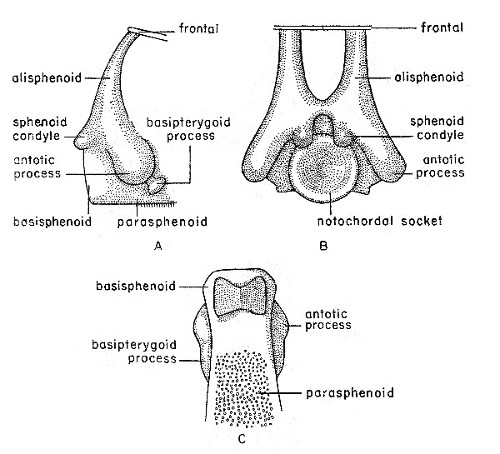
 Fig. 1. Synaptotylus newelli (Hibbard). Restoration of
the basisphenoid, based on K. U. no. 9939, × 5. A, lateral view, B,
posterior view, C, ventral view.
Fig. 1. Synaptotylus newelli (Hibbard). Restoration of
the basisphenoid, based on K. U. no. 9939, × 5. A, lateral view, B,
posterior view, C, ventral view.
The Project Gutenberg EBook of A New Genus of Pennsylvania Fish (Crossoperygii, Coelacanthiformes) from Kansas, by Joan Echols This eBook is for the use of anyone anywhere at no cost and with almost no restrictions whatsoever. You may copy it, give it away or re-use it under the terms of the Project Gutenberg License included with this eBook or online at www.gutenberg.org Title: A New Genus of Pennsylvania Fish (Crossoperygii, Coelacanthiformes) from Kansas Author: Joan Echols Release Date: August 28, 2010 [EBook #33560] Language: English Character set encoding: ISO-8859-1 *** START OF THIS PROJECT GUTENBERG EBOOK NEW GENUS OF PENNSYLVANIA FISH *** Produced by Chris Curnow, Joseph Cooper, Josephine Paolucci and the Online Distributed Proofreading Team at https://www.pgdp.net.
University of Kansas Publications
Museum of Natural History
Volume 12, No. 10, pp. 475-501, 7 figs.
October 25, 1963
University of Kansas
Lawrence
1963
University of Kansas Publications, Museum of Natural History
Editors: E. Raymond Hall, Chairman, Henry S. Fitch,
Theodore H. Eaton, Jr.
Volume 12, No. 10, pp. 475-501, 7 figs.
Published October 25, 1963
University of Kansas
Lawrence, Kansas
PRINTED BY
JEAN M. NEIBARGER, STATE PRINTER
TOPEKA, KANSAS
1963
In 1931 and 1932, H. H. Lane, C. W. Hibbard and W. K. McNown collected the specimens that Hibbard (1933) described and made the basis of two new species. These were from the Rock Lake shale member of the Stanton formation, six miles northwest of Garnett, Anderson County, Kansas. In 1954, from a locality (KAn-1/D, see page 480) approximately one fourth mile southwest of the first locality, specimens were quarried by F. E. Peabody, R. W. Wilson and R. Weeks. In 1955 R. R. Camp collected additional blocks of Rock Lake shale from this second locality. Study of all of the materials from the above mentioned localities reveals the existence of an hitherto unrecognized genus of coelacanth. It is named and described below.
I wish to thank Prof. Theodore H. Eaton, Jr., for suggesting the project and for much helpful advice. I am indebted to Dr. E. I. White of the British Museum (Natural History) for furnishing a cast of the endocranium of Rhabdoderma elegans (Newberry) for comparison, and to Drs. Donald Baird (Princeton University), Bobb Schaeffer (American Museum of Natural History) and R. H. Denison (Chicago Natural History Museum) for loans and exchanges of specimens for comparison. I am grateful to Dr. Bobb Schaeffer for advice on the manuscript. Mr. Merton C. Bowman assisted with the illustrations. The study here reported on was made while I was a Research Assistant supported by National Science Foundation Grant G-14013.[Pg 478]
Suborder DIPLOCERCIDOIDEI
Family Diplocercidae
Subfamily Rhabdodermatinae, new subfamily
Type genus.—Rhabdoderma Reis, 1888, Paleontographica, vol. 35, p. 71.
Referred genus.—Synaptotylus new, described below.
Horizon.—Carboniferous.
Diagnosis.—Sphenethmoid region partly ossified, and consisting of basisphenoid, parasphenoid, and ethmoid ossifications; paired basipterygoid process and paired antotic process on basisphenoid; parasphenoid of normal size, and closely associated with, or fused to, basisphenoid; ethmoids paired in Rhabdoderma (unknown in Synaptotylus).
Discussion.—Because of the great differences in endocranial structure between the Devonian and Pennsylvanian coelacanths, they are here placed in new subfamilies. The two proposed subfamilies of the family Diplocercidae are the Diplocercinae and the Rhabdodermatinae. The Diplocercinae include those coelacanths having two large unpaired bones in the endocranium (at present this includes Diplocercides Stensiö, Nesides Stensiö and Euporosteus Jaekel). The subfamily Rhabdodermatinae is composed of coelacanths having reduced endocranial ossification, as described in detail above, and now including Rhabdoderma Reis and Synaptotylus n. g.
Members of this subfamily differ from those of the subfamily Diplocercinae in having several paired and unpaired elements in the sphenethmoid region of the endocranium, instead of only one larger ossification. They differ from those of the suborder Coelacanthoidei in the retention of basipterygoid processes.
Synaptotylus is more closely related to Rhabdoderma than to the Diplocercines because the anterior portion of the endocranium contains only a basisphenoid, parasphenoid, and probably ethmoids. The sphenethmoid region was certainly not a large, unpaired unit as in the Diplocercines. Probably the posterior part, the otico-occipital region (not known in Synaptotylus), was much more nearly like that of Rhabdoderma, which consisted of unpaired basioccipital and supraoccipital, and paired prootics, exoccipitals, and anterior and posterior occipital ossifications (Moy-Thomas, 1937:[Pg 479] figs. 3, 4). Moy-Thomas (1937:389) points out that in Rhabdoderma the occipital region is "considerably more ossified" than in any coelacanths other than the Devonian forms. Berg (1940:390) thought that the Carboniferous coelacanths should be placed in a separate family because they did not have two large, unpaired bones in the endocranium. Rhabdoderma and Synaptotylus represent another stage in evolution of the endocranium in coelacanths, and, if classification is to be based on endocranial structure, then this stage (represented by the two genera) may later be given family rank as Berg suggested. Because Rhabdoderma and Synaptotylus have only part of the sphenethmoid region ossified and because they retain basipterygoid processes, they are considered to be related and are included in the subfamily Rhabdodermatinae.
Synaptotylus, new genus
Type species.—Synaptotylus newelli (Hibbard).
Horizon.—Rock Lake shale member, Stanton formation, Lansing group, Missouri series, Upper Pennsylvanian.
Diagnosis.—Late Pennsylvanian fishes of small size, having the following combination of characters: on basisphenoid, knoblike antotic processes connected by a low ridge to basipterygoid processes; entire ventral surface of parasphenoid toothed; anterior margin of parasphenoid notched and no evidence of hypophyseal opening. Dermal bones of skull smooth or with low, rounded tubercles and striae; fronto-ethmoid shield incompletely known but having one pair of large rectangular frontals with posterolaterally slanting anterior margins; intertemporals large, the lateral margins curving laterally; postorbital triangular, apex downward; subopercular somewhat triangular; squamosal carrying sensory canal that curves down posteriorly and extends onto a ventral projection; opercular generally triangular; supratemporals elongate, curving to fit lateral margin of intertemporals; circumorbital plates lightly ossified. Palatoquadrate complex consisting of endopterygoid and ectopterygoid (both toothed on medial surface), quadrate, and metapterygoid, the latter smooth and having widened border for articulation on anterodorsal margin. Pectoral girdle consisting of cleithrum and clavicle (supracleithrum not seen); small projection on medial surface of posterior portion of cleithrum; horizontal medial process on clavicle. Pelvic plate bearing three anteriorly diverging apophyses, and one denticulate ventromedian process for articulation to opposite plate. Lepidotrichia jointed distally,[Pg 480] but not tuberculated. Scales oval, having posteriorly converging ridges on posterior exposed parts.
The name refers to the most distinctive character of the genus, the connected antotic and basipterygoid processes on the basisphenoid, and is derived from Greek, synaptos—joined, tylos (masc.)—knob, projection.
Synaptotylus is excluded from the advanced suborder Coelacanthoidei by the retention of basipterygoid processes on the basisphenoid. Synaptotylus differs from Rhabdoderma in several characters of the basisphenoid, the most important being: knoblike antotic processes (those of Rhabdoderma are wider, more flattened and more dorsal in position); small, lateral basipterygoid processes (in Rhabdoderma these are larger and farther ventral in position).
Synaptotylus newelli (Hibbard)
Coelacanthus newelli Hibbard, 1933, Univ. Kansas Sci. Bull., 21:280, pl. 27, figs. 2, 3.
Coelacanthus arcuatus Hibbard, 1933, Univ. Kansas Sci. Bull., 21:282, pl. 26, fig. 8; pl. 27, fig. 1.
Rhabdoderma elegans Moy-Thomas, 1937 (in part), Proc. Zool. Soc. London, 107(ser. B, pt. 3):399.
Type.—K. U. no. 786F.
Diagnosis.—Same as for the genus.
Horizon.—Rock Lake shale member, Stanton formation, Lansing group, Missouri series, Upper Pennsylvanian.
Localities.—The specimens studied by Hibbard (K. U. nos. 786F, 787F, 788) and no. 11457 were taken from the Bradford Chandler farm, from the original quarry in SW-1/4, SE-1/4, sec. 32, T.19S, R.19E. The remainder were collected from University of Kansas Museum of Natural History locality KAn-1/D, a quarry in sec. 5, T.19S, R.19E. Both of these are approximately six miles northwest of Garnett, Anderson County, Kansas.
Referred specimens.—K. U. nos. 786F, 787F, 788, 9939, 11424, 11425, 11426, 11427, 11428, 11429, 11430, 11431, 11432, 11433, 11434, 11449, 11450, 11451, 11452, 11453, 11454, 11455, 11457.
Preservation.—Preservation of many of the specimens is good, few are weathered, but most of the remains are fragmentary and dissociated. One specimen (the type, no. 786F) and half of another were nearly complete. Specimens are found scattered throughout the Rock Lake shale (see p. 498).
Morphology.—Terminology used for bones of the skull is that of Moy-Thomas (1937) and Schaeffer (1952).
Fig. 1. Synaptotylus newelli (Hibbard). Restoration of
the basisphenoid, based on K. U. no. 9939, × 5. A, lateral view, B,
posterior view, C, ventral view.
The basisphenoid (see fig. 1) has been observed in only one specimen (K. U. no. 9939) in posterodorsal and ventral views. The basisphenoid, although somewhat crushed, appears to be fused to the parasphenoid. Both antotic and basipterygoid processes are[Pg 481] present, and are connected by a low, rounded ridge. The antotic processes are large, bulbar projections. These processes in Rhabdoderma are wider and more flattened (Moy-Thomas, 1937:figs. 3, 4). The antotic processes are at mid-point on the lateral surface, not dorsal as in Rhabdoderma, and both the processes and the ridge are directed anteroventrally. The basipterygoid processes are smaller, somewhat vertically elongated projections, situated at the end of the low connecting ridge extending anteroventrally from the antotic processes, and are not basal as are those of Rhabdoderma. The sphenoid condyles, seen in posterior view, issue from the dorsal margin of the notochordal socket. The margins of the socket are rounded, and slope down evenly to the center. A slight depression situated between and dorsal to the sphenoid condyles is supposedly for the attachment of the intercranial ligament (Schaeffer and Gregory, 1961:fig. 1). The alisphenoids extend[Pg 482] upward, anterodorsally from the region above the sphenoid condyles, and may connect to ridges on the ventral surface of the frontals. The lateral laminae are not preserved, and their extent is unknown.
In viewing the changes in the endocranium of Carboniferous and Permian coelacanths, it would be well to consider the mechanical relationship of the loss of the basipterygoid processes to the effect on swallowing prey. Evidently many of the coelacanths, Latimeria for example, are predators (Smith, 1939:104); to such fishes a more efficient catching and swallowing mechanism would be an adaptive improvement. Stensiö (1932:fig. 14) presents a cross section of the ethmosphenoid moiety of the endocranium of Diplocercides kayseri (von Koenen) showing the metapterygoid of the palatoquadrate loosely articulated to both the antotic and basipterygoid processes. According to Tchernavin (1948:137) and Schaeffer and Rosen (1961:190) the swallowing of large prey depends on the ability of the fish to expand its oral cavity by allowing the posteroventral portion of the palatoquadrate and the posterior end of the mandible to swing outward. Where the palatoquadrate articulates with the basisphenoid at the antotic and basipterygoid processes, as in the Devonian coelacanths, it can not swing so far laterally as where it articulates with only the dorsal, antotic process. Perhaps the loss of the basipterygoid articulation reflects the development of a more efficient mechanism for swallowing prey in these fishes. Schaeffer and Rosen (1961:191, 193) show that in the evolution of the actinopterygians several changes improved the feeding mechanism: some of these changes are: (1) freeing of the maxilla from the cheek, giving a larger chamber for the action of the adductor mandibulae; (2) development of a coronoid process on the mandible; and (3) increase in torque around the jaw articulation. In coelacanths, at least some comparable changes occurred, such as: (1) loss of the maxillary, thus increasing the size of the adductor chamber; (2) development of the coronoid bone, affording a greater area for muscle attachment; (3) development of an arched dorsal margin on the angular; (4) modification of the palatoquadrate complex, with resultant loss of the basipterygoid processes. In Synaptotylus the basipterygoid processes are small, not basally located, and perhaps not functional. A more efficient feeding mechanism developed rapidly during the Carboniferous and has remained almost unaltered.[Pg 483]
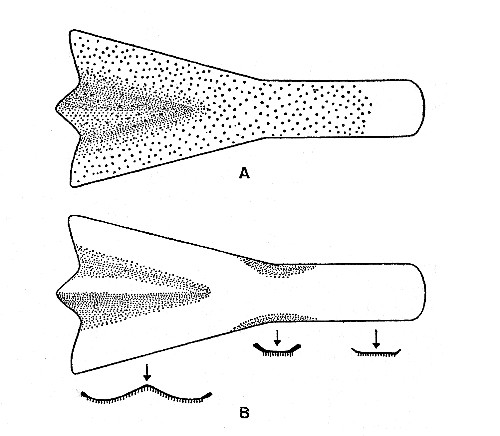 Fig. 2. Synaptotylus newelli (Hibbard). Restoration of
the parasphenoid, based on K. U. nos. 9939, 11451, × 5. A, ventral view,
B, dorsal view and cross sections.
Fig. 2. Synaptotylus newelli (Hibbard). Restoration of
the parasphenoid, based on K. U. nos. 9939, 11451, × 5. A, ventral view,
B, dorsal view and cross sections.
The parasphenoid (see fig. 2) is a shovel-shaped bone having a wide anterior portion and a narrower posterior portion of nearly uniform width. Most of the ventral surface is covered with minute granular teeth. The anterior margin is flared and curved posteromedially from the lateral margin to a median triangular projection. The lateral margins curve smoothly from the greatest anterior width to the narrow central portion, where the margins become somewhat thickened and turned dorsally. Posterior to this the lateral margins are probably nearly straight. The external surface of the anterior section is nearly flat and has a central depressed area the sides of which slope evenly to the center. The internal surface is smooth and centrally convex. Because of the fragmentary nature of all four observed specimens, total length was not measured but is estimated to be 15 to 20 mm. The opening of the hypophyseal canal was not present, possibly because of crushing. Ethmoidal ossifications were not preserved in any of the specimens studied. The parasphenoid differs from that of Rhabdoderma elegans (Newberry) in being more flared and widened anteriorly and more concave centrally.[Pg 484]
Various portions of the cranial roof are preserved in several specimens (see fig. 3). For comparisons with Rhabdoderma elegans, see Moy-Thomas (1937:fig. 1).
The premaxillaries and rostral elements are not preserved in any of the specimens. Only one pair of relatively large frontals have been observed; they are 5.5 to 9.0 mm. long and 2.0 to 3.5 mm. wide. These are nearly flat bones, with the greatest width posteriorly 0.1 to 1.0 mm. wider than the anterior portion. The midline suture is straight, the lateral margins are nearly straight, the anterior margin slopes evenly posterolaterally, and the posterior margin is slightly convex to straight. The anterior margin in R. elegans is essentially straight. Ornamentation consists of sparse, unevenly spaced, coarse tubercles or short striae. In one specimen both bones have small clusters of tubercles near the lateral margins and about 2.0 mm. from the posterior margin. None of these bones has alisphenoids or ridges on the ventral surface, as Stensiö (1921:65, 97) described for Wimania and Axelia.
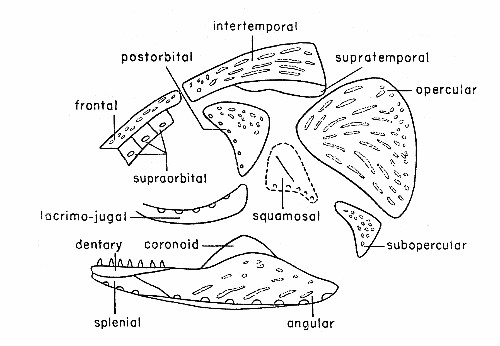 Fig. 3. Synaptotylus newelli (Hibbard). Diagram of the
dermal bones of the skull, in lateral view, based on K. U. nos. 788 and
11432. × 2-1/2 approximately.
Fig. 3. Synaptotylus newelli (Hibbard). Diagram of the
dermal bones of the skull, in lateral view, based on K. U. nos. 788 and
11432. × 2-1/2 approximately.
Only six supraorbitals have been preserved (see fig. 3). These are nearly square, flat, thin bones lying nearly in place adjacent to[Pg 485] a frontal on K. U. no. 788. The smallest is anterior; the margins of all are nearly straight. The bones are unornamented. Each bears a pore of the supraorbital line just below the midline. The supraorbitals of R. elegans have a triangular outline and do not bear pores.
Intertemporals (fig. 3) on several specimens vary from approximately 9.0 to 15.0 mm. in length, 2.0 to 2.7 mm. in anterior width, and increase to 4.5 to 8.0 mm. in maximum posterior width. The midline suture is straight, the anterior margin is concave and the lateral margin proceeds laterally in a concave curve to the widest portion. In R. elegans only the anterior half of the corresponding margin is concave. The posterior margin is slightly rounded and slopes anteriorly toward the lateral margin. Ornamentation is usually of randomly oriented tubercles and striae, although striae are more common in the posterior third and may be longitudinal, whereas tubercles occur mainly on the anterior section. No evidence of sensory pores, as seen on the intertemporal of R. elegans, has been found.
The supratemporals were observed on only one specimen (K. U. no. 788), (fig. 3). Sutures were difficult to distinguish but the medial margin is presumed to curve to fit and to articulate with the lateral margins of the intertemporals. Lateral margins are smoothly curved but the anterior and posterior margins were broken off. There appears to be no ornamentation on this bone. The supratemporals are much more elongated and curving than those in R. elegans.
The cheek region is nearly complete in one specimen (K. U. no. 788), and scattered parts occur in a few others (see fig. 3). The lacrimojugal of no. 788 is elongate, with both ends curving dorsally. It differs from the lacrimojugal in R. elegans, in which the anterior end extends anteriorly and is not curved dorsally. The posterior and anterior margins are not preserved; the greatest height appears to be posterior. Pores of the suborbital portion of the infraorbital sensory canal are seen on the dorsal surface of the bone. In R. elegans the pores are on the lateral surface. A section of the lacrimojugal on specimen no. 11425, broken at both ends, shows a thin layer of bone perforated by the pores and covering a groove for the canal within the dorsal margin of the bone. Both specimens are unornamented.
A nearly complete postorbital (fig. 3) on specimen no. 788 is nearly triangular, with the apex ventral. The concave anterior[Pg 486] margin bears pores of the postorbital part of the infraorbital line. Ornamentation consists of widely spaced, coarse tubercles.
Part of one squamosal is preserved. It is somewhat triangular and its apex is ventral. This bone is associated with the postorbital, subopercular and lacrimojugal on no. 788. The preopercular sensory line passes down the curving ventral margin of this bone, and extends ventrally onto a narrow projection. A low ridge, nearly vertical, passes dorsally from about mid-point of the canal to the dorsal portion. The anterior margin is nearly straight, the ventral margin is concave, and the dorsal margin is convex dorsally but may be incomplete. Perhaps the squamosal and preopercular are fused. The surface appears smooth; the view may be of the medial side. The squamosal of R. elegans is nearly triangular and notably different from that of Synaptotylus newelli.
The subopercular (fig. 3) shows closely spaced tubercles on the lateral surface. The bone is an elongated, irregular triangle with the apex pointing anterodorsally. The margins are incomplete, except for the concave, curving anterior margin.
Numerous operculars (fig. 3) occur in the suite of specimens, both isolated and nearly in place. Each is subtriangular; the apex of the triangle is ventral. A slight convexity projects from the anterodorsal border. The posterior margin is broadly but shallowly indented. Otherwise the margins are smooth. Maximum height ranges from 8.0 to 11.0 mm., and maximum width from 8.0 to 13.0 mm. Ornamentation varies from a few widely spaced, randomly oriented tubercles to closely spaced tubercles merging posteriorly into striae. On some specimens these are parallel to the dorsal border, and oblique in the central portion. On the posterior margins of several operculars the striae break up into tubercles. A few operculars have closely spaced tubercles over much of the surface. The internal surface is smooth.
The palatoquadrate complex, best seen on K. U. no. 9939 (fig. 4), consists of endopterygoid, ectopterygoid, metapterygoid and quadrate. No trace of epipterygoids, dermopalatines or autopalatines, such as Moy-Thomas (1937:392, fig. 5) described for Rhabdoderma, has been observed.
The endopterygoid has a long, ventral, anteriorly-directed process, and an anterodorsal process that meets the metapterygoid in forming the processus ascendens. The suture between the endopterygoid[Pg 487] and metapterygoid, seen in lateral view, is distinct in some specimens and has an associated ridge; these bones appear to be fused in others, without regard to size. This suture curves dorsally from a point anterior to the quadrate and passes anterodorsally to the extremity of the processus ascendens. The suture is visible on the medial side only near the processus ascendens, for it is covered by a dorsal, toothed extension of the endopterygoid. The endopterygoid has a smooth lateral surface; the medial surface is covered with tiny granular teeth, in characteristic "line and dot" arrangement. The teeth extend onto the ventral surface of the ventral process.
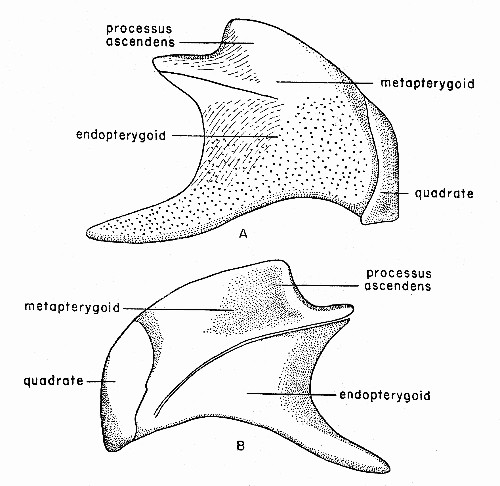 Fig. 4. Synaptotylus newelli (Hibbard). Restoration of
the palatoquadrate complex, based on K. U. no. 9939, × 5. A, medial
view, B, lateral view.
Fig. 4. Synaptotylus newelli (Hibbard). Restoration of
the palatoquadrate complex, based on K. U. no. 9939, × 5. A, medial
view, B, lateral view.
Two long, narrow, splintlike bones covered on one surface with granular teeth are interpreted as ectopterygoids. These are 13.0 and 16.0 mm. long and each is 1.5 mm. wide. Orientation of these is unknown, but they probably fitted against the ventral surface[Pg 488] of the ventral process of the endopterygoid (Moy-Thomas, 1937:fig. 5).
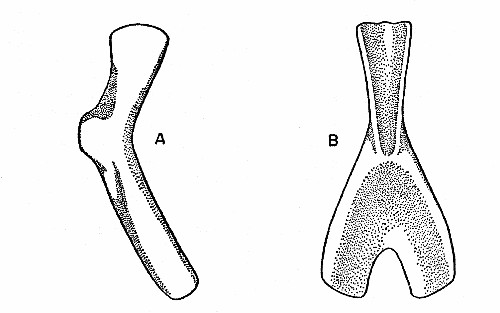 Fig. 5. Synaptotylus newelli (Hibbard). A, ceratohyal,
lateral (?) view, based on K. U. nos. 11429 and 11457, × 5. B, urohyal,
based on K. U. no. 11457, × 5.
Fig. 5. Synaptotylus newelli (Hibbard). A, ceratohyal,
lateral (?) view, based on K. U. nos. 11429 and 11457, × 5. B, urohyal,
based on K. U. no. 11457, × 5.
The metapterygoid has a smooth surface in both views. The dorsal edge has a thickened, flared margin that presumably articulated with the antotic process of the basisphenoid. No articular surface for the basipterygoid process has been observed.
The quadrate is distinct and closely applied to the posteroventral margin of the complex. In medial view the margin is nearly straight and continues to the ventral edge. The ventral surface is thickened and forms a rounded, knoblike articular surface. In lateral view the surface is smooth; the anterior margin is irregular (or perhaps broken on all specimens), and proceeds in an irregular convex curve from the posterior to the ventral margin.
The general shape of the palatoquadrate complex is most nearly like that of Rhabdoderma elegans (Moy-Thomas, 1937:fig. 5). The orientation of the complex in the living fish was probably oblique, with the processus ascendens nearly vertical, the quadrate oblique, and the ventral process of the endopterygoid extending dorsoanteriorly and articulating with the parasphenoid.
Of the hyoid arch only the ceratohyals (see fig. 5A) are preserved in several specimens. These are long, curved bones with a posteroventral process and widened, flaring posterior margin. The medial (?) surface is concave in one specimen. The lateral (?) surface displays a distinct ridge on several specimens, arising on the dorsal[Pg 489] surface opposite the posteroventral process and extending diagonally to the anteroventral end of the anterior limb. The impression of one other specimen appears to have a central ridge because of greater dorsal thickness and narrowness. Both surfaces are unornamented.
The urohyal (see fig. 5B) is an unornamented, Y-shaped bone, with the stem of the Y pointing anteriorly. Orientation with respect to dorsal and ventral surfaces is uncertain. In one view a faint ridge, also Y-shaped, occurs on the expanded posterior portion, and the surface is convex. The anterior process has a convex surface, sloping evenly off to the lateral margin; the opposite side of the process has a concave surface. The posterior portion has a slightly depressed area (see fig. 5B) at the junction of the "arms" of the Y.
The five branchial arches are represented by the ceratobranchials, several of which are preserved on K. U. no. 11431. These are long bones with anteriorly curving ventral ends. The medial surfaces are partly covered with minute granular teeth; only the dorsal part is without teeth. The dorsal articular surface is convex dorsally and rounded.
The mandible (fig. 3), the best specimens of which are K. U. nos. 788 and 11425, is seen only in lateral and ventral views, with only angular, splenial and dentary visible.
The angular forms the main body of the mandible, and is similar to that of Spermatodus. The dorsal margin of the angular is expanded in the central region, with some variation. One specimen has an expanded portion slightly anterior to that of the opposite angular. The articular surface near the posterior end has not been observed; the posterior end of the angular slopes off abruptly. The anterior sutures are seen in only two specimens, K. U. nos. 788, 11425. The dentary meets the angular in a long oblique suture; the dentary gradually tapers posterodorsally and ends on the dorsal surface of the angular. The splenial fits into a posteriorly directed, deep V-shaped notch on the ventral surface. The lateroventral surface of the angular contains sensory pores of the mandibular line. The ventral surface extends medially into a narrow shelf, approximately 1.0 mm. wide, which extends the full length of the bone; the external surface of this shelf is smooth and slightly concave dorsally. Ornamentation of the angular consists of tubercles and longitudinal or oblique striae, occurring mostly on the expanded portion. The medial surface is not seen. Several broken[Pg 490] specimens show a central canal filled with a rod of calcite; in one of these the sensory pores are also calcite-filled and appear to be connected to the rod. Thus the pores originally opened into a central canal.
The dentary is an unornamented bone with the anterior half curving medially; the greatest height is anterior. This bone in specimen K. U. no. 11425 bears irregularly spaced, simple, recurved, conical teeth; nine were counted, but there is space for many others. One other specimen, no. 11429, seems to have tiny tubercles on the surface. The dentary meets the splenial dorsally in a straight suture.
The splenial also curves medially, and as stated, meets the dentary in a straight suture. Ornamentation on this bone was not observed. The posterior margin is V-shaped and fits the notch in the angular. The ventral surface bears three or more sensory pores of the mandibular line.
The gular plates are oval. The medial margin is straight to slightly curved, the lateral margin curved crescentically, the posterior end is blunt, and the anterior end somewhat rounded. Ornamentation varies greatly; some bones show only a few tubercles, whereas others exhibit an almost concentric pattern of closely spaced striae. Typically there are some tubercles in the anterior quarter or third of the total length; these pass into longitudinally oriented striae in the posterior section. A few have only randomly oriented, widely-spaced striae. The internal surface is smooth.
The coronoid (K. U. no. 11428) is a triangular bone, with the apex pointing dorsally. The lateral surface is smooth; no teeth were observed. Moy-Thomas (1937:292, 293) mentions several tooth-bearing coronoids in Rhabdoderma, but as yet these have not been seen in Synaptotylus.
Only three specimens (K. U. nos. 786F, 787F, 11450) show parts of the vertebral column, but isolated neural and haemal arches are numerous. All are of the coelacanth type, having Y-shaped neural and haemal arches, without centra. A total count of 38 was obtained, but this was incomplete; the actual number was probably near 50. Counts of 10 and 16 haemal arches were obtained in two of the specimens. Total height of neural arches ranges from 7.5 to 12.0 mm., and of haemal arches, from 9.0 to 12.0 mm. The shorter arches are anterior and the height increases gradually to a maximum in the caudal region. Height of the spines varies from[Pg 491] 4.0 to 9.0 mm., or from twice the height of the arch in the anterior to three times the height in the caudal region. Total width of the base, measured in isolated specimens because lateral views in other specimens prevented measuring width, ranges from 0.7 to 4.2 mm. The short, broad arches having short spines occur at the anterior end of the spinal column; the narrower arches having tall spines occur toward the caudal end. Broken neural and haemal arches show a thin covering of bone with a central, calcite-filled cavity, which in life may have been filled with cartilage (Stensiö, 1932:58, fig. 20).
No ossified ribs have been observed, either isolated or in place.
For further description of the axial skeleton, see Hibbard (1933).
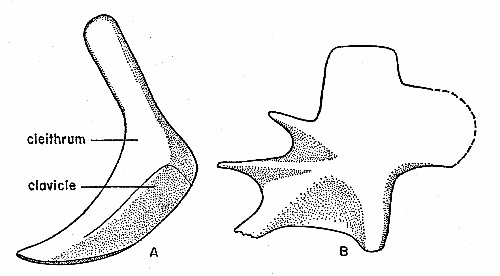 Fig. 6. Synaptotylus newelli (Hibbard). Paired fin
girdles. A, pectoral girdle, lateral view, based on K. U. no. 11433, ×
3.5. B, pelvic girdle basal plate, medial (?) view, based on K. U. no.
788, × 8. Anterior is toward the left.
Fig. 6. Synaptotylus newelli (Hibbard). Paired fin
girdles. A, pectoral girdle, lateral view, based on K. U. no. 11433, ×
3.5. B, pelvic girdle basal plate, medial (?) view, based on K. U. no.
788, × 8. Anterior is toward the left.
A nearly complete pectoral girdle on specimen K. U. no. 11433 (see fig. 6A) has only a cleithrum and clavicle. No evidence of an extracleithrum or supracleithrum has been observed, but the extracleithrum may be fused to the cleithrum. The two bones form a boot-shaped unit, with the anteroventral part turned medially to form a horizontal process which meets the opposite half of the girdle. In lateral view the surface is unornamented, and convex in the ventral half. The suture between the cleithrum and clavicle begins on the expanded posterior portion, the "boot-heel," at a point immediately below the greatest width on the posterior margin,[Pg 492] passes anteriorly, then turns sharply and parallels the anterior margin. The shape of the cleithrum resembles that in Rhabdoderma and the internal surface is not ridged (see Moy-Thomas, 1937:fig. 9). The exact orientation in the fish is uncertain, but if the median extension is really horizontal, then the posterior expansion is directed caudally. The medial surface is concave, steepest near the anterior margin, and then slopes outward evenly. In medial view one specimen (K. U. no. 11426) shows a small, caudally directed projection of bone, evidently for articulation of the fin-skeleton, at the widest portion of the cleithrum. Sutures on several specimens were indistinct. Broken specimens show sutural faces, but many nearly complete specimens show little or no indication of sutures, without regard to size of the girdles. The internal structure of the fin was not observed.
Numerous isolated basal plates of the pelvic girdle have revealed details of structure but no information on the orientation. Presumably the basal plates of Synaptotylus had essentially the same orientation as those of other coelacanths (Moy-Thomas, 1937:395). The most complete basal plate is K. U. no. 788 (see fig. 6B). The three apophyses diverge anteriorly; the horizontal one is best developed and the dorsal one is least well developed. A median process (Schaeffer, 1952:49), denticulate on several specimens, articulates with the corresponding process of the opposite plate. The expanded part that articulates with the skeleton of the fin extends caudally. The posterior expanded part is nearly square in outline, resembling the dorsal, rectangular projection. One side bears ridges leading to the extremities of the apophyses, and faint crenulations on the median process. This may be the medial view. The other view displays a smooth surface, usually without indication of the ridges seen in the reverse view. These specimens differ somewhat from the basal plates of Rhabdoderma and appear to be intermediate between Rhabdoderma and Coelacanthus (Moy-Thomas, 1937:fig. 10A, B). The apophyses are not free as in Rhabdoderma but webbed with bone almost to their extremities, as in Coelacanthus.
The pelvic fin is seen in only two specimens (K. U. nos. 786F, 788). That on no. 788 is lobate and has 25 lepidotrichia, jointed for approximately the distal half, and 2.5 to 13.0 mm. in length. Total length of the fin is 25.0 mm. There is no trace of the internal skeletal structure or of the articulation to the basal plate in either specimen. For a description of the fin on no. 786F, see Hibbard (1933:281).[Pg 493]
A few isolated bones on K. U. no. 788 (fig. 7) are interpreted as basal plates of the unpaired fins. For additional description of the unpaired fins on the type, K. U. no. 786F, see Hibbard (1933).
Two of these bones are flat, smooth and oblong, bearing a diagonal ridge that extends in the form of a projection. Orientation is completely unknown. These may be basal plates of the anterior dorsal fin. The fin on no. 786F that Hibbard (1933:281) interpreted as the posterior dorsal fin is now thought to be the anterior dorsal fin.
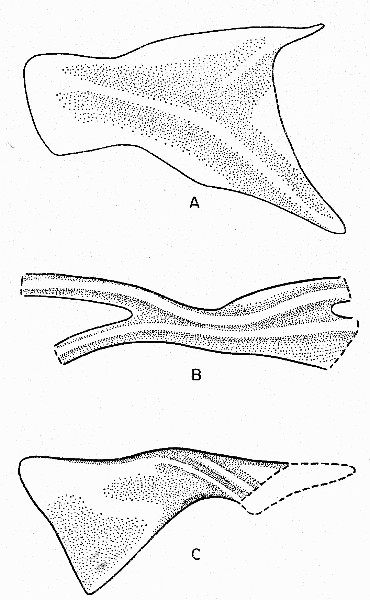 Fig. 7. Synaptotylus newelli (Hibbard). Basal plates of
unpaired fins. A, anterior dorsal fin, based on K. U. no. 788, × 10. B,
posterior dorsal fin, based on K. U. no. 788, × 12. C, anal fin, based
on K. U. no. 11450, × 5. Anterior is toward the left.
Fig. 7. Synaptotylus newelli (Hibbard). Basal plates of
unpaired fins. A, anterior dorsal fin, based on K. U. no. 788, × 10. B,
posterior dorsal fin, based on K. U. no. 788, × 12. C, anal fin, based
on K. U. no. 11450, × 5. Anterior is toward the left.
One distinctive bone may represent the basal plate of the posterior dorsal fin. This incomplete specimen shows two projecting curved processes, bearing low but distinct ridges, which diverge, probably anteriorly. The central portion is narrow. The two ridges continue onto the posterior portion. This has been broken off, but shows that the ridges diverge again. The surface is smooth, except for the ridges. As before, orientation is uncertain. On no. 786F this fin was interpreted by Hibbard (1933:281) as the anal fin.
Only part of one basal plate of the anal fin was preserved on K. U. no. 11450. That plate is oblong and has an expanded anterior end. The narrow, constricted part bears two oblique ridges and a few tubercles. The posterior part has nearly straight margins (represented by impressions) and the posterior margin is oblique, sloping anteroventrally. The flared anterior part has a smooth surface. This basal plate is more nearly like those of Coelacanthus, according to the descriptions given by Moy-Thomas (1937:399). The basal plate is associated with seven apparently unjointed, incomplete lepidotrichia. The anal fin on no. 786F is interpreted as the anterior dorsal fin (Hibbard, 1933:281).
The caudal fins are preserved on K. U. nos. 786F, 787F, and have a total of 24 lepidotrichia, 12 above and 12 below. These are jointed for the distal half or two-thirds, and are up to 16.0 mm. in length. In specimen no. 787F the supplementary caudal fin has at least seven lepidotrichia, the longest of which is 11.0 mm. but incomplete. Anterior lepidotrichia appear unjointed but the posterior ones are jointed for the distal two-thirds (?) (these are broken off). The supplementary caudal fin is approximately 1.5 mm. long and 8.0 mm. or more wide. The supplementary caudal fin on K. U. no. 786F described by Hibbard (1933:281) could not be observed; this part of the caudal fin is missing.
In the suite of specimens isolated scales are numerous, but patches of scales are rare. Only two specimens (K. U. nos. 786F, 787F) are complete enough for scale counts, but preservation permits only partial counts. In general the scales resemble those of Rhabdoderma elegans (Newberry).
The scales are oval. The exposed posterior part of each bears posteriorly converging ridges; the anterior part is widest and shows a fine fibrillar structure. There are at least six scale-rows on either[Pg 495] side of the lateral line. Lateral line scales show no pores, and except for slight irregularities in the orientation and length of the posterior ridges, closely resemble the others. Central ridges on the lateral line scales are shorter and tend to diverge from the center of the impression of the canal. The lateral line canal shows only as the impression of a continuous canal 0.7 mm. in diameter. Preservation is poorest in scales along the line of the neural and haemal arches; therefore lateral line scales are rarely preserved. Isolated scales are of two types: those on which the posterior ridges converge sharply and form the gothic arch configuration mentioned by Hibbard (1933:282), and those which do not. Both types of scales can be present on one fish, as shown by specimen no. 788. This is not apparent on nos. 786F and 787F; all of the scales on these specimens appear to be much alike. Both Moy-Thomas (1937:385) and Schaeffer (1952:51, 52) have remarked on the variation of the scales on different parts of the same fish. Because the number of ridges and amount of convergence of the ridges is not related to size of the scale, it is concluded that these characters are not of taxonomic significance.
The strong resemblance of the scales of the Garnett specimens to those of Rhabdoderma elegans (Newberry) caused Moy-Thomas (1937:399) to add Hibbard's two species to the synonymy of R. elegans. But at that time only the scales could be adequately described. If the shape of the scale and the number and pattern of ridges can vary with age, size and shape of the scale, it follows that assignment of isolated scales to a species should not be attempted. Assignment to genus should be made only with caution.
Discussion.—The relationship of Synaptotylus to other coelacanths is obscure at present. The knoblike antotic processes on the basisphenoid are unlike those of any other known coelacanth. The palatoquadrate complex is shaped like that of Rhabdoderma elegans but consists of fewer bones, probably because of fusion. The scales resemble those of Rhabdoderma. With regard to general shape of fin girdles, the pectoral girdle resembles that of Eusthenopteron more than that of Rhabdoderma, but the cleithrum is more nearly like the cleithrum of Rhabdoderma. The pelvic girdle appears to be midway between those of Rhabdoderma and Coelacanthus in general appearance. Regarding the basal plates of the remaining fins, those of Synaptotylus appear to resemble basal plates of both Rhabdoderma and Coelacanthus. Considering the structure of the sphenethmoid region of the braincase, Synaptotylus is probably[Pg 496] more closely related to Rhabdoderma than to other known coelacanth genera.
Classification of Carboniferous coelacanths has been difficult, partly because the remains are commonly fragmentary, and significant changes in anatomy did not become apparent in early studies. In general, coelacanths have been remarkably stable in most characters, and it has been difficult to divide the group into families. As Schaeffer (1952:56) pointed out, definition of coelacanth genera and species has previously been made on non-meristic characters, and the range of variation within a species has received little attention. For example, Reis (1888:71) established the genus Rhabdoderma, using the strong striation of the scales, gular plates and posterior mandible as the main characters of this Carboniferous genus. Moy-Thomas (1937:399-411) referred all Carboniferous species to Rhabdoderma, redescribed the genus and compared it to Coelacanthus, the Permian genus. He cited as specific characters the ornamentation of the angulars, operculars and gular plates (Moy-Thomas, 1935:39; 1937:385). Individual variation in some species has rendered ornamentation a poor criterion. This variation is apparent in Synaptotylus newelli (Hibbard), some specimens having little or no ornamentation; others having much more. The number of ridges and pattern of ridges on the scales also varies. Schaeffer (1952:56) has found this to be true of Diplurus also. Moy-Thomas (1935:40; 1937:385) realized that the type of scale is a poor criterion for specific differentiation. In the search for features useful in distinguishing genera of coelacanths, Schaeffer and Gregory (1961:3, 7) found the structure of the basisphenoid to be distinctive in known genera, and thought it had taxonomic significance at this level. Higher categories should have as their basis characters that display evolutionary sequences. A recent classification (Berg, 1940), followed in this paper, reflects two evolutionary trends in endocranial structure of coelacanths: reduction of endocranial ossification and loss of the basipterygoid processes. Because there has been little change in other structures in coelacanths, Berg's classification is the most useful. Berg (1940:390) includes Rhabdoderma in the suborder Diplocercidoidei because of the presence of the basipterygoid processes, and in the single family, Diplocercidae, but remarks that because of the reduced amount of endocranial ossification the Carboniferous[Pg 497] Diplocercidae "probably constitute a distinct family." In considering this concept of classification, the subfamilies Diplocercinae and Rhabdodermatinae of the family Diplocercidae are proposed above. The subfamily Rhabdodermatinae includes at present Rhabdoderma and Synaptotylus. The principal characters of the subfamily Rhabdodermatinae, named for the first known genus, are the retention of the basipterygoid processes and the reduction of endocranial ossification. Application of this classification based upon endocranial structure would probably change existing groupings of species of Carboniferous coelacanths; the entire complex of Carboniferous genera should be redescribed and redefined. It will be necessary to consider endocranial structure in any future classification.
The greater part of the evolution previously mentioned appears to have been accomplished during the Carboniferous; thereafter coelacanth structure became stabilized. The trend progressed from Devonian coelacanths which had two large unpaired bones in the endocranium, and both antotic and basipterygoid processes on the basisphenoid, to Carboniferous fishes in which ossification was reduced to a number of paired and unpaired bones embedded in cartilage, and retaining both processes, and then post-Carboniferous kinds with reduced ossification and no basipterygoid processes. The Pennsylvanian was evidently the time of greatest change for the coelacanths, and they have not changed significantly since, in spite of the fact that since the Jurassic they have shifted their environment from shallow, fresh water to moderate depth in the sea (Schaeffer, 1953:fig. 1). The changes in endocranial structure appear to be significant, and are perhaps related to higher efficiency of the mouth parts in catching and swallowing prey (see p. 482).
The coelacanth fishes from the Rock Lake shale are part of the varied fauna collected from Garnett. Peabody (1952:38) listed many elements of the fauna and flora, and concluded that the deposits are of lagoonal origin. In addition to numerous invertebrates (including microfossils) and arthropods, a number of vertebrates other than coelacanths have been found. These include at least one kind of shark, Hesperoherpeton garnettense Peabody, one or more kinds of undescribed labyrinthodonts and the reptiles Petrolacosaurus kansensis Lane, Edaphosaurus ecordi Peabody, and Clepsydrops (undescribed species). This is indeed a rich vertebrate[Pg 498] fauna, and the earliest known reptilian fauna. Much of the rock contains plant remains. The flora that has been identified is adapted to growing in a well-drained soil; although it contains some elements considered characteristic of the Permian, it is of Pennsylvanian age (Moore et al., 1936). Peabody (1952:38-39) discusses the features of these lagoonal sediments. Much of the fauna and flora suggests continental origin, but the many marine invertebrates at some horizons indicate that at least some of the sediments were of marine origin.
Little can be said about the actual environment of the living fishes of the genus Synaptotylus. Remains of these fishes occur in layers containing marine invertebrates, as well as in those containing plant remains and vertebrate skeletal parts, and in those nearly completely composed of dark carbonaceous material. Most of the remains are fragmentary and consist of isolated bones, isolated scales, and dissociated skulls; only one specimen and half of another are nearly complete. Many published statements on Rhabdoderma, a related genus, indicate both marine and fresh-water environments. Wehrli (1931:115) regarded Rhabdoderma elegans (Newberry) as a euryhaline species, and cited its occurrence with both marine and fresh-water fossils. Aldinger (1931:199) also found this to be the case with other species, and Fiege (1951:17) quotes others as giving the same information. Keller (1934:913) thought that few Carboniferous fishes were exclusively marine, and stated that the majority of them became adapted to fresh water during the late Carboniferous. Later, Schaeffer (1953:175) stated that all Carboniferous and Permian coelacanths were fresh-water fishes, and that many were from swamp deposits. If Keller is correct, then members of the genus Synaptotylus may have inhabited the lagoon, the adjacent sea, or the streams draining into the lagoon. Perhaps these fishes swam upstream, as modern salmon and tarpon do, although there is no direct evidence for this. Possibly they lived in the lagoon at times of scant rainfall and little runoff, when the salinity of lagoon water approached normal marine values or the fishes may have lived in the streams, and after death were washed into the lagoon. As numerous remains of land plants and animals were washed in, perhaps this best accounts for the presence of the fish in nearly all layers of the deposits, not only the marine strata.[Pg 499]
A new genus of Pennsylvanian coelacanths, Synaptotylus, is described and a previously named species, Coelacanthus newelli Hibbard, 1933 (C. arcuatus Hibbard, 1933, is a junior synonym), is referred to this genus. All specimens of Synaptotylus newelli (Hibbard) were collected from the Rock Lake shale member of the Stanton formation, Lansing group, Missouri series, six miles northwest of Garnett, Anderson County, Kansas. Synaptotylus is distinguished from all other coelacanths by a basisphenoid having large, knoblike antotic processes each connected by a low ridge to a small basipterygoid process. Synaptotylus is most closely related to Rhabdoderma, but is intermediate between Rhabdoderma and Coelacanthus in shape of the fin girdles and basal plates. Two new subfamilies, Diplocercinae and Rhabdodermatinae, of the family Diplocercidae, are proposed. Synaptotylus and Rhabdoderma are included in the subfamily Rhabdodermatinae, because both exhibit reduced ossification in the endocranium and retain basipterygoid processes.
Loss of the basipterygoid processes in post-Carboniferous coelacanths may reflect the development of a more efficient feeding mechanism, by allowing the palatoquadrate complex and mandible to swing farther laterally and expand the oral cavity.
Synaptotylus newelli (Hibbard) may have occupied either the sea or fresh water; these fishes occur in lagoonal deposits with reptiles and amphibians, arthropods, marine invertebrates and remains of land plants.
Because scale patterns on Synaptotylus and Rhabdoderma are so nearly similar and vary with size of the scale and its location on the fish, it is recommended that isolated scales not be assigned to a species, and to a genus only with great caution.[Pg 500]
Aldinger, H.
1931. Ueber karbonische Fische aus Westfälen. Paleont. Zeit., 13:186-201.
Berg, L. S.
1940. Classification of fishes, both Recent and fossil. Moscow and Leningrad, 1940 (J. W. Edwards, Ann Arbor, Michigan, 1947, offset reproduction, pp. 1-345, 197 figs., plus English translation of text, pp. 346-517, 1947.)
Fiege, K.
1951. Eine Fisch-Schwimmspur aus dem Culm bei Waldeck. Neues Jahrb. Geol. and Paläont. Jahrgang 1951:9-31.
Hibbard, C. W.
1933. Two new species of Coelacanthus from the middle Pennsylvanian of Anderson County, Kansas. Kansas Univ. Sci. Bull., 21:279-287.
Keller, G.
1934. Fischreste aus dem oberkarbon des Ruhrgebiets. Gluckauf, 70:913-917.
Moore, R. C., Elias, M. K., and Newell, N. D.
1936. A "Permian" flora from the Pennsylvanian rocks of Kansas. Jour. Geol., 44:1-31.
Moy-Thomas, J. A.
1935. A synopsis of the coelacanth fishes of the Yorkshire Coal Measures. Ann. Mag. Nat. Hist., 15 (ser. 10): 37-46.
1937. The Carboniferous coelacanth fishes of Great Britain and Ireland. Proc. Zool. Soc. London, 107 (B): 383-415.
Peabody, F. E.
1952. Petrolacosaurus kansensis Lane, a Pennsylvanian reptile from Kansas. Kansas Univ. Paleont. Contrib., 1:1-41.
Reis, O. M.
1888. Die Coelacanthinen mit besonderen Berücksichtigung der im Weissen Jura Bayerns verkommenden Arten. Palaeontographica, 35:1-96.
Schaeffer, B.
1952. The Triassic coelacanth fish Diplurus, with observations on the evolution of the Coelacanthini. Bull. Amer. Mus. Nat. Hist., 99:art. 2, 29-78.
1953. Latimeria and the history of the coelacanth fishes. New York Acad. Sci. Trans., (2) 15:170-178.
Schaeffer, B., and Gregory, J. T.
1961. Coelacanth fishes from the continental Triassic of the western United States. Amer. Mus. Novitates, 2036:1-18.
Schaeffer, B., and Rosen, D. E.
1961. Major adaptive levels in the evolution of the actinopterygian feeding mechanism. Am. Zool., 1:187-204.
Smith, J. L. B.
1939. A living coelacanthid fish from South Africa. Trans. Roy. Soc. South Africa, 28:1-106.
Stensiö, E. A.
1921. Triassic fishes from Spitzbergen. Part I. Vienna, Adolf Holzhausen: 1-307.
1932. Triassic fishes from East Greenland. Meddel. om Grønland, 38:1-305.[Pg 501]
Tchernavin, V. V.
1948. On the mechanical working of the head of bony fishes. Proc. Zool. Soc. London, 118:129-143.
Wehrli, H.
1931. Die Fauna der Westfälischen Stufen A und B der Bochumer Mulde zwischen Dortmund und Kamen (Westfälen). Palaeontographica, 74:93-134.
End of the Project Gutenberg EBook of A New Genus of Pennsylvania Fish
(Crossoperygii, Coelacanthiformes) from Kansas, by Joan Echols
*** END OF THIS PROJECT GUTENBERG EBOOK NEW GENUS OF PENNSYLVANIA FISH ***
***** This file should be named 33560-h.htm or 33560-h.zip *****
This and all associated files of various formats will be found in:
https://www.gutenberg.org/3/3/5/6/33560/
Produced by Chris Curnow, Joseph Cooper, Josephine Paolucci
and the Online Distributed Proofreading Team at
https://www.pgdp.net.
Updated editions will replace the previous one--the old editions
will be renamed.
Creating the works from public domain print editions means that no
one owns a United States copyright in these works, so the Foundation
(and you!) can copy and distribute it in the United States without
permission and without paying copyright royalties. Special rules,
set forth in the General Terms of Use part of this license, apply to
copying and distributing Project Gutenberg-tm electronic works to
protect the PROJECT GUTENBERG-tm concept and trademark. Project
Gutenberg is a registered trademark, and may not be used if you
charge for the eBooks, unless you receive specific permission. If you
do not charge anything for copies of this eBook, complying with the
rules is very easy. You may use this eBook for nearly any purpose
such as creation of derivative works, reports, performances and
research. They may be modified and printed and given away--you may do
practically ANYTHING with public domain eBooks. Redistribution is
subject to the trademark license, especially commercial
redistribution.
*** START: FULL LICENSE ***
THE FULL PROJECT GUTENBERG LICENSE
PLEASE READ THIS BEFORE YOU DISTRIBUTE OR USE THIS WORK
To protect the Project Gutenberg-tm mission of promoting the free
distribution of electronic works, by using or distributing this work
(or any other work associated in any way with the phrase "Project
Gutenberg"), you agree to comply with all the terms of the Full Project
Gutenberg-tm License (available with this file or online at
https://gutenberg.org/license).
Section 1. General Terms of Use and Redistributing Project Gutenberg-tm
electronic works
1.A. By reading or using any part of this Project Gutenberg-tm
electronic work, you indicate that you have read, understand, agree to
and accept all the terms of this license and intellectual property
(trademark/copyright) agreement. If you do not agree to abide by all
the terms of this agreement, you must cease using and return or destroy
all copies of Project Gutenberg-tm electronic works in your possession.
If you paid a fee for obtaining a copy of or access to a Project
Gutenberg-tm electronic work and you do not agree to be bound by the
terms of this agreement, you may obtain a refund from the person or
entity to whom you paid the fee as set forth in paragraph 1.E.8.
1.B. "Project Gutenberg" is a registered trademark. It may only be
used on or associated in any way with an electronic work by people who
agree to be bound by the terms of this agreement. There are a few
things that you can do with most Project Gutenberg-tm electronic works
even without complying with the full terms of this agreement. See
paragraph 1.C below. There are a lot of things you can do with Project
Gutenberg-tm electronic works if you follow the terms of this agreement
and help preserve free future access to Project Gutenberg-tm electronic
works. See paragraph 1.E below.
1.C. The Project Gutenberg Literary Archive Foundation ("the Foundation"
or PGLAF), owns a compilation copyright in the collection of Project
Gutenberg-tm electronic works. Nearly all the individual works in the
collection are in the public domain in the United States. If an
individual work is in the public domain in the United States and you are
located in the United States, we do not claim a right to prevent you from
copying, distributing, performing, displaying or creating derivative
works based on the work as long as all references to Project Gutenberg
are removed. Of course, we hope that you will support the Project
Gutenberg-tm mission of promoting free access to electronic works by
freely sharing Project Gutenberg-tm works in compliance with the terms of
this agreement for keeping the Project Gutenberg-tm name associated with
the work. You can easily comply with the terms of this agreement by
keeping this work in the same format with its attached full Project
Gutenberg-tm License when you share it without charge with others.
1.D. The copyright laws of the place where you are located also govern
what you can do with this work. Copyright laws in most countries are in
a constant state of change. If you are outside the United States, check
the laws of your country in addition to the terms of this agreement
before downloading, copying, displaying, performing, distributing or
creating derivative works based on this work or any other Project
Gutenberg-tm work. The Foundation makes no representations concerning
the copyright status of any work in any country outside the United
States.
1.E. Unless you have removed all references to Project Gutenberg:
1.E.1. The following sentence, with active links to, or other immediate
access to, the full Project Gutenberg-tm License must appear prominently
whenever any copy of a Project Gutenberg-tm work (any work on which the
phrase "Project Gutenberg" appears, or with which the phrase "Project
Gutenberg" is associated) is accessed, displayed, performed, viewed,
copied or distributed:
This eBook is for the use of anyone anywhere at no cost and with
almost no restrictions whatsoever. You may copy it, give it away or
re-use it under the terms of the Project Gutenberg License included
with this eBook or online at www.gutenberg.org
1.E.2. If an individual Project Gutenberg-tm electronic work is derived
from the public domain (does not contain a notice indicating that it is
posted with permission of the copyright holder), the work can be copied
and distributed to anyone in the United States without paying any fees
or charges. If you are redistributing or providing access to a work
with the phrase "Project Gutenberg" associated with or appearing on the
work, you must comply either with the requirements of paragraphs 1.E.1
through 1.E.7 or obtain permission for the use of the work and the
Project Gutenberg-tm trademark as set forth in paragraphs 1.E.8 or
1.E.9.
1.E.3. If an individual Project Gutenberg-tm electronic work is posted
with the permission of the copyright holder, your use and distribution
must comply with both paragraphs 1.E.1 through 1.E.7 and any additional
terms imposed by the copyright holder. Additional terms will be linked
to the Project Gutenberg-tm License for all works posted with the
permission of the copyright holder found at the beginning of this work.
1.E.4. Do not unlink or detach or remove the full Project Gutenberg-tm
License terms from this work, or any files containing a part of this
work or any other work associated with Project Gutenberg-tm.
1.E.5. Do not copy, display, perform, distribute or redistribute this
electronic work, or any part of this electronic work, without
prominently displaying the sentence set forth in paragraph 1.E.1 with
active links or immediate access to the full terms of the Project
Gutenberg-tm License.
1.E.6. You may convert to and distribute this work in any binary,
compressed, marked up, nonproprietary or proprietary form, including any
word processing or hypertext form. However, if you provide access to or
distribute copies of a Project Gutenberg-tm work in a format other than
"Plain Vanilla ASCII" or other format used in the official version
posted on the official Project Gutenberg-tm web site (www.gutenberg.org),
you must, at no additional cost, fee or expense to the user, provide a
copy, a means of exporting a copy, or a means of obtaining a copy upon
request, of the work in its original "Plain Vanilla ASCII" or other
form. Any alternate format must include the full Project Gutenberg-tm
License as specified in paragraph 1.E.1.
1.E.7. Do not charge a fee for access to, viewing, displaying,
performing, copying or distributing any Project Gutenberg-tm works
unless you comply with paragraph 1.E.8 or 1.E.9.
1.E.8. You may charge a reasonable fee for copies of or providing
access to or distributing Project Gutenberg-tm electronic works provided
that
- You pay a royalty fee of 20% of the gross profits you derive from
the use of Project Gutenberg-tm works calculated using the method
you already use to calculate your applicable taxes. The fee is
owed to the owner of the Project Gutenberg-tm trademark, but he
has agreed to donate royalties under this paragraph to the
Project Gutenberg Literary Archive Foundation. Royalty payments
must be paid within 60 days following each date on which you
prepare (or are legally required to prepare) your periodic tax
returns. Royalty payments should be clearly marked as such and
sent to the Project Gutenberg Literary Archive Foundation at the
address specified in Section 4, "Information about donations to
the Project Gutenberg Literary Archive Foundation."
- You provide a full refund of any money paid by a user who notifies
you in writing (or by e-mail) within 30 days of receipt that s/he
does not agree to the terms of the full Project Gutenberg-tm
License. You must require such a user to return or
destroy all copies of the works possessed in a physical medium
and discontinue all use of and all access to other copies of
Project Gutenberg-tm works.
- You provide, in accordance with paragraph 1.F.3, a full refund of any
money paid for a work or a replacement copy, if a defect in the
electronic work is discovered and reported to you within 90 days
of receipt of the work.
- You comply with all other terms of this agreement for free
distribution of Project Gutenberg-tm works.
1.E.9. If you wish to charge a fee or distribute a Project Gutenberg-tm
electronic work or group of works on different terms than are set
forth in this agreement, you must obtain permission in writing from
both the Project Gutenberg Literary Archive Foundation and Michael
Hart, the owner of the Project Gutenberg-tm trademark. Contact the
Foundation as set forth in Section 3 below.
1.F.
1.F.1. Project Gutenberg volunteers and employees expend considerable
effort to identify, do copyright research on, transcribe and proofread
public domain works in creating the Project Gutenberg-tm
collection. Despite these efforts, Project Gutenberg-tm electronic
works, and the medium on which they may be stored, may contain
"Defects," such as, but not limited to, incomplete, inaccurate or
corrupt data, transcription errors, a copyright or other intellectual
property infringement, a defective or damaged disk or other medium, a
computer virus, or computer codes that damage or cannot be read by
your equipment.
1.F.2. LIMITED WARRANTY, DISCLAIMER OF DAMAGES - Except for the "Right
of Replacement or Refund" described in paragraph 1.F.3, the Project
Gutenberg Literary Archive Foundation, the owner of the Project
Gutenberg-tm trademark, and any other party distributing a Project
Gutenberg-tm electronic work under this agreement, disclaim all
liability to you for damages, costs and expenses, including legal
fees. YOU AGREE THAT YOU HAVE NO REMEDIES FOR NEGLIGENCE, STRICT
LIABILITY, BREACH OF WARRANTY OR BREACH OF CONTRACT EXCEPT THOSE
PROVIDED IN PARAGRAPH 1.F.3. YOU AGREE THAT THE FOUNDATION, THE
TRADEMARK OWNER, AND ANY DISTRIBUTOR UNDER THIS AGREEMENT WILL NOT BE
LIABLE TO YOU FOR ACTUAL, DIRECT, INDIRECT, CONSEQUENTIAL, PUNITIVE OR
INCIDENTAL DAMAGES EVEN IF YOU GIVE NOTICE OF THE POSSIBILITY OF SUCH
DAMAGE.
1.F.3. LIMITED RIGHT OF REPLACEMENT OR REFUND - If you discover a
defect in this electronic work within 90 days of receiving it, you can
receive a refund of the money (if any) you paid for it by sending a
written explanation to the person you received the work from. If you
received the work on a physical medium, you must return the medium with
your written explanation. The person or entity that provided you with
the defective work may elect to provide a replacement copy in lieu of a
refund. If you received the work electronically, the person or entity
providing it to you may choose to give you a second opportunity to
receive the work electronically in lieu of a refund. If the second copy
is also defective, you may demand a refund in writing without further
opportunities to fix the problem.
1.F.4. Except for the limited right of replacement or refund set forth
in paragraph 1.F.3, this work is provided to you 'AS-IS' WITH NO OTHER
WARRANTIES OF ANY KIND, EXPRESS OR IMPLIED, INCLUDING BUT NOT LIMITED TO
WARRANTIES OF MERCHANTIBILITY OR FITNESS FOR ANY PURPOSE.
1.F.5. Some states do not allow disclaimers of certain implied
warranties or the exclusion or limitation of certain types of damages.
If any disclaimer or limitation set forth in this agreement violates the
law of the state applicable to this agreement, the agreement shall be
interpreted to make the maximum disclaimer or limitation permitted by
the applicable state law. The invalidity or unenforceability of any
provision of this agreement shall not void the remaining provisions.
1.F.6. INDEMNITY - You agree to indemnify and hold the Foundation, the
trademark owner, any agent or employee of the Foundation, anyone
providing copies of Project Gutenberg-tm electronic works in accordance
with this agreement, and any volunteers associated with the production,
promotion and distribution of Project Gutenberg-tm electronic works,
harmless from all liability, costs and expenses, including legal fees,
that arise directly or indirectly from any of the following which you do
or cause to occur: (a) distribution of this or any Project Gutenberg-tm
work, (b) alteration, modification, or additions or deletions to any
Project Gutenberg-tm work, and (c) any Defect you cause.
Section 2. Information about the Mission of Project Gutenberg-tm
Project Gutenberg-tm is synonymous with the free distribution of
electronic works in formats readable by the widest variety of computers
including obsolete, old, middle-aged and new computers. It exists
because of the efforts of hundreds of volunteers and donations from
people in all walks of life.
Volunteers and financial support to provide volunteers with the
assistance they need are critical to reaching Project Gutenberg-tm's
goals and ensuring that the Project Gutenberg-tm collection will
remain freely available for generations to come. In 2001, the Project
Gutenberg Literary Archive Foundation was created to provide a secure
and permanent future for Project Gutenberg-tm and future generations.
To learn more about the Project Gutenberg Literary Archive Foundation
and how your efforts and donations can help, see Sections 3 and 4
and the Foundation web page at https://www.pglaf.org.
Section 3. Information about the Project Gutenberg Literary Archive
Foundation
The Project Gutenberg Literary Archive Foundation is a non profit
501(c)(3) educational corporation organized under the laws of the
state of Mississippi and granted tax exempt status by the Internal
Revenue Service. The Foundation's EIN or federal tax identification
number is 64-6221541. Its 501(c)(3) letter is posted at
https://pglaf.org/fundraising. Contributions to the Project Gutenberg
Literary Archive Foundation are tax deductible to the full extent
permitted by U.S. federal laws and your state's laws.
The Foundation's principal office is located at 4557 Melan Dr. S.
Fairbanks, AK, 99712., but its volunteers and employees are scattered
throughout numerous locations. Its business office is located at
809 North 1500 West, Salt Lake City, UT 84116, (801) 596-1887, email
business@pglaf.org. Email contact links and up to date contact
information can be found at the Foundation's web site and official
page at https://pglaf.org
For additional contact information:
Dr. Gregory B. Newby
Chief Executive and Director
gbnewby@pglaf.org
Section 4. Information about Donations to the Project Gutenberg
Literary Archive Foundation
Project Gutenberg-tm depends upon and cannot survive without wide
spread public support and donations to carry out its mission of
increasing the number of public domain and licensed works that can be
freely distributed in machine readable form accessible by the widest
array of equipment including outdated equipment. Many small donations
($1 to $5,000) are particularly important to maintaining tax exempt
status with the IRS.
The Foundation is committed to complying with the laws regulating
charities and charitable donations in all 50 states of the United
States. Compliance requirements are not uniform and it takes a
considerable effort, much paperwork and many fees to meet and keep up
with these requirements. We do not solicit donations in locations
where we have not received written confirmation of compliance. To
SEND DONATIONS or determine the status of compliance for any
particular state visit https://pglaf.org
While we cannot and do not solicit contributions from states where we
have not met the solicitation requirements, we know of no prohibition
against accepting unsolicited donations from donors in such states who
approach us with offers to donate.
International donations are gratefully accepted, but we cannot make
any statements concerning tax treatment of donations received from
outside the United States. U.S. laws alone swamp our small staff.
Please check the Project Gutenberg Web pages for current donation
methods and addresses. Donations are accepted in a number of other
ways including including checks, online payments and credit card
donations. To donate, please visit: https://pglaf.org/donate
Section 5. General Information About Project Gutenberg-tm electronic
works.
Professor Michael S. Hart was the originator of the Project Gutenberg-tm
concept of a library of electronic works that could be freely shared
with anyone. For thirty years, he produced and distributed Project
Gutenberg-tm eBooks with only a loose network of volunteer support.
Project Gutenberg-tm eBooks are often created from several printed
editions, all of which are confirmed as Public Domain in the U.S.
unless a copyright notice is included. Thus, we do not necessarily
keep eBooks in compliance with any particular paper edition.
Most people start at our Web site which has the main PG search facility:
https://www.gutenberg.org
This Web site includes information about Project Gutenberg-tm,
including how to make donations to the Project Gutenberg Literary
Archive Foundation, how to help produce our new eBooks, and how to
subscribe to our email newsletter to hear about new eBooks.