infer that this would be the case, from
the analogy of many other birds, which are dark whilst
young, and when adult are white; and more especially
from the case of the Ardea gularis, the colours of which
are the reverse of those of A. asha, for the young are
dark-coloured and the adults white, the young having
retained a former state of plumage. It appears therefore
that the progenitors in their adult condition of the
Ardea asha, the Buphus, and of some allies, have undergone,
during a long line of descent, the following changes
of colour: firstly a dark shade, secondly pure white,
and thirdly, owing to another change of fashion (if I
may so express myself), their present slaty, reddish, or
golden-buff tints. These successive changes are intelligible
only on the principle of novelty having been
admired by birds for the sake of novelty.
Summary of the Four Chapters on Birds.—Most male
birds are highly pugnacious during the breeding-season,
and some possess weapons especially adapted for fighting
with their rivals. But the most pugnacious and the
best-armed males rarely or never depend for success
solely on their power to drive away or kill their rivals,233
but have special means for charming the female. With
some it is the power of song, or of emitting strange
cries, or of producing instrumental music, and the males
in consequence differ from the females in their vocal
organs, or in the structure of certain feathers. From
the curiously diversified means for producing various
sounds we gain a high idea of the importance of this
means of courtship. Many birds endeavour to charm
the females by love-dances or antics, performed on the
ground or in the air, and sometimes at prepared places.
But ornaments of many kinds, the most brilliant tints,
combs and wattles, beautiful plumes, elongated feathers,
top-knots, and so forth, are by far the commonest
means. In some cases mere novelty appears to have
acted as a charm. The ornaments of the males must
be highly important to them, for they have been acquired
in not a few cases at the cost of increased danger
from enemies, and even at some loss of power in fighting
with their rivals. The males of very many species
do not assume their ornamental dress until they
arrive at maturity, or they assume it only during the
breeding-season, or the tints then become more vivid.
Certain ornamental appendages become enlarged, turgid,
and brightly-coloured during the very act of courtship.
The males display their charms with elaborate care and
to the best effect; and this is done in the presence of
the females. The courtship is sometimes a prolonged
affair, and many males and females congregate at an
appointed place. To suppose that the females do not
appreciate the beauty of the males is to admit that
their splendid decorations, all their pomp and display,
are useless; and this is incredible. Birds have fine
powers of discrimination, and in some few instances it
can be shewn that they have a taste for the beautiful.
The females, moreover, are known occasionally to234
exhibit a marked preference or antipathy for certain
individual males.
If it be admitted that the females prefer, or are
unconsciously excited by the more beautiful males, then
the males would slowly but surely be rendered more
and more attractive through sexual selection. That it
is this sex which has been chiefly modified we may infer
from the fact that in almost every genus in which the
sexes differ, the males differ much more from each other
than do the females; this is well shewn in certain closely-allied
representative species in which the females can
hardly be distinguished, whilst the males are quite distinct.
Birds in a state of nature offer individual differences
which would amply suffice for the work of sexual
selection; but we have seen that they occasionally present
more strongly-marked variations which recur so
frequently that they would immediately be fixed, if
they served to allure the female. The laws of variation
will have determined the nature of the initial changes,
and largely influenced the final result. The gradations,
which may be observed between the males of
allied species, indicate the nature of the steps which
have been passed through, and explain in the most
interesting manner certain characters, such as the
indented ocelli of the tail-feathers of the peacock, and
the wonderfully-shaded ocelli of the wing-feathers of
the Argus pheasant. It is evident that the brilliant
colours, top-knots, fine plumes, &c., of many male
birds cannot have been acquired as a protection;
indeed they sometimes lead to danger. That they
are not due to the direct and definite action of the
conditions of life, we may feel assured, because the
females have been exposed to the same conditions, and
yet often differ from the males to an extreme degree.
Although it is probable that changed conditions acting235
during a lengthened period have produced some definite
effect on both sexes, the more important result will have
been an increased tendency to fluctuating variability or
to augmented individual differences; and such differences
will have afforded an excellent groundwork for
the action of sexual selection.
The laws of inheritance, irrespectively of selection,
appear to have determined whether the characters acquired
by the males for the sake of ornament, for producing
various sounds, and for fighting together, have
been transmitted to the males alone or to both sexes,
either permanently or periodically during certain seasons
of the year. Why various characters should sometimes
have been transmitted in one way and sometimes
in another is, in most cases, not known; but the period
of variability seems often to have been the determining
cause. When the two sexes have inherited all characters
in common they necessarily resemble each other;
but as the successive variations may be differently transmitted,
every possible gradation may be found, even
within the same genus, from the closest similarity to
the widest dissimilarity between the sexes. With many
closely-allied species, following nearly the same habits
of life, the males have come to differ from each other
chiefly through the action of sexual selection; whilst
the females have come to differ chiefly from partaking
in a greater or lesser degree of the characters thus
acquired by the males. The effects, moreover, of the
definite action of the conditions of life, will not have
been masked in the females, as in the case of the males,
by the accumulation through sexual selection of strongly-pronounced
colours and other ornaments. The individuals
of both sexes, however affected, will have been
kept at each successive period nearly uniform by the
free intercrossing of many individuals.
236With the species, in which the sexes differ in colour,
it is possible that at first there existed a tendency to
transmit the successive variations equally to both sexes;
and that the females were prevented from acquiring the
bright colours of the males, on account of the danger to
which they would have been exposed during incubation.
But it would be, as far as I can see, an extremely difficult
process to convert, by means of natural selection, one
form of transmission into another. On the other hand
there would not be the least difficulty in rendering a
female dull-coloured, the male being still kept bright-coloured,
by the selection of successive variations, which
were from the first limited in their transmission to the
same sex. Whether the females of many species have
actually been thus modified, must at present remain
doubtful. When, through the law of the equal transmission
of characters to both sexes, the females have
been rendered as conspicuously coloured as the males,
their instincts have often been modified, and they have
been led to build domed or concealed nests.
In one small and curious class of cases the characters
and habits of the two sexes have been completely transposed,
for the females are larger, stronger, more vociferous
and brightly-coloured than their males. They
have, also, become so quarrelsome that they often fight
together like the males of the most pugnacious species.
If, as seems probable, they habitually drive away rival
females, and by the display of their bright colours or
other charms endeavour to attract the males, we can
understand how it is that they have gradually been rendered,
by means of sexual selection and sexually-limited
transmission, more beautiful than the males—the latter
being left unmodified or only slightly modified.
Whenever the law of inheritance at corresponding
ages prevails, but not that of sexually-limited trans237mission,
then if the parents vary late in life—and we
know that this constantly occurs with our poultry, and
occasionally with other birds—the young will be left
unaffected, whilst the adults of both sexes will be
modified. If both these laws of inheritance prevail and
either sex varies late in life, that sex alone will be
modified, the other sex and the young being left unaffected.
When variations in brightness or in other
conspicuous characters occur early in life, as no doubt
often happens, they will not be acted on through sexual
selection until the period of reproduction arrives; consequently
they will be liable to be lost by the accidental
deaths of the young, and if dangerous will be eliminated
through natural selection. Thus we can understand
how it is that variations arising late in life have chiefly
been preserved for the ornamentation and arming of the
males, the females and the young being left almost unaffected,
and therefore like each other. With species
having a distinct summer and winter plumage, the males
of which either resemble or differ from the females
during both seasons or during the summer alone, the
degrees and kinds of resemblance between the young
and the old are exceedingly complex; and this complexity
apparently depends on characters, first acquired
by the males, being transmitted in various ways and
degrees, as limited by age, sex, and season.
As the young of so many species have been but little
modified in colour and in other ornaments, we are
enabled to form some judgment with respect to the
plumage of their early progenitors; and we may infer
that the beauty of our existing species, if we look to the
whole class, has been largely increased since that period
of which the immature plumage gives us an indirect
record. Many birds, especially those which live much
on the ground, have undoubtedly been obscurely co238loured
for the sake of protection. In some instances
the upper exposed surface of the plumage has been thus
coloured in both sexes, whilst the lower surface in the
males alone has been variously ornamented through
sexual selection. Finally, from the facts given in these
four chapters, we may conclude that weapons for battle,
organs for producing sound, ornaments of many kinds,
bright and conspicuous colours, have generally been
acquired by the males through variation and sexual
selection, and have been transmitted in various ways
according to the several laws of inheritance—the females
and the young being left comparatively but little
modified.287
239
CHAPTER XVII.
Secondary Sexual Characters of Mammals.
The law of battle—Special weapons, confined to the males—Cause
of absence of weapons in the female—Weapons common to both
sexes, yet primarily acquired by the male—Other uses of such
weapons—Their high importance—Greater size of the male—Means
of defence—On the preference shewn by either sex in the
pairing of quadrupeds.
With mammals the male appears to win the female
much more through the law of battle than through the
display of his charms. The most timid animals, not
provided with any special weapons for fighting, engage
in desperate conflicts during the season of love. Two
male hares have been seen to fight together until one
was killed; male moles often fight, and sometimes with
fatal results; male squirrels “engage in frequent contests,
and often wound each other severely;” as do
male beavers, so that “hardly a skin is without scars.”288
I observed the same fact with the hides of the guanacoes
in Patagonia; and on one occasion several were so
absorbed in fighting that they fearlessly rushed close by
me. Livingstone speaks of the males of the many animals
in Southern Africa as almost invariably shewing
the scars received in former contests.
The law of battle prevails with aquatic as with ter240restrial
mammals. It is notorious how desperately male
seals fight, both with their teeth and claws, during the
breeding-season; and their hides are likewise often
covered with scars. Male sperm-whales are very jealous
at this season; and in their battles “they often
lock their jaws together, and turn on their sides and
twist about;” so that it is believed by some naturalists
that the frequently deformed state of their lower jaws is
caused by these struggles.289
All male animals which are furnished with special
weapons for fighting, are well known to engage in fierce
battles. The courage and the desperate conflicts of stags
have often been described; their skeletons have been
found in various parts of the world, with the horns inextricably
locked together, shewing how miserably the
victor and vanquished had perished.290 No animal in the
world is so dangerous as an elephant in must. Lord
Tankerville has given me a graphic description of the
battles between the wild bulls in Chillingham Park,
the descendants, degenerated in size but not in courage,
of the gigantic Bos primigenius. In 1861 several contended
for mastery; and it was observed that two of
the younger bulls attacked in concert the old leader
of the herd, overthrew and disabled him, so that he was
believed by the keepers to be lying mortally wounded
in a neighbouring wood. But a few days afterwards one
of the young bulls singly approached the wood; and
241then the “monarch of the chase,” who had been lashing
himself up for vengeance, came out and, in a short
time killed his antagonist. He then quietly joined the
herd, and long held undisputed sway. Admiral Sir
B. J. Sulivan informs me that when he resided in the
Falkland Islands he imported a young English stallion,
which, with eight mares, frequented the hills near Port
William. On these hills there were two wild stallions,
each with a small troop of mares; “and it is certain
that these stallions would never have approached each
other without fighting. Both had tried singly to fight
the English horse and drive away his mares, but had
failed. One day they came in together and attacked
him. This was seen by the capitan who had charge of
the horses, and who, on riding to the spot, found one
of the two stallions engaged with the English horse,
whilst the other was driving away the mares, and had
already separated four from the rest. The capitan
settled the matter by driving the whole party into the
corral, for the wild stallions would not leave the
mares.”
Male animals already provided with efficient cutting
or tearing teeth for the ordinary purposes of life, as
in the carnivora, insectivora, and rodents, are seldom
furnished with weapons especially adapted for fighting
with their rivals. The case is very different with the
males of many other animals. We see this in the horns
of stags and of certain kinds of antelopes in which
the females are hornless. With many animals the
canine teeth in the upper or lower jaw, or in both, are
much larger in the males than in the females; or are
absent in the latter, with the exception sometimes of a
hidden rudiment. Certain antelopes, the musk-deer,
camel, horse, boar, various apes, seals, and the walrus,
offer instances of these several cases. In the females242
of the walrus the tusks are sometimes quite absent.291
In the male elephant of India and in the male dugong292
the upper incisors form offensive weapons. In the male
narwhal one alone of the upper teeth is developed into
the well-known, spirally-twisted, so called horn, which is
sometimes from nine to ten feet in length. It is believed
that the males use these horns for fighting together; for
“an unbroken one can rarely be got, and occasionally
one may be found with the point of another jammed
into the broken place.”293 The tooth on the opposite
side of the head in the male consists of a rudiment about
ten inches in length, which is embedded in the jaw. It
is not, however, very uncommon to find double-horned
male narwhals in which both teeth are well developed.
In the females both teeth are rudimentary. The male
cachalot has a larger head than that of the female, and
it no doubt aids these animals in their aquatic battles.
Lastly, the adult male ornithorhynchus is provided with
a remarkable apparatus, namely a spur on the foreleg,
closely resembling the poison-fang of a venomous snake;
its use is not known, but we may suspect that it serves
as a weapon of offence.294 It is represented by a mere
rudiment in the female.
When the males are provided with weapons which
the females do not possess, there can hardly be a doubt
that they are used for fighting with other males, and
that they have been acquired through sexual selection.
243
It is not probable, at least in most cases, that the females
have actually been saved from acquiring such weapons,
owing to their being useless and superfluous, or in some
way injurious. On the contrary, as they are often used
by the males of many animals for various purposes,
more especially as a defence against their enemies, it is
a surprising fact that they are so poorly developed or
quite absent in the females. No doubt with female deer
the development during each recurrent season of great
branching horns, and with female elephants the development
of immense tusks, would have been a great
waste of vital power, on the admission that they were
of no use to the females. Consequently variations in
the size of these organs, leading to their suppression,
would have come under the control of natural selection,
and if limited in their transmission to the female offspring
would not have interfered with their development
through sexual selection in the males. But how
on this view can we explain the presence of horns in the
females of certain antelopes, and of tusks in the females
of many animals, which are only of slightly less size
than in the males? The explanation in almost all cases
must, I believe, be sought in the laws of transmission.
As the reindeer is the single species in the whole
family of Deer in which the female is furnished with
horns, though somewhat smaller, thinner, and less
branched than in the male, it might naturally be
thought that they must be of some special use to her.
There is, however, some evidence opposed to this view.
The female retains her horns from the time when they
are fully developed, namely in September, throughout
the winter, until May, when she brings forth her young;
whilst the male casts his horns much earlier, towards the
end of November. As both sexes have the same requirements
and follow the same habits of life, and as the male244
sheds his horns during the winter, it is very improbable
that they can be of any special service to the female at
this season, which includes the larger proportion of the
time during which she bears horns. Nor is it probable
that she can have inherited horns from some ancient
progenitor of the whole family of deer, for, from the fact
of the males alone of so many species in all quarters of
the globe possessing horns, we may conclude that this
was the primordial character of the group. Hence it
appears that horns must have been transferred from the
male to the female at a period subsequent to the divergence
of the various species from a common stock; but
that this was not effected for the sake of giving her any
special advantage.295
We know that the horns are developed at a most
unusually early age in the reindeer; but what the cause
of this may have been is not known. The effect, however,
has apparently been the transference of the horns
to both sexes. It is intelligible on the hypothesis of
pangenesis, that a very slight change in the constitution
of the male, either in the tissues of the forehead or in
the gemmules of the horns, might lead to their early
development; and as the young of both sexes have
nearly the same constitution before the period of reproduction,
the horns, if developed at an early age in the
male, would tend to be developed equally in both sexes.
In support of this view, we should bear in mind that the
horns are always transmitted through the female, and
that she has a latent capacity for their development, as
we see in old or diseased females.296 Moreover the females
245of some other species of deer either normally or occasionally
exhibit rudiments of horns; thus the female of
Cervulus moschatus has “bristly tufts, ending in a knob,
instead of a horn;” and “in most specimens of the
female Wapiti (Cervus Canadensis) there is a sharp
bony protuberance in the place of the horn.”297 From
these several considerations we may conclude that the
possession of fairly well-developed horns by the female
reindeer, is due to the males having first acquired them
as weapons for fighting with other males; and secondarily
to their development from some unknown cause at an
unusually early age in the males, and their consequent
transmission to both sexes.
Turning to the sheath-horned ruminants: with antelopes
a graduated series can be formed, beginning with
the species, the females of which are completely destitute
of horns—passing to those which have horns so
small as to be almost rudimentary, as in Antilocapra
Americana—to those which have fairly well-developed
horns, but manifestly smaller and thinner than in the
male, and sometimes of a different shape,298 and ending
with those in which both sexes have horns of equal size.
As with the reindeer, so with antelopes there exists a
relation between the period of the development of the
horns and their transmission to one or both sexes; it
246is therefore probable that their presence or absence in
the females of some species, and their more or less perfect
condition in the females of other species, depend,
not on their being of some special use, but simply on
the form of inheritance which has prevailed. It accords
with this view that even in the same restricted
genus both sexes of some species, and the males alone
of other species, are thus provided. It is a remarkable
fact that, although the females of Antilope bezoartica
are normally destitute of horns, Mr. Blyth has seen no
less than three females thus furnished; and there was
no reason to suppose that they were old or diseased.
The males of this species have long straight spirated
horns, nearly parallel to each other, and directed backwards.
Those of the female, when present, are very
different in shape, for they are not spirated, and
spreading widely bend round, so that their points are
directed forwards. It is a still more remarkable fact
that in the castrated male, as Mr. Blyth informs me, the
horns are of the same peculiar shape as in the female,
but longer and thicker. In all cases the differences
between the horns of the males and females, and of
castrated and entire males, probably depend on various
causes,—on the more or less complete transference of
male characters to the females,—on the former state
of the progenitors of the species,—and partly perhaps on
the horns being differently nourished, in nearly the same
manner as the spurs of the domestic cock, when inserted
into the comb or other parts of the body, assume various
abnormal forms from being differently nourished.
In all the wild species of goats and sheep the horns
are larger in the male than in the female, and are sometimes
quite absent in the latter.299 In several domestic
247breeds of the sheep and goat, the males alone are furnished
with horns; and it is a significant fact, that in
one such breed of sheep on the Guinea coast, the horns
are not developed, as Mr. Winwood Reade informs me,
in the castrated male; so that they are affected in
this respect like the horns of stags. In some breeds,
as in that of N. Wales, in which both sexes are properly
horned, the ewes are very liable to be hornless. In
these same sheep, as I have been informed by a trustworthy
witness who purposely inspected a flock during
the lambing-season, the horns at birth are generally
more fully developed in the male than in the female.
With the adult musk-ox (Ovibos moschatus) the horns of
the male are larger than those of the female, and in the
latter the bases do not touch.300 In regard to ordinary
cattle Mr. Blyth remarks: “In most of the wild bovine
animals the horns are both longer and thicker in the
bull than in the cow, and in the cow-banteng (Bos
sondaicus) the horns are remarkably small, and inclined
much backwards. In the domestic races of
cattle, both of the humped and humpless types, the
horns are short and thick in the bull, longer and
more slender in the cow and ox; and in the Indian
buffalo, they are shorter and thicker in the bull, longer
and more slender in the cow. In the wild gaour
(B. gaurus) the horns are mostly both longer and
thicker in the bull than in the cow.”301 Hence with
most sheath-horned ruminants the horns of the male
are either longer or stronger than those of the female.
With the Rhinoceros simus, as I may here add, the
horns of the female are generally longer but less powerful
than in the male; and in some other species of
248rhinoceros they are said to be shorter in the female.302
From these various facts we may conclude that horns
of all kinds, even when they are equally developed in
both sexes, were primarily acquired by the males in
order to conquer other males, and have been transferred
more or less completely to the female, in relation
to the force of the equal form of inheritance.
The tusks of the elephant, in the different species or
races, differ according to sex, in nearly the same manner
as the horns of ruminants. In India and Malacca the
males alone are provided with well-developed tusks.
The elephant of Ceylon is considered by most naturalists
as a distinct race, but by some as a distinct
species, and here “not one in a hundred is found with
tusks, the few that possess them being exclusively
males.”303 The African elephant is undoubtedly distinct,
and the female has large, well-developed tusks,
though not so large as those of the male. These differences
in the tusks of the several races and species of
elephants—the great variability of the horns of deer,
as notably in the wild reindeer—the occasional presence
of horns in the female Antilope bezoartica—the
presence of two tusks in some few male narwhals—the
complete absence of tusks in some female walruses;—are
all instances of the extreme variability of secondary
sexual characters, and of their extreme liability to
differ in closely-allied forms.
Although tusks and horns appear in all cases to have
been primarily developed as sexual weapons, they often
serve for other purposes. The elephant uses his tusks
249in attacking the tiger; according to Bruce, he scores
the trunks of trees until they can be easily thrown
down, and he likewise thus extracts the farinaceous
cores of palms; in Africa he often uses one tusk, this
being always the same, to probe the ground and thus
to ascertain whether it will bear his weight. The
common bull defends the herd with his horns; and
the elk in Sweden has been known, according to Lloyd,
to strike a wolf dead with a single blow of his great
horns. Many similar facts could be given. One of the
most curious secondary uses to which the horns of any
animal are occasionally put, is that observed by Captain
Hutton304 with the wild goat (Capra ægagrus) of the
Himalayas, and as it is said with the ibex, namely, that
when the male accidentally falls from a height he
bends inwards his head, and, by alighting on his massive
horns, breaks the shock. The female cannot thus
use her horns, which are smaller, but from her more
quiet disposition she does not so much need this strange
kind of shield.
Each male animal uses his weapons in his own peculiar
fashion. The common ram makes a charge and
butts with such force with the bases of his horns, that I
have seen a powerful man knocked over as easily as a
child. Goats and certain species of sheep, for instance
the Ovis cycloceros of Afghanistan,305 rear on their hind
legs, and then not only butt, but “make a cut down
and a jerk up, with the ribbed front of their scimitar-shaped
horn, as with a sabre. When the O. cycloceros
attacked a large domestic ram, who was a noted
bruiser, he conquered him by the sheer novelty of his
250mode of fighting, always closing at once with his
adversary, and catching him across the face and nose
with a sharp drawing jerk of his head, and then
bounding out of the way before the blow could be
returned.” In Pembrokeshire a male goat, the master
of a flock which during several generations had run
wild, was known to have killed several other males in
single combat; this goat possessed enormous horns,
measuring 39 inches in a straight line from tip to tip.
The common bull, as every one knows, gores and tosses
his opponent; but the Italian buffalo is said never to
use his horns, he gives a tremendous blow with his
convex forehead, and then tramples on his fallen enemy
with his knees—an instinct which the common bull does
not possess.306 Hence a dog who pins a buffalo by
the nose is immediately crushed. We must, however,
remember that the Italian buffalo has long been domesticated,
and it is by no means certain that the wild
parent-form had similarly shaped horns. Mr. Bartlett
informs me that when a female Cape buffalo (Bubalus
caffer) was turned into an enclosure with a bull of
the same species, she attacked him, and he in return
pushed her about with great violence. But it was
manifest to Mr. Bartlett that had not the bull shewn
dignified forbearance, he could easily have killed her
by a single lateral thrust with his immense horns. The
giraffe uses his short hair-covered horns, which are
rather longer in the male than in the female, in a
curious manner; for with his long neck he swings his
head to either side, almost upside down, with such
force, that I have seen a hard plank deeply indented
by a single blow.
Fig. 61. Oryx leucoryx, male (from the Knowsley Menagerie).
251
With antelopes it is sometimes difficult to imagine
how they can possibly use their curiously-shaped horns;
thus the spring-boc (Ant. euchore) has rather short upright
horns, with the sharp points bent inwards almost
at a right angle, so as to face each other; Mr. Bartlett
does not know how they are used, but suggests that
they would inflict a fearful wound down each side of
the face of an antagonist. The slightly-curved horns of
the Oryx leucoryx (fig. 61) are directed backwards, and
are of such length that their points reach beyond the
middle of the back, over which they stand in an almost
parallel line. Thus they seem singularly ill-fitted for
fighting; but Mr. Bartlett informs me that when two
of these animals prepare for battle, they kneel down,
with their heads between their front legs, and in this
attitude the horns stand nearly parallel and close to
the ground, with the points directed forwards and a
little upwards. The combatants then gradually approach
each other and endeavour to get the upturned
points under each other’s bodies; if one succeeds in252
doing this, he suddenly springs up, throwing up his
head at the same time, and can thus wound or perhaps
even transfix his antagonist. Both animals always kneel
down so as to guard as far as possible against this
manœuvre. It has been recorded that one of these
antelopes has used his horns with effect even against a
lion; yet from being forced to place his head between
the forelegs in order to bring the points of the horns
forward, he would generally be under a great disadvantage
when attacked by any other animal. It is,
therefore, not probable that the horns have been modified
into their present great length and peculiar position, as
a protection against beasts of prey. We can, however,
see that as soon as some ancient male progenitor of the
Oryx acquired moderately long horns, directed a little
backwards, he would be compelled in his battles with
rival males to bend his head somewhat inwards or downwards,
as is now done by certain stags; and it is not
improbable that he might have acquired the habit of
at first occasionally and afterwards of regularly kneeling
down. In this case it is almost certain that the
males which possessed the longest horns would have
had a great advantage over others with shorter horns;
and then the horns would gradually have been rendered
longer and longer, through sexual selection, until
they acquired their present extraordinary length and
position.
With stags of many kinds the branching of the horns
offers a curious case of difficulty; for certainly a single
straight point would inflict a much more serious wound
than several diverging points. In Sir Philip Egerton’s
museum there is a horn of the red-deer (Cervus elaphus)
thirty inches in length, with “not fewer than
fifteen snags or branches;” and at Moritzburg there
is still preserved a pair of antlers of a red-deer, shot in253
1699 by Frederick I., each of which bears the astonishing
number of thirty-three branches. Richardson
figures a pair of antlers of the wild reindeer with twenty-nine
points.307 From the manner in which the horns
are branched, and more especially from deer being
known occasionally to fight together by kicking with
their fore-feet,308 M. Bailly actually came to the conclusion
that their horns were more injurious than useful
to them! But this author overlooks the pitched battles
between rival males. As I felt much perplexed about
the use or advantage of the branches, I applied to Mr.
McNeill of Colinsay, who has long and carefully observed
the habits of red-deer, and he informs me that
he has never seen some of the branches brought into
action, but that the brow-antlers, from inclining downwards,
are a great protection to the forehead, and their
points are likewise used in attack. Sir Philip Egerton
also informs me in regard both to red-deer and fallow-deer,
that when they fight they suddenly dash together,
and getting their horns fixed against each other’s bodies
a desperate struggle ensues. When one is at last
forced to yield and turn round, the victor endeavours
to plunge his brow-antlers into his defeated foe. It
thus appears that the upper branches are used chiefly
or exclusively for pushing and fencing. Nevertheless
with some species the upper branches are used as
weapons of offence; when a man was attacked by a
254Wapiti deer (Cervus Canadensis) in Judge Caton’s park
in Ottawa, and several men tried to rescue him, the stag
“never raised his head from the ground; in fact he kept
his face almost flat on the ground, with his nose nearly
between his fore-feet, except when he rolled his head
to one side to take a new observation preparatory to
a plunge.” In this position the terminal points of
the horns were directed against his adversaries. “In
rolling his head he necessarily raised it somewhat,
because his antlers were so long that he could not
roll his head without raising them on one side, while
on the other side they touched the ground.” The
stag by this procedure gradually drove the party of
rescuers backwards, to a distance of 150 or 200 feet;
and the attacked man was killed.309
Although the horns of stags are efficient weapons,
there can, I think, be no doubt that a single point
would have been much more dangerous than a branched
antler; and Judge Caton, who has had large experience
with deer, fully concurs in this conclusion. Nor
do the branching horns, though highly important as a
means of defence against rival stags, appear perfectly
well adapted for this purpose, as they are liable to
become interlocked. The suspicion has therefore crossed
my mind that they may serve partly as ornaments.
That the branched antlers of stags, as well as the
elegant lyrated horns of certain antelopes, with their
graceful double curvature, (fig. 62), are ornamental
in our eyes, no one will dispute. If, then, the horns,
like the splendid accoutrements of the knights of old,
add to the noble appearance of stags and antelopes,
they may have been partly modified for this purpose,
255though mainly for actual service in battle; but I have
no evidence in favour of this belief.
Fig. 62. Strepsiceros Kudu (from Andrew Smith’s ‘Zoology of South Africa’).
An interesting case has lately been published, from
which it appears that the horns of a deer in one district
in the United States are now being modified through
sexual and natural selection. A writer in an excellent256
American Journal310 says, that he has hunted for the
last twenty-one years in the Adirondacks, where the
Cervus Virginianus abounds. About fourteen years ago
he first heard of spike-horn bucks. These became from
year to year more common; about five years ago he
shot one, and subsequently another, and now they are
frequently killed. “The spike-horn differs greatly
from the common antler of the C. Virginianus. It
consists of a single spike, more slender than the antler,
and scarcely half so long, projecting forward from the
brow, and terminating in a very sharp point. It gives
a considerable advantage to its possessor over the
common buck. Besides enabling him to run more
swiftly through the thick woods and underbrush
(every hunter knows that does and yearling bucks
run much more rapidly than the large bucks when
armed with their cumbrous antlers), the spike-horn
is a more effective weapon than the common antler.
With this advantage the spike-horn bucks are gaining
upon the common bucks, and may, in time, entirely
supersede them in the Adirondacks. Undoubtedly
the first spike-horn buck was merely an accidental
freak of nature. But his spike-horns gave him an
advantage, and enabled him to propagate his peculiarity.
His descendants, having a like advantage,
have propagated the peculiarity in a constantly
increasing ratio, till they are slowly crowding the
antlered deer from the region they inhabit.”
Male quadrupeds which are furnished with tusks
use them in various ways, as in the case of horns.
The boar strikes laterally and upwards; the musk-deer
with serious effect downwards.311 The walrus,
257though having so short a neck and so unwieldy a body,
“can strike either upwards, or downwards, or sideways,
with equal dexterity.”312 The Indian elephant
fights, as I was informed by the late Dr. Falconer, in a
different manner according to the position and curvature
of his tusks. When they are directed forwards and
upwards he is able to fling a tiger to a great distance—it
is said to even thirty feet; when they are short and
turned downwards he endeavours suddenly to pin the
tiger to the ground, and in consequence is dangerous
to the rider, who is liable to be jerked off the
hoodah.313
Very few male quadrupeds possess weapons of two
distinct kinds specially adapted for fighting with rival
males. The male muntjac-deer (Cervulus), however,
offers an exception, as he is provided with horns and
exserted canine teeth. But one form of weapon, has
often been replaced in the course of ages by another
form, as we may infer from what follows. With ruminants
the development of horns generally stands
in an inverse relation with that of even moderately
well-developed canine teeth. Thus camels, guanacoes,
chevrotains and musk-deer, are hornless, and they
have efficient canines; these teeth being “always of
smaller size in the females than in the males.” The
Camelidæ have in their upper jaws, in addition to
their true canines, a pair of canine-shaped incisors.314
Male deer and antelopes, on the other hand, possess
horns, and they rarely have canine teeth; and these
when present are always of small size, so that it is
258doubtful whether they are of any service in their
battles. With Antilope montana they exist only as
rudiments in the young male, disappearing as he
grows old; and they are absent in the female at
all ages; but the females of certain other antelopes
and deer have been known occasionally to exhibit
rudiments of these teeth.315 Stallions have small canine
teeth, which are either quite absent or rudimentary
in the mare; but they do not appear to be used in
fighting, for stallions bite with their incisors, and do
not open their mouths widely like camels and guanacoes.
Whenever the adult male possesses canines now
in an inefficient state, whilst the female has either none
or mere rudiments, we may conclude that the early
male progenitor of the species was provided with efficient
canines, which had been partially transferred to
the females. The reduction of these teeth in the males
seems to have followed from some change in their
manner of fighting, often caused (but not in the case
of the horse) by the development of new weapons.
Tusks and horns are manifestly of high importance to
their possessors, for their development consumes much
organised matter. A single tusk of the Asiatic elephant,—one
of the extinct woolly species,—and of the
African elephant, have been known to weigh respectively
150, 160, and 180 pounds; and even greater weights
have been assigned by some authors.316 With deer, in
259which the horns are periodically renewed, the drain
on the constitution must be greater; the horns, for
instance, of the moose weigh from fifty to sixty pounds,
and those of the extinct Irish elk from sixty to seventy
pounds,—the skull of the latter weighing on an average
only five and a quarter pounds. With sheep, although
the horns are not periodically renewed, yet their development,
in the opinion of many agriculturists, entails
a sensible loss to the breeder. Stags, moreover,
in escaping from beasts of prey are loaded with
an additional weight for the race, and are greatly
retarded in passing through a woody country. The
moose, for instance, with horns extending five and a
half feet from tip to tip, although so skilful in their
use that he will not touch or break a dead twig
when walking quietly, cannot act so dexterously whilst
rushing away from a pack of wolves. “During his
progress he holds his nose up, so as to lay the
horns horizontally back; and in this attitude cannot
see the ground distinctly.”317 The tips of the horns
of the great Irish elk were actually eight feet apart!
Whilst the horns are covered with velvet, which lasts
with the red-deer for about twelve weeks, they are
extremely sensitive to a blow; so that in Germany
the stags at this time change their habits to a certain
extent, and avoid dense forests, frequenting young
woods and low thickets.318 These facts remind us, that
male birds have acquired ornamental plumes at the
cost of retarded flight, and other ornaments at the cost
of some loss of power in their battles with rival males.
260
With quadrupeds, when, as is often the case, the
sexes differ in size, the males are, I believe, always
larger and stronger. This holds good in a marked
manner, as I am informed by Mr. Gould, with the marsupials
of Australia, the males of which appear to
continue growing until an unusually late age. But
the most extraordinary case is that of one of the
seals (Callorhinus ursinus), a full-grown female weighing
less than one-sixth of a full-grown male.319 The
greater strength of the male is invariably displayed,
as Hunter long ago remarked,320 in those parts of the
body which are brought into action in fighting with
rival males,—for instance, in the massive neck of the
bull. Male quadrupeds are also more courageous and
pugnacious than the females. There can be little
doubt that these characters have been gained, partly
through sexual selection, owing to a long series of victories
by the stronger and more courageous males over
the weaker, and partly through the inherited effects of
use. It is probable that the successive variations in
strength, size, and courage, whether due to so-called
spontaneous variability or to the effects of use, by the
accumulation of which male quadrupeds have acquired
these characteristic qualities, occurred rather late in
life, and were consequently to a large extent limited
in their transmission to the same sex.
Under this point of view I was anxious to obtain
information in regard to the Scotch deerhound, the
sexes of which differ more in size than those of any
other breed (though blood-hounds differ considerably),
or than in any wild canine species known to me.
261
Accordingly, I applied to Mr. Cupples, a well-known
breeder of these dogs, who has weighed and measured
many of his own dogs, and who, with great kindness, has
collected for me the following facts from various sources.
Superior male dogs, measured at the shoulder, range
from twenty-eight inches, which is low, to thirty-three,
or even thirty-four inches in height; and in weight
from eighty pounds, which is low, to 120, or even more
pounds. The females range in height from twenty-three
to twenty-seven, or even to twenty-eight inches;
and in weight from fifty to seventy, or even eighty
pounds.321 Mr. Cupples concludes that from ninety-five
to one hundred pounds for the male, and seventy for
the female, would be a safe average; but there is reason
to believe that formerly both sexes attained a greater
weight. Mr. Cupples has weighed puppies when a
fortnight old; in one litter the average weight of four
males exceeded that of two females by six and a half
ounces; in another litter the average weight of four
males exceeded that of one female by less than one
ounce; the same males, when three weeks old, exceeded
the female by seven and a half ounces, and at the age
of six weeks by nearly fourteen ounces. Mr. Wright
of Yeldersley House, in a letter to Mr. Cupples, says:
“I have taken notes on the sizes and weights of puppies
of many litters, and as far as my experience goes,
dog-puppies as a rule differ very little from bitches
till they arrive at about five or six months old; and
then the dogs begin to increase, gaining upon the
262bitches both in weight and size. At birth, and for
several weeks afterwards, a bitch-puppy will occasionally
be larger than any of the dogs, but they are
invariably beaten by them later.” Mr. McNeill, of
Colinsay, concludes that “the males do not attain
their full growth till over two years old, though
the females attain it sooner.” According to Mr.
Cupples’ experience, male dogs go on growing in
stature till they are from twelve to eighteen months
old, and in weight till from eighteen to twenty-four
months old; whilst the females cease increasing in
stature at the age of from nine to fourteen or fifteen
months, and in weight at the age of from twelve to
fifteen months. From these various statements it is
clear that the full difference in size between the
male and female Scotch deerhound is not acquired
until rather late in life. The males are almost exclusively
used for coursing, for, as Mr. McNeill informs
me, the females have not sufficient strength and weight
to pull down a full-grown deer. From the names used
in old legends, it appears, as I hear from Mr. Cupples,
that at a very ancient period the males were the most
celebrated, the females being mentioned only as the
mothers of famous dogs. Hence during many generations,
it is the male which has been chiefly tested
for strength, size, speed, and courage, and the best
will have been bred from. As, however, the males
do not attain their full dimensions until a rather
late period in life, they will have tended, in accordance
with the law often indicated, to transmit
their characters to their male offspring alone; and
thus the great inequality in size between the sexes
of the Scotch deerhound may probably be accounted
for.
The males of some few quadrupeds possess organs or263
parts developed solely as a means of defence against
the attacks of other males. Some kinds of deer use,
as we have seen, the upper branches of their horns
chiefly or exclusively for defending themselves; and
the Oryx antelope, as I am informed by Mr. Bartlett,
fences most skilfully with his long, gently curved horns;
but these are likewise used as organs of offence. Rhinoceroses,
as the same observer remarks, in fighting
parry each other’s sidelong blows with their horns,
which loudly clatter together, as do the tusks of boars.
Although wild boars fight desperately together, they
seldom, according to Brehm, receive fatal blows, as
these fall on each other’s tusks, or on the layer of
gristly skin covering the shoulder, which the German
hunters call the shield; and here we have a part specially
modified for defence. With boars in the prime
 Fig. 63. Head of common wild boar, in prime
of life (from Brehm).
of life (see fig. 63) the
tusks in the lower jaw
are used for fighting
but they become in
old age, as Brehm
states, so much curved
inwards and upwards,
over the snout, that
they can no longer be
thus used. They may,
however, still continue
to serve, and even in
a still more effective
manner, as a means of defence. In compensation for
the loss of the lower tusks as weapons of offence, those
in the upper jaw, which always project a little laterally,
increase so much in length during old age, and
curve so much upwards, that they can be used as a
means of attack. Nevertheless an old boar is not so264
dangerous to man as one at the age of six or seven
years.322
Fig. 63. Head of common wild boar, in prime
of life (from Brehm).
of life (see fig. 63) the
tusks in the lower jaw
are used for fighting
but they become in
old age, as Brehm
states, so much curved
inwards and upwards,
over the snout, that
they can no longer be
thus used. They may,
however, still continue
to serve, and even in
a still more effective
manner, as a means of defence. In compensation for
the loss of the lower tusks as weapons of offence, those
in the upper jaw, which always project a little laterally,
increase so much in length during old age, and
curve so much upwards, that they can be used as a
means of attack. Nevertheless an old boar is not so264
dangerous to man as one at the age of six or seven
years.322
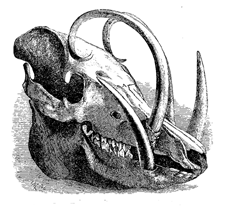
Fig. 64. Skull of the Babirusa Pig (from Wallace’s ‘Malay Archipelago’).
In the full-grown male Babirusa pig of Celebes
(fig. 64), the lower tusks are formidable weapons, like
those of the European boar in the prime of life, whilst
the upper tusks are so long and have their points so
much curled inwards, sometimes even touching the
forehead, that they are utterly useless as weapons of
attack. They more nearly resemble horns than teeth,
and are so manifestly useless as teeth that the animal
was formerly supposed to rest his head by hooking them
on to a branch. Their convex surfaces would, however,
265if the head were held a little laterally, serve as an
excellent guard; and hence, perhaps it is that in
old animals they “are generally broken off, as if by
fighting.”323 Here, then, we have the curious case of
the upper tusks of the Babirusa regularly assuming
during the prime of life, a structure which apparently
renders them fitted only for defence; whilst in the European
boar the lower and opposite tusks assume in a less
degree and only during old age nearly the same form,
and then serve in like manner solely for defence.
Fig. 65. Head of Æthiopian Wart-hog, from ‘Proc. Zool. Soc.’ 1869. (I now find that
this drawing represents the head of a female, but it serves to shew, on a reduced
scale, the characters of the male.)
In the wart-hog (Phacochœrus æthiopicus, fig. 65)
the tusks in the upper jaw of the male curve upwards
during the prime of life, and from being pointed,
serve as formidable weapons. The tusks in the lower
jaw are sharper than those in the upper, but from their
shortness it seems hardly possible that they can be used
as weapons of attack. They must, however, greatly
266strengthen those in the upper jaw, from being ground
so as to fit closely against their bases. Neither the
upper nor the lower tusks appear to have been specially
modified to act as guards, though, no doubt, they
are thus used to a certain extent. But the wart-hog is
not destitute of other special means of protection, for
there exists, on each side of the face, beneath the eyes,
a rather stiff, yet flexible, cartilaginous, oblong pad
(fig. 65), which projects two or three inches outwards;
and it appeared to Mr. Bartlett and myself, when viewing
the living animal, that these pads, when struck from
beneath by the tusks of an opponent, would be turned
upwards, and would thus protect in an admirable manner
the somewhat prominent eyes. These boars, as I
may add on the authority of Mr. Bartlett, when fighting
together, stand directly face to face.
Lastly, the African river-hog (Potamochoerus penicillatus)
has a hard cartilaginous knob on each side of
the face beneath the eyes, which answers to the flexible
pad of the wart-hog; it has also two bony prominences
on the upper jaw above the nostrils. A boar of this
species in the Zoological Gardens recently broke into
the cage of the wart-hog. They fought all night-long,
and were found in the morning much exhausted, but
not seriously wounded. It is a significant fact, as
shewing the purpose of the above-described projections
and excrescences, that these were covered with blood,
and were scored and abraded in an extraordinary
manner.
The mane of the lion forms a good defence against
the one danger to which he is liable, namely the attacks
of rival lions: for the males, as Sir. A. Smith
informs me, engage in terrible battles, and a young
lion dares not approach an old one. In 1857 a tiger
at Bromwich broke into the cage of a lion, and a267
fearful scene ensued; “the lion’s mane saved his neck
and head from being much injured, but the tiger at
last succeeded in ripping up his belly, and in a few
minutes he was dead.”324 The broad ruff round the
throat and chin of the Canadian lynx (Felis Canadensis)
is much longer in the male than in the female; but
whether it serves as a defence I do not know. Male
seals are well known to fight desperately together, and
the males of certain kinds (Otaria jubata)325 have great
manes, whilst the females have small ones or none.
The male baboon of the Cape of Good Hope (Cynocephalus
porcarius) has a much longer mane and larger
canine teeth than the female; and the mane probably
serves as a protection, for on asking the keepers
in the Zoological Gardens, without giving them any
clue to my object, whether any of the monkeys especially
attacked each other by the nape of the neck, I
was answered that this was not the case, excepting with
the above baboon. In the Hamadryas baboon, Ehrenberg
compares the mane of the adult male to that
of a young lion, whilst in the young of both sexes and
in the female the mane is almost absent.
It appeared to me probable that the immense woolly
mane of the male American bison, which reaches
almost to the ground, and is much more developed
in the males than in the females, served as a protection
to them in their terrible battles; but an experienced
hunter told Judge Caton that he had never
observed anything which favoured this belief. The
268stallion has a thicker and fuller mane than the mare;
and I have made particular inquiries of two great
trainers and breeders who have had charge of many
entire horses, and am assured that they “invariably
endeavour to seize one another by the neck.” It
does not, however, follow from the foregoing statements,
that when the hair on the neck serves as a
defence, that it was originally developed for this purpose,
though this is probable in some cases, as in that
of the lion. I am informed by Mr. McNeill that the
long hairs on the throat of the stag (Cervus elephas)
serve as a great protection to him when hunted, for
the dogs generally endeavour to seize him by the
throat; but it is not probable that these hairs were
specially developed for this purpose; otherwise the
young and the females would, as we may feel assured,
have been equally protected.
On Preference or Choice in Pairing, as shewn by either
sex of Quadrupeds.—Before describing, in the next chapter,
the differences between the sexes in voice, odour
emitted, and ornamentation, it will be convenient here
to consider whether the sexes exert any choice in their
unions. Does the female prefer any particular male,
either before or after the males may have fought together
for supremacy; or does the male, when not a polygamist,
select any particular female? The general impression
amongst breeders seems to be that the male
accepts any female; and this, owing to his eagerness,
is, in most cases, probably the truth. Whether the
female as a general rule indifferently accepts any male
is much more doubtful. In the fourteenth chapter,
on Birds, a considerable body of direct and indirect
evidence was advanced, shewing that the female selects
her partner; and it would be a strange anomaly if269
female quadrupeds, which stand higher in the scale of
organisation and have higher mental powers, did not
generally, or at least often, exert some choice. The
female could in most cases escape, if wooed by a male
that did not please or excite her; and when pursued,
as so incessantly occurs, by several males, she would
often have the opportunity, whilst they were fighting
together, of escaping with, or at least of temporarily
pairing with, some one male. This latter contingency
has often been observed in Scotland with female red-deer,
as I have been informed by Sir Philip Egerton.326
It is scarcely possible that much should be known
about female quadrupeds exerting in a state of nature
any choice in their marriage unions. The following
very curious details on the courtship of one of the
eared seals, Callorhinus ursinus, are given327 on the
authority of Capt. Bryant, who had ample opportunities
for observation. He says, “Many of the females on
their arrival at the island where they breed appear
desirous of returning to some particular male, and
frequently climb the outlying rocks to overlook the
rookeries, calling out and listening as if for a familiar
voice. Then changing to another place they do the
same again.... As soon as a female reaches the
shore, the nearest male goes down to meet her, making
meanwhile a noise like the clucking of a hen to her
chickens. He bows to her and coaxes her until he
gets between her and the water so that she cannot
escape him. Then his manner changes, and with a
270harsh growl he drives her to a place in his harem.
This continues until the lower row of harems is
nearly full. Then the males higher up select the
time when their more fortunate neighbours are off
their guard to steal their wives. This they do by
taking them in their mouths and lifting them over
the heads of the other females, and carefully placing
them in their own harem, carrying them as cats do
their kittens. Those still higher up pursue the same
method until the whole space is occupied. Frequently
a struggle ensues between two males for the possession
of the same female, and both seizing her at once pull
her in two or terribly lacerate her with their teeth.
When the space is all filled, the old male walks around
complacently reviewing his family, scolding those
who crowd or disturb the others, and fiercely driving
off all intruders. This surveillance always keeps him
actively occupied.”
As so little is known about the courtship of animals in
a state of nature, I have endeavoured to discover how far
our domesticated quadrupeds evince any choice in their
unions. Dogs offer the best opportunity for observation,
as they are carefully attended to and well understood.
Many breeders have expressed a strong opinion on this
head. Thus Mr. Mayhew remarks, “The females are
able to bestow their affections; and tender recollections
are as potent over them as they are known to
be in other cases, where higher animals are concerned.
Bitches are not always prudent in their
loves, but are apt to fling themselves away on curs
of low degree. If reared with a companion of vulgar
appearance, there often springs up between the pair a
devotion which no time can afterwards subdue. The
passion, for such it really is, becomes of a more than
romantic endurance.” Mr. Mayhew, who attended271
chiefly to the smaller breeds, is convinced that the
females are strongly attracted by males of large size.328
The well-known veterinary Blaine states329 that his own
female pug became so attached to a spaniel, and a
female setter to a cur, that in neither case would they
pair with a dog of their own breed until several weeks
had elapsed. Two similar and trustworthy accounts
have been given me in regard to a female retriever
and a spaniel, both of which became enamoured with
terrier-dogs.
Mr. Cupples informs me that he can personally vouch
for the accuracy of the following more remarkable case,
in which a valuable and wonderfully-intelligent female
terrier loved a retriever, belonging to a neighbour, to
such a degree that she had often to be dragged away
from him. After their permanent separation, although
repeatedly shewing milk in her teats, she would never
acknowledge the courtship of any other dog, and to the
regret of her owner, never bore puppies. Mr. Cupples
also states that a female deerhound now (1868) in his
kennel has thrice produced puppies, and on each
occasion shewed a marked preference for one of the
largest and handsomest, but not the most eager, of four
deerhounds living with her, all in the prime of life.
Mr. Cupples has observed that the female generally
favours a dog whom she has associated with and
knows; her shyness and timidity at first incline her
against a strange dog. The male, on the contrary,
seems rather inclined towards strange females. It
appears to be rare when the male refuses any particular
female, but Mr. Wright, of Yeldersley House,
272a great breeder of dogs, informs me that he has known
some instances; he cites the case of one of his own
deerhounds, who would not take any notice of a particular
female mastiff, so that another deerhound had
to be employed. It would be superfluous to give other
cases, and I will only add that Mr. Barr, who has carefully
bred many blood-hounds, states that in almost
every instance particular individuals of the opposite
sex shew a decided preference for each other. Finally
Mr. Cupples, after attending to this subject for another
year, has recently written to me, “I have had full confirmation
of my former statement, that dogs in breeding
form decided preferences for each other, being
often influenced by size, bright colour, and individual
character, as well as by the degree of their previous
familiarity.”
In regard to horses, Mr. Blenkiron, the greatest
breeder of race-horses in the world, informs me that
stallions are so frequently capricious in their choice,
rejecting one mare and without any apparent cause
taking to another, that various artifices have to be
habitually used. The famous Monarque, for instance,
would never consciously look at the dam of Gladiateur,
and a trick had to be practised. We can partly see the
reason why valuable race-horse stallions, which are in
such demand, should be so particular in their choice.
Mr. Blenkiron has never known a mare to reject a
horse; but this has occurred in Mr. Wright’s stable,
so that the mare had to be cheated. Prosper Lucas330
quotes various statements from French authorities, and
remarks, “On voit des étalons qui s’éprennent d’une
jument, et négligent toutes les autres.” He gives, on
the authority of Baëlen, similar facts in regard to bulls.
273
Hoffberg, in describing the domesticated reindeer of
Lapland, says, “Fœmina majores et fortiores mares
præ cæteris admittunt, ad eos confugiunt, a junioribus
agitatæ, qui hos in fugam conjiciunt.”331 A clergy-man,
who has bred many pigs, assures me that sows
often reject one boar and immediately accept another.
From these facts there can be no doubt that with
most of our domesticated quadrupeds strong individual
antipathies and preferences are frequently exhibited,
and much more commonly by the female than by the
male. This being the case, it is improbable that the
unions of quadrupeds in a state of nature should be
left to mere chance. It is much more probable that
the females are allured or excited by particular males,
who possess certain characters in a higher degree than
other males; but what these characters are, we can
seldom or never discover with certainty.
274
CHAPTER XVIII.
Secondary Sexual Characters of Mammals—continued.
Voice—Remarkable sexual peculiarities in seals—Odour—Development
of the hair—Colour of the hair and skin—Anomalous
case of the female being more ornamented than the male—Colour
and ornaments due to sexual selection—Colour acquired
for the sake of protection—Colour, though common to both
sexes, often due to sexual selection—On the disappearance of
spots and stripes in adult quadrupeds—On the colours and
ornaments of the Quadrumana—Summary.
Quadrupeds use their voices for various purposes,
as a signal of danger, as a call from one member of
a troop to another, or from the mother to her lost
offspring, or from the latter for protection to their
mother; but such uses need not here be considered.
We are concerned only with the difference between the
voices of the two sexes, for instance between that of
the lion and lioness, or of the bull and cow. Almost
all male animals use their voices much more during
the rutting-season than at any other time; and some,
as the giraffe and porcupine,332 are said to be completely
mute excepting at this season. As the throats (i.e. the
larynx and thyroid bodies333) of stags become periodically
enlarged at the commencement of the breeding-season,
it might be thought that their powerful voices
must be then in some way of high importance to them;
but this is very doubtful. From information given to
me by two experienced observers, Mr. McNeill and Sir
275P. Egerton, it seems that young stags under three
years old do not roar or bellow; and that the old ones
begin bellowing at the commencement of the breeding-season,
at first only occasionally and moderately, whilst
they restlessly wander about in search of the females.
Their battles are prefaced by loud and prolonged bellowing,
but during the actual conflict they are silent.
Animals of all kinds which habitually use their voices,
utter various noises under any strong emotion, as when
enraged and preparing to fight; but this may merely
be the result of their nervous excitement, which leads
to the spasmodic contraction of almost all the muscles of
the body, as when a man grinds his teeth and clenches
his hands in rage or agony. No doubt stags challenge
each other to mortal combat by bellowing; but it is
not likely that this habit could have led through
sexual selection, that is by the loudest-voiced males
having been the most successful in their conflicts, to
the periodical enlargement of the vocal organs; for the
stags with the most powerful voices, unless at the same
time the strongest, best-armed, and most courageous,
would not have gained any advantage over their rivals
with weaker voices. The stags, moreover, which had
weaker voices, though not so well able to challenge other
stags, would have been drawn to the place of combat as
certainly as those with stronger voices.
It is possible that the roaring of the lion may be
of some actual service to him in striking terror into
his adversary; for when enraged he likewise erects his
mane and thus instinctively tries to make himself appear
as terrible as possible. But it can hardly be supposed
that the bellowing of the stag, even if it be of
any service to him in this way, can have been important
enough to have led to the periodical enlargement
of the throat. Some writers suggest that the276
bellowing serves as a call to the female; but the
experienced observers above quoted inform me that
female deer do not search for the male, though the
males search eagerly for the females, as indeed might
be expected from what we know of the habits of
other male quadrupeds. The voice of the female,
on the other hand, quickly brings to her one or more
stags,334 as is well known to the hunters who in wild
countries imitate her cry. If we could believe that
the male had the power to excite or allure the female
by his voice, the periodical enlargement of his vocal
organs would be intelligible on the principle of sexual
selection, together with inheritance limited to the same
sex and season of the year; but we have no evidence
in favour of this view. As the case stands, the loud
voice of the stag during the breeding season does not
seem to be of any special service to him, either during
his courtship or battles, or in any other way. But may
we not believe that the frequent use of the voice, under
the strong excitement of love, jealousy, and rage, continued
during many generations, may at last have
produced an inherited effect on the vocal organs of
the stag, as well as of other male animals? This
appears to me, with our present state of knowledge,
the most probable view.
The male gorilla has a tremendous voice, and when
adult is furnished with a laryngeal sack, as is likewise
the adult male orang.335 The gibbons rank amongst the
noisiest of monkeys, and the Sumatra species (Hylobates
syndactylus) is also furnished with a laryngeal sack; but
Mr. Blyth, who has had opportunities for observation,
277does not believe that the male is more noisy than the
female. Hence, these latter monkeys probably use their
voices as a mutual call; and this is certainly the case
with some quadrupeds, for instance with the beaver.336
Another gibbon, the H. agilis, is highly remarkable,
from having the power of emitting a complete and
correct octave of musical notes,337 which we may reasonably
suspect serves as a sexual charm; but I shall have to
recur to this subject in the next chapter. The vocal
organs of the American Mycetes caraya are one-third
larger in the male than in the female, and are wonderfully
powerful. These monkeys, when the weather is
warm, make the forests resound during the morning and
evening with their overwhelming voices. The males
begin the dreadful concert, in which the females, with
their less powerful voices, sometimes join, and which
is often continued during many hours. An excellent
observer, Rengger,338 could not perceive that they were
excited to begin their concert by any special cause; he
thinks that like many birds, they delight in their own
music, and try to excel each other. Whether most of the
foregoing monkeys have acquired their powerful voices
in order to beat their rivals and to charm the females—or
whether the vocal organs have been strengthened
and enlarged through the inherited effects of long-continued
use without any particular good being gained—I
will not pretend to say; but the former view, at
least in the case of the Hylobates agilis, seems the most
probable.
I may here mention two very curious sexual peculiarities
occurring in seals, because they have been sup278posed
by some writers to affect the voice. The nose of
the male sea-elephant (Macrorhinus proboscideus), when
about three years old, is greatly elongated during the
breeding-season, and can then be erected. In this state
it is sometimes a foot in length. The female at no
period of life is thus provided, and her voice is different.
That of the male consists of a wild, hoarse,
gurgling noise, which is audible at a great distance,
and is believed to be strengthened by the proboscis.
Lesson compares the erection of the proboscis, to the
swelling of the wattles of male gallinaceous birds, whilst
they court the females. In another allied kind of seal,
namely, the bladder-nose (Cystophora cristata), the head
is covered by a great hood or bladder. This is internally
supported by the septum of the nose, which is
produced far backwards and rises into a crest seven
inches in height. The hood is clothed with short hair,
and is muscular; it can be inflated until it more than
equals the whole head in size! The males when rutting
fight furiously on the ice, and their roaring “is
said to be sometimes so loud as to be heard four
miles off.” When attacked by man they likewise roar
or bellow; and whenever irritated the bladder is inflated.
Some naturalists believe that the voice is thus
strengthened, but various other uses have been assigned
to this extraordinary structure. Mr. R. Brown thinks
that it serves as a protection against accidents of all
kinds. This latter view is not probable, if what the
sealers have long maintained is correct, namely, that
the hood or bladder is very poorly developed in the
females and in the males whilst young.339
279
Odour.—With some animals, as with the notorious
skunk of America, the overwhelming odour which they
emit appears to serve exclusively as a means of defence.
With shrew-mice (Sorex) both sexes possess abdominal
scent-glands, and there can be little doubt, from the
manner in which their bodies are rejected by birds and
beasts of prey, that their odour is protective; nevertheless
the glands become enlarged in the males during
the breeding-season. In many quadrupeds the glands
are of the same size in both sexes;340 but their use is
not known. In other species the glands are confined
to the males, or are more developed in them than in
the females; and they almost always become more
active during the rutting-season. At this period the
glands on the sides of the face of the male elephant
enlarge and emit a secretion having a strong musky
odour.
The rank effluvium of the male goat is well known,
and that of certain male deer is wonderfully strong
and persistent. On the banks of the Plata I have perceived
the whole air tainted with the odour of the male
Cervus campestris, at the distance of half a mile to
leeward of a herd; and a silk handkerchief, in which I
carried home a skin, though repeatedly used and washed,
retained, when first unfolded, traces of the odour for
one year and seven months. This animal does not emit
its strong odour until more than a year old, and if cas280trated
whilst young never emits it.341 Besides the general
odour, with which the whole body of certain ruminants
seems to be permeated during the breeding-season, many
deer, antelopes, sheep, and goats, possess odoriferous
glands in various situations, more especially on their
faces. The so-called tear-sacks or suborbital pits come
under this head. These glands secrete a semi-fluid
fetid matter, which is sometimes so copious as to stain
the whole face, as I have seen in the case of an antelope.
They are “usually larger in the male than in
the female, and their development is checked by castration.”342
According to Desmarest they are altogether
absent in the female of Antilope subgutturosa.
Hence, there can be no doubt that they stand in some
close relation with the reproductive functions. They
are also sometimes present, and sometimes absent, in
nearly-allied forms. In the adult male musk-deer
(Moschus moschiferus), a naked space round the tail
is bedewed with an odoriferous fluid, whilst in the
adult female, and in the male, until two years old, this
space is covered with hair and is not odoriferous. The
proper musk-sack, from its position, is necessarily confined
to the male, and forms an additional scent-organ.
It is a singular fact that the matter secreted by this
latter gland does not, according to Pallas, change in
consistence, or increase in quantity, during the rutting-season;
nevertheless this naturalist admits that its presence
is in some way connected with the act of repro281duction.
He gives, however, only a conjectural and
unsatisfactory explanation of its use.343
In most cases, when during the breeding-season the
male alone emits a strong odour, this probably serves
to excite or allure the female. We must not judge
on this head by our own taste, for it is well known
that rats are enticed by certain essential oils, and
cats by valerian, substances which are far from agreeable
to us; and that dogs, though they will not eat
carrion, sniff and roll in it. From the reasons given
when discussing the voice of the stag, we may reject
the idea that the odour serves to bring the females
from a distance to the males. Active and long-continued
use cannot here have come into play, as in the case of
the vocal organs. The odour emitted must be of considerable
importance to the male, inasmuch as large
and complex glands, furnished with muscles for everting
the sack, and for closing or opening the orifice,
have in some cases been developed. The development
of these organs is intelligible through sexual selection,
if the more odoriferous males are the most successful in
winning the females, and in leaving offspring to inherit
their gradually-perfected glands and odours.
Development of the Hair.—We have seen that male
quadrupeds often have the hair on their necks and
shoulders much more developed than in the females;
and many additional instances could be given. This
sometimes serves as a defence to the male during his
battles; but whether the hair in most cases has been
specially developed for this purpose is very doubtful.
We may feel almost certain that this is not the case,
282when a thin and narrow crest runs along the whole
length of the back; for a crest of this kind would
afford scarcely any protection, and the ridge of the back
is not a likely place to be injured; nevertheless such
crests are sometimes confined to the males, or are
much more developed in them than in the females.
Two antelopes, the Tragelaphus scriptus344 (see fig. 68,
p. 300) and Portax picta, may be given as instances.
The crests of certain stags and of the male wild goat
stand erect, when these animals are enraged or terrified;345
but it can hardly be supposed that they have
been acquired for the sake of exciting fear in their
enemies. One of the above-named antelopes, the Portax
picta, has a large well-defined brush of black hair on
the throat, and this is much larger in the male than in
the female. In the Ammotragus tragelaphus of North
Africa, a member of the sheep-family, the front-legs
are almost concealed by an extraordinary growth of
hair, which descends from the neck and upper halves
of the legs; but Mr. Bartlett does not believe that this
mantle is of the least use to the male, in whom it is
much more developed than in the female.
Male quadrupeds of many kinds differ from the
females in having more hair, or hair of a different
character, on certain parts of their faces. The bull
alone has curled hair on the forehead.346 In three
closely-allied sub-genera of the goat family, the males
alone possess beards, sometimes of large size; in two
other sub-genera both sexes have a beard, but this
283disappears in some of the domestic breeds of the common
goat; and neither sex of the Hemitragus has a
beard. In the ibex the beard is not developed during
the summer, and is so small at other seasons that it
may be called rudimentary.347 With some monkeys the
beard is confined to the male, as in the Orang, or is
much larger in the male than in the female, as in the
Mycetes caraya and Pithecia satanas (fig. 66). So it is
with the whiskers of some species of Macacus,348 and, as
we have seen, with the manes of some species of baboons.
But with most kinds of monkeys the various tufts of
hair about the face and head are alike in both sexes.

Fig. 66. Pithecia Satanas, male (from Brehm).
284
The males of various members of the Ox family
(Bovidæ), and of certain antelopes, are furnished with
a dewlap, or great fold of skin on the neck, which is
much less developed in the female.
Now, what must we conclude with respect to such
sexual differences as these? No one will pretend that
the beards of certain male-goats, or the dewlap of the
bull, or the crests of hair along the backs of certain
male antelopes, are of any direct or ordinary use to
them. It is possible that the immense beard of the
male Pithecia, and the large beard of the male Orang,
may protect their throats when fighting; for the keepers
in the Zoological Gardens inform me that many monkeys
attack each other by the throat: but it is not probable
that the beard has been developed for a distinct
purpose from that which the whiskers, moustache,
and other tufts of hair on the face serve; and no one
will suppose that these are useful as a protection. Must
we attribute to mere purposeless variability in the male
all these appendages of hair or skin? It cannot be denied
that this is possible; for with many domesticated
quadrupeds, certain characters, apparently not derived
through reversion from any wild parent-form, have appeared
in, and are confined to, the males, or are more
largely developed in them than in the females,—for instance
the hump in the male zebu-cattle of India, the
tail in fat-tailed rams, the arched outline of the forehead
in the males of several breeds of sheep, the mane in the
ram of an African breed, and, lastly, the mane, long
hairs on the hinder legs, and the dewlap in the male
alone of the Berbura goat.349 The mane which occurs in
285the rams alone of the above-mentioned African breed of
sheep, is a true secondary sexual character, for it is not
developed, as I hear from Mr. Winwood Reade, if the
animal be castrated. Although we ought to be extremely
cautious, as shewn in my work on ‘Variation
under Domestication,’ in concluding that any character,
even with animals kept by semi-civilised people, has
not been subjected to selection by man, and thus augmented;
yet in the cases just specified this is improbable,
more especially as the characters are confined
to the males, or are more strongly developed in them
than in the females. If it were positively known that
the African ram with a mane was descended from the
same primitive stock with the other breeds of sheep,
or the Berbura male-goat with his mane, dewlap, &c.,
from the same stock with other goats; and if selection
has not been applied to these characters, then
they must be due to simple variability, together with
sexually-limited inheritance.
t would appear reasonable to extend
the same view to the many analogous characters
occurring in animals under a state of nature. Nevertheless
I cannot persuade myself that this view is
applicable in many cases, as in that of the extraordinary
development of hair on the throat and forelegs
of the male Ammotragus, or of the immense beard of
the male Pithecia. With those antelopes in which the
male when adult is more strongly-coloured than the
female, and with those monkeys in which this is likewise
the case, and in which the hair on the face is of a
different colour from that on the rest of the head, being
arranged in the most diversified and elegant manner,
it seems probable that the crests and tufts of hair have
286been acquired as ornaments; and this I know is the
opinion of some naturalists. If this view be correct,
there can be little doubt that they have been acquired,
or at least modified, through sexual selection.
Colour of the Hair and of the Naked Skin.—I will
first give briefly all the cases known to me, of male
quadrupeds differing in colour from the females. With
Marsupials, as I am informed by Mr. Gould, the sexes
rarely differ in this respect; but the great red kangaroo
offers a striking exception, “delicate blue being
the prevailing tint in those parts of the female,
which in the male are red.”350 In the Didelphis opossum
of Cayenne the female is said to be a little more
red than the male. With Rodents Dr. Gray remarks:
“African squirrels, especially those found in the tropical
regions, have the fur much brighter and more
vivid at some seasons of the year than at others, and
the fur of the male is generally brighter than that
of the female.”351 Dr. Gray informs me that he
specified the African squirrels, because, from their unusually
bright colours, they best exhibit this difference.
The female of the Mus minutus of Russia is of
a paler and dirtier tint than the male. In some few
bats the fur of the male is lighter and brighter than
in the female.352
The terrestrial Carnivora and Insectivora rarely exhibit
sexual differences of any kind, and their colours
are almost always exactly the same in both sexes. The
287ocelot (Felis pardalis), however, offers an exception, for
the colours of the female, compared with those of the
male, are “moins apparentes, le fauve étant plus terne,
le blanc moins pur, les raies ayant moins de largeur
et les taches moins de diamètre.”353 The sexes of
the allied Felis mitis also differ, but even in a less
degree, the general hues of the female being rather
paler than in the male, with the spots less black.
The marine Carnivora or Seals, on the other hand,
sometimes differ considerably in colour, and they present,
as we have already seen, other remarkable sexual
differences. Thus the male of the Otaria nigrescens
of the southern hemisphere is of a rich brown shade
above; whilst the female, who acquires her adult tints
earlier in life than the male, is dark-grey above, the
young of both sexes being of a very deep chocolate
colour. The male of the northern Phoca groenlandica
is tawny grey, with a curious saddle-shaped dark mark
on the back; the female is much smaller, and has a
very different appearance, being “dull white or yellowish
straw-colour, with a tawny hue on the back;” the
young at first are pure white, and can “hardly be distinguished
among the icy hummocks and snow, their
colour thus acting as a protection.”354
With Ruminants sexual differences of colour occur
more commonly than in any other order. A difference
of this kind is general with the Strepsicerene antelopes;
thus the male nilghau (Portax picta) is bluish-grey
and much darker than the female, with the square white
patch on the throat, the white marks on the fetlocks,
288and the black spots on the ears, all much more distinct.
We have seen that in this species the crests and
tufts of hair are likewise more developed in the male
than in the hornless female. The male, as I am
informed by Mr. Blyth, without shedding his hair,
periodically becomes darker during the breeding-season.
Young males cannot be distinguished from young
females until above twelve months old; and if the
male is emasculated before this period, he never, according
to the same authority, changes colour. The importance
of this latter fact, as distinctive of sexual colouring,
becomes obvious, when we hear355 that neither the red
summer-coat nor the blue winter-coat of the Virginian
deer is at all affected by emasculation. With most or
all of the highly-ornamented species of Tragelaphus the
males are darker than the hornless females, and their
crests of hair are more fully developed. In the male
of that magnificent antelope, the Derbyan Eland, the
body is redder, the whole neck much blacker, and the
white band which separates these colours, broader,
than in the female. In the Cape Eland also, the male
is slightly darker than the female.356
In the Indian Black-buck (A. bezoartica), which belongs
to another tribe of antelopes, the male is very dark, almost
black; whilst the hornless female is fawn-coloured. We
have in this species, as Mr. Blyth informs me, an exactly
parallel series of facts, as with the Portax picta, namely in
the male periodically changing colour during the breed289ing
season, in the effects of emasculation on this change,
and in the young of both sexes being undistinguishable
from each other. In the Antilope niger the male is
black, the female as well as the young being brown; in
A. sing-sing the male is much brighter coloured than the
hornless female, and his chest and belly are blacker;
in the male A. caama, the marks and lines which occur
on various parts of the body are black instead of as
in the female brown; in the brindled gnu (A. gorgon)
“the colours of the male are nearly the same as those
of the female, only deeper and of a brighter hue.”357
Other analogous cases could be added.
The Banteng bull (Bos sondaicus) of the Malayan
archipelago is almost black, with white legs and buttocks;
the cow is of a bright dun, as are the young
males until about the age of three years, when they
rapidly change colour. The emasculated bull reverts
to the colour of the female. The female Kemas goat
is paler, and the female Capra ægagrus is said to be
more uniformly tinted than their respective males.
Deer rarely present any sexual differences in colour.
Judge Caton, however, informs me that with the males
of the Wapiti deer (Cervus Canadensis) the neck, belly,
and legs are much darker than the same parts in the
female; but during the winter the darker tints gradually
fade away and disappear. I may here mention that
Judge Caton has in his park three races of the Virginian
deer, which differ slightly in colour, but the
differences are almost exclusively confined to the blue
290winter or breeding coat; so that this case may be
compared with those given in a previous chapter of
closely-allied or representative species of birds which
differ from each other only in their nuptial plumage.358
The females of Cervus paludosus of S. America, as
well as the young of both sexes, do not possess the
black stripes on the nose, and the blackish-brown line
on the breast which characterise the adult males.359
Lastly, the mature male of the beautifully coloured and
spotted Axis deer is considerably darker, as I am informed
by Mr. Blyth, than the female; and this hue
the castrated male never acquires.
The last Order which we have to consider—for I am
not aware that sexual differences in colour occur in
the other mammalian groups—is that of the Primates.
The male of the Lemur macaco is coal-black, whilst
the female is reddish-yellow, but highly variable in
colour.360 Of the Quadrumana of the New World, the
females and young of Mycetes caraya are greyish-yellow
and alike; in the second year the young male
becomes reddish-brown, in the third year black, excepting
the stomach, which, however, becomes quite
black in the fourth or fifth year. There is also a
strongly-marked difference in colour between the sexes
in Mycetes seniculus and Cebus capucinus; the young
of the former and I believe of the latter species resembling
the females. With Pithecia leucocephala the
young likewise resemble the females, which are brownish-291black
above and light rusty-red beneath, the adult
males being black. The ruff of hair round the face
of Ateles marginatus is tinted yellow in the male and
white in the female. Turning to the Old World, the
males of Hylobates hoolock are always black, with the
exception of a white band over the brows; the females
vary from whity-brown to a dark tint mixed with
black, but are never wholly black.361 In the beautiful
Cercopithecus diana the head of the adult male is of an
intense black, whilst that of the female is dark grey; in
the former the fur between the thighs is of an elegant
fawn-colour, in the latter it is paler. In the equally
beautiful and curious moustache monkey (Cercopithecus
cephus) the only difference between the sexes is that
the tail of the male is chesnut and that of the female
grey; but Mr. Bartlett informs me that all the hues
become more strongly pronounced in the male when
adult, whilst in the female they remain as they were
during youth. According to the coloured figures given
by Solomon Müller, the male of Semnopithecus chrysomelas
is nearly black, the female being pale brown.
In the Cercopithecus cynosurus and griseo-viridis one
part of the body which is confined to the male sex is of
the most brilliant blue or green, and contrasts strikingly
with the naked skin on the hinder part of the body,
which is vivid red.
Fig. 67. Head of male Mandrill (from Gervais, ‘Hist. Nat des Mammifères’).
Lastly, in the Baboon family, the adult male of Cynocephalus
hamadryas differs from the female not only by
his immense mane, but slightly in the colour of the hair
and of the naked callosities. In the drill (Cynocephalus
292leucophœus) the females and young are much paler-coloured,
with less green, than the adult males. No
other member of the whole class of mammals is coloured
in so extraordinary a manner as the adult male mandrill
(Cynocephalus mormon). The face at this age becomes
of a fine blue, with the ridge and tip of the nose of the
most brilliant red. According to some authors the face
is also marked with whitish stripes, and is shaded in parts
with black, but the colours appear to be variable. On
the forehead there is a crest of hair, and on the chin a
293yellow beard. “Toutes les parties supérieures de leurs
cuisses et le grand espace nu de leurs fesses sont
également colorés du rouge le plus vif, avec un
mélange de bleu qui ne manque réellement pas
d’élégance.”362 When the animal is excited all the naked
parts become much more vividly tinted. Several authors
have used the strongest expressions in describing these
resplendent colours, which they compare with those of
the most brilliant birds. Another most remarkable
peculiarity is that when the great canine teeth are fully
developed, immense protuberances of bone are formed
on each cheek, which are deeply furrowed longitudinally,
and the naked skin over them is brilliantly-coloured, as
just described. (Fig. 67.) In the adult females and in
the young of both sexes these protuberances are scarcely
perceptible; and the naked parts are much less brightly
coloured, the face being almost black, tinged with blue.
In the adult female, however, the nose at certain regular
intervals of time becomes tinted with red.
In all the cases hitherto given the male is more
strongly or brightly coloured than the female, and differs
in a greater degree from the young of both sexes.
But as a reversed style of colouring is characteristic of
the two sexes with some few birds, so with the Rhesus
monkey (Macacus rhesus) the female has a large surface
of naked skin round the tail, of a brilliant carmine red,
which periodically becomes, as I was assured by the
keepers in the Zoological Gardens, even more vivid,
and her face is also pale red. On the other hand with
294the adult male and with the young of both sexes, as I
saw in the Gardens, neither the naked skin at the
posterior end of the body, nor the face, shew a trace
of red. It appears, however, from some published
accounts, that the male does occasionally, or during
certain seasons, exhibit some traces of the red. Although
he is thus less ornamented than the female, yet in the
larger size of his body, larger canine teeth, more developed
whiskers, more prominent superciliary ridges,
he follows the common rule of the male excelling the
female.
I have now given all the cases known to me of a difference
in colour between the sexes of mammals. The
colours of the female either do not differ in a sufficient
degree from those of the male, or are not of a suitable
nature, to afford her protection, and therefore cannot be
explained on this principle. In some, perhaps in many
cases, the differences may be the result of variations
confined to one sex and transmitted to the same sex,
without any good having been thus gained, and therefore
without the aid of selection. We have instances
of this kind with our domesticated animals, as in the
males of certain cats being rusty-red, whilst the females
are tortoise-shell coloured. Analogous cases occur
under nature; Mr. Bartlett has seen many black varieties
of the jaguar, leopard, vulpine phalanger and
wombat; and he is certain that all, or nearly all, were
males. On the other hand, both sexes of wolves,
foxes, and apparently of American squirrels, are occasionally
born black. Hence it is quite possible that
with some mammals the blackness of the males, especially
when this colour is congenital, may simply be the
result, without the aid of selection, of one or more
variations having occurred, which from the first were295
sexually limited in their transmission. Nevertheless
it can hardly be admitted that the diversified, vivid,
and contrasted colours of certain quadrupeds, for instance
of the above-mentioned monkeys and antelopes,
can thus be accounted for. We should bear in mind
that these colours do not appear in the male at birth,
as in the case of most ordinary variations, but only at
or near maturity; and that unlike ordinary variations,
if the male be emasculated, they never appear or subsequently
disappear. It is on the whole a much more
probable conclusion that the strongly-marked colours
and other ornamental characters of male quadrupeds
are beneficial to them in their rivalry with other males,
and have consequently been acquired through sexual
selection. The probability of this view is strengthened
by the differences in colour between the sexes occurring
almost exclusively, as may be observed by going
through the previous details, in those groups and subgroups
of mammals, which present other and distinct
secondary sexual characters; these being likewise due
to the action of sexual selection.
Quadrupeds manifestly take notice of colour. Sir
S. Baker repeatedly observed that the African elephant
and rhinoceros attacked with special fury white or grey
horses. I have elsewhere shewn363 that half-wild horses
apparently prefer pairing with those of the same colour,
and that herds of fallow-deer of a different colour, though
living together, have long kept distinct. It is a more
significant fact that a female zebra would not admit the
addresses of a male ass until he was painted so as to
resemble a zebra, and then, as John Hunter remarks,
“she received him very readily. In this curious fact,
296we have instinct excited by mere colour, which had
so strong an effect as to get the better of everything
else. But the male did not require this, the
female being an animal somewhat similar to himself,
was sufficient to rouse him.”364
In an early chapter we have seen that the mental
powers of the higher animals do not differ in kind,
though so greatly in degree, from the corresponding
powers of man, especially of the lower and barbarous
races; and it would appear that even their taste for the
beautiful is not widely different from that of the Quadrumana.
As the negro of Africa raises the flesh on his
face into parallel ridges “or cicatrices, high above the
natural surface, which unsightly deformities, are considered
great personal attractions;”365—as negroes, as
well as savages in many parts of the world, paint their
faces with red, blue, white, or black bars,—so the
male mandrill of Africa appears to have acquired his
deeply-furrowed and gaudily-coloured face from having
been thus rendered attractive to the female. No doubt
it is to us a most grotesque notion that the posterior
end of the body should have been coloured for the
sake of ornament even more brilliantly than the face;
but this is really not more strange than that the
tails of many birds should have been especially decorated.
e do not at present possess any evidence
that the males take pains to display their charms
before the female; and the elaborate manner in which
this is performed by male birds, is the strongest argument
in favour of the belief that the females admire,
297or are excited by, the ornaments and colours displayed
before them. There is, however, a striking parallelism
between mammals and birds in all their secondary sexual
characters, namely in their weapons for fighting with
rival males, in their ornamental appendages, and in their
colours. In both classes, when the male differs from the
female, the young of both sexes almost always resemble
each other, and in a large majority of cases resemble the
adult female. In both classes the male assumes the
characters proper to his sex shortly before the age for
reproduction; if emasculated he either never acquires
such characters or subsequently loses them. In both
classes the change of colour is sometimes seasonal, and
the tints of the naked parts sometimes become more
vivid during the act of courtship. In both classes the
male is almost always more vividly or strongly coloured
than the female, and is ornamented with larger crests
either of hair or feathers, or other appendages. In a
few exceptional cases the female in both classes is
more highly ornamented than the male. With many
mammals, and at least in the case of one bird, the
male is more odoriferous than the female. In both
classes the voice of the male is more powerful than that
of the female. Considering this parallelism there can be
little doubt that the same cause, whatever it may be,
has acted on mammals and birds; and the result, as far
as ornamental characters are concerned, may safely be
attributed, as it appears to me, to the long-continued
preference of the individuals of one sex for certain individuals
of the opposite sex, combined with their success
in leaving a larger number of offspring to inherit
their superior attractions.
Equal transmission of ornamental characters to both
sexes.—With many birds, ornaments, which analogy leads298
us to believe were primarily acquired by the males, have
been transmitted equally, or almost equally, to both
sexes; and we may now enquire how far this view
may be extended to mammals. With a considerable
number of species, especially the smaller kinds, both
sexes have been coloured, independently of sexual selection,
for the sake of protection; but not, as far as I can
judge, in so many cases, nor in nearly so striking a
manner as in most of the lower classes. Audubon remarks
that he often mistook the musk-rat,366 whilst sitting
on the banks of a muddy stream, for a clod of earth, so
complete was the resemblance. The hare on her form
is a familiar instance of concealment through colour;
yet this principle partly fails in a closely-allied species,
namely the rabbit, for as this animal runs to its burrow,
it is made conspicuous to the sportsman and no doubt
to all beasts of prey, by its upturned pure-white tail.
No one has ever doubted that the quadrupeds which
inhabit snow-clad regions, have been rendered white to
protect them from their enemies, or to favour their
stealing on their prey. In regions where snow never
lies long on the ground a white coat would be injurious;
consequently species thus coloured are extremely
rare in the hotter parts of the world. It deserves notice
that many quadrupeds, inhabiting moderately cold regions,
although they do not assume a white winter dress,
become paler during this season; and this apparently
is the direct result of the conditions to which they
have long been exposed. Pallas367 states that in Siberia
a change of this nature occurs with the wolf, two
species of Mustela, the domestic horse, the Equus he299mionus,
the domestic cow, two species of antelopes, the
musk-deer, the roe, the elk, and reindeer. The roe,
for instance, has a red summer and a greyish-white
winter coat; and the latter may perhaps serve as a
protection to the animal whilst wandering through the
leafless thickets, sprinkled with snow and hoar-frost.
If the above named animals were gradually to extend
their range into regions perpetually covered with snow,
their pale winter-coats would probably be rendered,
through natural selection, whiter and whiter by degrees,
until they became as white as snow.

Fig. 68. Tragelaphus scriptus, male (from the Knowsley Menagerie).

Fig. 69. Damalis pygarga, male (from the Knowsley Menagerie).
Although we must admit that many quadrupeds have
received their present tints as a protection, yet with a
host of species, the colours are far too conspicuous and
too singularly arranged to allow us to suppose that they
serve for this purpose. We may take as an illustration
certain antelopes: when we see that the square
white patch on the throat, the white marks on the fetlocks,
and the round black spots on the ears, are all
more distinct in the male of the Portax picta, than in
the female;—when we see that the colours are more
vivid, that the narrow white lines on the flank and
the broad white bar on the shoulder are more distinct
in the male Oreas derbyanus than in the female;—when
we see a similar difference between the sexes
of the curiously-ornamented Tragelaphus scriptus (fig.
68),—we may conclude that these colours and various
marks have been at least intensified through sexual
selection. It is inconceivable that such colours and
marks can be of any direct or ordinary service to these
animals; and as they have almost certainly been intensified
through sexual selection, it is probable that they
were originally gained through this same process, and
then partially transferred to the females. If this view
be admitted, there can be little doubt that the equally300
singular colours and marks of many other antelopes,
though common to both sexes, have been gained and
transmitted in a like manner. Both sexes, for instance,
of the Koodoo (Strepsiceros Kudu, fig. 62) have narrow
white vertical lines on their hinder flanks, and an
elegant angular white mark on their foreheads. Both
sexes in the genus Damalis are very oddly coloured; in
D. pygarga the back and neck are purplish-red, shading
on the flanks into black, and abruptly separated from the301
white belly and a large white space on the buttocks;
the head is still more oddly coloured, a large oblong
white mask, narrowly-edged with black, covers the face
up to the eyes (fig. 69); there are three white stripes
on the forehead, and the ears are marked with white.
The fawns of this species are of a uniform pale yellowish-brown.
In Damalis albifrons the colouring of the
head differs from that in the last species in a single
white stripe replacing the three stripes, and in the ears
being almost wholly white.368 After having studied to
302the best of my ability the sexual differences of animals
belonging to all classes, I cannot avoid the conclusion
that the curiously-arranged colours of many antelopes,
though common to both sexes, are the result of sexual
selection primarily applied to the male.
The same conclusion may perhaps be extended to the
tiger, one of the most beautiful animals in the world,
the sexes of which cannot be distinguished by colour,
even by the dealers in wild beasts. Mr. Wallace
believes369 that the striped coat of the tiger “so assimilates
with the vertical stems of the bamboo, as to
assist greatly in concealing him from his approaching
prey.” But this view does not appear to me satisfactory.
We have some slight evidence that his beauty
may be due to sexual selection, for in two species of
Felis analogous marks and colours are rather brighter
in the male than in the female. The zebra is conspicuously
striped, and stripes on the open plains of South
Africa cannot afford any protection. Burchell370 in describing
a herd says, “their sleek ribs glistened in the
sun, and the brightness and regularity of their striped
coats presented a picture of extraordinary beauty, in
which probably they are not surpassed by any other
quadruped.” Here we have no evidence of sexual
selection, as throughout the whole group of the Equidæ
the sexes are identical in colour. Nevertheless he who
attributes the white and dark vertical stripes on the
flanks of various antelopes to sexual selection, will probably
extend the same view to the Royal Tiger and
beautiful Zebra.
We have seen in a former chapter that when young
animals belonging to any class follow nearly the same
303habits of life with their parents, and yet are coloured
in a different manner, it may be inferred that they have
retained the colouring of some ancient and extinct
progenitor. In the family of pigs, and in the genus
Tapir, the young are marked with longitudinal stripes,
and thus differ from every existing adult species in
these two groups. With many kinds of deer the
young are marked with elegant white spots, of which
their parents exhibit not a trace. A graduated series
can be followed from the Axis deer, both sexes of
which at all ages and during all seasons are beautifully
spotted (the male being rather more strongly
coloured than the female)—to species in which neither
the old nor the young are spotted. I will specify
some of the steps in this series. The Mantchurian
deer (Cervus Mantchuricus) is spotted during the whole
year, but the spots are much plainer, as I have seen
in the Zoological Gardens, during the summer, when the
general colour of the coat is lighter, than during the
winter, when the general colour is darker and the horns
are fully developed. In the hog-deer (Hyelaphus porcinus)
the spots are extremely conspicuous during the
summer when the coat is reddish-brown, but quite disappear
during the winter when the coat is brown.371
In both these species the young are spotted. In the
Virginian deer the young are likewise spotted, and
about five per cent. of the adult animals living in
Judge Caton’s park, as I am informed by him, temporarily
exhibit at the period when the red summer
coat is being replaced by the bluish winter coat, a row
of spots on each flank, which are always the same in
304number, though very variable in distinctness. From
this condition there is but a very small step to the
complete absence of spots at all seasons in the adults;
and lastly, to their absence at all ages, as occurs with
certain species. From the existence of this perfect
series, and more especially from the fawns of so many
species being spotted, we may conclude that the now
living members of the deer family are the descendants
of some ancient species which, like the Axis deer, was
spotted at all ages and seasons. A still more ancient
progenitor probably resembled to a certain extent the
Hyomoschus aquaticus—for this animal is spotted, and
the hornless males have large exserted canine teeth,
of which some few true deer still retain rudiments. It
offers, also, one of those interesting cases of a form
linking together two groups, as it is intermediate in
certain osteological characters between the pachyderms
and ruminants, which were formerly thought to be
quite distinct.372
A curious difficulty here arises. If we admit that
coloured spots and stripes have been acquired as ornaments,
how comes it that so many existing deer, the
descendants of an aboriginally spotted animal, and
all the species of pigs and tapirs, the descendants of
an aboriginally striped animal, have lost in their adult
state their former ornaments? I cannot satisfactorily
answer this question. We may feel nearly sure that
the spots and stripes disappeared in the progenitors of
our existing species at or near maturity, so that they were
retained by the young and, owing to the law of inheritance
at corresponding ages, by the young of all succeeding
generations. It may have been a great advantage to
305the lion and puma from the open nature of the localities
which they commonly haunt, to have lost their stripes,
and to have been thus rendered less conspicuous to their
prey; and if the successive variations, by which this
end was gained, occurred rather late in life, the young
would have retained their stripes, as we know to be the
case. In regard to deer, pigs, and tapirs, Fritz Müller
has suggested to me that these animals by the removal
through natural selection of their spots or stripes would
have been less easily seen by their enemies; and they
would have especially required this protection, as soon
as the carnivora increased in size and number during
the Tertiary periods. This may be the true explanation,
but it is rather strange that the young should
not have been equally well protected, and still more
strange that with some species the adults should have
retained their spots, either partially or completely,
during part of the year. We know, though we cannot
explain the cause, that when the domestic ass varies and
becomes reddish-brown, grey or black, the stripes on the
shoulders and even on the spine frequently disappear.
Very few horses, except dun-coloured kinds, exhibit
stripes on any part of their bodies, yet we have good
reason to believe that the aboriginal horse was striped
on the legs and spine, and probably on the shoulders.373
Hence the disappearance of the spots and stripes in our
adult existing deer, pigs, and tapirs, may be due to a
change in the general colour of their coats; but whether
this change was effected through sexual or natural selection,
or was due to the direct action of the conditions
of life, or some other unknown cause, it is impossible
to decide. An observation made by Mr. Sclater well
illustrates our ignorance of the laws which regulate the
306appearance and disappearance of stripes; the species of
Asinus which inhabit the Asiatic continent are destitute
of stripes, not having even the cross shoulder-stripe,
whilst those which inhabit Africa are conspicuously
striped, with the partial exception of A. tæniopus, which
has only the cross shoulder-stripe and generally some
faint bars on the legs; and this species inhabits the almost
intermediate region of Upper Egypt and Abyssinia.374

Fig. 70. Head of Semnopithecus rubicundus. This and the following figures (from
Prof. Gervais) are given to shew the odd arrangement and development of the hair on
the head.
|
|
Fig. 71. Head of Semnopithecus comatus. |
Fig. 72. Head of Cebus capucinus. |
|
|
Fig. 73. Head of Ateles marginatus. |
Fig. 74. Head of Cebus vellerosus. |
Quadrumana.—Before we conclude, it will be advisable
to add a few remarks to those already given on the
307
ornamental characters of monkeys. In most of the
species the sexes resemble each other in colour, but
in some, as we have seen, the males differ from the
females, especially in the colour of the naked parts of
the skin, in the development of the beard, whiskers,
and mane. Many species are coloured either in so extraordinary
or beautiful a manner, and are furnished
with such curious and elegant crests of hair, that we
can hardly avoid looking at these characters as having
been gained for the sake of ornament. The accompanying
figures (figs. 70 to 74) serve to shew the308
arrangement of the hair on the face and head in several
species. It is scarcely conceivable that these crests of
hair and the strongly-contrasted colours of the fur and
skin can be the result of mere variability without the
aid of selection; and it is inconceivable that they can
be of any ordinary use to these animals. If so, they
have probably been gained through sexual selection,
though transmitted equally, or almost equally, to both
sexes. With many of the Quadrumana, we have additional
evidence of the action of sexual selection in
the greater size and strength of the males, and in the
greater development of their canine teeth, in comparison
with the females.
With respect to the strange manner in which both
sexes of some species are coloured, and of the beauty
of others, a few instances will suffice. The face of the
Cercopithecus petaurista (fig. 75) is black, the whiskers
and beard being white, with a defined, round, white
spot on the nose, covered with short white hair, which
gives to the animal an almost ludicrous aspect. The
Semnopithecus frontatus likewise, has a blackish face
with a long black beard, and a large naked spot on
the forehead of a bluish-white colour. The face of
Macacus lasiotus is dirty flesh-coloured, with a defined
red spot on each cheek. The appearance of Cercocebus
æthiops is grotesque, with its black face, white
whiskers and collar, chesnut head, and a large naked
white spot over each eyelid. In very many species, the
beard, whiskers, and crests of hair round the face are of
a different colour from the rest of the head, and when
different, are always of a lighter tint,375 being often pure
309white, sometimes bright yellow, or reddish. The whole
face of the South American Brachyurus calvus is of a
“glowing scarlet hue;” but this colour does not appear
310until the animal is nearly mature.376 The naked skin
of the face differs wonderfully in colour in the various
species. It is often brown or flesh-colour, with parts
perfectly white, and often as black as that of the
most sooty negro. In the Brachyurus the scarlet tint
is brighter than that of the most blushing Caucasian
damsel. It is sometimes more distinctly orange than
in any Mongolian, and in several species it is blue,
passing into violet or grey. In all the species known
to Mr. Bartlett, in which the adults of both sexes have
strongly-coloured faces, the colours are dull or absent
during early youth. This likewise holds good with the
Mandrill and Rhesus, in which the face and the posterior
parts of the body are brilliantly coloured in one sex
alone. In these latter cases we have every reason to
believe that the colours were acquired through sexual
selection; and we are naturally led to extend the same
view to the foregoing species, though both sexes when
adult have their faces coloured in the same manner.

Fig. 75. Cercopithecus petaurista (from Brehm)
Although, according to our taste, many kinds of
monkeys are far from beautiful, other species are universally
admired for their elegant appearance and
bright colours. The Semnopithecus nemæus, though
peculiarly coloured, is described as extremely pretty;
the orange-tinted face is surrounded by long whiskers
of glossy whiteness, with a line of chesnut-red over the
eyebrows; the fur on the back is of a delicate grey, with
a square patch on the loins, the tail and the fore-arms
all of a pure white; a gorget of chesnut surmounts the
chest; the hind thighs are black, with the legs chesnut-red.
I will mention only two other monkeys on account
of their beauty; and I have selected these as they present
slight sexual differences in colour, which renders it
311in some degree probable that both sexes owe their
elegant appearance to sexual selection. In the moustache-monkey
(Cercopithecus cephus) the general colour
of the fur is mottled-greenish, with the throat white; in
the male the end of the tail is chesnut; but the face is
the most ornamented part, the skin being chiefly bluish-grey,
shading into a blackish tint beneath the eyes,
with the upper lip of a delicate blue, clothed on the
lower edge with a thin black moustache; the whiskers
312are orange-coloured, with the upper part black, forming
a band which extends backwards to the ears, the latter
being clothed with whitish hairs. In the Zoological
Society’s Gardens I have often overheard visitors admiring
the beauty of another monkey, deservedly called
Cercopithecus Diana (fig. 76); the general colour of the
fur is grey; the chest and inner surface of the forelegs
are white; a large triangular defined space on the hinder
part of the back is rich chesnut; in the male the inner
sides of the thighs and the abdomen are delicate fawn-coloured,
and the top of the head is black; the face and
ears are intensely black, finely contrasted with a white
transverse crest over the eyebrows and with a long
white peaked beard, of which the basal portion is
black.377

Fig. 76. Cercopithecus Diana (from Brehm).
In these and many other monkeys, the beauty and
singular arrangement of their colours, and still more the
diversified and elegant arrangement of the crests and
tufts of hair on their heads, force the conviction on my
mind that these characters have been acquired through
sexual selection exclusively as ornaments.
Summary.—The law of battle for the possession of the
female appears to prevail throughout the whole great
class of mammals. Most naturalists will admit that
the greater size, strength, courage, and pugnacity of the
male, his special weapons of offence, as well as his
special means of defence, have all been acquired or
modified through that form of selection which I have
313called sexual selection. This does not depend on any
superiority in the general struggle for life, but on
certain individuals of one sex, generally the male sex,
having been successful in conquering other males, and
on their having left a larger number of offspring to
inherit their superiority, than the less successful males.
There is another and more peaceful kind of contest,
in which the males endeavour to excite or allure the
females by various charms. This may be effected by
the powerful odours emitted by the males during the
breeding-season; the odoriferous glands having been
acquired through sexual selection. Whether the same
view can be extended to the voice is doubtful, for the
vocal organs of the males may have been strengthened
by use during maturity, under the powerful excitements
of love, jealousy, or rage, and transmitted to the same
sex. Various crests, tufts, and mantles of hair, which
are either confined to the male, or are more developed
in this sex than in the females, seem in most cases to
be merely ornamental, though they sometimes serve as
a defence against rival males. There is even reason to
suspect that the branching horns of stags, and the
elegant horns of certain antelopes, though properly
serving as weapons of offence or of defence, have been
partly modified for the sake of ornament.
When the male differs in colour from the female he
generally exhibits darker and more strongly-contrasted
tints. We do not in this class meet with the splendid
red, blue, yellow, and green colours, so common with
male birds and many other animals. The naked parts,
however, of certain Quadrumana must be excepted; for
such parts, often oddly situated, are coloured in some
species in the most brilliant manner. The colours of
the male in other cases may be due to simple variation,314
without the aid of selection. But when the colours are
diversified and strongly pronounced, when they are not
developed until near maturity, and when they are lost
after emasculation, we can hardly avoid the conclusion
that they have been acquired through sexual selection for
the sake of ornament, and have been transmitted exclusively,
or almost exclusively, to the same sex. When
both sexes are coloured in the same manner, and the
colours are conspicuous or curiously arranged, without
being of the least apparent use as a protection, and
especially when they are associated with various other
ornamental appendages, we are led by analogy to the
same conclusion, namely, that they have been acquired
through sexual selection, although transmitted to both
sexes. That conspicuous and diversified colours, whether
confined to the males or common to both sexes, are as
a general rule associated in the same groups and subgroups
with other secondary sexual characters, serving
for war or for ornament, will be found to hold good if
we look back to the various cases given in this and
the last chapter.
The law of the equal transmission of characters to
both sexes, as far as colour and other ornaments are
concerned, has prevailed far more extensively with
mammals than with birds; but in regard to weapons,
such as horns and tusks, these have often been transmitted
either exclusively, or in a much higher degree
to the males than to the females. This is a surprising
circumstance, for as the males generally use their
weapons as a defence against enemies of all kinds,
these weapons would have been of service to the female.
Their absence in this sex can be accounted for,
as far as we can see, only by the form of inheritance
which has prevailed. Finally with quadrupeds the315
contest between the individuals of the same sex, whether
peaceful or bloody, has with the rarest exceptions been
confined to the males; so that these have been modified
through sexual selection, either for fighting with
each other or for alluring the opposite sex, far more
commonly than the females.
316
CHAPTER XIX.
Secondary Sexual Characters of Man.
Differences between man and woman—Causes of such differences
and of certain characters common to both sexes—Law of
battle—Differences in mental powers—and voice—On the
influence of beauty in determining the marriages of mankind—Attention
paid by savages to ornaments—Their ideas of
beauty in woman—The tendency to exaggerate each natural
peculiarity.
With mankind the differences between the sexes are
greater than in most species of Quadrumana, but not
so great as in some, for instance, the mandrill. Man
on an average is considerably taller, heavier, and
stronger than woman, with squarer shoulders and more
plainly-pronounced muscles. Owing to the relation
which exists between muscular development and the
projection of the brows,378 the superciliary ridge is generally
more strongly marked in man than in woman.
His body, and especially his face, is more hairy, and
his voice has a different and more powerful tone. In
certain tribes the women are said, whether truly I know
not, to differ slightly in tint from the men; and with
Europeans, the women are perhaps the more brightly
coloured of the two, as may be seen when both sexes
have been equally exposed to the weather.
Man is more courageous, pugnacious, and energetic
than woman, and has a more inventive genius. His
317brain is absolutely larger, but whether relatively to the
larger size of his body, in comparison with that of
woman, has not, I believe been fully ascertained. In
woman the face is rounder; the jaws and the base of the
skull smaller; the outlines of her body rounder, in parts
more prominent; and her pelvis is broader than in man;379
but this latter character may perhaps be considered
rather as a primary than a secondary sexual character.
She comes to maturity at an earlier age than man.
As with animals of all classes, so with man, the distinctive
characters of the male sex are not fully developed
until he is nearly mature; and if emasculated they
never appear. The beard, for instance, is a secondary
sexual character, and male children are beardless,
though at an early age they have abundant hair on
their heads. It is probably due to the rather late
appearance in life of the successive variations, by
which man acquired his masculine characters, that
they are transmitted to the male sex alone. Male
and female children resemble each other closely, like
the young of so many other animals in which the adult
sexes differ; they likewise resemble the mature female
much more closely, than the mature male. The female,
however, ultimately assumes certain distinctive
characters, and in the formation of her skull, is said to be
intermediate between the child and the man.380 Again,
as the young of closely allied though distinct species do
not differ nearly so much from each other as do the
adults, so it is with the children of the different races of
man. Some have even maintained that race-differences
318cannot be detected in the infantile skull.381 In regard to
colour, the newborn negro child is reddish nut-brown,
which soon becomes slaty-grey; the black colour being
fully developed within a year in the Sudan, but not
until three years in Egypt. The eyes of the negro are
at first blue, and the hair chesnut-brown rather than
black, being curled only at the ends. The children of
the Australians immediately after birth are yellowish-brown,
and become dark at a later age. Those of the
Guaranys of Paraguay are whitish-yellow, but they
acquire in the course of a few weeks the yellowish-brown
tint of their parents. Similar observations have
been made in other parts of America.382
I have specified the foregoing familiar differences between
the male and female sex in mankind, because they
are curiously the same as in the Quadrumana. With
these animals the female is mature at an earlier age than
the male; at least this is certainly the case with the
Cebus azaræ.383 With most of the species the males are
larger and much stronger than the females, of which
fact the gorilla offers a well-known instance. Even in
so trifling a character as the greater prominence of the
superciliary ridge, the males of certain monkeys differ
from the females,384 and agree in this respect with mankind.
In the gorilla and certain other monkeys, the
319cranium of the adult male presents a strongly-marked
sagittal crest, which is absent in the female; and Ecker
found a trace of a similar difference between the two
sexes in the Australians.385 With monkeys when there
is any difference in the voice, that of the male is the
more powerful. We have seen that certain male monkeys,
have a well-developed beard, which is quite deficient,
or much less developed in the female. No instance
is known of the beard, whiskers, or moustache
being larger in a female than in the male monkey.
Even in the colour of the beard there is a curious
parallelism between man and the Quadrumana, for
when in man the beard differs in colour from the hair
of the head, as is often the case, it is, I believe, invariably
of a lighter tint, being often reddish. I have
observed this fact in England, and Dr. Hooker, who
attended to this little point for me in Russia, found
no exception to the rule. In Calcutta, Mr. J. Scott,
of the Botanic Gardens, was so kind as to observe with
care the many races of men to be seen there, as well
as in some other parts of India, namely, two races in
Sikhim, the Bhoteas, Hindoos, Burmese, and Chinese.
Although most of these races have very little hair on
the face, yet he always found that when there was any
difference in colour between the hair of the head and
the beard, the latter was invariably of a lighter tint. Now
with monkeys, as has already been stated, the beard
frequently differs in a striking manner in colour from
the hair of the head, and in such cases it is invariably
of a lighter hue, being often pure white, sometimes
yellow or reddish.386
320
In regard to the general hairiness of the body, the
women in all races are less hairy than the men, and in
some few Quadrumana the under side of the body of
the female is less hairy than that of the male.387 Lastly,
male monkeys, like men, are bolder and fiercer than
the females. They lead the troop, and when there is
danger, come to the front. We thus see how close is
the parallelism between the sexual differences of man
and the Quadrumana. With some few species, however,
as with certain baboons, the gorilla and orang,
there is a considerably greater difference between the
sexes, in the size of the canine teeth, in the development
and colour of the hair, and especially in the
colour of the naked parts of the skin, than in the case
of mankind.
The secondary sexual characters of man are all highly
variable, even within the limits of the same race or
sub-species; and they differ much in the several races.
These two rules generally hold good throughout the
animal kingdom. In the excellent observations made
on board the Novara,388 the male Australians were found
to exceed the females by only 65 millim. in height,
whilst with the Javanese the average excess was 218
millim., so that in this latter race the difference in height
321between the sexes is more than thrice as great as with
the Australians. The numerous measurements of various
other races, with respect to stature, the circumference
of the neck and chest, and the length of the back-bone
and arms, which were carefully made, nearly all shewed
that the males differed much more from each other than
did the females. This fact indicates that, as far as these
characters are concerned, it is the male which has been
chiefly modified, since the races diverged from their
common and primeval source.
The development of the beard and the hairiness of
the body differ remarkably in the men belonging to
distinct races, and even to different families in the same
race. We Europeans see this amongst ourselves. In
the island of St. Kilda, according to Martin,389 the men
do not acquire beards, which are very thin, until the
age of thirty or upwards. On the Europæo-Asiatic
continent, beards prevail until we pass beyond India,
though with the natives of Ceylon they are frequently
absent, as was noticed in ancient times by Diodorus.390
Beyond India beards disappear, as with the Siamese,
Malays, Kalmucks, Chinese, and Japanese; nevertheless
the Ainos,391 who inhabit the northernmost islands
of the Japan archipelago, are the most hairy men in the
world. With negroes the beard is scanty or absent, and
they have no whiskers; in both sexes the body is almost
destitute of fine down.392 On the other hand, the Pa322puans
of the Malay archipelago, who are nearly as black
as negroes, possess well-developed beards.393 In the
Pacific Ocean the inhabitants of the Fiji archipelago
have large bushy beards, whilst those of the not-distant
archipelagoes of Tonga and Samoa are beardless; but
these men belong to distinct races. In the Ellice group
all the inhabitants belong to the same race; yet on
one island alone, namely Nunemaya, “the men have
splendid beards;” whilst on the other islands “they
have, as a rule, a dozen straggling hairs for a beard.”394
Throughout the great American continent the men
may be said to be beardless; but in almost all the tribes
a few short hairs are apt to appear on the face, especially
during old age. With the tribes of North America,
Catlin estimates that eighteen out of twenty men are
completely destitute by nature of a beard; but occasionally
there may be seen a man, who has neglected to
pluck out the hairs at puberty, with a soft beard an inch
or two in length. The Guaranys of Paraguay differ
from all the surrounding tribes in having a small beard,
and even some hair on the body, but no whiskers.395 I
am informed by Mr. D. Forbes, who particularly attended
to this subject, that the Aymaras and Quechuas of the
Cordillera are remarkably hairless, yet in old age a few
straggling hairs occasionally appear on the chin. The
men of these two tribes have very little hair on the
various parts of the body where hair grows abundantly
323in Europeans, and the women have none on the corresponding
parts. The hair on the head, however, attains
an extraordinary length in both sexes, often reaching
almost to the ground; and this is likewise the case with
some of the N. American tribes. In the amount of
hair, and in the general shape of the body, the sexes
of the American aborigines do not differ from each
other so much as with most other races of mankind.396
This fact is analogous with what occurs with some allied
monkeys; thus the sexes of the chimpanzee are not as
different as those of the gorilla or orang.397
In the previous chapters we have seen that with
mammals, birds, fishes, insects, &c., many characters,
which there is every reason to believe were primarily
gained through sexual selection by one sex alone, have
been transferred to both sexes. As this same form of
transmission has apparently prevailed to a large extent
with mankind, it will save much useless repetition if
we consider the characters peculiar to the male sex
together with certain other characters common to both
sexes.
Law of Battle.—With barbarous nations, for instance
with the Australians, the women are the constant cause
of war both between the individuals of the same tribe
and between distinct tribes. So no doubt it was in
ancient times; “nam fuit ante Helenam mulier teterrima
belli causa.” With the North American Indians,
the contest is reduced to a system. That excellent ob324server,
Hearne,398 says:—“It has ever been the custom
among these people for the men to wrestle for any
woman to whom they are attached; and, of course, the
strongest party always carries off the prize. A weak
man, unless he be a good hunter, and well-beloved,
is seldom permitted to keep a wife that a stronger
man thinks worth his notice. This custom prevails
throughout all the tribes, and causes a great spirit
of emulation among their youth, who are upon all
occasions, from their childhood, trying their strength
and skill in wrestling.” With the Guanas of South
America, Azara states that the men rarely marry till
twenty or more years old, as before that age they
cannot conquer their rivals.
Other similar facts could be given; but even if we
had no evidence on this head, we might feel almost
sure, from the analogy of the higher Quadrumana,399
that the law of battle had prevailed with man during
the early stages of his development. The occasional
appearance at the present day of canine teeth which
project above the others, with traces of a diastema or
open space for the reception of the opposite canines, is
in all probability a case of reversion to a former state,
when the progenitors of man were provided with these
weapons, like so many existing male Quadrumana. It
was remarked in a former chapter that as man gradually
became erect, and continually used his hands
and arms for fighting with sticks and stones, as well as
for the other purposes of life, he would have used his
325jaws and teeth less and less. The jaws, together with
their muscles, would then have become reduced through
disuse, as would the teeth through the not well understood
principles of correlation and the economy of
growth; for we everywhere see that parts which are
no longer of service are reduced in size. By such steps
the original inequality between the jaws and teeth in
the two sexes of mankind would ultimately have been
quite obliterated. The case is almost parallel with
that of many male Ruminants, in which the canine
teeth have been reduced to mere rudiments, or have
disappeared, apparently in consequence of the development
of horns. As the prodigious difference between
the skulls of the two sexes in the Gorilla and Orang,
stands in close relation with the development of the
immense canine teeth in the males, we may infer that
the reduction of the jaws and teeth in the early male
progenitors of man led to a most striking and favourable
change in his appearance.
There can be little doubt that the greater size and
strength of man, in comparison with woman, together
with his broader shoulders, more developed muscles,
rugged outline of body, his greater courage and pugnacity,
are all due in chief part to inheritance from
some early male progenitor, who, like the existing
anthropoid apes, was thus characterised. These characters
will, however, have been preserved or even
augmented during the long ages whilst man was still
in a barbarous condition, by the strongest and boldest
men having succeeded best in the general struggle for
life, as well as in securing wives, and thus having left a
large number of offspring. It is not probable that the
greater strength of man was primarily acquired through
the inherited effects of his having worked harder than
woman for his own subsistence and that of his family;326
for the women in all barbarous nations are compelled
to work at least as hard as the men. With civilised
people the arbitrament of battle for the possession of
the women has long ceased; on the other hand, the men,
as a general rule, have to work harder than the women
for their mutual subsistence; and thus their greater
strength will have been kept up.
Difference in the Mental Powers of the two Sexes.—With
respect to differences of this nature between man
and woman, it is probable that sexual selection has
played a very important part. I am aware that some
writers doubt whether there is any inherent difference;
but this is at least probable from the analogy of the
lower animals which present other secondary sexual
characters. No one will dispute that the bull differs
in disposition from the cow, the wild-boar from the
sow, the stallion from the mare, and, as is well known
to the keepers of menageries, the males of the larger
apes from the females. Woman seems to differ from
man in mental disposition, chiefly in her greater tenderness
and less selfishness; and this holds good even
with savages, as shewn by a well-known passage in
Mungo Park’s Travels, and by statements made by
many other travellers. Woman, owing to her maternal
instincts, displays these qualities towards her infants
in an eminent degree; therefore it is likely that she
should often extend them towards her fellow-creatures.
Man is the rival of other men; he delights in competition,
and this leads to ambition which passes too
easily into selfishness. These latter qualities seem to
be his natural and unfortunate birthright. It is generally
admitted that with woman the powers of intuition,
of rapid perception, and perhaps of imitation, are more
strongly marked than in man; but some, at least, of327
these faculties are characteristic of the lower races, and
therefore of a past and lower state of civilisation.
The chief distinction in the intellectual powers of
the two sexes is shewn by man attaining to a higher
eminence, in whatever he takes up, than woman can
attain—whether requiring deep thought, reason, or
imagination, or merely the use of the senses and
hands. If two lists were made of the most eminent
men and women in poetry, painting, sculpture, music,—comprising
composition and performance, history,
science, and philosophy, with half-a-dozen names under
each subject, the two lists would not bear comparison.
We may also infer, from the law of the deviation of
averages, so well illustrated by Mr. Galton, in his
work on ‘Hereditary Genius,’ that if men are capable
of decided eminence over women in many subjects, the
average standard of mental power in man must be
above that of woman.
The half-human male progenitors of man, and men
in a savage state, have struggled together during many
generations for the possession of the females. But mere
bodily strength and size would do little for victory,
unless associated with courage, perseverance, and determined
energy. With social animals, the young males
have to pass through many a contest before they win a
female, and the older males have to retain their females
by renewed battles. They have, also, in the case of
man, to defend their females, as well as their young,
from enemies of all kinds, and to hunt for their joint
subsistence. But to avoid enemies, or to attack them
with success, to capture wild animals, and to invent
and fashion weapons, requires the aid of the higher
mental faculties, namely, observation, reason, invention,
or imagination. These various faculties will thus
have been continually put to the test, and selected328
during manhood; they will, moreover, have been
strengthened by use during this same period of life.
Consequently, in accordance with the principle often
alluded to, we might expect that they would at least
tend to be transmitted chiefly to the male offspring
at the corresponding period of manhood.
Now, when two men are put into competition, or a
man with a woman, who possess every mental quality
in the same perfection, with the exception that the
one has higher energy, perseverance, and courage,
this one will generally become more eminent, whatever
the object may be, and will gain the victory.400
He may be said to possess genius—for genius has been
declared by a great authority to be patience; and
patience, in this sense, means unflinching, undaunted
perseverance. But this view of genius is perhaps
deficient; for without the higher powers of the imagination
and reason, no eminent success in many subjects
can be gained. But these latter as well as the former
faculties will have been developed in man, partly
through sexual selection,—that is, through the contest of
rival males, and partly through natural selection,—that
is, from success in the general struggle for life; and as
in both cases the struggle will have been during
maturity, the characters thus gained will have been
transmitted more fully to the male than to the female
offspring. Thus man has ultimately become superior to
woman. It is, indeed, fortunate that the law of the
equal transmission of characters to both sexes has commonly
prevailed throughout the whole class of mammals;
otherwise it is probable that man would have
329become as superior in mental endowment to woman, as
the peacock is in ornamental plumage to the peahen.
It must be borne in mind that the tendency in characters
acquired at a late period of life by either sex,
to be transmitted to the same sex at the same age, and
of characters acquired at an early age to be transmitted
to both sexes, are rules which, though general, do not
always hold good. If they always held good, we might
conclude (but I am here wandering beyond my proper
bounds) that the inherited effects of the early education
of boys and girls would be transmitted equally to both
sexes; so that the present inequality between the sexes
in mental power could not be effaced by a similar
course of early training; nor can it have been caused
by their dissimilar early training. In order that woman
should reach the same standard as man, she ought,
when nearly adult, to be trained to energy and perseverance,
and to have her reason and imagination exercised
to the highest point; and then she would probably
transmit these qualities chiefly to her adult
daughters. The whole body of women, however, could
not be thus raised, unless during many generations
the women who excelled in the above robust virtues
were married, and produced offspring in larger numbers
than other women. As before remarked with respect
to bodily strength, although men do not now fight for
the sake of obtaining wives, and this form of selection
has passed away, yet they generally have to undergo,
during manhood, a severe struggle in order to maintain
themselves and their families; and this will tend to keep
up or even increase their mental powers, and, as a consequence,
the present inequality between the sexes.401
330
Voice and Musical Powers.—In some species of Quadrumana
there is a great difference between the adult
sexes, in the power of the voice and in the development
of the vocal organs; and man appears to have inherited
this difference from his early progenitors. His vocal
cords are about one-third longer than in woman, or
than in boys; and emasculation produces the same effect
on him as on the lower animals, for it “arrests that prominent
growth of the thyroid, &c., which accompanies
the elongation of the cords.”402 With respect to the
cause of this difference between the sexes, I have nothing
to add to the remarks made in the last chapter on the
probable effects of the long-continued use of the vocal
organs by the male under the excitement of love, rage,
and jealousy. According to Sir Duncan Gibb,403 the
voice differs in the different races of mankind; and
with the natives of Tartary, China, &c., the voice of
the male is said not to differ so much from that of the
female, as in most other races.
The capacity and love for singing or music, though
not a sexual character in man, must not here be passed
over. Although the sounds emitted by animals of all
kinds serve many purposes, a strong case can be made
out, that the vocal organs were primarily used and perfected
in relation to the propagation of the species.
Insects and some few spiders are the lowest animals
which voluntarily produce any sound; and this is generally
effected by the aid of beautifully constructed
331stridulating organs, which are often confined to the
males alone. The sounds thus produced consist, I believe
in all cases, of the same note, repeated rhythmically;404
and this is sometimes pleasing even to the ears of man.
Their chief, and in some cases exclusive use appears to
be either to call or to charm the opposite sex.
The sounds produced by fishes are said in some cases
to be made only by the males during the breeding
season. All the air-breathing Vertebrata necessarily
possess an apparatus for inhaling and expelling air, with
a pipe capable of being closed at one end. Hence when
the primeval members of this class were strongly excited
and their muscles violently contracted, purposeless
sounds would almost certainly have been produced;
and these, if they proved in any way serviceable, might
readily have been modified or intensified by the preservation
of properly adapted variations. The Amphibians
are the lowest Vertebrates which breathe air; and
many of these animals, namely, frogs and toads, possess
vocal organs, which are incessantly used during the
breeding-season, and which are often more highly
developed in the male than in the female. The male
alone of the tortoise utters a noise, and this only during
the season of love. Male alligators roar or bellow
during the same season. Every one knows how largely
birds use their vocal organs as a means of courtship;
and some species likewise perform what may be called
instrumental music.
In the class of Mammals, with which we are here
more particularly concerned, the males of almost all the
species use their voices during the breeding-season
much more than at any other time; and some are abso332lutely
mute excepting at this season. Both sexes of other
species, or the females alone, use their voices as a love-call.
Considering these facts, and that the vocal organs
of some quadrupeds are much more largely developed
in the male than in the female, either permanently or
temporarily during the breeding season; and considering
that in most of the lower classes the sounds produced
by the males, serve not only to call but to excite or allure
the female, it is a surprising fact that we have not as yet
any good evidence that these organs are used by male
mammals to charm the females. The American Mycetes
caraya perhaps forms an exception, as does more probably
one of those apes which come nearer to man,
namely, the Hylobates agilis. This gibbon has an
extremely loud but musical voice. Mr. Waterhouse
states,405 “It appeared to me that in ascending and
descending the scale, the intervals were always exactly
half-tones; and I am sure that the highest note was
the exact octave to the lowest. The quality of the
notes is very musical; and I do not doubt that a good
violinist would be able to give a correct idea of the
gibbon’s composition, excepting as regards its loudness.”
Mr. Waterhouse then gives the notes. Professor
Owen, who is likewise a musician, confirms the
foregoing statement, and remarks that this gibbon
“alone of brute mammals may be said to sing.” It
appears to be much excited after its performance. Unfortunately
its habits have never been closely observed
in a state of nature; but from the analogy of almost
all other animals, it is highly probable that it utters its
musical notes especially during the season of courtship.
333
The perception, if not the enjoyment, of musical
cadences and of rhythm is probably common to all animals,
and no doubt depends on the common physiological
nature of their nervous systems. Even Crustaceans,
which are not capable of producing any
voluntary sound, possess certain auditory hairs, which
have been seen to vibrate when the proper musical notes
are struck.406 It is well known that some dogs howl
when hearing particular tones. Seals apparently appreciate
music, and their fondness for it “was well
known to the ancients, and is often taken advantage
of by the hunters at the present day.”407 With all
those animals, namely insects, amphibians, and birds,
the males of which during the season of courtship
incessantly produce musical notes or mere rhythmical
sounds, we must believe that the females are able to
appreciate them, and are thus excited or charmed;
otherwise the incessant efforts of the males and the
complex structures often possessed exclusively by them
would be useless.
With man song is generally admitted to be the basis
or origin of instrumental music. As neither the enjoyment
nor the capacity of producing musical notes are
faculties of the least direct use to man in reference
to his ordinary habits of life, they must be ranked
amongst the most mysterious with which he is endowed.
They are present, though in a very rude and as it
appears almost latent condition, in men of all races,
even the most savage; but so different is the taste of
the different races, that our music gives not the least
pleasure to savages, and their music is to us hideous
and unmeaning. Dr. Seemann, in some interesting
334remarks on this subject,408 “doubts whether even amongst
the nations of Western Europe, intimately connected
as they are by close and frequent intercourse, the
music of the one is interpreted in the same sense
by the others. By travelling eastwards we find that
there is certainly a different language of music.
Songs of joy and dance-accompaniments are no longer,
as with us, in the major keys, but always in the minor.”
Whether or not the half-human progenitors of man possessed,
like the before-mentioned gibbon, the capacity
of producing, and no doubt of appreciating, musical
notes, we have every reason to believe that man possessed
these faculties at a very remote period, for
singing and music are extremely ancient arts. Poetry,
which may be considered as the offspring of song, is
likewise so ancient that many persons have felt astonishment
that it should have arisen during the earliest
ages of which we have any record.
The musical faculties, which are not wholly deficient
in any race, are capable of prompt and high development,
as we see with Hottentots and Negroes, who have
readily become excellent musicians, although they do
not practise in their native countries anything that we
should esteem as music. But there is nothing anomalous
in this circumstance: some species of birds
which never naturally sing, can without much difficulty
be taught to perform; thus the house-sparrow has learnt
the song of a linnet. As these two species are closely
allied, and belong to the order of Insessores, which
includes nearly all the singing-birds in the world, it is
quite possible or probable that a progenitor of the spar335row
may have been a songster. It is a much more
remarkable fact that parrots, which belong to a group
distinct from the Insessores, and have differently-constructed
vocal organs, can be taught not only to speak,
but to pipe or whistle tunes invented by man, so that
they must have some musical capacity. Nevertheless
it would be extremely rash to assume that parrots are
descended from some ancient progenitor which was a
songster. Many analogous cases could be advanced
of organs and instincts originally adapted for one purpose,
having been utilised for some quite distinct
purpose.409 Hence the capacity for high musical development,
which the savage races of man possess, may
be due either to our semi-human progenitors having
practised some rude form of music, or simply to their
having acquired for some distinct purposes the proper
vocal organs. But in this latter case we must assume
that they already possessed, as in the above instance of
the parrots, and as seems to occur with many animals,
some sense of melody.
Music affects every emotion, but does not by itself excite
in us the more terrible emotions of horror, rage, &c.
It awakens the gentler feelings of tenderness and love,
which readily pass into devotion. It likewise stirs up in
us the sensation of triumph and the glorious ardour for
war. These powerful and mingled feelings may well give
rise to the sense of sublimity. We can concentrate, as
336
Dr. Seemann observes, greater intensity of feeling in a
single musical note than in pages of writing. Nearly the
same emotions, but much weaker and less complex, are
probably felt by birds when the male pours forth his full
volume of song, in rivalry with other males, for the sake
of captivating the female. Love is still the commonest
theme of our own songs. As Herbert Spencer remarks,
music “arouses dormant sentiments of which we had not
conceived the possibility, and do not know the meaning;
or, as Richter says, tells us of things we have not seen
and shall not see.”410 Conversely, when vivid emotions
are felt and expressed by the orator or even in common
speech, musical cadences and rhythm are instinctively
used. Monkeys also express strong feelings in different
tones—anger and impatience by low,—fear and pain by
high notes.411 The sensations and ideas excited in us
by music, or by the cadences of impassioned oratory,
appear from their vagueness, yet depth, like mental reversions
to the emotions and thoughts of a long-past age.
All these facts with respect to music become to a
certain extent intelligible if we may assume that
musical tones and rhythm were used by the half-human
337progenitors of man, during the season of courtship,
when animals of all kinds are excited by the
strongest passions. In this case, from the deeply-laid
principle of inherited associations, musical tones would
be likely to excite in us, in a vague and indefinite manner,
the strong emotions of a long-past age. Bearing in
mind that the males of some quadrumanous animals
have their vocal organs much more developed than in
the females, and that one anthropomorphous species
pours forth a whole octave of musical notes and may be
said to sing, the suspicion does not appear improbable
that the progenitors of man, either the males or females,
or both sexes, before they had acquired the power
of expressing their mutual love in articulate language,
endeavoured to charm each other with musical notes
and rhythm. So little is known about the use of the
voice by the Quadrumana during the season of love, that
we have hardly any means of judging whether the habit
of singing was first acquired by the male or female
progenitors of mankind. Women are generally thought
to possess sweeter voices than men, and as far as this
serves as any guide we may infer that they first acquired
musical powers in order to attract the other sex.412 But
if so, this must have occurred long ago, before the progenitors
of man had become sufficiently human to treat
and value their women merely as useful slaves. The
impassioned orator, bard, or musician, when with his
varied tones and cadences he excites the strongest
emotions in his hearers, little suspects that he uses the
same means by which, at an extremely remote period,
his half-human ancestors aroused each other’s ardent
passions, during their mutual courtship and rivalry.
338
On the influence of beauty in determining the marriages
of mankind.—In civilised life man is largely, but
by no means exclusively, influenced in the choice of
his wife by external appearance; but we are chiefly
concerned with primeval times, and our only means of
forming a judgment on this subject is to study the habits
of existing semi-civilised and savage nations. If it can
be shewn that the men of different races prefer women
having certain characteristics, or conversely that the
women prefer certain men, we have then to enquire
whether such choice, continued during many generations,
would produce any sensible effect on the race, either
on one sex or both sexes; this latter circumstance
depending on the form of inheritance which prevails.
It will be well first to shew in some detail that savages
pay the greatest attention to their personal appearance.413
That they have a passion for ornament is notorious;
and an English philosopher goes so far as to maintain
that clothes were first made for ornament and not for
warmth. As Professor Waitz remarks, “however poor
and miserable man is, he finds a pleasure in adorning
himself.” The extravagance of the naked Indians of
South America in decorating themselves is shewn “by
a man of large stature gaining with difficulty enough
by the labour of a fortnight to procure in exchange
339the chica necessary to paint himself red.”414 The
ancient barbarians of Europe during the Reindeer period
brought to their caves any brilliant or singular objects
which they happened to find. Savages at the present
day everywhere deck themselves with plumes, necklaces,
armlets, earrings, &c. They paint themselves in
the most diversified manner. “If painted nations,” as
Humboldt observes, “had been examined with the same
attention as clothed nations, it would have been perceived
that the most fertile imagination and the most
mutable caprice have created the fashions of painting,
as well as those of garments.”
In one part of Africa the eyelids are coloured black;
in another the nails are coloured yellow or purple. In
many places the hair is dyed of various tints. In different
countries the teeth are stained black, red, blue,
&c., and in the Malay Archipelago it is thought shameful
to have white teeth like those of a dog. Not one
great country can be named, from the Polar regions in
the north to New Zealand in the south, in which the
aborigines do not tattoo themselves. This practice was
followed by the Jews of old and by the ancient Britons.
In Africa some of the natives tattoo themselves, but it
is much more common to raise protuberances by rubbing
salt into incisions made in various parts of the body;
and these are considered by the inhabitants of Kordofan
and Darfur “to be great personal attractions.” In the
Arab countries no beauty can be perfect until the cheeks
or temples have been gashed.”415 In South America,
as Humboldt remarks, “a mother would be accused of
340culpable indifference towards her children, if she did
not employ artificial means to shape the calf of the leg
after the fashion of the country.” In the Old and New
World the shape of the skull was formerly modified
during infancy in the most extraordinary manner, as is
still the case in many places, and such deformities are
considered ornamental. For instance, the savages of
Colombia416 deem a much flattened head “an essential
point of beauty.”
The hair is treated with especial care in various
countries; it is allowed to grow to full length, so as to
reach to the ground, or is combed into “a compact
frizzled mop, which is the Papuan’s pride and glory.”417
In Northern Africa “a man requires a period of from
eight to ten years to perfect his coiffure.” With other
nations the head is shaved, and in parts of South America
and Africa even the eyebrows are eradicated. The
natives of the Upper Nile knock out the four front
teeth, saying that they do not wish to resemble brutes.
Further south, the Batokas knock out the two upper
incisors, which, as Livingstone418 remarks, gives the face
a hideous appearance, owing to the growth of the lower
jaw; but these people think the presence of the incisors
most unsightly, and on beholding some Europeans, cried
out, “Look at the great teeth!” The great chief Sebituani
tried in vain to alter this fashion. In various parts
of Africa and in the Malay Archipelago the natives file
the incisor teeth into points like those of a saw, or pierce
them with holes, into which they insert studs.
341
As the face with us is chiefly admired for its beauty,
so with savages it is the chief seat of mutilation. In
all quarters of the world the septum, and more rarely
the wings of the nose are pierced, with rings, sticks,
feathers, and other ornaments inserted into the holes.
The ears are everywhere pierced and similarly ornamented,
and with the Botocudos and Lenguas of South
America the hole is gradually so much enlarged that
the lower edge touches the shoulder. In North and
South America and in Africa either the upper or lower
lip is pierced; and with the Botocudos the hole in the
lower lip is so large that a disc of wood four inches in
diameter is placed in it. Mantegazza gives a curious
account of the shame felt by a South American native,
and of the ridicule which he excited, when he sold his
tembeta,—the large coloured piece of wood which is
passed through the hole. In central Africa the women
perforate the lower lip and wear a crystal, which, from
the movement of the tongue, has “a wriggling motion
indescribably ludicrous during conversation.” The
wife of the chief of Latooka told Sir S. Baker419 that his
“wife would be much improved if she would extract
her four front teeth from the lower jaw, and wear the
long pointed polished crystal in her under lip.” Further
south with the Makalolo, the upper lip is perforated,
and a large metal and bamboo ring, called a pelelé, is
worn in the hole. “This caused the lip in one case to
project two inches beyond the tip of the nose; and
when the lady smiled the contraction of the muscles
elevated it over the eyes. ‘Why do the women wear
these things?’ the venerable chief, Chinsurdi, was
asked. Evidently surprised at such a stupid question,
he replied, ‘For beauty! They are the only beautiful
342things women have; men have beards, women have
none. What kind of a person would she be without
the pelelé? She would not be a woman at all with a
mouth like a man, but no beard.’”420
Hardly any part of the body, which can be unnaturally
modified, has escaped. The amount of suffering
thus caused must have been wonderfully great, for
many of the operations require several years for their
completion, so that the idea of their necessity must be
imperative. The motives are various; the men paint
their bodies to make themselves appear terrible in battle;
certain mutilations are connected with religious
rites; or they mark the age of puberty, or the rank
of the man, or they serve to distinguish the tribes.
As with savages the same fashions prevail for long
periods,421 mutilations, from whatever cause first made,
soon come to be valued as distinctive marks. But
self-adornment, vanity, and the admiration of others,
seem to be the commonest motives. In regard to
tattooing, I was told by the missionaries in New Zealand,
that when they tried to persuade some girls to give up
the practice, they answered, “We must just have a few
lines on our lips; else when we grow old we shall be
so very ugly.” With the men of New Zealand, a most
capable judge422 says, “to have fine tattooed faces was
the great ambition of the young, both to render themselves
attractive to the ladies, and conspicuous in war.”
A star tattooed on the forehead and a spot on the chin
343are thought by the women in one part of Africa to be
irresistible attractions.423 In most, but not all parts of
the world, the men are more highly ornamented than
the women, and often in a different manner; sometimes,
though rarely, the women are hardly at all ornamented.
As the women are made by savages to perform the
greatest share of the work, and as they are not allowed
to eat the best kinds of food, so it accords with the characteristic
selfishness of man that they should not be
allowed to obtain, or to use, the finest ornaments.
Lastly it is a remarkable fact, as proved by the foregoing
quotations, that the same fashions in modifying
the shape of the head, in ornamenting the hair, in
painting, tattooing, perforating the nose, lips, or ears,
in removing or filing the teeth, &c., now prevail and
have long prevailed in the most distant quarters of
the world. It is extremely improbable that these practices
which are followed by so many distinct nations are
due to tradition from any common source. They rather
indicate the close similarity of the mind of man, to whatever
race he may belong, in the same manner as the
almost universal habits of dancing, masquerading, and
making rude pictures.
Having made these preliminary remarks on the
admiration felt by savages for various ornaments, and
for deformities most unsightly in our eyes, let us see
how far the men are attracted by the appearance
of their women, and what are their ideas of beauty.
As I have heard it maintained that savages are quite
indifferent about the beauty of their women, valuing
them solely as slaves, it may be well to observe that
this conclusion does not at all agree with the care which
the women take in ornamenting themselves, or with
344their vanity. Burchell424 gives an amusing account of
a Bushwoman, who used so much grease, red ochre,
and shining powder, “as would have ruined any but a
very rich husband.” She displayed also “much vanity
and too evident a consciousness of her superiority.”
Mr. Winwood Reade informs me that the negroes of
the West Coast often discuss the beauty of their women.
Some competent observers have attributed the fearfully
common practice of infanticide partly to the desire felt
by the women to retain their good looks.425 In several
regions the women wear charms and love-philters to
gain the affections of the men; and Mr. Brown enumerates
four plants used for this purpose by the women of
North-Western America.426
Hearne,427 who lived many years with the American
Indians, and who was an excellent observer, says, in
speaking of the women, “Ask a Northern Indian what
is beauty, and he will answer, a broad flat face, small
eyes, high cheek-bones, three or four broad black lines
across each cheek, a low forehead, a large broad chin,
a clumsy hook nose, a tawny hide, and breasts hanging
down to the belt.” Pallas, who visited the northern
parts of the Chinese empire, says “those women are
preferred who have the Mandschú form; that is to say,
a broad face, high cheek-bones, very broad noses, and
enormous ears;”428 and Vogt remarks that the obliquity
of the eye, which is proper to the Chinese and Japanese,
345is exaggerated in their pictures for the purpose, as
“it seems, of exhibiting its beauty, as contrasted with
the eye of the red-haired barbarians.” It is well
known, as Huc repeatedly remarks, that the Chinese of
the interior think Europeans hideous with their white
skins and prominent noses. The nose is far from being
too prominent, according to our ideas, in the natives of
Ceylon; yet “the Chinese in the seventh century, accustomed
to the flat features of the Mogul races, were
surprised at the prominent noses of the Cingalese; and
Thsang described them as having ‘the beak of a bird,
with the body of a man.’”
Finlayson, after minutely describing the people of
Cochin China, says that their rounded heads and faces
are their chief characteristics; and, he adds, “the
roundness of the whole countenance is more striking
in the women, who are reckoned beautiful in proportion
as they display this form of face.” The Siamese
have small noses with divergent nostrils, a wide mouth,
rather thick lips, a remarkably large face, with very
high and broad cheek-bones. It is, therefore, not wonderful
that “beauty, according to our notion is a stranger
to them. Yet they consider their own females to be
much more beautiful than those of Europe.”429
It is well known that with many Hottentot women
the posterior part of the body projects in a wonderful
manner; they are steatopygous; and Sir Andrew Smith
is certain that this peculiarity is greatly admired by the
men.430 He once saw a woman who was considered a
346beauty, and she was so immensely developed behind, that
when seated on level ground she could not rise, and had
to push herself along until she came to a slope. Some of
the women in various negro tribes are similarly characterised;
and, according to Burton, the Somal men “are
said to choose their wives by ranging them in a line,
and by picking her out who projects farthest a tergo.
Nothing can be more hateful to a negro than the
opposite form.”431
With respect to colour, the negroes rallied Mungo
Park on the whiteness of his skin and the prominence
of his nose, both of which they considered as “unsightly
and unnatural conformations.” He in return praised
the glossy jet of their skins and the lovely depression of
their noses; this they said was “honey-mouth,” nevertheless
they gave him food. The African Moors, also,
“knitted their brows and seemed to shudder” at the
whiteness of his skin. On the eastern coast, the negro
boys when they saw Burton, cried out “Look at the
white man; does he not look like a white ape?” On
the western coast, as Mr. Winwood Reade informs me,
the negroes admire a very black skin more than one of
a lighter tint. But their horror of whiteness may be
partly attributed, according to this same traveller, to
the belief held by most negroes that demons and spirits
are white.
The Banyai of the more southern part of the continent
are negroes, but “a great many of them are of a light
coffee-and-milk colour, and, indeed, this colour is considered
handsome throughout the whole country;” so
that here we have a different standard of taste. With the
347Kafirs, who differ much from negroes, “the skin, except
among the tribes near Delagoa Bay, is not usually
black, the prevailing colour being a mixture of black
and red, the most common shade being chocolate.
Dark complexions, as being most common are naturally
held in the highest esteem. To be told that he
is light-coloured, or like a white man, would be deemed
a very poor compliment by a Kafir. I have heard of
one unfortunate man who was so very fair that no
girl would marry him.” One of the titles of the
Zulu king is “You who are black.”432 Mr. Galton, in
speaking to me about the natives of S. Africa, remarked
that their ideas of beauty seem very different from
ours; for in one tribe two slim, slight, and pretty girls
were not admired by the natives.
Turning to other quarters of the world; in Java, a
yellow, not a white girl, is considered, according to
Madame Pfeiffer, a beauty. A man of Cochin-China
“spoke with contempt of the wife of the English
Ambassador, that she had white teeth like a dog,
and a rosy colour like that of potato-flowers.” We
have seen that the Chinese dislike our white skin, and
that the N. Americans admire “a tawny hide.” In
S. America, the Yura-caras, who inhabit the wooded,
damp slopes of the eastern Cordillera, are remarkably
pale-coloured, as their name in their own language
expresses; nevertheless they consider European women
as very inferior to their own.433
348
In several of the tribes of North America the hair
on the head grows to a wonderful length; and Catlin
gives a curious proof how much this is esteemed, for
the chief of the Crows was elected to this office from
having the longest hair of any man in the tribe, namely
ten feet and seven inches. The Aymaras and Quechuas
of S. America, likewise have very long hair; and this,
as Mr. D. Forbes informs me, is so much valued for
the sake of beauty, that cutting it off was the severest
punishment which he could inflict on them. In both
halves of the continent the natives sometimes increase
the apparent length of their hair by weaving into
it fibrous substances. Although the hair on the head
is thus cherished, that on the face is considered by
the North American Indians “as very vulgar,” and
every hair is carefully eradicated. This practice prevails
throughout the American continent from Vancouver’s
Island in the north to Tierra del Fuego in the
south. When York Minster, a Fuegian on board the
“Beagle” was taken back to his country, the natives told
him he ought to pull out the few short hairs on his face.
They also threatened a young missionary, who was left
for a time with them, to strip him naked, and pluck
the hairs from his face and body, yet he was far from
a hairy man. This fashion is carried to such an extreme
that the Indians of Paraguay eradicate their eyebrows
and eyelashes, saying that they do not wish to
be like horses.434
It is remarkable that throughout the world the races
349which are almost completely destitute of a beard dislike
hairs on the face and body, and take pains to eradicate
them. The Kalmucks are beardless, and they are well
known, like the Americans, to pluck out all straggling
hairs; and so it is with the Polynesians, some of the
Malays, and the Siamese. Mr. Veitch states that the
Japanese ladies “all objected to our whiskers, considering
them very ugly, and told us to cut them off, and
be like Japanese men.” The New Zealanders are
beardless; they carefully pluck out the hairs on the
face, and have a saying that “There is no woman for a
hairy man.”435
On the other hand, bearded races admire and greatly
value their beards; among the Anglo-Saxons every part
of the body, according to their laws, had a recognised
value; “the loss of the beard being estimated at twenty
shillings, while the breaking of a thigh was fixed at
only twelve.”436 In the East men swear solemnly by
their beards. We have seen that Chinsurdi, the chief
of the Makalolo in Africa, evidently thought that
beards were a great ornament. With the Fijians in
the Pacific the beard is “profuse and bushy, and is his
greatest pride;” whilst the inhabitants of the adjacent
archipelagoes of Tonga and Samoa are “beardless,
and abhor a rough chin.” In one island alone of the
Ellice group “the men are heavily bearded, and not a
little proud thereof.”437
350
We thus see how widely the different races of man
differ in their taste for the beautiful. In every nation
sufficiently advanced to have made effigies of their gods
or of their deified rulers, the sculptors no doubt have
endeavoured to express their highest ideal of beauty
and grandeur.438 Under this point of view it is well to
compare in our mind the Jupiter or Apollo of the Greeks
with the Egyptian or Assyrian statues; and these with
the hideous bas-reliefs on the ruined buildings of Central
America.
I have met with very few statements opposed to the
above conclusion. Mr. Winwood Reade, however, who
has had ample opportunities for observation, not only
with the negroes of the West Coast of Africa, but with
those of the interior who have never associated with
Europeans, is convinced that their ideas of beauty are
on the whole the same as ours. He has repeatedly found
that he agreed with negroes in their estimation of
the beauty of the native girls; and that their appreciation
of the beauty of European women corresponded
with ours. They admire long hair, and use artificial
means to make it appear abundant; they admire
also a beard, though themselves very scantily provided.
Mr. Reade feels doubtful what kind of nose is
most appreciated: a girl has been heard to say, “I
do not want to marry him, he has got no nose;” and
this shews that a very flat nose is not an object of admiration.
We should, however, bear in mind that the
depressed and very broad noses and projecting jaws of
the negroes of the West Coast are exceptional types
with the inhabitants of Africa. Notwithstanding the
foregoing statements, Mr. Reade does not think it pro351bable
that negroes would ever prefer the “most beautiful
European woman, on the mere grounds of physical
admiration, to a good-looking negress.”439
The truth of the principle, long ago insisted on by
Humboldt,440 that man admires and often tries to exaggerate
whatever characters nature may have given him,
is shewn in many ways. The practice of beardless races
extirpating every trace of a beard, and generally all the
hairs on the body, offers one illustration. The skull has
been greatly modified during ancient and modern times
by many nations; and there can be little doubt that this
has been practised, especially in N. and S. America, in
order to exaggerate some natural and admired peculiarity.
Many American Indians are known to admire a
head flattened to such an extreme degree as to appear
to us like that of an idiot. The natives on the nort-hwestern
coast compress the head into a pointed cone;
and it is their constant practice to gather the hair
into a knot on the top of the head, for the sake, as
Dr. Wilson remarks, “of increasing the apparent elevation
of the favourite conoid form.” The inhabitants
of Arakhan “admire a broad, smooth forehead, and in
order to produce it, they fasten a plate of lead on the
heads of the newborn children.” On the other hand,
352“a broad, well-rounded occiput is considered a great
beauty” by the natives of the Fiji islands.441
As with the skull, so with the nose; the ancient Huns
during the age of Attila were accustomed to flatten
the noses of their infants with bandages, “for the sake
of exaggerating a natural conformation.” With the
Tahitians, to be called, long-nose is considered as an
insult, and they compress the noses and foreheads of
their children for the sake of beauty. So it is with the
Malays of Sumatra, the Hottentots, certain Negroes,
and the natives of Brazil.442 The Chinese have by
nature unusually small feet;443 and it is well known
that the women of the upper classes distort their feet
to make them still smaller. Lastly, Humboldt thinks
that the American Indians prefer colouring their bodies
with red paint in order to exaggerate their natural tint;
and until recently European women added to their naturally
bright colours by rouge and white cosmetics; but
I doubt whether many barbarous nations have had any
such intention in painting themselves.
In the fashions of our own dress we see exactly the
same principle and the same desire to carry every point
to an extreme; we exhibit, also, the same spirit of
emulation. But the fashions of savages are far more
permanent than ours; and whenever their bodies are
353artificially modified this is necessarily the case. The
Arab women of the Upper Nile occupy about three days
in dressing their hair; they never imitate other tribes,
“but simply vie with each other in the superlativeness
of their own style.” Dr. Wilson, in speaking of the
compressed skulls of various American races, adds, “such
usages are among the least eradicable, and long survive
the shock of revolutions that change dynasties
and efface more important national peculiarities.”444
The same principle comes largely into play in the art of
selection; and we can thus understand, as I have elsewhere
explained,445 the wonderful development of all the
races of animals and plants which are kept merely for
ornament. Fanciers always wish each character to be
somewhat increased; they do not admire a medium
standard; they certainly do not desire any great and
abrupt change in the character of their breeds; they
admire solely what they are accustomed to behold, but
they ardently desire to see each characteristic feature
a little more developed.
No doubt the perceptive powers of man and the
lower animals are so constituted that brilliant colours
and certain forms, as well as harmonious and rhythmical
sounds, give pleasure and are called beautiful; but why
this should be so, we know no more than why certain
bodily sensations are agreeable and others disagreeable.
It is certainly not true that there is in the mind of
man any universal standard of beauty with respect to
the human body. It is, however, possible that certain
tastes may in the course of time become inherited,
though I know of no evidence in favour of this belief;
354and if so, each race would possess its own innate ideal
standard of beauty. It has been argued446 that ugliness
consists in an approach to the structure of the
lower animals, and this no doubt is true with the more
civilised nations, in which intellect is highly appreciated;
but a nose twice as prominent, or eyes twice
as large as usual, would not be an approach in structure
to any of the lower animals, and yet would be
utterly hideous. The men of each race prefer what
they are accustomed to behold; they cannot endure
any great change; but they like variety, and admire
each characteristic point carried to a moderate extreme.447
Men accustomed to a nearly oval face, to
straight and regular features, and to bright colours,
admire, as we Europeans know, these points when
strongly developed. On the other hand, men accustomed
to a broad face, with high cheek-bones, a depressed
nose, and a black skin, admire these points
strongly developed. No doubt characters of all kinds
may easily be too much developed for beauty. Hence a
perfect beauty, which implies many characters modified
in a particular manner, will in every race be a prodigy.
As the great anatomist Bichat long ago said, if every
one were cast in the same mould, there would be no such
thing as beauty. If all our women were to become as
beautiful as the Venus de Medici, we should for a time
be charmed; but we should soon wish for variety; and
as soon as we had obtained variety, we should wish to
see certain characters in our women a little exaggerated
beyond the then existing common standard.
355
CHAPTER XX.
Secondary Sexual Characters of Man—continued.
On the effects of the continued selection of women according to a
different standard of beauty in each race—On the causes which
interfere with sexual selection in civilised and savage nations—Conditions
favourable to sexual selection during primeval
times—On the manner of action of sexual selection with
mankind—On the women in savage tribes having some power
to choose their husbands—Absence of hair on the body, and
development of the beard—Colour of the skin—Summary.
We have seen in the last chapter that with all barbarous
races ornaments, dress, and external appearance are
highly valued; and that the men judge of the beauty
of their women by widely different standards. We
must next inquire whether this preference and the
consequent selection during many generations of those
women, which, appear to the men of each race the
most attractive, has altered the character either of the
females alone or of both sexes. With mammals the
general rule appears to be that characters of all kinds
are inherited equally by the males and females; we
might therefore expect that with mankind any characters
gained through sexual selection by the females
would commonly be transferred to the offspring of both
sexes. If any change has thus been effected it is almost
certain that the different races will have been differently
modified, as each has its own standard of beauty.
With mankind, especially with savages, many causes
interfere with the action of sexual selection as far as the
bodily frame is concerned. Civilised men are largely356
attracted by the mental charms of women, by their
wealth, and especially by their social position; for men
rarely marry into a much lower rank of life. The men
who succeed in obtaining the more beautiful women,
will not have a better chance of leaving a long line
of descendants than other men with plainer wives, with
the exception of the few who bequeath their fortunes
according to primogeniture. With respect to the opposite
form of selection, namely of the more attractive
men by the women, although in civilised nations women
have free or almost free choice, which is not the case
with barbarous races, yet their choice is largely influenced
by the social position and wealth of the men;
and the success of the latter in life largely depends on
their intellectual powers and energy, or on the fruits of
these same powers in their forefathers.
There is, however, reason to believe that sexual
selection has effected something in certain civilised and
semi-civilised nations. Many persons are convinced, as
it appears to me with justice, that the members of our
aristocracy, including under this term all wealthy families
in which primogeniture has long prevailed, from
having chosen during many generations from all classes
the more beautiful women as their wives, have become
handsomer, according to the European standard of
beauty, than the middle classes; yet the middle classes
are placed under equally favourable conditions of life
for the perfect development of the body. Cook remarks
that the superiority in personal appearance
“which is observable in the erees or nobles in all the
other islands (of the Pacific) is found in the Sandwich
islands;” but this may be chiefly due to their better
food and manner of life.
The old traveller Chardin, in describing the Persians,
says their “blood is now highly refined by frequent357
intermixtures with the Georgians and Circassians,
two nations which surpass all the world in personal
beauty. There is hardly a man of rank in Persia
who is not born of a Georgian or Circassian mother.”
He adds that they inherit their beauty, “not from their
ancestors, for without the above mixture, the men of
rank in Persia, who are descendants of the Tartars,
would be extremely ugly.”448 Here is a more curious
case: the priestesses who attended the temple of Venus
Erycina at San-Giuliano in Sicily, were selected for their
beauty out of the whole of Greece; they were not
vestal virgins, and Quatrefages,449 who makes this statement,
says that the women of San-Giuliano are famous
at the present day as the most beautiful in the island,
and are sought by artists as models. But it is obvious
that the evidence in the above cases is doubtful.
The following case, though relating to savages, is well
worth giving from its curiosity. Mr. Winwood Reade
informs me that the Jollofs, a tribe of negroes on the
west coast of Africa, “are remarkable for their uniformly
fine appearance.” A friend of his asked one of
these men, “How is it that every one whom I meet is
so fine-looking, not only your men, but your women?”
The Jollof answered, “It is very easily explained: it
has always been our custom to pick out our worse-looking
slaves and to sell them.” It need hardly
be added that with all savages female slaves serve as
concubines. That this negro should have attributed,
whether rightly or wrongly, the fine appearance of his
tribe, to the long-continued elimination of the ugly
358women, is not so surprising as it may at first appear;
for I have elsewhere shewn450 that negroes fully appreciate
the importance of selection in the breeding of
their domestic animals, and I could give from Mr. Reade
additional evidence on this head.
On the Causes which prevent or check the Action of
Sexual Selection with Savages.—The chief causes are,
firstly, so-called communal marriages or promiscuous
intercourse; secondly, infanticide, especially of female
infants; thirdly, early betrothals; and lastly, the low
estimation in which women are held, as mere slaves.
These four points must be considered in some detail.
It is obvious that as long as the pairing of man, or
of any other animal, is left to chance, with no choice
exerted by either sex, there can be no sexual selection;
and no effect will be produced on the offspring by
certain individuals having had an advantage over others
in their courtship. Now it is asserted that there exist
at the present day tribes which practise what Sir J.
Lubbock by courtesy calls communal marriages; that
is, all the men and women in the tribe are husbands and
wives to each other. The licentiousness of many savages
is no doubt astonishingly great, but it seems to me
that more evidence is requisite before we fully admit
that their existing intercourse is absolutely promiscuous.
Nevertheless all those who have most closely studied
the subject,451 and whose judgment is worth much more
359than mine, believe that communal marriage was the
original and universal form throughout the world, including
the intermarriage of brothers and sisters. The
indirect evidence in favour of this belief is extremely
strong, and rests chiefly on the terms of relationship
which are employed between the members of the same
tribe, implying a connection with the tribe alone, and
not with either parent. But the subject is too large
and complex for even an abstract to be here given, and
I will confine myself to a few remarks. It is evident in
the case of communal marriages, or where the marriage-tie
is very loose, that the relationship of the child to its
father cannot be known. But it seems almost incredible
that the relationship of the child to its mother should
ever have been completely ignored, especially as the
women in most savage tribes nurse their infants for a
long time. Accordingly in many cases the lines of
descent are traced through the mother alone, to the
exclusion of the father. But in many other cases the
terms employed express a connection with the tribe
alone, to the exclusion even of the mother. It seems
possible that the connection between the related members
of the same barbarous tribe, exposed to all sorts of
danger, might be so much more important, owing to the
need of mutual protection and aid, than that between
the mother and her child, as to lead to the sole use of
terms expressive of the former relationships; but Mr.
Morgan is convinced that this view of the case is by no
means sufficient.
The terms of relationship used in different parts of
360the world may be divided, according to the author just
quoted, into two great classes, the classificatory and
descriptive,—the latter being employed by us. It is
the classificatory system which so strongly leads to the
belief that communal and other extremely loose forms
of marriage were originally universal. But as far as
I can see, there is no necessity on this ground for believing
in absolutely promiscuous intercourse. Men and
women, like many of the lower animals, might formerly
have entered into strict though temporary unions for
each birth, and in this case nearly as much confusion
would have arisen in the terms of relationship as in
the case of promiscuous intercourse. As far as sexual
selection is concerned, all that is required is that choice
should be exerted before the parents unite, and it
signifies little whether the unions last for life or only
for a season.
Besides the evidence derived from the terms of relationship,
other lines of reasoning indicate the former
wide prevalence of communal marriage. Sir J. Lubbock
ingeniously accounts452 for the strange and widely-extended
habit of exogamy,—that is, the men of one
tribe always taking wives from a distinct tribe,—by
communism having been the original form of marriage;
so that a man never obtained a wife for himself
unless he captured her from a neighbouring and hostile
tribe, and then she would naturally have become his
sole and valuable property. Thus the practice of capturing
wives might have arisen; and from the honour
so gained might ultimately have become the universal
habit. We can also, according to Sir J. Lubbock,452
thus understand “the necessity of expiation for mar361riage
as an infringement of tribal rites, since, according
to old ideas, a man had no right to appropriate
to himself that which belonged to the whole tribe.”
Sir J. Lubbock further gives a most curious body of
facts shewing that in old times high honour was bestowed
on women who were utterly licentious; and this,
as he explains, is intelligible, if we admit that promiscuous
intercourse was the aboriginal and therefore
long revered custom of the tribe.453
Although the manner of development of the marriage-tie
is an obscure subject, as we may infer from
the divergent opinions on several points between the
three authors who have studied it most closely, namely,
Mr. Morgan, Mr. M’Lennan, and Sir J. Lubbock, yet
from the foregoing and several other lines of evidence it
seems certain that the habit of marriage has been gradually
developed, and that almost promiscuous intercourse
was once extremely common throughout the world.
Nevertheless from the analogy of the lower animals,
more particularly of those which come nearest to man
in the series, I cannot believe that this habit prevailed
at an extremely remote period, when man had hardly
attained to his present rank in the zoological scale.
Man, as I have attempted to shew, is certainly descended
from some ape-like creature. With the existing Quadrumana,
as far as their habits are known, the males of
some species are monogamous, but live during only a
part of the year with the females, as seems to be the
case with the Orang. Several kinds, as some of the
Indian and American monkeys, are strictly monogamous,
and associate all the year round with their wives.
Others are polygamous, as the Gorilla and several
362American species, and each family lives separate.
Even when this occurs, the families inhabiting the
same district are probably to a certain extent social:
the Chimpanzee, for instance, is occasionally met with
in large bands. Again, other species are polygamous,
but several males, each with their own females, live
associated in a body, as with several species of Baboons.454
We may indeed conclude from what we know of the
jealousy of all male quadrupeds, armed, as many of them
are, with special weapons for battling with their rivals,
that promiscuous intercourse in a state of nature is
extremely improbable. The pairing may not last for
life, but only for each birth; yet if the males which are
the strongest and best able to defend or otherwise assist
their females and young offspring, were to select the
more attractive females, this would suffice for the work
of sexual selection.
Therefore, if we look far enough back in the stream
of time, it is extremely improbable that primeval men
and women lived promiscuously together. Judging from
the social habits of man as he now exists, and from
most savages being polygamists, the most probable
view is that primeval man aboriginally lived in small
communities, each with as many wives as he could
support and obtain, whom he would have jealously
guarded against all other men. Or he may have lived
with several wives by himself, like the Gorilla; for
all the natives “agree that but one adult male is
seen in a band; when the young male grows up, a
contest takes place for mastery, and the strongest, by
363killing and driving out the others, establishes himself
as the head of the community.”455 The younger males,
being thus expelled and wandering about, would, when
at last successful in finding a partner, prevent too close
interbreeding within the limits of the same family.
Although savages are now extremely licentious, and
although communal marriages may formerly have
largely prevailed, yet many tribes practise some form
of marriage, but of a far more lax nature than with
civilised nations. Polygamy, as just stated, is almost
universally followed by the leading men in every tribe.
Nevertheless there are tribes, standing almost at the
bottom of the scale, which are strictly monogamous.
This is the case with the Veddahs of Ceylon: they
have a saying, according to Sir J. Lubbock,456 “that
death alone can separate husband and wife.” An
intelligent Kandyan chief, of course a polygamist,
“was perfectly scandalized at the utter barbarism of
living with only one wife, and never parting until
separated by death.” It was, he said, “just like the
Wanderoo monkeys.” Whether savages who now
enter into some form of marriage, either polygamous or
monogamous, have retained this habit from primeval
times, or whether they have returned to some form of
marriage, after passing through a stage of promiscuous
intercourse, I will not pretend to conjecture.
Infanticide.—This practice is now very common
throughout the world, and there is reason to believe
that it prevailed much more extensively during former
times.457 Barbarians find it difficult to support them364selves
and their children, and it is a simple plan to kill
their infants. In South America some tribes, as Azara
states, formerly destroyed so many infants of both sexes,
that they were on the point of extinction. In the Polynesian
Islands women have been known to kill from four
or five to even ten of their children; and Ellis could not
find a single woman who had not killed at least one.
Wherever infanticide prevails the struggle for existence
will be in so far less severe, and all the members of
the tribe will have an almost equally good chance of
rearing their few surviving children. In most cases
a larger number of female than of male infants are
destroyed, for it is obvious that the latter are of most
value to the tribe, as they will when grown up aid in defending
it, and can support themselves. But the trouble
experienced by the women in rearing children, their
consequent loss of beauty, the higher estimation set on
them and their happier fate, when few in number, are
assigned by the women themselves, and by various observers,
as additional motives for infanticide. In
Australia, where female infanticide is still common, Sir
G. Grey estimated the proportion of native women to
men as one to three; but others say as two to three.
In a village on the eastern frontier of India, Colonel
Macculloch found not a single female child.458
When, owing to female infanticide, the women of a
tribe are few in number, the habit of capturing wives
from neighbouring tribes would naturally arise. Sir J.
Lubbock, however, as we have seen, attributes the practice
in chief part, to the former existence of communal
marriage, and to the men having consequently captured
365women from other tribes to hold as their sole property.
Additional causes might be assigned, such as the communities
being very small, in which case, marriageable
women would often be deficient. That the habit of
capture was most extensively practised during former
times, even by the ancestors of civilised nations, is
clearly shewn by the preservation of many curious
customs and ceremonies, of which Mr. M’Lennan has
given a most interesting account. In our own marriages
the “best man” seems originally to have been
the chief abettor of the bridegroom in the act of capture.
Now as long as men habitually procured their
wives through violence and craft, it is not probable that
they would have selected the more attractive women;
they would have been too glad to have seized on any
woman. But as soon as the practice of procuring wives
from a distinct tribe was effected through barter, as now
occurs in many places, the more attractive women would
generally have been purchased. The incessant crossing,
however, between tribe and tribe, which necessarily
follows from any form of this habit would have tended
to keep all the people inhabiting the same country
nearly uniform in character; and this would have
greatly interfered with the power of sexual selection in
differentiating the tribes.
The scarcity of women, consequent on female infanticide,
leads, also, to another practice, namely polyandry,
which is still common in several parts of the world, and
which formerly, as Mr. M’Lennan believes, prevailed
almost universally; but this latter conclusion is doubted
by Mr. Morgan and Sir J. Lubbock.459 Whenever two
366or more men are compelled to marry one woman, it is
certain that all the women of the tribe will get married,
and there will be no selection by the men of the more
attractive women. But under these circumstances the
women no doubt will have the power of choice, and
will prefer the more attractive men. Azara, for instance,
describes how carefully a Guana woman bargains
for all sorts of privileges, before accepting some one
or more husbands; and the men in consequence take
unusual care of their personal appearance.460 The very
ugly men would perhaps altogether fail in getting a
wife, or get one later in life, but the handsomer men,
although the most successful in obtaining wives, would
not, as far as we can see, leave more offspring to inherit
their beauty than the less handsome husbands of the
same women.
Early Betrothals and Slavery of Women.—With
many savages it is the custom to betroth the females
whilst mere infants; and this would effectually prevent
preference being exerted on either side according to
personal appearance. But it would not prevent the
more attractive women from being afterwards stolen
or taken by force from their husbands by the more
powerful men; and this often happens in Australia,
America, and other parts of the world. The same consequences
with reference to sexual selection would to a
certain extent follow when women are valued almost
exclusively as slaves or beasts of burden, as is the case
with most savages. The men, however, at all times
would prefer the handsomest slaves according to their
standard of beauty.
We thus see that several customs prevail with savages
367which would greatly interfere with, or completely stop,
the action of sexual selection. On the other hand, the
conditions of life to which savages are exposed, and
some of their habits, are favourable to natural selection;
and this always comes into play together with sexual
selection. Savages are known to suffer severely from
recurrent famines; they do not increase their food by
artificial means; they rarely refrain from marriage,461
and generally marry young. Consequently they must
be subjected to occasional hard struggles for existence,
and the favoured individuals will alone survive.
Turning to primeval times when men had only doubtfully
attained the rank of manhood, they would probably
have lived, as already stated, either as polygamists or
temporarily as monogamists. Their intercourse, judging
from analogy, would not then have been promiscuous.
They would, no doubt, have defended their females to
the best of their power from enemies of all kinds, and
would probably have hunted for their subsistence, as
well as for that of their offspring. The most powerful
and able males would have succeeded best in the
struggle for life and in obtaining attractive females. At
this early period the progenitors of man, from having
only feeble powers of reason, would not have looked
forward to distant contingencies. They would have
been governed more by their instincts and even less
by their reason than are savages at the present day.
They would not at that period have partially lost one
of the strongest of all instincts, common to all the lower
animals, namely the love of their young offspring; and
368consequently they would not have practised infanticide.
There would have been no artificial scarcity of women,
and polyandry would not have been followed; there
would have been no early betrothals; women would
not have been valued as mere slaves; both sexes, if the
females as well as the males were permitted to exert
any choice, would have chosen their partners, not for
mental charms, or property, or social position, but almost
solely from external appearance. All the adults would
have married or paired, and all the offspring, as far as
that was possible, would have been reared; so that the
struggle for existence would have been periodically
severe to an extreme degree. Thus during these primordial
times all the conditions for sexual selection
would have been much more favourable than at a later
period, when man had advanced in his intellectual
powers, but had retrograded in his instincts. Therefore,
whatever influence sexual selection may have had in
producing the differences between the races of man, and
between man and the higher Quadrumana, this influence
would have been much more powerful at a very remote
period than at the present day.
On the Manner of Action of Sexual Selection with
mankind.—With primeval men under the favourable
conditions just stated, and with those savages who at the
present time enter into any marriage tie (but subject to
greater or less interference according as the habits of
female infanticide, early betrothals, &c., are more or
less practised), sexual selection will probably have
acted in the following manner. The strongest and most
vigorous men,—those who could best defend and hunt
for their families, and during later times the chiefs or
head-men,—those who were provided with the best
weapons and who possessed the most property, such as369
a larger number of dogs or other animals, would have
succeeded in rearing a greater average number of offspring,
than would the weaker, poorer and lower
members of the same tribes. There can, also, be no
doubt that such men would generally have been able
to select the more attractive women. At present the
chiefs of nearly every tribe throughout the world succeed
in obtaining more than one wife. Until recently,
as I hear from Mr. Mantell, almost every girl in New
Zealand, who was pretty, or promised to be pretty,
was tapu to some chief. With the Kafirs, as Mr. C.
Hamilton states,462 “the chiefs generally have the pick
of the women for many miles round, and are most
persevering in establishing or confirming their privilege.”
We have seen that each race has its own
style of beauty, and we know that it is natural to man
to admire each characteristic point in his domestic animals,
dress, ornaments, and personal appearance, when
carried a little beyond the common standard. If then
the several foregoing propositions be admitted, and I
cannot see that they are doubtful, it would be an inexplicable
circumstance, if the selection of the more
attractive women by the more powerful men of each
tribe, who would rear on an average a greater number
of children, did not after the lapse of many generations
modify to a certain extent the character of the tribe.
With our domestic animals, when a foreign breed
is introduced into a new country, or when a native
breed is long and carefully attended to, either for use or
ornament, it is found after several generations to have
undergone, whenever the means of comparison exist, a
greater or less amount of change. This follows from
unconscious selection during a long series of generations—that
370is, the preservation of the most approved individuals—without
any wish or expectation of such a result
on the part of the breeder. So again, if two careful
breeders rear during many years animals of the same
family, and do not compare them together or with
a common standard, the animals are found after a
time to have become to the surprise of their owners
slightly different.463 Each breeder has impressed, as
Von Nathusius well expresses it, the character of his
own mind—his own taste and judgment—on his
animals. What reason, then, can be assigned why
similar results should not follow from the long-continued
selection of the most admired women by those
men of each tribe, who were able to rear to maturity
the greater number of children? This would be unconscious
selection, for an effect would be produced,
independently of any wish or expectation on the part
of the men who preferred certain women to others.
Let us suppose the members of a tribe, in which
some form of marriage was practised, to spread over an
unoccupied continent; they would soon split up into
distinct hordes, which would be separated from each
other by various barriers, and still more effectually by
the incessant wars between all barbarous nations. The
hordes would thus be exposed to slightly different conditions
and habits of life, and would sooner or later
come to differ in some small degree. As soon as this
occurred, each isolated tribe would form for itself a
slightly different standard of beauty;464 and then un371conscious
selection would come into action through the
more powerful and leading savages preferring certain
women to others. Thus the differences between the
tribes, at first very slight, would gradually and inevitably
be increased to a greater and greater degree.
With animals in a state of nature, many characters
proper to the males, such as size, strength, special
weapons, courage and pugnacity, have been acquired
through the law of battle. The semi-human progenitors
of man, like their allies the Quadrumana, will
almost certainly have been thus modified; and, as
savages still fight for the possession of their women, a
similar process of selection has probably gone on in a
greater or less degree to the present day. Other characters
proper to the males of the lower animals, such
as bright colours and various ornaments, have been
acquired by the more attractive males having been
preferred by the females. There are, however, exceptional
cases in which the males, instead of having been
the selected, have been the selectors. We recognise
such cases by the females having been rendered more
highly ornamented than the males,—their ornamental
characters having been transmitted exclusively or
chiefly to their female offspring. One such case has
been described in the order to which man belongs,
namely, with the Rhesus monkey.
Man is more powerful in body and mind than woman,
and in the savage state he keeps her in a far more
abject state of bondage than does the male of any other
animal; therefore it is not surprising that he should
have gained the power of selection. Women are everywhere
conscious of the value of their beauty; and when
they have the means, they take more delight in decorating
themselves with all sorts of ornaments than do372
men. They borrow the plumes of male birds, with which
nature decked this sex in order to charm the females.
As women have long been selected for beauty, it is
not surprising that some of the successive variations
should have been transmitted in a limited manner; and
consequently that women should have transmitted their
beauty in a somewhat higher degree to their female
than to their male offspring. Hence women have become
more beautiful, as most persons will admit, than
men. Women, however, certainly transmit most of
their characters, including beauty, to their offspring
of both sexes; so that the continued preference by
the men of each race of the more attractive women,
according to their standard of taste, would tend to
modify in the same manner all the individuals of both
sexes belonging to the race.
With respect to the other form of sexual selection
(which with the lower animals is much the most common),
namely, when the females are the selectors, and
accept only those males which excite or charm them
most, we have reason to believe that it formerly acted
on the progenitors of man. Man in all probability owes
his beard, and perhaps some other characters, to inheritance
from an ancient progenitor who gained in this
manner his ornaments. But this form of selection may
have occasionally acted during later times; for in utterly
barbarous tribes the women have more power in
choosing, rejecting, and tempting their lovers, or of afterwards
changing their husbands, than might have been expected.
As this is a point of some importance, I will give
in detail such evidence as I have been able to collect.
Hearne describes how a woman in one of the tribes
of Arctic America repeatedly ran away from her husband
and joined a beloved man; and with the
Charruas of S. America, as Azara states, the power of373
divorce is perfectly free. With the Abipones, when
a man chooses a wife he bargains with the parents
about the price. But “it frequently happens that the
girl rescinds what has been agreed upon between the
parents and the bridegroom, obstinately rejecting the
very mention of marriage.” She often runs away,
hides herself, and thus eludes the bridegroom. In the
Fiji Islands the man seizes on the woman whom he
wishes for his wife by actual or pretended force; but
“on reaching the home of her abductor, should she not
approve of the match, she runs to some one who can
protect her; if, however, she is satisfied, the matter is
settled forthwith.” In Tierra del Fuego a young
man first obtains the consent of the parents by doing
them some service, and then he attempts to carry off
the girl; “but if she is unwilling, she hides herself
in the woods until her admirer is heartily tired of
looking for her, and gives up the pursuit; but this
seldom happens.” With the Kalmucks there is a
regular race between the bride and bridegroom, the
former having a fair start; and Clarke “was assured
that no instance occurs of a girl being caught, unless
she has a partiality to the pursuer.” So with the
wild tribes of the Malay archipelago there is a similar
racing match; and it appears from M. Bourien’s account,
as Sir J. Lubbock remarks, that “the race ‘is not to
the swift, nor the battle to the strong,’ but to the
young man who has the good fortune to please his
intended bride.”
Turning to Africa: the Kafirs buy their wives, and
girls are severely beaten by their fathers if they will
not accept a chosen husband; yet it is manifest from
many facts given by the Rev. Mr. Shooter, that they
have considerable power of choice. Thus very ugly,
though rich men, have been known to fail in getting374
wives. The girls, before consenting to be betrothed,
compel the men to shew themselves off, first in front
and then behind, and “exhibit their paces.” They
have been known to propose to a man, and they not
rarely run away with a favoured lover. With the
degraded bushwomen of S. Africa, “when a girl has
grown up to womanhood without having been betrothed,
which, however, does not often happen, her
lover must gain her approbation, as well as that of
the parents.”465 Mr. Winwood Reade made inquiries
for me with respect to the negroes of Western Africa,
and he informs me that “the women, at least among
the more intelligent Pagan tribes, have no difficulty
in getting the husbands whom they may desire, although
it is considered unwomanly to ask a man to
marry them. They are quite capable of falling in
love, and of forming tender, passionate, and faithful
attachments.”
We thus see that with savages the women are not
in quite so abject a state in relation to marriage as has
often been supposed. They can tempt the men whom
they prefer, and can sometimes reject those whom they
dislike, either before or after marriage. Preference
on the part of the women, steadily acting in any one
direction, would ultimately affect the character of the
tribe; for the women would generally choose not merely
the handsomer men, according to their standard of taste,
375but those who were at the same time best able to defend
and support them. Such well-endowed pairs would
commonly rear a larger number of offspring than the
less well endowed. The same result would obviously
follow in a still more marked manner if there was selection
on both sides; that is if the more attractive, and
at the same time more powerful men were to prefer,
and were preferred by, the more attractive women.
And these two forms of selection seem actually to have
occurred, whether or not simultaneously, with mankind,
especially during the earlier periods of our long history.
We will now consider in a little more detail, relatively
to sexual selection, some of the characters which
distinguish the several races of man from each other
and from the lower animals, namely, the more or less
complete absence of hair from the body and the colour
of the skin. We need say nothing about the great
diversity in the shape of the features and of the skull
between the different races, as we have seen in the last
chapter how different is the standard of beauty in these
respects. These characters will therefore probably have
been acted on through sexual selection; but we have no
means of judging, as far as I can see, whether they
have been acted on chiefly through the male or female
side. The musical faculties of man have likewise been
already discussed.
Absence of Hair on the Body, and its Development on
the Face and Head.—From the presence of the woolly
hair or lanugo on the human fœtus, and of rudimentary
hairs scattered over the body during maturity, we may
infer that man is descended from some animal which
was born hairy and remained so during life. The loss
of hair is an inconvenience and probably an injury to
man even under a hot climate, for he is thus exposed376
to sudden chills, especially during wet weather. As
Mr. Wallace remarks, the natives in all countries are
glad to protect their naked backs and shoulders with
some slight covering. No one supposes that the nakedness
of the skin is any direct advantage to man, so
that his body cannot have been divested of hair through
natural selection.466 Nor have we any grounds for believing,
as shewn in a former chapter, that this can be
due to the direct action of the conditions to which man
has long been exposed, or that it is the result of correlated
development.
The absence of hair on the body is to a certain extent
a secondary sexual character; for in all parts of the
world women are less hairy than men. Therefore we
may reasonably suspect that this is a character which
has been gained through sexual selection. We know
that the faces of several species of monkeys, and large
surfaces at the posterior end of the body in other species,
have been denuded of hair; and this we may
safely attribute to sexual selection, for these surfaces
are not only vividly coloured, but sometimes, as with
the male mandrill and female rhesus, much more
377vividly in the one sex than in the other. As these
animals gradually reach maturity the naked surfaces,
as I am informed by Mr. Bartlett, grow larger, relatively
to the size of their bodies. The hair, however,
appears to have been removed in these cases, not for
the sake of nudity, but that the colour of the skin
should be more fully displayed. So again with many
birds the head and neck have been divested of feathers
through sexual selection, for the sake of exhibiting the
brightly-coloured skin.
As woman has a less hairy body than man, and as
this character is common to all races, we may conclude
that our female semi-human progenitors were
probably first partially divested of hair; and that this
occurred at an extremely remote period before the
several races had diverged from a common stock. As
our female progenitors gradually acquired this new
character of nudity, they must have transmitted it in
an almost equal degree to their young offspring of both
sexes; so that its transmission, as in the case of many
ornaments with mammals and birds, has not been
limited either by age or sex. There is nothing surprising
in a partial loss of hair having been esteemed
as ornamental by the ape-like progenitors of man, for
we have seen that with animals of all kinds innumerable
strange characters have been thus esteemed, and have
consequently been modified through sexual selection.
Nor is it surprising that a character in a slight degree
injurious should have been thus acquired; for we know
that this is the case with the plumes of some birds, and
with the horns of some stags.
The females of certain anthropoid apes, as stated in a
former chapter, are somewhat less hairy on the under
surface than are the males; and here we have what
might have afforded a commencement for the process378
of denudation. With respect to the completion of the
process through sexual selection, it is well to bear in
mind the New Zealand proverb, “there is no woman
for a hairy man.” All who have seen photographs of
the Siamese hairy family will admit how ludicrously
hideous is the opposite extreme of excessive hairiness.
Hence the king of Siam had to bribe a man to marry
the first hairy woman in the family, who transmitted
this character to her young offspring of both sexes.467
Some races are much more hairy than others, especially
on the male side; but it must not be assumed that
the more hairy races, for instance Europeans, have retained
a primordial condition more completely than
have the naked races, such as the Kalmucks or Americans.
It is a more probable view that the hairiness
of the former is due to partial reversion, for characters
which have long been inherited are always apt to return.
It does not appear that a cold climate has been
influential in leading to this kind of reversion; excepting
perhaps with the negroes, who have been reared
during several generations, in the United States,468 and
possibly with the Ainos, who inhabit the northern
379islands of the Japan archipelago. But the laws of inheritance
are so complex than we can seldom understand
their action. If the greater hairiness of certain
races be the result of reversion, unchecked by any form
of selection, the extreme variability of this character,
even within the limits of the same race, ceases to be
remarkable.
With respect to the beard, if we turn to our best
guide, namely the Quadrumana, we find beards equally
well developed in both sexes of many species, but in
others, either confined to the males, or more developed
in them than in the females. From this fact, and from
the curious arrangement, as well as the bright colours, of
the hair about the heads of many monkeys, it is highly
probable, as before explained, that the males first
acquired their beards as an ornament through sexual
selection, transmitting them in most cases, in an equal or
nearly equal degree, to their offspring of both sexes.
We know from Eschricht469 that with mankind, the
female as well as the male fœtus is furnished with much
hair on the face, especially round the mouth; and this
indicates that we are descended from a progenitor, of
which both sexes were bearded. It appears therefore
at first sight probable that man has retained his beard
from a very early period, whilst woman lost her beard
at the same time when her body became almost completely
divested of hair. Even the colour of the beard
with mankind seems to have been inherited from an
ape-like progenitor; for when there is any difference
in tint between the hair of the head and the beard, the
latter is lighter coloured in all monkeys and in man.
There is less improbability in the men of the bearded
380races having retained their beards from primordial
times, than in the case of the hair on the body; for
with those Quadrumana, in which the male has a larger
beard than that of the female, it is fully developed
only at maturity, and the later stages of development
may have been exclusively transmitted to mankind.
We should then see what is actually the case, namely,
our male children, before they arrive at maturity, as
destitute of beards as are our female children. On the
other hand the great variability of the beard within
the limits of the same race and in different races indicates
that reversion has come into action. However
this may be, we must not overlook the part which
sexual selection may have played even during later
times; for we know that with savages, the men of the
beardless races take infinite pains in eradicating every
hair from their faces, as something odious, whilst the
men of the bearded races feel the greatest pride in their
beards. The women, no doubt, participate in these
feelings, and if so sexual selection can hardly have
failed to have effected something in the course of later
times.470
It is rather difficult to form a judgment how the long
381hair on our heads became developed. Eschricht
471 states
that in the human fœtus the hair on the face during
the fifth month is longer than that on the head; and
this indicates that our semi-human progenitors were not
furnished with long tresses, which consequently must
have been a late acquisition. This is likewise indicated
by the extraordinary difference in the length of the hair
in the different races; in the negro the hair forms a
mere curly mat; with us it is of great length, and with
the American natives it not rarely reaches to the
ground. Some species of Semnopithecus have their
heads covered with moderately long hair, and this probably
serves as an ornament and was acquired through
sexual selection. The same view may be extended to
mankind, for we know that long tresses are now and
were formerly much admired, as may be observed in the
works of almost every poet; St. Paul says, “if a woman
have long hair, it is a glory to her;” and we have seen
that in North America a chief was elected solely from
the length of his hair.
Colour of the Skin.—The best kind of evidence that
the colour of the skin has been modified through sexual
selection is wanting in the case of mankind; for the
sexes do not differ in this respect, or only slightly and
doubtfully. On the other hand we know from many
facts already given that the colour of the skin is regarded
by the men of all races as a highly important
element in their beauty; so that it is a character which
would be likely to be modified through selection, as has
occurred in innumerable instances with the lower animals.
It seems at first sight a monstrous supposition
that the jet blackness of the negro has been gained
382through sexual selection; but this view is supported by
various analogies, and we know that negroes admire
their own blackness. With mammals, when the sexes
differ in colour, the male is often black or much darker
than the female; and it depends merely on the form of
inheritance whether this or any other tint shall be transmitted
to both sexes or to one alone. The resemblance
of Pithecia satanas with his jet black skin, white rolling
eyeballs, and hair parted on the top of the head, to a
negro in miniature, is almost ludicrous.
The colour of the face differs much more widely in
the various kinds of monkeys than it does in the races
of man; and we have good reason to believe that
the red, blue, orange, almost white and black tints of
their skin, even when common to both sexes, and the
bright colours of their fur, as well as the ornamental
tufts of hair about the head, have all been acquired
through sexual selection. As the newly-born infants of
the most distinct races do not differ nearly as much in
colour as do the adults, although their bodies are completely
destitute of hair, we have some slight indication
that the tints of the different races were acquired subsequently
to the removal of the hair, which, as before
stated, must have occurred at a very early period.
Summary.—We may conclude that the greater size,
strength, courage, pugnacity, and even energy of man,
in comparison with the same qualities in woman, were
acquired during primeval times, and have subsequently
been augmented, chiefly through the contests of rival
males for the possession of the females. The greater
intellectual vigour and power of invention in man is
probably due to natural selection combined with the
inherited effects of habit, for the most able men will
have succeeded best in defending and providing for383
themselves, their wives and offspring. As far as the
extreme intricacy of the subject permits us to judge,
it appears that our male ape-like progenitors acquired
their beards as an ornament to charm or excite the
opposite sex, and transmitted them to man as he now
exists. The females apparently were first denuded of
hair in like manner as a sexual ornament; but they
transmitted this character almost equally to both sexes.
It is not improbable that the females were modified in
other respects for the same purpose and through the
same means; so that women have acquired sweeter
voices and become more beautiful than men.
It deserves particular attention that with mankind
all the conditions for sexual selection were much more
favourable, during a very early period, when man had
only just attained to the rank of manhood, than during
later times. For he would then, as we may safely conclude,
have been guided more by his instinctive passions,
and less by foresight or reason. He would not then
have been so utterly licentious as many savages now are;
and each male would have jealously guarded his wife or
wives. He would not then have practised infanticide;
nor valued his wives merely as useful slaves; nor have
been betrothed to them during infancy. Hence we may
infer that the races of men were differentiated, as far as
sexual selection is concerned, in chief part during a very
remote epoch; and this conclusion throws light on the
remarkable fact that at the most ancient period, of which
we have as yet obtained any record, the races of man
had already come to differ nearly or quite as much as
they do at the present day.
The views here advanced, on the part which sexual
selection has played in the history of man, want scientific
precision. He who does not admit this agency in
the case of the lower animals, will properly disregard384
all that I have written in the later chapters on man.
We cannot positively say that this character, but not
that, has been thus modified; it has, however, been
shewn that the races of man differ from each other and
from their nearest allies amongst the lower animals, in
certain characters which are of no service to them in
their ordinary habits of life, and which it is extremely
probable would have been modified through sexual selection.
We have seen that with the lowest savages the
people of each tribe admire their own characteristic
qualities,—the shape of the head and face, the squareness
of the cheek-bones, the prominence or depression
of the nose, the colour of the skin, the length of the
hair on the head, the absence of hair on the face and
body, or the presence of a great beard, and so forth.
Hence these and other such points could hardly fail to
have been slowly and gradually exaggerated from the
more powerful and able men in each tribe, who would
succeed in rearing the largest number of offspring, having
selected during many generations as their wives the
most strongly characterised and therefore most attractive
women. For my own part I conclude that of all
the causes which have led to the differences in external
appearance between the races of man, and to a certain
extent between man and the lower animals, sexual
selection has been by far the most efficient.
385
CHAPTER XXI.
General Summary and Conclusion.
Main conclusion that man is descended from some lower form—Manner
of development—Genealogy of man—Intellectual and
moral faculties—Sexual selection—Concluding remarks.
A brief summary will here be sufficient to recall to the
reader’s mind the more salient points in this work.
Many of the views which have been advanced are highly
speculative, and some no doubt will prove erroneous;
but I have in every case given the reasons which have
led me to one view rather than to another. It seemed
worth while to try how far the principle of evolution
would throw light on some of the more complex problems
in the natural history of man. False facts are
highly injurious to the progress of science, for they often
long endure; but false views, if supported by some
evidence, do little harm, as every one takes a salutary
pleasure in proving their falseness; and when this is
done, one path towards error is closed and the road to
truth is often at the same time opened.
The main conclusion arrived at in this work, and now
held by many naturalists who are well competent to
form a sound judgment, is that man is descended from
some less highly organised form. The grounds upon
which this conclusion rests will never be shaken, for the
close similarity between man and the lower animals in
embryonic development, as well as in innumerable
points of structure and constitution, both of high and
of the most trifling importance,—the rudiments which386
he retains, and the abnormal reversions to which he
is occasionally liable,—are facts which cannot be disputed.
They have long been known, but until recently
they told us nothing with respect to the origin of
man. Now when viewed by the light of our knowledge
of the whole organic world, their meaning is
unmistakeable. The great principle of evolution stands
up clear and firm, when these groups of facts are considered
in connection with others, such as the mutual
affinities of the members of the same group, their
geographical distribution in past and present times,
and their geological succession. It is incredible that
all these facts should speak falsely. He who is not
content to look, like a savage, at the phenomena of
nature as disconnected, cannot any longer believe that
man is the work of a separate act of creation. He will
be forced to admit that the close resemblance of the
embryo of man to that, for instance, of a dog—the construction
of his skull, limbs, and whole frame, independently
of the uses to which the parts may be put, on
the same plan with that of other mammals—the occasional
reappearance of various structures, for instance
of several distinct muscles, which man does not normally
possess, but which are common to the Quadrumana—and
a crowd of analogous facts—all point in
the plainest manner to the conclusion that man is the
co-descendant with other mammals of a common progenitor.
We have seen that man incessantly presents individual
differences in all parts of his body and in his
mental faculties. These differences or variations seem
to be induced by the same general causes, and to obey
the same laws as with the lower animals. In both
cases similar laws of inheritance prevail. Man tends to
increase at a greater rate than his means of subsistence;387
consequently he is occasionally subjected to a severe
struggle for existence, and natural selection will have
effected whatever lies within its scope. A succession
of strongly-marked variations of a similar nature are
by no means requisite; slight fluctuating differences
in the individual suffice for the work of natural selection.
We may feel assured that the inherited effects
of the long-continued use or disuse of parts will have
done much in the same direction with natural selection.
Modifications formerly of importance, though no longer
of any special use, will be long inherited. When one
part is modified, other parts will change through the
principle of correlation, of which we have instances in
many curious cases of correlated monstrosities. Something
may be attributed to the direct and definite
action of the surrounding conditions of life, such as
abundant food, heat, or moisture; and lastly, many
characters of slight physiological importance, some indeed
of considerable importance, have been gained
through sexual selection.
No doubt man, as well as every other animal, presents
structures, which as far as we can judge with
our little knowledge, are not now of any service to
him, nor have been so during any former period of his
existence, either in relation to his general conditions of
life, or of one sex to the other. Such structures cannot
be accounted for by any form of selection, or by the
inherited effects of the use and disuse of parts. We
know, however, that many strange and strongly-marked
peculiarities of structure occasionally appear in our
domesticated productions, and if the unknown causes
which produce them were to act more uniformly, they
would probably become common to all the individuals
of the species. We may hope hereafter to understand
something about the causes of such occasional modifications,388
especially through the study of monstrosities:
hence the labours of experimentalists, such as those of
M. Camille Dareste, are full of promise for the future.
In the greater number of cases we can only say that the
cause of each slight variation and of each monstrosity
lies much more in the nature or constitution of the
organism, than in the nature of the surrounding conditions;
though new and changed conditions certainly
play an important part in exciting organic changes of
all kinds.
Through the means just specified, aided perhaps by
others as yet undiscovered, man has been raised to his
present state. But since he attained to the rank of
manhood, he has diverged into distinct races, or as they
may be more appropriately called sub-species. Some
of these, for instance the Negro and European, are so
distinct that, if specimens had been brought to a naturalist
without any further information, they would undoubtedly
have been considered by him as good and
true species. Nevertheless all the races agree in so
many unimportant details of structure and in so many
mental peculiarities, that these can be accounted for
only through inheritance from a common progenitor;
and a progenitor thus characterised would probably
have deserved to rank as man.
It must not be supposed that the divergence of
each race from the other races, and of all the races
from a common stock, can be traced back to any one
pair of progenitors. On the contrary, at every stage
in the process of modification, all the individuals which
were in any way best fitted for their conditions of life,
though in different degrees, would have survived in
greater numbers than the less well fitted. The process
would have been like that followed by man, when
he does not intentionally select particular individuals,389
but breeds from all the superior and neglects all the
inferior individuals. He thus slowly but surely modifies
his stock, and unconsciously forms a new strain.
So with respect to modifications, acquired independently
of selection, and due to variations arising from
the nature of the organism and the action of the surrounding
conditions, or from changed habits of life, no
single pair will have been modified in a much greater
degree than the other pairs which inhabit the same
country, for all will have been continually blended
through free intercrossing.
By considering the embryological structure of man,—the
homologies which he presents with the lower
animals,—the rudiments which he retains,—and the
reversions to which he is liable, we can partly recall
in imagination the former condition of our early progenitors;
and can approximately place them in their
proper position in the zoological series. We thus learn
that man is descended from a hairy quadruped, furnished
with a tail and pointed ears, probably arboreal
in its habits, and an inhabitant of the Old World.
This creature, if its whole structure had been examined
by a naturalist, would have been classed amongst the
Quadrumana, as surely as would the common and still
more ancient progenitor of the Old and New World
monkeys. The Quadrumana and all the higher mammals
are probably derived from an ancient marsupial
animal, and this through a long line of diversified
forms, either from some reptile-like or some amphibian-like
creature, and this again from some fish-like animal.
In the dim obscurity of the past we can see that the
early progenitor of all the Vertebrata must have been
an aquatic animal, provided with branchiæ, with the two
sexes united in the same individual, and with the most
important organs of the body (such as the brain and390
heart) imperfectly developed. This animal seems to
have been more like the larvæ of our existing marine
Ascidians than any other known form.
The greatest difficulty which presents itself, when
we are driven to the above conclusion on the origin of
man, is the high standard of intellectual power and of
moral disposition which he has attained. But every one
who admits the general principle of evolution, must see
that the mental powers of the higher animals, which
are the same in kind with those of mankind, though
so different in degree, are capable of advancement.
Thus the interval between the mental powers of one
of the higher apes and of a fish, or between those
of an ant and scale-insect, is immense. The development
of these powers in animals does not offer any
special difficulty; for with our domesticated animals,
the mental faculties are certainly variable, and the
variations are inherited. No one doubts that these
faculties are of the utmost importance to animals in a
state of nature. Therefore the conditions are favourable
for their development through natural selection.
The same conclusion may be extended to man; the
intellect must have been all-important to him, even at
a very remote period, enabling him to use language,
to invent and make weapons, tools, traps, &c.; by
which means, in combination with his social habits,
he long ago became the most dominant of all living
creatures.
A great stride in the development of the intellect
will have followed, as soon as, through a previous considerable
advance, the half-art and half-instinct of language
came into use; for the continued use of language
will have reacted on the brain, and produced an inherited
effect; and this again will have reacted on the391
improvement of language. The large size of the brain
in man, in comparison with that of the lower animals,
relatively to the size of their bodies, may be attributed
in chief part, as Mr. Chauncey Wright has well remarked,472
to the early use of some simple form of
language,—that wonderful engine which affixes signs
to all sorts of objects and qualities, and excites trains
of thought which would never arise from the mere impression
of the senses, and if they did arise could not
be followed out. The higher intellectual powers of
man, such as those of ratiocination, abstraction, self-consciousness,
&c., will have followed from the continued
improvement of other mental faculties; but
without considerable culture of the mind, both in the
race and in the individual, it is doubtful whether these
high powers would be exercised, and thus fully attained.
The development of the moral qualities is a more
interesting and difficult problem. Their foundation
lies in the social instincts, including in this term the
family ties. These instincts are of a highly complex
nature, and in the case of the lower animals give
special tendencies towards certain definite actions; but
the more important elements for us are love, and the
distinct emotion of sympathy. Animals endowed with
the social instincts take pleasure in each other’s company,
warn each other of danger, defend and aid each
other in many ways. These instincts are not extended
to all the individuals of the species, but only to those
of the same community. As they are highly beneficial
to the species, they have in all probability been acquired
through natural selection.
A moral being is one who is capable of comparing
392his past and future actions and motives,—of approving
of some and disapproving of others; and the fact that
man is the one being who with certainty can be thus
designated makes the greatest of all distinctions between
him and the lower animals. But in our third
chapter I have endeavoured to shew that the moral
sense follows, firstly, from the enduring and always
present nature of the social instincts, in which respect
man agrees with the lower animals; and secondly, from
his mental faculties being highly active and his impressions
of past events extremely vivid, in which respects
he differs from the lower animals. Owing to this condition
of mind, man cannot avoid looking backwards and
comparing the impressions of past events and actions.
He also continually looks forward. Hence after some
temporary desire or passion has mastered his social instincts,
he will reflect and compare the now weakened
impression of such past impulses, with the ever present
social instinct; and he will then feel that sense of dissatisfaction
which all unsatisfied instincts leave behind
them. Consequently he resolves to act differently for
the future—and this is conscience. Any instinct which
is permanently stronger or more enduring than another,
gives rise to a feeling which we express by saying that
it ought to be obeyed. A pointer dog, if able to reflect
on his past conduct, would say to himself, I ought (as
indeed we say of him) to have pointed at that hare
and not have yielded to the passing temptation of
hunting it.
Social animals are partly impelled by a wish to aid
the members of the same community in a general
manner, but more commonly to perform certain definite
actions. Man is impelled by the same general wish to
aid his fellows, but has few or no special instincts.
He differs also from the lower animals in being able393
to express his desires by words, which thus become
the guide to the aid required and bestowed. The
motive to give aid is likewise somewhat modified in
man: it no longer consists solely of a blind instinctive
impulse, but is largely influenced by the praise or
blame of his fellow men. Both the appreciation and
the bestowal of praise and blame rest on sympathy;
and this emotion, as we have seen, is one of the most
important elements of the social instincts. Sympathy,
though gained as an instinct, is also much strengthened
by exercise or habit. As all men desire their own
happiness, praise or blame is bestowed on actions and
motives, according as they lead to this end; and as
happiness is an essential part of the general good, the
greatest-happiness principle indirectly serves as a nearly
safe standard of right and wrong. As the reasoning
powers advance and experience is gained, the more
remote effects of certain lines of conduct on the
character of the individual, and on the general good,
are perceived; and then the self-regarding virtues,
from coming within the scope of public opinion, receive
praise, and their opposites receive blame. But with the
less civilised nations reason often errs, and many bad
customs and base superstitions come within the same
scope, and consequently are esteemed as high virtues,
and their breach as heavy crimes.
The moral faculties are generally esteemed, and with
justice, as of higher value than the intellectual powers.
But we should always bear in mind that the activity of
the mind in vividly recalling past impressions is one of
the fundamental though secondary bases of conscience.
This fact affords the strongest argument for educating
and stimulating in all possible ways the intellectual
faculties of every human being. No doubt a man with
a torpid mind, if his social affections and sympathies are394
well developed, will be led to good actions, and may
have a fairly sensitive conscience. But whatever renders
the imagination of men more vivid and strengthens
the habit of recalling and comparing past impressions,
will make the conscience more sensitive, and may even
compensate to a certain extent for weak social affections
and sympathies.
The moral nature of man has reached the highest
standard as yet attained, partly through the advancement
of the reasoning powers and consequently of a just
public opinion, but especially through the sympathies
being rendered more tender and widely diffused through
the effects of habit, example, instruction, and reflection.
It is not improbable that virtuous tendencies may
through long practice be inherited. With the more
civilised races, the conviction of the existence of an
all-seeing Deity has had a potent influence on the
advancement of morality. Ultimately man no longer
accepts the praise or blame of his fellows as his chief
guide, though few escape this influence, but his habitual
convictions controlled by reason afford him the
safest rule. His conscience then becomes his supreme
judge and monitor. Nevertheless the first foundation
or origin of the moral sense lies in the social instincts,
including sympathy; and these instincts no doubt were
primarily gained, as in the case of the lower animals,
through natural selection.
The belief in God has often been advanced as not
only the greatest, but the most complete of all the
distinctions between man and the lower animals. It is
however impossible, as we have seen, to maintain that
this belief is innate or instinctive in man. On the
other hand a belief in all-pervading spiritual agencies
seems to be universal; and apparently follows from a395
considerable advance in the reasoning powers of man,
and from a still greater advance in his faculties of imagination,
curiosity and wonder. I am aware that the
assumed instinctive belief in God has been used by many
persons as an argument for His existence. But this
is a rash argument, as we should thus be compelled to
believe in the existence of many cruel and malignant
spirits, possessing only a little more power than man;
for the belief in them is far more general than of a
beneficent Deity. The idea of a universal and beneficent
Creator of the universe does not seem to arise in
the mind of man, until he has been elevated by long-continued
culture.
He who believes in the advancement of man from
some lowly-organised form, will naturally ask how does
this bear on the belief in the immortality of the soul.
The barbarous races of man, as Sir J. Lubbock has
shewn, possess no clear belief of this kind; but arguments
derived from the primeval beliefs of savages are,
as we have just seen, of little or no avail. Few persons
feel any anxiety from the impossibility of determining
at what precise period in the development of the individual,
from the first trace of the minute germinal
vesicle to the child either before or after birth, man
becomes an immortal being; and there is no greater
cause for anxiety because the period in the gradually
ascending organic scale cannot possibly be determined.473
I am aware that the conclusions arrived at in this
work will be denounced by some as highly irreligious;
but he who thus denounces them is bound to shew why
it is more irreligious to explain the origin of man as
a distinct species by descent from some lower form,
396
through the laws of variation and natural selection,
than to explain the birth of the individual through the
laws of ordinary reproduction. The birth both of the
species and of the individual are equally parts of that
grand sequence of events, which our minds refuse to
accept as the result of blind chance. The understanding
revolts at such a conclusion, whether or not we
are able to believe that every slight variation of structure,—the
union of each pair in marriage,—the dissemination
of each seed,—and other such events, have
all been ordained for some special purpose.
Sexual selection has been treated at great length in
these volumes; for, as I have attempted to shew, it has
played an important part in the history of the organic
world. As summaries have been given to each chapter,
it would be superfluous here to add a detailed summary.
I am aware that much remains doubtful, but I
have endeavoured to give a fair view of the whole case.
In the lower divisions of the animal kingdom, sexual
selection seems to have done nothing: such animals
are often affixed for life to the same spot, or have the
two sexes combined in the same individual, or what is
still more important, their perceptive and intellectual
faculties are not sufficiently advanced to allow of the
feelings of love and jealousy, or of the exertion of choice.
When, however, we come to the Arthropoda and Vertebrata,
even to the lowest classes in these two great Sub-Kingdoms,
sexual selection has effected much; and it
deserves notice that we here find the intellectual faculties
developed, but in two very distinct lines, to the
highest standard, namely in the Hymenoptera (ants,
bees, &c.) amongst the Arthropoda, and in the Mammalia,
including man, amongst the Vertebrata.
In the most distinct classes of the animal kingdom,397
with mammals, birds, reptiles, fishes, insects, and even
crustaceans, the differences between the sexes follow
almost exactly the same rules. The males are almost
always the wooers; and they alone are armed with special
weapons for fighting with their rivals. They are
generally stronger and larger than the females, and are
endowed with the requisite qualities of courage and pugnacity.
They are provided, either exclusively or in a
much higher degree than the females, with organs for
producing vocal or instrumental music, and with odoriferous
glands. They are ornamented with infinitely
diversified appendages, and with the most brilliant or
conspicuous colours, often arranged in elegant patterns,
whilst the females are left unadorned. When the sexes
differ in more important structures, it is the male which
is provided with special sense-organs for discovering the
female, with locomotive organs for reaching her, and
often with prehensile organs for holding her. These
various structures for securing or charming the female
are often developed in the male during only part of the
year, namely the breeding season. They have in many
cases been transferred in a greater or less degree to
the females; and in the latter case they appear in
her as mere rudiments. They are lost by the males
after emasculation. Generally they are not developed
in the male during early youth, but appear a short
time before the age for reproduction. Hence in most
cases the young of both sexes resemble each other;
and the female resembles her young offspring throughout
life. In almost every great class a few anomalous
cases occur in which there has been an almost complete
transposition of the characters proper to the two sexes;
the females assuming characters which properly belong
to the males. This surprising uniformity in the laws
regulating the differences between the sexes in so many398
and such widely separated classes, is intelligible if we
admit the action throughout all the higher divisions
of the animal kingdom of one common cause, namely
sexual selection.
Sexual selection depends on the success of certain
individuals over others of the same sex in relation to
the propagation of the species; whilst natural selection
depends on the success of both sexes, at all ages, in relation
to the general conditions of life. The sexual
struggle is of two kinds; in the one it is between the
individuals of the same sex, generally the male sex, in
order to drive away or kill their rivals, the females
remaining passive; whilst in the other, the struggle is
likewise between the individuals of the same sex, in
order to excite or charm those of the opposite sex,
generally the females, which no longer remain passive,
but select the more agreeable partners. This latter
kind of selection is closely analogous to that which man
unintentionally, yet effectually, brings to bear on his
domesticated productions, when he continues for a long
time choosing the most pleasing or useful individuals,
without any wish to modify the breed.
The laws of inheritance determine whether characters
gained through sexual selection by either sex shall
be transmitted to the same sex, or to both sexes; as
well as the age at which they shall be developed. It
appears that variations which arise late in life are commonly
transmitted to one and the same sex. Variability
is the necessary basis for the action of selection,
and is wholly independent of it. It follows from this,
that variations of the same general nature have often
been taken advantage of and accumulated through
sexual selection in relation to the propagation of the
species, and through natural selection in relation to the
general purposes of life. Hence secondary sexual cha399racters,
when equally transmitted to both sexes can be
distinguished from ordinary specific characters only by
the light of analogy. The modifications acquired through
sexual selection are often so strongly pronounced that
the two sexes have frequently been ranked as distinct
species, or even as distinct genera. Such strongly-marked
differences must be in some manner highly important;
and we know that they have been acquired in
some instances at the cost not only of inconvenience,
but of exposure to actual danger.
The belief in the power of sexual selection rests
chiefly on the following considerations. The characters
which we have the best reason for supposing to have
been thus acquired are confined to one sex; and this
alone renders it probable that they are in some way
connected with the act of reproduction. These characters
in innumerable instances are fully developed only
at maturity; and often during only a part of the year,
which is always the breeding-season. The males (passing
over a few exceptional cases) are the most active in
courtship; they are the best armed, and are rendered
the most attractive in various ways. It is to be especially
observed that the males display their attractions
with elaborate care in the presence of the females;
and that they rarely or never display them excepting
during the season of love. It is incredible that all this
display should be purposeless. Lastly we have distinct
evidence with some quadrupeds and birds that the individuals
of the one sex are capable of feeling a strong
antipathy or preference for certain individuals of the
opposite sex.
Bearing these facts in mind, and not forgetting the
marked results of man’s unconscious selection, it seems
to me almost certain that if the individuals of one sex
were during a long series of generations to prefer pair400ing
with certain individuals of the other sex, characterised
in some peculiar manner, the offspring would
slowly but surely become modified in this same manner.
I have not attempted to conceal that, excepting when
the males are more numerous than the females, or when
polygamy prevails, it is doubtful how the more attractive
males succeed in leaving a larger number of offspring
to inherit their superiority in ornaments or other
charms than the less attractive males; but I have shewn
that this would probably follow from the females,—especially
the more vigorous females which would be the first
to breed, preferring not only the more attractive but at
the same time the more vigorous and victorious males.
Although we have some positive evidence that birds
appreciate bright and beautiful objects, as with the
Bower-birds of Australia, and although they certainly
appreciate the power of song, yet I fully admit that it
is an astonishing fact that the females of many birds
and some mammals should be endowed with sufficient
taste for what has apparently been effected through
sexual selection; and this is even more astonishing in
the case of reptiles, fish, and insects. But we really
know very little about the minds of the lower animals.
It cannot be supposed that male Birds of Paradise or
Peacocks, for instance, should take so much pains in
erecting, spreading, and vibrating their beautiful plumes
before the females for no purpose. We should remember
the fact given on excellent authority in a former chapter,
namely that several peahens, when debarred from
an admired male, remained widows during a whole
season rather than pair with another bird.
Nevertheless I know of no fact in natural history
more wonderful than that the female Argus pheasant
should be able to appreciate the exquisite shading of
the ball-and-socket ornaments and the elegant patterns401
on the wing-feathers of the male. He who thinks that
the male was created as he now exists must admit that
the great plumes, which prevent the wings from being
used for flight, and which, as well as the primary
feathers, are displayed in a manner quite peculiar to
this one species during the act of courtship, and at no
other time, were given to him as an ornament. If so,
he must likewise admit that the female was created and
endowed with the capacity of appreciating such ornaments.
I differ only in the conviction that the male
Argus pheasant acquired his beauty gradually, through
the females having preferred during many generations
the more highly ornamented males; the æsthetic
capacity of the females having been advanced through
exercise or habit in the same manner as our own taste
is gradually improved. In the male, through the fortunate
chance of a few feathers not having been modified,
we can distinctly see how simple spots with a little
fulvous shading on one side might have been developed
by small and graduated steps into the wonderful
ball-and-socket ornaments; and it is probable
that they were actually thus developed.
Everyone who admits the principle of evolution, and
yet feels great difficulty in admitting that female
mammals, birds, reptiles, and fish, could have acquired
the high standard of taste which is implied by the
beauty of the males, and which generally coincides with
our own standard, should reflect that in each member
of the vertebrate series the nerve-cells of the brain are
the direct offshoots of those possessed by the common
progenitor of the whole group. It thus becomes intelligible
that the brain and mental faculties should be
capable under similar conditions of nearly the same
course of development, and consequently of performing
nearly the same functions.
402The reader who has taken the trouble to go through
the several chapters devoted to sexual selection, will be
able to judge how far the conclusions at which I have arrived
are supported by sufficient evidence. If he accepts
these conclusions, he may, I think, safely extend them
to mankind; but it would be superfluous here to repeat
what I have so lately said on the manner in which
sexual selection has apparently acted on both the male
and female side, causing the two sexes of man to differ
in body and mind, and the several races to differ from
each other in various characters, as well as from their
ancient and lowly-organised progenitors.
He who admits the principle of sexual selection will
be led to the remarkable conclusion that the cerebral
system not only regulates most of the existing functions
of the body, but has indirectly influenced the progressive
development of various bodily structures and of certain
mental qualities. Courage, pugnacity, perseverance,
strength and size of body, weapons of all kinds, musical
organs, both vocal and instrumental, bright colours,
stripes and marks, and ornamental appendages, have
all been indirectly gained by the one sex or the other,
through the influence of love and jealousy, through the
appreciation of the beautiful in sound, colour or form,
and through the exertion of a choice; and these powers
of the mind manifestly depend on the development of
the cerebral system.
Man scans with scrupulous care the character and
pedigree of his horses, cattle, and dogs before he
matches them; but when he comes to his own marriage
he rarely, or never, takes any such care. He is impelled
by nearly the same motives as are the lower animals
when left to their own free choice, though he is in so far
superior to them that he highly values mental charms403
and virtues. On the other hand he is strongly attracted
by mere wealth or rank. Yet he might by selection
do something not only for the bodily constitution and
frame of his offspring, but for their intellectual and
moral qualities. Both sexes ought to refrain from
marriage if in any marked degree inferior in body or
mind; but such hopes are Utopian and will never be
even partially realised until the laws of inheritance are
thoroughly known. All do good service who aid towards
this end. When the principles of breeding and of inheritance
are better understood, we shall not hear ignorant
members of our legislature rejecting with scorn a plan
for ascertaining by an easy method whether or not consanguineous
marriages are injurious to man.
The advancement of the welfare of mankind is a most
intricate problem: all ought to refrain from marriage
who cannot avoid abject poverty for their children; for
poverty is not only a great evil, but tends to its own
increase by leading to recklessness in marriage. On the
other hand, as Mr. Galton has remarked, if the prudent
avoid marriage, whilst the reckless marry, the inferior
members will tend to supplant the better members of
society. Man, like every other animal, has no doubt advanced
to his present high condition through a struggle
for existence consequent on his rapid multiplication;
and if he is to advance still higher he must remain
subject to a severe struggle. Otherwise he would soon
sink into indolence, and the more highly-gifted men
would not be more successful in the battle of life than
the less gifted. Hence our natural rate of increase,
though leading to many and obvious evils, must not
be greatly diminished by any means. There should be
open competition for all men; and the most able should
not be prevented by laws or customs from succeeding
best and rearing the largest number of offspring. Im404portant
as the struggle for existence has been and even
still is, yet as far as the highest part of man’s nature is
concerned there are other agencies more important. For
the moral qualities are advanced, either directly or indirectly,
much more through the effects of habit, the
reasoning powers, instruction, religion, &c., than through
natural selection; though to this latter agency the social
instincts, which afforded the basis for the development
of the moral sense, may be safely attributed.
The main conclusion arrived at in this work, namely
that man is descended from some lowly-organised form,
will, I regret to think, be highly distasteful to many
persons. But there can hardly be a doubt that we are
descended from barbarians. The astonishment which
I felt on first seeing a party of Fuegians on a wild
and broken shore will never be forgotten by me,
for the reflection at once rushed into my mind—such
were our ancestors. These men were absolutely
naked and bedaubed with paint, their long hair was
tangled, their mouths frothed with excitement, and
their expression was wild, startled, and distrustful.
They possessed hardly any arts, and like wild animals
lived on what they could catch; they had no government,
and were merciless to every one not of their own
small tribe. He who has seen a savage in his native
land will not feel much shame, if forced to acknowledge
that the blood of some more humble creature flows
in his veins. For my own part I would as soon be
descended from that heroic little monkey, who braved
his dreaded enemy in order to save the life of his
keeper; or from that old baboon, who, descending from
the mountains, carried away in triumph his young
comrade from a crowd of astonished dogs—as from a
savage who delights to torture his enemies, offers up405
bloody sacrifices, practises infanticide without remorse,
treats his wives like slaves, knows no decency, and is
haunted by the grossest superstitions.
Man may be excused for feeling some pride at having
risen, though not through his own exertions, to the
very summit of the organic scale; and the fact of his
having thus risen, instead of having been aboriginally
placed there, may give him hopes for a still higher
destiny in the distant future. But we are not here
concerned with hopes or fears, only with the truth as
far as our reason allows us to discover it. I have given
the evidence to the best of my ability; and we must
acknowledge, as it seems to me, that man with all his
noble qualities, with sympathy which feels for the most
debased, with benevolence which extends not only to
other men but to the humblest living creature, with his
god-like intellect which has penetrated into the movements
and constitution of the solar system—with all
these exalted powers—Man still bears in his bodily
frame the indelible stamp of his lowly origin.
406
INDEX.
-
A.
-
Abbott, C., on the battles of seals, ii. 240.
-
Abductor of the fifth metatarsal, presence of, in man, i. 128.
-
Abercrombie, Dr., on disease of the brain affecting speech, i. 58.
-
Abipones, marriage-customs of the, ii. 373.
-
Abou-Simbel, caves of, i. 217.
-
Abortion, prevalence of the practice of, i. 134.
-
Abstraction, i. 62.
-
Acalles, stridulation of, i. 384.
-
Acanthodactylus capensis, sexual differences of colour in, ii. 36.
-
Accentor modularis, ii. 198.
-
Acclimatisation, difference of, in different races of men, i. 216.
-
Achetidæ, stridulation of the, i. 352, 353, 355;
-
rudimentary stridulating organs in female, i. 359.
-
Acilius sulcatus, elytra of the female, i. 343.
-
Acomus, development of spurs in the female of, ii. 162.
-
Acridiidæ, stridulation of the, i. 352, 356;
-
rudimentary stridulating organs in female, i. 359.
-
Acting, i. 232.
-
Actiniæ, bright colours of, i. 322.
-
Admiral butterfly, i. 392.
-
Adoption of the young of other animals by female monkeys, i. 41.
-
Advancement in the organic scale, Von Baer’s definition of, i. 211.
-
Aeby, on the difference between the skulls of man and the quadrumana, i. 190.
-
Æsthetic faculty, not highly developed in savages, i. 64.
-
Affection, maternal, i. 40;
-
manifestation of, by animals, i. 40;
-
parental and filial, partly the result of natural selection, i. 81;
-
shown by birds in confinement, for certain persons, ii. 110;
-
mutual, of birds, ii. 108.
-
Africa, probably the birthplace of man, i. 199;
-
South, crossed population of, i. 225;
-
South, retention of colour by the Dutch in, i. 242;
-
South, proportion of the sexes in the butterflies of, i. 310;
-
tattooing practised in, ii. 339;
-
Northern, coiffure of natives of, ii. 340.
-
Agassiz, L., on conscience in dogs, i. 78;
-
on the coincidence of the races of man with zoological provinces, i. 218;
-
on the number of species of man, i. 226;
-
on the courtship of the land-snails, i. 324;
-
on the brightness of the colours of male fishes during the breeding season, ii. 13;
-
on the frontal protuberance of the males of Geophagus and Cichla, ii. 13, 20;
-
on the slight sexual differences of the South Americans, ii. 323;
-
on the tattooing of the Amazonian Indians, ii. 342.
-
Age, in relation to the transmission of characters in birds, ii. 183;
-
variation in accordance with, in birds, ii. 213.
-
Agelæus phœniceus, ii. 116.
-
Ageronia feronia, noise produced by, i. 387.
-
Agrion, dimorphism in, i. 363.
-
Agrion Ramburii, sexes of, i. 362.
-
Agrionidæ, difference in the sexes of, i. 362.
-
Agrotis exclamationis, i. 369.
-
Ague, tertian, dog suffering from, i. 13.
-
Aïthurus polytmus, young of, ii. 220.
-
Ainos, hairiness of the, ii. 321.
-
Alca torda, young of, ii. 217.
407
Alces palmata, ii. 259.
-
Alder and Hancock, MM., on the nudibranch mollusca, i. 326.
-
Algen, Mr., on the stridulation of Scolytus, i. 379.
-
Allen, J. A., on the relative size of the sexes of Callorhinus ursinus, ii. 260;
-
on the mane of Otaria jubata, ii. 267;
-
on the pairing of seals, ii. 279;
-
on sexual differences in the colour of bats, ii. 286.
-
Allen, S., on the habits of Hoplopterus, ii. 48;
-
on the plumes of herons, ii. 82;
-
on the vernal moult of Herodias bubulcus, ii. 84.
-
Alligator, courtship of the male, i. 272, ii. 29;
-
roaring of the male, i. 331.
-
Amadavat, pugnacity of male, ii. 49.
-
Amadina Lathami, display of plumage by the male, ii. 95.
-
Amadina castanotis, display of plumage by the male, ii. 95.
-
Amazons, butterflies of the, i. 309;
-
America, variation in the skulls of aborigines of, i. 108;
-
wide range of aborigines of, i. 218;
-
lice of the natives of, i. 220;
-
general beardlessness of the natives of, ii. 322.
-
America, North, butterflies of, i. 309;
-
Indians of, women a cause of strife among the, ii. 324;
-
Indians of, their notions of female beauty, ii. 344, 347.
-
America, South, character of the natives of, i. 216;
-
population of parts of, i. 225;
-
piles of stones in, i. 233;
-
extinction of the fossil horse of, i. 239;
-
desert-birds of, ii. 224;
-
slight sexual difference of the aborigines of, ii. 323;
-
prevalence of infanticide in, ii. 361.
-
American languages, often highly artificial, i. 112.
-
Americans, wide geographical range of, i. 112;
-
and negroes, difference of, i. 247;
-
aversion of, to hair on the face, ii. 348;
-
native, variability of, i. 226.
-
Ammophila, on the jaws of, i. 342.
-
Ammotragus tragelaphus, hairy forelegs of, ii. 282, 285.
-
Amphibia, affinity of, to the ganoid fishes, i. 204;
-
vocal organs of the, ii. 331.
-
Amphibians, i. 213, ii. 24;
-
breeding whilst immature, ii. 215.
-
Amphioxus, i. 204.
-
Amphipoda, males sexually mature while young, ii. 215.
-
Amunoph III., negro character of features of, i. 217.
-
Anal appendages of insects, i. 342.
-
Analogous variation in the plumage of birds, ii. 74.
-
Anas, ii. 180.
-
Anas acuta, male plumage of, ii. 84.
-
Anas boschas, male plumage of, ii. 84.
-
Anas histrionica, ii. 214.
-
Anastomus oscitans, sexes and young of, ii. 217;
-
white nuptial plumage of, ii. 228.
-
Anatidæ, voices of, ii. 60.
-
Anax junius, difference in the sexes of, i. 362.
-
Anglo-Saxons, estimation of the beard among the, ii. 349.
-
Animals, cruelty of savages to, i. 94;
-
domesticated, more fertile than wild, i. 132;
-
characters common to man and, i. 185;
-
domestic, change of breeds of, ii. 369.
-
Annelida, i. 327.
-
Annulosa, i. 327.
-
Anolis cristatellus, male, crest of, ii. 32;
-
pugnacity of the male, ii. 32;
-
throat-pouch of, ii. 33.
-
Anobium tessellatum, sounds produced by, i. 384.
-
Anser canadensis, ii. 116.
-
Anser cygnoides, ii. 114;
-
knob at the base of the beak of, ii. 129.
-
Anser hyperboreus, whiteness of, ii. 228.
-
Antelope, prong-horned, horns of, i. 289.
-
Antelopes, generally polygamous, i. 267;
-
horns of, i. 289, ii. 245;
-
canine teeth of some male, ii. 241;
-
use of horns of, ii. 251;
-
dorsal crests in, ii. 282;
-
dewlaps of, ii. 284;
-
winter change of two species of, ii. 299;
-
peculiar markings of, ii. 299.
-
Antennæ, furnished with cushions in the male of Penthe, i. 343.
-
Anthidium manicatum, large male of, i. 347.
408
-
Anthocharis cardamines, i. 388, 393, 394;
-
sexual difference of colour in, i. 409.
-
Anthocharis genutia, i. 393.
-
Anthocharis sara, i. 393.
-
Anthophora acervorum, large male of, i. 347.
-
Anthophora retusa, difference of the sexes in, i. 366.
-
Anthus, moulting of, ii. 83.
-
Anthropidæ, i. 195.
-
Antigua, Dr. Nicholson’s observations on yellow fever in, i. 245.
-
Antics of birds, ii. 68.
-
Antilocapra americana, horns of, i. 289, ii. 245.
-
Antilope bezoartica, horned females of, ii. 246, 248;
-
sexual difference in the colour of, ii. 288.
-
Antilope Dorcas and euchore, ii. 245.
-
Antilope euchore, horns of, ii. 251.
-
Antilope montana, rudimentary canines in the young male of, ii. 258.
-
Antilope niger, sing-sing, caama, and gorgon, sexual differences in the colours of, ii. 289.
-
Antilope oreas, horns of, i. 289.
-
Antilope saiga, polygamous habits of, i. 267.
-
Antilope strepsiceros, horns of, i. 289.
-
Antilope subgutturosa, absence of suborbital pits in, ii. 280.
-
Antipathy, shown by birds in confinement, to certain persons, ii. 110.
-
Ants, i. 186;
-
playing together, i. 39;
-
memory in, i. 45;
-
intercommunication of, by means of the antennæ, i. 58;
-
large size of the cerebral ganglia in, i. 145;
-
soldier-, large jaws of, i. 155;
-
difference of the sexes in, i. 365;
-
recognition of each other by, after separation, i. 365.
-
Ants, White, habits of, i. 364.
-
Anura, ii. 25.
-
Apatania muliebris, male unknown, i. 314.
-
Apathus, difference of the sexes in, i. 366.
-
Apatura Iris, i. 386, 388.
-
Apes, anthropomorphous, i. 196;
-
difference of the young, from the adult, i. 13;
-
building platforms, i. 53;
-
probable speedy extermination of the, i. 201;
-
Gratiolet on the evolution of, i. 230;
-
semi-erect attitude of some, i. 142;
-
mastoid processes of, i. 143;
-
influence of the jaw-muscles on the physiognomy of, i. 144;
-
female, destitute of large canines, i. 156;
-
imitative faculties of, i. 161;
-
canine teeth of male, ii. 241;
-
females of some, less hairy beneath than the males, ii. 377.
-
Apes, long-armed, their mode of progression, i. 143.
-
Apis mellifica, large male of, i. 347.
-
Apollo, Greek statues of, ii. 350.
-
Apoplexy in Cebus Azaræ, i. 12.
-
Approbation, influence of the love of, i. 86, 92, 164, 165.
-
Appendages, anal, of insects, i. 342.
-
Aprosmictus scapulatus, ii. 174.
-
Aquatic birds, frequency of white plumage in, ii. 229.
-
Aquila chrysaëtos, ii. 105.
-
Arab women, elaborate and peculiar coiffure of, ii. 353.
-
Arabs, gashing of cheeks and temples among the, ii. 339.
-
Arachnida, i. 337.
-
Arakhan, artificial widening of the forehead by the natives of, ii. 351.
-
Arboricola, young of, ii. 190.
-
Archeopteryx, i. 204.
-
Arctiidæ,, coloration of the, i. 396.
-
Ardea asha, rufescens, and cærulea, change of colour in, ii. 231, 232.
-
Ardea cærulea, breeding in immature plumage, ii. 214.
-
Ardea gularis, change of plumage in, ii. 232.
-
Ardea herodias, love-gestures of the male, ii. 68.
-
Ardea ludoviciana, age of mature plumage in, ii. 213;
-
continued growth of crest and plumes in the male of, ii. 216.
-
Ardea nycticorax, cries of, ii. 51.
-
Ardeola, young of, ii. 190.
-
Ardetta, changes of plumage in, ii. 179.
-
Argenteuil, i. 29.
-
Argus pheasant, ii. 72, 97, 181;
-
display of plumage by the male, ii. 91;
-
ocellated spots of the, ii. 134, 141;
-
gradation of characters in the, ii. 141.
409
-
Argyll, Duke of, the fashioning of implements peculiar to man, i. 52;
-
on the contest in man between right and wrong, i. 104;
-
on the physical weakness of man, i. 156;
-
on the primitive civilisation of man, i. 181;
-
on the plumage of the male Argus pheasant, ii. 91;
-
on Urosticte Benjamini, ii. 151;
-
on the nests of birds, ii. 167;
-
on variety as an object in nature, ii. 230.
-
Argynnis aglaia, colouring of the lower surface of, i. 396.
-
Aricoris epitus, sexual differences in the wings of, i. 345.
-
Aristocracy, increased beauty of the, ii. 356.
-
Arms, proportions of, in soldiers and sailors, i. 116;
-
direction of the hair on the, i. 192.
-
Arms and hands, free use of, indirectly correlated with diminution of canines, i. 144.
-
Arrest of development, i. 121, 122.
-
Arrow-heads, stone, general resemblance of, i. 233.
-
Arrows, use of, i. 232.
-
Arteries, variations in the course of the, i. 108.
-
Artery, effect of tying, upon the lateral channels, i. 116.
-
Arthropoda, i. 328.
-
Arts practised by savages, i. 234.
-
Ascension, coloured incrustation on the rocks of, i. 326.
-
Ascidia, affinity of the lancelet to, i. 205;
-
tadpole-like larvæ of, i. 205.
-
Ascidians, i. 324;
-
bright colours of some, i. 322.
-
Asinus, Asiatic and African species of, ii. 306.
-
Asinus tæniopus, ii. 306.
-
Ass, colour-variations of the, ii. 305.
-
Ateles, effects of brandy on an, i. 12;
-
absence of the thumb in, i. 140.
-
Ateles beelzebuth, ears of, i. 23.
-
Ateles marginatus, colour of the ruff of, ii. 291;
-
hair on the head of, ii. 109;
-
on the recognition of a dog by a turkey, ii. 110;
-
on the selection of a mate by female birds, ii. 307.
-
Ateuchus, stridulation of, i. 384.
-
Ateuchus cicatricosus, habits of, i. 376.
-
Athalia, proportions of the sexes in, i. 314.
-
Attention, manifestations of, in animals, i. 44.
-
Audouin, V., on a hymenopterous parasite with a sedentary male, i. 273.
-
Audubon, J. J., on the pugnacity of male birds, ii. 43, 48;
-
on Tetrao cupido, ii. 50;
-
on Ardea nycticorax, ii. 51;
-
on Sturnella ludoviciana, ii. 51;
-
on the vocal organs of Tetrao cupido, ii. 56;
-
on the drumming of the male Tetrao umbellus, ii. 61;
-
on sounds produced by the nightjar, ii. 63;
-
on Ardea herodias and Cathartes jota, ii. 68;
-
on the spring change of colour in some finches, ii. 86;
-
on Mimus polyglottus, ii. 116;
-
on the turkey, ii. 119, 121;
-
on variation in the male scarlet tanager, ii. 126;
-
on the habits of Pyranga æstiva, ii. 167;
-
on local differences in the nests of the same species of birds, ii. 171;
-
on the habits of woodpeckers, ii. 175;
-
on Bombycilla carolinensis, ii. 180;
-
on young females of Tanagra æstiva acquiring male characters, ii. 180;
-
on the immature plumage of thrushes, ii. 185;
-
on the immature plumage of birds, ii. 186 et seq.;
-
on birds breeding in immature plumage, ii. 214;
-
on the growth of the crest and plumes in the male Ardea ludoviciana, ii. 216;
-
on the change of colour in some species of Ardea, ii. 231;
-
on the speculum of Mergus cucullatus, ii. 291;
-
on the musk-rat, ii. 298.
-
Audubon and Bachman, MM., on squirrels fighting, ii. 239;
-
on the Canadian lynx, ii. 267.
-
Austen, N. L., on Anolis cristatellus, ii. 32, 33.
-
Australia, half-castes killed by the natives of, i. 220;
-
lice of the natives of, i. 220;
-
not the birthplace of man, i. 199;
-
prevalence of female infanticide in, ii. 364.
-
Australia, South, variation in the skulls of aborigines of, i. 108.
-
410
Australians, colour of newborn
children of, ii. 318;
-
height of the sexes of, ii. 320;
-
women a cause of war among the, ii. 323.
-
Axis deer, sexual difference in the colour of the, ii. 290.
-
Aymaras, measurements of the, i. 119;
-
no grey hair among the, ii. 320;
-
hairlessness of the face in the, ii. 322;
-
long hair of the, ii. 348.
-
Azara, on the proportion of men and women among the Guaranys, i. 302;
-
on Palamedea cornuta, ii. 48;
-
on the beards of the Guaranys, ii. 322;
-
on strife for women among the Guanas, ii. 324;
-
on infanticide, ii. 344, 364;
-
on the eradication of the eyebrows and eyelashes by the Indians of Paraguay, ii. 348;
-
on polyandry among the Guanas, ii. 366;
-
celibacy unknown among the savages of South America, ii. 367;
-
on the freedom of divorce among the Charruas, ii. 372.
-
B.
-
Babbage, C., on the greater proportion of illegitimate female births, i. 302.
-
Babirusa, tusks of the, ii. 264.
-
Baboon, employing a mat for shelter against the sun, i. 53;
-
manifestation of memory by a, i. 45;
-
protected from punishment by its companions, i. 78;
-
rage excited in, by reading, i. 42.
-
Baboon, Cape, mane of the male, ii. 267;
-
Hamadryas, mane of the male, ii. 267.
-
Baboons, effects of intoxicating liquors on, i. 12;
-
ears of, i. 23;
-
manifestation of maternal affection by, i. 41;
-
using stones and sticks as weapons, i. 51;
-
co-operation of, i. 75;
-
silence of, on plundering expeditions, i. 79;
-
diversity of the mental faculties in, i. 110;
-
hands of, i. 139;
-
habits of, i. 141;
-
variability of the tail in, i. 150;
-
apparent polygamy of, i. 266;
-
polygamous and social habits of, ii. 362.
-
Bachman, Dr., on the fertility of mulattoes, i. 221.
-
Baer, K. E. von, on embryonic development, i. 14.
-
Bagehot, W., on the social virtues among primitive men, i. 93;
-
on the value of obedience, i. 162;
-
on human progress, i. 166;
-
on the persistence of savage tribes in classical times, i. 239.
-
Bailly, E. M., on the fighting of stags, ii. 252;
-
on the mode of fighting of the Italian buffalo, ii. 250.
-
Bain, A., on the sense of duty, i. 71;
-
aid springing from sympathy, i. 77;
-
on the basis of sympathy, i. 82;
-
on love of approbation, &c., i. 86;
-
on the idea of beauty, ii. 354.
-
Baird, W., on a difference in colour between the males and females of some Entozoa, i. 321.
-
Baker, Mr., observation on the proportion of the sexes in pheasant-chicks, i. 306.
-
Baker, Sir S., on the fondness of the Arabs for discordant music, ii. 67;
-
on sexual difference in the colours of an antelope, ii. 289;
-
on the elephant and rhinoceros attacking white or grey horses, ii. 295;
-
on the disfigurements practised by the negroes, ii. 296;
-
on the gashing of the cheeks and temples practised in Arab countries, ii. 339;
-
on the coiffure of the North Africans, ii. 340;
-
on the perforation of the lower lip by the women of Latooka, ii. 341;
-
on the distinctive characters of the coiffure of central African tribes, ii. 342;
-
on the coiffure of Arab women, ii. 353.
-
“Balz” of the Black-cock, ii. 45, 100.
-
Bantam, Sebright, i. 259, 294.
-
Banteng, horns of, ii. 247;
-
sexual differences in the colours of the, ii. 289.
-
Banyai, colour of the, ii. 346.
-
Barbarism, primitive, of civilised nations, i. 181.
-
Barbs, filamentous, of the feathers, in certain birds, ii. 74.
-
Barr, Mr., on sexual preference in dogs, ii. 272.
-
Barrington, Daines, on the language of birds, i. 55;
-
on the clucking of the hen, ii. 51;
-
on the object of the song of birds, ii. 52;
411
-
on the singing of female birds, ii. 54;
-
on birds acquiring the songs of other birds, ii. 55;
-
on the muscles of the larynx in song-birds, ii. 55;
-
on the want of the power of song by female birds, ii. 163.
-
Barrow, on the widow-bird, ii. 98.
-
Bartlett, A. D., on the tragopan, i. 270;
-
on the development of the spurs in Crossoptilon auritum, i. 290;
-
on the fighting of the males of Plectropterus gambensis, ii. 46;
-
on the knot, ii. 82;
-
on display in male birds, ii. 87;
-
on the display of plumage by the male Polyplectron, ii. 89;
-
on Crossoptilon auritum and Phasianus Wallichii, ii. 93;
-
on the habits of Lophophorus, ii. 121;
-
on the colour of the mouth in Buceros bicornis, ii. 129;
-
on the incubation of the cassowary, ii. 204;
-
on the Cape Buffalo, ii. 250;
-
on the use of the horns of antelopes, ii. 251;
-
on the fighting of male wart-hogs, ii. 266;
-
on Ammotragus tragelaphus, ii. 282;
-
on the colours of Cercopithecus cephus, ii. 291;
-
on the colours of the faces of monkeys, ii. 310;
-
on the naked surfaces of monkeys, ii. 377.
-
Bartram, on the courtship of the male alligator, ii. 29.
-
Basque language, highly artificial, i. 61.
-
Bate, C. S., on the superior activity of male crustacea, i. 272;
-
on the proportions of the sexes in crabs, i. 315;
-
on the chelæ of crustacea, i. 330;
-
on the relative size of the sexes in crustacea, i. 332;
-
on the colours of crustacea, i. 335.
-
Bates, H. W., on variation in the form of the head of Amazonian Indians, i. 111;
-
on the proportion of the sexes among Amazonian butterflies, i. 309;
-
on sexual differences in the wings of butterflies, i. 345;
-
on the field-cricket, i. 353;
-
on Pyrodes pulcherrimus, i. 367;
-
on the horns of Lamellicorn beetles, i. 370, 371;
-
on the colours of Epicaliæ, &c., i. 388;
-
on the coloration of tropical butterflies, i. 391;
-
on the variability of Papilio Sesostris and Childrenæ, i. 402;
-
on male and female butterflies inhabiting different stations, i. 403;
-
on mimickry, i. 411;
-
on the caterpillar of a Sphinx, i. 416;
-
on the vocal organs of the umbrella-bird, ii. 58;
-
on the toucans, ii. 227;
-
on Brachyurus calvus, ii. 309.
-
Batokas, knocking out two upper incisors, ii. 340.
-
Batrachia, ii. 25;
-
eagerness of male, i. 272.
-
Bats, sexual differences in the colour of, ii. 286.
-
Battle, law of, i. 182;
-
among beetles, i. 375;
-
among birds, ii. 40;
-
among mammals, ii. 239 et seq.;
-
in man, ii. 323.
-
Beak, sexual difference in the forms of the, ii. 39;
-
in the colour of the, ii. 72.
-
Beaks, of birds, bright colours of, ii. 227.
-
Beard, development of, in man, ii. 317;
-
analogy of the, in man and the quadrumana, ii. 319;
-
variation of the development of the, in different races of men, ii. 321;
-
estimation of, among bearded nations, ii. 349;
-
probable origin of the, ii. 379.
-
Beards, in monkeys, i. 192;
-
Beautiful, taste for the, in birds, ii. 108;
-
in the quadrumana, ii. 296.
-
Beauty, sense of, in animals, i. 63;
-
appreciation of, by birds, ii. 111;
-
influence of, ii. 338, 343;
-
variability of the standard of, ii. 370.
-
Beavan, Lieut., on the development of the horns in Cervus Eldi, i. 288.
-
Beaver, instinct and intelligence of the, i. 37, 38;
-
voice of the, ii. 277;
-
castoreum of the, ii. 279.
-
Beavers, battles of male, ii. 239.
-
Bechstein, on female birds choosing the best singers among the males, ii. 52;
-
on rivalry in song-birds, ii. 53;
-
on the singing of female birds, ii. 54;
-
on birds acquiring the songs of other birds, ii. 55;
-
on pairing the canary and siskin, ii. 115;
-
on a sub-variety of the monk pigeon, ii. 132;
412
-
on spurred hens, ii. 162.
-
Beddoe, Dr., on causes of difference in stature, i. 115.
-
Bee-eater, ii. 56.
-
Bees, i. 73;
-
destruction of drones and queens by, i. 82;
-
pollen-baskets and stings of, i. 155;
-
female, secondary sexual characters of, i. 254;
-
difference of the sexes in, i. 365.
-
Beetle, luminous larva of a, i. 345.
-
Beetles, i. 366;
-
size of the cerebral ganglia in, i. 145;
-
dilatation of the fore tarsi in male, i. 343;
-
blind, i. 367;
-
stridulation of, i. 378.
-
Belgium, ancient inhabitants of, i. 237.
-
Bell, Sir C, on emotional muscles in man, i. 5;
-
“snarling muscles,” i. 127;
-
on the hand, i. 141.
-
Bell, T., on the numerical proportion of the sexes in moles, i. 305;
-
on the newts, ii. 24;
-
on the croaking of the frog, ii. 27;
-
on the difference in the coloration of the sexes in Zootoca vivipara, ii. 36;
-
on moles fighting, ii. 239.
-
Bell-bird, sexual difference in the colour of the, ii. 79.
-
Bell-birds, colours of, ii. 228.
-
Benevolence, manifested by birds, ii. 109.
-
Bennett, A. W., on the habits of Dromœus irroratus, ii. 205.
-
Bennett, Dr., on birds of paradise, ii. 89.
-
Bernicla antarctica, colours of, ii. 228.
-
Bernicle gander pairing with a Canada goose, ii. 114.
-
Bettoni, E., on local differences in the nests of Italian birds, ii. 171.
-
Bhoteas, colour of the beard in, ii. 319.
-
Bhringa, disciform tail-feathers of, ii. 83.
-
Bibio, sexual differences in the genus, i. 349.
-
Bichat, on beauty, ii. 354.
-
Bile, coloured, in many animals, i. 323.
-
Bimana, i. 190.
-
Birds, imitations of the songs of other birds by, i. 44;
-
dreaming, i. 46;
-
language of, i. 55;
-
sense of beauty in, i. 63;
-
pleasure of, in incubation, i. 79;
-
male, incubation by, i. 210;
-
and reptiles, alliance of, i. 213;
-
sexual differences in the beak of some, i. 255;
-
migratory, arrival of the male before the female, i. 259;
-
apparent relation between polygamy and marked sexual differences in, i. 270;
-
monogamous, becoming polygamous under domestication, i. 270;
-
eagerness of male in pursuit of the female, i. 272;
-
wild, numerical proportion of the sexes in, i. 306;
-
secondary sexual characters of, ii. 38;
-
difference of size in the sexes of, ii. 43;
-
fights of male, witnessed by females, ii. 49;
-
display of male, to captivate the females, ii. 50;
-
close attention of, to the songs of others, ii. 52;
-
acquiring the song of their foster-parents, ii. 55;
-
brilliant, rarely good songsters, ii. 56;
-
love-antics and dances of, ii. 68;
-
coloration of, ii. 74 et seqq.;
-
moulting of, ii. 80 et seqq.;
-
unpaired, ii. 103;
-
male, singing out of season, ii. 106;
-
mutual affection of, ii. 108;
-
in confinement, distinguish persons, ii. 110;
-
hybrid, production of, ii. 113;
-
European, number of species of, ii. 124;
-
variability of, ii. 124;
-
gradation of secondary sexual characters in, ii. 135;
-
obscurely coloured, building concealed nests, ii. 169;
-
young female, acquiring male characters, ii. 180;
-
breeding in immature plumage, ii. 214;
-
moulting of, ii. 214;
-
aquatic, frequency of white plumage in, ii. 229;
-
vocal courtship of, ii. 331;
-
naked skin of the head and neck in, ii. 377.
-
Birgus latro, habits of, i. 334.
-
Birkbeck, Mr. on the finding of new mates by Golden Eagles, ii. 105.
-
Birthplace of man, i. 199.
-
Births, numerical proportions of the sexes in, in animals and man, i. 263, 265;
-
male and female, numerical proportion of, in England, i. 300.
-
Bischoff, Prof., on the agreement between the brains of man and of the Orang, i. 11;
413
-
figure of the embryo of the dog, i. 15;
-
on the convolutions of the brain in the human fœtus, i. 16;
-
on the difference between the skulls of man and the quadrumana, i. 190.
-
Bishop, J., on the vocal organs of frogs, ii. 28;
-
on the vocal organs of corvine birds, ii. 55;
-
on the trachea of the Merganser, ii. 60.
-
Bison, American, mane of the male, ii. 267.
-
Bitterns, dwarf, coloration of the sexes of, ii. 179.
-
Biziura lobata, musky odour of the male, ii. 38;
-
large size of male, ii. 43.
-
Blackbird, sexual differences in the, i. 268;
-
proportion of the sexes in the, i. 307;
-
acquisition of a song by a, ii. 55;
-
colour of the beak in the sexes of the, ii. 72, 227;
-
pairing with a thrush, ii. 113;
-
colours and nidification of the, ii. 170;
-
young of the, ii. 219;
-
sexual difference in coloration of the, ii. 226.
-
Black-buck, Indian, sexual difference in the colour of the, ii. 288.
-
Blackcap, arrival of the male, before the female, i. 259;
-
Black-cock, polygamous, i. 269;
-
proportion of the sexes in the, i. 306;
-
pugnacity and love-dance of the, ii. 45;
-
call of the, ii. 60;
-
moulting of the, ii. 83;
-
duration of the courtship of the, ii. 100;
-
sexual difference in coloration of the, ii. 226;
-
crimson eye-cere of the, ii. 227;
-
and pheasant, hybrids of, ii. 113.
-
Black-grouse, characters of young, ii. 185, 194.
-
Blackwall, J., on the speaking of the magpie, i. 59;
-
on the desertion of their young by swallows, i. 84;
-
on the superior activity of male spiders, i. 272;
-
on the proportion of the sexes in spiders, i. 314;
-
on sexual variation of colour in spiders, i. 337;
-
on male spiders, i. 338.
-
Bladder-nose Seal, hood of the, ii. 278.
-
Blaine, on the affections of dogs, ii. 270.
-
Blair, Dr., on the relative liability of Europeans to yellow fever, i. 243.
-
Blake, C. C., on the jaw from La Naulette, i. 126.
-
Blakiston, Capt., on the American snipe, ii. 64;
-
on the dances of Tetrao phasianellus, ii. 69.
-
Blasius, Dr., on the species of European birds, ii. 124.
-
Bledius taurus, hornlike processes of male, i. 374.
-
Bleeding, tendency to profuse, i. 292.
-
Blenkiron, Mr., on sexual preference in horses, ii. 272.
-
Blennies, crest developed on the head of male, during the breeding season, ii. 12.
-
Blethisa multipunctata, stridulation of, i. 379.
-
Bloch, on the proportions of the sexes in Fishes, i. 308.
-
Blood, arterial, red colour of, i. 323.
-
Blood-pheasant, number of spurs in, ii. 46.
-
Bluebreast, red-throated, sexual differences of the, ii. 195.
-
Blumenbach, on Man, i. 111;
-
on the large size of the nasal cavities in American aborigines, i. 119;
-
on the position of man, i. 190;
-
on the number of species of man, i. 226.
-
Blyth, E., observations on Indian crows, i. 77;
-
on the structure of the hand in species of Hylobates, i. 140;
-
on the ascertainment of the sex of nestling bullfinches by pulling out breast-feathers, ii. 24;
-
on the pugnacity of the males of Gallinula cristata, ii. 41;
-
on the presence of spurs in the female Euplocamus erythropthalmus, ii. 46;
-
on the pugnacity of the amadavat, ii. 49;
-
on the spoonbill, ii. 60;
-
on the moulting of Anthus, ii. 83;
-
on the moulting of bustards, plovers, and Gallus bankiva, ii. 84;
-
on the Indian honey-buzzard, ii. 126;
-
on sexual differences in the colour or the eyes of hornbills, ii. 129;
-
on Oriolus melanocephalus, ii. 178;
-
on Palæornis javanicus, ii. 179;
-
on the genus Ardetta, ii. 179;
414
-
on the peregrine falcon, ii. 180;
-
on young female birds acquiring male characters, ii. 180;
-
on the immature plumage of birds, ii. 185;
-
on representative species of birds, ii. 190;
-
on the young of Turnix, ii. 202;
-
on anomalous young of Lanius rufus and Colymbus glacialis, ii. 211;
-
on the sexes and young of the sparrows, ii. 212;
-
on dimorphism in some herons, ii. 214;
-
on orioles breeding in immature plumage, ii. 214;
-
on the sexes and young of Buphus and Anastomus, ii. 217;
-
on the young of the blackcap and blackbird, ii. 219;
-
on the young of the stonechat, ii. 220;
-
on the white plumage of Anastomus, ii. 229;
-
on the horns of Antilope bezoartica, ii. 246;
-
on the horns of Bovine animals, ii. 247;
-
on the mode of fighting of Ovis cycloceros, ii. 249;
-
on the voice of the Gibbons, ii. 276;
-
on the crest of the male wild goat, ii. 282;
-
on the colours of Portax picta, ii. 287;
-
on the colours of Antilope bezoartica, ii. 288;
-
on the development of the horns in the Koodoo and Eland antelopes, i. 289;
-
on the colour of the Axis deer, ii. 290;
-
on sexual difference of colour in Hylobates hoolock, ii. 291;
-
on the hog-deer, ii. 303;
-
on the beard and whiskers in a monkey becoming white with age, ii. 319.
-
Boar, wild, polygamous in India, i. 267;
-
use of the tusks by the, ii. 256;
-
fighting of, ii. 263.
-
Boitard and Corbié, MM., on the transmission of sexual peculiarities in pigeons, i. 283;
-
on the antipathy shown by some female pigeons to certain males, ii. 118.
-
Bold, Mr., on the singing of a sterile hybrid canary, ii. 53.
-
Bombet, on the variability of the standard of beauty in Europe, ii. 370.
-
Bombus, difference of the sexes in, i. 366.
-
Bombycidæ, coloration of, i. 394;
-
Bombycilla carolinensis, red appendages of, ii. 179.
-
Bombyx cynthia, i. 346;
-
proportion of the sexes in, i. 309, 313;
-
pairing of, i. 401.
-
Bombyx mori, difference of size of the male and female cocoons of, i. 346;
-
Bombyx Pernyi, proportion of sexes of, i. 313.
-
Bombyx Yamamai, i. 346;
-
M. Personnat on, i. 310;
-
proportion of sexes of, i. 313.
-
Bonaparte, C. L., on the call-notes of the wild turkey, ii. 60.
-
Bond, F., on the finding of new mates by crows, ii. 104.
-
Bone, implements of, skill displayed in making, i. 138.
-
Boner, C., on the transfer of male characters to an old female chamois, ii. 245;
-
on the antlers of the red deer, ii. 252;
-
on the habits of stags, ii. 259;
-
on the pairing of red deer, ii. 269.
-
Bones, increase of, in length and thickness, when carrying a greater weight, i. 116.
-
Bonnet monkey, i. 192.
-
Boomerang, i. 183.
-
Boreus hyemalis, scarcity of the male, i. 314.
-
Bory St. Vincent, on the number of species of man, i. 226;
-
on the colours of Labrus pavo, ii. 16.
-
Bos gaurus, horns of, ii. 247.
-
Bos primigenius, ii. 240.
-
Bos sondaicus, horns of, ii. 247;
-
Botocudos, i. 181;
-
mode of life of, i. 247;
-
disfigurement of the ears and lower lip of the, ii. 341.
-
Boucher de Perthes, J. C. de, on the antiquity of man, i. 3.
-
Bourbon, proportion of the sexes in a species of Papilio from, i. 310.
-
Bourien, on the marriage-customs of the savages of the Malay Archipelago, ii. 373.
-
Bovidæ, dewlaps of, ii. 284.
-
Bower-birds, ii. 102;
-
habits of the, ii. 69;
-
ornamented playing-places of, i. 63, ii. 112.
-
Bows, use of, i. 232.
415
-
Brachiopoda, i. 329.
-
Brachycephalic structure, possible explanation of, i. 148.
-
Brachyscelus, second pair of antennæ in the male, i. 337.
-
Brachyura, i. 332.
-
Brachyurus calvus, scarlet face of, ii. 309.
-
Brain, of man, agreement of the, with that of lower animals, i. 10;
-
convolutions of, in the human fœtus, i. 16;
-
larger in some existing mammals than in their tertiary prototypes, i. 51;
-
relation of the development of the, to the progress of language, i. 57;
-
disease of the, affecting speech, i. 58;
-
influence of development of mental faculties upon the size of the, i. 145;
-
influence of the development of, on the spinal column and skull, i. 146;
-
difference in the convolutions of, in different races of men, i. 216.
-
Brakenridge, Dr., on the influence of climate, i. 115.
-
Braubach, Prof., on the quasi-religious feeling of a dog towards his master, i. 68;
-
on the self-restraint of dogs, i. 78.
-
Brauer, F., on dimorphism in Neurothemis, i. 363.
-
Brazil, skulls found in caves of, i. 218;
-
population of, i. 225;
-
compression of the nose by the natives of, ii. 352.
-
Break between man and the apes, i. 200.
-
Bream, proportion of the sexes in the, i. 308.
-
Breeding, age of, in birds, ii. 214.
-
Breeding season, sexual characters making their appearance in the, in birds, ii. 80.
-
Brehm, on the effects of intoxicating liquors on monkeys, i. 12;
-
on the recognition of women by male Cynocephali, i. 13;
-
on revenge taken by monkeys, i. 40;
-
on manifestations of maternal affection by monkeys and baboons, i. 41;
-
on the instinctive dread of monkeys for serpents, i. 42;
-
on a baboon using a mat for shelter from the sun, i. 53;
-
on the use of stones as missiles by baboons, i. 51;
-
on the signal-cries of monkeys, i. 57;
-
on sentinels posted by monkeys, i. 74;
-
on co-operation of animals, i. 75;
-
on an eagle attacking a young Cercopithecus, i. 76;
-
on baboons in confinement protecting one of their number from punishment, i. 78;
-
on the habits of baboons when plundering, i. 79;
-
on the diversity of the mental faculties of monkeys, i. 110;
-
on the habits of baboons, i. 141;
-
on polygamy in Cynocephalus and Cebus, i. 266;
-
on the numerical proportion of the sexes in birds, i. 306;
-
on the love-dance of the Black-cock, ii. 45;
-
on Palamedea cornuta, ii. 48;
-
on the habits of the Black-grouse, ii. 49;
-
on sound produced by Birds of Paradise, ii. 63;
-
on assemblages of grouse, ii. 101;
-
on the finding of new mates by birds, ii. 106;
-
on the fighting of wild boars, ii. 263;
-
on the habits of Cynocephalus hamadryas, ii. 362.
-
Brent, Mr., on the courtship of fowls, ii. 117.
-
Breslau, numerical proportion of male and female births in, i. 301.
-
Bridgman, Laura, i. 57.
-
Brimstone butterfly, i. 393;
-
sexual difference of colour in the, i. 409.
-
British, ancient, tattooing practised by, ii. 339.
-
Broca, Prof., on the occurrence of the supra-condyloid foramen in the human humerus, i. 28;
-
on the capacity of Parisian skulls at different periods, i. 146;
-
on the influence of natural selection, i. 152;
-
on hybridity in man, i. 220;
-
on human remains from Les Eyzies, i. 237;
-
on the cause of the difference between Europeans and Hindoos, i. 240.
-
Brodie, Sir B., on the origin of the moral sense in man, i. 71.
-
Bronn, H. G., on the copulation of insects of distinct species, i. 342.
-
Bronze period, men of, in Europe, i. 160.
-
Brown, R., sentinels of seals generally females, i. 74;
-
on the battles of seals, ii. 240;
-
on the narwhal, ii. 242;
416
-
on the occasional absence of the tusks in the female walrus, ii. 242;
-
on the bladder-nose seal, ii. 278;
-
on the colours of the sexes in Phoca grœnlandica, ii. 287;
-
on the appreciation of music by seals, ii. 333;
-
on plants used as love-philters, by North American women, ii. 344.
-
Brown-Séquard, Dr., on the inheritance of the effects of operations by guinea pigs, ii. 380.
-
Bruce, on the use of the elephant’s tusks, ii. 249.
-
Brulerie, P. de la, on the habits of Ateuchus cicatricosus, i. 376;
-
on the stridulation of Ateuchus, i. 384.
-
Brünnich, on the pied ravens of the Feroe islands, ii. 126.
-
Bryant, Capt., on the courtship of Callorhinus ursinus, ii. 269.
-
Bubas bison, thoracic projection of, i. 372.
-
Bucephalus capensis, difference of the sexes of, in colour, ii. 29.
-
Buceros, nidification and incubation of, ii. 169.
-
Buceros bicornis, sexual differences in the colouring of the casque, beak, and mouth in, ii. 129.
-
Buceros corrugatus, sexual difference in the beak of, ii. 72.
-
Büchner, L., on the origin of man, i. 4;
-
on the want of self-consciousness, &c., in low savages, i. 62;
-
on the use of the human foot as a prehensile organ, i. 142;
-
on the mode of progression of the apes, i. 142.
-
Buckland, F., on the numerical proportion of the sexes in rats, i. 305;
-
on the proportion of the sexes in the trout, i. 308;
-
on Chimæra monstrosa, ii. 12.
-
Buckland, W., on the complexity of crinoids, i. 61.
-
Buckler, W., proportion of sexes of Lepidoptera reared by, i. 313.
-
Buckinghamshire, numerical proportion of male and female births in, i. 300.
-
Bucorax abyssinicus, inflation of the neck-wattle of the male, during courtship, ii. 72.
-
Budytes Raii, i. 260.
-
Buffalo, Cape, ii. 250.
-
Buffalo, Indian, horns of the, ii. 247.
-
Buffalo, Italian, mode of fighting of the, ii. 250.
-
Buffon, on the number of species of man, i. 226.
-
Bugs, i. 349.
-
Buist, R., on the proportion of the sexes in salmon, i. 308;
-
on the pugnacity of the male salmon, ii. 3.
-
Bulbul, pugnacity of the male, ii. 41;
-
display of under tail-coverts by the male, ii. 96.
-
Bull, mode of fighting of the, ii. 250;
-
curled frontal hair of the, ii. 282.
-
Bullfinch, sexual differences in the, i. 269;
-
piping, ii. 52;
-
female, singing of the, ii. 54;
-
courtship of the, ii. 94;
-
widowed, finding a new mate, ii. 105;
-
attacking a reed-bunting, ii. 111;
-
nestling, sex ascertained by pulling out breast-feathers, ii. 214.
-
Bullfinches distinguishing persons, ii. 110;
-
rivalry of female, ii. 121.
-
Bulls, two young, attacking an old one, i. 75;
-
wild, battles of, ii. 240.
-
Bull-trout, male, colouring of, during the breeding season, ii. 14.
-
Bunting, reed, head feathers of the male, ii. 95;
-
attacked by a bullfinch, ii. 111.
-
Buntings, characters of young, ii. 184.
-
Buphus coromandus, sexes and young of, ii. 217;
-
change of colour in, ii. 231, 232.
-
Burchell, Dr., on the zebra, ii. 302;
-
on the extravagance of a Bushwoman in adorning herself, ii. 344;
-
celibacy unknown among the savages of South Africa, ii. 367;
-
on the marriage-customs of the Bushwomen, ii. 374.
-
Burke, on the number of species of man, i. 226.
-
Burmese, colour of the beard in, ii. 319.
-
Burton, Capt., on negro ideas of female beauty, ii. 346;
-
on a universal ideal of beauty, ii. 351.
-
Bushmen, i. 157.
417
-
Bushwoman, extravagant ornamentation of a, ii. 344.
-
Bushwomen, hair of, i. 216;
-
marriage-customs of, ii. 374.
-
Busk, Prof. G., on the occurrence of the supra-condyloid foramen in the human humerus, i. 28.
-
Bustard, throat-pouch of the male, ii. 58;
-
humming noise produced by a male, ii. 65;
-
Indian, ear-tufts of a, ii. 73.
-
Bustards, occurrence of sexual differences and of polygamy among the, i. 269;
-
love-gestures of the male, ii. 68;
-
double moult in, ii. 81, 83.
-
Butler, A. G., on sexual differences in the wings of Aricoris epitus, i. 345;
-
>on the colouring of the sexes in species of Thecla, i. 389;
-
on the resemblance of Iphias glaucippe to a leaf, i. 394;
-
on the rejection of certain moths and caterpillars by lizards and frogs, i. 417.
-
Butterfly, noise produced by a, i. 387;
-
Emperor, i. 386, 388;
-
meadow brown, instability of the ocellated spots of, ii. 132.
-
Butterflies, proportion of the sexes in, i. 309;
-
forelegs atrophied in some male, i. 344;
-
sexual difference in the neuration of the wings of, i. 345;
-
pugnacity of male, i. 386;
-
protective resemblances of the lower surface of, i. 392;
-
display of the wings by, i. 396;
-
white, alighting upon bits of paper, i. 400;
-
attracted by a dead specimen of the same species, i. 400;
-
courtship of, i. 400;
-
male and female, inhabiting different stations, i. 403.
-
Buxton, C., observations on macaws, i. 76;
-
on an instance of benevolence in a parrot, ii. 109.
-
Buzzard, Indian honey-, variation in the crest of, ii. 126.
-
C.
-
Cabbage butterflies, i. 393.
-
Cachalot, large head of the male, ii. 242.
-
Cadences, musical, perception of, by animals, ii. 333.
-
Cæcum, i. 27;
-
large, in the early progenitors of man, i. 206.
-
Cairina moschata, pugnacity of the male, ii. 43.
-
Callianassa, chelæ of, figured, i. 330.
-
Callionymus lyra, characters of the male, ii. 7.
-
Callorhinus ursinus, relative size of the sexes of, ii. 260;
-
Calotes nigrilabris, sexual difference in the colour of, ii. 36.
-
Cambridge, O. Pickard, on the sexes of spiders, i. 315.
-
Camel, canine teeth of male, ii. 241, 257.
-
Campbell, J., on the Indian elephant, i. 267, 268;
-
on the proportion of male and female births in the harems of Siam, i. 303.
-
Campylopterus hemileucurus, i. 307.
-
Canaries distinguishing persons, ii. 110.
-
Canary, polygamy of the, i. 270;
-
change of plumage in, after moulting, i. 294;
-
female, selecting the best singing male, ii. 52;
-
sterile hybrid, singing of a, ii. 53;
-
female, singing of the, ii. 54;
-
selecting a greenfinch, ii. 115;
-
and siskin, pairing of, ii. 115.
-
Canestrini, G., on rudimentary characters and the origin of man, i. 4;
-
on rudimentary characters, i. 17;
-
on the movement of the ear in man, i. 20;
-
on the variability of the vermiform appendage in man, i. 27;
-
on the abnormal division of the malar bone in man, i. 124;
-
on abnormal conditions of the human uterus, i. 124;
-
on the persistence of the frontal suture in man, i. 125;
-
on the proportion of the sexes in silk-moths, i. 309, 311.
-
Canine teeth in man, i. 126;
-
diminution of, in man, i. 144;
-
diminution of, in horses, i. 144;
-
disappearance of, in male ruminants, i. 144;
-
large, in the early progenitors of man, i. 206.
418
Canines, and horns, inverse development of, ii. 257.
-
Canoes, use of, i. 137, 234.
-
Cantharis, difference of colour in the sexes of a species of, i. 367.
-
Capercailzie, proportion of the sexes in the, i. 306;
-
pugnacity of the male, ii. 45;
-
pairing of the, ii. 49;
-
autumn meetings of the, ii. 54;
-
call of the, ii. 61;
-
duration of the courtship of, ii. 100;
-
behaviour of the female, ii. 121;
-
inconvenience of black colour to the female, ii. 154;
-
sexual difference in coloration of the, ii. 226;
-
crimson eye-cere of the male, ii. 227;
-
polygamous, i. 269.
-
Capital, i. 169.
-
Capitonidæ, colours and nidification of the, ii. 171.
-
Capra ægagrus, ii. 249;
-
crest of the male, ii. 282;
-
sexual difference in the colour of, ii. 289.
-
Capreolus Sibiricus subecaudatus, ii. 298.
-
Caprice, common to man and animals, i. 65.
-
Caprimulgus, noise made by the males of some species of, with their wings, ii. 62.
-
Caprimulgus virginianus, pairing of, ii. 49.
-
Carabidæ, bright colours of, i. 367.
-
Carbonnier, on the natural history of the pike, i. 308;
-
the relative size of the sexes in fishes, ii. 7.
-
Carcineutes, sexual difference of colour in, ii. 173.
-
Carcinus mænas, i. 331, 333.
-
Carduelis elegans, sexual differences of the beak in, ii. 39.
-
Carnivora, marine, polygamous habits of, i. 268;
-
sexual differences in the colours of, ii. 286.
-
Carp, numerical proportion of the sexes in the, i. 308.
-
Carr, R., on the peewit, ii. 48.
-
Carrier pigeon, late development of the wattle in the, i. 293.
-
Carrion beetles, stridulation of, i. 378.
-
Carus, Prof. V., on the development of the horns in merino sheep, i. 289.
-
Cassowary, sexes and incubation of the, ii. 204.
-
Castoreum, ii. 279.
-
Casuarius galeatus, ii. 204.
-
Cat, convoluted body in the extremity of the tail of a, i. 30;
-
sick, sympathy of a dog with a, i. 77.
-
Cataract in Cebus Azaræ, i. 12.
-
Catarrh, liability of Cebus Azaræ to, i. 11.
-
Catarrhine monkeys, i. 196.
-
Caterpillars, bright colours of, i. 415.
-
Cathartes aura, ii. 116.
-
Cathartes jota, love-gestures of the male, ii. 68.
-
Catlin, G., on the development of the beard among North American Indians, ii. 322;
-
on the great length of the hair in some North American tribes, ii. 348.
-
Caton, J. D., on the development of the horns in Cervus virginianus and strongyloceros, i. 288;
-
on the presence of traces of horns in the female wapiti, ii. 245;
-
on the fighting of deer, ii. 252;
-
on the crest of the male wapiti, ii. 282;
-
on the colours of the Virginian deer, ii. 288;
-
on sexual differences of colour in the wapiti, ii. 289;
-
on the spots of the Virginian deer, ii. 303.
-
Cats, dreaming, i. 46;
-
tortois-eshell, i. 283, 285, 293;
-
enticed by valerian, ii. 281;
-
colours of, ii. 299.
-
Cattle, domestic, sexual differences of, late developed, i. 293;
-
rapid increase of, in South America, i. 135;
-
domestic, lighter in winter in Siberia, i. 282;
-
horns of, i. 289, ii. 247;
-
numerical proportion of the sexes in, i. 305.
-
Caudal vertebræ, number of, in macaques and baboons, i. 150;
-
basal, of monkeys, imbedded in the body, i. 151.
-
Cebus, maternal affection in a, i. 40;
-
gradation of species of, i. 227.
-
Cebus Azaræ, liability of, to the same diseases as man, i. 11;
-
distinct sounds produced by, i. 53;
-
early maturity of the female, ii. 318.
-
Cebus capucinus, polygamous, i. 266;
-
sexual differences of colour in, ii. 290;
-
hair on the head of, ii. 307.
-
Cebus vellerosus, hair on the head of, ii. 307.
419
-
Cecidomyidæ, proportions of the sexes in, i. 314.
-
Celibacy, unknown among the savages of South Africa and South America, ii. 367.
-
Centipedes, i. 339.
-
Cephalopoda, absence of secondary sexual characters in, i. 325.
-
Cephalopterus ornatus, ii. 58, 59.
-
Cephalopterus penduliger, ii. 59.
-
Cerambyx heros, stridulant organ of, i. 380.
-
Ceratophora aspera, nasal appendages of, ii. 34.
-
Ceratophora Stoddartii, nasal horn of, ii. 34.
-
Cerceris, habits of, i. 364.
-
Cercocebus æthiops, whiskers, &c., of, ii. 308.
-
Cercopithecus, young, seized by an eagle and rescued by the troop, i. 76;
-
definition of species of, i. 227.
-
Cercopithecus cephus, sexual difference of colour in, ii. 291, 311.
-
Cercopithecus cynosurus and griseo-viridis, colour of the scrotum in, ii. 291.
-
Cercopithecus Diana, sexual differences of colour in, ii. 291, 311, 312.
-
Cercopithecus griseo-viridis, i. 75.
-
Cercopithecus petaurista, whiskers, &c., of, ii. 308.
-
Ceres, of birds, bright colours of, ii. 227.
-
Ceriornis Temminckii, swelling of the wattles of the male during courtship, ii. 72.
-
Cervulus, weapons of, ii. 257.
-
Cervulus moschatus, rudimentary horns of the female, ii. 245.
-
Cervus alces, i. 288.
-
Cervus campestris, odour of, ii. 279.
-
Cervus canadensis, traces of horns in the female, ii. 245;
-
attacking a man, ii. 253;
-
sexual difference in the colour of, ii. 289.
-
Cervus elaphus, battles of male, ii. 240;
-
horns of, with numerous points, ii. 252.
-
Cervus Eldi, i. 288.
-
Cervus mantchuricus, ii. 303.
-
Cervus paludosus, colours of, ii. 290.
-
Cervus strongyloceros, i. 288.
-
Cervus virginianus, i. 288;
-
horns of, in course of modification, ii. 255.
-
Ceryle, male black-belted in some species of, ii. 173.
-
Cetacea, nakedness of, i. 148.
-
Ceylon, frequent absence of beard in the natives of, ii. 321.
-
Chaffinch, proportion of the sexes in the, i. 306, 307;
-
courtship of the, ii. 94.
-
Chaffinches, ii. 53;
-
new mates found by, ii. 105.
-
Chalcophaps indicus, characters of young, ii. 185.
-
Chalcosoma atlas, sexual differences of, i. 368.
-
Chamæleon, sexual differences in the genus, ii. 34.
-
Chamæleon bifurcus, ii. 34, 35.
-
Chamæleon Owenii, ii. 34, 36.
-
Chameleons, ii. 32.
-
Chamois, danger-signals of, i. 74;
-
transfer of male characters to an old female, ii. 245.
-
Chamæpetes unicolor, modified wing-feather in the male, ii. 64.
-
Chapuis, Dr., on the transmission of sexual peculiarities in pigeons, i. 283;
-
on streaked Belgian pigeons, i. 294, ii. 157.
-
Char, male, colouring of, during the breeding season, ii. 14.
-
Characters, male, developed in females, i. 280;
-
natural, artificial exaggeration of, by man, ii. 351;
-
secondary sexual, transmitted through both sexes, i. 279.
-
Charadrius hiaticula and pluvialis, sexes and young of, ii. 216.
-
Chardin on the Persians, ii. 356.
-
Charms, worn by women, ii. 344.
-
Charruas, freedom of divorce among the, ii. 372.
-
Chasmorhynchus, difference of colour in the sexes of, ii. 79;
-
Chastity, early estimation of, i. 96.
-
Chatterers, sexual differences in, i. 269.
-
Cheiroptera, absence of secondary sexual characters in, i. 268.
-
Chelæ of crustacea, i. 330, 336.
-
Chelonia, sexual differences in, ii. 28.
-
Chenalopex ægyptiacus, wing-knobs of, ii. 46.
-
Chera progne, ii. 84, 120.
420
-
Chest, proportions of, in soldiers and sailors, i. 117;
-
large, of the Quechua and Aymara Indians, i. 119.
-
Chevrotains, canine teeth of, ii. 257.
-
Chiasognathus, stridulation of, i. 384.
-
Chiasognathus Grantii, mandibles of, i. 377.
-
Children, legitimate and illegitimate, proportion of the sexes in, i. 302.
-
Chiloe, lice of the natives of, i. 220;
-
Chimæra monstrosa, bony process on the head of the male, ii. 12.
-
Chimæroid fishes, prehensile organs of male, ii. 1.
-
Chimpanzee, ii. 323;
-
ears of the, i. 21;
-
representatives of the eyebrows in the, i. 25;
-
platforms built by the, i. 36;
-
cracking nuts with a stone, i. 51;
-
hands of the, i. 139;
-
absence of mastoid processes in the, i. 143;
-
direction of the hair on the arms of the, i. 192;
-
supposed evolution of the, i. 230;
-
polygamous and social habits of the, ii. 362.
-
China, North, idea of female beauty in, ii. 344.
-
China, Southern, inhabitants of, i. 246.
-
Chinese, use of flint tools by the, i. 183;
-
difficulty of distinguishing the races of the, i. 215;
-
colour of the beard in, ii. 319;
-
general beardlessness of the, ii. 321;
-
opinions of the, on the appearance of Europeans and Cingalese, ii. 345, 347;
-
compression of the feet of, ii. 352.
-
Chinsurdi, his opinion of beards, ii. 341, 349.
-
Chlamydera maculata, ii. 70.
-
Chloëon, pedunculated eyes of the male of, i. 341.
-
Chloephaga, coloration of the sexes in, ii. 178.
-
Chlorocœlus Tanana (figured), i. 355.
-
Chorda Dorsalis, i. 207.
-
Chough, red beak of the, ii. 227.
-
Chromidæ, frontal protuberance in male, ii. 13;
-
sexual differences in colour of, ii. 20.
-
Chrysemys picta, long claws of the male, ii. 28.
-
Chrysococcyx, characters of young of, ii. 185.
-
Chrysomela cerealis, bright colours of, i. 367.
-
Chrysomelidæ, stridulation of, i. 379.
-
Cicada pruinosa, i. 351.
-
Cicada septendecim, i. 351.
-
Cicadæ, songs of the, i. 350;
-
rudimentary sound-organs in females of, i. 359.
-
Cicatrix of a burn, causing modification of the facial bones, i. 147.
-
Cichla, frontal protuberance of male, ii. 13.
-
Cimetière du Sud, Paris, i. 28.
-
Cincloramphus cruralis, large size of male, ii. 43.
-
Cinclus aquaticus, ii. 170.
-
Cingalese, Chinese opinion of the appearance of the, ii. 345.
-
Cirripedes, complemental males of, i. 255.
-
Civilisation, effects of, upon natural selection, i. 170;
-
influence of, in the competition of nations, i. 239.
-
Clanging of Geese, &c., ii. 51.
-
Claparède, E., on natural selection applied to man, i. 137.
-
Clarke, on the marriage-customs of the Kalmucks, ii. 373.
-
Classification, i. 188.
-
Claus, C., on the sexes of Saphirina, i. 336.
-
Cleft-palate, inherited, i. 121.
-
Climacteris erythrops, sexes of, ii. 206.
-
Climate, i. 115;
-
cool, favourable to human progress, i. 167;
-
power of supporting extremes of, by man, i. 237;
-
want of connexion of, with colour, i. 241.
-
Cloaca, existence of a, in the early progenitors of man, i. 207.
-
Cloacal passage existing in the human embryo, i. 16.
-
Club, origin of the, i. 234.
-
Clucking of fowls, ii. 51.
-
Clythra 4-punctata, stridulation of, i. 379.
-
Cobra, ingenuity of a, ii. 31.
-
Coccus, i. 186.
-
Coccyx, i. 29, 30;
-
in the human embryo, i. 16;
-
convoluted body at the extremity of the, i. 30;
-
imbedded in the body, i. 151.
-
Cochin-China, notions of beauty of the inhabitants of, ii. 345, 347.
421
-
Cock, game, killing a kite, ii. 44;
-
blind, fed by its companions, i. 77;
-
comb and wattles of the, ii. 98;
-
preference shown by the, for young hens, ii. 121;
-
game, transparent zone in the hackles of a, ii. 136.
-
Cock of the rock, ii. 100.
-
Cockatoos, ii. 226, 228, 230;
-
nestling, ii. 109;
-
black, immature plumage of, ii. 188.
-
Cœlenterata, absence of secondary sexual characters in, i. 321.
-
Coffee, fondness of monkeys for, i. 12.
-
Cold, supposed effects of, i. 116;
-
power of supporting, by man, i. 237.
-
Coleoptera, i. 366;
-
stridulant organs of, discussed, i. 381.
-
Collingwood, C., on the pugnacity of the butterflies of Borneo, i. 386;
-
on butterflies being attracted by a dead specimen of the same species, i. 400.
-
Colombia, flattened heads of savages of, ii. 340.
-
Colonists, success of the English as, i. 179.
-
Coloration, protective, in birds, ii. 223.
-
Colour, supposed to be dependent on light and heat, i. 115;
-
correlation of, with immunity from certain poisons and parasites, i. 242;
-
purpose of, in lepidoptera, i. 399;
-
relation of, to sexual functions, in fishes, ii. 14;
-
difference of, in the sexes of snakes, ii. 29;
-
sexual differences of, in lizards, ii. 36;
-
influence of, in the pairing of birds of different species, ii. 115;
-
relation of, to nidification, ii. 167, 172;
-
sexual differences of, in mammals, ii. 286, 294;
-
recognition of, by quadrupeds, ii. 295;
-
of children, in different races of man, ii. 318;
-
of the skin in man, ii. 381.
-
Colours, admired alike by man and animals, i. 64;
-
bright, due to sexual selection, i. 322;
-
bright, among the lower animals, i. 322, 323;
-
bright, protective to butterflies and moths, i. 395;
-
bright, in male fishes, ii. 7, 13;
-
transmission of, in birds, ii. 159.
-
Colquhoun, example of reasoning in a retriever, i. 48.
-
Columba passerina, young of, ii. 188.
-
Colymbus glacialis, anomalous young of, ii. 211.
-
Comb, development of, in fowls, i. 295.
-
Combs and wattles in male birds, ii. 98.
-
Community, preservation of variations useful to the, by natural selection, i. 155.
-
Compositæ, gradation of species among the, i. 227.
-
Comte, C., on the expression of the ideal of beauty by sculpture, ii. 380.
-
Conditions of life, action of changed, upon man, i. 113;
-
influence of, on plumage of birds, ii. 196.
-
Condor, eyes and comb of the, ii. 129.
-
Conjugations, origin of, i. 61.
-
Conscience, i. 91, 104;
-
absence of, in some criminals, i. 92.
-
Constitution, difference of, in different races of men, i. 216.
-
Consumption, liability of Cebus Azaræ to, i. 12;
-
connexion between complexion and, i. 244.
-
Convergence, i. 230.
-
Cooing of pigeons and doves, ii. 60.
-
Cook, Capt., on the nobles of the Sandwich Islands, ii. 356.
-
Cope, E. D., on the dinosauria, i. 204;
-
on the origin of genera, ii. 215.
-
Cophotis ceylanica, sexual differences of, ii. 32, 36.
-
Copris, i. 370.
-
Copris Isidis, sexual differences of, i. 369.
-
Copris lunaris, stridulation of, i. 380.
-
Corals, bright colours of, i. 322.
-
Coral-snakes, ii. 31.
-
Cordylus, sexual difference of colour in a species of, ii. 36.
-
Corfu, habits of the chaffinch in, i. 307.
-
Cornelius, on the proportions of the sexes in Lucanus Cervus, i. 313.
-
Corpora Wolffiana, i. 207;
-
agreement of, with the kidneys of fishes, i. 16.
422
-
Correlated variation, i. 130.
-
Correlation, influence of, in the production of races, i. 247.
-
Corse, on the mode of fighting of the elephant, ii. 257.
-
Corvus corone, ii. 104.
-
Corvus graculus, red beak of, ii. 227.
-
Corvus pica, nuptial assembly of, ii. 102.
-
Corydalis cornutus, large jaws of the male, i. 342.
-
Cosmetornis, ii. 181.
-
Cosmetornis vexillarius, elongation of wing-feathers in, ii. 73, 97.
-
Cotingidæ, sexual differences in, i. 269;
-
coloration of the sexes of, ii. 177;
-
resemblance of the females of distinct species of, ii. 192.
-
Cottus scorpius, sexual differences in, ii. 9.
-
Counting, origin of, i. 181;
-
limited power of, in primeval man, i. 234.
-
Courage, variability of, in the same species, i. 40;
-
universal high appreciation of, i. 95;
-
importance of, i. 162;
-
a characteristic of men, ii. 328.
-
Courtship, greater eagerness of males in, i. 272;
-
of fishes, ii. 2;
-
of birds, ii. 50, 100.
-
Cow, winter change of the, ii. 299.
-
Crab, devil, i. 332.
-
Crab, shore, habits of, i. 331.
-
Crabro cribrarius, dilated tibiæ of the male, i. 343.
-
Crabs, proportions of the sexes in, i. 315.
-
Cranz, on the inheritance of dexterity in seal-catching, i. 117.
-
Crawfurd, on the number of species of man, i. 226.
-
Crenilabrus massa and C. melops, nests built by, ii. 19.
-
Crest, origin of, in Polish fowls, i. 284.
-
Crests, of birds, difference of, in the sexes, ii. 189;
-
dorsal hairy, of mammals, ii. 282.
-
Cricket, field-, stridulation of the, i. 353;
-
pugnacity of male, i. 360.
-
Cricket, house-, stridulation of the, i. 352, 354.
-
Crickets, sexual differences in, i. 361.
-
Crioceridæ, stridulation of the, i. 379.
-
Crinoids, complexity of, i. 61.
-
Croaking of frogs, ii. 27.
-
Crocodiles, musky odour of, during the breeding season, ii. 29.
-
Crocodilia, ii. 28.
-
Crossbills, characters of young, ii. 184.
-
Crosses in man, i. 225.
-
Crossing of races, effects of the, i. 241.
-
Crossoptilon auritum, ii. 93, 166, 196;
-
adornment of both sexes of, i. 290;
-
sexes alike in, ii. 178.
-
Crotch, G. R., on the stridulation of beetles, i. 379, 382;
-
on the stridulation of Heliopathes, i. 383;
-
on the stridulation of Acalles, i. 384.
-
Crow Indians, long hair of the, ii. 348.
-
Crow, young of the, ii. 209.
-
Crows, ii. 226;
-
vocal organs of the, ii. 55;
-
living in triplets, ii. 106.
-
Crows, carrion, new mates found by, ii. 104.
-
Crows, Indian, feeding their blind companions, i. 77.
-
Cruelty of savages to animals, i. 94.
-
Crustacea, amphipod, males sexually mature while young, ii. 215;
-
parasitic, loss of limbs by female, i. 255;
-
prehensile feet and antennæ of, i. 256;
-
male, more active than female, i. 272;
-
parthenogenesis in, i. 315;
-
secondary sexual characters of, i. 328;
-
auditory hairs of, ii. 333.
-
Crystal worn in the lower lip by some Central African women, ii. 341.
-
Cuckoo fowls, i. 294.
-
Culicidæ, i. 254, 349.
-
Cullen, Dr., on the throat-pouch of the male bustard, ii. 58.
-
Cultivation of plants, probable origin of, i. 167.
-
Cupples, Mr., on the numerical proportion of the sexes in dogs, sheep, and cattle, i. 304, 305;
-
on the Scotch deerhound, ii. 261;
-
on sexual preference in dogs, ii. 271, 272.
-
Curculionidæ, sexual difference in length of snout in some, i. 255;
-
hornlike processes in male, i. 374;
-
musical, i. 378, 379.
-
Curiosity, manifestations of, by animals, i. 42.
423
-
Curlews, double moult in, ii. 80.
-
Cursores, comparative absence of sexual differences among the, i. 269.
-
Curtis, J., on the proportion of the sexes in Athalia, i. 314.
-
Cuvier, F., on the recognition of women by male quadrumana, i. 13.
-
Cuvier, G., views of, as to the position of man, i. 190;
-
on instinct and intelligence, i. 37;
-
on the number of caudal vertebræ in the mandrill, i. 150;
-
on the position of the seals, i. 190;
-
on Hectocotyle, i. 325.
-
Cyanecula suecica, sexual differences of, ii. 195.
-
Cyanalcyon, sexual difference in colours of, ii. 173;
-
immature plumage of, ii. 188.
-
Cychrus, sounds produced by, i. 382.
-
Cycnia mendica, sexual difference of colour in, i. 398.
-
Cygnus ferus, trachea of, ii. 59.
-
Cygnus olor, white young of, ii. 211.
-
Cyllo Leda, instability of the ocellated spots of, ii. 133.
-
Cynanthus, variation in the genus, ii. 125.
-
Cynipidæ, proportions of the sexes in, i. 314.
-
Cynocephalus, difference of the young, from the adult, i. 13;
-
male, recognition of women by, i. 13;
-
polygamous habits of species of, i. 266.
-
Cynocephalus chacma, i. 41.
-
Cynocephalus gelada, i. 51.
-
Cynocephalus hamadryas, i. 51;
-
sexual difference of colour in, ii. 291.
-
Cynocephalus leucophœus, colours of the sexes of, ii. 292.
-
Cynocephalus mormon, colours of the male, ii. 292, 296, 310.
-
Cynocephalus porcarius, mane of the male, ii. 267.
-
Cypridina, proportions of the sexes in, i. 315.
-
Cyprinidæ, proportion of the sexes in the, i. 308.
-
Cyprinidæ, Indian, ii. 17.
-
Cyprinodontidæ, sexual differences in the, ii. 7, 9.
-
Cyprinus auratus, ii. 16.
-
Cyprinus phoxinus, spawning of, ii. 15.
-
Cypris, relations of the sexes in, i. 315.
-
Cystophora cristata, hood of, ii. 278.
-
D.
-
Dacelo, sexual difference of colour in, ii. 174.
-
Dacelo Gaudichaudi, young male of, ii. 188.
-
Dal-ripa, a kind of ptarmigan, i. 306.
-
Damalis albifrons, peculiar markings of, ii. 301.
-
Damalis pygarga, peculiar markings of, ii. 300.
-
Dampness of climate, supposed influence of, on the colour of the skin, i. 116, 242.
-
Danaidæ, i. 387.
-
Dances of birds, ii. 68.
-
Dancing, i. 232.
-
Daniell, Dr., his experience of residence in West Africa, i. 245.
-
Darfur, protuberances artificially produced in, ii. 339.
-
Darwin, F., on the stridulation of Dermestes murinus, i. 379.
-
Dasychira pudibunda, sexual difference of colour in, i. 398.
-
Davis, A. H., on the pugnacity of the male stag-beetle, i. 375.
-
Davis, J. B., on the capacity of the skull in various races of men, i. 146;
-
on the beards of the Polynesians, ii. 322.
-
Death-rate higher in towns than in rural districts, i. 175.
-
Death-tick, i. 384.
-
De Candolle, Alph., on a case of inherited power of moving the scalp, i. 20.
-
Declensions, origin of, i. 61.
-
Decoration in birds, ii. 71.
-
Decticus, i. 355.
-
Deer, spots of young, ii. 184, 303;
-
horns of, ii. 243, 248;
-
use of horns of, ii. 252, 263;
-
size of the horns of, ii. 259;
-
female, pairing with one male, whilst others are fighting for her, ii. 269;
-
male, attracted by the voice of the female, ii. 276;
-
male, odour emitted by, ii. 279;
-
development of the horns in, i. 288;
424
-
horns of a, in course of modification, ii. 255.
-
Deer, Axis, sexual, difference in the colour of the, ii. 290.
-
Deer, fallow, different coloured herds of, ii. 295.
-
Deer, Mantchurian, ii. 303.
-
Deer, Virginian, ii. 303;
-
colour of the, not affected by castration, ii. 288;
-
colours of, ii. 289.
-
Deerhound, Scotch, greater size of the male, i. 293, ii. 260.
-
Defensive organs of mammals, ii. 263.
-
De Geer, C., on a female spider destroying a male, i. 339.
-
Dekay, Dr., on the bladder-nose seal, ii. 278.
-
Demerara, yellow fever in, i. 243.
-
Dendrocygna, ii. 185.
-
Dendrophila frontalis, young of, ii. 220.
-
Denny, H., on the lice of domestic animals, i. 219.
-
Dermestes murinus, stridulation of, i. 379.
-
Descent traced through the mother alone, ii. 359.
-
Deserts, protective colouring of animals inhabiting, ii. 224.
-
Desmarest, on the absence of suborbital pits in Antilope subgutturosa, ii. 280;
-
on the whiskers of Macacus, ii. 283;
-
on the colour of the opossum, ii. 286;
-
on the colours of the sexes of Mus minutus, ii. 286;
-
on the colouring of the ocelot, ii. 287;
-
on the colours of seals, ii. 287;
-
on Antilope caama, ii. 289;
-
on the colours of goats, ii. 290;
-
on sexual difference of colour in Ateles marginatus, ii. 291;
-
on the mandrill, ii. 293;
-
on Macacus cynomolgus, ii. 318.
-
Desmoulins, on the number of species of man, i. 226;
-
on the musk-deer, ii. 281.
-
Desor, on the imitation of man by monkeys, i. 44.
-
Despine, P., on criminals destitute of conscience, i. 92.
-
Development, embryonic, of man, i. 14, 16;
-
Devil, not believed in by the Fuegians, i. 67.
-
Devil-crab, i. 332.
-
Devonian, fossil insect from the, i. 360.
-
Dewlaps, of cattle and antelopes, ii. 284.
-
Diadema, sexual differences of colouring in the species of, i. 388.
-
Diadema anomala, mimickry by the female of, i. 413.
-
Diadema bolina, i. 413.
-
Diamond-beetles, bright colours of, i. 367.
-
Diastema, occurrence of, in man, i. 126.
-
Diastylidæ, proportion of the sexes in, i. 315.
-
Diodorus, on the absence of beard in the natives of Ceylon, ii. 321.
-
Dicrurus, racket-shaped feathers in, ii. 73;
-
nidification of, ii. 167.
-
Dicrurus macrocercus, change of plumage in, ii. 179.
-
Didelphis opossum, sexual difference in the colour of, ii. 286.
-
Differences, comparative, between different species of birds of the same sex, ii. 192.
-
Digits, supernumerary, more frequent in men than in women, i. 276;
-
supernumerary, inheritance of, i. 285;
-
supernumerary, early development of, i. 292.
-
Dimorphism in females of water-beetles, i. 343;
-
in Neurothemis and Agrion, i. 363.
-
Dipelicus Cantori, sexual differences of, i. 369.
-
Diplopoda, prehensile limbs of the male, i. 340.
-
Dipsas cynodon, sexual difference in the colour of, ii. 29.
-
Diptera, i. 348.
-
Disease, generated by the contact of distinct peoples, i. 239.
-
Diseases common to man and the lower animals, i. 11;
-
difference of liability to, in different races of men, i. 216;
-
new, effects of, upon savages, i. 238;
-
sexually limited, i. 292.
-
Display, coloration of Lepidoptera for, i. 395;
-
of plumage by male birds, ii. 86, 96.
-
Distribution, wide, of man, i. 137;
425
-
geographical, as evidence of specific distinctness in man, i. 218.
-
Disuse, effects of, in producing rudimentary organs, i. 18;
-
and use of parts, effects of, i. 116;
-
of parts, influence of, on the races of men, i. 247.
-
Divorce, freedom of, among the Charruas, ii. 372.
-
Dixon, E. S., on the habits of the guinea-fowl, i. 270;
-
on the pairing of different species of geese, ii. 114;
-
on the courtship of peafowl, ii. 121.
-
Dobrizhoffer, on the marriage-customs of the Abipones, ii. 374.
-
Dogs, suffering from Tertian ague, i. 13;
-
memory of, i. 45;
-
domestic, progress of, in moral qualities, i. 50;
-
distinct tones uttered by, i. 54;
-
parallelism between his affection for his master and religious feeling, i. 68;
-
sociability of the, i. 74;
-
sympathy of, with a sick cat, i. 77;
-
sympathy of, with his master, i. 77;
-
possible use of the hair on the forelegs of the, i. 193;
-
races of the, i. 229;
-
diverging when drawing sledges over thin ice, i. 40;
-
dreaming, i. 46, 158;
-
exercise of reasoning faculties by, i. 48;
-
their possession of conscience, i. 78;
-
numerical proportion of male and female births in, i. 304;
-
sexual affection between individuals of, ii. 270;
-
howling at certain notes, ii. 333;
-
rolling in carrion, ii. 281.
-
Dolichocephalic structure, possible cause of, i. 148.
-
Dolphins, nakedness of, i. 148.
-
Domestic animals, races of, i. 229;
-
change of breeds of, ii. 369.
-
Domestication, influence of, in removing the sterility of hybrids, i. 222.
-
D’Orbigny, A., on the influence of dampness and dryness on the colour of the skin, i. 242;
-
on the Yura-caras ii. 347.
-
Dotterel, ii. 203.
-
Doubleday, E., on sexual differences in the wings of butterflies, i. 345.
-
Doubleday, H., on the proportion of the sexes in the smaller moths, i. 311;
-
on the attraction of the males of Lasiocampa quercus and Saturnia carpini by the female, i. 312;
-
on the proportion of the sexes in the Lepidoptera, i. 312;
-
on the ticking of Anobium tessellatum, i. 385;
-
on the structure of Ageronia feronia, i. 387;
-
on white butterflies alighting upon paper, i. 400.
-
Douglas, J. W., on the sexual differences of the Hemiptera, i. 349;
-
on the colours of British Homoptera, i. 352.
-
Down, of birds, ii. 80.
-
Draco, gular appendages of, ii. 33.
-
Dragonet, Gemmeous, ii. 7.
-
Dragon-flies, caudal appendages of male, i. 344;
-
relative size of the sexes of, i. 347;
-
difference in the sexes of, i. 361;
-
want of pugnacity by the male, i. 364.
-
Drake, breeding plumage of the, ii. 84.
-
Dreams, i. 46;
-
a possible source of the belief in spiritual agencies, i. 66.
-
Drill, sexual difference of colour in the, ii. 291.
-
Dromœus irroratus, ii. 204.
-
Dromolæa, Saharan species of, ii. 172.
-
Drongo shrike, ii. 179.
-
Drongos, racket-shaped feathers in the tails of, ii. 73, 83.
-
Dryness, of climate, supposed influence of, on the colour of the skin, i. 242.
-
Dryopithecus, i. 199.
-
Duck, harlequin, age of mature plumage in the, ii. 213;
-
breeding in immature plumage, ii. 214.
-
Duck, long-tailed, preference of male, for certain females, ii. 122.
-
Duck, pintail, pairing with a wigeon, ii. 114.
-
Duck, voice of the, ii. 60;
-
pairing with a shield-drake, ii. 114;
-
immature plumage of the, ii. 188.
-
Duck, wild, sexual differences in the, i. 268;
-
speculum and male characters of, i. 291;
-
pairing with a pintail drake, ii. 115.
-
Ducks, dogs and cats recognised by, ii. 110;
-
wild, becoming polygamous under partial domestication, i. 270.
426
-
Dugong, tusks of, ii. 242;
-
Dujardin, on the relative size of the cerebral ganglia in insects, i. 145.
-
Duncan, Dr., on the fertility of early marriages, i. 174.
-
Dupont, M., on the occurrence of the supra-condyloid foramen in the humerus of man, i. 29.
-
Durand, J. P., on causes of variation, i. 113.
-
Dureau de la Malle, on the songs of birds, i. 55;
-
on the acquisition of an air by blackbirds, ii. 55.
-
Dutch, retention of their colour by the, in South Africa, i. 242.
-
Duty, sense of, i. 70.
-
Duvaucel, female Hylobates washing her young, i. 40.
-
Dyaks, pride of, in mere homicide, i. 94.
-
Dynastes, large size of males of, i. 347.
-
Dynastini, stridulation of, i. 381.
-
Dytiscus, dimorphism of females of, i. 343;
-
grooved elytra of the female, i. 343.
-
E.
-
Eagle, young Cercopithecus rescued from, by the troop, i. 75.
-
Eagle, white-headed, breeding in immature plumage, ii. 214.
-
Eagles, golden, new mates found by, ii. 105.
-
Ear, motion of the, i. 20;
-
external shell of the, useless in man, i. 21;
-
rudimentary point of the, in man, i. 22.
-
Ears, piercing and ornamentation of the, ii. 341.
-
Echidna, i. 201.
-
Echini, bright colours of some, i. 322.
-
Echinodermata, absence of secondary sexual characters in, i. 321.
-
Ecker, figure of the human embryo, i. 15;
-
on sexual differences in the pelvis in man, ii. 317;
-
on the presence of a sagittal crest in Australians, ii. 319.
-
Edentata, former wide range of, in America, i. 219;
-
absence of secondary sexual characters in, i. 268.
-
Edolius, racket-shaped feathers in, ii. 73.
-
Edwards, Mr., on the proportion of the sexes in North American species of Papilio, i. 309.
-
Egerton, Sir P., on the use of the antlers of deer, ii. 252;
-
on the pairing of red deer, ii. 269;
-
on the bellowing of stags, ii. 275.
-
Eggs, hatched by male fishes, ii. 20.
-
Egret, Indian, sexes and young of, ii. 217.
-
Egrets, breeding plumage of, ii. 82;
-
Ehrenberg, on the mane of the male Hamadryas baboon, ii. 267.
-
Ekström, M., on Harelda glacialis, ii. 122.
-
Elachista rufocinerea, habits of male, i. 311.
-
Eland, development of the horns of the, i. 289.
-
Elands, sexual differences of colour in, ii. 288.
-
Elaphomyia, sexual differences in, i. 349.
-
Elaphrus uliginosus, stridulation of, i. 379.
-
Elaps, ii. 31.
-
Elateridæ, proportions of the sexes in, i. 313.
-
Elaters, luminous, i. 345.
-
Elephant, i. 200;
-
nakedness of the, i. 148;
-
rate of increase of the, i. 135;
-
Indian, polygamous habits of the, i. 267;
-
pugnacity of the male, ii. 240;
-
tusks of, ii. 242, 243, 248, 249, 258;
-
Indian, mode of fighting, of the, ii. 257;
-
male, odour emitted by the, ii. 279;
-
attacking white or grey horses, ii. 295.
-
Elevation of abode, modifying influence of, i. 120.
-
Elimination of inferior individuals, i. 172.
-
Elk, ii. 249;
-
winter change of the, ii. 299.
-
Elk, Irish, horns of the, ii. 259.
-
Ellice Islands, beards of the natives, ii. 322, 349.
-
Elliot, R., on the numerical proportion of the sexes in young rats, i. 305;
-
on the proportion of the sexes in sheep, i. 305.
427
-
Elliott, D. G., on Pelecanus erythrorhynchus, ii. 80.
-
Elliott, Sir W., on the polygamous habits of the Indian wild boar, i. 267.
-
Ellis, on the prevalence of infanticide in Polynesia, ii. 364.
-
Elphinstone, Mr., on local differences of stature among the Hindoos, i. 115;
-
on the difficulty of distinguishing the native races of India, i. 215.
-
Elytra, of the females of Dytiscus, Acilius, Hydroporus, i. 343.
-
Emberiza, characters of young, ii. 184.
-
Emberiza miliaria, ii. 185.
-
Emberiza schœniculus, ii. 111;
-
head-feathers of the male, ii. 95.
-
Embryo of man, i. 14, 15;
-
Embryos of mammals, resemblance of the, i. 32.
-
Emigration, i. 172.
-
Emotions experienced by the lower animals in common with man, i. 39;
-
manifested by animals, i. 42.
-
Emperor moth, i. 398.
-
Emulation of singing-birds, ii. 53.
-
Emu, sexes and incubation of, ii. 204.
-
Endurance, estimation of, i. 95.
-
Energy, a characteristic of men, ii. 328.
-
England, numerical proportion of male and female births in, i. 300.
-
Engleheart, Mr., on the finding of new mates by starlings, ii. 106.
-
English, success of, as colonists, i. 179.
-
Engravers, short-sighted, i. 118.
-
Entomostraca, i. 332.
-
Entozoa, difference of colour between the males and females of some, i. 321.
-
Eocene, possible divergence of man during the, i. 200.
-
Eolidæ, colours of, produced by the biliary glands, i. 323.
-
Epeira, i. 337.
-
Epeira nigra, small size of the male of, i. 338.
-
Ephemeræ, i. 341.
-
Ephemeridæ, i. 361.
-
Ephemerina, proportions of the sexes in, i. 314.
-
Ephippiger vitium, stridulating organs of, i. 354, 358.
-
Epicalia, sexual differences of colouring in the species of, i. 388.
-
Equus hemionus, winter change of, ii. 298.
-
Erateina, coloration of, i. 397.
-
Erect attitude of man, i. 141, 142.
-
Eschricht, on the development of hair in man, i. 24;
-
on a lanuginous moustache in a female fœtus, i. 25;
-
on the want of definition between the scalp and the forehead in some children, i. 192;
-
on the arrangement of the hair in the human fœtus, i. 193;
-
on the hairiness of the face in the human fœtus of both sexes, ii. 379, 380.
-
Esmeralda, difference of colour in the sexes of, i. 368.
-
Esox lucius, i. 308.
-
Esox reticulatus, ii. 14.
-
Esquimaux, i. 157, 167;
-
their belief in the inheritance of dexterity in seal-catching, i. 117;
-
mode of life of, i. 246.
-
Estrelda amandava, pugnacity of the male, ii. 49.
-
Eubagis, sexual differences of colouring in the species of, i. 389.
-
Euchirus longimanus, sound produced by, i. 381.
-
Eudromias morinellus, ii. 203.
-
Eulampis jugularis, colours of the female, ii. 168.
-
Euler, on the rate of increase in the United States, i. 131.
-
Eumomota superciliaris, racket-shaped feathers in the tail of, ii. 73.
-
Eupetomena macroura, colours of the female, ii. 168.
-
Euphema splendida, ii. 174.
-
Euplocamus erythropthalmus, possession of spurs by the female, ii. 46.
-
Euplœa midamas, mimickry of, by the female of Diadema anomala, i. 413.
-
Europe, ancient inhabitants of, i. 237.
-
Europeans, difference of, from Hindoos, i. 240;
-
hairiness of, probably due to reversion, ii. 378.
-
Eurostopodus, sexes of, ii. 206.
-
Eurygnathus, different proportions of the head in the sexes of, i. 344.
428
-
Eustephanus, sexual differences of species of, ii. 39;
-
Exaggeration of natural characters by man, ii. 351.
-
Exogamy, ii. 360, 364.
-
Expression, resemblances in, between man and the apes, i. 191.
-
Extinction of races, causes of, i. 238.
-
Eye, destruction of the, i. 116;
-
change of position in, i. 147;
-
obliquity of, regarded as a beauty by the Chinese and Japanese, ii. 345.
-
Eyebrows, elevation of, i. 19;
-
development of long hairs in, i. 25;
-
in monkeys, i. 192;
-
eradicated in parts of South America and Africa, ii. 340;
-
eradication of, by the Indians of Paraguay, ii. 348.
-
Eyelids, coloured black, in part of Africa, ii. 339.
-
Eyelashes, eradication of, by the Indians of Paraguay, ii. 348.
-
Eyes, difference in the colour of, in the sexes of birds, ii. 128;
-
pillared, of the male of Chloëon, i. 341.
-
Eyton, T. C., observations on the development of the horns in the fallow-deer, i. 288.
-
Eyzies, Les, human remains from, i. 237.
-
F.
-
Fabre, M., on the habits of Cerceris, i. 364.
-
Facial bones, causes of modification of the, i. 147.
-
Faculties, mental, variation of, in the same species, i. 36;
-
diversity of, in the same race of men, i. 109;
-
inheritance of, i. 110;
-
diversity of, in animals of the same species, i. 110;
-
of birds, ii. 108.
-
Fakirs, Indian, tortures undergone by, i. 96.
-
Falco leucocephalus, ii. 214.
-
Falco peregrinus, ii. 104, 179.
-
Falco tinnunculus, ii. 109.
-
Falcon, peregrine, new mate found by, ii. 104.
-
Falconer, H., on the mode of fighting of the Indian elephant, ii. 257;
-
on canines in a female deer, ii. 258;
-
on Hyomoschus aquaticus, ii. 304.
-
Falkland islands, horses of, i. 236.
-
Fallow-deer, different coloured herds of, ii. 295.
-
Famines, frequency of, among savages, i. 333.
-
Farr, Dr., on the structure of the uterus, i. 123;
-
on the effects of profligacy, i. 173;
-
on the influence of marriage on mortality, i. 175.
-
Farrar, F. W., on the origin of language, i. 56;
-
on the crossing or blending of languages, i. 60;
-
on the absence of the idea of God in certain races of men, i. 65;
-
on early marriages of the poor, i. 173;
-
on the middle ages, i. 178.
-
Fashions, long prevalence of, among savages, ii. 343, 352.
-
Faye, Prof., on the numerical proportion of male and female births in Norway and Russia, i. 301;
-
on the greater mortality of male children at and before birth, i. 302.
-
Feathers, modified, producing sounds, ii. 63 et seqq., 163;
-
elongated, in male birds, ii. 72, 97;
-
racket-shaped, ii. 73;
-
barbless and with filamentous barbs in certain birds, ii. 74;
-
shedding of margins of, ii. 85.
-
Feeding, high, probable influence of, in the pairing of birds of different species, ii. 115.
-
Feet, modification of, in man, i. 141;
-
thickening of the skin on the soles of the, i. 118.
-
Felis canadensis, throat-ruff of, ii. 267.
-
Felis pardalis and F. mitis, sexual differences in the colouring of, ii. 287.
-
Female, behaviour of the, during courtship, i. 273.
-
Female birds, differences of, ii. 193.
-
Females, presence of rudimentary male organs in, i. 208;
-
preference of, for certain males, i. 262;
-
pursuit of, by males, i. 272;
-
occurrence of secondary sexual characters in, i. 276;
-
development of male characters by, i. 280.
-
Females and males, comparative mortality of, while young, i. 264, 276;
-
Femur and tibia, proportions of, in the Aymara Indians, i. 119.
-
Ferguson, Mr., on the courtship of fowls, ii. 118.
-
Fertilization, phenomena of, in plants, i. 273;
-
in the lower animals, i. 274.
-
Fevers, immunity of Negroes and Mulattoes from, i. 243.
-
Fiber zibethicus, protective colouring of, ii. 298.
-
Fidelity of savages to one another, i. 95;
-
Field-slaves, difference of, from house-slaves, i. 246.
-
Fijians, burying their old and sick parents alive, i. 77;
-
estimation of the beard among the, ii. 349;
-
admiration of, for a broad occiput, ii. 352.
-
Fiji Islands, beards of the natives, ii. 322, 349;
-
marriage-customs of the, ii. 373.
-
Filial affection, partly the result of natural selection, i. 81.
-
Filum terminale, i. 30.
-
Finch, racket-shaped feathers in the tail of a, ii. 73.
-
Finches, spring change of colour in, ii. 85;
-
British, females of the, ii. 193.
-
Fingers, partially coherent, in species of Hylobates, i. 140.
-
Finlayson, on the Cochin Chinese, ii. 345.
-
Fire, use of, i. 137, 183, 234.
-
Fischer, on the pugnacity of the male of Lethrus cephalotes, i. 376.
-
Fish, proportion of the sexes in, i. 307;
-
eagerness of male, i. 272.
-
Fishes, kidneys of, represented by Corpora Wolffiana in the human embryo, i. 16;
-
male, hatching ova in their mouths, i. 210;
-
receptacles for ova possessed by, i. 254;
-
relative size of the sexes in, ii. 7;
-
freshwater, of the tropics, ii. 17;
-
protective resemblances in, ii. 18;
-
nest-building, ii. 19;
-
spawning of, ii. 19;
-
sounds produced by, ii. 23, 331;
-
continued growth of, ii. 216.
-
Flexor pollicis longus, similar variation of, in man, i. 129.
-
Flint tools, i. 183.
-
Flints, difficulty of chipping into form, i. 138.
-
Florida, Quiscalus major in, i. 307.
-
Flounder, coloration of the, ii. 18.
-
Flower, W. H., on the abductor of the fifth metatarsal in apes, i. 128;
-
on the position of the Seals, i. 190;
-
on the throat-pouch of the male Bustard, ii. 58.
-
Fly-catchers, colours and nidification of, ii. 170.
-
Fœtus, human, woolly covering of the, i. 25;
-
arrangement of the hair on, i. 193.
-
Food, influence of, upon stature, i. 115.
-
Foot, prehensile, in the early progenitors of man, i. 206;
-
prehensile power of the, retained in some savages, i. 142.
-
Foramen, supra-condyloid, exceptional occurrence of in the humerus of man, i. 28, 130;
-
in the early progenitors of man, i. 206.
-
Forbes, D., on the Aymara Indians, i. 119;
-
on local variation of colour in the Quechuas, i. 246;
-
on the hairlessness of the Aymaras and Quechuas, ii. 322;
-
on the long hair of the Aymaras and Quechuas, ii. 320, 348.
-
Forel, F., on white young swans, ii. 211.
-
Formica rufo, size of the cerebral ganglia in, i. 145.
-
Fossils, absence of, connecting man with the apes, i. 201.
-
Fowl, occurrence of spurs in the female, i. 280;
-
game, early pugnacity of, i. 295;
-
Polish, early development of cranial peculiarities of, i. 295;
-
variations in plumage of, ii. 74;
-
examples of correlated development in the, ii. 130;
-
domestic, breeds and sub-breeds of, ii. 178.
-
Fowls, spangled Hamburgh, i. 281, 294;
-
sexual peculiarities in, transmitted only to the same sex, i. 283;
-
loss of secondary sexual characters by male, i. 284;
-
inheritance of changes of plumage by, i. 281;
-
Polish, origin of the crest in, i. 284;
-
period of inheritance of characters by, i. 294;
430
-
cuckoo-, i. 294;
-
development of the comb in, i. 295;
-
numerical proportion of the sexes in, i. 306;
-
courtship of, ii. 117;
-
mongrel, between a black Spanish cock and different hens, ii. 131;
-
pencilled Hamburgh, difference of the sexes in, ii. 158;
-
Spanish, sexual differences of the comb in, ii. 158;
-
spurred, in both sexes, ii. 162.
-
Fox, W. D., on some half-tamed wild ducks becoming polygamous, and on polygamy in the guinea-fowl and canary-bird, i. 270;
-
on the proportion of the sexes in cattle, i. 305;
-
on the pugnacity of the peacock, ii. 46;
-
on a nuptial assembly of magpies, ii. 102;
-
on the finding of new mates by crows, ii. 104;
-
on partridges living in triplets, ii. 107;
-
on the pairing of a goose with a Chinese gander, ii. 114.
-
Foxes, wariness of young, in hunting districts, i. 50;
-
France, numerical proportion of male and female births in, i. 301.
-
Francesco, B., on the Simian resemblances of man, i. 4.
-
Fraser, C., on the different colours of the sexes in a species of Squilla, i. 335.
-
Fringilla cannabina, ii. 86.
-
Fringilla ciris, age of mature plumage in, ii. 213.
-
Fringilla cyanea, age of mature plumage in, ii. 213.
-
Fringilla leucophrys, young of, ii. 217.
-
Fringilla spinus, ii. 115.
-
Fringilla tristis, change of colour in, in spring, ii. 85;
-
Fringillidæ, resemblance of the females of distinct species of, ii. 192.
-
Frogs, ii. 25;
-
male, temporary receptacles for ova possessed by, i. 254;
-
ready to breed before the females, i. 260;
-
vocal organs of, ii. 28.
-
Frontal bone, persistence of the suture in, i. 124.
-
Fruits, poisonous, avoided by animals, i. 36.
-
Fuegians, i. 167, 181;
-
mental capacity of the, i. 34;
-
quasi-religious sentiments of the, i. 67;
-
power of sight in the, i. 118;
-
skill of, in stone-throwing, i. 138;
-
resistance of the, to their severe climate, i. 156, 237;
-
difference of stature among the, i. 115;
-
mode of life of the, i. 246;
-
resemblance of, in mental characters, to Europeans, i. 232;
-
aversion of, to hair on the face, ii. 348;
-
said to admire European women, ii. 351.
-
Fulgoridæ, songs of the, i. 351.
-
Fur, whiteness of, in arctic animals, in winter, i. 282.
-
Fur-bearing animals, acquired sagacity of, i. 50.
-
G.
-
Gallicrex, sexual difference in the colour of the irides in, ii. 128.
-
Gallicrex cristatus, red caruncle occurring in the male during the breeding-season, ii. 80.
-
Gallinaceæ, frequency of polygamous habits and of sexual differences in the, i. 269;
-
love-gestures of, ii. 68;
-
decomposed feathers in, ii. 74;
-
stripes of young, ii. 184;
-
comparative sexual differences between the species of, ii. 192, 194;
-
plumage of, ii. 195.
-
Gallinaceous birds, weapons of the male, ii. 44;
-
racket-shaped feathers on the heads of, ii. 73.
-
Gallinula chloropus, pugnacity of male, ii. 40.
-
Gallinula cristata, pugnacity of the male, ii. 41.
-
Galloperdix, spurs of, ii. 46;
-
development of spurs in the female, ii. 162.
-
Gallophasis, young of, ii. 190.
-
Gallus bankiva, ii. 158;
-
Gallus Stanleyi, pugnacity of the male, ii. 44.
-
Galls, i. 152.
-
Galton, Mr., on the struggle between the social and personal impulses, i. 104;
-
on hereditary genius, i. 111;
-
on the effects of natural selection on civilised nations, i. 168;
431
-
on the sterility of sole daughters, i. 170;
-
on the degree of fertility of people of genius, i. 171;
-
on the early marriages of the poor, i. 173;
-
on the ancient Greeks, i. 177;
-
on the Middle Ages, i. 178;
-
on the progress of the United States, i. 179;
-
on South African notions of beauty, ii. 347.
-
Gammarus, use of the chelæ of, i. 331.
-
Gammarus marinus, i. 334.
-
Gannets, white only when mature, ii. 228.
-
Ganoidei, i. 204.
-
Ganoid fishes, i. 212.
-
Gaour, horns of the, ii. 247.
-
Gap between man and the apes, i. 200.
-
Gaper, sexes and young of, ii. 217.
-
Gardner, on an example of rationality in a Gelasimus, i. 334.
-
Garrulus glandarius, ii. 104.
-
Gärtner, on sterility of hybrid plants, i. 223.
-
Gasteropoda, i. 324;
-
pulmoniferous, courtship of, i. 324.
-
Gasterosteus, i. 271;
-
Gasterosteus leiurus, ii. 2, 14, 20.
-
Gasterosteus trachurus, ii. 2.
-
Gastrophora, wings of, brightly coloured beneath, i. 397.
-
Gauchos, want of humanity among the, i. 101.
-
Gaudry, M., on a fossil monkey, i. 197.
-
Gavia, seasonal change of plumage in, ii. 228.
-
Geese, clanging noise made by, ii. 51;
-
pairing of different species of, ii. 114;
-
Canada, selection of mates by, ii. 116.
-
Gegenbaur, C., on the number of digits in the Ichthyopterygia, i. 125;
-
on the hermaphroditism of the remote progenitors of the vertebrata, i. 207.
-
Gelasimus, use of the enlarged chela of the male, i. 331;
-
pugnacity of males of, i. 333;
-
proportions of the sexes in a species of, i. 315;
-
rational actions of a, i. 334;
-
difference of colour in the sexes of a species of, i. 336.
-
Gemmules, sexual selection of, i. 285.
-
Genesis, i. 318.
-
Genius, ii. 328;
-
Genius, fertility of men and women of, i. 171.
-
Geoffroy-Saint-Hilaire, Isid., on the recognition of women by male quadrumana, i. 13;
-
on the occurrence of a rudimentary tail in man, i. 29;
-
on monstrosities, i. 113;
-
on animal-like anomalies in the human structure, i. 125;
-
on the correlation of monstrosities, i. 130;
-
on the distribution of hair in man and monkeys, i. 149;
-
on the caudal vertebræ of monkeys, i. 150;
-
on correlated variability, i. 151;
-
on the classification of man, i. 186;
-
on the long hair on the heads of species of Semnopithecus, i. 192;
-
on the hair in monkeys, i. 194;
-
on the development of horns in female deer, ii. 244;
-
and F. Cuvier, on the mandrill, ii. 293;
-
on Hylobates, ii. 318, 320.
-
Geographical distribution, as evidence of specific distinctions in man, i. 218.
-
Geometræ, brightly coloured beneath, i. 397.
-
Geophagus, frontal protuberance of male, ii. 13, 20;
-
eggs hatched by the male, in the mouth or branchial cavity, ii. 200.
-
Georgia, change of colour in Germans settled in, i. 246.
-
Geotrupes, stridulation of, i. 380, 382.
-
Gerbe, M., on the nest-building of Crenilabrus massa and C. melops, ii. 19.
-
Gerland, Dr., on the prevalence of infanticide, i. 94; ii. 344, 364;
-
on the extinction of races, i. 237, 238.
-
Gervais, P., on the hairiness of the gorilla, i. 149;
-
on the mandrill, ii. 293.
-
Gesture-language, i. 232.
-
Ghost-moth, sexual difference of colour in the, i. 399, 402.
-
Gibb, Sir D., on differences of the voice in different races of men, ii. 330.
432
-
Gibbon, Hoolock, nose of, i. 192.
-
Gibbons, voice of, ii. 276.
-
Giraffe, mute, except in the rutting season, ii. 274;
-
its mode of using the horns, ii. 250.
-
Giraud-Teulon, on the cause of short sight, i. 118.
-
Glanders, communicable between man and the lower animals, i. 11.
-
Glands, odoriferous, in mammals, ii. 279, 281.
-
Glareola, double moult in, ii. 80.
-
Glomeris limbata, difference of colour in the sexes of, i. 340.
-
Glowworm, female, apterous, i. 255;
-
luminosity of the, i. 345.
-
Gnats, dances of, i. 349.
-
Gnu, sexual differences in the colour of the, ii. 289.
-
Goat, male, wild, falling on his horns, ii. 249;
-
male, odour emitted by, ii. 279;
-
male, wild, crest of the, ii. 282;
-
Berbura, mane, dewlap, &c., of the male, ii. 284;
-
Kemas, sexual difference in the colour of the, ii. 289.
-
Goats, sexual differences in the horns of, i. 283;
-
horns of, i. 289, ii. 246;
-
domestic, sexual differences of, late developed, i. 293;
-
beards of, ii. 282;
-
mode of fighting of, ii. 249, 250.
-
Goat-sucker, Virginian, pairing of the, ii. 49.
-
Gobies, nidification of, ii. 20.
-
God, want of the idea of, in some races of men, i. 65.
-
Godron, M., on variability, i. 112;
-
on difference of stature, i. 115;
-
on the want of connexion between climate and the colour of the skin, i. 241;
-
on the odour of the skin, i. 248;
-
on the colour of infants, ii. 318.
-
Goldfinch, ii. 56, 85;
-
proportion of the sexes in the, i. 307;
-
sexual differences of the beak in the, ii. 39;
-
courtship of the, ii. 95.
-
Goldfinch, North American, young of, ii. 216.
-
Gold-Fish, ii. 16.
-
Gomphus, proportions of the sexes in, i. 314;
-
difference in the sexes of, i. 362.
-
Gonepteryx Rhamni, i. 393;
-
sexual difference of colour in, i. 409.
-
Goodsir, Prof., on the affinity of the lancelet to the ascidians, i. 205.
-
Goosander, young of, ii. 189.
-
Goose, Antarctic, colours of the, ii. 228.
-
Goose, Canada, pairing with a Bernicle gander, ii. 114.
-
Goose, Chinese, knob on the beak of the, ii. 129.
-
Goose, Egyptian, ii. 46.
-
Goose, Sebastopol, plumage of, ii. 74.
-
Goose, Snow-, whiteness of the, ii. 228.
-
Goose, Spur-winged, ii. 46.
-
Gorilla, ii. 323;
-
semi-erect attitude of the, i. 142;
-
mastoid processes of the, i. 143;
-
direction of the hair on the arms of the, i. 192;
-
supposed evolution of the, i. 230;
-
polygamy of the, i. 266, ii. 361, 362;
-
voice of the, ii. 276;
-
cranium of, ii. 318;
-
fighting of male, ii. 324.
-
Gosse, P. H., on the pugnacity of the male Humming-birds, ii. 40.
-
Gosse, M., on the inheritance of artificial modifications of the skull, ii. 380.
-
Gould, B. A., on variation in the length of the legs in man, i. 108;
-
measurements of American soldiers, i. 114, 116;
-
on the proportions of the body and capacity of the lungs in different races of men, i. 216;
-
on the inferior vitality of mulattoes, i. 221.
-
Gould, J., on the arrival of male snipes before the females, i. 260;
-
on the numerical proportion of the sexes in birds, i. 306;
-
on Neomorpha, ii. 39;
-
on the species of Eustephanus, ii. 39;
-
on the Australian Musk-duck, ii. 39;
-
on the relative size of the sexes in Biziura lobata and Cincloramphus cruralis, ii. 43;
-
on Lobivanellus lobatus, ii. 48;
-
on the habits of Menura Alberti, ii. 56;
-
on the rarity of song in brilliant birds, ii. 58;
-
on Selasphorus platycercus, ii. 65;
-
on the Bower-birds, ii. 69, 102;
-
on the ornamental plumage of the Humming-birds, ii. 78;
-
on the moulting of the ptarmigan, ii. 83;
-
on the display of plumage by the male Humming-birds, ii. 86;
-
on the shyness of adorned male birds, ii. 97;
433
-
on the decoration of the bowers of Bower-birds, ii. 112;
-
on the decoration of their nests by Humming-birds, ii. 112;
-
on variation in the genus Cynanthus, ii. 125;
-
on the colour of the thighs in a male parrakeet, ii. 126;
-
on Urosticte Benjamini, ii. 151, 152;
-
on the nidification of the Orioles, ii. 168;
-
on obscurely-coloured birds building concealed nests, ii. 169;
-
on Trogons and Kingfishers, ii. 173;
-
on Australian parrots, ii. 174;
-
on Australian pigeons, ii. 175;
-
on the moulting of the ptarmigan, ii. 181;
-
on the immature plumage of birds, ii. 186 et seq.;
-
on the Australian species of Turnix, ii. 201;
-
on the young of Aithurus polytmus, ii. 220;
-
on the colours of the bills of Toucans, ii. 227;
-
on the relative size of the sexes in the Marsupials of Australia, ii. 260;
-
on the colours of the Marsupials, ii. 286.
-
Goureau, on the stridulation of Mutilla europæa, i. 366.
-
Gout, sexually transmitted, i. 292.
-
Graba, on the Pied Ravens of the Feroe Islands, ii. 126;
-
on the Bridled Guillemot, ii. 127.
-
Gradation of secondary sexual characters in birds, ii. 135.
-
Grallatores, absence of secondary sexual characters in, i. 270;
-
double moult in some, ii. 81.
-
Grallina, nidification of, ii. 169.
-
Grasshoppers, stridulation of the, i. 356.
-
Gratiolet, Prof., on the anthropomorphous apes, i. 196;
-
on the evolution of the anthropomorphous apes, i. 230.
-
Gray, Asa, on the gradation of species among the Compositæ, i. 227.
-
Gray, J. E., on the caudal vertebræ of monkeys, i. 150;
-
on the presence of rudiments of horns in the female of Cervulus moschatus, ii. 245;
-
on the horns of goats and sheep, ii. 246;
-
on the beard of the Ibex, ii. 283;
-
on the Berbura goat, ii. 285;
-
on sexual differences in the coloration of Rodents, ii. 286;
-
on the colours of the Elands, ii. 288;
-
on the Sing-sing antelope, ii. 289;
-
on the colours of goats, ii. 290;
-
on the Hog-deer, ii. 303.
-
“Greatest happiness principle,” i. 97, 98.
-
Greeks, ancient, i. 177.
-
Green, A. H., on beavers fighting, ii. 239;
-
on the voice of the beaver, ii. 277.
-
Greenfinch, selected by a female canary, ii. 115.
-
Greg, W. R., on the early marriages of the poor, i. 173;
-
on the Ancient Greeks, i. 178;
-
on the effects of natural selection on civilised nations, i. 167.
-
Grenadiers, Prussian, i. 112.
-
Grey, Sir G., on female infanticide in Australia, ii. 364.
-
Greyhounds, numerical proportion of the sexes in, i. 263, 265;
-
numerical proportion of male and female births in, i. 304.
-
Grouse, red, monogamous, i. 269;
-
pugnacity of young male, ii. 48;
-
producing a sound by scraping their wings upon the ground, ii. 61;
-
duration of courtship of, ii. 100;
-
colours and nidification of, ii. 170.
-
Grube, Dr., on the occurrence of the supra-condyloid foramen in the humerus of man, i. 28.
-
Grus americanus, age of mature plumage in, ii. 213;
-
breeding in immature plumage, ii. 214.
-
Grus virgo, trachea of, ii. 60.
-
Gryllus campestris, i. 353;
-
pugnacity of male, i. 360.
-
Gryllus domesticus, i. 354.
-
Grypus, sexual differences in the beak in, ii. 39.
-
Guanacoes, battles of, ii. 239;
-
canine teeth of, ii. 257.
-
Guanas, strife for women among the, ii. 324;
-
polyandry among the, ii. 366.
-
Guanche skeletons, occurrence of the supra-condyloid foramen in the humerus of, i. 29.
-
Guaranys, proportion of men and women among, i. 302;
-
colour of newborn children of the, ii. 318;
-
beards of the, ii. 322.
434
-
Guenée, A., on the sexes of Hyperythra, i. 310.
-
Guilding, L., on the stridulation of the LOCUSTIDÆ, i. 352.
-
Guillemot, variety of the, ii. 127.
-
Guinea, sheep of, with males only horned, i. 289.
-
Guinea-fowl, monogamous, i. 269;
-
occasional polygamy of the, i. 270;
-
markings of the, ii. 134.
-
Guinea-pigs, inheritance of the effects of operations by, ii. 380.
-
Gull, instance of reasoning in a, ii. 108.
-
Gulls, seasonal change of plumage in, ii. 228;
-
Günther, Dr., on hermaphroditism in Serranus, i. 208;
-
on male fishes hatching ova in their mouths, i. 210, ii. 20;
-
on mistaking infertile female fishes for males, i. 308;
-
on the prehensile organs of male Plagiostomous fishes, ii. 2;
-
on the pugnacity of the male salmon and trout, ii. 3;
-
on the relative size of the sexes in fishes, ii. 7;
-
on sexual differences in fishes, ii. 8 et seqq.;
-
on the genus Callionymus, ii. 9;
-
on a protective resemblance in a Pipe-fish, ii. 18;
-
on the genus Solenostoma, ii. 22;
-
on Megalophrys montana, ii. 26;
-
on the coloration of frogs and toads, ii. 26;
-
on sexual differences in the Ophidia, ii. 29;
-
on differences of the sexes of lizards, ii. 32 et seqq.
-
Gynanisa Isis, ocellated spots of, ii. 132.
-
Gypsies, uniformity of, in various parts of the world, i. 242.
-
H.
-
Habits, bad, facilitated by familiarity, i. 101;
-
variability of the force of, i. 183.
-
Häckel, E., on the origin of man, i. 4;
-
on rudimentary characters, i. 17;
-
on the canine teeth in man, i. 126;
-
on death caused by inflammation of the vermiform appendage, i. 28;
-
on the steps by which man became a biped, i. 142;
-
on man as a member of the Catarrhine group, i. 199;
-
on the position of the Lemuridæ, i. 202;
-
on the genealogy of the Mammalia, i. 203;
-
on the lancelet, i. 204;
-
on the transparency of pelagic animals, i. 323;
-
on the musical powers of women, ii. 337.
-
Hagen, H., and Walsh, B. D., on American neuroptera, i. 314.
-
Hair, development of, in man, i. 24;
-
character of, supposed to be determined by light and heat, i. 116;
-
distribution of, in man, i. 149, ii. 375;
-
possibly removed for ornamental purposes, i. 149;
-
arrangement and direction of, i. 192;
-
of the early progenitors of man, i. 206;
-
different texture of, in distinct races, i. 216;
-
and skin, correlation of colour of, i. 248;
-
development of, in mammals, ii. 281;
-
management of, among different peoples, ii. 340;
-
great length of, in some North American tribes, ii. 348;
-
elongation of the, on the human head, ii. 380.
-
Hairiness, difference of, in the sexes, in man, ii. 320;
-
variation of, in races of men, ii. 321.
-
Hairs and excretory pores, numerical relation of, in sheep, i. 248.
-
Hairy family, Siamese, ii. 378.
-
Hamadryas baboon, turning over stones, i. 75;
-
mane of the male, ii. 267.
-
Hamilton, C., on the cruelty of the Kafirs to animals, i. 94;
-
on the engrossment of the women by the Kafir chiefs, ii. 369.
-
Hammering, difficulty of, i. 138.
-
Hancock, A., on the colours of the nudibranch mollusca, i. 326.
-
Hands, larger at birth, in the children of labourers, i. 117;
-
structure of, in the quadrumana, i. 139;
-
and arms, freedom of, indirectly correlated with diminution of canines, i. 144.
-
Handwriting, inherited, i. 58.
-
Harcourt, E. Vernon, on Fringilla cannabina, ii. 86.
-
Harelda glacialis, ii. 122.
-
Hare, protective colouring of the, ii. 298.
435
Hares, battles of male, ii. 239.
-
Harlan, Dr., on the difference between field- and house-slaves, i. 246.
-
Harris, J. M., on the relation of complexion to climate, i. 245.
-
Harris, T. W., on the Katy-did locust, i. 353;
-
on the stridulation of the grasshoppers, i. 357;
-
on Œcanthus nivalis, i. 361;
-
on the colouring of Lepidoptera, i. 396;
-
on the colouring of Saturnia Io, i. 398.
-
Harry-long-legs, pugnacity of male, i. 349.
-
Hartman, Dr., on the singing of Cicada septendecim, i. 351.
-
Haughton, S., on a variation of the flexor pollicis longus in man, i. 129.
-
Hawks, feeding orphan nestling, ii. 107.
-
Hayes, Dr., on the diverging of sledge-dogs on thin ice, i. 46.
-
Head, altered position of, to suit the erect attitude of man, i. 143;
-
hairiness of, in man, i. 149;
-
processes of, in male beetles, i. 370;
-
artificial alterations of the form of the, ii. 351.
-
Hearne, on strife for women among the North American Indians, ii. 324;
-
on the North American Indians’ notion of female beauty, ii. 344;
-
repeated elopements of a North American woman, ii. 372.
-
Heart, in the human embryo, i. 16.
-
Heat, supposed effects of, i. 116.
-
Hectocotyle, i. 325.
-
Hedge-warbler, ii. 198;
-
Heel, small projection of, in the Aymara Indians, i. 120.
-
Hegt, M., on the development of the spurs in peacocks, i. 290.
-
Heliconidæ, i. 387;
-
mimickry of, by other butterflies, i. 411.
-
Heliopathes, stridulation peculiar to the male, i. 383.
-
Heliothrix auriculata, young of, ii. 188, 189.
-
Helix pomatia, example of individual attachment in, i. 325.
-
Hellins, J., proportions of sexes of Lepidoptera reared by, i. 313.
-
Helmholtz, on the vibration of the auditory hairs of crustacea, ii. 333.
-
Hemiptera, i. 349.
-
Hemitragus, beardless in both sexes, ii. 283.
-
Hepburn, Mr., on the autumn song of the water-ouzel, ii. 54.
-
Hepialus humuli, sexual difference of colour in the, i. 399, 402.
-
Herbs, poisonous, avoided by animals, i. 36.
-
Hermaphroditism of embryos, i. 207.
-
Herodias bubulcus, vernal moult of, ii. 84.
-
Heron, love-gestures of a, ii. 68.
-
Heron, Sir R., on the habits of peafowl, ii. 119, 120, 152.
-
Herons, decomposed feathers in, ii. 74;
-
breeding plumage of, ii. 82, 83;
-
young of the, ii. 208;
-
sometimes dimorphic, ii. 214;
-
continued growth of crest and plumes in the males of some, ii. 216;
-
change of colour in some, ii. 231.
-
Hetærina, difference in the sexes of, i. 362;
-
proportion of the sexes in, i. 314.
-
Heterocerus, stridulation of, i. 379.
-
Hewitt, Mr. on a game-cock killing a kite, ii. 44;
-
on the recognition of dogs and cats by ducks, ii. 110;
-
on the pairing of a wild duck with a pintail drake, ii. 115;
-
on the courtship of fowls, ii. 117;
-
on the coupling of pheasants with common hens, ii. 122.
-
Hindoo, his horror of breaking his caste, i. 99, 103.
-
Hindoos, local difference of stature among, i. 115;
-
difference of, from Europeans, i. 240;
-
colour of the beard in, ii. 319.
-
Hipparchia Janira, instability of the ocellated spots of, ii. 132.
-
Hipparchiæ, i. 387.
-
Hippocampus, development of, i. 210;
-
marsupial receptacles of the male, ii. 21.
-
Hippopotamus, nakedness of, i. 148.
-
Hips, proportions of, in soldiers and sailors, i. 117.
-
Hodgson, S., on the sense of duty, i. 71.
-
Hoffberg, on the horns of the reindeer, ii. 244;
436
-
on sexual preferences shown by reindeer, ii. 273.
-
Hog, wart-, ii. 265;
-
Hog-deer, ii. 303.
-
Holland, Sir H., on the effects of new diseases, i. 238.
-
Homologous structures, correlated variation of, i. 130.
-
Homoptera, i. 350;
-
stridulation of the, and orthoptera, discussed, i. 360.
-
Honduras, Quiscalus major in, i. 307.
-
Honey-buzzard of India, variation in the crest of, ii. 126.
-
Honey-suckers, moulting of the, ii. 83;
-
Australian, nidification of, ii. 169.
-
Honour, law of, i. 99.
-
Hooker, Jos., on the colour of the beard in man, ii. 319.
-
Hoolock Gibbon, nose of, i. 192.
-
Hoopoe, ii. 56;
-
sounds produced by the male, ii. 62.
-
Hoplopterus armatus, wing-spurs of, ii. 48.
-
Hornbill, African, inflation of the neck-wattle of the male during courtship, ii. 72.
-
Hornbills, sexual difference in the colour of the eyes in, ii. 129;
-
nidification and incubation of, ii. 169.
-
Horne, C., on the rejection of a brightly-coloured locust by lizards and birds, i. 361.
-
Horns, of deer, ii. 243, 248, 259;
-
and canine teeth, inverse development of, ii. 257;
-
sexual differences of, in sheep and goats, i. 283;
-
loss of, in female merino sheep, i. 284;
-
development of, in deer, i. 288;
-
development of, in antelopes, i. 289;
-
from the head and thorax, in male beetles, i. 370.
-
Horse, polygamous, i. 267;
-
canine teeth of male, ii. 241;
-
winter change of the, ii. 298;
-
fossil, extinction of the, in South America, i. 239.
-
Horses, dreaming, i. 46;
-
rapid increase of, in South America, i. 135;
-
diminution of canine teeth in, i. 144;
-
of the Falkland Islands and Pampas, i. 236;
-
numerical proportion of the sexes in, i. 263, 265;
-
lighter in winter in Siberia, i. 282;
-
sexual preferences in, ii. 272;
-
pairing preferentially the same colour, ii. 295;
-
numerical proportion of male and female births in, i. 303;
-
formerly striped, ii. 305.
-
Hottentot women, peculiarities of, i. 225.
-
Hottentots, lice of, i. 220;
-
readily become musicians, ii. 334;
-
notions of female beauty of the, ii. 345;
-
compression of nose by, ii. 352.
-
House-slaves, difference of, from field-slaves, i. 246.
-
Huber, P., on ants playing together, i. 39;
-
on memory in ants, i. 45;
-
on the intercommunication of ants, i. 58;
-
on the recognition of each other by ants after separation, i. 365.
-
Huc, on Chinese opinions of the appearance of Europeans, ii. 345.
-
Human kingdom, i. 186.
-
Human sacrifices, i. 68.
-
Humanity, unknown among some savages, i. 94;
-
deficiency of, among savages, i. 101.
-
Humboldt, A. von, on the rationality of mules, i. 48;
-
on a parrot preserving the language of a lost tribe, i. 236;
-
on the cosmetic arts of savages, ii. 339, 340;
-
on the exaggeration of natural characters by man, ii. 351;
-
on the red painting of American Indians, ii. 352.
-
Hume, D., on sympathetic feelings, i. 85.
-
Humming-bird, racket-shaped feathers in the tail of a, ii. 73;
-
display of plumage by the male, ii. 86.
-
Humming-birds, ornament their nests, i. 63, ii. 112;
-
polygamous, i. 269;
-
proportion of the sexes in, i. 307, ii. 221;
-
sexual differences in, ii. 39, 40, 151;
-
pugnacity of male, ii. 40;
-
modified primaries of male, ii. 65;
-
coloration of the sexes of, ii. 78;
-
young of, ii. 220;
-
nidification of the, ii. 168;
-
colours of female, ii. 168.
-
Humphreys, H. N., on the habits of the Stickle-back, i. 271, ii. 2.
-
Hunger, instinct of, i. 89.
437
-
Huns, ancient, flattening of the nose by the, ii. 352.
-
Hunter, J., on the number of species of man, i. 226;
-
on secondary sexual characters, i. 253;
-
on the general behaviour of female animals during courtship, i. 273;
-
on the muscles of the larynx in song-birds, ii. 55;
-
on the curled frontal hair of the Bull, ii. 282;
-
on the rejection of an ass by a female zebra, ii. 295.
-
Hunter, W. W., on the recent rapid increase of the Santali, i. 133;
-
Hussey, Mr., on a partridge distinguishing persons, ii. 110.
-
Hutchinson, Col., example of reasoning in a retriever, i. 48.
-
Hutton, Capt., on the male wild goat falling on his horns, ii. 249.
-
Huxley, T. H., on the structural agreement of man with the apes, i. 3;
-
on the agreement of the brain in man with that of lower animals, i. 10;
-
on the adult age of the Orang, i. 13;
-
on the embryonic development of man, i. 14;
-
on the origin of man, i. 4, 17;
-
on variation in the skulls of the natives of Australia, i. 108;
-
on the abductor of the fifth metatarsal in apes, i. 128;
-
on the position of man, i. 191;
-
on the sub-orders of primates, i. 195;
-
on the Lemuridæ, i. 202;
-
on the Dinosauria, i. 204;
-
on the amphibian affinities of the Ichthyosaurians, i. 204;
-
on variability of the skull in certain races of man, i. 226;
-
on the races of man, i. 229.
-
Hybrid birds, production of, ii. 113.
-
Hydrophobia communicable between man and the lower animals, i. 11.
-
Hydroporus, dimorphism of females of, i. 343.
-
Hyelaphus porcinus, ii. 303.
-
Hygrogonus, ii. 21.
-
Hyla, singing species of, ii. 27.
-
Hylobates, maternal affection in a, i. 40;
-
absence of the thumb in, i. 140;
-
upright progression of some species of, i. 143;
-
direction of the hair on the arms of species of, i. 192;
-
females of, less hairy below than males, ii. 320.
-
Hylobates agilis, i. 140;
-
hair on the arms of, i. 193;
-
musical voice of the, ii. 277;
-
superciliary ridge of, ii. 318;
-
voice of, ii. 332.
-
Hylobates hoolock, sexual difference of colour in, ii. 291.
-
Hylobates lar, i. 140;
-
hair on the arms of, i. 193.
-
Hylobates leuciscus, i. 140.
-
Hylobates syndactylus, i. 140;
-
laryngeal sac of, ii. 276.
-
Hymenoptera, i. 364;
-
large size of the cerebral ganglia in, i. 145;
-
classification of, i. 188;
-
sexual differences in the wings of, i. 345;
-
aculeate, relative size of the sexes of, i. 347.
-
Hymenopteron, parasitic, with a sedentary male, i. 272.
-
Hyomoschus aquaticus, ii. 304.
-
Hyperythra, proportion of the sexes in, i. 310.
-
Hypogymna dispar, sexual difference of colour in, i. 398.
-
Hypopyra, coloration of, i. 397.
-
I.
-
Ibex, male, falling on his horns, ii. 249;
-
Ibis, scarlet, young of the, ii. 208;
-
white, change of colour of naked skin in, during the breeding season, ii. 80.
-
Ibis tantalus, age of mature plumage in, ii. 213;
-
breeding in immature plumage, ii. 214, 215.
-
Ibises, decomposed feathers in, ii. 74;
-
white, ii. 228, and black, ii. 230.
-
Ichneumonidæ, difference of the sexes in, i. 365.
-
Ichthyopterygia, i. 125.
-
Ichthyosaurians, i. 204.
-
Ideas, general, i. 62.
-
Idiots, microcephalous, imitative faculties of, i. 57;
-
microcephalous, their characters and habits, i. 121.
-
Iguana tuberculata, ii. 32.
-
Iguanas, ii. 32.
-
Illegitimate and legitimate children, proportion of the sexes in, i. 302.
-
Imagination, existence of, in animals, i. 45.
-
Imitation, i. 39;
-
of man by monkeys, i. 44;
438
-
tendency to, in monkeys, microcephalous idiots and savages, i. 56;
-
influence of, i. 161.
-
Immature plumage of birds, ii. 183, 187.
-
Implacentata, i. 202.
-
Implements, employed by monkeys, i. 51;
-
fashioning of, peculiar to man, i. 52.
-
Impregnation, period of, influence of, upon sex, i. 303.
-
Improvement, progressive, man alone supposed to be capable of, i. 49.
-
Incisor teeth, knocked out or filed by some savages, ii. 340.
-
Increase, rate of, i. 131;
-
necessity of checks in, i. 135.
-
Indecency, hatred of, a modern virtue, i. 96.
-
India, difficulty of distinguishing the native races of, i. 215;
-
Cyprinidæ of, ii. 17;
-
colour of the beard in races of men of, ii. 319.
-
Indian, North American, honoured for scalping a man of another tribe, i. 93.
-
Individuality, i. 62.
-
Individuation, i. 318.
-
Indopicus carlotta, colours of the sexes of, ii. 175.
-
Infanticide, prevalence of, i. 94, 134;
-
supposed cause of, ii. 344;
-
prevalence and causes of, ii. 363 et seq.
-
Inferiority, supposed physical, of man, i. 156.
-
Inflammation of the bowels, occurrence of, in Cebus Azaræ, i. 12.
-
Inheritance, i. 110;
-
of effects of use of vocal and mental organs, i. 58;
-
of moral tendencies, i. 102, 104;
-
of long and short sight, i. 118;
-
laws of, i. 279;
-
sexual, i. 285;
-
sexually limited, ii. 154.
-
Inquisition, influence of the, i. 179.
-
Insanity, hereditary, i. 111.
-
Insect, fossil, from the Devonian, i. 360.
-
Insectivora, ii. 286;
-
absence of secondary sexual characters in, i. 268.
-
Insects, relative size of the cerebral ganglia in, i. 145;
-
male, appearance of, before the females, i. 260;
-
pursuit of female, by the males, i. 272;
-
period of development of sexual characters in, i. 291;
-
secondary sexual characters of, i. 341;
-
stridulation of, ii. 331.
-
Insessores, vocal organs of, ii. 55.
-
Instep, depth of, in soldiers and sailors, i. 117.
-
Instinct and intelligence, i. 37.
-
Instinct, migratory, vanquishing the maternal, i. 83, 90.
-
Instinctive actions, the result of inheritance, i. 80.
-
Instinctive impulses, difference of the force of, i. 87, 89;
-
and moral impulses, alliance of, i. 88.
-
Instincts, i. 36;
-
complex origin of, through natural selection, i. 38;
-
possible origin of some, i. 38;
-
acquired, of domestic animals, i. 79;
-
variability of the force of, i. 83;
-
difference of force between the social and other, i. 89, 104;
-
utilised for new purposes, ii. 335.
-
Instrumental music of birds, ii. 61, 66.
-
Intellect, influence of, in natural selection in civilised society, i. 171.
-
Intellectual faculties, their influence on natural selection in man, i. 158;
-
probably perfected through natural selection, i. 160.
-
Intelligence, Mr. H. Spencer on the dawn of, i. 37.
-
Intemperance, no reproach among savages, i. 96;
-
its destructiveness, i. 172.
-
Intoxication in monkeys, i. 12.
-
Iphias glaucippe, i. 394.
-
Iris, sexual difference in the colour of the, in birds, ii. 72, 128.
-
Ischio-pubic muscle, i. 127.
-
Ithaginis cruentus, number of spurs in, ii. 46.
-
Iulus, tarsal suckers of the males of, i. 340.
-
J.
-
Jackals learning to bark from dogs, i. 44.
-
Jack-snipe, coloration of the, ii. 226.
-
Jacquinot, on the number of species of man, i. 226.
439
-
Jaeger, Dr., on the difficulty of approaching herds of wild animals, i. 74;
-
on the increase of length in bones, i. 116;
-
on the deposition of a male Silver pheasant on account of spoiled plumage, ii. 120.
-
Jaguars, black, ii. 294.
-
Janson, E. W., on the proportions of the sexes in Tomicus villosus, i. 314;
-
on stridulant beetles, i. 379.
-
Japan, encouragement of licentiousness in, i. 134.
-
Japanese, general beardlessness of the, ii. 321;
-
aversion of the, to whiskers, ii. 349.
-
Jardine, Sir W., on the Argus pheasant, ii. 72, 97.
-
Jarrold, Dr., on modifications of the skull induced by unnatural position, i. 147.
-
Javanese, relative height of the sexes of, ii. 320;
-
notions of female beauty, ii. 347.
-
Jaw, influence of the muscles of the, upon the physiognomy of the apes, i. 144.
-
Jaws, smaller in the same ratio with the extremities, i. 117;
-
influence of food upon the size of, i. 118;
-
diminution of, in man, i. 144;
-
in man, reduced by correlation, ii. 325.
-
Jay, young of the, ii. 209;
-
Canada, young of the, ii. 209.
-
Jays, new mates found by, ii. 104;
-
distinguishing persons, ii. 110.
-
Jeffreys, J. Gwyn, on the form of the shell in the sexes of the Gasteropoda, i. 324;
-
on the influence of light upon the colours of shells, i. 326.
-
Jelly-fish, bright colours of some, i. 322.
-
Jenner, Dr., on the voice of the rook, ii. 61;
-
on the finding of new mates by magpies, ii. 103;
-
on retardation of the generative organs in birds, ii. 107.
-
Jenyns, L., on the desertion of their young by swallows, i. 84;
-
on male birds singing after the proper season, ii. 107.
-
Jerdon, Dr., on birds dreaming, ii. 46;
-
on the pugnacity of the male bulbul, ii. 41;
-
on the pugnacity of the male Ortygornis gularis, ii. 44;
-
on the spurs of Galloperdix, ii. 46;
-
on the habits of Lobivanellus, ii. 48;
-
on the spoonbill, ii. 60;
-
on the drumming of the Kalij pheasant, ii. 63;
-
on Indian bustards, ii. 65;
-
on Otis bengalensis, ii. 69;
-
on the ear-tufts of Sypheotides auritus, ii. 73;
-
on the double moults of certain birds, ii. 82;
-
on the moulting of the honey-suckers, ii. 83;
-
on the moulting of bustards, plovers, and drongos, ii. 84;
-
on display in male birds, ii. 86;
-
on the spring change of colour in some finches, ii. 86;
-
on the display of the under tail-coverts by the male bulbul, ii. 96;
-
on the Indian honey-buzzard, ii. 126;
-
on sexual differences in the colour of the eyes of hornbills, ii. 129;
-
on the markings of the Tragopan pheasant, ii. 134;
-
on the nidification of the Orioles, ii. 168;
-
on the nidification of the hornbills, ii. 169;
-
on the Sultan yellow-tit, ii. 174;
-
on Palæornis javanicus, ii. 180;
-
on the immature plumage of birds, ii. 186 et seq.;
-
on representative species of birds, ii. 190;
-
on the habits of Turnix, ii. 202;
-
on the continued increase of beauty of the peacock, ii. 216;
-
on coloration in the genus Palæornis, ii. 231.
-
Jevons, W. S., on the migrations of man, i. 135.
-
Jews, ancient, use of flint tools by the, i. 183;
-
uniformity of, in various parts of the world, i. 242;
-
numerical proportion of male and female births among the, i. 301;
-
ancient, tattooing practised by, ii. 339.
-
Johnstone, Lieut., on the Indian elephant, i. 268.
-
Jollofs, fine appearance of the, ii. 357.
-
Jones, Albert, proportion of sexes of Lepidoptera, reared by, i. 313.
-
Juan Fernandez, humming-birds of, ii. 221.
-
Junonia, sexual differences of colouring in species of, i. 389.
-
Jupiter, Greek statues of, ii. 350.
440
-
K.
-
Kafir skull, occurrence of the diastema in a, i. 126.
-
Kafirs, their cruelty to animals, i. 94;
-
lice of the, i. 220;
-
colour of the, ii. 347;
-
engrossment of the handsomest women by the chiefs of the, ii. 369;
-
marriage-customs of the, ii. 373.
-
Kalij pheasant, drumming of the male, ii. 62;
-
Kallima, resemblance of, to a withered leaf, i. 392.
-
Kalmucks, aversion of, to hairs on the face, ii. 349;
-
marriage-customs of the, ii. 373.
-
Kangaroo, great red, sexual difference in the colour of, ii. 286.
-
Kant, Imm., on duty, i. 70;
-
on self-restraint, i. 86;
-
on the number of species of man, i. 226.
-
Katy-did, stridulation of the, i. 352.
-
Keller, Dr., on the difficulty of fashioning stone implements, i. 138.
-
Kestrels, new mates found by, ii. 104.
-
Kidney, i. 116.
-
King, W. R., on the vocal organs of Tetrao cupido, ii. 56;
-
on the drumming of grouse, ii. 63;
-
on the reindeer, ii. 244;
-
on the attraction of male deer by the voice of the female, ii. 276.
-
King and Fitzroy, on the marriage-customs of the Fuegians, ii. 374.
-
King-crows, nidification of, ii. 167.
-
Kingfisher, ii. 56;
-
racket-shaped feathers in the tail of a, ii. 73.
-
Kingfishers, colours and nidification of the, ii. 171, 173, 176;
-
immature plumage of the, ii. 188, 190;
-
young of the, ii. 209.
-
King Lory, ii. 174;
-
immature plumage of the, ii. 188.
-
Kingsley, C., on the sounds produced by Umbrina, ii. 23.
-
Kirby and Spence, on the courtship of insects, i. 272;
-
on sexual differences in the length of the snout in curculionidæ, i. 255;
-
on the elytra of Dytiscus, i. 343;
-
on peculiarities in the legs of male insects, i. 344;
-
on the relative size of the sexes in insects, i. 345;
-
on the luminosity of insects, i. 345;
-
on the Fulgoridæ, i. 351;
-
on the habits of Termites, i. 364;
-
on difference of colour in the sexes of beetles, i. 367;
-
on the horns of the male lamellicorn beetles, i. 371;
-
on hornlike processes in male curculionidæ, i. 374;
-
on the pugnacity of the male stag-beetle, i. 375.
-
Kite, killed by a game-cock, ii. 44.
-
Knot, retention of winter plumage by the, ii. 82.
-
Knox, R., on the semilunar fold, i. 23;
-
on the occurrence of the supra-condyloid foramen in the humerus of man, i. 28;
-
on the features of the young Memnon, i. 217.
-
Koala, length of the cæcum in, i. 27.
-
Kölreuter, on the sterility of hybrid plants, i. 223.
-
Kobus ellipsiprymnus, proportion of the sexes in, i. 305.
-
Koodoo, development of the horns of the, i. 289;
-
markings of the, ii. 300.
-
Köppen, F. T., on the migratory locust, i. 352.
-
Kordofan, protuberances artificially produced in, ii. 339.
-
Kowalevsky, A., on the affinity of the Ascidia to the Vertebrata, i. 205.
-
Kowalevsky, W., on the pugnacity of the male Capercailzie, ii. 45;
-
on the pairing of the Capercailzie, ii. 49.
-
Krause, on a convoluted body at the extremity of the tail in a Macacus and a cat, i. 30.
-
Kuppfer, Prof., on the affinity of the Ascidia to the Vertebrata, i. 205.
-
L.
-
Labidocera Darwinii, prehensile organs of the male, i. 329.
-
Labrus, splendid colours of the species of, ii. 16.
-
Labrus mixtus, sexual differences in, ii. 9.
-
Labrus pavo, ii. 16.
-
Lacertilia, sexual differences of, ii. 32.
441
-
Lafresnaye, M. de, on Birds of Paradise, ii. 78.
-
Lamarck, on the origin of man, i. 4.
-
Lamellibranchiata, i. 324.
-
Lamellicorn beetles, hornlike processes from the head and thorax of, i. 370, 373;
-
analogy of, to Ruminants, i. 373;
-
influence of sexual selection on, i. 377.
-
Lamellicornia, stridulation of, i. 380.
-
Lamont, Mr., on the tusks of the Walrus, ii. 242;
-
on the use of its tusks by the Walrus, ii. 257.
-
Lampornis porphyrurus, colours of the female, ii. 168.
-
Lancelet, i. 204, 212.
-
Landois, H., on the production of sound by the Cicadæ, i. 351;
-
on the stridulating organ of the Crickets, i. 354;
-
on Decticus, i. 355;
-
on the stridulating organs of the Acridiidæ, i. 356;
-
on the presence of rudimentary stridulating organs in some female Orthoptera, i. 359;
-
on the stridulation of Necrophorus, i. 378;
-
on the stridulant organ of Cerambyx heros, i. 380;
-
on the stridulating organs in the Coleoptera, i. 382;
-
on the ticking of Anobium, i. 385;
-
on the stridulant organ of Geotrupes, i. 380.
-
Language an art, i. 55;
-
articulate, origin of, i. 56;
-
relation of the progress of, to the development of the brain, i. 57;
-
effects of inheritance in production of, i. 58;
-
complex structure of, among barbarous nations, i. 61;
-
natural selection in, i. 61;
-
gesture, i. 232;
-
primeval, i. 235;
-
of a lost tribe preserved by a parrot, i. 236.
-
Languages, presence of rudiments in, i. 60;
-
classification of, i. 60;
-
variability of, i. 60;
-
crossing or blending of, i. 60;
-
complexity of, no test of perfection or proof of special creation, i. 62;
-
resemblance of, evidence of community of origin, i. 189.
-
Languages and species, identity of evidence of their gradual development, i. 59.
-
Lanius, ii. 180;
-
characters of young, ii. 185.
-
Lanius rufus, anomalous young of, ii. 211.
-
Lankester, E. R., on comparative longevity, i. 168, 171;
-
on the destructive effects of intemperance, i. 173.
-
Lanugo, of the human fœtus, i. 25; ii. 375.
-
Lapponian language, highly artificial, i. 61.
-
Lark, proportion of the sexes in the, i. 307;
-
female, singing of the, ii. 54.
-
Larks, attracted by a mirror, ii. 112.
-
Lartet, E., on the size of the brain in mammals, i. 51;
-
comparison of cranial capacities of skulls of recent and tertiary mammals, i. 146;
-
on Dryopithecus, i. 199.
-
Larus, seasonal change of plumage in, ii. 228.
-
Larva, luminous, of a Brazilian beetle, i. 345.
-
Larynx, muscles of the, in song-birds, ii. 55.
-
Lasiocampa quercus, attraction of males by the female, i. 311;
-
sexual difference of colour in, i. 398.
-
Latham, R. G., on the migrations of man, i. 136.
-
Latooka, perforation of the lower lip by the women of, ii. 341.
-
Laurillard, on the abnormal division of the malar bone in man, i. 124.
-
Lawrence, W., on the superiority of savages to Europeans in power of sight, i. 118;
-
on the colour of negro infants, ii. 318;
-
on the fondness of savages for ornaments, ii. 338;
-
on beardless races, ii. 349;
-
on the beauty of the English aristocracy, ii. 357.
-
Layard, E. L., on an instance of rationality in a Cobra, ii. 30;
-
on the pugnacity of Gallus Stanleyi, ii. 44.
-
Laycock, Dr., on vital periodicity, i. 12.
-
Leaves, decaying, tints of, i. 323.
-
Lecky, Mr., on the sense of duty, i. 71;
-
on suicide, i. 94;
-
on the practice of celibacy, i. 96;
-
his view of the crimes of savages, i. 97;
-
on the gradual rise of morality, i. 103.
442
-
Leconte, J. L., on the stridulant organ in the Coprini and Dynastini, i. 381.
-
Lee, H., on the numerical proportion of the sexes in the trout, i. 308.
-
Leg, calf of the, artificially modified, ii. 340.
-
Legitimate and illegitimate children, proportion of the sexes in, i. 302.
-
Legs, variation of the length of the, in man, i. 108;
-
proportions of, in soldiers and sailors, i. 116;
-
fore, atrophied in some male butterflies, i. 344;
-
peculiarities of, in male insects, i. 344.
-
“Lek” of the black-cock and capercailzie, ii. 100.
-
Lemoine, Albert, on the origin of language, i. 56.
-
Lemur macaco, sexual difference of colour in, ii. 290.
-
Lemuridæ, i. 195;
-
their origin, i. 213;
-
position and derivation of the, i. 202;
-
ears of the, i. 23;
-
variability of the muscles in the, i. 128.
-
Lemurs, uterus in the, i. 123;
-
tailless species of, i. 194.
-
Leopards, black, ii. 294.
-
Lepidoptera, i. 386;
-
numerical proportions of the sexes in the, i. 309;
-
colouring of, i. 387;
-
ocellated spots of, ii. 132.
-
Lepidosiren, i. 204, 212.
-
Lenguas, disfigurement of the ears of the, ii. 341.
-
Leptorhynchus angustatus, pugnacity of male, i. 375.
-
Leptura testacea, difference of colour in the sexes of, i. 367.
-
Lequay, on the occurrence of the supra-condyloid foramen in the humerus of man, i. 29.
-
Leroy, on the wariness of young foxes in hunting-districts, i. 50;
-
on the desertion of their young by swallows, i. 84.
-
Lesse, valley of the, i. 29.
-
Lesson, on the Birds of Paradise, i. 269, ii. 98;
-
on the sea-elephant, ii. 278.
-
Lestis bombylans, difference of the sexes in, i. 366.
-
Lethrus cephalotes, pugnacity of the males of, i. 371, 376.
-
Leuckart, R., on the vesicula prostatica, i. 31;
-
on the influence of the age of parents on the sex of offspring, i. 302.
-
Levator claviculæ muscle, i. 128.
-
Libellula depressa, colour of the male, i. 363.
-
Libellulidæ, relative size of the sexes of, i. 347;
-
difference in the sexes of, i. 361.
-
Lice of domestic animals and man, i. 219.
-
Licentiousness, prevalence of, among savages, i. 96;
-
a check upon population, i. 134.
-
Lichtenstein, on Chera progne, ii. 120.
-
Life, inheritance at corresponding periods of, i. 280, 285.
-
Light, supposed effects of, i. 116;
-
influence of, upon the colours of shells, i. 326.
-
Lilford, Lord, the ruff attracted by bright objects, ii. 111.
-
Limosa lapponica, ii. 204.
-
Linaria, ii. 180.
-
Linaria montana, i. 307.
-
Linnæus, views of, as to the position of man, i. 190.
-
Linnet, numerical proportion of the sexes in the, i. 307;
-
crimson forehead and breast of the, ii. 86;
-
courtship of the, ii. 94.
-
Linyphia, i. 337.
-
Lion, polygamous, i. 268;
-
mane of the, defensive, ii. 266;
-
roaring of the, ii. 275.
-
Lions, stripes of young, ii. 183.
-
Lips, piercing of the, by savages, ii. 341.
-
Lithobius, prehensile appendages of the female, i. 340.
-
Lithosia, coloration in, i. 396.
-
Littorina littorea, i. 324.
-
Livingstone, Dr., on the influence of dampness and dryness on the colour of the skin, i. 242;
-
on the liability of negroes to tropical fevers after residence in a cold climate, i. 243;
-
on the spur-winged goose, ii. 47;
-
on weaver-birds, ii. 63;
-
on an African nightjar, ii. 73, 97;
-
on the battle-scars of South African male mammals, ii. 239;
443
-
on the removal of the upper incisors by the Batokas, ii. 340;
-
on the perforation of the upper lip by the Makalolo, ii. 342;
-
on the Banyai, ii. 347.
-
Livonia, numerical proportion of male and female births in, i. 301.
-
Lizards, relative size of the sexes of, ii. 32;
-
gular pouches of, ii. 33.
-
Lloyd, L., on the polygamy of the capercailzie and bustard, i. 269;
-
on the numerical proportion of the sexes in the capercailzie and blackcock, i. 306;
-
on the salmon, ii. 5;
-
on the colours of the sea-scorpion, ii. 9;
-
on the pugnacity of male grouse, ii. 45;
-
on the capercailzie and black-cock, ii. 49, 54;
-
on the call of the capercailzie, ii. 61;
-
on assemblages of grouse and snipes, ii. 101;
-
on the pairing of a shield-drake with a common duck, ii. 114;
-
on the battles of seals, ii. 240;
-
on the elk, ii. 249.
-
Lobivanellus, wing-spurs in, ii. 48.
-
Local influences, effect of, upon stature, i. 114.
-
Lockwood, Mr., on the development of Hippocampus, i. 210.
-
Locust, bright-coloured, rejected by lizards and birds, i. 361.
-
Locust, migratory, i. 352.
-
Locustidæ, stridulation of the, i. 352, 354;
-
Longicorn beetles, difference of the sexes of, in colour, i. 367;
-
Lonsdale, Mr., on an example of personal attachment in Helix pomatia, i. 325.
-
Lophobranchii, marsupial receptacles of the male, ii. 21.
-
Lophophorus, habits of, ii. 121.
-
Lophorina atra, sexual difference in coloration of, ii. 226.
-
Lophornis ornatus, ii. 76.
-
Lord, J. K., on Salmo lycaodon, ii. 5.
-
Lory, King, ii. 174;
-
immature plumage of the, ii. 188.
-
Love-antics and dances of birds, ii. 68.
-
Lowne, B. T., on Musca vomitoria, i. 145, 349.
-
Loxia, characters of young of, ii. 184.
-
Lubbock, Sir J., on the antiquity of man, i. 3;
-
on the origin of man, i. 4;
-
on the mental capacity of savages, i. 34;
-
on the origin of implements, i. 52;
-
on the simplification of languages, i. 62;
-
on the absence of the idea of God among certain races of men, i. 65;
-
on the origin of the belief in spiritual agencies, i. 66;
-
on superstitions, i. 69;
-
on the sense of duty, i. 71;
-
on the practice of burying the old and sick among the Fijians, i. 77;
-
non-prevalence of suicide among the lowest barbarians, i. 94;
-
on the immorality of savages, i. 97;
-
on Mr. Wallace’s claim to the origination of the idea of natural selection, i. 137;
-
on the absence of remorse among savages, i. 164;
-
on the former barbarism of civilised nations, i. 181;
-
on improvements in the arts among savages, i. 182;
-
on resemblances of the mental characters in different races of men, i. 232;
-
on the power of counting in primeval man, i. 234;
-
on the arts practised by savages, i. 234;
-
on the prehensile organs of the male Labidocera Darwinii, i. 329;
-
on Chloëon, i. 341;
-
on Smynthurus luteus, i. 348;
-
on strife for women among the North American Indians, ii. 324;
-
on music, ii. 334;
-
on the ornamental practices of savages, ii. 338;
-
on the estimation of the beard among the Anglo-Saxons, ii. 349;
-
on artificial deformation of the skull, ii. 352;
-
on “communal marriages,” ii. 358, 360;
-
on exogamy, ii. 360, 364;
-
on the Veddahs, ii. 363;
-
on polyandry, ii. 365.
-
Lucanidæ, variability of the mandibles in the male, i. 376.
-
Lucanus, large size of males of, i. 347.
-
Lucanus cervus, numerical proportion of sexes of, i. 313;
-
weapons of the male, i. 375.
-
Lucanus elaphus, use of mandibles of, i. 377;
-
large jaws of male, i. 342.
-
Lucas, Prosper, on sexual preference in horses and bulls, ii. 272.
-
Lunar periods, i. 212.
444
-
Lund, Dr., on skulls found in Brazilian caves, i. 218.
-
Lungs, enlargement of, in the Quechua and Aymara Indians, i. 119;
-
a modified swim-bladder, i. 207;
-
different capacity of in races of man, i. 216.
-
Luminosity in insects, i. 345.
-
Luschka, Prof., on the termination of the coccyx, i. 30.
-
Lust, instinct of, i. 89.
-
Luxury, comparatively innocuous, i. 171.
-
Lycæna, sexual differences of colouring in species of, i. 390.
-
Lyell, Sir C., on the antiquity of man, i. 3;
-
on the origin of man, i. 4;
-
on the parallelism of the development of species and languages, i. 59;
-
on the extinction of languages, i. 60;
-
on the Inquisition, i. 178;
-
on the fossil remains of vertebrata, i. 201;
-
on the fertility of mulattoes, i. 221.
-
Lynx, Canadian, throat-ruff of the, ii. 267.
-
Lyre-bird, assemblies of, ii. 101.
-
M.
-
Macacus, ears of, i. 23;
-
convoluted body in the extremity of the tail of, i. 30;
-
variability of the tail in species of, i. 150;
-
whiskers of species of, ii. 283.
-
Macacus cynomolgus, superciliary ridge of, ii. 318;
-
beard and whiskers of, becoming white with age, ii. 319.
-
Macacus inornatus, i. 151.
-
Macacus lasiotus, facial spots of, ii. 308.
-
Macacus radiatus, i. 192.
-
Macacus rhesus, sexual difference in the colour of, ii. 293, 310.
-
Macalister, Prof., on variations of the palmaris accessorius muscle, i. 109;
-
on muscular abnormalities in man, i. 128, 129;
-
on the greater variability of the muscles in men than in women, i. 275.
-
Macaws, Mr. Buxton’s observations on, i. 76;
-
McCann, J., on mental individuality, i. 63.
-
McClelland, J., on the Indian cyprinidæ, ii. 17.
-
Macculloch, Col., on an Indian village without any female children, ii. 364.
-
Macculloch, Dr., on tertian ague in a dog, i. 13.
-
Macgillivray, W., on the vocal organs of birds, i. 59;
-
on the Egyptian goose, ii. 48;
-
on the habits of woodpeckers, ii. 63;
-
on the habits of the snipe, ii. 64;
-
on the whitethroat, ii. 69;
-
on the moulting of the snipes, ii. 82;
-
on the moulting of the anatidæ, ii. 85;
-
on the finding of new mates by magpies, ii. 103;
-
on the pairing of a blackbird and thrush, ii. 113;
-
on pied ravens, ii. 126;
-
on the guillemots, ii. 127;
-
on the colours of the tits, ii. 174;
-
on the immature plumage of birds, ii. 186 et seqq.
-
Machetes, sexes and young of, ii. 216.
-
Machetes pugnax, numerical proportion of the sexes in, i. 306;
-
supposed to be polygamous, i. 270;
-
pugnacity of the male, ii. 41;
-
double moult in, ii. 81.
-
Mackintosh, on the moral sense, i. 70.
-
MacLachlan, R., on Apatania muliebris and Boreus hyemalis, i. 314;
-
on the anal appendages of male insects, i. 342;
-
on the pairing of dragon-flies, i. 347;
-
on dragon-flies, i. 362, 363;
-
on dimorphism in Agrion, i. 363;
-
on the want of pugnacity in male dragon-flies, i. 364;
-
on the ghost-moth in the Shetland Islands, i. 402.
-
McLennan, Mr., on the origin of the belief in spiritual agencies, i. 66;
-
on the prevalence of licentiousness among savages, i. 96, ii. 358;
-
on infanticide, i. 134, ii. 363;
-
on the primitive barbarism of civilised nations, i. 181;
-
on traces of the custom of the forcible capture of wives, i. 182, ii. 365;
-
on polyandry, ii. 365.
-
McNeill, Mr., on the use of the antlers of deer, ii. 252;
-
on the Scotch deerhound, ii. 261;
-
on the long hairs of the throat of the stag, ii. 268;
445
-
on the bellowing of stags, ii. 274.
-
Macrorhinus proboscideus, structure of the nose of, ii. 278.
-
Magpie, power of speech of, i. 59;
-
stealing bright objects, ii. 112;
-
nuptial assemblies of, ii. 102;
-
new mates found by, ii. 103;
-
young of the, ii. 209;
-
coloration of the, ii. 230.
-
Magpies, vocal organs of the, ii. 55.
-
Maillard, M., on the proportion of the sexes in a species of Papilio from Bourbon, i. 310.
-
Maine, Mr., on the absorption of one tribe by another, i. 159;
-
on the want of a desire for improvement, i. 166.
-
Makalolo, perforation of the upper lip by the, ii. 341.
-
Malar bone, abnormal division of, in man, i. 124.
-
Malay, Archipelago, marriage-customs of the savages of the, ii. 373.
-
Malays, line of separation between the Papuans and the, i. 218;
-
general beardlessness of the, ii. 321;
-
staining of the teeth among, ii. 339;
-
aversion of some, to hairs on the face, ii. 349.
-
Malays and Papuans, contrasted characters of, i. 216.
-
Male animals, struggles of, for the possession of the females, i. 259, 260;
-
eagerness of, in courtship, i. 272, 273;
-
generally more modified than female, i. 272, 275;
-
differ in the same way from females and young, i. 285.
-
Male characters, developed in females, i. 280;
-
transfer of, to female birds, ii. 193.
-
Male, sedentary, of a hymenopterous parasite, i. 272.
-
Malefactors, i. 172.
-
Males, presence of rudimentary female organs in, i. 208.
-
Males and females, comparative mortality of, while young, i. 264, 276;
-
comparative numbers of, i. 261, 263.
-
Malherbe, on the woodpeckers, ii. 174.
-
Malthus, T., on the rate of increase of population, i. 131, 132, 134.
-
Maluridæ, nidification of the, ii. 169.
-
Malurus, young of, ii. 216.
-
Mammæ, i. 254;
-
rudimentary, in male mammals, i. 17, 30, 208, 209, 210;
-
supernumerary, in women, i. 125;
-
of male human subject, i. 130.
-
Mammalia, Prof. Owen’s classification of, i. 187;
-
genealogy of the, i. 203.
-
Mammals, secondary sexual characters of, ii. 239;
-
weapons of, ii. 241;
-
recent and tertiary, comparison of cranial capacity of, i. 146;
-
relative size of the sexes of, ii. 260;
-
pursuit of female, by the males, i. 272;
-
parallelism of, with birds in secondary sexual characters, ii. 297;
-
voices of, used especially during the breeding season, ii. 331.
-
Man, variability of, i. 108;
-
erroneously regarded as more domesticated than other animals, i. 111;
-
definitive origin of, i. 235;
-
migrations of, i. 135;
-
wide distribution of, i. 137;
-
causes of the nakedness of, i. 149;
-
supposed physical inferiority of, i. 156;
-
numerical proportions of the sexes in, i. 264;
-
a member of the Catarrhine group, i. 198;
-
early progenitors of, i. 206;
-
secondary sexual characters of, ii. 316;
-
primeval condition of, ii. 367.
-
Mandans, correlation of colour and texture of hair in the, i. 248.
-
Mandible, left, enlarged in the male of Taphroderes distortus, i. 344.
-
Mandibles, use of the, in Ammophila, i. 342;
-
large, of Corydalis cornutus, i. 342;
-
large, of male Lucanus elaphus, i. 342.
-
Mandrill, number of caudal vertebræ in the, i. 150;
-
Mantegazza, Prof., on the ornaments of savages, ii. 338 et seqq.;
-
on the beardlessness of the New Zealanders, ii. 349;
-
on the exaggeration of natural characters by man, ii. 351.
-
Mantell, W., on the engrossment of pretty girls by the New Zealand chiefs, ii. 369.
446
-
Mantis, pugnacity of species of, i. 360.
-
Marcus Aurelius, on the origin of the moral sense, i. 71;
-
on the influence of habitual thoughts, i. 101.
-
Mareca penelope, ii. 114.
-
Marks, retained throughout groups of birds, ii. 131.
-
Marriage, influence of, upon morals, i. 96;
-
restraints upon, among savages, i. 133;
-
influence of, on mortality, i. 175;
-
development of, ii. 361.
-
Marriages, communal, ii. 358, 360;
-
Marshall, Mr., on the brain of a Bushwoman, i. 216.
-
Marsupials, i. 202;
-
possession of nipples by, i. 209;
-
their origin from Monotremata, i. 213;
-
uterus of, i. 122;
-
development of the nictitating membrane in, i. 23;
-
abdominal sacks of, i. 254;
-
relative size of the sexes of, ii. 260;
-
colours of, ii. 286.
-
Marsupium, rudimentary, in male marsupials, i. 208.
-
Martin, W. C. L., on alarm manifested by an orang at the sight of a turtle, i. 43;
-
on the hair in Hylobates, i. 194;
-
on a female American deer, ii. 258;
-
on the voice of Hylobates agilis, ii. 277;
-
on Semnopithecus nemæus, ii. 312.
-
Martin, on the beards of the inhabitants of St. Kilda, ii. 321.
-
Martins deserting their young, i. 84.
-
Martins, C., on death caused by inflammation of the vermiform appendage, i. 28.
-
Mastoid processes in man and apes, i. 143.
-
Maudsley, Dr., on the influence of the sense of smell in man, i. 24;
-
on Laura Bridgman, i. 58;
-
on the development of the vocal organs, i. 59.
-
Mayers, W. F., on the domestication of the goldfish in China, ii. 17.
-
Mayhew, E., on the affection between individuals of different sexes in the dog, ii. 270.
-
Maynard, C. J., on the sexes of Chrysemys picta, ii. 28.
-
Meckel, on correlated variation of the muscles of the arm and leg, i. 130.
-
Medicines, effect produced by, the same in man and in monkeys, i. 12.
-
Medusæ, bright colours of some, i. 322.
-
Megalithic structures, prevalence of, i. 233.
-
Megalophrys montana, sexual differences in, ii. 26, 27.
-
Megapicus validus, sexual difference of colour in, ii. 174.
-
Megasoma, large size of males of, i. 347.
-
Meigs, Dr. A., on variation in the skulls of the natives of America, i. 108.
-
Meinecke, on the numerical proportion of the sexes in butterflies, i. 309.
-
Meliphagidæ, Australian, nidification of, ii. 169.
-
Melita, secondary sexual characters of, i. 331.
-
Meloë, difference of colour in the sexes of a species of, i. 367.
-
Memory, manifestations of, in animals, i. 45.
-
Memnon, young, i. 217.
-
Mental characters, difference of, in different races of men, i. 216.
-
Mental faculties, variation of, in the same species, i. 36, 110;
-
diversity of, in the same race of men, i. 109;
-
inheritance of, i. 110;
-
similarity of the, in different races of man, i. 232;
-
of birds, ii. 108.
-
Mental powers, difference of, in the two sexes in man, ii. 326.
-
Menura Alberti, ii. 102;
-
Menura superba, ii. 101, 102;
-
long tails of both sexes of, ii. 164.
-
Merganser, trachea of the male, ii. 60.
-
Mergus cucullatus, speculum of, i. 291.
-
Mergus merganser, young of, ii. 189.
-
Merganser serrator, male plumage of, ii. 85.
-
Metallura, splendid tail-feathers of, ii. 152.
447
-
Methoca ichneumonides, large male of, i. 347.
-
Meves, M., on the drumming of the snipe, ii. 63.
-
Mexicans, civilisation of the, not foreign, i. 183.
-
Meyer, on a convoluted body at the extremity of the tail in a Macacus and a cat, i. 30.
-
Meyer, Dr. A., on the copulation of phryganidæ of distinct species, i. 342.
-
Migrations of man, effects of, i. 135.
-
Migratory instinct of birds, i. 79;
-
vanquishing the maternal, i. 83, 90.
-
Mill, J. S., on the origin of the moral sense, i. 71;
-
on the “greatest happiness principle,” i. 97;
-
on the difference of the mental powers in the sexes of man, ii. 328.
-
Millipedes, i. 339.
-
Milne-Edwards, H., on the use of the enlarged chela of the male Gelasimus, i. 331.
-
Milvago leucurus, sexes and young of, ii. 205.
-
Mimickry, i. 411.
-
Mimus polyglottus, ii. 109.
-
Mind, difference of, in man and the highest animals, i. 104;
-
similarity of the, in different races, i. 232.
-
Minnow, proportion of the sexes in the, i. 308, 309.
-
Minnows, spawning habits of, ii. 15.
-
Mirror, larks attracted by, ii. 112.
-
Mivart, St. George, on the reduction of organs, i. 18;
-
on the ears of the lemuroidea, i. 23;
-
on variability of the muscles in lemuroidea, i. 128, 136;
-
on the caudal vertebræ of monkeys, i. 150;
-
on the classification of the primates, i. 196;
-
on the orang and on man, i. 197;
-
on differences in the lemuroidea, i. 198;
-
on the crest of the male newt, ii. 24.
-
Mocking-thrush, partial migration of, ii. 109;
-
Modifications, unserviceable, i. 153.
-
Moles, numerical proportion of the sexes in, i. 305;
-
battles of male, ii. 239.
-
Mollienesia petenensis, sexual difference in, ii. 9.
-
Mollusca, beautiful colours and shapes of, i. 326;
-
absence of secondary sexual characters in the, i. 324.
-
Molluscoida, i. 205, 324.
-
Monacanthus scopas and M. Peronii, sexual differences in, ii. 12.
-
Mongolians, perfection of the senses in, i. 119.
-
Monkey, protecting his keeper from a baboon, i. 78, 87;
-
bonnet-, i. 192;
-
rhesus, sexual difference in colour of the, ii. 293, 310;
-
moustache-, colours of the, ii. 291.
-
Monkeys, liability of, to the same diseases as man, i. 11;
-
male, recognition of women by, i. 13;
-
revenge taken by, i. 40;
-
maternal affection in, i. 40;
-
variability of the faculty of attention in, i. 44;
-
using stones and sticks, i. 51;
-
imitative faculties of, i. 56;
-
signal-cries of, i. 57;
-
sentinels posted by, i. 74;
-
diversity of the mental faculties in, i. 110;
-
mutual kindnesses of, i. 75;
-
hands of the, i. 139, 140;
-
breaking hard fruits with stones, i. 140;
-
basal caudal vertebræ of, imbedded in the body, i. 151;
-
human characters of, i. 191;
-
gradation of species of, i. 227;
-
beards of, ii. 283;
-
ornamental characters of, ii. 306;
-
analogy of sexual differences of, with those of man, ii. 318;
-
different degrees of difference in the sexes of, ii. 323;
-
expression of emotions by, ii. 336;
-
generally monogamous habits of, ii. 361;
-
polygamous habits of some, ii. 362;
-
naked surfaces of, ii. 376;
-
American, manifestation of reason in, i. 47;
-
American, direction of the hair on the arms of some, i. 192.
-
Monogamy, not primitive, i. 182.
-
Monogenists, i. 228.
-
Mononychus pseudacori, stridulation of, i. 382.
-
Monotremata, i. 202;
-
development of the nictitating membrane in, i. 23;
-
lactiferous glands of, i. 209;
-
connecting mammals with reptiles, i. 213.
-
Monstrosities, analogous, in man and lower animals, i. 113;
-
caused by arrest of development, i. 121;
448
-
correlation of, i. 130;
-
transmission of, i. 224.
-
Montagu, G., on the habits of the black and red grouse, i. 269;
-
on the pugnacity of the ruff, ii. 41;
-
on the singing of birds, ii. 52;
-
on the double moult of the male pintail, ii. 84.
-
Monteiro, Mr., on Bucorax abyssinicus, ii. 72.
-
Montes de Oca, M., on the pugnacity of male Humming-birds, ii. 40.
-
Monticola cyanea, ii. 172.
-
Monuments, as traces of extinct tribes, i. 237.
-
Moose, battles of, ii. 240;
-
horns of the, an incumbrance, ii. 259.
-
Moral and instinctive impulses, alliance of, i. 88.
-
Moral faculties, their influence on natural selection in man, i. 158.
-
Moral rules, distinction between the higher and lower, i. 100.
-
Moral sense, origin of the, i. 102;
-
so-called, derived from the social instincts, i. 97, 98.
-
Moral tendencies, inheritance of, i. 102.
-
Morality, supposed to be founded in selfishness, i. 97;
-
test of, the general welfare of the community, i. 98;
-
gradual rise of, i. 103;
-
influence of a high standard of, i. 166.
-
Morgan, L. H., on the Beaver, i. 37;
-
on the reasoning powers of the Beaver, i. 46;
-
on the forcible capture of wives, i. 182;
-
on the castoreum of the beaver, ii. 279;
-
marriage unknown in primeval times, ii. 359;
-
on Polyandry, ii. 365.
-
Morris, F. O., on hawks feeding an orphan nestling, ii. 107.
-
Mortality, comparative, of females and males, i. 264, 276, 302.
-
Morton, on the number of species of man, i. 226.
-
Moschus moschiferus, odoriferous organs of, ii. 280.
-
Motacillæ, Indian, young of, ii. 190.
-
Moths, i. 394;
-
absence of mouth in some male, i. 254;
-
apterous female, i. 255;
-
male, prehensile use of the tarsi by, i. 256;
-
male, attracted by females, i. 311;
-
coloration of, i. 397;
-
sexual differences of colour in, i. 398.
-
Motmot, racket-shaped feathers in the tail of a, ii. 73.
-
Moult, double, ii. 181;
-
double annual, in birds, ii. 80.
-
Moulting of birds, ii. 214.
-
Moults, partial, ii. 83.
-
Moustache-monkey, colours of the, ii. 291, 311.
-
Moustaches, in monkeys, i. 192.
-
Mud-turtle, long claws of the male, ii. 28.
-
Mulattoes, persistent fertility of, i. 221;
-
immunity of, from yellow fever, i. 243.
-
Mule, sterility and strong vitality of the, i. 221.
-
Mules, rational, i. 48.
-
Müller, Ferd., on the Mexicans and Peruvians, i. 183.
-
Müller, Fritz, on astomatous males of Tanais, i. 255;
-
on the disappearance of spots and stripes in adult mammals, ii. 305;
-
on the proportions of the sexes in some crustacea, i. 315;
-
on secondary sexual characters in various crustaceans, i. 328 et seqq.;
-
on the luminous larva of a beetle, i. 345;
-
musical contest between male Cicadæ, i. 351;
-
on the sexual maturity of young amphipod crustacea, ii. 215.
-
Müller, J., on the nictitating membrane and semilunar fold, i. 23.
-
Müller, Max, on the origin of language, i. 56;
-
struggle for life among the words, &c., of languages, i. 60.
-
Müller, S., on the Banteng, ii. 290;
-
on the colours of Semnopithecus chrysomelas, ii. 291.
-
Muntjac-deer, weapons of the, ii. 257.
-
Murie, J., on the reduction of organs, i. 18;
-
on the ears of the Lemuroidea, i. 23;
-
on variability of the muscles in the Lemuroidea, i. 128, 136;
-
basal caudal vertebræ of Macacus inornatus imbedded in the body, i. 151;
-
on differences in the Lemuroidea, i. 198;
449
-
on the throat-pouch of the male Bustard, ii. 58;
-
on the mane of Otaria jubata, ii. 267;
-
on the suborbital pits of Ruminants, ii. 280;
-
on the colours of the sexes in Otaria nigrescens, ii. 287.
-
Murray, A., on the Pediculi of different races of men, i. 219.
-
Murray, T. A., on the fertility of Australian women with white men, i. 220.
-
Mus coninga, i. 50.
-
Mus minutus, sexual difference in the colour of, ii. 286.
-
Musca vomitoria, i. 145.
-
Muscicapa grisola, ii. 170.
-
Muscicapa luctuosa, ii. 170.
-
Muscicapa ruticilla, breeding in immature plumage, ii. 214.
-
Muscle, ischio-pubic, i. 127.
-
Muscles, rudimentary, occurrence of, in man, i. 19;
-
variability of the, i. 109;
-
effects of use and disuse upon, i. 116;
-
animal-like abnormalities of, in man, i. 127;
-
correlated variation of, in the arm and leg, i. 130;
-
variability of, in the hands and feet, i. 136;
-
of the jaws, influence of, on the physiognomy of the Apes, i. 144;
-
habitual spasms of, causing modifications of the facial bones, i. 147;
-
of the early progenitors of man, i. 206;
-
greater variability of the, in men than in women, i. 275.
-
Musculus sternalis, Prof. Turner on the, i. 19.
-
Music, i. 232;
-
of birds, ii. 51;
-
discordant, love of savages for, ii. 67;
-
different appreciation of, by different peoples, ii. 333;
-
origin of, ii. 333, 337;
-
effects of, ii. 335.
-
Musical cadences, perception of, by animals, ii. 333;
-
powers of man, ii. 330 et seqq.
-
Musk-deer, canine teeth of male, ii. 241, 256, 257;
-
male, odoriferous organs of the, ii. 280;
-
winter change of the, ii. 299.
-
Musk-duck, Australian, ii. 38;
-
large size of male, ii. 43;
-
of Guiana, pugnacity of the male, ii. 43.
-
Musk-ox, horns of, ii. 247.
-
Musk-rat, protective resemblance of the, to a clod of earth, ii. 298.
-
Musophagæ, colours and nidification of the, ii. 171;
-
both sexes of, equally brilliant, ii. 177.
-
Mussels opened by monkeys, i. 140.
-
Mustela, winter change of two species of, ii. 298.
-
Mutilations, healing of, i. 13.
-
Mutilla europæa, stridulation of, i. 366.
-
Mutillidæ, absence of ocelli in female, i. 341.
-
Mycetes caraya, polygamous, i. 266;
-
vocal organs of, ii. 277;
-
beard of, ii. 283;
-
sexual differences of colour in, ii. 290;
-
voice of, ii. 332.
-
Mycetes seniculus, sexual differences of colour in, ii. 290.
-
Myriapoda, i. 339.
-
N.
-
Nägeli, on the influence of natural selection on plants, i. 152;
-
on the gradation of species of plants, i. 227.
-
Nails, coloured yellow or purple in part of Africa, ii. 339.
-
Naples, greater proportion of female illegitimate children in, i. 301.
-
Narwhal, tusks of the, ii. 242, 248.
-
Nasal cavities, large size of, in American aborigines, i. 119.
-
Nascent organs, i. 18.
-
Nathusius, H. von, on the improved breeds of pigs, i. 230;
-
on the breeding of domestic animals, ii. 370.
-
Natural selection, its effects on the early progenitors of man, i. 136;
-
influence of, on man, i. 151, 154;
-
limitation of the principle, i. 152;
-
influence of, on social animals, i. 155;
-
Mr. Wallace on the limitation of, by the influence of the mental faculties in man, i. 158;
-
influence of, in the progress of the United States, i. 179.
-
Natural and sexual selection contrasted, i. 278.
-
Naulette, jaw from, large size of the canines in, i. 126.
-
Neanderthal skull, capacity of the, i. 146.
-
Neck, proportion of, in soldiers and sailors, i. 117.
450
-
Necrophorus, stridulation of, i. 378, 382.
-
Nectarinia, young of, ii. 190.
-
Nectariniæ, nidification of, ii. 169;
-
Negro, resemblance of a, to Europeans, in mental characters, i. 232.
-
Negro-women, their kindness to Mungo Park, i. 95.
-
Negroes, character of, i. 216;
-
lice of, i. 220;
-
blackness of, i. 224, ii. 381;
-
variability of, i. 225, 226;
-
immunity of, from yellow fever, i. 243;
-
difference of, from Americans, i. 247;
-
disfigurements of the, ii. 296;
-
colour of newborn children of, ii. 318;
-
comparative beardlessness of, ii. 321;
-
readily become musicians, ii. 334;
-
appreciation of beauty of their women by, ii. 344, 346;
-
idea of beauty among, ii. 350;
-
compression of the nose by some, ii. 352.
-
Neolithic period, i. 183.
-
Neomorpha, sexual difference of the beak in, ii. 39.
-
Nephila, i. 337.
-
Nests, made by fishes, ii. 19;
-
decoration of, by Humming-birds, ii. 112.
-
Neumeister, on a change of colour in pigeons after several moultings, i. 294.
-
Neuration, difference of, in the two sexes of some butterflies and hymenoptera, i. 345.
-
Neuroptera, i. 314, 361.
-
Neurothemis, dimorphism in, i. 363.
-
New Zealand, expectation by the natives of, of their extinction, i. 240;
-
practice of tattooing in, ii. 342;
-
aversion of natives of, to hairs on the face, ii. 349;
-
pretty girls engrossed by the chiefs in, ii. 369.
-
Newton, A., on the throat-pouch of the male bustard, ii. 58;
-
on the difference between the females of two species of Oxynotus, ii. 193;
-
on the habits of the phalarope, dotterel, and godwit, ii. 204.
-
Newts, ii. 24.
-
Nicholson, Dr., on the non-immunity of dark Europeans from yellow fever, i. 245.
-
Nictitating membrane, i. 23, 207.
-
Nidification, of fishes, ii. 19;
-
relation of, to colour, ii. 167, 172;
-
of British birds, ii. 169.
-
Night-heron, cries of the, ii. 51.
-
Nightingale, arrival of the male before the female, i. 259;
-
object of the song of the, ii. 52.
-
Nightingales, new mates found by, ii. 105.
-
Nightjar, selection of a mate by the female, ii. 116;
-
Australian, sexes of, ii. 206;
-
coloration of the, ii. 226.
-
Nightjars, noise made by some male, with their wings, ii. 62;
-
elongated feathers in, ii. 73, 97.
-
Nilghau, sexual differences of colour in the, ii. 287.
-
Nilsson, Prof., on the resemblance of stone arrow-heads from various places, i. 233;
-
on the development of the horns in the reindeer, i. 288.
-
Nipples, absence of, in Monotremata, i. 209.
-
Nitzsch, C. L., on the down of birds, ii. 80.
-
Noctuæ, brightly-coloured beneath, i. 397.
-
Noctuidæ, coloration of, i. 394.
-
Nordmann, A., on Tetrao urogalloides, ii. 100.
-
Nomadic habits, unfavourable to human progress, i. 167.
-
Norway, numerical proportion of male and female births in, i. 301.
-
Nose, resemblance of, in man and the apes, i. 192;
-
piercing and ornamentation of the, ii. 341;
-
flattening of the, ii. 352;
-
very flat, not admired in negroes, ii. 350.
-
Nott and Gliddon, on the features of Rameses II., i. 217;
-
on the features of Amunoph III., i. 218;
-
on skulls from Brazilian caves, i. 218;
-
on the immunity of negroes and mulattoes from yellow fever, i. 243;
-
on the deformation of the skull among American tribes, ii. 352.
-
Nudibranch mollusca, bright colours of, i. 326.
-
Numerals, Roman, i. 182.
-
Nunemaya, natives of, bearded, ii. 322, 349.
451
-
O.
-
Obedience, value of, i. 162.
-
Observation, powers of, possessed by birds, ii. 109.
-
Occupations, sometimes a cause of diminished stature, i. 115;
-
effect of, upon the proportions of the body, i. 116.
-
Ocelli, absence of, in female Mutillidæ, i. 341.
-
Ocelli of birds, formation and variability of the, ii. 132.
-
Ocelot, sexual differences in the colouring of the, ii. 287.
-
Ocyphaps lophotes, ii. 96.
-
Odonata, i. 314.
-
Odonestis potatoria, sexual difference of colour in, i. 398.
-
Odour, correlation of, with colour of skin, i. 248;
-
emitted by snakes in the breeding-season, ii. 30;
-
of mammals, ii. 278.
-
Œcanthus nivalis, difference of colour in the sexes of, i. 361.
-
Oidemia, ii. 226, 227.
-
Olivier, on sounds produced by Pimelia striata, i. 385.
-
Omaloplia brunnea, stridulation of, i. 381.
-
Onitis furcifer, processes of anterior femora of the male, and on the head and thorax of the female, i. 372.
-
Onthophagus, i. 370.
-
Onthophagus rangifer, sexual differences of, i. 369;
-
variation in the horns of the male, i. 370.
-
Ophidia, sexual differences of, ii. 29.
-
Opossum, wide range of, in America, i. 219.
-
Optic nerve, atrophy of the, caused by destruction of the eye, i. 116.
-
Orang-Outan, ii. 323;
-
Bischoff on the agreement of the brain of the, with that of man, i. 11;
-
adult age of the, i. 13;
-
ears of the, i. 21;
-
vermiform appendage of, i. 27;
-
platforms built by the, i. 36;
-
alarmed at the sight of a turtle, i. 43;
-
using a stick as a lever, i. 51;
-
using missiles, i. 52;
-
using the leaves of the Pandanus as a night covering, i. 53;
-
hands of the, i. 139;
-
absence of mastoid processes in the, i. 143;
-
direction of the hair on the arms of the, i. 192;
-
its aberrant characters, i. 197;
-
supposed evolution of the, i. 230;
-
voice of the, ii. 276;
-
monogamous habits of the, ii. 361;
-
male, beard of the, ii. 284.
-
Oranges, treatment of, by monkeys, i. 139.
-
Orange-tip butterfly, i. 388, 393, 394.
-
Orchestia Darwinii, dimorphism of males of, i. 332.
-
Orchestia Tucuratinga, limbs of, i. 330, 331, 337.
-
Ordeal, i. 68.
-
Oreas canna, colours of, ii. 288.
-
Oreas Derbyanus, colours of, ii. 288, 299.
-
Organs, prehensile, i. 256;
-
utilised for new purposes, ii. 335.
-
Organic scale, von Baer’s definition of progress in, i. 211.
-
Orioles, nidification of, ii. 167.
-
Oriolus, species of, breeding in immature plumage, ii. 214, 215.
-
Oriolus melanocephalus, coloration of the sexes in, ii. 178.
-
Ornaments, prevalence of similar, i. 233;
-
fondness of savages for, ii. 338;
-
of male birds, ii. 50.
-
Ornamental characters, equal transmission of, to both sexes, in mammals, ii. 297;
-
Ornithoptera crœsus, i. 310.
-
Ornithorhynchus, i. 200;
-
spur of the male, ii. 242;
-
reptilian tendency of, i. 204.
-
Orocetes erythrogastra, young of, ii. 219.
-
Orrony, Grotto of, i. 28.
-
Orsodacna atra, difference of colour in the sexes of, i. 368.
-
Orthoptera, i. 352;
-
metamorphosis of, i. 292;
-
stridulating, auditory apparatus of, i. 353;
-
colours of, i. 360;
-
rudimentary stridulating organs in female, i. 359;
-
stridulation of the, and Homoptera, discussed, i. 360.
-
Ortygornis gularis, pugnacity of the male, ii. 44.
-
Oryctes, stridulation of, i. 381;
-
sexual differences in the stridulant organs of, i. 383.
452
-
Oryx leucoryx, use of the horns of, ii. 251, 263.
-
Osphranter rufus, sexual difference in the colour of, ii. 286.
-
Ostrich, African, sexes and incubation of the, ii. 205.
-
Ostriches, stripes of young, ii. 184.
-
Otaria jubata, mane of the male, ii. 267.
-
Otaria nigrescens, difference in the coloration of the sexes of, ii. 287.
-
Otis bengalensis, love-antics of the male, ii. 68.
-
Otis tarda, polygamous, i. 269;
-
throat-pouch of the male, ii. 58.
-
Ouzel, ring-, colours and nidification of the, ii. 179.
-
Ouzel, water-, colours and nidification of the, ii. 170.
-
Ovibos moschatus, horns of, ii. 247.
-
Ovipositor of insects, i. 254.
-
Ovis cycloceros, mode of fighting of, ii. 249.
-
Ovule of man, i. 14.
-
Owen, Prof., on the Corpora Wolffiana, i. 16;
-
on the great toe in man, i. 16;
-
on the nictitating membrane and semilunar fold, i. 23;
-
on the development of the posterior molars in different races of man, i. 26;
-
on the length of the cæcum in the Koala, i. 27;
-
on the coccygeal vertebræ, i. 29;
-
on rudimentary structures belonging to the reproductive system, i. 31;
-
on abnormal conditions of the human uterus, i. 123;
-
on the number of digits in the Ichthyopterygia, i. 125;
-
on the canine teeth in man, i. 126;
-
on the walking of the chimpanzee and orang, i. 139;
-
on the mastoid processes in the higher apes, i. 143;
-
on the hairiness of elephants in elevated districts, i. 149;
-
on the caudal vertebræ of monkeys, i. 150;
-
classification of mammalia, i. 187;
-
on the hair in monkeys, i. 194;
-
on the piscine affinities of the Ichthyosaurians, i. 204;
-
on polygamy and monogamy among the antelopes, i. 267;
-
on the horns of Antilocapra americana, i. 289;
-
on the musky odour of crocodiles during the breeding season, ii. 29;
-
on the scent-glands of snakes, ii. 30;
-
on the Dugong, Cachalot and Ornithorhynchus, ii. 242;
-
on the antlers of the red deer, ii. 252;
-
on the dentition of the camelidæ, ii. 257;
-
on the tusks of the Mammoth, ii. 258;
-
on the horns of the Irish elk, ii. 259;
-
on the voice in the giraffe, porcupine, and stag, ii. 274;
-
on the laryngeal sac of the gorilla and orang, ii. 276;
-
on the odoriferous glands of mammals, ii. 279, 280;
-
on the effects of emasculation on the vocal organs of men, ii. 330;
-
on the voice of Hylobates agilis, ii. 332;
-
on American monogamous monkeys, ii. 362.
-
Owls, white, new mates found by, ii. 105.
-
Oxynotus, difference of the females of two species of, ii. 193.
-
P.
-
Pachydermata, i. 268.
-
Paget, on the abnormal development of hairs in man, i. 25;
-
on the thickness of the skin on the soles of the feet of infants, i. 118.
-
Painting, i. 232.
-
Palæmon, chelæ of a species of, i. 331.
-
Palæornis, sexual differences of colour in, ii. 231.
-
Palæornis Javanicus, colour of beak of, ii. 179.
-
Palæornis rosa, young of, ii. 188.
-
Palamedea cornuta, spurs on the wings, ii. 47.
-
Paleolithic period, i. 183.
-
Palestine, habits of the chaffinch in, i. 307.
-
Pallas, on the perfection of the senses in the Mongolians, i. 119;
-
on the want of connexion between climate and the colour of the skin, i. 241;
-
on the polygamous habits of Antilope Saiga, i. 267;
-
on the lighter colour of horses and cattle in winter in Siberia, i. 282;
-
on the tusks of the musk-deer, ii. 256, 258;
-
on the odoriferous glands of mammals, ii. 279;
-
on the odoriferous glands of the musk-deer, ii. 280;
453
-
on winter changes of colour in mammals, ii. 298;
-
on the ideal of female beauty in North China, ii. 344.
-
Palmaris accessorius muscle, variations of the, i. 109.
-
Pampas, horses of the, i. 236.
-
Pangenesis, hypothesis of, i. 280, 284.
-
Panniculus carnosus, i. 19.
-
Papilio, sexual differences of colouring in species of, i. 389;
-
proportion of the sexes in North American species of, i. 309;
-
coloration of the wings in species of, i. 396.
-
Papilio ascanius, i. 389.
-
Papilio Sesostris and Childrenæ, variability of, i. 402.
-
Papilio Turnus, i. 310.
-
Papilionidæ, variability in the, i. 402.
-
Papuans, line of separation between the, and the Malays, i. 218;
-
beards of the, ii. 322;
-
hair of, ii. 340.
-
Papuans and Malays, contrast in characters of, i. 216.
-
Paradise, Birds of, ii. 100, 181;
-
supposed by Lesson to be polygamous, i. 260;
-
rattling of their quills by, ii. 61;
-
racket-shaped feathers in, ii. 73;
-
sexual differences in colour of, ii. 76;
-
decomposed feathers in, ii. 74, 97;
-
display of plumage by the male, ii. 88.
-
Paradisea apoda, barbless feathers in the tail of, ii. 74;
-
plumage of, ii. 78;
-
and P. papuana, divergence of the females of, ii. 192.
-
Paradisea rubra, ii. 75, 78.
-
Paraguay, Indians of, eradication of eyebrows and eyelashes by, ii. 348.
-
Parrakeet, Australian, variation in the colour of the thighs of a male, ii. 126.
-
Parallelism of development of species and languages, i. 59.
-
Parasites on man and animals, i. 12;
-
as evidence of specific identity or distinctness, i. 219;
-
immunity from, correlated with colour, i. 242.
-
Parental affection, partly a result of natural selection, i. 81.
-
Parents, age of, influence upon sex of offspring, i. 302.
-
Parinæ, sexual difference of colour in, ii. 174.
-
Park, Mungo, negro-women teaching their children to love the truth, i. 95;
-
his treatment by the negro-women, i. 95, 326;
-
on negro opinions of the appearance of white men, ii. 346.
-
Parrot, racket-shaped feathers in the tail of a, ii. 73;
-
instance of benevolence in a, ii. 109.
-
Parrots, imitative faculties of, i. 44;
-
change of colour in, i. 152;
-
living in triplets, ii. 106;
-
affection of, ii. 108;
-
colours of, ii. 223;
-
sexual differences of colour in, ii. 231;
-
colours and nidification of the, ii. 171, 174, 176;
-
immature plumage of the, ii. 188;
-
musical powers of, ii. 335.
-
Parthenogenesis in the Tenthredinæ, i. 314;
-
in Cynipidæ, i. 314;
-
in crustacea, i. 315.
-
Partridge, monogamous, i. 269;
-
proportion of the sexes in the, i. 306;
-
female, ii. 194.
-
“Partridge-dances,” ii. 68.
-
Partridges, living in triplets, ii. 106;
-
spring coveys of male, ii. 107;
-
distinguishing persons, ii. 110.
-
Parus cæruleus, ii. 174.
-
Passer, sexes and young of, ii. 212.
-
Passer brachydactylus, ii. 212.
-
Passer domesticus, ii. 170, 212.
-
Passer montanus, ii. 170, 212.
-
Patagonians, self-sacrifice by, i. 88.
-
Patterson, Mr., on the AGRIONIDÆ, i. 362.
-
Paulistas of Brazil, i. 225.
-
Pavo cristatus, i. 290; ii. 136.
-
Pavo muticus, i. 290, ii. 136;
-
possession of spurs by the female, ii. 46, 162.
-
Pavo nigripennis, ii. 120.
-
Payaguas Indians, thin legs and thick arms of the, i. 117.
-
Payan, Mr., on the proportion of the sexes in sheep, i. 305.
-
Peacock, polygamous, i. 269;
-
sexual characters of, i. 290;
-
pugnacity of the, ii. 46;
-
rattling of the quills by, ii. 61;
-
elongated tail-coverts of the, ii. 72, 97;
-
love of display of the, ii. 68, 87, 135;
-
ocellated spots of the, ii. 135;
454
-
inconvenience of long tail of the, to the female, ii. 154, 164, 165;
-
continued increase of beauty of the, ii. 216.
-
Peacock-butterfly, i. 392.
-
Peafowl, preference of females for a particular male, ii. 120;
-
first advances made by the female, ii. 120.
-
Pediculi of domestic animals and man, i. 219.
-
Pedigree of man, i. 213.
-
Pedionomus torquatus, sexes of, ii. 201.
-
Peewit, wing-tubercles of the male, ii. 48.
-
Pelagic animals, transparency of, i. 323.
-
Pelecanus erythrorhynchus, horny crest on the beak of the male, during the breeding season, ii. 80.
-
Pelecanus onocrotalus, spring plumage of, ii. 85.
-
Pelelé, ii. 341.
-
Pelican, blind, fed by his companions, i. 77;
-
young, guided by old birds, i. 77;
-
pugnacity of the male, ii. 43.
-
Pelicans, fishing in concert, i. 75.
-
Pelobius Hermanni, stridulation of, i. 380, 382.
-
Pelvis, alteration of, to suit the erect attitude of man, i. 143;
-
differences of the, in the sexes in man, ii. 317.
-
Penelope nigra, sound produced by the male, ii. 64.
-
Pennant, on the battles of seals, ii. 240;
-
on the bladder-nose seal, ii. 278.
-
Penthe, antennal cushions of the male, i. 343.
-
Perch, brightness of male, during breeding season, ii. 13.
-
Peregrine Falcon, new mate found by, ii. 104.
-
Period of variability, relation of, to sexual selection, i. 296.
-
Periodicity, vital, Dr. Laycock on, i. 12.
-
Periods, lunar, followed by functions in man and animals, i. 12, 212.
-
Periods of life, inheritance at corresponding, i. 280, 285.
-
Perisoreus canadensis, young of, ii. 209.
-
Peritrichia, difference of colour in the sexes of a species of, i. 367.
-
Periwinkle, i. 324.
-
Pernis cristata, ii. 126.
-
Perseverance, a characteristic of man, ii. 328.
-
Persians, said to be improved by intermixture with Georgians and Circassians, ii. 357.
-
Personnat, M., on Bombyx Yamamai, i. 310.
-
Peruvians, civilisation of the, not foreign, i. 183.
-
Petrels, colours of, ii. 230.
-
Petrocincla cyanea, young of, ii. 219.
-
Petronia, ii. 212.
-
Pfeiffer, Ida, on Javanese ideas of beauty, ii. 347.
-
Phacochœrus æthiopicus, tusks and pads of, ii. 265.
-
Phalanger, Vulpine, black varieties of the, ii. 294.
-
Phalaropus fulicarius, ii. 203.
-
Phalaropus hyperboreus, ii. 203.
-
Phanæus, i. 373.
-
Phanæus carnifex, variation of the horns of the male, i. 370.
-
Phanæus faunus, sexual differences of, i. 369.
-
Phanæus lancifer, i. 370.
-
Phasgonura viridissima, stridulation of, i. 354, 356.
-
Phasianus Sœmmerringii, ii. 157.
-
Phasianus versicolor, ii. 89.
-
Phasianus Wallichii, ii. 93, 196.
-
Phasmidæ, mimickry of leaves by the, i. 414.
-
Pheasant, polygamous, i. 269;
-
production of hybrids with the common fowl, ii. 122;
-
and black grouse, hybrids of, ii. 113;
-
immature plumage of the, ii. 188.
-
Pheasant, Argus, ii. 72, 181;
-
display of plumage by the male, ii. 91;
-
ocellated spots of the, ii. 134, 141;
-
gradation of characters in the, ii. 141.
-
Pheasant, Blood-, ii. 46.
-
Pheasant, Cheer, ii. 93, 195.
-
Pheasant, Eared, i. 290, ii. 93, 195;
-
sexes alike in the, ii. 178;
-
length of the tail in the, ii. 166.
455
-
Pheasant, Golden, display of plumage by the male, ii. 89;
-
sex of young, ascertained by pulling out head-feathers, ii. 214;
-
age of mature plumage in the, ii. 213.
-
Pheasant, Kalij, drumming of the male, ii. 62.
-
Pheasant, Reeve’s, length of the tail in, ii. 166.
-
Pheasant, Silver, sexual coloration of the, ii. 228;
-
triumphant male, deposed on account of spoiled plumage, ii. 120.
-
Pheasant, Sœmmerring’s, ii. 156, 166.
-
Pheasant, Tragopan, ii. 72;
-
display of plumage by the male, ii. 91;
-
markings of the sexes of the, ii. 134.
-
Pheasants, period of acquisition of male characters in the family of the, i. 290;
-
proportion of sexes in chicks of, i. 306;
-
length of the tail in, ii. 156, 164, 166.
-
Philodromus, i. 337.
-
Philters, worn by women, ii. 344.
-
Phoca grœnlandica, sexual difference in the coloration of, ii. 287.
-
Phœnicura ruticilla, ii. 105.
-
Phosphorescence of insects, i. 345.
-
Phryganidæ, copulation of distinct species of, i. 342.
-
Phryniscus nigricans, ii. 25.
-
Physical inferiority, supposed, of man, i. 156.
-
Pickering, on the number of species of man, i. 226.
-
Picton, J. A., on the soul of man, ii. 395.
-
Picus auratus, ii. 43.
-
Pieridæ, mimickry by female, i. 413.
-
Pieris, i. 393.
-
Pigeon, carrier, late development of the wattle in, i. 293;
-
domestic, breeds and sub-breeds of, ii. 178;
-
pouter, late development of the crop in, i. 293;
-
female, deserting a weakened mate, i. 262.
-
Pigeons, nestling, fed by the secretion of the crop of both parents, i. 210;
-
changes of plumage in, i. 281;
-
transmission of sexual peculiarities in, i. 283;
-
changing colour after several moultings, i. 294;
-
numerical proportion of the sexes in, i. 306;
-
cooing of, ii. 60;
-
variations in plumage of, ii. 74;
-
display of plumage by male, ii. 96;
-
local memory of, ii. 109;
-
antipathy of female, to certain males, ii. 118;
-
pairing of, ii. 118, 119;
-
profligate male and female, ii. 119;
-
wing-bars and tail-feathers of, ii. 131;
-
supposititious breed of, ii. 155;
-
pouter and carrier, peculiarities of predominant in males, ii. 158;
-
nidification of, ii. 168;
-
immature plumage of the, ii. 188;
-
Australian, ii. 175;
-
Belgian, with black-streaked males, i. 285, 293; ii. 157.
-
Pigs, origin of the improved breeds of, i. 230;
-
numerical proportion of the sexes in, i. 305;
-
stripes of young, ii. 184, 303;
-
sexual preference shown by, ii. 273.
-
Pike, American, brilliant colours of the male, during the breeding season, ii. 14.
-
Pike, male, devoured by females, i. 308.
-
Pike, L. O., on the psychical elements of religion, i. 68.
-
Pimelia striata, sounds produced by the female, i. 385.
-
Pintail Drake, plumage of, ii. 84;
-
pairing with a wild duck, ii. 115.
-
Pintail Duck, pairing with a Wigeon, ii. 114.
-
Pipe-fish, filamentous, ii. 18;
-
marsupial receptacles of the male, ii. 21.
-
Pipits, moulting of the, ii. 83.
-
Pipra, modified secondary wing-feathers of male, ii. 65.
-
Pipra deliciosa, ii. 65, 66.
-
Pirates stridulus, stridulation of, i. 350.
-
Pithecia leucocephala, sexual differences of colour in, ii. 290.
-
Pithecia Satanas, beard of, ii. 283, 284, 285;
-
resemblance of, to a negro, ii. 381.
-
Pits, suborbital, of Ruminants, ii. 280.
-
Pittidæ, nidification of, ii. 167.
-
Placentata, i. 202.
-
Plagiostomous fishes, ii. 1.
-
Plain-wanderer, Australian, ii. 201.
456
-
Planariæ, bright colours of some, i. 322.
-
Plantain-eaters, colours and nidification of the, ii. 171;
-
both sexes of, equally brilliant, ii. 177.
-
Plants, cultivated, more fertile than wild, i. 132;
-
Nägeli, on natural selection in, i. 152;
-
male flowers of, mature before the female, i. 260;
-
phenomena of fertilisation in, i. 273;
-
relation between number and size of seeds in, i. 317.
-
Platalea, ii. 60;
-
change of plumage in, ii. 179.
-
Platyblemnus, i. 361.
-
Platycercus, young of, ii. 209.
-
Platyphyllum concavum, i. 352, 356.
-
Platyrrhine monkeys, i. 196.
-
Platysma myoides, i. 19.
-
Plecostomus, head-tentacles of the male of a species of, ii. 10.
-
Plecostomus barbatus, peculiar beard of the male, ii. 10.
-
Plectropterus gambensis, spurred wings of, ii. 46.
-
Ploceus, ii. 54.
-
Plovers, wing-spurs of, ii. 48;
-
Plumage, changes of, inheritance of, by fowls, i. 281;
-
tendency to analogous variation in, ii. 74;
-
display of, by male birds, ii. 86, 96;
-
changes of, in relation to season, ii. 180;
-
immature, of birds, ii. 183, 187;
-
colour of, in relation to protection, ii. 223.
-
Plumes on the head in birds, difference of, in the sexes, ii. 164.
-
Pneumora, structure of, i. 357.
-
Podica, sexual difference in the colour of the irides of, ii. 128.
-
Poeppig, on the contact of civilised and savage races, i. 239.
-
Poison, avoidance of, by animals, i. 49.
-
Poisonous fruits and herbs avoided by animals, i. 36.
-
Poisons, immunity from, correlated with colour, i. 242.
-
Polish fowls, origin of the crest in, i. 284.
-
Pollen and van Dam, on the colours of Lemur macaco, ii. 290.
-
Polyandry, ii. 365;
-
in certain cyprinidæ, i. 309;
-
among the elateridæ, i. 313.
-
Polydactylism in man, i. 125.
-
Polygamy, influence of, upon sexual selection, i. 265;
-
superinduced by domestication, i. 270;
-
supposed increase of female births by, i. 303;
-
in the stickleback, ii. 2.
-
Polygenists, i. 228.
-
Polynesia, prevalence of infanticide in, ii. 364.
-
Polynesians, aversion of, to hairs on the face, ii. 349;
-
wide geographical range of, i. 112;
-
difference of stature among the, i. 115;
-
crosses of, i. 225;
-
variability of, i. 225;
-
heterogeneity of the, i. 241.
-
Polyplectron, display of plumage by the male, i. 89;
-
number of spurs in, ii. 46;
-
gradation of characters in, ii. 137;
-
female of, ii. 194.
-
Polyplectron chinquis, ii. 90, 138, 139.
-
Polyplectron Hardwickii, ii. 138, 139.
-
Polyplectron malaccense, ii. 139, 140.
-
Polyplectron Napoleonis, ii. 138, 140.
-
Polyzoa, i. 324.
-
Pontoporeia affinis, i. 329.
-
Porcupine, mute, except in the rutting season, ii. 274.
-
Pores, excretory, numerical relation of, to the hairs in sheep, i. 248.
-
Porpitæ, bright colours of some, i. 322.
-
Portax picta, dorsal crest and throat-tuft of, ii. 282;
-
sexual differences of colour in, ii. 287, 299.
-
Portunus puber, pugnacity of, i. 332.
-
Potamochoerus penicillatus, tusks and facial knobs of the, ii. 266.
-
Pouchet, G., on the ratio of instinct and intelligence, i. 37;
-
on the instincts of ants, i. 187;
-
on the caves of Abou-Simbel, i. 217;
-
on the immunity of negroes from yellow fever, i. 243.
-
Pouter pigeon, late development of the large crop in, i. 293.
-
Power, Dr., on the different colours of the sexes in a species of Squilla, i. 335.
-
Powys, Mr., on the habits of the chaffinch in Corfu, i. 307.
-
Pre-eminence of man, i. 137.
457
-
Preference for males by female birds, ii. 113, 122;
-
shown by mammals, in pairing, ii. 268.
-
Prehensile organs, i. 256.
-
Presbytis entellus, fighting of the male, ii. 324.
-
Preyer, Dr., on supernumerary mammæ in women, i. 125.
-
Prichard, on the difference of stature among the Polynesians, i. 115;
-
on the connection between the breadth of the skull in the Mongolians and the perfection of their senses, i. 119;
-
on the capacity of British skulls of different ages, i. 146;
-
on the flattened heads of the Colombian savages, ii. 340;
-
on Siamese notions of beauty, ii. 345;
-
on the beardlessness of the Siamese, ii. 349;
-
on the deformation of the head among American tribes and the natives of Arakhan, ii. 352.
-
Primary sexual organs, i. 254.
-
Primates, i. 190;
-
sexual differences of colour in, ii. 290.
-
Primogeniture, evils of, i. 170.
-
Primula, relation between number and size of seeds in, i. 317.
-
Prionidæ, difference of the sexes in colour, i. 367.
-
Proctotretus multimaculatus, ii. 26, 37.
-
Proctotretus tenuis, sexual difference in the colour of, ii. 37.
-
Profligacy, i. 173.
-
Progenitors, early, of man, i. 206.
-
Progress, not the normal rule in human society, i. 166;
-
Prong-horn, horns of, i. 289.
-
Proportions, difference of, in distinct races, i. 216.
-
Protective colouring in butterflies, i. 392;
-
Protective nature of the dull colouring of female Lepidoptera, i. 403, 405, 414.
-
Protective resemblances in fishes, ii. 18.
-
Protozoa, absence of secondary sexual characters in, i. 321.
-
Pruner-Bey, on the occurrence of the supra-condyloid foramen in the humerus of man, i. 29;
-
on the colour of negro infants, ii. 318.
-
Prussia, numerical proportion of male and female births in, i. 301.
-
Psocus, proportions of the sexes in, i. 314.
-
Ptarmigan, monogamous, i. 269;
-
summer and winter plumage of the, ii. 81, 83;
-
nuptial assemblages of, ii. 101;
-
triple moult of the, ii. 181;
-
protective coloration of, ii. 198.
-
Puff-birds, colours and nidification of the, ii. 171.
-
Pugnacity of fine-plumaged male birds, ii. 93.
-
Pumas, stripes of young, ii. 183.
-
Puppies learning from cats to clean their faces, i. 44.
-
Pycnonotus hæmorrhous, pugnacity of the male, ii. 41;
-
display of under tail coverts by the male, ii. 96.
-
Pyranga æstiva, male aiding in incubation, ii. 167.
-
Pyrodes, difference of the sexes in colour, i. 367.
-
Q.
-
Quadrumana, hands of, i. 139;
-
differences between man and the, i. 190;
-
dependence of, on climate, i. 218;
-
sexual differences of colour in, ii. 290;
-
ornamental characters of, ii. 306;
-
analogy of sexual differences of, with those of man, ii. 318;
-
fighting of males for the females, ii. 324;
-
monogamous habits of, ii. 361;
-
beards of the, ii. 378.
-
Quain, R., on the variation of the muscles in man, i. 109.
-
Quatrefages, A. de, on the occurrence of a rudimentary tail in man, i. 29;
-
on the moral sense as a distinction between man and animals, i. 70;
-
on variability, i. 112;
-
on the fertility of Australian women with white men, i. 221;
-
on the Paulistas of Brazil, i. 225;
-
on the evolution of the breeds of cattle, i. 230;
-
on the Jews, i. 242;
-
on the liability of negroes to tropical fevers after residence in a cold climate, i. 243;
458
-
on the difference between field- and house-slaves, i. 246;
-
on the influence of climate on colour, i. 246;
-
on the Ainos, ii. 321;
-
on the women of San-Giuliano, ii. 357.
-
Quechua Indians, i. 119;
-
local variation of colour in the, i. 246;
-
no grey hair among the, ii. 320;
-
hairlessness of the, ii. 322;
-
long hair of the, ii. 348.
-
Querquedula acuta, ii. 114.
-
Quiscalus major, proportions of the sexes of, in Florida and Honduras, i. 307.
-
R.
-
Rabbit, white tail of the, ii. 298.
-
Rabbits, danger-signals of, i. 74;
-
domestic, elongation of the skull in, i. 147;
-
modification of the skull in, by the lopping of the ear, i. 147;
-
numerical proportion of the sexes in, i. 305.
-
Races, distinctive characters of, i. 215;
-
or species of man, i. 217;
-
crossed, fertility or sterility of, i. 220;
-
of man, variability of the, i. 225;
-
of man, resemblance of, in mental characters, i. 232;
-
formation of, i. 235;
-
of man, extinction of, i. 236;
-
effects of the crossing of, i. 240;
-
of man, formation of the, i. 240;
-
of man, children of the, ii. 318;
-
beardless, aversion of, to hairs on the face, ii. 349.
-
Raffles, Sir S., on the Banteng, ii. 290.
-
Rafts, use of, i. 137, 234.
-
Rage, manifested by animals, i. 40.
-
Raia batis, teeth of, ii. 6.
-
Raia clavata, female spined on the back, ii. 2;
-
sexual difference in the teeth of, ii. 6.
-
Raia maculata, teeth of, ii. 6.
-
Rails, spur-winged, ii. 48.
-
Ram, mode of fighting of the, ii. 249;
-
African, mane of an, ii. 284;
-
fat-tailed, ii. 284.
-
Rameses II., i. 217.
-
Ramsay, Mr., on the Australian Musk-duck, ii. 38;
-
on the Regent-bird, ii. 113;
-
on the incubation of Menura superba, ii. 165.
-
Rana esculenta, vocal sacs of, ii. 28.
-
Rat, common, general dispersion of, a consequence of superior cunning, i. 50;
-
supplantation of the native, in New Zealand, by the European rat, i. 240;
-
common, said to be polygamous, i. 268;
-
numerical proportion of the sexes in, i. 305.
-
Rats, enticed by essential oils, ii. 281.
-
Rationality of birds, ii. 108.
-
Rattle-snakes, difference of the sexes in the, ii. 29;
-
said to use their rattles as a sexual call, ii. 30.
-
Raven, vocal organs of the, ii. 55;
-
stealing bright objects, ii. 112;
-
pied, of the Feroe Islands, ii. 120.
-
Rays, prehensile organs of male, ii. 1.
-
Razor-bill, young of the, ii. 217.
-
Reade, Winwood, on the Guinea sheep, i. 289;
-
non-development of horns in castrated male Guinea sheep, ii. 247;
-
on the occurrence of a mane in an African ram, ii. 285;
-
on the negroes’ appreciation of the beauty of their women, ii. 344;
-
on the admiration of negroes for a black skin, ii. 346;
-
on the idea of beauty among negroes, ii. 350;
-
on the Jollofs, ii. 357;
-
on the marriage-customs of the negroes, ii. 374.
-
Reason, in animals, i. 46.
-
Redstart, American, breeding in immature plumage, ii. 214.
-
Redstarts, new mates found by, ii. 105.
-
Reduvidæ, stridulation of, i. 350.
-
Reed-bunting, head-feathers of the male, ii. 95;
-
attacked by a bullfinch, ii. 111.
-
Reefs, fishes frequenting, ii. 17.
-
Regeneration, partial, of lost parts in man, i. 13.
-
Regent-bird, ii. 112.
-
Reindeer, antlers of, with numerous points, ii. 252;
-
sexual preferences shown by, ii. 273;
-
horns of the, i. 288;
-
winter change of the, ii. 299;
-
battles of, ii. 240;
-
horns of the female, ii. 243.
459
-
Relationship, terms of, ii. 360.
-
Religion, deficiency of, among certain races, i. 65;
-
psychical elements of, i. 68.
-
Remorse, i. 91;
-
deficiency of, among savages, i. 164.
-
Rengger, on the diseases of Cebus Azaræ, i. 11;
-
on maternal affection in a Cebus, i. 40;
-
revenge taken by monkeys, i. 40;
-
on the reasoning powers of American monkeys, i. 47;
-
on the use of stones by monkeys for cracking hard nuts, i. 51;
-
on the sounds uttered by Cebus Azaræ, i. 54;
-
on the signal-cries of monkeys, i. 57;
-
on the diversity of the mental faculties of monkeys, i. 110;
-
on the Payaguas Indians, i. 117;
-
on the inferiority of Europeans to savages in their senses, i. 118;
-
on the polygamous habits of Mycetes caraya, i. 266;
-
on the voice of the howling monkeys, ii. 277;
-
on the odour of Cervus campestris, ii. 279;
-
on the beards of Mycetes caraya and Pithecia Satanas, ii. 283;
-
on the colours of Felis mitis, ii. 287;
-
on the colours of Cervus paludosus, ii. 290;
-
on sexual differences of colour in Mycetes, ii. 291;
-
on the colour of the infant Guaranys, ii. 318;
-
on the early maturity of the female of Cebus azaræ, ii. 318;
-
on the beards of the Guaranys, ii. 322, 323;
-
on the emotional notes employed by monkeys, ii. 336;
-
on American polygamous monkeys, ii. 362.
-
Representative species, of birds, ii. 190, 191.
-
Reproduction, unity of phenomena of, throughout the mammalia, i. 13;
-
period of, in birds, ii. 214.
-
Reproductive system, rudimentary structures in the, i. 30;
-
accessory parts of, i. 207.
-
Reptiles, ii. 28.
-
Reptiles and birds, alliance of, i. 213.
-
Resemblances, small, between man and the apes, i. 191.
-
Retrievers, exercise of reasoning faculties by, i. 48.
-
Revenge, manifested by animals, i. 40.
-
Reversion, i. 122;
-
perhaps the cause of some bad dispositions, i. 173.
-
Rhagium, difference of colour in the sexes of a species of, i. 367.
-
Ramphastos carinatus, ii. 227.
-
Rhinoceros, nakedness of, i. 148;
-
horns of, ii. 247;
-
horns of, used defensively, ii. 263;
-
attacking white or grey horses, ii. 295.
-
Rhynchæa, sexes and young of, ii. 202.
-
Rhynchæa australis, ii. 203.
-
Rhynchæa bengalensis, ii. 203.
-
Rhynchæa capensis, ii. 202.
-
Rhythm, perception of, by animals, ii. 333.
-
Richard, M., on rudimentary muscles in man, i. 19.
-
Richardson, Sir J., on the pairing of Tetrao umbellus, ii. 49;
-
on Tetrao urophasianus, ii. 58;
-
on the drumming of grouse, ii. 63;
-
on the dances of Tetrao phasianellus, ii. 69;
-
on assemblages of grouse, ii. 101;
-
on the battles of male deer, ii. 240;
-
on the reindeer, ii. 244;
-
on the horns of the musk-ox, ii. 247;
-
on antlers of the reindeer with numerous points, ii. 252;
-
on the moose, ii. 259.
-
Richardson, on the Scotch deerhound, ii. 261.
-
Richter, Jean Paul, on imagination, i. 45.
-
Riedel, on profligate female pigeons, ii. 119.
-
Ring-ouzel, colours and nidification of the, ii. 170.
-
Ripa, Father, on the difficulty of distinguishing the races of the Chinese, i. 215.
-
Rivalry, in singing, between male birds, ii. 53.
-
River-hog, African, tusks and knobs of the, ii. 266.
-
Rivers, analogy of, to islands, i. 204.
-
Roach, brightness of male during breeding-season, ii. 13.
-
Robbery, of strangers, considered honourable, i. 94.
-
Robertson, Mr., remarks on the development of the horns in the roebuck and red-deer, i. 288.
460
-
Robin, pugnacity of the male, ii. 40;
-
autumn song of the, ii. 54;
-
female, singing of the, ii. 54;
-
attacking other birds with red in their plumage, ii. 111;
-
young of the, ii. 208.
-
Robinet, on the difference of size of the male and female cocoons of the silk-moth, i. 346.
-
Rodents, uterus in the, i. 123;
-
absence of secondary sexual characters in, i. 268;
-
sexual differences in the colours of, ii. 286.
-
Roe, winter change of the, ii. 299.
-
Rolle, F., on the origin of man, i. 4;
-
on a change in German families settled in Georgia, i. 246.
-
Roller, ii. 56.
-
Romans, ancient, gladiatorial exhibitions of the, i. 101.
-
Rook, voice of the, ii. 61.
-
Rössler, Dr., on the resemblance of the lower surface of butterflies to the bark of trees, i. 392.
-
Rostrum, sexual difference in the length of, in some weevils, i. 255.
-
Rudimentary organs, i. 17;
-
Rudiments, presence of, in languages, i. 60.
-
Rudolph, on the want of connexion between climate and the colour of the skin, i. 241.
-
Ruff, supposed to be polygamous, i. 270;
-
proportion of the sexes in the, i. 306;
-
pugnacity of the, ii. 41, 48;
-
double moult in, ii. 81, 84;
-
duration of dances of, ii. 100;
-
attraction of the, to bright objects, ii. 111.
-
Ruminants, male, disappearance of canine teeth in, i. 144, ii. 325;
-
generally polygamous, i. 266;
-
analogy of Lamellicorn beetles to, i. 373;
-
suborbital pits of, ii. 280;
-
sexual differences of colour in, ii. 287.
-
Rupicola crocea, display of plumage by the male, ii. 87.
-
Rüppell, on canine teeth in deer and antelopes, ii. 258.
-
Russia, numerical proportion of male and female births in, i. 301.
-
Ruticilla, ii. 180.
-
Rütimeyer, Prof., on the physiognomy of the apes, i. 149;
-
on the sexual differences of monkeys, ii. 323.
-
Rutlandshire, numerical proportion of male and female births in, i. 301.
-
S.
-
Sachs, Prof., on the behaviour of the male and female elements in fertilisation, i. 274.
-
Sacrifices, Human, i. 182.
-
Sagittal crest in male apes and Australians, ii. 319.
-
Sahara, birds of the, ii. 172;
-
animal inhabitants of the, ii. 224.
-
Sailors, growth of, delayed by conditions of life, i. 114;
-
Sailors and soldiers, difference in the proportions of, i. 116.
-
St. John, Mr., on the attachment of mated birds, ii. 108.
-
St. Kilda, beards of the inhabitants of, ii. 321.
-
Salmo eriox, and S. umbla, colouring of the male, during the breeding season, ii. 14.
-
Salmo lycaodon, ii. 4.
-
Salmo salar, ii. 4.
-
Salmon, leaping out of fresh water, i. 83;
-
male, ready to breed before the female, i. 260;
-
proportion of the sexes in, i. 308;
-
male, pugnacity of the, ii. 3;
-
male, characters of, during the breeding season, ii. 3, 14;
-
spawning of the, ii. 19;
-
breeding of immature male, ii. 215.
-
Salvin, O., on the Humming-birds, i. 269, ii. 168;
-
on the numerical proportion of the sexes in Humming-birds, i. 307, ii. 221;
-
on Chamæpetes and Penelope, ii. 64;
-
on Selasphorus platycercus, ii. 65;
-
on Pipra deliciosa, ii. 66;
-
on Chasmorhynchus, ii. 79.
-
Samoa Islands, beardlessness of the natives of, ii. 322, 349.
-
Sand-skipper, i. 334.
-
Sandwich Islands, variation in the skulls of the natives of the, i. 108;
461
-
superiority of the nobles in the, ii. 356.
-
Sandwich Islanders, lice of, i. 219.
-
San-Giuliano, women of, ii. 357.
-
Santali, recent rapid increase of the, i. 133;
-
Mr. Hunter on the, i. 241.
-
Saphirina, characters of the males of, i. 335.
-
Sarkidiornis melanonotus, characters of the young, ii. 185.
-
Sars, O., on Pontoporeia offinis, i. 329.
-
Saturnia carpini, attraction of males by the female, i. 311.
-
Saturnia Io, difference of coloration in the sexes of, i. 398.
-
Saturniidæ, coloration of the, i. 396, 398.
-
Savage, Dr., on the fighting of the male gorillas, ii. 324;
-
on the habits of the gorilla, ii. 363.
-
Savage and Wyman, on the polygamous habits of the gorilla, i. 266.
-
Savages, imitative faculties of, i. 57, 161;
-
causes of low morality of, i. 97;
-
uniformity of, exaggerated, i. 111;
-
long-sighted, i. 118;
-
rate of increase among, usually small, i. 132;
-
retention of the prehensile power of the feet by, i. 142;
-
tribes of, supplanting one another, i. 160;
-
improvements in the arts among, i. 182;
-
arts of, i. 234;
-
fondness of, for rough music, ii. 67;
-
attention paid by, to personal appearance, ii. 338;
-
relation of the sexes among, ii. 363.
-
Saw-fly, pugnacity of a male, i. 364.
-
Saw-flies, proportions of the sexes in, i. 314.
-
Saxicola rubicola, young of, ii. 220.
-
Scalp, motion of the, i. 20.
-
Scent-glands in snakes, ii. 30.
-
Schaaffhausen, Prof., on the development of the posterior molars in different races of man, i. 26;
-
on the jaw from La Naulette, i. 126;
-
on the correlation between muscularity and prominent supra-orbital ridges, i. 130;
-
on the mastoid processes of man, i. 143;
-
on modifications of the cranial bones, i. 147;
-
on human sacrifices, i. 182;
-
on the probable speedy extermination of the anthropomorphous apes, i. 201;
-
on the ancient inhabitants of Europe, i. 237;
-
on the effects of use and disuse of parts, i. 247;
-
on the superciliary ridge in man, ii. 316;
-
on the absence of race-differences in the infant skull in man, ii. 318;
-
on ugliness, ii. 354.
-
Schaum, H., on the elytra of Dytiscus and Hydroporus, i. 343.
-
Schelver, on dragon-flies, i. 363.
-
Schiodte, on the stridulation of Heterocerus, i. 379.
-
Schlegel, F. von, on the complexity of the languages of uncivilised peoples, i. 61.
-
Schlegel, Prof., on Tanysiptera, ii. 190.
-
Schleicher, Prof., on the origin of language, i. 56.
-
Schleiden, Prof., on the rattle-snake, ii. 30.
-
Schomburgk, Sir R., on the pugnacity of the male musk-duck of Guiana, ii. 43;
-
on the courtship of Rupicola crocea, ii. 87.
-
Schoolcraft, Mr., on the difficulty of fashioning stone implements, i. 138.
-
Sclater, P. L., on modified secondary wing-feathers in the males of Pipra, ii. 65;
-
on elongated feathers in nightjars, ii. 73;
-
on the species of Chasmorhynchus, ii. 79;
-
on the plumage of Pelecanus onocrotalus, ii. 85;
-
on the plantain-eaters, ii. 177;
-
on the sexes and young of Tadorna variegata, ii. 206;
-
on the colours of Lemur macaco, ii. 290;
-
on the stripes in asses, ii. 305.
-
Scolecida, absence of secondary sexual characters in, i. 321.
-
Scolopax frenata, tail-feathers of, ii. 64.
-
Scolopax gallinago, drumming of, ii. 63.
-
Scolopax javensis, tail-feathers of, ii. 64.
-
Scolopax major, assemblies of, ii. 101.
-
Scolopax Wilsonii, sound produced by, ii. 64.
462
-
Scolytus, stridulation of, i. 379.
-
Scoter-duck, black, sexual difference in coloration of the, ii. 226;
-
bright beak of male, ii. 227.
-
Scott, J., on the colour of the beard in man, ii. 319.
-
Scrope, on the pugnacity of the male salmon, ii. 3;
-
on the battles of stags, ii. 240.
-
Scudder, S. H., imitation of the stridulation of the Orthoptera, i. 353;
-
on the stridulation of the ACRIDIIDÆ, i. 356;
-
on a Devonian insect, i. 360;
-
on stridulation, ii. 331.
-
Sculpture, expression of the ideal of beauty by, ii. 350.
-
Sea-anemonies, bright colours of, i. 322.
-
Sea-bear, polygamous, i. 268.
-
Sea-elephant, male, structure of the nose of the, ii. 278;
-
Sea-lion, polygamous, i. 268.
-
Seal, bladder-nose, ii. 278.
-
Seals, their sentinels generally females, i. 74;
-
evidence furnished by, on classification, i. 190;
-
sexual differences in the coloration of, ii. 287;
-
appreciation of music by, ii. 333;
-
battles of male, ii. 240;
-
canine teeth of male, ii. 241;
-
polygamous habits of, i. 268;
-
pairing of, ii. 269;
-
sexual peculiarities of, ii. 277.
-
Sea-scorpion, sexual differences in, ii. 9.
-
Season, changes of colour in birds, in accordance with the, ii. 80;
-
changes of plumage of birds in relation to, ii. 180.
-
Seasons, inheritance at corresponding, i. 282.
-
Sebituani, ii. 340.
-
Sebright Bantam, i. 294.
-
Secondary sexual characters, i. 253;
-
relations of polygamy to, i. 266;
-
gradation of, in birds, ii. 135;
-
transmitted through both sexes, i. 279.
-
Sedgwick, W., on hereditary tendency to produce twins, i. 133.
-
Seemann, Dr., on the different appreciation of music by different peoples, ii. 334;
-
on the effects of music, ii. 335.
-
Selasphorus platycercus, acuminate first primary of the male, ii. 65.
-
Selby, P. J., on the habits of the black and red grouse, i. 269.
-
Selection, double, i. 276.
-
Selection of male by female birds, ii. 99, 122.
-
Selection, methodical, of Prussian grenadiers, i. 112.
-
Selection, sexual, influence of, on the colouring of Lepidoptera, i. 403;
-
Selection, sexual and natural, contrasted, i. 278.
-
Self-command, habit of, inherited, i. 92;
-
Self-consciousness, i. 62.
-
Self-preservation, instinct of, i. 89.
-
Self-sacrifice, by savages, i. 88;
-
Semilunar fold, i. 23.
-
Semnopithecus, i. 197;
-
long hair on the heads of species of, i. 192; ii. 380.
-
Semnopithecus chrysomelas, sexual differences of colour in ii. 291.
-
Semnopithecus comatus, ornamental hair on the head of, ii. 307.
-
Semnopithecus frontatus, beard, &c., of, ii. 308.
-
Semnopithecus nasica, nose of, i. 192.
-
Semnopithecus nemæus, colouring of, ii. 310.
-
Semnopithecus rubicundus, ornamental hair on the head of, ii. 306.
-
Senses, inferiority of Europeans to savages in the, i. 118.
-
Sentinels, i. 74, 82.
-
Serpents, instinctively dreaded by apes and monkeys, i. 37, 42.
-
Serranus, hermaphroditism in, i. 208.
-
Sex, inheritance limited by, i. 282.
-
Sexes, relative proportions of, in man, i. 300, ii. 320;
-
probable relation of the, in primeval man, ii. 362.
-
Sexual characters, secondary, i. 253;
-
relations of polygamy to, i. 266;
-
transmitted through both sexes, i. 279; gradation of, in birds, ii. 135.
463
-
Sexual and natural selection, contrasted, i. 278.
-
Sexual characters, effects of the loss of, i. 284;
-
Sexual differences in man, i. 14.
-
Sexual selection, explanation of, i. 256, 260, 271;
-
influence of, on the colouring of Lepidoptera, i. 403;
-
action of, in mankind, ii. 368.
-
Sexual similarity, i. 277.
-
Sharks, prehensile organs of male, ii. 1.
-
Sharpe, R. B., on Tanysiptera sylvia, ii. 165;
-
on Ceryle, ii. 173;
-
on the young male of Dacelo Gaudichaudi, ii. 188.
-
Shaw, Mr., on the pugnacity of the male salmon, ii. 3.
-
Shaw, J., on the decorations of birds, ii. 71.
-
Sheep, danger-signals of, i. 74;
-
sexual differences in the horns of, i. 283;
-
horns of, i. 289, ii. 246, 259;
-
domestic, sexual differences of, late developed, i. 293;
-
numerical proportion of the sexes in, i. 304;
-
mode of fighting of, ii. 249;
-
arched foreheads of some, ii. 284.
-
Sheep, Merino, loss of horns in females of, i. 284;
-
Shells, difference in form of, in male and female Gasteropoda, i. 324;
-
beautiful colours and shapes of, i. 326.
-
Shield-drake, pairing with a common duck, ii. 114;
-
New Zealand, sexes and young of, ii. 206.
-
Shooter, J., on the Kafirs, ii. 347;
-
on the marriage-customs of the Kafirs, ii. 373.
-
Shrew-mice, odour of, ii. 279.
-
Shrike, Drongo, ii. 179.
-
Shrikes, characters of young, ii. 185.
-
Shuckard, W. E., on sexual differences in the wings of Hymenoptera, i. 345.
-
Shyness of adorned male birds, ii. 97.
-
Siagonium, proportions of the sexes in, i. 314;
-
dimorphism in males of, i. 374.
-
Siam, proportion of male and female births in, i. 303.
-
Siamese, general beardlessness of the, ii. 321;
-
notions of beauty of the, ii. 345;
-
hairy family of, ii. 378.
-
Siebold, C. T. von, on the auditory apparatus of the stridulant orthoptera, i. 353.
-
Sight, inheritance of long and short, i. 118.
-
Signal-cries of monkeys, i. 57.
-
Silk-moth, difference of size of the male and female cocoons of the, i. 346;
-
pairing of the, i. 401;
-
male, fertilising two or three females, i. 406;
-
proportion of the sexes in, i. 309, 311;
-
Ailanthus, Prof. Canestrini, on the destruction of its larvæ by wasps, i. 311.
-
Simiadæ, i. 195;
-
their origin and divisions, i. 213.
-
Similarity, sexual, i. 277.
-
Singing of the Cicadæ and Fulgoridæ, i. 351;
-
of tree-frogs, ii. 27;
-
of birds, object of the, ii. 52.
-
Sirenia, nakedness of, i. 148.
-
Sirex juvencus, i. 365.
-
Siricidæ, difference of the sexes in, i. 365.
-
Siskin, ii. 85;
-
pairing with a canary, ii. 115.
-
Sitana, throat-pouch of the males of, ii. 33, 36.
-
Size, relative, of the sexes of insects, i. 345.
-
Skin, movement of the, i. 19;
-
nakedness of, in man, i. 148;
-
colour of the, i. 241.
-
Skin and hair, correlation of colour of, i. 248.
-
Skull, variation of, in man, i. 108;
-
cubic contents of, no absolute test of intellect, i. 145;
-
Neanderthal, capacity of the, i. 146;
-
causes of modification of the, i. 147;
-
difference of, in form and capacity, in different races of men, i. 216;
-
variability of the shape of the, i. 226;
-
differences of, in the sexes in man, ii. 317;
-
artificial modifications of the shape of, ii. 340.
-
Skunk, odour emitted by the, ii. 279.
-
Slavery, prevalence of, i. 94;
-
Slaves, difference between those of field and house, i. 246.
-
Smell, sense of, in man and animals, i. 23.
-
Smith, Adam, on the basis of sympathy, i. 82.
-
Smith, Sir A., on the recognition of women by male Cynocephali, i. 13;
-
on an instance of memory in a baboon, i. 45;
-
on the retention of their colour by the Dutch in South Africa, i. 242;
-
on the polygamy of the South African antelopes, i. 267;
-
on the proportion of the sexes in Kobus ellipsiprymnus, i. 305;
-
on Bucephalus capensis, ii. 29;
-
on South African lizards, ii. 37;
-
on fighting gnus, ii. 240;
-
on the horns of rhinoceroses, ii. 248;
-
on the fighting of lions, ii. 266;
-
on the colours of the Cape Eland, ii. 288;
-
on the colours of the gnu, ii. 289;
-
on Hottentot notions of beauty, ii. 345.
-
Smith, F., on the Cynipidæ and Tenthredinidæ, i. 314;
-
on the relative size of the sexes of Aculeate Hymenoptera, i. 347;
-
on the difference between the sexes of ants and bees, i. 365;
-
on the stridulation of Trox sabulosus, i. 380;
-
on the stridulation of Mononychus pseudacori, i. 382.
-
Smynthurus luteus, courtship of, i. 348.
-
Snakes, sexual differences of, ii. 29;
-
male, ardency of, ii. 30.
-
“Snarling muscles,” i. 127.
-
Snipe, drumming of the, ii. 63;
-
coloration of the, ii. 226.
-
Snipe, painted, sexes and young of, ii. 202.
-
Snipe, solitary, assemblies of, ii. 101.
-
Snipes, arrival of male before the female, i. 260;
-
pugnacity of male, ii. 43;
-
double moult in, ii. 80.
-
Snow-goose, whiteness of the, ii. 228.
-
Social animals, affection of, for each other, i. 76;
-
defence of, by the males, i. 83.
-
Sociability, the sense of duty connected with, i. 71;
-
impulse to, in animals, i. 79, 80;
-
manifestations of, in man, i. 84;
-
instinct of, in animals, i. 86.
-
Sociality, probable, of primeval men, i. 155;
-
influence of, on the development of the intellectual faculties, i. 160;
-
origin of, in man, i. 161.
-
Soldiers, American, measurements of, i. 114.
-
Soldiers and sailors, difference in the proportions of, i. 116.
-
Solenostoma, bright colours and marsupial sack of the females of, ii. 22.
-
Song of male birds appreciated by their females, i. 63;
-
want of, in brilliant plumaged birds, ii. 94;
-
of birds, ii. 163.
-
Sorex, odour of, ii. 279.
-
Sounds admired alike by man and animals, i. 64;
-
produced by fishes, ii. 23;
-
produced by male frogs and toads, ii. 27;
-
instrumentally produced by birds, ii. 63 et seqq.
-
Spain, decadence of, i. 178.
-
Sparassus smaragdulus, difference of colour in the sexes of, i. 337, 338.
-
Sparrow, pugnacity of the male, ii. 40;
-
acquisition of the Linnet’s song by a, ii. 55;
-
coloration of the, ii. 198;
-
immature plumage of the, ii. 188.
-
Sparrow, white-crowned, young of the, ii. 217.
-
Sparrows, house- and tree-, ii. 170.
-
Sparrows, new mates found by, ii. 105.
-
Sparrows, sexes and young of, ii. 212;
-
learning to sing, ii. 334.
-
Spathura Underwoodi, ii. 77.
-
Spawning of fishes, ii. 15, 19.
-
Spear, origin of the, i. 234.
-
Species, causes of the advancement of, i. 172;
-
distinctive characters of, i. 214;
-
or races of man, i. 217;
-
sterility and fertility of, when crossed, i. 122;
-
supposed, of man, i. 226;
-
gradation of, i. 227;
-
difficulty of defining, i. 228;
-
representative, of birds, ii. 190, 191;
-
of birds, comparative differences between the sexes of distinct, ii. 192.
465
-
Spectre-insects, mimickry of leaves by, i. 414.
-
Spectrum femoratum, difference of colour in the sexes of, i. 361.
-
Speech, connection between the brain and the faculty of, i. 58.
-
“Spel” of the black-cock, ii. 60.
-
Spencer, Herbert, on the dawn of intelligence, i. 37;
-
on the origin of the belief in spiritual agencies, i. 66;
-
on the origin of the moral sense, i. 101;
-
on the influence of food on the size of the jaws, i. 118;
-
on the ratio between individuation and genesis, i. 318;
-
on music, ii. 336.
-
Sperm-whales, battles of male, ii. 240.
-
Sphingidæ, coloration of the, i. 396.
-
Sphinx, Humming-bird, i. 399.
-
Sphinx, Mr. Bates on the caterpillar of a, i. 416.
-
Spiders, i. 337;
-
male, more active than female, i. 272;
-
proportion of the sexes in, i. 314;
-
male, small size of, i. 338.
-
Spilosoma menthrasti, rejected by turkeys, i. 398.
-
Spine, alteration of, to suit the erect attitude of man, i. 143.
-
Spirits, fondness of monkeys for, i. 12.
-
Spiritual agencies, belief in, almost universal, i. 65.
-
Spoonbill, ii. 60;
-
Chinese, change of plumage in, ii. 179.
-
Spots, retained throughout groups of birds, i. 131;
-
disappearance of, in adult mammals, ii. 303.
-
Sprengel, C. K., on the sexuality of plants, i. 260.
-
Spring-boc, horns of the, ii. 251.
-
Sproat, Mr., on the extinction of savages in Vancouver Island, i. 239;
-
on the eradication of facial hair by the natives of Vancouver Island, ii. 348;
-
on the eradication of the beard by the Indians of Vancouver Island, ii. 380.
-
Spurs, occurrence of, in female fowls, i. 280, 284;
-
development of, in various species of Phasianidæ, i. 290;
-
of Gallinaceous birds, ii. 44, 46;
-
development of, in female Gallinaceæ, ii. 162.
-
Squilla, different colours of the sexes of a species of, i. 335.
-
Squirrels, battles of male, ii. 239;
-
African, sexual differences in the colouring of, ii. 286;
-
black, ii. 294.
-
Stag, long hairs of the throat of, ii. 268;
-
horns of the, i. 279, 282;
-
battles of, ii. 240;
-
horns of the, with numerous branches, ii. 252;
-
bellowing of the, ii. 274;
-
crest of the, ii. 282.
-
Stag-beetle, large size of male, i. 347;
-
weapons of the male, i. 375;
-
numerical proportion of sexes of, i. 313.
-
Stainton, H. T., on the numerical proportion of the sexes in the smaller moths, i. 310;
-
habits of Elachista rufocinerea, i. 311;
-
on the coloration of moths, i. 397;
-
on the rejection of Spilosoma menthrasti, by turkeys, i. 398;
-
on the sexes of Agrotis exclamationis, i. 399.
-
Stallion, mane of the, ii. 268.
-
Stallions, two, attacking a third, i. 75;
-
fighting, ii. 241;
-
small canine teeth of, ii. 258.
-
Stansbury, Capt., observations on pelicans, i. 77.
-
Staphylinidæ, hornlike processes in male, i. 374.
-
Starfishes, bright colours of some, i. 322.
-
Stark, Dr., on the death-rate in towns and rural districts, i. 175;
-
on the influence of marriage on mortality, i. 176;
-
on the higher mortality of males in Scotland, i. 302.
-
Starling, American field, pugnacity of male, ii. 51.
-
Starling, red-winged, selection of a mate by the female, ii. 116.
-
Starlings, three, frequenting the same nest, i. 269, ii. 106;
-
new mates found by, ii. 105.
-
Statues, Greek, Egyptian, Assyrian, &c., contrasted, ii. 350.
-
Stature, dependence of, upon local influences, i. 114.
-
Staudinger, Dr., his list of Lepidoptera, i. 312;
-
on breeding Lepidoptera, i. 311.
466
-
Staunton, Sir G., hatred of indecency a modern virtue, i. 96.
-
Stealing of bright objects by birds, ii. 112.
-
Stebbing, T. R., on the nakedness of the human body, ii. 375.
-
Stemmatopus, ii. 278.
-
Stenobothrus pratorum, stridulating organs of, i. 357.
-
Sterility, general, of sole daughters, i. 170;
-
when crossed, a distinctive character of species, i. 214.
-
Sterna, seasonal change of plumage in, ii. 228.
-
Stickle-back, polygamous, i. 271;
-
male, courtship of the, ii. 2;
-
male, brilliant colouring of, during the breeding season, ii. 14;
-
nidification of the, ii. 20.
-
Sticks used as implements and weapons by monkeys, i. 51.
-
Sting in bees, i. 254.
-
Stokes, Capt., on the habits of the great Bower-bird, ii. 70.
-
Stonechat, young of the, ii. 220.
-
Stone implements, difficulty of making, i. 138;
-
as traces of extinct tribes, i. 237.
-
Stones, used by monkeys for breaking hard fruits and as missiles, i. 140;
-
Stork, black, sexual differences in the bronchi of the, ii. 60;
-
red beak of the, ii. 227.
-
Storks, ii. 226, 230;
-
sexual difference in the colour of the eyes of, ii. 128.
-
Strange, Mr., on the Satin Bower-bird, ii. 69.
-
Stretch, Mr., on the numerical proportion in the sexes of chickens, i. 306.
-
Strepsiceros kudu, horns of, ii. 255;
-
Stridulation, by males of Theridion, i. 339;
-
of the Orthoptera and Homoptera discussed, i. 360;
-
of beetles, i. 378.
-
Stripes, retained throughout groups of birds, ii. 131;
-
disappearance of, in adult mammals, ii. 303.
-
Strix flammea, ii. 105.
-
Structure, existence of unserviceable modifications of, i. 153.
-
Struggle for existence, in man, i. 180, 185.
-
Struthers, Dr., on the occurrence of the supra-condyloid foramen in the humerus of man, i. 28.
-
Sturnella ludoviciana, pugnacity of the male, ii. 51.
-
Sturnus vulgaris, ii. 105.
-
Sub-species, i. 227.
-
Suffering, in strangers, indifference of savages to, i. 94.
-
Suicide, i. 172;
-
formerly not regarded as a crime, i. 94;
-
rarely practised among the lowest savages, i. 94.
-
Suidæ, stripes of young, ii. 184.
-
Sumatra, compression of the nose by the Malays of, ii. 352.
-
Sumner, Archb., man alone capable of progressive improvement, i. 49.
-
Sun-birds, nidification of, ii. 169.
-
Superstitions, i. 182;
-
Superstitious customs, i. 68.
-
Superciliary ridge in man, ii. 316, 318.
-
Supernumerary digits, more frequent in men than in women, i. 276;
-
inheritance of, i. 285;
-
early development of, i. 292.
-
Supra-condyloid foramen in the early progenitors of man, i. 206.
-
Suspicion, prevalence of, among animals, i. 39.
-
Sulivan, Sir B. J., on two stallions attacking a third, ii. 241.
-
Swallow-tail Butterfly, i. 393.
-
Swallows deserting their young, i. 84, 90.
-
Swan, black, red beak of the, ii. 227;
-
black-necked, ii. 230;
-
white, young of, ii. 211;
-
wild, trachea of the, ii. 59.
-
Swans, ii. 226, 230;
-
Swaysland, Mr., on the arrival of migratory birds, i. 259.
-
Swinhoe, R., on the common rat in Formosa and China, i. 50;
-
on the sounds produced by the male Hoopoe, ii. 62;
-
on Dicrurus macrocercus and the Spoonbill, ii. 179;
-
on the young of Ardeola, ii. 190;
-
on the habits of Turnix, ii. 202;
-
on the habits of Rhynchæa bengalensis, ii. 203;
-
on Orioles breeding in immature plumage, ii. 214, 215.
467
-
Sylvia atricapilla, young of, ii. 219.
-
Sylvia cinerea, aerial love-dance of the male, ii. 68.
-
Sympathy, i. 168;
-
among animals, i. 77;
-
its supposed basis, i. 82.
-
Sympathies, gradual widening of, i. 100.
-
Syngnathous fishes, abdominal pouch in male, i. 210.
-
Sypheotides auritus, acuminated primaries of the male, ii. 64;
-
T.
-
Tabanidæ, habits of, i. 254.
-
Tadorna variegata, sexes and young of, ii. 206.
-
Tadorna vulpanser, ii. 114.
-
Tahitians, i. 183;
-
compression of the nose by the, ii. 352.
-
Tail, rudimentary, occurrence of, in man, i. 29;
-
convoluted body in the extremity of the, i. 30;
-
absence of, in man and the higher apes, i. 150, 194;
-
variability of, in species of Macacus and in baboons, i. 150;
-
presence of, in the early progenitors of man, i. 206;
-
length of, in pheasants, ii. 156, 164, 166;
-
difference of length of the, in the two sexes of birds, ii. 164.
-
Tait, Lawson, on the effects of natural selection on civilised nations, i. 168.
-
Tanager, scarlet, variation in the male, ii. 126.
-
Tanagra æstiva, ii. 180;
-
age of mature plumage in, ii. 213.
-
Tanagra rubra, ii. 126;
-
Tanais, absence of mouth in the males of some species of, i. 255;
-
relations of the sexes in, i. 315;
-
dimorphic males of a species of, i. 328.
-
Tankerville, Earl, on the battles of wild bulls, ii. 240.
-
Tanysiptera, races of, determined from adult males, ii. 190.
-
Tanysiptera sylvia, long tail-feathers of, ii. 165.
-
Taphroderes distortus, enlarged left mandible of the male, i. 344.
-
Tapirs, longitudinal stripes of young, ii. 184, 303.
-
Tarsi, dilatation of front, in male beetles, i. 343.
-
Tarsius, i. 200.
-
Tasmania, half-castes killed by the natives of, i. 220.
-
Tattooing, i. 232;
-
universality of, ii. 339.
-
Taste, in the Quadrumana, ii. 296.
-
Taylor, G. on Quiscalus major, i. 307.
-
Tea, fondness of monkeys for, i. 12.
-
Tear-sacks, of Ruminants, ii. 280.
-
Teebay, Mr., on changes of plumage in spangled Hamburgh fowls, i. 281.
-
Teeth, rudimentary incisor, in Ruminants, i. 17;
-
posterior molar, in man, i. 25;
-
wisdom, i. 26;
-
diversity of, i. 108;
-
canine, in the early progenitors of man, i. 206;
-
canine, of male mammals, ii. 241;
-
in man, reduced by correlation, ii. 325;
-
staining of the, ii. 339;
-
front, knocked out or filed by some savages, ii. 340.
-
Tegetmeier, Mr., on the abundance of male pigeons, i. 306;
-
on the wattles of game-cocks, ii. 98;
-
on the courtship of fowls, ii. 117;
-
on dyed pigeons, ii. 118.
-
Tembeta, ii. 341.
-
Temper, in dogs and horses, inherited, i. 40.
-
Tench, proportions of the sexes in the, i. 308, 309;
-
brightness of male, during breeding season, ii. 13.
-
Tenebrionidæ, stridulation of, i. 379.
-
Tennent, Sir J. E., on the tusks of the Ceylon Elephant, ii. 248, 258;
-
on the frequent absence of beard in the natives of Ceylon, ii. 321;
-
on the Chinese opinion of the aspect of the Cingalese, ii. 345.
-
Tennyson, A., on the control of thought, i. 101.
-
Tenthredinidæ, proportions of the sexes in, i. 314;
-
fighting habits of male, i. 364;
-
difference of the sexes in, i. 365.
468
-
Tephrodornis, young of, ii. 190.
-
Terai, i. 237.
-
Termites, habits of, i. 364.
-
Terns, white, ii. 228;
-
Terns, seasonal change of plumage in, ii. 228.
-
Terror, common action of, upon the lower animals and man, i. 39.
-
Testudo nigra, ii. 28.
-
Tetrao cupido, battles of, ii. 50;
-
sexual difference in the vocal organs of, ii. 56.
-
Tetrao phasianellus, dances of, ii. 68;
-
duration of dances of, ii. 100.
-
Tetrao scoticus, ii. 170, 185, 194.
-
Tetrao tetrix, ii. 170, 185, 194;
-
pugnacity of the male. ii. 45.
-
Tetrao umbellus, pairing of, ii. 49;
-
battles of, ii. 50;
-
drumming of the male, ii. 61.
-
Tetrao urogalloides, dances of, ii. 100.
-
Tetrao urogallus, pugnacity of the male, ii. 45.
-
Tetrao urophasianus, inflation of the œsophagus in the male, ii. 57.
-
Thamnobia, young of, ii. 190.
-
Thaumalea picta, display of plumage by the male, ii. 89.
-
Thecla, sexual differences of colouring in species of, i. 389.
-
Thecla rubi, protective colouring of, i. 392.
-
Theridion, i. 337;
-
stridulation of males of, i. 339.
-
Theridion lineatum, variability of, i. 338.
-
Thomisus citreus, and T. floricolens, difference of colour in the sexes of, i. 337.
-
Thompson, J. H., on the battles of sperm-whales, ii. 240.
-
Thompson, W., on the colouring of the male char during the breeding season, ii. 14;
-
on the finding of new mates by magpies, ii. 103;
-
on the finding of new mates by Peregrine falcons, ii. 104.
-
Thorax, processes of, in male beetles, i. 370.
-
Thorell, T., on the proportion of the sexes in spiders, i. 315.
-
Thornback, difference in the teeth of the two sexes of the, ii. 6.
-
Thoughts, control of, i. 101.
-
Thrush, pairing with a blackbird, ii. 113;
-
colours and nidification of the, ii. 170.
-
Thrushes, characters of young, ii. 185, 269.
-
Thug, his regrets, i. 94.
-
Thumb, absence of, in Ateles and Hylobates, i. 140.
-
Thury, M., on the numerical proportion of male and female births among the Jews, i. 301.
-
Thylacinus, possession of the marsupial sack by the male, i. 208.
-
Thysanura, i. 348.
-
Tibia, dilated, of the male Crabro cribrarius, i. 343.
-
Tibia and femur, proportions of, in the Aymara Indians, i. 119.
-
Tierra del Fuego, marriage-customs of, ii. 373.
-
Tiger, colours and markings of the, ii. 302.
-
Tigers, depopulation of districts by, in India, i. 134.
-
Tillus elongatus, difference of colour in the sexes of, i. 368.
-
Timidity, variability of, in the same species, i. 40.
-
Tineina, proportion of the sexes in, i. 310.
-
Tipula, pugnacity of male, i. 349.
-
Tits, sexual difference of colour in, ii. 174.
-
Toads, ii. 25;
-
male, treatment of ova by some, i. 210;
-
male, ready to breed before the female, i. 260.
-
Toe, great, condition of, in the human embryo, i. 17.
-
Tomtit, blue, sexual difference of colour in the, ii. 174.
-
Tonga Islands, beardlessness of the natives of, ii. 322, 349.
-
Tooke, Horne, on language, i. 55.
-
Tools, flint, i. 183;
-
used by monkeys, i. 51;
-
use of, i. 137.
-
Top-knots in birds, ii. 74.
-
Tomicus villosus, proportion of the sexes in, i. 314.
-
Tortoise, voice of the male, ii. 331.
-
Tortures, submitted to by American savages, i. 95.
469
-
Totanus, double moult in, ii. 81.
-
Toucans, colours and nidification of the, ii. 171;
-
beaks and ceres of the, ii. 227.
-
Towns, residence in, a cause of diminished stature, i. 115.
-
Toynbee, J., on the external shell of the ear in man, i. 21.
-
Trachea, convoluted and imbedded in the sternum, in some birds, ii. 59;
-
structure of the, in Rhynchæa, ii. 203.
-
Trades, affecting the form of the skull, i. 147.
-
Tragelaphus, sexual differences of colour in, ii. 288.
-
Tragelaphus scriptus, dorsal crest of, ii. 282;
-
Tragopan, i. 270;
-
swelling of the wattles of the male, during courtship, ii. 72;
-
display of plumage by the male, ii. 91;
-
markings of the sexes of the, ii. 134.
-
Tragops dispar, sexual difference in the colour of, ii. 30.
-
Training, effect of, on the mental difference between the sexes of man, ii. 329.
-
Transfer of male characters to female birds, ii. 193.
-
Transmission, equal, of ornamental characters, to both sexes in mammals, ii. 297.
-
Traps, avoidance of, by animals, i. 49;
-
Treachery, to comrades, avoidance of, by savages, i. 88.
-
Tremex columbæ, i. 365.
-
Tribes, extinct, i. 160;
-
Trichius, difference of colour in the sexes of a species of, i. 368.
-
Trimen, R., on the proportion of the sexes in South African butterflies, i. 310;
-
on the attraction of males by the female of Lasiocampa quercus, i. 312;
-
on Pneumora, i. 358;
-
on difference of colour in the sexes of beetles, i. 367;
-
on moths brilliantly coloured beneath, i. 397;
-
on mimickry in butterflies, i. 412;
-
on Gynanisa Isis, and on the ocellated spots of Lepidoptera, ii. 132;
-
on Cyllo Leda, ii. 133.
-
Tringa, sexes and young of, ii. 216.
-
Tringa canutus, ii. 82.
-
Triphæna, coloration of the species of, i. 395.
-
Tristram, H. B., on unhealthy districts in North Africa, i. 244;
-
on the habits of the chaffinch in Palestine, i. 307;
-
on the birds of the Sahara, ii. 172;
-
on the animals inhabiting the Sahara, ii. 224.
-
Triton cristatus, ii. 24.
-
Triton palmipes, ii. 24.
-
Triton punctatus, ii. 24, 25.
-
Troglodytes vulgaris, ii. 198.
-
Trogons, colours and nidification of the, ii. 171, 173.
-
Tropic-birds, white only when mature, ii. 228.
-
Tropics, freshwater fishes of the, ii. 17.
-
Trout, proportion of the sexes in, i. 308;
-
male, pugnacity of the, ii. 3.
-
Trox sabulosus, stridulation of, i. 380.
-
Truth, not rare between members of the same tribe, i. 95;
-
more highly appreciated by certain tribes, i. 100.
-
Tulloch, Major, on the immunity of the negro from certain fevers, i. 243.
-
Tumbler, almond, change of plumage in the, i. 294.
-
Turdus merula, ii. 170;
-
Turdus migratorius, ii. 185.
-
Turdus musicus, ii. 170.
-
Turdus polyglottus, young of, ii. 219.
-
Turdus torquatus, ii. 170.
-
Turkey, swelling of the wattles of the male, ii. 72;
-
variety of, with a top-knot, ii. 74;
-
recognition of a dog by a, ii. 110;
-
wild, pugnacity of young male, ii. 48;
-
wild, notes of the, ii. 60;
-
male, wild, acceptable to domesticated females, ii. 119;
-
wild, first advances made by older females, ii. 121;
-
wild, breast-tuft of bristles of the, ii. 179.
-
Turkey-cock, scraping of the wings of, upon the ground, ii. 61;
-
wild, display of plumage by, ii. 87;
-
fighting habits of, ii. 98.
470
-
Turner, Prof. W., on muscular fasciculi in man referable to the panniculus carnosus, i. 19;
-
on the occurrence of the supra-condyloid foramen in the human humerus, i. 28;
-
on muscles attached to the coccyx in man, i. 29;
-
on the filum terminale in man, i. 30;
-
on the variability of the muscles, i. 109;
-
on abnormal conditions of the human uterus, i. 123;
-
on the development of the mammary glands, i. 209;
-
on male fishes hatching ova in their mouths, i. 210.
-
Turnix, sexes of some species of, ii. 201, 207.
-
Turtle-dove, cooing of the, ii. 60.
-
Tuttle, H., on the number of species of man, i. 226.
-
Tylor, E. B., on emotional cries, gestures, &c., of man, i. 54;
-
on the origin of the belief in spiritual agencies, i. 66;
-
on the primitive barbarism of civilised nations, i. 181;
-
on the origin of counting, i. 181;
-
on resemblances of the mental characters in different races of man, i. 232.
-
Type of structure, prevalence of, i. 211.
-
Typhœus, stridulating organs of, i. 378;
-
Twins, tendency to produce, hereditary, i. 133.
-
Twite, proportion of the sexes in the, i. 307.
-
U.
-
Ugliness, said to consist in an approach to the lower animals, ii. 354.
-
Umbrella-bird, ii. 58, 59.
-
Umbrina, sounds produced by, ii. 23.
-
United States, rate of increase in, i. 131;
-
influence of natural selection on the progress of, i. 179;
-
change undergone by Europeans in the, i. 246.
-
Upupa epops, sounds produced by the male, ii. 62.
-
Uraniidæ, coloration of the, i. 396.
-
Uria troile, variety of, (= U. lacrymans), ii. 127.
-
Urodela, ii. 24.
-
Urosticte Benjamini, sexual differences in, ii. 151.
-
Use and disuse of parts, effects of, i. 116;
-
influence of, on the races of man, i. 247.
-
Uterus, reversion in the, i. 123;
-
more or less divided, in the human subject, i. 123, 130;
-
double, in the early progenitors of man, i. 206.
-
V.
-
Vaccination, influence of, i. 168.
-
Vancouver Island, Mr. Sproat on the savages of, i. 239;
-
natives of, eradication of facial hair by the, ii. 348.
-
Vanellus cristatus, wing tubercles of the male, ii. 48.
-
Vanessæ, i. 387;
-
resemblance of lower surface of, to bark of trees, i. 392.
-
Variability, causes of, i. 111;
-
in man, analogous to that in the lower animals, i. 112;
-
of the races of man, i. 225;
-
greater in men than in women, i. 275;
-
period of, relation of the, to sexual selection, i. 296;
-
of birds, ii. 124;
-
of secondary sexual characters in man, ii. 320.
-
Variation, correlated, i. 30;
-
laws of, i. 113;
-
in man, i. 185;
-
analogous, i. 194;
-
analogous, in plumage of birds, ii. 74.
-
Variations, spontaneous, i. 131.
-
Varieties, absence of, between two species, evidence of their distinctness, i. 215.
-
Variety, an object in nature, ii. 230.
-
Variola, communicable between man and the lower animals, i. 11.
-
Vauréal, i. 29.
-
Veddahs, monogamous habits of, ii. 363.
-
Veitch, Mr., on the aversion of Japanese ladies to whiskers, ii. 349.
-
Vengeance, instinct of, i. 89.
-
Venus Erycina, priestesses of, ii. 357.
-
Vermes, i. 327.
-
Vermiform appendage, i. 27.
471
-
Verreaux, M., on the attraction of numerous males by the female of an Australian Bombyx, i. 312.
-
Vertebræ, caudal, number of, in macaques and baboons, i. 150;
-
of monkeys, partly imbedded in the body, i. 151.
-
Vertebrata, ii. 1;
-
common origin of the, i. 203;
-
most ancient progenitors of, i. 212;
-
origin of the voice in air-breathing, ii. 331.
-
Vesicula prostatica, the homologue of the uterus, i. 31, 208.
-
Vibrissæ, represented by long hairs in the eyebrows, i. 25.
-
Vidua, ii. 181.
-
Vidua axillaris, i. 269.
-
Villerme, M., on the influence of plenty upon stature, i. 115.
-
Vinson, Aug., on the male of Epeira nigra, i. 338.
-
Viper, difference of the sexes in the, ii. 29.
-
Virey, on the number of species of man, i. 226.
-
Virtues, originally social only, i. 93;
-
gradual appreciation of, i. 165.
-
Viscera, variability of, in man, i. 109.
-
Viti Archipelago, population of the, i. 225.
-
Vlacovich, Prof., on the ischio-pubic muscle, i. 127.
-
Vocal music of birds, ii. 51.
-
Vocal organs of man, i. 58;
-
of birds, i. 59; ii. 163;
-
of frogs, ii. 28;
-
of the Insessores, ii. 55;
-
difference of, in the sexes of birds, ii. 56;
-
primarily used in relation to the propagation of the species, ii. 330.
-
Vogt, Carl, on the origin of species, i. 1;
-
on the origin of man, i. 4;
-
on the semilunar fold in man, i. 23;
-
on the imitative faculties of microcephalous idiots, i. 57;
-
on microcephalous idiots, i. 121;
-
on skulls from Brazilian caves, i. 218;
-
on the evolution of the races of man, i. 230;
-
on the formation of the skull in women, ii. 317;
-
on the Ainos and negroes, ii. 321;
-
on the increased cranial difference of the sexes in man with race-development, ii. 329;
-
on the obliquity of the eye in the Chinese and Japanese, ii. 344.
-
Voice in mammals, ii. 274;
-
in monkeys and man, ii. 319;
-
in man, ii. 330;
-
origin of, in air-breathing vertebrates, ii. 331.
-
Von Baer, definition of advancement in the organic scale, i. 211.
-
Vulpian, Prof., on the resemblance between the brains of man and of the higher apes, i. 11.
-
Vultures, selection of a mate by the female, ii. 116;
-
colours of, ii. 229.
-
W.
-
Waders, young of, ii. 217.
-
Wagner, R., on the occurrence of the diastema in a Kafir skull, i. 126;
-
on the bronchi of the black stork, ii. 60.
-
Wagtail, Ray’s, arrival of the male before the female, i. 260.
-
Wagtails, Indian, young of, ii. 190.
-
Waist, proportions of, in soldiers and sailors, i. 117.
-
Waitz, Prof., on the number of species of man, i. 226;
-
on the colour of Australian infants, ii. 318;
-
on the beardlessness of negroes, ii. 321;
-
on the fondness of mankind for ornaments, ii. 338;
-
on the liability of negroes to tropical fevers after residence in a cold climate, i. 243;
-
on negro ideas of female beauty, ii. 346;
-
on Javanese and Cochin Chinese ideas of beauty, ii. 347.
-
Walckenaer and Gervais, on the Myriapoda, i. 340.
-
Waldeyer, M., on the hermaphroditism of the vertebrate embryo, i. 207.
-
Wales, North, numerical proportion of male and female births in, i. 301.
-
Walker, Alex., on the large size of the hands of labourers’ children, i. 117.
-
Walker, F., on sexual differences in the diptera, i. 348.
-
Wallace, Dr. A., on the prehensile
-
472
-
use of the tarsi in male moths, i. 256;
-
on the rearing of the Ailanthus silk-moth, i. 311;
-
on breeding Lepidoptera, i. 311;
-
proportion of sexes of Bombyx cynthia, B. yamamai, and B. Pernyi, reared by, i. 313;
-
on the development of Bombyx cynthia and B. yamamai, i. 346;
-
on the pairing of Bombyx cynthia, i. 401;
-
on the fertilisation of moths, i. 406.
-
Wallace, A. R., on the origin of man, i. 4;
-
on the power of imitation in man, i. 39;
-
on the use of missiles by the orang, i. 52;
-
on the varying appreciation of truth among different tribes, i. 100;
-
on the limits of natural selection in man, i. 137, 158;
-
on the occurrence of remorse among savages, i. 165;
-
on the effects of natural selection on civilised nations, i. 168;
-
on the use of the convergence of the hair at the elbow in the orang, i. 193;
-
on the contrast in the characters of the Malays and Papuans, i. 216;
-
on the line of separation between the Papuans and Malays, i. 218;
-
on the sexes of Ornithoptera Crœsus, i. 310;
-
on protective resemblances, i. 322;
-
on the relative sizes of the sexes of insects, i. 346;
-
on Elaphomyia, i. 349;
-
on the Birds of Paradise, i. 269;
-
on the pugnacity of the males of Leptorhynchus angustatus, i. 375;
-
on sounds produced by Euchirus longimanus, i. 381;
-
on the colours of Diadema, i. 388;
-
on Kallima, i. 392;
-
on the protective colouring of moths, i. 394;
-
on bright coloration as protective in butterflies, i. 395;
-
on variability in the Papilionidæ, i. 402;
-
on male and female butterflies inhabiting different stations, i. 403;
-
on the protective nature of the dull colouring of female butterflies, i. 403, 405, 414;
-
on mimickry in butterflies, i. 412;
-
on the mimickry of leaves by Phasmidæ, i. 414;
-
on the bright colours of caterpillars, i. 416;
-
on brightly-coloured fishes frequenting reefs, ii. 17;
-
on the coral snakes, ii. 31;
-
on Paradisea apoda, ii. 74, 78;
-
on the display of plumage by male Birds of Paradise, ii. 88;
-
on assemblies of Birds of Paradise, ii. 101;
-
on the instability of the ocellated spots in Hipparchia Janira, ii. 132;
-
on sexually limited inheritance, ii. 155;
-
on the sexual coloration of birds, ii. 166, 196, 197, 200, 206;
-
on the relation between the colours and nidification of birds, ii. 166, 171;
-
on the coloration of the Cotingidæ, ii. 177;
-
on the females of Paradisea apoda and papuana, ii. 193;
-
on the incubation of the cassowary, ii. 204;
-
on protective coloration in birds, ii. 223;
-
on the hair of the Papuans, ii. 340;
-
on the Babirusa, ii. 264;
-
on the markings of the tiger, ii. 302;
-
on the beards of the Papuans, ii. 322;
-
on the distribution of hair on the human body, ii. 375.
-
Walrus, development of the nictitating membrane in the, i. 23;
-
tusks of the, ii. 241, 248;
-
use of the tusks by the, ii. 257.
-
Walsh, B. D., on the proportion of the sexes in Papilio Turnus, i. 310;
-
on the Cynipidæ and Cecidomyidæ, i. 314;
-
on the jaws of Ammophila, i. 342;
-
on Corydalis cornutus, i. 342;
-
on the prehensile organs of male insects, i. 342;
-
on the antennæ of Penthe, i. 343;
-
on the caudal appendages of dragon-flies, i. 344;
-
on Platyphyllum concavum, i. 356;
-
on the sexes of the Ephemeridæ, i. 361;
-
on the difference of colour in the sexes of Spectrum femoratum, i. 361;
-
on sexes of dragon-flies, i. 361;
-
on the difference of the sexes in the Ichneumonidæ, i. 365;
-
on the sexes of Orsodacna atra, i. 368;
-
on the variation of the horns of the male Phanæus carnifex, i. 370;
-
on the coloration of the species of Anthocharis, i. 393.
-
Wapiti, battles of, ii. 240;
-
traces of horns in the female, ii. 245;
-
attacking a man, ii. 253;
-
crest of the male, ii. 282;
-
sexual difference in the colour of the, ii. 289.
-
Warbler, Hedge-, ii. 198;
-
young of the, ii. 209.
473
-
Warblers, Superb, nidification of, ii. 169.
-
Wariness, acquired by animals, i. 50.
-
Warington, R., on the habits of the sticklebacks, ii. 2, 20;
-
on the brilliant colours of the male stickleback during the breeding season, ii. 14.
-
Wart-hog, tusks and pads of the, ii. 265.
-
Watchmakers, short-sighted, i. 118.
-
Water-hen, ii. 40.
-
Waterhouse, C. O., on blind beetles, i. 367;
-
on difference of colour in the sexes of beetles, i. 367.
-
Waterhouse, G. R., on the voice of Hylobates agilis, ii. 332.
-
Water-ouzel, autumn song of the, ii. 54.
-
Waterton, C., on the pairing of a Canada goose with a Bernicle gander, ii. 114;
-
on hares fighting, ii. 239;
-
on the Bell-bird, ii. 79.
-
Wattles, disadvantageous to male birds in fighting, ii. 98.
-
Wealth, influence of, i. 169.
-
Weale, J. Mansel, on a South African caterpillar, i. 416.
-
Weapons, employed by monkeys, i. 51;
-
use of, i. 137;
-
offensive, of males, i. 257;
-
of mammals, ii. 241 et seq.
-
Weaver-bird, ii. 54.
-
Weaver-birds, rattling of the wings of, ii. 62;
-
Webb, Dr., on the wisdom teeth, i. 25.
-
Wedgwood, Hensleigh, on the origin of language, i. 56.
-
Weevils, sexual difference in length of snout in some, i. 255.
-
Weir, Harrison, on the numerical proportion of the sexes in pigs and rabbits, i. 305;
-
on the sexes of young pigeons, i. 306;
-
on the songs of birds, ii. 52;
-
on pigeons, ii. 109;
-
on the dislike of blue pigeons to other coloured varieties, ii. 118;
-
on the desertion of their mates by female pigeons, ii. 119.
-
Weir, J. Jenner, on the nightingale and blackcap, i. 259;
-
on the relative sexual maturity of male birds, i. 261;
-
on female pigeons deserting a feeble mate, i. 262;
-
on three starlings frequenting the same nest, i. 269;
-
on the proportion of the sexes in Machetes pugnax and other birds, i. 306, 307;
-
on the coloration of the Triphænæ, i. 395;
-
on the rejection of certain caterpillars by birds, i. 417;
-
on sexual differences of the beak in the goldfinch, ii. 40;
-
on a piping bullfinch, ii. 52;
-
on the object of the nightingale’s song, ii. 52;
-
on song-birds, ii. 53;
-
on the pugnacity of male fine-plumaged birds, ii. 93;
-
on the courtship of birds, ii. 94;
-
on the finding of new mates by Peregrine-falcons and Kestrels, ii. 104;
-
on the bullfinch and starling, ii. 105;
-
on the cause of birds remaining unpaired, ii. 107;
-
on starlings and parrots living in triplets, ii. 107;
-
on recognition of colour by birds, ii. 110;
-
on hybrid birds, ii. 113;
-
on the selection of a greenfinch by a female canary, ii. 115;
-
on a case of rivalry of female bullfinches, ii. 121;
-
on the maturity of the Golden pheasant, ii. 213.
-
Weisbach, Dr., measurement of men of different races, i. 216;
-
on the greater variability of men than of women, i. 275;
-
on the relative proportions of the body in the sexes of different races of man, ii. 320.
-
Welcker, M., on Brachycephaly and Dolichocephaly, i. 148;
-
on sexual differences in the skull in man, ii. 317.
-
Wells, Dr., on the immunity of coloured races from certain poisons, i. 243.
-
Westring, on the stridulation of Reduvius personatus, i. 350;
-
on the stridulating organs of the Coleoptera, i. 382;
-
on sounds produced by Cychrus, i. 382;
-
on the stridulation of males of Theridion, i. 339;
-
on the stridulation of beetles, i. 379;
-
on the stridulation of Omaloplia brunnea, i. 381.
-
Westphalia, greater proportion of female illegitimate children in, i. 301.
474
-
Westropp, H. M., on the prevalence of certain forms of ornamentation, i. 233.
-
Westwood, J. O., on the classification of the Hymenoptera, i. 188;
-
on the Culicidæ and Tabanidæ, i. 254;
-
on a Hymenopterous parasite with a sedentary male, i. 272;
-
on the proportions of the sexes in Lucanus cervus and Siagonium, i. 313;
-
on the absence of ocelli in female mutillidæ, i. 341;
-
on the jaws of Ammophila, i. 342;
-
on the copulation of insects of distinct species, i. 342;
-
on the male of Crabro cribrarius, i. 343;
-
on the pugnacity of male Tipulæ i. 349;
-
on the stridulation of Pirates stridulus, i. 350;
-
on the Cicadæ, i. 351;
-
on the stridulating organs of the crickets, i. 354;
-
on Pneumora, i. 357;
-
on Ephippiger vitium, i. 355, 358;
-
on the pugnacity of the Mantides, i. 360;
-
on Platyblemnus, i. 361;
-
on difference in the sexes of the Agrionidæ, i. 362;
-
on the pugnacity of the males of a species of Tenthredinæ, i. 364;
-
on the pugnacity of the male stag-beetle, i. 375;
-
on Bledius taurus and Siagonium, i. 375;
-
on lamellicorn beetles, i. 378;
-
on the coloration of Lithosia, i. 396.
-
Whale, Sperm-, battles of male, ii. 240.
-
Whales, nakedness of, i. 148.
-
Whately, Archb., language not peculiar to man, i. 53;
-
on the primitive civilisation of man, i. 181.
-
Whewell, Prof., on maternal affection, i. 40.
-
Whiskers, in monkeys, i. 192.
-
White, Gilbert, on the proportion of the sexes in the partridge, i. 306;
-
on the house-cricket, i. 352;
-
on the object of the song of birds, ii. 52;
-
on the finding of new mates by white owls, ii. 105;
-
on spring coveys of male partridges, ii. 107.
-
Whiteness, a sexual ornament in some birds, ii. 232;
-
of mammals inhabiting snowy countries, ii. 298.
-
White-throat, aerial love-dance of the male, ii. 68.
-
Widow-bird, polygamous, i. 269;
-
breeding plumage of the male, ii. 84, 97;
-
female, rejecting the unadorned male, ii. 120.
-
Widows and widowers, mortality of, i. 176.
-
Wigeon, pairing with a pintail duck, ii. 114.
-
Wilckens, Dr., on the modification of domestic animals in mountainous regions, i. 120;
-
on a numerical relation between the hairs and excretory pores in sheep, i. 248.
-
Wilder, Dr. Burt, on the greater frequency of supernumerary digits in men than in women, i. 276.
-
Williams, on the marriage-customs of the Fijians, ii. 374.
-
Wilson, Dr., on the conical heads of the natives of North-Western America, ii. 351;
-
on the Fijians, ii. 352;
-
on the persistence of the fashion of compressing the skull, ii. 353.
-
Wing-spurs, ii. 162.
-
Wings, differences of, in the two sexes of butterflies and Hymenoptera, i. 345;
-
play of, in the courtship of birds, ii. 95.
-
Winter, change of colour of mammals in, ii. 298.
-
Witchcraft, i. 68.
-
Wives, traces of the forcible capture of, i. 182.
-
Wolf, winter change of the, ii. 298.
-
Wolff, on the variability of the viscera in man, i. 109.
-
Wollaston, T. V., on Eurygnathus, i. 344;
-
on musical curculionidæ, i. 378;
-
on the stridulation of Acalles, i. 384.
-
Wolves learning to bark from dogs, i. 44;
-
Wolves, black, ii. 294.
-
Wombat, black varieties of the, ii. 294.
-
Women distinguished from men by male monkeys, i. 13;
-
preponderance of, in numbers, i. 302;
-
effects of selection of, in accordance with different standards of beauty, ii. 355;
-
practice of capturing, ii. 360, 364;
-
early betrothals and slavery of, ii. 366;
-
selection of, for beauty, ii. 372;
475
-
freedom of selection by, in savage tribes, ii. 372.
-
Wonder, manifestations of, by animals, i. 42.
-
Wonfor, Mr., on sexual peculiarities in the wings of butterflies, i. 345.
-
Woolner, Mr., observations on the ear in man, i. 22.
-
Wood, J., on muscular variations in man, i. 109, 128, 129;
-
on the greater variability of the muscles in men than in women, i. 275.
-
Wood, T. W., on the colouring of the orange-tip butterfly, i. 394;
-
on the habits of the Saturniidæ, i. 398;
-
on the habits of Menura Alberti, ii. 56;
-
on Tetrao cupido, ii. 56;
-
on the display of plumage by male pheasants, ii. 89;
-
on the ocellated spots of the Argus pheasant, ii. 144;
-
on the habits of the female Cassowary, ii. 204.
-
Woodcock, coloration of the, ii. 226.
-
Woodpecker, selection of a mate by the female, ii. 116.
-
Woodpeckers, ii. 56;
-
tapping of, ii. 62;
-
colours and nidification of the, ii. 171, 174, 223;
-
characters of young, ii. 185, 199, 209.
-
Wormald, Mr., on the coloration of Hypopyra, i. 397.
-
Wounds, healing of, i. 13.
-
Wren, ii. 198;
-
Wright, C. A., on the young of Orocetes and Petrocincla, ii. 220.
-
Wright, Chauncey, on correlative acquisition, ii. 335;
-
on the enlargement of the brain in man, ii. 391.
-
Wright, Mr., on the Scotch deerhound, ii. 261;
-
on sexual preference in dogs, ii. 271;
-
on the rejection of a horse by a mare, ii. 272.
-
Wright, W. von, on the protective plumage of the Ptarmigan, ii. 81.
-
Writing, i. 182.
-
Wyman, Prof., on the prolongation of the coccyx in the human embryo, i. 16;
-
on the condition of the great toe in the human embryo, i. 17;
-
on variation in the skulls of the natives of the Sandwich Islands, i. 108;
-
on the hatching of the eggs in the mouths and branchial cavities of male fishes, i. 210, ii. 20.
-
X.
-
Xenarchus, on the Cicadæ, i. 350.
-
Xenorhynchus, sexual difference in the colour of the eyes in, ii. 129.
-
Xiphophorus Hellerii, peculiar anal fin of the male, ii. 9, 10.
-
Xylocopa, difference of the sexes in, i. 366.
-
Y.
-
Yarrell, W., on the habits of the Cyprinidæ, i. 309;
-
on Raia clavata, ii. 2;
-
on the characters of the male salmon during the breeding season, ii. 4, 14;
-
on the characters of the rays, ii. 6;
-
on the gemmeous dragonet, ii. 8;
-
on the spawning of the salmon, ii. 19;
-
on the incubation of the Lophobranchii, ii. 21;
-
on rivalry in song-birds, ii. 53;
-
on the trachea of the swan, ii. 60;
-
on the moulting of the anatidæ, ii. 85;
-
on an instance of reasoning in a gull, ii. 108;
-
on the young of the waders, ii. 217.
-
Yellow fever, immunity of negroes and mulattoes from, i. 243.
-
Youatt, Mr., on the development of the horns in cattle, i. 284.
-
Yura-caras, their notions of beauty, ii. 347.
-
Z.
-
Zebra, rejection of an ass by a female, ii. 295;
-
Zebus, humps of, i. 284.
-
Zigzags, prevalence of, as ornaments, i. 233.
-
Zincke, Mr., on European emigration to America, i. 179.
-
Zootoca vivipara, sexual difference in the colour of, ii. 36.
-
Zygænidæ, coloration of the, i. 396.
THE END.
LONDON:
PRINTED BY WILLIAM CLOWES AND SONS, DUKE STREET, STAMFORD STREET,
AND CHARING CROSS.
1
50a, Albemarle Street, London
January, 1871.
MR. MURRAY’S
LIST OF STANDARD WORKS.
AIDS TO FAITH; a Series of Theological Essays. By various
Writers. Seventh Edition. 8vo. 9s.
CONTENTS:
Miracles.—Dean Mansel.
Evidences of Christianity.—Bishop of Killaloe.
Prophecy—and the Mosaic Record of Creation.—Rev. Dr. M’Caul.
Ideology and Subscription.—Canon F. C. Cook.
The Pentateuch.—Rev. George Rawlinson.
Inspiration.—Bishop of Ely.
Death of Christ.—Archbishop of York.
Scripture and its Interpretation.—Bishop of Gloucester and Bristol.
AUSTIN’S (John) LECTURES ON JURISPRUDENCE; or, The
Philosophy of Positive Law. Third Edition. Revised by ROBERT
CAMPBELL. 2 vols. 8vo. 32s.
——— (Sarah) POLITICAL AND ECCLESIASTICAL
HISTORY OF THE POPES OF ROME. Translated from the German of
Leopold Ranke. Fourth Edition. With a Preface by Dean Milman. 3 vols.
8vo. 30s.
BARROW’S (Sir John) AUTOBIOGRAPHICAL MEMOIR,
including Reflections, Observations, and Reminiscences at Home and Abroad.
From Early Life to Advanced Age. Portrait. 8vo. 15s.
——— VOYAGES OF DISCOVERY AND RESEARCH
WITHIN THE ARCTIC REGIONS, since 1818. Abridged and Arranged
from the Official Narratives. 8vo. 15s.
BARRY’S (Alfred, D.D.) MEMOIR OF THE LIFE AND
WORKS OF SIR CHARLES BARRY, R.A. Second Edition. With Portrait
and 40 Illustrations. Medium 8vo. 15s.
BELCHER’S (Lady) MUTINEERS OF THE ‘BOUNTY,’ AND
THEIR DESCENDANTS; in Pitcairn and Norfolk Islands. With
Illustrations. Post 8vo. 12s.
BELL’S (Sir Chas.) FAMILIAR LETTERS. With Portrait.
Crown 8vo. 12s.
BERTRAM’S (Jas. G.) HARVEST OF THE SEA; A Contribution
to the Natural and Economic History of the British Food
Fishes. Second Edition. With 50 Illustrations. 12s.
BIBLE COMMENTARY; THE HOLY BIBLE, according to
the Authorized Version, a.d. 1611. With an Explanatory and Critical
Commentary and a Revision of the Translation. By BISHOPS and
other CLERGY of the ANGLICAN CHURCH. Edited by F. C. COOK, M.A.,
Canon of Exeter. Vol. I. The Pentateuch. PARTS 1 & 2. Medium 8vo. 30s.
BIRCH’S (Samuel) HISTORY OF ANCIENT POTTERY AND
PORCELAIN. Egyptian, Assyrian, Greek, Etruscan, and Roman. With
coloured Plates and 200 Woodcuts. 2 vols. Medium 8vo. 42s.2
BANKES’ (George) STORY OF CORFE CASTLE, including
the Private Memoirs of a Family resident there in the time of the Civil Wars,
together with Unpublished Correspondence of the Ministers and Court of
Charles I. at York and Oxford. With Woodcuts. Post 8vo. 10s. 6d.
BISSET’S (Andrew) HISTORY OF THE COMMONWEALTH
OF ENGLAND, from the Death of Charles the First to the Expulsion
of the Long Parliament by Cromwell. From MSS. in the State Paper
Office, &c. 2 vols. 8vo. 30s.
BYRON’S (Lord) POETICAL WORKS. Edited with Notes.
Library Edition. With Portrait. 6 vols. 8vo. 45s.
—————— With Notes and Illustrations.
Cabinet Edition. With Plates. 10 vols. Fcap. 8vo. 30s.
—————— With Portrait and
Illustrations. One Volume. Royal 8vo. 9s.
——— LIFE. With his Letters and Journals. By THOMAS
MOORE. With Notes and Illustrations. Cabinet Edition. With Plates.
6 vols. Fcap. 8vo. 18s.
—————— With Portraits. One Volume. Royal 8vo. 9s.
BLUNT’S (Rev. J. J.) LECTURES ON THE RIGHT USE OF
THE EARLY FATHERS. Third Edition. 8vo. 9s.
——— UNDESIGNED COINCIDENCES IN THE OLD
AND NEW TESTAMENTS: an Argument of their Veracity. With an
Appendix, containing Undesigned Coincidences between the Gospels, Acts,
and Josephus. Ninth Edition. Post 8vo. 6s.
——— CHRISTIAN CHURCH DURING THE FIRST
THREE CENTURIES. Fourth Edition. Post 8vo. 6s.
——— PARISH PRIEST: His Duties, Acquirements, and
Obligations. Fifth Edition. Post 8vo. 6s.
——— PLAIN SERMONS PREACHED TO A COUNTRY
CONGREGATION. Fifth Edition. 2 vols. Post 8vo. 12s.
BONAPARTE’S (Napoleon) CONFIDENTIAL CORRESPONDENCE
WITH HIS BROTHER JOSEPH, KING OF SPAIN. Selected
and Translated with Explanatory Notes. 2 vols. 8vo. 26s.
BORROW’S (George) GYPSIES OF SPAIN; their Manners,
Customs, Religion and Language. Third Edition. 2 vols. Post 8vo. 18s.
——— BIBLE IN SPAIN; or, The Journeys, Adventures,
and Imprisonments of an Englishman in an attempt to circulate the Scriptures
in the Peninsula. Fourth Edition. 3 vols. Post 8vo. 27s.
——— LAVENGRO; The Scholar—The Gipsy—and The
Priest. With Portrait. 3 vols. Post 8vo. 30s.
——— ROMANY RYE; A Sequel To Lavengro. 2 vols.
Post 8vo. 21s.
BOSWELL’S (James) LIFE OF SAMUEL JOHNSON, LL.D.;
including the TOUR to the HEBRIDES. By the Rt. Hon. J. W. CROKER.
With Portraits. Royal 8vo. 10s.
BRAY’S (Mrs.) REVOLT OF THE PROTESTANTS IN THE
CEVENNES. With some Account of the Huguenots in the Seventeenth
Century. Post 8vo. 10s. 6d.
——— LIFE OF THOMAS STOTHARD, R.A. With Personal
Reminiscences. With Portrait and Illustrations. 8vo. 21s.3
BROGDEN’S (Rev. Jas.) ILLUSTRATIONS OF THE
LITURGY AND RITUAL OF THE CHURCH OF ENGLAND AND
IRELAND; selected from the Works of eminent Divines of the 17th
Century. 3 vols. Post 8vo. 27s.
——— CATHOLIC SAFEGUARDS against the Errors,
Corruptions, and Novelties of the Church of Rome. 3 vols. 8vo. 42s.
BULGARIA; Notes on the Resources and Administration of
Turkey—the Condition, Character, Manners, Customs, and Language of
the Christian and Mussulman Populations, &c. By S. G. B. ST. CLAIR
and CHARLES A. BROPHY. 8vo. 12s.
CAMPBELL’S (Lord) LIVES OF THE LORD CHANCELLORS
AND KEEPERS OF THE GREAT SEAL OF ENGLAND, from
the Earliest Times to the Reign of George the Fourth. 10 vols. Post 8vo.
60s.
—————— LIVES OF LORDS LYNDHURST
AND BROUGHAM. 8vo. 16s.
——— (Sir Neil) JOURNAL OF OCCURRENCES,
and Notes of Conversations with Napoleon at Fontainbleau and Elba in
1814-15. With a Memoir of that Officer. By his Nephew, REV. A. N. C.
MACLACHLAN. With Portrait. 8vo. 15s.
——— (George) MODERN INDIA. A Sketch of the
System of Civil Government. With some Account of the Natives and Native
Institutions. Second Edition. 8vo. 16s.
——— INDIA AS IT MAY BE. An Outline of a
Proposed Government and Policy. 8vo. 12s.
CASTLEREAGH’S (Viscount) MEMOIRS. CORRESPONDENCE,
AND DESPATCHES. Edited by THE MARQUIS OF LONDONDERRY.
12 vols. 8vo. 14s. each.
CATHCART’S (Sir George) COMMENTARIES ON THE
WAR IN RUSSIA AND GERMANY, 1812-13. With Plans. 8vo. 14s.
——— MILITARY OPERATIONS IN KAFFRARIA.
Second Edition. 8vo. 12s.
CHALMERS’ (George) POETICAL REMAINS OF SOME OF
THE SCOTTISH KINGS. Post 8vo. 10s. 6d.
CHURCH AND THE AGE. Essays on the Principles and
Present Position of the Anglican Church. By various Writers. Second
Edition. 8vo. 14s.
CONTENTS:
Anglican Principles.—Dean Hook.
Modern Religious Thought.—Bishop of Gloucester and Bristol.
The State, The Church, and Synods.—Rev. Dr. Irons.
Religious Use of Taste.—Rev. R. St. John Tyrwhitt.
Place of the Laity.—Professor Burrows.
The Parish Priest.—Rev. Walsham How.
Divines of 16th and 17th Centuries.—Rev. A. W. Haddan.
Liturgies and Ritual.—Rev. M. F. Sadler.
The Church and Education.—Rev. Dr. Barry.
Indian Missions.—Sir Bartle Frere.
The Church and the People.—Rev. W. D. Maclagan.
Conciliation and Comprehension.—Rev. A. Weir.
CHURTON AND JONES’ (Archdeacon) NEW TESTAMENT.
With a Plain Explanatory Commentary for Families and General Readers;
with more than 100 Illustrations of Scripture Scenes, from Photographs
and Sketches taken on the Spot. 2 vols. 8vo. 21s.4
CICERO’S LIFE and TIMES, with his Character as a Statesman,
Orator, and Friend; and a Selection from his Correspondence and
Orations. By WILLIAM FORSYTH. Third Edition. With 40 Illustrations.
8vo. 10s. 6d.
CLODE’S (C. M.) HISTORY OF THE ADMINISTRATION
AND GOVERNMENT OF THE BRITISH ARMY from the Revolution,
1688. 2 vols. 8vo. 42s.
COLCHESTER’S (Lord) DIARY AND CORRESPONDENCE
WHILE SPEAKER OF THE HOUSE OF COMMONS, 1802-1817. Edited
by HIS SON. With Portrait. 3 vols. 8vo. 42s.
CORNWALLIS’S (Marquis) CORRESPONDENCE DURING
THE AMERICAN WAR: Administrations in India,—Union with Ireland,
and Peace of Amiens. Edited by CHARLES ROSS. Second Edition. With
Portrait. 3 vols. 8vo. 63s.
COWPER’S (Countess) DIARY WHILE LADY OF THE
BEDCHAMBER TO CAROLINE, PRINCESS OF WALES, 1714-20.
Edited by Hon. Spencer Cowper. Second Edition. Portrait. 8vo. 10s. 6d.
CRABBE’S (Rev. George) POETICAL WORKS; with his Life,
Letters, and Journals. By HIS SON. With Notes and Illustrations.
Cabinet Edition. With Plates, 8 vols., Fcap. 8vo, 24s.; or, with Illustrations,
one volume, Royal 8vo. 7s.
CROKER’S (Rt. Hon. J. W.) WORKS OF ALEXANDER
POPE. With Introductions and Notes by Rev. Whitwell Elwin. Vols. I.
to III. With Portraits. 8vo. 10s. 6d. each.
——— BOSWELL’S LIFE OF SAMUEL JOHNSON, D.D.,
including their Tour to the Hebrides. Edited with Notes. With Portraits.
1 vol. Royal 8vo. 10s.
——— ESSAYS ON THE FRENCH REVOLUTION.
8vo. 16s.
CROWE AND CAVALCASELLE’S HISTORY OF PAINTING
IN ITALY, from the Second to the Sixteenth Century. With 100
Illustrations. 3 vols. 8vo. 63s.
—————— HISTORY OF PAINTING IN NORTH
ITALY. With Illustrations. 2 vols. 8vo.
—————— EARLY FLEMISH PAINTERS. With
Illustrations. Post 8vo. 12s.
CUNNINGHAM’S (Peter) GOLDSMITH’S WORKS. Printed
from the last Edition, revised by the Author, and edited, with Notes. With
Vignettes. 4 vols. 8vo. 30s.
——— JOHNSON’S LIVES OF THE MOST
EMINENT ENGLISH POETS. With Critical Observations on their Works.
Edited, with Notes. 3 vols. 8vo. 22s. 6d.
——— (J. D.) HISTORY OF THE SIKHS, from
the Origin of the Nation to the Battles of the Sutlej. Second Edition. With
Maps. 8vo. 15s.
CUST’S (Sir Edward) ANNALS OF THE WARS OF THE
18th and 19th CENTURIES, 1700-1815. With Maps. 9 vols. Fcap. 8vo.
5s. each.
——— LIVES OF THE WARRIORS OF THE 17th CENTURY—The
Thirty Years’ War—The Civil Wars of France and England—The
Commanders of Fleets and Armies before the Enemy. 1604-1704. 6 vols.
Post 8vo. 50s.5
DARWIN’S (Charles) Journal of Researches into the Natural
History of the Countries visited during a Voyage round the World. Tenth
Edition. Post 8vo. 9s.
——— ORIGIN of SPECIES by MEANS of NATURAL
SELECTION; or, the Preservation of Favoured Races in the Struggle for Life.
Fifth Edition. Post 8vo. 14s.
——— FERTILIZATION OF ORCHIDS THROUGH
INSECT AGENCY, and as to the good of Intercrossing. With Woodcuts.
Post 8vo. 9s.
——— VARIATION OF ANIMALS AND PLANTS
UNDER DOMESTICATION. With Illustrations. 2 vols. 8vo. 28s.
——— DESCENT OF MAN, and on SELECTION in
RELATION to SEX. With Illustrations. 2 vols. Crown 8vo. 24s.
DELEPIERRE’S (Octave) HISTORY OF FLEMISH LITERATURE
FROM THE TWELFTH CENTURY. 8vo. 9s.
DENISON’S (E. B.) LIFE OF BISHOP LONSDALE. With
Portrait. Crown 8vo. 10s. 6d.
DERBY’S (Earl of) HOMER’S ILIAD RENDERED INTO
ENGLISH BLANK VERSE. Seventh Edition. 2 vols. Post 8vo. 10s.
DE ROS’ (Lord) MEMORIALS OF THE TOWER OF LONDON.
Second Edition. With Illustrations. Crown 8vo. 12s.
DEVEREUX’S (W. B.) LIVES OF THE EARLS OF ESSEX
IN THE REIGNS OF ELIZABETH, JAMES I., AND CHARLES I.
Portraits. 2 vols. 8vo. 30s.
DOUGLAS’ (Sir Howard) LIFE AND ADVENTURES. By
S. W. FULLOM. 8vo. 14s.
——— TREATISE ON GUNNERY. Fifth Edition.
Woodcuts. 8vo. 21s.
——— CONSTRUCTION OF MILITARY BRIDGES
and the Passage of Rivers in Military Operations. Plates. 8vo. 21s.
DUCANGE’S MEDIÆVAL LATIN-ENGLISH DICTIONARY.
Illustrated and enlarged by numerous additions, derived from patristic
and scholastic authors, the works of the Record Commission, Mediæval
Histories, Charters, Glossaries, &c., &c. By E. A. DAYMAN, B.D. 4to.
[In Preparation.
DUDLEY’S (Earl of) LETTERS TO BISHOP COPLESTONE.
Second Edition. Portrait. 8vo. 10s. 6d.
DYER’S (Thos. H.) HISTORY OF MODERN EUROPE, from
the Taking of Constantinople by the Turks to the Close of the War in the
Crimea, 1453-1857. With an Index. 4 vols. 8vo. 42s.
——— LIFE AND LETTERS OF JOHN CALVIN. Compiled
from authentic Sources. With Portrait. 8vo. 15s.
EASTLAKE’S (Sir Charles) CONTRIBUTIONS TO THE
LITERATURE OF THE FINE ARTS. Second Edition. 8vo. 12s.
——— MEMOIR; With Selections from his Correspondence,
and Additional Contributions to the Literature of the Fine
Arts. By LADY EASTLAKE. 8vo. 12s.
——— ITALIAN SCHOOLS OF PAINTING. From
the German of Kugler. Edited, with Notes. Sixth Edition. With 100
Illustrations. 2 vols. Post 8vo. 30s.6
EGYPTIANS (ANCIENT): Their Manners and Customs. By
SIR J. GARDNER WILKINSON. Fourth Edition. with illustrations.
2 vols. Post 8vo. 12s.
——— (MODERN): Their Manners and Customs. By
E. W. LANE. Fifth Edition. With Illustrations. 2 vols. Post 8vo. 12s.
ELLESMERE’S (Lord) ESSAYS ON HISTORY, BIOGRAPHY,
GEOGRAPHY, and ENGINEERING. 8vo. 12s.
ELPHINSTONE’S (Mount Stuart) HISTORY OF INDIA.
The Hindu and Mahometan Periods. Fifth Edition. With Notes and
Additions by PROFESSOR COWELL. With Map. 8vo. 18s.
ELWIN’S (Rev. Whitwell) WORKS OF ALEXANDER POPE.
With Introductions and Notes, and many original Letters now for the first
time published. With Portrait. Vols. I to III. 8vo. 10s. 6d. each.
ENGEL’S (Carl) MUSIC OF THE MOST ANCIENT
NATIONS; particularly of the Assyrians, Egyptians, and Hebrews; with
Special Reference to the Discoveries in Western Asia and in Egypt. Second
Edition. With 100 Illustrations. 8vo. 10s. 6d.
FARRAR’S (Rev. A. S.) CRITICAL HISTORY OF FREE
THOUGHT IN REFERENCE TO THE CHRISTIAN RELIGION. 8vo.
16s.
FEATHERSTONHAUGH’S (G. W.) TOUR THROUGH THE
SLAVE STATES OF NORTH AMERICA, from the River Potomac, to
Texas and the Frontiers of Mexico. 2 vols. 8vo. 26s.
FERGUSSON’S (James) HISTORY OF ARCHITECTURE IN
ALL COUNTRIES. From the Earliest Times. With 1200 Illustrations.
VOLS. I. & II. 8vo. 42s. each.
——— Vol. III. The Modern Styles. With 312 Illustrations.
8vo. 31s. 6d.
FERRIER’S (T. P.) CARAVAN JOURNEYS IN PERSIA,
AFFGHANISTAN, HERAT, TURKISTAN, AND BELOOCHISTAN, with
Descriptions of Meshed, Balk, and Candahar, and Sketches of the Nomade
Tribes of Central Asia. Second Edition. With Map. 8vo. 21s.
——— HISTORY OF THE AFFGHANS. With Map.
8vo. 21s.
FORSTER’S (John) HISTORY OF THE GRAND REMONSTRANCE,
1641. With an Introductory Essay on English Freedom under
Plantagenet and Tudor Sovereigns. Second Edition. 8vo. 12s.
——— LIFE OF SIR JOHN ELIOT, 1590-1632. With
Portrait. 2 vols. 8vo. 30s.
——— CROMWELL, DEFOE, STEELE, CHURCHILL,
FOOTE.—Biographies. Post 8vo. 12s.
FORSYTH’S (William) LIFE AND TIMES OF CICERO.
With Selections from his Correspondence and Orations. Third Edition.
With Illustrations. 8vo. 10s. 6d.
FOSS’ (Edward) JUDGES OF ENGLAND. With Sketches of
their Lives, and Notices of the Courts at Westminster, from the Conquest to
the Present Time. 9 vols. 8vo. 126s.
——— BIOGRAPHICAL DICTIONARY OF THE JUDGES
OF ENGLAND, FROM THE CONQUEST TO THE PRESENT TIME, 1066-1870.
Condensed from the above work. Medium 8vo. 21s.7
GEORGE THE THIRD’S CORRESPONDENCE WITH LORD
NORTH, 1769-82. Edited, with Notes and Introduction, by W. BODHAM
DONNE. 2 vols. 8vo. 32s.
GIBBON’S (Edward) HISTORY OF THE DECLINE AND
FALL OF THE ROMAN EMPIRE. With Notes by DEAN MILMAN
and M. GUIZOT. A new Edition. Edited, with additional Notes incorporating
the Researches of recent writers, by WM. SMITH, D.C.L. With
Portrait and Maps. 8 vols. 8vo. 60s.
GOLDSMITH’S (Oliver) WORKS. Edited, with Notes, by
PETER CUNNINGHAM, F.S.A. With Portrait and Vignettes. 4 vols. 8vo.
30s.
GRENVILLE’S (George) PUBLIC AND PRIVATE CORRESPONDENCE
WITH HIS FRIENDS AND CONTEMPORARIES,
during a period of Thirty Years. Including his Diary of Political Events
while First Lord of the Treasury. Edited, with Notes, by W. J. SMITH.
4 vols. 8vo. 16s. each.
GREY’S (Earl) CORRESPONDENCE WITH KING WILLIAM
IV. and SIR HERBERT TAYLOR, from November, 1830, to the Passing of
the Reform Act in 1832. Edited by HIS SON. 2 vols. 8vo. 30s.
GROTE’S (George) HISTORY of GREECE, from the Earliest
Period to the Close of the Generation contemporary with Alexander the Great.
Fourth Edition. With Portrait, Maps, and Plans. 8 vols. 8vo. 112s.
—————— Cabinet Edition. With Portrait and
Plans. 12 vols. Post 8vo. 6s. each.
——— PLATO AND THE OTHER COMPANIONS OF
SOCRATES. Second Edition. 3 vols. 8vo. 45s. ***. 2s. 6d.
GRUNER’S (Lewis) TERRA-COTTA ARCHITECTURE OF
NORTH ITALY. From careful Drawings and Restorations. With Illustrations,
engraved and printed in Colours. Small folio. 5l. 5s.
GUIZOT’S (M.) MEDITATIONS ON CHRISTIANITY, AND
ON THE RELIGIOUS QUESTIONS OF THE DAY. 2 vols. Post 8vo. 20s.
GURWOOD’S (Col.) SELECTIONS FROM THE WELLINGTON
DESPATCHES AND GENERAL ORDERS. Intended as a convenient
Manual for Officers while Travelling or on Service. 8vo. 18s.
GUSTAVUS VASA (Life of). His Exploits and Adventures.
With Extracts from his Correspondence. With Portrait. 8vo. 10s. 6d.
HALLAM’S (Henry) CONSTITUTIONAL HISTORY OF
ENGLAND, from the Accession of Henry VII. to the Death of George II.
Eighth Edition. 3 vols. 8vo. 30s.
——— HISTORY OF THE STATE OF EUROPE
DURING THE MIDDLE AGES. Eleventh Edition. 3 vols. 8vo. 30s.
——— LITERARY HISTORY OF EUROPE. Fourth
Edition. 3 vols. 8vo. 36s.
——— HISTORICAL WORKS. With the Author’s latest
Corrections and Additions. Containing HISTORY OF ENGLAND—EUROPE
DURING THE MIDDLE AGES—LITERARY HISTORY OF EUROPE.
Cabinet Edition. 10 vols. Post 8vo. 6s. each.
*** The public are cautioned against editions of Hallam’s Histories recently
advertised, which are merely reprints of old editions, which the author himself
declared to be full of errors, and do not contain the additional notes, &c.
The only correct editions are published by John Murray.
8
HAMILTON’S (James) WANDERINGS IN NORTHERN
AFRICA, BENGHAZI, CYRENE, THE OASIS OF SIWAH, &c. Second
Edition. With Woodcuts. Post 8vo. 12s.
——— (W. J.) RESEARCHES IN ASIA MINOR,
PONTUS, AND ARMENIA; with some Account of the Antiquities and
Geology of those Countries. With Map and Plates. 2 vols. 8vo. 38s.
HANDBOOK TO THE CATHEDRALS OF ENGLAND; a
Concise History of each See, with Biographical Notices of the Bishops. By
RICHARD J. KING, B.A. With 300 Illustrations. 6 vols. Post 8vo.
Containing:—
Southern Division; Winchester, Salisbury, Exeter, Wells,
Rochester, Canterbury, and Chichester. With 110 Illustrations.
2 vols. Crown 8vo. 24s.
Eastern Division; Oxford, Peterborough, Ely, Norwich,
and Lincoln. With 90 Illustrations. Crown 8vo. 18s.
Western Division; Bristol, Gloucester, Hereford, Worcester,
and Litchfield. With 60 Illustrations. Crown 8vo. 16s.
Northern Division; York, Ripon, Durham, Carlisle, Chester,
and Manchester. With 60 Illustrations. Crown 8vo. 2 vols. 21s.
HANNAH’S (Rev. Dr.) DIVINE AND HUMAN ELEMENTS
IN HOLY SCRIPTURE. 8vo. 10s. 6d.
HATHERLEY’S (Lord) CONTINUITY OF SCRIPTURE, as
declared by the Testimony of our Lord and of the Evangelists and Apostles.
Fourth Edition. Crown 8vo. 6s.
HEAD’S (Sir F. B.) ROYAL ENGINEER, and the Royal
Establishments at Woolwich and Chatham. With Illustrations. 8vo. 12s.
——— DEFENCELESS STATE OF GREAT BRITAIN.
Contents—1. Military Warfare. 2. Naval Warfare. 3. The Invasion of
England. 4. The Capture of London by a French Army. 5. The Treatment
of Women in War. 6. How to Defend Great Britain. Post 8vo. 12s.
——— FAGGOT OF FRENCH STICKS; or, Description of
Paris in 1851. 2nd Edition. 2 vols. Post 8vo. 12s.
——— DESCRIPTIVE ESSAYS. Contributed to the ‘Quarterly
Review.’ 2 vols. Post 8vo. 18s.
HERODOTUS: A New English Version. Edited, with copious
Notes, from the most Recent Sources of Information. By GEORGE
RAWLINSON, M.A. Assisted by Sir Henry Rawlinson and Sir Gardner
Wilkinson. Second Edition. With Maps and Woodcuts. 4 vols. 8vo.
48s.
HESSEY’S (Rev. Dr.) SUNDAY: its Origin, History, and
Present Obligations. Second Edition. Post 8vo. 9s.
HILL (Frederick) ON CRIME: its Amount, Causes, and
Remedies. 8vo. 12s.
HOMER’S ILIAD, rendered into English Blank Verse. By the
EARL OF DERBY. Seventh Edition. 2 vols. Small 8vo. 10s.
HOOK’S (Dean) CHURCH DICTIONARY: a Manual of Reference
for the Clergy—Students—and General Readers. Tenth Edition. 8vo. 16s.
HORACE. A New Edition of the Text. Edited by DEAN
MILMAN. With 100 Illustrations. Small 8vo. 7s. 6d.
——— LIFE. By Dean Milman. With Illustrations. 8vo. 9s.9
JAMESON’S (Mrs.) LIVES OF THE EARLY ITALIAN
PAINTERS—and the Progress of Painting in Italy from Cimabue to Bassano.
Tenth Edition. With 50 Portraits. Post 8vo. 12s.
JOHNSON’S (Samuel) LIFE. By JAMES BOSWELL. Including
the Tour to the Hebrides. Edited by the Rt. Hon. J. W. Croker.
With Portraits. Royal 8vo. 10s.
——— LIVES OF THE MOST EMINENT ENGLISH
POETS, with Critical Observations on their Works. Edited, with Notes,
by Peter Cunningham, F.S.A. With Portrait. 3 vols. 8vo. 22s. 6d.
JOHNSTON’S (Wm.) ENGLAND AS IT IS: Political, Social,
and Industrial, in the Nineteenth Century. 2 vols. Post 8vo. 18s.
JONES AND CHURTON’S (Archdeacon) NEW TESTAMENT.
Edited, with a Plain Practical Commentary for the use of
Families and General Readers. With 100 Panoramic and other Views
from Sketches and Photographs made on the Spot. 2 vols. Crown 8vo. 21s.
JUNIUS; the Handwriting of Junius professionally investigated.
By MR. CHABOT, Expert. With Preface and Collateral Evidence, by the
Hon. Edward Twisleton. With Facsimiles, Woodcuts, &c. 4to.
KEN’S (Bishop) LIFE. Second Edition. With Portrait. 2 vols.
8vo. 18s.
KERR’S (Robert) GENTLEMAN’S HOUSE; or, How to Plan
English Residences, from the Parsonage to the Palace. Third Edition.
With Views and Plans. 8vo. 24s.
——— (R. Malcolm) BLACKSTONE’S COMMENTARIES,
adapted to the present state of the Law. Fourth Edition. 4 vols. 8vo.
[In the Press.
KING’S (Rev. C. W.) ANTIQUE GEMS; their Origin, Use, and
Value, as Interpreters of Ancient History, and as illustrative of Ancient Art.
Second Edition. With Illustrations. 8vo. 24s.
KIRK’S (J. Foster) HISTORY OF CHARLES THE BOLD,
DUKE OF BURGUNDY. With Portraits. 3 vols. 8vo. 45s.
KORFF’S (Baron) ACCESSION OF NICHOLAS I., compiled
by special command of the Emperor Alexander II. Translated from the
Russian. 8vo. 10s. 6d.
KUGLER’S (Franz) HISTORY OF PAINTING (The Italian
Schools). Edited, with Notes, by SIR CHARLES EASTLAKE. Sixth
Edition. With Illustrations. 2 vols. Post 8vo. 30s.
————————— (German, Dutch, and Flemish Schools).
Edited, with Notes, by DR. WAAGEN. Second Edition. With Illustrations.
2 vols. Post 8vo. 24s.
LANE’S (Edw. W.) ACCOUNT OF THE MANNERS AND
CUSTOMS OF THE MODERN EGYPTIANS. Fifth Edition. With Woodcuts.
2 vols. Post 8vo. 12s.
LAYARD’S (A. H.) TRAVELS AND RESEARCHES AT
NINEVEH AND BABYLON. With an Account of the Manners and Arts
of the Ancient Assyrians; being the Narrative of a First and Second Expedition
to the Ruins of Assyria. With Maps and Illustrations. 3 vols. 8vo.
57s.
LENNEP’S (H. Van) TRAVELS IN ASIA MINOR. With
Illustrations of Biblical Literature and Archæology. With Maps and Illustrations.
2 vols. Post 8vo. 24s.10
LEWIS’ (Sir G. C.) ESSAY ON THE GOVERNMENT OF
DEPENDENCIES. 8vo. 12s.
LEXINGTON (The) PAPERS; or, Some Account of the Courts
of London and Vienna at the end of the 17th Century. Edited by HON. H.
MANNERS SUTTON. 8vo. 14s.
LIDDELL’S (Dean) HISTORY OF ROME: from the Earliest
Times to the Establishment of the Empire. With Chapters on the History
of Literature and Art. 2 vols. 8vo. 28s.
LINDSAY’S (Lord) LIVES OF THE LINDSAYS; or, a
Memoir of the Houses of Crawford and Balcarres. 3 vols. 8vo. 24s.
LOWE’S (Sir Hudson) HISTORY OF THE CAPTIVITY OF
NAPOLEON AT ST. HELENA. Edited by WILLIAM FORSYTH. With
Portrait. 3 vols. 8vo. 45s.
LYELL’S (Sir Charles) PRINCIPLES OF GEOLOGY; or,
the Ancient Changes of the Earth and its Inhabitants, as illustrated by Geological
Monuments. Tenth Edition. With Illustrations. 2 vols. 8vo. 32s.
——— ANTIQUITY OF MAN FROM GEOLOGICAL
EVIDENCES. With Remarks on Theories of the Origin of Species by
Variation. Third Edition. With Illustrations. 8vo. 14s.
LYTTON’S (Lord) LOST TALES OF MILETUS. Second
Edition. Post 8vo. 7s. 6d.
——— POEMS. A New Edition. Post 8vo. 10s. 6d.
MACDOUGALL’S (Col.) MODERN WARFARE AS INFLUENCED
BY MODERN ARTILLERY. With Plans and Woodcuts.
Post 8vo. 12s.
MACGREGOR’S (John) CRUISE IN THE ‘ROB ROY’
CANOE ON THE JORDAN, THE NILE, THE RED SEA, LAKE OF
GENNESARETH, &c. Third Edition. With Maps and Illustrations. Crown
8vo. 12s.
MAETZNER’S (Professor) COPIOUS ENGLISH GRAMMAR.
A Methodical, Analytical, and Historical Treatise on the Orthography,
Prosody, Inflections, and Syntax of the English Tongue. With numerous
authorities, cited in the order of historical development. 3 vols. 8vo.
[In the Press.
MAHON (Lord). See STANHOPE (Earl of).
MAINE’S (H. Sumner) ANCIENT LAW; its Connection with
the Early History of Society, and its relation to Modern Ideas. Fourth
Edition. 8vo. 12s.
MANSEL’S (Dean) LIMITS OF RELIGIOUS THOUGHT
EXAMINED. Fifth Edition. Post 8vo. 8s. 6d.
MARCO POLO’S TRAVELS. A New English Version. Illustrated
by the Light of Modern Travels and Oriental Writers. By COL.
YULE, C.B. With Maps and Illustrations. 2 vols. Medium 8vo.
MARRYAT’S (Joseph) HISTORY OF MEDIÆVAL AND
MODERN POTTERY AND PORCELAIN Third Edition. With Coloured
Plates and 240 Woodcuts. Medium 8vo. 42s.11
MILMAN’S (Dean) HISTORY OF THE JEWS, from the
Earliest Period, continued to Modern Times, with a new Preface and
Notes. 3 vols. Post 8vo. 18s.
—————— OF CHRISTIANITY, from the
Birth of Christ to the Abolition of Paganism in the Roman Empire.
3 vols. Post 8vo. 18s.
—————— LATIN CHRISTIANITY;
and of the Popes down to Nicholas V. 9 vols. Post 8vo. 54s.
——— CHARACTER AND CONDUCT OF THE APOSTLES
CONSIDERED AS AN EVIDENCE OF CHRISTIANITY. 8vo.
10s. 6d.
——— ANNALS OF ST. PAUL’S CATHEDRAL. Second
Edition. With Portrait and Illustrations. 8vo. 18s.
——— SAVONAROLA, ERASMUS, and other LITERARY
ESSAYS. 8vo. 15s.
——— HISTORICAL WORKS; containing the ‘History
of the Jews,’ ‘Early Christianity,’ and ‘Latin Christianity.’ With
the Author’s latest Additions and Corrections. Cabinet Edition. 15 vols.
Post 8vo. 6s. each.
——— POETICAL WORKS; containing ‘Samor,’ ‘Fall of
Jerusalem,’ ‘Belshazzar,’ ‘Martyr of Antioch,’ ‘Anne Boleyn,’ &c. With Plates.
3 vols. Fcap. 8vo. 18s.
——— AGAMEMNON OF ÆSCHYLUS AND THE
BACCHANALS OF EURIPIDES. With Passages from the Lyric and Later
Poets of Greece. With Illustrations. Crown 8vo. 12s.
——— HORACE; a New Edition of the Text. With 100
Woodcuts. Small 8vo. 7s. 6d.
—————— LIFE OF. With Illustrations. 8vo. 9s.
MOLTKE’S (Baron) RUSSIAN CAMPAIGNS ON THE
DANUBE AND THE PASSAGE OF THE BALKAN, 1828-9. With Plans.
8vo. 14s.
MONGREDIEN’S (A.) TREES AND SHRUBS FOR ENGLISH
PLANTATION. A Selection and Description of the most Ornamental
which will flourish in the open air. With Classified Lists. With 30 Illustrations.
8vo. 16s.
MOORE’S (Thomas) LIFE OF LORD BYRON; his Letters
and Journals. With Notes and Illustrations. Cabinet Edition. With Plates.
6 vols. Fcap. 8vo. 18s.
————————— With Portraits.
One Volume. Royal 8vo. 9s.
MOTLEY’S (J. L.) HISTORY OF THE UNITED NETHERLANDS,
from the Death of William the Silent to the Twelve Years’ Truce:
with a full view of the English-Dutch struggle against Spain, and of the origin
and destruction of the Spanish Armada. Fourth Edition. With Portraits.
4 vols. 8vo. 60s.; or, Cabinet Edition, 4 vols., Post 8vo, 6s. each.
MOZLEY’S (Rev. J. B.) TREATISE ON THE AUGUSTINIAN
DOCTRINE OF PREDESTINATION. 8vo. 14s.
——— PRIMITIVE DOCTRINE OF BAPTISMAL REGENERATION.
8vo. 7s. 6d.
MURCHISON’S (Sir Roderick) SILURIA: a History of the
Oldest Rocks in the British Isles and other Countries. With a Sketch of
the Distribution of Native Gold. Fourth Edition. With Plates and Woodcuts.
8vo. 30s.12
NAPIER’S (Sir Charles) LIFE AND OPINIONS; chiefly
derived from his Journals and Letters. Second Edition. With Portraits.
4 vols. Post 8vo. 48s.
——— (Sir William) ENGLISH BATTLES AND SIEGES
IN THE PENINSULA. Extracted from his History of the Peninsular
War. Fifth Edition. With Portrait. Post 8vo. 9s.
——— LIFE. With Portraits. 2 vols. Post 8vo. 30s.
NELSON’S (Robert) LIFE AND TIMES. By Rev. C. T.
Secretan, M.A. With Portrait. 8vo. 12s.
NEWBOLD’S (Lieut.) STRAITS OF MALACCA, PENANG,
AND SINGAPORE. 2 vols. 8vo. 26s.
NEW TESTAMENT. With a Plain Explanatory Commentary
for General Readers. By ARCHDEACON CHURTON, M.A., and
ARCHDEACON BASIL JONES, M.A. With 110 authentic Views of
Scripture Sites, &c., from Sketches and Photographs taken on the Spot.
2 vols. 8vo. 21s.
NICHOLAS’ (Sir Harris) HISTORIC PEERAGE OF ENGLAND.
Exhibiting the Origin, Descent, and Present State of every Title of
Peerage which has existed in this Country since the Conquest. A New
Edition. Edited by W. COURTHOPE. 8vo. 30s.
NICHOLLS’ (Sir George) HISTORY OF THE ENGLISH,—IRISH,—AND
SCOTCH POOR LAWS. 4 vols. 8vo. 54s.
NORTH’S (Lord) CORRESPONDENCE WITH KING GEORGE
THE THIRD, 1769-82. Edited, with Notes and Introduction, by W. BODHAM
DONNE. 2 vols. 8vo. 32s.
OLD LONDON; its Archæology and Antiquities; A Series
of Papers read at the Meeting of the Archæological Institute, July, 1866. By
Various Writers. 8vo. 12s.
CONTENTS:
Archæology in its Religious Aspect.—Dean Stanley.
An Address.—A. J. Beresford Hope, M.P.
Chapter House of Westminster Abbey.—G. G. Scott, R.A.
Sculpture in Westminster Abbey.—R. Westmacott, R.A.
Westminster Hall.—E. Foss, F.S.A.
Architectural History of the Tower.—G. T. Clark.
Public Record Office.—Joseph Burtt.
London and her Election of Stephen.—Rev. J. R. Green.
Royal Picture Galleries.—G. Scharf, F.S.A.
OWEN’S (Lieut.-Col.) PRINCIPLES AND PRACTICE OF
MODERN ARTILLERY, including Artillery Material, Gunnery, and
Organization and Use of Artillery in Warfare. With Illustrations.
8vo.
PARKMAN’S (Fras.) DISCOVERY OF THE GREAT WEST;
or, The Valleys of the Mississippi and the Lakes of North America. An
Historical Narrative. With Map. 8vo. 10s. 6d.
PEEL’S (Sir Robert) MEMOIRS. I. Roman Catholic Relief
Bill, 1828-9. II. Formation of the New Government in 1834-5.
III. Repeal of the Corn Laws in 1845-6. Edited by EARL STANHOPE
and Rt. Hon. EDWARD CARDWELL. 2 vols. Post 8vo. 15s.13
PERCY’S (John) METALLURGY: or, The Art of Extracting
Metals from their Ores, and Adapting them to Various Purposes
of Manufacture. With numerous Illustrations. 5 vols. 8vo.
I.—Fuel, Wood, Peat. Coal, Charcoal, Coke. Fire-Clays. Copper,
Zinc, and Brass. 30s.
II.—Iron and Steel. 42s.
III.—Lead, including Desilverization and Cupellation. 30s.
IV.—Gold, Silver, and Mercury.
[In the Press.
V.—Platinum, Tin, Nickel, Cobalt, Antimony, Bismuth, Arsenic, &c.
[In the Press.
PHILLIP’S (John) RIVERS, MOUNTAINS, AND SEA-COAST
OF YORKSHIRE; with Essays on the Climate, Scenery, and
Ancient Inhabitants. With 36 Plates. 8vo. 15s.
POPE’S (Alexander) WORKS. Collected in part by the late
Rt. Hon. J. W. CROKER. Edited, with Introductions and Notes, by Rev.
Whitwell Elwin. With Portraits. Vol. I. to III. 8vo. 10s. 6d. each.
POTTERY (ANCIENT): Egyptian, Assyrian, Greek, Etruscan,
and Roman. By SAMUEL BIRCH, F.S.A. With Coloured Plates and 200
Woodcuts. 2 vols. Medium 8vo. 42s.
——— (MEDIÆVAL AND MODERN). By JOSEPH
MARRYAT. Third Edition. With Coloured Plates and 300 Woodcuts.
Medium 8vo. 42s.
——— NOTES ON VENETIAN CERAMICS. By W. R.
DRAKE, F.S.A. A Supplement to ‘Marryat’s Pottery.’ Medium 8vo. 4s.
PRINCIPLES AT STAKE. Essays on Church Questions of the
Present Day. By various Writers. Second Edition. 8vo. 12s.
CONTENTS:
Ritualism and Uniformity.—Benjamin Shaw, M.A.
Increase of the Episcopate.—Bishop of Bath and Wells.
Powers and Duties of the Priesthood.—Canon Payne Smith.
National Education.—Rev. Alex. R. Grant.
Doctrine of the Eucharist.—Rev. G. H. Sumner.
Scripture and Ritual.—Canon T. D. Bernard.
The Church in South Africa.—Arthur Mills, M.A.
Schismatical Tendency of Ritualism.—Rev. Dr. Salmon.
Revisions of the Liturgy.—Rev. W. G. Humphry.
Parties and Party Spirit.—Dean of Chester.
RANKE’S (Leopold) HISTORY OF THE POPES OF ROME:
Political and Ecclesiastical. Translated from the German, by MRS. AUSTIN.
Fourth Edition. With a Preface by Dean Milman. 3 vols. 8vo. 30s.
RASSAM’S (Hormuzd) NARRATIVE OF THE BRITISH
MISSION TO ABYSSINIA. With Notices of the Countries from Massowah,
through the Soodan, and back to Annesley Bay, from Magdala.
With Map and Illustrations. 2 vols. 8vo. 28s.
RAWLINSON’S (Rev. George) MONARCHIES OF THE
ANCIENT WORLD; or, The History, Geography, and Antiquities of
Chaldæa, Media, Assyria, Babylonia, and Persia. With Maps and Illustrations.
3 vols. 8vo. 42s.
——— HERODOTUS. A New English Version.
Edited, with Notes and Essays, Historical, Ethnographical, and Geographical.
By SIR GARDNER WILKINSON and SIR HENRY RAWLINSON.
Second Edition. With Maps and Woodcuts. 4 vols. 8vo. 48s.14
REED’S (E. J.) SHIPBUILDING IN IRON AND STEEL;
a Practical Treatise, giving full details of Construction, Processes of Manufacture,
and Building Arrangements. With Plans and Woodcuts. 8vo. 30s.
——— IRON-CLAD SHIPS; their Qualities, Performances,
and Cost. With Chapters on Turret Ships, Iron-clad Rams, &c. With Illustrations.
8vo. 12s.
REYNOLDS’ (Sir Joshua) LIFE. With Notices of Hogarth,
Wilson, Gainsborough, and other Artists, his Contemporaries. Commenced
by C. R. LESLIE, R.A., and continued by TOM TAYLOR. With
Portraits. 2 vols. 8vo. 42s.
ROBERTSON’S (Canon) HISTORY OF THE CHRISTIAN
CHURCH, from the Apostolic Age to the End of the Fifth Council of the
Lateran. 4 vols. 8vo.
Vol. I., A.D. 64-590. 18s.
Vol. II., A.D. 590-1122. 20s.
Vol. III., A.D. 1122-1303. 18s.
Vol. IV., A.D. 1303-1517. [In the Press.
ROBINSON’S (Rev. Edward) BIBLICAL RESEARCHES IN
PALESTINE AND THE ADJACENT REGIONS; a Journal of Travels in
1838 and 1852. Third Edition. Maps. 3 vols. 8vo. 42s.
——— PHYSICAL GEOGRAPHY OF THE HOLY
LAND. Post 8vo. 10s. 6d.
SCOTT’S (Gilbert) REMARKS ON GOTHIC ARCHITECTURE,
SECULAR AND DOMESTIC, PRESENT AND FUTURE. 8vo. 9s.
SIMMONS (Capt. T. F.) ON THE CONSTITUTION AND
PRACTICE OF COURTS MARTIAL; with a Summary of the Law of
Evidence, &c. Sixth Edition, revised and corrected. 8vo.
[In the Press.
SMILES’S (Samuel) LIVES OF BRITISH ENGINEERS.
From the Earliest Times down to the Death of Robert Stephenson, with an
Account of their Principal Works, a History of Inland Communication in
Britain, and the Introduction and Invention of the Steam Engine. With
Portraits and Illustrations. 4 vols. 8vo. 21s. each.
SMITH’S (Dr. Wm.) DICTIONARY OF THE BIBLE: its
Antiquities, Biography, Geography, and Natural History. By various
Writers. With Illustrations. 3 vols. Medium 8vo. 5l. 5s.
——— CONCISE BIBLE DICTIONARY, condensed from
the above work. With Maps and 300 Illustrations. Medium 8vo. 21s.
——— DICTIONARY OF CHRISTIAN BIOGRAPHY AND
ANTIQUITIES: from the times of the Apostles to the age of Charlemagne.
With Illustrations. 2 vols. Medium 8vo.
[In the Press.
—————— GREEK AND ROMAN ANTIQUITIES.
With Woodcuts. Royal 8vo. 21s.
—————— GREEK AND ROMAN BIOGRAPHY
AND MYTHOLOGY. With Woodcuts. 3 vols. Royal 8vo. 63s.
—————— GREEK AND ROMAN GEOGRAPHY.
With Woodcuts. 2 vols. Royal 8vo. 42s.
——— LATIN-ENGLISH DICTIONARY. With Tables
of the Roman Calendar, Measures, Weights, and Money. Eighth Edition
(1250 pp.). Medium 8vo. 21s.
——— ENGLISH-LATIN DICTIONARY. Compiled from
Original Sources. By WM. SMITH, D.C.L., and THEOPHILUS D.
HALL, M.A. (964 pp.) Medium 8vo. 21s.15
SOMERVILLE’S (Mary) PHYSICAL GEOGRAPHY. Sixth
Edition, revised. By H. W. BATES. With Portrait. Post 8vo. 9s.
——— SCIENCES. Ninth Edition. With Portrait and
Woodcuts. Post 8vo. 9s.
——— MOLECULAR AND MICROSCOPIC
SCIENCE. With Illustrations. 2 vols. Post 8vo. 21s.
STANHOPE’S (Earl) HISTORY OF ENGLAND, from the
Peace of Utrecht to the Peace of Versailles, 1713-83. Library Edition.
7 vols. 8vo. 93s. Or, Cabinet Edition, 7 vols., Post 8vo, 35s.
——— REIGN OF QUEEN ANNE UNTIL THE
PEACE OF UTRECHT, 1701-1713. 8vo. 16s.
——— SPAIN UNDER CHARLES THE SECOND.
Second Edition. 8vo. 6s. 6d.
——— LIFE OF BELISARIUS. 8vo. 10s. 6d.
—————— WILLIAM PITT. With Portraits.
4 vols. 8vo. 24s.
—————— MISCELLANIES. 8vo. 5s. 6d.
STANLEY’S (Dean) SINAI AND PALESTINE IN CONNECTION
WITH THEIR HISTORY. Eleventh Edition. With Map.
8vo. 14s.
——— EPISTLES OF ST. PAUL TO THE CORINTHIANS.
With Dissertations and Notes. Third Edition. 8vo. 18s.
——— HISTORY OF THE EASTERN CHURCH. Fourth
Edition. Plans. 8vo. 12s.
—————— JEWISH CHURCH. From
Abraham to the Captivity. Third Edition. 2 vols. 8vo. 24s.
——— MEMORIALS OF CANTERBURY. Fourth
Edition. With Woodcuts. Post 8vo. 7s. 6d.
—————— WESTMINSTER ABBEY.
Third Edition. With Illustrations. 8vo. 21s.
——— ESSAYS ON CHURCH AND STATE. 8vo. 16s.
——— SERMONS PREACHED IN THE EAST
DURING A TOUR WITH H.R.H. THE PRINCE OF WALES. With
Notices of the Localities Visited. 8vo. 9s.
——— ADDRESSES AND CHARGES OF BISHOP
STANLEY. With Memoir. 8vo. 10s. 6d.
STREET’S (G. E.) GOTHIC ARCHITECTURE IN SPAIN.
Second Edition. With Illustrations. Medium 8vo. 30s.
STYFFE (Knut) ON THE STRENGTH OF IRON AND
STEEL. Translated from the Swedish. By CHRISTER P. SANDBERG.
With Preface, by DR. PERCY. With Lithographic Plates. 8vo. 12s.
SYBEL’S (Von) HISTORY OF EUROPE DURING THE
FRENCH REVOLUTION, 1789-1795. From Secret Papers and Documents
in the Archives of Germany, &c. Translated by W. C. Perry. 4 vols. 8vo.
48s.
TAIT’S (Archbishop) DANGERS AND SAFEGUARDS; or,
Suggestions to the Theological Student under present Difficulties. 2nd
Edition. 8vo. 12s.
THOMSON’S (A. S.) STORY OF NEW ZEALAND: PAST
AND PRESENT—SAVAGE AND CIVILIZED. With Map and Illustrations.
2 vols. Post 8vo. 24s.16
THOMSON’S (Archbishop) LIFE IN THE LIGHT OF GOD’S
WORD. 2nd Edition. Post 8vo. 5s.
TOZER’S (H. F.) RESEARCHES IN THE ISLANDS OF
TURKEY, ALBANIA, MONTENEGRO, &c. With Notes on the Classical
Superstitions of the Modern Greek. With Map and Illustrations. 2 vols.
Crown 8vo. 24s.
TYLOR’S (E. B.) RESEARCHES INTO THE EARLY HISTORY
OF MANKIND, and the Development of Civilization. Second
Edition. With Illustrations. 8vo. 12s.
——— PRIMITIVE CULTURE; Researches into the Development
of Mythology, Philosophy, Religion, Art, and Custom. 2 vols. 8vo.
UBICINI’S (M. A.) TURKEY AND ITS INHABITANTS.
The Moslems, Greeks, Armenians, &c.—The Reformed Institutions, Army,
&c., described. 2 vols. Post 8vo. 21s.
WAAGEN’S (DR.) TREASURES OF ART IN GREAT
BRITAIN. Being an Account of the Chief Collections of Paintings,
Sculptures, Drawings, MSS., Miniatures, &c. 4 vols. 8vo. 54s.
WELLINGTON’S (Duke of) DESPATCHES DURING HIS
VARIOUS CAMPAIGNS. Edited by Col. Gurwood. 8 vols. 8vo. 8l. 8s.
——— SUPPLEMENTARY DESPATCHES. Edited
by HIS SON. 12 vols. 8vo. 20s. each.
——— CIVIL AND POLITICAL CORRESPONDENCE.
Edited by HIS SON. Vols. I. to III. 8vo. 20s. each.
——— SPEECHES ON VARIOUS OCCASIONS.
2 vols. 8vo. 42s.
WHYMPER’S (Edward) SCRAMBLES AMONG THE ALPS,
1860-9. Including the First Ascent of the Matterhorn. With Observations
on Glacier Phenomena in the Alps and in Greenland. With 100 Maps
and Illustrations. Medium 8vo. 21s.
WILKINSON’S (Sir Gardner) ANCIENT EGYPTIANS;
their Private Life, Manners, and Customs, Fourth Edition. With Illustrations.
2 vols. Post 8vo. 12s.
WILLIAM THE FOURTH’S CORRESPONDENCE WITH
SIR HERBERT TAYLOR AND EARL GREY, from Nov., 1830, to the
Passing of the Reform Act in 1832. 2 vols. 8vo. 30s.
WILSON’S (Sir Robert) SECRET HISTORY OF EVENTS
DURING THE INVASION OF RUSSIA AND RETREAT OF THE
FRENCH ARMY, 1812. Second Edition. With Map and Plans. 8vo. 15s.
WORDSWORTH’S (Bishop) GREECE—Pictorial, Historical,
and Descriptive. With an Essay on Greek Art, by George Scharf, F.S.A.
Fourth Edition. With 600 Illustrations. Royal 8vo. 21s.
——— TOUR IN ATHENS AND ATTICA.
Fourth Edition. With Plates. Post 8vo. 5s.
YULE’S (Col. Henry) MARCO POLO’S TRAVELS. Illustrated
by the light of Modern Travels and Oriental Writers. With Maps
and Illustrations. 2 vols. Medium 8vo. 42s.
JOHN MURRAY, 50A, ALBEMARLE STREET.
LONDON: PRINTED BY WILLIAM CLOWES AND SONS, STAMFORD STREET,
AND CHARING CROSS.
***END OF THE PROJECT GUTENBERG EBOOK THE DESCENT OF MAN AND SELECTION IN RELATION TO SEX***
******* This file should be named 36520-h.txt or 36520-h.zip *******
This and all associated files of various formats will be found in:
http://www.gutenberg.org/3/6/5/2/36520
Updated editions will replace the previous one--the old editions
will be renamed.
Creating the works from public domain print editions means that no
one owns a United States copyright in these works, so the Foundation
(and you!) can copy and distribute it in the United States without
permission and without paying copyright royalties. Special rules,
set forth in the General Terms of Use part of this license, apply to
copying and distributing Project Gutenberg-tm electronic works to
protect the PROJECT GUTENBERG-tm concept and trademark. Project
Gutenberg is a registered trademark, and may not be used if you
charge for the eBooks, unless you receive specific permission. If you
do not charge anything for copies of this eBook, complying with the
rules is very easy. You may use this eBook for nearly any purpose
such as creation of derivative works, reports, performances and
research. They may be modified and printed and given away--you may do
practically ANYTHING with public domain eBooks. Redistribution is
subject to the trademark license, especially commercial
redistribution.
*** START: FULL LICENSE ***
THE FULL PROJECT GUTENBERG LICENSE
PLEASE READ THIS BEFORE YOU DISTRIBUTE OR USE THIS WORK
To protect the Project Gutenberg-tm mission of promoting the free
distribution of electronic works, by using or distributing this work
(or any other work associated in any way with the phrase "Project
Gutenberg"), you agree to comply with all the terms of the Full Project
Gutenberg-tm License (available with this file or online at
http://www.gutenberg.org/license).
Section 1. General Terms of Use and Redistributing Project Gutenberg-tm
electronic works
1.A. By reading or using any part of this Project Gutenberg-tm
electronic work, you indicate that you have read, understand, agree to
and accept all the terms of this license and intellectual property
(trademark/copyright) agreement. If you do not agree to abide by all
the terms of this agreement, you must cease using and return or destroy
all copies of Project Gutenberg-tm electronic works in your possession.
If you paid a fee for obtaining a copy of or access to a Project
Gutenberg-tm electronic work and you do not agree to be bound by the
terms of this agreement, you may obtain a refund from the person or
entity to whom you paid the fee as set forth in paragraph 1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be
used on or associated in any way with an electronic work by people who
agree to be bound by the terms of this agreement. There are a few
things that you can do with most Project Gutenberg-tm electronic works
even without complying with the full terms of this agreement. See
paragraph 1.C below. There are a lot of things you can do with Project
Gutenberg-tm electronic works if you follow the terms of this agreement
and help preserve free future access to Project Gutenberg-tm electronic
works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the Foundation"
or PGLAF), owns a compilation copyright in the collection of Project
Gutenberg-tm electronic works. Nearly all the individual works in the
collection are in the public domain in the United States. If an
individual work is in the public domain in the United States and you are
located in the United States, we do not claim a right to prevent you from
copying, distributing, performing, displaying or creating derivative
works based on the work as long as all references to Project Gutenberg
are removed. Of course, we hope that you will support the Project
Gutenberg-tm mission of promoting free access to electronic works by
freely sharing Project Gutenberg-tm works in compliance with the terms of
this agreement for keeping the Project Gutenberg-tm name associated with
the work. You can easily comply with the terms of this agreement by
keeping this work in the same format with its attached full Project
Gutenberg-tm License when you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern
what you can do with this work. Copyright laws in most countries are in
a constant state of change. If you are outside the United States, check
the laws of your country in addition to the terms of this agreement
before downloading, copying, displaying, performing, distributing or
creating derivative works based on this work or any other Project
Gutenberg-tm work. The Foundation makes no representations concerning
the copyright status of any work in any country outside the United
States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other immediate
access to, the full Project Gutenberg-tm License must appear prominently
whenever any copy of a Project Gutenberg-tm work (any work on which the
phrase "Project Gutenberg" appears, or with which the phrase "Project
Gutenberg" is associated) is accessed, displayed, performed, viewed,
copied or distributed:
This eBook is for the use of anyone anywhere at no cost and with
almost no restrictions whatsoever. You may copy it, give it away or
re-use it under the terms of the Project Gutenberg License included
with this eBook or online at www.gutenberg.org
1.E.2. If an individual Project Gutenberg-tm electronic work is derived
from the public domain (does not contain a notice indicating that it is
posted with permission of the copyright holder), the work can be copied
and distributed to anyone in the United States without paying any fees
or charges. If you are redistributing or providing access to a work
with the phrase "Project Gutenberg" associated with or appearing on the
work, you must comply either with the requirements of paragraphs 1.E.1
through 1.E.7 or obtain permission for the use of the work and the
Project Gutenberg-tm trademark as set forth in paragraphs 1.E.8 or
1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted
with the permission of the copyright holder, your use and distribution
must comply with both paragraphs 1.E.1 through 1.E.7 and any additional
terms imposed by the copyright holder. Additional terms will be linked
to the Project Gutenberg-tm License for all works posted with the
permission of the copyright holder found at the beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm
License terms from this work, or any files containing a part of this
work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this
electronic work, or any part of this electronic work, without
prominently displaying the sentence set forth in paragraph 1.E.1 with
active links or immediate access to the full terms of the Project
Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary,
compressed, marked up, nonproprietary or proprietary form, including any
word processing or hypertext form. However, if you provide access to or
distribute copies of a Project Gutenberg-tm work in a format other than
"Plain Vanilla ASCII" or other format used in the official version
posted on the official Project Gutenberg-tm web site (www.gutenberg.org),
you must, at no additional cost, fee or expense to the user, provide a
copy, a means of exporting a copy, or a means of obtaining a copy upon
request, of the work in its original "Plain Vanilla ASCII" or other
form. Any alternate format must include the full Project Gutenberg-tm
License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying,
performing, copying or distributing any Project Gutenberg-tm works
unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing
access to or distributing Project Gutenberg-tm electronic works provided
that
- You pay a royalty fee of 20% of the gross profits you derive from
the use of Project Gutenberg-tm works calculated using the method
you already use to calculate your applicable taxes. The fee is
owed to the owner of the Project Gutenberg-tm trademark, but he
has agreed to donate royalties under this paragraph to the
Project Gutenberg Literary Archive Foundation. Royalty payments
must be paid within 60 days following each date on which you
prepare (or are legally required to prepare) your periodic tax
returns. Royalty payments should be clearly marked as such and
sent to the Project Gutenberg Literary Archive Foundation at the
address specified in Section 4, "Information about donations to
the Project Gutenberg Literary Archive Foundation."
- You provide a full refund of any money paid by a user who notifies
you in writing (or by e-mail) within 30 days of receipt that s/he
does not agree to the terms of the full Project Gutenberg-tm
License. You must require such a user to return or
destroy all copies of the works possessed in a physical medium
and discontinue all use of and all access to other copies of
Project Gutenberg-tm works.
- You provide, in accordance with paragraph 1.F.3, a full refund of any
money paid for a work or a replacement copy, if a defect in the
electronic work is discovered and reported to you within 90 days
of receipt of the work.
- You comply with all other terms of this agreement for free
distribution of Project Gutenberg-tm works.
1.E.9. If you wish to charge a fee or distribute a Project Gutenberg-tm
electronic work or group of works on different terms than are set
forth in this agreement, you must obtain permission in writing from
both the Project Gutenberg Literary Archive Foundation and Michael
Hart, the owner of the Project Gutenberg-tm trademark. Contact the
Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable
effort to identify, do copyright research on, transcribe and proofread
public domain works in creating the Project Gutenberg-tm
collection. Despite these efforts, Project Gutenberg-tm electronic
works, and the medium on which they may be stored, may contain
"Defects," such as, but not limited to, incomplete, inaccurate or
corrupt data, transcription errors, a copyright or other intellectual
property infringement, a defective or damaged disk or other medium, a
computer virus, or computer codes that damage or cannot be read by
your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right
of Replacement or Refund" described in paragraph 1.F.3, the Project
Gutenberg Literary Archive Foundation, the owner of the Project
Gutenberg-tm trademark, and any other party distributing a Project
Gutenberg-tm electronic work under this agreement, disclaim all
liability to you for damages, costs and expenses, including legal
fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT
LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE
PROVIDED IN PARAGRAPH 1.F.3. YOU AGREE THAT THE FOUNDATION, THE
TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE
LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR
INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH
DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a
defect in this electronic work within 90 days of receiving it, you can
receive a refund of the money (if any) you paid for it by sending a
written explanation to the person you received the work from. If you
received the work on a physical medium, you must return the medium with
your written explanation. The person or entity that provided you with
the defective work may elect to provide a replacement copy in lieu of a
refund. If you received the work electronically, the person or entity
providing it to you may choose to give you a second opportunity to
receive the work electronically in lieu of a refund. If the second copy
is also defective, you may demand a refund in writing without further
opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth
in paragraph 1.F.3, this work is provided to you 'AS-IS,' WITH NO OTHER
WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT LIMITED TO
WARRANTIES OF MERCHANTIBILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied
warranties or the exclusion or limitation of certain types of damages.
If any disclaimer or limitation set forth in this agreement violates the
law of the state applicable to this agreement, the agreement shall be
interpreted to make the maximum disclaimer or limitation permitted by
the applicable state law. The invalidity or unenforceability of any
provision of this agreement shall not void the remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the
trademark owner, any agent or employee of the Foundation, anyone
providing copies of Project Gutenberg-tm electronic works in accordance
with this agreement, and any volunteers associated with the production,
promotion and distribution of Project Gutenberg-tm electronic works,
harmless from all liability, costs and expenses, including legal fees,
that arise directly or indirectly from any of the following which you do
or cause to occur: (a) distribution of this or any Project Gutenberg-tm
work, (b) alteration, modification, or additions or deletions to any
Project Gutenberg-tm work, and (c) any Defect you cause.
Section 2. Information about the Mission of Project Gutenberg-tm
Project Gutenberg-tm is synonymous with the free distribution of
electronic works in formats readable by the widest variety of computers
including obsolete, old, middle-aged and new computers. It exists
because of the efforts of hundreds of volunteers and donations from
people in all walks of life.
Volunteers and financial support to provide volunteers with the
assistance they need are critical to reaching Project Gutenberg-tm's
goals and ensuring that the Project Gutenberg-tm collection will
remain freely available for generations to come. In 2001, the Project
Gutenberg Literary Archive Foundation was created to provide a secure
and permanent future for Project Gutenberg-tm and future generations.
To learn more about the Project Gutenberg Literary Archive Foundation
and how your efforts and donations can help, see Sections 3 and 4
and the Foundation web page at http://www.gutenberg.org/fundraising/pglaf.
Section 3. Information about the Project Gutenberg Literary Archive
Foundation
The Project Gutenberg Literary Archive Foundation is a non profit
501(c)(3) educational corporation organized under the laws of the
state of Mississippi and granted tax exempt status by the Internal
Revenue Service. The Foundation's EIN or federal tax identification
number is 64-6221541. Contributions to the Project Gutenberg
Literary Archive Foundation are tax deductible to the full extent
permitted by U.S. federal laws and your state's laws.
The Foundation's principal office is located at 4557 Melan Dr. S.
Fairbanks, AK, 99712., but its volunteers and employees are scattered
throughout numerous locations. Its business office is located at
809 North 1500 West, Salt Lake City, UT 84116, (801) 596-1887, email
business@pglaf.org. Email contact links and up to date contact
information can be found at the Foundation's web site and official
page at http://www.gutenberg.org/about/contact
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
gbnewby@pglaf.org
Section 4. Information about Donations to the Project Gutenberg
Literary Archive Foundation
Project Gutenberg-tm depends upon and cannot survive without wide
spread public support and donations to carry out its mission of
increasing the number of public domain and licensed works that can be
freely distributed in machine readable form accessible by the widest
array of equipment including outdated equipment. Many small donations
($1 to $5,000) are particularly important to maintaining tax exempt
status with the IRS.
The Foundation is committed to complying with the laws regulating
charities and charitable donations in all 50 states of the United
States. Compliance requirements are not uniform and it takes a
considerable effort, much paperwork and many fees to meet and keep up
with these requirements. We do not solicit donations in locations
where we have not received written confirmation of compliance. To
SEND DONATIONS or determine the status of compliance for any
particular state visit http://www.gutenberg.org/fundraising/pglaf
While we cannot and do not solicit contributions from states where we
have not met the solicitation requirements, we know of no prohibition
against accepting unsolicited donations from donors in such states who
approach us with offers to donate.
International donations are gratefully accepted, but we cannot make
any statements concerning tax treatment of donations received from
outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation
methods and addresses. Donations are accepted in a number of other
ways including checks, online payments and credit card donations.
To donate, please visit: http://www.gutenberg.org/fundraising/donate
Section 5. General Information About Project Gutenberg-tm electronic
works.
Professor Michael S. Hart is the originator of the Project Gutenberg-tm
concept of a library of electronic works that could be freely shared
with anyone. For thirty years, he produced and distributed Project
Gutenberg-tm eBooks with only a loose network of volunteer support.
Project Gutenberg-tm eBooks are often created from several printed
editions, all of which are confirmed as Public Domain in the U.S.
unless a copyright notice is included. Thus, we do not necessarily
keep eBooks in compliance with any particular paper edition.
Each eBook is in a subdirectory of the same number as the eBook's
eBook number, often in several formats including plain vanilla ASCII,
compressed (zipped), HTML and others.
Corrected EDITIONS of our eBooks replace the old file and take over
the old filename and etext number. The replaced older file is renamed.
VERSIONS based on separate sources are treated as new eBooks receiving
new filenames and etext numbers.
Most people start at our Web site which has the main PG search facility:
http://www.gutenberg.org
This Web site includes information about Project Gutenberg-tm,
including how to make donations to the Project Gutenberg Literary
Archive Foundation, how to help produce our new eBooks, and how to
subscribe to our email newsletter to hear about new eBooks.
EBooks posted prior to November 2003, with eBook numbers BELOW #10000,
are filed in directories based on their release date. If you want to
download any of these eBooks directly, rather than using the regular
search system you may utilize the following addresses and just
download by the etext year.
http://www.gutenberg.org/dirs/etext06/
(Or /etext 05, 04, 03, 02, 01, 00, 99,
98, 97, 96, 95, 94, 93, 92, 92, 91 or 90)
EBooks posted since November 2003, with etext numbers OVER #10000, are
filed in a different way. The year of a release date is no longer part
of the directory path. The path is based on the etext number (which is
identical to the filename). The path to the file is made up of single
digits corresponding to all but the last digit in the filename. For
example an eBook of filename 10234 would be found at:
http://www.gutenberg.org/dirs/1/0/2/3/10234
or filename 24689 would be found at:
http://www.gutenberg.org/dirs/2/4/6/8/24689
An alternative method of locating eBooks:
http://www.gutenberg.org/dirs/GUTINDEX.ALL
*** END: FULL LICENSE ***